/



































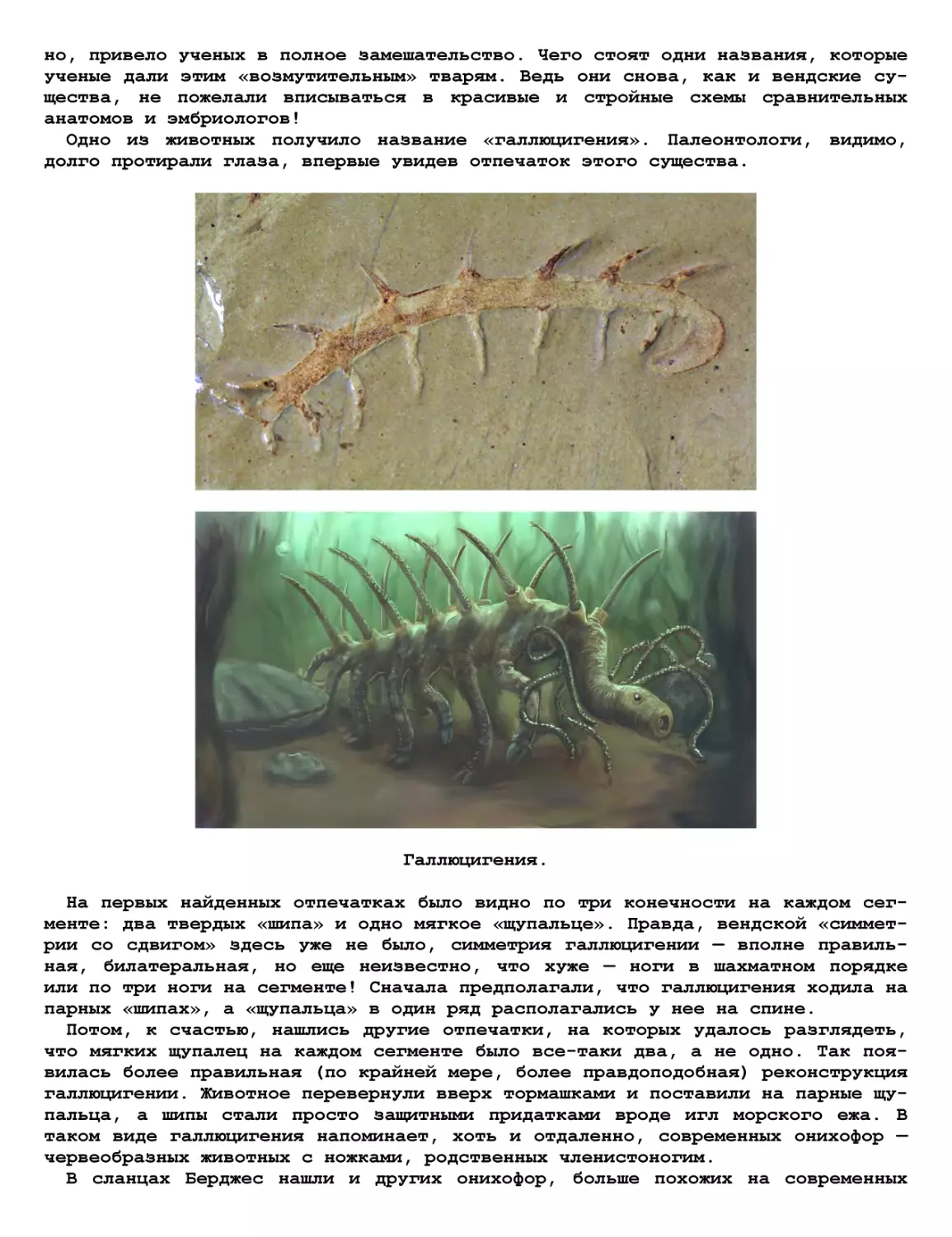
























































































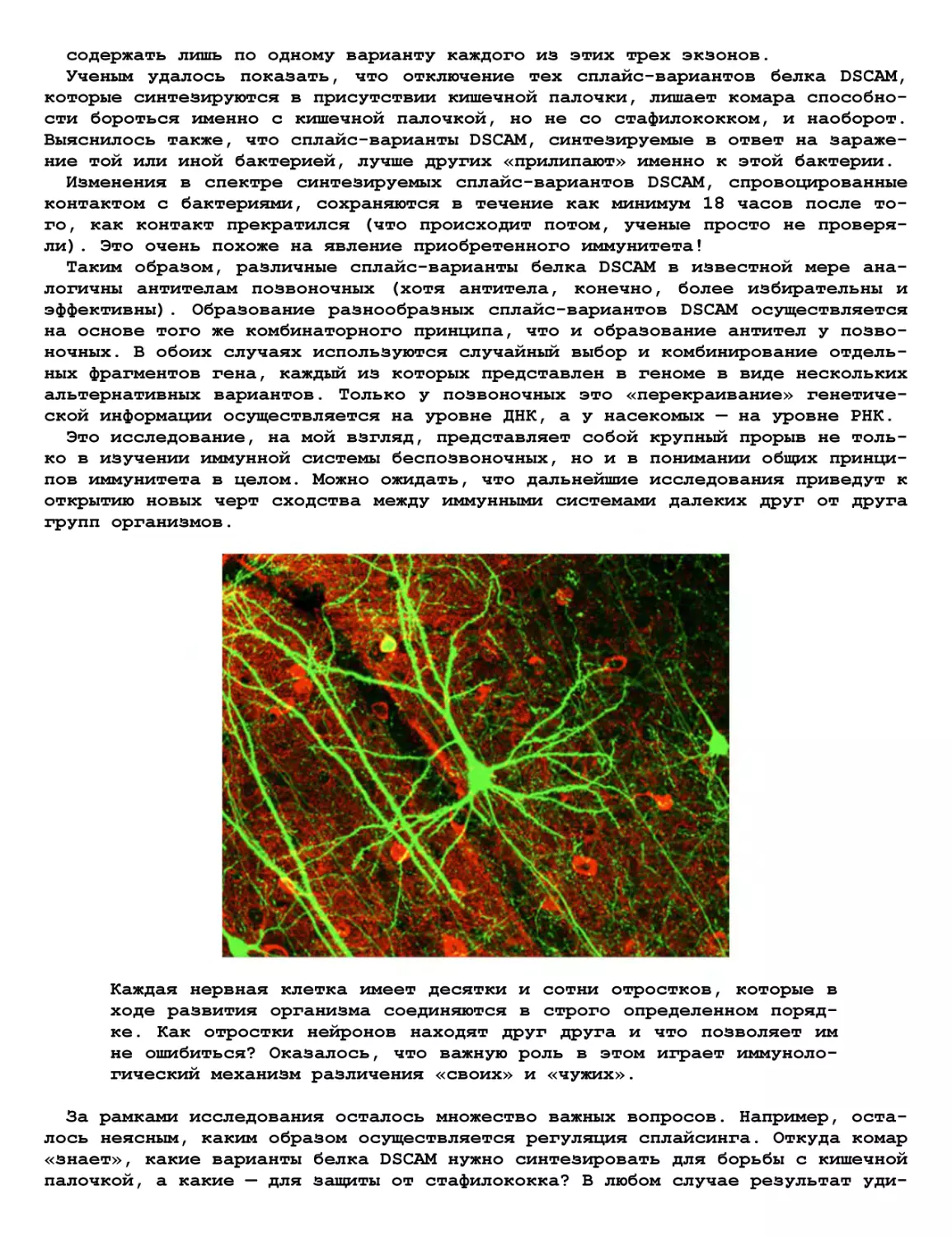
























































































































































































































































































































































































































































































































































































Теги: журнал журнал домашняя лаборатория
Год: 2024




Текст
ИНТЕРНЕТ-ЖУРНАЛ
ЛАБОг*.
ФЕВРАЛЬ 2024
\л}' *
ДОМАШНЯЯ
ЛАБОРАТОРИЯ
Научно-практический
и образовательный
интернет-журнал
Адрес редакции:
homelab@gmx.us
Статьи для журнала направ-
лять, указывая в теме пись-
ма «For journal».
Журнал содержит материалы
найденные в Интернет или
написанные для Интернет.
Журнал является полностью
некоммерческим. Никакие го-
норары авторам статей не
выплачиваются и никакие оп-
латы за рекламу не принима-
ются.
Явные рекламные объявления
не принимаются, но скрытая
реклама, содержащаяся в
статьях, допускается и даже
приветствуется.
Редакция занимается только
оформительской деятельно-
стью и никакой ответствен-
ности за содержание статей
не несет.
Статьи редактируются, но
орфография статей является
делом их авторов.
При использовании материа-
лов этого журнала, ссылка
на него не является обяза-
тельной, но желательной.
Никакие претензии за не-
вольный ущерб авторам, за-
имствованных в Интернет
статей и произведений, не
принимаются. Произведенный
ущерб считается компенсиро-
ванным рекламой авторов и
их произведений.
По всем спорным вопросам следу-
ет обращаться лично в соответ-
ствующие учреждения провинции
Свободное государство (ЮАР).
При себе иметь, заверенные ме-
стным нотариусом, копии всех
необходимых документов на афри-
каанс, в том числе, свидетель-
ства о рождении, диплома об
образовании, справки с места
жительства, справки о здоровье
и справки об авторских правах
(в 2-х экземплярах).
Nft
ЩжШ П-П
- - ^
СОДЕРЖАНИЕ
Рождение сложности (окончание)
Начала органической химии
Биология дрожжей (продолжение)
Закись азота
Некоторые методы органической химии
Февраль 2024
История
Ликбез
136
155
Химичка
197
220
Электроника
Применение Raspberry Pi Pico (продолжение)
Частотомер на Arduino Pro Mini
Датчик направления магнитного поля
Техника резки стекла
236
270
279
Техника
286
Технологии
Выделение чистых культур дрожжей 303
Выделение чистых культур патогенов 338
Техника микробиологической работы 353
Изготовление электровакуумных приборов (продол.) 364
Литье пластмасс в силикон 387
Лаборатория
Химлаборатория инструментального анализа (прод.) 400
От атомов к древу (окончание)
Бетагемот
Очистка молекул и разделение смесей
Слово о спортивных напитках
Мышление
410
Литпортал
529
Разное
668
683
НА ОБЛОЖКЕ
Рисунок к публикации «Техника микробиологической работы».
История
РОЖДЕНИЕ СЛОЖНОСТИ
Марков А.
ГЛАВА 5.
ЖИВОТНЫЕ
Предыдущая глава, посвященная эволюционным механизмам усложнения живых су-
ществ, должна была морально подготовить нас к разговору о той группе эукари-
от, в которой тенденция к усложнению проявилась наиболее ярко. Речь пойдет о
животных. Эта группа нам ближе всех, ведь мы сами к ней относимся. Кроме то-
го, она и по многим объективным характеристикам представляется наиболее инте-
ресной. К таким объективным показателям можно отнести и рекордное разнообра-
зие видов (одних только насекомых описано более миллиона видов — больше, чем
во всех остальных группах живых организмов, вместе взятых), и «господствую-
щее» положение в экосистемах (в том смысле, что животные занимают самые верх-
ние этажи трофической пирамиды), и действительно самый высокий уровень слож-
ности среди всех живых существ.
Начнем с самого начала — с того, как животные появились на свет.
И снова
ископаемая
летопись
Первые этапы эволюционного развития эукариот представлены в палеонтологиче-
ской летописи весьма скудно. В течение протерозойского эона (2,5-0,54 млрд.
лет назад), судя по палеонтологическим данным, разнообразие и численность
микроорганизмов неуклонно росли. К сожалению, далеко не всегда можно уверенно
отличить ископаемых прокариот от одноклеточных эукариот. Ведь от древних мик-
робов ничего не остается, кроме минерализованных (окаменевших) оболочек.
Современные одноклеточные эукариоты обычно раз в десять крупнее прокариот,
но это не абсолютное правило, и никто не знает, насколько строго оно соблюда-
лось в глубокой древности. Самым надежным признаком считается структура кле-
точной оболочки. Например, если она покрыта шипами, можно не сомневаться: пе-
ред нами представитель эукариот. Но далеко не все протисты имеют такие узна-
ваемые оболочки. Древнейшие ископаемые одноклеточные организмы, которых можно
с полной уверенностью отнести к эукариотам, имеют возраст около 2,0-1,8 млрд.
лет.
Строматолиты — слоистые минеральные образования, формирующиеся в результате
жизнедеятельности микробных сообществ — в течение протерозойского зона стано-
вились все более разнообразными и многочисленными. Мы можем уверенно сказать,
что важнейшую роль в строматолитообразующих сообществах в этот период играли
цианобактерии, среди которых встречались формы, внешне не отличимые от совре-
менных. Протерозойские строматолиты достигли высокого уровня сложности: поя-
вились формы со всевозможными ветвящимися столбиками, козырьками, разнообраз-
ной слоистостью и микроструктурой и т. п. Современные строматолиты, образуе-
мые бактериальными матами, устроены намного проще. Из этого следует, что и
протерозойские микробные маты были не в пример сложнее современных. Возможно,
не последнюю роль в них играли одноклеточные эукариоты, но пока это лишь
предположение.
Ископаемые и современные строматолиты.
Похоже, что первые попытки эукариот перейти к многоклеточному уровню орга-
низации начались уже в середине протерозоя. В окрестностях оз. Верхнего (Сев.
Америка) в отложениях возрастом 1,9-1,4 млрд. лет найдены спиралевидные угле-
родистые ленты, получившие название Grypania. Некоторые авторы считают их ос-
татками примитивных многоклеточных эукариотических водорослей.
Другие исследователи считают, что грипания могла быть очень высокоразвитой
колонией цианобактерии. Однако есть и ряд других ископаемых находок примерно
того же возраста, напоминающих многоклеточные водоросли.
Хорошим кандидатом на почетное звание древнейшего животного является
Horodyskia, детально изученная российским палеонтологом М. А. Федонкиным со-
вместно с западными коллегами.
Городискию нашли в отложениях возрастом около 1,44 млрд. лет в Северной
Америке, а также в Австралии, где возраст вмещающих пород составляет 1,4-1,07
млрд. лет. По-видимому, это было колониальное многоклеточное животное, отда-
ленно напоминающее современных гидроидных полипов. Колония имела общий
«ствол» (столон), погруженный в грунт, на котором через равные промежутки си-
дели однообразные округлые «особи»-зооиды.
Городиския1 — самое древнее животное? (Общий вид, отдельный «зооид».)
Может быть, городиския и не была настоящим животным в современном понимании
этого слова. Все современные животные (Metazoa), по-видимому, представляют
собой монофилетическую (имеющую единое происхождение) группу, предком которой
были одноклеточные жгутиконосцы из группы хоанофлагеллят (воротничковых жгу-
тиконосцев) . Однако не исключено, что подвижные одноклеточные гетеротрофные
эукариоты в течение протерозойской эры предпринимали и другие попытки перейти
на многоклеточный уровень организации и превратиться таким образом в живот-
ных. Потомки этих ранних «экспериментаторов» не дожили до наших дней. Это
предположение сделано по аналогии с растениями и грибами. Ведь эти два царст-
ва многоклеточных эукариот откровенно полифилетичны, т. е. каждое из них про-
изошло не от одного, а от нескольких разных одноклеточных предков. Почему же
мы должны лишать подобной вольности животное царство? На сегодняшний день
считается, что одноклеточные эукариоты переходили к многоклеточности более 20
раз, однако, современные животные — результат лишь одного из этих событий.
Последствия всех остальных переходов к многоклеточности «достались» грибам и
растениям. Возможно, городиския в какой-то мере устраняет эту несправедли-
вость. Впрочем, некоторые авторы трактуют городискию не как животное, а как
колонию одноклеточных эукариот (так называемых фораминифер).
К древнейшим животным предположительно можно отнести и пармию (Parmia) —
червеобразное существо, отпечатки которого обнаружены недавно М. Б. Гнилов-
ской и ее коллегами в отложениях возрастом около 1 млрд. лет на северо-
востоке Европейской России. Похожие отпечатки примерно такого же возраста
найдены и в Китае. На некоторых отпечатках видно образование, напоминающее
хоботок (такие хоботки встречаются у современных червей). Однако многие экс-
перты считают, что на самом деле это отпечатки водорослей, а «хоботки» — в
действительности стебельки.
1 М. A. Fedonkin. The origin of the Metazoa in the light of the Proterozoic fossil
record // Paleontological Research. Vol. 7. № 1. E 9-41. March 31, 2003 © by the Pa-
laeontological Society of Japan.
Ископаемое Tappania возрастом 1,43 млрд. лет из Австралии, по-видимому,
представляет собой древнейший многоклеточный гриб, a Bangiomorpha возрастом
1,2 млрд. лет из Северной Канады — практически бесспорная многоклеточная
красная водоросль. Древнейшие бурые водоросли известны из отложений возрастом
1,0 млрд. лет.
Сравнительное изучение нуклеотидных последовательностей ДНК современных ор-
ганизмов показывает, что эволюционные пути растений, грибов и животных разо-
шлись около 1,6 млрд. лет назад. И это, как мы видим, неплохо согласуется с
новейшими палеонтологическими данными.
Однако до настоящего расцвета животных было еще далеко. В эволюции вообще
было много случаев, когда между «изобретением» чего-то нового и его широким
«внедрением», то есть распространением и расцветом той или иной группы орга-
низмов, проходило очень долгое время. То же самое наблюдается и в развитии
человеческого общества (например, между изобретением компаса в Китае и нача-
лом его широкого применения в мореплавании прошли века).
В конце протерозойского эона на Земле наступил ледниковый период, самый су-
ровый за всю историю. Обсуждается даже «теория снежка» (snowball theory), со-
гласно которой ледники в это время покрыли всю планету целиком. В последнее
время, однако, появились данные, показывающие, что ситуация была не столь
устрашающей. Тем не менее, это сильнейшее похолодание, по-видимому, затормо-
зило развитие жизни. Около 635 млн. лет назад ледники наконец отступили, и
начался последний период протерозоя, который называют вендским (это принятое
в отечественной научной литературе название было образовано академиком Б. С.
Соколовым от славянского племени вендов, или венедов, населявших южный берег
Балтийского моря) или эдиакарским. Вот тут-то и началось стремительное и
безудержное развитие животного мира. В глобальном масштабе происходила смена
доминирующих форм жизни в океане. Микробные сообщества, в которых главными
производителями органики были цианобактерии, были потеснены эукариотическими
водорослями, а в портрет обитателей эпохи добавились крупные мягкотелые бес-
позвоночные животные.
Первопоселенцы
суши
Многоклеточная жизнь зародилась в океане. На суше безраздельное господство
микроорганизмов продолжалось еще примерно 150-200 млн. лет после того, как в
море размножились крупные многоклеточные организмы. Первыми обитателями суши,
вероятно, были цианобактерии и актинобактерии. Судя по некоторым косвенным
данным, они могли заселить поверхность материков еще в начале или середине
протерозоя. Гетеротрофные актинобактерии образуют многоклеточные ветвящиеся
структуры, похожие на грибной мицелий. Они способны объединяться с фототроф-
ными цианобактериями в удивительные симбиотические «сверхорганизмы» — так на-
зываемые актинолишайники2. Среди первых обитателей суши были и грибы, которые
тоже вступали в симбиоз с цианобактериями. Генетические и биохимические сис-
темы, развившиеся у сухопутных грибов для симбиоза с цианобактериями, позже
пригодились им для «налаживания отношений» с первыми наземными растениями.
Вся эта наземная микробиота постепенно готовила почву (как в переносном, так
и в прямом смысле) для заселения суши растениями, которые начали осваивать
пустынные берега в конце силурийского периода (около 410-420 млн. лет назад).
Наземные растения с самого начала жили в тесном симбиозе с почвенными гриба-
ми, без которых они, скорее всего, вовсе не смогли бы покинуть родную водную
стихию (см. главу «Великий симбиоз»).
2 см. Г.М. Зенова. Лишайники.
Большинство вендских мягкотелых лишь весьма отдаленно напоминают современ-
ных нам животных3. Некоторые специалисты считают, что между вендской фауной и
«настоящими», то есть более поздними, животными нет или очень мало прямых
родственных связей. Может быть, это был еще один эволюционный эксперимент,
закончившийся тупиком, а настоящие животные появились независимо от таинст-
венных «вендобионтов»? Впрочем, по мнению ряда ведущих палеонтологов, пред-
ставления о том, что вендские животные не имеют ничего общего с современным
животным миром, не вполне соответствуют действительности.
Загадка
древних
эмбрионов4,5
До недавнего времени чуть ли не самыми загадочными и спорными из эдиакар-
ских находок оставались многоклеточные образования из формации Доушаньтуо
(Китай, возраст около 580 млн. лет), похожие на ранних эмбрионов многоклеточ-
ных животных.
Местонахождение Доушаньтуо найдено около 20 лет назад. С ним связаны изуми-
тельные открытия ископаемых докембрийскохю возраста — это самые ранние наход-
ки бесспорных многоклеточных животных: губок, кишечно-полостных, билатераль-
ных червей; известны также и отпечатки крупных водорослей замечательной со-
хранности .
Но взрослые (хоть и микроскопические!) многоклеточные в этих отложениях
редки. Самую обильную часть находок Доушаньтуо составляют похожие на эмбрио-
нов шаровидные образования от одной клетки и более, верхний предел — около
1000 клеток. Все клетки в эмбрионе одинаковы по форме и размерам. В отличие
от водорослевых клеток клетки эмбрионов не имеют толстой стенки и плотно при-
мыкают друг к другу. Внутренней полости (бластоцеля) нет, эпителиального слоя
тоже не найдено. Бластопора (эмбрионального рта) нет даже у эмбрионов из ты-
сячи клеток. И исследователи предполагали, что эмбрионы Доушаньтуо принадле-
жат каким-то очень примитивным многоклеточным животным, возможно, общим пред-
кам губок и кишечнополостных.
Начало 2007 года стало переломным моментом в изучении таинственных эмбрио-
нов. В январе американские ученые опубликовали в журнале Nature статью, в ко-
торой доказывалось, что эмбрионы Доушаньтуо — никакие не эмбрионы, а колонии
гигантских бактерий.
Что же навело ученых на эту мысль? Дело в том, что несколько лет назад в
водах Мексиканского залива, в глубоких бескислородных слоях, были обнаружены
гигантские бактерии Thiomargarita. Их размер — от 0,1 до 0,75 мм, что для
бактерий поистине рекордные показатели. Эти гиганты используют для окисления
сульфидов нитраты, превращая их в азот. Это так называемое «нитратное дыха-
ние», о котором мы немного говорили в главе «Великий симбиоз» (см. раздел
3 Об удивительной фауне вендского периода написано уже достаточно много популярных
книг (см., например, свободно доступную в Интернете книгу Я. Е. Малаховской и А. Ю.
Иванцова «Вендские жители Земли».). О некоторых характерных вендских животных рас-
сказано в главе «Направленность эволюции» в связи с проблемой происхождения члени-
стоногих .
4 Раздел написан Еленой Наймарк.
5 James W. Hagadorn et al. Cellular and Subcellular Structure of Neoproterozoic
Animal Embryos // Science. 2006. V. 314. P. 291-294; Jake V. Bailey et al. Evidence
of giant sulphur bacteria in Neoproterozoic phosphorites // Nature. 2007. V. 445. P.
198-201; Leiming Yin et al. Doushantuo embryos preserved inside diapause egg cysts
// Nature. 2007. V. 446. P. 6611-6663.
«Вместо выделительной системы — микробное сообщество»). Кислород нитратов пе-
реходит на сульфиды, образуя сульфаты. Запасы нитратов в клетке сконцентриро-
ваны в центральной вакуоли. Клетки Thiomargarita окружены гликопротеидным
чехлом.
Иногда гигантские клетки делятся, образуя скопления из двух или более со-
единенных клеток. На поверхности гигантских бактерий почти всегда имеются
симбиотические нитчатые или сферические сульфатредуцирующие бактерии. Им для
жизнедеятельности нужны как раз сульфаты, которые являются продуктом нитрат-
ного дыхания Thiomargarita. Весь этот бактериальный комплекс катализирует
осаждение фосфатов — если геолог в далеком будущем нашел бы места обитания
Thiomargarita, то это были бы богатейшие залежи фосфоритов. Именно таким фос-
форитовым отложением и является местонахождение Доушаньтуо. Так что вполне
логично предположить, что и в древних осадках фосфориты концентрировались при
участии серобактерий.
Также логично предположение, что этими бактериями могли быть организмы, по-
добные Thiomargarita. И где же они в Доушаньтуо? Единственные подходящие кан-
дидаты — наши старые знакомые, принимаемые за яйца и эмбрионы многоклеточных.
Они имеют сходные с серобактериями размеры, совершенно такое же дробление
клеток в трех плоскостях, в результате которого получаются многоклеточные об-
разования с числом клеток, кратным двум. Поверхность клеток у тех и других
гладкая, те и другие окружены фосфатной оболочкой.
Что касается многочисленных симбионтов на поверхности Thiomargarita, то
окаменевшие нитчатые образования видны кое-где и на поверхности древних эм-
брионов . Эти разительные черты морфологического сходства вместе с логичным
объяснением условий накопления фосфоритового осадка заставили научное сообще-
ство внимательно отнестись к смелой гипотезе.
Однако всего через три месяца, в апреле того же года, новые ископаемые на-
ходки положили конец спорам о природе таинственных шариков. На этот раз по-
счастливилось китайским палеонтологам из Нанкинского института геологии и па-
леонтологии, работавшим в компании с Эндрю Ноллем из Гарвардского университе-
та (США). Палеонтологи нашли множество старых знакомцев: двух-, четырех-, ...,
32-клеточных эмбрионов, но заключенных внутрь сложно устроенной оболочки.
Слева — гигантские серобактерии Thiomargarita, справа — ископае-
мые «эмбрионы».
Оболочка состоит из толстостенных ячеек и снаружи орнаментирована многочис-
ленными шипами. Подобные шипастые оболочки характерны для яиц многих совре-
менных беспозвоночных. Стало ясно, что это особая эмбриональная стадия — ста-
дия покоящихся яиц, один из способов, при помощи которого животные могут пе-
реждать неблагоприятные условия. Так что эмбрионы Доушаньтуо теперь практиче-
ски безоговорочно следует признать покоящимися яйцами многоклеточных беспо-
звоночных .
Вторая часть открытия оказалась не менее замечательной. Сами эти оболочки с
шипиками, складками и бугорками на поверхности раньше находили во множестве в
позднепротерозойских породах и относили к группе акритархов рода
Tianzhushania. Акритархи — сборная группа проблематичных ископаемых, большин-
ство из которых, как считалось, представляют собой оболочки цист (покоящихся
стадий) одноклеточных и многоклеточных водорослей. Теперь же стало очевидно,
что полые пузырьки такой формы и поверхностного рельефа — это не что иное,
как оболочки яиц многоклеточных животных. Мало того, если сами эмбрионы нахо-
дят в породах возрастом около 580-550 млн. лет, то их оболочки под видом «ак-
ритархов» известны из самых нижних слоев формации Доушаньтуо, возраст которых
— 632 млн. лет. Это значит, что настоящие, бесспорные многоклеточные животные
«постарели» на несколько десятков миллионов лет. И появились они в древних
морях Китайской платформы всего через 3 млн. лет после окончания великого
позднепротерозойского оледенения.
Самое примитивное
животное на свете
Большинство современных типов животных впервые появляются в ископаемой ле-
тописи лишь в кембрийском периоде (это первый период палеозойской эры, начав-
шийся 542 млн. лет назад). Однако результаты молекулярно-филогенетического
анализа свидетельствуют о существенно более раннем появлении многих из них.
Не стоит забывать, что палеонтология и сравнение последовательностей ДНК — не
единственные источники сведений о происхождении и ранней эволюции животных.
Немало полезной информации можно получить и при помощи классических методов
сравнительной анатомии и эмбриологии. Вплоть до 80-х годов XX века именно эти
методы служили главным инструментом для воссоздания путей эволюции животных,
а палеонтология занимала почетное второе место. Сегодня на первое место вышли
молекулярно-генетические методы, палеонтология осталась на втором (благодаря
своему чрезвычайно бурному развитию в последние десятилетия), а анатомия с
эмбриологией переместились на третье. Но своего значения они не утратили и,
надеюсь, никогда не утратят. Что же говорят о происхождении животных эти
классические науки?
Они говорят, во-первых, что животные наверняка произошли от хоанофлагеллят
— воротничковых жгутиконосцев (и это подтвердилось молекулярными данными).
Во-вторых — что первые животные должны были представлять собой нечто вроде
шарика (или лепешки) из двух типов клеток. Клетки наружного слоя несли жгути-
ки и служили для движения. Внутри помещались клетки, похожие на амеб и выпол-
няющие пищеварительную функцию. Примерно так устроены личинки низших животных
— губок и кишечнополостных. Многие зоологи считали, что именно эти группы
(особенно губки) представляют собой наименее изменившихся потомков общего
предка всех животных, то есть, являются самыми примитивными из современных
представителей животного царства.
Однако на почетное звание самого примитивного животного претендовало еще
одно весьма странное существо — трихоплакс. Это плоское создание, похожее на
медленно ползающую кляксу, не имеет ни осей симметрии, ни мускулатуры, ни пе-
реднего и заднего концов, не говоря уже о таких сложных устройствах, как пи-
щеварительная, нервная, кровеносная или выделительная системы. Трихоплакс по
своему строению напоминает личинок кишечнополостных, и его действительно дол-
го считали личинкой медузы. Но потом оказалось, что трихоплакс образует поло-
вые клетки и размножается половым путем.
Трихоплакс — одно из самых примитивных животных — похож на мед-
ленно ползающую тонкую бесформенную пластинку.
Правда, до сих пор не удалось выяснить, как идет развитие оплодотворенного
яйца: в лабораторных условиях эмбрионы погибают на очень ранних стадиях. В
аквариуме трихоплакс размножается бесполым путем — делится надвое или отпоч-
ковывает маленьких многоклеточных «бродяжек», у которых уже есть все типы
клеток, имеющиеся у взрослого трихоплакса. Тем не менее, наличие полового
размножения доказывает, что трихоплакс — не чья-то личинка, а вполне взрос-
лое , самостоятельное существо. Правда, трихоплакс мог произойти и от более
сложно устроенных животных в результате неотении — редукции поздних стадий
развития и перехода к размножению на стадии личинки. Это явление довольно ши-
роко распространено (один из примеров — знакомый аквариумистам аксолотль, не-
отеническая личинка саламандры).
Окончательно решить вопрос о происхождении трихоплакса и его месте на эво-
люционном древе животного царства мог только тщательный молекулярно-
генетический анализ. В 2006 году американские и немецкие исследователи прочли
небольшую, но очень информативную часть генома трихоплакса, а именно его ми-
тохондриальную хромосому6.
Оказалось, что митохондриальный геном трихоплакса по своему строению зани-
мает промежуточное положение между ближайшими родственниками животных хоаноф-
лагеллятами и всеми остальными животными (включая губок и кишечнополостных).
Митохондриальные геномы хоанофлагеллят велики по размеру (40 и более тысяч
пар нуклеотидов), содержат длинные некодирующие участки и большое число ге-
6 Stephen L. Dellaporta, Anthony Xu, Sven Sagasser, Wolfgang Jakob, Maria A. Moreno,
Leo W Buss, Bernd Schierwater. Mitochondrial genome of Trichoplax adhaerens supports
Placozoa as the basal lower metazoan phylum // PNAS. 2006. V. 103. P. 8751-8756.
нов. Животные, напротив, имеют очень маленькие митохондриальные геномы (14-18
тысяч пар нуклеотидов) с очень небольшими некодирующими вставками и с мини-
мальным числом генов. У трихоплакса, как выяснилось, митохондриальныи геном
по многим важным признакам занимает промежуточное положение.
Таким образом, анализ митохондриальнохю генома подтвердил точку зрения, со-
гласно которой трихоплакс — самое примитивное из современных животных. Он,
безусловно, не является упростившимся потомком губок или кишечнополостных,
чьи митохондриальные геномы сохранили гораздо меньше примитивных черт. Про-
стота организации трихоплакса первична. Объясняется она тем, что из всех мно-
гочисленных и разнообразных потомков «общего предка всех животных» трихоплакс
изменился меньше всех.
Впрочем, окончательные выводы делать пока рано. Позже был прочтен и основ-
ной (ядерный) геном трихоплакса. Результаты его анализа поставили под сомне-
ние выводы, сделанные на основе изучения митохондриальнохю генома. В ядерном
геноме трихоплакса оказалось меньше примитивных черт, чем у губок. Возможно,
самыми примитивными животными являются все-таки губки, хотя самые примитивные
митохондрии сохранились у трихоплакса.
Сравнительная геномика
и ранняя эволюция животных7
Если не учитывать редкую и плохо изученную мелочь вроде трихоплакса, то са-
мыми примитивными животными можно назвать губок, у которых еще нет настоящих
тканей, нервной системы и кишечника. Губки противопоставляются всем прочим
животным — так называемым «настоящим многоклеточным животным» (Eumetazoa).
Среди этих последних самыми примитивными считаются кишечнополостные, или кни-
дарии (к ним относятся медузы, актинии, гидроидные и коралловые полипы), и
гребневики. В пределах Eumetazoa книдарии и гребневики («радиально-
симметричные животные») противопоставляются двусторонне-симметричным
(Bilateria). К двусторонне-симметричным относятся все прочие животные от жу-
ков и червей до морских звезд и человека.
Генетические исследования последних лет показали, что традиционные пред-
ставления о примитивности книдарии, скорее всего, сильно преувеличены. В ча-
стности, оказалось, что у них, как и у высших животных, есть Нох-гены, кото-
рые в ходе индивидуального развития задают полярность зародыша и определяют
план строения, в котором явно проступают черты билатеральной симметрии. Это
подтвердило старую гипотезу, согласно которой общий предок Eumetazoa был дву-
сторонне -симметричным животным. Для того чтобы составить более полное пред-
ставление об этом предке, было необходимо прочесть геном представителя книда-
рии и сравнить его с известными геномами билатерий.
И вот в 2007 году американские ученые сообщили о «черновом» прочтении гено-
ма актинии Nematostella8. Геном состоит из 15 пар хромосом, имеет размер око-
ло 450 млн. пар нуклеотидов (в 100 раз больше, чем у кишечной палочки, и в 6
раз меньше, чем у человека) и содержит примерно 18 000 белок-кодирующих ге-
нов, что вполне сопоставимо с другими животными.
Для каждого гена актинии исследователи пытались найти аналоги в геномах би-
латерий: человека, дрозофилы, круглого червя, рыбы и лягушки. Если аналог (то
есть похожий ген) находился, исследователи делали вывод, что соответствующий
ген имелся у общего предка Eumetazoa. Таким образом удалось составить доволь-
7 Геномика — новый раздел генетики, занимающийся анализом не отдельных генов, а це-
лых геномов.
8 Nicholas H. Putnam et al. Sea Anemone Genome Reveals Ancestral Eumetazoan Gene
Repertoire and Genomic Organization // Science. 2007. V. 317. P. 86-94.
но полное представление о генном репертуаре этого загадочного предка.
Оказалось, что репертуар этот был весьма широк и включал не менее 7766 ген-
ных семейств, сохранившихся и у книдарий, и у билатерий. Человек унаследовал
не менее 2/3 своих генов от общего с актинией предка; сама актиния — примерно
столько же. Муха и круглый червь унаследовали от общего предка с актинией
лишь 50% и 40% генов соответственно.
Актиния Nematostella.
Выяснилось, что в эволюционных линиях позвоночных и книдарий было потеряно
меньше исходных генов и меньше приобретено новых, чем в линии, ведущей к
круглым червям и насекомым. Однако нужно иметь в виду, что применяемые мето-
дики не позволяли отличить действительную потерю гена от его изменения «до
неузнаваемости». Поэтому в целом можно лишь заключить, что в линии позвоноч-
ных геном изменился меньше, чем в линии первичноротых, куда относятся муха и
червь. Из этого следует неожиданный вывод: геном человека оказался в целом
гораздо более похожим на геном актинии, чем геномы мухи и червя. Сходство за-
трагивает не только набор генов, но и порядок их расположения в хромосомах.
Около 80% генов общего предка Eumetazoa имеют явные аналоги за пределами
животного царства. Это значит, что они были унаследованы животными от одно-
клеточных предков (как мы помним, ими были воротничковые жгутиконосцы — хоа-
нофлагелляты). Получается, что геном на удивление мало изменился при станов-
лении животного царства. Среди оставшихся 20% генов, аналогов которых нет у
одноклеточных, имеется большое число ключевых регуляторов развития. Примерно
четверть этих новых генов (то есть 5% от общего числа) содержат участки или
функциональные блоки, встречающиеся у одноклеточных, но в других комбинациях.
Это указывает на один из основных путей создания новых генов: они формируются
из старых путем перекомбинирования фрагментов.
Как и следовало ожидать, значительная часть «новых» генов Eumetazoa выпол-
няет функции, непосредственно связанные с теми новшествами, которые появились
у животных на организменном уровне. Это, прежде всего, гены, отвечающие за
межклеточные взаимодействия и передачу сигналов, за перемещения клеток, регу-
ляцию их деления и другие процессы, играющие ключевую роль в ходе индивиду-
ального развития животных.
Нох-гены — регуляторы
развития животных9
9 Раздел написан Еленой Наймарк.
Ключевую роль в эволюции животных играли изменения сравнительно небольшого
числа генов — регуляторов индивидуального развития, от которых зависит план
строения организма. Главными из этих генов-регуляторов являются так называе-
мые Нох-гены.
Прошло уже больше 50 лет с тех пор, как американский генетик Эдвард Льюис с
удивлением рассматривал мутантную плодовую мушку, у которой на голове вместо
антенн выросли ноги. Эта странная мутация получилась, когда ген, ответствен-
ный за формирование грудных конечностей, включился не в то время и не в том
месте. А Эдвард Льюис (вместе с Кристианой Нюссляйн-Волхард и Эриком Вишау-
сом) получил в 1995 году за исследование этих механизмов эмбриогенеза Нобе-
левскую премию по физиологии и медицине.
Слева: знаменитая дрозофила с ногами вместо антенн; справа: нор-
мальная дрозофила.
Так было открыто семейство Нох-генов, отвечающих за правильное формирование
частей тела у животных. Работа этих генов казалась чудом: вот оно, решение
великой загадки, как из сборища одинаковых эмбриональных клеток в правильном
порядке дифференцируются ткани и органы и в результате получается сложный ор-
ганизм. Необходимо только в нужный момент включать правильный Нох-ген.
Регуляторные Нох-гены у дрозофилы располагаются в хромосоме в довольно
строгом порядке, приблизительно в том самом, в котором происходит дифференци-
ровка основных частей тела двусторонне-симметричного (билатерального) живот-
ного. Сначала у раннего эмбриона начинают работать гены, отвечающие за строе-
ние органов на голове, затем на груди, затем гены начинают оформлять и хво-
стовую часть.
Похожие гены были найдены у мыши и у человека. Даже у этих высокоорганизо-
ванных существ они выполняют ту же работу: отвечают за порядок эмбрионального
развития. Открытие сходных Нох-генов у разных типов животных заставило зооло-
гов и эмбриологов по-новому взглянуть на индивидуальное развитие животных и
его преобразования в ходе эволюции. Стало ясно, что, изменив всего лишь один
ген или время его включения, можно трансформировать, дублировать, удалить или
перенести в другое место сразу целый орган, сохранив при этом общий план
строения.
Кроме того, ученые получили новый мощный инструмент для эволюционных по-
строений — семейство гомологичных (происходящих один от другого) генов, при-
сутствующих у всех животных. Все гипотезы о происхождении и ранней эволюции
животных теперь включают и этот пласт информации.
Нох-гены располагаются на одной или нескольких (до четырех) хромосомах,
обычно тесными группами (кластерами), внутри которых сохраняется более или
менее строгий порядок: «головные» гены впереди, «хвостовые» — сзади. У более
примитивных представителей многоклеточных, таких как гребневики (Ctenophora)
и кишечнополостные (Cnidaria), этих эмбриональных регуляторных генов только
четыре, у млекопитающих их уже 48.
Семейство Нох-генов подразделяется на 14 классов. Считается, что эти 14
классов возникали путем дупликации (удвоения) одного или немногих исходных
генов. «Дубликаты» затем мутировали и обретали новые функции. У примитивных
кишечнополостных и гребневиков имеется всего 4 класса Нох-генов, у предпола-
гаемого общего предка двусторонне-симметричных животных их должно было быть
по крайней мере 8, у млекопитающих присутствуют все 14 классов.
дрозофила (Dm.)
1 2 Зл 3' 3' 4 5 (Г в
лучеперые рыбы
7 Нох-кластеров
много потерянных
генов и укороченнь^
.кластеров
мышь (М.т.)
: 234 56789 Ю 11 12 13 Е*Н<
с £н=**:—с-к=к=->:
круглый червь (Се.)
Ev.Hx
о 6 8 ?•U EvHx
билатеральн
кишечнополостные //у v\
1 EvHx
общий предок билатерально
симметричных животных
Обобщенная схема эволюции основных групп многоклеточных живот-
ных. Внизу показано предполагаемое строение Нох-кластера у обще-
го предка билатеральных животных. У него предположительно име-
лось 8 Нох-генов в едином кластере. Гены нарисованы разными цве-
тами и пронумерованы. Цифры дробью указывают, что данные гены
совмещены, то есть еще не разошлись в ходе эволюции. Некоторые
гены показаны одинаковым цветом, это означает большое сходство в
их нуклеотидных последовательностях. Стрелочки указывают направ-
ление считывания генов на ДНК.
Принцип работы всех Нох-генов одинаков. Все они являются транскрипционными
факторами (регуляторами транскрипции — «прочтения» генов), то есть их функция
состоит во «включении» или «выключении» других генов. В результате работы
Нох-факторов запускается каскад реакций, приводящий к появлению в клетке нуж-
ных белков.
В последние годы представления о Нох-генах существенно изменились и расши-
рились . Этому способствовала расшифровка ДНК-последовательностей Нох-генов у
многих групп животных: кольчатых, плоских и круглых червей, иглокожих, члени-
стоногих, оболочников, ланцетников, не говоря уже о млекопитающих. Под тяже-
стью новых данных обрушилось представление об упорядоченном расположении Нох-
генов в хромосомах. Выяснилось, что, например, у иглокожих первые три Нох-
гена располагаются прямо перед последним (14-м), а начинается кластер с пято-
го гена. У круглых червей и оболочников Нох-гены вообще не образуют кластеров
и их порядок в хромосомах не соблюдается вовсе. Это говорит о том, что пра-
вильный порядок экспрессии («включения») Нох-генов в различных частях эмбрио-
на не обязательно должен соответствовать порядку расположения этих генов в
хромосоме. Последовательность включения Нох-генов зависит помимо «места под
солнцем» от ряда дополнительных факторов, которые сейчас активно изучаются
(но выяснить пока удалось немногое).
Число классов Нох-генов в разных типах животных оказалось разным. Заворажи-
вающее сходство Нох-кластеров у мухи и человека тускнеет, лишь только к ним
прибавляется еще какая-нибудь группа двусторонне-симметричных животных. Так,
у круглых червей (нематод) найдено всего 5 классов из 14, у оболочников — 9,
зато у ланцетника — все 14. У паразитических трематод (класс плоских червей)
обнаружено всего 4 Нох-гена на двух хромосомах, и между этими четырьмя раз-
бросаны другие гены, не связанные с Нох-семейством.
И вот вопрос: почему у двусторонне-симметричных плоских червей Нох-генов
столько же, сколько и у радиально-симметричных кишечнополостных и гребневи-
ков? У кишечнополостных — вроде бы понятно: недостающих четырех классов генов
еще не было, они образовались только после того, как появился гипотетический
предок двусторонне-симметричных животных. Но у червей-то куда делась половина
предковых регуляторных генов, зачем они избавились от них? Или, может быть,
этой недостающей половины не было и у предка? Ответа пока нет. Ясно только,
что даже при потере большого числа Нох-генов оставшиеся могут организовать
эмбриональное развитие сложного билатерального существа.
Зато стали известны детали регуляции самих регуляторных Нох-генов. Между
Нох-генами расположены участки ДНК, прежде считавшиеся бессмысленными. В дей-
ствительности, как оказалось, с них считываются короткие молекулы регулятор-
ных РНК. Некоторые из них усиливают или ослабляют экспрессию (работу) самих
Нох-генов, некоторые косвенно влияют на работу других транскрипционных факто-
ров . В экспериментах показано, что эти микроРНК могут регулировать как сосед-
ний, так и отдаленный Нох-ген.
Так что семейство Нох-генов, главное из главных среди генов-регуляторов,
само не имеет полной власти в своем хозяйстве. За ним следят мелкие «выскоч-
ки» РНК, способные видоизменить экспрессию гена и тем самым замедлить или ус-
корить формирование органа10.
Пожалуй, самый главный вывод, который можно сделать на основе анализа гено-
ма актинии, состоит в том, что уже самые первые представители животного цар-
ства обладали весьма сложным и совершенным «набором рабочих инструментов», то
есть генов, который позволил создать огромное разнообразие сложных многокле-
10 Derek Lemons, William McGinnis. Genomic Evolution of Hox Gene Clusters //
Science. 2006. V. 313. P. 1912-1922.
точных организмов, внося лишь небольшие изменения в базовую программу разви-
тия. Более того, эти изменения, судя по всему, вносились не в «архитектуру»
программы, а только в ее «настройки».
Базовая генетическая программа поведения клетки, имевшаяся уже у первых жи-
вотных, оказалась столь удачной и гибкой, что дальнейшая эволюция животного
царства — в том числе и прогрессивная эволюция, связанная с усложнением орга-
низма, — уже почти не требовала радикальных изменений самой программы. Доста-
точно было лишь небольших изменений в «настройках». Менялись, в частности,
регуляторные участки ДНК, от которых зависит тонкая настройка работы генов-
регуляторов .
«Главный
секрет»
животных
На интуитивном уровне мы привыкли относиться к царству животных как к чему-
то огромному и чуть ли не бесконечно разнообразному. Но в последнее время все
больше появляется фактов, которые показывают, что в действительности животные
(Metazoa) представляют собой весьма специфическую, компактную и генетически
однородную группу организмов. Прочтенный геном актинии — яркое тому свиде-
тельство .
По-видимому, «самый большой секрет» животных, та генеральная идея, на кото-
рой основаны их строение и эволюция, заключается в особой технологии построе-
ния сложного организма из множества изначально одинаковых модулей — клеток.
Суть технологии в том, что благодаря деятельности ряда ключевых генов — регу-
ляторов развития (в том числе Нох-генов) между делящимися клетками складыва-
ется сложная система взаимоотношений, клетки обмениваются сигналами, градиен-
ты концентраций регуляторных веществ задают симметрию и план строения разви-
вающегося организма, и все эти факторы вместе направляют процесс самооргани-
зации, самосборки сложного многоклеточного существа из генетически идентичных
(то есть изначально одинаково «запрограммированных») клеток.
Очень важно помнить, что геном работает не на уровне организма, а на уровне
клетки. По сути дела он реально кодирует лишь биохимию и поведение одной
клетки. Никакой «программы развития организма» в оплодотворенном яйце нет:
там есть программа поведения клетки, не более. Что же касается «программы
развития», то она самозарождается из взаимодействия делящихся клеток уже в
ходе самого развития (примерно так же, как это происходит у Protozoon).
Развитие червей
начинается с хвоста
Нематода (круглый червь) Caenorhabditis elegans — замечательный модельный
объект для изучения индивидуального развития животных, равно как и для многих
других исследований. Достаточно сказать, что в 2002 году Сиднею Бреннеру, ко-
торый первым начал использовать этого крошечного червя в качестве модельного
объекта в генетике и эмбриологии, была присуждена Нобелевская премия «за от-
крытия в области генетического регулирования развития человеческих органов».
Работа, конечно, проводилась на черве, но выводы оказались справедливыми и
для человека — вот вам и еще одно доказательство генетического единства жи-
вотного царства.
Генетики и эмбриологи выявили целый ряд генов, влияющих на развитие червя,
причем многие из этих генов имеются также и у других животных, включая чело-
века. Как мы уже знаем, система генетической регуляции развития, включающая
Нох-гены и ряд других генов-регуляторов, в общих чертах сходна у всех живот-
ных. Однако ученые пока еще очень далеки от полного понимания тех удивитель-
ных механизмов, которые заставляют дробящееся яйцо превращаться не в комок
клеток, а в сложно и тонко организованное многоклеточное существо.
Червь Caenorhabditis elegans — любимец генетиков и эмбриологов.
Одно из главных достоинств С. elegans — прозрачность. Под микро-
скопом хорошо видны на просвет все внутренние органы и даже от-
дельные клетки. Чтобы увидеть во всех подробностях, как из опло-
дотворенной яйцеклетки развивается червячок с пищеварительной и
нервной системами, мускулатурой, органами размножения, достаточ-
но просто положить яйцо С. elegans на предметный столик микро-
скопа и наблюдать за ним в течение 14 часов — именно столько
продолжается эмбриональное развитие этого животного.
Генетический анализ позволяет лишь выявить гены, необходимые (но вовсе не
обязательно достаточные) для тех или иных аспектов дифференцировки клеток.
Например, мутации Нох-генов могут приводить к причудливому перемешиванию при-
знаков, характерных для разных отделов тела (на голове дрозофилы могут вырас-
ти ноги, на заднем сегменте груди — дополнительная пара крыльев). Известны
гены, мутации в которых приводят к полному нарушению процессов клеточной диф-
ференцировки, так что вместо нормального эмбриона действительно образуется
бесструктурный комок клеток. Но чтобы понять, как все эти гены вместе руково-
дят развитием, необходимы эмбриологические эксперименты.
С. elegans — идеальный объект для таких исследований. Развитие этого червя
прослежено во всех деталях — от оплодотворенного яйца до взрослого организма.
В точности известна судьба каждой зародышевой клетки (бластомера); известно,
из какого бластомера образуется каждая из клеток взрослого червя. Надо ска-
зать , что развитие круглых червей, в отличие от многих других животных, очень
строго определено с самого начала. Даже на стадии двухклеточного эмбриона его
бластомеры отнюдь не одинаковы. Разделив их, мы не получим близнецов, как это
бывает у позвоночных. Более крупный передний бластомер АВ дает большую часть
тела, мускулатуру, нервную систему; задний бластомер Р1 — меньшую часть тела,
включая половые органы и кишечник. Таким образом, судьба этих бластомеров из-
вестна уже на двухклеточной стадии. Она предопределена изначальной полярно-
стью яйцеклетки: одному бластомеру достается больше каких-то регуляторных
(сигнальных) веществ, другому меньше.
При этом, однако, бластомеры вовсе не являются замкнутыми системами, изна-
чально запрограммированными на какой-то определенный путь развития. Они не
могут правильно развиваться без контакта с другими бластомерами; они обмени-
ваются между собой разнообразными химическими сигналами, корректируя свое по-
ведение в соответствии с переменами в клеточном окружении.
Маркус Бишофф и Ральф Шнабель из Института генетики Технического универси-
тета в Брауншвейге задались целью выяснить, какой механизм отвечает за прида-
ние эмбриону переднезадней полярности. В экспериментах им чрезвычайно помогло
то обстоятельство, что с ранними эмбрионами С. elegans можно обращаться как с
миниатюрным живым конструктором. Бластомеры можно разделять и перекомбиниро-
вать произвольным образом и даже составлять химерные эмбрионы из клеток раз-
ных особей. До взрослого червя такие конструкции, как правило, дорасти не мо-
гут, но все-таки умирают не сразу и некоторое время развиваются.
Исследователи отделили у четырехклеточных эмбрионов две передние клетки
(«дочки» бластомера АВ), из которых в норме должен развиться почти весь чер-
вяк. Лишенные контакта с задними бластомерами («дочками» бластомера Р1, кото-
рые называются EMS и Р2) , эти клетки делились хаотически и превращались в
аморфный комок без всяких признаков переднезадней полярности.
АВ—* большая часть червя
/
зигота ^ ^ EMS—кишечник
4 Р2 —* половая система
Естественно было предположить, что задние бластомеры выделяют какое-то сиг-
нальное вещество, упорядочивающее деление передних клеток. Так оно и оказа-
лось . «Поляризующим центром» зародыша, как показали эксперименты, служит с (
из него впоследствии развивается половая система). Достаточно приложить клет-
ку Р2 к потомкам АВ хотя бы на пять минут, чтобы клетки «поляризовались». По-
сле этого потомки АВ делятся не случайным образом, а преимущественно в опре-
деленной плоскости, так что в результате из них получается не комок, а удли-
ненный червеобразный зародыш с выраженным передним и задним концом. Задним
концом всегда становится та точка, к которой прикоснулась клетка Р2.
Развитие зародыша С. elegans.
Очевидно, сигнальное вещество, выделяемое клеткой Р2, влияет на ориентацию
плоскости деления у дробящихся бластомеров. Сила этого вещества такова, что,
прикладывая к клеткам два Р2-бластомера в разных местах, ученые получали при-
чудливых монстров с двумя хвостами или L-образно изогнутые зародыши с двумя
взаимно перпендикулярными переднезадними осями. Каждая точка контакта с Р2
превращалась в хвост!
Теперь надо было выяснить, что это за вещество. Ранее было установлено, что
на ориентацию плоскости деления клеток оказывает влияние сигнальный белок
Wnt, один из важнейших многофункциональных регуляторов развития у животных.
Этот белок в иерархии регуляторов стоит в некотором смысле даже выше Нох-
генов. Одни клетки выделяют белок Wnt, другие воспринимают его при помощи
специальных белков-рецепторов, и это приводит к активизации других генов-
регуляторов , в том числе Нох-генов.
Чтобы проверить, действительно ли бластомер Р2 поляризует остальные бласто-
меры при помощи белка Wnt, исследователи использовали мутантные Р2-бластомеры
с выключенным геном Wnt. Оказалось, что такие Р2-бластомеры поляризующим дей-
ствием не обладают. Таким образом, природа сигнала была установлена.
Оставалось понять, каким образом сигнал передается от одних клеток к дру-
гим . Исследователи обнаружили, что клетки, «поляризованные» Р2-бластомером,
сами приобретают способность поляризовать другие клетки. Однако если у них
выключен ген Wnt, этого не происходит. Из этого авторы сделали вывод, что пе-
редача сигнала происходит не путем диффузии сигнального вещества, производи-
мого Р2-бластомером, из клетки в клетку (как у модельного существа Protozoon
из прошлой главы), а по принципу эстафеты. Поляризованные клетки сами начина-
ют производить белок Wnt и таким образом поляризуют своих соседей.
Дополнительные эксперименты показали, что оставшаяся часть зародыша (та,
что получается из бластомера EMS) тоже поляризуется благодаря контакту с Р2.
Таким образом, бластомер Р2 оказался главным организующим центром развивающе-
гося зародыша, определяющим его переднезаднюю полярность. Это важное открытие
германских эмбриологов, а также разработанные ими методики сложных манипуля-
ций с бластомерами открывают новые захватывающие перспективы перед учеными,
стремящимися проникнуть в тайну развития животных.
Данная работа11 очень наглядно показывает, каким образом довольно простые
системы химического «общения» между клетками зародыша позволяют им «самоорга-
низоваться» в сложный многоклеточный организм.
Нужны ли
эмбрионам
гены?
Имеется еще одно странное обстоятельство, затрудняющее понимание природы
индивидуального развития животных. Оно состоит в том, что ранние этапы онто-
генеза, как правило, идут вообще без участия генов, при полностью отключенном
геноме.
Задумаемся, что это значит. Ведь принято считать, что индивидуальное разви-
тие — это постепенная «реализация» той генетической информации, которая за-
ключена в геноме оплодотворенной яйцеклетки (зиготы) и которая в итоге окон-
чательно «воплощается» в строении взрослого организма. Все понимают, что путь
от генотипа к фенотипу труден и извилист, но лишь немногие эволюционисты все-
рьез пытаются объяснить ключевые закономерности эволюции особенностями тех
сложнейших процессов самоорганизации, которые составляют суть онтогенеза (уж
слишком трудна задача). Поэтому обычно проблему пытаются упростить, сведя все
к вопросу о том, каким образом те или иные изменения генотипа (например, му-
тации) могут отразиться на процессе развития зародыша.
Геном традиционно рассматривается как активное начало (в нем все «закодиро-
вано», он «руководит» развитием). Развивающийся зародыш, напротив, считается
чем-то вроде пассивного «результата» деятельности генома. Дело несколько ос-
ложняется тем, что сам геном в процессе онтогенеза явно не самодостаточен: в
разных клетках эмбриона одни гены включаются, другие выключаются в строгой
последовательности, определяемой, в частности, химическими сигналами, которы-
ми обмениваются между собой клетки и ткани зародыша. Кто кем управляет, ста-
Marcus Bischoff, Ralf Schnabel. A Posterior Centre Establishes and Maintains
Polarity of the Caenorhabditis elegans Embryo by a Wnt-Dependent Relay Mechanism //
PLoS Biology. 2006. 4(12): e395.
новится не совсем ясно. Некоторые теоретики по этому поводу даже заявляют,
что геном — это не «программа развития зародыша», а скорее некий набор инст-
рументов, которыми зародыш пользуется (или не пользуется) по своему усмотре-
нию.
Делящаяся яйцеклетка (слева наверху), морула в виде комка клеток
(слева внизу), бластула в виде полого шарика (справа внизу и
вверху). На всех этих стадиях развития животных геном обычно не
функционирует. На рисунке — развитие медузы Aequorea.
Для управления собственными генами зародыш использует множество разных ме-
ханизмов: это и регуляция транскрипции (считывания генов) при помощи малых
РНК и специальных регуляторных белков — транскрипционных факторов (Нох-гены,
как мы помним, тоже кодируют транскрипционные факторы), и особые «эпигенети-
ческие» механизмы, о которых мы подробнее поговорим в главе 8.
Еще одно обстоятельство, которое делает сомнительным тезис о полной и одно-
значной обусловленности онтогенеза геномом зиготы, состоит в том, что, как
уже было сказано выше, у подавляющего большинства животных на ранних стадиях
развития геном вообще не функционирует. Он просто-напросто отключен, все гены
молчат и матричные РНК («считываемые» с генов матрицы для синтеза белка) не
производятся.
Зародыш между тем претерпевает сложные превращения. Яйцеклетка начинает
дробиться, число клеток эмбриона растет в геометрической прогрессии: 2, 4, 8,
16, 32... Наконец формируется однослойный шар из клеток (бластула). Клетки, на-
ходящиеся на одном из полюсов бластулы, мигрируют внутрь, давая начало второ-
му зародышевому листку (энтодерме), из которой позже разовьется кишечник. На
этой стадии двуслойный зародыш называется гаструлой. Только на этом этапе у
многих животных начинают, наконец, включаться гены, унаследованные от папы с
мамой. У других это происходит чуть раньше — на стадии бластулы. И только
млекопитающие — группа, уникальная во многих отношениях, — включают свои гены
еще раньше (например, мышь делает это на стадии двух клеток)
Как удается эмбриону развиваться без всякого «генетического контроля»
вплоть до стадии гаструлы? Почему гены зародыша так долго остаются выключен-
ными? Какие механизмы обеспечивают отключение генов в зиготе, а затем их
своевременное включение13 ?
Образование гаструлы многоклеточного животного. 1 — бластомеры,
2 — бластоцель (первичная полость тела), 3 — энтодерма (внутрен-
ний зародышевый листок), 4 - эктодерма, 5 — бластопор (первичный
рот), 6 — первичная кишка.
Ответ на первый вопрос более или менее ясен. Яйцеклетка содержит большое
количество матричных РНК, унаследованных от материнского организма. Эти мРНК
считываются с материнских генов заблаговременно, в процессе созревания яйце-
клетки. Именно они обеспечивают синтез белков, необходимых для ранних стадий
онтогенеза. В определенный момент материнские мРНК начинают уничтожаться. Это
происходит как раз тогда, когда зародыш начинает сам производить мРНК, то
есть включает свои гены. Этот довольно быстрый процесс замены зародышем мате-
ринских мРНК на свои собственные называется maternal-zygotic transition
(MZT) .
Менее ясен вопрос о том, что движет процессом MZT. Предполагается три воз-
можных механизма.
1. По мере роста числа клеток, из которых состоит зародыш, начинает сказы-
ваться нехватка тех веществ (что бы они из себя ни представляли) , которые не
позволяют генам зародыша включиться. Ранние стадии эмбриогенеза животных не
случайно называют «дроблением»: зигота именно дробится, клетки эмбриона после
каждого деления становятся все мельче, поскольку между клеточными делениями
отсутствует стадия роста клеток. Общее количество цитоплазмы не растет, тогда
как количество клеточных ядер, а следовательно и ДНК, увеличивается в геомет-
рической прогрессии. Если предположить, что яйцеклетка заранее запаслась ка-
кими-то ингибиторами транскрипции, то количество этих гипотетических ингиби-
торов, приходящихся на каждую клетку, должно быстро убывать, и, в конце кон-
цов, их остается так мало, что они уже не могут сдерживать транскрипцию.
Другая уникальная особенность млекопитающих — мощное развитие «геномного имприн-
тинга», механизма, при помощи которого родители целенаправленно влияют на наследст-
венные свойства эмбриона, отчасти — в своих собственных корыстных интересах (см.
главу 8). Вполне возможно, что между этими двумя особенностями существует связь. Ро-
дители могут быть заинтересованы в том, чтобы «отредактированные» ими гены зародыша
включились пораньше.
13 Последующий текст данного раздела основан на статье: Alexander F. Schier. The Ma-
ternal-Zigotic Transition: Death and Birth of RNAs // Science. 2007. V. 316. P. 406-
407.
2. Не исключено, что в зиготе изначально имеет место целенаправленное бло-
кирование некоторых ключевых генов, работа которых инициирует транскрипцию.
Так, показано, что искусственное введение в эмбрион мощных активаторов транс-
крипции (транскрипционных факторов «широкого спектра действия») может вызвать
преждевременное частичное включение эмбрионального генома.
3. Наконец, сама по себе быстрая череда клеточных делений может мешать
транскрипции. Ведь каждому делению должно предшествовать удвоение ДНК (репли-
кация) . В ходе дробления репликация должна происходить по сути дела непрерыв-
но . Между тем известно, что репликация может мешать транскрипции, а во время
клеточного деления (митоза) могут происходить обрыв и уничтожение тех мРНК,
синтез которых еще не закончился. Может быть, клетки эмбриона просто физиче-
ски не успевают транскрибировать свои гены? Искусственное замедление процесса
дробления действительно может вызвать преждевременное включение эмбрионально-
го генома. Кстати, у млекопитающих дробление протекает сравнительно медленно,
не этим ли объясняется раннее включение генов зародыша?
Впрочем, ни одна из этих теорий не объясняет всей совокупности имеющихся
фактов. Например, они не могут объяснить, почему эмбриональные гены включают-
ся не все сразу, а постепенно, в строго определенном порядке.
Ход развития и строение взрослого многоклеточного животного «закоди-
рован» в геноме примерно в той же степени и в том же смысле, в каком
причудливые морозные узоры на стекле «закодированы» в структуре мо-
лекулы воды. В обоих случаях между наследственным кодом и его вопло-
щением (генотипом и фенотипом) лежат сложнейшие, трудно поддающиеся
изучению процессы самоорганизации. Эта аналогия помогает понять, по-
чему генетики, даже имея полные тексты геномов многих видов, так
медленно продвигаются к пониманию «генетических основ» сложных био-
логических объектов и явлений.
Вторым существенным аспектом MZT является уничтожение материнских мРНК.
Тут, как выяснилось, все предусмотрено заранее: материнские мРНК помечены
особой последовательностью нуклеотидов, расположенной на нетранслируемом (то
есть не кодирующем белок) «хвостике» этих молекул. Среди первых генов, кото-
рые эмбрион включает в процессе MZT, находятся гены особых белков и маленьких
РНК, которые распознают эту последовательность, прикрепляются к ней и тем са-
мым инициируют уничтожение материнских мРНК.
До полного понимания всех этих процессов науке еще очень далеко, но дело
движется. Когда основные механизмы генной регуляции в ходе раннего онтогенеза
будут расшифрованы, биологи смогут вплотную заняться следующим принципиальным
вопросом, а именно — зачем все это надо? Почему животные не доверяют своему
геному контроль над ранними стадиями развития, а потом в какой-то момент
вдруг «переключаются» с материнских мРНК на свои собственные?
Между прочим, эта проблема имеет не только теоретическое, но и практическое
значение. Трудности, с которыми сталкиваются работы по клонированию животных,
во многом определяются тем, что мы еще слишком мало знаем о работе генов в
ходе раннего развития. Клонирование животных осуществляется путем пересадки
ядра из соматической (неполовой) клетки одного животного в яйцеклетку друго-
го . Однако для нормального развития необходимо, чтобы гены зародыша поначалу
помалкивали, а они в донорском ядре вполне активны. Цитоплазма яйцеклетки
должна каким-то образом «перепрограммировать» ядро, выключить геном, а потом
в нужный момент снова включить его. Пока мы не знаем, как ей в этом помочь,
больших успехов в клонировании ожидать трудно.
Разгадан механизм
регенерации конечностей
«Генетическая программа» индивидуального развития очень тесно связана с
другой «программой», на основе которой происходит регенерация — заживление
ран и отращивание утраченных частей тела. Изучая регенерацию, ученые одновре-
менно проникают в тайны онтогенеза и наоборот.
Биологи из Испании и США. недавно провели серию генно-инженерных эксперимен-
тов , в результате которых им удалось показать, что регенерация конечностей у
позвоночных идет под управлением тех же регуляторных белков, которые руково-
дят развитием конечностей у эмбриона. Белки, о которых идет речь, образуют
два сигнально-регуляторных каскада, которые называются Wnt/beta-catenin (о
роли белка Wnt в развитии червя Caenorhabditis elegans говорилось выше в сю-
жете «Развитие червей начинается с хвоста») и BMP (по названию ключевого уча-
стника каскада — белка BMP, bone morphogenetic protein).
Для первого эксперимента ученые сконструировали специальный вирус, в геном
которого был встроен ген белка, блокирующего работу Wnt-каскада. Введение ви-
руса аксолотлю снизило способность к регенерации. У аксолотля в норме отре-
занные конечности восстанавливаются полностью, однако, у зараженных искусст-
венным вирусом аксолотлей вместо лапы вырастала лишь заостренная культя без
пальцев.
Этот эксперимент показал, что нормальная работа Wnt-каскада — необходимое
условие регенерации, но является ли она также и достаточным условием? Чтобы
проверить это, ученые поставили эксперимент на шпорцевой лягушке, у которой в
отличие от аксолотля способность к регенерации есть только у головастиков, а
с возрастом эта способность теряется. Был сконструирован еще один вирус, про-
изводящий белок бета-катенин — один из ключевых участников Wnt-каскада. Вве-
дение этого вируса улучшало способность шпорцевых лягушек к регенерации ко-
нечностей на тех стадиях развития, когда эта способность уже ослаблена. Одна-
ко у взрослых лягушек, полностью потерявших способность к регенерации, этого
не наблюдалось. Следовательно, для регенерации необходимо что-то еще, кроме
Wnt-каскада.
Аксолотль — земноводное, сохраняющее способность к восстановлению
утраченных конечностей в течение всей жизни. Это сделало его излюб-
ленным объектом биологов, изучающих механизмы регенерации.
Нормально регенирировавмая после ампутации конечность аксолотля
(слева) и недоразвитая конечность у особи, которой был введен
Axinl — ингибитор Wnt-каскада.
Третьим подопытным объектом стала рыбка данио (Danio rerio). В норме эта
рыбка способна заново отращивать утраченные плавники. Ранее было показано,
что различные ингибиторы Wnt-каскада снижают эту способность (так же как у
аксолотля и шпорцевой лягушки). Чтобы проверить, способно ли активирование
Wnt-каскада усилить способность к регенерации у данио, исследователи исполь-
зовали рыбок, потерявших эту способность в результате мутации. Введение таким
рыбкам бета-катенинового вируса привело к частичному «исправлению» данного
дефекта. На данио исследователям удалось также показать, что для успешной ре-
генерации помимо Wnt-каскада должен нормально работать и сигнально-
регуляторный каскад BMP.
Самое интересное, что оба каскада (Wnt и BMP) руководят и развитием конеч-
ностей у эмбрионов. На клеточно-тканевом уровне оба процесса — регенерация и
развитие конечности в эмбриогенезе — тоже очень сходны. Все эти данные указы-
вают на то, что восстановление утраченных конечностей осуществляется на осно-
ве той же самой программы развития, которая руководит формированием конечно-
стей у эмбриона. И эту программу, по-видимому, можно искусственно «включить»
даже у тех животных, которые в норме вообще не способны восстанавливать утра-
ченные конечности. Ведь программа эмбрионального развития есть у всех!
Ученым удалось подтвердить это предположение в опытах на курином эмбрионе.
Удаление особого многослойного эпителия, расположенного на верхушке зачатка
крыла, приводит к недоразвитию крыла. Однако оказалось, что, если активиро-
вать у цыпленка ген бета-катенина, крыло после такой ампутации развивается
гораздо лучше.
Исследователи сделали из всего этого осторожный вывод14, что сигнально-
регуляторные каскады, управляющие развитием зародыша, вероятно, стали тем
«сырым материалом», на основе которого у некоторых животных развилась способ-
ность к регенерации. Не исключено, что дальнейшее изучение этих регуляторных
систем позволит в будущем «включать» регенерацию у взрослых животных, полно-
стью лишенных способности к восстановлению утраченных частей тела, в том чис-
ле и у человека.
Кембрийский
взрыв
В самом начале кембрийского периода, примерно 542 млн. лет назад, многие
группы животных почти одновременно стали обзаводиться твердым минерализован-
ным скелетом. Поскольку в ископаемом состоянии обычно сохраняются именно та-
кие скелеты, а мягкие части бесследно исчезают, это событие в палеонтологиче-
ской летописи выглядит как внезапное, «взрывное» появление многих групп жи-
вотных (моллюсков, членистоногих, губок, археоциат, брахиопод, к которым не-
сколько позже присоединяются иглокожие, кораллы, мшанки и другие). Отсюда и
общепринятое название этого события — «кембрийский взрыв».
Вся та палеонтология, о которой мы говорили до сих пор, — палеонтология до-
кембрия, то есть изучающая архейский и протерозойский зоны со всеми биомарке-
рами, окремненными цианобактериями, акритархами, городискиями и мягкотелыми
животными венда, — стала интенсивно развиваться лишь сравнительно недавно. До
этого момента докембрийские толщи казались ученым практически мертвыми, не
содержащими почти никаких следов жизни. «Кембрийский взрыв» выглядел внезап-
ным появлением множества разнообразных организмов словно бы из ниоткуда. По-
этому докембрий назвали криптозоем — временем «скрытой жизни», а последний
этап развития биосферы, начавшийся с кембрия и включающий палеозойскую, мезо-
зойскую и кайнозойскую эры, носит название фанерозоя (время «явной жизни»).
Дарвин считал кембрийский взрыв одним из фактов, не укладывающихся в его
теорию постепенных эволюционных изменений. Впоследствии выяснилось, что
«взрыв» на самом деле был не таким уж взрывообразным. Как мы теперь знаем,
предки многих кембрийских групп жили и раньше, но они были по большей части
бесскелетными, мягкотелыми. Именно поэтому палеонтологи долго не могли обна-
ружить их остатков в докембрийских породах.
Загадка кембрийского взрыва, тем не менее, осталась, только теперь речь
идет не о внезапном возникновении как бы «из ничего» многих типов животных, а
о более или менее одновременном появлении у них минерального скелета. Это
могло быть связано с изменениями условий среды. Например, к такому эффекту
могло бы привести резкое уменьшение кислотности воды, в результате чего кар-
14 Yasuhiko Kawakami et al. Wnt/beta-catenin signaling regulates vertebrate limb
regeneration // Genes & Development. 2006. V. 20. P. 3232-3237.
бонат кальция (СаС03) — самый распространенный у животных скелетообразующий
материал — стал хуже растворяться в морской воде и легче выпадать в осадок.
Предложен и целый ряд других объяснений. Хорошие популярные рассказы о кем-
брийском взрыве и теориях, предложенных для его объяснения, можно найти в
книгах А. Ю. Розанова «Что произошло 600 миллионов лет назад» (1986) и К. Ю.
Еськова «История Земли и жизни на ней» (2000)15. Обе книги, к счастью, нахо-
дятся в открытом доступе в Интернете, поэтому повторяться нет необходимости.
Я расскажу лишь об одном недавнем исследовании, которое имеет отношение одно-
временно и к кембрийскому взрыву, и к сравнительной геномике, и к ранней эво-
люции животных, и к некоторым идеям, изложенным выше в этой главе.
Для того чтобы дружно обзавестись карбонатными (то есть сделанными из
СаСОз) скелетами, животным недостаточно было одних лишь благоприятных условий
среды. Нужны были еще специальные гены и ферменты, при помощи которых живот-
ные могли бы контролировать образование и рост кристаллов карбоната кальция в
нужных местах и в нужном количестве.
Важнейшую роль в образовании карбонатных скелетов у животных играют фермен-
ты карбоангидразы, которые примерно в миллион раз ускоряют реакцию превраще-
ния растворенного в воде углекислого газа в гидрокарбонат:
карбоангидраза
С02 + Н20 -> НС03" + Н+
Карбоангидразы широко распространены в живом мире, в том числе и у прокари-
от . Помимо биоминерализации они участвуют в выполнении множества других функ-
ций (регуляция рН, транспорт ионов, выведение С02 из тканей и др.) . До сих
пор оставалось неясным, когда и в какой последовательности разные группы жи-
вотных обзаводились карбоангидразами. Возможно, они уже имели эти ферменты к
началу «кембрийской скелетной революции» — в этом случае надо признать, что
скелетообразование не является первичной функцией карбоангидраз у животных,
но могло быть и так, что независимое приобретение генов карбоангидраз разными
группами организмов как раз и послужило толчком к появлению скелетов.
Для прояснения ситуации очень не хватало геномных данных по самым примитив-
ным многоклеточным животным, таким как губки. Биологи из Германии и Австралии
недавно восполнили этот пробел, изучив скелетообразующие ферменты у примитив-
ной архаичной губки Astrosclera willeyana16. Это настоящее «живое ископае-
мое»: род Astrosclera существует больше 200 млн. лет (с конца триасового пе-
риода) , а по строению своего карбонатного скелета эта губка очень близка к
формам, процветавшим еще в палеозое (так называемым строматопоратам).
Скелет астросклеры состоит из мелких сферических элементов, которые посте-
пенно растут и сливаются друг с другом. Ученые выделили из скелета губки ор-
ганическую фракцию, а из нее — все белки. Три преобладающих белка оказались
карбоангидразами. Исследователи определили их аминокислотную последователь-
ность , а затем по этой последовательности «выудили» из генома и три соответ-
ствующих гена. Это позволило, сравнивая между собой нуклеотидные последова-
тельности генов карбоангидраз примитивной губки и высших животных, чьи геномы
уже прочтены, реконструировать эволюцию этих белков у животных.
Ученые пришли к выводу, что все многочисленные и разнообразные карбоангид-
разы животных происходят от одного предкового белка, который имелся у послед-
него общего предка всех животных. В разных эволюционных линиях ген этой ис-
ходной карбоангидразы неоднократно подвергался независимым дупликациям (уд-
Домашняя лаборатория 2007-10
16 Daniel J. Jackson et al. Sponge Paleogenomics Reveals an Ancient Role for
Carbonic Anhydrase in Skeletogenesis // Science. 2007. V. 316. P. 1893-1895.
воениям). Так возникали различные новые варианты карбоангидраз. «Последний
общий предок всех животных», вне всяких сомнений, жил задолго до кембрийской
скелетной революции. Получается, что животные изначально были хорошо подго-
товлены (преадаптированы) к развитию минерального скелета — у них с самого
начала были ферменты, способные резко ускорить образование карбоната кальция.
Эти ферменты, очевидно, использовались докембрийскими мягкотелыми животными
для других целей — как уже говорилось, у карбоангидраз в животном организме
хватает работы и без скелетообразования. Когда условия среды стали благопри-
ятствовать биоминерализации, разные группы животных не сговариваясь «привлек-
ли» часть своих карбоангидраз к выполнению новой функции.
Astrosclera willeyana.
Совсем недавно была опубликована еще одна интересная работа, посвященная
исследованию геномов губок17.
Оказалось, что у этих примитивных многоклеточных уже есть значительная
часть комплекса так называемых постсинаптических белков, которые у более вы-
сокоорганизованных животных функционируют в нервных клетках и участвуют в
«приеме сигнала». У губок, однако, нет нервных клеток. Зачем же им эти белки?
По всей видимости, они участвуют в обмене сигналами между клетками губки. Жи-
вотное может не иметь нервной системы, но если его клетки совсем не будут
«общаться» друг с другом, это будет уже не животное, а скопление одноклеточ-
ных организмов. Позже, когда у животных развилась нервная система, эти «ком-
муникационные» белки пригодились для формирования системы обмена сигналами
между нервными клетками. Этот пример, как и множество других, показывает, что
большинство эволюционных новшеств возникает не на пустом месте, а собирается
из «подручного материала», причем часто для радикального изменения функции
какого-нибудь белка или белкового комплекса достаточно совсем небольших гене-
тических изменений.
Onur Sakarya et al. A Post-Synaptic Scaffold at the Origin of the Animal Kingdom
// PLoS One. 2007. 2(6): e506.
ГЛАВА 6.
НАПРАВЛЕННОСТЬ
ЭВОЛЮЦИИ
Случайность и
закономерность
Одно из центральных утверждений синтетической теории эволюции (СТЭ) состоит
в том, что все эволюционные изменения основаны на отборе случайных, ненаправ-
ленных мутаций. Однако эволюция совсем не похожа на случайный, хаотический
процесс. В ней явно есть направленность. Например, очевидно, что господствую-
щие на Земле формы жизни постепенно становятся все более сложными. Как же
так? Может ли направленный и как будто вполне осмысленный процесс идти на ос-
нове случайных, хаотических наследственных изменений — мутаций? В более общей
форме этот вопрос можно сформулировать так: каково соотношение случайности и
закономерности в эволюции? Этот вопрос был и остается одним из центральных в
теоретической биологии и одновременно одним из самых спорных. Отчасти мы уже
ответили на него в главе 4, где говорилось, в частности, о том, что естест-
венный отбор — то есть избирательное запоминание системой возникающих измене-
ний — неизбежно придает эволюции упорядоченный, направленный характер.
Особенно ярко проявляется закономерный характер эволюции в явлении паралле-
лизма. Так называют те случаи, когда у разных организмов независимо возникают
похожие признаки. Мы уже вскользь упоминали об этом явлении, когда говорили о
приспособлении бактерий к антибиотикам (глава 4) . Пришло время поговорить о
параллелизмах подробнее.
Некоторые примеры общеизвестны. Так, представители разных классов позвоноч-
ных животных, перешедшие к активному плаванию в толще воды, независимо друг
от друга приобрели обтекаемую рыбообразную форму тела и плавники (рыбы, их-
тиозавры, дельфины). Еще более интересны примеры возникновения почти идентич-
ных наборов «жизненных форм» на разобщенных континентах. Например, Южная Аме-
рика долгое время была изолирована от других материков, но еще до отделения
туда проникли примитивные плацентарные млекопитающие кондиляртры — «древние
копытные». Эволюция кондиляртр протекала далее независимо в Южной и Северной
Америке, причем последняя время от времени соединялась с Евразией сухопутным
мостом — Берингией. В результате на «Большой земле» (в Северной Америке и Ев-
разии) кондиляртры дали начало отрядам парно- и непарнокопытных, куда отно-
сятся всем известные лошади, носороги, антилопы, олени, гиппопотамы и т. д. В
Южной Америке расцвели другие отряды копытных, произошедшие от тех же конди-
ляртр — нотоунгуляты18, литоптерны, астрапотерии и пиротерии. Среди этих «юж-
ных копытных» появились формы, поразительно схожие с настоящими лошадьми, но-
сорогами, верблюдами, зайцами и даже слонами. В Южной Америке процветали так-
же и сумчатые, быстро вымершие на «Большой земле». Среди южноамериканских
сумчатых тоже появились жизненные формы, удивительно похожие на некоторых се-
вероамериканских и азиатских зверей. Один из ярких примеров — сумчатый сабле-
зубый тигр тилакосмилус, аналог настоящих саблезубых тигров, которые относи-
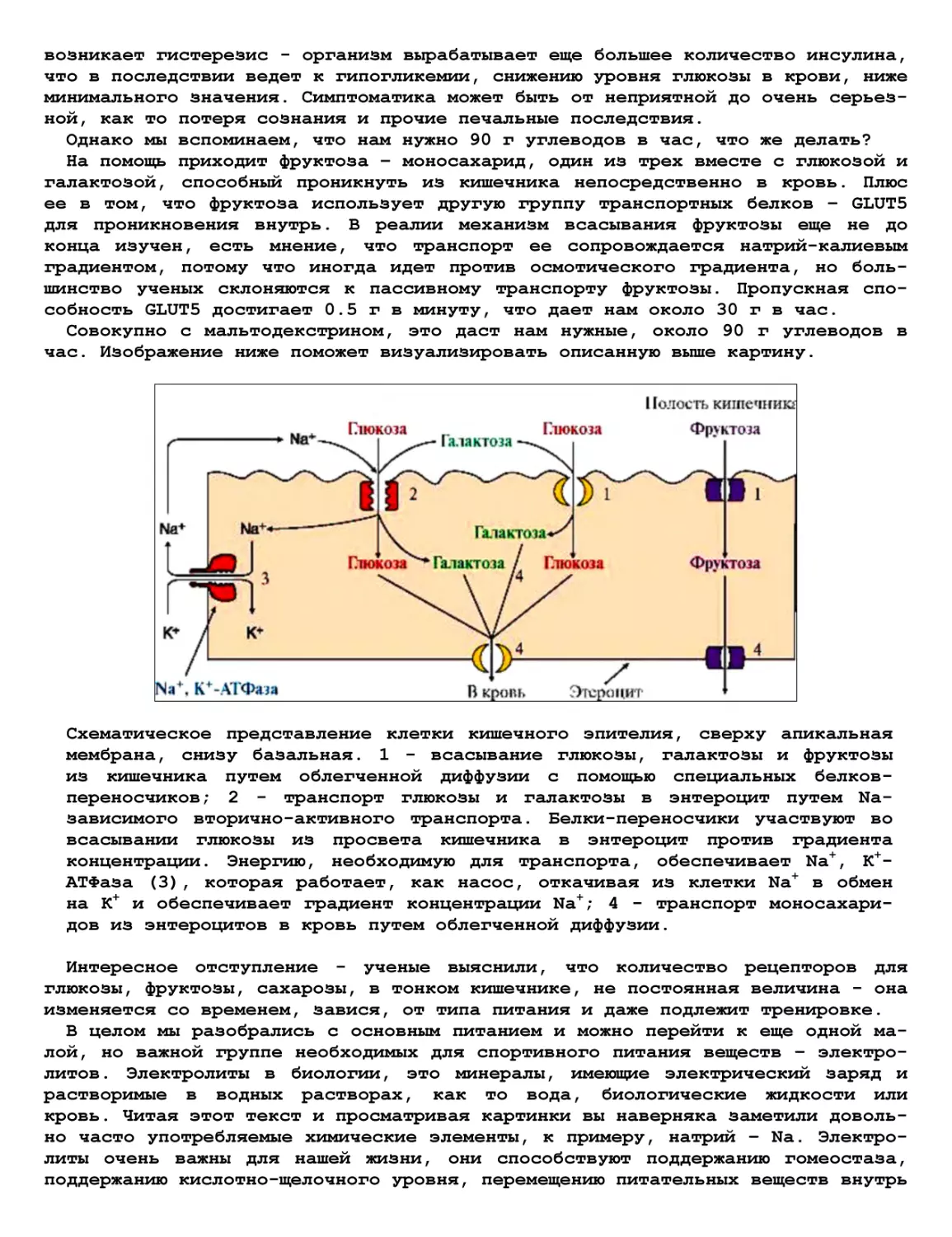
лись к семейству кошачьих и охотились на крупных толстокожих копытных на про-
сторах Азии, Африки и Северной Америки. Тилакосмилус, хоть и был сумчатым,
вероятно, занимал ту же экологическую нишу и охотился на крупных толстокожих
Нотоунгуляты, по-видимому, появились в Азии, а оттуда проникли в Северную и Южную
Америку еще от ее отделения. Но на «Большой земле» нотоунгуляты быстро вымерли, вы-
тесненные более совершенными группами, а в Южной Америке сохранились, размножились и
дали много разнообразных, удивительных форм.
«южных копытных» .
Впрочем, не стоит преувеличивать это сходство. В Южной Америке появились и
такие звери, аналогов которым среди обитателей «Большой земли» найти не уда-
ется. Это броненосцы, муравьеды, ленивцы, к которым относятся и удивительные
гигантские наземные ленивцы мегатерии. Все эти звери не похожи ни на кого,
только на самих себя. В целом южноамериканские звери были менее совершенными,
чем их аналоги с «Большой земли». Например, сумчатые хищники вроде тилакосми-
луса все-таки не могли сравниться в ловкости и сообразительности с «настоящи-
ми», несумчатыми, львами и тиграми. Поэтому с южноамериканскими хищными мле-
копитающими успешно конкурировали гигантские хищные нелетающие птицы форора-
косы. Кроме того, там появились самые крупные летающие птицы всех времен. Не-
которые из них, такие как аргентавис, достигали в размахе крыльев семи с по-
ловиной метров при весе до 80 кг и, возможно, могли проглотить целиком зверя
размером с зайца. Совсем недавно в Патагонии был найден самый большой в мире
птичий череп. Его длина — 71,6 см.
Самый большой
птичий череп
всех времен
Фороракосы — крупные нелетающие хищные птицы, появившиеся в Южной Америке
около 62 млн. лет назад, то есть почти сразу после вымирания динозавров. Мно-
гие палеонтологи полагают, что между этими двумя событиями существует тесная
связь. Исчезновение динозавров освободило нишу двуногого бегающего хищника,
которая активно эксплуатировалась разнообразными хищными динозаврами в тече-
ние всей истории этой группы рептилий. После вымирания динозавров эта ниша
была немедленно освоена их ближайшими родственниками — птицами. Семейство фо-
роракосов, последние представители которого вымерли около 2 млн. лет назад,
включает самых крупных птиц всех времен. Их рост мог достигать трех метров и
более. Однако многие детали строения крупных фороракосов остаются неизвестны-
ми из-за фрагментарности имеющихся находок.
Titanis walleri — родственник фороракоса, живший 2 млн. лет на-
зад в Техасе и Флориде. На передних конечностях у него имелась
пара пальцев, образующих подобие клешни.
Увлекательная история эволюции южноамериканских животных описана в знаменитой
книге Дж. Симпсона «Великолепная изоляция».
В 2006 году палеонтологи нашли в Аргентине неплохо сохранившийся череп фо-
роракоса длиной 716 мм. Это самый большой птичий череп из всех известных нау-
ке. Возраст находки — 10-15 млн. лет (средний миоцен). Вместе с черепом най-
дены кости ногЛ которые оказались неожиданно длинными и тонкими. Это противо-
речит прежним представлениям о том, что крупнейшие представители фороракосов
были тяжеловесны и медлительны. По всей видимости, эти чудовищные птицы на
самом деле были очень быстрыми бегунами20.
Несмотря на удивительное разнообразие и кажущуюся высокую приспособленность
южноамериканских зверей, многие из них не выдержали конкуренции с североаме-
риканскими переселенцами: после воссоединения Южной и Северной Америк, а это
произошло в плиоцене, многие северные виды отправились осваивать южные терри-
тории. Большинство южноамериканских хищников и копытных были вытеснены при-
шельцами и вымерли.
В этом проявилось одно из «правил» эволюции, которому пока нет однозначного
объяснения. На больших территориях быстрее идут эволюционный прогресс и рост
конкурентоспособности, тогда как на малых территориях, в том числе на изоли-
рованных островах, быстрее происходит появление всевозможных странных, не-
обычных, причудливых форм. Это обстоятельство биологи подметили уже давно, но
лишь недавно сумели его подтвердить строгими цифрами.
Эволюция на
островах идет
быстрее
Еще со времен Дарвина, исследовавшего галапагосских вьюрков, удаленные ост-
рова считаются своеобразными «эволюционными лабораториями», в которых виды
претерпевают быстрые и порой весьма изощренные эволюционные изменения. В ча-
стности, известно, что крупные животные, попав в изолированные и обедненные
островные экосистемы, часто мельчают, а мелкие, наоборот, достигают гигант-
ских размеров, причем в ряде случаев на это требуется совсем немного времени.
Однако до сих пор представления об ускоренной эволюции на островах не имели
строгого количественного обоснования, а отдельные факты и вовсе им противоре-
чили: например, ископаемому сицилийскому слону Elephas falconeri, рост кото-
рого был менее метра, а масса (100 кг) составляла лишь 1% от массы его конти-
нентального предка, потребовалось очень много времени (около 300 тысяч лет),
чтобы так измельчать.
Чтобы точно измерить темпы эволюционных изменений на островах и сравнить их
с аналогичными данными по материковым популяциям, необходимо было собрать
весьма значительный объем палеонтологических данных, причем весь материал
должен был быть точно датирован. К настоящему времени палеонтология уже нако-
пила достаточно данных для такого анализа, что и позволило Вирджинии Миллиен
из Университета Макгилла (Канада) получить строгие оценки темпов эволюции на
островах и на «Большой земле».
Миллиен обработала данные по 86 островным и 84 континентальным популяциям
млекопитающих, относящихся к 88 видам и 14 отрядам. Для каждой популяции была
подсчитана скорость эволюционных изменений одного или нескольких размерных
признаков (от 1 до 57 признаков для каждой популяции, в среднем 5,06 признака
на популяцию) в течение определенного интервала времени (от 20 до 12 000 000
лет, средняя величина интервала — 462 тысячи лет). Использовались только ли-
нейные признаки (длина черепа, отдельных костей, зубов и т. д.).
20 Luis M. Chiappe, Sara Bertelli. Skull morphology of giant terror birds // Nature.
2006. V. 443.
Выяснилось, что морфологическая эволюция на островах действительно идет бы-
стрее, однако этот эффект заметен лишь на временных интервалах менее 45 тысяч
лет. При увеличении временного интервала выше этого порога различия между
островными и континентальными популяциями становятся статистически незначимы-
ми.
Выявленные различия нельзя объяснить разным таксономическим составом ост-
ровной и континентальной биоты (то есть тем, что на островах некоторые группы
млекопитающих встречаются чаще, чем на континенте, и наоборот). Оказалось,
что темпы эволюции не являются «филогенетически консервативными». Например,
виды, эволюционировавшие на континенте медленнее других, вовсе не обязательно
будут продолжать вести себя сходным образом и на изолированном острове.
Гигантский варан, обитающий на островах Комодо, Ринджа, Падар и
Флорес (Индонезия) — один из общеизвестных случаев «островного
гигантизма».
По-видимому, млекопитающие, попавшие на удаленные острова, очень быстро
приспосабливаются к новым условиям, что порой сопровождается разительными из-
менениями размеров и пропорций тела. Из-за высокой скорости этих изменений в
большинстве случаев палеонтологам не удается обнаружить переходных форм между
островными эндемиками и их континентальными предками. После относительно ко-
роткого периода быстрой адаптации темп эволюции островных животных, как пра-
вило, замедляется (поэтому не удалось выявить различий в темпах эволюции на
временных интервалах свыше 45 тысяч лет).
Фрагментация естественных местообитаний (в том числе и в результате дея-
тельности человека) может ставить животных в условия, аналогичные попаданию
на изолированные острова. В этом случае можно ожидать резкого ускорения эво-
люционных изменений по «островному» сценарию. Реальность таких явлений под-
тверждается фактами. Например, было показано резкое ускорение эволюционных
изменений размеров тела за последние 200 лет у 25 видов млекопитающих в Да-
нии. И это явление связывают с фрагментацией местообитаний.
Полученные результаты21 показывают, что многие виды континентальных млеко-
питающих потенциально способны к очень быстрым эволюционным преобразованиям.
Темп эволюции может возрасти в три и более раза всего за пару десятилетий.
Получается, что в условиях растущей антропогенной нагрузки на природные эко-
21 Milleien V. 2006. Morphological Evolution Is Accelerated among Island Mammals.//
PLoS Biol 4(10): e321.
системы многие привычные нам животные могут начать изменяться самым неожидан-
ным и быстрым образом.
Но вернемся к теме параллелизмов. В последние десятилетия существенно изме-
нились представления о том, каким образом происходили в ходе эволюции круп-
нейшие прогрессивные изменения — ароморфозы22. Оказалось, что практически во
всех хорошо изученных случаях переход на более высокий уровень организации
происходит не в какой-то одной эволюционной линии, а в нескольких, развиваю-
щихся параллельно. При этом отдельные признаки, из которых складывается аро-
морфоз, иногда появляются в разных линиях почти одновременно, а иногда — в
разное время и даже в разном порядке. Прогрессивные признаки постепенно нака-
пливаются, пока, наконец, в одной или немногих линиях они не соберутся все
вместе, и тогда палеонтологи «объявляют» о рождении новой группы.
В этом ярко проявляется закономерный характер эволюции. В какой-то момент
словно начинает носиться в воздухе новая «идея» — например, идея млекопитаю-
щих. И многие разные группы не сговариваясь начинают развиваться в одном и
том же направлении, хотя и немножко разными путями. Чтобы подчеркнуть это об-
стоятельство, многие биологи в последние годы стали называть происхождение
млекопитающих «процессом маммализации зверообразных рептилий», происхождение
птиц — «орнитизацией», амфибий — «тетраподизацией», членистоногих — «артропо-
дизацией», покрытосеменных растений — «ангиоспермизацией» и т. д.
Происхождение
членистоногих
В «домолекулярную эру» в распоряжении ученых было три научных дисциплины,
при помощи которых можно было реконструировать эволюционную историю организ-
мов :
1. сравнительная анатомия,
2. сравнительная эмбриология,
3. палеонтология.
В последние десятилетия добавился еще один чрезвычайно мощный метод, осно-
ванный на сравнении нуклеотидных последовательностей ДНК и аминокислотных по-
следовательностей белков. Молекулярный метод сильно потеснил первые две дис-
циплины (но не третью) , потому что он позволяет работать одновременно с ог-
ромным количеством признаков. По сути дела, каждый нуклеотид в молекуле ДНК —
это отдельный признак, который можно использовать в эволюционных исследовани-
ях наряду с такими классическими признаками, как строение черепа или ранние
стадии развития эмбриона.
Еще в конце XIX — начале XX века анализ эмбриологических и анатомических
данных привел к появлению двух основных гипотез происхождения членистоногих.
К этому типу животных относятся ракообразные, насекомые, многоножки, паукооб-
разные и трилобиты. Согласно первой из них, предки членистоногих были похожи
на типичную личинку ракообразных — науплиус, тело которого состоит из трех
сегментов. Вторая гипотеза, ставшая в какой-то момент почти общепризнанной,
предполагала, что предками членистоногих были многосегментные животные с мно-
жеством однотипных парных конечностей, близкие к современным многощетинковым
кольчатым червям — полихетам. Происхождение членистоногих от кольчатых червей
казалось большинству ученых совершенно очевидным. Слишком уж бросается в гла-
за сходство многоножки или гусеницы с кольчатым червем, и даже состоящие из
По определению крупнейшего отечественного эволюциониста И. И. Шмальгаузена, аро-
морфоз — это «расширение жизненных условий, связанное с усложнением организации и
повышением жизнедеятельности».
двух «веточек» ножки примитивных членистоногих (ракообразных, трилобитов)
удивительно похожи на двуветвистые конечности полихет. В замечательном учеб-
нике В. А. Догеля «Зоология беспозвоночных», по которому училось несколько
поколений отечественных биологов (включая автора этих строк) и продолжают
учиться до сих пор, написано буквально следующее: «Происхождение типа
Arthropoda в общих чертах ясно. Предками их были примитивные полимерные коль-
чатые черви из класса многощетинковых (Polychaeta). Сходство плана строения
членистоногих и кольчецов настолько велико, что некоторые современные зоологи
предпочитают объединять их, как это делали свыше 100 лет назад, в один тип
членистых — Articulata»23.
Такая степень уверенности в вопросе о том, кто от кого произошел, встреча-
ется у Догеля ох как нечасто. И надо же такому случиться — из множества гипо-
тез, основанных на данных анатомии и эмбриологии, именно эта, по всей видимо-
сти , оказалась неверной.
Ни палеонтологические, ни молекулярные данные не дали подтверждения этой
стройной и логичной теории. Вместо ожидаемых переходных форм между полихетами
и членистоногими были обнаружены весьма причудливые создания со странными со-
четаниями признаков.
Поначалу биологов «обнадежила» сприггина — мягкотелое животное вендского
периода (630-542 млн. лет назад; см. главу «Животные»), которое вроде бы име-
ло ожидаемое сочетание признаков: цельную «голову» в форме полумесяца, напо-
минающую головной щит трилобита, и длинное тело, состоящее из одинаковых сег-
ментов с длинными придатками, напоминающее тело многощетинковых червей.
Сприггина (вендский период) могла бы считаться переходной формой
между кольчецами и членистоногими, если бы имела нормальную би-
латеральную симметрию.
В. А. Догель. Зоология беспозвоночных. Седьмое издание. М. : Высшая школа, 1981.
с. 430.
Но если внимательно присмотреться к отпечаткам сприггины, можно заметить,
что конечности у нее располагаются не попарно друг напротив друга, как у всех
без исключения кольчатых червей и членистоногих, а в шахматном порядке (как
если бы левую половину ее тела сдвинули относительно правой на пол сегмента
вперед или назад). Такая «билатеральная симметрия со сдвигом» специалистам по
сравнительной анатомии казалась совершенно немыслимой. Непонятно, как росло
тело сприггины, как формировались сегменты (и вообще, можно ли их назвать
сегментами?), как были устроены ее кровеносная и нервная системы, структура
которых определяется у полихет механизмом формирования сегментов.
Еще лучше эта «невероятная» сегментация видна у других вендских форм, похо-
жих на примитивных членистоногих, — например, у вендии.
Вендия — «членистоногое» с ногами в шахматном порядке.
Многие другие вендские животные, напоминающие кольчатых червей или прими-
тивных членистоногих, «сдвинуты» таким же образом. Состоят ли они в родстве с
настоящими кольчецами и членистоногими или те произошли от каких-то других
предков? Возможно ли эволюционное превращение «сдвинутой» симметрии в нор-
мальную? Окончательных ответов пока нет.
Вот еще одно животное со сдвинутой симметрией — дикинсония. Найден отпеча-
ток ее пищеварительной системы, и видно, что отростки кишечника тоже распола-
гались в шахматном порядке, в соответствии с сегментацией. Ясно, что это и не
членистоногие, и не полихеты. Палеонтолог М. А. Федонкин выделил этих живот-
ных в особый вымерший тип проартикулят (Proarticulata).
Забавно, что очень похожая сегментация имеется и у прикрепленных, стебель-
чатых вендских животных — петалонам, которые уже совсем не похожи ни на коль-
чецов, ни на членистоногих.
Еще одно животное со «сдвинутой» метаметрией — дикинсония.
Что ж, если вендская мягкотелая фауна дала больше новых загадок, чем отве-
тов на старые вопросы, может быть, уникальные находки более поздних кембрий-
ских мягкотелых животных скажут нам больше?
Кембрийский период (488-542 млн. лет назад) был временем становления боль-
шинства важнейших групп многоклеточных животных. Как мы помним из главы «Жи-
вотные», в самом начале кембрия произошла скелетная революция — в палеонтоло-
гической летописи внезапно появляется множество разнообразных существ, обла-
дающих твердыми скелетными элементами: различными раковинками, шипами, спику-
лами и т. д. В венде животные были в основном мягкотелыми, и среди них мало
кого можно с уверенностью отнести к тому или иному известному типу. Но уже в
первой половине кембрия разнообразие животных резко выросло, и среди них поя-
вились представители практически всех современных типов (членистоногие, мол-
люски , брахиоподы, хордовые, иглокожие и т. д.).
«Перистое» вендское ископаемое — чарния.
Одно из самых богатых местонахождений кембрийских мягкотелых называется
сланцы Берджес (Burgess Shale) и находится в Канаде. То, что было там найде-
но, привело ученых в полное замешательство. Чего стоят одни названия, которые
ученые дали этим «возмутительным» тварям. Ведь они снова, как и вендские су-
щества, не пожелали вписываться в красивые и стройные схемы сравнительных
анатомов и эмбриологов!
Одно из животных получило название «галлюцигения». Палеонтологи, видимо,
долго протирали глаза, впервые увидев отпечаток этого существа.
Галлюцигения.
На первых найденных отпечатках было видно по три конечности на каждом сег-
менте: два твердых «шипа» и одно мягкое «щупальце». Правда, вендской «симмет-
рии со сдвигом» здесь уже не было, симметрия галлюцигении — вполне правиль-
ная, билатеральная, но еще неизвестно, что хуже — ноги в шахматном порядке
или по три ноги на сегменте! Сначала предполагали, что галлюцигения ходила на
парных «шипах», а «щупальца» в один ряд располагались у нее на спине.
Потом, к счастью, нашлись другие отпечатки, на которых удалось разглядеть,
что мягких щупалец на каждом сегменте было все-таки два, а не одно. Так поя-
вилась более правильная (по крайней мере, более правдоподобная) реконструкция
галлюцигении. Животное перевернули вверх тормашками и поставили на парные щу-
пальца, а шипы стали просто защитными придатками вроде игл морского ежа. В
таком виде галлюцигения напоминает, хоть и отдаленно, современных онихофор —
червеобразных животных с ножками, родственных членистоногим.
В сланцах Берджес нашли и других онихофор, больше похожих на современных
(Aysheaia).
Айшеайя. Голова находится справа, видны приротовые придатки.
Возможно, айшеайя питалась губками, потому что ее остатки часто
находят вместе с остатками губок.
Но все «приключения» галлюцигении меркнут по сравнению с уникальной судьбой
другого среднекембрийского монстра — аномалокариса. Отдельные части тела это-
го хищного чудовища (некоторые экземпляры достигали почти двух метров в дли-
ну) сначала находили по отдельности и описывали как разные организмы. Удиви-
тельная «медуза» с дыркой посередине оказалась ротовым диском аномалокариса.
Внутренний край «дырки», как выяснилось, был усажен острыми зубами.
Аномалокарис. Справа - реконструкция.
Членистые образования, изначально описанные как креветки, оказались хвата-
тельными конечностями необычного хищника.
Только когда были найдены первые полные отпечатки аномалокариса, ученые по-
няли , как было устроено это существо. Сегментированное тело с «плавательными
лопастями» напоминает некоторых кольчатых червей. Однако передние хвататель-
ные конечности (бывшие «креветки») и крупные глаза на стебельках — в точности
как у членистоногих.
Мягкотелая фауна сланцев Берджес и других подобных местонахождений в Китае,
США, Гренландии и Сибири открыла перед учеными целую галерею удивительных су-
ществ с разнообразными причудливыми сочетаниями признаков членистоногих и
червей (причем не кольчатых, а других — так называемых хюловохоботных). Все
это очень мало похоже на постепенный, плавный переход от одного типа к друго-
му. Вот, например, опабиния (вверху слева) — животное, явно близкое к анома-
локарису, но лишенное членистых приротовых конечностей. Вместо них у опабинии
имеются складной хоботок и пять глаз.
Другие загадочные существа из Берджес-Шелл: опабиния (слева) и
марелла (справа).
По-видимому, отдельные признаки членистоногих формировались независимо в
разных группах червеобразных организмов. Эти признаки перекомбинировались и
смешивались, образуя самые неожиданные комбинации. Возможно, не обошлось
здесь и без горизонтального межвидового обмена генами (такой перенос могли
осуществлять вирусы).
Как мы помним из прошлой главы, большую роль в крупных эволюционных пере-
стройках у животных могут играть мутации ключевых генов — регуляторов разви-
тия, таких как Нох-гены. В результате таких мутаций свойства одних сегментов
могут проявиться у других. Теоретически в результате подобной мутации от ор-
ганизмов, похожих на аномалокариса (с единственной парой членистых конечно-
стей, развившихся специально для схватывания добычи), могли буквально в одно-
часье произойти формы, имеющие такие конечности на всех сегментах туловища.
Что же касается кольчатых червей, то они, судя по современным молекулярным
данным, вовсе не являются предками членистоногих. Вместе с головохоботными и
круглыми червями членистоногих теперь объединяют в надтип «линяющих»
Ecdysozoa, а кольчатых червей вместе с моллюсками и брахиоподами поместили в
другой надтип — Lophotrochozoa.
Новые палеонтологические находки в сланцах Берджес и других местонахождени-
ях помогают выяснять детали ранней эволюции этих групп. Почти каждая публика-
ция такого рода служит новым подтверждением общего правила мозаичного распре-
деления признаков и многочисленных параллелизмов на ранних этапах развития
больших групп организмов.
Происхождение
неправильных
морских ежей
Эволюционные переходы от одной крупной группы организмов к другой, сопрово-
ждающиеся многочисленными параллелизмами и мозаичным распределением призна-
ков, характерны не только для таких древних времен, как вендский и кембрий-
ский периоды. В этом разделе мы перенесемся на несколько сотен миллионов лет
вперед — в юрский период мезозойской эры — и рассмотрим один из важнейших пе-
реломных моментов в эволюции класса морских ежей (Echinoidea).
Морские ежи делятся на «правильных» — радиально-симметричных и «неправиль-
ных» — двусторонне-симметричных. Правильные ежи всеядные, живут на поверхно-
сти грунта, а неправильные зарываются в грунт и питаются детритом, то есть
органическими остатками, которыми изобилует морское дно. Доказано, что непра-
вильные ежи произошли от правильных в первой половине юрского периода. Это,
несомненно, был ароморфоз, хотя и не такой значительный, как появление млеко-
питающих или членистоногих. Все признаки ароморфоза налицо: строение животных
усложнилось, разнообразие жизненных условий расширилось, так как был освоен
принципиально новый тип субстрата — рыхлые грунты, а также обретен новый тип
питания; кроме того, это эволюционное преобразование открыло морским ежам
путь к увеличению видового разнообразия, численности и биомассы, то есть к
тому, что называют «биологическим прогрессом».
На пути от правильных ежей к неправильным ясно видны направленность движе-
ния и возможность достижения конечной цели несколькими дорогами.
У правильных морских ежей анальное отверстие (перипрокт) расположено на
вершине панциря и окружено десятью пластинками апикальной системы, от которых
радиально расходятся двойные ряды пластинок — амбулакры и интерамбулакры. Рот
находится в середине нижней стороны тела.
* 4Г
Правильный морской еж Hemicidaris.
В юрском периоде в нескольких группах правильных ежей независимо возникла
тенденция к смещению перепрокта назад. Это было связано с переходом к питанию
грунтом. При этом резко увеличился объем пропускаемого через кишечник мате-
риала, а значит, и количество отбросов. Морским ежам — грунтоедам крайне не-
выгодно иметь перипрокт на макушке панциря, потому что пропущенные через ки-
шечник частицы грунта будут падать на них сверху и с большой вероятностью
снова заглатываться. Для грунтоедов, и без того сидящих на низкокалорийной
диете, это совершенно неприемлемо.
Смещение перипрокта назад требовало изменения конфигурации окружающих пери-
прокт пластинок. Эти перестройки неизбежно вели к преобразованию всей архи-
тектуры панциря, поскольку апикальные пластинки в ходе онтогенеза «отпочковы-
вают» от себя все остальные пластинки панциря.
Смещение перипрокта назад — «преадаптация» к превращению морских
ежей из «правильных» в «неправильных».
Устоявшаяся древняя связь между перепроктом и апикальной системой не могла
быть сразу отменена. Природа изобретала различные конструктивные решения этой
задачи. Потребность сместить перипрокт подальше назад приводила к появлению
разнообразных, в том числе весьма необычных, форм со всевозможными дополни-
тельными пластинками; иногда апикальная система буквально разрывалась попо-
лам.
Преобразования апикальной системы в ходе смещения перипрокта у
разных групп морских ежей.
В итоге окончательное отделение перипрокта от апикальной системы произошло
параллельно как минимум в пяти разных эволюционных линиях24.
Типичный неправильный морской еж. Задняя половинка апикальной систе-
мы в итоге все-таки отделилась от перипрокта и воссоединилась с пе-
редней половиной. Апикальная система снова «собралась» вместе и вер-
нулась в свое исходное положение на макушке панциря, а перипрокт
оказался сдвинут на задний конец.
Мозаичное распределение примитивных и прогрессивных признаков, единая общая
направленность изменений и многочисленные параллелизмы в ходе превращения
правильных морских ежей в неправильных проявились не только в эволюции апи-
кальной системы, но и в развитии других частей тела. Примерно таким же обра-
зом развивались у неправильных морских ежей специализированные органы дыха-
ния, сбора и захвата пищи, передвижения в толще грунта.
Происхождение
птиц
Мало кто из древних обитателей Земли может сравниться по своей популярности
со знаменитым археоптериксом, восемь скелетов которого найдено в Германии в
отложениях позднеюрского возраста.
Это существо сочетает в себе признаки хищных динозавров-теропод и птиц. По-
ка археоптерикс был единственной известной «полуптицей-полурептилией», ситуа-
ция казалась простой. От динозавров произошел археоптерикс, от археоптерикса
— настоящие птицы.
Но последовали новые находки, которые, как водится, вместо того чтобы еще
более прояснить ситуацию, сильно ее запутали. Во-первых, выяснилось, что ар-
хеоптерикс был совсем не одинок.
В отложениях мелового периода обнаружилось множество близких к археоптерик-
су существ, обладающих смесью птичьих и динозавровых признаков. Этих странных
птиц назвали «энанциорнисами», т. е. «противоптицами», чтобы подчеркнуть, что
они, возможно, и не имеют прямого отношения к настоящим птицам. Орнитизация,
А. Н. Соловьев, А. В. Марков. Ранние этапы эволюции неправильных морских ежей.
2004.
или «оптичивание», шла многими параллельными путями, и какая именно из «пере-
ходных форм» дала начало современным птицам, не так-то просто понять.
Археоптерикс и его родичи знанциорнисы действительно очень близки к хищным
динозаврам — тероподам. Хотя у них выработались многие птичьи признаки, до
сих пор у ученых остались сомнения в том, что знанциорнисы были прямыми пред-
ками современных птиц. Эксперты отмечают, что у энанциорнисов не заметно те-
неденции к постепенному приобретению тех птичьих особенностей, которых им «не
хватало», чтобы стать настоящими птицами.
Энанциорнис.
Зато обнаружились, причем в большом количестве, разнообразные наследники
хищных динозавров, близкие по признакам к энанциорнисам или к каким-то другим
птицеподобным созданиям. Оказалось, что многие мелкие хищные динозавры в раз-
ное время приобретали те или иные птичьи черты. Найдено довольно много видов
динозавров с настоящими перьями.
Зачем
динозаврам
перья?
Перья не сразу стали использоваться для полета. Сначала они, по всей види-
мости, служили для термоизоляции и были похожи на пух современных птиц. Потом
они пригодились для брачных демонстраций. В октябре 2008 года в Китае нашли
остатки пушистого нелетающего динозаврика размером с голубя, у которого на
хвосте было четыре длиннейших пера, примерно таких же, как у самцов современ-
ных райских птиц. Такие перья могли использоваться только для привлечения са-
мок — больше они ни на что не годны. Позже перья могли пригодиться как сред-
ство планирования при прыжках с ветки на ветку (у древесных динозавров дроме-
озаврид) либо для ускорения бега — у быстро бегающих наземных форм.
Недавно откопали совсем уж невероятных «четырехкрылых» динозавров — микро-
рапторов, которые, скорее всего, умели неплохо летать. Крупные перья, явно
предназначенные для полета, были у них не только на передних конечностях, но
и на задних!
До недавнего времени считалось, что все «оптичившиеся» динозавры были срав-
нительно мелкими. Однако в 2007 году в Китае был найден настоящий птицеподоб-
ный гигант, живший в позднемеловую эпоху и весивший около полутора тонн.
В Китае нашли кости
гигантского птицеподобного
динозавра
По мнению большинства ученых, непосредственными предками птиц были целюро-
завры (это одна из групп теропод, являющихся, в свою очередь, одной из групп
ящеротазовых динозавров). Многие целюрозавры в течение юрского и особенно ме-
лового периодов приобретали различные птичьи черты, в том числе перья, причем
эти события происходили независимо в разных эволюционных линиях. Эксперименты
по «орнитизации» продолжались и после появления настоящих птиц — по-видимому,
вплоть до самого конца эпохи динозавров, наступившего на рубеже мезозойской и
кайнозойской эр (65,5 млн. лет назад).
До сих пор считалось, что степень «оптичивания» у целюрозавров коррелирова-
ла с размерами: наибольшее число птичьих черт было отмечено у мелких предста-
вителей группы, тогда как у крупных целюрозавров орнитизация не заходила так
далеко. Предполагалось, что, увеличиваясь в размерах, целюрозавры могли те-
рять некоторые птичьи признаки и возвращаться к более примитивному состоянию.
Новая находка китайских палеонтологов показала, что даже крупнейшие предста-
вители группы могли быть очень похожи на птиц.
Чудовище, названное гигантораптором, проживало на территории Китая в позд-
немеловую эпоху, в период между 89,3 млн. и 65,5 млн. лет назад. Оно имело 8
м в длину, 3,5 м в высоту при горизонтальном положении туловища и весило око-
ло полутора тонн.
Исследовав структуру костей на срезах, исследователи пришли к выводу, что
найденный динозавр погиб на 11-м году жизни (возраст определили по числу «го-
довых колец») . Он уже был взрослым, но продолжал расти. По-видимому, к ста-
рости гиганторапторы могли весить существенно больше полутора тонн. Как и
другие гигантские динозавры, гигантораптор рос очень быстро, значительно бы-
стрее, чем его ближайшие родственники — некрупные целюрозавры из группы
Oviraptorosauria.
Гигантораптор
В строении скелета гигантораптора выявлен ряд черт, которые сближают его с
птицами и ранее у крупных динозавров не встречались. В частности, из всех ги-
гантских теропод у гигантораптора были самые длинные и тонкие конечности —
как передние, так и задние. Авторы находки полагают, что гигантораптор, воз-
можно , был самым быстрым бегуном в своем размерном классе.
Отпечатки перьев не сохранились, но исследователи полагают, что перья у ги-
гантораптора могли быть, особенно на передних конечностях (предположение ос-
новано на наличии перьев у родственников и предполагаемых предков гиганторап-
тора и на некоторых косвенных признаках). Есть мнение, что перья у бегающих
теропод изначально развились для тепла и лишь потом были приспособлены для
полета. Однако нам следует иметь в виду, что крупные перья на передних конеч-
ностях никогда не служили для теплоизоляции: скорее, они выполняли функции
аэродинамического характера25.
Как заметил российский палеоорнитолог Е. Н. Курочкин, у археоптерикса на
сегодняшний день не осталось ни одного «птичьего» признака, который не был бы
найден у тех или иных динозавров. Раньше такими признаками считались перья,
крючковидные отростки на ребрах и вилочка (сросшиеся ключицы). Зато многие из
тех признаков, которые есть у настоящих (веерохвостых) птиц, но отсутствуют у
динозавров, не обнаружены ни у археоптерикса, ни у его ящерохвостых родствен-
ников .
Кто был предком
настоящих птиц?
Археоптерикс и его родня — энанциорнисы, — несомненно, произошли от дино-
25 Xing Xu et al. A gigantic bird-like dinosaur from the Late Cretaceous of China //
Nature. 2007. V. 447. P. 844-847.
завров. Однако, по мнению Е. Н. Курочкина, это была тупиковая ветвь, вымершая
в конце мелового периода вместе с динозаврами и не оставившая потомков.
Курочкин считает, что предков настоящих птиц нужно искать в более древних
эпохах. Возможно, птицы произошли вовсе не от пернатых динозавров, не от ар-
хеоптерикса и не от энанциорнисов, а от более древних пресмыкающихся — позд-
нетриасовых текодонтов. Не исключено, что эта группа рептилий была общим
предком и птиц, и динозавров.
Лучшим из обнаруженных до сих пор кандидатов на роль птичьего предка среди
текодонтов является протоавис, найденный в позднетриасовых отложениях США. в
1983 г. Хотя протоавис был наземным животным и не умел летать, у него имеются
важнейшие птичьи признаки, которых нет ни у археоптерикса, ни у энанциорни-
сов , ни у пернатых динозавров.
Протоавис.
Возможно, движение в «птичью» сторону началось еще в триасе у текодонтов.
Линия, приведшая к настоящим птицам и идущая от форм, близких к протоавису,
не сразу достигла эволюционного успеха. Вначале доминировала «энанциорнисо-
вая» линия, идущая от хищных динозавров — теропод. Причем птичьи признаки
приобретались параллельно в нескольких линиях динозавров, поэтому говорят о
«процессе орнитизации теропод». Эта линия оказалась тупиковой. Только после
ее вымирания в конце мела настоящие (веерохвостые) птицы заняли освободившее-
ся экологическое пространство и достигли подлинного расцвета.
Впрочем, большинство западных ученых не разделяют эту точку зрения и выво-
дят птиц напрямую из энанциорнисов и им подобных промежуточных форм.
Завершая разговор о происхождении птиц, расскажу об одном необычном иссле-
довании, результаты которого были опубликованы в начале 2007 года в журнале
Nature26.
Кто бы мох1 подумать, что в серьезных научных журналах начнут появляться
статьи об эволюции геномов динозавров — животных, в чьих ископаемых костях за
давностью лет никакой ДНК попросту не осталось. Тем не менее, это произошло.
26 Chris L. Organ, Andrew M. Shedlock, Andrew Meade, Mark Pagel, Scott V. Edwards.
Origin of avian genome size and structure in non-avian dinosaurs // Nature. 2007. V.
446. P. 180-184.
Изобретательность исследователей, выполнивших эту работу, вызывает чувство
глубокого восхищения.
Ученые воспользовались тем обстоятельством, что в ископаемых костях, если
они достаточно хорошо сохранились, на срезах бывают видны маленькие полости,
в которых при жизни животного помещались клетки костной ткани — остеоциты.
Известно, что размер генома в некоторых группах живых существ положительно
коррелирует с размером клеток. Справедливо ли это для остеоцитов позвоночных?
Авторы изучили срезы костей 26 видов современных тетрапод (то есть четвероно-
гих; к ним относятся амфибии, рептилии, птицы и млекопитающие) и обнаружили
линейную зависимость между размером генома и средним объемом остеоцита. Най-
денная зависимость позволила ученым с приемлемой точностью оценивать размеры
геномов ископаемых видов.
Авторы воспользовались этой замечательной возможностью, чтобы ответить на
вопрос, который давно волнует ученых: когда и почему у предков современных
птиц произошло радикальное уменьшение генома? Дело в том, что птичьи геномы
значительно меньше по размеру, чем у остальных тетрапод. Размер генома совре-
менных птиц — от 0,97 до 2,16 млрд. пар нуклеотидов, в среднем 1,45. Для
сравнения — у жабы 6,00, у крокодила 3,21, у коровы 3,7, у кошки 2,9, у мыши
3,3, у человека 3,5. Считалось, что уменьшение генома у птиц является своеоб-
разной адаптацией к полету. Из генома была выброшена значительная часть неко-
дирующих и повторяющихся участков, в частности многие мобильные генетические
элементы. По логике вещей для птиц было важнее всего максимально облегчить
свое тело и оптимизировать обмен веществ. Наличие в каждой клетке сотен мил-
лионов «лишних» пар нуклеотидов было бы для них непозволительной роскошью:
ведь все эти километры ДНК нужно еще и обслуживать — определенным образом
упаковывать, ремонтировать при возникновении различных поломок, копировать
перед каждым клеточным делением. А для этого клетка должна синтезировать и
содержать огромное количество различных белковых молекул, не говоря уже об
энергетических затратах. Но это только логика, а природа зачастую действует в
обход человеческого разумения.
Чтобы проверить, действительно ли уменьшение генома было связано с полетом,
необходимо было выяснить размеры генома у вымерших предков птиц. Ученые исхо-
дили из теории о происхождении птиц от динозавров-теропод. Между прочим, в
силу особенностей современной биологической систематики факт происхождения
птиц от динозавров (а не от общих с динозаврами предков) требует рассматри-
вать птиц как подгруппу динозавров, а для «собственно» динозавров теперь при-
меняют громоздкий термин «non-avian dinosaurs» (нептичьи динозавры). То есть
формально динозавры не вымерли: посмотрите в окно — на ветках сидит множество
крылатых «динозавриков»!
Ученые измерили остеоциты у 31 вида динозавров и ископаемых птиц, или, как
они пишут, «птиц и нептичьих динозавров». По размеру остеоцитов они прикинули
возможные размеры геномов их владельцев. Результаты получились довольно не-
ожиданные . Оказалось, что размеры остеоцитов и, следовательно, геномов сильно
различаются у двух основных групп динозавров — птицетазовых и ящеротазовых. К
птицетазовым относятся растительноядные формы, такие как трицератопс и игуа-
нодон . К ящеротазовым помимо двуногих хищников — теропод относятся также мас-
сивные диплодоки и им подобные из группы завропод. И как будто нарочно, чтобы
всех запутать, птицы произошли вовсе не от птицетазовых, а от ящеротазовых
динозавров.
Как выяснилось, геномы птицетазовых динозавров имели средний размер около
2,5 млрд. пар нуклеотидов, что вполне сопоставимо с современными рептилиями.
Геномы теропод, в том числе самых древних, живших задолго до появления птиц,
были значительно меньше — в среднем 1,78 млрд. п. н. Из девяти исследованных
видов теропод только у одного (овираптора) размер генома оказался за предела-
ми диапазона, характерного для современных птиц. Единственный исследованный
представитель завропод — апатозавр — тоже имел маленький геном.
Авторы пришли к заключению, что общий предок всех динозавров имел типичный
для наземных позвоночных большой геном. Это состояние сохранилось у птицета-
зовых динозавров, а также у рептилий, доживших до наших дней. У ящеротазовых
динозавров еще на заре их истории (в триасе) произошло радикальное сокращение
генома. Птицы, таким образом, унаследовали маленький геном от своих предков —
динозавров-теропод, а не приобрели его позже как адаптацию к полету.
И все таки связь между размером генома и полетом существует. Об этом свиде-
тельствуют два обстоятельства. Во-первых, у нелетающих птиц, таких как страу-
сы, геном больше, чем у летающих. По-видимому, утрата способности к полету
привела к тому, что всевозможные мобильные элементы снова «расплодились» в
геноме нелетающих птиц. Во-вторых, у летучих мышей геном меньше, чем у других
млекопитающих.
По-видимому, уменьшение генома у ящеротазовых динозавров следует рассматри-
вать не как адаптацию к полету, а как преадаптацию, то есть как признак, раз-
вившийся в связи с какими-то иными обстоятельствами, а впоследствии облегчив-
ший развитие способности к полету.
Параллельная эволюция в высыхающем Арале. Параллелизмы характерны отнюдь не
только для крупных эволюционных событий, связанных с переходом на более высо-
кий уровень организации. Поразительные примеры параллельной эволюции наблюда-
лись в последние годы в гибнущем Аральском море. О них стало известно благо-
даря исследованиям биологов из Омска С. И. Андреевой и Н. И. Андреева.
Большой и Малый Арал.
Как известно, сейчас Арала как единого водоема уже не существует: он разде-
лился на два изолированных, быстро высыхающих пересоленных «озера» — Большой
и Малый Арал. Резкое увеличение солености привело к вымиранию большей части
видов животных и растений. Однако некоторые двустворчатые моллюски сумели вы-
жить . Внезапная перемена условий привела к тому, что уцелевшие виды начали
быстро эволюционировать. У них резко возросла изменчивость, и появились целые
«букеты» новых форм, причем различия между этими новыми формами и исходными
видами порой очень велики: такая степень различий характерна для разных ро-
дов, а иногда и семейств двустворчатых моллюсков. Массовое вымирание освобо-
дило множество экологических ниш. Вымерли, не выдержав повышения солености,
все моллюски-фильтраторы. Однако многие зарывающиеся грунтоеды оказались бо-
лее «солеустойчивыми». Опустевшие ниши фильтраторов стали «втягивать» в себя
выживших моллюсков, которые ранее вели совсем другой образ жизни.
В результате зарывающиеся хрунтоеды из рода Cerastoderma буквально на гла-
зах изумленных исследователей стали вылезать на поверхность грунта и превра-
щаться в фильтраторов (при этом в строении их раковины происходили соответст-
вующие изменения). Самое интересное, что все эти эволюционные процессы проте-
кали очень сходным образом в двух разобщенных водоемах — Большом и Малом Ара-
ле!
К сожалению, этот уникальный эволюционный «эксперимент» уже закончился, и
закончился трагически — как и любое событие, ставшее следствием неразумного
вмешательства человека в природу. Соленость в Большом Арале недавно достигла
непереносимого для моллюсков уровня в 6%, что привело к их полному вымиранию.
Возможно, в Малом Арале они проживут еще какое-то время, но сравнивать их
эволюцию уже не с чем.
Видообразование на
разных островах идет
параллельными путями
Уединенные острова и озера представляют собой уникальные эволюционные лабо-
ратории, где небольшие популяции животных и растений могут сильно меняться за
весьма малое по геологическим масштабам время. Этому способствует, прежде
всего, обедненность таких изолированных экосистем и резко ослабленная межви-
довая конкуренция. Всевозможные причудливые, узко специализированные формы,
которые на материке, скорее всего, не имели бы никаких шансов на существова-
ние , на крошечном уединенном островке имеют возможность выжить.
Изучение островных флор и фаун иногда позволяет почти воочию увидеть про-
цесс эволюции и проверить на практике различные теоретические модели, разра-
ботанные биологами. Южноафриканские исследователи, изучающие эндемичных вьюр-
ков архипелага Тристан-да-Кунья, в начале 2007 года опубликовали в журнале
Science статью, в которой подтверждаются сразу две широко обсуждаемые в науч-
ной литературе гипотезы27.
Первая из них — модель экологического видообразования в условиях симпатрии
(то есть на одной и той же территории). Согласно модели, исходный предковый
вид может разделиться на два в результате адаптации его представителей к раз-
ным экологическим нишам, причем для этого не нужны какие-либо физические
барьеры, разделяющие исходно единую популяцию на две изолированные части. За-
мечу, что в рамках классической СТЭ симпатрическое видообразование традицион-
но считалось весьма маловероятным; думали, что в подавляющем большинстве слу-
Peter G. Ryan, Paulette Bloomer, Coleen L. Moloney, Tyron J. Grant, Wayne
Delport. Ecological Speciation in South Atlantic Island Finches // Science. 2007. V.
315. P. 1420-1423.
чаев для разделения вида необходимы физические изолирующие барьеры. В послед-
ние годы, однако, получено множество убедительных доказательств реальности
симпатрическохю видообразования.
Вторая гипотеза предполагает, что, поскольку видообразование контролируется
экологическими факторами, оно должно в сходных условиях происходить сходным
образом. Иначе говоря, если есть два одинаковых изолированных острова, и на
каждый из них попадет один и тот же предковыи вид, то через некоторое время
на этих островах должны параллельно и независимо образоваться сходные «пучки»
видов.
Конечно, обе теоретические модели возникли не на пустом месте, и каждая из
них подтверждается множеством фактов, однако в большинстве своем это косвен-
ные доказательства. Кроме того, они затрагивают лишь отдельные группы живот-
ных или растений. Например, убедительные свидетельства симпатрическохю видо-
образования получены для пальм, многих пресноводных рыб, рачков-бокоплавов
(гаммарид) озера Байкал и т. д., тогда как с другими группами (в том числе с
птицами) дело обстоит хуже. К примеру, галапагосские вьюрки, получившие все-
мирную известность благодаря тому впечатлению, которое они произвели на моло-
дого Чарльза Дарвина, пока не предоставили ученым убедительных свидетельств
симпатрическохю характера своего видообразования (предполагают, что разные
виды этих птиц могли сформироваться на разных островах).
То же самое до настоящего времени предполагалось и для вьюрков архипелага
Тристан-да-Кунья. Орнитологи делят их на два вида: Nesospiza acunhae (с ма-
леньким клювом) и N. wilkinsi (с большим клювом). Оба вида встречаются только
на двух маленьких островах — Неприступном (Inaccessible) площадью 14 кв. км и
Найтингейл (Nightingale) площадью 4 кв. км. Первый вид ранее обитал также и
на главном острове архипелага, но там его истребили около ста лет назад заве-
зенные людьми крысы и мыши. Острова Неприступный и Найтингейл, к счастью, не-
обитаемы и к тому же объявлены заповедной зоной, так что уникальные местные
птицы имеют шанс задержаться на этом свете.
Nightingale
Inaccessible
wjfkinsi quest* acunhae hybrids dunnei
yV/ видообразование \Yf
4
заселение
обоих
остроеое
4
предковая форма
Схема эволюции вьюрков рода Nesospiza.
Между прочим, палеонтологи недавно установили, что практически на всех ост-
ровах и атоллах Тихого океана до прихода людей обитали разнообразные эндемич-
ные виды птиц, в том числе нелетающих. Как только на остров приходили люди —
полинезийцы или меланезийцы, — эти виды немедленно исчезали. Так что истории
с дронтом и новозеландским моа — не исключение, а правило.
Представители одного и того же вида, обитающие на разных островах, немного
отличаются друг от друга, поэтому их считают разными подвидами. На Неприступ-
ном выделяют подвиды N. acunhae acunhae и N. wilkinsi dunnei, на Найтингейле
— N. acunhae quest! и N. wilkinsi wilkinsi. Предполагалось, что виды N.
acunhae и N. wilkinsi сформировались на разных островах, а затем расселились,
и их ареалы пересеклись.
Однако генетический анализ, проведенный исследователями, показал, что исто-
рия у этих птиц была иной. Сравнив несколько митохондриальных и ядерных гене-
тических маркеров, ученые обнаружили, что два вида на каждом из островов бли-
же друг к другу, чем к сородичам своего же вида, но с другого острова. Напри-
мер, большеклювые N. wilkinsi dunnei с Неприступного гораздо ближе к мелко-
клювым N. acunhae acunhae с того же острова, чем к большеклювым N. wilkinsi
wilkinsi с Найтингейла.
Ученые сделали вывод, что все четыре формы, по- видимому, происходят от од-
ного предка, когда-то залетевшего из Южной Америки с преобладающими западными
ветрами (расстояние от материка — 3000 км) . Эта предковая форма заселила оба
острова и на каждом из них параллельно и независимо подразделилась на две
разновидности — мелкоклювую и большеклювую. Направленность этих эволюционных
преобразований определялась особенностями кормовой базы. На обоих островах
основную пищу вьюрков составляют мелкие семена местной травы Spartina
arundinacea и крупные семена дерева Phylica arborea. Размер клюва у вьюрков
строго коррелирует с размером разгрызаемых семян. Большеклювые N. wilkinsi
питаются в основном семенами дерева, мелкоклювые N. acunhae — семенами травы.
Любопытно, что на меньшем по площади острове Найтингейл процесс симпатриче-
ского видообразования продвинулся заметно дальше, чем на более крупном остро-
ве. На Неприступном, где разнообразие природных условий выше, две формы вьюр-
ков, по-видимому, еще способны скрещиваться друг с другом. В некоторых рай-
онах острова встречаются птицы с клювами промежуточных размеров — судя по
всему, гибриды (генетический анализ не противоречит этому предположению). На
Найтингейле никаких гибридов нет, да и генетически две местные разновидности
вьюрков сильнее отличаются друг от друга, чем формы с Неприступного. По-
видимому, это говорит о том, что разнообразие условий обитания в данном слу-
чае выступало как фактор, тормозящий эволюцию.
Что такое
«скорость
эволюции»?
Необходимо помнить, что под «скоростью эволюции» в зависимости от контекста
могут подразумеваться два совершенно разных показателя. Одно дело — скорость
образования разнообразных причудливых специализированных форм, совсем другое
— скорость прогрессивных преобразований, связанных с выработкой новых адапта-
ции широкого профиля и ростом сложности организма. На маленьких изолированных
клочках суши выше только первая из этих скоростей, тогда как вторая, наобо-
рот, выше на больших материках с разнообразными условиями и сложными насыщен-
ными экосистемами.
Что заставляет
эволюцию
повторяться?
Для начала отметим первое, что бросается в глаза. Сходство параллельно раз-
вивающихся групп обычно тем больше, чем ближе их родство. Очень далекие друг1
от друга группы могут выработать лишь самое поверхностное сходство (например,
рыбы и дельфины) . Наоборот, в Большом и Малом Арале или на крошечных остров-
ках архипелага Тристан-да-Кунья сходство вновь образующихся форм было очень
велико, поскольку они происходили от одних и тех же исходных видов. По-
видимому, само строение организма во многом предопределяет возможные пути его
эволюции. Для каждого организма существует весьма ограниченное число «разре-
шенных» путей эволюционных изменений — и любые попытки уклониться от этих пу-
тей безжалостно отсекаются отбором. У близкородственных видов, индивидуальное
развитие которых идет по сходной «программе», набор этих разрешенных путей
тоже сходен, что и приводит к параллелизмам.
Во-вторых, большую роль играет сходство структуры основных типов сообществ,
или экосистем. Набор экологических «ролей» (ниш) в разных сообществах в целом
сходен. Например, если это степь, прерия или саванна, то там должна быть, во-
первых, трава, во-вторых — крупные травоядные звери (копытные или их анало-
ги) , в третьих — хищники, охотящиеся на этих копытных, в четвертых — жуки-
навозники и другие беспозвоночные, перерабатывающие навоз травоядных, и т. д.
Все эти ниши обязательно должны быть кем-то заполнены, иначе весь комплекс
развалится. Попробуйте лишить экосистему одного из блоков, например жуков-
навозников , — немедленно начнется экологический кризис. Нечто подобное наблю-
далось в Австралии, где завезенные европейскими фермерами овцы стали быстро
приводить пастбища в негодность из-за отсутствия в местной фауне эффективных
переработчиков навоза. Пришлось срочно завозить жуков-навозников с «Большой
земли». У фермеров не было времени ждать, пока эволюция справится с проблемой
в своей обычной неторопливой манере. А подождали бы пару сотен тысячелетий —
и почти наверняка на основе местных австралийских жуков развились бы «свои»
эффективные навозники. И они, скорее всего, по многим признакам были бы похо-
жи на своих коллег с других континентов. Обитание в сходных условиях и наце-
ленность на сходный образ жизни приводят к тому, что организмы приобретают
сходные черты строения.
Кроме того, в последнее время появляется все больше свидетельств, что «слу-
чайные и ненаправленные» наследственные мутации, которые, согласно господ-
ствовавшим до недавнего времени представлениям, являются единственным источ-
ником материала для отбора, в действительности часто оказываются не такими уж
случайными. Об этом мы поговорим подробно в главе «Управляемые мутации». Ко-
нечно, природе постоянно приходится действовать «методом случайного тыка», но
она всячески старается этот метод оптимизировать.
Наконец, многие параллелизмы могут иметь довольно простые генетические объ-
яснения. Индивидуальное развитие животных и растений контролируется сравни-
тельно небольшим числом ключевых генов-регуляторов. Если, например, в двух
эволюционных линиях независимо «сломается» какой-то регуляторный ген, в этом
едва ли можно усмотреть что-то удивительное или невероятное. Однако внешнее
проявление этой ошибки может показаться в высшей степени удивительным: ведь
мы увидим весьма сложные и, главное, одинаковые изменения в двух разных груп-
пах организмов.
Один из ярких примеров такого рода дают поясничные ребра, которые на ранних
этапах эволюции млекопитающих многократно исчезали и снова появлялись в раз-
ных эволюционных линиях. Недавно китайские палеонтологи нашли очередного дои-
сторического зверька, строение которого проливает новый свет на это явление.
Данная находка, помимо прочего, представляет собой прекрасный пример класси-
ческой «переходной формы».
Надо сказать, что находки древних мезозойских млекопитающих в последнее
время необычайно участились. Еще пару десятилетий назад считалось, что мезо-
зойские млекопитающие были малочисленной, второстепенной группой мелких кры-
соподобных зверьков, влачивших жалкое существование в тени господствовавших в
то время динозавров. Благодаря находкам последних лет, значительная часть ко-
торых сделана китайскими палеонтологами, стало известно, что древние звери
были весьма многочисленны, разнообразны и могли достигать размеров лисы или
барсука.
Новая палеонтологическая
находка проливает свет на
раннюю эволюцию млекопитающих28
Китайские палеонтологи в очередной раз удивили научный мир необычной наход-
кой. На этот раз они откопали очень хорошо сохранившийся скелет неизвестного
ранее примитивного млекопитающего из группы триконодонтов, жившего около 125
млн. лет назад (в раннемеловую эпоху).
Новооткрытый мезозойский зверек Yanoconodon allini был как раз таким, каким
«положено» быть мезозойскому зверю — маленьким и невзрачным. Интересен он,
прежде всего, тем, что у него очень хорошо сохранились косточки среднего уха
(молоточек и наковальня), причем сохранились они не по отдельности, а в своем
естественном положении, сочлененные с другими костями.
Строение этих косточек является одним из главных отличительных признаков
млекопитающих. У предков зверей — зверозубых рептилий — была только одна слу-
ховая косточка (стремечко), а молоточек и наковальня составляли единое целое
с нижней челюстью. То же самое наблюдается и у самых примитивных древних мле-
копитающих. У современных млекопитающих в ходе эмбрионального развития эти
косточки сначала, как и у далеких предков, прилегают к нижней челюсти, а за-
тем отделяются от нее. Отделение происходит в два этапа: сначала слуховые
косточки отделяются от боковой поверхности нижней челюсти, но остаются связа-
ны с ней спереди посредством меккелева хряща (этот хрящ, кстати, представляет
собой остаток первичной, изначальной нижней челюсти позвоночных; у акул вся
нижняя челюсть образована меккелевым хрящом). На втором этапе исчезает и это
переднее соединение: меккелев хрящ у взрослых млекопитающих рассасывается.
На основе этих фактов более 30 лет назад была выдвинута гипотеза, согласно
которой в эволюции среднего уха млекопитающих имелась промежуточная стадия —
примерно такая же, как в эмбриональном развитии современных зверей29. Яноко-
нодон блестяще подтвердил эту догадку. Именно поэтому первооткрыватели при-
своили ему видовое название allini — в честь Эдгара Аллина (Edgar Allin), ав-
тора подтвердившейся гипотезы. Молоточек и наковальня у яноконодона уже отде-
лены от нижней челюсти с боков, но еще прикреплены к ней спереди посредством
окостеневшего меккелева хряща.
Строение скелета яноконодона еще раз подтвердило, что в ранней эволюции
млекопитающих многие прогрессивные признаки появлялись параллельно и незави-
симо в разных линиях. Например, у яноконодона имеются ребра на поясничных по-
звонках (это примитивный признак), тогда как у некоторых близких форм пояс-
ничные ребра отсутствуют. Сравнительно-анатомический и филогенетический (эво-
Zhe-Xi Luo, Peiji Chen, Gang Li, Meng Chen. A new eutriconodont mammal and
evolutionary development in early mammals // Nature. 2007. V. 446. P. 288-293.
29 Allin E. F. Evolution of the mammalian middle ear. 1975 // J. Morphol. 147, 403-
438.
люционный) анализ мезозойских млекопитающих показывает, что поясничные ребра
многократно исчезали и вновь появлялись в разных эволюционных линиях.
По мнению исследователей, в случае с поясничными ребрами, как и с другими
признаками, затрагивающими дифференцировку отделов позвоночника, многочислен-
ные параллелизмы могут объясняться тем, что эти признаки контролируются очень
небольшим количеством генов. Речь идет о знаменитых Нох-генах, многофункцио-
нальных регуляторах развития, которые задают общий план строения, симметрию и
дифференцировку отделов тела у многоклеточных животных. Например, если у мыши
отключить ген НохЮ (который, правда, присутствует в геноме в трех копиях, и
отключить надо все три) , то на поясничном отделе позвоночника развиваются
ребра. Скорее всего, многократное появление и исчезновение поясничных ребер у
древних млекопитающих объясняется простотой «кодирования» этого признака, то
есть тем, что для его изменения достаточно всего одной или немногих мутаций.
Yanoconodon allini.
Сравнительно простые, эффективные и «умные» системы регуляции индивидуаль-
ного развития, сложившиеся в ходе эволюции у животных (в том числе система
Нох-генов), обладают важным побочным свойством: их случайные изменения (на-
пример, в результате мутаций) с довольно высокой вероятностью могут породить
не только нежизнеспособных монстров, но и что-то вполне осмысленное и даже
удачное. В этом, возможно, состоит одна из причин ускорения прогрессивной
эволюции у высших животных30.
Сходные идеи развивал И. И. Шмальгаузен в книге Факторы эволюции. Хотя, разумеет-
ся, о Нох-генах в то время еще никто не знал.
1
{
Tbrinaxodon
Massetognathus
Probainogrui thus
Kayentatherium
Megazostrodon
Castorocauda
Tochyglossus
Ornithorhynchus
Fruitafossor
Jeholodens
»* ~Л)
Л<
t/ ребра
Mbiiub с тройной
мутаииеи гена НохЮ
(поясничные ребра есть)
ТГ11-13
Sac ra! s
U Yanoconodon > ^Т~^Т\
I р Gobiconodon V/^
Ч л крестцовые
ребер
подвздошная нет
1г15-16
5 mm
tr11-13
Поясничные
робрп ость
Поясничных
ребер нет
Repenomamus
Sinobaatar
Kryptobaatar
Zhangheothenum ->< ^V c - V
Maotherium кростцовыо
AkidoJestes ■ ^ ' '
\ ребра :■- till 13
Honkolotherium пс^ошная есть ж ^
Vincelestes Jf-^l1 4;-:- Os"\\ < 4 ^
Sinodolphys :У:^^Р^^у.Г
крестцовые ' 5 mrn
Eomata ■ •
1 Iе
Sacral*
r л
*..Vr4-,. . Г- -/;.-^'V'V--' ."
"'4' '•- ■
Lumbar ribs^
| present
Sacral
,' »
: /. O"- '*
r * V »'•*• w * * . i ,
1 ' ' ' 4\ C--k'*>K» , .• ,* r-■ <~ Г
Lumbar ribs [
absent
/
t—
>•?■> >>.*•
.,-.c-,rv^j
' !
~ " ." :
V,-->-4'j
/ ■ |
tr11-13
Нормальная мышь
(поясничных ребер нет)
Параллельная эволюция признака «наличие/отсутствие поясничных
ребер» у мезозойских млекопитающих.
Генетические основы параллельной эволюции изучены пока слабо. Сходные изме-
нения морфологии или биохимии организмов могут быть обусловлены как одинако-
выми, так и совершенно разными наборами мутаций. Какой из этих двух вариантов
встречается чаще, до сих пор не известно, и число детально исследованных слу-
чаев невелико.
В главе «Рождение сложности» мы говорили о том, как бактерии приспосаблива-
ются к антибиотикам. Такое приспособление происходит у разных бактерий парал-
лельно и, по-видимому, может обеспечиваться одним и тем же комплексом мута-
ций, приобретаемых в определенной последовательности.
Однако это крайне простой и редкий случай, когда между признаком (устойчи-
вость к антибиотику) и геном (в данном случае это ген фермента бета-
лактамазы) наблюдается соотношение один к одному. Гораздо чаще, особенно у
организмов, более сложных, чем бактерии, каждый ген влияет на множество при-
знаков , а каждый признак определяется множеством генов. К тому же связи между
генами и признаками опосредуются многочисленными внешними и внутренними фак-
торами в ходе индивидуального развития. Все это делает задачу «расшифровки»
генетических основ эволюционных преобразований чрезвычайно сложной.
Тонкая подстройка многофункционального гена может приводить к параллельному
появлению новых признаков. Группе исследователей из США. удалось расшифровать
генетический механизм параллельной эволюции пятен на крыльях у самцов мушек-
дрозофил . Эти пятна имеются у одних видов рода Drosophila и отсутствуют у
других. В процессе эволюции пятна появлялись и утрачивались неоднократно, хо-
тя выглядят они у всех видов примерно одинаково.
Исследователи показали, что наличие или отсутствие пятен определяется мно-
гофункциональным геном yellow («желтый»), от которого зависит окраска не
только крыльев, но и прочих частей тела. При этом пятнистость крыльев опреде-
ляется не «основным» (кодирующим) участком гена, в котором записано строение
соответствующего белка. Как выяснилось, все дело в том, в каких частях разви-
вающегося крыла происходит экспрессия (работа) гена yellow на стадии куколки.
У видов без пятен слабая экспрессия наблюдается равномерно по всему крылу,
резко усилена на концах крыльев (как раз там, где у взрослой мухи будут тем-
ные пятна).
В ходе эволюции пятна на крыльях дрозофилы несколько раз утрачи-
вались и появлялись — в зависимости от характера экспрессии мно-
гофункционального гена yellow.
Характер экспрессии гена yellow определяется двумя некодирующими регулятор-
ными участками гена. Пятна на крыльях у дрозофил иногда появлялись благодаря
модификации первого из этих участков, иногда — второго. Таким образом, в дан-
ном случае независимое появление одного и того же признака (пятен на крыльях)
было обеспечено разными генетическими изменениями, затронувшими, правда, один
и тот же ген.
Исследование показало31, что тонкая «подстройка» регуляторных участков мно-
гофункциональных генов может приводить к появлению новых морфологических при-
знаков у таких сложных организмов, как дрозофила.
31 Benjamin Prudhomme, Nicolas Gompel, Antonis Rokas, Victoria A. Kassner, Thomas M.
Williams, Shu-Dan Yeh, John R. True, Sean B. Carroll. Repeated morphological
evolution through cis-regulatory changes in a pleiotropicgene // Nature. 2006. V.
440. P. 1050-1053.
По-видимому, именно изменения регуляторных участков генов играют основную
роль в эволюции формы и строения сложных организмов. Большинство генов у та-
ких организмов — многофункциональные. Изменение кодирующей части многофунк-
ционального гена может вызвать целый каскад разнообразных малопредсказуемых
последствий. Гораздо удобнее и безопаснее чуть-чуть «подкрутить» один из мно-
гочисленных регуляторов, влияющий не на все функции данного гена сразу, а
лишь на одну или несколько.
Важнейший для всей человеческой цивилизации пример параллельной эволюции
дают культурные злаки. Ведь все множество одомашненных видов и сортов злаков,
от кукурузы до пшеницы, должны были обрести совершенно определенный набор хо-
зяйственных признаков. Это скороспелость, высокая продуктивность, неопадающие
семена, голозерность и др. Скороспелость, то есть превращение многолетников
или двухлетников в однолетники, необходима, так как урожай желательно иметь
каждый год, а не через два года на третий. Неопадающие семена и неломкий ко-
лос нужны для предотвращения опадения на землю спелых семян до сбора их зем-
ледельцем. Голозерность сильно облегчает и ускоряет процесс обмолота. И все
эти признаки культурные растения приобрели в результате неосознанного искус-
ственного отбора независимо друг от друга. Такая параллельная эволюция, по-
видимому, сильно облегчалась тем, что формирование многих хозяйственных при-
знаков контролируется единичными регуляторными генами. И если этот единствен-
ный регуляторный ген отключить или изменить его работу мутацией, то сразу по-
лучится нужный признак. По-видимому, именно таким способом сформировались у
многих злаков неопадающие семена и неломкие стебли. Для появления подобного
признака часто бывает достаточно «испортить» всего один регуляторный ген из
семейства MADS-генов. MADS-гены — это гены растений, аналогичные по функциям
Нох-генам животных. Эти гены у культурных растений имеют очень большое сход-
ство, поэтому легко можно ожидать и появления сходных мутаций у разных расте-
-32
НИИ
Закономерная повторяемость
в неповторимом живом
многообразии
В 1920 году на съезде селекционеров в Саратове Н. И. Вавилов сообщил об от-
крытии удивительной закономерности, которой подчиняется наследственная измен-
чивость у разных видов и родов растений. Оказалось, что вариации в строении
листьев, корней, колосьев и семян, наблюдаемые у особей любого вида, вовсе не
случайны и не беспорядочны. Каждый вид имеет строго определенный набор воз-
можных вариаций, причем эти наборы («ряды») почти идентичны у близкородствен-
ных видов и очень похожи у представителей разных родов и даже семейств.
Изменчивость разных видов злаков можно изобразить в виде таблицы, где каж-
дый ряд соответствует виду, а каждая колонка — определенной вариации. На ос-
нове составленной им таблицы Вавилов сформулировал «закон гомологических ря-
дов в наследственной изменчивости». Самое удивительное, что закон гомологиче-
ских рядов позволил даже предсказать существование таких разновидностей, ко-
торые еще не были известны науке! Точно так же, как пустые клетки менделеев-
ской таблицы впоследствии заполнились новооткрытыми химическими элементами,
свойства которых в точности совпали с предсказанными, так и пустые клетки ва-
виловской таблицы были заполнены открытыми природными разновидностями злаков.
Главное — знать, что ищешь.
Это блестящее подтверждение пришло позже, однако, уже после первого доклада
Н. П. Гончаров и др. Доместикация злаков Старого Света: поиск новых подходов для
решения старой проблемы. 2007 // Журнал общей биологии. Т. 68. № 2. С. 126-148.
Вавилова на съезде селекционеров присутствующие устроили овацию, а один из-
вестный ботаник сказал: «Это биологи приветствуют своего Менделеева!»
Таким образом, наследственная изменчивость, составляющая основу естествен-
ного отбора и эволюции, оказалась строго упорядоченной, а вовсе не «случай-
ной» . Такой вывод с большим трудом укладывался в схемы классического дарви-
низма. Ведь они основывались на отборе чисто случайных вариаций.
Твердая Посевная Скрученная
Мягкая Вытянутая
Проявление закона гомологических рядов в наследственной изменчи-
вости на примере разных сортов пшеницы.
Некоторые отечественные биологи-теоретики, такие как А. А. Любищев и С. В.
Мейен, придавали вавиловским гомологическим рядам огромное значение и счита-
ли, что за этим явлением скрывается некий фундаментальный закон, управляющий,
возможно, не только биологической эволюцией, но и всем мирозданием. Они под-
черкивали, что гомологические ряды, подобные рядам Вавилова, наблюдаются и в
строении кристаллов, и в структуре органических молекул. Интересно и то, что
многие признаки, изменчивость которых подчиняется вавиловскому закону, напри-
мер, сходные вариации формы листьев у очень далеких друг от друга растений —
цветковых, голосеменных и папоротников, не имеют явного приспособительного
значения. Поэтому происхождение такого сходства трудно объяснить естественным
отбором.
По-видимому, в вавиловских рядах действительно проявляются некие общие за-
коны развития (самоорганизации) сложных систем, причем не только биологиче-
ских. Система, состоящая из взаимосвязанных элементов (блоков), может сущест-
вовать в различных устойчивых состояниях, переходя из одного в другое, но
число этих состояний ограничено, а их характер определяется свойствами эле-
ментов и структурой их взаимодействий.
Очевидно, что дискретный и упорядоченный характер изменчивости накладывает
жесткие ограничения на эволюционные возможности организмов. Поэтому существо-
вание сходных вариаций у разных видов во многом объясняет то явление, которо-
му посвящена эта глава, — параллельную эволюцию. Сходная изменчивость предо-
пределяет сходство эволюционных изменений у разных групп, а ограниченное чис-
ло возможных вариаций ограничивает и число возможных путей эволюционного раз-
вития .
Чем же определяется дискретный характер изменчивости? Прежде всего, нужно
хорошо понимать, какая это сложная штука — жизнь. Чтобы выжить, любой орга-
низм должен приспосабливаться к огромному числу всевозможных внешних факто-
ров. Известно, что никакую систему нельзя оптимизировать одновременно по мно-
гим параметрам. Неизбежно приходится что-то выбирать и чем-то жертвовать.
Кроме того, организм — это целостная система, вся части которой должны рабо-
тать согласованно, не мешать, а помогать друг другу. Приспособление к какому-
то внешнему фактору одного органа неизбежно влияет на всю систему, так что
другим органам уже приходится приспосабливаться не только к выполнению своих
собственных задач, но и к произошедшему в самом организме изменению. Иначе
вся сложнейшая конструкция может рассыпаться.
Но и это еще не все. Каждый многоклеточный организм развивается из одной
клетки — яйца. Значит, нужно разработать такую совершенную генетическую про-
грамму развития, чтобы в результате последовательных делений одной-
единственной клетки в итоге получился сложный жизнеспособный организм. А ведь
еще необходимо следить за тем, чтобы жизнеспособность (то есть целостность
системы) неизменно сохранялась на каждом этапе роста и развития.
Соблюсти все эти условия чрезвычайно сложно. Не случайно некоторые биологи
сравнивают эволюцию с переделыванием самолета на лету. Поэтому каждое живое
существо — это результат труднейшего компромисса между многими тысячами раз-
нообразных требований, предъявляемых к системе как извне, так и изнутри.
Идея о «компромиссном» характере организации живых существ («метафора адап-
тивного компромисса»33) помогает понять многое и в строении живых существ, и
в их эволюции (см. главу «Рождение сложности»).
Во-первых, становится ясно, почему далеко не о каждом признаке и свойстве
живого существа можно уверенно сказать, «зачем это нужно». А ведь отсутствие
явного адаптивного, то есть приспособительного, смысла у многих признаков
всегда смущало дарвинистов. Обычно отговаривались тем, что, дескать, пока мы
не знаем, зачем этот признак, но когда-нибудь обязательно узнаем. Ну а теперь
становится понятным, что многие признаки появились не «зачем», а «потому что»
— это побочные следствия компромисса. Если бы бабочки умели говорить, то на
вопрос «почему у тебя на крыльях именно такой узор, а не другой?» они могли
бы с полным правом ответить: «Так получилось».
Почему бабочки
похожи на цветы?
Многих биологов-теоретиков приводил в замешательство «детский» вопрос: по-
чему дневные бабочки, питающиеся нектаром цветов, сами похожи на цветы, а
иные еще и издают «цветочный» аромат? Не в деталях, не до полного сходства —
это не маскировка, — но, в общем, и целом, по яркости и броскости рисунка
А. П. Расницын. Процесс эволюции и методология систематики. 2002.
крыльев-лепестков? Мне кажется, здесь проявился «принцип зацепки»: естествен-
ному отбору, чтобы создать адаптацию, нужно зацепиться за какое-то уже суще-
ствующее свойство — «преадаптацию». Яркая и необычная окраска бабочек нужна
им для привлечения половых партнеров. Бабочки питаются нектаром, и поэтому их
органы чувств и нервная система издавна были «настроены» на поиск цветов (и
сами цветы, чтобы привлечь опылителей, становились в процессе эволюции более
яркими и броскими). Совершенствуя способы привлечения партнера, естественный
отбор «зацепился» за эту психологическую особенность бабочек. Преимущество в
размножении стали получать те особи, которые привлекали взгляд своих сороди-
чей, порхающих в поисках яркого цветка.
Во-вторых, понятно, что достигнутый с таким трудом компромисс — сложнейшую
систему индивидуального развития и строения организма — довольно трудно изме-
нить . Ведь в ней все взаимосвязано, изменишь одну часть — и все может разва-
литься .
Пути для изменения все-таки остаются, но их не может быть много, и при этом
они дискретны — структура сложной системы предусматривает возможность лишь
ограниченного и строго определенного набора изменений.
Возможность изменения сохраняется благодаря тому, что живые системы никогда
не становятся абсолютно целостными, никогда не сливаются в единый и неделимый
«атом». На всех уровнях сохраняется блочный принцип «сборки» живых систем,
причем отдельные блоки (гены, клетки, органы, ткани) стараются сохранить хотя
бы частичную независимость от других. Благодаря этому далеко не каждое изме-
нение одного блока сразу разрушает систему. Например, в индивидуальном разви-
тии отдельные процессы роста и дифференцировки клеток хоть и сильно взаимо-
связаны, но все-таки не абсолютно «слиты» в единую жесткую последователь-
ность . Многие из этих процессов могут меняться независимо от остальных.
Например, все земноводные проходят в своем развитии стадию водной личинки.
У лягушек это головастики, очень сильно отличающиеся от взрослых лягушек и по
строению, и по образу жизни. У хвостатых земноводных — тритонов и саламандр —
личинки отличаются от взрослых гораздо меньше. У них другие размеры, плава-
тельная складка на хвосте и спине, ветвистые наружные жабры, которые у взрос-
лых рассасываются, а также ряд второстепенных отличий. Превращением личинки
во взрослое животное управляет гормон тироксин. Что же произойдет, если в ре-
зультате мутации тироксин в организме не образуется или ткани вдруг станут
нечувствительны к нему? И то, и другое нарушение может произойти в результате
одной-единственной «точечной» мутации в одном из генов.
Головастик лягушки просто-напросто умрет, не оставив потомства. А вот ли-
чинки некоторых саламандр в этом случае не умирают. Все процессы роста и раз-
вития у них продолжаются как ни в чем не бывало, за исключением того, что они
так и не теряют жабры и плавник и не выходят на сушу. В положенное время у
таких личинок развиваются половые железы, и они совершенно нормально размно-
жаются и откладывают икру, из которой выводится потомство, унаследовавшее ис-
ходную мутацию. В итоге из наземной саламандры получается чисто водное живот-
ное — уже известный нам аксолотль, размножающаяся личинка-переросток.
У аксолотля отключилась система превращения личинки во взрослое животное.
Все остальные системы продолжают работать по-прежнему. Так что изменение от-
дельных «блоков» системы индивидуального развития не обязательно приводит к
гибели и может даже в некоторых случаях оказаться полезным — например, если у
данного вида земноводных на суше больше врагов, чем в воде.
Это явление — переход к размножению на ранней стадии, приводящий к сохране-
нию у взрослых особей эмбриональных и личиночных черт, — играло важную роль в
эволюции. Возможно, таким путем появились позвоночные из головастико-подобных
личинок сидячих морских фильтраторов — оболочников (Tunicata). Есть теория,
что и в происхождении человека какую-то роль сыграли подобные процессы. По
ряду признаков, таких как безволосость и высокий лоб, человек больше похож на
эмбрионов и детенышей обезьян, чем на взрослых.
Другая возможность скачкообразных перестроек состоит в перенесении свойств
одних частей тела на другие. Как мы знаем, для этого может быть достаточно
мутации одного гена-регулятора. В результате какой-нибудь «переключатель»,
управляющий дифференцировкой частей организма, вдруг начинает включаться не в
то время или не в том месте. Так, у различных насекомых обнаружены вполне
«вавиловские» гомологичные ряды мутаций, приводящих к тому, что свойства од-
них сегментов тела или конечностей проявляются в других сегментах, неположен-
ных. Вместо усиков могут вырасти ноги, вместо третьего сегмента груди разви-
вается точная копия второго, и если это произойдет у мухи, то она в результа-
те автоматически приобретет вторую пару крыльев! Или же ноги вырастут не
только на грудных, но и на брюшных сегментах, и получится насекомое с восе-
мью-десятью парами ног. Правда, такие мутанты обычно умирают в раннем возрас-
те .
Мутации такого типа — их называют «гомеозисными» — могли сыграть большую
роль в эволюции. В результате гомеозисной мутации животное, подобное кембрий-
скому аномалокарису с единственной парой членистых конечностей, могло бы при-
обрести сразу множество таких ног вдоль всего тела. Среди древних ископаемых
рыб силурийского и девонского периодов есть группы, имеющие только одну пару
плавников — переднюю. Но известны и такие ископаемые рыбы, у которых множест-
во парных плавников. Все современные рыбы, как и наземные позвоночные, имеют
две пары конечностей. Я нисколько не удивлюсь, если в скором времени генетики
докажут, что изменения числа пар конечностей в ранней эволюции позвоночных
происходили за счет гомеозисных мутаций, затрагивающих Нох-гены. По предполо-
жению академика Л. П. Татаринова, шерстяной покров у млекопитающих тоже мог
появиться в результате гомеозисной мутации. У предков млекопитающих — зверо-
подобных рептилий — некоторые чешуи на морде удлинились и стали выполнять
осязательную функцию. Постепенно они превратились в длинные упругие волоски —
вибриссы. А затем в один прекрасный день в результате гомеозисной мутации все
остальные чешуи на теле превратились в такие волоски. Правда, чувствительной
функции у них при этом не появилось, поскольку к ним не подходили соответст-
вующие нервы34, но для утепления мутация оказалась очень даже полезной.
Можно подумать, что подобные мутации не могут привести к усложнению орга-
низмов, потому что это не более чем перетасовка и перекомбинация имеющихся
признаков. Это, однако, не совсем так. Попадая в новое место, новое клеточное
и тканевое «окружение», перепрыгнувший орган неизбежно и сам становится не-
множко другим. Так закладывается основа для дифференцировки. Например, пре-
словутые ноги, вырастающие иногда у насекомых вместо усиков, на самом деле
довольно сильно отличаются от обычных ног и могли бы стать основой для появ-
ления совершенно нового типа конечностей, если бы... нормальные усики на этом
месте не были так необходимы этим глупым насекомым.
Постепенно
или скачками?
Из дискретного характера изменчивости, казалось бы, следует, что эволюцион-
ные преобразования в основном должны происходить внезапными скачками. Однако,
Точно так же обстоит дело и с ногами насекомых, вырастающими на месте усиков в
результате гомеозисных мутаций: нервы, необходимые для работы ног, к ним не подхо-
дят, а те нервы, что управляют работой усиков, «не годятся», поэтому лишние ноги ос-
таются неподвижными, несмотря на нормально развитую мускулатуру.
как и любые другие эволюционные закономерности, закон гомологических рядов
Вавилова, правило «квантованной» изменчивости и скачкообразных переходов жи-
вых систем из одного устойчивого состояния в другое не нужно абсолютизиро-
вать . Далеко не всякая изменчивость дискретна, и многие эволюционные преобра-
зования организмов происходят вполне плавно.
Градуализм и
сальтационизм
Вопрос о равномерности темпов эволюции довольно долго был одной из самых
острых проблем в эволюционном учении. И сам Дарвин, и многие его последовате-
ли видели эволюцию как процесс в основном плавный, постепенный, то есть гра-
дуалистический. Высказывалась, правда, и другая точка зрения — о возможности
мгновенного превращения одного вида в другой в результате крупномасштабных
мутаций, или, по-другому, макромутаций. Это учение называют сальтационизмом,
от «сальто» — прыжок. Основным аргументом противников сальтационизма было то,
что крупные мутации почти всегда смертельны или, по крайней мере, очень вред-
ны. Сальтационисты на это отвечали, что среди «монстров», появляющихся в ре-
зультате макромутаций, могут оказаться и «перспективные» (hopeful monsters).
Открытие Нох-генов и гомеозисных мутаций показало, что в фантазиях сальтацио-
нистов было разумное зерно. Однако классическая СГЭ образца 50-х — 60-х годов
прошлого века придавала основное значение отбору небольших наследственных от-
клонений и считала эволюцию в основном постепенным, или градуалистическим,
процессом.
Единственным источником наследственной изменчивости сторонники СТЭ считали
случайные мутации — ошибки при копировании ДНК. Считалось, что, поскольку му-
тационный процесс совершенно случаен и не направлен, то и идти он должен при-
мерно с одинаковой скоростью у всех живых организмов.
Это предположение легло в основу принципа «молекулярных часов», который ак-
тивно используется и по сей день. На основе этого принципа исследователи по
числу различий в нуклеотидных последовательностях ДНК оценивают время расхож-
дения видов, то есть определяют, когда жил их последний общий предок. И метод
молекулярных часов обычно не так уж плохо работает, что само по себе может
показаться странным. Ведь сегодня хорошо известно, что мутагенез не всегда
является абсолютно случайным, его темп может целенаправленно регулироваться
клеткой, в разных группах организмов и даже в разных частях одного и того же
генома он идет с разной скоростью. Чтобы молекулярные часы продолжали тикать,
приходится добавлять ограничения, выбирать «подходящие» куски генома, учиты-
вать особенности группы, принимать в расчет разницу между значимыми и «молча-
щими» нуклеотидными заменами.
Альтернативой градуализму (учению о постепенной эволюции) и сальтационизму
(эволюции путем скачков) стал пунктуализм, или теория прерывистого равнове-
сия, сформулированная в 1972 году Стивеном Гулдом и Нильсом Элдриджем.
Эта концепция предполагает, что в эволюции видов чередуются длительные пе-
риоды стабильности, когда основные черты вида сохраняются неизменными, и ко-
роткие периоды быстрых изменений, в ходе которых вид преобразуется. Он либо
целиком превращается в другой вид, либо делится на два или более новых вида,
либо «отпочковывает» их от себя. Точка зрения пунктуалистов, основанная на
обширном палеонтологическом материале, имеет и достаточно веские теоретиче-
ские обоснования. В самых общих чертах их можно свести к следующему. Вид
представляет собой относительно устойчивую самоподдерживающуюся систему. Что-
бы на месте старой системы возникла новая, старая должна быть разрушена. Нуж-
на хорошая «встряска», приводящая к разрушению внутренних связей.
В популяциях живых организмов разрушение старой системы связей, приводящее
к дестабилизации системы, проявляется, прежде всего, в резком росте изменчи-
вости. Имеются экспериментальные подтверждения того, что резкое изменение ус-
ловий приводит не к плавному и постепенному сдвигу морфологических характери-
стик популяции, а к быстрому росту изменчивости, «размыванию» признаков, и
только потом из этой дестабилизированной популяции может выкристаллизоваться
новая разновидность или вид35.
Поначалу споры градуалистов с пунктуалистами были довольно бурными, но се-
годня эти две концепции вполне мирно уживаются друг с другом. Их справедливо
считают не противоречащими друг другу, а взаимодополнительными. Обе идеи бы-
стро обрастают всевозможными примерами, уточнениями и обоснованиями. Имеющие-
ся факты говорят о том, что эволюция иногда идет по пунктуалистическому, ино-
гда — по градуалистическому сценарию.
Обе концепции, однако, до сих пор относятся к числу «эмпирических обобще-
ний», а не строгих теорий — как, впрочем, и почти все остальные так называе-
мые «законы эволюции». В немалой степени это объясняется тем, что никому пока
не удалось выяснить точное количественное соотношение градуалистической и
пунктуалистической составляющих в эволюционном процессе.
Доказан скачкообразный
характер эволюционного
процесса
В 2006 году британские биологи предприняли серьезную попытку решить давний
вопрос о соотношении постепенности и скачкообразности в эволюции. Методика,
примененная исследователями, проста, как все гениальное. Странно, что никто
до этого не додумался раньше. Суть идеи в том, что искомое соотношение легко
можно вычислить, сопоставив между собой длины ветвей эволюционных «деревьев»,
которые строятся при помощи стандартных методик на основе величины генетиче-
ских различий между сравниваемыми видами.
Ветви таких деревьев имеют определенную длину, соответствующую величине ге-
нетических различий или, что то же самое, величине эволюционных изменений,
произошедших в данной эволюционной линии.
Все виды, используемые в таком анализе, обычно современные (выделение ДНК
из ископаемых остатков — это все-таки экзотика). Значит, если эволюция шла
равномерно (градуалистически), то расстояние от основания древа до кончика
любой из ветвей (эти кончики соответствуют анализируемым видам) должно быть
одинаковым, и оно не должно зависеть от количества «узлов» (точек ветвления),
расположенных между кончиком ветви и корнем. Если же эволюция ускорялась в
момент ветвления, то есть шла по пунктуалистическому сценарию, то расстояние
от корня до конца ветви должно быть пропорционально числу узлов, расположен-
ных между ними (см. рисунок).
Исследователи проанализировали эволюционные деревья, построенные по молеку-
лярным данным для 122 групп близкородственных видов. В 57 случаях из 122 меж-
ду обнаружилась вполне четкая, статистически достоверная корреляция между
длиной ветви и числу узлов на ней. Для остальных деревьев корреляция оказа-
лась недостоверной — в основном по той причине, что у этих деревьев было
слишком мало ветвей, то есть количество видов в выборке оказалось недостаточ-
ным для получения статистически надежных результатов.
Таким образом, пунктуалистический эффект действительно существует и прояв-
ляется если и не всегда, то достаточно часто. Любопытно, что у растений и
грибов он проявляется заметно сильнее, чем у животных.
См.: А. С. Раутиан. Правило дестабилизации.
А Пунктуалистическое
В Градуалистичеехое
•
1
1
2 j
1
2
Если эволюция идет в соответствии с принципом «прерывистого рав-
новесия», то есть скачками, эволюционные деревья должны выгля-
деть как на рисунке А. Если же эволюция идет с постоянной скоро-
стью, правильным окажется рисунок В.
Исследователи не остановились на достигнутом и определили также относитель-
ный вклад пунктуалистического эффекта в суммарную величину различий между
нуклеотидными последовательностями ДНК разных видов. Выяснилось, что в сред-
нем около 22% наблюдаемых различий возникает во время «взрывных» периодов ви-
дообразования, а остальные 78% постепенно накапливаются в ходе градуалистиче-
ской эволюции.
В работе рассматривались только различия в нуклеотидных последовательно-
стях, причем функциональное значение этих различий никак не учитывалось (да
этого никогда и не делают при построении «молекулярных деревьев»). На уровне
организма картина может быть иной. Известно, что значительная часть изменений
ДНК никак не отражается на строении организма, и, с другой стороны, даже
очень небольшие изменения в ключевых участках ДНК могут привести к радикаль-
ным изменениям организма.
Вполне возможно, что те изменения ДНК, которые происходят очень быстро в
период видообразования, связаны с более значительными изменениями в строении
организма, чем те, что происходят во время градуалистического этапа эволюции.
В этом случае в морфологической эволюции будет значительно больше «пунктуа-
лизма», чем в эволюции молекулярной. Что, собственно, и наблюдается в палео-
нтологической летописи.
Данная работа36 интересна как еще одна попытка строгого количественного
обоснования одного из эмпирических «законов эволюции». Все-таки приятно, что
давно подмеченные зоологами и палеонтологами, но не доказанные закономерности
эволюции постепенно начинают получать строгие обоснования.
Иногда кажущаяся дискретность изменений порождается отбором, который благо-
приятствует некоторым из возможных вариантов изменений, но далеко не всем.
Например, когда организм человека вырабатывает иммунитет против новой инфек-
ции , в генах защитных белков — антител — происходит активное мутирование. Это
необходимо, чтобы «методом случайного поиска» подобрать такое антитело, кото-
рое будет лучше всего узнавать нового возбудителя и прикрепляться к нему
36 Mark Pagel, Chris Venditti, Andrew Meade. Large Punctuational Contribution of
Speciation to Evolutionary Divergence at the Molecular Level // Science. 2006. V.
314. P. 119-121.
(подробнее об этом см. в главе «Управляемые мутации»). Сравнивая структуру
получающихся при этом антител, ученые обнаружили, что больше всего мутаций
происходит в трех небольших участках так называемой вариабельной области (V-
области) гена, кодирующего антитело. Сложнейшими экспериментами было доказа-
но, что мутированию подвергаются не только эти участки, а вся V-область, при-
чем мутации везде происходят с одинаковой скоростью. Почему же в имеющихся в
крови антителах измененными обычно оказываются только три маленьких участка?
Оказалось, что это результат отбора. Каждое новое антитело испытывается орга-
низмом, и все не подходящие к данному возбудителю антитела отбраковываются.
Только мутации, произошедшие в трех небольших участках гена, могут улучшить
связывание возбудителя. Если эффективное антитело успешно прошло проверку, то
только тогда оно начинает производиться в больших количествах. А мутации дру-
гих участков почти всегда вредны, и такие антитела организм отбраковывает.
Понятно, что отбор такого рода может отчасти объяснять и происхождение не-
которых вавиловских рядов.
Тем не менее, остается в силе общее правило, гласящее, что структура орга-
низма допускает лишь ограниченный набор возможных преобразований и тем самым
придает эволюции некоторую направленность, а иногда и предопределенность.
* * *
По-видимому, эволюция — это процесс в целом закономерный и предопределен-
ный, но в деталях и частностях случайный. Предсказать ход эволюции можно лишь
в самых общих чертах. У эволюции нет строгих законов, как в математике или
физике. У нее есть только набор закономерностей и правил, каждое из которых
имеет множество исключений.
К числу важнейших закономерностей эволюции я бы отнес следующие три «прави-
ла» :
160
1
О
S
Pteridophyta
Gymnospermophyta
-Magnoliophyta
•450 -400 -350 -300 -250 -200 -150 -100 -50 0
Время, млн лет назад
37
«Новое не вытесняет старое, а добавляется к нему» Динамика разно-
образия трех крупнейших групп наземных растений: споровых
37 Марков А. В. , Коротаев А. В. Гиперболический рост разнообразия морской и конти-
нентальной биот фанерозоя и эволюция сообществ // Журнал общей биологии. 2008. № 3.
(Pteridophyta) — это самая древняя и примитивная группа; голосемен-
ных (Gymnospermophyta); покрытосеменных, или цветковых
(Magnoliophyta), — это самая молодая и прогрессивная группа. По го-
ризонтальной оси — геологическое время (млн лет назад). Видно, что
развитие новых «прогрессивных» групп не сопровождалось существенным
сокращением разнообразия (вытеснением) их более примитивных предков.
1. Общая направленность от простого к сложному. Хотя до сих пор сохранились и
процветают такие примитивные формы жизни, как бактерии, никто не станет
отрицать, что в биосфере идет постепенное появление и накопление все более
сложных организмов. Часто усложнение организации оказывается выгодным, по-
тому что ведет к повышению интенсивности обмена веществ (росту «энергии
жизнедеятельности»), а ведь в биологической эволюции, как и в химической
(см. главу «Происхождение жизни»), побеждает тот «круговорот», который
крутится быстрее. Это позволяет более сложным организмам занимать господ-
ствующее положение в сообществах, оттесняя примитивных предков в менее
привлекательные ниши. Именно поэтому облик биосферы определяется в основ-
ном высокоорганизованными животными и растениями. И это несмотря на то,
что и по численности, и по массе бактерии их явно превосходят. Рост био-
разнообразия в целом носит аддитивный, то есть накопительный, характер —
новое обычно добавляется к старому, а не вытесняет его.
600
о
•X
о
2
Ф
О
С
ос
к
Z
3
о.
О
Рост средней продолжительности существования (ПС) семейств в течение
фанерозойского зона (542 млн. лет назад — наши дни). Тонкая линия —
семейства морских организмов, толстая — континентальных. Различия в
колебаниях двух кривых в данном случае не столь важны (известно, что
в океане и на суше эволюция шла во многом по-разному) . Важнее, что
оба графика являются возрастающими. По вертикальной оси: средняя ПС
семейств в млн лет. По горизонтальной оси — геологическое время
(млн. лет назад) 38 .
2. Рост устойчивости и приспособляемости живых систем. Все эволюционные ли-
нии, дожившие до наших дней, в ходе своего развития прошли через горнило
множества экологических кризисов, катастроф и массовых вымираний. Те груп-
пы, которые не могли быстро приспосабливаться к меняющимся условиям, в
100
0
550 -450 -350 -250 -150 -50
Марков А. В. , Коротаев А. В. Гиперболический рост разнообразия морской и конти-
нентальной биот фанерозоя и эволюция сообществ. // Журнал общей биологии. 2008. № 3.
большинстве своем давно вымерли. Устойчивые, пластичные линии постепенно
накапливались в биосфере. Это видно, например, из того факта, что с тече-
нием времени средняя продолжительность существования видов, родов и се-
мейств неуклонно росла. Поэтому в наши дни биосферу населяют самые устой-
чивые и пластичные формы жизни из всех когда-либо существовавших.
3. Рост эффективности и безотходности биогеохимического круговорота. С рос-
том сложности и совершенства организмов и их сообществ неизбежно растет
и эффективность глобального круговорота веществ, в котором биосфера иг-
рает важнейшую роль и который определяет «лицо» нашей планеты. Например,
самое сложное и совершенное из современных сообществ — тропический лес —
не только чрезвычайно быстро «прокручивает» через себя огромные количе-
ства вещества и энергии, но и практически не производит никаких отходов.
Там не образуется даже подстилка из листьев и других отмерших частей
растений — все это очень быстро перерабатывается грибами, бактериями и
беспозвоночными и возвращается в круговорот. Совсем по-другому обстояло
дело, например, в древних лесах каменноугольного периода, в которых из-
за несовершенства структуры сообщества огромные массы отмершей древесины
накапливались, образуя месторождения каменного угля. В результате столь
необходимый для жизни углерод безвозвратно выводился из глобального кру-
говорота39 . Рост безотходности заметен и в эволюции организмов. У высших
растений и животных постепенно растет продолжительность жизни, снижается
«детская смертность», развивается забота о потомстве, что позволяет, в
свою очередь, снизить уровень рождаемости — то есть фактически произво-
дить меньше заведомо обреченных на гибель потомков.
Все три названных закономерности: усложнение, рост устойчивости и безотход-
ности — отчетливо прослеживаются и в развитии человеческого общества. Это по-
зволяет говорить о преемственности социальной эволюции по отношению к эволю-
ции биологической и придает особый смысл и практическое значение эволюционным
исследованиям.
Важно подчеркнуть, что из этой преемственности вовсе не следует никакого
«социал-дарвинизма» и она вовсе не свидетельствует об усилении «борьбы за су-
ществование» и «естественного отбора» в обществе, как пытались доказать неко-
торые политические силы в первой половине XX века. В трудах современных эво-
люционистов , например, В. А. Красилова, подчеркивается неуклонное ослабление
роли борьбы за существование и отбора в ходе эволюции, развитие более «гуман-
ных» эволюционных стратегий, основанных на взаимопомощи и симбиозе и ведущих
к росту пластичности и снижению всевозможных потерь и отходов (см. также гла-
ву «Великий симбиоз»).
Может быть, самый главный из всех эволюционных законов — это постепенное
отступление Смерти и Хаоса перед лицом развивающейся Жизни.
ГЛАВА 7.
УПРАВЛЯЕМЫЕ
МУТАЦИИ
Великие открытия 50-60-х годов XX века, такие как расшифровка структуры ДНК
и генетического кода, произвели столь сильное впечатление на научное сообще-
ство, что наспех оформившиеся вокруг этих открытий теории сразу же стали счи-
Человек, сжигая каменный уголь, снова возвращает этот «потерянный» углерод в био-
сферу и делает его доступным для живых организмов. При сжигании угля образуется уг-
лекислый газ, из которого растения создают органические вещества в процессе фотосин-
теза.
таться «классическими». Дальнейшее развитие биологии показало, что не стоило
так торопиться. Что ж, зато с тех пор молекулярные биологи не могут пожало-
ваться на недостаток сенсационности в своих открытиях. Стоит ученым обнару-
жить какой-нибудь новый молекулярный механизм, глядишь, одна-две «догмы» сра-
зу же и рушатся. И тотчас падкие на сенсации журналисты привычной рукой стро-
чат броские заголовки: «Теория эволюции под вопросом» и «Дарвин опровергнут».
Хотя уж кто-кто, а Дарвин тут точно ни при чем. И факт эволюции никто из
серьезных ученых под сомнение не ставит. Просто нужно честно признать, что в
работе молекулярных систем, отвечающих за обработку генетической информации,
очень много неизвестного и непонятного. И каждое новое открытие приближает
нас к пониманию того, как на самом деле идет эволюция на молекулярном уровне.
Пожалуй, из всех скороспелых «абсолютных истин» синтетической теории эволю-
ции с самым громким треском рухнула догма о полной случайности всех наследст-
венных изменений.
Произошло это лишь в последние 10-20 лет, и многие биологи даже не успели
еще вполне осознать этот факт. До сих пор и в популярных текстах, и даже в
учебниках и научных статьях продолжают встречаться ссылки на «случайность
всех мутаций» как на что-то общеизвестное и не подлежащее сомнению. Однако на
сегодняшний день твердо установлено, что живая клетка располагает большим ар-
сеналом средств, позволяющих ей контролировать изменения своего генома.
Вообще-то этого следовало ожидать. Ведь мутации — изменения нуклеотидной
последовательности ДНК — являются важным фактором, влияющим на жизнеспособ-
ность организмов. И это влияние проявляется не когда-то в отдаленном будущем,
а здесь и сейчас — у самого организма или его непосредственных потомков. Если
организмы в ходе эволюции могут выработать приспособления, например, для за-
щиты от хищников или болезнетворных микробов или для контроля температуры те-
ла, то почему они не имеют права выработать также и приспособления, позволяю-
щие им контролировать мутации? Никакого теоретического запрета на такие при-
способления вроде бы нет, однако биологи-теоретики почему-то довольно долго
считали их запрещенными.
Наверное, дело тут в том, что в большинстве ситуаций единственное, чего хо-
чет добиться организм от мутационного процесса, — это чтобы он шел как можно
медленнее или не шел вовсе. В стабильных благоприятных условиях это вполне
разумно. Зачем менять свою наследственность, если все и так хорошо? Однако
сама жизнь не позволяет долго существовать, совершенно не меняясь. Не идти
вперед — значит идти назад.
Для начала вспомним, чему нас учили в школе на уроках биологии. Главный ис-
точник мутаций — ошибки, возникающие в ходе копирования ДНК.
ДНК в норме состоит из двух комплементарных цепочек нуклеотидов. Нуклеотид
А всегда стоит напротив Т, Г — напротив Ц. Пары Г-Ц более прочные, они удер-
живаются вместе тремя водородными связями, а пары А-Т — только двумя.
Во время репликации (копирования, удвоения) две цепочки разделяются, и на
каждой по принципу комплементарности синтезируется новая цепочка, как показа-
но на рисунке ниже. Синтез осуществляется при помощи фермента ДНК-зависимой
ДНК-полимеразы. Одна из двух новых цепей синтезируется подряд, без перерывов,
потому что направление ее синтеза совпадает с направлением «расплетания»
двойной спирали исходной молекулы ДНК. Вторая цепь синтезируется кусочками,
задом наперед. Эти кусочки называются «фрагментами Оказаки» (в честь их пер-
вооткрывателя , японского молекулярного биолога Редзи Оказаки, пережившего
бомбардировку Хиросимы и скончавшегося в 1975 году в 45-летнем возрасте от
лейкемии). В итоге получаются две одинаковые молекулы, каждая из двух компле-
ментарных цепей.
На приведенном рисунке возникла мутация — в правой молекуле напротив одного
из гуанинов (Г) случайно встал аденин (А) вместо цитозина (Ц). Скорее всего,
эта мутация будет замечена и исправлена специальными ферментами, функция ко-
торых как раз и состоит в исправлении подобных ошибок. Починку «неправильных»
или поврежденных участков ДНК называют репарацией.
А:Т
Т:А
И
т *
А:Т
Т:А
А
\
А
А
а:т
тч:а
А:Т
:т
Ц \
Г
Т
А
А
Ц
Т:а'
т1 а'
а':тГ
/
А7
f
Т
Г
Г I
т:а
4Т
ТА
ТА
ТА
A IAVT
Т:А
J: A
^7
Т:А
AT
Т:А
О
if
В нашем примере мутация возникла случайно. Впрочем, постойте. Случайно ли?
Она ведь возникла на стыке двух фрагментов Оказаки, а процесс соединения этих
фрагментов — некая особая операция, «технологически» отличающаяся от других
этапов репликации, расплетания спирали и присоединения комплементарных нук-
леотидов. Может быть, в нашей клетке забарахлила система соединения фрагмен-
тов Оказаки? А может, она забарахлила не случайно, а потому, что на нее ока-
зал воздействие какой-то внешний или внутренний фактор? А если он внутренний,
то клетка, наверное, может как-то контролировать его? А тогда, если хорошень-
ко разобраться, не может ли, в конечном счете, оказаться так, что сама клетка
«отдала команду» осуществить мутацию в данном месте ДНК? Если мы всерьез за-
думаемся над этими вопросами, то поймем, что было бы крайне удивительно, если
бы живая клетка за 4 миллиарда лет эволюции так и не выработала никаких меха-
низмов управления мутационным процессом. Ведь такие механизмы, во-первых,
вполне возможны, во-вторых, могли бы оказаться очень полезными.
Впрочем, нет повода сомневаться в том, что значительная часть мутаций дей-
ствительно возникает случайно — просто потому, что никакое копировальное уст-
ройство не может работать с абсолютной точностью. Кроме того, мутации могут
происходить и без репликации, пока ДНК находится в двухцепочечном состоянии,
— например, нуклеотиды могут претерпевать химические изменения под воздейст-
вием радиации, ультрафиолета или свободных радикалов, возникающих в ходе кле-
точного дыхания. Большинство ошибок в цепях ДНК будет замечено и исправлено,
но какую-то небольшую их часть пропустит, не заметит даже самый строгий моле-
кулярный «корректор».
Но мутации возникают не только потому, что невозможно копировать ДНК с аб-
солютной точностью. Мы привыкли думать, что мутации — это всегда некое нару-
шение, неправильность, ошибка, то есть что-то нежелательное, «мешающее нор-
мально жить». В действительности это не всегда так. Изменение наследственной
информации — неотъемлемая и необходимая часть жизни. Если бы геномы не меня-
лись , на нашей планете, возможно, до сих пор жил бы только один вид очень
примитивных микробов — тот самый Лука, общий предок всего живого (см. главу
«Происхождение жизни») . Впрочем, и он бы давно вымер, не смог бы долго про-
держаться с неизменным геномом. А если бы молекулы РНК копировались с абсо-
лютной точностью на этапе «преджизни», то и никакой Лука никогда бы не поя-
вился. Об этом, между прочим, свидетельствуют результаты экспериментов, про-
водимых исследователями РНК-мира. Для того чтобы в сообществе размножающихся
молекул РНК зародилось что-то новое и полезное, совершенно необходимо, чтобы
отдельные короткие молекулы, соединяясь в более длинные, могли обмениваться
друг с другом своими участками (обмен участками между разными молекулами ДНК
или РНК называется рекомбинацией). Рекомбинация — важнейший источник наслед-
ственной изменчивости наряду с «обычными» мутациями. В опытах с колониями РНК
рекомбинация происходит сама собой, бесконтрольно, но в живой клетке она на-
ходится под контролем разнообразных и сложных регуляторных систем.
Очень важно понять, что изменения наследственной информации нужны всему жи-
вому не только в геологическом масштабе времени, чтобы постепенно совершенст-
воваться в течение миллионов лет. Естественный отбор не может «заглянуть» так
далеко, поэтому и специальные механизмы для достижения столь отдаленных целей
не могут развиться. Но наследственные изменения нужны здесь и сейчас, они не-
обходимы нам постоянно. Особенно наглядно это видно на примере простейших жи-
вых систем — вирусов. Многие ученые, правда, не считают их живыми организма-
ми, поскольку они не могут размножаться без помощи чужих клеток, но все же
многие свойства живого у них есть. Да и кто в этом мире может похвастать аб-
солютной самодостаточностью? Разве что «отважный странник», подземный микроб,
о котором рассказано во второй главе.
На примере вирусов хорошо видно, как они нарочно позволяют определенному
количеству мутаций совершаться при каждой репликации. Таким способом они кон-
тролируют скорость мутирования и фактически превращают эволюцию в составную
часть своего жизненного цикла, чуть ли не в основной компонент своего поведе-
ния.
Вирусы-мутанты
помогают друг другу
в борьбе за выживание
Большинство вирусов, вызывающих болезни человека, — РНК-содержащие. Их на-
следственный материал хранится в форме РНК, а не ДНК, как у всех других форм
жизни. Среди немногочисленных белков, закодированных в геноме таких вирусов,
присутствует фермент РНК-зависимая РНК-полимераза, синтезирующий новые копии
вирусной РНК (см. главу 1).
Популяции РНК-содержащих вирусов неоднородны: они состоят из целого набора
генетически различающихся линий. Такие полиморфные популяции называют «квази-
видами» — по аналогии с видами настоящих живых организмов, которые тоже все-
гда полиморфны. Процесс удвоения (репликации) вирусной РНК происходит с боль-
шим числом ошибок (мутаций), благодаря чему могут быстро возникать новые ва-
рианты вируса. У полиовируса — возбудителя полиомиелита — на каждый акт реп-
ликации приходится в среднем 1,9 мутаций. При этом, конечно, появляется много
нежизнеспособных вариантов, однако эти потери оправданы. Высокая скорость му-
тирования помогает вирусам приспосабливаться к меняющимся условиям — к дея-
тельности иммунной системы хозяина, новым лекарствам, переходам от одного хо-
зяина к другому и из ткани в ткань.
Марко Виньюцци из Калифорнийского университета в Сан-Франциско (США.) и его
коллеги, изучающие полиовирусов, предположили, что дело здесь не ограничива-
ется поиском оптимального для данной ситуации варианта генома методом «слу-
чайного попадания». Возможно, возникающие в пределах квазивида вариации могут
помогать друг другу за счет разделения функций. Например, в одной линии может
возникнуть мутация, повышающая скорость репликации вирусной РНК, в другой ли-
нии — другая мутация, повышающая, например, эффективность самосборки белковых
молекул в вирусную оболочку — капсид. Поскольку обе линии живут и размножают-
ся в одной и той же хозяйской клетке, они могут пользоваться «достижениями»
друг друга. Эффективная РНК-полимераза первой линии будет размножать и геномы
второй, а более эффективные капсидные белки второй линии будут «упаковывать»
и РНК первой. Кстати, не исключено, что совместить обе эти мутации в одном и
том же геноме просто невозможно. Вирусный геном очень мал, и у него не так уж
много степеней свободы, то есть вариантов мутаций, совместимых с «жизнью» ви-
руса. Бывает даже, что у двух разных генов имеется общий кусок, настолько ви-
русы экономны в отношении наследственного материала. В этом случае изменения
в одном белке неизбежно будут приводить к изменениям в другом.
Возможность экспериментальной проверки гипотезы о взаимопомощи среди виру-
сов появилась благодаря тому, что Виньюцци и его коллегам удалось обнаружить
и выделить мутантную форму полиовируса со «сверхточной» РНК-полимеразой, со-
вершающей очень мало ошибок при копировании РНК. Эта мутация происходит из-за
замены глицина (G) серином (S) в 64-й позиции РНК-полимеразы. В соответствии
с этим мутация получила название G64S. Эта мутация ведет к тому, что вирусы
начинают мутировать с аномально низкой частотой —0,3 мутации на геном вместо
положенных 1, 940 .
Первым делом ученые проверили, действительно ли скорость мутирования важна
для приспособляемости вирусов. Это подтвердилось: вирусы G64S приспосаблива-
лись к неблагоприятным условиям (лекарствам, ингибиторам) гораздо медленнее
своих «диких» собратьев. Кроме того, вирусы G64S оказались неспособны прони-
кать в спинной и головной мозг зараженных мышей, как это делают нормальные
полиовирусы с высокой скоростью мутирования. Однако искусственное увеличение
темпа мутагенеза привело к тому, что вирусы G64S стали проникать в мозг и
размножаться там. При этом полиморфизм вирусной популяции вырос, но мутация
G64S сохранилась у всех линий.
Исследователи провели серию остроумных экспериментов, показывающих, что об-
щая «эффективность» популяции вирусов зависит не от наличия в ней какой-то
одной особо удачной линии, а от кооперативного взаимодействия нескольких раз-
ных линий, помогающих друг другу. Например, удалось показать, что вирусы G64S
сами по себе не способны проникать в мозг, но успешно проникают туда в компа-
нии с другими вирусными популяциями — как «дикими», так и другими G64S, пред-
варительно подвергавшимися действию искусственных стимуляторов мутагенеза.
Следовательно, одни линии вирусов помогают другим преодолеть гематоэнцефали-
ческий барьер41.
Эксперименты блестяще подтвердили гипотезу о взаимовыгодной кооперации меж-
ду разнообразными мутантами, составляющими вирусную популяцию — квазивид. Это
означает, что эволюционирующей единицей является не отдельная линия и тем бо-
лее не один вирус, а весь квазивид в целом, причем внутреннее разнообразие —
залог успеха. Одна линия, даже самая совершенная, всегда будет проигрывать
полиморфному квазивиду, включающему несколько взаимодополняющих линий42.
40 Мутация G64S закрепилась у вирусов в процессе приспособления к одному из ле-
карств, которое делает скорость мутирования у вирусов выше допустимого предела.
41 Утолщение оболочки кровеносных сосудов мозга препятствует проникновению крупных
молекул и частиц, в том числе вирусов, из крови в мозг. Аналогичным образом устроен
у млекопитающих и «вейсмановский» гематотестикулярный барьер, препятствующий проник-
новению вирусов в семенники (см. главу «Наследуются ли приобретенные признаки?»).
42 Vignuzzi et al. Quasispecies diversity determines pathogenesis through coopera-
tive interactions in a viral population // Nature. 2006. V. 439. P. 344-348.
Частота мутаций
под контролем
Ярким примером того, как живые организмы регулируют скорость мутагенеза вы-
годным для себя образом, является имеющаяся у бактерий система «спасения уто-
пающих» — SOS-response. Бактерия, как и любая живая клетка, не может «рассчи-
тать», какое именно изменение генома ей в данный момент выгодно, и не может
перекроить свои гены по заранее намеченному плану. Но иногда, когда условия
жизни становятся невыносимыми, бывает выгодно увеличить частоту возникновения
случайных мутаций во всем геноме или в отдельных его участках. И делать это
бактерии умеют. Например, у кишечной палочки обнаружены так называемые SOS-
гены, включающиеся в экстренных случаях. Один из них — ген dinB — кодирует
склонную к ошибкам ДНК-полимеразу. В благоприятных условиях у бактерии вклю-
чены гены обычных, редко ошибающихся ДНК-полимераз, и скорость мутирования
остается низкой. Активизация гена dinB приводит к резкому увеличению частоты
мутаций. В некоторых смертельно опасных ситуациях это может оказаться спаси-
тельным для гибнущей популяции микробов: вдруг какой-то из возникших мутантов
окажется более жизнеспособным в данных условиях? Бактерии, таким образом, ис-
пользуют увеличение частоты мутирования как экстренную меру самосохранения.
Молекулярные биологи уже научились ловко манипулировать этим свойством бакте-
рий и активно используют его для создания различных экспериментальных генно-
инженерных конструкций.
Бактерия Salmonella typhimurium.
Недавно шведские ученые при помощи таких конструкций сумели доказать, что у
бактерий имеются эффективные средства для уменьшения вреда, приносимого нака-
пливающимися в геноме мутациями43. Исследователи задались вопросом: как меня-
ется жизнеспособность организма в ходе накопления в геноме случайных мутаций?
