/
























































































































































































































Автор: Барышников Г.Ф.
Теги: палеонтология млекопитающие млекопитающие евразии
ISBN: 0206-0477
Год: 1996




Текст
ISSN 0206-0477
РОССИЙСКАЯ АКАДЕМИЯ НАУК
ЗООЛОГИЧЕСКИЙ ИНСТИТУТ
ЧЕТВЕРТИЧНЫЕ И СОВРЕМЕННЫЕ
МЛЕКОПИТАЮЩИЕ
ЕВРАЗИИ
I
РОССИЙСКАЯ АКАДЕМИЯ НАУК
ТРУДЫ ЗООЛОГИЧЕСКОГО ИНСТИТУТА, ТОМ 270
Выпускаются с 1932 года
ЧЕТВЕРТИЧНЫЕ И СОВРЕМЕННЫЕ
МЛЕКОПИТАЮЩИЕ ЕВРАЗИИ ut
Под редакцией Г Ф. БАРЫШНИКОВА
САНКТ-ПЕТЕРБУРГ
1996
RUSSIAN ACADEMY OF SCIENCES
PROCEEDINGS OF THE ZOOLOGICAL INSTITUTE
VOL. 270
QUATERNARY AND RECENT MAMMALS OF EURASIA
Editor Gennady BARYSHNIKOV
No part of this book may be reproduced in any form by photostat, microfilm or any other
means, without written permission from publisher
Главный редактор-
директор Зоологического института РАН
член-кор. РАН А.Ф.Алимов
Редакционная коллегия:
Я.И.Старобогатов (редактор серии), Ю. С. Балашов, Л.Я.Боркин, И.С.Даревский,
В.А.Заславский, И.М.Кержнер, В.А. Тряпицын, И.М. Фокин, В.В.Хлебович (зам. редактора),
С.Я.Цалолихин
Рецензенты:
Ф.Н. Голенищев, Р.Л. Потапов
Сборник содержит статьи по эволюции, систематике и морфологии палеогеновых,
плейстоценовых и современных млекопитающих из отрядов хищных, хоботных, грызунов и
зайцеобразных. Дан обзор четвертичных гиен России, подробно рассмотрены молочные зубы
современных и вымерших виверр и мангустов. Представляет интерес для научных работников в
области зоологии, палеонтологии и геологии.
На обложке - палеолитическое изображение крупного хищника (пещерной гиены или
медведя) из грота Шовэ, Франция, ориньяк (по Шовэ, Брюнель-Дешампу и Иллеру).
2002000000-05
М------------- Без объявления © Зоологический институт РАН, 1996
055(02)3-96
Никакая часть этой книги не может быть воспроизведена ни фотоспособом, ни
микрофильмированием и каким-либо другим методом без письменного разрешения
Зоологического института РАН.
ПРЕДИСЛОВИЕ
С 16 по 21 октября 1995 г. в Зоологическом институте Российской Ака-
демии наук в Санкт-Петербурге прошел Первый международный симпозиум
по изучению мамонтов и мамонтовой фауны. В его работе приняли участие
более 100 специалистов из Великобритании, Франции, Нидерландов, Герма-
нии, Австрии, Польши, Финляндии, Украины, Грузии, США, Японии и Рос-
сии.
Идея проведения международного симпозиума по мамонтовой тематике
принадлежала покойному академику О.А.Скарлато, который на протяжении
последних 8 лет был председателем Комитета по изучению мамонтов и ма-
монтовой фауны РАН (Мамонтовый комитет). Традиции изучения мамонтов
и их спутников по фауне ледникового периода - шерстистого носорога, пер-
вобытных овцебыка и бизона, пещерных хищников, заложены участником
великих “академических” путешествий XVIII века академиком П.С.Палласом
и первым директором Зоологического музея Императорской Академии наук
академиком Ф.Брандтом. Особое место в деятельности Российской академии
и Зоологического музея в частности занимает организация в начале нашего
века раскопок, а затем и комплексного изучения замерзшего полного трупа
мамонта с р.Березовка в Восточной Сибири. Позднее, в связи с находкой ске-
лета мамонта на п-ове Таймыр, был организован Мамонтовый комитет, кото-
рый, во многом благодаря усилиям его председателей - акад. Е.Н.Павлов-
ского, акад. Б.Е.Быховского, проф. Н.К.Верещагина и акад. О.А.Скарлато,
стал признанным центром по изучению фауны ледникового периода.
Изучение скелетов таймырского и других мамонтов, мерзлой тушки мо-
лодого мамонта с р.Киргилях, мумифицированных частей лошади с р.Селе-
рикан, гигантского скопления костей мамонта с р.Берелех, многотысячных
остатков плейстоценовых млекопитающих и птиц из палеолитических стоя-
нок разных областей России принесли Мамонтовому комитету мировое
признание.
Мамонтовым комитетом были проведены шесть Всесоюзных коорди-
национных совещаний по изучению мамонтов и мамонтовой фауны, органи-
зованы несколько палеозоологических экспедиций, преимущественно в Си-
бирскую Арктику, опубликованы 17 тематических сборников о четвертичной
фауне России и прилежащих территорий.
Во время Международного симпозиума был обновлен состав Мамон-
тового комитета. Почетным председателем выбран проф. Н.К.Верещагин,
председателем комитета стал Директор Зоологического института, член-кор.
РАН А. Ф.Алимов. Заместителями его утверждены Г.Ф.Барышников (Санкт-
Петербург) и П.А.Лазарев (Якутск), ученым секретарем И.Е.Кузьмина
(Санкт-Петербург). В состав Мамонтового комитета также вошли В.Е.Га-
з
рутт, акад. А.П.Деревянко, А.К.Маркова, проф. В.М.Михельсон, проф.
Р.Л.Потапов, Н.Д.Праслов, проф. С.В.Томирдиаро, проф. В.В.Украинцева,
И.М.Фокин и И.В. Форонова. Иностранными членами избраны проф.
Л.Агенброуд (США), г-н К.Араи (Япония), Б.Батыров (Узбекистан), проф.
Г.Бозинский (Германия), проф. А.Давид (Молдова), Л.Рековец (Украина) и
д-р Дж.Сондерс (США).
Настоящий сборник продолжает серию публикаций Зоологического ин-
ститута и Мамонтового комитета по фауне четвертичного (ледникового) пе-
риода. Он включает статьи авторов, участвовавших в работе Первого
Международного мамонтового симпозиума, и является началом реализации
программы по четвертичной палеозоологии, которая была намечена на
симпозиуме. Основное внимание уделено изменчивости вымерших и
современных млекопитающих - географической, геохронологической, поло-
вой и возрастной. Изучение ее представляет собой одну из наиболее актуаль-
ных задач, стоящих перед исследователями плейстоценовых видов.
Мы глубоко признательны г-ну К.Араи, директору Палеонтологичес-
кого музея Нагано, Япония, за участие в работе симпозиума и за финансовую
поддержку, оказанную для публикации настоящей книги.
Председатель Комитета по изучению
мамонтов и мамонтовой фауны
член-кор. РАН, проф. А.Ф.Алимов
PREFACE
The First International Symposium on Mammoths and Mammoth Faunas was
held at the Zoological Institute of the Russian Academy of Sciences in Saint
Petersburg on October 16-21, 1995. More than 100 scientists from Great Britain,
France, Netherlands, Germany, Austria, Poland, Finland, Ukraine, Georgia, USA,
Japan and Russia participated.
The idea for such a symposium was first proposed by the late Academician
O.A.Scarlato, who headed the Committee on the Study of Mammoths and
Mammoth Faunas of the Russian. Academy of Sciences (Mammoth Committee) for
the past 8 years. There is a long tradition in Russia of studying mammoths and
other Ice Age mammals - woolly rhinoceros, prehistory musk ox and bison, cave
carnivores. It was initiated in XVIII century by Academician P. S. Pallas, the
organiser and a participant in the Great Academic expeditions, and by
Academician F. Brandt, the first director of the Zoological Museum of the
Imperial Russian Academy of Sciences.
The discovery, excavation, and multidisciplinary investigation of the com-
plete, frozen carcass of the Berezovka River mammoth in Eastern Siberia occupies
a special place in the history of the Russian Academy of Sciences and Zoological
Museum. Later, after discovery of a mammoth skeleton on the Taimir Peninsula,
the Mammoth Committee was established. This Committee became the recognised
centre for the study of the Ice Age fauna, primarily through the outstanding efforts
of its leaders: Academicians E.N.Pavlovskyi, B.E.Bykhovskyi, and O.A.Scarlato,
and Professor N.K.Vereschagin. The Mammoth Committee became recognised
world-wide because of its investigations of the Taimir and other mammoth skele-
tons, the recovery and study of the frozen carcass of a mammoth baby from Kir-
gilakh River and the mummified remains of a horse from the Selerikan River, the
excavation and study of the extensive mammoth bone accumulation on the Bere-
lekh River, plus the recovery of countless other Pleistocene fossil mammal and
bird bones from a host of Palaeolithic sites around Russia.
Six All-Union co-ordination workshops on the study of mammoths and mam-
moth faunas were organised by the Mammoth Committee. Several palaeo-
zoological expeditions, predominantly to Arctic Siberia, were conducted by this
organisation. The results of these expeditions and other studies devoted to Quater-
nary faunas of Russia and adjacent territories were published in 17 volumes.
During the recent International Symposium, the Mammoth Committee was
reorganised: Prof. N. K. Vereschagin was elected Honorary President; the Director
of the Zoological Institute, Corresponding Member of the Russian Academy of
Sciences, Prof. A.F.Alimov became President; Dr. G.F.Baryshnykov (Saint Peters-
burg) and Dr. P.A.Lazarev (Yakutsk) were elected Vice-Presidents; Dr. I.E.Kuz-
mina (Saint Petersburg) was elected Scientific Secretary; Dr. W.E.Garutt, Acade-
mician A.P.Derevyanko, Dr.I.V.Foronova, Dr. I.M.Fokin, Dr. A.K.Markova, Prof.
V.M.Mikhelson, Prof. R.L.Potapov, Dr. N.D.Praslov, Prof. S.V.Tomirdiaro, and
5
Prof. V.V.Ukraintseva, were elected members of the Committee; and Prof. L.
Agenbroad (USA), Mr. K.Arai (Japan), Dr. B.Batyrov (Uzbekistan), Prof. H.Bo-
sinski (Germany), Prof. A.David (Moldova), Dr. L.Rekovetz (Ucraine), and Dr.
J.Saunders (USA) become foreign members of the Committee.
This volume continues the series of publications of the Zoological Institute
and Mammoth Committee on the fauna of the Quaternary Period (Ice Age). It cob-
tains articles of by authors who participated in this First International Mammoth
Symposium. This volume is a first step in the realisation of a program in Quater-
nary palaeozoology that was begun at the Symposium. In this volume special
attention is given to questions of biogeography and biochronological, and to me-
thods and studies for determining sex and age related variability in extinct and
extant mammals. Evaluating variability in Pleistocene species is one of the major
problems facing investigators.
The efforts of Mr. K.Arai, Director of the Palaeontological Museum in Naga-
no, Japan, in helping to conduct the symposium and the financial support of his
museum in the publication of this volume are gratefully acknowledged.
The President of the Mammoth Committee,
Corresponding Member of the
Russian Academy of Sciences,
Prof. A. F. Alimov
ISSN 0206-0477 ЧЕТВЕРТИЧНЫЕ И СОВРЕМЕННЫЕ
МЛЕКОПИТАЮЩИЕ ЕВРАЗИИ
СПб., 1996 (ТРУДЫ ЗООЛОГИЧЕСКОГО ИНСТИТУТА РАН, Т. 270)
RUSSIAN ACADEMY OF SCIENCES
PROCEEDINGS OF THE ZOOLOGICAL INSTITUTE
St. PETERSBURG, 1996, VOL. 270
УДК 569.742.6: 551.791
Г.Ф.Барышников, H.К.Верещагин
Зоологический институт РАН, Санкт-Петербург
КРАТКИЙ ОБЗОР ЧЕТВЕРТИЧНЫХ ГИЕН (СЕМЕЙСТВО
HYAENIDAE) РОССИИ И СОПРЕДЕЛЬНЫХ ТЕРРИТОРИЙ
Рассмотрены находки четвертичных гиен с территории бывшего СССР, датируемые
второй половиной плейстоцена (начиная с кромера). Они принадлежат 3 видам:
Pliocrocuta perrieri (Украина, Закавказье и Средняя Азия), Pachycrocuta brevirostris (За-
кавказье, Казахстан, Таджикистан и Забайкалье) и Crocuta spelaea. Последний вид был
широко распространен от Молдовы и Кавказа на западе и до Приморского края на восто-
ке. Приведены данные по геохронологической и географической изменчивости С. spelaea.
Из позднего плейстоцена Приморского края (пещера Географического общества) описан
новый подвид С. spelaea ussurica subsp. nov., отличавшийся от номинативного более круп-
ными размерами коренных зубов. Достоверные остатки Hyaena hyaena известны только из
голоценовых местонахождений Закавказья и Средней Азии, что объясняется поздним рас-
селением вида на север.
Семейство гиеновых (Hyaenidae) - одно из наиболее обычных семейств
хищных млекопитающих в составе плейстоценовых фаунистических комп-
лексов Евразии. Крупные и массивные костедробящие зубы и мощные
челюсти гиен хорошо сохраняются в ископаемом состоянии и часто при-
сутствуют в костеносных местонахождениях, особенно пещерных. Пещеры
использовались гиенами в качестве убежищ или для рождения детенышей,
поэтому палеозоологическая литература с описанием плейстоценовых остат-
ков гиен из захоронений Европы и Китая очень обширна, но материалы с
территории России и сопредельных стран публиковались весьма споради-
чески. Даже по такому широко распространенному виду, как пещерная гие-
на, Crocuta spelaea (Gold.), данных немного, что до сих пор не позволяло
судить об ее географической изменчивости и внутривидовой таксономии.
7
Данный очерк является первым кратким обзором находок четвертич-
ных гиен с территории бывшего Советского Союза, как опубликованных, так
и ранее не публиковавшихся. Во временном отношении мы ограничиваемся
рассмотрением гиеновых из среднего и позднего плейстоцена, проводя ниж-
нюю границу по кромерскому (морозовскому) межледниковью (в широком
его понимании) - времени первого появления в геологической летописи уме-
ренных широт Евразии гиен рода Crocuta.
МАТЕРИАЛ И МЕТОДИКА
В ходе проведенного исследования нами были изучены коллекции Зоологического
института РАН в Санкт-Петербурге (ЗИН), а также некоторые сборы из Палеонтоло-
гического института РАН в Москве (ПИН), Института археологии и этнографии СО РАН
в Новосибирске (ИАЕ), Центрального научно-исследовательского геолого-разведочного
музея в Санкт-Петербурге (ЦНИГР), Геолого-минералогического музея Казанского уни-
верситета (ГМК), Естественно-исторического музея АН Азербайджана в Баку (ЕИМ) и
Института археологии АН Украины в Киеве (ИАУ). Для сравнения были использованы
коллекции лаборатории четвертичной геологии (Laboratoire de Geologic du Quatemaire,
Marseille-Luminy) в Марселе, Франция (LGQ), университета Пантеон-Сорбонна (University
Pantheon-Sorbonne) в Париже, Франция (UPS), университета Клода Бернара (University
Clodt Bemar Lyon 1) в Лионе, Франция (UCB) и Института систематики и эволюции
животных (Institute of Systematics and Evolution of Animals) в Кракове, Польша (ISE).
В общей сложности были исследованы остатки 3 видов плейстоценовых гиен из 20
местонахождений. Основное внимание уделено описанию коллекции из пещеры Геогра-
фического общества, выполненного впервые.
Pliocrocuta perrieri
Франция: Лунель-Вьель (Lunel-Viel) (LGQ).
Pachycrocuta brevirostris
Россия: Тологой (ЗИН).
Crocuta spelaea
Россия: р.Казанка (ГМК), Ильская 1 (ЗИН), пещера Носорога (ЗИН), пещера
Смеловская 2 (ЗИН), Красный Яр (ЗИН), Усть-Канская пещера (ЗИН), Денисова пещера
(ИАЕ), Заиграево (ЦНИГР), пещера Географического общества (ЗИН).
Украина: Пролом 2 (ИАУ), Аювул-Коба (ЗИН).
Азербайджан: Бинагады (ЗИН, ЕИМ).
Казахстан: Железинское (ПИН), Бухтарминская пещера (ЗИН).
Узбекистан: Аман-Кутан (ЗИН).
Франция: Лунель-Вьель (Lunel-Viel)(LGQ), Жоранс (Jaurens) (UCB), Арси-сюр-Кюр
(Arcy-sur-Cure) (UPS).
Польша: Голобец (Golabiec), Вержховска Гурна (Wierzchowska Goma) (ISE).
Объем таксонов и синонимия гиеновых приведены в соответствии с последней
ревизией Hyaenidae (Werdelin, Solounias, 1991), за исключением выделения в отдельные
семейства Protelidae и Ictitheriidae.
При измерении зубов использованы следующие промеры: L - наибольшая передне-
задняя длина коронки (у резцов и клыков бралась по нижнему краю эмали), Lm - длина
метастиля Р4/, Lp - длина главного бугорка Р2/2 от его заднего края до передней стенки
коронки, Lpa - длина паракона Р4/, Lpr - длина протоконида (главного бугорка) на Р/4, Ltr
8
длина тригонида Л//1, IV наибольшая поперечная ширина коронки, Wa передняя
ширина /4/ на уровне протокона, Wb - задняя ширина Р4/ на уровне смыкания паракона с
метастилем.
ОБЗОР ТАКСОНОВ
СЕМЕЙСТВО HYAENIDAE GRAY, 1821
ТРИБА HYAENINI S. STR.
Род Hyaena Brunnich, 1771
Типовой вид. Canis hyaena Linnaeus, 1758, современность,
Ларистан (южный Иран).
Диагноз. Размеры средние. Твердое небо сравнительно узкое. Шов
между носовым отростком межчелюстной кости (processus nasalis ossis prae-
maxillare) и носовым отростком лобной кости (processus nasalis ossis frontalis)
отсутствует. Р4/ с умеренно длинным метастилем, который не длиннее
паракона. ММ относительно крупный, с 3 корнями. Передний дополнитель-
ный бугорок на /74 маленький. М/\ с метаконидом и умеренно укороченным
талонидом, который несет 2-3 бугорочка.
Состав рода. Монотипичен.
Сравнение. Род Hyaena может рассматриваться как один из наи-
более примитивных представителей семейства (в принятом здесь объеме).
Для него типичны одиночный образ жизни (иногда встречаются небольшие
семьи), собирательство, падалеядение, реже активное хищничество. В зубной
системе Hyaena характерны сравнительно небольшие размеры хищнических
зубов, укороченный метастиль на Р4/, крупный ММ и М/\ с хорошо разви-
тым метаконидом и относительно длинным талонидом. Примитивные при-
знаки прослеживаются также в морфологии молочных коренных (Барыш-
ников, Аверьянов, 1995). Коренные зубы Hyaena приспособлены для дробле-
ния костей, а уровень развития зубных признаков приблизительно соответ-
ствует предковому для Hyaenidae (без иктитериев) состоянию. Зубная
система гиен эволюционировала от этого исходного состояния в нескольких
направлениях: в сторону возрастания костедробящей функции премоляров
(Pliocrocuta) или усиления режущей роли хищнических зубов (Adcrocuta,
Chasmaporthetes), или же параллельного развития обеих функций (Pachy-
crocuta, Crocuta).
Замечания. К роду Hyaena в течение долгого времени относили всех
ископаемых гиен, в том числе четвертичные виды Европы и Азии. В настоящее время
можно считать, что гиены среднего и позднего плейстоцена внетропической зоны Евразии
принадлежали к другой трибе Crocutini. Что касается рода Hyaena, то он известен в
Африке с начала виллафранка (//. hyaena makapani Toerien, 1952), но в Евразию проник
очень поздно, и достоверные древние находки его здесь неизвестны, (Werdelin, Solounias,
1991).
9
Hyaena hyaena (Linnaeus, 1758)
Canis hyaena Linnaeus, 1758: 40.
Распространение. Полное отсутствие находок плейстоце-
новых остатков Hyaena hyaena (L.) в Крыму, на Русской равнине, Кавказе, в
Сибири и Казахстане говорит о том, что этот вид не входил в состав мамон-
товой фауны. Не было его и в плейстоцене Китая (Huang, 1989). Будучи юж-
ным по своему происхождению, Н. hyaena, очевидно, лишь в голоцене засе-
лил северные части своего азиатского ареала. Небольшое число голоценовых
находок имеется в Закавказье (Баиловская пещерка близ Баку, аллювиальные
суглинки в низовьях р.Сумгаит на Апшеронском полуострове, Гиеновая
пещера на Коцахурском хребте в районе Цители-Цкаро) и в Средней Азии
(Верещагин, 1959; Векуа и др., 1980; Ишунин, Тетюхин, 1989). В Узбекис-
тане остатки Н. hyaena определены на стоянке Сазаган 2 близ Самарканда,
слои которой датируются от мезолита до эпохи бронзы (Батыров, 1985).
Субфоссильный череп Н. hyaena с р.Сумгаит (кол. МЕИ) не отличается
по величине от черепов современной полосатой гиены из Закавказья и Турк-
мении (см. Огнев, 1931). Он имеет следующие размеры (мм): общая длина
черепа 238, кондилобазальная длина 211, длина твердого неба 107, длина
ряда С1/-Р4/ 87, длина Р4/ 31.6, ширина в клыках 55.5, межглазничная
ширина 48, ширина височного сужения 38.2, ширина твердого неба 94, длина
нижней челюсти 164; M/1 L=22A, W=\1.0.
ТРИБА CROCUTINI BARYSHNIKOV ЕТ AVERIANOV, 1995
Род Pliocrocuta Kretzoi, 1938
Типовой вид. Hyaena perrieri Croizet et Jobert, 1828, ранний
виллафранк, Этуэр (Перрье), Франция.
Диагноз. Размеры средние. Шов между носовыми отростками
межчелюстной кости и лобной кости имеется. Метастиль Р4/ длинный, пре-
вышающий длину паракона. ММ умеренно крупный. Передний дополни-
тельный бугорок на Р/4 большой. Л//1 с метаконидом (иногда без него) и
удлиненным двухбугорковым талонидом.
Состав рода. Монотипичен.
Сравнение. Плиокрокуты изрестны в Европе, начиная с русциния
(Werdelin, Solounias, 1991). Представляют собой раннюю ступень в эволюции
гиен трибы Crocutini. Зубная характеристика плиокрокут показывает пре-
обладание дробящей функции коренных, специализированных на раска-
лывание костей и поедание различной падали. Обнаруживают морфоло-
гическое сходство с современным африканским родом Parahyaena Hendey,
1974 (Kurtdn, 1956; Galiano, Frailey, 1977).
10
Pliocrocutaperrieri (Croizet et Jobert, 1828)
Hyaena perrieri Croizet et Jobert, 1828: 178; Hyaena arvernensis Croizet et Jobert,
1828: 180; Hyaena monspessulana Christol et Bravard, 1828: 368 (nom. nud.); Hyaena prisca
Serres, Dubreuil et Jeanjean, 1828: 269; Hyaena topariensis Forsyth Major, 1883: 2; Hyaena
arvernensis var. pyrenaica Deperet, 1890: 112; Hyaena brunnea Thunberg - Верещагин, 1951:
64-65; Crocuta sivalensis (Falconer et Cautley) - Яцко, 1956: 335; Hyaena donnezani Viret,
1954: 52; Hyaena arambourgi Ozansoy, 1965: 40-41; Hyaena perrieri kuruksaensis Sharapov,
1986: 52; Pachycrocuta perrieri (Croizet et Jobert) - Сотникова, 1989: 37-41; Hyaena sp.
Давид и др., 1990: 68; Pachycrocuta pyrenaica (Deperet) - Калмыков, 1992: 22-24; Hyaena
prisca de Serres - Барышников, Батыров, 1994: 32-34.
Распространение. В Западной Европе Р perrieri просле-
живается вплоть до миндель-рисского (голштейнского) межледниковья. Он
известен, например, из голштейнского местонахождения Лунель-Вьель во
Франции, где был определен как Hyaenaprisca (Bonifay, 1971).
На территории бывшего СССР вид найден у южной государственной
границы (рис.1). Он указан для русциния Украины (Одесские катакомбы),
Забайкалья (Удунга) и для среднего виллафранка Таджикистана (Куруксай,
точка Наврухо и точка 97) (Яцко, 1956; Howell, Petter, 1980; Шарапов, 1986;
Сотникова, 1989; Калмыков, 1992). В Средней Азии Р perrieri жил и
значительно позже. В частности, к данному виду принадлежат остатки гиены
из отложений Пенджикентского карьера (Таджикистан), датируемые послед-
ним межледниковьем, и, вероятно, из ашельских слоев пещеры Сель-Унгур в
Киргизии (Барышников, Батыров, 1984). Клык гиены из слоев среднего па-
леолита (мустье) стоянки Кульбулак в Ташкентской области, по величине не
отличающийся от клыка Hyaena hyaena (Худайбердиев, 1973), тоже может
быть отнесен к Р perrieri.
Скорее всего, к данному виду принадлежит фрагмент нижней челюсти
Hyaena sp. из местонахождения Чортков, Западная Украина, который был
найден совместно с Blarina ucrainica, Mimomys sp., Allophajomys pliocaenicus,
Bison priscus и другими видами среднего плейстоцена (Давид и др., 1990). Во
всяком случае по длине Р/4 (£=23.0 мм) украинская гиена не отличается от Р.
perrieri (= Н. prisca) из Лунель-Вьеля (табл. 1).
К Р. perrieri, вероятно, относятся также череп и фрагменты зубов гие-
ны из строительного известняка древнекаспийских плиоценовых террас близ
Баладжар в Азербайджане. Первоначально они были определены как Hyaena
hyaena (Богачев, 1938), но позднее Верещагин (1951) сближал их с предками
Hyaena brunnea. Верхние коренные зубы РЗ/ (£=22.5, 23.0; ИМ 5.0, 15.0 мм)
и Р4/ (£=34.5 мм) крупные, их величина вполне типична для европейских
находок Р. perrieri (Howell, Petter, 1980).
Приуроченность поздних четвертичных находок Р. perrieri к межледни-
ковым местонахождениям Европы и Центральной Азии, очевидно, указывает
на проникновение вида во внетропические широты только в теплые эпохи
плейстоцена, когда он расселялся сюда из областей с субтропическим клима-
том.
11
Судя по относительно более узким (по сравнению с Crocuta crocuta)
хоанам на черепе Р perrieri из Лунель-Вьеля, можно предполагать, что евро-
пейская плиокрокута не обладала высокими скоростными способностями.
Вероятнее всего, она была менее быстрым бегуном, чем пятнистая гиена, и
не использовала групповую охоту. По образу жизни Р perrieri был похож,
вероятнее всего, на современные африканские виды Parahyaena brunnea и
Hyaena hyaena, являясь, в первую очередь, падалеядом, собирателем мелких
позвоночных и, в меньшей степени, активным охотником на молодых и
ослабевших копытных. Размеры коренных зубов указывают на способность
вида к потреблению крупной падали.
Замечания. Сотникова (1989, с.39) отмечает, что у Р perrieri из Куруксая все
нижние хищнические зубы без метаконида и с относительно длинным талонидом.
Отмеченные признаки сближают куруксайскую гиену с Р perrieri из среднего
виллафранка Европы (Сен-Валье, Пуэбла де Вальверде). По значениям относительной
длины тригонида А//1 (см. Сотникова, 1989; Барышников, Батыров, 1994) плиокрокута из
Куруксая (82.4-83.52-84.4%, и=6) превосходит гиен из Лунель-Вьеля (80.1, 80.6, л=2) и
Пенджикента (78.0, 81.3, п=2). Метаконид имеется на одном из двух изученных Л/71 в
Лунель-Вьеле и на обоих экземплярах из Пенджикента.
Рис. 1. Места находок остатков Pliocrocuta perrieri (а) и Pachycrocuta brevirostris (b)
Fig. 1. Map showing the localities with findings of Pliocrocuta perrieri (a) and Pachycrocuta
brevirostris (b)
12
Таблица 1. Размеры нижних коренных зубов у гиен из родов
Pliocrocuta, Pachycrocuta и Hyaena
Table 1. Measurements (mm) of lower cheeck teeth in the genus Pliocrocuta,
Pachycrocuta and Hyaena
Промеры (мм) и индексы (%) Pliocrocuta perrieri Лунель-Вьель кол. LGQ Pachycrocuta brevirostris Тологой ЗИН Hyaena hyaena современность кол. ЗИН
10010 1 3716 | 2002 34476 n | lim | x
Длина 107.1 111.2 6 90.9-100.1 96.10
PI2-MIX 79.1 76.2 6 97.9-70.8 69.55
Р/2 L 15.1 16.3 17.5 6 13.5-15.0 14.18
Lp 11.1 11.1 14.6 6 10.3-12.0 11.03
W 9.6 10.8 13.6 6 8.2-10.0 8.93
Р/3 L 22.5 22.3 6 17.1- 19.5 18.52
FT 13.7 15.1 6 10.7- 12.8 11.62
Р/4 L 23.5 23.3 26.7 6 18.9-20.6 19.83
Lpt 11.6 11.9 13.7 6 9.2-10.2 9.82
W 13.8 13.9 18.0 6 10.7-12.9 11.77
М/1 L 24.1 23.2 29.1 6 19.4-21.4 20.65
Ltr 19.3 18.7 25.6 6 16.1- 17.3 16.68
FT 12.8 12.8 15.8 6 9.6-11.3 10.48
Wta 8.9 9.0 9.8 6 7.9 - 9.3 8.90
Индексы
Р/2 W.L 63.6 66.3 77.7 6 59.0- 66.7 62.95
Р/3 W\L 60.9 67.7 6 58.8- 67.8 62.77
Р/4 W:L 58.7 59.6 67.4 6 56.6- 62.6 59.28
М!\ Ltr.L 80.1 80.6 88.0 6 79.4- 83.0 80.82
Р!4 L.M!\L 97.6 91.7 6 90.7- 98.5 96.01
Таким образом, виллафранкские плиокрокуты выглядят более эволюционно
продвинутыми, специализированными, чем гиены четвертичного возраста. Однако у более
древней плиокрокуты из русциния Забайкалья (Удунга) индекс относительной длины Л//1
равен 79.8%, метаконид присутствует на всех экземплярах (Калмыков, 1992). Отмеченные
признаки, вероятнее всего, варьировали в истории плиокрокут. Их развитие могло
зависеть от увеличения или уменьшения давления естественного отбора на различные
популяции. Условия межледниковых эпох были, очевидно, благоприятными для
плиокрокут, и уменьшение давления отбора приводило к появлению признаков,
утраченных предками в эпохи ухудшения природной обстановки. Данные факты вполне
согласуются с высказанной ранее гипотезой о цикличности в морфологической эволюции
гиеновых (Барышников, Аверьянов, 1995), в которой фазы специализации сменялись
фазами деспециализации.
13
Род Pachycrocuta Kretzoi, 1938
Типовой вид. Hyaena brevirostris Aymard, 1846, поздний вилла-
франк, Франция.
Диагноз. Размеры очень крупные. Шов между носовыми отрост-
ками межчелюстной и лобной костей имеется. Метастиль Р4/ длинный,
превышающий длину паракона. Л/1/ сранительно небольшой. Р/2 и Р/4
относительно более широкие, чем у Pliocrocuta. Передний дополнительный
бугорок на Р/4 крупный. На М/\ метаконид редуцирован, вплоть до полного
его исчезновения; талонид короткий, однобугорковый.
Состав рода. Монотипичен.
Замечание. Сотникова (1989), вслед за Хоуэллом и Петтер (Howell, Petter,
1980), принимает более широкий объем рода, включая в него Pliocrocuta, и выделяет в
нем 4 вида.
Сравнение. Высоко специализированные гиены. Филогенети-
чески связаны с Pliocrocuta и представляют собой дальнейшее развитие
эволюционной линии, развивавшейся в направлении увеличения премоляров
для разгрызания толстостенных трубчатых костей крупных млекопитающих,
с одновременным усилением режущей функции хищнических зубов.
Pachycrocuta brevirostris (Aymard, 1846)
Hyaena brevirostris Aymard, 1846: 153; Hyaena sinensis Owen, 1870: 422-424; Hyaena
robusta Weithofer, 1889: 46; Hyaena bathygnata Dubois, 1908: 1265; Hyaena zdanskyi Pei,
1934: 110-116; Hyaena licenti Pei, 1934: 120-121; Crocuta cf. sinensis Zdansky - Векуа, 1962:
38-42; Crocuta sp. - Байгушева, 1971: 12; Pachycrocuta brevirostris (Aymard) - Сотникова,
1989:41-44.
Описание. Имеющийся в нашем распоряжении обломок левой
нижнечелюстной кости молодого зверя (ЗИН 34476) происходит из местона-
хождения Тологой в Забайкалье. На поверхности кости нанесены обозна-
чения "Т-2, Р-П, С-2а", которые позволяют привязать находку к верхней
части средней толщи осадочных образований. Фауна из этого горизонта
относится к тологойскому фаунистическому комплексу (Вангенгейм, 1977).
Он является вероятным хроностратиграфическим аналогом тираспольского
териокомплекса Восточной Европы и может быть датирован нижним
средним плейстоценом (MNQ 20-21). Ранее Р brevirostris уже отмечалась в
видовом списке фауны Тологоя (Калмыков, 1990), но морфологическое
описание тологойской гиены нем не известно.
На челюсти сохранился ряд зубов Р/2-Л//1, при этом Р/3 полностью не
прорезался (рис. 2, а, вклейка). Видна также коронка режущегося клыка. Длина
PI2-MIX составляет 89.2 мм, высота челюсти за Р/2 равна 43 мм. Данные
параметры очень близки к тем, которые приводит Сотникова (1989) для
14
молодой особи Р brevirostris из местонахождения Лахути 2 в Таджикистане.
Подбородочное отверстие одно, крупное, расположено под Р/2.
Клык относительно небольших размеров, его длина по основанию
эмали равна 18.5 мм. По внутренней стенки коронки проходит мощный
вертикальный гребень, почти достигающий вершины клыка.
Коренные зубы очень крупные, высокие (см. табл. 1). Р/2 и Р/4
абсолютно и относительно широкие, островершинные. На Р/2 задний допол-
нительный бугорок маленький, передний вообще невыражен. Р/4 трех-
бугорчатый, по высоте не уступает Р/3. Оба дополнительных бугорка его
слабо разнятся по величине. В задний части коронки хорошо развит ворот-
ничок, заходящий на лингвальную сторону зуба. А7/1 с рудиментарным мета-
конидом, коротким и широким талонидом. Края последнего приподняты, в
центре талонида расположен небольшой гребневидный бугорок.
Вероятно, из одновозрастных отложений Забайкалья происходит череп
Р. brevirostris sinensis (Owen), выставленный в экспозиции Палеонтологи-
ческого музея РАН в Москве. Судя по сильной изношенности жевательной
поверхности коронок, череп принадлежал старому зверю. Промеры черепа,
любезно выполненные по нашей просьбе Е.Н.Мащенко, следующие (мм):
общая длина 334, кондилобазальная длина 300, межглазничная ширина 93,
длина PXI-P4I 93; промеры сильно истертого Р4/: Z=47, Zw=29, Wa=22,
Wb=\5.
Замечания. По строению и размерам нижних коренных зубов пахикрокута
из Тологоя ничем существенным не отличается от Р. brevirostris из местонахождения
Лахути 2 (Сотникова, 1989), за исключением присутствия у тологойской гиены на М/1
рудиментарного метаконида. Длина верхнего хищнического зуба у забайкальского черепа
близка к максимальному значению ее у Р. brevirostris из Чжоукоудяня 1 (Howell, Petter,
1980).
Очень близка по величине зубов к пахикрокуте правая нижнечелюстная кость гиены
из Ливенцовского карьера в Ростовской области, отнесенная к Crocuta sp. Байгушева
(1971), изучавшая челюсть, уже указала на сходство между ливенцовской формой и
гиеной из Тологоя. Она отметила также наличие на талониде Л//1 двух зубцов и
присутствие точечной альвеолы Р/\. Байгушева приводит следующие промеры зубов: Р/4
Z=25.4, ИМ 7.0; А//1 Z=28, ИМ 5 мм.
К Р brevirostris принадлежат также остатки гиены из местонахождения Ахалкалаки
в южной Грузии (ранний средний плейстоцен), которые были ранее определены как
Crocuta cf. sinensis (Векуа, 1962). Высота нижнечелюстной кости за Л/71 (47.5 мм) и
размеры А/71 (1=29.0, ИМ5.5 мм) вполне типичны для Р. brevirostris. Талонид короткий
(4.5 мм), однобугорковый, метаконид отсутствует.
Крупные и массивные предкоренные зубы у Pachycrocuta были приспособлены для
раскалывания костей, представляя собой дальнейшее развитие костедробящих премо-
ляров Pliocrocuta. В тоже время нижний хищнический зуб у Pachycrocuta имеет более
выраженный режущий тип, дробящие структуры которого сильно редуцированы.
Следовательно, мы видим у Pachycrocuta по сравнению с Pliocrocuta специализацию
зубного аппарата как для дробления костей, так и для разрезания мясной пищи. Она
проявляется и в морфологии молочных коренных (Барышников, Аверьянов, 1995).
Распространение. Р. brevirostris имеет, вероятно, африкан-
ское происхождение и появляется в Евразии в позднем виллафранке (Howell,
15
Petter, 1980). Вид был широко распространен в Европе, Китае, Индии и
вплоть до Явы, доживая до среднего плейстоцена в Китае, где он определен
из Чжоукоудяня (Pei, 1934). В Европе Р brevirostris прослеживается до кро-
мера (Странска Скала, Бактон Форест Бед, Вестбари, Гомбасцог, Зюссен-
борн, Петралона), но в Мауере и Мосбахе отсутствует (Kurt6n, Poulianos,
1977).
В границах бывшего СССР находки Р. brevirostris (см. рис.1) известны
для фауны раннего плейстоцена Приазовья (Ливенцовка) и Забайкалья (Засу-
хино), нижнего среднего плейстоцена Закавказья (Ахалкалаки), Казахстана
(Актогай на р.Чарын, Алма-Атинская обл.; илийский териокомплекс), Тад-
жикистана (Лахути 2 на р.Обимазар) и Забайкалья (Тологой на р.Селенга)
(Векуа,1962; Байгушева, 1971; Алексеева, 1977; Иметхенов, Калмыков, 1988;
Сотникова, 1989; Калмыков, 1990; Тлеубердина и др., 1990).
По образу жизни пахикрокута, вероятно, напоминала современную
пятнистую гиену (Crocuta crocuta), но вела, скорее всего, одиночный образ
жизни. Крупные размеры и мощные зубы позволяли ей как разгрызать
останки павших гигантских толстокожих, так и самостоятельно охотиться на
различных копытных, успешно конкурируя с другими гиенами и большими
кошками, пока ее не вытеснил Crocuta spelaea.
Род Crocuta Каир, 1828
Типовой вид. Canis crocuta Erxleben, 1777, современность, Аф-
рика.
Диагноз. Размеры средние и крупные. Твердое небо относительно
широкое. Шов между носовыми отростками межчелюстной и лобной костей
отсутствует. Р4/ с длинным метастилем, который значительно длиннее пара-
кона. М1\ очень маленький, с 2 корнями, и с возрастом часто выпадает.
Передний дополнительный бугорок на Р/4 небольшой. Л//1 с метаконидом
или без него, талонид очень короткий и обычно без бугорков или с одним
режущим бугорочком (у архаичных форм с 1-2 бугорками).
Состав рода. Включает 3-4 вида, хотя Верделин и Солунис (Wer-
delin, Solounias, 1991) объединяют их в один вид.
Сравнение. Наиболее эволюционно продвинутый род трибы
Crocutini. Активные хищники с хорошо организованными приемами коллек-
тивной охоты. Адаптивные изменения в зубной системе характеризуют
усиление режущей функции хищнических зубов (относительное удлинение
метастиля Р4/, исчезновение метаконида и редукция талонида на Л//1),
сочетающееся с возрастанием размеров и гипсодонтии РЗ/З как приспо-
собления для раскалывания крупных костей. Подобный "дуалистинный"
характер адаптации коренных зубов наблюдается также у рода Pachycrocuta
(см. выше), но происхождение и развитие его в обоих родах было незави-
симым, конвергентным. Возможно, что незавимое развитие линии Crocuta и
16
линии Pliocrocuta Pachycrocuta указывает на их весьма удаленные род-
ственные связи и на правомерность выделения для двух последних родов
отдельной трибы Parahyaenini, вместе с современным родом Parahyaena.
Crocuta spelaea (Goldfuss, 1823)
Hyaena spelaea Goldfuss, 1823: 456-462; Hyaena ultima Matsumoto, 1915: 2-3;
Crocuta crocuta spelaea (Goldf.) - Верещагин, 1951: 55-65; Crocuta crocuta spelaea (Goldf.)
- Татаринов, 1958: 797-800; Crocuta spelaea (Goldf.) - Кожамкулова, 1969: 34-35; Crocuta
spelaea (Goldf.) - Алексеева, 1980: 35-36; Crocuta spelaea (Goldf.) - Давид, 1980: 68-71;
Crocuta spelaea (Goldf.) Давид и др., 1990: 68-73; Crocuta crocuta spelaea (Goldf.) -
Барышников, Батыров, 1994: 34; Crocuta spelaea (Goldf.) - Барышников, 1985: 3-45.
Типовая местность. Пещера Гайленройтер (Gailenreuther
Hdhle=Zoolithen Hohle), южная Германия, поздний плейстоцен.
Описание. Из находок пещерной крокуты в фаунистических комп-
лексах среднего плейстоцена России в первую очередь следует отметить
крупный череп с р.Казанка в Татарстане (ГМК 191), который был детально
изучен одним из нас (Барышников, 1992). По общим размерам черепа и
зубов (табл. 2, 3, 4) и по пропорциям хищнических зубов (относительно
укороченный метастиль Р4/, несколько удлиненный талонид А7/1) гиена из
Татарстана оказалась близкой к С. spelaea praespelaea из кромерско-мин-
дельской фауны Европы (Schutt, 1971). От другого крупного среднеплей-
стоценового подвида С. 5. petralona (Kurtdn et Poulianos, 1977) она отлича-
ется менее широкими премолярами.
Аналогичные параметры (см. табл. 2) имеет череп (без нижней челюс-
ти) из местонахождения Железинское на р.Иртыш в Казахстане (кол. ПИН).
Эта находка также может быть отнесена к подвиду С. spelaea praespelaea,
ареал которого, скорее всего, охватывал не только всю Европу, но и юг
Западной Сибири и Казахстан.
Еще один фрагмент черепа без зубов крупной крокуты найден в
бассейне р.Уда близ ст.Заиграево в Западном Забайкалье (ЦНИГР 3922). Он
был определен Рябининым (1932) как Hyaena crocuta Егх. var. spelaea Goldf.
Сделанные промеры (мозговая длина 173, межглазничная ширина 68, шири-
на височного сужения 45, слуховая ширина 103, высота в области bullae
osseae 128 мм) не позволяют точно определить таксономическую принадлеж-
ность данного экземпляра. Вполне возможно, что он принадлежит -к С. Spe-
laea ultima (Matsumoto, 1915).
Серийный материал (рис.З, 4, вклейка) по небольшой пещерной крокуте
С. spelaea из асфальтовых песков Бинагады возле Баку в Азербайджане (кол.
ЕИМ, ЗИН 22382, 22384, 24407) был монографически опубликован ранее
(Верещагин, 1951). Величиною черепа и нижнечелюстных костей (табл. 2, 5)
бинагадинская крокута заметно уступала С. 5. praespelaea (кромер-миндель)
и С. s. spelaea (вюрм), но в то же время была крупйег^чем С. 5. intermedia из
17
Таблица 2. Размеры черепа пещерной гиены (Crocuta spelaea)
Table 2. Measurements (mm) of cranium in Crocuta spelaea
Промеры, мм Сл. praespelaea С. .V. spelaea С. s. ussurica ssp.n.
р.Казанка, Поволжье ГММ 191 Жслезин- ское, Казахстан кол. ПИН Бинагады, Закавказье ЗИН 24407 пещера Географического общества, Приморье ЗИН 34478, голотип
Длина общая (1) 315.0 296.0 287.0
кондилобазальная (2) 268.5 257.0 250.0
твердого неба (11) 147.0 130.0 128.0
горизонтальной
части небной кости са63.5 50.0 53.0
С1/-Р4/ 111.0 112.0 105.5
Ширина мастоидная (18) 110.0 104.6 112.0
“ мозговой коробки (22) 85.0 86.3 87.5
в мыщелках (19) 60.8 57.0 52.5
скуловая (23) 196.0 201.0 180.6
в надглазничных
отростках (24) 95.0 87.0 саЮЗ.6 са77.0
межглазничная (25) 70.5 67.0 64.9 64.3
твердого неба (26) 129.8 133.0 111.5 127.0
в клыках (27) 75.7 72.0 69.2
в хоанах 43.6 36.3 35.4
между овальными
отверстиями 55.2 33.4 40.6
височная(28) 52.0 46.0 49.6 48.5
между подглазничными
отверстиями (29) 69.7 67.0 69.4
Примечание. Цифры в круглых скобках соответствуют номерам промеров в работе von den
Driesch (1976).
миндель-рисса Европы (Kurtdn, Poulianos, 1977). По кондилобазальной длине
она близка к европейской крокуте из фауны последнего оледенения (рисс-
вюрма) Англии. Сближают их также длина хищнического зуба Р4/ и индекс
относительной длины метастиля. Последний показатель в выборке из Бинага-
дов оказался в среднем заметно меньше, чем у С. spelaea dorotheae, описан-
ной из пещеры Зуттиех (Zuttiyeh Cave) в Галилее, Палестина, содержащую
ябрудийскую и леваллуа-мустьерскую индустрию (Kurtdn, 1965; Коробков,
1978). М/] у бинагадинской крокуты относительно очень широкий. Метако
нид присутствует в 6 случаях из 11 (см. рис. 4, а). Талонид короткий, одно-
бугорковый, но у 1 экз. он несет 2 мелких бугорочка.
18
Таблица 3. Размеры резцов и клыков пещерной гиены (Crocuta spelaea)
Table 3. Measurements (mm) of incisors and canines in Crocuta spelaea
Промеры, мм С. spelaea praespelaea р.Казанка, Поволжье ГММ 191 С.spelaea ussurica ssp.n. пещера Географического общества, Приморье кол. ЗИН
/1/ L 8.9 9.0
IV 6.4 6.3
/2/ L 11.0 10.7
W 7.8 7.5
13/ L 12.8 14.3
W 11.0 12.0
С1/ L 19.2 18.8, 20.7
IV 13.6 13.4, 15.0
7/2 L 8.7 9.4
IV 5.6 5.9
7/3 L 10.0 10.5
IV 9.5 10.2
С/\ L 18.2, 18.4
IV 15.1, 15.2
Заслуживает внимания серия из зубов и обломков костей С. spelaea из
мустьерских слоев пещеры Аман-Кутан в Узбекистане (сборы Д.Н.Льва,
1957; кол. ЗИН 29058). По длине нижнего ряда кореных крокута из Аман-
Кутана близка к гиене из Бинагадов и из мустьерских памятников Крыма
(Барышников, 1995). Большинство зубов очень сильно истерты. Единствен-
ный целый Р4/ очень короткий (табл. 6), его длина меньше минимальных
значений длины даже у наиболее мелкого подвида С. spelaea dorotheae.
Укороченность данного экземпляра связана с сильной стертостью коронки,
поскольку два других обломка Р4/, судя по длине метастиля (16.1, 16.2 мм),
были нормальных размеров. Индекс длины метастиля (41.7 %) также обычен
для С. spelaea. Два имеющихся М/1 небольшие, в обоих случаях с метакони-
дом. Обращает на себя внимание сильно укороченный талонид. Серия
верхних резцов 13/ (А=10.0, 10.8, 11.0, 12.7; 1Г=9.6, 10.3, 10.3, 9.4 мм) по ве-
личине не отличается от резцов пещерной гиены из мустьерских стоянок
Крыма (Барышников, 1995).
19
Таблица 4. Размеры верхних коренных зубов среднеплейстоценовой
пещерной гиены Crocuta spelaeapraespelaea
Table 4. Measurements (mm) of upper cheek teeth in Crocuta spelaea praespelaea
from the Middle Pleistocene
Промеры, мм р.Казанка, Поволжье ГММ 191 Железинское, Казахстан кол. ПИН
Р\/ L 8.4
W 7.8
Р2/ L 17.9
Lp 11.3
W 13.4
РЗ/ L 25.3 27.0
IV 18.5 21.5
Р4/ L 42.7 43
Lm 18.3
Wa 23.3 24
Wb 12.7
В нашем распоряжении имеется небольшая коллекция остатков С.
spelaea из пещер Южного Урала. Левая нижнечелюстная кость из пещеры
Носорога (ЗИН 34477, сборы А.В.Рюмина), судя по заметной истертости
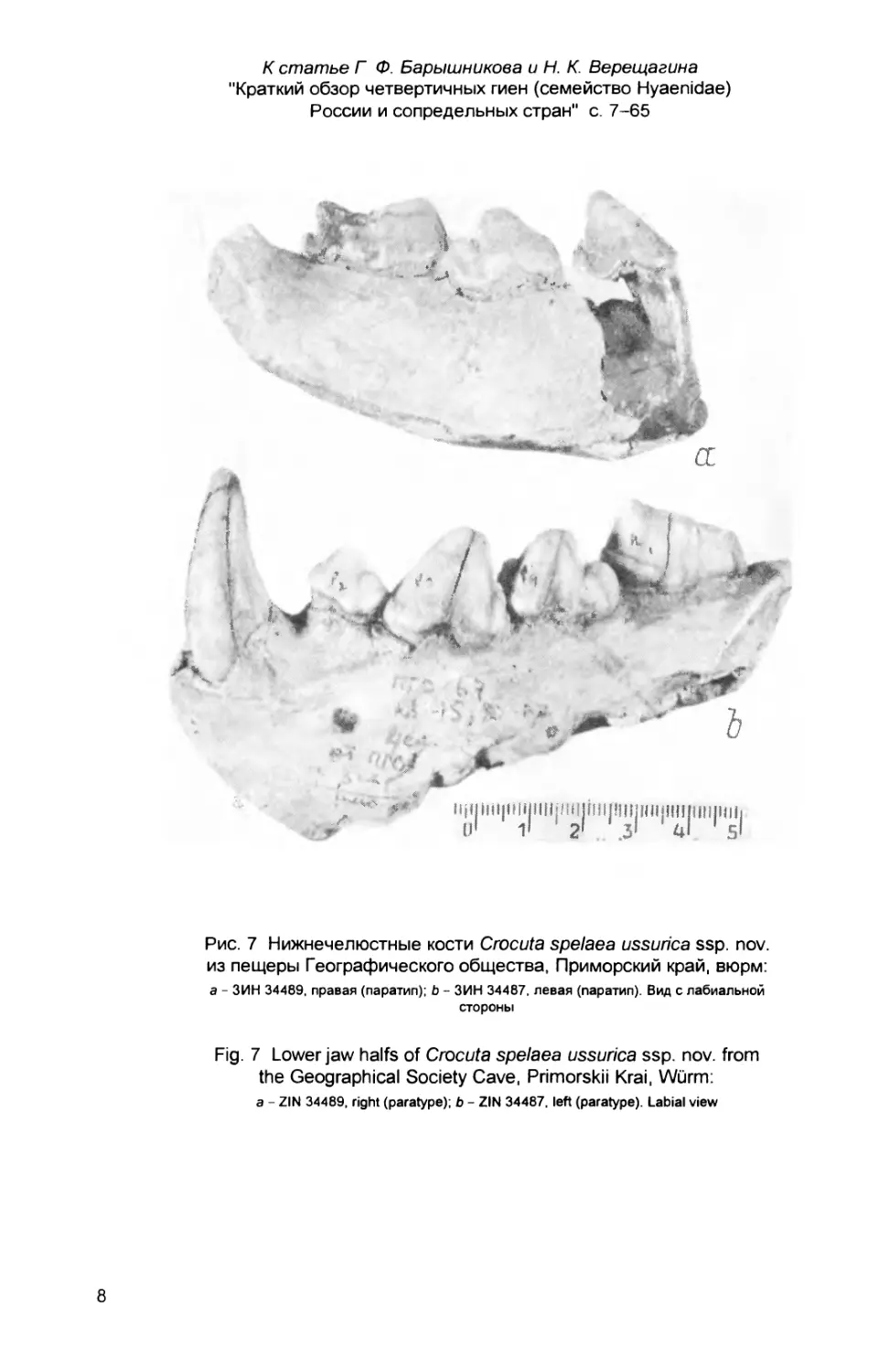
коренных зубов, принадлежала старой особи (см. рис. 2, Ь, вклейка). Клык ее
крупный (Л=17.9, ИМ4.7 мм). Зубной ряд С/1-Л//1 более короткий, чем на
экземплярах из Сибири (см. табл. 5), и по значению длины близок к таковому
у гиены из палеолита Крыма (Барышников, 1995). М/\ также небольшой, с
отчетливым метаконидом и маленьким однобугорковым талонидом.
Несколько изолированных зубов происходят из пещеры Смеловская 2 (ЗИН
34491, сборы О.Н.Бадера). Их промеры (мм): Р2/А=17.8, ^=12.8; РЗ/ L=25.3,
^=18.0; Р/3 А=22.7, 24.6, ИМ7.1, 18.5; Р4/ А=23.0, 23.6, Lpr=\3.3, 15.3,
ИМ 5.0, 15.3. По величине зубов гиена из Смеловской практически не отли
чалась от крокуты из пещеры Носорога. Более крупные размеры (см. табл. 5)
имеет левая нижнечелюстная кость старой гиены из местонахождения Крас-
ный Яр на р.Обь (ЗИН 32752, сборы Э.В.Алексеевой) (см. рис. 2, с, вклейка).
Клык большой (А=18.7, ИМ5.1 мм). М/\ с коротким талонидом без бугор-
ков; на месте метаконида наблюдается вздутие эмали (рудиментарный мета-
конид ?). По величине коренных зубов С. spelaea из Красного Яра (табл. 7)
сходна с С. spelaea praespelaea, но более короткий метаконид М/\ сближает
ее с номинативным подвидом из позднего плейстоцена Европы.
20
Таблица 5. Размеры нижней челюсти пещерной гиены (Crocuta spelaea)
Table 5. Measurements (mm) of mandible in Crocuta spelaea
Промеры, MM С. spelaea spelaea С. 5. ussurica ssp. n.
Бинагады, Закавказье ЗИН 22382 пещера Носорога, Южный Урал ЗИН 34477 Красный Яр, Западная Сибирь ЗИН 32752 пещера Географического общества, Приморье кол. ЗИН
” 1 lim | X и = 1 и = 1 " 1 lim | X | а
Длина общая 4 181.0-192.6 185.7
" С/1-Л//1 5 105.1-116.0 110.6 116.7 125.7 2 117.4, 120.0
РП-М/Х 5 82.4-92.3 85.6 86.8 96.6 3 91.5-98.3 93.83
PI2-PI4 4 55.2-64.4 58.5 62.1 64.0 6 59.7-66.9 62.77 2.83
PI3-PI4 4 41.0-48.7 44.3 46.5 47.0 10 43.2-50.6 47.73 2.21
Высота венечного отростка 4 89.7-96.7 92.6
Высота перед Р/2 5 35.1-41.6 38.2 39.4 4 42.5-47.4 44.60 1.78
Таблица 6. Размеры верхних коренных зубов позднеплейстоценовой пещерной гиены
(Crocuta spelaea)
Table 6. Measurements (mm) of upper cheek teeth in Crocuta spelaea from the Late Pleistocene
C. spelaea spelaea С. spelaea ussurica ssp.n.
Промеры, Бинагады, Аман-Куган, пещера Географического общества,
MM Закавказье Узбекистан Приморье
ЗИН 22382,24407 ЗИН 29058 кол. ЗИН
n lim | X n 1 lim | X п 1 Нт | * 1 ст
PM L 1 7.0 1 7.2 4 6.9-8.2 7.55 0.56
W 1 6.8 1 7.5 4 6.2-8.5 7.57 0.84
PH L 4 15.4-17.6 16.75 3 15.2-16.8 16.03 5 16.0-20.9 18.72 1.76
Lp 4 13.2-13.8 13.47 2 11.4, 13.4 5 13.0-15.7 14.68 1.09
W 4 11.4-13.4 12.57 3 11.2-12.1 11.63 5 11.5-15.0 13.42 1.31
РЗ/ L 1 22.2 1 20.7 7 23.0-28.9 27.17 1.94
W 1 16.4 1 15.4 7 16.3-21.7 19.74 1.71
PM L 8 36.6-42.2 39.64 1 33.3 4 40.7-46.5 44.27 2.53
Lm 8 14.7-18.0 16.85 3 13.9-16.2 15.40 4 17.4-21.8 19.95 1.87
Wa 8 18.7-23.9 21.76 1 18.8 5 19.8-27.9 24.08 3.03
Wb 8 11.3-14.5 12.69 1 10.0 3 13.8-15.0 14.53 0.64
Изолированные коренные зубы и отдельные кости пещерной крокуты
были добыты в ходе археологических раскопок на Алтае. В Бухтарминской
пещере (ЗИН 34403) пригодных для измерения образцов немного (Р/2
Z=17.3, Zp=13.5, ИМЗ.З; Р/4 Z=24.6, Zpr=14.0, JF=16.2). Также малочислен-
ны целые коренные зубы в Денисовой пещере (кол. ИАЭ). Их промеры
(мм): Р/2 1=18.3, £р=14.4, ^=13.2; Р/3 ^=16.6; М/\ Л=35.6, И7г=31.9,
ИМ5.4. Целый М/l из слоя 17 в Денисовой пещере очень крупный, с метако-
нидом. Талонид сравнительно длинный, и индекс относительной длины три-
гонида лежит в области минимальных значений его у С. spelaea spelaea из
Европы.
Серийный материал по пещерной крокуте извлечен из верхнечетвер-
тичных отложений в пещере Географического общества на р.Партизанской
(Сучан) в Приморском крае. Имеется почти полный осевой череп (ЗИН
34478), передняя часть морды, носовые кости и скуловые дуги у которого
отсутствуют (рис.5, вклейка). Частично обломаны затылочные мыщелки, са-
гиттальный гребень, надглазничные отростки и верхнечелюстные кости. Че-
реп местами окатан, местами несет следы погрызов крупных хищников (ги-
ен?). Из зубов сохранился правый Р2/ и обломок правого РЗ/.
По общим размерам данный экземпляр несколько превосходит череп
С. spelaea из Бинагадов, но заметно меньше черепа с р.Казанка (см. табл.2).
Обращает на себя внимание, что череп из Приморского края очень широкий
в лицевой части. По ширине твердого неба и расстоянию между подглазнич-
ными отверстиями гиена из пещеры Географического общества не только
превосходит зверей из Бинагадов, но не уступает С. spelaea praespelaea из
Татарстана.
Нижнечелюстные кости из пещеры Географического общества (ЗИН
34484-34489) высокие, с одним подбородочным отверстием (рис. 6, 7, вклей-
ка). Зубы посажены плотно, кулисообразно. Небольшая диастема между С/1
и Р/2 достигает в длину 8 мм. В целом длина нижнего зубного ряда вполне
типична для С. spelaea из миндельских и вюрмских фаун Европы (см. табл.
5). По соотношению длины четырех нижних коренных зубов Р/2 М/\
(16.35-23.85-25.1-34.7%, п=2) гиена из Приморского края сходна с С. spelaea
из Красного Яра и из Крыма (Барышников, 1995), за исключением относи-
тельно более короткого у нее Р/2. По длине Р/2-Л//1 к уссурийской гиене
близка крокута из позднеплейстоценовых местонахождений Китая - Верх-
няя пещера Чжоукоудяня и Вурчжи (Pei, 1940; Lu et al., 1986).
Резцы и клыки обычной для С. spelaea величины (см. табл.З). Среди
верхних коренных Р1/ небольшой, но остальные зубы, особенно Р/3 и Р4/,
очень длинные (см. табл. 6). Если по длине и ширине Р2/ изученная выборка
не выходит за пределы европейских популяций С. 5. spelaea, то соответству-
ющие параметры РЗ/ у гиены из Приморья заметно превышают средние по
казатели, известные для Европы, за исключением С. 5. petralona, у которой
РЗ/ еще более широкий (Kurtdn, Poulianos, 1977). Большинство РЗ/уссурий-
23
Таблица 7. Размеры нижних коренных зубов пещерной гиены
(Crocuta spelaea)
Table 7. Measurements (mm) of lower cheek teeth in Crocuta spelaea
Промеры, C. spelaea praespelaea p. Казанка, Поволжье ГММ 191 C spelaea spelaea
Красный Яр, Западная Сибирь ЗИН 32752 пещера Носорога, Южный Урал ЗИН 34477
мм
Р/2 L 17.4 16.9 16.8
Lp 12.1 12.5
W 12.5 13.4 13.0
Р/3 L 22.8 23.1 22.8
IV 16.8 18.0 17.1
Р/4 L 24.8 24.4 24.8
Lpr 13.6 13.1 11.1
IV 15.1 15.6 15.8
Л//1 L 33.7 33.8 29.6
Ltr 31.2 27.3
Lpr 11.9 12.6 11.8
IV 14.0 13.6 12.6
ской крокуты имеет сильно истертую коронку, что делает невозможным
определить степень гипсодонтии зуба (отношение высоты коронки к ее
длине). Единственный нестертый экземпляр РЗ/ очень высокий (высота
коронки 30 мм), но по индексу гипсодонтии (106%) он вполне соответствует
С. s. spelaea и заметно уступает среднеплейстоценовому подвиду С. spelaea
ultima из Китая (Kurtdn, 1956).