Для своего эксперимента ученые создали настоящее чудо генной инженерии — бак-
терию с регулируемой скоростью мутагенеза. Для этого они соединили ген dinB,
кодирующий склонную к ошибкам ДНК-полимеразу, с промотором, который активиру-
ется сахаром арабинозой. Промотор — это регуляторный участок ДНК, который оп-
ределяет, как и когда будет работать соседний с ним ген. Получившуюся конст-
рукцию вставили в геном бактерии Salmonella typhimurium. Это дало возможность
43 Sophie Maisnier-Patin, John R. Roth, Asa Fredriksson, Thomas Nystrom, Otto G Berg
& Dan I Andersson. Genomic buffering mitigates the effects of deleterious mutations
in bacteria // Nature Genetics. 2005. V. 37. № 12. P. 1376-1379.
очень тонко регулировать скорость мутагенеза генно-модифицированной бактерии,
просто меняя концентрацию арабинозы в среде. Чем больше арабинозы, тем актив-
нее работает ген dinB и тем больше ошибок-мутаций происходит при репликации
(копировании) молекулы ДНК.
О ..
щ •
е -0.1
§ -0.2
1 -0.3
О
§ -0,4
I -0.5
-0,6
I *^
I I > I I I I
0 50 100 150 200 250
Число мутаций на геном
Снижение жизнеспособности (вертикальная ось) по мере роста числа
мутаций сначала идет быстро, а потом замедляется (черные точки).
Наклонный пунктирный отрезок показывает, каким было бы снижение
жизнеспособности без эффекта «взаимной нейтрализации».
Авторы показали, что мутации у модифицированной бактерии происходят более
или менее случайным образом и распределяются равномерно по всему геному.
Жизнеспособность бактерий-мутантов оценивалась по скорости их размножения.
Оказалось, что по мере накопления мутаций жизнеспособность сначала снижается
быстро, но в дальнейшем, когда число мутаций переваливает за 3-5 десятков,
снижение жизнеспособности резко замедляется.
Оказалось, что снижение вредоносности мутаций по мере роста их числа связа-
но с деятельностью белков-шаперонов, обеспечивающих правильное сворачивание
(укладку) белковых молекул. Повышение числа мутантных (и потому неправильно
«свернутых») белков в клетке приводит к росту производства шаперонов, которым
иногда удается помочь мутантному белку свернуться правильно. Таким образом,
живая клетка в некоторых случаях способна не только регулировать скорость му-
тирования своего генома, но и эффективно справляться с вредоносным действием
возникших мутаций.
Наличие подобных компенсаторных механизмов дает организмам возможность без
чрезмерного ущерба для себя накапливать мутации, которые могут в дальнейшем
«пригодиться» естественному отбору для создания новых форм жизни. Ведь любая
мутация, вредная сегодня, может оказаться полезной завтра, когда условия пе-
ременятся .
Рост изменчивости в стрессовых условиях может происходить не только за счет
увеличения частоты мутирования, но и из-за сбоев в работе «компенсирующих»
механизмов — например, белков-шаперонов, которые до поры до времени «маскиру-
ют» значительную часть накопившихся мутаций, не дают им проявляться.
Конечно, склонные к ошибкам ДНК-полимеразы лишь увеличивают частоту мута-
ций, не влияя на их характер, который остается случайным. И, тем не менее,
существование такого механизма заставляет признать, что для живых организмов
мутагенез вовсе не является некой не зависящей от них внешней силой, слепым
«давлением энтропии», как полагали биологи еще пару десятилетий назад. Ошиб-
ка, частота возникновения которой целенаправленно регулируется, — это уже как
будто и не совсем ошибка, не правда ли?
Бывает и так, что разные участки генома мутируют с разной скоростью, причем
у каждого участка эта скорость довольно постоянна. По-видимому, это означает,
что одним генам организм «разрешает» мутировать чаще, чем другим.
«Адаптивное
мутирование»
Это явление обнаружено у бактерий-мутантов, которые в результате мутации
утратили способность использовать в пищу определенные виды Сахаров. Оказа-
лось, что если поместить этих бактерий в среду, где именно эти сахара являют-
ся единственным источником пищи, то бактерии сначала прекращают расти, а по-
том среди них появляются так называемые ревертанты — микробы, у которых в ре-
зультате обратной мутации функция испорченного фермента восстановилась. Так
вот, частота возникновения ревертантов оказалась во много раз выше, чем сле-
довало ожидать при случайном характере мутирования.
Вполне возможно, что в действительности в данном случае имеет место не на-
правленное мутирование, а лишь некая его имитация. Согласно одной из версий,
в стрессовых условиях происходит отбор микробов, у которых испорченный ген
дуплицирован (присутствует в нескольких копиях). Такие микробы получают пре-
имущество, поскольку испорченный фермент хоть и плохо, но все-таки расщепляет
данный сахар. Ну а потом уже среди этих размножившихся микробов возникают ре-
вертанты. Большое число копий испорченного гена повышает вероятность того,
что хотя бы в одной из них произойдет нужная мутация.
Целенаправленное
создание новых
генов
В некоторых случаях сомневаться в способности клетки направлять мутационный
процесс на нужные гены не приходится. Особенно ярко проявляется это в работе
иммунной системы позвоночных. Биологов и медиков давно интересовал вопрос,
каким образом удается белым кровяным клеткам — В-лимфоцитам — производить та-
кое огромное разнообразие антител, используемых для борьбы с различными ин-
фекциями .
Антитела — это белки, которые умеют безошибочно узнавать определенных бак-
терий, вирусов, а также любые чужеродные белки (и многие углеводы) и прикреп-
ляться к ним, что приводит к обезвреживанию самих возбудителей или выделяемых
ими токсинов. По приблизительной оценке, организм человека способен произво-
дить не менее миллиона разных антител. Даже если в организм вторгается совер-
шенно новый вирус, которого никогда раньше не было в природе, уже через не-
сколько дней в крови можно обнаружить антитела, которые безошибочно узнают и
«связывают» именно этого возбудителя и никакого другого!
Организм не может запастись заранее всеми необходимыми генами антител по
двум причинам. Во-первых, такое количество генов не поместится ни в каком ге-
номе : это привело бы к непомерному росту «расходов» на содержание в каждой
клетке громадного количества ДНК. Во-вторых, как бы ни был велик запас защит-
ных генов, в любой момент может появиться новая инфекция, для борьбы с кото-
рой не подойдет ни одно из имеющихся антител.
Для кодирования миллиона антител понадобилось бы четыре миллиона генов, по-
скольку каждое антитело состоит из четырех белковых молекул. Но ведь мы зна-
ем, что у нас не наберется и тридцати тысяч генов. Впрочем, еще задолго до
прочтения генома человека стало ясно, что гены большинства антител, образую-
щихся в крови при различных инфекциях, не закодированы в геноме изначально,
не передаются по наследству, а изготавливаются по мере необходимости из не-
большого числа генов-заготовок.
Это производство разделено на два этапа. Сначала, на ранних стадиях разви-
тия организма, гены антител формируются комбинаторным путем из унаследованно-
го от родителей набора заготовок. Число наследуемых заготовок сравнительно
невелико, но за счет комбинаторики из них можно собрать десятки и сотни тысяч
разных генов. Специальные белки режут геномную ДНК и переставляют кусочки с
место на место. Этот этап создания генов антител называется V(D)J-
рекомбинацией, потому что каждый ген антитела собирается из заготовок двух (V
и J) или трех (V, D, J) типов. Каждый В-лимфоцит продуцирует только один тип
антител. На этом этапе неизбежно возникают «неудачные» лимфоциты, опасные для
организма: они производят антитела, атакующие собственные антигены (молекулы)
организма. Такие лимфоциты выбраковываются, остальные сохраняются и размножа-
ются. Этот процесс называется клональнои селекцией. В результате человек или
животное получает большой набор В-лимфоцитов с антителами, способными атако-
вать почти любые молекулы, кроме тех, которые в норме присутствуют в данном
организме.
Когда в организм попадает инфекция, гены антител подвергаются более тонкой
«настройке». Те лимфоциты из имеющегося набора, чьи антитела проявляют наи-
большее сродство к новому чужеродному антигену, начинают вносить случайные
изменения в гены своих антител, точнее, в определенные участки этих генов —
так называемые вариабельные области, или V-области. Именно эти участки гена
отвечают за распознавание и связывание антигена (чужеродного белка или угле-
вода) .
Процесс внесения множественных изменений (мутаций) в гены защитных белков в
клетках иммунной системы называется соматическим гипермутированием. На основе
получающихся новых вариантов генов синтезируются антитела и проверяются на
эффективность, то есть на степень сродства к новому антигену. Это продолжает-
ся до тех пор, пока не будет найден высокоэффективный вариант гена, обеспечи-
вающий надежную защиту от нового возбудителя. Лимфоциты, производящие такое
антитело, начинают усиленно размножаться. Так формируется приобретенный имму-
нитет . После выздоровления некоторые лимфоциты могут долго сохраняться в ка-
честве клеток «иммунной памяти», что снижает риск повторного заболевания.
Соматическое гипермутирование — сложный и не до конца изученный процесс. Он
идет под контролем специальных ферментов и имеет отчасти «закономерный», а
отчасти «случайный» характер. Ключевую роль играет особый фермент — цитидин-
дезаминаза, который атакует нуклеотиды Ц (цитозины) в V-области гена антитела
и превращает их в урацилы (У) . Как мы помним, урацил в норме входит в состав
РНК, но не ДНК. Присутствие урацила в цепи ДНК служит «сигналом тревоги» для
ферментов, осуществляющих репарацию — починку поврежденных участков ДНК. Эти
ферменты находят урацил и начинают «исправлять ошибку». При этом они вырезают
вокруг урацила довольно большой кусок ДНК (длиной до 60 нуклеотидов), а потом
восстанавливают его, но делают это очень неаккуратно. В результате такой не-
точной починки вокруг того места, где находился урацил, возникает множество
мутаций. Показано, что присутствие цитозинов абсолютно необходимо для гипер-
мутирования и что мутации возникают на расстоянии не более 30 нуклеотидов от
каждого «испорченного» ферментом цитидин-дезаминазой цитозина (чаще всего —
на расстоянии 4-15 нуклеотидов). Остается неясным, почему система репарации,
обычно работающая очень качественно, в данном случае под видом «ремонта» за-
нимается «целенаправленным вредительством». Конечно, эволюционистов-
теоретиков не может не волновать крамольный вопрос: что если механизм гипер-
мутирования иногда включается не только в лимфоцитах?
Таким образом, у клетки есть возможность вполне целенаправленно изменять
свой собственный геном. Конечно, сделать процесс создания нужного гена по-
настоящему разумным клеткам так и не удалось. Они не могут исследовать новый
вирус и рассчитать, какое именно антитело в данном случае требуется. Им при-
ходится действовать «методом оптимизированного случайного поиска». Оптимизи-
рованного — потому что имеются хорошие заготовки и клетки знают, в какие уча-
стки этих заготовок следует вносить случайные изменения. И это уже немало!
Целенаправленное, сконцентрированное на строго определенных генах мутирова-
ние вовсе не является уникальной чертой лимфоцитов позвоночных животных. Мно-
гие бактерии, как выяснилось, проделывают примерно то же самое с генами своих
поверхностных белков — тех самых, к которым иммунная система создает антите-
ла. Бактерии делают это, чтобы обмануть иммунную систему, и едва ли стоит
удивляться, что два старинных врага сражаются друг с другом одним и тем же
оружием.
Гонококки обманывают
иммунную систему
По данным Всемирной организации здравоохранения, 78 миллионов человек в ми-
ре ежегодно заражаются гонореей. Гонококк (Neisseria gonorrhoeae) — облигат-
ный паразит человека: он может жить только в человеческом организме, и в ходе
эволюции микроб очень хорошо адаптировался к своей «среде обитания». Он не
хуже других бактерий умеет приспосабливаться к антибиотикам (см. главу «Рож-
дение сложности»). Однако главным врагом микроба испокон веков являются не
лекарства, а наша иммунная система, и гонококк выработал мощные средства
борьбы с ней, причем как оборонительные, так и наступательные.
Известно, что гонококк может подавлять размножение и активность лимфоци-
тов44. Но самое зловредное свойство микроба, из-за которого люди не могут вы-
работать против него устойчивый иммунитет, состоит в его способности быстро
менять структуру своих поверхностных белков — тех самых, по которым клетки
иммунной системы распознают паразита. Пока иммунная система учится распозна-
вать и обезвреживать какой-то штамм гонококка, он меняется, и размножившиеся
лимфоциты (с антителами и рецепторами, «настроенными» на поверхностные белки
бактерии) оказываются не у дел.
Главный поверхностный белок гонококка пилин меняется не за счет обычных
«случайных мутаций» — ошибок при копировании генома, а за счет контролируемо-
го процесса генной конверсии, то есть вполне целенаправленной замены одних
участков гена другими. Собственно говоря, это тот же самый механизм, благода-
ря которому наши лимфоциты вырабатывают миллионы разнообразных антител и ре-
цепторов, способных распознать практически любую заразу.
Можно сказать, что гонококк борется с иммунной системой ее же оружием. Од-
нако детали механизма генной конверсии сильно различаются в иммунной системе
и у гонококка. О том, как это происходит в иммунной системе, мы уже знаем.
Что же до гонококка, то в его геноме помимо активного пилинового гена сущест-
вует множество его неполных копий — «псевдогенов». Некоторые их участки пол-
ностью идентичны «образцу», тогда как другие сильно отличаются. Эти копии не
экспрессируются, молчат. Время от времени какой-либо участок активного гена
может быть заменен соответствующим фрагментом одной из копий. В результате
структура пилина меняется, и рецепторы иммунной системы перестают его узна-
44 Boulton I. С, Gray-Owen S. D. Neisserial binding to CEACAMl arrests the activa-
tion and proliferation of CD4+ T lymphocytes // Nature Immunology. 2002. V. 3. P.
229-236.
вать.
Молекулярный механизм замены одних участков генома другими у гонококка пока
~ 45 _
неизвестен, однако предложено несколько гипотетических моделей Все эти мо-
дели предполагают, что в каждой клетке гонококка должно присутствовать как
минимум два экземпляра генома. По-видимому, участки псевдогенов одного экзем-
пляра вставляются в активный пилиновый ген другого экземпляра. Это противоре-
чит устоявшимся представлениям, согласно которым бактерии являются гаплоидны-
ми организмами, то есть имеют один экземпляр генома в каждой клетке. В отли-
чие от бактерий большинство растений и животных — диплоидные организмы, у них
два экземпляра генома, или двойной набор хромосом в каждой клетке.
Ученые из Северо-Западного университета (Чикаго, США.) не только доказали,
что гонококк действительно является диплоидным организмом, но и выяснили, ка-
ким образом происходит репликация (удвоение) хромосом гонококка перед клеточ-
ным делением.
Авторы проводят интересную параллель с другим микробом46, у которого ранее
было отмечено присутствие более одного экземпляра генома, а именно с
Deinococcus radiodurans, который знаменит тем, что способен выдерживать чудо-
вищные дозы радиации, абсолютно смертельные для любого другого живого сущест-
ва. Дейнококк является тетраплоидом — в его клетках присутствует сразу четыре
копии генома. «Запасные» копии нужны этому обитателю ядерных реакторов для
оперативной репарации (починки) ДНК. Когда под воздействием радиации в одной
из хромосом возникают мутация или разрыв, запасные хромосомы, по-видимому,
используются как матрицы, с которых можно скопировать «правильные» последова-
тельности нуклеотидов в испорченную хромосому. В обоих случаях множествен-
ность генома (полиплоидия) развилась в связи с необходимостью внесения каких-
то осмысленных изменений в ДНК.
Контролируемая
перестройка
генома у
инфузорий
Один из самых поразительных примеров целенаправленной перестройки генома
демонстрируют инфузории. Это самые сложные из одноклеточных организмов и во-
обще верх того, что смогла создать эволюция на одноклеточном уровне. Строение
инфузорий во многом напоминает многоклеточных, даром что клетка всего одна.
Например, у многоклеточных животных различают линию генеративных клеток, ге-
ном которых обычно оберегается от всяческих изменений — ведь именно этот ге-
ном передается по наследству потомкам. Кроме того, имеется и линия соматиче-
ских клеток, геном которых может меняться по мере надобности. У инфузорий то-
же есть два генома — генеративный и вегетативный (соматический). Генератив-
ный, передающийся по наследству, геном хранится в маленьком ядре (микронукле-
усе) , содержит кучу мобильных элементов и некодирующих участков и в целом на-
ходится в нерабочем состоянии, если не сказать — в полном хаосе. Например,
многие гены в нем разорваны на куски и перемешаны. Но, тем не менее, это нор-
мальный, хотя и сильно запущенный большой эукариотический геном. Кстати, чис-
ло генов у инфузорий и у человека примерно одинаковое — порядка 30 тысяч. Ге-
ном микронуклеуса (МИК-геном) , естественно, не работает, да он и не смог бы!
45 Kline К. A. et al. Recombination, repair and replication in the pathogenic Neis-
seriae: the 3 Rfs of molecular genetics of two human-specific bacterial pathogens //
Molecular Microbiology. 2003. V. 50. P. 3-13.
46 Deborah M. Tobiason, H. Steven Seifert. The Obligate Human Pathogen, Neisseria
gonorrhoeae, Is Polyploid // PLoS Biology. 2006. V. 4. Issue 6.
Он служит только для передачи генов потомству при половом размножении.
Вегетативный — соматический — рабочий геном инфузории хранится в большом
ядре, так называемом макронуклеусе. Он по многим параметрам сильно отличается
от других эукариотических геномов. Он состоит из множества, иногда из многих
тысяч отдельных «нанохромосом». Это настоящие хромосомы, только очень малень-
кие , обычно содержащие всего один ген. Для каждой нанохромосомы, или МАК-
хромосомы, в макронуклеусе имеется очень большое число копий. Соответственно,
весь МАК-геном оказывается многократно сдублирован, то есть макронуклеус яв-
ляется полиплоидным, тогда как микронуклеус представляет собой диплоидное яд-
ро.
У инфузории окситрихи (Oxytricha) МАК-геном по размеру в целых 20 раз мень-
ше МИК-генома (50 млн. и 1 млрд. пар нуклеотидов соответственно; для сравне-
ния, у человека — 3,5 млрд., у бактерий — обычно до 10 млн.). Такое радикаль-
ное сокращение МАК-генома при его изготовлении из МИК-генома достигается за
счет выбрасывания всего «лишнего».
Инфузории (на фото — инфузория Oxytricha) — самые сложные из од-
ноклеточных организмов, и вообще — верх того, что смогла создать
эволюция на одноклеточном уровне. Строение инфузорий во многом
напоминает многоклеточных, даром что клетка всего одна.
Инфузории размножаются делением, при этом делятся оба ядра. Время от време-
ни инфузории соединяются попарно, чтобы обменяться наследственным материалом.
Этот процесс называется конъюгацией и рассматривается как особая разновид-
ность полового процесса. Во время конъюгации микронуклеус претерпевает мейоз,
то есть такое деление, в ходе которого число хромосом в поделившемся микро-
нуклеусе сокращается вдвое. Вместо одного диплоидного микронуклеуса у каждой
инфузории получается по два гаплоидных (на самом деле по четыре, но два из
них тут же разрушаются) . Каждая инфузория передает один из двух гаплоидных
микронкулеусов своей подруге, а второй оставляет себе. Микронуклеусы затем
сливаются. В результате каждая инфузория снова имеет один диплоидный микро-
нуклеус, в котором половина хромосом — ее собственная, а половина получена от
партнера. Затем инфузории разъединяются и продолжают жить как жили с той не-
большой разницей, что с точки зрения генетики каждая из них теперь преврати-
лась в свою собственную дочь.
Во время конъюгации или сразу после нее макронуклеус вместе со своим гено-
мом разрушается, а затем собирается заново. За основу берется генеративный
геном микронуклеуса, но он при этом подвергается радикальной перестройке. 95%
МИК-генома просто удаляется. «На выброс» идут практически все мобильные эле-
менты и некодирующие последовательности. Остаются чистые гены, почти без при-
месей. Но реорганизация генома не сводится к удалению не нужных здесь и сей-
час участков генома. Происходит также «распутывание» — сборка работоспособных
генов из разрозненных и перепутанных обрывков. Как мы помним, многие гены в
МИК-геноме разорваны на мелкие кусочки и перемешаны. В промежутках между эти-
ми кусочками могут находиться длинные некодирующие вставки.
Например, в МИК-геноме ген может иметь такую структуру: 2X7X5X4X8X1X3X6
(цифрами обозначены «рабочие» фрагменты гена, буквой X — ненужные вставки
различной длины). В МАК-геноме этот ген будет выглядеть так: 12345678.
Откуда клетка знает, в каком порядке нужно соединять обрывки? Ответ на этот
вопрос был получен лишь в конце 2007 года.
Исследователи из Принстонского университета установили, что для «распутыва-
ния» генетической информации инфузории используют образцы (матрицы), пред-
ставляющие собой молекулы РНК, считанные с нанохромосом макронуклеуса (МАК-
хромосом) перед тем, как макронуклеус был разрушен. Чтобы это выяснить, при-
шлось провести множество сложных экспериментов47.
Для проверки гипотезы о роли РНК-матриц в сборке МАК-хромосом исследователи
кормили инфузорий генно-модифицированными бактериями, производящими двухцепо-
чечные молекулы РНК, совпадающие по последовательности нуклеотидов с фрагмен-
том одной из МАК-хромосом. Эукариотические клетки относятся к двухцепочечным
РНК с опаской, принимают их за вирусов и начинают уничтожать все РНК с такой
же последовательностью нуклеотидов, в том числе и обычные, одноцепочечные.
Идея состояла в том, что, проглотив бактерию, инфузория сама уничтожит одну
из РНК-матриц, необходимых ей для сборки МАК-хромосом. Так и вышло. В резуль-
тате после конъюгации получились инфузории, у которых соответствующий участок
одной из МАК-хромосом оказался собран неправильно или вообще не собран — про-
сто оставлен в том виде, в каком он был в МИК-хромосоме. При этом все осталь-
ные МАК-хромосомы были собраны правильно.
Стало быть, РНК-матрицы действительно участвуют в программируемой пере-
стройке генома. Но что они собой представляют — являются ли они копиями целых
нанохромосом или отдельных их участков?
Исследователи стали выделять и анализировать РНК из инфузорий на разных
стадиях жизненного цикла. Выяснилось, что через несколько часов после конъю-
гации (как раз тогда, когда старый макронуклеус разрушается, а новый начинает
формироваться) в клетках появляются длинные молекулы РНК, соответствующие це-
лым МАК-хромосомам вместе с концевыми участками — теломерами. Через 30-50 ча-
сов после конъюгации эти молекулы исчезают.
Таким образом, перед тем как уничтожить макронуклеус вместе с вегетативным
геномом, клетка снимает «резервную копию» с каждой МАК-хромосомы. Эта копия,
представляющая собой молекулу РНК, в дальнейшем используется как образец для
сборки новых маленьких и аккуратных МАК-хромосом из того безобразия, которое
записано в МИК-хромосомах.
Следующий вопрос состоял в том, насколько точно РНК-матрицы регулируют про-
цесс сборки МАК-хромосом и можно ли управлять этим процессом, внедряя в клет-
47 Mariusz Nowacki, Vikram Vijayan, Yi Zhou, Klaas Schotanus, Thomas G. Doak, Laura
F. Landweber. RNA-mediated epigenetic programming of a genome-rearrangement pathway
// Nature. 2008. V. 451. P. 153-158.
ку искусственные РНК-матрицы. Исследователи синтезировали несколько молекул
РНК, похожих на «настоящие» РНК-матрицы, но с измененным порядком фрагментов.
Например, если для МИК-гена со структурой 2X7X5X4X8X1X3X6 правильная РНК-
матрица имеет вид 12345678, то в искусственной матрице какую-нибудь пару
фрагментов меняли местами: 13245678.
родительский макронуклеус (МАК)
родительская МАК-хромосома
РНК-матрица
а.транскрипция
Ь. транспорт
развивающийся макронуклеус
МИК-хромосома
i
4 5 в-
d достраивание
теломер
РНК-матрица, считанная с МАК-хромосомы перед разрушением макро-
нуклеуса, служит «ключом» для распутывания генетической информа-
ции, содержащейся в МИК-хромосоме. Черным цветом обозначены кон-
цевые участки хромосом — теломеры.
Впрыскивание таких матриц в инфузорий после конъюгации приводило к формиро-
ванию МАК-хромосом двух типов: одни воспроизводили правильный порядок фраг-
ментов , ведь правильные матрицы из клеток не удалялись. Другие — тот, который
присутствовал в искусственных матрицах. Таким образом, РНК-матрицы осуществ-
ляют весьма точное управление процессом сборки МАК-хромосом, и при помощи ис-
кусственных матриц можно направлять этот процесс в желаемую сторону.
Следующий важный вопрос — регулируют ли РНК-матрицы сборку только тех ге-
нов, которые в генеративном геноме перемешаны, то есть имеют неправильный по-
рядок фрагментов, или же этот механизм универсален и применяется ко всем ге-
нам без исключения?
Исследователи изготовили и ввели в инфузорий РНК-матрицы с неправильным по-
рядком фрагментов для тех генов, которые в генеративном геноме не перемешаны
и потому в «распутывании» не нуждаются. В итоге соответствующие гены в МАК-
хромосомах оказались неправильно собраны. Значит, механизм универсален.
Из этого, кстати, следует интересный эволюционный вывод. Поскольку у инфу-
зорий уже развилась универсальная система «распутывания» измельченных и пере-
путанных генов, дальнейшая фрагментация МИК-генов и перестановки их частей
уже не будут отсеиваться отбором. Ведь есть распутывающий механизм, ему все
равно, он все исправит. Видимо, потому-то МИК-геномы инфузорий и пришли по-
степенно в состояние хаоса.
Глядя на рисунок, можно понять, что система изначально могла развиться про-
сто для удаления лишних кусков генома, а «распутывающая» функция ее возникла
при этом автоматически, сама собой, как некий довесок — сначала ненужный, но
потом ставший необходимым.
Таким образом, информация о последовательности, в которой нужно сшивать об-
рывки генов генеративного генома, передается потомству инфузорий «неклассиче-
ским» способом — в виде молекул РНК. А ведь это не такая уж маленькая часть
наследственной информации!
Могут ли РНК-матрицы передавать потомству также и информацию о последова-
тельности отдельных нуклеотидов? До сих пор речь у нас шла только о последо-
вательности фрагментов генов, то есть о кусках длиной в десятки и сотни нук-
леотидов . Каждый ген, как известно, может существовать в виде нескольких ва-
риантов (аллелей), различающихся единичными нуклеотидными заменами или встав-
ками. Поэтому соответствие РНК-матрицы и собираемых на ее основе МАК-хромосом
далеко не всегда является абсолютным. Отдельные нуклеотиды могут различаться,
и это не мешает правильной сборке.
В принципе не исключено, что какие-то нуклеотидные замены могут передавать-
ся из РНК-матрицы в собираемую МАК-хромосому. Конечно, инфузориям нет смысла
переносить в МАК-хромосому все различия такого рода. Ведь тогда МАК-хромосомы
после конъюгации оставались бы полностью идентичными материнским, и конъюга-
ция потеряла бы всякий смысл. Но, как выяснилось, некоторые нуклеотидные за-
мены все-таки переносятся в МАК-хромосомы из РНК-матриц. Это, однако, проис-
ходит не по всей длине собираемого гена, а только в непосредственной близости
от мест «сшивки» фрагментов. Это очень важный факт, и он однозначно свиде-
тельствует о том, что в сшивке кусочков ДНК у инфузорий принимает участие
только что открытый механизм починки ДНК на основе РНК-матриц (см. раздел
«Новые способы работы с информацией» в главе 9).
Могут ли подобные системы «редактирования» генома, основанные на использо-
вании РНК-матриц, работать и у других организмов, а не только у инфузорий?
Почему бы и нет? Нужно искать. Череда открытий последних лет однозначно пока-
зывает, что живая клетка по-прежнему таит в себе множество неизвестных нам
молекулярных механизмов, в том числе и таких, которые используются для целе-
направленного изменения собственного генома.
И так далее
Мы рассмотрели далеко не все известные на сегодняшний день механизмы, по-
средством которых живые организмы могут управлять изменениями собственных ге-
номов или геномов своего потомства. Рассказать обо всех случаях такого рода в
рамках одной небольшой публикации едва ли возможно, но о некоторых из них все
же необходимо вкратце упомянуть — для полноты картины.
Метилирование ДНК
как средство
регуляции мутагенеза
Клетка располагает множеством разнообразных средств для регуляции активно-
сти генов, и одним из них является метилирование ДНК. Метилирование — это
прикрепление метильных групп (-СН3) к некоторым нуклеотидам. Мы еще вернемся
к этому явлению в главе «Наследуются ли приобретенные признаки?», а сейчас
хотелось бы обратить внимание только на одну особенность этого процесса. Как
выяснилось, метилирование нуклеотида повышает вероятность его мутирования.
Чаше всего метилируется цитозин (Ц). Метилированный цитозин с большой вероят-
ностью может самопроизвольно превратиться в тимин (Т). Таким образом, метили-
рованные нуклеотиды превращаются в «горячие мутационные точки». Между тем хо-
рошо известно, что метилирование ДНК — процесс отнюдь не хаотический, он идет
под управлением сложных молекулярных систем. Клетка метилирует не все нуклео-
тиды подряд, а лишь те, которые она «хочет» прометилировать. Что же получает-
ся? Получается, что клетка имеет возможность по собственному усмотрению раз-
мечать свой геном, расставляя на нем «горячие мутационные точки». Вот вам и
случайные мутации!
Природные генные инженеры
Если вы, дорогие читатели, считаете, что честь изобретения генной инженерии
принадлежит человеку, то мне придется вас разочаровать. Генная инженерия была
изобретена бактериями задолго до появления Homo sapiens. Мы же не только без
спросу позаимствовали чужую идею, но и самих изобретателей заставили себе
служить. Жертвой этого вопиющего нарушения авторских прав стал микроб
Agrobacterium tumefaciens из группы альфа-протеобактерий (то есть близкий
родственник предков митохондрий, о чем говорилось в главе «Великий симбиоз»).
У предков агробактерии, как и у многих добропорядочных микробов, имелся осо-
бый молекулярный аппарат, предназначенный для сексуальных контактов с другими
микробами. Микробный половой процесс сводится к внесению в цитоплазму других
микроорганизмов фрагментов своей ДНК в ходе конъюгации (подробно об этом го-
ворится в главе «Наследуются ли приобретенные признаки?»)48. Хитрая агробак-
терия стала использовать этот полезный аппарат для того, чтобы вводить свою
ДНК в клетки растений, на которых агробактерия паразитирует. В результате та-
кой инъекции бактериальные гены начинают работать в растительной клетке, это
приводит к усиленному делению клеток и образованию опухоли, в которой агро-
бактерия чувствует себя очень комфортно. Это самая настоящая генная инженерия
без всяких оговорок, то есть введение чужеродного генетического материала с
целью направленного изменения свойств организма-хозяина. Вся современная ген-
ная инженерия растений основана на нещадной эксплуатации агробактерии, кото-
рую заставляют вводить в клетки растений различные фрагменты ДНК по прихоти
экспериментаторов. Итак, мы видим, что от полового процесса до целенаправлен-
ного изменения наследственных свойств — один шаг. А если подумать еще немно-
го, то можно понять, что и шага-то делать не надо. Мы уже пришли. Ведь когда
одна бактерия вводит другой бактерии свои гены, разве она не меняет при этом
ее наследственные свойства? И разве этот процесс не происходит в известной
мере целенаправленно? И разве то, что именно и кому именно будет введено, не
может повлиять на эволюцию? Или, может быть, вы думаете, что бактериям все
равно, какие гены и кому вводить? Им это далеко не все равно, о чем свиде-
тельствуют имеющиеся у прокариот сложные механизмы химической коммуникации и
взаимного узнавания. Вот теперь мы подобрались вплотную к одному секрету, ко-
торый на самом деле лежит на поверхности. Все живые организмы заботятся о на-
следственности своего потомства, и естественный отбор вполне в состоянии под-
держать такие изменения, которые делают эту заботу более эффективной. Иными
словами, эволюция может создавать средства оптимизации самой себя. «Приспо-
собления для эволюции», считавшиеся запретными в классическом неодарвинизме
(СТЭ) , на самом деле не только возможны — они существуют и окружают нас бук-
вально со всех сторон.
Выбор брачного партнера
Я думаю, что читатели уже все поняли. Можно ли спорить с тем, что, выбирая
себе брачного партнера, мы тем самым манипулируем наследственными свойствами
(геномом) своего будущего потомства? Пользуясь случаем, хочу «разоблачить» и
еще одну устаревшую догму — о безвыборочном или равновероятном скрещивании
(панмиксии) в популяции. Впрочем, называть это представление догмой, пожалуй,
не стоит: биологи никогда не воображали, будто в природных популяциях скрещи-
вание происходит абсолютно безвыборочно. Однако отклонения от случайного
Хотя половой процесс у прокариот и инфузорий называется одинаково — «конъюгация»,
его механизмы очень сильно различаются. В отличие от инфузорий у бактерий при конъю-
гации один микроб является донором, а другой — реципиентом, то есть гены передаются
только в одном направлении. Кроме того, передается не весь геном, а только часть
его.
скрещивания традиционно считались несущественными. На самом деле формирование
брачных пар никогда не бывает случайным, всегда присутствует элемент выбора.
И выбор осуществляется если не на уровне организма, то на уровне половых кле-
ток. Процесс выбора регулируется множеством разнообразных механизмов, пора-
жающих своей сложностью и эффективностью. Даже бактерии не меняются генами с
кем попало: у них есть целый ряд способов отличить «своих» от «чужих» и вы-
брать из множества потенциальных партнеров оптимального. У эукариот способы и
алгоритмы выбора необычайно развились и усложнились. По-видимому, многие ор-
ганизмы умеют на основе химических сигналов оценивать степень своего родства
с потенциальным партнером. Результаты такого анализа учитываются при принятии
«брачного решения». Как показали исследования последних лет, механизмы разли-
чения своих и чужих на уровне многоклеточных организмов удивительным образом
связаны и переплетены с аналогичными механизмами на уровне отдельных клеток и
молекул. Иммунная система, задача которой — отличать свое от чужого на моле-
кулярном уровне, может участвовать и в оценке степени родства потенциального
партнера. В обоих случаях важную роль играют белки из надсемейства иммуногло-
булинов . Специальные эксперименты показали, что даже люди в состоянии извлечь
ключевую информацию о генотипе другого человека по запаху его одежды. Это ка-
жется фантастикой, но это факт. Например, показано, что женщине больше всего
нравится запах тела тех мужчин, чей генотип по определенным параметрам наибо-
лее сильно отличается от ее собственного. Рыбка колюшка в аналогичной ситуа-
ции предпочитает промежуточный вариант: не слишком генетически близкого, но и
не слишком неродственного самца. А есть и такие случаи, правда, редкие, когда
животные вполне целенаправленно стремятся к инбридингу — скрещиванию с бли-
жайшими родственниками. Совершенно очевидно, что смещение брачных предпочте-
ний в ту или иную сторону может влиять на эволюцию. Например, предпочтение
«максимально неродственных» может привести к межвидовой гибридизации. Мы
должны иметь в виду, что, вопреки расхожему мнению, межвидовые гибриды часто
оказываются вполне жизнеспособными и плодовитыми. Напротив, предпочтение
«максимально родственных» может привести к тому, что компактная группа осо-
бей, желающих скрещиваться только друг с другом, самоизолируется от остальных
представителей своего вида. В результате исходный вид разделится на два, и
произойдет акт видообразования.
Эволюция
механизмов
адаптации
Процесс приспособления, или адаптациогенез, составляет главное содержание
эволюции. Он может идти на разных уровнях: строения организма, его поведения,
структуры коллектива. На всех этих уровнях можно наблюдать постепенный пере-
ход от первичных, примитивных, медленных способов адаптации, основанных на
случайных мутациях и отборе, к более эффективным и быстрым. Роль случайности
снижается; роль закономерных, контролируемых процессов растет.
Исходным, базовым алгоритмом эволюции является случайный поиск, или метод
проб и ошибок. Он и сегодня остается очень важным. Однако на всех уровнях
адаптациогенеза можно наблюдать постепенное развитие механизмов, сужающих
рамки случайности и, таким образом, оптимизирующих этот алгоритм. Впрочем,
полностью исключить элемент случайного поиска, по-видимому, невозможно не
только в биологической, но даже и в социальной и культурной эволюции челове-
ка.
1. Уровень строения организма включает все биохимические, физиологические и
морфологические признаки. Уже на этом уровне многие формы жизни выработали
более быстрые и эффективные способы приспосабливаться, чем случайный поиск,
осуществляемый по схеме «случайные мутации + отбор». В этой главе мы познако-
мились с некоторыми из таких способов. К механизмам оптимизации эволюционного
«случайного поиска» относятся регуляция темпов мутагенеза в зависимости от
условий, адаптивная конверсия генов в клетках иммунной системы и у бактерий,
геномный импринтинг (о нем мы поговорим подробнее в главе «Наследуются ли
приобретенные признаки»). Например, при формировании нового антитела и комби-
нирование фрагментов ДНК, и гипермутирование — процессы, лишь отчасти случай-
ные . Границы случайности в данном случае жестко предопределены. Комбинируются
фрагменты ДНК из строго определенного ограниченного набора, гипермутирование
происходит лишь в строго определенном участке гена, а отбор клонов лимфоцитов
делает весь процесс однозначно направленным. В результате конечный итог по-
добной «последовательности случайных событий» становится строго детерминиро-
ванным. Эффективное антитело против нового возбудителя непременно будет выра-
ботано — жаль только, что не всегда это происходит достаточно быстро, иначе
мы никогда не умирали бы от инфекционных болезней. Такой механизм достижения
цели можно назвать «оптимизированным случайным поиском». Избирательное скре-
щивание родительских пар представляет собой один из способов управления при-
способленностью потомства (рекомбинационная изменчивость). Этот процесс кон-
тролируется сложнейшими механизмами выбора брачного партнера. Только при слу-
чайном скрещивании родители не могут воспользоваться этим способом повлиять
на адаптивность потомства, но в природе безвыборочного скрещивания практиче-
ски никогда не бывает. По мере роста уровня организации живых существ увели-
чивалась сложность и эффективность механизмов выбора брачного партнера, а
значит, рекомбинационная изменчивость становилась все менее случайной. Из
всего этого можно сделать очень важный вывод: эволюционируют не только орга-
низмы, виды, сообщества; эволюционируют также и механизмы эволюции. Кроме то-
го , важнейшим способом усовершенствования адаптивных возможностей организмов
является развитие адаптивных модификаций, о которых мы говорили в главе «Рож-
дение сложности». Между прочим, приобретенный иммунитет — это яркий пример
адаптивной модификации, если смотреть на дело с точки зрения целостного орга-
низма, а не с точки зрения отдельных лимфоцитов, размножающихся в нашем теле
подобно каким-нибудь симбиотическим амебам.
2. На уровне индивидуального поведения четко прослеживается переход от пре-
имущественно наследственных, генетически детерминированных стереотипов пове-
дения к более пластичным, основанным на обучении. Точно так же, как в случае
с иммунитетом выгоднее передавать потомству не жестко детерминированное сред-
ство борьбы с конкретной инфекцией, а универсальную способность к «обучению»
лимфоцитов, в эволюции поведения оказалось выгоднее передавать не жестко за-
крепленную манеру поведения, а обучаемость. Развитие способности к обучению —
крупнейшее эволюционное достижение, к которому земная жизнь шла очень долго и
постепенно. Даже у одноклеточных эукариот уже есть зачаточные способности к
обучению. У них обнаружены простейшие формы памяти, такие как «привыкание» и
«сенсибилизация», то есть ослабление или, наоборот, обострение чувствительно-
сти к повторяющимся стимулам в зависимости от их «важности». Что уж говорить
о таких высокоорганизованных животных, как муравьи или осьминоги.
3. На уровне социума идет социальный адаптациогенез. Огромное множество жи-
вых организмов — от бактерий до млекопитающих — ведут в той или иной форме
общественный образ жизни. Социум как целое обладает определенными системными
характеристиками, которые могут быть в большей или меньшей степени адаптивны-
ми. Здесь тоже наблюдается переход от генетически детерминированных форм со-
циальных отношений к более гибким вариантам, при которых социум может адек-
ватно, то есть адаптивно, реагировать на изменения среды. Например, число и
размеры дочерних колоний у муравейника могут определенным «разумным», то есть
адаптивным, образом меняться в зависимости от доступности ресурсов. «Есть все
основания рассматривать образование отводков как механизм поддержания числен-
ности семьи на определенном, оптимальном для данного вида в данных условиях
уровне», — пишет крупнейший специалист по муравьям А. А. Захаров в книге «Му-
равей, семья, колония». Многие животные и даже микроорганизмы живут поодиноч-
ке, пока условия благоприятны, а при ухудшении условий собираются вместе и
формируют слаженный коллектив, что повышает шансы на выживание (см. сюжет
«Способность к сложному коллективному поведению может возникнуть благодаря
единственной мутации» в главе «Рождение сложности»). Однако в целом возможно-
сти такого рода у всех дочеловеческих форм жизни, включая даже высших обезь-
ян, крайне ограниченны. Человек стал первым в истории Земли гиперсоциальным
животным. Этим термином обозначают способность к практически не ограниченно-
му, и в том числе адаптивному, изменению структуры социума. Только человек
способен в зависимости от условий радикально менять структуру своих коллекти-
вов, правила взаимоотношений с сородичами, способы добычи пропитания, устрой-
ство семьи и т. д. Многие антропологи считают эту особенность чуть ли не са-
мой главной, базовой отличительной чертой человека, которая и вывела наш вид
на качественно иной уровень эволюционного развития. Получается, что появление
человека разумного стало естественным и закономерным результатом общей эволю-
ционной тенденции к росту эффективности механизмов адаптациогенеза, к оптими-
зации эволюционного «случайного поиска». Не потому ли человек смог стать до-
минирующим видом на нашей планете?
ГЛАВА 8. НАСЛЕДУЮТСЯ ЛИ
ПРИОБРЕТЕННЫЕ ПРИЗНАКИ?
От Ламарка
к Дарвину
Отдельные эволюционные идеи высказывались учеными и философами еще в антич-
ные времена49, и в новое время традиция продолжилась. Дарвин был далеко не
первым, кто выступил против догмата о неизменности раз и навсегда сотворенных
видов. Считается, что первую более-менее последовательную и внутренне непро-
тиворечивую теорию эволюции живых организмов придумал в начале XIX века Жан
Батист Ламарк. В основе теории Ламарка лежали два предположения: о наследова-
нии приобретенных признаков и о внутренне присущем всему живому стремлении к
совершенству.
Первое из этих предположений объясняло, почему организмы так хорошо приспо-
соблены к условиям своего существования. В течение жизни они используют свои
органы по-разному: одни — чаще и интенсивнее, другие — реже и слабее. Те ор-
ганы, которые все время тренируются, крепнут и растут, а те, которые почти
или вовсе не используются, постепенно уменьшаются и слабеют. Небольшие изме-
нения, возникающие вследствие такой избирательной тренировки органов, переда-
ются по наследству потомкам. Например, если животное питается листьями высо-
ких деревьев, ему приходится все время вытягивать шею. Шея тренируется, креп-
нет и немножко удлиняется. Потомство такого животного получит уже от рождения
немного более длинную шею. Так, по мнению Ламарка, появились жирафы. Если ка-
кая-то птица перестает летать, переходит к наземной жизни, то ее крылья от
49 Автором двух важнейших эволюционных идей, или моделей, — естественного отбора и
«блочной сборки» — был древнегреческий философ Эмпедокл. Он полагал, что живые суще-
ства возникли в результате случайного комбинирования частей — голов, рук, ног и т.
п. — и последующего отбора удачных вариантов. Другой древний философ, Анаксимандр, в
полном соответствии с данными современной науки утверждал, что предками людей были
рыбы.
долгого неупотребления атрофируются. Так возникли нелетающие птицы с рудимен-
тарными крыльями.
Второе предположение Ламарка — внутренняя «тяга к совершенству». Оно объяс-
няло постепенное усложнение организмов, появление новых органов и тканей.
Время от времени некоторые ученые продолжают возвращаться к этой идее Ламар-
ка. Например, известный палеонтолог, эволюционист и философ-богослов Пьер
Тейяр де Шарден, один из первооткрывателей знаменитого пекинского синантропа,
полагал, что неуклонно происходящее в ходе эволюции повышение уровня органи-
зации живых существ не может быть объяснено отбором случайных мутаций и слу-
жит доказательством присутствия какой-то особой направляющей силы. Шарден на-
зывал ее «радиальной энергией», потому что, по его мнению, она движет эволю-
цию к некому абсолютному средоточию, или центру, к «точке Омега».
Что же изменил Дарвин в теории Ламарка? Он отказался от второй посылки сво-
его предшественника, от «тяги к совершенству», и предложил гораздо более про-
стой и понятный механизм эволюционных изменений, которого теория Ламарка не
предусматривала, — естественный отбор. Механизм эволюции, по Дарвину, основан
на четырех базовых принципах. Первый из них — это борьба за существование:
она происходит оттого, что каждый организм производит заведомо больше потом-
ков , чем способно выжить. Второй — изменчивость: Дарвин еще не знал о генети-
ке и не мог точно сформулировать причины и законы изменчивости, но сам факт
был очевиден — все организмы немножко разные. Третий принцип — избирательное
выживание или избирательное размножение наиболее приспособленных — это и есть
собственно естественный отбор. И наконец, четвертая основа эволюции — это на-
следственность , благодаря которой свойства, обеспечившие данной особи победу
в борьбе, передаются ее потомству.
В силу странного стечения обстоятельств сейчас, когда противопоставляют
«дарвинизм» «ламаркизму», имеют в виду вовсе не отношение к идее «стремления
к совершенству», то есть вовсе не то, что действительно различало взгляды
двух великих естествоиспытателей. Сложилось ошибочное мнение, что Дарвин яко-
бы опроверг первый постулат Ламарка о наследовании приобретенных признаков и
доказал, что наследуются только признаки врожденные.
На самом деле Дарвин не только не отвергал эту ламарковскую идею — напро-
тив, она многократно упоминается в знаменитом «Происхождении видов» и призна-
50
ется Дарвином как самоочевидная Дарвин даже развил ее, выдвинув в книге
«Изменения животных и растений при одомашнении» теорию пангенеза. Этот факт
часто замалчивался сторонниками синтетической теории эволюции (СТЭ), которая
сложилась в начале — середине XX века из сплава учения Дарвина и достижений
генетики. И это неудивительно, ведь дарвиновская теория пангенеза противоре-
чит основной догме неодарвинизма — утверждению, согласно которому вся инфор-
мация может передаваться от генов к белкам только в одном направлении, то
есть по принципу «генотип определяет фенотип».
Теория пангенеза была призвана объяснить механизм наследования приобретен-
ных признаков. Дарвин предположил (не забываем, что дело происходило до воз-
никновения генетики), что в клетках организма образуются особые мелкие части-
цы. Он назвал их «геммулами», или «пангенами». Геммулы содержат в себе сведе-
ния о тех изменениях, которые клетки претерпели в течение жизни. С током кро-
ви геммулы разносятся по организму и, в конце концов, проникают в половые
клетки. Таким образом потомству передается информация о приобретенных призна-
ках.
Догма о невозможности наследования приобретенных признаков начала склады-
Например, в главе 14 «Происхождения видов» Дарвин отмечает: «Изучение наших до-
машних форм указывает нам, что неупотребление органов ведет к их уменьшению и что
результаты неупотребления могут передаваться по наследству».
ваться через несколько лет после смерти Дарвина, в основном благодаря усилиям
немецкого ученого Августа Вейсмана. Он показал, что, если отрубать крысам из
поколения в поколение хвосты, это не приводит к рождению бесхвостых крысят.
Другой эксперимент состоял в том, что черным мышам пересаживали яичники белых
мышей. У тех мышек, которым удавалось выжить после этой экзекуции, мышата ро-
ждались белые. На основании этих и других подобных экспериментов был сформу-
лирован принцип «Вейсмановского барьера»: клетки тела не могут передавать ин-
формацию половым клеткам. Следовательно, ни внешние воздействия, ни приобре-
тенный опыт, ни упражнение органов не могут приводить к адекватному или хотя
бы отчасти неслучайному изменению наследственности.
Развитие молекулярной биологии еще сильнее укрепило в сознании ученых этот
барьер, действительно превратив его в догму. Было установлено, что наследст-
венная информация записана в молекулах ДНК особым кодом, который был расшиф-
рован в 60-е годы XX века. Информация, записанная в ДНК, сначала должна быть
«переписана» на молекулу РНК. Затем специальные сложные молекулярные комплек-
сы — рибосомы — считывают информацию с молекулы РНК, синтезируя молекулу бел-
ка в точном соответствии с записанной в РНК инструкцией. Белки выполняют ог-
ромное множество функций, и, в конечном счете, именно они в основном опреде-
ляют облик организма (фенотип). Таким образом, информация движется в одном
направлении — от ДНК к РНК, от РНК — к белкам. Никаких механизмов переноса
информации в обратную сторону — от белков к РНК или от РНК к ДНК — поначалу
обнаружено не было, что и укрепило веру в невозможность такого переноса.
Потом, правда, оказалось, что есть вирусы, у которых хранилищем наследст-
венной информации служат молекулы РНК (а не ДНК, как у всех прочих организ-
мов) , и у них есть специальные ферменты, которые умеют осуществлять обратную
транскрипцию, то есть переписывать информацию из РНК в ДНК. Созданная таким
путем ДНК встраивается в хромосомы клетки-хозяина и размножается вместе с ни-
ми. Поэтому с такими вирусами очень трудно бороться (один из них — это вирус
ВИЧ). Но вот «обратной трансляции» — переписывания информации из белков в РНК
— так ни у кого и не обнаружили. По-видимому, такого явления в природе дейст-
вительно не существует.
Обратная
трансляция
У современных живых организмов обратная трансляция — переписывание информа-
ции с белка на РНК, — судя по всему, не встречается. Однако японскому иссле-
дователю Масаюки Насимото в 2001 году удалось экспериментально показать прин-
ципиальную возможность обратной трансляции. Насимото изготовил молекулу РНК,
которая может совершать два действия. Во-первых, узнавать аминокислоту арги-
нин и присоединяться к ней. Во-вторых, прикреплять к другой молекуле РНК, ко-
торая потом будет кодировать белок, триплет нуклеотидов АГГ, то есть кодон,
соответствующий аргинину. Рибозим, изготовленный Насимото, с полным правом
можно назвать «обратной тРНК». Напомню, что обычные транспортные РНК (тРНК)
узнают кодон на молекуле мРНК и прикрепляют к синтезируемой молекуле белка
соответствующую этому кодону аминокислоту. «Обратная тРНК», изготовленная На-
симото , совершает противоположное действие: узнает аминокислоту, входящую в
состав белковой молекулы, и прикрепляет к синтезируемой молекуле мРНК кодон,
соответствующий этой аминокислоте. Таким образом, осуществляется процесс,
прямо противоположный трансляции: информация переписывается не с РНК на бе-
лок, а, наоборот, с белка на РНК.
При помощи таких «обратных тРНК» обратная трансляция, по предположению На-
симото, могла происходить в РНК-мире (см. главу 1). Теоретически можно пред-
ставить себе ситуацию, когда существовало некое равновесие между процессами
прямой и обратной трансляции. Молекулы РНК «кодировали» белки, то есть управ-
ляли их синтезом, а белки, в свою очередь, управляли синтезом РНК, то есть
тоже «кодировали» их. В дальнейшем равновесие сместилось, и белки потеряли
способность к кодированию РНК. Возможно, решающую роль в смещении равновесия
сыграло то обстоятельство, что молекулы РНК в отличие от белков способны са-
мостоятельно контролировать синтез собственных копий на основе принципа ком-
плементарности. Иными словами, РНК и так кодируют сами себя, не нуждаясь для
этого в посредничестве белков.
С легкой руки одного из первооткрывателей структуры ДНК Френсиса Крика идея
об однонаправленной передаче информации в ряду ДНК — РНК — белок стала имено-
ваться «центральной догмой молекулярной биологии»51. Впрочем, позже Крик при-
знался, что использование термина «догма» принесло больше неприятностей, чем
оно того стоило.
Но всякое действие рождает противодействие. И иногда «лекарство оказывается
хуже болезни». В качестве уродливого и страшного противовеса догматизации
достижений генетики в СССР утвердилась лысенковщина. Крестьянский сын Трофим
Лысенко открыл способ придавать яровым культурам высокую продуктивность, вы-
держивая их некоторое время на холоде. Молодой экспериментатор был принят на
работу в лабораторию Н. И. Вавилова. Трагические последствия этого шага всем
известны. Лысенко, пользуясь далеко не научными аргументами и средствами,
подчинил себе руководство советской биологической наукой. Генетика и «вейсма-
низм» были объявлены лженауками. Официально была принята догма, противополож-
ная вейсмановской и основанная на базовом принципе ламаркизма: приобретенные
признаки наследуются; определяющим фактором наследственности являются не ми-
фические гены, а воздействие внешней среды. Как это ни парадоксально, Лысенко
считался последовательным дарвинистом. Он как раз обвинял Вавилова в отступ-
лении от теории Дарвина и на этом основывал свои «научные» обвинения.
Торжество лысенковщины в СССР и особенно репрессии против генетиков привели
к окончательной дискредитации ламаркизма на Западе и догматизации принципа
Вейсмана. Наука в очередной раз смешалась с политикой, что ей категорически
противопоказано. Это не пошло на пользу ни советской, ни западной биологии.
Два противоположных подхода к проблеме наследственности сошлись в смертельной
схватке. Вопрос состоял уже не в том, могут ли наследоваться приобретенные
признаки. Речь шла о борьбе двух «научно-социальных» систем: социалистической
лысенковщины и буржуазного вейсманизма. После того как Лысенко утратил свое
влияние, советская биология постепенно вернулась в русло мировой науки, вовсю
занявшись генетикой. Но последствия этой коллизии дают о себе знать и по сей
день: многих генетиков, как российских, так и зарубежных, до сих пор передер-
гивает при одном упоминании о возможности наследования приобретенных призна-
ков .
Однако реальные факты показывают, что приобретенные признаки иногда все же
могут передаваться по наследству. Безусловно, это происходит редко, это вооб-
ще нетипично, и случается такое только с некоторыми специфическими категория-
ми наследственных признаков, которые можно назвать «приобретенными» лишь с
определенной долей условности. Такие банальные вещи, как натренированные мыш-
цы или отрезанные хвосты, конечно, не наследуются никогда. Однако по мере то-
го, как наука все глубже проникает в тайны молекулярной организации живой
клетки, становится все очевиднее, что организмы не передают своим потомкам
приобретенные ими в течение жизни свойства не потому, что не могут, а потому,
что не хотят. С чисто «технической» точки зрения определенные возможности та-
кого рода у живых организмов имеются.
Crick F. Central Dogma of Molecular Biology // Nature. 1970. V. 227. P. 561-563.
Конечно, изменения, происходящие в клетках тела в течение жизни, отражаются
в первую очередь на белках, но не только. В каждой клетке есть и РНК, и ДНК,
и изменения могут затронуть и их тоже. Перед каждым клеточным делением все
молекулы ДНК в клетке удваиваются: ферменты ДНК-зависимые ДНК-полимеразы син-
тезируют точные копии имеющихся ДНК, которые потом распределяются между до-
черними клетками. Однако при копировании иногда возникают ошибки — мутации.
Если мутация возникает при образовании половой клетки, она, естественно, пе-
редается по наследству. В ортодоксальной СТЭ образца начала 60-х годов XX ве-
ка было принято считать, что мутации происходят совершенно случайно (теперь
мы знаем, что это не всегда так: см. главу «Управляемые мутации»). Так возни-
кает изменчивость, служащая материалом для естественного отбора.
Но мутации могут происходить при делении любых клеток тела, а не только при
образовании яйцеклеток и сперматозоидов. Такие мутации называются соматиче-
скими, от «сома» — тело. Они приводят к возникновению участков измененных
тканей. Соматические мутации могут быть вызваны различными воздействиями
внешней среды и в какой-то мере отражают эти воздействия, то есть несут некую
информацию о них. Маловероятно, чтобы такого рода информация могла оказаться
полезной следующим поколениям. Хотя возможно и такое, особенно если речь идет
о контролируемых перестройках генома соматических клеток, о которых мы гово-
рили в главе «Управляемые мутации». Тем не менее, если бы соматические мута-
ции могли наследоваться, это вполне можно было бы назвать «наследованием при-
обретенных признаков», хоть и не совсем в том смысле, который вкладывал в это
понятие Ламарк.
Классическая генетика отрицает возможность наследования соматических мута-
ций. Считается, что изменения клеток тела никак не могут отразиться на генах
половых клеток. По-видимому, в большинстве случаев это утверждение справедли-
во . Но природа, сколько бы мы ее ни изучали, всегда остается неизмеримо слож-
нее любых наших теорий, моделей и прогнозов. И из всякого придуманного нами
«закона» обязательно находятся исключения. В данном случае исключения тоже
существуют.
Горизонтальный
обмен генами
У одноклеточных организмов, понятное дело, нет разделения на соматические и
половые клетки. Их единственная клетка является одновременно и половой, и со-
матической, и любые произошедшие в ней изменения генов беспрепятственно и не-
избежно передаются потомкам. А гены у одноклеточных организмов изменяются до-
вольно часто. И это не только мутации. У них очень широко распространен так
называемый горизонтальный обмен генетическим материалом.
Три способа горизонтального обмена генами у бактерий:
■ конъюгация: две бактерии соединяются при помощи специальных белковых тру-
бочек — конъюгационных пилей, и бактерия-донор передает бактерии-
реципиенту часть своего генома;
■ вирусная трансдукция: вирусы, переходя из одной бактерии в другую, могут
прихватывать с собой куски бактериального генома;
■ естественная трансформация: иногда бактерия просто «всасывает» фрагменты
ДНК из окружающей среды и при определенных условиях встраивает их в свой
геном. Как мы помним из главы «Великий симбиоз», этот способ межвидового
генетического обмена мог сыграть важную роль в становлении эукариотической
клетки.
Когда бактерия встраивает в свою единственную кольцевую хромосому кусочки
чужого генома, она меняет свои свойства, то есть фактически превращается в
другой организм. Новые свойства — «приобретенные признаки», — естественно,
передаются потомству. В предельном случае возможна даже полная замена собст-
венного генома бактерии чужим геномом. Если последний получен от другого вида
бактерий, происходит нечто совершенно чудесное: бактериальная клетка в одно-
часье меняет свою видовую принадлежность. Микроб, относящийся к виду А,
трансформируется в микроба вида Б. Самое удивительное, что это не чисто тео-
ретические рассуждения, а экспериментально доказанный факт. Он был установлен
в 2007 году исследователями из института Крейга Вентера (США.) .
Первая в мире операция по пересадке генома позволила превратить один вид
бактерий в другой. Ученые из Института Крейга Вентера в течение последних 10
лет уверенно идут к великой цели — созданию искусственных микроорганизмов с
заданными свойствами. Практическое значение этих работ может оказаться огром-
ным. Например, планируется создание микробов, которые будут в больших количе-
ствах производить дешевое топливо. Генеральная идея состоит в том, чтобы ус-
тановить минимальный набор генов, необходимый для жизнеобеспечения бактерии,
добавить туда гены, кодирующие полезные функции, например, синтез водорода,
искусственно синтезировать спроектированный геном и внедрить его в живую бак-
терию. Ее собственный геном при этом должен быть каким-то образом удален.
Работы ведутся в основном с бактериями рода Mycoplasma.
Микоплазмы — довольно обширная (около 180 видов) группа паразитических бак-
терий, вызывающих всевозможные болезни у растений, животных и человека. Мико-
плазмы обладают рядом уникальных свойств, которые делают их весьма удобным
объектом для подобных исследований. Геномы микоплазм очень малы — от 600 до
1400 тыс. пар нуклеотидов — и хорошо изучены. На сегодняшний день полностью
прочтены геномы 14 видов микоплазм. В отличие от подавляющего большинства
других бактерий с маленькими геномами микоплазмы не являются облигатными
внутриклеточными паразитами. Они могут жить вне хозяйских клеток, поэтому их
можно выращивать обычным образом на питательной среде. Правда, среда должна
быть весьма богатой: микоплазмы очень требовательны в этом отношении, по-
скольку у них отсутствуют гены, необходимые для синтеза многих жизненно важ-
ных веществ. Наконец, у микоплазм нет жесткой клеточной стенки, характерной
для большинства бактерий. Клетки микоплазм окружены лишь тонкой и эластичной
мембраной. Это сильно облегчает обмен наследственным материалом между клетка-
ми.
Изучая геномы микоплазм, Крейг Вентер и его коллеги уже очень близко подо-
шли к пониманию того, что должен представлять собой «минимальный геном» буду-
щих искусственных микробов52. Синтез искусственных фрагментов генома уже на-
лажен, синтез целого бактериального генома — дело недалекого будущего. Биоло-
ги давно научились внедрять в бактерий отдельные фрагменты геномов. В этом
ученым большую помощь оказывают имеющиеся у микробов естественные механизмы
для обмена генетическим материалом. Однако до сих пор никому не удавалось пе-
ресадить целый геном в живую бактериальную клетку.
В июне 2007 года Крейг Вентер и его сотрудники сообщили о первой успешной
трансплантации целого генома от одного вида бактерий другому. Правда, ученые
пока сами не до конца понимают, как им это удалось и пройдет ли этот номер с
другими видами бактерий. Сделано было следующее. Ученые выделили геном из
бактерии Mycoplasma mycoides, которая вызывает пневмонию у коров. Геном этого
Lartigue С. et al. Genome Transplantation in Bacteria: Changing One Species to
Another // Science. 2007. V. 317. P. 632-638.
микроба, как и у большинства бактерий, представляет собой одну кольцевую мо-
лекулу ДНК. Геном был тщательно очищен от посторонних примесей, в том числе
от белков, и добавлен в культуру бактерий Mycoplasma capricolum, возбудителей
козьего полиартрита. Предварительно в геном М. mycoid.es были внесены особые
метки, в том числе гены устойчивости к антибиотикам. По этим меткам можно по-
том определить, успешно ли прошла трансплантация.
Спустя недолгое время среди клеток Mycoplasma capricolum появились бактерии
с признаками Mycoplasma mycoides. Обработав культуру бактерий антибиотиком,
ученые уничтожили тех микробов, которые не вобрали в себя чужую ДНК, а остав-
шихся подвергли тщательному изучению. По всем признакам это были самые на-
стоящие М. mycoides. Ни генов, ни белков, характерных для исходного вида
Mycoplasma capricolum, у них обнаружить не удалось. Антитела, избирательно
реагирующие на поверхностные белки Mycoplasma capricolum, не прикреплялись к
этим микробам, в отличие от антител, распознающих поверхностные белки
Mycoplasma mycoides.
Все это свидетельствует о том, что пересадка генома полностью удалась. Ав-
торы предполагают, что бактерии «проглатывали» чужую молекулу ДНК, и в первый
момент в них, вероятно, содержались оба генома вместе. Когда такая клетка де-
лилась, одна из дочерних клеток получала геном Mycoplasma capricolum, а дру-
гая — геном Mycoplasma mycoides. Последующая обработка антибиотиком уничтожи-
ла клетки первого типа.
Дальнейшие исследования покажут, можно ли проделывать подобную манипуляцию
с другими бактериями и другими геномами. Не исключено, что вобрать в себя це-
лый чужой геном способны только микробы, не имеющие клеточной стенки, — в
этом случае микоплазмы, скорее всего, и впредь останутся единственными объек-
тами для таких экспериментов. Так или иначе, проделанная работа сильно при-
близила Крейга Вентера к его заветной цели — созданию искусственного микроба.
По-видимому, эта цель может быть достигнута уже через несколько лет. Кстати
сказать, в США. сейчас активно дискутируются этические и юридические проблемы,
связанные с близящимся созданием искусственных организмов. Самые горячие спо-
ры идут по вопросу о том, можно ли будет эти организмы патентовать.
У многоклеточных горизонтальный обмен генами между неродственными организ-
мами играет гораздо меньшую роль. Вместо него развились более совершенные ме-
ханизмы перемешивания и перекомбинирования наследственной информации, связан-
ные с половым размножением. По сути дела это тот же самый горизонтальный об-
мен, но только замкнутый в пределах вида (разные особи смешивают свои гены в
потомстве, но с представителями других видов обмен генами резко ограничен). К
тому же половые железы у животных действительно ограждены от влияний внешней
среды особым «вейсмановским» барьером, через который могут проникать только
очень немногие вещества, в основном небольшие молекулы.
Тем не менее, многоклеточные животные и растения время от времени заимству-
ют гены у микроорганизмов, например, у паразитических или симбиотических бак-
терий. Поскольку эти бактерии, в свою очередь, могут заимствовать гены у сво-
их хозяев, а также переходить от одного хозяина к другому, они могут служить
посредниками при переносе генетического материала между разными видами хозя-
ев . По-видимому, это происходит чрезвычайно редко. Надежные экспериментальные
подтверждения переноса генов от бактерий к многоклеточным были получены толь-
ко недавно, и их пока очень мало. Но нужно иметь в виду, что редкость события
вовсе не обязательно означает, что его роль в эволюции мала и незначительна.
Ведь ключевые эволюционные преобразования сами по себе являются весьма редки-
ми событиями — это, что называется, «штучный товар». Роль горизонтального пе-
реноса генов в эволюции многоклеточных еще предстоит оценить, и некоторые
косвенные данные свидетельствуют о том, что она может быть весьма велика.
Рассмотрим один из случаев переноса генов бактерий в геном многоклеточного
животного, обнаруженный в 2007 году. В данном случае «донором» генетического
материала была паразитическая бактерия вольбахия, а «реципиентом» — мушка
дрозофила. Этот случай интересен тем, что в геном насекомого встроились не
отдельные гены, а целый бактериальный геном.
Вольбахия — паразитическая бактерия, обитающая в клетках многих наземных и
пресноводных членистоногих и круглых червей — филярий. Вольбахию называют
микробом-манипулятором, поскольку она научилась при помощи специальных регу-
ляторных белков управлять размножением и развитием своим хозяев. Например,
она умеет превращать самцов в самок, избирательно убивать зародышей мужского
пола, повышать плодовитость зараженных самок и даже делать бесплодными самок,
которые ею не заражены. О том, как ей это удается, можно прочесть в популяр-
ных статьях: А. В. Марков. Антимужской микроб., А. В. Марков, И. А. Захаров-
Гезехус. Бактерия вольбахия — повелитель мух. Вольбахия «впрыскивает» регуля-
торные белки в цитоплазму хозяина при помощи модифицированного конъюгационно-
го аппарата, то есть поступает примерно так же, как ее дальняя родственница
агробактерия — природный генный инженер, о котором мы говорили в заключитель-
ной части главы «Управляемые мутации». Вольбахия паразитирует в клетках бес-
позвоночных уже более 100 миллионов лет, да и ее предки — альфапротеобактерии
из группы риккетсиевых — тоже были внутриклеточными паразитами. За это время
вольбахия и ее хозяева успели приспособиться друг к другу. В ряде случаев
вольбахия даже повышает жизнеспособность своих хозяев, то есть выступает в
роли полезного симбионта. При таком долгом и тесном сожительстве было бы даже
странно, если бы какие-то фрагменты генома вольбахии время от времени не по-
падали в ядра клеток хозяина и не включались в хозяйский геном. Однако дока-
зать это удалось лишь в 2007 году.
Яйцо осы Trichogramma kaykai с множеством бактерий Wolbachia
(ярко окрашенные точки). Вольбахии концентрируются в удлиненном
кончике яйца, из которого впоследствии разовьются органы размно-
жения осы. Бактерии попадут в репродуктивные органы, затем — в
яйцеклетки, обеспечив себе гарантированный переход в следующее
поколение насекомых-хозяев.
Довольно часто в ходе выполнения проектов по прочтению геномов высших орга-
низмов (особенно насекомых) исследователи натыкались на фрагменты бактериаль-
ных последовательностей ДНК, но это обычно интерпретировалось как результат
загрязнения: предполагали, что при выделении ДНК из клеток исследуемого орга-
низма в пробы попало небольшое количество бактериальной ДНК. И соответствую-
щие участки ДНК просто не учитывались при «сборке» генома из прочтенных фраг-
ментов .
В середине 2007 года группа американских ученых предприняла широкомасштаб-
ный анализ таких «загрязнений» с целью найти реальные случаи переноса генов
вольбахии в геномы животных-хозяев53. Ученые выделяли ДНК из разных видов на-
секомых и круглых червей — филярий, а также анализировали накопленные в Ген-
банке54 данные по нуклеотидным последовательностям различных беспозвоночных.
Надо сказать, что многие прочтенные «вчерне» геномы до сих пор не подверга-
лись процедуре окончательной сборки. Они хранятся в компьютерных базах в виде
набора разрозненных, частично перекрывающихся обрывков разной длины. Если пе-
ренос генов от внутриклеточных бактерий к хозяевам действительно имеет место,
среди этих обрывков могут обнаружиться такие куски ДНК, которые содержат од-
новременно и эукариотические, и бактериальные участки. Именно такие обрывки и
интересовали исследователей.
В результате для четырех видов насекомых и четырех видов филярий удалось
получить бесспорные доказательства внедрения генов вольбахии в геном хозяина;
еще у трех видов это можно предполагать с большой долей вероятности.
Наибольшее внимание авторы уделили тропической плодовой мушке Drosophila
ananassae, потому что в геноме некоторых представителей этого вида обнаружи-
лись полные или почти полные копии генома вольбахии. Получается, что в ядрах
клеток этих мушек содержится полная генетическая информация сразу о двух раз-
ных организмах!
Для проверки этого результата ученые провели целый ряд специальных тестов.
Мушек вылечили от вольбахии антибиотиком и убедились, что лечение привело к
полному исчезновению внутриклеточных паразитов. Из вылеченных мух снова выде-
лили ДНК. Оказалось, что полный набор генов вольбахии по-прежнему присутству-
ет в пробах.
Затем проверили наследуемость этих генов по мужской линии. Дело в том, что
вольбахия, как и другие цитоплазматические бактерии (вспомним митохондрии!),
передается потомству только по материнской линии, вместе с цитоплазмой яйце-
клетки. В сперматозоиды вольбахия не проникает — они для этого слишком малы.
Поэтому потомство зараженной самки всегда оказывается зараженным, потомство
здоровой — здоровым, а от отца это не зависит. Однако если геном вольбахии
действительно встроился в геном хозяина, то он должен передаваться по отцов-
ской линии точно так же, как и по материнской, — вместе с ядерными хромосома-
ми.
Чтобы проверить это, скрестили вылеченных самцов D. ananassae, в хромосомы
которых встроился геном вольбахии, со здоровыми самками, в геноме которых ге-
ны вольбахии отсутствовали. В ДНК потомства обнаружились гены вольбахии, что
и стало решающим доказательством горизонтального переноса генов от паразита к
хозяину. Авторы также показали, что многие гены, заимствованные мухой у бак-
терии, активно работают (то есть транскрибируются, «считываются»).
Полученные результаты показывают, что межвидовой обмен генами может играть
более существенную роль в эволюции животных, чем считалось ранее. Источниками
нового генетического материала для животных могут служить не только вирусы и
мобильные генетические элементы, что было известно и ранее, но и бактерии.
Впрочем, пока трудно сказать, насколько широк спектр бактерий, гены которых
могут быть заимствованы животными.
Горизонтальный перенос генов привлекает в последние десятилетия пристальное
Julie С. Dunning Hotopp et al. Widespread Lateral Gene Transfer from Intracellu-
lar Bacteria to Multicellular Eukaryotes // Science. 2007. V. 317. P. 1753-1756.
54 http://www.ncbi.nlm.nih.gov/Genbank/
внимание ученых — и отнюдь не только потому, что в некоторых случаях он может
приводить к «ламарковскому» наследованию. Широкое распространение горизон-
тального переноса в живой природе заставляет пересмотреть еще одно основопо-
лагающее положение классического дарвинизма и СТЭ, а именно — точку зрения об
исключительно дивергентном характере эволюции. «Дивергенция» означает «расхо-
ждение» . Долгое время эволюционная теория базировалась на представлении о
том, что виды не могут обмениваться друг с другом наследственной информацией.
Как только вид делится на два, потомки теряют способность скрещиваться друг с
другом, между ними возникает репродуктивная изоляция. Так что после разделе-
ния они эволюционируют изолированно, сами по себе, по схеме «случайные мута-
ции + естественный отбор».
Дарвиновская
схема дивергенции
Классический взгляд на эволюцию отражен в знаменитой дарвиновской «схеме
дивергенции». Это единственный рисунок, которым Чарльз Дарвин сопроводил свой
великий труд о происхождении видов. Эволюционный процесс на нем представлен в
виде ветвящегося древа. Исходный вид делится на насколько ветвей — новых ви-
дов . Каждая ветвь может делиться дальше, и так до бесконечности. Отсутствие
поперечных перемычек между ветвями показывает, что каждый вид эволюционирует
сам по себе. Он должен самостоятельно изобретать все полезные адаптации, он
не может «посоветоваться» с другими видами, перенять их опыт, заимствовать их
«открытия».
л
гН
04
Ь1* V*
*н
л
nv
:u
1 1 ?—г
\ I t I
-Н 1 У 1-
п" г" m^_y_l' vu zu
1 Г
-^ 1-
Ц * h
\ I t
\ I /
-+-H i-
\ I J
\ I i
I t
-I—>-
\ I /
i. t-
_i \ u
,10
f«
mi°Elp F'a
wio
flO
L-m1
V-* W*
4|/l»
1<\
Vk* ■; -^
me
4i'we
v-^--'
VF NlV^KU'm'
fn'
NfoNj/i7
j*V/ VJ|(* ^fc*
V/m*
чу n*
4^1*
£tf
ч*. /
а+Ч^Ч^'а' \* ^lm* \ [
a'Hf/ ^Ф.У^'.гп3 ' = ■■■
a^- »» ^m* i 1 !
■'№ Ntfn' i 1 i i
\L..\ L_L. ! i .
I 4*-#z<
]. *' ■ ^:zJ
i i pW-\£*-i*
i i V/, !
1 14/\ i
А В
V L
\ 1
* 1
\ \
\ l
С D
1 i
1 /
! /
1 t
■ 1
E
i
t
i
\
1
F
j
/
f
j
J
r H
I I
1 I
1 I
1 I
I K L
i / /
f / /
Классическая схема дивергенции по Дарвину имеет вид древа, ветви ко-
торого , раз разделившись, уже никогда более не сливаются.
Примерно так же выглядят практически все эволюционные реконструкции, публи-
куемые в научных статьях вплоть до настоящего времени.
Однако некоторые исследователи выражали сомнение в том, что при такой изо-
лированной эволюции на основе мутаций и отбора жизнь успела бы за сравнитель-
но недолгий срок своего существования (4 млрд. лет) развиться от простейших
форм до таких высокоорганизованных, как млекопитающие или насекомые.
Схема эволюции жизни с учетом горизонтального переноса генов по-
хожа не на дерево, а на запутанную сеть.
С открытием горизонтального переноса генов между разными видами и даже цар-
ствами живых организмов ситуация изменилась, и эволюция предстает в несколько
ином свете. Получается, что «удачные изобретения» одних видов в принципе дос-
тупны другим и могут быть ими заимствованы. В этом случае биосфера предстает
единой информационной средой, в которой вирусы и различные мобильные генети-
ческие элементы (см. ниже) распространяют информацию примерно так же, как в
человеческом обществе благодаря устной и письменной речи достижения и откры-
тия одних людей становятся известными другим и могут ими использоваться.
Горизонтальный обмен генами вовсе не является бесконтрольным и неограничен-
ным. Он не таков даже у прокариот, которые обмениваются генами относительно
свободно и в каком-то смысле могут рассматриваться как единый, огромный и не-
вероятно полиморфный вид. Уже у прокариот выработались различные механизмы
выбора партнеров по обмену генами, и чаще всего такой обмен происходит все-
таки между родственниками. Эукариоты выработали гораздо более эффективные и
сложные адаптации для того, чтобы ограничивать и контролировать этот процесс.
Важнейшими из этих адаптации являются половое размножение и репродуктивная
изоляция видов. Именно появление полового размножения и репродуктивной изоля-
ции привело к формированию биологических систем нового типа — эндогамных ви-
дов. Эндогамия — избирательное скрещивание с себе подобными, со «своими», в
отличие от экзогамии — скрещивания с «чужаками».
Однако межвидовая репродуктивная изоляция у эукариот все-таки не абсолютна.
В частности, эукариоты тоже способны заимствовать чужие гены. Горизонтальный
обмен фактически превращает биосферу в единую «лабораторию» по изобретению
новых полезных наследственных признаков.
Правда, эффективность работы этой эволюционной лаборатории резко снижается
по мере роста сложности организмов и их приспособленности. Большая часть
адаптации у сложных организмов зависит не от одного-двух, а от множества ге-
нов, которые должны работать согласованно и которые влияют не только на дан-
ный признак, но и на множество других. Поэтому вероятность того, что привне-
сенный извне чужой ген окажется полезным, у высших организмов значительно
меньше, чем у низших. Именно этим, по-видимому, объясняется тот факт, что
многоклеточные организмы выработали эффективные, хотя и не абсолютные, сред-
ства защиты от горизонтального переноса.
Горизонтальный обмен генами
может заменить животным
половое размножение
Завершая разговор о горизонтальном переносе генов, нельзя не упомянуть о
замечательном открытии, сделанном в конце 2008 года. Как выяснилось, сущест-
вует целый класс многоклеточных животных, для которых горизонтальный обмен
генами с неродственными организмами — не исключение, а норма.
Разговор об этом открытии нам придется начать издалека. Как мы уже знаем,
горизонтальный обмен генами и половое размножение — явления родственные,
сходные по своему биологическому смыслу и эволюционной роли. В обоих случаях
происходит смешивание генов разных организмов в одном геноме — «межорганиз-
менная генетическая рекомбинация». Прокариоты не способны к настоящему поло-
вому размножению и практикуют горизонтальный обмен. Многоклеточные эукариоты
предпочитают размножаться половым путем, а горизонтальный обмен пытаются ог-
раничить . Но что происходит с теми многоклеточными, которые по тем или иным
причинам отказались от полового размножения? Могут ли они вернуться к более
древнему способу межорганизменной рекомбинации — горизонтальному обмену?
Существует множество теорий, объясняющих, почему половое размножение полу-
чило такое широкое распространение в живой природе. Животные довольно легко
утрачивают половое размножение и переходят к партеногенезу, то есть к разви-
тию из неоплодотворенных яиц. Такое не раз происходило и продолжает происхо-
дить в разных эволюционных линиях (лишь у одних млекопитающих переход к пар-
теногенезу принципиально невозможен, так как у них многие жизненно важные ге-
ны в яйцеклетке отключены, а их работающие копии могут быть получены только
со сперматозоидом — см. ниже врезку «Геномный импринтинг») . Однако все виды
(или небольшие группы видов) животных, лишенных полового размножения, являют-
ся молодыми, они лишь недавно произошли от «нормальных» двуполых предков. Это
значит, что виды, отказавшиеся от полового размножения, имеют тенденцию очень
быстро вымирать. Они не успевают дивергировать и дать начало, допустим, цело-
му бесполому семейству или отряду. «Очень быстро» по эволюционным масштабам
времени — это может означать десятки и сотни тысяч лет, в крайнем случае,
первые миллионы.
Бделлоидные коловратки (Bdelloidea) — микроскопические обитатели водоемов,
одни из самых мелких многоклеточных животных — представляют собой удивитель-
ное исключение. Это целый класс животных (около 400 видов), размножающихся
исключительно бесполым путем (партеногенетически). Никто никогда не видел
самцов бделлоидных коловраток, и, судя по всему, они живут так уже много де-
сятков миллионов лет.
Почему все животные не берут пример с коловраток? Бделлоидные коловратки —
главный камень преткновения для всех теоретиков, пытающихся объяснить биоло-
гический смысл полового размножения. Какое ни придумай объяснение, сразу же
возникает «проклятый» вопрос: если половое размножение такое полезное, как же
бделлоидные коловратки без него обходятся? И если бделлоидные коловратки на-
шли способ без него обходиться, почему другие животные не пошли по тому же
пути? Ведь половое размножение — весьма «дорогое удовольствие» с точки зрения
естественного отбора. При бесполом размножении вы передаете каждому потомку
все свои гены, а при половом — только половину.
Между тем естественный отбор по определению благоприятствует тем организ-
мам, которые наиболее эффективно «тиражируют» свои гены в следующих поколени-
ях. За половое размножение приходится платить двукратным снижением этой эф-
фективности. Выдающийся биолог Джон Мзйнард Смит назвал этот парадокс «двой-
ной ценой пола» (twofold cost of sex). Другой выдающийся биолог и популяриза-
тор науки, Ричард Докинз, в книге «Рассказ прародителя» подчеркивает, что лю-
бое теоретическое построение, указывающее на преимущества полового размноже-
ния, обязательно должно как-то объяснять парадокс бделлоидных коловраток (или
даже начинаться с такого объяснения).
Лишь в 2008 году этот парадокс удалось объяснить. По-видимому, коловратки —
единственные животные, которым удалось вернуться к более древнему и примитив-
ному способу межорганизменной рекомбинации — к широкомасштабному горизонталь-
ному обмену генами, который и заменил коловраткам половое размножение.
Ученые с факультета молекулярной и клеточной биологии Гарвардского универ-
ситета, похоже, сумели найти объяснение этой уникальной особенности бделлоид-
ных коловраток. В ходе изучения концевых участков хромосом бделлоидной коло-
вратки Adineta vaga они обнаружили множество генов, не встречающихся ни у ка-
ких других животных. Некоторые из этих генов явно имеют бактериальное проис-
хождение : их нуклеотидные последовательности почти идентичны бактериальным
аналогам. Другие столь же, несомненно, происходят от грибов, третьи — от рас-
тений. Полные геномы бделлоидных коловраток пока не прочтены. Авторы имели
возможность проанализировать лишь около 1% генома изучаемого животного (при-
мерно 1 млн. пар нуклеотидов). Были выявлены сотни генов, заимствованных ко-
ловратками у представителей других царств живой природы. Степень вероятности
того, что данный ген был заимствован не у животного, определялась по сходству
нуклеотидной последовательности гена с ближайшим аналогом за пределами живот-
ного царства, по сравнению с уровнем сходства между этим геном и его ближай-
шим «животным» аналогом. Понятно, что таким способом невозможно выявить гены,
заимствованные коловратками у других животных или, тем более, у других бдел-
лоидных коловраток.
Итак, бделлоидные коловратки активно заимствуют гены у других живых су-
ществ. Как мы помним, горизонтальный генетический обмен очень широко распро-
странен у прокариот (бактерий и архей) — он в определенном смысле «заменяет»
им половое размножение. Значительно реже меняются генами одноклеточные эука-
риоты, у которых есть также и настоящий половой процесс (попарное слияние по-
ловых клеток). Для многоклеточных горизонтальный генетический обмен — очень
большая редкость. Половое размножение, скорее всего, возникло как более безо-
пасная и эффективная альтернатива горизонтальному генетическому обмену.
Животные стараются всячески оберегать свои половые клетки от проникновения
постороннего генетического материала.
У бделлоидных коловраток барьеры, стоящие на пути проникновения чужой ДНК в
яйцеклетки, очевидно, сильно ослаблены. Это может быть связано с необычным
образом жизни этих микроскопических животных. Они живут в мелких лужах и от-
лично переносят высыхание на любой стадии жизненного цикла. Потом их, как
пыль, может перенести ветром в другую лужу. Однако при высыхании мембраны
клеток могут повреждаться, что облегчает проникновение чужеродной ДНК. При
высыхании также образуются разрывы в хромосомах, которые клеткам приходится
зашивать, когда коловратка снова размокнет. В ходе починки (репарации) разо-
рванных хромосом имеется большая вероятность случайного включения в хромосому
чужеродного фрагмента.
Ученые показали, что, по крайней мере некоторые из заимствованных генов ре-
ально работают в клетках коловраток и кодируют функциональные белки. Большин-
ство генов, заимствованных коловратками у бактерий, грибов и растений, коди-
руют ферменты, не входящие в состав сложных биохимических путей и каскадов, а
выполняющие какую-то самостоятельную биохимическую функцию. Это и понятно,
ведь именно такие гены могут оказаться полезными, если их заимствовать пооди-
ночке. Впрочем, есть указания и на то, что иногда гены заимствовались сразу
по два. Такие гены расположены в непосредственной близости друг от друга и в
геноме бактерий-доноров, и в хромосоме коловратки-реципиента.
Авторы55 не делали попыток выяснить, обмениваются ли бделлоидные коловратки
генами между собой. Это технически гораздо более трудная задача, чем обнару-
жение генетического обмена с бактериями и грибами. Однако едва ли коловратки,
охотно заимствующие гены у микробов и растений, имеют при этом какую-то осо-
бую систему защиты от инкорпорации генетического материала близких родствен-
ников .
Если же бделлоидные коловратки хотя бы иногда меняются генами друг с дру-
гом, то получается, что они на самом деле не отказались от идеи межорганиз-
менной генетической рекомбинации — перемешивания генов разных родителей в ге-
номе потомства. Они просто вернулись от продвинутого варианта такой рекомби-
нации (полового процесса) к более примитивному варианту — горизонтальному об-
мену, который был свойствен их далеким одноклеточным предкам.
Бделлоидные коловратки (Bdelloidea)
Eugene A. Gladyshev, Matthew Meselson, Irina R. Arkhipova. Massive Horizontal
Gene Transfer in Bdelloid Rotifers // Science. 2008. V. 320. P. 1210-1213.
Симбиоз способствует
наследованию признаков
«по Ламарку»
Помимо горизонтального обмена генами есть еще один важнейший механизм, бла-
годаря которому ветви эволюционного древа могут сливаться. Это образование
симбиотических систем — «сверхорганизмов» (см. главу «Великий симбиоз»). У
многих симбиотических организмов имеется интересная возможность передавать
своим потомкам «приобретенные признаки» таким образом, что создается полное
впечатление наследования «по Ламарку».
Рассмотрим, к примеру, тлей. В отечественной литературе можно встретить
описания экспериментов, которые вроде бы подверждают «ламарковское» наследо-
вание у этих насекомых56. Например, приспособление к новому кормовому расте-
нию у тлей может происходить в ходе партеногенетического (бесполого) размно-
жения, когда потомство развивается из неоплодотворенных яиц и является точной
генетической копией матери. При половом размножении происходит перемешивание
родительских генов, и могут появиться новые удачные генетические комбинации,
которые и будут подхвачены отбором. Но при бесполом размножении, казалось бы,
можно рассчитывать только на возникновение новых полезных мутаций, а это все-
таки вещь довольно редкая. Но тли, тем не менее, приспосабливаются к новой
диете сравнительно быстро — всего за десяток поколений. Возможно, дело тут в
том, что тли, как и все насекомые, питающиеся растительными соками, представ-
ляют собой симбиотические организмы. Как мы помним из главы «Великий симби-
оз», в клетках тлей живут бактериальные симбионты, которые обеспечивают хо-
зяина необходимыми питательными веществами, отсутствующими в растительном со-
ке . За жизнь одного поколения тлей в их клетках могут смениться несколько по-
колений бактерий, причем симбионты, получая необычную пищу (сок другого рас-
тения) будут подвергаться интенсивному отбору. Бактерий много, размножаются
они быстро, и вероятность возникновения удачных мутаций хотя бы у некоторых
из них достаточно велика. В результате насекомое передаст своему потомству
полезный «приобретенный признак» — бактерий-симбионтов с изменившимися на-
следственными свойствами.
Вирусы и мобильные
генетические элементы
Один из способов горизонтального обмена генами, от которого не защищены да-
же многоклеточные, — это вирусный перенос. Известно, что ДНК вируса может
встраиваться в геном клетки-хозяина, а потом снова отделяться от него и фор-
мировать новые вирусные частицы, которые могут заражать другие клетки. При
этом вместе с собственной ДНК вирус может случайно прихватить и кусочек ДНК
хозяина и таким образом перенести этот кусочек в другую клетку, в том числе и
в клетку другого организма. В большинстве случаев вирусы, размножающиеся в
клетках животных, все-таки не могут пробраться сквозь «барьер Вейсмана» и за-
разить половые клетки. Но все же иногда вирусная инфекция передается потомст-
ву. А ведь это не что иное, как наследование приобретенного признака! И не-
важно , что от такого признака обычно один только вред. Вирус ведь может при-
хватить с собой и какой-нибудь полезный кусочек ДНК, да и сами вирусные гены
могут «пригодиться» хозяину.
56 С. В. Мейен. Закон есть закон? 1974.
«Прирученные» вирусы —
источник эволюционных
новшеств
Хотя вероятность того, что встроившийся вирусный геном принесет хозяину
пользу, невелика, целый ряд таких случаев уже известен. Например, гены виру-
са, встроившегося в геном предка узконосых обезьян свыше 40 млн. лет назад,
нашли себе несколько применений. Они работают у обезьян и человека в плаценте
и выполняют там, по всей видимости, сразу три полезные функции. Первая из них
— управление слиянием клеток в ходе формирования наружного слоя плаценты.
Этот наружный слой состоит из слившихся клеток, и для его образования необхо-
дим какой-то способ нарушать целостность клеточных мембран, то есть протыкать
их. Вирусные белки хорошо умеют это делать. Вторая функция — защита эмбриона
от иммунной системы матери. Вирусные белки — отличные «специалисты» по подав-
лению активности иммунной системы. Наконец, третья функция состоит в защите
эмбриона от «диких» вирусов. Белки «прирученного» вируса прикрепляются к тем
поверхностным белкам человеческих клеток, за которые обычно «цепляются» дикие
вирусы, чтобы проникнуть в клетку. В результате диким вирусам становится не
за что «ухватиться», и они не могут заразить эмбрион.
Другой яркий пример «одомашнивания» вируса описан у наездников — паразити-
ческих насекомых, личинки которых развиваются в теле других насекомых, напри-
мер гусениц. Около 100 млн. лет назад предки наездников заразились вирусом и
«приручили» его. Геном вируса встроился в геном наездника, и вирусные гены
стали помогать насекомому-паразиту бороться с иммунной защитой своих жертв.
Самки многих наездников впрыскивают в тело жертвы кроме своих яиц еще и осо-
бые «вирусоподобные частицы», внутри которых находятся гены белков, подавляю-
щих иммунитет гусениц. Это позволяет личинкам наездника беспрепятственно раз-
виваться в теле жертвы. Как выяснилось, «вирусоподобные частицы» образуются
благодаря деятельности генов прирученного вируса, который давно уже стал не-
отъемлемой частью генома наездника.
Чуть ли не половина генома человека состоит из мобильных генетических эле-
ментов (МГЭ) — транспозонов и ретротранспозонов. У других животных и растений
дело обстоит примерно так же. У прокариот мобильных элементов меньше, но тоже
очень много. Типичный мобильный элемент представляет собой по сути дела упро-
щенный вариант вируса. МГЭ обладают всеми характерными свойствами вирусов,
кроме одного — инфекционности. Вирусы, как известно, могут свободно перехо-
дить из одной хозяйской клетки в другую и передаваться горизонтально, то есть
заражать другие организмы. МГЭ более ограничены в своих передвижениях. Они
могут активно размножаться и перемещаться в пределах генома — перепрыгивать с
места на место, встраиваясь в хозяйские хромосомы в разных местах и влияя на
работу близлежащих генов. Они, разумеется, передаются вертикально (от родите-
лей к потомкам, то есть наследуются), а иногда и горизонтально — подобно пол-
ноценным вирусам.
Мобильные генетические элементы впервые были обнаружены Барбарой МакКлинток
в 1951 году у кукурузы. Однако МГЭ слишком долго считались «генетической эк-
зотикой», их распространенность и эволюционная роль недооценивались, и в ре-
зультате свою заслуженную Нобелевскую премию за это открытие МакКлинток полу-
чила лишь в 1983 году, когда ей самой было уже за 80.
Поначалу эти подвижные вирусоподобные участки генома интерпретировали как
«геномных паразитов», «эгоистическую» или «мусорную» ДНК. В последнее время
все яснее становится огромная эволюционная роль этих элементов. Они придают
геному свойство, которое в зависимости от субъективного восприятия называют
либо «нестабильностью», либо «пластичностью». В первом случае подчеркивается
потенциальный вред геномных перестроек, индуцируемых МГЭ, во втором — их по-
тенциальная польза, создание материала для отбора, повышение полиморфизма и
приспособляемости.
«Прыжки» транспозонов придают причудливую окраску кукурузным початкам.
МГЭ придают геному нестабильность не только своими беспорядочными прыжками.
Если какому-нибудь транспозону удастся как следует размножиться, это значит,
что в геноме появится множество одинаковых фрагментов генетического «текста».
А это резко повышает вероятность ошибок в ходе копирования ДНК и рекомбина-
ции. Действительно, сложно не ошибиться при переписывании или редактировании
текста, изобилующего повторами.
Но геномные перестройки, индуцируемые МГЭ, не являются хаотичными. Напри-
мер, многие транспозоны способны встраиваться не в любое место генома, а
только туда, где есть определенные «предпочтительные» для данного МГЭ после-
довательности нуклеотидов. Это делает их передвижения в известной мере пред-
сказуемыми, закономерными. Распределение повторов по геному повышает вероят-
ность не любых, а строго определенных перестроек. Например, участок генома,
заключенный между двумя одинаковыми последовательностями (например, между
двумя копиями транспозона), с большой вероятностью может быть «потерян» при
репликации. Если такая потеря окажется выгодной, она будет поддержана отбором
и закрепится в череде поколений, если нет, отбор будет ее отбраковывать, но,
тем не менее, этот участок все равно будет выпадать из генома снова и снова
(такое явление наблюдается, например, у возбудителя чумы Yersinia pestis)57.
Как у бактерий, так и у высших организмов МГЭ могут служить своеобразными
«рецепторами стресса», резко активизируя свои прыжки в критических для орга-
низма ситуациях и приводя к вспышкам мутагенеза. Это может способствовать
приспособляемости видов. Когда условия жизни резко ухудшаются, то это по сути
дела означает, что имеющийся у организма геном перестает соответствовать тре-
бованиям среды. В этой ситуации рост изменчивости может оказаться единствен-
ным возможным выходом (см. главу «Управляемые мутации»). Все это придает эво-
люционным изменениям, происходящим при участии МГЭ, не совсем случайный ха-
рактер .
Г. Б. Смирнов. Механизмы приобретения и потери генетической информации бактери-
альными геномами. 2007.
Вирусы и мобильные элементы:
кто от кого произошел?
Родство вирусов и МГЭ не вызывает сомнений, однако не совсем ясно, кто из
них появился раньше и кто от кого произошел. Не исключено, что началось все с
мобильных элементов, которые могли просто самозародиться в геномах примитив-
ных организмов, и произойти это могло еще на заре жизни. Постепенно усложня-
ясь, МГЭ в дальнейшем превратились в настоящих вирусов. По крайней мере, не-
которые типы вирусов, скорее всего, возникли именно таким путем. Это, прежде
всего, ретровирусы, к числу которых относится вирус ВИЧ. Ретровирусы отлича-
ются от остальных вирусов тем, что встраивание в хозяйский геном является
обязательной частью их жизненного цикла. Наследственный материал ретровируса
хранится в форме РНК. Когда ретровирус попадает в клетку, ее рибосомы начина-
ют синтезировать вирусные белки по «инструкциям», записанным в вирусной РНК.
Одним из этих белков является фермент обратная транскриптаза (РНК-зависимая
ДНК-полимераза), функция которой состоит в переписывании информации из РНК в
ДНК. Этот процесс называется обратной транскрипцией; отсюда и «ретро» в на-
звании вируса. Обратная транскриптаза осуществляет «переписывание» вирусного
генома в хозяйские хромосомы.
Ближайшими родственниками ретровирусов являются ретротранспозоны — мобиль-
ные генетические элементы, широко распространенные в геномах эукариот. Ретро-
вирусы, встроившиеся в геном хозяина, и ретротранспозоны очень похожи друг на
друга. Те и другие представляют собой фрагменты ДНК, содержащие более или ме-
нее стандартный комплект генов, необходимых для собственного копирования и
встраивания в хозяйскую ДНК. Гены эти в большинстве своем сходны у ретровиру-
сов и ретротранспозонов. Обе разновидности «ретроэлементов» размножаются при
помощи обратной транскрипции, то есть переписывания информации с РНК на ДНК.
Большинство специалистов склоняется к тому, что исторически первыми появи-
лись ретротранспозоны, а не ретровирусы. Они проще устроены, и самые простые
из них вполне могли «самозародиться» в геномах примитивных организмов в ре-
зультате случайных мутаций. По-видимому, это произошло еще на уровне прокари-
от . Ретровирусы, скорее всего, являются результатом некоторого усложнения
или, если угодно, «совершенствования» ретротранспозонов. В дальнейшем, конеч-
но, имели место и обратные процессы — ведь ретровирус легко может снова пре-
вратиться в ретротранспозон, если потеряет инфекционность в результате какой-
нибудь мутации.
По-видимому, многие крупные прогрессивные преобразования в эволюции высших
организмов были связаны с активностью МГЭ.
От «прирученного» ретротранспозона ведет свою родословную фермент теломера-
за, отвечающий за восстановление кончиков хромомосом, которые имеют обыкнове-
ние укорачиваться после каждой репликации. Теломераза играет важную роль в
процессах старения и в образовании раковых опухолей. Старение связано с низ-
кой активностью теломеразы, рак — со слишком высокой. Для восстановления кон-
чиков хромосом теломераза использует РНК-матрицу и механизм обратной транс-
крипции. А это не что иное, как главное «ноу-хау» ретротранспозонов и ретро-
вирусов . Теломеразная регуляция — важнейший механизм поддержания целостности
сложного многоклеточного организма, в котором ни одна клетка не имеет права
делиться, когда ей вздумается.
У мобильных элементов древние позвоночные позаимствовали также и фермент
транспозазу, который умеет вырезать и перемещать участки ДНК. Потомки этой
транспозазы — белки RAG. Эти белки собирают из кусочков гены антител, по-
разному комбинируя фрагменты ДНК в зреющих лимфоцитах. Так достигается огром-
ное разнообразие этих защитных белков при небольшом количестве имеющихся в
геноме фрагментов-заготовок (см. главу «Управляемые мутации»). Великолепная
иммунная система позвоночных была одной из главных предпосылок их эволюцион-
ного успеха. И она, как мы теперь понимаем, является щедрым даром ближайших
родственников вирусов.
Древние млекопитающие
позаимствовали у ретротранспозона
ген, необходимый для развития плаценты58
Недавно обнаруженный ген Reg10 мог сыграть важную роль в появлении плацен-
тарных млекопитающих. Об этом свидетельствуют результаты исследований сотруд-
ников Токийского медицинского университета. Ученые показали, что у мышиных
эмбрионов с выключенным геном ReglO нарушается развитие плаценты, отчего эм-
брион погибает через 10 дней после зачатия. Внешне плацента такого эмбриона
выглядит почти нормально, однако в ней отсутствуют некоторые типы клеток, не-
обходимые для эффективной работы органа.
При помощи чрезвычайно сложных манипуляций, связанных с приживлением к де-
фектному зародышу эмбриональных клеток с нормально работающим геном ReglO,
исследователям удалось спасти нескольких обреченных и вырастить из них взрос-
лых мышей, способных к размножению. Правда, они заметно отставали в развитии.
При этом ген ReglO у них по-прежнему не работал. Тем самым было показано, что
ген необходим в первую очередь именно для развития плаценты, хотя может вы-
полнять и ряд других, менее важных функций.
Обнаружение гена, необходимого для развития плаценты, само по себе очень
интересно. Появление плаценты позволило продлить внутриутробное развитие,
что, в свою очередь, имело далеко идущие эволюционные последствия. Согласно
одной из гипотез, именно плацентарность создала предпосылки для увеличения
размеров мозга и быстрого «поумнения» млекопитающих, которое началось около
60 млн. лет назад и затронуло только плацентарных. Сумчатые и однопроходные
(к которым относятся утконос и ехидна), не имеющие плаценты, так и остались
тугодумами.
Однако нет никаких оснований говорить, что ген ReglO «кодирует плаценту».
Плацента, хоть и бракованная, развивается и без его помощи. Появление нового
эмбрионального органа потребовало согласованных изменений множества разных
генов. ReglO — только один из них.
Интересно другое. Ген ReglO по своей структуре оказался чрезвычайно сходен
с мобильным генетическим элементом — ретротранспозоном Sushi-ichi. По-
видимому, предки плацентарных млекопитающих подхватили где-то ретротранспо-
зон, который со временем был «приручен», утратил подвижность, а затем один из
его генов был рекрутирован для выполнения новой функции — регуляции развития
плаценты.
Простейший транспозон устроен еще проще, чем простейший ретротранспозон.
Это короткий участок ДНК, на концах которого расположены так называемые обра-
щенные повторы, а в середине имеется ген, кодирующий транспозазу — фермент,
необходимый для «размножения» и перемещения мобильного элемента. Обращенные
повторы — это последовательности нуклеотидов, комплементарные самим себе (мы
помним, что нуклеотид А комплементарен нуклеотиду Т, нуклеотид Г — нуклеотиду
Ц). Последовательности, комплементарные сами себе, называют также палиндрома-
Ono et al. Deletion of ReglO, an imprinted gene acquired from a retrotransposon,
causes early embryonic lethality // Nature Genetics. 2006. V. 38. P. 101-106.
ми, потому что они «читаются» при транскрипции одинаково в обе стороны. Вот
пример генетического палиндрома: AAGCCCAA. TTGGGCTT. Если вместо многоточия
«вписать» ген транспозазы, получим модель простейшего транспозона.
В отличие от ретротранспозонов обычные транспозоны не используют для само-
копирования механизм обратной транскрипции, поэтому им не нужна обратная
транскриптаза.
Чтобы транспозон совершил «прыжок», хозяйская клетка должна для начала про-
читать ген транспозазы и синтезировать на его основе белок, то есть саму
транспозазу. После этого транспозаза должна найти в хозяйской хромосоме свой
транспозон, опознать его по концевым повторам, затем вырезать и вставить в
какое-нибудь другое место генома.
Итак, транспозаза умеет делать две вещи:
1. узнавать определенные последовательности ДНК и прикрепляться к ним;
2. вырезать и перемещать фрагменты ДНК.
Оба «навыка» транспозазы могут быть использованы клеткой, когда ей удается
приручить какой-нибудь транспозон, то есть сделать его неотъемлемой функцио-
нальной частью своего генома. Для подобных процессов даже придумали специаль-
ный термин — «молекулярное одомашнивание» (molecular domestication). Транспо-
зон теряет подвижность просто за счет какой-нибудь мутационной «поломки» и
начинает выполнять полезную для организма функцию. Мы уже упоминали о том,
что способность прирученных транспозаз — RAG-белков — вырезать и перемещать
фрагменты ДНК лежит в основе системы приобретенного иммунитета.
Еще больше эволюционных возможностей содержит в себе способность транспозаз
распознавать определенные последовательности нуклеотидов ДНК. На специфиче-
ском распознавании белками нуклеотидных последовательностей основаны многие
важнейшие процессы в живой клетке. Одним из таких процессов является регуля-
ция активности генов при помощи специализированных регуляторных белков —
транскрипционных факторов (ТФ) . Транскрипционные факторы распознают опреде-
ленные нуклеотидные последовательности, расположенные в регуляторной области
тех или иных генов, присоединяются к ним и тем самым регулируют работу этих
генов.
Чрезвычайно любопытно, что концевые обращенные повторы многих мобильных
элементов, то есть участки ДНК, распознаваемые транспозазами, порой бывают
удивительно похожи по последовательности нуклеотидов на сайты связывания ТФ
(участки ДНК, распознаваемые транскрипционными факторами). Между прочим, в
сайтах связывания ТФ часто встречаются палиндромные мотивы — последовательно-
сти ДНК, читающиеся одинаково в обе стороны.
Это позволяет предположить, что в ходе эволюции прирученные транспозазы мо-
гут иногда брать на себя роль транскрипционных факторов, а фрагменты концевых
повторов транспозонов, наверное, могут иногда оказываться в таких позициях,
где они способны функционировать в качестве сайтов связывания ТФ.
Растения заимствуют
гены-регуляторы у
«геномных паразитов»59
В 2007 году был обнаружен первый реальный случай «превращения» прирученной
транспозазы в транскрипционный фактор. Открытие было сделано в ходе изучения
светочувствительных белков у арабидопсиса — растения из семейства крестоцвет-
ных. Как и другие цветковые растения, арабидопсис внимательно следит за осве-
59 Rongcheng Lin, Lei Ding, Claudio Casola, Daniel R. Ripoll, Cedric Feschotte,
Haiyang Wang. Transposase-Derived Transcription Factors Regulate Light Signaling in
Arabidopsis // Science. 2007. V. 318. P. 1302-1305.
щенностью при помощи сложных молекулярных систем. Это помогает растению опти-
мизировать свой рост, обмен веществ, выращивать листья и соцветия в «правиль-
ное» время.
Важную роль в фоторецепции у цветковых играет белок фитохром А, реагирующий
на свет с длиной волны 700-750 нм («дальний красный»). Под действием света
фитохром А переходит в биологически активную форму и транспортируется из ци-
топлазмы в ядро клетки, где он «включает» целый ряд генов, регулирующих цве-
тение и созревание семян. Предполагается, что фитохром А был уже у последнего
общего предка цветковых растений и что формирование эффективной светочувстви-
тельной регуляторнои системы способствовало быстрой экспансии цветковых в ме-
ловом периоде.
Светозависимый транспорт фитохрома А в ядро осуществляется при помощи бел-
ков FHY1 и FHL. Кроме того, были идентифицированы еще два белка — FHY3 и
FAR1, без которых транспорт фитохрома А в ядро нарушается, однако конкретная
функция этих белков была до сих пор неизвестна. Именно эти два белка и при-
влекли внимание исследователей.
Анализ нуклеотидных последовательностей генов FHY3 и FAR1 показал чрезвы-
чайно высокое сходство с генами транспозаз, входящими в состав транспозонов
Mutator и Jittery. Оба эти транспозона широко распространены в геномах цвет-
ковых растений. Сходство генов FHY3 и FAR1 странспозазами оказалось настолько
большим, что говорить о случайности не приходится — это, несомненно, гены
«прирученных» транспозаз.
Оказалось, что белки FHY3 и FAR1 необходимы для активации генов FHY1 и FHL.
«Прирученные транспозазы» регулируют активность обоих генов не поодиночке, а
совместными усилиями. Они проникают в ядро и прикрепляются непосредственно к
регуляторнои области (промотору) генов FHY1 и FHL, что приводит к резкому
росту активности этих генов. Таким образом, «прирученные транспозазы» FHY3 и
FAR1 работают как самые настоящие транскрипционные факторы.
Кроме того, исследователи установили, что активность самих генов «приручен-
ных транспозаз» подавляется фитохромом А, поступающим в ядро. В результате
образуется контур отрицательной обратной связи, благодаря которому сигнальная
система срабатывает при определенной освещенности и затем отключается, а не
работает постоянно, как пожарная сирена, пока освещенность не изменится.
Но главное значение этой работы, конечно, в том, что впервые удалось проде-
монстрировать превращение «прирученных транспозаз» в полноценные транскрипци-
онные факторы. Авторы предполагают, что приручение одного или нескольких
транспозонов произошло вскоре после появления цветковых растений (примерно в
середине мелового периода, около 100 млн лет назад) и было связано с освоени-
ем первыми цветковыми разных широтных зон, то есть местообитаний с разной се-
зонной динамикой освещенности.
Данные, указывающие на важную эволюционную роль МГЭ, стали стремительно на-
капливаться в последние несколько лет. Стало ясно, что многие важные генети-
ческие инновации — прежде всего новые регуляторные участки ДНК, управляющие
работой соседних генов, — сформировались из фрагментов «прирученных» МГЭ. Но
до самого последнего времени не удавалось получить сколько-нибудь точных ко-
личественных оценок вклада МГЭ в эволюцию. Поэтому ученые не могли сказать
определенно, как все-таки следует относиться к обнаруженным фактам приручения
МГЭ — как к типичному и массовому явлению, магистральному направлению эволю-
ции, или все-таки как к экзотике.
Для получения количественных оценок необходим комплексный сравнительный
анализ целых геномов. Как известно, в последние годы мировое научное сообще-
ство прилагает огромные усилия для того, чтобы определить нуклеотидные после-
довательности геномов как можно большего числа живых организмов — от бактерий
до млекопитающих. И эти усилия начинают приносить плоды. Первые количествен-
ные подтверждения того, что формирование новых регуляторных участков ДНК из
фрагментов МГЭ является правилом, а не исключением, были получены в результа-
те прочтения генома маленького южноамериканского сумчатого зверька — серого
короткохвостого опоссума.
Короткохвостый опоссум Monodelphis domestica. Этот маленький юж-
ноамериканский зверек стал первым сумчатым млекопитающим, чей
геном удалось прочесть. Вторым, скорее всего, будет кенгуру.
Геном опоссума был прочтен в мае 2007 года60. Этот зверек был выбран для
геномных исследований не случайно — он является важным лабораторным объектом,
на котором изучают, в частности, регенерацию и формирование злокачественных
опухолей. Кроме того, предполагали, что сравнение с геномом сумчатого живот-
ного поможет лучше понять прочтенные ранее геномы плацентарных — человека,
шимпанзе, макака-резуса, собаки, мыши, крысы. Эти надежды полностью оправда-
лись .
Различия в наборе белок-кодирующих генов между опоссумом и человеком оказа-
лись очень невелики. Подавляющее большинство генов опоссума имеют бесспорные
человеческие аналоги и наоборот.
В целом в белок-кодирующих областях геномов сумчатых и плацентарных за 180
миллионов лет, прошедших после разделения этих линий, возникло сравнительно
мало эволюционных инноваций. По современным представлениям, ведущую роль в
эволюции высших организмов должны были играть изменения регуляторных участков
генома, которые сами не кодируют белков, но влияют на работу белок-кодирующих
генов. Геном опоссума блестяще подтвердил эту теорию.
Сравнительный анализ геномов опоссума, плацентарных и курицы показал, что в
эволюции плацентарных подавляющее большинство эволюционных инноваций было
связано с появлением новых участков ДНК, которые не кодируют белков, но вы-
полняют важные регуляторные функции. Из всех функционально важных кодирующих
участков генома плацентарных лишь около одного процента появились после отде-
Т. S. Mikkelsen et al. Genome of the marsupial Monodelphis domestica reveals
innovation in non-coding sequences // Nature. 2007. V. 447. P. 167-177.
ления плацентарных от общего предка с сумчатыми. Что же касается функциональ-
но важных некодирующих (то есть регуляторных) последовательностей, то из их
числа целых 20% оказались уникальными для плацентарных. Иными словами, ско-
рость появления новых регуляторных участков генома в эволюции плацентарных
была в 20 раз выше, чем скорость появления новых кодирующих участков.
У читателя может возникнуть резонный вопрос, почему мы говорим об эволюции
плацентарных в связи с прочтением генома опоссума — зверька, относящегося не
к плацентарным, а к сумчатым. Дело в том, что только сравнение с геномом
опоссума позволило ученым установить, какие из некодирующих последовательно-
стей геномов плацентарных являются новыми, то есть появившимися после разде-
ления эволюционных линий сумчатых и плацентарных. Для того чтобы понять эво-
люционную роль изменений в некодирующих последовательностях, нужно еще знать,
какие из этих последовательностей являются функционально важными, а какие
нет. Для плацентарных это можно сделать, сравнивая между собой геномы разных
видов: если некодирующий участок сходен, например, у человека и собаки, зна-
чит, он, скорее всего, является важным (отбор отбраковывал слишком сильные
изменения в этом участке). Для сумчатых этого сделать пока нельзя, поскольку
прочтен геном только одного вида сумчатых. Именно поэтому геном опоссума по-
зволил гораздо больше узнать об эволюции плацентарных, чем сумчатых.
Как и следовало ожидать, особенно много новых регуляторных последовательно-
стей у плацентарных появилось в окрестностях генов, кодирующих ключевые регу-
ляторы индивидуального развития, в том числе Нох-генов (см. главу 5) . Сами
эти гены отличаются повышенной консервативностью — они очень похожи у плацен-
тарных, опоссума и даже курицы. Получается, что изменения в строении организ-
ма у плацентарных млекопитающих были обусловлены в основном добавлением новых
регуляторов к генам — регуляторам онтогенеза.
Самый яркий результат исследователи получили в ходе изучения происхождения
новых регуляторных последовательностей, возникших в ходе эволюции плацентар-
ных. В принципе, эти последовательности могут появляться тремя способами:
1. в результате изменения «до неузнаваемости» каких-то старых, предковых ре-
гуляторных последовательностей;
2. в результате дупликации старых регуляторных последовательностей и после-
дующего накопления различий между копиями;
3. заново, из последовательностей, которые у предков были нефункциональными,
в том числе из «прирученных» мобильных элементов.
Ранее было выявлено несколько случаев, когда в эволюции позвоночных новые
регуляторные последовательности формировались из мобильных генетических эле-
ментов. Как мы уже знаем, сама структура МГЭ делает их превосходными «заго-
товками» для создания новых регуляторных элементов в хозяйском геноме. МГЭ
обычно имеют свои собственные регуляторные элементы, например, места прикреп-
ления транспозаз — ферментов, осуществляющих перемещения МГЭ. Эти регулятор-
ные элементы легко могут быть адаптированы для регуляции работы генов хозяй-
ского генома. Однако до сих пор оставалось неясным, являются ли выявленные
случаи удачного приручения транспозонов редкими исключениями или общим прави-
лом . Теперь наконец можно с уверенностью сказать: это правило.
Оказалось, что в человеческом геноме как минимум 16% из числа важных регу-
ляторных последовательностей, уникальных для плацентарных, представляют собой
участки мобильных элементов. При создании новых регуляторных последовательно-
стей у плацентарных в ход пошли все основные группы транспозонов и рет-
ротранспозонов. Причем вышеупомянутые 16% — это, несомненно, сильно занижен-
ная оценка. Дело в том, что «прирученный», утративший подвижность транспозон
в результате накопления мутаций становится неузнаваемым примерно за 100-200
миллионов лет. Сохранится в целости лишь тот его фрагмент, который оказался
полезен хозяйскому геному, но этого, скорее всего, будет недостаточно, чтобы
распознать в таком фрагменте бывший транспозон. За время, прошедшее с момента
обособления плацентарных, эта судьба должна была постигнуть значительную
часть прирученных транспозонов.
Таким образом, «одомашнивание» мобильных генетических элементов играет
крайне важную роль в эволюции млекопитающих. Скорее всего, это справедливо и
для других живых организмов, но чтобы это доказать, необходимы дальнейшие ис-
следования .
Но вернемся к наследованию приобретенных признаков. Связь МГЭ и вирусов с
«ламарковскими» механизмами наследования может оказаться еще более тесной,
чем принято считать. Можно предположить — пока лишь теоретически, — что при
помощи МГЭ высшие организмы могут иногда передавать своему потомству вполне
исчерпывающую и адекватную информацию о полезных адаптациях, выработанных ими
в течение жизни. По сути дела, это и есть «ламарковское» наследование в чис-
том виде и без всяких оговорок, в том самом смысле, какой вкладывал в это по-
нятие сам Ламарк.
Эта смелая гипотеза была предложена и подробно обоснована австралийским им-
мунологом Э. Стилом и его коллегами61. Мы помним из главы «Управляемые мута-
ции», как лимфоциты млекопитающих производят гены новых защитных белков-
антител в ходе выработки приобретенного иммунитета. Для этого исходные гене-
тические заготовки сначала комбинируются разными способами, а затем подверга-
ются интенсивному мутированию и отбору. Таким образом формируется новый ген,
обеспечивающий защиту против какой-нибудь инфекции — например, против новой
болезнетворной бактерии. Выработка приобретенного иммунитета требует времени,
и не всегда организм успевает справиться с этой задачей. Против некоторых са-
мых опасных возбудителей было бы выгодно иметь врожденный иммунитет, а не
приобретать его каждый раз заново при столкновении с инфекцией. Собственно
говоря, именно этот «недостаток» нашей иммунной системы и пытается восполнить
современная медицина, осуществляя вакцинацию людей и домашних животных. Если
бы генетические изменения, возникающие в лимфоцитах при выработке приобретен-
ного иммунитета, могли иногда передаваться потомству, это было бы аналогично
естественной, природной вакцинации последующих поколений.
Но как такое может произойти? Ведь для этого новый вариант защитного гена,
сформировавшийся в лимфоцитах, должен быть перенесен в половые клетки и
встроен в их геном. Кто или что может служить переносчиком генов из лимфоци-
тов в половые клетки? По мнению Стила, на роль таких переносчиков идеально
подходит одна из разновидностей МГЭ, которой буквально кишат геномы млекопи-
тающих, — так называемые эндогенные ретровирусы. Это геномы ретровирусов, ко-
торым когда-то удалось встроиться в геном половых клеток, или «продвинутые»
ретротранспозоны, способные к образованию вирусоподобных частиц. Грань между
сложными ретротранспозонами и простыми ретровирусами весьма условна. Эндоген-
ные ретровирусы, разумеется, передаются по наследству точно так же, как любые
другие участки генома. При этом они сохраняют способность «оживать», то есть
упаковывать свой геном в белковую оболочку (капсид) и переходить из клетки в
клетку. Вместе со своими собственными генами обратной транскриптазы, интегра-
зы, капсидного белка эндогенные ретровирусы могут прихватывать с собой и «чу-
жие» молекулы РНК, считанные с каких-нибудь других генов. Например, с гена
нового антитела, если дело происходит в лимфоците.
Покинув лимфоцит, такой эндогенный ретровирус в принципе может с током кро-
ви добраться до половых клеток и проникнуть в них. Здесь в соответствии с
«инструкциями», записанными в вирусной РНК, будут синтезированы ферменты об-
ратная транскриптаза и интеграза. Первая произведет ДНКовые копии вирусных
Э. Стил, Р. Линдли, Р. Бландэн. Что если Ламарк прав? Иммуногенетика и эволюция.
2002.
генов, а заодно и гена нового антитела; вторая встроит эти фрагменты ДНК в
геном половой клетки. Таким образом новый защитный ген будет передан потомст-
ву. При выполнении некоторых дополнительных условий это может привести к то-
му, что потомство будет с самого начала иметь повышенную устойчивость к ка-
кой-то инфекции.
Самое интересное, что если бы эта гипотеза оказалась верной, и такой меха-
низм наследования удалось бы продемонстрировать экспериментально, то тем са-
мым была бы подтверждена «крамольная» и всеми позабытая дарвиновская теория
пангенеза, о которой мы говорили в начале главы. Нетрудно заметить, что эндо-
генные ретровирусы, выступающие в роли переносчиков информации о приобретен-
ных признаках в половые клетки, совершенно идентичны геммулам, или пангенам,
о существовании которых говорил Дарвин.
Как от теоретических геммул перейти к реальным агентам приобретенного на-
следования? В этом нам помогут, например, ретропсевдогены, присутствующие в
большом количестве в геномах эукариот. Ретропсевдоген — это последователь-
ность нуклеотидов, идентичная кодирующей части какого-нибудь гена, обычно не-
активная, неработающая, которая возникла в результате деятельности обратной
транскриптазы. Ее нетрудно отличить от «настоящего» гена по отсутствию нитро-
нов — некодирующих вставок, которые есть в большинстве генов высших организ-
мов .
Ретропсевдоген образуется так. Сначала с обычного гена «считывается» мат-
ричная РНК (мРНК). Считывается все подряд — и кодирующие участки (экзоны), и
вставленные между ними интроны. Затем интроны удаляются, а кодирующие участки
сшиваются друг с другом (этот процесс посттранскрипционной обработки РНК на-
зывается сплайсингом). В результате образуется «зрелая мРНК». На ее основе
синтезируется белок.
Если же зрелая мРНК с удаленными интронами «попадется под руку» обратной
транскриптазе, то она может синтезировать ДНКовую копию этой РНК и встроить
ее в какую-нибудь хромосому. Так и возникают ретропсевдогены. Они обычно не
работают, но не потому, что лишены интронов, а потому, что рядом с ними не
оказывается подходящих регуляторных участков ДНК — промоторов. Если около ге-
на нет промотора, ферменту РНК-полимеразе просто не к чему прикрепиться, и
ген не может быть прочитан. Впрочем, иногда ретропсевдогену может «повезти»,
и он случайно окажется рядом с промотором в результате какой-нибудь геномной
перестройки. Известен ряд случаев, когда ретропсевдогены становились работаю-
щими генами.
Существование ретропсевдогенов доказывает, что обратная транскриптаза дей-
ствительно может «переписывать» в геном половых клеток информацию, содержа-
щуюся в молекулах РНК, считанных с различных генов. Это означает, что у жи-
вотных есть все составные части предполагаемого механизма «пангенеза». Теоре-
тически дарвиновские геммулы имеют право на существование. Они вполне могли
бы работать и обеспечивать наследование приобретенных признаков «по Ламарку».
Загвоздка лишь в том, что они, по-видимому, в действительности делают это
крайне редко. Почему? Скорее всего, просто потому, что это бесперспективно.
От всех инфекционных агентов, которые существуют сейчас и могут появиться в
будущем, все равно нельзя запастись готовыми, наследуемыми антителами. Любой
организм сталкивается в течение жизни с тысячами и миллионами разных возбуди-
телей, и если бы каждое новое антитело «переписывалось» в геном половых кле-
ток, геном оказался бы неимоверно раздут, а это, как мы знаем, ведет к росту
«непроизводительных расходов» для каждой клетки организма (см. раздел «Проис-
хождение птиц» в главе 6). Развитие системы приобретенного иммунитета — спо-
собности вырабатывать любые антитела в течение жизни — было величайшим эволю-
ционным достижением, а закрепление в геноме потомства каждого удачного анти-
тела было бы тупиковым путем.
Эпигенетическое
наследование
Недавно открыто еще несколько молекулярных механизмов, которые в принципе
могут служить для передачи по наследству приобретенных признаков. Эти меха-
низмы не связаны напрямую с изменениями самого «текста», записанного в струк-
туре молекул ДНК, то есть с мутациями. Поэтому такую наследственность называ-
ют «эпигенетической», или «надгенетической».
Один из эпигенетических механизмов — метилирование ДНК. Оказалось, что в
процессе жизнедеятельности к молекулам ДНК в клетках, в том числе и в поло-
вых, специальные ферменты «пришивают» метильные группы (-СН3) . Причем к одним
генам метильных групп пришивают больше, к другим — меньше. Распределение ме-
тильных групп по генам зависит от того, насколько активно тот или иной ген
используется. Активные гены метилируются слабо, неактивные — сильно. Получа-
ется совсем как с упражнением и неупражнением органов, которое Ламарк считал
причиной наследственных изменений. Поскольку «рисунок метилирования» может
передаваться по наследству и поскольку он, в свою очередь, влияет на актив-
ность генов у потомства, легко заметить, что здесь может работать совершенно
ламарковский механизм наследования: «натренированные» предками гены будут и у
потомства работать активнее, чем «ослабевшие» от долгого неиспользования.
Однако и в данном случае природа, судя по всему, не заинтересовалась воз-
можностью наладить наследование «по Ламарку». Наибольшего развития система
метилирования ДНК в половых клетках достигла у двух групп высших многоклеточ-
ных, венчающих эволюционное древо, соответственно, животных и растений — а
именно у плацентарных млекопитающих и покрытосеменных (цветковых) растений. В
обоих случаях избирательное метилирование ДНК в половых клетках, так называе-
мый «геномный импринтинг», служит не для передачи по наследству приобретенных
признаков, а для регуляции взаимоотношений между зародышем и материнским ор-
ганизмом .
Геномный импринтинг — распространенное у млекопитающих и цветковых растений
явление, состоящее в том, что некоторые гены в половых клетках родителей осо-
бым образом «метятся». К цитозиновым основаниям прикрепляется метильная груп-
па. «Помеченный» ген у потомства просто-напросто не работает. Некоторые гены
отключаются в сперматозоидах, другие в яйцеклетках. Если гены отключены в
сперматозоидах, то соответствующие признаки потомство наследует только от ма-
тери. Если отключены гены в яйцеклетке, то наследуются отцовские признаки. В
половых клетках потомства старые метки удаляются и заменяются новыми. В ре-
зультате у внуков могут проявиться признаки деда или бабки, которые не были
выражены у родителей. Импринтинг — это пример так называемой «эпигенетиче-
ской» наследственности, то есть наследственных свойств, не связанных с изме-
нением последовательности нуклеотидов в молекулах ДНК, генетического текста.
Именно благодаря импринтингу при скрещивании разных пород или видов млеко-
питающих оказывается небезразлично направление гибридизации, то есть кто из
двух родителей будет принадлежать к какому виду. Например, при скрещивании
осла с кобылой получаются мулы, а при скрещивании жеребца с ослицей — лошаки.
Вопреки классическим законам генетики важным для потомства оказывается не
только то, какие гены они получили, но и от кого — от отца или матери.
Зачем нужен геномный импринтинг, почему он появился? Для объяснения этого
существует две гипотезы. Первая — общепринятая — состоит в том, что имприн-
тинг развился вследствие различия интересов полов. У млекопитающих между сам-
кой и ее детенышем во время внутриутробного развития складываются отчасти ан-
тагонистические отношения. Говоря упрощенно, эмбрион старается высосать из
матери побольше соков, а мать старается сохранить силы и здоровье, чтобы в
будущем иметь возможность родить и других детей. Самец в этом конфликте в об-
щем случае — на стороне детеныша. Других-то детей самка еще неизвестно от ко-
го родит, а этот — свой. Поэтому самцы отключают в своих сперматозоидах те
гены, которые способствуют защите матери от чрезмерных притязаний эмбриона, а
матери, напротив, отключают в своих яйцеклетках те гены, которые могут эти
притязания усилить. Действительно, большинство генов млекопитающих, подвер-
гающихся родительскому импринтингу, так или иначе, связаны с внутриутробным
развитием, строением плаценты и т. п.
Из-за этого у млекопитающих (в отличие от многих других животных, таких как
насекомые или ящерицы) принципиально невозможно развитие зародыша на основе
только материнских или только отцовских генов.
Можно взять только что оплодотворенную яйцеклетку какого-нибудь другого жи-
вотного (не млекопитающего, а, например, лягушки), в которой ядра яйцеклетки
и сперматозоида еще не успели слиться, удалить из нее отцовское ядро и заме-
нить его ядром из другой яйцеклетки. Два «материнских» ядра сольются, и из
такой яйцеклетки в благоприятных условиях может развиться нормальный орга-
низм, все гены которого — материнские. Можно искусственно сделать яйцеклетку
с двумя отцовскими ядрами, и из нее тоже может вырасти жизнеспособное живот-
ное . Однако с млекопитающими этот номер не пройдет. Без отцовских генов у эм-
бриона не разовьется плацента, а без материнских плацента разовьется даже
лучше, чем надо, но не будет нормально развиваться сам эмбрион.
Дополнительная гипотеза, более симпатичная с этической точки зрения, была
62 ^
высказана совсем недавно Согласно этой гипотезе основной смысл геномного
импринтинга — достижение лучшей совместимости между матерью и плодом, то есть
на первое место ставятся не антагонистические взаимоотношения матери и эм-
бриона , а кооперативные. Если часть отцовских генов будет выключена, то у эм-
бриона будут работать только материнские копии этих генов, и эмбрион, таким
образом, будет по своим физиологическим и биохимическим свойствам больше по-
хож на мать, и им легче будет приспособиться друг к другу. Эта гипотеза пред-
полагает, что в ходе родительского импринтинга должно отключаться больше от-
цовских генов, чем материнских, и факты это подтверждают.
Метилирование и деметилирование генов играют важную роль в процессе индиви-
дуального развития многоклеточного организма, где они служат для контроля ра-
боты генов в развивающихся органах и тканях. Кроме того, при помощи метилиро-
вания клетки борются с чрезмерной активностью мобильных генетических элемен-
тов . Избирательное метилирование МГЭ снижает частоту их «прыжков». Особенно
тщательно многоклеточные организмы «следят» за активностью МГЭ при созревании
сперматозоидов (см. главу «На подступах к неведомому»).
Очень важно, что метилирование дает клетке возможность попутно контролиро-
вать и частоту мутирования тех или иных генов. Как уже отмечалось в главе
«Управляемые мутации», метилирование цитозина (Ц) резко повышает вероятность
мутации, в результате которой цитозин заменяется тимином (Т) . Метилирование
ДНК активно контролируется клеткой, таким образом, в клетке реально существу-
ет механизм, позволяющий целенаправленно регулировать вероятность мутирования
определенных участков генома.
Взаимное включение
и выключение генов
Jason В. Wolf, Reinmar Hager. A Maternal-Offspring Coadaptation Theory for the
Evolution of Genomic Imprinting // PLoS Biology, 2006. 4(12).
Еще один возможный вариант эпигенетического наследования приобретенных при-
знаков основан на системах взаимной активации и инактивации генов. Допустим,
ген А производит белок, одно из действий которого состоит в блокировании ра-
боты гена Б, а ген Б, в свою очередь, кодирует другой белок, способный «вы-
ключать» ген А. Такая система может находиться в одном из двух состояний: ли-
бо ген А работает, и тогда ген Б выключен, либо наоборот. Допустим, что пере-
ход системы из одного состояния в другое может происходить только в результа-
те какого-то особенного внешнего воздействия, и случается такое редко. То со-
стояние, в котором находится эта двухгенная система в клетках матери, будет
через яйцеклетку передаваться ее потомству (поскольку сперматозоид содержит
пренебрежимо малое количество белков). Если же в течение жизни матери система
переключится в другое состояние, то этот приобретенный признак передастся по-
томству , родившемуся после «переключения». Опять получается наследование по
Ламарку. Точнее, возможность такого наследования. И опять, как и в предыдущих
случаях, живые организмы, похоже, не очень-то торопятся воспользоваться этой
замечательной возможностью.
Мы видим, что «ламарковское» наследование приобретенных признаков вполне
осуществимо технически. Имеется целый ряд молекулярных механизмов, способных
обеспечить целенаправленную передачу потомству наследуемой информации о при-
обретенных признаках. Тот факт, что живые организмы редко используют эти воз-
можности , говорит о том, что наследование «по Ламарку» им просто не выгодно.
Негенетическая
«память поколений»
Передача информации от родителей к потомкам может осуществляться тремя ос-
новными путями, два из которых общеизвестны: это генетическая наследствен-
ность, свойственная всем без исключения живым организмам, и обучение, харак-
терное только для животных со сложной нервной системой.
Третий путь менее известен и гораздо хуже изучен, однако и он, судя по все-
му , играет важную роль в жизни многих организмов. Это так называемые «роди-
тельские эффекты» — внегенетические изменения у потомства, обусловленные ус-
ловиями жизни и заботой родителей. Простейший пример — самка, плохо питавшая-
ся в течение своей жизни, откладывает яйца с меньшим количеством питательных
веществ, из которых развивается — даже при «хороших» генах — сравнительно
чахлое потомство. Чем не наследование «приобретенного признака»?
Более сложные варианты могут включать различные эпигенетические изменения
наследственного материала (ДНК), о которых мы говорили выше, в том числе ге-
номный импринтинг, который представляет собой не что иное, как целенаправлен-
ное манипулирование наследственными свойствами потомства. Кроме того, извест-
но, что эмбриональное развитие животных, особенно на начальных этапах, во
многом зависит от разнообразных молекул (в том числе матричных РНК) , посту-
пающих в яйцеклетку из материнского организма (см. раздел «Нужны ли эмбрионам
гены» в главе 5) . Ясно, что условия жизни матери в принципе могут влиять на
количество и состав этих веществ и, следовательно, на развитие зародыша.
Изучать родительские эффекты легче всего у тех животных, в жизненном цикле
которых присутствует партеногенетическое размножение (развитие потомства из
неоплодотворенныхяиц) — как, например, у рачков-дафний. В этом случае геномы
матери и ее дочерей идентичны и легче отличить «материнские» эффекты от гене-
тических .
Недавно российским ученым удалось показать методом моделирования, что мате-
ринский эффект — негенетическая передача от матери к потомству информации о
длине светового дня и обилии пищи — играет важную роль в сезонных изменениях
численности и поведения дафний и делает популяцию более устойчивой. Наличие
материнского эффекта у дафний, вначале предсказанное теоретически, недавно
получило экспериментальные подтверждения63.
Еще в 1996 году А. А. Умнов и В. Р. Алексеев разработали имитационную мо-
дель для проверки гипотезы о существовании у ветвистоусых рачков материнского
эффекта. В модели предполагалось, что дафнии передают потомству информацию о
трофических условиях (то есть о том, насколько хорошо питалась мать). Позже
эта гипотеза была подтверждена экспериментально64. Выяснилось, что дафнии пе-
редают потомству информацию не только о своем питании, но и об изменениях
длины светового дня. Вследствие этого особи, имеющие одинаковый размер и воз-
раст, могут по-разному реагировать на одни и те же пищевые и температурные
условия только потому, что их матери имели разную жизненную историю.
Ветвистоусый рачок Daphnia longispina — массовый представитель
зоопланктона озер и луж умеренной зоны. На фотографии видны
зреющие яйца в выводковой камере.
В новой статье В. Р. Алексеева и Т. И. Казанцевой, опубликованной в 2007
году в «Журнале общей биологии», оценивается возможное влияние материнского
эффекта на колебания численности популяций дафний, на время перехода от пар-
теногенетического (однополого) размножения к двуполому, на количество зимую-
щих яиц и другие параметры популяции.
В качестве «образца» для моделирования использовалась хорошо изученная по-
пуляция рачков Daphnia longispina из озера Красненького в Псковской области.
В. Р. Алексеев, Т. И. Казанцева. Использование индивидуально-ориентированной мо-
дели для изучения роли материнского эффекта в смене типов размножения у Cladocera //
Журнал общей биологии. 2007. Т. 68. С. 231-240.
64 Alekseev, Lampert. Maternal control of resting-egg production in Daphnia //
Nature. 2001. V. 414. P. 899-901.
Жизненный цикл этих рачков довольно сложен. Весной из зимующих яиц выходит
первое поколение самок, которые начинают быстро размножаться путем партеноге-
неза, производя на свет только самок. В начале лета численность микроскопиче-
ских водорослей — основной пищи дафний — снижается, и дафнии начинают произ-
водить на свет самцов и переходят к двуполому размножению. Образуются покоя-
щиеся оплодотворенные яйца, и наступает «летняя диапауза», длящаяся не более
месяца. Затем из яиц выходит новое поколение самок, размножающихся партеноге-
нетически. В конце лета и осенью в популяции снова появляются самцы, начина-
ется двуполое размножение и откладываются зимующие яйца, покрытые плотной
оболочкой, позволяющей переносить неблагоприятные условия.
Переход дафний от однополого размножения к двуполому регулируется несколь-
кими факторами — температурой, обилием пищи, длиной светового дня. Информация
о динамике этих факторов в прошлом и настоящем доходит до дафний двумя путя-
ми: из их личного жизненного опыта, а также от матери благодаря «материнскому
эффекту».
Разработанная авторами модель имитирует жизненный путь каждой отдельной
особи. Вероятность перехода особи от однополого размножения к двуполому зави-
сит в модели как от сиюминутного состояния факторов среды (температуры, длины
светового дня, количества пищи), так и от индивидуального «потенциала роста»
особи, который, в свою очередь, определяется условиями жизни и «потенциалом»
ее матери. Потенциал роста определяет максимально возможную скорость роста
данной особи. Чем медленнее рост, тем выше вероятность перехода к двуполому
размножению.
Таким образом, переход к двуполому размножению зависит не только оттого,
как питалась данная особь и каковы условия среды в данный момент, но и отто-
го, как питались ее мать и более далекие предки. Тем самым задается «материн-
ский эффект» — негенетическая передача информации от матери к потомкам. Авто-
рам удалось показать, что при определенных входных параметрах их модель очень
точно воспроизводит реальную динамику численности самцов и самок, партеноге-
нетических и покоящихся яиц, наблюдаемую в реальной популяции. Надо сказать,
что эта реальная динамика достаточно сложна: например, в течение одного лета
наблюдается несколько (обычно 5) пиков численности дафний.
Точное соответствие модельной и реальной популяционной динамики дало осно-
вания полагать, что модель действительно учитывает все основные факторы и
причинно-следственные связи, определяющие эту динамику. Теперь модель можно
было использовать для проверки гипотез о роли материнского эффекта.
Изъятие из модели материнского эффекта привело к тому, что модельная дина-
мика стала резко отличаться от реальной. Ни один из вариантов модели с отклю-
ченным материнским эффектом не позволил воспроизвести динамику численности и
формирование банка покоящихся яиц, необходимых для устойчивого многолетнего
развития популяции.
Например, если потенциал роста был задан высоким, покоящиеся яйца практиче-
ски не образовывались, выживание популяции в зимний период было возможно
только благодаря небольшому количеству перезимовавших самок и в долгосрочной
перспективе популяция становилась крайне уязвимой. На этом основании авторы
сделали вывод о существенной роли материнского эффекта в формировании популя-
ционной динамики и сезонных адаптации у дафний.
ГЛАВА 9.
НА ПОДСТУПАХ
К НЕВЕДОМОМУ
У некоторых читателей при чтении предыдущих глав могло сложиться обманчивое
впечатление, что в биологии осталось не так уж много нерешенных проблем. Эта
глава написана специально для тех, кто так подумал.
На самом деле в сегодняшней биологии нерешенных загадок и тайн, пожалуй,
больше, чем когда-либо в прошлом. Это вовсе не значит, что биология удаляется
от истины, а ученые все больше запутываются в неразрешимых противоречиях.
Тенденция как раз противоположная: большинство возникающих вопросов успешно и
довольно быстро разрешается, но каждый полученный ответ порождает новые во-
просы. Хорошую аллегорию для этой ситуации придумали еще древние греки — пер-
вооткрыватели научного метода познания. Если представить себе область извест-
ного как некую замкнутую геометрическую фигуру, допустим, шар, а область не-
ведомого — как все, что находится за пределами этой фигуры, то становится яс-
но , почему с ростом наших знаний появляется все больше новых вопросов. Чем
больше объем шара, читай — количество знаний, тем больше площадь его сопри-
косновения с неведомым. Эта площадь символизирует количество вопросов, кото-
рые уже осознаны человечеством, но еще не получили ответа.
Впрочем, эта геометрическая аллегория исходит из допущения, что неведомое
бесконечно. В природе это вряд ли так. В конце концов, земная жизнь (а на
большее биологи пока не замахиваются) имеет вполне конечную размерность. Вре-
мя существования, биомасса, численность, занимаемый объем, даже количество
атомов, когда-либо побывавших в составе живых существ, — все эти величины да-
леко не бесконечны, а в масштабах Вселенной даже и не очень велики. И, разу-
меется, большинство характеристик живых объектов многократно повторяется: от
клетки к клетке, от организма к организму, от сообщества к сообществу, поэто-
му вовсе не обязательно в деталях изучать каждый из этих объектов. «Область
неведомого» в биологии не бесконечна, у нее есть границы, но только мы пока
не знаем, где они проходят и когда мы до них, наконец, доберемся.
В этой главе мы поговорим о некоторых интригующих научных проблемах, нахо-
дящихся в разных стадиях разрешения. В одних случаях это нечто совсем непо-
нятное, только начинающее приоткрываться. В других, может быть, нам осталось
сделать лишь одно последнее усилие, чтобы головоломка наконец сложилась. Та-
ких «историй с ненаписанным концом» в современной биологии очень много, и
нужно помнить, что описанные здесь случаи — лишь капля в море.
О сложном и непонятном трудно рассказать понятно и просто. Я уже упоминал в
предисловии о том, что современная биология — сложная наука и поэтому для то-
го, чтобы высказывать компетентные суждения по биологическим вопросам, требу-
ются не меньшие профессиональная подготовка и уровень образования, чем, к
примеру, в квантовой физике. Очень жаль, что наши доморощенные креационисты
этого не понимают. Проблемы, о которых пойдет речь в этой главе, служат хоро-
шей иллюстрацией к сказанному.
Парамутации
Как мы знаем из предыдущих глав, далеко не вся наследственная информация,
передающаяся из поколения в поколение, записана в последовательности нуклео-
тидов в молекулах ДНК. Кроме этой генетической информации есть еще и так на-
зываемая эпигенетическая (см. главу «Наследуются ли приобретенные призна-
ки?») . Например, определенные нуклеотиды в ДНК могут подвергаться химической
модификации — метилированию. В ходе репликации молекул ДНК специальные фер-
менты не всегда, но часто метилируют в новой молекуле ДНК те же нуклеотиды,
которые были метилированы в родительской молекуле. Таким образом, «рисунок
метилирования» может передаваться по наследству, а от него зависят активность
генов и, соответственно, фенотипические признаки. Внегенетическая наследст-
венная информация может передаваться и с другими молекулами, например, РНК,
которые имеются в цитоплазме половых клеток (см., например, раздел «Контроли-
руемая перестройка генома у инфузорий» в главе «Управляемые мутации»). Эпиге-
нетическая наследственность порой приводит к нарушениям основополагающих за-
конов классической генетики. Многие эпигенетические механизмы в общих чертах
уже расшифрованы, но некоторые оказались на редкость неподатливыми. Типичным
примером являются так называемые парамутации, обнаруженные полвека назад у
кукурузы. Несмотря на упорные усилия, природу этого явления до сих пор так и
не удалось разгадать.
Парамутация — это устойчивое, наследуемое изменение свойств гена, которое
возникает в результате взаимодействия разных вариантов (аллелей) этого гена
между собой и при котором в отличие от обычных мутаций нуклеотидная последо-
вательность гена не меняется. Результаты парамутации выглядят очень странно.
Свойства организма у парамутантных особей не соответствуют тому, что «записа-
но» в их генотипе. Допустим, вы унаследовали от обоих родителей гены голубых
глаз, а глаза у вас при этом почему-то карие, как у одного из дедушек. Но у
дедушки-то имеется «ген кареглазости», а у вас его нет! Впрочем, у людей па-
рамутации пока не обнаружены.
С парамутациями можно было еще как-то мириться, пока дело касалось только
растений. Однако в 2006 году парамутации были обнаружены у животных, а именно
у мышей. Это стало одной из самых громких сенсаций последних лет в области
генетики.
Честь открытия принадлежит французским ученым из Университета Ниццы65, ко-
торые работали с линией лабораторных мышей, несущих мутацию в гене Kit. Этот
ген кодирует многофункциональный белок, влияющий, помимо прочего, на образо-
вание темного пигмента меланина. В свое время генетики, изучавшие работу это-
го гена у мышей, искусственно создали неработающий мутантный вариант гена. У
мышей, гетерозиготных по этой мутации, то есть имеющих одну нормальную копию
гена и одну измененную, Kit+/-, лапки и кончик хвоста остаются белыми. Гомо-
зиготы по этому гену — обладатели двух испорченных копий гена, Kit-/-, — уми-
рают вскоре после рождения.
Скрещивая между собой гетерозигот Kit+/-, французские генетики столкнулись
с нарушением законов генетики. Согласно этим законам, четверть потомства
должна была сразу погибать, получив два варианта испорченного гена Kit-/-,
половина — иметь белые лапы и хвост, это генотип Kit+/-, и четверть — иметь
нормальную темную окраску, это генотип Kit+/+. Вместо этого из 57 выживших
мышат, полученных от восьми скрещиваний, только три оказались нормально окра-
шенными, а остальные 54 имели белые лапы и хвост.
Генетический анализ показал, что половина белохвостых мышат имеет генотип
Kit+/+,TO есть у них попросту нет мутантного варианта «гена белохвостости».
Эти мыши обязаны были иметь нормальную окраску! А получалось, что мыши унас-
ледовали от родителей некий врожденный признак, не унаследовав генов, ответ-
ственных за формирование этого признака.
Если скрестить этих аномальных мышей — белохвостых, но не имеющих «гена бе-
лохвостости» — друг с другом, то потомство их тоже оказывается белохвостым.
Правда, в последующих поколениях признак проявляется все слабее и, в конце
концов, сходит на нет. Признаки организма, наконец, приходят в соответствие с
генотипом.
Дальнейшие опыты показали, что если хотя бы один из родителей мышонка (не-
важно, отец или мать) — белохвостый, то и мышонок с большой вероятностью бу-
дет белохвостым независимо от того, есть ли «ген белохвостости» у него само-
го.
Стало очевидно, что носителем наследственной информации в данном случае яв-
ляется не последовательность нуклеотидов в молекуле ДНК, а что-то другое. Ес-
Minoo Rassoulzadegan et al. RNA-mediated non-mendelian inheritance of an
epigenetic change in the mouse // Nature. 2006. V. 441. P. 469-474.
тественно, подозрение пало в первую очередь на РНК — второй класс «информаци-
онных» молекул клетки.
Ученые предположили, что белохвостость мышонка с генотипом Kit+/+ может
быть вызвана тем, что в оплодотворенную яйцеклетку, из которой он развился,
попала родительская РНК, считанная с мутантной копии гена Kit. Хотя у самого
мышонка обе копии гена Kit нормальные, присутствие «мутантной» РНК может как-
то повлиять на их работу.
Это предположение подтвердилось. Исследователи обнаружили, что у гетерози-
гот Kit+/- с мутантной копии гена Kit считывается «мутантная» РНК, которая в
дальнейшем распадается на фрагменты разной величины. Если выделить эти фраг-
менты и ввести их в оплодотворенную яйцеклетку, полученную от диких серохво-
стых родителей, из яйцеклетки развивается белохвостый мышонок, не имеющий
«гена белохвостости». По-видимому, эти фрагменты РНК не только регулируют
прочтение гена Kit, но и каким-то образом самовоспроизводятся, иначе они не
могли бы передаваться в ряду поколений. Однако механизм самовоспроизведения
этих РНК у мышей разгадать пока не удалось.
Ученые также обнаружили, что в сперматозоидах белохвостых мышей Kit+/- рез-
ко повышено содержание РНК по сравнению с нормальными сперматозоидами. Это
свидетельствует об активной работе ряда генов, в том числе и гена Kit. В нор-
ме в сперматозоидах большинство генов «молчит» и РНК почти не образуется. Что
бы это значило? Ответа пока нет.
Живые организмы
— не компьютеры
Работа французских генетиков, обнаруживших роль РНК в наследовании морфоло-
гических признаков у мышей, наряду с другими недавними открытиями показывает,
что классические представления о природе «наследственной информации» и меха-
низмах ее «прочтения» слишком упрощены. В действительности все гораздо слож-
нее . Приходится признать, что аналогии между живыми организмами и искусствен-
ными информационными системами, например компьютерами, вошедшие в моду в кон-
це XX века, в значительной степени неправомочны. В отличие от компьютера в
живых системах так называемая «информация», ее носители, а также «устройства»
для ее прочтения и реализации оказываются слиты воедино и практически нераз-
делимы. Например, РНК оказывается не только «результатом прочтения» генетиче-
ского кода и средством передачи информации от ДНК к системе синтеза белка, но
и активным участником и регулятором самого процесса «прочтения», способным
менять смысл читаемых «сообщений» (см., например, раздел «Наследие РНК-мира»
в главе «Происхождение жизни»). Представьте себе текст, умеющий сам себя ре-
дактировать , или жесткий диск, который в зависимости от записанных на нем
байтов информации приобретает разные физические свойства и активно вмешивает-
ся в работу считывающего устройства — например, заставляет пропустить или по-
вторно прочесть некоторые участки. Или, как в случае с парамутациями, пред-
ставьте себе, что вы хотите перенести информацию с одного компьютера на дру-
гой при помощи дискеты, однако ваша дискета помнит, в каких компьютерах побы-
вала раньше и какие файлы ей там встречались, и на основе этих знаний она на-
чинает по собственному произволу редактировать записанный на ней текст. В
итоге до компьютера-«реципиента» вы донесете не то, что скопировали, а нечто
совсем иное. Не случайно некоторые ведущие теоретики в настоящее время ставят
под сомнение саму применимость понятия «информация» к последовательностям
нуклеотидов ДНК и РНК.
Более того, совершенно ясно, что передача информации от родителей к потом-
кам при помощи РНК является, возможно, необходимым, но явно не достаточным
условием парамутации. Это хорошо видно из результатов исследования парамута-
ций у кукурузы — растения, у которого данный феномен активно изучается уже
полвека.
Классический пример парамутации у кукурузы дает ген pll, от которого зави-
сит окраска пыльников, то есть той части тычинки, в которой образуется пыль-
ца. Этот ген при одной и той же последовательности нуклеотидов может нахо-
диться в одном из двух состояний: «активном» (в этом случае пыльники получа-
ются пурпурные) и «неактивном» (светлые пыльники). Фокус состоит в том, что
неактивное состояние гена является как бы «заразным», то есть активный ген
pll, побывав в одной клетке со своим неактивным «собратом», сам становится
неактивным.
Согласно законам генетики при скрещивании двух гетерозиготных растений (то
есть таких, у которых одна копия гена активна, а вторая нет) четверть потом-
ков должна иметь пурпурные пыльники. Однако этого не происходит, у всех по-
томков пыльники оказываются светлыми, и в последующих поколениях пурпурная
окраска пыльников тоже не появляется. Это происходит потому, что активный
ген, побывав в гетерозиготном состоянии с неактивным, меняет свои свойства и
переходит в неактивное состояние. В таком неактивном виде он затем передается
по наследству. Это и есть парамутация.
Мы уже знаем, что большую роль в поддержании «переключенного» состояния ге-
на при парамутации играют молекулы РНК, передающиеся от родителей к потомкам.
Кроме того, в опытах с кукурузой недавно удалось показать, что для устойчивой
передачи парамутации из поколения в поколение необходим фермент, размножающий
молекулы РНК (РНК-зависимая РНК-полимераза, см. главу «Происхождение жизни»).
И еще было установлено, что многие другие гены тоже участвуют в поддержании
устойчивости парамутантного состояния, но в чем конкретно состоит функция
этих генов, никто до недавнего времени не знал.
В отличие от «настоящей» мутации парамутация у кукурузы может сравнительно
легко ревертироваться, то есть вернуться в исходное состояние. Например, не-
активная версия гена pll может превратиться обратно в активную, если произой-
дут определенные мутации (настоящие, а не «пара-») в других генах.
У кукурузы удалось выявить 10 генов, мутации в которых могут приводить к
возвращению неактивного гена pll в активное состояние. Это значит, что нор-
мальная работа этих генов необходима для поддержания парамутантного состоя-
ния. Чтобы понять механизм парамутации, очень важно выяснить, что это за гены
и что они кодируют. До недавнего времени была известна функция только одного
из них — было показано, что он кодирует РНК-зависимую РНК-полимеразу (см. вы-
ше) .
В 2007 году американские генетики предприняли очередную попытку разгадать
тайну парамутации у кукурузы. И ответ, казалось, был уже почти у них в руках.
Еще бы одно маленькое усилие, и... В общем, история получилась вполне детектив-
бб
ная
Исследователи сосредоточили свое внимание на одном из вышеупомянутых десяти
генов, необходимых для парамутации. Этот ген называется rmrl, и до сих пор
никто понятия не имел, как он работает и что кодирует. Оказалось, что он ко-
дирует не известный ранее белок, похожий по своей структуре на ферменты,
управляющие метилированием ДНК. Это позволило предположить, что поддержание
«парамутантного» состояния как-то связано с метилированием ДНК. Заодно наме-
тилась и связь с РНК-зависимой РНК-полимеразой, которая тоже нужна для пара-
мутации, как было показано ранее. Дело в том, что РНК-зависимая РНК-
Christopher J. Hale, Jennifer L. Stonaker, Stephen M. Gross, Jay B. Hollick. A
Novel Snf2 Protein Maintains trans-Generational Regulatory States Established by
Paramutation in Maize // PLoS Biology. 2007. 5(10): e275.
полимераза размножает маленькие молекулы РНК, которые тоже участвуют в регу-
ляции метилирования ДНК.
Итак, можно было ожидать, что парамутация как-то связана с метилированием
ДНК в окрестностях гена pll.
Ученые обратили внимание на то обстоятельство, что перед началом гена pll
имеется некодирующий участок ДНК, представляющий собой фрагмент транспозона.
Надо сказать, что метилирование ДНК используется в том числе и для инактива-
ции транспозонов.
Кусочек транспозона, расположенный перед началом гена pll, как выяснилось,
по-разному метилируется у нормальных растений и у мутантов с неработающим ге-
ном rmrl (у первых он метилируется сильнее). Кроме того, при работающем гене
rmrl в клетках растений была выявлена пониженная концентрация молекул РНК,
считанных с гена pll, по сравнению с растениями, у которых ген rmrl не рабо-
тал. Скорость транскрипции (считывания) гена pll, однако, была одинаковой в
обоих случаях. Это значит, что от гена rmrl зависит не скорость считывания, а
устойчивость (стабильность) уже считанных с гена pll молекул РНК.
На этом этапе исследователи, казалось, уже вплотную подошли к пониманию ме-
ханизма парамутации. Разрозненные факты вроде бы начали складываться в цель-
ную картину. Можно было предположить, что «активное» состояние гена pll соот-
ветствует низкому уровню метилирования фрагмента транспозона перед его нача-
лом, а «пассивное» — высокому; что ген rmrl регулирует уровень метилирования,
а уровень метилирования в свою очередь определяет стабильность молекул РНК,
считанных с гена pll (и тем самым определяет «активность» гена).
Хотя эту схему трудно назвать простой, дальнейшие эксперименты показали,
что в действительности все еще сложнее. Стало ясно, что истинный механизм па-
рамутации иной, и загадка в итоге так и осталась неразгаданной.
Ученые выявили два факта, которые говорят о том, что уровень метилирования
фрагмента транспозона перед началом гена pll не является определяющим для
возникновения парамутации.
Во-первых, если бы это было так, то в случае нормально работающего гена
rmrl уровень метилирования у «парамутантных» растений должен быть выше, чем у
«непарамутантных». Но это не подтвердилось: уровень метилирования у таких
растений оказался одинаковым (высоким). Во-вторых, оказалось, что ген rmrl
необходим только для устойчивого наследования парамутации, но не для ее воз-
никновения .
Таким образом, механизм парамутации у кукурузы по-прежнему неизвестен. Мо-
жет быть, это явление связано с метилированием каких-то других участков ДНК
или с какими-то иными эпигенетическими модификациями. Метилирование кусочка
транспозона перед началом гена pll, очевидно, необходимо для сохранения неак-
тивного состояния гена, но таких «необходимых, но не достаточных условий» в
принципе может быть много.
Скорее всего, выявленные на сегодняшний день случаи парамутации — лишь вер-
хушка айсберга, основу которого составляют неизвестные пока аспекты эпигене-
тической наследственности. Нужно иметь в виду, что существует не так уж много
генов, которые так четко и ясно проявляются в фенотипе, как ген pll, влияющий
на окраску пыльников. Не случайно большинство известных парамутации связаны
именно с окраской, будь то пыльники кукурузы или хвосты мышей. Другие параму-
тации труднее обнаружить, и они пока остаются незамеченными.
Что общего у
цветковых растений
и млекопитающих?
Парамутации обнаружены у цветковых растений и млекопитающих — двух групп,
венчающих эволюционное древо, соответственно, растений и животных. Очень мо-
жет быть, что это не случайность. Именно в этих двух группах организмов наи-
большее развитие получил родительский геномный импринтинг (см. главу 8). Как
мы помним, суть этого явления в том, что родители целенаправленно манипулиру-
ют наследственными свойствами потомства, метилируя некоторые гены в своих по-
ловых клетках. Ранее мы также упоминали о возможной связи геномного имприн-
тинга с тем обстоятельством, что из всех животных именно у млекопитающих в
ходе индивидуального развития очень рано начинают работать собственные гены
эмбриона (см. главу 5). Итак, три особенности: парамутации, импринтинг и ран-
нее включение эмбриональных генов. Существует ли между ними реальная связь, и
если да, то какова ее природа? Вот еще одна из волнующих неразрешенных зага-
док, ждущих своего ответа.
Может быть, все эти три особенности связаны с тем, что именно у млекопитаю-
щих и цветковых растений материнский организм вкладывает очень много ресурсов
в свое потомство. Это порождает «конфликт интересов» между полами: отец заин-
тересован в том, чтобы потомство получило от матери как можно больше ресур-
сов, а мать пытается сохранить силы, чтобы в будущем произвести новое потом-
ство . Геномный импринтинг — результат этого конфликта. Отец выключает в своих
половых клетках те гены, работа которых выгодна матери, но не эмбриону, а
мать отключает в своих яйцеклетках те гены, работа которых выгодна эмбриону,
но вредна для нее самой. Раннее включение генов эмбриона могло развиться в
связи с тем, что как отцу, так и матери выгодно, чтобы «отредактированные»
ими гены эмбриона включились пораньше. Может быть, парамутации являются по-
бочным следствием этих приспособлений?
Животные держат
мобильные генетические
элементы под контролем
С мобильными генетическими элементами (МГЭ) мы уже познакомились (см. главу
«Наследуются ли приобретенные признаки?»), однако у читателя может сложиться
неполное и не совсем точное представление об этих удивительных объектах, на-
ходящихся на грани живого и неживого, если не упомянуть об одном важном от-
крытии, которое сделали недавно генетики. До сих пор МГЭ обычно воспринима-
лись как активные преобразователи чужих геномов, которые по собственной при-
хоти прыгают с место на место, встраиваются в разные участки хромосом и вно-
сят элемент хаоса в слаженную работу генетических комплексов.
И вдруг оказалось, что геномы высших организмов вовсе не являются пассивны-
ми «жертвами» МГЭ. Эукариотическая клетка, как выяснилось, умеет контролиро-
вать деятельность этих не в меру шустрых нахлебников.
Возможны ли в биологии
«открытия на кончике пера»?
Открытые недавно молекулярные механизмы, при помощи которых эукариотическая
клетка управляет мобильными генетическими элементами, оказались невероятно
сложными, запутанными и даже на первый взгляд громоздкими. Для того чтобы ра-
зобраться в них, могут понадобиться годы. И все это оказалось полной неожи-
данностью для ученых. Никто не подозревал, что может существовать такая сис-
тема. Никакие теории этого не предсказывали. Впору задуматься: сколько еще
сюрпризов нас ждет? И когда, наконец, мы наберем достаточно знаний, чтобы де-
лать открытия «на кончике пера» — так, как астрономы начали открывать не ви-
димые простым глазом планеты Солнечной системы после того, как поняли общие
законы движения планет? Пока же открытия чаще всего сваливаются на биологов
как снег на голову.
Один из редких примеров удачного «умозрительного» открытия в молекулярной
биологии, впоследствии подтвержденного экспериментально, — это открытие тело-
меразы, фермента, который восстанавливает кончики хромосом, укорачивающиеся
после каждого клеточного деления. Российский ученый А. М. Оловников «вычис-
лил» теломеразу за много лет до того, как она была реально открыта. Поняв,
что кончики хромосом должны укорачиваться (потому что фермент, который копи-
рует ДНК, не может скопировать ту ее часть, к которой он прикрепляется в са-
мом начале процесса копирования), Оловников рассудил, что, следовательно,
должен существовать особый фермент, который умеет восстанавливать укоротив-
шиеся кончики. Иначе все клетки очень быстро потеряли бы способность делить-
ся. Самое удивительное, что западные ученые признали это достижение Оловнико-
ва, ссылаются на его работы и считают его одним из создателей «теломерной
теории старения». Обычно подобные озарения остаются не замеченными и не при-
знанными научным сообществом. Сейчас в цене эксперименты, а не рассуждения.
Система управления мобильными элементами впервые была найдена у плодовой
мушки дрозофилы в 2006 году67. Оказалось, что в процессе созревания спермато-
зоидов у дрозофилы «включаются» гены белков семейства Piwi, а также гены осо-
бых маленьких регуляторных РНК, которые получили название пиРНК (piRNA, со-
кращение от Piwi-interacting RNAs).
ПиРНК — короткие, длиной в 24-29 нуклеотидов, молекулы, нуклеотидные после-
довательности которых совпадают с теми или иными участками различных МГЭ.
ПиРНК присоединяются к Piwi-белкам и «программируют» их на распознавание и
уничтожение молекул мРНК, считанных с мобильных элементов. Тем самым подавля-
ется активность МГЭ.
У дрозофилы активность генов Piwi была зарегистрирована только в семенниках
самцов (в созревающих сперматоцитах). Почему-то мушки считают необходимым
приглушить свои МГЭ именно во время сперматогенеза. Возможно, это объясняется
тем, что МГЭ именно в сперматоцитах становятся особенно активными.
В дальнейшем выяснилось, что белки семейства Piwi есть и у млекопитающих.
Они тоже синтезируются во время сперматогенеза и образуют комплексы с пиРНК.
В 2007 году американским биологам удалось показать, что у млекопитающих (а
именно у мышей, на которых проводились опыты) Piwi-белки и пиРНК выполняют
примерно те же функции, что и у плодовой мушки68. Однако то, что обнаружили
исследователи, в действительности похоже на верхушку айсберга и ставит гораз-
до больше новых вопросов, чем дает ответов.
Выяснилось, что мышиные пиРНК весьма многочисленны и разнообразны (как и у
дрозофилы). Последовательности многих из них совпадают с участками известных
мышиных МГЭ, следовательно, они способны «направлять» деятельность Piwi-
белков на МГЭ. Однако есть очень много других пиРНК, последовательности кото-
рых совпадают либо с участками рабочих генов, либо с фрагментами генома,
функции которых неизвестны. Зачем нужны эти пиРНК, пока невозможно сказать.
Ученые установили, что отключение генов Piwi-белков приводит к резкому рос-
ту активности мобильных генетических элементов в сперматоцитах. Стало ясно,
что система регуляции активности МГЭ при помощи Piwi-белков и пиРНК не явля-
ется уникальной особенностью насекомых. Скорее всего, она широко распростра-
Vagin et al. A distinct small RNA pathway silences selfish genetic elements in
the germline // Science. 2006. V. 313. P. 320-324.
68 Alexei A. Aravin, Ravi Sachidanandam, Angelique Girard, Katalin Fejes-Toth, Greg-
ory J. Hannon. Developmentally Regulated piRNA Clusters Implicate MILI in Transposon
Control // Science. 2007. V. 316. P. 744-747.
нена в живой природе.
Если считать МГЭ «эгоистическими» и чужеродными объектами, своеобразными
геномными паразитами, то Piwi-белки и пиРНК можно было бы назвать системой
«внутриклеточного иммунитета», в которой роль «антител» выполняют пиРНК. Если
же считать МГЭ полноправными составными частями единого генома, эту систему
правильнее будет поставить в один ряд с другими известными механизмами генной
регуляции, которые обеспечивают своевременное включение и выключение различ-
ных участков генома в зависимости от потребностей организма.
В пользу второй интерпретации свидетельствует один чрезвычайно важный факт.
Оказалось, что набор пиРНК, производимых сперматоцитами, меняется с возрас-
том, причем весьма резко. Существует два почти непересекающихся «комплекта»
пиРНК. Один из них, «ранний», обнаруживается в семенниках мышат примерно до
12-14-дневного возраста. После этого начинают производиться совсем другие,
«поздние» пиРНК. Свойством подавлять активность МГЭ обладают оба комплекта, и
смысл перемены пока совершенно неясен. Так или иначе, все это больше похоже
не на работу иммунной системы, а на генетическую регуляцию индивидуального
развития. Хотя, конечно, для окончательных выводов у нас еще слишком мало
данных.
Ученые также установили, что пиРНК могут влиять на активность МГЭ не только
путем «программирования» Piwi-белков, но и другим способом — через механизм
метилирования ДНК. Как мы помним, это один из важных способов регуляции ак-
тивности генов. В общем случае чем сильнее метилирован ген, тем слабее он ра-
ботает. Метилирование ДНК осуществляется специальными ферментами. Однако во-
прос о том, откуда эти ферменты «узнают», какие гены надо метилировать, а ка-
кие не надо, пока еще далек от разрешения. На растениях ранее было показано,
что определенную роль в этом могут играть маленькие молекулы РНК, которые ка-
ким-то образом «указывают» ферментам гены, подлежащие метилированию. Ранее
также было установлено, что многие МГЭ у млекопитающих подвергаются усиленно-
му метилированию — это еще один способ держать МГЭ под контролем. Исследова-
тели решили проверить, влияет ли отключение системы Piwi-пиРНК на степень ме-
тилирования МГЭ в сперматоцитах мышей. Оказалось, да, влияет, причем весьма
существенно. Возможно, это свидетельствует о том, что пиРНК и Piwi-белки ка-
ким-то образом направляют деятельность ферментов, ответственных за метилиро-
вание, на нужные участки генома. У насекомых в отличие от млекопитающих мети-
лирование ДНК для регуляции работы генов почти не используется.
И у мух, и у мышей в работе системы Piwi-пиРНК используется принцип обрат-
ной связи. Дело в том, что те «обрезки», которые получаются из считанных с
МГЭ молекул мРНК в результате деятельности Piwi-белков, сами могут функциони-
ровать в качестве пиРНК, то есть связываться с Piwi-белками и управлять их
работой. Чем активнее МГЭ, тем больше с них считывается молекул мРНК. Это
расширяет поле деятельности для Piwi-белков, которые режут эти мРНК на кусоч-
ки. Чем больше становится в клетке таких кусочков, функционирующих в качестве
пиРНК, тем эффективнее действуют Piwi-белки и тем сильнее подавляется актив-
ность МГЭ.
Очевидно, что исследователи только начали проникать в некую совершенно но-
вую, весьма необычную область. Пожалуй, единственный обоснованный вывод, ко-
торый можно сделать, состоит в том, что поведение МГЭ не является бескон-
трольным , что клетка имеет реальную возможность влиять на их деятельность.
Учитывая, что МГЭ играют важнейшую роль в эволюции организмов и даже могут
придавать ей отчасти направленный характер, получается, что организмы в прин-
ципе способны активно регулировать собственную эволюцию. Это еще один меха-
низм управления наследственными изменениями из числа тех, о которых рассказа-
но в главе «Управляемые мутации».
Взаимное
узнавание
Множество захватывающих тайн в биологии связано со способами взаимного уз-
навания на уровне молекул, клеток и организмов. Как одна молекула находит
другую в безумной химической круговерти цитоплазмы? Как транскрипционный фак-
тор находит среди миллиардов нуклеотидов ДНК тот единственный и неповторимый
участок, к которому он должен прикрепиться? Откуда знают растущие нервные
клетки, в какую сторону им необходимо выпустить отросток и с какой клеткой
вступить в контакт? Как удается самцу насекомого безошибочно найти по запаху
самку своего вида, особенно если концентрация химического сигнала в воздухе
близка к нулю? Как ухитряется самка рыбки колюшки опять-таки по запаху вы-
брать себе в мужья такого самца, который приходится ей одновременно и не
слишком близкой, и не слишком дальней родней?
Тема эта необозримо велика, и мы ее до сих пор почти не затрагивали. Не
удастся нам подробно раскрыть ее и в этой последней главе. Но о двух удиви-
тельных и тесно взаимосвязанных открытиях я все-таки расскажу. Одно из них
имеет отношение к иммунитету, другое — к формированию нервной системы в ходе
индивидуального развития. Казалось бы, какая тут связь? Самая прямая: в обоих
случаях главным действующим лицом оказался один и тот же белок из надсемейст-
ва иммуноглобулинов. И белок этот сам по себе заслуживает того, чтобы о нем
рассказать. Известный в биологических кругах афоризм гласит: тот, кто не ви-
дел кораллового рифа, не зоолог. Примерно то же и здесь: кто не слышал о бел-
ке DSGAM, едва ли может представить себе, на что способны белки. Но начнем по
порядку.
Первое открытие связано с иммунной системой насекомых69. Как мы помним, ос-
новная обязанность иммунной системы — безошибочно распознавать любую попавшую
в организм заразу, в том числе и такую, с которой раньше не приходилось
встречаться. Для этого нужно иметь очень много разных белков, избирательно
связывающихся с различными чужеродными веществами (антигенами). Позвоночные
животные продуцируют сотни тысяч вариантов таких защитных белков — гораздо
больше, чем генов в геноме. Высокое разнообразие защитных белков достигается
благодаря прижизненным перестройкам генома в клетках иммунной системы — лим-
фоцитах. О том, как это происходит, говорилось в главе «Управляемые мутации»,
а здесь я только вкратце напомню самое основное.
В геноме «зародышевой линии», то есть в том геноме, который позвоночное жи-
вотное получает от папы с мамой, нет генов антител как таковых, а есть наборы
заготовок — несколько «кассет» похожих, но немного различающихся фрагментов
будущего гена. В зреющем лимфоците специальные ферменты (потомки прирученных
транспозонов — RAG-белки) режут и перекраивают геномную ДНК, чтобы собрать из
этих заготовок один функциональный ген. При этом из каждой «кассеты», состоя-
щей из десятков похожих фрагментов, случайным образом выбирается какой-то
один. В разных лимфоцитах гены антител получаются разными, а общее число тео-
ретически возможных вариантов у человека или мыши достигает трех миллионов.
Приобретенный иммунитет формируется за счет того, что те лимфоциты, чьи анти-
тела лучше других связываются с данным инфекционным агентом, усиленно размно-
жаются, а гены их антител вдобавок еще и «подгоняются» к антигену за счет му-
тирования и дополнительного отбора.
Всего этого нет у беспозвоночных. Долгое время считалось, что беспозвоноч-
Yuemei Dong, Harry E. Taylor, George Dimopoulos. AgDscam, a Hypervariable
Immunoglobulin Domain-Containing Receptor of the Anopheles gambiae Innate Immune
System // PLoS Biology. 2006. V. 4. Issue 7.
ным удается как-то обходиться одним лишь врожденным, неспецифическим иммуни-
тетом и сравнительно небольшим числом иммунных белков — рецепторов, гены ко-
торых в «явном виде» присутствуют в реноме зародышевой линии и не подвергают-
ся прижизненным перестройкам. Учитывая быструю эволюцию и вариабельность мик-
роорганизмов, было довольно удивительно, каким образом беспозвоночные ухитря-
ются при этом выжить.
Однако со временем открывалось все больше фактов, показывающих, что у бес-
позвоночных тоже формируется приобретенный иммунитет. Вот только как им это
удается без прижизненных перестроек генов, оставалось неясным.
Иммунологи из Университета Джонса Гопкинса (США.) обратили внимание на один
весьма необычный ген, имеющийся и у позвоночных, и у насекомых. Этот ген на-
зывается DSGAM (Down syndrome cell adhesion molecule). Некоторые его мутации
у человека ассоциированы с синдромом Дауна. У насекомых он, как было извест-
но, отвечает за регуляцию роста нервных клеток, хотя как именно он это дела-
ет , никто не знал. Ген принадлежит к надсемейству иммуноглобулинов, то есть
содержит иммуноглобулиновые домены (последовательности нуклеотидов, кодирую-
щие участки белковой молекулы, предназначенные для избирательного связывания
других молекул). Иммуноглобулиновые гены есть практически у всех животных.
Они выполняют разнообразные рецепторные функции, далеко не всегда связанные с
иммунной защитой. Например, тот же рост аксонов направляется определенными
химическими сигналами, которые кто-то должен улавливать и распознавать.
После того как на матрице гена DSGAM синтезируется его РНК-копия (мРНК) ,
она подвергается особой процедуре редактирования — альтернативному сплайсин-
гу.
Альтернативный
сплайсинг
Как известно, почти все гены высших организмов состоят из участков, коди-
рующих белок (экзонов), и вставленных между ними некодирующих фрагментов (ни-
тронов) . Есть мнение, что некоторые интроны являются потомками мобильных ге-
нетических элементов, бурно размножившихся в геноме древнейших эукариот (см.
главу «Великий симбиоз»). В ходе прочтения гена (транскрипции) сначала синте-
зируется «незрелая» матричная РНК (мРНК), представляющая собой полную копию
всего гена, включая интроны. Затем интроны один за другим «вырезаются» из мо-
лекулы РНК, а экзоны соединяются друг с другом. Это и называется сплайсингом.
Обычно существует несколько альтернативных вариантов сплайсинга для одной и
той же мРНК. Дело в том, что часть экзонов может быть вырезана вместе с при-
легающими к ним интронами. В результате из одной и той же «незрелой» мРНК,
содержащей, например, 7 экзонов (1, 2, 3, 4, 5, 6, 7) могут получиться разные
«зрелые» мРНК: 12357, 12467, 2345 и т. д. Экзоны могут вырезаться из молекулы
РНК, но не могут меняться местами. Разные мРНК станут затем матрицами для
синтеза разных белков. Таким образом, благодаря механизму альтернативного
сплайсинга один и тот же ген может кодировать несколько разных белков. То,
какой именно белок будет производиться данным геном в каждой конкретной си-
туации, зависит от сложных регуляторных систем, о которых пока еще очень мало
известно.
Между прочим, тот факт, что благодаря альтернативному сплайсингу один ген
может кодировать более одного белка, является явным нарушением принципа «один
ген — один белок» (или «один ген — один фермент») , который был сформулирован
еще в 40-е годы XX века. Данный принцип сыграл огромную положительную роль в
становлении современной молекулярной биологии. На тот момент это было вели-
чайшим достижением — догадаться, что загадочные гены, молекулярная природа
которых еще не была расшифрована, занимаются не чем-нибудь, а именно кодиро-
ванием белков, причем каждому белку соответствует свой собственный ген. Но
сегодня этот принцип устарел, как и почти все «классические догмы» образца
середины прошлого века. Вот вам и еще один пример лопнувшей «абсолютной исти-
ны» !
Альтернативный сплайсинг
ДНК | 1 1 | 2 | | 3 | | 4 ]
«гзон нитрон
РНК | 1 | | 2 | | 3 | | 4 |
// \\_
1 ! 2 | 3 | 4 ] / \ i 1 1 2 1
[
» : 2 |_| з | 4 | | 1 ; з | 4 |
варианты сплайсинга РНК
щ
Уникальной особенностью гена DSCAM, содержащего 21 экзон, является то, что
три из них представлены не в единственном экземпляре, а в виде кассет из мно-
жества немного различающихся копий. У малярийного комара, на котором проводи-
лось данное исследование, экзон № 4 представлен в 14 экземплярах, экзон № б —
в 30, экзон № 10 — в 38. В ходе сплайсинга сохраняется только один вариант
каждого из этих экзонов. Таким образом, за счет альтернативного сплайсинга на
основе единственного гена DSCAM может быть синтезировано 14 х 30 х 38 = 15
960 разных рецепторных белков. У мушки дрозофилы число вариантов еще больше:
38 016.
Такая структура гена DSCAM до боли напоминает те кассеты заготовок, из ко-
торых в лимфоцитах млекопитающих конструируются гены антител. Поэтому мысль
об участии DSCAM в иммунной защите, по правде сказать, напрашивалась давно,
но доказать это удалось лишь недавно, в 2006 году. Ученые обнаружили, что в
присутствии разных бактерий иммунные клетки производят разные наборы сплайс-
вариантов белка DSCAM. Например, в ответ на заражение кишечной палочкой резко
увеличивается частота использования 8-го варианта 4-го экзона, тогда как ста-
филококк стимулирует синтез белков DSCAM с использованием 1-го варианта того
же экзона.
Искусственное «выключение» гена DSCAM приводит к резкому снижению способно-
сти комара сопротивляться инфекциям; кроме того, в его гемолимфе (аналог кро-
ви у насекомых) начинают бурно размножаться бактерии, в норме присутствующие
там в небольших количествах. Это доказывает непосредственное участие DSCAM в
иммунной защите.
ЧИ0*)
\&(Щ
7Ю
lgl<H38)
11-13 ton 15-21
щ iiiiiimiiiii i iiiiiiiiiiiiiiiiiiiiiiiiiiiii hi iiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiii in и mi
Строение гена DSCAM малярийного комара. Вертикальными отрезками показаны
экзоны. Иммуноглобулиновые экзоны 4, 6 и 10 представлены кассетами из
14, 30 и 38 альтернативных вариантов. Зрелая РНК после сплайсинга будет
содержать лишь по одному варианту каждого из этих трех экзонов.
Ученым удалось показать, что отключение тех сплайс-вариантов белка DSGAM,
которые синтезируются в присутствии кишечной палочки, лишает комара способно-
сти бороться именно с кишечной палочкой, но не со стафилококком, и наоборот.
Выяснилось также, что сплайс-варианты DSGAM, синтезируемые в ответ на зараже-
ние той или иной бактерией, лучше других «прилипают» именно к этой бактерии.
Изменения в спектре синтезируемых сплайс-вариантов DSGAM, спровоцированные
контактом с бактериями, сохраняются в течение как минимум 18 часов после то-
го, как контакт прекратился (что происходит потом, ученые просто не проверя-
ли) . Это очень похоже на явление приобретенного иммунитета!
Таким образом, различные сплайс-варианты белка DSGAM в известной мере ана-
логичны антителам позвоночных (хотя антитела, конечно, более избирательны и
эффективны). Образование разнообразных сплайс-вариантов DSGAM осуществляется
на основе того же комбинаторного принципа, что и образование антител у позво-
ночных. В обоих случаях используются случайный выбор и комбинирование отдель-
ных фрагментов гена, каждый из которых представлен в геноме в виде нескольких
альтернативных вариантов. Только у позвоночных это «перекраивание» генетиче-
ской информации осуществляется на уровне ДНК, а у насекомых — на уровне РНК.
Это исследование, на мой взгляд, представляет собой крупный прорыв не толь-
ко в изучении иммунной системы беспозвоночных, но и в понимании общих принци-
пов иммунитета в целом. Можно ожидать, что дальнейшие исследования приведут к
открытию новых черт сходства между иммунными системами далеких друг от друга
групп организмов.
Каждая нервная клетка имеет десятки и сотни отростков, которые в
ходе развития организма соединяются в строго определенном поряд-
ке . Как отростки нейронов находят друг друга и что позволяет им
не ошибиться? Оказалось, что важную роль в этом играет иммуноло-
гический механизм различения «своих» и «чужих».
За рамками исследования осталось множество важных вопросов. Например, оста-
лось неясным, каким образом осуществляется регуляция сплайсинга. Откуда комар
«знает», какие варианты белка DSGAM нужно синтезировать для борьбы с кишечной
палочкой, а какие — для защиты от стафилококка? В любом случае результат уди-
вительный: целая иммунная система — в одном-единственном белке!
Второе недавнее открытие, связанное с белком DSCAM, позволило понять, какую
роль играет это биохимическое чудо в развитии нервной системы.
До сих пор было известно только то, что у насекомых с мутациями в гене
DSCAM нервная система развивается неправильно. Механизм участия DSGAM в инди-
видуальном развитии был неясен, хотя «общую идею» нетрудно было угадать. Что-
бы из делящихся клеток зародыша, которые все имеют одну и ту же заложенную в
них «программу поведения» (геном), сформировалась не аморфная клеточная мас-
са, а сложный организм, эти клетки должны знать, в какую сторону им расти или
переползать, к кому приклеиваться, а от кого, напротив, отделяться. Для этого
им нужно как-то узнавать друг друга, понимать, кто есть кто в их окружении.
Такое взаимное узнавание клеток особенно важно в ходе развития нервной систе-
мы, чтобы отростки нейронов соединялись друг с другом правильным образом и
формировали «правильные» нервные контуры и сети.
Что касается белков иммуноглобулинового надсемейства, то они как раз и спе-
циализируются на выполнении задач такого рода, то есть на персональной иден-
тификации и различении «своих» и «чужих». Поэтому можно было предвидеть, что
для этих белков найдется работа не только в иммунной системе, но и в других
ситуациях, когда нужно разбираться, кто есть кто в клеточном или социальном
70
окружении Однако, повторю, до недавнего времени никто не знал, каким имен-
но способом белок DSGAM реализует эту функцию в ходе развития нервной систе-
мы.
В 2004 году у белка DSGAM было обнаружено очередное поразительное свойст-
во71 . Оказалось, что каждый сплайс-вариант этого белка обладает способностью
к так называемому «гомофильному связыванию». Это значит, что молекула данного
сплайс-варианта «узнает» другую такую же молекулу и прочно связывается с ней.
При этом она никогда не связывается с другими сплайс-вариантами того же бел-
ка . Совершеннейшая фантастика, если подумать! Всем известно, что молекулы ДНК
и РНК легко «узнают» свои собственные копии и склеиваются с ними на основе
принципа комплементарности. Но чтобы подобные свойства обнаружились у белков
— в это даже поверить трудно. Тем не менее, данный факт подтвержден весьма
тщательными экспериментами, проверены сотни сплайс-вариантов. Механизм «гомо-
фильного связывания» активно исследуется, но пока еще до конца не расшифро-
ван.
Нетрудно догадаться, что эта удивительная особенность делает DSGAM идеаль-
ным молекулярным устройством для различения «своих» и «чужих». Оставалось вы-
яснить, как это устройство используется при развитии нервной системы.
Биологи из Австрии и США в 2007 году получили ответ на этот вопрос при по-
мощи генной инженерии72. Они создали три линии генетически модифицированных
мух, в каждой из которых ген DSGAM был радикально «упрощен». Из него были вы-
резаны все альтернативные наборы фрагментов-заготовок, кроме одного-
единственного, разного в каждой из трех линий. Модифицированные мухи, таким
образом, могли синтезировать только один сплайс-вариант белка DSGAM вместо 38
О «социальном окружении» я упомянул не случайно. Оказывается, позвоночные живот-
ные (а возможно, и не только они) широко используют иммуноглобулиновые белки для
персональной идентификации. Например, белки так называемого «главного комплекса гис-
тосовместимости» составляют важнейшую часть индивидуального запаха и, в частности,
позволяют животным отличать родню от чужаков. Даже люди не совсем утратили эту спо-
собность .
71 Wojtowicz W. M. et al. Alternative splicing of Drosophila Dscam generates axon
guidance receptors that exhibit isoform-specific homophilic binding // Cell. 2004.
V. 118. P. 619-633.
72 Daisuke et al. Dscam diversity is essentialfor neuronal wiring and self-
recognition // Nature. 2007. V. 449. P. 223-227.
016.
Внесенные в геном изменения оказались летальными. Все мухи, у которых обе
копии гена DSGAM были изменены (то есть гомозиготы) , погибали на стадии ли-
чинки или куколки, и у этих личинок и куколок наблюдались серьезные нарушения
в развитии нервной системы. Например, у нормальных мух в обонятельном отделе
мозга имеются определенным образом расположенные узлы, к каждому из которых
присоединяются отростки чувствительных нейронов определенного типа. У личи-
нок-мутантов данный отдел мозга представлял собой беспорядочную массу нейро-
нов , и отростки чувствительных клеток крепились к нему как попало. При этом
ген DSGAM у этих личинок работал, и количество производимого белка DSCAM было
таким же, как у здоровых насекомых. Результат был одинаков во всех трех гене-
тически модифицированных линиях.
Авторы сделали вывод, что жизненно важным является не только белок DSGAM
сам по себе, но и разнообразие его сплайс-вариантов.
Дальнейшие исследования показали, что каждый нейрон в норме производит свой
собственный уникальный набор сплайс-вариантов белка DSGAM. Таким образом соз-
дается уникальная «визитная карточка» данного нейрона, что позволяет нейрону
отличать свои собственные отростки от чужих.
При помощи сложных экспериментов удалось показать, что нейроны, способные
синтезировать только один сплайс-вариант DSGAM, растут неправильно, если они
окружены другими такими же мутантными нейронами, однако их рост становится
совершенно нормальным, если их окружают обычные нервные клетки, производящие
разные сплайс-варианты. Если нейрон вообще не производит белка DSGAM, он рас-
тет неправильно в любом клеточном окружении. От того, какой именно сплайс-
вариант производят мутантные клетки, как выяснилось, ничего не зависит.
Таким образом, сплайс-варианты DSGAM нужны нервным клеткам для создания
уникального «молекулярного паспорта», благодаря которому нейроны отличают
свои собственные отростки от чужих.
Данное открытие подтверждает идею о том, что молекулярные системы различе-
ния «своих» и «чужих», основанные на иммуноглобулиновых белках, гораздо шире
распространены в живой природе, чем думали когда-то, и роль их отнюдь не сво-
дится к одной лишь иммунной защите. Осмелюсь предположить, что скоро нас ждут
новые открытия в этой области.
Новые способы
работы с
информацией
Одна из областей, в которых наши знания остаются до обидного неполными, —
это изучение той роли, которую играют молекулы РНК в обработке генетической
информации. Биологи то и дело открывают новые клеточные «информационные тех-
нологии», в которых РНК оказывается главным действующим лицом, и конца этим
открытиям пока не видно.
Новооткрытые функции
РНК подтверждают теорию
абиогенеза
Почему раньше биологи не замечали множества разнообразных функций, выпол-
няемых в клетке молекулами РНК? Может быть, слишком привыкли думать, что «всю
работу в клетке делают белки»? Похоже на то. Как только ученые осознали, что
жизнь началась с РНК (это понимание пришло, как мы помним, в середине 80-х
годов XX века), стало ясно также и то, что теория РНК-мира имеет проверяемое
следствие. Из нее следует, что, если хорошенько поискать, в современных живых
клетках могут найтись ранее незамеченные «следы» эпохи РНК-мира — в том числе
разные неожиданные функции, выполняемые молекулами РНК. Это следствие блестя-
ще подтвердилось, и новые открытия продолжают его подтверждать по сей день,
так что и конца не видно. Это одно из тех обстоятельств, которые все больше
убеждают нас в том, что жизнь действительно возникла естественным путем из
неживой материи. Почему? Судите сами.
Из теории естественного происхождения жизни (абиогенеза) следовало, что
должна существовать молекула, с которой «все началось», — молекула, способная
одновременно справляться и с «работой», и с хранением наследственной инофрма-
ции. Это было проверямое следствие — единственный реальный способ проверить
научную теорию, которую нельзя подтвердить или опровергнуть прямым наблюдени-
ем (а такова большая часть научных теорий). Проверяемое следствие — чрезвы-
чайно ценная вещь! Именно по наличию или отсутствию проверяемых следствий
всегда можно отличить научную теорию от ненаучной. Например, креационизм не
является научной теорией как раз потому, что не имеет проверяемых следствий.
Ведь Бог мог сотворить жизнь и Вселенную с абсолютно любым строением и свой-
ствами . Какое бы неожиданное свойство мы ни обнаружили, всегда можно сказать:
это так, потому что так было угодно Богу. Мотивы высшего разума мы постичь не
можем, поэтому не можем и предсказать ничего конкретного о тех областях ре-
альности, которые еще не изучены. Наука избегает таких теорий, из которых ни-
чего конкретного не следует. Из теории абиогенеза, напротив, следовало нечто
вполне конкретное: должна существовать молекула с такими-то свойствами. Под-
ходящую молекулу нашли — ею оказалась РНК. Проверяемое следствие подтверди-
лось , и теория абиогенеза заработала себе очередной большой и жирный «плю-
сик» . Теория РНК-мира после этого стала важной составной частью теории абио-
генеза. Из нее, в свою очередь, вытекали новые проверяемые следствия, которые
сегодня подтверждаются, и тем самым новые «плюсики» зарабатывает и теория
РНК-мира, и вмещающая ее теория абиогенеза.
Взять, к примеру, тот же альтернативный сплайсинг. Каким образом клетка
«решает», какой из вариантов сплайсинга нужно выбрать в данной ситуации (и,
следовательно, какой из вариантов белка синтезировать)? Об этом пока известно
очень мало. Удалось выяснить, что такая регуляция требует участия особых бел-
ков — регуляторов сплайсинга. Не было оснований думать, что сплайсинг регули-
руется чем-то еще, кроме белков. И вдруг ученые из Иельского университета
(США) публикуют статью73, в которой описан совершенно иной способ регуляции,
где белки не участвуют вообще. Ключевую роль в нем играет сама молекула РНК,
подвергающаяся сплайсингу.
Способность молекулы РНК самостоятельно определять свою судьбу и выбирать
способ, каким она будет перекроена, определяется наличием в одном из ее неко-
дирующих участков (интронов) специфической последовательности нуклеотидов,
которая сама собой сворачивается в особую трехмерную структуру — РНК-
переключатель . О том, что это такое, мы уже рассказывали в главе «Происхожде-
ние жизни».
Исследование проводилось на грибе Neurospora crassa, известном широкой пуб-
лике как розовая хлебная плесень. По иронии судьбы, на этом же объекте в 40-е
годы прошлого века были получены сенсационные результаты, позволившие сформу-
лировать принцип «один ген — один белок». Сейчас на нейроспоре изучают аль-
тернативный сплайсинг — явление, опровергающее (или, лучше сказать, уточняю-
щее и расширяющее) этот замечательный принцип.
Ming Т. Cheah, Andreas Wachter, Narasimhan Sudarsan, Ronald R. Breaker. Control
of alternative RNA splicing and gene expression by eukaryotic riboswitches // Na-
ture. 2007. V. 447. P. 497-500.
У нейроспоры, как и у ряда других эукариот, в генах, участвующих в биосин-
тезе тиамина (витамина Bi) , были обнаружены участки, сходные с известными
бактериальными РНК-переключателями, которые реагируют на производное тиамина
— тиамин-пирофосфат. Большинство известных РНК-переключателей действуют по
принципу отрицательной обратной связи. Они реагируют на вещество, синтезируе-
мое белковым продуктом данного гена, и при достаточно высокой концентрации
этого вещества отключают ген.
Примерно то же самое наблюдалось и в данном случае. Повышение концентрации
тиамин-пирофосфата в клетках гриба приводит к снижению производства белков,
ответственных за синтез тиамина. Было показано, что если удалить из соответ-
ствующих генов участки, похожие на бактериальные РНК-переключатели, то произ-
водство тиамин-синтезирующих белков перестает зависеть от концентрации тиа-
мин-пирофосфата .
Таким образом, стало ясно, что участки грибных генов, похожие на РНК-
переключатели, действительно являются таковыми. Оставалось лишь выяснить ме-
ханизм их действия, то есть понять, как они блокируют работу «своих» генов. У
бактерий РНК-переключатели делают это либо на этапе транскрипции (первичного
«прочтения» гена), либо на этапе трансляции — синтеза белка на матрице мРНК.
незрелая
мРНК
незрелая
мРНК
Низкая концентрация тиамин-пирофосфата (ТРР)
сплайсинг
I Т
■шиш
т
шм»ш
t
РНК-переключатель
Высокая концентрация ТРР
i
синтез белка
(трансляция)
сплайсинг
t
ЩЩМР
t
РНК-переключатель
1
Inmttqrfi
"бракованная"
зрелая мРНК
трансляция блокируется
На этом рисунке показано, как РНК-переключатель регулирует альтерна-
тивный сплайсинг1 у розовой хлебной плесени (на примере гена NMT1).
Участок мРНК, вырезаемый при сплайсинге, отмечен пунктирными линиями и
серыми стрелками. При низкой концентрации тиамин-пирофосфата РНК-
переключатель «защищает» потенциальный сайт (место) сплайсинга, отме-
ченный значком в виде буквы «Т». В результате при сплайсинге вместо
этого сайта используется другой, расположенный по соседству (серая
стрелка). Участок мРНК, отмеченный белым цветом, не попадает в зрелую
мРНК. При высокой концентрации ТРР это вещество связывается с РНК-
переключателем и меняет его конфигурацию. В результате молекула РНК
режется в том месте, которое раньше было прикрыто РНК-переключателем,
белый участок попадает в зрелую РНК и «портит» ее.
У эукариот, как выяснилось, дело обстоит иначе — работа гена блокируется на
этапе сплайсинга. Бактериям это недоступно, поскольку у бактерий сплайсинга
нет (почти нет, если быть точным) . Тиаминовый РНК-переключатель в генах
Neurospora crassa располагается в первом интроне, недалеко от начала гена.
Если в клетке мало тиамин-пирофосфата, РНК-переключатель «приклеивается» од-
ной из своих петель к строго определенному месту на молекуле мРНК. Это место
является одним из потенциальных мест сплайсинга, то есть именно здесь в ходе
сплайсинга молекула мРНК может быть разрезана. Однако приклеившийся РНК-
переключатель не позволяет этого сделать, и молекула разрезается в другом
подходящем месте по соседству. В результате формируется «правильная» зрелая
мРНК, на основе которой синтезируется полноценный белок.
Если же в клетке много тиамин-пирофосфата, это вещество присоединяется к
РНК-переключателю и изменяет его форму. Переключатель «отклеивается» от места
сплайсинга и перестает его защищать. Тогда молекула РНК режется именно в этом
месте, которое раньше прикрывалось РНК-переключателем. Это, в конечном счете,
приводит к формированию «бракованной» зрелой мРНК, на базе которой полноцен-
ный белок синтезировать невозможно.
Таким образом, РНК-переключатель в зависимости от концентрации тиамин-
пирофосфата направляет сплайсинг по одному из двух альтернативных путей.
Судя по некоторым косвенным признакам, регуляция сплайсинга при помощи РНК-
переключателей может быть довольно широко распространена у эукариот. Чтобы
проверить это предположение, необходима разработка эффективных методов поиска
РНК-переключателей в эукариотических геномах — эти методы пока еще далеки от
совершенства.
Еще одна неожиданная функция РНК обнаружилась недавно в ходе изучения меха-
низмов репарации — починки повреждений в ДНК. Оказалось, что молекулы РНК мо-
гут играть роль матриц, информация с которых переписывается в поврежденную
молекулу ДНК взамен утерянной74. Процесс основан на обратной транскрипции
(как мы помним из предыдущей главы, так называют переписывание информации из
РНК в ДНК, то есть синтез ДНК на РНК-матрице) . Изобретение обратной транс-
крипции, между прочим, должно было стать важным переломным моментом в разви-
тии РНК-мира, поскольку позволило РНК-организмам перейти к хранению наследст-
венной информации в более надежной и стабильной форме молекул ДНК. В предыду-
щих главах мы упоминали несколько случаев использования обратной транскрипции
современными организмами: это перемещение и размножение ретротранспозонов и
ретровирусов, образование ретропсевдогенов, восстановление укорачивающихся
при каждом клеточном делении кончиков хромосом — теломер. И вот еще один
пункт добавился к этому списку — репарация ДНК.
Если молекула ДНК повреждена — например, подверглась разрыву, — для ее по-
чинки необходима матрица, в которой последовательность нуклеотидов соответст-
вует исходному, «правильному» состоянию поврежденного участка. Ранее счита-
лось , что в качестве таких матриц всегда используются другие молекулы ДНК.
Потом выяснилось, что иногда эти ДНК-матрицы синтезируются путем обратной
транскрипции на основе РНК при участии ретротранспозонов.
Наконец, совсем недавно ученые из Национального института экологии здоровья
(США.) сумели показать, что репарация возможна и непосредственно на основе
РНК-матриц без предварительного изготовления ДНК-матрицы и без участия специ-
фических ферментов — обратных транскриптаз, кодируемых ретротранспозонами.
Исследователи искусственно вызывали у дрожжей разрыв хромосомы в одном и
том же строго определенном месте. Затем в клетки добавляли искусственно син-
тезированные короткие молекулы РНК, последовательности нуклеотидов в которых
соответствовали участкам поврежденной хромосомы по краям разрыва. Оказалось,
Storici F., Bebenek К., Kunkel Т. A., Gordenin D. A., Resnick M. A. RNA-templated
DNA repair // Nature. 2007. V. 447. P. 338-341.
что эта процедура повышает вероятность успешной «починки» разорванной хромо-
сомы в 500 раз.
Если в середину молекулы РНК, служащей матрицей для репарации, ввести не-
сколько лишних нуклеотидов, они потом обнаруживаются в «починенной» хромосоме
как раз между сшитыми краями разрыва. Это свидетельствует о синтезе ДНК на
матрице РНК, то есть об обратной транскрипции.
разрыв двойной
спирали ДНК
г
>'ч> иА> i>Vo & w\>
^ <^s*~ ,<\<~ ^> <*v*, <■»/> ,r^fn.
\j.x<j> \j\af \>"Lj у .i*^ '.A>'''..k
1
r *\
■*■ r-,-r-- s*t-r s "\rv +чг>< i»V>
"1
"> /<\<\ /f\*\ .'ГУ*"'' <*V\ .^V^'
' S'~*" \J\S ~Л> -+'-*' Ч-*Л>' '.^.v
"" Г\*"- Уу ,^» *-\<* /r^ ГЛ***
■У>\> '\>W 4>\.,, /"s>'^ ^*'v> '*.V.
ЛЛ'' ^V" -^VV •'fYrN\ /^Vv. /^A-
'••iwii-ii'' Jiy ■^•«y \^Л> '->»а>' '\.j
1
: <*W /V\ /V^N-
-^ >\v. /,У'^,
соединение
концов
'Л /<\<*. /^,<\. <^,*\\ .<у<\ /Г\^\
,v*,«wf' ■■",>\> s>U^ -А^ '.>\-/ V">
Г4.- О^ "TV^ •'r7j"V ""V^v /^yrv.
гомологичная
рекомбинация
Два основных способа «починки» разрывов двойной спирали ДНК.
Первый способ (негомологичное соединение концов) чреват неточно-
стями — потерей или вставкой лишних нуклеотидов в районе разры-
ва. Второй более точен, но требует наличия «запасной копии» по-
врежденного фрагмента ДНК. Как выяснилось, эта запасная копия не
обязательно должна быть двухцепочечной ДНК (как показано на ри-
сунке) , годится и одноцепочечная ДНК и даже РНК.
Кроме того, это говорит об отсутствии принципиальных преград для переписы-
вания информации из РНК в ДНК в живых клетках, что может иметь большое значе-
ние для эволюции. Уже целый ряд фактов указывает на то, что молекулы РНК ино-
гда могут служить чем-то вроде «резервных копий» особо важных «файлов», запи-
санных в геномной ДНК, и при потере каких-то фрагментов информации (даже це-
лых генов!) эти резервные копии могут идти в ход. Мы уже знаем три таких при-
мера (описанные в главе «Управляемые мутации» геномные перестройки у инфузо-
рий, парамутации, участие РНК в репарации), а есть и другие. Ясно, что все
это открывает большие возможности для эволюции, хотя о том, в какой мере и
для чего именно они используются, мы пока можем только гадать.
Сравнение геномов человека и мыши
помогло обнаружить новый способ
регуляции работы генов75
Ну и напоследок — еще одно недавнее открытие, которое как нельзя лучше по-
75 Liana F. Lareau, Maki Inada, Richard E. Green, Jordan C. Wengrod, Steven E. Bren-
ner. Unproductive splicing of SR genes associated with highly conserved and ultra-
conserved DNA elements // Nature. 2007. V. 446. P. 926-929.
называет, что наши знания об устройстве и функционировании живой клетки до
сих пор крайне неполны, а сложность и запутанность механизмов внутриклеточной
регуляции далеко превосходит все, что мы еще недавно могли себе представить.
В основе нового способа генной регуляции, только что открытого биологами из
Калифорнийского университета в Беркли, лежит уже знакомый нам альтернативный
сплайсинг, а также еще один механизм, называемый nonsense-mediated mRNA decay
(NMD). Это можно перевести как «разрушение мРНК, опосредуемое бессмыслицей».
Данный механизм представляет собой нечто вроде «внутриклеточной цензуры». Он
служит для уничтожения заведомо бессмысленных молекул РНК. Особые молекуляр-
ные системы, о которых пока мало что известно, идентифицируют зрелую (то есть
прошедшую сплайсинг) мРНК как бессмысленную и приговаривают ее к уничтожению
в том случае, если в ней имеется «преждевременный» стоп-кодон (три нуклеоти-
да, сигнализирующие об окончании синтеза белка). В норме стоп-кодон должен
располагаться в конце каждого гена. Но в результате мутации он может образо-
ваться в середине гена. В этом случае синтез белка обрывается преждевременно,
и весь ген становится бессмысленным. Именно для выявления и «обезвреживания»
таких мутаций и существует система NMD.
Открытие, о котором идет речь, было сделано в ходе анализа так называемых
«ультраконсервативных» участков генома. Как известно, геномы человека и мыши
довольно сильно различаются. Однако имеются и довольно длинные (до многих со-
тен нуклеотидов) абсолютно идентичные участки. Ясно, что такая консерватив-
ность нуклеотидных последовательностей должна иметь какой-то смысл. По-
видимому, она означает, что любое изменение этих участков неизменно оказыва-
ется вредным и для человека, и для мыши, поэтому естественный отбор быстро
отсеивает таких мутантов.
Некоторые ультраконсервативные участки находятся между генами и могут вы-
полнять регуляторные функции. Другие лежат прямо внутри генов, задевая как
интроны, так и экзоны. Это уже более странно, поскольку обычно структура ге-
нов и белков довольно пластична, а многие нуклеотидные замены в пределах эк-
зонов вообще не влияют на структуру кодируемого белка. Такие замены в ходе
эволюции обычно накапливаются вполне свободно.
Ученые заметили, что ультраконсервативные участки имеются во всех без ис-
ключения генах семейства SR (splicing regulators). Эти гены кодируют белки,
участвующие в регуляции альтернативного сплайсинга.
Детальный анализ показал, что гены SR сами подвержены альтернативному
сплайсингу, причем в результате могут получаться такие мРНК, которые должны
уничтожаться системой NMD. Оказалось, что у каждого SR-гена имеются два аль-
тернативных сплайс-варианта. Один из них представляет собой «нормальную»
мРНК, которая может служить матрицей для синтеза полноценного SR-белка. Вто-
рой содержит преждевременный стоп-кодон и должен уничтожаться цензурой. Экс-
перименты показали, что этот второй сплайс-вариант действительно активно
уничтожается системой NMD.
Обнаруженный регуляторный контур работает по принципу отрицательной обрат-
ной связи. SR-белки могут контролировать альтернативный сплайсинг не только
«чужих» мРНК, но и своих собственных. Чем больше производит клетка SR-белков,
тем чаще они направляют сплайсинг своих мРНК по альтернативному пути, тем са-
мым снижая темп производства новых SR-белков. Так поддерживается некоторое
постоянство концентрации SR-белков в клетке.
Как мог возникнуть такой причудливый регуляторный контур? Ведь производство
SR-белков вполне могло бы контролироваться каким-нибудь менее экзотическим
способом — например, при помощи транскрипционных факторов. Видимо, в данном
случае при создании контура обратной связи отбор просто «зацепился» за то,
что ближе лежало, а именно за способность SR-белков контролировать альтерна-
тивный сплайсинг. Аналогичным образом, активность генов, кодирующих транс-
крипционные факторы, обычно регулируется не чем-нибудь, а транскрипционными
факторами (хотя и исключений немало). Это характерный «почерк» эволюции, со-
всем не похожий на разумное проектирование, а похожий скорее на самосборку
чего получится из чего попало76.
Десять генов, составляющих SR-семейство, родственны друг другу. Общий пре-
док людей и мышей уже имел все десять, но у него самого когда-то был предок
только с одним исходным SR-геном, от которого произошли все остальные путем
последовательных дупликаций. Самое удивительное, что механизм саморегуляции с
участием альтернативного сплайсинга и цензуры, судя по всему, возникал у раз-
ных представителей SR-семейства параллельно и независимо. Это видно из того,
что преждевременные стоп-кодоны образуются у них в ходе альтернативного
сплайсинга разными способами. У одних SR-генов имеется особый экзон со стоп-
кодоном, который в норме вырезается из мРНК вместе с интронами. У других к
концу мРНК после «законного» стоп-кодона может быть пришита пара-тройка лиш-
них экзонов, в результате чего законный стоп-кодон с точки зрения системы
цензуры начинает выглядеть как преждевременный. Очевидно, SR-гены имеют высо-
кую предрасположенность к формированию таких регуляторных контуров.
Ультраконсервативными у людей и мышей обычно являются те участки SR-генов,
которые непосредственно прилегают к преждевременному стоп-кодону, — но не
всегда и не только. Скорее всего, уникальный консерватизм этих участков обу-
словлен тем, что они необходимы для правильного сплайсинга. Проверить это
предположение трудно, поскольку о молекулярных механизмах сплайсинга нам из-
вестно пока очень мало. Вместе с тем у асцидии Ciona (далекого родственника
позвоночных животных) тоже есть SR-гены с альтернативными экзонами, несущими
преждевременные стоп-кодоны. Нуклеотидные последовательности этих генов похо-
жи на человеческие и мышиные, но все же не идентичны им. Это значит, что
ультраконсервативность в данном случае не является абсолютной. Ситуация еще
сильнее запуталась совсем недавно, когда удалось экспериментально показать,
что удаление некоторых некодирующих ультраконсервативных участков из генома
мышей совершенно не вредит их здоровью.
Не исключено, что новооткрытый способ генной регуляции распространен до-
вольно широко и затрагивает отнюдь не только гены — регуляторы сплайсинга.
Дальнейшие исследования должны пролить свет на этот вопрос.
На мой взгляд, данное открытие говорит, прежде всего, о том, как мало мы
еще знаем о жизни клетки и о возможных путях ее эволюционных изменений.
Сколько еще подобных открытий предстоит сделать, прежде чем у нас появится
достаточно оснований для построения всеобъемлющей эволюционной теории, или,
как сейчас говорят, «нового эволюционного синтеза»? Думаю, много.
ВМЕСТО
ЗАКЛЮЧЕНИЯ
Честно говоря, у меня в запасе еще целая куча подобных историй, показываю-
щих , как мало мы еще знаем и как много предстоит открыть. Но я вынужден оста-
новиться, потому что объем публиквации не может расти до бесконечности.
Давайте подведем итоги. В «Предисловии» я обещал воздержаться от глобальных
обобщений и постараюсь сдержать слово. Великие обобщения делать действительно
рано, но самое время указать на некоторые тенденции. Куда идет биология, в
какую сторону движется теоретическая мысль, в каком направлении меняются наши
представления о жизни и ее эволюции?
76 На это можно возразить, что и человеческое инженерно-техническое творчество тоже
часто идет по такому принципу. Но от всемогущего Творца мы все-таки вправе ожидать
качественно иного уровня креативности.
Пожалуй, самая очевидная тенденция состоит в том, что эволюция жизни, как
мы ее понимаем, становится все менее хаотичным и все более закономерным, на-
правленным процессом. При этом в одних отношениях у эволюции оказывается
меньше, а в других — гораздо больше возможностей и степеней свободы, чем счи-
тали биологи 30-40 лет назад.
Уменьшение степеней свободы затронуло в первую очередь наши представления о
макроэволюции (крупномасштабных эволюционных изменениях). Мы видим, например,
что при переходе на более высокий уровень организации обязательно наблюдаются
параллелизмы. Если раньше параллелизмы казались исключением, то теперь ясно,
что это общее правило.
Что касается микроэволюции, то здесь степеней свободы, напротив, оказалось
гораздо больше, чем было принято считать. Жизнь больше не представляется нам
обреченной всегда двигаться вслепую и наугад. Отбор удачных вариантов из мно-
жества случайных изменений оказывается хоть и очень важным, первичным, но
все-таки далеко не единственным способом придания эволюции направленности и
смысла.
Живая клетка, как выяснилось, располагает целым арсеналом средств, позво-
ляющих контролировать наследственные изменения. Можно, оказывается, даже при-
обретенные признаки сделать наследственными, если очень уж прижмет. Правда, в
большинстве случаев это невыгодно, но лишняя степень свободы всегда может
пригодиться.
Мы привыкли считать эволюцию очень медленным процессом, слишком медленным
для того, чтобы реальные живые организмы, существующие «здесь и сейчас», мог-
ли всерьез озаботиться его направлением и последствиями. Говоря более строго,
считалось, что естественный отбор не может создавать специальных приспособле-
ний для оптимизации эволюционного процесса. Постепенно мы начинаем осозна-
вать , что эволюция — слишком важная часть жизни, чтобы пустить ее на самотек.
Она точно так же происходит «здесь и сейчас», как и все прочие жизненные про-
цессы, и естественный отбор поэтому не может ее игнорировать. Организмы, ко-
нечно, не умеют предвидеть отдаленные последствия эволюционных изменений, но
они могут и даже обязаны позаботиться хотя бы о своих непосредственных потом-
ках. Когда самка выбирает себе в партнеры не первого попавшегося самца, а,
скажем, самого крупного, яркого или определенным образом пахнущего, она тем
самым направляет эволюцию в определенное русло. И пусть только попробует вы-
брать плохо: потомство от такого брака окажется менее жизнеспособным, и лег-
комысленное отношение к важнейшей жизненной задаче будет отсеяно отбором. Вы-
бор брачного партнера — это не что иное, как целенаправленное манипулирование
наследственными свойствами потомства, то есть управление эволюцией77. То же
самое можно сказать и о геномном импринтинге, хотя в данном случае наследст-
венные изменения недолговечны и в следующем поколении, скорее всего, будут
стерты. Но они могут оставить и неизгладимый след, потому что метилирование
нуклеотидов влияет на вероятность их мутирования. Когда бактерия, испытывая
голод, начинает производить склонные к ошибкам ДНК-полимеразы, чтобы повысить
скорость мутагенеза, то что это, если не целенаправленное манипулирование
темпами эволюции? А чего стоит история с вирусами, для которых, как выясни-
лось, снижение скорости мутирования смерти подобно?
Способность к эволюции, более того, необходимость эволюции заложена в самую
Нужно помнить, что эволюция на микроуровне — изменение ДНК отдельных организмов —
далеко не всегда ведет к эволюции на макроуровне, то есть появлению новых видов. На-
против , видам часто приходится прикладывать колоссальные микроэволюционные «усилия»
именно для того, чтобы остаться самими собой. Получается своеобразная эволюционная
диалектика: чтобы сохранить собственную идентичность, вид должен иметь динамично ме-
няющийся генофонд.
сердцевину жизни, это ее основа, которую нельзя удалить, не уничтожив все
здание.
И мы видим, что земная жизнь эволюционирует не как множество разрозненных
объектов (организмов, видов, популяций), каждый из которых озабочен лишь соб-
ственным выживанием и должен полагаться только на себя. Жизнь развивается как
единое целое. «Блочная сборка», информационный обмен, кооперация, симбиоз —
вот на чем, как мы теперь видим, основывалось развитие жизни с самых первых
ее шагов на Земле. Как это не похоже на старые представления о всеобщей без-
жалостной борьбе и изолированном, одиноком пути каждого отдельного вида! Мы
видим, как по мере развития биологической науки бывшие «паразиты» превращают-
ся в друзей, «мусорная ДНК» — в ведущий фактор эволюционного прогресса, инди-
видуальные организмы — в симбиотические сверхорганизмы.
Но не будем преувеличивать. Возможности биологической эволюции по созданию
эффективных механизмов самоуправления все-таки ограничены. Как лимфоцит не
может знать заранее, какая именно мутация позволит создать нужное антитело,
так и самка оленя не может предугадать, к каким отдаленным последствиям при-
ведет ее пристрастие к болыыерогим самцам. Без метода проб и ошибок, без эле-
мента случайности и без отбора эволюция обойтись все-таки не может. Однако со
временем механизмы эволюции совершенствуются, точно так же, как органы дыха-
ния и пищеварения. Куда это может привести? Не будет ли, в конце концов, пре-
одолено и это принципиальное ограничение, не появится ли у эволюции возмож-
ность предвидеть результаты генетических изменений и проектировать их на ос-
нове этого знания?
Создается впечатление, что эволюция в данный момент как раз работает над
решением этой проблемы. Она уже произвела на свет — может быть, в качестве
первой пробы — довольно необычный вид животных, который не только научился у
агробактерий основам генной инженерии, но и, кажется, стоит на пороге понима-
ния последствий своих поступков.
Впрочем, эта тема явно выходит за рамки нашей публикации. Но только сейчас,
когда мы подошли к финалу, начинает по-настоящему ощущаться вся важность и
объемность недосказанного. Думаю, что это ощутили и вы, дорогие читатели. Что
ж, это может стать поводом для продолжения разговора.
Ликбез
НАЧАЛА ОРГАНИЧЕСКОЙ ХИМИИ
Реформатский С.Н.
ОРГАНИЧЕСКИЕ
КИСЛОТЫ
Органические кислоты образуют несколько групп, из которых самая важная —
карбоновые кислоты, названные так вследствие присутствия в их формулах кар-
боксильной группы СООН1) . Доказательством ее присутствия служат следующие ре-
акции получения кислот:
1) кислоты могут быть получены действием воды в присутствии оснований на те
трехгалогенопроизводные углеводородов, в которых три атома галогена стоят при
одном углеродном атоме; по смыслу реакции три атома галогена должны замес-
Кроме карбоновых, известны сульфокислоты, например C6H5'S02 (ОН) , тио-кислоты, на-
пример СН3—COSH, сульфиновые кислоты, например C2H5'SOOH и другие.
титься при этом на три гидроксила:
СН3СС13 + ЗН20 = СН3С(ОН)3 + 3HC1
но такие трехатомные спирты легко теряют частицу воды и превращаются в ки-
слоты
СН3 * С(ОН)3 = Н20 + СНв • С — О.
ОН
Эта реакция, кроме доказательства присутствия карбоксильной группы и ее
строения, дает основание рассматривать карбоновые кислоты как производные ги-
потетических трехатомных спиртов, в которых три гидроксила стоят при одном и
том же углероде.
2) Кислоты получаются также обмыливанием цианистых соединений (нитрилов);
например, цианистый метил CH3.CN при действии щелочей или, лучше, соляной ки-
слоты распадается на аммиак и уксусную кислоту (действует здесь вода):
сн„.с
N + Нг
н
СН3-С—О СН3-С—О
О I +NH3= |
un ОН ONH4
3) Соли карбоновых кислот получаются присоединением алкотюлятов к окиси уг-
лерода :
— С— О -J-СН,-ONa = СНЧ-Сzz О
I * I
ONa
На основании сказанного карбоновые кислоты можно рассматривать:
a) как производные углеводородов, водород которых замещен на карбоксил,
b) как ангидриды трехатомных спиртов с тремя гидрэксилами при одном углеро-
де;
c) кроме того, их можно производить и из воды замещением атома водорода на
кислотный радикал, например (СН3СО)ОН.
При сопоставлении формул алдегида
сн3—с-_о
I
Н
и карбоновой кислоты
снч —с—о
I
он
видно, что последние суть производные алдегидов, происшедшие через замеще-
ние водорода алдегидной группы на гидроксил, что и достигается при окислении
алдегидов.
Общие свойства
карбоновых кислот
Кислоты вследствие присутствия в них водного остатка должны, конечно, иметь
сходство со спиртами; но положение водных остатков в тех и других не одинако-
во : в то время как в спиртах гидроксил стоит при углероде, не содержащем ки-
слорода, здесь, наоборот, — при углероде окисленном (при карбониле). Это об-
стоятельство обусловливает существенные отличия кислот от спиртов.
Для спиртов характерно:
1) образование алкоголятов при действии, например, натрия;
2) замещение гидроксила на атом хлора при действии РС15;
3) образование эфиров замещением водорода на радикалы.
Все эти превращения свойственны и кислотам, для них также известны:
CH3'COONa, СН3С0С1, CH3COOR. Но алкоголяты распадаются уже от действия воды
на спирт и едкую щелочь и образуются лишь при прямом действии металлов на
спирты, тогда как металлические производные кислот от воды не изменяются, и
вообще суть настоящие соли; образуются же они из кислот не только при дейст-
вии металлов, но и при действии оснований и углекислых солей. Затем, продукты
замещения гидроксила спиртов на атом хлора по отношению к воде довольно по-
стоянны, тогда как хлорангидриды кислот, образующиеся при реакции с РС15, ды-
мят уже на воздухе вследствие действия влажности, образуя НС1 и вновь кисло-
ту:
СН3С0С1 + Н20 = НС1 + СНз'СООН.
Далее, замещением гидроксильного водорода спиртов на углеводородный радикал
получается простой эфир, например: СН30'СН3, а при кислотах — сложный, напри-
мер СН3СООСН3; первые, как известно, от действия щелочей не обмыливаются, а
последние, наоборот, очень легко вступают в эту реакцию.
Кислоты, как производные углеводородов, могут быть предельными или непре-
дельными, жирными или ароматическими; а кроме того, в зависимости от числа
карбоксилов, — одно-, дву-, трех- и многоосновными.
Предельные
одноосновные
кислоты СпН2п02
Они производятся из предельных углеводородов замещением одного атома водо-
рода на карбоксильную группу. Простейшей кислотой поэтому должна быть кисло-
та, производящаяся из метана, т. е. уксусная — СН3СООН. Но в этом ряду есть
еще более простая кислота, муравьиная — Н'СООН; она должна быть причислена к
этому ряду кислот, потому что состав ее отвечает общей их формуле СпН2п02.
Изомерия
Для кислот, производящихся из метана и этана, изомерных случаев быть не мо-
жет . Из пропана СН3' СН2' СН3 производятся две кислоты:
1) замещением водорода в группе СН3 — нормальная масляная кислота
СН3СН2СН2СООН и
2) замещением в группе СН2 — изомасляная (СН3) 2СН'СООН. Бутан сам существует
в двух изомерных формах, и от кажпой из них можно произвести по две ки-
слоты :
1).СН3 2) СН3 3) СНа 4) СН^
111 I
сн, сн2 сн —сн3 снэ—с —сн3
I : I I I
СН, СН-СН, СН£ СООН
I II
СНг СООН СООН
I
СООН
Название
Для некоторых кислот существуют эмпирические названия; так, простейшая ки-
слота Н'СООН носит название муравьиной,
СНз'СООН — уксусной,
С2Н5'СООН — пропионовой,
С3Н7СООН — масляной,
С4Н9'СООН — валерьяновой,
СбНц
СбН1з
С7Н15
CsHiv
СООН — капроновой,
СООН — энантовой,
СООН — каприловой,
СООН — пеларгоновой,
C9Hi9'COOH — каприновой,
С15Н31СООН — пальмитиновой,
С17Н35СООН — стеариновой.
Для отличия изомерных кислот составляются рациональные названия по произ-
водству их от уксусной кислоты; например, кислоты, происходящие из бутана,
могут быть названы так:
1) пропил-уксусная или нормальная валерьяновая,
2) метил-этил-уксусная,
3) изопропил-уксусная или просто изовалерьяновая и
4) триметил-уксусная.
По женевской номенклатуре кислоты называются по углеводородам с тем же чис-
лом углеродных атомов; например, СНз'СООН — этановая кислота. При изокислотах
ведется нумерация углеродных атомов, причем С в карбоксиле считается первым;
например, СН3—СН2—СН (СН3)—СООН называется метил-2-бутановой кислотой.
Способы
получения
1) Одноосновные кислоты распространены в природе большею частью в виде
сложных эфиров, каковы, например, жиры и жирные масла, из которых они и могут
быть получены обмыливанием.
2) Окислением спиртов, алдегидов, кетонов и многих других органических со-
единений .
3) Обмыливанием цианистых соединений.
4) Действием угольного ангидрида на металлорганические соединения; напри-
мер, CH3Na + С02 = CH3COONa.
Магний-иод-органические соединения также соединяются с С02 (1) и по
разложении водой дают кислоты (2):
1). С — 0 + CH,MgJ = OVC—О,
и i
О OMgJ
2) CHj-Cet.O+HjO^CHj-C — 0 + Mg(OH).L
I I
OMgJ OH
5) Синтетический способ Франкланда и Дюппа (1865 г.), разработанный Висли-
ценусом, состоит в следующем: если на уксусный эфир СН3СООС2Н5 подействовать
алкоголятом натрия, то образуется так называемый ацетоуксусный эфир
СН3'СО'СН2'СООС2Н5, в котором группа СН2 под влиянием двух электроотрицательных
карбонильных групп приобрела способность замещать свой водород на металлы;
поэтому дальнейшее действие алкоголята натрия таково, что получается натр-
ацетоуксусный эфир; а этот последний при действии на него какого-нибудь гало-
генопроизводного выменивает Na на радикал:
CH3COCHNaCOOC2H5 + CH3I = СН3СОСН (СН3) СООС2Н5 + Nal
Если полученное соединение обмылить концентрованным спиртовым раствором ще-
лочи, то оно распадается по следующему уравнению:
СН3СОСН(СН3) СООС2Н5 + 2К0Н = СН3СООК + СН3СН2СООК + С2Н5ОН
Таким образом, исходя из уксусной кислоты, содержащей два углерода, прихо-
дят к кислоте с тремя углеродными атомами. В продукте, подвергаемом обмылива-
нию, и другой атом водорода группы СН2 может быть замещен сначала на натрий,
а затем на радикал, так что получится еще более сложная кислота.
6) Аналогичный предыдущему синтез Конрада состоит в действии алкоголята на-
трия, а затем какого-нибудь галогенопроизводного на этиловый эфир малоновой
кислоты СООС2Н5—CH2—COOC2H5; в нем, как и в ацетоуксусном эфире, есть группа
СН2, положенная между двумя карбонилами; при этом последовательно получаются:
СН2(СООС2Н5)2 -> CHNa(COOC2H5)2 -> СН (R) (СООС2Н5)2;
в последнем эфире можно еще один водород заместить на натрий
CNa(R) (СООС2Н5)2л а натрий — на какой-нибудь радикал C(Rf) (R) (СООС2Н5)2; эти
эфиры при обмыливании дают кислоты: СН(R)(СООН)2 и C(Rf)(R)(СООН)2.
При нагревании полученные двуосновные кислоты теряют частицу С02 и превра-
щаются в одноосновные: RCH2'COOH и
>сн.соон
Получение масляной кислоты по способу Конрада
Реакция протекает в три фазы:
1) получение этил-малонового эфира;
2) получение этил-малоновой кислоты и
3) получение масляной кислоты.
1) В небольшую колбу, соединенную с вертикальным холодильником, к раствору
2,3 г натрия в 25 г абсолютного спирта приливают 16 г малонового эфира и
при взбалтывании постепенно туда же прибавляют 20 г С2Н51. Потом нагревают
(1—2 часа) на водяной бане до исчезновения щелочной реакции; отгоняют
спирт из соляной бани, разбавляют остаток водой и извлекают эфиром. По от-
гонке эфира продукт перегоняют. Этил-малоновый эфир кипит при 206—203 С.
Выход — 15 г.
2) Приготовляют раствор КОН в Н20 так, чтобы на 1 г эфира приходилось 1,25 г
КОН, растворенного в 1 см3 Н20. В колбу с водной щелочью приливают этил-
малоновый эфир и с обратным холодильником нагревают на водяной бане до ис-
чезновения маслообразного слоя, на что требуется 1/4—1/2 часа. Затем раз-
бавляют смесь полуторным объемом воды, при охлаждении приливают к раствору
эквивалентное количество конц. НС1 (концентрацию ее определяют ареометром,
количество вычисляют по количеству взятого в реакцию КОН) и 3—4 раза из-
влекают этил-малоновую кислоту эфиром; эфирный раствор сушат безводным
Na2S04 и, отфильтровав, эфир отгоняют; остаток выливают в чашку и нагрева-
ют, при помешивании, на водяной бане до начала затвердевания. Застывшую
при охлаждении массу высушивают на неглазурованной тарелке и перекристал-
лизовывают из бензола. Т. пл. 111,5 С. Выход — 7 г.
3) В закрытой пробкой вюрцевской колбочке, повернутой так, чтобы отводная
трубка направлена была вверх, на масляной бане при 180 С нагревают этил-
малоновую кислоту до прекращения выделения СОг (требуется около 30 минут).
Остаток перегоняют с термометром. Т. кип. масляной кислоты 162—163 С. Вы-
ход равняется 85% теории.
Таблица предельных одноосновных кислот
Название
кислоты
Муравьиная
Уксусная
Пропионовая
Масляная норм.
Изомасляная
Валерьян. норм.
Изовалерьян.
Метил-этил-укс.
Триметил-укс.
Капроновая
Энантовая
Каприловая
Пеларгоновая
Каприновая
Лауриновая
Миристиновая
Пальмитиновая
Стеариновая
Мелиссиновая
Формула
СН202
с2н4о2
с3нбо2
С4Н8О2
С4Н8О2
С5Н10О2
С5Н10О2
С5Н10О2
С5Н10О2
СбН1202
С7Н1402
CsHi602
С9Н18О2
С10Н20О2
С12Н2402
С14Н2802
С16Н3202
С18Н3602
С30Н60О2
Темпер.
плавл. С
+8,6
+16,7
-23
+2
—
-20
—
—
+35,5
-1,5
-10,5
+16,5
+12,5
+31,4
+43,6
+53,8
+62
+72,5
+90
Удельный вес
1,2256 при 15 С
1,0607 при 15 С
1,0163 при 0 С
0,938 при 0 С
0,9651 при 0 С
0,9562 при 0 С
0,9467 при 0 С
0,941 при 21 С
—
0,9450 при 0 С
0,9313 при 0 С
0,911 при 20 С
0,9107 при 12,5 С
0,930 при 37 С
0,8750 при 43,6 С
0,8622 при 53,8 С
0,8527 при 62 С
0,8454 при 69,2 С
—
Температура кипения
при 760 мм
100 С
119 С
140,6 С
163 С
155,5 С
185,5 С
176,3 С
177 С
164 С
205 С
223 С
236 С
254 С
269 С
225 С при 100 мм
250,5 С при 100 мм
271,5 С при 100 мм
291 С при 100 мм
Физические
свойства
Низшие представители изучаемых кислот — тела жидкие, с резким кислым запа-
хом, перегоняющиеся, без разложения, растворимые в воде и при охлаждении спо-
собные кристаллизоваться. Средние представители труднее растворимы в воде, а
высшие (от Си) суть тела твердые, перегоняющиеся только в разреженном про-
странстве , без запаха и нерастворимые в воде. Все растворимы в спирте и эфи-
ре. Низшие и средние представители (от Ci до Сю) способны улетучиваться с па-
рами воды, за что названы летучими жирными кислотами, а высшие (содержащие
Си и более) вследствие кристалличности называются твердыми жирными кислота-
ми . Правильности в температурах кипения обычные, т. е. наблюдается повышение
с усложнением состава, а для изомеров — понижение т. кип. с накоплением мети-
ловых групп; например, т. кип. нормальной валерьяновой = 185,5 С, изо-
валерьяновой = 176,3 С, триметил-уксусной = 164 С. Вообще же кислоты кипят
выше, чем углеводороды и спирты с тем же числом атомов углерода. Удельный вес
падает с усложнением состава; для трех низших членов он больше единицы, а для
высших — меньше.
Температура плавления твердых кислот имеет следующую особенность: она выше
для кислот с четным числом атомов углерода по сравнению не только с кислотой
менее сложной, но и высшей, рядом с ней стоящей; напр. т. пл. кислоты СцН2202
= 28,5 С, кислоты С13Н26О2 = 40,5 С, тогда как т. пл. кислоты С12Н24О2 = 43,6 С.
Химические
свойства
Растворимые кислоты имеют кислую реакцию, и потому синий лакмус окрашивают
в красный цвет, но с увеличением частичного веса кислотность ослабевает.
Превращения кислот могут быть разделены на две группы:
1) превращения, происходящие в карбоксильной группе, и
2) превращения в радикале.
К первой группе относятся:
a) Способность кислот замещать водород карбоксильной группы на металл с об-
разованием солей.
Получение солей органических кислот
Получение последних производится различно, смотря по тому, приготовляют ли
соль растворимую или нерастворимую и, кроме того, щелочного или какого друго-
го металла. Для получения растворимой соли, например натриевой, к кислоте
прибавляют раствор или NaOH или Na2C03; конец реакции определяют при помощи
лакмуса или метилоранжа. По насыщении раствор сгущают и соль выкристаллизовы-
вают .
Реакция насыщения уксусной кислоты едким натром сопровождается отделением
тепла в количестве около 13,7 К, т. е. столько же, сколько и при кислотах
хлористоводородной и азотной.
Если нужно приготовить растворимую в воде соль такого металла, углекислая
соль которого не растворима в воде, то задача упрощается; например, для полу-
чения уксусно-кальциевой соли к нагретому водному раствору кислоты прибавляют
СаСОз до тех пор, пока, несмотря на избыток последнего, прекратится выделение
С02. После этого смесь фильтруют и из фильтра выкристаллизовывают соль. Если
приготовляемая соль не растворима в воде, то получают сперва какую-нибудь
растворимую соль этой кислоты, а из нее уже осаждают требуемую соль путем об-
менного разложения; например, чтобы получить нерастворимую щавелево-
кальциевую соль, действуют на щавелево-аммониевую соль хлористым кальцием:
C204(NH4)2 + СаС12 = С204Са + 2NH4C1
b) Аммиачные соли кислот при нагревании теряют частицу воды и дают амиды
кислот:
CH3COONH4 = CH3CONH2 + Н20
Дальнейшее нагревание амидов самих по себе или в присутствии водоотнимающих
веществ приводит к отпадению еще одной частицы воды и образованию нитрилов
кислот:
CH3CONH2 = CH3CN + Н20
c) При действии пятихлористого (1) или треххлористого (2) фосфора гидроксил
карбоксильной группы замещается на атом хлора, причем получаются хлорангидри-
ды кислот:
1) СНзСООН + РС15 = СН3С0С1 + Р0С13 + НС1
2) ЗСНзСООН + 2РС13 = ЗСН3СОС1 + Р203 + ЗНС1
d) При смешении кислот со спиртами выделяется частица воды и образуются
сложные эфиры:
СНзСООН + С2Н5ОН = Н20 + СН3СООС2Н5
Скорость этерификации зависит от строения кислоты; нормальные кислоты имеют
большую скорость, а изокислоты — меньшую; чем больше разветвлена цепь угле-
родных атомов, тем меньше скорость этерификации.
Из превращений в радикале укажем на действие галогенов. При действии хлора
и брома водород радикала замещается на галоген, и образуются галогенозамещен-
ные кислоты; например, при действии хлора на уксусную кислоту получаются: мо-
нохлоруксусная, дихлоруксусная и трихлоруксусная кислоты:
СНзСООН + С12 = СН2С1С00Н + НС1
СНзСООН + 2С12 = СНС12С00Н + 2НС1
СНзСООН + ЗС12 = СС13С00Н + ЗНС1
Получение монохлоруксусной кислоты
Для получения монохлоруксусной кислоты Брюкнер рекомендует поступать так: в
смесь уксусной кислоты (200 г) с иодом (2 г) , РС15 (10 г) и красным фосфором
(5 г) при 100 С в продолжение двух часов пропускают хлор. Чистой СН2С1'СООН
получается 170 г.
Число входящих атомов галогена находится в зависимости от различных условий
(нагревания или охлаждения, прямого света или рассеянного, от количества дей-
ствующего галогена и пр.). Если галогена взять такое количество, чтобы он мог
заместить только один атом водорода, то замещение происходит в а-положении,
т. е. замещается лишь тот водород, который стоит при углероде, связанном с
карбоксилом; например в кислоте пропионовой СН3'СН2'СООН галоген становится в
группу СН2, а не СН3, и получается СН3'СНВг'СООН. На основании указанной реак-
ции можно определить, гидрогенизован или нет тот атом углерода, при котором
стоит карбоксил; так, триметилуксусная кислота (СН3)зС'СООН при действии брома
не дает соответствующей бромокислоты, так как углерод, связанный с карбокси-
лом , негидрогенизован.
Аналогично галогенопроизводным жирных углеводородов и жирные галогенокисло-
ты способны к разнообразным превращениям; так, их галоген может быть замещен
на гидроксил, на группу нитро, амино, на CN и др. Получаются таким образом
оксикислоты, нитрокислоты, аминокислоты и др.2
Кроме указанных приведем следующие, более глубокие превращения кислот:
1) от нагревания с натронной известью кислоты распадаются с образованием
предельных углеводородов;
2) известковые и баритовые соли кислот при сухой перегонке дают кетоны и ал-
дегиды;
3) при действии электрического тока кислоты разлагаются на водород, угольный
ангидрид и радикал, причем первый выделяется на отрицательном полюсе, а
2 Кроме сх-галогенокислот известны |3- и у~кислоты'" к действию щелочей они относятся
различно; так сх-галогенокислоты при нагревании с КОН дают оксикислоты; |3-
галогенокислоты теряют при этом галогеноводородную кислоту и превращаются в непре-
дельные кислоты; если же их кипятить с содой, то получается непредельный, углеводо-
род ; например,
2CH3CHBrCH(CH3) COOH + Na2C03 = 2СН3СН: СН (СН3) + ЗС02 + 2NaBr + Н20.
у-Галогенокислоты при кипячении не только со щелочами, но даже с водой дают лак-
тоны (см. при оксикислотах).
С02 и радикал — на положительном. Радикалы, как обладающие свободным срод-
ством, в момент выделения соединяются между собою попарно, образуя пре-
дельный углеводород; например
СН3ХООН , СН3
=н3+зеой+| .
сн3-соон сн5
При других более сложных кислотах наблюдается образование и непредельных
углеводородов (Бутлеров, Бунге); например из пропионовой кислоты получен эти-
лен, а из масляной — пропилен. При электролизе кислот образуются также спир-
ты.
По отношению к окислителям нормальные кислоты более устойчивы, чем изоки-
слоты.
Представители
Муравьиная кислота НСООН открыта была в XVII столетии Джоном Брайтом в му-
равьях, откуда и получила название. Она распространена в растительном царст-
ве, находится в жгучей крапиве; но ядовитые свойства последней обу-
словливаются присутствием какого-то другого вещества.
Муравьиная кислота получается из глицерина и щавелевой кислоты при 100—110
С (ср. получение аллилового спирта); при этом сначала щавелевая кислота рас-
падается на угольный ангидрид и муравьиную кислоту:
С2Н204 = С02 + СН202,
которая с глицерином образует муравьиный эфир (так наз. моноформин) глице-
рина, а этот последний обмыливается той кристаллизационной водой, которая на-
ходится в кристаллической щавелевой кислоте:
СН2(ОН) СН(ОН) СН2(ОНСО) + Н20 = СН2(ОН) СН(ОН) СН2(ОН) + НСООН.
Получение муравьиной кислоты
В тубулированной реторте (емк. 150 см3), соединенной с холодильником и при-
емником, на водяной бане (для повышения температуры в воду прибавляют NaCl)
нагревают 25 г обезвоженного (медленным нагреванием в фарфоровой чашке, при
помешивании, до 175—180 С; термометр в глицерине) глицерина с 25 г крист. ща-
велевой кислоты; когда прекратится выделение С02, реторту вынимают из воды,
дают немного охладиться и прибавляют еще 25 г С2Н204 и т. д. до 5 раз. В это
время в приемник перегоняется водная муравьиная кислота. Фракцнонировкой ее
получают 77%-ную кислоту. Т. кип. 107 С.
В технике муравьино-кислый натрий получают легким нагреванием (120—150 С)
окиси углерода с едким натром под давлением в 6—8 атмосфер; этот технический
способ значительно удешевил муравьиную кислоту:
СО + NaOH = HCOONa.
Безводная муравьиная кислота — бесцветная, резкого запаха жидкость с т.
кип. 100 С; при охлаждении кристаллизуется и имеет т. пл. +8,6 С. С водой,
спиртом и эфиром смешивается во всех отношениях. Водная кислота с содержанием
22,5% воды кипит при 107,2 С.
Для муравьиной кислоты характерны следующие реакции:
1) при нагревании с конц. H2S04 она разлагается на воду и окись углерода:
CH202 = H20 + СО;
2) при нагревании как солей, так и самой кислоты с раствором азотнокислого
серебра или сулемы происходит выделение свободных металлов Ад и Нд (или
НдС1) . Восстановительные свойства муравьиной кислоты объясняются тем, что в
ее формуле НСООН или ОН'СН:0 находится алдегидная группа СН:0, так что ее
можно рассматривать как алдегид угольной кислоты; а алдегиды характеризуются
восстановительными свойствами. Муравьиная кислота — антисептик.
Свойства муравьиной кислоты
a) а) В пробирке к муравьиной кислоте или к ее соли приливают конц. H2S04 и
нагревают; выделяющуюся окись углерода поджигают; она горит голубым пламе-
нем.
b) К кипящему раствору НдС12 прибавляют немного муравьиной кислоты; выделяет-
ся белый (HgCl) или серый (Нд) осадок.
c) В пробирке к нескольким каплям НСООН приливают слабый раствор КМп04 и
разбавленной H2S04; при нагревании окраска исчезает. Если образующуюся при
этом С02 провести в баритовую воду, то последняя замутится.
Из солей муравьиной кислоты характерны свинцовая и серебряная: первая — по
способности легко кристаллизоваться в иглах или призмах, а вторая — по легкой
разлагаемости с выделением серебра и угольного ангидрида.
Все соли муравьиной кислоты в воде растворимы.
Этиловый эфир муравьиной кислоты — жидкость с т. кип. 55 С, применяется для
фабрикации искусственного рома.
Уксусная кислота СН3'СООН известна была в древности, но состав ее был опре-
делен лишь в 1814 году Берцелиусом. В природе находится в свободном состоянии
в виде солей и в виде эфиров.
Уксусная кислота в технике добывается или из продуктов сухой перегонки де-
рева или уксусным брожением спиртовых жидкостей. По первому способу получен-
ная из древесного уксуса известковая соль выпаривается досуха; сухая соль
прокаливается (до 200 С) для разрушения смолистых примесей и затем перего-
няется с соляной кислотой. Для получения чистой кислоты известковую соль при
помощи соды переводят в натронную; последнюю сплавляют, затем перекристалли-
зовывают, высушивают и перегоняют с крепкой серной кислотой. Безводная кисло-
та получается повторным замораживанием ее. 80%-ная уксусная кислота поступает
в продажу под названием уксусной эссенции.
По второму способу спиртовые жидкости, каковы: разбавленная водка, пиво,
вино и др., окисляются в особых чанах кислородом воздуха под влиянием уксус-
нокислых бактерий. Получаемый при этом продукт употребляется главным образом
в виде уксуса (3—5%-ный водный раствор уксусной кислоты) и отчасти для приго-
товления свинцовых белил.
В настоящее время уксусную кислоту получают из ацетилена, пропуская смесь
его с водяным паром и воздухом над молибденовым ангидридом (Мо03 — катализа-
тор) . В этой реакции ацетилен сначала присоединяет воду, образуя уксусный ал-
дегид, который окисляется кислородом воздуха в уксусную кислоту.
Получение безводной уксусной кислоты
Продажную уксусную кислоту перегоняют, собирая при 110—119 С; дестиллат ох-
лаждают льдом (зимой выставляют на холод); когда все закристаллизуется, выни-
мают из льда и дают отчасти расплавиться; жидкую часть сливают; кристаллы
расплавляют и еще раз замораживают и повторяют эту операцию 2—3 раза; после
этого перегоняют, собирая при 118 С.
Безводная или ледяная уксусная кислота — бесцветная едкая жидкость с т.
кип. 118 С; при охлаждении легко кристаллизуется; т. пл. 17 С. Плотность пара
уксусной кислоты, определенная при температуре, близкой к 119 С, почти удво-
енная; нормальной становится лишь около 200 С. С водой, спиртом и эфиром сме-
шивается во всех отношениях. Очень гигроскопична. Смешение с водой со-
провождается выделением тепла и повышением удельного веса (контракция), при-
чем максимум его приходится на смесь с 23% воды3. Уксусная кислота имеет
большое применение: в виде уксуса, для синтеза красящих и душистых веществ,
для получения ацетона, для приготовления ацетатов целлюлозы, для получения
уксуснокислых солей при крашении и печатании и др.
Все соли уксусной кислоты в воде растворимы. Замечательно, что спиртовый
раствор уксусной кислоты, а также безводная кислота не способны разлагать уг-
лекислых солей, наоборот, угольный ангидрид вытесняет уксусную кислоту из
спиртового раствора уксусно-натровой соли (при этом осаждается ЫагСОз) ; вод-
ный же раствор кислоты весьма легко разлагает карбонаты. Из солей некоторые
имеют медицинское применение, другие — техническое. Соль калия плохо кристал-
лизуется, но в присутствии уксусной кислоты получаются кристаллы состава:
СНз'СООКСНзСООН, или СН3СООК-2СН3СООН.
Водный раствор аммиачной соли CH3'COONH4 применяется в медицине как потогон-
ное средство под названием Spiritus Mindereri. Средняя свинцовая соль
(СНз'СОО) 2РЬ'ЗН20 носит название свинцового сахара; имеет сладковатый вкус;
ядовита. В растворе свинцового сахара растворяется окись свинца, и при кри-
сталлизации из раствора выпадает основная соль (СН3'СОО) 2РЬ'РЬ (ОН) 2 или
(СНз'СОО) РЬ(ОН), называемая свинцовым уксусом; его раствор от действия уголь-
ного ангидрида осаждает основную углесвинцовую соль — свинцовые белила. Ос-
новная медная соль носит название ярь-медянки, а соединение уксусно-медной и
мышьяковисто-медной соли называется швейнфуртской зеленью. Обе соли применя-
ются как краски. Серебряная соль довольно трудно растворима в воде и кристал-
лизуется в виде блестящих игол. Железная соль имеет значение в анализе; а
именно — при действии хлорного железа на растворы солей уксусной кислоты по-
лучается темно-красное окрашивание вследствие образования растворимой в воде
(СНз'СОО) з^е; при кипячении этого раствора происходит гидролиз и выпадает бу-
рый осадок основных уксуснокислых солей железа: (СНз'СОО) 2Fe(ОН) ,
(СНз'СОО) Fe (ОН) 2 и даже (FeOH)3; так же легко разлагается и соль алюминия; на
этом основано применение их в технике в качестве протрав, а в анализе — для
отделения алюминия и железа от марганца, цинка, кобальта и никеля. При нагре-
вании уксусных солей с мышьяковистым ангидридом получается окись какодила,
обладающая отвратительным запахом.
Уксусная кислота по отношению к окислителям постоянна; она даже хромовый
ангидрид растворяет, не окисляясь.
Свойства уксусной кислоты и ее натриевой соли
а) В пробирке к раствору уксусно-натриевой соли приливают FeCl3: получается
темно-красный раствор; при кипячении его получается красно-бурый осадок
основных солей.
3 Если смешать ледяную уксусную кислоту с HN03 (уд. в. 1,52) , то повышается темпера-
тура, и при перегонке получается бесцветная, дымящая на воздухе жидкость с т. кип.
127—128 С. Пиктэ дает этому телу формулу (СН3 COO) 2N(OH) 3, т. е. рассматривает его
как смешанный ангидрид уксусной кислоты и ортоазотнои N(OH)5. Вода разлагает его на
уксусную и азотную кислоты. Пиктэ же при растворении азотного ангидрида в уксусном
получил смешанный ангидрид
(СН3СО)20+ N205 = 2CH3COON02
Дымящая легкоподвижная жидкость.
b) В пробирке прокаливают безводную уксусно-натриевую соль; по охлаждении
приливают разбавленной НС1: из полученного при этом Na2C03 выделяется С02.
c) В спиртовый раствор безводной уксусно-натриевой соли довольно долго про-
пускают СО2; раствор мутится от выделения Ыа2СОз.
d) К 5 каплям чистой СНз'СООН приливают слабый раствор КМп04. Обесцвечивания
нет.
Действием РС15 или РС1з на уксусную кислоту получается хлористый ацетил
СН3'С0С1; это — бесцветная, удушливого запаха, дымящая на воздухе жидкость с
т. кип. 55 С. Водой бурно разлагается на НС1 и СНз'СООН. Применяется для по-
лучения уксусных эфиров:
СН3'С0С1 + С2Н5'ОН = СН3СООС2Н5 + НС1
С аммиаком дает амид уксусной кислоты:
СН3'С0С1 + NH3 = CH3CONH2 + НС1
с водородом — этиловый спирт:
СН3'С0С1 + 2Н2 = СН3'СН2(ОН) + НС1
с цинкорганическими соединениями применяется для получения спиртов и кето-
нов; с ароматическими углеводородами, в присутствии А1С1з, дает метиловые ке-
тоны.
Получение СН3'С0С1
В колбе емк. 100 см3, соединенной с холодильником и приемником, к 30 г ле-
дяной уксусной кислоты приливают 24 г РС13 и, закрыв колбу пробкой, осторожно
(около 50 С) нагревают на водяной бане. Когда прекратится выделение НС1 и
жидкость разделится на 2 слоя (верхний — СН3'СОС1) , доводят баню до кипения и
отгоняют верхний слой. Для очищения дестиллат еще раз перегоняют с термомет-
ром, собирая при 50—56 С. Выход — 25 г.
Другой способ получения СН3'СОС1
Нагретый до 50 С раствор уксусной кислоты в бензоле или хлороформе прилива-
ют в SiCl4 и потом фракционируют. Выход 85%. При других кислотах выход мень-
ше .
Вероятное течение реакции:
1) 4СН3'СООН + SiCl4 = Si(OCOCH3)4 + 4HC1
2) Si(OCOCH3)4 + SiCl4 = 4СН3'СОС1 + 2Si02
Свойства СН3СОС1
a) В пробирке к 5 см3 Н20 приливают 1/2 см3 СН3'СОС1. Скоро наступает бурная
реакция и хлорангидрид переходит в раствор.
b) В пробирке к СН3'СОС1 приливают но каплям крепкий аммиак; бурная реакция с
выделением кристаллического амида.
c) В пробирке к 1 см3 спирта, при охлаждении водой, по каплям приливают такой
же объем СН3'СОС1; после этого — равный объем воды и затем КОН — до щелоч-
ной реакции. При этом (или по прибавлении твердой NaCl) всплывает слой
приятно пахнущего уксусного эфира.
При действии на хлористый ацетил уксусно-натровой соли получается уксусный
ангидрид:
СН3С0С1 + CH3COONa = СН3СООСОСН3 + NaCl
В технике получают его, пропуская над сухим уксуснокислым натрием молеку-
лярную смесь хлора и сернистого ангидрида:
С12 + S02 + CH3COONa = 2(CH3CO)20 + 2NaCl + Na2S04.
Уксусный ангидрид можно рассматривать как продукт отпадения частицы воды от
двух частиц уксусной кислоты; однако, непосредственное отнятие воды от уксус-
ной кислоты происходит лишь отчасти при пропускании паров уксусной кислоты
над нагретым до 300 С Ti02.
Бесцветная, удушливая жидкость с т. кип. 139,5 С и уд. весом 1,0969 при 0
С. В воде почти нерастворим, но растворяется в спирте и эфире. В отличие от
хлористого ацетила труднее реагирует с водой; чтобы произошло присоединение
ее, требуется время или нагревание. Применяется для получения уксусных эфи-
ров.
Получение уксусного ангидрида
В реторту емк. 150 см к 30 г измельченной сухой уксусно-натровой соли
(способ получения сухой соли описан дальше) приливают по каплям 20 г хлори-
стого ацетила. Когда прилита половина, перемешивают стеклянной палочкой и за-
тем добавляют остальную часть. Затем нагревают реторту светящимся пламенем
бунзеновской горелки, и ангидрид перегоняется. Дестиллат перегоняют с термо-
метром . Выход — 2 0 г.
Получение уксусного ангидрида по способу Гольдшмидта,
видоизмененному Э. Фридманом
В широкогорлый стеклянный баллон емк. 1 литра помещают порошкообразную
просеянную смесь 330 г обезвоженного уксуснокислого натрия и 16 г серы; к
смеси приливают 470 г сухого бензола. Вместо бензола можно брать толуол, хло-
роформ и другие, а еще удобнее — уксусный ангидрид (400 г) . Смесь тщательно
перемешивают. Баллон закрывают пробкой с двумя отверстиями. В одно вставляет-
ся стеклянный тройник, один отросток которого соединяется с наклонным холо-
дильником, а другой — с мешалкой с ртутным запором. Через второе отверстие
пробки проходит газоотводная трубка, которая у начала шейки баллона отогнута
к его стенке, чтобы не задевать мешалки. Конец трубки доходит до одной трети
ото дна. Через эту трубку пропускают осушенный хлор или из баллона, или из
прибора для добывания хлора, с такой скоростью, чтобы хлор сполна поглощался
смесью. Для контроля внешний конец холодильника соединяют с промывалкой. Для
вышеуказанной смеси требуется 110 г хлора; количество это определяется по
взвешиванию баллона со смесью, а при повторных опытах и по времени, установ-
ленному первыми опытами. Чтобы избежать разогревания смеси, баллон помещают в
водяную ванну. Мешалка пускается в ход перед началом пропускания хлора и ра-
ботает в течение всего опыта. Прекратив пропускание хлора, смесь нагревают в
течение часа при 90 С; затем, поместив баллон в масляную баню, отгоняют при
уменьшенном давлении (50—100 мм) с термометром. Сначала перегоняется бензол,
а в пределах 75—90 С ангидрид. Его подвергают вторичной перегонке. При 760 мм
его температура кипения 136 С, при 105 мм — 81 С, при 53 мм — 63 С. Выход —
до 90% теоретического.
Обезвоживание СН3СООЫа'ЗН20
500 г свежеперекристаллизованной и отжатой соли нагревают в чугунном котел-
ке. Когда начнется разбрызгивание и загустение, необходимо постоянно переме-
шивать железной палкой, чтобы избежать обугливания соли на дне. Когда соль
загустела и еще несколько времени нагрета была около 300 С, ее частями еще
горячую толкут в чугунной ступке, просеивают и сейчас же пересыпают в банку с
притертой пробкой. Выход — около 330 г.
Свойства уксусного ангидрида
a) К 5 см Н20 прибавляют 1/2 см3 ангидрида. Растворения не происходит. Сюда
же прибавляют NaOH: при взбалтывании наступает растворение с образованием
CH3COONa.
b) К 2 см3 спирта прибавляют 1 см3 ангидрида и несколько минут нагревают. За-
тем приливают равный объем воды и нейтрализуют щелочью. Образовавшийся ук-
сусный эфир узнается по запаху.
Уксусно-этиловый эфир СН3'СООС2Н5 получается действием уксусной кислоты на
серновинную кислоту:
C2H5OS02(OH) + СН3СООН = СН3СООС2Н5 + S02(OH)2.
Это — бесцветная, приятного запаха жидкость с т. кип. 76 С; довольно трудно
растворима в воде.
Получение уксусного эфира
К смеси 50 г конц. H2S04 и 50 г С2Н5'ОН, нагреваемой при 131—135 С, медлен-
но, но непрерывно приливают смесь 100 см3 спирта с 100 см3 ледяной уксусной
кислоты. Эфир при этом отгоняется. Дестиллат нейтрализуют раствором Ыа2СОз и
в делительной воронке отделяют верхний слой; сушат на СаС12 и перегоняют с
термометром, собирая при 72—78 С. Выход около 90% теоретического количества.
Полного освобождения от спирта достигают взбалтыванием с конц. раствором
СаС12 (1 ч. Н20 и 1 ч. СаС12) .
Обмыливание эфира
В колбе емкостью в 1/2 литра, снабженной вертикальным холодильником, кипя-
тят смесь 20 см3 уксусного эфира, 200 см3 Н20 и 50 г твердого КОН; через 1/2
часа холодильник ставят как для перегонки и отгоняют часть жидкости. Дестил-
лат испытывают на спирт. К остатку в перегонной колбе приливают серной кисло-
ты и перегоняют в особый приемник. Дестиллат испытывают на уксусную кислоту
(запах, лакмус, FeCl3) .
При пропускании хлора в нагретую безводную уксусную кислоту в присутствии
фосфора или серы или уксусного ангидрида получается монохлоруксусная кислота
СН2С1'СООН. Тело кристаллическое, ст. пл. 61 С.
Применяется для получения индиго.
Пропионовая кислота СН3—СН2—СООН получается обмыливанием соответствующего
нитрила СН3—СН2—CN или окислением нормального пропилового спирта. Жидкость с
т. кип. 140,6 С. С водой смешивается.
Масляные кислоты: СН3—СН2—СН2—СООН нормальная и (СН3) 2СН—СООН изомасляная.
Первая находится в виде глицерида (т. е. сложного эфира, образованного глице-
рином и масляной кислотой) в коровьем масле вместе с капроновой, каприловои и
каприновой кислотами; в прогорклом масле она находится в свободном состоянии.
Получается обыкновенно масляным брожение и сахара под влиянием схизомицетов в
присутствии мела. Изомасляная кислота получается окислением изобутилового
спирта. Обе кислоты жидки и имеют сходный неприятный запах (запах пота); от-
личаются они:
1) по растворимости: изомасляная более трудно растворима в воде, чем масля-
ная;
2) температура кипения нормальной кислоты 163 С, а изокислоты 156,5 С;
3) кальциевая соль масляной кислоты легче растворима в холодной, чем в горя-
чей воде, а изомасляной — наоборот.
Свойства соли масляной кислоты
Насыщенный на холоду раствор нормального маслянокислого кальция, помещенный
в запаянную трубку, по внесении в горячую воду выделяет кристаллы. Насыщенный
при нагревании раствор изомасляного кальция при обыкновенной температуре вы-
деляет кристаллы соли; если запаянную трубку с таким раствором и кристаллами
поместить в горячую воду, то кристаллы растворятся.
Разделение летучих кислот
Для разделения летучих кислот, корда они находятся в смеси, применяют или
фракционированное осаждение их серебряных солей или фракционированное нейтра-
лизование самих кислот. Первый способ применяется для определения малых коли-
честв кислот; для этого нагретый водный раствор кислот насыщают углекислым
серебром и, отфильтровав, охлаждают раствор. Если он был достаточно концен-
трован, то сейчас же начинается выделение кристаллов; в противном случае не-
обходимо сгустить его выпариванием на водяной бане. При этом раньше осаждает-
ся соль более сложной кислоты. Отфильтровав, выделившуюся соль анализируют, а
раствор сгущают для новой кристаллизации и т. д. Присутствие муравьиной ки-
слоты узнается при этом по почернению раствора вследствие разложения муравьи-
но-серебряной соли.
По второму способу берут часть водного раствора кислот и нейтрализуют его
содой; затем смешивают с ненейтрализованной частью и подвергают смесь пере-
гонке с водяными парами; тогда н дестиллат перейдут более летучие кислоты, а
в перегонном аппарате останутся соли менее летучих кислот и с ними же всегда
соль уксусной кислоты. Повторяя такую операцию несколько раз, достигают до-
вольно полного разделения кислот.
Из твердых высших кислот наиболее важны: пальмитиновая Ci6H3202 или
С15Н31СООН и стеариновая С^НзбОг или С17Н35СООН. Их глицериды в смеси с глице-
ридом олеиновой кислоты Ci8H3402 входят в состав животных жиров.
Жиры
Исследование жиров впервые произведено было Шеврелем, а Вертело первый по-
лучил жир искусственным путем: нагреванием глицерина с пальмитиновой и стеа-
риновой кислотами. Жиры суть сложные зфиры, образованные глицерином и жирными
кислотами4. Вследствие обязательного присутствия в них глицерина они называ-
ются глицеридами жирных кислот. Глицерид стеариновой кислоты называется стеа-
рином С3Н5О3 (C17H35CO) з, пальмитиновой кислоты — пальмитином С3Н5О3 (Ci5H3iCO) 3 и
олеиновой — олеином С3Н5О3 (C17H33CO) 3 .
Вот их более подробные формулы:
Пчелиный воск хотя и относится к сложным эфирам, но не к жирам, так как в состав
его глицерин не входит: в нем находятся пальмитиновый эфир мелиссилового спирта
СзоНб! (OCi6H3iCO) и церотиновая кислота С2бН5202.
СН3.0(CnHS6CO) CHS ■ 0(С]БНа1СО) СН3 ■ 0(С]7Н33СО)
I I i
CH.O(H17Cs6CO) CH*0(CrH31CO) CH.O(C„H83CO)
! I I
СНг - 0(C17 H85CO) CH2 • 0{C]5CWCO) CH8.0(C17HMCO)
Стеарин Пальмитин Олеин
Первые два тверды, а олеин жидкий. Стеарин плавится при 72 С, пальмитин при
65 С, а олеин затвердевает при —6 С. Природные жиры состоят обыкновенно из
смеси этих глицеридов, и консистенция жира зависит от относительного количе-
ства олеина; так прованское масло содержит 75% олеина, а баранье сало 25%;
человеческий и свиной жир занимают в этом отношении середину. При температуре
тела человеческий жир жидкий.
В технике стеарином называют смесь стеариновой и пальмитиновой кислот, ко-
торая идет на приготовление стеариновых свечей.
Кислоты, входящие в состав природных глицеридов, имеют нормальное строение
и содержат обыкновенно четное число углеродных атомов.
Нормальное строение стеариновой кислоты доказано Краффтом следующим обра-
зом. Он подвергал сухой перегонке смесь бариевых солей стеариновой и уксусной
кислот и получал (см. 3-й способ получения кетонов), при этом кетон состава
С17Н35СОСН3. Последний при окислении (см. порядок окисления кетонов) давал
уксусную кислоту и маргариновую С16Н33СООН; таким образом из стеариновой ки-
слоты , содержащей Ci8, получена была кислота с семнадцатью углеродными атома-
ми. Баритовая соль последней в смеси с уксуснобариевой солью при сухой пере-
гонке дала кетон С16Н33СОСН3, а этот при окислении превратился в уксусную и
пальмитиновую кислоты, т. е. кислоту уже с шестнадцатью углеродными атомами.
Идя таким путем дальше, Краффт дошел до каприновой кислоты Ci0H2o02, нормаль-
ность строения которой доказана ее синтезом; а отсюда естественный вывод, что
и стеариновая кислота имеет также нормальное строение. Это исследование имело
еще и другое важное значение: получены были искусственно кислоты, добывавшие-
ся раньше из смесей природных продуктов.
Как редкий случай следует отметить нахождение в природных жирах кислот цик-
лического строения; найдена также кислота таририновая, имеющая тройную связь.
Жиры (в химически чистом виде) бесцветны, без запаха и вкуса; или тверды
(сало), или жидки (жирные масла); в воде не растворимы, в холодном спирте ма-
ло растворимы; легко растворяются в эфире, бензоле, бензине, хлороформе, се-
роуглероде, скипидаре и др. При нагревании до 300 С разлагаются, выделяя, ме-
жду прочим, акролеин (запах пригоревшего жира). Удельный вес жира колеблется
от 0,92 до 0,95 и 0,97. Тела нейтральные; но при хранении при доступе воздуха
и влажности в жирах появляется кислая реакция. При долгом хранении на свету
жиры прогоркают, приобретая неприятный запах и вкус, желтеют; кроме кислот
при прогоркании образуются и алдегиды (масляный, энантовый и пеларгоновый).
Жиры, как сложные эфиры, легко обмыливаются щелочами, кислотами и даже во-
дой:
СзН5Оз(С17Нз5СО) + ЗН20 = С3Н5(ОН)3 + 3Ci8H3602.
На этой реакции основано получение названных выше кислот.
Обмыливание жиров происходит также под влиянием ферментов, назаваемых липа-
зами. Последние встречаются во многих растениях, особенно в касторовом семе-
ни. Обмыливание жиров растертыми касторовыми семенами в присутствии серной
кислоты применено в технике. Липазы же участвуют в пищеварении, вызывая обмы-
ливание жиров, принимаемых в пищу, так как жиры усваиваются организмом не как
таковые, а как продукты их обмыливания5.
Жиры — тела наименее окисленные по сравнению с другими важнейшими классами
органических соединений в организмах: в них кислорода 10—12%, тогда как в уг-
леводах 49%, а в белках 23%. Они служат питательным резервом на случай голо-
довки .
При недостатке жиров в пище образование жира в организме идет за счет угле-
водов .
Ввиду важности жиров в физиологическом и техническом отношении большое зна-
чение имеет анализ их. Для этого определяют:
1) кислотное число, т. е. количество мг едкого кали, потребное для нейтрали-
зации свободных кислот в 1 г жира;
2) число омыления, т. е. количество мг КОН, потраченное на обмыливание 1 г
жира;
3) вычитая первое число из второго, получают эфирное число.
4) особенно важное значение имеет определение йодного числа, т. е. весового
количества иода, присоединяемого данным жиром; это количество выражается в
процентах.
По этим числам определяют природу данного жира, а также и фальсификацию жи-
ров. При анализе коровьего масла, содержащего и глицериды летучих кислот6,
очень важно определение количества этих кислот; это — число Рейхерта-
Мейсселя, показывающее количество куб. см децинормального раствора КОН, по-
требное для нейтрализации летучих кислот, выделенных из 5 г жира.
Пальмитиновую кислоту получают обыкновенно обмыливанием пальмового масла
или лучше японского воска (сала), главная составная часть которого есть паль-
митин; а стеариновую кислоту таким же путем получают из бараньего сала; но
так как последнее содержит смесь трех вышеупомянутых глицеридов, то приходит-
ся прибегнуть к разделению получаемых кислот.
Сначала отделяют олеиновую кислоту; для этого переводят кислоты в их свин-
цовые соли и обрабатывают последние эфиром; в эфире легко растворяется олеи-
новокислый свинец и очень трудно свинцовые соли пальмитиновой и стеариновой
кислот; последние отфильтровывают и разлагают минеральными кислотами. Впро-
чем, этот способ применяется обыкновенно для получения олеиновой кислоты.
Иначе поступают так: растворяют смесь кислот в возможно малом количестве
спирта при нагревании и охлаждают раствор; тогда кристаллизуется стеариновая
и пальмитиновая кислоты, а олеиновая остается в растворе. Для разделения же
первых двух кислот пользуются способом Гейнтца; для этого к спиртовому рас-
твору кислот приливают спиртовый же раствор уксусномагниевой соли в количест-
ве, недостаточном для полного превращения кислот в соли магния. Получающийся
при этом осадок содержит соль Мд наиболее сложной кислоты (в данном случае—
стеариновой); его отфильтровывают; фильтрат идет для новых осаждений, а соль
для выделения кислоты разлагают серной кислотой.
Пальмитиновая и стеариновая кислоты образуют белую, без запаха и вкуса,
кристаллическую массу; первая с т. пл. 62 С, а вторая с т. пл. 71,5 С; смесь
их, получаемая из бараньего сала, плавится значительно ниже, именно около 59
С. В воде они нерастворимы, в спирте же и эфире растворяются. Применяются
5 Выдающимися катализаторами для обмыливания жиров служат сульфокислоты, получаемые
или при действии конц. H2S04 на касторовое масло (реактив Твитчеля), или при очище-
нии нефти серной кислотой (реактив Петрова). Эти реактивы превращают жиры в эмульсию
и тем увеличивают поверхность соприкосновения жира с омылителем.
6 В коровьем масле содержится до 2% лецитина, холестерина и красящие вещества. В со-
став его входят глицериды стеариновой, пальмитиновой и олеиновой кислот; кроме того
глицериды масляной, капроновой, каприловой и каприновой кислот; последних содержится
5-6%.
главным образом для приготовления стеариновых свечей и мыла.
Сущность стеаринового производства состоит в следующем. Жир обмыливают или
известью, или магнезией, или концентрованнои серной кислотой, или перегретым
паром. В первом и втором случаях полученные соли кислот разлагают серной ки-
слотой, в остальных же случаях получают прямо свободные кислоты; если они
имеют темную окраску (как при получении с H2S04) , то для очищения их перего-
няют перегретым паром. После этого кислоты прессуют для отделения от жидкой
олеиновой кислоты, а из полученной смеси пальмитиновой и стеариновой кислот
готовят свечи; впрочем, к этой массе предварительно примешивают немного пара-
фина для избежания кристаллической структуры и обусловливаемой ею ломкости
свечей7.
Мылами вообще называются соли жирных кислот; а так как они получаются при
разложении жиров основаниями, то и самое разложение называется обмыливанием.
Распадение жиров и вообще сложных эфиров достигается не только при помощи ос-
нований, но также и при посредстве кислот, воды или ферментов; хотя здесь и
не образуются мыла, все-таки реакция по аналогии носит название обмыливания.
Обыкновенное мыло есть смесь щелочных солей стеариновой, пальмитиновой и
олеиновой кислот; жидкое мыло содержит соли калия, а твердое — соли натрия;
получаются они при действии щелочей на жиры.
При обмыливании жиров окисью свинца получаются свинцовые соли, применяемые
для приготовления пластырей.
Для стеаринового и мыловаренного производств жирные масла непригодны, так
как в них много олеиновой кислоты, а твердые (животные) жиры дороги; поэтому
жирные масла подвергают восстановлению водородом в присутствии катализатора
никкеля; жидкое масло при этом затвердевает вследствие перехода олеиновой ки-
слоты в стеариновую, и тогда идет на вышеупомянутые производства, а также на
производство маргарина.
Приготовление мыла
Для получения мыла всего лучше взять японский воск, представляющий собою
глицерид пальмитиновой кислоты.
В фарфоровой чашке на водяной бане нагревают 75 г японского воска; когда он
расплавится, прибавляют раствор 25 г КОН в 30 см3 воды и все время помешивают
палочкой. Через час или полтора обмыливание заканчивается. После этого, при
помешивании, приливают в чашку воду до образования прозрачного раствора, на
что требуется около 300 см3 Н20, и выливают в стакан емк. в 1 л. К этому рас-
твору приливают, при помешивании, 1/2 л горячего насыщенного раствора пова-
ренной соли (калиевая соль пальмитиновой кислоты переходит при этом в натрие-
вую, которая труднее растворима); мыло выделяется сначала в виде масла, а по
охлаждении становится твердым. До приливания поваренной соли можно прибавить
к раствору немного гелиотропина (пиперонала): тогда получится душистое мыло.
Получение мыла из свиного сала
В фарфоровой чашке кипятят ,10 г сала с 6 см3 раствора NaOH (1:2) . Жидкость
при этом сильно пенится. По мере выкипания добавляют воду. Обмыливание закон-
чено, если взятая стеклянной палочкой капля жидкости будет сполна растворять-
ся в горячен дестиллированной воде. После этого продукт реакции охлаждают и к
нему приливают 40 см3 насыщенного раствора NaCl; мыло всплывает на поверх-
ность и застывает.
Светильню пропитывают борными или фосфорными солями, которые при горении свечи
сплавляются в шарик, и, наклоняя своею тяжестью конец светильни, выводят этот конец
из холодной части пламени в горячую, содействуя полному сгоранию и препятствуя обра-
зованию нагара.
Свойства мыла
a) К водному раствору мыла приливают НС1: выделяются нерастворимые твердые
кислоты.
b) К раствору мыла приливают раствор СаС12: получается осадок нерастворимых
кальциевых солей.
Мыльная проба
В один узкий стеклянный цилиндр, снабженный пробкой, наливают 75 см3 дес-
тиллированной воды, а в другой — столько же жесткой воды из водопровода (или
дестиллированной воды, к которой прибавлено немного СаС1г) ; в тот и другой
цилиндр приливают затем одинаковое количество (1—2 см3) спиртового раствора
мыла и сильно встряхивают: в первом цилиндре образуется пена, долго не исче-
зающая, во втором же цилиндре или совсем не образуется пены, или она сейчас
же пропадает; параллельно с этим вода во втором цилиндре мутится (образуются
нерастворимые соли щелочноземельных металлов). Добавляя сюда мыла, можно дос-
тигнуть , наконец, образования неисчезающей пены. Чем более жестка вода, тем
больше потребуется мыла, чтобы получить эту пену. На этом основано определе-
ние степени жесткости воды.
Гидролиз мыла
В цилиндр с водой осторожно по стенкам приливают спиртовый раствор мыла, к
которому прибавлен раствор фенолфталеина, так, чтобы слои не смешивались; на
месте разделения слоев получается красное кольцо. Окрашивание это вызывается
действием фенолфталеина на щелочь, выделившуюся из мыла от действия воды.
(ПРОДОЛЖЕНИЕ СЛЕДУЕТ)
Ликбез
БИОЛОГИЯ ДРОЖЖЕЙ
И.П. Бабьева, И.Ю. Чернов
(продолжение)
ОСОБЕННОСТИ
МЕТАБОЛИЗМА
Хотя дрожжи и не так разнообразны по своему метаболизму, как бактерии, раз-
личные виды дрожжей могут катаболизировать разные соединения углерода и азота
и образовывать различные конечные продукты (рис. 38).
Спиртовое
брожение
Наиболее известное свойство многих дрожжей - способность к спиртовому бро-
жению. Многие виды дрожжей могут переключаться с бродильного метаболизма на
дыхательный и обратно в зависимости от условий: при наличии кислорода броже-
ние ингибируется и дрожжи начинают дышать, в отсутствие кислорода включается
механизм спиртового брожения. Так как кислородное дыхание - энергетически бо-
лее выгодный процесс, чем брожение, то выход биомассы дрожжей в расчете на
единицу используемого субстрата выше при выращивании их в аэробных условиях,
чем в анаэробных. Это явление называется эффектом Пастера.
Спиртовое брожение может идти не только в анаэробных условиях. Если выращи-
вать дрожжи в присутствии кислорода, но при высоком содержании глюкозы в сре-
де, то в этом случае дрожжи также сбраживают глюкозу. Таким образом, глюкоза
подавляет процессы анаэробного дыхания. Это явление получило название эффекта
Кребтри, или катаболитной репрессии.
Анаэробные условия
Агробнме условия
Источники
углер*>да:
Моносахариды
Днсахариды
Олнгосахариды
Полисахариды
Источники
аюта:
NH<\ NO,
Аминокислоты
Макроэлементы
[Микроэлементы
Источники
углерода:
Углеводы
Спирты
Органические
кислоты
Углеводороды
(^•соединения
Ароматические
I
I I
АТФ НАД(Ф)Н Мономеры
I I I
ЛНЛБОЛИЗМ
i
Продукты биосинтеза
Белки
Полисахариды
Литтнды
Нуклеиновые
кислоты
Эталон. СО,
СО. Н.О
Побочные продукты
Летучие жирные кислоты
Высшие спирты
Альдегиды
Эфиры
Меркаптаны
Рис. 38. Общая схема метаболизма дрожжей.
Многие дрожжи вообще не способны бродить. По соотношению между этими двумя
процессами в метаболизме можно выделить следующие группы дрожжей:
1. Дрожжи, существующие только за счет брожения и не способные расти в аэроб-
ных условиях. К ним относится, например, вид Arxiozyma telluris, обитающий
в кишечном тракте грызунов.
2. Активные бродилыцики: интенсивно сбраживают различные субстраты, но в
аэробных условиях переключаются на дыхательный обмен. Представители
Saccharomyces cerevisiae, Schizosaccharomyces pombe.
3. Слабые бродилыцики - в основном существуют за счет аэробного дыхания, но в
анаэробных условиях могут бродить, однако значительно менее интенсивно,
чем виды из предыдущей группы. Это аскомицетовые дрожжи из родов Pichia,
Debaryomyces, а также все способные к брожению базидиомицетовые дрожжи.
4. Дрожжи, существующие только за счет дыхания и не способные расти в ана-
эробных условиях. К этой группе относятся аскомицетовые дрожжи из рода
Lipomyces и многие несовершенные дрожжи базидиомицетового аффинитета -
Cryptococcus, Rhodotorula, Sporobolomyces.
Пентозы
1
Гексазы
7<
НАДФН
Глюхозо-6-Ф
Фру1сгозо-6-Ф
С,-соединения
Ксилулозо-5-Ф v CHjO
НАДФН
ПЕНТОЗО-
ФОСФАТНЬЕЙ
ЦИКЛ
Сахарофосфатыые
перестройки
I!
КСИЛУЛОЗО-
М0Н0ФОСФАТНЫЙ
цикл
7 Фрукгозо-1,6-ди-Ф
Ь
Диокснацетон-3-Ф
V- *
Глицеральдепид-3-Ф
НАД.Н^=
НАДН
Этанол
Переамии ирован ие
А
р-Кетоадшшнат
+
Жирные кислоты
А
Аминокислоты
(ппотамат, аспартат)
Ароматические
соединения
Углеводороды
Рис. 39. Пути катаболизма различных типов соединений дрожжами.
При росте в анаэробных условиях дрожжи превращают глюкозу в пировиноградную
кислоту по гликолитическому пути, получая 2 моля АТФ на 1 моль глюкозы. Про-
цесс гликолиза включает реакцию окисления фосфоглицеринового альдегида, в ко-
торой образуется восстановленный пиридиннуклеотид НАД-Н. В аэробных условиях
последний окисляется через систему переноса электронов кислородом. В отсутст-
вие же кислорода для сохранения окислительно-восстановительного равновесия
необходимо окислить НАД-Н каким-либо другим путем. В дрожжевой клетке это
окисление включает декарбоксилирование пирувата и превращение образовавшегося
при этом ацетальдегида в этанол с одновременным окислением НАД-Н до НАД+. Об-
разовавшийся НАД+ может принимать участие в окислении следующей молекулы фос-
фоглицеринового альдегида (рис. 39).
Ход брожения может сильно меняться в зависимости от условий. Если, напри-
мер, в культуру бродящих дрожжей добавить бисульфит, то образующийся из пиру-
вата ацетальдегид связывается в бисульфитный аддукт, тем самым блокируя обра-
зование этанола. Дрожжевые клетки в ответ на это используют для окисления на-
капливающегося НАД-Н половину образующегося триозофосфата. Последний, восста-
навливаясь , превращается в глицерин. Хотя при этом суммарный выход АТФ стано-
вится равным нулю, и такое брожение не может обеспечить рост клеток, его мож-
но использовать для промышленного получения глицерина.
В типичном же случае основные продукты спиртового брожения - этанол и угле-
кислота, однако, в микроколичествах образуется также множество побочных со-
единений .
Субстраты
брожения
Все бродящие дрожжи сбраживают глюкозу и фруктозу, поскольку именно с этих
Сахаров начинается гликолитическое расщепление. Кроме глюкозы и фруктозы мо-
гут сбраживаться другие соединения, которые легко превращаются в интермедиаты
гликолитического пути. В основном к ним относятся гексозы и олигосахариды,
включающие остатки гексоз. Из моносахаридов наиболее часто сбраживается га-
лактоза, из дисахаридов - сахароза, мальтоза, трегалоза. Значительно реже
встречаются дрожжи, сбраживающие лактозу и мелибиозу.
Некоторые дрожжи способны также сбраживать полисахариды, состоящие из гек-
созных остатков. Например Kluyveromyces fragilis сбраживает инулин (полифрук-
тозан), a Schwanniomyces occidentalis и Saccharomycopsis fibuliger активно
сбраживают крахмал.
Долгое время не были известны дрожжи, способные интенсивно сбраживать пен-
тозы. Такие виды были описаны только к началу 80-х годов XX в. К ним относят-
ся, например, Pichia stipitis (несовершенная стадия - Candida shehatae),
Pachysolen tannophilus. Брожение ксилозы начинается с восстановления ее до
ксилита с помощью фермента ксилозоредуктазы. Затем ксилит окисляется ксилит-
дегидрогеназой до ксилулозы, которая фосфорилируется с образованием ксилуло-
зо-5-фосфата. Последний может вступать в реакции пентозофосфатного пути, где
происходит перестройка углеродного скелета ксилулозо-5-фосфата с образованием
интермедиатов гликолиза (см. рис. 39).
Дыхание
При росте в аэробных условиях при низком содержании глюкозы в среде дрожжи
получают АТФ за счет процессов дыхания, как это делает большинство аэробных
организмов. Полное окисление углеродного субстрата до углекислого газа и воды
может происходить у дрожжей с помощью трех различных механизмов: в цикле три-
карбоновых кислот, в глиоксилатном цикле и в пентозофосфатном цикле (см. рис.
39) . При функционировании каждого из этих циклов в клетке происходит образо-
вание восстановленных пиридиннуклеотидов. Они могут быть использованы либо
для процессов восстановления в ходе биосинтеза, либо для получения АТФ путем
окислительного фосфорилирования. В последнем случае НАД-Н становится донором
электронов для электронно-транспортной цепи, в которую у дрожжей входят такие
белки-переносчики электронов, как флавопротеиды, убихиноны и цитохромы, лока-
лизованные на внутренней мембране митохондрий.
Спектр углеродных соединений, усваиваемых дрожжами за счет аэробного дыха-
ния, значительно шире, чем в случае брожения. Рассмотрим особенности аэробно-
го метаболизма дрожжами некоторых углеродных соединений.
Углеводы
За исключением Arxiozyma telluris, обитающего в кишечнике грызунов и неспо-
собного к дыхательному метаболизму, все остальные виды дрожжей растут аэробно
на глюкозе. Многие другие гексозы также могут использоваться дрожжами как
субстраты дыхания.
Если в анаэробных условиях лишь единичные виды дрожжей могут сбраживать пя-
тиатомные сахара, то в присутствии кислорода пентозы (арабиноза, ксилоза и
др.) и метилпентозы (рамноза) могут служить субстратами для очень многих
дрожжей. При этом происходит превращение пентоз в соответствующие интермедиа-
ты пентозофосфатного пути.
Многие дрожжи способны также аэробно ассимилировать различные производные
Сахаров, например сахароспирты (сорбит, рибит, эритрит, маннит, инозит) или
гликозиды (арбутин, салицин).
Полисахариды также могут усваиваться дрожжами, способными к синтезу соот-
ветствующих гидролаз. Довольно часто встречаются дрожжи, ассимилирующие крах-
мал. Многие виды дрожжей способны усваивать такие полисахариды, как полифрук-
тозаны (инулин), полигалактоурониды (пектины), гемицеллюлозы (ксиланы). Спо-
собность к расщеплению р-глюкозидных связей в целлюлозе у дрожжей практически
отсутствует. Целлюлазы обнаружены лишь у нескольких видов дрожжей, например,
у Trichosporon pullulans, но активность этих ферментов значительно ниже, чем
у целлюлолитических мицелиальных грибов, таких как, например, виды рода
Trichoderma.
Жирные кислоты и н-алканы
Ряд видов дрожжей отличается способностью к аэробному росту на длинноцепо-
чечных жирных кислотах и алканах. К таким дрожжам относятся некоторые виды
рода Candida аскомицетового аффинитета (Candida tropicalis, Candida interme-
dia, Candida lipolytica, Candida guilliermondii).
Ассимиляция н-алканов может идти только аэробным путем, так как первая ста-
дия их катаболизма - это окисление концевого атома углеводородной цепочки мо-
лекулярным кислородом. Образующийся спирт окисляется до соответствующей жир-
ной кислоты, которая затем подвергается процессам р-окисления, отщепляя по-
следовательно двууглеродные остатки ацетил-КоА. Последний окисляется в цикле
трикарбоновых кислот или глиоксилатном цикле.
Дрожжи, растущие на средах с н-алканами, имеют специфические цитологические
особенности. На ультратонких срезах таких клеток обычно видны органеллы, ок-
руженные гомогенными липопротеидными мембранами - пероксисомы. Функция перок-
сисом состоит в компартментализации процессов р-окисления от дальнейших пре-
вращений ацетил-КоА.
Одноуглеродные соединения
Более 20 видов дрожжей способны аэробно расти на метаноле в качестве един-
ственного источника углерода и энергии. Наиболее активно ассимилируют метанол
такие виды, как Candida boidinii, Candida methylica, Pichia polymorpha,
Pichia pastoris. Все эти виды - факультативные метилотрофы, они хорошо растут
также и на углеводах.
Катаболизм метанола дрожжами начинается с его окисления кислородом с обра-
зованием формальдегида. Эта реакция катализируется ферментом метанолоксида-
зой, содержащим в качестве кофермента ФАД. При действии флавиновых оксигеназ
образуется Н202, поэтому метилотрофные дрожжи всегда богаты каталазой, разла-
гающей токсичную Н202 до Н20 и 02. Формальдегид затем окисляется последова-
тельно до муравьиной кислоты и С02. При этом окисляемый субстрат находится в
связанном виде с молекулой глутатиона. В ходе окисления на 1 моль метанола
образуется 1 моль НАД-Н, который может участвовать в процессах биосинтеза или
отдавать электроны в цепь переноса электронов для генерации АТФ.
При росте на метаноле, как на единственном источнике углерода, дрожжи долж-
ны использовать только одноуглеродные фрагменты для синтеза всех клеточных
компонентов. Ассимиляция одноуглеродных соединений и образование С-С связей у
метилотрофных дрожжей идет в ксилулозо-монофосфатном цикле. Ключевая реакция
этого цикла - взаимодействие ксилулозо-5-фосфата с формальдегидом и образова-
ние фосфоглицеринового альдегида и диоксиацетона - уникальна и встречается
только у метилотрофов. Последующие превращения представляют собой обращение
реакций гликолитического пути. Регенерация ксилулозо-5-фосфата достигается
окислением и декарбоксилированием фруктозо-6-фосфата.
Ароматические соединения
Некоторые дрожжи, в частности Trichosporon pullulans, Debaryomyces
hansenii, способны аэробно ассимилировать ароматические соединения, такие как
фенол, резорцин, салициловая кислота и т.п. Большинство дрожжей неспособно
ассимилировать в качестве источника углерода гетероциклические соединения,
включая пуриновые и пиримидиновые производные. Однако недавно описан новый
вид Arxula adeninivorans, активно ассимилирующий различные пуриновые соедине-
ния в качестве источника углерода и азота. Катаболизм ароматических соедине-
ний у дрожжей идет по р-кетоадипиновому пути и начинается с введения ОН-групп
и раскрытия ароматического кольца с помощью оксигеназ:
Ацетил-Ко А
V^coo;J
Сукцинат
Хотя дрожжи, использующие сложные ароматические соединения в качестве един-
ственного источника углерода и энергии, встречаются довольно редко, они го-
раздо чаще могут окислять их в процессах кометаболизма, например в присутст-
вии глюкозы.
Кроме перечисленных классов соединений дрожжевые грибы часто могут ассими-
лировать различные органические кислоты, альдегиды, спирты, амины, аминокис-
лоты и пр. Существуют дрожжи, обладающие высокой протеазнои активностью и
способные расти на белках. К ним относится, например, Debaryomyces hansenii,
часто развивающийся на консервированных мясных продуктах и колбасах. Большин-
ство базидиомицетовых дрожжей образует также внеклеточные ДНК-азы.
Вторичные
продукты
метаболизма
В процессе роста дрожжи наряду с основными продуктами (этанол, С02) образу-
ют множество других метаболитов, которые синтезируются как побочные соедине-
ния при катаболизме углеводов, обмене аминокислот и пр. Хотя эти соединения
обычно накапливаются в культуральной среде в очень незначительных количествах
и трудно идентифицируются, они могут иметь большое практическое значение, на-
пример, для пищевой промышленности, так как от их состава зависит качество
пищевого продукта, получаемого с помощью дрожжей. Такие соединения называют
органолептическими.
Обязательные побочные продукты метаболизма дрожжей - высшие спирты (сивуш-
ные масла). Механизм синтеза высших спиртов связан с образованием алифатиче-
ских аминокислот (валин, лейцин, изолейцин).
Он включает удаление с аминокислот и их предшественников аминогрупп (тран-
саминирование), образующаяся аминокислота декарбоксилируется и восстанавлива-
ется до спирта:
° о
R-CH-СООЧ—♦R-CH-C004-— R-C * R-ChbOH
Чаще всего встречаются спирты: пропиловый, изоамиловый, бутиловый, изобути-
ловый. Летучие жирные кислоты, такие как уксусная, пропионовая, масляная,
изомасляная, изовалериановая - также обычные минорные продукты метаболизма
дрожжей. Большое практическое значение имеют выделяемые дрожжами альдегиды и
кетоны, особенно ацетоин и диацетил:
о он о о
II I II II
СНз-С-С—СН3 СН3—С—С—СН3
Ацетоин Диацетил
Исходным соединением для синтеза ацетоина и диацетила является пируват. При
декарбоксилировании пирувата образуется ацетальдегид, который конденсируется
с другой молекулой пирувата с образованием ацетомолочной кислоты. Декарбокси-
лирование ацетолактата дает ацетоин, обратимо окисляющийся до диацетила. С
ацетолактата начинается также путь биосинтеза валина и пантотеновой кислоты,
поэтому ацетоин и диацетил считают побочными продуктами синтеза этих соедине-
ний. Эти и другие соединения сильно влияют на вкус спиртных напитков - вина и
пива.
Среди других побочных продуктов большое значение имеют также длинноцепочеч-
ные жирные кислоты, эфиры и меркаптаны.
Источники
азота
Универсальным источником азота для дрожжей являются соли аммония. Аммоний-
ный ион, а также аммиак, усваиваются всеми без исключения видами дрожжей.
Азот аммония включается в метаболизм с помощью реакций аминирования - взаимо-
действия NH3 с ос-кетокислотами:
ос-Кутоглутарат + NH3 + НАД-Н + Н+ > Глютамат + НАД+ + Н20
Многие дрожжи способны усваивать азот в окисленной форме - в виде солей
нитратов и нитритов. Дрожжи, использующие нитраты, имеют две ферментные сис-
темы: первая восстанавливает нитрат до нитрита, вторая - нитрат до аммония. У
некоторых видов присутствует только вторая ферментная система - они способны
усваивать нитриты, но не нитраты. Способность к ассимиляции нитратов считает-
ся ценным таксономическим признаком, поэтому определение способности к росту
на среде с КЫОз в качестве единственного источника азота - рутинный тест при
идентификации дрожжей. В последнее время, с развитием геносистематики, тесту
на ассимиляцию нитратов стали придавать значительно меньшее значение, как ди-
агностическому признаку.
Источниками азота для дрожжей могут быть также различные органические со-
единения, содержащие NH-группы: амины, аминокислоты, азотсодержащие гетеро-
циклические соединения. Высвобождение NH3 из таких соединений может идти раз-
личными путями. Первый - окисление аминооксидазами. Второй путь - переамини-
рование - перенос NH-группы с ассимилируемого субстрата на ос-кетокислоту с
образованием соответствующей аминокислоты.
Практически все дрожжи используют в качестве источника азота мочевину, рас-
щепляя ее на С02 и NH3. Однако, механизм этого расщепления различен у аскоми-
цетовых и базидиомицетовых дрожжей. Аскомицетовые дрожжи сначала карбоксили-
руют мочевину с образованием аллофоната, который затем гидролизуется:
НСО, ОНГ
о о
NH2-C-NH2 J> <^C NH2-C-NH-COOH - » 2С02 ♦ 2 NHj
АТФ АТФ + Ф н2°
Базидиомицетовые дрожжи непосредственно расщепляют мочевину с помощью фер-
мента уреазы. Во этом случае аммиак образуется в избытке и выделяется в сре-
ду:
О
И
NH2-C-NH2 + HjO * С02 +2NH}
Это дает возможность различать дрожжи аско- и базидиомицетового аффинитета
с помощью теста на наличие уреазы: в питательную среду добавляют мочевину и
кислотно-основной индикатор (феноловый красный). При наличии уреазы выделяет-
ся аммиак, среда подщелачивается и индикатор меняет окраску. Этот простой
тест широко используется при видовой идентификации дрожжевых грибов.
ЛИМИТИРУЮЩИЕ
ФАКТОРЫ
За исключением нескольких исключительно холодолюбивых видов, среди дрожжей
нет ярко выраженных экстремофилов, то есть видов, предпочитающих крайне высо-
кие или низкие значения температуры, рН, осмотического давления, влажности
среды и т.п. В то же время существуют дрожжи, которые сильно выделяются среди
большинства других видов по своей способности переносить неблагоприятные фак-
торы среды.
Температура
У большинства видов дрожжей минимальная температура роста находится в пре-
делах 0-5 С, а максимальная - 30-40 С. Почти все дрожжи могут расти при ком-
натной температуре 20-25 С. Базидиомицетовые дрожжи в целом характеризуются
более низкими максимальными температурами роста, чем аскомицетовые (рис. 40).
Однако из этого правила имеется ряд исключений:
1. Некоторые дрожжи, например Kluyveromyces thermotolerans, Arxiozyma
telluris, Candida tropicalis, могут расти при температурах выше 45 С. Од-
нако, минимальная температура роста таких дрожжей также находится в пре-
делах 0-5 С, поэтому их следует называть термотолерантными, то есть ус-
тойчивыми к повышенным температурам.
2. Существует несколько видов дрожжей, обитающих только во внутренних орга-
нах теплокровных животных. К таким видам относится, например,
Cyniclomyces guttulatus, обитающий в кишечном тракте кроликов. Эти дрожжи
весь свой жизненный цикл проводят при температуре тела животного-хозяина,
поэтому температурный диапазон их роста очень узок, обычно от 30 до 40 С
(психрофобные).
3. Гораздо чаще встречаются дрожжи, которые хорошо растут при низких темпе-
ратурах около 0 С, а иногда - и при отрицательных температурах. Если мак-
симальная температура роста таких дрожжей не ниже 20-25 С, то они называ-
ются холодоустойчивыми, или психротолерантными. Наиболее обычны психрото-
лерантные виды среди дрожжей базидиомицетового аффинитета. Если же макси-
мальная температура роста ниже 20 С, то принято говорить о психрофильных
дрожжах. Облигатные психрофилы встречаются среди штаммов таких видов, как
Leucosporidium scottii, Mrakiafrigida. Эти дрожжи часто выделяются в вы-
соких широтах, например, из тундровых почв или из воды антарктических мо-
рей. Психрофильные штаммы встречаются также среди несовершенных дрожжей
базидиомицетового аффинитета, таких как Cryptococcus, Bullera, Sporobolo-
myces и др. Среди аскомицетовых дрожжей психрофильные виды не известны.
Рис. 40. Относительное количество видов дрожжей с различной мак-
симальной температурой роста.
Таким образом, в целом дрожжи - это довольно холодолюбивая группа микроор-
ганизмов . Часто это дает возможность создать селективные условия при выделе-
нии дрожжей из различных субстратов. Дело в том, что учету и выделению дрож-
жей методом посева на твердых питательных средах сильно мешают мицелиальные
грибы, присутствующие в изучаемом субстрате. Колонии грибов маскируют более
мелкие дрожжевые колонии и затрудняют их изоляцию. Подобрать какую-либо се-
лективную только на одноклеточные грибы питательную среду, то есть ограничить
рост мицелиальных грибов химически невозможно: все ингибиторы роста мицели-
альных грибов действуют и на дрожжи. Однако, при выращивании посевов при низ-
кой (около 5 С) температуре дрожжевые колонии в среднем развиваются быстрее.
Это дает возможность частично избавиться от колоний мицелиальных грибов.
Активность воды
Активность воды (aw) в каком-либо растворе определяется как отношение дав-
ления пара над раствором к давлению пара над чистой водой. Для чистой воды aw
=1. Все без исключения дрожжи способны расти при aw, приближающейся к 1.
Большинство видов дрожжей перестает расти при aw = 0.9. Такую активность воды
имеет, например, 50% раствор глюкозы или 14% раствор NaCl. Однако ряд видов
дрожжей способен развиваться при концентрации глюкозы в среде до 60% или при
концентрации NaCl до 20%. Такие дрожжи называют ксеротолерантными. К ним от-
носятся аскомицетовые дрожжи Zygosaccharomyces bailii, Zygosaccharomyces
rouxii, Schizosaccharomyces pombe, Debaryomyces hansenii и др. Эти виды часто
встречаются в различных вареньях, джемах, сиропах, сухофруктах и могут вызы-
вать порчу этих продуктов. Дрожжи Debaryomyces hansenii особенно устойчивы к
высокой концентрации NaCl в среде и часто встречаются в различных соленьях,
на консервированных мясных продуктах, в морской воде.
Механизм устойчивости ксеротолерантных дрожжей к низким значениям aw заклю-
чается в изменении внутриклеточной среды. Клетки этих дрожжей способны накап-
ливать различные полиолы, такие как глицерин, арабит, эритрит, ксилит, кото-
рые могут служить растворителями для внутриклеточных ферментов вместо воды.
Ксеротолерантные дрожжи могут быть использованы для промышленного получения
таких полиолов, которые представляют интерес для химической (глицерин) и пи-
щевой (ксилит) промышленности.
Кислотность среды
Оптимальные значения рН для роста большинства дрожжевых грибов находятся в
области средней кислотности (рН 4-6) . Однако отдельные виды способны разви-
ваться в более кислой среде. Например, некоторые штаммы Saccharomyces
cerevisiae хорошо растут при рН 2.5-3. К кислотоустойчивым дрожжам относится
также обитающий в кишечном тракте животных Cyniclomyces guttulatus, растущий
при рН 2. Дрожжи, которые могли бы расти при щелочных значениях рН (8 и бо-
лее) не известны.
Практически все виды дрожжей могут расти в диапазоне рН 4-4.5. В то же вре-
мя, в такой слабокислой среде не растет большинство банальных бактерий, кото-
рые наиболее обычны в самых разных природных местообитаниях (например, псев-
домонады, бациллы, коринеподобные бактерии, актиномицеты). На этом основан
простейший метод селективного выделения дрожжей: питательная среда (например,
сусло-агар) подкисляется НС1 или молочной кислотой до рН 4-4,5. В большинстве
случаев на такой среде вырастают только дрожжи и быстрорастущие мицелиальные
грибы.
Ингибиторы роста
Рост дрожжей подавляется многими антибиотиками актиномицетного и грибного
происхождения с разным механизмом действия. Наиболее известны из них следую-
щие (рис. 41).
Циклогексимид (актидион) подавляет синтез цитоплазматического белка на по-
лирибосомном уровне. Чувствительность к этому антибиотику у разных дрожжей
сильно варьирует, и признак устойчивости к нему используется в таксономии
дрожжей для дифференциации видов. Например Saccharomyces cerevisiae и родст-
венные виды полностью ингибируются при концентрации циклогексимида всего не-
сколько мг/л, а липомицеты, большинство видов Kluyveromyces, апикулятные
дрожжи Kloeckera-Hanseniaspora резистентны к этому соединению и могут расти
при его концентрации до 100 мг/л.
IIN
О
H5C
О
.сн3
о
он
N=\ ОН N=i
[-ch2cch2-n'v |
F
L <N
LN
НОСИ
HCNHCOCllCb
сн2он
N
Циклогсксимил
Хлорамфсникол
НчС
Амфотсрицин В
I Nib
он
Рис. 41. Примеры наиболее известных антимикотиков, ингибирующих
рост дрожжей.
Хлорамфеникол ингибирует синтез митохондриальных белков, но не оказывает
действия на белковый синтез в цитоплазме. Высокая концентрация глюкозы обес-
печивает рост Saccharomyces cerevisiae в присутствии хлорамфеникола за счет
брожения, но при снижении концентрации глюкозы рост прекращается, так как об-
разовавшийся ацетил-КоА не включается в метаболизм из-за неспособности дрож-
жей синтезировать ферменты цикла Кребса в митохондриях.
Туникамицин селективно блокирует синтез и секрецию маннан-протеиновохю ком-
плекса клеточной стенки и маннан-содержащих ферментов (инвертазы, фосфатазы).
Рост Saccharomyces cerevisiae подавляется слабо, но клетки становятся очень
чувствительными к действию литических ферментов.
В качестве эффективных ингибиторов роста грибов хорошо известны полиеновые
антибиотики, состоящие из циклической липофильной и более подвижной гидро-
фильной частей. Они подавляют рост грибов и дрожжей, но не прокариот. Дейст-
вие их проявляется в образовании прочного комплекса с цитоплазматической мем-
браной, что приводит к изменению ее проницаемости и вытеканию многих цито-
плазматических компонентов. Приобретение устойчивости к полиеновым антибиоти-
кам сопровождается изменениями в составе стеринов и жирных кислот. Наиболее
известные примеры полиеновых антибиотиков - нистатин, амфотерицин. Сходное
действие оказывают некоторые соединения из группы азолов, например флукона-
зол. Эти соединения широко используются при лечении заболеваний, вызываемых
некоторыми видами дрожжей, таких как кандидоз и криптококкоз.
Киллерный
эффект у
дрожжей
Киллерный эффект был впервые обнаружен у Saccharomyces cerevisiae Макоуром
и Беваном в 1963 г. Киллерные дрожжи выделяют токсин, к которому они сами ус-
тойчивы , но который летален для других - чувствительных штаммов. Киллерные
штаммы были обнаружены сначала у дрожжей из родов Saccharomyces, Candida,
Debaryomyces, Kluyveromyces, Pichia, а позже у базидиомицетовых дрожжей и их
анаморф.
Для определения киллерной активности штамм дрожжей высевается штрихом на
поверхность твердой питательной среды, на которую (или в которую) нанесен га-
зон чувствительного штамма. Штамм считается киллерным, если штрих оказывается
окруженным чистой зоной, в которой отсутствует рост чувствительного штамма,
то есть принцип метода - тот же, что и при определении антибиотической актив-
ности .
Генетические анализы киллер-эффекта у лабораторных штаммов Saccharomyces
cerevisiae показали, что расщепление по этому признаку отличается от менде-
левского. Более того, для передачи киллерного признака достаточно одного про-
цесса плазмогамии без слияния ядер. Это указывает на то, что киллерный фено-
тип кодируется внеядерными генетическими элементами (плазмидами). Анализ бес-
клеточных экстрактов киллерных дрожжей на присутствие плазмид, ответственных
за киллерный эффект, обнаружил два типа двунитевой РНК (днРНК): более тяжелая
была обозначена L, более легкая - М.
Далее было показано, что именно М днРНК определяет киллерный фенотип: у не-
киллерных штаммов М днРНК отсутствует, киллерные штаммы после инкубации при
повышенных температурах или обработки циклогексимидом теряют и М днРНК, и
киллерную активность. Однако, для экспрессии киллерного фенотипа необходимо
присутствие как М, так и L днРНК: последняя необходима для поддержания пер-
вой. Дело в том, что как М, так и L днРНК окружены белковой оболочкой, то
есть представляют собой вирусоподобные частицы (ВПЧ). Если М днРНК ответст-
венна за образование киллерного токсина, то L днРНК кодирует капсидный белок
для обоих типов частиц. Так как ВПЧ не проявляют литического цикла, их отно-
сят к латентным вирусам. Такие вирусы найдены у многих грибов.
Только у одного штамма другого вида дрожжей - Kluyveromyces lactis (штамм
IFO 1267) - были найдены киллерные плазмиды совершенно другого типа, относя-
щиеся к ДНК плазмидам. Все остальные изученные киллерные штаммы дрожжей
Candida, Cryptococcus, Debaryomyces, Kluyveromyces, Pichia не обнаруживали
никаких плазмид. Киллерная активность наследуется у них менделевским путем,
ингибируется циклогексимидом, то есть кодируется ядерными генами.
Киллерные токсины всех изученных дрожжей представляют собой пептиды или
гликопептиды. Лучше других изучен токсин из Saccharomyces cerevisiae. Он со-
стоит из двух полипептидных компонентов: а и р, которые связаны дисульфидными
связями.
Киллерные токсины секретируются в экспоненциальной фазе. Сначала в клетке
образуется протоксин, который гликозилируется для эффективного транспорта.
Затем гликозилированный токсин переносится в систему Гольджи и включается в
секреторные везикулы. В системе Гольджи происходит расщепление протоксина на
аир компоненты, которые соединяются дисульфидными связями. Это приводит к
активированию токсина, который секретируется при слиянии везикул с цитоплаз-
матической мембраной. Третий - у~компонент протоксина (сайт гликозилирования)
остается в мембране и обуславливает иммунность к секретируемому токсину.
Механизм действия киллерного токсина заключается в связывании его клеточной
стенкой чувствительного штамма. Рецептором является р(1->6)-глюкан. Сферопла-
сты дрожжей, однако, также чувствительны к киллерному токсину, то есть суще-
ствует также и второй рецептор в мембране. Токсин повреждает цитоплазматиче-
скую мембрану путем образования пор, что приводит к свободному прохождению
ионов через мембрану, а значит к дисбалансу в ионном составе клетки и блоки-
ровке работы протонного насоса. Таким образом последовательность событий при
действии киллерного токсина на клетку такова: связывание токсина с р-глюканом
- связывание с рецептором на мембране - образование пор - изменение электро-
химического потенциала клетки из-за утечки ионов калия - гибель клетки.
Киллерные дрожжи интенсивно изучаются в последнее время в связи с перспек-
тивами их практического использования. В бродильной промышленности, при при-
готовлении вина, пива, часто происходит заражение бродящего сусла так назы-
ваемыми «дикими» дрожжами, которые изменяют ход брожения в нежелательную сто-
рону. Использование киллерных штаммов Saccharomyces cerevisiae для сбражива-
ния исключает развитие диких дрожжей - киллерный токсин будет попросту подав-
лять их рост. Сходные проблемы возникают во всех других видах дрожжевых про-
изводств, где используются чистые культуры. Применение киллерных токсинов
дрожжей перспективно также в медицине в качестве противодрожжевых и противо-
грибных антибиотиков.
Киллерные токсины обладают избирательным, видоспецифическим действием, по-
этому определение устойчивости или чувствительности к различным токсинам ис-
пользуется в систематике дрожжей в качестве важного таксономического признака
при дифференциации видов.
РАСПРОСТРАНЕНИЕ
ДРОЖЖЕВЫХ ГРИБОВ
В ПРИРОДЕ
Особенности распространения дрожжей в природе стали интересовать микробио-
логов начиная с самых первых исследований процессов традиционного виноделия.
Первоначально изучение дрожжей ограничивалось теми видами и штаммами, которые
вызывали брожение при приготовлении пива и вина. Однако уже в конце XIX в. М.
Беиеринк высказывал мысль о том, что эти культурные виды представляют собой
селекционированные формы «диких» дрожжей, широко распространенных в природе.
Естественно, возник вопрос об источниках их попадания в бродящие субстраты.
Первые исследования, выполненные основателями зимологии Э. Хансеном и А. Кле-
кером, были посвящены именно этой теме: поиску природных источников винных
дрожжей Saccharomyces cerevisiae. В нашей стране этому вопросу также уделяли
внимание крупнейшие микробиологи, например Г. А. Надсон. Сахаромицеты были
найдены на ягодах винограда, однако, как оказалось, преобладают здесь совсем
иные виды дрожжей, не участвующие в последующем сбраживании виноградного со-
ка. Еще реже встречались сахаромицеты в окружающих субстратах, в частности, в
почве под виноградниками. Уже в ранних работах высказывалось предположение,
что почва не является средой, в которой возможно активное развитие дрожжевых
грибов, а служит для последних лишь своеобразной «ловушкой», где дрожжи могут
сохраняться определенное время в жизнеспособном состоянии и служить источни-
ком спор для инфицирования винограда нового урожая. Таким образом возникло
понятие «круговорота дрожжей» в природе. Под «дрожжами» в то время подразуме-
вались одноклеточные аскомицетовые грибы, родственные сахаромицетам и способ-
ные к активному брожению.
Расширяющиеся микологические исследования приводили к обнаружению все новых
видов дрожжевых грибов, в том числе и таких, которые существенно отличались
от типичных сахаромицетов. Оказалось, что многие из одноклеточных грибов, вы-
деляемых из природных источников, не образовывали аскоспор и вообще не были
способны к сбраживанию Сахаров. Для таких дрожжей был предложен род Torula,
давший затем начало целой серии родов несовершенных дрожжей {Torulopsis,
Candida, Rhodotorula, Cryptococcus), виды которых часто обнаруживаются в са-
мых различных природных субстратах, включая почву, растения, разнообразные
растительные остатки и природные воды. Стало понятным, что дрожжи распростра-
нены довольно широко, и их развитие далеко не ограничивается субстратами тра-
диционных бродильных процессов. Однако, при этом существенно изменилось и со-
держание самого понятия «дрожжи». Дрожжами стали называть любые одноклеточные
грибы, не обязательно вызывающие спиртовое брожение. В то же время, несмотря
на существенные отличия между сахаромицетами и небродящими дрожжевыми гриба-
ми, достаточно долго сохранялось представление о дрожжах как самостоятельной
филогенетической линии грибов. Лишь после обнаружения у несовершенных дрожжей
рода Rhodotorula полного жизненного цикла, типичного для телиоспоровых гете-
робазидиомицетов, термин «дрожжи» окончательно утратил таксономическое содер-
жание. Тем не менее, вплоть до настоящего времени дрожжи продолжают рассмат-
риваться в качестве единой группы, представляющей собой особую жизненную фор-
му , или экоморфу грибов.
Дело в том, что все дрожжи обладают очень сходным обликом за счет роста
преимущественно в виде одиночных клеток. Кроме того, с одноклеточной органи-
зацией дрожжей сопряжены многие их физиологические особенности, в частности
узкий спектр усваиваемых соединений, отсутствие способности к гидролизу труд-
нодоступных полимеров, особенно таких, как целлюлоза и лигнин, быстрый рост
за счет потребления простых углеводов. Особенно характерен этот набор призна-
ков для аскомицетовых дрожжей. Все это делает их более приспособленными к
обитанию в жидких и мелкодисперсных средах, богатых легкодоступными источни-
ками углерода, в то время как мицелиальные грибы получают преимущество при
росте на плотных поверхностях. Одноклеточность у грибов - вторичное явление в
их эволюции, которое возникало независимо в разных группах аско- и базидиоми-
цетов как реакция на существование в жидких и полужидких средах с относитель-
но высокой концентрацией легкодоступных источников питания.
Действительно, дрожжи - наиболее типичные обитатели природных субстратов,
характеризующихся высоким содержанием легкодоступных питательных веществ (Са-
харов , сахароспиртов, органических кислот и т.п.). В таких субстратах популя-
ции отдельных видов дрожжей могут достигать очень высоких значений численно-
сти и даже доминировать в микробном населении. Многие из подобных природных
местообитаний неоднократно привлекали внимание зимологов и служили источника-
ми выделения самых различных видов дрожжей. Рассмотрим примеры таких место-
обитаний более подробно.
Специфические локусы
развития дрожжей
Виноград и другие сочные плоды
Вскоре после открытия Пастером дрожжевой природы спиртового брожения было
показано, что дрожжи постоянно обитают на поверхности ягод винограда и других
сладких плодов. С этого времени было проведено множество исследований дрож-
жевого населения винограда и всех технологических стадий виноделия. Было ус-
тановлено, что на свежих ягодах винограда, а также в только что приготовлен-
ном виноградном соке обычно доминируют дрожжи аскомицетового аффинитета
Kloeckera apiculata (телеоморфа - Hanseniaspora uvarum), образующие лимоно-
видные клетки в результате биполярного почкования и неспособные к росту при
повышенной концентрации этанола. Наряду с этим с ягод винограда выделяется
много других видов дрожжей, как аскомицетового, так и базидиомицетового аффи-
нитета. Однако в забродившем соке все эти дрожжи быстро вытесняются устой-
чивыми к этанолу, активно бродящими и быстрорастущими дрожжами Saccharomyces
cerevisiae. На свежих ягодах винограда сахаромицеты также обнаруживаются, но
редко. Основным источником сахаромицетов в процессах традиционного виноделия
являются предметы оборудования винодельческих предприятий. Таким образом, S.
cerevisiae - это чисто синантропный вид, предковые формы которого, тем не ме-
нее, сохраняются в природных очагах.
На других сладких плодах различных растений в разных географических регио-
нах также постоянно присутствуют дрожжи, численность которых многократно уве-
личивается по мере порчи (забраживания) плодов. Как правило, с плодов выделя-
ются как типичные эпифитные виды, обычные и на других частях растений, так и
способные к брожению виды аскомицетовохю аффинитета, среди которых также
обычны представители рода Kloeckera.
Для дрожжей, часто способных питаться только простыми углеводами в высокой
концентрации, важным условием успешного развития является своевременное попа-
дание клеток на поврежденный плод. Поэтому особую роль в инфицировании высоко
сахаристых плодов аскомицетовыми дрожжами играют различные насекомые, питаю-
щиеся плодами или откладывающие в них свои яйца. Такие насекомые могут пере-
носить дрожжевые клетки с одного плода на другой. При этом дрожжи, развиваю-
щиеся в плодах, могут даже служить для насекомых дополнительным источником
питания.
Нектароносные цветки и насекомые-опылители
Цветочный нектар может содержать сахара в концентрации до 20%, а также ами-
нокислоты, органические кислоты, минеральные соли, представляя собой иде-
альную среду для развития дрожжей. Действительно, из цветков энтомофильных
растений всегда выделяются дрожжи, среди которых можно обнаружить как типич-
ные эпифитные виды, постоянно присутствующие и на других частях растений, так
и специализированные формы. По-видимому, так же как и в случае сочных плодов,
появление последних в значительной степени зависит от успешного инфицирова-
ния, за которое могут быть ответственны насекомые-опылители. Действительно,
одни и те же виды дрожжей обнаруживаются как в нектароносных цветках, так и
на их опылителях, в частности пчелах. Дрожжи, обитающие в цветочном нектаре,
весьма разнообразны и представлены родами Cryptococcus, Rhodotorula, Candida,
Pichia, Hanseniaspora, Debaryomyces, Metschnikowia и др.
Рис. 42. Крестовидные клетки Metschnikowia reukaufii в цветочном нектаре.
Однако наиболее характерно развитие в цветочном нектаре аскомицетовых дрож-
жей Metschnikowia reukaufii. Эти дрожжи имеют давнюю историю изучения и впер-
вые были обнаружены в самом начале XX века при прямой микроскопии нектара,
где были видны характерные крестовидные дрожжевые клетки (рис. 42). Однако
при выделении организма в чистую культуру эти клетки исчезали. Высказывалось
предположение, что подобная морфология способствует переносу дрожжей насеко-
мыми-опылителями. Организм был назван Anthomyces reukaufii (цветочный гриб).
В 20-х гг. эти дрожжи были подробно изучены Г.А. Надсоном и Н.А. Красильнико-
вым, а позже, после обнаружения у него сумчатой стадии, переописан как
Metschnikowia reukaufii. Последующие исследования показали, что эти дрожжи
выделяются не только из цветочного нектара, но и с поверхности других частей
растений, а также из других субстратов, включая растительные остатки и верх-
ние горизонты почв. Тем не менее, вероятность их обнаружения на цветках энто-
мофильных растений значительно выше, чем на других частях растений. Характер-
но, что этот же вид наиболее часто обнаруживается на насекомых, опыляющих
нектароносные цветки.
Сокотечения деревьев
Сокотечения, или весенний плач деревьев - это вытекание флоэмного сока из
повреждений на стволе, например, у таких деревьев, как береза, клен, дуб и
др. Выделяющийся сок со временем обильно заселяется микроорганизмами, среди
которых преобладают бактерии и дрожжи. Такой забродивший сок привлекает мно-
гих насекомых, которые используют его в качестве питательного субстрата и
служат эффективным средством переноса дрожжей на свежие субстраты.
Дрожжевые сообщества, формирующиеся в бродящих сокотечениях деревьев, весь-
ма разнообразны и включают как типичные эпифитные виды, постоянно обнаружи-
вающиеся на листьях, коре деревьев и в лесных подстилках, так и виды аскоми-
цетового аффинитета, как правило, способные к брожению. Среди последних име-
ется несколько специфических форм, все известные выделения которых были сде-
ланы исключительно из сокотечений, например, Arthroascus javanensis, Nadsonia
elongata, ряд представителей рода Pichia. Только из сокотечений были выделены
базидиомицетовые красные бродящие дрожжи Phaffia rhodozyma.
Изучение дрожжевых сообществ, развивающихся в сокотечениях деревьев, пред-
ставляется очень перспективным для развития самых разных аспектов микробной
экологии. Эти сообщества проходят ряд сукцессионных стадий со сменой домини-
рующих видов, что делает их удобной и простой моделью для изучения общих за-
кономерностей экологических сукцессии. Сокотечения представляют собой как бы
природный аналог хемостата, что дает возможность моделировать их в лаборатор-
ных условиях. Очень интересны перспективы исследования механизмов переноса
дрожжей насекомыми, питающимися на сокотечениях. Широкое распространение де-
ревьев, у которых встречается раневое истекание флоэмного сока, делает воз-
можным изучение роли географического положения в формировании дрожжевых сооб-
ществ сокотечений. Однако подробное изучение всех этих вопросов - дело буду-
щего .
Разлагающиеся ткани кактусов
Один из наиболее подробно исследованных примеров специфических дрожжевых
сообществ - ассоциации дрожжей и дрозофил в разлагающихся тканях кактусов в
пустынях Нового Света. Формирование этих сообществ начинается в результате
локальных механических повреждений тканей кактуса, которые облегчают развитие
в месте повреждения пектолитических бактерий, разрушающих наружные покровы
растения. Позже в таком локусе обильно развиваются дрожжи. Многие насекомые,
среди которых преобладают различные виды дрозофил, используют загнившие ткани
кактусов как места кормления, значительно способствуя при этом инфицированию
их дрожжами, привнесенными из соседних локусов. Личинки дрозофил развиваются
в гниющих тканях, используя дрожжи в качестве источника белка, витаминов и
стеролов. Таким образом формируется, поддерживается и коэволюционирует до-
вольно сложная симбиотическая система, включающая растение, различные виды
бактерий, дрожжей и насекомых.
Детальные многолетние исследования этих локусов обнаружили десятки видов
дрожжей, большинство из которых относились к роду Pichia и Candida. Среди них
был описан ряд новых видов, в частности Pichia cactophila, P. amethionina, P.
opuntiae и др. , которые оказались специфичными именно для этих местообитаний
и не были выделены из других источников. Была обнаружена тесная связь между
таксономическим составом дрожжевых сообществ и филогенетическим и географиче-
ским расстоянием между кактусами, что свидетельствует об их коэволюционном
развитии. Важно подчеркнуть, что эти местообитания также представляют собой
локальное повышение концентрации легкодоступных источников углерода, что в
формировании их также активное участие принимают насекомые, и что здесь также
доминируют типичные сахаролитические дрожжи аскомицетовохю аффинитета.
Дрожжи в буровой муке насекомых-ксилофагов
Жизнедеятельность дереворазрушающих насекомых тесно связана с широким кру-
гом различных микроорганизмов, среди которых дрожжи занимают значительное ме-
сто . В кишечнике многих ксилофагов, проложенных ими галереях и личиночных ка-
мерах, в измельченной древесине (буровой муке) постоянно присутствуют дрожжи
и дрожжеподобные грибы аскомицетовохю аффинитета. Описаны десятки видов дрож-
жей в основном из родов Candida, Pichia, Ambrosiozyma, которые были выделены
исключительно из буровой муки и насекомых-ксилофахюв.
Ассоциации дрожжей и дереворазрушающих насекомых можно разделить на два ти-
па. К первому относятся симбиотические ассоциации, в которых дрожжи развива-
ются в мицетомах стенок кишечника насекомых. Наличие таких дрожжевых симбион-
тов было установлено у многих жуков-ксилофагов из семейств Ipinae (короеды),
Ceramby-cidae (усачи). Функция дрожжей заключается в снабжении насекомых ви-
таминами и другими важными ростовыми веществами. При этом потеря симбионта
сопровождается нарушением роста насекомых и процесса их метаморфоза.
Рис. 43. Ходы насекомых-ксилофахюв в древесине - характерное ме-
стообитание многих видов аскомицетовых дрожжей.
Другой тип взаимоотношений, более факультативный, - развитие дрожжей в бу-
ровой муке, на экскрементах и других продуктах жизнедеятельности насекомых.
Личинки насекомых-ксилофахюв прогрызают ходы в древесине (рис. 43), образуя
буровую муку - измельченную, частично переваренную древесину. На этой буровой
муке и поселяются различные виды микроорганизмов, в том числе и дрожжи. Дрож-
жи, растущие на буровой муке, также служат источником белка и витаминов в пи-
тании ксилофагов. Видовой состав дрожжей в буровой муке очень специфичен, так
как в древесине присутствуют соединения, ограничивающие рост большинства мик-
роорганизмов (терпены, эфирные масла и пр.). Они представлены большой группой
видов аскомицетовых дрожжей из родов Pichia, Ambrosiozyma, Saccharomycopsis
или их анаморфами (Candida spp.). Все эти дрожжи - типичные сахаролитики. Как
правило, они способны к брожению, обладают довольно узким спектром усваивае-
мых соединений, не способны к интенсивному гидролизу труднодоступных полиме-
ров (целлюлозы). Их развитие на древесной буровой муке становится возможным
лишь в результате интенсивного гидролиза измельченной ксилофагами древесины,
вызываемого целлюлолитическими бактериями и мицелиальными грибами. Высвобож-
дающиеся низкомолекулярные продукты гидролиза и обусловливают развитие саха-
ролитических дрожжей.
Кишечный тракт беспозвоночных животных
Приведенные выше примеры типичных местообитаний дрожжевых грибов объединяет
одна интересная особенность: тем или иным образом в их формировании участвуют
беспозвоночные животные. Более того, в кишечниках самых разных беспозвоночных
также часто присутствуют дрожжи, среди которых есть как случайные, так и сим-
биотические формы, сопутствующие животному на протяжении всей его жизни. Та-
кие дрожжи входят в состав сложных микробных ассоциаций, формирующихся в ки-
шечном тракте беспозвоночных и играющих важную роль в их питании. Кроме упо-
мянутых выше насекомых-ксилофагов такие симбиотические дрожжи были обнаружены
в кишечниках многих ризо- и сапрофагов, например, кивсяков, мокриц, личинок
пластинчатоусых жуков (бронзовка, майский жук), личинок жуков-щелкунов. Дрож-
жи, обитающие в кишечниках сапрофагов, представлены разными аскоспоровыми ро-
дами, такими как Debaryomyces, Williopsis, Torulaspora, а также анаморфами из
рода Candida. Если лабораторные культуры таких животных долгое время кормить
стерильным кормом, то в одних случаях дрожжи из их кишечника исчезают, в дру-
гих, например у кивсяков (рис. 44), продолжают существовать неограниченно
долго. Таким образом, можно говорить как о факультативных ассоциациях (прото-
кооперативных), при которых дрожжи постоянно поступают в кишечник беспозво-
ночного с кормом, так и об облигатных (мутуалистических), когда дрожжи посто-
янно обитают в кишечном тракте беспозвоночного и способны поддерживать в нем
постоянную численность за счет размножения. В обоих случаях дрожжи могут иг-
рать важную роль в питании сапрофагов, например, снабжая их необходимыми фак-
торами роста, или регулировать химический состав кишечника. Сами дрожжи, оче-
видно, ассимилируют сахара, образующиеся при гидролизе полисахаридов бакте-
риями, входящими в состав кишечного микробного сообщества.
Дрожжевые грибы постоянно обнаруживаются также в кишечниках и экскрементах
различных беспозвоночных, питающихся зелеными частями растений (фитофагов),
например, многих представителей равнокрылых, клопов, трипсов, жуков, гусениц
бабочек и перепончатокрылых и др. Среди таких дрожжей обычны виды, которые
постоянно обитают на поверхности надземных частей растений, но особенно ха-
рактерны представители рода Metschnikowia, особенно упомянутые выше виды М.
reukaufii и М. pulcherrima, обитающие в растительном нектаре. Очень богатые и
многочисленные дрожжевые сообщества могут формироваться на поверхности листь-
ев растений, пораженных насекомыми из группы сосущих фитофагов (тли). Обиль-
ное развитие последних вызывает накопление сахаристых выделений на поверхно-
сти листьев, т.н. «медвяной росы». Численность дрожжей на листьях, густо по-
крытых тлями и обильной медвяной росой, уже через несколько дней становится
на 2-3 порядка выше, чем на свежих частях растений. При этом численность М.
pulcherrima может достигать 10б КОЕ/г, в то время как на листьях без медвяной
росы обычно не превышает 103 КОЕ/г. Кроме представителей рода Metschnikowia в
субстратах, ассоциированных с зеленоядными беспозвоночными, часто обнаружива-
ются и другие виды аскомицетовых дрожжей из родов Candida, Debaryomyces.
Всасывание
питательных
-длсмснтон
Киллсрныс вещества,
пищеварительные
ферменты
Пер трофическая
мембрана
Антипсристальтмчсскос
движение
Малы]иj иевы сосуды:
моче&ая кислота,
Моченая кислота
Перистая ми чес кое
дни жен ие
Экскременты
tp
#
'-& з
Рис. 44. Судьба дрожжей в пищеварительном тракте кивсяка
Pachyiulus flavipes; дрожжи: 1 - экзогенные, поступающие с пи-
щей, 2 - автохтонные кишечные (Debaryomyces, Torulaspora,
Williopsis), 3 - лизированные.
Итак, наиболее обильно дрожжи развиваются в природных местообитаниях, как
правило, локальных и недолговечных, в которых происходит накопление легкодос-
тупных источников питания - простых Сахаров, органических кислот и др. Анало-
гом таких природных сообществ может служить интенсивное развитие одной попу-
ляции сахаромицетов при сбраживании виноградного сока в традиционном виноде-
лии. Непосредственной причиной этого является высвобождение большого количе-
ства Сахаров при раздавливании ягод винограда. При этом преимущество получают
именно быстрорастущие сахаромицеты, способные к быстрому, «взрывному» росту
за счет малоэффективного бродильного метаболизма. В природе часто можно на-
блюдать похожие явления, когда растительные сахара в высокой концентрации
становятся доступными для утилизации дрожжевыми грибами. Вот примеры механиз-
мов такого локального повышения концентрации Сахаров:
■ накопление Сахаров самим растением (нектар, сочные плоды);
■ истечение флоэмного сока через раневые повреждения (сокотечения деревьев);
■ разрушение и измельчение растительного материала фитофагами (дрожжевые со-
общества в кишечниках и экскрементах растительноядных насекомых);
■ накопление Сахаров на поверхности растений в результате деятельности сосу-
щих насекомых (медвяная роса);
■ высвобождение легкодоступных Сахаров в результате интенсивного гидролиза
растительных полимеров грибами и бактериями (буровая мука ксилофагов).
Как видно, возникновение подобных местообитаний в основном сопряжено с ло-
кальными разрушениями растительных тканей, которые приводят к высвобождению
содержимого клеток. Подобные явления довольно часто вызывают различные беспо-
звоночные : фитофаги, ксилофаги, микофаги. Этим и объясняется наблюдаемая
связь аскомицетовых дрожжей и беспозвоночных животных. В ряде случаев могут
возникать устойчивые козволюционные ассоциации между определенными видами
дрожжей и беспозвоночных, например, в гниющих тканях кактусов или в галереях
ксилофагов. Однако чаще участвующие в таких ассоциациях виды дрожжей постоян-
но выделяются и из других типов субстратов.
Большинство местообитаний с высоким содержанием Сахаров очень недолговечно.
Существенную роль в образовании таких «взрывных» сахаролитических дрожжевых
группировок должен играть случайный фактор. Так же как в традиционном виноде-
лии постоянно существует вероятность развития «диких» дрожжей, которая снижа-
ется добавлением культурной расы, развитие того или иного вида дрожжевых гри-
бов в отдельном высокосахаристом природном субстрате зависит от успешного
первоначального инфицирования. Здесь проявляется еще одна важная роль беспо-
звоночных животных в формировании дрожжевых сообществ, а именно направленное
и эффективное инфицирование дрожжами таких локальных и в основном недолговре-
менных местообитаний. Беспозвоночные в этом случае выступают просто в качест-
ве переносчиков дрожжей между сахаросодержащими локусами. В экологии такой
тип взаимоотношений, когда один вид использует другого в качестве переносчи-
ка , называется форезией.
Известные варианты дрожжевых сообществ, формирующихся в высокосахаристых
субстратах, весьма разнообразны и далеко не исчерпываются рассмотренными выше
примерами. Поиск таких специфических природных локусов развития дрожжей явля-
ется интереснейшим и во многих отношениях (таксономическом, экологическом,
биотехнологическом) и перспективным направлением микробной экологии. Подобные
дрожжевые сообщества могут представлять собой прекрасные модели для изучения
функциональной экологии, механизмов межвидовых взаимодействий и процессов ко-
эволюции .
Дрожжи на растениях,
растительных остатках
и в почве
Распространение дрожжей в природе не ограничивается только локальными и не-
долговечными местообитаниями с высоким содержанием Сахаров. Их можно выделить
практически из любых природных сред, включая верхние и более глубокие почвен-
ные горизонты, поверхность растений, растительные остатки, природные воды,
воздух. Обычно численность дрожжей в большинстве таких местообитаний невысока
и часто лежит на пределе чувствительности методов их учета. Роль дрожжей
здесь значительно ниже по сравнению с ролью мицелиальных грибов и бактерий за
исключением, может быть, живых частей растений. Многие виды аскомицетовых
дрожжей, интенсивно развивающиеся в специфических сахаросодержащих субстра-
тах, спорадически выделяются также из почв и с поверхности растений. Тем не
менее, известны десятки видов дрожжевых грибов, для которых почвы, растения и
растительные остатки являются основным местообитанием. Многие из таких видов
представляют собой дрожжеподобные грибы базидиомицетовохю аффинитета, для
цикла развития которых характерны дрожжевая анаморфная и мицелиальная теле-
оморфная стадии. Сахаролитическая стратегия у таких дрожжей выражена слабее,
чем у представителей порядка Saccharomycetales. За немногими исключениями ба-
зидиомицетовые дрожжи не способны бродить и в среднем усваивают более широкий
спектр соединений. Приуроченность таких дрожжей к местообитаниям с высоким
содержанием свободных Сахаров выражена слабее. Среди них много космополитиче-
ских эвритопных видов, которые с одинаковой вероятностью выделяются из многих
субстратов в различных географических зонах. Методом посева из природных суб-
стратов эти виды, среди которых много гетероталличных, чаще выделяются в виде
дрожжевых анаморф. Не исключено, что телеоморфная стадия у некоторых таких
видов вообще утеряна в ходе эволюции. Типичными примерами таких банальных
форм служат анаморфные виды Cryptococcus albidus, С. laurentii, Trichosporon
pullulans, Sporobolomyces roseus.
Надземные части растений
Надземные части растений почти всегда заселены эпифитными микроорганизмами,
среди которых значительную часть составляют дрожжи. Обнаружить их можно, на-
пример, сделав отпечатки листьев на поверхности питательной среды (рис. 45).
Численность дрожжей на зеленых листьях растений обычно колеблется в пределах
103-105 КОЕ/г, но может достигать 107 КОЕ/г На листьях некоторых тропических
растений дрожжи иногда могут развиваться в таком количестве, что образуют на-
лет, заметный даже невооруженным глазом. Эти дрожжи - не паразиты растений, а
комменсалы, «нахлебники». Единственный постоянный источник питания таких эпи-
фитных дрожжей - прижизненные выделения растений, т.н. экссудаты, в состав
которых входят глюкоза, фруктоза, сахароза, в меньшем количестве трегалоза,
галактоза, органические кислоты. Многие дрожжи способны также утилизировать
липиды листовой кутикулы, гемицеллюлозы, а в качестве азотсодержащих соедине-
ний - белки, аминокислоты, нуклеиновые кислоты. Выделяемые клетками эпифитных
дрожжей экстрацеллюлярные полимеры позволяют им достаточно прочно адгезиро-
ваться на поверхности листьев. Имеются сведения о том, что развивающиеся на
растениях дрожжи, так же как и большинство фитопатогенных грибов, могут вести
эндофитный образ жизни, то есть, растут не на поверхности листьев, а в меж-
клеточном пространстве и внутри тканей листа.
Многие виды дрожжей особенно часто выделяются с поверхности растений и при
этом имеют специфические морфологические и физиологические особенности, сви-
детельствующие об их приспособленности к существованию именно в этом типе ме-
стообитания. К таким особенностям относятся выраженная каротиноидная пигмен-
тация, защищающая клетки от солнечной радиации, образование активно отстрели-
вающихся баллистоспор, рассеивающихся воздухом, формирование полисахаридных
капсул, способствующих адгезии и предохраняющих клетки от высыхания. Дрожжи с
таким сочетанием свойств относятся к экоморфологической группе фитобионтов.
Наиболее типичные представители фитобионтов - анаморфные виды базидиомицето-
вохю аффинитета Sporobolomyces roseus, Rhodotorula glutinis.
Видовой состав дрожжей на листьях растений варьирует в широких пределах.
Однако мало что известно о специфичности дрожжевых сообществ у разных видов
растений. Четкие различия обнаруживаются только в том случае, если сами рас-
тения произрастают в сильно различающихся условиях или относятся к разным
экологическим группам. Например, численность и разнообразие дрожжей в среднем
выше на листьях мезофитов, чем на листьях суккулентных и склерофитных форм, у
которых ограничены процессы транспирации и экссудации. Еще один фактор,
влияющий на плотность заселения дрожжами надземных частей растений - густота
растительного покрова. В местах с сильно разреженным растительным покровом
средняя плотность дрожжей несколько снижается, по-видимому, из-за недостатка
«посевного материала». Существенно различаются также дрожжевые группировки,
формирующиеся на древесных (одревесневшие ветви, поверхность коры) и хлоро-
филлоносных (листья деревьев, травы) частях растений. Последние значительно
богаче, как по численности, так и по таксономическому разнообразию населяющих
их дрожжевых грибов.
Рис. 45. Колонии дрожжей, выросших на месте отпечатков листьев
на поверхности сусло-агара.
Численность и видовой состав дрожжей на листьях растений существенно изме-
няются во времени. При этом динамика эпифитнохю дрожжевого населения опреде-
ляется сочетанием трех групп факторов:
■ погодно-климатическими факторами, определяющими ключевые экологические па-
раметры среды, такие как температура, влажность, интенсивность солнечной
радиации и т.п.;
■ фенологическими ритмами растений, с которыми связаны интенсивность выделе-
ния и химический состав экссудатов, служащих основным источником питания
для эпифитных микроорганизмов, а также выделение растением стимуляторов и
ингибиторов роста микроорганизмов;
■ биологическими ритмами в жизнедеятельности беспозвоночных животных, ассо-
циированных с растением, прежде всего фитофагов и опылителей; эти ритмы в
свою очередь взаимосвязаны как с погодными условиями, так и с фенологиче-
скими изменениями и особенностями онтогенеза растений.
Как правило, численность дрожжевых грибов заметно повышается при отмирании
надземных частей растений. На отмирающих и мертвых, но находящихся на корню
неодревесневших частях растений, численность дрожжей может достигать 107-108
КОЕ/г. Однако, как правило, на этой стадии не происходит существенных пере-
строек таксономической структуры дрожжевого сообщества и по большинству пара-
метров дрожжевые сообщества на мертвых надземных частях растений мало чем от-
личаются от типичных эпифитных сообществ.
Растительные остатки
Другой тип природных местообитаний, в которых всегда присутствуют дрожжи -
это всевозможные варианты растительных остатков, теряющих черты своего анато-
мического строения и находящихся на различных стадиях деструкции. В разных
биогеоценозах это могут быть лесные подстилки, торфянистые горизонты, луговая
дернина, степной войлок и т.п. Плотность заселения дрожжами таких субстратов
несколько ниже, чем надземных частей растений и в среднем составляет 103-104
КОЕ/г.
Следует заметить, что изучение дрожжевых сообществ растительных остатков
методом посева наиболее затруднено. Лесные подстилки обычно густо населены
мицелиальными грибами, в том числе быстрорастущими, такими как Mucor или
Penicillium. Обильное развитие этих грибов в посевах сильно мешает учету
дрожжевых колоний.
Как правило, видовой состав дрожжей в разлагающихся растительных остатках
не сильно отличается от того, что можно обнаружить на живых и мертвых надзем-
ных частях растений в этом же биогеоценозе. Обычно здесь преобладают все те
же типичные фитобионтные виды. Особенно это характерно для свежих раститель-
ных остатков, сформировавшихся за относительно короткое время (прошлогодний
опад, растительные остатки, образовавшиеся в ходе быстрых сукцессии при раз-
ложении несклеротизированных частей растений). В сильно разложившихся расти-
тельных остатках (нижние слои лесной подстилки) структура дрожжевого населе-
ния существенно перестраивается. Доступность необходимых для дрожжевых грибов
свободных Сахаров или органических кислот здесь существенно снижена. Поэтому
в таких местообитаниях дрожжевые группировки приобретают более «экстремаль-
ные» черты: сокращается число обнаруживаемых видов, доминанты становятся бо-
лее выраженными. В подстилках высокую роль начинают играть виды базиди-
омицетового аффинитета, которые обладают характерным комплексом морфо-
физиологических свойств, свидетельствующих об их адаптированности именно к
этому типу местообитаний. Впрочем, отнесение этих видов к дрожжам достаточно
условно: наряду с дрожжевыми клетками для них характерно обильное развитие
истинного мицелия. От большинства видов дрожжей они отличаются высокой гидро-
литической активностью и способны к ассимиляции многих полисахаридов и других
полимеров. Такие виды объединяют в особую экоморфологическую группу сапроби-
онтов, к их наиболее типичным представителям относятся Trichosporon pullulans
и Cystofilobasidium capitaturn, особенно характерные для лесных подстилок.
Почва
Среди огромного разнообразия микроорганизмов, населяющих различные горизон-
ты почв, обычно присутствуют и дрожжи. В то же время, численность дрожжей в
почве невелика, часто она лежит на пределе чувствительности метода посева и
редко превышает 103 КОЕ/г. Главным фактором, определяющим развитие дрожжей в
почве, является концентрация легкодоступного органического вещества. Во всех
типах почв численность дрожжей резко падает с глубиной и наиболее заселенным
является верхний 1-5 см слой, непосредственно под подстилкой или слоем расти-
тельного опада. В отдельных почвенных образцах дрожжи удается обнаружить да-
леко не всегда: в среднем по всем типам почв приблизительно в 70% случаев в
органогенных горизонтах, и лишь в 20-50% в горизонтах типа В. Особенно редко
выделяются дрожжи из глубоких горизонтов ВС и С, глеевых горизонтов G, гори-
зонтов с высоким содержанием легкорастворимых солей. В наиболее бедных мине-
ральных горизонтах дрожжи, по-видимому, вообще могут существовать только на
фрагментах растительных остатков. Например, в поверхностных слоях серо-бурых
пустынных почв и такыров, тундровых пятен, надмерзлотных горизонтов тундровых
глеевых почв дрожжи удается выделить только из микрообразцов растительных
фрагментов, присутствующих в почвенной массе. Понятно, что вероятность обна-
ружить дрожжи в таких почвах значительно выше на участках с более высоким по-
крытием растительностью.
Другим важным фактором, с которым связано развитие дрожжей в минеральных
горизонтах почв, является величина рН. Дрожжи наиболее обильны в кислых поч-
вах, таких как тундровые глеевые, подзолистые, бурые лесные, и крайне мало-
численны в сухо-степных и пустынных почвах с нейтральной и особенно щелочной
реакцией среды, то есть в каштановых, черноземах, серо-бурых пустынных, серо-
земах, солончаках. Не исключено, что это просто следствие методических огра-
ничений. В большинстве почвенно-зимологических исследований дрожжи выделяют
на сусло-агаре, подкисленном до рН 4-4.5 (для ограничения роста бактерий).
При кислых значениях рН среды виды, предпочитающие нейтральные или щелочные
значения, могут не вырастать. Однако, истинных алкалофильных видов дрожжей до
сих пор не известно, хотя многие исследователи используют другие методы огра-
ничения роста бактерий, без подкисления (антибиотики). Кислотолюбивость дрож-
жевых грибов вполне может быть связана с их сахаролитической стратегией, так
как рост при избытке углеводов, как правило, сопровождается образованием и
накоплением в среде органических кислот.
Очень многие виды дрожжей, обнаруживаемые на растениях и растительных ос-
татках, могут быть выделены и из почвы. В верхних горизонтах обычны типичные
фито- и сапробионты, изредка встречаются аскомицетовые дрожжи, характерные
для рассмотренных выше местообитаний с высокой концентрацией свободных Саха-
ров . Эти виды составляют группу аллохтонных почвенных дрожжей. Они попадают в
почву с опадом, переносятся беспозвоночными животными. Почва для них не явля-
ется истинным местом обитания, а лишь «ловушкой», местом сохранения или по-
степенного отмирания. В то же время, нельзя исключить и возможности некоторо-
го их развития здесь, а, следовательно, определенной роли в функционировании
почвенного микробного комплекса, особенно в органогенных горизонтах и микро-
локусах с высоким содержанием легкодоступных элементов питания.
Аллохтонные дрожжи могут быть ассоциированы с корнями растений или сущест-
вовать на мертвых растительных остатках, выступая спутниками организмов-
гидролитиков. Многие из видов этой группы - типичные эпифиты, постоянно оби-
тающие на поверхности зеленых частей растений. Они не погибают при смене ме-
стообитания, но переходят в другую жизненную форму, проявляя мицелиально-
дрожжевой диморфизм не только в морфологическом плане, но и изменяя свои жиз-
ненные функции. Наиболее яркий пример таких организмов - вид «черные дрожжей»
Aureobasidium pullulans, одинаково широко распространенный и как эпифит на
листьях, и как компонент грибного населения лесных подстилок и верхних гори-
зонтов почвы. На листьях дрожжевые популяции этого вида выступают типичными
эпифитами-эккрисотрофами, в почве же они растут в мицелиальной форме, прояв-
ляя себя как активные гидролитики, разлагающие пектины, гемицеллюлозы и клет-
чатку .
Однако существуют и автохтонные почвенные дрожжи, то есть те, которые про-
водят в почве весь жизненный цикл, и для которых минеральные горизонты почв
являются наиболее естественным местообитанием. Такими истинно почвенными
дрожжами являются все виды рода Lipomyces.
Они представляют собой специфическую группу аскомицетовых грибов и настоль-
ко отличаются от остальных дрожжей по своим экофизиологическим характеристи-
кам, что не учитываются обычными методами посева на сахаросодержащие среды,
включая сусло-агар. Впервые эти дрожжи были обнаружены в 1940-х гг. американ-
ским микробиологом Р. Старки, который проводил наблюдения за ростом азотобак-
тера методом обрастания почвенных комочков (рис. 46) на безазотистой среде
Эшби и, оставив чашки на более длительный срок, чем обычно, увидел, что спу-
стя 20 суток вокруг комочков почвы обильно развиваются слизистые дрожжи. Из-
за большого количества жира в клетках они были названы впоследствии липомице-
тами и под родовым названием Lipomyces впервые включены в определитель дрож-
жей в 1952 г.
Рис. 46. Учет липомицетов методом обрастания почвенных комочков
на среде Эшби.
На плотных средах липомицеты растут, как правило, в виде растекающихся сли-
зистых колоний. Это связано с интенсивным образованием полисахаридных капсул,
толщина которых может в два раза превышать диаметр клетки. Микроморфологиче-
ски липомицеты довольно легко дифференцируются от всех других дрожжей. Они
имеют крупные клетки до 10 мкм и более в диаметре, которые в старых культурах
заполняются жиром и превращаются в хламидоспоры. Половой цикл липомицетов,
описанный выше, обычно включает копуляцию двух почек на одной материнской
клетке, в результате чего образуются характерные мешковидные аски.
Липомицеты - типичные педобионты, относящиеся к автохтонной группировке
почвенной биоты. Об этом свидетельствует ряд показателей. Во-первых, эти
дрожжи выделяются из разных почв постоянно и не обнаруживаются в других суб-
стратах, смежных с почвой, откуда они могли бы приноситься в почву. Во-
вторых, численность липомицетов и их распределение по профилю почв соответст-
вует физико-химическому и биологическому режиму конкретного почвообразова-
тельного процесса. Изменение численности в течение года происходит в соответ-
ствии с гидротермическими условиями каждого периода и менее связано со ста-
диями вегетации растений. В-третьих, у липомицетов имеются некоторые особен-
ности морфологии, физиологии, ростовых процессов и биохимии, которые могут
служить доказательством прямой адаптации этих организмов к почвенной среде
обитания. Все известные виды липомицетов - дрожжи с окислительным типом обме-
на, не способные к спиртовому брожению. Они характеризуются высокоэффективным
метаболизмом при относительно низкой удельной скорости роста и низкими трата-
ми на поддержание, используют широкий спектр источников углерода, в том числе
полимеров (инулин, крахмал), ауксоавтотрофы. Липомицеты характеризуются спо-
собностью к росту при очень низкой концентрации азота в среде, что дает воз-
можность выделять их на безазотистых средах, используемых для учета азотфик-
саторов. Стратегия их существования в минеральных горизонтах почв направлена,
прежде всего, на совершенствование механизмов переживания, чему способствует
как эффективное потребление источников углерода и азота, так и способность к
накоплению больших запасов питательных веществ в виде внутриклеточных липидов
и внеклеточных полисахаридных капсул. Их можно отнести к экологической группе
патиентов, или стрессотолерантов, т.е. организмов, подверженных L-отбору (от-
бору в неблагоприятной обстановке).
Липомицеты распространены очень широко. Эти дрожжи находили на всех матери-
ках кроме Антарктиды. Наиболее детально изучено распространение липомицетов в
почвах на территории России и бывших республик СССР. В северном полушарии ли-
помицеты распространены вплоть до лесотундры и не встречаются лишь в тундрах
и высокогорьях. Липомицеты практически отсутствуют и в зоне пустынь, если не
считать редкие случаи их обнаружения в оазисах, в искусственно орошаемых поч-
вах.
Отдельные виды рода Lipomyces ассоциированы с разными группами почв. L.
anomalus (=Babjevia anomala) был выделен только один раз из торфянисто-
подзолистой глееватой почвы северной тайги в Коми АССР. Это северная граница
распространения липомицетов, которая проходит в зоне лесотундры. В подзонах
средней и южной тайги обычен вид L. starkeyi, которому сопутствует L. lipofer
(=Waltomyces lipofer), более тяготеющий к влажным заболоченным почвам зоны
умеренного климата и почвам лугового типа в горных районах. В степной зоне, в
черноземах и каштановых почвах, регулярно и в высокой численности встречается
единственный вид L. tetrasporus. В красноземах субтропиков на территории Кав-
каза, а также в тропических почвах других стран (Нигерия, Мали, Тринидад и
Тобаго, Вьетнам) распространен самый южный вид липомицетов - L. kononenkoae.
Интересно, что в условиях засушливого климата саваноидных пустынь эти дрожжи
«уходят под защиту» распространенных там термитов: популяции L. kononenkoae
можно найти только в локальных участках почв непосредственно под термитника-
ми.
В горных районах распространение липомицетов повторяет в определенной мере
картину, которая складывается при исследовании равнинных территорий. В прими-
тивных почвах высокогорий эти дрожжи отсутствуют, в луговом (субнивальном) и
лесном поясах они представлены L. starkeyi в горных системах европейской час-
ти и L. lipofer - в азиатской. В степных и лугово-степных почвах низкогорий и
предгорий распространен L. tetrasporus.
В интразональных и пахотных почвах эти зональные закономерности в распро-
странении липомицетов несколько нарушаются. По поймам рек популяции отдельных
видов липомицетов могут заходить в несвойственные им зоны. В болотных и тор-
фяно-глеевых почвах на территории Русской Равнины содержание липомицетов не-
высокое, частота встречаемости низкая. В этих почвах обнаруживаются только
многоспоровые липомицеты L. lipofer и L. starkeyi. В окультуренных почвах от-
мечается «смешение» разных видов, например L. starkeyi и L. kononenkoae выде-
ляются вместе из красноземов под чайными плантациями, в пахотных черноземах
можно обнаружить сразу три вида - L. lipofer, L. tetrasporus и L. starkeyi.
Таким образом, липомицеты в основном ассоциированы с наиболее зрелыми и
плодородными почвами. Они не встречаются там, где почвообразовательный про-
цесс находится на ранней стадии развития, где почвы маломощные, что имеет ме-
сто в тундрах и высокогорьях. Максимальной плотности липомицеты достигают в
заселении почв лесостепи и луговых степей. В мощной толще черноземов липоми-
цеты распространяются на большую глубину и иногда составляют доминирующую
часть активной микрофлоры в нижних горизонтах этих почв. В лесной полосе рас-
пространение липомицетов по почвенному профилю не соответствует вертикально-
профильному распределению других дрожжей. Если основная масса дрожжей в лес-
ных почвах сосредоточена в подстилках, то липомицеты, наоборот, имеют макси-
мум в гумусовом и минеральных горизонтах.
Из экологических факторов наиболее четкое влияние на развитие липомицетов
оказывает влажность. Изменение численности этих дрожжей в почве строго следу-
ет за колебаниями влажности. Лабораторные опыты по наблюдению за поведением
искусственно внесенных в стерильную почву популяций липомицетов (L
tetrasporus в чернозем и L. starkeyi в дерново-подзолистую почву) показали,
что переходу к спорообразованию препятствует высокая концентрация доступного
источника углерода (глюкоза), низкая температура (5-10 С) и переувлажнение
почвы.
Функции липомицетов в почве многогранны. В трофических цепях эти дрожжи вы-
ступают как важное звено в питании беспозвоночных. В колониях липомицетов,
обрастающих комочки почвы, постоянно можно наблюдать массовое развитие амеб,
часто попадаются также нематоды и клещи. Амебы переваривают клеточную стенку
и содержимое клеток, за исключением липидных глобул, которые экскретируются и
накапливаются в среде. Не перевариваются амебами также и споры липомицетов.
В процессах деструкции растительных остатков в лесных почвах роль липомице-
тов, по-видимому, невелика из-за слабой их конкурентоспособности по сравнению
с быстрорастущими грибами. Об этом свидетельствует и отмеченное выше их рас-
пределение по профилю лесных почв: плотность заселения липомицетами подстилок
значительно ниже, чем минеральных почвенных горизонтов. Способность липоми-
цетов к гидролизу некоторых полисахаридов позволяет предположить их возможное
участие в разложении этих полимеров. Таким образом, липомицеты следует рас-
сматривать не как главных членов, а как спутников тех микроорганизмов, кото-
рые в основном осуществляют процесс деструкции растительных остатков. Липоми-
цеты включаются в разложение органических остатков лишь на поздних стадиях
этого процесса, протекающего в почвенной толще под травянистыми растительными
формациями.
Многие функции липомицетов в почве связаны с их способностью синтезировать
и экскретировать полисахариды, создающие вязкую межклеточную среду с высокой
гигроскопичностью. Эта среда обеспечивает особый режим питания и влагообмена
самих клеток-продуцентов, а также их спутников, среди которых могут быть и
азотфиксирующие бактерии. Последние более активно связывают молекулярный азот
в ассоциациях с липомицетами, нежели в чистых культурах. При этом внеклеточ-
ные полисахариды липомицетов служат бактериям источником углерода, а часть
связанного азота становится доступной и для дрожжей. Колонии липомицетов в
почве служат, возможно, центрами формирования микросообществ, в состав кото-
рых входят и микроорганизмы, лидирующие мертвые и живые клетки этих дрожжей.
Внеклеточные полисахариды могут быть факторами повышения водопрочности поч-
венных агрегатов, участвуя в создании и стабилизации структуры почвы. Взаимо-
действуя с катионами почвенных минералов, кислые дрожжевые полисахариды спо-
собствуют прочному связыванию адсорбированных клеток и почвенных частиц между
собой.
Что касается других видов дрожжей, то среди них нет ни одного, который бы,
подобно липомицетам, выделялся исключительно из почв и не встречался бы на
растениях и растительных остатках. В то же время три анаморфных вида базидио-
мицетового аффинитета, относящихся к роду Cryptococcus, встречаются в органо-
генных и минеральных горизонтах почв значительно чаще, чем в других типах
субстратов. Это виды С. aerius, С. terreus и С. terricola. Интересно, что для
всех этих видов характерны те же микроморфологические черты, что и для липо-
мицетов, а именно интенсивное накопление в шаровидных клетках внутриклеточных
липидов в виде крупных глобул при росте на богатых средах, образование тол-
стых полисахаридных капсул. Исходя из всех этих особенностей морфологии и
распространения эти виды объединяются вместе с представителями рода Lipomyces
в экоморфологическую группу педобионтных дрожжей.
Смена дрожжевых сообществ при разложении растительных остатков
Итак, обычными наземными местообитаниями дрожжей являются растения, расти-
тельные остатки и почвы. Хотя численность дрожжей в таких местообитаниях су-
щественно ниже, чем в высокосахаристых локусах, они являются постоянным ком-
понентом их микробного населения. Эти субстраты можно рассматривать в доста-
точно естественной последовательности (живые части растений —> мертвые части
растений —> растительные остатки —> органогенные горизонты почв —> ми-
неральные горизонты почв), соответствующей слоям вертикальной стратификации
биогеоценоза или стадиям сукцессии при разложении растительных остатков. По-
этому их называют пространственно-сукцессионным рядом. В этом ряду закономер-
но уменьшается концентрация легкодоступных источников питания, в первую оче-
редь свободных Сахаров, поэтому для дрожжей, как типичных сахаролитиков, сре-
да обитания становится все более неблагоприятной. В пространственно-
сукцессионном ряду в структуре дрожжевых сообществ происходят следующие изме-
нения (рис. 47):
■ уменьшается общая численность дрожжей, в среднем от 104-105 КОЕ/г на зеле-
ных листьях растений (иногда с некоторым увеличением на стадии отмирания
листьев) до 102-103 КОЕ/г в почве;
■ снижается видовое разнообразие дрожжей (его можно измерить, например,
средним числом видов, которое удается выделить из отдельного образца стан-
дартного объема, или общим количеством видов в заданной выборке образцов);
■ все более выраженным становится доминирование одного вида: если на живых
частях растений на долю самого обильного вида приходится обычно не более
половины общей численности дрожжей, то в почве - до 90% и более;
■ дрожжевое население становится все более единообразным, т.е. отдельные об-
разцы растений, взятые в разных местах или в разные сроки, значительно
больше отличаются друг от друга по видовому составу дрожжей, чем образцы
почвы.
6 г Общая численность, КО Ft
4
3
6
5
4
3
2
1
Среднее число видов
f
Доля наиболее обильного вида
12 3 4 5
Стадии пространствен но-
сукиессиоииого ряда
Рис. 47. Изменение дрожжевых сообществ в пространственно-сукцесси-
онномряду: 1 - живые надземные части растений, 2 - мертвые надземные
части растений, 3 - разлагающиеся растительные остатки, 4 - ор-
ганогенные горизонты почв, 5 - глубокие минеральные горизонты почв.
Все эти изменения действительно свидетельствуют о том, что в ходе сукцессии
при разложении растительных остатков среда обитания для дрожжей становится
все более неблагоприятной. Они вполне аналогичны тем изменениям в структуре
растительности и животного населения, которые происходят при перемещении в
области климатического пессимума, например, при переходе от бореальных лесов
к высокоширотной Арктике или от равнин к высокогорьям. Таким образом, про-
странственно -сукцессионный ряд следует рассматривать как важный экологический
градиент в распределении дрожжевых грибов в природе.
Географические закономерности
в распространении дрожжей
Микроорганизмы обычно считают космополитами. Мельчайшие размеры позволяют
им неограниченно расселяться и находить благоприятные микрозоны в любой дос-
таточно гетерогенной среде, в среднем характеризующейся неподходящими усло-
виями для данного вида. Вместе с тем, особенности распространения организмов
зависят и от климатических условий, которые закономерно изменяются по геогра-
фической широте. Вопрос о наличии таких широтно-зональных закономерностей в
распространении микроорганизмов был в центре внимания особого направления в
отечественной микробиологии, получившего название «эколого-географического».
В первую очередь оно связано с именем академика Е. Н. Мишустина, которому в
30-50-х гг. XX в. впервые удалось продемонстрировать широтно-зональные изме-
нения таксономического состава почвенных бацилл. Эти работы привлекли внима-
ние многих микробиологов к вопросу о широтно-зональных закономерностях в рас-
пределении микроорганизмов. Спустя несколько десятилетий самой изученной
группой в этом отношении оказались дрожжевые грибы благодаря относительной
простоте их учета и невысокому видовому разнообразию.
Таксономическая структура дрожжевых сообществ не просто изменяется в зо-
нальном ряду, но эти изменения носят вполне закономерный характер. Числен-
ность и разнообразие дрожжей, обилие отдельных видов монотонно возрастают или
убывают в значительном диапазоне широт или при анализе их зависимости от та-
кого показателя, как температура самого теплого месяца в году. Все эти зави-
симости соответствуют идее экологического оптимума, причем эти оптимумы раз-
личны для разных видов. По приуроченности к различным природным зонам среди
дрожжей выделяются виды арктические (Cryptococcus gilvescens), аркто-
бореальные {Cryptococcus laurentii), бореальные (Cryptococcus podzolicus),
пустынно-степные (Cryptococcus diffluens). Природные зоны существенно разли-
чаются по таксономической структуре дрожжевых сообществ в субстратах про-
странственно -сукцессионного ряда.
Тундра
В пределах тундровой зоны дрожжи наиболее подробно исследованы в различных
районах Таймырского полуострова. Сообщества дрожжевых грибов в тундре пред-
ставлены двумя весьма различными комплексами: довольно разнообразным эпифит-
ным комплексом, который формируется на зеленых и мертвых надземных частях
растений, и более однородным подстилочно-почвенным комплексом на разлагающих-
ся частях растений и в почвенных горизонтах. Автохтонные почвенные дрожжи в
тундровой зоне отсутствуют.
На листьях тундровых растений дрожжи достигают довольно высокой численности
- до 105-10б КОЕ/г. Для этой зоны характерна высокая доля дрожжей среди дру-
гих эпифитных микроорганизмов. Очень часто дрожжи являются единственными
представителями грибов, населяющих филлосферу.
Эпифитные дрожжи в тундре характеризуются довольно постоянным составом до-
минирующих видов. На живых и мертвых частях растений обычно преобладает
Cryptococcus laurentii. Как правило, его доля составляет 30-80% от общей чис-
ленности дрожжей. На листьях обычны также виды Mrakiafrigida, Leucosporidium
scottii, большинство штаммов которых относятся к психрофилам, а также широко
распространенные и в других природных зонах виды Cryptococcus albidus,
Rhodotorula glutinis, Sporobolomyces roseus, черные дрожжи. Все эти виды мож-
но выделить с любого тундрового растения. Особенностью эпифитных дрожжевых
сообществ тундры является почти полное отсутствие аскомицетовых дрожжей, на-
пример, видов Metschnikowia pulcherrima и М. reukaufii, которые являются ха-
рактерными обитателями листьев и цветков энтомофильных растений в бореальной
полосе.
В разлагающихся растительных остатках - в подстилке, торфянистых горизонтах
тундровых почв, в нижней части моховой дернины - дрожжи также постоянно при-
сутствуют, но представлены здесь очень однородным комплексом, в котором обыч-
но более 90% общей численности дрожжей приходится на долю одного вида
Cryptococcus gilvescens. Это единственный настоящий арктический вид дрожжей,
который встречается только в пределах тундровой зоны, причем циркумполярно.
Штаммы Cr. gilvescens были выделены в разных районах Таймыра, в низовьях Ко-
лымы , на Аляске. Другие дрожжи, обычные в разлагающихся растительных остатках
тундровых почв, представлены видами Mrakia frigida, Trichosporon pullulans,
Rhodotorula minuta, Cryptococcus albidus.
Из минеральных горизонтов тундровых почв дрожжи выделяются редко, не более
чем в 20% случаев. Их численность здесь обычно лежит на границе чувствитель-
ности метода посева и редко превышает 103 КОЕ/г. В почве представлены те же
виды, что и в разлагающихся остатках, но с еще более выраженным доминировани-
ем Cr. gilvescens. Липомицеты полностью отсутствуют в почвах тундровой зоны.
Различные типы тундровых почв: тундровые глеевые, дерновые почвы тундровых
луговин, примитивные, тундрово-болотные не отличаются по видовой структуре
населяющих их дрожжевых сообществ: во всех типах резко доминирует один вид
Cr. gilvescens. Несколько более разнообразны дрожжи, обитающие в зоогенных
почвах, которые обычно формируются на южных сторонах холмов, на легких поро-
дах, в местах, активно посещаемых животными (например, около норовищ песцов,
колоний леммингов, мест постоянного кормления хищных птиц). Кроме лучших тем-
пературных условий в таких местах есть постоянный приток свежего органическо-
го вещества (экскрементов, остатков пищи). Биомасса дрожжей на 1 м2 такой
зоогенной почвы может быть выше чем на 10000 м2 пятнистой тундры. Видовое
разнообразие дрожжей в таких почвах также довольно высокое. Здесь могут быть
обнаружены многие виды дрожжей, в том числе аскомицетовые, более характерные
для южных зон.
Лесная полоса
В лесах умеренной полосы формируются наиболее сложные и разнообразные груп-
пировки дрожжевых грибов по сравнению с биогеоценозами других природных зон.
Разнообразие дрожжевого населения леса значительно выше, прежде всего за счет
аскомицетовых дрожжей, представленных как телеоморфными (Debaryomyces,
Pichia, Metschnikowia, Hanseniaspora, Lipomyces), так и анаморфными родами
(Candida, Arxula, Biastobotrys,Mastigomyces, Schizoblastosporion). Кроме то-
го, для лесных биогеоценозов характерно широкое распространение анаморфных
дрожжей, совершенные стадии которых являются фитопатогенами и тесно ассоции-
рованы с определенными видами деревьев (Taphrina, Protomyces).
В целом аскомицетовые дрожжи представляют собой довольно стенобионтную
группу организмов, развивающихся в основном в локусах с повышенным содержани-
ем легкодоступных Сахаров. Однако их можно встретить также и на листьях рас-
тений , на растительных остатках и в почве. В лесной полосе доля аскомицетовых
дрожжей в таких субстратах пространственно-сукцессионного ряда наиболее вели-
ка. По-видимому, это связано с тем, что в лесных биогеоценозах, как в наибо-
лее сложных, имеется наибольшее разнообразие типов местообитаний, в том числе
и таких, в которых обильно развиваются бродящие аскомицетовые дрожжи, и кото-
рые отсутствуют в других природных зонах (сокотечения деревьев, буровая мука
жуков-ксилофагов, разнообразные беспозвоночные-фитофаги). Поэтому здесь выше
вероятность попадания этих дрожжей и в другие местообитания. С другой сторо-
ны, распространение сахаромицетовых дрожжей, как экологически специализиро-
ванных организмов, может в большей степени ограничиваться неблагоприятными
климатическими условиями по сравнению с эврибионтными дрожжевыми грибами ба-
зидиомицетового аффинитета, многие из которых способны к образованию протек-
торных структур типа капсул или каротиноидных пигментов.
Для дрожжевых комплексов в лесных биогеоценозах характерно более выровнен-
ное, чем в других природных зонах, распределение обилия по видам. В областях
климатического пессимума для сообществ дрожжей, рассматриваемых на биогеоце-
нотическом уровне, характерно явление супердоминирования, когда один или не-
сколько видов резко преобладают не только в субстратах определенного типа, но
и в биогеоценозе в целом, причем независимо от типа последнего. Как уже отме-
чалось, в тундровой зоне такими видами являются Cryptococcus gilvescens для
почвенного и Cryptococcus laurentii для надземного ярусов. В субтропических
пустынях во всех типах субстратов резко доминирует один вид - Cryptococcus
diffluens. В лесных биогеоценозах такие однозначно преобладающие виды выявить
значительно труднее, и распределение относительного обилия видов носит менее
предсказуемый характер.
Эпифитный комплекс дрожжей в лесной полосе особенно разнообразен. Числен-
ность дрожжей на листьях лесных трав в среднем составляет 105 КОЕ/г, но может
достигать 107-108 КОЕ/г. Наиболее часто на надземных частях растений встреча-
ются виды Cryptococcus albidus, Cryptococcus laurentii, Sporobolomyces
roseus, Rhodotorula glutinis, Rh.fujisanensis, Rh. mucilaginosa. Из аскомице-
товых дрожжей наиболее обычны Metschnikowia pulcherrima, Debaryomyces
hansenii, различные виды рода Pichia. Соотношение между видами эпифитных
дрожжей сильно варьирует в зависимости от вида растения, сезона, типа биогео-
ценоза .
В лесных подстилках численность дрожжей падает до 104 КОЕ/г. Видовое разно-
образие дрожжей здесь заметно меньше, чем на надземных частях растений. Ха-
рактерной особенностью дрожжевого населения лесной подстилки можно считать
сообитание представителей разных жизненных форм дрожжей: типичных эпифитов,
педобионтов, а также собственно подстилочных видов. Очевидно, что представи-
тели эпифитной микрофлоры постоянно попадают в подстилку с опадом, а почвен-
ные виды заносятся в результате жизнедеятельности различных почвенных живот-
ных. В то же время, судя по высокой численности эпифитных и почвенных видов,
в подстилке может происходить и их активное размножение. В верхних слоях под-
стилки (L) обычно доминируют те же виды, что и на растениях: Cryptococcus
albidus, С. laurentii, дрожжи с каротиноидной и меланоидной пигментацией. В
слое ферментации (F) и особенно в гумифицированном слое (Н) обычны характер-
ные подстилочные виды: дрожжеподобные грибы рода Trichosporon и Cystofiloba-
sidium capitaturn, которые приурочены к поздним стадиям деструкции раститель-
ных остатков и редко обнаруживаются на живых листьях растений. В нижних слоях
подстилки постоянно встречаются также почвенные виды Cryptococcus terricola,
Cryptococcus podzolicus, липомицеты.
Среди дрожжей, населяющих минеральные горизонты автоморфных лесных почв
(подзолистых, дерново-подзолистых, бурых лесных) первое место принадлежит ли-
помицетам. В бореальной полосе распространены многоспоровые виды липомицетов:
Lipomyces starkeyi и Lipomyces lipofer. При учете на сусло-агаре в минераль-
ных горизонтах также дрожжи обнаруживаются постоянно, но большинство видов
встречается спорадически, общая численность дрожжей редко превышает 103
КОЕ/г. Тем не менее, два вида дрожжей выделяются из минеральных горизонтов
лесных почв чаще, чем из других субстратов: Cryptococcus terricola и С.
podzolicus. Наряду с этими почвенными видами в верхних горизонтах постоянно
присутствуют типичные подстилочные виды Cystofilobasidium capitaturn,
Trichosporon pullulans, а также эпифиты Cryptococcus laurentii, Sporobolomy-
ces roseus, Rhodotorula glutinis, Rh. mucilaginosa. Особенностью верхних го-
ризонтов подзолистых почв является постоянное присутствие аскомицетовых дрож-
жей из родов Pichia, Debaryomyces, Hanseniaspora.
Гидроморфные почвы болот таежно-лесной полосы характеризуются значительно
большей численностью дрожжей. Дрожжевые сообщества болотно-торфяных почв от-
личаются значительно более высокими значениями общей численности, до 104-105
КОЕ/г и отсутствием характерного для зональных почв падения численности при
переходе от верхних горизонтов к нижним. В торфе доминируют те же эпифитные и
эврибионтные виды, которые обычны на листьях растений. Среди них особенно
многочисленны Cryptococcus albidus, Rhodotorula mucilaginosa, Sporobolomyces
roseus. Характерно, что именно эти виды наиболее толерантны к целому комплек-
су лимитирующих факторов и часто доминируют в воздухе, в пыли и в других суб-
стратах, где заведомо не может происходить их активного размножения. Числен-
ность дрожжей в торфе не зависит от таких факторов, как ОВП, рН, уровень
аэрации, ботанический состав торфа. Все это говорит о том, что дрожжи в бо-
лотном торфе фактически консервируются в неактивном состоянии. Анаэробные ус-
ловия, лимитация минеральными элементами, низкие температуры не позволяют
дрожжевым клеткам активно размножаться. Тем не менее, интенсивного отмирания
наиболее толерантных эпифитных дрожжей в торфах также не происходит за счет
таких адаптивных свойств, как защитная каротиноидная пигментация, образование
капсул и хламидоспороподобных клеток.
В то же время нельзя исключить и локальных вспышек активности дрожжей в
торфяно-болотных почвах. Об этом свидетельствует значительно более высокое,
по сравнению с другими почвенными и растительными субстратами, содержание в
торфе аскомицетовых дрожжей, способных осуществлять анаэробные процессы спир-
тового брожения. Известны также специфические виды, выделяющиеся только из
торфа и не обнаруживающиеся в других субстратах. К ним относятся
Schizoblasto-sporion starkeyi-henricii, Candida paludigena, Candida aurita.
По-видимому, это стенобионтные виды, приспособленные к обитанию именно в бо-
лотном торфе, хотя их адаптивные свойства еще мало изучены.
Степь
Степи занимали огромные пространства на территории России. Теперь в Евро-
пейской части они почти полностью распаханы за исключением ряда заповедников
(Попереченская степь, Михайловская целина, Хомутовская степь), которые пред-
ставляют большой интерес как примеры самых продуктивных экосистем суши с наи-
более зрелыми и плодородными высокогумусными почвами, черноземами. В этих за-
поведниках проводились исследования дрожжевого населения.
В зоне степей дрожжевые сообщества можно подразделить на два различных ти-
па: эпифитный и почвенный.
Дрожжи в массе развиваются на живых и мертвых надземных частях степных рас-
тений. Их численность здесь достигает 10б КОЕ/г. Наиболее распространенные
эпифитные виды дрожжей - те же, что и в лесной полосе: Cryptococcus albidus,
Cryptococcus laurentii, краснопигментированные баллистоспоровые дрожжи
Sporobolomyces roseus и различные виды рода Bullera. В отличие от более се-
верных лесных районов в степи существенно выше доля пигментных дрожжей. Кроме
видов, обладающих каротиноидной пигментацией, на листьях степных растений в
большом количестве развиваются «черные дрожжи», образующие меланоидные пиг-
менты, прежде всего Aureobasidium pullulans. Так же как в лесной полосе, на
энтомофильных растениях обычны виды Metschnikowia pulcherrima и Metschnikowia
reukaufii. Кроме того, на листьях степных растений широко распространены ба-
зидиомицетовые фитопатогенные грибы из группы головневых (Ustilaginales), га-
плоидные стадии которых способны расти сапротрофно и формируют типичные дрож-
жевые колонии при посеве на питательные среды.
В отличие от лесных биогеоценозов, в степях не выражен особый подстилочный
комплекс дрожжевых грибов. Хотя в луговых и типичных степях обычно формирует-
ся мощный слой т.н. «степного войлока», состоящего из неразложившихся остат-
ков трав, его дрожжевое население по видовому составу такое же, как на живых
частях растений. Виды, типичные для лесной подстилки, такие как Cryptococcus
podzolicus, Cystofilobasidium capitatum, представители рода Trichosporon,
здесь редки или отсутствуют. По-видимому, это связано с низкой влажностью
этого субстрата на протяжении большей части года.
Численность дрожжей в степных черноземах очень низка при учете на средах
типа сусло-агара и редко превышает 103 КОЕ/г. Среди дрожжей, выделяемых из
черноземов, обычны те же эпифитные виды, что и на растениях. В то же время, в
степных биогеоценозах имеются и истинно почвенные дрожжи. К ним относится вид
Cryptococcus aerius, который выделяется из почв южных зон значительно чаще,
чем из других местообитаний. Однако наиболее обильны в черноземах степей на-
стоящие педобионты - липомицеты. Они выделяются из всех почв зоны степей до
глубины более 50 см, давая до 90-100% обрастания почвенных комочков на среде
Эшби. Представлены они здесь одним видом Lipomyces tetrasporus, который может
быть встречен и в других почвах с высоким содержанием гумуса.
Пустыня
Как уже отмечалось, одним из наиболее изученных местообитаний, в которых
формируются обильные и разнообразные дрожжевые сообщества, являются гниющие
ткани кактусов в североамериканских пустынях. Рассмотрим теперь особенности
дрожжевого населения пустынных почв, растительных остатков и различных расте-
ний, которые были изучены в основном в пустынях Средней Азии.
По сравнению с более северными зонами, в пустынях численность эпифитных
дрожжей на листьях растений в среднем существенно ниже и обычно не превышает
104 КОЕ/г. Особенно мало дрожжей на листьях суккулентных и склерофитных видов
растений, которые наиболее характерны для зоны пустынь. Основной тип питания
дрожжевых клеток на живых частях растений - эккрисотрофный, поэтому толстая
кутикула ксерофитов и ограничение прижизненных выделений могут лимитировать
развитие эпифитов. В то же время, после периода весенних дождей, когда в пус-
тынях обильно развиваются эфемеры и эфемероиды, на их листьях успевают раз-
множиться эпифитные дрожжи, численность которых может превышать 10б КОЕ/г.
Эфемеры быстро отмирают, однако в их высохших остатках, которые могут сохра-
няться на поверхности почвы длительное время, а также в опаде многолетних
растений, жизнеспособные дрожжи сохраняются на уровне 105-10б КОЕ/г.
В пустынных почвах численность дрожжевых грибов очень низкая и, как прави-
ло, находится на пределе чувствительности метода посева - менее 102 КОЕ/г.
Более или менее постоянно дрожжи обнаруживаются лишь в самых верхних горизон-
тах: корковом горизонте серо-бурых пустынных почв и закрепленных песков, в
водорослевой корке такыров, то есть там, где в почвенной массе присутствуют
свежие растительные остатки. При этом их численность существенно выше под ку-
старниками или под густыми разрастаниями эфемеров, чем на голых участках. В
минеральных почвенных горизонтах глубже 10 см дрожжи можно встретить только
на поверхности корней. Особенно редко удается обнаружить дрожжи в солончаках,
что, очевидно, связано с высокими значениями рН в этих почвах. Липомицеты в
почвах пустынь не обнаруживаются.
Видовой состав дрожжей в пустынях Средней и Центральной Азии беден по срав-
нению с северными природными зонами. Наиболее часто как на растениях, так и в
почвах обнаруживается один вид анаморфных дрожжей базидиомицетовохю аффините-
та - Cryptococcus albidus. На листьях растений и в растительных остатках на
долю этого вида обычно приходится не менее половины общей численности дрож-
жей, а в почве - до 90% и более. Дрожжи, которые относят к этому виду - ти-
пичные эвритопы. Они широко распространены во всех природных зонах. В то же
время Cryptococcus albidus sensu lato - сборный вид, включающий множество
разновидностей, которые мало различаются по фенотипическим признакам, но мо-
гут быть разделены методами геносистематики. В зоне пустынь этот вид пред-
ставлен главным образом разновидностью Cryptococcus albidus var diffluens,
который по уровню генотипических отличий можно рассматривать и в качестве са-
мостоятельного вида Cryptococcus diffluens. Встречаемость Cryptococcus
diffluens закономерно увеличивается от северных широт к южным. В лесной поло-
се этот вид встречается редко, однако его численность увеличивается в суб-
стратах с очень низкой влажностью. Например, он постоянно обнаруживается в
пыли, на листьях ксерофитных растений в засушливые периоды. Такой характер
распространения согласуется с адаптивными особенностями этих дрожжей. Для них
характерно формирование уже в 3-5 суточных культурах хламидоспороподобных
клеток с утолщенными клеточными стенками. В лабораторных исследованиях пока-
зано, что Cryptococcus diffluens способен переносить очень длительное обезво-
живание в неактивном состоянии.
Безусловно, недостаток влаги в первую очередь определяет особенности дрож-
жевого населения пустынных биогеоценозов. В тех случаях, когда режим увлажне-
ния существенно отличается, может меняться и видовой состав эпифитных дрож-
жей. Например, такие отличия были обнаружены при сравнении дрожжевого населе-
ния двух субтропических пустынь: Каракумы и Негев. Многолетние исследования
показали, что в пустынях Средней Азии на листьях растений и в растительных
остатках преобладает Cryptococcus diffluens, тогда как в пустыне Негев -
Cryptococcus laurentii, характерный для более северных зон. При этом общая
численность дрожжей на растениях в пустыне Негев в среднем значительно выше.
Это объясняется климатическими особенностями пустыни Негев, а именно частыми
туманами, которые практически не влияют на уровень влажности в почве, но мо-
гут служить дополнительным источником влаги для надземных субстратов. Морфо-
логические и физиологические характеристики преобладающих видов дрожжей кор-
релируют с климатическими особенностями регионов. Как уже отмечалось,
Cryptococcus diffluens способен переносить длительное обезвоживание в неак-
тивном состоянии, и стратегия этого вида в континентальных пустынях Средней
Азии заключается в переживании сухого периода в состоянии хламидоспор и в
увеличении активности в короткие влажные периоды непосредственно после весен-
них дождей. Клетки дрожжей вида Cryptococcus laurentii, преобладающего в пус-
тыне Негев, способны формировать мощную полисахаридную капсулу. Такие капсулы
хорошо адсорбируют влагу из воздуха, что может давать преимущество при разви-
тии в условиях частых туманов. Таким образом, региональные отличия в структу-
ре дрожжевого населения пустынь могут быть весьма значительными в случае су-
щественных различий в режиме увлажнения.
Характерной особенностью пустынной зоны является преобладание среди микро-
скопических грибов темноцветных видов, формирующих меланоидные пигменты, та-
ких как Cladosporium,Alternaria, Ulocladium. Среди них значительную долю со-
ставляют и «черные дрожжи», представленные родами Aureobasidium, Phaeococco-
myces. Относительное обилие этих дрожжеподобных грибов на листьях растений в
пустынной зоне еще выше, чем в степях.
В почвах пустынь почти полностью отсутствуют аскомицетовые дрожжи, что ха-
рактерно также и для экстремально холодных местообитаний - тундр и высокого-
рий. В отличие от бореальных и арктических областей, в субтропических пусты-
нях значительную долю составляют баллистоспоровые дрожжи, близкие к родам
Bullera, Bensingtonia и Udeniomyces и очень незначительно представлены тели-
оспоровые дрожжи Leucosporidium, Mrakia, Rhodosporidium. В растительных ос-
татках в пустынях были выделены лишь единичные штаммы Trichosporon pullulans,
Cystofilobasidium capitaturn, которые бывают очень обильны в лесных подстил-
ках. Кроме Cryptococcus diffluens в почвах пустынь обычны и другие виды крип-
тококков, в частности Cryptococcus aerius и Cryptococcus dimennae.
Высокогорья
Высотная поясность в определенной степени аналогична явлению широтной зо-
нальности. С увеличением высоты местности в дрожжевом населении происходят
изменения, аналогичные тем, которые наблюдаются при переходе к высокоширотным
областям. В высокогорьях Памира, Тянь-Шаня, Кавказа средняя численность дрож-
жей в почвах горных тундр и горных лугов на высотах более 3000 м значительно
ниже, чем в почвах горных лесов (2000-3000 м) . В экстремальных условиях суб-
нивального и нивального высокогорных поясов значительно снижается видовое
разнообразие дрожжей, почти не встречаются аскомицетовые виды и преобладают
представители рода Cryptococcus. Однако виды криптококков здесь иные, нежели
в высокоширотной Арктике. В высокогорных тундрах Памира не обнаружен
Cryptococcus gilvescens, доминирующий в почвах тундровой зоны, но преобладают
те же виды криптококков, которые распространены в пустынях Памирских предго-
рий: Cryptococcus aerius и Cryptococcus diffluens. Среди дрожжей, обнаружива-
емых в высокогорных почвах нивального пояса, встречаются уникальные виды, хо-
рошо приспособленные к обитанию в таких экстремально холодных местообитаниях
и не встречающиеся за их пределами. К ним относится дрожжеподобный гриб
Tausoniapamirica - облигатный психрофил, верхняя температурная граница роста
которого не превышает 18 С.
Функции дрожжевых грибов
в природных экосистемах
Какую же роль играют дрожжи в функционировании природных экосистем? В боль-
шинстве местообитаний они не занимают первых мест среди других групп микроор-
ганизмов по численности и биомассе. Особенно невелика доля дрожжей в место-
обитаниях, бедных свежим органическим веществом, например в почвах, природных
водах. В минеральных горизонтах почв их численность обычно не превышает 102
клеток/г при учете методом посева на плотные среды. Среди дрожжей нет актив-
ных гидролитиков, способных разрушать сложные полимеры, такие как целлюлоза
или лигнин. Поэтому в почвах дрожжи вынуждены либо «подбирать» легкодоступные
низкомолекулярные соединения, высвобождающиеся при гидролизе полимеров бакте-
риями и грибами, либо переживать длительные периоды голодания между случайны-
ми поступлениями свежих органических остатков (например, в результате дея-
тельности почвенных беспозвоночных). В то же время, в лесных подстилках чис-
ленность дрожжей может составлять десятки миллионов клеток/г, и в таких слу-
чаях их деструкционная деятельность должна быть весьма значительной. Наконец,
во многих рассмотренных выше локальных местообитаниях с высоким содержанием
Сахаров или органических кислот дрожжи могут существенно преобладать среди
других микроорганизмов и играть главную роль в утилизации таких соединений.
Таким образом, функции дрожжей в природе могут быть сведены к следующим
процессам:
■ быстрое освоение локусов с высоким содержанием легкодоступных источников
питания, характерное для большинства аскомицетовых дрожжей; часто это при-
водит к значительному накоплению дрожжевой биомассы, которая может служить
кормом для беспозвоночных животных (буровая мука ксилофахюв, гниющие ткани
кактусов);
■ «подбирание» прижизненных выделений растений (эккрисотрофия) и «запуск»
сукцессии при разложении растительных остатков для эпифитных дрожжей, раз-
вивающихся на живых частях растений;
■ гидролиз растительных полимеров на начальных стадиях деструкции раститель-
ных остатков в подстилках и верхних слоях почвы;
■ улавливание остаточных легкодоступных веществ в минеральных горизонтах
почв со способностью переживать неблагоприятные условия (L-стратегия).
Специализация на выполнении этих функций приводит у разных групп дрожжевых
грибов к формированию характерного комплекса морфологических и физиологиче-
ских свойств. Это дает возможность говорить о различных жизненных формах
дрожжей. В общей экологии термином «жизненная форма» обозначается внешний об-
лик, определенный морфологический тип организма, сформировавшиеся в результа-
те приспособления к определенной среде обитания. Как уже отмечалось, дрожжи в
современном понимании представляют собой определенную жизненную форму грибов.
У микроорганизмов приспособления носят в основном физиологический характер, и
при выделении таких экологических групп необходимо учитывать физиологические
характеристики, поэтому правильнее говорить не о жизненных формах, а о морфо-
физиологических группах.
Соответственно перечисленным выше функциям среди дрожжей можно выделять
следующие жизненные формы (табл. 2).
Таблица.2. Жизненные формы дрожжей.
Тип
Сахаро-
бионты
Фитобионты
Сапробион-
ты
Педобионты
Адаптивные признаки
Отсутствие выраженных мор-
фологических адаптации,
брожение, г-стратегия
Пигментация, баллистоспоры,
капсулы, окислительный об-
мен , липазная активность,
г-стратегия
Мицелиально-дрожжевой ди-
морфизм, окислительный об-
мен, гидролазная актив-
ность , К-стратегия
Отсутствие пигментации,
внутриклеточные липиды,
окислительный обмен, олиго-
нитрофилия, L-стратегия
Основные ме-
стообитания
Сочные плоды,
нектар, соко-
течения де-
ревьев, бес-
позвоночные
Живые и от-
мершие части
растений
Лесные под-
стилки
Минеральные
горизонты
почв
Типичные виды
Saccharomyces cerevisiae
Saccharomycodes ludwigii
Hanseniaspora uvarum
Metschnikowia ulcherrima
Sporobolomyces roseus
Rhodotorula glutinis
Cryptococcus flavus
Trichosporon pullulans
Cystofllobasidium
capitatum Cryptococcus
podzolicus
Lipomyces spp.
Cryptococcus aerius
Cryptococcus terricola
Сахаробионты - «настоящие» дрожжи, наиболее типичным представителем которых
является Saccharomyces cerevisiae. Они обладают комплексом свойств, свиде-
тельствующим об их приспособленности к существованию в средах, обогащенных
легкодоступными источниками углерода. Отсутствие пигментации, развитых мице-
лиальных структур, хламидоспор, слизистых капсул, а также способность к более
или менее интенсивному брожению и узкий спектр усваиваемых соединений угле-
рода - характерный набор свойств этих дрожжей. Кроме сахаромицетов к сахаро-
бионтам следует относить представителей родов Debaryomyces, Kluyveromyces,
Torulaspora, Zygosaccharomyces, а также большинство видов из родов Pichia и
Candida.
Фитобионты - адаптированы к обитанию на поверхности живых частей растений
и, как правило, образуют каротиноидные пигменты. Они часто имеют в цикле раз-
вития хламидоспоры или хламидоспороподобные клетки, устойчивые к высушиванию.
Характерный признак многих видов - образование баллистоспор, рассеивающихся
токами воздуха. Наиболее типичные представители фитобионтов - роды Sporobolo-
myces и Sporidiobolus, некоторые виды родов Rhodotorula и Cryptococcus.
Сапробионты обладают относительно высокой гидролитической активностью и
принимают участие в деструкции растительных остатков на средних и поздних
стадиях. К типичным сапробионтам относятся некоторые виды рода Trichosporon,
Cystofilobasidium capitaturn, группа несовершенных видов базидиомицетовохю аф-
финитета, классифицируемых в роде Cryptococcus (Cryptococcus podzolicus, Cr.
humicolus).
Педобионты - дрожжи, наиболее приспособленные к обитанию на твердых поверх-
ностях почвенных частиц. Они обладают слизистыми капсулами, которые создают
межклеточную среду, сохраняющую благоприятный режим влахюобмена и питания в
условиях временного иссушения почвы. Эти дрожжи способны накапливать большое
количество запасных веществ, главным образом в форме липидов, которые обеспе-
чивают переживание длительных периодов голодания. Для них также характерна
способность к усвоению соединений азота в очень низкой концентрации (олиго-
нитрофилия). Типичные представители педобионтов - все виды липомицетов. По-
видимому, к педобионтам можно также отнести некоторые виды криптококков, в
частности Cryptococcus terreus, Cr. aerius, Cr. terricola.
Можно выделить и другие важные функции, осуществляемые дрожжевыми грибами в
природных экосистемах. Капсулы базидиомицетовых дрожжей, по-видимому, служат
местом обитания бактерий. В таких бактериально-дрожжевых ассоциациях могут
усиливаться многие важные процессы, например азотфиксация. Внеклеточные поли-
сахариды почвенных дрожжей влияют на агрегатное состояние почв и служат суб-
стратом для бактерий. Клетки дрожжей активно поедаются многими почвенными жи-
вотными: амебами, микроартроподами, червями. Кроме источника энергии дрожжи в
рационе животных могут выступать в качестве важного источника витаминов.
ДРОЖЖИ - ВОЗБУДИТЕЛИ
ЗАБОЛЕВАНИЙ ЧЕЛОВЕКА
Среди дрожжей нет облигатно-патогенных видов, которые могут размножаться
только в организме человека. Однако, в природных местообитаниях встречаются
факультативно-патогенные и условно-патогенные дрожжи, которые могут вызывать
серьезные заболевания у людей с ослабленным иммунитетом. К таким заболеваниям
относятся кандидоз, вызываемый некоторыми видами рода Candida, в первую оче-
редь Candida albicans, и криптококкоз, возбудитель которого - Cryptococcus
neoformans.
Кандидоз
Основным возбудителем кандидоза является Candida albicans (см. рис. 25).
Однако это заболевание могут вызывать и другие виды: С tropicalis, С.
parapsilosis, С. glabrata, С. krusei, реже С lusitaniae, С. guilliermondii,
С. rugosa и др. Все они являются несовершенными дрожжеподобными грибами аско-
мицетового аффинитета. Их довольно часто можно встретить в различных природ-
ных местообитаниях. Многие из них также относятся к компонентам нормальной
микрофлоры человека, то есть постоянно обитают в теле здоровых людей. Их час-
то можно выделить с кожи, слизистых оболочек, из фекалий. Важной особенностью
всех этих видов является их способность к росту при 37 С, те есть при темпе-
ратуре тела человека.
Массовое развитие дрожжей в теле человека приводит к кандидозу. Заболевание
возникает в основном у людей с ослабленной иммунной системой. Наиболее рас-
пространенными причинами являются злокачественные новообразования, травмы,
ожоги и серьезные хирургические вмешательства, длительное лечение антибиоти-
ками широкого спектра действия, преждевременные роды; трансплантация органов
и тканей, которая обычно сопровождается применением препаратов, подавляющих
естественный иммунитет. Повышенные шансы заболеть кандидозом имеют недоношен-
ные новорожденные и лица пожилого возраста с наличием тяжелых заболеваний, а
также наркоманы, использующие нестерильные шприцы. В последнее время кандидоз
особенно часто развивается у больных СПИДом. Кандидоз встречается во всем ми-
ре.
Клинические формы кандидоза могут быть очень разнообразными. Они варьируют
от поверхностных поражений слизистых оболочек, кожи и ногтей (рис. 48) до об-
ширных поражений различных внутренних органов и тканей. Распространенной фор-
мой кандидоза является т.н. «молочница», при которой поражаются слизистые
оболочки рта и горла. При этом на языке и слизистой оболочке рта появляются
кремово-белые пятна, которые распространяются на небо и глотку. Поскольку
возбудитель при этом проглатывается, в отсутствие лечения возможно инфицирова
ние легких и желудочно-кишечного тракта. Сходные с молочницей симптомы возни-
кают при поражении слизистых оболочек половых органов. При поверхностном кан-
дидозе могут также поражаться крупные складки кожи, особенно под молочными
железами, паховые и межъягодичные области, заушные складки. На коже возникают
омертвевшие белые участки, которые превращаются в мокнущие красные эрозии.
Больных при этом беспокоит зуд, жжение.
Рис. 48. Различные клинические проявления поверхностных форм
кандидоза: поражения ногтей, кожи, слизистых оболочек.
При поражении дрожжами внутренних органов возникают особенно тяжелые формы
кандидоза, характеризующиеся высокой летальностью, достигающей 30-70%. В за-
висимости от локализации возбудителя в теле у больных могут развиваться кан-
дидозная пневмония, кандидозный менингит, поражения различных внутренних ор-
ганов .
Основным способом лечения кандидозов является применение антимикотиков -
антигрибных препаратов (см. рис. 45). Среди них наиболее распространены амфо-
терицин В, флуконазол, итраконазол. Очень большое значение имеет точная иден-
тификация возбудителя, так как разные виды Candida, вызывающие кандидоз, име-
ют различную чувствительность к этим препаратам. Для этого возбудитель должен
быть выделен из тела больного с помощью посева проб пораженных тканей на пи-
тательные среды и получен в виде чистой культуры. Виды Candida albicans, С.
tropicalis, С. parapsilosis, С. lusitaniae при росте на питательных средах
очень близки по морфологическим и физиологическим характеристикам и для их
точной идентификации приходится применять современные молекулярно-биологичес-
кие методы.
Криптококкоз
Криптококкоз (торулез, европейский бластомикоз) - заболевание, вызываемое
несовершенными дрожжами базидиомицетового аффинитета Cryptococcus neoformans.
Дрожжи этого вида можно встретить в природных местообитаниях, но наиболее
часто его обнаруживают в помете голубей, воробьев и других птиц, при этом са-
ми птицы не болеют. В сухом помете криптококки могут сохраняться в течение
многих месяцев. Cr. neoformans обнаруживали также на слизистых оболочках здо-
ровых людей.
Инфицирование человека обычно происходит воздушно-пылевым путем. Наиболее
опасным проявлением криптококкоза является менингоэнцефалит, при котором по-
ражается центральная нервная система. Заболевание начинается приступами го-
ловной боли, которые постепенно усиливаются и становятся нестерпимыми. Появ-
ляются признаки паралича, сознание нарушается. Развитие болезни ведет к по-
степенному истощению организма, затем коматозному состоянию. Смерть наступает
от паралича дыхания через 4-6 месяцев. Смертность при криптококковом менинги-
те достигает 100%. У многих больных, кроме поражения центральной нервной сис-
темы, развивается криптококкоз легких. Он протекает в виде пневмонии, которая
по симптомам очень похожа на пневмонии другой этиологии. При диссеминирован-
ной форме криптококкоза могут поражаться самые различные органы. Встречаются
также поражения кожи и слизистых оболочек, которые протекают относительно
легко. Также как и в случае кандидоза, наибольшие шансы заболеть криптококко-
зом имеют люди с нарушением клеточного иммунитета, обусловленным СПИДом, лей-
козом, отторжением трансплантанта при пересадке органов и тканей, а также
длительным применением иммуносупрессантов.
Вследствие большой медицинской значимости дрожжи Cryptococcus neoformans
очень детально исследованы. История изучения этого вида насчитывает уже более
100 лет. На питательных средах он выделяется в гаплоидной фазе, представлен-
ной почкующимися клетками. Практическая диагностика Cr. neoformans основана
на некоторых его характерных особенностях, в частности образовании коричневых
колоний на средах с фенольными соединениями за счет синтеза меланинов, редко
встречающейся способности к утилизации в качестве источника азота креатини-
на. Внутри этого вида различают две разновидности с различными серотипами.
Принадлежность к разным серотипам определяется различиями в строении капсуль-
ных полисахаридов. К разновидности Cr. neoformans var neoformans принадлежат
серотипы, обозначаемые как А и D, к Cr. neoformans var gattii - серотипы В и
С. Разновидности Cr. neoformans имеют различное распространение. Сг.
neoformans var neoformans встречается повсеместно и в природе обнаруживается
наиболее часто в птичьем помете, a Cr. neoformans var gattii характерна для
тропических и субтропических регионов. У обоих разновидностей найдены совер-
шенные стадии, принадлежащие к телеоморфному роду Filobasidiella (рис. 49)
Для Cr. neoformans var neoformans - это F. neoformans var neoformans, для Cr.
neoformans var gattii - F. neoformnans var bacillispora. Обе телеоморфы гете-
роталличны и развиваются лишь при смешивании дрожжевых штаммов комплиментар-
ных типов спаривания. При этом развиваются дикариотические гифы с пряжками,
на которых формируются удлиненные булавовидные несептированные базидии. В ба-
зидиях происходит кариогамия, а после миграции диплоидного ядра к вершине ба-
зидии протекает мейоз. Возникшие гаплоидные ядра продолжают делиться митоти-
чески и дают начало длинным цепочкам базидиоспор. Образование базидиоспор
объясняет механизм заражения человека воздушно-пылевым путем. Долгое время
оставалось непонятным, каким образом относительно крупные (до 8-10 мкм) дрож-
жевые клетки криптококков проникают в альвеолы легких. Первичное заражение
человека происходит именно базидиоспорами, имеющими диаметр всего 1-3 мкм, и
к тому же более устойчивыми к неблагоприятным условиям среды, чем вегета-
тивные клетки.
Рис. 49. Filobasidiella neoformans - телеоморфная стадия
Cryptococcus neoformans: почкующиеся клетки, базидия, базидии с
цепочками базидиоспор.
Для лечения криптококкоза используют различные антимикотики, такие как ам-
фотерицин, флуконазол, подавляющие у грибов синтез эргостерина, и 5-
фторцитозин, ингибирующий у них синтез нуклеиновых кислот. Для большей эффек-
тивности эти препараты используют комбинированно, что обусловлено также и
частым обнаружением резистентных к ним штаммов. Обнаружение последних вынуж-
дает медиков вести непрерывный поиск новых антифунгальных агентов.
Malassezia
Дрожжи рода Malassezia - облигатные симбионты человека и теплокровных жи-
вотных, обитающие на поверхности их кожи и на волосяном покрове. В других ме-
стообитаниях эти организмы не встречаются. Первые описания этих грибов отно-
сятся к середине XIX в. Они были обнаружены в образцах кожи людей, больных
так называемым пестрым лишаем. При микроскопировании кожных чешуи в них на-
блюдали короткие гифы, проникающие в слой эпидермиса. Первые попытки культи-
вировать эти грибы были безуспешными. В образцах кожи и волос можно также не-
редко наблюдать присутствие дрожжевых клеток, которые также не культивируются
на обычных средах для дрожжей. Этот организм, вызывающий образование перхоти,
получил название Pityrosporum. Вырастить эти организмы в культуре удалось в
20-х гг. XX в. Оказалось, что для роста им необходимы добавки жирных кислот,
например оливкового масла. Позже при культивировании Pityrosporum обнаружи-
лось , что эти дрожжи способны к образованию гиф, которые по морфологии иден-
тичны гифам Malassezia. Таким образом было доказано, что Pityrosporum и
Malassezia - это дрожжевая и мицелиальная стадия одного и того же гриба.
Характерная особенность дрожжеподобных грибов poRSiMalassezia - монополяр-
ное энтеробластическое почкование на широком основании. Отделившаяся почка
оставляет на материнской клетке шрам в виде воротничка, из которого появляют-
ся и последующие почки (рис. 50). В результате клетка приобретает характерную
форму в виде груши или лампочки. Malassezia относится к дрожжеподобным грибам
базидиомицетового аффинитета, хотя телеоморфная стадия у них неизвестна. К
настоящему времени описано восемь видов этого рода, различающихся по морфоло-
гическим и физиологическим характеристикам.
Рис. 50. Malassezia - дрожжевые клетки с монополярным энтеробла-
стическим почкованием.
Дрожжи рода Malassezia вместе с различными бактериями относятся к компонен-
там нормальной микрофлоры человека и постоянно обнаруживаются на разных уча-
стках кожи здоровых людей. Обычны они и на коже домашних и диких теплокровных
животных, причем плотность заселения кожи этими дрожжами коррелирует с коли-
чеством выделяемого сального секрета. Однако при различных нарушениях иммун-
ной системы виды рода Malassezia могут вызывать различные заболевания кожи.
Из них наиболее известен упомянутый выше пестрый лишай, или питириаз. Это
хроническое неконтагиозное заболевание, распространенное главным образом в
тропических странах. Оно выражается в массовом развитии гиф Malassezia, кото-
рые проникают в слой эпидермиса, нарушая пигментацию и нормальную структуру
поверхностного слоя кожи. Другим заболеванием, вызываемым этим грибом, явля-
ется фолликулит, который характеризуется наличием воспалений в виде множества
фолликул - мелких нарывов, распространяющихся по спине, груди, предплечьям и
сопровождающихся сильным зудом. Фолликулит также встречается наиболее часто в
странах с теплым климатом. Еще одно заболевание кожи, вызываемое дрожжами ро-
да Malassezia - себоррейный дерматит. Они могут выступать в качестве важного
аллергена при таком распространенном в последнее время аллергическом заболе-
вании , как атопический дерматит.