По среднему значению длины верхнего хищнического зуба Р4/ пещер-
ная гиена из Приморья превосходит почти все изученные выборки С. spelaea
как позднего, так и среднего палеолита (см. табл. 6), однако метастиль Р4/
сравнительно короткий, и индекс его длины (в среднем 44.27%) вполне типи-
чен, например, для большинства выборок из позднего плейстоцена Франции
(Clot, 1980).
Нижнекоренные зубы также выделяются крупными размерами (табл. 8).
Особенно велико среднее значение длины ЛУ/1, которое превышает аналогич-
ные параметры у номинативного подвида и у С. spelaea ultima и сходно со
средним значением длины М/1 у С. spelaea praespelaea из Ополья (Татари-
нов, 1958). В тоже время относительная длина тригонида М/\ уссурийской
гиены (в среднем 91.86%) такая же, как у С. spelaea spelaea из грота Жоранс
во Франции (Ballesio, 1979). Короткий талонид ЛУ/1 с одним режущим бугор-
24
ком- метаконид встречен на 2 экз. из 7 (29% случаев). По длине и ширине
нижних коренных зубов от уссурийской гиены не отличается крокута из
позднего плейстоцена северо-восточного Китая (Верхняя пещера Чжоу-
коудяня, Вурчжи, Ксиалатуо) (Pei, 1940; Zong, Huang, 1985; Lu et al., 1986).
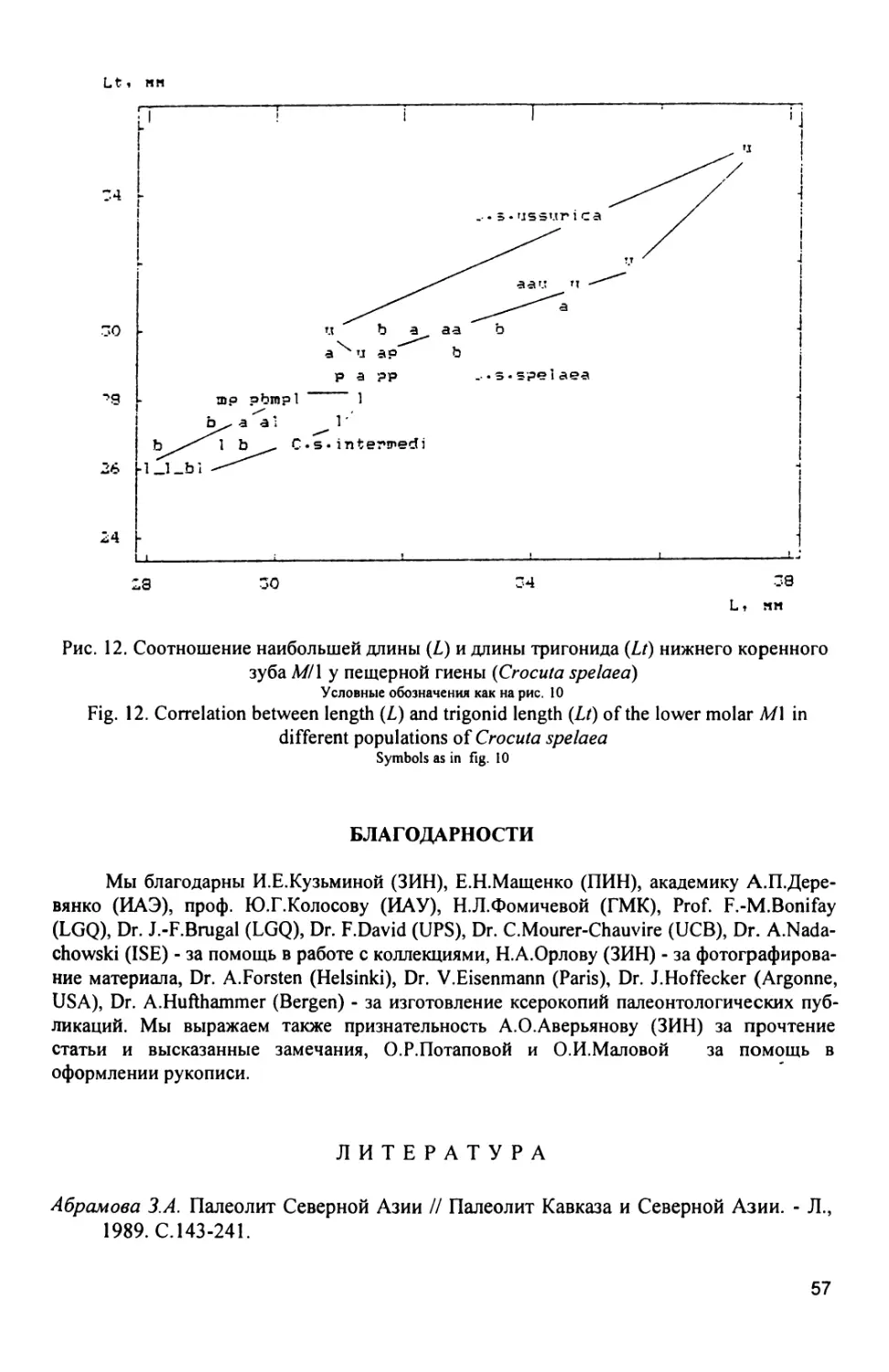
Морфометрические особенности пещерной гиены из позднеплейсто-
ценовой фауны пещеры Географического общества позволяют выделить ее в
отдельный подвид С. spelaea ussurica subsp. nov. (см. систематическую
часть).
распространение. В России находки ископаемых костей
гиены стали известны еще в первой половине XIX века благодаря раскопкам
на Алтае пещер по рекам Чарыш и Ханхара. Академик Брандт (Brandt, 1871),
просмотревший впоследствии собранную коллекцию, подтвердил присут-
ствие пещерной крокуты, которую он называет спутницей мамонта и
шерстистого носорога. В дальнейшем остатки С. spelaea определяли из
Украины, Крыма, Кавказа, Средней Азии, Урала, других частей южной
Сибири. Большинство ранних материалов не имеет, к сожалению, страти-
графической привязки, и поэтому история ареала С. spelaea на территории
бывшего СССР может быть прослежена только в самых общих чертах.
Наиболее древняя регистрация в Европе гиен из рода Crocuta происхо-
дит из эпивиллафранка Италии (Домеглиара близ Вероны; Azzaroli, 1983).
Палеарктическую крокуту можно рассматривать как потомка С. sivalensis
(Falconer et Cautley, 1868), известного из виллафранка Индии (Пинджор) и
являющегося предковым видом не только по отношению к евроазиатскому
С. spelaea, но и к африканскому С. crocuta (Kurtdn, 1956; Барышников, 1995).
Средний плейстоцен. В течение кромера (в широком его
понимании) С. spelaea распространилась в Палеарктике и сукцессионно
вытеснила преобладавшие здесь ранее виды гиен {Pliocrocuta perrieri,
Pachycrocuta brevirostris). В Европе пещерная гиена определена из многих
местонахождений нижнего среднего плейстоцена (кромер и ранний миндель)
и представлена крупным подвидом С. spelaea praespelaea Schutt, 1971. Она
найдена в Гомбасцоге (Kretzoi, 1938), Зюссенборне (Kurtdn, 1969), Странска
Скала (Kurtdn, 1972), в основной фауне Мосбаха (Schutt, 1971), в Петралоне
(Petralona-Crenian и PetroIonian Layer) (Kurtdn, Poulianos, 1981: 38-42),
Шарцфельде (Schutt, 1971: 66), Фойгтштедте (Kurtdn, 1986), а также на ряде
местонахождений Британских островов (Westbury-Sub-Mendip, Upper Fresh-
water Bed) (Newton, 1883; Bishop, 1982).
Черепа крупной крокуты, морфометрически близкой к С. spelaea
praespelaea, происходят из Татарстана (р. Казанка) и из Казахстана (Желе-
зинское на р.Иртыш, Павлодарская область) (Распопов, 1925; Беляева, 1933;
Кожамкулова, 1969; Барышников, 1992) и, возможно, из Забайкалья (Заигра-
ево) (рис. 8). К гиенам этого же эволюционного уровня принадлежат находки
из Молдовы (Колкотова Балка) и Западной Украины (Синяково 1, Девичьи
Скалы, хутор Винява) (Татаринов, 1958; Верещагин, Давид, 1971; Давид и
др., 1990) и, скорее всего, обломок нижнечелюстной кости, извлеченный из
25
cn
Таблица 8. Размеры нижних коренных зубов позднеплейстоценовой пещерной гиены
(Crocuta spelaea)
Table 8. Measurements (mm) of lower cheek teeth in Crocuta spelaea from the Late Pleistocene
C. spelaea spelaea C. s. ussurica ssp. n.
Промеры, MM Бинагады, Закавказье ЗИН 22382 Аман-Кутан, Узбекистан ЗИН 29058 пещера Географического общества, Приморье кол. ЗИН
" 1 lim 1 * 1 ° " 1 lim | X « 1 lim | * 1 ст
PH L 7 15.0-18.5 16.44 1.36 3 15.2-17.6 16.73 10 15.5-18.3 17.12 0.98
Lp 7 11.2-15.0 12.33 1.32 1 11.7 10 11.4-13.6 12.72 0.80
W 7 10.4-13.0 11.63 1.06 3 10.9-12.0 11.60 10 10.1-14.0 12.77 1.13
P/3 L 7 19.5-23.3 21.30 1.48 3 20.0-21.8 21.00 13 21.6-26.0 23.95 1.30
W 7 13.8-16.7 15.29 1.20 3 15.7-16.4 16.03 13 15.2-18.8 17.11 1.17
p/4 L 5 21.4-25.9 23.68 1.71 3 21.2-22.8 22.23 13 22.1-26.4 24.62 1.39
Lpr 5 11.7-15.0 13.02 1.49 1 12.2 13 13.1-14.5 13.67 0.40
W 5 12.6-15.4 14.28 1.33 3 12.6-14.5 13.83 13 13.8-17.1 15.01 1.01
M!\ L 10 28.1-33.5 30.19 1.75 2 29.1,30.0 8 30.8-37.4 34.11 2.17
Ltr 10 25.9-29.6 27.61 1.27 2 27.4,27.8 7 28.9-34.7 31.37 1.94
Lpr 10 10.4-12.8 11.85 0.72 1 12.2 7 11.4-14.2 12.84 1.11
W 10 12.4-15.3 13.41 0.92 3 12.5-12.9 12.67 8 13.6-15.1 14.31 0.52
аллювия VI террасы р.Дунай возле Нагорного (Алексеева, 1977). К С. spelaea
praespelaea близка также крупная крокута из раннеашельской фауны
Треугольной пещеры (слой 7а, абсолютная дата 583 000 лет) на Северном
Кавказе (Барышников, 1993) и, по-видимому, из пещеры Азых (слой 6) в
Азербайджане (Гаджиев, Алиев, 1966; Сулейманов, 1982).
Другой среднеплейстоценовый подвид С. spelaea petralonae Kurtdn
1977, отличающийся от С. spelaea praespelaea большой величиной клыков и
очень широкими верхними премолярами, происходит из верхней части
отложений (возможно, миндельских) в пещере Петралона (Post Petralonian) в
Греции (Kurten, Poulianos, 1977). Очень крупные размеры этой пещерной
гиены позволяют думать о том, что она едва ли была исходной формой для
более мелких подвидов конца среднего и позднего плейстоцена. Поэтому
Куртен и Пулианос (Kurten, Poulianos, 1981) предполагали ’’локальное”
вымирание крокут в Европе в течение эльстерской (миндельской) эпохи и за-
тем новую миграцию их из азиатского региона. Отсутствие остатков С. spe-
laea в миндельских отложениях грота Араго, Тутавель на юге Франции
(Cregut, 1980) как будто бы подтверждает такую гипотезу (Kahlke, 1994).
Рис. 8. Места находок довюрмских остатков Crocuta spelaea'.
а - кромер и миндель, b - рисе, с - рисс-вюрм
Fig. 8. Prewiirmian localities with remains of Crocuta spelaea'.
a - Cromerian and Mindel, b - Riss, c - Riss-Wurm
27
Чрезвычайно многочисленные остатки мелкой С. spelaea intermedia
Bonifay, 1971 были найдены в отложениях теплой голштейнской (миндель-
рисской) межледниковой эпохи на юге Франции (пещера Лунель-Вьель;
Bonifay, 1971). Морфологически крокута из Лунель-Вьеля занимает проме-
жуточную позицию между С. spelaea praespelaea из раннего среднего плей-
стоцена и позднеплейстоценовым С. spelaea spelaea.
В границах бывшего СССР убедительных доказательств присутствия
пещерной крокуты в миндельских и миндель-рисских местонахождениях по-
ка нет. Указание на существование С. spelaea в ашельской фауне пещеры Ку-
даро 1 на Большом Кавказе (Верещагин, 1959) в дальнейшем не подтверди-
лось (Барышников, 1987). Возможно, что к данному геологическому отрезку
(сингильский териокомплекс) принадлежит обломок зуба С. spelaea из осыпи
разреза в пещере Сель-Унгур в Киргизии (Барышников, Батыров, 1994).
Для рисских фаун Западной Европы сведений о пещерной крокуте не-
много. Она определена, в частности, из нескольких пещерных костеносных
скоплений во Франции (Шатильон-Сен-Жан, Гаргас; Clot, 1980). К данному
возрастному срезу относятся, вероятно, остатки С. spelaea из среднеплейсто-
ценовых гротов Молдовы - Старые Дуруиторы (нижний слой 5) и Выхватин-
цы (слой 3) (Давид, 1978, 1980; Анисюткин, Кетрару, 1982), к сожалению
детально не изученные. Не исключено, однако, что Выхватинцы имеет более
молодой, рисс-вюрмский возраст (Черныш, 1965; Алексеева, 1990).
Возможно, что аналогичную датировку имеют фрагменты костей С.
spelaeus из нижней части среднеплейстоценовых серо-зеленых суглинков
Кузнецкой котловины в Сибири, где гиена найдена совместно с Mammuthus
chosaricus (Форонова, 1982).
Поздний плейстоцен. С. crocuta из местонахождений
последнего межледниковья достоверно известна для нескольких пещер Анг-
лии (Klein, Scott, 1989) и, возможно, для пещеры Линденталь в Германии
(Soergel, 1937). Вместе с тем пещерные отложения Линденталь могут быть и
более древними, рисскими (см. Ballesio, 1979).
На территории бывшего СССР последним межледниковьем датируется
фауна из асфальтовой ловушки Бинагады близ Баку в Азербайджане, заклю-
чающая обильные остатки С. spelaea (Богачев, 1939; Верещагин, 1951). Для
верхнехазарских отложений, подстилающих Бинагаинское местонахождение,
опубликованы абсолютные датировки (76 000, 81 000 по слою ракушечника с
р.Шура-Озень в Дагестане), которые позволяют хронологически сопоставить
верхнехазарскую трансгрессию Каспийского моря с периодом высокого
уровня океана во время рисс-вюрмского межледниковья и последующих ран-
невюрмских межстадиалов (Арсланов и др., 1978). Зубаков (1986) заключает
бинагадинский комплекс млекопитающих во временные рамки между перио-
дами формирования VIII раннехвалынской (53 000 лет) и IX хазарской
террасе (130 000 лет).
Вполне вероятно, что рисс-вюрмский возраст имеют остатки С. spelaea
из пещеры Ильинка в Одесской области (Пидопличко, 1951, 1956), в фауни-
28
стическом спектре которой найден мелкий дикобраз Hystrix vinogradovi
(Барышников, Баранова, 1983). Данный вид мелкого дикобраза характерен в
Средней Европе для последнего межледниковья (Mottl, 1967), но геологи-
ческая документация по его находкам из Ильинки не опубликована.
Рисс-вюрмский (микулинский) возраст вполне могут иметь остатки С.
spelaea из древнейших слоев в палеолитических пещерах Крыма (Киик-Коба,
Заскальная 5 и, возможно, Пролом 2) и Северного Кавказа (Ильская 1)
(Hoffecker et al., 1990).
В центральных областях Русской равнины в это время пещерная
крокута, очевидно, не встречалась или была редка, как, впрочем, и в конце
среднего плейстоцена (одинцовское межледниковье и московское оледе-
нение). Во всяком случае, Алексеева (1990) в своем обзоре основных
местонахождений хазарской и шкурлатовской териофауны Поволжья и
Подонья С. spelaea не упоминает. Впрочем, к данному временному отрезку,
возможно, приурочено скопление костей С. spelaea и других млекопитаю-
щих, обнаруженное в суглинке овражного делювия на глубине 8-12 м около
с.Шубное в Воронежской области. В выносах оврага были встречены также
грубо оббитые рубящие орудия (Громов, 1935).
Рисс-вюрмским возрастом датирует Васильев (1995) комплекс крупных
млекопитающих, включающий С. spelaea, найденный им в разрезе Красный
Яр на правом берегу р.Обь в 17 км ниже Новосибирска.
Наиболее широко на территории Евразии представлены остатки С.
spelaea из отложений первой половины вюрмской (валдайской) эпохи (рис.
9). К ней мы относим все находки гиен из археологических памятников, слои
которых содержат мустьерские каменные изделия. В Европе в это время С.
Spelaea была распространена от Испании и Пиренеев до Британских остро-
островов и области Северного моря на западе и до Италии, включая Сици-
лию, и Греции (Пелопоннес) на юге (Kahlke, 1994).
Вюрмские крокуты относятся к номинативному подвиду С. spelaea
spelaea (Goldfuss), который был описан по двум целым черепам и
фрагментам нижних челюстей из пещеры Гайленройтер во Французской
Юре в Германии (Goldfuss, 1823, Tab. XV, XVI: 1-3, XVII:3). К номина-
тивному подвиду мы относим также упомянутые выше остатки гиен из
отложений рисской и рисс-вюрмской эпох, которые имели несколько
меньшие в среднем размеры. По мере накопления нового, более надежно
стратифицированного материала станет понятным, заслуживают ли эти,
более древние формы пещерных гиен обособления на подвидовом уровне.
В России и других странах бывшего СССР остатки С. 5. spelaea опреде-
лены из целого ряда местонахождений Молдовы (Старые Дуруиторы (слои
3-4), Бутешты, Буздужаны 1, Тринка 3, Сахарна) и континентальной Украи-
ны (Старый Кодак в Днепропетровской области; 1 кость), но особенно мно-
гочисленны они в Крыму (Киик-Коба (слои 3-6), Пролом 2 (слои 2-4), За-
скальная 5, Волчий грот, Кош-Коба, Аджи-Коба (слой 3), Чокурча 1, Шай-
29
Рис. 9. Места находок вюрмских остатков Crocuta spelaea-.
а -С. s. spelaea, b -С. s. ussurica
Fig. 9. Wiirmian localities with remains of Crocuta spelaea-.
a - C. s. spelaea, b - C. s. ussurica
тан-Коба, Староселье, Аювул-Коба) (Громов, 1948; Пидопличко, 1951; Да
вид, 1980; Барышников, 1995).
На Северном Кавказе С. spelaea известен как из мустьерских стоянок
открытого типа - Ильская 1, Ильская 2, так и из пещерных - Даховская, Бара-
каевская (Верещагин, 1959; Hoffecker et al., 1990; Барышников, 1991; Barysh-
nikov Hoffecker, 1994). Две плечевые кости С. spelaea найдены в отложениях
карстового колодца на Лагонакском плато в верховьях р.Белая (Алексеева,
Ломизе, 1960). В Закавказье остатки пещерной гиены определены из мусть-
ерских отложений Верхней пещеры (1 кость) близ Кутаиси в Грузии и из
Тагларской пещеры (слой 2) на р.Куручай в Азербайджане (Векуа, 1978;
Джафаров, 1983).
Для Средней Азии существование С. spelaea отмечено в фаунистичес-
ких комплексах мустьерских стоянок в гроте Тешик-Таш (1 клык) в долине
р.Турган-Дарья и в пещере Аман-Кутан, расположенной в западных отрогах
Зеравшанского хребта в Узбекистане (Верещагин, Батыров, 1967). Остатки
гиены из пещеры Аман-Кутан довольно обильны (более 55 экз.), но они не
имеют привязки к какому либо стратиграфическому горизонту. С позиций
геоморфологии мустье обеих стоянок не древнее середины позднеилякского
этапа развития рельефа (Ранов, Несмеянов, 1973), что соответствует второй
половине раннего вюрма.
В Казахстане отдельные кости и зубы пещерной гиены найдены на
стоянке Актас в Кокчетавской области и в карьере у г.Зыряновска, Восточно-
Казахстанская область (Кожамкулова, 1969; Кожамкулова, Пак, 1988).
В позднем плейстоцене Северного и Среднего Урала пещерная крокута,
по всей видимости, отсутствовала (Дуброво, 1966; Кузьмина, 1975, 1982), но
она встречалась на Южном Урале. Здесь остатки С. spelaea определены из
местонахождений на р.Ай, из пещеры Смеловская 2 на р.Малый Кизыл (кол.
ЗИН), Усть-Катавской пещеры на р.Сим, Ключевой пещеры на р.Юрюзань (1
зуб) и из пещеры Носорога (Громов, 1948). Кости гиены из Усть-Катавской
пещеры принадлежат нескольким молодым и взрослым особям. Большин-
ство находок не имеет, к сожалению, надежной стратиграфической привязки,
но Громов (1948) полагал, что по облику фауны Усть-Катавская пещера вы-
глядит древнее стоянки Талицкого, которая, скорее всего, приурочена к сред-
невалдайскому мегаинтерстадиалу (Рогачев, Аникович, 1984). В пещере
Смеловская 2 остатки гиены (45 штук) происходят из слоя 5 (опесчаненная
глина), подстилающего суглинистые отложения, в которых найдены поздне-
палеолитические изделия (Бадер, 1971). Из верхних горизонтов пещеры
Л.Д.Сулержицким (ГИН РАН, Москва) были получены по костям мамонта и
лошади три радиоуглеродные даты - 25 000±600 (ГИН-8403), 31 000±1 500
(ГИН-8401)и41 000±1 800 (ГИН-8402).
Для Западной Сибири С. spelaea приведен в фаунистических списках
материала, собранного на бечевнике р.Обь у с.Красный Яр Томской области
(Алексеева, 1980). Фрагмент нижней челюсти и кости конечностей из Крас-
ного Яра не имеют точной геологической привязки и датированы позднем
31
плейстоценом предположительно, на основе морфологии зубов. Одн^
метакарпальная кость найдена в верхнечетвертичных глинах второй террась
р. Томь у с.Салтымаково (Алексеева, 1980).
К первой половине вюрма (зырянское оледенение и каргинский интер-
стадиал) могут быть отнесены остатки гиены из мустьерских пещер Алтая
Усть-Канская, Страшная, Денисова и Окладникова (Руденко, 1960; Галкина.
Оводов, 1975; Оводов, 1980). Цейтлин (1974, 1979), изучавший геологию
Усть-Канской пещерной стоянки, находящейся в верховьях р.Чарыщ,
разделил плейстоценовые отложения пещеры на две пачки. К нижней пачке
он отнес залегание мустьерских материалов, и связал ее формирование с тегь
лыми условиями каргинского времени. Фаунистический материал из Усть-
Канской пещеры, к сожалению, не расчленен по слоям.
В пещере Страшная, расположенной в среднем течении р.Иня в бас-
сейне р.Чарыш, пещерные слои нарушены. Кости С. spelaea встречаются,
начиная с глубины 180 см, в совместном залегании с палеолитическими на-
ходками, которым присуща леваллуазская техника расщепления камня (Ок-
ладников и др., 1973). Древние отложения с глубины 4-6 м имеют возраст
значительно древнее 45 000 лет (Деревянко, Зенин, 1992), и Цейтлин (1979)
относит их к каргинскому межледниковому комплексу. В расположенной по
соседству со Страшной пещере Логово Гиены культурных остатков не найде-
но, однако для этого костеносного местонахождения имеется радиоуглерод-
ная дата 32 700 лет, выполненная по костям бизона, собранных с глубины 40-
60 см (Оводов, 1980).
Несколько абсолютных датировок опубликовано для палеолитических
слоев Денисовой пещеры на р. Ануй (Деревянко и др., 1993). Наиболее древ-
няя из них (282 000 для низов слоя 22) отвечает чаганской межледниковой
свите среднего плейстоцена. Кости С. spelaea извлечены в Денисовой пеще-
ре преимущественно из мустьерских слоев 12-20 ее внутренней камеры. Воз-
раст костей определяется первой половиной последнего оледенения, что под-
твердилось позднее полученной термолюминисцентной датой из слоя 14 в 69
000 лет (устное сообщение М.В.Шунькова, Новосибирск).
В пещере Окладникова (Сибирячиха) кости пещерной гиены собраны в
отложениях мустьерского времени (слои 1-6), абсолютный возраст которых
колеблется от 33 300 до 42 500 лет (Деревянко, Маркин, 1992).
Остатки С. spelaea приводятся также для нескольких пещер Кузнецкого
Алатау - грот Проскурякова (дата более 46 000 лет), Тохзасский грот, пещера
Фанатиков, Николаевская пещера, а также для мустьерского грота Двуглазка
(слои 5-7) в бассейне р.Толчея в Хакассии (Агеева и др., 1978; Оводов, 1980).
В гроте Двуглазка гиена (Crocuta sp.) найдена в фаунистическом комплексе,
свидетельствующем об относительно теплом и сухом климате с преоблада-
ием степных ассоциаций (Абрамова, 1989). С. spelaea упоминается также в
списках фауны из верхнечетвертичных отложений Кузнецкой котловины
(Форонова, 1982).
32
Для более восточных районов юга Средней Сибири сведения о
гаходках С. spelaea крайне скудны. Вид отсутствует в Нижнеудинской и
1ругих известных пещерах Саян и на палеолитических стоянках в долинах
)ек Енисей и Ангара (Ермолова, 1978). Мозговая часть черепа гиены найдена
/ с Торгашинское близ Красноярска (Рябинин, 1932), а несколько костей
лещерной гиены были собраны на глинистом полу Унылкской пещеры в
10Лине р.Уда в бассейне Ангары (Оводов, 1980). В позднем плейстоцене
Забайкалья и в Якутии остатки С. spelaea пока не найдены (Мочанов, 1977;
Лазарев, Томская, 1987; Иметхенов, Калмыков, 1988; Абрамова, 1989).
Скудость данных о пещерной крокуте на юге Сибири может объясняться как
суровостью плейстоценового климата, так и молодым возрастом палеолити-
ческих местонахождений, содержащих фаунистические материалы (в
основном не древнее средней поры верхнего палеолита).
Во второй половине вюрма (валдая), в эпоху верхнего палеолита
численность С. spelaea в фаунистических комплексах резко уменьшается.
Эта тенденция прослеживается по всему ареалу вида. Кости и зубы С.
spelaea, например, вполне обильны в мустьерских отложениях много-
слойных пещерных памятников Арси-сюр-Кюр (Arcy-sur-Cure) во Франции,
но в слоях верхнего палеолита они становятся здесь очень редки (Dr.F.David,
Paris, устное сообщение). Остаточные популяции С. spelaea в Западной
Европе все же прослеживаются до конца плейстоцена. Одни из наиболее
поздних находок связаны, в частности, со стоянками позднего мадлена на
юго-западе Франции (Delpech, 1983).
В Восточной Европе остатки С. spelaea, датируемые поздним валдаем,
отмечены для Молдовы и Крыма. В Молдове они указаны для стоянок
Брынзены 1 (нижний слой 3), Старые Дуруиторы (верхний слой 2), Тринка
(верхний слой), но во всех памятниках малочисленны (Давид, 1980). Стоянки
в Молдове, скорее всего, относятся к ранней поре верхнего палеолита
(Рогачев, Аникович, 1984).
На Крымских стоянках позднего палеолита единичные кости и зубы С.
spelaea встречены в Аджи-Кобе 1 (3 кости в слое 2) и в Сюрень 1 (Громова,
Громов, 1937). В гроте Сюрень 1 они присутствуют только в самом нижнем
слое 4 (4 кости) (Громов, 1948; Векилова, 1957), который явно древнее 20
ООО лет, поскольку залегает значительно ниже слоя 2 с радиоуглеродной
датой 17 100±700 (ГИН-8081).
На памятниках верхнего палеолита центра Русской равнины (Костенки
1-12, Юдиново, Елисеевичи, Сунгирь, Авдеево; серия радиоуглеродных
датировок от 32 700 до 12 300 лет) остатки С. spelaea не найдены (Вереща-
гин, Кузьмина, 1977; Алексеева, 1990). Нет их и на стоянках бассейнов рек
Днестр (Молодово 5, Кормань 4; даты от 25 140 до 16 750) и Днепр
(Пушкари, Новгород-Северский, Чулатов, Мезин, Гонцы, Журавка, Меже-
рич) (Корниец, 1962; Пидопличко, 1976; Татаринов, 1977; Алексеева, 1987).
Старые определения гиены в фаунистических комплексах позднепалео-
Заказ 255(a)
33
литических стоянок Кирилловская в г.Киеве и Боршево 2 на р.Дон вызывают
обоснованные сомнения (см. Громов, 1948) и требуют подтверждения.
В фауне позднего палеолита Кавказа пещерная крокута, скорее всего,
уже не встречалась. Имеется, правда, определение С. spelaea из верхнепалео-
литической стоянки Окуми 1 в Абхазии (Бурчак-Абрамович, 1981). Здесь
отмечены 8 костей от 1 особи, но точная стратиграфическая приуроченность
их не указана. Скорее всего, кости гиены происходят из нижних слоев, в
индустрии которых присутствуют следы мустьерской техники обработки
камня (Любин, 1989), и в этом случае данная находка вполне может иметь
мустьерский возраст. Также неясна достоверность присутствия пещерной
гиены на стоянке Сагварждиле в Западной Грузии (Векуа, 1978).
Далее к востоку кости С. spelaea из верхневюрмских отложений
собраны на Южном Урале - Первая и Вторая Серпиевская пещера и местона-
хождение Аша 1 на берегу р.Сим (по 1 кости), навес Устиново в верхнем
течении р.Миасс (2 кости в слое 2) (Смирнов и др., 1990). Поздний возраст
для этих сборов недостаточно доказан. Лучше других датированы материалы
из Игнатиевской пещеры на р.Сим, где костные фрагменты С. spelaea
происходят с поверхности пола (1 кость), а также из слоя 2а в раскопе II (2
кости) и слоев 2-8 в раскопе V (15 костей) (Смирнов и др., 1990). Ряд радио-
углеродных дат, полученных по обломкам крупных костей для слоя 2а (14
038) и слоев 3-8 (более 27 500 лет), как будто бы свидетельствует о том, что
пещерная гиена на Южном Урале могла сохраняться, по крайней мере, до
максимальной стадии поздневалдайского оледенения.
На Алтае несколько обломков костей С. spelaea встречено в Денисовой
пещере совместно с костяными и каменными изделиями верхнего палеолита
(слои 9-11). Более обширная коллекция (18 остатков, преимущественно от
молодых и полувзрослых животных) происходит из палеолитической
пещеры в устье р.Бухтарма в Восточно-Казахстанской области (Верещагин,
Мельникова, 1958). По характеру археологического инвентаря стоянка в
Бухтарминской пещере отнесена к поздней поре палеолита (Гохман, 1957).
На позднепалеолитических стоянках Средней и Восточной Сибири С.
spelaea отсутствует, однако многочисленные остатки ее (около 170 облом-
ков) собраны в пещере Географического общества на юге Приморского края
(Верещагин, Оводов, 1968; Оводов, 1977). Пещера Географического об-
щества расположена на правом берегу р.Партизанской (Сучан). Раскапывал
ее Н.Д.Оводов под руководством Н.К.Верещагина; часть костного материала
была собрана местным краеведом Е.Г.Лешком. Мощность вскрытых рыхлых
отложений достигает 5 м. Выделены 6 литологических слоев, при этом
основная масса каменных отщепов и галечных нуклеусов верхнепалео-
литического облика найдена в слое 4 (Окладников и др., 1968). Остатки
пещерной гиены, хранящихся в коллекции ЗИН, не имеют четкой
стратиграфической привязки; значительная часть их происходит из слоя 4, на
большинстве других отмечена лишь глубина их залегания (80-140 см).
Опубликованный фаунистический список (см. Оводов, 1977) также приведен
34
суммарно для плейстоценовых слоев 4-5, что не позволяет проследить
количественное распределение костей С. spelaea в разрезе пещерных отло-
жений. Цейтлин (1979) относит культурный горизонт стоянки к началу сар-
танской эпохи, но, скорее всего, с учетом присутствия на глубине 1.3-1.7 м
прослоя с пыльцой хвойных и широколиственных пород, костеносный слой
пещеры имеет более древний, каргинский возраст (Алексеев, 1978). Это под-
тверждает и полученная по фрагментам костей лошади и мамонта с глубины
60-80 м радиоуглеродная дата 32 570 лет (Герасимов и др., 1983).
Крокута из плейстоцена Китая, которую обычно определяют как С.
crocuta ultima (Kurt£n, 1956) или как С. ultima (Huand, 1989), представляет
собой, несомненно, не более чем подвид пещерной гиены - С. spelaea ultima.
Эта форма была обозначена по очень незначительному материалу (один Р4/)
из мергелей Сычуаня в сочетании с фауной, которая коррелирует с фауной
синантропа из Чжоукоудяня (Kurten, 1956). С. spelaea ultima указывают для
среднего и позднего плейстоцена северного и северо-восточного Китая и для
северной Кореи (Kurten, 1956; Sun, 1988; Zhou et al., 1990). Имеются очень
поздние находки его во Внутренней Монголии - Вурджи (Wurji) с абсолют-
ной датировкой 19 800 (Lu et al., 1986), и в Сычуане - Ксиалатуо (Xialatuo) с
датой 11 550 (Zong, Huang, 1985). Судя по крупным размерам нижних корен-
ных зубов из верхней пещеры Чжоукоудяня (Pei, 1940) и Вурджи (Lu et al.,
1986), позднеплейстоценовая гиена северо-восточного Китая принадлежала,
скорее всего, к тому же подвиду, что и гиена с юга Приморского края, т.е. к
С. spelaea ussurica ssp. nov. Уссурийскую крокуту можно считать потомком
С. s. ultima, стратиграфически сменившим его в позднем плейстоцене.
Возможно, что в Китае пещерная гиена дожила вплоть до неолита (Kahlke,
1994).
Заключая обзор находок С. spelaea, следует сказать, что ареал вида
нигде в Евразии не пересекал параллель 58° с.ш. (рис. 8, 9). Наиболее север-
ные из известных находок в России показывают, что северная граница ареала
располагалась на одной широте как в среднем (р.Казанка в Татарстане), так и
в позднем плейстоцене (Красный Яр на р.Обь). По мере продвижения на
восток она все более и более сдвигалась к югу, демонстрируя тем самым
зависимость распространения С. spelaea от климатических условий. Будучи
относительно теплолюбивой, пещерная гиена не смогла освоить суровые
просторы плейстоценовой Сибири, за исключением наиболее южных ее
частей.
Климатическую обусловленность расселения С. spelaea наглядно
Демонстрирует пульсация северной границы видового ареала, которая опре-
делялась чередованием теплых и холодных эпох в течение плейстоцена. Пер-
в°е, в геологическом смысле чрезвычайно быстрое распространение С. spe-
laea (вплоть до Англии, Татарстана и юга Западной Сибири), связано с бла-
гоприятными условиями кромерского межледниковья. Затем область распро-
странения вида (особенно в ее восточной части), очевидно, сократилась, хотя
отдельные продвижения к северу могли иметь место в теплые фазы второй
35
половины среднего плейстоцена и в последнее межледниковье. Волна широ-
кой экспансии С. spelaea зафиксирована для теплой эпохи внутривалдай-
ского (каргинского) интерстадиала, когда гиены обитали на юге Русской
равнины, на Южном Урале, в Горном Алтае и в Приморском крае. Суровые
условия поздней стадии валдаского (сартанского) оледения привели к рез-
кому сокращению численности вида и дроблению ареала, в результате чего
только некоторые популяции С. spelaea смогли сохраниться в горных усло-
виях Урала и Алтая вплоть до стадии главного климатического минимума,
наступившей 18 000 лет тому назад.
В более южных частях ареала пещерная гиена непрерывно обитала на
обширной территории от Средиземноморья до Восточного Китая, хотя обра-
щает на себя внимание почти полное исчезновение С. spelaea к началу верх-
него палеолита из фауны Молдовы, Крыма, Кавказа и Средней Азии, т.е.
задолго до наступления наиболее холодной фазы последнего оледенения.
Образ жизни. Недавно один из нас (Барышников, 1995) рекон-
струировал образ жизни С. spelaea как активного хищника, жившего боль-
шими группами и самостоятельно добывавшего различной величины копыт-
ных зверей. Реконструкция эта основывалась на большей хищнической спе-
циализации зубного аппарата у С. spelaea по сравнению с современным
африканским видом С. crocuta. Именно питание свежим мясом (и, в
меньшей, мере падалью) позволило пещерной гиене освоить северные
регионы и войти в состав мамонтового биома, не обладая специальными
приспособлениями для выживания в климате с продолжительным морозным
периодом.
Дополнительные доказательства в поддержку такой точки зрения дают
сравнительная этология и тафономия. Еще со времен Геккеля для выяснения
истории органического мира используют сравнительно-анатомические дан-
ные, группируя их в эволюционные ряды форм, которые можно рассмат-
ривать как конечный результат конкретных филогенезов. Построение таких
морфологических рядов представляет собой частный случай более общего
сравнительно-биологического подхода, в пределах которого можно получать
и другие сравнительные ряды - экологические, этологические и т.д.
Применительно к современным и позднечетвертичным видам гиен мы
имеем следующее распределение их (морфологический ряд) согласно
степени преобразования зубной системы и координационно связанных с нею
частей черепа в сторону повышения эффективности переработки мясной
пищи (усиления хищнической специализации): Hyaena hyaena - Parahyaena
brunnea - Crocuta crocuta - C. spelaea. Аналогичным образом может быть
построен и этологический ряд, отражающий социальное поведение гиен (см.
Gittleman, 1989) с точки зрения филогенетической перспективы: Н. hyaena
(одиночное поведение) - Р brunnea (семейные пары) - С. crocuta (формиро-
вание групп). Логичным продолжением данного этологического ряда являет-
ся допущение того, что плейстоценовый С. spelaea жил и охотился больши-
36
ми группами, кланами, имевшими сложную иерархическую социальную
структуру.
Подтверждение этому дают те тафономические различия, которые
можно наблюдать между костными скоплениями, образованными гиенами
Африки и плейстоценовой гиеной Европы (Brugal et al., 1993). В европейских
местонахождениях всегда высок процент остатков самих гиен, которые
иногда принимают форму самостоятельных крупных скоплений, что не
наблюдается ни в современных, но в ископаемых скоплениях Африки. При
этом в выборках костей С. spelaea доминируют старые и молодые
животные, включая новорожденных, которые накапливались в результате
устройства" в пещерах беременными самками родильных логовищ. В
современных логовищах Африки, напротив, остатки молодых гиен весьма
немногочисленны.
Подобная тафономическая картина может быть объяснена разным
поведением С. spelaea и С. crocuta. В Европе она является результатом
жизнедеятельности больших кланов пещерных гиен, которые в скальных
укрытиях устраивали временные убежища и коллективные "родильные
дома" Крупные скопления гиеновых костей известны из многих пещер
Англии (Киркдаль, Торньютон, Кентские пещеры), Германии (Линденталь),
Австрии (Тойфельслукен) и Чехии (Шведов Стул) (Soergel, 1937; Ehrenberg,
1938-1940; Musil, 1962; Sutcliffe, 1969), и отчасти из пещер Крыма (грот
Пролом 2), Алтая (Логово Гиены), Приморья (пещера Географического
общества) и Китая (Верхняя пещера Чжоукоудяня) (Pei, 1940; Оводов, 1980;
Барышников, 1995). Увеличение численности кланов и усложнение
социальной внутриклановой структуры у С. spelaea могло способствовать
как успеху охоты на стадных копытных, так и противодействию конкуренции
в поисках пищи и удобных убежищ со стороны пещерных львов, медведей,
волков и первобытных людей, особо обострявшейся в зимние месяцы.
ГЕОХРОНОЛОГИЧЕСКАЯ И ГЕОГРАФИЧЕСКАЯ ИЗМЕНЧИВОСТЬ
CROCUTA SPELAEA
Пещерная гиена представляет собой классический пример увеличения
или уменьшения величины тела в зависимости от температурных флуктуаций
на разных этапах плейстоцена. Их реконструкция основана на изменениях в
размерах черепа и коренных зубов, которые достоверно различаются у
животных ледниковых и межледниковых эпох (Kurt£n, Poulianos, 1977; Klein,
Scott, 1989). Опубликованные данные относятся преимущественно к Европе,
но мы имеем возможность проследить изменчивость пещерной гиены по
всему ее ареалу, что позволило уточнить внутривидовую структуру С.
spelaea.
Кондилобазальная длина черепа. Европейские
популяции С. spelaea показывают (см. Kurt£n, Poulianos, 1977) возрастание
37
кондилобазальной длины черепа от большой в кромере и раннем минделе (С.
spelaea praespelaea) до очень большой в минделе (С. spelaea petralona)b
затем следует резкое уменьшение ее значений до минимальных в миндель-
рисское межледниковье (С. spelaea intermedia), новое незначительное увели-
чение в риссе, очередное уменьшение в рисс-вюрмское межледниковье и,
наконец, возрастание промеров длины вплоть до максимальных ее показа-
телей в вюрмское оледенение (С. spelaea spelaea). Единичные данные по
Кавказу, Казахстану и Китаю вполне укладываются в эту схему (табл. 9).
Длина зубного ряда. Изменения значений длины верхнего
и нижнего зубных рядов С. spelaea на разных этапах плейстоценовой исто-
рии вида происходило, как и следовало ожидать, аналогично изменениям
величины черепа (табл. 10, И). При этом размеры образцов из Сибири не
уступают наиболее крупным европейским экземплярам.
Несколько иная картина наблюдается в том случае, если рассматривать
распределение во времени и пространстве суммарной величины средних
значений длины трех верхних коренных. Она приблизительно равна у С. s.
praespelaea (86.4мм, и=7-9), С. s. petlalonae (88.2, и=1) и С. s. ultima (84.5,
и=4-5) и заметно меньше у С. s. intermedia (74.9, л=20-32). Ее значения
увеличиваются в рисской выборке из Гаргаса (82.4, л=6-8) и вновь умень-
шаются в материале из Бинагадов (78.6, л=1-8). Близкое значение к послед-
ней имеет подвид С. s. dorotheae (79.0, и=7-9). В вюрмских выборках Европы
данная величина меняется незначительно: Жерд (83.9, /7=9-13), Рипа (85.6,
и=7), Крымские местонахдения (81.6, /7=10-25). Наибольший показатель
среди изученных коллекций имеет выборка из пещеры Географического
общества (90.2, и=4-7). Учитывая общие размеры уссурийской крокуты
можно заключить, что она была относительно наиболее крупнозубой.
Вариабельность РЗ/. Этот зуб, взаимодействуя при окклюзии
Р/3, играет важную роль в дроблении гиенами крупных костей. В европей-
ских популяциях С. spelaea основная тенденция изменения размеров и про-
порций коронки РЗ/ сводится к тому, что у гиен межледниковий она была
более короткой и широкой, чем у гиен ледниковых эпох (Kurt6n, Poulianos,
1977). Относительное увеличение третьего премоляра (по сравнению с вели-
чиною хищнических зубов) наблюдается у С. s. dorotheae из Леванта (Kurten,
1965). Что касается С. s. ultima из среднего плейстоцена Китая, то для этого
подвида была характерна очень высокая коронка РЗ/ (Kurt£n, 1956). Подоб-
ная степень гипсодонтии не отмечена в других популяциях С. spelaea, в том
числе и в коллекции из пещеры Географического общества.
Соотношение промеров длины и ширины коронки в изученных нами
выбоках приведены на рис. 10. Обращает на себя внимание относительно ши-
рокий РЗ/ из Лунель-Вьеля, а также вытянутость коронки в пещере Геогра-
фического общества при общих крупных ее размерах. Первый показатель
может рассматриваться как усиление РЗ/ при возростании дробящей функ-
ции зубного аппарата, второй - как его ослабление. Подобное варьирование
зубных адаптаций могло происходить независимо в разных локальных попу-
38
Таблица 9. Географическая изменчивость кондилобазальной длины черепа (мм)
у пещерной гиены {Crocuta spelaea)
Table 9. Geographical variability of condylobasal length (mm) in Crocuta spelaea
“ Местонахождения | n | lim | х
Средний плейстоцен Кромер и миндель С. spelaea praespelaea Германия Мосбах (Schutt, 1971) 1 267
Поволжье р.Казанка (Барышников, 1992) 1 268.5
Казахстан Железинское (кол. ПИН) 1 257
С. spelaea petralonae Греция Петралона (Kurten, Poulianos, 1977) 2 274.0-285.0 279.5
С. spelaea ultima Китай (Colbert, Hooiler, 1953) 1 260
Миндель-рисс C. spelaea intermedia Франция Лунель-Вьель (Bonifay, 1971) 8 240-251 242.8
Рисе С. spelaea spelaea Франция Шатийон-Сен-Жан (Kurt6n, Poulianos, 1977) 1 260
Гаргас (Gaudry, Boule, 1892) 1 245
Поздний плейстоцен Рисс-вюрм С. spelaea spelaea Англия, главным образом Баррингтон (Kurten, Poulianos, 1977) 3 236.0-262.0 248.7
Закавказье Бинагады (кол. ЕИМ и ЗИН) 3 243.0-258.0 250.7
39
Продолжение табл. 9
Местонахождения I и I lim | х ~~
В ю р м
Франция
Жоранс (Ballesio, 1979) 2 262.0-272.0 267.0
Комб Капелль (Gaudry, Boule, 1892) 1 294
Монако
Гримальди (Boule, 1919) 1 285
Европа (K.urt£n, Poulianos, 1977) 12 240.0-288.0 264.2
Таблица 10. Географическая изменчивость длины верхнего зубного ряда
Р1/-Р4/ (мм) у пещерной гиены (Crocuta spelaea)
Table 10. Geographical variability of length of Р1/-Р4/ (mm) in Crocuta spelaea
Местонахождения | и | lim 1*1 Q
Средний плейстоцен
Кромер и миндель
С. spelaea praespelaea
Германия
Мосбах (Schutt, 1971) Поволжье р.Казанка (Барышников, 1$ 4 >92) 1 87.0-96.0 92.5 89.3 3.87
С. spelaea intermedia Франция Лунель-Вьель (кол.ЬСР) Миндель-рисс 7 81.1-85.6 83.39 1.79
С. spelaea spelaea Франция Taprac(Clot, 1980) Рисе 3 86.4-89.6 87.7
С. spelaea spelaea Закавказье Бинагады (кол. ЗИН) Поздний плейстоцен Рисс-вюрм 3 79.7-84.5 82.07
40
Продолжение табл. 10
Местонахождения
| л | lim 1*1 G
В ю р м
С. spelaea spelaea
Франция Жерд (Clot, 1980) Буадер (Clot, 1980) 1 4 85.8-91.0 91.4 87.45 2.40
Жоранс (кол. UCB) 2 84.4-89.8 87.15 2.90
С. spelaea ussurica Приморье пещера Географического общества (кол. ЗИН) 1 87.8
Таблица 11. Географическая изменчивость длины нижнего зубного ряда
Р/2-М/1 (мм) у пещерной гиены (Crocuta spelaea)
Table 11. Geographical variability of length of P/2-M/1 (mm) in Crocuta spelaea
Местонахождения
Средний плейстоцен
Кромер и миндель
С. spelaea praespelaea
Германия
Мосбах (Schutt, 1971) 2 94.0-99.0 96.5
Греция Петралона (Kurt£n, Poulianos, 1977) Поволжье р.Казанка (Барышников, 1992) 4 1 85.0-93.0 88.25 92.5 3.59
Миндель
С. spelaea petralonae Греция Петралона (Kurt£n, Poulianos, 1977) 1 96.0
Миндел ь-рисс
С. 'spelaea intermedia Франция Лунель-Вьель (колл. LGQ) 9 83.3-88.4 85.77 1.55
41
Продолжение табл. 11
Местонахождения 1п | lim 1 - 1 с
Рисе
С. spelaea spelaea
Франция
Гаргас (Clot, 1980) 2 86.8-87.0 86.9
Поздний плейстоцен
Рисс-вюрм
С. spelaea spelaea
Франция
Лерм (Clot, 1980) 5 85.4-88.0 86.7 0.97
Закавказье
Бинагады (кол. ЗИН) 6 82.2-92.1 86.02 3.51
В ю р м
С. spelaea spelaea
Франция
Арси-сюр-Кюр (кол. UPS) 4 85.7-94.1 90.05 3.13
Жерд(С1о1, 1980) 5 88.5-93.0 91.2 2.08
Пэр-но-Пэр (Clot, 1980) 6 82.5-94.4 89.4 4.37
Жоранс (Ballesio, 1979) 9 84.5-92.0 88.9 2.31
Польша
Облазова и Вержховска Гурна (кол. ISE) 2 92.9-95.2 94.05
Румыния
Рипа (Coloman, 1978) 3 86.5-98.5 92.5
Молдова
разные пещеры (Давид, 1980) 5 89.0-92.0 90.5
Южный Урал
пещера Носорога (кол. ЗИН) 1 86.8
Западная Сибирь
Красный Яр (кол. ЗИН) 1 96.6
С. spelaea ussurica
Приморье
пещера Географического общества (кол. ЗИН) 3 91.2-98.3 93.83
Китай
Верхняя пещера Чжоукоудяня (Pei, 1940) 1 94.0
Вурджи (Lu et al., 1986) 1 90.2
ляциях. Были, по крайней мере, три направления модификации РЗ/ у С. spe-
laea: увеличение абсолютной (С. s. petralona) и относительной (С. s. inter-
media) ширины, увеличение высоты коронки (С. 5. ultima) (костедробящая
42
адаптация) и возрастание общих размеров зуба с относительным его
удлинением (С. 5. ussurica) (хищнеческая адаптация). У С. s. spelaea относи-
тельная ширина коронки РЗ/ несколько больше, чем у С. 5. ussurica.
Вариабельность Р4/. Для С. spelaea предполагалось уве-
личение размеров Р4/ к позднему плейстоцену (Kurt6n, Poulianos, 1977),
однако наши данные показывают, что длина зуба в течение плейстоцена
заметно варьировала в соответствии с изменениями величины черепа,
достигая более крупных размеров в ледниковые эпохи (табл. 12). Наиболее
длинными оказались зубы в выборке из пещеры Географического общества.
Изменения относительной длины метастиля также связаны с климати-
ческими условиями, в которых обитали животные. Данный индекс достигал
наибольших значений в миндельскую и вюрмскую эпохи и наименьших - в
миндель-рисскую и рисс-вюрмскую (табл. 13). Удлинение метастиля Р4/
было обусловлено усилением хищнической специализации зубного аппарата
С. spelaea, которая ярче проявлялась в популяциях, обитавших в холодном
климате ледниковых эпох. Среди крокут вюрмского оледенения более
продвинутыми по этому признаку были звери из северо-западных частей
ареала (рис. 11).
Вариабельность М/\. Длина нижнего хищнического зуба С.
spelaea также изменялась на протяжение плейстоцена в зависимости от
температурного фактора (табл. 14). Кроме того, в выборках вюрмской эпохи
отчетливо видно, что длина М/\ возрастала по мере продвижения с юга на
север.
В процессе эволюции С. spelaea талонид М/\ становился все более
коротким (специализация к миофагии), и у подвидов позднего плейстоцена
длина его в среднем не превышала 10% от общей длины зуба (табл. 15). У
вюрмских гиен величина талонида находилась в прямой зависимости от
размеров самого зуба (рис. 12).
Присутствие на М/\ маленького метаконида значительно варьирует и,
как уже отмечалось в литературе (Kretzoi, 1938; Kurt£n, 1956), не является
надежным систематическим признаком. Частота встречаемости метаконида,
по нашим данным, изменялась от 23-29% в кромере и минделе до 50% в
миндель-риссе, затем она уменьшалась в риссе, возрастала в рисс-вюрме
(Бинагады) и вновь уменьшалась до 40-43% в вюрме (табл. 16). По другим
данным (Kurten, Poulianos, 1977) частота присутствия метаконида в выборках
из рисс-вюрмских местонахождений, например, в Европе, очень мала (11%).
Подобное варьирование в проявлении признака может быть поставлено в
связи не только с общими изменениями климата и характера питания живот-
ных, но и с особенностями локальных популяций, в частности с ослаблением
межпопуляционного обмена генами при оседлом образе жизни или ограни-
чении межклановых контактов.
Обсуждение. В геологическом масштабе времени практически
мгновенно С. spelaea расселился в кромерскую эпоху по всей Европе,
Казахстану и Китаю. Такое парадаксальное распространение крокут на север
43
Таблица 12. Географическая изменчивость длины Р4/ у пещерной гиены
(Crocuta spelaea)
Table 12. Geographical variability of length of Р4/ in Crocuta spelaea
Местонахождения | п 1 | lim 1 X 1 с
Средний плейстоцен
Кромер и миндель
С. spelaea praespelaea
Англия
Форест-Бед (Kurten, 1969) 1 39.7
Германия
Зюссенборн (Kurten, 1969) 1 40.0
Мосбах (Schutt, 1971) 7 39.4-42.5 41.3
Чехия
Странска Скала (Kurten, 1972) 1 36.2
Украина
хутор Винява (Давид и др., 1990) 1 42.4
Поволжье
р.Казанка (Барышников, 1992) 1 42.7
С. spelaea petralonae
Греция
Петралона (Kurten, Poulianos, 1977) 2 43.0-43.5 43.25
С. spelaea ultima
Китай (Kurten, 1956) 5 37.2-46.4 41.80 2.43
Миндель-рисс
С. spelaea intermedia
Франция
Лунель-Вьель (кол. LGQ) 10 35.7-39.5 37.53 1.24
Рисе
С. spelaea spelaea
Франция
Шатийон-Сен-Жан (Clot, 1980) 5 39.7-43.5 41.4
Гаргас (Clot, 1980) 8 39.0-42.4 40.3 1.23
Поздний плейстоцен
Рисс-вюрм
С. spelaea spelaea
Англия
Торньютон (Kurten, 1963) 63 37.7-43.5 41.2
44
Продолжение табл. 12
Местонахождения 1 « 1 | lim 1 X 1 о
Германия
Линденталь (Soergel, 1937) 5 37.7-40.1 38.2
Закавказье
Бинагады (кол. ЗИН) 8 36.6-42.2 39.63 1.95
В ю р м
С. spelaea dorotheae
Палестина (Kurten, 1965) 9 35.2-39.7 38.4 1.3
С. spelaea spelaea
Англия
Кентские пещеры (Kurten, 1956) 39 36.7-45.0 40.83 1.28
Франция
Арси-сюр-Кюр (кол. UPS) 12 38.8-43.3 40.86 1.38
Жерд(С1оГ 1980) 13 39.2-43.1 41.3 1.20
Буадер (Clot, 1980) 12 39.6-42.3 40.65 0.95
Эйшель (Clot, 1980) 9 37.5-44.6 41.7 1.91
Пэр-но-Пэр (Clot, 1980) 37 38.5-45.8 41.6 1.63
Жоранс (Ballesio, 1979) 9 37.7-41.0 39.9 1.82
Польша
Голобец (кол. ISE) 3 40.6-42.1 41.3
Румыния
Рипа (Coloman, 1978) 7 40.0-45.0 42.07 1.59
Молдова
разные пещеры (Давид, 1980) 9 39.2-43.0 41.4
Крым
Пролом 2 (Барышников, 1995) 13 37.4-43.9 40.38 1.90
Узбекистан
Аман-Кутан (кол. ЗИН) 1 33.3
С. spelaea ussurica
Приморье
пещера Географического общества (кол. ЗИН) 4 40.7-46.5 44.27 2.53
находит свое логическое объяснение в том, что предковый вид С. sivalensis
достиг в это время стадии биологического расцвета, и плотность населения
его в тропической Азии возросла настолько, что сделалось возможным заво-
евание новых территорий (Вангенгейм, 1978). Эволюция С. sivalensis в стра-
нах с умеренным климатом сопровождалась увеличением размеров живот-
ных и становлением вида С. spelaea.
Крупный подвид C.s.praespelaea был распространен от Англии до Ка-
захстана, не проявляя больших различий в величине черепа и зубов. Хищни-
45
Таблица 13. Географическая изменчивость относительной длины метастиля Р4/
к общей длине зуба (%) у пещерной гиены (Crocuta spelaea)
Table 13. Geographical variability of ratio of Р4/ metastyle length to teeth length (%)
in Crocuta spelaea
Местонахождения | п | lim 1 * 1 о
Средний плейстоцен Кромер и миндель
С. spelaea praespelaea Англия Форест Бед (Kurten, 1969) Германия Зюссенборн (Kurt£n, 1969) Мосбах (Schutt, 1971) Чехия Странска Скала (Кш1ёп, 1972) Украина хутор Винява (Давид и др., 1990) Поволжье р.Казанка (Барышников, 1992) 1 1 7 1 1 1 40.8-46.0 46.8 43.7 43.0 44.5 46.2 42.9
С. spelaea petralonae Греция Петралона (Кш1ёп, Poulianos, 1977) 1 42.8
Миндель-рисс
С. spelaea intermedia Франция Лунель-Вьель (кол. LGQ) 10 35.7-39.5 37.53
Рисе
С. spelaea spelaea Франция Faprac(Clot, 1980) 8 42.9-46.1 44.45 1.0
Поздний плейстоцен Рисс-вюрм
С. spelaea spelaea Англия Торньютон (Kurten, 1963) Закавказье Бинагады (кол. ЗИН) 63 7 37.7-43.5 41.0-44.4 41.2 42.82 1.18
46
Продолжение табл. 13
Местонахождения 1 п | lim 1 х 1 с
В ю р м
С. spelaea dorotheae Палестина (Kurten, 1965) 9 44.09
С. spelaea spelaea Англия
Кентские пещеры (Kurten, 1962) 100 37.7-45.6 41.06
" (Kurten, 1956) 39 40.83 3.08
Франция
Арси-сюр-Кюр (кол. UPS) 12 38.8-43.3 40.86 2.93
Жерд (Clot, 1980) 13 42.9-48.5 5.9 1.86
Буадер (Clot, 1980) 12 42.5-47.5 5.2 1.83
Эйшель(С1о1, 1980) 8 42.1-46.6 4.8 1.41
Пэр-но-Пэр (Clot, 1980) 37 42.9-49.1 46.25 1.48
Жоранс (Ballesio, 1979) 9 37.7-41.0 39.9 3.48
Польша Голобец (кол. ISE) Румыния 3 42.5-44.3 43.57
Рипа (Coloman, 1978) 7 40.0-45.0 42.07 1.79
Молдова разные пещеры (Давид, 1980) Крым 9 41.1
Пролом 2 (Барышников, 1995) 13 40.1-44.6 42.62 1.41
Узбекистан
Аман-Кутан (кол. ЗИН) 1 41.7
С. spelaea ussurica Приморье
пещера Географического общества (кол. ЗИН) 4 40.7-46.5 44.98 1.76
ческие адаптации этих гиен были несколько ослаблены, по сравнению с
гиенами геологически более поздних эпох. Другой крупный подвид C.s.petra-
lona, определенный пока только из Греции, имел широкие верхние премо-
ляры и являлся, очевидно, локальной формой, питавшейся преимущественно
падалью. В еще большей степени костедробящие адаптации наблюдаются в
зубной системе C.s.ultima из среднего плейстоцена Восточного Китая.
В миндельскую эпоху в северных областях своего распространения
пещерная крокута исчезла или же уменьшила свои размеры под
воздействием неблагоприятных для своего существования условий. Мелкий
подвид С. 5. intermedia характеризовался относительно широким РЗ/ и
большой частотой встречаемости метаконида на М/\. Эта мелкая гиена была,
47
очевидно, не только охотником, но и собирателем и потребителем падали
Начиная с рисского оледенения, ее сменил подвид С. s. spelaea, который
встречался в Европе, на Кавказе, в Средней Азии и на юге Сибири. У
номинативного подвида хищническая специализация зубного аппарата
достигла высшего своего проявления для пещерных гиен. Наблюдалось
увеличение размеров животных по направлению к эпохе последнего
оледенения, а внутри ареала - от южных областей к северным в пределах
одного временного среза. Среди южных гиен самые мелкие принадлежали к
подвиду С. 5. dorotheae из восточных районов Средиземного моря (Левант).
На Дальнем Востоке развитие пещерных гиен происходило аналогич-
ным образом - от С. 5. ultima через развитие режущих черт хищнических
зубов к С. 5. ussurica, который отличался, кроме того, особой крупнозу-
бостью и относительно узким РЗ/. Крупные размеры зубов уссурийской кро-
куты, возможно, связаны с обилием в Восточной Азии крупных травоядных.
Ш, мм
мм
Рис. 10. Соотношение длины (L) и ширины (JV) верхнего коренного зуба РЗ/ у пещерной
гиены (Crocuta spelaea)'.
а - Арси-сюр-Кюр, Франция; b - Бинагады, Закавказье; g - Голобец, Польша; / - Лунель-Вьель, Франция; т - Аман-
Кутан, Узбекистан;/? - Пролом 2, Крым; и - пещера Географического общества, Приморский край
Fig. 10. Correlation between length (L) and width (IF) of the upper premolar РЗ/ in different
populations of Crocuta spelaea'.
a- Arcy-sur-Cure, France; b- Binagady, Transcaucasia; g- Golabiec, Poland; /- Lunel-Viel, France; m- Aman-Kutan,
Uzbekistan; p- Prolom 2, Crimea; u- Geographical Society Cave, Primorskii Krai, Russia
48
l.m t , мм
Рис. 11. Соотношение длины (L) и длины метастиля (Lmt) верхнего коренного зуба ?4/ у
пещерной гиены (Crocuta spelaea)
Условные обозначения как на рис. 10
Fig. 11. Correlation between length (L) and metastyle length (Lmt) of the upper premolar Р4/ in
different populations of Crocuta spelaea
Symbols as in fig. 10
Приведенные выше особенности плейстоценовой истории С. spelaea
свидетельствуют о том, что размеры животных увеличивались в холодные
эпохи и уменьшались в теплые, подчиняясь тем самым общей зоогеогра-
фической закономерности, известной как ’’правило Бергмана” Эта же
закономерность проявлялась и в изменениях размеров гиен в пределах
видового ареала.