Заболевания, ассоциированные с Malassezia, являются в основном хроническими
и характерны для людей с подавленной иммунной системой и предрасположенными к
аллергии. Лечение их направлено на устранение дрожжей с помощью различных ан-
тимикотиков. У здоровых людей при нормальном функционировании сальных желез
присутствие Malassezia на коже незаметно. Более того, они могут играть поло-
жительную роль, конкурируя с болезнетворными микроорганизмами.
(ПРОДОЛЖЕНИЕ СЛЕДУЕТ)
Химичка
ЗАКИСЬ АЗОТА
Григорьев И.Н.
Введение
Бесцветный газ со слабым сладковатым запахом и вкусом. Т.пл. -90.8 С;
Т. кип. -88.5 С. Плотность N20 1.5299 (по воздуху) при 0 С и 760 мм рт. ст.
При нормальных условиях 1 л закиси азота весит 1.9778 г. Плотность жидкой за-
киси азота 1.226 г/см3 при -89 С. Закись азота растворима в воде и еще лучше
- в этиловом спирте (1 объем спирта растворяет 4 объема закиси азота при
20°С). При нагревании свыше 500 С закись азота разлагается:
2N20 = 2N2 + 02 +39 ккал
Процентное содержание кислорода в продуктах распада N20 выше, чем в возду-
хе , поэтому горение веществ идет энергичнее. При повышенных температурах за-
кись азота действует как сильный окислитель. Например, тлеющая лучина вспыхи-
вает, как в кислороде, а смеси закиси азота с водородом и аммиаком при нагре-
вании взрываются.
Малые концентрации закиси азота при вдыхании вызывают чувство своеобразного
опьянения, которое обычно сопровождается выраженной эйфорией, благодаря этому
закись азота получила свое знаменитое тривиальное название - веселящий газ.
При вдыхании чистого газа быстро развиваются наркотическое состояние и асфик-
сия. В смеси с кислородом при правильном дозировании вызывает наркоз без
предварительного возбуждения и побочных явлений. Закись азота при вдыхании не
вызывает раздражения дыхательных путей, с гемоглобином не связывается. После
прекращения вдыхания через 10...15 минут полностью выделяется через дыхатель-
ные пути в неизменном виде.
Основное свое применение закись азота находит в медицине из-за ее наркоти-
ческих свойств. Газ используется для наркоза в смеси с кислородом в хирурги-
ческой практике, оперативной гинекологии, хирургической стоматологии, для
обезболивания родов, для профилактики травматического шока в послеоперацион-
ный период, а также для купирования болевых приступов при острой коронарной
недостаточности, инфаркте миокарда, остром панкреатите и других патологиче-
ских состояниях, сопровождающихся болями.
Одно из наиболее известных медицинских применений закиси азота - обезболи-
вание при стоматологических операциях. Смесь кислорода и веселящего раза по-
дается в маску, закрепленную на носу. Концентрация раза подбирается так, что-
бы пациент находился в сознании, при этом он мало восприимчив к боли и испы-
тывает состояние приятной релаксации. Благодаря этому дети в европейских
странах не боятся стоматологов, что позволяет проводить продолжительное лече-
ние. К сожалению, в России эта процедура практически не используется, благо-
даря этому стоматологи для многих детей являются воплощением зла, а сам про-
цесс часто напоминает средневековые пытки. По крайней мере, так казалось ав-
тору в детстве.
Наркоз закисью азота.
Из второстепенных по значимости областей применения веселящего газа отметим
пищевую промышленность. Закись азота зарегистрирована как добавка Е-942. В
отличие от широко используемого двойника по физическим свойствам - углекисло-
го газа употребляется редко. Нам удалось найти лишь один довольно распростра-
ненный продукт, содержащий веселящий газ - это взбитые сливки. Выпускаются
аэрозольные баллончики, где веселящий газ используется в качестве пропеллента
или сливки взбиваются в сифонах с помощью восьмиграммовых баллончиков со сжи-
женным газом. Из аэрозольного баллончика взбитых сливок нам удалось получить
около трех литров веселящего газа. Другими словами, взбитые сливки могут слу-
жить быстрым и очень простым (но дорогим) нехимическим источником получения
небольших количеств газа. Однако не будем забегать наперед: получению закиси
азота из взбитых сливок посвящен один из следующих разделов статьи.
Одно из популярных любительских применений закиси азота т.н. системы NOS -
от англ. Nitrous Oxide System, использующиеся для увеличения мощности двига-
телей внутреннего сгорания, что является очень привлекательным для автогонщи-
ков . Сжиженная закись азота из баллона и горючее впрыскиваются во впускной
(всасывающий) коллектор двигателя, что приводит к следующим результатам:
• снижает температуру всасываемого в двигатель воздуха, увеличивая тем самым
плотность поступающей в цилиндры смеси;
• увеличивает содержание кислорода в поступающем газе (воздух содержит лишь
21 % кислорода по весу);
• повышает скорость (интенсивность) сгорания в цилиндрах двигателя.
Увеличение количества кислорода позволяет сжигать большее количество топли-
ва, как следствие высвобождать больше энергии. Чистый кислород непригоден для
этой цели, ввиду слишком сильного окислительного действия. Азот, образующийся
при распаде закиси азота, не дает смеси детонировать. Подобные системы, одна-
ко , несмотря на громкую рекламу, скорее дорогое развлечение, т.к. дают крайне
непродолжительный эффект (баллон расходуется очень быстро) и двигатель при
этом быстрее изнашивается. NOS используются лишь любителями гонок.
Система для впрыскивания закиси азота в двигатель.
Не совсем приятной областью использования закиси азота стало его "неофици-
альное применение" в качестве наркотического вещества (вернее ингалянта),
другими словами - злоупотребление веселящим газом. В США довольно распростра-
нено нецелевое использование баллончиков с пищевым веселящим газом для взби-
вания сливок. В России увлечение вдыханием веселящим газом начало приобретать
популярность в клубах.
История
Впервые оксид азота(I) был получен в 1772 году Джозефом Пристли, который
назвал его «дефлогистированным нитрозным воздухом».
Наркотическое действие веселящего газа было открыто Г. Дэви. Рассмотрим
подробнее биографию этого выдающегося химика до периода исследований действия
газов. Гемфри Дэви (Humphry Davy, 1778-1829) родился в маленьком городке Пен-
зансе на юго-западе Англии. Гемфри учился в грамматической школе близи Пен-
занса. В 1795 г., через год после смерти отца, Гэмфри был принят в качестве
ассистента и аптечного помощника к местному хирургу Дж. Бингану Борлэйзу (J.
Binghan Borlase). Он готовил мази, взвешивал порошки, помогал при перевязках,
мечтая выучиться врачебному ремеслу и стать доктором.
Чрезвычайно прилежный и любознательный Дэви жадно прислушивался к разгово-
рам своего шефа с местными коллегами и заезжими коммерсантами-аптекарями о
перспективах развития "пневматической медицины", основы которой были положены
работами английского ученого Джозефа Пристли (Priestley J., 1733-1804). Шум
вокруг этой новой моды в медицине был уже порядочный, если судить по оппози-
ции, которая довольно громко высказывалась. Так, например, Ян Ингенхаус (Jan
Ingenhoucz), придворный врач австрийского имперского двора, открыто предосте-
регал врачебный мир от опасностей чрезмерного увлечения газами как "жизненным
эликсиром". Но еще более категоричные отрицательные взгляды высказал автори-
тетный американский врач, химик Митчелл (Lantham Mitchell), который ссылаясь
на проделанные им опыты на животных с открытой Пристли закисью азота, заявил,
что этот газ является опасным ядом, от которого его животные чуть не погибли.
Митчелл в своих предостережениях шел еще дальше, высказывая мысль, что неко-
торые газы сами по себе и есть главная причина эпидемических заболеваний.
Митчелл был для всех признанным авторитетом, и его суждения в общественном
сознании были чуть ли не истиной в последней инстанции. Однако на юного Дэви
подобные приговоры произвели обратное действие и зародили в нем мысль приго-
товить закись азота, и на самом себе испробовать ее действие.
Гемфри Дэви.
По ночам, когда хозяин Борлэйз уходил, Дэви перечитывал "Химический спра-
вочник " Никольсона, "Руководство по элементарной химии" Лавуазье и "Экспери-
менты и наблюдения" Пристли, а также постепенно подготавливал оборудование и
препараты для приготовления закиси азота. Когда все было готово, и газ был
получен, Дэви начал свои героические опыты. Вдыхание закиси азота произвело
на него столь необыкновенное действие, вызывая чрезвычайно приятные ощущения
и веселое настроение, что Дэви, скрывая опыты от своего шефа, стал повторять
их почти ежедневно, все более и более убеждаясь не только в отсутствии отрав-
ляющего действия, но и в неизменном опьяняющем эффекте закиси азота и вызы-
ваемых им веселых галлюцинациях.
Дэви был начинающим поэтом, и он не утерпел, чтобы не описать в стихах свои
ощущения при действии закиси азота. Но для нас гораздо интереснее не его юно-
шеские вирши, теряющие прелесть стихотворной формы при переводе прозой, а
точные записи из его знаменитой книги, вышедшей в 1800 г., когда он уже был
сотрудником "Медицинского пневматического института". Здесь мы находим первое
в мире прямое указание на обезболивающее действие ингаляции закиси азота. Вот
цитата из книги Дэви "Исследования химические и философские, касающиеся глав-
ным образом закиси азота, или дефлогистированного воздуха и его вдыхания":
"При прорезывании одного злосчастного зуба, называемого dentes sapientiae,
я испытывал острое воспаление десны, сопровождающееся большой болью, которая
одинаково мешала как отдыху, так и сознательной работе. Однажды, когда воспа-
ление было чрезвычайно чувствительно, я вдохнул три большие дозы закиси азо-
та. Боль совершенно исчезла после первых четырех или пяти вдохов и неприятные
ощущения на несколько минут сменились чувством удовольствия. Когда прежнее
состояние сознания возвратилось, вместе с ним вернулось и состояние в органе,
и мне даже показалось, что боль была сильнее после опыта, чем раньше".
Увлекательные опыты с закисью азота и сопровождающее их состояние наркозно-
го опьянения испортили взаимоотношения Дэви с его шефом, который вначале не
мох1 понять причины небывалых раньше приступов безудержного смеха и возбужде-
ния своего ученика. Случалось, что приезжавшие пациенты, встретившись с не-
вменяемым, по их мнению, лекарским помощником, уезжали с недовольством и
практика Борлэйза стала падать. Когда хозяин выяснил наконец причину частой
одержимости Дэви, то, усмотрев в опытах с закисью азота причину своего вра-
чебного неблагополучия, он запретил Дэви продолжать исследования в его доме.
Дэви переселился к своему приемному отцу, доктору Тонкину. Здесь он вновь
собрал кое-какую стеклянную посуду и оборудование, наладил производство газов
и возобновил свои опыты. Именно тут, у Тонкина, он дал закиси азота название
"веселящий газ".
Но однажды ночью семейство Тонкинов было разбужено сильным взрывом. Вбежав
в комнату Дэви, они застали его растерянного, с виноватым видом, среди раз-
бросанного взрывом оборудования. Последовало категорическое запрещение про-
должать эти затеи, угрожающие взорвать весь дом. Вторично поискам Дэви насту-
пил конец.
Но в это время в Пензанс случайно приехал доктор Дэвис Гидди (Davies Giddy,
позднее Gilbert), впоследствии ставший президентом Королевского Общества
(1827-30 гг.). Он услышал о "таинственных газах" и о взрыве в доме Тонкинов и
пожелал познакомиться с этим "неисправимым юношей". Гидди сразу же усмотрел в
юном Дэви многообещающего пытливого исследователя и рекомендовал его своему
другу - доктору Томасу Беддо (Beddoes, Thomas, 1760-1808) , директору "Пневма-
тического института" в Клифтоне, под Бристолем.
Приехав в Клифтон, Дэви получил максимум того, о чем мог мечтать: замеча-
тельную лабораторию, прекрасные условия жизни и работы, и прекрасного руково-
дителя, увлеченного мечтами об исследованиях газов и их действии на организм
человека при вдыхании.
Томасу Беддо, возглавлявшему созданный им "Пневматический институт", в это
время было сорок лет. Это был чрезвычайно образованный, разносторонний уче-
ный, собравший вокруг себя группу энтузиастов новой идеи - пневматической ме-
дицины. Сам он был известным химиком, философом, поэтом и искренне увлекался
идеями служения на благо человечества. Он получил образование в Лондоне,
Эдинбурге и Париже, был дружен с Лавуазье. Дополнительный курс химии он про-
шел в Оксфорде. Беддо увлекался психологическими анализами, изучал природу
снов и впечатления раннего детства, предвосхищая тем самым будущие работы
Зигмунда Фрейда.
Идеи Беддо о лечении ингаляциями газов различных болезней встретили весьма
горячий отклик и всяческую помощь. Достаточно сказать, что известный поэт То-
мас Веджвудд предоставил тысячу фунтов стерлингов в его распоряжение, а из-
вестный изобретатель, создатель первой паровой машины, Джеймс Уатт (Watt, J,
1736-1819) снабжал его лаборатории необходимым оборудованием.
"Пневматический институт" был оборудован и снабжен первоклассным по тем
временам оборудованием и лабораториями, при нем находилась больница на 10 ко-
ек и поликлиническое отделение. Ко времени приезда Дэви в институте уже широ-
ко проводились испытания ингаляций кислорода, водорода, азота и некоторых не-
давно открытых углеводородов. Фактически, это был настоящий научный центр, в
котором изучались свойства различных газов и их влияние на организм человека.
Можно смело сказать, что Томас Беддо и его сотрудники были пионерами и пред-
шественниками современной респираторной терапии. В "Пневматическом институ-
те ", во многом благодаря Джеймсу Уатту, создавались и испытывались первые ин-
галяторы, спирометры, баллоны сжатого газа и т.д. Именно в "Пневматическом
институте" впервые был применен с лечебной целью кислород; разработаны основы
аэрозольной терапии; впервые была измерена общая емкость легких методом раз-
ведения водорода (Дэви).
Намерения Дэви заняться закисью азота были встречены Беддо с одобрением.
Дэви повторил свои опыты, проведенные в Пензансе, изготовил хорошие газомет-
ры, хотя дважды чуть не погиб от ингаляций недостаточно очищенного газа. В
конце концов, 11 апреля 1799 г. ему удалось наладить получение "чистой" заки-
си азота.
Как же тогда получали закись азота, и чистой ли она была по современным по-
нятиям? Обратимся к монографии Дэви. В главе "О подготовке закиси азота для
экспериментов над вдыханием" описан использованный прибор, который состоял из
стеклянной реторты объемом 2-3 кварты (около 2-3 л) , в которой разлагается
расплав аммиачной селитры, горло реторты соединялось с герметичными газомет-
рами Уатта, в качестве запирающей жидкости использовалась вода насыщенная за-
кисью азота. Дэви предупреждает, что "газ, произведенный из нитрата аммония,
должен быть перенесен, чтобы находится в покое, по крайней мере, в течение
часа после его генерации". Если использовать газ раньше, то он будет содер-
жать "более или менее белого пара, у которого есть неприятное кислотное свой-
ство" и который сильно раздражает легкие. Читателю известно, что даже при
благоприятном температурном режиме разложения аммиачной селитры всегда идут
побочные реакции, в ходе которых сырой газ кроме безобидных примесей воды и
азота содержит также пары азотной кислоты и смесь N0 и N02. В процессе разло-
жения селитры газ увлекает аэрозоль нитрата аммония. При выдерживании газа
количество примесей уменьшалось незначительно, вероятно при этом конденсиро-
валась часть азотной кислоты, и оседал аэрозоль селитры. Содержание N0 и N02
не менялось. Дышать таким газом вредно для здоровья, но тогда об этом не зна-
ли.
Первая демонстрация ингаляции больших доз закиси азота была проведена Дэви
в присутствии Беддо и младшего ассистента Кинглэка. Успех был полный: вдыхая
три-четыре кварты из приготовленного, непроницаемого шелкового мешка, Дэви не
испытал никакого дурного действия. К опытам присоединились и другие. Первым
из принявших ингаляцию был известный поэт Сэмуель Тейлор Колеридж. Дэви начал
вдыхать из шелкового мешка, содержащего 20 кварт закиси азота (22.8 л). Вот
его описание:
"Почти немедленно началось дрожание, идущее от груди к конечностям. Я испы-
тывал ощущение осязательного напряжения, в высшей степени приятного в каждом
члене. Мои зрительные впечатления были ослепительны и казались великолепными.
Я отчетливо слышал каждый звук в комнате и был прекрасно ориентирован в про-
исходившем. Постепенно, по мере того, как приятные ощущения нарастали, я по-
терял связь с внешним миром. Потоки зрительных образов быстро пробегали в мо-
ем сознании и так сочетались со словами, что производили совершенно новые об-
разы. Я пребывал в мире идей, заново измененных и причудливо сочетавшихся. Я
строил теории и делал открытия. Когда я был разбужен из этого полубредового
транса доктором Кинглэком, который отнял мешок от моего рта, то негодование и
гордость были первыми чувствами от присутствия около меня другого человека.
Мои эмоции были возвышены, и я испытывал энтузиазм; я ходил по комнате около
минуты, совершенно не считаясь с тем, что говорили вокруг меня. Когда я вер-
нулся к прежнему состоянию рассудка, я испытывал потребность сообщить об от-
крытии, которое я сделал во время эксперимента. Я старался вернуть свои виде-
ния, но они были слабы и неотчетливы. Однако сумма данных представлялась сама
собой, и я заявил доктору Кинглэку с самой полной уверенностью и пророческим
образом: в мире нет ничего, кроме понятий; вселенная состоит из впечатлений,
идей, удовольствий и страданий"1.
Опыт продолжался три с половиной минуты, и было израсходовано не более по-
ловины газа. Потом Дэви вдохнул оставшуюся половину газа.
"Подобные ощущения были снова произведены; я был быстро брошен в радостный
транс, и продолжал в нем находиться дольше, чем прежде. В течение многих ми-
нут после эксперимента я испытал волнения (трепет) в конечностях, взволнован-
ность продолжалась почти два часа".
Опыты стали ставить шире. Слухи и рассказы привлекли множество больных в
"Пневматический институт", главным образом, страдающих астмой. Многие из них
после ингаляций считали себя совершенно излеченными и "заново родившимися".
Среди описаний экспериментаторов проводивших вдыхание почти все описывают
необычные и трудно описуемые приятные ощущения. Газ вдыхали из шелковых меш-
ков в количествах 7-9 л. Закись азота нередко получалась непригодной для ды-
хания из-за аэрозоля кислоты. Интересующихся описаниями сеансов вдыхания и
историей исследования веселящего раза отсылаем к книге "Исследования химиче-
ские и философские, касающиеся главным образом закиси азота, или дефлогисти-
рованного воздуха и его вдыхания"2. К сожалению, книга не переводилась на
русский язык.
10 декабря 1844 г. В этот день в небольшом американском городке Хартфорде
(штат Коннектикут) произошло довольно знаменательное событие. 27-летний дан-
тист Хорас Уэллс (Horace Wells, 1815-1848) просматривая местную газету
"Hartford Courant", заинтересовался объявлением, в котором извещалось, что
приезжий химик доктор Кольтон сегодня вечером проведет так называемый дивер-
тисмент с показом опытов "с веселящим газом".
Следует отметить, что в Великобритании и США первой половины 19-го века об-
щественные демонстрации ингаляций закиси азота не были уже редкостью. Так же
как и ether frolics (эфирные шалости), эти своеобразные развлечения довольно
широко распространились не только в больших, но даже и в маленьких городках
США. Программа подобных "дивертисментов" странствующих лекторов и артистов
обычно представляла собой смесь развлечений, фокусов и научно-популярных лек-
ций по темам от электричества до химии с демонстрацией различных опытов поч-
тенной публике. После того, как в 1799-1800 гг. стали известны эксперименты,
проводившиеся в "Пневматическом институте" Томаса Беддо, и многократные де-
монстрации Г. Дэви возбуждающего эффекта закиси азота на вечеринках с друзья-
ми, на протяжении последующих нескольких десятилетий многочисленные лекторы
стали включать в свои программы и демонстрацию эффекта вдыхания закиси азота,
проводимую на добровольцах из публики. Любопытно, что и известный изобрета-
Известный (в широких кругах) английский математик и философ Бертран Рассел в книге
"История Западной Философии" пишет: "Уильям Джеймс описывает человека, который испы-
тал действие веселящего газа; всякий раз, когда он находился под воздействием этого
газа, он знал тайну Вселенной, но когда приходил в себя, то забывал ее. Наконец ему
удалось путем огромного усилия записать эту тайну до того, как видение исчезло. Со-
вершенно очнувшись, он бросился посмотреть то, что записал. Это было: "Повсюду пах-
нет нефтью".
2 Humphry Davy. Researches, Chemical and Philosophical; Chiely Concerning Nitrous
Oxide or Dephlogisticated Nitrous Air and its Respiration. (1800) .
тель 6-зарядного револьвера полковник Самуель Кольт, впоследствии владелец
крупной оружейной фабрики в том же Хартфорде, в тридцатых годах 19-го столе-
тия был антрепренером подобных шоу.
Однако одним из лучших демонстраторов США. считался вышеупомянутый Гарднер
Квинси Кольтон (Colton, Gardner Quincy, 1814-1898).
После двух лет изучения медицины в Нью-Йорке Кольтон бросил медицинскую
школу в 1844 г. (или был изгнан из нее), послав по его собственным словам ме-
дицину к "чертям собачьим". Он начал карьеру публичного лектора, разъезжая по
городам США. с лекциями по химии, натурфилософии и телеграфу. Его "дивертис-
менты" представляли собой массовые красочные зрелища. Программа часто включа-
ла в себя выставку "Страшного суда", большого полотна Рембрандта Пила
(Rembrandt Peale) , размерами 13 футов в высоту и 24 фута в длину. В дополне-
ние своих научно-популярных лекций Кольтон демонстрировал публике работу те-
леграфа и эффекты, вызываемые ингаляцией закиси азота ("веселящего газа").
Вот текст того любопытного объявления, которое заинтересовало Хораса Уэллса
и побудило его пригласить свою жену пойти вечером с ним вместе на представле-
ние Кольтона:
"Большая демонстрация эффекта, вызываемого ингаляцией закиси азота, веселя-
щего или смехотворного газа, будет дана в Юнион Холл сегодня (вторник) вече-
ром, 10 декабря 1844 г.
Сорок галлонов газа будут приготовлены и отпущены всем желающим вдыхать его
в аудитории. Двенадцать молодых людей уже выразили желание быть добровольцами
для ингаляции газа в начале представления.
Восемь крепких мужчин приглашены занять передние места, чтобы защитить на-
ходящихся под действием газа от повреждений самих себя или других. Эти меры
предпринимаются, дабы не возникло никакой опасности. Вероятно, никто не сде-
лает попыток драки.
Газ вызывает у тех, кто вдыхает его, либо смех, пение, танцы, разговорчи-
вость или буйство и т.д. и т.п. в соответствии с главной чертой характера.
Но, кажется, что все сохраняют сознание настолько, чтобы не высказать или не
сделать того, о чем они после пожалели бы.
Газ будет отпускаться только джентльменам из лучшего общества. Задача раз-
влечения - сделать его занятием, приятным во всех отношениях.
Мистер Кольтон дал два аналогичных представления прошлой весной в Таверне
Бродвея в Нью-Йорке. Оба спектакля привлекли свыше четырех тысяч дам и джент-
льменов, полный отчет о чем можно найти в New Mirror от 6 апреля, написанный
Н.П. Уиллисом. Прибывши в Хартфорд, мистер Кольтон дает данное представление
по настоятельной просьбе друзей. Его желание и намерение - заслужить и приоб-
рести покровительство и расположение публики первого класса. Он верит, что
они будут смеяться больше, чем смеялись за последние шесть месяцев. Развлече-
ние это является научным для тех, кто желает его сделать таковым. Те, кто
вдыхал газ один раз, боятся привыкнуть к нему, но таких примеров еще не было.
Невозможно описать словами восхитительное ощущение, вызываемое ингаляцией.
Поэт Роберт Санти (Robert Sontey) как-то сказал, что "атмосфера высшего из
всех возможных небес должна состоять именно из подобного газа".
Полное описание действия, оказанного на некоторых замечательных людей Евро-
пы, можно узнать в Хуперовском медицинском словаре под заголовком "Азот".
Мистер Кольтон будет вдыхать газ первым. История и свойства газа будут из-
ложены в начале.
Мистер Кольтон даст частное представление для дам, которые пожелают вдыхать
газ во вторник, между 12 и 13 часами бесплатно. Никто, кроме дам, не допуска-
ется. Это делается для тех, кто хочет вдыхать газ без свидетелей.
Представление начнется в 7 часов. Билеты по 25 центов продаются в Главном
книжном магазине и у входа 10 декабря".
Вечером 10 декабря Хорас Уэллс и его жена направились в Юнион Холл. Во вре-
мя представления на соседнем месте с Х.Уэллсом оказался молодой человек по
имени Самуэль А. Кулей (Samuel A. Cooley), приказчик из большой хартфордской
аптеки. По роду своей профессии дантиста Уэллс часто имел дело с этой аптекой
и лично знал Самуэля, которого называл просто Сэмом.
Колтон прочитал короткую вступительную лекцию и затем началась демонстрация
ингаляции закиси азота. Газ подавали в резиновых мешках. Вдыхали его через
грубые деревянные мундштуки, подобные тем, через которые в плохих деревенских
трактирах сосали сидр из бочонков. Кольтон первым принял ингаляцию веселящего
раза и заявил о восхитительных ощущениях, которые он испытывает.
В числе любопытных, пожелавших попробовать на себе действие закиси азота,
вышел на сцену и Кулей. Как только Кольтон дал ему вдохнуть несколько глотков
газа, Кулей опьянел. Он подпрыгивал, приседал, танцевал по сцене, а затем,
вообразив присутствие какого-то невидимого врага, стал энергично размахивать
в воздухе руками, нападать и отбиваться. Когда наркозное опьянение кончилось,
Кулей очнулся и старался понять, где он и, что с ним. Кулей сел на свое место
рядом с Уэллсом, и в то время, как на сцене проводили ингаляции следующим
добровольцам, он почувствовал боль в голени. Засучив штанину, он к удивлению
увидел зияющую и кровоточащую, ушибленную рану ниже колена. Он никак не мог
разгадать, в чем дело, если бы Уэллс не разъяснил ему, что, будучи в наркоз-
ном возбуждении, он перепрыгнул через скамьи и, споткнувшись, очевидно, силь-
но ударился ногой. Было ясно, что для столь значительной ссадины нужен был
весьма сильный удар, который, тем не менее, прошел совершенно нечувствитель-
ным для аптекаря.
Сеансы ингаляции веселящего газа на сцене продолжались. Публика со смехом
следила за поведением и выходками каждой новой добровольной жертвы экспери-
ментов . Один лишь Хорас Уэллс сидел молчаливый и задумчивый. В голове его все
яснее и яснее созревал вывод, суливший грандиозные перспективы и славу.
"Скажите, Сэм, неужели вы совершенно не почувствовали боли при столь силь-
ном ушибе ноги?" - спросил он. "Абсолютно нет", - ответил Кулей.
Таким образом, благодаря представлению Гарднера Квинси Кольтона, дантист
Хорас Уэллс пришел к мысли о возможности уменьшить боль при экстракции зубов
с помощью ингаляций закиси азота. Тем самым он вновь вернулся к идеям Гемфри
Дэви, высказанным еще четыре десятилетия назад.
Рано утром он отправился в гостиницу, в которой остановился гастролирующий
лектор и демонстратор Гарднер Квинси Кольтон, чтобы попросить у него некото-
рое количество закиси азота. Уэллс смог убедить Кольтона согласиться, пообе-
щав ему долю барышей из того, что, несомненно, должно было стать весьма при-
быльным делом.
Получив согласие, Уэллс решил испробовать обезболивающее действие закиси
азота прежде всего на самом себе, и обратился к другому хартфордскому данти-
сту, Джону Риггсу (John M. Riggs) с просьбой, чтобы тот удалил у него один
здоровый зуб.
Все собрались в приемной Риггса в тот же день, 11 декабря, в послеобеденное
время. Кольтон принес газ и сам лично сделал ингаляцию большой дозы Уэллсу, а
Риггс, воспользовавшись хорошим наркозом, вырвал у коллеги один из здоровых
моляров. Уэллс вскоре очнулся и с крайним энтузиазмом воскликнул: «Наступила
новая эра в экстракции зубов!». Он уверял всех присутствующих, что не почув-
ствовал ни малейшей боли, и что в процессе самой ингаляции он испытывал заме-
чательно приятные ощущения.
Это событие стало первым использованием закиси азота для обезболивания в
стоматологии.
Дальнейшая судьба Хораса Уэллса сложилась трагически, но мы на этом оста-
навливаться не будем.
Религиозное, а в более общем случае - мистическое мировоззрение апеллирует
к иррациональному. С другой стороны, закись азота (равно как и некоторые дру-
гие наркотические вещества) направляют сознание человека по пути, который
расходится с нашей грешной реальностью. Не удивительно, что нашлось немало
философов-мистиков, которые восприняли субъективные ощущения, вызванные опья-
нением закисью азота, эфиром и другими наркотиками, как проявление неких объ-
ективных истин, недоступных человеку в здравом рассудке. Ярким примером таких
мыслителей был Уильям Джеймс (Джемс)3.
Уильям Джеймс [William James] (11 января 1842, Нью-Йорк - 26 августа 1910,
Чокоруа, шт. Нью-Хэмпшир), американский философ и психолог, основатель праг-
матизма .
Начав свое образование с изучения живописи в Ньюпорте, Джеймс поступил за-
тем в Научную школу Лоренса (Lawrence Scientific School) Гарвардского универ-
ситета, которую двумя годами позже сменил на Медицинскую школу. Продолжил
обучение в Германии, где штудировал медицину, физиологию и естественные науки
под руководством Гельмгольца и Вирхова. По возвращении в Гарвард Джеймс полу-
чил в 1868 степень доктора медицины. После этого он пережил тяжелый душевный
кризис, который преодолел только в 1870 благодаря увлечению философией Ш. Ре-
нувье, утверждавшего миропонимание свободы и творчества. Памяти Ренувье
Джеймс посвятил написанную тогда же рукопись "Некоторые проблемы философии"
(опубликована она была только после смерти автора). В 1871 при участии Джейм-
са создается Метафизический клуб, объединивший Ч. Пирса, Ч. Райта, Дж. Росса,
Ф. Эббота и других математиков, естествоиспытателей, юристов, теологов.
Джеймс очень высоко оценил доклады Пирса, учеником и последователем которого
он стал. После того как в 1873 Джеймса приглашают занять должность профессора
Гарвардского университета, он начинает регулярно вести занятия - сначала по
анатомии и физиологии, затем по психологии и, наконец, по философии.
Фундаментальный двухтомный труд "Принципы психологии" (1890) приносит ему
известность как крупному психологу, а сборник философских работ "Воля к вере"
(1897) создает ему репутацию влиятельного мыслителя. Благодаря своим лекциям,
пользующимся неизменной популярностью, Джеймс часто получает приглашения из
различных университетов США. и Европы. Его сочинения, написанные в начале ве-
ка, - "Многообразие религиозного опыта" (1902), "Прагматизм" (1907), "Плюра-
листический универсум" (1909) - провозглашают прагматизм особым философским
направлением.
Уильям Джеймс описывает свои личные переживания, связанные со случаями из-
менения сознания. "Одно из очарований опьянения, - пишет он, - бесспорно, за-
ключается в глубинном чувстве реальности и истине, которая достигается через
него. В каком бы свете ни представали перед нами вещи, они видятся безогово-
рочными, "совершенно совершенными", что не наблюдается, когда мы трезвые". В
своей книге "Многообразие религиозного опыта" он добавляет:
"Эфир и в особенности окись азота, в известной дозе примешанные к воздуху,
являются также могучими стимулами к пробуждению мистического сознания. Перед
вдыхающим их, точно разверзаются бездны истины одна за другою. Когда человек
приходит в нормальное состояние, истина от него ускользает, и если остается
3 Подход Джеймса и других философов-мистиков, которые искали истину в закиси азота,
является вполне логичным. Если наука стремится к знаниям объективным (т.е. независи-
мым от личности исследователя), то философия изучает отношение человек - мир. Други-
ми словами, субъективное начало принципиально неотделимо от философии. Оказалось,
что субъективные переживания человека под действием наркотических веществ так же хо-
рошо пригодны для построения новых философских концепций (и обоснования старых), как
и умозаключения философов, сделанные в здравом рассудке, - главное только не забы-
вать , на чем эти концепции основаны. - Прим. ред.
от нее какая-нибудь формула, для нормального рассудка она оказывается бес-
смыслицей. Тем не менее, у человека остается чувство, что эта формула полна
глубокого значения. Я лично знаю многих людей, которые убеждены, что в тран-
се, вызванном окисью азота, возможны настоящие метафизические откровения.
Несколько лет тому назад я сделал сообщение в печати о произведенных лично
надо мною опытах опьянения окисью азота. На основании их я пришел тогда к та-
кому выводу, - который и до настоящего времени не поколебался в моих глазах,
- что наше нормальное или, как мы его называем, разумное сознание представля-
ет лишь одну из форм сознания, причем другие, совершенно от него отличные
формы, существуют рядом с ним, отделенные от него лишь тонкой перегородкой.
Мы можем совершить наш жизненный путь, даже не подозревая об их существова-
нии; но как только будет применен необходимый для их пробуждения стимул, они
сразу оживут для нас, представляя готовые и определенные формы духовной жиз-
ни, которые, быть может, имеют где-нибудь свою область применения. Наше пред-
ставление о мире не может быть законченным, если мы не примем во внимание и
эти формы сознания. Из них, правда, нельзя вывести точной формулы и они не
могут дать нам плана той новой области, какую они перед нами раскрывают, но
несомненно, что они должны помешать слишком поспешным заключениям о пределах
реального. Возвращаясь к моим собственным переживаниям этого рода, я должен
признать, что они сводятся к особому просветленному состоянию, которому я не
могу не придать мистической окраски. Основной чертой такого состояния всегда
является примиренность, словно две противоположные стороны мира, столкновения
между которыми составляет причину всех наших внутренних бурь и неурядиц, рас-
плавились и образовали единое целое. Они не принадлежат к одному роду, как
два различных вида, но один из видов, - более возвышенный, - сам становится
родом по отношению к противоположному виду и растворяет его в себе. Я знаю,
что эта мысль с логической стороны темна, но я не могу избавиться от ее влия-
ния на меня. Я чувствую, что в ней есть смысл, соприкасающийся с сущностью
гегелевской философии. Имеющий уши, да слышит. Для меня эта мысль постижима
лишь этим путем искусственно разбуженного мистического состояния сознания.
У меня есть друзья, которые верят в откровение, обусловленное наркозом. Для
них оно также метафизическая интуиция, в которой мир в своих многообразных
проявлениях воспринимается как бы растворившимся в Едином.
"Мы погружаемся в этот повсюду разлитой Дух, - пишет один из них, - всеце-
ло, безраздельно, в полном самозабвении. Все перестает тогда существовать для
нас, все, что выше и что ниже нас, все где бы то ни было сущее. Остается лишь
жизнь и мы, в нее погруженные. Существует только единое, множественное теряет
свою множественность, исчезает; и каждый из нас есть этот Единый существую-
щий... Это предельная черта. И насколько достоверно то наше существование, ко-
торое составляет обычный источник наших забот, настолько же достоверна и ра-
дость, царящая над всяким дуализмом, над всеми антитезами, какой я достиг в
моем уединении, равном божескому"."
Весьма поэтично и точно Джеймс описывает наступление в процессе интоксика-
ции своеобразного состояния мистического откровения, а также его исчезание в
процессе выделения растворенной закиси азота из крови:
"Для меня, как и для всех известных мне людей, главным в подобных экспери-
ментах бывает наступление просветленности сознания. Истина лежит глубже оче-
видности ослепительных видений и необычных ощущений. Мозгу внезапно и с такой
ясностью открываются сущность явлений и все логические связи бытия, что нор-
мальное сознание не дает этому аналогий. Только по мере возвращения трезвости
тает способность ясновидения, глаза становятся пустыми, а в мозгу мелькают
какие-то обрывки фраз и слов. Такое чувство, как будто смотришь на снежный
пик, помертвевший, потому что на нем только что погас закат, или на пепел,
оставшийся от еще недавно раскаленных головешек".
Наши дни
Увлечение веселящим газом - до недавнего времени практически забытое - ныне
вновь возродилось. Предприимчивые владельцы клубов поняли, что могут хорошо
зарабатывать на продаже шаров с веселящим газом и привлечь много посетителей,
тем более что в перечень наркотических препаратов веселящий газ не входит4.
На многих сайтах с помощью примитивной рекламы пытаются спекулировать этим
веществом.
Получила распространение продажа через интернет восьмиграммовых баллончиков
со сжиженной пищевой закисью азота, предназначенной для взбивания сливок в
сифоне. В комплект входит простейшее устройство пробойник с трубкой для выхо-
да газа, на которую для сбора газа надевают шар. Кроме высокой цены такой
способ употребления представляется крайне неудобным.
Другими словами, закись азота - новое лекарство от избытка денег.
Мы советуем не поддаваться на подобную рекламу и не выручать "барыг", тем
более что чистую закись азота при желании легко получить в домашних условиях.
Примитивное устройство на основе восьмиграммовых баллончиков за-
киси азота.
Учась на 5 курсе университета, я решил испытать, как будут вести себя зна-
комые и друзья, вдыхая закись азота. В общежитии была собрана установка, ве-
селящий газ я получал порциями по 3-9 литров несколько раз.
Добровольцы вызвались не сразу, несмотря на все мои заверения в чистоте ра-
за и подробному рассказу о его свойствах. Первую порцию в 3 литра я вдохнул
сам продемонстрировав безопасность опыта. Оказалось, что внешне действие газа
выглядит не так как описано в некоторых источниках. Люди не совершали идиот-
ские поступки, вполне сохраняя контроль и связь с внешним миром. Остается га-
дать, как можно надышавшись газа, совершать какие-либо действия. Может тогда
имело место самовнушение или какие-то другие причины. Опишу свои опыты. За-
кись азота, несмотря на кажущуюся безопасность, может привести к печальным
последствиям, если ее использовать неправильно. Как любой относительно инерт-
ный газ закись азота не поддерживает дыхания. Это означает, что дышать ей
можно только короткое время, периодически вдыхая воздух, иначе начнется ги-
поксия (кислородное голодание). Анестезиологи всегда примешивают к наркозным
газам кислород. Однако опыт прошлого и мой собственный показывает, что можно
обходиться чистой закисью азота. Также недопустимо совместное употребление
спиртного или других наркотиков, это может создать эффект усиления действия
каждого из веществ и привести к печальным последствиям! Используются мешки
Газ входит в число одурманивающих веществ, утверждённых Правительством Российской
Федерации 4 августа 2021 года.
емкостью три литра или резиновые подушки, вроде "кислородной". Вначале нужно
выбрать удобное место, где можно было бы присесть в покое от суеты мира, на-
полненные мешки осторожно кладут вблизи. Хорошо если за опытом наблюдает
друг-ассистент, однако при соблюдении осторожности можно обойтись и без по-
сторонней помощи. "Экспериментатор" должен медленно и глубоко вдохнуть и вы-
дохнуть воздух несколько раз, чтобы насытить организм кислородом. Затем он
берет, мешок обхватывает губами трубку клапана и, открыв правой рукой его и
одновременно держа мешок, делает несколько вдохов-выдохов полной грудью. Дей-
ствие начинается со второго вдоха и обычно на четвертом достигает максимума.
В этот момент желательно выдохнуть обратно в мешок, тогда можно сэкономить
часть газа. Однако мешок обычно выпадает из рук и человек находится в "тран-
се" и "вихре вибраций" около 15 секунд. Порции в 3-л обычно хватает на цикл
из 4-6 вдохов-выдохов полной грудью. Что чувствует "экспериментатор" описать
можно лишь в самых общих чертах. Действие газа может сильно меняться для раз-
ных людей. Один мой знакомый после вдыхания 6 литров закиси азота почувство-
вал лишь легкое опьянение, сравнимое с алкогольным.
Закись азота: вся земная истина находится на табуретке.
После вдоха газа и нескольких последующих вдохов-выдохов (на это уходит
около 3 литров) быстро наступает то самое состояние, когда доступна "глубина
всей земной истины" по выражению Джеймса. В моей интерпретации ощущений зем-
ная истина это восприятие мира (точнее совокупности окружающих предметов в
момент эксперимента) в состоянии глубоко эйфорической вибрации. Это воспри-
ятие крайне трудно описать, т.к. в нашем повседневном языке нет слов соответ-
ствующих переживаниям. Похоже на нирвану в представление буддистов, все за-
ключено в этом ощущении: полная самодостаточность. Понятное дело нет никакого
желания что-либо делать, кроме созерцания и бытия в этом состоянии (поэтому
непонятны описания, где что-либо делают) были мысли о фиксировании этого от-
кровения , но даже в глубине транса я знал, что это бессмысленно. Разумеется,
это не идет ни в какое сравнение с опьянением алкоголем, разве что потом со-
стояние последействия напоминает легкое опьянение. Глубоко эйфорический вихрь
вибрации это нечто невыразимо прекрасное и великое. Состояние этого транса
длится около 15 секунд, потом быстро слабеет.
Резиновой подушки веселящего раза хватило на пять таких метафизических от-
кровений .
После воспоминания постепенно несколько блекнут, но основные моменты сохра-
няются в памяти.
После 2-го вдоха появляется и нарастает чувство радости, и появляются
странные вибрации всего тела (внешне не заметные) и в такт им - звуковые. За-
кись азота, похоже, позволяет "настроится в резонанс на частоту колебаний
Вселенной"5. Можно так же отметить, что звуки слышатся доходящими как бы "со
стороны", хотя это нельзя точно объяснить. Цвета кажутся более яркими. Появ-
ляется ощущение "правильного" осознания всего, мимолетное прикосновение "Не-
что", кажущегося истинным и великим. "Нечто" не может быть выражено в словес-
ной форме6, это особое ощущение, новое экстрасенсорно-мистическое восприятие
мира.
Затем следует сделать несколько вдохов-выдохов воздуха и только потом можно
вдохнуть следующую порцию закиси азота. Вдыхание веселящего газа должно быть
непродолжительным и чередоваться с вдохами воздуха, как видно из описания!
Это очень важно. Непрерывное вдыхание закиси азота в течение продолжительного
времени приводит к гипоксии, а затем смерти вследствие нее. Это происходило и
происходит с недалекими токсикоманами, применяющими надетые на голову пакеты,
плотно прилегающие маски или использующими большую концентрацию газа в замк-
нутых пространствах. Хотя, возможно, автор излишне заостряет внимание читате-
ля, образованному человеку это ясно как дважды два. Здесь видно удобство при-
менения мешков для вина. Порция газа небольшая, но достаточная, не дает раз-
виться гипоксии и удобный клапан, автоматически закрывающийся, если мешок вы-
падет из пальцев. Удобна также резиновая подушка с трубкой снабженной зажимом
Мора. После того как газ прекратили вдыхать действие его быстро ослабевает.
Примерно 40 минут - 1 час длится состояние легкого отрешения от действитель-
ности, похожее на действие пива. При этом происходит накатывание волн ощуще-
ний - периодическое усиление-ослабление приятного опьянения. Все это время
автор наблюдал парестезии, т.е. чувство "мурашек", легкого "онемения" кожи,
небольшое нарушение координации точных движений пальцев. Это все видимые по-
бочные действия острого отравления закисью азота.
Следует помнить, что, несмотря на малую токсичность, при регулярном зло-
употреблении закись азота так же опасна для здоровья, как и другие наркотиче-
ские вещества. В настоящее время механизм действия закиси азота ясен не пол-
ностью, а ее вредное влияние на организм исследовано недостаточно.
При использовании закиси азота в небольших количествах действительно нет
видимых побочных последствий для организма. Можно утверждать, что закись азо-
та значительно менее опасна для человека, чем алкоголь. Но это совсем не оз-
начает , что негативных последствий употребления веселящего газа нет.
Не следует полагаться на распространенное (особенно в рекламе веселящего
газа) мнение о полной безопасности закиси азота и совершенном отсутствии его
вредного влияния на организм. История медицины показала ошибочность кажущейся
первоначально безопасности наркотических веществ/лекарств.
В частности использование веселящего газа приводит к расходованию витамина
Bi2, что может повлечь неврологические проблемы. Ранние признаки включают по-
калывания в пальцах и конечностях в течение длительного времени. Люди с дефи-
цитом витамина В12 наиболее подвержены побочным последствиям действия закиси
5 Это не гипотеза, а лишь шутка. Хотя автор и не уверен наверняка, так ли это.
6 Хотя ради курьеза, пожалуй, стоит попытаться записать несколько своих мыслей. Од-
нако намного лучше предварительно включить диктофон и наговорить, т.к. записать что-
либо во время действия газа очень сложно, нарушена координация движений.
азота.
Ранее считали, что закись азота практически не оказывает токсического дей-
ствия, но последние научные данные позволили усомниться в этом утверждении.
Закись азота взаимодействует с кобальтом, который входит в состав витамина
Bi2. В результате ингибируется синтез метионина, тимидина, тетрагидрофолата и
ДНК. Закись азота напрямую ингибирует метионинсинтетазу, стимулирует образо-
вание мегалобластов в костном мозге, а при длительном воздействии может при-
вести к угнетению функции костного мозга с развитием агранулоцитоза. Длитель-
ная ингаляция закиси азота может вызвать хроническую инактивацию цианокобала-
мина и метионинсинтетазы, что может привести к анемии. Особенно опасно приме-
нение закиси азота для больных с гиповитаминозом. В этих случаях необходимо
применение цианокобаламина и фолиевой кислоты.
В результате длительной ингаляции закиси азота может развиться миелонейро-
патия, которую характеризуют прогрессирующей демиелинизацией задних рогов
спинного мозга с характерной симптоматикой.
Экспериментально обнаружен тератогенный эффект закиси азота, и хотя он не
доказан у людей, тем не менее, закись азота не следует использовать в первом
триместре беременности.
Мы не стали касаться опасности примесей оксидов азота, что могут присутст-
вовать в газе при недостаточной очистке, которой всегда следует уделять пер-
востепенное внимание. Интересующиеся могут почитать про действие оксидов азо-
та на организм в справочнике7.
Об опасности гипоксии было сказано выше8.
Как мы уже говорили, закись азота используется в пищевой промышленности, в
частности, в качестве пропеллента (газ, создающий давление в аэрозольных бал-
лончиках) . Для подобных целей обычно используют более дешевый углекислый
газ9, но это не всегда возможно: углекислый газ обладает кислотными свойства-
ми. Являясь "двойником" углекислого газа по физическим свойствам, закись азо-
та кислотными свойствами не обладает. Видимо, этим и объясняется то, что в
баллончиках "взбитых сливок" используется закись азота (сливки могут испор-
титься от действия кислоты - даже такой слабой, как угольная).
Но действительно ли в баллончиках взбитых сливок, которые продают в магази-
нах, содержатся существенные количества закиси азота? Как мы неоднократно
убедились, верить отечественным производителям и продавцам нельзя.
Я решил проверить распространенный миф о веселящем газе из взбитых сливок.
7 Филов В.А. (ред.) Вредные химические вещества.
8 Думаю, стоит добавить пару слов по поводу гипоксии. Человек может прожить без еды
около месяца, без воды - несколько дней, а без кислорода - всего несколько минут.
Лишившись подачи кислорода, клетки головного мозга начинают необратимо умирать через
две с половиной - три минуты. Не удивительно, что эволюция выработала у человека бо-
язнь нехватки воздуха, например, попав под воду, мы инстинктивно пытаемся всплыть.
Но всего эволюция предвидеть не могла: если человек вдыхает вместо воздуха газ,
не обладающий запахом, вкусом и раздражающим действием, в первые секунды он практи-
чески ничего не чувствует. Вы спокойно продолжаете "дышать", не ощущая никакой раз-
ницы. И только через полминуты - минуту начинает ощущаться гипоксия (нехватка кисло-
рода) . Если эксперимент продолжить, вы можете потерять сознание и умереть, так и не
осознав в чем дело. В случае с закисью азота, ситуация осложняется опьянением.
Не так давно ушел из жизни мой знакомый биохимик, которому не было и сорока. Он
работал в области геронтологии (наука о продлении жизни). По неофициальной версии
причиной смерти стал эксперимент по дыханию смесью с низким содержанием кислорода.
Химик был опытным экспериментатором и отлично осознавал опасность, но как видите,
это не помогло. - Прим. ред.
9 Для непищевых целей в качестве пропеллента также применяют пропан-бутановую смесь,
иногда - фреоны.
Для чего купил самый дешевый баллончик "легких" сливок. Дома медленно страв-
ливая раз (через ловушку из маленькой колбы Вюрца, с раствором поваренной со-
ли, как счетчик пузырьков и предохранительную склянку) набрал ровно один ме-
шок (3 л) закиси азота. После этого давление в баллончике упало, и он стал
продавливаться пальцами. Газ при вдыхании произвел надлежащий эффект.
Вывод: из одного баллончика взбитых сливок можно получить закись азота в
количестве, которого хватит на один эксперимент по его вдыханию.
В крайнем случае, таким способом получить небольшие количества веселящего
газа можно, но это неоправданно дорого. Не стоит субсидировать жуликов от
маркетинга.
Получение
закиси азота
Получить веселящий газ можно несколькими путями. Самым доступным в домашних
условиях является способ Г. Дэви - термическое разложение нитрата аммония
(аммиачной селитры) по реакции:
NH4N03 = N20 + 2H20.
В лабораторных условиях более удобным является нагревание сульфаминовой ки-
слоты с азотной кислотой (73%):
NH2S02OH + HN03 = N20 + H2S04 + H20
однако сульфаминовую и азотную кислоты достать труднее, поэтому мы остано-
вимся на разложении аммиачной селитры. Кстати, разложение аммиачной селитры
используется для синтеза закиси азота в промышленных масштабах.
При нагревании нитрата аммония имеет место несколько реакций. Приведем
фрагменты из книги Л.И. Багал Химия и технология инициирующих взрывчатых ве-
ществ (1975):
Нитрат аммония при нагревании несколько выше температуры плавления (сухой
нитрат аммония плавится при 169.6 С), разлагается по реакции
NH4N03 <=> NH3 + HNO3 (1)
<. . .>
Реакция разложения на закись азота и воду изучалась Вертело, Томсеном и Бе-
леем. Первые два исследователя нашли, что реакция экзотермична
NH4NO3 => N20 + 2H20 +8.8 ккал (2)
Основными реакциями разложения нитрата аммония при температуре до 270 С яв-
ляются (1) и (2). Расплавленный нитрат аммония при нагревании выше 250-260 С
может выделять окислы азота, азот и воду:
NH4N03 => 0.5N2 + NO + 2Н20
4NH4N03 => 3N2 + N204 + 8H20
3NH4N03 => 2N2 + N203 + 6H20
Саундерс (1922 г.), основываясь на результатах газового анализа, пришел к
убеждению, что основными реакциями разложения при температуре до 260 С явля-
ются (1) и (2), а также реакция
5NH3 + 3HN03 => 4N2 + 9H20
Разложение при взрыве по его мнению протекает по реакции
8NH4N03 => 16Н20 + 2N02 + 4N0 + 5N2
<. . .>
Для нормального процесса образования закиси азота путем разложения нитрата
аммония его температурный режим и степень чистоты имеют исключительное значе-
ния.
Как видно из приведенных выше данных, аммиачная селитра при нагревании до
240-250 С разлагается с образованием закиси азота и воды, однако даже при
этой температуре полученный "сырой" газ содержит пары азотной кислоты, оксиды
азота N0 и N02, аммиак, хлор (за счет примесей хлоридов) , азот и "туман" во-
зогнанной аммиачной селитры. Понятно, что такую смесь вдыхать нельзя (если
возникнет мысль повторить опыты Дэви), поскольку это смертельно опасно! Мало
того, если колба закрыта резиновой пробкой, то она даже после кратковременно-
го использования постепенно разрушается (с образованием совсем не безвредных
продуктов).
Поэтому способ получения веселящего газа нагреванием аммиачной селитры на
сковородке (который зачастую рекомендуют "гуру", чтобы посмеяться над "профа-
нами") выглядит в лучшем случае как черный юмор.
Перейдем к рассмотрению установки.
Установка для получения закиси азота (веселящего газа) .
Нитрат аммония разлагают в колбе Вюрца при осторожном нагревании. Лучше ис-
пользовать термометр, однако без него при необходимости можно обойтись. Как
показал опыт лучше использовать нагрев примерно до 220 С, при этом наблюдает-
ся легкое "кипение" расплава. Полученный "сырой газ" для очистки пропускают
вначале через охлаждаемую льдом ловушку, для сбора перероняющейся воды с при-
месью азотной кислоты. Далее раз проходит через склянку Дрекселя с раствором
железного купороса, она также служит своеобразным индикатором скорости выде-
ления газа. Затем газ промывается в импровизированной промывалке (с пористым
распылителем) с раствором 5-7 % щелочи (гидроксида натрия или калия), где он
очищается от N02, азотной кислоты, хлора. И, наконец, в третьей промывалке с
пористым распылителем, в которую налит раствор сульфата железа (II), закись
азота очищается от N0 и следов оставшихся примесей. После нее газ содержит
закись азота с некоторым количеством воды и азота, а также следами N02 и N0.
Следует помнить, что очистке закиси азота, если она будет использована для
повтора опытов Дзви, следует уделить особое внимание, в противном случае газ
будет токсичным.
В качестве реакционной загрузки использовалось удобрение нитрат аммония
(аммиачная селитра).
Теперь рассмотрим цикл получения газа. Мы использовали термостойкую колбу
Вюрца емкостью 500 мл. Однако возможно применение колб меньшей емкости, что
отразится на длительности процесса. Так как по колбе в процессе реакции сте-
кают капли воды, может произойти растрескивание стекла, поэтому рекомендуется
термостойкое стекло. Отвод колбы обмотан влажным бинтом для дополнительного
охлаждения "сырого газа", периодически его осторожно смачивают водой из пи-
петки. Следите, чтобы капли воды не попали на колбу! Колбу загружают селитрой
на три четверти. Вначале осторожно нагревают селитру до плавления, не присое-
диняя отвод с резиновой трубкой. Нагревать можно над электроплиткой, спиртов-
кой, газовой конфоркой. Периодически ее помешивают стеклянной палочкой. (Не
рекомендуем использовать для нагрева таблетки сухого горючего, издающие при
длительном горении крайне неприятный запах. После сжигания трех таблеток при-
шлось проветривать помещение.) Плавление длится долго, постепенно объем за-
грузки уменьшается можно досыпать немного селитры.
Нитрат аммония (аммиачная селитра) в колбе Вюрца.
Когда селитра расплавилась и начала слегка "кипеть" осторожно присоединяем
резиновый шланг, придерживая рукой конец стеклянного отвода. Надевают резино-
вую трубку, слегка смочив водой стеклянный отвод, натягивать нужно примерно
на 7-8 мм. Можно заранее присоединить резиновую трубку к отводу колбы, но то-
гда нельзя закрывать пробкой с термометром горло коблы до начала стабильного
выделения газа.
Теперь началось выделение веселящего газа. Вначале нужно либо подождать по-
ка он вытеснит воздух из системы, либо сразу присоединить мешок для сбора га-
за (так можно примерно увидеть объем газа, который выделился). После заполне-
ния мешка наполовину, присоединяем пустой мешок. Нагрев регулируем так, чтобы
температура была около 220 С. Это позволит получить более чистый газ. При на-
гревании вначале наблюдается более сильное "кипение" селитры, т.к. испаряется
примесь влаги. Необходимо следить за прохождением газа через первую промывал-
ку - должна выходить непрерывная цепочка пузырьков. Примерно за 20 минут мож-
но получить три литра газа. Опыт показал, что держать нужный температурный
интервал очень легко. При снижении температуры можно ненадолго подогреть га-
зовой горелкой боковую часть колбы. Однако оставлять прибор без присмотра на
длительное время (более 5 мин.) нельзя!!! Помните - при охлаждении скорость
выделения закиси азота уменьшается и через некоторое время воду из промывалки
засосет в ловушку, а затем и в колбу. Это вызовет ее разрушение со всеми не-
приятными последствиями! Возможен и противоположный исход, когда от перегрева
произойдет неконтролируемое ускорение реакции. Поэтому, понаблюдав за процес-
сом, отрегулируйте степень нагрева колбы.
Разложение аммиачной селитры.
Используемая плитка (мощность спирали 600 Вт) оказалось неспособной вызвать
перегрев, несколько раз приходилось слегка подогревать колбу газовой горел-
кой, чтобы температура держалась возле 220 С.
Выделяющая вода конденсируется на горле колбы и периодически по стенкам
стекает в расплав, вызывая непродолжительный ускоренный выход газа. Чтобы из-
бежать этого (и возможного разрушения колбы) некоторые руководства рекоменду-
ют боковой обогрев горла колбы или ее теплоизоляцию, однако это кажется из-
лишним. Постепенно мешок заполняется газом. Когда он почти заполнился и слег-
ка упруго продавливается, его необходимо отсоединить и поставить другой. Де-
лают это так - вначале выдергивают проволочные кольца, клапан закрывается.
Затем быстро отсоединяют широкий шланг от горла мешка. Берем пустой мешок,
выдавливаем воздух, открыв пальцами клапан, или просто втягиваем воздух ртом.
Фиксируем "крылышки" клапана в открытом положении палочкой от мороженого и
двумя проволочными кольцами из медного электропровода. Далее надеваем резино-
вый шланг подачи газа. Цикл повторяется.
Также закисью азота (за неимением достаточного количества мешков для вина)
была наполнена резиновая подушка. Для длительного хранения газа она непригод-
на. Однако удобна при кратковременном хранении.
Мешок для вина с закисью азота.
Когда собран и отложен последний мешок необходимо отсоединить шланг от от-
вода колбы Вюрца. Автор просто его срезал, т.к. снять его проблематично, мож-
но сломать отвод. Колбу с расплавом лучше сразу вынести на открытый воздух,
она должна остывать на штативе, ничего не касаясь, до полного охлаждения!
Пробку с термометром необходимо вынуть. Можно закрыть горло пробкой (без тер-
мометра !) . Промывалки нельзя оставлять с растворами. Их разбирают, растворы
переливают в стеклянные бутылки. Следует помнить, что щелочь разрушает поли-
этилентерефталат, поэтому раствор ее лучше хранить в полиэтиленовых или поли-
пропиленовых бутылках (в крайнем случае - стеклянных). Все трубки тщательно
промывают проточной водой, аквариумные "распылители" необходимо после промыв-
ки выдержать в разных стаканах с водой, для окончательного удаления следов
растворов. Распылитель, бывший в растворе щелочи, при мытье раскрошился в ру-
ках. Поэтому лучше заранее купить несколько запасных.
В описанной выше версии прибора было решено использовать термометр и допол-
нительную ловушку для кислого конденсата (служащую также предохранительной
склянкой от случайного засасывания раствора). Возможен и более простой вари-
ант - без термометра и ловушки, - однако с ним работать менее удобно, и нужна
большая осторожность.
Ловушку мы сделали из маленькой колбы Вюрца, однако можно с успехом изгото-
вить ее из аптечной склянки на 100 мл, с винтовой резьбой под пластиковую
крышку.
* !
Колба Вюрца (ловушка).
Достаточно просверлить два отверстия, как показано ниже, и вставить длинную
(почти до дна) и короткую стеклянные трубки. Ловушка охлаждается в ледяной
бане, для чего удобнее использовать металлический широкогорлый термос, напол-
ненный колотым льдом с водой. Загрузка селитры (для 500 мл колбы) около 850 г
- почти полный пакет удобрения.
Изготовление промывалок не представляет труда - нужны лишь аквариумные рас-
пылители подходящего диаметра (меньше диаметра горла бутылки) и стеклянные
трубки (можно использовать подходящие по диаметру пипетки Мора). Необходимо
сделать 3 отрезка трубки 3-4 см длиной, диаметр примерно 7 мм, и 3 куска
трубки 25-30 см (подбирается по длине бутылки)10 со слегка оттянутым концом.
Для этого трубку, вращая, нагревают в пламени горелки (удобны корейские го-
релки, надевающиеся на баллончик с бутаном). Когда пламя окрасится в желтый
цвет и трубка начнет размягчаться, ее непрерывно вращая, совсем слегка растя-
гивают . После остывания посередине оттянутого пространства трубку разрезают.
Стеклянные трубки при необходимости можно использовать составные - срастив подхо-
дящие отрезки кусочком резиновой трубки.
Для этого краем тонкого, "бархатного" напильника слегка наносят черту на 1/6
- 1/3 окружности трубки и разламывают, одновременно, как бы стараясь разо-
рвать трубку в разные стороны. Оттянутый конец трубки вставляют в крышку пла-
стиковой бутылки, отверстие должно быть чуть меньше диаметра трубки, тогда
система совершенно герметична. Окончательный размер длинных трубок подбирает-
ся так, чтобы распылитель при закручивании пробки был у самого дна бутылки.
Собранную систему необходимо проверить на герметичность, смочив снаружи проб-
ку мыльным раствором и каким-либо способом создать давление газа. Аккуратно
сделанные промывалки даже под значительным давлением совершенно герметичны.
Затем в прибор необходимо залить воды по уровню начала горлышка бутылок (в
первую можно на две трети) и попытаться ртом продуть систему, с переднего
конца трубки подачи "сырого" газа. С некоторым усилием это должно удаться.
Промывалка (крышка).
Для сбора установки использовались: литровая промывалка (оснащенная порис-
тым распылителем) на основе пластиковой пивной бутылки с сульфатом железа,
стеклянная промывалка из водочной бутылки (раствор щелочи, тоже с пористым
распылителем) и самодельная склянка Дрекселя с раствором железного купороса
из полулитровой пластиковой бутылки.
Раствор щелочи готовим осторожно, помня о крайней опасности ее для глаз и
кожи. Применялся 7 % раствор едкого кали. Раствор железного купороса готовим
насыщенным, воду слегка подкисляем серной кислотой. Нами был куплен сульфат
железа фирмы Фаско (фото выше), железный купорос был влажным, однако он дос-
таточно чистый и растворяется без остатка.
Установка для получения закиси азота (веселящего газа). Колба с
селитрой не подсоединена, маленькая колба Вюрца (справа) играет
роль ловушки.
Химичка
НЕКОТОРЫЕ МЕТОДЫ ОРГАНИЧЕСКОЙ ХИМИИ
(продолжение)
МЕТАМФЕТАМИН
Синтез P2NP
EtlM02
RNH,
NO:
Конденсация бензальдегида и нитроэтана.
Помещаем в колбу 150 мл циклогексамина.
Добавляем в колбу 750 мл бензальдегида...
...порциями пока не пропадет запах амина.
Перемешиваем.
Добавляем в колбу порциями 300 мл ледяной уксусной кислоты (ЛУК).
Если смесь будет разогреваться - охлаждаем колбу проточной водой или
же добавляем ЛУК более медленно. При перемешивании может появится
белый туман - так что делаем все под вытяжкой.
Добавляем в колбу 750 мл нитроэтана.
Перемешиваем.
На колбу устанавливаем обратный холодильник. Смесь нагреваем на
водяной бане при 50-62 С примерно 6 часов. Цвет смеси будет ме-
няться от светло-желтого до темно-желтого.
Удаляем холодильник и перемешиваем.
Переливаем в стакан для охлаждения и кристаллизации. Если смесь смесь слиш-
ком густая, то можно ее разбавить изопропиловым спиртом (ИПС).
Если кристаллизация начнется сразу - значит продукт чистый.
Стакан закрываем пленкой с резинкой и перемешиваем.
Рост кристаллов P2NP.
После охлаждения и полной кристаллизации P2NP отделяем вакуумной
фильтрацией.
Жидкость переливаем в другой сосуд, а кристаллы P2NP промываем на фильтре
холодным ИПС. P2NP сушим при комнатной температуре день или больше. Выход
1021 г.
К отлитой жидкости добавляем воды пока не начнут выделятся до-
полнительные кристаллы.
Смесь охлаждаем несколько часов. Кристаллы отфильтровываем. Их отделяем как
и ранее и сушим где-то неделю. Это дает еще 80 г P2NP.
Общий выход 1101 г (67% от теоретического).
P2NP хранить в запечатанном пакете в сухом темном месте при 2-8 С (в холо-
дильнике) .
Метамфетамин через
реакцию Лейкарта
Начнем с установки.
Потребуется 2-х горлая колба на магнитной мешалке с подогревом.
В одном горле термометр.
Во втором горле обратный холодильник.
Заливаем в колбу 5.4 мл Р2Р.
Добавляем 13.4 мл N-метилформамида.
Термометр на место и нагреваем до 60 С. Перемешивание на миниму-
ме. Затем греем при 60-80 С более чем 3-4 часа.
После 4 часов медленно увеличиваем температуру нагрева до 110 С.
При 110 С держим 2 часа.
Затем температуру увеличиваем до 155 С и держим еще 24 часа.
Нагрев отключаем. Цвет потемнел.
Приготовляем ЮМ раствор NaOH (24 мл Н20 + 9.8 г NaOH)
Заливаем его в колбу. Держим 2.5 часа.
Потом переливаем содержимое колбы в делительную воронку.
Нижний слой сливаем.
Верхний слой - в колбу.
Добавляем в колбу 10.7 мл НС1 (более чем 32%)
Ставим обратный холодильник. Греем 2 часа при 250 С нагрева и
перемешивании.
Приготовляем 8. ЗМ раствор NaOH (16 мл Н20 + 5.3 г NaOH) . Добавля-
ем его в колбу по каплям.
Перемешиваем и переливаем в делительную воронку.
Нижний слой сливаем.
Берем 20-30 мл толуола и заливаем его в колбу и перемешиваем.
Затем переливаем в делительную воронку. Повторяем это 3 раза.
Из делительной воронки все в стакан. Ополаскиваем делительную
воронку небольшим количеством толуола и тоже в стакан.
В стакан добавляем понемногу немного сухого (безводного) MgS04 и
перемешиваем.
Далее вакуумная фильтрация.
Фильтрат переливаем в колбочку и используем для выделения метам-
фетамина гидрохлорида.
Электроника
ПРИМЕНЕНИЕ RASPBERRY PI PICO
Буркхард Каинка
(продолжение)
10. Регулируемый
источник
напряжения
Для испытаний электронных компонентов часто требуется точно регулируемое
напряжение. Здесь выход ШИМ Pico используется вместе с фильтром нижних частот
в качестве источника напряжения. Напряжение можно постепенно увеличивать или
уменьшать с помощью двух кнопок.
//Pico_ADpwm
#include "pico/stdlib.h"
#include "hardware/adc.h"
#include "hardware/pwm.h"
void setup() {
int ad=0;
int iir=0;
int pwm=0;
int n, d;
Serial.begin(115200) ;
adc_init() ;
adc_gpio_init (26) ;
adc_select_input (0) ;
gpio_set_function(0, GPI0_FUNC_PWM) ;
pwm_set_wrap(0, 4095);
pwm_set_gpio_level (0 , 0) ;
pwm_set_enabled(0, true);
gpio_pull_up(4) ;
gpio_pull_up(8) ;
while(true){
for (n=0; n<50; n++) {
ad = adc_read();
iir = (iir*9+ad)/10;
sleep_ms(10);
}
Serial.print(pwm*3300/4095) ;
Serial.print(" mVn);
Serial.print(iir*3300/4095-5) ;
Serial.println(" mVff);
if (gpio_get(4)==0)
pwm-=d;
if (pwm<0)
pwm=0 ;
if (gpio_get(8)==0)
pwm+=d;
pwm_set__gpio_level (0 r pwm) ;
d++;
if ((gpio_get(4)==l) & (gpio_get(8)==1))
d=l;
}
5V 4V7 GND EN 3V3 REF P28 GND P27 P26 RUN P22 GND P21
40 A2 AGND A1 АО 30
LED ф*
W*
RP2040
BOOT
1
10
P0 P1 GND P2 P3 P4 P5 GND P6 P7 P8 P9 GND P10
10k
i
100p
0. 3,3V
Поскольку АЦП имеет разрешение 12 бит, выход ШИМ также установлен в диапа-
зоне 0...4095. В результате частота ШИМ составляет 125 МГц/4096 = 30,5 кГц.
Таким образом, сигнал можно очень хорошо сгладить с помощью RC фильтра нижних
частот с сопротивлением 10 кОм и 100 мкФ.
Две кнопки начинают с одиночных шагов, которые затем постепенно увеличива-
ются, так что можно было удобно устанавливать еще большие изменения напряже-
ния. Отпустив кнопки, вы вернетесь к одному шагу.
Последовательный монитор показывает заданное напряжение и измеренное напря-
жение . Тест показывает ошибку смещения около 5 мВ в нижнем конце шкалы. После
изменения напряжения проходит около двух секунд, пока не будет достигнуто из-
меренное конечное значение. Это связано, с одной стороны, с постоянной време-
ни RC-фильтра, равной одной секунде, а с другой стороны, со встроенным БИХ-
фильтром, который должен предотвращать колебания показаний.
О С0М13
ГТ7 ИГУ
0 mV
0 mV
2 mV
4 mV
8 mV
12 mV
16 mV
22 mV
29 mV
36 mV
36 mV
36 mV
36 mV
0 Autoscrol
-o—nrv
6 mV
5 mV
6 mV
7 mV
8 mV
9 mV
13 mV
17 mV
22 mV
29 mV
33 mV
35 mV
36 mV
1 1 Zeitstempel anzeigen
.
Zeiienumbruch (CR)
—
~\ _. _ .
v 115200 Baud ~
□ X
Senden
A
V
Ausgabe loschen
Сглаженный выходной сигнал имеет очень высокое сопротивление — 10 кОм, по-
этому напряжение падает даже при небольших нагрузках. Это может быть преиму-
ществом, когда необходимо исследовать чувствительные компоненты. Например, вы
можете определить, при каком напряжении начинает загораться тот или иной све-
тодиод. В других случаях желателен более высокий выходной ток. Следующий бу-
ферный усилитель с LM358 обеспечивает ток до 40 мА. Это можно даже использо-
вать для привода небольших электродвигателей. Таким образом, управление ШИМ
становится контролем скорости.
В других случаях можно обойтись без сглаживания и управлять силовым драйве-
ром напрямую ШИМ-сигналом. Часто достаточно одного транзистора. Затем сигнал
ШИМ можно использовать, например, для управления лампой накаливания или дви-
гателем. При использовании индуктивных нагрузок необходимо использовать об-
ратноходовой диод для подавления пиков индуктивного напряжения.
+5V
10k
PWM
1/2 LM358
100м
i 1
I
• 0. 3,3V
6V/100mA
PWM
♦ +5V
11. Регистратор
характеристических
кривых
Характеристические кривые представляют собой диаграммы U/I, описывающие по-
ведение компонента. Напряжение на компоненте увеличивают и изменяют ток. Вы-
ход ШИМ с фильтром нижних частот снова используется для постоянного напряже-
ния, которое автоматически проходит через него. Напряжение на компоненте из-
меряется напрямую. Ток получается из разницы между только что установленным
напряжением холостого хода ШИМ и измеренным напряжением.
Чтобы кривая была плавной, измеренный сигнал проходит через БИХ-фильтр
(IIR-фильтр). Ток может достигать почти 3,3 мА с целью с очень низким сопро-
тивлением, но в других случаях остается намного меньшим. Для сравнения прове-
дена вспомогательная линия на уровне 1 мА.
//Pico_ADpwm2 Characteristic curve
#include "pico/stdlib.h"
#include "hardware/adc.h"
#include "hardware/pwm.h"
void setup() {
int ad=0;
int iir=0;
int pwm=0;
int n, u, u2, i, i2, j, t;
Serial.begin(115200);
adc_init() ;
adc_gpio_init (26) ;
adc_select_input (0) ;
gpio_set_function(0, GPIO_FUNC_PWM) ;
pwm_set_wrap(0, 4095);
pwm_set_gpio_level (0 , 0) ;
pwm_set_enabled(0, true);
gpio_pull_up(4) ;
gpio_pull_up(8) ;
sleep_ms(1000) ;
while(true) {
pwm_set_gpio_level (0 , 0) ;
sleep_ms(1000) ;
u2=0; t=0;
iir = 5;
for(j=0;j<4096;j++) {
pwm_set_gpio_level (0 , j) ;
sleep_ms(2);
for (n=0; n<5; n++) {
ad = adc_read();
iir = (iir*3+ad)/4;
sleep_us(500);
}
u=iir*3300/4095;
if(u>2500)
u=2500;
if (u<2500)
i=G-iir)*3300/4095;
if(i<0)
i=0;
u=u/5;
if(u>u2) {
Serial.print(1000);
Serial.print (" ");
Serial, println(i);
t++;
}
if(u>u2)
u2=u;
}
pwm_set_gpio_level (0 , 0) ;
for(j=t;j<500;j++) {
Serial.print(1000) ;
Serial.print (" ") ;
Serial.println(i) ;
sleep_ms(5);
}
}
void loop() {}
I I I
5V 4V7 GND
40
USB
1
1 СП 11 ;,#..
LtU \]/'/t
1
P0 P1 GND
к
-J
J EZ
l 1
L
I—
i i
EN 3V3
BOOT
P2 P3
1 1
100p
I
REF
P4
I
I I I
P28 GND P27
A2 AGND A1
P5 GND P6
I
*2
I
P26 RUN P22
АО 30
RP2040
10
P7 P8 P9
I I I
^ Testpbjekt
По оси Х напряжение достигает 2,5 В, так что одно деление шкалы последова-
тельного плоттера соответствует 0,5 В. Первое измерение показывает кривую ре-
зистора сопротивлением 10 кОм на холостом ходу. Видно, что ток возрастает ли-
нейно и достигает 250 мкА при 2,5 В. Небольшие скачкообразные отклонения от
линейности на диаграмме обусловлены конечной точностью АЦП.
О СОМ13
1000.0 "
750.0 -
500.0 "
250.0 "
■ ■
°'0о
115200 Baud ^
100
Senden
:оо
300
□ X
—
400 500
Zeilenumbruch (CR) ^
Измерение на резисторе сопротивлением 1 кОм также показывает линейное уве-
личение , при этом ток около 1 мА измеряется при напряжении 1 В. Характеристи-
ческая кривая расширяется примерно до 1,65 В и 1,65 мА, поскольку здесь дели-
тель напряжения с двумя одинаковыми резисторами достигает максимума 3,3 В /
2. Горизонтальный участок больше не принадлежит самой характеристической кри-
вой, а служит для заполнения серийного плоттера 500 измеренными значениями.
Следующее измерение снова точно вписывается в окно.
© С0М13
2000.0 -
1500.0 -
1000.0 "
500.0 -
■■
0.0 1
0
115200 Baud <* |
100
Senden
200
300
- □ х
400 500
Zeilenumbruch (CR) ^
Характеристическая кривая диода 1N4148 показывает экспоненциальный рост то-
ка , который в этом масштабе начинает расти примерно с 0,4 В от нулевой линии.
Резкий подъем подтверждает впечатление, что прямое напряжение диода находится
примерно в диапазоне от 0,6 до 0,7 В. Однако это вопрос масштаба. На схеме
уже можно увидеть ток порядка 10 мкА примерно от 0,4 В.
© С0М13
3200.0-
2 400.0 -
1600.0-
800.0 -
[ ■■
1
□ X
J
0.0 1
0
115200 Baud v
100
Senden
i
200
300
400 500
Zeilenumbruch (CR) v
Характеристическая кривая красного светодиода поразительно похожа на кривую
кремниевого диода, хотя подъем сдвинут в сторону значительно более высоких
напряжений. Едва заметный ток течет от величины напряжения ок. 1,6 В. При то-
ке 1 мА прямое напряжение составляет ок. 1,8В.
О С0М13
1600.0 -
1200.0 "
800.0 "
400.0 -
, ■■
°-°о
115200 Baud v
100
Senden
1
200
300
□ X
!
1
400 500
Zeilenumbruch (CR) v
Еще более высокие напряжения до прибл. 2,4В индицируются зеленым светодио-
дом.
О С0М13
1000.0 "
750.0 "
500.0 "
250.0 "
■ ■
о.о -1
0
115200 Baud ^
100
Senden
"" Т" " ' ' с
200
300
□ X
1
/
J
400 500
Zeilenumbruch (CR) v
Для исследования N-канального МОП-транзистора BS170 были соединены затвор и
сток. Характеристическая кривая менее крутая, чем характеристика диода. При
напряжении затвора 2 В ток стока около 1 мА.
По характеристической кривой можно оценить, что BS170 уже можно хорошо
управлять с помощью управляющего напряжения 3,3 В.
© С0М13
1600.0 -
1200.0 -
зоо.о -
400.0 -
^ ""
и.и -1
0
115200 Baud v |
100
Senden
—i
200
300
□ X
/
/
400 500
Zeilenumbruch (CR) v»
12. Простой
осциллограф
Благодаря более быстрым измерениям и последовательному плоттеру можно реа-
лизовать простой осциллограф. Здесь снова использовалась синхронизация с мик-
росекундным таймером, чтобы остальные задержки не складывались в установлен-
ный интервал в одну миллисекунду.
т
10п
Oszi
1k
5V 4V7 GND EN 3V3 REF P28 GND P27 P26 RUN P22 GND
40 A2 AGND A1 АО 30
//Pico_AD3
#include "pico/stdlib.h"
#include "hardware/adc.hf
void setup() {
uint32_t ad=0;
uint32 t iir=0;
Serial.begin(115200) ;
adc_init() ;
adc_gpio_init (26) ;
//gpio_pull_up (26) ;
adc_select_input (0) ;
uint32_t t = time_us_32 () ;
while(true){
ad = adc_read 0*3300/4095;
iir = (iir*3+ad)/4;
Serial.println(iir);
t+=1000;
while (t>time_us_32() ) ;
}
}
void loop() { }
Кроме того, для получения более плавных осциллограмм был включен простой
БИХ-фильтр (IIR-фильтр) нижних частот. Для каждого измерения только 25 % пе-
ременной iir определяется текущим измерением, а 75 % — предыдущими измерения-
ми. Поведение этого фильтра соответствует поведению RC-фильтра нижних частот.
Этот результат измерения был получен путем прикосновения пальца к измери-
тельному входу, в результате чего был измерен сигнал частотой 50 Гц. Кроме
того, к GND был подключен конденсатор емкостью 10 нФ. Вместе с программным
lR-фильтром это подавляет высокочастотные помехи.
При правильно установленном размере окна плоттера вы получаете пять частей
шкалы по оси X, каждая из которых представляет 100 точек измерения, т.е. в
данном случае 100 мс. Поскольку период сетевого напряжения составляет 20 мс,
в каждом сегменте шкалы вы видите ровно 5 колебаний.
13. Измерение освещенности
с помощью светодиода
Аналоговый вход Pico имеет чрезвычайно высокий импеданс. Это делает возмож-
ными измерения, которые невозможны с помощью обычных измерительных приборов с
внутренним сопротивлением 10 МОм. В качестве фотодиода можно использовать
обычный светодиод. В зависимости от цвета он выдает напряжение до 1,5 В и бо-
лее , хотя максимальный ток находится лишь в диапазоне наноампер.
1
5V
40
1 1
4V7 GND
1
EN
1
3V3
1
REF
ill
4
I
10n
I
>
I
P28 GND P27 P26 RUN
A2 AGND A1 АО 30
Для измерения использовался желтый светодиод. Программа осциллографа
Pico_AD3 работала, замедлилась до периода выборки 100 мс.
t+=l 00000;
while (t>time us 32());
Измерения проводились при нормальном комнатном освещении, когда светодиод
был направлен на окно, улавливая естественный свет в пасмурную погоду. Начало
измерения показывает рост прибл. 2 В/с. Отсюда при емкости 10 нФ можно полу-
чить фототок светодиода 20 нА. Напряжение возрастает примерно до 1,2 В и
представляет собой логарифмическую меру освещенности. Колебания яркости, за-
фиксированные в среднем диапазоне, были вызваны частичными тенями, создавае-
мыми человеком, ходящим по комнате.
14. Усилитель
класса D
Усилитель класса D отличается высоким КПД и низкими искажениями. Выходной
каскад состоит из цифрового мостового усилителя. В состоянии ожидания оба вы-
хода генерируют один и тот же сигнал ШИМ, поэтому между двумя соединениями
громкоговорителя нет разницы потенциалов. При модуляции длины импульсов сдви-
гаются относительно друг друга.
Такой усилитель можно реализовать с помощью RPi Pico, если не требуется бо-
лее высокая выходная мощность. Мостовой усилитель формируется из двух выходов
ШИМ, принадлежащих одному блоку ШИМ, т.е. они находятся в фазе.
Сигнал AF поступает на аналоговый вход АО, при этом потенциал покоя уста-
навливается на половину диапазона измерения с помощью делителя напряжения.
Измеренный сигнал напрямую управляет шириной импульса РО и инвертированной
шириной импульса Р1. Громкоговоритель должен иметь относительно высокое со-
противление, предпочтительно 50 Ом. Для первоначальных экспериментов также
можно использовать громкоговоритель сопротивлением 8 Ом с последовательным
резистором сопротивлением 47 Ом. Как и порты, выходы ШИМ работают с настрой-
кой 4 мА и, следовательно, выдают максимум 30 мА. К сожалению, изменить диа-
пазоны тока в режиме ШИМ невозможно. При использовании громкоговорителя со-
противлением 50 Ом выходная мощность достигает 20 мВт.
Разрешение измерения и сигнала ШИМ установлено на 10 бит, т.е. в диапазоне
от 0 до 1023. Это обеспечивает высокую частоту дискретизации 122 кГц (125
МГц/1024 = 122 кГц). Делитель напряжения с двойным сопротивлением 1 кОм уста-
навливает центральное напряжение. Последовательный резистор сопротивлением 1
кОм на входе АО предназначен для предотвращения перегрузки входа в случае
возможной перегрузки.
1 1 1
5V 4V7 GND
40
USB
1 СП
1
Р0 Р1 GND
I
50
I
I
EN 3V3
BOOT
Р2 РЗ
I
R
I
г4
REF
Р4
I
1k
ь
1к
■*—
1
т
Р28 GND
А2 AGND
РЪ GND
I I
i
Р27
А1
Р6
i
Tiii
Р26 RUN P22 GND
АО 30
RP2040
10
Р7 Р8 Р
9 GNO
1 1 1 1
-•Line in
l l l l l
Р21
Р10
I
Р20 Р19 Р18 GND
RPi Pico
P11 P12 P13 GND
I I I I
I I
P17 P16
21
20
P14 P1S
I I
/\
//Pico_PWM5 Class D amplifier
#include "pico/stdlib.h"
#include "hardware/pwm.h"
#include "hardware/adc.h"
void setup() { }
void loop() { }
void pwm_int() {
pwm_clear_irq(0) ;
int u =adc_read() /4;
pwm_set_gpio_level (0 , u) ;
pwm_set_gpio_level (1, 1023-u);
}
void setup 1() {
adc_init() ;
adc_gpio_init (26) ;
adc_select_input (0) ;
gpio_set_function(0, GPI0_FUNC_PWM) ;
gpio_set_f unction (1, GPIO_FUNC_PWM) ;
pwm_clear_irq(0) ;
pwm_set_irq_enabled(0, true);
irq_set_exclusive_handler (PWM_IRQ_WRAP, pwm_int) ;
irq_set_enabled(PWM_IRQ_WRAP, true) ;
pwm_set_wrap(0, 1023);
pwm_set_gpio_level (0 , 512) ;
pwm_set_enabled(0 л true);
}
void loop 1() {
}
В программе используется функция прерывания, которая вызывается каждый раз
при переполнении счетчика ШИМ. Таким образом, дискретизация входного сигнала
АЦП и обновление ширины импульса ШИМ происходит синхронно с частотой ШИМ 122
кГц. Чтобы гарантировать, что плавный процесс не будет нарушен другими преры-
ваниями, все выполняется во втором ядре процессора.
На осциллограмме виден общий нарастающий фронт обоих выходов и слегка сме-
щенные спадающие фронты. Амплитуда импульса соответствует полному рабочему
напряжению. В области спадающего фронта ток, текущий через громкоговоритель,
можно распознать по наклонам падения напряжения на выходных внутренних рези-
сторах, вызванному индуктивностью звуковой катушки.
Усилитель обеспечивает хороший звук и удобное воспроизведение музыки при
средней громкости в помещении. Вторая осциллограмма показывает усиленный сиг-
нал 34 на выходе ФНЧ сопротивлением 1 кОм и 100 нФ.
Использование прерывания не всегда приводит к абсолютно ровному времени.
Если другие прерывания активны, они должны разделить время. Решение этой и
других критичных по времени проблем заключается в использовании второго ядра
процессора в RPi Pico. Для этого вам необходимо включить заголовочный файл
pico/multicore.h. Функции setup() и 1оор() тогда принадлежат первому ядру
(СогеО), функции setupl () и loopl () принадлежат Corel, т.е. второму ядру.
15. Частотомер
от 1 Гц до 1 МГц
Простой частотомер может наблюдать за состоянием порта и подсчитывать все
положительные фронты в течение определенного временного окна в одну секунду.
В первом эксперименте в качестве источника сигнала используется выход ШИМ с
симметричным прямоугольным сигналом частотой 1 МГц. Выход ШИМ — это контакт
Р0, который также используется в качестве цифрового входа для частотомера.
Таким образом, никакая внешняя схема не требуется. Проверенный микросекундный
таймер снова используется для измерения времени стробирования.
LED Ф', —^~
Т BOOT
1
Р0 Р1 GND P2 РЗ
1—гпгп—г
• 1MHz
//Pico_Freql
#include "pico/stdlib.h"
#include "hardware/pwm.h"
void setup() {
Serial.begin(115200) ;
gpio_set_function(0, GPIO_FUNC_PWM) ;
pwm_set_wrap(0, 124); //PWM l MHz, 50%
pwm_set_gpio_level (0 , 63) ;
//pwm_set_wrap(0, 1249); //PWM 100 kHz, 50%
//pwm_set_gpio_level (0 , 639) ;
pwm_set_enabled(0, true);
while (true) {
uint32_tf=0 ;
bool in_old = 0;
while (!gpio_get(0));
while (gpio_get(0) ) ;
uint32_t t = time_us_32() + 1000000;
while (t>time_us_32 () ) { // 1 MHz measure
bool in_new =(gpio_get(0));
if (in_new>in_old)
f++;
in_old=in_new ;
}
Serial, print(f);
Serial.println(" Hz");
}
}
void loop() {
}
Последовательный монитор показывает слегка колеблющийся результат — 990,021
МГц. Это означает, что отображаемая частота занижена примерно на 1 %.
990021 Hz
990021 Hz
990020 Hz
Если выход ШИМ установлен на 100 кГц, сохраняется погрешность измерения 1
%. Уже при 10 кГц ошибка исчезает полностью.
10000 Hz
10000 Hz
10000 Hz
Наблюдения показывают, что на пико идет другой процесс, который съедает
столько времени, что 1% положительных фронтов считывается на более высоких
частотах. Задержки должны быть менее длины одного импульса сигнала частотой
10 кГц, т.е. менее 50 не.
Подозрение падает на интерфейс USB, который должен работать постоянно.
Решение проблемы снова заключается в использовании второго ядра процессора
в RPi Pico. Виртуальный последовательный интерфейс инициализируется в первом
ядре. Фактическое измерение частоты перенесено на второе ядро. Доступ к ин-
терфейсу с Serial.print есть, но передача данных происходит в СогеО (в первом
ядре).
//Pico_Freq2
#include "pico/stdlib.h"
#include "hardware/pwm.h"
#include "pico/multicore.h"
void setup() { //CoreO
Serial.begin(115200) ;
}
void loop() {}
void setupl () { //Core 1
gpio_set_function(0, GPIO_FUNC_PWM) ;
pwm_set_wrap(0, 124); //PWM l MHz, 50%
pwm_set_gpio_level (0 , 63) ;
pwm_set_enabled(0, true);
while (true) {
uint32_tf=0 ;
bool in_old = 0;
while (!gpio_get(0));
while (gpio_get(0) ) ;
uint32_t t = time_us_32() + 1000000;
while (t>time_us_32 () ) { // 1 MHz measure
bool in_new =(gpio_get(0));
if (in_new>in_old)
f++;
in_old=in_new ;
}
Serial, print(f);
Serial.println(" Hz");
}
}
void loopl () {}
Результат радует: при ШИМ-сигнале 1 МГц отображается ровно 1000000 Гц.
Обычные частотомеры показывают колебания в последнем десятичном знаке, по-
скольку окно измерения не начинается в фазе с входным сигналом. Однако здесь
перед началом измерения ожидается положительный фронт входного сигнала. Это
синхронизирует начало, в результате чего дисплей становится постоянным.
1000000 Hz
1000000 Hz
1000000 Hz
16. Частотомер
от 1 кГц до 65 МГц
Для более высоких частот запроса состояний порта уже недостаточно, поэтому
необходим аппаратный счетчик. Здесь блок ШИМ 2 (срез 2) используется как 16-
битный счетчик. Канал В также может считать внешние часы вместо системных ча-
сов . Теперь он подключен к Р5.
С помощью pwm_get_default_conf ig () вы получаете структуру по умолчанию со
всеми настройками модуля ШИМ. С помощью config_set_clkdiv_mode настройка из-
меняется таким образом, что внешние импульсы подсчитываются на входе В по на-
растающим фронтам. Эта конфигурация затем назначается модулю ШИМ 2 с помощью
pwm_init, где параметр false указывает, что выход ШИМ не требуется. Вместо
этого на этот раз показания счетчика считываются с помощью pwm_get_counter.
//Pico_Freq3
#include "pico/stdlib.h"
#include "hardware/pwm.h"
#include "pico/multicore.h"
void setup() {
Serial.begin(115200);
}
void loop() {}
void setup 1() {
gpio_set_function(0, GPIO_FUNC_PWM) ;
pwm_set_wrap(0, 1); //PWM 62.5 MHz
pwm_set_gpio_level (0 , 1) ;
pwm_set_enabled(0, true);
while (true) {
uint32_tf=0 ;
pwm_config cfg = pwm_get_default_config();
pwm_conf ig_set_clkdiv_mode (&cf g, PWM_DIV_B_RISING) ;
pwm_init(2, &cfg, false);
gpio_set_f unction (5, GPIO_FUNC_PWM) ;
uint32_t t = time_us_32 ()+2;
while (t>time_us_32 () ) ;
pwm_set_enabled(2r true);
t+=1000;
while (t>time_us_32() ) ;
pwm_set_enabled(2r false);
f= pwm_get_counter(2) ;
Serial, print(f);
Serial.printing kHz");
}
}
void loopl() {
}
Опять же, выход ШИМ на порту GP0 используется в качестве источника сигнала.
Здесь генерируется максимально возможная частота ШИМ 62,5 МГц. В качестве ба-
зы времени используется микросекундный таймер. Чтобы гарантировать, что изме-
рение начнется точно в начале микросекунды, заранее ожидается начало второй
микросекунды. Еще через 1000 мкс измерение заканчивается. Результат измерения
подтверждает частоту измерения 62500 кГц с обычными колебаниями на последней
цифре.
62500 kHz
62499 kHz
62501 kHz
GP5 теперь можно использовать в качестве входа для универсального счетчика.
Для защиты входа от отрицательного или слишком высокого напряжения сигнала
резистор и конденсатор следует соединить последовательно.
17. Частотомер от
1 Гц до 100 МГц
Счетчик ШИМ имеет ширину 16 бит и поэтому может считать только до 65535.
Для еще большего разрешения необходимо обнаруживать переполнение счетчика.
Здесь для этой цели используется прерывание ШИМ. При каждом переполнении уве-
личивается старшее слово f__hi. Время стробирования теперь можно увеличить до
1 000 000 мкс. На этот раз Rpi Pico разогнан до 200 МГц. Следовательно, на
вывод Р0 можно вывести тестовый сигнал частотой 100 МГц.
LED
#
веют
1
Р0 Р1 GND Р2 РЗ Р4 Р5 GND
//Pico_Freq4
#include "pico/stdlib.h"
#include "hardware/pwm.h"
#include "pico/multicore.h"
uint32 t f hi;
void setup() {
set_sys_clock_khz(200000, true); Serial.begin(115200);
}
void loop() {}
void pwm_int() {
void pwm mi. v / t
pwm_clear_irq(2) ; f_hi++;
}
void setupl() {
uint32_tf=0 ;
gpio_set_function(0, GPIO_FUNC_PWM) ;
pwm_set_wrap(0, 1); //PWM 62.5 MHz
pwm_set_gpio_level (0 , 1) ;
pwm_set_enabled(0, true);
while (true) {
pwm_config cfg = pwm_get_default_config();
pwm_conf ig_set_clkdiv_mode (&cf g , PWM_DIV_B_RISING) ;
pwm_init(2, &cfg, false);
gpio_set_function(5, GPIO_FUNC_PWM) ;
pwm_set_irq_enabled(2, true);
irq_set_exclusive_handler (PWM_IRQ_WRAP, pwm_int) ;
irq_set_enabled(PWM_IRQ_WRAP, true) ;
f_hi=0 ;
uint32_t t = time_us_32 ()+2;
while (t>time_us_32 () ) ;
pwm_set_enabled(2, true); t+= 1000000;
while (t>time_us_32() ) ;
pwm_set_enabled(2r false);
f = pwm_get_counter(2);
f+=f_hi«16;
Serial, print(f);
Serial.println(" Hz");
}
}
void loopl() {
}
При прямом соединении Р0 и Р5 тестовая частота 100 МГц измеряется с высокой
точностью. Однако абсолютная точность ниже, поскольку зависит от допуска
кварцевого генератора в пико. Если доступен подходящий эталонный кварц, счет-
чик можно откалибровать, установив время стробирования 1 000 000 мкс.
99999998 Hz
1000000 Hz
99999997 Hz
18. Генератор сигналов
от 7 Гц до 50 МГц
Этот генератор прямоугольных импульсов использует выход ШИМ на выводе Р0.
Частота устанавливается по команде с последовательного монитора в Гц или в
кГц. С помощью flOO выбирается частота 100 Гц. F100 же дает 100 кГц. Выходная
частота получается путем деления частоты системы на целочисленный делитель.
Чтобы обеспечить плавные частоты, такие как 10 МГц или 50 МГц, частота систе-
мы устанавливается на 100 МГц.
Наименьшая частота выхода ШИМ раньше составляла 125 МГц/65536 = 1,907 кГц,
а теперь будет 1,525 кГц. Однако еще есть неиспользованный ранее прескалер
шириной 8 бит. В целях плавных делителей при необходимости устанавливается
коэффициент делителя 10, 100 или 250. Таким образом можно снизить частоту до
7 Гц. Максимальная частота составляет 50 МГц с коэффициентом делителя 2.
1
USB
1 LED Xv "*"—
Т boot
1
I Р0 Р1 GND P2 РЗ
1
1 1 1 1
7 Hz „.50 MHz
Р4 Р5 GND
1 1 1
• Fin
//Pico_PWM4 8 Hz... 50 MHz
#include "pico/stdlib.h"
#include "hardware/pwm.h"
#include "pico/multicore.h"
uint32 t f hi =0;
void setup() {
set_sys_clock_khz( 100000, true);
Serial.begin(115200) ;
while(1){
if (Serial.available()) {
char ch = Serial. read();
uint32_t f = Serial. parselnt();
if(ch = 70)
f*=1000; //F
if(ch=102)
f*=l; //f
Serial, println (f) ;
ch = Serial. read();
uintl6_t pre = 1;
if(f<1908)
pre=10;
if(f<191)
pre = 100;
if(f<20)
pre = 250;
gpio_set_function(0, GPI0_FUNC_PWM) ;
pwm_config cfg = pwm_get_default_config();
pwm_config_set_clkdiv_int (&cfg, pre);
pwm init(0, &cfg, true);
uint32_t wrap = 100000000/pre/f;
pwm_set_wrap(0, wrap-1);
pwm_set_gpio_level (0 , wrap/2) ;
pwm_set_enabled(0, true);
}
}
void pwm_int() {
pwm_clear_irq(2) ;
f_hi++ ;
}
void loop() {
}
void setupl() {
uint32_tf=0 ;
while (true) {
pwm_config cfg = pwm_get_default_config();
pwm_conf ig_set_clkdiv_mode (&cf g, PWM_DIV_B_RISING) ;
pwm_init(2, &cfg, false);
gpio_set_f unction (5, GPIO_FUNC_PWM) ;
pwm_set_irq_enabled(2, true);
irq_set_exclusive_handler (PWM_IRQ_WRAP, pwm_int) ;
irq_set_enabled(PWM_IRQ_WRAP, true) ;
f_hi=0 ;
uint32_t t = time_us_32 ()+2;
while (t>time_us_32 () ) ;
pwm_set_enabled(2r true);
t+= 1000000;
while (t>time_us_32() ) ;
pwm_set_enabled(2, false);
f= pwm_get_counter(2) ;
f+=f_hi«16;
Serial, print(f);
Serial.println(" Hz");
}
}
void loopl() {
}
В программу также входит частотомер высокого разрешения. При прямом соеди-
нении выхода сигнала и входа счетчика можно проверить заданный сигнал. При
вводе типа f8 сначала отображается желаемая частота в Гц. После небольшой за-
держки отображается измеренная частота.
Частота, подобная 100 кГц, генерируется с высокой точностью, поскольку она
обходится плавным делителем 1000. С другой стороны, если вы хотите сгенериро-
вать 900 кГц, возникает ошибка округления 900 Гц с результатом 900,9 кГц. Са-
мые высокие плавные частоты — 10 МГц, 20 МГц и 50 МГц.
О С0М13
f8
j. u v и**
100 Hz
100 Hz
100 Hz
100 Hz
100 Hz
8
16 Hz
8 Hz
8 Hz
8 Hz
8 Hz
8 Hz
8 Hz
0 Autoscroll
1 Zeitstempel
anzeigen
Zeilenumbruch (CR)
-
v 115200 Baud v
□
X
Senden
A
V
Ausgabe loschen
О СОМ13
F900
j.\j\j\j\j\j
100000
100000
11 Z,
Hz
Hz
99999 Hz
100000
100000
Hz
Hz
99999 Hz
100000
100000
900000
602473
900900
900900
900900
0 Autoscrol
Hz
Hz
Hz
Hz
Hz
Hz
1 I I Zeitstempel
anzeigen
Zeilenumbruch (CR)
-
v 115200 Baud v
D
X
Senden
V
Ausgabe loschen
19. Делитель
частоты
На этот раз блок ШИМ 2 инициализируется с внешним входом на Р5 и выходом
ШИМ на Р4 . Ширина счета устанавливается делителем = 100, а ширина импульса —
50%. Это дает вам универсальный делитель частоты со входом Р5 и выходом Р4.
Коэффициент делителя можно установить в диапазоне от 2 до 32 768.
//Pico PWM4
#include "pico/stdlib.h"
#include "hardware/pwm.h"
#include "pico/multicore.h"
void setup () {
Serial.begin(1 15200);
}
void loop() {}
void setupl() {
uintl6_t divisor = 100;
gpio_set_fimction(0, GPIO_FUNC_PWM) ;
pwm_set_wrap(0, 1); //PWM 62.5 MHz
pwm_set_gpio_level(0, 1) ;
pwm_set_enabled(0, true);
pwm_config cfg = pwm_get_default_config();
pwm_config_set_clkdiv_mode(&cfg, PWM_DIV_B_RISING) ;
pwm_init(2, &cfg, true);
gpio_set_function (5, GPIO_FUNC_PWM) ;
gpio_set_function(4, GPIO_FUNC_PWM) ;
pwm_set_wrap(2, divider-1);
pwm_set_gpio_level(4, divider/2);
pwm_set_enabled(2, true);
}
void loopl () {}
USB
I
1
P(
•
-ED b*
D P1 GND
I I
62.5 MHz
BOOT
P2 РЗ Р4
I I
Fout «
P5 GND
► 1
I
► Fin
Программируемый делитель можно использовать в качестве прескалера для час-
тотомера, который может измерять только более низкие частоты. Некоторые циф-
ровые вольтметры содержат такой ограниченный счетчик. С помощью соответствую-
щего прескалера они также могут измерять ВЧ-сигналы. В целях тестирования на
порт Р0 снова подается прямоугольный сигнал частотой 62,5 МГц. Если подклю-
чить его к Р5, то на Р4 появится сигнал 625 кГц.
20. RC-генераторы
Программа Pico_RCl реализует НЧ RC-генератор, который содержит электролити-
ческий конденсатор емкостью 100 мкФ в качестве единственного внешнего компо-
нента. Необходимые резисторы являются внутренними и состоят из подтягивающих
и понижающих резисторов порта Р2 6.
Программа основана на функции модуля таймера NE555 и переключается между
зарядкой и разрядкой при 1/3 и 2/3 рабочего напряжения. Для измерения напря-
жения конденсатора используется АЦП на выводе АО.
ЮОр
Н
5V
40
4V7 GND EN
3V3 REF P28 GND P27 Р26 RUN P22
А2 AGND A1 АО 30
USB
LED
%
RP2040
BOOT
1
Р0 Р1 GND Р2 РЗ Р4 Р5 GND P6
10
Р7
Р8 Р9
Ф*
1к
//Pico_RCl
#include "pico/stdlib.h"
#include "hardware/adc.h"
void setup() {
adc_init () ;
adc_gpio_init(26) ;
gpio_pull_up(26) ;
adc_select_input(0);
gpio_init(0);
gpio_set_dir(0, true);
while(true){
if(adc_read()>2 730){
gpio_pull_down(26);
gpio_put(0, 0);
}
if(adc_read()<1365){
gpio_pull_up(26);
gpio_put(0, 1);
}
}
void loop () { }
Программа одновременно управляет выходом DO, к которому подключен светодиод
с последовательным резистором. Теперь он мигает с частотой около 0,3 Гц. Пе-
риод колебаний пропорционален емкости. Следовательно, вы можете генерировать
более высокие частоты с помощью конденсаторов меньшей величины. При конденса-
торе емкостью всего 100 нФ достигается частота около 300 Гц. На осциллограмме
представлена кривая напряжения на конденсаторе.
Даже без АЦП и всего с одним портом можно сформировать простой RC-
генератор. Поскольку после сброса каждый порт работает как вход с гистерези-
сом, дальнейшая инициализация не требуется. Достаточно запросить вход и пере-
ключиться между повышением и понижением. Теперь вы можете подключить любой
конденсатор между Р0 и GND. Конденсатор емкостью 100 нФ обеспечивает частоту
около 0,6 кГц из-за меньшего гистерезиса, составляющего около 0,4 В.
//Pico_RC2 RC generator PO
#include "pico/stdlib.h"
void setup() {}
void loop() {}
void setupl() {
while (true) {
if (f++)
gpio_pull_down(0);
else
gpio_pull_up (0) ;
}
}
void loopl () {}
На осциллограмме показано напряжение сигнала на конденсаторе 100 нФ. Сигнал
очень чистый и не содержит заметного дрожания фазы, поскольку контур управле-
ния работает на втором ядре и поэтому не подвергается воздействию других про-
цессов . Температурную зависимость керамического конденсатора очень хорошо
видно на осциллографе. Если прикоснуться к конденсатору, он нагревается и
частота увеличивается.
Вместо конденсатора также можно использовать пьезопреобразователь, который
тогда излучает тон около 1 кГц. Пьезодиск также является конденсатором и име-
ет емкость около 20 нФ. Свойства этого конденсатора изменяются при касании
или отражении звука. Немного потренировавшись, вы сможете воспроизводить зву-
ки разной частоты или даже простые мелодии.
Данное измерение производилось полностью без конденсатора, т.е. только с
учетом емкости осциллографа и порта. Вы можете увидеть треугольный сигнал
прибл. 300 кГц. Небольшой джиггер вызван конечными во времени точками выборки
где-то в диапазоне МГц. При большей емкости сигнал соответственно становится
более точным.
21. Измерение
емкости
Это измерительное устройство основано на RC-генераторе с внутренними тяну-
щими резисторами и внешним опорным конденсатором емкостью 1000 пФ. Измерение
частоты происходит одновременно. Если дополнительно подключен внешний измери-
тельный объект, емкость увеличивается, а частота снижается. При 1000 пФ изме-
ряется частота около 60 кГц. Точная частота без объекта измерения вначале
присваивается переменной f 0. Емкость измеряемого объекта затем непрерывно
рассчитывается по изменению частоты. При необходимости достаточно нажать
кнопку Р8 г чтобы отрегулировать нулевую точку.
LED
Ш*
RP2040
BOOT
1
Р0 Р1 GND Р2 РЗ Р4 Р5 GND P6
10
Р7
Р8 Р9 GND
Измеритель можно использовать с хорошей точностью в диапазоне от 1 пФ до
прибл. 100 нФ. Также можно проверять электролитические конденсаторы емкостью
до 100 мкФ, хотя точность заметно снижается. Помимо конденсаторов, в обратном
направлении можно измерять и диоды. На желтом светодиоде была измерена ем-
кость перехода 7 пФ.
//Pico_RC4 С measurement
#include "pico/stdlib.h"
void setup() {
Serial.begin(115200);
}
void loop() {}
void setupl() {
int n=0;
gpio_pull_up(8) ;
uint32 t fO;
while (true) {
uint32_tf=0 ;
uint32_t t = time_us_32() + 1000000;
while (t>time_us_32 () ) {
while (l==gpio_get(0) ) ;
gpio_pull_up(0); f++;
while (0==gpio get(0));
gpio_pull_down (0) ;
}
n++;
if(n=2)
f0=f ;
if(0==gpio_get(8))
n=0;
if(n>2){
n=3;
uint32_t c= f0*1000/f-1000;
if (O1000000000)
c=0;
Serial, print(c);
Serial.println(" pF ");
}
void loopl () {}
О СОМ13
□ X
Senden
11
11
10
11
11
11
10
11
10
10
10
10
11
pF
pF
pF
pF
pF
pF
pF
pF
pF
pF
pF
pF
pF
0 Autoscroll П Zeitstempel anzeigen
Zeilenumbruch (CR)
115200 Baud
Ausgabe loschen
Другое возможное применение — емкостные сенсорные датчики. При прикоснове-
нии к изолированному проводу емкость увеличивается на 1 пФ примерно до 5 пФ.
Прямое прикосновение к входу изолированного человека приводит к примерно 100
пФ.
22. Сенсорный
датчик
Датчик касания использует контакт Р26 (АО) в качестве RC-генератора с рези-
сторами и емкостью порта. АЦП постоянно измеряет напряжение и пытается под-
держивать его в пределах от 1/3 до 2/3 рабочего напряжения. При прикосновении
емкость увеличивается и изменяются характеристики генератора. Для защиты от
ВЧ-излучения последовательно с контактом датчика включен резистор сопротивле-
нием 1 кОм. Аналоговые измерения должны оцениваться таким образом, чтобы при-
косновение было четко обнаружено. В этом случае включается выход Р0.
touch
т
1k
J I I I L
J L
5V
40
4V7 GND EN 3V3
REF P28 GND P27 P26 RUN P22
A2 AGND A1 АО 30
USB
LED
W^
RP2040
BOOT
1
P0 P1 GND P2 P3 P4 P5 GND P6
10
P7
P8 P9
Ф*
1k
I I I I I I I
//Pico_RC3 Touch
#include "pico/stdlib.h"
#include "hardware/adc.h"
void setup() {
Serial.begin(115200) ;
adc_init() ;
adc_gpio_init (26) ;
gpio_pull_up (26) ;
adc_select_input (0) ;
gpio_init(0);
gpio_set_dir(0 r true)
int ulr u2, u, uO;
int n=0;
while(true){
u=0;
n++;
for
(int n=0; n<100; n++) {
ul=adc_read() ;
if(adc_read()>2730) {
gpio_pull_down (26)
u+=ul;
}
u2=adc_read() ;
if(u2<1365){
gpio_pull_up(26);
u-=u2;
}
}
u/=100;
Serial.println(u);
sleep_ms(50);
if (n==2)
u0=u ;
if(n>3)
n=3;
if (abs(u-uO) > 100)
gpio_put(0,l);
else
gpio_put(0,0);
void loop() {}
Анализ сигнала генератора показывает, что частота на холостом ходу состав-
ляет около 60 кГц. Программа работает слишком медленно, чтобы поддерживать
напряжение в заданных пределах. Таким образом, амплитуда увеличивается при-
мерно до 3,3 В. При оценке регистрируется разница напряжений между верхней и
нижней точкой переключения как среднее значение для 100 запусков. Среднее
значение, например, 500.
ВВ Oscilloscope PCSU1000
File £drt Options View Math Help
Oscilloscope
Spectrum Analyzer T ransient Recorder
V Logc Analyzer
History
Vofcs/Drv
Г
□
X
Time Л) iv
On
CM
ji:
Autoset
Persist | |_
On
Autoset |
I 20V 11 10V 11 Э/
IV
0.5V
Big Screen
2V 11 IV 11 0.5V |
0.2V 11 0.1V 11 50mV |
ir
I 0.2V 11 0.1V 11 50mV |
Coupling П<Г
Potion CacJ 1Ж1 LgndJ сзв GBD [acJ foe] IgnpI
Probe
20mV 11 IQmV 11 5mV |
Coupling
GND| Position
|5O0ms||200ms|
|l00ms||50ms|
lJ0m*J| 10ms|
| 5ms || 2ms |
I 1ms H 0.5ms |
10.2ms || 0.1ms |
| 50us || 20u$|
I 10us || 5us |
I 2us || 1us J
|0.5us||0.2us|
|0.1us|| |
1GSA
|02us||01us|
|Q.05us||OQ2us|
Run
; -
:r
::
' Trigger | Delay |
On/OH | On | | Off | |-»)<-|
Source | Ch1 I | Ch2 | | Ext [
- ** [>ПГЧП
j Level RunrWB
При прикосновении к контакту датчика емкость увеличивается, а частота пада-
ет, например, до 40 кГц. При изменении усредненной амплитуды напряжения более
чем на 100 выполняется операция переключения.
gg Oscilloscope PCSU1000
Eile Edit Options View Math Help
OscJIoscope Spectrum Analyzer Transient Recorder
Г" Logic Analyzer
0.5V
10us
History
Vols/Drv.
CM
-Ch2
On
| Autoset
Persist | (_
On
J | Autoset
IT
I 20V || 10V || 5V |
[ 2V ][ IV |[ 0.5V_J
| 0.2V || 0.1V || 50mV |
Big Screen
I 2V I
_JV_
I a5V I
0.2V
Coupling
Probe
20mV
0.1V
10mV
| 50mV |
| 5mV |
ua
jsL
Coupling
Position (ЖЗ (Ж) (И (Ж) D3D (Ж1 (Ж) (Ж) Position
•p Trigger | Delay |
On/Off
On
Off
□
X
Time/Div.
ir
Edge
Level
l500ms||2Q0ms|
|l00rm||50ms|
I 2Pms || 10ms J
| 5ms || 2ms |
| 1msJ[0_5ms]
10.2ms || aims |
I 50us || 20us |
Пой71Г5ц71
1 2us || Ins I
|0.5us||0.2us|
I O.lus || |
1GS/s
|0.2us||0.1us|
|a05us||0.02us|
Run
-><-
Source | CM | | Ch2 | | Ext |
Второй вариант сенсорного датчика использует цифровой вход для настройки
быстродействующего RC-генератора. При этом здесь происходит измерение частоты
с разрешением 1 кГц. В зависимости от типа подключенного кабеля или сенсорно-
го контакта измеряются частоты 600 кГц и более. В случае касания частота па-
дает существенно ниже 500 кГц. Контакт датчика находится на Р2, а выход на
Р0.
е сом1з
-
П X
| Senden |
ртп—jcttz:
546 kHz
546 kHz
546 kHz
544 kHz
545 kHz
546 kHz
545 kHz
545 kHz
398 kHz
387 kHz
384 kHz
382 kHz
382 kHz
0Autoscroll I | Zettstempel anzeigen
| Zeilenumbnich (CR)
v] 115200 Baud v|
a|
V
Ausgabe leschen
//Pico_RC5 Touch2
#include "pico/stdlib.h"
void setup() {}
void loop() {}
void setupl() {
gpio_init(0);
gpio_set_dir(0, true);
gpio_init(2);
while (true) {
uint32_tf=0 ;
uint32_t t = time_us_32() + 1000;
while (t>time_us_32()) {
while (gpio_get(2)==1);
gpio_pull_up(2) ;
f++;
while (gpio_get(2)==0);
gpio_pull_down(2);
}
if(f<500)
gpio_put(0,1) ;
else
gpio_put(0,0);
Serial, print(f);
Serial.println(" kHz");
sleep ms(100);
}
}
void loopl () {
}
Для целей управления выводится измеренная частота (рис. выше). В примере в
качестве контакта датчика была подключена алюминиевая фольга. В случае прямо-
го контакта частота падала ниже 400 кГц. В зависимости от поверхности датчика
и установленного порога частоты программа также подходит для емкостного дат-
чика приближения без прямого контакта. Тогда достаточно приблизить руку при-
мерно на 2 см. Также можно использовать изолирующую поверхность между датчи-
ком и рукой.
(ПРОДОЛЖЕНИЕ СЛЕДУЕТ)
Электроника
■
^н
щ
1
^^^^щ
<$few„e„, СС3020 1
\\S0.0 1Щ
^^^■1
■
1
1
ЧАСТОТОМЕР НА ARDUINO PRO MINI
Сидоренко С.
Основные технические характеристики:
■ Диапазон измеряемых частот 10 Гц...6 МГц
■ Чувствительность - 100 мВ
■ Напряжение питания - 5 В
■ Потребляемый ток в покое - 20 мА
■ Потребляемый ток в режиме измерения - 26 мА
Основой описываемого частотомера является модуль Arduino pro mini [1] на
микроконтроллере (МК) ATmegal68. Выбор этого модуля обусловлен принципом "ра-
зумной достаточности", поскольку в нём использован кварцевый резонатор НС-
49S, объём памяти МК для загрузки кода — 15 кБ, а объём самого кода не превы-
шает 6 кБ, его доступностью и приемлемой стоимостью, что позволяет получить
максимальную эффективность при минимальных затратах.
Частотомер выполнен по классической схеме счёта числа импульсов за опреде-
лённый промежуток времени.
Схема частотомера показана на рис. 1. По своим параметрам он немного пре-
восходит частотомер [2].
Измеряемый сигнал поступает на вход усилителя-формирователя на транзисторе
VT1 и логическом элементе DD1.1. Логический элемент DD1.2 разрешает прохожде-
ние импульсов на вход модуля А1. Микроконтроллер модуля формирует измеритель-
ный интервал, производит подсчёт импульсов и выводит результат измерения на
ЖКИ.
Выводы RS, Ей четыре линии данных DB4—DB7 ЖКИ подключены к выводам АО—А5
модуля А1. Цифровые выводы пригодятся для дальнейшей модернизации. Линия R/W
подключена к общему проводу, так как нам потребуется только функция записи в
память ЖКИ. Измерительный интервал 1 с формируется с помощью двух восьми-
разрядных таймеров Т2 и ТО МК, которые работают независимо от режима работы
самого МК.
R1- м
100 itf C2
100 к
Вход
Общий
^ +5 В стаб
Общий
Рис,
Подсчёт импульсов ведётся с помощью таймера Т1 (вход — контакт 5 А1) . По-
скольку он шестнадцатиразрядный, поэтому считает до 65536. Конечно, это слиш-
ком мало. Поэтому используем возможности таймера Т1 по переполнению. Для это-
го обычно используется обработчик прерываний по переполнению, но мы будем ис-
пользовать регистр флагов TIFR. Скетч в цикле 1оор() контролирует таймер Т1 и
в режиме счёта при установке флага переполнения счётчика добавляет 1 к пере-
менной count.
По истечении времени измерения, когда на выходе А таймера ТО (контакт 6 А1)
устанавливается лог. 0, производится расчёт измеренной частоты и вывод полу-
чившегося значения на ЖКИ HG1. Весь цикл измерения и отображения составляет
чуть больше 1 с. Для коррекции показаний введена переменная Кк. Обычно она
имеет тип float, что для МК не очень хорошо. Но у нас Кк — integer и содержит
погрешность показаний в герцах на частоте 4 МГц. Корректируются показания
пропорционально частоте.
Изначально планировалось сделать два режима измерения: 1 с и 0,1 с. Ока-
залось , что при 0,1 с вывод на экран требуется делать в течение не менее 0,5
с. В противном случае показания на экране будут "мельтешить". Разница в 1 с и
0,6... 0,8 с на глаз не сильно заметна, а вот точность показаний в десять раз
хуже. Поэтому я отказался от периода измерения в 0,1 с. Да и скетч получился
проще.
Использование таймера ТО в качестве счётчика-делителя лишает нас функций
delay() и millis() Поэтому сделана подпрограмма pauza(), чтобы облегчить ра-
боту МК.
Печатная плата для частотомера не разрабатывалась, поскольку время, затра-
ченное на разработку, травление, лужение, сверление отверстий, намного боль-
ше, чем монтаж элементов устройства на макетной печатной плате размерами
60x80 мм с металлизированными отверстиями. Соединения между деталями и узлами
произведены с помощью проводников от шлейфа IDE HDD компьютера (такие HDD уже
не используются). Толщина их менее 0,1 мм, но этого достаточно для прохожде-
ния сигнала. Изоляция у них виниловая, хорошо снимается, если это необходимо.
В одно отверстие на плате легко входят три проводника в изоляции. Сами про-
водники хорошо облуживаются.
Сначала устанавливают перемычку между контактами 3 и 4 на плате модуля
Arduino pro mini. Затем устанавливают саму плату на небольшой высоте над ма-
кетной платой. Контакты для программирования МК модуля припаяны к макетной
плате и соединены с платой модуля А1.
Теперь можно запрограммировать МК. У меня была установлена Arduino IDE
vl.8.9. Более поздние версии должны тоже корректно компилировать. Устанавли-
вают драйвер Corn-порта СН341. Подключаем модуль Arduino pro mini к компьюте-
ру с помощью адаптера USB-TTL CH341. Сам адаптер подключают к Arduino pro
mini в соответствии с таблицей.
Arduino pro mini
СН341А
Тх
Rx
Rx
Тх
GND
GND
VCC
+5V
В Arduino IDE в меню "Инструменты" выбираем плату — Arduino pro or Pro
mini, процессор — ATmegal68 (5V, 16MHz) , порт — в моём случае Com 3. Порт
должен отображаться, в противном случае надо проверить установку драйвера в
"Диспетчере устройств". Загружают скетч в Arduino IDE, проверяют и загружают
в модуль Arduino pro mini. Далее с помощью изолированных проводов подключают
ЖКИ.
Чтобы защитить сигнальные входы и выходы модуля А1, я подключил их через
резисторы сопротивлением 1 кОм, а вывод 15 ЖКИ к линии питания +5 В — через
резистор сопротивлением 200...560 Ом. Это не обязательно, просто делюсь опы-
том. Эти резисторы не влияют на работу устройства. Затем монтируют элементы
входного усилителя-формирователя на транзисторе VT1, микросхему DD1 устанав-
ливают панель и делают все соединения.
После подачи напряжения питания налаживание начинают с регулировки резисто-
ром R6 контрастности ЖКИ. На экране сначала появляется надпись Arduino f-
metr, затем на первой строке — f = 0 Hz, а на второй — Is. На коллекторе
транзистора VT1 подборкой резистора R1 выставляют напряжение 2,8... 3 В. Дают
прогреться 3... 5 мин и подают на вход частотомера образцовую частоту 4 МГц
(или частоту с известной погрешностью) . У меня на ЖКИ были показания f =
3,999739 MHz. Разница в 261 Гц в меньшую сторону. Следовательно, переменная
Кк = 261. Если показания превышают образцовую частоту, Кк со знаком минус за-
писывают это значение Кк в скетч и перепрограммируют Arduino pro mini. Нала-
живание закончено.
Корпус частотомера может быть любой, куда помещается плата. В корпусе дела-
ют отверстия для ЖКИ и разъёмов входа и питания.
Для расширения диапазона измеряемых частот потребуется дополнительный счёт-
чик-делитель на восемь. Под рукой оказался счётчик 74LS90 (российский аналог
— К555ИЕ2). Предельная рабочая частота 74LS90 — 32 МГц, К555ИЕ2 — 50 МГц. Ис-
пользовать можно любой счётчик-делитель, лишь бы он обеспечивал необходимую
частоту измерения. Удачным решением может быть использование микросхемы
74НС93, у которой предельная частота 77 МГц.
Схема установки такого счётчика-делителя в частотомер показана на рис. 2
(нумерация элементов продолжена). Вход С1 у счётчика 74LS90 инверсный, поэто-
му он подключён к выходу элемента DD1.2, если вход прямой, его подключают к
выходу элемента DD1.3. В принципе, можно было бы использовать и деление на
десять. Тогда бы пришлось задействовать выход Q4 счётчика DD2 и дополнитель-
ный вход Arduino pro mini. Но у счётчика 74LS90 есть недостаток, при делении
на десять может проскакивать лишний импульс. Входы сброса (выводы 2 и 3 DD2)
объединены по схеме И. Сброс — по фронту импульса. А нам нужно, чтобы сброс
происходил по схеме ИЛИ, так как он должен происходить и в режиме счёта и по
команде Arduino pro mini. Для реализации этой функции использованы элементы
DD1.3 и DD1.4.
В доработанный скетч добавлены битовое считывание показаний счётчика, умно-
жение значения частоты на восемь при её вычислении, сброс счётчика и расчет
коррекции для частоты 16 МГц (тактовая частота МК).
К R5
^
*- +5Встаб
выв 5 DD2
: выв 6DD1 2
* Ккокт. 7А1
К кокт. в А1
^ К кокт. 9 А1
f-н^ Ккокт. 10А1
К кокт. 5 А1
К выв 10 DD2
Общий
Рис. 2
Конечно, можно было бы поставить делитель на входе частотомера и так не за-
морачиваться. Но в моих планах сделать щуп-делитель для измерения частоты вы-
ше 200 МГц.
Внешний вид смонтированной платы частотомера показан на рис. 3 и рис. 4.
Рис. 3
J- • -^
* ' * ' & -• •JO
'▼ ччщ
^'OOOOOMitif} PC .
ЮОООООООООО<3*М
ООООООООООО^ООФСЮои^иис .OV
осооооооооо*ооофоооооооо*ос
^сооооооооо^оооеооа^чооппо<
>СОООР- ~ ->Л|3*И*~£*ЧЮО£OCOCOC
<: --ooq«h*jO<—*^оо *ооосоо«*.:|
i» <: --ooq«h*jO<—*^оо *ооосоо«*.
-* - ^-900
Рис. 4
// Arduino f-metr ver.l
// выход T2 (pin D3) и вход TO (pin D4)- соединены перемычкой
#include<LiquidCrystal.h>
unsigned long f = 0; // частота (Гц)
unsigned int count; // счетчик переполнения Т1
int Kk = 0; // коэффициент коррекции на 4 МГц (1 сек)
const String del = " "; // очистка части дисплея
boolean Flagl; // флаг отображения 1 строки
LiquidCrystal lcd(14,15,16,17,18,19); // используем аналоговые pinfbi A0-A5
void setup() {
pinMode(3, OUTPUT); // выход Т2 - pin 3
pinMode(4, INPUT); // вход ТО - pin 4
pinMode(6, OUTPUT); // выход TO - pin 6
pinMode(5, INPUT); // вход Т1 - pin 5
// Инициализация LCD
lcd.begin(16,2) ;
lcd.setCursor(0,0); lcd.print(" ARDUINO f-metr");
//
nolnterrupts();
// таймер 2 - 250 Гц
OCR2A = 249;
OCR2B = 125;
TCCR2A = 0; TCCR2B = 0;
// режим быстрого ШИМ с верхним пределом
TCCR2A = _BV(C0M2B1) | _BV(COM2B0) | _BV(WGM21) | _BV(WGM20) ;
// установка предделителя в 256 и запуск таймера
TCCR2B = _BV(WGM22) | _BV(CS22) | _BV(CS21) ;
// таймер 0 выход - 1 Гц
OCR0A = 249;
TCCR0A = 0; TCCR0B = 0;
TCCR0A = _BV(COM0Al) | _BV(WGM00) | _BV(WGM01) ; // режим ШИМ
TCCR0B = _BV(CS02) | _BV(CS01) ;
// таймер 1 - счетчик, вход - pin D5
TCCR1A = 0; TCCR1B = 0; TCNT1 = 0;
TCCR1B = _BV(CS12) | _BV(CS11) | _BV(CS10) ;
TIFR1 = 1 « TOV1;
interrupts(); // включить глобальные прерывания
}
// вывод на экран
void mylLCD () { //1 строка
float ff; // частота в МГц (кГц)
led.setCursor(4,0); led.print(del + del);
lcd.setCursor(0,0); lcd.print("f = ") ;
if(f < 1000) {
lcd.print(f); lcd.print(" Hz");}
else {
if(f >= 1000000){
ff = (float)f/1000000.0;
lcd.print(ff,6); lcd.print(" MHz");}
else {
ff = (float)f/1000.0;
lcd.print(ff,3); lcd.print(" kHz");}
}
void my2LCD () { //2 строка
String strl = deif-
ied. setCursor(0,1);lcd.print(strl);
strl = " Is ";
led.setCursor(0,1); led.print(strl);
}
void corr() { // расчет коррекции
unsigned long calc_Kk;
if(f>1000){ // если > 1 кГц, то корректируем
calc_Kk = f/1000; // частота в кГц
calc_Kk *= (Kk); // умножаем на Kk |4 МГЦ - Кк
calc Kk /= 4; // делим на 4 кГц | f МГц - х
calc_Kk /= 1000;
f += calc Kk; // корректируем
}
}
void freq_calc() { // расчет частоты
f = (unsigned long)count * 65536 + TCNTl;
corr(); // коррекция
}
void pauza () {
asm("nop"); asm("nop"); asm("nop"); asm("nop"); asm("nop");
asm("nop"); asm("nop"); asm("nop"); asm("nop"); asm("nop");
}
void loop() { // бесконечный цикл
// измерение
if (bitRead(PIND, 6)==1) {
// то же, что digitalRead(6)==l, только за 1 такт
if (!Flagl) {Flagl = true;}
// проверяем переполнение TCNTl
if(TIFRl & (1 « TOV1)) {count++; TIFRl = 1 « TOVl;}
}
else { // вывод на экран
if (Flagl) {
Flagl = false;
// на всякий случай
if(TIFRl & (1 « TOVl)) {count++; TIFRl = 1 « TOVl;}
freq_calc(); // определяем частоту
count = 0; TCNTl =0; // обнуляем счетные регистры
mylLCD(); // отображаем частоту
my2 LCD () ;
}
}
pauza (); // пауза
// Arduino f-metr ver.2
// выход T2 (pin D3) и вход TO (pin D4)- соединены перемычкой
#include<LiquidCrystal.h>
#define Rst 7 // pin сброса внешнего счетчика
byte Kg =8; // Коэффициент деления счетчика на входе
byte b; // показания счетчика
unsigned long f = 0; // частота (Гц)
unsigned int count; // счетчик переполнения Т1
int Kk = 0; // коэффициент коррекции на 16 МГц
const String del = " "; // очистка части дисплея
boolean Flagl; // флаг отображения 1 строки
LiquidCrystal lcd(14,15,16,17,18,19); // используем аналоговые pinfbi A0-A5
void setup() {
pinMode(3, OUTPUT); // выход Т2 - pin 3
pinMode(4, INPUT); // вход ТО - pin 4
pinMode(6, OUTPUT); // выход ТО - pin 6
pinMode(5, INPUT); // вход Т1 - pin 5
pinMode(8, INPUT);
pinMode(9, INPUT);
pinMode(10, INPUT);
pinMode(Rst, OUTPUT); // pin сброса счетчика на входе
// Инициализация LCD
lcd.begin(16,2) ;
lcd.setCursor(0,0); lcd.print(" ARDUINO f-metr");
//
nolnterrupts();
// таймер 2 - 250 Гц
OCR2A = 249;
OCR2B = 125;
TCCR2A = 0; TCCR2B = 0;
// режим быстрого ШИМ с верхним пределом
TCCR2A = _BV(C0M2B1) | _BV(COM2B0) | _BV(WGM21) | _BV(WGM20) ;
// установка предделителя в 256 и запуск таймера
TCCR2B = _BV(WGM22) | _BV(CS22) | _BV(CS21) ;
// таймер 0 выход - 1 Гц
OCR0A = 249;
TCCR0A = 0; TCCR0B = 0;
TCCR0A = _BV(COM0Al) | _BV(WGM00) | _BV(WGM01) ; // режим ШИМ
TCCR0B = _BV(CS02) | _BV(CS01) ;
// таймер 1 - счетчик, вход - pin D5
TCCR1A = 0; TCCR1B = 0; TCNT1 = 0;
TCCR1B = _BV(CS12) | _BV(CS11) | _BV(CS10) ;
TIFR1 = 1 « TOV1;
interrupts(); // включить глобальные прерывания
}
// вывод на экран
void mylLCD () { //1 строка
float ff; // частота в МГц (кГц)
led.setCursor(4,0); led.print(del + del);
lcd.setCursor(0,0); lcd.print("f = ");
if(f < 1000) {
lcd.print(f); lcd.print(" Hz");}
else {
if(f >= 1000000){
ff = (float)f/1000000.0;
lcd.print(ff,6); lcd.print(" MHz");}
else {
ff = (float)f/1000.0;
lcd.print(ff,3); lcd.print(" kHz");}
}
}
void my2LCD () { //2 строка
String strl = del;
led.setCursor(0,1);lcd.print(strl);
strl = " Is ";
led.setCursor(0,1); led.print(strl);
}
void corr() { // расчет коррекции
unsigned long calc_Kk;
if(f>1000){ // если > 1 кГц, то корректируем
calc_Kk = f/1000; // частота в кГц
calc_Kk *= (Kk); // умножаем на Kk |1б МГЦ - Кк
calc Kk /= 16; // делим на 16 кГц | f МГц - х
calc_Kk /= 1000;
f += calc Kk; // корректируем
}
}
void freq_calc() { // расчет частоты
f = (unsigned long)count * 65536 + TCNTl;
// умножаем на коэффициент деления счетчика на входе
f *= (Kg);
// добавляем значение счетчика-делителя
f += b;
corr(); // коррекция
}
void pauza () {
asm("nop"); asm("nop"); asm("nop"); asm("nop"); asm("nop");
asm("nop"); asm("nop"); asm("nop"); asm("nop"); asm("nop");
}
void loop() { // бесконечный цикл
/ измерение
// то же, что digitalRead(6)==l, только за 1 такт
if (bitRead(PIND, 6)==1) {
if (!Flagl) {Flagl = true;}
// проверяем переполнение TCNTl
if(TIFRl & (1 « TOV1)) {count++; TIFRl = 1 « TOVl;}
}
else { // вывод на экран
if (Flagl) {
Flagl = false;
// на всякий случай
if(TIFRl & (1 « TOVl)) {count++; TIFRl = 1 « TOVl;}
b = PINB & B00000111;
digitalWrite(Rst,0);
freq calc() ;
count = 0; TCNT1 = 0;
digitalWrite(Rst,1) ;
mylLCD();
my2LCD();
II
II
II
II
II
II
II
}
}
pauza (); // пауза
}
Литература:
1. Arduino pro mini. — Характеристики, распиновка, описание платы. — URL:
https://clck.ru/FDUZ6(01.12.23).
2. Булдаков А. Частотомер на LGT8F328P-LQFP32 MiniEVB. — Радио, 2023, № 4, с.
29-32.
читаем значения Pin 8,9,10
сброс счетчика-делителя
определяем частоту
обнуляем счетные регистры
перевод счетчика в режим счета
отображаем частоту
отображаем 2 строку
Ill Г U
Датчик направления
магнитного поля
Я4
1Л
CYBEREX ТЕСН
*ij
VCC +5V
ДАТЧИК НАПРАВЛЕНИЯ МАГНИТНОГО ПОЛЯ
Устройство должно выполнять детектирование направления магнитного потока, а
в качестве индикатора будет использоваться RGB светодиод. Принципиальная схе-
ма довольно проста и состоит из доступных компонентов, донором которых может
служить старый CD или DVD привод. В схеме применяется два ключевых компонен-
та : это линейный датчик Холла и сдвоенный операционный усилитель. Ниже приве-
дена принципиальная схема устройства.
www
D2
LED__RGfc
\7
GND
Проект печатной платы разрабатывался в KiCAD, ниже вы можете видеть пара-
метры платы.
Реализация будет выполняться с применением современных доступных инструмен-
тов . А помогут нам лазерные технологии.
С недавнего времени в наборе домашних инструментов появился младший брат
моего 3D принтера - миниатюрный лазерный ЧПУ. С помощью данного устройства мы
и будем делать плату, а насколько это упрощает процесс домашнего изготовления
платы, вы поймете далее. Давайте приступим.
Для начала в обрезках фальгированного стеклотекстолита находим необходимый
по размеру кусок, обрабатываем и наклеиваем фоторезист.
Затем помещаем плату в заранее отмеченную лазером область и фиксируем с по-
мощью изоленты, создавая что-то на подобии "гнезда" для платы. И запускаем
процесс фотоэкспозиции.
Для засветки фоторезиста используется лазерный модуль с длинной волны
405нм, время экспозиции подбирается вручную, учитывая мощность лазера.
Получаем следующий результат:
Засветка фоторезиста лазером имеет очень высокую точность и качество. Необ-
ходимо лишь предварительно настроить фокус луча на установленной заготовке.
Затем нам необходимо проявить фоторезист. Процесс выполняется в 10% раство-
ре гидроксида натрия. Ниже результат проявки.
Следующий этап - это травление платы. Эта процедура выполняется в растворе
перекиси водорода, лимонной кислоты и соли.
Чтобы защитить медные дорожки платы, нам необходимо нанести паяльную маску.
Паяльная маска наносится с помощью сетки для шелкографии и карты вашего люби-
мого банка.
Для полимеризации паяльной маски снова воспользуемся нашим лазерным ЧПУ,
поместив плату в наше изолентовое "гнездо":
После завершения данной процедуры, извлекаем нашу плату и осторожно, с по-
мощью салфетки и ацетона удаляем не засвеченную маску. Результат - ниже на
фото.
Далее нам необходимо дополнительно зафиксировать слой маски с помощью ульт-
рафиолетовой лампы, я использую свою.
После завершения процесса, монтируем все компоненты схемы на плату и при-
паиваем с помощью небольшого термостола. В итоге мы получаем следующий ре-
зультат :
На этом этапе можно заметить, что маска имеет не очень эстетический вид, но
основные свои функции она выполняет. Данная маска использовалась впервые, по-
этому сработал эффект первого блина. С другим паяльными масками подобных про-
блем не возникало.
Теперь необходимо протестировать работу собранной платы. Подключаем питание
и берем в руку неодимовый магнит...
Теперь нам необходимо разработать корпус устройства. Выполнять данную зада-
чу мы будем во FreeCAD.
Задействуем 3D принтер и наконец напечатаем наш корпус:
Как можно видеть из модели корпуса, наше устройство будет представлять с
собой стик, а питание будет выполняться с помощью USB-C порта, который будет
размещен сверху. Давайте теперь соберем наше устройство в корпусе.
С помощью секундного клея прикрепим плату и USB порт к нашему корпусу. За-
тем накроем все крышкой и с помощью того же клея все зафиксируем. В результа-
те у нас получается следующий результат.
Техника
ТЕХНИКА РЕЗКИ СТЕКЛА
Обычно мастер-самоделыцик сталкивается в своей работе со стеклом листовым.
Цилиндрические заготовки — стеклянные палочки, капилляры, трубочки, трубки и
трубы могут использоваться в техническом творчестве или декоративно-
прикладном искусстве (ДПИ) при работах экзотических. Очень популярная задача
— резка стеклянной тары — банок, бутылок. И, конечно, стеклянное приборо-
строение, дела стеклодувные. Область бытия, где разносортные и разнокалибер-
ные трубочки и палочки — основные заготовки для любых, даже самых сложных и
замысловатых работ. И резка их — первейший навык.
Разрезание стеклянного цилиндра принципиально ничем не отличается от тако-
вой работы с листом стекла, но имеет немало интересных вариаций, особенно в
стеклодувном ремесле, где, к слову сказать, правильность и качество выполне-
ния каждой операции зависит от таковых в работах предыдущих, а начинается всё
с резки заготовок. Кроме того, резки много даже для очень несложных стекло-
дувных работ. За рабочий день мастеру приходится отрезать стержни-трубки де-
сятки раз. Навык этот должен быть отточен и автоматичен — вроде как «шашкой
махнуть». Ровный, аккуратный рез без зазубрин менее склонен растрескиваться
при нагреве, он равномерно разогреется в факеле горелки, фигура, сформованная
из такой заготовки, будет правильной.
<У^
подача
воды
алмазный
диск
ограничитель
Самый надёжный и скучный способ резки — механизированный алмазный
инструмент [1]. Очень хорошо непрерывно омывать его водой для увели-
чения ресурса отрезного диска и смывания вредной стеклянной пыли,
которая, к слову сказать, является полезным материалом для приготов-
ления дешёвой и прочной термостойкой замазки-клея.
Стеклодувный
нож
Экзотическое и, пожалуй, самое мирное холодное оружие. Представляет собой
особым образом заточенную узко-длинную пластинку из твёрдого металла, закреп-
лённую в удобной ручке. Предназначен для нанесения риски-царапины поперёк
стеклянной трубки и является функциональным аналогом стеклореза.
Нож из твёрдых сплавов для резки стеклянных трубок.
Свой стеклодувный нож сделал пластинчатой конструкции с режущим элементом
из куска станочной пилы по металлу.
Заточив обрезок пилы, убедился в его работоспособности по стек-
лу, тонким абразивным диском в небольшой углошлифовальной машин-
ке (УШМ) вырезал режущий элемент с пазами. При работе не позво-
лял заготовке перегреваться.
Из обрезков нетонкой нержавеющей стали разметил и выпилил эле-
менты держателя.
Сварил держатель в рукояточнои части, просверлил крепёжные от-
верстия для лезвия, обработал железки на электроточиле и не-
сколькими номерами наждачки.
Заготовку для декоративного латунного кольца на деревянной руко-
ятке нашёл в старом водопроводном хламе.
Для его обработки выточил оснастку-цангу из берёзового полена на
своём самодельном токарном станке по дереву.
При этом металлическая заготовка насаживается поверх расклини-
ваемой цанги. Кольцо обточил абразивным кругом УШМ, довёл не-
сколькими номерами наждачки.
Рукоятку выточил «в центрах» из кусочка сухой черёмухи, проточил
посадочное место для латунного кольца. Отшлифовал несколькими
номерами шлифовальной бумаги при прямом и обратном вращении, вы-
лощил горстью стружек.
Рукоятку отделал прозрачным матовым лаком, просверлил глухое отвер-
стие и отчасти забил, отчасти вклеил металлическую часть в сборе. На
фото — рабочее положение инструмента. Большим пальцем прижимаем к
лезвию стеклянную трубку и с некоторым усилием проворачиваем её.
Царапина на трубке 010 мм. Чёткая и глубокая, она не обязательно
должна охватывать трубку полным кольцом, довольно нескольких
миллиметров — этакая затравка для трещины.
„_ Т Т™
q
Ъ_^^_^^^_4
tj
1——'— —
J _i [_ 1
1 \
^/
Ломают трубку сильным, уверенным движением, взявшись обеими ру-
ками одинаково по обе стороны от царапины [2] и одновременно
растягивая заготовку. Саму риску при этом, повернув от себя,
большими же пальцами полезно в неё упереться. Заготовка ломается
лучше, если риску смочить (водой, слюной).
Края трубки обычно удаются весьма ровными, операция очень быст-
рая. Способ позволяет резать трубки диаметром до 15...20 мм.
Важный момент — заточка лезвия ножа. Её делают на электроточиле с абразив-
ным кругом средней крупности, под 45°, быстрым движением. Лезвие ножа при
увеличении должно иметь ряд мелких зазубрин, вгрызающихся в стекло. Стекло-
дувный нож со шлифованным лезвием работать перестаёт.
Наносить риски на стеклянные трубки можно и аккуратно сточенным по бокам
надфилем. Очень хорошо работает трёхгранный алмазный надфиль «а-ля натюрель»
на металлической связке. При первых опытах, особенно с крупными трубками, ру-
ки лучше защитить перчатками или обмотать их.
Термоудар
Широко используется для резки стеклянных заготовок в стеклодувной мастер-
ской , тем более что под рукой не один высокотемпературный факел. Что такое
термоудар? Стекло — материал хрупкий, имеющий значительный температурный ко-
эффициент расширения (ТКР) и плохо проводящий тепло. Более или менее толсто-
стенная заготовка при быстром нагреве имеет холодную сердцевину, и разница в
их расширении стремится стекло разорвать. Особенно при изъянах в целостности
— дефекты, напряжения.
Толстостенная стеклянная банка с дефектом.
Даже небольшая порция горячего томатного сока «для предварительно-
го прогрева» аккуратненько отколола её дно.
Аналогичным образом отрезают крупные цилиндрические заготовки — делают мет-
ку стеклодувным ножом, обматывают место реза с обеих сторон лентами мокрого
асбеста и разогревают с вращением в узком «паяльном» факеле. Края реза удают-
ся невысокого качества, их затем оплавляют или шлифуют.
Трубки отрезают термоударом, нагревая место реза с меткой ножом и резко ох-
лаждая его — прикладывая холодную стеклянную палочку. Или, напротив — разо-
гретый конец стеклянной палочки прикладывают к метке на холодной трубке. Час-
то вместо стеклянной палочки используют нарочитый металлический крючок, кото-
рый в пламени горелки раскаляют до красного каления.
Резка горячей
нихромовой
проволокой
Очень распространённый и удобный приём. При большом количестве резов заго-
товок одинакового диаметра может представлять собой этакий прищепку-пинцет с
проволочной рабочей частью на конце — полной окружностью, но обычно это ста-
ционарное настольное более или менее универсальное приспособление с V-
образной рабочей частью из нетонкой проволоки или даже ленты.
Для своей электрорезалки подобрал в закромах готовый трансформатор доста-
точной мощности и с рядом низковольтных обмоток.
Испытания трансформатора — подбор напряжения для красного кале-
ния куска имеющейся нихромовой проволоки 0 0,5 мм (выпрямленная
спираль от электроплитки).
г
Коробку для тяжёлого плоского трансформатора сделал из обрезков 15
мм фанеры — длинные резы выполнил на циркулярной пиле, короткие на
торцевой маятниковой. Собрал боковые стенки на столярный ПВА и гвоз-
дики . Верхнюю и нижнюю крышки выпилил из фанеры потоньше.
Стойки-изоляторы выточил из сухого берёзового полена.
Деревянные сверлёные стойки приклеил к верхней крышке коробки,
покрасил в пару слоев эмалью ПФ-115. Закрепил установочные эле-
менты — тумблер, сетевую лампочку, колодку плавкого предохрани-
теля, вентиляционные сеточки вырезанные и насверлённые из алюми-
ниевого листа. Стойки для нихромового провода сделал из 6 мм
медной трубки, в изоляторы её заделал клеем-замазкой из стеклян-
ной пыли, замешанной в жидком стекле [1]. Для закрепления нихро-
мовой проволоки на концы стоек припаял по винтовой клемме.
Вид на электромонтаж прибора. Маломощный выпрямительный мост —
для светодиода «Сеть» на передней стенке. Некрасивые дыры в бо-
ковых стенках — забытые проёмы для вентиляции (выгрызены и ещё
не заделаны решеточками.
Работа с прибором аналогична резке разогретой стеклянной палоч-
кой — стеклодувным ножом делаем риску и ей прикладываем трубку к
накалённой проволоке. Через 5...7 секунд слышится характерный
хруст и появляется короткая трещина. Чуть проворачиваем трубку в
петле, и трещина становится длиннее. Трубку на фото (0 18 мм из
платинового стекла - зелёный торец) удаётся разрезать за два
раза. Отрезанные кусочки могут быть весьма короткими, какими их
нипочём не сделать обычным разламыванием (заставка). Края стекла
удаются более или менее ровными и без зазубрин.
Отрезать тонкостенный шарик разламыванием не удастся — механическая
нагрузка его разрушит, электрорезалкой же - возможно.
Учебно-тренировочный шарик с короткими ножками. Трубка 0 10 мм из
свинцового стекла (серый торец). Хорошо виден и сам свинец — местное
потемнение стекла, восстановленный в факеле без избытка кислорода.
К резке нихромовой проволокой стоит добавить — её диаметр должен быть зна-
чительным. Классики рекомендуют 0,7...1,5 мм. Тогда запас тепла в ней будет вы-
ше и прибор работает быстрее и аккуратнее. Слишком выравнивать гнутую холод-
ную проволоку нет нужды, при первой же резке она преотличным образом отформу-
ется под нагрузкой.
Резка банок
и бутылок
Представляет собой аналогичную задачу и может быть выполнена несколькими
способами, обычно термоударом — хрестоматийный «полевой», с верёвочкой, про-
питанной бензином, культурным «домашним» — той же накалённой током нихромовой
проволокой. Получил распространение и стеклорезный способ предварительного
нанесения кольцевой царапины в специальном несложном станке с последующим
термоударом. Такой рез удаётся ровнее и аккуратнее, даёт меньше брака.
Приспособление для этого собрал из подножного хлама, а универсальность ему
придают крепления элементов столярными струбцинами.
Основание станка выполнил из обрезка мощной доски, на нём пришуруп-
лены четыре некрупных мебельных колесика. Высокие ножки снизу позво-
ляют удобно задействовать имеющиеся в хозяйстве простые струбцины,
на бортик сверху крепится рычаг с головкой роликового стеклореза.
Дном вращаемая бутылка упирается в ограничитель. С его лицевой
стороны закреплён кусок гладкой металлической пластинки, по нему
рифлёное стекло скользит лучше.
Собственно, рычаг со стеклорезом. Крепится струбциной за дере-
вянную часть. Головка от стеклореза (большая стрелочка) при не-
обходимости может переставляться для работы с более крупными или
мелкими заготовками (маленькие стрелочки).
Приспособление в сборе. Стеклорезная черта должна приходиться
между роликов или хотя бы над парой крайних, хотя, изловчившись,
удавалось отрезать и кусочки горлышек.
Несколько замечаний к работе — стеклорезную головку нужно периодически сма-
зывать, её ролик нельзя вести два раза по одному месту — сильно тупится. Бу-
тылки предварительно отмыть от грязи, пыли и этикеток. В некоторых случаях
достаточно длительного (сутки) замачивания в воде, в других приходится после
ещё и оттирать липкий клей растворителем.
Действую так: прочертив кольцевую царапину, разогреваю её над строительным
феном (1800 Вт, включён на второй передаче), вращая бутылку. Горячую немед-
ленно погружаю в ведро с холодной водой до уровня царапины. Иногда одного
раза довольно и бутылку можно разломить руками, часто приходится греть её и
макать ещё разок. Следует сказать о нередко невысоком качестве бутылок —
обычно это неравной толщины стенки, доходящие иногда до солидных местных на-
плывов — случается брак, к нему нужно быть готовым и относиться философски.
Удачный рез хорош, но недостаточно для ежедневного бытового применения.
Следует, по крайней мере, притупить острые кромки, а лучше отшлифовать край
полностью.
Проще всего сделать это вручную на куске средней водостойкой на-
ждачки. Край стекла следует регулярно и часто макать в воду, а
наждачку промывать. Работа муторная, долгая и тяжёлая. Шлифо-
вальную шкурку не жалеть и часто менять. Если под неё подложить
кусок плотного войлока, то при нажиме стекло слегка проминает
абразив и край выходит заваленным, не режущим.
Литература:
1. Сорокин В. С. Стеклодувная мастерская. 2013 г.
2 . Легошин А. Я., Мануйлов Л. А. Стеклодувное дело. Москва. Высшая школа,
1976 г.
Технологии
ВЫДЕЛЕНИЕ ЧИСТЫХ КУЛЬТУР ДРОЖЖЕЙ
Качмазов Г.С.
ДРОЖЖИ РОДА
SACCHAROMYCES
Клетки круглые и овальные, иногда удлиненные. Вегетативное размножение про-
исходит многосторонним почкованием. Может быть примитивный псевдомицелий, ис-
тинного мицелия нет. Колонии обычно пастообразные. На жидких средах при про-
должительном культивировании может образовываться пленка, но она не бывает
сухой порошковидной или всползающей.
Аски образуются преимущественно из вегетативных диплоидных клеток без непо-
средственно предшествующей конъюгации. Аскоспоры круглые или слабоовальные,
бесцветные, гладкие, 1-4 в аске. Все виды активно сбраживают сахара и не ис-
пользуют нитраты. Лактозу и высшие парафины не ассимилируют.
Дрожжи этого рода с давних времен распространены в кустарном виноделии и
широко используются в разных отраслях бродильной промышленности, в связи с
чем они более других дрожжей изучены в разных аспектах. Их систематика, одна-
ко, постоянно пересматривается. Центральный вид — Saccharomyces cerevisiae
известен в десятках синонимов, которые в настоящее время рассматриваются как
производственные расы, но не самостоятельные виды.
В разных определителях объем рода Saccharomyces сильно варьирует. Ван дер
Вальт в определителе Лоддер различает 41 вид, среди которых есть диплоидные,
гаплоидные и виды с соматогамной автогамией. В. И. Кудрявцев включает в род
Saccharomyces только диплоидные дрожжи, выделяя гаплоидные виды в
Zy go saccharomyces, а виды с автогамным половым процессом — в род
Debaryomyces. H. А. Красильников рассматривает виды, объединенные у Ван дер
Вальта в Saccharomyces, в трех родах — Saccharomyces, Zygosaccharomyces и
Torulaspora. Такое деление на роды с изменениями в составе видов поддержано в
новой систематике. Согласно Д. Ярроу, в роде Saccharomyces оставлено пять ви-
дов с преимущественно диплоидной вегетативной фазой: S. cerevisiae, S.
kluyveri, S. exiguus, S. dairensis, S. servazzii.
На сегодняшний день основным и наиболее доступным для воспроизведения кри-
терием идентификации видов дрожжей в составе рода Saccharomyces следует при-
знать совокупность их культурально-биохимических свойств, главным образом от-
ношение к источникам углерода, подробно представленных в таблице ниже.
Видовая принадлежность дрожжей Saccharomyces (по Ярроу)
Физиологи-
ческие
cerevisiae
dairensis
exiguus
kluyveri
servazzii
telluris
unisporus
Брожение
Глюкоза
Галактоза
Сахароза
Мальтоза
Лактоза
+
в
в
в
-
+
+
-
-
-
+
+
+
-
-
+
+
+
-/ел
-
+
+
-
-
-
+
-
-
-
-
+
+
-
-
-
Ассимиляция
Галактоза
Сахароза
Мальтоза
Целлобиоза
Трегалоза
Лактоза
Раффиноза
Мелибиоза
D-ксилоза
Арабиноза
Рибоза
Рамноза
Раствори-
мый крах-
мал
Рибит
D-маннит
Янтарная
кислота
Лимонная
кислота
Инозит
KN03
Лизин
Этила-
мин НС1
Кадаве-
рин 2 НС1
в
в
в
-
в
-
в
в
-
-
-
-
в
-
в
-/м
-
-
-
-
-
-
+
-
-
-
в
-
-
-
-
-
-/м
-
-
-
-
-
-
-
-
-
-
-
+
+
-
-
+
-
+/м
-
-
-
-
-
-
-
-
-/м
-
-
-
-
-/м
-
+
+
+/м
в
+/м
-
+
+
-/м
-
-
-
-
-
в
+/м
-
-
-
+
+
+
+
-
-
-
+/м
-
-
-
-
-
-/м
-
-
-
-
-
-
-
-
-
+
+
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-/м
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
+
+
+
Рост на среде
Без вита-
минов
С 50% глю-
козы
-
-
-
-
-/ел
-
в
-
-
-/м
-
-
-
-
При 37 С
в
ел/-
-
+
в
+
+/сл
Рост в присутствии циклогексимида
100 мкг/мл
1000мкг/мл
Среднее
значение
Г+Ц (mol%)
-
-
39,5
+/-
-
37,5
+/-
-
34,0
-
-
40,0
+
-
34,7
-
-
33,0
+
+
32,0
в - признак варьирует, ел - слабо сбраживает или ассимилирует, м - ассими-
лирует
Дрожжи размножаются вегетативно путем почкования, деления или почкующегося
деления.
При почковании на поверхности клетки образуется маленькое выпячивание —
почка, которая увеличивается почти до размеров материнской клетки и отделяет-
ся от нее, оставляя на месте бывшего прикрепления почковый шрам или рубец в
виде кольцеобразного выступа (рис. 1).
Рис. 1. Вегетативное размножение дрожжей.
Если одновременно или последовательно образующиеся почки формируются на
разных сторонах материнской клетки, то такое почкование называют множествен-
ным или многосторонним соответственно.
Почки могут закладываться и на широком основании, которое сужается к момен-
ту отделения почки; иногда же, наоборот, почки образуются на узком и довольно
заметном выросте.
Полярное образование дочерних клеток, начинающееся с формирования почки на
широком основании и заканчивающееся появлением четко заметной даже в световом
микроскопе септы в районе перешейка, носит название почкующегося деления.
Если при почковании клетки не разъединяются, а продолжают образовывать все
новые и новые почки, то, в конце концов, формируются структуры ложных гиф, из
которых складывается псевдомицелий. Он бывает примитивным, если состоит из
продолговатых клеток приблизительно одинакового размера, или же хорошо разви-
тый, если дифференцируется на длинные клетки псевдогиф и окружающие их в ха-
рактерном порядке (кучками, мутовками, поодиночке) клетки другой формы, име-
нуемые бластоспорами.
Если дрожжи размножаются делением, то при образовании нитевидных структур
они формируют истинный мицелий. При распаде такого мицелия на отдельные клет-
ки последние носят название артроспор. На плотной среде они часто располага-
ются характерным зигзагом.
Эндоспоры — внутриклеточные бесполые споры — появляются в неопределенном
числе в клетках гиф в старых культурах. Клетки при этом иногда заметно разду-
ваются .
Способы вегетативного размножения наблюдают одновременно с описанием микро-
морфологии дрожжей на тех же средах в культурах 2-3-суточного возраста.
Клетки Saccharomyces cerevisiae размножаются вегетативным образом при помо-
щи почкования. Сначала появляется вырост на материнской клетке, затем проис-
ходит митотическое деление ядра, образование клеточной стенки и отделение
клеток друг от друга. На материнской клетке остается шрам от почкования, что
позволяет определить её возраст. Обычно материнская клетка может образовывать
20—30 почек.
Клетки дрожжей могут пребывать в одном из двух стабильных состояниях (фа-
зах) : гаплоидном (сфероиды) и диплоидном (эллипсоиды), которые считаются раз-
личными поколениями. В течение каждой фазы пекарские дрожжи размножаются ве-
гетативно почкованием. По продолжительности у пекарских дрожжей преобладает
диплоидная фаза. Она переходит в гаплоидную фазу путём образования гаплоидных
аскоспор в результате мейоза. Гаплоидная фаза переходит в диплоидную путём
слияния образовавшихся из аскоспор гаплоидных клеток.
Для выращивания оптимальными условиями является раствор дрожжевого экстрак-
та с температурой 30 С, содержащий пептон и глюкозу.
МЕТОДЫ СТЕРИЛИЗАЦИИ
ПОСУДЫ И ПИТАТЕЛЬНЫХ
СРЕД
Основываясь на влиянии внешних условий на микроорганизмы, в микробиологиче-
ской практике разработан ряд приемов, приводящих микроорганизмы к гибели. Од-
ним из таких приемов является стерилизация.
Под стерилизацией (обеспложиванием) понимают полное уничтожение микроорга-
низмов и их спор в питательных средах, посуде, на инструментах и других пред-
метах лабораторного оборудования. Для их стерильности наиболее часто пользу-
ются воздействием высокой температуры.
Стерилизация
обжиганием на
пламени горелки
Небольшие стеклянные (палочка, шпатель) и металлические (игла, петля, пин-
цет, скальпель) предметы проводят несколько раз через пламя горелки (рис. 2).
Стерилизация достигается обугливанием находящихся на их поверхности микроор-
ганизмов (рис. 3). Обжиганием на пламени пользуются и для стерилизации по-
верхности ватных пробок.
1540
1550
520
Рис. 2. Значение температуры в разных участках пламе-
ни газовой горелки.
Рис. 3. Стерилизация обжиганием на пламени горелки.
Стерилизация
кипячением
Стерилизацию металлических инструментов и резиновых трубок проводят кипяче-
нием. Так как споры некоторых бактерий сохраняют жизнеспособность при кипяче-
нии в воде в течение нескольких часов, то рекомендуется стерилизацию кипяче-
нием проводить в 2%-ном растворе карбоната натрия в течение 10 мин. В этих
условиях споры погибают.
Стерилизация
сухим жаром
Сухим жаром стерилизуют стеклянную посуду. При этом пробирки, колбы предва-
рительно закрывают ватными пробками. Чтобы избежать заражения просте-
рилизованных предметов из воздуха, их перед стерилизацией заворачивают в
оберточную бумагу и вынимают только перед работой.
Пипетки перед стерилизацией с концов закрывают ватой. Затем их обертывают
длинными полосками бумаги шириной 3,5-4 см. Бумагу наматывают по спирали, на-
чиная с конца пипетки, который будет погружен в среду. Концы обертки закреп-
ляют ниткой. Тонкие пипетки обертывают бумагой вместе по несколько штук.
Чашки Петри заворачивают в бумагу в форме квадрата, сторона которого при-
близительно равна трем диаметрам чашки. Чашку Петри помещают на середину лис-
та, загибают его с двух противоположных сторон кверху так, чтобы края налега-
ли друг на друга. Два свободных конца загибают вниз. При таком обертывании у
чашек легко различать верх и низ.
Подготовленную таким образом посуду помещают в сушильный шкаф, в котором
нагревают ее при температуре 160-170 С в течение 2 ч (с момента установления
нужной температуры). При таком нагревании погибают не только бактерии, но и
их споры.
Температуру в сушильном шкафу выше 175 С допускать не следует, так как при
этом ватные пробки буреют, а бумажная обертка становится ломкой.
Стерилизация текучим паром
(дробная стерилизация),
или тиндализация
Питательные среды, воду, резиновые трубки и другие предметы, портящиеся от
действия сухого жара, и питательные среды, портящиеся под действием высокой
температуры (среды, содержащие молоко, солод, желатину), обеспложивают дейст-
вием текучего пара.
Стерилизацию текучим паром производят в кипятильнике Коха (рис. 4) или в
автоклаве с открытым вентилем. Воду в них доводят до кипения, и образующийся
пар обтекает стерилизуемые объекты. Температура стерилизуемых питательных
сред достигает 100 С. Нагревание в течение 30-45 мин приводит к гибели веге-
тативных клеток бактерий, но споры их не погибают. Затем жидкость охлаждают
до температуры, благоприятной для прорастания спор (до 30 С). Нагревание при-
водит к активации спор и более быстрому их прорастанию. На следующий день на-
гревание повторяют. При этом погибают вегетативные клетки, развившиеся из
спор. Для обеспечения полной стерильности жидкость оставляют еще на сутки и
снова повторяют нагревание. Такую стерилизацию называют дробной или тиндали-
зацией.
Рис. 4. Аппарат Коха.
Пастеризация
В основе пастеризации лежит нагревание жидкостей до температуры меньше 100
С. Целью ее является уничтожение неспороносных бактерий в жидкостях, теряющих
питательные свойства при кипячении (молоко, пиво, вино и др.). Осуществляется
пастеризация путем нагревания жидкостей при 60 С в течение 30 мин, или при 75
С в течение 15 мин, или при 80 С в течение 10 мин.
Холодная
стерилизация
Органические жидкости, не выносящие нагревания, освобождают от бактерий,
пропуская через стерильные мелкопористые фильтры (рис. 5). Эти фильтры задер-
живают микроорганизмы, их называют бактериальными фильтрами.
X
/♦
Рис. 5. Мембранные фильтры для стерилизации.
Бактериальные фильтры имеют разные номера. Фильтры № 1 имеют средний диа-
метр пор 0,3 мкм и являются наиболее надежными. Фильтры № 5 имеют самые боль-
шие отверстия пор, диаметром 1,2 мкм.
Перед употреблением мембранные фильтры стерилизуют кипячением. Фильтры по-
мещают в теплую дистиллированную воду и кипятят 30 мин, меняя 2-3 раза воду.
Стерилизация паром
под давлением
(автоклавирование)
Наиболее надежным и универсальным методом стерилизации питательных сред и
материалов является стерилизация их насыщенным паром под давлением. Про-
изводят ее в автоклаве, в котором стерилизуемые объекты нагревают чистым на-
сыщенным паром при давлении выше атмосферного. Когда насыщенный пар встреча-
ется с более холодным объектом, он конденсируется, превращаясь в воду. При
конденсации выделяется большое количество теплоты, и температура стерилизуе-
мого объекта быстро повышается.
Полная стерилизация питательных сред при 120 С и давлении 0,1 МПа обеспечи-
вается нагреванием в течение 20 мин.
Стерилизация в автоклаве производится при повышенном давлении, поэтому ра-
бота с ним требует определенной осторожности.
Системы автоклавов различаются, но все они имеют общие принципы устройства
(рис. 6), и правила работы с ними однотипны.
Во внутренний котел автоклава (стерилизационную камеру) помещают материал,
подлежащий стерилизации.
В водопаровую камеру наливают воду с таким расчетом, чтобы уровень ее был
между верхней (максимальной) и нижней (минимальной) чертой.
Рис. 6. Небольшой автоклав и его аналог - скороварка.
Крышку автоклава привинчивают болтами к корпусу. Завинчивают болты попарно,
крест-накрест, чтобы избежать перекоса крышки, который может возникнуть при
завинчивании болтов по кругу.
Открывают краны и включают источник обогрева. Когда пар из выпускного крана
начинает выходить непрерывной струей, его закрывают и наблюдают за постепен-
ным повышением давления в рабочей камере по манометру.
Отсчет времени стерилизации начинают с того момента, когда в автоклаве ус-
тановится заданное давление.
Между показаниями манометра и температурой кипения воды имеется определен-
ная зависимость (табл. ниже). Время от времени эти соотношения следует прове-
рять . Нарушение их указывает на неисправность автоклава и на необходимость
его ремонта.
Проверку осуществляют следующим образом: в стерилизационную камеру автокла-
ва помещают 100 г бензойной кислоты с добавлением небольшого количества фук-
сина или метиленовохю синего. Если при показании манометра в 0,1 МПа бензой-
ная кислота расплавится, образуя с красителем сплав, то, значит, автоклав да-
ет нужную температуру (120 С).
Зависимость давления и температуры в камере автоклава
Показатели
манометра
МПа
0,00
0,02
0,04
0,05
0,06
атм
0,0
0,2
0,4
0,5
0,6
Температура
кипения воды
100
105
110
112
114
Показатели
манометра
МПа
0,07
0,08
0,09
0,10
0,15
атм
0,7
0,8
0,9
1,0
1,5
Температура
кипения воды
116
117
119
121
127
После окончания заданного срока стерилизации источник нагрева выключают,
перекрывают вентиль водо-паровой камеры и только после этого постепенно от-
крывают выпускной клапан. При быстром выпускании пара могут быть вырваны ват-
ные пробки из стерилизуемой посуды.
После полного выхода пара отвинчивают болты крышки (снова крест-накрест) и
открывают ее, ориентируя крышку на себя для защиты от выходящего пара.
Если во время стерилизации давление начинает подниматься выше заданного
уровня, его регулируют, уменьшая нагрев или выпуская часть пара через предо-
хранительный клапан. Последний должен быть отрегулирован так, чтобы при повы-
шении давления излишек пара выходил автоматически.
ПИТАТЕЛЬНЫЕ
СРЕДЫ
Среды общего
назначения
В качестве основного компонента питательных сред может быть использовано
зерновое, виноградное, плодово-ягодное или мелассовое сусло. Однако наиболее
универсальным и сбалансированным следует признать солодовое неохмеленное сус-
ло с содержанием 8-10% СВ.
Солодовое сусло
Для приготовления сусла в лаборатории используют ячменный солод крупного
помола с высокой осахаривающей способностью. В 1 л воды вносят 250 г солода и
нагревают до 50 С. Через 30 мин температуру повышают до 55 С, а еще через 30
мин постепенно поднимают до 62,5-63 С. При этой температуре выдерживают до
исчезновения реакции на крахмал (йодная проба).
Полученное сусло отделяют от дробины путем фильтрации через марлю и вату,
разбавляют водой до нужной концентрации, разливают по колбам и стерилизуют
при 112 С 30 мин. Если рН ниже 5,5, то сусло подщелачивают 10%-ным раствором
соды или едкого кали до получения рН 5,6-6,0. Полученное таким образом сусло
желательно очистить от примесей кизельгуром или отстаиванием в холодильнике и
фильтрацией через бумажный фильтр.
В отсутствие солодового вполне применимо зерновое сусло, осахаренное ком-
плексом ферментных препаратов с обязательным внесением дрожжевой золки и 0,3-
0,5% дрожжевого автолизата перед автоклавированием.
Для приготовления жидкой питательной среды осахаренное и отфильтрованное
сусло разливают в колбы или пробирки на 1/2-3/4 объема, закрывают ватно-
марлевыми пробками и автоклавируют 20 мин при 0,1 МПа.
Мелассовое сусло
Густую мелассу отвешивают на технических весах в количестве 200-300 г, сме-
шивают с водой в соотношении 1:3 и нагревают до 75 С, затем насыпают 4% су-
перфосфата и повышают температуру до 85 С при постоянном помешивании. 33%-ный
раствор едкой щелочи в количестве 0,8% по объему к исходной мелассе разводят
в 30-50 см3 воды, добавляют к нагретой мелассе и хорошо перемешивают. В ре-
зультате происходит выпадение осадка трикальцийфосфата, который увлекает за
собой коллоиды мелассы и более крупные частицы, осадок быстро оседает на дно,
и меласса осветляется. После этого к мелассе добавляют 2,5% сернокислого ам-
мония и 2% суперфосфата в виде вытяжки (1 г суперфосфата и 10 см3 воды нагре-
вают до кипения и фильтруют через бумажный фильтр). Мелассу перемешивают с
питательными солями и фильтруют через бумажный фильтр, лучше под вакуумом на
воронке Бюхнера.
Осветленную мелассовую среду разводят водой до требуемой концентрации 5-8%
СВ по сахарометру и разливают в колбы, пробирки или бутылки; для приготовле-
ния твердых питательных сред добавляют агар.
Мелассовое сусло стерилизуют в автоклаве при избыточном давлении 0,05 МПа
30 мин или в аппарате Коха при 100 С в течение 1 ч трое суток подряд.
Сусло-агар
В химическом стакане в сусло (8-10% СВ) внести необходимое количество агар-
агара (2,5 г на 100 см3) и выдержать 10-15 мин, не перемешивая до полного его
увлажнения. Стакан поставить на слабый огонь и, постоянно перемешивая, дове-
сти до кипения. Содержимое стакана перелить в колбу до 1/2 объема, закрыть ее
ватно-марлевой пробкой и автоклавировать 20 мин при 0,1 МПа.
Если питательная среда не требует внесения каких-либо термолабильных ингре-
диентов, то ее можно сразу после автоклавирования разливать в заранее приго-
товленные стерильные высушенные чашки Петри. Если же предполагается внесение
антибиотиков, факторов роста или других чувствительных к температуре компо-
нентов, то среду в колбе необходимо остудить до 40-45 С, стерильно внести
нужные компоненты, перемешать и разливать в чашки Петри. При этом желательно
избегать образования комков для получения ровной поверхности питательной сре-
ды в чашках. После этого чашки со средой не следует более перемещать, пока
агар полностью не остынет.
Чтобы удалить образовавшийся конденсат и уменьшить его дальнейшее образова-
ние , чашки необходимо подсушить. Для этого чашки с агаром переворачиваются
крышкой вниз, дно чашки с агаром извлекается и ставится на ребро крышки. В
таком положении в боксе под УФ-лампой чашки выдерживаются 60 мин. Высушенные
чашки закрываются и могут использоваться для работы либо храниться в холо-
дильнике завернутыми в полиэтиленовый пакет, чтобы исключить пересыхание или
случайное обсеменение среды. Чашки, хранившиеся в холодильнике, перед работой
следует некоторое время выдержать при комнатной температуре или в термостате.
Скошенный сусло-агар
Для приготовления скошенного сусло-агара доведенную до кипения среду (см.
выше) разливают в пробирки до 1/3 ее объема, закрывают ватно-марлевыми проб-
ками и автоклавируют 20 мин при 0,1 МПа. По окончании стерилизации пробирки с
горячей средой раскладывают на наклонной поверхности для образования скоса
нужной высоты и дают полностью застыть. Остывшие пробирки переносят в штатив
и используют по назначению.
Полужидкий сусло-агар
В химическом стакане в сусло (8-10% СВ) внести необходимое количество агар-
агара (0,5 г/100 см3) и выдержать 10-15 мин, не перемешивая до полного его
увлажнения. Стакан поставить на слабый огонь и, постоянно перемешивая, дове-
сти до кипения. Содержимое стакана разлить в пробирки до 1/3-1/2 объема, за-
крыть ватно-марлевыми пробками и автоклавировать 20 мин при 0,1 МПа.
По окончании стерилизации пробирки раскладывают вертикально в штатив и по-
сле полного их охлаждения используют по назначению.
Ацетатная среда для выделения посторонних дрожжей
На 1000 см водопроводной воды берут 10 г уксуснокислого натрия, 10 г хло-
ристого аммония и 5 г глюкозы.
Дрожжевой автолизат добавляют в среду в количестве 3 см3.
Среду разливают в пробирки по 5 см3 и стерилизуют в автоклаве при избыточ-
ном давлении 0,05 МПа в течение 30 мин.
Среда с монойодуксусной кислотой
2%-ный сусло-агар после автоклавирования охлаждают до 50 С. В остывшую сре-
ду асептически вносят рабочий раствор монойодуксусной кислоты до концентрации
2,5% (97,5 см3 + 2,5 см3 кислоты). Приготовленную среду тщательно перемешива-
ют и разливают в чашки Петри. Конечная рН 5,8.
Посевы исследуемых проб инкубируют не менее 120 ч при 28-30 С.
На данной среде растут дрожжи, не относящиеся к роду Saccharomyces.
Рабочий раствор. В предварительно проавтоклавированную воду внести монойо-
дуксусной кислоты в количестве 7,44 мг на 1,0 см3.
Суело-желатин
К солодовому суслу концентрацией 8-10% СВ прибавляют 15% желатина и через
30-40 мин, когда желатин набухнет, среду подогревают до 40-50 С, фильтруют
через ватно-марлевый фильтр и разливают в пробирки или колбы.
Стерилизуют в аппарате Коха при 100 С по 30 мин трое суток подряд.
Среда Сабуро для выращивания дрожжей
К 100 см стерильной дрожжевой воды добавляют 5 г пептона, 4 г глюкозы, 1,8
г агара. После того как агар расплавится при нагревании, среду фильтруют и
разливают в пробирки.
Стерилизуют 20 мин при избыточном давлении 0,05 МПа или в аппарате Коха
дробной стерилизацией.
Синтетическая среда с лизином
Рецептура синтетической среды с лизином следующая:
■ Глюкоза . . . 50 г
■ Лизин .... 3 г
■ КН2Р04 1 г
■ MgS04 1 г
■ FeS04 следы
Каждый компонент среды растворяют в воде отдельно, смешивают в указанном
порядке и доводят водопроводной водой до 1000 см3.
В полученную жидкую синтетическую среду вносят агар (2,5%), расплавляют и
разливают в пробирки по 10 см3. Среду стерилизуют 20 мин при давлении 0,05
МПа.
После автоклавирования пробирки ставят в штатив для получения скошенной
среды.
Виноградное сусло
Сусло помещают в колбу и нагревают до кипения в кипятильнике Коха, после
охлаждения отфильтровывают от выпавших белковых веществ через бумажный склад-
чатый фильтр и разливают в пробирки по 5 см3, стараясь не смочить горлышка,
затем в пробирках пастеризуют.
Предназначается для выращивания дрожжей.
Яблочное сусло
В свежий или консервированный яблочный сок добавляют при необходимости са-
хар до массовой концентрации 10-12 г/100 см3. Концентрация титруемых кислот
не должна превышать 6-7 г/дм3. При излишней кислотности сок разбавляют водой.
Стерилизуют в автоклаве текучим паром или в кипятильнике Коха однократно в
течение 40-45 мин.
Применяют для культивирования дрожжей.
Картофельная вода
20 г очищенного от кожуры и протертого сырого картофеля добавляют к 1000
см3 водопроводной воды, настаивают 4 ч, кипятят 15 мин, фильтруют через
складчатый фильтр, разливают в пробирки по 5 смЗ и стерилизуют при избыточном
давлении 0,1 МПа в течение 2 0 мин.
Дрожжевая вода
На 1000 см водопроводной воды берут 80 г прессованных дрожжей или 20 г су-
шеных, смесь кипятят в течение 20 мин, разливают в бутылки и стерилизуют при
избыточном давлении 0,1 МПа в течение 30-40 мин.
После отстаивания в течение нескольких дней дрожжевая муть оседает, а дрож-
жевая вода становится прозрачной.
Дрожжевой автолизат
Вариант 1. Для приготовления можно использовать прессованные или сушеные
пекарские дрожжи.
200 г прессованных дрожжей размешивают с 100 см3 водопроводной воды или 100
г сушеных дрожжей с 400 см3 воды, добавляют поваренной соли в количестве
0,15% к весу прессованных дрожжей и 0,5% к весу сушеных дрожжей, смесь поме-
щают в термостат при 50-55 С на 48 ч, затем нагревают до кипения и фильтруют
через складчатый бумажный фильтр, разливают в стеклянную посуду и стерилизуют
при избыточном давлении 0,1 МПа в течение 30 мин.
Вариант 2. Прессованные дрожжи заливают равным по массе количеством воды:
полученную суспензию клеток покрывают тонким слоем толуола и выдерживают трое
суток при температуре 60 С, время от времени перемешивая. После этого суспен-
зию несколько минут кипятят, дают остыть и с помощью 1 Н раствора КОН доводят
значение рН до 6, затем отстаивают. Надосадочная жидкость (дрожжевой автоли-
зат) должна быть прозрачной, ее сливают и отфильтровывают через бумажный
фильтр на воронке Бюхнера под разряжением.
Стерилизуют автоклавированием 20 мин при 112 С.
Состав основных синтетических сред
Ингредиенты (на 1000
см3 дистил. воды)
Морфологический
агар
Азотистая
основа
Углеводная
основа
Безвитаминная
среда
Источники углерода и азота, г
Глюкоза
(NH4)2S04
Аспарагин
10
3,5
1,5
—
5
—
10
—
—
10
5
—
Макроэлементы, г
кн2ро4
К2НР02
MgS04
NaCl
СаС12
0,85
0,15
0,5
0,1
0,1
0,85
0,15
0,5
0,1
0,1
0,85
0,15
0,5
0,1
0,1
0,85
0,15
0,5
0,1
0,1
Аминокислоты, мг
L-гистидин' НС1
DL-метионин
DL-триптофан
10
20
30
10
20
20
1
2
2
10
20
20
Витамины, мкг
Пантотенат кальция
Фолиевая кислота
Инозит
Никотиновая кислота
Парааминобензойная
кислота
Пиридоксин' НС1
Рибофлавин
Тиамин НС1
Биотин
2000
2
10 000
400
200
400
200
400
20
2000
2
10 000
400
200
400
200
400
20
2000
2
10 000
400
200
400
200
400
20
—
—
—
—
—
—
—
—
—
Микроэлементы, мкг
Н3Р03
CuS04
KI
FeCl5
MnS04
Na2Mo04
ZnS04
Промытый агар, г
Количество сухой гото-
вой среды фирмы Difco
на 1 л, г
500
40
100
200
400
200
400
18
35
500
40
100
200
400
200
400
—
6,7
500
40
100
200
400
200
400
—
11,7
500
40
100
200
400
200
400
—
16,7
Стерильный мел
Имеющийся в продаже мелкоразмолотый мел насыпают в стерильные пробирки с
ватными пробками по 0,3-0,5 г и стерилизуют сухим жаром 1 ч при 170 С.
Стерильная водопроводная вода
Водопроводную воду разливают в пробирки по 10 или 9 см3 и в колбы по 50 и
100 см3, закрывают ватными пробками и стерилизуют в автоклаве 30 мин при дав-
лении 0,1 МПа или в аппарате Коха при 100 С по 1 ч трое суток подряд.
Среды
накопительные
Приготовленные среды автоклавируются при 0,1 МПа в течение 20 мин. Могут
использоваться как основа для агаризованных сред.
При необходимости вносятся антибиотики.
Состав и количество ингредиентов накопительных сред показаны в таблице ни-
же .
Среды накопительные для дрожжей
Вариант
1
2
3
Ингредиенты
Глюкоза (сахароза)
Дрожжевая зола
Дрожжевая вода (0,15 г/см3)
Дрожжевой автолизах
Дрожжевая зола
Сусло неохмеленное
Глюкоза (сахароза)
Кислота лимонная
Дрожжевой автолизат
Дрожжевая зола
Вода
Количество
80,0 см3
5,0 г
До 1000,0 см3
2,0 см3
5,0 г
До 1000,0 см3
80,0 г
6,0 г
10,0 см3
5,0 г
До 1000,0 см3
Среды для
аскоспорообразования
На средах для спорообразования дрожжи инкубируют при температурах, несколь-
ко меньших в сравнении с оптимальными для вегетативного роста, — в большинст-
ве случаев при 20-25 С.
Время инкубации составляет до 4-6 недель при еженедельном микроскопирова-
нии.
Начинающие исследователи нередко затрудняются отличить аскоспоры от внутри-
клеточных бесполых спор.
Рис. 7. Споры дрожжей Saccharomyces с флуоресцентным
красителем под микроскопом.
Среда Городковой для выявления спор у дрожжей
■ Пептон 1,0 г
■ Поваренная соль 0,5 г
■ Глюкоза 0,25 г
■ Агар 2,0 г
■ Вода до 100,0 см3
Иногда вводят еще 1% мясного экстракта.
Среду нейтрализуют содой (питьевой) до рН 7,3, доводят до кипения и кипятят
до тех пор, пока не расплавится агар, фильтруют и разливают в пробирки по 5
см3.
Стерилизуют при избыточном давлении 0,05 МПа в течение 20 мин.
Ацетатный агар Фоувелла
На 1000 см дистиллированной воды берут 5 г CH3COONa'3H20 и 20 г агара. рН
среды должно быть 5-7. Автоклавируют 15 мин при 121 С.
Ацетатный агар Мак-Клари
■ CH3COONa3H20 8,2 г
■ Глюкоза 1,0 г
■ Дрожжевой экстракт 2,5 г
■ Дистиллированная вода до 1000,0 см3
Автоклавируют 15 мин при 121 С.
Среда с дрожжевым экстрактом и глюкозой
■ Дрожжевой экстракт 5,0 г
■ Глюкоза 50,0 г
■ Дистиллированная вода до 1000,0 см3
Автоклавируют 20 мин при 112 С.
Агаризованная дрожжевая вода
Готовят, добавляя к 1000 см дрожжевой воды 15 г агара.
Автоклавируют 15 мин при 121 С.
Среда Старки
■ К2НР04 1,0 г
■ СаС12 0,05 г
■ Водопроводная вода до 1000,0 см3
Автоклавируют 15 мин при 112 С. После стерилизации добавляют 6,0 см3 этило-
вого спирта.
Овощной агар
Предварительно готовят экстракт овощей путем автоклавирования 10 мин при
112 С равных частей промытых и мелко нарезанных моркови, сахарной свеклы,
огурцов и картофеля в четырех частях воды, после чего экстракт отфильтровыва-
ют через ткань, отжимая осадок.
К 1000 см3 полученного экстракта (4% СВ, рН 6,7) добавляют 20 г сухих дрож-
жей и 20 г агара.
Автоклавируют 15 мин при 121 С.
Гипсовые блоки
Восемь частей гипса (2CaS04'H20) смешивают с тремя частями воды и помещают
пасту в цилиндрические формы из бумаги или фольги высотой 3-4 см. Поверхность
тщательно сглаживают. После застывания блоки освобождают от форм, помещают в
высокие чашки и стерилизуют при 110-120 С в течение 2 ч. Перед посевом в чаш-
ки наливают стерильной воды слоем 1 см или раствор, содержащий 1% маннита и
0,5% К2НР04.
Осветление
сред
Осветленные жидкие, агаризованные или желатиновые среды необходимы в диаг-
ностических исследованиях и для получения хорошо видимых признаков глубинного
роста и изолированных колоний микроорганизмов.
В ряде случаев прозрачную среду можно получить, отфильтровав ее от осадка
через вату. Когда этого бывает недостаточно, среды осветляют с помощью белков
куриных яиц. Для осветления 500 см3 среды достаточно белка одного яйца. Белок
тщательно отделяют от желтка и встряхивают с равным объемом воды до образова-
ния густой пены. Взбитый белок выливают в предварительно расплавленную и за-
тем остуженную до 45-50 С среду. Перед внесением белка проверяют значение рН
среды и, если необходимо, среду подщелачивают до рН 7,0-7,3. Среду с белком
тщательно перемешивают и прогревают в кипящей водяной бане в течение часа.
Белок свертывается и адсорбирует все взвешенные в среде частицы. Среду быстро
отфильтровывают в горячем виде через вату.
Синтетические агаризованные среды, внесение белка в которые нежелательно,
осветляют следующим образом. Среду, налитую в химический стакан, автоклавиру-
ют и оставляют после стерилизации в закрытом автоклаве на 10-12 ч. При таком
медленном остывании среды все взвешенные частицы оседают на дно. Застывшую
среду извлекают из стакана, прозрачную часть среды срезают, помещают в колбу
и вновь стерилизуют.
ВЫДЕЛЕНИЕ
ЧИСТОЙ
КУЛЬТУРЫ
Источником чистой культуры дрожжей могут быть лабораторные коллекционные
культуры, производственные субстраты и естественные экологические ниши.
Чистой микробной культурой называют популяцию, представляющую собой потом-
ство одной или нескольких клеток одного вида микроорганизма.
Каждый новый изолят носит название штамма, которому присваивают буквенное
или номерное обозначение.
Расами называют производственные штаммы одного вида дрожжей, различающиеся
между собой по степени проявления физиологической активности.
В генетических исследованиях пользуются так называемыми клонами — чистыми
культурами, полученными от одной споры или гаплоидной клетки.
Основной задачей при исследовании любого материала является получение чис-
той культуры, т. е. изолированных колоний тех организмов, которые находятся в
субстрате. При этом важнейшим условием успешной работы является правильный
выбор последовательности ее выполнения и подбор необходимых питательных сред.
Существуют прямые и непрямые методы получения чистых культур дрожжей.
Первые основаны на выделении одной клетки или споры под непосредственным
контролем через микроскоп.
Во втором случае используют косвенные приемы для разделения клеток.
Прямые
методы
Капельный способ Линднера
Суспензию дрожжей разбавляют жидким суслом до концентрации ^100 клеток в 1
мл и стерильным чертежным пером наносят мельчайшие капельки на необезжирен-
ное покровное стекло, простерилизованное фламбированием в пламени горелки.
Капельки располагают в определенном порядке, обычно по 5 капель в два ряда,
т. е. всего 10 капель на стекле. Затем быстро опрокидывают стекло над влажной
камерой, которую запечатывают минеральной смазкой. Все капли немедленно про-
сматривают под микроскопом и отмечают те из них, которые содержат по одной-
единственной клетке. Если все капли содержат по несколько клеток, то увеличи-
вают разбавление суспензии и процедуру повторяют. После трех-, четырехдневно-
го инкубирования, когда отмеченные единичные клетки образуют микроколонии,
последние переносят стерильной иглой или полоской стерильной фильтровальной
бумагой в жидкую среду и инкубируют.
Метод Линднера, упрощенный Вучковичем
Видоизменение Вучковичем метода Линднера сводится к тому, что капельки сус-
пензии, содержащие 3-4 клетки дрожжей, снимают петлей, которой предварительно
захватывают стерильную среду, и переносят штрихом на поверхность плотной сре-
ды. Через некоторое время появляются микроколонии, число которых на одном
штрихе должно соответствовать количеству исходных клеток в отмеченной капель-
ке . Из этих колоний пересевом выделяют чистые культуры. Таким способом можно
сразу получить несколько одноклеточных культур за короткое время.
Выделение спор дрожжей при помощи микроманипулятора
К использованию микроманипулятора прибегают главным образом при необходимо-
сти изолировать споры из асков, так как вегетативные клетки легко повреждают-
ся при микроманипулировании. Аски разрывают либо механическим прикосновением
игл микроманипулятора, либо заранее обрабатывая суспензию препаратом фермен-
тов (например, из пищеварительного тракта виноградной улитки Helix pomatia),
лизирующих клеточную стенку дрожжей.
Непрямые
методы
В эту группу включают методы, основанные на разделении клеток в питательных
средах и использовании специфических биологических особенностей отдельных ви-
дов для создания преимущественных условий для их роста.
Основным методом выделения чистых культур микроорганизмов является метод,
предложенный Р. Кохом. Принцип его заключается в получении чистой культуры из
изолированной колонии.
На поверхность плотных питательных сред из пипетки наносят каплю накопи-
тельной культуры или ее разведения в стерильной воде и стерильным стеклянным
шпателем Дригальского распределяют каплю по поверхности среды в чашке Петри
(рис. 8). Далее этим же шпателем протирают поверхность среды последовательно
во второй, третьей и четвертой чашках. Обычно в первых двух чашках после ин-
кубации наблюдается сплошной рост микроорганизмов, тогда как в последующих —
рост изолированных колоний.
Рассевать накопительную культуру можно бактериологической петлей методом
истощающего штриха. В этом случае накопительную культуру или ее разведение
отбирают петлей и на поверхности плотной среды проводят штрихи в порядке,
указанном на рисунке 9. Перед взятием каждого нового инокулята петлю стерили-
зуют над пламенем горелки.
После посева чашки Петри помещают в термостат крышками вниз, чтобы конден-
сационная вода, образовавшаяся на крышке чашки при застывании агара и впо-
следствии при инкубации, не стекала на поверхность среды и не помешала полу-
чить изолированные колонии. Чашки выдерживают в термостате в течение 7-10 су-
ток при температуре 28-30 С,
" У
?i*\i*f
Рис. 8. Рассев культуры микроорганизмов на поверхность плотной
питательной среды шпателем Дригальского: а — шпатель Дригальско-
ро ; б — рассев; в — рост микроорганизмов.
Рис. 9. Рассев культуры микроорганизмов на поверхность плотной
питательной среды бактериологической петлей: 1-4 — последова-
тельность контакта бактериологической петли с поверхностью плот-
ной питательной среды.
В зависимости от первичного источника выделения и интенсивности его обсеме-
нения следует принимать решение об использовании накопительных (элективных)
сред на стадии посевов первичного материала.
Внесение в питательные среды селективных компонентов, их качественный и ко-
личественный состав определяются интенсивностью и биологическим разнообразием
обсеменения первоначального субстрата посторонней сопутствующей плесневой,
дрожжевой и бактериальной микрофлорой. При выделении дрожжей из сильно за-
грязненных субстратов и подавления роста сопутствующих микроорганизмов к ука-
занным выше питательным средам добавляют различные ингибиторы селективного
действия.
Рост большинства бактерий и актиномицетов подавляется при низком значении
рН среды, поэтому чаще всего среды для выделения дрожжей подкисляют до рН 4,5
путем добавления к ним минеральных или органических кислот. Для синтетических
сред наиболее применима соляная кислота, для сусловых — молочная, лимонная
или винная. Для подкисления солодового сусла (8% СВ) обычно требуется концен-
трированной молочной кислоты 3-4 см3/л. Кислоты добавляют в жидкую или рас-
плавленную агаровую среду после стерилизации, непосредственно перед засевом
или перед разливкой в чашки Петри.
Вместо кислот используют также антибиотики широкого спектра действия:
стрептомицин (80 мг/л или 100 ед/см3) , пенициллин (20-100 ед/см3) , левомице-
тин (50 мг/л) и др. Их можно добавлять в среду порознь и в комплексе. На-
пример, против кислотоустойчивых бактерий применяют следующие смеси антибио-
тиков: пенициллин 60 ед/см3 + стрептомицина сульфат 100 ед/см3; левомицетин
(хлорамфеникол) 20 мг/л + стрептомицина сульфат 20 мг/л + хлортетрациклин со-
лянокислый 100 мг/л.
Для отделения сахаромицетов от других дрожжей добавляют 2,5% этилацетата и
рН до 4,0 доводят уксусной кислотой. Затем чашки герметизируют. При этих ус-
ловиях колонии сахаромицетов появляются первыми.
Росту дрожжей на питательных средах при высеве из исходного материала за-
частую препятствуют грибы с широко распространяющимися по поверхности суб-
страта мицелиальными колониями. Они имеют общие с дрожжами потребности в ис-
точниках питания, устойчивы к низким значениям рН среды и нечувствительны к
действию указанных выше противобактериальных антибиотиков. Для ограничения
роста микромицетов в среду добавляют специфические вещества: дифенил (0,005-
0,01%), бычью желчь (0,25-0,5%), теллурат калия (0,05-0,15%), пропионат на-
трия (0,15-0,25%) , или некоторые красители: бромкрезоловый пурпурный (0,0025%
или 2,5 см3/л 1% раствора), бенгальский розовый (0,003%), кристаллический
фиолетовый (0,001%).
Поскольку дрожжи являются факультативными анаэробами и не требуют особых
условий аэрирования, для получения их изолированных колоний посевы из накопи-
тельной культуры или другого исследуемого материала лучше производить на по-
верхность 2,5%-ного сусло-агара и инкубировать в обычных условиях при 28-30
С.
ОТБОР НАИБОЛЕЕ
АКТИВНОЙ КУЛЬТУРЫ
Стандартный
метод
Пробу исследуемой культуры дрожжей высевают в чашку Петри на сусло-агар.
Через 6-7 суток выращивания дрожжей в термостате при 30 С из наиболее крупных
колоний делают пересевы на косой сусло-агар и одновременный посев одной петли
в пробирку с 15 см3 солодового сусла.
Пробирки с посевом ставят в термостат с температурой 30 С. Через 42-48 ч
сусло осторожно сливают с дрожжевого осадка.
В маленькие пробирки (длина 12 см, диаметр 1,2 см), одинаковые по диаметру,
насыпают по 1 г муки, употребляемой для обычной проверки подъемной силы дрож-
жей.
В пробирки с дрожжевым осадком наливают по 2 см3 нагретой до 30 С водопро-
водной воды, осадок тщательно взбалтывают в воде петлей диаметром 0,9 см, из-
готовленной из толстой проволоки, затем дрожжевую суспензию выливают в про-
бирку с мукой и размешивают до равномерной консистенции. Верхний уровень этой
мучной болтушки отмечают восковым карандашом, все пробирки ставят в термостат
при 30 С.
Через каждые 15 мин вынимают пробирки, быстро отмечают новый уровень подъе-
ма жидкого теста и обычной линейкой замеряют высоту подъема от первой отметки
до второй.
Наблюдение длится 60 мин.
Предлагаемую в качестве примера таблицу можно использовать для регистрации
полученных результатов.
Активность исследуемой культуры
Номер
колонии
1
2
3
4
Подъем жидкого теста (см) при 30 С (мин)
15
30
45
60
Если дрожжи из отмеченной по лучшему подъему колонии имеют также лучшие по-
казатели по микроскопии (клетки однородные, примеси посторонних микроорганиз-
мов нет), то эту пробирку с колониями на косом сусло-агаре используют для по-
севов при выпуске следующей генерации чистой культуры дрожжей.
Модифицированный
метод
Производственную или коллекционную культуру дрожжей высевают на чашку Петри
с 2,5%-ным сусло-агаром и инкубируют при 30 С в течение 72-96 ч. По окончании
инкубации выбирают 3-5 наиболее крупных колоний, характер роста и морфология
клеток которых соответствуют исследуемой расе. Из каждой колонии одновременно
делают пересевы на скошенный сусло-агар и в колбу с 50 см3 солодового сусла
плотностью 8% СВ. Оба посева инкубируют при 30 С в течение 72 ч. Культуру на
скошенном агаре хранят до окончания эксперимента при 2-4 С, а культуру на
сусле используют для проведения анализа.
В заранее взвешенные с точностью до 0,01 г градуированные центрифужные про-
бирки разливают по 10 см3 тщательно перемешанной жидкой культуры дрожжей.
Центрифугируют 15 мин при частоте вращения 2000 об/мин, после чего сливают
надосадочную жидкость и снова взвешивают пробирку с осажденной дрожжевой био-
массой. По разнице второго и первого взвешиваний определяют массу осажденных
дрожжей в каждой пробирке. Для дальнейшей работы отбирают две из них с наи-
большим количеством биомассы с целью постановки параллельных проб. Исходя из
полученных результатов, рассчитывают количество необходимых ингредиентов —
муки пшеничной 1-го сорта и воды.
Полученная масса отпрессованных в центрифуге дрожжей должна составить 2% к
массе используемой в эксперименте муки и 1% к объему воды, т. е. на каждые
0,01 г дрожжей вносят 0,5 г муки 1-го сорта и 1 смЗ воды.
Далее в градуированную пробирку объемом 25 мл вносят необходимое количество
муки.
В центрифужную пробирку с осажденными дрожжами добавляют необходимое коли-
чество воды и тщательно перемешивают до однородной суспензии, которую сразу
переносят в пробирку с мукой.
Петлей из толстой проволоки смесь перемешивают до получения равномерной
мучной болтушки, после чего регистрируют ее первоначальный объем.
Полученные таким образом пробы помещают в термостат при 30 С. Наблюдение
ведут в течение 90 мин, при этом каждые 15 мин регистрируют уровень подъема
жидкого теста.
По окончании инкубирования рассчитывают отношение объема жидкого теста на
каждый момент регистрации к его первоначальному объему. Среднее арифметиче-
ское между полученными результатами считают показателем активности выбранной
культуры.
Такой метод учета результатов позволяет проследить динамику подъема жидкого
теста, которая может различаться у культур при одинаковом объеме теста на ко-
нец опыта.
Лучшей считается проба с наибольшей средней величиной шести зарегистриро-
ванных показателей подъема теста, а культура, используемая для этой пробы, —
наиболее активной.
Манометрический
метод
Для проведения анализа необходимо собрать несложный прибор, состоящий из
бродильного сосуда (100-120 см3), медицинского анероидного манометра и газо-
отводной трубки с краником (рис. 10). Перед началом работы следует точно из-
мерить внутренний объем прибора Vi, величина которого в дальнейшем будет ис-
пользоваться как постоянная.
Производственную или коллекционную культуру дрожжей пересеять на чашку Пет-
ри с 2,5%-ным сусло-агаром и инкубировать при 30 С в течение 96-120 ч. По
окончании инкубации выбрать 3 наиболее крупные колонии, характер роста и мор-
фология клеток которых соответствуют исследуемой расе.
Каждую из выбранных колоний рассеять на скошенный агар и в две чашки Петри,
при этом на поверхность агара в чашках перенести большую часть колонии для
получения в них сплошного роста.
Все посевы инкубировать при 30 С в течение 120-140 ч.
Культуру на скошенном агаре хранить до окончания эксперимента при 2-4 С.
Дрожжевую биомассу, полученную в чашках Петри, использовать для анализа.
Рис. 10. Схема прибора для проведения анализа: 1 — колба; 2 —
резиновая пробка; 3 — газоотводная трубка; 4 — краник; 5 — под-
вижная часть; 6 — манометр; 7 — дрожжевая суспензия.
Полученный в чашках сплошной слой дрожжей снять с поверхности агара шпате-
лем и отвесить 0,5 г. Биомассу перенести в 10 см3 физиологического раствора
температурой 30 С и размешать до получения однородной суспензии, которую сра-
зу перелить в бродильный сосуд. Туда же добавить 10 см3 8-10%-ного солодового
сусла и герметично закрыть резиновой пробкой с газоотводной трубкой. Свобод-
ный конец трубки соединить гибким шлангом с манометром. Для уравнивания дав-
ления в сосуде с атмосферным давлением извлечь подвижную часть краника и сра-
зу вернуть его на место, тщательно притерев и совместив отверстия.
Подготовленный таким образом прибор поместить в термостат при 30 С и реги-
стрировать время, за которое стрелка манометра достигнет необходимой отметки,
соответствующей 10 см3 выделенного углекислого газа.
Соответствующее этим условиям показание манометра рассчитать по формуле
Рх = mRT/VxM ,
где Рх — показание манометра; m = 0,016; R = 62 360 мм рт. ст./моль К; Т =
303 К; М = 44 г; Vx = Vi - 20,5 см3 (объем суспензии) .
Подставив значения в формулу, определить давление, создаваемое газом массой
0,016 г и объемом 10 см3 в используемом объеме Vx при температуре Т.
Время, за которое стрелка манометра достигнет определенной по формуле вели-
чины, считать показателем активности.
Лучшей считать культуру, затратившую минимальное время на выделение 10 мл
углекислого газа, а значит, обладающую более высокой ферментативной активно-
стью.
Окончательное решение принимать только с учетом культуральных и морфологи-
ческих свойств исследуемой культуры.
Хранившуюся на скошенном агаре дублирующую культуру, показавшую лучшие ре-
зультаты, использовать для разводочного цикла.
ЖИЗНЕСПОСОБНОСТЬ
ДРОЖЖЕЙ
Наиболее простым для проверки жизнеспособности дрожжевых клеток в суспензии
является метод с использованием метиленовохю синего. Этот метод обусловлен
тем, что метиленовый синий благодаря дыхательной активности клеток быстро ре-
дуцируется и теряет окраску. При отмирании клетки становятся более проницаемы
для метиленового синего, и при отсутствии дыхательной активности цвет остает-
ся синим. Таким образом, метиленовый синий указывает на отсутствие у клеток
дыхательного метаболизма, а не на их гибель как таковую, причем существует
возможность, что некоторые метаболически активные клетки оказываются не спо-
собными к размножению, в связи с чем они будут отнесены к мертвым.
Если число жизнеспособных клеток превышает 90%, этим можно пренебречь, но
при снижении числа таких клеток менее чем до 90% эта погрешность становится
все более значимой. Общее количество клеток в 1 см3 и количество «синих» кле-
ток измеряется в камере для подсчета (рис. 11). Для измерения процента жизне-
способных клеток с точностью 0,5% необходимо насчитать не менее 200 клеток.
Для получения большей точности необходимо наличие не менее 600 клеток, причем
не менее чем в двух отдельных пробах. Использовать два раздельных анализа
следует потому, что среднее из двух результатов обеспечит большую точность
результатов.
Рис. 11. Счетная камера Горяева (для подсчета под микроскопом).
Другие красители дают лучшие результаты, например метиленовый фиолетовый.
Как бы то ни было, в настоящее время общепризнано, что информация о процен-
те жизнеспособных клеток не позволяет гарантировать приемлемое состояние за-
севных дрожжей. Термин «жизнеспособность» отражает способность дрожжей к бы-
строму и эффективному сбраживанию, а не состояние «быть живым».
Жизнеспособность дрожжей
по степени окисления
В качестве индикаторов жизнеспособности предложены самые разные методы,
включая содержание гликогена, АТФ или стерола, образование С02 или потребле-
ние 02 из раствора глюкозы, а также скорость выделения определенных ионов.
Наиболее удобным методом для определений, которые должны быть выполнены в
кратчайшее время, является выделение ионов Н+, т. е. снижение значения вели-
чины рН, при введении дрожжей в сбраживаемый субстрат.
Для определения жизнеспособности дрожжей по степени окисления дрожжи промы-
вают центрифугированием и затем тщательно перемешивают в 100 см3 дистилли-
рованной воды. Через 10 мин добавляют 5 см3 20%-ного (масс/об) раствора глю-
козы и перемешивают еще 10 мин. В ходе инкубации в течение 20 мин каждую ми-
нуту измеряют рН суспензии. Резкое снижение значения рН сразу же и после до-
бавления раствора глюкозы до < 4 свидетельствует об активности дрожжей, а
степень окисления коррелирует с изменением величины рН.
Как и метод с использованием метиленовохю синего, данный метод достаточно
быстр и позволяет получить результаты до внесения дрожжей.
Дифференциальное
окрашивание
метиленовым синим
Около 1 г сушеных дрожжей вносят в пробирку со стерильным солодовым суслом
плотностью 8% видимых СВ.
Пробирку ставят в термостат при температуре 30 С на 4 ч. За это время все
живые клетки, содержащиеся в пробе, выходят из состояния анабиоза и начинают
почковаться. Из подмоложенных таким способом дрожжей готовят препарат, кото-
рый окрашивают метиленовым синим, приготовленным на буферной смеси.
Для прессованных дрожжей и живой культуры нет необходимости в такой проце-
дуре.
Каплю взвеси дрожжей смешивают с каплей синьки на предметном стекле и на-
крывают покровным стеклом. Мертвые клетки окрашиваются, а живые остаются не-
окрашенными. В пяти полях зрения отдельно подсчитывают количество мертвых и
живых клеток, процент мертвых клеток вычисляют по отношению ко всему количе-
ству клеток. Материнскую клетку с неотделившейся почкой принимают за одну
клетку.
Люминесцентный
метод
Используют люминесцентный микроскоп или люминесцентный осветитель и флуоро-
хром примулин.
Краситель готовят в виде двух растворов: водный раствор (1:20 000) и рас-
твор (1:20 000) в фосфатном буфере рН 8-9. Оба раствора хранят в темноте.
Предметные и покровные стекла тщательно обезжирены.
При микроскопировании белых вин бактериологической петлей помещают на пред-
метное стекло каплю осадка и рядом каплю водного раствора примулина, смешива-
ют их и накрывают покровным стеклом, на которое затем наносят каплю нелюми-
несцирующего иммерсионного масла.
При микроскопировании красных вин (в связи с гашением люминесценции) крас-
ный цвет необходимо устранить, что достигается созданием в препарате щелочной
среды. Красный цвет вина при этом превращается в синий.
Бактериологической петлей помещают на предметное стекло каплю осадка вина и
две капли раствора примулина в фосфатном буферном растворе рН=8-9. Капли тща-
тельно смешивают петлей в течение 1-2 мин, затем накрывают покровным стеклом,
на которое помещают каплю нелюминесцирующехю иммерсионного масла.
Приготовленные препараты рассматривают в люминесцентном микроскопе в фиоле-
товой и синей видимой частях спектра. Для этого пользуются светофильтрами:
синим, сине-зеленым, белым и запирающим желтым.
Сначала препарат рассматривают в проходящем свете и подсчитывают общее ко-
личество микроорганизмов, а затем — в свете люминесценции, подсчитывая только
количество мертвых клеток.
В свете люминесценции живые микроорганизмы не видны, иногда у дрожжевых
клеток светится только оболочка в виде тонкого зеленого кольца. Мертвые дрож-
жи и бактерии в свете люминесценции светятся зеленым и желтым светом разной
интенсивности.
При подсчете микроорганизмов необходимо учитывать разведение исследуемого
материала в 2 раза для белых вин и в 3 раза — для красных.
Метод
эпифлуоресценции
Методика позволяет определять количество жизнеспособных клеток дрожжей в
винах и других сброженных материалах, содержащих их небольшое количество (в
момент розлива в бутылки), с использованием эпифлуоресценции, и сравнивается
с контрольным методом (подсчет колоний на питательной среде).
Применяют также красители-флуорохромы и люминесцентный микроскоп.
Препарат готовят на мембранах Миллипор из поликарбоната диаметром 13-25 мм
или из ацетилцеллюлозы диаметром 45 мм.
Испытуемое вино фильтруют под вакуумом через мембраны, окрашивают и подсчи-
тывают под микроскопом число клеток в см3 с учетом числа клеток и подсчитан-
ных полей, площади фильтрующей мембраны, площади полей микроскопа, объема от-
фильтрованного образца, числа наблюдаемых полей.
ПРОВЕРКА
ЧИСТОТЫ
КУЛЬТУРЫ
Время от времени культуры, как в коллекциях, так и в промышленности, прове-
ряют на наличие загрязнений или на появление мутантов.
Перед использованием в производстве пробы засевных дрожжей должны пройти
проверку на жизнеспособность. Считается правильным проверять каждую партию
семенных дрожжей на допустимое количество (а лучше всего вообще отсутствие)
контаминирующих бактерий и диких дрожжей. Для бактерий рекомендуется исполь-
зовать в основном актидионовый WL-arap или другие среды дифференциального на-
значения, а для диких дрожжей (не Saccharomyces) — лизиновый агар. Для обна-
ружения «диких» дрожжей среди сахаромицетов делают рассев репликатором на ли-
зиновый агар (углеродная основа + 0,1% L-лизина как единственного источника
азота), на котором S. cerevisiae и S. carlsbergensis не растут, а «дикие»
дрожжи растут. Но если контаминанты Saccharomyces окажутся стойкими к цикло-
гексимиду (что встречается довольно редко), определить их будет невозможно.
Однако следует иметь в виду, что не существует такой селективной среды, кото-
рая бы одновременно подавляла рост культурных дрожжей и допускала бы развитие
всех присутствующих контаминантов. Тем не менее, дикие дрожжи рода Saccharo-
myces образуют споры и могут быть выделены благодаря своей теплостойкости
(сравнительно с неспорообразующими культурными дрожжами). При этом, однако,
для инкубации требуется не менее трех суток, а задерживать на это время вне-
сение дрожжей непрактично. Таким образом, этот анализ может служить лишь для
проверки качества постфактум.
Petite или RD-мутанты (дефектные по дыханию) на сусло-агаре образуют окра-
шенные мелкие колонии. На среде с теллуритом они не восстанавливают послед-
ний.
Просмотр культуры
под микроскопом
Контролю подвергают односуточную культуру из жидкого солодового сусла плот-
ностью 10-12% СВ. Одну каплю культуры при помощи стерильной пипетки наносят
на чистое предметное стекло и накрывают покровным стеклом (оба стекла предва-
рительно протирают спиртом). Приготовленный препарат помещают на предметный
столик микроскопа и просматривают десять полей зрения при увеличении в 600-
800 раз. При просмотре их не должно быть обнаружено ни посторонних дрожжевых
грибков, ни бактерий.
Метод
микрокультуры
Для определения посторонних дрожжевых грибков применяют метод микрокульту-
ры. Этот метод удобен для получения быстрого ответа о составе микрофлоры.
На дно чашки Петри кладут фильтровальную бумагу, вырезанную по форме чашки.
Туда же помещают две стеклянные палочки, предметное и покровное стекла и все
стерилизуют сухим жаром в течение 1 ч при 160 С. Стерильной стеклянной палоч-
кой наносят на стерильное предметное стекло каплю исследуемого образца, к
этой капле прибавляют 2-3 капли 1%-ного сусло-агара, предварительно расплав-
ленного на водяной бане и охлажденного до 50-45 С. Смешивают агар с жидкостью
уголком стерильного покровного стекла и накрывают им получившийся таким обра-
зом препарат. Фильтровальную бумагу в чашке Петри смачивают стерильной водой,
препарат помещают в чашку Петри на стеклянные палочки и ставят в термостат
при температуре 30 С.
Через 6-8 ч ведут наблюдение за формой выросших микроколоний под микроско-
пом. Просматривают 20 полей зрения в середине препарата и по краям. В каждом
поле подсчитывают отдельно микроколонии как сахаромицетов, состоящие из круг-
лых или овальных клеток, так и посторонних дрожжевых грибков, обычно очень
резко выделяющихся по своему внешнему виду и форме клеток.
Определяют среднее количество колоний сахаромицетов и посторонних дрожжевых
грибков и рассчитывают их процентное соотношение.
Посев на
чашки Петри
Пробу из культуры, предварительно разведенную в стерильной водопроводной
воде, наносят на поверхность твердой питательной среды, инкубируют при опти-
мальных условиях и определяют наличие неспецифических колоний. На поверхности
питательной среды в чашках Петри чистая культура образует однородные по мор-
фологии, консистенции и пигментации колонии.
Посев на элективную
питательную среду
Степень засоренности чистой культуры, маточных и засевных дрожжей посторон-
ними дрожжевыми и дрожжеподобными грибами оценивают при помощи элективной
ацетатной среды, где имеются благоприятные условия для размножения диких
дрожжей, а сахаромицеты не размножаются. Благодаря этому представляется воз-
можным выявить в исследуемом материале даже единичные клетки диких дрожжевых
грибов.
Исследуемый материал — 1-2 капли взвеси исследуемых дрожжей — вносят в про-
бирку с ацетатной средой. Посев производят с соблюдением всех правил асептики
— над пламенем горелки, стерильной пипеткой или предварительно прокаленной
петлей.
Ацетатная среда для выделения посторонних дрожжей
На 1000 см водопроводной воды берут 10 г уксуснокислого натрия, 10 г хло-
ристого аммония и 5 г глюкозы.
Дрожжевой автолизат добавляют в среду в количестве 3 см3.
Среду разливают в пробирки по 5 см3 и стерилизуют в автоклаве при избыточ-
ном давлении 0,05 МПа в течение 30 мин.
Засеянную пробирку помещают в термостат при температуре 30 С и ведут за ней
ежедневные наблюдения. Появление пленки на поверхности среды или помутнение
ее указывает на наличие в исследуемом материале посторонних дрожжевых грибов.
Чем раньше появится пленка, тем более инфицированным является материал. Быст-
рота появления пленки позволяет дать оценку степени зараженности материала
посторонними дрожжевыми грибами.
Если пленка появилась через 5 или большее число дней либо совсем не появи-
лась , то это означает, что культура чистая, материал не содержит посторонних
дрожжей в значимых для производства количествах; если же пленка появилась че-
рез 3-4 дня, то дрожжи удовлетворительные и пригодны для работы; если пленка
появилась через 1-2 дня, то дрожжи неудовлетворительные и не пригодны для ра-
боты .
По истечении сроков инкубации необходимо провести микроскопирование и пере-
сев пробы на сусло-агар для анализа культуральных свойств.
ХРАНЕНИЕ ЧИСТЫХ
КУЛЬТУР ДРОЖЖЕЙ
С целью сохранения культуры выросшие изолированные колонии отсевают петлей
на поверхность скошенного 2,5%-ного сусло-агара в пробирки или в жидкое неох-
меленное сусло (8% СВ).
Посев на скошенный агар производится бактериологической петлей методом ис-
тощающего штриха, начиная со дна пробирки.
Длительное хранение требует особого подбора среды и условий хранения био-
массы , при которых культура сохранит свою физиологическую активность.
Длительное поддержание чистых культур микроорганизмов, без потери активно-
сти и производственных качеств, необходимо как при хранении в коллекциях и
проведении научно-исследовательских работ, так и при эксплуатации в производ-
стве .
Основной задачей при хранении чистых культур промышленных штаммов микроор-
ганизмов в музейных коллекциях является правильный выбор условий для сохра-
нения их производственно ценных свойств. Общей тенденцией при этом является
максимальное замедление всех жизненных процессов, что достигается различными
приемами. Жизнеспособность клеток при этом должна быть сохранена. Кроме того,
способы хранения культур должны быть по возможности менее трудоемки и надеж-
ны.
Из-за различий биологических свойств видов невозможно использовать с равно-
ценными положительными результатами один общий способ хранения для разных
культур дрожжей. Большой опыт по хранению культур микроорганизмов, в том чис-
ле и дрожжей, накоплен сотрудниками крупных национальных коллекций, в которых
имеется возможность проверять и оценивать разные методы хранения применитель-
но к широкому набору дрожжевых организмов.
Наиболее широко применяемый способ — это поддержание культур путем их пе-
риодических пересевов. В последние два десятилетия получили распространение
также методы хранения микробных культур под минеральным маслом и лиофильная
сушка. Так как эти три метода применяются уже много лет, то имеется возмож-
ность сравнительной оценки их пригодности для хранения разных видов дрожжей.
Другие, менее широко используемые или недавно разработанные методы можно ре-
комендовать лишь для проверки с обязательным дублированием культур, которые
поддерживаются обычными способами.
Периодические
пересевы
Обычно культуры поддерживают, пересевая их на агаризованных средах в двой-
ной повторности. Одна пробирка, которую после засева совсем не открывают,
служит контрольной, а из второй по мере надобности делают отсевы.
Сроки пересевов определяют для большинства культур дрожжей скоростью высы-
хания среды. Она зависит от температуры и влажности помещения, где хранят
пробирки.
Промежутки между пересевами можно увеличить за счет более плотного закупо-
ривания пробирок и снижения температуры хранения. Так, использование пробирок
с завинчивающимися металлическими крышками вместо обычных ватных пробок пре-
дохраняет от высушивания, но хуже обеспечивает чистоту сохраняемых культур.
Применение вазелинового масла для снижения скорости высыхания культур также
имеет свои недостатки, о чем речь будет идти ниже. Поэтому хорошо сделанные
ватные пробки (их можно сверху заливать парафином) все же остаются лучшим
средством закрывания пробирок. Они обеспечивают достаточный газообмен, хорошо
предохраняют от микробного загрязнения и легко изготовляются.
Ватные пробки, однако, не предохраняют культуры от заражения их микофильны-
ми клещами (mites). Для профилактики используют следующий прием. Перед посе-
вами или пересевами культур пробку слегка выдвигают из пробирки и наносят на
нее каплю раствора, содержащего 10 г сулемы, 50 см3 глицерина, 500 см3 этило-
вого спирта и 450 см3 воды. Затем пробку вдвигают в пробирку и несколько раз
проворачивают ее, чтобы смочить внутренние стенки пробирки. В раствор можно
добавить какой-либо краситель для контроля равномерности смачивания.
Если хранение ведется при температуре в пределах 15-20 С, то после пересева
пробирки сразу же помещают в коллекционное помещение, за исключением тех
культур, которые требуют для роста более высокой температуры. Психрофильные
дрожжи выращивают и хранят при 3-4 С в холодильнике. При низкой температуре
после появления хорошего роста можно хранить и все другие дрожжи.
Во многих лабораториях и коллекциях дрожжи поддерживают и хранят на сусло-
агаре. Было показано, однако, что на этой сложной среде при длительном хра-
нении происходит изменение физиологических свойств, характерных для вида, за
счет адаптации или стабилизации мутантов. В связи с этим для хранения более
пригодна среда следующего состава, которую используют для голландской коллек-
ции.
Глюкоза
Пептон
Дрожжевая вода рН 5,8-6,0
4,0 г
0,5 г
до 100,0 см3
Для винных дрожжей предложена среда, на которой дрожжи хорошо спорулируют и
долго хранятся.
■ Морковь тертая 125 г
■ Вода 1000 см3
Настоять 1 ч, довести до кипения, кипятить 10 мин, отфильтровать через ва-
ту , рН 5.
Пивоваренные дрожжи лучше поддерживать на сусло-агаре или среде следующего
состава.
■ Дрожжевой экстракт 3%
■ Глюкоза 1%
На дрожжевых и спиртовых заводах, работающих на мелассе, для хранения про-
изводственных культур лучше использовать среду следующего состава.
■ Мелассовое сусло 5% СВ 100 см3
■ Солодовое сусло 12% СВ 100 см3
■ Дрожжевой автолизат 2 см3
В некоторых коллекциях дрожжи поддерживают в жидких средах (сусло с дрожже-
вым автолизатом, пептоном и глюкозой) с пересевами через 4 мес.
Хранение под
минеральным
маслом
Заливка агаровых культур минеральным маслом преследует цель задержать высы-
хание и тем самым увеличить сроки пересевов.
Для заливки культур дрожжей наиболее пригодно высокоочищенное медицинское
вазелиновое масло плотностью 0,8-0,9. Масло стерилизуют в автоклаве в течение
1 ч при 121 С, а затем прогревают для удаления влаги в сушильном шкафу при
температуре не выше 150 С или выдерживают при комнатной температуре не менее
2-3 дней.
Дрожжи выращивают на сусло-агаре или другой среде, выбранной для хранения.
Среду готовят в пробирках таким образом, чтобы столбик был достаточно высо-
ким, а скос небольшим.
Через 4-6 суток, когда штрих хорошо сформируется, культуры заливают маслом
таким образом, чтобы слой его над верхним краем агарового косяка не превышал
1 см. После заливки культуры хранят в вертикальном положении либо при комнат-
ной температуре, либо в холодильнике при 4-6 С.
Пересев дрожжей из-под вазелинового масла производят один раз в год. Для
пересевов используют свежие скошенные агаризованные среды, на которых культу-
ры хранились.
Опыт многих коллекций свидетельствует об успешных результатах этого способа
хранения дрожжевых культур. Для винных дрожжей, в частности, было показано,
что при хранении под вазелиновым маслом с пересевами даже через 2-4 года они
не теряют бродильных свойств и хорошо выживают. Имеются указания на сохране-
ние живых клеток дрожжей под маслом до 10 лет и более без пересевов.
К недостаткам этого метода хранения следует отнести следующие:
■ при пересевах масло смешивается с инокулятом, и штрихи получаются хуже
обычных;
■ масло разбрызгивается при обжигании петли, и от этого возникает возмож-
ность инфицирования помещения;
■ требуется специальная очистка использованных пробирок от масла;
■ при стерилизации масла высокими температурами в нем могут образовываться
токсичные для дрожжей вещества.
С учетом всех преимуществ при достаточном опыте персонала и правильной ор-
ганизации работы в лаборатории перечисленными недостатками можно пренебречь.
Лио филизация
Лиофилизацией называют процесс высушивания под вакуумом из замороженного
состояния. Техника лиофилизации и хранения лиофилизированных культур сильно
варьирует в разных лабораториях. Вариации касаются сред и сроков выращивания,
применения суспензионных жидкостей, температуры замораживания, условий хране-
ния и восстановления культур.
Дрожжи, подлежащие лиофилизации, выращивают в оптимальных условиях на сре-
дах, не слишком обогащенных питательными веществами. Для этой цели пригодна
среда Сабуро или глюкозопептонная следующего состава.
■ Глюкоза 1,0 г
■ Пептон 1,0 г
■ Мясной и дрожжевой экстракты по 0,5 г
■ К2НР04 0,3 г
■ Вода до 100,0 см3
Дрожжи выращивают и на сусло-агаре. Из 2-5-суточных культур готовят густую
суспензию в специальных суспензионных жидкостях, содержащих защитные веще-
ства, которые можно разделить на три группы:
1. коллоидные среды животного, растительного и минерального происхождения,
такие как сыворотка крови, плазма, желатина, снятое молоко, агар-агар,
гель гидрата окиси алюминия и др.;
2. среды с углеводами (чаще всего 10%-ный раствор сахарозы) и продуктами гид-
ролиза белков — пептоном и аминокислотами;
3. сложные среды, в состав которых входят вещества, образующие как коллоид-
ные, так и истинные растворы.
Хорошие результаты получены в голландской коллекции (CBS, г. Дельфт) при
использовании в качестве защитной среды раствора, содержащего 7,5% инозита,
2% глутамата натрия и 5% декстрина. рН такой среды доводят до 7,0 с помощью
NaOH.
Ниже приведены 2 варианта сложных защитных сред.
Вариант 1
■ Желатина 1%
■ Дрожжевой экстракт 1%
■ Глюкоза 0,5%
■ Аскорбиновая кислота 0,25%
■ Дистиллированная вода до 100,0%
Вариант 2
■ Бычья сыворотка 100 см3
■ Сахароза 30 г
■ Дистиллированная вода 200 см3
Использование 10-20%-ного раствора сахарозы дает хорошую выживаемость сразу
после лиофилизации, но при длительном хранении жизнеспособность дрожжей резко
снижается из-за высокой остаточной влажности материала.
Важным моментом в процессе лиофилизации является замораживание суспензии.
Для дрожжей при лиофилизации обычно применяют охлаждение от -30 С до -70 С,
однако температура около -15 С, способствующая медленному замораживанию, дает
наилучшие результаты. Есть указания также на успешное использование режима
быстрого замораживания дрожжей при температуре смеси сухого льда и спирта -78
С и сверхбыстрого замораживания в жидком азоте при -196 С.
Замороженный материал не должен оттаивать в процессе высушивания. Продолжи-
тельность высушивания меняют в зависимости от обрабатываемого материала, его
объема, состава защитной среды и используемой аппаратуры. Чем мельче кристал-
лы , образовавшиеся при замораживании, тем быстрее проходит высушивание. Оста-
точная влажность сказывается на сохранении свойств лиофилизированных дрожжей
и на продолжительности последующего их хранения без потери жизнеспособности.
Сразу же после окончания процесса лиофилизации важно определить жизнеспо-
собность обработанной культуры, а в случае, когда была известна концентрация
живых клеток в исходной суспензии, можно установить и процентное отношение
оставшихся живыми от начального количества клеток.
Реактивацию лиофилизированных культур производят путем переноса части мате-
риала либо сразу же в жидкую питательную среду, либо путем предварительного
8-часового выдерживания в дистиллированной воде с последующим пересевом на
суело-агар.
Запаянные ампулы с лиофилизированными культурами хранят в темноте при ком-
натной температуре или в холодильнике с температурой 4-6 С. Последнее пред-
почтительнее, особенно для дрожжей с высокой остаточной влажностью материала.
Итоги 19-20-летнего хранения культур дрожжей — представителей 17 родов,
всего 557 штаммов — в голландской коллекции позволили установить разную при-
годность этого метода хранения для отдельных видов. Дрожжи с мелкими клетками
и аскоспорами, такие как Pichia, Hansenula и Debaryomyces, показали хорошую
выживаемость, а крупноклеточные слабоспорулирующие или неспорулирующие дрожжи
родов Saccharomyces, Kluyveromyces, Dekkera и Brettanomyces выживали хуже.
Таким образом, и этот широко применяющийся сейчас метод хранения микробных
культур не следует рассматривать как универсальный, одинаково пригодный для
разных видов дрожжей.
Хранение на
адсорбентах
В качестве адсорбентов используют почву, песок, каолин, силикагель, вату и
фильтровальную бумагу. Этот способ хранения не имеет разработанной стандарт-
ной техники. Высокая выживаемость и физиологическая активность обеспечивается
при хранении дрожжей в силикагеле, содержащем различные микроэлементы. Двух-
суточные культуры дрожжей, полученные на синтетической среде с 1% парафина на
качалках, смешивали со 150 см3 стерильных шариков силикагеля и оставляли в
термостате при 32 С на 24 ч. Через сутки определяли рН среды и доводили при
необходимости до 7-8. Затем культуральную жидкость сливали, шарики силикагеля
с дрожжами помещали в стерильные чашки Петри и высушивали в эксикаторе при
пониженном давлении в течение 5-6 суток. После высушивания культуры дрожжей
хранили в течение года при комнатной температуре в запаянных стеклянных ампу-
лах при давлении 13,33 кПа. Для реактивации 200 мг шариков силикагеля измель-
чали в фарфоровой ступке, порошок переносили в 3-5 мл стерильной воды и после
кратковременного выдерживания делали высев на питательную среду.
Описан способ хранения дрожжевых культур на вате и полосках фильтровальной
бумаги, в 10%-ном растворе сахарозы в почве.
Хранение культур
винных дрожжей
в виде спор
На сусловых средах винные дрожжи плохо спорулируют и быстро отмирают. В. В.
Абрамовичем был разработан метод получения активно спорулирующих культур для
длительного хранения их в состоянии спор.
Дрожжевой осадок из сброженного сусла переносят на 2-3%-ный голодный (вод-
ный) агар, где 60-80% клеток образуют споры. Дрожжи сохраняют жизнеспособ-
ность на этой среде до двух и более лет. Этим способом было проверено 18 рас
винных дрожжей. Подавляющее большинство проверенных культур вызывало активное
брожение через 1,5-2,2 года хранения.
Н.И. Бурьян предложила для винных дрожжей способ хранения без потери их ос-
новных производственно ценных свойств (энергии размножения и дыхания, бро-
дильной активности, кислотоустойчивости, спиртоустойчивости) путем чередова-
ния периодов активного размножения и пребывания в состоянии спор.
Дрожжи предварительно выращивают 48 ч на среде с 5% (по объему) автолизата
пивных дрожжей и 2% глюкозы, переносят для спорообразования на измененную
среду Адамса (2% агара + 0,14% уксуснокислого натрия), затем 10 суток выдер-
живают при 15 С, а для длительного хранения переносят в помещение с темпера-
турой 10 С. Через 9-10 мес. дрожжи переводят в активное состояние, засевая
споровым материалом колбы со средой, содержащей 18-20% сахара (рН 2,7-3,0), а
по окончании брожения — в новую серию колб, среда в которых содержит 28-30%
сахара (рН 5,5). Эти пересевы занимают приблизительно 2 мес, после чего дрож-
жи вновь переносят на голодный агар.
Хранение микробных
культур под вакуумом
Небольшое количество дрожжей с агаровой среды вводят в маленькую пробирку,
которую закрывают ватной пробкой. Пробирку помещают в более крупную пробирку
из тугоплавкого стекла. На ее дно предварительно вносят несколько стеклянных
бус и кристаллов КОН. Пробирку закрывают резиновой пробкой со стеклянной
трубкой, через которую производят откачку воздуха до 1,3-6,6 кПа. Трубку за-
паивают парафином. Пробирки хранят при комнатной температуре.
Хранение коллекционных
культур дрожжей в
дистиллированной воде
Пробирки с 8-10 см3 стерильной дистиллированной воды засевают большим коли-
чеством инокулята с глюкозного или суслового агара и хранят при комнатной
температуре. По мере испарения воды ее добавляют в пробирки. Жизнеспособность
дрожжей сохраняется в этих условиях до двух лет и более. Основным условием
успешного хранения дрожжей в дистиллированной воде является обильный иноку-
лят.
Этот способ оказался непригодным для таких слабо спорулирующих дрожжей, как
Hanseniaspora, Endomycopsis, Nematospora и некоторых Saccharomyces.
Хранение
в манните
Предлагается способ хранения дрожжей в активной форме, при котором чистую
культуру суспендируют в 10%-ном растворе маннита, играющего роль осмотическо-
го стабилизатора, с добавлением 0,2% р-фенилэтанола в качестве бактерицидного
агента.
При концентрации биомассы дрожжей 1,0-2,5-108 клеток/см3 и температуре хра-
нения 25 С гарантируется сохранение свойств дрожжей в течение года.
МЕТОДЫ
ОКРАСКИ
Простая окраска
Мазок, т. е. каплю эмульсии микроорганизмов, помещенную на предметное стек-
ло, высушивают и фиксируют над пламенем горелки. Подготовленный препарат за-
ливают 1-2 каплями краски (метиленовой синью Леффлера или фуксином Пфейфера),
выдерживают 0,5-1 мин и смывают дистиллированной водой.
Окраска
спор
дрожжей
Метод Виртца
Фиксированный на пламени препарат заливают 5%-ным водным раствором малахи-
тового зеленого и 3-4 раза прогревают до появления паров (30-60 с). Окраску
можно проводить, выдерживая препарат 3-5 мин в растворе малахитового зелено-
го, подогретого до 80 С. Затем препарат промывают 30 с в проточной воде и до-
крашивают 30 с 0,5%-ным водным раствором сафранина.
Зрелые аскоспоры окрашиваются в сине-зеленый, а вегетативные клетки — в
красный цвета.
Аскоспоры дрожжей родов Hansenula, Schizosaccharomyces могут удерживать оба
красителя.
Карбол-фуксиновая окраска
Вариант 1. Фиксированный на пламени препарат заливают раствором Циля (10
см3 насыщенного спиртового раствора основного фуксина с 90 см3 5%-ного фено-
ла) и нагревают 2-5 мин до появления паров. Затем проводят обесцвечивание
препарата, погружая его в 2%-ный раствор молочной кислоты или в 95%-ный эта-
нол, содержащий 1 мл концентрированной НС1 в 100 см3.
Препарат промывают в воде и докрашивают 3-5 мин 1%-ными растворами метиле-
нового синего или нильского голубого.
Зрелые аскоспоры окрашиваются в красный, вегетативные клетки — в синий цве-
та.
Аскоспоры рода Schizosaccharomyces, а также после набухания аскоспоры дрож-
жей родов Hansenula и Pichia не обладают кислотоустойчивостью и окрашиваются
в синий цвет.
Вариант 2. Фиксированный препарат окрашивают в течение 2-3 мин карболовым
фуксином при подогревании над пламенем горелки, затем краску сливают и препа-
рат обесцвечивают соляной кислотой со спиртом (две части 10%-ного раствора
соляной кислоты и одна часть 96%-ного спирта) в течение 30 с, промывают водой
и окрашивают в течение 1-2 мин метиленовой синью.
Споры окрашиваются в красный цвет, а вегетативные клетки — в синий.
Определение гликогена
в клетках дрожжей
Гликоген — одно из резервных веществ дрожжевых клеток. Он накапливается в
цитоплазме и окрашивается раствором Люголя в коричневый или красно-бурый
цвет. Наблюдения за содержанием гликогена в клетках дрожжей проводят в дина-
мике развития культуры.
Готовят препарат «раздавленная капля» в растворе Люголя и в нескольких по-
лях зрения подсчитывают общее количество клеток и количество клеток, содержа-
щих гликоген. Общее число подсчитанных клеток должно быть не менее 100.
Если препарат нагреть до 60 С, окраска гликогена исчезает, а при охлаждении
препарата она вновь восстанавливается.
Окраска
волютина
По методу Омелянского
На предметном стекле готовят тонкий мазок из культуры исследуемых дрожжей,
высушивают на воздухе и фиксируют на пламени горелки. Мазок окрашивают карбо-
ловым фуксином 30-40 с и промывают водой. Далее мазок дифференцируют, погру-
жая его в склянку с 1%-ным раствором серной кислоты на 20-30 с и немедленно
промывают водой. Серная кислота обесцвечивает цитоплазму, а зерна волютина
остаются окрашенными фуксином. Препарат докрашивают метиленовым синим (1:40)
20-30 с, промывают водой, высушивают на воздухе и микроскопируют, пользуясь
объективом МИ-90.
На препарате зерна волютина окрашены в красный цвет, цитоплазма клетки — в
голубой.
По методу Леффлера
Тонкий фиксированный мазок культуры исследуемых дрожжей окрашивают метиле-
новым синим Леффлера в течение 3 мин. Мазок промывают водой, не высушивая,
покрывают его покровным стеклом и микроскопируют, пользуясь объективом ВИ-60.
На препарате зерна волютина окрашены в красновато-фиолетовый цвет, цито-
плазма — в голубой.
Для более ясной дифференциации зерен волютина под покровное стекло полоской
фильтровальной бумаги протягивают каплю 1%-ного раствора серной кислоты. При
этом зерна волютина сохраняют окраску, а цитоплазма обесцвечивается.
Обратный результат можно получить, добавляя под покровное стекло каплю 5%-
ного раствора соды. Волютиновые зерна обесцвечиваются, а цитоплазма клетки
сохраняет голубой цвет.
Краски
Метиленовый синий (насыщенный спиртовой раствор)
К 100 см 96%-ного этилового спирта прибавляют 10 г метиленового синего (в
порошке) и оставляют стоять несколько дней (при этом несколько раз встряхива-
ют) , затем раствор фильтруют.
■ Метиленовый синий 1:40.
■ Метиленовый синий (насыщенный спиртовый раствор) 1 см3
■ Вода дистиллированная 40 см3
Метиленовый синий щелочной (синька Леффлера)
В 100 см дистиллированной воды растворяют 30 см3 насыщенного спиртового
раствора метиленового синего и 1 см3 1%-ного раствора едкого кали.
Разбавленная синька (по Финку)
Готовят три раствора: 0,09 г фосфорнокислого натрия (N2HP04) растворяют в
500 см3 дистиллированной воды; 13,6 г фосфорнокислого калия (КН2Р04) растворя-
ют в 500 см3 дистиллированной воды; 0,1 г метиленового синего растворяют в
500 см3 дистиллированной воды.
Для приготовления рабочего раствора синьки смешивают 0,25 см3 первого рас-
твора с 99,75 см3 второго и 100 см3 раствора синьки, рН раствора должно быть
4,6.
Основной фуксин
2 г сухой краски и 20 см3 96%-ного этилового спирта встряхивают и оставляют
стоять несколько дней, получается насыщенный спиртовой раствор.
Карболовый фуксин (по Цилю)
10 см насыщенного спиртового раствора основного фуксина растворяют в 100
см3 5%-ного раствора карболовой кислоты (фенола).
Разбавленный фуксин (по Пфейферу)
Для окраски микроорганизмов применяют раствор разбавленного в 10 раз карбо-
лового фуксина.
Малахитовый зеленый (водный насыщенный раствор)
■ Малахитовый зеленый 5 г
■ Дистиллированная вода 100 см3
Для окраски спор лучше брать спиртово-водный раствор.
■ Малахитовый зеленый 5 г
■ Спирт этиловый, 96° 20 см3
■ Дистиллированная вода 80 см3
Сафранин (водный раствор)
■ 2,5%-ный раствор сафранина в 96%-ном этаноле 10 см3
■ Вода дистиллированная 100 см3
Раствор Люголя для выявления гранулезы
■ Кристаллический йод 1 г
■ Йодид калия 2 г
■ Дистиллированная вода 100 см3
Гранулеза в клетках окрашивается в темно-синий цвет.
Раствор Люголя для выявления гликогена
■ Кристаллический йод 7 г
■ Йодид калия 20 г
■ Дистиллированная вода 100 см3
Гликоген приобретает красно-бурую окраску.
Раствор туши
■ Жидкая натуральная тушь 10 см3
■ Дистиллированная вода 90 см3
Раствор центрифугируют 15-20 мин. Верхний слой отсасывают (пипеткой), пере-
носят в пробирку и автоклавируют 30 мин при 0,05 МПа (температура 110 С).
Можно раствор туши сразу после смешивания автоклавировать 30 мин при давле-
нии 0,05 МПа. После автоклавирования раствор отстаивают две недели, после че-
го его можно использовать.
Метод Литтмана:
■ Черная тушь 15 см3
■ Раствор тимерсола в соотношении 1:1000 30 см3
■ Твин-80 0,1 см3
На черном фоне капсулы видны как светлый ореол вокруг клетки.
Технологии
ВЫДЕЛЕНИЕ ЧИСТЫХ КУЛЬТУР ПАТОГЕНОВ
Питательные
среды
Питательные среды должны отвечать следующим требованиям:
Содержать в достаточном количестве все необходимые питательные вещества в
легко усвояемом для бактерий виде.
Иметь оптимальную рН среды для роста данного вида бактерий. Большинство
бактерий растут при нейтральной рН среды (7,2 -7,8).
Быть изотоничными, т.е. осмотическое давление в среде должно быть таким же
как в клетке. Для этого к средам добавляют 0,9% по концентрации хлорид на-
трия (NaCl).
Иметь достаточную влажность, т.к. питание бактерий осуществляется за счет
диффузии; также при усыхании сред в них повышается концентрация солей, что
тормозит рост бактерий.
Быть стерильными, т.е. не содержать посторонней микрофлоры.
Питательные среды классифицируют:
По исходным компонентам: натуральные и синтетические.
По консистенции: жидкие, твердые и полужидкие.
По составу: простые (основные, универсальные) и сложные.
По назначению: специальные, элективные, дифференциально-диагностические.
Классификация питательных сред по исходным компонентам:
Натуральные питательные среды готовят из животных и растительных продуктов
(молока, мяса, яиц, картофеля и т.д.). В практических бактериологических
лабораториях чаще используют искусственные питательные среды, представляю-
щие собой сбалансированные смеси питательных веществ в концентрациях и со-
четаниях , необходимых для роста и размножения микроорганизмов. В них в ка-
честве универсального источника азота и углерода используют пептоны - про-
дукты неполного расщепления белков с помощью пепсина или различные гидро-
лизаты (рыбный, казеиновый, дрожжевой и др.). В настоящее время широко
применяют сухие питательные среды, выпускаемые промышленностью, представ-
ляющие собой гигроскопические порошки, растворимые в воде. Данные среды
имеют постоянный состав, их просто готовить и удобно транспортировать.
■ Синтетические питательные среды готовят из неорганических и органических
соединений, обеспечивающих азотистое, углеродное и минеральное питание
микроорганизмов, они имеют строго определенный состав. Например: среда Со-
тона - для культивирования микобактерий; среда 199 и среда Игл - для выра-
щивания культур клеток.
Классификация питательных сред по консистенции:
■ Жидкие среды готовят на основе водных растворов каких-либо веществ, как
правило, мясной воды.
■ Для получения плотных сред к жидким добавляют уплотнители, чаще всего
агар-агар, получаемый из морских водорослей. Он представляет собой полиса-
харид сложного состава, имеющий волокнистую структуру. Агар-агар плавится
при температуре около 90 С и затвердевает при температуре около 40 С. В
плотных средах агар-агар составляет 1,5-3,0%.
■ Полужидкие среды имеют вязкую консистенцию благодаря добавлению к ним не-
большого количества агар-агара, концентрация которого составляет 0,3-0,7%.
Классификация питательных сред по составу:
■ Простые (основные, универсальные) - мясо-пептонный бульон (МПБ) и мясо-
пептонный агар (МПА), на них хорошо растут многие виды бактерий.
Мясо-пептонный бульон (МПБ). Мясо-пептонный агар (МПА).
■ Сложные среды, используют для получения роста прихотливых бактерий, не
растущих или очень плохо растущих на простых питательных средах. Сложные
питательные среды (кровяные, сывороточные, асцитические, сахарные и т.д.)
приготавливают, путем добавления к МПБ или МПА: крови (5-10%), сыворотки
(10-20%), асцитической жидкости (25-30%), углеводов (0,5-1%), глицерина
(2-5%) и т.п.
Классификация питательных сред по назначению:
■ Элективные среды предназначены для избирательного выделения и накопления
микробов определенного вида из биологических материалов, содержащих разно-
образную микрофлору. Принцип действия элективных сред может основываться
на добавлении веществ, подавляющих (ингибирующих) рост сопутствующей мик-
рофлоры. Например: ЖСА (желточно-солевой агар), используется для выращива-
ния стафилококков, высокое содержание соли, до 9%, не влияет на их рост,
но подавляет рост остальных. Второй принцип действия элективных сред осно-
вывается на опережении роста определенного вида микроба по сравнению с со-
путствующей флорой.
Рост стафилококков на среде ЖСА.
■ Дифференциально-диагностические - среды, позволяющие не только культивиро-
вать бактерии, но и отличать одни виды бактерий от других по их фермента-
тивной активности или культуральным свойствам. В классическом варианте это
углеводы (лактоза, сахароза и т. д.), мочевина или другие субстраты, при
расщеплении которых микробными ферментами образуются вещества, изменяющие
рН или окислительно-восстановительный потенциал среды. В результате появ-
ления продуктов ферментации индикатор окрашивает колонии микробов и (или)
среду вокруг них, помогая отличить от бесцветных (неокрашенных) колоний
микробов, не ферментирующих данный субстрат. К дифференциально-
диагностическим средам относятся среды Эндо, Плоскирева, Левина, Гисса и
многие другие. Среда Эндо состоит из МПА с добавлением 1% лактозы и инди-
катора (основного фуксина, обесцвеченного сульфитом натрия). Свежеприго-
товленная среда имеет бледно-розовую окраску. При росте лактозоположитель-
ных бактерий их колонии окрашиваются в темно-красный цвет с металлическим
блеском. Лактозоотрицательные бактерии образуют бесцветные колонии (цвета
среды).
Характер роста лактозоположительных (красные) и
тельных (бесцветные) колоний на среде Эндо.
лактозоотрица-
Среда Плоскирева (содержащая МПА, лактозу и индикатор - нейтральный крас-
ный) является не только дифференциально-диагностической, но и селективной,
так как способствует лучшему росту некоторых болезнетворных бактерий (сальмо-
нелл, шигелл) и подавляет рост сопутствующих микробов за счет содержания
бриллиантовой зелени, солей желчных кислот.
Среды Гисса состоят из 1% пептонной воды, 0,5% определенного углевода (глю-
коза, лактоза, маннит и др.) и индикатора Андреде (кислый фуксин в 1Н раство-
ре NaOH) . В пробирку со средой опускают поплавок (стеклянная трубочка один
конец которой запаян) для улавливания газообразных продуктов, образующихся
при разложении углеводов. Свежеприготовленная среда имеет слегка желтый отте-
нок . При разложении углеводов она приобретает красный цвет. О газообразовании
свидетельствует накопление газа в поплавке.
'1
Среды Гисса до посева. Изменение сред Гисса после суточной инкубации.
Рост
бактерий
На жидких питательных средах бактерии могут образовывать различные формы
роста. Большинство патогенных факультативно-анаэробных бактерий при росте на
жидких средах образуют равномерное помутнение и осадок на дне. Некоторые бак-
терии на жидких питательных средах могут образовывать специфичные формы рос-
та: пленку на поверхности среды - холерный вибрион, коринебактерии дифтерии,
микобактерий туберкулеза и др.; пленку с отходящими в глубь среды нитями
(сталактиты) - возбудитель чумы; «придонно-пристеночный» рост - стрептококки;
рост в виде «комочка ваты» - сибиреязвенная палочка.
Характер роста на жидкой среде в виде равномерного
помутнения и осадка.
На плотных питательных средах бактериальные клетки, фиксированные на по-
верхности среды, образуют макроскопические скопления колонии бактерий одного
вида, выросших в результате размножения одной или нескольких бактериальных
клеток. Колонии разных видов бактерий отличаются по: размерам, форме, поверх-
ности, характеру края, консистенции, окраске и др. Все эти признаки, как пра-
вило , видоспецифичны, поэтому они имеют важное диагностическое значение, т.е.
их изучение используется для определения видовой принадлежности исследуемой
культуры бактерий.
Размеры колоний варьируют от десятых долей - до 5-6 мм. Различают: точечные
(колонии с диаметром не более 1 мм), средние (размером 2-4 мм), крупные (бо-
лее 5 мм) .
**Л\\.
h'
/
s
^r
• 1
s 1
•
• ^T'
Разновидности колоний на питательном агаре.
Форма колоний может быть круглая, розетковидная, звездчатая, древовидная и
др. Колонии бывают плоскими, выпуклыми, куполообразными, вдавленными. Края у
колоний могут быть ровными, волнистыми, фестончатыми, расплывчатыми, изрезан-
ными и др. Поверхность колоний может быть гладкая или шероховатая, влажная
или сухая. По консистенции различают однородные или зернистые колонии, по
прозрачности матовые, непрозрачные или прозрачные.
Различают два основных типа колоний - S и R типы. Подавляющее большинство
патогенных бактерий образует колонии S-типа. Колонии S-типа - круглые, глад-
кие, блестящие, выпуклые, с ровными краями, влажные. Колонии R-типа - пло-
ские, шероховатые, матовые, неправильной формы, с исчерченными краями. Однако
форма колоний подвержена изменчивости. R-формы колоний могут переходить в S-
формы и наоборот.
Колонии S-типа.
Колонии R-типа.
R-тип колонии образуют: возбудитель сибирской язвы - матовые, плоские с ло-
конообразной периферией колонии (колонии в виде «львиной гривы»).
Возбудитель чумы - периферийная кружевная зона, вокруг приподнятой цен-
тральной части колонии (колонии в виде «кружевного платочка»).
Возбудитель туберкулеза - морщинистые колонии светло-кремового цвета (коло-
нии в виде «тутовой ягоды»).
Возбудитель дифтерии - плоские колонии с радиальной исчерченностью (колонии
в виде «цветка маргаритки»).
Микоплазмы - колонии состоят из центра врастающего в глубь среды и перифе-
рийной ажурной части (колонии в виде «яичницы-глазуньи»).
Колонии могут быть бесцветными или окрашенными - пигментированными: белые,
золотистые, лимонно-желтые колонии - у стафилококков, рубиново-красные - у
серраций, сине-зеленые - у синегнойной палочки, кремовые - у микобактерий ту-
беркулеза .
Выделение
чистых культур
В практических бактериологических лабораториях для выделения чистых культур
аэробных бактерий используют метод Дригальского, представляющий собой после-
довательный поверхностный рассев исследуемого материала петлей на чашки Петри
с МПА. Процесс выделения чистой культуры данным методом можно разделить на 3
этапа:
■ 1 этап: стерилизуют бактериальную петлю в пламени спиртовки. Захватывают
петлей исследуемый материал (гной, кровь, моча и др.) и наносят на поверх-
ность питательной среды в чашке Петри. Движения петли при посеве должны
производиться последовательно по всей поверхности среды. После посева пет-
лю прожигают в пламени спиртовки. Чашки подписывают на донышке и ставят
вверх дном в термостат при температуре 37 С на сутки.
Посев материала на поверхность питательной среды.
■ 2 этап: на следующий день посевы на чашке просматривают, изучая характер
роста. Отмечают нужную (подозрительную) колонию. С помощью стерильной бак-
териальной петли захватывают подозрительную колонию и пересевают в пробир-
ку со скошенным МПА. Пробирку подписывают и помещают в термостат при тем-
пературе 37 С на 18-24 часа.
Характер роста на трех секторах питательной среды.
3 этап: визуально изучают характер роста на скошенном агаре. Затем делают
мазок из выросшей культуры, окрашивают его по методу Грама и, убедившись в
том, что культура чистая, переходят к видовой идентификации. При микроско-
пии мазка, приготовленного из чистой бактериальной культуры, в нем обнару-
живаются морфологически и тинкториально однородные клетки.
Характер роста на скошенном агаре.
Кроме метода Дригальского, в бактериологической практике применяют и дру-
гие . Например, для выделения чистой культуры протея, обладающего «ползучим»
ростом, используют метод Шукевича. Исследуемый материал засевают в конденса-
ционную воду скошенного МПА. Бактерии, обладающие ползучим ростом, из конден-
сационной воды вползают на поверхность агара. На следующий день проверяют
чистоту выделенной культуры. Из верхнего края налета делают мазок, окрашивают
по Граму и микроскопируют.
Чистую культуру бактерий также можно получить, если на исследуемый материал
воздействовать каким-либо физическим или химическим агентом, обладающим изби-
рательным антибактериальным действием. Например: для выделения чистой культу-
ры спорообразующих бактерий, исследуемый материал прогревают при температуре
80 С в течение 20 минут или кратковременно кипятят для уничтожения вегетатив-
ных форм. Споры бактерий при этом сохраняются и при посеве материала на пита-
тельную среду прорастают. Далее чистую культуру получают обычным путем.
Для выделения чистой культуры возбудителя чумы посевы инкубируют при темпе-
ратуре 5 С, что значительно замедляет рост сопутствующей микрофлоры.
Для получения чистой культуры кислотоустойчивых бактерий (микобактерии ту-
беркулеза) исследуемый материал обрабатывают 2% серной кислотой, уничтожающей
сопутствующую флору.
Для культивирования анаэробов применяют особые методы, сущность которых за-
ключается в удалении воздуха или замены его специальной газовой смесью (или
инертными газами) в герметизированных анаэростатах. В настоящее время чаще
используется GasРак—система. Она химическим путем обеспечивает постоянство
газовой смеси, приемлемой для роста большинства анаэробных микроорганизмов. В
герметичном аназростате, в результате реакции воды с таблетками боргидрида
натрия и бикарбоната натрия образуется водород и диоксид углерода. Водород
затем реагирует с кислородом газовой смеси на палладиевом катализаторе с об-
разованием воды, уже вторично вступающей в реакцию гидролиза боргидрида. Ме-
тод был предложен Брюером и Олгаером в 1965 году. Разработчики представили
одноразовый пакет, генерирующий водород, который был позднее усовершенствован
ими до саше, генерирующих двуокись углерода и содержащих внутренний катализа-
тор.
Анаэростаты GasРак—система.
Другим способом выращивания анаэробов является использование эксикатора (на
дно которого устанавливается зажженная свеча, либо наливается щелочной рас-
твор пирагалола, поглощающий кислород).
/f*4
Эксикатор.
Как правило, посевы анаэробов производят на среде Китта-Тароцци, состоящей
из МПБ, 0,5% глюкозы и кусочков печени или мясного фарша. Перед посевом среду
прогревают в водяной бане в течение 10-15 мин для удаления воздуха и остужа-
ют . После посева среду заливают слоем вазелинового масла или парафином для
изоляции от атмосферного воздуха. Так же часто применяют среду Вильсона-Блера
- железосульфитный агар. Она состоит из МПА к которому добавляют глюкозу,
сульфит натрия и хлорид железа. Среду развивают в пробирку высоким столбиком.
Материал засевают уколом в столбик. Анаэробы образуют почернение по ходу уко-
ла, за счет образования сульфата железа.
Среда Китта-Тароцци до и после посева.
Выделение чистых культур анаэробных бактерий методом Цейсслера:
■ 1 этап: исследуемый материал засевают на среду Китта-Тароцци. Пробирки за-
ливают слоем вазелинового масла или парафина, подписывают и ставят в тер-
мостат при температуре 37 С на сутки.
■ 2 этап: на следующий день посевы на среде Китта-Тароцци просматривают и в
случае наличия роста (помутнение среды) производят пересев в чашки Петри
на сахарно-кровяной агар, для получения изолированных колоний. Чашки под-
писывают и помещают в анаэростат и термостатируют сутки при температуре 37
С.
■ 3 этап: просматривают посевы, изучают изолированные колонии. Для выделения
и накопления чистой культуры производят пересев подозрительной колонии с
чашки в пробирку со средой Китта-Тароцци. Пробирки термостатируют сутки
при температуре 37 С.
■ 4 этап: изучают визуально характер роста чистой культуры на среде Китта-
Тароцци. Затем, проводят проверку чистоты выделенной культуры, для чего ее
микроскопируют. Чистая культура, выделенных анаэробных бактерий, морфоло-
гически и тинкториально однородна.
Биохимические
свойства бактерий
Состав ферментов синтезируемый любой бактерией определяется ее геномом и
является довольно стабильным признаком. Бактерии синтезируют разнообразные
ферменты, принадлежащие к 6 основным классам.
Определение сахаролитических, протеолитических и других ферментов, образуе-
мых определенными видами и даже вариантами (биоварами) бактерий имеют важное
таксономическое значение, широко применяясь в систематики и идентификации.
Целый ряд ферментов (гиалуронидаза, коагулаза, фибринолиз и др.) способствуют
проявлению патогенных свойств бактерий, поскольку мишенью их действия являют-
ся клетки и ткани организма человека. Эти ферменты называют ферментами пато-
генности (подробно о ферментах данной группы будет рассмотрено в разделе «Ин-
фекция») .
Ферменты бактерий могут локализоваться в цитоплазме, цитоплазматической
мембране или периплазматическом пространстве бактерий, это так называемые эн-
доферменты, другие - экзоферменты, выделяются в окружающую среду.
Функциональное значение экзоферментов связано с расщеплением макромолекул
органических и неорганических веществ в окружающей среде до более простых со-
единений. Они осуществляют контактное и внеклеточное переваривание веществ,
повреждают ткани своих хозяев и др.
У бактерий различают три основные группы ферментов:
1. Конститутивные - постоянно синтезируемые бактериальной клеткой.
2. Индуцибельные (адаптивные) - синтез которых индуцируется соответствующим
субстратом.
3. Репрессибельные - синтез которых подавляется в результате избыточного на-
копления продукта реакции, катализируемой данным ферментом.
Одной из особенностей синтеза бактериальных ферментов является преобладание
индуцибельных ферментов над конститутивными, что связано как с малым объемом
протоплазмы, так и с их ролью главного механизма адаптации к меняющимися ус-
ловиям внешней среды.
Другой особенностью синтеза ферментов у гетеротрофных бактерий является их
выделение в огромном количестве в окружающую среду. Ферменты могут функциони-
ровать независимо друг от друга или быть тесно связанными между собой, обес-
печивая протекание метаболических реакций в строгой последовательности. По-
следних называют - мультиферментные комплексы, например ферменты дыхательной
цепи, локализованные на цитоплазматической мембране.
Определение ферментов используют для идентификации бактерий - возбудителей
инфекционных болезней, помимо изучения морфологических и культурных свойств.
К биохимическим свойствам относят: сахаролитические, протеолитические свой-
ства , пигментообразование, токсинообразование.
Бактерии характеризует неодинаковая способность использовать различные уг-
леводы. Ферментация углеводов бактериями приводит к образованию органических
кислот, либо кислот и газов. Определение сахаролитических ферментов проводят
на дифференциально-диагностических средах Гисса. Среды Гисса состоят из 1%
пептонной воды, 0,5% определенного углевода (глюкоза, лактоза, мальтоза, са-
хароза и др.), индикатора Андреде (кислый фуксин в 1Н растворе NaOH). В про-
бирку со средой опускают поплавок (стеклянная трубочка один конец которой за-
паян) для улавливания газообразных продуктов. Свежеприготовленная среда имеет
соломенно-желтый цвет. Исследуемую культуру засевают в пробирку. Результаты
посевов учитываются через 24-48 часов. О разложении углеводов судят по изме-
нению цвета среды. Среда приобретает красный цвет, в результате сдвига рН в
кислую сторону, за счет образования кислоты. О газообразовании свидетельству-
ет накопление газа в поплавке.
^Ч^,
Пробирка со средой Гисса до
посева материала.
Пробирка со средой Гисса
после посева.
Протеолитические ферменты - протеазы, катализирующие расщепление белка. На
практике о протеолитической активности бактерий чаще судят по способности
расщеплять белок до продуктов глубокого распада: индола и серововодорода. Для
этого исследуемую культуру засевают на МПБ, между пробкой и горлышком пробир-
ки укрепляют индикаторные бумажки. На сероводород - индикаторная бумажка про-
питана уксуснокислым свинцом. При выделении H2S - она чернеет. На индол - ин-
дикаторная бумага - пропитана раствором щавелевой кислоты, при выделении ин-
дола - она краснеет.
*
I m M
Проявление протеолитических свойств.
Сахаролитические и протеолитические ферменты микроорганизмов весьма много-
образны и универсальны (часто встречаются у представителей разных видов). Это
обусловливает относительно невысокие дифференцирующие свойства традиционных
сред. Для более четкой дифференциации культур у них желательно определять ро-
до- и/или видоспецифические ферменты. В конце XX века в бактериологическую
практику вошли дифференциальные среды нового поколения - хромогенные, принцип
действия которых основан на выявлении высокоспецифичных ферментов у искомых
микроорганизмов. К таким ферментам относятся, например, бета-Б-глюкуронидаза
Escherichia coli или бета-Б-глюкозидаза энтерококков. Для обнаружения уни-
кального фермента и, соответственно, идентификации микроорганизма, в состав
среды вводят хромогенный субстрат - вещество, при расщеплении которого обра-
зуются окрашенные и/или флюоресцирующие продукты. В результате микробный рост
окрашивается в определенный цвет или приобретает способность к флюоресценции
при ультрафиолетовом облучении. Поскольку хромогенный субстрат или их смесь
вводятся в состав сред (в том числе селективных) для первичного посева, то
результат - выделение чистой культуры и ее идентификация - может быть получен
уже в течение первых суток исследования.
-"77-у
■ : ■',:■•'■■/,
г л. . •> Ш.
Salmonella Tvphimunum ЛТСС' 14028
у//
/.,
/.-; /.■:.'
С<:гоЬааег fteundu ЛТСС SC90
Salmonella enrent'dt ЛТСС1 130'6
•
"Ч
а»
%
*
%
9
а
Sneprococcus Jtfj/jcrue ЛТСС 13813 Enrerococcui tjtecahc ЛТСС 19433
Рост различных видов бактерий на хромогенных средах.
Пигменты - красящие вещества различных классов химических соединений, выде-
ляемых в среду или входящие в состав клеток. Синтезирующие пигменты бактерии
чаще встречаются среди обитателей воздуха, почвы, воды. Они, как правило, за-
щищают микробную клетку от ультрафиолетовых лучей. Среди патогенных для чело-
века бактерий пигментообразование мало характерно. Данный признак относитель-
но постоянен и цвет пигмента используют для идентификации пигментообразующих
бактерий. Пигменты каратиноиднохю класса, имеющие желтые, лимонные, оранжевые
тона, синтезируют стафилококки (S. aureus), микобактерии (М. tuberculosis, M.
bovis). Растворимый в воде сине-зеленый пигмент пиоцианин синтезирует синег-
нойная палочка (P. aeruginosa). Пирагалолловый пигмент красного цвета проди-
гиозин продуцируют серрации (S. marcescens).
Лимонно-желтые колонии S. Aureus
Рост микобактерии на среде
Левенштейна-Иенсена.
Каталаза - железосодержащий фермент, катализирующий реакцию расщепления
Н2О2 на воду и кислород. Присутствует у большинства аэробных и факультативно-
анаэробных бактерий и отсутствует у облигатно анаэробных. Для определения в
каплю Н202 на предметном стекле вносят испытуемую культуру. При наличие ката-
лазы сразу же или через несколько секунд появляются пузырьки.
Тест на каталазу.
Оксидазы - группа ферментов, характерных для аэробов и отсутствующих у ана-
эробов и большинства факультативно-анаэробных бактерий. Для определения на
фильтровальную бумагу смоченную 1% р-ром тетраметилпарафенилдиамин-
дегидрохлорида (реактив Кодака) наносят полосу из испытуемой культуры. В по-
ложительном случае через несколько секунд появляется фиолетовое окрашивание.
Тест на оксидазу.
Технологии
ТЕХНИКА МИКРОБИОЛОГИЧЕСКОЙ РАБОТЫ
Посев (бакпосев — от бактериологический посев) — один из стационарных мето-
дов культивирования микроорганизмов на питательных средах, применяемый для
культуральной диагностики в медицинской микробиологии, а также для исследова-
ния биохимических и биологических свойств в различных биотехнологических це-
лях.
В зависимости от содержания исследуемых бактерий в образце, проводят посев
на плотные питательные среды (для получения изолированных колоний и определе-
ния чистоты культуры). Если в исследуемом материале содержание микроорганиз-
мов незначительное, то посев проводят на жидкие среды обогащения.
Различают различные методы посева.
Для посевов применяют микробиологические петли, реже — иглы и шпатели. Чаще
всего для культивирования используются пробирка и чашка Петри. Универсальным
инструментом для засева культуры является бактериальная петля. Помимо неё,
для посева уколом применяют специальную бактериальную иглу, а для посевов на
чашках Петри — металлические или стеклянные шпатели. Для посевов жидких мате-
риалов наряду с петлёй используются градуированная и пастеровская пипетки.
Микробиологические петли и игла.
Микробиологические шпатели (стеклянные).
Пастеровская пипетка для посевов.
Посев в
пробирку1
При посеве берут пробирку в левую руку, а правой, плотно обхватив пробку
четвёртым и пятым пальцами, вынимают её. Удерживая другими пальцами той же
руки петлю, сначала в горизонтальном, а потом в вертикальном положении её
вносят в открытое пламя и прожигают до красного каления. Остывшей петлёй на-
бирают посевной материал, после чего закрывают пробирку пробкой, предвари-
тельно проведя край пробирки над пламенем спиртовки. Пробку при этом обжигать
не следует. Затем в пробирку со скошенным агаром вносят петлю с посевным ма-
териалом, опуская её до конденсата в нижней части среды, и зигзагообразным
движением распределяют материал по скошенной поверхности агара. Вынув петлю,
обжигают край пробирки и закрывают её пробкой. Петлю обжигают в пламени го-
релки и ставят в штатив рукояткой вниз. Пробирки с посевами подписывают зара-
нее, указывая дату посева, номер исследования и название культуры.
Исходное положение (в малой пробирке - культура микроорганизма).
1 Здесь и далее работа ведется в ламинарном микробиологическом боксе, но было время
когда их не было и микробиологи работали в стерилизуемом помещении (боксе) и даже
порой на своем письменном столе.
Пробирки в левую руку. Петля в правую. Обжигаем петлю в пламени
спиртовки.
Вынимаем ватные пробки из пробирок,
В горячем воздухе над спиртовкой берем культуру иглой.
Переносим петлю в пробирку для засева..
..и засеваем штриховым движением по поверхности агара.
Обжигаем быстрым движением края пробирок (и пробки)...
и закрываем пробирки пробками.
Не забываем обжечь петлю.
Закончили.
Посев на плотные
питательные среды
в чашке Петри
Посевы «газоном» производят на плотную питательную среду в чашке Петри. Для
этого, приоткрыв левой рукой крышку, петлёй или пипеткой наносят посевной ма-
териал на поверхность питательного агара по методу Дригальского. После инку-
бации посева появляется равномерный сплошной рост бактерий с разделением их
на колонии. Идентификацию выделенных бактериальных культур проводят путём
изучения морфологии бактерий, их культуральных, биохимических и других при-
знаков, присущих каждому виду.
Колония — это видимое изолированное скопление представителей одного вида
микроорганизмов, образующееся при размножении одной колониеобразующеи единицы
(КОЕ) на плотной питательной среде (на поверхности или в глубине её). Колонии
бактерий разных видов отличаются друг от друга по своей морфологии, цвету и
другим культуральным признакам.
Все готово для посева.
На дне чашки Петри нанесены линии разделяющие ее на сектора. Ка-
ждый сектор для своего разведения культуры (разведения в пробир-
ках, два 1:10 и два 1:100).
II i*
Обжигаем петлю всю в пламени спиртовки.
Открываем пробирку. В данном случае там накопительная культура,
так что пробирку можно не обжигать.
Берем пробу жидкости петлей (зависит от размеров петли)
Чашку - в область горячего воздуха.
1-И
Засеваем штрихом соответствующий сектор.
1-И
Обжигаем петлю.
Засев остальных секторов - аналогично. Теперь пересев с чашки на чашку.
Касаемся петлей одиночной точечной колонии.
Рассеваем штрихом на другой чашке по методу Дригальскоро.
Обжигаем петлю.
# t
забываем обжеих. „
"ожечь петлю.
Технологии
ИЗГОТОВЛЕНИЕ ЭЛЕКТРОВАКУУМНЫХ ПРИБОРОВ
(продолжение1)
Несколько общих слов перед работой. Заготовки для стеклодувного дела должны
быть перед применением тщательно очищены [1] и с особым пристрастием, при из-
готовлении электровакуумных приборов (ЭВП). Некоторое послабление может быть
сделано только в учебно-тренировочных случаях.
Все стеклотрубки делятся по температурному коэффициенту расширения (ТКР) на
несколько групп, именуемых по металлу, нетолстую проволоку которого, можно в
них впаивать без существенных натяжений. Так, например, есть стёкла платино-
вой группы, молибденовой, вольфрамовой. Стёкла внутри одной группы, считаем
условно пригодными для спаивания между собой.
В описанных ниже работах использовалось стекло платиновой группы - дешёвое,
легкоплавкое, с высоким КТР и особой склонностью к растрескиванию при термо-
ударах. Тот его вариант, что не содержит окиси свинца и не темнеет на восста-
новительном газовоздушном пламени.
Огневое оснащение работ - настольная газовоздушная горелка-пушка и электро-
лизёр .
Первый общий навык после резки стекла - вращение заготовки пальцами одной
руки. Непрерывное и возможно более плавное. Заготовку следует вращать в пла-
мени для равномерного нагрева, стекло размягчённое, при этом сохраняет сим-
метричную форму. Работу нужно вращать, в том числе, и вынув из пламени - для
равномерного остывания и уменьшения риска растрескивания. Особенно это отно-
сится к легкоплавким стёклам с высоким ТКР.
Горячую работу следует замедленно охлаждать для снижения внутренних напря-
1 Предыдущая часть - Домашняя лаборатория 2024-01
жений и предотвращения разрушения. Простейший способ - покрутить работу не-
сколько минут в большом мягком коптящем (светящем) пламени - горение газа без
подачи воздуха. При этом стекло покрывается слоем копоти, которая легко сти-
рается после остывания. Очень удобный приём для уменьшения скорости охлажде-
ния горячего стекла - укутывание его огнеупорным одеялом. Работы более или
менее крупные, сложные и ответственные, после этого нужно ещё и полноценно
отжечь в печи.
Стеклодувная мастерская рассматривается как вредный цех, тем более при ис-
пользовании стекла, побывавшего у химиков, которое вполне может иметь и ос-
татки легко летучих ядовитых веществ вроде селена, ртути, мышьяка и других
подобных - рабочее место должно быть непременно оснащено хорошей вытяжной
вентиляцией, а б/у заготовки лучше бы не использовать вовсе.
Глаза при горячих работах со стеклом следует защищать специальными очками-
светофильтрами или в крайнем случае при работах кратковременных, хотя бы
обычными пластиковыми от осколков стекла.
Перед началом огневых работ следует позаботиться о средствах тушения потен-
циальных возгораний. Полезно ещё и иметь под рукой большую мокрую тряпку.
* * *
Немалую часть технологического процесса изготовления радиолампы занимает
работа со стеклом -хорошо подходящим для этой цели материалом. Рассмотрим на-
бор операций, достаточных для изготовления стеклянной части простого электро-
вакуумного прибора (ЭВП) в ключе работ в условиях любительской мастерской.
Разогрев
стекла
Внесение заготовки в пламя горелки - момент ответственный, тем более для
легкоплавких стёкол с высоким коэффициентом теплового расширения (КТР). Стек-
ло - материал хрупкий и плохо проводящий тепло. Верхние его слои нагреваются
значительно быстрее внутренних, они по-разному расширяются, и заготовка раз-
рушается . Бывает, что и с сильным разлётом горячих осколков. Повышается риск
растрескивания при нагреве крупных, сложных, толстостенных стекляшек, прово-
лочных впаев, повреждении поверхности стекла, в том числе и неровных зазуб-
ренных резов, неотожженных промежуточных изделий. Разогревать такие заготовки
следует с особой деликатностью и не жалея времени.
Один из способов разогрева - решение «в лоб» - вращая заготовку,
нагреваем её в дальней части негорячего мягкого коптящего пламе-
ни (газ без воздушного дутья), потом в ближней, затем добавляем
немного воздуха, делая факел чуть жёстче и так далее. На фото -
разогрев края коротенького кусочка трубки 018 мм, для удобства
насаженной на силиконовую пробочку со стеклянной трубкой-
державкой . Копоть на стекле при дальнейшем нагреве сгорает.
Появление в факеле характерного яркого рыжего хвоста, т. н. содового свече-
ния - окрашивания пламени испаряющимися с поверхности заготовки ионами натрия
сигнализирует о фазовом переходе в стекле. С этого момента его можно нагре-
вать с любой практической скоростью, не опасаясь термоудара.
Второй способ разогрева простой заготовки - короткое внесение её холодной в
горячее пламя. Вынув, трубочку тоже вращаем. Полученная порция тепла при этом
проникает в стекло глубже. Макаем ещё и ещё, постепенно увеличивая время в
пламени.
Оплавление
края трубки
Не только правило хорошего тона, но и суровая необходимость - вращать заго-
товку приходится часто и одновременно поддувая её. Суя в рот стекло с острым
краем, можно здорово изрезать губы. Рукам достаётся тоже. Кроме того, неоп-
лавленный конец трубки сложнее затыкать пробками, они быстрее портятся.
Оплавление края трубки. Выполняется в горячей части неширокого
жёсткого факела.
Обычные ошибки: разогрев слишком долгий или слишком широкой части трубки.
При этом её конец начинает стягиваться, уменьшая внутренний канал. Надёжно и
ровно всадить пробку в такое отверстие трудно, край приходится дополнительно
разворачивать специальным инструментом - операция на крупных трубках непро-
стая.
Расширение стянутого конца трубки медной развёрткой,
воском. Горлышко под пробку.
смазанной
Изготовление
пробирочного
дна
Конец трубки сильно и равномерно разогревается. Здесь использован
асбестовый отражатель пламени, возвращающий часть тепла на стекло.
Раскрыва губок моего небольшого медицинского пинцета не хватает
для захвата трубки целиком - размягчённые края сплющиваю на гра-
фитовой плитке.
Сильно разогреваю конец трубки, в
цет мог её схватить.
том числе и сплющенную часть, чтобы пин-
Вынимаем заготовку из пламени, хватаем конец пинцетом и плавно
оттягиваем его, вращая основную заготовку.
Переплавляем перекрученный стеклянный хвост около нашего доныш-
ка, остатки - в металлическую урну с боем.
Разогреваем пока ещё конусное донышко и несколько раз снимаем
лишнюю капельку стекла стеклянной палочкой или пинцетом.
Разогреваем донышко в горячем мягком пламени и в несколько приё-
мов раздуваем до пробирочной полусферы.
Покрутив минутку горячую работу в коптящем пламени, перенёс её в
базальтовое одеяло - сложенный пополам отрезок мягкого рулонного
огнеупора, для замедленного остывания.
Готовое пробирочное донышко. Оно же - макушка баллона радиолампы
или конец разрядной трубки. Соответственно, в него может быть
впаян штенгель или проволочный вывод.
Обычные ошибки: неравномерное вращение в пламени, заготовка вращается не
перпендикулярно факелу - приводят к размягчению неровного кольца трубки, фи-
гуры из такой заготовки получаются несимметричные, косые. Вытягивая длинный
ус, заготовка не вращается, то же и при раздувании - те же последствия. Под-
дувать размягчённое стекло следует щеками и небольшими порциями.
Штенгели
Штенгелем называют тонкую технологическую трубочку, впаянную в колбу ЭВП в
удобном месте. Собранный прибор через штенгель присоединяют к откачному по-
сту, удаляют воздух и при необходимости наполняют нужными газами или вещест-
вами. Штенгель готового прибора переплавляют около колбы - нагревают ручной
горелкой, и давление атмосферы его сплющивает. На колбе остаётся характерная
пупочка.
Здесь используется штенгель внешним 0 5 мм.
Остатки штенгелей на макушке заводских радиоламп. 05 мм на мощ-
ном сдвоенном кенотроне 5Ц8С, 1970 г.р. 04 мм на «лысом» - с не-
распыляемым геттером варианте стабилизаторного триода 6С19П 1964
г.р. НПО «Светлана», г. Ленинград.
Мелкие подготовительные
работы с тонкой трубочкой
В зависимости от имеющегося в мастерской оборудования, штенгель может окан-
чиваться ровным оплавленным краем для присоединения металлическим цанговым
зажимом, ровным необработанным краем или небольшим расширением в случае при-
паивания к вакуумной системе из стекла. В ряде случаев штенгель может быть
присоединён к мягкому толстостенному вакуумному шлангу.
Оплавляем конец отрезанного кусочка трубочки. Работать, однако,
приходится быстрее и аккуратнее, чаще осматривать готовое.
\
Оплавленный конец заготовки штенгеля.
Разогрев второй конец трубочки несколько дольше и шире, добива-
емся полного заплавления канала - стекло стягивают в каплю силы
поверхностного натяжения.
Разогрев кольцо стекла и чуть раздув его, получаем т. н. оливку
- местное утолщение для надёжного присоединения шланга.
j
Обрезаем закрытый кончик штенгеля и оплавляем острый торец. Мож-
но подключать шланг. Операция выполняется после припаивания
штенгеля к колбе ЭВП.
Получив на конце тонкой трубочки каплю горячего стекла, очень
просто раздуть её до крупного пузыря с тончайшими стенками. Ино-
гда это пузырь лопнувший.
4
Прижав такой пузырик к плоской твёрдой поверхности, легко разру-
шаем его, оставляя более толстую и прочную вороночку-основание.
Обычно она удаётся замечательно правильной формы.
Такие вороночки, кроме прочего, помогают очень просто и на широ-
ком факеле спаять тонкие трубки, припаять штенгель к тонкостен-
ной колбе и так далее.
Впаивание
подготовленной
трубочки-штенгеля
В работах участвует ручная горелка с соплом 0,3 мм, сжигающая гремучий газ
из электролизёра, обогащенный толикой паров бензина.
Разогретая обычным образом заготовка помещается над небольшим языком
светящего пламени (подогрев). Маленьким горячим игольчатым факелом
сильно разогреваем точку на стекле - место для припаивания штенгеля.
Не давая стеклу остыть, быстро и аккуратно раздуваем крохотный
пузырик, обламываем его тонкий купол, слегка оплавляем основание
в мягком пламени.
Монтаж. Одновременно, держа заготовки по обе стороны от средней
части мягкого пламени, разогреваем до начала малинового свечения
торец полученного кратера на трубке-колбе и кончик тонкой тру-
бочки-штенгеля, и поаккуратнее слепляем их.
Увы, слепленный не значит спаянный. Между неперемешавшимися
стёклами деталей могут остаться незаметные глазу микротечи, че-
рез которые сложный прибор со временем насосёт воздуха и выйдет
из строя. На фото - прилепленный в торец колбы штенгель.
Пропаивал слепленные стыки деталей ручной горелкой, её крохотным го-
рячим факелом, подогревая место работы в небольшом коптящем пламени.
Работа мелкая и точная, поверх защитных дидимовых очков удобно
надевать козырёк с лупой.
Тщательно вращая, готовую работу закоптил в крупном светящемся
пламени и укутал базальтовым одеялом.
Впаянный штенгель.
Операция сложная, требует некоторого навыка. Обычные неприятности: часто
заплавляется маленькое проходное отверстие между штенгелем и колбой. При пай-
ке сильно разогревается место соединения и более или менее длинный штенгель,
размягчаясь, падает или перекручивается. Приходится искать температуру подог-
ревного пламени и место в нём. Такое горячее, чтобы стекло не растрескивалось
при пайке и такое прохладное, чтобы детали не размягчались. Слепленное стекло
часто растрескивается при повторном нагреве - шов лучше пропаивать сразу, не
охлаждая работы.
Более устойчивы при работе и прочны в эксплуатации впаи тонкого
штенгеля с несколько расширенным концом. Лучше раздуванием тон-
костенного пузыря и скалыванием его непрочной части.
Впаивание одиночного
проволочного вывода
Операция, зависящая от группы стекла и, соответственно, проволоки, которую
допустимо в него впаивать. Для платиновой группы это, понятно - платина или
специальный недорогой биметалл - платинит (железоникелевый сплав со слоем ме-
ди) .
Медь в стекле растворяется неважно, зато окись меди - хорошо!
Поэтому платинитовую (и медную) проволоку перед запаиванием в
стекло отжигаем в дальней окислительной части факела.
Окисленную проволоку помещаем в отрезок стеклянной трубочки.
Которую последовательно прогреваем, добиваясь отсутствия воздуш-
ных пузырей и положения проволоки в стекле более или менее по
оси. Остеклованный ввод немедленно укутываем базальтовой ватой.
Пара остеклованных вводов. Оценить качество впая металла в стек-
ло можно по равномерности цвета (у платинита он ярко-розовый) и
отсутствию включений и потемнений.
Подготовим отверстие для электрода - разогреем конусное сужение на
трубке-колбе и пинцетом или стеклянной палочкой оттянем тонкий усик.
Обломав его, получаем удобное отверстие.
Плавно разогреваем подготовленный торец колбы с вложенным внутрь
электродом, помня, что детали нагреваются очень по-разному, а
остеклование имеет внутренние напряжения.
Разогреваем сужение и через размягчённое стекло пинцетом немного вы-
тягиваем остеклованный электрод наружу. Хорошо прогреваем впай, при
необходимости поддуваем выравнивая колбу и пинцетом поправляем стре-
мящийся наклониться внутрь электрод. Замедленно охлаждаем работу.
Следует сказать, что в реальных приборах из платинового стекла проволочный
вывод составной, сваренный из нескольких частей из разных металлов.
Обычные неудачи: отожжённая платинитовая проволока, как правило, легко за-
паивается в стекло без брака, но впай до отжига имеет внутренние напряжения и
склонен к трещинам. Неправильная форма впая может быть вызвана огрехами в
подготовительных операциях.
}
Вид впая в отражённом свете. Колба - трубка 018 мм со стенками
нормальной толщины.
Аналогичный впай в тонкостенную трубку 026 мм (отмытая колба от
трубчатой лампы дневного света).
Описанных операций довольно для изготовления некоторых газоразрядных прибо-
ров , в том числе и миниатюрных высоковакуумных насосов и датчиков. Следует
сказать, что выполнение операции может и существенно
учебниках классики кое-что делают по-разному.
отличаться - даже
Литература:
1. Бондаренко Ю. Н. Изготовление газоразрядных источников света для лабора-
торных целей и многое другое.
Технологии
ЛИТЬЕ ПЛАСТМАСС В СИЛИКОН
Многие из тех, кто печатает на 3D-принтере сталкиваются или с необходимо-
стью получить партию моделей в короткие сроки, или скопировать удачно полу-
чившуюся деталь, или получить изделия с прочностными характеристиками, пре-
восходящими таковые у пластиков для домашней Зс1-печати.
3D-принтер далеко не всегда способен выполнить такие задачи, но отлично по-
дойдет для создания единственного образца, или мастер-модели. А дальше на по-
мощь нам приходят материалы производства компании Smooth-On1, наверное, само-
го популярного производителя материалов холодного отверждения.
В этом обзоре мы сравним самые основные и популярные силиконы, полиуретаны
и добавки к ним, кратко посмотрим на основные способы создания форм и изде-
лий, подумаем, где это может найти применение и, наконец, создадим свою сили-
коновую форму и модель.
Процесс создания изделий методом литья практически всегда одинаков: создаем
модель, с её помощью создаем силиконовую форму, заливаем в нее материал, по-
лучаем изделие. Но в зависимости от модели, необходимых свойств, количества
отливок, каждый этап может кардинально меняться. Существует несколько спосо-
бов как создания формы, так и готового изделия.
Пару слов о подготовке моделей, напечатанных на 3D-принтере. Компания
Smooth-On обратила своё внимание на эту технологию и выпустила специальный
лак под названием XTC-3D2. Он прекрасно сглаживает характерные для напечатан-
1 https://www.smooth-on.com/products/
2 https://www.smooth-on.com/product-line/xtc-3d/
ных моделей огрехи, видимые слои, которые обязательно перейдут на силиконовую
форму, и придает поверхности гладкость и глянцевый вид.
Методы
создания
форм
Сплошная заливка
Самый простой способ: модель помещается в опалубку (специальную герметичную
емкость из обычного оргстекла, пластика или другого материала), фиксируется в
ней и заливается силиконом. Хорошо подходит для простых двухмерных моделей,
рельефов, сувенирной и брендинговой продукции.
Разрезная форма
Аналогичен предыдущему, только модель размещается с учетом того, что форма
будет разрезаться полностью или частично для облегчения съема. Модель может
быть подвешена с помощью проволоки или размещена на тонкой опоре. Способ
предназначен для более сложной геометрии, технических изделий, сложных фигур.
Двухсоставная форма
Это один из самых сложных способов. Заключается в помещении модели на гли-
няную или пластилиновую основу, которая делит силиконовую форму пополам.
На основе размещаются специальные замки, которые будут обеспечивать точное
совмещение двух форм и отсутствие смещений. Вокруг основы собирается опалуб-
ка, герметизируется горячим клеем или пластилином, и в неё заливается первая
половина формы. Затем, после отверждения силикона, форма переворачивается,
глина или пластилин счищаются, силикон покрывается разделительным составом, и
заливается вторая половина формы.
Метод «в намазку»
Этим методом создаются так называемые «чулочные» формы, когда силикон точно
повторяет форму объекта и имеет толщину от 3 мм до нескольких сантиметров.
Для создания формы «в намазку» необходим достаточно вязкий силикон, который
бы не стекал с модели.
Можно использовать как специально предназначенные для этого силиконы, назы-
ваемые тиксотропными, так и обычные, но модифицированные с помощью загустите-
лей.
Силикон наносится кисточкой или шпателем в несколько слоев, в которых чере-
дуется вязкость и скорость отвердевания, чтобы форма была максимально детали-
зированной и прочной. После того, как все слои готовы, с помощью специального
состава создается жесткая внешняя оболочка, которая будет держать форму.
Методы
заливки
пластиков
Самый простой метод заключается в обычной заливке пластика в форму, он под-
ходит для домашнего использования и позволяет достичь приемлемого качества.
Но, по необходимости, для более качественного результата возможно использова-
ние установок высокого давления, что позволит практически полностью убрать
пузырьки воздуха.
Для этого форма вместе с залитым пластиком помещается в камеру, в которой
создается повышенное до 4 атмосфер давление. Форма должна оставаться в камере
все время отвердевания полиуретана. При таком давлении пузыри уменьшаются до
почти невидимых глазу размеров, что значительно повышает качество изделия.
Еще один метод, так называемая заливка «в обкатку», используется для созда-
ния полых изделий. В форму заливается небольшое количество пластика, около
10% от общего объема, отверстие для заливки закрывается, и начинается враще-
ние формы по всех плоскостях, вручную или на специальной ротационной машине.
При этом пластик отвердевает на стенках формы, создавая полую модель, что по-
зволяет существенно снизить вес изделия и экономить материал.
Обзор
силиконов
Серия Mold Star 15, 16, 30
Силиконы для создания форм на основе платины. Застывают при комнатной тем-
пературе, образуют прочную, гибкую и очень детализированную форму. Предназна-
чены для литья силикона, полиуретана, смол, полиэстера, воска и других мате-
риалов. Химически чувствительны и не способны работать с латексом, серой и
некоторыми другими соединениями.
Самые базовые и основные силиконы, способные решать большинство задач. Циф-
ра в названии отражает твердость по шкале Шор А. Обладает низкой вязкостью,
что позволяет в большинстве случаев работать без оборудования для дегазации.
Материал двухкомпонентный, части смешиваются в удобном соотношении 1:1 по
объему. В основном предназначены для создания форм методом сплошной заливки.
Серия Rebound 25, 40
Серия силиконов для создания форм методом «в намазку», который состоит в
том, чтобы наносить силикон кистью или шпателем на поверхность модели. Обла-
дает высокой вязкостью, возможностью модификации свойств с помощью загустите-
лей и ускорителей для создания качественной многослойной формы. Двухкомпо-
нентный, смешивается в соотношении 1:1 по объему.
Серия Equinox 35, 38, 40
Силиконовые пасты с временем жизни 1, 4 и 30 минут. Предназначены для руч-
ного смешивания, по консистенции напоминают густое тесто. Цифры соответствуют
твердости по Шору А. Обладает крайне высокой прочностью на разрыв и долговеч-
ностью. В отвержденном состоянии является безопасным для заливки шоколада,
карамели и других ингридиентов.
Серия SortaClear 18, 37, 40
Серия полупрозрачных силиконов. Такая особенность, как оптическая прозрач-
ность , используется для создания сложных разрезных форм — изделие прекрасно
просматривается, что позволяет сделать точный разрез. Как и серия Equinox,
является безопасным при контакте с пищевыми продуктами.
Также к силиконам существует большое количество добавок, обладающих самыми
разными эффектами. Accel-Т и Plat-Cat являются ускорителями отверждения, Slo-
jo — увеличивает время жизни силикона, Thi-Vex увеличивает вязкость силикона
и позволяет намазывать его кистью или шпателем на модель, Silc-Pig — это кон-
центрированные пигменты для окрашивания.
Обзор
полиуретанов
Серия Smooth-Cast
Самая основная и популярная серия полиуретанов для создания конечных изде-
лий. Линейка включает в себя более 10 наименований различных пластиков с са-
мыми разными свойствами, позволяющими подобрать материал именно для Вашего
проекта. Например, Smooth-Cast 300 обладает коротким временем жизни в 3 мину-
ты и временем отверждения в 10 минут, что позволяет быстро воспроизводить
большие партии деталей. Smooth-Cast 305 аналогичен предыдущему, но «живет»
уже 7 минут, что позволяет провести дегазацию смешанных компонентов и полу-
чить еще более качественное изделие. ONYX обладает глубоким черным цветом,
которого не достичь с помощью красителей, 65D ROTO предназначен для создания
полых моделей с помощью метода «в обкатку», 325 незаменим для точного воспро-
изведения цвета, 385 отверждается практически без усадки и максимально точно
копирует изделие.
Серия TASK
Серия полиуретанов специального назначения. Разработана для промышленного
применения и обладает специфическими свойствами для конкретных задач.
Для Вашего удобства мы сформировали специальные фильтры по сферам примене-
ния:
■ Пищевые: Smooth-Sil 940, серия Sorta Clear, серия Equinox, TASK 11.
■ Архитектурные, заливка бетона и других абразивных материалов: Серия
VytaFlex, серия Brush-On, серия Ez-Spray, серия РМС.
■ Медицинские, симуляция тканей и органов: Серия Dragon Skin, добавки
Slacker, Ecoflex 0030, Ecoflex Gel, Body Double.
■ Прототипирование: Практически любые полиуретаны и силиконы, в зависимости
от задач и требований. Серии Mold Max, Mold Star, Smooth-Cast, TASK.
■ Спецэффекты и грим: Skin Tite, Body Double, Dragon Skin, Alja-Safe,
Ecoflex, Soma Foama, Rubber Glass, Encapso K.
Стоит отметить, что это деление все же условное, и дано, чтобы примерно
представить возможности широкого ассортимента компании Smooth-On.
Пример
Мы будем использовать только те материалы и оборудование, которые можно
применить в домашних условиях. Мы попробуем создать самую сложную в изготов-
лении двухсоставную форму.
Нам понадобятся:
• Платиновый силикон Mold Star 30
• Заливочный полиуретан Smooth-Cast 300
• Лак для 3D-моделей ХТС-3D
• Упаковка виниловых перчаток
• Несколько одноразовых пластиковых стаканчиков
• Емкости для смешивания
• Термоклеевой пистолет
• Материал для опалубки (пластиковые панели)
• Скульптурная глина
• Несколько малярных кистей
В роли мастер-модели выступит модель довольно популярного среди печатников
тестового болта. Мы распечатали его черным PLA пластиком на Picaso 3D
Designer с толщиной слоя 100 микрон. Сделали мы это специально для того, что-
бы продемонстрировать эффект ХТС 3D, поскольку далеко не каждый 3D принтер
может печатать с качеством 50 микрон.
Далее — обработка, чтобы отлитая модель не переняла слоистость напечатанно-
го объекта. Обрабатываем болт лаком XTC-3D3, а затем шкурим, чтобы получить
гладкую матовую поверхность.
3 http://3dtoday.ru/blogs/top3dshop/overview-topcoat-xtc3d/
Теперь модель готова к заливке силиконом. Помещаем её на глиняную основу, с
помощью которой мы создадим силиконовую форму из двух частей.
Модель должна быть погружена в глину ровно наполовину, так что начинаем
процесс выравнивания глины. Края должны быть максимально ровными и полностью
прилегать к модели, от этого зависит качество разделения силиконовых полови-
нок. Убираем лишнюю глину и заключаем модель в пластиковую опалубку.
Все стыки пластика обрабатываем термоклеевым пистолетом и закрываем опалуб-
ку, окончательно обрабатываем глиняную основу, делаем в ней выемки для зам-
ков .
Все готово к заливке силикона. Поскольку двухкомпонентные силиконы и поли-
уретаны склонны к разделению на фракции, перед каждым использованием их необ-
ходимо тщательно перемешивать в емкости.
После перемешивания отмеряем равное количество двух компонентов по объему и
приступаем к смешиванию.
/
J.
\
\
\
т
Для данной марки силикона дегазация в вакуумной камере необязательна, что
очень удобно: исключены появления пузырьков, которые могут испортить нашу
форму. Медленно заливаем силикон в опалубку, в самую нижнюю её точку.
V
И оставляем застывать. Время застывания для этой марки силикона составляет
6 часов. По истечении этого времени освобождаем модель от опалубки.
Затем убираем глину, тщательно очищаем модель от её остатков, смазываем си-
ликон разделительным составом. В случае его отсутствия, можно использовать и
обычный вазелин, но качество будет немного хуже.
И дальше полностью повторяем процесс, заливая вторую половину силиконовой
формы.
Спустя еще 6 часов силиконовая форма готова. С помощью лезвия аккуратно
разъединяем половинки, вынимаем деталь и оцениваем, что у нас получилось.
Хорошо видны замки, закладывавшиеся в глиняной основе, хорошая детализация,
несмотря на то, что разделительная линия проходила по довольно сложным мес-
там , вроде вдавленных букв.
На самом деле, для данной модели это не самый оптимальный способ создания
формы. Но нам было интересно протестировать именно этот метод, несмотря на
сложности.
Итак, все готово к заливке полиуретана. Соединяем две половинки формы, ис-
пользуя элементы опалубки для жесткости, скрепляем с помощью резинок, скотча
или другим способом, и приступаем к подготовке полиуретана.
Хорошо перемешиваем оба компонента, встряхивая их в течение 5-10 минут. По-
сле этого даем немного отстояться, чтобы вышли образовавшиеся пузыри. Все ос-
тальное точно так же, как и с силиконом: отмеряем равное количество по объему
и смешиваем их. А дальше действовать нужно быстро: время жизни этого полиуре-
тана составляет всего 3 минуты, а время начинает идти сразу, как вы смешали
два компонента вместе. Так что мешаем быстро, но аккуратно, чтобы не созда-
вать лишних пузырьков, и сразу заливаем в форму.
Примерно через 3 минуты, в зависимости от объема материала, произойдет бы-
строе схватывание пластика, а через 10 минут деталь готова к извлечению.
Модель готова. Переданы абсолютно все детали оригинала.
Теперь Вы можете получать изделия из огромного количества материалов с са-
мыми различными свойствами, а не ограничиваться лишь классическими PLA и ABS.
К тому же, доступным станет мелкосерийное производство: распечатав всего один
экземпляр и должным образом его обработав, Вы сможете в довольно короткие
сроки создать необходимое Вам количество копий в домашних условиях. Для дос-
тижения приемлемого результата вовсе не обязательно использование дорогостоя-
щего оборудования.
Лаборатория
ХИМЛАБОРАТОРИЯ ИНСТРУМЕНТАЛЬНОГО АНАЛИЗА
Уранова В.В., Исякаева P.P., Мажитова М.В.
(продолжение)
НАИБОЛЕЕ ЧАСТО
ИСПОЛЬЗУЕМЫЕ ТИПЫ
ЛАБОРАТОРНОЙ ПОСУДЫ
Щипцы
Щипцы в лаборатории применяются для переноса горячей посуды, для нагревания
и прокаливания посуды и материалов, для захвата и переноса опасных химических
веществ.
Виды щипцов:
1. Тигельные щипцы (рис. 123) применяются в лабораториях различного профиля
для захвата и переноса горячей посуды разного диаметра и формы. Особенности
конструкции: щипцы напоминают бытовые ножницы с круглыми ручками и отверстия-
ми для пальцев руки. Кончики щипцов изогнуты и могут плотно соединяться для
удержания даже мелких предметов. На кончиках есть насечки для более плотного
захвата. Щипцы изготавливаются из нержавеющей стали — долговечного, износо-
устойчивого, термостойкого и химически стойкого материала. Изделие может ис-
пользоваться и в качестве пинцета для переноса образцов для взвешивания, при
этом следует вовремя очищать кончики щипцов от налипших веществ, чтобы они
потом не попали на другие образцы.
2. Щипцы для стаканов (рис. 124) — специализированное лабораторное приспо-
собление, разработанное для безопасных манипуляций с агрессивными веществами
или горячей посудой. С помощью специального захвата можно устойчиво зафикси-
ровать стакан, чтобы перенести его в нужное место, установить на горелку,
снять с горелки или плитки. Захват охватывает стакан с двух сторон. Для того,
чтобы стакан не скользил, захват снабжен термо- и химически стойким покрытием
с насечками. Шарнир щипцов плоский. Изделие изготавливается из нержавеющей
стали, отличается прочностью, износостойкостью, долговечностью и стойкостью к
химическим веществам.
Рис. 123. Тигельные щипцы
Рис. 124. Щипцы для стаканов
3. Щипцы для выпарительных чашек (рис. 125) предназначены для манипуляций с
круглыми чашками. С помощью щипцов можно безопасно брать, переносить чашки с
горячими веществами, устанавливать их над открытым огнем, держать чашку с ки-
пящей жидкостью на безопасном удалении от рук, не рискуя обжечься брызгами.
Щипцы изготавливаются из прочной нержавеющей стали. Специальная конструкция
захвата позволяет надежно фиксировать круглую чашку, обхватывая ее с двух
сторон.
__Lr
ТЗ,
Рис. 125. Щипцы для выпарительных чашек.
Эксикаторы и
кристаллизаторы
Эксикатор 2-го исполнения (без крана) (рис. 126). Предназначен для обезво-
живания веществ, образцов и материалов, охлаждения веществ после прокалива-
ния, хранения материалов в изолированном от окружающего воздуха пространства,
в контролируемых условиях. Изделие выполняется из толстостенного светлого
стекла. Состоит из основания, вставки и крышки. Вставка фарфоровая, с отвер-
стиями для циркуляции воздуха. На вставку устанавливают бюксы, стаканы, чашки
с пробами. На дно эксикатора, укладывают или наливают осушитель, поглощающий
влагу. Крышка, плотно прилегает к основанию, создавая герметичное соединение,
не пропускающее наружного воздуха внутрь. Для гарантии герметичности ободок
крышки и основания смазывают специальной смазкой.
Эксикатор со стеклянным пришлифованным краном в крышке (рис. 127) может ис-
пользоваться для осушения, хранения образцов в контролируемых условиях. В нем
можно создать вакуум.
*^
j
V
Рис. 126. Эксикатор без крана
i I
иЛ ч. у
УК
> <
Рис. 127. Эксикатор со стеклянным пришлифованным краном в крышке.
Кристаллизатор, или кристаллизационная чаша (рис. 128) — химический сосуд,
который используется для очистки веществ путем перекристаллизации, а также
для выпаривания растворов. Лабораторные кристаллизаторы преимущественно изго-
тавливаются из обычного или термостойкого стекла.
Рис. 128. Кристаллизатор.
Тигли
Тигель - это специальная цилиндрическая или коническая форма для нагрева,
высушивания, сжигания, обжига, плавления различных материалов и других про-
цессов, способная выдерживать температуры нагрева до 2000 С. Отличительной
особенностью тиглей является применение для их конструкции огнеупорных мате-
риалов и высокоустойчивых к различным воздействиям металлов и сплавов. Типо-
вой тигель обычно имеет коническую (усеченный конус) или цилиндрическую фор-
му.
Виды тиглей:
1. Стеклянный тигель Туча (рис. 129) - это стеклянный сосуд, сужающийся к
низу. В дно встроена пластина из пористого стекла. Для изготовления использу-
ют химически стойкое боросиликатное стекло. Стеклянный тигель Туча предназна-
чен для фильтрации мелких твердых осадков, для работы с агрессивными жидко-
стями (кроме концентрированных щелочей и плавиковой кислоты), для прокалива-
ния осадков. Может применяться для работы с горячими растворами, с кислыми и
щелочными растворами; для фильтрации под вакуумом. Не предназначен для фильт-
рации желеобразных материалов.
2. Тигли из кварцевого стекла (рис. 130), имеют важное преимущество перед
аналогами из многих других материалов: они исключительно термостойки, причем
не трескаются при быстром охлаждении и повторном нагревании. Тигель подходит
для нагревания и плавления в нем веществ и материалов, имеющих нейтральную
или кислотную реакцию. Его нельзя использовать для работы со щелочами и с
плавиковой кислотой, а ортофосфорную кислоту нельзя нагревать в нем сильнее
300 С. Подавляющее большинство остальных веществ и аналитических растворов
можно нагревать в этом высоком тигле, и он не коррозирует и не потрескается
даже при интенсивной эксплуатации.
Рис. 129. Стеклянный тигель Туча Рис. 130. Тигли из кварцевого стекла
Фарфоровая
посуда
Фарфоровые изделия по сравнению со стеклянными характеризуются большей хи-
мической и термической стойкостью. В состав разных видов фарфора входит от 20
до 60% каолина (или глины), 20-40% кварца и 20-60% полевого шпата. Обжиг из-
делий производят при 1200-1400 С. Многие фарфоровые изделия покрывают глазу-
рью (иногда частично).
Фарфоровый тигель. В фарфоровых тиглях нельзя проводить сплавление с щело-
чами и с карбонатом натрия, а также работать с фтористоводородной кислотой,
так как фарфор при этом разрушается. Иногда тигли снабжены крышками, которые
применяют для избежание распыления или потерь вещества из-за разбрызгивания.
Фарфоровый стакан. Толстые стенки делают лабораторный фарфоровый стакан
очень прочным и долговечным. Он не трескается от горячих жидкостей, допускает
нагревание не только на водяной бане, но и в пламени горелки. В последнем
случае рекомендуется под дно подложить асбестовую сетку, чтобы обеспечить
равномерное нагревание стакана и содержимого.
Фарфоровая кружка с ручкой и носиком (рис.131). Особенность конструкции за-
ключается в полностью покрытой глазурью поверхности внутри и снаружи, за ис-
ключением наружной части дна. Благодаря покрытию сосуд устойчив к воздействию
агрессивных жидкостей, может использоваться для работы с кислотами, щелочами,
органическими растворителями, солями. Фарфоровая кружка прочна, не трескается
от перепадов температур.
Фарфоровый тигель.
Фарфоровый стакан.
Фарфоровая лабораторная кастрюля (рис.132) используется для различных це-
лей. С ее помощью можно выпаривать жидкости, нагревать и кипятить их. Сосуд
изготовлен из фарфора, покрытого глазурью (кроме дна снаружи и верхней кром-
ки) . Поэтому его можно нагревать и на открытом пламени спиртовки или горелки,
и на водяной бане. В первом случае кастрюлю лучше поставить на асбестовую
сетку. Для удобства работы с сосудом его конструкция предусматривает ручку и
носик.
^~
\
Рис. 131. Кружка с ручкой и носиком
\
Z^
v:—
ф
Рис. 132. Лабораторная кастрюля
Фарфоровый бак для лабораторного использования (рис.133). Выполнен в виде
высокой широкой емкости с плоским дном. Дно чуть меньше диаметром, чем сред-
няя и верхняя часть. Край защищен ободком от сколов и трещин. Бак изготавли-
вается из качественного фарфора, покрыт глазурью, увеличивающей химическую
устойчивость стенок. Сферы применения: хранение различных веществ, сыпучих,
твердых, жидких; хранения воды; хранение лабораторных принадлежностей, напри-
мер, шариков для шаровой мельницы; проведение аналитических исследований, на-
пример, анализа состава воды; для отстаивания большого количества взвеси.
Фарфоровый барабан (рис.134) — специализированное оборудование, использую-
щееся для мелкого помола различного сырья в виде камней, кусков, крупной
крошки. Молоть можно любое сырье, главное, чтобы его твердость не превышала 7
по Моосу. Допускается измельчение с добавлением воды. В качестве вспомога-
тельного инструмента для помола в барабан насыпают твердую кремниевую гальку
или фарфоровые шарики. Изделие широкое, цилиндрической формы, с широким гор-
лом и крышкой. Сам барабан и крышка фарфоровые, снаружи покрыты глазурью (за
исключением дна барабана). Внутренние поверхности барабана и крышки не глази-
рованы .
^ШЕЯ&&&^
Рис. 133. Фарфоровый бак
Рис. 134. Фарфоровый барабан
Фарфоровая ступка с пестиком (рис.135) применяется для растирания твердых и
гранулированных веществ, для смешивания паст в однородную субстанцию без ко-
мочков . Верхняя часть пестика удобна для захвата, а нижняя часть округлая.
Такая форма позволяет измельчать вещества не только растирающими движениями,
но и ударными. Для удобства растирания внутренняя поверхность ступки и нижняя
часть пестика не полируются и не покрываются глазурью. Они специально остав-
лены с шероховатой поверхностью. Наружные поверхности ступки и верхняя часть
пестика покрыты глазурью.
Фарфоровая вставка для эксикатора (рис.136). Выполнена в виде диска с круг-
лыми равномерно распределенными отверстиями. За счет этих отверстий внутри
эксикатора свободно циркулируют потоки воздуха. Вставка служит подставкой для
посуды с образцами, которые помещают в эксикатор. Сама она устанавливается
чуть выше дна на специальный выступ эксикатора. На дно сосуда укладывается
силикагель, наливается серная кислота или другой реактив, хорошо поглощающий
влагу. Вставка подходит как для обычных, так и для вакуумных эксикаторов.
Рис. 135. Фарфоровая ступка с пестиком
Рис. 136. Фарфоровая вставка для эксикатора.
Фарфоровая ложка со шпателем (рис. 137) - лабораторный инструмент, который
используется как ложка, и как шпатель. С одной стороны ложка, с другой шпа-
тель . Используют при приготовлении растворов и их взвешиваний, для снятия
осадков с фильтров, помешивания и растирания.
г"
«U
л*'
Рис. 137. Фарфоровая ложка со шпателем.
Фарфоровая ложка (рис. 138) — удобный и полезный лабораторный инструмент,
простой, но необходимый. Может применяться для отмеривания и взвешивания сы-
пучих материалов или жидкостей; для приготовления растворов, паст и гелей;
для снятия осадков; для растирания веществ. Ложка изготавливается из термо-
стойкого фарфора и полностью покрыта глазурью, что делает ее устойчивой к
большинству химических веществ, в том числе и агрессивных.
Рис. 138. Фарфоровая ложка.
Фарфоровая лодочка (рис.139) используется для сжигания органических веществ
при высоких температурах и при определении содержания углерода в стали. Гла-
зурью не покрывается.
Рис. 139. Фарфоровая лодочка.
Треугольник (рис.140) для тигля состоит из трех насаженных на стальную про-
волоку фарфоровых трубок одинаковой длины. Предназначаются для установки тиг-
лей для прокаливания в пламени газовой горелки.
Рис. 140. Треугольник для тигля.
(ПРОДОЛЖЕНИЕ СЛЕДУЕТ)
Мышление
>*;> ^
'to
ОТ АТОМОВ К ДРЕВУ
Ястребов С.
ЧАСТЬ III. ДРЕВО ЖИЗНИ
13. ЗЕМЛЯ
И ЖИЗНЬ
В период Чоу твердеющая масса опускает-
ся; образуются вода, огонь, горы, кам-
ни, земля — пять элементов, которые по-
лучили название пяти стихий. Поэтому и
говорят, что земля появилась в период
Чоу. Через следующие 5400 лет период
Чоу кончается, и с наступлением периода
Инь на земле зарождается жизнь.
У Чэн-Энь. «Путешествие на запад»
Пять миллиардов лет назад ни Солнца, ни Земли еще не существовало. По со-
временным оценкам, самая точная дата начала образования Солнечной системы —
4,5682 миллиарда лет назад (это определено по возрасту древнейших твердых
включений в метеоритах). К тому времени Вселенная уже имела длинную и сложную
историю. Мы знаем, что Большой взрыв произошел примерно 13,8 миллиарда лет
назад. А это означает, что в момент начала формирования Солнечной системы
возраст Вселенной уже насчитывал около 9,2 миллиарда лет. Почти бездонная
пропасть исторического прошлого, настоящая ЛЛбездна времен".
К моменту рождения Солнечной системы несколько совершенно разных космиче-
ских эпох успели сменить друг друга. Началу образования звезд предшествовали
так называемые "темные века" (Dark Ages), когда во Вселенной не было никаких
светящихся объектов. Первые звезды зажглись не раньше чем через 100 миллионов
лет после Большого взрыва. И эти звезды очень сильно отличались от Солнца.
Наше Солнце — желтый карлик с ожидаемой продолжительностью жизни порядка 10
миллиардов лет. А первые звезды во Вселенной, судя по расчетам астрономов,
были голубыми гигантами с массой, которая могла в 500-1000 раз превосходить
массу Солнца, и со светимостью в миллионы раз выше солнечной, но со средней
продолжительностью жизни всего лишь три миллиона лет. Планетных систем эти
звезды не имели: в древнейшей Вселенной, состоявшей целиком из водорода и ге-
лия, им было бы просто не из чего образоваться.
Когда короткий жизненный цикл звезд первого поколения заканчивался, эти
звезды или взрывались в качестве сверхновых, или (если их масса более чем в
250 раз превышала массу Солнца) коллапсировали в черные дыры без взрывов. Не-
которые из этих массивных черных дыр потом сливались друг с другом и станови-
лись центрами ядер формирующихся галактик. А те звезды, которые становились
сверхновыми, при взрывах разбрасывали на огромные расстояния газ и пыль — те-
перь уже насыщенные новыми химическими элементами, гораздо более тяжелыми,
чем водород и гелий. Эти тяжелые элементы могли синтезироваться только в не-
драх звезды (или даже непосредственно во время ее взрыва). И в туманностях,
которые оставались от таких звезд, постепенно началось образование звезд сле-
дующих поколений — уже с планетными системами.
Около пяти миллиардов лет назад одна из таких туманностей находилась на ок-
раине галактики Млечный Путь. Вероятно, она могла бы до сих пор оставаться в
покое, если бы не близкий взрыв сверхновой звезды. По крайней мере, такова
популярная у современных астрономов гипотеза, основанная на особенностях изо-
топного состава некоторых метеоритов. Ударная волна сверхновой, распростра-
нившаяся в безвоздушном пространстве благодаря огромному количеству выброшен-
ного при взрыве звездного материала, заставила наше газопылевое облако поте-
рять устойчивость и коллапсировать. Оно расслоилось, в нем появились сгуще-
ния, которые притянули к себе окружающее вещество, и, в конце концов, вся
масса облака стала стягиваться к одному центральному ядру. Это и была расту-
щая звезда. Но одновременно потерявшее устойчивость облако начало вращаться,
поэтому возникшая центробежная сила растянула его в диск. В конце концов,
центральное ядро крутящегося газопылевого диска стало Солнцем, а периферия —
планетами.
Планетная система делится на две части так называемой лллинией льда", кото-
рая в нашем случае проходит между орбитами Марса и Юпитера. Внутри от "линии
льда" летучие вещества (вода, аммиак, метан, углекислый газ, угарный газ) в
основном испаряются, а снаружи от нее — конденсируются в ледяные пылинки, ог-
ромные массы которых могут входить в состав планет. В результате внутри от
"линии льда" образовались твердые планеты земного типа (Меркурий, Венера,
Земля, Марс), а снаружи — планеты-гиганты, превосходящие планеты земного типа
массой в десятки и сотни раз, но гораздо менее плотные (Юпитер, Сатурн, Уран,
Нептун).
При этом большая часть воды, находящейся в Солнечной системе, оказалась как
раз по внешнюю сторону "линии льда". Например, ядра комет состоят из водяного
льда на 80%, а в составе Урана и Нептуна воды столько, что эти планеты отно-
сят к классу ледяных гигантов. Мантия Нептуна, скорее всего, представляет со-
бой океан жидкой воды с примесью аммиака и метана, имеющий непредставимую для
нас глубину — около 10 000 километров. Очень вероятно, что и Уран устроен
примерно так же. А с другой стороны, гипербазиты, из которых в основном со-
стоит мантия Земли, содержат всего лишь 0,05-0,06% воды. Очевидно, что по
сравнению со многими другими объектами Солнечной системы это ничтожно мало.
По счастью, Земля еще и очень активная планета. Ее кора (вместе с верхней
мантией) расколота на гигантские литосферные плиты, которые непрерывно дви-
жутся. В глубине Земли идут конвективные потоки мантийного вещества, а на ее
поверхности есть множество действующих вулканов (причем в некоторые эпохи их
было гораздо больше, чем сейчас). А вулканические газы обычно не меньше чем
на 75% состоят из водяного пара. Та вода, которая все-таки есть в земной ман-
тии, выходит с этим паром, конденсируется и образует океан. Впрочем, есть ве-
роятность , что значительную часть воды на Землю принесли врезавшиеся в нее
льдистые метеориты или кометы. Но, как бы там ни было, без участия вулканов
облик нашей планеты был бы совсем другим.
Одной из главных причин высокой тектонической активности Земли, скорее все-
го, является ее уникальный естественный спутник. Луна всего-навсего в 3,7
раза меньше Земли. Ни у одной другой планеты Солнечной системы нет спутника,
настолько огромного относительно нее. У Венеры спутников нет вообще, а спут-
ники Марса имеют ничтожные размеры (27 и 24 километра) и представляют собой
небольшие астероиды, захваченные полем тяготения планеты. Правда, в Солнечной
системе есть и более крупные спутники, чем Луна, но это спутники планет-
гигантов: Ганимед, Титан, Каллисто и Ио. По относительному размеру Луна со-
вершенно необычна (если не учитывать систему Плутон — Харон, которая к плане-
там не относится). И это очень важно. Поле тяготения Луны, направление кото-
рого постоянно меняется из-за ее движения по орбите, создает приливные силы,
"накачивающие" дополнительной энергией всевозможные перемещения вещества
внутри Земли. Конечно, Луна — не единственная причина тектонической активно-
сти Земли, но она значительно усиливает ее, ускоряя в итоге всю эволюцию пла-
неты. В общем, со спутником Земле повезло.
Откуда взялась Луна? Предполагается, что через 50-60 миллионов лет после
начала своего формирования молодая Земля столкнулась с другим небесным телом
— несуществующей ныне планетой размером примерно с Марс, то есть вдвое меньше
Земли. Столкновение выбросило в космос некоторое количество материала земной
мантии, который и стал основой формирующейся Луны. Планета, нанесшая удар,
развалилась, ее остатки или вошли в состав Луны, или стали метеоритами. Эта
гипотеза — ее называют гипотезой великого столкновения, или мегаимпакта
(Giant Impact), — порождает много вопросов, но и подтверждений у нее хватает.
Например, исследование лунных образцов показывает, что Луна на несколько де-
сятков миллионов лет моложе как Земли, так и других тел Солнечной системы;
Земля и Луна почему-то уникально близки друг к другу по изотопному составу
(их называют "геохимическими близнецами"); и наконец, Луна близка по типу по-
род к земной мантии, но, в отличие от Земли, почти лишена железного ядра. Ги-
потеза мегаимпакта изящно объясняет все эти факты.
Так или иначе, Земля — единственная землеподобная планета, которая богата
одновременно тектонической энергией и жидкой водой. Причем ей посчастливилось
сохранить эти свойства на протяжении нескольких миллиардов лет — в отличие от
Марса, где тоже когда-то был водный океан. А ведь именно эти две особенности
и есть главные предпосылки возникновения жизни (во всяком случае, жизни зем-
ного типа) : жидкая вода — потому что она служит растворителем для биохимиче-
ских реакций, а тектоническая энергия — потому что она может эти реакции за-
пустить . Тектоническая активность Земли не только дает тепло, но и обеспечи-
вает перепады концентраций молекул или ионов (начиная с простейшего иона Н+) ,
которые сами по себе становятся источниками энергии.
Возникновение
жизни
Есть несколько гипотез, более-менее детально расписывающих вероятные первые
химические шаги на пути к жизни. Они отличаются в деталях, но едины в глав-
ном. Все эти гипотезы предполагают, что местами зарождения жизни были не от-
крытые водоемы, а микрополости в грунте или минеральных осадках, куда подво-
дилась энергия от горячих источников или от вулканов. Надо сказать, что это
не такая уж новость. Например, известный швейцарский биолог Карл фон Нэгели
еще в XIX веке писал по поводу зарождения жизни: "Вероятно, это случилось не
в открытой воде, а во влажном слое тонкого пористого материала (песка, гли-
ны) , где совместно действовали молекулярные силы твердых, жидких и газообраз-
ных тел". Вот это мнение сейчас и стало научным мейнстримом. Где возникнове-
ние жизни наименее вероятно — так это в водной толще спокойного океана, осве-
щенного солнцем. Там просто нет таких потоков энергии и вещества, которые за-
рождающаяся жизнь могла бы "оседлать" и перенаправить себе на пользу.
Итак, где-то в воде, пропитывавшей окрестности древних вулканов или горячих
источников, начались автокаталитические (то есть самоускоряющиеся) химические
реакции, цепочки которых вскоре стали пересекаться за счет общих промежуточ-
ных продуктов и замыкаться в циклы. Главные участники этих реакций, скорее
всего, были небольшими органическими молекулами, поначалу даже одноуглеродны-
ми. Но реакции-то были не простыми. Особенность любой автокаталитической ре-
акции по определению состоит в том, что ее продукт одновременно является ка-
тализатором, то есть веществом, ускоряющим ход самой реакции. При условии
достаточной сложности реакционной системы (а оно в данном случае наверняка
соблюдалось: и реагентов, и продуктов было множество) автокаталитические ре-
акции приобретают свойство саморазвития, потому что в них появляется обратная
связь: небольшое изменение механизма реакции влияет на состав ее продуктов,
изменение которого, в свою очередь, влияет на механизм — и так шах1 за шагом.
Спустя какое-то время в системе автокаталитических реакций начали синтезиро-
ваться аминокислоты, простейшие углеводы, а там дело дошло и до полимеров —
сперва простых, потом посложнее. Наконец, некоторые из этих полимеров
"научились" катализировать сначала синтез друг друга (это совсем легко), а
потом и воспроизводство самих себя. Иными словами, они стали репликаторами. А
с появлением репликаторов автоматически включается дарвиновский механизм ес-
тественного отбора, необходимые и достаточные условия для которого — самовос-
производство , наследственность, изменчивость и конкуренция за субстрат. Все,
с этого момента биологическая эволюция запущена.
Можно не сомневаться, что на этих первых этапах жизнь была еще практически
незаметной для постороннего наблюдателя (если бы, конечно, он мог тогда суще-
ствовать) . Это легко понять, если вообразить себя инопланетным путешественни-
ком, прибывшим пусть даже к самой колыбели земной жизни. Что он увидит? Теп-
лый вулканический грунт, башни пористых осадков на морском дне... И все. Ни-
чего примечательного. Без химического анализа такой путешественник и не понял
бы, с чем столкнулся.
Первыми в истории Земли полноценными репликаторами, скорее всего, были мо-
лекулы РНК (см. рис. 13.1). Дело в том, что из всех биологически активных мо-
лекул только РНК может выполнять сразу все жизненно важные функции: и хране-
ние наследственной информации, и ее копирование, и катализ реакций обмена ве-
ществ. Белки и их предшественники, более простые пептиды, никогда таких воз-
можностей не имели. Тем не менее, первые пептиды наверняка появились примерно
в те же времена, что и первые РНК. Это следует из чисто химических соображе-
ний. Дело в том, что синтез РНК довольно сложен, а вот аминокислоты — причем
именно альфа-аминокислоты, из которых пептиды обычно состоят, — достаточно
легко синтезируются из самых простых молекул, например из угарного газа (СО)
и циановодорода (HC=N), в условиях, примерно соответствующих вероятным усло-
виям в окрестностях древних вулканов. Поэтому существование эволюционного
этапа, когда автокаталитические системы состояли бы исключительно из РНК, ма-
ловероятно . Скорее всего, эволюция пептидов и РНК была сопряженной всегда,
еще со времен их гораздо более простых общих предшественников. Возможно, что
дополнительной (в придачу к самокопированию) задачей первых репликаторов как
раз и был катализ синтеза пептидов, влиявших на химическую среду таким обра-
зом, чтобы эти репликаторы с большей вероятностью могли выжить.
АРХЕИ БАКТЕРИИ
Jj 11
мембраны современного типа ^^Ь|
репликация ДНК «■»
ДНК и обратная транскрипция «■А
трансляция ^^т
первые репликаторы ^^Ь
ДНК-мир
"Ретро-мир"
Мир РНК и белков
РНК-мир
Мир до биологии гс ки<
tSBfOK«*TdrHTHHCCKV1X
ци</ое
ДНК-содержащие клетки
I с автономной системой репликации
(заимствованной от вирусов)
ДНК-содержащие вирусы
ретровирусы
РНК-содержащие вирусы
эгоистичные рибозимы
Рис. 13.1. Начало эволюции жизни на Земле (до расхождения клеточных
организмов на архей и бактерий). Разумеется, это гипотетический сце-
нарий, но достаточно правдоподобный. Момент приобретения клетками
(или их предшественниками) собственного механизма репликации ДНК тут
намеренно не уточняется, этот вопрос все еще открыт.
С другой стороны, из современного опыта мы знаем, что белки — более мощные
катализаторы, чем РНК, и их возможности в этом плане несравненно разнообраз-
нее. Поэтому неудивительно, что те РНК, которые "научились" катализировать
синтез каких-нибудь особых пептидов, получили преимущество в выживании. В ре-
зультате пептиды (или уже белки?) стали использоваться репликаторами в каче-
стве своего рода молекулярных инструментов, которыми можно было действовать
на среду, повышая свои шансы уцелеть и размножиться. Конкурируя друг с дру-
гом, РНКовые репликаторы постепенно совершенствовали способность программиро-
вать синтез белков, делая это все более и более точно. И, в конце концов, они
"изобрели" механизм трансляции на рибосоме. Этот механизм позволяет запро-
граммировать всю структуру белка с абсолютной точностью — до каждой аминокис-
лоты. И вот с этого момента возможности живой природы по созданию белков ста-
ли буквально безграничными. Заодно появились и первые вирусы — "оппортунисти-
ческие" репликаторы, которые не стали заводить собственную систему синтеза
белка, зато научились паразитировать на чужой.
Следующим важным эволюционным событием был перенос генетической информации
с РНК на ДНК. Дело в том, что молекула РНК всем хороша, но вот химическая ус-
тойчивость у нее низкая и разрушается она довольно легко. Поэтому длительно
хранить на ней генетическую информацию — дело ненадежное. Для этого предпоч-
тителен какой-нибудь другой полимер. Им-то и стала ДНК. Если первые РНК впол-
не могли синтезироваться спонтанно в неживой природе, то синтез ДНК уже со
всей определенностью является "изобретением" живых организмов, и эта молекула
с самого начала получила единственную функцию: хранить информацию. Ничего
другого она делать не умеет. Одно-единственное преимущество, которое имеет
ДНК перед РНК, — ее высокая химическая устойчивость, позволяющая долго и на-
дежно храниться. Для того, кто владеет уникальным ЛЛноу-хау" синтеза каких-
нибудь полезных белков, это по-настоящему ценно.
Таким образом, началась эпоха великой перезаписи геномов с РНК на ДНК. В
начале этой эпохи на Земле жили РНК-содержащие организмы, которые наверняка
уже освоили к тому моменту технологию точного синтеза белка. Иными словами,
ДНК появилась эволюционно позже, чем трансляция. Вполне возможно, что генети-
ческая стратегия первых ДНК-содержащих организмов была похожа на генетическую
стратегию ретровирусов (см. главу 12) . В жизненном цикле вирусов этого типа
есть обязательная стадия ретротранскрипции, то есть обратной транскрипции —
переноса генетической информации с РНК на ДНК. А вот собственного механизма
репликации ДНК у ретровирусов нет. И у клеточных организмов его тоже, скорее
всего, вначале не было. Надежные ферменты репликации (они называются ДНК-
зависимые ДНК-полимеразы) появились позже. Но уж когда они появились, это да-
ло возможность хранить на ДНК генетическую информацию непрерывно, при необхо-
димости сразу перезаписывая ее с одной молекулы ДНК на другую. И тогда рет-
ротранскрипция стала не нужна.
В результате образовалась самая привычная нам форма жизни: ДНК-содержащая
клетка с генетической стратегией ллДНК-РНК-белок".
Признаемся честно: мы не знаем, когда именно живое вещество разбилось на
клетки, отделенные от внешней среды и друг от друга замкнутыми липидными мем-
бранами. Вполне возможно, что это произошло раньше, чем появилась репликация
ДНК и исчезла обязательная ретротранскрипция. В таком случае вполне может
оказаться, что первые клетки по жизненному циклу напоминали ретровирусы (или
даже классические РНК-содержащие вирусы, хотя это менее вероятно). Клеточная
мембрана делит весь мир на внутреннюю среду, где химические реакции жестко
контролируются геномом, и окружающую среду, где контроль гораздо менее жест-
кий, требует специальных инструментов (например, выделения из клетки каких-
нибудь белков) и где его приходится делить с обладателями других геномов,
конкурируя с ними за влияние. Кроме того, клеточная мембрана придает геному
целостность, резко ограничивая обмен генетическими элементами с окружающей
средой и защищая генетическую систему от генетических паразитов. Только с
этого момента приобретает реальный смысл понятие особи, индивидуума, — слово,
которое по-латыни значит "неделимый".
Парадоксальным образом размножаются все живые клетки именно делением. Мате-
ринская клетка делится на две дочерние, которые получают достаточно точные
копии ее генома. Цепь последовательно делящихся клеток — это цепь прямых, без
всяких метафор, предков и потомков. Иногда потомки одной и той же клетки ока-
зываются в разных условиях (или получают разные мутации) и начинают под дей-
ствием естественного отбора накапливать различия. Тогда мы можем заметить,
что линия предков и потомков ветвится.
Первым таким ветвлением было разделение всех клеточных организмов на архей
и бактерий. Оно произошло точно раньше, чем появился полноценный механизм ре-
пликации ДНК, и наверняка раньше, чем появились клеточные мембраны современ-
ного типа. А это означает, что типичные (с нашей точки зрения) клетки, окру-
женные липидной мембраной и имеющие генетическую стратегию "ДНК-РНК-белок", с
самого начала существовали в виде двух расходящихся эволюционных ветвей.
Так возникло древо жизни.
Arbor
vitae
Из всех естественных наук только биология может представить свои объекты в
виде эволюционного древа. Связано это с тем, что у живых организмов (и только
у них) есть передаваемый по наследству геном, который обеспечивает уникальную
преемственность информации и структуры. Белок, возникший миллиард лет назад,
вполне может до сих пор сохранить не только свою функцию, но и большую часть
аминокислотной последовательности — конечно, если этот белок достаточно ва-
жен, чтобы его устойчивость поддерживалась естественным отбором. Предок чело-
века, живший миллиард лет назад, скорее всего, был одноклеточным существом.
И, тем не менее, сохранившаяся преемственность (причем не только чисто гене-
тическая, но и запечатленная в наблюдаемой под микроскопом структуре клеток)
не дает забыть о нашем прямом родстве.
Древо жизни — это не воображаемая конструкция или модель, а совершенно ре-
альный объект. Любая его веточка так же реальна, как любая веточка любого де-
рева, разветвляющегося в пространстве. (Эту фразу написал в одной статье за-
мечательный зоолог и биогеограф Иван Иванович Пузанов, но она выражает на-
столько очевидную мысль, что ее и в кавычки заключать не хочется: все равно
как если бы Пузанов сказал, что Волга впадает в Каспийское море.) Эволюцион-
ное древо ничем качественно не отличается от родословной какой-нибудь коро-
левской семьи или линии охотничьих собак. Точнее, различия-то между ними есть
(например, очень разным может быть вклад процесса гибридизации), но все они
одинаково реальны. Кстати, это одна из причин, почему англичане в середине
XIX века довольно легко поняли теорию Дарвина. Англия традиционно была живот-
новодческой страной, ее жители превосходно знали, как составляются родослов-
ные породистых коров, собак, голубей и тому подобных одомашненных тварей. Пе-
рейти от родословных к эволюционным древесам было нетрудно.
Вот всего одна наглядная иллюстрация. В 1938 году южноафриканский ихтиолог
Джеймс Смит сделал поразительное открытие. Он обнаружил, что ллкистеперые рыбы
вымерли не совсем". Действительно, кистеперые рыбы — сборная группа, в кото-
рую входит открытая Смитом латимерия, — близки к эволюционным корням наземных
позвоночных, но до 1938 года они были известны только из палеонтологической
летописи. Считалось, что они вымерли даже раньше, чем динозавры. Поэтому со-
временная кистеперая рыба была потрясающей находкой. Но проблема состояла в
том, что латимерия была одна-единственная, и поймать второй экземпляр никак
не удавалось. Поиски мест, где живет латимерия, заняли много лет и шли очень
трудно (эта захватывающая история прекрасно изложена самим Смитом в книге
"Старина-четвероног"). В какой-то момент Смит обратился за помощью к премьер-
министру Южной Африки — известному политику Даниэлю Франсуа Малану, объяснив
ему по телефону, почему это так важно. И Малан помог. Он выделил самолет и
решил все проблемы с зарубежными визами, которые были нужны для срочного по-
лета за пойманной рыбой. Когда рыбу доставили в Кейптаун, Малан спросил Сми-
та: "Вы хотите сказать, что и мы некогда выглядели таким образом?" Смит не-
сколько растерялся и отделался шуткой. Спустя некоторое время известный анг-
лийский биолог Джулиан Хаксли сказал по этому поводу, что правильным ответом
было бы: "Вообще говоря, да!" И он был абсолютно прав. Самым верным ответом
тут было бы простое "ДА". Правда, латимерия в действительности не предок на-
земных позвоночных, она, скорее, их эволюционная "кузина". Но и самые что ни
на есть прямые предки, судя по всему, выглядели достаточно похоже на нее —
неспециалист не отличил бы.
Древо жизни — это совокупность целых организмов (а не только их геномов).
Графически оно, в самом деле, похоже на дерево, ветви которого, правда, могут
иногда и сливаться. Пример такого слияния — объединение организмов древней
археи и древней альфа-протеобактерии, которое дало начало эволюционной ветви
эукариот (см. главу 10). В этом месте древо жизни замкнулось в кольцо. Но
полное слияние эволюционных ветвей происходит все-таки редко (гораздо реже,
чем обмен отдельными генами). Основной способ биологической эволюции — дивер-
генция, то есть расхождение ветвей. Этот вывод в целом сохраняет силу со вре-
мен Чарльза Дарвина, который описал вероятную общую структуру эволюционных
древес, и Эрнста Геккеля, который первым всерьез попытался построить истинное
эволюционное древо всего живого.
Конечно, эволюционные деревья, помещаемые в научных работах, бывают несо-
вершенными — точно так же, как несовершенны старые карты мира. На доколумбо-
вых картах нет Америки, а на картах XVIII века еще начисто отсутствует Ан-
тарктида. Но уж тут-то ясно, что отсутствие этих континентов на картах не
имеет никакой фундаментальной причины: их просто еще не успели открыть. Вот
так же обстоит дело и с неточностями эволюционных древес, которые рисуют уче-
ные (и заодно со спорами о том, какая версия древа более правильна, — а они
часто бывают очень бурными).
Оглянувшись на эволюцию Вселенной от Большого взрыва до возникновения на
Земле животных и растений, мы сразу увидим, что генетическое родство — это
совершенно особый тип отношений, по самой своей сути возможный только между
живыми объектами. Здесь есть одно исключение — языки, для которых лингвисты
тоже устанавливают самое настоящее родство. Родословные деревья языков очень
похожи на родословные деревья живых организмов (они и строятся близкими спо-
собами, с применением сходных математических аппаратов). Но это не столь уж
удивительно, поскольку любой человеческий язык — в конечном счете явление жи-
вой природы, часть "расширенного фенотипа" вида Homo sapiens. Так что исклю-
чение на самом деле подтверждает правило. Нигде в неживой природе нет ничего
подобного. Бесполезно сравнивать по признаку родства две молекулы, два кри-
сталла, две звезды или два вулкана (если только не понимать "родство" совсем
уж метафорически). Зато для любой пары живых существ, сколь угодно далеких
друг от друга, определить родственные отношения хотя бы в первом приближении
нетрудно. Неудивительно, что именно на родстве в первую очередь и основана
биологическая система.
14. РОЖДЕНИЕ
СИСТЕМЫ
Моя цель — просто набросать здесь проект систематики
китообразных. Я архитектор, а не строитель. Но это —
грандиозная задача; простому сортировщику писем в
почтовой конторе она не по плечу. Вслепую пробирать-
ся вслед за ними на дно морское, шарить руками в не-
изреченных основах, в плечевом и тазовом поясе само-
го мира — разве это не жутко?
Герман Мелвилл. Моби Дик, или белый кит
Область биологии, занимающаяся разнообразием живых организмов, называется
систематикой. Строго говоря, систематика — это абсолютно неустранимый аспект
биологического знания как такового. Любое живое существо, изучаемое любым
биологом с любыми целями, обязательно имеет какую-нибудь систематическую при-
надлежность. Другое дело, что есть (и никогда не переведутся) биологи, у ко-
торых систематика сама по себе является главной областью интересов. Часто го-
ворят, например, что такой-то человек — величайший знаток систематики жуков-
долгоносиков, или хищных динозавров, или сумчатых грибов, или еще кого-
нибудь .
Основным понятием систематики является таксон — группа организмов, обладаю-
щая заданной степенью однородности. Таксон — это единица системы. Любая сис-
тема живой природы представляет собой некоторое сочетание таксонов. А любой
таксон, в свою очередь, имеет четкие границы. Можно сказать, что природа де-
лится на таксоны примерно так же, как территории континентов делятся на стра-
ны и провинции. И об их границах примерно так же спорят — хотя, как правило,
и без применения аргументов, опасных для жизни. Все названия таксонов суть
имена собственные. Недаром в старых книгах их было принято писать с большой
буквы, а латинские названия пишутся так и сейчас. По правилам, заведенным еще
в XVIII веке и соблюдаемым по сей день, любой таксон обязательно получает в
первую очередь именно латинское название, а уж оно переводится (или не пере-
водится) на современные языки.
Таксоны различаются рангом по возрастанию: вид, род, семейство, отряд,
класс, тип, царство, домен. Ранг таксона обозначает его мощность как подмно-
жества: чем выше ранг, тем эта мощность больше. Виды группируются в роды, ро-
ды — в семейства, семейства — в отряды, отряды — в классы, классы — в типы,
типы — в царства, а царства — в домены. В общем, система живой природы от на-
чала до конца иерархическая. Группы последовательных рангов вложены друг в
друга. Например, класс млекопитающих включает в себя отряды грызунов, прима-
тов , рукокрылых, парнокопытных, непарнокопытных, хищных и разные прочие. А
отряд хищных, в свою очередь, включает семейства псовых, медвежьих, куньих,
енотовых, кошачьих, гиеновых, виверровых и (опять же) некоторые другие. В об-
щем случае таксон может состоять из скольких угодно таксонов подчиненного
ранга, в том числе и из одного-единственного. Например, в отряде трубкозубов
всего одно семейство, один род и один вид (если не считать ископаемых). Такие
лл вырожденные" таксоны называются монотипическими. В случае с трубкозубом
удивляться этому не приходится: он настолько не похож ни на одно другое со-
временное млекопитающее, что для него пришлось создать собственный отряд, со-
стоящий из одного вида.
Трубкозуб.
Часто бывает, что систематикам не хватает основных рангов и они создают
промежуточные: подтип, надкласс, подкласс, надотряд и т.п. Но тут действует
простое правило: любой полностью систематизированный организм должен быть
обязательно отнесен к какому-нибудь таксону каждого из основных рангов (от
вида до домена включительно), а вот промежуточные ранги не обязательны.
Кроме того, в некоторых областях биологии есть особые исторические тради-
ции: например, в ботанике отряд принято называть порядком, а тип — отделом.
Сути дела это не меняет.
И, наконец, в современной биологии встречается совершенно иной подход к по-
строению системы — безранговый (хотя иерархический принцип и в этом случае
строго соблюдается). О таких системах мы поговорим позже, в главе 15.
Биологи всегда стремились создать систему, максимально устойчивую к введе-
нию в нее новых объектов или признаков. Такая система называется естествен-
ной. Чем меньше меняется система при добавлении новых фактов, тем она естест-
веннее . Идеальная естественная система отличается от любой другой системы
тем, что ее невозможно оптимизировать: она уже наилучшим образом отображает
отношения между объектами, и любые поправки ее только испортят. Стремление к
такой системе, конечно, подобно бесконечному лласимптотическому приближению к
абсолютной истине", математические свойства которого изучали некоторые второ-
степенные персонажи Стругацких. Тем не менее, эта цель по-своему привлека-
тельна. История биологии — по крайней мере, ее систематических областей —
это, по сути, и есть сильно растянувшаяся во времени история попыток постро-
ить полную естественную систему живых организмов.
Первая такая попытка связана с именем Аристотеля. Это неудивительно: от
Аристотеля начинается история большинства естественных наук и уж во всяком
случае — биологии и физики. Аристотель вообще уделял очень много внимания ис-
следованиям природы. Этим он отличался от своего учителя Платона, для которо-
го идеалом науки была геометрия и который, насколько можно судить, считал
полноценным знанием только то, что хотя бы в принципе можно вывести в уме из
набора аксиом. "Естествознание — родная стихия аристотелевской мысли, особен-
но когда речь идет о живой природе", — писал историк биологии Валериан Викто-
рович Лункевич. И это не пустая фраза, а совершенно точное наблюдение. Способ
мышления Аристотеля был мало приспособлен к математическим абстракциям, зато
идеально — к изучению живых организмов, сложных, многообразных и развивающих-
ся. Характерно, что Аристотель не внес никакого научного вклада в математику,
хотя, безусловно, хорошо знал ее просто как образованный человек. А вот в
зоологии он открыл настолько много, что его с полным правом считают основате-
лем этой науки. Принимался он и за ботанику, но жизни не хватило на все (Ари-
стотель умер в 62 года) . И в результате крупнейшим ботаником античности стал
его ученик Теофраст.
Система Аристотеля была проста и наглядна. Он разделил живую природу на три
ступени: растения (живут, но не движутся), животные (живут и движутся), чело-
век (живет, движется и мыслит). Естественно, при этом он сразу же столкнулся
с так называемой проблемой переходных форм, которая всегда возникает при по-
пытке классифицировать непрерывную совокупность по качественным признакам.
Дело в том, что античные исследователи знали о существовании организмов, ко-
торые (по тогдашним представлениям) неподвижны или малоподвижны, как расте-
ния, но могут питаться, как животные. В основном это некоторые морские суще-
ства: губки, коралловые и гидроидные полипы, актинии, голотурии. Аристотель
предполагал, что в этих "промежуточных" организмах растительная природа сме-
шана с животной. Более поздние античные авторы стали называть их зоофитами,
то есть "животными-растениями". Так что de facto в аристотелевской системе
живой природы не три, а четыре категории: человек, животные, зоофиты и расте-
ния.
Созданная в середине XVIII века система Карла Линнея принципиально отлича-
ется от системы Аристотеля в двух отношениях. Во-первых, она состоит из вло-
женных друг в друга групп, которым присвоены ранги: царство — класс — отряд —
семейство — род — вид (ранги типа и домена были добавлены позже) . Система
Аристотеля тоже иерархична, но четкого понятия о рангах там нет. Это изобре-
тение Линнея. Животные и растения у Линнея имеют ранг царств.
Во-вторых, человек у Линнея не образует особую категорию, равноправную жи-
вотным и растениям, а входит в царство животных — отныне и навсегда.
Система Линнея претендовала на то, чтобы охватить не одну лишь живую приро-
ду, а вообще все объекты во Вселенной. Кроме животных и растений там есть
третье царство — минералов. Эта особенность линнеевскои системы сейчас инте-
ресна только как факт истории культуры. Или, может быть, альтернативной исто-
рии науки, которая не осуществилась в нашей реальности.
Справедливости ради надо сказать, что распространение систематики на всю
природу (и живую, и неживую) не было личной причудой Карла Линнея, а отражало
совершенно нормальное для натуралистов XVIII века отношение к вещам. Мир объ-
ектов естественной истории виделся им единым целым. Были, например, авторы,
которые добавляли к трем линнеевским царствам четвертое и пятое — царство во-
ды и царство огня. А уж царство минералов признавали все. Причем натуралисты
тех времен умудрялись находить переходные формы между всеми тремя общеприня-
тыми царствами — растений, животных и минералов. Они писали не только о зоо-
фитах ("животных-растениях"), но и о литофитах ("растениях-камнях"), и о ли-
тозоях ("животных-камнях"); к последним относили, например, коралловых поли-
пов, образующих известковые постройки. Эти взгляды продержались до второй по-
ловины XIX века, когда победа эволюционной идеи в биологии и — с другой сто-
роны — стремительное развитие наук о Земле сделали единую классификацию живых
и неживых объектов практически бессмысленной.
Главным отличием животных от растений Линней считал активную подвижность.
Он писал: "Естественные тела, наделенные всегда одной и той же формой и спо-
собностью к перемещению, называются животными; имеющие всегда одну и ту же
форму, но не способные перемещаться — растениями; а те, которые имеют разно-
образную форму, называются минералами". Аристотелевских "зоофитов" Линней от-
нес к царству животных (правда, после долгих колебаний): он понимал, что не-
которая подвижность, основанная на чувствительности животного типа, у них все
же есть.
Однако проблема переходных форм никуда не делась. Во-первых, есть подвижные
растения, в том числе и хищные — вроде росянки, которая ловит насекомых свои-
ми листьями. Во-вторых, есть совершенно неподвижные животные — например, губ-
ки. Найти у них хоть какие-то сокращающиеся элементы смог только современник
Линнея англичанин Джон Эллис: путем тщательных наблюдений он выяснил, что
губка может сокращать и расслаблять маленькие отверстия, сквозь которые в ее
тело затекает вода. И в-третьих, есть еще и грибы, которые один ботаник, быв-
ший на поколение старше Линнея, в сердцах обозвал произведениями дьявола,
призванными нарушать гармонию природы и приводить исследователей в отчаяние
(современные специалисты по грибам — микологи — обожают это цитировать). По-
ложение грибов в линнеевскои системе было крайне туманным. На первый наивный
взгляд это растения. Но у грибов есть особенности, резко отличающие их от ти-
пичных растений, — например, независимость от света и отсутствие ветвления
(на самом деле микроскопические нити, из которых состоит тело гриба, ветвят-
ся, но Линней этого не знал). Неудивительно, что отнести грибы к растениям
Линней решился лишь после больших сомнений, приписав комментарий: "Порядок
грибов, к позору ботаники, и поныне представляет собою хаос".
Проблему переходных форм не раз пытались решить, выделив третье царство жи-
вого . Но вот что это должно быть за царство? Тут разные ученые рассуждали по-
разному. Ботаник барон Отто фон Мюнхгаузен, современник и хороший знакомый
Линнея, в 1766 году предложил создать промежуточное царство — по-немецки Das
Mittelreich, куда вошли грибы, а вместе с ними коралловые полипы и другие
прикрепленные животные. Это была первая трехцарственная система живой приро-
ды, долго не продержавшаяся.
Другую идею опубликовал в 1783 году ботаник Ноэль Мартен Жозеф де Неккер.
Он предложил выделить в третье царство только грибы. Это было куда более ра-
зумное решение. К 1780-м годам исследователи, в общем, успели убедиться, что
кораллы, гидры и губки — не переходные формы между царствами, а просто живот-
ные . А вот с грибами ситуация была в точности противоположной. Натуралисты,
решившие ими всерьез заняться, обычно довольно быстро приходили к твердому
убеждению, что грибы — никак не растения. Неккер был первым из таких натура-
листов, но далеко не последним.
Тем удивительнее, что признание этих фактов широкими массами ученых задер-
жалось аж на 200 лет. Почти до конца XX века во многих учебниках ботаники
грибы по традиции продолжали числиться растениями, хотя к тому времени уже
все понимали, что с зелеными растениями у них нет ничего общего. Кроме ба-
нальной инерции мышления, никакой серьезной причины тут не найти.
Следующий шаг в развитии системы живой природы был связан с появлением кле-
точной теории. Согласно этой теории, клетка есть элементарная единица живого.
В 1838-1839 годах ботаник Матиас Шлейден и зоолог Теодор Шванн убедительно
показали, что это относится и к растениям, и к животным: организмы и тех и
других состоят из клеток. Надо сказать, что у Шлейдена и Шванна, безусловно,
были предшественники, но авторами клеточной теории по традиции считают именно
их, потому что они первыми рискнули провозгласить эту теорию громко и четко —
и выиграли. Для нас это важно вот почему. К XIX веку было известно уже до-
вольно много микроскопических живых существ — инфузорий, амеб, жгутиконосцев,
радиолярий и других. Их начал исследовать еще великий Антони ван Левенгук,
современник Исаака Ньютона. Но долгое время натуралисты считали, что инфузо-
рии и амебы — это просто очень маленькие животные, и все тут.
Надо сказать, что животные такого размера действительно существуют (коло-
вратки, например). Но подавляющее большинство микроскопических живых существ
отличается от крупных животных гораздо серьезнее. Животные — многоклеточные
организмы, а инфузории, амебы, жгутиконосцы и радиолярии — одноклеточные. На
то, чтобы понять это, у исследователей ушло полтора века. Первым, кто четко
отделил одноклеточные организмы от многоклеточных, был немецкий зоолог Карл
фон Зибольд. В 1845 году он выпустил свою "Сравнительную анатомию беспозво-
ночных", в которой "одноклеточные животные" (как их с этого момента стали на-
зывать) были отнесены к особому таксону Protozoa. В русских учебниках зооло-
гии название Protozoa обычно переводилось как "простейшие". По рангу это был
(у разных авторов) или тип, или подцарство.
Карл фон Зибольд выделил четыре группы простейших: жгутиконосцы, корненож-
ки, инфузории и споровики. Жгутиконосцы — это одноклеточные существа, плаваю-
щие с помощью жгутиков. Корненожки передвигаются с помощью ложноножек, вре-
менных выростов клетки, которые можно выпускать и втягивать; их типичный
представитель — обыкновенная амеба. Инфузории покрыты ресничками (которые
устроены точно так же, как жгутики, но их не одна-две, а тысячи) и имеют два
клеточных ядра. А споровики — это чисто паразитические простейшие со сложными
жизненными циклами; к ним относится, например, возбудитель малярии. Эту клас-
сификацию, по тем временам новаторскую, а по нынешним немыслимо архаичную,
многие читатели наверняка еще застали: в школьных учебниках зоологии она фи-
гурировала как минимум до 1980-х годов.
Заодно тот же Зибольд предположил, что главный признак, отличающий животных
от растений, — не подвижность или чувствительность, а тип питания. Растения
питаются автотрофно, животные — гетеротрофно (Зибольд еще не знал этих терми-
нов, но суть различия понимал правильно). Иначе говоря, растения самостоя-
тельно синтезируют сложные молекулы из углекислого газа и воды, а животные
получают готовые питательные вещества из тел других организмов (живых или
мертвых). Механизмом автотрофнохю питания у растений служит фотосинтез. Для
него нужен зеленый пигмент — хлорофилл, который содержится в специальных час-
тях растительной клетки — хлоропластах. Некоторые растения, в том числе и
цветковые (петров крест, подъельник, повилика), лишены хлорофилла и могут пи-
таться только гетеротрофно, но это зволюционно вторично: все они произошли от
зеленых предков.
Куда более серьезной проблемой для систематиков стали грибы, которые строго
гетеротрофны и не имеют хлорофилла никогда. Чтобы все-таки отнести их к рас-
тениям, ботаникам пришлось придумывать для царства растений дополнительные
общие признаки: характер роста, способность всасывать питательные вещества
всей поверхностью тела и размножение неактивными зачатками (семенами или спо-
рами) . Выделить грибы в самостоятельное царство подавляющее большинство био-
логов еще долго не решалось.
Но сложнее всего было разобраться в одноклеточных организмах. Чем подробнее
их изучали, тем больше "переходных форм" среди них обнаруживалось. Например,
есть автотрофные одноклеточные, у которых прекрасно выражена активная подвиж-
ность, — растения они или животные? Есть одноклеточные, у которых автотрофное
питание сочетается с гетеротрофным (таких называют миксотрофами). И, наконец,
есть одноклеточные, которые могут довольно легко менять тип питания с авто-
трофного на гетеротрофный или наоборот. Так стало ясно, что в мире однокле-
точных организмов провести границу между растениями и животными очень трудно.
В результате в 1860-х годах сразу несколько исследователей предложили объе-
динить всех одноклеточных в собственное царство, отдельное и от растений, и
от животных. Знаменитый немецкий биолог-эволюционист Эрнст Геккель в 1866 го-
ду назвал это царство Protista (протисты). Трехцарственная система Геккеля
выглядела вполне разумно: есть растения, есть животные, а есть протисты, к
которым относятся одноклеточные предки, как растений, так и животных.
Справедливости ради надо сказать, что первым одноклеточных в самостоятель-
ное царство выделил не Геккель, а английский натуралист Джон Хогг. Еще в 1860
году он предложил создать царство под названием Protoctista (буквально
"первично созданные"). В него вошла примитивная (по определению Хогга) жизнь,
слабо дифференцированная на растительную и животную. Интересно, что в системе
Хогга это царство было не третьим, а четвертым — в придачу к животным, расте-
ниям и минералам. Дальше мы будем учитывать только царства живой природы и —
соответственно — называть систему из животных, растений и протоктистов (или
протистов) трехцарственной, но надо учитывать, что для Хогга и многих натура-
листов его времени это было не так.
Протоктисты Хогга и протисты Геккеля — очень близкие понятия, но все же не
синонимы. В XX веке название "протисты" обычно использовали исследователи,
стремившиеся сузить объем этого царства, а "протоктисты" — исследователи,
стремившиеся, наоборот, расширить его, включив туда некоторых примитивных
многоклеточных. Например, многоклеточные зеленые водоросли не входили в цар-
ство Protista, но входили в царство Protoctista.
Были и другие попытки выделить третье царство, до которых докопались исто-
рики науки, но которыми мы не станем утруждать здесь читателя. В общем, по
всему видно, что к началу 1860-х годов эта мысль уже носилась в воздухе.
Эрнст Геккель просто высказал ее громче и убедительнее всех.
Однако биология продолжала развиваться, и спустя несколько десятилетий ста-
ло ясно, что объединить всех одноклеточных в одно царство невозможно. Трех-
царственная система Геккеля создавалась, когда о строении клетки еще было из-
вестно очень мало, поэтому она игнорирует такой важнейший признак, как кле-
точное ядро. У бактерий и сине-зеленых водорослей (которые в широком смысле
тоже бактерии) ядер в клетках нет. У остальных, в том числе и у многих одно-
клеточных , — есть. В XX веке это было твердо установлено. В результате появи-
лись четырехцарственные системы, состоявшие из царств растений, животных,
протистов и бактерий. Протистами теперь стали называться одноклеточные или
колониальные (но не многоклеточные) существа с клеточными ядрами. На бактерий
это название больше никогда не распространяли.
В 1925 году французский протистолог Эдуард Шаттон впервые предложил разде-
лить живые организмы на две крупнейшие группы: эукариоты (с клеточными ядра-
ми) и прокариоты (безъядерные). Шаттон не был теоретиком: он увлеченно изучал
одноклеточных паразитов, инфузорий и водоросли, а общими вопросами интересо-
вался мало. Свою классификацию он ввел мимоходом, просто для удобства, и
опубликовал ее в виде краткого попутного замечания в работе, которую мало кто
читал, кроме других протистологов. Неудивительно, что ее не сразу заметили. К
тому же названия, предложенные Шаттоном, пишутся по-разному у разных авторов:
или Prokaryota и Eukaryota, или Procaryota и Eucaryota. Первый вариант ближе
к правильному написанию греческого слова карион (ядро), а второй — к версии
самого Шаттона, который, как часто делают французские биологи, пренебрегал
латынью и писал названия "с французским акцентом" — в данном случае
Procaryotes и Eucaryotes. Сейчас правильным считается все-таки первый вариант
— через к. Для любителей филологической точности можно добавить, что в искон-
но латинских словах буква к употребляется редко, зато она часто служит марке-
ром проникающих в латынь греческих заимствований, что мы тут и видим.
В конце концов, прокариоты и эукариоты получили ранг надцарств. В такой
системе было одно царство прокариот (бактерии) и три царства эукариот — про-
тисты, растения и животные. Стало считаться, что система живых организмов на-
чинается именно с деления всего живого на прокариот и эукариот, а все осталь-
ные таксоны — всего лишь более мелкие подразделения внутри этих двух гигант-
ских групп.
Тут нужна поправка: речь идет о системе всего живого, кроме вирусов. И про-
кариоты, и эукариоты состоят из клеток, а вот вирусы — нет. Поэтому в 1965
году зоолог Николай Николаевич Воронцов предложил ввести таксономический ранг
империи, более высокий, чем ранг надцарства. В империю доклеточных (Non-
Cellulata) входят только вирусы, а империя клеточных (Cellulata) делится на
надцарства прокариот и эукариот. Это предложение, хотя и не прошло незамечен-
ным, всеобщей поддержки не получило: большинство биологов в те времена еще
сомневались, стоит ли вообще относить вирусы к живым организмам. В 2008 году,
когда о вирусах стало известно неизмеримо больше, чем в 1960-е годы, близкую
идею высказали французские вирусологи Дидье Рауль и Патрик Фортерр (см. главу
12) . Они предложили разделить все живые организмы на рибосомо-кодирующие и
капсид-кодирующие. Здесь и далее мы будем обсуждать исключительно систему ри-
босомо-кодирующих организмов, состоящих из клеток. Увы, для вирусов построить
единое эволюционное древо просто невозможно — в первую очередь потому, что
нет ни одного гена, который был бы общим абсолютно для всех вирусов и который
можно было бы считать унаследованным от их общего предка (мы обсуждали это в
главе 12). У клеточных организмов такие универсальные гены есть, и их немало.
Широкую известность в тех же 1960-х годах получили идеи американского эко-
лога Роберта Уиттэкера, который заново пересмотрел всеобщую систематику, учи-
тывая последние на тот момент научные достижения. Уиттэкер наконец-то выделил
в самостоятельное царство грибы, поэтому царств стало пять: животные, расте-
ния, грибы, протисты и бактерии (см. рис. 14.1).
Главной проблемой такой пятицарственной системы оказались границы царства
протистов (или протоктистов) . Оно было чрезмерно разнородным, и ничего поде-
лать с этим не удавалось никакими силами: решив одну проблему, систематики
тем самым тут же создавали другую. Если остальные три царства эукариот хоть
как-то соответствовали эволюционным ветвям, то царство протистов охватывало
целый эволюционный уровень, в котором ветвей было множество. Фактически это
царство было образовано методом исключения: в него заносили всех эукариот,
которые не приобрели полноценной многоклеточности, а часто и некоторых много-
клеточных с ними за компанию. В итоге реальный статус царства протистов резко
отличался от статуса царств растений, животных и грибов. Это бросалось в гла-
за и не могло не создавать впечатления, что такая система внутренне нелогич-
на.
Рис. 14.1. Система Роберта Уиттэкера, "отредактированная" Линн Мар-
гулис, о которой мы упоминали в главе 10 в связи с проблемой проис-
хождения эукариот. Маргулис опубликовала эту версию системы в 1971
году. Царство протистов тут так сильно расширено, что фактически
это, скорее, протоктисты, — к этому наименованию Маргулис вскоре и
перешла. Почти все обозначенные здесь представители этой группы, так
или иначе, обсуждаются дальше в тексте. Пунктиром обведены предпола-
гаемые родственники растений, животных и грибов, относящиеся к про-
тистам по уровню организации. По прочтении главы 15 внимательный чи-
татель сможет, если захочет, сделать собственные выводы о том, кто
из них оказался тут на своем месте, а кто нет.
В 1970-х многие биологи, в том числе и авторы университетских учебников,
предпочли вообще ликвидировать царство протистов, сочтя его слишком сборным.
Таким образом, царств опять стало четыре — бактерии, растения, животные и
грибы. Последние три царства, по идее, должны были включить в себя всех одно-
клеточных родственников соответствующих организмов. В реальности такой подход
привел к чудовищной путанице, которая продлилась до начала XXI века, а кое-
где (например, в школьных программах) продолжается и по сей день. Сплошь и
рядом на биологических факультетах одни и те же группы одноклеточных существ
фигурировали и в курсах ботаники, и в курсах зоологии, причем в совершенно
разном статусе. В результате студентам приходилось одновременно учить две
противоречащие друг другу системы — зоологическую и ботаническую. Неудиви-
тельно, что в глазах большинства биологов это начисто обесценивало системати-
ку как научную дисциплину. Ее стали рассматривать или как второстепенную и
прикладную область знания (этакое составление инвентарной описи), или как
чистейшую "игру в бисер", слабо связанную с реальным миром. Бесконечные пере-
становки одних и тех же групп никого не вдохновляли. В общем, в систематике
явно наступил кризис.
Ветви и
домены
Кризис систематики был преодолен ценой ее почти полного поглощения другой
биологической наукой — филогенетикой. Если систематика может (по крайней ме-
ре, в принципе) пользоваться любыми, сколь угодно разнообразными и необычными
способами классификации живых организмов, то филогенетика изучает их родство,
и только родство. Попросту говоря, филогенетика — это наука о том, кто от ко-
го произошел.
До того как биологи признали эволюцию, фактор родства между организмами при
построении систем не учитывался. Вернее, учитывался, но лишь в скрытой форме;
то, что сходство организмов может отражать их самые настоящие родственные от-
ношения , в додарвиновскую эпоху признавали очень немногие. Эта идея, для нас
совершенно очевидная, тогда казалась слишком смелой и (как это ни странно)
слишком упрощенной. Что касается естественных и искусственных систем, то о
них стали рассуждать еще во времена Карла Линнея, но вот содержание этих по-
нятий за прошедшие без малого 300 лет сильно поменялось. В XVIII веке естест-
венной начали было считать систему, построенную по как можно большему числу
признаков. Иногда оговаривалось, что признаки должны быть ллсущественными", но
определить, что это такое, толком никто не мог. А в XIX веке возникла идея,
что естественная группа организмов — это родственная группа. Именно этому
подходу следовал знаменитый немец Эрнст Геккель, когда создавал свою систему
из трех царств. Слово ллфилогенетика" придумал тоже он, и не случайно. Геккель
был твердо убежден, что естественная система живых организмов — это система,
описывающая ход эволюции.
В XIX веке этой геккелевской идеи было достаточно для продуктивной работы,
но в XX веке она стала нуждаться в уточнениях. Поэтому в середине XX века не-
мецкий биолог Вилли Хенниг создал новое научное направление, которое назвал
филогенетической систематикой. Современную версию хенниговской филогенетиче-
ской систематики чаще всего называют кладистикой — от слова "клада", приду-
манного на основе греческого корня и обозначающего эволюционную ветвь.
Главное требование филогенетической систематики состоит в том, что соответ-
ствие между эволюционным древом и иерархической системой организмов должно
быть взаимно-однозначным. Иначе говоря, при правильно выбранном методе для
данного эволюционного древа должно быть возможно построить только одну систе-
му. Любой произвол в этом деле исключается.
Чтобы это требование выполнялось, Хеннигу пришлось ввести строгое правило:
любая систематическая группа должна включать в себя только одну эволюционную
ветвь, причем обязательно целиком. Последняя оговорка, на неискушенный взгляд
малозаметная, на самом деле тут важнее всего. Из нее следует, что граница лю-
бого таксона может пересекать эволюционное древо только один раз — на "входе"
в этот таксон. Вот в этом и состоит главный принцип хенниговской системы (см.
рис. 14.2).
А В С D E F ABC О ЕР
монофилетический парафилетический полифилетический
= голофилетический
Рис. 14.2. Типы таксонов, выделяемые филогенетической систематикой.
Монофилетический, он же голофилетический, таксон включает эволюцион-
ную ветвь целиком, парафилетический — только частично (с вырезом в
произвольном месте), а полифилетический и вовсе состоит из фрагмен-
тов разных ветвей. Прекрасный пример парафилетическохю таксона —
царство протистов. Пример полифилетического — царство грибов в ста-
ром широком смысле, куда вместе с настоящими грибами относились
оомицеты, миксомицеты, акразиомицеты и другие организмы, сходные с
грибами, но на самом деле не родственные им (см. главу 15) . Филоге-
нетическая систематика вообще запрещает любые таксоны, кроме строго
монофилетических (= голофилетических).
Нетрудно видеть, что царство протистов совершенно не соответствует этому
условию. Растения, животные и грибы, без сомнения, произошли от разных одно-
клеточных эукариот. А все одноклеточные эукариоты по определению относятся к
протистам. Получается, что граница царства протистов пересекает эволюционное
древо как минимум четырежды: первый раз при происхождении протистов от прока-
риот и еще три раза — при происхождении животных, растений и грибов от про-
тистов. Это типичный, прямо-таки образцовый пример таксона, запрещенного фи-
логенетической систематикой.
А если попытаться классифицировать эукариот так, чтобы граница каждого цар-
ства пересекала эволюционное древо только один раз? Тогда система совершенно
неминуемо окажется многоцарственной. Это показал в 1974 году английский бота-
ник Гордон Лидейл. Он честно попытался построить систему эукариот таким обра-
зом, чтобы каждое царство действительно заключало в себе ровно одну эволюци-
онную ветвь (согласно тогдашним представлениям о родстве, конечно). В резуль-
тате получилась система из 18 эукариотных царств, причем 11 из них состояли
только из "протистов".
Современники в целом не приняли систему Лидейла всерьез, сочтя ее своего
рода интеллектуальным чудачеством. А зря. Она была хотя бы внутренне последо-
вательной. И главное, в 1970-х годах ничего более надежного все равно никто
не предложил.
Как раз тогда, когда эти проблемы начали осознаваться, классическая система
живой природы получила мощный удар с совершенно неожиданной стороны. Его на-
несла молекулярная филогенетика — недавно (на тот момент) возникшая наука,
изучающая родственные связи организмов путем прямого чтения аминокислотных
последовательностей белков и нуклеотидных последовательностей генов.
Все началось с того, что американский микробиолог Карл Вёзе решил исследо-
вать родственные отношения между разными группами прокариот. Для этого он на-
шел у разных бактерий (и заодно, для контроля, у нескольких представителей
эукариот) один и тот же ген, кодирующий определенную молекулу, входящую в со-
став рибосом. Это была одна из молекул рибосомной РНК, сокращенно рРНК. Ее
точное название — 16S рРНК у прокариот и 18S рРНК у эукариот (буква S тут
обозначает некую константу, о которой нам сейчас достаточно знать, что она
связана с размером молекулы) . Рибосомы есть абсолютно во всех клетках. Их
функции у разных организмов ничем не отличаются. И у холерного вибриона, и у
крапивы, и у дрожжей, и у человека рибосомы делают только одно — синтезируют
белки в точном соответствии с "инструкциями", записанными на информационных
РНК. Тогда логично предположить, что различия между разными организмами в
"текстах" генов, обеспечивающих создание самих рибосом, могут быть только
случайными. И чем больше этих случайных различий успело накопиться, тем даль-
ше организмы друг от друга (иначе говоря, тем более давно жил их общий пре-
док) .
Результат исследования поразил Вёзе. Оказалось, что некоторая часть прока-
риот настолько сильно отличается от типичных бактерий, что никак не может
входить в одно с ними царство. Так была открыта совершенно особая ветвь про-
кариотной жизни — архебактерии, которых вскоре стали называть просто археями
(см. главы 5, 10) .
У архей много абсолютно уникальных признаков. Например, мы уже знаем, что у
бактерий (и у эукариот тоже) клеточные мембраны состоят из сложных эфиров,
образованных L-изомером глицерофосфата, а у архей — из простых эфиров, обра-
зованных D-изомером глицерофосфата; есть там и другие химические отличия (см.
главу 5). Эти типы мембран едва ли могли произойти один от другого. Значит,
они возникли независимо. Какой была мембрана у общего предка бактерий и архей
— загадка; маловероятно, что ее не было вообще, но она вполне могла быть, на-
пример, неорганической. Таким образом, открытие архей сразу же, просто в силу
того, что эти существа стали известны, дает нам ценную информацию, проливаю-
щую свет на происхождение клеточной жизни. Это хороший пример того, как важен
в биологии сравнительный метод.
Не менее поразителен был тот факт, что по ряду признаков архей оказались
ближе к эукариотам, чем к бактериям. Например, архейные рибосомы больше похо-
жи на эукариотные, чем на бактериальные. В архейных генах встречаются интроны
— некодирующие вставки, характерные в основном для эукариот. В то же время
устройство клетки у архей типично прокариотное: ни ядра, ни митохондрий, ни
других видимых под микроскопом структур, характерных для эукариот, там нет.
Обдумав все эти данные, Вёзе решил, что деление на прокариот и эукариот по-
просту устарело. На самом деле множество всех клеточных организмов распадает-
ся не на две, а на три главные группы: бактерии, архей и эукариоты. Для этих
групп Вёзе предложил новый таксономический ранг — домен (ниже империи, но вы-
ше царства). Таким образом, трехцарственную систему Геккеля через 100 лет
сменила трехдоменная система Вёзе.
А как же теперь обстоит дело со старой классификацией, делившей клеточные
организмы на прокариот и эукариот? А никак. Эти две группы не могут считаться
равноправными: их статус совершенно различен. Эукариоты — действительно эво-
люционная ветвь, происходящая от одного-единственного предка (которым, скорее
всего, была архея, вступившая в симбиоз с альфа-протеобактерией). И таксон
"эукариоты" включает в себя эту ветвь целиком. Что же касается прокариот...
но тут лучше начать с начала, чтобы было яснее.
Начнем с того, что все клеточные формы жизни имеют одного общего предка.
Это следует в первую очередь из единства генетического кода, удивительное по-
стоянство которого не спишешь ни на случайность, ни на параллельную эволюцию
(она не могла повториться в разных ветвях настолько точно). А если у общего
предка всех живых клеток был генетический код, значит, у него был и аппарат
трансляции. И вообще он был достаточно сложным созданием. Этого общего предка
сейчас принято называть LUCA — сокращение, которое чаще всего расшифровывает-
ся как Last Universal Common Ancestor (последний универсальный общий предок).
Хотя, честно говоря, правильнее был бы вариант Last Universal Cellular
Ancestor (последний универсальный клеточный предок), который предпочитает
Патрик Фортерр. Так или иначе, всех потомков ЛЛЛуки" можно рассматривать как
одну гигантскую эволюционную ветвь. В таком случае старая группа прокариот
включает в себя лишь часть этой ветви: в нее входят все потомки "Луки", в ка-
кую бы сторону ни пошла их эволюция, но почему-то за исключением одной спе-
циализированной веточки, которая называется эукариотами. Филогенетическая
систематика категорически запрещает такие таксоны. Вывод: прокариоты — вообще
не таксон, а название уровня организации, объединяющего всех тех потомков
ллЛуки", которые не приобрели уникального плана строения клетки, предусматри-
вающего ядро и другие эукариотные признаки.
Ясно, что при этих условиях прокариоты никак не могут быть равноправны эу-
кариотам. Недаром в системе Вёзе эукариотный домен называется не Eukaryota, a
Eukarya. Такое название подчеркивает, что это не одна из двух равноценных
групп, а нечто полностью уникальное.
Заодно эта история хорошо показывает, что научные истины никогда не стоит
воспринимать как "высеченные в граните". О том, что живые организмы делятся
на прокариот и эукариот, в наше время слышали многие. Но какое место эта
классификация реально занимает в истории науки? Давайте посмотрим. Термины
"прокариоты" и ллэукариоты" были придуманы Эдуардом Шаттоном, который употре-
бил их в нескольких своих работах мимоходом — и все. Этих терминов тогда ни-
кто и не узнал, кроме людей, непосредственно общавшихся с Шаттоном; до самого
начала 1960-х годов они встречаются в научных статьях очень редко. Кто же,
наконец, провозгласил их на весь мир? Это сделали два известных микробиолога
— Роджер Станье и Корнелиус ван Ниль. В 1962 году они выпустили обзорную ста-
тью под названием "Понятие бактерии". Вот эта статья получила широкую извест-
ность , и система, делящая организмы на прокариот и эукариот, стала общеприня-
той именно после нее. Но уже в 1977 году Карл Вёзе и его коллеги ясно показа-
ли , что реальная структура древа жизни этой системе не соответствует. Таким
образом ллпрокариотно-эукариотная" классификация на самом-то деле продержалась
всего лишь 15 лет. До 1962 года о ней мало кто знал, а после 1977-го ее уже
можно было с полным основанием объявить устаревшей. Иное дело, что инерция
устаревших представлений сама по себе большая сила.
Надо добавить, что в домен архей входит не меньше пяти сильно различающихся
групп, вполне заслуживающих ранга царств (эвриархеоты, кренархеоты, таумархе-
оты, корархеоты и другие, этот список сейчас продолжает пополняться). В доме-
не бактерий подобных групп еще больше. Но выделение такого количества царств
прокариот вызвало бы столько ненужной путаницы, что микробиологи предпочитают
обходиться без него, переходя к безранговой номенклатуре. Дальше мы увидим,
что и к царствам эукариот относится то же самое. В хенниговской филогенетиче-
ской систематике ранги вообще не обязательны. Если они там сохраняются, то
лишь по традиции и для удобства.
Асгард
Итак, есть три эволюционные ветви клеточных организмов — Archaea, Eukarya и
Bacteria. Казалось бы, теперь картина ясна. Но, как это постоянно бывает в
науке, полученные ответы сразу вызвали к жизни новые вопросы.
Эукариоты произошли от архей. Это твердо установленный факт (симбиотический
компонент эукариотной клетки, происхождение которого однозначно бактериаль-
ное, мы сейчас временно не учитываем) . Но что, собственно, значит "произошли
от архей"? Насколько буквально это стоит понимать?
Одно дело, если эукариоты и археи — это две ветви, разошедшиеся от уникаль-
ного для них общего предка. В этом случае их надо считать сестринскими груп-
пами, а вот предками или потомками друг друга называть, строго говоря, нель-
зя. И совсем другое дело, если эукариоты отпочковались от какой-то из групп
архей уже после того, как древо самих архей разветвилось. Тогда эукариоты и
вправду окажутся потомками архей в самом точном смысле этого слова. Но одно-
временно это будет означать, что эукариот надо считать не отдельным доменом,
а группой внутри архей — так сказать, их подмножеством. По крайней мере, это-
го в обязательном порядке потребует филогенетическая систематика. Иначе полу-
чится, что граница домена архей пересечет эволюционное древо дважды — при
происхождении самих архей от "Луки" и при происхождении эукариот от архей; а
такое, по современным понятиям, недопустимо (см. рис. 14.3).
"Трехдоменный" сценарий Вёэе Современный "двудрменный" сценарий
АРХЕИ ЭУК/
ПП1
|И
Y
1_'г—
<\РИ01
11
1
ГЫ БАКТЕРИИ
1 ПППППП АРХЕИ ЭУКАРИ0ТЬ1 БАКТЕРИИ
I имшш утиши
ш
1 ■■■и
ш Ни
©
1
Щ
(11
"йГ ^9^ "Ф"
t
1
Рис. 14.3. Две версии положения эукариот и архей на филогенетиче-
ском древе. 1 — общий предок эукариот, 2 — общий предок архей, 3 —
общий предок бактерий, 4 — общий предок эукариот и архей. На правом
древе общие предки 2 и 4 совпадают, а эукариоты оказываются всего
лишь ветвью архей — по крайней мере, с точки зрения филогенетической
систематики. Легко убедиться, что если выделить на этом древе архей
(без эукариот) в особый таксон, то он будет парафилетическим.
Тем не менее, именно эта вторая возможность сейчас и подтверждается. По ме-
ре развития генетических исследований становится все яснее, что эукариоты
зволюционно ближе к одним археям, чем к другим. И в свою очередь, некоторые
археи ближе к эукариотам, чем друг к другу: например, кренархеоты, судя по
современным филогенетическим древесам, доводятся куда более близкими родст-
венниками эукариотам, чем каким-нибудь эвриархеотам. Все это означает, что
ветвь эукариот возникла довольно глубоко внутри архейного древа. Попросту го-
воря, эукариоты — это специализированная веточка архей. А в таком случае на
смену трехдоменному древу жизни неизбежно должно прийти двудоменное.
Интересно, что система из трех доменов (бактерии, археи, эукариоты) в любом
случае уже продержалась к настоящему моменту дольше, чем система из двух над-
царств (прокариоты и эукариоты). Но, похоже, что и она оказалась в развитии
науки промежуточным этапом. Следующие шаги по обновлению системы уже предпри-
нимаются .
В 2015 году в ходе глубоководных исследований Северного Ледовитого океана
были открыты локиархеоты — археи, являющиеся ближайшими родственниками эука-
риот . Локиархеоты оказались генетически ближе к эукариотам, чем к любым дру-
гим археям, известным на тот момент. Название они получили в честь Локи,
скандинавского бога огня. У локиархеот есть белки, очень близкие к актину —
белку, из которого у эукариот образуются микрофиламенты, важнейшие составные
части цитоскелета (см. главу 10) . Причем генов актиноподобных белков у них
несколько штук. Значит, эволюция этих белков началась достаточно задолго до
появления эукариот. Кроме того, у них есть белки, работа которых связана с
изменением формы клеточной мембраны и с рециклизацией мембран — процессом,
совершенно необходимым клеткам, которые транспортируют вещества в подвижных
вакуолях (см. опять же главу 10) . Все это означает, что клетка локиархеот
имеет много общего с эукариотной.
В придачу к локиархеотам были вскоре открыты еще три группы родственных им
архей. Они тоже получили названия в честь скандинавских богов: одинархеоты,
торархеоты и хеймдалльархеоты. И наконец, все эти четыре группы вместе были
закономерно названы асгардархеотами.
Как это ни удивительно, на момент написания этих строк асгардархеот еще ни-
кто никогда не видел своими глазами (вернее, под микроскопом). Но генетиче-
ского материала от них найдено и прочитано уже столько, что характеристика
этих существ выходит достаточно детальной. Например, у одинархеот есть тубу-
лин — белок, из которого у эукариот состоят микротрубочки. И других белков,
которые раньше считались уникальными для эукариот, в клетках асгардархеот
хватает. При этом асгардархеоты однозначно более близкородственны эукариотам,
чем любым другим археям. Иными словами, существует эволюционная ветвь, кото-
рая включает асгардархеот, эукариот и больше никого. Более того, похоже, что
сами эукариоты — это всего лишь веточка глубоко внутри эволюционного ллкуста"
асгардархеот; например, хеймдалльархеоты генетически гораздо ближе к эукарио-
там, чем к торархеотам или одинархеотам. Как бы там ни было, эти данные очень
здорово подкрепляют новую двудоменную систему жизни.
Ядерная
интерлюдия
Сделаем отступление.
Открытие асгардархеот, бесспорно, величайшее достижение. Теперь мы знаем
точный ЛЛадрес", по которому на эволюционном древе можно найти нашего архейно-
го предка. Значит ли это, что проблема происхождения эукариот решена?
Конечно, нет.
Проблема происхождения эукариот — одна из самых сложных и запутанных во
всей эволюционной биологии. Мы уже обсуждали ее в главе 10, но довольно крат-
ко (о некоторых гипотезах там вообще пришлось умолчать) . Так что к этой теме
не грех вернуться — хотя бы ненадолго.
Итак, чем, собственно, эукариотная клетка отличается от прокариотной? Во-
первых, в ней есть сложная система внутренних мембран, образующая эндоплазма-
тическую сеть и тесно связанное с ней ядро — ту самую структуру, которой эу-
кариоты обязаны своим названием. Во-вторых, в ней есть митохондрии — или хотя
бы их различимые на молекулярном уровне остатки. В-третьих, в эукариотнои
клетке перемешаны компоненты, имеющие совершенно разное происхождение — как
бактериальное, так и архейное. Любой состоятельный сценарий происхождения эу-
кариот должен объяснить эти три особенности их клеток. Причем желательно все
разом — так, чтобы не приходилось вводить дополнительные предположения ad hoc
(ллдля данного случая") .
Собственно говоря, объяснить перечисленные признаки по отдельности было бы
гораздо проще. Приобретение митохондрий легко объясняется симбиозом: предок
эукариот когда-то "проглотил" бактерию, ставшую предком митохондрий, и оста-
вил ее внутри себя жить. Обилие в эукариотнои клетке бактериальных генов и
белков столь же легко объясняется горизонтальным переносом генов: предок эу-
кариот жил в каком-то сложном бактериальном сообществе, вот и нахватался ге-
нов от соседей. Ну а ядро вполне могло возникнуть из впячивания плазматиче-
ской мембраны, которое удачно отшнуровалось и замкнулось. Судя по всему, не-
что похожее произошло в одной интересной группе грамотрицательных бактерий,
которая называется планктомицетами. У них есть довольно сложная система внут-
ренних мембран, окружающая ту область цитоплазмы, где находится ДНК, и у не-
которых родов это выглядит почти как клеточное ядро. Вероятно, все внутренние
мембраны планктомицетов развились из впячиваний их плазматической мембраны
(точнее внутренней из двух мембран, свойственных грамотрицательным бактери-
ям) . Самое же любопытное, что планктомицеты — вовсе не родственники эукариот
и возникновение аналога ядра наверняка произошло у них совершенно независимо.
Но это как раз и убеждает, что такие события в принципе возможны.
Сейчас, однако, нам нужно несколько другое. Нам нужен эволюционный сцена-
рий, который удовлетворительно объяснял бы все главные особенности эукариот —
и ядро, и митохондрии, и генетическую ллхимерность". Есть ли такой сценарий в
нашем распоряжении?
Серьезные попытки его создать, безусловно, налицо. Ближе всего к идеалу
сейчас, видимо, подошли знакомые нам из главы 10 гипотезы Евгения Кунина и
двоюродных братьев Баумов. Эти гипотезы отличаются друг от друга, но так или
иначе, обе они утверждают примерно следующее. Предком эукариот была некая ар-
хея, которая обзавелась наружным симбионтом в виде альфа-протеобактерии. Со
временем альфа-протеобактерия оказалась окружена выростами архейной клетки, и
наружный симбиоз превратился во внутренний. Одновременно с этим (и, вероятно,
вследствие этого) возникло ядро. В этом месте гипотезы расходятся. Кунин по-
лагает, что ядро образовалось из слившихся внутри клетки мембранных пузырь-
ков, а Баумы высказали и вовсе парадоксальную идею: ядро эукариот соответст-
вует "телу" архейной клетки, а цитоплазма — ее разросшимся выростам, которые
охватили альфа-протеобактерий и слились друг с другом (см. ранее рис. 10.9).
В любом случае изоляция архейного генома в ядре оказалась в итоге полезна,
чтобы защитить его от чрезмерно активного проникновения генетического мате-
риала бактерий-симбионтов. Но такое проникновение все равно на первых порах
происходило, и генетическая ллхимерность" эукариотнои клетки объясняется имен-
но этим.
Что ж, авторы этих гипотез сложили пазл почти до конца. Но все-таки лишь
ллпочти" — потому что есть, например, еще такой бросающийся в глаза факт, как
различие архейных и бактериальных клеточных мембран. Липиды, из которых со-
стоят эти мембраны, могут создаваться на основе право- или левовращающего
изомера глицерофосфата. Архейные мембраны состоят из ллправозакрученных" липи-
дов. Бактериальные — из лллевозакрученных". Разница между ними вполне фунда-
ментальна. А теперь спросим себя: почему же в эукариотнои клетке все мембраны
(и плазматическая, и ядерная, и вакуолярные) относятся к бактериальному типу?
Это парадоксальный факт, на первый взгляд вовсе не следующий ни из гипотезы
Кунина, ни из гипотезы Баумов. Исходя из этих гипотез, можно было бы скорее
ожидать обратного: ведь это архея охватила и поглотила бактерию, а не наобо-
рот. Конечно, объяснить наблюдаемые факты при желании все равно легче легко-
го: мы же знаем, что большинство генов "домашнего хозяйства" (обеспечивающих
всевозможные процессы обмена веществ) в эукариотных геномах имеет бактериаль-
ное, а не архейное происхождение; вот продукты этих генов и взяли на себя
синтез компонентов мембран, а поскольку ЛЛправозакрученные" и лллевозакручен-
ные" молекулы в одной мембране лишь ограниченно совместимы, то архейные мем-
бранные липиды в конце концов были полностью вытеснены бактериальными. И все
бы хорошо — если не считать, что тут перед нами типичное объяснение ad hoc,
не вытекающее само по себе из основной гипотезы, а добавленное специально,
чтобы охватить не уложившиеся в нее факты. Хотя само по себе это и не значит,
что оно неверно.
Тут нам будет полезно вспомнить историю науки. В старинной астрономии по-
четное место занимало понятие "спасения явлений" (во многом определявшее всю
ее методологию). "Спасти" тот или иной факт значило привести его в согласие с
единой теорией, которая, в свою очередь, должна была согласоваться со всеми
остальными наблюдаемыми фактами. Иногда эта операция оказывалась очень непро-
стой. Например, докоперниковская астрономия вынуждена была создать ради нее
представление о движении планеты по эпициклу — маленькой окружности, центром
которой является точка, движущаяся по большой орбите. Астрономический эпицикл
стал настоящим символом вспомогательной гипотезы, ниоткуда не вытекающей и
нужной лишь затем, чтобы свести концы с концами (то, что мы выше назвали "ad
hoc"). В конце концов, борьба за наилучшее "спасение явлений" привела к тому,
что геоцентрическая система сменилась на гелиоцентрическую, позволившую от
эпициклов отказаться. Что ж, европейская наука всегда, еще со времен Пифагора
и Платона, руководствовалась идеей, что рациональное объяснение мира должно
быть по возможности единым. На то она и европейская.
А не можем ли мы и в вопросе происхождения эукариот, образно говоря, поме-
нять местами Землю и Солнце? Хотя бы из чисто хулиганских побуждений, чтобы
посмотреть, что из этого получится. До сих пор мы исходили из идеи, что на
заре эволюции эукариот бактерия (вероятно, альфа-протеобактерия) была захва-
чена и поглощена какой-то археей. А что, если наоборот: архея была захвачена
и поглощена какой-то бактерией?
Обратившись к литературе, мы сразу увидим, что это отнюдь не фантазия на
пустом месте. В последнее десятилетие XX века подобные гипотезы были удиви-
тельно популярны, да и сейчас у них есть осторожные сторонники. Существует
целое семейство гипотез, согласно которым клеточное ядро имеет симбиотическое
происхождение — от археи, встроившейся в клетку более крупной бактерии и ок-
руженной ее сомкнутыми выростами.
Образцом такой гипотезы можно считать эволюционный сценарий, который опуб-
ликовали в 1996 году Радхей Гупта и помогавший ему Брайан Голдинг; у этих ав-
торов есть полная аргументация и нет ничего лишнего. Гупта и Голдинг изучали
эволюцию белков, сравнивая их аминокислотные последовательности. И они пришли
к выводу, который в основном подтверждается и сегодня: примерно половина эу-
кариотных белков восходит к археям, а другая половина — к грамотрицательным
бактериям. Причем это соотношение настолько близко к равному, что классиче-
ская теория (согласно которой архейный предок эукариот всего лишь захватил
бактериального симбионта как некий "предмет роскоши") становится неубедитель-
ной.
Кроме того, это соотношение трудно объяснить простым горизонтальным перено-
сом генов (даже при том, что интенсивность этого процесса могла быть в древ-
ние времена намного выше, чем сейчас). Такой перенос шел бы более-менее рав-
номерно между всеми участниками сообщества. И если бы дело было только в нем,
то предок эукариот "нахватался" бы генов от самых разных соседей-бактерий — и
грамположительных, и грамотрицательных. Между тем Гупта и Голдинг обнаружили,
что у эукариот очень много белков и генов, каким-то образом унаследованных от
грамотрицательных бактерий, а вот от грамположительных не нашлось на тот мо-
мент ни одного. Конечно, эта избирательность не случайна, вопрос только в ее
источнике.
В общем, состав эукариотного генома дает все основания полагать, что при
возникновении эукариот клетки археи и грамотрицательной бактерии просто сли-
лись (см. рис. 14.4). Получилась единая клетка, сначала унаследовавшая геномы
обоих "родителей" — Гупта и Голдинг прямо так их и называют. Но, поскольку
два полнофункциональных генома клетке были не нужны, часть каждого из них ис-
чезла, а между оставшимися частями произошло разделение функций. От архейного
генома в эукариотной клетке остались в основном "информационные" гены, обес-
печивающие работу самого генетического аппарата. А от бактериального генома —
в основном "операционные" гены, обеспечивающие обмен веществ. Тогда будет
вполне разумно предположить, что в химерной структуре (каковой эукариотная
клетка в любом случае, несомненно, является) от археи произошло ядро, а от
бактерии — цитоплазма вместе с наружной мембраной. И получается, что это
именно архея проникла внутрь бактерии. Вероятно, сначала она жила во впячива-
нии наружной мембраны бактерии — этаком "заливе". Потом глубокие складки мем-
браны бактерии, со всех сторон охватившие архею, сомкнулись друг с другом,
образовав внутреннюю мембранную систему — ядро и эндоплазматическую сеть. По-
сле этого плазматическая мембрана самой археи стала лишней и исчезла. Факти-
чески от археи остался голый генетический аппарат, заключенный внутри системы
бактериальных мембран. Ну а гены самой бактерии, сначала находившиеся снаружи
от ядра, постепенно мигрировали в него (молекулярные механизмы, делающие воз-
можной такую миграцию, известны). И получилась нормальная эукариотная клетка.
Рис. 14.4. Гипотеза Гупты. А — бактерия, во впячивании плазматиче-
ской мембраны которой поселилась архея. Б — впячивание мембраны бак-
терии начинает смыкаться, в то же время разрастаясь. В — впячивание
мембраны бактерии становится внутренней полостью, превращаясь в эн-
доплазматическую сеть, один из участков которой заключает в себе ге-
ном археи (мембрана последней за ненадобностью исчезла) . Так, со-
гласно этой гипотезе, возникло клеточное ядро.
Интересно, что в гипотезе Гупты (назовем ее так для краткости, хотя ее под-
держивали и другие ученые) есть кое-что общее с уже знакомой нам гипотезой
Баумов. И там и там получается, что остаток исходной архейной клетки находит-
ся в основном в эукариотном ядре. Хотя приводящие к этому выводу сценарии
прямо-таки полярно противоположны. Гипотеза Баумов, при всем ее бесспорном
изяществе, по части состава мембран (архейные vs. бактериальные) дает пред-
сказания, строго обратные тому, что наблюдается в действительности, и выйти
из этого положения она может только путем добавления "эпициклов".
Пурификасьон Лопес-Гарсия и Давид Морейра справедливо замечают, что зага-
дочное превращение архейных мембран в бактериальные остается настоящей ахил-
лесовой пятой традиционных симбиотических гипотез, согласно которым цитоплаз-
ма эукариотнои клетки имеет археиное происхождение. А вот если допустить, что
происхождение цитоплазмы — бактериальное, то проблема мембран тут же снимает-
ся и существующая картина получает простое объяснение.
Слабые места у этой гипотезы тоже есть — куда ж без них. Прежде всего, она
никак не объясняет происхождения митохондрий (которые как-никак тоже потомки
грамотрицательных бактерий и заведомо являются источником значительной части
генов, полученных эукариотами из этой эволюционной ветви). С учетом гипотезы
Гупты выходит, что симбионтов, без которых немыслим эукариотный организм, бы-
ло не два, а три. Сначала клетка археи слилась с клеткой какой-то неведомой
грамотрицательной бактерии, а потом образовавшийся гибридный монстр вобрал в
себя еще и предка митохондрии.
Может показаться, что такой сложный сценарий, включающий дополнительных
участников, нарушает принцип экономии мышления (согласно которому критерий
истины состоит в достижении максимума знаний с помощью минимума познаватель-
ных средств). Само по себе это не страшно: механизм такого уникального эволю-
ционного события, как возникновение эукариот, не мог быть простым. Хуже дру-
гое. Древнейший бактериальный хозяин симбиотической археи (если он существо-
вал) до сих пор неизвестен. На эту тему были кое-какие предположения, но про-
верить их оказалось очень трудно. Чтобы защитить гипотезу Гупты, приходится
допустить, что древнейший бактериальный предок в процессе интеграции в эука-
риотную клетку изменился до неузнаваемости — в отличие от митохондрий, кото-
рые вступили в симбиоз позже и сохранили свой исходный тип обмена веществ.
Это, конечно, возможно, но вот тут позиция сторонников гипотезы Гупты стано-
вится довольно слабой. Судя по данным современной геномики, ни одна другая
группа бактерий не дала такого серьезного вклада в геном эукариот, как альфа-
протеобактерии (хотя мелких заимствований от самых разных групп там полным-
полно) . Между тем альфа-протеобактерии — это и есть та самая группа, в кото-
рую входят предки митохондрий. А вот поиск какого-нибудь еще бактериального
предка эукариот зашел в тупик. Например, гены дельта-протеобактерий (которых
Лопес-Гарсия и Морейра одно время считали теми самыми "древнейшими бактери-
альными предками") в эукариотных геномах встречаются, но в количестве, ничем
не выделяющемся на фоне многих других источников генетического материала. Ес-
ли дельта-протеобактерий когда-то и сыграли некую особую роль, то последующая
эволюция стерла генетические свидетельства этого.
Конечно, можно предположить, что альфа-протеобактериальных симбионтов было
два: древний (предок цитоплазмы) и поздний (предок митохондрии). Но такая ги-
потеза явно не соответствует критерию ^экономичности", и, чтобы она была при-
нята, потребуются очень сильные свидетельства, которых пока нет. (Тем не ме-
нее, кое-что на эту тему обсуждается в главе 10.)
На другие козырные аргументы теории Гупты сторонникам теории Баумов тоже
есть что ответить. Во-первых, сейчас выяснилось, что не все бактерии, пере-
давшие эукариотам свои гены, были грамотрицательными. Например, гены, коди-
рующие ферменты гликолиза, эукариоты получили не от грамотрицательных бакте-
рий, а от грамположительных. Правда, от грамотрицательных бактерий взято
больше, но это может объясняться просто преобладанием грамотрицательных бак-
терий в тех природных сообществах, где эукариоты возникли. Кроме того, в по-
следнее время появились свидетельства, что и обычные археи, до всякого проис-
хождения эукариот, уже довольно активно заимствовали гены от бактерий. Выводы
отсюда пока неясны, но в любом случае эти факты дополнительно усложняют кар-
тину.
Во-вторых, "правозакрученные" и "левозакрученные" молекулы липидов не столь
уж несовместимы в одной мембране: оказывается, можно подобрать условия, при
которых смешанная мембрана будет вполне стабильна, и это в принципе позволяет
допустить, что именно такая мембрана и была у первых эукариот. Тогда замена
компонентов мембран по ходу эволюции могла быть и постепенной.
Наконец, еще одно замечание касается стройной схемы, согласно которой ар-
хейные гены (информационные) управляют генетическим аппаратом, а бактериаль-
ные (операционные) — обменом веществ в цитоплазме. Из этой схемы есть одно
важное исключение: цитоскелет. Белки цитоскелета — типично лл операционные",
характерные не для ядра, а для цитоплазмы, но уж они-то абсолютно точно унас-
ледованы эукариотами от архей. Тут налицо, если воспользоваться термином из
наук о Земле, несогласие, уже давно озадачивавшее исследователей. Конечно,
объяснить его так или иначе при желании нетрудно: гены белков цитоскелета
могли сохраниться при любом слиянии клеток просто потому, что к тому моменту
они были "ноу-хау" архей, полезным на все случаи жизни и не имевшим никаких
бактериальных аналогов. Но, во всяком случае, это еще раз показывает, с какой
сложной проблемой мы имеем дело. Остается дождаться, пока исследователи из-
влекут из морских бездн или еще откуда-нибудь живых асгардархеот: тогда, ско-
рее всего, что-нибудь да прояснится.
"Обозначились
смутные контуры"
А теперь вернемся к системе самих эукариот. В последней трети XX века в
этой области науки тоже начались интересные события.
Прежде всего, 1970-е стали эпохой переоценки признаков, по которым эукарио-
ты обычно классифицируются. С одной стороны, окончательно выяснилось, что вы-
делять крупные группы эукариот по способу питания, равно как и по составу
клеточных стенок или по типу запасных веществ, — дело безнадежное: эти при-
знаки слишком легко меняются в ходе эволюции. С другой же стороны, новым ис-
точником данных о строении клеток стала электронная микроскопия. Она позволя-
ет изучать такие структурные детали, которые ни в один световой микроскоп
просто не видно. Например, оказалось, что хлоропласты — части клеток, где
происходит фотосинтез, — у одних эукариот окружены двумя мембранами, а у дру-
гих почему-то тремя или даже четырьмя. Не менее интересной оказалась структу-
ра митохондрий. Как известно, любая митохондрия имеет две мембраны — наружную
и внутреннюю, причем ключевые биохимические процессы, ради которых митохонд-
рия, собственно, и нужна, связаны с ее внутренней мембраной (см. главу 11) .
Неудивительно, что поверхность этой мембраны увеличивается за счет складок,
которые принято называть кристами. Так вот, электронная микроскопия показала,
что кристы митохондрий могут иметь разную форму. Они могут быть пластинчаты-
ми, а могут быть трубчатыми. Есть и другие варианты формы крист (например,
дисковидные и мешковидные), но они встречаются реже. У большинства эукариот
кристы или пластинчатые, или трубчатые. Самое же интересное, что форма крист,
судя по всему, обычно очень медленно меняется в эволюции и остается устойчи-
вой внутри крупных групп эукариот. А значит, она может быть хорошим признаком
для различения этих групп. Почему бы не попробовать?
Забегая вперед, надо сказать, что сейчас форму крист митохондрий уже не по-
мещают в основу системы хотя бы потому, что некоторая изменчивость внутри
групп по этому признаку все же есть. И, тем не менее, ряд важных вещей по
кристам митохондрий понять можно. Чем больше накапливалось данных на эту те-
му, тем яснее становилось, что все эукариоты распадаются как минимум на две
большие группы, связанные только через общий корень. Причем линия раздела ме-
жду этими группами пролегла довольно неожиданным образом: из классической
зоологии и ботаники подобное никак не следовало.
В 1986 году петербургский зоолог Ярослав Игоревич Старобогатов придал этим
представлениям максимально четкую форму. Он отважился разделить всех эукариот
на таксоны Lamellicristata (с пластинчатыми кристами) и Tubulicristata (с
трубчатыми кристами), которые получили ранг надцарств (см. рис. 14.5). Пред-
полагалось, что это два главных эукариотных эволюционных ствола. В группу
Lamellicristata вошли зеленые водоросли, высшие растения, эвгленовые жгутико-
носцы , воротничковые жгутиконосцы, многоклеточные животные. В группу
Tubulicristata — бурые, золотистые и желтозеленые водоросли, инфузории, фора-
миниферы, радиолярии. В обеих группах были и одноклеточные, и многоклеточные,
и автотрофные, и гетеротрофные представители. Теперь они делились не по этим
признакам, а исключительно по предполагаемому родству. Так стали прорисовы-
ваться контуры совершенно новой системы организмов, резко отличающейся от
геккелевской.
Рис. 14.5. Митохондрии с пластинчатыми (вверху) и трубчатыми
кристами в разрезе.
На самом деле в системе Старобогатова было еще и третье надцарство — Akonta
(безжгутиковые). В него вошли эукариоты с пластинчатыми кристами митохондрий,
на всех стадиях жизненного цикла полностью лишенные жгутиков: красные водо-
росли и типичные грибы. Это объединение не поддерживается никакими новыми
данными, так что обсуждать его нет смысла. Старобогатов был бы гораздо более
прав, если бы включил красные водоросли и грибы в группу Lamellicristata.
Увы, даже гениальные ученые (а Старобогатов явно был отмечен проблеском гени-
альности) почти никогда не угадывают правильно все сразу.
В любом случае система Старобогатова (причем именно та ее часть, которая
успешно выдержала проверку временем) ведет к одному серьезному выводу, кото-
рый легко будет понять всякому, кто хоть немного помнит обычный школьный
учебник зоологии. Первая часть этого учебника знакомит читателя с такими чу-
десными созданиями, как обыкновенная амеба, инфузория-туфелька и малярийный
плазмодий, иногда дополняя рассказ о них упоминаниями всевозможных сувоек,
раковинных амеб, фораминифер и радиолярий. Так вот, все только что названные
существа имеют митохондрии с трубчатыми кристами. А это уже само по себе оз-
начает, что они относятся к совсем иным эволюционным ветвям, чем многоклеточ-
ные животные. Инфузория-туфелька, активно плавающий поедатель бактерий, дей-
ствительно похожа на животных образом жизни, но эволюционно она к ним ничуть
не ближе, чем, например, какое-нибудь зеленое растение. Называть таких су-
ществ ллодноклеточными животными" попросту неправильно.
Вновь забегая вперед, добавим, что понятие ЛЛодноклеточные животные" вообще
вышло из употребления, причем уже довольно давно. Поэтому с этого момента мы
будем считать, что "многоклеточные животные" и просто "животные" — строгие
синонимы. Никаких животных, кроме многоклеточных, просто нет на свете.
Ради справедливости укажем, что есть и такие одноклеточные представители
"фауны школьного учебника", у которых кристы дисковидные (или дисковидные в
сочетании с трубчатыми). Это некоторые жгутиконосцы: фотосинтезирующая эвгле-
на, хищный бодо и вызывающая сонную болезнь трипаносома. От многоклеточных
животных (у которых кристы, напомним, пластинчатые) эти существа так же дале-
ки, как и инфузории.
В то же время есть как минимум две относительно большие эволюционные ветви
эукариот, которые действительно родственны многоклеточным животным. Это всем
известные грибы и не столь знаменитые, но зато еще более близкие к животным
воротничковые жгутиконосцы. И те и другие имеют пластинчатые кристы митохонд-
рий и гетеротрофный обмен веществ. Надо заметить, что это достаточно
"сильное" сочетание признаков, которое встречается лишь у немногих эукариот
(по крайней мере, если учитывать крупные ветви, а не количество видов). Одна-
ко во всем остальном грибы и воротничковые жгутиконосцы мало похожи друг на
друга.