Основная тенденция эволюции С. spelaea связана с усилением хищни-
ческой специализации зубного аппарата. Именно переход к миофагии, к
активной охоте позволил С. spelaea освоить территории с продолжитель-
ными холодными зимами. Однако развитие режущей функции зубов не было
однонаправленным, аккумулятивным: за фазами хищнической специали-
зации, приуроченным к ледниковьям, следовали фазы деспециализации в
межледниковья, во время которых наблюдался как бы возврат к предыдуще-
му состоянию и развитие не режущей, а дробящей функции зубного аппара-
та. Такое варьирование в характере озубления С. spelaea может объясняться
ослаблением давления естественного отбора на популяции пещерной гиены в
49
Таблица 14. Географическая изменчивость длины М/1 (мм) у пещерной гиены
(Crocuta spelaea)
Table 14. Geographical variability of length of M/1 (mm) in Crocuta spelaea
Местонахождения | n 1 lim I 1 X 1 CT
Средний плейстоцен Кромер и миндель С. spelaea praespelaea Германия Зюссенборн (Kurten, 1969) 1 Мосбах (Schutt, 1971) 3 33.0-34.0 26.5 33.6
Греция Петралона (Kurten, Poulianos, 1977) 6 28.8-33.0 31.12 1.51
Украина Ополье (Татаринов, 1958) 7 32.3-35.9 34.21
Поволжье р.Казанка (Барышников, 1992) С. spelaea petralonae Греция Петралона (Kurten, Poulianos, 1977) 1 2 33.0-33.1 33.7 33.05
С. spelaea ultima Китай (Kurten, 1956) 6 27.3-39.3 32.78 2.01
Миндель-рисс С. spelaea intermedia Франция Лунель-Вьель (кол. LGQ) 9 28.0-31.4 29.58 1.12
Рисе С. spelaea spelaea Франция Шатийон-Сен-Жан (Clot, 1980) 10 29.5-34.4 31.7
Поздний плейстоцен Рисс-вюрм С. spelaea spelaea Англия Киркдаль (Klein, Scott, 1989) i 21 30.0-34.4 31.50 1.01
Баррингтон (Klein, Scott, 1989) 10 30.3-33.7 31.77 1.15
Торкорт (Klein, Scott, 1989) 17 29.5-33.6 31.38 1.19
Торньютон (Klein, Scott, 1989) 90 28.7-34.6 31.60 1.30
Германия Линденталь (Soergel, 1936) 11 30.9-33.5 31.9
50
Продолжение табл. 14
Местонахождения 1 ” | lim 1 * 1 °
Закавказье
Бинагады (кол. ЗИН) 10 28.1-33.5 30.19 1.75
В ю р м
С. spelaea dorotheae
Палестина (Кш1ёп, 1965) 6 27.3-30.6 29.0 1.3
С. spelaea spelaea
Англия
Пин Хоул (Klein, Scott, 1989) 44 30.0-40.7 32.77 2.21
Коиган (Klein, Scott, 1989) 16 29.5-34.8 32.14 1.58
Пикенс Хоул (Klein, Scott, 1989) 25 30.0-34.7 32.29 1.42
Кентские пещеры (Kurten, 1956) 41 28.9-36.3 32.61 1.14
Франция
Арси-сюр-Кюр (кол. UPS) 12 29.5-34.5 32.11 1.52
Жерд(СкН, 1980) 23 30.5-34.3 32.8 1.20
Эйшель (Clot, 1980) 19 29.3-34.3 32.7 1.51
Пэр-но-Пэр (Clot, 1980) 59 28.7-38.2 32.7 1.69
Жоранс (Ballesio, 1979) 9 30.8-34.6 32.5 2.69
Польша
Голобец (кол. ISE) 6 29.7-33.7 31.2 1.64
Вержховска Гурна (кол. ISE) 4 31.2-33.6 32.52 1.09
Молдова
разные пещеры (Давид, 1980) 11 31.2-34.0 32.8
Крым
Пролом 2 (Барышников, 1995) 9 29.4-34.2 31.02 1.58
Аювул-Коба (кол. ЗИН) 2 32.7-33.3 33.0
Северный Кавказ
Ильская 1 (кол. ЗИН) 4 30.5-32.8 31.47 0.97
Узбекистан
Аман-Кутан (кол. ЗИН) 2 29.1-30.0 29.55
Южный Урал
пещера Носорога (кол. ЗИН) 1 29.6
Западная Сибирь
Красный Яр (кол. ЗИН) 1 33.8
Алтай
Усть-Канская пещера (кол. ЗИН) 2 29.6-33.5 31.55
Денисова пещера (кол. ИАЭ) 1 35.6
С. spelaea ussurica
Приморье
пещера Географического общества (кол. ЗИН) 8 30.8-37.4 34.11 2.17
Китай
Верхняя пещера Чжоукоудяня (Pei, 1940) 1 35.5
Вурджи (Lu et al., 1986) 1 34.2
51
4
Таблица 15. Географическая изменчивость относительной длины тригонида
М/1 (%) у пещерной гиены (Crocuta spelaea)
Table 15. Geographical variability of ratio of M/1 trigonide length to teeth length (%)
in Crocuta spelaea
Местонахождения | п | lim J ' 1 2
Средний плейстоцен Кромер и мин дель С. spelaea praespelaea Германия Зюссенборн (Kurten, 1969) 1 Мосбах (Schutt, 1971) 3 87.1-88.8 92.1 88.1
Греция Петралона (Kurt6n, Poulianos, 1977) 6 86.8-90.1 88.61 1.16
Поволжье р. Казанка (Барышников, 1992) С. spelaea petralonae Греция Петралона (Kurten, Poulianos, 1977) С. spelaea ultima Китай (Kurten, 1956) 1 1 5 88.7 86.1 89.90 1.47
Миндель-рисс С. spelaea intermedia Франция Лунель-Вьель (кол. LGQ) 9 87.6-91.8 89.78 1.36
Поздний плейстоцен Рисс-вюрм С. spelaea spelaea Закавказье Бинагады (кол. ЗИН) । 10 88.4-93.9 91.53 2.00
В ю р м С. spelaea spelaea Франция Арси-сюр-Кюр (кол. UPS) 12 87.8-93.8 91.44 1.49
Жоранс (Ballesio, 1979) 4 89.9-91.6 90.52 0.75
Польша Вержховска Гурна (кол. ISE) 4 88.8-92.0 90.42 1.61
Крым Пролом 2 (Барышников, 1995) 9 89.0-93.2 91.38 1.38
Аювул-Коба (кол. ЗИН) 2 91.1-91.9 91.5
52
Продолжение табл. 15
Местонахождения 1 " 1 Нт | 1 £ 1 ст
Северный Кавказ
Ильская 1 (кол. ЗИН) 3 91.0-92.5 91.54
Узбекистан
Аман-Кутан (кол. ЗИН) 2 91.3-95.5 93.4
Южный Урал пещера Носорога (кол. ЗИН) Западная Сибирь 1 92.2
Красный Яр (кол. ЗИН) 1 92.3
Алтай
Усть-Канская пещера (кол. ЗИН) 2 93.1-93.9 93.5
Денисова пещера (кол. ИАЭ) 1 89.6
С. spelaea ussurica Приморье
пещера Географического общества (кол. ЗИН) 7 89.9-95.8 91.86 2.03
Таблица 16. Географическая изменчивость частоты встречаемости метаконида
М /1 у пещерной гиены (Crocuta spelaea)
Table 16. Geographical variability of frequency of M/1 metaconid in populations of
Crocuta spelaea
Местонахождения Всего зубов Зубы с метаконидом п | %
Средний плейстоцен
Кромер и миндель
С. spelaea praespelaea
Германия
Зюссенборн (Soergel, 1936) 1
Мосбах (Schutt, 1971) 3 2 67
Румыния
Гомбасцог (Kretzoi, 1938) 1
Греция
Петралона (Kurt£n, Poulianos, 1977) 7 1 14
Поволжье
р.Казанка (Барышников, 1992) 1 1
С. spelaea petrolonae
Греция
Петралона (Kurten, Poulianos, 1977) 1
53
Продолжение табл. 16
Местонахождения Всего зубов Зубы с метаконидом п | %
Кромер и миндель, всего
Kurten, Poulianos, 1977 наши данные 13 14 3 4 23.1 28.6
Миндель-рисс С. spelaea intermedia Франция Лунель-Вьель (Bonifay, 1971) нижний уровень 19 14 73
верхний уровень 11 1 10
Миндель-рисс, всего 30 15 50
Рисе С. spelaea spelaea Франция Шатийон-Сен-Жан (Ballesio, 1979) 8 3 38
Поздний плейстоцен Рисс-вюрм С. spelaea spelaea Германия Линденталь (Soergel, 1937) 13 13 100
Закавказье 'Бинагады (кол. ЗИН) 11 6 55
Рисс-вюрм, всего Kurt£n, Poulianos, 1977 160 18 11.2
наши данные 24 19 79.2
В ю р м С. spelaea spelaea Англия Кентские пещеры (Kurt£n, 1956) 37 17 46
Франция Арси-сюр-Кюр (кол. UPS) 12 3 25
Жоранс (Ballesio, 1979) 11 4 36
Крым Пролом 2 (Барышников, 1995) 9 2 22
Аювул-Коба (кол. ЗИН) 2 1
54
Окончание табл. 16
Местонахождения Всего зубов Зубы с метаконидом
п 1 %
Северный Кавказ
Ильская 1 (кол. ЗИН) 3 3 100
Узбекистан
Аман-Кутан (кол. ЗИН) 2 2
Южный Урал
пещера Носорога (кол. ЗИН) 1 1
Западная Сибирь
Красный Яр (кол. ЗИН) 1
Алтай
Усть-Канская пещера (кол. ЗИН) 2
Денисова пещера (кол. ИАЭ) 1 1
С. spelaea ussurica
Приморье
пещера Географического общества (кол. ЗИН) 7 2 29
Вюрм, всего
Kurten, Poulianos, 1977 230 100 43.5
наши данные 88 28 40.9
теплые межледниковые эпохи (благоприятные климатические условия,
обилие пищи, поедание падали) и усилением его в холодные эпохи оледене
ний (неблагоприятные климатические условия, потребность в калорийном
свежем мясе, увеличение охотничьей активности, обострение конкуренции за
пищу со стороны других крупных хищников и человека).
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
ОТРЯД CARNIVORA BOWDICH, 1821
СЕМЕЙСТВО HYAENIDAE GRAY, 1821
ТРИБА CROCUTINI BARYSHNIKOV ET AVERIANOV, 1995
Род Crocuta Каир, 1828
Crocuta spelaea (Goldfuss, 1823)
Crocuta spelaea ussurica Baryshnikov et Vereshchagin subsp. nov.
Название. Дано по нахождению в Уссурийском крае (от р.Ус-
сури).
55
Голотип. ЗИН № 0.34478, фрагмент черепа, пещера Геогра-
фического общества, долина р.Партизанская (Сучан), юг Приморского края;
поздний плейстоцен (каргинский интерстадиал).
П а р а т и п ы. ЗИН № 0.34479, левая верхнечелюстная кость с Р1 /-def.M/ (сл.4,
глубина 80-100 см, кв. 15, 1967); ЗИН № 0.34480, правая верхнечелюстная кость с РХ/-Р4/
(сл.4, глубина 80-100 см, кв. 15, 1967); ЗИН № 0.34481, обломок левой верхнечелюстной
кости с РЗ/-Р4/ (сл.4, кв. 17, 1967); ЗИН № 0.34482, обломок правой верхнечелюстной
кости с РЗ/-Р4/ (глубина 80-100 см, кв. 15, 1967); ЗИН № 0.34483, обломок левой
верхнечелюстной кости с РЗ/-Р4/ (левый ход, Лешок, 1972); ЗИН 0.34484, нижняя
челюсть с PI2-MIX ; ЗИН № 0.34486, нижняя челюсть с P/2-def.A7/l; ЗИН № 0.34487, ле-
вая нижнечелюстная кость с С/1-Л//1 (сл.4, кв. 15, 1967); ЗИН № О. 34489, правая нижне-
челюстная кость с Р/З-М/Х (шурф 3, 1970). Местонахождение и возраст как у голотипа.
Диагноз. Пещерная гиена средних размеров, с относительно
широким твердым небом и очень крупными коренными зубами. Средние
значения длины РЗ/ (27.17, и=7) и длины Р4/ (44.27, и=4) превышают
таковые у всех известных подвидов.
Размеры типовой серии (мм). Промеры голотипа даны в табл.2.;
сохранившийся Р2/ ZM8.6, Zp=14.4, ИМ 3.1. Верхнечелюстные кости: ЗИН №№ 34479,
(Р/3 Z=27.9, ИМ9.3), 34480 (РХ/-Р4/ 87.8, РЗ/-Р4/ 64.3, РЗ/ L=ca23.0, ИМ6.3, Р4/
£=ся40.7, ИЪМ9.8), 34481 (РЗ/-Р4/ 72.1, РЗ/ Z=28.9, ИМ1.7, Р4/ А=46.5, Zw=21.8,
ИЪ=27.9), 34482 (РЗ/-Р4/ 71.6, РЗ/ ZM8.3, ИМ 1.0, Р4/ ZM5.5, Z/tMO.8, ИЪ=25.3), 34483
(РЗ/-Р4/ 70.0, РЗ/ ZM7.5, ИМО.О, Р4/ L=44.4, Lm=X9.8, №а=24.7); нижнечелюстные кости:
ЗИН №№ 34486 (Р2/-М/1 98.3, Р/3 L=25.2, ИМ 8.0, А//1 L=37.4, Lt=34.7, ИМ4.7), 34484
(С/1-Л//1 120.0, Р2/-Л//1 92.0, Р/2-Р/4 59.7, Р/3 ZM2.8, ИМ7.0, М/Х L=34.\, Lt=30.9,
ИМ3.6), 34487 (С/1-Л//1 117.4, Р/2-М/Х 91.2, Р/2-Р/4 61.6, Р/3 L=24.0, ИМ7.0, Л//1
Z=ca33.7, ИМа14.7), 34489 (Л//1 L=31.4, Z./M8.9, ИМ3.8).
Сравнение. От одновозрастного номинативного подвида отли-
чается абсолютно и относительно более крупными верхними коренными
зубами. Так, различия по длине РЗ/ в сравнении с выборками из Франции -
Жерд (/=3.11, и=13), Буадер (/=3.70, л=11), Пэр-но-Пэр (/=3.09, и=42),
Жоранс (/=2.61, и=11), из Польши Голобец (/=2.67, и=9) и из Крыма -
Пролом 2 (/=3.54, и=19), оказались достоверно значимыми (Р<0.02). Также
достоверны (Р<0.05) различия по длине Р4/: Жерд (/=2.56, и=13), Пэр-но-Пэр
(/=2.37, и=37), Жоранс (/=3.48, и=9), Аси-сюр-Кюр (/=2.93, и=12), Пролом 2
(/=3.21, и=13). Еще сильнее по длине хищнического зуба выражены различия
с очень мелким С. s. dorotheae (/=5.03, Р<0.001). От среднеплейстоценового
подвида С. 5. ultima, помимо более длинной в среднем коронки РЗ/ (t=3.56,
п=5; Р<0.0Х) и верхнего хищнического зуба (/=2.43, и=5; Р<0.05), отличался
меньшей гипсодонтией РЗ/, относительно более длинным метастилем Р4/ и
укороченным талонидом Л//1.
Распространение и возраст. Южная часть Примор-
ского края, северо-восточной и восточный Китай и, очевидно, север Кореи;
поздний плейстоцен (вюрм).
Материал. Помимо типовой серии еще около 160 костных остат-
ков из пещеры Географического общества (ЗИН № 0.34485, 34488, 34490).
56
38
мм
Рис. 12. Соотношение наибольшей длины (Z) и длины тригонида (Lt) нижнего коренного
зуба Л771 у пещерной гиены (Crocuta spelaea)
Условные обозначения как на рис. 10
Fig. 12. Correlation between length (L) and trigonid length (Lt) of the lower molar M\ in
different populations of Crocuta spelaea
Symbols as in fig. 10
БЛАГОДАРНОСТИ
Мы благодарны И.Е.Кузьминой (ЗИН), Е.Н.Мащенко (ПИН), академику А.П.Дере-
вянко (ИАЭ), проф. Ю.Г.Колосову (ИАУ), Н.Л.Фомичевой (ГМК), Prof. F.-M.Bonifay
(LGQ), Dr. J.-F.Brugal (LGQ), Dr. F.David (UPS), Dr. C.Mourer-Chauvire (UCB), Dr. A.Nada-
chowski (ISE) - за помощь в работе с коллекциями, Н.А.Орлову (ЗИН) - за фотографирова-
ние материала, Dr. A.Forsten (Helsinki), Dr. V.Eisenmann (Paris), Dr. J.Hoffecker (Argonne,
USA), Dr. A.Hufthammer (Bergen) - за изготовление ксерокопий палеонтологических пуб-
ликаций. Мы выражаем также признательность А.О.Аверьянову (ЗИН) за прочтение
статьи и высказанные замечания, О.Р.Потаповой и О.И.Маловой за помощь в
оформлении рукописи.
ЛИТЕРАТУРА
Абрамова З А. Палеолит Северной Азии И Палеолит Кавказа и Северной Азии. - Л.,
1989. С.143-241.
57
Агеева Е.А. Гричан Ю.В., Оводов Н.Д. Плейстоценовая гиена в каменной ловушке //
Природа, 1978. № 12. С. 114-115.
Алексеев М.Н. Антропоген Восточной Азии: Стратиграфия и корреляция. М.,
1978. 208 с.
Алексеева Л.И. Териофауна раннего антропогена Восточной Европы. М., 1977.
214 с. (Труды Геол, ин-та АН СССР, вып. 300).
Алексеева Л. И. Териофауна многослойной стоянки Молодова 5 И Многослойная па-
леолитическая стоянка Молодова 5. Люди каменного века и окружающая среда.
-М., 1987. С.153-162.
Алексеева Л.И. Териофауна верхнего плейстоцена Восточной Европы (крупные мле-
копитающие). - М., 1990. 109 с. (Труды Геол, ин-та АН СССР, вып. 455).
Алексеева Л. И., Ломизе М.Г О находке плейстоценовой фауны млекопитающих в
верховьях реки Белой (Северный Кавказ) И Изв. Высш, учебн. завед. Геол, и
разведка, 1960. № 2. С.29-33.
Алексеева Э.В. Млекопитающие плейстоцена юго-востока Западной Сибири (хищ-
ные, хоботные, копытные). - М., 1980. 188 с.
Анисюткин Н.К., Кетрару И.А. Грот Выхватинцы и проблема домустьерских
памятников Молдавии И Проблемы антропогена Молдавии. Кишинев, 1982.
С.120-132.
Арсланов Х.А., Герасимова С.А., Леонтьев О.К. и др. О возрасте плейстоценовых и
голоценовых отложений Каспийского моря (по данным радиоуглеродного и
уран-иониевого методов датирования) И Бюл. Комис, по изуч. четвертичн.
периода, 1978. Т.48. С.39-48.
Бадер О.Н. Смеловская 2 палеолитическая стоянка в степях Южного Урала И
Матер, и исслед. по археол. СССР, 1971. № 173. С.200-208.
Байгушева В.С. Ископаемая териофауна Ливенцовского карьера (северо-восточное
Приазовье) И Труды Зоол. ин-та АН СССР, 1971. Т.49. С.5-29.
Барышников ГФ. Млекопитающие Кавказа в эпоху раннего палеолита И Труды Зоол.
ин-та АН СССР, 1987. Т.168. С.3-20.
Барышников ГФ. Позвоночные Баракаевской мустьерской стоянки на Северном
Кавказе//Труды Зоол. ин-та АН СССР, 1991 (1992). Т.238. С. 139-166.
Барышников ГФ. Плейстоценовые хищные (Carnivora) волжской фауны из
коллекции Казанского университета И Волжская фауна плейстоценовых млеко-
питающих в Геолого-минералогическом музее Казанского университета.
Казань, 1992. С. 107-140.
Барышников ГФ. Крупные млекопитающие ашельской стоянки в пещере
Треугольная на Северном Кавказе И Труды Зоол. ин-та РАН, 1993 (1994). Т.249.
С.3-47.
Барышников ГФ. Пещерная гиена, Crocuta spelaea (Carnivora, Hyaenidae) из
палеолитической фауны Крыма И Труды Зоол. ин-та РАН, 1995. Т.263. С.3-45.
Барышников Г.Ф., Аверьянов А.О. Молочные зубы хищных млекопитающих (отряд
Carnivora). Часть V Семейства Protelidae и Hyaenidae И Труды Зоол. ин-та РАН,
1995. Т.263. С.46-84.
Барышников Г.Ф., Баранова Г.И. Hystrix vinogradovi в плейстоцене Европы и Кавказа
И Грызуны: Материалы VI Всесоюзного совещания, Ленинград, 25-28 января
1984 г. - Л., 1983. С.39-41.
Барышников Г.Ф., Батыров Б.Х. Среднеплейстоценовые хищные млекопитающие
(Carnivora, Mammalia) Средней Азии И Труды Зоол. ин-та РАН, 1994. Т.256.
С.3-43.
58
Батыров Б.Х. Костные остатки млекопитающих из стоянки Сазаган 2 И Вопросы
археологии, древней истории и этнографии. - Самарканд, 1985. С.40-44.
Беляева Е.И. Новые находки четвертичных млекопитающих с р. Иртыша И Природа,
1933. № 10. С.62.
Бируля А.А. Предварительное сообщение о хищниках (Carnivora) из четвертичных
отложений Крыма//Докл. АН СССР, 1930. № 6. С. 139-144.
Богачев В.В. Палеонтологические заметки. 11. Гиены из окрестностей Баку И Труды
Азерб. фил. АН СССР. Геол, сер., 1938. Т. IX (39). С. 18-22.
Богачев В.В. Бинагады кладбище четвертичной фауны на Апшеронском полу-
острове. - Баку, 1939. 84 с.
Бурчак-Абрамович Н.И. Ископаемые позвоночные верхнепалеолитической стоянки
пещеры Окуми в Южной Абхазии И Пещеры Грузии, 1981. № 9. С. 123-134.
Вангенгейм Э.А. Палеонтологическое обоснование стратиграфии антропогена
Северной Азии (по млекопитающим). - М., 1977. 172 с.
Вангенгейм Э.А. Некоторые вопросы палеозоогеографии антропогена И Бюл. Комис,
изуч. четвертичн. периода, 1978. № 48. С.3-14.
Васильев С. Фауна млекопитающих последнего межледниковья Новосибирского
Приобья И Первое Между нар. мамонт, совещ. 16-22 октября 1995, Санкт-
Петербург. Тез. докл. - СПб, 1995. С.604- 605.
Векилова Е.А. Стоянка Сюрень 1 и ее место среди палеолитических местонахождений
Крыма и ближайших территорий // Матер, и исслед. по археол. СССР, 1957. №
59. С.235-323.
Векуа А.К. Ахалкалакская нижнеплейстоценовая фауна млекопитающих. - Тбилиси,
1962. 207 с. + XIX табл, (на груз. яз. с русск. рез.)
Векуа А.К. Ископаемые позвоночные Цуцхватских пещер И Изучение пещер
Колхиды. Цуцхватская многоярусная карстовая пещерная система. Тбилиси,
1978. С.94-126.
Векуа А.К., Габелая Ц.Д., Квавадзе Е.В., Чхиквадзе В.М. Гиеновая пещера И Пещеры
Грузии, 1980. Т.8. С.110-119.
Верещагин Н.К. Хищные (Carnivora) из бинагадинского асфальта И Труды Естеств.-
истор. музея АН АзербССР, 1951. Вып.4. С.28-140.
Верещагин Н.К. Млекопитающие Кавказа. История формирования фауны. - М.-Л.,
1959. 704 с.
Верещагин Н.К., Батыров Б.Х. Фрагменты истории териофауны Средней Азии И
Бюл. Моск, о-ва испыт. природы. Отд. биол., 1967. Т.72. № 4. С. 104-115.
Верещагин Н.К., Давид А. И. Млекопитающие эпохи отложений тираспольского
гравия //Труды Зоол. ин-та АН СССР, 1971. Т.49. С.30-43.
Верещагин Н.К, Кузьмина И.Е. Остатки млекопитающих из палеолитических
стоянок на Дону и Верхней Десне И Труды Зоол. ин-та АН СССР, 1977. Т.72.
С.77-110.
Верещагин Н.К., Мельникова Н.Н. Зоогеографические открытия археологов в
Восточном Казахстане и Алтайском крае И Изв. Всес. географ, о-ва, 1958. Т.90.
Вып.4. С.385-387.
Верещагин Н.К, Оводов Н.Д. История фауны Приморья И Природа, 1968. № 9. С.42-
49.
Еаджиев Д.В., Алиев С.Д. Некоторые представители хищных млекопитающих
(Mammalia, Carnivora) из палеолитических отложений Азыхской пещеры И
Учен. зап. Азерб. гос. мед. ин-та, 1966. Т.23. С.7-13.
59
Галкина Л.И, Оводов Н.Д. Антропогеновая териофауна пещер Западного Алтая //
Систематика, фауна, зоогеография млекопитающих и их паразитов.
Новосибирск, 1975. С. 165-180.
Герасимов И.П. Чичагова О. А., Черкинский А.Е. и др. Радиоуглеродные исследо-
вания радиометрической лаборатории Института географии АН СССР. Сообщ.
5 // Бюл. Комис, по изуч. четвертичн. периода АН СССР, 1983. № 52. С.205-211.
Гохман И.И. Палеолитическая стоянка "Пещера" на Бухтарме //.Краткие сообщ. Ин-
та истории матер, культуры АН СССР, 1957. Вып.6-7. С.54-58.
Громов В.И. Некоторые новые данные о фауне и геологии палеолита Восточной
Европы и Сибири // Изв. Акад, истории матер, культуры, 1935. Вып. 118. С.246-
270.
Громов В. И. Палеонтологическое и археологическое обоснование стратиграфии
континентальных отложений четвертичного периода на территории СССР
(млекопитающие, палеолит) // Труды Ин-та геол, наук, 1948. Вып. 64. Геол,
серия. № 17. 521 с.
Громова В., Громов В.И. Материалы к изучению палеолитической фауны Крыма в
связи с некоторыми вопросами четвертичной стратиграфии // Труды сов. секции
междунар. ассоц. изучен, четвертичн. периода, 1937. Вып. 1. С. 52-96.
Давид А.И. Остатки пещерной гиены из плейстоценовых отложений Молдавии //
Фауна позднего кайнозоя междуречья Днестр - Прут. - Кишинев, 1978. С.99-
104.
Давид А И. Териофауна плейстоцена Молдавии. - Кишинев, 1980. 188 с.
Давид А.И., Татаринов К.А., Свистун В.И. Хищные, хоботные и копытные раннего
плейстоцена юго-запада СССР. - Кишинев, 1990. 132 с.
Деревянко А.П., Зенин А.Н. Археологические исследования пещеры Страшная //
Altaica, 1992. № 1. С. 13-18.
Деревянко А.П., Маркин С.В. Мустье Горного Алтая (по материалам пещеры им.
Окладникова). - Новосибирск, 1992. 225 с.
Деревянко А.П., Шуньков М.В., Нэш Д., Ли Х.-Д. Исследования палеолита Денисовой
пещеры // Altaica, 1993. № 2. С.6-10.
Джафаров А.К. Мустьерская культура Азербайджана (по материалам Тагларской
пещеры). - Баку, 1983. 98 с.
Дуброво И.А. Плейстоценовые млекопитающие Среднего и Южного Урала и их
стратиграфическое значение // Научн. труды Пермского политехи, ин-та, 1966.
Сб.20. С.125-135.
Ермолова Н.М. Териофауна долины Ангары в позднем антропогене. - Новосибирск,
1978. 223 с.
Зубаков В.А. Глобальные климатические события плейстоцена. - Л., 1986. 288 с.
Иметхенов А.Б., Калмыков Н.П. Фауна и флора кайнозоя Бурятии. - Улан-Удэ, 1988.
181 с.
Ишунин Г.И., Тетюхин Г.Ф. Вероятные пути образования фауны млекопитающих на
территории Узбекистана. - Ташкент, 1989. 80 с.
Калмыков Н.П. Фауна крупных млекопитающих плейстоцена Прибайкалья и
западного Забайкалья. - Улан-Удэ, 1990. 116 с.
Калмыков Н.П. Биостратиграфия и фауна млекопитающих плиоцена Забайкалья.
Новосибирск, 1992. 97 с.
Кожамкулова Б.С. Антропогеновая ископаемая териофауна Казахстана. - Алма-Ата,
1969. 149 с. + XXXIV табл.
60
Кожамкулова Б.С., Пак Т.К. Позднеплейстоценовая териофауна стоянки Актас
(Кокчетавская область) и ее сопоставление с одновозрастной фауной
Восточного Казахстана // Материалы по истории фауны и флоры Казахстана.
Т. 10. - Алма-Ата, 1988. С. 121 -154.
Корниец НЛ. Про причини вимирання мамонта на територй Украши // Викопж
фауни Украши i сум!жних територш. Вип.1. - Кшв, 1962. С. 91-169.
Коробков И. И. Палеолит Восточного Средиземноморья // Палеолит Ближнего и
Среднего Востока. - Л., 1978. С.7-185.
Кузьмина И.Е. Некоторые данные о млекопитающих Среднего Урала в позднем
плейстоцене//Бюл. Комис, по изуч. четвертичн. периода, 1975. № 43. С.63-77.
Кузьмина И.Е. Видовой состав и относительная численность млекопитающих
Среднего Урала в позднем плейстоцене // Труды Зоол. ин-та АН СССР, 1982.
Т.111. С.44-48.
Лазарев П.А., Томская А.И. Млекопитающие и биостратиграфия позднего кайнозоя
Северной Якутии. - Якутск, 1987. 172 с.
Любин В.П. Палеолит Кавказа // Палеолит Кавказа и Северной Азии. - Л., 1989. С.7-
142.
Мочанов Ю.А. Древнейшие этапы заселения человеком Северо-Врсточной Азии.
Новосибирск, 1977. 263 с.
Оводов Н.Д. Позднеантропогеновая фауна млекопитающих (Mammalia) юга
Уссурийского края // Фауна и систематика позвоночных Сибири.
Новосибирск, 1977. С. 157-177.
Оводов Н.Д. Пещерные местонахождения остатков млекопитающих Сибири и
Дальнего Востока (краткий обзор) // Карст Дальнего Востока и Сибири.
Владивосток, 1980. С. 154-163.
Огнев С.И. Звери Восточной Европы и Северной Азии. Т.2. Хищные млекопитающие.
-М.-Л., 1931. 776 с.
Окладников А.П., Верещагин Н.К., Оводов Н.Д. Открытие пещерного палеолита в
Приморье // Вестник Акад, наук СССР, 1968. № 10. С.54-63.
Окладников А.П., Муратов В.М., Оводов Н.Д., Фриденберг Э.О. Пещера Страшная -
новый памятник палеолита Алтая // Матер, по археологии Сибири и Дальнего
Востока. 4.2. - Новосибирск, 1973. С.3-54.
Пидопличко И.Г О ледниковом периоде. Вып.2. - Киев, 1951. 264 с.
Шдогинчко 1.Г Матер1али до вивчення минулих фаун УРСР. Вип.2. Кшв, 1956.
236 с.
Пидопличко И.Г Межиричские жилища из костей мамонта. - Киев, 1976. 240 с.
Ранов В.А., Несмеянов С.А. Палеолит и стратиграфия антропогена Средней Азии.
Душанбе, 1973. 163 с.
Распопов Г Остатки гиены, найденные в Казани // Труды О-ва естествоиспыт. при
Казанск. ун-те, 1925. Т.51. Вып.1. С. 1-8.
Рогачев А.Н., Аникович М.В. Поздний палеолит Русской равнины и Крыма //
Археология СССР. Палеолит СССР. - М., 1984. С. 162-271.
Руденко СИ. Усть-Канская пещерная палеолитическая стоянка// Матер, и исслед. по
археол. СССР, 1960. № 79. С. 104-125.
Рябинин А.Н. Об остатках пещерных льва и гиены из четвертичных отложений
Сибири // Бюл. Информ, бюро Ассоц. по изуч. четвертичн. отложений Европы
при ВГРО, 1932. №2. С. 1-14.
Савинов П.Ф. Общие результаты палеобиологических исследований Павлодарского
Прииртышья // Териология. Т. 1. - Новосибирск, 1972. С. 131 -142.
61
Смирнов Н.Г Большаков В.Н., Косинцев П.А., Панова Н.К и др. Историческая
экология животных гор Южного Урала. - Свердловск, 1990. 245 с.
Сотникова М.В. Хищные млекопитающие плиоцена - раннего плейстоцена. М.,
1989. 123 с. (Труды Геол, ин-та АН СССР. вып.440).
Сулейманов М.Б. Среда обитания первобытного человека на юго-востоке Малого
Кавказа (по данным палеолитических пещер Азых и Таглар): Автореф. канд.
дис. -М., 1982. 23 с.
Татаринов КА. Печерш гиени з четвертинних вщклад!в Отлля // Доповш АН
УРСР, 1958. № 7. С.797-800.
Татаринов КА. Фауна позвоночных стоянки Кормань IV // Многослойная палео-
литическая стоянка Кормань IV на Среднем Днестре. - М., 1977. С. 112-118.
Тлеубердина П.А., Кожамкулова Б.С., Кондратенко ГС. Каталог кайнозойских
млекопитающих Казахстана. - Алма-Ата, 1990. 159с.
Форонова ИВ. Новые находки млекопитающих в плейстоценовых отложениях
Кузнецкой котловины //Труды Зоол. ин-та АН СССР, 1982. Т. 111. С.50-57.
Худайбердиев Т.Н. Материалы по териофауне северо-восточного Узбекистана в
позднем антропогене: Автореф. канд. дис. - Ташкент, 1973. 24 с.
Цейтлин СМ. Геология пещерных палеолитических стоянок Алтая (бассейн
р.Чарыш) // Бюл. Комис, по изуч. четвертичн. периода, 1974. № 42. С. 108-115.
Цейтлин С.М. Геология палеолита Северной Азии. - М., 1979. 287 с.
Черныш А.П. Ранний и средний палеолит Приднестровья. - М., 1965. 139 с. (Труды
Комис, по изуч. четвертичн. периода, вып. 25).
Шарапов Ш. Куруксайский комплекс позднеплиоценовых млекопитающих Афгано-
Таджикской депрессии. - Душанбе, 1986. 272 с.
Яцко И.Я. О находках гиен в древних карстовых пещерах г.Одессы // Ежегодник
Всес. палеонтол. о-ва, 1956. Вып. 15. С.335-340.
Aymard A. Communication sur le gisement de Sainzelles // Annals Soc. Agricul. Sci. Arts
Commerce Puy, 1846. T.13. P.153-155.
Azzaroli A. Quaternary mammals and the "End-Villafranchian" dispersal event - a turning
point in the histofy of Eurasia// Palaeogeogr. Palaeoclim. Palaeoecol., 1983. Vol.44.
P.117-139.
Ballesio R. Le gisement Pl£istocene sup£rieur de la grotte de Jaurens a Nespouls, Correze,
France: les Carnivores (Mammalia, Carnivora). I. Canidae et Hyaenidae // Nouv.
Arch. Mus. Hist. nat. Lyon, 1979. Fasc.17. P.25-55 + V Pl.
Baryshnikov G., Hoffecker J. Mousterian hunters of the NW Caucasus: Preliminary results
of recent investigations//}. Field Archaeol., 1994. Vol.21. P.1-14.
Bishop M.J. The mammal fauna of the early Middle Pleistocene cavern infill site of
Westbury-sub-Mendip, Somerset// Spec. Pap. Palaeontol., 1982. Vol.28. P. 1-108.
Bonifay M.-F. Carnivores quaternaires du sud-est de la France // Мёт. Mus. Nat. Hist.
Natur., 1971. Ser.C. T.21. P.43-377+27 Pl.
Boule M. Les grottes de Grimaldi (Baousse-Rousse). G£ologie et paleontologie. T.l. Fasc.4
- Carnassiers. Monaco, 1919. P.237-362 + XXX-XLI Pl.
Brandt J.F. Neue Untersuchungen Ober die in den Altaischen Hohlen aufgefundenen
Saugethierreste: ein Beitrag zur quaternaren Fauna des Russischen Reiches // Bull.
Acad. Sci. St.Petersb., 1871. T.15. N 2. S.147-202.
Brugal J.-Ph., Fosse Ph., Guadelli J.-L. Comparative study of bone assemblages made by
Recent and Pleistocene hyenids // Paper for "Eigth meeting of working group on bone
modification" Hot Springs (SD), 1993.
62
Christol J., Bravard A. M£moires sur de nouvelles especes d'hyenes fossiles decouvertes
dans la caverne de Lunel-Viel pres Montpellier // Mem. Soc. Hist. Natur. Paris,
1828. T.4. P.368-378.
Clot A. La grotte de la Carriere (Gerde, Ht. Pyrenees). Stratigraphic et paleontologie des
carnivores. - Toulouse, 1980. 239 pp.+ 19 P1.+ 131 Tabl.
Colbert E., Hooijer D. Pleistocene mammals from the Limestone fissures of Szechwan,
China// Bull. Amer. Mus. Natur. Hist., 1953. Vol. 102. Art. 1. P. 1-134 + 40 pl.
Colotnan C. Crocuta spelaea (Goldfuss) in pleistocenul superior de la Ripa (bazinul Beius)
//Nymphaea, 1978. N 6. P.183 -204.
Cregut E. La faune de mammiferes du gisement Pleistocene moyen anterissien de la Caune
de 1'Arago (Tautavel, Pyren£es-Orientales, France) // C. R. Acad. Sc. Paris. Ser.D.,
1980. T.290. P.751-754.
Croizet J.B., Jbbert A. Recherches sur les ossemens fossiles du departement du Puy-de-
Dome. Paris, 1828. 224 pp.
Delpech F. Les faunes du Paleolithique sup£rieur dans le sud-ouest de la France // Cahiers
Quatern. Paris, 1983. T.6. P. 1-453.
Deperet C.J. Les animaux Pliocenes de Roussilion // Мёт. Soc. G6ol. France, 1890. T.3.
P.1-164.
Dubois E. Das geologische Alter der Kendeng- oder Trinilfauna // Tijdschrift Kon.
Nederland. Aardrijksk. Genootschap, 1908. T.25. S. 1235-1270.
Ehrenberg K. Die Fuchs- oder Teufelslukken bei Eggenburg Niederdonau // Abh. Zool.
Bot. Ges. Wien, 1938-1940. Bd.17. Hf.l. S. 1-130; Hf.2. S. 131-301.
Falconer H. Palaeontological memoirs. I. Fauna antiqua sivalensis. London, 1868. 590 pp.
Forsyth Major C.I. Die Tyrrhenis. Studien liber geographische Verbreitung von Thieren
und Pflanzen im westlichen Mittelmeergebiet // Kosmos, 1883. Bd.13. S. 1-17, 81-
106.
Galiano H, Frailey D. Chasmaporthetes kani, new species from China, with remarks on
phylogenetic relationships of genera within the Hyaenidae (Mammalia, Carnivora) //
Amer. Mus. Novitates, 1977. N 2632. P.1-16.
Gaudry A., Boule M. Les oubliettes de Gargas // Mater, histor. temps quatern. Paris, 1892.
Fasc.4. P. 105-130 + XX-XXIV Pl.
Gittleman J.L (ed.). Carnivora behavior, ecology and evolution. - London, 1989. 620 pp.
Goldfuss A. Osteologische Beitrage zur Kenntnis verschiedener SSugetiere der Vorwelt //
Nova Acta Phys.-Med. Acad. Cesar. Leopold.-Carol., 1823. Bd. 21. S. 449-490.
Hoffecker J., Baryshnikov G., Potapova O. Vertebrate remains from the Mousterian site of
Ilskaya 1 (Northern Caucasus, USSR): New analysis and interpretation // Journ.
Archaeol. Sci., 1991. Vol. 18. P.l 13-147.
Hoyveil C., Petter G. The Pachycrocuta and Hyaena lineages (PlioPleistocene and extant
species of the Hyaenidae). Their relationships with Miocene ictitheres: Palhyaena and
Hyaenictitherium//Geobios, 1980. N 13. Fasc.4. P.579-623.
Huang Wanpo. Taxonomy of the Hyaenidae (Hyaena and Crocuta) of the Pleistocene in
China // Vertebrata Palasiatica, 1989. Vol.27. N 3. P. 197-204 (на кит. яз. с англ,
рез.).
Kahlke R.-D. Die Entstehungs-, Entwicklungs- und Verbreitungsgeschichte des
oberpleistozanen Mammuthus-Coelodonta-Faunenkomplexes in Eurasien (Gross-
sSuger) // Abh. Senckenberg. naturforsch. Ges., 1994. Bd.546. S. 1-164.
Klein R., Scott K. Glacial/Interglacial size variation in fossil spotted hyenas (Crocuta
crocuta) from Britain // Quaternary Research, 1989. Vol.32. P.88-95.
63
Kretzoi Л/. Die Raubtiere von GombaszOg nebst einer Ubersicht der Gesamtfauna. (Ein
Beitrag zur Stratigraphic des Altquartaers) // Ann. Mus. Nat. Hung. Min. Geol,
Palaeontol., 1938. Bd.31. S.88-157.
Kurten B. The status and affinities of Hyaena sinensis Owen and Hyaena ultima Matsumoto
// Amer. Mus. Novitates, 1956. N 1764. P. 1-48.
Kurten B. The spotted hyena (Crocuta crocuta) from the middle Pleistocene of Mosbach at
Wiesbaden, Germany //Comment, biol. Soc. Scient. Fennica, 1962. T.24. N 3. P.1-9.
Kurten B. The cave hyena, an essay in statistical analysis // (Brothwell D. & Higgs E.S
eds.) Science in Archaeology. - London, 1963. P.217-223.
Kurten B. The Carnivora of the Palestine caves // Acta Zool. Fennica, 1965. N 107. 74 pp.
Kurten B. Die Carnivoren-Reste aus den Kiesen von Siissenborn bei Weimar // PalSontol.
Abh., 1969. Bd.3. Hf.3/4. S.735-756.
Kurten B. Fossil Hyaenidae from the excavation at Stranska Skdla // Anthropos (Brno),
1972. T.20 (n.s. 12). P.113-120.
Kurten B. Crocuta (Hyaenidae) from the Pleistocene of Voigtstedt, Thtlringia (G.D.R.) //
Quartarpalaontologie, 1986. Bd. 6. S.99-100.
Kurten B., Poulianos N. New stratigraphic and faunal material from Petralona Cave with
special reference to the Carnivora // Anthropos (Athen), 1977. Vol.4. N 1-2. P.47-
130.
Kurten B., Poulianos N. Fossil Carnivora of Petralona Cave: status of 1980 // Anthropos
(Athen), 1981. Vol.8. P.9-56.
Linnaeus C. Systema naturae. Ed. X. T.l. - Stockholm, 1758, 824 pp.
Lu Youquan, Li Yi, Jin Changzhu. Mammalian remains from the Late Pleistocene of Wurji,
Nei Mongol // Vertebrata Palasiatica, 1986. Vol.24. N 2. P. 152-162 (на кит. яз. с
англ, рез.)
Matsumoto Н. On some fossil mammals from Sze-chuan, China // Sci. Reports Tohoku
Imper. University. Ser.2 (Geol.), 1915. T.3. P.1-28.
Mottl M. Neuer Beitrag zum Hystrix-Horizont Europas // Ann. Naturhistor. Mus. Wien,
1967. Bd.71. S.305-327.
Musil R. Die Hohle "Sveduv Stul" Ein typischer Hohlenhyanenhorst // Anthropos (Brno),
1962. T.13. S.97-260.
Newton E.T. On the occurrence of the cave hyaena in the "Forest Bed" at Corton Cliff,
Suffolk//Geol. Mag., 1883. Vol.10. P.404-406.
Owen R. On fossil remains of mammals found in China // Quarterly J. Geol. Soc. London,
1870. Vol.26. P.417-434.
Ozansoy F. Etude des gisements continentaux et des mammiferes du C£nozoique de
Turquie//Мёт. Soc. Geol. France, 1965. T.l02. P.1-92.
Pei W.C. On the Carnivora from Locality 1 of Choukoutien // PaleontoL Sinica. Ser.C.,
1934. Vol.8. Facs.l.P. 1-166.
Pei W.C. The Upper Cave Fauna of Choukoutien // Palaeont. Sin. N. Ser. C., 1940. Vol.
10. P.1-84.
Schutt G. Die Hyftnen der Mosbacher Sande (Altpleistozan, Wiesbaden/Hessen) mit einem
Beitrag zur Stammesgeschichte der Gattung Crocuta // Mainz Naturw. Arch., 1971.
Bd.10. S.29-76.
Serres M. de, Dubreuil J., Jeanjean B. Memoires sur des especes d'hyenes fossiles
dёcouvertes dans les cavernes de Lunel-Viel // Мёт. Mus. Nat. Hist. Natur., 1828.
T.l7. P.269 -312.
64
Soergel И7 Hyaena brevirostris Aymard und Hyaena ex aff. crocutta Erxl. aus den Kiesen
von Suessenborn // Zeitschr. Deutschen Geol. Gesell., 1936. Bd 88. S. 525-539 +
4 Taf.
Soergel W. Die Stellung der Hyaena spelaea Goldf. aus der Lindentaler HySnenhohle bei
Gera//Beitr. Geol. Thiir., 1937. Bd.4. Hf.5. S. 171-189.
Sun J.P The palaeoenvironmental changes and macromammal evolution during the
Pleistocene in East Asia//Korean J. Quatern. Res., 1988. Vol.2. N 1. P.51-86.
Sutcliffe A.J. Adaptations of spotted hyaenas to living in the British Isles // Bull. Mammal.
Soc., 1969. Vol.31. P.1-4.
Viret J. Le loess a bancs dur?is de Saint-Vallier (Drome) et sa faune de mammiferes
Villafranchiens//Nouv. Arch. Mus. Hist. Natur. Lyon, 1954. T.4. P. 1-200.
von den Driesch A. A giude to the measurement of animal bones from archaeological sites
// Peabody Mus. Bull., 1976. N 1. 136 pp.
Weithofer A. Die fossilen Hyanen des Arnotales in Toskana // Denkschriften Akad.
Wissenschaften Wien, 1889. Bd.55. S.337 -360.
Werdelin L., Solounias N. The Hyaenidae: taxonomy, systematics and evolution // Fossils
and Strata, 1991. N 30. P. 1-104.
Zhou X., Sun Y Wang Z., Wang H. Gulongshan Cave site. A Upper Paleolithic site at
Dalian City. Beijing, 1990. 94 pp. (на кит. яз. с англ, рез.)
Zong Guanfu, Huang Xueshi. Note on some mammalian fossils from the Latest Pleistocene
of Xialatuo, Luhuo, Sichuan // Vertebrata Palasiatica, 1985. Vol.23. N 4. P.287-294
(на кит. яз. с англ, рез.)
Summary
G.F.Baryshnikov, N.K.Vereshchagin
A BRIEF REVIEW OF QUATERNARY HYENAS (HYAENIDAE)
OF RUSSIA AND ADJOINING REGIONS
Discoveries of Quaternary hyenas in the former Soviet Union dated to the second half of
the Pleistocene (beginning from the Cromerian) are considered. These are assigned to three
species: Pliocrocuta perrieri (Middle to Late Pleistocene, Ukraine, Transcaucasia, and Middle
Asia), Pachycrocuta brevirostris (Middle Pleistocene, Transcaucasia, Kazakhstan, Tajikistan,
and Transbaikalia), and Crocuta spelaea (Middle to Late Pleistocene). The last species had a
wide distribution, extending from Moldova and the Crimea in the west up to the Primorski Krai
in the east. C. spelaea penetrated northwards in the Middle and Late Pleistocene but never
migrated further than 58 degrees N. In the Late Pleistocene the most extensive expansion of the
species occurred in the inter-Wurm (Bryanskian, Karginian) interstadial. In the Crimea and in
the Caucasus C. spelaea became extinct by the beginning of the Upper Paleolithic, but in the
southern Urals and in the Altai it survived until the main climatic minimum of the Late
Pleistocene. Data on geochronological and geographic variation of C.spelaea are presented. The
main evolutionary trend was the development of carnivorous specialization of the dentition,
which strengthened in the glacial epochs and became weaker in the interglacials. C spelaea
ussurica subsp. nov. was described from the Upper Paleolithic layer in Geographic Society Cave
in southern Primorski Krai. It differs from the nominative subspecies by larger sizes of molars.
Significant remains of Hyaena hyaena are only known from Holocene deposits from the
Transcaucasia and Middle Asia, which is attributed to the late expansion of distribution of the
species northwards.
5 Зака.,
ISSN 0206-0477 ЧЕТВЕРТИЧНЫЕ И СОВРЕМЕННЫЕ
МЛЕКОПИТАЮЩИЕ ЕВРАЗИИ
СПб., 1996 (ТРУДЫ ЗООЛОГИЧЕСКОГО ИНСТИТУТА РАН, Т. 270)
RUSSIAN ACADEMY OF SCIENCES
PROCEEDINGS OF THE ZOOLOGICAL INSTITUTE
St. PETERSBURG, 1996, VOL. 270
УДК 599.742.5
Г.Ф.Барышников, А.О.Аверьянов
Зоологический институт РАН, Санкт-Петербург
МОЛОЧНЫЕ ЗУБЫ ХИЩНЫХ МЛЕКОПИТАЮЩИХ
(ОТРЯД CARNIVORA)
Часть VI. СЕМЕЙСТВА VIVERRIDAE И HERPESTIDAE
Исследованы молочные зубы 30 современных и 1 вымершего вида виверровых
(Viverridae) и мангустовых (Herpestidae) мировой фауны. Зубы обоих семейств построены
по единому плану и имеют единую зубную формулу, исходную для Camivora. Среди
Viverridae выделяются группы, различающиеся по направлению и уровню эволюционных
преобразований молочной зубной системы. Главные из них - примитивно-насекомоядные
Hemigalinae и Viverrinae (наблюдается продвинутость в миофагии Viverricula - Viverra,
Prionodon Genetta), насекомоядные Euplerinae и всеядные Paradoxurinae (усиление
растениеядности Arctogalidia - Arctictis, Paguma). Обособленное положение занимают
Cynogale (питание рыбой и водными беспозвоночными) и Nandinia (специализированный
миофаг, показывающий черты сходства с Felidae). В семействе Herpestidae различаютсяся
две группы - Galidiinae (более специализированные миофаги) и Herpestinae. У последних
развитие шло от примитивного уровня (At Пах, Bdeogale) в сторону усиления миофагии
(Icheumia, Herpestes) или же энтомофагии (Mungos, Crossarchus, Cynictis, Rhynchogale). В
целом Herpestidae эволюционно более продвинуты по строению молочных зубов, чем
Viverridae.
Данная работа продолжает серию исследований молочной зубной
системы хищных млекопитающих (Барышников, Аверьянов, 1990, 1992,
1993а, б, 1995). Аелуроидные хищники близких семейств Viverridae и
Herpestidae наиболее разнообразная и богатая видами группа в отряде
Camivora, сравнимая по разнообразию пищевых адаптаций лишь с
Mustelidae. В целом это примитивные хищники, характеризующиеся, в
частности, длительным периодом функционирования молочной зубной
системы. Благодаря последнему обстоятельству молодые экземпляры виверр
66
и мангустов довольно обычны в музеях мира, однако описания их молочных
зубов в литературе редки (Mivart, 1882; Leche, 1915; Butler, 1946).
Палеонтологическая летопись виверровых и мангустовых, как и
большинства других тропических и субтропических млекопитающих, весьма
фрагментарна. Несмотря на это, имеются характеристики молочных зубов
для ряда вымерших форм (Dehm, 1950; Petter, 1967; Beaumont, 1967, 1974;
Petter, Heintz, 1970; Ginsburg, 1977; Fejfaretal., 1987).
МАТЕРИАЛ
Изучены 105 черепов с молочными зубами 30 современных видов и 2 экз. 1
вымершего вида виверр и мангустов из коллекций Зоологического института РАН в
Санкт-Петербурге (далее ЗИН), Зоологического музея Московского государственного
университета в Москве (ЗММУ), Естественно-исторического музея (Naturhistoriska
Riksmuseet) в Стокгольме (NRM), Зенкенбергского музея (Senckenberg Museum) во
Франкфурте-на-Майне (SM), университета Клод-Бернар (Universite Claude-Bernard Lyon I)
в Лионе (UCB), Национального музея естественной истории (Museum National d'Histoire
Naturelle) в Париже (MNHN) и Музея естественной истории Фильда (Field Museum of
Natural History) в Чикаго (FMNH).
Nandinia binotata, NRM A 580146 ? , Мазизи (Masisi), Конго; MNHN №№ 1887-449
и 1962-1704, оба без местонахождения (б/м).
Viverricula indica, ЗИН №№ 270 и 273, о. Ява, Индонезия, 8190, о. Тайвань, Китай;
ЗММУ №№ 53328 ?, б/м, S-132771, Вьетнам, S-137605 ? и S-144515 сГ Южный Вьетнам;
NRM №№ А 582015 и А 595030, б/м, А 584194, Сикара (Sikara), Мадагаскар и А 590436
сГ, о. Ява.
Civettictis civetta, NRM №№ А 580147 сГ, Мазизи, Конго и А 580331, Какамега
(Kakamega forest, N.Kavirondo).
Viverra zibetha, ЗИН №№ 2283 и 5800, г.Фу-Чао, Южный Китай, 5340, б/м; ЗММУ
S-133456 $, Северный Вьетнам; NRM №№ А 584026, б/м, А 591160 ?, Тонкин (N.Tonkin
Chapa) и А 591702 ? Китай.
Viverra modica, UCB №№ 65569 и 65571, изолированные D/4, средний миоцен
(астараций, MN 7), Ла Грив-Сан-Альбан (La Grive-Saint-Alban), Франция.
Genetta servalina, NRM №№ A 580217 сГ, Бени (Beni), Конго, A 580220 сГ, Люленга
(Lulenga, Kivu District, foot of Mikeno), A 581145 сГ, Мазизи, Конго; MNHN 1983-092,
Экваториальная Гвинея.
Genetta tigrina, ЗИН 1363, Вост. Африка; NRM №№ 580288 и 580326 - Какамега,
А 590085 и А 601201 - Кибоното (Kibonoto) [г. Килиманджаро] и А 590144 сГ, Мазизи
(Mazizi), Конго.
Genetta dongolana, ЗИН №№ 8185, Египет и 14684, Зап. Африка.
Pseudogenetta villiersi, MNHN 1977-694, Сенегал.
Eupleres goudoti, MNHN 1880-2542, Мадагаскар.
Paradoxurus hermaphroditic, ЗИН №№ 5801, Калькутта, Индия, 264, 8193 и 8195-
8197, о. Ява, Индонезия, 34288, зоопарк; ЗММУ №№ S-115212 ? и S-115213 сГ, пров. Ха-
Шон-Бинь, Вьетнам; NRM №№ А 581114 ? и А-590121 ?, Хат Санук (Hat Sanuk,
Siamenska Malacca), A 582714, о. Ява, A 594555 и A 604734, б/м, A 606098 сГ, Аннам
(C.Annam Lao-Bao).
5’
67
Paguma larvata, ЗИН №№ 5793 ?, 8186 ? и 8188 сД ущ. Хо-Цзи-Тоу, Китай; ЗММУ
№№ S-93390, Гонконг и S-133458 ?, Северный Вьетнам; NRM А 595230, Саравак
(Sarawak).
Arctogalidia trivirgata, ЗИН 8198 сД о. Борнео, Индонезия; FMNH №№ 98650 ? и
98651 сД Паханг (Pahang, Bentong), Малайзия.
Arctictis binturong, NRM A 582332, Талок Бетонг (Talok Belong), о. Суматра и А
595046 ?, б/м; MNHN 1884-229, Мархе (Marche), Филиппинские о-ва.
Hemigalus derbyanus, ЗИН 5802, о. Борнео, Индонезия; MNHN 1962-1603, Пуло-
Пинанг (Poulo-Pinang), Малайзия.
Galidia elegans, ЗИН 1628 ?, Мароваре (Marovare), Мадагаскар; MNHN 1-1278,
Мадагаскар.
Galidictis vittata, MNHN 1-1248, "Южная Африка" [Мадагаскар].
Salanoia unicolor, MNHN 1962-326, Мадагаскар.
Atilax paludinosus, NRM A 592005, Донга Сабу к (Donga Sabuk), бывшая Брит. Вост.
Африка.
Rhynchogale melleri, NRM A 585397, б/м. [Африка]
Herpestes ichneumon, NRM №№ A 580319, A 580322, A 580334, A 590300, A 590333,
Какамега, A 580640 ?, p. Яла (Yala River, N.Kavirondo), A 581062 и A 591152, Кибоното,
близ г. Килиманджаро [Кения]; MNHN CG 1974-173 ?, Букоко (Boukoko, La Maboke)
[Экваториальная Африка].
Herpestes edwardsi, ЗИН №№ 8173 сД 8174 ? и 8179, г.Бадман, Иран.
Herpestes javanicus, ЗММУ S-144510, Южный Вьетнам.
Herpestes sanguineus, NRM №№ А 581295 ?, г. Магали (Magalies Berg, Transvaal),
ЮАР и А 590729 сД бывш. Брит. Вост Африка.
Suricata suricatta, ЗИН 283, Южная Африка; SM 19421, зоопарк [Южная Африка].
Bdeogale crassicauda, NRM А 580166, Кисмагу (Kismagu), бывшая Брит. Вост.
Африка.
Bdeogale nigripes, MNHN 1885-220, Габон.
Crossarchus obscurus, ЗИН 8700, оз. Виктория, Вост. Африка; NRM №№ А 580519,
Либерия и А 590346, Камерун.
Mungos mungo, ЗММУ S-58223 сД б/м; NRM №№ А 581985 Дамарсаландет
(Damarsalandet), А 591985 сД Дамараланд (Damaraland) и А 590231 ?, Бени (Beni), Конго.
Ichneumia albicauda, ЗИН №№ 5795, б/м и 14687, Зап. Африка; NRM №№ А 580271
сД Кикуйу (Kikuyu), Кения, А 580412 сД Рутсхуру (Rutshuru) и А 581000, Какамега;
MNHN №№ 1892-2052, Хаут Кемо (Haut Кето) и 1962-2077, б/м.
Cynictis penicillata, NRM 581303 ?, б/м. [Южная Африка]
МЕТОДИКА
Состав и объем таксонов современных виверровых принят нами
согласно системе, предложенной Михелис (Michaelis, 1972), с некоторыми
изменениями (см. Wenzel, Haltenorth, 1972 и др.). Вслед за Михелис и
другими авторами мы исключаем из виверровых мадагаскарскую фоссу
(Cryptoprocta ferox\ но делим виверровых на два семейства - Viverridae и
Herpestidae. В семействе Viverridae мы принимаем 5 современных
подсемейств, в семействе Herpestidae - 2 (см. Wozencraft, 1993; Павлинов и
68
др., 1995). Русские названия мангустов и виверр приведены в основном по
Соколову (1979).
Номенклатура бугорков и схема промеров молочных зубов опублико-
ваны нами ранее (Барышников, Аверьянов, 1990, рис. 2, 3). Для промеров в
таблицах использованы следующие обозначения: Н- высота, L- длина, Lmts -
длина метастиля, Lt - длина талона, Ltl - длина талонида, IV - ширина, Wt -
ширина талона, JVtl - ширина талонида, IVtг - ширина тригонида. Все промеы
в тексте и таблицах даны в мм. Промеры молочных коренных зубов виверр и
мангустов обрабатывались одним из методов многомерной статистики
анализом главных компонент. Расчеты производились на IBM PC AC386SX с
помощью модулей DATA, FACTOR и GRAPH из пакета статистических
программ SYSTAT.
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
ОТРЯД CARNIVORA BOWDICH, 1821
СЕМЕЙСТВО VIVERRIDAE GRAY, 1821
ПОДСЕМЕЙСТВО INSERTAE SEDIS
Род Herpestides de Beaumont, 1967
Herpestides antiquus (de Blainville, 1842)
Описание (no Beaumont, 1967, p.93; pl.2, fig.7a,b). Коронка D/3
трехзубчатая, с крупными передним и задним добавочными бугорками.
Последний относительно ниже, чем у Herpestidae. Коронка в задней части не
расширена, без ’’талонида”
D/4 мощный, почти вдвое длиннее D/3. Тригонид режущий, метаконид
очень небольшой. Все три бугорка талонида мощные, выше метаконида (у
Herpestidae ниже). Бассейн талонида небольшой.
Замечания. Описанный материал происходит из Montaigu-le-BIin, Франция,
нижний миоцен, агений (MN 2). На основании изучения признаков базикраниума
Herpestides помещен в основание ствола Viverridae (Hunt, 1991).
ПОДСЕМЕЙСТВО NANDINIINAE РОСОСК, 1921
Род Nandinia Gray, 1843
Nandinia binotata (Gray, 1830) - Пальмовая цивета
Описание (рис. 1; изображения молочных зубов имеются также в
работе Leche, 1915, fig. 4-7). Молочные резцы мелкие. ^/2/игольчатый. dI3!
69
Рис. 1. Молочные зубы Nandinia binotata, NRM А 580146:
а, г - снаружи; б, д - со стороны жевательной поверхности; в, е - изнутри
Fig. 1. Deciduous teeth of Nandinia binotata, NRM A 580146:
а, г - lateral; б, d - occlusal; в, e - labial views
70
крупнее, со слабо расширенной приостренной коронкой, dI/2 игольчатый.
dI/З немного шире, с латеральным бугорком.
Корни верхних клыков почти одной толщины с основанием коронки,
нижних - немного мощнее. dCM острый, вершина его направлена вниз. dC/\
слабо изогнутый, островершинный, без четких валиков и бугорков.
D2/ простой, довольно крупный, очень узкий зуб, с одной вершиной и
без дополнительных элементов. Длиннее £>4/. Сближен с D3/. Корней 2.
D3/ хищнического типа. Передний отдел коронки довольно крупный, с
четкими, но невысокими парастилями А и В (на экземпляре, изображенном
Лехе, оба парастиля отсутствуют). Метастиль режущий, умеренно длинный
(36% общей длины), но невысокий. В основании метастиля имеется
короткий цингулюм, несущий 3 сравнительно крупных бугорочка. Талон
маленький, короткий и узкий, без бугорка протокона (MNHN 1962-1704;
Leche, 1915, fig.5), либо более крупный, расширенный в лингвальной части
(NRM А 580146). Смещен вперед от вершины амфикона. Корней 3.