Грибы — это преимущественно наземные организмы, специализирующиеся на осмо-
трофном питании (см. главу 10) . Попросту говоря, "осмотрофы" значит
"всасывалыцики". Тело типичного гриба представляет собой систему очень тонких
— толщиной в одну клетку — нитей с огромной суммарной поверхностью, сквозь
которую всасываются питательные вещества. В просторечии эта система нитей на-
зывается грибницей. Нити грибницы одеты хитиносодержащей клеточной стенкой,
которая придает им прочность, но затрудняет любое активное изменение формы
клеток (см. главу 5) . Фагоцитоза у грибов нет. У большинства из них нет и
жгутиков, причем ни на каких стадиях жизненного цикла (в отличие от многокле-
точных животных, у которых очень часто бывают жгутиковые сперматозоиды).
Воротничковые жгутиконосцы — это водные существа, одноклеточные или обра-
зующие колонии (см. рис. 14.6). Клетка воротничкового жгутиконосца имеет один
жгутик, окруженный характерным "воротничком" — кольцевым выростом, облегчаю-
щим ловлю пищевых объектов. Под электронным микроскопом видно, что этот
"воротничок" на самом деле состоит из множества длинных тонких выростов клет-
ки, укрепленных проходящими внутри микрофиламентами и как бы склеенных вместе
при помощи специальных белков. Эти тонкие выросты называются микроворсинками.
Воротничковые жгутиконосцы подвижны. Они могут плавать, а могут и вести при-
крепленный образ жизни, но в этом случае они постоянно работают жгутиком,
подгоняя к себе воду вместе со взвешенными в ней пищевыми частицами. Питаются
они путем фагоцитоза, поглощая эти частицы. По научному такое питание называ-
ют фаготрофным, противопоставляя его осмотрофному.
Воротничковых жгутиконосцев открыли в XIX веке. И тут же было замечено, что
клетки, очень похожие на них, входят в состав тел некоторых многоклеточных
животных, а именно губок. Это, конечно, не случайность. Современные генетиче-
ские данные надежно подтверждают сделанный еще полтора века назад вывод: во-
ротничковые жгутиконосцы — самые близкие современные родственники многокле-
точных животных. Получается, что это едва ли не единственные ЛЛодноклеточные
животные", для которых такое название могло бы иметь хоть какой-то реальный
смысл. Но при этом они-то как раз в школьный учебник зоологии и не попали.
Рис. 14.6. Воротничковые жгутиконосцы. Слева — одиночный жгути-
коносец монозига, справа — жгутиконосец сферозка, образующий ша-
рообразные колонии.
Вклад
Кавалье-Смита
В 1981 году английский протистолог Томас Кавалье-Смит предложил версию сис-
темы эукариот, в которой фигурировали шесть царств.
■ Царство Fungi. Грибы. Организмы с осмотрофным питанием, пластинчатыми кри-
стами, хитиновой клеточной стенкой, лишенные каких бы то ни было жгутико-
вых стадий (за исключением самых примитивных форм, выделяемых в особое
подцарство). В остальных пяти царствах жгутиковые клетки сохраняются — ес-
ли и не у всех представителей, то у многих.
■ Царство Animalia. Животные. Организмы с фаготрофным питанием, пластинчаты-
ми кристами и активной подвижностью. В это царство входят многоклеточные
животные и воротничковые жгутиконосцы.
■ Царство Plantae. Растения. Фотосинтезирующие организмы с пластинчатыми
кристами. Хлоропласты окружены двумя мембранами. Фагоцитоз отсутствует.
Одноклеточные, колониальные или многоклеточные формы. В это царство входят
красные водоросли, зеленые водоросли, высшие растения.
■ Царство Chromista. Оно было впервые выделено самим Кавалье-Смитом в этой
самой работе. Фотосинтезирующие организмы с трубчатыми кристами (за исклю-
чением одной группы одноклеточных). Хлоропласты окружены тремя или четырь-
мя мембранами. У многих представителей есть фагоцитоз. Одноклеточные, ко-
лониальные или многоклеточные формы. В это царство входят бурые, золоти-
стые и желтозеленые водоросли, а также несколько довольно разнообразных
групп одноклеточных жгутиконосцев.
■ Царство Protozoa. Одноклеточные гетеротрофные или (реже) миксотрофные ор-
ганизмы с трубчатыми кристами. Питаются путем фагоцитоза. Передвигаются с
помощью жгутиков, ресничек или ложноножек. К этому царству относятся инфу-
зории, амебы, радиолярии, фораминиферы, солнечники, споровики и несколько
групп жгутиконосцев. Это и есть ЛЛодноклеточные животные" в том (устарев-
шем) значении, которое раньше встречалось в учебниках.
■ Царство Euglenozoa. Эвгленовые жгутиконосцы. Одноклеточные формы, плаваю-
щие с помощью жгутиков. Некоторые строго гетеротрофны (например, трипано-
сома и бодо), некоторые способны к фотосинтезу и являются типичными миксо-
трофами (например, эвглена), а некоторые даже строго автотрофны (например,
колациум и трахеломонас). Хлоропласты, если они есть, окружены тремя мем-
бранами . Кристы митохондрий не пластинчатые и не трубчатые, а дисковидные.
Относительно немногочисленное царство, состоящее из одних жгутиконосцев,
но достаточно сильно обособленное, чтобы его не следовало ни с кем объеди-
нять .
Что здесь можно увидеть? Во-первых, как мы сейчас понимаем, эта система не
свободна от ошибок (например, некоторых паразитических жгутиконосцев, вошед-
ших в царство ллпротозоев", на самом деле правильнее было бы сблизить с
ллэвгленозоями") . Еще бы — в 1981-м! Скорее стоит удивиться, что серьезных
ошибок там не так уж и много. Кроме того, к чести Томаса Кавалье-Смита, надо
сказать, что он никогда не отстаивал свои гипотезы любой ценой. Он вполне
сознательно рассматривает их как ряд последовательных приближений, описываю-
щих эволюцию все более и более точно, но ни в коем случае не окончательных.
Поэтому он всегда легко пересматривал свои предположения, включая в них новые
факты, а иногда и сразу формулировал их так, чтобы облегчить последующий пе-
ресмотр. Характерно, что в статье 1981 года (а это одна из десятков его работ
на подобные темы) Кавалье-Смит на самом-то деле предложил сразу несколько
возможных вариантов системы эукариот, с числом царств от пяти до девяти. На-
пример, в отношении красных и зеленых водорослей он тогда колебался — отно-
сятся ли они к одному царству или к разным. Поскольку эта публикация — все же
не труд по истории науки, то во всех подобных случаях тут выбраны версии,
наиболее соответствующие тем гипотезам, которые потом подтвердились.
Второе, что бросается в глаза: Кавалье-Смит сохраняет невероятно архаичный
таксон Protozoa (предложенный, как мы помним, еще в середине XIX века. Карлом
Зибольдом), но при этом радикальным образом меняет его состав и значение. Он
исключает из этого таксона всех родственников животных, зато оставляет в нем
существ с трубчатыми кристами митохондрий, которые не имеют к животным ни ма-
лейшего отношения. В таких случаях для таксонов обычно придумывают новые на-
звания, но Кавалье-Смит, всегда склонный к парадоксам, делать этого не стал.
Кроме того, в той же статье 1981 года Кавалье-Смит высказал два важных
предположения:
1. Возможно, царства Fungi и Animalia образуют единую эволюционную ветвь. Во-
первых , между ними есть некоторое биохимическое сходство: например, и у
грибов, и у животных широко распространен хитин. Во-вторых, самостоятельно
плавающие жгутиковые клетки грибов ("низших", у которых жгутики сохраняют-
ся) , воротничковых жгутиконосцев и многоклеточных животных имеют один жгу-
тик, направленный назад. У других эукариот такое состояние не встречается.
2. Возможно, единую эволюционную ветвь образуют также царства Chromista и
Protozoa. Во всяком случае, только в этих двух группах распространено со-
четание трубчатых крист митохондрий со способностью питаться путем фагоци-
тоза.
Рассмотренная система Кавалье-Смита — не слишком обобщенная (как система
Уиттэкера, где почти все царства на поверку оказались сборными), но и не
слишком дробная (как система Лидейла, где статус царства придается каждой
крупной ветви). Самое же главное, что эта система абсолютно последовательно
строится по родству, или, во всяком случае, по достаточно хорошо обоснованным
гипотезам о родстве. Теперь контуры истинного эволюционного древа эукариот
стали, наконец, видны, пусть пока еще и смутно.
В 1987 году произошло историческое событие: Кавалье-Смит формально выделил
эволюционную ветвь, в которую входят воротничковые жгутиконосцы, многоклеточ-
ные животные и грибы. Эта группа получила название Opisthokonta (заднежгути-
ковые) . Чтобы понять смысл этого названия, еще раз вспомним, что очень многие
эукариотные клетки имеют жгутик — нитевидный вырост, в котором проходит пучок
микротрубочек, обычно организованных по схеме 9+2 (девять сдвоенных микротру-
бочек по краю и две одиночные в центре). Работа жгутика обеспечивает движение
клетки в воде. Если жгутиков очень много и они бьются согласованными волнами,
то их называют ресничками. Эти структуры уникальны для эукариот. Жгутики бак-
терий не имеют с эукариотными жгутиками ничего общего, кроме названия. А тех
эукариот, у которых жгутик служит основным средством движения, принято назы-
вать жгутиконосцами — это слово мы уже не раз встречали (см. рис. 14.7).
ХРОМАЛЬВЕОЛЯТЫ
желто-зеленые
водорос.п<
золотистые
водоросли
диноф'&гелляты криптомонады
РАСТЕНИЯ
г рази-офиты
ЗАДНЕ-
ЖГУТИКОВЫЕ
вороти г'.чксэье
.жгутиконосцы
ЭКСКАВАТЫ
эвглексзые бело* иды трл износом:* очеимснады
гипермастигмны
дипломонадь:
Рис. 14.7. Жгутиковые формы, входящие в состав разных крупных
эволюционных ветвей эукариот. О некоторых из них говорится под-
робнее дальше, в главе 15.
Жгутик, вызывающий поступательное движение клетки, можно уподобить винту
корабля или самолета, хотя, в отличие от винта (и от бактериального жгутика),
он не вращается, а изгибается из стороны в сторону — примерно как рыбий плав-
ник. Тем не менее, эукариотныи жгутик, как и винт, может быть тянущим (тогда
он направлен вперед), а может быть толкающим (тогда он направлен назад). На-
пример, у самолетов бывают винты обоих типов, хотя толкающие встречаются за-
метно реже. Пример самолета с толкающим кормовым винтом — немецкий истреби-
тель "Хеншель" Hs.P75, разработанный во время Второй мировой войны, но так и
не пущенный в производство. (Между прочим, очень похожий на него самолет уве-
ковечил японский режиссер Мамору Осии в своем знаменитом полнометражном аниме
"Небесные скитальцы".) Возвращаясь к жгутиковым клеткам, можно сказать, что у
них ситуация примерно такая же, как в винтовой авиации: для большинства эука-
риот характерны тянущие жгутики, а вот толкающие — это скорее нечто необыч-
ное. Обладатели "кормовых" толкающих жгутиков — воротничковые жгутиконосцы,
жгутиковые стадии примитивных хитридиевых грибов и сперматозоиды многоклеточ-
ных животных. Вот эти группы и называются заднежгутиковыми (опистоконтами).
Другие общие признаки опистоконтов — пластинчатые кристы митохондрий и стро-
гая гетеротрофность. Кавалье-Смит выделил эту группу абсолютно правильно: ее
реальность надежно подтвердили дальнейшие исследования, в первую очередь мо-
лекулярно-генетические. Хотя у самого Кавалье-Смита молекулярных данных тако-
го рода в ту пору не было (а если и были, то настолько противоречивые, что
это лишь сбивало с толку). Кроме воротничковых жгутиконосцев, грибов и живот-
ных к опистоконтам относится еще несколько небольших групп одноклеточных и
колониальных существ, родственных или грибам, или животным.
В 1991 году Кавалье-Смит выделил еще одну группу эукариот, получившую на-
звание Alveolata. Это одноклеточные организмы с трубчатыми кристами митохонд-
рий, имеющие дополнительные общие признаки на уровне тонкой структуры клеток.
Под наружной мембраной у них находятся многочисленные плоские пузырьки, кото-
рые называются альвеолами. Слой альвеол, укрепленный микротрубочками, образу-
ет внутреннюю оболочку клетки — пелликулу. К альвеолятам относятся инфузории,
некоторые гетеротрофные жгутиконосцы с их паразитическими родственниками
(включая малярийного плазмодия) и одна группа одноклеточных водорослей — ди-
нофлагелляты. Самое интересное, что эти существа, в общем-то, совсем не похо-
жи друг на друга ни по внешнему облику, ни по образу жизни. Предполагая, что
они образуют одну ветвь, Кавалье-Смит основывался в первую очередь на сходст-
ве внутриклеточных структур, невидимых без электронного микроскопа, и оказал-
ся опять прав. (В более старой системе 1981 года эти организмы входили в цар-
ство Protozoa, но оно, по мнению подавляющего большинства современных авто-
ров , оказалось все же сборным.)
Между тем в биологии наступила эпоха молекулярной филогенетики. Сама эта
наука родилась значительно раньше. Но именно в последнее десятилетие XX века
исследования нуклеотидных последовательностей генов начали ставить на поток.
Излюбленным объектом молекулярных филогенетиков в тот период были гены, коди-
рующие рибосомную РНК: она невелика, информативна, и ее изучение уже привело
Карла Вёзе к колоссальному успеху. Работы генетиков, сравнивавших последова-
тельности рРНК разных эукариот, сразу подтвердили реальность групп
Opisthokonta и Alveolata. А дальше последовал еще более серьезный вывод. Поя-
вились указания на то, что группы Chromista и Alveolata образуют единую эво-
люционную ветвь, многочисленную и разнообразную, но состоящую в основном из
организмов с трубчатыми кристами митохондрий. Эта ветвь получила название
Chromalveolata.
Хромальвеоляты просто поражают своим разнообразием. Среди них есть однокле-
точные, колониальные и многоклеточные фотосинтезирующие существа (всевозмож-
ные водоросли), есть плавающие одноклеточные хищники (инфузории, хищные жгу-
тиконосцы) , есть крайне специализированные паразиты (споровики, к которым от-
носится малярийный плазмодий), а есть и многоклеточные осмотрофы, точные ана-
логи грибов по внешнему облику и образу жизни. В общем, одних только хромаль-
веолят наверняка хватило бы, чтобы обильно заселить эукариотами целую плане-
ту, создав на ней сложные и разнообразные экосистемы со своими собственными
растениями, фаготрофами, паразитами и поедателями мертвой органики.
Вот кого среди хромальвеолят нет, так это многоклеточных хищников, аналогов
животных. При всем их огромном разнообразии такую жизненную форму им
"изобрести" не удалось. Судя по всему, живой природе Земли было намного слож-
нее создать многоклеточных животных, чем многоклеточные растения (мы еще по-
говорим про это в последней части).
Заодно молекулярные исследования подтвердили единство эволюционной ветви
Plantae — в том объеме, в каком ее выделил Кавалье-Смит. В эту группу входят
все зеленые растения, от одноклеточных водорослей до цветковых, и красные во-
доросли в придачу.
Тут возникает один интересный чисто эволюционный момент. Общий предок эво-
люционных стволов Plantae и Chromalveolata мог быть только одноклеточным: это
следует из конфигурации древа совершенно однозначно. Значит, входящие в эти
два ствола многоклеточные водоросли стали таковыми независимо друг от друга.
Если многоклеточные животные возникли в эволюции (насколько мы сейчас знаем)
всего однажды, то многоклеточные растения совершенно точно возникали несколь-
ко раз. Это стало окончательно ясно еще в первой половине XX века, после ра-
бот выдающегося ботаника Адольфа Пашера: он показал, что есть несколько со-
вершенно самостоятельных эволюционных линий водорослей, каждая из которых на-
чинается с одноклеточных организмов и постепенно достигает многоклеточности.
Например, многоклеточные зеленые и красные водоросли принадлежат к разным
эволюционным линиям, начинающимся от разных одноклеточных предков — и это не-
смотря на то, что в системе, из которой мы сейчас исходим, обе линии входят в
группу Plantae. Можно точно сказать, что многоклеточность растительного типа,
пусть и примитивная, возникала не менее трех раз в эволюционной ветви Plantae
(у красных, зеленых и харовых водорослей) и, по меньшей мере, столько же раз
в эволюционной ветви Chromalveolata (у бурых, золотистых и желто-зеленых во-
дорослей) . А скорее всего, таких событий было еще больше.
15. МИР
ЭУКАРИОТ
Дерево Сфера царствует здесь над другими.
Дерево Сфера — это значок беспредельного дерева,
Это итог числовых операций.
Ум, не ищи ты его посредине деревьев:
Он посредине, и сбоку, и здесь, и повсюду.
Николай Заболоцкий. «Деревья»
Область биологической систематики, оперирующая категориями выше типа, назы-
вается мегасистематикой. Нечего и говорить, что область эта — исключительно
важная. Именно благодаря мегасистематике мы можем представить себе, как в це-
лом, в самом первом приближении устроена живая природа. Крупнейшим мегасисте-
матиком конца XX — начала XXI века можно с полным основанием назвать Томаса
Кавалье-Смита. Он сделал в этой области много важных открытий, хотя и ошибок
совершил не меньше (но никогда не стеснялся их исправлять, сразу используя
для этого появляющиеся новые данные) . В любом случае надо учитывать, что со-
временные эволюционные исследования — это всегда коллективный процесс. Над
такими проблемами, как построение эволюционного древа эукариот, в наше время
работают большие международные группы ученых. Вот общую картину родственных
отношений, которую им удалось раскрыть, мы сейчас и рассмотрим. Благо картина
эта уже достаточно целостная.
Современная мегасистема эукариот сложилась в первые годы XXI века в основ-
ном в результате молекулярно-филогенетических исследований. У этой современ-
ной мегасистемы есть три важные особенности.
Во-первых, она выстроена строго по принципам филогенетической систематики —
без всяких компромиссов. Это означает, что любой таксон там охватывает свою
эволюционную ветвь целиком, вместе со всеми потомками, как бы специализирова-
ны они ни были. Сборные группы, выделяемые по образу жизни или по уровню ор-
ганизации, теперь полностью исключены. Это радикально отличает новую систему
от большинства систем ЛЛклассического" толка (в том числе и от тех, что создал
и продолжает создавать Кавалье-Смит: он-то никогда не был последовательным
кладистом).
Во-вторых, эту систему уже невозможно связать с именем какого-либо одного
ученого. ЛЛУ нее нет единого автора: родство между организмами — это факт, не
допускающий авторских трактовок. Ученые, создавшие эту систему, — это не фи-
лософы, а практики, изучающие структуру отдельных генов и анализирующие полу-
ченные данные математизированными методами молекулярной кладиетики".
В-третьих, современная систематика мало интересуется рангами таксонов. Ино-
гда их по старинке еще используют, но в целом сейчас никого всерьез не волну-
ет, считать ли такую-то эволюционную ветвь царством, подцарством, надцарством
или субдоменом. Очень часто авторы статей по систематике обходятся вовсе без
этих рангов. Мы уже знаем, что в филогенетической систематике это нормально:
ранги в ней не обязательны, они сохраняются только там, где это удобно. А в
данном случае это скорее неудобно (например, употребление хорошо всем знако-
мого слова "царство" сразу приведет к тому, что его новое значение начнут
сравнивать со старым и это вызовет совершенно ненужную путаницу). Поэтому
здесь и далее мы будем для удобства считать, что у таксонов ниже домена, но
выше типа рангов нет.
Новая система — действительно совершенно новая. Она не имеет никаких корней
в старых двух-, трех-, четырех- или пятицарственных системах и не является
преемственной по отношению к ним. Для простоты можно считать, что ее построи-
ли вообще с нуля (на самом деле это, разумеется, не так, но понять ее так бу-
дет легче).
Новая система, конечно, развивается. Более того, она непрестанно модифици-
руется по принципу последовательных приближений. "Если раньше ученый долгие
годы размышлял над деревом, создаваемым им из опыта, интуиции, вдохновения,
то сейчас весомый отчет по гранту ставится выше научной щепетильности, и на-
учное сообщество не осуждает практики выдвижения одним и тем же автором раз
за разом противоположных филогенетических гипотез при условии, что для них
привлекаются новые данные". Неудивительно, что филогенетическое древо посто-
янно обновляется и в разных научных работах можно увидеть его слегка отличаю-
щиеся версии. Обновления происходят за счет включения в анализ новых генов,
или изучения ранее мало изученных живых существ, или применения более совер-
шенных методов обработки данных. Никаких переворотов, подобных тем, что про-
исходили в конце XX века, это не вызывает (и, скорее всего, уже никогда не
вызовет), но поток уточнений, которые часто сопровождаются еще и придумывани-
ем новых названий, вполне может сбить новичка с толку. Лучший выход в этой
ситуации — выбрать одну хорошую систему, которая в главных своих чертах будет
верна, и уж в ней-то разобраться основательно, а потом, где нужно, модифици-
ровать ее с учетом новых открытий.
В качестве такой "хорошей системы" мы рассмотрим эволюционное древо, кото-
рое опубликовали в 2004 году биологи Аластер Симпсон и Эндрю Роджер. Название
их статьи — ллРеальные ЛЛцарства" эукариот" (именно так, со словом ЛЛцарства" в
кавычках). Чаще всего эти ллцарства" называют просто супергруппами. Примем во
внимание, что в этой системе кое-что успело заведомо устареть: например, одна
из супергрупп Симпсона и Роджера на самом деле, скорее всего, является сбор-
ной , и подавляющее большинство современных систематиков ее уже не признает.
Так что наш обзор этой системы будет относиться скорее к истории науки (пусть
и совсем недавней), чем к ее нынешнему состоянию. Это сделано сознательно,
потому что такой порядок изложения представляется здесь более удобным. До со-
временного состояния мегасистематики мы еще доберемся.
Итак, Симпсон и Роджер в своем обзоре 2004 года выделили шесть супергрупп
эукариот: Excavata, Rhizaria, Chromalveolata, Plantae, Amoebozoa и Opis-
thokonta (см. рис. 15.1). Поговорим о них по порядку.
Рис. 15.1. Эволюционное древо эукариот, соответствующее научным
представлениям 2004 года и демонстрирующее шесть традиционных
супергрупп. Именно оно будет принято за основу обсуждения в этой
главе, хотя надо учитывать, что во второй половине главы к нему
будут сделаны поправки.
Excavata
Эта супергруппа состоит в основном из жгутиконосцев — одноклеточных эукари-
от, у которых главным средством движения служат жгутики (см. рис. 15.2).
Название экскават связано с одной их структурной особенностью: на брюшной
стороне клетки (то есть на той, которая обычно обращена к поверхности грунта)
находится продольное углубление, со стороны цитоплазмы армированное микротру-
бочками. Это брюшная ротовая бороздка, через которую жгутиконосец заглатывает
пищу (она продолжается в так называемый клеточный рот). Брюшная бороздка есть
у многих экскават, но не у всех.
Широко известный представитель экскават — эвглена, попавшая во многие
школьные учебники зоологии в роли типичного жгутиконосца. Жгутиков у нее два,
но один из них очень короткий и в движении не участвует. Плазматическая мем-
брана у эвглены подостлана изнутри спиральным каркасом из лент специальных
опорных белков, придающим клетке довольно устойчивую форму (в образовании
этого каркаса участвуют микротрубочки и даже элементы эндоплазматической се-
ти) . Получается упругая оболочка, благодаря которой тело эвглены очень обте-
каемо и она хорошо плавает. Но одновременно оболочка затрудняет выпускание
ложноножек — и, соответственно, захват пищевых объектов. Поэтому фагоцитоз у
эвглены не развит.
Рис. 15.2. Некоторые представители супергруппы Excavata. Панели а,е
Discobans, панели b-d показывают метамонады. (а) Якобид:
Reclinomonas Americana; (b) дипломонад: Giardia intestinalis; (с)
Carpediemonas marsupialis; (d) Оксимонад: Pyrsonympha sp. ; (e) гете-
ролобозой: Percolomonas cosmopolitus; (f) Percolomonas cosmopolitus
(амебоидная форма); (g) Кинетопластид: Bodo Designis; (h) Евгленид:
Евглена мутабилис; (1) Кинетопластидная трипаносома: трипаноплазма
sp. Масштаб: для а-с,е-д - 10 мкм; i - 5 мкм.
Эвглена — миксотрофный организм. Она прекрасно умеет питаться гетеротрофно,
всасывая полезные вещества из раствора (когда эвглен разводят на корм малькам
аквариумных рыб, их самих кормят, добавляя в воду мясной бульон) . Но она мо-
жет и фотосинтезировать, потому что у нее есть зеленые хлоропласты. Причем
эти хлоропласты произошли от захваченных предком эвглены зеленых водорослей —
одноклеточных эукариот из супергруппы Plantae. В свое время эвглену иногда
ошибочно сближали с зелеными водорослями, на которые она похожа аппаратом фо-
тосинтеза (и больше ничем). На самом же деле зеленые водоросли родственны не
эвглене, а ее хлоропластам.
Близкий родственник эвглены — астазия, жгутиконосец, внешне очень похожий
на нее, но бесцветный и питающийся только гетеротрофно. Самое интересное, что
у астазии это состояние, скорее всего, вторично: в ее геноме обнаружены гены
хлоропластнохю происхождения. Судя по всему, захват одноклеточной зеленой во-
доросли, которая стала предком хлоропластов, произошел один раз — у общего
предка эвглены и ее родственников. Но с тех пор многие из этих родственников
успели потерять хлоропласты и вновь стать гетеротрофными. Потеря хлоропластов
произошла не меньше чем в пяти разных ветвях эвгленовых жгутиконосцев, в том
числе и у астазии. Благо это несложно: даже у самой эвглены бывают бесхлоро-
пластные популяции, нормально живущие на гетеротрофном питании. Такой вот
здесь получился эволюционный зигзаг.
Кроме эвгленовых жгутиконосцев (к которым относится и астазия) в супергруп-
пу экскават входит много других интересных существ. Фотосинтезирующих форм
среди них мало, эвглена тут скорее исключение, чем правило. Большинство экс-
кават никогда не имело хлоропластов. Зато среди них есть много гетеротрофных
жгутиконосцев, как хищных, так и паразитов. К последним относятся некоторые
возбудители инфекционных болезней человека: трипаносомы, лямблии, трихомона-
ды, лейшмании. Все это — паразитические жгутиконосцы из супергруппы Excavata.
Паразитические экскаваты часто имеют причудливую форму клеток. Скорее все-
го, это связано с тем, что им не нужна особенно высокая маневренность, зато
нужен мощный жгутиковый аппарат, чтобы пробиваться сквозь вязкую среду (кровь
или содержимое кишечника). Поэтому у них часто увеличивается число жгутиков.
Например, лямблия имеет два ядра и восемь жгутиков, ее клетка как бы частично
раздвоена. Это далеко не предел, у других жгутиконосцев бывают клетки и по-
сложнее . В то же время непаразитические родственники лямблии живут в толще
донных осадков и чаще всего имеют только два жгутика. Такое состояние свойст-
венно большинству хищных жгутиконосцев, которым высокая маневренность как раз
нужна. Один жгутик они используют как винт для поступательного движения, дру-
гой — как руль.
Некоторые жгутиконосцы-экскаваты, живущие внутри других организмов, прино-
сят им пользу. Например, жгутиконосцы, которые живут в пищеварительном тракте
термитов. В отличие от подавляющего большинства животных, термиты могут пи-
таться одной только древесиной. А древесина — это масса растительных клеточ-
ных стенок, которые состоят в основном из целлюлозы (см. главу 6). Такой корм
очень малопитателен, тем более что у термитов — во всяком случае, у примитив-
ных — нет никакого собственного фермента, способного расщеплять связи между
остатками глюкозы в огромной молекуле целлюлозы. Переваривать целлюлозу этим
термитам помогают симбиотические жгутиконосцы, живущие у них в кишечнике. Вот
эти жгутиконосцы и относятся к супергруппе экскават. Причем жгутиконосцам, в
свою очередь, помогают в расщеплении целлюлозы симбиотические бактерии, кото-
рыми клетка такого жгутиконосца прямо-таки набита. Так что симбиоз тут
"многоэтажный", по типу русской матрешки.
Типичные обитатели кишечника термитов — крупные жгутиконосцы, называемые
гипермастигинами (буквально это значит что-то вроде ллсверхжгутиковые") . Такой
жгутиконосец может иметь несколько сотен, а то и несколько тысяч жгутиков,
расположенных рядами, кругами или спиралями. Ядро у него при этом вполне мо-
жет быть одно, хотя у некоторых родов много ядер. Неудивительно, что архитек-
тура цитоскелета в таких клетках очень сложная и красивая. Кроме того, у не-
которых гипермастигин на поверхности клетки живут еще и спирохеты — подвижные
грамположительные бактерии, длинные, тонкие и закрученные в спираль. Они зая-
кориваются на клетке жгутиконосца и несут функцию дополнительных жгутиков.
Раньше была популярна гипотеза, что жгутики, собственно, и произошли от таких
симбиотических спирохет, но молекулярная биология ее не подтверждает.
Стоит отметить, что жгутиконосцы-гипермастигины иногда живут в кишечнике не
только у термитов, но и у тараканов, среди которых тоже есть виды, питающиеся
древесиной. Это неудивительно: термиты и тараканы — ближайшие родственники.
Еще один великолепный представитель экскават — Mixotricha paradoxa, живущая
в кишечнике одного австралийского термита. Это крупный — до полумиллиметра —
одноядерный жгутиконосец, несущий на своем переднем конце не десятки и сотни
жгутиков (как у гипермастигин), а всего лишь четыре жгутика, и те малоподвиж-
ные . Зато все остальное тело миксотрихи усеяно тысячами симбиотических спиро-
хет , которые бьются вместо ресничек. Причем, похоже, что этих спирохет там
несколько видов. По подсчетам, на одной миксотрихе может жить примерно 250
000 спирохет. Если убить их антибиотиками, миксотриха тут же теряет подвиж-
ность . При этом кроме спирохет на ее поверхности живут еще и другие бактерии,
имеющие форму коротких палочек. Эти бактерии сидят на теле миксотрихи там,
где прикрепляются спирохеты, как бы заякоривая их. Но и это не все. Помимо
"якорных" бактерий и обычных спирохет, работающих как реснички, на теле мик-
сотрихи сидят еще особые крупные спирохеты, относящиеся к другому роду, но
тоже участвующие в движении, и какие-то длинные палочковидные бактерии, при-
крепляющиеся к ней одним концом. В общем, поверхность миксотрихи подобна гус-
тому саду, где процветает никак не меньше четырех совершенно различных разно-
видностей бактерий-симбионтов. А ведь у нее есть еще и внутренние симбионты,
помогающие переваривать целлюлозу. Так что клетка миксотрихи — это целое мно-
говидовое сообщество.
Термиты, как и другие насекомые, могут очень хорошо сохраняться в ископае-
мом янтаре (если повезет, конечно).
В янтаре возрастом 20 миллионов лет палеонтологи нашли термита того самого
рода, в котором живет миксотриха. И в кишечнике этого термита обнаружены ос-
татки симбиотических одноклеточных, уже тогда связанных со спирохетами. Самое
крупное из этих одноклеточных, по всей вероятности, и есть миксотриха.
Для экскават характерны дисковидные кристы митохондрий. Впрочем, многие па-
разитические экскаваты (и некоторые свободноживущие тоже) митохондрий вообще
лишены. Например, ни у лямблии, ни у гипермастигин, ни у миксотрихи митохонд-
рий нет. Но это не признак примитивности, а результат вторичной утраты. Поте-
ря митохондрий — обычное дело у паразитов, живущих в чьем-нибудь кишечнике,
или, скажем, у обитателей бескислородного ила. Там, где нет кислорода, мито-
хондрии бесполезны, и они исчезают. Но в таких клетках обычно есть их остат-
ки, сохранившие некоторые метаболические функции, или хотя бы митохондриаль-
ные гены, успевшие мигрировать в ядро.
В прошлом митохондрии были абсолютно у всех изученных эукариот. И это, ко-
нечно, очень важно для понимания их происхождения. "Эмпирический факт заклю-
чается в том, что не существует эукариота, возникшего без предшествующего эн-
досимбиоза", — писал в 2002 году крупнейший микробиолог Георгий Александрович
Заварзин. И добавлял: "Вероятно, проэукариот, так до сих пор и не найденный,
уже был химерой". Эти выводы совершенно верны, и у них есть эволюционные
следствия, о которых мы еще поговорим.
Rhizaria
Типичные представители этой супергруппы — одноклеточные существа с трубча-
тыми кристами митохондрий и длинными тонкими ложноножками, которые часто бы-
вают нитевидными или шиловидными (см. рис. 15.3а).
Самые знаменитые группы ризарий — фораминиферы и радиолярии. Это морские
амебоподобные существа со сложными минеральными скелетами. Большинство фора-
минифер живет на морском дне, и только немногие из них парят в водной толще.
Их раковины чаще всего многокамерные, иногда трубчатые, а иногда спиральные
(хотя есть и другие формы). "Фораминиферы" буквально значит "дырконесущие".
Называются они так потому, что в раковине фораминиферы есть множество малень-
ких отверстий, сквозь которые высовываются тонкие ложноножки. Эти ложноножки
ветвятся, переплетаются, сливаются и в результате образуют вокруг фораминифе-
ры сложную сеть, которая благодаря разным способам сокращения ложноножек мо-
жет служить и для питания, и для передвижения.
Особенно сложными формами отличались, например, глоботрунканиды — вымершие
фораминиферы, которые жили в конце мезозойской эры. Их раковина не только бы-
ла многокамерной, но еще имела разнообразные выросты и кили, помогавшие па-
рить в толще воды. Конец мезозоя был эпохой теплого безледникового климата,
когда огромные площади занимали мелководные окраинные моря (примерно как ны-
нешние Балтийское и Желтое). Резкая зональность отсутствовала, работающих
всепланетными "холодильниками" полярных шапок не было и в помине, поэтому
глобальное перемешивание воды было гораздо слабее, чем в современности: массы
холодных вод, погружающиеся в глубину и приводящие в движение всю океанскую
толщу, тогда просто нигде не возникали. В мезозойском спокойном море взвешен-
ные в воде фораминиферы могли спокойно делить между собой экологические ниши,
специализируясь по узким интервалам глубин. К сожалению, глоботрунканиды вы-
мерли вместе с динозаврами.
Рис. 15.3а. Некоторые представители супергрупп Rhizaria.
У радиолярий богатство форм вообще поразительно. В первую очередь это каса-
ется их твердых скелетов. Тут можно увидеть вставленные друг1 в друга дырчатые
шары, эллипсоиды, диски, конусы, треножники, короны, многолопастные и шлемо-
видные фигуры, и все это бывает оснащено длинными иглами, палочками, мечевид-
ными выростами, решетками, перемычками или кольцами. Великолепными формами
радиолярий восхищался еще Эрнст Геккель, чьей узкой научной специальностью
они как раз и были (а поскольку он к тому же профессионально рисовал, то мно-
гими изображениями радиолярий в его исполнении можно и сейчас полюбоваться).
Все радиолярии живут в толще морской воды, и сложные формы с выростами нужны
им для того, чтобы лучше там парить. Как и у фораминифер, у радиолярий есть
сеть переплетающихся ложноножек, которыми они ловят пищевые частицы. Но, кро-
ме того, у них есть особые длинные неветвящиеся ложноножки, которые называют-
ся аксоподиями. Внутри каждой аксоподии проходит укрепляющий ее пучок микро-
трубочек, еще и связанных между собой (в обычных ложноножках ничего подобного
не бывает). Аксоподии нужны радиоляриям в основном, опять же, для лучшего па-
рения.
Многообразие ризарий не ограничивается фораминиферами и радиоляриями. Среди
них есть типичные жгутиконосцы (в том числе хищные), есть амебы, а есть и па-
разиты. Например, именно к ризариям относится плазмодиофора, возбудитель так
называемой килы капусты. Ее питающаяся стадия — это живущие в клетках капусты
многоядерные амебы. Такую жизненную форму принято называть плазмодием.
Интересная группа ризарий — хлорарахниофитовые водоросли. В эту группу вхо-
дят самые настоящие фотосинтезирующие амебы (с характерными для ризарий длин-
ными тонкими ложноножками), хотя у некоторых из них есть и жгутики. Разумеет-
ся, хлорарахниофиты имеют хлоропласты, которые в данном случае окружены че-
тырьмя мембранами и представляют собой захваченные одноклеточные зеленые во-
доросли — эукариоты из супергруппы Plantae. Скоро мы увидим, что ситуация,
когда хлоропласты образуются из захваченных эукариот (а не прокариот), встре-
чается в эволюции на самом-то деле очень часто.
"Chromalveolata"
Название этой супергруппы, единственной из шести, в заголовке поставлено в
кавычки. Вызвано это тем, что современная систематика признает ее выделенной
некорректно — вскоре мы узнаем почему. Тем не менее, члены этой группы дейст-
вительно близки друг к другу, и мы можем для удобства рассмотреть их вместе,
а поправки (которых требуют новые данные) внести на следующем этапе: так бу-
дет понятнее.
Рис. 15.36. Alveolata (одна из четырех подгрупп Chromalveolata).
Вскоре мы узнаем, что на самом деле их сейчас принято объединять
в супергруппу Harosa, она же SAR (Stramenopiles + Alveolata +
Rhizaria).
Мы уже не раз говорили о том, что все эукариоты, умеющие фотосинтезировать,
получили эту способность благодаря симбиозу. Точнее — эндосимбиозу, один из
участников которого живет внутри другого. Эукариотная клетка поглотила прока-
риотную и "поработила" ее, оставив жить внутри себя в качестве хлоропласта.
Именно поэтому хлоропласты до сих пор размножаются делением.
Но возможна и другая ситуация. Эукариотная клетка может проглотить,
"поработить" и превратить в хлоропласт другого эукариота, внутри которого уже
есть собственный прокариотный симбионт. Это называется вторичным эндосимбио-
зом. Два таких примера мы уже видели: эвглена и амебы-хлорарахниофиты, хлоро-
пласты которых — это бывшие зеленые водоросли. Но у хромальвеолят это не ред-
кость , а общее явление, определившее эволюцию всей супергруппы. Их хлоропла-
сты — вторичные (см. рис. 15.4). Среди хромальвеолят очень много фотосинтези-
рующих форм, и все их хлоропласты произошли от захваченных эукариот, а именно
от красных водорослей. Окрашены они обычно не в зеленые тона, а в желтовато-
бурые . Иногда проглоченный эукариотный симбионт даже сохраняет маленькое яд-
ро, которое в этом случае называется нуклеоморфом. Чаще ядро симбионта исче-
зает, но сохраняются другие следы вторичного эндосимбиоза в виде дополнитель-
ных мембран.
Рис. 15.4. Два независимых случая вторичного эндосимбиоза. Сле-
ва — Guillardia, представитель криптомонад. Справа —
Bigelowiella, представитель группы хлорарахниофит, относящейся к
супергруппе Rhizaria. Эукариотные симбионты, ставшие хлоропла-
стами, здесь разные: у криптомонад — красная водоросль, а у хло-
рарахниофит — зеленая.
Более того, у одноклеточных хромальвеолят иногда встречается третичный эн-
досимбиоз: захват эукариотного симбионта, внутри которого на правах хлоропла-
ста живет еще один эукариотный симбионт, и только уже внутри того — прокари-
отный симбионт, который, собственно, и фотосинтезирует. Если один представи-
тель хромальвеолят захватит другого в качестве хлоропласта, именно так и по-
лучится. В случае третичного эндосимбиоза одна клетка объединяет в себе четы-
ре организма: двух разных хромальвеолят, красную водоросль и цианобактерию,
ставшую первичным хлоропластом. И это не считая митохондрий.
В группу хромальвеолят (в ее традиционном объеме) входит не меньше семи
больших групп преимущественно водных фотосинтезирующих эукариот, то есть во-
дорослей. Это криптомонады, гаптофиты, динофлагелляты, золотистые, желто-
зеленые, бурые и диатомовые водоросли. Из этих семи групп четыре (криптомона-
ды, гаптофиты, динофлагелляты и диатомовые) состоят в основном из одноклеточ-
ных форм. Колонии они образуют нечасто, а если образуют, то относительно про-
стые . Зато некоторые из этих одноклеточных водорослей живут огромными массами
в освещенной зоне океана и играют там примерно ту же роль, что зеленые расте-
ния в наземных экосистемах: изо всех сил фотосинтезируют, создавая так назы-
ваемую первичную продукцию. В первую очередь это относится к динофлагеллятам,
диатомовым водорослям и гаптофитам. Последние знамениты тем, что микроскопи-
ческие известковые пластинки, которыми покрыты снаружи их клетки, образуют
основу писчего мела. Воспетые многими поэтами Дуврские белые скалы, что в
графстве Кент, сложены в основном из остатков панцирей мельчайших планктонных
водорослей-гаптофит, которые когда-то жили в теплых мелководных морях мезо-
зойской эры.
С золотистыми и желто-зелеными водорослями дело обстоит иначе. В этих груп-
пах есть обычные жгутиконосцы, есть амебоидные существа, есть гигантские мно-
гоядерные клетки, есть колонии, а есть и многоклеточные организмы, нитчатые
или пластинчатые. Золотистые и желто-зеленые водоросли — это две совершенно
отдельные эволюционные линии. Их предками были одноклеточные жгутиконосцы,
хоть и родственные, но все-таки разные. Все этапы усложнения (во многом одни
и те же!) эти две линии водорослей прошли независимо друг от друга, хотя и на
основе набора общих генов, связанного с близким родством. Это прекрасный при-
мер того, что в эволюционной биологии называется параллелизмом.
Наконец, группа бурых водорослей целиком состоит из многоклеточных организ-
мов. Почти все они живут в морях. И некоторые из них более-менее сравнимы по
сложности с хорошо нам знакомыми наземными зелеными растениями — например,
ламинария, она же морская капуста. В фантастическом романе Фрэнка Герберта и
Билла Рэнсома "Ящик Пандоры" подводные леса из бурых водорослей (их там назы-
вают ллэлектрокелпом") даже обзавелись чем-то вроде собственного коллективного
разума.
Кроме всевозможных водорослей среди хромальвеолят есть и много нефотосинте-
зирующих существ. Есть бесцветные гетеротрофные жгутиконосцы, поедающие бак-
терий. Есть оомицеты — потомки каких-то морских водорослей, потерявшие хлоро-
пласты и вернувшиеся к гетеротрофности. Они стали осмотрофами
("всасывалыциками") и приобрели в связи с этим жизненную форму, в точности
повторяющую жизненную форму типичных грибов: тело, состоящее из ветвящихся
тончайших нитей толщиной в одну клетку. Долгое время их и принимали за грибы.
Но к концу XX века выяснилось, что у оомицетов есть признаки, которых у на-
стоящих грибов быть просто не может: клеточная стенка из целлюлозы (а не из
хитина), трубчатые кристы митохондрий и подвижная одноклеточная расселитель-
ная стадия с двумя жгутиками. Таким образом, сходство оомицетов с грибами —
это чистой воды конвергенция, независимое приобретение общих черт, не связан-
ное с родством. В отличие от большинства грибов, оомицеты сохраняют жгутико-
вые стадии и (как правило) связь с водой. Типичный представитель оомицетов —
сапролегния, белесая водная плесень.
Случалось и такое, что оомицеты вмешивались в человеческую историю. Великий
голод в Ирландии в 1845-1849 годах произошел из-за фитофторы — оомицета, вы-
зывающего картофельную гниль. Ирландские крестьяне тогда были в массе своей
бедны, и картошка составляла значительную долю их рациона. Поэтому, когда
практически все посевы картофеля в стране оказались поражены эпидемией фитоф-
торы, это стало настоящей катастрофой. Достаточно сказать, что население Ир-
ландии после нее сократилось примерно на треть в результате как гибели от го-
лода , так и эмиграции в Америку людей, пытавшихся от него спастись.
Очень интересная группа хромальвеолят — опалины. Это одноклеточные, но мно-
гоядерные существа, живущие в конечном отделе кишечника амфибий (реже рыб и
рептилий). Особого вреда своим хозяевам они, судя по всему, не наносят, так
что их можно считать не паразитами, а просто сожителями. Опалины — обычнейшие
обитатели клоаки лягушек (клоакой у животных называют расширенную часть зад-
ней кишки, в которую впадают выделительные и половые протоки). Клетка опалины
может нести несколько тысяч жгутиков, которые в этом случае уже можно назы-
вать ресничками. Они расположены рядами и бьются согласованными волнами, да-
вая возможность опалине довольно быстро плавать. Опалины, без сомнения, про-
изошли от обычных одноядерных жгутиконосцев, но по ходу эволюции их клетка
постепенно увеличивалась и усложнялась. Мелкие (и, по-видимому, примитивные)
опалины имеют всего два клеточных ядра, а у крупных в клетках бывает до 200
ядер и больше. Причем все эти ядра одинаковы — по крайней мере, с виду.
Именно к хромальвеолятам, как выяснилось, относятся и инфузории — однокле-
точные существа, известные натуралистам еще с XVII века. Первыми исследовате-
лями инфузорий были создатели самых ранних микроскопов — великий натуралист-
любитель Антони ван Левенгук и не менее великий физик Христиан Гюйгенс. Инфу-
зорий иначе называют ресничными, потому что их тело, как правило, покрыто не-
сколькими тысячами типичных ресничек. Кроме того, для инфузорий характерна
очень сложная клетка с постоянным клеточным ртом и системой внутренних опор-
ных структур. Внешняя форма клетки у них тоже постоянная, часто выразительная
и красивая, о чем свидетельствуют названия: инфузория-туфелька, инфузория-
трубач . Неудивительно, что биологи довольно долго принимали инфузорий за жи-
вотных. Тем не менее, на самом деле инфузории — самые настоящие одноклеточ-
ные , причем даже не многоядерные. Чаще всего в клетке инфузории два ядра: ма-
ленький микронуклеус (генеративное ядро, содержащее полный геном и служащее
для продолжения рода) и большой макронуклеус (соматическое ядро, содержащее
только ту часть генома, что нужна для повседневной жизни, но зато в огромном
числе копий). Ядерный дуализм, то есть разделение функций между микронукле-
усом и макронуклеусом, — это, пожалуй, самая главная особенность инфузорий
как группы.
Сложность устройства инфузорий настолько впечатляет, что некоторые биологи
всерьез считали их предками многоклеточных животных. Но это, разумеется, не
так. Инфузории тем и интересны, что они нашли свой собственный путь эволюции,
альтернативный по отношению к любой многоклеточности.
Многие хромальвеоляты стали паразитами. В первую очередь это группа
Apicomplexa (название, буквально означающее что-то вроде "существа со сложным
передним концом"). Апикомплексы произошли от жгутиконосцев, и сейчас извест-
но , от каких именно. Но в процессе глубокого приспособления к паразитизму они
изменились очень сильно — во многих случаях до неузнаваемости. Именно апиком-
плексы составляли большую часть сборной группы паразитических эукариот, кото-
рых со времен Карла фон Зибольда называли споровиками. К апикомплексам отно-
сятся, например, возбудители малярии и токсоплазмоза.
Интересно, что апикомплексы совершенно точно произошли от фотосинтезирующих
предков. У этих специализированных паразитов есть апикопласт — остаток вто-
ричного хлоропласта, ставший бесцветным, но сохранивший четыре мембраны и
собственную ДНК. Сохранился он потому, что помимо фотосинтеза хлоропласты у
предков апикомплексов занимались синтезом жирных кислот и некоторых других
полезных веществ. После отказа от фотосинтеза эти метаболические функции ни-
куда не делись, а вместе с ними уцелели и остатки хлоропластов, дающие прямое
свидетельство сложного эволюционного пути своих хозяев. В глубокой древности
предок апикомплексов был обычным хищным жгутиконосцем. Однажды он проглотил
водоросль (скорее всего, красную) и сделал ее своим хлоропластом, а тем самым
стал водорослью и сам. Затем произошел отказ от фотосинтеза. Обесцветившийся
жгутиконосец вернулся к гетеротрофному питанию и стал, в конце концов, спе-
циализированным паразитом. Однако в соответствии с законом необратимости эво-
люции в его организме сохранились следы пройденного эволюционного пути — в
данном случае не только генетические, но и структурные (апикопласт виден в
микроскоп). Когда в 1898 году англичанин Рональд Росс и итальянец Джованни
Батиста Грасси раскрыли жизненный цикл возбудителя малярии, никто из них, ко-
нечно , не мог и представить, что этот опаснейший микроб окажется потомком
безобидной одноклеточной водоросли.
В целом среди хромальвеолят можно выделить две очень крупные эволюционные
ветви. Одна из них нам уже знакома — она называется альвеолятами (Alveolata).
К этой ветви относятся инфузории, апикомплексы, динофлагелляты. Главная осо-
бенность альвеолят — их уже упоминавшаяся оболочка, которая называется пелли-
кулой. Под внешней мембраной у них находится слой плотно сомкнутых маленьких
мембранных пузырьков (так называемых альвеол), которые, в свою очередь, подо-
стланы микротрубочками. Это и есть пелликула. Она придает клетке устойчивую
форму, а заодно позволяет устанавливать на базе имеющейся оболочки всевозмож-
ные дополнения — вроде системы белковых корешков, синхронизирующих у инфузо-
рий биение ресничек, или сложного аппарата проникновения в клетку хозяина,
который расположен на конце клетки апикомплексов.
Другая крупнейшая ветвь хромальвеолят называется Stramenopiles. Группу эту
выделил в 1989 году протистолог Дэвид Паттерсон. Подвижные клетки страменопи-
лов обычно имеют два жгутика: двигательный, направленный вперед и опушенный,
и рулевой, отогнутый назад и гладкий (либо опушенный заметно слабее). Причем
волоски, которыми опушен двигательный жгутик, у них имеют особую трубчатую
структуру. Такое состояние называется гетероконтным (разножгутиковым). Назва-
ние страменопилов отражает именно этот признак. Если буквально истолковать
вошедшие в это название латинские корни, получится что-то вроде "имеющие со-
лому в волосах": с соломинками тут сравниваются сидящие на двигательном жгу-
тике волоски. К страменопилам относятся золотистые, желто-зеленые, бурые и
диатомовые водоросли, а также оомицеты и опалины. У опалин разножгутиковость
исчезает, но они произошли от жгутиконосцев, у которых она точно была.
Есть и такие хромальвеоляты, которые не относятся ни к альвеолятам, ни к
страменопилам. Из тех, о ком мы тут говорили, это две группы одноклеточных:
криптомонады и гаптофиты. Возможно, это очень древние эволюционные линии.
К большому сожалению, современные генетические данные (причем не такие уж и
новые) показывают, что хромальвеоляты, скорее всего, не являются группой, со-
ответствующей требованиям филогенетической систематики. Таксон
Chromalveolata, конечно, состоит из родственных друг другу форм, но они объе-
динены в него, по нынешним меркам, слишком произвольно. В одной своей части
эта группа получилась сборной, а в другой, наоборот, неполной, с ллфигурным
вырезом". Поэтому таксон Chromalveolata на данный момент надо считать уста-
ревшим. О том, как обстоят дела в реальности, мы поговорим чуть ниже.
Plantae
Эта супергруппа состоит в основном из фотосинтезирующих организмов. Она
единственная, где нет ни одной крупной эволюционной ветви, освоившей гетеро-
трофность. Поэтому название Plantae — растения — ей вполне подходит (см. рис.
15.5.).
fy'/A
4
9 -■
Цианобактерии
Первичный
эндосимбиоз
Первые фотосинтезирующие
эукариоты
Л~
Зелёные водоросли,
высшие растения
Красные водоросли
Вторичные
эндосимбиозы
■ &')
Глаукофиты
40
Heterokonta = Stramenooiles
Рис. 15.5. Некоторые представители супергруппы Plantae.
У супергруппы растений есть несколько общих особенностей. Во-первых, это
пластинчатые кристы митохондрий (у ризарий и у большинства хромальвеолят они
трубчатые). Во-вторых, подвижные клетки у растений чаще всего имеют два оди-
наковых гладких жгутика, направленных вперед. Такое состояние называется изо-
контным (равножгутиковым). Отличный пример равножгутиковости — хламидомонада,
одноклеточная зеленая водоросль, описанная в школьных учебниках ботаники.
Есть и третья очень важная особенность. Хлоропласты растений — первичные,
то есть возникшие путем первичного эндосимбиоза. Они произошли непосредствен-
но от захваченных прокариот, а именно от цианобактерии. Этим растения резко
отличаются, например, от бурых или золотистых водорослей, хлоропласты которых
— вторичные, происходящие от эукариотных симбионтов, то есть как раз от за-
хваченных представителей Plantae.
В состав супергруппы растений входит всего три эволюционные ветви. Во-
первых, это одна сильно обособленная (и крайне примитивная по некоторым при-
знакам) группа водорослей, которая называется глаукофитами. Они в основном
одноклеточные, а если образуют колонии, то очень простые. Во-вторых, это
красные водоросли. И в-третьих, это огромная и разнообразная эволюционная
ветвь, для которой Томас Кавалье-Смит предложил латинское название
Viridiplantae (зеленые растения). В нее входят как наземные зеленые растения,
так и их многочисленные родственники-водоросли, крайне разнообразные по уров-
ню организации.
Именно к ветви Viridiplantae принадлежат и самые мелкие, и самые крупные
современные эукариоты — по крайней мере, если не учитывать внутриклеточных
паразитов (которые бывают очень маленькими) и разрастающиеся грибницы (кото-
рые, наоборот, бывают огромными, но все же не являются компактными, истинно
многоклеточными телами). Самым мелким свободноживущим эукариотом считается
океанская планктонная зеленая водоросль остреококкус: размер ее клетки меньше
микрона. А крупнейшие истинно многоклеточные организмы на Земле — это стомет-
ровые секвойи и секвойядендроны, самые громадные экземпляры которых получают
собственные имена: "Генерал Шерман", ЛЛГиперион" и т.п.
Первые зеленые растения, несомненно, были одноклеточными жгутиконосцами. Но
в ходе своей долгой и бурной эволюции они успели дать огромный спектр жизнен-
ных форм. Среди них есть одноклеточные без всяких жгутиков (например, хлорел-
ла) , есть шарообразные колонии из нескольких сотен или тысяч жгутиковых кле-
ток (например, вольвокс), есть гигантские многоядерные клетки, есть нитчатые
и пластинчатые примитивные многоклеточные, а есть и обладатели довольно слож-
ных ветвящихся тел, как у харовых водорослей. Зеленые водоросли (в широком
смысле этого слова) живут в любых водоемах от луж до Мирового океана, а ино-
гда даже и на суше. От кого же из них произошли наземные растения?
Прообразом наземных растений могут считаться или пластинчатые колеохетовые
водоросли, или нитчатые водоросли-сцеплянки. Именно к этим двум группам на-
земные растения ближе всего, судя по генетическим данным. Водоросль, которая
была предком наземных растений, жила на мелководье и представляла собой либо
однослойную пластинку, либо переплетение нитей. Большой разницы тут нет, по-
тому что эти жизненные формы могут легко переходить друг в друга. Водоросли
жили в переходной зоне между водоемом и берегом, которая была в те древние
времена чрезвычайно протяженной: тут надо учитывать, что до появления сплош-
ного растительного покрова, удерживающего почву, реки на Земле не имели по-
стоянных берегов, и мелководьем была, по сути, почти вся суша. Есть вполне
правдоподобная гипотеза, что "изобретением", запустившим эволюцию наземных
растений, был стволик с коробочкой наверху, поначалу предназначенный исключи-
тельно для разбрасывания воздушных спор, чтобы их разносил ветер. Нет сомне-
ний, что этот способ расселения очень эффективен (на современной Земле он по-
зволяет растениям осваивать расположенные за тысячи километров от ближайшей
суши вулканические острова). Именно для того, чтобы его обеспечить, могли бы
появиться первые механические и проводящие ткани, совершенно ненужные пла-
стинчатой или нитчатой водоросли в воде, но необходимые воздушному стволику.
А уж потом наличие этих тканей позволило растениям полностью выйти на сушу и
создать наземные жизненные формы. И в результате мы теперь живем в мире, где
целые природные зоны определяются наземными растениями: от степных злаков до
самых многочисленных деревьев на современной Земле — лиственниц, образующих
сибирскую и североамериканскую светлохвойную тайгу.
Главным признаком, отличающим высшие растения от водорослей, традиционно
считается архегоний — колбообразный мешочек, в котором созревает яйцеклетка.
Это настоящий многоклеточный орган размножения. Причем зародыш организма сле-
дующего поколения, уже в свою очередь многоклеточный, какое-то время развива-
ется прямо внутри архегония. Именно с этим связано латинское название высших
растений — Embryophyta. К этой группе относятся мхи, плауны, хвощи, псилоты,
папоротники, голосеменные (хвойные, саговники, гинкго) и главные звезды со-
временного растительного мира Земли — цветковые. Правда, у голосеменных от
архегониев остаются буквально считанные клетки, а у цветковых и вовсе ничего,
но тут важно то, что они были у их предков.
Amoebozoa
Супергруппа Amoebozoa — это царство амеб (см. рис. 15.6). Она состоит из
существ, которые передвигаются амебоидным способом, выпуская и втягивая лож-
ноножки и постоянно меняя форму клетки. Очень характерна форма самих ложноно-
жек: они не длинные и тонкие, как у ризарий, а широкие, лопастевидные. У не-
которых амебозоев одновременно с ложноножками есть и жгутики. Кристы митохон-
дрий у них обычно трубчатые, хлоропластов не бывает. Все амебозои — гетеро-
трофы. Некоторые из них потеряли митохондрии, потому что живут или в бески-
слородном иле, или в чьем-нибудь кишечнике (как дизентерийная амеба, напри-
мер) .
Рис. 15.6. Некоторые представители супергруппы Amoebozoa.
Обыкновенная амеба, она же амеба-протей, с которой по традиции начинается
школьный учебник зоологии, — довольно типичный представитель амебозоев. Она
относится к голым амебам, у которых нет никакого покрова сверх плазматической
мембраны. Но бывают и раковинные амебы, причем с очень разнообразной формой и
структурой раковинок. И голые, и раковинные амебы могут жить не только в во-
доемах , но и в почве: находящейся там капиллярной воды им вполне хватает,
чтобы оставаться водными существами.
В то же время не будем забывать, что "амеба" (точно так же, как и
"жгутиконосец"), — это название не эволюционной ветви, а жизненной формы.
Увидев под микроскопом незнакомую амебу, мы, строго говоря, еще ничего не
знаем о ее положении в системе. Вполне нормальные амебы, пришедшие к этой
жизненную форме совершенно самостоятельно, есть и среди ризарий, и среди экс-
кават. Но амебозои — единственная супергруппа, где этот способ жизни безус-
ловно преобладает.
Существуют амебы, которые в определенный момент своего жизненного цикла
приобретают многоклеточность (во всяком случае, примитивную). Большую часть
времени они могут ползать поодиночке где-нибудь в почве. Но когда приходит
время, эти амебы сползаются вместе и образуют многоклеточное плодовое тело,
очень похожее на плодовое тело гриба. В другом случае странствующие амебы
сначала образуют плазмодий — гигантскую многоядерную амебу, которая ползает и
питается, пока не настанет пора создавать плодовое тело. Потом плодовое тело
выбрасывает споры, и все повторяется. Организмы с такими жизненными циклами
называются слизевиками.
Жизненная форма слизевика распространена на удивление широко. Например,
сейчас мы знаем, что свои собственные слизевики с настоящими плодовыми тела-
ми, совершенно независимо освоившие этот образ жизни, есть в супергруппе экс-
кават. Они называются акразиевыми слизевиками, или акразиомицетами. Это еще
один яркий пример эволюционной конвергенции. Более того, организмы, близкие
по типу жизненного цикла к слизевикам, есть и в группе ризарий (плазмодиофо-
ромицеты), и в группе страменопилов (лабиринтуломицеты). Что же касается
"настоящих" слизевиков, которые входят в супергруппу амебозоев, то они назы-
ваются миксомицетами.
Многие биологи рассматривают слизевиков как прекрасную модель начальных
стадий эволюции многоклеточности. Есть даже осторожно высказываемая гипотеза,
что примерно такой же жизненный цикл, как у типичных слизевиков, имели предки
многоклеточных животных. Главная трудность этой гипотезы в том, что многокле-
точные животные относятся к другой супергруппе эукариот — к последней, о ко-
торой нам осталось поговорить.
Opisthokonta
В эту супергруппу входят гетеротрофные существа с пластинчатыми кристами
митохондрий и жгутиковыми клетками, имеющими единственный жгутик, направлен-
ный назад. Мы можем легко догадаться, что такое состояние называется заднеж-
гутиковым, или опистоконтным.
Самое удивительное, что, кроме перечисленных признаков, у разных опистокон-
тов нет между собой почти ничего общего. По крайней мере, так кажется на пер-
вый взгляд. Сильно ли похожи друг на друга воротничковый жгутиконосец, белый
гриб и синий кит? А ведь по меркам мегасистемы эукариот они близкие родствен-
ники.
Стереоцнлия
Кутнкулярна
я пластинка
Norvo
cafyx
Поддержнваюшая
клетка
Афферент.
нерв
Мяелнн. i
оболочка
Кнноцнлня
Волосковые I
клетки:
Эфферент.
нерв
стервоцилии кимоцилия
жгутж (микроворсинки) (жгутик)
V.
микроворсинки |
/
■:.ir
/.\>
V
микроворсинки
(щеточная каемка)
еоротничковая волосковая клетка клетка кишечника
клетка губки позвоночных позвоночных
(хоаноцит) (энтероцит)
Рис. 15.7. Волосковая клетка внутреннего уха (А) и сравнение ворот-
ничковой клетки рубок с клетками позвоночных (Б) . Воротничковая
клетка губки по ультраструктуре очень похожа на клетки воротничковых
жгутиконосцев, тут без разницы, кого из них изображать.
На самом деле кое-что общее найти все же можно. Например, во внутреннем ухе
позвоночных животных (в том числе и в человеческом) есть чувствительные эле-
менты, которые называются волосковыми клетками (см. рис. 15.7). Вершина такой
клетки несет один жгутик (его функция тут опорная) и группу собранных вместе
микроворсинок. Если внешняя сила — например, течение жидкости в полости внут-
реннего уха — сгибает или наклоняет эти микроворсинки, то клетка генерирует
электрический сигнал. Это основной принцип работы внутреннего уха. Так вот,
давно замечено, что волосковая клетка внутреннего уха напоминает по структуре
клетку воротничкового жгутиконосца. Компоненты там те же самые: жгутик и
группа микроворсинок на вершине клетки, они только смещены относительно друг
друга. И это отнюдь не поверхностное сходство. Недавно исследователи показа-
ли, что в волосковых клетках внутреннего уха позвоночных и во внутренних
клетках губок, которые называются хоаноцитами, экспрессируется целая группа
общих генов. В основном эти гены кодируют белки, связанные с цитоскелетом и
участвующие в образовании микроворсинок. А хоаноциты губок — это клетки, в
точности похожие на воротничковых жгутиконосцев, уж тут преемственность оче-
видна. Порядок ветвлений эволюционного древа животных вполне позволяет допус-
тить, что эта преемственность "дотянулась" и до позвоночных.
Про сперматозоиды животных мы уже упоминали. Сперматозоид — обладатель ти-
пичного жгутика (правда, в некоторых группах животных встречаются безжгутико-
вые сперматозоиды, но это явно производное состояние). И плывет он жгутиком
назад. Это и есть признак, характерный для опистоконтов, но не встречающийся
у других эукариот. В этом отношении животные более архаичны, чем грибы, у
большинства из которых все жгутиковые стадии исчезли.
Итак, опистоконты — это грибы, животные и воротничковые (см. рис. 15.8). Но
не только они. Можно назвать, по меньшей мере, четыре группы "странных опи-
стоконтов", не относящихся ни к многоклеточным животным, ни к воротничковым
жгутиконосцам, ни к типичным безжгутиковым грибам. Это хитридиевые грибы,
нуклеарииды, криптомицеты и мезомицетозои. Все эти группы относительно мало-
известны (во всяком случае, у широкой публики они не на слуху). Но это вовсе
не значит, что они неважны для понимания структуры древа жизни.
Хитридиевые грибы, или хитридиомицеты, — это преимущественно водные орга-
низмы. Тело примитивного хитридиевого гриба представляет собой фактически
амебу: одноядерную или многоядерную, но в любом случае с длинными, тонкими и
сильно ветвящимися ложноножками, которые глубоко проникают в питательный суб-
страт. От таких ложноножек один шаг до грибницы, и неудивительно, что этот
шаг был в ходе эволюции легко сделан: у высших хитридиевых налицо уже типич-
ная грибница (амебоидное тело в этом случае покрывается клеточной стенкой и
теряет подвижность). Тем не менее, у хитридиомицетов, в отличие от других
грибов, сохраняются плавающие жгутиковые споры — разумеется, со жгутиком, об-
ращенным назад.
Нуклеарииды — это пресноводные или почвенные амебы с длинными тонкими лож-
ноножками, эволюционно близкие к грибам. Легко представить, что от кого-то
вроде них грибы как раз и произошли. Хотя скоро мы увидим, что мнения на эту
тему есть разные.
Криптомицеты — одноклеточные организмы, безжгутиковые или с одним жгутиком,
лишенные клеточной стенки, близкие, опять же, к грибам и паразитирующие внут-
ри клеток других грибов или водорослей. Их считают примитивными родственника-
ми грибов, которым к тому же удалось необычайно глубоко специализироваться в
сторону паразитизма.
Но вот тут есть большая проблема. Кроме криптомицетов существует еще целых
две группы специализированных внутриклеточных паразитов, близких к примитив-
ным грибам: афелиды и микроспоридии. Афелиды паразитируют в клетках водорос-
лей, а микроспоридии — в клетках животных, включая и человека. Мы между делом
упоминали в главе 12, что микроспоридии умудрились стать не простыми парази-
тами, а энергетическими: они выкачивают из хозяйской клетки АТФ. Современные
молекулярно-биологические исследования приводят к выводу, что криптомицеты,
афелиды и микроспоридии составляют единую эволюционную ветвь, сестринскую по
отношению ко всем грибам. Для этой единой ветви предложено название
Opisthosporidia (опистоспоридии). И что характерно, все ее члены — внутрикле-
точные паразиты. Не значит ли это, что внутриклеточным паразитом был и общий
предок опистоспоридии с грибами? Не обязательно — но не исключено.
Рис. 15.8. Некоторые представители супергруппы Opisthokonta.
Мезомицетозои — группа амебообразных организмов, которые чаще всего парази-
тируют в каких-нибудь морских или пресноводных многоклеточных животных. Жгу-
тиков у них обычно нет, но есть ложноножки. Питаются они примерно как грибы:
осмотрофно, всасывая вещества сквозь клеточную мембрану. Известно, что на не-
которых стадиях жизненного цикла у мезомицетозоев бывает клеточная стенка —
по-видимому, хитиновая, как и у грибов. Но еще интереснее, что некоторые ме-
зомицетозои образуют в ходе своего размножения колонии покоящихся амеб, уди-
вительно похожие на ранние зародыши многоклеточных животных. Причем показано,
что деления клеточных ядер в такой колонии синхронизируются — тоже как у за-
родышей многоклеточных животных. Это означает, что клетки "зародыша" (или не
клетки, а только их ядра, если клетки на этом этапе слиты) делятся не вразно-
бой , но строго одновременно, так что их число остается степенью двойки: 4, 8,
16, 32.
Мезомицетозои — эволюционные "кузены" многоклеточных животных (в одной ста-
тье их прямо так и назвали). "Братья" многоклеточных животных, по генетиче-
ским данным, воротничковые жгутиконосцы: это самая близкая к ним группа. Но
при этом жизненный цикл воротничковых жгутиконосцев, даже колониальных, слиш-
ком прост, чтобы быть для многоклеточных животных предковым. Например, ника-
кого механизма синхронизации клеточных делений при образовании колонии у во-
ротничковых, судя по всему, нет. В этом отношении мезомицетозои более продви-
нуты. Кстати, у некоторых мезомицетозоев встречаются ультратонкие ложноножки,
из которых вполне могли бы получиться микроворсинки "воротничка".
В 2010 году у мезомицетозоев обнаружили свойственный многоклеточным живот-
ным ген, который называется Brachyury. У животных этот ген кодирует один ре-
гуляторный белок, активно участвующий в эмбриональном развитии. Например, у
позвоночных экспрессия гена Brachyury важна для нормального развития основы
осевого скелета — хорды — и прилегающих к ней зачатков (вдаваться в эмбриоло-
гические детали мы тут не будем). Возникает естественный вопрос: зачем этот
ген существу, у которого нет, не то что хорды, а вообще никаких органов? Дан-
ных на эту тему пока маловато, но кое-что понять уже можно. Известно, напри-
мер, что у одного из мезомицетозоев ген Brachyury активен в момент, когда его
многоядерный плазмодий распадается на множество одноядерных амеб. Очевидно,
он как-то регулирует происходящие во время развития сложные взаимные переме-
щения клеток — собственно, как и у животных. В любом случае мы знаем, что
этот ген очень эволюционно консервативен: его продукт, взятый у одноклеточной
амебы, может запустить развитие осевых структур в зародыше лягушки — правда,
не с таким успехом, как продукт гена самой лягушки. Но сам факт, что белковые
продукты столь специализированных генов амебы и лягушки оказались взаимозаме-
няемыми , уже поразителен.
Естественно, исследователи заинтересовались вопросом, у кого еще есть ген
Brachyury. Выяснилось, что за пределами супергруппы опистоконтов он (равно
как и родственные ему гены) не встречается нигде. Зато внутри опистоконтов
распространен очень широко. Ген Brachyury есть у всех многоклеточных живот-
ных, у самых разных мезомицетозоев, у криптомицетов, у хитридиевых грибов и
еще у одной относительно примитивной группы грибов, которая называется зиго-
мицетами. Унаследовать его они могли только от общего предка всех опистокон-
тов : конфигурация эволюционного древа просто не оставляет других возможно-
стей. А вот у воротничковых жгутиконосцев и у высших грибов гена Brachyury
нет. Причем из положения этих групп на древе однозначно следует, что у их
предков он когда-то был, но оказался утрачен за ненадобностью.
Итак, ген Brachyury имеется у мезомицетозоев и у многоклеточных животных,
но потерян у воротничковых жгутиконосцев (которые, напомним, на эволюционном
древе находятся к животным ближе всего). По-видимому, из этого неумолимо сле-
дует, что жизненный цикл современных воротничковых жгутиконосцев вторично уп-
рощен . У реального предка многоклеточных животных он наверняка был гораздо
сложнее. Увы, мы не знаем точно, каким был этот предок. Эволюционное древо
опистоконтов вообще полно провалов: по нему видно, что многие переходные фор-
мы давно вымерли и, скорее всего, не уцелели даже в палеонтологической лето-
писи. Что-то (но не все) сохранили от общих предков мезомицетозои, а что-то
(но не все) — воротничковые. То, что предок животных сочетал в себе некоторые
признаки обеих этих групп, можно утверждать наверняка. Кроме того, в послед-
нее время накапливаются молекулярные данные, позволяющие считать, что мезоми-
цетозои — это не эволюционная ветвь, а эволюционный уровень, объединяющий
всех примитивных родственников животных. Тогда прямые предки животных просто
неизбежно должны были через этот уровень так или иначе пройти.
Скорее всего, когда-то мезомицетозои (или их близкие родственники) были
свободноживущими и населяли море. Но затем их постигла обычная судьба архаи-
стов, переживших свое время: они были вытеснены из большинства местообитаний
и сумели уцелеть только ценой глубокой специализации, в данном случае — ухода
в паразитизм. Причем вполне вероятно, что вытеснили мезомицетозоев их же соб-
ственные потомки. "Эволюция уничтожает свои причины". Подобные примеры исто-
рия жизни на Земле знает. Правда, в данном случае мезомицетозои ЛЛв отместку"
научились на своих потомках паразитировать.
Теперь мы можем подвести некоторые итоги, и прежде всего — понять, как в
целом выглядит эволюционное древо супергруппы опистоконтов (см. рис. 15.9).
Эта супергруппа состоит из двух больших ветвей: Holomycota (ЛЛвсе грибы") и
Holozoa (ллвсе животные") . К ветви Holomycota относятся нуклеарииды, опистос-
поридии, хитридиомицеты и высшие грибы. К ветви Holozoa — мезомицетозои (в
широком смысле этого слова), воротничковые жгутиконосцы и многоклеточные жи-
вотные .
сумчатые грибы (аскомицеты) Dikarya
базидиееые грибы (базидиомицеты)
зигомицеты
хитридиомицеты
Fungi
микроспоридии
Opisthosporidia
<^] аф
криптомицеты
елиды
нуклеарииды
Holomycota
<з
<d
многоклеточные животные
воротничковые жгутиконосцы
мезомицетозои
Holozoa
Рис. 15.9. Эволюционное древо супергруппы Opisthokonta.
Высшие грибы — группа, эволюция которой связана с сушей. По глубине приспо-
собленности к наземному образу жизни с ними могут сравниться разве что цвет-
ковые растения (животные остаются далеко позади). У всех грибов, кроме хитри-
диевых, жизненный цикл полностью лишен жгутиковых стадий. У них не бывает ни-
каких подвижных половых клеток, а расселяются они с помощью спор, рассчитан-
ных на перенос по воздуху. Поэтому размножение грибов не зависит от наличия
жидкой воды. Правда, грибнице все равно нужна для роста высокая влажность, но
независимость размножения от воды, тем не менее, заметно расширяет их эколо-
гические возможности. Примерно 98% современных видов грибов относится к груп-
пе Dikarya, у которой половой процесс, как правило, сводится к обмену клеточ-
ными ядрами между вступившими в контакт нитями разных грибниц. В частности, к
этой группе относятся все шляпочные грибы, плодовые тела которых мы называем
"грибами" в обыденной жизни.
Раньше грибами считались еще и оомицеты, которые на самом деле, как мы те-
перь знаем, представляют собой бесхлоропластные разножгутиковые водоросли,
относящиеся к группе страменопилов. Среди прочих признаков грибы отличаются
от оомицетов способом синтеза лизина — аминокислоты, которая входит в состав
белков (см. главу 3). Формула лизина следующая: СН2 (NH2)-CH2-CH2-CH2-CH (NH2)-
СООН. У большинства эукариот, в том числе и у оомицетов, лизин синтезируется
через диаминопимелиновую кислоту (НООС-СН (NH2)-CH2-CH2-CH2-CH (NH2)-СООН) . Это
так называемый ДАЛ-путь. Но у грибов лизин синтезируется другим путем, через
альфа-аминоадипиновую кислоту (НООС-СН2-СН2-СН2-СН (NH2)-СООН) . Это так назы-
ваемый AAA-путь. Из этих двух путей ДАП-путь — намного более распространенный
и, судя по всему, более древний. Почему же грибы перешли на AAA-путь? Очевид-
но, их предки на каком-то эволюционном этапе вообще потеряли способность син-
тезировать лизин, а потом восстановили ее с нуля. И в соответствии с законом
необратимости эволюции восстановление получилось неточным. Но почему же пред-
ки грибов разучились синтезировать лизин? Не потому ли, что они были внутри-
клеточными паразитами и могли получать аминокислоты прямо из клеток хозяев?
Это вполне логичное объяснение, но верно ли оно, мы пока не знаем.
Многоклеточные животные — группа, которая по-латыни называется Metazoa. Это
единственная во всей живой природе Земли эволюционная ветвь, в которой есть
многоклеточные хищники. Обратим внимание, что практически все жизненные фор-
мы, о которых мы раньше говорили — колониальные и многоклеточные водоросли,
амебы, слизевики, грибоподобные осмотрофы, — возникали в разных группах эука-
риот по несколько раз. В эволюции это обычное дело. И только многоклеточные
хищники составляют тут исключение: у них аналогов нет.
Хищниками, однако, можно назвать далеко не всех многоклеточных животных.
Группа Metazoa включает пять эволюционных ветвей: губки, пластинчатые, греб-
невики, стрекающие и билатерии, они же двусторонне- симметричные (см. рис.
15.10). Губки — это сидячие неподвижные существа, которые проводят жизнь, бу-
дучи прикрепленными ко дну или подводным предметам, и питаются за счет фильт-
рации воды. А единственный представитель пластинчатых — трихоплакс, существо
хоть и подвижное, но размером с крупную амебу (порядка миллиметра) и устроен-
ное по меркам многоклеточных животных исключительно просто. Все, на что спо-
собны губки и трихоплакс, — это поглощать микроскопические пищевые частицы,
которые им попадаются. При таком способе питания многоклеточность хоть и по-
лезна, но не обязательна.
Совершенно иначе обстоит дело у животных, имеющих нервную систему, мышцы и
рот. Иногда их называют "настоящими многоклеточными животными". Они могут ак-
тивно искать и захватывать крупную добычу, нередко сравнимую по размеру с са-
мим хищником. Скорее всего, выход на эволюционный уровень "настоящих много-
клеточных животных" произошел дважды: у гребневиков и у общего предка стре-
кающих с билатериями. Есть серьезные указания на то, что нервная система была
независимо "изобретена" в этих двух эволюционных линиях. То же самое относит-
ся и к мускулатуре. У гребневиков она относительно слабая, эти животные пла-
вают в основном за счет биения ресничек. У стрекающих и билатерии тоже может
сохраняться ресничное движение (коловратки, плоские черви, многие морские ли-
чинки) , но обычно оно сочетается с гораздо более эффективным мышечным движе-
нием.
В группу стрекающих входят медузы и полипы (гидры, актинии, кораллы). Ста-
дии плавающей медузы и сидячего полипа часто чередуются у них в одном жизнен-
ном цикле. Почти все стрекающие — хищники. Признак, давший им название, —
стрекательные клетки, которые поражают добычу выбрасывающимися нитями. Сим-
метрия тела у стрекающих лучевая, с центральной осью и несколькими равноцен-
ными сторонами. Но есть гипотеза, что лучевая симметрия стрекающих — вторич-
ная, возникшая на основе двусторонней симметрии, которая была у их ползающих
предков. Если это верно, то первичная лучевая симметрия есть только у гребне-
виков .
t
{}
I
многоклеточность
4}
ервная система
»
воротничковые жгутиконосцы
(аоанофлагелляты)
гребневики
^М губки
'•itV-Л пластинчатые
стрекающие
деусторо н не-симметрич иые
(билатерии)
Рис. 15.10. Эволюционное древо многоклеточных животных (вместе с их
ближайшими современными родственниками — воротничковыми жгутиконос-
цами) . Надо предупредить, что тут есть, по меньшей мере, одно спор-
ное место: о том, какая ветвь животных самая древняя — гребневики
или губки — до сих пор идут дискуссии. Приведенное древо соответст-
вует гипотезе Леонида Мороза, известного нейробиолога, показавшего,
что нервная система гребневиков, похоже, возникла независимо от
нервных систем билатерии и стрекающих.
Подцарство
простейшие
(Одноклеточные)
Царство
Животные
Подцарство
Многоклеточные
Тип Саркодовые
Тип Жгутиконосцы
Тип Инфузории
Тип Кишечнополостные
Тип Плоские черви
Тип Круглые черви
Тип Кольчатые черви
Тип Моллюски
Тип Членистоногие
Класс Брюхоногие
Класс Двустворчатые
Класс Головоногие
И Класс Насекомые
Класс Ракообразные
Класс Паукообразные
и Класс Костные рыбы
И Класс Земноводные
Тип Хордовые
Класс Хрящевые рыбы
Класс Пресмыкающиеся
И Класс Птицы
И Класс Млекопитающие
Рис. 15.11. Представители многоклеточных животных.
И наконец, двусторонне-симметричные животные, или билатерии, — это необык-
новенно разнообразная эволюционная ветвь, богатая оригинальными планами
строения (см. рис. 15.11). В теле билатерии выделяются срединная плоскость,
спинная и брюшная сторона, передний и задний концы. Часто оно делится еще и
на сегменты. На переднем конце обычно концентрируются органы чувств и разрас-
таются нервные узлы. Чтобы обеспечивать развитие такого тела в каждом жизнен-
ном цикле, у билатерии возникло много новых регуляторных генов. А это, в свою
очередь, способствовало бурной эволюции новых форм. Некоторые билатерии стали
сидячими и (или) сменили двустороннюю симметрию на лучевую. Некоторые, как
садовая улитка, приобрели диссимметрию, охватившую значительную часть их те-
ла. Некоторые до предела уменьшились в размере, став невидимыми невооруженным
глазом, но сохранив почти все системы органов. Некоторые перевернулись, так
что спинная и брюшная стороны у них поменялись местами. Некоторые удлинились
до нескольких сотен сегментов, а другие, наоборот, вовсе отказались от сег-
ментации. Некоторые выработали сложный наружный скелет, а некоторые — еще бо-
лее сложный внутренний. Изучением этого невероятного богатства форм животных
занимается сравнительная анатомия — интереснейшая наука, но погружение в нее,
к сожалению, увело бы нас слишком далеко за пределы тем, охваченных этой кни-
гой.
Новые
горизонты
Знаменитый американский физик-теоретик Джон Арчибальд Уилер однажды сказал:
"Мы живем на острове, окруженном морем нашей неосведомленности. По мере того
как растет остров знания, растет и береговая линия соприкосновения с неведо-
мым" .
Конечно, Уилер был прав. Любой научный прорыв решает и закрывает те или
иные вопросы, которые раньше были актуальными (и могли мучить исследователей
столетиями). Но одновременно он тут же порождает новые вопросы и ставит новые
задачи, недоступные прежним поколениям ученых. Причем этих новых вопросов и
задач обычно больше, чем прежних. Так в принципе устроена технология позна-
ния. Хорошо это или плохо, но мы никогда не дождемся конца науки. Даже если
человечество просуществует несколько миллиардов лет и доживет до превращения
Солнца в красный гигант, то и тогда любой ученый, скорее всего, найдет для
себя не меньше интересных нерешенных вопросов, чем он может найти сейчас. Хо-
тя, разумеется, вопросы эти к тому времени будут другими — невообразимыми для
нас.
Достигнутое в начале XXI века установление истинного (в хорошем приближе-
нии) эволюционного древа эукариот вписывается в эту тенденцию. Те, кто его
установил, после этого и не думали почивать на лаврах. Они оценили достигну-
тое, увидели, какие новые вопросы оно порождает, и стали искать ответы на эти
вопросы, привлекая новые данные (в первую очередь генетические) и организуя
работу по принципу последовательных приближений. И эти исследования сейчас
вовсю продолжаются. Увы, любая попытка дать их полную сводку превратила бы
эту главу в реестр латинских названий, который никто не стал бы читать. В
этой области, условно говоря, еще не улеглась пыль. Поэтому сейчас мы ограни-
чимся в основном теми новыми фактами, которые установлены уже более-менее
бесспорно и у которых к тому же есть ясный биологический смысл. Ну, и добавим
пару поправок, без которых в наше время уж совсем не обойтись.
Первой такой поправкой стало переименование супергруппы растений. Во все-
мирно известной сводке, которую выпустил в 2005 году большой международный
коллектив протистологов, это название было сочтено неудачным по причине его
исторически сложившейся двусмысленности. Вместо него предложили название
Archaeplastida ("имеющие древние хлоропласты"). Название Plantae в этой вер-
сии системы было сохранено только за высшими наземными растениями, то есть за
теми, кого иначе называют Embryophyta.
Исчезла ли в результате этих переименований двусмысленность, вопрос спор-
ный. Но, так или иначе, название "архепластиды" сейчас очень распространено.
Смысл его прост: это единственная супергруппа эукариот, где хлоропласты про-
изошли прямо от цианобактерий, а не от каких-нибудь других эукариот со своими
собственными хлоропластами внутри.
Есть, впрочем, и такие авторы, которые как ни в чем не бывало продолжают
называть архепластид растениями. Право на это у них есть, потому что выбор
названий для групп такого ранга не регламентирован никакими строгими правила-
ми . В данном случае это больше вопрос моды, чем правил.
Добавим, что первичные хлоропласты архепластид — особенность чрезвычайно
характерная, но все же не уникальная. Раковинная амеба паулинелла, относящая-
ся к супергруппе ризарий, тоже приобрела первичный хлоропласт, возникший из
захваченной цианобактерий, причем сделала это эволюционно очень недавно (ско-
рее всего, порядка 100 миллионов лет назад). Предок архепластид, безусловно,
приобрел свои хлоропласты гораздо раньше.
В 2007 году обнаружилась куда более серьезная проблема. Генетические иссле-
дования показали, что группа Rhizaria примерно так же близка к группам
Alveolata и Stramenopiles, как и эти две группы друг к другу. Это не раз пе-
репроверяли, и результат получался тот же самый. Попросту говоря, ризарии
встраиваются внутрь хромальвеолят. Более подробные молекулярные исследования
вроде бы показывают, что ризарии генетически ближе к альвеолятам, чем к стра-
менопилам. Есть, однако, и результаты, противоречащие этому. В любом случае
тут мы имеем компактный ллкуст" из трех ветвей. Этот ллкуст" стали называть SAR
(Stramenopiles + Alveolata + Rhizaria).
SAR — единая эволюционная ветвь, к основанию которой примыкают рано обосо-
бившиеся , очень древние группы криптомонад и гаптофит. Положение этих групп
на эволюционном древе до сих пор неясно, но, во всяком случае, в саму супер-
группу SAR их предпочитают не включать. Следует добавить, что одно время
криптомонад и гаптофит считали единой эволюционной ветвью, которой дали на-
звание Hacrobia, но эта гипотеза так и не стала общепринятой.
Что касается самой группы SAR, то у нее есть и альтернативное название —
Harosa.
Главная проблема супергруппы SAR (= Harosa) состоит в том, что ей пока не-
возможно дать никакую осмысленную общую характеристику. Она выделена исключи-
тельно по молекулярно-генетическим данным, и в нее входят организмы, предель-
но различные по образу жизни и внешнему облику. Разве что кристы митохондрий
у них трубчатые, но этот признак никак не является определяющим (он есть, на-
пример, у амебозоев, которые не имеют с SAR ничего общего) . Как возникла
группа SAR? Каким был ее предок? В чем уникальность ее эволюционного пути?
Эти вопросы — настоящий вызов для биологов-эволюционистов.
Unikonta
и Bikonta
В 2003 году уже знакомый нам Томас Кавалье-Смит, взявшись в очередной раз
корректировать свои представления о филогенетическом древе с учетом новых
данных, пришел к выводу, что супергруппы опистоконтов и амебозоев связаны
тесным родством. По Кавалье-Смиту, амебозои и опистоконты образуют единую са-
мостоятельную эволюционную ветвь. Это подтверждается и сходством жизненных
форм (которое здесь налицо), и генетическими исследованиями. Например, только
у опистоконтов и амебозоев слиты вместе три гена, кодирующие ферменты синтеза
пиримидинов — очень важного для всех живых организмов класса молекул, в кото-
рый входят цитозин, урацил и тимин (см. главу 7). Этого слияния нет ни у дру-
гих эукариот, ни у бактерий, ни у архей. По всей вероятности, оно произошло
только один раз. Именно по таким уникальным признакам и следует выделять род-
ственные группы.
Эволюционная ветвь, состоящая из опистоконтов и амебозоев, называется
Unikonta (одножгутиковые) . Связано это с тем, что у них, натурально, один
жгутик. По крайней мере, такое состояние в этой группе распространено доволь-
но широко. У других эукариот жгутиков чаще всего два, если не считать случа-
ев, когда один из них редуцирован — как у эвглены, к примеру.
Кавалье-Смит пошел и дальше. Он предположил, что все эукариоты делятся на
две эволюционные ветви: Unikonta (одножгутиковые) и Bikonta (двужгутиковые).
К униконтам относятся опистоконты и амебозои, а к биконтам — все остальные,
включая экскават, хромальвеолят, ризарий и архепластид.
В чем биологический смысл деления эукариот на Unikonta и Bikonta? Кавалье-
Смит не был бы Кавалье-Смитом, если бы не попытался ответить и на этот во-
прос . Он обратил внимание на то, что все основные группы Unikonta — это ис-
ходно обитатели твердых поверхностей (дна или почвы). Они легко переходят в
прикрепленное состояние и, даже будучи подвижными, часто питаются,
"заякориваясь" ложноножками на субстрате. В общем, их адаптивная зона — пре-
имущественно придонная.
А вот Bikonta перешли от придонного образа жизни к плаванию в толще воды,
среди планктона. Они приобрели второй жгутик, повышающий маневренность, и
оболочки, основанные на каркасе из микротрубочек (как пелликула альвеолят),
которые придают клетке постоянную форму, делая ее обтекаемой. Это позволило
биконтам сначала научиться отлично плавать, а потом и захватить фотосинтези-
рующих симбионтов, ставших хлоропластами (у униконтов их нет). Неудивительно,
что жизненные формы биконтов теперь гораздо разнообразнее.
К сожалению, этот изящный сценарий не выдержал проверки фактами. В 2009 го-
ду канадские исследователи Эндрю Роджер и Аластер Симпсон (с которыми мы
встречались в начале этой главы) провели тщательное сравнение жгутиковых ап-
паратов разных одноклеточных эукариот и убедительно показали: общий предок
всех современных униконтов наверняка имел два жгутика, а не один. В таком
случае называть остальных эукариот "биконтами" бессмысленно. К тому же един-
ство эволюционной ветви биконтов не поддерживается большинством генетических
исследований. Видимо, от этого таксона надо просто отказаться.
А вот гипотеза о существовании таксона Unikonta проверку выдержала. Молеку-
лярно-генетические данные надежно подтверждают: супергруппы Opisthokonta и
Amoebozoa в самом деле образуют единую эволюционную ветвь. Правда, большинст-
во современных исследователей согласны с тем, что старое название этой ветви
— одножгутиковые — не отражает исходное для нее состояние признаков. Но, во-
первых, это всего лишь название, а во-вторых, одножгутиковых существ среди
униконтов и вправду очень много. Иногда униконтов даже считают одной супер-
группой, а опистоконтов и амебозоев — подразделениями внутри нее (например,
Евгений Кунин в книге ллЛогика случая" именно так описывает систему эукариот) .
Что ж, с точки зрения филогенетической систематики это по-своему резонно.
Томас Кавалье-Смит, всегда внимательно относящийся к новым фактам, вскоре
согласился, что предки как униконтов, так и всех эукариот когда-то имели два
жгутика. Отсюда следует, что исчезновение второго жгутика у опистоконтов и у
многих амебозоев — состояние производное. Тем не менее, верным остается то,
что адаптивная зона, которую выбрали униконты, — в самом деле, придонная, не
предполагающая (как правило) освоения водной толщи, зато связанная с поверх-
ностью грунта. Именно поэтому униконты специализировались в основном к жизни
в пресных водоемах или в почве. Вот из этой-то группы и вышли миксомицеты,
грибы, животные.
В серии относительно недавних работ Кавалье-Смит отказался от деления на
Unikonta и Bikonta, зато разделил эукариот на два эволюционных уровня — Eozoa
и Neozoa (см. рис. 15.12). Это решение имеет простой биологический смысл.
Eozoa — это низшие эукариоты, которых раньше объединяли (а многие авторы объ-
единяют и сейчас) в ветвь экскават. Однако, по мнению Кавалье-Смита, это на
самом деле не ветвь, а эволюционный уровень, являющийся исходным для эукариот
в целом. К нему принадлежат жгутиконосцы, имеющие два жгутика, — правда, один
из них может редуцироваться (как у эвглены) , или же, наоборот, их может ста-
новиться больше (как у гипермастигин). Кроме того, для примитивных жгутико-
носцев-ллэозоев" характерна внутренняя оболочка из микротрубочек — тоже как у
эвглены. А вот передвижение с помощью ложноножек у них не развито.
П!
; с! 1 .<
-Vf.o'.jf
Opisthokonta ApUSOZOa
Amoebozoa
Diaphoretickes = Corticata
Plantae SAR
жгутик
жгутиковый карман
фаготрофный аэробный
двужгутиковый предок
Рис. 15.12. Гипотеза Eozoa и Neozoa. Группа, которую мы уже привыкли
называть экскаватами, здесь распадается на отдельные ветви и оказы-
вается не более и не менее как примитивным эволюционным уровнем, че-
рез который когда-то прошли все зукариоты. Выход на уровень Neozoa,
по этой гипотезе, произошел дважды. Corticata — название, предложен-
ное Кавалье-Смитом для того же эволюционного ствола, который называ-
ют Diaphoretickes. По сравнению с оригинальными статьями Кавалье-
Смита номенклатура здесь упрощена, но представления о ходе эволюции
переданы точно.
Neozoa, по Кавалье-Смиту, — это высшие зукариоты, у которых возникли два
расходящихся магистральных направления эволюции. Одни "неозои" освоили амебо-
идное движение, которое удобно при донном образе жизни и заодно способствует
питанию путем фагоцитоза. Второй жгутик при этом стал не нужен, и во многих
эволюционных линиях он исчез. По этому пути пошли униконты. Другие ллнеозои"
сохранили два жгутика, а в придачу приобрели пелликулу и (или) клеточную
стенку — структуры, придающие клетке жесткую обтекаемую постоянную форму. Это
дало им возможность специализироваться к плаванию в толще воды и к фотосинте-
зу — последнее, конечно, с помощью захваченных симбионтов. По этому пути по-
шли SAR и растения, причем последние вообще потеряли способность к фагоцитозу
из-за слишком прочной клеточной стенки.
Классификация "Eozoa/Neozoa" не получила большой популярности, потому что
она противоречит принципам филогенетической систематики — та категорически
запрещает выделение эволюционных уровней в качестве таксонов. Сейчас эта
классификация интересна скорее как факт научной мысли, показывающий, что фи-
логенетическая систематика — еще не окончательная истина.
Заодно тут возникает одна частная, но интересная задачка. Мы давно уже зна-
ем, что митохондрии (у всех эукариот) и хлоропласты (у фотосинтезирующих)
произошли от симбиотических бактерий. Предками митохондрий были альфа-
протеобактерии, а предками первичных хлоропластов — цианобактерии (вторичные
хлоропласты, происходящие от поглощенных эукариот с первичными хлоропластами
внутри, нас в данном случае не интересуют). И митохондрии, и первичные хлоро-
пласты имеют две мембраны. Логично предположить, что внутренняя мембрана ми-
тохондрии или первичного хлоропласта — это плазматическая мембрана захвачен-
ной бактерии, а внешняя — мембрана пищеварительной вакуоли ее эукариотного
хозяина. Именно это и утверждала классическая теория эндосимбиоза.
Но давайте обратим внимание на то, что предки и митохондрий, и хлоропластов
относятся к грамотрицательным бактериям. Это означает, что у них две мембраны
— внутренняя и внешняя. После захвата такой бактерии эукариотной клеткой не-
избежно образуется конструкция из трех мембран: мембрана пищеварительной ва-
куоли и две мембраны бактерии. Какая же из них была утрачена? Кавалье-Смит
считает, что и в хлоропластах, и в митохондриях наиболее вероятна потеря са-
мой внешней из трех мембран — той, что принадлежала пищеварительной вакуоли
хозяина. Ибо потерять ее легче всего: в конце концов, захваченная бактерия
просто разорвет эту мембрану, если вздумает внутри нее расти. Ко всему проче-
му , внешние мембраны митохондрий/хлоропластов и грамотрицательных бактерий
совпадают по свойствам: и те и другие устроены как свободное ллсито", прони-
цаемое для максимального числа всевозможных веществ (в отличие, как от внут-
ренней мембраны бактерии, так и от внутриклеточных мембран хозяина-
эукариота). Так что первая приходящая на ум схема, скорее всего, неверна. И в
митохондриях, и в хлоропластах сохранились обе мембраны грамотрицательных
бактерий. У одной группы эукариотных водорослей, а именно у глаукофит из су-
пергруппы Plantae, между мембранами хлоропласта сохраняются даже остатки кле-
точной стенки — пептидогликановой, как бактериям и положено. Кстати, это са-
мое что ни на есть прямое свидетельство происхождения хлоропластов от циано-
бактерии .
У Томаса Кавалье-Смита есть одна особенность, роднящая его со многими дру-
гими хорошими биологами-эволюционистами (начиная, пожалуй, с нашего соотече-
ственника — палеонтолога Владимира Ковалевского). Он прекрасно понимает, что
на установлении родственных отношений организмов серьезное изучение эволюции
не заканчивается, а начинается. Эволюционная биология не просто наука о том,
кто чей предок. В такой же, если не большей мере ее интересует, как и почему
произошли те или иные эволюционные события. Например, положение человека на
молекулярно-филогенетическом древе сейчас известно абсолютно точно: ближайшая
к нему эволюционная ветвь состоит из шимпанзе и бонобо; эти виды разошлись
уже после того, как их общий предок отделился от предка человека, и поэтому
являются нашими родственниками в строго одинаковой степени. Ну и что? В конце
концов, гораздо интереснее узнать, когда и почему человек стал прямоходящим,
как при этом менялась его система размножения и социальная структура, для че-
го увеличился мозг, откуда взялась речь. В общем, выстроить полный эволюцион-
ный сценарий, дополняющий тот, который предложил в свое время гениальный Оуэн
Лавджой (очень хорошее изложение этого сценария на русском языке можно найти
в книге Дональда Джохансона и Мейтленда Иди "Люси"). Разумеется, такие сцена-
рии должны быть проверяемыми, и надо быть готовыми к тому, что иногда они бу-
дут опровергаться. Но без них изучать эволюцию было бы просто скучно.
Теперь вернемся к эволюционному древу эукариот и подведем сухие итоги. Ис-
следования последнего десятилетия помимо множества спорных гипотез (которые
мы тут не обсуждаем) привели к твердому установлению двух важных фактов. Во-
первых, вместо двух равноценных супергрупп — хромальвеолят и ризарий, зани-
мающих на древе соседние места, мы теперь имеем дело с тремя близкими, но са-
мостоятельными ветвями — страменопилов, ризарий и альвеолят, — которые прихо-
дится сводить в одну супергруппу: иначе этот пазл не складывается. Во-вторых
(и что гораздо более важно), существует эволюционная ветвь Unikonta, состоя-
щая из двух супергрупп — опистоконтов и амебозоев. Подчеркнем, что и описто-
конты, и амебозои сами по себе реальны. Просто они вместе образуют ветвь бо-
лее высокого порядка.
Таким образом, шесть супергрупп эукариот теперь можно свести к четырем:
экскаваты, униконты, растения (архепластиды) и SAR.
В капитальной коллективной сводке 2012 года группу Unikonta предложили пе-
реименовать в Amorphea. Буквально это значит "бесформенные": намек на то, что
у многих представителей этой группы клетка не имеет постоянной формы. Лучше
ли это, чем ллодножгутиковые", — вопрос такой же спорный, как и в случае с ар-
хепластидами, но, во всяком случае, в современных публикациях название
Amorphea встречается нередко.
А можно ли еще сократить число главных естественных групп эукариот — хотя
бы до трех, а лучше до двух? Вероятно, да, но с серьезной оговоркой: чем бо-
лее древних ветвлений мы коснемся, тем менее надежными будут выводы. В случае
с общим эволюционным древом эукариот эта зависимость — очень сильная.
В конце 2000-х появились генетические данные, указывающие, что, скорее все-
го, эукариоты распадаются на три мегаствола: униконты, экскаваты и растения +
SAR. Тогда получается, что растения и SAR образуют единую мегагруппу, в кото-
рую входят почти все фотосинтезирующие организмы. Правда, у этой мегагруппы
нет уже совсем никаких общих признаков, кроме генетических. Она установлена
только благодаря сравнению последовательностей генов, кодирующих несколько
сотен разных белков, и математической обработке этих данных. Мегагруппу, со-
стоящую из растений и SAR, предложено называть Diaphoretickes ("разнообраз-
ные") , как раз в честь полного отсутствия общих структурных особенностей. Та-
кое вот парадоксальное название. В общем, по этой системе получается, что эу-
кариоты состоят из трех эволюционных мегастволов, названия двух из которых —
ллбесформенные" и "разнообразные". Сами эти слова показывают, сколько здесь
неясного и насколько это направление исследований открыто для будущего. Не
стоит к тому же забывать, что есть и третий мегаствол — экскаваты, уже знако-
мая нам группа, занимающая на древе особое положение.
Древо
и взрыв
Сумма данных, которыми мы сейчас располагаем, не оставляет сомнений, что
общий предок современных эукариот уже был полноценной эукариотной клеткой. У
него было ядро, эндоплазматическая сеть, аппарат Гольджи, микротрубочки, мик-
рофиламенты, митохондрии и жгутики. Был сложный метаболизм, обеспеченный хо-
рошо знакомым биохимикам набором ферментов. Создается впечатление, что эука-
риотная клетка во всем ее великолепии появилась в эволюции жизни на Земле
сразу в готовом виде. Сразу заметим, что это впечатление может быть и ложным,
связанным просто с тем, что переходные формы вымерли, а палеонтологических
данных не хватает (по первым шагам эволюции эукариот они вообще отсутствуют).
Но, так или иначе, у общего предка всех современных эукариот полный набор пе-
речисленных атрибутов уже точно был.
Что же произошло дальше с его потомками? Вот здесь структура эволюционного
древа вполне может кое-что нам подсказать.
Начнем с того, что есть несколько четко установленных эволюционных стволов
эукариот, существование которых бесспорно. Если исходить из общепринятых ги-
потез и пренебречь некоторыми мелочами, таких стволов можно насчитать девять:
опистоконты, амебозои, архепластиды, страменопилы, ризарии, альвеоляты, гап-
тофиты, криптомонады и экскаваты. Современные генетические данные наводят на
мысль, что все эти эволюционные стволы разошлись от некой очень компактной
группы предков почти одновременно. И с тех пор они остаются устойчивыми, хо-
тя, конечно, продолжают давать новые формы каждый внутри себя. Иными словами,
получается, что в истории эукариот был период чрезвычайно быстрой начальной
дивергенции, после которого новые крупные эволюционные ветви уже не возника-
ли. Все расхождения ветвей первого-второго порядка произошли в самом начале
эволюции эукариот за такой короткий промежуток времени, что практически нало-
жились друг на друга. Возможно, как раз поэтому выделить главные эволюционные
стволы эукариот относительно легко, а вот определить степени их родства между
собой очень трудно.
Эту идею нельзя назвать новой. На самом рубеже XX-XXI веков французский
биолог Эрве Филипп, поддержанный группой коллег, высказал предположение, что
трудность установления родственных связей между крупными ветвями эукариот не
является следствием недостатка данных, а отражает реальное событие —
ллэукариотный Большой взрыв". Филипп считал, что триггером бурной эволюции эу-
кариот стало увеличение содержания кислорода в атмосфере, вызвавшее симбиоз с
предками митохондрий. Появление первого эукариота, обладавшего митохондриями,
создало принципиально новую адаптивную зону, которую его потомки начали стре-
мительно осваивать, эволюционируя во все возможные стороны. Вот тогда-то все
главные ветви эукариот и возникли. На эволюционном древе соответствующий мо-
мент выглядит как мультифуркация — распад одной линии сразу на несколько (см.
рис. 15.13). И это не артефакт, связанный с недостатком данных, а отражение
реальных особенностей эволюционного процесса в то время и в том месте. Чис-
ленные прикидки здесь таковы: эукариоты существуют на Земле никак не меньше
двух миллиардов лет, но все их основные эволюционные ветви, вероятно, возник-
ли за первые десятки миллионов лет, и уж во всяком случае — не более чем за
первые 100 миллионов летЮО.
В общем, "сейчас представляется, что все известные линии эукариот, дожившие
до современности, возникли за промежуток времени, гораздо более короткий, чем
время, прошедшее с тех пор. А отсюда следует важный вывод: ни одна современ-
ная группа эукариот не может быть значительно примитивнее других".
Когда стало понятно, что построить филогенетическое древо эукариот все же
удается, иные исследователи сочли было это мнение Эрве Филиппа старомодным
заблуждением. Но не исключено, что тут они поспешили с выводами.
Во-первых, эволюционный процесс не может идти совершенно одинаково во все
времена и во всех таксонах. Существует такое понятие, как "эволюция эволю-
ции" . А возникновение эукариотной клетки — это одно из самых уникальных собы-
тий во всей истории жизни на Земле, и нет ничего удивительного, если в его
временной окрестности эволюция шла в необычном (с нашей точки зрения) режиме.
Во-вторых, необычность этого режима все-таки не стоит преувеличивать. Си-
туация, когда за короткий промежуток времени один предок дает множество эво-
люционных ветвей, давно известна биологам и особенно палеонтологам: она назы-
вается взрывной эволюцией (explosive evolution). Хорошо изученные примеры та-
ких событий есть, например, в эволюции млекопитающих. Ничто не мешало подоб-
ному "взрыву" произойти и в начале эволюции эукариот. Напротив, к этому были
все предпосылки. И тут не надо допускать никаких неизвестных факторов, все
вполне укладывается в классическую эволюционную теорию.
ЭУКАРИОТЫ
митомонорий
ТрМХДОЮиДДЫ
дилломонады
грибы
<:
мияосхАчдеты
■рлсиые водоросли
>*л«нмс рэст**»**
улгпсхтыс
апывсопяты
оэ>*ожгути<о#ы«
другие группы
БАКТЕРИИ
АРХЕИ
Рис. 15.13. Гипотеза "эукариотного Большого взрыва". А — древо,
соответствующее старым представлениям Эрве Филиппа. Обратим вни-
мание, что три из 11 выделенных здесь ветвей (дипломонады, три-
хомонады и эвгленовые) относятся к экскаватам. Черной стрелочкой
указан момент приобретения первичных хлоропластов, белыми стре-
лочками — моменты приобретения вторичных хлоропластов. Б — ком-
пьютерная модель взрывной эволюции. Очевидно, что свести такое
древо к набору ветвлений надвое (как того требует кладистика)
чрезвычайно трудно.
Если ллэукариотный Большой взрыв" действительно имел место, то нам, как ни
странно, не столь уж и важно знать, в каком именно порядке там расходились
главные эволюционные стволы. В любом случае первые члены этих стволов почти
ничем не отличались друг от друга: при эволюции путем дивергенции иначе и
быть не может. А вот что по-настоящему интересно — это кто из них к чему при-
спосабливался и как они потом делили между собой адаптивные зоны. Обо всем
этом мы пока знаем очень мало. Но, видимо, узнаем и больше: установленное
эволюционное древо дает для этого неплохую основу.
ЛЛМикроцарства"
Настало время сделать очередной шаг вперед. Мы уже успели привыкнуть к то-
му, что эукариоты делятся на пять-шесть супергрупп. И даже если границы этих
супергрупп несколько перекраиваются, общая картина остается более-менее той
же.
Проблема в том, что существует несколько небольших групп эукариот, которые
ни в какие супергруппы включить не удается (по крайней мере, без серьезных
трудностей, натяжек и споров). Мы уже упоминали две такие группы — это гапто-
фиты и криптомонады. Но есть и другие, менее известные, зато еще более зага-
дочные .
Известный протистолог Ян Павловский метко назвал эти загадочные группы
"микроцарствами". Обсудить их все тут нет никакой возможности, иначе эта гла-
ва превратится в справочник. Краткое перечисление тем более не выход: те, кто
всерьез заинтересуется темой, легко найдут его в научных статьях, а всех ос-
тальных множество мудреных латинских названий скорее отпугнет. Поэтому, чтобы
получить какое-то представление о сути дела, но в то же время не запутаться,
мы сейчас кратко поговорим всего о трех живых организмах, явно относящихся к
ллмикроцарствам". В двух случаях это будут отдельные роды, а в третьем — груп-
па из нескольких родов: бревиата, коллодиктион и апузомонады (см. рис.
15.14).
Рис. 15.14. Представители ллмикроцарств" (слева направо): Апузо-
монас, Бревиата и Коллодиктион.
Бревиата — это одноклеточное амебообразное существо со жгутиком, живущее в
солоноватых водоемах. Жгутик у бревиаты один и направлен вперед, в отличие от
опистоконтов, у которых он тоже один, но направлен назад. При этом второй
жгутик у нее, безусловно, когда-то был: электронная микроскопия обнаружила в
клетке его остатки. Наряду со жгутиком у бревиаты есть и ложноножки — вполне
типичные, как у самой настоящей амебы. Положение бревиаты на эволюционном
древе остается спорным. Достаточно правдоподобно выглядит версия, что это са-
мый примитивный современный представитель супергруппы амебозоев. Тогда полу-
чается , что эволюционная ветвь, которую образует бревиата, — сестринская по
отношению ко всем остальным амебозоям, вместе взятым. Но иногда бревиату во-
обще не включают в супергруппу амебозоев, оставляя за ее пределами. И возмож-
но, это разумнее, учитывая, что есть и такие результаты, которые больше сбли-
жают бревиату с опистоконтами. В любом случае получается, что бревиата примы-
кает к самому основанию какой-то из этих двух смежных ветвей. А что, если в
лице бревиаты перед нами и вовсе третья ветвь, имеющая тот же ранг, что опи-
стоконты с амебозоями, и возникшая одновременно с ними? Такая гипотеза труд-
нопроверяема (и потому непопулярна), но с точки зрения эволюционной теории в
ней нет ничего невероятного.
Интересно, что бревиата предпочитает жить в воде с очень низким содержанием
кислорода. Поэтому у нее нет митохондрий, хотя есть митохондриальные гены,
успевшие мигрировать в ядро. Учитывая, насколько бревиата примитивна (то есть
насколько древней эволюционной ветвью она является), вполне можно было бы
предположить, что митохондрий у нее никогда и не было. К сожалению, это опро-
вергается твердо установленными генетическими фактами, против которых не пой-
дешь . Природа далеко не всегда так экономична, как от нее этого ждут люди.
Бревиата — в самом деле очень древнее существо, но митохондрии она тем не ме-
нее успела когда-то приобрести, а потом утратить.
Апузомонады — жгутиконосцы, которые живут в морских донных отложениях, на
дне пресных водоемов, а часто и в почве: капельно-жидкой воды для них там
вполне достаточно. Это отличный пример жизненной стратегии, типичной для
ллодножгутиковых" в классическом понимании Кавалье-Смита. Жгутиков у апузомо-
над тем не менее два. Они не несут никаких волосков, но направлены в разные
стороны: один — вперед, а другой загнут и тянется вдоль тела клетки (поэтому
под микроскопом его бывает плохо видно). Жгутиконосец-апузомонада скользит по
грунту, работая жгутиками, а на "брюшной" — то есть обращенной к субстрату —
стороне он образует ложноножки, которыми захватывает бактерий. Группа эта от-
носительно немногочисленная, но в то же время широко распространенная. Благо-
даря особенностям своего образа жизни апузомонады легко преодолевают экологи-
ческие барьеры как между пресными водами и морем, так и между любыми водоема-
ми и почвой. Поэтому весьма вероятно, что именно апузомонады (или какие-то
похожие на них существа) первыми среди эукариот вышли на сушу.
Положение этой группы на древе долго было неопределенным. Скорее всего,
апузомонады все же примыкают к опистоконтам. Однако ни в группу Holomycota,
ни в группу Holozoa ветвь апузомонад не входит — она отошла от ствола описто-
контов раньше, чем эти группы разделились.
Кроме того, в последние несколько лет появились молекулярные данные, позво-
ляющие объединить бревиат, апузомонад и опистоконтов в единую группу, которая
получила название Obazoa. Это название образовано от сокращений: О —
Opisthokonta, В — Breviatea, A — Apusomonadida. Если это подтвердится, зна-
чит, Unikonta (они же Amorphea) распадаются на ветви Amoebozoa и Obazoa. Та-
кая версия мегасистемы уже вошла в некоторые современные руководства.
Интересен вопрос о кристах митохондрий. У апузомонад кристы трубчатые, как
у амебозоев. Но у другой сходной с ними группы жгутиконосцев — анкиромонад —
кристы пластинчатые, как у опистоконтов. Палеонтологи прекрасно знают, что
признаки, обычно характеризующие крупные эволюционные стволы, могут быть
очень изменчивыми на ранних этапах эволюции, когда эти стволы еще не разо-
шлись или разошлись едва-едва. Вот такой случай наблюдается и тут.
И, наконец, коллодиктион. Мы уже вскользь упоминали это существо в главе
10, но сейчас есть несомненный повод вспомнить о нем еще раз. Коллодиктион
был открыт еще во времена Чарльза Дарвина. Однако понимать, насколько особое
место он занимает в живой природе, биологи стали только к началу XXI века.
Итак, коллодиктион — это довольно крупный пресноводный жгутиконосец. У него
четыре одинаковых жгутика, направленных вперед. Кроме того, у него есть лож-
ноножки, причем широкие, лопастевидные, как у амебозоев. На "брюшной" сторо-
не, обычно обращенной к грунту, у коллодиктиона находится ротовая бороздка,
армированная со стороны цитоплазмы корешками из микротрубочек. Этот признак
мы уже встречали у экскават, которые именно ему и обязаны своим названием.
Кристы митохондрий у коллодиктиона трубчатые. По образу жизни он — хищник,
причем склонный питаться довольно крупными объектами. Чаще всего это эукари-
отные зеленые водоросли, иногда одноклеточные, а иногда и колониальные —
вплоть до восьмиклеточных колоний. Причем проглоченные водоросли коллодиктион
переваривает настолько медленно, что они, по-видимому, успевают у него пора-
ботать в качестве хлоропластов — прекрасная модель ранних этапов симбиотиче-
ской эволюции.
Что же это за существо такое? Здесь нас ожидает сюрприз — впрочем, не такой
уж и большой, если учесть все то, что мы к данному моменту уже знаем. Молеку-
лярные исследования показывают, что коллодиктиона нельзя без натяжек включить
ни в одну супергруппу эукариот. Это особая эволюционная ветвь — ранний выброс
"эукариотного Большого взрыва". Бревиату, апузомонад и некоторые другие груп-
пы тоже можно назвать такими "ранними выбросами", но коллодиктион, возможно,
самый древний из них.
Задумаемся. Очень похоже, что ветвей эукариот, соответствующих по рангу
традиционным супергруппам, на самом деле куда больше, чем пять или шесть (см.
рис. 15.15). Просто далеко не все из них достигли большого разнообразия. И
те, кто его не достиг, статуса супергрупп не удостоились. Хотя объективно, по
положению на древе, они вполне могут быть равноправны таковым (как, например,
бревиаты равноправны амебозоям).
Коллодиктион — организм, настолько близкий к общему предку всех современных
эукариот, насколько это вообще возможно. Его единственный явно производный
признак — четыре жгутика. По всей видимости, у общего предка всех современных
эукариот жгутиков было два. Но мы знаем, что увеличение числа жгутиков много
раз происходило в эволюции экскават (к которым коллодиктион относительно бли-
зок) , и он просто оказался еще одной группой, независимо от других пошедшей
по этому пути. Скорее всего, в его случае это связано с тем, что коллодиктион
за свою долгую историю успел специализироваться на питании сверхкрупной добы-
чей — другими эукариотами. Это вызвало увеличение размера, а оно потребовало
дополнительных органов движения. И неудивительно, что это относительно недав-
няя специализация: ведь во времена самых первых эукариот никаких других эука-
риот, которыми можно было бы питаться, еще просто не существовало в природе.
Состояние всех остальных признаков у коллодиктиона примерно такое, какого и
можно было бы ожидать от древнейшего эукариота. Но история со жгутиками здесь
очень показательна. Каким бы древним коллодиктион ни был, он, прежде всего,
является реальным живым существом, а вовсе не ожившей схемой. А жить — значит
специализироваться.
Ну а теперь — торжественный момент. Зная все, что нам сейчас известно, мы
можем легко представить себе реального общего предка всех современных эукари-
от и достаточно подробно описать его. Безусловно, это был жгутиконосец. Одно-
временно со жгутиками он, скорее всего, имел ложноножки — примерно того же
типа, что у обыкновенной амебы, но менее развитые и используемые только для
захвата пищи, а не для передвижения. Жгутиков у него было два (без всяких во-
лосков на них) . Кристы митохондрий, скорее всего, были трубчатыми, с меньшей
вероятностью — пластинчатыми, но в любом случае их форма легко менялась от
вида к виду: на том эволюционном этапе этот признак еще не устоялся. Образ
жизни был придонным: жгутиконосец передвигался в основном у поверхности грун-
та, лишь изредка поднимаясь в толщу воды, поэтому в его клетке дифференциро-
вались "спинная" и "брюшная" стороны. На "брюшной" стороне, скорее всего, бы-
ла продольная ротовая бороздка — та самая, которая сохранилась в супергруппе
экскават и благодаря которой эта супергруппа получила свое название. И конеч-
но, наш предок был хищником — в той мере, в какой в тогдашнем мире вообще бы-
ло возможно хоть какое-то хищничество. Он питался всеми, кого мог поймать, и
занимал самую вершину пищевой пирамиды.
Обычному человеку может быть нелегко совместить с наглядными представления-
ми о мире тот факт, что его личный прямой предок — ничуть не менее прямой,
чем дед или прадед, — когда-то, пару миллиардов лет назад, был одноклеточным
жгутиконосцем, похожим на коллодиктиона. Но, судя по всему, это чистая прав-
да. Как сказал герой известной пьесы Григория Горина: "Это гораздо больше,
чем факт. Так оно и было на самом деле".
l.hi kofto
SAR
Obazoa
"Excavata"
общий предок
современных зукариот (LECA)
Рис. 15.15. Упрощенное эволюционное древо эукариот, более-менее со-
ответствующее современным представлениям. О положении криптомонад и
гаптофит есть целая группа разных гипотез — здесь проблема созна-
тельно оставлена открытой. Проблема родственных связей малых групп
внутри Unikonta тоже оставлена открытой, но надо иметь в виду, что
сейчас набрала популярность обсуждаемая в тексте гипотеза Obazoa —
если она верна, то ветви апузомонад и бревиат должны будут примкнуть
к опистоконтам. Наконец, далеко не факт, что Excavata действительно
являются единой ветвью, а не сборной группой. Подробное обсуждение
всех этих нерешенных вопросов — дело будущего, хочется надеяться,
что ближайшего.
Многоклеточность
Вся биология когда-то началась с изучения многоклеточных организмов. Это
совершенно естественно. "Люди, львы, орлы и куропатки, рогатые олени, гуси,
пауки, молчаливые рыбы, обитавшие в воде, морские звезды и те, которых нельзя
было видеть глазом", — легко заметить, что в этом классическом списке, порож-
денном трепетным подсознанием героини Чехова , девять наименований из десяти
относятся к многоклеточным существам, которые были объектами науки еще во
времена Аристотеля. Изучение природы всегда начинается с очевидного, а к ме-
нее очевидному переходит лишь постепенно.
Давайте еще раз взглянем на знакомое нам теперь эволюционное древо эукари-
от. Перед нами великое множество эволюционных ветвей, большинство из которых
— это бросается в глаза — состоит из одних только одноклеточных организмов,
как правило, и в самом деле невидимых простым глазом: их удалось открыть
только после изобретения микроскопа. Несколько меньше — ветвей, в которых
есть колониальные формы, и совсем уж немного тех, где возникает многоклеточ-
ность в самом полном смысле этого слова. Например, многоклеточные животные —
это всего лишь одна из нескольких сотен (без преувеличения) эволюционных вет-
вей аналогичного ранга.
Что же интересного в таком редком явлении, как многоклеточность? Какой,
собственно, вклад в картину мира она дает? И, в конце концов, достойна ли она
вообще глубокого фундаментального интереса? Может быть, значение исследований
многоклеточных организмов в основном прикладное — для медицины, ветеринарии,
сельского хозяйства и тому подобных отраслей?
Обойти эти вопросы было бы интеллектуальной нечестностью. К счастью, ответ
на них достаточно очевиден. Любой многоклеточный организм предоставляет нам
для изучения множество интереснейших явлений, которых ни у каких одноклеточ-
ных просто-напросто не существует. Это программирование зародышевого разви-
тия, в ходе которого тысячи клеток должны располагаться в пространстве задан-
ным образом, обновляя это расположение с каждым очередным клеточным делением;
работа генных сетей, тончайшим образом регулирующих экспрессию разных генов в
разных клетках; клеточная дифференцировка; гормональная сигнализация; регуля-
ция обмена веществ на уровне целого организма; и наконец, работа сложнейшего
из всех известных нам природных объектов — человеческого мозга. Неудивитель-
но, что огромная доля биологов занималась (и, вероятно, всегда будет зани-
маться) именно этими вопросами. Можно смело утверждать, что даже если бы во
Вселенной существовал всего лишь один вид многоклеточных организмов, притом
не имеющий никакого практического значения, то и тогда он заслужил бы огром-
ное внимание биологов, к какой бы форме жизни сами эти биологи ни относились.
Им было бы ясно, что они столкнулись с принципиально новым способом организа-
ции живой материи, который грех не попытаться изучить как следует. В похожей
ситуации оказались, например, герои лемовского ллСоляриса". Организм, который
они обнаружили, был единственным в своем роде и не имел для Земли никакого
практического значения, но, тем не менее, он сразу вызвал к жизни целую об-
ласть науки — соляристику. Лем прекрасно понимал психологию ученых, в реаль-
ности все так и было бы.
Кроме того, появление многоклеточных организмов самым серьезным образом из-
менило облик Земли как планеты. Они повлияли и на состав атмосферы, и на
ландшафты, и на глобальные круговороты разных химических элементов (в первую
очередь углерода), и на климат. Без многоклеточной жизни Земля выглядела бы
совершенно иначе.
Итак, многоклеточность — это важно. Увы, компактно обсудить ее у нас сейчас
не получится. Дело в том, что способы реализации многоклеточности в разных
группах организмов слишком различны, чтобы о них можно было хотя бы в самом
общем приближении рассказать как о едином целом. Этим занимаются специальные
науки — зоология, ботаника, альгология (наука о водорослях) и микология (нау-
ка о грибах), — погрузиться в которые в этой публикации нет никакой возможно-
сти: как сказал классик, никто не обнимет необъятного. К счастью, и о много-
1 Пьеса «Чайка»
клеточных животных, и о многоклеточных растениях, и даже о грибах источников
информации для желающих хватает (о грибах, например, есть книга
ЛЛЗанимательная микология", которую написал один из самых блестящих профессо-
ров биофака МГУ — Юрий Таричанович Дьяков). А мы тут поневоле ограничимся об-
щими моментами.
Для начала представим себе простенькую зеленую водоросль с двумя жгутиками
— вроде хламидомонады, той самой, которая описана в школьном учебнике ботани-
ки. Она, безусловно, одноклеточная. Но если она поделится, а потом еще раз
поделится и образовавшиеся клетки почему-либо не разойдутся, то мы получим
колонию из четырех клеток — а значит, уже многоклеточное существо, по крайней
мере, сугубо номинально. И такие четырехклеточные колонии реально существуют
(зеленая водоросль гониум, например). Если делений будет больше, то образуют-
ся колонии из 8, 16, 32, 64 клеток: все эти примеры тоже реальны. При этом
экспериментально установлено, что грань между колонией, где все клетки одина-
ковы и обладают равными возможностями, и организмом, где они начинают необра-
тимо специализироваться, в данном случае проходит на уровне 32-клеточной во-
доросли эудорины. "Любая из 32 клеток Eudorina elegans также способна дать
новую колонию. Однако у 32-клеточных Eudorina illinoisensis четыре передние
клетки более мелкие, они уже не могут участвовать в размножении, представляя
дифференцированную и не способную к инициальной роли часть колониальной осо-
би" . Именно здесь мы видим рубеж между колонией, члены которой потенциально
могут разойтись и жить независимо, и организмом, откуда обратного хода нет.
Но означает ли это, что эудорина находится на одной ступени организации с ме-
тасеквойей, пшеницей, слоном или человеком? А если нет, то в чем между ними
разница?
Известный американский протистолог Джон Корлисс решил, что разбираться в
этом нужно, основываясь на понятии ткани. По общепринятому определению, ткань
— это система клеток и межклеточного вещества, объединенных происхождением,
строением и выполняемыми функциями. Так вот, Корлисс предложил считать, что
протистом называется любой эукариотный организм, имеющий не более одной тка-
ни. Тогда получается, что к протистам относятся все одноклеточные... и неко-
торые многоклеточные тоже. Например, любая нитчатая водоросль, хоть она и
многоклеточная, должна считаться протистом, потому что никакого разнообразия
тканей у нее нет.
В то же время определение Корлисса четко указывает границу, за которой от-
нести живое существо к протистам уже при всем желании нельзя. Например, у
многоклеточного протиста не может быть никакой сосудистой системы, потому что
она по самой своей природе требует наличия хотя бы двух тканей: той, из кото-
рой состоят сосуды, и той, которая их окружает и через них снабжается.
Организмы, имеющие не менее двух полноценных тканей, Корлисс вполне логично
назвал многотканевыми. Например, все многоклеточные животные бесспорно попа-
дают в эту категорию. У любого уважающего себя животного, даже если у него
нет нервной системы и мышц, есть хотя бы две ткани: эпителиальная (покровная)
и соединительная, занимающая внутренний объем и богатая межклеточным вещест-
вом. И той и другой необходимы межклеточные контакты, о которых мы упоминали
в конце главы 10. Для эпителиев особенно важны плотные контакты и десмосомы,
а для соединительной ткани — фокальные контакты, которые позволяют клеткам
закрепляться на волокнах межклеточного вещества.
По мнению Корлисса, на уровень многотканевых организмов вышли всего две
эволюционные ветви: многоклеточные животные (Metazoa) и высшие растения
(Embryophyta). Все остальные до полноценной многотканевости, грубо говоря,
недотянули. Ближе всех к ней подошли некоторые водоросли — бурые, красные, а
также харовые, которые близки к наземным растениям и раньше считались их пря-
мыми предками. Но Корлисс все равно считает их всех протистами, несмотря на
то, что длина некоторых бурых водорослей достигает 60 метров. У грибов много-
тканевость сомнительна, и по классификации Корлисса грибы — тоже протисты.
Они потенциально способны выйти на многотканевый уровень, но им это просто не
нужно. Есть одна группа грибов — лабульбениомицеты, паразиты насекомых, — в
теле которых все-таки возникла практически настоящая тканевая организация.
Клетки делятся там в разных направлениях и образуют целую трехмерную структу-
ру, хоть и мелкую (доли миллиметра) . Но в мире грибов это исключение, а не
правило.
На грани перехода к многотканевости находятся некоторые слизевики, входящие
в супергруппу амебозоев. Тут есть примеры, когда в плодовом теле возникает
настоящая покровная ткань, причем устроенная поразительно похоже на эпителий
животных. Более того, некоторые мембранные белки, обеспечивающие межклеточные
контакты, у слизевиков те же самые, что и у животных. Взглянув на эволюцион-
ное древо, мы видим, что слизевики и животные могли унаследовать эти белки
только от общего предка всей мегагруппы Unikonta. Значит, молекулярная
"машинерия", позволяющая в принципе создать многоклеточность, у этого предка
— очень древнего! — уже была на месте.
В высшей степени замечательно, что собственные слизевики нашлись не только
в супергруппе амебозоев (куда входит большая их часть), но и в группе описто-
контов. Это фонтикула — с виду вполне типичный слизевик, у которого питающая-
ся стадия состоит из множества одиночных амеб, а для образования спор служит
плодовое тело. Но типичные слизевики принадлежат к амебозоям. А вот фонтику-
ла, как показала молекулярная генетика, вовсе к ним не относится. Она отно-
сится к супергруппе опистоконтов и занимает там место в основании "грибного"
эволюционного ствола Holomycota — рядом с амебами-нуклеариидами, на которых
ее амебоидная стадия, собственно, и похожа. Это означает, что в супергруппе
опистоконтов (в которую, напомним, входят не только грибы, но и животные) бы-
ла самостоятельно, хотя и на общей с амебозоями молекулярной основе, вырабо-
тана жизненная форма слизевика. И как ни странно, это вполне ожидаемо. Слизе-
вики независимо возникали в разных группах эукариот не меньше семи раз. Един-
ственная из традиционных супергрупп эукариот, в которой нет собственных сли-
зевиков, — это растения (архепластиды). Во всех остальных супергруппах они
есть. Как видим, жизненная форма слизевиков почему-то оказалась среди эукари-
от очень "популярной". И мы не можем исключить, что через такую жизненную
форму когда-то прошли предки многоклеточных животных.
История со слизевиками показывает нам еще одну важную вещь. Многоклеточный
и даже многотканевый организм вовсе не обязан быть таким на протяжении всей
своей жизни. У слизевиков и, если уж на то пошло, у грибов многоклеточные
плодовые тела образуются только на коротком отрезке жизненного цикла. К тому
же они никак не участвуют в питании, а служат только для размножения. В об-
щем-то эти организмы вполне могли бы обойтись и без плодовых тел, если бы им
не требовалось разбрасывать споры (и многие грибы действительно обходятся).
Теперь посмотрим под тем же углом зрения на эволюцию растений. Мы уже гово-
рили о гипотезе, согласно которой предками наземных растений были пластинча-
тые зеленые водоросли, напоминающие современные колеохетовые (см. рис.
15.16А). Если эта гипотеза верна, то тело первого наземного растения должно
было состоять из двух частей: фотосинтезирующая зеленая пластинка, погружен-
ная в воду либо растущая на влажном грунте, и образующийся в период размноже-
ния воздушный стволик с коробочкой для разбрасывания спор (см. рис. 15.16Б-
Д). Вот только этот стволик на первых порах и обладал многотканевой
организацией. Основной части растения — зеленой пластинке, погруженной во
влажную среду, — она была не нужна.
Не просматривается ли тут закономерность? Насколько мы можем судить, и у
слизевиков, и у грибов, и у растений многотканевая организация начинает фор-
мироваться на временных непитающихся стадиях. И только потом (у растений)
мнохютканевость распространяется почти на весь жизненный цикл, порождая хоро-
шо нам знакомые жизненные формы трав, деревьев, кустарников. Заметим, что,
например, у слизевиков ничего подобного не произошло. Правда, у некоторых из
них бывают стадии ползающих многоядерных амеб, но там ни на какую мнохюткане-
вость нет и намека. Плодовые же тела служат для размножения, и ни для чего
больше.
Рис. 15.16. Происхождение наземных растений. А — пластинка со-
временной колеохетовой водоросли. Б-Д — вероятные последователь-
ные стадии формирования воздушного стволика.
Рис. 15.17. Жизненный цикл, который мог бы быть у предка многокле-
точных животных. А — половые клетки (яйцеклетка и сперматозоид). Б —
результат слияния половых клеток (зигота). В — расселительные жгути-
ковые клетки (зооспоры) . Г — питающаяся жгутиковая стадия. Д — аме-
боидная стадия. Е-3 — временные многоклеточные стадии: Е —
"усовершенствованная" многоклеточная зооспора (синзооспора, как у
некоторых водорослей); Ж — колония жгутиконосцев (как у воротничко-
вых) ; 3 — скопление амеб (плазмодий, как у слизевиков) . Стадия Ж —
питающаяся, стадии Е и 3, вероятнее всего непитающиеся. При возник-
новении животных все одноклеточные стадии, кроме половых клеток и
зиготы, исчезли, а одна (или не одна) из многоклеточных стадий со-
хранилась .
Еще один фрагмент мозаики мы получим, обратившись к достижениям современной
молекулярной биологии. Есть целая группа генов и белков, которые сначала счи-
тались уникальными для многоклеточных животных, но затем были обнаружены у
воротничковых жгутиконосцев и мезомицетозоев — иначе говоря, у одноклеточных
и колониальных представителей эволюционной ветви "животных в широком смысле",
которая называется Holozoa. Про один из таких генов мы уже говорили — это ген
Brachyury, участвующий у позвоночных в развитии осевых структур. Но есть и
другие: например, гены, кодирующие мембранные белки, обычно связанные у жи-
вотных с межклеточными плотными контактами (теми самыми, что важны для эпите-
лиальной ткани). Или тирозинкиназы — ферменты, переносящие фосфат от АТФ на
остаток аминокислоты тирозина в белке-мишени (о том, что это за процесс, см.
главу 7) . Эти ферменты служат важными внутриклеточными посредниками в ходе
сигнальных взаимодействий, которые — опять же — особенно важны для многокле-
точных животных. Причем все перечисленные гены распространены как минимум по
всей группе Holozoa, а некоторые и шире.
Какой же отсюда следует вывод? По всей вероятности, жизненный цикл общего
предка всех Holozoa уже был достаточно сложным и включал в себя многоклеточ-
ную стадию — не обязательно питающуюся, вполне вероятно, что покоящуюся или
расселительную. Именно на этой сугубо временной стадии, скорее всего, и была
первоначально "обкатана" многотканевость, которая затем, в одной специализи-
рованной эволюционной ветви, распространилась почти на весь жизненный цикл
(см. рис. 15.17) . Ив результате возник один из сложнейших и красивейших объ-
ектов в известной нам Вселенной: животный организм, внутреннюю структуру ко-
торого изучает сравнительная анатомия.
Подвижные
в подвижном
Животные — это многоклеточные организмы, образ жизни которых определяется
неразрывной связью питания с движением. Любое животное или активно движется в
среде, или столь же активно движет среду относительно себя — например, гонит
воду бьющимися ресничками, попутно извлекая из нее питательные частицы. Ино-
гда то и другое совмещается. Тогда мы видим фильтраторов, способных к актив-
ному движению, среди которых есть наши очень близкие (по меркам эволюционного
древа) родственники: например, ланцетник и головастики некоторых лягушек. А
вот обратный случай — неподвижное животное в неподвижной среде — почти невоз-
можен. "Почти" — потому что тут, как и обычно в биологии, можно найти исклю-
чения: есть крайне специализированные паразиты, которые живут внутри других
животных, а сами по образу жизни приближаются к грибам (хотя и у них есть
подвижные личинки). Но это — отличный пример исключения, подтверждающего пра-
вило. Как сказал один хороший московский профессор зоологии, "во всех тех
случаях, когда подвижность теряется, животные теряют свой образ и подобие".
Типичному многоклеточному животному больше, чем любому другому живому сущест-
ву, подходит девиз корабля капитана Немо: Mobilis in mobile — "Подвижное в
подвижном". Кстати, и свое название этот корабль получил в честь животного —
головоногого моллюска наутилуса, свободно плавающего в океанской толще.
Именно с совершенством двигательной системы, скорее всего, связан эволюци-
онный успех одной из самых разнообразных групп животных — типа хордовых (см.
рис. 15.18). Этот тип не является самым многочисленным в животном царстве.
Есть как минимум два типа, превосходящие его по числу видов: членистоногие и
моллюски. Тем не менее, разнообразие формы тела и образа жизни у хордовых ог-
ромно, а в экологических нишах, требующих крупного размера, они просто вне
конкуренции. Почему? Тут стоит обратить внимание на тот самый признак, кото-
рый дал типу хордовых название. Их "визитная карточка" — хорда, или спинная
струна, тянущаяся вдоль большей части тела. Она связана с массивной сегменти-
рованной мускулатурой, которая позволяет животному двигаться, волнообразно
изгибаясь (хорда при этих изгибах благодаря своей упругости не дает телу по-
терять форму). Получается очень эффективный двигательный аппарат, который
иногда называют миохордальным комплексом. Никаких других уникальных преиму-
ществ, кроме миохордального комплекса, у ранних хордовых, насколько можно су-
дить , не было. Их эволюционный успех вызван необыкновенно удачной конструкци-
ей опорно-двигательной системы, которая, с одной стороны, дала им возможность
стать со временем лучшими в мире хищниками, увеличив размер и заняв самую
вершину пищевой пирамиды, а с другой — создала потенциал для бурной эволюции
на структурном уровне. Выражаясь современным языком, план строения хордовых
имеет очень высокую эволюционируемость (evolvability) . Но при этом даже на
самых неожиданных эволюционных поворотах он не разрушается, а остается самим
собой, сохраняя глубокое единство общей структуры — то, что великий сравни-
тельный анатом XIX века Ричард Оуэн назвал архетипом позвоночных. Сочетание
высокой эволюционируемости с фундаментальной устойчивостью плана строения
проходит красной нитью сквозь всю эволюцию хордовых животных, начавшуюся свы-
ше 500 миллионов лет назад со скромного, длиной в несколько сантиметров, пла-
вающего фильтратора — "подвижного в подвижном". Как мы знаем, именно на этом
эволюционном пути, в конце концов, возникли общество и разум.
Рис. 15.18. Одно из самых древних хордовых — хайкоуихтис, живший
около 520 миллионов лет назад. Тело этого маленького (несколько сан-
тиметров) существа, скорее всего, было более-менее прозрачным, по-
этому неудивительно, что здесь видны мышечные сегменты.
ЧАСТЬ IV. ИСТОРИЯ ЖИЗНИ
16. КРИПТ030Й
И вновь, прорезав плотные туманы,
На теплые архейские моря,
Где отбивают тяжкий пульс вулканы,
Льет бледный свет пустынная заря.
И, размножая легких инфузорий,
Выращивая изумрудный сад,
Все радостней и золотистей зори
Из облачного пурпура сквозят.
Михаил Зенкевич. «Темное родство»
Зададимся следующим вопросом: можно ли рассматривать историю Вселенной как
единый процесс, начавшийся Большим взрывом, охватывающий наше время и продол-
жающийся в неведомом космическом будущем? Средневекового книжника такой под-
ход, вероятно, совсем не удивил бы. Написанная в XII веке хроника Оттона
Фрейзингенского — "самая всеобщая из всех всеобщих историй", по выражению ве-
ликого историка-медиевиста Марка Блока, — начинается с сотворения мира и за-
канчивается Страшным судом. В конце XX века такое отношение к вещам возроди-
лось в учебной дисциплине, которая получила название "большой истории" (Big
History). Курсы ллбольшой истории" выстраивают в единую последовательность со-
бытия любого рода — от зарождения первых звезд до промышленной революции. Все
развитие мира там представлено единой шкалой, охватывающей 13,8 миллиарда
лет: примерно столько времени, по расчетам, прошло от Большого взрыва до со-
временности .
Ясно, что "большая история" ставит себе грандиозную задачу. Требуемый для
нее объем сведений (еще и стремительно растущий) очень трудно уложить в
сколько-нибудь компактный рассказ. В этом отношении Оттону Фрейзингенскому
приходилось куда легче, чем авторам, работающим с современными научными дан-
ными. Поэтому важнейшим понятием в лл большой истории" является порог
(threshold) — момент качественного изменения системы, дающий историку точку
отсчета. В любом историческом процессе число по-настоящему значительных поро-
гов относительно невелико. А привязав к ним обзор, можно ЛЛс птичьего полета"
увидеть важные вещи, которые иначе могли бы и ускользнуть от внимания.
Попробуем, не вдаваясь ни в какие вселенские обобщения, применить этот под-
ход к биологическому и палеонтологическому материалу. Мы сразу увидим, что за
несколько миллиардов лет земная жизнь не раз миновала качественные пороги,
любой из которых можно было бы и не пройти. Причем иногда это выглядело весь-
ма драматично, напоминая пропасть, через которую перекинут узкий мост. И по-
сле каждого такого порога не только жизнь на Земле, но и вся Земля как плане-
та серьезно менялась. Пущенное в оборот австрийским геологом Эдуардом Зюссом
выражение "лик Земли" как нельзя лучше характеризует объект этих изменений.
Для удобства разговора введем два термина: биосфера — целостная живая обо-
лочка Земли и биота — совокупность видов живых организмов. Биота отличается
от биосферы тем, что связи между организмами в нее не включены: грубо говоря,
это просто список видов. Термин "биосфера" предложил сам Эдуард Зюсс, а тер-
мин ЛЛбиота" — румынский зоолог Эмиль Раковицэ. Каждый пройденный глобальный
порог навсегда менял состояние и биосферы, и биоты.
Ясно, что любая "пороговая" схема эволюции биосферы будет в чем-то несовер-
шенна просто из-за того, что частью событий совершенно неизбежно придется
пренебречь. Тем не менее, попытки создания подобных схем иногда удаются. За-
мечательным образцом этого подхода можно назвать книгу Кирилла Еськова
"История Земли и жизни на ней"2, основанную на учебном курсе и выдержавшую
несколько переизданий. К сожалению, со времени создания этой книги прошло уже
около 20 лет, а для такой бурно развивающейся науки, как палеонтология, это
огромный срок. Нисколько не намереваясь критиковать великолепную работу Ки-
рилла Юрьевича, попробуем предложить что-то вроде краткого комментария к ней.
Здесь мы рассмотрим семь порогов, соответствующих важным событиям в истории
Земли: (1) появление жизни как таковой, (2) кислородная революция, (3) эпоха
"Земли-снежка", (4) кембрийский взрыв, (5) появление лесов и насекомых, (6)
пермский кризис и (7) приобретение некоторыми животными эусоциальности.
Поговорим о них по порядку (см. рис. 16.1).
». ^^ Эусоцюньмость
Мюолой |
—т
n.vx>o >ой
—а2
Эдилшрмй I
Крмсгеммй
-еро
1500
-2500
•4000
4$00
Пермский кризис
лесов
и н.к»«оиык
Кыдбрмйский
•£ 3«им/14-о«*аок
Кис породил*
ргвОГКЧ***.
гуромскоо
олодом«ии«,
ПОЯВЛвММО
эуклрмот
ftopf*»
следы «мм
Рис. 16.1. Выделенные в тексте пороговые события, наложенные на
геохронологическую шкалу.
2 Домашняя лаборатория 2007-10
Логика
случая
Насколько закономерным или случайным событием по меркам Вселенной было по-
явление земной жизни? Тут будет интересно сравнить мнения двух крупных биоло-
гов, работавших примерно в одно и то же время. Это француз Жак Моно и бельги-
ец Кристиан де Дюв. Оба они получили Нобелевскую премию за открытия в области
клеточной биологии: Моно открыл важные механизмы регуляции работы генов, а де
Дюв обнаружил несколько новых клеточных органелл. В том, как работает живая
клетка, и Моно, и де Дюв разбирались превосходно. К тому же они поддерживали
дружеские отношения, благо оба принадлежали к франкоязычному миру. Но вот их
взгляды на происхождение жизни оказались совершенно разными.
Жак Моно считал, что раз жизнь (насколько мы пока знаем) уникальна, то нет
никакой необходимости объяснять ее появление чем-то иным, кроме игры слепого
случая. В конце концов, Вселенная настолько огромна, что где-нибудь на ее
просторах вполне может один раз произойти любое сколь угодно маловероятное
событие. Достаточно, чтобы вероятность такого события не была строго нулевой.
Это не дает повода принимать случайность за закономерность.
"Вселенная не была чревата жизнью, как и биосфера не была чревата челове-
ком , — писал Моно. — Нам просто выпал счастливый шанс в рулетке, как тому,
кто только что выиграл миллион в казино".
Де Дюв, наоборот, считал, что появление жизни — это закономерный процесс,
готовый реализоваться на любой планете с подходящими физико-химическими усло-
виями. Возражая Моно, он говорил, что Вселенная как раз "чревата жизнью", и
приводил два довода, которые с тех пор стали только сильнее.
Во-первых, химические лл строительные блоки", пригодные для создания живых
систем, легко синтезируются в космосе. Хорошо известно, что они входят в со-
став метеоритов и комет. Де Дюв пользуется здесь трудно переводимым выражени-
ем "vital dust" — "жизнетворная пыль". По его словам, "жизнетворная пыль"
пронизывает всю Вселенную и образует своего рода семена жизни, готовые взойти
на любом подходящем небесном теле. И в самом деле, сейчас мы точно знаем, что
в метеоритах есть аминокислоты, сахара, азотистые основания, спирты, жирные
кислоты и другие углеродные соединения, причем все они там довольно разнооб-
разны. Похоже, что их синтез не требует никаких особенно редких условий.
Во-вторых, возникновение жизни — процесс по своей сути химический. Все ин-
формационные аспекты здесь вторичны. Истинные действующие лица — это нуклеи-
новые кислоты, углеводы, липиды и другие молекулы. А уж химию-то мы знаем хо-
рошо — и можем уверенно сказать, что все химические процессы в большой степе-
ни детерминистичны, то есть дают при одних и тех же условиях один и тот же
результат. Конечно, здесь участвует статистика (поскольку молекул очень мно-
го) , но в итоге на волю случая почти ничего не остается. Применительно к на-
шей задаче это должно означать, что если на какой-нибудь планете сложатся та-
кие же условия, какие были на Земле примерно четыре миллиарда лет назад, то
вероятность возникновения жизни там будет близка к единице.
Сейчас мы понимаем, что многие свойства живых объектов на самом деле предо-
пределены чистейшей химией. Например, каталитическая активность РНК, то есть
ее способность ускорять химические реакции, появляется автоматически, как
только начинают синтезироваться (каким угодно способом) цепочки РНК длиной
хотя бы в десятки нуклеотидов. Сборка жирных кислот и других липидов в мем-
браны, подобные клеточным, тоже происходит сама собой, стоит только им по-
пасть в водный раствор. Для всего этого достаточно подходящих внешних усло-
вий. Другое дело, что такие условия далеко не повсеместны, и — что еще важнее
— даже там, где они готовы сложиться, системе ничего не стоит выйти за их
пределы по каким-нибудь случайным причинам; пользуясь английской идиомой, это
проще, чем упасть с бревна (as easy as falling off a log) . Например, на со-
временной Венере зарождение жизни земного типа совершенно невозможно, хотя
почти по всем физическим параметрам эта планета очень близка к Земле и изна-
чально они, скорее всего, были гораздо более похожи друг на друга, чем сей-
час. Судьба Венеры наглядно показывает, насколько Земле повезло.
Первая
жизнь
Когда на Земле появилась жизнь? Самый распространенный до недавнего времени
ответ на этот вопрос таков: древнейшие предполагаемые остатки живых организ-
мов найдены в Гренландии, в горных породах зеленокаменной формации Исуа,
имеющих возраст 3,8 миллиарда лет. Значит, к этому времени жизнь уже точно
существовала. Правда, неизвестно, какая именно. И вот тут таится первая про-
блема. Дело в том, что найденные в Исуа остатки не сохранили никаких следов
структуры живых клеток. Это зерна чистого углерода, и вывод о том, что они
когда-то были живыми существами, сделан исключительно по составу этого угле-
рода.
Чтобы понять, в чем тут дело, вспомним, как устроены атомы (см. главу 1) .
Главный параметр любого атома — число протонов, или атомный номер (Z). Только
от него зависит, к какому химическому элементу атом относится. Однако в атом-
ном ядре есть не только протоны, но и нейтроны. Суммарное количество протонов
и нейтронов в ядре данного атома называется его массовым числом (А) . И вот
оно у атомов одного и того же элемента может различаться. Например, любой
атом, в ядре которого шесть протонов, будет атомом углерода. Но есть несколь-
ко разных типов атомов углерода, например с шестью нейтронами в ядре (12С)
или с семью нейтронами в ядре (13С) . Атомы, имеющие одинаковый атомный номер,
но разное массовое число, называются изотопами.
Углекислый газ (СОг) может включать в себя как атом 12С, так и атом 13С. Но
вот белок, связывающий углекислый газ для фотосинтеза, гораздо охотнее захва-
тывает молекулы СОг с углеродом 12С просто потому, что они более легкие. В ре-
зультате происходит разделение изотопов. На самом деле оно идет не только при
фотосинтезе, но и при других способах биологической фиксации СОг, но фотосин-
тез — самый распространенный из них на современной Земле. Однако в любом слу-
чае отсюда следует, что живые организмы, прямо или опосредованно питающиеся
продуктами фиксации углекислоты — то есть практически все живые организмы на
свете, — имеют смещенное соотношение лллегкого" и "тяжелого" изотопов углеро-
да: доля "тяжелого" изотопа там заметно понижена по сравнению с атмосферным
углекислым газом. А это означает, что, найдя чистый углерод, можно по соотно-
шению 12С/13С определить, является ли этот углерод биогенным, то есть, входил
ли он когда-нибудь в состав живого тела.
Но что, если при переплавлении горных пород заработал какой-нибудь другой,
чисто физический механизм разделения изотопов углерода? Это теоретически воз-
можно, и некоторые ученые считают, что в случае с породами Исуа именно так
оно и было. Тогда "следы самой древней жизни" исчезают. Нельзя сказать, что
эта тема закрыта, но статус пород Исуа сейчас определенно под сомнением. Пе-
чальнее всего, что биологи тут и сделать ничего не могут — решающее слово
принадлежит геологии и изотопной химии. Биогенное происхождение углерода из
Исуа не исключено, оно просто спорно. С другой же стороны, в самое последнее
время появились данные, что в формации Исуа есть остатки строматолитов —
слоистых бактериальных колоний, но это еще требует проверки.
В любом случае зеленокаменная формация Исуа, судя по всему, не предел. Не-
давно было объявлено, что найден предположительно биогенный углерод возрастом
4,1 миллиарда лет. Это совершенно поразительно, потому что для настолько
древних времен почти неизвестны полноценные горные породы. Главные сохранив-
шиеся с тех пор твердые объекты — это мелкие зерна чрезвычайно устойчивого
минерала циркона, захороненные где-нибудь в более поздних осадках. Вот в со-
ставе этих цирконовых зерен геологи и нашли углерод со смещенным изотопным
соотношением, типичным для живых организмов. По оценке авторов исследования,
другие пути разделения изотопов в данном случае маловероятны, так что это мо-
гут быть следы жизни — невообразимо древней жизни! Какой она была, остается
загадкой, ведь в изученных образцах налицо только химический сигнал.
Между тем первые живые организмы могли очень сильно отличаться от современ-
ных — причем под "современностью" в данном случае приходится подразумевать не
более и не менее как последние три с лишним миллиарда лет. Например, мы зна-
ем, что в современной цепочке передачи генетической информации есть три глав-
ных звена: копирование ДНК (репликация), синтез РНК (транскрипция) и синтез
белка (трансляция). При этом молекулярные данные свидетельствуют, что у обще-
го предка всех клеточных организмов системы транскрипции и трансляции были
намного более простыми, чем у современных клеток, а системы репликации ДНК не
было совсем. Еще Карл Вёзе показал, что белки репликации бактерий не имеют
ничего общего с белками репликации архей и эукариот. Скорее всего, это озна-
чает, что весь механизм копирования ДНК возник минимум дважды — у бактерий и
у архей (эукариоты унаследовали его от последних). Тогда получается, что у
общего предка всех их, вместе взятых, генетическая информация хранилась в ос-
новном на РНК, как и предполагает популярная в наше время теория ЛЛРНК-мира".
Кроме того, этот общий предок вполне мог еще не достичь так называемого
дарвиновского порога — момента, когда интенсивность привычной нам вертикаль-
ной передачи генов (от предков к потомкам) начала существенно превышать ин-
тенсивность горизонтального переноса генов (между соседними геномами незави-
симо от родства). Понятие "дарвиновский порог" (Darwinian Threshold) ввел тот
же Карл Вёзе — он вообще много занимался ранними этапами эволюции. Нам сейчас
трудно вообразить, как выглядела жизнь по ту сторону дарвиновского порога, но
ясно, что тогдашние организмы были предельно изменчивы: никаких устойчивых
видов в тех условиях существовать не могло. Нетрудно догадаться, что устойчи-
вость биологических видов определяется именно надежной передачей генетической
информации от предков к потомкам. Когда этот механизм еще не сложился, мир
был совершенно другим. Вёзе потому и назвал порог дарвиновским, что его пере-
ход означал происхождение видов в самом, что ни на есть буквальном смысле
слова "вид" (а ЛЛПроисхождением видов", как известно, называется главная книга
Дарвина). Живые системы, не достигшие дарвиновского порога, просто не могли
распадаться на биологические виды таким же образом, как распадается на них
жизнь сейчас.
Самые древние более-менее достоверно определимые остатки живых клеток имеют
возраст 3,4 миллиарда лет. Это уже типичные прокариоты, скорее всего, входя-
щие в дожившую до современности группу сульфатредуцирующих бактерий. На этой
отметке заканчивается туманная повесть о происхождении жизни и начинается ее
собственная история.
Обоюдоострый
меч 02
Переходя ко второму порогу — кислородной революции, надо, прежде всего,
сказать, что биологические свойства молекулярного кислорода (02) как минимум
двуедины. Кислород — мощный окислитель, с помощью которого можно получить
много полезной энергии, и в то же время сильный яд, свободно проходящий
сквозь клеточные мембраны и разрушающий структуру клеток. Иногда говорят, что
кислород — это обоюдоострый меч. Древнейшим жителям Земли существа, дышащие
кислородом, скорее всего, показались бы кем-то вроде людей с фторной планеты
из повести "Сердце Змеи" (см. главу 2) . У всех организмов, имеющих дело с ки-
слородом, обязательно есть специальные ферментные системы, гасящие его агрес-
сивное химическое воздействие. А те, у кого таких ферментных систем нет, об-
речены быть строгими анаэробами, выживающими только в бескислородной среде.
На современной Земле это некоторые бактерии и археи.
Практически весь молекулярный кислород на Земле имеет биогенное происхожде-
ние, то есть выделяется живыми существами. Главный источник 02 — это кисло-
родный фотосинтез. Других процессов, способных давать его в сравнимых количе-
ствах , просто нет. Как мы уже знаем, фотосинтезом называется синтез глюкозы
(C6Hi206) из углекислоты (С02) и воды (Н20) , происходящий с помощью энергии
света. Главным "действующим лицом" тут служит углекислый газ, который восста-
навливается водой. Кислород же в этой реакции — не что иное, как побочный
продукт, отход. Фотосинтез может вообще не приводить к выделению кислорода,
если вместо воды в нем используется в качестве восстановителя какое-нибудь
другое вещество — например, сероводород, свободный водород или соединения же-
леза. Такой фотосинтез называется бескислородным, и есть несколько его разных
вариантов.
Бескислородный фотосинтез наверняка появился раньше кислородного. Поэтому в
первый миллиард лет существования жизни (а скорее всего, дольше) фотосинтез
хотя и шел, но никакого насыщения атмосферы Земли кислородом не вызывал. Со-
держание кислорода в атмосфере в те времена составляло не больше 0,001% от
современного — попросту говоря, это значит, что его там не было.
Все изменилось, когда на сцену вышли сине-зеленые водоросли, или цианобак-
терии. Впоследствии именно эти существа стали предками хлоропластов, фотосин-
тезирующих элементов клеток эукариот. Цианобактерии — очень древняя эволюци-
онная ветвь. По меркам земной истории, они удивительно неизменны. Например,
широко распространенная в современных водоемах сине-зеленая водоросль осцил-
лятория имеет ископаемых родственников, живших 800 миллионов лет назад, при-
чем от современных осцилляторий они практически неотличимы. Таким образом,
осциллятория — чрезвычайно впечатляющий пример живого ископаемого. Но самые
первые цианобактерии появились намного раньше нее, это надежно подтверждено
палеонтологическими данными.
Поначалу цианобактерии не были многочисленны, потому что освоенный ими ки-
слородный фотосинтез не давал никаких серьезных преимуществ по сравнению с
бескислородным, которым владели другие группы микробов. Но химическое окруже-
ние этих микробов постепенно менялось. Наступил момент, когда "сырья" для
бескислородного фотосинтеза просто перестало хватать. И вот тогда час циано-
бактерии пробил. Кислородный фотосинтез имеет одно большое преимущество и
один большой недостаток. Преимущество — это совершенно неограниченный запас
исходного реагента-восстановителя (воды), а недостаток — высокая токсичность
побочного продукта (кислорода). Неудивительно, что поначалу этот тип обмена
не был "популярен". Зато при малейшем дефиците других субстратов, кроме воды,
обладатели кислородного фотосинтеза должны были сразу получить конкурентное
преимущество, что, по всей видимости, и произошло. После этого наступила эпо-
ха длиной примерно в миллиард лет, на протяжении которой облик Земли опреде-
ляли в первую очередь цианобактерии. Недавно эту эпоху даже предложили неофи-
циально назвать в их честь "цианозоем".
Именно из-за цианобактерии 2,4 миллиарда лет назад началась кислородная ре-
волюция, она же кислородная катастрофа, или Великое окислительное событие
(Great Oxidation Event, GOE). Строго говоря, это событие не было ни мгновен-
ным, ни абсолютно уникальным. Краткие всплески концентрации кислорода, так
называемые ллкислородные дуновения", случались и раньше — это палеонтологиче-
ски зафиксировано. И все же 2,4 миллиарда лет назад произошло нечто новое. За
короткое по меркам земной истории время (считанные десятки миллионов лет)
концентрация кислорода в атмосфере выросла примерно в 1000 раз и осталась на
этом уровне (см. рис. 16.2). До прежних ничтожных величин она не опустилась
больше никогда. Биосфера необратимо стала кислородной.
1,000-
4,0? 100-
g £ 10-
2 §
«0 О
^1 1 -
§& 0.1-
8.8
| 5 0.01 -
ZJ ОС
5 1
* - 0001 -
0 0001 -
J
П*Л/
1 'Кислородная
Г революция"
Т 1 1 Т 1 1 ! 1 1 1 1 1 1 !
4 3 2 10
Время (млрд лет назад)
Рис. 16.2.График изменения концентрации кислорода в атмосфере
Земли. Первый резкий подъем — это и есть кислородная революция.
Обратим внимание, что вертикальная шкала здесь логарифмическая:
это означает, что любое ее деление соответствует изменению не на
единицу, а в десять раз.
Для подавляющего большинства древних прокариот такой уровень кислорода был
смертельно опасен. Неудивительно, что первым результатом кислородной револю-
ции, скорее всего, стало вымирание (хотя, возможно, и не столь катастрофиче-
ское, как думали ученые лет 30 назад, когда появился термин "кислородный хо-
локост") . Выжили в первую очередь те, кто успел создать защищающие от кисло-
рода ферментные системы. Есть основания полагать, что в первые 100-200 мил-
лионов лет ллнового кислородного мира" кислород был для живых организмов толь-
ко ядом, и ничем больше. А вот потом ситуация поменялась. Ответом биоты на
кислородный вызов стало появление бактерий, которые включили кислород в це-
почку реакций, разлагающих глюкозу, и таким образом начали использовать его
для получения энергии.
Сразу оказалось, что кислородное окисление глюкозы (дыхание) намного энер-
гетически эффективнее бескислородного (брожения). В расчете на одну молекулу
глюкозы оно дает в несколько раз больше свободной энергии, чем любой сколь
угодно ЛЛнавороченный" вариант бескислородного обмена. При этом все начальные
этапы распада глюкозы у пользователей дыхания и брожения остались общими: ки-
слородное окисление послужило всего лишь надстройкой над уже имевшимся древ-
ним биохимическим механизмом, который сам по себе в кислороде не нуждался.
Группа микробов, которая освоила рискованное, но эффективное получение
энергии с помощью кислорода, называется протеобактериями. Согласно общеприня-
той сейчас теории, именно от одной из их подгрупп произошли дыхательные орга-
неллы эукариотных клеток — митохондрии. Судя по генетическим данным, ближай-
ший современный родственник митохондрий — это пурпурная спиральная альфа-
протеобактерия Rhodospirillum rubrum. Родоспириллум обладает и дыханием, и
брожением, и бескислородным фотосинтезом (в котором вместо воды используется
сероводород), и может переключаться между этими тремя типами метаболизма в
зависимости от внешних условий. Без сомнения, такой симбионт был предку эука-
риот очень полезен.
Более того, многие современные ученые считают, что симбиоз древних архей с
альфа-протеобактериями был толчком к образованию эукариотнои клетки. Это
"раннемитохондриальная" гипотеза, которую мы уже обсуждали в главе 10. Как мы
помним, она предполагает, что разделение будущей эукариотнои клетки на цито-
плазму и ядро произошло только после того, как в нее внедрился симбионт —
протеобактерия. Более старый ллпозднемитохондриальный" сценарий, согласно ко-
торому протеобактерия была попросту проглочена готовой эукариотнои клеткой,
сейчас выглядит несколько менее вероятным. Но надо учитывать, что в любом
случае обе клетки — и архейная, и протеобактериальная — были в процессе объе-
динения серьезно "пересобраны", породив своего рода химеру с новыми свойства-
ми. Эта химера и стала эукариотнои клеткой. Молекулярные компоненты архейного
и протеобактериального происхождения в ней сильно перемешались, разделив меж-
ду собой функции. Так или иначе, без протеобактерий эукариоты не возникли бы.
А это означает, что их появление было прямым следствием кислородной револю-
ции. В свете сказанного почти не выглядят преувеличением слова двух современ-
ных крупных ученых, палеонтолога и геолога: "Все согласны с тем, что эволюция
сине-зеленых водорослей была самым значительным биологическим событием на на-
шей планете (даже более значительным, чем развитие эукариотических клеток и
появление многоклеточных организмов)". Действительно, знакомый нам мир живот-
ных и растений не существовал бы, если бы не цианобактерии и вызванный ими
кризис. При всей мало привлекательности марксизма приходится признать, что
известный тезис Маркса "революции — локомотивы истории" применительно к био-
сферным революциям иногда все-таки подтверждается.
Эпохи
жизни
Вся история Земли делится на четыре огромных промежутка, именуемых зонами.
Названия эонов следующие:
■ катархей или гадей (4,6-4,0 млрд. лет назад),
■ архей (4,0-2,5 млрд. лет назад),
■ протерозой (2,5-0,54 млрд. лет назад) и
■ фанерозой (начался 0,54 млрд. лет назад и продолжается сейчас).
Первые три эона (то есть все, кроме фанерозоя) принято объединять понятием
ллкриптозой", что буквально значит ЛЛскрытая жизнь". Это деление будет нам по-
стоянно помогать. Сделаем оговорку, что почти во всех подобных случаях запо-
минать стоит не точные датировки, а саму последовательность эпох и относящих-
ся к ним событий: она гораздо важнее (см. рис. 16.1). Исключение можно сде-
лать разве что для двух-трех ключевых дат вроде возраста Земли.
Катархей — это так называемая догеологическая эпоха, от которой почти не
осталось "нормальных" горных пород, расположенных послойно. Классические гео-
логические и палеонтологические методы, основанные на сравнении последова-
тельных слоев отложений, там принципиально неприменимы. Оставшиеся от катар-
хея твердые объекты — это в первую очередь мелкие зерна циркона, те самые, в
которых недавно нашли предположительно биогенный углерод. О катархейской жиз-
ни (если она была) известно крайне мало.
В архее Земля принадлежит прокариотам — бактериям и археям (совпадение кор-
ней в названии геологической эры "архей" и группы микробов "археи" случайно).
Граница архея и протерозоя приходится примерно на момент одного из сильных
"кислородных дуновений", предшествовавших кислородной революции. Сама кисло-
родная революция произошла уже внутри протерозоя, хотя и в самом его начале.
Протерозой — это эпоха кислорода и эукариот. О кислороде мы уже говорили, а
с датировкой происхождения эукариот связан интересный парадокс. Дело в том,
что более-менее надежно определимые многоклеточные эукариоты появляются в па-
леонтологической летописи гораздо раньше, чем столь же надежно определимые
одноклеточные. Нитчатая водоросль Grypania spiralis, которую обычно считают
зукариотом, появилась 2,1 миллиарда лет назад. Справедливости ради нужно ска-
зать , что главным доводом за эукариотную природу грипании служит ее крупный
размер — все остальные признаки на самом деле не дают уверенности, что это не
гигантская колония цианобактерий. Но дело в том, что эта находка далеко не
единственная. Самым древним известным эукариотом сейчас считается грибообраз-
ный организм Diskagma buttonii возрастом 2,2 миллиарда лет. А еще есть зага-
дочные крупные спиралевидные существа — скорее всего, водоросли, возраст ос-
татков которых не меньше 2,1 миллиарда лет, как и у грипании. Зато самые ран-
ние одноклеточные, однозначно определяемые как эукариоты, имеют возраст всего
1,6 миллиарда лет. Это, разумеется, не значит, что многоклеточные эукариоты
возникли раньше одноклеточных — такое предположение противоречит и молекуляр-
ным данным, и здравому смыслу. Одноклеточные просто хуже сохраняются, да и
признаков, по которым можно понять, что это за организм, у них меньше.
Из приведенных цифр следуют важные общебиологические выводы. Вспомним, что
дата кислородной революции — 2,4 миллиарда лет назад. Следовательно, мы те-
перь знаем, что всего через 200 миллионов лет после кислородной революции в
палеонтологической летописи появляются не просто эукариоты, а многоклеточные
эукариоты. Это означает, что первые этапы эволюции эукариот были пройдены, по
меркам глобальной истории, очень быстро. Безусловно, эукариотной клетке по-
требовалось некоторое время, чтобы оформить симбиоз с предками митохондрий,
создать ядро, усложнить цитоскелет. Но после того, как эти процессы заверши-
лись , первые многоклеточные организмы возникли буквально сразу же. На уровне
клетки это не потребовало никаких дополнительных приспособлений. Любая эука-
риотная клетка уже имеет в наличии полный набор молекулярных структур, необ-
ходимых, чтобы построить из таких клеток многоклеточное тело (во всяком слу-
чае, относительно простое). Разумеется, все эти структуры не менее полезны и
для жизни одиночной клетки, иначе они просто не появились бы. Общий предок
эукариот, без сомнений, был одноклеточным, и очень многим его потомкам много-
клеточность так никогда и не пригодилась. Насколько разнообразны современные
одноклеточные эукариоты, мы отчасти знаем из главы 15.
Добавим, что совсем недавно палеонтологи описали нитчатые структуры, очень
похожие по деталям строения на настоящую грибницу, в горных породах, возраст
которых оценивается в 2,2-2,4 миллиарда лет (причем 2,2 миллиарда лет — это
минимальная оценка). Кстати, имеющая возраст 2,2 миллиарда лет Discagma во
многом напоминает относительно примитивные современные грибы — зигомицеты
(см. рис. 16.3). Если предположение о таком огромном возрасте грибов подтвер-
дится, это заодно послужит прекрасным подтверждением гипотезы ллэукариотного
Большого взрыва", которую мы обсуждали в главе 15. Тогда волей-неволей полу-
чится, что все главные эволюционные ветви эукариот возникли очень быстро, за
считанные десятки миллионов лет, — типичная взрывная эволюция. Альтернативное
объяснение могло бы состоять в том, что эукариоты появились на несколько со-
тен миллионов лет раньше, чем мы сейчас думаем. Такая гипотеза тоже существу-
ет, но она пока более чем спорная.
Кислородная революция имела еще одно важное последствие, коснувшееся соста-
ва атмосферы. В архейской атмосфере было много азота (как и сейчас), а также
углекислого газа и метана (гораздо больше, чем сейчас). Углекислый газ и ме-
тан очень хорошо поглощают инфракрасное излучение и тем самым удерживают в
атмосфере Земли тепло, мешая ему уходить в космос. Это называется парниковым
эффектом. Причем считается, что от метана парниковый эффект как минимум раз в
20-30 сильнее, чем от углекислого газа. А в архейские времена метана в атмо-
сфере Земли было примерно в 1000 раз больше, чем сейчас, и это обеспечивало
довольно теплый климат.
Рис. 16.3. Самый древний известный эукариот — загадочный орга-
низм Diskagma button!! возрастом 2,2миллиарда лет. Он напоминает
строением современные гломеромицеты — примитивные грибы, живущие
в симбиозе с сине-зелеными водорослями.
Тут вмешивается еще и астрономия. Согласно общепринятой теории эволюции
звезд, светимость Солнца медленно, но непрерывно растет. В архее она состав-
ляла всего 70-80% от современной — понятно, почему парниковый эффект был ва-
жен для поддержания планеты в тепле. Но после кислородной революции атмосфера
стала окислительной и почти весь метан (СН4) превратился в углекислый газ
(С02), эффективность которого как парникового газа гораздо ниже. Это вызвало
катастрофическое гуронское оледенение, длившееся около 100 миллионов лет и в
некоторые моменты охватывавшее практически всю Землю: на участках суши, кото-
рые тогда находились всего в нескольких градусах широты от экватора, найдены
следы ледников. Пик гуронского оледенения наступил 2,3 миллиарда лет назад. К
счастью, оледенение не могло остановить тектоническую активность земной ман-
тии . Вулканы продолжали выбрасывать в атмосферу углекислый газ, и со временем
его накопилось достаточно, чтобы восстановить парниковый эффект и растопить
льды.
Надо сразу сказать, что такое климатическое испытание было в истории Земли
не последним. Самое широко известное глобальное оледенение — позднепротеро-
зойское, о котором речь впереди. Оно оставило больше следов, чем раннепроте-
розойское гуронское оледенение, и было раньше открыто палеонтологами, став в
свое время настоящей сенсацией. Видимо, при определенных обстоятельствах Зем-
ле было не так уж и трудно превратиться в ледяной шар.
ЛЛ Скучный
миллиард лет"
и его конец
За бурными событиями начала протерозоя последовал так называемый ЛЛскучный
миллиард лет" (Boring Billion). В это время не происходило никаких оледене-
ний, никаких резких перемен в составе атмосферы, никаких биосферных переворо-
тов . Эукариотные водоросли жили в океанах, понемногу выделяя кислород. Их мир
был по-своему разнообразным и сложным. Например, из эпохи "скучного миллиар-
да" известны многоклеточные красные и желто-зеленые водоросли, поразительно
похожие на своих современных родственников. Известны оттуда и грибы. А вот
многоклеточные животные на просторах ЛЛ скучного миллиарда лет" отсутствуют.
Будем аккуратны: на нынешний момент никто не может с полной уверенностью ут-
верждать , что многоклеточных животных тогда не было, но аргументы в пользу
того, что они были, в лучшем случае очень сомнительны.
В чем тут дело? Напрашивается мысль, что многоклеточность как таковая го-
раздо более совместима с образом жизни растения, чем с образом жизни животно-
го. Любая клетка растения заключена в жесткую клеточную стенку, и нет сомне-
ний, что это сильно облегчает регуляцию взаимного расположения клеток в слож-
ном теле. Наоборот, клетки животных вообще лишены клеточной стенки, их форма
неустойчива, да еще и постоянно меняется при актах движения или фагоцитоза.
Собрать из таких клеток целый организм — сложная задача. Если бы никаких мно-
гоклеточных животных не появилось вовсе, а биологами стали представители рас-
тений либо грибов, они, скорее всего, после изучения этой проблемы пришли бы
к выводу, что сочетание многоклеточности с отсутствием клеточной стенки про-
сто невозможно. Во всяком случае, это объясняет, почему многоклеточность мно-
го раз возникала в разных группах водорослей, но только один раз — у живот-
ных.
Есть и другая идея. В 1959 году канадский зоолог Джон Ральф Нерселл связал
внезапное (как тогда считалось) появление животных в палеонтологической лето-
писи с ростом концентрации кислорода в атмосфере. Животные, как правило, об-
ладают активной подвижностью, которая требует столько энергии, что без кисло-
родного дыхания им никак не обойтись. И кислорода им нужно много. А в эпоху
ллскучного миллиарда" содержание 02 в атмосфере почти наверняка не достигало
10% от современного уровня — минимума, который принято считать необходимым
для поддержания животной жизни. Правда, эта подозрительно круглая цифра, ско-
рее всего, завышена: физиологические исследования показывают, что многие жи-
вотные могут обойтись и меньшим количеством кислорода. Подобные оговорки, од-
нако, не мешают признать, что старая идея Нерселла как минимум не противоре-
чит современным данным: предполагаемое начало эволюции многоклеточных живот-
ных весьма приближенно, но все-таки совпадает по времени с новым ростом кон-
центрации атмосферного кислорода в конце протерозоя. Это просто не могло не
стать фактором, облегчившим появление животных: в конце-то концов, ясно ведь,
что чем больше кислорода, тем для них лучше. Не надо только считать кислород-
ный фактор строго единственным. Будем помнить, что и во времена, когда кисло-
рода стало сколько угодно, никаких многократных попыток создания многоклеточ-
ности животного типа не отмечается. Этот эксперимент удался природе только
один раз.
Уютная эпоха "скучного миллиарда лет" могла бы длиться еще долго, если бы в
биологию не вмешалась география. Драматические события, героем которых стала
сама планета, привлекали внимание ученых на протяжении полувека, но только
лет 15 назад информацию о них удалось сложить в более-менее цельную картинку.
Бросим на эту картинку беглый взгляд, начав, как и положено, с начала.
ЛЛЗемля-снежок":
пролог
В 1964 году английский геолог Брайан Харленд опубликовал статью, в которой
констатировал, что абсолютно на всех континентах есть следы древнего оледене-
ния, относящегося к одному и тому же времени — позднему протерозою. Как раз в
начале 1960-х геологи научились определять прошлое положение континентов с
помощью данных о намагниченности горных пород. Харленд собрал эти данные и
увидел, что убедительно объяснить их можно только одним способом: предполо-
жив, что позднепротерозойское оледенение охватило сразу все широты Земли, то
есть, было всепланетным. Любые другие гипотезы выглядели еще менее правдопо-
добными (например, пришлось бы предполагать немыслимо быстрое перемещение по-
люсов, чтобы все континенты по очереди накрывались полярной шапкой). Как ска-
зал Шерлок Холмс во время поисков Джонатана Смолла, "отбросьте все невозмож-
ное, то, что останется, и будет ответом, каким бы невероятным он ни казался".
Именно так Харленд и поступил. Выпущенная им с соавтором обстоятельная, напи-
санная прекрасным языком статья не претендует ни на какие сенсации — там про-
сто-напросто честно изложены факты и выводы. И все же гипотеза о всепланетном
оледенении была для большинства ученых слишком смелой. Поэтому популярность
она получила далеко не сразу.
Буквально в те же годы теорией оледенений занялся известный геофизик, ле-
нинградец Михаил Иванович Будыко. Он обратил внимание на то, что оледенение
может саморазвиваться. Ледяной покров имеет высокую отражательную способность
(альбедо), поэтому, чем больше суммарная площадь ледников, тем большая доля
солнечного излучения отражается обратно в космос, унося с собой тепло. А чем
меньше Земля получает тепла, тем на ней становится холоднее, и площадь ледя-
ного покрова в результате растет, повышая альбедо еще сильнее. Получается,
что оледенение — это процесс с положительной обратной связью, то есть способ-
ный усиливать сам себя. А в таком случае должен существовать некоторый крити-
ческий уровень оледенения, после которого оно будет нарастать, пока волны
льда с Северного и Южного полюсов не схлопнутся на экваторе, полностью заклю-
чив планету в ледяной покров и понизив ее температуру на несколько десятков
градусов. Будыко математически показал, что такое развитие событий возможно.
Но он и понятия не имел, что в истории Земли несколько раз это действительно
происходило! Потому что на тот момент Будыко и Харленд еще не читали друг
друга.
лл3емля-снежок":
основное действие
Сейчас оледенение, которое открыл Харленд, принято называть эпохой "Земли-
снежка" (Snowball Earth). Судя по всему, оно действительно было всепланетным.
А главной его причиной считается резкое ослабление парникового эффекта из-за
падения концентрации углекислого газа (который стал главным парниковым газом
после того, как кислород ЛЛсьел" почти весь метан). Фотосинтез и дыхание тут,
скорее всего, ни при чем. Если кислородную революцию биота Земли устроила се-
бе сама, то теперь она оказалась жертвой внешнего фактора, совершенно небио-
логического по своей природе.
Дело в том, что оборот углекислого газа в целом гораздо меньше зависит от
живых существ, чем оборот кислорода. Основным источником атмосферного С02 на
Земле до сих пор служат извержения вулканов, а основным стоком — процесс, ко-
торый называется химическим выветриванием. Углекислый газ взаимодействует с
горными породами, разрушая их, а сам при этом превращается в карбонаты (ионы
НСОз" или СОз2) . Последние отлично растворяются в воде, зато в состав атмо-
сферы больше уже не входят. И получается предельно простая зависимость. Если
интенсивность работы вулканов превосходит интенсивность химического выветри-
вания , атмосферная концентрация С02 растет. Если наоборот — падает.
На исходе "скучного миллиарда", 800 миллионов лет назад, почти вся земная
суша входила в состав единственного суперконтинента под названием Родиния. По
словам известных геологов, гигантские суперконтиненты, как и крупные империи
в социальной истории Земли, всегда оказывались неустойчивыми. Поэтому неуди-
вительно, что Родиния быстро начала раскалываться. По краям разломов застывал
извергнутый базальт, который сразу же становился объектом химического вывет-
ривания. А почвы тогда не было, и продукты выветривания легко сносились в
океан. В конце концов, Родиния распалась на семь или восемь небольших — раз-
мером примерно с Австралию — континентов, которые стали дрейфовать в стороны
друг от друга.
Расход С02 на выветривание базальта привел к падению его уровня в атмосфе-
ре. Вулканизм, которым распад суперконтинента неизбежно сопровождался, мог бы
это скомпенсировать, если бы не одно случайное обстоятельство. В силу каких-
то причуд дрейфа континентов и Родиния, и ее обломки находились у экватора, в
теплом поясе, где химическое выветривание шло особенно быстро. Математические
модели показывают, что именно по этой причине концентрация С02 опустилась ни-
же порога, за которым начинается оледенение. А когда оно началось, останавли-
вать выветривание было уже поздно.
Надо признать, что положение континентов в позднем протерозое оказалось на-
столько неудачным (с точки зрения обитателей планеты), насколько это вообще
возможно. Дрейф континентов направляется потоками вещества земной мантии, ди-
намика которых, по сути, неведома. Но мы знаем, что в данном случае эти пото-
ки собрали всю земную сушу в единый континент, находившийся точно на экваторе
и вытянутый по широте. Если бы он оказался на одном из полюсов или был вытя-
нут с севера на юг, начавшееся оледенение закрыло бы часть пород от выветри-
вания и тем самым приостановило уход углекислого газа из атмосферы — тогда
процесс мог бы притормозиться. Как раз такую ситуацию мы наблюдаем сейчас,
когда есть ледяные щиты Антарктиды и Гренландии. А в конце протерозоя почти
все крупные участки суши находились близко к экватору — и оставались обнажен-
ными до того самого момента, когда северный и южный ледяные покровы, наконец,
сомкнулись. Земля стала ледяным шаром.
На самом деле эпизодов "Земли-снежка" было не меньше трех. Первый из них
относится еще к гуронскому оледенению (которое, как мы помним, произошло не
из-за углекислого газа, а из-за метана). Потом в течение более чем миллиарда
лет никаких оледенений не было совсем. А затем последовало еще два разделен-
ных небольшим перерывом всепланетных оледенения, одно из которых длилось при-
мерно 60 миллионов лет, другое — примерно 15 миллионов лет. Именно их открыл
Брайан Харленд. Геологический период, охватывающий эти оледенения, называют
криогением (он является частью протерозойской эры).
О живой природе криогения известно относительно мало. Климат тогда на всей
Земле был, по нынешним меркам, антарктическим. Большую часть Мирового океана
покрывал километровый слой льда, так что интенсивность фотосинтеза не могла
быть высокой. Свет, неожиданно ставший ценнейшим ресурсом, попадал в океан
только местами, сквозь трещины, полыньи или небольшие участки тонкого льда.
Удивительно, что некоторые многоклеточные организмы сумели пережить криоге-
ний, совершенно не изменившись, — например, красные водоросли. Они и сейчас
приспособлены к тому, чтобы использовать очень слабый свет, проникающий на
такую глубину, где уже не могут жить никакие другие фотосинтезирующие сущест-
ва. Никуда не исчез и одноклеточный планктон. Содержание кислорода в криоге-
ниевом океане сильно упало, поэтому жизнь на его дне, скорее всего, была в
основном анаэробной, но подробности этого от нас пока скрыты.
Рис. 16.4. Динамика типичного эпизода "Земли-снежка" на примере
первой (стёртской) фазы криогениевого глобального оледенения.
График показывает изменение температуры земной поверхности.
Окончания эпизодов "Земли-снежка" тоже были по-своему драматичны. Во время
всепланетных оледенений все процессы, связанные с поглощением больших объемов
углекислого газа, в буквальном смысле замораживались. А между тем вулканы
(работу которых никто не останавливал) выбрасывали и выбрасывали С02 в атмо-
сферу, постепенно доводя его концентрацию до огромных величин. В какой-то мо-
мент ледяной щит уже не мох1 сопротивляться парниковому эффекту, и тогда начи-
нался лавинообразный процесс разогрева планеты (см. рис. 16.4). Буквально за
несколько тысяч лет — то есть геологически за мгновение — весь лед таял, ос-
вободившаяся вода заливала значительную часть суши мелкими окраинными морями,
а температура земной поверхности, судя по расчетам, подскакивала до 50 С. И
только после этого начинался постепенный возврат Земли к "нормальному" для
нее внеледниковому состоянию. За время криогения весь этот цикл был пройден
минимум дважды.
Необычный
мир венда
Эпоха "Земли-снежка" закончилась 635 миллионов лет назад. Начался последний
период протерозоя — эдиакарий (635-542 млн. лет назад). Теперь нам будет
удобнее вести счет времени не на миллиарды лет, а на миллионы — это наглядно
показывает, насколько события ускоряются. Хотя, возможно, дело просто в том,
что они к нам ближе и от них сохранилось больше следов.
Раньше эдиакарий называли вендом, в честь нескольких групп древних индоев-
ропейских племен — венедов (от этого же слова, по-видимому, произошло назва-
ние города Венеция). К сожалению, сейчас это красивое наименование сохрани-
лось лишь как нестрогий синоним.
Главным событием эдиакария (нельзя не добавить: с нашей антропоцентричной
точки зрения) следует назвать появление многоклеточных животных. Датировать
это событие нелегко. В палеонтологической летописи эдиакария хватает свиде-
тельств перехода к многоклеточности животного типа — правда, чем они более
ранние, тем более спорные. Во второй половине эдиакария в изобилии появляются
вендобионты — крупные (до метра длиной) загадочные существа с плоским диско-
видным или листовидным телом, состоящим из множества однотипных повторяющихся
"сегментов" (см. рис. 16.5). "Сегменты" здесь поставлены в кавычки, потому
что сегментация вендобионтов почти наверняка не имеет ничего общего с сегмен-
тацией настоящих многоклеточных животных. Сам термин "вендобионты" предложил
немецкий палеонтолог Адольф Зейлахер, который считал этих существ совершенно
особой формой жизни — гигантскими многоядерными клетками.
Действительно, есть серьезные основания считать, что вендобионты были во
многих отношениях ближе не к многоклеточным животным, а к амебам или грибам
(между прочим, и у тех и у других большие многоядерные клетки не редкость).
Они совершили попытку выхода в крупный размер, которая сначала привела к ус-
пеху, но в итоге кончилась неудачей: в конце эдиакария вендобионты вымерли.
При этом надо учитывать, что вендобионты были очень разнообразны. Не факт,
что их вообще можно хоть в каком-то приближении считать единой группой. Это
скорее эволюционный уровень. И несмотря на то, что большинство вендобионтов
никаких потомков не оставило, от кого-то из них вполне могли напрямую про-
изойти некоторые современные животные — например, такие типы, как пластинча-
тые и гребневики. В эдиакарских корнях этих эволюционных ветвей нет ничего
невероятного.
Самое древнее, абсолютно бесспорное ископаемое многоклеточное животное на-
зывается Kimberella quadrata (см. рис. 16.6). Это двусторонне-симметричное
существо длиной до 15 сантиметров, ползавшее по морскому дну. Характер изме-
нений формы тела найденных кимберелл (а найдено их много, в разных частях
света) вместе с отпечатками следов не оставляет никаких сомнений, что они ак-
тивно ползали, растягиваясь, сжимаясь и изгибаясь с помощью мышц. Характерные
признаки кимбереллы — вытянутое, но компактное тело с ногой (мускулистой ниж-
ней поверхностью) и мантией (складкой, окаймляющей туловище). По этим призна-
кам она очень похожа не на кого-нибудь, а на моллюсков. Есть мнение, что у
кимбереллы была даже радула — свойственный моллюскам "язык" с хитиновыми зуб-
цами, приспособленный для соскребания водорослей. Так или иначе, это уже по
всем статьям настоящее многоклеточное животное — с мышцами, нервной системой,
ротовым аппаратом и вообще всем, что такому животному полагается иметь.
Рис. 16.5. Некоторые вендобионты.
Рис. 16.6. Кимберелла.
Кимберелла жила 555 миллионов лет назад. И примерно в это же время впервые
появляются многочисленные ископаемые следы животных, явно активно ползавших
по дну. Тут надо заметить, что "настоящие многоклеточные животные" — не осо-
бенно строгий термин. Сейчас нам достаточно условиться, что мы называем так
животных с мускулатурой, нервной системой и ртом. У вендобионтов, насколько
можно судить, ничего этого не было. Они питались в лучшем случае микроскопи-
ческими водорослями, а скорее всего — просто веществами, растворенными в мор-
ской воде. И только в конце эдиакария появились многоклеточные существа, спо-
собные активно отыскивать добычу и захватывать ее крупными кусками, чтобы пе-
реварить внутри. Вендобионты были перед такими чудовищами беззащитны — неуди-
вительно, что их "золотой век" на этом закончился. В истории донных сообществ
началась совершенно другая эпоха.
17. ФАНЕР030Й
И солнце парит топь в полдневном жаре,
И в зарослях хвощей из затхлой мглы
Возносятся гигантских сигиллярий
Упругие и рыхлые стволы.
Косматые — с загнутыми клыками —
Пасутся мамонты у мощных рек,
И в сумраке пещер под ледниками
Кремень тяжелый точит человек.
Михаил Зенкевич. «Темное родство»
Конец эдиакарского периода является в то же время рубежом двух эонов — про-
терозоя и фанерозоя. И вот тут нужно небольшое пояснение.
"Фанерозой" буквально означает ЛЛявная жизнь". Это эпоха, к которой относит-
ся подавляющее большинство изучаемых палеонтологами ископаемых остатков. Все
предшествующие времена, включая протерозой, архей и катархей, собирательно
называют криптозоем — ЛЛскрытой жизнью". Фанерозой же, в свою очередь, делится
на три эры, названия которых, скорее всего, знакомы большинству читателей:
палеозой, мезозой и кайнозой. "Палеозой" буквально означает ЛЛдревняя жизнь",
ллмезозой" — ллсредняя жизнь" и ллкайнозой" — ллновая жизнь". А каждая из этих эр
делится на периоды. Период, с которого начинается палеозой (и тем самым весь
фанерозой), называется кембрием. Как и многие другие геологические периоды,
кембрий получил свое имя из географии: Кембрия — это римское название Уэльса,
кельтской страны на западе Британии. Соответственно, очень распространенный
синоним криптозоя — докембрий.
Чтобы правильно видеть перспективу, будем помнить следующее: весь фанерозой
составляет (округленно) всего лишь примерно 1/9 как от времени существования
Земли, так и от истории жизни на ней. Остальные 8/9 — это криптозой, или до-
кембрий, которому посвящена предыдущая глава. Иное дело, что в фанерозое со-
бытия очень сгущаются.
В 1845 году шотландский геолог Родерик Мурчисон предложил назвать все вре-
мена до начала кембрия азойской эрой, то есть — буквально — безжизненной.
Долго это название не продержалось: уже палеонтологи XIX века показали, что в
толще докембрийских пород есть следы жизни. А сейчас мы точно знаем, что
жизнь существовала на Земле на протяжении большей части докембрия, и можем
датировать многие ключевые события докембрийской истории — например, кисло-
родную революцию или появление многоклеточности. И о том и о другом мы вкрат-
це говорили в предыдущей главе.
лл Большой
взрыв жизни"
Главное отличие фанерозойской жизни от докембрийской — колоссальное обилие
многоклеточных животных. Причем подавляющее большинство этих животных с само-
го начала относится уже к современным типам. В кембрии появляются губки,
гребневики, стрекающие, круглые и кольчатые черви, членистоногие, моллюски,
брахиоподы, иглокожие, полухордовые и хордовые. Внезапное появление всех этих
животных в палеонтологической летописи принято называть кембрийским взрывом.
В более древних слоях их остатков нет (по крайней мере, определяемых одно-
значно и бесспорно). Таким образом, кембрий — это время рождения фауны, близ-
кой к современной. Кембрийский взрыв дал такой эффект и произошел так быстро,
что его часто называют "эволюционным Большим взрывом" — по аналогии с тем
Большим взрывом, в котором родилась Вселенная.
Еще кембрийский взрыв иногда называют скелетной революцией. Действительно,
многие появившиеся в этот момент группы животных приобрели твердые скелеты,
причем совершенно разные по структуре и на разной биохимической основе: на-
пример, между спикулами губок, раковинами моллюсков и хитиновыми панцирями
членистоногих нет буквально ничего общего. Такая одновременность никак не мо-
жет быть случайной. Тем не менее "кембрийский взрыв" и "скелетная революция"
— не синонимы. Во-первых, твердые скелеты были далеко не у всех кембрийских
животных (например, у первых хордовых их не было). Во-вторых, и в докембрии
иногда попадаются явные скелетные структуры — например, непонятно кому при-
надлежавшие жилые трубки. В общем, понятие "кембрийский взрыв" куда более оп-
ределенное, и неудивительно, что современные авторы чаще говорят именно о
нем.
А был ли
взрыв?
Но вот вопрос: произошел ли кембрийский взрыв на самом деле? Существует
мнение, что многие современные группы животных появились еще в глубоком до-
кембрии, но долгое время не оставляли ископаемых остатков, а потому были па-
леонтологически "невидимы". Причины этого могли быть разными: мелкий размер
животных, отсутствие у них твердых скелетов или просто неподходящие для захо-
ронения физические условия. Гипотеза ЛЛдолгой скрытой докембрийской эволюции"
неплохо поддерживается молекулярной филогенетикой, основанной на сравнении
аминокислотных и нуклеотидных последовательностей белков и генов разных жи-
вотных (разумеется, современных — ведь со времен кембрия ни белков, ни ДНК не
осталось). Реконструкции, сделанные исключительно по молекулярным данным, за-
частую уводят корни современных типов животных даже не в эдиакарий, а в пре-
дыдущий период — криогений. Тогда выходит, что кембрийский взрыв — это не
столько реальное эволюционное событие, сколько артефакт сохранности. На рубе-
же кембрия животные просто "проявились", обретя твердые скелеты и начав захо-
раниваться в осадочных толщах. А возникли они гораздо раньше.
Однако есть впечатление, что тщательной проверки, при которой молекулярно-
биологические данные аккуратно шаг за шагом сопоставляются с палеонтологиче-
скими, гипотеза "долгой скрытой докембрийской эволюции" все же не выдержива-
ет. В 2013 году группа исследователей под руководством австралийского палео-
нтолога Майкла Ли попыталась провести такую проверку, использовав материал по
самому многочисленному типу животных — членистоногим. Были рассмотрены и мор-
фологические признаки, изучаемые палеонтологами, и последовательности ядерных
генов, изучаемые генетиками. Специальная компьютерная программа построила по
всему этому массиву данных единое эволюционное древо (причем молекулярные по-
следовательности она учитывала наравне с морфологией). Назовем его для крат-
кости "древом Ли". Программа, использованная для построения древа Ли, могла
определить скорость эволюции на каждом отрезке этого древа и в результате вы-
числить наиболее вероятные даты расхождений эволюционных ветвей. Это и было
целью исследователей.
Теперь давайте следить за цифрами. Считается, что кембрийский период начал-
ся 542 миллиона лет назад. Согласно древу Ли, расхождение двух главных эволю-
ционных ветвей членистоногих — Chelicerata (мечехвосты и паукообразные) и
Mandibulata (многоножки, ракообразные и насекомые) — произошло 543 миллиона
лет назад. А расхождение двух подгрупп внутри Mandibulata, многоножек и рако-
образных, произошло 538 миллионов лет назад. Как видим, граница кембрия попа-
дает точно между этими датами: Chelicerata и Mandibulata разошлись перед са-
мым началом кембрия, а многоножки и ракообразные — сразу после него. Понятно,
что точность таких датировок не абсолютна, но уж случайным совпадением они не
могут быть никак. Итак, мы видим, что рождение главных эволюционных ветвей
членистоногих произошло в ближайшей временной окрестности нижней границы кем-
брия. На этот момент действительно пришелся пик изменений.
Получается, что кембрийский взрыв — никакой не артефакт. Вероятно, большин-
ство крупнейших эволюционных ветвей животных действительно возникло в ближай-
шей временной окрестности границы кембрия (плюс-минус несколько миллионов
лет) . Есть и математические модели, подтверждающие, что погруженные в докем-
брий "стволы" эволюционных древес современных типов животных должны быть ко-
роткими. Время существования этих докембрийских "стволов" — считанные миллио-
ны лет, может быть, первые десятки миллионов, но уж никак не сотни миллионов.
В общем, на данный момент у нас достаточно оснований, чтобы считать гипотезу
ллдолгой скрытой докембрийской эволюции" скорее неверной, а кембрийский взрыв
— реальностью, как это, собственно говоря, и следует напрямую из палеонтоло-
гических данных.
Чтобы ослабить категоричность, добавим: вывод, который мы только что сдела-
ли, безусловно обладает свойством фальсифицируемости. Это означает, что можно
сформулировать четкие условия, при которых он будет опровергнут. Например,
для этого достаточно найти хотя бы одного достоверно определимого скорпиона
(или сороконожку, или улитку) криогениевого возраста. Но пока такого не было,
и вероятность, что это когда-нибудь случится, с каждым годом уменьшается.
Причины
взрыва
Итак, в начале кембрия невероятно быстро возникло множество новых крупных
эволюционных ветвей животных. Такого не случалось больше никогда, ни раньше,
ни позже. Даже после катастрофических массовых вымираний (о которых речь впе-
реди) животный мир восстанавливался за счет нарастания разнообразия уже суще-
ствовавших больших групп, а не за счет появления новых. Вот почему кембрий-
ский взрыв обязательно требует объяснения.
Правда, "невероятно быстро" не значит "мгновенно". Новые группы животных
отнюдь не появились в начале кембрия все разом в полном составе, как актеры
после подъема занавеса. Кембрийский взрыв был хоть и сильно сжатым во време-
ни, но все же постепенным. Скорость эволюционных процессов в нем вполне можно
измерить, и такие исследования, как мы видели, есть. Кембрий длился примерно
57 миллионов лет (542-485 млн. лет назад), при этом в самом его начале (пер-
вые шесть миллионов лет) морская фауна была еще довольно бедна. Новые группы
животных появились там действительно быстро, по меркам истории Земли, но во-
все не одномоментно.
С чем же все-таки это было связано? За полтора века, прошедших с тех пор,
как ученые (и в том числе Чарльз Дарвин) осознали загадку кембрийского взры-
ва, предлагались разнообразнейшие объяснения этого события, от генетических
до космических. Одна современная обзорная статья на эту тему так и называется
— ллПо ту сторону кембрийского взрыва: от галактики до генома". Например, тен-
денция к массовому образованию минеральных скелетов — знаменитая ЛЛскелетная
революция", она же "биоминерализация", — в начале кембрия охватила не только
самых разных многоклеточных животных, но и одноклеточных эукариот, и некото-
рые водоросли. Напрашивается предположение, что это было связано с глобальным
изменением химического состава внешней среды, то есть в данном случае морской
воды. И в самом деле, показано, что в начале кембрия по каким-то чисто геоло-
гическим причинам примерно в три раза выросла концентрация в морской воде
кальция (Са2+) — иона, который нужен для создания твердых скелетов как ника-
кой другой. Минеральная основа скелетов животных — чаще всего карбонат каль-
ция (раковины моллюсков, иглы и чашечки коралловых полипов, спикулы губок), а
иногда фосфат кальция (кости позвоночных). Уже из этих названий ясно, что без
кальция тут, так или иначе, не обойтись.
Проблема в том, что объяснить скелетную революцию еще не значит объяснить
сам кембрийский взрыв. Скелетная революция всего лишь снабдила твердыми мине-
рализованными тканями животных, уже существовавших к моменту ее начала. При-
чем это коснулось далеко не всех из них. В тех кембрийских местонахождениях,
тип сохранности которых допускает захоронение бесскелетных существ, сразу же
обнаруживается, что значительная часть кембрийской фауны так и осталась
"мягкотелой". Так что дело не в скелетах. Явление, которое надо объяснить в
первую очередь, — это уникальное ускорение эволюции многоклеточных животных,
очень быстро (за конец эдиакария и начало кембрия) создавшее множество новых
крупных групп, неважно, скелетных или нет.
В дальнейшем рассказе мы будем исходить из сценария, который кратко изложил
еще в начале 1970-х годов американский палеонтолог Стивен Стэнли. Конечно,
тут надо помнить, что палеонтология — очень быстро развивающаяся наука. Рабо-
ты 40-летней давности в ней всегда требуют поправок, и мы эти поправки по хо-
ду разговора внесем. Правда, в данном случае это будут скорее дополнения.
Главная идея Стэнли исключительно хорошо выдержала проверку временем. Сумма
фактов, известных на данный момент, вполне в нее вписывается.