D4/ небольшой, треугольный. Эктофлексус и стилярная полка хорошо
развиты, парастилярная лопасть сильно выдается лабиально, либо почти не
развита (Leche, 1915, fig.5). Паракон и метакон небольшие, невысокие, при-
мерно равновеликие (MNHN 1962-1704), либо метакон заметно меньше
(NRM А 580146). Талон маленький, слабо моделированный, либо заметно
более крупный. Несет небольшой зубец протокона.
D/2 небольшой, одновершинный или же сравнительно крупный, с
дополнительными передним и задним зубцами и острым бугорком на
"талониде" (MNHN 1962-1704 и NRM А 580146; Leche, 1915, fig.6, 7).
Корней 2.
D/3 лишь немногим меньше D/4. Зуб сравнительно широкий, особенно
в заднем отделе (экз. MNHN), либо существенно более узкий (экз. NRM).
Главная вершина высокая и острая. Впереди нее расположен дополнитель-
ный острый зубец. Иногда еще один более мелкий зубчик присутствует на
задней стенке главного бугорка; на экземпляре NRM А 580146 и изобра-
женном Лехе этот зубец ("метаконид") очень крупный и сравнительно высо-
кий. Один из изученных образцов из MNHN без "талонида" На другом эк-
земпляре и на экземпляре NRM А 580146 в задней части коронки имеется
"талонид", в который с главного бугорка спускается гребень. Лингвальнее
последнего находится небольшой бассейн. По заднему краю "талонида" про-
ходит гребешок. Корней 2.
D/4 сравнительно небольшой, массивный, относительно широкий,
похож на М/\, В основании параконида имеется короткий прецингулид, об-
разующий спереди крупный острый зубец. Метаконид небольшой, низкий,
слабо смещен назад и отделен от протоконида долинкой. Талонид короткий,
с маленьким бассейном, без четко различимых бугорков (экз. MNHN), либо с
отчетливыми гипоконидом (наиболее крупный), гипоконулидом и энтокони-
Дом (самый маленький) (NRM А 580146).
71
ПОДСЕМЕЙСТВО VIVERRINAE S. STR.
Род Viverricula Hodgson, 1838
Viverricula indica (Desmarest, 1804) - Малая цивета
Описание (рис. 2). Молочные зубы довольно изменчивы по раз-
мерам (табл. 1). Резцы мелкие, простого строения, без лингвального пояска.
dI3l клыкоподобный. На dH\-3 имеется очень слабо развитый латеральный
зубчик.
Клыки невысокие, слабо изогнутые. Лингвальный поясок на dC/\ ко-
роткий, образует спереди бугорок.
D2/2 сравнительно высокие, но короткие, их коронки слабо расширены
в задней части. На D2I имеются едва различимые передний и задний до-
бавочный зубчик (задний может быть заметно крупнее: экз. ЗИН 8190 и
Leche, 1915, fig. 10), на D/2 передний зубчик более отчетливый, задний
зубчик часто отсутствует. Коронка D/2 наклонена вперед, гребень, спускаю-
щийся с главной вершины назад, образует выпуклость в основании коронки.
Корней 2.
D3/ обычного хищнического типа. Область парастиля А очень короткая,
хотя сам парастиль обычно хорошо развит в виде острого зубца; реже пред-
ставлен 2-3 мелкими бугорками (NRM А 584194). Парастиль В хорошо раз-
вит, намного выше парастиля А и отделен от амфикона глубокой бороздкой.
В основании метастиля имеются короткие лабиальный и лингвальный цингу-
люмы. Метастиль длинный (39-46% от общей длины зуба, Л/=42.0±0.8%,
л=9). Талон очень небольшой, но хорошо обособлен от основания амфикона.
Имеются отчетливый протокон в виде невысокого продольно вытянутого бу-
горка, и короткая препротокриста, идущая до основания парастиля В. Прото-
кон иногда двухзубчатый (ЗММУ S-132771).
D4/ треугольной формы, варьирует по размерам (на экземпляре ЗИН
8393 очень короткий, ширина зуба меньше длины). Эктофлексус неглубокий.
Стилярная полка широкая. Парастилярная лопасть хорошо развита, заметно
выдается лабиально. Паракон хорошо развит, высокий. Метакон заметно
меньше и ниже, гребневидный. Талон сравнительно небольшой, с малень-
ким, но отчетливым бассейном. Бугорок протокона крупный, но невысокий.
D/3 трехзубчатый, сравнительно длинный (77-97% от длины £>/4,
Л/=82.3±2.1%, и=9) и очень узкий, лишь в задней части коронка слегка
расширена. Передний и задний добавочные зубцы относительно крупные,
примерно равные по размерам и по высоте, отделены от основной вершины
глубокими бороздками. От основания главной вершины к "талониду" по
лабиальному краю тянется отчетливый добавочный гребневидный бугорок
(иногда очень высокий: ЗММУ S-53328). С лингвальной стороны на "тало-
ниде" постцингулид образует продольно вытянутый острый зубчик.
D/4 обычного хищнического типа. Режущие гребни протоконида и па-
раконида хорошо развиты, разделены очень глубокой бороздкой. Параконид
72
Рис. 2. Молочные зубы Viverricula indica, NRM А 584194:
а, г - снаружи; о - со стороны жевательной поверхности; в, е - изнутри
Fig. 2. Deciduous teeth of Viverricula indica, NRM A 584194:
а, г - lateral; б, о - occlusal; «, e - labial views
73
Таблица 1. Размеры молочных зубов у Nandinia, Eupleres и некоторых Viverrinae
Table 1. Measurements of deciduous dentition in Nandinia, Eupleres and some Viverrinae
Промеры, Nandinia bmotata Viverricula indica Civet ticli.s civella Pseudogenetta thierryi Eupleres goudotii
MM «=1-3 | M±m n 1 lim | М±П! n=2 n=l n= 1
Верхние зубы
IM L 7 0.6-1.5 0.9±0.12 1.4, 1.7 0.4
W 7 0.5-1.9 0.9±0.18 1.3, 1.4 0.6
H 7 1.0-2.1 1.4±0.14 2.5, 3.0 0.7
72/ L 0.7, 0.9 8 0.8-1.9 l.l±0.13 1.7, 1.9 0.8 0.5
W 0.6, 0.9 8 0.6-2.4 1.1 ±0.20 1.7, 2.0 0.7 0.7
H 1.2, 1.3 8 1.2-3.1 1.7±0.21 3.0, 3.2 1.1 1.0
73/ L 1.2, 1.2 9 1.1-2.0 1.3±0.09 2.2, 2.8 1.2 0.8
W 1.2, 1.2 9 1.2-3.1 1.6±0.26 2.3, 2.3 1.4 0.5
H 1.6, 2.0 9 2.2-3.6 2.5±0.16 3.4, 5.8 2.1 0.7
CM L 2.2, 2.7 11 2.1-3.7 2.6±0.14 3.6, 4.0 2.3 0.9
W 1.6, 2.4 11 1.3-2.5 1.8±0.10 2.6, 2.8 1.5 0.8
Продолжение табл. 1
Промеры, мм Nandmia bmotata Viverncula indica Civet lid is civelta Pseudogenetta thierryi Eupleres goudolii
/1=1-3 | M±m " 1 lim | Mdm n=2 n=l n=l
Н 5.3, 5.4 11 5.7-10.0 7.1±0.45 9.0, 11.2 5.0 1.1
DU L 3.4, 3.7 9 3.1-6.1 4.1 ±0.31 6.3, 6.5 3.9 2.4
W 1.0, 1.8 9 1.1-2.3 1.4±0.13 2.4, 3.0 1.2 0.8
н 2.1, 2.2 9 2.5-4.1 3.1±0.17 5.1. 6.0 2.4 1.2
D3/ L 5.2, 6.4 9 5.7-9.6 6.9±0.39 9.7, 11.0 5.8 3.2
Lmts 1.9, 2.6 9 2.5-4.1 2.9±0.17 3.6, 4.1 2.0 0.9
Lt 1.1, 1.1 9 1.2-2.0 1.5±0.09 2.5, 3.0 1.5 0.7
W 2.9, 2.9 9 2.8-3.8 3.3±0.11 5.2, 5.6 3.5 1.7
Wt 1.1. 1.3 9 0.7-1.5 l.l±0.10 1.5, 2.3 1.6 0.4
H 2.8, 3.3 9 3.4-5.2 3.9±0.26 5.0, 6.1 3.0 1.5
D4/ L 3.0, 3.0 9 3.2-6.2 4.1±0.28 7.9, 7.9 3.6 3.0
Ж 3.7, 4.3 9 4.1-6.4 4.8±0.22 7.8, 9.4 4.0 2.2
H 1.6, 1.9 9 1.7-2.4 2.1±0.07 3.1, 3.8 1.9 1.1
сл
Промеры, Nandima bmotata
мм n=l-3 | M±m n
7/1 L 0.1 6
W 0.2 6
77 0.5 6
7/2 L 0.3, 0.8 8
W 0.4, 0.7 8
77 0.6, 0.8 8
7/3 L 0.7, 0.9 9
W 0.5, 1.0 9
77 1.2, 1.7 9
C/1 L 2.6, 3.0 11
W 1.4-1.8 1.6±0.11 11
77 5.1-5.3 5.2±0.44 9
Продолжение табл. 1
Viverricula indica Ci vein cl is civelta Pseudogenet ta th terry i Eupleres goudolii
1 l''n 1 Mini n=2 n=l n=l
Нижние зубы
0.4-1.0 0.7±0.11 0.9, 1.0
0.4-1.2 0.7±0.13 0.8, 1.0
0.8-1.4 1.2±0.10 1.9, 2.5
0.6-1.5 0.8±0.11 1.4, 1.5 0.6
0.6-2.0 0.9±0.17 1.4, 1.7 0.6
0.7-2.3 1.2±0.17 2.3, 2.8 0.8
0.7-1.7 l.l±0.12 1.8, 2.2 1.0
1.1-2.5 1.5±0.16 2.1,3.1 1.0
1.2-3.1 1.6±0.19 3.3, 4.3 1.2
2.2-3.5 2.5±0.12 4.2, 4.2 1.9
1.2-2.3 1.6±0.09 2.3, 2.9 1.3
4.8-7.7 6.1±0.60 8.0, 9.9 4.0
Окончание табл. А
Промеры, Nandmia bmotata 1'iverricula indica Civettictis civelta Pseudogenetta thierryi Eupleres goudotii
мм /1=1-3 M±m » 1 lim | M±m n=2 n=l n=l
£)/2 L 3.0-3.6 3.4±0.20 8 3.4-6.0 4.0±0.30 6.1, 6.9 3.8
W 1.1-1.8 1.5±0.21 8 1.0-2.1 1.3±0.12 2.3, 2.7 1.3
Н 2.0-2.6 2.2±0.20 8 3.1-4.5 3.7±0.14 5.0, 5.8 2.5
D/3 L 3.6-5.2 4.4±0.46 9 4.6-7.4 5.5±0.27 8.4, 8.7 5.0
W 1.5-2.3 1.8±0.23 9 1.6-2.8 1.9±0.12 3.2. 3.5 1.6
Н 2.0-3.5 2.7±0.45 9 3.1-5.0 4.1±0.17 5.0, 6.7 3.2
£)/4 L 4.4-5.8 5.2±0.43 10 5.7-10.0 6.7±0.39 10.5, 12.0 5.4
Ltl 1.0-1.5 1.2±0.15 10 2.1-4.2 2.6±0.20 4.5. 5.9 1.4
Wtr 2.1-2.6 2.3±0.14 10 2.4-3.9 2.8±0.13 4.9, 5.4 2.4
Wtl 1.9-2.1 2.0±0.06 10 2.3-3.9 2.7±0.14 5.0, 5.5 2.2
Н 2.5-3.5 2.9±0.32 10 4.0-5.9 4.5±0.18 5.9, 6.2
довольно низкий, в его основании с лабиальной стороны имеется хорошо
развитый прецингулид (на конечной стадии окклюзии в этот прецингулид
упирается вершина амфикона). Метаконид высокий, иногда даже выше пара-
конида, хорошо обособлен от протоконида. Талонид варьирует по длине (34-
46% от длины зуба, Л/=38.9±1.0%, л=10), с хорошо развитыми бугорками и
небольшим бассейном. Энтоконид очень высокий, шиловидный, часто на-
много выше других бугорков талонида (у других виверр энтоконид обычно
ниже гипоконида), либо такой же высоты, как гипоконид. Гипоконулид в ви-
де небольшого бугорка, сближен с гипоконидом.
Род Civettictis Pocock, 1915
Civettictis civetta (Schreber, 1776) - Африканская цивета
Описание (по Mivart, 1882, fig.4). Молочные резцы сравнительно
крупные, с простыми конусовидными коронками. <7/3/3 лишь немного круп-
нее предыдущих резцов.
dC\/\ крупные, со слегка изогнутыми коронками, без лингвальных
поясков в основании коронки.
D2/2 крупные, с массивными главными вершинами и небольшими до-
бавочными зубчиками. Коронка D/2 слегка расширена в задней части.
Корней 2.
D3/ хищнический зуб. Коронка почти симметричная: талон расположен
в центре лингвальной стороны зуба, напротив амфикона; парастилярная об-
ласть длинная, равна по величине метастилярной части зуба. Лабиальный
край коронки вогнутый. Парастиль В, скорее всего, небольшой. Талон корот-
кий, видимо, без отчетливого протокона.
D4/ крупный, в форме равнобедренного треугольника. Эктофлексус не-
глубокий. Стилярная полка короткая. Паракон немного крупнее метакона.
Имеются сравнительно очень крупные параконуль и метаконуль. Протокон
крупный.
D/3 длинный, с высоким передним добавочным зубцом и гораздо более
низким задним. Коронка расширена в задней части, где задний цингулид об-
разует "талонид"
D/4 хищнический зуб с мощными, хорошо развитыми бугорками. На
протокониде и паракониде хорошо выражены режущие гребни. Метаконид
крупный, почти равен по величине паракониду. В основании лабиальной
стороны параконида имеется отчетливый прецингулид. Талонид с обширным
бассейном и мощными гипоконидом и энтоконидом.
78
Род Viverra L., 1758
Viverra zibetha L., 1758 - Большая цивета
Описание (рис. 3, вклейка). Молочные резцы сравнительно
крупные, без лингвального пояска и добавочных зубчиков. dI3/ крупнее
других резцов, клыкоподобный.
dC\/\ с невысокой сильно изогнутой коронкой. В основании коронки
dC/\ имеется лингвальный поясок.
D2/2 сравнительно крупные (табл. 2), вытянутые в длину, двухкорне-
вые. Коронка D2/ без добавочных зубчиков, в форме равностороннего тре-
угольника. На D/2 имеются очень маленькие передний и задний добавочные
зубчики.
D3/ обычного хищнического типа. Область парастиля А сравнительно
короткая, но сам бугорок довольно крупный. Парастиль В отчетливый, за-
метно ниже парастиля А. Метастиль относительно длинный (37.9-44.4% от
общей длины зуба, Л/=42.5±0.86%, л=7). Талон очень небольшой, без прото-
кона, опоясан цингулюмом.
Коронка DA/ треугольной формы. Эктофлексус практически не выра-
жен. Имеется лабиальный цингулюм, иногда несущий два маленьких бугорка
(NRM А 591160). Парастилярная лопасть хорошо развита, сильно выдается
вперед и лабиально. Паракон и метакон в виде низких продольных гребней,
первый из них заметно длиннее. Талон сравнительно крупный, с обширным
бассейном. Протокон невысокий, гребневидный. Иногда имеются отчетли-
вые парастиль и метаконуль (ЗММУ S-133456).
D/3 сравнительно низкий и короткий (70.0-77.8% от длины £>/4,
Л/=74.8±1.09%, п=7). Передний добавочный зубец хорошо развит, отделен от
главной вершины глубокой бороздкой. Задний добавочный зубец обычного
типа, крупный, либо едва заметен и сливается с задним краем основной вер-
шины (NRM А 591160). В основании коронки сзади на лабиальной стороне
обычно имеется дополнительная вершина, в виде полого поднимающегося
гребня (среди других Viverridae отмечен еще только для Viverricula). Пост-
цингулид очень короткий.
DIA обычного хищнического типа. Режущие гребни протоконида и па-
раконида хорошо развиты. Параконид очень низкий, ниже метаконида. В
основании параконида с лабиальной стороны имеется короткий прецингулид.
Метаконид сравнительно небольшой, не достигает половины высоты прото-
конида. Талонид с тремя невысокими, примерно равными по размерам бу-
горками, и с отчетливым бассейном. Гипоконид и энтоконид несколько вытя-
нуты продольно. Энтоконид может быть намного выше гипоконида (экз.
ЗММУ S-133456). Гипоконулид отчетливый, выдается назад.
Сравнение. Изолированные D/4 из среднемиоценового (астара-
Ний, MN 7) местонахождения Ла-Грив-Сант-Альбан, Франция, отнесенные
79
Таблица 2. Размеры молочных зубов у Viverra, Paradoxurus и Hemigalus
Table 2. Measurements of deciduous dentition in Viverra, Paradoxurus and Hemigalus
Промеры, Viverra zibelha Viverra modica Paradoxurus hermaphroditus Hemigalus derbyanus
MM n | lim | Mun n=2 n | lim | Msm n=\-2
Верхние з у 6 i >1
71/ L 5 1.0-1.7 1.3±0.12 9 1.0-1.1 1.0±0.02 0.8,1.1
W 5 1.2-1.5 1.4±0.06 9 0.8-1.3 l.l±0.06 0.9,1.0
77 5 1.4-2.2 1.8±0.13 9 1.2-1.9 1.5±0.07 1.2,1.3
72/ L 7 1.4-2.0 1.7±0.08 11 1.1-1.3 1.2±0.02 1.0,1.1
W 7 1.5-2.6 2.0±0.12 11 1.0-1.4 1.2±0.04 1.2,1.3
77 7 1.5-3.0 2.3±0.17 11 1.3-2.1 1.8±0.09 1.4,1.4
73/ L 7 1.6-2.5 2.1±0.11 14 1.4-2.0 1.7±0.04 1.2,1.4
W 7 2.0-3.2 2.4±0.14 14 1.3-2.0 1.7±0.05 1.7,1.9
H 7 2.8-3.6 3.3±0.12 14 2.3-3.4 2.7±0.09 1.8,2.0
С1/ L 7 3.4-4.4 3.8±0.13 14 2.6-3.4 3.0±0.07 2.8,3.1
W 7 2.0-2.9 2.4±0.11 14 1.2-2.3 1.9±0.07 1.6,1.8
77 7 8.1-9.5 8.7±0.21 13 5.3-8.0 6.6±0.23 5.1,5.8
DU L 7 5.1-6.8 5.9±0.24 12 3.1-5.0 4.0±0.16 4.4,5.2
W 7 1.9-2.4 2.H0.06 12 1.4-2.1 1.7±0.06 1.6,1.9
77 7 4.1-4.5 4.3±0.06 12 2.3-3.8 3.2±0.12 3.1,3.3
D3/ L 7 9.0-10.6 10.0±0.24 13 6.3-7.5 6.9±0.10 6.2,7.1
Lmts 7 3.9-4.5 4.2±0.09 13 2.3-3.2 2.7±0.07 1.8,2.0
Lt 7 1.4-2.5 2.0±0.14 13 1.7-2.5 2.0±0.06 1.4,2.6
W 7 4.2-4.9 4.4±0.10 13 3.6-4.9 4.0±0.10 3.6,4.0
Wt 7 0.8-2.1 1.5±0.18 13 1.3-2.2 1.6±0.07 1.4,1.7
77 7 4.6-6.1 5.3±0.20 13 3.3-4.4 3.8±0.11 3.3,4.0
80
Продолжение табл. 2
Промеры, мм Viverra zibetha Viverra modica Paradoxurus hermaphrodilus Hemigalus derbyanus
п | lim | Mun n=2 n | lim | M^m n=l-2
D4/ L 7 5.6-7.2 6.2±0.19 12 4.8-6.0 5.5±0.11 5.0,5.5
W 7 6.0-8.4 7.3±0.29 12 4.8-6.3 5.6±0.13 4.8,5.0
H 7 1.4-3.3 2.6±0.25 12 1.5-3.1 2.3±0.13 2.7,2.7
Нижние зубы
7/1 L 5 0.8-1.1 0.9±0.05 8 0.4-0.8 0.6±0.05 0.7
W 5 0.6-1.3 0.9±0.12 8 0.4-0.8 0.6±0.05 0.9
H 5 1.2-2.0 1.4±0.15 8 0.5-1.7 l.l±0.13 1.2
7/2 L 7 1.1-1.6 1.3±0.06 9 0.7-0.9 0.8±0.02 1.1
W 7 0.7-1.8 1.3±0.13 9 0.7-1.1 0.9±0.05 1.4
H 7 1.6-2.4 2.0±0.12 9 1.0-1.4 1.2±0.04 1.7
7/3 L 7 1.6-2.0 1.8±0.06 10 1.1-1.3 1.2±0.02 1.3
W 7 1.4-2.6 2.0±0.14 10 1.2-1.8 1.5±0.06 2.0
H 7 1.7-2.7 2.2±0.16 10 1.5-2.7 2.0±0.11 2.5
C/1 L 7 3.0-3.9 3.4±0.12 12 2.7-3.6 3.1±0.07 2.2
W 7 2.0-2.5 2.3±0.06 12 1.6-2.0 1.8±0.04 1.8
H 7 3.4-8.2 6.7±0.61 12 4.3-6.3 5.4±0.17 5.1
D/2 L 7 5.0-6.4 5.8±0.18 10 3.3-4.5 4.1±0.11 5.0,5.4
W 7 1.7-2.3 2.0±0.09 10 1.4-2.2 1.7±0.07 1.4,1.6
H 7 3.7-5.0 4.1±0.17 10 2.0-4.3 3.H0.19 3.1,4.0
D/3 L 7 7.0-8.1 7.6±0.18 11 5.3-6.3 5.8±0.09 6.1,7.2
W 7 2.4-3.0 2.8±0.08 11 2.3-3.0 2.7±0.08 1.7,2.2
H 7 4.7-5.4 5.1±0.10 11 2.9-4.1 3.5±0.11 3.3,3.9
6
81
Окончание табл. 2
Промеры, мм Viverra zibetha Vi verra modica Paradoxurux hermaphroditus Hemtgalus der by anus
n lim | M-m n=l " 1 1 1™ | Mrm n=\-2
D/4 L 7 9.0-10.9 10.3±0.23 5.6, 6.1 12 7.7-9.3 8.6±0.13 8.5,8.6
Ltl 7 4.1-4.4 4.3±0.04 1.9, 2.2 12 3.0-4.2 3.5±0.09 3.7,3.9
Wtr 7 3.8-4.2 4.0±0.06 2.3, 2.4 12 3.3-4.3 3.8±0.10 2.9,3.4
Wtl 7 3.6-4.0 3.9±0.06 2.3, 2.4 12 3.2-4.4 4.0±0.11 3.0,3.4
Н 7 5.0-6.3 6.0±0.18 3.2, 3.6 12 2.9-4.2 3.5±0.11 3.7,3.7
при описании к Viverra (Viverrictis) modica Gaillard, 1899 (рис. 4; Beaumont,
1974, pl.II, fig.9, 10), по строению и размерам (см. табл. 2) больше напомина-
ют D/4 Viverricula indica, чем представителей рода Viverra. Между режущи-
ми гребнями параконида и протоконида имеется глубокая вырезка, энтоко-
нид шиловидный, выше гипоконида.
Род Semigenetta Helbing, 1927
Semigenetta? elegans Dehm, 1950
Описание (no Dehm, 1950, Abb.225-227). D3/ с крупным пара-
стилем А и, видимо, без парастиля В. Метастиль сравнительно длинный. Та-
лон не широкий, как у Genetta, без протокона. Длина £>3/ 7.1, ширина - 3.5.
На D/3 передний добавочный зубчик очень небольшой, задний - более
крупный и высокий (у Genetta оба зубчика примерно равны по размерам).
Длина D/3 - 5.6.
£>/4 с тригонидом обычного строения (метаконид сравнительно неболь-
шой). Талонид низкий, из бугорков имеется, по-крайней мере, энтоконид.
Длина D/4 - 6.2-6.7, Л/=6.36±0.09, п=5.
Замечания. Материал происходит из местонахождения Wintershof-West,
Германия (по Dehm, 1950 - бурдигал, по Шевырева, 1983 и Вислобокова, 1990 - аквитан
[агений]; нижний миоцен, MN 3). Не исключено, что молочные зубы принадлежат
представителю семейства Herpestidae (для них больше характерна разница в высоте
добавочных бугорков на D/3).
82
Рис. 4. Z?/4 Viverra modica, UCB 65569:
a - снаружи, б - co стороны жевательной поверхности, в - изнутри, г - сзади
Fig. 4. D/4 Viverra modica, UCB 65569:
a - labial, 6 - occlusal, в - lingual, г - posterior views
6’
83
Род Genetta G.Cuvier, 1816
Genetta servalina Pucheran, 1855 - Серваловая генета
Описание (рис. 5). Верхние резцы мелкие. dl\/, d!2/ игольчатые.
d!3/ несколько крупнее, без боковых зубчиков. Нижние резцы очень мелкие,
с двухзубчатой коронкой.
Клыки островершинные, их корни значительно толще коронки (MNHN
1983-092), либо такой же толщины (NRM А 581145). dCM уплощен с боков,
без дополнительных гребней и зубцов. dC/\ почти такой же величины, слабо
изогнутый.
D2/ расположен значительно ближе к 2)3/, чем к dCM. Коронка вытя-
нутая, узкая, с одной вершиной и небольшим бугорком на заднем цингулю-
ме. Корней 2.
2)3/ - крупный хищнический зуб обычного строения. Передний отдел
коронки удлинен, несет сравнительно очень крупный парастиль А. У основа-
ния амфикона с лингвальной стороны расположен четкий зубчик парастиля
В. Талон маленький, без протоконового бугорка. Он лежит несколько кпере-
ди от уровня вершины амфикона. Метастиль режущий, относительно длин-
ный (36-41%, А/=37.8±1.2%, п=4, от общей длины зуба). У его основания с
лингвальной стороны имеется короткий цингулюм, иногда несущий мелкие
бугорки (NRM 581145). Мелкие бугорочки имеются также в передней и
задней частях лабиального цингулюма.
Е>4/ обычного для подсемейства Viverrinae вида. Короче 2)2/. Эктофле-
ксус резкий и сравнительно глубокий, моделированный им передний край
коронки (парастилярная лопасть) заметно выступает лабиально. Стилярная
полка хорошо выражена. Паракон и метакон небольшие, но четкие, пример-
но одинаковые по размерам. Протокон имеет вид островершинного зубца.
D/2 относительно длинный, вытянутый. Высокий главный бугорок
несет едва намечающийся зубчик на задней стенке. В основании коронки
спереди и сзади имеется по небольшому бугорку. Корней 2.
D/3 крупный, трехвершинный, лишь немногим меньше Е>/4. Передний и
задний дополнительные зубцы относительно высокие; задний немного выше
переднего. Оба зубца отделены от главной вершины глубокой щелью. Корон-
ка в заднем отделе слегка расширена и образует короткий "талонид”, кото-
рый несет 2 небольших бугорка (NRM 581 145), либо посередине его прохо-
дит продольный гребешок (MNHN 1983-092).
D/4 умеренно длинный, типичного хищнического вида. Параконид и
протоконид режущие, разделены очень глубокой щелью. В основании пара-
конида спереди и с боковых сторон имеется мощный цингулид. Метаконид
довольно крупный, прислонен к задне-лингвальной стенке протоконида.
Талонид уже тригонида. Он несет крупный гипоконид и небольшой энтоко-
нид (MNHN 1983-092), либо оба бугорка примерно равны по размерам (NRM
84
Рис. 5. Молочные зубы Genetta servalina, NRM А 581145:
а, г - снаружи; б, д - со стороны жевательной поверхности; в, е - изнутри
Fig. 5. Deciduous teeth of Genetta servalina, NRM A 581145:
а, г • lateral; б, d - occlusal; в, e - labial views
85
A 581145). Гипоконид и энтоконид связаны гребнем, тянущимся по заднему
краю коронки. Имеется небольшой внутренний бассейн талонида, смещен-
ный лингвально.
Сравнение. У пятнистой генеты, С tigrina (Schreber, 1776) на D3/
лабиальный цингулюм не выражен, на лингвальном цингулюме в основании
метастиля нет бугорков, метастиль сравнительно длиннее (42-47%,
А/=43.8±1.0%, п=5, от общей длины зуба). На 2)4/ эктофлексус менее глубо-
кий, талон сравнительно короче. На D/2 на задней стенке главной вершины
добавочный бугорок отсутствует.
У обыкновенной генеты, G. genetta (L. 1758) молочные зубы крупнее
(табл. 3). У египетской генеты, G. dongolana (Hemprich et Ehrenberg, 1832) на
D3/ сильнее развиты талон и протокон.
Из среднего миоцена (MN 7) Бени Мелал (Марокко) описан фрагмент
нижней челюсти Genetta sp. cf. G. tigrina c 2)4 (Ginsburg, 1977, pl.Ill, fig. 1).
По небольшим размерам 2)4 (длина 5.2, ширина тригонида 2.25) этот экзем-
пляр ближе к G. tigrina, чем к G. servalina или G. genetta (см. табл. 3). От
современных генет отличается относительно более крупными бугорками
талонида 2)/4.
Из отложений начала позднего плейстоцена (ранний вюрм) грота Геула,
Израиль известен фрагмент нижней челюсти Genetta sp. с D/3,4 (Petter,
Heintz, 1970, pl.I, fig. 11). От современных генет этот экземпляр отличается
менее обособленным задним добавочным зубчиком на D/3. По относительно
крупным размерам зубов (длина DI3 5.5, D/4 6.6) близок к G. genetta (см.
табл. 3). Экземпляр принадлежит, видимо, данному виду, распространенному
ныне в Средиземноморье.
Род Pseudogenetta Dekeyser, 1949
Pseudogenetta thierryi Matschie, 1902 - Ложная генета
Описание. Верхние молочные резцы маленькие, ложковидные.
dI3/ заметно шире dI2l, с внутренним воротничком у основания коронки.
Нижние резцы мелкие, со слабо расширенными коронками. На dI/З хорошо
различим латеральный зубец.
У клыков корни толще коронок. dCM тонкий, острый, дугообразно изо-
гнутый, без дополнительных зубцов и валиков. Вершина dC!\ незначительно
отогнута вверх.
D2/ сближен с D3/. Он узкий, длиннее 2)4/. Основная вершина высокая,
острая. Имеются слабые расширения коронки в передней и задней ее частях.
Задний отдел коронки более длинный, несет продольный режущий эмалевый
тяж. D/2 узкий, относительно длинный зуб, сближен с D/3. Основная вер-
шина его высокая, острая. Имеется дополнительный зубчик на переднем
конце коронки и более слабо развитый - на задней стенке основного конуса.
Корней 2.
86
Таблица 3. Размеры молочных зубов у Genetta
Table 3. Measurements of deciduous dentition in Genetta
Промеры, Genetta tigrina Genetta genetta (jenetta servahna Genetta dongolana
MM n | lim M±m /1=1 n I lim | /7=1
Верхние зубы
/1/ 7 1 0.6 2 0.4,0.6
FT 1 0.5 2 0.4,0.4
77 1 1.0 2 1.0,1.2
72/ 7 1 0.7 1.0 2 0.8,0.8
W 1 0.5 0.9 2 0.6,0.6
H 1 1.1 1.3 2 1.0,1.2
73/ L 5 1.0-1.2 1.Ы0.04 1.5 3 0.9-1.1 1.0±0.06 1.1
W 5 1.0-1.2 l.l±0.04 1.1 3 0.9-1.1 1.0±0.06 1.3
H 5 1.4-2.0 1.6±0.12 1.9 3 1.3-1.7 1.5±0.12 1.7
С1/ L 5 2.0-2.5 2.2±0.08 2.5 3 2.3-2.6 2.4±0.10 2.7
W 6 1.4-2.2 1.6±0.13 1.5 3 1.3-1.5 1.4±0.06 1.5
H 5 5.5-6.5 6.0±0.20 5.6 3 5.7-7.0 6.5±0.39 53
7)2/ L 5 3.2-3.7 3.4±0.15 4.3 2 3.2,3.6 43
W 5 0.9-1.6 l.l±0.13 1.1 2 1.0,1.5 1.3
H 5 2.0-2.5 2.3±0.08 1.7 2 1.3,2.5 2.3
7)3/ L 6 5.1-6.5 6.H0.22 7.6 3 6.3-6.9 6.6±0.18 7.0
Lmts 6 2.4-2.9 2.6±0.08 2.8 3 2.4-2.6 2.5±0.07 2.S
Lt 6 1.0-1.2 1.Ы0.04 1.1 3 1.1-1.3 1.2±0.07 1.4
W 6 2.8-3.3 3.0±0.08 3.3 3 2.8-3.2 3.0±0.12 4.0
Wt 6 0.5-1.5 l.l±0.15 1.4 3 0.6-1.2 0.9±0.17 1_8
H 6 2.5-3.4 3.1±0.15 3.9 3 3.2-3.4 3.3±0.07 3 .8
87
Продолжение табл. 3
Промеры, Genetta tigrina Genetta genetta Genetta .servahna Genetta dongolana
мм п | lim М±т п=\ п I lim | М^т /7=1 ~
£4/ L 3 3.1-3.3 3.2±0.07 4.0 2 3.0,3.4 3.7
W 3 4.2-4.5 4.3±0.10 5.0 2 4.1,4.4 5.0
Н 3 1.7-2.0 1.9±0.09 2.3 2 1.8,2.2 1.7
Нижние зубы
//1 L 1 0.3 0.7 3 0.3-0.6 0.4±0.09
W 1 0.3 0.8 3 0.4-0.4 0.4±0.00
Н 1 0.9 0.9 3 0.6-1.2 0.9±0.18
Z/2 L 1 0.6 3 0.5-0.9 0.7±0.12
W 1 0.4 3 0.4-0.7 0.6±0.10
Н 1 0.6 3 0.6-1.2 0.9±0.18
//3 L 5 0.6-0.9 0.7±0.05 0.9 3 0.7-0.8 0.7±0.03
W 5 0.7-1.0 0.8±0.06 1.1 3 0.6-1.1 0.8±0.15
Н 5 1.0-1.4 1.2±0.08 1.4 3 0.8-1.6 1.2±0.23
С/1 L 6 2.0-2.5 2.2±0.08 2.5 3 2.1-2.1 2.1±0.00
W 6 1.2-1.5 1.3±0.05 1.4 3 1.2-1.3 1.3±0.03
Н 6 4.7-5.8 5.2±0.17 4.7 3 5.3-5.8 5.6±0.15
D/2 L 6 3.0-4.1 3.7±0.18 4.8 3 3.4-4.4 3.8±0.31
W 6 0.9-1.7 1.Ы0.12 1.7 3 1.1-1.3 1.2±0.06
Н 6 2.0-3.3 2.8±0.18 2.8 3 2.7-4.2 3.3±0.45
D/3 L 6 4.4-5.3 4.9±0.13 6.2 3 5.3-5.3 5.3±О.ОО
W 6 1.2-2.3 1.5±0.16 2.1 3 1.3-1.6 1.5±0.09
Н 6 3.0-3.8 3.4±0.14 4.0 3 3.7-3.8 3.7±0.03
88
Окончание табл. 3
Промеры, мм Genetta tigrina Genetta genetta Genetta servahna Genetta dongolana
n | lim Л/i m /7=1 n | lim | M m n=l
Р/4 L 6 5.2-5.9 5.5±0.10 6.8 3 5.7-6.0 5.8±0.10
Ltl 6 1.4-2.2 1.7±0.11 1.6 3 1.4-2.0 1.7±0.18
Wtr 6 2.1-2.5 2.3±0.09 2.8 3 2.1-2.2 2.2±0.03
Wtl 6 1.8-2.2 1.9±0.09 2.4 3 1.7-2.0 1.8±0.10
Н 6 2.8-4.5 4.0±0.25 5.0 3 4.0-4.1 4.0±0.03
D3/ хищнического типа. Зубцы и режущие гребни острые. Передний
отдел зуба сравнительно длинный. Присутствуют парастиль А и парастиль В.
От последнего в сторону протокона отходит препротокриста. Метастиль
небольшой (34% длины зуба), лезвиевидный, четко отделен от амфикона.
Талон короткий, но широкий, расположен несколько впереди от вершины
амфикона. Несет четкий бугорок протокона.
£>4/ с треугольной в плане коронкой. Эктофлексус и стилярная полка
развиты хорошо. Паракон и метакон небольшие, но четкие, островершин-
ные; из них первый несколько крупнее. Талон небольшой, поскольку
лабиальные бугорки сильно смещены лингвально. Имеется острый протокон.
D/3 крупный, островершинный. Выделяются три зубца, причем передий
и задний дополнительные зубцы сопоставимы по высоте. Коронка расшиена
в заднем отделе, где имеется острый продольный гребень.
D/4 - типичный хищнический зуб, лишь незначительно длиннее D/3.
Метаконид довольно крупный, острый, прижат к задней стенке протоконида.
Талонид широкий, с небольшим бассейном. Он несет гипоконид и энтоонид,
которые смещены назад и расположены на одном уровне. Оба бугорка соеди-
нены высоким гребнем, проходящим по заднему краю талонида (гребневид-
ный гипоконулид).
Род Pseudocivetta Petter, 1967
Pseudocivetta ingens Petter, 1967
Описание (no Petter, 1967, fig.3). Молочные зубы сравнительно
очень крупные. D/2 двухкорневой, добавочные зубчики почти не выражены.
На D/3 имеется небольшой передний добавочный зубчик, задний добавочный
89
зубец очень низкий и продольно вытянутый (не характерно для других Viver-
ridae). Длина £3/ 14, £>4/ 11, D/2 12, D/3 16.
Замечания. Вид описан из слоя I ущелья Олдувай, Танзания (виллафранк).
Крупная виверра, постоянные зубы которой несут черты адаптации ко всеядности.
Род Prionodon Horsfield, 1823
Prionodon linsang (Hardwicke, 1821) - Полосатый линзанг
Описание (по Leche, 1915, fig.22, 23). D2/2 сравнительно крупные,
с узкими трехзубчатыми коронками, двухкорневые. -
D3/ обычного типа, с небольшим парастилем А и без парастиля В. Ме-
тастиль сравнительно длинный. Талон крупный, расположен у основания ам-
фикона.
£)4/ крупный, по размерам почти равен D3/. Эктофлексус небольшой,
стилярная полка имеется. Парастилярная и метастилярная лопасти примерно
равны по размерам, не выдаются лабиально. Паракон и метакон высокие,
почти равных размеров. Талон крупный, с бассейном.
D/3 крупный, по длине и высоте не уступает D/4. Коронка с крупными,
примерно равными по размерам передним и задним добавочным зубцам и
отчетливым ’’талонидом” в виде острого бугорка.
D/4 с режущим тригонидом, гребни протоконида и параконида разделе-
ны глубокой бороздкой. Метаконид крупный, по высоте лишь немного усту-
пает протокониду. Талонид очень короткий, видимо, представлен одним ост-
рым бугорком.
ПОДСЕМЕЙСТВО Е U PL ERIN А Е CHENU, 1852
Род Eupleres Doyere, 1835
Eupleres goudotii Doyere, 1835 - Мелкозубый фаналук
Описание. На оригинальном материале представлены только
верхние молочные зубы, нижние молочные зубы описаны по Лехе (Leche,
1915, fig. 26, 27).
Резцы очень маленькие, посажены редко. Между dI3/3 и dC\/\ нет
большой диастемы. Размеры резцов заметно возрастают от dlM к <У/3/,
последний почти вдвое крупнее первого, либо все резцы примерно одина-
ковы (Leche, 1915, fig.24, 25). dl\/ мелкий, со слабо расширенной коронкой,
которая сильнее выдается латерально. dI2/ имеет латеральный и медиальный
выступы коронки, но не оформленных в виде боковых зубчиков. dI3/ клыко-
образный, сближен с клыком, но меньше его. Его острая вершина отогнута
90
назад. Имеется внутренний воротничок. Нижние резцы меньше верхних,
примерно одинаковые по размерам.
Клык dCM маленький, слабый, по величине сопоставим с Р1/. Его
коронка крючковидно изогнута, поэтому вершина направлена не только
вниз, но и несколько назад. dC/\ еще меньше, почти равен J//3 по величине.
D2/2 отделены большой диастемой от JC1/1 и несколько меньшей
диастемой от 7)3/3. Зубы узкие, островершинные, сравнительно крупные. На
заднем конце D2/ заметно слабое возвышение эмали. Корней 2.
D3/ очень слабый, лишь немногим длиннее D4/. Внешняя стенка корон-
ки вогнутая. Передний отдел зуба без бугорков. В него спускается с вершины
амфикона эмалевый гребешок. Амфикон высокий и острый, слабо обособ-
ленный от короткого режущего метастиля. Длина последнего составляет не
более 30% от всей длины коронки. Талон маленький, слабо выступающий,
без протокона. Лежит немного кпереди от уровня вершины амфикона.
D4/ более длинный, чем широкий. Отдаленно напоминает D3/. Экто-
флексус не в виде входящего угла, как и других виверр, но похож на пологую
вогнутую дугу. Маленькая стилярная полка имеется. Паракон и метакон не-
большие, острые. Для них характерны продольные кили, отходящие от
вершин. Особенно удлинен задний киль метакона, переходящий в метастиль
(по длине не уступает метастилю D3I). Талон маленький, короткий, смещен
вперед на уровень заднего отдела паракона. Несколько напоминает типич-
ный талон D3/. На его лингвальном конце расположен маленький, но четкий
зубец протокона.
D/3 сравнительно крупный; главная вершина высокая, шиловидная; пе-
редние и задние добавочные зубцы очень низкие. Имеется очень небольшой
"талонид” - выступ на заднем цингулиде.
D/4 сравнительно небольшой. Бугорки тригонида хорошо развиты,
высокие, приостренные, но без режущих гребней. Метаконид почти равен
паракониду по высоте. Талонид очень короткий и узкий, без четко выражен-
ных бугорков (имеется только возвышение на гипоконида), но с небольшим
бассейном.
ПОДСЕМЕЙСТВО PARADOXURINAE GRAY, 1865
ТРИБА PARADOXURINI S. STR.
Род Paradoxurus F.Cuvier, 1821
Paradoxurus hermaphroditus (Pallas, 1777) - Обыкновенный мусанг
Описание (рис. 6). J/1,2/1,2 игольчатые, dI3/3 намного крупнее,
простого строения (табл. 2).
91
г
е
Рис. 6. Молочные зубы Paradoxurus hermaphroditus, NRM А 582714:
а, г - снаружи; б, о - со стороны жевательной поверхности; в, е - изнутри
Fig. 6. Deciduous teeth of Paradoxurus hermaphroditus, NRM A 582714:
а, г - lateral; 6, () - occlusal; в, e - labial views
92
JC1/1 небольшие, слабо изогнутые. На dCM вдоль заднего края с линг-
вальной стороны идет неглубокая щель. На месте лингвального пояска на
нижнем клыке имеется слабый бугорок.
В строении всех молочных коренных присутствуют признаки, связан-
ные с адаптацией к всеядному типу питания: бугорки низкие, притупленные,
режущие структуры не развиты. D2/2 сравнительно высокие, одновершин-
ные, двухкорневые. Коронка D2I слабо расширена в задней части (NRM А
582714), либо очень узкая, без расширения.
На D3/ имеется крупный парастиль А, его основание вытянуто в виде
поперечного гребня. Парастиль В отсутствует, но вместо него от парастиля А
к вершине амфикона поднимается короткий валик. Метастиль очень корот-
кий (34.3-44.1% от общей длины зуба, Л/=39.2±0.92%, /7=13). Талон длинный,
но неширокий. Протокон хорошо развит, в виде острого бугорка (NRM А
582714), либо почти не развит (ЗММУ S-115213).
7)4/ в плане имеет вид почти равностороннего треугольника; очертания
коронки довольно изменчивы. Эктофлексус практически не развит; стиляр-
ная полка широкая. Иногда в глубине эктофлексуса имеется маленький
добавочный бугорок (NRM А 581114). Паракон и метакон очень низкие,
гребневидные, почти равны по размерам. Протокон в виде широкого про-
дольного гребня. Бассейн талона в виде небольшой, но глубокой овальной
долинки.
D/3 относительно очень короткий (59.1-74.0% от длины 7)/4,
Л/=66.8±1.47%, /7=11), трехзубчатый, главная вершина очень низкая,
притупленная. Передний и задний добавочный зубцы почти равны по высоте
(у других Viverridae задний зубец обычно намного выше). Имеется короткий,
поперечно ориентированный постцингулид.
7)/4 вытянутый в длину. Бугорки тригонида очень низкие, притуплен-
ные. Параконид и метаконид примерно равны по высоте. Имеется отчетли-
вый передний цингулид в основании параконида. Талонид короткий (35.5-
46.2% от общей длины зуба, Л/=40.5±0.96%, и=12), но широкий, обычно
шире тригонида. Гипоконид и энтоконид в виде очень низких, притуплен-
ных, продольно вытянутых бугорков, приближены к тригониду. Гипоко-
нулид в виде низкого поперечного гребня. Бассейн талонида сравнительно
большой.
Род Paguma Gray, 1831
Paguma larvata (Hamilton Smith, 1827) - Гималайская цивета
Описание (рис. 7). Молочные зубы гималайской циветы очень
близки по строению к таковым у обыкновенного мусанга. Отличия касаются
следующих признаков: молочные клыки относительно более мелкие, dCM
без щели; на dC/\ лингвальный бугорок либо отсутствует (NRM А 595230),
93
a
Рис. 7. Молочные зубы Paguma larvata, NRM A 595230:
а, г - снаружи; б, <) - co стороны жевательной поверхности; в, е - изнутри
Fig. 7. Deciduous teeth of Paguma larvata, NRM A 595230:
а, г - lateral; б, О - occlusal; в, e - labial views
либо очень крупный (ЗММУ S-133458); на D3/ область парастиля А относи-
тельно более длинная, иногда отогнута лабиально; метастиль более длинный
(38.6-51.6% от длины зуба, Л/=43.7±1.95%, п=6; табл. 4); на 7)4/ лучше развит
лабиальный цингулюм, имеется отчетливый парастиль, паракон существенно
крупнее метакона; D/3 такой же длины (62.7-71.4% от длины D/4,
Л/=66.5±1.16%, п=6); на £>/4 гипоконулид лучше обособлен, выдается назад,
талонид такой же длинный (31.0-46.8% от длины зуба, Л/=38.1 ±2.49%, п=6).
ТРИБА ARCTOGALIDIINI SIMPSON, 1945
Род Arctogalidia Merriam, 1897
Arctogalidia trivirgata (Gray, 1832) - Мелкозубая цивета
Описание (рис. 8). Верхние и нижние резцы довольно крупные.
<//1-2/ со слабо расширенной притупленной коронкой. dI3/ заостренный, с
заметным задним воротничком. <///1-3 слабо расширенные, с небольшим
внутренним воротничком.
Клыки острые. dCM слабо изогнутый. Коронка dC/\ имеет у своего
основания с внутренней стороны слабое вздутие эмали.
D2/ отделен небольшой диастемой от D3/. Зуб относительно крупный, с
высокой и острой основной вершиной и с двумя добавочными вершинами.
D/2 сближен с D/3. Длина зуба составляет почти половину длины /9/4. Он
высокий, остроконечный. Перед главным бугорком имеется небольшой
острый добавочный зубец. Корней 2.
D3/ хищнический зуб, с широким передним отделом. Имеется крупный
йарастиль А, парастиль В отсутствует. Амфикон высокий, острый. Метастиль
рчень короткий (22% всей длины коронки), имеет вид невысокого бугорка.
Талон длинный, неширокий, слабо смещен кзади. Протокон маленький.
Мощная препротокриста соединяет протокон с парастилем А.
D4/ крупный, почти равный по длине и ширине. Эктофлексус слабый.
Стилярная полка не выражена, эктоцингулюм отчетливый. Лабиальные бу-
горки большие, высокие, примерно равны по размерам. Талон длинный и
Широкий, с продольно вытянутой внутренней долинкой. Бугорок протокона
выражен слабо.
D/3 сравнительно широкий, особенно в заднем отделе коронки. Основ-
ная вершина высокая, острая. Впереди нее находится небольшой дополни-
тельный зубчик. Кроме того, по заднему краю коронки идет поперечный гре-
бень.
/9/4 крупный, широкий. Тригонид состоит из трех широко расстав-
ленных бугорков. Режущие гребни на тригониде не выражены. Протоконид
Низкий, ниже лингвальных бугорков. Метаконид хорошо обособлен от про-
т°конида, почти не уступает ему по величине, сильно сдвинут лингвально.
95
Рис. 8. Молочные зубы Arctogalidia trivirgata, ЗИН 8198:
а, г - снаружи; б, о - со стороны жевательной поверхности; в, е - изнутри
Fig. 8. Deciduous teeth of Arctogalidia trivirgata, ZIN 8198:
а, г - lateral; 6, <) - occlusal; в, e - labial views
96
Таблица 4. Размеры молочных зубов у Paguma, Arctogalidia и Arctictis
Table 4. Measurements of deciduous dentition in Paguma, Arctogalidia and Arctictis
Промеры, Paguma larvata Arctogalidia tnvirgala, n=3 Arctictis bint urong
MM "1 lim | M i m lim | Л/t m " 1 lim | M tin
Верхние зубы
/1/ L 4 1.0-1.4 1.2±0.09 1.2-1.5 1.3±0.10 1 1.3
W 4 0.5-1.3 l.l±0.19 1.0-1.4 1.2±0.12 1 1.2
H 4 1.3-2.2 1.7±0.20 1.7-2.1 1.9±0.12 1 2.1
12/ L 5 1.1-2.0 1.4±0.16 1.1-1.5 1.3±0.12 3 1.4-1.6 1.5±0.07
W 5 0.8-2.0 1.5±0.19 1.2-1.4 1.3±0.06 3 1.4-1.9 1.7±0.16
H 5 1.5-2.7 2.0±0.21 1.7-2.2 2.0±0.15 3 1.9-2.9 2.3±0.32
13/ L 5 1.7-2.4 2.0±0.12 1.5-1.7 1.6±0.07 3 1.8-2.3 2.1±0.15
W 5 1.6-2.4 2.2±0.15 1.4-1.7 1.5±0.10 3 2.1-2.5 2.3±0.12
H 5 2.8-3.6 3.U0.14 2.2-2.7 2.4±0.15 3 3.0-3.4 3.2±0.12
CM L 5 2.3-3.5 3.U0.22 3.0-3.4 3.3±0.13 3 3.7-4.6 4.2±0.26
W 5 1.3-2.5 2.1±0.21 2.1-2.1 2.1±0.00 3 2.4-2.6 2.5±0.06
H 5 5.4-11.4 7.1±1.11 5.2-6.7 5.7±0.48 3 5.5-11.3 9.3±1.89
DU L 4 3.7-5.0 4.U0.31 3.5-4.1 3.8±0.17 3 3.0-4.0 3.4±0.30
W 4 1.5-2.0 1.7±0.12 1.7-1.9 1.8±0.06 3 1.8-2.1 1.9±0.10
H 4 2.5-3.4 3.H0.20 2.7-4.0 3.2±0.42 3 3.6-4.1 3.8±0.15
D3! L 6 5.4-6.4 6.0±0.15 4.7-5.4 5.0±0.22 3 5.2-5.5 5.3±0.09
Lmts 6 2.1-3.3 2.6±0.18 1.2-2.2 1.6±0.31 3 1.5-1.8 1.6±0.10
Lt 6 1.7-2.1 1.9±0.06 1.6-2.0 1.8±0.12 3 1.8-2.4 2.1±0.17
W 6 3.3-4.4 3.7±0.17 2.7-3.0 2.8±0.10 3 3.3-4.0 3.7±0.20
Wt 6 0.6-1.5 1.0±0.14 0.6-0.6 0.6±0.00 3 0.8-1.3 l.l±0.15
H 6 2.6-4.1 3.2±0.20 3.0-3.4 3.U0.13 3 3.0-3.6 3.3±0.18
97
Продолжение табл. 4
Paguma larvata Arctogalidia trivirgata, Arctictis binturong
Промеры, П=5
мм п 1 lim | М'.т lim | М'.т " 1 lim | М±т
D4/ L 5 5.3-6.7 5.9±0.24 4.6-5.0 4.7±0.13 2 5.0,5.3
W 5 5.3-5.8 5.5±0.09 3.7-4.4 4.1±0.22 2 4.8,5.0
Н 5 2.5-3.2 2.8±0.13 2.3-2.7 2.5±0.12 2 3.0,3.2
Нижние з у б ы
Z/1 L 4 0.5-0.9 0.7±0.09 1.0-1.3 l.lzbO.10 3 0.5-1.6 1.0±0.32
W 4 0.3-0.9 0.7±0.13 0.6-1.2 0.9±0.18 3 0.9-1.4 1.U0.17
Н 4 0.8-1.3 llzbO.ll 1.0-1.9 1.3±0.30 3 1.6-2.1 1.9±0.17
1/2 L 5 0.8-1.4 1.1±0.10 1.0-1.2 1.1±0.07 2 1.3,1.6
W 5 0.6-1.5 1.1±0.15 1.0-1.2 1.1±0.07 2 1.3,1.4
Н 5 1.2-1.9 1.5±0.16 1.3-2.0 1.5±0.23 2 1.3,2.1
1/3 L 4 1.1-1.6 1.3d=o.ro 1.2-1.7 1.4±0.15 3 1.5-2.1 1.8±0.18
W 4 0.8-2.3 1.8±0.33 1.2-1.4 1.3±0.07 3 2.0-2.4 2.2±0.12
Н 4 1.8-2.5 2.2±0.16 1.4-2.6 2.0±0.35 3 2.1-2.8 2.4±0.22
С/\ L 5 2.4-3.6 3.3±0.23 2.7-3.4 3.1±0.21 3 3.5-4.9 4.3±0.41
W 5 1.4-2.3 2.0±0.16 1.8-1.9 1.8±0.03 3 2.1-2.4 2.3±0.09
Н 5 4.5-9.3 5.9±0.86 4.7-5.4 5.1±0.21 3 5.4-6.5 5.9±0.32
D/2 L 5 3.9-4.5 4.1±0.11 3.4-3.6 3.5±0.06 2 4.0,4.5
W 5 1.5-2.1 1.8±0.11 1.7-1.8 1.7±0.03 2 1.7,2.3
Н 5 2.4-3.4 2.7±0.19 3.2-3.5 3.3±0.09 2 3.2,3.4
D/3 L 6 4.8-55 53dbO.ll 4.3-4.5 4.4±0.07 3 4.8-5.7 5.3±0.28
W 6 2.5-3.7 2.9±0.19 2.1-2.3 2.2±0.07 3 2.3-3.1 2.6±0.25
Н 6 2.4-4.4 3.1±0.34 3.2-3.8 3.4±0.20 3 3.0-3.1 3.0±0.03
98
',К0ПЧа1,Ие табл. 4
Промеры, мм Pagurna larvata A rctogalidta tn virgcUa, л=3 Arctictis btnturonK
n 1 lim | Л/ m lim | M m J2_l lim 1 M 7>i
D/4 L 6 7.2-8.4 7.9±0.18 6.8-7.1 7.0±0.09 3 6.7-7.3 7.1 ±0.19
Ltl 6 2.6-3.6 3.0±0.18 2.0-2.8 2.3±0.24 3 2.0-2.8 2.4±0.23
Wtr 6 3.3-4.0 3.7±0.13 3.1-3.4 3.2±0.10 3 3.0-3.7 3.5±0.23
Wtl 6 3.4-4.6 4.1±0.20 3.1-3.3 3.2±0.06 3 2.9-3.5 3.2±0.18
Н 6 2.6-3.0 2.8±0.07 2.9-3.3 3.1±0.12 3 2.5-3.4 3.0±0.26
Талонид большой, с широким бассейном. Гипоконид и энтоконид крупные
бугорки, хорошо обособленные. От первого из них по заднему краю зуба
идет поперечный гребень.
ТРИБА ARCTICTIINI GRAY. 1865
Род Arctictis Temminck, 1824
Arctictis binturong (Raffles, 1821) - Бинтуронг
Описание (рис. 9). Верхние молочные резцы небольшие. Как и у
других виверр, коронки их слабо расширены, с хорошо развитым задним
воротничком. Нижние резцы маленькие, слабо расширенные, с одним лате-
ральным (J//2) или медиальным (J//3) зубцами (на нижних постоянных рез-
цах развиты оба боковых зубца). Внутренний воротничок скошен, резцы на-
поминают таковые у Canidae и Ursidae.
Верхний клык dCM простой, с острой вершиной, направленной вниз.
Ширина и длина коронки у основания такая же, как у корня. Клык не изо-
гнут, его задняя стенка почти прямая, либо слабо изогнута (NRM А 582332).
dC/\ имеет слабо изогнутую либо почти прямую коронку, вершина которой
направлена вверх. По его передней и задней сторонам, ближе к внутреннему
краю, проходят гребни.
D2/ довольно крупный, одновершинный, с высокой коронкой. Зуб упло-
щен с боков, без дополнительных бугорочков, со слабым пояском в осно-
вании коронки с лингвальной стороны. Корень 1.
D3/ похож на соответствующий зуб других всеядных хищников. Режу-
щие гребни развиты слабо. Передний отдел коронки развит очень слабо, без
парастиля А и без площадки (имеется у большинства Viverridae). Амфикон
99
7’
Рис. 9. Молочные зубы Arctictis binturong, NRM А 582332:
а, г - снаружи; б, д - со стороны жевательной поверхности; в, е - изнутри
Fig. 9. Deciduous teeth of Arctictis binturong, NRM A 582332:
а, г - lateral; б, c) - occlusal; в, e - labial views
100
притупленный, занимает почти всю площадь коронки. Его передний гребень
доходит до переднего края зуба. Метастиль слабый (его длина составляет 29-
33% от общей длины зуба), в виде притупленного бугорка. Талон небольшой,
сравнительно массивный, без бугорка протокона, со слабой препротокрис-
той. Корней 3.
D4/ треугольной формы, заметно крупнее D3/, по строению несколько
напоминает D4/ у Ргосуоп. Эктофлексус и стилярная полка отсутствуют;
лабиальный край коронки выпуклый; имеется слабый лабиальный цингулюм.
Наиболее крупный бугорок зуба паракон; его основание занимает почти
половину коронки. Метакон заметно меньше, но хорошо обособлен. Прото-
кон крупный, в виде вытянутого продольного бугорка. Бассейн талона резко
уменьшен, представлен узкой продольной долинкой между параконом и
протоконом. На стертом экземпляре (см. рис. 9) образуется гребень, идущий
от вершины паракона к небольшому парастилю и затем к продольному
гребню протокона. Корней 3.
D/2 короткий, но высокий зуб, с островершинной коронкой. В основа-
нии коронки сзади имеется небольшой бугорок. Корней 2.
D/3 простой, одновершинный зуб, сжатый с боков. Задний отдел его
немного расширен. Имеется передний добавочный бугорок. По середине
задней стенки главного бугорка проходит продольный эмалевый гребень,
доходящий до заднего края коронки. Корней 2.
D/4 по форме напоминает Л//1, но меньше его и относительно более
узкий. Протоконид в виде невысокого тупого конуса. Параконид и метако-
нид - небольшие притупленные бугорки, прислоненные к лингвальной стенке
протоконида. Талонид короткий, с крупным, но низким гипоконидом и
небольшим бассейном. Вдоль средней части талонида тянется эмалевый
гребень, несколько смещенный лабиально и доходящий до гипоконида.
Корней 2.
Замечания. Вершины £>3/, £>4/, D/3 и D/4 на обоих исследованных
экземплярах существенно стерты.
ТРИБА CYNOGALINI GRAY, 1865
Род Cynogale Gray, 1837
Cynogale bennettii Gray, 1837 - Выдровая цивета
Описание (по рисункам в работах Майварта (Mivart, 1882, fig. 12)
и Лехе (Leche, 1915, fig.55-58). Резцы мелкие, без лингвального пояска. dl3/
крупнее других верхних резцов. dl/З с небольшим латеральным зубчиком.
dC\/ с высокой прямой коронкой. Коронка dC/\ ниже, изогнута, напра-
влена вперед, с отчетливым зубчиком на месте лингвального пояска.
D2/2 очень крупные, выше других молочных коренных, с острой треу-
гольной коронкой, двухкорневые. На D2I имеется очень слабый задний доба-
101
вочный зубчик, коронка слегка расширена в задней части. На D/2 присут-
ствует крупный передний добавочный зубец.
D3/ отличается по строению от хищнического зуба других виверр.
Коронка трехзубчатая, боковые зубцы образуют парастиль А и метастиль,
которые примерно равны по высоте. Основание парастиля А длиннее основа-
ния метастиля (для других виверр характерно обратное соотношение). Пара-
стиль В отсутствует. Талон представлен протяженным лингвальным цингу-
люмом, несущим несколько мелких бугорков. Его центр смещен относи-
тельно середины вершины амфикона назад. Зуб, возможно, двухкорневой.
D4/ превышает по длине и ширине £)3/, треугольной формы. Лабиаль-
ный край коронки прямой, без эктофлексуса, с цингулюмом в передней
части, несущим мелкие бугорочки. Из лабиальных бугорков имеются от-
четливый парастиль, очень крупный паракон (его основание занимает почти
половину лабиального края зуба) и небольшой метакон (метастиль?), связан-
ный с параконом продольным гребнем, несущим мелкие зубчики. Ряд лаби-
альных бугорков максимально придвинут к краю коронки (нет никакой сти-
лярной полки), отделен от лингвального края коронки обширным и глубоким
бассейном. Протокон небольшой или не выражен в виде самостоятельного
бугорка, вдоль всего лингвального края коронки расположены 7 зубчиков,
примерно равных по размерам. В основании лингвального края талона име-
ется четкий цингулюм, несущий мелкие бугорочки.
D/3 крупный, с четырехзубчатой коронкой: позади обычной для виверр
задней добавочной вершины, расположенной на заднем крае основной вер-
шины, имеется еще один дополнительный зубец. Коронка узкая, без расши-
рения в задней части. Постцингулюм короткий, образует маленький зубчик.
D/4 длинный, но невысокий, необычного для виверр строения. Триго-
нид намного короче и уже талонида, состоит только из вытянутых в одну ли-
нию параконида и протоконида (второй из них вдвое выше первого) (Mivart,
1882, fig. 12), либо имеется также небольшой метаконид (Leche, 1915, fig.57).
В основании тригонида с лабиальной стороны имеется длинный прецингу-
лид, несущий мелкие бугорочки. На талониде хорошо выражены двойные
гипоконид и энтоконид, разделенные обширным бассейном. Спереди от
гипоконида и энтоконида имеются ряды мелких зубчиков, протянувшихся до
тригонида.
ПОДСЕМЕЙСТВО HEMIGALINAE GRAY, 1865
ТРИБА HEMIGALINI S. STR.
Род Hemigalus Jourdan, 1837
Hemigalus derbyanus (Gray, 1837) - Полосатая цивета
Описание (рис. 10). Верхние резцы посажены дугообразно. Есть
обычная большая диастема между d!3/ и dCM. Коронки резцов слабо расши-
102
Рис. 10. Молочные зубы Hemigalus derbyanus, ЗИН 5802:
а, г - снаружи; б, <) - со стороны жевательной поверхности; в, е - изнутри
Fig. 10. Deciduous teeth of Hemigalus derbyanus, ZIN 5802:
а, г - lateral; б. о - occlusal; в, e - labial views
103
ренные, асимметричные, ложковидные. На dl\-3l хорошо выражен внутрен-
ний воротничок. dI3/ заметно крупнее и шире других верхних резцов.
Нижние резцы небольшие (табл. 2). dU\ почти игольчатый. Коронки dI/2-З
более широкие. Сильнее развит латеральный отдел, где намечается дополни-
тельный боковой зубец.
Клыки обычного типа, корень несколько толще коронки или же одной с
ней толщины. dCM умеренно высокий, у основания коронки с внутренней
стороны виден слабый валик эмали. dC/\ с изогнутой коронкой, вершина
которой направлена вверх. С внутренней стороны нижнего клыка имеется
слабый продольный эмалевый валик.
D2I крупный, узкий, очень высокий. Вершина одна. Вдоль заднего края
бугорка тянется режущий гребень. Корней 2, широко расставленных. D/2
крупный, узкий, сближен с D/3. Главная вершина острая, с длинным задним
гребнем. Перед ней находится слабый зубчик. Корней 2.
D3/ крупный, сложный зуб. Он совсем не похож на Р4/ (последний
широкий, с высоким парастилем, массивным амфиконом, без метастиля, но с
большим талоном и крупным протоконом); зуб несколько напоминает ММ.
Передний отдел большой, с крупным парастилем А и хорошо обособленным
парастилем В. Амфикон высокий и острый. Метастиль имеет вид не ре-
жущего лезвия, обычного для Carnivora, а единичного островершинного бу-
горка. Он очень короткий (28% всей длины зуба). Талон крупный, несет
высокий острый протокон, который сопоставим по высоте с парастилем В и
соединен с ним препротокристой. По лабиальной стороне вдоль всей корон-
ки тянется цингулюм, особенно хорошо выраженный в переднем и заднем
отделах зуба. Лингвальный постцингулюм также имеется.
D4/ треугольной в плане формы. Эктофлексус выражен слабо. Стиляр-
ная полка практически отсутствует, но лабиальные бугорки окаймлены по
наружному краю четким цингулюмом. Паракон и метакон высокие, остро-
конечные, широко расставленные. Талон массивный, с внутренним бассей-
ном. Протокон также массивный, к нему по переднему и заднему краям
талона идут гребни, причем задний гребень непосредственно за протоконом
образует подобие бугорка (метаконуль?). Вдоль лингвальной стенки прото-
кона идет едва заметный эмалевый валик (аналог хорошо выраженного
цингулюма у ММ).
D/3 крупный, похож на D/4. Имеется острый бугорок впереди главного
конуса ("параконид") и хорошо моделированный высокий острый бугорок у
его заднего внутреннего края ("метаконид"). Задний отдел коронки расширен
и образует "талонид" Последний несет более крупный внешний бугорок
("гипоконид") и небольшой, смещенный назад "энтоконид" Между обоими
бугорками "талонида" находится маленькая площадка (аналог бассейна). D/3
моляризован в существенно большей степени, чем у остальных известных
нам виверрид.
D/4 очень крупный, широкий, с большим талонидом. Напоминает Л//1,
но у последнего талонид развит еще сильнее. Параконид и протоконид остро-
104
вершинные. Метаконид высокий, острый, немногим мельче протоконида,
либо выше его (ЗИН 5802) и отделен от него глубокой долинкой. Талонид
широкий, с обширным бассейном, который обрамляют бугорки гипоконида,
энтоконида и гипоконулида. Перед гипоконидом и энтоконидом расположе-
ны мелкие дополнительные бугорочки. Энтоконид более высокий и сильнее
смещен назад, чем гипоконид, открывая бассейн талонида лингвально.
ПОДСЕМЕЙСТВО LOPHOCYONINAE
FEJFAR ET SCHMIDT-KITTLER, 1987
Род Lophocyon Fejfar et Schmidt-Kittler, 1987
Lophocyon carpathicus Fejfar et Schmidt-Kittler, 1987
, Описание (no Fejfar et al., 1987, Text-fig. 11). Молочные коренные
l (известны только D/3 и фрагмент D/4) переходного от туберкуло-секториаль-
I ного к лофодонтному строения, сравнительно крупные.
г D/3 полностью моляризован, с трехбугорчатым тригонидом (не отмеча-
ется для других Carnivora; даже у Nasua, молочные зубы которых в значи-
тельной степени специализированы к поеданию растительной пищи, D/3
моляризован в существенно меньшей степени). Паралофид длиннее метало-
< фида. Параконид, видимо, был очень низкий. Метаконид ниже протоконида;
. оба бугорка разделены бороздкой на металофиде (протокристиде). Вершины
протоконида и метаконида приострены. Прецингулид в основании лабиаль-
' ной стороны параконида четкий и очень длинный, доходит до основания
i протоконида. Талонид заметно шире тригонида, несет очень сильно
развитый cristid obliqua, образующий высокий гребень, идущий от задней
стенки метаконида до гипоконулида, почти параллельно металофиду.
Гребень разделен на 2 примерно равные по длине части. Гипоконулид в виде
Очень длинного гребня, идущего параллельно cristid obliqua, но гораздо более
^низкий. Все 3 параллельных гребня (металофид, cristid obliqua и гипокону-
лид) несут следы стирания. Длина D/3 8.2, ширина - 5.7.
От D/4 сохранились только талонид и задняя часть тригонида. Cristid
'Obliqua, видимо, более короткий, чем на D/3, но также сильно развит, ср
^следами стирания. Гипоконид и энтоконид соединены поперечным гребнем,
’который заметно стерт. Гипоконулид отсутствует. Ширина талонида D/4 5.9.
Замечания. Описанные молочные зубы присутствуют на голотипе L.
Carpathicus, происходящем из среднемиоценовых отложений (сармат?) Косице-Банков,
Словакия (Fejfar et al., 1987). Постоянные зубы L. carpathicus характеризуются наиболее
Сраженной среди Lophocyoninae (включает также Sivanasua Pilgrim, 1931 и Euboictis
Fejfar et Schmidt-Kittler, 1984 из раннего-среднего миоцена Европы) специализацией к
растительноядению. По морфологии коренных зубов лофоционины конвергентно сходны
с зубами ранних Equidae, нижние зубы с зубами Coryphodon из Pantodonta. Эти
Животные питались, видимо, мягкой растительной пищей. Lophocyon дтнесен к Viverridae
Иа основании строения слуховой области (Fejfar et al., 1987).
105
СЕМЕЙСТВО HERPESTIDAE BONAPARTE, 1845
ПОДСЕМЕЙСТВО G A L I D I I N A E GRAY, 1865
Род Galidia I.Geoffroy, 1837
Galidia elegans I.Geoffroy, 1837 - Кольцехвостый мунго
Описание (рис. 11). Верхние молочные резцы длинные и узкие,
нижние - небольшие, узкие. dCM и dC/\ относительно крупные (табл. 5).
D2I сближен с D3/. Это маленький одноконусный зуб. Корней 2.
D3/ обычного хищнического типа. Передний отдел коронки сравни-
тельно длинный, с крупным парастилем А. Парастиль В отсутствует. Амфи-
кон высокий, острый. Метастиль относительно длинный (41% общей длины
зуба). Талон маленький, без ясно выраженного зубца протокона.
D4/ в плане почти треугольной формы. Хорошо развиты эктофлексус и
стилярная полка; парастилярная лопасть сильно выдается лабиально. Пара-
кон и метакон небольшие; первый бугорок немного крупнее. Талон относи-
тельно большой, длинный, с четкой вершиной протокона.
D/2 маленький, одновершинный, сближен с D/3. Корней 2.
D/3 довольно крупный (77-83% от длины D/4); коронка его расширена в
заднем отделе. Перед главной вершиной имеется небольшой зубчик. Еще
один дополнительный зубец расположен на задней стенке главного конуса.
Позади него различим маленький "талонид"
D/4 хищнический зуб обычного облика. Метаконид сравнительно
крупный, приближен к заднему краю протоконида. Талонид довольно круп-
ный, но уже тригонида. Несет гипоконид и энтоконид, разделенные неболь-
шим бассейном.
Замечание. Верхние и нижние молочные резцы и клыки на исследованном
экземпляре сильно стертые, что указывает на их участие в обработке пищи. Молочные
резцы прорезаются на восьмой день, коренные - на 21-й день (Larkin, Roberts, 1979).
Род Galidictis I.Geoffroy, 1839
Galidictis fasciata (Gmelin, 1788) - Узкополосый мунго
Описание. Изучены только верхние молочные зубы. Резцы
сравнительно крупные, плотно прилегают друг к другу. Длина коронка почти
равна ее ширине. Боковых зубцов нет. dI3/ заметно крупнее остальных
резцов.
dCM небольшой, слабо изогнутый, с острой вершиной, направленной
вниз.
D2I очень маленький, простой, по-видимому, с одним корнем (такая
степень редукции D2/ не наблюдается у других изученных виверр и
мангустов). Сближен с D3/.
106
Рис. 11. Молочные зубы Galidia elegans, ЗИН 1628:
а, г - снаружи; б, д - со стороны жевательной поверхности; в, е - изнутри
Fig. 11. Deciduous teeth of Galidia elegans, ZIN 1628:
а, г - lateral; d - occlusal; в, e - labial views '
107
Таблица 5. Размеры молочных зубов у Galidiinae и некоторых Herpestinae
Table 5. Measurements of deciduous dentition in Galidiinae and some Herpestinae
Galidia Salanoia Galidictis Atilax Rhyne ho gale
Промеры, мм elegans concolox fasciata paludinosus mellen
n=l-2 «=1 n=l n=l
Верхние зубы
IM L 1.0 0.5 1.1 1.0
W 0.3 0.6 0.9 1.0
H 1.2 0.7 1.2 1.0
/2/ L 1.1 0.9 1.1 1.3
W 0.7 0.6 1.2 1.1
H 1.4 1.0 1.2 1.7
13/ L 1.4 1.1 1.5 2.0
W 1.1 1.0 1.6 2.0
H 2.4 2.0 2.8 3.1
CM L 2.2,2.3 2.0 1.6 3.7
W 1.7,1.8 1.1 1.2 2.8
H 5.5 3.6 3.1 8.4
DU L 2.0,2.4 1.7 1.7 4.2
FT 1.0,1.1 0.7 1.0 3.0
H 1.0,1.3 1.0 0.7 3.8
D3I L 5.0,5.4 4.7 4.9 8.2
Lmts 2.1,2.2 2.0 1.5 3.4
Lt 1.0,1.0 0.5 0.0 2.9
W 2.3,2.6 2.2 1.6 5.9
Wt 0.9,1.0 0.5 0.0 1.8
H 2.7,3.0 2.6 2.6 5.1
108
Продолжение табл. 5
Промеры, мм Galidia elegans п=\-2 Salanoia concolor Л1=1 Gahdictis fasciata Л=1 Atilax paludinosus Л1=1 Rhynchogale melleri n=l
7)4/ L 3.1,3.! 2.9 3.6 5.0
W 3.2,4.1 3.8 3.9 7.2
Н 1.0,1.8 1.5 1.5 1.8
Нижние зубы
7/1 L 0.5 0.4 1.0
W 0.4 0.5 0.7
H 0.7 0.9 1.0
7/2 L 1.0 0.4 1.1
W 0.5 0.7 0.9
H 0.7 0.9 1.1
7/3 L 1.3 0.8 1.6
W 1.1 1.1 2.0
H 1.9 1.2 2.1
C/1 L 2.5,2.6 2.1 4.1
W 1.5,1.9 1.3 2.8
H 5.3,5.3 3.5 8.2
D/2 L 2.0,2.1 2.1 3.8
W 0.9,1.0 0.8 2.5
H 1.0,1.4 2.1 3 1
D/3 L 3.2,3.7 3.2 5.9
W 1.4,1.5 1.4 3.6
H 1.8,2.7 2.1 4.4
109
Окончание табл. 5
Промеры, Gahdia elegans n=\-2 Salanoia concolox n=l Gahdictis fasciata n=l Ar i lax paludinosus n=\ Rhynchogale mellen n=l
D/4 L 4.4,4.9 4.5 8.4 5.8
Ltl 1.1,1.2 1.3 3.1 2.8
Wtr 2.1,2.2 2.0 4.4 2.7
Wtl 1.6,1.8 1.8 3.8 3.0
Н 2.9,3.1 2.5 5.4 3.1
D3/ небольших размеров, узкий, с острым амфиконом. Перед амфико-
ном находится небольшой зубец (парастиль А?). Метастиль очень короткий
(31%), режущий. Талон не выражен. Имеются лишь его следы на уровне
амфикона.
D4I обычного строения. Эктофлексу с и стилярная полка выражены хо-
рошо. Паракон и метакон небольшие, приостренные, слабо различаются по
величине. Протокон в виде зубца, к которому идут два гребня, обрамляющих
внутренний бассейн талона.
Род Salanoia Gray, 1865
Salanoia concolor (I.Geoffroy, 1837) - Одноцветный мунго
Описание. Верхние резцы относительно длинные. Коронка их
более длинная, чем широкая. На tZ/2-З/ развит внутренний воротничок. Ниж-
ние tZZ/1-2 очень мелкие, со слабо расширенной коронкой. dl/З немного круп-
нее; его коронка ассиметрична из-за сильного развития латерального края.
Клыки простые, острые. Вершина dCM направлена вниз. По его пе-
редне-внутреннему краю проходит вертикальный эмалевый гребень. d& с
более мощным корнем. Коронка его при взгляде сбоку крючковидная; вер
шина клыка отогнута вверх и назад.
D2I маленький узкий зуб, с одной округлой вершиной. Сближен с D3
Корней 2. D/2 маленький, узкий, сближен с D/3. Коронка его одновершинная,
но на переднем и заднем конце ее и на заднем ребре главного бугорка наме'
чаются мелкие зубчики.
D3/ хищнического типа, небольших размеров. Передний отдел зуба уз-
кий, несет высокий парастиль А. Парастиль В не выражен. Амфикон высо-
110
кий, острый. Метастиль лезвиеобразный, длинный (43% от общей длины).
Талон очень маленький, без зубца протокона. Расположен на уровне верши-
ны амфикона.
D4/ сравнительно крупный, с треугольной коронкой. Эктофлексус и
особенно стилярная полка развиты хорошо. Паракон и метакон невысокие,
но четкие, острые, примерно равные по величине. Талон небольшой. Острый
протокон отделен от лабиальных бугорков продольной долинкой.
D/3 крупный, трехвершинный. Все вершины острые, четкие. Главный
бугорок высокий. В заднем отделе коронки зуб несколько расширен и
образует ’’талонид", в который с вершины главного бугорка спускается про-
дольный гребешок.
D/4 обычный хищнический зуб маленьких размеров (немного короче,
чем у Galidia). Метаконид острый, обособлен от протоконида глубокой
щелью. Талонид сравнительно большой, с тремя четкими бугорками гипоко-
нида, энтоконида и гипоконулида, между которыми лежит бассейн. Гипоко-
нид и энтоконид смещены кзади в равной степени.
ПОДСЕМЕЙСТВО HERPESTINAE S. STR.
ТРИБА HERPESTINI S. STR.
( Род Atilax F.Cuvier, 1826
j Atilaxpaludinosus (G.Cuvier, 1829) - Водяной мангуст
l Описание (рис. 12). Резцы сравнительно крупные, постепенно уве-
гличиваются в размерах от мезиального зуба к дистальному. Коронки tZZ/1,2
|,двухзубчатые. В основании коронок d!3/ и dl/\-3 с лингвальной стороны
|имеется поясок.
* Клыки простого строения. В основании d&\ с лингвальной стороны
Вимеются небольшой бугорок и короткий поясок.
| D2/2 сравнительно крупные, с овальной в плане одновершинной корон-
ной, двухкорневые. На D2I присутствует короткий "талон" (лингвальный
Цингулюм).
г Хищнический зуб D3I сравнительно короткий. Парастиль А крупный,
Но невысокий; парастиль В отсутствует. Имеется лабиальный цингулюм с
относительно крупными бугорочками. Метастиль длинный (42% от общей
Длины зуба), не лезвиевидный. Талон крупный, расположен напротив амфи-
кона. Протокон в виде низкого гребня. На постпротокристе выражено греб-
невидное поднятие.
D4/ крупный, по размерам сопоставим с £>3/, треугольной в плане
формы. Лабиальный край короткий; эктофлексус неглубокий, hq парастиляр-
Кая лопасть хорошо развита. Паракон и метакон небольшие, примерно равны
По размерам, далеко отставлены от лабиального края. Талон большой, с от-
111
a
e
Рис. 12. Молочные зубы Atilaxpaludinosus, NRM A 592005:
а, г- снаружи; б, д - со стороны жевательной поверхности; в, е - изнутри
Fig. 12. Deciduous teeth of Atilax paludinosus, NRM A 592005:
а, г - lateral; б, d - occlusal; в, e - labial views
112
четливым бассейном. Гребень протокона низкий. Имеются отчетливые пара-
конуль и метаконуль.
D/3 сравнительно короткий (70% от длины 2)/4), коронка его резко рас-
ширена-в задней части. Задняя добавочная вершина высокая, тесно прижата
к заднему краю основной вершины. В задне-лингвальном углу коронки рас-
положен короткий постцингулид.
D/4 приближается по строению к "всеядному" типу. Бугорки тригонида
хорошо развиты, без четко выраженных режущих гребней. По высоте мета-
конид равен паракониду, но по величине основания уступает паракониду и
протокониду. Имеется четкий прецингулид в основании параконида. На та-
лониде бугорки плохо различимы; более или менее выделяется только гипо-
конид (низкий гребень); бассейн хорошо развит.
Род Herpestes liliger, 1811
Herpestes ichneumon (L., 1758) - Египетский мангуст
Описание (рис. 13). Резцы мелкие, t/21/l игольчатые. На <У72,3/
имеется лингвальный поясок, на tZZ/2,3- лабиальный бугорок.
Клыки невысокие, слабо изогнутые, простого строения. Лингвальный
поясок развит только на d&\.
D2/2 сравнительно крупные зубы с одновершинной коронкой, двух-
корневые. Коронка D2I несколько расширена в задней части и несет неболь-
шой добавочный зубчик. На D/2 имеются задний добавочный зубчик и воз-
вышение на гребне, идущем от главной вершины к этому зубчику.
2)3/ обычного строения. Парастиль А крупный, но невысокий. На лаби-
альном цингулюме, примыкающем к парастилю Л, находятся 2-3 небольших
бугорка. Парастиль В отсутствует, но к месту его расположения тянется от-
четливый гребень препротокристы. Метастиль сравнительно длинный (37.5-
43.5% от общей длины зуба, А/=40.2±0.79%, и=9), его лезвие прямое. Талон
короткий, но сравнительно широкий. Протокон сравнительно крупный, в
виде острого зубца; его основание занимает только лингвальную половину
талона.
2)4/ треугольной в плане формы. Эктофлексус неглубокий. Парасти-
лярная лопасть сильно выдается вперед. Стилярная полка широкая. Паракон
и метакон небольшие, гребневидные; первый из них несколько крупнее.
Талон относительно небольшой, но с бассейном. Протокон низкий, гребне-
видный.
2)/3 высокий, но сравнительно короткий (69.2-78.9% от длины 2)/4,
Л/=73.6±1.11%, «=9), с трехзубчатой коронкой. Передний зубец очень ма-
ленький; задний сравнительно невысокий, не достигает половины высоты
главной вершины. Коронка расширена в задней части. На лабиальном цингу-
8 Заказ 255(
113
Рис. 13. Молочные зубы Herpestes ichneumon, NRM А 580334:
а, г - снаружи; б, д - со стороны жевательной поверхности; в, е - изнутри
Fig. 13. Deciduous teeth of Herpestes ichneumon, NRM A 580334:
а, г - lateral; б, d - occlusal; в, e - labial views
114
лиде сзади и на поперечном постцингулиде обычно имеются маленькие буго-
рочки.
D/4 обычного хищнического типа. Режущие гребни параконида и
протоконида хорошо развиты. Параконид невысокий, ниже метаконида. По-
следний маленький, не достигает половины высоты протоконида, прижат к
его лингвальной стенке. Талонид сравнительно короткий (31.7-41.0% от об-
щей длины зуба, Л/=35.8± 1.12%, и=9); его бугорки практически не выра-
жены; бассейн отсутствует. Можно распознать только очень невысокий греб-
невидный гипоконид.
Сравнение. У стройного мангуста, Н sanguineus (Riippel, 1836)
молочные зубы заметно более мелкие (табл. 6); лингвальный поясок в осно-
вании dC/\ выражен намного слабее; на D3/ нет лабиального цингулюма, ме-
тастиль длиннее (в среднем 42.7% от общей длины зуба), талон существенно
менее широкий; на D4/ талон относительно более крупный; на D/3 передний
и задний добавочный зубцы относительно намного выше; на D/4 тригонид
относительно более короткий.
У яванского мангуста, Н. javanicus (Е. Geoffroy, 1818) на D3/ нет
лабиального цингулюма; талон не такой широкий; метастиль такой же длин-
ный, 40.0% от длины зуба; на D4/ парастилярная лопасть развита слабее;
паракон и метакон примерно равны по размерам; на D/3 задняя добавочная
вершина практически отсутствует.
У обыкновенного мангуста, Н edwardsii (Е. Geoffroy, 1818) на D3/ про-
токой очень маленький (рис. 14, вклейка)’, длина метастиля 38.2-38.9% от
длины зуба (А/=39.4±0.87%, /7=3); на D4/ метастилярная лопасть по величине
почти равна парастилярной; на D/4 талонид сравнительно более короткий
(см. табл. 6).
У короткохвостого мангуста, Н brachyurus Gray, 1837 на D/4
(Beaumont, 1974, pl.I, fig. 10) метаконид заметно более крупный, а талонид
короче и уже, чем у других известных нам Herpestes. В целом по морфологии
напоминает моляр мезозойских дельтатероидов.
Молочные зубы Herpestes (Leptoplesictis) aurelianensis Beaumont, 1974
из среднего миоцена (астараций, MN 7) Ла Грив-Сан-Албан, Франция (Beau-
mont, 1974, pl.I, fig.7) по размерам (длина D/4 4.2) меньше, чем у совре-
менных Herpestes (см. табл. 6). На D/3 боковые зубцы относительно высокие.
D/4 не отличается по строению от зубов других Herpestes.
Род Rhynchogale Thomas, 1894
Rhynchogale melleri (Gray, 1865) - Умби
Описание (рис. 15). Исследован только D/4. Зуб "всеядного" типа,
с шиловидными параконидом и метаконидом и очень низким протоконидом
8*
115
Рис. 15. Z)/4 Rhynchogale melleri, NRM A 585397:
a - снаружи, б - co стороны жевательной поверхности, в - изнутри
Fig. 15. D/4 of Rhynchogale melleri, NRM A 585397:
a - lateral, 6 - occlusal, в - labial views
(на MIX, 2 протоконид также ниже параконида и метаконида). На бугорках
тригонида режущие гребни не выражены. Талонид заметно шире тригонида,
с продольно вытянутым бассейном. Гипоконид низкий, гребневидный.
Энтоконид выше гипоконида. Гипоконулид гребневидный.
ТРИБА SURICATINI THOMAS, 1882
Род Suricata Desmarest, 1804
Suricata suricatta (Schreber, 1776) - Суриката
Описание (рис. 16). Верхние и нижние резцы простые. dl\-2l и
tZZ/1-2 игольчатые. Коронка d!3/ более расширена, со слабым внутренним
воротничком. dI/З с хорошо заметным латеральным зубчиком.
Клыки острые, слабо изогнутые. dCl/ без валика в основании коронки.
dC/\ менее широкий, со слабым валиком эмали на внутренней стороне.
116
D2I примыкает к Л)3/, от клыка отделен диастемой. По длине почти
равен £>4/. Простой зуб с высокой остроконечной коронкой и с небольшими
выступами на переднем и заднем концах коронки. D/2 очень высокий,
островершинный, сближен с £>/3. Коронка, как и у других нижних коренных,
наклонена вперед. Имеется небольшой задний добавочный зубчик. Корней 2.
D3/ маленький хищнический зуб. Внешняя стенка его слабо вогнута.
Передний отдел коронки крупный, несет крупный парастиль А. Парастиль В
отсутствует. Амфикон высокий, острый. Метастиль короткий (34% от общей
длины зуба). Талон умеренно длинный, широкий, расположен на уровне
вершины амфикона. Протокон в виде маленького зубца (SM 19421), либо
сравнительно крупный (ЗИН 283). Корней 3.
D4/ заметно более широкий, чем длинный. Эктофлексус почти не выра-
жен. Стилярная полка широкая. Паракон и метакон небольшие, но высокие,
сближенные. Талон имеет высокую препротокристу и значительно более низ-
кую постпротокристу. Они смыкаются у протокона, который по высоте сопо-
ставим с лабиальными бугорками. Корней 3.
D/3 очень высокий, небольшой (65-67% от длины D/4). К главному ко-
нусу на уровне половины его высоты сзади прижат крупный бугорок. Спере-
ди и сзади от них имеются более мелкие дополнительные бугорки. Зуб нем-
ного расширен в задней части (SM 19421), либо одинаковой ширины по всей
длине (ЗИН 283). Корней 2.
D/4 - хищнический зуб обычного строения. Тригонид с 3 четкими и
хорошо обособленными бугорками. Метаконид лишь немногим ниже прото-
конида (SM 19421), либо даже выше последнего (ЗИН 283). Талонид имеет
глубокий бассейн. Бассейн окружен гребнем, на котором можно различить
вершины гипоконида и энтоконида (на экз. ЗИН 283 имеется также отчет-
ливый гипоконулид). Корней 2.
ТРИБА MUNGOTINI РОСОСК, 19016
Род Bdeogale Peters, 1850
Bdeogalle crassicauda Peters, 1852 - Пушистохвостый мангуст
Описание (рис. 17). Верхние молочные резцы относительно круп-
ные, размеры их умеренно возрастают от dl\l к dI3i (табл. 7). Коронка почти
квадратная в плане, с хорошо развитым задним воротничком. Нижние резцы
также сравнительно крупные, узкие. Латеральный зубчик имеется на dI/2,3.
D2I сравнительно крупный, прижат к D3/. Зуб островершинный, расши-
рен в задней части. Имеется довольно крупный талон, поддерживаемый вы-
ростом заднего корня. Корней 2.
D3/ крупный, широкий, "всеядного" облика. Амфикон массивный,
Умеренно высокий. Перед ним расположен сравнительно небольшой пара-
117
Рис. 16. Молочные зубы Suricata suricatta, ЗИН 283:
, г - снаружи; б, с) - со стороны жевательной поверхности; в, е - изнутри
Fig. 16. Deciduous teeth of Suricata suricatta, ZIN 283:
а, г - lateral; б, 0 - occlusal; в, e - labial views
118
Таблица 6. Размеры молочных зубов у Herpestes
Table 6. Measurements of deciduous dentition in Herpestes
Промеры, Н. edwardsii H. ichneumon H. javanicus H. sanguineus
мм п | lim | Mun n | lim | M^m n=l n=\-2
Верхние зубы
IM L 1 0.3 6 0.4-1.0 0.6±0.08 0.6
W 1 0.6 6 0.3-0.8 0.5±0.07 0.3
H 1 1.0 6 0.9-1.4 l.l±0.08 0.4
in L 2 0.5,0.5 7 0.7-1.5 0.8±0.11 0.6
W 2 0.4,0.6 7 0.5-1.1 0.6±0.07 0.3
H 2 1.2,1.2 7 0.9-1.6 1.2±0.09 0.5
13/ L 3 0.9-1.0 1.0±0.03 8 1.1-1.9 1.3±0.08 0.8
3 0.8-1.1 1.0±0.09 8 1.1-1.8 1.3±0.08 0.6
H 3 1.5-2.2 1.8±0.21 8 1.5-2.5 2.1±0.13 2.0
CM L 3 1.8-2.1 2.0x0.09 9 2.1-3.4 2.5±0.12 2.1 1.7,1.8
W 3 1.3-1.5 1.4±0.06 9 1.6-2.4 1.8±0.07 1.7 1.2,1.3
H 3 4.6-5.7 5.2±0.33 9 5.4-7.4 6.1±0.19 6.7 3.6,4.5
D2/ L 3 3.1-3.6 3.3±0.17 9 3.8-4.4 4.1±0.07 2.3,2.4
W 3 1.2-1.3 1.3±0.03 9 1.4-2.1 1.8±0.08 1.0,1.0
H 3 2.5-3.4 2.9±0.26 9 3.1-4.1 3.8±0.11 2.3,2.3
D3/ L 3 5.4-5.6 5.5±0.06 9 6.3-7.9 6.9±0.20 5.0 4.4,4.5
Lmts 3 2.1-2.3 2.2±0.07 9 2.4-3.3 2.8±0.10 2.0 1.8,2.0
Lt 3 1.2-1.4 1.3±0.06 9 1.0-2.2 1.4±0.11 1.3 0.6,1.0
W 3 3.1-4.1 3.5±0.30 9 3.7-4.7 4.1±0.10 3.4 2.5,2.7
Wt 3 1.3-1.5 1.4±0.07 9 1.4-2.1 1.7±0.07 1.4 1.1,1.3
H 3 3.3-3.4 3.4±0.03 9 4.1-4.8 4.5±0.11 2.4 3.0,3.0
119
Продолжение табл. 6
Промеры, Н. edwardsii Н. ichneumon Н. javanicus Н. sanguineus
мм п | lim | М~т п | lim | М'.т и=1 п=\-2
D4/ L 3 2.7-3.2 3.0±0.15 9 3.0-5.1 3.7±0.21 3.1 lA.l.'l
И7 3 4.3-4.4 4.3±0.03 9 5.0-6.2 5.6±0.15 4.4 3.8,4.1
Н 3 1.7-2.1 1.8±0.13 9 2.3-2.9 2.5±0.06 1.3 1.0,1.5
Нижние зубы
//! L 1 0.3 5 0.4-0.8 0.5±0.06 0.2
IV 1 0.3 5 0.3-0.6 0.5±0.05 0.2
Н 1 0.4 5 0.8-1.3 1.1±0.08 0.6
//2 L 2 0.4,0.4 7 0.5-1.0 0.7±0.06 1.1 0.4
IV 2 0.4,0.4 7 0.4-1.4 0.7±0.13 0.9 0.3
Н 2 0.7,1.0 7 0.8-1.4 1.1±0.09 1.2 0.8
1/3 L 3 0.7-0.8 0.7±0.03 8 1.0-1.3 1.1±0.03 1.2 0.5
W 3 0.8-0.9 0.8±0.03 8 1.0-1.8 1.2±0.08 1.2 0.7
н 3 1.0-1.2 1.1=4=0.07 8 1.3-2.0 1.6±0.10 1.7 1.7
С/\ L 3 1.7-2.6 2.2±0.26 9 2.4-3.8 2.8±0.14 2.5 2.1,2.2
W 3 1.3-1.4 1.3±0.03 9 1.5-2.3 1.8±0.07 1.9 1.1,1.2
н 3 4.1-5.2 4.7±0.32 9 4.0-5.8 4.8±0.19 4.9 3.6,3.6
D/2 L 3 2.3-2.7 2.5±0.12 9 3.0-3.7 3.3±0.08 2.7 2.1
W 3 1.1-1.2 1.2±0.03 9 1.3-2.1 1.6±0.08 1.3 1.0
Н 3 1.7-2.8 2.2±0.32 9 3.0-4.3 3.6±0.14 1.7 2.2
D/3 L 3 3.5-3.7 3.6±0.06 9 4.3-5.4 4.7±0.12 4.0 2.7,3.3
W 3 1.4-1.7 1.5=1=0.10 9 1.7-2.8 2.1±0.10 2.0 1.2,1.4
Н 3 3.4-4.1 3.7±0.20 9 4.0-5.0 4.4±0.09 2.7 2.8,3.2
120
Окончание табл. 6
Промеры, мм Н. edwardsii H. ichneumon H. javanicus H. sanguineus
lim М tm n 1 lisj 1 n=l n=\-2
Г>/4 L 3 4.4-4.9 4.6±0.15 9 5.7-7.7 6.4±0.19 4.5 4.3,4.4
Ltl 3 1.7-2.1 1.9±0.12 9 2.0-2.8 2.3±0.08 1.7 1.4,2.1
Wtr 3 2.1-2.2 2.U0.03 9 2.4-3.8 2.7±0.14 2.5 1.8,2.0
Wtl 3 1.2-1.8 1.5±0.17 9 1.9-3.3 2.3±0.13 2.0 1.4,1.5
Н 3 3.4-4.1 3.8±0.20 8 4.1-4.6 4.4±0.07 3.0 3.0,3.3
стиль А. Парастиля В нет, но препротокриста, идущая к прото кону, хорошо
различима. Метастиль сравнительно длинный (51% от общей длины), задний
конец его лабиально отогнут. Талон массивный, длинный и широкий, с
островершинным протоконом. Лежит несколько кзади относительно верши-
ны амфикона, загнут назад. По краям талона идут гребни, окаймляющие
небольшой внутренний бассейн.
D4/ треугольной формы, по длине немногим уступает D3/. Эктофлексус
почти не выражен. Имеется стилярная полка. Паракон и метакон небольшие,
лежат на одной продольной линии. Паракон заметно меньше метакона. Про-
токон крупный, островершинный; хорошо выражены пре- и постпрото-
криста, ограничивающие бассейн талона.
DI2 крупный, прижат к D/3. Корона одновершинная, с небольшим
добавочным зубчиком сзади, расширена в заднем отделе. Корней 2.
D/3 относительно широкий, особенно в заднем отделе (67% от длины
D/4). Позади его главной вершины находится массивный бугорок, достигаю-
щий половины высоты последней. Имеются короткие талонид и передний
цингулид, по середине которого проходит продольный гребень.
7)/4 крупный, массивный, широкий. Бугорки тригонида широко рас-
ставлены, невысокие, с притупленными вершинками. Бугорки тригонида со-
поставимы по величине, параконид несколько массивнее. Протоконид и
метаконид разделены глубокой долинкой. Талонид занимает почти половину
Коронки зуба. Он шире тригонида, с обширным бассейном. Имеются четко
обозначенные гипоконид, энтоконид и гипоконулид, которые по высоте при
Приближаются к бугоркам тригонида. Энтоконид более сдвинут кзади, чем
гипоконид.
Сравнение. У черноногого мангуста, В. nigripes (Pucheron, 1855)
На ctf/2,3 нет латерального зубчика. D2I без талона. На 7)3/ метастиль более
Короткий (34% от общей длины зуба). DI2 без заднего добавочного зубчика.
121
Рис. 17. Молочные зубы Bdeogale crassicauda, NRM А 580166:
а, г - снаружи; б, д - со стороны жевательной поверхности; в, е - изнутри
Fig. 17. Deciduous teeth of Bdeogale crassicauda, NRM A 580166:
а, г - lateral; б, d - occlusal; e, e - labial views
122
Таблица 7. Размеры молочных зубов у Suricata, Bdeogale и Crossarchus
Table 7. Measurements of deciduous dentition in Suricata, Bdeogale and Crossarchus
Промеры, Suricata suricatta Bdeogale crassicauda Bdeogale nigripes Crossarchus obscurus
MM n=\-2 n=\ n=\ n | lim | M±m
Верхние зубы
/1/ L 0.4,0.5 1.0 1.6 2 0.6,0.6
IV 0.3.0.4 0.7 1.6 2 0.4,0.4
H 0.8,0.8 1.5 1.8 2 0.9,1.2
/2/ L 0.5,0.6 1.2 1.7 2 0.7,0.8
IV 0.6,0.7 1.2 1.9 2 0.6,0.8
H 0.9,1.1 1.7 2.5 2 1.1,1.6
73/ L 1.0 1.8 1.9 2 1.1,1.1
IV 1.0 1.7 2.5 2 1.0,1.3
H 1.7 2.9 2.8 2 1.5,2.1
CM L 1.4,2.0 2.5 2 1.7,1.7
IV 1.3,1.4 1.7 2 1.0,1.3
H 3.4 4.5 2 4.4,4.9
DU L 2.5,2.7 2.8 4.8 3 2.6-2.9 2.7±0.09
IV 1.1,1.6 1.8 2.8 3 1.1-1.2 l.l±0.03
H 2.1,3.2 2.7 3 1.5-2.7 2.3±0.39
D3I L 3.7,4.1 4.3 6.7 3 4.0-4.1 4.0±0.03
Lmts 1.4,1.5 2.2 2.3 3 1.5-1.8 1.7±0.10
Lt 1.1,1.2 1.7 2.7 3 1.0-1.2 l.l±0.06
IV 2.5,2.8 3.6 5.0 3 2.6-3.1 2.8±0.17
Wt 1.0,1.2 1.4 1.7 3 1.2-1.3 1.2±0.03
H 2.8,3.2 3.7 3.5 3 1.8-3.2 2.6±0.41
123
Продолжение табл. 7
Промеры, мм Suricata suricalta Bdeogale crassicauda Bdeogale nigripes C'rossarchus obscurus
п=\-2 п=\ n=\ n | lim | Mun
Г>4/ L 2.7,3.1 3.5 5.5 2 2.8,3.6
И7 4.0,4.2 4.8 6.7 2 4.1,4.1
И 1.6,2.5 1.1 3.1 2 1.3,1.6
Нижние зубы
7/1 L 0.6 0.5 1.2 2 0.4,0.4
PK 0.6 0.5 0.9 2 0.3,0.5
H 0.6 1.2 1.5 2 0.5,1.1
7/2 L 0.5,0.7 1.0 1.5 2 0.5,0.6
PK 0.5,0.7 1.0 1.8 2 0.5,1.0
H 0.8,0.8 1.9 1.5 2 0.8,1.2
Z/3 L 0.7,0.8 1.3 1.8 2 0.7,0.8
PK 0.7,1.0 1.6 2.7 2 0.8,1.2
H 1.2,1.4 2.3 2.0 2 1.2,2.5
C/1 L 1.6,1.9 2.7 2 1.5,1.7
IV 1.1,1.2 1.6 2 1.2,1.3
H 3.3,4.0 4.5 2 3.4,3.6
D/2 L 2.3,2.4 2.9 4.5 2 2.0,2.7
IV 1.2,1.2 1.2 2.1 2 1.1,1.2
H 2.3,3.3 2.5 2.1 2 2.6,3.2
D/3 L 3.0,3.1 3.4 5.7 2 3.0,3.2
IV 1.4,1.7 2.1 2.8 2 1.2,1.6
H 2.7,3.5 3.0 3.3 2 2.7,3.5
124
Окончание табл. 7
Промеры, мм Suricata suricalta Bdeogale crassicauda Bdeogale nigripcs Crossarchus obscurus
n=\-2 n=l n=\ n | lim |
Z)/4 L 4.5,4.7 5.2 7.8 3 4.1-4.5 4.3±0.12
Ltl 1.5,2.2 2.1 3.1 3 1.3-1.7 1.5±0.12
Wtr 2.1,2.2 2.3 3.5 3 1.9-2.4 2.1±0.15
Wtl 2.0,2.0 2.4 3.7 3 1.6-2.3 1.9±0.21
Н 3.0,3.7 2.6 3.6 3 2.4-3.2 2.9±0.24
На 2)/4 между параконидом и метаконидом расположен дополнительный
бугорочек.
Род Crossarchus F.Cuvier, 1825
Crossarchus obscurus F.Cuvier, 1825 - Длинноносый кузиманзе
Описание (рис. 18). Резцы сравнительно крупные. dl\!\,2 с двух-
зубчатой коронкой. На сИЪ/ имеется лингвальный поясок, на dI/З - хорошо
выраженный латеральный бугорок.
dC\/\ невысокие, простого строения, с очень слабыми лингвальными
поясками.
7)2/2 сравнительно крупные, с одновершинной коронкой, похожи друг
на друга (имеется небольшой задний добавочный зубчик и бугорок на пояске
позади него). На экземпляре ЗИН 8700 2)2/2 без задних добавочных зуб-
чиков. На 7)2/ различим очень короткий лингвальный цингулюм ("талон").
7)3/ сравнительно короткий, с крупным талоном. Парастиль А может
быть двойным (NRM А 590346), парастиль В отсутствует. Метастиль очень
низкий, но длинный (44-45% от общей длины зуба). Талон крупный; его
лингвальную половину, развернутую назад, занимает основание протокона.
Протокон в виде высокого острого зубца. Препротокриста отчетливая.
7)4/ сравнительно крупный, заметно больше 7)3/. Эктофлексус неглу-
бокий, смещен к переднему краю зуба (в отличие от большинства других
Мангустовых и виверровых, метастилярная лопасть заметно крупнее парасти-
лярной). Стилярная полка большая. Паракон и метакон в виде очень низких
продольных гребней, первый из них короче. Талон большой, но без выражен-
ного бассейна. Протокон большой, гребневидный.
125
Рис. 18. Молочные зубы Crossarchus obscurus, NRM А 590346:
а, г - снаружи; д - со стороны жевательной поверхности; в. е - изнутри
Fig. 18. Deciduous teeth of Crossarchus obscurus, NRM A 590346:
а, г - lateral; б, d - occlusal; в, e - labial views
126
D/3 - высокий зуб с трехвершинной коронкой, основание которой резко
расширено в задней части, 72% от длины D/4. Передний и задний доба-
вочные зубцы отделены от главной вершины глубокими бороздками; задний
зубец намного выше переднего, достигает половины высоты главной верши-
ны. В основании коронки имеется поперечный постцингулид, несущий в
лингвальной части 2 бугорочка. Другой бугорочек ("метаконид") расположен
в основании косого гребня, поднимающегося с лингвальной стороны на глав-
ную вершину.
D/4 с высокими шиловидными бугорками тригонида. Режущие гребни
на протокониде и паракониде слабо выражены. Метаконид немного выше па-
раконида и приближается по высоте к протокониду. Талонид без выраженно-
го бассейна; большую его часть занимает основание высокого гипоконида.
Энтоконид и гипоконулид в виде маленьких бугорочков, сближенных друг с
другом.
Род Mungos E.Geoffroy et F.Cuvier, 1795
Mungos mungo (Gmelin, 1788) - Полосатый мангуст
Описание (рис. 19). Резцы небольшие; tZZT ,2/ игольчатые; dI3/
более крупный, без добавочных бугорков и пояска; коронки dI/\,2 могут
;быть двухзубчатыми; dI/З с латеральным добавочным зубчиком.
dC\l\ небольшие, простого строения. В основании dC/\ идет очень
^слабый лингвальный поясок.
’ D2/2 сравнительно крупные, с продольно вытянутой одновершинной
коронкой. На D2/ имеется небольшой передний, а на D/2 задний доба-
вочный бугорок.
D3/ легкого строения, с относительно крупным талоном. Парастиль А
Относительно крупный, парастиль В отсутствует. Метастиль низкий. Талон
Короткий, но широкий, расположен напротив вершины амфикона. Протокон
в виде высокого острого зубца. Отчетливо выражена препротокриста.
£>4/ по ширине превышает D3/. Эктофлексус очень слабый. Стилярная
Полка широкая. Парастилярная лопасть иногда выдается лабиально (ЗММУ
S-58223). Паракон и метакон примерно равны по размерам, низкие, гребне-
видные. Талон короткий и широкий, без бассейна, весь занят основанием
Протокона. Протокон высокий, гребневидный.
D/3 сравнительно узкий и короткий (45-58% от длины Z)/4). Задняя
Добавочная вершина очень низкая, передняя - отчетливая. Имеется короткий
Поперечный постцингулид.
D/4 с ослабленными признаками "хищнического" зуба. Бугорки триго-
Нида относительно мелкие, шиловидные, низкие (иногда даже ниже вершины
экз. NRM А 591985), со слабо выраженными режущими гребнями. Ме-
127
Рис. 19. Молочные зубы Mungos mungo, NRM А 591985:
а, г - снаружи; б, с) - со стороны жевательной поверхности; в, е - изнутри
Fig. 19. Deciduous teeth of Mungos mungo, NRM A 591985:
а, г - lateral; б, d - occlusal; в, e - labial views
128
таконид такой же высокий, как и параконид, либо несколько ниже (ЗММУ S -
58223). Талонид без бассейна. Гипоконид и энтоконид очень слабо выра-
жены (в виде низких гребневидных поднятий). Гипоконулид, напротив, от-
четливый и сравнительно высокий, выше остальных бугорков талонида.
Род Ichneumia I.Geoffroy, 1837
Ichneumia albicauda (G.Cuvier, 1829) - Белохвостый мангуст
Описание (рис. 20). Размеры верхних и нижних резцов заметно
увеличиваются от dIM к <7/3/3 (табл. 8). Коронка верхних резцов ложко-
видная, с хорошо выраженным внутренним воротничком. Среди нижних рез-
цов dU\ игольчатый, dI/2 со слабо расширенной коронкой, а dI/З имеет хоро-
шо моделированный латеральный зубчик.
Клыки довольно слабые. На dCM имеется короткий гребешок в основа-
нии коронки с лингвальной стороны. Вершина клыка направлена вниз. На
dC/\ корень несколько толще коронки. Последняя крючковидно изогнутая;
ее вершина направлена вверх и несколько назад. С внутренней стороны у
переднего края основания коронки прослеживается утолщение эмали.
D2/2 высокие, островершинные. Спереди и сзади коронки D2/ нахо-
дятся добавочные зубчики. У переднего края основной вершины D/2 распо-
ложен маленький острый зубчик, смещенный лингвально. Иногда присут-
ствует задний добавочный зубчик и зубцевидный "талонид" Корней 2.
D3/ - крупный хищнический зуб обычного строения. Лабиальный край
зуба прямой. Передний отдел коронки небольшой, с парастилем А. Пара-
стиль В очень небольшой или отсутствует, иногда двойной (ЗИН 5795). Ме-
тастиль умеренно длинный (36-44% общей длины, Л/=39.1±1.1%, и=7). Талон
довольно крупный, с высоким острым зубцом протокона. Лежит на уровне
вершины амфикона.
£)4/ с треугольной в плане коронкой, относительно крупный. Экто-
флексус не развит. Стилярная полка большая, часто имеется бугорок в глу-
бине эктофлексуса ("бугорок С"). Лабиальные бугорки мелкие, острые; их
продольные кили образуют единый режущий гребень. Размеры их разли-
чаются незначительно, но паракон обычно несколько крупнее. Талон круп-
ный, с глубоким бассейном, который ограничен спереди и сзади гребешками.
Передний из этих гребешков более мощный. Протокон в виде массивного
зубца.
D/3 - высокий, крупный зуб (72% от длины D/4). Передний дополни-
тельный бугорок на цингулиде небольшой; задний, расположенный за основ-
ной вершиной, высокий и хорошо обособленный, иногда распадается на 2
зубчика (ЗИН 5795). Имеется небольшой "талонид" с бассейном, который
моделирован гребнем, проходящим по заднему краю коронки.
D/4 - крупный и широкий хищнический зуб. Параконид по высоте мало
Уступает протокониду. Метаконид высокий, обособленный, но заметно мень-
129
9 Заказ 255(а'
Рис. 20. Молочные зубы Ichneumia albicauda, NRM А 580271:
а, г - снаружи; б, д - со стороны жевательной поверхности; в, е - изнутри
Fig. 20. Deciduous teeth of Ichneumia albicauda, NRM A 580271:
а, г - lateral; () - occlusal; в, e - labial views
130
Таблица 8. Размеры молочных зубов у Mungos, Ichneumia и Cynictis
Table 8. Measurements of deciduous dentition in Mungos, Ichneumia and Cynictis
Промеры, Mungos mungo Ichneumia albicauda Cynictis penicillata
MM n | lim | Mim n 1 bm | M~m n=l
Верхние зубы
/1/ L 2 0.5,0.6 3 0.9-1.0 1.0±0.05 0.5
IV 2 0.4,0.5 3 0.6-0.7 0.7±0.03 0.3
H 2 1.0,1.2 3 0.9-1.5 l.l±0.19 0.6
in L 2 0.6,0.7 5 1.1-1.3 l.l±0.04 0.5
IV 2 0.5,0.7 5 1.0-1.2 1.U0.02 0.4
H 2 1.0,1.5 5 1.2-2.1 1.6±0.17 0.7
13/ L 4 1.0-1.5 1.2±0.11 7 1.4-1.9 1.7±0.06 1.0
IV 4 1.0-1.1 l.l±0.03 7 1.6-2.2 1.7±0.08 0.7
H 4 1.4-2.9 2.4±0.33 7 1.9-3.5 2.8±0.25 1.7
CM L 3 2.1-2.5 2.2±0.13 7 2.3-2.9 2.6±0.08 2.2
IV 3 1.1-1.7 1.4±0.17 7 1.8-2.1 2.0±0.03 1.3
H 3 3.8-6.7 5.2±0.84 7 4.9-6.1 5.6±0.21 4.4
D2/ L 3 2.7-3.7 3.U0.31 7 4.4-5.1 4.7±0.09 3.0
IV 3 1.1-1.4 1.2±0.09 7 1.7-2.0 1.9±0.05 1.1
H 3 2.2-2.7 2.4±0.17 7 3.1-4.1 3.7±0.17 2.7
D3/ L 3 4.2-6.3 4.9±0.70 7 6.5-7.2 6.9±0.09 5.1
Lmts 3 1.8-2.8 2.2±0.32 7 2.4-3.0 2.7±0.09 2.1
Lt 3 1.2-1.7 1.4±0.15 7 1.6-2.0 1.9±0.07 1.2
IV 3 2.6-3.3 3.0±0.22 7 4.0-5.3 4.5±0.15 3.2
Wt 3 1.2-2.0 1.5±0.24 7 1.5-3.1 2.0±0.20 1.3
H 3 2.4-3.6 2.9±0.36 7 3.8-4.6 4.U0.12 3.2
9-
131
Продолжение табл. 8
Промеры, Mungos mungo Ichneumia albicauda Cymctis penicillata
мм л | lim | M±m n| lim | M+m n=l
DM L 3 3.0-3.5 3.3±0.17 7 5.1-5.5 5.2±0.06 4.3
W 3 4.2-5.0 4.5±0.27 7 6.3-6.8 6.6±0.07 3.3
Н 3 1.3-1.8 1.6±0.15 7 2.0-3.4 2.6±0.16 1.3
Нижи iи e зубы
Z/l L 2 0.3,0.4 2 0.7,0.8 0.3
W 2 0.3,0.4 2 0.7,0.8 0.3
н 2 0.4,1.1 2 1.1,1.2 0.7
Z/2 L 2 0.5,0.6 4 0.9-1.1 1.0±0.05 0.5
W 2 0.4,0.5 4 1.1-1.4 1.2±0.06 0.5
н 2 0.4,1.2 4 1.3-2.2 1.7±0.22 0.7
ИЗ L 3 0.7-0.9 0.8±0.06 5 1.1-1.3 1.2±0.03 0.7
W 3 0.9-1.4 l.l±0.15 5 1.5-2.0 1.8±0.08 0.8
н 3 1.3-2.0 1.7±0.20 5 1.9-3.0 2.5±0.21 1.3
а\ l 3 1.9-2.5 2.2±0.18 5 2.7-3.3 2.9±0.13 2.4
W 3 1.2-1.5 1.3±0.10 5 1.8-1.9 1.9±0.02 1.2
н 3 3.5-6.0 4.5±0.76 5 4.3-5.3 4.8±0.19 3.4
DU L 3 2.4-2.8 2.6±0.12 5 3.7-4.3 4.1±0.10 2.3
W 3 1.0-1.4 l.l±0.13 5 1.6-2.0 1.7±0.07 1.0
Н 3 2.1-2.8 2.4±0.22 5 3.0-4.3 3.6±0.25 2.5
D/3 L 3 3.1-4.0 3.5±0.26 5 5.0-5.7 5.3±0.14 3.4
W 3 1.4-2.0 1.6±0.20 5 2.2-2.7 2.5±0.09 1.5
Н 3 3.0-3.4 3.2±0.12 5 3.7-4.6 4.2±0.16 3.4
132
Окончание табл. 8
Промеры, мм Mungos mungo Ichneumia alhicauda Cynictis penicillata
n lim | M-m n 1 lim |
£)/4 L 3 4.8-6.4 5.4±0.52 5 7.1-7.4 7.3±0.07 5.0
Ltl 3 2.0-2.2 2.H0.06 5 2.5-3.0 2.9±0.11 2.5
Wtr 3 2.1-2.4 2.2±0.09 5 3.1-3.4 3.2±0.06 2.2
Wtl 3 1.9-2.1 2.0±0.07 5 3.3-3.4 3.4±0.02 2.1
H 3 3.1-4.0 3.5±0.28 5 4.0-4.5 4.2±0.10 3.1
ше других бугорков тригонида. По его внутреннему краю и по внутреннему
краю протоконида спускаются навстречу друг к другу эмалевые гребешки.
Талонид крупный, с глубоким бассейном. Хорошо выражены гипоконид и
более мелкий энтоконид. На месте гипоконулида идет короткий гребешок.
Род Cynictis Ogilby, 1833
Cynictispenicillata (G.Cuvier, 1829) - Желтый мангуст
Описание (рис. 21). Резцы мелкие, <У/1,2/1 игольчатые, dI3/ с
лингвальным пояском, <7//2,3 с пояском и латеральным бугорком.
Клыки короткие, с изогнутой коронкой. На dCM имеется слабый, а на
dC/\ более отчетливый лингвальный поясок.
D2/2 - сравнительно крупные, вытянутые в длину зубы. D2I с высокой
главной вершиной и маленькими зубчиками в основании коронки спереди и
сзади. D/2 с более низкой коронкой; передний добавочный зубчик распола-
гается намного выше заднего.
03/ обычного хищнического типа, со сравнительно крупным талоном.
Парастиль А в виде низкого продольного гребня, парастиль В отсутствует.
Метастиль длинный (41% от общей длины зуба). Талон широкий; большую
его часть занимает основание протокона. Протокон низкий.
£>4/ сравнительно крупный, треугольной в плане формы. Эктофлексус
неглубокий. Метастилярная лопасть развита больше парастилярной. Стиляр-
ная полка широкая. Паракон и метакон в виде продольно вытянутых неболь-
ших бугорков, примерно равных по размерам. Талон с небольшим бассей-
ном, вершина протокона невысокая.
133
Рис. 21. Молочные зубы Cynictispenicillata, NRM А 581303:
а, г- снаружи; б, д - со стороны жевательной поверхности; в, е - изнутри
Fig. 21. Deciduous teeth of Cynictispenicillata, NRM A 581303:
а, г - lateral; б, d - occlusal; в, e - labial views
134
D/3 с высокой трехвершинной коронкой, но задний добавочный зубец
сравнительно слабо развит (68% от длины £>/4). Имеется короткий поперед
ный постцингулид, заходящий на лингвальную сторону коронки.
D/4 обычного типа. Режущие гребни протоконида и параконида хороню
развиты. Метаконид высокий, выше параконида. Талонид с обширным бас-
сейном. Гипоконид в виде невысокого пологого бугорка. Энтоконид очень
маленький. Гипоконулид шиповидный, выше других бугорков талонида.
ОБСУЖДЕНИЕ
Молочные зубы Viverridae и Herpestidae построены по единому плану,
варьирующему в деталях у ряда аберрантных групп и являющемуся исход-
ным для Carnivora, наряду с планом строения молочных зубов Canidae. У
Canidae молочные зубы меняются в пределах семейства в меньшей степени и
отличаются следующими существенными чертами: 1) резцы всегда крупные,
2-3-зубчатые; 2) парастиль В на D3/ редуцированный, с развитыми гребнями,
идущими к парастилю А и к протокону; 3) задняя добавочная вершина на D/3
и метаконид на D/4 почти всегда уменьшены. Полярность первого признака
неясна; возможно, что это - аутапоморфия для Canidae. Признаки 2 и 3 четко
характеризуют развитие режущей специализации молочных зубов канид. По
этим признакам семейство Canidae в целом более продвинуто, чем семейства
Viverridae и Herpestidae, хотя среди последних имеются отдельные предста-
вители, молочные зубы которых приспособлены для разрезания мясной
пищи в существенно большей степени, чем у канид.
Зубная формула. Молочная зубная формула dI\-3/\-3 dC\/\
D2-4/2-4 сохраняется неизменной у всех виверровых и мангустовых и исход-
на для отряда Carnivora. То же характерно для Canidae и, видимо, для Ргосуо-
nidae (Барышников, Аверьянов, 1993а, б). У Ursidae и Mustelidae среди
Arctoidea и у Felidae из Aeluroidea наблюдается уменьшение числа зубов в
различной степени. У примитивных млекопитающих, в частности у миацид
Miacidae (Springhom, 1985), Р1/1, будучи по происхождению молочными зу-
бами (см. Барышников, Аверьянов, 1990), функционируют одновременно с
другими молочными зубами (смена зубов происходит довольно поздно в
онтогенезе) и у более взрослых животных обычно выпадают. Для изученных
виверр (Nandinia, Viverricula, Civettictis, Viverra, Genetta, Paradoxurus, Pagu-
ma, Cynogale, Hemigalus, видимо, также для Prionodon, Eupleres, Arctogalidia
и Arctictis} и для некоторых мангустов (Bdeogale и, возможно, Ichneumia)
также характерно ранние прорезание и выпадение Р1/1, функционирующих
вместе с молочными зубами. У большинства мангустов (Galidia, Atilax,
Herpestes, Suricata, Crossarchus, Mungos и Cynictis) прорезание P\/\ задер-
живается, и эти зубы функционируют уже вместе с постоянными зубами. По
Данному признаку мангустовые в целом более продвинуты, чем виверровые.
135
Резцы. Молочные резцы у изученных виверр и мангустов сравни-
тельно крупные, уменьшены в размерах обычно лишь d!\/\. У большинства
видов верхние резцы имеют простую коническую коронку, иногда расширен-
ную дистально (Nandinia). Нижние резцы у Viverridae, как правило, простые,
с конической коронкой, иногда со следами двухзубчатости (явно двух-
зубчатые dU\-3 наблюдаются у Genetta). У мангустов нижние резцы чаще
всего отчетливо двухзубчатые. Последний признак, видимо, является более
продвинутым (по аналогии с Canidae). Для миацид были характерны неболь-
шие молочные резцы с простой конусовидной коронкой (Springhom, 1985). В
целом, по строению молочных резцов, Herpestidae кажутся более специали-
зированными, чем Viverridae.
Клыки. Молочные клыки виверровых и мангустовых обычно круп-
ные, со слабо изогнутой коронкой (в большей степени изогнуты нижние клы-
ки). Соотносительные размеры молочных клыков, коренных и резцов у
Viverridae и Herpestidae аналогичны таковым у Miacidae (Springhom, 1985).
Наиболее относительно крупными клыками обладает Genetta. Верхние клы-
ки обычно без лингвального пояска (слабый поясок имеется у Crossarchus и
Cynictis). На нижних клыках лингвальный поясок в основании коронки чаще
всего присутствует. У ряда форм (некоторые Paradoxurinae и Genetta) вместо
пояска имеется лишь отчетливый передний бугорок (состояние, обычное для
Felidae). Поясок на нижних клыках полностью отсутствует у некоторых
мангустов (Suricata) и у Nandinia. Степень редукции пояска, видимо, связана
с усилением режущей функции молочных зубов (у Paradoxurinae, возможно,
редуцируется по другой причине). В этом отношении Nandinia более
специализирована, чем даже Felidae.
Коренные. D2/2 сравнительно крупные, двухкорневые зубы, обыч-
но со слабо развитыми добавочными зубчиками (хорошо развиты у Nandinia,
Arctogalidia, Herpestes, Crossarchus, Mungos, Ichneumia, Cynictis и др.).
Иногда коронки D2/2 довольно высокие (Viverricula, Arctictis) или усложне-
ны, образуя подобие талона и талонида (Bdeogale).
D/3 крупный зуб, с небольшими зубчиками спереди и сзади коронки и с
крупным добавочным зубцом позади главной вершины, достигающим неред-
ко половины высоты последней (примитивное состояние, характерное для
миацид (Springhom, 1985) и большинства виверрид и герпестид). Задний
добавочный зубец в различной степени редуцирован у Viverra, Arctogalidia,
Arctictis, Mungos и Ichneumia. Значение редукции этого зубца у виверр и
мангустов не совсем понятно. У Canidae, Felidae и других Camivora эта ре-
дукция связана с усилением режущей функции молочных зубов.
D3I устроен принципиально сходным образом у большинства Viverridae
и Herpestidae. Это типичный хищнический зуб с парастилем А, крупным ам-
фиконом, метастилем различной длины и, как правило, с талоном. Схема
строения D3/ меняется лишь у абберантных виверровых: у Cynogale из
Paradoxurinae и Eupleres из Euplerinae. У Cynogale парастиль А удлинен, а
метастиль укорочен и уступает по длине первому. Талон заметно преобра-
136
зован, имеет вид пояска, несущего несколько мелких зуочиков. У Eupleres
D3/, как и другие молочные зубы, заметно упрощен (нет парастиля В
метастиль короткий, талон небольшой), но в целом сохраняет исходное для
Viverridae строение. Такое упрощение объясняется, видимо, энтомофагией
Eupleres. Редукция метастиля D3/ наблюдается и у известных авторам
Paradoxurinae {Arctictis, Arctogalidia, Paguma, Paradoxurus) и у Hemigalus,
что может объяснятся всеядностью последних. У наиболее продвинутых
миофагов (Genetta, Nandinia) метастиль D3/, напротив, заметно удлинен.