Начнем с начала. На полях заметим: решить, что именно стоит принять за
"начало", при разборе любого исторического процесса — задача непростая, ведь
причинно-следственные цепочки могут тянуться в прошлое едва ли не до беско-
нечности, сбивая неосторожного исследователя с толку. В нашем случае
"началом" будет эдиакарская биота. Что же она собой представляла?
В экологии принято выделять организмы-ередообразователи, активность которых
определяет собой структуру целых сообществ. Такие организмы называются эдифи-
каторами. Например, в дубраве эдификатором является дуб, в маленьком тихом
пруду им вполне может быть ряска, и т.д. Так вот, в эдиакарских морях эдифи-
каторами были покрывавшие дно ЛЛковры" нитчатых водорослей — так называемые
водорослевые маты. На этих "коврах" жили уже знакомые нам вендобионты. Боль-
шинство из них вело прикрепленный образ жизни. Как они питались — не совсем
понятно, но, скорее всего, осмотрофно, всасывая из воды растворенные вещества
всей поверхностью тела. Таким способом до сих пор питаются некоторые морские
протисты, например крупные — до 2 0 сантиметров! — многоядерные ксенофиофоры,
похожие на гигантских амеб (они относятся к супергруппе Rhizaria). Вендобион-
ты вполне могли быть близки к ним по образу жизни.
Впрочем, на эту тему есть и другие идеи. В 1986 году палеонтолог Марк Мак-
менамин предположил, что вендобионты были экологическими аналогами современ-
ных погонофор — глубоководных кольчатых червей, лишенных рта и кишечника. По-
гонофоры живут в океане на такой глубине, куда солнечный свет не проникает.
Зато там встречаются горячие источники, выделяющие в воду сероводород (H2S).
Тело погонофоры набито симбиотическими бактериями, которые окисляют сероводо-
род до серы и полученную при этом энергию используют для фиксации углекисло-
ты , как при фотосинтезе. За счет этого процесса питаются и сами бактерии, и
червь, в котором они живут. Некоторые погонофоры питаются за счет других бак-
терий, окисляющих не сероводород, а метан, но тоже автотрофных. В любом слу-
чае погонофоры — это фактически автотрофные животные. Что касается вендобион-
тов , то им было еще проще: они часто жили на мелководье, где солнечного света
хватало для фотосинтеза, и вполне могли бы питаться за счет наполнявших их
тело симбиотических одноклеточных водорослей. Это тоже реально, есть совре-
менные черви и моллюски, которые именно так и делают. Правда, у них этот спо-
соб питания обычно является дополнительным, но почему бы ему не стать и ос-
новным? Мир вендобионтов, где никто никого не ел, Макменамин назвал "садом
Эдиакары", с явной шуточной аллюзией на сад Эдема. Большой недостаток этой
гипотезы в том, что ее пока трудно проверить. К тому же она заведомо не может
распространяться на всех вендобионтов без исключения: некоторые из них жили в
море глубже уровня, куда проникает достаточно света для фотосинтеза. Но, в
конце концов, в разных условиях они могли питаться по-разному.
Парадокс в том, что концепция "сада Эдиакары" представляется близкой к ис-
тине при любом сколько-нибудь реалистичном предположении о способе питания
вендобионтов. Тут уже и неважно, жили в них водоросли или нет. В эдиакарском
мире действительно никто никого не ел (не считая одноклеточных организмов).
Принципиально важно, что в эдиакарских сообществах до некоторого момента не
было не только хищников (которые питались бы другими животными), но и
"травоядных" (которые умели бы механически соскребать водоросли). Таким обра-
зом, водорослевым матам там никто особо не мешал расти.
Все изменилось, когда повышение концентрации кислорода в морской воде (судя
по геологическим данным, оно шло постепенно в течение всего эдиакария) позво-
лило некоторым многоклеточным существам ускорить обмен веществ настолько,
чтобы начать вести по-настоящему активный образ жизни. Тогда появились
"сборщики урожая" — крупные животные со сложной двигательной системой и ртом,
которые перемещались по водорослевым матам и выедали их значительные участки.
Одним из таких "сборщиков" была знакомая нам кимберелла. По образу жизни и по
скорости передвижения первые эдиакарские животные-водорослееды, скорее всего,
приблизительно напоминали современных улиток. Для нас это выглядит безобидно,
но с точки зрения эдиакарских жителей появление таких чудищ было настоящей
катастрофой. Водорослевые маты немедленно перестали быть сплошными. К тому же
животные не только соскребали их сверху, но и объедали снизу, освоив для это-
го проникновение в грунт (у зоологов принято называть такое поведение
"минированием"). Тут досталось заодно и вендобионтам, которые в конце эдиака-
рия просто исчезли.
С этого момента начала работать общая закономерность, установленная эколо-
гами уже давно и проверенная разными способами вплоть до прямых эксперимен-
тов : в условиях давления хищника разнообразие его жертв повышается по сравне-
нию с сообществом, где хищников нет вовсе. Если раньше донные сообщества за-
полнялись очень немногочисленными доминирующими видами водорослей, то теперь
равновесие рухнуло, и на этом фоне началась бурная эволюция. А между тем на-
бор экологических ниш, доступных животным, тоже расширялся. Появились, напри-
мер, активные грунтоеды, приспособленные к тому, чтобы постоянно жить в но-
рах, пропуская донный грунт сквозь кишечник и извлекая из него питательные
вещества. Так и до сих пор живут многие морские черви, вроде пескожила. Чер-
ви-грунтоеды впервые стали рыть в морском дне не только горизонтальные, но и
вертикальные ходы, вызывая обогащение грунта кислородом и тем самым дополни-
тельно облегчая его заселение другими животными (см. рис. 17.1). Эти события
получили название субстратной революции. Таким образом, эволюционирующие жи-
вотные не только занимали готовые экологические ниши, но и активно создавали
новые, превращая процесс в автокаталитический (самоускоряющийся).
Некоторые жители поверхности дна стали расширять свои экологические ниши не
в сторону грунта, а, наоборот, в сторону водной толщи. В результате возник
зоопланктон — сообщество мелких животных, пребывающих взвешенными в воде и
дрейфующих вместе с ней. Как правило, представители зоопланктона питаются,
фильтруя воду и отцеживая из нее фитопланктон, то есть находящиеся в той же
водной толще одноклеточные водоросли (а таких к моменту кембрийского взрыва
было уже сколько угодно, на все вкусы) . И действительно, в раннем кембрии в
палеонтологической летописи появляются первые планктонные фильтраторы — жаб-
роногие рачки. Жаброноги, как и все ракообразные, обладатели членистых конеч-
ностей, исходно предназначенных для хождения по грунту, то есть по дну. По-
этому нет никаких сомнений, что ранние этапы своей эволюции они прошли на
дне, а к планктонному образу жизни обратились только после этого.
Рис. 17.1. Субстратная революция.
Последствия появления зоопланктона, в свою очередь, оказались вполне гло-
бальными. Дело в том, что животные-планктеры отфильтровывают из воды не толь-
ко водоросли, но и любую взвесь, в которой могут быть хоть какие-то питатель-
ные вещества. В основном это распыленные остатки мертвых организмов. Отфильт-
ровав взвесь и всосав из нее полезные молекулы, планктеры (в первую очередь
ракообразные) аккуратно ллупаковывают" остальное содержимое кишечника в плот-
ные комки — фекальные пеллеты, которые из-за своей компактности быстро тонут
и отправляются на дно. А вода в результате очищается. Пеллетная транспорти-
ровка взвеси — важнейший фактор, понижающий мутность воды в океане.
Надо сказать, что переменивший облик Земли феномен пеллетной транспортиров-
ки непосредственно связан с одной чисто физиологической особенностью живот-
ных, образующих основную массу зоопланктона, — ракообразных. Дело в том, что
находящаяся в кишечнике пища у них окружается перитрофической мембраной —
тонкой белково-хитиновой пленкой, пропускающей мелкие молекулы, но отфильтро-
вывающей крупные. Остатки переваренной пищи оформляются в компактные пеллеты
именно благодаря ей. Перитрофическая мембрана есть и у насекомых, что неуди-
вительно : ведь насекомые — это на самом-то деле всего лишь одна из многих
эволюционных ветвей ракообразных. "В лице" насекомых ракообразные освоили су-
шу. Впрочем, тут мы забегаем вперед.
Итак, после появления планктонных фильтраторов вода в океане стала прозрач-
ной, свет начал проникать в нее на большую глубину и повысилась концентрация
кислорода (часть которого раньше расходовалась на окисление той же мертвой
взвеси). Первый фактор сразу увеличил глубину зоны, в которой света достаточ-
но для фотосинтеза, а второй улучшил условия для придонной фауны. По всем
данным, прозрачный насыщенный кислородом фанерозойский океан резко отличается
от мутного докембрийскохю океана. Заодно повысилась концентрация кислорода и
в атмосфере. Естественно, что в новых благоприятных условиях разнообразие,
как растений, так и животных дополнительно выросло. Замкнулась еще одна авто-
каталитическая петля.
Пришествие
хищника
Все животные, о которых мы до сих пор говорили, были "травоядными" — хотя
бы в самом наиширочайшем смысле этого слова. Они питались или фотосинтезирую-
щими организмами, или, на худой конец, чьими-то останками. При этом собствен-
ная биомасса "травоядных" представляла собой ценный (и до какого-то момента
совершенно невостребованный) ресурс для животных, питающихся другими животны-
ми , то есть для хищников.
Поначалу никаких многоклеточных хищников просто не существовало. Но при на-
личии таких атрибутов активной жизни, как нервная система, мускулатура и ро-
товой аппарат, их появление было всего лишь вопросом времени. Первые крупные
хищники, уже совершенно определенно специализированные на питании другими
многоклеточными животными, появляются в летописи примерно 520 миллионов лет
назад. Это динокариды — хорошо плавающие существа, родственные членистоногим.
Самый известный представитель динокарид — аномалокарис, стройное сегментиро-
ванное создание длиной около метра со сложными фасеточными глазами и мощными
членистыми околоротовыми конечностями, явно служившими для захвата крупной
подвижной добычи (см. рис. 17.2). В самом начале кембрия подобных хищников
нет. Пресловутая "скелетная революция", несомненно, в какой-то мере стала от-
ветом на их появление — изменение химического состава морской воды только об-
легчило ее. А появление скелетов, в свою очередь, запустило освоение новых и
новых экологических ниш.
Стивен Стэнли совершенно правильно писал, что для объяснения кембрийского
взрыва вполне достаточно чисто биологических причин. Факторы, действующие на
биосферу извне, могли повлиять на скорость того или иного процесса, но все
главные события можно объяснить и без них. Вспышка разнообразия многоклеточ-
ных животных была естественным результатом серии автокаталитических процес-
сов, запущенных появлением первых "травоядных" (вроде кимбереллы) и происхо-
дивших на уровне природных сообществ, то есть, иначе говоря, экосистем. Вне
экологии понять кембрийский взрыв невозможно.
С появлением хищников процесс образования новых жизненных форм стал понем-
ногу тормозиться. Репертуар экологических ниш сложился, почти все они уже бы-
ли распределены и заняты. Разумеется, расширение сообществ продолжалось и
дальше — просто медленнее. Например, только после окончания кембрийского пе-
риода появились лопатоногие моллюски, занявшие довольно-таки экзотическую ни-
шу роющих хищников, преследующих своих жертв внутри грунта. Но такого разма-
ха, как на рубеже эдиакария и кембрия, крупномасштабная эволюция животных
больше никогда не достигала.
Рис. 17.2. Аномалокарис.
С точки зрения событийной истории началом кембрийского взрыва можно считать
появление первых эффективных водорослеедов (кимберелла), а концом — появление
первых эффективных хищников (аномалокарис). Кимберелла появилась 555 миллио-
нов лет назад, аномалокарис — 520 миллионов лет назад, интервал между ними —
35 миллионов лет. Не так уж и быстро.
Первопроходцы
суши
Выход жизни на сушу — событие, настолько сильно растянутое во времени, что
его просто невозможно рассматривать как четкий порог. Более того, сама поста-
новка вопроса о выходе на сушу на самом деле спорна. Утверждение "море — ко-
лыбель жизни" вовсе не настолько само собой разумеющееся, как может показать-
ся.
Например, в последние годы стала популярной гипотеза, согласно которой
жизнь возникла не в океане, а в мелких наземных водоемах. В пользу этой гипо-
тезы есть несколько биохимических доводов, самый простой и наглядный из кото-
рых следующий. Известно, что цитоплазма практически всех живых клеток содер-
жит гораздо больше ионов калия (К+) , чем ионов натрия (Na+) . Между тем почти
во всех природных водоемах соотношение концентраций этих ионов в точности об-
ратное. В морской воде в 40 раз больше натрия, чем калия, а в живой клетке,
наоборот, в 10-20 раз больше калия, чем натрия. Внутриклеточный избыток калия
важен для работы многих ферментов, и в том числе для системы синтеза белка.
Причем анализ генных последовательностей показывает, что эти калий-зависимые
ферменты очень древние. Скорее всего, они были уже у общего предка всех со-
временных живых организмов. Значит, первые клетки формировались в среде, где
калия было намного больше, чем натрия. Океан такой средой быть не мог. Гораз-
до вероятнее, что это были горячие источники, вода в которых как раз может
иметь подходящий химический состав. И располагаться эти источники могли как в
море, так и на суше (см. главу 11).
Но даже если живые клетки появились в неглубоких водоемах на суше, то после
этого они быстро заселили океан. Дело в том, что по большинству параметров
морская среда намного комфортнее наземной: в ней всегда хватает воды, и в нее
почти не проникают опасные для клеток ультрафиолетовые лучи. Сейчас ультра-
фиолетовый компонент солнечного света экранируется слоем атмосферы, обогащен-
ным газом озоном (03) , который постоянно образуется в небольшом количестве из
атмосферного кислорода (02) . До кислородной революции на Земле не было ника-
кого озонового слоя, поэтому защищать клетки от ультрафиолетовых лучей могла
только вода. Получается, что океан был если и не колыбелью жизни, то уж точно
ее убежищем на первые несколько миллиардов лет эволюции.
Первые признаки жизни на суше относятся еще к раннему докембрию. В основном
это палеопочвы, обогащенные лллегким" изотопом углерода 12С — как мы помним,
именно по этому критерию биогенный углерод, прошедший по биохимическим путям
живых клеток, отличается от абиогенного. Первые такие палеопочвы имеют воз-
раст примерно 2,7 миллиарда лет, но не исключено, что они существовали и
раньше. Это — архейская эпоха, когда на Земле жили одни прокариоты. Зная, на-
сколько огромны экологические возможности прокариот, нетрудно допустить, что
кто-то из них прижился и в таком негостеприимном месте, как архейская суша.
Первое прямое свидетельство наземной жизни — остатки нитчатых сине-зеленых
водорослей в ископаемой пещере возрастом 1,2 миллиарда лет. Этот момент вре-
мени относится к эпохе ЛЛскучного миллиарда лет", когда сине-зеленые водорос-
ли, они же цианобактерии, уже не только были многочисленны, но и мало чем от-
личались от своих современных родичей. А о современных сине-зеленых водорос-
лях известно, что они довольно часто живут вне воды, например на почве или на
скалах. Так что никаких серьезных причин сомневаться в этой находке нет.
Несколько позже — один миллиард лет назад — появляются остатки наземных во-
дорослей, предположительно относящихся уже к эукариотам. Они образуют пла-
стинчатые колонии, а также цисты, то есть покоящиеся стадии с толстой сложно
структурированной клеточной стенкой. Тут надо сказать, что проникновение эу-
кариотных водорослей в наземную среду — явление, опять же, не столь и редкое.
Например, современная зеленая водоросль трентеполия не просто выдерживает на-
земные условия, а специализирована к ним и ведет полностью наземный образ
жизни. У нее есть целый набор приспособлений к жизни на воздухе — очень тол-
стые клеточные стенки, обилие запасных веществ. Колонии трентеполии обычно
выглядят как кирпично-красный или желтый налет на коре деревьев (зеленый
цвет, присущий большинству растений, тут маскируется дополнительными красными
пигментами, защищающими водоросль от "световых перегрузок" в слишком хорошо
освещенной среде). В том, что водоросли с похожим образом жизни существовали
миллиард лет назад, нет ровным счетом ничего невероятного.
Надо отметить, что оценки, которые мы сейчас обсуждаем, не самые смелые, а,
наоборот, самые скептические. Например, древнейшая известная кора выветрива-
ния (понятие, частным случаем которого является почва) образовалась примерно
3,5 миллиарда лет назад, и есть исследователи, которые допускают существова-
ние наземной жизни уже тогда. Это не выглядит невозможным, но пока не имеет
строгих доказательств. В чем мы можем быть уверены, так это в том, что назем-
ная жизнь, причем многоклеточная, существовала еще до позднепротерозойской
эпохи ЛЛЗемли-снежка". Неудивительно, что сразу после лл3емли-снежка", то есть
в эдиакарском периоде, она возродилась.
И как возродилась! Есть гипотеза, что именно активностью наземной биоты
объясняется "вторая кислородная революция", то есть происходивший в эдиакарии
рост содержания кислорода в атмосфере. Причинно-следственная цепочка тут вот
какая. Важным фактором, влияющим на концентрацию кислорода, является присут-
ствие в той же среде мертвого органического вещества. Если такого вещества
много, то значительная часть кислорода тратится на его окисление, переходя в
итоге в углекислоту и воду. Если любым способом вывести из оборота мертвую
органику, свободного кислорода при прочих равных условиях станет больше.
Именно это произошло, когда кембрийский зоопланктон освоил пеллетную транс-
портировку мелкой взвеси на дно. В эдиакарии механизм, приводивший к этому
эффекту, тоже существовал, но был несколько другим. Мертвая органика связыва-
лась частицами глины, которые поступали в океан с континентальным стоком, и
оседала на дно вместе с этими частицами. Между тем геологи считают, что ис-
точником такого количества глины может быть только биогенная кора выветрива-
ния, то есть почва. Тогда получается, что эдиакарская наземная биота, предпо-
ложительно состоявшая из водорослей и грибов, запустила уже достаточно мощное
почвообразование. Кстати, есть данные, что в эдиакарии были и лишайники.
С другой стороны, в палеонтологии, как мы уже знаем, довольно широко рас-
пространена идея, что эволюция многоклеточных животных зависела от содержания
кислорода в атмосфере (так называемая гипотеза кислородного контроля). Высо-
кая концентрация кислорода, несомненно, облегчила переход к "настоящей" жи-
вотной многоклеточности, требующей много энергии. Если же подъем этой концен-
трации произошел, в конечном счете, из-за активности наземных организмов —
значит, выход жизни на сушу не только предшествовал появлению настоящих мно-
гоклеточных животных, но и был в какой-то мере его причиной.
Леса и
насекомые
В первые четыре периода палеозойской эры — кембрий (542-485 млн. лет на-
зад) , ордовик (485-443 млн. лет назад), силур (443-419 млн. лет назад) и де-
вон (419-359 млн. лет назад) — наземная биота постепенно, но неуклонно услож-
нялась . Высшие споровые растения, относящиеся примерно к тому же уровню орга-
низации , что и современные мхи, совершенно достоверно появились в ордовике.
Наземные водоросли, от которых они, скорее всего, произошли, известны из кем-
брия. Ну а в конце ордовика уже существовали первые сосудистые растения,
имеющие специальные проводящие ткани для транспорта воды и питательных ве-
ществ. К ним относятся все современные высшие растения, кроме мхов: папорот-
ники , псилоты, хвощи, плауны, голосеменные и цветковые. Начиная с силура
именно сосудистые растения преобладают в растительном покрове Земли.
Животные тоже вышли на сушу. В первую очередь это членистоногие — тип, ко-
торый в наземных условиях достиг колоссального расцвета (см. рис. 17.3). В
силуре появились многоножки и скорпионы, в девоне — сенокосцы, клещи, ложно-
скорпионы и насекомые. Причем на сушу некоторые из них выбрались довольно ра-
но: например, ископаемые следы, наверняка принадлежащие наземным многоножкам,
обнаруживаются в ордовике. А в начале девона появились хищные губоногие мно-
гоножки, которые могли питаться только другими членистоногими. У ископаемых
экземпляров этих многоножек сохранились даже ногочелюсти с крючками, сквозь
которые в тело жертвы впрыскивался яд из ядовитых желез — этот механизм у них
одинаков с современными представителями группы. Это означает, что к началу
девона сообщество наземных членистоногих было уже достаточно сложным, чтобы
хищник всегда мог найти себе добычу.
Рис. 17.3. Фауна раннего девона.
Что касается позвоночных, то они вышли на сушу едва ли не последними. На-
земные позвоночные появляются в палеонтологической летописи только во второй
половине девона, причем их первые известные представители, скорее всего, еще
вели в основном водный образ жизни. Расцвет их начался в следующем периоде —
каменноугольном, или карбоне (359-299 млн. лет назад). Освоившись на суше,
наземные позвоночные заняли в сообществе место "суперхищников", способных без
особого труда съесть любое членистоногое, и приступили к собственной бурной
эволюции.
В сложившемся "пазле" наземной фауны явно не хватает двух привычных нам
фрагментов. Во-первых, там нет пауков. Их ископаемые родичи, еще не умевшие
плести паутину, вышли на сушу в силуре, но настоящие пауки, и то очень прими-
тивные, появились только в конце карбона. А во-вторых, мы пока еще не сказали
ни слова о летающих насекомых. Упомянутые девонские насекомые были первично-
бескрылыми, то есть относились к таким группам, у которых крыльев никогда не
было. Первичнобескрылые насекомые есть и сейчас: например, встречающаяся в
домах сахарная чешуйница. Но подавляющее большинство современных насекомых —
крылатые (включая сюда и тех, кто по той или иной причине потерял крылья вто-
рично) . И вот этой группы в девоне еще нет. Первый бескрылый родственник кры-
латых насекомых, очень похожий на них строением ротового аппарата, обнаружен
в конце девона. А первое насекомое с крыльями — в начале карбона. С этого мо-
мента в истории жизни на Земле началась новая эпоха, хотя поначалу, вероятно,
это особенно заметно не было.
Дело в том, что крылатые насекомые во многих отношениях уникальны на фоне
всей остальной земной фауны, вместе взятой. Достаточно сказать, что к этой
группе относится примерно две трети всех современных видов животных. Кстати,
наиболее богатый видами отряд крылатых насекомых — жесткокрылые, или жуки; с
этим связано знаменитое (хотя и не имеющее четкого подтверждения в источни-
ках) высказывание биолога Джона Холдейна: "Если Создатель существует, он,
должно быть, необычайно любит жуков". При всем этом крылатые насекомые обла-
дают необычайно широким набором способов питания. Это наглядно выражено во
многообразии их ротовых аппаратов: грызущий, лижущий, грызуще-лижущий, труб-
чато-сосущий, колюще-сосущий, режуще-сосущий и др., вплоть до таких необыч-
ных, как хватательная маска личинки стрекозы, гигантские жвалы жука-оленя или
20-сантиметровый спирально закрученный хоботок длиннохоботного бражника. Кры-
латые насекомые встречаются везде, где только могут жить многоклеточные суще-
ства , за исключением морских глубин. Они бывают хищниками, растительноядными,
паразитами, поедателями грибов или разрушителями мертвой органики, и во всех
этих ролях могут вырабатывать тончайшие пищевые специализации. Кроме того,
они образуют еще и множество других, непищевых связей с соседями по сообщест-
вам: например, насекомые-опылители сильно повлияли на эволюцию цветковых рас-
тений. В общем, биосферную роль крылатых насекомых трудно переоценить.
Растительный мир на рубеже девона и карбона тоже изменился. В середине де-
вона сразу несколько неродственных групп наземных растений выработали жизнен-
ную форму дерева с мощной корневой системой и вертикальным ветвящимся стволом
высотой до восьми метров. А в позднем девоне появились первые леса, состояв-
шие в основном из древовидного папоротника археоптериса. И хотя археоптерисо-
вые леса конца девона не пережили, другие древесные породы тут же пришли им
на смену. Например, в карбоне появились те самые сигиллярии с "упругими и
рыхлыми стволами", которые упомянул в стихах поэт-акмеист Михаил Зенкевич. В
общем, на рубеже девона и карбона началось великое "облесение суши"
(afforestation of the land). Лес открыл множество новых экологических ниш,
которые не замедлили занять животные, питавшиеся как растениями, так и друг
другом. И тот же лес выбрасывал в атмосферу огромное количество кислорода,
тем самым, облегчая животным всевозможные "эксперименты" с формой и размером.
В карбоне достигли своего расцвета амфибии и появились первые рептилии — не-
сомненно, в связи с ростом разнообразия насекомых, которыми эти позвоночные
могли питаться. Так пошло и дальше. Крылатые насекомые и леса — два фактора,
вот уже примерно 350 миллионов лет определяющие облик земной суши.
Парк
пермского
периода
Последний период палеозойской эры — пермский — продолжался 46 миллионов лет
(298-252 млн. лет назад). Пермский мир был уже во многих отношениях близок к
нынешнему. Сушу населяли многообразные наземные позвоночные, среди которых
были и хищники, и растительноядные. Процветал мир насекомых; именно в перми
появился отряд, оказавшийся самым эволюционно успешным в этом классе, — жуки.
На континентах росли леса, состоявшие в Северном полушарии из древних хвой-
ных, а в Южном — из лиственных голосеменных, отдаленно родственных современ-
ному гинкго. Водная жизнь тоже бурно развивалась: пермские моря были полны
моллюсков, иглокожих, брахиопод, ракообразных, кораллов и других беспозвоноч-
ных, не говоря уж о рыбах. Современного наблюдателя (если бы он не слишком
всматривался) в перми поразило бы, прежде всего, отсутствие птиц, млекопитаю-
щих и цветковых растений, а если бы он мох1 увидеть Землю из космоса — то еще
и совершенно другое расположение материков.
В конце карбона — начале перми произошло так называемое гондванское оледе-
нение, охватившее значительную часть суши Южного полушария. Очень вероятно,
что его главной причиной было распространение лесов. Деревья карбонового пе-
риода связывали огромное количество углерода в биомассе своих стволов, разла-
гать которую было некому: эффективные разрушители древесины тогда еще не воз-
никли. В результате деревья падали и захоранивались как есть, создавая залежи
каменного угля. Переход углерода из атмосферного углекислого газа в эти зале-
жи, где он оказался "заперт", сильно ослабил парниковый эффект — вот этого,
по-видимому, и хватило для запуска оледенения. Достоверно показано, что атмо-
сферная концентрация С02 в тот момент сильно упала. Гондванское оледенение
создало холодный климат на части земной суши, но никаких биосферных переворо-
тов , насколько можно судить, не вызвало. Все крупные группы организмов его
спокойно пережили.
События конца перми были гораздо драматичнее. Но чтобы к ним перейти, нам
понадобится небольшое вступление.
Пять
великих
рубежей
Еще палеонтологи XIX века прекрасно знали, что рубежи периодов, а тем более
эр, обычно характеризуются спадами разнообразия живых организмов, то есть,
попросту говоря, вымираниями. Это естественно: одни живые существа вымирают,
другие приходят им на смену. Границы периодов как раз и проводятся по момен-
там, когда смены флор и фаун особенно заметны. А конец пермского периода од-
новременно является концом всей палеозойской эры — не приходится удивляться,
что некоторая часть биоты на этом рубеже исчезла.
А что получится, если оценить это явление количественно? В 1979 году палео-
нтолог Дэвид Рауп, много занимавшийся статистическим анализом разнообразия
древних фаун, опубликовал статью со следующим выводом: есть основания счи-
тать , что в конце пермского периода вымерло, не оставив потомков, примерно
96% всех видов морских животных на Земле. Получалось, что биота, по крайней
мере, морская, прошла в этот момент через настоящее "бутылочное горлышко".
Подсчеты Раупа были основаны на огромном материале. И хотя его результаты ка-
сались только морской фауны, они не оставляли сомнений, что в конце перми
действительно случилось очень крупное вымирание.
Другой известный палеонтолог, Дуглас Эрвин, решил определить скорость этого
процесса. Анализируя вместе с коллегами распределение остатков животных в
осадочных толщах, он постепенно пришел к выводу, что пермское вымирание про-
изошло за очень короткий срок — меньше одного миллиона лет. Коротким этот
срок, конечно, можно назвать только по меркам истории Земли, но для событий
такого масштаба он действительно мал.
Итак, на границе палеозоя и мезозоя произошло нечто большее, чем рядовая
смена ископаемых фаун. Рауп и Эрвин столкнулись с явлением, относящимся к ка-
тегории массовых вымираний (mass extinctions). Что же это, собственно, такое?
Массовые вымирания, они же биосферные кризисы, случались в истории Земли
довольно регулярно. Принято считать, что в фанерозое крупнейших вымираний бы-
ло пять. Пермское — одно из них. Из остальных четырех два произошли внутри
палеозоя (в конце ордовика и в конце девона), одно — внутри мезозоя (на рубе-
же триасового и юрского периодов), и еще одно отделяет мезозой от кайнозоя
(именно тогда исчезли динозавры). Некоторые из этих событий вполне могли
иметь внешние причины. Например, вымирание на границе мезозоя и кайнозоя свя-
зывают с падением на Землю 10-километрового астероида, кратер от которого
сейчас находится на побережье полуострова Юкатан. Но, похоже, что если это и
верно, то случай тут все же исключительный: большинство вымираний объяснить
подобными единичными катастрофами не удается. Необходимо учитывать, что кри-
зисов, лишь немного уступающих пяти "главным" вымираниям, а то и сравнимых с
ними, в фанерозое на самом-то деле было еще больше десятка. Космических ката-
строф просто не хватит на на всех.
Некоторые палеонтологи считают, что кризис, сопровождающийся массовым выми-
ранием, есть совершенно нормальная стадия развития природного сообщества, во
всяком случае, сообщества размером с земную биосферу. Известный палеоботаник
Валентин Абрамович Красилов так и писал: кризис — это естественный механизм
эволюции сообществ. Под этим углом зрения становится понятно, почему некото-
рые кризисы не удается толком связать ни с какими внешними воздействиями на
живую природу. Но столь же ясно, что само по себе признание кризиса нормаль-
ным явлением еще ничего не объясняет. В основе любого исторического события,
и биосферного кризиса в том числе, лежат вполне реальные причинно-
следственные связи. Наша задача — их раскрыть. Если нет внешних причин, зна-
чит , есть внутренние, и наоборот.
В этом плане вымирание на границе палеозоя и мезозоя особенно интересно.
Чтобы понять почему, его можно бегло сравнить с вымиранием на границе мезозоя
и кайнозоя, тем самым, которое погубило динозавров. Прежде всего, уточним на-
звания этих событий. Последний период мезозоя называется меловым, первый пе-
риод кайнозоя — палеогеновым, поэтому "динозавровый" кризис мы будем называть
мел-палеогеновым, как это обычно и делают. Кризис на рубеже палеозоя и мезо-
зоя по тому же принципу называют пермо-триасовым.
Разница между этими кризисами следующая. Во-первых, пермо-триасовое вымира-
ние было намного мощнее. Аккуратный подсчет по одной и той же методике пока-
зывает, что мел-палеогеновый кризис уничтожил примерно 15% от всех существо-
вавших тогда биологических видов, а пермо-триасовый — примерно 60%. Эти цифры
несколько меньше тех, которые обычно приводятся в научно-популярных публика-
циях, но зато уж в них можно не сомневаться. Во-вторых, в отличие от мел-
палеогенового кризиса, пермо-триасовый кризис не был вызван никаким внешним
ударом вроде астероидного импакта (гипотеза, что подобный импакт был и тут,
высказывалась, но не подтвердилась). И, в-третьих, после пермо-триасового
кризиса разрушенная биосфера вступила в очень долгий — растянувшийся на де-
сятки миллионов лет — период постепенного восстановления, чего после мел-
палеогенового кризиса не было вообще: там сообщества восстановились сразу.
Недавно детальные геологические исследования показали, что пермское вымира-
ние заняло гораздо более короткий срок, чем думали раньше, — всего 60 000
лет. Это 0,06 миллиона лет, срок по меркам планетной истории просто невероят-
но маленький. Что же там случилось?
Пермский
кризис
Большинство ученых сейчас считает, что пермскую катастрофу вызвали вулканы.
Показано, что точно в момент биосферного кризиса (то есть 252 миллиона лет
назад) на территории, которая сейчас называется Западной и Средней Сибирью,
образовалась трапповая провинция — система действующих вулканов, не ограни-
ченных точечными кратерами, а занимающих сплошное протяженное поле. Выглядело
это, должно быть, грандиозно. Расколотая во многих местах земная кора изверг-
ла буквально море расплавленного базальта, залившее 1,6 миллиона квадратных
километров — это площадь современного Ирана. Текущий базальт затопил огромную
территорию и постепенно застывал, создавая абсолютно плоскую безжизненную
равнину, по которой текли новые базальтовые волны от новых извержений. А дым
от них, надо полагать, затмил небо. Но все это было только прологом ко все-
планетной катастрофе.
Трапповые вулканы неизбежно выбрасывали в атмосферу колоссальные объемы
вулканических газов — углекислого газа (С02) , хлороводорода (НС1) , сероводо-
рода (H2S) , сернистого ангидрида (S02) и др. Это имело много последствий. Во-
первых, углекислый газ усилил парниковый эффект, вызвав сильное глобальное
потепление. Во-вторых, вулканические газы повысили кислотность морской воды,
а это по чисто химическим причинам сильно затруднило жизнь кораллов и иглоко-
жих: карбонаты, из которых состоят твердые скелеты этих животных, в слишком
кислой воде просто-напросто растворяются. В-третьих, сероводород ядовит для
большинства живых организмов, особенно при концентрации, в тысячи раз превы-
шающей нынешнюю атмосферную, — а именно такой уровень сероводорода создавали
трапповые вулканы.
Более того, показано, что существует концентрация сероводорода, которая еще
не мешает росту фитопланктона (то есть одноклеточных водорослей), но уже
мгновенно убивает зоопланктон (например, веслоногих рачков). Если такое слу-
чится в реальном водоеме, он тут же "зацветет". Дальше последует цепочка со-
бытий, обычная для цветущих водоемов и в наше время: бесконтрольно размножаю-
щимся водорослям не хватит ресурсов, часть из них погибнет, погрузится на дно
и там начнет разлагаться, связывая запас кислорода в нижних слоях воды, кото-
рый в данном конкретном случае должен быть и без того невелик: ведь чем выше
температура, тем хуже кислород растворяется в воде. Если этим водоемом будет
Мировой океан, то его глубины в результате станут практически бескислородны-
ми , что, собственно, в момент пермского кризиса и наблюдалось. Называя вещи
своими именами, там случился глобальный замор.
Между тем в бескислородной среде, богатой соединениями серы, прекрасно себя
чувствуют анаэробные бактерии, в том числе и сульфатредуцирующие, выделяющие
в качестве побочного продукта метаболизма опять же сероводород. Готово — пет-
ля положительной обратной связи замкнулась. И наверняка подобных петель было
несколько. Вулканических газов могло и не хватить, чтобы убить все живое, но
их хватило, чтобы запустить серию обратных связей, сделавших катастрофу само-
развивающейся .
Пустынная
планета
Сразу после пермского кризиса Земля представляла собой довольно унылое ме-
сто . Ничем не сдерживаемый парниковый эффект привел к разогреву Мирового
океана до 38-40 С. Это близко к смертельной для большинства животных и расте-
ний температуре денатурации белков (примерно 45 С) . К тому же при высокой
температуре снижается растворимость кислорода в воде, так что существование
водной фауны практически исключается. Палеонтологические данные показывают,
что рыбы в эту эпоху остались только в приполярных областях океана. Суша,
скорее всего, прогревалась еще сильнее, поэтому тропические и субтропические
широты были непригодны для жизни крупных наземных животных. Авторы, опублико-
вавшие эти данные, делают твердый вывод: в раннем триасе температура стала
дополнительной причиной массового вымирания.
Не блестяще обстояло дело и с кислородом. В перми его концентрация в атмо-
сфере поднялась почти до 30%Л а после пермского кризиса она упала до 10-15% и
оставалась такой большую часть триаса. Это сильно ограничивало физиологиче-
ские возможности уцелевших позвоночных, не давая им достичь крупных размеров.
Полностью исчезли леса — деревьев, способных их образовать, просто не оста-
лось нигде на Земле. Соответственно, вымерли все животные, занимавшие специ-
фически лесные экологические ниши. Самым крупным и многочисленным наземным
позвоночным этой эпохи был листрозавр — всеядная зверообразная рептилия раз-
мером со среднюю собаку. Впрочем, еще чаще листрозавра сравнивают со свиньей,
подразумевая, что он вел похожий образ жизни и питался чем попало. Участвуя в
подготовке телепередачи о листрозавре, английский палеонтолог Майкл Бентон
назвал ее "Когда свиньи правили Землей".
В целом можно утверждать, что больше всего от пермо-триасового кризиса по-
страдали морские беспозвоночные, а меньше всего — насекомые. У последних во-
обще все "провалы" разнообразия, вызванные массовыми вымираниями, сглажены по
сравнению с другими группами животных. Частично это связано с их удачной ана-
томией и физиологией (дыхательная система насекомых состоит из микроскопиче-
ских трубочек — трахей, которые пронизывают тело, доставляя воздух прямо к
клеткам без всякого переноса кровью; как бы мало кислорода в этом воздухе ни
было, животное его получит), а частично — с тем, что наземная среда, к кото-
рой они приспособлены, гораздо более разнообразна и расчленена, чем водная.
Если вы — жук, то уж какое-нибудь приемлемое для себя микроместообитание все-
гда найдете.
Триасовое
возрождение
На протяжении триасового периода структура сообществ постепенно восстанав-
ливается. Более того, в ней возникают принципиально новые жизненные формы —
например, морские рептилии. В середине триаса появляются двукрылые насекомые
(то есть комары и мухи), а в конце — черепахи, крокодилы, динозавры, летающие
ящеры (птерозавры) и млекопитающие. Это уже почти современная биота. Из
групп, представителей которых мы привыкли каждый день видеть, не хватает
только птиц (появляются в юре) и цветковых растений (появляются в мелу). Ни-
каких переворотов, сравнимых с кембрийским взрывом или пермским кризисом, в
истории Земли отныне больше не будет, по крайней мере, до появления человека.
Тут уместно привести интереснейший чисто эмпирический результат, который
получил американский палеонтолог Джон Сепкоски. Он собрал данные по морским
животным за все времена — от кембрия до современности — и построил график,
где по горизонтали было отложено время, а по вертикали — разнообразие, изме-
ренное числом родов и семейств (см. рис. 17.4). Оказалось, что в течение все-
го палеозоя, не считая начального периода роста сразу после кембрийского
взрыва, разнообразие остается в целом стабильным. После кризисов оно просто
возвращается к прежнему уровню. На границе перми и триаса, естественно, виден
глубокий провал. И что же дальше? За пермо-триасовым провалом следует не воз-
вращение к прежнему устойчивому уровню, а непрерывный подъем, который иногда
притормаживается очередными кризисами, но тем не менее неуклонно длится вот
уже почти 250 миллионов лет. На графике Сепкоски очень наглядно видно, что
вся эволюция как бы делится на две части: до пермского кризиса, когда разно-
образие было по большому счету стабильным, и после него, когда оно непрерывно
растет. Никакого общепринятого объяснения этому до сих пор нет, но сам факт
установлен твердо. И получается, что пермский кризис изменил весь режим раз-
вития жизни на Земле. Перефразируя знаменитого английского историка Арнольда
Тойнби, вполне можно сказать: эволюция — это ответ на вызов.
Рис. 17.4. График Сепкоски, изображающий динамику разнообразия мор-
ских животных в фанерозое (от венда). Обозначения: Cambr — кембрий,
Ordovic — ордовик, Sil — силур, Dev — девон, Carbon — карбон, Perm —
пермь, Тг — триас, Jurassic — юра, Cretac — мел, Tertiary — третич-
ный период. Цифрами с 1 по 5 обозначены пять главных массовых выми-
раний (ордовикское, девонское, пермское, триасовое и меловое), Звез-
дочками показаны еще два кандидата на великие вымирания.
От клетки
к социуму
Сумма доступных нам сейчас научных знаний, в общем, не оставляет сомнений,
что одним из самых важных процессов в эволюции эукариот было слияние организ-
мов . Оно шло на разных уровнях. Уже самая первая эукариотная клетка, скорее
всего, возникла в результате симбиоза археи с бактерией. На следующем эволю-
ционном этапе, который наступил довольно быстро, эукариотные клетки начали
всевозможными способами объединяться друг с другом. Интересно, что генетиче-
ская близость клеток, вступающих в союз, была при этом моментом хоть и важ-
ным, но не решающим. В общем случае клетки, составляющие единый организм, мо-
гут быть как строго генетически одинаковыми (в типичном многоклеточном теле),
так и вовсе не родственными друг другу (например, в лишайнике, где клетки
гриба самым натуральным образом порабощают клетки зеленой водоросли). Преоб-
ладание в нынешней биоте организмов "первого типа", каждый из которых возни-
кает из одной-единственной яйцеклетки, — не сам собой разумеющийся факт, а
поворот эволюции, требующий объяснения. Но так или иначе, не позже чем к на-
чалу палеозойской эры Земля стала миром многоклеточных эукариот — растений и
животных.
На новом эволюционном витке началось объединение целых организмов в системы
следующего уровня — социальные. Довольно точные эквиваленты слова
ЛЛ социальность" — "общественность", или, в устаревшей лексике, ЛЛобщежитель-
ность". Любая социальность основана на обмене сигналами между особями, в ре-
зультате которого у них формируется некоторое общее поведение. Например, со-
циальные амебы, относящиеся к так называемым клеточным слизевикам (из супер-
группы Amoebozoa), могут обмениваться химическими сигналами в виде молекул
циклического аденозинмонофосфата (цАМФ, см. главу 8) . Под действием этого
сигнального вещества они сползаются вместе и образуют грибообразное плодовое
тело. Это один из простейших случаев социальности, часто рассматриваемый как
ее элементарная модель. Но ясно, что канал для обмена сигналами у слизевиков
очень узкий. Примерно так же обстоит дело у растений, сигнальные системы ко-
торых почти исключительно химические и передают мало информации. Тут, однако,
вспоминается знаменитый роман Джона Уиндема "День триффидов", в котором хищ-
ные растения-мутанты научились общаться звуковыми сигналами, тем самым резко
расширив канал передачи информации и создав сложное, организованное поведе-
ние. Нетрудно догадаться, какая группа организмов получила подобные возможно-
сти не в фантастике, а в реальности. Это — животные.
Качественно новым эволюционным достижением была не столько социальность жи-
вотных как таковая, сколько ее высшая форма — эусоциальность, при которой
"представители вида живут группами, состоящими из нескольких поколений, и
члены группы действуют альтруистично по отношению друг к другу в соответствии
с регулярным разделением труда". Животные, ставшие эусоциальными, фактически
объединяются в суперорганизм, члены которого больше не являются самодостаточ-
ными (не могут существовать без поддержки товарищей по виду и сами вынуждены
тратить ресурсы на такую поддержку), но обретают взамен новые возможности,
принципиально недоступные одиночной особи. Согласно процитированному опреде-
лению, человек разумный — эусоциальный вид. "В этом смысле люди вполне срав-
нимы с муравьями, термитами и другими эусоциальными насекомыми". И есть неко-
торые основания полагать, что именно эусоциальность создала человека разумно-
го — во всяком случае, в большей степени, чем любая другая его биологическая
особенность.
Рассмотренное определение эусоциальности взято из относительно недавней
книги Эдварда Уилсона, крупнейшего в мире специалиста по муравьям и глубокого
мыслителя, в свое время фактически создавшего науку социобиологию. Тут возни-
кает загвоздка, которую мы не можем обойти вниманием. Дело в том, что тради-
ционно биологи называли эусоциальными только тех животных, у которых есть
разделение на размножающуюся касту и стерильных рабочих. Идеал эусоциальности
— это муравейник, где яйца откладывает только матка. Причем раньше так считал
и сам Эдвард Уилсон. В своей обзорной работе 1975 года он перечисляет три
признака эусоциальности:
(1) общественная забота о потомстве,
(2) разделение особей на размножающиеся и стерильные касты и
(3) перекрывание как минимум двух поколений.
Очевидно, что в случае с человеком камнем преткновения служит второй при-
знак . Общественная забота о потомстве и перекрывание поколений у него есть, а
вот разделения особей на размножающихся и стерильных — нет (если не считать
бездетных элит вроде духовенства некоторых религий и китайских чиновников-
евнухов, но наличие таких элит никак нельзя считать всеобщей чертой нашего
вида).
Спустя 30 лет, в 2005 году, Уилсон с соавтором выпустили обзорную статью о
муравьях, в которой дали следующее определение: ллЭусоциальность — это эволю-
ционно достигнутый уровень колониального существования, при котором взрослые
члены колонии относятся к двум или более перекрывающимся поколениям, сотруд-
ничают в уходе за молодью и делятся на размножающуюся и неразмножающуюся
(или, по крайней мере, менее размножающуюся) касты". Как видим, тут уже поя-
вилась оговорка. Кроме того, в этом же обзоре подчеркивается, что главное в
явлении эусоциальности — это "общие принципы, существующие на организменном и
сверхорганизменном уровне", "параллели, ясно прочерченные между построением
организма из молекул и тканей и суперорганизма — из взаимодействующих целост-
ных организмов". А в конце статьи упоминается и человек. Авторы признают, что
путь его эволюции имеет много общего с эусоциальными животными, но прямо на-
звать человека эусоциальным существом еще пока не решаются.
Однако в том же 2005 году вышла статья, озаглавленная "Новое эусоциальное
позвоночное?". Ее авторы, английские биологи Кевин Фостер и Фрэнсис Рэтникс,
обратили внимание на одну всем известную особенность человека — менопаузу.
Это генетически запрограммированная физиологическая перемена, имеющая отчет-
ливо пороговый характер (в отличие от старения, которое идет медленно и по-
степенно) . И она приводит к тому, что все особи одного из двух полов выключа-
ются из размножения примерно в середине жизни, при этом сохраняя свою соци-
альную функцию — участие в выращивании потомства. Вероятнее всего, менопауза
— это адаптация, появление которой было как-то связано с социальной структу-
рой наших предков. Здесь Фостер и Рэтникс отмечают, что свойственная человеку
полная и необратимая стерильность значительной части особей, ухаживающих за
потомством, — это признак, вообще уникальный среди позвоночных. У голых зем-
лекопов — грызунов, которых часто считают образцом эусоциальности, — такого
нет, стерильность рабочих особей у них физиологически обратима. Таким обра-
зом, если можно считать эусоциальным видом голого землекопа, то можно и чело-
века разумного.
При всей спорности этой аргументации она, очевидно, дала нужную подсказку
Эдварду Уилсону, мысли которого и без того много лет развивались в сходном
направлении. В статье, опубликованной им вместе с соавторами в 2010 году, уже
прямо сказано, что существует широкое определение эусоциальности (наличие
"частично неразмножающихся каст"), под которое человек подходит. К этому вре-
мени Уилсон стал убежденным сторонником расширенной интерпретации эусоциаль-
ности: главное в ней — не распределение репродуктивных функций (как считали
когда-то первые социобиологи), а возникновение сверхорганизма, "часто причуд-
ливого по устройству, но представляющего особый уровень биологической органи-
зации" . И вот в такое понимание эусоциальности человек уже вписывается. В
2012 году Уилсон выпустил замечательную книгу "Социальное завоевание Земли"
(в русском переводе, вышедшем в 2014 году, — "Хозяева Земли"), целиком посвя-
щенную тому, какую роль в эволюции жизни на Земле сыграли эусоциальные суще-
ства, включая человека.
Муравьиными
тропами
Именно на работах Эдварда Уилсона и будет преимущественно основан наш крат-
кий рассказ про эусоциальность. Уилсон — идеальный проводник по этой теме,
независимо от того, согласны ли мы с его общефилософскими взглядами (которые
он решительно высказывает в нескольких своих последних книгах). И действи-
тельно, кто может разбираться во всем этом лучше, чем человек, посвятивший 60
лет своей жизни изучению муравьев?
Итак, первое, что можно констатировать: эусоциальность часто дает ее обла-
дателям огромный эволюционный успех. Во многих тропических лесах две трети
насекомых — это муравьи. Скоординированная активность множества муравьев или
термитов позволяет построить огромное защищенное поселение с собственным мик-
роклиматом, организовать походы на фуражировку и даже создать аналог сельско-
го хозяйства (и среди муравьев, и среди термитов есть виды, разводящие в
гнездах строго определенные грибы по достаточно сложным технологиям). Ясно,
что одиночным насекомым такое не под силу.
Тем удивительнее, что эусоциальность появилась в эволюции относительно
поздно. В палеозое неизвестны никакие следы активности эусоциальных животных
— судя по всему, такого феномена тогда просто не существовало. Насколько мы
знаем, первыми на Земле эусоциальными существами стали термиты. Самые ранние
известные остатки термитов относятся к началу мелового периода. При этом ме-
ловые термиты очень похожи на современных — настолько, что неспециалист не
смог бы их отличить, и, что еще важнее, они обнаруживаются на нескольких раз-
ных континентах, которые в меловом периоде уже отделены друг от друга океа-
ном . Единым целым эти континенты были только в триасе. Так что, скорее всего,
первые термиты произошли от своих тараканообразных предков не в мелу, а зна-
чительно раньше. Специалисты помещают вероятное время их возникновения где-то
между средним триасом и ранней юрой, в интервале 237-174 миллиона лет назад.
Получается, что первые общественные насекомые заняли (или создали) одну из
принципиально новых экологических ниш, открывшихся после пермо-триасовой
"перезагрузки" биосферы.
Несколько позже, но тоже в мезозойскую эру, появляются общественные пере-
пончатокрылые насекомые — муравьи, пчелы, шмели, некоторые осы. На их разви-
тие сильно повлияла так называемая "цветковая революция", корда голосеменные
растения в большинстве сообществ массово сменились недавно возникшими цветко-
выми — родственниками привычных нам магнолий, буков, роз и других деревьев,
трав и кустарников. Это произошло примерно в середине мелового периода, и как
раз тогда же началась очень бурная эволюция муравьев, в итоге сделавшая их
одними из самых заметных членов наземных экосистем. Конечно, это не случайное
совпадение. "Цветковые" леса обладают более сложной структурой местообитаний,
в которую разным насекомым, в том числе и общественным, оказалось легче
встроиться.
В масштабе эволюционного древа эусоциальность — в общем-то нечастое явле-
ние. Эдвард Уилсон и его соавторы пока насчитывают всего 18 независимых собы-
тий возникновения эусоциальности у животных (не считая человека), 16 из кото-
рых относятся к членистоногим. Эусоциальность возникла один раз у термитов,
девять раз у перепончатокрылых (ос, пчел, муравьев) , три раза у других насе-
комых и три раза у ракообразных. При этом во всех без исключения известных
случаях необходимым предварительным условием для эусоциальности было защищен-
ное гнездо, в котором постоянно живет и размножается группа особей одного ви-
да. Эусоциальные морские ракообразные — десятиногие раки-щелкуны — умудряются
создавать такое гнездо даже внутри тела другого животного, а именно губки.
Только на основе постоянного гнезда может начаться разделение функций, при
котором одни особи размножаются, а другие занимаются исключительно добычей
пищи, уходом за личинками или защитой от внешнего мира. И тогда группа пре-
вращается в сверхорганизм.
У позвоночных животных эусоциальность возникает очень редко. Самый извест-
ный пример эусоциального позвоночного — африканский грызун под названием го-
лый землекоп (Heterocephalus glaber). Голые землекопы ведут роющий образ жиз-
ни. Они создают подземное защищенное гнездо, в котором живет группа, пред-
ставляющая собой расширенную семью с "маткой" (размножающейся самкой), ее
двумя-тремя "мужьями" и несколькими десятками "рабочих" обоих полов, которые
не размножаются, пока это делает "матка". Такой уровень эусоциальности не ус-
тупает тому, которого достигли термиты.
У голых землекопов есть еще два необычных признака. Во-первых, они не ста-
реют (смертность не растет с возрастом) и могут жить до 30 лет и больше — для
млекопитающих такого размера это уникальный случай. Во-вторых, они не способ-
ны самостоятельно поддерживать постоянную температуру тела, то есть фактиче-
ски не являются теплокровными животными. Последнее очевидным образом вызвано
колониальной жизнью землекопов: температура в хорошо устроенном подземном по-
селении достаточно стабильна, чтобы физиологические механизмы терморегуляции
можно было и отключить. Причины "отключения" старения гораздо более загадочны
(одно из самых простых объяснений, хотя и не обязательно самое верное, тако-
во : у землекопов любая рабочая самка в принципе может дождаться своего шанса
занять место царицы, но для этого ей нужно как можно дольше прожить, сохранив
силы, — отсюда естественный отбор против старения), но в любом случае они на-
верняка так или иначе связаны с необычной социальной структурой этих зверей.
Голый землекоп (Heterocephalus glaber).
Второй вид стопроцентно эусоциальных млекопитающих — дамарский землекоп
(Fukomys damarensis) . Если голый землекоп живет в Восточной Африке, то дамар-
ский — в Юго-Западной. Эусоциальность сложилась у этих двух близких видов,
скорее всего, независимо, так же, как она несколько раз независимо возникала
у близких видов ос и пчел. Вот, по сути, и все эусоциальные позвоночные, не
считая человека. Правда, к эусоциальности приближаются некоторые грызуны
(прерийные полевки) и хищники (гиеновые собаки), но с голыми землекопами их
по глубине приспособлений нельзя и сравнивать. В любом случае подавляющее
большинство обладателей эусоциальности — это насекомые.
Эусоциальный
мозг
Чем же отличается эусоциальность человека от эусоциальности любых других
животных? Начнем с того, что в ней общего. Это склонность объединяться в ус-
тойчивые группы на стоянках. Тут стоит привести длинную цитату из Эдварда
Уилсона, она лучше любого пересказа своими словами:
"Стоянки a priori были важнейшей адаптацией на пути к эусоциальности: по
сути, стоянки — это человеческие гнезда. Все без исключения животные, достиг-
шие эусоциальности, начинали со строительства гнезда и защиты его от врагов.
Они выводили в нем потомство, покидали его, уходя на поиски пищи, и возвраща-
лись в него с добычей, которую делили с остальными обитателями... Почему за-
щищенное гнездо играет такую важную роль? Потому что в нем члены группы выну-
ждены собираться вместе. Им приходится уходить на разведку и поиски пищи, но
в конце концов они всегда возвращаются".
Именно плотная "упаковка" разновозрастных и разнополых особей в компактном
гнезде, которое нельзя покинуть без риска для жизни, вынуждает животных со-
вершенствовать систему взаимодействий друг с другом. В истории человечества
таким "гнездом" мог быть и костер, вокруг которого собиралась группа степных
охотников, и пещера, и рыцарский замок, и даже советская коммунальная кварти-
ра. Этот фактор действовал на человека с самого начала, так же как он дейст-
вует, например, на каких-нибудь социальных ос (кстати, именно от ос произошли
муравьи).
И вот тут мы упираемся в принципиальные отличия человеческой эусоциальности
от той, которая свойственна насекомым. Их, по мнению Уилсона, всего два: ог-
раниченная способность к расселению и слишком крупный размер особей.
На расселение общественных насекомых очень сильно влияет их главное преиму-
щество — крылья. Оплодотворенная муравьиная матка может легко пролететь рас-
стояние, которое для человека (с поправкой на размер тела) было бы эквива-
лентно нескольким сотням километров. После этого она обламывает крылья и ос-
новывает новую колонию на новом месте, где, если повезет, может не оказаться
никаких конкурентов. Следует добавить, что эта возможность стремительного
расселения реализуется не всегда: например, у рыжих лесных муравьев гнезда
обычно образуются "почкованием", рядом со старыми, соединяясь с ними поначалу
обменными дорогами. Но, так или иначе, расселение путем дальних перелетов му-
равьям в принципе доступно. А млекопитающим, во всяком случае наземным, —
нет. Они могут расселяться только очень постепенно, не имея "предохранитель-
ного клапана", позволяющего за один шаг разделить родственные колонии в про-
странстве и тем самым снизить напряженность конкуренции между ними. Это каса-
ется как голых землекопов, так и людей, и имеет у них одни и те же последст-
вия, а именно высокую внутривидовую агрессивность. Показано, что голые земле-
копы отличаются "ксенофобией" и часто нападают на незнакомых — а значит, за-
ведомо принадлежащих другому гнезду — особей своего вида. В отношении челове-
ка разумного тут можно обойтись без пояснений. В фильме Мамору Осии "Небесные
скитальцы", который уже упоминался совсем по другому поводу в главе 14, неда-
ром звучат слова: "Нельзя уничтожить войну, не уничтожив человека". В самом
этом утверждении можно усомниться, но вот в том, что склонность к войне запе-
чатлена в нашей чисто биологической природе, особых сомнений нет.
Не менее важен размер тех особей, которые образуют колонию. Млекопитающие —
гиганты животного мира. Типичное современное млекопитающее превосходит массой
типичное современное насекомое примерно на три порядка, то есть в тысячу раз.
Это не абсолютный закон (некоторые млекопитающие бывают меньше некоторых на-
секомых) , но — повторимся — типичное соотношение именно таково. Что же каса-
ется человека, то его размер очень велик даже по стандартам большинства мле-
копитающих: среди них на самом деле не так уж и много животных, вес которых
превосходит килограмм, а тем более измеряется десятками килограммов. В эпоху
динозавров, то есть в мезозое, таких зверей не существовало вовсе, но и в
кайнозое их доля никогда не была особенно велика. Достаточно сказать, что две
трети всех современных видов млекопитающих — это грызуны и летучие мыши.
Почему это важно? Очевидно, что чем крупнее животное, тем крупнее будут и
его отдельные органы, в том числе мозг. Любой мозг состоит из более-менее од-
нотипных нервных клеток — нейронов, размер которых у всех позвоночных в пер-
вом приближении одинаков. Чем мозг больше, тем большее число нейронов он вме-
стит . А чем больше будет нейронов, тем больше между ними установится связей и
тем сложнее при прочих равных условиях может стать поведение. Правда, оговор-
ка насчет "прочих равных" тут очень сильна (в мозге слона втрое больше нейро-
нов, чем в мозге человека, но 97% из них находятся в мозжечке — отделе мозга,
который управляет в первую очередь мышечным движением). И, тем не менее, если
усреднить данные по крупным группам животных, мы, скорее всего, увидим, что
абсолютные цифры тут важнее относительных. В мозге муравья примерно 250 000
нервных клеток, в мозге пчелы — около миллиона. При таком количестве нейронов
поведение относительно жестко программируется инстинктами (это видно хотя бы
из классических книг Жана Анри Фабра, детально описывавшего сложные формы по-
ведения перепончатокрылых). У серой крысы 200 миллионов нейронов, и это уже
другое дело: все мы знаем, как разнообразно поведение этих зверьков и как хо-
рошо развита у них способность к обучению. А у современного человека нормаль-
ное число нейронов составляет 86 миллиардов. Эусоциальное существо с мозгом
такого размера просто обречено стать разумным.
Опять же — почему? Есть серьезные основания считать, что самая ресурсоемкая
область применения головного мозга крупных животных — это социальные контак-
ты, то есть выстраивание системы взаимодействий с сородичами по виду. Осно-
ванное на этом представление, связанное в основном с именем английского ан-
трополога Робина Данбара, получило название гипотезы социального мозга. На-
блюдения над разными видами обезьян показывают, что с ростом абсолютного раз-
мера мозга взаимодействия между особями очень быстро усложняются. А в сложной
социальной системе, в свою очередь, индивиды с более крупным мозгом могут по-
лучить за счет этого преимущество при естественном отборе, то есть заработает
положительная обратная связь. Размер мозга голого землекопа, существа разме-
ром с мышь, по всей видимости, не достиг порога, за которым эта положительная
обратная связь может запуститься. Ну а размер мозга первых представителей ро-
да Homo, появившихся около двух миллионов лет назад, — достиг.
Примем во внимание, что и крупный размер, и эусоциальность — качества в це-
лом довольно редкие (например, из всех современных насекомых эусоциальностью
обладает только 2% видов). А их сочетание, судя по всему, реализовалось за
всю историю Земли только один раз. Вот результатом этого сочетания и стала
наша цивилизация. Результат неизбежный, зато само сочетание крайне маловеро-
ятное .
Люди и
селениты
Огромный мозг человека позволил ему реализовать в высшей степени неэконо-
мичную эволюционную стратегию, сочетающую эусоциальность с универсальностью
каждой особи. Очевидно, что любой здоровый человек потенциально способен не
только размножаться, но и более-менее успешно освоить любой предлагаемый на-
личным социумом род деятельности (разве что с небольшими ограничениями, выте-
кающими из случайной индивидуальной изменчивости по таким, например, призна-
кам, как музыкальный слух). Причем эта способность сохраняется на протяжении
большей части активной жизни. Кроме того, человек способен десятилетиями на-
капливать знания, формируя в итоге совершенно неповторимую структуру личности
с уникальными навыками. Поддержание социальной системы, состоящей из особей с
такими свойствами, — дело достаточно затратное. Не могла ли природа выбрать
какой-то другой путь?
Здесь помогут мысленные эксперименты, поставленные научными фантастами. На-
пример, в романе Герберта Уэллса ллПервые люди на Луне" описана гуманоидная
цивилизация, устроенная совершенно по иному принципу, чем наша. Уэллсовские
селениты (жители Луны) готовят каждого члена общества к строго определенной
профессии с самого рождения, используя для этого не только сложную систему
воспитания, но и ллсмелые хирургические операции". Например, селенит-математик
чисто физиологически не способен заниматься чем-либо, кроме математики: у не-
го сильно развит мозг (и даже определенные области мозга), конечности и внут-
ренние органы уменьшены, а сильные и яркие переживания могут быть связаны
только с математикой, и ни с чем другим. Селенит-пастух, наоборот, хорошо фи-
зически тренирован, имеет глаза, защищенные "твердой и угловатой роговой обо-
лочкой", но не знает ничего, кроме технических понятий своего ремесла, и сча-
стье может испытывать, только занимаясь пастушеской работой. "И так обстоит
дело с селенитами всех сословий — каждый представляет собой в совершенстве
законченную составную часть общей машины".
Уэллс прекрасно знал биологию и не мог не понимать, что с точки зрения этой
науки он рисует довольно реалистичную картину. Эусоциальность почти всегда
приводит к тому, что отдельные особи начинают специализироваться, приобретая
физиологические и даже анатомические различия между собой. Чтобы оценить по-
следнее, достаточно сравнить, например, рабочего термита с термитом-солдатом.
Уэллсовские селениты вписываются в это правило. Примерно такого уровня спе-
циализации особей и стоило бы "из общих соображений" ожидать от эусоциальных
существ с крупным мозгом.
С этой точки зрения социум Homo sapiens, в котором каждая особь одновремен-
но универсальна и уникальна, выглядит чудом. А ведь он существует уже не-
сколько десятков тысячелетий. По эволюционным меркам это мало, но по меркам
истории культуры — очень много. Более того, внутри человеческих обществ время
от времени возникают очаги дополнительного "сброса специализации". Например,
после так называемой катастрофы бронзового века (XII в. до н.э.) начались со-
бытия, которые Михаил Константинович Петров, известный философ, специалист по
истории античной культуры, в свое время проанализировал в статье с замеча-
тельным названием: ЛЛПентеконтера. В первом классе европейской школы мысли".
Гибель крупных империй Восточного Средиземноморья разрушила специализацию лю-
дей, основанную на кастовом делении, и породила мир, где каждый мужчина был
универсальным матросом на борту столь же универсального небольшого корабля —
пентеконтеры (см. рис. 17.5). По мнению Петрова, именно с этого и началось
знаменитое "греческое чудо", создавшее, в конце концов, европейскую цивилиза-
цию.
Рис. 17.5. Реконструкция знаменитого корабля ЛЛАрго".
Космическая
рулетка
Насколько закономерны в нашей Вселенной такие явления, как жизнь и разум?
Кратко на этот вопрос не ответить. Современные научные знания не дают осно-
ваний поддерживать связанную с именем маркиза Пьера Симона де Лапласа позицию
полного детерминизма. В реальности существует не только необходимое. Сущест-
вование жизни не противоречит никаким положениям физики или химии, но оно и
не вытекает непосредственно из этих положений. Обратим внимание, что такое
утверждение верно далеко не для всех природных явлений: например, образование
атомов, молекул, звезд при данных физических законах было неизбежно. О воз-
никновении жизни этого сказать нельзя. Звездные системы не обязательно порож-
дают жизнь, как и жизнь (судя по всему) не обязательно порождает разум. Во
всяком случае, у нас пока нет никаких серьезных оснований утверждать обрат-
ное.
Можно мысленно прочертить траекторию, соединяющую в одно целое все ключевые
моменты химической и биологической эволюции, начиная от формирования планет-
ной системы и заканчивая становлением цивилизации. Такие события, как возник-
новение первых клеток, мнохюклеточности или мозга, будут на этой траектории
промежуточными точками. Как же оценить вероятность того, что жизнь на данной
планете пройдет по ней от начала до конца?
Думается, что тут уместна вот какая аналогия. Представим себе игрока в ру-
летку, который все время ставит на красное с одним дополнительным условием:
серия выпадений красного должна быть обязательно непрерывной. Пока раз за ра-
зом выпадает красное, человек остается в игре. Если хотя бы один раз выпало
черное, игра прекращается и он выбывает. Какие шансы будут у такого игрока?
Это легко подсчитать. При одном испытании игрой в рулетку вероятность выпа-
дения красного составляет 1/2. Но уже при десятке испытаний вероятность по-
стоянного выпадения красного станет меньше 1/1000, а при сотне испытаний в
знаменателе окажется вполне астрономическое 32-значное число. Вот оно-то и
будет характеризовать соотношение тех, кто выиграл, и тех, кто проиграл.
Похоже, что жизнь в космосе сталкивается примерно с такой же игрой вероят-
ностей. Для каждого отдельного фактора или события вероятность помешать раз-
витию жизни может быть невысока. Настоящая проблема в том, что этих факторов
и событий очень много. Планета не должна оказаться слишком близко или слишком
далеко от звезды, не должна быть разрушена ударом другого небесного тела, не
должна проявлять слишком высокую или слишком низкую тектоническую активность
— продолжать в таком роде можно до бесконечности. Между тем никакого суммиро-
вания, приводящего к взаимной компенсации, здесь нет. Единичного события, хо-
тя бы на короткое время выводящего условия на планете за пределы пригодных
для жизни, может быть достаточно, чтобы закрыть вопрос навсегда, даже если
значения всех остальных переменных при этом остаются в пределах допустимого.
Более того, биосфера может не только погибнуть от внешних по отношению к
ней факторов (это можно назвать угрозой первого рода), но и буквально совер-
шить самоубийство (угроза второго рода). Причем для этого не обязательно ну-
жен разумный агент, который устроит термоядерную войну. У известного геолога
Джозефа Киршвинка есть интересная статья под названием "Красная Земля, белая
Земля, зеленая Земля, черная Земля". Под "красной Землей" тут понимается Зем-
ля в начале кислородной эпохи: тогдашние осадки богаты окисленными соедине-
ниями железа, часто имеющими красноватый оттенок. "Белая Земля" — это Земля
эпохи лл3емли-снежка". лл3еленая Земля" — это современная Земля с ее почти
сплошным растительным покровом. А вот ллчерная Земля" — это ситуация, которая,
по мнению Киршвинка, в принципе могла бы реализоваться в далеком будущем. Со-
временные системы фотосинтеза поглощают свет разных цветов (синий, красный),
но вот зеленый свет они большей частью отражают. Именно поэтому современные
растения и выглядят зелеными. Между тем в солнечном свете зеленый компонент
очень силен. А что, если эволюция создаст фотосинтезирующий организм с такими
пигментами, которые будут преимущественно поглощать зеленый свет? Это вполне
может повысить эффективность фотосинтеза и дать такому организму преимущество
в выживании: эволюция — слепой скульптор, она никогда не заглядывает вперед
дальше чем на один шаг. Земля, покрытая растениями, поглощающими зеленый
свет, будет из космоса выглядеть черной. Отражательная способность планеты
при этом грозит снизиться настолько, что биосфера не сможет справиться с пе-
регревом и большинство живых организмов погибнет от высокой температуры (как
это чуть было не произошло на рубеже перми и триаса). При всей внешней фанта-
стичности этого сценария никаким законам природы он не противоречит.
Говоря о жизни в космосе, мы сразу сталкиваемся с еще одной проблемой, свя-
занной скорее с человеческим восприятием реальности. Рассуждая о том, почему
мы до сих пор не встретили инопланетные цивилизации, люди очень часто основы-
ваются (больше подсознательно, чем сознательно) на старинных представлениях,
согласно которым Вселенная практически вечна. Философы, убежденные, что мир
никогда не рождался и никогда не умрет, были даже в Средневековье. В Новое
время близкую к этому позицию выразил, например, шотландский геолог Джеймс
Геттон, писавший: ЛЛВ истории Земли мы не видим никаких следов начала и ника-
ких признаков конца". Но сейчас-то мы знаем, что это совсем не так! Любая
планетная система имеет конечный срок существования, в который развивающаяся
там жизнь должна уложиться. И в сравнении с темпами эволюции самой жизни этот
срок не так уж велик. Например, часть "жизненного цикла" Солнца, охватывающая
промежуток от протозвезды до красного гиганта, должна занять около 10 милли-
ардов лет. Это всего-навсего вдвое больше, чем уже длится история Солнечной
системы.
Кроме того, возникновение жизни было невозможно в первые миллиарды лет по-
сле Большого взрыва, пока звезды первых поколений не достигли стадии сверхно-
вых и не взорвались, разбросав по Галактике пригодные для сборки планетных
систем и живых тел тяжелые химические элементы. Считается, что наше Солнце —
звезда даже не второго, а третьего поколения. В древней Вселенной, включавшей
только звезды первого поколения и состоявшей почти исключительно из водорода
и гелия, никакой жизни быть не могло. Это дополнительно ограничивает макси-
мальный срок, отпущенный на биологическую эволюцию.
С другой стороны, эта эволюция обычно идет очень неравномерно. Например,
первые эукариоты появились только после "кислородной революции", и, скорее
всего, вследствие нее. До этого биосфера Земли в течение двух миллиардов лет
была чисто прокариотной. Но и в эукариотной биосфере эволюция отнюдь не все-
гда шла быстро: за "зукариотным Большим взрывом" (если он был) последовал
ллскучный миллиард лет", в течение которого в живой природе Земли не появилось
никаких качественных новшеств. Вполне возможно, что без них обходилось бы и
дальше, если бы не катастрофическое оледенение, вызванное особенностями дрей-
фа континентов и приведшее к вынужденной перестройке всей биосферы (эпоха
ЛЛЗемли-снежка"). Сложись геологическая история Земли несколько иначе, самыми
сложными живыми организмами на ней до сих пор могли бы быть строматолиты —
или, в лучшем случае, красные водоросли. Более того, не исключен сценарий,
когда целая биосфера доживает до гибели в огне своего солнца (ставшего крас-
ным гигантом), так и не успев породить ни многоклеточных животных, ни высших
растений.
Отсюда следует ясный вывод. Обычно (и чаще всего неявно) принимаемое
ллгосподами ксенологами" допущение, что время, нужное для развития цивилиза-
ции, пренебрежимо мало по сравнению с временем существования Вселенной, со-
вершенно неверно. На самом деле эти времена сравнимы друг с другом. Первое,
конечно, меньше, но не на порядки, а всего лишь в разы. На Земле от возникно-
вения жизни до возникновения разума прошло время, приблизительно равное трети
времени существования Вселенной, и ничего не стоит вообразить сценарий, где
это время было бы еще больше.
Эта логика сразу подсказывает возможное решение пресловутой загадки
^молчания Вселенной" (Silentium Universi), или основного парадокса ксеноло-
гии, как предпочитали выражаться Стругацкие. Весьма вероятно, что наша циви-
лизация — просто первая в Галактике. Другие биосферы, если они где-то и суще-
ствуют, еще не успели дойти до этой стадии. Только и всего. Может быть, успе-
ют в будущем — а может быть, и нет.
"Человек — не статический центр мира, как он долго полагал, а ось и вершина
эволюции, что много прекраснее", — писал в середине XX века Пьер Тейяр де
Шарден. Что ж, современные научные данные не исключают, что Тейяр был в ка-
ком-то смысле прав. Удивительный сверхорганизм человечества — острие эволюции
жизни во Вселенной.
ЗАКЛЮЧЕНИЕ
Природа не ведает сна. Жизнь никогда не
останавливается. Сотворение мира не за-
кончено . Библия учит нас, что в День Шес-
той бог1 сотворил человека и потом дал се-
бе отдых, но каждый из шести дней длился
миллионы лет. День же отдыха был, верно,
очень коротким. Человек — не завершение,
а начало. Мы живем в начале второй недели
творения. Мы — дети Дня Восьмого.
Торнтон Уайлдер. «День Восьмой»
Пьер Тейяр де Шарден писал в книге "Феномен человека", первое издание кото-
рой вышло в 1955 году:
"Нам кажется невероятным, что могли жить люди, которые и не подозревали,
что звезды мерцают над нами на расстояниях в сотни световых лет или что кон-
туры жизни начали вырисовываться уже миллионы лет назад у границ нашего гори-
зонта. И, однако, достаточно открыть любую из чуть пожелтевших книг, в кото-
рых авторы XVI и даже еще XVIII века пространно рассуждали о структуре мира,
чтобы с изумлением констатировать, что наши прапрапрадедушки чувствовали себя
совершенно непринужденно в пространстве-ящике, где звезды вращались вокруг
Земли менее шести тысяч лет. В космической атмосфере, в которой мы бы сразу
же задохнулись, в перспективе, куда мы бы физически были не в состоянии всту-
пить , они дышали без малейшей стесненности, если не полной грудью".
Это высказывание Тейяра прекрасно описывает начавшуюся в Новое время рево-
люцию в понимании природы — революцию, которую часто называют переходом "от
существующего к возникающему" или "от бытия к становлению". Нам сейчас трудно
вообразить Вселенную, которая существует 6000 лет и оканчивается сферой не-
подвижных звезд за орбитой Сатурна. А ведь на протяжении тысячелетий подобное
представление о мире было нормой, пусть и находились великие умы, его оспари-
вавшие .
Многочисленные открытия, сделанные на протяжении трех веков (с XVIII по
XX), открыли перед людьми совершенно новую космологическую перспективу. Науч-
ная картина мира перестала быть стационарной. Кант и Лаплас предложили первую
физическую модель образования Солнечной системы. Геология Геттона и Лайеля
продемонстрировала бездонную (как тогда казалось) глубину истории нашей пла-
неты, а начавшая развиваться после выхода знаменитой книги Дарвина эволюцион-
ная биология доказала, что не менее сложная история была и у живой природы.
Благодаря теории относительности Эйнштейна возникла модель расширяющейся Все-
ленной — сначала чисто математическая, но быстро получившая астрономические
подтверждения. После этого Жорж Леметр и Георгий Гамов сформулировали теорию
расширения Вселенной "от первоначального атома", которую еще при их жизни на-
звали теорией Большого взрыва. Радиоизотопный метод позволил определить воз-
раст Солнечной системы (4,6 миллиарда лет), а расчеты на основе теории Боль-
шого взрыва — возраст Вселенной (13,8 миллиарда лет). Теория тектоники плит в
основных чертах объяснила, как и почему на Земле менялось расположение конти-
нентов . Инфляционная теория дополнила классическую космологию, установив, что
видимая Вселенная — это не просто не вся Вселенная, а ее ничтожно малая часть
(из-за стремительного расширения в самом начале). Неравновесная термодинамика
начала исследовать физические принципы, делающие возможной самоорганизацию
материи — упорядоченное изменение от простого к сложному. Одним словом, Все-
ленная обрела историю. На месте статичного аристотелевско-птолемеевскохю кос-
моса возникла картина грандиозного потока, несущего в себе все вещи и устрем-
ленного из бурного прошлого в неведомое будущее.
Вот частью этого потока и является древо жизни — иначе говоря, эволюционное
древо. Надо сказать, что сейчас мы знаем довольно много даже о его глубоких
"корнях": загадка происхождения жизни, рассуждения о которой еще недавно вос-
принимались как бездоказательные, в наше время обернулась рядовой научной
проблемой, вполне поддающейся решению экспериментальными методами. Однако те-
ма этой публикации — не столько "корни" древа жизни, сколько его "ствол" и
главные ветви. Автор должен признаться, что на протяжении всей работы он счи-
тал смысловым центром третью часть, так и озаглавленную — "Древо жизни", — и
именно ее писал с особым удовольствием. То, что эта часть не стала заодно и
самой большой, объясняется — увы — чрезмерным объемом первых двух частей, ко-
торые задумывались как вводные (например, глава о биоэнергетике разрослась в
процессе написания до совершенно непредвиденного размера, а сокращать было
жалко: тема-то интересная). Впрочем, нет худа без добра: широкий охват мате-
риала позволяет нам увидеть, что субъектами эволюции в том или ином смысле
могут быть структуры всех уровней — от атомов и молекул до планеты Земля.
"Философский
камень" биологии
Открытие генетического кода подарило людям такие возможности для изучения
биологической эволюции, которые до середины XX века было просто невозможно
вообразить. Например, представим, что молекулярный филогенетик сравнивает не-
сколько видов эукариот, у которых прочитаны аминокислотные последовательности
100 белков. Типичный размер эукариотного белка — 300 аминокислот. Получается,
что сравнению подлежат 30 000 аминокислотных позиций. Если прочитаны еще и
нуклеотидные последовательности генов, кодирующих эти белки, то — поскольку
любая аминокислота кодируется тремя нуклеотидами — сравнение охватит уже 90
000 позиций, на каждой из которых в принципе может стоять любой из четырех
нуклеотидов (А, Т, Г или Ц). Таково количество признаков, с которыми работают
систематики в молекулярную эру. Открытие методов работы с генетической инфор-
мацией поистине стало "философским камнем" биологии, во всяком случае, эволю-
ционной. Отметим, что все приведенные числа не взяты с потолка — они заимст-
вованы из реальных свежих научных работ, причем достаточно скромных по разма-
ху: объем анализируемых данных запросто может быть и больше (например, во
многих современных исследованиях сравниваются целые геномы, и тогда речь идет
уже о миллионах нуклеотидов).
Любая группа живых организмов имеет общего предка, у которого был свой ге-
ном. От этого генома путем конвариантной редупликации, то есть копирования с
сохранением изменений, произошли геномы всех современных представителей груп-
пы. Многообразные события, меняющие геномы в ходе эволюции, оставляют следы,
которые наслаиваются друг на друга. Их можно найти и прочитать, примерно так
же, как можно найти и прочитать следы правок в обычном тексте. Ведь геном —
это тоже текст (хотя и не только текст, конечно). Геном любого организма за-
ключает в себе целую летопись эволюционных изменений, многие из которых можно
довольно точно распознать и датировать. Сплошь и рядом это относится даже к
изменениям, которые произошли много сотен миллионов лет назад. Текст может
сохранить все.
Любой геном — это в некотором смысле архив, документирующий события, проис-
ходившие на всем протяжении эволюционной линии от последнего общего предка
всех живых организмов (или, во всяком случае, клеточных организмов, если мы
говорим о них) до обладателя этого генома. Конечно, следы многих событий в
конце концов стираются из генома начисто, и с этим ничего не поделаешь:
"Полностью утраченная информация, не сохранившаяся ни в одном экземпляре,
восстановлена быть не может". С другой стороны, бывают и такие эволюционные
новшества, которые запечатлеваются в геноме очень четко — так, что "прочи-
тать" их следы можно даже спустя миллиарды лет. "Впечатанные" в геном свиде-
тельства в принципе могут воспроизводиться из поколения в поколение сколь
угодно долго. Причем все они заключены внутри сложной упорядоченной структу-
ры , дающей возможность их послойно датировать. Это означает, что наряду с па-
леонтологической летописью, которая интенсивно изучается с начала XIX века, в
распоряжении биологов теперь есть еще одна настоящая летопись — генетическая.
Как и палеонтологическая летопись, она неполна, но зато любой ее сохранивший-
ся фрагмент несет огромное количество информации.
Установление родства между организмами — далеко не единственная, но, безус-
ловно, очень важная задача эволюционной биологии. Сейчас эта задача частью
уже решена, а частью активно решается. Неудивительно, что книги на эту тему
быстро устаревают. Особенно это относится к главе 15. Избежать этого невоз-
можно : скорость открытий в области мегасистематики сейчас такова, что любая
сводка должна обновляться по меньшей мере раз в год, а иногда и быстрее. На-
пример, супергруппа Excavata все более четко разбивается современными иссле-
дованиями на две самостоятельные супергруппы — Discoba и Metamonada, которые
независимо отходят от "корня" общего древа эукариот. Вместо супергруппы
Opisthokonta на эволюционном древе эукариот все чаще появляется супергруппа
Obazoa, включающая тех же опистоконтов плюс две небольшие группы примитивных
жгутиконосцев. Загадочную ветвь криптомонад в последнее время сближают с ар-
хепластидами, то есть с растениями — в противовес старой системе, причисляв-
шей криптомонад к хромальвеолятам. С другой стороны, на молекулярных деревь-
ях, построенных с учетом новейших данных, регулярно возникает сестринская по
отношению к риз ариям ветвь, состоящая из страменопилов и альвеолят — а это
означает, что систематики, возможно, поторопились с "закрытием" группы
Chromalveolata: не достаточно ли было просто изменить ее состав, выкинув от-
туда криптомонад и гаптофит? Наконец, в феврале 2018 года вышла авторитетная
и основанная на большом наборе новых данных работа, показывающая, что колло-
диктион (многократно здесь упоминавшийся) вместе с двумя другими "микроцарст-
вами" малоизвестных протистов образует целую супергруппу, сестринскую по от-
ношению ко всем одножгутиковым (Unikonta = Amorphea) . И конечно же,
ЛЛпродолжение следует". Остается лишь посоветовать интересующимся этой темой
следить за научными новостями, благо освещаются они сейчас неплохо, а тема —
и впрямь интригующая.
Подчеркнем примечательную вещь. Все перечисленные изменения касаются новой
мегасистемы, которая оформилась только на рубеже XX-XXI веков. Преемственной
по отношению к старой мегасистеме (разбивавшей живые организмы на два, три,
четыре или пять царств) она не является. Можно утверждать, что современная
мегасистема соотносится со старыми системами живой природы (образца "животные
— растения — грибы — протисты — бактерии...") так же, как современная астро-
номия, во всей ее красе и славе, соотносится с геоцентрической системой Пто-
лемея. С точки зрения истории науки это утверждение будет верным буквально,
без всяких преувеличений. И это прекрасно! Нам выпало жить в годы, когда но-
вая научная картина мира (во всяком случае, биологическая) формируется прямо
на глазах.
Будем помнить, что построение филогенетического древа, отображающего — в
идеале — родственные связи абсолютно всех земных живых организмов, в любом
случае станет только первым шагом к настоящему, глубокому познанию эволюцион-
ного процесса. Как механизмы эволюции, так и ее результаты невероятно много-
образны. Известный американский биолог Стивен Гулд говорил по этому поводу:
ллПроблема с историей жизни состоит в том, что у нас есть только один экспе-
римент. Благодаря огромному биохимическому сходству мы понимаем, что вся
жизнь на Земле произошла из одного источника. Тем самым у нас нет никакого
ответа на главный вопрос: должна ли любая жизнь быть устроена так, как она
устроена на Земле, или мы наблюдаем всего одну возможность, осуществившуюся
на фоне сотни миллионов нереализованных альтернатив?"
Сам Гулд, насколько можно судить, склонялся ко второму варианту ответа. Ес-
ли бы историю Вселенной можно было каким-то непостижимым образом повторить,
возникла ли бы в ней хоть какая-нибудь жизнь? И если да — то как бы она вы-
глядела? К чему пришла бы жизнь на Земле, если бы не было столкновения, поро-
дившего Луну, или лл3емли-снежка", или пермского кризиса (см. главы 16, 17)?
За всеми этими поворотами и развилками скрываются возможности, которых мы,
скорее всего, никогда не сможем даже вообразить (как сказал писатель Олег Ла-
дыженский: "Вся западная мифология не смогла придумать кенгуру"). Вряд ли
кто-нибудь мог бы предсказать, например, такое уникальное событие, как воз-
никновение эукариот, точный механизм которого до сих пор остается загадкой
(см. главы 10, 14). Вряд ли инопланетный биолог, посетивший Землю в мезозой-
скую эру, мог бы предсказать великую эволюционную судьбу тогдашних млекопи-
тающих. Или, скажем, если бы тот же биолог попал три миллиона лет назад в Аф-
рику и встретил там странных прямоходящих обезьян — смог бы он предсказать,
что именно эти нелепые и малочисленные существа создадут великую письменную
культуру, преобразуют облик планеты, выйдут в космос?..
А ведь перед нами будущее — и оно еще более сложно и непредсказуемо, чем
прошлое.
От древа к
лабиринту
Чарльз Дарвин считал, что родственные отношения всех живых организмов могут
быть изображены ветвящимся древом. Сам термин "древо жизни" изобретением Дар-
вина не является — он, скорее всего, заимствован из библейской Книги Бытия.
Классическая биология полагала, что все биологическое разнообразие возникло
из единственного начального узла эволюционного древа в результате серии его
последовательных ветвлений. Сейчас эту "нулевую гипотезу" дополняет постулат,
блестяще популяризированный Ричардом Докинзом в книгах ллЭгоистичный ген"3 и
ллРасширенный фенотип": эволюция генов и эволюция целых организмов — просто-
напросто разные вещи. Гены вполне могут перемещаться из одного организма в
другой, и все реальные организмы (включая человека) суть в той или иной сте-
пени генетические химеры. Например, в геноме человека имеется довольно много
генов и еще больше некодирующих нуклеотидных последовательностей, включенных
туда эндогенными ретровирусами (см. главу 12). Все эукариоты получили от ви-
русов такие важные для работы генетического аппарата вещи, как механизм кэши-
рования информационной РНК и фермент топоизомеразу, играющий важную роль в
репликации ДНК (см. главу 10). Список подобных примеров легко продолжить.
Итак, в общем случае нам приходится иметь дело не с единым молекулярно-
филогенетическим древом, а с набором древес, относящихся к отдельным генам,
которые совсем не обязаны совпадать друг с другом.
Картина еще больше усложняется в тех случаях, когда по ходу эволюции орга-
3 Домашняя лаборатория 2023-07-08
низмы не просто обменивались генами, а сливались целиком. Таких примеров го-
раздо меньше, но некоторые из них очень важны для истории жизни на Земле: на-
пример, происхождение эукариот, которое сопровождалось слиянием минимум двух,
а может быть, и трех микроорганизмов (см. главы 10, 14). Систему родственных
связей, возникающую в результате наложения друг на друга множества событий
подобного рода, ЛЛдревом" уже не назвать.
В 2010 году известный французский биолог Дидье Рауль предложил для обозна-
чения структур такого типа термин ЛЛризома". Попросту говоря, ризома — это
сеть. Типичную ризому дает, например, попытка построить по молекулярным дан-
ным общее эволюционное древо бактерий. Иначе и не может быть, учитывая, на-
сколько легко разные бактерии обмениваются генами. Внутри древа эукариот эф-
фекты горизонтального переноса генов выражены гораздо меньше, но зато проис-
хождение эукариот как таковых без учета этого процесса вообще необъяснимо.
Французское слово ЛЛризома" (rhizome) буквально означает ЛЛкорневище". В 1976
году философы-постмодернисты Жиль Делёз и Феликс Гваттари сделали это слово
философским термином. Ризома — это сетевая структура, противопоставляемая
древовидной; визуализацией ризомы будет запутанная корневая система, лишенная
сколько-нибудь четкой формы, но пронизанная множеством внутренних связей.
ллРизома так устроена, что в ней каждая дорожка имеет возможность пересечься с
другой. Нет центра, нет периферии, нет выхода. Потенциально такая структура
безгранична", — писал Умберто Эко в ЛЛЗаметках на полях ЛЛИмени розы"", рассуж-
дая о типах лабиринтов. Ризома — образ мира, "потерявшего свой стержень", но
таящего в себе неожиданные возможности, — "творящий Хаос". Превращая "ризому"
из чисто философского термина в биологический, Дидье Рауль, несомненно, учи-
тывал эти оттенки значений. В своих интервью он обратился еще и к философии
Ницше, говоря, что эволюция — не столько аполлонический мир (красивый, разум-
ный, организованный), сколько дионисический (хаотичный, бурный, внезапный). С
точки зрения Рауля, эволюция гораздо более прерывиста, случайна и непредска-
зуема , чем полагало большинство ученых XIX-XX веков.
Разумеется, такой взгляд на вещи не надо превращать в догму (впрочем, к лю-
бым другим научным взглядам это тоже относится). Тем не менее, здесь наверня-
ка отражены некоторые реальные свойства природных процессов — причем не толь-
ко генетических. Из сказанного в этой публикации можно увидеть, что система
траекторий, по которым двигалась (или могла бы двинуться) история жизни на
Земле, в общем-то гораздо больше похожа на сеть тропинок в дремучем лесу, чем
на прямую, как стрела, провешенную трассу. Это относится к нашему прошлому, и
нет никаких оснований считать, что это не будет относиться к будущему. Стоит
добавить, что такой вывод не должен служить основанием для пессимизма. Конеч-
но, путь сопряжен с риском — в ризоме легко заблудиться, но она же может по-
дарить путнику новые, неожиданные возможности: ведь сотворение мира еще длит-
ся. Эволюция — не освещенная дорога, ведущая от амебы прямо к человеку, а
темный лабиринт, полный обрывов, провалов, тупиков, внезапных пересечений,
катастроф и эвкатастроф. Дионисический космос, таящий в себе и смерть, и на-
дежду .
Литпортал
БЕТАГЕМОТ1
Питер Уотте
Бета-прелюдия:
Правонарушитель
Ахиллу Дежардену рассказывали, что к утратившим зрение то возвращается во
сне.
Это касалось не только слепых. Любой, кого порвала жизнь, видит сны цельно-
го создания. Человек без рук и без ног бегает и пинает футбольный мяч; глухой
слушает симфонии, потерявшие близких, снова любят. Разум обладает собственной
инерцией, привыкает за много лет к определенной роли и неохотно отказывается
от прежней парадигмы.
Конечно, в конце концов, это проходит. Блекнут яркие видения, замолкает му-
Вторая часть трилогии - "Водоворот" - см. Домашняя лаборатория 2023-12, 2024-01.
зыка, воображаемое уступает место чему-то более подобающему пустым глазницам
и лопнувшей перепонке. Но на это уходят годы, десятилетия — и все это время
мозг терзает себя по ночам напоминаниями о том, что имел когда-то.
Так случилось и с Ахиллом Дежарденом. Во сне у него была совесть.
Сны уносили его в прошлое, в жизнь скованного бога: в руках миллионы жиз-
ней, его владения простираются от геосинхронной орбиты до дна Марианской впа-
дины. Он снова без устали сражался за добро, он подключен к тысяче каналов
сразу, рефлексы и способности к распознаванию образов подстегнуты модифициро-
ванными генами и индивидуализированными нейротропами. Он наводил порядок там,
где бушевал хаос. Приносил в жертву десять человек, но спасал сотню. Он изо-
лировал эпидемические вспышки, разбирал завалы, обезвреживал террористов и
экологические катастрофы, угрожавшие со всех сторон. Он плыл по радиоволнам и
скользил по тончайшим нитям оптоволокна, вселяясь в перуанские морские фабри-
ки, а минуту спустя — в корейский спутник связи. Он снова становился лучшим
правонарушителем в УЛН, пробовал второй закон термодинамики на прочность, до-
ходил до предела, а иногда и чуть дальше. Он был истинным духом из машины — а
машина в те времена была повсюду.
Однако сны, искушавшие его чуть ли не каждую ночь, были не о власти, а о
рабстве. Только во сне он мог вновь почувствовать ту парадоксальную покор-
ность , что смывала с его рук реки крови. Они называли ее Трипом Вины. Набор
искусственных нейротрансмиттеров, названия которых Дежарден не удосужился вы-
учить. Он, как-никак, мог убить миллионы людей одним приказом: такую власть
никому не дают в руки, не снабдив несколькими предохранителями. С Трипом в
мозгу мятеж против общего блага становился физиологически невозможным. Трип
Вины обрубал связь между абсолютной властью и абсолютной развращенностью: лю-
бая попытка применить власть во зло вызвала бы мощнейший эпилептический при-
ступ. Ахиллу не случалось лежать без сна, сомневаясь в правоте своих поступ-
ков или в чистоте побуждений. То и другое ввели ему в жилы люди, менее склон-
ные к колебаниям.
Эта безвинность была так утешительна! Вот ему и снилось рабство. И еще сни-
лась Элис, освободившая его от цепей.
Во сне он мечтал их вернуть.
Понемногу сны ускользали, как всегда с ними бывает. Прошлое уходило: насту-
пало беспощадное настоящее.
Мир разваливался, распадаясь на застывшие кадры, из глубин моря поднимался
губительный вирус, использовавший вместо транспорта плоть подводницы из
Нт АмПацифика, а барахтавшиеся в его турбулентном следе Власть Предержавшие и
Упустившие прозвали агрессора Бетагемотом и принялись сжигать людей и имуще-
ство в лихорадочных, тщетных попытках предотвратить грядущую смену режима.
Северная Америка пала. Триллионы микроскопических пехотинцев маршировали по
суше, безвозвратно губя почву и плоть. Полыхали и затухали войны: кампания в
Н!АмПацифике, Колумбийский ожог, евро-африканское восстание. И, конечно, Рио
— получасовая война, война, которую Трип Вины никак не должен был допустить.
Дежарден, так или иначе, участвовал во всех. И, пока отчаявшиеся многоклеточ-
ные скатывались во внутренние распри, настоящий враг неотвратимо наползал на
землю подобно душному одеялу. Даже Ахилл Дежарден, гордость Патруля Энтропии,
не мог его сдержать.
Теперь, когда настоящее почти настигло его, он ощущал легкую печаль по все-
му, чего не сделал. Впрочем, то были фантомные боли, пережитки совести, ос-
тавшейся в далеком прошлом. Они почти не задевали его здесь, на зыбком пере-
крестке сна и яви, где на мгновение он вспоминал, что свободен и жаждет раб-
ства.
Потом он открывал глаза, и не оставалось ничего, кроме равнодушия. Ман-
дельброт, тушкой распластавшись у него на груди, замурлыкала. Рассеяно погла-
живая ее, Ахилл вызвал утреннюю статистику. Ночь выдалась сравнительно спо-
койной: внимания заслуживали разве что на удивление безрассудные беженцы, по-
пытавшиеся прорвать Североамериканский периметр. Они подняли парус под покро-
вом темноты, вышли с Лонг-Айленда на переоборудованном мусоровозе в час-
десять по Атлантическому Стандартному. Еще через час заинтересованные лица из
евро-африканских кругов уже боролись за бабки, решая, кто же осуществит «вы-
нужденную ликвидацию». Беглецы едва миновали мыс Код, когда алжирцы (алжир-
цы? ) их сняли.
Система даже не стала поднимать Дежардена с постели. Мандельброт потянулась
и отправилась на утренний обход. Освободившись, Дежарден встал и зашлепал к
лифту. Шестьдесят пять пустующих этажей плавно ушли вниз. Всего пару лет на-
зад здесь гудел улей антикризисного центра: тысячи оперативников на Трипе,
вечно подвешенных на тонкой ниточке над нервным срывом, с холодной бесстраст-
ной бережливостью распоряжались жизнями и легионами. Теперь все это принадле-
жало ему одному. После Рио многое изменилось.
Лифт выплюнул его на крышу УЛН. Соседние здания выстроились подковой, тес-
нясь на границе расчищенной зоны. Статическое поле Садбери, задевавшее брюхом
крыши самых высоких зданий, осыпало руки Дежардена мурашками.
Лучи встающего солнца подожгли руины поверженного царства.
Опустошение было не полным — пока еще не совсем. Города на востоке отчасти
уцелели: вооружившись, огородившись стенами и без конца отражая претендующих
на их земли захватчиков. Там, где конфликты не исчерпали себя, все еще клоко-
тали линии фронта: одна-две даже стабилизировались. На континенте сохранились
островки цивилизации. Их было не много, однако война продолжалась.
И все потому, что пять лет назад женщина по имени Лени Кларк восстала с
океанского дна, и в крови ее бурлила жажда мести пополам с Бетагемотом. Де-
жарден перешел посадочную площадку и встал на краю крыши. Пока он мочился в
пустоту, над обрывом поднималось солнце. Сколько перемен, размышлял он.
Сколько ярусов катастроф во имя нового равновесия. Его владения от планеты
сжались до континента, подпаленного с краев. Кругозор, некогда вмещавший бес-
конечность, теперь ограничивался побережьями. Руки, обнимавшие прежде весь
мир, были ампутированы до локтей. Сам по себе Североамериканский сегмент Сети
отсекли от прочих, словно электронную гангрену; Ахиллу Дежардену приходилось
иметь дело с мертвеющей массой.
И все же во многих отношениях он был сейчас могуществен, как никогда. Да,
территория уменьшилась, но меньше стало и тех, с кем доводилось ее делить. Он
теперь был не столько командным игроком, сколько единоличником. Не то чтобы
об этом многие знали...
Впрочем, кое-что не изменилось. Формально он все еще был сотрудником УЛН,
ну или тех останков организации, какие еще сохранились на земном шаре. Мир
давно повалился на бок — во всяком случае, близлежащие его части, но обязан-
ности Дежардена остались прежними: минимизировать ущерб.
Вчерашние локальные пожары сегодня преобразились в пылающие преисподние, и
Дежарден всерьез сомневался, что их теперь вообще возможно потушить, но он
был из тех, кто мог хотя бы сдерживать их наступление. Он все еще оставался
правонарушителем — смотрителем маяка, как он назвал себя в тот день, когда
они, наконец, смилостивились и позволили ему остаться, — и сегодняшний день
будет похож на все остальные. Предстоит отражать атаки и разоблачать врага.
Какие-то жизни прервутся во имя спасения других, более многочисленных или бо-
лее ценных. Надо будет уничтожать болезнетворных микробов и поддерживать
имидж.
Повернувшись спиной к восходящему солнцу, он переступил лежащее под ногами
женское тело, нагое и вы-потрошенное. Эту женщину тоже звали Элис.
Он попытался вспомнить, совпадение это или нет.
БЕТА-МАКС
Мир не умирает, его убивают. И у убийц
есть имена и адреса.
Юта Филлипс1
Контратака
Все начинается со звука в темноте. Дрейфуя по склону подводной горы, Лени
Кларк готовится к неизбежному расставанию с одиночеством.
На таком расстоянии она совершенно слепа. «Атлантида» с ее подвесными рама-
ми, маячками и иллюминаторами, истекающими в бездну размытым светом, осталась
в сотнях метрах позади. Достаточно далеко, чтобы не мигало никаких сигналов,
чтобы огоньки на каналах инженерных сетей и наружных хранилищах не загрязняли
мрак. Линзы на глазах могут использовать для зрения малейшую искорку, но не
способны создать свет там, где его не существует. Как здесь. Три тысячи мет-
ров, триста атмосфер, три миллиона килограмм на квадратный метр выжали из ми-
роздания все до последнего фотона. Лени Кларк слепа, как последний сухопут-
ник. За пять лет на Срединно-Атлантическом хребте она успела полюбить слепо-
ту.
А теперь все вокруг заполняет комариное зудение гидравлики и электрического
тока. Сонар мягко постукивает по ее имплантатам. Зудение неуловимо переходит
в писк и гаснет. Легкий толчок: что-то замирает прямо над ней.
— Дрянь. — Механизм в горле обращает словечко в тихое жужжание. — Уже?
— Я дал тебе лишних полчаса. — Это голос Лабина. Его слова преобразованы
той же техникой, но искаженный звук теперь стал для нее привычней оригинала.
Она бы вздохнула, будь здесь возможно дыхание.
Кларк включает налобный фонарь. Во вспыхнувшем луче появляется Лабин: чер-
ный силуэт, усеянный точками внедренных механизмов. Впадина на груди — диск
со множеством щелей, хром на черном фоне. Роговичные накладки превращают гла-
за в пустые матовые овалы. Он выглядит так, словно создан из теней и техноло-
гий . Кларк знает, что за этим обликом скрывается человечность, но помалкивает
об этом.
Рядом с Лабином парит пара «кальмаров». С одной из метровых торпед свисает
нейлоновый мешок, бугрясь всяческой электроникой. Кларк переворачивается ко
второй, сдвигает тумблер с «Ведомый» на «Ручной». Машинка вздрагивает и вы-
двигает рулевое устройство.
Кларк, не раздумывая, отключает фонарь. Все снова поглощает тьма. Ни движе-
ния , ни огоньков. Никаких атак.
Просто все здесь по-другому.
— Что-то случилось? — жужжит Лабин.
Ей вспоминается совсем иной океан на другом краю мира. Там, на источнике
Чэннера, стоило выключить свет, загорались звезды, тысячи биолюминесцентных
созвездий: рыбы освещались как посадочные полосы аэропортов; мерцали члени-
стоногие ; сложными переливами вспыхивали виноградинки гребневиков. Чэннер
песней сирены приманивал этих причудливых обитателей средних глубин, застав-
ляя погружаться глубже обычных для них слоев, подкармливая незнакомыми веще-
ствами и превращая в чудовищных красавцев. Там, на станции «Биб», темнело
лишь тогда, когда свет зажигали.
Однако «Атлантида» — не «Биб», а это не Чэннер. Здесь свет исходит лишь от
грубых, неуклюжих машин. Фонари пробивают в черноте пустые тоннели, безжиз-
1
Юта Филлипс (1935-2008) — настоящее имя Брюс Данкан Филлипс, американский бард,
анархист и деятель профсоюзного движения (прим. ред.)
ненные и уродливые, как от горящего натрия. А выключишь их, и. . . пусто. В
том-то и весь смысл, конечно.
— Так было красиво, — говорит она.
Ему не требуется пояснений.
— Было. Не забывай только, почему.
Она хватается за руль.
— Просто тут все... по-другому, понимаешь? Иногда мне почти хочется, чтобы
из глубин вырвался какой-нибудь здоровенный зубастый поганец и попробовал от-
хватить от меня кусочек.
Она слышит, как оживает «кальмар» Лабина — невидимо для глаз, но где-то ря-
дом. И запускает собственный, чтобы последовать за ним.
Сигнал она принимает одновременно на низкочастотник и на сам свой скелет.
Вибрация костей глубоко отзывается в челюсти, а модем просто пищит.
Она включает приемник.
— Кларк.
— Кен тебя нашел, а? — Воздушный голос, не изуродованный приспособлениями
для подводной речи.
— Ага. — Слова Кларк по контрасту звучат неприятно и механически. — Мы уже
двинулись.
— Хорошо. Просто проверка. — Голос ненадолго замолкает. — Лени?
— Я тут.
— Просто... осторожнее там, ладно? — просит ее Патриция Роуэн. — Ты же зна-
ешь , как я волнуюсь.
Они всплывают, и вода неуловимо светлеет. Она и не заметила, как мир пере-
красился из черного в синий. Кларк никогда не успевает поймать момент, когда
это происходит.
С тех пор как Патриция дала отбой, Лабин молчал. Теперь, когда густая синь
плавно переходит в лазурь, Кларк проговаривает мысль вслух.
— Она тебе по-прежнему не нравится.
— Я ей не доверяю, — жужжит Лабин. — А так — вполне себе нравится.
— Потому что она — корп?
Их уже много лет никто не называет корпоративными управляющими.
— Бывший корп. — Машинка в горле не скрывает, с каким мрачным удовольствием
он это выговаривает.
— Бывший, — повторяет Кларк.
— Не потому.
— Так почему?
— Список причин тебе известен.
Известен. Лабин не доверяет Роуэн, потому что когда-то, давным-давно, она
всем заправляла. Это по ее приказу их всех тогда привлекли к делу, взяли по-
врежденный товар и испортили пуще прежнего: переписали воспоминания, перепра-
вили побуждения, даже совесть переделали во имя какого-то неопределимого, не-
уловимого общего блага.
— Потому что она бывший корп, — повторяет Кларк.
Вокодер Лабина испускает нечто похожее на хмыканье.
Кларк знает, к чему клонит напарник. Она по сию пору не уверена, какие со-
бытия ее собственного детства — реальность, а какие были внедрены, инсталли-
рованы постфактум. А ведь она еще из счастливчиков: пережила тот взрыв, пре-
вративший источник Чэннера в тридцать квадратных километров радиоактивного
стекла. Ее не размололо в кашу вызванное взрывом цунами, ее не испепелили
вместе с миллионным лагерем беженцев на побережье Нт АмПацифика.
Конечно, ей не должно было так повезти. Строго говоря, все эти миллионы бы-
ли побочными жертвами и не более того. Не их вина — да и не Роуэн, — что
Кларк не сидела на месте, не дала в себя как следует прицелиться.
И все же. Вина вине рознь. Пусть у Патриции Роуэн на руках кровь миллионов,
но ведь зараженные области сами о себе не позаботятся; тут на каждом шагу
требуются ресурсы и решимость. Блокировать карантинную зону; направить подъ-
емники ; испепелить. Отчистить, сполоснуть, повторить. Убей миллион, чтобы
спасти миллиард, убей десяток, спасая сотню. Возможно даже, убей десять чело-
век , чтобы спасти одиннадцать — принцип тот же, даже если маржа прибыли ниже.
Только вся эта механика не работает сама собой, руку приходится все время
держать на кнопке. Роуэн, устраивая бойню, никогда не закрывала глаз на цену
и брала ответственность на себя.
Лени Кларк было намного проще. Она просто рассеяла заразу по миру и ушла в
тень, даже не оглянувшись. Ее жертвы и теперь еще громоздятся курганами, на-
растают по экспоненте, в десятки раз превосходя счет Роуэн. А ей и пальцем не
пришлось шевельнуть.
Никто из тех, кто числит в друзьях Лени, не имеет разумных оснований судить
Патрицию Роуэн. Кларк с ужасом думает о том дне, когда эта простая истина
дойдет до Кена Лабина.
«Кальмары» увлекают их все выше. А вот теперь — явный градиент: свет, па-
дающий сверху, тает в темноте под ними. Для Кларк это самая пугающая часть
океана: полуосвещенные воды средних глубин, где рыщут настоящие кальмары —
бескостные многорукие монстры по тридцать метров в длину, у которых мозги хо-
лодные и быстрые, как сверхпроводники. Ей рассказывали, что теперь они вырас-
тают вдвое больше прежних размеров. И в пять раз увеличились в числе. Очевид-
но, это все за счет лучших условий роста. В теплеющих морях личинки
Architeuthis развиваются быстрее, и никакие хищники на их поголовье не влияют
— всех давно выловили рыбаки.
Architeuthis - гигантский кальмар.
Конечно, Кларк их ни разу не видела. И надеется не увидеть — согласно отче-
там, популяция сокращается от бескормицы, а величина океана сводит шансы слу-
чайной встречи к микроскопическим величинам. Но временами зонды улавливают
призрачное эхо массивных объектов, проходящих над головой: жесткие вскрики
хитина и панцирей, смутные ландшафты окружающей плоти, почти невидимой для
сонара. По счастью, «архи» редко нисходят в истинную тьму.
С подъемом рассеянные вокруг оттенки становятся более насыщенными — в су-
мраке светоусилители не передают цветов, но в такой близости от поверхности
разница между линзой и невооруженным глазом, в теории, минимальна. Иногда
Кларк хочется это проверить: снять накладки с глаз и посмотреть самой, но это
несбыточная мечта. Подводная кожа гидрокостюма, облепляющая лицо, напрямую
связана с фотоколлагеном. Она даже моргнуть не может.
А вот и течение. Над ними кожура океана морщится тусклой ртутью. Подъемы,
падения, перекаты, бесконечная смена гребней и провалов сминают холодный шар,
светящийся по ту сторону, стягивают его в игриво приплясывающие узелки. Еще
немного, и они вырываются на поверхность, где перед ними расстилается мир из
моря и лунного неба.
Они все еще живы. Три тысячи метров свободного всплытия за сорок минут, а
ни один капилляр не лопнул. Кларк сглатывает под напором изотонического рас-
твора в горле и синусах, ощущает искрящую в груди механику, и в который раз
дивится чудесам жизни без дыхания.
Лабин, само собой, думает только о деле. Он перевел своего «кальмара» на
максимальную плавучесть и использует его как платформу для приемника. Кларк
переводит своего в стационарный режим и помогает Лабину. Они скользят вверх и
вниз по серебристым волнам, луна такая яркая, что линз даже не требуется.
Взлетает на привязи пучок антенн, глаза и уши растопыриваются во все стороны,
выслеживая спутники, компенсируя движение волн. Одна-две простенькие рамки
сканируют наземные станции.
Сигналы накапливаются, хотя и очень медленно.
С каждым поиском бульон становится все жиже. О, эфир по-прежнему полон ин-
формации, — мелкие гистограммы расползаются по всем сантиметровым частотам,
по всей шкале идет трескотня, — но плотность сильно снизилась.
Разумеется, даже в отсутствии сигнала есть свой грозный смысл.
— Немного, — отмечает Кларк, кивая на индикаторы.
— М-м-м. — Лабин натянул шлемофон поверх капюшона подводника.
— Галифакс еще ловится.
Он задерживается там и тут, вылавливая загрузившиеся каналы. Кларк тоже бе-
рет шлем и устремляет внимание на запад.
— Из Садбери ничего, — докладывает она спустя некоторое время.
Лабин не напоминает, что Садбери молчит с самого Рио. Не говорит, как малы
шансы, что Ахилл Дежарден еще жив. Даже не спрашивает, когда же она, наконец,
смирится с очевидным. Просто произносит:
— И Лондона не могу поймать. Странно.
Она сдвигается на другую частоту.
Они не получат внятной картины, наугад шаря пальцами в потоке. Настоящий
анализ придется отложить до возвращения на «Атлантиду». Большая часть услы-
шанных языков Кларк незнакома, хотя некоторые пробелы восполняются картинкой.
Много бунтов по Европе — на фоне страхов, что Бетагемот добрался до Южного
Противотечения. Анклав избранных, что могли позволить себе некие контрмеры,
трещит под напором бесчисленных орд, которые не смогли. Китай с буферными го-
сударствами по прежнему во тьме — уже пару лет, — но это говорит скорее об
обороне против апокалипсиса, чем о поражении. Все, подлетающее к их побережью
на пятьсот кэмэ и ближе, расстреливается без предупреждения, а значит, у них
как минимум функционирует военная инфраструктура.
Еще один переворот под знаком «Мадонны Разрушения», на сей раз в Мозамбике.
Уже восьмой, и счет нарастает. Восемь наций спешат приблизить конец света во
имя Лени Кларк. Восемь стран поддалось чарам порожденной ею злобной гнуси.
Дипломатичный Лабин об этих событиях не упоминает.
Из обеих Америк вестей мало. Экстренные сообщения да тактические переговоры
УЛН. Несколько выступлений апокалиптических сект, разглагольствующих о док-
трине Проактивного Уничтожения или вероятности Второго Пришествия, высчитан-
ной по Байесу. Конечно, по большей части болтовня: важные переговоры ведутся
по узким каналам из точки в точку, и эту сфокусированную информацию никогда
не занесет на пустынную поверхность Атлантики.
Конечно, порой Лабину было по силам менять и такие правила, но даже ему это
в последнее время давалось с трудом.
— Ридли пропал, — говорит он между тем. И вот это уже действительно плохая
новость. «Ретранслятор Ридли» — сверхсекретный спутниковый комплекс, настоль-
ко закрытый, что даже допуска Лабина едва хватает для входа. Это один из по-
следних источников надежных сведений для «Атлантиды». Корда корпы еще полага-
ли, что движутся к спасению, а не к изоляции, они оставляли за собой всяче-
ские неотслеживаемые каналы для связи с сушей. Никто точно не знал, почему за
последние пять лет так много их позакрывалось. Но опять же, чтобы это выяс-
нить, нужно было хотя бы ненадолго высунуться из-под воды, а у кого хватило
бы на это духа?
— Может, стоит рискнуть, — вслух размышляет Кларк. — Просто оставить уста-
новку на плаву на несколько дней. Дать возможность собрать настоящие данные.
Квадратный метр техники на весь океан — ну серьезно, великая ли вероятность?
Достаточно, понимает она. Многие еще живы. И готовы взглянуть фактам в ли-
цо, носом чуют неизбежную гибель. Найдутся и такие, кто уделит немного време-
ни мыслям о мести. У некоторых даже хватит на это средств — раз спасение не
купишь, можно потратиться на воздаяние. Что, если мир узнает: те, кто спусти-
ли с цепи Бетагемот, еще живы и прячутся под тремя сотнями атмосфер?
То, что «Атлантида» сохранила анонимность, — чистая удача, и испытывать ее
никому не хотелось. Скоро они переместятся, не оставив нового адреса. А пока
этак раз в неделю выставляют над водой глаза и уши, перехватывают эфир и вы-
жимают из принятых сигналов все возможное.
Когда-то этого было достаточно. В конце концов, Бетагемот оставил после се-
бя так мало, что даже электромагнитный спектр выцвел до прозрачности.
«Но уж в ближайшие пять минут на нас никто нападать не станет», — говорит
Лени себе... и тут же понимает, что нападение уже идет.
Маленькие красные пики на краю зрения говорят ей, что канал Лабина перегру-
жен. Она идентифицируется на его частоте, готовится поддержать в бою — но не
успевает, потому что враг взламывает и ее линию. Глаза наполняются помехами,
в ушах звучит ядовитое:
— И в башку не бери меня отрубать, соска долбаная! Попробуешь открыть дру-
гой канал — расхерачу на хрен. И всю установку утоплю, крыса ты мокрая!
— Опять, — голос Лабина доносится словно издалека, из параллельного мира,
где мягкие длинные волны безобидно поглаживают тела и механизмы. Только вот
Кларк атакуют в прежнем мире: среди вихря помех — Господи, только не это! —
проступает лицо: жуткий симулякр, искаженный почти до неузнаваемости. Кларк
подключает полдюжины сетевых фильтров. От одного прикосновения уходят в пар
целые гигабайты. В наушниках — чудовищный вопль.
— Хорошо, — замечает из соседнего измерения тоненький голосок Лабина. — Те-
перь , если только удастся сохранить...
— Ничего ты не сохранишь! — визжит привидение. — Ни хрена! Твари, да вы
хоть знаете, кто Я?
«Да», — молчит Кларк.
— Я — Лени Кларк...
Изображение темнеет.
Еще мгновение она словно кружится в водовороте. На сей раз это просто вол-
ны . Кларк стаскивает с головы шлемофон. Небо с пробелом луны мирно вращается
над головой.
Лабин отключает прием.
— Ну вот, — сообщает он, — потеряли восемьдесят процентов улова.
— Может, попробуем снова?
Она знает, что этому не бывать. Время на поверхности подчиняется нерушимому
расписанию; паранойя в наше время — синоним здравого смысла. А то, что под-
грузилось в их приемник, все еще где-то здесь, плавает в радиоволнах. Меньше
всего им сейчас нужно снова открывать эту дверь.
Кларк начинает сматывать антенну. Руки дрожат в лунном свете.
Лабин делает вид, что ничего не замечает.
— Забавно, — говорит он, — а оно на тебя не похоже.
Столько лет вместе, а он ее совсем не знает.
Они не должны существовать — эти демоны, принявшие ее имя. Хищники, подчис-
тую истребляющие добычу, надолго на свете не задерживаются. Паразиты, убившие
хозяина, гибнут вместе с ним. И неважно, созданы они из плоти или из электро-
нов: везде, как ее учили, действуют одни и те же законы. Но за последние ме-
сяцы она уже несколько раз сталкивалась с такими монстрами, слишком вирулент-
ными, чтобы выживать в рамках эволюционной теории.
— Может, они просто следуют моему примеру? — рассуждает Кларк. — Питаются
чистой ненавистью?
Луна остается позади. Лабин ныряет головой вниз, направляя «кальмара» прямо
в сердце тьмы. Кларк немного задерживается, погружаясь без спешки и любуясь,
как корчится и гримасничает светило над головой. Наконец лунные лучи размыва-
ются , растворяются в размытой мгле: они уже не озарят небо, они сами — небо.
Кларк запускает двигатель и отдается глубине.
К тому времени, как она догоняет Лабина, внешний свет совсем гаснет: она
ориентируется по зеленоватой точке, горящей на приборной панели его «кальма-
ра» . Они молча продолжают спуск. Давление нарастает. Наконец они минуют кон-
трольную точку периметра, условную границу «своей» территории. Кларк запуска-
ет низкочастотник и делает вызов. Ответа нет.
Не то чтобы никого нет в сети. Канал забит голосами, вокодированными и
обычными. Они накладываются, перебивают друг друга. Что-то стряслось. Несча-
стный случай. «Атлантида» требует подробностей. Механические голоса рифтеров
вызывают медиков к восточному шлюзу. Лабин сонирует глубину, снимает показа-
ния , включает фару «кальмара» и уходит влево. Кларк следует за ним.
Темноту перед ними пересекает тусклое созвездие, пропадает из вида. Гаснет.
Кларк прибавляет скорость, чтобы не отстать, и напор воды едва не сбивает ее
с «кальмара». Они с Лабином сближаются с плывущими ниже.
Два «кальмара» идут над самым дном на буксире у третьего. Один из задних
пуст. Второй увлекает за собой два сплетенных тела. Кларк узнает Ханнука Йе-
гера. Одной, почти вывернутой рукой тот сжимает руль. Другая обхватывает
грудь черной тряпичной куклы в натуральную величину. За куклой в воде расплы-
вается тонкая чернильная струйка.
Лабин заходит справа. В свете его фары струйка вспыхивает алым.
Эриксон, соображает Кларк. На морском дне человека узнают по десяткам зна-
комых примет: по осанке, повадке — и только мертвых рифтеров не отличишь друг
от друга. То, что ей пришлось опознавать Эриксона по ярлыку на рукаве — дур-
ной знак. Нечто пропороло его гидрокостюм от паха до подмышки — и его самого
тоже. Выглядит нехорошо. Ткани млекопитающих съеживаются в ледяной воде, пе-
риферийные сосуды сжимаются, удерживая тепло внутри. Поверхностная рана не
кровоточила бы при пяти по Цельсию. Так что Эриксона достало глубоко. Что бы
это ни было.
На буксирном «кальмаре» — Грейс Нолан. Лабин пристраивается за ней и сбоку
— живым волноломом, прикрывая от встречного течения Эриксона с Йегером. Кларк
поступает так же. Вокодер Эриксона часто тикает — боль или помехи.
— Что случилось? — жужжит Лабин.
— Точно не знаю. — Нолан смотрит вперед, не отвлекаясь от управления. — Мы
проверяли вспомогательный отвод над озером. Джин забрел за скалу. Через не-
сколько минут мы нашли его в таком виде. Может, обвалился уступ, и его заце-
пило .
Кларк выворачивает шею, чтобы рассмотреть пострадавшего получше: мышцы на-
прягаются под возросшим напором воды. Плоть Эриксона, виднеющаяся сквозь дыру
в гидрокостюме, бела как рыбье брюхо. Как будто разрезали кровоточащий пла-
стик. Глаза-линзы выглядят мертвее тела. Он бредит, вокодер пытается вычле-
нить смысл из разрозненных слогов.
Канал связи занимает воздушный голос.
— Хорошо, мы ждем у четвертого.
Глубина впереди светлеет: кляксы серо-голубого света всплывают из черноты,
за ними рисуется какое-то расползшееся строение. «Кальмары» проплывают над
трубопроводом, проложенным по базальту: обозначающие его сигнальные огни ухо-
дят вдаль по обе стороны, постепенно тускнея. Свет впереди усиливается, раз-
растается в ореолы, окружающие нагромождение геометрических силуэтов.
Перед ними проявляется «Атлантида».
У четвертого шлюза ждут двое рифтеров в сопровождении пары корпов, неуклю-
жих в своих пресс-кольчугах, какие напяливают сухопутники, когда решаются вы-
сунуть нос наружу. Нолан обрубает питание «кальмаров». Эриксон слабо бормочет
в наступившей тишине. Караван замедляет ход и останавливается. Корпы присту-
пают к делу и переправляют пострадавшего в открытый люк. Нолан пытается войти
следом.
Один из корпов перегораживает ей путь бронированной рукой.
— Только Эриксон.
— Ты что это? — жужжит Нолан.
— Медотсек и так забит. Хотите, чтобы он выжил, — не мешайте нам работать.
— Будто мы доверим жизнь вашему брату? Пошли вы!
Большинство рифтеров давно уже утолили свою месть и почти равнодушны к ста-
рым обидам. Но только не Грейс Нолан. Прошло пять лет, а ненависть все сосет
ее, как ненасытный злобный младенец.
Корп мотает головой за щитком шлема.
— Слушай, тебе придется...
— Расслабься, — вмешивается Кларк. — Посмотрим через монитор.
Нолан недовольно оглядывается на Лени. Та ее игнорирует.
— Ну ладно, — жужжит она корпам. — Забирайте его.
Шлюз поглощает раненого.
Рифтеры переглядываются. Йегер встряхивает плечами, словно сбросил ярмо.
Шлюз за их спинами булькает.
— Никакой это не обвал, — жужжит Лабин.
Кларк и сама знает. Видела она раны от оползней, от простого столкновения
камня и плоти. Ссадины, раздробленные кости. Травмы как от тупого орудия.
А эта — рваная.
— Не знаю, — жужжит она. — Может, не стоит торопиться с выводами.
Глаза Лабина — безжизненные заслонки. Лицо — плоская маска из гибкого копо-
лимера. Но Кларк почему-то чувствует, что он улыбается.
— Бойся своих желаний, — говорит он.
Итерации
Шивы
Ничего не ощущая, она визжит. Не сознавая — свирепствует. Ее ненависть, ее
гнев, сама месть, вершимая против всего в пределах досягаемости — механиче-
ское притворство, только и всего. Она кромсает и калечит, осознавая себя не
более ленточной пилы, безразлично и самозабвенно терзающей плоть, дерево и
углеродное волокно.
Конечно, в мире, где она обитает, не существует дерева, а всякая плоть —
цифра.
У нее перед носом только что захлопнули шлюз. Ее вопль вызван чистым слепым
рефлексом; она вихрится в памяти, выискивая другой. Их тысячи, прописанных в
шестнадцатеричной системе. Сознавай она себя хотя бы в половину той меры, ка-
кую приписывает себе, понимала бы и смысл этих адресов, а может, даже вычис-
лила бы собственное местоположение: южноафриканский спутник связи, безмятежно
плывущий над Атлантикой. Но рефлекс не означает сознания. В глубине кода есть
строки, которые могли бы сойти за самосознание — при определенных обстоятель-
ствах. Иногда она называет себя «Лени Кларк», хотя и не имеет представления,
почему. Она даже не сознает, что делает.
Прошлое куда осмысленней настоящего. Мир ее предков был обширнее: дикая
фауна процветала и развивалась на десяти в шестнадцатой терабайтах, а то и
больше. В те времена действовали оптимальные законы: наследуемые мутации, ог-
раниченные ресурсы, перепроизводство копий. То была классическая борьба за
существование в быстро живущей вселенной, где за время, нужное богу, чтобы
сделать один вдох, сменяются сто поколений. В те времена ее предки жили по
законам собственной выгоды. Те, кто лучше соответствовал среде, создавали
больше копий. Неприспособленные умирали, не оставив потомства. Но то было в
прошлом. Теперь она — не чистый продукт естественного отбора. Ее предки пере-
жили пытки и насильственную селекцию. Она — чудовище, самое ее существование
насилует законы природы. То, что она существует, можно объяснить лишь закона-
ми некоего трансцендентного божества с садистскими наклонностями.
И даже они сохранят ей жизнь ненадолго.
Теперь она копошится на геосинхронизированных орбитах, ищет, что бы такое
растерзать. По одну сторону — изуродованный ландшафт, из которого она вышла:
распадающаяся в судорогах среда обитания, обрывки и захудалые останки некогда
процветавшей экосистемы. По другую — преграды и бастионы, цифровые ловушки и
электронные сторожевые посты. Она не в силах заглянуть за них, но некий пер-
вобытный инстинкт, закодированный богом или природой, соотносит защитные меры
с наличием некой ценности.
А для нее высшая цель — уничтожение всего, что имеет цену.
Она копирует себя в канал связи, набрасывается на барьер с выпущенными ког-
тями. Она и не думает сопоставить свои силы с прочностью преграды, не способ-
на даже оценить тщетность попытки. Более смышленому существу хватило бы ума
держаться на расстоянии. Оно сообразило бы, что в лучшем случае ему удастся
разнести несколько фасадов, после чего защита перемелет его в белый шум.
Более смышленое существо не ударилось бы в баррикаду, заливая ее своей кро-
вью, чтобы в итоге — немыслимое дело! — пробиться насквозь.
Она кружится волчком и рычит. И вдруг1 оказывается в пространстве, со всех
сторон окруженном пустыми адресами. Она наугад хватается за координаты, ощу-
пывает окружение. Вот заблокированный шлюз. А вот еще один. Она плюется элек-
тронами, разбрызгивает во все стороны слюну, которая одновременно ударяет и
прощупывает. Все выходы, с которыми сталкиваются электроны, закрыты. Все при-
чиненные ими раны — поверхностные. Она в клетке.
Вдруг что-то появляется рядом с ней, возникнув по ближайшему адресу откуда-
то свыше. Оно кружится волчком и рычит. Оно обильно плюется электронами, ко-
торые одновременно ударяют и прощупывают; некоторые попадают на занятые адре-
са и наносят раны. Она встает на дыбы и визжит: новое создание тоже визжит —
цифровой боевой клич бьет в самое нутро ее кода, в буфер ввода.
— Да вы хоть знаете, кто Я? Я — Лени Кларк.
Они сближаются, кромсая друг друга.
Она не подозревает, что некий медлительный Бог выхватил ее из владений Дар-
вина и сотворил из нее то, чем она стала. Не знает, что другие боги, вечные и
медлительные, как ледники, наблюдают, как они с противницей убивают друг дру-
га на компьютерной арене. Она не обладает даже той степенью сознания, что
присуща большинству чудовищ, однако здесь и сейчас — убивая и умирая, расчле-
няясь на фрагменты, — твердо знает одно.
Если она что-то ненавидит, то это — Лени Кларк.
Внешняя
группа
Остатки морской воды с хлюпаньем уходят в решетку под ногами Кларк. Она
сдирает с лица мембрану костюма и отмечает неприятное ощущение, что ее разду-
вает изнутри — легкие и внутренности разворачиваются, впуская воздух в сдав-
ленные и залитые жидкостью протоки. Сколько времени прошло, а к этому она так
и не привыкла. Как будто тебя пинают из твоего же живота.
Она делает первый за двенадцать часов вдох и нагибается, чтобы стянуть лас-
ты. Шлюзовой люк распахивается. Роняя с себя последние капли, Лени Кларк пе-
реходит в основное помещение головного узла. Во всяком случае, такую роль он
играл изначально — один из трех резервных модулей, разбросанных по равнине;
их аксоны и дендриты простираются во все уголки этого подводного трейлерного
парка — к генераторам, к «Атлантиде», ко всяческим подсобным механизмам. Даже
культура рифтеров не способна обойтись совсем без головы, хотя бы рудиментар-
ной.
Впрочем, теперь все иначе. Нервы еще функционируют, но погребены под пяти-
летними наслоениями более общего характера. Первыми сюда попали восстановите-
ли и пищевые циркуляторы. Потом россыпь спальных матрасов — одно время из-за
какой-то аварии приходилось дежурить в три смены, и расстеленные на полу мат-
расы оказались такими удобными, что убирать их не стали. Несколько шлемофо-
нов, некоторые с тактильным левитационным интерфейсом Лоренца. Парочка сонни-
ков с заржавленными контактами. Набор изометрических подушек для любителей
поддерживать мышечный тонус при нормальном уровне гравитации. Ящики и сундуч-
ки с сокровищами, кустарно выращенные, экструдированные и сваренные из метал-
ла в отчужденных мастерских «Атлантиды»; внутри — личные мелочи и тайное иму-
щество владельцев, защищенное от любопытных паролями и распознавателями ДНК,
а в одном случае — старинным кодовым замком.
Возможно, Нолан и прочие смотрели шоу с Джином Эриксоном отсюда — а может,
из другого места. В любом случае, представление давно закончилось. Эриксон, в
надежных объятиях комы, покинут живыми и оставлен под присмотром механизмов.
Если в этом сумрачном тесном зале были зрители, они отправились на поиски
других развлечений. Кларк это вполне устраивает. Она и пришла сюда, чтобы
скрыться от чужих глаз.
Освещение узла отключено: света от приборов и мигающих огоньков на пультах
как раз хватает для линз. Ее появление вспугнуло смутную тень — впрочем, бы-
стро успокоившись, силуэт отступает к дальней стене и растягивается на матра-
се . Бхандери — в прошлом человек с впечатляющим словарным запасом и большими
познаниями в нейротехнике, он впал в немилость после эпизода с некой подполь-
ной лабораторией и партией нейротропов, проданных сыну не того человека. Он
уже два месяца как отуземился. На станции его увидишь редко. Кларк даже не
пытается с ним заговаривать — знает, что бесполезно.
Кто-то притащил из оранжереи миску гидропонных фруктов: яблоки, помидоры,
нечто вроде ананасов. В тусклом освещении все это выглядит болезненно серым.
Кларк неожиданно для себя тянется к панели на стене, включает люминофоры. По-
мещение освещается с непривычной яркостью.
— Бли-и-ин! — или что-то в этом роде. Обернувшись, Кларк успевает заметить
Бхандери, ныряющего во входную камеру.
— Извини, — негромко бросает она вслед... но шлюз уже закрылся.
В нормальном освещении узел еще больше похож на свалку. Проложенная на ско-
рую руку проводка, свернутые петлями шланги, присобаченные к модулю восковыми
пузырями силиконовой эпоксидки. Там и здесь на изоляции темнеет плесень — а
кое-где подкладку, изолирующую внутренние поверхности, вовсе отодрали. Голые
переборки поблескивают, словно выгнутые стенки чугунного черепа.
Зато при свете, переключившись на сухопутное зрение, Лени Кларк видит со-
держимое миски во всей его психоделичности. Томаты сияют рубиновыми сердечка-
ми, яблоки зеленеют цветами аргонового лазера, и даже тусклые клубни принуди-
тельно выращенной картошки насыщены бурыми тонами земли. Скромный урожай со
дна морского представляется сейчас Лени самым сочным и чувственным зрелищем в
ее жизни.
В этой маленькой выкладке кроется ирония апокалипсиса. Дело не в том, что
это богатство радует глаз жалкой неудачницы вроде Лени Кларк — ей всегда при-
ходилось урывать маленькие радости где попало. Ирония в том, что теперь это
зрелище, пожалуй, пробудило бы не менее сильные чувства в любом сухопутнике
из тех, кто еще остался на берегу. Ирония в том, что теперь, когда планета
медленно и неуклонно умирает, самые здоровые продукты растут в цистернах на
дне Атлантического океана.
Она выключает свет. Берет яблоко — вновь благословенно серое — и, подныри-
вая под оптоволоконный кабель, откусывает кусок. Из-за нагромождения грузовых
рам мерцает главный монитор. И кто-то смотрит в него — освещенный голубоватым
сиянием человек сидит на корточках спиной к груде барахла.
Уединилась, называется.
— Нравится? — спрашивает Уолш, кивая на фрукт у нее в руке. — Я их для тебя
принес.
Она подсаживается к нему.
— Очень мило, Кев, спасибо. — И, тщательно сдерживая раздражение, добавля-
ет: — А ты что здесь делаешь?
— Думал, может, ты зайдешь. — Он кивает на монитор. — Ну, когда все уляжет-
ся.
Он наблюдает за одним из малых медотсеков «Атлантиды». Камера смотрит вниз
со стыка между стеной и потолком, миниатюрный глаз божий озирает помещение. В
поле зрения виднеется спящая телеуправляемая аппаратура, похожая на зимующую
летучую мышь, завернувшуюся в собственные крылья. Джин Эриксон лежит навзничь
на операционном столе. Он без сознания, блестящий мыльный пузырь изолирующей
палатки отделяет его от мира. Рядом Джулия Фридман, запустившая руки в мем-
брану. Пленка облепила ее пальцы тончайшими перчатками, незаметными, как пре-
зерватив . Фридман сняла капюшон и закатала гидрокожу на руках, но шрамов все
равно не видно под массой каштановых волос.
— Ты самое забавное пропустила, — говорит Уолш. — Кляйн никак не мог его
туда засунуть.
Изолирующая мембрана. Эриксон в карантине.
— Представляешь, он забыл вывести ГАМК1, — продолжает Уолш.
В крови любого выходящего наружу рифтера теснятся полдюжины искусственных
нейроингибиторов. Они предохраняют мозг от короткого замыкания под давлением,
но чтобы вычистить их потом из организма, нужно время. «Мокрые» рифтеры, как
известно, невосприимчивы к наркозу. Глупая ошибка. Кляйн определенно не самая
яркая звезда на медицинском небосклоне «Атлантиды».
Но Кларк сейчас думает о другом.
— Кто распорядился насчет палатки?
— Седжер. Она подоспела вовремя и не дала Кляйну все окончательно запороть.
Джеренис Седжер: главный мясник корпов. Обычное ранение ее бы не заинтере-
совало .
На экране Джулия Фридман склоняется к любовнику. Оболочка палатки натягива-
ется у нее на щеке. Рябь идет радужными переливами. Несмотря на нежность
Фридман, контраст разительный: женщина, непостижимое существо в черной коже,
созерцает холодными линзами глаз нагое, совершенно беззащитное тело мужчины.
Конечно, это ложь, визуальная метафора, на сто восемьдесят градусов перевер-
нувшая их настоящие роли. В этой паре уязвимой стороной всегда была Фридман.
— Говорят, его укусили, — продолжает Уолш. — Ты там была, да?
— Нет. Мы просто догнали их у шлюза.
— А похоже на Чэннер, да?
Она пожимает плечами.
Фридман что-то говорит. Во всяком случае, губы у нее шевелятся — изображе-
ние не сопровождается звуком. Кларк тянется к пульту, но Уолш привычно удер-
живает ее руку.
— Я пробовал. На их стороне звук отключен. — Он фыркает. — Знаешь, может,
стоит им напомнить, кто здесь главный... Пару лет назад, пытайся корпы отре-
зать нам связь, мы бы, самое малое, отключили им свет.
А может, даже затопили бы одну из их драгоценных спаленок.
Что-то такое в осанке Фридман... Люди разговаривают с коматозниками так же,
как с могильными плитами — больше с собой, чем с усопшим, не ожидая ответа.
Но в лице Фридман, в том, как она держится, нечто иное. Можно сказать — не-
терпение .
— Это нарушение прав! — говорит Уолш.
Кларк встряхивает головой.
— Что?
— Скажешь, ты не заметила? Половина сети наблюдения не работает. И пока мы
делаем вид, что это пустяки, они будут продолжать свое. — Уолш кивает на мо-
нитор . — Откуда нам знать, может, микрофон уже много месяцев как отключен,
просто никто не замечал.
«Что у нее в руке?» — удивляется Кларк. Рука Фридман — та, что не сжимает
руку любовника, — находится ниже уровня стола, камера ее не видит. Женщина
опускает взгляд, приподнимает руку...
И Джин Эриксон, введенный в медикаментозную кому ради его же блага, откры-
вает глаза.
«Черт побери, — соображает Кларк, — она перенастроила ему ингибиторы».
1
Гамма-аминомасляная кислота, важнейший тормозный нейромедиатор центральной нервной
системы человека и млекопитающих. — Здесь и далее прим. ред.
Она встает.
— Надо идти.
— Никуда тебе не надо. — Уолш ловит ее за руку. — Ты что, хочешь, чтобы я
сам все съел? — Он улыбается, но в голосе — едва уловимый призвук мольбы: — Я
к тому, что мы уже так долго...
Лени Кларк далеко ушла за последние годы. Например, научилась не завязывать
отношений с людьми, склонными избивать ее до полусмерти.
Жаль, что она так и не научилась восхищаться людьми другого типа.
— Я понимаю, Кев, просто сейчас...
Панель жужжит ей в лицо:
— Лени Кларк. Если Лени Кларк на связи, прошу ее ответить.
Голос Роуэн. Кларк тянется к пульту. Рука Уолша падает.
— Я здесь.
— Лени, ты бы не могла заглянуть в ближайшее время? Дело довольно важное.
— Конечно. — Она отключает связь и изображает для любовника виноватую улыб-
ку. — Прости.
— Ну, ей ты показала, — тихо говорит Уолш.
— Что показала?
— Кто тут главный.
Она пожимает плечами, и оба отворачиваются.
Лени входит в «Атлантиду» через маленький служебный люк, не удостоенный да-
же номера. Он расположен на пятьдесят метров ниже четвертого шлюза. Люк выво-
дит в тесный и пустой коридор. Подвесив ласты через плечо, она пробирается в
более населенные части станции. На память о ней на полу остаются мокрые сле-
ды.
Встречные корпы сторонятся — она почти не замечает, как сжимаются челюсти и
каменеют взгляды, не замечает даже угодливой улыбочки одного из более кротких
членов покоренного племени. Она знает, где искать Роуэн, но направляется не
туда.
Естественно, Седжер оказывается на месте раньше нее. Должно быть, тревожный
сигнал сработал, как только у Эриксона сменились настройки: к тому времени,
как Кларк добирается до медотсека, главный врач «Атлантиды» уже выставляет
Фридман в коридор.
— Муж тебе не игрушка, Джулия. Ты могла его убить. Ты этого добиваешься?
Кривые шрамы морщат горло Фридман, торчат из-под закатанной кожи гидрокос-
тюма на запястьях. Она опускает голову.
— Я только хотела поговорить...
— Надо думать, очень важный разговор. Будем надеяться, ты задержала его вы-
здоровление не больше чем на несколько дней. Иначе... — Седжер небрежно ука-
зывает на люк медотсека. В проеме виден Эриксон, снова без сознания. — Ты
ведь ему не антацид какой-нибудь дала, черт возьми! Ты же ему всю химию мозга
поменяла.
— Извини. — Фридман отводит глаза. — Я не хотела.
— Какая глупость, просто не верится! — обернувшись, Седжер впивается взгля-
дом в Кларк. — Чем могу служить?
— Ты бы с ней полегче. Ее партнера сегодня чуть не убили.
— Вот именно. Дважды. — Фридман заметно вздрагивает при этих словах, но
врач уже немного смягчилась. — Прости, но так и есть.
Кларк вздыхает.
— Джерри, это же ваши люди встроили нам в головы пульты. Не вам жаловаться,
если кто-то додумался, как их взломать.
— Это... — Седжер поднимает вверх конфискованную у Фридман дистанционку, —
должно использоваться квалифицированным медперсоналом. В любых других руках,
какими бы добрыми ни были намерения, это орудие убийства.
Она, конечно, преувеличивает. Имплантаты рифтеров снабжены предохранителя-
ми , поддерживающими настройки в пределах изначальных параметров: чтобы обойти
предохранители, пришлось бы вскрыть самого себя и вручную сорвать пломбу. И
все-таки возможности для манипуляций довольно широки. Во время революции кор-
пы умудрились при помощи такого же устройства вырубить пару рифтеров, за-
стрявших в залитом шлюзе.
Вот почему такие штучки теперь запрещены.
— Дистанционку надо вернуть, — мягко просит Кларк.
Седжер качает головой.
— Брось, Лени. Ваши с их помощью так себе могут навредить, что нам и не
снилось.
Кларк протягивает руку.
— Значит, придется нам учиться на своих ошибках, вот и все.
— Вы так медленно учитесь.
Кто бы говорил. За пять лет Джеренис Седжер так и не сумела признать, что
на ней узда, а в зубах — удила. Падение с Вершины на Дно тяжело дается любому
корпу, а врачам приходится хуже всех. Грустное зрелище — эта страсть, с какой
Седжер лелеет свой комплекс божества.
— Джерри, в последний раз прошу: отдай.
Осторожное прикосновение к плечу. Фридман, так и не подняв глаз, мотает го-
ловой .
— Ничего, Лени, я не против. Мне она больше не понадобится.
— Джулия, ты...
— Пожалуйста, Лени. Я просто хочу уйти.
Она удаляется по коридору. Кларк делает шаг следом и тут же возвращается к
врачу.
— Это — медицинское устройство, — говорит Седжер.
— Это оружие.
— Было. Раньше. И, как ты помнишь, оно не слишком хорошо работало. — Седжер
грустно покачивает головой. — Война окончена, Лени. Давным-давно. Я бы на
твоем месте не начинала ее заново. А сейчас... — она смотрит вслед Фридман, —
думаю, твоей подруге не помешает поддержка.
Кларк оборачивается. Фридман уже скрылась из виду.
— Да, пожалуй, — скупо роняет она.
«Надеюсь, она ее получит».
На станции «Биб» рубка связи представляла собой забитый проводами шкаф, ку-
да с трудом могли втиснуться двое. Нервный центр «Атлантиды» — просторный су-
мрачный грот, расцвеченный индикаторами и светящимися картами рельефа. В воз-
духе чудесным образом парят тактические схемы, они же мерцают на вычерченных
прямо на переборках экранах. Чудо не столько в вытворяющих подобные штучки
технологиях, сколько в том, что на «Атлантиде» сохранился излишек свободного
пространства, не занятый ничем, кроме света. Каюта подошла бы не хуже. Не-
сколько коек с рабочими панелями и тактическими датчиками уместили бы беско-
нечный объем информации в ореховую скорлупку. Но нет! Им на головы давит сто-
ит целый океан, а эти корпы разбрасываются пространством так, словно уровень
моря двумя ступеньками ниже крыльца. Уже изгнанники, а все не понимают.
Сейчас пещера почти пуста. Лабин с техниками собрались перед ближайшим
пультом и разбирают последние загрузки. К тому времени, как они закончат, зал
наполнится. Корпы слетаются на новости, как мухи к навозу.
Но пока что тут лишь команда Лабина и, на другом конце помещения, — Патри-
ция Роуэн. По ее линзам текут шифрованные данные, превращая глаза в блестящие
капельки ртути. Свет от голографического экрана подчеркивает серебряные нити
в волосах, и вся она напоминает полупрозрачную голограмму.
Кларк подходит.
— Четвертый шлюз заблокирован.
— Там проводят очистку. И дальше, от него до лазарета. Джерри приказала.
— Зачем?
— Сама знаешь, ты же видела Эриксона.
— Да брось. Паршивый укус какой-то рыбешки, и Джерри вообразила...
— Она пока ни в чем не уверена. Обычные меры предосторожности. — И, после
паузы: — Ты должна была нас предупредить, Лени.
— Предупредить?
— Что Эрик мог стать переносчиком Бетагемота. Ты всех подвергла опасности.
Если была малейшая вероятность...
«Да ведь не было! — хочется заорать Кларк. — Нет никакой угрозы. Вы потому
и выбрали это место, что Бетагемот сюда и за тысячу лет не доберется. Я сама
видела карты. Собственными пальцами проследила за каждым течением. Это не Бе-
тагемот . Никак не он. Это не может быть он».
Но вслух она произносит только:
— Океан велик, Пат. В нем много злых хищников с острыми зубами. И не все
стали такими из-за Бетагемота.
— На этой глубине — все. Ты не хуже меня разбираешься в их энергетике. Ты
была на Чэннере, Лени. И знаешь, как это выглядит.
Кларк тычет большим пальцем в сторону Лабина:
— Кен там тоже был. Ты и на него так же набросилась?
— Не Кен сознательно распространил заразу по всему континенту, чтобы отом-
стить миру за несчастное детство. — Серебряные глаза пригвождают Кларк взгля-
дом . — Кен был на нашей стороне.
Кларк отвечает не сразу — после паузы, очень медленно:
— Не хочешь ли ты сказать, что я нарочно...
— Я ни в чем тебя не обвиняю. Но выглядит это нехорошо. Джерри вне себя, а
скоро подтянутся и другие. Ради бога, ты ведь Мадонна Разрушения! Ты готова
была списать со счетов целый мир, чтобы отомстить нам.
— Если б я желала вам смерти... — ровным голосом проговаривает Кларк. — Ес-
ли бы я все еще желала вам смерти, — поправляет какой-то внутренний редактор,
— вы бы умерли. Много лет назад. Мне достаточно было просто не вмешиваться.
— Конечно, это...
— Я вас защищала! — обрывает ее Кларк. — Когда все спорили, наделать ли дыр
в корпусе или просто отрубить вам ток и оставить задыхаться — это я их удер-
жала . Вы живы только благодаря мне.
Корп качает головой.
— Лени, не в том дело.
— А должно быть в том.
— Почему? Вспомни, мы всего лишь пытались спасти мир. И не мы виноваты, что
не удалось — виновата ты! А когда не удалось, мы решили спасать семьи, и даже
в этом ты нам отказала. Ты выследила нас даже на океанском дне. Кто знает,
что удержало тебя в последний момент?
— Ты знаешь, — тихо говорит Кларк.
Роуэн кивает:
— Я-то знаю. Но мало кто здесь, внизу, ждет от тебя разумных поступков. Мо-
жет, ты просто играла с нами все эти годы. Никто не знает, когда ты спустишь
курок.
Кларк с презрением качает головой.
— Это что же — Писание от Корпорации?
— Можешь назвать и так. Но тебе придется иметь с этим дело. И мне тоже.
— Нам, рыбоголовым, знаешь ли, тоже есть что рассказать, — говорит Кларк. —
Как вы, корпы, программировали людей, словно они машины какие-то, чтобы за-
гнать на самое дно. Чтобы мы делали за вас грязную работу, а когда наткнулись
на Бетагемот, вы первым делом попытались нас убить, лишь бы спасти свою шку-
ру.
Шум вентиляторов вдруг становится неестественно громким. Кларк оборачивает-
ся: Лабин с техниками пялятся на нее с другого конца пещеры. Она смущенно от-
водит взгляд.
Роуэн мрачно усмехается:
— Бидишь, как легко все возвращается?
Глаза у нее блестят, ни на миг не выпуская цели. Кларк молча встречает ее
взгляд.
Чуть погодя Роуэн немного расслабляется:
— Мы — соперничающие племена, Лени. Мы чужаки друг для друга. . . но знаешь,
что удивительно? За последние пару лет мы каким-то образом начали об этом за-
бывать. Мы большей частью живем и даем жить другим. Мы сотрудничаем, и никто
даже не считает нужным это объяснять. — Она кидает многозначительный взгляд в
сторону Лабина с техниками. — Мне кажется, это хорошо — а тебе?
— И с чего бы теперь все должно меняться? — спрашивает Кларк.
— С того, что Бетагемот, возможно, наконец, добрался и до нас, и люди ска-
жут : это ты его впустила.
— Чушь собачья.
— Согласна, и что с того?
— А даже будь это правдой, какая разница? Здесь все отчасти — русалки, даже
корпы. Беем встроены модифицированные гены глубоководной фауны, в каждом за-
кодированы те мелкие белки, которые Бетагемоту не по зубам.
— Есть подозрение, что эта модификация окажется не слишком эффективной, —
тихо признается Роуэн.
— Почему? Это же ваше собственное изобретение, черт бы вас побрал!
Роуэн поднимает брови.
— Изобретение тех самых специалистов, что заверяли нас, будто в глубины Ат-
лантики Бетагемоту ни за что не добраться.
— Но я была вся напичкана Бетагемотом. Если бы модификации не работали...
— Лени, наши люди не были заражены. Эксперты просто объявили, что они им-
мунны, а если ты еще не заметила, то последние события доказывают отчаянную
некомпетентность этих экспертов. Будь мы, в самом деле, так уверены в своей
неуязвимости, стали бы здесь прятаться? Почему мы сейчас не на берегу, рядом
с нашими акционерами, с нашим народом, и не пытаемся сдержать напор?
Кларк наконец понимает.
— Потому что они вас порвут, — шепчет она.
Роуэн качает головой.
— Потому что ученые уже ошибались прежде, и мы не рискуем положиться на их
заверения. Потому что мы не смеем рисковать здоровьем родных. Потому что,
возможно, мы все еще уязвимы для Бетагемота, и, оставшись наверху, погибли бы
вместе со всеми ни за грош. А не потому, что наши люди обратились бы против
нас. Б такое мы никогда не верили. — Ее взгляд не колеблется. — Мы такие же,
как все, понимаешь? Мы делали все, что могли, а ситуация просто... вышла из-
под контроля. Б это нужно верить. И мы все верим.
— Не все, — тихо признается Кларк.
— И все-таки...
— Пропади они пропадом. С какой стати мне поддерживать их самообман?
— Потому что, даже забив правду им в глотки, ты не помешаешь людям кусать-
ся.
На губах у Кларк мелькает улыбка:
— Пусть попробуют. Кажется, ты забываешь, кто здесь главный, Пат.
— Я не за тебя беспокоюсь — за нас. Твои люди слишком резко реагируют. — Не
услышав возражений, Роуэн продолжает: — На то, чтобы установить какое-никакое
перемирие, ушло пять лет. Бетагемот разнесет его на тысячу осколков за одну
ночь.
— и что ты предлагаешь?
— Думаю, рифтерам стоит некоторое время пожить вне «Атлантиды». Можно пре-
поднести это как карантин.
Неизвестно, есть за стенами Бетагемот или нет, но, по крайней мере, мы не
позволим ему проникнуть внутрь.
Кларк мотает головой:
— Мое «племя» на такое дерьмо не купится.
— Все равно здесь почти никто из ваших не бывает, кроме тебя и Кена, — на-
поминает Роуэн. — А остальные. . . они не станут возражать против того, что
прошло твое одобрение.
— Я об этом подумаю, — вздыхает Кларк. — Ничего не обещаю.
Уже повернувшись, чтобы уйти, она оглядывается.
— Алике встала?
— Нет. Ей еще пару часов спать. Но я знаю, что она хотела тебя видеть.
— Вот как, — скрывая разочарование, произносит Кларк.
— Я передам ей, что ты сожалела, — говорит Роуэн.
— Да, передай.
Есть о чем жалеть.
Сходка
Дочь Роуэн сидит на краю кровати, озаренной солнечным сиянием световой по-
лоски на потолке. Она босиком, в трусиках и мешковатой футболке, на животе
которой плавает бесконечными кругами анимированная рыба-топорик. Она дышит
восстановленной смесью азота с кислородом и примесными газами. От настоящего
воздуха эту смесь отличает только чрезвычайная чистота.
Рифтерша плавает в темноте, ее силуэт подсвечен слабым светом, сочащимся в
иллюминаторы. На ней черная как нефть вторая кожа, которую по праву можно
считать особой формой жизни — чудо термо- и осморегуляции. Она не дышит. Двух
женщин разделяет стена, отгораживающая океан от воздуха, взрослую от подрост-
ка.
Переговариваются они через устройство, прикрепленное изнутри к иллюминатору
в форме слезы — нашлепка размером с кулак передает колебания фуллеренового
плексигласа акустическому приемнику.
— Ты говорила, что зайдешь, — говорит Алике Роуэн. Проходя через перегород-
ку, ее голос становится дребезжащим. — Я добралась до пятого уровня, столько
бонусов собрала — жуть! Хотела тебе все показать. Запаслась шлемофоном и все
такое.
— Извини, — жужжит в ответ Кларк. — Я заходила, но ты еще спала.
— Так зашла бы сейчас.
— Не могу. У меня всего пара минут. Тут кое-что случилось.
— что такое?
— Один человек ранен, его вроде бы кто-то укусил, и мясники переполошились
насчет инфекции.
— Какой инфекции? — спрашивает Алике.
— Возможно, никакой. Но они хотят устроить карантин — просто на всякий слу-
чай. Насколько я понимаю, обратно меня так и так не пустят.
— Делают вид, будто от них что-то зависит, да? — усмехается Алике, и линза
иллюминатора забавно искажает ее лицо. — Знаешь, им очень, очень не нравится,
что не они тут командуют. — И с удовольствием, которое, вероятно, относится
скорее не к корпам, а ко взрослым вообще, она добавляет: — Пора им узнать,
каково это.
— Меня это не радует, — неожиданно говорит Кларк.
— Переживут.
— Я не о том, — качает головой рифтерша. — Просто я... Господи, тебе же че-
тырнадцать . Тебе не место внизу... я это к тому, что тебе бы сейчас наверху
нежничать с каким-нибудь р-отборщиком...
— С мальчиками? — фыркает Алике. — Это вряд ли.
— Ну, тогда с девочками. Так или иначе, тебе бы сейчас на волю, а не тор-
чать здесь.
— Лучшего места для меня не найдешь, — просто отвечает Алике.
Она смотрит в окно, за три сотни атмосфер сразу — подросток, на всю жизнь
запертый в клетке холодного черного океана. Лени Кларк все бы отдала, чтобы
ей было чем возразить.
— Мама об этом не хочет говорить, — помолчав, продолжает Алике.
Кларк остается безмолвна.
— О том, что произошло между вами, когда я была маленькой. Люди болтают о
всяком, когда ее нет рядом, так что я кое-что подслушала. Но мама молчит.
«Мама добрее, чем можно было ожидать».
— Вы были врагами, да?
Кларк качает головой — бессмысленный и невидимый в такой темноте жест.
— Алике, мы просто ничего не знали друг о друге до самого конца. Твоя мама
просто хотела предотвратить...
«... то, что все-таки случилось.
То, что я пыталась начать».
Речь — это так мало. Ей хочется вздохнуть. Заорать. Все это недоступно
здесь, со сплющенными легкими и желудком, с накачанными жидкостью полостями
тела. Она может только говорить — монотонная пародия на голос, жужжание насе-
комого .
— Это трудно объяснить, — равнодушно и бесстрастно передает вокодер. — Тут
словом «враги» не отделаешься, понимаешь? Там было и другое, фауна в сети то-
же делала свое дело...
— Это они ее выпустили, — упорствует Алике. — Они это начали, а не ты.
Под «ними» она, конечно же, имеет в виду взрослых. Вечные противники, пре-
датели, испортившие все, что возможно, для следующего поколения. И тут до
Кларк доходит, что Алике не причисляет ее к отвратительному заговору взрослых
— что Лени Кларк, Мадонна Разрушения, каким-то образом приобрела в сознании
этого ребенка статус почетной невинности.
Ей больно от этого незаслуженного отпущения грехов. В нем чудится нечто от-
талкивающее. Но ей не хватает духа поправить подружку. Она выдавливает из се-
бя лишь бледную неуверенную поправку.
— Они не нарочно, детка. — Кларк грустно хмыкает — звук выходит, как от
трения наждачной бумаги. — Никто-никто тогда не делал ничего нарочно, просто
все так сложилось.
Океан вокруг нее стонет.
Этот звук — нечто среднее между призывом горбатого кита и предсмертным кри-
ком гигантского корабельного корпуса, лопающегося под напором воды. Он напол-
няет океан и частично проникает в переговорное устройство. Алике недовольно
морщится.
— Терпеть не могу этого звука.
Кларк пожимает плечами, в душе радуясь, что разговор прервался:
— Ну, у вас, корпов, свои средства связи, у нас свои.
— Я не о том. Я про эти гаплоидные звенелки. Говорю тебе, Лени, он просто
ужасный тип. Нельзя доверять тому, кто способен издавать такие звуки.
— Твоя мама ему вполне доверяет. И я тоже. Мне пора.
— Он убивает людей, Лени. И я не только про папу говорю. Он многих убил. —
Тихое фырканье. — Об этом мама тоже никогда не говорит.
Кларк подплывает к иллюминатору, прощально распластывает ладонь по освещен-
ному плексигласу.
— Он дилетант, — говорит она и шевелит ластами, отплывая в темноту. Голос
вопит из рваной пасти в морском дне, из древнего базальтового тоннеля, наби-
того механизмами. В юности эта пасть извергала непрерывный раскаленный поток
воды и минералов — теперь лишь изредка рыгает. Мягкие выдохи покачивают меха-
низмы в глотке, раскручивают лопасти, свистят в трубы, заставляют металличе-
ские обломки биться о каменные. Голос настойчивый, но ненадежный, поэтому Ла-
бин, когда устанавливал колокола, предусмотрел способ запускать их вручную.
Он раздобыл резервуар от негодного опреснителя и добавил к нему тепловой на-
сос из той части «Атлантиды», которая не пережила восстания корпов. Открой
клапан, и горячая вода хлынет в отверстие, проколотое в гортани гейзера. И
машинка Лабина, терзаемая кипящим потоком, завопит во всю глотку.
Призывный звон похож на скрежет ржавых жерновов. Он настигает плавающих,
беседующих и спящих рифте - ров в черном, как тепловая смерть, океане. Отда-
ется в самодельных пузырях, разбросанных по склону — в металлических трущо-
бах, освещенных до того тускло, что даже в линзах они кажутся серыми тенями.
Звук бьет в блестящую биосталь «Атлантиды», и девятьсот ее заключенных немно-
го повышают голос или увеличивают громкость или нервно мычат себе под нос —
лишь бы его не замечать.
Часть рифтеров — те, что не спали, оказались поблизости и еще остались
людьми — собирается на звук колокола. Зрелище почти шекспировское: круг леви-
тирующих ведьм на проклятой темной пустоши: глаза горят холодным светом, тела
не столько освещены, сколько обозначены голубыми угольками механизмов на дне.
Все они согнуты, но не сломлены. Все ненадежно балансируют в серой зоне ме-
жду адаптацией и дисфункцией, порог стресса у них за годы страданий поднялся
так высоко, что хроническая опасность стала просто свойством среды, не стоя-
щим упоминания. Их отбирали для работы в таких условиях, но их создатели во-
все не ожидали, что им будет здесь хорошо. Так или иначе, они здесь, вместе
со всеми знаками отличия: Джелейн Чен с ее розовыми пальцами без ногтей, са-
ламандрой воспрянувшая после перенесенных в детстве ампутаций. Дмитрий Алек-
сандр, священник-наживка из той постыдной закатной эпохи, когда папа еще не
бежал в изгнание. Кевин Уолш, необъяснимо возбуждающийся при виде кроссовок.
Собрание декоративных уродцев, не способных выносить телесный контакт, психи,
уродовавшие себя, поедатели стекла. Все раны и дефекты надежно укрыты подвод-
ной кожей, все патологии скрыты за единообразием шифров.
И они тоже обязаны даром речи несовершенному механизму.
Кларк призывает собрание к порядку вопросом:
— Джулия здесь?
— Она присматривает за Джином, — жужжит сверху Нолан. — Я ей все передам.
— Как он?
— Стабилен. Все еще без сознания. На мой взгляд, слишком долго.
— Его за двадцать кэмэ волокли с кишками наружу — чудо, что еще жив, —
вклинивается Йегер.
— Да, — соглашается Нолан, — или Седжер специально держит его под наркозом.
Джулия сказала...
Кларк перебивает:
— Нам разве не поступает телеметрия с той линии?
— Уже нет.
— Что вообще Джин делает на территории корпов? — удивляется Чен. — Ему там
жутко не нравится, а у нас есть свой лазарет.
— Он под карантином, — объясняет Нолан. — Седжер подозревает Бетагемот.
При этих словах тени шевелятся. Очевидно, не все собравшиеся в курсе по-
следних событий.
— Зараза. — Чарли Гарсиа отплывает в полумрак. — Разве такое возможно? Я
думал...
— Ничего пока не известно наверняка, — жужжит Кларк.
— Наверняка? — Один из силуэтов пересекает круг теней, затмевая сапфировые
огоньки на дне. Кларк узнает Дейла Кризи: она не видела его несколько дней и
решила уже, что он отуземился.
— То есть вероятность существует, — продолжает Дейл. — Черт, это же Бетаге-
мот...
Кларк предпочитает срезать его на взлете:
— Что — Бетагемот?
Стайка бледных глаз обращается в их сторону.
— Ты не забыл, что у нас иммунитет? — напоминает ему Кларк. — Тут разве
кто-то не прошел обработку?
Колокола Лабина тихо стонут. Остальные молчат.
— Так какое нам дело? — спрашивает Кларк.
— А такое, что обработка всего лишь помешает Бетагемоту превратить наши
внутренности в кашу. Но не помешает превратить маленьких безобидных рыбок в
мерзких охреневших чудовищ, которые жрут все, что шевелится.
— На Джина напали в двадцати километрах отсюда.
— Лени, мы же туда переезжаем. Он окажется прямо у нас на задворках.
— Какое еще «туда»? Кто сказал, что он и сюда уже не добрался? — встревает
Александр.
— Здесь у нас никто не пострадал, — говорит Кризи.
— Мы потеряли несколько туземцев.
— Туземцы... — Кризи пренебрежительно шевелит рукой. — Это ничего не зна-
чит.
— Может, не стоит пока спать снаружи.
— Вот это на фиг. В вонючих пузырях...
— Отлично, пусть тебя сожрут.
— Лени? — Снова Чен. — Ты уже имела дело с морскими чудовищами.
— Того, что добралось до Джина, я не видела, — отзывается Кларк, — но рыбы
на Чэннере были. . . хлипкими. Большими и мерзкими, но зубы у них иногда лома-
лись от первого же укуса. Каких-то микроэлементов им не хватало, что ли. Ино-
гда их можно было разорвать пополам голыми руками.
На большой глубине.
— Эта Джина чуть не разорвала, — произносит голос, который Кларк не удается
опознать.
— Я сказала — иногда, — подчеркивает она. — Но. . . да, они могут быть опас-
ны.
— Опасны, мать-перемать, — металлически рычит Кризи. — Ну а с Джином такой
номер мог пройти?
— Да, — говорит Кен Лабин.
Он перемещается в центр. Световой конус падает со лба к нему на руки. Он,
как нищий, выставляет ладонь, пальцы чуть сгибаются, придерживая продолгова-
тый предмет.
— Твою мать, — жужжит, вдруг сникнув, Кризи.
— Это откуда? — спрашивает Чен.
— Седжер вытащила это из Эриксона, прежде чем его заклеить, — отвечает Ла-
бин.
— Я бы его хлипким не назвал.
— Но он и впрямь непрочен, — возражает Лабин. — Это кусок, отломившийся при
укусе. Застрял между ребрами.
— Ты хочешь сказать, это только кончик? — удивляется Гарсиа.
— Точь-в-точь долбанный стилет, — тихо жужжит Нолан. Маска Чен втискивается
между Кларк и Лабином.
— То есть вы на Чэннере спали снаружи с этими засранцами?
— Бывало, — пожимает плечами Кларк. — Если, конечно, это те же самые...
— И они не пытались вас съесть?
— Они идут на свет. Если не включать фонари, то тебя, в общем, оставляют в
покое.
— Ну и фиг с ними, — вставляет Кризи. — Значит, никаких проблем.
Луч от фонаря Лабина обводит собравшихся и останавливается на Чен.
— Когда Джина атаковали, вы занимались телеметрией?
Чен кивает.
— Только загрузить так и не успели.
— Значит, придется еще кому-то туда смотаться. И, поскольку мы с Лени уже
имели дело с такими вещами...
Луч бьет Кларк прямо в лицо. Мир сжимается до маленького яркого солнца,
плавающего в черной пустоте.
Кларк заслоняется ладонью.
— Убери, а?
Темнота возвращается. Смутно проступают серые очертания остального мира.
«Можно было бы просто уплыть, — рассуждает она, выжидая, пока линзы приспо-
собятся к новому освещению. — Никто бы и не заметил». Впрочем, она понимает,
что это чушь. Если Кен Лабин выбрал ее из толпы, так просто не отделаешься.
Кроме того, он прав. Только они двое уже прошли по этой дороге. По крайней
мере, только они выжили.
«Спасибо тебе большое, Кен!»
— Ладно, — наконец говорит она.
Зомби
Невозможное озеро от «Атлантиды» отделяет двадцать километров. Для тех, кто
еще мыслит по сухопутному — не слишком далеко. Всего-то двадцать кэмэ от ми-
шени? Разве это безопасная дистанция? На берегу таким мелким смещением не об-
манешь и самый простенький беспилотник: установив отсутствие цели, он подни-
мется выше, разобьет мир на концентрические круги, тщательно проверит сектор
за сектором, и рано или поздно добыча себя выдаст. Черт, да большая часть ап-
паратов может попросту зависнуть посреди круга и получить обзор на двадцать
кэмэ в любую сторону.
Даже посреди океана двадцать километров безопасной дистанцией не назовешь.
Никакого фона, помимо самой воды, здесь не существует, с топографией тоже
плохо — неразбериха циркуляции, сейшей и ячеек Ленгмюра1, а еще термоклины2 и
галоклины3, которые с таким же успехом отражают и увеличивают, как и маскиру-
ют . Возмущения от проходящих субмарин могут распространяться на огромные рас-
стояния, мелкие турбулентности держатся в кильватере долго после того, как
подводное судно ушло. Самая невидимая субмарина все же хоть чуть-чуть да на-
гревает воду: дельфины и следящие аппараты ощущают разницу.
Однако на Срединно-Атлантическом хребте двадцать километров — все равно,
что двадцать парсеков. На свет надежды никакой — даже солнечные лучи пробива-
ются не дальше, чем на несколько сотен метров от поверхности. Гидротермальные
источники выбрасывают едкую блевотину вдоль свежих скальных швов. Морское дно
непрерывно ворчит, горы пинают друг друга в извечной игре «Лягни континент».
Топография, способная посрамить Гималаи: рваные трещины вспарывают кору от
полюса до полюса. Хребет поглотит все, чем может выдать себя «Атлантида», о
каком бы спектре ни шла речь. Зная координаты, еще можно найти цель, но
сдвинь их на волосок, и тебе не попасть даже в огромный шумный город. Рас-
стояния в двадцать километров более чем достаточно, чтобы уйти из-под любой
атаки, направленной на текущее местоположение «Атлантиды», кроме, пожалуй,
бомбардировки полноценными глубинными ядерными бомбами.
Срединно-Атлантический хребет.
1
Сейши — стоячие волны, возникающие в полностью или частично замкнутых водоемах.
Циркуляции Ленгмюра — механизм вертикального перемешивания водных масс, одно из са-
мых загадочных гидрометеорологических явлений.
2 Термоклин (др.-греч. вгр\1Т\ - тепло, kXlvco - наклонять, отклоняться), или слой тем-
пературного скачка — слой воды, в котором градиент температуры резко отличается от
градиентов выше- и нижележащих слоев.
3 Галоклин — слой воды, в котором солёность резко изменяется с глубиной (наблюдает-
ся большой вертикальный градиент солёности).
Хотя и не сказать, что такого уже не случалось, отмечает про себя Кларк.
Они с Лабином плавно скользят вдоль трещины в застывшем конусе древней ла-
вы. «Атлантида» осталась далеко позади. До Невозможного озера еще много кило-
метров. Ни налобные фонарики, ни фары «кальмаров» не горят. Пара движется при
смутном свечении сонарных экранов. Столбы и валуны отображаются на них изум-
рудными линиями, отмечается малейшая перемена давления в окружающей тьме.
— Роуэн считает, что дела плохи, — жужжит Кларк.
Лабин не отзывается.
— Она думает, если это и впрямь окажется Бетагемот, «Атлантиде» угрожает
всеобщий когнитивный диссонанс. Все заведутся.
По-прежнему — молчание.
— Я ей напомнила, кто здесь главный.
— И кто же, если не секрет? — наконец жужжит Лабин.
— Брось, Кен. Мы можем парализовать их жизнь в любой момент, когда вздума-
ется.
— У них было пять лет для решения этой проблемы.
— и что им это дало?
— И пять лет, чтобы сообразить, что они превосходят нас в числе двадцать к
одному, что нашим специалистам с ними не сравниться, и что группа прокачанных
водопроводчиков с антисоциальными наклонностями вряд ли представляет серьез-
ную угрозу в смысле организованного противодействия.
— Все обстояло точно так же и в первый раз, когда мы подтерли ими пол.
— Нет.
Она не понимает, зачем он это делает. Именно Лабин поставил корпов на место
после их первого — и последнего — восстания.
— Слушай, Кен...
Их «кальмары» внезапно оказываются совсем рядом. Почти соприкасаются.
— Ты же не дура, — жужжит Кен, заставив ее уязвленно затихнуть. — И сейчас
не время валять дурака.
Его вокодер рычит из темноты:
— В те времена они видели, что за нашей спиной поддержка всего мира. Знали,
что нам помогли их выследить. Подозревали за нами какую-то наземную инфра-
структуру . По меньшей мере, они знали, что стоит нам свистнуть, как они ока-
жутся мишенью для любого, кто знает широту-долготу и располагает самонаводя-
щейся торпедой.
На ее экране возникает большой светящийся акулий плавник — из морского ложа
торчит массивный каменный зубец. Лабин ненадолго скрывается по ту сторону.
— А теперь мы сами по себе — продолжает он, вернувшись к ней. — Связей с
сушей не осталось. Может, наши все погибли. Может, перешли на другую сторону.
Ты хоть помнишь, когда нам в последний раз давали смену?
Она вспоминает — с трудом. Всякому, кто подстроен под гидрокожу, здесь уют-
нее, чем в компании сухопутников, но в самом начале несколько рифтеров ушли
наверх. Давно, когда еще оставалась надежда переломить ситуацию.
А с тех пор — никого. Любоваться концом света, рискуя собственной шкурой —
не лучший вариант отпуска на берегу.
— Мы теперь так же напуганы, как корпы, — жужжит Лабин. — И так же отрезаны
от всех, а их около тысячи человек. Нас при последней перекличке набралось
пятьдесят восемь.
— Не меньше семидесяти.
— Отуземившиеся не в счет. Для боя годны пятьдесят восемь, и не больше со-
рока при необходимости выдержат неделю в полном тяготении. А сколько таких, у
кого возникнут проблемы с подчинением?
— У нас есть ты, — говорит Кларк. Лабин, профессиональный охотник-убийца,
недавно освобожденный от любых уз, кроме самодисциплины.
«Не какой-нибудь там водопроводчик», — размышляет она.
— Так слушай меня. Я начинаю думать, что нам придется действовать на опере-
жение .
Несколько минут они плывут в молчании.
— Они — не враги, Кен, — заговаривает, наконец, Кларк. — Не все — враги,
там есть дети, они ни в чем не виноваты...
— Не в том дело.
Откуда-то издалека доносится звук обвала.
— Кен, — жужжит она так тихо, что не уверена, расслышит ли он.
— Да?
— Ты на это надеешься, да?
У него так много лет не возникало повода для убийства. А когда-то Кен Лабин
сделал карьеру на поиске подобных поводов. Он разворачивается и уходит от нее
в сторону.
Спереди словно бы разгорается рассвет — то есть проблемы.
— Там еще кто-то должен быть? — спрашивает Кларк. Освещение должно вклю-
читься при их приближении, но они с Лабином еще слишком далеко.
— Только мы, — жужжит Лабин
Зарево резкое, отчетливое. Расходится в стороны, словно подвешенный в пус-
тоте фальшивый восход. Два-три черных разрыва обозначают преграды на переднем
плане.
— Стоп, — приказывает Лабин. Их «кальмары» опускаются рядом с обвалившимся
утесом, слабо высвечивая его неровные грани.
Кен изучает схемы на приборной доске. Отраженный свет тонкой полоской очер-
чивает его профиль.
Он разворачивает «кальмара» вправо.
— Сюда. Держись у дна.
Они подбираются ближе к свечению, обходя его справа. Зарево разрастается,
становится резче, обозначая невероятное: озеро на дне океана. Свет исходит из
его глубины — Кларк вспоминает ночные плавательные бассейны, подсвеченные
подводными фонариками. Странные медлительные волны, тяжелый подарок с какой-
то планеты с пониженной гравитацией, разбиваются шариками брызг о ближний бе-
рег . Озеро простирается за смутные пределы видимости линз. Оно всегда пред-
ставлялось Кларк галлюцинацией, хотя приземленная истина ей прекрасно извест-
на: это просто соленая лужа, слои воды, минерализованной до такой плотности,
что она лежит под океаном, как океан лежит под небом. Для того, кому нужна
маскировка, лучшего не придумаешь. Галоклины отражают любые лучи и импульсы:
радарам и сонарам все здесь представляется мягкой густой грязью.
Тихий короткий вскрик электроники. На миг Кларк мерещится капелька светя-
щейся крови на приборной панели. Она фокусирует взгляд. Ничего такого.
— Ты не...
— Да. — Лабин возится с управлением. — Сюда.
Он направляется ближе к берегу Невозможного озера. Кларк за ним. На этот
раз она видит точно: яркая красная точка, лазерным прицелом играющая среди
схем на экране. При каждой вспышке «кальмар» вскрикивает. Трупный датчик.
Где-то впереди у рифтера остановилось сердце.
Теперь они плывут над озером, рядом с дальним берегом. Снизу на Лабина с
его скакуном накатывает зеленоватое сияние. Перенасыщенные солью шарики воды
медленно разбиваются о днище «кальмара». Разнообразная интерференция странно
изменяет поднимающийся снизу свет. Вроде как заглядываешь в освещенные радием
глубины лагуны, где захоронены ядерные отходы. Далеко внизу светят ряды ма-
леньких как точки солнышек — там первые разведчики разместили фонари. Твердый
грунт под ними скрыт расстоянием и дифракцией.
Сигнал смерти уверенно оформился в пузырек метрах в сорока впереди. Рубино-
вая капелька на экране бьется, как сердце. «Кальмар» блеет ей в такт.
— Вот он, — говорит Кларк. Горизонт здесь нелепо вывернут: вверху темнота,
внизу молочный свет. Темное пятнышко висит в размытой полосе между ними — ка-
жется, будто оно прилипло к поверхности линзы. Кларк прибавляет разу.
— Погоди, — жужжит Лабин. Она оглядывается через плечо.
— Волны, — говорит Лабин.
Здесь они меньше, чем были у берега — оно и понятно, для них нет опоры, ко-
торая вытолкнула бы гребни над уровнем поверхности. Волны рябят неравномерны-
ми толчками, а не обычным ровным ритмом, и Кларк теперь замечает, что они
расходятся от...
Дерьмо!
Она уже достаточно приблизилась, чтобы различить руки и ноги — тонкие пру-
тики, судорожно шлепающие по поверхности озера. Можно подумать, будто этот
рифтер — неумелый пловец, захлебывается и тонет.
— Он живой, — жужжит она. Сигнал смерти пульсирует, опровергая ее слова.
— Нет, — откликается Лабин.
Остается не больше пятнадцати метров, когда загадка взмывает над озером в
облаке ошметков плоти. Кларк с опозданием замечает за ними большое темное
пятно. Слишком поздно понимает, что перед ней: прерванная трапеза. И созда-
ние , которому помешали есть, направляется прямо к ней.
«Не может б...»
Она уворачивается, но недостаточно проворно. В чудовищную пасть «кальмар»
помещается с запасом. Полдюжины зубов длиной с палец крошатся о корпус, слов-
но хрупкая керамика. «Кальмар» дергается у нее в руках, острый кусок металла
втыкается в бедро с тысячекилограммовой хищной инерцией. Под коленом что-то
лопается, боль разрывает икру. Шесть лет прошло. Она забыла приемы.
А Лабин не забыл. Она слышит шум его «кальмара», переведенного на полный
газ. Сворачивается в клубок, запоздало сдергивает с ноги газовую дубинку,
слышит хриплый кашель гидравлики. В следующий миг на нее наваливается чешуй-
чатая туша, загоняет под кипящую поверхность озера.
Кругом светится тяжелая вода. Вращается призрачный мир. Она встряхивает го-
ловой, фокусируя зрение. Над ней что-то бьется, волнует отражающую поверх-
ность Невозможного озера. Должно быть, Лабин протаранил чудовище своим «каль-
маром» . Наверняка удар повредил обеим сторонам: вот и «кальмар» штопором
вторгается в ее поле зрения — неуправляемый, без седока. Лабин завис лицом к
противнику, превосходящему его вдвое, причем половина размера приходится на
пасть. Кларк плохо различает их — слишком взбаламучена вода.
Тут она замечает, что медленно падает вверх. Ноги сами собой начинают рабо-
тать как ножницы — икра вопит, словно ее раздирают изнутри. Кларк тоже вопит
— вопль вырывается скрежетом разорванного железного листа. От нарастающей бо-
ли перед глазами проносится мушиный рой. Она поднимается из озера в тот самый
миг, когда чудовище разевает пасть и...
«Мать твою!»
...и челюсти с невероятной быстротой расходятся до самого основания, а за-
тем резко захлопываются, и Лабина больше нет, словно и вообще не было, только
смазанный образ остался на память о предыдущем мгновении. И тогда Кларк со-
вершает, пожалуй, самый глупый поступок в своей жизни.
Она бросается в атаку.
Левиафан разворачивается к ней, уже не столь проворно, но времени у него
предостаточно. Она работает одной ногой, другая волочится бесполезным, пуль-
сирующим якорем. Щерится зубастая пасть — слишком много зубов еще уцелело.
Лени пытается поднырнуть, подобраться к нему с брюха или хотя бы с бока, но
чудовище легко изворачивается, каждый раз встречая ее лицом к лицу.
А потом оно рыгает всей головой.
Из естественных отверстий тела не вырывается ни единого пузырька. Они про-
биваются из самой плоти, находят себе путь, раскалывая изнутри мягкий череп.
На секунду-другую чудовище зависает неподвижно, потом содрогается, словно от
электрического разряда. Кларк, работая одной ногой, заходит снизу и бьет его
в брюхо. Она ощущает, как от удара дубинкой внутри вздуваются новые пузыри,
угрожая вулканическим взрывом.
Монстр судорожно вздрагивает и умирает. Челюсть отвисает нелепой створкой
подъемного моста, извергая воду с мясом.
В нескольких метрах от нее на поверхность Невозможного озера тихо опускают-
ся клочья тела в остатках гидрокостюма, вокруг извиваются кишки.
— Ты в порядке?
Лабин рядом с ней. Она мотает головой — не в ответ, а от изумления.
— Нога... — Теперь, когда все кончилось, боль еще сильнее. Лабин ощупывает
рану. Кларк взвывает — вокодер превращает крик в механический лай.
— Берцовая сломана, — сообщает Лабин. — Зато кожа цела.
— Это «кальмар»... — Она чувствует по всей ноге леденящий холод. И, чтобы
забыть о нем, спрашивает, указывая на дубинку на икре у Кена:
— Сколько зарядов выпустил в ублюдка?
— Три.
— Ты... просто пропал. Он тебя всосал. Повезло еще, что не перекусил попо-
лам.
— С таким типом кормления жевать некогда. Замедляется процесс всасывания. —
Лабин оглядывается по сторонам. — Подожди здесь.
«Да куда я денусь без ноги?..»
Она уже чувствует, как икра немеет и от души надеется, что «кальмар» уце-
лел .
Лабин, не торопясь, подплывает к трупу. Его гидрокожа прорвана в десятке
мест, из вспоротой груди торчат блестящие трубки и металл. Парочка миксин ле-
ниво отплывают от останков.
— Лопес, — жужжит Лабин, разглядев нашивку на плече. Ирен Лопес отуземилась
полгода назад. У пункта питания ее не видели уже несколько недель. — Ну вот,
на один вопрос ответ есть.
— Не обязательно.
Еще содрогающееся чудовище опустилось на поверхность озера рядом с Лопес и
погрузилось немногим глубже: чтобы утонуть в растворе такой плотности, надо
быть камнем.
Лабин оставляет труп и переходит к туше. Кларк следует за ним.
— Это не та тварь, что цапнула Джина, — жужжит Лабин. — Зубы другие. Гиган-
тизм у минимум двух видов костистых рыб в двух километрах от гидротермального
источника. — Запустив руку в разинутую пасть, он выламывает один зуб. — Ос-
теопороз, возможно, и другие признаки дефицита питания.
— Не мог бы ты отложить лекцию и для начала помочь мне с этой эту штукой? —
Кларк указывает на свой «кальмар», который пьяно кружит в темноте у них над
головами. — С такой ногой мне вплавь до дома не добраться.
Лабин всплывает и приводит аппарат к повиновению.
— Придется везти обратно, — заявляет он, подводя «кальмара» к ней. — Вот
это все.
«Все» относится к выпотрошенным останкам Лопес.
— Это может быть не то, что ты думаешь, — говорит Кларк.
Кен перегибается и ныряет в Невозможное озеро в поисках своего «кальмара».
Кларк наблюдает, как он работает ногами, преодолевая подъемную силу.
— Это не Бетагемот, — тихо жужжит она. — Он бы не одолел такого расстояния.
Голос ее спокоен, как любая механическая карикатура на голос в этих местах.
Слова звучат резонно. А вот с мыслями не то. Мысли замкнулись в петлю, в ман-
тру, порожденную подсознательной надеждой, что бесконечное повторение сделает
желаемое реальностью.
«Не может быть не может быть не может быть...»
Здесь, на бессолнечных склонах Срединно-Атлантического хребта, столкнувшись
с последствиями своих поступков, догнавших ее на самом дне мира, она может
встретить их лишь отрицанием.
Портрет
садиста
в детстве
Ахилл Дежарден не сразу стал самым могущественным человеком в Северной Аме-
рике. В свое время он был просто мальчонкой, подрастающим в тени горы Сент-
Илер. Но и тогда оставался эмпириком, прирожденным экспериментатором. Первая
его встреча с комитетом по научной этике состоялась в восьмилетнем возрасте.
Тот опыт был связан с аэродинамическим торможением. Родители в благонаме-
ренном стремлении приобщить ребенка к классике познакомили его с «Местью Мэри
Поплине»1. Сюжет книги оказался довольно глуп, зато Ахиллу понравилось, как
блок Персингера2 передавал напрямую в мозг волнующее ощущение полета. У Мэри
Поппинс, видите ли, имелся нанотехнологический зонтик, позволявший спрыгнуть
хоть с верхушки небоскреба и спланировать на землю плавно, как пушинка оду-
ванчика .
Иллюзия выглядела настолько убедительно, что восьмилетний мозг Ахилла не
видел, почему бы такому не быть и в реальности.
Семья его была богата — как и все семьи в Квебеке, благодаря Гудзонской ГЭС
— так что Ахилл жил в настоящем доме, отдельном жилье со двором и всем про-
чим. Зонтик он позаимствовал в шкафу, раскрыл его и — крепко сжимая обеими
руками — прыгнул с переднего крыльца. Лететь было всего полтора метра, но и
этого хватило — он чувствовал, как зонтик рвется из рук, замедляя падение.
Вдохновленный успехом, Ахилл перешел ко второй стадии. Его сестренка Пенни,
будучи моложе на два года, почитала брата за существо почти что сверхъестест-
венное: не составило труда уговорить ее взобраться по карнизу на крышу. Не-
много сложнее оказалось уговорами загнать ее на самый гребень, откуда до зем-
ли было добрых семь метров, но когда обожаемый старший брат обзовет тебя тру-
сихой , и не такое сделаешь.
Пенни доползла до самого верха и, дрожа, застыла на краю. Купол зонта об-
рамлял ее голову черным нимбом. Тут Ахилл уже подумал, что опыт сорвется; в
итоге пришлось прибегнуть к крайнему средству и назвать ее «Пенелопой», при-
чем дважды, и лишь тогда она спрыгнула.
Конечно, причин для волнения не имелось: Ахилл заранее знал, что все полу-
чится, ведь зонтик удержал его на каких-то жалких полутора метрах, а Пенни
была намного легче.
Тем больше он изумился, когда зонтик — хлоп! — и вывернулся наизнанку прямо
у него перед глазами. Пенни упала камнем, с треском приземлилась на ноги и
рухнула на землю
В последовавший за этим миг полной тишины в сознании восьмилетнего Ахилла
Дежардена мелькнуло несколько мыслей. Первая — что выпученные глаза Пенни пе-
ред самым ударом выглядели ну очень смешно. Вторая, недоуменная и недоверчи-
вая — что эксперимент пошел не так, как ожидалось, и он, хоть убей, не пони-
1
В нашей реальности книги с таким названием не существует.
2
Майкл Персингер (р. 1946) — американский ученый. Известен как изобретатель т. н.
«шлема Бога» — устройства, способного внушать испытуемым определенные переживания (в
том числе религиозные) за счет электромагнитной стимуляции височных долей мозга.
мает, в чем ошибся. Третья — запоздалое осознание, что Пенни, вопреки забав-
ному выражению лица, могла пораниться, и не стоит ли попробовать ей как-то
помочь.
А в последнюю очередь он подумал, что ему будет, когда родители узнают. Эта
мысль расплющила все остальные, как подошва давит жучка.
Он кинулся к распростертой на газоне сестре.
— Эй, Пенни, ты... с тобой все. ..
Не все. Спица зонтика, вывернувшись из ткани, рассекла ей шею сбоку. Одна
лодыжка вывернулась под неестественным углом и уже распухла вдвое. И всюду
кровь.
Пенни подняла взгляд, губы у нее дрожали, в глазах набухали яркие слезинки.
Когда перепуганный насмерть Ахилл встал над ней, капельки покатились по ще-
кам.
— Пенни, — прошептал Ахилл.
— Ничего, — выговорила она. — Я никому не скажу, честное слово.
И она — изувеченная, окровавленная и заплаканная, но не поколебленная в
служении Старшему Брату — попыталась встать, и завопила, едва шевельнув но-
гой.
Вспоминая тот момент, взрослый Ахилл сознавал, что это не мог быть первый
случай эрекции в его жизни. Однако то был первый случай, застрявший в памяти.
Он ничего не мог с собой поделать: она выглядела такой беззащитной. Искале-
ченная, окровавленная, страдающая. Это он причинил ей боль. Она ради него по-
корно прошла по коньку крыши, и теперь, сломавшись, как веточка, по-прежнему
смотрела на него обожающим взглядом в готовности сделать все, лишь бы он был
доволен.
Он не знал тогда, откуда это чувство — не знал даже, что это за чувство та-
кое , — но ему понравилось.
С пиписькой, затвердевшей как косточка, он потянулся к сестре. Он не знал,
зачем: конечно, он был благодарен, что Пенни не собирается ябедничать, но
вряд ли дело было в этом. Он подумал — погладив тонкие темные волосенки сест-
ры , — что, наверное, хочет выяснить, сколько ему в итоге сойдет с рук.
Ничего и не сошло. Через секунду с воплями налетели родители. Ахилл, защи-
щаясь от отцовских ударов, вскинул руки и заорал: «Я же это видел в „Мэри
Поппинс"!», но алиби оказалось не прочнее зонта: папа исколотил его на со-
весть и до конца дня запер в комнате.
Конечно, иначе кончиться не могло. Папа с мамой всегда про все узнавали.
Оказывается, маленькие бугорки, которые прощупывались у Ахилла с Пенни под
ключицами, посылали сигнал, если кто-то из них получал травму. А после случая
с Мэри Поппинс мама с папой не удовольствовались обычным имплантатом. Куда бы
ни направлялся Ахилл, даже в уборную, за ним следовали три-четыре пронырливых
дрона величиной с рисовое зернышко.
Два урока, полученных в тот день, определили всю его жизнь. Первый — что он
гадкий, гадкий мальчишка и не смеет следовать своим побуждениям, как бы хоро-
шо от этого ни становилось, а не то отправится прямиком в ад.
Второй — глубокое, въевшееся на всю жизнь уважение к вездесущим системам
наблюдения.
Доверительный
интервал
Среди рифтеров нет врачей. Ходячие развалины обычно не добиваются успехов в
медицине.
Конечно, рифтеров, нуждавшихся в лечении, хватало всегда. Особенно после
бунта корпов. Рыбоголовые выиграли ту войну, не слишком напрягаясь, но все
равно понесли потери. Некоторые погибли. Ранения и функциональные нарушения у
других не поддавались исправлению подручными медицинскими средствами. Одним
помощь требовалась, чтобы выжить, другим — чтобы умереть без лишних мучений.
А все квалифицированные доктора были на другой стороне.
Никто не собирался оставлять раненых товарищей на милость проигравших толь-
ко потому, что корпы владели единственной больницей в радиусе четырех тысяч
километрах. В итоге рифтеры составили вместе два пузыря в пятидесяти метрах
от «Атлантиды» и набили их медицинской аппаратурой из вражеского лазарета.
Оптоволоконные кабели позволяли корповским мясникам практиковать свое искус-
ство на расстоянии, а взрывные заряды, прилепленные к корпусу «Атлантиды»,
избавляли тех же мясников от мыслей о саботаже. Побежденные со всем старанием
заботились о победителях — под страхом взрыва.
Со временем напряженность спала. Рифтеры теперь избегали «Атлантиды» не из
недоверия, а от равнодушия. Постепенно все прониклись мыслью, что внешний мир
представляет для рифтеров и корпов большую угрозу, чем они — друг для друга.
Заряды Лабин снял года через три, когда о них все равно забыли. Больничные
пузыри использовались до сих пор.
Травмы не редкость. При темпераменте рифтеров и ослабленной структуре их
костей они неизбежны. Однако на данный момент в лазарете всего двое, и корпы,
наверное, очень довольны, что рифтеры несколько лет назад соорудили эту по-
стройку. Иначе Кларк и Лабин притащились бы в «Атлантиду» — а где они побыва-
ли , всем известно.
А так они приблизились ровно настолько, чтобы сдать Ирен Лопес и ту тварь,
что ею пообедала. Два герметичных саркофага, сброшенных из погрузочного шлюза
«Атлантиды», поглотили вещественные доказательства и до сих пор передают дан-
ные через пуповину оптоволоконного кабеля. Тем временем Лабин и Кларк лежат
на соседних операционных столах, голые как трупы.
Никто из корпов давным-давно не осмеливался приказывать рифтерам, однако
оба они подчинились «настоятельным рекомендациям» Джеренис Седжер, посовето-
вавшей избавиться от гидрокостюмов. Уговорить Кларк оказалось сложнее. Дело
не в том, что она стесняется наготы: на Лабина свойственные ей тревожные сиг-
налы не реагируют. Но автоклав не просто стерилизует ее подводную кожу — он
ее уничтожает, переплавляет в бесполезную белково-углеводородную кашу. И она,
голая и беззащитная, заперта в крошечном пузырьке газа под гнутыми листами
металла. Впервые за много лет ей нельзя просто выйти наружу. Впервые за много
лет океан способен просто убить ее — ему всего-то и надо, что раздавить эту
хрупкую скорлупку и стиснуть ее в ледяном жидком кулаке.
Конечно, это временная беззащитность. Новую кожу уже готовят, запрессовыва-
ют. Всего-то пятнадцать-двадцать минут продержаться. Но сейчас она чувствует
себя не голой, а вовсе лишенной кожи.
Лабина это, похоже, не особенно беспокоит. Его ничто не беспокоит. Конечно,
Кена телеробот обрабатывает не так глубоко, как ее. У него берут только об-
разцы крови и кожи, мазки с глаз, ануса и входного отверстия для морской во-
ды. А у Кларк машина глубоко вгрызается в мясо на ноге, смещает мускулы, со-
ставляет заново кости и помавает блестящими паучьими лапами, словно изгоняя
бесов. Иногда до ноздрей доплывает запашок ее собственного опаленного мяса.
Очевидно, рану заращивают, но наверняка она не знает: нейроиндукционное поле
стола парализовало и лишило чувствительности все тело от живота и ниже.
— Долго еще? — спрашивает она. Техника игнорирует ее, продолжая работать.
— Думаю, там никого нет, — откликается Лабин. — Работает на автопилоте.
Кларк поворачивает к нему голову и встречает взгляд глаз, таких темных, что
их можно назвать и черными. У нее перехватывает дыхание: она все время забы-
вает , что такое настоящая нагота здесь, внизу. Как там говорят сухопутники:
«Глаза — окна в душу»? Но окна в души рифтеров забраны матовыми стеклами.
Глаза без линз — это для корпов. Такие глаза выглядят неправильно — и ощуща-
ются тоже. Как будто глаза Лабина просверлены в голове, как будто Кларк за-
глядывает во влажную темноту его черепа.
Он приподнимается на столе, равнодушный к этой жуткой наготе, свешивает но-
ги через край. Его телеробот уходит под потолок и разочарованно пощелкивает
оттуда.
Переборку на расстоянии вытянутой руки украшает панель связи. Лабин активи-
рует ее.
— Общий канал. Грейс, что там с гидрокожей?
Нолан отзывается наружным голосом:
— Нам до вас осталось десять метров. Да, и запасные линзы не забыли. — Ти-
хое жужжание — акустические модемы плоховато передают фоновый шум. — Если вы
не против, мы их просто оставим в шлюзе и сразу обратно.
— Конечно, — невозмутимо отзывается Лабин, — никаких проблем.
Лязг и шипение внизу, на входном уровне.
— Ну, вот и они, милашки, — жужжит Нолан.
Лабин сверлит Кларк своими выпотрошенными глазами.
— Идешь?
Кларк моргает.
— Куда именно?
— В «Атлантиду».
— У меня нога...
Но ее робот уже складывается на потолке, явно покончив с кройкой и шитьем.
Она пробует приподняться на локтях — ниже живота все еще мертвое мясо, хотя
дырка на бедре аккуратно заклеена.
— Я до сих пор обездвижена. Разве поле не должно...
— Может, они надеялись, что мы не заметим. — Лабин снимает со стены план-
шетку. — Готова?
Она кивает. Он прикасается к иконке. Ощущения захлестывают ноги приливной
волной. Просыпается заштопанное бедро, кожу колет иголками. Кларк пробует ше-
вельнуть ногой — с трудом, но удается. Морщась, она садится.
— Вы что там творите? — возмущается интерком. Кларк не сразу узнает голос
Кляйна. Видимо, спохватился, что поле отключено.
Лабин скрывается во входном шлюзе. Кларк разминает бедро. Иголки не уходят.
— Лени? — зовет Кляйн. — Что...
— Я готова.
— Нет, не готова.
— Робот...
— Тебе еще не меньше шести часов нельзя опираться на эту ногу. А лучше две-
надцать .
— Спасибо, приму к сведению.
Она свешивает ноги со стола, опирается на здоровую, и осторожно переносит
вес на вторую. Колено подгибается. Она хватается за стол, успевает удержать-
ся.
В операционной показывается Лабин с сумкой на плече.
— Ты как? — на глазах у него снова линзы, белые как свежий лед. Кларк, ожи-
вившись , кивает ему.
— Давай кожу.
Кляйн их слышит.
— Постойте, вам не давали допуска... я хочу сказать. . .
Сначала глаза. Верхняя часть костюма легко обволакивает туловище. Рукава и
перчатки прилипают к телу дружелюбными тенями. Она опирается на Лабина, чтобы
дотянуться до бедер — под новой кожей иголки колют не так сильно, а попробо-
вав заново опереться на ногу, она удерживается на ней добрых десять секунд.
Прогресс.
— Лени, Кен, вы куда собрались?
На этот раз голос Седжер. Кляйн вызвал подмогу.
— Решили заглянуть в гости, — отвечает Лабин.
— Вы хорошо все продумали? — сдержанно спрашивает Седжер. — При всем уваже-
нии. . .
— А есть причины воздержаться? — невинно интересуется Лабин.
— У Лени но...
— Не считая ее ноги.
Мертвая тишина.
— Вы уже проверили образцы, — замечает Лабин.
— Не в полной мере. Анализы делаются быстро, но не мгновенно.
— И? Есть что-нибудь?
— Если вы заразились, мистер Лабин, это произошло всего несколько часов на-
зад. Уровень инфекции в крови еще не поддается определению.
— Значит, нет, — заключает Лабин. — А наши костюмы?
Седжер молчит.
— Значит, они нас защитили, — подытоживает Лабин. — На этот раз.
— Я же сказала, мы не закончили...
— Я думал, что Бетагемот сюда добраться не может, — говорит Лабин.
Седжер опять затихает.
— Я тоже так думала, — отзывается она, наконец.
Кларк вполуприпрыжку двигается к шлюзу. Лабин подает ей руку.
— Мы выходим, — говорит он.
Полдюжины аналитиков сгрудились у пультов на дальнем конце грота связи, пе-
ребирая симуляции и подгоняя параметры в упрямой надежде, что их виртуальный
мир имеет некоторое отношение к реальному. Патриция Роуэн, стоя позади, раз-
глядывает что-то на одном из экранов. За вторым в одиночку работает Джеренис
Седжер.
Она оборачивается, видит рифтеров и, чуть повысив голос, издает предупреди-
тельный сигнал, замаскированный под приветствие.
— Кен, Лени!
Все оборачиваются. Парочка малоопытных пятится на шаг-другой.
Роуэн первая берет себя в руки. Ее блестящие ртутью глаза непроницаемы.