Важным в систематическом отношении признаком представляется на-
личие или отсутствие на D3/ парастиля В. Этот бугорок имеется у боль-
шинства изученных Viverrinae (кроме Prionodon), у Nandinia (Nandiniinae) и
у Hemigalus (Hemigaliinae). Парастиль В отсутствует у Eupleres (Euplerinae), у
всех изученных Paradoxurinae, Galidiinae и Herpestinae (сильно уменьшенный
парастиль В имеется только у Ichneumia). Распределение этого признака
показывает, что редукция парастиля В происходила независимо в разных
линиях виверрид и герпестид, и последние более продвинуты в этом отноше-
нии. Редукция парастиля В, очевидно, связана с усилением миофагии. У
Nandinia парастиль В занимает нетипичное для виверровых и мангустовых
положение: этот бугорок сдвинут лабиально и находится почти на одной
линии с вершинами парастиля А и амфикона. Такое состояние типично для
Felidae. Трудно судить, является ли данный признак синапоморфией Nan-
diniinae и Felidae, свидетельствующей о филогенетической близости этих
групп, либо он развился у Nandinia независимо, как результат глубоко зашед-
шей адаптации к миофагии, параллельной таковой у Felidae.
Присутствие парастиля В на D3/ является, возможно, синапоморфией
для всех Fissipedia современного типа (без Miacoidea). Изолированный D3/
из нижнего палеоцена (пуэркий) Канады (Fox, Youzwyshyn, 1994, figs.9,10),
принадлежавший, видимо, Viverravidae, лишен парастиля В и препрото-
кристы, зато имеет сильно развитую постпротокристу. Метастиль очень ко-
роткий, окружен лабиальным и лингвальным цингулюмами; талон большой,
с крупным протоконом. Возможно, что основными режущими структурами
молочных зубов миацид семейства Viverravidae были постпротокриста D3/ -
протокристид £>/4, а не метастиль D3/ паракристид D/4, как у Fissipedia
современного типа. Подобная редущая структура менее эффективна для
разрезания мясной пищи; обладание ею может исключать Viverravidae из
числа возможных предков остальных Fissipedia.
Другой признак, отражающий степень развития хищнических
адаптаций в молочной зубной системе, - редукция протокона на D3/. Хорошо
развитый протокон имеется у примитивных Viverrinae (Civettictis, Prionodon,
Pseudogenetta, Viverricula) и отсутствует у более продвинутых членов
Подсемейства (Genetta и Viverra)', среди Hemigaliinae он есть у Hemigalus, нет
У Nandinia и Eupleres, большинства Paradoxurinae (иногда может быть у
Paradoxurus), у всех Galidiinae, но имеется у всех изученных нами Herpes-
tinae (у последних протокон обычно располагается на широком и коротком
137
(спереди назад) талоне, не характерном для Viverridae). Редукция протокона
£>3/ происходила, очевидно, независимо и параллельно в разных филогене-
тических линиях виверровых и мангустовых.
Е>4/ устроен принципиально сходным образом у всех изученных Viver-
ridae и Herpestidae, варьируя лишь в относительных размерах и степени вы-
раженности главных бугорков. Наиболее преобразовано строение £>4/ у
Cynogale, где лабиальные бугорки сильно сдвинуты к краю зуба, паракон
увеличен, метакон сильно уменьшен, протокон разбит на серию мелких
бугорков и развит крупный бассейн талона. Параконуль и метаконуль на £>4/
имеются среди изученных форм только у Civettictis, Hemigalus, Galidia и
Atilax, т.е. соответственно у представителей подсемейств Viverrinae, Hemi-
galiinae, Galidiinae и Herpestidae. У более продвинутых членов этих подсе-
мейств и у Euplerinae и Paradoxurinae оба бугорка отсутствуют. Для некото-
рых характерна крупная парастилярная лопасть, сильно выступающая ла-
биально (плезиоморфный признак, широко распространенный у меловых
Eutheria на постоянных молярах): для Viverra, Viverricula, Pseudogenetta и
Genetta из Viverrinae, Nandinia, Galidiinae, и для Atilax и Herpestes из Herpes-
tinae. Редукция подобного состояния, очевидно, происходила независимо в
разных линиях обоих семейств. Интересно, что данный признак отмечается
для наиболее специализированных миофагов: Genetta, Nandinia, а также для
Hyaenidae и Felidae (но не для Mustelidae). Усиление парастилярной лопасти
(возможно, вторичное) связано с развитием у миофагов мощного поперечно
ориентированного режущего гребня парастиля £>4/, участвующего в разреза-
нии мяса, и могло развиваться независимо в разных линиях.
Строение £>/4 у Viverridae и Herpestidae мало варьирует, сохраняя при-
митивное для Camivora состояние. Прецингулид £>/4 (при окклюзии связан с
парастилем В D3/) имеется у Viverrinae (кроме Prionodon), Nandinia, Para-
doxurinae (кроме Arctictis), Hemigallinae и у Atilax и Bdeogale из Herpestinae.
Отсутствие прецингулида £>/4 характеризует также Galidiinae. У Arctictis,
Cynogale, Salanoia, Mungos, Suricata главные вершины тригонида Z)/4 при-
остренные, без резко выраженных режущих гребней, что может быть об-
условлено в основном насекомоядным характером питания этих форм. У
остальных изученных виверровых и мангустовых параконид и протоконид
D/4 образуют более или менее выраженный лезвиевидный режущий гребень
(паракристид), даже у Paradoxurinae, имеющих всеядный тип питания.
У примитивных Viverrinae (Civettictis), Paradoxurinae (Arctictis и
Paradoxurus), Hemigaliinae (Hemigalus), Galidiinae (Salanoia) и у большинства
Herepstinae (кроме Herpestes и Ichneumia) метаконид D/4 крупный, примерно
равный по размерам паракониду. У остальных изученных родов метаконид в
той или иной степени уменьшен, что связывается с усилением адаптации к
миофагии (включая наиболее специализированных миофагов Genetta и
Nandinia). Тенденция усиления адаптации к миофагии прослеживается у дрУ'
гих Feliformia. Так, уменьшенный метаконид D/4 характерен для всех Hyaeni-
138
dae и Protelidae; еще больше он редуцируется, часто до полного исчезнове-
ния, у Felidae.
По строению талонида £>/4 изученные рода виверровых и мангустовых
мало варьируют. Талонид всегда крупный, очень часто с бассейном и в той
или иной степени выраженными бугорками. То же характерно для Ictitheri-
idae. У Hyaenidae талонид £>/4 всегда уменьшен; у Felidae он либо очень ма-
ленький, в виде небольшого бугорка, либо вовсе отсутствует.
По размерам и пропорциям верхних молочных коренных зубов в ана-
лизе главных компонент (табл. 9; рис. 22) наиболее различаются подсе-
мейства Viverrinae, Herpestinae и Galidiinae (в обоих факторных простран-
ствах, FlxF2 и FlxF3). Paradoxurinae в пространстве FlxF2 располагаются, в
основном, внутри Herpestinae, в пространстве FlxF3 внутри Viverrinae.
Следовательно, по размерам и пропорциям D3/ (F2 в большей степени связан
с длиной, длиной метастиля и высотой £>3/) Paradoxurinae ближе к Herpesti-
nae, а по размерам £>4/ и промерам ширины £>2/, £>3/, талона D3/ (F3) - к
Viverrinae.
При анализе главных компонент промеров нижних молочных коренных
зубов Herpestinae образуют более широкое "облако" (рис. 23), включающее
Hemigalus, почти всех Paradoxurinae и близко подходящее к Galidia. "Обла-
ко" Viverrinae в пространстве FlxF2 частично перекрывается с Paradoxurinae,
в пространстве FlxF3 (F3 связан с длиной талона £>/4) четко отделено от
Paradoxurinae и других групп. Hemigalus в обоих пространствах по верхним и
нижним молочным коренным близок к Paradoxurinae. Анализ главных
компонент свидетельствует о различии, по крайней мере, Viverrinae и Para-
doxurinae по пропорциям молочных коренных зубов.
Замечания о филогении Viverridae и Herpe-
stidae. Распределение признаков молочных зубов среди Viverridae и
Herpestidae не позволяет выделить четко охарактеризованные монофилети-
ческие группы (подсемейства). Наиболее обособленными являются Paradoxu-
rinae (бугорки на молочных коренных притуплены), в меньшей степени
Galidiinae. Уникальные черты имеют некоторые терминальные таксоны
(Cynogale, Eupleres), что, естественно, не дает информации об их родствен-
ных связях.
Среди Viverridae и Herpestidae можно выделить несколько групп (град)
по уровню эволюционной продвинутое™ строения молочных зубов. Низшую
граду образуют примитивные Viverrinae (например, Viverricula), характери-
зующиеся в целом исходно хищническим (насекомоядным) типом строения
Молочных зубов. Однако, некоторые представители подсемейства более про-
двинуты в направлении приспособления к миофагии (Viverra, Prionodon, и, в
существенно большей степени, Genetta). Paradoxurinae представляют со-
бой, по видимому, филогенетически единую группу (ветвь), развившуюся из
Примитивных Viverrinae (типа Viverricula) в направлении адаптации ко все-
яДению (наиболее продвинуты Arctictis и Paguma, наименее - Arctogalidia).
139
FACTORS)
Рис. 22. Анализ главных компонент по промерам верхних молочных коренных зубов у
различных Viverridae и Herpestidae:
а - Arctictis binturong, b - Arctogalidia trivirgata, c - A tilax paludinosus, d - Bdeogale crassicaudata, e - Bdeogale nigripes,
f- Crossarchus obscurus, g - Cynictis penicillata, h - Galidia elegans, I - Galidictis vitattaj - Genetta donglana, к - Genetta
genetta, I - Genetta servalina, m - Genetta tigrina, n - Hemigalus derbyanus, о - Herpestes edwardsi, p - Herpestes ichneumon,
q - Herpestes sanguineus, r - Ichneumia albicauda, s - Mungos mungo, t - Nandinia binotata, и - Paguma larvala,
v - Paradoxurus hermaphroditus, w - Pseudogenetta villiersi, x - Suricata suricata, у - Civettictis civetta, z - Viverra zibetha,
Viverricula indica
Fig. 22. Principal component analysis based on the measurements of upper deciduous cheek
teeth in some Viverridae and Herpestidae:
a - Arctictis binturong, b - Arctogalidia trivirgata, c - Atilax paludinosus, d - Bdeogale crassicaudata, e - Bdeogale nigripes,
f- Crossarchus obscurus, g - Cynictis penicillata, h - Galidia elegans, i - Galidictis vitattaj - Genetta donglana, к - Genetta
genetta, I - Genetta servalina, m - Genetta tigrina, n - Hemigalus derbyanus, о - Herpestes edwardsi, p - Herpestes ichneumon,
q - Herpestes sanguineus, r - Ichneumia albicauda, s - Mungos mungo, I - Nandinia binotata, и - Paguma larvala,
v - Paradoxurus hermaphroditus, w - Pseudogenetta villiersi, x - Suricata suricata, у - Civettictis civetta, z - Viverra zibetha,
@ - Viverricula indica.
140
Рис. 23. Анализ главных компонент по промерам нижних молочных коренных зубов у
различных Viverridae и Herpestidae:
а - Arctictis binturong, b - Arctogalidia trivirgata, c - Atilaxpaludinosus, d - Bdeogale crassicaudata, e - Bdeogale nigripes,
f- Crossarchus obscurus, g - Cynictis penicillata, h - Galidia elegans, i - Genetta genetta, J - Genetta servalina, к - Genetta
tigrina, I - Hemigalus derbyanus, m - Herpestes edwardsi, n - Herpestes ichneumon, о - Herpestes javanicus, p - Herpestes
sanguineus, q - Ichneumia albicauda, r - Mungos mungo, s - Nandinia bmotata, t - Paguma larvata, и - Paradoxurus
hermaphroditus, v - Pseudogenetta villiersi, w - Suricata suricata, x - Civettictis civetta, у - Viverra zibetha, z - Viverricula
indica
Fig. 23. Principal component analysis based on the measurements of lower deciduous cheek teeth
in some Viverridae and Herpestidae:
° - Arctictis binturong, b - Arctogalidia trivirgata, c - Atilaxpaludinosus, d- Bdeogale crassicaudata, e - Bdeogale nigripes,
f' Crossarchus obscurus, g - Cynictis penicillata, h - Galidia elegans, i - Genetta genetta, J - Genetta servalina, к - Genetta
tigrina, I - Hemigalus derbyanus, m - Herpestes edwardsi, n - Herpestes ichneumon, о - Herpestes javanicus, p - Herpestes
sanguineus, q - Ichneumia albicauda, r - Mungos mungo, s - Nandinia binotata, t - Paguma larvata, и - Paradoxurus
hermaphroditus, v - Pseudogenetta villiersi, w - Suricata suricata, x - Civettictis civetta, у - Viverra zibetha, z - Viverricula
indica.
141
Таблица 9. Матрица факторных нагрузок первых трех факторов
анализа главных компонент
Table 9. Correlations of characters with the first three principal components
Промеры, мм | П I /•2 1 л
Верхние зубы
D2/ L 0.921 0.089 0.153
W 0.870 -0.293 -0.149
H 0.892 -0.319 -0.077
D3I L 0.861 0.381 0.285
Lmts 0.785 0.568 0.124
Lt 0.878 -0.318 -0.241
W 0.935 0.009 -0.309
И7 0.686 0.202 -0.590
H 0.888 0.340 0.035
DM L 0.861 -0.252 0.306
W 0.954 0.038 0.036
H 0.764 -0.401 0.380
Дисперсия, % 74.159 9.558 7.412
Нижние зубы
D/2 L 0.914 0.003 0.185
W 0.897 0.317 -0.062
H 0.805 -0.341 0.215
D/3 L 0.929 -0.027 0.177
W 0.868 0.399 -0.144
H 0.843 -0.484 0.055
142
Продолжение табл. 9
Промеры, мм | /1 1 /•2 | /3
£>/4 L 0.970 0.095 0.002
Ltl 0.502 -0.391 -0.759
Wtr 0.949 0.247 -0.038
Wtl 0.906 0.331 -0.096
н 0.791 -0.449 0.165
Дисперсия, % 74.151 10.414 6.847
Молочные зубы Cynogale специализированы для потребления, видимо,
рыбы и водных беспозвоночных. Обычно у рыбоядных хищных млекопи-
тающих (ластоногие и древние средеэоценовые киты Pakicetus (см. Ginge-
rich, Russell, 1990), потомки мезонихид) молочные зубы упрощаются, корон-
ки коренных становятся высокими, без добавочных бугорков или с неболь-
шим числом последних, треугольной формы. Характер молочных зубов Cy-
nogale позволяет предполагать преобладание в пище беспозвоночных (раз-
виты дробящие поверхности, в частности, очень крупный талон на 7)4/, этот
зуб вообще очень крупный, крупнее D3/ - аналогичная картина наблюдается
и у Enchydra из Lutrinae, также питающегося водными беспозвоночными, см.
Барышников, Аверьянов, 1990). Значительные отличия по строению молоч-
ных зубов Cynogale и Hemigalus, ранее часто относимых к одному подсе-
мейству (Michaelis, 1972; Соколов, 1979 и др.), подтверждают мнение Тэйлор
(Taylor, 1988) об искусственности такого объединения. Cynogale, в частнос-
ти, резко отличается от Hemigalus отсутствием парастиля В и протокона на
W По мнению Тэйлор Hemigaliinae должно включать только типовой род.
По характеру специализации Hemigalus близок к примитивным Viverrinae.
Наиболее продвинутым миофагом среди Viverridae является Nandinia.
Очевидно, что эта форма заслуживает выделение в особое подсемейство (фи-
логенетическая ветвь) Nandiniinae. Пальмовая цивета в строении молочных
3Убов показывает явно сходные черты с Felidae (положение парастиля В на
^3/, которое не отмечается у других виверровых и мангустовых, редукция
Метаконида Z?/4 и др.). Однако трудно однозначно интерпретировать эти при-
знаки как синапоморфии с Felidae. Скорее всего, они развились параллельно
(возникли независимо?) в обоих группах как результат продвинутой адапта-
к миофагии. Тем не менее, даже параллельное развитие указанных при-
знаков само по себе свидетельствует о филогенетической близости данных
143
групп. Интересно отметить, что пальмовая цивета, как и кошачьи, имеет
втяжные когти и по строению скелета задних конечностей приспособлена к
древесному образу жизни (Taylor, 1976). Другие наиболее специализирован-
ные миофаги из виверровых, например, Genetta, также лазают по деревьям.
Возможно, миофагия в какой-то степени скоррелирована в данном случае с
лазающим образом жизни. В частности, Miacis? kessleri Springhom, 1982 из
среднего эоцена Германии единственный миацид, у которого известно
строение молочных зубов (Springhom, 1985) также был, скорее всего,
древолазающей формой и, наряду с этим, он показывает некоторые черты
усиления хищнической адаптации в молочных зубах (на Z?/4 параконид
уменьшен, талонид заметно укорочен). Остальные виверровые и мангусто-
вые - в основном наземные собиратели (Taylor, 1976).
Herpestidae в целом более продвинуты по строению молочных зубов,
чем Viverridae. Среди них Galidiinae являются более специализированными
миофагами, чем большинство Herpestinae. У мангустов молочные зубы в
основном насекомоядного облика, видимо, в болыиейчггепени, чем у прими-
тивных Viverrinae (талон на D3/, как правило, более длинный в поперечном
направлении; главные бугорки тригонида D/4 приостренные). Среди мангус-
товых наиболее примитивными формами являются Atilax и Bdeogale\ не-
сколько более продвинуты в направлении миофагии Ichneumia и Herpestes
(метаконид D/4 уменьшен); Mungos, Crossarchus, Cynictis и Rhynchogale
более продвинуты в направлении энтомофагии (бугорки тригонида Z?/4
приострены, метаконид большой, талонид крупный, с обширным бассейном).
В строении молочных зубов нет общих эволюционно-продвинутых при-
знаков, объединяющих вместе Galidiinae и Herpestinae. В пользу объедине-
ния этих групп в одно семейство свидетельствуют другие признаки, в част-
ности, их пальцехождение (Taylor, 1988).
В целом по строению молочных зубов изученные виверровые и ман-
густовые распределяются на филогенетических ветвях приблизительно в
соответствии со схемой эволюции группы, представленной Михелис (Micha-
elis, 1972, S.92), за исключением следующих моментов, касающихся оценки
уровня эволюционной продвинутое™ отдельных форм: 1) среди Viverrinae
род Genetta наиболее продвинут, а не является исходной группой для
подсемейства; 2) Nandinia наиболее продвинута среди Viverridae и пред-
ставляет особую филогенетическую ветвь; 3) среди Herpestinae роды В de а
gale и Ichneumia не наиболее продвинуты, а одни из самых примитивных
Распределение изученных родов виверрид и герпестид по адаптивным зонам
характеризующим специализацию молочной зубной системы, представлено
на рис. 24.
144
ЗН ГОМОФАГ ИЯ I
Рис. 24. Распределение изученных родов Viverridae и Herpestidae по степени
специализации молочной зубной системы внутри адаптивных зон
Fig. 24. Distribution of studied genera of Viverridae and Herpestidae according to the deciduous
dentition specializations in adaptive zones
БЛАГОДАРНОСТИ
Авторы благодарны И.Я.Павлинову (Москва), Dr. J.Englund и Mr. A.Stanczak
(Стокгольм), Dr. V.Eisenmann (Париж), Dr. C.Mourer-Chauvire и Md. Sirven (Лион), Fr.
J.Altmann (Франкфурт-на-Майне) и Miss M.Morales (Чикаго) за содействие при работе с
коллекциями. Фотоработы выполнил Н.А.Орлов. Мы признательны Prof. J.D.Archibald
(Сан-Диего, США) за редактирование английского абстракта. Статья выполнена при
финансовой поддержке по программе "Биологическое разнообразие".
ЛИТЕРАТУРА
Барышников ГФ., Аверьянов А.О. Молочные зубы хищных млекопитающих (отряд
Carnivora). Часть I. Семейство Mustelidae // Труды Зоол. ин-та АН СССР, 1990.
Т.212. С.73-119.
Барышников Г.Ф., Аверьянов А. О. Молочные зубы хищных млекопитающих (отряд
Carnivora). Часть II. Семейство Ursidae И Труды Зоол. ин-та АН СССР, 1992
(1991). Т.238. С.68-99.
10
Заказ 255(
145
Барышников ГФ., Аверьянов А.О. Молочные зубы хищных млекопитающих (отряд
Carnivora). Часть III. Семейство Procyonidae // Труды Зоол. ин-та АН СССР,
1993а (1992). Т.246. С.103-128.
Барышников Г.Ф., Аверьянов А.О. Молочные зубы хищных млекопитающих (отряд
Carnivora). Часть IV Семейства Amphicyonidae и Canidae // Труды Зоол. ин-та
РАН, 19936. Т.249. С. 158-197.
Барышников Г.Ф., Аверьянов А.О. Молочные зубы хищных млекопитающих (отряд
Carnivora). Часть V Семейства Protelidae и Hyaenidae // Труды Зоол. ин-та РАН,
1995. Т.263. С. 46-84.
Вислобокова И.А. Ископаемые олени Евразии // Труды Палеонтол. ин-та АН СССР,
1990. Т.240. 208 с.
Павлинов И.Я., Борисенко А.В., Крускоп С. В., Яхонтов Е.Л. Млекопитающие
Евразии. II. Non-Rodentia: систематико-географический справочник. - М., 1995.
336 с.
Соколов В.Е. Систематика млекопитающих (китообразные, хищные, ластоногие,
трубкозубые, хоботные, даманы, сирены, парнокопытные, мозоленогие,
непарнокопытные). - М., 1979. 528 с.
Шевырева Н.С. Грызуны (Rodentia, Mammalia) неогена Евразии и Северной Африки -
эволюционная основа плейстоценовой и современной фауны грызунов
Палеарктики // История и эволюция современной фауны грызунов (неоген -
современность). - М., 1983. С.9-145.
Beaumont G. de. Observations sur les Herpestinae (Viverridae, Carnivora) de 1’Oligocene
superieur avec quelques remarques sur des Hyaenidae du Neogene // Arch. Sci.
Geneve, 1967. T.20. F.l. P.79-108.
Beaumont G. de. Contribution a 1’etude des Viverrides (Carnivora) du Miocene d’Europe П
Arch. Sci. Geneve, 1974. T.26. F.3. P.285-296.
Butler P.M. The evolution of carnassial dentitions in the Mammalia 11 Proc. Zool. Soc.
London, 1946. Vol. 116. Pt.2. P. 198-220.
Dehm R. Die Raubtiere aus dem Mittel-Miocan (Burdigalium) von Wintershof-West bei
Eichstatt in Bayern // Abh. Bayer Akad. Wiss., Math.-Naturw. KI., Neue Folge, 1950.
Hf.58. S.9-141.
Fejfar O., Schmidt-Kittler N., Zacharov M. Lophocyon carpathicus n. gen. n. sp. aus dem
Jungtertiar der Ostslowakei und eine neue unterfamilie der Schleichkatzen
(Viverridae) // Palaeontographica, Abt. A., 1987. Bd.199. Lfg.1-3. S. 1-22.
Fox R.C., Youzwyshyn G.P New primitive carnivorans (Mammalia) from the Paleocene of
Western Canada, and their bearing on relationships of the order // J. Vert. Paleontol.,
1994. Vol.14.N3. P.382-404.
Gingerich P.D., Russell D.E. Dentition of Early Eocene Pakicetus (Mammalia, Cetacea) //
Contrib. Mus. Paleontol. Univ. Michigan, 1990. Vol.28. N 1. P.1-20.
Ginsburg L. Les Carnivores du Miocene de Beni Mellal (Maroc) // Geol. Mediterr., 1977.
T.4, N3. P.225-240.
Hunt R.M., Jr. Evolution of the Aeluroid Carnivora: viverrid affinities of the Miocene
camivoran Herpestides H Amer. Mus. Novit., 1991. № 3023. P.1-34.
Larkin P., Roberts M. Reproduction in the ring-tailed mongoose Galidia elegans at the
National Zoological Park, Washington П Int. Zoo Yearb., 1979. Vol. 19. P. 188-193.
Leche Ж Zur Frage nach der stammesgeschichtlichen Bedeutung des Milchgebiss bei den
Saugetieren. II.Viverridae, Hyaenidae, Felidae, Mustelidae, Creodonta // Zool.
Jahrb., Syst., 1915. Bd.38. S.275-370.
146
Michaelis В. Die Schleichkatzen (Viverriden) Afrikas П Saugetierk. Mitteil., 1972. Bd.2O
Hf.1-2. S.l-110.
Mivart S.G. On the classification and distribution of the Aeluroidea // Proc. Zool. Soc
London, 1882. P. 135-208.
Petter G. Petits carnivores villafranchiens du Bed I d’Oldoway (Tanzanie) // Problems
actuels de Paleontologie (Evolution des Vertebres). Paris, 1967. P.529-538.
Petter G., Heintz E. Mammiferes quaternaires de la grotte de Geula (Nord d’Haifa, etat
d’Israel) // Bull. Mus. Nat. Hist. Natur., ser.2, 1970. T.41. N 5. P. 1292-1298.
Springhorn R. Zwei neue Skelette von Miacis? kessleri (Mammalia, Carnivora) aus den
lutetischen Olschiefern der ’’Grube Messel” H Senckenberg. lethaea, 1985. Bd. 66. N
1/2. S.121-144.
Taylor M.E. The functional anatomy of the hindlimb of some African Viverridae // J.
Morphol., 1976. Vol. 148. N 2. P.227-252.
Taylor M.E. Foot structure and phylogeny in the Viverridae // J. Zool., 1988. Vol.216. N 1.
P.131-139.
Wenzel E., Haltenorth T. System der Schleichkatzen (Viverridae) // Saugetierk. Mittl.,
1972. Bd.20.N 1-2. S.l 10-127.
Wozencraft W.C. Order Carnivora // D.E.Wilson, D.M.Reeder (eds.). Mammal species of
the world. A taxanomic and geographic reference (2nd ed.). Washington, 1993.
P.279-348.
Summary
G.F.Baryshnikov, A.O.Averianov
DECIDUOUS TEETH OF CARNIVOROUS MAMMALS (ORDER
CARNIVORA).
PART VI. FAMILIES VIVERRIDAE AND HERPESTIDAE
Milk dentition in 30 recent and one extinct species of Viverridae and Herpestidae were
analyzed. Both families have the same general dental morphology and dental formula primitive
for the order. Among Viverridae the groups which are differs by direction and level of
specialization of the deciduous teeth generation can be recognized. The main groups are
insectivorous Hemigalinae and Viverrinae (Viverricula - Viverra and Prionodon - Genetta are
miophagous), insectivorous Euplerinae and omnivorous Paradoxurinae (with more plant-eating
specializations in Arctogalidia - Arctictis and Paguma). Cynogale, which eats fish and water
invertebrates and Nandinia, which is specialized for miophagy and show some similarities with
Felidae take the side branch position. Two groups in the family Herpestidae can be recognized:
Galidiinae and Herpestinae. The former is more specialized for miophagy. The latter shows
trends for miophagy (Ichneumia and Herpestes) or entomophagy (Mungos, Crossarchus,
Cynictis, Rhynchogale). As a whole, Herpestidae has a more derived deciduous dentition than
Viverridae.
10*
ISSN 0206-0477. ЧЕТВЕРТИЧНЫЕ И СОВРЕМЕННЫЕ
МЛЕКОПИТАЮЩИЕ ЕВРАЗИИ
СПб., 1996 (ТРУДЫ ЗООЛОГИЧЕСКОГО ИНСТИТУТА РАН, Т. 270)
RUSSIAN ACADEMY OF SCIENCES
PROCEEDINGS OF THE ZOOLOGICAL INSTITUTE
St. PETERSBURG, 1996, VOL. 270
УДК 569.325
А.О.Аверьянов
Зоологический институт РАН, Санкт-Петербург
О СИСТЕМАТИЧЕСКОМ ПОЛОЖЕНИИ КРОЛИКА
"CAPROLAGUS" BRACHYPUS YOUNG, 1927
(LAGOMORPHA, LEPORIDAE) ИЗ ВИЛЛАФРАНКА КИТАЯ
Кролик “Caprolagus" brachypus Young, 1927, описанный из позднего виллафранка
Китая, выделен в новый род Sericolagus gen. nov., отличающийся от других лепорид соче-
танием алилепусного и некролагусного вариантов строения Р/3 с далеко идущим назад
нижним резцом (до заднего края Р/3). Этот вид был, видимо, в значительной степени
приспособлен к рытью.
В 1927 г. китайский палеонтолог Ян Чжунь-чжень (Young, 1927, SS.63-
65, Taf.III, figs. 14-18) описал по двум нижнечелюстным костям и нескольким
элементам посткраниального скелета из виллафранкских отложений окрест-
ностей Пекина новый вид лепорид Caprolagus brachypus. При описании он
отметил, что, судя по относительно коротким костям конечностей, новый
вид относится к примитивным зайцам и, возможно, близок к ископаемому С.
sivalensis Forsyth Major, 1899 из сиваликской фауны Индии (известен только
по зубам) и современным С. hispidus (Pearson, 1839) (щетинистый заяц или
ассамский кролик) и С. netscheri (Schlegel et Jentink, 1899) (полосатый заяц,
ныне относится к роду Nesolagus Forsyth Major, 1899).
Позднее к С. brachypus были отнесены 4 фрагмента нижнечелюстных
костей и несколько посткраниальных костей из "средней зоны красноватых
глин" (виллафранк) местонахождения Шуангыйнцы (Shuangyinzi, =Jingle,
=Locality 2), пров. Хэбэй, Китай (Teilhard de Chardin, Young, 1931, pp.29-30,
Pl.V, figs.25-29 [экземпляр, изображенный на рис. 26, принадлежит пищухе,
см. Qiu, 1987, р.383]) и фрагмент нижнечелюстной кости из "нижних
саньменьских" отложений (поздний виллафранк) близ г.Шоуянь (Locality 20),
148
Пров. Хенань, Китай (Young, 1935, Text-fig.2, Pl.I, fig.3). К характеристике
вида было добавлено, что он принадлежит к группе мелких южноазиатских
зайцев с ’’плотными" конечностями и особой формой нижней челюсти, с
сильно расходящимися ветвями и утолщением под резцом и на симфизе
(Teilhard de Chardin, Young, 1931, p.29).
Систематическое положение C. brachypus было пересмотрено Шрой-
дером (Schreuder, 1936), который отметил, что этот вид, имеющий хорошо
развитые задне-наружную (гипофлексид) и задне-внутреннюю (мезофлексид)
складки эмали на Р/3, должен относиться к подсемейству Palaeolaginae, тогда
как оба вида Caprolagus (hispidus и sivalensis) имеют Р/3 только с одной,
задне-наружной складкой, идущей до лингвального края зуба, и, поэтому,
относятся к Leporinae. Кроме того, для современного и ископаемого
Caprolagus характерно сильное развитие вторичных складочек эмали на
тригониде Р/3 и на передней стенке талонида Р/3-Л//2, что не наблюдается у
С. brachypus [на самом деле, на передней стенке талонида в зубном ряду Р/3-
А//2 С. brachypus имеются складочки, но не такие сильные, как у капрола-
гусов (см. Bohlin, 1939-1943, р. 133; pers. obs.)].
По последним данным (Steensma, Hussain, 1992) С. sivalensis происхо-
дит из слоев формации Самвал (верхнесиваликская подгруппа) штата Каш-
мир, Пакистан, датируемых 1.8-2.0 млн лет, что соответствует верхнему вил-
лафранку (зона MQ 18). Таким образом, С. sivalensis и "Caprolagus" bra-
chypus одновозрастны, и последний вид не может быть предковым для
морфологически однородной группировки С. sivalensis - C.hispidus.
Шройдер (Schreuder, 1936) отнес С. brachypus к роду Hypolagus Dice,
1917 (последний часто относится к подсемейству Archaeolaginae, а не Palaeo-
laginae, представители которого имеют Р/3 без мезофлексида и с гипофлек-
сидом, доходящим только до половины ширины коронки). Точку зрения
Шройдера поддерживали, в основном, авторы, не разделявшие подсемейства
Palaeolaginae и Archaeolaginae (Teilhard de Chardin, 1940, 1942 и др.). В
последней работе П.Тейяр-де-Шарден отнес к Hypolagus cf. brachypus около
12 нижнечелюстных костей и поврежденный череп мелкого зайца (мельче
одновозрастного Hypolagus schreuderi Teilhard de Chardin, 1940) из верхнего
горизонта местонахождения Юше (III зона виллафранка; Юше-Ш по Qiu,
1987; видимо, формация Мажегоу, 2.9-3.4 млн лет, см. Flynn et al., 1991). Из
трех изображенных Р/3 из этой выборки (Teilhard de Charden, 1942, fig.53, a-
c) два имеют типичный гиполагусный морфотип, третий некролагусный
морфотип (мезофлексид замкнут в эмалевый островок). Позднее экземпляры
с гиполагусными Р/3 были отнесены к Н. schreuderi (Cai, 1989, р.170), к
которому, возможно, принадлежат все экземпляры из Юше (Qiu, 1987; Flynn
et al., 1991).
Болин (Bohlin, 1939-1943, p.133) также считал, что "Caprolagus"
brachypus не относится к роду Caprolagus, но, по его мнению, включение
этого вида в род Hypolagus настолько же сомнительно. По Болину, Р/3 у "С.”
brachypus имеет почти такую же структуру, как и у рода Alilepus. Гуреев
149
(1964, с. 124), Даусон (Dawson, 1967, р.ЗОЗ) и Кай Баокван (Cai, 1989, р. 180)
относили "С." brachypus к роду Alilepus. Даусон, а также Цю Чжань-Сян и
Хань Дэфен (Qiu, Han, 1986, р.53) считали, что экземпляры Р/3 A. brachypus
с замкнутым мезофлексидом, возможно, представляют промежуточную
стадию эволюции лепорид от типичных Alilepus Lepus. Экземпляр Р/3 с
замкнутым мезофлексидом, отнесенный к Н. brachypus, был описан также из
среднего плейстоцена Хешуй (Heshui), пров. Ганьсу, Китай (Zheng, 1976,
fig.4a). По мнению Цю Чжань-Сяна (Qiu, 1987, р.383) родовая принадлеж-
ность "Caprolagus" brachypus неясна; этот вид может относиться к особому
роду, более продвинутому, чем Alilepus (аналогично североамериканскому
роду Pratilepus Hibbard, 1939).
Как видно из приведенного выше обзора, систематическое положение
"Caprolagus" brachypus трактуется разными авторами неоднозначно, что
может объясняться отчасти малочисленностью и фрагментарностью мате-
риала по этому виду. В этой ситуации решению проблемы может способ-
ствовать привлечение для анализа новых признаков, в частности строение
известных посткраниальных элементов. Такие признаки были обнаружены
автором в ходе изучения типового материала по "Caprolagus" brachypus,
хранящимся в Палеонтологическом музее г.Уппсала, Швеция.
МАТЕРИАЛ И МЕТОДИКА
Изучена типовая коллекция остатков "Caprolagus" brachypus из Палеонтологичес-
кого музея в Уппсале, Швеция (Paleontologiska Museet, Uppsala, далее PMU): PMU M
15010a, фрагмент правой нижнечелюстной кости с 773-Л//3 (рис. 1, а; рис. 2, a-в, вклейка)',
PMU М 15010b, фрагмент левой нижнечелюстюстной кости с Р/3-Л//2 (рис. 1, б; рис.2, а,
г, d); PMU М 15011, фрагмент правой тазовой кости; PMU М 15013, фрагмент второго(?)
поясничного позвонка (рис. 2, к-u); PMU М 15015, правая бедренная кость (рис. 2, е-з,
вклейка). Все остатки, возможно, принадлежат одному индивидууму и происходят из
Санджиадьян (Sanjiadian, =Sanchiatien, =Locality 60) (нумерация Lagrelius Collection, см.
Mateer, Lucas, 1985), Хуи-ю (Hui-Yu, =Huiy), Ваньпинь Цянь (Wangping Xian,
=Wangpinghsien), Beijing Shi (Пекинский округ, Китай); поздний виллафранк (ранний
плейстоцен). Экземпляр PMU М 15010 обозначен здесь как лектотип вида, остальные
экземпляры - паралектотипы.
В качестве сравнительного материала изучены следующие экземпляры
современных лепорид:
Pentalagus furnessi (Stone, 1900): Зоологический институт РАН, Санкт-Петербург,
ЗИН 69275, ? , скелет.
Caprolagus hispidus (Pearson, 1839): Музей естественной истории Швеции
(Naturhistoriska Riksmuseet, Stockholm), NRM A 848, посткраниальный скелет,- NRM A
585848,череп.
Nesolagus netscheri (Schlegel et Jentink, 1880): Британский музей естественной
истории (Natural History Museum, London), BMNH 1921.1.18.1, скелет.
Oryctolagus cuniculus (L., 1758): ЗИН 15180, ? , скелет..
Sylvilagus bachmani (Waterhouse, 1838): ЗИН 14470, скелет.
Lepus americanus Erxleben, 1777: ЗИН 40772, скелет.
150
Рис. 1. Жевательная поверхность
Р/3 Sericolagus brachypus (лекто-
тип). Поздний виллафранк, Китай:
а - правый, PMU М 15010а, б - левый,
PMU М 15010b
Fig. 1. Occlusal surface of P/3 of
Sericolagus brachypus (lectotype).
Late Villafranchian, China:
«-right, PMU M 15010a, 6 - left,
PMU M 15010b
Lepus europaeus Pallas, 1778: ЗИН 23659, ? , скелет.
Lepus mandshuricus Radde, 1861: ЗИН 13439, ? , скелет.
Lepus oiostolus Hodgson, 1840: ЗИН 13445, ? , скелет.
Lepus timidus L., 1758: ЗИН 20732, сГ , скелет.
Промеры бедренной кости обрабатывались одним из методов многомерной
статистики - анализом главных компонент. Расчеты производились на IBM PC AC386SX с
помощью модулей DATA, FACTOR и GRAPH из пакета статистических программ
SYSTAT.
РЕЗУЛЬТАТЫ
Зубы. Нижний резец изогнут в большей степени, чем у других
лепорид. Р/3 с маленьким, сравнительно узким тригонидом, без передней
складки (рис. 1). Передне-наружная складка хорошо развита, заполнена
цементом. Задне-наружная складка доходит до половины ширины коронки,
ее задняя стенка может быть слабо складчатая. Задне-внутренняя складка
короткая, со складчатой передней стенкой. Передняя стенка талонида Р/4-
Л//2 слабо складчатая. Л//3 сравнительно крупный, состоит из двух конидов.
Нижнечелюстная кость. Кость массивная, с короткой
диастемой и круто поднимающимся вверх резцом (рис. 2, а-д, вклейка',
табл. 1). Левая и правая нижнечелюстные кости расходятся под сравнительно
большим углом. Задний конец нижнего резца идет назад дальше, чем у
большинства других лепорид, почти достигая заднего конца Р/4 (то же
наблюдается у Caprolagus hispidus', у Caprolagus sivalensis резец доходит до
заднего конца Р/3 (Steensma, Hussain, 1992); у остальных лепорид задний ко-
нец резца обычно не доходит даже до переднего края Р/3; строение нижне-
челюстной кости у Nesolagus netscheri автору неизвестно).
Посткраниальный скелет. На втором (?) поясничном
позвонке (рис. 2, и-л, вклейка) поперечные отростки обломаны, но, видимо,
они были небольшими (характерно для плохо бегающих лепорид).
Шейка подвздошной кости слабо изогнута в дорзо-вентральной плос-
кости. Вертлужная впадина сужена вентрально-дорзально. На крыле под-
вздошной кости гребни прикрепления ягодичных мышц почти не выражены.
151
Таблица 1. Размеры зубов, нижнечелюстной кости, второго(?) поясничного
позвонка и тазовой кости у Sericolagus brachypus
Table 1. Measurements of teeth, mandible, second(?) lumbar vertebra and innominate
bone of Sericolagus brachypus
Промеры, мм | Экземпляры
Нижнечелюстная кость PMU М 15010а PMU М 15010b
Длина резца (//1) 2.4 2.5
Ширина резца (//1) 2.5 2.6
Длина Р/3 2.8 3.0
Длина Р/З-М/З альвеолярная 15.3
Длина диастемы 14.0 14.6
Высота кости перед Р/3 12.4 12.0
То же, после М/3 14.1
Второй(?) поясничный позвонок PMUM 15013
Длина тела 11.3
Ширина тела спереди 8.4
Тазовая кость PMUM 15011
Передне-задний диаметр вертлужной впадины 10.1
Высота наименьшая шейки подвздошной кости 5.5
Бедренная кость сравнительно короткая, плотная (рис. 2, е-з, вклейка).
По пропорциям близка к бедренным костям таких примитивных кроликов,
как Oryctolagus, Nesolagus и Sylvilagus (табл. 2, 3). Распределение промерен-
ных бедренных костей в факторном пространстве (рис. 3) показывает, что
значение фактора 1 (связан с длиной и шириной проксимального конца
кости) для этих примитивных форм меньше, чем у бегающих Lepus, роющих
Caprolagus и Trischizolagus и лазающих Pentalagus. По значению фактора 2
примитивные кролики близки к Caprolagus и Trischizolagus и отличаются от
Pentalagus и продвинутых Lepus, которые не роют нор. Среди изученных
лепорцд, бедренная кость "Caprolagus" brachypus выделяется очень сильным
развитием гребня (linea pectinea), идущим вдоль медиального края кости под
152
Таблица 2. Размеры бедренной кости у Sericolagus brachypus и некоторых других
лепорид
Table 2. Measurements of femur of Sericolagus brachypus and some other leporids
Виды Промеры, мм
GL CL I И7Ч | WP2 1 DC- | WD1< | | WD1S
Sericolagus brachypus 68.3 65.5 14.3 11.0 5.8 5.7 11.9
Nesolagus netscheri 76.2 72.6 13.3 14.0 5.9 6.0 11.8
Oryctolagus cuniculus 72.6 70.0 13.2 12.7 5.9 5.5 11.8
Silvilagus bachmani 62.2 60.4 11.7 11.5 4.9 5.0 10.0
Caprolagus hispidus 98.1 91.6 17.8 16.4 8.0 7.2 1.6.3
t Pentalagus furnessi 93.0 87.8 19.8 19.4 8.1 8.6 17.5
| Trischizolagus dumitrescuae 103.3 99.2 20.0 19.8 8.5 8.3 17.1
^Lepus americanus 95.1 93.6 17.0 15.0 7.0 5.8 14.1
Lepus mandshuricus 105.8 101.7 18.2 17.0 7.3 6.9 14.5
Lepus oiostolus 113.0 107.4 23.0 21.0 9.6 8.3 16.6
Lepus europaeus 133.3 126.0 25.6 25.0 10.8 9.8 19.3
Lepus timidus 129.5 124.8 25.9 23.7 10.1 8.9 20.0
Примечание:^ - длина наибольшая; CL - расстояние от проксимального края головки бедра
До дистального конца кости; И7Ч - ширина проксимального конца кости от края большого трохантера до
головки бедра; WP2 - то же, на уровне третьего и малого трохантеров; DC - передне-задний диаметр головки
бедра; WDF - ширина диафиза наименьшая; WD1S - ширина дистального эпифиза.
Малым трохантером. К этому гребню крепится гребешковая мышца (т.
pectineus), относящаяся к ягодичной группе мышц, приводящих (разгибаю-
щих) бедро в тазобедренном суставе. Незначительное усиление ягодичных
мышц наблюдается у хорошо приспособленных к бегу лагоморф (Гамбарян,
1972), однако, у "Caprolagus” brachypus эти мышцы, видимо, были развиты в
существенно большей степени. Это может свидетельствовать о далеко за-
Щедшей адаптации данного вида к рытью, при котором земля из норы выбра
сывалась задними ногами. Такая трактовка не противоречит другим
Известным особенностям "Caprolagus” brachypus (свидетельствующим об
отсутствии развитой способности к быстрому бегу), хотя у наземных земле-
153
Таблица 3. Матрица факторных нагрузок первых трех факторов
анализа главных компонент
Table 3. Correlations of characters with the first three principal components
Промеры бедренной кости I Fl j 1 F2 1 F3
GL 0.977 -0.206 0.004
CL 0.969 -0.241 -0.002
WP\ 0.991 -0.018 -0.010
WP2 0.988 0.057 0.113
DC 0.992 0.017 0.015
WDF 0.959 0.267 0.064
WDIS 0.973 0.127 -0.185
Дисперсия, % 95.747 2.736 0.731
Примечание. Обозначения промеров как в табл. 2.
роев в задних конечностях, несущих небольшую нагрузку при рытье, фос-
сориальные признаки обычно не развиваются или маскируются другими
адаптациями (Гамбарян, 1960).
ОБСУЖДЕНИЕ
Как видно из приведенной характеристики, "Caprolagus" brachypus от-
личается от всех известных родов лепорид (кроме Caprolagus) далеко иду-
щим назад нижним резцом, от Caprolagus - строением коренных зубов. Этот
кролик по пропорциям бедренной кости близок к небольшим примитивным
роющим лепоридам (Qryctolagus, Nesolagus, Sylvilagus), но отличается от
них существенно менее продвинутым строением Р/3 (алилепусный и некро
лагусный морфотипы, у Oryctolagus и Sylvilagus - лепусный морфотип, у
Nesolagus - еще более продвинутый педоморфный морфотип). От вымерших
кроликов рода Hypolagus, также приспособленных к рытью, "Caprolagus'
brachypus резко отличается строением Р/3 (у последних никогда не на-
блюдается мезофлексид). Возможно, что по особенностям посткраниального
скелета он был ближе всего к представителям рода Alilepus (скелет послед-
них обнаружен только недавно (см. Winkler, 1992) и до сих пор не описан)
154
FACTOR С1)
FACTOR(2)
Рис. 3. Анализ главных компонент по 7 промерам бедренной кости у различных Leporidae
а - Lepus amencanus, b - Sericolagus brachypus, c - Caprolagus hispidus, e - Lepus europaeus, I - Lepus timidus, m - Lepus
mandshuricus, n - Nesolagus nelscheri, о - Lepus oiosiolus, p - Pentalagus furnesst, r - Oryctolagus cunt cuius, s - Sylvilagus
bachmant, t - Trischizolagus dumitrescuae
Fig. 3. Principal component analysis based on 7 measurements of femur in some Leporidae
a - Lepus amencanus, b - Sericolagus brachypus, c - Caprolagus hispidus, e - Lepus europaeus, I - Lepus timidus, m - Lepus
mandshuricus, n - Nesolagus nelscheri, о - Lepus oiosiolus, p - Pentalagus jurnesst, r - Oryctolagus cuntculus, s -
Sylvilagus bachmani, I - Trischizolagus dumitrescuae
Приведенных отличий достаточно, чтобы выделить ’’Caprolagus" brachypus в
самостоятельный род.
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
СЕМЕЙСТВО LEPORIDAE FISHER, 1817
ПОДСЕМЕЙСТВО LEPORINAE S. STR.
ТРИБА ALILEPINI GUREEV, 1964
155
Род Sericolagus Averianov, gen. nov.
Название рода от лат. "sericum" - шелк ("Seres" в Древнем Риме - назва-
ние для китайцев и Китая).
Типовой вид Caprolagus brachypus Young, 1927, нижний и
средний (?) плейстоцен, Китай.
Диагноз. Небольшие лепориды с алилепусными и некролагусными
вариантами строения Р/3. Тригонид Р/3 без передней складки. Задний конец
нижнего резца достигает заднего края Р/4.
Видовой состав. Только типовой вид.
БЛАГОДАРНОСТИ
Автор благодарен Dr. S.Stuenes (PMU) за помощь в изучении типовой коллекции
"Caprolagus” brachypus, Drs. S.Parfitt, D.Hills и R.Sabin (BMNH) за присылку промеров
бедренной кости Nesolagus netscheri, Г.Ф.Барышникову (ЗИН) - за прочтение рукописи и
высказанные замечания.
ЛИТЕРАТУРА
Гамбарян П.П. Приспособительные особенности органов движения роющих млеко-
питающих. - Ереван, 1960. 195 с.
Гамбарян П.П. Бег млекопитающих. - М, 1972. 334 с.
Гуреев А.А. Фауна СССР. Зайцеобразные (Lagomorpha). - М.-Л., 1964. 276 с. (Фауна
СССР. Млекопитающие. ТЛИ, Вып.Ю).
Bohlin В. A revision of the fossil Lagomorpha in the Paleontological Museum, Upsala И
Bull. geol. Inst. Upsala, 1939-1943. Vol.30. P.l 17-154.
Cai B. Fossil Lagomorphs from the late Pliocene of Yangyuan and Yuxian, Hebei //
VertebrataPalasiatica, 1989. Vol.27. N 3. P.l70-181.
Dawson M.R. Lagomorph history and the stratigraphic record // Teichert, C & Yochelson.
E.L., (eds.) Essays in paleontology and stratigraphy, Raymond C. Moore Comme-
morative volume. Univ. Kansas Dept. Geol. Spec. Publ., 1967. Vol. 2. P.287-316.
Flynn L.J., Tedford R.H., Qiu Z. Enrichment and stability in the Pliocene mammalian fauna
of North China//Paleobiology, 1991. Vol.17.N3. P.246-265.
Mateer N.J., Lucas S.G. Swedish vertebrate paleontology in China: a history of the Lagreli-
us Collection // Bull. Geol. Inst. Univ. Uppsala, new ser., 1985. Vol.l 1. P.1-24.
Qiu Z. The Neogene mammalian faunas of Ertemte and Harr Obo in Inner Mongolia (Nei
Mongol), China. 6. Hares and pikas - Lagomorpha: Leporidae and Ochotonidae //
Senckenberg. Leth., 1987. Bd.67. Hf.5/6. S.375-399.
Qiu Z., Han D. Fossil Lagomorpha from the Hominoid locality of Lufeng, Yunnan H Acta
Anthropol. Sinica, 1986. Vol.5. N 1. P.41-53.
Schreuder A. Hypolagus from the Tegelen Clay; with a note on recent Nesolagus // Arch
Neerland. Zool., 1936. T.2. Liv.2-3. P.225-239.
156
Steensma К.J., Hussain S.T. Caprolagus sivalensis (Lagomorpha, Mammalia) from the
Upper Siwalik Subgroup of Azad Kashmir, Pakistan H N. Jb. Geol. PaleontoL Mh.,
1992. Hf.7. S.429-434.
Teilhard de Chardin P The fossils from Locality 18 near Peking П PaleontoL Sinica, new
ser.C, 1940. Vol.9. 94 pp.
Teilhard de Chardin P New rodents of the Pliocene and Lower Pleistocene of North China
// Inst. Geo-BioL, Pekin, 1942. N 9. 101 pp.
Teilhard de Chardin P., Young C.C. Fossil mammals from the Late Cenozoic of Northern
China//PaleontoL Sinica, ser. C, 1931. Vol.9. Fasc.l. 66 pp.
Winkler A.J. Small mammals from the Late Miocene Lothagam locality, West Turkana,
Kenya//J. Vert. PaleontoL, 1992. Vol. 12. N 3, SuppL P.60A.
Young C.C. Fossile Nagetiere aus Nord-China // PaleontoL Sinica, ser. C., 1927. Vol.5.
Fasc.3. 82 pp.
Young C.C. Miscellaneous mammalian fossils from Shansi and Honan // PaleontoL Sinica,
ser. C, 1935. VoL9. Fasc.2. 42 pp.
Zheng S. Small Mammals of Middle Pleistocene in Heshui, Gansu 11 Vertebrata Palasiatica,
1976. Vol.14.N2. P.112-119.
Summary
A.O.Averianov
ON THE SYSTEMATIC POSITION OF RABBIT
“CAPROLAGUS” BRACHYPUS YOUNG, 1927 (LAGOMORPHA,
LEPORIDAE) FROM THE VILLAFRANCHIAN OF CHINA
“Caprolagus” brachypus Young, 1927 described from the Late Villafranchian of China
is assigned to the new genus Sericolagus, which differs from other leporids by combination of
Alilepus- and Nekrolagus-\\ke structures of /73, lower incisor protruding far posteriorly at the
level of posterior margin of P/4. In the proportions of the femur this rabbit is close to modem
small species of Orydolagus, Nesolagus and Sylvilagus. Sericolagus brachypus is characterized
by very strong development of the linea pectinea on the femur. This feature and proportions of
the femur may indicate fossor ial adaptations of this rabbit.
ISSN 0206-0477. ЧЕТВЕРТИЧНЫЕ И СОВРЕМЕННЫЕ
МЛЕКОПИТАЮЩИЕ ЕВРАЗИИ
СПб., 1996 (ТРУДЫ ЗООЛОГИЧЕСКОГО ИНСТИТУТА РАН, Т. 270)
RUSSIAN ACADEMY OF SCIENCES
PROCEEDINGS OF THE ZOOLOGICAL INSTITUTE
St. PETERSBURG, 1996, VOL. 270
УДК 569.32:551.781 (517.3)
А.О.Аверьянов
Зоологический институт АН России, Санкт-Петербург
О ВОЗРАСТНЫХ ИЗМЕНЕНИЯХ ЗУБНОЙ СИСТЕМЫ
TSAGANOMYS ALTAICUS MATTHEW ET GRANGER, 1923
(RODENTIA, TSAGANOMYIDAE) ИЗ ОЛИГОЦЕНА
ЦЕНТРАЛЬНОЙ АЗИИ
Остатки грызуна Tsaganomys altaicus из олигоценовых отложений Монголии и
Китая существенно различаются по микроскопическому строению коренных зубов. По
мнению Шевыревой (1971 и др.), эти отличия имеют филогенетическое значение. Форма,
характеризующаяся более эволюционно-продвинутым строением коренных зубов, был t
выделена ею в особый род и вид Beatomus bisus Shevyreva, 1972. Здесь предлагается
альтернативная гипотеза, согласно которой различия в строении зубов Tsaganomys
"Beatomus" связаны с онтогенетическими изменениями зубного аппарата грызунов одного
вида, причем зубы типа "Beatomus” характерны для молодых животных. Название
Beatomus bisus Shevyreva, 1972, исходя из этого предположения, а также из сравнения
голотипа этого таксона с опубликованным описанием типовых экземпляров Т. altaicus.
следует рассматривать как младший субъективный синоним названия Tsaganomys altaicus
Matthew et Granger, 1923.
Своеобразный высоко специализированный роющий грызун Tsagano-
mys altaicus известен по многочисленным остаткам из среднеолигоценовог
свиты Шанд-Гол и ее аналогов в Монголии и средне-верхнеолигоценовых
отложений провинций Внутренняя Монголия и Ганьсу в Китае (Matthev
Granger, 1923; Teilhard de Chardin, 1926; Bohlin, 1937; Mellet, 1968; Дашзэвэг
1970; Kowalski, 1974; Шувалов, Решетов, 1974; Шевырева, 1976; Wang et al
1981). По общей форме и пропорциям черепа этот грызун поразительно
похож на современных африканских землекоповых (семейство Bathyergidae)-
к которым он и был отнесен при первоописании (Matthew, Granger, 1923)
Впоследствии большинство исследователей относило Tsaganomys к выме[’
158
тему семейству Cylindrodontidae (олигоцен Азии и Северной Америки).
Сравнительно недавно Tsaganomys вместе с близкими азиатскими родами
был выделен Вудом (Patterson, Wood, 1982) в особое семейство Tsagano-
myidae*, близкое, по его мнению, к современным Bathyergidae (см. также
Wood, 1985). Как Tsaganomyidae, так и Cylindrodontidae имели свой набор
общих эволюционно-продвинутых признаков (Jaeger, 1988, fig. 7.2), что
говорит о самостоятельности этих ветвей. В последнее время накапливается
все больше данных, свидетельствующих против родства Tsaganomyidae с
Bathyergidae. Последние развились, видимо, из каких-то Thryonomyoidea,
вторично утратив гистрикоморфию (Lavocat, 1973; Sarich, 1985; Maier,
Schrenk, 1987).
Внешнее сходство Tsaganomys и близких форм с батиергидами объ-
ясняется сходной специализацией к роющему образу жизни. Многочислен-
ные признаки этой специализации в строении посткраниального скелета
цаганомиид были подмечены Виноградовым и Гамбаряном (1952), которые
реконструировали мускулатуру Т. altaicus. Из анализа морфобиологических
особенностей скелета авторы сделали вывод, что Tsaganomys "в основном
был наземным животным, но он рыл более или менее сложные и глубокие
норы, служившие ему преимущественно убежищем, и добывал пищу в значи-
тельной степени посредством рытья. Вполне вероятно, что это животное
обитало в местностях с резкими летними засухами, когда вся травянистая
растительность выгорала и животное вынуждено было переходить на
питание корнями, клубнями и луковицами, для добывания которых ему
приходилось рыть лунки" (там же, с.41).
Ряд исследователей (Teilhatd de Chardin, 1926; Bohlin, 1937; Вино-
градов, Гамбарян, 1952**) отмечали неоднородность остатков, относимых к
Т. altaicus, что обычно объяснялось большой индивидуальной изменчи-
востью. По мнению Шевыревой (1971; 1972; 1974а, б, в; 1976а, б), эта не-
однородность обусловлена не большим диапазоном изменчивости, а
качественными различиями самого материала. У одних экземпляров в щеч-
ных зубах имеются стержни с ячеисто-волокнистой структурой (репара-
тивный дентин), коренные конической формы, Р/4 с рудиментом доба-
вочного корня, ростральная часть черепа относительно длинная, лобноно-
совой шов выступает углом назад, дорсальная поверхность черепа упло-
щенная. За этими экземплярами Шевырева предложила закрепить назва ние
£ altaicus*. Другие экземпляры, с относительно более высокими призма-
тическими щечными зубами без стержня, с изогнутым профилем дорсальной
стороны черепа и сильно выступающими вперед концами лобных костей,
* Это всего лишь новый ранг для подсемейства Tsaganomyinae Matthew et Granger, 1923
Ф. 4) - старшего синонима среди названий семейственной группы для Tsaganomys.
Б-С.Виноградов считал, что в материале представлены 2 вида, однако приводимые им
Различия в форме нижнечелюстной кости, скорее всего, связаны с возрастными
ИзМеНениями.
159
она отнесла к новому роду и виду Beatomus bisus Shevyreva, 1972. Ковалы
ский (Kowalski, 1974, рр. 157, 158), просмотревший материал Шевыревой, не
согласился с тем, что различия в гистологической структуре коренных зубов
достаточны для выделения нового таксона и вида и свел В. bisus в синонимы
Т. altaicus. На большую вероятность синонимии В. bisus и Т. altaicus недавно
указал также Брайент (Bryant, 1994). Как нам представляется, признаки,
приводимые Шевыревой для различения родов Tsaganomys и Beatomus,
столь существенны, что простое сведение Beatomus в синоним Tsaganomys.
без объяснения этих различий, выглядит неубедительным. Тем не менее,
отличия между "цаганомисами" и "беатомусами", видимо, могут быть
объяснены, если попытаться реконструировать онтогенетические изменения
зубной системы этих своеобразных грызунов.
Ключевым моментом для такой реконструкции является, видимо,
выяснение биологического значения особых стержней ячеисто-волокнистой
ткани в коренных зубах Tsaganomys. По мнению Шевыревой (19746, с.47),
эти стержни являются образованием, компенсирующим при стирании слабый
эмалевый рельеф жевательной поверхности, слабость которого характерна
для цилиндродонтид. Причем стержни имеются только у Т altaicus, Cyclo-
mylus lohensis Matthew et Granger, 1923 и Cyclomylus turgaicus (Vinogradov et
Gambarjan, 1952), но их нет у В. bisus (у которого эмалевый рельеф жева-
тельной поверхности коренных зубов столь же рудиментарен), так как пер-
вые три вида имеют другой тип полости зубной пульпы. Таким образом,
Шевырева придает описанным ею стержням филогенетическое значение: это
структуры, появившиеся в эволюции для компенсации исчезновения эмале-
вых гребней на поверхности коронок коренных зубов. Но у цилиндродонтид
эмалевые гребни на жевательной поверхности зубов смогли исчезнуть толь-
ко потому, что функция резания стала осуществляться полукруглыми полос-
ками эмали (имеющимися только на передне-внутренней стороне верхних и
задне-внутренней стороне нижних зубов), направленных на противолежащих
зубах вогнутостью друг к другу и действующих наподобие ножниц. Следова-
тельно, развилась более мощная, по сравнению с гребнями, режущая струк-
тура жевательной поверхности, и исчезновение гребней вряд ли нужно было
компенсировать. О филогенетическом значении различий в строении полос-
ти пульпы у Tsaganomys и Beatomus Шевырева не упоминает.
Ближе к истине, как нам кажется, замечание Шевыревой (19746, с.47) о
том, что "картина присутствия стержня и в нем участков дентина ^совокуп-
* Формально это неправильно: не опубликовано никаких доказательств присутствия
стержней ячеисто волокнистой ткани в зубах голотипа Т altaicus, хранящегося
Американском музее естественной истории (AMNH № 19019); паратипы этого вида
(AMNH №№ 19033, 19031, см. Matthew, Granger, 1923, fig. 4) не имеют таких стержне!’
которые "обычно хорошо видны невооруженным взглядом" (Шевырева, 1972, с. 143)
поэтому в новый таксон следовало бы выделить форму со стержнями в зубах.
160
цости есть не что иное, как явление "заместительного" дентина при некото-
рых повреждениях, например, кариесе у млекопитающих" Здесь же она
совершенно справедливо проводит аналогию с "пломбированием" дентином
поврежденных пластинок панциря у девонских бесчелюстных Psammosteidae.
При стирании контактирующих с грунтом пластинок псаммостеид или иных
их механических повреждений вскрывались заполненные соединительной
тканью межбалочные полости. В таких местах появлялись одонтобласты и
заполняли дентином ("пломбировали") поврежденные участки (Быстров,
1955), т. е. такая пломбировка имеет регенариционное значение. Но ведь этот
процесс происходил в течение жизни одной особи псаммостеида! По-
видимому, и образование стержня ячеисто-волокнистой ткани на зубах
цаганомисов могло иметь отношение, скорее, к онтогенезу зубной системы,
чем к филогенетическим преобразованиям в линии цаганомиид.
Если сравнивать продольные разрезы нижнечелюстной кости у
Tsaganomys и "Beatomus" (рис. 1, а, в; см. также Шевырева, 19766, рис. 13, г,
м; табл. XII, 2 и 3), то можно отметить два существенных момента: 1) у
"Beatomus" "корни" коренных зубов и "корень" резца широко открыты (зубы
призматические), коронки относительно высокие, полость пульпы достигает
лишь половины высоты коронки, 2) у Tsaganomys "корни" коренных зубов и
резца замыкаются (зубы конические), коронки относительно меньше по
высоте, полость пульпы открывается на жевательной поверхности коронки, и
в этом месте она "запломбирована" репаративным дентином. Из этого
сравнения следует важный вывод - полость пульпы заполнялась дентином у
цаганомисов медленнее, чем коронка стиралась. Следовательно, в какой-то
момент, на относительно поздней стадии онтогенеза, полость пульпы
вскрывалась стиранием. Тогда должен был включаться регенерационный
механизм, и зубы "пломбировались" дентинобластами, работавшими по
измененной генетической программе образовывались стержни ячеисто-
волокнистой ткани. То, что происходило в зубах предков цаганомиид при
болезненных изменениях и механических повреждениях, у Т altaicus стало
онтогенетической нормой (может быть, не у всех особей вида?), регенера-
ционный механизм закрепился в генетической программе, став своеобразной
адаптацией к потреблению чрезвычайно жестких кормов. Проблема снаши-
ваемости зубов актуальна для всех роющих грызунов, и каждая группа выхо-
дит из положения по своему. У представителей семейства Bathyergidae, на-
пример, функционирование зубов разделено во времени, и поэтому, не имея
последовательной серии черепов молодых и взрослых животных, можно
(ошибиться в определении их зубной формулы (Wood, 1985; Denys, 1988).
Приняв это предположение, можно гипотетически реконструировать
онтогенез зубной системы Т altaicus. Молочные зубы Т. altaicus кратко
описаны Мэтью и Грэнжером* (Matthew, Granger, 1923, р.4), но они нигде не
были изображены. Видимо, они существенно не отличались от молочных
"Зубов близкого вида Cyclomylus lohensis (Kowalski, 1974, Pl.XLVI, 2); по
последним данным (Bryant, 1994), оба таксона конспецифичны. Нестертые
постоянные зубы имеют гипселодонтное строение, эмалевая поверхность
коронок очень быстро стирается в онтогенезе (Wood, 1989). В развитии
постоянных щечных зубов можно восстановить следующие изменения: эмаль
утолщается, полость пульпы постепенно заполняется дентином, вскрывается
стиранием (рис. 1, б) и "пломбируется" ячеисто-волокнистой тканью (рис. 1,
б, в - стадия “Tsaganomys”), стержень ячеисто-волокнистой ткани увеличи-
вается в размерах (рис. 1, в, г), рост зубов замедляется и совсем прекраща-
ется, поэтому коронки постепенно уменьшаются в высоте за счет стирания
(рис. \, б, в, г). "Корень" резца постепенно закрывается, его рост замедляется
и прекращается (рис. 1, а-г).
Таким образом, вариант строения зубов цаганомиид типа "Beatomus"
является, по моему мнению, лишь одной из онтогенетических стадий
развития зубной системы Т altaicus (наблюдается у молодых животных, рис.
1, а) и, следовательно, Beatomus bisus является младшим синонимом
названия Tsaganomys altaicus.
Следует отметить, что гипотеза Шевыревой (1971; 1972; 1974а, б, в;
1976а, б), согласно которой строение зубов типа "Beatomus” (истинная гипсо-
донтия) представляет собой более эволюционно-продвинутую филогенети-
ческую стадию по сравнению с зубами типа Tsaganomys, содержит рацио-
нальное зерно: более молодая онтогенетическая стадия развития зубов
выглядит более эволюционно-продвинутой, так как прогрессивная эволюция
зубной системы у грызунов (приобретение истинной гипсодонтией и т.п.)
происходила путем фетализации, задержки на все более позднее время в
онтогенезе ювенильного состояния.
О геологической разновозрастности Beatomus и Tsaganomys свидетель-
ствует только указание Шевыревой (19746, с.50), что "на основании сборов,
проведенных in situ, можно предположить, что Beatomus bisus связан с
кровлей толщи [свиты Шанд-Гол - А.А.], где совместно с ним пока не был
обнаружен Tsaganomys altaicus" Однако, не сказано, сколько экземпляров В.
bisus найдено in situ в кровле толще (различия могут быть случайными) и
найдены ли экземпляры В. bisus in situ в основании толщи. Догадка о том,
что "оба вида существовали в различное время" (Шевырева, 19716 с. 144) - Т
altaicus в среднем олигоцене, а В. bisus в позднем - пока не подтверждена.
Для обоснования синонимии В. bisus и Т altaicus следует рассмотреть
также другие признаки, которые, по мнению Шевыревой, отличают обе
формы:
1) На Р/4 у Tsaganomys имеется рудимент дополнительного корня или
ребро вместо него. Как известно, у грызунов с высококоронковыми зубами
корни формируются на относительно поздней стадии онтогенеза. Видимо, и
в данном случае у молодых животных (стадия "Beatomus") корни еше
отсутствовали, у более поздних формировались, и у старых (стадия "Tsaga-
♦В первоописании Т altaicus ясно упомянут dP3l, а не РЗ/, как пишут Виноградов и
Гамбарян (1952, с.23) и Шевырева (19766, с.57).
162
Рис. 1. Гипотетическая последовательность онтогенетических преобразований в зубной
системе Tsaganomys altaicus (продольные срезы нижнечелюстной кости с Р/4 - Л/72):
а - стадия ''Beatomus" б - промежуточная стадия, в - стадия "Tsaganomy , г - конечная стадия (а и в по Шевыревой,
19766, рис 13, г, м; в - модифицировано).
() - денгин, пн - полость пульпы, j - эмаль, явс - ячеисто-волокнистая структура
Fig. 1. A hypothetic scheme of the ontogenetic changes in the dentition of Tsaganomys altaicus
(sections of the mandible fragments with P/4 - Л//2):
- "Beatomus" stage, 6 - intermediate stage, в - "Tsaganomys" stage, г - final stage (a and a after Shevyreva, 19766, fig 13, г,
.v, « - modifyed)
о - dentyne, nn - pulp cavity, j - enamel, явс - cellular-filamentous structure
nomys,r) при стирании коронки они выходили на жевательную поверхность.
Возможно, дополнительный корень Р/3 формировался не у всех особей.
2) Внутренние края альвеол верхнекоренных зубов у Beatomus сходятся
под углом к Р4/, а у Tsaganomys не сходятся. Значение признака непонятно.
3) Гребни, идущие от передней стенки Р4/ к резцовым отверствиям у
Tsaganomys почти слиты. У голотипа В. bisus (Шевырева, 19766, табл. XII, 4)
11*
163
эти гребни слиты не меньше, чем у изображенных там же экземпляров
Т. altaicus.
4) Ростральная часть черепа более удлинена у Tsaganomys. Так как зубы
типа "Tsaganomys " соответствуют более поздней онтогенетической стадии Т
altaicus, то, естественно, что взрослые особи этого вида будут иметь более
удлиненную ростральную часть черепа, чем молодые, имеющие зубы типа
"Beatomus ".
5) У Beatomus профиль дорсальной стороны черепа вогнутый. Признак,
скорее всего, также связан с возрастной изменчивостью.
6) У Tsaganomys латеральные и медиальные края носовых костей вза-
имно параллельны, доходят до краев передних резцовых отверствий. При-
знак, возможно, индивидуально изменчив.
7) У Tsaganomys угол лобно-носового шва выступает кзади (показаны 2
экз. Шевырева, 19766, табл. XI, 1в, Зв), у Beatomus концы лобных костей
сильно выступают вперед (показаны 2 экз.; Шевырева, 19766, табл. ХШ, 1,
2). Различия в положении носо-лобного шва, видимо, связаны с индивиду-
альной изменчивостью. В коллекции Зоологического института РАН в
Санкт-Петербурге (ЗИН) имеется фрагментарный череп с выступающим кза-
ди углом лобно-носового шва (рис. 2, б, вклейка; признак Tsaganomys), ассо-
циированный с ветвями нижней челюсти, несущими зубы без стержня (рис
3, 7 и 2, вклейка; признак "Beatomus").
Размах возрастной (и половой?) изменчивости Т. altaicus виден даже на
примере небольшой выборки остатков этого грызуна, хранящейся в кол-
лекции ЗИН (Монголия, Заалтайская Гоби, Гоби-Алтайский аймак, Цогтсо-
мон, гора Улан-Ганг, 20 км западнее озера Цзохой-Нур). Здесь представлены,
в частности, нижнечелюстная кость молодой особи со следами нестертой
эмалевой поверхности на PI4-MIX (см. рис. 3, 3, вклейка), череп, фрагмент
верхнечелюстной кости и 2 нижнечелюстные кости более взрослых особей,
еще сохранявших зубы без стержня (рис. 3, 4, рис. 4, вклейка) и ростральная
часть черепа и изолированные резцы заметно более крупных особей (старые
самцы?, рис. 5, 1,4 и 5, вклейка).
Таким образом, кажется достаточно убедительным полагать, что корен-
ные зубы цилиндродонтид типа "Beatomus" принадлежали молодым особям
цаганомисов, и название Beatomus bisus Shevyreva, 1972 следует считать
младшим синонимом Tsaganomys altaicus Matthew et Granger, 1923.
БЛАГОДАРНОСТИ
Автор благодарен| Н.С.Шевыревой], И.М.Громову, Г.Ф.Барышникову, | Л.А.Несову]
и А.К.Агаджаняну за чтение рукописи и высказанные замечания.
164
ЛИТЕРАТУРА
Быстров А.П. Микроструктура панциря бесчелюстных позвоночных силура и девона
И Сб. памяти акад. Л.С.Берга. - М.-Л., 1955. С.472-523.
Виноградов Б.С., Гамбарян П.П. Олигоценовые цилиндродонтиды Монголии и
Казахстана//Труды Палеонтол. ин-та АН СССР, 1952. Т.41. С. 13-42.
Дашзэвэг Д. Новые данные о стратиграфии среднеолигоценовых отложений МНР И
Труды Совм. сов.-монгол. палеонтол. эксп., 1970. Вып.2. С.37-43.
Шевырева Н.С. К вопросу о систематическом положении "Tsaganomys altaicus"
(Rodentia, Mammalia) И Бюлл. Моск, о-ва испыт. природы. Отд. геол., 1971.
Т.46. №6. С. 144.
Шевырева Н.С. Новые грызуны из палеогена Монголии и Казахстана // Палеонтол.
журн., 1972. №3. С.134-145.
Шевырева Н.С. Историческое развитие некоторых грызунов подотряда Sciuromorpha
в Азии И Палеонтол. журн., 1974а. № 1. С.90-100.
Шевырева Н.С. О систематическом положении "Tsaganomys altaicus" на основе
микроскопического строения коренных зубов некоторых Cylindrodontidae
(Rodentia, Mammalia) // Труды Совм. сов.-монгол. палеонтол. эксп., 19746.
Вып. 1. С.46-59.
Шевырева Н.С. Гипсодонтия цилиндродонтид - показатель климата и ландшафтов
палеогена Азии // Первый Междунар. териол. конгресс, 1974 г. - М., 1974в. Т.П.
С.336.
Шевырева Н.С. О параллельном развитии некоторых типов зубной системы у
грызунов (Rodentia, Mammalia) // Труды Зоол. ин-та АН СССР, 1976а. Т.60. С.4-
47.
Шевырева Н.С. Палеогеновые грызуны Азии (семейства Paramyidae, Sciuravidae,
Ischyromyidae, Cylindrodontidae) // Труды Палеонтол. ин-та АН СССР, 19766.
Т.158. 115 с.
Шувалов В.Ф., Решетов В.Ю. Новое местонахождение млекопитающих среднего
олигоцена на юге МНР // Труды Совм. сов.- монгол, палеонтол. эксп., 1974.
Вып. 1. С.326-329.
Bohlin В. OberoligozSne SSugetiere aus dem Shargaltein-Tal (Western Kansu) //
Palaeontol. Sinica, ser. C, 1937. N 3. 66 pp.
Bryant J.D. Tsaganomys altaicus from the Early Oligocene Hsanda Gol Formation,
Mongolia: the oldest hystricognath rodent known from a complete skull // J. Vert.
Paleontol., 1994. Vol.14. N 3. Suppl. P.18A.
Denys C. Nouvelles observation de la structure dentaire de specimens juvenilles de
Cryptomys hottentotus (Rongeurs, Bathyergidae) // Mammalia, 1988. T.52. N 2.
P.292-294.
Jaeger J.-J. Rodent phylogeny: new data and old problems 11 Benton M.J. (ed.). The
Phylogeny and Classification of the Tetrapods. Vol. 2. Mammals. - Oxford, 1988.
P.l 77-199.
Kowalski K. Middle Oligocene rodents from Mongolia// Paleontol. Polonica, 1974. N 30.
P.147-178.
Lavocat R. Les rongeurs du Miocene d'Afrique Orientale. 1. Miocene inferieur И Мёт.
Inst. Montpellier, 1973. T.l. 284 pp.
Maier IF Schrenk F. The hystricomorphy of the Bathyergidae as determined from
ontogenetic evidence //Z. SSugetierk., 1987. Bd.52. Hf.3. P.l56-164.
165
Matthew W.D., Granger W New Bathyergidae from the Oligocene of Mongolia П Amer.
Mus. Novitates, 1923. N 101. 5 pp.
Mellet J.S. The Oligocene Hsanda Gol Formation, Mongolia: a revised faunal list // Amer.
Mus. Novitates, 1968. N 2318. 16 pp.
Patterson B., Wood A.E. Rodents from the Deseadan Oligocene of Bolivia and the
Relationships of the Caviomorpha // Bull. Mus. Compar. Zool., 1982. Vol. 149. N 7.
P.371-543.
Sarich V.M. Rodent macromolecular systematics // Luckett W.P, Hartenberger J.L. (eds.).
Evolutionary Relationships among Rodents, a Multidisciplinary Analysis. New York,
1985. P.423-452.
Teilhard de Chardin P Description de mammiferes tertiares de China et de Mongolia //
Ann. Paleontol., 1926. T.15. P. 1-179.
Wang B.-Y., Chang J., MengX.-l, ChenJ.-R. Stratigraphy of the Upper Middle Oligocene
of Qianlishan District, Nei Mongol (Inner Mongolia) П Vertebr. Palasiat., 1981.
Vol. 19. N 1. P.25-34.
Wood A.E. The relationships, origin and dispersal of the hysrticognathous Rodents //
Luckett W.P, Hartenberger J.L. (eds.). Evolutionary Relationships among Rodents, a
Multidisciplinary Analysis. New York, 1985. P.475-513.
Wood A.E. Unworn cheek teeth of the Mongolian Oligocene rodent Tsaganomys: their
significance for the relationships 11 J. Vert. Paleontol., 1989. Vol.9. N 3. Suppl.
P.46A.
Summary
A.O.Averianov
ONTOGENETIC CHANGES IN THE DENTITION OF
TSAGANOMYS ALTAICUS MATTHEW ET GRANGER, 1923
(RODENTIA, TSAGANOMYIDAE) FROM THE OLIGOCENE OF
CENTRAL ASIA
Remains of the rodent Tsaganomys altaicus from Oligocene deposits of Mongolia and
China vary greatly in the microstructure of their cheek teeth. According to Shevyreva (1971 and
others), these differences are of phylogenetic significance. The form characterized by a more
derived structure of the cheek teeth was assigned by her to a distinct genus and species,
Beatomus bisus Shevyreva, 1972. In this paper an alternative hypothesis, explaining the
differences in the teeth structure in Tsaganomys and ^Beatomus" by ontogenetic changes of the
dentition of animals from one species, is proposed. The teeth of "Beatomus" type are
characteristic of young individuals. Based on this analysis, Beatomus bisus Shevyreva, 1972 is
considered junior subjective synonym of Tsaganomys altaicus Matthew et Granger, 1923.
SSN 0206-0477. ЧЕТВЕРТИЧНЫЕ И СОВРЕМЕННЫЕ МЛЕКОПИТАЮЩИЕ
ЕВРАЗИИ
_ СПб., 1996 (ТРУДЫ ЗООЛОГИЧЕСКОГО ИНСТИТУТА РАН, Т. 270)
- RUSSIAN ACADEMY OF SCIENCES ’
PROCEEDINGS OF THE ZOOLOGICAL INSTITUTE
St. PETERSBURG, 1996, VOL. 270
УДК 569.614
А.Н.Тихонов
Зоологический институт РАН, Санкт-Петербург
МАМОНТ, MAMMUTHUS PRIMIGENIUS (ELEPHANTIDAE,
PROBOSCIDEA) С ОСТРОВА БОЛЬШОЙ ЛЯХОВСКИЙ,
ВОСТОЧНАЯ СИБИРЬ
Проведен сравнительный анализ скелета и зубов взрослого самца мамонта,
найденного в 1906 г. на о. Большой Ляховский (Новосибирские острова) и находящегося в
экспозиции Национального музея естественной истории в Париже. Индивидуальный
возраст зверя около 25-30 лет>В челюстях идет смена Л/1 на Л/2, большинство эпифизов
трубчатых костей еще не приросло. По размерам бивней и некоторых костей конечностей
ляховский мамонт не уступает более взрослым (по генерации коренных зубов) мамонтам-
самцам. Геологические и тафономические условия залегания скелета свидетельствуют о
том, что гибель животного произошла в каргинское межледниковье (45-25 тыс.лет назад),
или же несколько раньше, в эпоху зырянского оледенения (более 45 тыс.лет назад).
ВВЕДЕНИЕ
Находки целых скелетов мамонта Mammuthus primigenius (Blumenbach,
1799) с остатками мягких тканей редки. Ценность подобных находок опре-
деляется тем, что позволяет воссоздать облик зверя, более подробно описать
их морфологию. Большинство скелетов, найденных в вечной мерзлоте Сиби-
ри, находятся в отечественных музеях, однако есть одно исключение - это
Мамонт, найденный на о. Б. Ляховский в 1906 г. В настоящее время его
скелет экспонируется в Национальном музее естественной истории в Париже
(Франция). Подробного описания экземпляра не было опубликовано до сих
Пор: отдельные статьи французских ученых посвящены в основном изуче-
нию мягких тканей, а также реконструкции и монтажу скелета (Boule, 1913,
Neuville, Gautrelet, 1914; Neuville, 1921); в отечественной же литературе име-
ется только краткие сведения об истории находки (Воллосович, 1914). Ля
167
ховский мамонт* один из немногих экземпляров, пол которого подтвержден
наличием гениталий. На данный момент - это единственный почти полный
скелет мамонта-самца, возраст которого не превышает 30 лет (смена корен-
ных зубов М\ на М2). Его изучение позволяет заполнить пробел, существу-
ющий в описании зверей данного возраста.
В работе использована схема промеров черепа, предложенная Дуброво
(1960). Для костей посткраниального скелета выбраны промеры из несколь-
ких работ разных авторов (Гарутт, 1972; Дуброво, 1982; Гарутт, 1992; Аверь-
янов, 1992). Номенклатура промеров длинных костей конечностей взята из
работы фон ден Дриш (von den Driesch, 1976): GL - наибольшая длина; Bp -
ширина проксимального отдела; Dp - поперечник проксимального отдела;
SD - ширина диафиза; Bd - ширина дистального отдела; Dd - поперечник
дистального отдела, HS - высота лопатки, Ld - дорзальная длина лопатки,
SLC - длина шейки лопатки, BG - ширина суставной впадины лопатки, LG
длина суставной впадины лопатки, SDO поперечник олекранона, DC
поперечник головки бедренной кости.
К сожалению, ряд промеров черепа и отдельных костей оказалось не-
возможно сделать на монтированном скелете. Измерения производились с
помощью рулетки с точностью в большинстве случаев до 5 мм. Здесь не при-
ведены промеры мелких костей дистальных частей конечностей, описанных
ранее (Neuville, 1944).
ИСТОРИЯ НАХОДКИ •
Необходимо кратко коснуться истории находки ляховского мамонта,
так как в отечественной и зарубежной литературе иногда неверно приводятся
дата и место находки. Например, Лазарев и Томская (1987, с.55) пишут, что
мамонт был найден в 1903 г. на южном берегу о. Большой Ляховский, а в
последней зарубежной монографии по мамонтам (Lister, Bahn, 1995, с. 159)
повторяется ошибка французских авторов, писавших, что мамонт был раско-
пан в 1901 и 1903 годах. История находки изложена в основном по публи-
кации Воллосовича (1914).
Летом 1906 г. промышленник А.Горохов обнаружил на дне ручья в
долине р. Этериканка, в 20 км выше места ее впадения в р.Большой Этери-
кан, на северо-востоке о. Б. Ляховский (рис. 1), голову мамонта в коже, с хо-
ботом и двухпудовым правым бивнем. Горохов выломал бивень и отрубил
кусок хобота (впоследствии они были утеряны). Информация о находке была
передана геологу К.А.Воллосовичу, участвовавшему в 1908 г. в раскопках
* В связи с редкостью находок полных скелетов мамонта, каждая из них получила в
литературе собственное имя, основанное на географическом положении скелета
ляховский мамонт, березовский, таймырский, моховской (Кутоманова) и т.д.
168
м.Мед«ежий
с ХННИКОв л
Рис. 1. Карта находок остатков мамонта (Mammuthus primigenius) с мягкими тканями
на о. Большой Ляховский
Крестиком отмечено место находки дяховского мамонта
Fig. 1. The map with finds of woolly mammoth (Mammuthus primigenius) remains with soft
tissues on Bolshoi Liakhowski Island
The place of find of liakhowski mammoth is marked by cross
п ’
О Л и *
санга-юряхского мамонта. Он предложил местным промышленникам отпра-
виться на о. Б. Ляховский летом 1908 г. и разыскать стоянку Горохова (к
тому времени умершего), возле которой, вероятно, и был обнаружен мамонт.
Уже в декабре 1908 г. Воллосович получил известие от поисковой группы,
что мамонт найден. Он обратился за финансовой помощью в Российскую
Академию наук к академику Ф.Б.Шмидту, который, однако, отказал ему под
тем предлогом, что интерес к мамонтам (после находок на рр. Березовка и
Санга-Юрях) ослабел и поэтому деньги на раскопки не получить. Тогда Вол-
лосович, понимавший, что найден прекрасно сохранившийся труп мамонта,
обратился к графу А.В.Стенбок-Фермору, который, не раздумывая, предоста-
вил деньги на необходимые изыскания. Летом 1909 г. и весной 1910 г.
169
группа промышленников, по поручению Воллосовича, раскопала остатки
мамонта. Были добыты четыре ноги, большая часть скелета, куски кожи (в
том числе с ухом, глазницей и хвостом), куски мышц с жиром и половой
член. Часть мягких тканей была оставлена в ледяном погребе около места на-
ходки, а наиболее ценные части трупа и кости были вывезены весной 1910 г.
в Якутск и далее в пригород Санкт-Петербурга, пос. Лахту, где была прове-
дена специальная обработка и препарирование этого материала, частично
пострадавшего в пути. В 1912 г. ляховский мамонт был подарен А.В.Стен-
бок-Фермором правительству Франции, за что он был удостоен ордена
“Почетного Легиона”, а российская наука и Зоологический музей Академии
Наук лишились ценнейшего экспоната. Скелет был смонтирован в Нацио-
нальном музее естественной истории в Париже, часть мягких тканей пере-
дана другим парижским музеям, а некоторые и вовсе утрачены (одна из
задних ног). Руководство Российской Академии наук, вероятно, осознало
допущенную ошибку и уже в 1913 г. профинансировало экспедицию Кутома-
нова на Таймыр за остатками мамонта, которые оказались намного худшей
сохранности.
Французские ученые поначалу проявили большой интерес к подарен-
ному уникальному экспонату, но позднее писали о нем только в связи с
перемонтажом скелета (Coppens, 1958). Интересно отметить, что примерно в
те же годы, когда был доставлен в Париж скелет ляховского мамонта, во
Франции был найден и смонтирован в Булонском музее (Boulogne Museum)
почти полный скелет мамонта из долины Аа (de ГАа) (Pontier, 1913).
ГЕОЛОГИЧЕСКИЕ И ТАФОНОМИЧЕСКИЕ УСЛОВИЯ ЗАХОРОНЕНИЯ
В отчете Воллосовича сказано, что остатки трупа залегали на дне ручья
в мерзлом илу на глубине в 2 сажени (более 4 м). Так как сам Воллосович не
был на месте находки, то точное геологическое описание отсутствует. По
словам промышленников, на противоположном берегу ручья возвышался
двумя уступами обрыв, высотой до 6 саженей, с ледяными жилами в верхней
части. Воллосович сравнивает этот обрыв с описанным им же обрывом око-
ло Малого Зимовья (южное побережье острова) и приходит к выводу, что
они идентичны. Обнажения около Малого Зимовья сложены в основном зы-
рянскими и каргинскими отложениями (Романовский, 1958, 1961). Эти же
отложения преобладают и в долине р. Б. Этерикан (А.А. Архангелов, устное
сообщение). Как и барон Толль (1897), Воллосович считал, что погребенные
льды в береговых обрывах острова - не что иное как остатки ископаемого ги-
гантского ледника. То обстоятельство, что ляховский мамонт был выкопан у
основания берега, а на дне ямы появился "чистый лед", свидетельствует, по
его мнению, о достаточно древнем геологическом возрасте скелета, который
вряд ли был переотложен.
170
Тафономические замечания Воллосовича сводятся к тому, что мамонт
погиб в грязевом потоке в начале лета, поскольку шерсть на левом боку
варьирует по цвету и длине, что, по его мнению, соответствует весенней
линьке. На глубине около 2 м были найдены куски шкуры и желудка,
разорванные хищниками (встречена шерсть песца и когти хищных птиц),
свидетельствующие о том, что уже в более позднее время труп обнажался и
был расчленен на отдельные части. Вероятнее всего, коренное залегание
трупа неоднократно подвергалось размыву ручьем, часто менявшим свое
русло, однако, остатки левой части туши с глубины около 4 м прекрасно
сохранились и не несут следов переотложения или иного воздействия потока.
Подробное геологическое изучение о. Б.Ляховский проводится в по-
следние годы экспедицией Московского университета под руководством
А.А.Архангелова. Удалось установить, что значительные скопления костей
млекопитающих приурочены к отложениям каргинского и, особенно, зырян-
ского времени. На восточном побережье острова, в 15 км к югу от устья р.
Б.Этерикан, имеются значительные костеносные слои зырянского возраста.
Именно в этих местах в последние годы найдены новые части туш мамонтов
(Лазарев и др., 1995). По мнению автора этих находок, А.А.Архангелова, они
происходят из слоев зырянского времени. Неподалеку отсюда располагается
место захоронения ляховского мамонта. Абсолютный возраст 32 100±900 лет
(МАГ-136), соответствующий эпохе каргинского межледниковья, определял-
ся для остатков мамонтов с южного побережья о. Б. Ляховский из основания
разреза едомы (Ложкин, 1977). По другим данным о. Большой Ляховский
сложен исключительно зырянскими отложениями, а более поздние - каргин-
ские и сартанские - полностью вытаяли (Томирдиаро, Черненький, 1987). С
определенной долей уверенности можно предположить, что ляховский
мамонт погиб в зырянское или каргинское время, так как практически все
остальные находки остатков мамонтов с мягкими тканями на Большом
Ляховском относятся к этому периоду.
ОПИСАНИЕ СКЕЛЕТА
Общие размеры смонтированного в парижском музее скелета, могут
быть определены лишь приблизительно, так как большая часть позвонков и
обе плечевые кости у него искусственные. Длина скелета от переднего конца
межчелюстных костей до основания хвоста составляет 321 см, что на 60 см
короче скелета березовского мамонта, а высота в холке (256 см) почти равна
таковой у березовского (265 см) и даже превышает высоту (250 см) более
длинного (403 см) таймырского. Приведенные данные по общим размерам,
плохо согласующиеся между собой, лишь подтверждают тот факт, что
любые выводы, основанные на промерах смонтированных скелетов, должны
приниматься с известной долей вероятности. Практически каждый скелет
имеет искусственные элементы и смонтирован в различных позах, придавае-
171
мых авторами монтажа. Более точные и сопоставимые данные можно полу-
чить по промерам отдельных костей скелета. Для сравнения использовались
измерения березовского мамонта, как наиболее близкого по биологическому
возрасту самца (смена зубов М2 на Л/3), и взрослого таймырского мамонта
(Л/3), являющегося неотипом вида Mammuthus primigenius. Большая часть
промеров этих двух скелетов заимствована из работ Заленского (1903) и
Гарутта (1992). В некоторых случаях для сравнения использованы данные по
юрибейскому (Дуброво, 1982) и моховскому (Аверьянов, 1994) мамонтам.
Череп. Сохранился почти полностью, повреждена только передняя
часть альвеолы левого бивня, вероятно, в результате выламывания бивня. По
своим размерам череп ляховского мамонта несколько меньше, чем у таймыр-
ского (табл. 1), хотя в целом, учитывая сравнительно молодой возраст жи-
вотного, размеры черепа вполне сопоставимы с более взрослым березовским
мамонтом. Практически все швы между костями черепа хорошо различимы.
Межчелюстные кости длинные и относительно широкие на уровне подглаз-
ничных отверстий. Ширина лицевой части уступает экземплярам более
взрослых мамонтов, носовое отверстие - с несколько опущенными вниз и
слегка заостренными латеральными краями. Скуловая дуга короткая и отно-
сительно массивная; ее минимальная высота такая же, как у таймырского
мамонта. Мозговая часть черепа широкая, затылок шириной 60 см, он не-
сколько разрушен в верхней части (протерлись тонкие стенки пневматизи-
рованной костной ткани). Высота черепа (от затылочных мыщелков) не усту-
пает высоте более взрослых экземпляров и даже превосходит этот промер у
таймырского мамонта, однако значение промера сильно варьирует (см. табл.
1) и, вероятно, связано со степенью развития бивней, тяжесть которых влия-
ет на размеры затылочной части черепа. Затылочные мыщелки крупные, их
высота наибольшая среди почти всех изученных экземпляров. Учитывая, что
по большинству промеров черепа ляховский мамонт уступает более взрос-
лым особям, можно предположить, что кости черепа (и, в первую очередь,
его лицевой части) еще не закончили свой рост. Исключение, возможно,
составляют некоторые кости мозгового отдела черепа, прекращающие свой
рост по достижении животным половой зрелости (Haynes, 1991). Обычно
размеры черепа не используются как критерий определения индивидуаль-
ного возраста, однако, сравнивая имеющиеся показатели (см. табл. 1), можно
предположить, что некоторые из них длина черепа и размеры скуловой
дуги, так же, как и степень сращения костей черепа, позволяют определить
приблизительный индивидуальный возраст при отсутствии коренных зубов и
бивней.
Нижняя челюсть. Кость длинная, за счет удлиненного под-
бородочного отростка, что является характерным признаком самцов (Аверья-
нов, 1994). Основные характерные особенности нижней челюсти мамонтов
приведены в описании юрибейского мамонта (Дуброво, 1982). Восходящая
ветвь нижней челюсти сравнительно широкая -37.7% от длины челюсти; у
172
Таблица 1. Размеры черепа мамонта (Mammuthus primigenius)
Table 1. Measurements (mm) of the skull of Mammuthus primigenius
Промеры, Юрибей- ский Ляхов- ский Березов- ский Таймыр- ский Мохов- ской Адыч- ский
М\ А/1-2 AV2-3 М3 М3 М3
Длина черепа 550 594 589 625 681
Высота черепа 708 720 786 852 825
। Расстояние от макушки до
$ переднего края j межчелюстных костей 1100 1198 1172 1279
Высота черепного свода 440 430 522 464
Ширина затылка 500 600 610 655 661
I Ширина в надглазничных [ отростках 500 620 715 661 617 723
। Ширина скуловая 600 590 660 676 625
Высота затылка 408 530 560 432 530 574
Ширина между мыщелками 195 205 239 233 220
Длина скуловой дуги 330 375 420 450
Минимальная высота скуловой дуги 40 43 39 45
Ширина межчелюстных на уровне подглазничных отверстий 220 320 375 312 385
Ширина в средней части 280 312 307 332 340
Ширина на концах альвеол 400 415 445 432 457
Ширина мыщелка 95 85 92 80 108
Высота мыщелка 80 75 84 78 79
Диаметр альвеолы наибольший 130 135 221 193 143
Диаметр альвеолы наименьший 118 120 115 147 140
173
таймырского мамонта этот показатель меньше - 36%. Высота кости у заднею
края альвеолы последнего зуба небольшая (табл. 2). Вероятно, изменение
этого промера связано с возрастом зверя и может служить критерием для
определения смены зубов при отсутствии таковых. У ляховского мамонта
Л//3 только начинает формироваться в альвеолярной полости, вследствие че-
го высота челюсти заметно меньше, чем у экземпляров с прорезавшимся
М/3.
Бивни. Правый бивень отсутствовал уже на месте находки. Левый
бивень светло-бурого цвета сохранился полностью. Его длина по большой
кривизне 251 см, диаметр у выхода из альвеолы: 130 мм - наибольший и 118
мм наименьший. Гетеронимная закрученность хорошо выражена, радиус
кривизны 57 см. Коэффициент радиуса кривизны 43.8 типичен для бивней
самцов (Верещагин, Тихонов, 1986). По своим параметрам бивень ляховско-
го мамонта должен принадлежать животному в возрасте около 40 лет
(Верещагин, Тихонов, 1990), однако методика определения возраста живот-
ного по количеству темных полос на бивне (конусов нарастания дентиновой
ткани), использованная в указанной выше работе, очевидно, неточна. Скорее
всего, в течение одного года могут формироваться несколько подобных
полос, что связано с лунным циклом, сменами сезонов года и минерализо-
ванностью кормов (Клевезаль, Клейненберг, 1967). Размеры бивней сильно
варьируют у современных слонов; например, самцы африканское слона
(Loxodonta africand) в возрасте 30 лет могут иметь бивни, различающиеся по
длине у разных особей в 1.5-2 раза (Haynes, 1991). Сравнивая бивень ляхов-
ского мамонта с бивнями взрослых мамонтов с более поздними сменами
коренных зубов, можно предположить, что изменения в размерах бивней у
мамонтов в основном коррелируют с возрастом, хотя размах индивидуаль-
ной изменчивости очень велик.
Коренные зубы. В более ранних работах, посвященных
ляховскому мамонту (Воллосович, 1914; Coppens, 1958), нигде не были
охарактеризованы его коренные зубы. Крупные бивни позволяли считать,
что, скорее всего, функционировали последние коренные зубы (А/3/3) и,
следовательно, возраст животного около 40-50 лет (Верещагин, Тихонов.
1990). Однако непосредственное изучение материала показало, что у ляхов-
ского мамонта заканчивалась смена Л/1/1 на Л/2/2. В верхней челюсти по
обеим сторонам сохранились остатки ММ с 8 эмалевыми пластинами и МП с
начинавшими стираться 7 эмалевыми пластинами. На правой стороне ниж-
ней челюсти М/\ уже отсутствует, а МП имеет 14 эмалевых пластин, из ко-
торых 11 затронуты стиранием; на левой стороне М/\ сохранился в виде не-
большого остатка с 2 пластинами. По методике, разработанной Шером и Га-
руттом (1985) для определения генерации коренных зубов слонов (отноше-
ние количества эмалевых пластин к ширине зуба), зубы ляховского мамонта
более всего соответствуют в верхней челюсти смене Р4/ на ММ, а в нижней
М/\ на М/2, Данная методика, вероятно, несовершенна в том случае, когда
один из зубов уже заканчивает свое функционирование. Сопоставляя данные
174
Таблица 2. Размеры нижнечелюстной кости мамонта (Mammuthus primigenius)
Table 2. Measurements (mm) of the mandible of Mammuthus primigenius
Промеры, Юрибей- ский Ляхов-ский Березов- ский Таймыр- ский Мохов- ской
М\ А/1-2 Л/2-3 М3 М3
Длина кости 352 650 620 628 582
расстояние между головками суставных отростков 420 415 494 470
Высота кости у заднего края альвеолы 120 139 135 135
Ширина восходящей ветви 205 245 230 226 255
Длина подбородочного выступа 240 221 188 240
по зубам ляховского мамонта с известной последовательностью смены зубов
у индийского слона, Elephas maximus (Roth, Shoshani, 1988), можно предпо-
ложить, что по своим размерным характеристикам и количеству пластин они
полностью соответствуют смене Л/1/1 на Л72/2 в обеих челюстях (рис. 2).
Такое же сравнение с данными по африканскому слону (Haynes, 1991) пока-
зывает, что по количеству эмалевых пластин зубы последних генераций не-
сопоставимы у мамонтов и африканских слонов, так как у последних
количество пластин в 1.5-2 раза меньше.
Осевой скелет. На смонтированном скелете имеются 29 шей-
ных, грудных и поясничных позвонков, подавляющее большинство которых
искусственные. В шейном отделе настоящие только последние 6-й и 7-й
позвонки, в грудном 8-й, 12-й, 15-й, в поясничном - 20-й и 21-й; крестец -
также муляж, состоящий из 5 позвонков. Всего у мамонтов насчитывается от
34 до 37 позвонков в перечисленных отделах и 20-22 хвостовых. Последние
на скелете ляховского мамонта отсутствуют, хотя исходно был найден хвост
в шкуре. Позвоночник смонтирован без учета межпозвонковых дисков, из-за
Чего скелет получился непропорционально коротким. Сохранившиеся насто-
ящие позвонки имеют заметные швы между суставными поверхностями и
Телом позвонка. Ребра в основном искусственные, настоящих только 10 ре-
бер с правой стороны и 3 с левой. Всего смонтированы 19 пар ребер.
Пояс передних конечностей. Отсутствуют обе пле-
чевых кости и правая локтевая.
Л о п а т к а. По общим размерам лопатка ляховского мамонта усту-
пает экземплярам более взрослых мамонтов, однако ширина шейки лопатки
175
mm
—I---------->---------1-----—I-------------1—
l-ll Il-Ill III—IV IV-V V-VI
300r
250 г
200^-
150^-
100 5
50 r
Qb-
Рис. 2. Положение зубов ляховского мамонта (показаны точками) на диаграмме
зависимости между длиной верхнего (вверху) и нижнего (внизу) зубных рядов и фазой
функционирования двух зубов одновременно у индийского слона, Elephas maximus
(по Roth, Shoshani, 1988, р. 582)
Порядковый номер зубов указан по горизонтальной оси римскими цифрами
Fig. 2. Position of teeth of Liakhowski mammoth (marked by dots) on the diagram of grinding
length for upper jaws (above) and lower jaws (below), with stage of function of two teeth
simultaneously in Indian elephant Elephas maximus (Roth, Shoshani, 1988, p. 582)
The number of tooth on the horizontal axe marked by rome numbers
и размеры суставной впадины отличаются у них незначительно. Это позволя-
ет говорить о том, что лопатка еще не закончила свой рост в длину (табл. 3)
Передний край лопатки прямой, хотя он выпуклый у юрибейского мамонта и
вогнутый у березовского.
Локтевая. Несколько короче, чем у других особей, но дистальны11
конец по своим размерам не уступает взрослым экземплярам. Диафиз сра^'
нительно массивный (см. табл.З). Ширина дистального отдела и массивность
диафиза с возрастом увеличиваются (Дуброво, 1982), хотя эта закона
мерность соблюдается, скорее всего, у полувзрослых животных. Мож1’1
176
определенно утверждать, что локтевая кость еще не достигла своей наиболь-
шей длины, так как оба эпифиза не приросли. По аналогии с африканским
слоном это соответствует возрасту до 30 лет, поскольку проксимальный
эпифиз прирастает в промежутке от 20 до 30 лет, а дистальный - от 30 до 40
(Haynes, 1991).
Лучевая. Массивная, слабо изогнутая, по длине уступает кости
таймырского и березовского мамонтов. Заметно, что увеличение параметров
лучевой кости идет только в сторону увеличения длины кости (см. табл.З).
Проксимальный эпифиз полностью прирос, дистальный еще нет. Прираста-
ние проксимального эпифиза у африканских слонов происходит до 30 лет
(Haynes, 1991).
Кости кисти. Ляховский мамонт - один из немногих экземпля-
ров, имеющих полный набор костей дистального отдела передней конеч-
ности. Все они были отпрепарированы и смонтированы на скелете, вследст-
вие чего промерить их оказалось невозможным.
Пояс задних конечностей. Имеются все элементы пояса.
Тазовые кости. Сохранились почти полностью, только крылья
подвздошных костей частично обломаны. По своим размерам таз очень
близок к таковому у березовского и таймырского экземпляров (табл. 4).
Вырезка вертлужной впадины открывается не в овальное отверстие, что
является диагностичным признаком для Mammuthus primigenius (Заленский,
1903; Дуброво, 1982). Согласно соотношениям в промерах таза, предложен-
ных для определения пола [длина и ширина входа в тазовую полость к
ширине таза в месте сращения лонной и седалищной кости (Lister, 1996)],
ляховский мамонт - типичный самец. Выбранные соотношения спорны, так
как при постоянном росте одного параметра, другой уже после 20-30 лет
жизни практически не меняется: кости таза продолжают расти за счет
крыльев подвздошных костей, увеличиваются и размеры входа в тазовую
полость; в то же время тело лонной и седалищной кости заканчивает свой
рост при достижении животным половой зрелости, что видно на примере
африканского слона. У ляховского мамонта ширина тазовых костей в лонно-
седалищном сращении даже больше, чем у более крупных и взрослых бере-
зовского, таймырского и ленского мамонтов. Несомненно, что тазовые кости
могут использоваться для определения пола у некоторых млекопитающих и,
в частности, у слонов, но неудачно выбранное Листером соотношение не
показательно для особей, чей возраст превышает 30-40 лет. Таз юрибей-
ского мамонта по своему внешнему виду больше соответствует самцу, неже-
ли самке, однако из за отсутствия промеров и пропорций нельзя оконча-
тельно установить пол этого экземпляра, который из-за его небольших раз-
меров принимался за самку (Верещагин, Тихонов, 1990).
Бедренная. Левая кость сохранилась полностью, дистальная
Часть правого бедра отсутствовала на месте находки. По длине она уступает
костям более взрослых мамонтов, хотя по размерам головки бедра, массив-
12 Заказ 2 S 177
Таблица 3. Размеры костей передней конечности мамонта
(Mammuthus primigenius)
Table 3. Measurements (mm) of bones from the forelimb
of Mammuthus primigenius
Промеры, мм | Юрибейский | | Ляховский 1 Березовский | Таймырский
Лопатка (scapula)
HS 574 710 795 778
Ld 530 600 665 690
SLC 173 210 221 207
BG 84 110 102 НО
LG 154 180 191 196
Локтевая (ulna)
GL 720 778 782
Bp 176 210 205 210
SDO 142 150 175 160
SD 72 99 103 91
Bd 114 200 175 178
Dd 130 140 145 161
Лучевая (radius)
GL 467 670 683 711
Bp 95 110 117 109
SD 33 40 32 36
Bd 109 145 130 135
Dd 83 110 по 116
178
Таблица 4. Размеры тазовой кости мамонта (Mammuthus primigenius)
Table 4. Measurements (mm) of the pelvis of Mammuthus primigenius
Промеры, мм | Ляховский | Березовский | Таймырский
щирина наибольшая 1300 1340 1250
Ширина тела подвздошной кости 210 183 194
Ширина крыла наибольшая 49 52 47
Ширина тазовой полости 43 41 43
Высота тазовой полости 32 33 41
ности диафиза и дистального эпифиза даже превосходит таймырского ма-
монта (табл. 5). Большой вертел расположен выше основания головки, как и
У юрибейского мамонта, который намного моложе. Как и у большинства
длинных костей конечностей, рост после полового созревания происходит
только в длину с очень незначительным увеличением эпифизов и массив-
ности диафиза. Положение большого вертела относительно головки бедра
сильно варьирует и не связано с возрастом или полом.
Размеры бедренной кости и сроки прирастания ее эпифизов некото-
рыми авторами используются для более или менее точного определения
возраста и пола у современных слонов (Roth, 1984; Haynes, 1991). У самцов
современного Loxodonta africana проксимальный эпифиз прирастает в воз-
расте от 30 до 40 лет, а дистальный - в возрасте 26-29 лет. У ляховского ма-
монта дистальный эпифиз прирастает или только что прирос, а прокси-
мальный - еще не приросший. По данным Хэйнеса (Haynes, 1991), длина бед-
ра у самцов и самок африканского слона заметно различается по достижении
20-летнего возраста, после чего бедренная кость самок практически не
растет в длину (в отличии от самцов). В этой же работе отмечается, что дли-
на бедра у самцов слонов коррелирует с массивностью диафиза, что нельзя
сказать про бедренные кости мамонтов.
Большая берцовая. Кость крупных размеров, по длине' и
Массивности не уступает кости таймырского мамонта (см. табл. 5). Оба эпи
эПифиза не приросли. По Хэйнесу (Haynes, 1991) прирастание эпифизов у
loxodonta africana наступает после 25-28 лет. Вероятнее всего, рост берцо-
вМх костей заканчивается раньше бедренных. Сходная ситуация наблюдается
11 в передней конечности при сравнении сроков роста плечевой кости и кос-
^Й предплечья.
12*
179
V
Таблица 5. Размеры костей задней конечности мамонта (Mammuthus primigenius^
Table 5. Measurements (mm) of bones from the hindlimb of Mammuthus primigenius
Промеры, мм | Юрибейский | Ляховский | Березовский | Таймырский
Бедренная (femur)
GL 1000 1050 1070
DC 136 150 153 144
SD 130 122 122
Bd 180 210 215 230
Dd 197 250 226 244
Большая берцовая (tibia)
GL 491 570 626 568
Bp 170 230 216 224
Dp 139 170 164 177
SD 72 90 88 86
Bd 140 160 169 166
Dd 113 120 135 130
Малая берцовая (fibula)
GL 460 560 585 556
Bp 47 55 52 69
Dp 35 40 36 47
Bd 53 50 77 78
Dd 84 110 95 106
Малая берцовая. По длине так же, как и предыдущая кость
не уступает костям более взрослых экземпляров (см. табл. 5). Эпифизы
приросшие. Промеры проксимального эпифиза были затруднены из-за при'
сохших сухожилий и мышц.
180
Кости стопы. Так же, как и на передних конечностях, проме-
рять их на смонтированной стопе не удалось.
КРАТКИЙ ОБЗОР МЯГКИХ ТКАНЕЙ
В отчете Воллосовича (1914) указывалось, что найдены два больших
куска шкуры (один с ухом и глазом, второй - с хвостом), а также ряд неболь-
ших обрывков кожи с шерстью. Вдоль спины шерсть темно-бурого цвета со-
хранилась небольшими участками, сменяясь на боку шерстью светло-рыжей,
а на голове почти черной. Различие в окраске шерсти может свидетель-
ствовать о линьке, что дало основание Воллосовичу предположить, что ма-
мрнт погиб в начале лета. Прекрасный кусок шкуры от основания хобота до
середины спины был разрезан французскими препараторами на несколько
частей, и кожа с головы с глазницей и ухом была смонтирована на муляж в
парижском музее.
Левые ноги мамонта сохранились намного лучше правых: они были в
шкуре и с мышцами. На правых ногах шкура осталась только около ступней,
подошвы которых сильно разрушились. Разрастания в виде пальцевидных
выростов на подошвах левых ног были приняты Воллосовичем за свидетель-
ство адаптации мамонта к обитанию на мягких грунтах влажной тундры. По-
добные аномалии встречаются и у полорогих, пасущихся на заливных лугах,
и. вряд ли имеют какое-либо адаптивное значение. На передних ногах
ляховского мамонта были три копытца, на задних - четыре. Кости ног были
отпрепарированы и затем смонтированы на скелете (Neuville, 1944).
Внутренние органы мамонта (часть желудка и кишечника) были остав-
лены промышленниками на острове, однако они не поленились захватить
полутораметровый половой член мамонта, который, по мнению Воллосови-
ча, своими крупными размерами указывает на то, что мамонт погиб от
Удушья при заносе его туши грязевыми наплывами.
ОБСУЖДЕНИЕ
На примере ляховского мамонта, имеющего точно установленный пол,
Подтверждаются все характерные признаки самцов у мамонтов (Аверьянов,
1994) - крупные размеры скелета, массивный череп, заостренные края носо-
вого отверстия, удлиненный симфизный отросток нижнечелюстной кости и
большие бивни. В зарубежных музеях полные скелеты М. primigenius, близ-
по возрасту и размерам, практически отсутствуют. Наиболее сходны по
^ПЭДивидуальному возрасту (смена М2 на М3) мамонты из Зигсдорфа (Siegs-
d°rf) в Германии (Mol et al., 1988) и из Пра-Роде (Praz-Rodet) в Швейцарии
Wiedemann, 1969), однако по размерам зубов и костей они намного превос-
*°Дят ляховский экземпляр. Скелеты мамонтов из Европейской России, со-
181
поставимые по возрасту, принадлежат, скорее всего, самкам (Севское место,
нахождение). Полные скелеты сибирских мамонтов заметно различаются по
своим размерам, но в целом поздние мамонты были мельче.
Индивидуальный возраст. При первоначальном описи,
нии считалось, что ляховский мамонт - взрослый самец с последней сменой
зубов (Л/2 на ЛУЗ). Эту ошибку повторяли в дальнейшем все авторы, писав-
шие об этом экземпляре. Причиной данной ошибки были, вероятно, крупные
размеры скелета и бивней. В действительности же не вызывает никакого со.
мнения, что у ляховского мамонта - предпоследняя смена зубов (Л/1 на Л/2),
причем в нижней челюсти она практически закончилась, так как в правой
челюстной кости Л/1 уже отсутствует. Следовательно, ляховский мамонт
самый молодой из известных полных скелетов половозрелых мамонтов.
Окончание функционирования М\ в челюстях азиатских и африканских сло-
нов наступает в возрасте от 22 до 28 лет.
Определение возраста по бивням дает противоречивые результаты. На-
пример, на левом бивне ляховского мамонта - 48 темных колец, что, казалось
бы, должно соответствовать 40-45 годам. Этот метод подвергался сомнению
и ранее (Haynes, 1991) и, вероятно, не может быть использован для опреде-
ления индивидуального возраста. Размеры и вес бивней также сильно варьи-
руют. У Loxodonta africana окружность бивня на выходе из черепа коррели-
рует с возрастом до 15 лет (время полового созревания); после 20 лет окруж-
ность и длина бивней никак не связана с возрастом. Например, у самцов аф-
риканских слонов из одной популяции размеры и вес бивней могут быть рав-
ными у 28 летнего и 38 летнего самцов (Haynes, 1991). Окружность бивня на
выходе из альвеолы у ляховского мамонта (40.5 см) почти равна окружности
бивня таймырского мамонта (40 см) и несколько уступает этому промеру \
березовского (44 см). Очевидно, что этот параметр никак не связан с возрас-
том.
Степень прирастания эпифизов на тубчатых костях конечностей также
дает информацию об индивидуальном возрасте слонов. В своем сравнении
мы опираемся на разработанные схемы возрастной изменчивости для совре
менных животных (Sikes, 1966, 1971; Roth, 1984; Haynes, 1991). На примере
ляховского мамонта видно, что биологический возраст по прирастанию эпи-
физов трубчатых костей можно определить с точностью почти до года. На
локтевой кости оба эпифиза не приросли, следовательно, возраст зверя мо
жет определяться до 30 лет - времени окончательного прирастания прокси
мального эпифиза. Лучевая кость, напротив, с приросшим проксимальным
эпифизом, что свидетельствует о возрасте свыше 25 лет. По костям пред
плечья получается возраст от 25 до 30 лет. Бедренная кость с обеими непри
росшими эпифизами показывает, что мамонту было не более 30 лет - время
прирастания дистального эпифиза. Наиболее точную картину для ляховского
мамонта устанавливает степень прирастания эпифизов большой берцовой
кости, прирастание которых происходит в возрасте не старше 28 лет. На на-
шем экземпляре проксимальный эпифиз еще только прирастает, а дистал»
182
ный - не приросший. Следовательно, возраст ляховского мамонта получается
от 25 до 28 лет.
В целом противоречий между определением возраста по зубам и
скелету нет. Суммируя эти данные, можно предположить, что биологический
возраст ляховского мамонта, с учетом его обитания в экстремальных услови-
ях Арктики, замедлявших развитие животного, будет в пределах 25-30 лет.
По сравнению с мамонтами из Берелеха и Севска (Барышников и др., 1977;
Мащенко, 1992), это очень крупный экземпляр для своего возраста, соответ-
ствующий самым крупным и старым представителям из указанных выше
местонахождений.
Геологический возраст и систематическое
положение. Отсутствие радиоуглеродных датировок остатков ляхов-
ского мамонта не позволяет точно определить геологический возраст наход-
ки. Отложения в долине р. Б. Этерикан датируются в основном зырянским и
каргинским временем. Сравнительно крупные для его индивидуального воз-
раста размеры костей скелета и бивней не противоречат этим датировкам,
так как мамонты сартанского времени (таймырский и юрибейский, а по-
следний, по нашему мнению, скорее, самец, нежели самка) в среднем мень-
ших размеров, чем мамонты зырянского и каргинского времени (ленский,
моховской, хатангский). Учитывая также то, что остатки трупов других эк-
земпляров, найденных в 10-15 км от места находки ляховского мамонта,
происходят из слоев зырянского или каргинского времени, с определенной
степенью вероятности можно предположить, что его геологический возраст
также зырянский или каргинский.
Подвидовая систематика мамонтов Сибири практически не разрабо-
тана. За исключением номинативного подвида М. primigenius primigenius
(Bltlmenbach, 1799), обитавшего на огромной территории Евразии, выделены
еще ряд подвидов, включая голоценового мелкого мамонта с о.Врангеля М.
primigenius vrangeliensis Garutt, Averianov at Vartanyan, 1993. Однако обраща-
ет на себя внимание то обстоятельство, что мамонты более позднего времени
были в целом мельче, так что не исключено, что в ближайшем будущем бо-
лее ранние зырянские и каргинские мамонты приобретут отдельный подви-
довой статус. К этому подвиду, вероятнее всего, и будет отнесен ляховский
мамонт.
БЛАГОДАРНОСТИ
Изучение скелета ляховского мамонта стало возможным благодаря поддержке моих
французских коллег Dr. Francine David (Laboratoire d'Ethnologie Prehistorique, Paris) и Prof.
Leonard Ginsburg (Laboratoire de Paleontologie, Paris). Большую помощь в работе оказал
А.О.Аверьянов, чьи советы и консультации были очень полезны. Я также признателен
Г.Ф.Барышникову за просмотр рукописи и сделанные замечания.
183
ЛИТЕРАТУРА
Аверьянов АО. Описание остатков мамонтов из Камского Устья И Волжская фауна
плейстоценовых млекопитающих в Геолого-минералогическом музее Казан-
ского университета. - Казань, 1992. С. 56-65.
Аверьянов АО. Мамонт Кутоманова // Труды Зоол. ин-та РАН, 1994. Т. 256. С. 111-
135.
Барышников Г.Ф., Кузьмина И.Е., Храбрый В.М. Результаты измерений трубчатых
костей мамонтов Берелехского "кладбища" // Труды Зоол. ин-та АН СССР,
1977. Т. 72. С. 58-67.
Верещагин Н.К., Тихонов А.Н. Исследования бивней мамонтов И Труды Зоол. ин-та
АН СССР, 1986. Т. 149. С. 3-14.
Верещагин Н.К, Тихонов А.Н. Экстерьер мамонта. - Якутск, 1990. 39 с.
Воллосович КА. Мамонт острова Большого Ляховского (Новосибирские острова)
Геологический очерк И Записки Имп. минерал, об-ва. Сер.2, 1914. 4.50. С. 305-
338.
Гарутт В.Е. Скелет хозарского мамонта Mammuthus cf. chosaricus Dubrovo из
среднеплейстоценовых отложений р.Орья (басе. р. Камы) И Вопросы
стратиграфии и корреляции плиоценовых и плейстоценовых отложений
северной и южной части Предуралья. - Уфа, 1972. С. 35-55.
Гарутт В.Е. Скелет мамонта Mammuthus primigenius (Blumenbach, 1799) из
окрестностей с.Козлово Чермезского района Пермской области // Волжская
фауна плейстоценовых млекопитающих в Геолого-минералогическом музее
Казанского университета. - Казань, 1992. С. 31-54.
Дуброво И.А. Древние слоны СССР. Вып. 1. // Труды Палеонтол. инта АН СССР,
1960. Т. 85. С. 1-78.
Дуброво И.А. Морфология скелета юрибейского мамонта И Юрибейский мамонт. -М.,
1982. С. 53-99.
Заленский В.В. Остеологические и одонтографические исследования над мамонтом
(Elephas primigenius Blum.) и слонами (El. indicus L. и El. africanus Blum.) // На-
учные результаты экспедиции, снаряженной Императорской Академией наук
для раскопки мамонта, найденного на р. Березовке в 1901 г. Т. 1. - СПб., 1903.
С. 1-124.
Клевезаль Г.А., Клейненберг С.Е. Определение возраста млекопитающих по слоистым
структурам зубов и кости. - М., 1967. 167 с.
Лазарев П.А., Тихонов А.Н., Абрамов А.В., Михайлов С.С. Новые находки остатков
мамонтов на о-ве Большой Ляховский И Первое Международное мамонтовое
совещание: Тез. докл. - СПб., 1995. С. 622-623. (Цитология, 1995. Т. 37. № 7.).
Лазарев П.А., Томская А.И. Млекопитающие и биостратиграфия позднего кайнозоя
Северной Якутии. - Якутск, 1987. 172 с.
Ложкин А. В. Радиоуглеродные датировки верхнеплейстоценовых отложений Ново-
сибирских островов и возраст едомной свиты северо -востока СССР И Докл. АН
СССР, 1977. Т.235. № 2. С. 435-437.
Мащенко Е.Н. Структура стада мамонтов из Севского позднеплейстоценового масто-
нахождения (Брянская область) // Труды Зоол. ин-та РАН, 1992. Т.246. С. 41-59.
Романовский НН. Палеогеографические условия образования четвертичных отло-
жений о. Большой Ляховский (Новосибирские острова) И Вопросы физической
географии полярных стран, Вып. 1. М., 1958. С. 80-88.
184
романовский Н.Н. Эрозионно-термокарстовые котловины на севере приморских
низменностей Якутии и Новосибирских островах // Мерзлотные исследования,
Вып. 1.М., 1961. С. 124-144.
рояль Э.В. Ископаемые ледники Новосибирских островов, их отношение к трупам
мамонтов и к ледниковому периоду И Зап. Рус. геогр. о-ва по общей географии,
1897. Т. 32, № 1.С. 130-152.
ромирдиаро С.В., Черненький Б.И. Криогенно-эоловые отложения Восточной Аркти-
ки и Субарктики. - М., 1987. 197 с.
Boule Л/. Un cadavre de mammouth siberien au museum 11 L'Anthropologie, 1913. T. 24.
*' p. 116-117.
Spppens Y. Le mammouth de 1'Atrikanova (Siberie) H Bull. Mus. Nat. Hist. Natur. Ser. 2,
I 1958. T. 30, N 4. P. 402-406.
Maynes G. Mammoths, mastodonts and elephants. Biology, behavior, and the fossil record.
p - Cambridge, 1991. 413 pp.
[ster A. Sexual dimorphism in the mammoth pelvis: an aid to gender determination // The
Proboscidea: Evolution and paleoecology of elephants and their relatives. - Oxford,
1996. P. 254-259.
Ister A., Bahn P Mammoths. - Boxtree, 1995. 168 pp.
rol D., Verhagen A., van Essen H. De mammoet en de prehistorische mens van Siegsdorf
11 Cranium, 1988. N 5. S. 57-64.
euville H. Remarques sur les rayons digitaux du mammouth de Siberie (Elephas
primigenius Blumenbach) // Bull. Mus. Nat. Hist. Natur. Ser. 2, 1944. T. 16. P. 353-
( 358.
euville H., Gautrelet J. Observations faites sur le sang du mammouth offert au museum
par le Comte Stenbocq-Fermor 11 Bull. Mus. Nat. Hist. Natur., 1914. T. 20. P.106-
109.
ontier G. Etude sur lemammouth de ГАа // Bull. Soc. Prehist. Fran., 1913. N 10. P. 621-
Г 646.
froth V.L. How elephant grow: Heterochrony and calibration of developmental stages in
£ some living and fossil species H J. Vertbr. Paleontol., 1984. Vol. 4, N 1. P. 126-145.
froth V.L, Shoshani J. Dental identification and age determination in Elephas maximus П J.
I Zool. London, 1988. Vol.214. P. 567-588.
Sikes S.K. The African elephant, Loxodonta africana: A field method for the estimation of
age П J. Zool. London, 1966. Vol. 150. P. 279-295.
Sikes S.K. The natural history of african elephant. - London, 1971. 397 pp.
von den Driesch A. A guide to the measurement of animal bones from archaeological sites
11 Bull. Peabody Mus., 1976. Vol.l. 137 pp.
Wiedmann Л/. Le mammouth de Praz-Rodet (Le Brassus, Vaud) // Bull. Soq. vaudoise Sci.
Natur., 1969. Vol. 70. P. 229-239.
185
Summary
A.N. Tikhonov
WOOLLY MAMMOTH, MAMMUTHUS PRIMIGENIUS
(PROBOSCIDEA, ELEPHANTIDAE) FROM BOLSHOY
LYAKHOVSKY ISLAND, EAST SIBERIA
A comparative analysis of the skeleton and teeth of an adult male mammoth found in 1906
on Bolshoi Lyakhovsky Island (New Siberian Islands) and exhibited in the Museum National
d'Histoire Naturelle, Paris was conducted. In the jaws, Л/l is being replaced by Ml and most
epiphyses of the limb bones are not yet fused, conditions that indicate an age of approximately
25-30 years. In the size of its tusks and some limb bones the Lyakhovsky mammoth is not smal-
ler than older mammoth males (determined on the basis of molar generation). The geological and
taphonomic conditions of the skeleton's occurrence suggest that the animal died in the Karginian
interglacial (45-25 thousand years ago) or somewhat earlier, in the epoch of the Zyryanian
glaciation (more than 45 thousand years ago).
ISSN 0206-0477. ЧЕТВЕРТИЧНЫЕ И СОВРЕМЕННЫЕ
МЛЕКОПИТАЮЩИЕ ЕВРАЗИИ
СПб., 1996 (ТРУДЫ ЗООЛОГИЧЕСКОГО ИНСТИТУТА РАН, Т. 270)
RUSSIAN ACADEMY OF SCIENCES
PROCEEDINGS OF THE ZOOLOGICAL INSTITUTE
St. PETERSBURG, 1996, VOL. 270
УДК 569.614
К.Араи, A.H.Тихонов, А.В.Абрамов
Палеонтологический музей Нагано, Мацумото, Япония; Зоологический институт
РАН, Санкт-Петербург
НАХОДКИ МАМОНТА, MAMMUTHUS PRIMIGENIUS
(PROBOSCIDEA, ELEPHANTIDAE) НА ОСТРОВЕ БОЛЬШОЙ
ЛЯХОВСКИЙ (ВОСТОЧНАЯ СИБИРЬ)
В результате работы российско-японских экспедиций на о. Бол. Ляховский в 1993-
1995 гг. были найдены остатки нескольких Особей мамонта (Mammuthus primigenius
Bliim.) с мягкими тканями. На восточном побережье у р. Бол. Этерикан обнаружены круп-
ный кусок шкуры взрослого животного с хорошо сохранившимся ухом, левая передняя
нога молодого зверя с остатками мягких тканей, часть кожи со спины и левая задняя нога
полувзрослого мамонта. На р.Зимовье (южное побережье) раскопана часть скелета и
задняя правая нога с остатками мягких тканей от крупного взрослого зверя. Все находки
происходят из слоев каргинского или зырянского времени.
В августе 1993 г. на о. Бол. Ляховский (Новосибирские острова, Яку-
тия) работала российско-японская геологическая экспедиция. 23 августа
участник экспедиции А.А.Архангелов (Москва) обнаружил на восточном по-
бережье острова, в 2 км к югу от устья р.Большой Этерикан, обрывки шкуры
мамонта, Mammuthus primigenius (Bliim.). В 500 м от первой находки
находилась передняя нога с куском шкуры. Несколько дней спустя на р.Зи-
мовье (южное побережье острова) были обнаружены части скелета еще од-
ного крупного мамонта, в том числе правая задняя нога с остатками мягких
тканей (Сайдзе и др., 1995). К сожалению, в тот год вывезти материал с ост-
рова не удалось. Для его транспортировки на материк в августе следующего
года была организована специальная совместная экспедиция Палеонтологи-
ческого музея Нагано (Япония), Всемирного Музея мамонта (Якутск, Рос-
сия) и Зоологического института РАН (Санкт-Петербург, Россия). Она увен-
чалась успехом - обе находки с восточного побережья благополучно прибы-
ли в Якутск (Лазарев и др., 1995).
187
Новая российско-японская экспедиция состоялась летом 1995 г. Непо-
далеку от прошлогодних находок в районе р. Бол. Этерикан был обнаружен
большой кусок кожи мамонта и задняя нога. Эти находки, а также найденные
еще в 1993 г. остатки скелета мамонта и задняя правая нога с р.Зимовье,
были успешно доставлены в Якутск.
В настоящее время перечисленные выше материалы находятся на вре-
менном хранении в Палеонтологическом музее Нагано.
ОПИСАНИЕ
Первая находка представляет собой большой кусок шкуры мамонта с
левой стороны головы и бока с областью глаза и ушной раковиной. Длина
шкуры - 219 см, наибольшая ширина - 92 см. Толщина кожи достигает на
холке 22 мм. Пряди рыжевато-бурой шерсти сохранились на холке и ухе.
Валиковидная ушная раковина имеет длину 30 см и ширину 10 см, диаметр
слухового отверстия - 4.5 см. Расстояние от уха до глазницы - 40 см, от глаз-
ницы до переднего края шкуры (основание хобота) - 38 см. Сопоставляя эти
промеры с данными по ленскому и хатангскому мамонтам, можно предпо-
ложить, что шкура принадлежала взрослой особи небольшого размера. Вали-
ковидная форма уха заметно отличается от обычной треугольной, известной
по предыдущим находкам мамонтов (Верещагин, Тихонов, 1990). С внут-
ренней стороны шкуры сохранился небольшой кусок скуловой кости черепа.
Левая передняя нога мамонта с куском шкуры с шеи и головы оказалась
сильно объеденной песцом. Сохранились небольшой участок кожи с височ-
ной части головы с отверстием для глаза и узкая лента кожи, соединяющая
шкуру с ногой, общей длиной около 1 м. С внутренней стороны к шкуре
прикреплены обломки черепа, включая альвеолу левого бивня диаметром 6
см. По расчетам, произведенным для бивней мамонтов и современных сло-
нов, возраст животного с такой альвеолой определяется в 15-20 лет (Вере-
щагин, Тихонов, 1986). На шкуре местами видна светло-рыжая шерсть, неко-
торые пряди отпавшей шерсти (с шеи или холки) имеют длину 50-54 см. От
ноги сохранились запястье и предплечье; последнее без шкуры и почти пол-
ностью без мышц. Присутствует также обломок дистального эпифиза плече-
вой кости. Длина предплечья - 53 см; такой размер у поздних мамонтов с р.
Берелех соответствует возрасту около 20 лет (Барышников и др., 1977).
Подошва ноги сильно растрескавшаяся, со следами четырех ногтеобразных
копытец. Ее ширина - 28 см, передне-задний диаметр - 25 см. Шерсть с ноги
полностью отвалилась. Предплечье и запястье сходятся почти под прямым
углом, что характерно для слонов, умерших своей смертью (Haynes, 1991). В
целом проведенный анализ позволяет определить индивидуальный возраст
зверя в 18-20 лет.
Трехметровый кусок шкуры со спины полувзрослого мамонта, найден-
ный в августе 1995 г. на морской литорали, был разорван в клочья морскими
188
волнами и соединялся с задней левой ногой тонкой полоской кожи. На шкуре
не было шерсти, и поэтому хорошо заметны характерные для мамонта склад-
ки кожи у основания хвоста. У ноги отсутствует подошва, но сохранились
некоторые кости заплюсны и обе большие берцовые кости. Судя по разме-
рам последних, остатки принадлежат небольшому по размерам животному,
возраст которого не превышает 20-25 лет.
Задняя правая нога взрослого мамонта, а также некоторые кости скеле-
та, найденные на р.Зимовье еще в 1993 г., на момент раскопок были почти
полностью без мягких тканей. В общей сложности собрано 18 костей (вклю-
чая нижнюю челюсть), 8 позвонков, 5 ребер, обломки таза и трубчатые кос-
ти. Их размеры с учетом того, что эпифизы всех без исключения трубчатых
костей полностью приросли, позволяют предположить, что это был очень
крупный старый мамонт. В нижней челюсти функционировали зубы послед-
ней генерации, причем стиранию подверглись все эмалевые пластины, а не-
сколько передних уже полностью стерты. Зверю было, вероятнее всего,
более 50 лет.
ТАФОНОМИЯ И ГЕОЛОГИЧЕСКИЙ ВОЗРАСТ
Материалы с восточного побережья о.Бол. Ляховский не имеют точно-
го местонахождения, поэтому не представляется возможным оценить усло-
вия их захоронения. Берег в этих местах представляет собой едому высотой
до 15 м с типичным байджераховым рельефом. Скелет на р.Зимовье обнару-
жен по нескольким костям, торчавшим из берегового речного обрыва. Рас-
копки позволили установить, что основное костное скопление залегает на
глубине около 4 м от поверхности тундры. Кости оказались расположенными
в мерзлоте не в анатомическом порядке, что, возможно, свидетельствует о
том, что скелет перемещался водой (Сайдзе и др., 1995).
В последние годы подробное геологическое изучение о.Бол. Ляховский
проводится экспедицией Московского университета под руководством
А.А.Архангелова. Удалось установить, что остатки плейстоценовых млеко-
питающих в основном приурочены к отложениям каргинского и зырянского
времени. На восточном побережье острова, в 15 км к югу от устья р. Бол.
Этерикан, где было найдено большинство из описанных выше находок, по
мнению геолога-мерзлотоведа А.А.Архангелова, преобладают отложения
каргинского времени. Это подтверждается полученной в Японии радио-
карбоновой датой 26.860±290 лет (TH-1786), выполненной в 1993 г. по
шкуре мамонта (Сайдзе и др., 1995).
Абсолютный возраст 32 100±900 лет (МАГ-136), соответствующий эпо-
хе каргинского межледниковья, имеют остатки мамонтов с южного побе-
режья о. Бол. Ляховский, происходящие из основания разреза едомы непо-
далеку от местонахождения на р.Зимовье (Ложкин, 1977). Береговые обрывы
р.Зимовье сложены в основном зырянскими и каргинскими отложениями
189
(Романовский, 1961). Вполне допустимо считать, что мамонт, найденный на
этой реке в 1993 г., скорее всего, погиб в зырянскую или каргинскую эпохи.
ЛИТЕРАТУРА
Барышников Г.Ф., Кузьмина И.Е., Храбрый В.М. Результаты измерений трубчатых
костей мамонтов Берелехского "кладбища" // Труды Зоол. ин-та АН СССР,
1977. Т. 72. С. 58-67.
Верещагин Н.К, Тихонов А.Н. Исследования бивней мамонтов // Труды Зоол. ин-та
АН СССР, 1986. Т. 149. С. 3-14.
Верещагин Н.К, Тихонов А.Н. Экстерьер мамонта. - Якутск, 1990. 39 с.
Лазарев П.А., Тихонов А.Н., Абрамов А.В., Михайлов С.С. Новые находки остатков
мамонтов на о-ве Большой Ляховский И Первое Международное мамонтовое
совещание: Тез. докл. - СПб., 1995. С. 622-623. (Цитология, 1995. Т. 37. № 7.).
Ложкин А. В. Радиоуглеродные датировки верхнеплейстоценовых отложений
Новосибирских островов и возраст едомной свиты северо-востока СССР И
Докл. АН СССР, 1977. Т.235. № 2. С. 435-437.
Романовский НН Эрозионно-термокарстовые котловины на севере приморских
низменностей Якутии и Новосибирских островах И Мерзлотные исследова-
ния. Вып. 1. - М., 1961. С. 124-144.
Сайдзе К, Нагаока Д., Фукуда М., Архангелов А.А., Куницкий В.В. Радиоуглеродное
определение возраста кожного покрова мамонта, обнаруженного на острове
Большой Ляховский в Заполярной Сибири И Quaternary Research, 1995.
Vol.34. N 4. Р.315-317 ( на япон. яз.).
Haynes G. Mammoths, mastodonts and elephants. Biology, behavior, and the fossil record.
- Cambridge, 1991. 413 pp.
Summary
K.Arai, A.Tikhonov, A.Abramov
FINDS OF WOOLLY MAMMOTH, MAMMUTHUS PRIMIGENIUS
(PROBOSCIDEA, ELEPHANTIDAE) FROM THE BOLSHOY
LYAKHOVSKY ISLAND (EAST SIBERIA)
As a result of two expeditions on Bolshoy Lyakhovsky Island in 1993-1995 remains of
some mammoth specimens (Mammuthus primigenius) with soft tissues were found. On the
eastern coast near Eterikan River were found a large piece of skin with ear of an adult animal
and the left forelimb with soft tissues and skin from the head of subadult mammoth. The skin
from the back with left hindlimb of young mammoth was also found. On Zimovye River
(southern coast), part of a skeleton and right hindlimb with soft tissues of a large adult male were
excavated. All finds originated from the layers of Karginski and Zyryanski epochs. These finds
add new information about the exterior appearance of mammoths. The comparative analysis of
these remains permited an estimation of the ontogenetic age of these individuals.
ISSN 0206-0477. ЧЕТВЕРТИЧНЫЕ И СОВРЕМЕННЫЕ
МЛЕКОПИТАЮЩИЕ ЕВРАЗИИ
СПб., 1996 (ТРУДЫ ЗООЛОГИЧЕСКОГО ИНСТИТУТА РАН, Т. 270)
RUSSIAN ACADEMY OF SCIENCES
PROCEEDINGS OF THE ZOOLOGICAL INSTITUTE
St. PETERSBURG, 1996, VOL. 270
УДК 569.614/940.25
К. Коэн
। Центр Александра Куайрэ, Национальный музей естественной истории,
Париж, Франция
♦
* СТО ЛЕТ (1695-1796) В ИСТОРИИ ИЗУЧЕНИЯ МАМОНТА:
j НАХОДКИ МАМОНТОВЫХ КОСТЕЙ И ИХ ОБЪЯСНЕНИЯ
В ЗАПАДНОЙ ЕВРОПЕ
Рассмотрен столетний период ранней истории изучения остатков мамонта в Европе,
I течение которого фантастические представления об ископаемых костях как ’’игре
трироды’’ заменяются данными о том, что они принадлежат слонам, некогда жившим на
1емле. Описание Э.Тенцелиусом ископаемого скелета из Тонны и изучение Й.Ф.Брей-
^иусом зубов мамонта, собранных в Сибири Д.Г.Мессершмиттом, положили начало
естественно-научному исследованию мамонта, для которого И.Ф.Блюменбах в конце
JfVIII века дал научное название Elephas primigenius.
i\
Л
] Период развития интеллектуальной жизни Европы, начавшийся в
680 г., характеризуется важным сдвигом в видении и интерпретации иско-
темых объектов Начиная со средних веков, ископаемые находки объяс-
шлись как "игра природы" или как объекты, производимые самой землей. За
>то лет до исследований Ж.Кювье ископаемых костей четвероногих, начав-
шихся в 1796 г. и в дальнейшем предоставивших мамонту почетное место в
[стории основания новой научной дисциплины 2), начали появляться объяс-
|ения остатков мамонта, многочисленныхне только в России, но и во многих
По вопросу объяснения ископаемых остатков в течение этого периода см. Martin
udwick. The Meaning of fosssils. Chicago University Press, 1972 (2nd ed. 1976); Paolo Rossi.
he Dark Abyss of Time. - Chicago: University Press, 1984.
- Georges Cuvier. "Mdmoire sur les especies d'£l£phants, tant vivantese que fossiles" // Magasin
Encyclop£dique. T. III. - Paris, 1796.
191
местах Западной Европы, согласно которым это были фактические останки
живых существ.
Данное исследование посвящено трем главным примерам ископаемых
находок, представляющим три основных этапа в ранней истории изучения и
толкования остатков мамонта в Западной Европе и характеризующим три ос-
новных этапа в построении самой палеонтологии как научной дисциплины.
ЭРНСТ ТЕНЦЕЛИУС и слон из тонны
В 1697 г. в Готе, Германия было опубликовано (рис. 1) на латинском
языке письмо, которое Эрнст Тенцелиус (Ernst Tentzelius), историограф
короля Саксонии, послал советнику эрцгерцога Тосканского Антонио Малья-
бечи (Antonio Magliabechi) 3). В письме содержалось свидетельство о не-
обыкновенной находке. "В декабре месяце 1695 г." в горе, расположенной
вблизи Тоннен, "деревни, находящейся недалеко от Эрфорда в Тюрингии,
являющейся частью курфюршества Саксонии", был обнаружен скелет "ока-
меневшего слона", глубоко погребенного в холме. Данное письмо не только
сугубо информативно, оно затрагивает научные и метафизические вопросы
огромной важности. Проблема заключалась в том, чтобы доказать, что "все
окаменелости, найденные в земле, не имеют минерального происхождения и
не являются игрой природы" Это означало доказать (в противоположность
традиционному подходу к природе, унаследованному от эпохи Возрожде-
ния), что кости, обнаруженные в земле, были костными остатками некогд^
существовавших живых существ. Тенцелиус решается утверждать здесь, чтй
данная находка, действительно, не "игра природы" или образование, появив|
шееся в земле, а скелет слона.
Для того, чтобы доказать это, Тенцелиус очень тщательно и точно опи;
сал обстоятельства ее обнаружения. Как это часто происходило в конце XVI
века, на находку наткнулись случайно: остатки животного были обнаружень
в песчаном карьере, где добывался "очень чистый и белый песок". Различные;
кости, составлявшие целый скелет, были извлечены в анатомической после!;
довательности. "Сперва кости огромных размеров, которые были частьк
задних ног животного; одна из них весом 19 фунтов". Затем другая кост
округлой формы, "сустав которой был крупнее человеческой головы и веси.
9 фунтов". После этого была обнаружена еще од на кость, крупнее предыду
щей, "принадлежащая бедру и весящая 32 фунта". Остатки можно было опре
делить как настоящие кости, среди которых различались не только разный
элементы задних конечностей, но и их суставы. ft
Первая находка была случайной, затем поиски приобрели организован^
ный характер. "В начале следующего года, после того как закончились сильй
f
De sceleto elephantino a celeberrimo Wilhelmo Tentzelio Historigrapho ducali saxonico,
quoque Testaceorum petrificationes defenduntur... - Urbini, 1697.
192
ИЛЛЮСТРАЦИИ
К статье Г Ф. Барышникова и Н. К. Верещагина
"Краткий обзор четвертичных гиен (семейство Hyaenidae)
России и сопредельных стран" с. 7-65
Рис. 2. Нижнечелюстные кости Pachycrocuta brevirostris {а) и Crocuta spelaea
spelaea (b, с); левые (а, с) и правые (Ь):
а - Тологой, Забайкалье (ЗИН 34476); b - пещера Носорога. Южный Урал (ЗИН 34477); с - Красный Яр.
р. Обь (ЗИН 32752). Вид с лабиальной (а, с) и лингвальной (Ь) сторон
Fig. 2. Lower jaw halts of Pachycrocuta brevirostris (a) and Crocuta spelaea
spelaea (b, c); left (a, c) and right (b):
a - Tologoi, Transbaikalia (ZIN 34476); b - Nosorog Cave, South Urals (ZIN 34477); c - Krasnyi Yar, Ob River
(ZIN 32752). Labial (a, c) and lingual (b) view
К статье Г Ф. Барышникова и Н. К. Верещагина
"Краткий обзор четвертичных гиен (семейство Hyaenidae)
России и сопредельных стран" с. 7-65
Рис. 3. Череп Crocuta spelaea spelaea из местонахождения Бинагады в
Азербайджане, рисс-вюрм (ЗИН 24407).
Вид сверху (а), снизу (Ь) и сбоку (с)
Fig. 3. Skull of Crocuta spelae spelae from Binagady site in Azerbaijan, Riss-Wurm
(ZIN 24407).
Dorsal (a), palatal (b), and lateral (c) views
Рис. 4. Нижнечелюстные кости Crocuta spelaea spelaea (а) и Crocuta crocuta (b). Хорошо различим метаконид
на М/1 у плейстоценового вида:
- Бинагады, Азербайджан, рисс-вюрм (ЗИН 22382-40); b - Южная Африка, современность (ЗИН 5779). Вид с линг
стороны
К статье Г Ф Барышникова и Н. К. Верещагина
краткий обзор четвертичных гиен (семейство Hyaenidae)
России и сопредельных стран" с. 7-65
Fig. 4. Lower jaw halfs of Crocuta spelaea spelaea (a) and Crocuta crocuta (b). Metaconid of M/1 is
distinguished in cave hyena:
m - Binagady, Azerbaijan, Riss-Wurm (ZIN 22382-40); b - South Africa, Recent (ZIN 5779). Lingual view
К статье Г Ф. Барышникова и Н. К. Верещагина
'Краткий обзор четвертичных гиен (семейство Hyaenidae)
России и сопредельных стран" с. 7 -65
Рис. 5. Череп Crocuta spelaea ussurica ssp. nov из пещеры Географического
общества, Приморский край, вюрм (ЗИН 34478, голотип)
Вид сверху (а), снизу (Ь) и сбоку (с)
Fig. 5. Skull of Crocuta spelaea ussurica ssp. nov. from the Geographical Society
Cave, Primorskii Krai, Wurm (ZIN 34478, holotype)
6 Dorsal (a), palatal (b) and lateral (c) views
Рис. 6. Нижние челюсти Crocuta spelaea ussurica ssp. nov. из пещеры Географического общества,
Приморский край, вюрм:
- ЗИН 34484 (паратип), b - 34486 (паратип). Вид сверху
Fig. 6. Mandibles of Crocuta spelaea ussurica ssp. nov.from the Geographical Society Cave, Primorskii Krai,
Wurm:
- ZIN 34484 (paratype), b - 34486 (paratype). Dorsal view
К статье Г Ф. Барышникова и Н. К. Верещагина
'Краткий обзор четвертичных гиен (семейство Hyaenidae)
России и сопредельных стран" с. 7-65
К статье Г Ф. Барышникова и Н. К. Верещагина
"Краткий обзор четвертичных гиен (семейство Hyaenidae)
России и сопредельных стран" с. 7-65
Рис. 7 Нижнечелюстные кости Crocuta spelaea ussurica ssp. nov.
из пещеры Географического общества, Приморский край, вюрм:
а - ЗИН 34489, правая (паратип): b - ЗИН 34487, левая (паратип). Вид с лабиальной
стороны
Fig. 7 Lower jaw halfs of Crocuta spelaea ussurica ssp. nov. from
the Geographical Society Cave, Primorskii Krai, Wurm:
a - ZIN 34489, right (paratype); b - ZIN 34487, left (paratype). Labial view
8
К статье Г Ф Барышникова и А. О. Аверьянова "Молочные зубы хищных
млекопитающих (отряд Carnivora). Часть VI.
Семейства Viverridae и Herpestidae", с. 66-147
Рис. 3. Молочные зубы Viverra zibetha, ЗИН 2283.
- снаружи, б. д - со стороны жевательной поверхности; в. е - изнутри
Fig. 3. Deciduous teeth of Viverra zibetha. ZIN 2283:
а. г - lateral, б d - occlusal, в. e - labial views
9
К статье Г Ф. Барышникова и А. О. Аверьянова "Молочные зубы хищных
млекопитающих (отряд Carnivora). Часть VI.
Семейства Viverridae и Herpestidae" с. 66-147
Рис. 14. Молочные зубы Herpestes edwardsi, ЗИН 8174:
а. г - снаружи; б.д - со стороны жевательной поверхности; в, е - изнутри
Fig. 14 Deciduous teeth of Herpestes edwardsi, ZIN 8174:
а, г - lateral; б, d - occlusal; в, e - labial views
10
-татье А. О. Аверьянова "О систематическом положении "Caprolagus" brachypus
Young, 1927 (Lagomorpha Leporidae) из виллафранка Китая", с. 148-157
Рис 2. Остатки Sericolagus brachypUs (типовая серия). Поздний виллафранк,
Китай:
а - нижнечелюстные кости (PMU М 15010а, Ь), вид сверху; б, в- правая нижнечелюстная кость (PMU М 15010а),
вид снаружи и изнутри; г. д - левая нижнечелюстная кость (PMU М 150106), вид снаружи и изнутри; е-з - правая
бедренная кость (PMU М 15015), вид с дорсальной стороны, с вентральной стороны и с проксимального конца;
и-л - второй (?) поясничный позвонок (PMU М 15013), вид сверху, сбоку и снизу
Fig. 2. Remains of Sericolagus brachypus (type series). Late Villafranchian, China:
a - mandibles (PMU M 15010a, b). occlusal view; б. в - right mandible (PMU M 15010a), medial and lateral
view; г, d - left mandible (PMU M 15010b), medial and lateral view; е-з - right femur (PMU M 15015), dorsal,
ventral and proximal views; и-л - second (?) lumbar vertebra (PMU M 15013), dorsal, ventral and lateral views
11
К статье А. О. Аверьянова "О возрастных изменениях в зубной системе
Tsaganomys altaicus Matthew et Granger, 1923 (Rodentia, Tsaganomyidae) из
олигоцена Центральной Азии" с. 158—166
Рис. 2. Деформированный череп Tsaganomys altaicus (ЗИН 78351а):
а — снизу, б — сверху, в — сбоку
Fig. 2. Damaged skull of Tsaganomys altaicus (ZIN 78351a):
a — ventral, 6 — dorsal, в — lateral viev
12
К статье А. О. Аверьянова "О возрастных изменениях в зубной системе
Tsaganomys altaicus Matthew et Granger, 1923 (Rodentia, Tsaganomyidae) из
олигоцена Центральной Азии" с. 158—166
Рис. 3. Нижнечелюстные кости (7а, 2а) и зубы (16, 26, 3, 4) Tsaganomys
altaicus:
ЗИН 783516; 2 - ЗИН 78351 в (а - кости сверху. 6 Р/4-М/3); 3 - ЗИН 78352, Р/4-М/2; 4 - ЗИН 78353,
Р4/-МЗ/
Fig. 3. Mandibles (1а, 2а) and teeth (16, 26, 3, 4) of Tsaganomys altaicus:
1 - ZIN 783516; 2 ZIN 78351b (a - dorsal view, б P/4-M/3); 3 - ZIN 78352, P/4-M/2; 4 - ZIN 78353.
Р4/-МЗ/
13
К статье А. О. Аверьянова "О возрастных изменениях в зубной системе
Tsaganomys altaicus Matthew et Granger, 1923 (Rodentia, Tsaganomyidae) из
олигоцена Центральной Азии" с. 158—166
Рис. 4. Левая (7, ЗИН 783516) и правая (2, ЗИН 78351 в) нижнечелюстные
кости Tsaganomys altaicus:
1а, 2а - с лабиальной стороны; 26 - с лингвальной стороны; 16, 2в - снизу
Fig. 4 Left ( 1, ZIN 783516) and right (2, ZIN 78351b) mandibles of Tsaganomys
altaicus:
1a, 2a - labial, 26 - lingual; 16, 2e - ventral views
14
К статье А. О. Аверьянова "О возрастных изменениях в зубной системе
Tsaganomys altaicus Matthew et Granger, 1923 (Rodentia, Tsaganomyidae) из
олигоцена Центральной Азии" с. 158—166
Рис. 5. Остатки Tsaganomys altaicus:
1 - ростральная часть черепа (ЗИН 78354), сверху (а) и снизу (б); 2 - дистальный фрагмент плечевой
кости (ЗИН 78358), сзади (а) и спереди (б); 3 - нижнечелюстная кость с Р/4-М/2 (ЗИН 78352), с лабиальной (а),
лингвальной (б) сторон и снизу (в); 4 и 5 - нижние резцы (ЗИН 78355 и 78357), вид спереди
Fig. 5. Remains of Tsaganomys altaicus:
1 - rostral part of the skull (ZIN 78354), dorsal (a) and ventral (6) views; 2 - distal part of the humerus
(ZIN 78358) posterior (a) and anterior (6) views; 3 - mandibles with P/4-M/2 (ZIN 78352), labial (a), lingual (6) and
ventral (e) views; 4 and 5 - lower i isors (ZIN 78355 and 78357), anterior view
15
I 3 Заказ 255(a 193
ZS'CIiTj. -.mi
SoLr^TrGLbiak
Cl.n ijfi nu*» if Saf itntiffinuri llrW-J
ANTONIVM MAGLIABECH1VM
BIBLIOTECARIUM M. D. E,
in Epinohm ci confcnpum
JOANNIS BAPTIST/F SCARAMVCCI
Primi Medici Urbini, ac cjusStatus Genculis
Prothomedici
MED ITAT IONES FAMILTARF.S
VRD1N!, L1TTERIS LEONARD!. M.DC.XCVH.
Рис. 1. Первая страница трактата Тенцелиуса о слоне из Тонны (1697 г.).
Fig. 1. Front page of Tentzelius' treatise on the Tonn Elephant (1697).
, rcdmtdirtf TBafirf gtreorffcnreirb/bnrdjbimitbemJaffer Ш ten
poosbifft vklQ3luf(il VOU fid)/ reit ЬйЙ Unicornu fbfli-
Jc aurf) rF>ur.
(f nblldjtn rea$ beflen Virrutes nnb TBittfungen anlengcf / fo ifl
jtrar pen brmftlbrn nod) nldjfd btfani/ aiiflh bag bit (finreofjncr in
Preftn Torntu baitjtnigt/'lo in pollanbgrfniibcii reorben/ ntif t*7ur;cit
rrtOcr Oa? l’6[f)QU(|ui btt 5tin0<r gtbraudjef (jaben ; Ifl abcc ftin
^n?ci|ffl/r!5 tutrbtfold)t< bialiincninit btm Unicornu foflilitbcnmuf*
|ig librrdn fommrn/ inbtm Orffcn nitrrflirf) abibrbictnbt Krafft to:
3ugtn i|l/ unb tebaburdjUt bbftn 3cucf)tlgMfcnbrt Ulbtf frbe cor-
ligirtn unbvtrbtljtrn fan, •*"
/ Фптфсйп?кЬ bit bamn offenbaren/bannbaiaxioma.
Dem&Natura mhi! faciunr fruftra ip nnb ЫгДи!rear/ Ьфс
btp bftfer $ tit gtnbti t au$ relr Ьей arofftn ®otf а ЭШтафе/ 0ЗД
{>;!! un b ©life an s feint m mit bantfbartm -tyr&tn
jrftnntn/uribjumQJrfalugfagcn
SCELETO
ELEPHANT1N0
X Ct liber rintf
W1LHELM0 ERNESTO TENTZELIO
HISTORIOGRAPH© DVCALf SAXONICO,
Vbi quexjue
Ш7ИС£0Я^Л/ PmiFACTIOKFS DEtEXDUVniR,
«
Aliqua fubtcrranta'phznometu tximlnl
(uVpcryntur.
ные холода, люди возобновили раскопки в том же месте”. Были постепенно
извлечены другие части скелета: "спинной хребет, к которому все еще при-
креплялись ребра", "две еще более крупные сферические кости, с костями
передних конечностей и плечевой костью, длиной четыре фута и шириной
два с половиной палма" (1 палм=7.6 см, прим. ред.). Далее шли позвонки
шеи и "заостренная кость, которая образует вершину или ее верхнюю часть".
И, наконец, огромных размеров голова "с четырьмя молярами, каждый весом
12 фунтов, и двумя большими зубами или рогами, торчащими из головы,
шириной два с половиной палма и длиной восемь футов".
Доводы Тенцелиуса были приняты одними учеными и отвергнуты
другими. В этот период на рубеже двух столетий продолжались споры отно-
сительно природы ископаемых объектов, и многие натуралисты придержи-
вались традиционного объяснения, согласно которому "ископаемые объек-
ты" следовало рассматривать как "игру природы" В 1696 г. ученые из Меди-
цинского колледжа Готы опубликовали брошюру (рис. 2), в которой остатки
определялись как принадлежащие "ископаемому единорогу" 4). Однако были
и просвещенные умы того времени, которые допускали правильность выво-
дов Тенцелиуса. В письме к Томасу Бурнету (Thomas Burnet) из Кимни, да-
тированном июлем 1696 г., великий немецкий философ Лейбниц, с которым
консультировались относительно найденных костей, писал, что они, вероят-
нее всего, принадлежали слону: "Я не сомневаюсь в том, что это ex regno
animali, и если они не принадлежат слону, то они должны принадлежать
аналогичному животному"5).
Ситуация вокруг находки слона в Тонне прекрасно отражает сложные
интеллектуальные искания последних лет XVII века. В противовес теориям
средних веков и Реннесанса об "игре природы" необходимо было доказы-
вать, что ископаемые находки являются остатками некогда существовавших
живых существ. Доказательства могли быть представлены путем сравнения
ископаемых остатков и последовательного изучения костей во время тща-
тельно проводившихся раскопок. Для опровержения тезиса, все еще сущест-
вовавшего в то время об ископаемых "гигантах" и "единорогах" 6), следовало
аргументированно показать, что исследователи имеют дело с остатками ре-
альных, а не мифических существ; применение/Сравнительной анатомии поз-
волило сделать это. Было необходимо также отвергнуть предположение о
том, что найденные кости были преднамеренно погребены в историческое
время ("некоторые люди утверждали, что это животное было привезено рим-
4) Des Unicomu fossilis oder gegrabenen Einhorn Welches in der herzshafft Tonna gefunden
morden Berfertiget von dem Coliegio Medico in Gotha den 14 febr. 1696.
5) Г.В.Лейбниц, письмо Томасу Бурнетту из Кимни (Ганновер, 17-27 июля 1696 г.) в труде
C.I.Gerhardt. Die philosophischen Schreiben von Gottfried W.Leibniz. Bd. III. - Berlin, 1875-
1890, p. 184.
6) Claudine Cohen. Le Destin du Mammouth. - Paris, Seuil, 1994. Chap. II et III, p. 47-86 (на
англ. яз. Princeton U.Press, in press).
194
г. •:><»<>• • c-:rrjpЭ-К>
- Hiftoriai.
\^3 ;.'>• Jutfnhttcrtfl abgm^cncnpflm/unbTfnfatig jcfcfleujfrr..
- 5 ' frfn rd'X’ttn^iM pat dn,t3apb«$rSbtr (n Ьц.вопапгп. ..
(д >' J d'tubcn/bfp7>j!rg'Zone^/tfl((ptgrp([f|<^tint gr*
fiinbfn/btrnidn!<roftdn^ithBdh/ ba<anbtrtroltrin .
.OiifTth!!i mi** finer 5? ugel/einett еггоафрепсп5Ясп|феп ^opfftgrog/eti,
l:dv aberivif cin Сгф|:1ЬегЬ!а1/’Xipptn/<Q3ru|lbdn/ Ktlefgrubfi/ unb
-Oal<grni,fi?.’(*4lcnrfr qffhlt geroeffii/unb пафЪспЛсFefrl&fl rotifer
паФдг(n6tf/fo iflcr и!Ь1ефгпапеиипдго|ргг( t»on. £(п|еп(ц 7. 6>ф|ф ;
Reifert 4opff/ drum (ilepbaffen Ropff п!фС tingle ф '^ntmen/ btfitn<'
.^unhufcnjcbcr j.S^ublang/iuib buifen f.Sdjiibrodf vontinunbtr
geflanben; ??cbrnbenbtn Лидел (?ubtn??orb-Oftroarfg $crau£geftbtn
jroectrt ^ibnf/оГггgcgtntinahtuцгЬоко;» «heer £>bnrr/(tf fornfpfc
fcig' bi nr г л aber aid tin jmenfpaitneg /)ol$ blrf/ #. €?фи(? lang/ unb ti*
neiicettfnerfdiroerqeroefeit. Qlon bem5fopffunbbtrtmd(ltn anbcrn
Q'einenM man Fein grivif ЭДаа£ ппЬ-(Ягго1фГ fcabrn ffcnnen/roeilen
fc(Aeimbtraudne(?menmtbrftifbeild jcrbrodxn/ unbbiel barwn mit
in ben Sanb fontmtn; abcr 1(1 rd/ bag (о1фгStueft jufammen
nr!ifentnergeroogen 'unbbniungeuitintr GMfitgerotftn. 2(ufbepbtn
Sdftnbf?.4cp(fd/uiibjroareiiirotnlgbarvon7 fraben 4. groffrQWr
pj!?nf gtlrgen/bertn febrveber t Spann brdf/cine Spanned?/unb
j.pollbiefgerotfen/aniQ5cn?id)tabrr finer roitbcranbtrt ii.'Pfunb gt«
госдеп. ' * ' : -
-Hitrben bafntainnfcnberbeifmargrnommen/bagfdndnigCMitb/
oulfer her ^opffuiibbiez. lunge ^abnt/anditanbtr де^аядеп/ fonbern
a lie geif jtun roenigflen cine gtirr /tunb breif 0апЬЬЙпго1|фт gtrotfen;
ДС'иmebrrflcn aua lautcrTaffelnоЬегОЯаКtrn/role bu^Jrautn (Jfig/
hri*Specularh obft felemtetfcuifl genunr/bepupben; (JtUd)f/d«bet
Scpff/dnhnger£abn unb ein’-Fein/ pop!/ unb mil Sanb obtrSop*
fdnauttgtfiilld/edi$( aber birfjfe unb boUfommm/ affc ofmt fonbtr»
baren
I ..
baren (!5сгиф unb ©efebmae?/In ber^rben grl(4fcftf/f<$irer Hub murbt
genufen/ аиф roann man baran gd reft Г/ n ld)f f Irben tr ollrn/ fo bulb fie
cberanbie (ii|jffomnKnimbfrtidcnmorbrn/pnb|u itidtfalldnlddjtet
uub barter/ fonbern аиф tony ebrr gra и trerben/ 1:11b fo bann allefamnit
fefir angtjogtn babrn.
Siie k£rbr/in пи1фгг(Ь1феСеГ:<Г turbcrgrngdaun/ifl alfobrfd;af«
frn; «Obrn (Id?rt егрИф 4. Sdjtib (кФ |фгост Samm-ff rbe/ bar-
cuf folgeerinrodjftie|7фг unbтйгЬей ©anb-C'eburgr/von 1J.Sd)»<
ben / nntre гоНфет tin l.Sebub/. in.ter Olt'ffen uber beffni jroep
СфиЬ ^Oiocolla unb barfe 2cp(lei:u ju f<btn fenn ; паф fcldjtn:
ftnitt fieb апвапЬ1ф((гигГеп obrr2t>oir/7. Se^uh (?o(f)/badnnrn
tintСф|фг von 1. 2оП/ unb unftn briefer abcrmal tin Sefjuh (?оф
оПккоПл nuf jrorp £ф1ф fanbldjfrnTbcti (1феп/пи1фгг рф im Sett*
frr/rooobigt Sfitrft gefiinbciiroorben/erlbigef. (ftfetcbtn and? Ьигф
bufn!iitrrfd)itbfncQ:rbni/SanbunbS(dntvoncl4niti bit‘Xirtfe bin*
tititev vidgrc([tunbddiic2(btrn/rofldu>itbif lEurrdnanb(fiQ4ii*
innt/ftrf) |'гх1спе»ггп/Ьаг5ГО||фгпvibl langtlirb гипЬг^фпе^епЬйеи
fer U’adjfrn/ncnroddbtnbif gr^cnrintrJ)afdiiii?grovftpn/bir(ldr.t*
Pen ubrr Ьигф ein Microkopiururdenttef tverbrntnuiirn. •
СиЬ1|фсп1ПЬ1ггЬгрпгф ingcbrnrfeii/ bag bit Olliorolh tn bet
Jf?e г.'Гфа(Ге Tonna f<$r gtmrin ftp/ uttb bag vor 11 Нфсп ЗаЬпп In fo gt«
uanfeit PolknbbtpWriftn^onna/roo vidTop ипЬЭДаисгрппгцг#
Ьгофеп rotrbtn/allbtrtff rin Unicornu foible in (tyflalf tint? ^irfcp*
Okrorpbdgffiinbfn/unbvonbfnUnftrtbantnmifytaftnrotbrrbit fal*
lente ё-пфг jur (tlbfn ^dt деЬгаифгt roorbtn.
Wann banit nun ubtr obgtmrUc auGgtgrabtnt T'firtt unb
^apnt uuf гг|фкЬепе •Sfrpnintgtn t nfpanbtn/inbtm rflldjt fo!rf)t fur ti*
nrn SXirftii/aiibcre fur dnttt (£ltpbanf tn rptr/unb поф anbrrt frir dn
LT nicor nu Tortile at puffen/fo puf her ©и г ф I a и фГ I g(l 15 »ГП « nb -Of»/
-0 tr t -0/ £cr£ og |u Sad?[rn/ 3 it UcbAS I r v r unb^rrg/
-еиф^пдггпппЬ2Gt(lpbaRn/ к.unfergnabigfltrftw0 unb/)rr?/iun
btit tigrntИфгпФсиПЬ brr ^сфг |u trfaprtnf und/аиф unfertrornlge
(Mutufrn banibtr jn trbffhrn/grtabigfl anbtfopltn/ n?dcf;rn ‘^rftpl
irir bfnnbfennntngeborfamfl^gdebef. ’?• '
Wicbulhn'abnbpfeir/ bag bltft О(Га, с ЬГГ Паф Georg. Horrtir
* )( i Opci.
i
i
i>
195
P„. 2. On»™ «3 трзкззтз М««—»Г» “ °™™ '““”“”ro
Fig. 2. Exm« «Г the treatise of the Goth. Medical Collee“3”en.reied Tentxeltas’ End as the remains of a "tell uniarn"
скими купцами, другие, что оно было привезено Аттилой, третьи припи-
сывали это Шарлеманю, четвертые - герцогу Глейхену, и все они последо-
вательно утверждали, что животное было похоронено в этом холме”) и найти
доказательства того, что они захоронены в земле с незапамятных времен.
Свидетельство их древнего возраста могло быть предоставлено в результате
изучения последовательности слоев земли и, в частности, в том месте, где
они были обнаружены. Наконец, оставалась загадка их присутствия в данном
месте: Священная история и эпизод о Всемирном потопе давали простой
ответ на этот вопрос 7)
После проведения этого прекрасного исследования, которое уже вклю-
чало сравнительно-анатомическое изучение, касалось функциональных свя-
зей и стратиграфии, окончательная концепция по-прежнему сводилась к биб-
лейскому рассказу.
БРЕЙНИУС И МЕССЕРШМИТТ: ОПРЕДЕЛЕНИЕ СЛОНА
Несколько десятилетий спустя другая серия публикаций показала, что в
определении ископаемых костей слона наступила новая фаза. Она явилась
результатом сотрудничества тех, кто проводил полевые исследования в Си-
бири и академические исследования в Западной Европе8).
В 1720 г. в Сибирь по указу Петра Великого была направлена важная
экспедиция, возглавляемая Даниелем Готтлибом Мессершмиттом (Daniel
Gottlib Messerschmitt), которая продлилась 7 лет. В архиве Российской Ака-
демии наук в Санкт-Петербурге до сих пор хранятся 7 неопубликованных
томов мемуаров и записей этой экспедиции 9). Мессершмит, который был
географом и натуралистом, знал, как и другие путешественники по Сибири, о
существовании таинственной "мамонтовой кости" Во время путешествия он
хотел понять ее природу и происхождение. В 1723 г. он писал одному из
своих помощников, Ивану Толстукову: "Если ты найдешь рога мамонта, ты
должен провести очень тщательный поиск других костей этого животного и
собрать их в неповрежденном состоянии все до последней" 10).
Еще в 1722 г. Мессершмитт направил из Сибири в Данциг свое му
другу голландскому натуралисту Йоханну Филиппу Брейниусу (Johann Philip
Breynius) два зуба таинственного "мамонтового животного", к которым была
приложена короткая записка (на латинском языке) следующего содержания:
"Моляры, вероятно, дилювиальные, принадлежат животному в настоящее
время неизвестному - русскому мамонту или, может быть, слону. Я прошу
7) С.С. Gillispie. Genesis and Geology. - Cambridge: U. Press, 1959.
8) Более подробный отчет о сибирских полевых исследованиях в XV11-XVIII веках
представлен у С. Cohen, op. cit., pp. 88-100.
9) П. Пекарский. Наука и литература в России при Петре Великом. Т. I. СПб., 1862.
С. 350-362.
,0) Там же
196
Вас решить, какому животному они принадлежат. Зубы были обнаружены на
вершине горы около реки Томас. Другой образец - кусок слоновой кости,
очень сходной с наружным зубом слона, был обнаружен среди других в горе
Томас" п).
Брейниус после тщательного изучения присланных материалов
представил Королевскому обществу в Данциге в 1728 г. свои идеи относи-
тельно ’’зубов и костей мамонта, найденных в земле’’ 12). Первый изученный
им объект был определен как ’’моляр (Dens molaris) или коренной зуб
(Grinder) шириной один фут, длиной в половину фута и толщиной три
дюйма, весом 8 фунтов и три унции, достаточно целый, если не считать того,
что он разломан на два куска и что концы корня испорчены’’. Другой эк-
земпляр - ’’кусок dens exertus, в некоторых местах не отличающийся от сло-
новой кости, но в других местах калцинированный подобно обычному иско-
паемому единорогу (Unicom Fossile)” На основе изучения остатков из Сиби-
ри Брейниус делает два главных вывода, давая, тем самым, ясный ответ на
просьбу Мессершмита относительно их определения. Он заключает:
”1) Зубы и кости мамонта являются настоящими естественными зубами
и костями, принадлежавшими очень крупным жившим животным, так как у
них есть не только наружное сходство, но и внутренняя структура, анало-
гичная таковой зубов и костей животных’’
2) Эти крупные животные были слонами, что следует из формы, строе-
ния и больших размеров зубов, которые точно соответствуют зубам, перети-
рающим пищу, а также бивням слонов’’
Таким образом, выводы Брейниуса оказались идентичными тем,
которые Ханс Слоун (Hans Sloan), президент Лондонского Королевского об-
щества, представил годом раньше этому уважаемому собранию, а также Ака-
демии наук в Париже.
По возвращении из экспедиции в Сибирь Мессершмитт в 1730 г. при-
был в Данциг, привезя с собой серию рисунков с изображением нескольких
частей скелета: ’’очень крупный череп, Dens exertus и моляры, Os femoris,
принадлежащие животному, обычно называемому мамонтом, и обнаружен-
ные в Сибири’’ Эти ’’любопытные наброски” показывают фронтальный и два
латеральных вида (справа и слева) черепа (рис. 3), один моляр, один бивень и
одно бедро. Комментарии на латинском языке указы вают на размеры
остатков и анализируют детали их морфологии. Присутствие в верхней че-
люсти одного моляра, который все еще находится в альвеоле, представляют
убедительное свидетельство того, что данный скелет, действительно, при-
надлежал слону, а не "химерическому Бегемоту" Библии.
John Philip Breynius. Observations, and a description of some Mammoth's Bones Dug up in
Siberia, Proving them to Have belonged to Elephants // Philosophical Transactions (1737, 38).
-London, 1741, p. 124-139.
,2) Там же
197
Рис. 3. Рисунок Мессершмитта, опубликованный Брейниусом в "Philosophical Transactions" (1741 г.).
Fig. 3. Messerschmitt's drawing, published by Breynius in the "Philosophical Transactions" (1741).
Более того, наряду с рисунками Мессершмитта, имелось письмо, напи-
санное по-латински "живым свидетелем" Михаилом Волоховичем и поме-
ченное: Иркутск, 10 февраля 1724 г. В нем сообщается о находке куска кожи,
покрытой густой коричневой шерстью:
"Эта голова была найдена русским солдатом по имени Василий Ерлов
на восточном берегу реки Индигирки, недалеко от устья речки Волоковой
ручей. После того как она была обнаружена, я, будучи не занят, присут-
ствовал и явился живым свидетелем раскопок этого скелета или костей. И
затем на другом берегу той же реки, называемом Станойяр (Sztanoijahr), я
увидел кусок сгнившей кожи, видневшейся из обрыва песчаного холма; он
был довольно большим, очень толстым и был покрыт длинной шерстью,
весьма густой и имевшей коричневый цвет, несколько напоминавшей козью
шерсть. Эту шкуру я посчитал не козьей, а шкурой Бегемота, поскольку я не
смог отнести ее ни к одному из известных мне животных" 13).
В 1738 г. Брейниус, ставший членом-корреспондентом Лондонского
Королевского общества, направил к Слоуну несколько документов, каса-
ющихся вопроса о сибирском мамонте, под названием "Зубы и кости ма-
монта - примечательная и удивительная находка в России" 14). В этой статье,
опубликованной так же, как и рисунки Мессершмитта в 1741 г. в "Philo-
sophical Transactions" (рис. 4), делается попытка объединить сведения, накоп-
ленные за несколько десятилетий в России и Западной Европе. Она впервые
представила прямые свидетельства, подтверждающие, что так называемый
мамонт принадлежит к слонам. Это явилось официальным актом опреде-
ления "сибирского мамонта" как слона за 70 лет до того, как Кювье пере-
смотрел это положение.
С мемуаром Брейниуса отождествлялись не только "наружные зубы"
"русского мамонта", которые местные жители Сибири использовали для
изготовления предметов из "мамонтовой кости", но также те фрагменты
"мамонтовой кости" (ископаемый единорог), которые можно найти под
землей в западно-европейских странах. Но проблема заключалась в том, как
объяснить присутствие этих остатков в Европе и Сибири. Последним
заключением Брейниуса было то, что эти остатки были принесены туда в
результате Библейского потопа.
"Эти зубы и кости слонов были принесены не человеком, а Потопом,
волнами и ветрами, и остались там после того, как воды вернулись в свои
берега, и были затем погребены под землею, даже около вершин высоких
гор. И так как мы ничего не знаем ни о каких необыкновенных потопах в
этих странах, а только о Всемирном потопе Ноя, который был описан Мои-
сеем, я думаю, что мы должны с наибольшей вероятностью отнести это
,3) John Philip Breynius. Observations, and a description of some Mammoth's Bones Dug up in
Siberia, Proving them to Have belonged to Elephants // Philosophical Transactions (1737, 38).
-London, 1741, p. 124-139.
,4) Там же
199
PHILOSOPHICAL
TRANSACTIONS.
GIVING SOME
ACCOUNT
О E T H E
Prefent Undertakings, Studies, and Labours,
I N G e'n I О U S,
Conliderable Parts of the WORLD.
VOL. XL. For the Years 1737, ‘1738.
With a Supplement, being the Croonean Lectures
on Mufcular Motion, for the Year 1738.
L О N <D О N:
Printed forT. Woodward, at the Half-Moon, between the
Temple-Gates in Fleetfreet; and C. Davis in Tater-
nofter-row; Printers to the Royal Society.
M.dcc.xuT
Рис. 4, Титульный лист тома "Philosophical Transactions", в котором была опубликована
работа Брейниуса (1741).
Fig. 4. Front page of the volume of the "Philosophical Transactions" in which Breynius’ paper
was published (1741).
200
странное явление к вышеупомянутому Потопу. Таким образом, не только
Священное писание может служить подтверждением естественной истории,
но и истина, изложенная в Священном писании, в котором говорится, что
потоп Ноя был всемирным и в существовании которого многие сомневаются,
вновь может быть подтверждена естественной историей" 15).
Приведенная цитата означает, что к середине XVIII века присутствие
остатков слонов в странах, удаленных друг от друга (Ирландия, Англия,
Франция, Италия, Германия, Польша и Сибирь), широко принималось во
всей Европе, как "естественное" свидетельство того, что Библейский Потоп
был всемирным.
В течение второй половины XVIII века подобное объяснение, заимство-
ванное из теорий о Земле английских дилювиалистов, таких как Бурнет
(Burnet) и Вудвард (Woodward), получило широкое распространение. Многие
европейские путешественники по Сибири (например, Гмелин 16) и Паллас 17))
также объясняли присутствие замороженных мамонтов в земле тем, что это
были остатки животных, пытавшихся спастись от потопа, убегая на север.
Но еще в 1749 г. многие другие натуралисты (например, Бюффон 18)) отка-
зались от религиозных объяснений естественных явлений. Бюффон в своем
трактате 1778 г. "Epochs of Nature" 19) объяснял присутствие слонов на севере
изменением климата вследствие постепенного охлаждения Земли, которое
вынудило животных мигрировать из полярного района (где когда-то был
теплый климат) в тропические страны с более мягким климатом.
БЛЮМЕНБАХ И УСТАНОВЛЕНИЕ ELEPHAS PRIMIGENIUS
В 1780-х гг. в работах великого немецкого натуралиста Иоганна Фрид-
риха Блюменбаха (Johann Friedrich Bliimenbach) был осуществлен важный
синтез различных находок и публикаций. Блюменбах связывал интерес к
находкам мамонта с общим вопросом о природе ископаемых: в эти послед-
ние годы XVIII столетия начали допускать идею "утраченного вида" Бюф-
фон принял ее, а Блюменбах в 6-м издании свего труда "Handbuch fiir Natur-
geschichte" 20), опубликованного в Геттингене в 1799 г., предложил классифи-
цировать "окаменелости" и "ископаемых" в соответствии с их степенью
,5) John Philip Breynius. Observations, and a description of some Mammoth's Bones Dug up in
Siberia, Proving them to Have belonged to Elephants // Philosophical Transactions (1737, 38).
-London, 1741, p. 124-139.
,6) Gtnelin. Voyages en Siberie, франц, пер. с нем. оригинала, 2 vol. - Paris 1767.
,7) Паллас. Комментарии Академии Санкт-Петербурга за 1772 г. Т. XVII, с. 572.
,8) Buffon. Theory of the Earth, in Hisoire Naturelle. T. 1. - Paris, 1749
,9) George Louis Leclerc de Buffon. Les Epoques de la Nature, 1778 (ed. by Jaques Roger.
Editions du Museum d'Histoire Naturelle. - Paris (1962), 2nd ed.
20) Johann Friedrich Bliimenbach. Handbuch fur Naturgeschichte. 1st German edit. - G6ttingen,
1779; 6th ed., 1799, N 262 sq.
201
сходства с животными, существующими в природе в настоящее время (рис.
5). Согласно его утверждению, ’’сравнение окаменелостей с организован-
ными существами настоящего Творения’’ приводит к необходимости про-
ведения различий между ископаемыми, аналоги которых все еще существу-
ют в природе (petrificata superstitorum), ископаемыми, аналоги которых
сомнительны (petrificata dubiorum), и ископаемыми, аналоги которых неиз-
вестны (petrifucata incognitorum). Так называемые "сомнительные" окамене-
лости ’’имеют некоторое сходство с ныне существующими созданиями: они
отличаются или своими чудовищно большими размерами или некоторыми
различиями; эти различия могут быть небольшими, но постоянными в неко-
торых их частях" К данной категории Блюменбах отнес, в частности, "остат-
ки слона огромных размеров... (это так называемые кости гигантов наших
добрых отцов)" 2 °. Он говорит, что такие кости можно найти не только во
всей Сибири, но также и в Германии. Блюменбах упоминает работы зоолога
Мерка, а также знаменитый скелет слона, который, как говорилось ранее,
был обнаружен в Бургтонне в 1695 г.
Таким образом, в конце XVIII века был сделан третий, самый важный и
решающий шаг в изучении костей мамонтов в Европе и того, что в работах
Кювье будет названо “ископаемыми слонами” В 6 издании своего труда
“Handbuch fiir Naturgeschichte”, опубликованном на немецком языке в
1799 г. 22) и переведенном на французский язык в 1803 г. 23), Блюменбах
впервые дал мамонту латинское название в соответствии с правилами номен-
клатуры Линнея - Elephas primigenius ("первый слон"). Под этим названием
он включил вид primigenius в род Elephas. В качестве главного примера
Блюменбах взял "экземпляр из Тонны", различные интерпретации которого
были им тщательно изучены. До недавнего времени он оставался типовым
экземпляром этого вида24). На первой странице сборника брошюр, опублико-
ванных в течение XVII века (но, вероятно, переплетенных вместе позднее),
примечания, выполненные почерком Блюменбаха (рис. 6), выявляют тот
глубокий интерес, который исследователь питал к этому ископаемому, и
показывает нам, что Блюменбах был знаком со всей библиографией (как
русской, так и западно-европейской), посвященной ископаемым костям
мамонта. Этот бесценный сборник очерков, собранных и прокомментиро-
ванных Блюменбахом, является сейчас собственностью Сти вена Джея Го-
улда, просвещенного библиофила и выдающегося палеонтолога, работаю-
щего в Гарвардском университете.
2,) Blumenbach, op. cit., N 267, p. 401.
22) Там же, p. 403.
23) Bliimenbach. Manuel d'Histoire Naturelie. - Metz, 1803.
24) В 1990 г. был предложен новый типовой экземпляр для рода Mammuthus’. см. У.Е.
Garutt, A. Gentry, et A. Lister. "'Mammuthus Brookes, 1828 (Mammalia, Proboscidea):
proposed Conservation, and Elephas primigenius Bliimenbach, 1799 (currently Mammuthus
primigemius)'. proposed Designation as the Type Species of Mammuthus, and Designation of
Neotype" // Bull. Zool. Nomenclature, 1990. N 47. P. 38-44.
202
Рис. 5. Сравнение Блюменбахом коренных зубов "совершенно неизвестных" (Ohio
Incognitum [А]), "сомнительных" (сибирский мамонт [В] и хорошо известных
(африканский слон [С]) животных (из "Handbuch fur Naturgeschichte", 6-е издание, 1799).
Fig. 5. Bliimenbach: Comparison of molars from "totally unknown" (the Ohio Incognitum [A]),
"dubious" (the Siberian Mammoth [B]) and well known (the African Elephant [C]) animals
(from "Handbuch fur Naturgeschichte, 6th edit., 1799).
203
204
Рис 6 "Рождение" Elephas primigenius: рукопись Блюменбаха с библиографическими заметками о "русском мамонте
Fig. 6. The "Birth-act" of Elephasprimigenius: Blumenbah manuscript about bibliographical notes on the "Russian Mammoth
В тексте Блюменбаха высказана идея об ископаемых видах, неизвест-
ных в современной природе, и идея о мирах, погибших в результате ката-
строф, а также идея стратиграфической последовательности ’’окаменелых”
остатков живых существ в слоях земли. Соотношение перечисленных идей
очень сходно со структурой тезисов, выдвинутых Кювье. Труд Кювье "Мё-
moire sur les especes d’eldphants, tant vivantes que fossiles" 25), представленный
французской Академии наук в 1796 г., стал краеугольным камнем в изучении
костей мамонта как остатков вымершего вида. В 1812 г. глава работы Кювье
"Recherches sur les ossements fossiles de quadrupedes", посвященная ископае-
мым слонам 26), дала полный обзор находок мамонтовых костей и их интер-
претаций в Европе и России от древних времен и до современной эпохи.
Можно сказать, что это исследование сыграло важную роль не только в
изучении мамонта, но также в создании новой научной дисциплины
палеонтологии позвоночных.
БЛАГОДАРНОСТИ
Я выражаю огромную благодарность Стивену Джею Гоулду (Prof. Stephen Jay
Gould), профессору Гарвардского университета, который показал мне свой личный экзем-
пляр сборника очерков Блюменбаха, посвященных экземпляру, обнаруженному в Тонне, и
любезно согласился предоставить мне право опубликовать копию бесценной рукописи
Блюменбаха. Я также хотела бы поблагодарить Вадима Гарутта, профессора из Зоологи-
ческого института, Санкт-Петербург, который помог мне в изучении ранних находок и
исследований остатков мамонта в России.
Summary
Cl. Cohen
A HUNDRED YEARS OF MAMMOTH BONE FINDS AND
INTERPRETATIONS IN WESTERN EUROPE (1695-1796)
European mammoths have been studied for over one hundred years. During this time, the
perception that fossils were merely "games of Nature" was replaced by scientifically gathered
data showing that these remains belonged to once living creatures. Fossil remains of extinct ele-
phants were even used to argue that these species still existed somewhere on the Earth. Two
early studies that helped this shift from superstition to scientific inquiry were the description of a
fossil elephant skeleton from Tonna, Gemany by E.Tentzelius and the study of mammoth molars
by J.Ph.Breynius that had been collected in Siberia by D.G.Messerschmitt. The idea that species
could become extinct, which was an unorthodox idea some 200 years ago, was becoming more
widely accepted through the description of animals that resembled living species but were
obviously different. Such was the case with the description of Elephas primigenius by J.F.Blu-
menbach near the end of the 18th century that established an extinct species of elephant distinct
from either living species.
25) См. прим. 2
26) George Cuvier. Recherches sur les Ossemens fossiles de Quadrupedes. T. II..- Paris, 1812.
СОДЕРЖАНИЕ
Предисловие 3
Барышников ГФ., Верещагин Н.К. Краткий обзор четвертичных гиен (семейство
Hyaenidae) России и сопредельных стран......................................... 7 V
Барышников ГФ., Аверьянов А.О. Молочные зубы хищных млекопитающих (отряд
Carnivora). Часть VI. Семейства Viverridae и Herpestidae 66'
Аверьянов А. О. О систематическом положении кролика "Caprolagus" brachypus
Young, 1927 (Lagomorpha, Leporidae) из виллафранка Китая 148v
Аверьянов А. О. О возрастных изменениях зубной системы Tsaganomys altaicus Mat-
thew et Granger, 1923 (Rodentia, Tsaganomyidae) из олигоцена Центральной Азии. 158V
Тихонов А.Н. Мамонт, Mammuthusprimigenius (Proboscidea, Elephantidae), с острова
Большой Ляховский, Восточная Сибирь........................................ 167tJ
Араи К., Тихонов А.Н., Абрамов А.В. Находки мамонта, Mammuthus primigenius
(Proboscidea, Elephantidae) на острове Большой Ляховский (Восточная Сибирь).. 187’*
Коэн К Сто лет (1695-1796) в истории изучения мамонта: находки мамонтовых
костей и их объяснения в Западной Европе (пер. с англ. яз. Т.Н.Платоновой). 191V
CONTENTS
Preface............................................................................ 5
Baryshnikov G., Vereshchagin N. A brief review of Quaternary hyenas (Hyaenidae) of
Russia and adjoining regions.......................................................... 7
Baryshnikov G., Averianov A. Deciduous teeth of carnivorous mammals (order
Carnivora). Part VI. Families Viverridae and Herpestidae............................. 66
Averianov A. On the systematic position of rabbit "Caprolagus" brachypus Young, 1927
(Lagomorpha, Leporidae) from the Villafranchian of China............................ 148
Averianov A. Ontogenetic changes in the dentition of Tsaganomys altaicus Mattew et
Granger, 1923 (Rodentia, Tsaganomyidae) from the Oligocene of Central Asia.......... 158
Tikhonov A. Woolly mammoth, Mammuthus primigenius (Proboscidea, Elephantidae)
from Bolshoy Lyakhovsky Island, East Siberia........................................ 167
Arai K., Tikhonov A., Abramov A. Finds of woolly mammoth, Mammuthus primigenius
(Proboscidea, Elephantidae) from Bolshoy Lyakhovsky Island (East Siberia) 187
Cohen. C. A hundred years of Mammoth bone finds and interpretations in Western
Europe (1695-1796) 191
ЧЕТВЕРТИЧНЫЕ И СОВРЕМЕННЫЕ МЛЕКОПИТАЮЩИЕ ЕВРАЗИИ
Труды Зоологического института РАН
Том 270
Утверждено к печати
редакционно-издательским советом
Зоологического института РАН
План 1996 г.
Редактор Т.А.Асанович
Художник О.И.Малова
Изготовитель оригинал-макета В.И.Квачко
Подписано к печати 25.11.96. Формат 60x90 */]б- Печать офсетная.
Бумага офсетная. Гарнитура литер. Печ. л. 13+1 п. л. вкл. Уч.-изд. л. 14.
Тираж 200 экз. Цена договорная.13 25‘-^:
Зоологический институт РАН, 199034, СПб., Университетская наб., 1
ГППП-3, 191104, СПб., Литейный пр., 55