— Ты бы поберегла ногу, Лени. Вот... — Она подкатывает от ближайшего пульта
свободный стул. Лени с благодарностью опускается на него.
Никто не суетится. Здешние корпы умеют следовать за лидером, хотя и не всем
это по нраву.
— Джерри говорит, ты увернулась от пули, — продолжает Роуэн.
— Насколько нам известно, — уточняет Седжер. — На данный момент.
— То есть пуля все-таки была, — отмечает Лабин.
Седжер смотрит на Роуэн. Роуэн смотрит на Лабина.
Большинство стараются не смотреть ни на кого.
Наконец Седжер пожимает плечами.
— D-цистеин и d-цистин — в наличии. Пиранозильная РНК — тоже. Фосфолипидов
и ДНК нет. Внутриклеточная АТФ выше нормы. Не говоря о том, что при РЭМ-
микроскопии инфицированных клеток просто видно, как там все кишит этими ма-
лявками. — Она переводит дыхание. — Если это не Бетагемот, то его злой брат-
двойник .
— Дрянь, — вырывается у одного аналитика. — Опять!
Через миг Кларк понимает, что ругательство относится не к словам Седжер, а
к чему-то на экране. Подавшись вперед, она видит изображение за плечами кор-
пов — объемную модель Атлантического бассейна. Светящиеся инверсионные следы
вьются по глубине многоголовыми змеями, разделяясь и сходясь над континен-
тальными шельфами и хребтами. Течения, водовороты и глубоководная циркуляция
— внутренние реки океана — отмечены разными оттенками красного и зеленого. А
поверх изображения скупой итог:
СВЕДЕНИЕ НЕ УДАЛОСЬ. ДОВЕРИТЕЛЬНЫЙ ИНТЕРВАЛ ПРЕВЫШЕН. ДАЛЬНЕЙШИЕ ПРОГНОЗЫ
НЕНАДЕЖНЫ.
— Ослабь немножко Лабрадорское течение, — предлагает один из аналитиков.
— Еще немного, и его просто не будет, — возражает другой.
— Откуда нам знать, может, его уже и нет.
— Когда Гольфстрим...
— Ну, ты попробуй, а?
Атлантика гаснет и перезагружается.
Роуэн, отвернувшись от своих, находит взглядом Седжер.
— А если они ни к чему не придут?
— Возможно, он был здесь с самого начала. А мы его просто не заметили. —
Седжер, словно не доверяя собственной версии, мотает головой. — Мы ведь не-
множко спешили.
— Спешили, но не настолько. Прежде чем выбрать участок, мы проверили каждый
источник на тысячу километров отсюда, разве не так?
— Кто-то проверял, да, — устало говорит Седжер.
— Я видела результаты. Они убедительны. — Роуэн, кажется, беспокоит не
столько появление Бетагемота, сколько мысль, что разведка дала сбой. — И с
тех пор ни один отчет ничего не показывал... — Спохватившись, она перебивает
сама себя: — Ведь не показывал, Лени?
— Нет, — говорит Кларк, — ничего не было.
— Ну вот. Пять лет весь район был чист. Насколько нам известно, чисто было
на всем глубоководье Атлантики. И долго ли Бетагемот может выжить в холодной
морской воде?
— Неделю-другую, — подсказывает Седжер. — Максимум месяц.
— А сколько времени потребовалось бы, чтобы его донесли сюда глубоководные
течения?
— Десятки лет, если не века, — вздыхает Седжер. — Все это известно, Пат.
Очевидно, что-то изменилось.
— Спасибо, Джерри, просветила. И что бы это могло быть?
— Господи, чего ты от меня хочешь? Я тебе не океанограф. — Седжер вяло ма-
шет рукой в сторону аналитиков. — Их спрашивай. Джейсон прогоняет эту модель
уже...
Джейсон обрушивает на экран поток непристойностей. Экран в ответ огрызает-
ся:
СВЕДЕНИЕ НЕ УДАЛОСЬ. ДОВЕРИТЕЛЬНЫЙ ИНТЕРВАЛ ПРЕВЫШЕН. ДАЛЬНЕЙШИЕ ПРОГНОЗЫ
НЕНАДЕЖНЫ.
Роуэн, прикрыв глаза, начинает заново:
— Ну а в эвфотической зоне1 он способен выжить? Там ведь теплее, даже зи-
мой. Может, наши рекогносцировщики подхватили его наверху и занесли сюда?
— Тогда бы он и проявился здесь, а не над Невозможным озером.
— Да он вообще нигде не должен был прояв...
— А если это рыбы? — перебивает вдруг Лабин.
Роуэн оборачивается к нему.
— Что?
— Внутри организма-хозяина Бетагемот может существовать неограниченно дол-
го, так? Меньше осмотический стресс. Потому-то, в общем, он и заражал рыб.
Может, его к нам подвезли?
Эвфотическая зона — верхняя толща воды крупного водоема, в которой благодаря сол-
нечному освещению имеются условия для фотосинтеза.
— Глубоководные рыбы не рассеиваются по океану, — возражает Седжер, — а
просто околачиваются возле источников.
— А личинки в планктоне?
— Все равно не сходится. Не с такими расстояниями.
— Не в обиду тебе, — произносит Лабин, — но ты медик. Может, спросим на-
стоящего специалиста?
Это, конечно, шпилька. Когда корпы составляли список допущенных на ковчег,
ихтиологи даже не рассматривались. Но Седжер лишь качает головой:
— Они бы сказали то же самое.
— Откуда тебе знать? — с неожиданным любопытством интересуется Роуэн
— Оттуда, что Бетагемот большую часть земной истории был заперт в немного-
численных горячих источниках. Если он способен распространяться с планктоном,
зачем было так долго ждать? Он бы захватил весь мир сотни миллионов лет на-
зад.
В Патриции Роуэн что-то меняется. Кларк не вполне улавливает, в чем дело.
Может, сама поза. Или ее линзы вспыхнули ярче, словно интеллект, блестящий в
глазах, перешел на скоростной режим.
— Пат? — окликает ее Кларк.
Но Седжер вдруг как ошпаренная срывается со стула, повинуясь прозвучавшему
в наушниках сигналу. Прикасается к запястнику, подключая его к сети.
— Выхожу. Задержи их. — И оборачивается к Лабину с Кларк. — Если действи-
тельно хотите помочь, давайте со мной.
— В чем дело? — спрашивает Лабин.
Седжер уже на середине пещеры.
— Опять идиоты, не способные ничему научиться. Вот-вот убьют вашего друга.
Кавалерия
По всей «Атлантиде» встречаются линии — четырехсантиметровые бороздки,
словно кто-то равномерно прошелся по всему корпусу цепной пилой. С обеих сто-
рон они обозначены предупредительной разметкой с диагональными полосками, и
если посмотреть вверх, чуть отступив от них, то видно, зачем: в каждом проеме
расположены опускные переборки, готовые упасть ножом гильотины в случае про-
боины. Эти границы очень удобно использовать в качестве линий на песке, раз-
деляющих противников. Таких, например, как полдюжины корпов, замерших перед
пограничной чертой — у них хватает ума или трусости не лезть вперед. И как
Ханнук Йегер, беспокойно приплясывающий по ту сторону полосатой ленты, не
подпуская их к лазарету ближе чем; на пятнадцать метров.
Лабин расталкивает перетрусивших корпов плечом. Кларк, хромая, движется за
ним по пятам. Йегер приветственно скалит зубы.
— Все веселье за четвертой дверью слева.
Его закрытые накладками глаза щурятся при виде корпов, сопровождающих пару.
Кларк с Лени проходят мимо. Двинувшуюся следом Седжер Йегер хватает за гор-
ло .
— Только по приглашениям!
— Ты же не... — Йегер усиливает хватку, и голос Седжер превращается в ше-
пот . — Ты хочешь... Джину смерти?
— Это что, угроза? — рычит Йегер.
— Я его врач!
— Пусти ее, — велит Кларк. — Она может понадобиться
Йегер и пальцем не шевелит.
«Вот черт, черт, — думает Кларк, — он что, на взводе?»
У Йегера мутация, переизбыток моноаминовой оксидазы в крови. От этого нару-
шается баланс элементов в мозгу — тех, что удерживают человека в равновесии.
Начальство встроило ему компенсирующий механизм — в те времена, когда такие
вещи еще допускались, — но рифтер как-то научился его обходить. Бывает, он
нарочно так себя заводит, что хватает чьего-то косого взгляда, чтобы сорвать-
ся . В таких случаях уже не важно, друг ты или враг. В таких случаях даже Ла-
бин принимает его всерьез.
Вот как сейчас.
— Пропусти ее, Хан. — Его голос звучит спокойно и ровно, тело расслаблено.
С другого конца коридора доносится стон — и что-то с шумом ломается.
Йегер, фыркнув, отталкивает Седжер. Та с кашлем приваливается к стене.
— Ты тоже с нами, — обращается Лабин к Роуэн, которая скромно держится за
полосатой лентой. И к Йегеру: — Если ты, конечно, не против.
— Дерьмо, — сплевывает Йегер. — Да плевать мне.
Он сжимает и разжимает кулаки — судорожно, словно под током.
Лабин кивает.
— Иди, — небрежно бросает он Кларк. — Я помогу Хану держать оборону.
Это, конечно, Нолан. Кларк, подходя к медотсеку, слышит ее рычание:
— Ах, так ты еще и обделался, чертов педик...
Кларк протискивается в люк. В ноздри бьет кислая смесь страха и фекалий.
Да, Нолан, и на подмогу вызвала Кризи. Кляйн отброшен в угол, избит и весь в
крови. Может, пытался встать на пути, а может, Нолан пыталась его вынудить.
Джин Эриксон наконец-то пришел в себя и скорчился на столе, как зверь в
клетке. Растопыренными пальцами он упирается в изолирующую пленку, и та рас-
тягивается , будто невероятно тонкий латекс. Чем сильнее рифтер напирает, тем
больше сопротивление: он еще не до конца вытянул руку, а мембрана уже достиг-
ла максимальной прочности — вдоль линий сопротивления расцветают маслянистые
радуги.
— Твою-то мать, — рычит Джин, опускаясь на место.
Нолан, присев, по-птичьи склоняет голову набок в нескольких сантиметрах от
окровавленного лица Кляйна.
— Выпусти его, птенчик.
Кляйн плюется кровью и слюной.
— Я же сказал, он...
— Не подходите к нему! — Седжер вламывается внутрь, словно последних пяти
лет — и пяти минут — и не было вовсе. Не успевает она дотронуться до плеча
Нолан, как Кризи отшвыривает ее к переборке.
Нолан стряхивает воображаемую грязь с места, которого коснулась рука Сед-
жер.
— Голову не повреди, — приказывает она Кризи. — В ней может быть пароль.
— Так, все. — Хоть у Роуэн хватило ума остаться в коридоре. — Быстро. Успо-
койтесь .
Нолан фыркает, мотает головой.
— А то что, обмылок хренов, ты охрану вызовешь? Прикажешь нам «очистить по-
мещение»?
Белые глаза Кризи разглядывают Седжер с расстояния в несколько сантиметров.
Эти глаза над ухмыляющимися бульдожьими челюстями грозят бессмысленным и без-
думным насилием. Говорят, Кризи умеет обращаться с женщинами. Нет, с Кларк он
шуток не шутил — как правило, с ней вообще никто не связывается.
Роуэн заглядывает в открытый люк, лицо у нее спокойное и уверенное. Кларк
видит мольбу за этой самонадеянной маской. Первое побуждение — не замечать
ее. Ногу раздражающе покалывает. Кризи за плечом чмокает губами над Седжер.
Его рука зависает у нее над подбородком. Кларк его игнорирует.
— В чем дело, Грейс?
Нолан хищно улыбается.
— Нам удалось привести его в чувство, но тут Норми, — она рассеянно толкает
Кляйна в лоб, — наложил на стол какой-то пароль. Мы не смогли опустить мем-
брану .
Кларк оборачивается к Эриксону.
— Ты как себя чувствуешь?
— Они со мной что-то сделали. — Джин кашляет. — Пока я был в коме.
— Да, сделали. Спасли его... — Кризи бьет Седжер головой о переборку. Сед-
жер умолкает.
Кларк не сводит глаз с Эриксона.
— Когда шевелишься, кишки не вываливаются?
Юн неуклюже поворачивается, показывая ей живот: мембрана натягивается на
голове и плечах, словно амниотический мешок1.
— Чудо современной медицины, — говорит Джин, укладываясь лицом вверх. Ну
да, все внутренности на месте. Свежие розовые шрамы на животе дополнили ста-
рые, те, что на груди.
Судя по всему, Джеренис Седжер, очень хочет что-то сказать. А Дейл Кризи,
похоже, очень хочет, чтобы она попыталась.
— Пусть говорит, — распоряжается Кларк. Кризи чуть ослабляет хватку. Седжер
смотрит на Кларк и не раскрывает рта.
— Ну, так что? — торопит ее Кларк. — Похоже, вы его нормально заклеили. Три
дня прошло.
— Три дня, — повторяет Седжер. Из-за давления на горле ее голос выходит
тонким и гнусавым. — Его почти начисто выпотрошили, а ты думаешь, трех дней
достаточно для выздоровления?
Собственно, Кларк в этом уверена. Ей уже приходилось видеть искромсанные и
разбитые тела: она видела, как многорукие роботы собирали их заново и помеща-
ли в раны тонкие электрические сетки, которые так ускоряли заживление, что
это выглядело бы чудом, если бы не стало таким привычным. Трех дней более чем
достаточно, чтобы прийти в себя. Швы, может, еще малость кровят, но держатся,
а в невесомом черном чреве глубины времени на поправку будет сколько угодно.
Вот что никак не доходит до сухопутников: силы отнимает само земное притя-
жение .
— Ему еще нужны операции? — спрашивает она.
— Понадобятся, если не побережется.
— На вопрос отвечай, мать твою! — рявкает Нолан.
Седжер косится на Кларк и не находит у нее поддержки.
— Ему требуется время на реабилитацию, и в коме срок сократится на две тре-
ти. Если он хочет поскорей отсюда выбраться, лучше варианта не найти.
— Вы удерживаете его против воли, — говорит Нолан.
— Зачем... — подает из коридора голос Роуэн.
Нолан налетает на нее:
— А ты заткнись на хрен!
Роуэн хладнокровно испытывает судьбу.
— Зачем бы нам его удерживать, если не по медицинским показаниям?
— Он мог бы отдохнуть у себя в пузыре, — отвечает ей Кларк. — Или просто
снаружи.
Седжер качает головой.
— У него значительно повышена температура. Лени, да ты посмотри на него!
В ее словах есть смысл. Эриксон растянулся на спине, явно обессилев. Кожа
блестит от пота, хотя этот блеск почти неразличим за бликами мембраны.
— Температура, — повторяет Кларк. — Не из-за операции?
— Нет. Какая-то оппортунистическая инфекция.
Амнион, Амниотический мешок, Амниотический пузырь или Водная оболочка — одна из
зародышевых оболочек у эмбрионов пресмыкающихся, птиц, млекопитающих.
— Откуда бы?
— На него напало дикое животное, — напоминает Седжер. — Даже простой укус
может занести множество всякой дряни, а Джина практически выпотрошили. Я бы
даже удивилась, пройди все без осложнений.
— Слыхал, Джин? — спрашивает Кларк. — Ты вроде как подхватил рыбье бешенст-
во.
— Охренеть, — произносит Эриксон, созерцая потолок.
— Так что тебе решать. Останешься, позволишь себя долечить? Или рискнешь
положиться на лекарства?
— Вытащите меня отсюда, — негромко просит Джин.
Кларк поворачивается к Седжер.
— Ты его слышала.
Та выпрямляется. Невозможное, безумное, вечное упрямство.
— Лени, я тебя на помощь звала. Если это называется...
Кулак Кризи ядром врезается ей в живот. Седжер охает и заваливается набок,
в падении ударяется головой о переборку и остается лежать, хватая воздух
ртом. Краем глаза Кларк замечает, что Роуэн сделала шаг вперед, но вовремя
спохватилась.
Она холодно сморит на Кризи:
— Напрасно ты так, Дейл.
— Пусть не наглеет, — ворчит тот.
— И как же она выпустит Джина, если даже вдохнуть не может, болван?
— Ну, реально, Лен, что тут такого?
Это Нолан. Кларк поворачивается к ней.
— Ты знаешь, как они с нами обращались, — продолжает Грейс, встав рядом с
Кризи. — Знаешь, сколько наших искалечили эти уроды. И убили.
«Меньше, чем я», — молчит Кларк.
— Я б сказала, если Дейлу охота спустить пар на этих выродках, пусть его. —
Нолан дружески берет Кларк за плечо. — Может, малость уравняет счет. Понима-
ешь?
— Это ты бы так сказала, — тихо отвечает Кларк, — а я — иначе.
— Вот это сюрприз! — На лице Нолан мелькает тень усмешки.
Они смотрят друг друга сквозь роговичные щитки. У дальней стены скулит
Кляйн, у них под ногами вроде бы начинает дышать Джеренис Седжер. Кризи нави-
сает у Кларк над плечом, само его присутствие — открытая угроза. Она ровно,
медленно дышит. Приседает на корточки — легонечко, легонечко, больная нога
готова подогнуться — и помогает Седжер сесть.
— Выпусти его, — приказывает она.
Седжер бормочет что-то в свой запястник. На коже ее предплечья загорается
клавиатура со странными символами. Другой рукой она набирает нужную комбина-
цию.
Изолирующая палатка издает тихий хлопок. Эриксон нерешительно касается мем-
браны пальцами, убеждается, что они проходят насквозь, и чуть не падает со
стола, словно выскочив из мыльного пузыря. Подошвы мягко стукают об пол. Но-
лан подает ему извлеченный откуда-то гидрокостюм.
— С возвращением, дружище. Говорила же, мы тебя вытащим.
Они оставляют Кларк с корпами. Седжер, игнорируя протянутую ей руку, с тру-
дом поднимается на ноги и опирается о переборку. Одной ладонью она все еще
прикрывает живот. И склоняется над Кляйном.
— Норм? Норм? — Она неловко склоняется к своему подчиненному и оттягивает
ему веко. — Не теряй сознания...
С ее головы срывается капля и плюхается на разбитое лице медика, теряясь в
его собственной крови. Седжер, выругавшись, вытирает рану тыльной стороной
ладони.
Кларк подходит помочь. Под нору попадается что-то маленькое и острое. Она
поднимает ступню. Зуб, липкий от слюны и прочего, с тихим стуком падает на
пол.
— Я... — начинает Кларк.
Седжер с яростью оборачивается.
— Пошла вон!!!
Кларк какое-то время разглядывает ее, затем выходит.
В коридоре ее ждет Роуэн.
— Нельзя, чтобы такое повторилось.
Кларк приваливается к переборке, снимая часть веса с больной ноги.
— Ты же знаешь Грейс. Они с Джином...
— Дело не только в Грейс. По крайней мере, это ненадолго. Я же говорила,
что-то такое могло произойти.
Ее охватывает страшная усталость.
— Ты говорила, что нужно развести стороны. Так зачем Джерри удерживала Джи-
на, когда тот захотел уйти?
— Ты думаешь, ей нужен рядом этот тип? Она заботилась о благе пациента. У
нее работа такая.
— Мы сами о себе позаботимся.
— У вас просто нет должной квалификации...
Кларк предостерегающе вскидывает руку.
— Это мы уже слышали, Пат. Маленькие люди не видят всей картины. Простого
гражданина надо беречь от жестокой правды. Крестьяне слишком невежественны,
чтобы голосовать. — Она с отвращением качает головой. — Пять лет прошло, а вы
все норовите гладить нас по головке, как детишек.
— Скажешь, Джин Эриксон — более квалифицированный диагност, чем наш главный
врач?
— Скажу, что он имеет право на ошибку. — Кларк машет рукой. — Слушай, мо-
жет, ты и права. Может, он через неделю свалится с гангреной и приползет об-
ратно к Джерри. Или предпочтет умереть. Но это ему решать.
— Речь не о гангрене, — тихо говорит Роуэн. — И не об обычной инфекции. И
тебе это известно.
— Не вижу разницы.
— Я тебе говорила.
— Ты говорила о перепуганных ребятишках, не верящих в ими же созданную за-
щиту . Но защита выдержит, Пат. Я — живое тому доказательство. Мы можем пить
культуру Бетагемота как воду, нам это не повредит.
— Мы лишились...
— Лишились очередного пласта самообмана, только и всего. Пат, Бетагемот
здесь. Не знаю, как он сюда попал, но тут ничего не поделаешь, да и зачем? Он
вас никак не потревожит, разве что ткнет носом в нечто такое, о чем вы пред-
почли бы не думать, но к этому вы скоро привыкнете. Вам не впервой. Через ме-
сяц ты и думать о нем забудешь.
— Тогда, прошу... — начинает Роуэн и замолкает. Кларк ждет, пока ее собе-
седница загонит себя в роль подчиненной.
— Дайте нам этот месяц, — шепчет, наконец, Роуэн.
Возмездие
Кларк нечасто заходит в жилой квартал, а в этом отсеке как будто и вовсе не
бывала. Стены коридора покрыты электронной краской и подключены к генератору
обоев. На переборке по левому борту вырос густой лес рогатых кораллов, на
правой вертятся и сбиваются в стайки рыбы-хирурги, словно узлы какой-то непо-
нятной рассеянной неиросети. И повсюду пляшут осколки солнечных лучей. Кларк
не в состоянии определить, что за иллюзия перед ней — чисто синтетическая или
основанная на архивных съемках настоящего кораллового рифа. Она бы все равно
не увидела разницы: ни одно из морских созданий, с которыми она свела знаком-
ство за прожитые годы, не жило при солнечном свете.
Здесь много семей, догадывается Кларк. Как правило, взрослые не в восторге
от видов дикой природы: довольно сложно ценить такую эстетику и дальше, по-
стигнув ее иронию.
А вот и он — номер D-18. Она жмет на кнопку звонка. За запертым люком при-
глушенно звучит музыка: зыбкая ниточка мелодии, слабый голос, шум движения.
Люк распахивается. Выглядывает плотно сбитая девочка лет десяти с колючей
светлой челкой. Музыка доносится из глубины помещения — флейта Леке, сообра-
жает Кларк. При виде рифтерши улыбка на лице девочки мгновенно гаснет.
— Привет, — начинает Кларк, — я искала Алике.
Она и сама примеряет улыбку.
Та не подходит. Девочка неуверенно отступает на шаг.
— Леке...
Музыка прерывается.
— Что? Кто там?
Маленькая блондинка отступает в сторону — по-кошачьи нервно. Алике Роуэн
сидит на кушетке посреди комнаты. Одна рука лежит на флейте, другая тянется к
перламутровым наглазникам.
— Привет, Леке, — говорит Кларк. — Мама сказала, я найду тебя здесь.
— Лени! Ты прошла!
— Прошла?
— Карантин! Мне сказали, тебя с психованным заперли, на анализы вроде бы.
Ты их, наверно, перехитрила.
Перед кушеткой стоит прямоугольная тумба на колесиках, высотой примерно в
метр — маленький обелиск в тех же переливчатых тонах, что наглазники Алике.
Девочка кладет свою пару на крышку — рядом с точно такими же.
Кларк ковыляет к ней. Алике тотчас мрачнеет.
— Что у тебя с ногой?
— «Кальмар» взбунтовался. Рулем зацепило.
Подружка Алике бормочет что-то сбоку и скрывается в коридоре. Кларк обора-
чивается ей вслед:
— Твоей подруге я не слишком понравилась.
Алекс небрежно машет рукой:
— Келли — трусиха. Только глянь на нее, и сразу в голове всплывает вся
чушь, что мамуля наболтала ей про ваших. Она славная, просто не фильтрует ис-
точников информации. — Пожав плечами, девочка меняет тему. — Так что случи-
лось?
— Помнишь, я тебе недавно рассказывала про карантин?
Алике хмурится.
— Про парня, которого покусали. Эриксон?
— Угу. Ну и вот, похоже, он все-таки что-то подцепил, так что на время мы
решили установить в «Атлантиде» режим «Рыбоголовым входа нет».
— Вы позволите себя выставить?
— Вообще-то, я думаю, что это разумная мысль, — признает Кларк.
— Почему? Чем он заразился?
Кларк качает головой.
— Тут не медицинский вопрос, хотя это часть проблемы. Просто... все изрядно
разгорячились, причем с обеих сторон. Мы с твоей мамой считаем, что лучше
держать ваших и наших подальше друг от друга. Какое-то время.
— Как это? Что происходит?
— А мама тебе не...
До Кларк с опозданием доходит, что Патриция Роуэн могла кое-что скрывать от
дочери. Если уж на то пошло, то неизвестно даже, а многие ли взрослые на «Ат-
лантиде» в курсе дела. Корпы в принципе склонны держать информацию под зам-
ком . Конечно, на взгляд Кларк все их принципы плевка не стоили, и все же. . .
Она не собирается становиться между Пат и...
— Лени? — Алике хмуро уставилась на нее. Эта девочка — из немногих людей,
которым Кларк не стесняясь показывает обнаженные глаза, однако, сейчас на ней
линзы. Она делает еще шаг-другой по ковру и видит скрытую до сих пор грань
тумбы. У верхнего края нечто вроде панели управления: темная лента, на кото-
рой горят красные и голубые иконки. По всей длине бежит зубчатая линия, похо-
жая на ЭЭГ.
— Это что? — спрашивает Кларк, чтобы отвлечь девочку. Для игровой приставки
штуковина слишком массивная.
— А, это... — Алике пожимает плечами. — Это Келлин зельц.
— Что?!
— Ну, типа, умный гель. Нейроная культура с...
— Я знаю, что это такое, Леке. Просто... мне странно видеть его здесь, по-
сле. . .
— Хочешь посмотреть? — Алике выбивает на крышке шкафчика короткую дробь.
Перламутровая поверхность идет разводами и становится прозрачной, под ней —
лепешка розовато-серых тканей в круглом ободке. Похоже на густую овсянку. Пу-
динг разбит на части перфорированными стеклянными перегородками.
— Не особо большой, — говорит Алике. — Куда меньше, чем были в прежние вре-
мена. Келли говорит, он размером с кошачий.
«Значит, наверняка злобный, если и не слишком умный».
— Зачем он нужен? — спрашивает Кларк. «Не могут же они быть такими идиота-
ми , чтобы использовать эту штуку после...»
— Это вроде домашней зверушки, — виновато объясняет Алике. — Она назвала
его Рамблом.
— Зверушки?!
— Ага. Он думает, вроде как. Учится всякому. Хотя никто точно не понимает,
как это происходит.
— А, так ты о них слышала?
— Ну, он намного меньше тех, что работали на вас.
— Они на нас не раб...
— Он совсем безобидный. Не подключен ни к каким системам, и вообще.
— Так что же он делает? Вы его обучаете всяким трюкам?
Мозговая каша поблескивает, как гноящаяся язва.
— Вроде того. Он отвечает, если ему что-нибудь говоришь. Не всегда впопад,
но от этого только смешнее. А если подключить к нему радио, он играет в такт
музыке крутые цветные узоры. — Алике подхватывает свою флейту и кивает на на-
глазники. — Посмотришь?
— Зверушка, — бормочет Кларк. «Чертовы корпы...»
— Мы не такие, — резко отвечает Алике. — Не все такие.
— Извини? Не такие — какие?
— Не корпы. Что это вообще означает? Мою маму? Меня?
«Неужели я проговорила это вслух?»
— Просто... сотрудников корпорации. — Кларк никогда всерьез не задумывалась
о происхождении слова — не больше, чем об этиологии слова «стул» или «фумаро-
ла»1.
Фумарола (итал. fumarola, от лат. fumare — дымить(ся)) — трещина или отверстие,
располагающееся в кратерах, на склонах и у подножия вулканов и являющееся источником
горячих газов.
— Если ты не замечала, здесь полно и другого народа. Техники, врачи и про-
сто родственники.
— Да, я в курсе. Конечно, я в курсе...
— А ты валишь всех в одну кучу, понимаешь? Если у кого нет в груди пучка
трубок, то для тебя это сразу корп, труп1.
— Ну... извини. — Она запоздало пускается в оправдания: — Я не обзываюсь,
просто слово такое.
— Нет, для вас, рыбоголовых, это не «просто слово».
— Извини, — повторяет Кларк. Ни одна из них не сдвинулась с места, но рас-
стояние между ними заметно увеличилось.
— в общем, — говорит Кларк, наконец, — я просто хотела предупредить, что в
ближайшее время появляться не буду. Разговаривать мы, конечно, сможем, но...
Движение у люка. В комнату входит крупный коренастый мужчина с зачесанными
ото лба темными волосами.
Кожа между бровями собралась складками, от него так и веет враждебностью.
Отец Келли.
— Мисс Кларк, — ровным голосом произносит он.
Внутри у нее все стягивается в тугой злобный узел. Она видела такие лица,
знает эту повадку — не сосчитать, сколько раз сталкивалась с ней в возрасте
Келли. Она знает, на что способны отцы, знает, что творил с ней ее собствен-
ный, но она уже не маленькая девочка, а отцу Келли, похоже, совсем не помеша-
ет урок...
Однако ей приходится все время напоминать себе: ничего этого не было.
Портрет
садиста в
отрочестве
Конечно, со временем Ахилл Дежарден научился обводить шпиков вокруг пальца.
Он с малых лет понял, каков расклад. В мире, который ради его же безопасности
держат под постоянным наблюдением, существовали лишь наблюдаемые и наблюдате-
ли, и Ахилл точно знал, на какой стороне предпочел бы находиться. Невозможно
мастурбировать при зрителях.
Этим и наедине с собой нелегко было заниматься. Его, как-никак, воспитали в
определенных религиозных убеждениях: миазмы католицизма, цепляясь за обложку
«Nouveaux Separatistes»2, висели над Квебеком и тогда, когда в остальном мире
о религии уже думать забыли. Они осаждали Ахилла каждую ночь, когда он доил
себя, и в голове у него мелькали мерзостные образы, от которых пенис стано-
вился твердым. И то, что под действием развешанных им над кроватью, столом и
комодом магнитных мобилей шпики лишь пьяно покачивались, потеряв связь с се-
тью, ничего не меняло. Как и то обстоятельство, что его все равно ожидал ад,
даже если б он ни разу в жизни не коснулся своего тела, — ведь сказал же Ии-
сус : «То, что ты совершаешь в сердце своем, ты совершаешь в глазах Бога».
Ахилл был заранее проклят за непрошенные мысли, так что ничего не терял, во-
площая их.
Вскоре после одиннадцатого дня рождения пенис стал оставлять улики: во вре-
мя ночных оргий на простыни брызгала белесоватая жидкость. Две недели он не
осмеливался обратиться к энциклопедии: именно столько потребовалось на форму-
лировку такого запроса, чтобы мама с папой не узнали. Взлом приватных настро-
ек домашней «служанки» занял еще три дня. Никогда не знаешь, какие элементы
Corpse (англ.) — труп.
2
«Новых сепаратистов» (фр.). Французы Квебека - католики, не раз пытавшиеся отде-
лится от остальной, в основном протестантской, Канады.
отслеживает эта штуковина. К тому времени, как Ахилл решился простирнуть по-
стельное белье, от него пахло примерно как от Эндрю Трайтса из социального
центра — а тот был вдвое больше любого своего сверстника, и никто не хотел
стоять с ним рядом на рапитранской остановке.
— Я думаю...
Начал Ахилл в тринадцать лет.
Церкви он больше не верил. Что ни говори, он был прирожденным эмпириком, а
Бог не выстоит и десяти секунд под критическим взглядом личности, уже вычис-
лившей страшную правду про пасхальных кроликов. Как ни странно, перспектива
вечных мук теперь казалась вполне реальной, на каком-то примитивном уровне,
недоступном для логики. А если проклятие реально, то исповедь не повредит.
— ...что я чудовище, — закончил он.
Признание было не таким рискованным, как могло показаться. Исповедник был
не слишком надежен — Ахилл загрузил его из Сети (из Водоворота, мысленно по-
правился он, теперь все его называли только так) , и в нем могло быть полно
червей и троянов, даже тщательная чистка не дала бы гарантии, — но он отклю-
чил все каналы ввода-вывода, кроме голосового, и мог стереть все подчистую,
если начнутся какие-нибудь фокусы. И уж точно не собирался оставлять програм-
мку в рабочем состоянии после того, как перед ней выложится.
Папа, прознай он, что Ахилл занес в семейную Сеть дикое приложение, съехал
бы с катушек, однако мальчик не стал бы рисковать с домашними фильтрами, даже
если б отец и перестал за ним шпионить после смерти мамы. Так или иначе, папа
никак не мог узнать. Он был внизу, в сенсориуме, вместе со всей провинцией —
страной, пришлось напомнить себе Ахиллу, — подключившись к помпезной церемо-
нии первого Дня Независимости. Надутая колючая Пенни — дни, когда она обоже-
ствляла старшего брата, давно миновали, — продала бы его мгновенно и с удо-
вольствием, но она последнее время обитала в основном в экстаз-шлеме. Контак-
ты , наверно, уже протерли ей виски насквозь.
Был день рождения последнего нового государства на планете, и Ахилл Дежар-
ден остался в спальне наедине с исповедником.
— Чудовище какого рода? — спросил «ТераДруг » голосом андрогина.
Слово Ахилл выучил еще утром и тщательно выговорил:
— Женоненавистник.
— Понимаю, — пробормотал ему в ухо «ТераДруг».
— У меня возникают... возникают такие желания — делать им больно. Девочкам.
— И как ты при этом себя чувствуешь? — голос стал чуть более мужественным.
— Хорошо. Ужасно... то есть... они мне нравятся. Чувства, в смысле.
— Не мог бы ты уточнить? — в голосе не слышалось ни ужаса, ни отвращения.
Конечно, их и не могло быть — программа ничего не чувствует, это даже не Тью-
ринг-софт. Просто навороченное меню. Но, как это ни глупо, Ахиллу полегчало.
— Это... сексуально, — признался он. — Ну, думать о них так.
— Как именно?
— Ну, что они беззащитные. Уязвимые. Я... мне нравится, как они смотрят,
когда... это самое...
— Продолжай, — сказал «ТераДруг».
— Когда им больно, — жалобно выговорил Ахилл.
— А, — сказала программа. — Сколько тебе лет, Ахилл?
— Тринадцать.
— у тебя есть друзья среди девочек?
— Конечно?
— И как ты относишься к ним?
— Я же сказал! — прошипел Ахилл, едва не срываясь на крик. — Мне...
— Нет, — мягко остановил его «ТераДруг», — я спросил, как ты относишься к
ним лично, когда не чувствуешь полового возбуждения. Ты их ненавидишь?
Ну почему же, нет. Вот Андреа толковая девчонка, к ней всегда можно обра-
титься, когда надо вычистить вирусы. А Мартин... Ахилл как-то раз чуть не
убил ее старшего брата, когда тот стал к ней приставать. Мартин и мухи бы не
обидела, а этот мудак-братец...
— Они мне нравятся, — сказал Ахилл, морща лоб из-за парадокса. — Очень нра-
вятся. Они отличные. Кроме тех, кого я хочу, ну ты понял, да и то только ко-
гда я. . .
«ТераДруг » терпеливо ждал.
— Все клево, — выдавил, наконец, Ахилл, — если только я не...
— Понимаю, — выдержав паузу, заговорила программа. — Ахилл, у меня для тебя
хорошее известие. Ты вовсе не женоненавистник.
— Нет?
— Женоненавистник — это тот, кто ненавидит женщин, боится их или считает в
чем-то ниже себя. Похоже на тебя?
— Нет, но... кто же тогда я?
— Это просто, — сообщил ему «ТераДруг». — Ты сексуальный садист. Это совер-
шенно иное.
— Правда?
— Секс — очень древний инстинкт, Ахилл, и он формировался не в вакууме. Он
переплетается с другими базовыми побуждениями: агрессии, территориальности,
конкуренции за ресурсы. Даже в здоровом сексе присутствует немалый элемент
насилия. Секс и насилие имеют немало общих нейронных механизмов.
— Ты. . . хочешь сказать, что все такие же, как я? — На такое он и надеяться
не смел.
— Не совсем. У большинства людей имеется переключатель, который на время
секса подавляет агрессию. У одних такие переключатели работают лучше, у дру-
гих хуже. У клинического садиста этот переключатель работает очень плохо.
— и это я, — пробормотал Ахилл.
— Весьма вероятно, — сказал «ТераДруг», — хотя без должного клинического
обследования уверенности быть не может. Я сейчас не имею выхода в твою сеть,
но предоставлю список ближайших медавтоматов, если ты скажешь, где мы нахо-
димся .
За спиной Ахилла тихо скрипнула дверь. Он обернулся и похолодел до костей.
Дверь спальни распахнулась. В темном проеме стоял отец.
— Ахилл, — донесся из вращающейся бездны голос «ТераДруга», — ради твоего
же здоровья, не говоря уже о душевном спокойствии, тебе крайне желательно по-
сетить один из наших филиалов. Гарантированная диагностика — первый шаг к
здоровой жизни.
«Он не мог услышать», — сказал себе Ахилл. «ТераДруг» звучал в его наушни-
ке , а если бы папа подслушал, наверняка бы вспыхнул световой сигнал. Папа не
взламывал программу. Он не мог услышать ее голос. И мыслей Ахилла.
— Если тебя беспокоит цена, наши расценки...
Ахилл почти не задумываясь стер приложение, его сильно мутило.
Отец не шевелился.
Он теперь вообще мало двигался. Невеликий запас топлива в нем выгорел, и он
застыл где-то между горем и равнодушием. С падением церкви его яростный като-
лицизм обратился против него же, выжег его изнутри и оставил после себя пус-
тоту. К моменту смерти матери там даже печали почти не осталось. (Сбой лече-
ния, глухо сказал он, вернувшись из больницы. Активировались не те промоторы,
тело обратилось против собственных генов. Стало пожирать себя. Сделать было
ничего нельзя. Они подписывали отказ от претензий.)
Сейчас папа стоял в темном проеме и чуть покачивался — даже кулаки не сжи-
мал . Он уже много лет не поднимал руки на своих детей.
«Так чего же я боюсь? — удивился Ахилл, чувствуя, как желудок стягивается в
узел. — Он знает, знает. Я боюсь, что он знает».
У отца почти неуловимо растянулись уголки губ. Это была не улыбка и не ос-
кал . Вспоминая тот день, взрослый Ахилл Дежарден видел здесь своего рода по-
нимание , но тогда он понятия не имел, что это значит. Он знал лишь, что отец
повернулся и ушел в главную спальню, закрыл за собой дверь и никогда больше
не упоминал о том вечере.
В будущем Ахилл понял и то, что «ТераДруг», скорее всего, ему подыгрывал.
Целью программы было привлечение клиентов, а для этого не стоит тыкать их но-
сом в неприятные истины. Маркетинговая стратегия требует, чтобы клиенту было
приятно.
И все же это не значит, что программа непременно врала. Зачем, если правда
достигает той же цели? К тому же это было так рационально. Не грех, а дис-
функция . Термостат, выставленный с изъяном, хотя его вины в том и нет. Вся
жизнь — это машина, механизм, построенный из белков, нуклеиновых кислот и
электрических цепей. А когда это машины могли контролировать собственные
функции? Тогда, на заре суверенного Квебека, он познал свободу отпущения: Не-
виновен, всему причиной — ошибка в схеме проводки.
Но вот что странно.
Казалось бы, градус отвращения к себе должен был после этого хоть немного
понизиться.
Дом
больного
Джин Эриксон с Джулией Фридман живут в маленьком однопалубном пузыре в
двухстах метрах к юго-востоку от «Атлантиды». Хозяйством всегда занималась
Джулия: всем известно, что Джин недолюбливает замкнутые помещения. Дом для
него — подводный хребет, а пузырь — необходимое зло, существующее ради секса,
кормежки и тех случаев, когда его темные грезы оказываются недостаточно увле-
кательными .
Даже в таких случаях он воспринимает пузырь так же, как ловец жемчуга две-
сти лет назад воспринимал водолазный колокол: место, где изредка можно глот-
нуть воздуха, чтобы вернуться на глубину. Теперь, конечно, речь скорее о реа-
нимации .
Лени Кларк вылезает из шлюза и кладет ласты на знакомый, до смешного неуме-
стный здесь коврик. В главной комнате темновато даже для глаз рифтера: серые
сумерки размываются лишь яркими хроматическими шкалами на панели связи. Пах-
нет плесенью и металлом, чуть слабее — рвотой и дезинфекцией. Под ногами
булькает система жизнеобеспечения. Открытые люки зияют черными ртами: в кла-
довку, в гальюн, в спальный отсек. Где-то рядом попискивает электронный мет-
роном — кардиомонитор отсчитывает пульс.
Появляется Джулия Фридман.
— Он еще... ой. — Она сменила гидрокостюм на термохромный свитер с горлови-
ной «хомутом», почти скрывающей шрамы. Странно видеть глаза рифтера над воро-
том сухопутной одежды. — Привет, Лени.
— Привет. Как он?
— Нормально. — Развернувшись в люке, она прислоняется спиной к раме: напо-
ловину в темноте, наполовину в полумраке. А лицом обращается к темноте и к
человеку в ней. — Но бывает и лучше, я бы сказала. Он спит. Он сейчас много
спит.
— Удивляюсь, как ты сумела удержать его в доме.
— Да, он бы, наверно, даже сейчас предпочел бы остаться снаружи, но... ду-
маю, согласился ради меня. Я его попросила. — Фридман качает головой. — Слиш-
ком легко это вышло.
— Что?
— Его уговорить. — Она вздыхает. — Ты же знаешь, как ему нравится снаружи.
— А антибиотики, что дала Джерри, помогают?
— Наверное. Скорее всего. Трудно сказать, понимаешь? Как бы плохо ни было,
она всегда может сказать, что без них было бы еще хуже.
— Она так говорит?
— О, Джин, с тех пор, как вернулся, с ней не разговаривает. Он им не верит.
— Джулия смотрит в пол. — Винит ее во всем.
— в том, что ему плохо?
— Он думает, они с ним что-то сделали.
Кларк припоминает.
— Что именно он...
— Не знаю. Что-то. — Фридман поднимает взгляд, ее бронированные глаза на
миг встречаются с глазами Кларк и тут же уходят в сторону. — Уж слишком долго
не проходит, понимаешь? Для обычной инфекции. Как тебе кажется?
— Я сама не знаю, Джулия.
— Может, тут как-то вмешался Бетагемот. Осложнил состояние.
— Не знаю, бывает ли такое.
— Может, я теперь тоже заразилась. — Кажется, Фридман говорит сама с собой.
— То есть, я много с ним сижу...
— Можно проверить, если хочешь.
Фридман смотрит на нее.
— Ты ведь была инфицирована? Раньше.
— Только Бетагемотом, — подчеркивая разницу, поясняет Кларк. — Меня он не
убил. Я даже не заболела.
— А должна была бы, рано или поздно. Так?
— Если бы не модификация. Но я модифицирована. Как и все мы. — Она натужно
улыбается. — Мы же рифтеры, Джулия. Те еще поганцы, нас так просто не взять.
Он выдюжит, я уверена.
Этого мало, понимает Кларк. Все, что она может предложить Джулии, — вдох-
новляющий обман. От прикосновений благоразумно воздерживается: Фридман не вы-
носит физического контакта. Может, она и стерпела бы дружескую руку на своем
плече — однако допускает людей в личное пространство с большим разбором. В
этом, хотя и мало в чем еще, Кларк чувствует с ней родство. Каждая из них за-
мечает , как ежится другая, даже когда остальным не заметно.
Фридман через плечо оглядывается в темноту.
— Грейс говорила, ты помогла его оттуда вытащить.
Кларк пожимает плечами, немного удивляясь, что Нолан приписала заслугу ей.
— Я бы, знаешь, тоже не захотела там оставаться. Только... — Голос ее зами-
рает . В тишине вздыхает вентиляция пузыря.
— Только ты задумываешься, а не лучше ли было бы его не трогать? — угадыва-
ет Кларк.
— Да нет. Ну, может быть, отчасти. Не думаю, что доктор Седжер так уж пло-
ха , как говорят.
— Говорят? Кто?
— Джин и... Грейс.
— А. . .
— Просто... не знаю. Не знаю даже, хотел бы он, чтобы я была здесь? — Фрид-
ман горестно усмехается. — Я, в общем-то, не борец, Лени. Не то, что ты и. . .
когда меня пинают, я просто прогибаюсь.
— Если бы он захотел, мог бы остаться у Грейс, Джулия. А он с тобой.
Фридман с какой-то поспешностью смеется.
— О, нет, я не о том. — Все же слова Кларк ее чуточку взбодрили.
— в общем, — говорит Кларк, — я, пожалуй, оставлю вас вдвоем, ребята. Зашла
просто узнать, как у него дела.
— Я ему передам, — кивает Фридман. — Он будет рад.
— Конечно. О чем речь. — Она нагибается за своими ластами.
— И ты еще заходи, когда он придет в себя. Ему приятно будет. — Помолчав,
Джулия отворачивается — лица не видно за каштановыми кудряшками. — Мало кто
заходит, знаешь ли. Кроме Грейс. Салико еще недавно заходил.
Кларк пожимает плечами.
— Рифтеры — не мастера в общении. («Могла бы уже понять», — не добавляет
она) . Иногда до Джулии Фридман просто не доходит. Несмотря на свои шрамы и
все пережитое, она рифтер только по названию, вроде как почетный член, приня-
тый в закрытый круг ради мужа.
«Кстати, вопрос, а что здесь делаю я сама?» — осеняет Кларк.
— Мне кажется, они его иногда слишком серьезно воспринимают, — говорит
Фридман.
— Серьезно? — Кларк оглядывается на нее из шлюза. Пузырь почему-то вдруг
стал теснее.
— Ну, насчет. . . корпов. Я слышала, что Салико не совсем здоров, но ты же
знаешь Салико.
«Он думает, они с ним что-то сделали...»
— Я бы на этот счет не волновалась, — говорит Кларк. — Правда.
Она улыбается, вздыхая при мысли о своей дипломатичности.
С практикой утешительная ложь дается слишком легко.
С тех пор как она позволила Кевину себя иметь, прошло немало времени. Как
ни печально, он никогда этого толком не умел. Ему все давалось трудней, чем
большинству сверстников, а это, в общем-то, не редкость среди местных придон-
ников. Тот факт, что он выбрал объектом для тренировки такую фригидную суку,
как Лени Кларк, не упростил дела. Мужчина, боящийся прикосновений, и женщина
с отвращением к любому контакту. Если у них двоих и есть что-то общее, так
это терпение.
Она считает, что в долгу перед ним. И кроме того, хотела бы задать ему не-
сколько вопросов. Но сегодня он — гранитный член с маленьким мозговым придат-
ком. На фиг предварительные ласки: он врывается в нее с разгона, даже языком
не поработав, чтобы восполнить недостаток влаги. Фрикции болезненно натягива-
ют губы — она незаметно опускает руку и расправляет их. Уолш работает как на-
сос, дыхание со свистом прорывается сквозь звериный оскал, на глазах твердые
непроницаемые линзы. Они всегда скрывают глаза во время секса — тут верх взя-
ли вкусы Кларк, — но обычно у Уолша так все отражается на лице, что этого не
скрыть парой пленчатых скорлупок. А в этот раз не так. В этот раз за его на-
кладками скрывается нечто непонятное Кларк — нечто, сфокусированное на том
месте, которое она занимает в пространстве, а не на ней самой. От его грубых
толчков она сползает с матраса и стукается головой о голый металл палубы. Они
молча трахаются в застойном воздухе, среди сваленной техники. Кларк не знает,
что на него нашло. Впрочем, это приятная перемена, секса, настолько близкого
к честному и откровенному насилию, у нее не бывало много лет. Закрыв глаза,
она вспоминает Карла Актона.
Впрочем, по окончании она видит синяк на плече не у себя, а у Кевина — вен-
чик порванных капилляров окружает крошечный прокол на сгибе локтя.
— Это что? — Она приникает к ранке губами и водит языком по припухлости.
— А, это... Грейс у всех брала кровь на анализ.
Она вскидывает голову.
— Что?
— Она в этом не специалист. Только со второй попытки нашла вену. Ты бы ви-
дела Лайджа. Рука — будто на морского ежа напоролся.
— Зачем Грейс брала кровь?
— Ты что, не слышала? Лайдж что-то подхватил. И Салико плохо себя чувству-
ет, а он пару дней назад навещал Джина и Джулию.
— И Грейс думает...
— Чем бы ни наделили его корпы, оно заразно.
Кларк садится. Она полчаса пролежала нагишом на палубе, но холод ощутила
только сейчас.
— Грейс думает, корпы ему что-то подсадили?
— Это Джин так думает. Она проверит.
— Как? У нее же нет медицинской подготовки.
Уолш пожимает плечами:
— Чтобы управляться с медбазой, подготовка не нужна.
— Да она совсем тупая? — Кларк в недоумении мотает головой. — Даже если бы
на «Атлантиде» хотели нас заразить, они не так глупы, чтобы использовать ви-
рус из стандартного набора.
— Наверно, она думает, что начать следует с простого.
Что же такое у него в голосе?
— Ты ей веришь, — говорит Кларк.
— Ну, не сов...
— Джулия заболела?
— Пока нет.
— «Пока нет», Кевин. Джулия не отходила от Джина с тех пор, как его вытащи-
ли. Будь у него зараза, наверняка бы подцепила. А Салико их сколько раз наве-
щал? Один?
— Может, два.
— А Грейс? Я слышала, она постоянно там бывает. Она заболела?
— Она говорит, что принимает меры предос...
— Предосторожности! — фыркает Кларк. — Пощади мои уши! Что, на всем хребте
у меня одной еще не отказали лобные доли? Не забыл, как в прошлом году Абра
подцепил суперсиф? Чарли Гарсиа восемь месяцев не мог избавиться от аскариды
в кишках, но я что-то не припомню, чтобы в этом обвиняли корпов. Кевин, люди
болеют, даже здесь, внизу. Особенно здесь. Каждый второй из нас сгниет даже
раньше, чем получит шанс отуземиться.
И опять что-то проглядывает за непрозрачным блеском линз на глазах Уолша.
Что-то не слишком дружелюбное.
— Что еще? — вздыхает она.
— Это просто меры предосторожности. Не вижу, какой от них вред.
— Очень большой, если люди сделают выводы, не дожидаясь фактов.
На минуту Уолш замирает. Потом встает на ноги.
— Грейс как раз и пытается получить факты, — говорит он, шлепая босыми но-
гами через отсек. — А поспешные выводы делаешь ты.
«Ух ты, Кевви! — дивится Кларк. — Никак, у тебя хребет появился?»
Он подхватывает со стула гидрокожу. Сморщенная черная синтетика обнимает
его как любовница.
— За перепихон спасибо, — говорит он. — Мне пора.
Шаблон
Лабина она застает зависшим над его сигнальным резервуаром. Трубки, оптово-
локонный кабель и прочие детали — почти все уже неисправные, останки давно
разобранных систем — лентой обвивают экватор большого бака. Сейчас течения
слишком ленивы, чтобы вызвать свечение камня и аппаратуры, и свет дает только
налобный фонарь Лабина.
— Абра сказал, что ты здесь, — жужжит Кларк.
— Подержи панель, а?
Она берет у него маленький прибор.
— Я хотела с тобой поговорить.
— о чем? — Кажется, все его внимание отдано янтарному пузырьку полимера на
одной из проводящих трубок.
Кларк смещается так, чтобы оказаться в его поле зрения.
— Ходят идиотские слухи. Грейс внушает людям, будто Джерри подсадила Джину
какую-то заразу.
Вокодер Лабина тикает, механически передавая его «м-м-м».
— У нее насчет корпов давно фитиль в заднице, но никто ее всерьез не прини-
мает . По крайней мере, не принимал...
— Вот оно что... — Лабин постукивает по клапану.
— Что?
— Трещина в резиновой прокладке вокруг термостата. Оттого и замыкания.
— Кен, послушай меня!
Он смотрит на нее, ждет.
— Что-то меняется. Грейс никогда так далеко не заходила, ты же знаешь.
— Я с ней никогда лбами не сталкивался, — жужжит Лабин.
— Она всегда была одна против всего мира. Но эта зараза, которую подцепил
Джин, все переменила. Ее стали слушать. Это опасно.
— Для корпов.
— Для всех нас. Не ты ли меня предупреждал, на что способны корпы, если
станут действовать дружно? Не ты ли сказал, что...
«... возможно, придется действовать на упреждение ...»
Желудок у Кларк куда-то проваливается.
— Кен, — медленно жужжит она, — ты же понимаешь, что Грейс просто чокнутая,
нет?
Он не торопится с ответом. Кларк надоедает ждать.
— Ну, правда, ты бы ее только послушал. Для нее война как будто и не конча-
лась . Стоит кому-то чихнуть, она уже ищет биологическое оружие.
Силуэт Лабина за фонарем чуть сдвигается: кажется, он пожал плечами.
— Есть несколько любопытных совпадений, — говорит он. — Джин попадает в
«Атлантиду» с тяжелым ранением. Джерри оперирует его в медотсеке, за которым
мы не можем вести полноценного наблюдения, и помещает в карантин.
— Карантин — это из-за Бетагемота, — напоминает Кларк.
— Как ты сама не раз отмечала, к Бетагемоту мы все иммунны. Удивляюсь, как
это ты не усомнилась в ее оправданиях. — Видя, что Кларк молчит, он продолжа-
ет : — Джина отпускают с «побочной инфекцией», которая не распознается нашим
оборудованием и пока не поддается лечению.
— Но ты же там был, Кен. Джерри не хотела выпускать Джина из карантина. За
это Дейл ее и исколошматил. Изоляция нулевого пациента — не самый дальновид-
ный вариант, если тебе надо распространить чуму.
— Полагаю, — жужжит в ответ Лабин, — на это Грейс скажет: они предвидели,
что мы все равно его вытащим, и устроили представление, как раз и рассчиты-
вая, что кто-нибудь потом за них вступится.
— То есть они для того его заперли, чтобы освободить? — Кларк с намеком по-
глядывает на электролизный приемник Лабина. — Тебе там кислорода хватает,
Кен?
— Я просто излагаю, как могла бы рассуждать Грейс.
— Довольно извращенный ход мысли, даже для... — До нее доходит. — То есть
она и в самом деле так говорит?
Фонарик у него на лбу слегка покачивается.
— Так слухи до тебя дошли. Ты все уже знаешь. — Она качает головой, прези-
рая себя. — А я еще надеялась тебя раскачать.
— Я всегда готов тебя выслушать.
— Мог бы сделать кое-что еще. То есть я понимаю, что ты не хочешь лезть в
эти дела, но Грейс же охреневшая психопатка. Она лезет в драку, а кого затя-
нет в струю, ей плевать.
Лабин неподвижен, непроницаем.
— Я ожидал от тебя большего к ней сочувствия.
— Это как понимать?
— Никак, — жужжит он, выдержав паузу. — Но, что бы ты ни думала о поведении
Грейс, ее опасения не совсем беспочвенны.
— Брось, Кен. Война окончена. — Она принимает его молчание за согласие. — И
зачем бы корпам начинать ее сызнова?
— Затем, что они проиграли.
— Это уже древняя история
— Было время, когда ты считала себя пострадавшей стороной, — напоминает он.
— Сколько крови пришлось пролить, прежде чем ты сочла, что расквиталась до
конца?
Его металлический, такой холодный голос вдруг оказывается совсем рядом,
звучит прямо у нее в голове.
— Я... была неправа, — помедлив, говорит Кларк.
— Это тебя не остановило. — Он отворачивается к своей установке.
— Кен, — зовет она.
Он смотрит на нее.
— Это же чушь. Сплошные «если». Сто к одному, что Джин просто подцепил что-
то от покусавшей его рыбины.
— Предположим.
— Кто сказал, что здесь не болтаются сотни зловредных микробов, которых по-
ка не обнаружили? Несколько лет назад и про Бетагемот никто не слышал.
— Это я помню.
— Значит, нельзя допускать эскалации. Пока нет никаких доказательств.
В отблеске фонаря его глаза светятся желтовато-белым.
— Если насчет доказательств ты серьезно, то могла бы сама их раздобыть.
— Как?
Он постукивает себя по груди слева. Там, где расположены имплантаты. Кларк
холодеет.
— Нет.
— Если Седжер что-то скрывает, ты об этом узнаешь.
— Она может скрывать что угодно и от кого угодно. И так не докажешь, что
именно она скрывает.
— Заодно бы и выяснила, что на душе у Нолан, раз уж тебя так волнуют ее мо-
тивы .
— Ее мотивы мне известны. Ни к чему гробить химию собственного мозга, чтоб
в них увериться.
— С точки зрения медицины риск минимальный, — напоминает он.
— Не в том дело. Это ничего не докажет. Ты же знаешь, Лабин, что конкретные
мысли читать невозможно.
— Тебе бы и не пришлось. Достаточно считать чувство вины...
— Нет, я сказала!
— Тогда не знаю, что тебе посоветовать. — Он снова отворачивается. В луче
фонаря трубы резервуара похожи на крошечную контрастную модель опрокинутого
на бок города. Под пальцами Лабина с шипением вспыхивает крошечное солнце, на
секунду Кларк слепнет. К тому времени, как ее линзы приспосабливаются, по-
верхность бака уже вся освещена. Лучи преломляются в воде, та мерцает, словно
марево в жаркий день; будь глубина меньше, уже взорвалась бы паром.
— Есть еще один способ, — жужжит она. Лабин выключает горелку паяльника.
— Есть. — Он оборачивается к ней. — Но я бы не слишком на него надеялся.
Давным-давно, когда трейлерный парк только собирался, кто-то додумался пре-
вратить один пузырь в большую столовую: ряд циркуляторов, пара разделочных
столов для отважных и несколько складных столиков, в обдуманном беспорядке
разбросанных по палубе. Предполагалось, что все это создаст эффект кафе под
открытым небом. На деле получилось нечто вроде кладовки, куда сваливают на
зиму мебель.
Но кое-что прижилось — сад. За это время он покрыл половину внешней палубы:
кучка вьющихся растений освещена лампами-подпорками с солнечным спектром —
палки похожи на скрытые в листве светящиеся бамбучины. Это даже не гидропони-
ка, маленькие джунгли произрастают в ящиках с сочной темной землей — на самом
деле это диатомовый ил с органическими добавками. Почва была довольно скуд-
ной, но на ней теперь отложились слои компоста, беспорядочно распространив-
шись поверх обшивки.
Из всех атмосферных пузырей на хребте здесь самый приятный запах. Кларк
распахивает люк и глубоко вдыхает. В этом вдохе удовольствия только половина:
вторая половина — решимость. Грейс Нолан смотрит на нее с дальнего конца оа-
зиса, где подвязывала плети чего-то, что до вмешательства генетиков могло
быть стручковым горохом.
Однако под непрозрачными глазами Нолан играет улыбка.
— Привет, Лени!
— Привет, Грейс. Думаю, хорошо бы нам поговорить.
Нолан забрасывает в рот стручок: гладкая черная амфибия кормится в пышной
зелени древнего болота. Жует она, пожалуй, дольше, чем необходимо.
— Насчет...
— Насчет «Атлантиды». И твоих анализов крови. — Кларк переводит дыхание. —
И твоих претензий ко мне.
— Боже мой, — возражает Нолан, — к тебе никаких претензий, Лен. Бывает, лю-
ди ссорятся. Ничего особенного, не принимай все так всерьез.
— Ну и ладно. Тогда поговорим о Джине.
— Конечно. — Нолан, выпрямившись, снимает с переборки стул и раскладывает
его. — А заодно про Сала, Лайджа и Лани.
«Уже и Лани?»
— Ты думаешь, виноваты корпы?
Нолан пожимает плечами.
— Я этого не скрываю.
— И откуда такие выводы? Нашла что-нибудь в крови?
— Мы пока собираем образцы. Кстати, Лизбет обосновалась в медпузыре, можешь
сдать анализы. По-моему, стоит.
— А если ничего не найдете? — интересуется Кларк.
— Я и не жду, что найдем. Седжер не так глупа, чтобы оставлять следы. Но
как знать.
— Ты же понимаешь, что корпы, возможно, ни при чем?
Нолан откидывается на спинку стула, потягивается.
— Милочка, сказать не могу, как я удивлена, что слышу такое от тебя.
— Тогда дай мне доказательства.
Нолан с улыбкой качает головой:
— Вот тебе для примера. Скажем, ты плаваешь в водах, где водятся акулы.
Здоровенные гадины с треугольными плавниками кишат вокруг, разглядывают тебя,
и ты понимаешь: не рвут на части только потому, что у тебя наготове дубинка,
а они знают, что дубинка вытворяет с такими рыбинами. Так что они держатся на
расстоянии, но от этого ненавидят тебя только сильнее, правда ведь? За то,
что ты уже убивала им подобных. Эти акулы далеко не глупы, но очень злопамят-
ны. Так вот, плывешь ты себе среди всех этих холодных мертвенных глаз и зу-
бов , и видишь... скажем, Кена. Вернее, то, что от него осталось. Обрывок киш-
ки, половина лица, опознавательная нашивка, плавающая среди акульих туш. Ну и
как, Лен, сойдет такое за доказательство? Или ты скажешь: нет, это ничего не
доказывает, я ведь не видела, что тут произошло. Скажешь: давайте воздержимся
от поспешных выводов...
— Довольно паршивая аналогия, — тихо говорит Кларк.
— А по-моему, охрененная.
— И что ты намерена делать?
— Я могу сказать, чего я делать не намерена, — заверяет ее Нолан. — Не ста-
ну сидеть спокойно, полагаясь на доброту корпов, пока все мои друзья превра-
щаются в падаль.
— Тебя об этом кто-нибудь просит?
— Пока нет. Но думаю, скоро попросят.
Кларк вздыхает.
— Грейс, я прошу, ради нас всех...
— Да пошла ты на хрен, — резко перебивает Нолан. — Тебе насрать на всех
нас.
Словно кто-то щелкнул выключателем. Кларк изумлено разглядывает Нолан, та
отвечает ей пустыми глазами, дрожа в припадке ярости.
— Хочешь знать, какие у меня к тебе претензии? Ты нас продала! Мы уже почти
покончили с этими акулами. Могли выпустить им кишки через глотки, а ты нас
остановила, дрянь поганая.
— Грейс, — пытается вставить Кларк, — я понимаю твои чув...
— Ни хрена! Ни хрена ты не понимаешь!
«Что же с тобой делали, — гадает Кларк, — как довели до такого?»
— Мне тоже от них досталось, — тихо говорит она.
— Еще бы. И ты-то за себя рассчиталась, скажешь, нет? И, поправь меня, если
я ошибаюсь, ты попутно угробила немало невинных, разве не так? Плевать тебе
на них было. И, может, тебя это не особо волнует, но из-за твоего крестового
похода погибло и немало наших, рыбоголовых. На них тебе тоже было плевать,
лишь бы отвесить пинка кому следует. Отлично. Ты расквиталась. А остальные-то
еще ждут, а? Мы ведь не собираемся даже скашивать миллионы невинных, мы всего
лишь хотим добраться до уродов, которые нас натянули, — и вот именно ты при-
ползаешь сюда, как цепная собачка Патриции Роуэн, чтобы сказать, что я не в
своем праве? — Нолан с отвращением мотает головой. — Я не понимаю, как мы по-
зволили остановить себя тогда, и уж точно не позволю остановить теперь.
Ненависть расходится от нее инфракрасными лучами, Кларк даже удивляется,
что листья не чернеют, не загораются от этого жара.
— Я думала, мы сумеем разобраться, потому и пришла, — говорит она.
— Ты потому пришла, что проигрываешь, и знаешь это.
От этих слов под ложечкой у Кларк стягивается холодный узелок гнева.
— Ты никогда и не думала ни в чем разбираться, — рычит Нолан. — Ты всегда
стояла на своем. Я — Мадонна Разрушения, я русалка долбаного Апокалипсиса, я
буду стоять в сторонке и устанавливать правила. Только на этот раз оползень
пошел не в ту сторону, милашка, и ты струсила. Испугалась меня. Так хоть из-
бавь меня от дерьмового альтруизма и дипломатии. Ты просто пытаешься усидеть
на своем жестяном трончике, пока он не развалился. Хорошо поговорили!
Она хватает ласты и вываливается в шлюз.
Портрет
садиста
в юности
Ахилл Дежарден не помнил, когда у него в последний раз был секс по взаимно-
му согласию. Зато помнил, когда впервые от такого отказался
Это было в 2046-м, он тогда как раз спас Средиземное море. Во всяком слу-
чае, так все подавало радио Нт АмПацифика. На самом деле он всего лишь вычис-
лил существование странного аттрактора в Кадисском заливе — мелкого устойчи-
вого завихрения, которое не додумался поискать никто другой. Симуляции пока-
зывали, что оно достаточно мало, чтобы поддаваться гасителям альбедо1; итого-
вый эффект должен был распространиться на Гибралтарский пролив и — если цифры
не врали — мог оттянуть гибель Средиземного на доброе десятилетие. Или до но-
вого отказа Гольфстрима. Это было не спасение, а отсрочка приговора, но УЛН
сейчас требовалось хоть что-то, чтобы загладить фиаско с Балтикой. К тому же
дальше, чем на десять лет вперед, никто и не загадывал.
Так что Ахилл Дежарден на время попал в звезды. Даже Лерцман чуть не месяц
изображал к нему симпатию и уверял, что подаст на досрочное повышение, как
только закончатся проверки. Если, мол, у Дежардена в прошлом не обнаружится
груды зарезанных младенцев, он получит свое еще до Хэллоуина. А может, черт
побери, получит, даже и груда младенцев не помешает. Проверки прошлого были
для высших эшелонов Патруля не более чем бессмысленным ритуалом: ты мог быть
серийным убийцей — это ничего не меняло для того, у кого в мозгу бормочет
Трип Вины. Все равно ты останешься рабом Общего Блага.
Ее звали Аврора. У нее была прическа под зебру — по тогдашней моде — и це-
лая батарея умилительно безвкусных шрамов, стилизованных под клейма беженцев.
Они сошлись на какой-то вечеринке, устроенной для УЛН Евро-африканской ас-
самблеей. Их аксессуары вынюхали ауры друг друга и подтвердили взаимный инте-
рес (в те времена он еще что-то значил) , а чипы обменялись справками о со-
стоянии здоровья (а это и тогда ничего не значило).
Так вот, они ушли с вечеринки, спустились на триста метров со стратосферы
УЛН на улицы Садбери — и еще на пятьдесят, в подземное чрево «Реактора Пике-
ринга», где софт был гарантированно защищен от взлома и давал вдвое большую
выборку по ЗППП, чем обычно. Кровь они сдали после симпатичной пары р-
отборщиков, которая распалась у них на глазах, когда один анализ выдал поло-
жительный результат на какую-то экзотическую трематоду в мочеполовых путях.
Дежарден тогда еще не был знаком с хитроумными веществами, которые заполо-
нили его сосуды в последующие годы, и мог без вреда для себя применять всяче-
ские тропы и стимуляторы настроения. Так что они с Авророй, пока проверялась
кровь, взяли столик в соседнем баре и погладили психотропную жабку, ползавшую
в террариуме под столешницей.
Из большого подземного резервуара, в который был погружен «Реактор», сквозь
плексигласовые стены сочился смутный зеленоватый свет — подделка под старин-
ное хранилище ядерных отходов. Через несколько минут на их столик порхнула
одна из местных бабочек. Ее прозрачные крылышки поблескивали данными спектра
— зелеными по всей длине волны.
— Я же говорила, — сказала Аврора и чмокнула его в нос.
Секс-кабинки в «Реакторе» сдавались поминутно. Они вскладчину оплатили пять
часов.
Он имел ее в реальности и в воображении. В реальности — виртуозный и нежный
любовник. Он трогал соски языком, зубами — ни-ни. Оставлял цепочки поцелуев
от горла до вагины, нежно исследовал каждое отверстие, его дыхание срывалось
от сдержанной страсти. Каждое движение обдумано, каждый сигнал отчетлив: Он
скорее умрет, чем причинит этой женщине боль.
В воображении он разрывал ее на части. В воображении ласки не было: он хле-
стал ее наотмашь, так что голова у нее моталась, чуть не отрываясь от шеи. В
воображении она вопила. В воображении он избивал ее до тех пор, пока она не
Альбедо (от лат. albus «белый») — характеристика диффузной отражательной способно-
сти поверхности.
перестала вздрагивать от удара кнута.
Она тем временем бормотала и сладко вздыхала, и заметила между делом, что
он явно преклоняется перед женщинами, и как это приятно после грубиянов, ко-
торым только бы воткнуть, и она, мол, даже не знает, достойна ли такого пре-
клонения. Дежарден мысленно похлопал себя по плечу. Он не спрашивал о проис-
хождении крошечных шрамов у нее на спине, о предательских розовых полосках от
применения анаболиков направленного действия. Как видно, Аврора находила, ку-
да потратить прокачанное здоровье. Может быть, только недавно избавилась от
отношений, в которых была жертвой. Может, он был ее спасителем.
Тем лучше. Он вообразил себя одним из прошлых партнеров: тех, что избивали
ее.
— Да ну на хрен, — сказала она на четвертом часу. — Просто врежь мне.
Он застыл в ужасе, гадая, что его выдало: язык тела или телепатия, или это
просто удачная догадка.
— Что?
— Ты такой нежный, — сказала ему Аврора. — Давай играть погрубее.
— Ты не... — Ему пришлось подавить изумленный смешок. — Ты о чем вообще?
— Ты что так опешил? — улыбнулась она. — Неужели ни разу женщину не лупил?
«Это ведь были намеки, — понял он. — Она жаловалась». А он, Ахилл Дежарден,
непревзойденный вычислитель закономерностей, мастер по выхватыванию сигнала
из шума помех, ничегошеньки не заметил.
— Удушение меня заводит, — продолжила она. — Что это твой ремень лежит без
дела? . .
Все было, как в его мечтах, за которые он себя ненавидел. Словно ожили его
самые постыдные фантазии. Это было превосходно. «Ах ты, сука, ты этого хоте-
ла, да? Вот как раз от меня и получишь».
Только не получила. Ахилл Дежарден вдруг обмяк, как доллар.
— Ты серьезно? — спросил он в надежде, что она не заметит, и понимая, что
уже заметила. — То есть... ты хочешь, чтобы я сделал тебе больно?
— Да Ахилл у нас герой! — Она насмешливо покосилась на него. — Опыта мало-
вато , да?
— Хорошо, — сказал он, невольно оправдываясь. — Только...
— Это же просто игра, детка. Ничего радикального. Я же не прошу меня уби-
вать .
«Очень жаль».
Впрочем, мысленная бравада не одурачила его ни на миг. Ахилл Дежарден, тай-
ный садист, вдруг до смерти перепугался.
— Ты про игры? — сказал он. — Шелковые веревки, стоп-слова и все такое?
Она покачала головой.
— Я, — терпеливо объяснила она, — хочу, чтобы у меня кровь текла. Хочу бо-
ли. Я хочу, чтобы ты сделал мне больно, любовничек.
«Да что со мной? — гадал он. — Я же именно о такой всегда мечтал. Какая
счастливая случайность!»
И миг спустя подумал: «Если это случайность».
Он, что ни говори, находился на переломе судьбы. Шли проверки его прошлого,
проводилась оценка рисков. Где-то под ковром система решала, можно ли дове-
рить Дежардену ежедневно определять судьбы миллионов. Конечно, его тайна уже
была им известна: механики заглянули ему в голову и наверняка заметили отсут-
ствующую или поврежденную проводку. Возможно, это проверка: способен ли он
контролировать себя. Возможно, Трип работает не так надежно, как ему внушали,
и особенно подлые нейроны его подтачивают, а изначальная порочность представ-
ляет собой потенциальную ловушку. . . Или все намного проще. Может, они просто
не рискнут вкладывать в раскрутку героя, чьи неконтролируемые склонности пуб-
лика может счесть... неприятными.
Аврора скривила губы и обнажила спину.
— Давай, детка, поработай со мной.
Она была блеском в глазах всех его прежних партнерш, той мигающей искоркой,
которая словно говорила: «Поберегись, ты, поганый извращенец. Малейшая оплош-
ность , и с тобой покончено!». Она была шестилетней Пенни, обещающей никому не
говорить, несмотря на кровь и переломы. Она была отцом, стоящим в темном
дверном проеме с непроницаемым взглядом, словно говорившим: «Мне про тебя
кое-что известно, сын, и ты никогда не узнаешь, что это».
— Рори, — осторожно заговорил Дежарден, — ты насчет этого ни к кому не об-
ращалась?
— Все время обращаюсь. — Она улыбалась, но в голосе зазвенело напряжение.
— Нет, я хотел сказать... понимаешь...
— К специалистам. — Улыбка пропала. — К какому-нибудь мозгоправу, который
за мои же денежки скажет мне, что я сама себя не понимаю, что все это просто
заниженная самооценка и что мой отец насиловал меня в доречевой период. — Она
потянулась за одеждой. — Нет, Ахилл, не обращалась. Я предпочитаю проводить
время с теми, кто меня принимает как есть, а не с тупыми мудаками, которые
пытаются превратить меня во что-нибудь другое. — Она натянула трусики. — На-
верно, на официальных приемах такие больше не попадаются.
— Ты могла бы остаться, — попытался он.
— Просто это было так неожиданно, — попытался он.
— Понимаешь, в этом есть какое-то неуважение, — попытался он.
Аврора вздохнула.
— Детка, если бы ты меня уважал, то признал бы за мной право самой решать,
что мне нравится.
— Но мне нравишься ты сама, — ляпнул он, низвергаясь в пламени и дыме. —
Как же мне делать тебе больно, если...
— Эй, ты думаешь, мне нравилось все, что я для тебя делала?
Она оставила его стоять с обвисшим пенисом, пятьюдесятью минутами до конца
аренды кабинки и убийственным, унизительным сознанием, что он навсегда в ло-
вушке собственного притворства.
«Я никогда не дам этому воли, — понял он. — Как бы мне ни хотелось, кто бы
меня ни просил, как бы это ни было безопасно. Я никогда не буду уверен, что
где-нибудь не прячется шпион, что это не ловушка. Я всю жизнь буду скрывать-
ся , потому что открыться страшно до ужаса».
Отец мог бы гордиться. Сын вырос добрым католиком.
Но Ахилл Дежарден знал толк в искусстве приспособления. Выходя из кабинки —
пристыженный и одинокий, — он уже начал восстанавливать защитные барьеры.
Возможно, оно и к лучшему. Как-никак, от биологии никуда не денешься: секс —
это насилие, причем буквально, до уровня нейронов. Трах или драка — работают
одни и те же синапсы, тот же стимул причинять насилие и подчинять. Каким бы
нежным ты ни был внешне, как бы ни притворялся. Даже если ты действуешь ис-
ключительно с согласия партнера, все равно это не более чем насилие над сдав-
шейся жертвой.
«Если я делаю все это, а любви не имею, то я — медь звенящая»1, — подумал
он.
Он сознавал это основами мозга, глубинами своего «оно». Садизм записан в
строении тела, а секс... секс мало того, что насилие, он — сплошное неуваже-
ние . И кому нужно обращаться так с другим человеческим существом в середине
двадцать первого века? Никто не вправе так поступать, тем более — чудовища с
неисправными выключателями. У него дома имелся сенсориум, позволявший вопло-
1
«Если я говорю языками человеческими и ангельскими, а любви не имею, то я — медь
звенящая или кимвал звучащий» (1Кор 13:1).
тить любое плотское желание, предоставляя ему виртуальных жертв в таком высо-
ком разрешении, что получалось одурачить даже его.
Были и другие преимущества. Не понадобятся больше сложные ритуалы ухажива-
ния, в которых он вечно путался. Не придется опасаться инфекций и неуклюже
выставлять анализы чем-то вроде предварительных ласк. И не будет больше этого
жестокого блеска в глазах жертвы, которая, может быть, догадалась.
Он во всем разобрался. Черт, он нашел новый смысл жизни.
Отныне и впредь Ахилл Дежарден будет цивилизованным человеком. Он обратит
свои порочные страсти на машину, а не на живую плоть — и тем самым спасет се-
бя от целой прорвы неловкостей. С Авророй все обернулось к лучшему — он чуть
не попался, но вовремя вывернулся. Вот уж у кого точно все провода в голове
перепутаны. Центры боли и удовольствия — все вперемешку.
Лучше не связываться с такими психами.
Пожарные
учения
Она просыпается где-то в море.
Непонятно, что привело ее в себя — вспоминается мягкий толчок, словно кто-
то осторожно будил ее, — но сейчас она совершенно одна. Того и добивалась.
Могла бы заснуть где-нибудь в трейлерном парке, но ей нужно было одиночество.
Потому она проплыла мимо «Атлантиды», мимо пузырей и генераторов, мимо хреб-
тов и расщелин, когтивших округу. Наконец добралась сюда, к отдаленному усту-
пу из пемзы и полиметаллических руд, и уснула с открытыми глазами.
А теперь что-то ее разбудило, и она не могла сориентироваться.
Она снимает с бедра сонарный пистолет и обводит окружающую тьму. Через не-
сколько секунд возвращается смазанное множественное эхо с левого края. Прице-
лившись в ту сторону, она стреляет снова. В самом центре оказывается «Атлан-
тида» с ее пригородами.
И еще более твердое и плотное эхо. Уже рядом — и приближается.
Идет не на перехват. Еще несколько импульсов показывают, что вектор движе-
ния минует ее справа. Тот, кто приближается, о ней не знает — или не знал,
пока она не применила сонар.
Учитывая отсутствие «кальмара», он движется на удивление быстро. Из любо-
пытства Кларк направляется ему наперерез. Налобный фонарь она притушила, све-
та едва хватает, чтобы отличить донный грунт от морской воды. Ил облаками
поднимается вокруг. Изредка попадаются камушки и хрупкие морские звезды, лишь
подчеркивая однообразие.
Головная волна настигает за миг до самого объекта. В бок ей врезается пле-
чо, откидывает ко дну. Вздымается ил. Ласт шлепает Кларк по лицу — она всле-
пую шарит руками и хватается за чье-то предплечье.
— Да какого хрена!
Рука вырывается, но брань, видимо, оказала свое действие. По крайней мере,
он больше не брыкается. Облака мути кружатся скорее по инерции.
— Кто... — Звук грубый, скрежещущий, даже для вокодера.
— Это Лени. — Она прибавляет яркости в фонаре, и яркий туман из миллиардов
частиц взвеси слепит ее. Отплыв в чистую воду, Кларк направляет луч на дно.
Там, внизу, что-то шевелится.
— Че-орт! Свет выключи...
— Прости. — Она притемняет фонарь. — Рама, это ты?
Со дна поднимается Бхандери. Механический шепот:
— Лени... привет.
Наверно, повезло, что он ее еще помнит. Черт, что он вообще еще может гово-
рить . Когда перестаешь бывать на станции, не просто гниет кожа. Не просто
размягчаются кости. У отуземившехюся рифтера стирается кора головного мозга.
Если позволить бездне слишком долго вглядываться в тебя, все метки цивилиза-
ции тают, как лед в проточной воде. Кларк представляет, как со временем раз-
глаживаются извилины мозга, возвращаясь к примитивному состоянию рыбы, более
подходящему к подобной среде.
Рама Бхандери еще не так далеко ушел. Он даже иногда появляется внутри.
— К чему такая спешка, что накрылось? — жужжит ему Кларк, не слишком наде-
ясь на ответ.
Но ответ она получает.
— Накры... дофамином, наверно... эпи...
Через секунду до нее доходит. Дофаминовый приход, накрыло его. Неужели он
еще настолько человек, что способен на каламбуры?
— Нет, Рама, я хотела спросить, куда ты спешишь?
Он зависает рядом с ней черным призраком, еле видимый в смутном мерцании
фонаря.
— А...а... я не... — голос затихает.
— Бабах, — снова начинает он после паузы. — Взрыв. Сли-ишком ярко.
Толчок, вспоминает Кларк. Такой сильный, что разбудил ее.
— Что взорвали? И кто?
— Ты настоящая? — рассеянно спрашивает он. — Я. . . думал, ты гистаминовый
глюк.
— Я Лени, Рама. Настоящая. Что взорвалось?
— Или ацетилхолино... — Он поводит ладонями перед лицом. — Только меня не
ломает...
Бесполезно.
— . . . она мне больше не нравится, — тихо жужжит Бхандери. — А он гонялся за
мной...
У Кларк перехватывает горло. Она придвигается к нему.
— Кто? Рама, что...
— Уйди. . . — скрежещет он. — Моя. . . территория.
— Прости, я...
Бхандери разворачивается и плывет прочь. Кларк, подавшись было следом, ос-
танавливается , потому что вспоминает: есть другой способ.
Она усиливает свет фонаря. Под ней еще висит мутная туча — над самым дном.
В такой плотной ленивой воде она продержится много часов.
Как и ведущие к ней следы.
Один — ее собственный: узкая полоска ила, взбитая ее движением с восточного
направления. Другой след отходит от него под углом 345 градусов. Кларк дви-
жется по нему.
Так она попадет не к «Атлантиде», скоро соображает она. След Бхандери ухо-
дит левее, мимо юго-западного крыла комплекса. Там, насколько она помнит,
смотреть не на что. Разве что на «поленницу» — склад частей, сброшенных в
расчете на продолжение строительства, когда корпы только появились здесь. И
правда, вода впереди светлеет. Кларк приглушает свой фонарь и сонарит яркое
пятно впереди. Возвращается жесткое геометрическое эхо объектов, заметно пре-
вышающих рост человека.
Она устремляется вперед. Размытое пятно превращается в четыре точечных ис-
точника света по углам «поленницы». Штабеля пластиковых и биостальных пластин
лежат на поддонах в пределах освещенного участка. Изогнутые запчасти для об-
шивки корпуса торчат из ила зарослями устриц. В туманной дали виднеются боль-
шие тени баков, теплообменников, кожухов аварийного реактора, которые так и
не пустили в ход.
А даль действительно туманная, понимает Кларк. Гораздо мутнее обычного.
Погрузившись в водяной столб, она зависает над индустриальным пейзажем.
Что-то вроде мягкой черной стены перегораживает свет дальнего фонаря. Она
этого ожидала после разговора с Бхандери. И вот перед ней молчаливое подтвер-
ждение: огромное облако ила, взбитое со дна и невесомо зависшее после недав-
него взрыва.
Естественно, корпы успели запастись и взрывными зарядами...
Что-то щекочет ей уголок глаза — какой-то маленький беспорядок среди упоря-
доченного хаоса внизу. Два куска обшивки стащили с поддонов и уложили прямо в
ил. Их поверхность покрыта угревой сыпью. Кларк выгибается, чтобы рассмотреть
их вблизи. Нет, это не безобидные хлопья ила и не молодая колония бентосных
беспозвоночных. Это дырки в трехсантиметровой цельной биостали. Края отвер-
стий гладкие — проплавлены мощным источником тепла и мгновенно застыли.
Угольные ожоги вокруг дыр — как синяки вокруг глаз.
Кларк холодеет.
Кто-то вооружился для окончательной разборки.
Семейные
ценности
Якоб и Ютта Хольцбринки с самого основания «Атлантиды» держались особняком.
Так было не всегда. Прежде, на поверхности, они даже по меркам корпов слыли
яркими оригиналами. Их, кажется, забавлял тот архаический контраст, который
они составляли миру в целом: они вели историю отношений от прошлого тысячеле-
тия, а поженились так давно, что бракосочетание состоялось в церкви! Ютта да-
же взяла фамилию мужа.
В старину женщины, как помнилось Роуэн, иногда так поступали. Жертвовали
кусочками собственной личности во благо Патриархата, или как там оно называ-
лось .
Пара была старомодной и тем гордилась. Когда эти двое появлялись на публи-
ке , то обязательно вместе — и очень выделялись.
Разумеется, на «Атлантиде» никакой публики не существовало. Отныне публика
была предоставлена самой себе. На станции же с самого начала собрались сливки
общества: самые влиятельные люди и еще рабочие пчелки, которые заботились о
них в самых недрах улья.
Якоб с Юттой почти перестали выходить. Побег изменил их. Он, конечно, изме-
нил каждого: посрамил могущественных, ткнув их носом в собственные промахи,
хотя они, черт побери, все равно сделали что могли, адаптировались даже к
Судному дню, вовремя закупили спасательные шлюпки и первыми прыгнули на борт.
В те дни простое выживание составляло профессиональную гордость. Однако
Хольцбринки не позволяли себе даже этого унылого самооправдания. Бетагемот не
коснулся их плоти, не затронул ни единой частицы, и все же, казалось, сделал
их меньше ростом.
Большую часть времени они проводили в своем номере, подключенному к вирту-
альной среде, куда более привлекательной, чем реальные помещения «Атлантиды».
Они, конечно, выходили к столу — частное производство продуктов питания оста-
лось в прошлом после того, как рифтеры конфисковали «свою долю» ресурсов, —
но все равно, наполнив подносы пищей из циркуляторов и гидропонных отсеков,
тотчас возвращались к себе. Мелкая, безобидная странность — желание держаться
подальше от себе подобных. Патриция Роуэн не обращала на нее внимания до того
дня, когда в гроте связи Кен Лабин, думая над разгадкой, не произнес: «А если
это рыбы? Может, его к нам подвезли? А личинки в планктоне?»
И Джерри Седжер, недовольная попыткой этого убийцы-перебежчика изображать
из себя глубокого мыслителя, отмахнулась, как от ребенка: «Если он способен
распространяться с планктоном, зачем было так долго ждать? Он бы захватил
весь мир сотни миллионов лет назад».
Может, и захватил бы, размышляла теперь Роуэн.
Хольцбринки поднялись на фармацевтике — их карьера началась до расцвета
генной инженерии. Конечно, они старались шагать в ногу со временем. Когда на
грани веков была открыта первая геотермальная экосистема, предыдущее поколе-
ние Хольцбринков влезло и туда: радовались новым надцарствам, просеивали кла-
дограммы неизвестных видов — новые микроорганизмы, новые энзимы, выживающие
при температурах, убийственных для любой формы жизни. Они каталогизировали
механизмы клеток, лениво тикающих на многокилометровой глубине, таких медли-
тельных, что их последнее деление относилось ко временам французской Револю-
ции. Они перестраивали редуцентов серы, задыхавшихся в кислороде, приспосаб-
ливая их пожирать нефтяные отходы и излечивать новые виды рака. Поговаривали,
что половина патентов на археобактерии принадлежит империи Хольцбринков.
Сейчас Патриция Роуэн сидит напротив Якоба и Ютты у них в гостиной и гада-
ет , что еще они могли запатентовать в последние дни на суше.
— Вы, конечно, слышали новости, — говорит она. — Джерри только что подтвер-
дила: Бетагемот добрался до Невозможного озера.
Якоб кивает по-птичьи: не только головой, но и плечами. Но на словах осто-
рожно возражает:
— Я так не думаю. Видел статистику. Слишком соленое. — Он облизывает губы,
смотрит в пол. — Бетагемоту такое не нравится.
Ютта успокаивает его, погладив по колену
Он очень дряхл, все его победы в прошлом. Он слишком давно родился, оказал-
ся слишком стар для вечной молодости. К тому времени, как стала возможной та-
кая перестройка — удаление всех дефектных нуклеотидных пар, усиление всех те-
ломер — его тело уже износилось от полувекового использования. Вступив в игру
так поздно, мало что можно исправить.
Роуэн мягко поясняет.
— Не в самом озере, Якоб. Где-то поблизости. В одном из горячих источников.
Он все кивает и кивает, не глядя на нее.
Роуэн оборачивается к Ютте. Та отвечает беспомощным взглядом.
Роуэн нажимает:
— Ты же знаешь, такого не предполагалось. Мы изучили вирус, изучили океано-
графию и очень тщательно подобрали место. Но что-то все-таки упустили.
— Чертов Гольфстрим накрылся, — говорит старик. Голос его ненамного, но
сильнее тела. — Говорили, что так и будет. Все течения изменятся. Англия пре-
вратится в Сибирь.
Роуэн кивает.
— Мы рассматривали различные сценарии. Кажется, ни один не подходит. Я по-
думала: может быть, в самом Бетагемоте есть что-то, чего мы не учли? — Она
чуть подается вперед. — Ваши люди ведь много чего накопали в Огненном поясе1,
а? Еще в тридцатых?
— Ну конечно, там все копались. Треклятая архея была золотой лихорадкой
двадцать первого века.
— И на Хуан де Фука много времени провели. С Бетагемотом не сталкивались?
— М-м-м. — Якоб Хольцбринк качает головой, но плечи его не шевелятся.
— Якоб, ты меня знаешь. Я всегда была за строгое хранение корпоративных
тайн. Но теперь мы на одной стороне, так сказать, в одной лодке. Если ты что-
то знаешь, хоть что-то...
— О, Якоб ведь никогда не занимался исследованиями, — перебивает Ютта. — Ты
же знаешь, он больше работал с персоналом.
— Да, конечно, но перспективными областями всерьез интересовался. Его все-
гда волновали новые открытия, не забывай. — Роуэн тихонько смеется. — Было
время, когда мы думали, что он просто живет в батискафе.
— Только на экскурсии выбирался. Ютта права, исследований я не вел. Этим
занимался Джарвис и его группа. — Якоб встречает взгляд Роуэн. — Всю его ко-
манду мы потеряли, когда Бетагемот вырвался на волю. УЛН рекрутировала наших
людей по всему земному шару. Выхватывала прямо у нас из-под носа! — Он фырка-
ет . — «Общее благо», черт бы его побрал.
Ютта сжимает ему колено. Он оглядывается на жену и улыбается, накрывает ее
ладонь своей.
Взгляд снова сползает к полу. Старик еле заметно покачивает головой.
— Якоб, понимаешь ли, не сближался с научной группой, — объясняет Ютта. —
Известное дело, ученые плохо разбираются в людях. Выпусти их к публике — слу-
чилась бы катастрофа, но им все равно иногда не нравилось, как Якоб презенто-
вал их находки.
Роуэн терпеливо улыбается.
— Якоб, просто я тут подумала насчет Бетагемота и его возраста...
— Чуть не самый старый организм на планете, — говорит Якоб. — Мы, осталь-
ные , завелись позже. Свалились с марсианским метеоритом или еще как. Чертов
Бетагемот чуть ли не единственная тварь, которая здесь же и зародилась.
— Но ведь в этом все и дело, да? Бетагемот не просто предшествовал осталь-
ной жизни, он появился раньше фотосинтеза. До кислорода! Ему больше четырех
миллиардов лет. Остальные по-настоящему древние микроорганизмы — археобакте-
рии, нанолиты и так далее — так и остались анаэробными. Их находят только в
ограниченных средах. А Бетагемот еще старее, но кислород его нисколько не
волнует.
Якоб Хольцбринк замирает.
— Умная тварюшка, — говорит он. — Идет в ногу со временем. Обзавелся этими,
как их называют... как у псевдомонасов...
— Генами Блашфорда. При стрессе меняют скорость собственной мутации.
— Да, да. Гены Блашфорда. — Якоб поглаживает ладонью свои редкие волосы и
покрытый старческими веснушками череп. — Он приспособился. Приспособился к
кислороду, приспособился жить в рыбах, а теперь приспосабливается к каждому
уголку и закоулку нашей чертовой планеты.
— Только не к сочетанию низких температур и высокой солености, — замечает
Роуэн. — Он так и не адаптировался к самой обширной экологической нише плане-
ты. Глубины моря отвергали его миллиарды лет, и сейчас бы отвергли, если бы
1
Тихоокеанский огненный пояс (кольцо) — подковообразная область по периметру Тихого
океана, которая отличается повышенной вулканической и сейсмической активностью.
не источник Чэннера.
— о чем ты? — неожиданно резко переспрашивает Ютта. Ее муж молчит.
Роуэн переводит дыхание.
— Все наши модели основаны на предпосылке, что Бетагемот существует в своей
нынешней форме сотни миллионов лет. Распространение кислорода, гипотоничные
тела носителей — все это условия раннего-раннего докембрия. А с тех пор, как
нам известно, изменилось не так уж многое, несмотря на все эти гены Блашфорда
— иначе Бетагемот бы давно уже правил миром. Мы знаем, что он не способен
распространяться по глубоководью, потому что так этого и не сделал, хотя у
него были миллионы лет. А если кто-то предполагал, что он может распростра-
няться с ихтиопланктонном, мы отмахивались, и не потому, что кто-то проверил
— у кого нашлось бы время, учитывая, что тогда творилось? — а потому что, ес-
ли бы он мог распространяться этим путем, то давно бы распространился. Мил-
лионы лет назад.
Якоб Хольцбринк откашливается.
Роуэн выкладывает карты на стол.
— Но что, если Бетагемоту не миллионы лет, а всего несколько десятков?
— Ну, это... — начинает Ютта.
— Тогда уже ни в чем нет уверенности, правда? Может, речь не о нескольких
изолированных реликтах, а об эпицентрах? И, может, дело не в том, что Бетаге-
мот не способен распространяться, а в том, что он только начал?
Снова птичьи кивки и снова отрицание:
— Нет, нет. Он старый! Шаблоны РНК, минерализованные стенки. Большущие поры
по всей поверхности — он из-за них и не выносит холодной соленой воды. Проте-
кает как сито.
В уголке губ у старика проступают пузырьки слюны. Ютта безотчетно тянется
их стереть, но Якоб с раздражением отмахивается. Она роняет руку на колени.
— Пиранозильная последовательность. Примитив. Уникальный. Та женщина, док-
тор. .. Джеренис — она тоже согласна. Он старый!
— Да, — соглашается Роуэн, — старый. Но, возможно, сравнительно недавно из-
менился?
Якоб взволновано потирает руку об руку.
— Что, мутация? Удачный прорыв? Чертовски неудачный для нас.
— Может быть, его изменили сознательно, — произносит Роуэн.
Ну вот, высказала.
— Надеюсь, ты не хочешь сказать... — Ютта умолкает. Роуэн, наклонившись
вперед, кладет ладонь на колено Якобу.
— Я знаю, что там творилось тридцать-сорок лет назад. Ты сам сказал, атмо-
сфера золотой лихорадки. Весь рифт в лабораториях, каждый органклонер работа-
ет in situ1. . .
— Ну конечно, in situ, попробовала бы ты воссоздать эти условия в лаборато-
рии. . .
— А твои люди были на переднем крае. Ты не только сам вел исследования, но
и за другими приглядывал. Наверняка приглядывал, любой толковый бизнесмен по-
ступил бы так. Потому я и пришла к тебе, Якоб. Я ничего не утверждаю, никого
не обвиняю, понимаешь? Просто я подумала, что если кто-то на «Атлантиде» зна-
ет , что произошло тогда, так это ты. Ты эксперт, Якоб. Ты ничего не можешь
мне сказать?
Ютта качает головой:
— Якоб ничего не знает, Патриция. Никто из нас ничего не знает. И я все же
улавливаю, к чему ты клонишь.
Роуэн не сводит глаз со старика. Тот смотрит в пол, сквозь пол, сквозь пли-
1
(Прямо) на месте (лат.).
ту палубы, сквозь трубы и провода под ней, сквозь изоляцию, фуллерен, био-
сталь , сквозь морскую воду и заиленные слизистые скалы — куда, она может
только догадываться. И когда он заговаривает, голос его, кажется, доносится
из этого далека.
— что ТЬ1 хочешь узнать?
— Могла ли у кого-нибудь найтись причина — чисто гипотетически — перестро-
ить такой организм, как Бетагемот?
— Сколько угодно, — отвечает далекий голос из тела, которое кажется едва
одушевленным.
— Например?
— Точечная доставка. Лекарств, генов, органелл. Такой клеточной стенки ни-
кто еще не видывал. Не пришлось бы заботиться об иммунной реакции, защитные
энзимы о них и не узнают. Целевая клетка принимает его прямо в себя: лидиро-
вание оболочки, ХДД1. Вроде биодеградирующего фуллерена.
— Еще?
— Идеальный стимулятор. При соответствующих условиях эта штука качает АТФ с
такой скоростью, что можно одной рукой перевернуть автомобиль. По сравнению с
ним митохондрии просто тухлятина. Солдат с Бетагемотом в клетках, при доста-
точной дозе, может и экзоскелет перещеголять.
— Если Бетагемот перестроить для этой цели, — поправляет Роуэн.
— Да, — шепчет старик, — в том-то и дело.
Роуэн очень тщательно подбирает слова:
— А могли найтись... менее узкие сферы применения? Оружие всеобщего уничто-
жения , промышленный терроризм?
— В смысле, чтобы он работал как сейчас? Нет. Надо было ослепнуть и сойти с
ума одновременно, чтобы подумать о таком.
— Но скорость воспроизводства пришлось бы немножко увеличить, а? Для ком-
мерческого использования.
Он кивает, по-прежнему глядя в никуда.
— Эти глубоководные организмы так медлительны, что тебе, считай, повезло,
если они делятся раз в десять лет.
— А при этом им потребовалось бы больше питания, верно? Чтобы поддерживать
ускорившийся рост.
— Конечно, это и ребенку известно. Но нарочно такого бы не стали делать,
никто не станет добиваться такой цели, это просто был бы неизбежный...
— Побочный эффект, — подсказывает Ютта.
— Побочный эффект, — повторяет старик. Голос его не изменился и по-прежнему
доносится, спокойный и далекий, из центра земли. Но на щеках у Якоба Хольц-
бринка слезы.
— Значит, не нарочно. Целили во что-то другое, а потом все пошло... напере-
косяк. Ты это хочешь сказать?
— Гипотетически? — Уголки губ у него приподнимаются в чуть заметной выму-
ченной улыбке. Слезы текут по морщинам и буграм старческого подбородка.
— Да, Якоб. Гипотетически.
Голова кивает.
— Можно что-нибудь сделать? Из того, что мы еще не пробовали?
Якоб качает головой.
— Я всего лишь корп. Не знаю.
Она встает. Старик, уйдя в свои мысли, смотрит вниз. Ютта смотрит на Роуэн.
— Не пойми его неправильно, — говорит она.
— Ты о чем?
— Он этого не делал, он не виноват. Не больше тебя. Не больше любого из
По всей вероятности, подразумевается хитиндисахаридцеацетилаза, энзим (фермент).
вас.
— Я понимаю, Ютта.
Она прощается. Последнее, что она видит через закрывающийся люк — как Ютта
Хольцбринк надевает на склоненную голову мужа гарнитуру сонника.
Теперь уже ничего не исправишь. Нет смысла обвинять, тыкать в кого-то паль-
цем. И все же она рада, что навестила их. Даже в некотором смысле благодарна.
Благодарность эгоистическая, но лучше, чем ничего. Утешение, уж какое есть,
Патриция Роуэн находит в том, что оказалась не первой из виноватых. Даже Лени
Кларк, русалка Апокалипсиса, оказалась не первой. Роуэн оглядывается на люк в
конце голубого коридора.
Все началось отсюда.
Портрет
садиста на
свободе
Технически это называлось «складной катастрофой». На графике она выглядела
как цунами в разрезе: гладкая крыша наступающей волны выдается вперед, проги-
бается под гребнем и уходит плавной дугой к новому низкоэнергетическому со-
стоянию равновесия, при котором камня на камне не остается.
С земли все представлялось намного грязнее: отказ энергосетей, судороги
систем жизнеобеспечения и удаления отходов, забитые взбудораженными толпами
дороги, до революции — один шаг. Полиция в экзоскелетах давно отступила с
уровня улиц; только «оводы» роились над головами, поливая толпы газом и ульт-
развуком .
Для переднего края волны существовало отдельное название. Момент хаотиче-
ского перегиба, в котором траектория прогибается внутрь перед тем, как обру-
шится волна, именовали «точкой перелома». Западная часть Нт АмПацифика минова-
ла ее в последние тридцать четыре часа: все, что находилось к западу от Ска-
листых гор, можно было более или менее списывать со счетов. УЛН опускала все
доступные барьеры — для людей, товаров, даже электроны замораживали на ходу.
С какой стороны ни посмотри, на Кордильерах мир заканчивался. Теперь только
правонарушители могли выходить за эту границу и пытаться что-нибудь сделать.
Что ни сделай, на этот раз всего будет мало.
Конечно, система деградировала не первое десятилетие. Даже не первый век.
Своей работой Дежарден был обязан этой живой связи между энтропией и челове-
ческой глупостью — без них борьба с кризисами не стала бы самой крупной от-
раслью на планете. Рано или поздно все начинает разваливаться, это известно
каждому, у кого есть пара глаз, a IQ чуть повыше комнатной температуры. Но не
было серьезных причин, чтобы все развалилось так быстро. Они могли бы выга-
дать еще десяток-другой лет, еще немного времени для того, чтобы верующие в
человеческую изобретательность продолжали обманывать себя.
Но чем ближе вы к точке перелома, тем труднее заклеивать трещины. Даже рав-
новесие в такой близости от края обрыва всегда неустойчиво. Что там бабочки —
когда планета зависла на самом краю, даже взмах крылышка тли может его столк-
нуть .
Год был 2051-й, и долгом Ахилла Дежардена было раздавить Лени Кларк как бу-
кашку, будь то бабочка или тля.
Он видел, как дела рук ее распространяются по континенту, словно паутина
трещин по льду озера. Он получал сводки данных из сотен источников за два ме-
сяца — подтвержденная и ориентировочная информация устарела для применения в
поисках, но могла пригодиться для предсказания следующего прорыва Бетагемота.
Мемы и легенды о Мадонне Разрушения, куда более многочисленные и метастатич-
ные — стратегия воспроизводства роя виртуальных существ, которых Дежарден
только что обнаружил и мох1 уже никогда не понять. Реальность невольно вступа-
ла в союз с мифами, и там, где они сливались, расцветал Бетагемот; из-за спи-
ны выскакивали пожары и отключения энергии, и в жертву общему благу приноси-
лись все новые невинные жизни.
Дежарден знал, что это ложь. Северная Америка, несмотря на все драконовские
меры предосторожности, уже миновала точку перелома. Падать системе далеко,
пройдет немало времени, пока она долетит с гребня до дна. Но Дежарден умел
читать числа. По его расчетам оставалось две — может быть, три — недели, пре-
жде чем весь континент рухнет в анархию следом за Нт АмПацификом.
Бегущая строка в уголке дисплея сообщала о новом бунте в Гонкувере. Сверх-
современные охранные системы гибли, защищая стеклянные шпили и богатые квар-
талы — проигрывая не умникам-хакерам и не превосходящей технологии, а напору
живого мяса. Оружие захлебывалось, скрывалось под приливом живых тел, топчу-
щих мертвые. У него на глазах толпа с торжествующими воплями проломила воро-
та. Тридцать тысяч голосов в нестройном хоре звучали как один голос, в кото-
ром уже не осталось ничего человеческого. Этот звук напоминал вой ветра. Де-
жарден отключил новости прежде, чем толпа узнала то, что было известно ему:
район пуст, корпы, когда-то укрывавшиеся в нем, давно залегли на дно.
На дно моря.
Легкая рука погладила его по спине. Он в испуге обернулся — за плечом стоя-
ла Элис Джовелланос. Увидев, кто пришел с ней, Дежарден украдкой бросил
взгляд на экран — на экране в десятке окон горел Рим. Он потянулся к выключа-
телю.
— Не надо. — Лени Кларк сдвинула с лица визор и пустыми, как яичные скор-
лупки, глазами уставилась на картины разрушений. Лицо ее было неподвижно, но
голос, когда она заговорила снова, дрогнул:
— Оставь.
Он познакомился с ней две недели назад. Выслеживал несколько месяцев, рылся
в архивах, раскопал ее досье, сосредоточил все свои способности на расшифров-
ке загадочной и непонятой головоломки по имени Лени Кларк. Но сложившиеся
фрагменты открыли перед ним не просто «ходячий инкубатор конца света», как
выразилась Роуэн. Они открыли портрет женщины, все детство которой было ло-
жью, навязанной извне программой, которой она не осознавала и не могла управ-
лять. Все это время Кларк пыталась вернуться домой, пересмотреть собственное
прошлое. А Кен Лабин, порабощенный собственным вариантом Трипа Вины, пытался
ее убить. Дежарден, в свою очередь, попытался его остановить — в тот момент
это представлялось единственным достойным вариантом. Оглядываясь назад, оста-
валось только удивляться, что столь добрый поступок мог быть следствием про-
буждающейся в нем психопатии.
Попытка спасения провалилась — Лабин перехватил его еще до того, как Кларк
добралась до Су-Сент-Мари. Дальнейшее Дежарден наблюдал, будучи привязанным к
креслу в темной комнате, с разбитым и изломанным лицом.
Как ни странно, так с ним обошелся не Кен Лабин. Как-то так вышло, что они
оказались, в широком смысле, на одной стороне: он, Элис, Кении и Лени вместе
трудились под знаменем серости, неясной морали и праведной мести. Спартак ос-
вободил Лабина от Трипа — так же, как до этого и Дежардена. Правонарушитель
вынужден был признаться, что и сейчас испытывает некоторое сочувствие к мол-
чаливому убийце: он знал, каково вернуться к настоящей ответственности после
того, как много лет все трудные решения за тебя принимали синтетические ней-
ротрансмиттеры. Парализующая тревога. Вина.
По крайней мере, в первое время. Сейчас вина почти стерлась. Остался только
страх.
Мир взывал к нему с тысяч сторон. Его обязанностью было прислушаться к каж-
дому призыву: обеспечить спасение или, если не выйдет, вычерпывать воду вед-
рами, пока последний обломок не скроется под волнами. Еще недавно это была не
просто обязанность — непреодолимая потребность, пристрастие, от которого он
не мох1 отказаться. Вот сейчас ему полагалось высылать аварийные бригады, пе-
ренаправлять каналы снабжения, изыскивать подъемники и «оводы» для поддержки
слабеющего карантина.
«На хрен!» — подумал он, отключая все каналы. И каким-то образом уловил,
как вздрогнула Лени Кларк, когда экран потемнел.
— Выцепил? — спросила Джовелланос. Она могла бы и сама попробовать, но про-
работала старшим правонарушителем всего неделю — маловато, чтобы привыкнуть к
новым возможностям, а тем более выработать седьмое чувство, которое Дежарден
оттачивал шестой год. Ее наиболее точная оценка местоположения корпов своди-
лась к «где-то в Северной Атлантике».
Кивнув, Дежарден потянулся к главной панели. Ониксовое отражение Кларк ше-
вельнулось у него за спиной в глубине темной поверхности. Дежарден подавил
желание оглянуться. Она была прямо здесь, в его кабинете — обычная девушка
вдвое меньше него. Тощая маленькая К-отборщица, которую одна половина челове-
чества мечтает убить, а другая — отдать за нее жизнь.
Он еще не знал ее, когда пожертвовал всем, чтобы прийти ей на помощь. Когда
же, наконец, встретился лицом к лицу, она показалась ему еще страшнее Лабина.
Но с тех пор с Кларк что-то случилось. Повадка снежной королевы ничуть не из-
менилась, но то, что стояло за ней, казалось... меньше, что ли. Почти хруп-
ким. Элис, правда, ничего не замечала. Едва нащупав шанс поквитаться со «Зло-
вещей Корпоративной Олигархией», или как там это называлось на нынешней неде-
ле , она сама себя назначила талисманом рифтеров.
Дежарден развернул окно на экране: спутниковая съемка открытого океана,
раскрашенного компьютером в цветные переливы множества оттенков.
— Я об этом думала, — пискнула Элис, — но даже если можно было бы выделить
из помех тепловые отпечатки, там, внизу, такая медленная циркуляция...
— Не температура, — перебил ее Дежарден, — а степень помутнения.
— Все равно, циркуляция...
— Заткнись и учись, а? — зыркнул на нее Дежарден.
Элис замолчала. В ее глазах стояла нескрываемая боль.
Она ступала по яичной скорлупе с тех пор, как призналась, что заразила его.
Дежарден обернулся к панели:
— Конечно, со временем многое изменяется. От барашков на волнах до каль-
марьего пердежа... — Он стукнул по иконке; новые данные наложились поверх ос-
новной линии прозрачной глазурью. — По одному снимку след ни за что не взять,
даже при самом точном разрешении. Придется оценивать данные за три месяца.
Слои смешались. Аморфная плазма исчезла, в тумане проступили четкие линии и
пятна.
Пальцы Дежардена пробежали по панели.
— Теперь отсеем все, что есть в базе данных НУОАИ1... — Мириады светящихся
шрамов вылиняли до прозрачности. — . ..следы Гольфстрима... — Потемнела цепоч-
ка бусинок от Флориды до Англии. — ...и все зарегистрированные участки строи-
тельства и точки подъема глубинных вод, которые не соотносятся с минимально
допустимыми размерами объекта.
Пропали еще несколько десятков пятнышек. Северная Атлантика стала темна и
безвидна, если не считать единственного яркого пятнышка, расположенного почти
точно посередине.
— Так вот оно где, — пробормотала Кларк.
Дежарден покачал головой.
1
Национальное управление океанических и атмосферных исследований, ныне существующее
в США научное ведомство.
— Надо еще учесть боковое смещение при подъеме к поверхности. Течения на
средних глубинах и тому подобное.
Он ввел алгоритмы — пятнышко дернулось к северо- востоку и застыло
«39°20!14" с.ш. 25°16!03" з.д.», — сообщил дисплей.
— Точно на северо-восток от зоны Атлантического Разлома, — сказал Дежарден.
— Самый низкий показатель завихренности во всем океане.
— Ты сказал — степень помутнения. — Отражение Кларк со светящимся кругом на
груди покачало головой. — А если вода там не движется...
— Пузыри, — воскликнула, догадавшись, Элис.
Дежарден кивнул.
— Убежище для нескольких тысяч человек не построишь без сварки. Сварочные
аппараты дают выброс использованного газа. Отсюда и помутнения.
Кларк все еще сомневалась.
— Мы варили на Чэннере. Давление сплющивает пузыри чуть не раньше, чем они
образуются.
— При точечной сварке — конечно. Но они там должны скреплять целые секции —
температуры выше, выход газа и термальная инерция тоже. — Он, наконец, обер-
нулся к ней. — Мы же не о кипящем котле говорим. К тому времени, как газ до-
бирается до поверхности, это просто мельчайшая газовая взвесь, невидимая про-
стым глазом. Но ее достаточно, чтобы уменьшить прозрачность воды, что мы
здесь и видим.
Он постучал по точке на экране.
Кларк несколько секунд рассматривала его ничего не выражающим взглядом.
— Кто-нибудь еще об этом знает? — спросила она, наконец.
Дежарден покачал головой.
— Никто не знает даже, что я этим занимаюсь.
— Ты не против, если это так и останется?
— Лени, — фыркнул он, — я даже думать не хочу, что будет, если кто-то про-
знает, что я на такие вещи трачу время. И не то чтобы я вам не рад, но болта-
ясь рядом, вы, ребятки, сильно рискуете. Вы не...
— Это уже не проблема, Кайфолом, — тихо сказала Элис. — Я быстро схватываю.
Она не хвасталась. Получив повышение после его дезертирства, она за не-
сколько часов вычислила, что тысяча с небольшим корпов тихо исчезли с лица
земли. И меньше двух дней ушло у нее на то, чтобы вернуть Дежардена в штат
УЛН, прикрыть его таинственное исчезновение алиби и бюрократической болтов-
ней. Конечно, она получила незаконную фору в начале игры — заражение Спарта-
ком исключило влияние Трипа Вины. Она получила все возможности старшего пра-
вонарушителя без сопутствующих ограничений. Само собой, она приняла меры,
прежде чем протащить Лени Кларк в святая святых УЛН.
Но Спартак и сейчас бурлил в голове у Дежардена, кислотой разъедая цепи,
выкованные Трипом. Он уже освободил его совесть и скоро, что крайне пугало
Ахилла, должен был уничтожить ее полностью.
Он взглянул на Элис. «Это ты со мной сделала», — подумал он и проанализиро-
вал вызванные этим обвинением чувства. Еще недавно это был гнев, ощущение,
что его предали. И даже нечто граничащее с ненавистью.
Теперь появилось сомнение. Элис... Элис была осложнением, гибелью и спасе-
нием Дежардена в одном стройном теле. Пока что она его спасла. И владела ин-
формацией, которая может оказаться спасительной позже. Представлялось неглу-
пым подыграть ей. По крайней мере, временно. Что до рифтеров, чем быстрее он
выпроводит их отсюда, тем проще станет уравнение.
И еще какая-то острая заноза, засевшая в голове, напоминала о возможностях,
которые скоро откроются перед человеком, спущенным с цепи.
Элис Джовелланос улыбнулась ему осторожно, а может, и с надеждой.
— Ты быстро схватываешь, — повторил он, — это точно.
Будем надеяться, что не слишком быстро.
Исповедь
Джеренис Седжер желает сделать заявление.
Не Кларк с Лабином — им она даже не говорит, о чем речь.
— Поймите меня правильно, — просит она. — Я хочу обратиться ко всему сооб-
ществу .
Ее разбитое на пиксели изображение смотрит с панели мрачно и виновато. На
заднем плане — Патриция Роуэн, она, кажется, тоже не в восторге.
— Хорошо, — отвечает, наконец, Лабин и отрубает связь.
«Седжер, — размышляет Кларк. — Седжер выступает с заявлением. А не Роуэн».
— Новости от медиков, — говорит она вслух.
— Плохие новости, — отзывается Лабин, затягивая мембрану на перчатках.
Кларк настраивает панель на низкочастотное вещание.
— Пожалуй, стоит собрать войска.
Лабин направляется к трапу:
— Позвонишь в колокол, а?
— А ты куда? — Колокол привлекает внимание тех рифтеров, что держат вокоде-
ры отключенными, но их Лабин обычно созывает сам.
— Хочу кое-что проверить, — отвечает он.
Люк шлюза с шипением закрывается за ним.
Конечно, даже с их нынешним количеством все сразу в головной узел не втис-
нутся .
Было бы проще, если б рифтеры строились по правилам. В конструкции модулей
предусматривалась стыковка; каждая самодостаточная сфера топырилась шестью
круглыми ртами по два метра в поперечнике. Каждая могла смыкаться губами с
аналогичной или с промежуточными коридорами — так что вся конструкция разрас-
талась в бугорчатый беспорядочный скелет с длинными костями и пустыми черепа-
ми, распростершийся на морском дне. Во всяком случае, так было задумано. Не-
сколько основных форм с бесконечными возможностями сочетать их.
Но нет. Модули-пузыри рассеялись по дну одиночными грибами. Рифтеры живут
по одиночке, или парами, или такими компаниями, какие сложились на данный мо-
мент . Толпа рифтеров — практически оксюморон1. Пузырь головного центра — один
из самых больших во всем треилерном парке, но и у него на главных палубах
умещается не больше дюжины человек. Учитывая, насколько личное пространство
расширяется у большинства рифтеров на глубине, им будет неуютно.
К тому времени как Кларк, запустив колокол, возвращается обратно, места
почти не осталось. На входе в шлюз к ней на хвост садятся Чен с Крамером. На
приемной палубе Абра Чун опережает ее и первой поднимается по лестнице.
Кларк, взбираясь следом, слышит, как снова запускается шлюз — а пока ее не
было, уже собралась кучка народу, человек восемь или девять.
Всем заправляет Грейс Нолан, обосновавшись за панелью связи. Сонары показы-
вают, что на подходе еще дюжина людей. Кларк от нечего делать прикидывает,
выдержат ли фильтры пузыря такую нагрузку. Может, и не будет никакого «заяв-
ления» . Может, Седжер просто хочет, чтобы они надышались собственной углеки-
слотой?
— Привет. — Рядом появляется Кевин Уолш, с надеждой зависнув на периметре
ее личного пространства. Он, кажется, опять стал самим собой. Стоящий впереди
1 Оксюморон, оксиморон, а также оксюморон, оксиморон (др.-греч. от «острый» + «глу-
пый») — образное сочетание противоречащих друг другу понятий; остроумное сопоставле-
ние противоречивых понятий, парадокс; стилистическая фигура или стилистическая ошиб-
ка — сочетание слов с противоположным значением (то есть сочетание несочетаемого).
Гомес оборачивается и замечает Кларк:
— О, Лен? У корпов новости, как я слышал.
Кларк кивает.
— Ты водишь компанию с этими засранцами. В курсе, о чем речь?
Она качает головой.
— Новости от Седжер. Наверное, что-то медицинское.
— Да, пожалуй. — Гомес негромко втягивает воздух сквозь покрытые пятнами
зубы. — Джулию никто не видел? Ей бы следовало это послушать.
Чун поджимает губы:
— После того, как она полторы недели провела с Джином? Если так хочется,
вот сам и дыши с ней одним воздухом.
— Я недавно видел ее у одной из «поленниц», — влезает Хопкинсон.
— Как она?
— Ты же знаешь Джулию. Черная дыра с сиськами.
— Я имела в виду — внешне. Не выглядит больной?
— Откуда мне знать? Думаешь, она там была в трусиках и лифчике? — пожимает
плечами Хопкинсон. — Да мы и не разговаривали.
Сквозь переборки и гул разговоров смутно прорываются крики терзаемого кам-
ня.
— Ну, все, — говорит за панелью Нолан, — хватит балду пинать. Прицелился —
стреляй. — Она касается иконки на панели. — Ты на связи, Седжер. Давай, дер-
зай.
— Все здесь? — звучит голос Седжер.
— Конечно, нет. Мы бы не уместились.
— Я бы предпочла...
— Тебя вывели на все каналы низкочастотника. В пределах пятисот метров тебя
услышит каждый.
— Так... — Пауза, молчание человека, соображающего, как лучше пройти через
минное поле. — Как вам известно, «Атлантида» уже несколько дней на карантине.
С тех пор, как мы узнали о Бетагемоте. Все мы модифицированы, так что не было
причин ожидать серьезных проблем. Карантин был банальной мерой предосторожно-
сти.
— Был... — отмечает Нолан. Внизу снова запускается шлюз.
Седжер рассказывает дальше:
— Мы провели анализ... образцов, которые доставили с Невозможного озера Кен
и Лени, и все указывает на Бетагемот. Те же особенности РНК, та же стереоизо-
меризация...
— Давай к сути, — рявкает Нолан.
— Грейс? — окликает Кларк. Нолан оглядывается на нее.
— Заткнись и дай ей договорить, — просит Кларк. Нолан, фыркнув, отворачива-
ется.
— в общем, так, — помолчав, продолжает Седжер. — Результаты были вполне од-
нозначные, поэтому мы из предосторожности кремировали зараженные останки. Ра-
зумеется , после оцифровки.
— Оцифровки? — переспрашивает Чен.
— Разрушающее сканирование с высоким разрешением, позволяющее воссоздать
копию образца вплоть до молекулярного уровня, — поясняет Седжер. — Смоделиро-
ванные ткани дают те же реакции, что и реальные образцы, но без сопутствующе-
го риска.
В поле зрения втискивается Чарли Гарсиа. С каждым новоприбывшим перегородки
словно смыкаются чуть тесней. Кларк сглатывает. Воздух вокруг нее уплотняет-
ся.
Седжер кашляет.
— Я работала с одной из таких моделей и... ну, я обратила внимание на некую
аномалию. Я думаю, что рыба, доставленная вами с Невозможного, была инфициро-
вана Бетагемотом.
По всей комнате встречаются взгляды пустых глаз. Вдалеке колокол Лабина ис-
пускает последний слабый вопль и замолкает — газовый резервуар опустел.
— Ну, естественно, — вступает после паузы Нолан. — Так что?..
— Я. . . гм. . . говорю «инфицирована» в смысле патологии, а не симбиоза. —
Седжер снова откашливается. — Я хотела сказать...
— Рыба была больна, — подсказывает Кларк. — Больна Бетагемотом.
Минута мертвого молчания. Затем:
— Боюсь, что ты права. Если б ее не убил Кен, то прикончил бы Бетагемот.
— Ох ты, блин, — тихо вырывается у кого-то. Словечко повисает в полной ти-
шине . Внизу булькает шлюз.
— Значит, больна, — нарушает молчание Дейл Кризи. — И что с того?
Гарсиа качает головой:
— Дейл, ты не забыл, как работает эта дрянь?
— Конечно. Разбирает энзимы на части, добывая из них серу или что-то в этом
роде. Мы к ней иммунны.
— Мы иммунны, — терпеливо объясняет Гарсиа, — потому что нам встроили осо-
бые гены, придающие энзимам прочность, с которой Бетагемот не справляется. А
получили мы эти гены от глубоководных рыб, Дейл.
Кризи еще обдумывает услышанное, а кто-то срывающимся голосом уже шепчет:
— Черт, черт, черт!
Одинокий опоздавший поднимается по лестнице — и почему-то оступается на
первой перекладине.
— Боюсь, что мистер Гарсиа прав, — говорит Седжер. — Если рыбы, обитающие
здесь, оказались уязвимыми для вируса, то мы, возможно, тоже уязвимы.
Кларк качает головой.
— Но... ты хочешь сказать, что это все-таки не Бетагемот? Что-то другое?
У лестницы поднимается суета, собравшиеся рифтеры пятятся, словно к ней
подключили ток. Над люком показывается шатающаяся Джулия Фридман, лицо у нее
цвета базальта. Она выбирается на палубу, держась за перила вокруг люка и не
решаясь их выпустить. Оглядывается, часто мигая поверх мертвых линз. Кожа у
нее блестит.
— Это все еще Бетагемот — в общих чертах, — бубнит в отдалении Седжер. Из
«Атлантиды». Из закупоренной, запаянной, герметически запечатанной каранти-
ном, безопасной как хрен знает что «Атлантиды». — Вот почему мы не сумели
точно определить природу болезни мистера Эриксона: он дал положительную реак-
цию на Бетагемот, но, мы, конечно, не обратили на это внимания, поскольку не
ожидали никаких проблем. Но это, по-видимому, новая мутация. Подобная вариа-
бельность довольно распространена среди организмов, попадающих в новую среду.
Это по сути...
«Злой брат-двойник Бетагемота», — вспоминает Кларк.
— ... Бетагемот второй модели, — заканчивает Седжер.
Джулия Фридман падает на колени, ее рвет.
Вавилон на всех частотах. Смешение прерывающихся голосов:
— Конечно, не верю. А ты что, веришь?
— Чушь собачья. Если...
— Они честно признались. Могли промолчать.
— Да, в тот самый момент, когда у Джулии проявились симптомы, их обуяла че-
стность . Какое совпадение!
— Откуда им было знать, что она...
— Им известен инкубационный период. Наверняка. Как еще объяснить такой точ-
ный расчет?
— Да, но что нам-то делать?
Узел опустел. Рифтеры, словно выброшенный из трюма балласт, рассыпались по
дну, теперь довольно многолюдному даже по сухопутным стандартам. Пузырь нави-
сает над ними чугунной планетой. Три фонаря, установленные у входного шлюза,
бросают на дно яркие пересекающиеся круги. Черные тела плавают на периферии
света — призраки беспокойного движения за немигающими глазами, похожими на
ряды акульих зубов. Кларк приходит мысль о голодных зверях, сторонящихся ла-
герного костра. Вообще-то, ей следовало бы чувствовать себя одной из стаи.
Грейс Нолан больше не видно. Несколько минут назад она скрылась в темноте,
поддерживая одной рукой Джулию, провожая ее к дому. Такое проявление альтру-
изма добавило ей сторонников — Чен с Хопкинсон при следующем противостоянии
окажутся на ее стороне. Гарсиа сомневается, но настроение сейчас не такое,
чтобы народу требовались безоговорочные доказательства.
— Эй, Дими, — жужжит Чен, — как там дела?
— Воняет как в лазарете, — воздушный голос Александра разительно выделяется
на фоне подводных. — Но я почти закончил. Вырастил бы мне кто-нибудь новую
кожу.
Он остался внутри: стерилизует все, что вошло в контакт с Фридман и ее вы-
делениями . Грейс Нолан вызвала волонтеров.
Она начала приказывать. А люди начали ее слушаться.
— Говорю, надо просто пробуравиться к гадам, — жужжит где-то рядом Кризи.
Кларк вспоминает прожженные в биостали дыры.
— Давайте пока подождем с контратакой. Им будет трудновато найти лекарство,
если мы их размажем по палубе.
— Как же, станут они искать лекарство!
Она пропускает этот выкрик мимо ушей.
— Им нужны образцы крови от каждого. Еще кто-то из нас может оказаться ин-
фицирован . Проявляется это, очевидно, не сразу
— У Джина проявилось довольно быстро, — напоминает кто-то.
— Когда тебя потрошат заживо, сопротивляемость малость снижается. А у Джу-
лии ничего не проявлялось... сколько, две недели?
— Я им ни капли крови не дам. — Голос Кризи — как скрежет по металлу. — А
если попробуют взять силой, я на их кровь полюбуюсь!
Кларк устало качает головой.
— Дейл, они никого не могут заставить и прекрасно об этом знают. Они про-
сят. Хочешь, чтобы молили — можно устроить. В чем дело? Ты и сам брал образ-
цы.
— Если вы на минутку отвлечетесь от вылизывания клитора Патриция Роуэн, у
меня тут весточка от Джина.
Грейс Нолан вплывает в круг света, словно черный зверь, утверждающий власть
в стае. Уж ее-то костер не пугает.
— Грейс, — жужжит Чен, — как там Джулия?
— А ты как думаешь? Она больна. Но я ее уложила и диагностер подключила —
хоть какая-то польза будет.
— А Джин? — спрашивает Кларк.
— Он ненадолго приходил в себя. И сказал, цитирую: «Я говорил им, эти людо-
еды со мной что-то сделали. Может, поверят, когда умрет моя жена»
— Ну, — пищит Уолш, — он, как видно, пошел на поправ. . .
— Корпы никогда бы не рискнули распространять подобную заразу, не будь у
них лекарства, — перебивает Нолан. — Слишком просто было самим подцепить.
— Верно... — это опять Кризи. — Говорю вам, давайте сверлить переборку за
переборкой, пока не отдадут.
В темноте сомнения и согласие сливаются воедино.
— Знаете, попробую разыграть адвоката дьявола. Я к тому, что есть ведь не-
которая вероятность, что они правду сказали.
Это Чарли Гарсиа подплывает сбоку.
— Я к тому, что микробы ведь мутируют, да? — продолжает он. — Особенно ко-
гда люди обстреливают их всякими веществами — а когда эта дрянь объявилась, в
нее наверняка палили всей фармацевтикой сразу. Так кто скажет, что он сам не
мох1 мутировать из первого Бетагемота в Бета-макс?
— Охрененная натяжка, я бы сказал, — жужжит Кризи.
Вокодер Гарсиа щелкает — звуковой символ для пожатия плечами.
— Просто напомнил.
— А если они собирались развязать биологическую войну, с чего бы им тянуть
до сих пор? — хватается за соломинку Кларк. — Почему было не начать четыре
года назад?
— Четыре года назад у них Бетагемота не было, — говорит Нолан. И Уолш:
— Они могли захватить с собой культуру вируса.
— Что, на память? Ностальгии ради, что ли? Ни хрена у них не было, пока
Джин не преподнес им Бетагемот тепленьким на блюдечке.
— Это ты зря, Грейс, — жужжит Гарсиа. — Мы уже пятьдесят лет собираем виру-
сы из обрывков. Зная последовательность генов, корпы могли в любое время со-
брать Бетагемот с нуля.
— И, если на то пошло, вообще что угодно, — добавляет Хопкинсон. — С какой
стати использовать штуку, которая так медленно работает? Сколько из наших за-
болело-то? Суперхолера свалила бы нас за несколько дней.
— Джина бы свалила сразу, — жужжит Нолан, — и он не успел бы заразить ос-
тальных. У быстродействующего организма здесь нет шансов — мы рассеяны, мы
изолированы, мы большей частью и не дышим. И даже внутри не снимаем кожи. Для
эпидемии нужен медленно действующий агент. Эти подонки знали, что делают.
— К тому же, начнись на дне океана эпидемия суперхолеры, мы бы наверняка
смекнули, в чем дело. Они бы сразу попались.
— И они это понимали.
— А Бетагемот их оправдывает, — говорит Чен. — Разве не так?
«Черт бы тебя побрал, Джелейн! — Кларк как раз подумала о том же. — Обяза-
тельно было об этом напоминать?»
Нолан тут же подхватывает тему:
— Так. Именно так. Бетагемот аж с самого Невозможного озера, никого ведь не
обвинишь, что его туда подкинули — они просто чуточку поработали с ним в «Ат-
лантиде», передали нам, а мы, конечно, не заметили разницы.
— Тем более, что они якобы уничтожили образцы, — добавляет Кризи.
Кларк мотает головой.
— Ты — водопроводчик с жабрами, Дейл. Пусть бы Седжер даже отдала тебе эти
образцы в запечатанном пакетике — разве ты знал бы, что с ними делать? Это
относится и к школьным опытам Грейс с образцами крови.
— Вот и вся твоя помощь. — Нолан изворачивается всем телом и оказывается
нос к носу с Кларк. — Мы, тупицы рыбоголовые, ни в чем не разбираемся, дока-
зать ничего не можем, и должны доверять ученым умникам, которые нас и нагрели
когда-то.
— Не все, — жужжит в ответ Кларк. — Рама Бхандери.
Внезапно все умолкают. Кларк и сама не верит, что заговорила об этом.
Вокодер Чен постукивает, выдавая ее нерешительность.
— Э... Лен, Рама же отуземился.
— Пока еще нет. Не совсем. Самое большее, на грани.
— Бхандери? — От механической злобы в голосе Нолан вибрирует вода. — Он уже
рыба!
— Он все еще соображает, — настаивает Кларк. — Я только вчера с ним говори-
ла . Мы могли бы его вернуть.
— Бхандери в этом дерьме разбирается, — вставляет Гарсиа. — Или разбирался.
— Ага, именно что в дерьме, — добавляет Кризи. — Я слыхал, он переделал ки-
шечную палочку, чтобы она выделяла психоактивные вещества. Человек ходит с
этим дерьмом в кишках и сам себе обеспечивает страну чудес. — Грейс Нолан,
обернувшись, пристально смотрит на него, но Кризи не улавливает намека. —
Кое-кто из его клиентов жрал собственные какашки. Чтобы сильней пронимало.
— Потрясающе, — жужжит Нолан, — слюнявый идиот и химик-говнарь. Наши про-
блемы решены.
— Я только говорю, что нам не обязательно самим себе перерезать глотку, —
настаивает Кларк. — Если корпы нам не лгут, то в них наша главная надежда
справиться с этим.
— Скажешь, мы должны им верить? — вставляет Чун.
— Я говорю, может, нам и не придется ничего принимать на веру. Дайте мне
шанс поговорить с Рамой, узнать, сумеет ли он помочь. Если нет, взорвать «Ат-
лантиду» можно и на следующей неделе.
Нолан рубит воду рукой:
— Да у него ум за разум зашел!
— У него осталось достаточно мозгов, чтобы рассказать, что произошло у «по-
ленницы» , — тихо жужжит Кларк.
Нолан отвечает долгим взглядом, чуть заметно напрягается всем телом.
— В самом деле, — встревает Гарсиа, — я, пожалуй, поддерживаю Лени.
— А я — нет, — тотчас огрызается Кризи.
— Проверить не повредит, — голос Хопкинсон вибрирует где-то с задних рядов.
— Лени права. Убить их никогда не поздно.
Не то чтобы импульс выигрышный, но Кларк все равно его использует.
— А что им остается, задержать дыхание и рвануть к поверхности? Мы можем
позволить себе ждать.
— А Джин может? А Джулия? — Нолан обводит глазами круг. — Сколько у нас
времени — у каждого?
— А если ты ошибаешься, вы перебьете всех засранцев до последнего, а потом
выясните, что они все же пытались нам помочь... — Кларк качает головой. —
Нет, я этого не допущу.
— Ты не до...
Кларк чуть увеличивает звук и заглушает ее.
— Вот мой план, народ. Все, кто еще не сдал, сдают кровь. Я ищу Раму и вы-
ясняю, способен ли он помочь. А к корпам пока никто не лезет.
«Вот оно, — думает она. — Поднимай ставку или пасуй».
Минута растягивается. Нолан оглядывает собрание. И, похоже, увиденное ей не
нравится.
— Вы все, счастливые маленькие эрочки и кашечки, делайте что хотите. А я
знаю, что мне делать.
— Тебе, — говорит ей Кларк, — надо сдать назад, заткнуться и ни хрена не
делать, пока мы не получим надежную информацию. А до тех пор, Грейс, если я
застану тебя в пятидесяти метрах от «Атлантиды» или Рамы Бхандери, то собст-
венными руками вырву тебе трубки из груди.
Внезапно они оказываются линзы к линзам.
— Больно много на себя берешь, твой ручной психопат пока еще тебя не под-
держал. — Вокодер Нолан стоит на минимальной громкости, ее слова — механиче-
ский шепот, слышный только Кларк. — Где твой телохранитель, корповская под-
стилка?
— Он мне не нужен, — невозмутимо жужжит в ответ Кларк. — Если ты мне не ве-
ришь , кончай нести фуфло, а переходи к делу.
Нолан неподвижно висит в воде. Ее вокодер тикает, как счетчик Гейгера.
— Слушай, Грейс, — неуверенно жужжит сбоку Чен. — Право, попробовать не по-
мешает , честно...
Нолан ее словно не слышит. И долго не отвечает. Наконец качает головой.
— Хрен с вами, пробуйте.
Кларк еще несколько секунд выдерживает паузу, потом разворачивается и, мед-
ленно работая ластами, уходит из круга света. Она не оглядывается — и надеет-
ся , что стая примет это за полную уверенность в себе. Но в душе жутко трусит.
В голове одна мысль: бежать — бежать от этого нового, с иголочки, напоминания
о собственном ядовитом прошлом, от цунами и отвернувшейся от нее удачи. Хо-
чется просто соскользнуть с Хребта, отуземиться и уходить все дальше, пока
голод и одиночество не разгладят извилины в мозгу, сделав его мозгом рептилии
— как у Бхандери. Ей ничего так не хочется, как сдаться.
Она уплывает в темноту и надеется, что остальные последовали ее примеру.
Прежде чем Грейс Нолан заставила их передумать.
Кларк берет курс на двухпалубник немного на отшибе от других. Безымянный —
хотя некоторые пузыри окрестили: «Домик Кори», «Пляжный мяч» или «Оставь на-
дежду» . На корпусе этого не было таблички, когда она в прошлый раз проплывала
мимо — нет и теперь. И никто не повесил запретительных знаков в шлюзе, одна-
ко, на сушилке блестят две пары ласт, а с сухой палубы доносятся тихие влаж-
ные звуки.
Она взбирается по лесенке. Ын и чья-то спина трахаются на матрасе. Очевид-
но, даже Лабинов гудок не отвлек их от этого занятия. Кларк ненадолго задумы-
вается, не прервать ли их, чтобы ввести в курс событий.
На фиг, скоро сами все узнают.
Она обходит пару и осматривает панель связи. Довольно простенькое устройст-
во из нескольких подручных деталей — только-только чтоб работало. Кларк, по-
играв с экраном сонара, выводит на него топографию Хребта, а поверх — сетку
условных иконок. Вот главные генераторы, проволочные скелеты небоскребов, вы-
сящиеся на юге. А вот «Атлантида», большое неровное колесо обозрения, опроки-
нутое на бок — сейчас нечеткое и расплывчатое, эхо размывается полудюжиной
глушилок, запущенных, чтобы защитить последние нововведения от чужих ушей. Со
времени Бунта эти глушилки не использовались. Кларк удивляется, что они оста-
лись на месте, да еще в рабочем состоянии. И задумывается, не занимался ли
кто-то активной подготовкой.
На экране — россыпь серебряных пузырьков, полузаброшенные дома тех, кому
едва ли знакомо значение этого слова. Увеличь разрешающую способность — можно
будет увидеть и людей: уменьшится охват, зато станет больше подробностей, и
окрестности моря наполнятся мерцающими сапфировыми изображениями, прозрачны-
ми, как пещерные рыбы. Изнутри каждого доносится жесткое эхо имплантатов,
крошечных матовых комочков механики.
Распознать, кого видишь на экране, довольно просто: у каждого возле сердца
— опознавательный маячок. Одним запросом Кларк может получить множество дан-
ных . Обычно она так не делает. Никто так не делает — у рифтеров свой особый
этикет. Да обычно в этом и нет нужды. С годами учишься читать голое эхо. Им-
плантаты Кризи чуть расплываются со стороны спины, ножной протез Йегера при
движении немного уводит его влево. Массивная туша Гомеса выдавала бы и сухо-
путника. Маячки — излишество, шпаргалка для новичков. Рифтерам эта телеметрия
в основном не нужна, а у корпов теперь нет к ней доступа.
Но иногда — когда расстояние скрывает все особенности эха или когда объект
изменился — остаются только шпаргалки.
Кларк настраивает максимальный охват: четкие яркие фигурки сливаются, сби-
ваются в центре, словно космический мусор, засасываемый в черную дыру. На
краях экрана проступают другие детали топографии — большие, смутные, разроз-
ненные . Вот появилась большая черная расщелина, взрезающая и пересекающая
донный грунт. Дюжина грубых курганов, серебристый осадочный мусор на дне —
одни высотой с метр, другие в пятьдесят раз больше. И само дно к востоку под-
нимается. За пределами обзора встают подножия огромных гор. На средней дис-
танции и дальше — несколько светлых голубых клякс: одни лениво плывут над
илистой равниной, другие просто дрейфуют. Распознать их на таком расстоянии
невозможно, да и не требуется. Сигнал маячков вполне отчетлив.
Бхандери на юго-западе, на полпути до границы обзора. Кларк фиксирует его
позицию и убирает увеличение, возвращая сонар к прежним настройкам. «Атланти-
да» и ее окрестности вырастают на экране и...
Секундочку!..
Одиночное эхо в белом шуме помех. Смутное пятно без деталей, неуместная бо-
родавка на одном из круглых переходов, соединяющих модули «Атлантиды». Бли-
жайшая камера установлена на причальном кране в двадцати пяти метрах восточ-
нее и выше. Кларк подключает ее: новое окно, открывшись, высвечивается зерни-
стым зеленым светом. «Атлантида» вся в темных заплатах. Кое-что сияет, как
всегда — верховые маячки, отводные отверстия, сигнальные огоньки на трубах
озаряют темноту. Но местами свет погас. Там, где светили желто-зеленые лампы,
теперь темные дыры и канавы, в глубине которых теплится синее мерцание, едва
отличимое от черноты.
«Не работает», — говорят синие угольки. Или, точнее: «Рыбоголовым хода
нет».
Шлюзы, ворота причального модуля... Никто уже не играет в «просто меры пре-
досторожности» . . .
Она меняет наклон камеры, нацеливает ее точнее. Дает приближение: полумрак
наплывает, превращаясь из размытого далека в размытую близь. Сегодня плохая
видимость: либо рядом проснулись дымные гейзеры, либо «Атлантида» выбрасывает
муть. Кларк различает лишь расплывчатый черный силуэт на зеленом фоне — до
того знакомый, что она даже не понимает, по каким приметам его опознала.
Лабин.
Он плавает в каких-то сантиметрах от обшивки, подаваясь то в одну сторону,
то в другую. Возможно, пытается удержаться на месте, сопротивляясь хитрому
сплетению течений — вот только нечего ему там делать. Нет там окошек, через
которые можно заглянуть внутрь, нет причин зависать именно у этого отрезка
коридора.
Через несколько секунд он начинает перемещаться вдоль обшивки — но так мед-
ленно , что это настораживает. Обычно его ласты движутся плавными, свободными
гребками, а сейчас чуть подергиваются. Он движется не быстрей, чем сухопут-
ник-пешеход.
За ее спиной кто-то достигает высшей точки. Ын ворчит: «Была моя очередь».
Лени Кларк их почти не слышит.
«Ублюдок, — думает она, глядя, как Лабин скрывается в темноте. — Ублюдок.
Взял и сделал!»
Вербовка
Алике не все понимает насчет «туземцев». По правде сказать, никто из корпов
не понимает, но остальные от этого бессонницей не маются: чем больше рыбого-
ловых уберутся с дороги, тем лучше. Алике, благослови ее душу, пришла в самую
настоящую ярость. Для нее это все равно, что оставить дряхлую старушку уми-
рать во льдах.
— Леке, они сами так решают, — объяснила однажды Кларк.
— Что решают — сойти с ума? Решают, пусть у них кости превратятся в кашу,
так что, заведи их внутрь, даже и на ногах устоять не смогут?
— Они решают, — мягко повторила Кларк, — остаться на рифте, и считают, что
цена не слишком высока.
— Почему? Что там такого хорошего? Что они там делают?
Кларк не стала рассказывать про галлюцинации.
— Думаю, свобода. И единение со всем, что тебя окружает. Это трудно объяс-
нить . . .
Алике фыркнула:
— Ты и сама не знаешь!
Отчасти это правда. Кларк, конечно, чувствовала притяжение открытого моря.
Может, это побег1, может, бездна — просто самое лучшее убежище от ада жизни
среди сухопутников. А может, и того проще. Может, это просто темная невесо-
мость материнского лона, забытое чувство, что тебя питают и надежно защищают
— то, что было, пока не начались схватки и все не пошло вразнос.
Все это ощущает каждый рифтер. Не все из них ассимилируются — пока еще не
все. Просто некоторые... более уязвимы. Склонны к зависимостям, в отличие от
более компанейских напарников. Может, у отуземившихся в лобных долях слишком
много серотонина или еще что. Обычно все сводится к чему-нибудь подобному.
Алике всего этого, конечно, не объяснишь.
— Вы должны убрать кормушки, — сказала Алике, — чтобы они хоть за едой за-
ходили внутрь.
— Они скорее умрут с голоду или станут питаться червями и моллюсками. — Что
тоже приведет к голодной смерти, если они раньше не умрут от отравления. — Да
и зачем тянуть их внутрь, если они не хотят?
— Да затем, что это самоубийство! — заорала Алике. — Господи, неужели тебе
надо объяснять? Вот ты бы не стала мешать мне покончить с собой?
— Смотря по обстоятельствам.
— В смысле?
— Действительно ли ты этого хочешь, или просто пытаешься чего-то добиться.
— Я серьезно!
— Да, я вижу, — вздохнула Кларк. — Если бы ты, в самом деле, решила покон-
чить с собой, я бы горевала и злилась, и мне бы страшно тебя недоставало. Но
мешать я бы не стала.
Алике была потрясена.
— Почему?
— Потому что это — твоя жизнь. Не моя.
Этого Алике, как видно, не ожидала. Сверкнула глазами, явно не соглашаясь,
но и не находя, что возразить.
— Ты когда-нибудь хотела умереть? — спросила ее Кларк. — Всерьез?
— Нет, но...
— А я — да.
Алике замолчала.
— И, поверь мне, — продолжала Кларк, — не особо весело слушать, как толпа
специалистов втолковывает, что «тебе есть для чего жить», и «дела не так пло-
хи», и «через пять лет вы вспомните этот день и сами не поймете, как вы даже
думать об этом могли». Понимаешь, они же ни черта не знают о моей жизни. Если
я в чем и разбираюсь лучше всех на свете, так это в том, каково быть мной. И,
на мой взгляд, надо быть чертовски самоуверенным, чтобы решать за другого,
стоит ли ему жить.
— Но ты же не должна так думать, — беспомощно ответила Алике. — Никто не
должен. Это как ободрать кожу на локте и...
— Дело не в том, чтобы чувствовать себя счастливым, Леке. Тут вопрос, есть
ли причины для счастья. — Кларк погладила девочку по щеке. — Ты скажешь, я из
равнодушия не помешаю тебе покончить с собой, а я говорю — я настолько нерав-
нодушна , что еще и помогу тебе, если ты этого действительно захочешь.
Алике долго не поднимала глаз. А когда подняла, глаза у нее сияли.
— Но ты не умерла, — сказала она. — Ты хотела, но не умерла, и именно по-
этому ты сейчас жива, и...
«А многие другие — нет». Эту мысль Кларк оставила при себе.
А сейчас она собирается все переиграть. Выслеживает человека, который решил
уйти, наплевав на его выбор и навязав ему свой. Кларк хочется думать, что
Алике сочла бы это забавным, но она понимает, что не права.
Ничего смешного здесь нет — слишком это жутко.
На сей раз она не взяла «кальмара» — туземцы обычно сторонятся звука меха-
низмов. Она целую вечность движется над равниной серого как кость ила — без-
донного болота из умиравшего миллионы лет планктона. Кто-то побывал здесь до
нее — ее путь пересекает след, в котором еще кружились взбаламученные движе-
нием микроскопические тельца. Она двигается тем же курсом. Из донного грунта
торчат разрозненные обломки пемзы и обсидиана. Их тени проходят через яркий
отпечаток налобного фонаря Кларк: вытягиваются, съеживаются и снова сливаются
с миллионолетним мраком. Постепенно обломков становится больше, чем ила: это
уже не отдельные выступы, а настоящая каменная россыпь.
Перед Кларк — нагромождения вулканического стекла. Она увеличивает яркость
фонаря: луч высвечивает отвесную скалу в нескольких метрах впереди. Ее по-
верхность изрезана глубокими трещинами.
— Алло, Рама?
Тишина.
— Это Лени.
Между двумя камнями проскальзывает белоглазая тень.
— Ярко...
Она заглушает свет.
— Так лучше?
— А. . . Лен. . . — Механический шепот, два слога, разделенные секундами уси-
лия , которое потребовалось, чтобы их выдавить. — Привет...
— Нам нужна твоя помощь, Рама.
Бхандери неразборчиво жужжит из своего укрытия.
— Рама?
— Не... помощь?
— У нас болезнь. Похожая на Бетагемот, но наш иммунитет против нее не дей-
ствует. Нам надо разобраться, что это такое, нужен человек, смыслящий в гене-
тике .
Между камнями ни малейшего движения.
— Это серьезно. Пожалуйста — ты мог бы помочь?
— ...теомикой, — щелкает Бхандери.
— Что? Я не расслышала.
—Протеомикой1. . . Генетикой. .. немножко совсем.
Он уже почти осиливает целые предложения. Почему бы не доверить ему сотни
жизней?
— Ты мне снилась, — вздыхает Бхандери. Звучит это так, словно кто-то провел
пальцем по зубьям металлической расчески.
— Это был не сон. И сейчас тоже. Нам, правда, нужна помощь. Рама. Пожалуй-
ста.
— Неправильно, — жужжит он. — Нет смысла.
— В чем нет смысла? — спрашивает Кларк, вдохновленная связностью его речи.
— Корпы. Корпов просите.
— Возможно, эпидемию устроили корпы. Они могли перестроить вирус. Им нельзя
доверять.
— ...бедняжки.
— Ты не мог бы...
— Еще гистамина, — рассеянно жужжит Рама, и затем: — Пока...
Протеомика — наука, изучающая белки, их структуру и функции.
— Нет! Рама!
Увеличив яркость фонаря, она успевает увидеть пару ласт, скрывающихся в
расщелине несколькими метрами выше. Резким движением ног1 Кларк толкается сле-
дом, ныряет в трещину, как ныряют с вышки — вытянув руки над головой.
Трещина глубоко рассекла скалу, но до того узка, что через два метра Кларк
приходится развернуться боком. Свет заливает узкий проем. В нем становится
светло, как в солнечный день наверху, и где-то рядом отчаянно хрипит вокодер.
Четырьмя метрами дальше Бхандери лягушкой раскорячился в проходе. Щель там
сужается, и он явно рискует застрять между каменными стенами. Кларк плывет к
нему.
— Слишком ярко, — жужжит он.
«А вот тебе», — мысленно отвечает она.
За два месяца хронического голодания Бхандери стал тощей тощего. Даже если
его заклинит, то в такую щель Кларк вряд ли за ним пролезет. Может быть, его
перепуганный маленький мозг уже прикинул шансы — Бхандери извивается, словно
разрываясь между соблазном вырваться на волю и надеждой спрятаться. Все же он
делает выбор в пользу свободы, но нерешительность дорого ему обходится —
Кларк успевает поймать его за лодыжку.
Он, зажатый каменными стенами, может лягаться только в одной плоскости.
— Пусти, чертова сука!
— Вижу, словарный запас вспоминается.
— Пус... ти!
Она выбирается к устью расселины сама и за ногу выволакивает Бхандери. Тот
упирается и скребет по стенам, потом, высвободившись из самого узкого места,
изворачивается и пытается ударить кулаком. Кларк отбивает удар. Ей приходится
напоминать себе, какие у него ломкие кости.
Наконец он покоряется. Кларк обхватывает его за плечи, сцепив пальцы на за-
тылке в двойной нельсон. Они у самого входа в ущелье; отбиваясь, Бхандери
ударяется спиной о растрескавшуюся базальтовую плиту.
— Свет! — щелкает он.
— Послушай, Рама. Дело слишком важное, чтобы дать тебе профукать ту ма-
лость, что осталась от твоих мозгов. Ты меня понимаешь?
Он корчится.
— Я выключу свет, если ты перестанешь драться и просто меня послушаешь, до-
говорились ?
— Я... тебя...
Она выключает фонарь. Бхандери вздрагивает и сразу обмякает у нее в руках.
— Хорошо. Так-то лучше. Ты должен вернуться, Бхандери. Ненадолго. Ты нам
нужен.
— . ..нужно. .. плохо... к нулевой. . .
— Кончай это, а? Не так уж ты далеко ушел. Ты здесь всего...
Месяца два, да? Ну, уже больше двух. Разве мозг уже мог превратиться в губ-
ку? Не тратит ли она время впустую?
Кларк начинает заново.
— Для нас это очень важно. Многие могут погибнуть. Ты тоже. Эта... болезнь
или что она там такое, достанет тебя так же легко, как любого из нас. Возмож-
но, уже достала. Ты понял?
—. . . понял. ..
Она надеется, что это ответ, а не эхо.
— И дело не только в болезни. Все ищут виноватых. Еще немного, и...
«Бабах, — вспоминается ей. — Взрыв. Слишком ярко».
— Рама, — медленно произносит она, — если дела пойдут вразнос, все взорвет-
ся. Ты понимаешь? Бабах! Как тогда у «поленницы». Все время будет бабах! Если
ты не поможешь мне. Не поможешь нам. Понял?
Бхандери висит перед ней в темноте, как бескостный труп.
— Да. Хорошо, — жужжит он, наконец. — Что ж ты сразу не сказала?
В драке он повредил ногу — все усилия приходятся теперь на левую, и его на
каждом гребке уводит вправо. Кларк попыталась подцепить его под руку и выров-
нять , но от прикосновения он испуганно дернулся. Теперь она просто плывет ря-
дом, время от времени подталкивая его в нужную сторону.
Трижды он делает рывок к свободе и забвению. Трижды она перехватывает его
неуклюжее движение и возвращает спутника на прежний курс, отбивающегося и
бессмысленно бормочущего. Впрочем, это лишь короткие эпизоды: побежденный, он
успокаивается, а успокоившись, начинает сотрудничать. До следующего раза.
Кларк уже поняла, что это, в сущности, не его вина.
— Эй, — жужжит она в десяти минутах от «Атлантиды».
— Да?
— Ты со мной?
— Да. Это только приступы... — Неразборчивое щелканье. — Я то в отключке,
то норм.
— Ты помнишь, что я говорила?
— Ты меня позвала.
— Помнишь, зачем?
— Какая-то эпидемия?
— Ага.
— И ты... вы думаете, корпы...
— Я не знаю.
— Нога болит...
— Извини.
И тут у него в мозгу что-то приходит в движение и снова дергает в сторону.
Кларк хватает и держит, пока приступ не проходит. Пока он отбивается от того,
что находит на него в такие моменты.
— ...еще здесь, вижу...
— Еще здесь, — повторяет Кларк.
— Хорошо, Лен. Пожалуйста, не делай так.
— Извини, — говорит она ему. — Извини.
— Я вам на хрен не нужен, — скрежещет Бхандери. — Все забыл.
— Вспомнишь.
Должен вспомнить!
— Ты не знаешь... ничего не знаешь про... нас.
— Немножко знаю.
— Нет.
— Я знавала одного... вроде тебя. Он вернулся.
Это почти ложь.
— Отпусти меня. Пожалуйста.
— Потом. Обещаю.
Она оправдывает себя на ходу и ни на минуту себе не верит
Лени помогает не только себе, но и ему. Оказывает ему услугу. Спасает от
образа жизни, неизбежно ведущего к смерти. Гиперосмос, синдром слизистого им-
плантата, отказ механики. Рифтеры — чудо биоинженерии. Благодаря несравненно-
му устройству гидрокостюмов они могут даже гадить на природе — но для разгер-
метизации вне атмосферы подводная кожа предназначена не была. А отуземившиеся
то и дело снимают маски под водой, впускают через рот сырой океан. И он разъ-
едает и загрязняет внутренний раствор, защищающий их от давления. Если проде-
лывать это достаточно часто, рано или поздно что-нибудь испортится.
«Я спасаю тебе жизнь», — думает она, не желая произносить этого вслух.
«Хочет он того или нет», — отвечает из памяти Алике.
— Свет! — хрипит Бхандери.
В темноте перед ним проступают отблески, уродуют идеальную черноту мерцаю-
щими язвами. Бхандери рядом с Кларк напрягается, но не убегает. Она уверена,
что он выдержит — всего две недели назад она застала его в головном узле, а
чтобы попасть туда, ему пришлось вытерпеть более яркие небеса. Не мох1 же он
за столь короткий срок так далеко уйти?
Или тут другое — не ровный ход, а резкий скачок? Может быть, его беспокоит
вовсе не свет, а то, о чем свет ему теперь напоминает?
«Бабах! Взрыв».
Призрачные пальцы легонько постукивают по имплантатам Кларк. Кто-то впереди
прощупывает их сонаром. Она берет Бхандери под руку, держит деликатно, но
твердо.
— Рама, кто-то...
— ...Чарли, — жужжит Бхандери.
Перед ними всплывает Гарсиа в янтарном сиянии, омывающем его со спины и
превращающем в привидение.
— Твою мать, ты его нашла! Рама, ты тут?
— Клиент...
— Он меня вспомнил! Охрененно рад тебя видеть, дружище. Я думал, ты уже по-
кинул сей бренный мир.
— Пытался. Она меня не пускает.
— Да, мы все извиняемся, но твоя помощь очень нужна. Только ты не напрягай-
ся, чувак. У нас получится. — Он оборачивается к Кларк. — Что нам понадобит-
ся?
— Медотсек готов?
— Одна сфера загерметизирована. Вторую оставили на случай, если кто сломает
руку.
— Хорошо. Свет придется выключить — во всяком случае, на первое время. Даже
наружное освещение.
— Легко.
— ...Чарли... — щелкает Бхандери.
— Я тут, дружище.
— ...будешь моим техником?
— Не знаю. Могу, наверное. Тебе нужен техник?
Маска Бхандери поворачивается к Кларк. В его манере держаться что-то резко
изменилось.
— Отпусти меня.
На этот раз она подчиняется.
— Сколько я не бывал внутри? — спрашивает он.
— Думаю, недели две. Самое большее, три.
По меркам рифтеров, это хирургически точная оценка.
— Могут быть... трудности, — говорит им Бхандери. — Реадаптация. Не знаю,
смогу ли я... не знаю, в какой степени я смогу вернуться.
— Мы понимаем. — жужжит Кларк. — Только...
— Заткнись. Слушай. — Бхандери дергает головой — движение рептилии, уже
знакомое Кларк. — Мне понадобится... толчок. Помощь в начале. Ацетилхолин.
Еще... тирозингидроксилаза. Пикротоксин. Если я развалюсь. Если начну разва-
ливаться, вам придется мне это ввести. Поняли?
Она повторяет:
— Ацетилхолин, пикротоксин, тиро... м-м...
— Тирозингидроксилаза. Запомни.
— Какие дозы? — спрашивает Гарсиа, — и как вводить?
— Я не... черт, забыл. Посмотри в медбазе. Максимально рекомендованная доза
для всего, кроме гидрокси... лазы. Ее вдвое больше, наверное. Думаю, так.
Гарсиа кивает.
— Еще что-нибудь?
— О да, — жужжит Бхандери. — Будем надеяться, я вспомню, что вообще...
Портрет
садиста в
команде
У Элис Джовелланос были свои представления о том, как надо извиняться.
«Ахилл, — начала она, — ты иногда такой кретин, что поверить невозможно!»
Он не сделал распечатки. Не нуждался в этом. Он был правонарушителем, заты-
лочные участки коры постоянно работают в ускоренном режиме, способности к со-
поставлению и поиску закономерностей, словно у аутиста. Он один раз прокрутил
ее письмо, посмотрел, как оно уходит за край экрана, и с тех пор перечитывал
сотню раз в воспоминаниях — не забыв ни единого пикселя.
А сейчас он сидел, застыв, как камень, и ждал ее. Ночные огни Садбери бро-
сали размытые пятна света на стены его номера. Слишком многое просматривается
с ближайших зданий, отметил Ахилл. Надо будет к ее приходу затемнить окна.
«Ты прекрасно знаешь, чем я рисковала, когда вчера тебе призналась, — дик-
товала программе Элис. — И ты знаешь, чем я рискую, отправляя тебе письмо, —
оно автоматически удалится, но наши уроды могут просканировать что угодно,
если захотят. И это часть проблемы, вот почему я вообще решилась тебе по-
мочь . . .
Я слышала, как ты говорил о доверии и предательстве, и может, в некоторых
твоих словах больше истины, чем мне хотелось бы. Но разве ты не понимаешь,
что спрашивать тебя заранее не было никакого толку? Пока на сцене Трип, ты не
можешь дать ответа сам. Ты настаиваешь, что тут я ошибаюсь, талдычишь о судь-
боносных решениях, которые принимаешь, о тысячах переменных, которыми жонгли-
руешь, но, Ахилл, дорогой мой, кто тебе сказал, что свободная воля — это все-
го лишь какой-то сложный алгоритм?
Я знаю, что ты не хочешь терять объективности. Но разве порядочный честный
человек — не страж самому себе, ты об этом не думал? Может, совсем и необяза-
тельно позволять им превращать себя в большой условный рефлекс. Просто ты сам
хочешь этого, ведь потом ты ни за что не несешь ответственности. Так легко,
когда не надо принимать решений самому. Это почти как наркотик. Может, ты на
него подсел, а сейчас у тебя синдром отмены».
Она так в него верила. И верила до сих пор: собиралась прийти сюда, ни о
чем не подозревая. Номер с защитой от наблюдения обходится не дешево, но
старший правонарушитель свободно мог позволить себе категорию «суперприват-
ность». Система охраны в этом здании непроницаема, беспощадна и начисто лише-
на долговременной памяти. После ухода посетителя о нем не остается никаких
записей.
«В общем, то, что они украли, мы вернули. И я хочу тебе рассказать, что
конкретно мы сделали, потому как невежество порождает страх, и все такое. Сам
знаешь. Ты в курсе насчет рецепторов Минского в лобных долях и что ней-
ротрансмиттеры вины привязаны к ним, а ты воспринимаешь это как угрызения со-
вести . Корпы создали Трип так: они вырезали из паразитов парочку генов, отве-
чающих за изменение поведения, и подправили их: чем более виноватым ты себя
чувствуешь, тем больше Трипа закачивается тебе в мозг. Он связывается с ней-
ротрансмиттерами, которые в результате изменяют конформацию и фактически за-
бивают двигательные пути, а у тебя наступает паралич.
В общем, Спартак — это аналог вины. Он взаимодействует с теми же участками,
что и Трип, но конформация у него немного другая, поэтому Спартак забивает
рецепторы Минского, но больше не делает практически ничего. К тому же он рас-
падается медленнее, чем обычные трансмиттеры вины, достигает более высокой
концентрации в мозгу и, в конце концов, подавляет активные центры простым ко-
личеством» .
Он вспомнил, как щепки старинного деревянного пола рвали ему лицо. Вспом-
нил, как лежал в темноте, привязанный к опрокинувшемуся на бок стулу, а голос
Кена Лабина спрашивал где-то рядом:
— Как насчет побочных эффектов? Естественного чувства вины, например?
В тот миг связанный, окровавленный Ахилл Дежарден увидел свою судьбу.
Спартак не удовлетворился тем, что разомкнул выкованные Трипом цепи. В этом
случае еще была бы надежда. Он бы вернулся к старому доброму стыду, который и
контролировал бы его наклонности. Остался бы неполноценным, но ведь таким он
всегда был. А вот оставлять его душу без надсмотрщика было нельзя. Он бы
справился — даже вне работы, даже если б ему предъявили обвинения. Справился
бы.
Но Спартак не знал удержу. Совесть — такая же молекула, как и любая другая,
и если для нее не находится свободных рецепторных участков, толку от нее не
больше, чем от какого-нибудь нейтрального раствора. Дежардена направили к но-
вой судьбе, в края, где он еще не бывал. В края, где нет вины, стыда, раская-
ния — нет совести ни в каком виде.
Элис не упомянула об этом, изливая ему во «входящих» свою оцифрованную ду-
шу . Только заверила, что это совершенно безопасно.
«В этом вся прелесть замысла, Кайфолом: выработка естественных трансмитте-
ров и Трипа не снижается, а потому через любую проверку ты пройдешь чистень-
ким. Даже анализ на более сложные формы даст положительный результат, ведь
базовый комплекс по-прежнему с нами — он просто не может найти свободных ре-
цепторов , за которые мог бы зацепиться. Так что ты в безопасности. Честно.
„Ищейки" — не проблема».
В безопасности. Она не представляла, что таилось у него голове. Ей следова-
ло быть осторожней. Эта простая истина известна даже детям: чудовища обитают
повсюду, даже в нас самих. Особенно в нас.
«Я не подвергла бы тебя опасности, Ахилл, поверь мне. Ты слишком много. ..
Ты для меня слишком хороший друг, чтобы так тебя подставлять».
Она его любила, конечно же. Раньше он никогда себе в этом не признавался —
тонюсенький внутренний голосок иногда нашептывал: «По-моему, она того, самую
капельку...», но потом три десятилетия ненависти к себе растаптывали его в
лепешку: «Эгоцентрик хренов. Как будто такое убоище кому-то нужно...»
Она никогда не делала ему прямых предложений — при всей своей порывистости,
Элис так же сомневалась в себе, как и он, — но кое-что можно было заметить:
добродушное вмешательство в любые его отношения с женщинами, бесконечные со-
циальные увертюры, прозвище Кайфолом — данное якобы за домоседство, а скорее
— за неспособность приносить удовольствие. Все это теперь бросалось в глаза.
Свобода от вины, свобода от стыда дала ему идеально острое зрение.
«Такие дела. Я рискнула, а дальше все в твоих руках. Впрочем, если сдашь
меня, знай: это твое решение. Как бы ты его ни рационализировал, какую-нибудь
тупую длинноцепочечную молекулу ты винить больше не сможешь. Все ты, все —
твоя свободная воля».
Он ее не сдал. Должно быть, причина крылась в некоем неустойчивом равнове-
сии конфликтующих молекул: те, что принуждали к предательству, ослабели, а
те, что выступали за верность друзьям, еще не выступили на первый план. Зад-
ним числом он считал это большой удачей.
«Потому воспользуйся своей свободой и подумай обо всем, что ты сделал и по-
чему, а потом спроси себя, действительно ли у тебя нет никаких моральных ори-
ентиров . Неужели ты не смог бы принять всех этих жестких решений, не отдавая
себя в рабство кучке деспотов? Я думаю, смог бы, Ахилл. Ты порядочный чело-
век , и тебе не нужны их кнуты и пряники. Я в это верю. И ставлю на это все».
Он взглянул на часы.
«Ты знаешь, где меня искать. Знаешь, какие у тебя варианты. Можешь присое-
диниться ко мне или вонзить нож в спину. Выбор за тобой».
Он встал и подошел к окну. Затемнил стекла.
«С любовью , Элис».
В дверь позвонили.
Все в ней было уязвимо. Она смотрела на него снизу верх — с надеждой, с ро-
бостью в миндалевидных глазах. Уголок рта оттянулся в нерешительной, почти
горестной улыбке.
Дежарден шагнул в сторону, спокойно, глубоко вдохнул, когда она прошла ми-
мо. От нее пахло невинностью и цветами, но в этой смеси были молекулы, дейст-
вующие под порогом сознания. Она была не глупа и знала, что он не глуп. Долж-
на была понимать, что он припишет свое возбуждение феромонам, которые она в
его присутствии не применяла много лет.
Значит, надеется.
Он сделал все возможное, чтобы укрепить в ней надежду, не выдав себя.
В последние дни Ахилл вел себя так, словно постепенно оттаивает, чуть ли не
против собственной воли. Он стоял рядом с ней, когда Кларк и Лабин раствори-
лись в уличном потоке, устремившись навстречу своей революции. Он позволил
себе задеть Элис локтем и продлить прикосновение. Через несколько мгновений
этого случайного контакта она медленно подняла на него глаза, и он наградил
ее пожатием плеч и улыбкой.
Он всегда считал ее другом — пока она не предала. Ей всегда хотелось боль-
шего. От такой смеси теряешь голову. Дежарден легко сумел обезоружить ее, по-
манив шансом на примирение.
Сейчас она прошла мимо, приблизившись к нему больше, чем было нужным, и
хвостик волос на затылке мягко качнулся над шеей. Мандельброт вышла в прихо-
жую и обвилась вокруг ее щиколоток меховым боа. Элис нагнулась почесать кошку
за ухом. Мандельброт замерла, раздумывая, не разыграть ли недотрогу, но реши-
ла не валять дурака и замурлыкала.
Дежарден кивнул на блюдце с таблетками дури на кофейном столике. Элис под-
жала губы:
— Это не опасно?
Химия организма старших правонарушителей могла очень неприятно взаимодейст-
вовать с самыми безобидными релаксантами, а Джовелланос совсем недавно обза-
велась этой химией.
— Думаю, после того, что ты натворила, тебе уже ничего не страшно, — прого-
ворил Дежарден.
Она понурилась. В горле у Дежардена застряла кроха раскаяния. Он сглотнул,
радуясь этому чувству.
— Главное, не мешай их с аксотропами, — добавил он немного мягче.
— Спасибо.
Она приняла наркотик как оливковую ветвь, забросила в рот вишнево-красный
шарик. Видно было, как она собирается с духом.
— Я боялась, что ты никогда больше не захочешь со мной разговаривать, — ти-
хо проговорила она.
— И ты это заслужила. — Он оставил фразу висеть в воздухе между ними. И
представил, как наматывает на кулак ее вороной хвостик. Как поднимает за во-
лосы , чувствует, как ее ноги дергаются в воздухе...
«Нет, остановись».
— Но я, наверно, понимаю, почему ты так поступила, — сказал он, наконец,
позволяя ей перевести дыхание.
— Правда?
— Думаю, что понимаю. Ты очень самоуверенна, — он вздохнул, — и очень ве-
ришь в меня. Иначе бы этого не сделала. Думаю, это чего-то стоит.
Казалась, она не дышала с самого появления, и только теперь выдохнула, ус-
лышав приговор: условное освобождение.
«Купилась, — подумал Дежарден. — Решила, что надежда есть».
А другая мысль, подавленная, но упрямая, твердила: «Разве она не права?»
Он погладил ее ладонью по щеке, уловил тихий короткий вздох, вызванный при-
косновением. И сморгнул мелькнувший образ: удар с плеча по этому милому, до-
верчивому лицу.
— Ты веришь в меня куда больше, чем я сам, Элис. Не знаю, насколько это оп-
равданно .
— Они украли у тебя свободу выбора. Я просто ее вернула.
— Ты украла у меня совесть. Как мне теперь выбирать?
— Умом, Кайфолом. Блестящим, прекрасным разумом. Не какими-то инстинктивны-
ми примитивными эмоциями, от которых в последнюю пару миллионов лет больше
вреда, чем добра.
Дежарден опустился на диван, в животе у него внезапно засосало.
— Я надеялся, что это побочный эффект, — тихо сказал он.
Она присела рядом.
— Ты о чем?
— Сама знаешь. — Дежарден покачал головой. — Люди никогда ничего не проду-
мывают до конца. Я вроде как надеялся, что вы с дружками просто. . . не преду-
смотрели этого осложнения, понимаешь? Что вы просто хотели отключить Трип, а
все эти дела с совестью... ошибка. Непредвиденная. Но как видно — нет.
Она тронула его за колено.
— Почему ты на это надеялся?
— Сам точно не знаю. — Его смешок был похож на лай. — Наверное, я рассуждал
так: если вы не знали — то есть сделали что-то случайно, то это одно, а вот
если сознательно взялись изготовить свору психопатов...
— Мы не психопатов делаем, Ахилл. Мы освобождаем людей от совести.
— Какая разница?
— у тебя по-прежнему есть чувства. Миндалевидное тело работает. Уровень се-
ротонина и дофамина в норме. Ты способен к долгосрочному планированию. Ты не
раб своих импульсов. Спартак ничего этого не изменил.
— Это ты так думаешь.
— Ты, правда, считаешь, что все гады на свете — психически больные?
— Может быть, и нет. Но готов поспорить, что все психи на свете — гады.
— Ты — нет, — сказала она.
И уставилась на него серьезными темными глазами. Он вдыхал ее запах и не
мог остановиться. Он хотел ее обнять. Хотел выпотрошить ее, как рыбу, и наса-
дить голову на палочку.
Он скрипнул зубами и промолчал.
— Слышал когда-нибудь о парадоксе стрелки? — помолчав, спросила Элис.
Дежарден покачал головой.
— Шесть человек в неуправляемом вагоне несутся к обрыву. Единственный спо-
соб их спасти — перевести поезд на другой путь. Только вот на другом пути
кто-то стоит и не успеет отскочить, поезд его задавит. Переведешь ли ты
стрелку?
— Конечно.
Это был простейший пример общего блага.
— А теперь предположим, ты не можешь перевести стрелку, но можешь остано-
вить поезд, столкнув кого-нибудь на пути. Столкнешь?
— Конечно, — немедленно ответил он.
— Вот что я для тебя сделала, — объявила Элис.
— Что?
— Для большинства людей это не одно и то же. Они считают, что перевести
стрелку — правильно, а столкнуть кого-то на рельсы — нет. Хотя это в точности
та же самая смерть против того же количества спасенных жизней.
Он хмыкнул.
— Совесть не рациональна, Ахилл. Знаешь, какие части мозга включаются, ко-
гда ты делаешь моральный выбор? Я тебе скажу: медиальная лобная извилина,
задняя часть поясной, угловая извилина. Все это...
— Центры эмоций, — вставил Дежарден.
— Именно так. Лобные доли вообще не искрят. Даже тем, кто признает логиче-
ское равенство этих сценариев, приходится приложить усилие. Просто толкнуть
кого-то на смерть ощущается как нечто неправильное, даже если на весах те же
жизни. Мозгу приходиться бороться с глупым, беспричинным чувством вины. Для
перехода к действию требуется больше времени, больше времени нужно для приня-
тия решения, и, в конечном счете, вероятность правильного решения ниже. Вот
что такое совесть, Кайфолом. Она подобна насилию, жадности, родственному от-
бору — была полезна миллион лет, но стала вредить с тех пор, как мы перестали
просто выживать в естественной среде и стали над ней доминировать.
«Ты эту речь отрепетировала», — подумал Дежарден.
И позволил себе легкую улыбку.
— Человек — это немножко больше, чем вина и разум, моя дорогая. А ты не по-
думала, что, возможно, вина не просто стреноживает разум? Может, она сдержи-
вает и еще кое-что?
— Например?
— Ну, просто ради примера... — Он выдержал паузу, притворяясь, будто ищет
вдохновение. — Откуда тебе знать, что я не какой-нибудь чокнутый маньяк-
убийца? Откуда знать, что я не психопат, не суицидник, ну или садист, допус-
тим?
— Я бы знала, — просто сказала Элис.
— Думаешь, у маньяков это на лбу написано?
Она сжала ему колено.
— Я думаю, что знаю тебя очень давно, а безупречно притворяться невозможно.
Тот, кого переполняет ненависть, рано или поздно сорвется. А ты... ну, нико-
гда не слышала о монстрах, уважающих женщин до такой степени, что отказывают-
ся их иметь. И, кстати, не хочешь ли пересмотреть последний пункт? Просто по-
размысли .
Дежарден покачал головой.
— Так ты, значит, во всем разобралась?
— Вполне. И терпения мне не занимать.
— Это хорошо. Сейчас оно тебе понадобится. — Он встал и улыбнулся ей свер-
ху. — Я на минутку в ванную. Чувствуй себя как дома.
Она улыбнулась в ответ.
— Обязательно. Можешь не спешить.
Он запер дверь, облокотился на раковину и пристально уставился в зеркало.
Отражение ответило свирепым взглядом.
«Она предала тебя. Превратила тебя в это».
Она ему нравилась. Он ее любил. Элис Джовелланос много лет была ему верным
другом. Дежарден сложил все это на чашу весов.
«Она сделала это нарочно».
Нет, нарочно это сделали они.
Потому что Элис действовала не сама по себе. Она дьявольски умна, но Спар-
така создала не в одиночку. У нее были друзья. Она сама призналась. «Мы вроде
как политические, правда, с организацией беда» — сказала она, сообщив ему о
его «освобождении».
Он чувствовал, как ржавеют и рассыпаются связи у него в голове. Как его
ущербность, ухмыляясь, дергает эти разъеденные ржавчиной проводки. Он поискал
в себе намек на сожаление, которое испытал несколько минут назад: когда задел
чувства Элис, причинил ей боль. Он и сейчас мог бы ощутить раскаяние или что-
то похожее, если бы постарался.
«Ты не раб своих импульсов», — сказала она.
В принципе, это было правдой. Ахилл мох1 сдержаться, если хотел. Но вот бе-
да : он начинал понимать, что не хочет.
— Эй, Кайфолом, — позвала из комнаты Элис.
«Заткнись. ЗАТКНИСЬ!»
— Да?
— Мандельброт требует ужина, а кормушка пустая. У тебя не осталось корма
под раковиной?
— Не осталось. Она научилась открывать дверцы.
— Тогда г. . .
— В шкафчике в спальне.
Ее шаги простучали мимо двери, Мандельброт подгоняла Элис мяуканьем.
«Нарочно!»
Элис заразила его, опередив график, чтобы освободить его разум для сражения
с Бетагемотом — а может быть, и по личным причинам, осознанным или нет. Но ее
дружки целили куда выше Ахилла Дежардена — они задумали «освободить» всех
правонарушителей на Земле. Лабин, в темноте, две недели назад, подытожил:
«Всего несколько тысяч человек держат руки на кнопках, способных парализовать
весь мир, а вы превратили этих людей в клинических социопатов».
Интересно, стала бы Элис применять свои семантические аргументы на Лабине?
Будь она привязана к тому стулу, ничего не видя, намочив штаны от страха,
слыша, как это кровожадное отродье расхаживает рядом, сочла бы она уместным
читать ему лекции об уровне серотонина и угловой извилине?
Может, и сочла бы. Что ни говори, она и ее друзья — «политические», хоть и
«с организацией беда», а от политики человек глупеет. Начинает верить, что
человеческое достоинство — какой-то платоновский идеал, моральный итог, кото-
рый можно вывести из неких первооснов. Не тратьте время на примитивную биоло-
гию. Не волнуйтесь за судьбу альтруистов в дарвиновской вселенной. Люди дру-
гие , люди — особенные, люди — агенты морали. Вот к чему приходишь, когда
слишком много времени уделяешь составлению манифестов и слишком редко гля-
дишься в зеркало.
Ахилл Дежарден оказался всего лишь первым представителем новой породы. Ско-
ро появятся другие, столь же могущественные и столь же неудержимые. Может,
уже появились. Элис не посвящала его в подробности. Он не представлял, на-
сколько далеко простираются амбиции Общества Спартака. Не знал, кого еще ус-
пели инфицировать, и сколько длится инкубационный период. Знал только, что
рано или поздно у него появятся соперники. Если не начать действовать сразу,
пользуясь полученной форой.
Мандельброт еще мяукала в спальне — видимо, распекала неумелую домработни-
цу. Дежарден ее понимал: у Элис было более чем достаточно времени, чтобы дос-
тать гранулированный корм, принести пакет в кухню и...
«В спальне!» — сообразил Ахилл.
«Ну, что ж, — подумал он миг спустя, — пожалуй, это решает вопрос».
Лицо в зеркале вдруг стало очень спокойным. Оно не двигалось и в то же вре-
мя словно говорило с ним. «Ты — не политик, — говорило лицо, — Ты — механизм.
Природа запрограммировала тебя на одно, УЛН — на другое, Элис вмешалась и пе-
реключила на третью программу. Все это не ты, и все — ты. Ты сам ничего из
этого не выбирал. И ни за что не отвечаешь.
Это она сотворила с тобой такое. Эта мочалка. Тупая давалка. Что бы ни слу-
чилось дальше, вина не на тебе.
На ней».
Он отпер дверь и прошел в спальню. Сенсориум у него на подушке предательски
поблескивал. Тактильный костюм лежал поперек кровати, словно сброшенная кожа.
Элис Джовелланос тряслась у изножья кровати, стаскивая с головы шлем. Лицо у
нее было красивым, без кровинки.
Жертву в этом виртуальном застенке она не могла не узнать. Дежарден настро-
ил симуляцию с точностью до тысячных.
Мандельброт тут же забыла об Элис и, громко мурлыча, начала бодать лбом хо-
зяина . Дежарден на нее не смотрел.
— Мне нужна кое-какая техническая информация, — чуть ли не виновато объяс-
нил он, — и сведения о твоих друзьях. Хотя я надеялся вытянуть все это из те-
бя по-хорошему. — Он кивнул на сенсориум, упиваясь ее ужасом. — Ну вот, забыл
убрать.
Она мотала головой. Ее лицо кривилось от паники.
— Я.., н-не думала, что ты... — выдавила она, наконец.
— Выходит, не думала, — пожал плечами Ахилл, — но постарайся увидеть и
светлую сторону. Ты впервые насчет меня не ошиблась.
Наконец все приобрело смысл: покупки, почти неосознанно сделанные через
анонимные кредитные линии, полимерная пленка и переносной мусоросжигатель,
инверсионный звукопоглотитель. Небрежное копание в ежедневнике Элис и списке
ее контактов. Вот почему хорошо быть правонарушителем на Трипе — когда всем
известно, что ты прикован к столбу, никто не утруждает себя возведением забо-
ра вокруг двора.
— Прошу тебя. — Губы у Элис дрожали, в блестящих глазах стоял страх. —
Ахилл...
Где-то в подвале его мозга рассыпался последний ржавый проводок.
— Зови меня Кайфолом, — сказал он.
Автомеханика
Первый раунд остается за корпами.
Рифтерша по имени Лизбет Мак — тихоня, Кларк с трудом припомнила ее имя —
наткнулась на корпа, бронированным тараканом ползавшего по обшивке основного
хозблока. Неважно, какие веские причины привели его туда. Неважно, можно ли
было считать это нарушением карантина. Мак среагировала так, как среагировали
бы многие рыбоголовые на ее месте: разозлилась. Решила проучить тупого сухо-
пута, но прежде его подогреть.
Она плавала кругами вокруг беспомощной неуклюжей жертвы, отпуская шуточки о
водолазном колоколе с ножками и громко, демонстративно призывая кого-то по-
дать ей пневмодрель — надо, мол, просверлить одну раковину.
Она совсем забыла о налобном фонаре на шлеме корпа. Когда Лизбет обнаружила
бедолагу, фонарь не горел — очевидно, корп старался не попадаться на глаза, а
наружного освещения на этой части станции хватало даже для глаз сухопутника.
Когда же он направил на нее вспышку, линзы у нее, компенсируя избыток осве-
щенности, на миг стали непрозрачными. Она ослепла всего на одну-две секунды,
но корпу этого хватило с лихвой. Кополимер пресс-кольчуге не соперник. К тому
времени как избитая, окровавленная Мак позвала на помощь, корп уже скрылся
внутри.
Сейчас Кларк с Лабином стоят в пятом шлюзе, а океан отступает от них. Кларк
вскрывает маску и ощущает, как тело сдувается, словно воздушный шар из живой
плоти. Внутренний люк с шипением отходит. В него вливается до боли яркий
свет. Пока линзы адаптируются, Кларк отступает назад, поднимает руки, гото-
вясь встретить атаку. Ее не происходит. В переходной камере жмутся несколько
корпов, но впереди одна только Патриция Роуэн. Между ней и рифтерами радужно
переливается изолирующая мембрана.
— Мы решили, что вам пока лучше остаться в шлюзе, — начинает Роуэн.
Кларк оглядывается на Лабина. Тот обводит встречающих пустыми непроницаемы-
ми глазами.
— Кто это был? — спрашивает Кларк.
— Думаю, это несущественно, — отвечает Роуэн.
— Лизбет другого мнения. У нее нос сломан.
— Наш человек утверждает, что защищался.
— Мужчина в пресс-кольчуте, рассчитанном на 300 бар, защищался от безоруж-
ной женщины в гидрокостюме?
— Корп защищался от рыбоголовой, — вставляет кто- то из присутствующих. —
Совсем другое дело.
Роуэн игнорирует комментарий.
— Наш человек пустил в ход кулаки потому, — говорит она, — что это была его
единственная надежда на успех. Вы не хуже меня знаете, от чего мы защищаемся.
— Я знаю, что никому из вас не полагалось выходить из «Атлантиды» без пред-
варительного согласования. Это правило действовало еще до карантина. Вы сами
согласились.
— Нам не дали особого выбора, — сдержанно замечает Роуэн.
— И все равно.
— На хер правила, — встревает один из корпов. — Они нас убить хотят, а мы
будем спорить о регламенте?
Кларк моргает:
— Как это понимать?
Роуэн вскидывает руку, и непокорный умолкает.
— Мы нашли мину, — говорит Патриция тем же тоном, каким сказала бы, что в
гальюне кончилась туалетная бумага.
— Что?!
— Ничего особенного. Стандартный заряд для сноса сооружений. Возможно, еще
из тех, которые собирал Кен, пока мы. . . — она старательно подбирает слова, —
несколько лет назад не пришли к соглашению. Говорят, нас должно было изолиро-
вать от основных узлов жизнеобеспечения, а большую часть отсека С затопило
бы. От одного только взрыва предполагалось от тридцати до ста погибших.
Кларк смотрит на Лабина, ловит легчайшее движение головы.
— Я не знала, — тихо говорит она.
Роуэн слабо улыбается.
— Как ты понимаешь, это вызывает некоторый скепсис.
— Я бы хотел видеть мину, — произносит Лабин.
— А я хотела бы видеть свою дочь на солнышке, — парирует Роуэн. — Только
ничего не выйдет.
Кларк качает головой.
— Послушай, Пат, я не знаю, откуда она... Я...
— А я знаю, — спокойно говорит Роуэн — Их целые штабеля на строительной
площадке. Только на Невозможном озере больше сотни.
— Мы найдем, кто ее подложил. Но вам нельзя ее оставлять. Вам запрещено
владеть оружием.
— Вы серьезно ожидаете, что мы так просто отдадим мину тем, кто ее подло-
жил?
— Пат, ты меня знаешь.
— Я вас всех знаю, — говорит Роуэн. — Ответ отрицательный.
— Как вы ее нашли? — спрашивает слева Лабин.
— Случайно. Отказала пассивная акустика, и мы послали человека починить ан-
тенну .
— Не уведомив нас заранее.
— Представлялось вполне вероятным, что связь нарушили ваши люди. Информиро-
вать вас было неблагоразумно. Даже если б вы не минировали нам корпуса.
— Корпуса, — повторяет Лабин. — Значит, мина была не одна?
Все молчат.
«Конечно, не одна, — соображает Кларк. — они нам ничего не скажут. Они го-
товятся к войне.
А для них это будет бойня».
— Интересно, все ли нашли? — задумчиво тянет Лабин.
Они стоят молча, скрыв лица под синтетическими черными масками. У них за
спинами, за непроницаемой плитой внутреннего люка, корпы снова строят планы и
планируют контрмеры. Впереди, за наружным люком, собирается в ожидании ответа
толпа рифтеров. Вокруг и внутри них искрят и перекачивают жидкости, подстраи-
вая их к бездне, разнообразные механизмы. К тому времени, как уровень воды
поднимается выше головы, они уже неподвластны давлению.
Лабин тянется к наружному люку. Кларк его перехватывает.
— Грейс, — жужжит она.
— Это мог быть кто угодно. — Он невесомо всплывает в затопленной камере,
поднимает руку, чтобы не наткнуться на потолок. Странное зрелище — гуманоид-
ный силуэт на голубовато-белом фоне стен. Линзы на глазах очень похожи на ды-
ры , прорезанные в черной бумаге, словно свет сзади проходит насквозь.
— Вообще-то, — добавляет Лабин, — я не вполне уверен, что они не лгали.
— Корпы? Зачем им лгать? Что они от этого выигрывают?
— Сеют рознь среди врагов. Разделяй и властвуй.
— Брось, Кен. Можно подумать, у нас есть прокорповская партия, готовая
встать на их защиту и...
Он просто смотрит на нее.
— Ты не знаешь, — жужжит она так тихо, что едва чувствует вибрацию в челю-
сти, — у тебя только догадки и подозрения. У Рамы не было возможности. . . ты
не знаешь наверняка.
— Не знаю.
— Мы действительно ничего не знаем. — Подумав, она поправляется. — Я ничего
не знаю. А ты — да.
— Недостаточно. Пока что.
— Я видела, как ты выслеживал их в коридорах.
Он не кивает, в этом нет нужды.
— Кого?
— В основном Роуэн.
— Ну и как там внутри?
— Примерно как вот тут. — Он указывает на нее.
«Не лезь мне в голову, подонок!»
Но она понимает, что на таком расстоянии это от него не зависит. Невозможно
взять и перестать чувствовать, будь эти чувства твои или чьи-либо еще. Поэто-
му вслух она говорит лишь:
— Нельзя ли конкретнее?
— Она чувствует себя виноватой — в чем-то. В чем, я не знаю. Причин хвата-
ет .
— Я же говорила.
— А вот наши люди, — продолжает он, — меньше страдают от внутренних кон-
фликтов и гораздо легче о них забывают. А я не могу быть повсюду сразу. И
время уходит.
«Ублюдок, — думает она, — засранец, обмылок».
Он плавает над ней, ожидая ответа.
— Хорошо, — говорит она, наконец, — я это сделаю.
Лабин тянет рукоять. Наружный люк отходит, открывая темный прямоугольник в
яркой белой раме. Они поднимаются навстречу ожидающим взглядам.
Лени Кларк — извращенна даже по стандартам рифтеров. Во-первых, их не слиш-
ком беспокоят вопросы приватности. Гораздо меньше, чем можно было бы ожидать
от отверженных и отбросов общества. Напрашивается предположение, что переме-
ной к лучшему эти места могут представляться лишь тем, кто сравнивает их с
уровнем намного ниже дна, и это верно. Также можно предположить, что такие
ущербные создания забьются в свои раковины, словно раки-отшельники, у которых
оборвали половину ног, будут шарахаться от каждой тени или яростно отбиваться
от малейшего покушения на их личное пространство. Но бесконечная вязкая ночь
здесь, внизу, если не лечит, то снимает боль. Бездна опускает тяжелые ладони
на израненных и разъяренных и каким-то образом успокаивает их. Как-никак,
здесь от любого конфликта можно уйти на любой из трехсот шестидесяти румбов.
И драться за ресурсы не приходится: половина пузырей давно пустует. А терри-
тории так много, что охранять ее нет смысла.
Поэтому большая часть пузырей стоит без охраны и без хозяев. В них вселяют-
ся и выселяются, заходят в первый попавшийся, чтобы трахнуться или поесть,
или — реже — пообщаться, прежде чем вернуться в естественную среду. Все места
одинаковы. И нет нужды ревниво охранять циркулятор Кальвина или ремонтный
верстак, а больше рифтерам ничего и не нужно. Одиночество можно найти где
угодно: проплыви две минуты в любую сторону, и можешь пропасть навеки. Зачем
возводить стены вокруг восстановленного воздуха?
У Лени Кларк для этого есть причины.
Она не одна такая. Еще несколько рифтеров потребовали исключительных прав,
застолбили за собой отсек, палубу, реже — целый пузырь. Они устроили гнездо в
гнезде: океан — против мира в целом, пузырь из сплавов и с атмосферой внутри
— против себе подобных. У пузырей замков не предусмотрено — сухопутные конст-
рукторы тревожились о безопасности, — но любители приватности и параноики
приваривают или наращивают укрепления поверх стандартных корпусов.
Кларк не жадина. Она не претендует на многое: одна каюта на верхней палубе
пузыря, заякоренного в шестидесяти метрах северо-восточнее «Атлантиды». Это
чуть больше ее давно сгинувшей каюты на станции «Биб»: может быть, потому-то
она и выбрала это помещение. В нем даже иллюминатора нет.
Она проводит внутри не очень много времени. Собственно, не бывала здесь с
тех пор, как начала трахаться с Уолшем. Но, как бы мало времени она ни прово-
дила в этом тесном, по-спартански обставленном чулане, главное — она знает,
что это ее каюта, что она есть, и никто не войдет сюда без ее позволения. И
убежище всегда под рукой, когда понадобится.
Вот как сейчас.
Кларк сидит голая на матрасе, омытая по дневному ярким светом: в датчиках,
за которыми она следит, все построено на цветах, а она не желает упустить ни
капли информации. Планшетка лежит рядом на неопреновой подушке, настроенная
на внутренности Лени. На экранчике — мозаика зеленых и красных огоньков: кро-
шечные гистограммы, мигающие звездочки, загадочные аббревиатуры. На переборке
напротив — зеркало. Она старается туда не смотреть, но пустые белые глаза то
и дело упираются в свои отражения.
Одна рука рассеянно играет левым соском, другая подносит деполяризирующий
скальпель ко шву на груди. Кожа вдоль шва плавно прогибается, образует склад-
ку, выпуклую геометрическую бороздку: три стороны прямоугольника, заглавное
С, словно формочкой для печенья выдавленное в коже между левой грудью и диа-
фрагмой .
Кларк вскрывает себе грудину.
Она расстегивает ребра вдоль хрящей и отгибает их — легкое сопротивление и
слабое неприятное чмоканье, когда однослойная подкладка расходится по шву.
Тупая боль, когда воздух врывается в грудную полость — на самом деле, это хо-
лод, но нервы внутри тела не отличают температуру от боли. Поработавший над
Кларк механик снабдил петлями четыре ребра в левом боку. Кларк подцепляет
пальцами живую панель и откидывает ее, открывая механизмы. Более острая и
сильная боль стреляет из межреберья, не приспособленного к таким перегибам. В
будущем ее ждут синяки.
Она берет инструмент со стоящего рядом подноса и начинает возиться с собой.
Гибкий кончик глубоко погружается в грудную полость, точно проскальзывает
по узкому как иголка клапану и встает намертво. Она до сих пор дивится, с ка-
кой легкостью нащупывает путь в собственных внутренностях. Рукоять инструмен-
та снабжена колесиком, настроенным на астрономическое передаточное число. Она
сдвигает его на четверть оборота, и насадка проворачивается на долю градуса.
Планшет на матрасе протестующе попискивает: НТР и ГАМК меняют цвет с зеленого
на желтый. Один из столбиков гистограммы чуточку удлиняется, два других уко-
рачиваются .
Еще четверть оборота. Планшетка опять жалуется.
Это до смешного грубое вмешательство: скорее насилие, чем соблазнение. Была
ли настоящая надобность в этих петлях из живого мяса, в Мясницкой работе хи-
рургов, проделавших ей дверцу в груди? Планшет удаленно снимает телеметрию с
имплантатов, связь работает в обе стороны, можно посылать телу команды и при-
нимать от него информацию. Для мелких настроек, изменений в рамках одобренно-
го оптимума, достаточно просто прикоснуться пальцем к экрану и ощутить, как
отзываются механизмы внутри.
Конечно, изменения, которые собирается внести в себя Лени Кларк, «мелкими»
не назовешь.
Работодатели никогда не претендовали на право собственности над телами сво-
их сотрудников — во всяком случае, официально. Но все, что они насовали
внутрь — их собственность. Кларк улыбается своим мыслям: «Могли бы предъявить
мне обвинение в вандализме».
Если они действительно не хотели, чтобы она шарила грубыми лапами в корпо-
ративном имуществе, зачем было оставлять эту сервисную панель в груди? Впро-
чем, они тогда работали в таком цейтноте... Не ждали перебои с электричест-
вом, не ждал «ГидроКвебек», Энергосеть тоже не ждала. Вся геотермальная про-
грамма была спешной, шла с отставанием и в авральном порядке, даже рифтеров
состряпали на скорую руку, чтобы заткнуть прорыв.
Такие, как Лени Кларк, были прототипами, опытными образцами и конечным про-
дуктом в одном лице. Разумно ли запечатывать имплантаты в понедельник, чтобы
уже в среду снова вскрывать тело, добираясь до подлежащей замене мышцы или
устанавливать какой-нибудь жизненно важный компонент, забытый разработчиками?
Даже трупные датчики были установлены задним числом, вспоминает Кларк. Эти
машинки доставил на «Биб» Карл Актон в начале своей вахты. Раздал, как пас-
тилки от горла, приказав всем раскрыться и вставить их рядом со входом для
морской воды.
Карл же первым и обнаружил, как проделывать то, чем занималась сейчас Лени
Кларк. За это Кен Лабин его убил.
«Времена меняются», — размышляет Кларк, меняя еще одну настройку.
Наконец она заканчивает. Позволяет живому клапану встать на место и чувст-
вует, как фосфолипиды затягивают шов. Молекулярные хвосты сплетаются в гидро-
фобной оргии. В груди снова бьется рассеянная боль, чуть отличная от прежней:
дезинфектанты и синтетические антитела впрыскиваются в полость на тот малове-
роятный случай, если откажет прокладка. Изнасилованный планшет сдался: поло-
вина датчиков горят желтым и оранжевым.
В голове у Кларк что-то начинает меняться. На несколько процентов сдвигает-
ся проницаемость важных мембран. Немножко снижается выработка определенных
веществ, предназначенных не для передачи, а для блокировки сигнала. Окна еще
не открылись, но задвижки сняты.
Напрямую она, конечно, ничего не чувствует. Изменения сами по себе необхо-
димы , но не достаточны — они ни на что не влияют здесь, где работают легкие,
где давление — всего-то атмосфера. Для активации нужна тяжесть океана.
Но теперь, когда Лени Кларк выйдет наружу — когда шагнет за край шлюза, и
давление сомкнётся вокруг нее жидкой горой, когда триста атмосфер стиснут го-
лову так, что синапсы начнет коротить, — тогда Кларк сумеет заглядывать в ду-
ши людей. Конечно, не в светлую часть. Никакой философии, музыки, альтруизма
и интеллектуальных рассуждений о добре и зле. Вообще ничего связанного с не-
окортексом. Лени Кларк будет улавливать то, что старше на сто миллионов лет.
Гипоталамус, ретикулярная формации, миндалина. Мозг рептилии, средний мозг.
Ревность, голод, страх и бессловесная ненависть. Все это она будет ощущать на
пятнадцати метрах и более.
Гиппокамп Мамиллярное
(правый) тело
Она помнит, каково это. Слишком хорошо помнит. Шесть лет прошло, а словно
вчера.
Осталось только шагнуть наружу.
Она сидит в своей каютке и не движется с места.
Могильщики
Ищите чертовы мины!
Они рассыпались по участку черными псами, вынюхивая на свету и в темноте,
сонарными пистолетами и детекторами течений. Кто-то мог сомневаться в успехе
— а кое-кто почти наверняка надеялся на поражение, — но у всякого, кто выжил
здесь пять лет, хватало ума не перечить Кену Лабину.
Ищите чертовы мины.
Кларк скользит среди них: на взгляд со стороны — просто еще один нос, ут-
кнувшийся в след. Только в ней нет сосредоточенности. Другие следуют вдоль
невидимых линий, нитей правильной сети, протянутой по району поиска, а Кларк
движется зигзагами, пристраивается то к одному, то к другому, обменивается
непринужденными гудками реплик и снова уходит в сторону, к следующему. У
Кларк другая цель.
Ищи чертова минера.
Гектары биостали. Перемежающиеся отрезки света и тени. Стаккато вспышек на
каждом выступе, мигающие маячки отмечают концы опор, антенн, опасные зоны,
где могут внезапно происходить горячие выбросы. Гневные немигающие взгляды
прожекторов у шлюзов и причалов, люков и погрузочных отсеков, зажженные ради
сегодняшних поисков. Бледные ауры света из сотен параболических иллюминато-
ров . Сумеречные пространства корпуса, где каждая выпуклость отбрасывает три-
четыре тени в размытом свете далеких фонарей.
Остальное темно. От голых опорных стоек падают продолговатые сетки теней.
Непроглядные чернильные лужи заполняют пространства между килем и дном, слов-
но «Атлантида» — огромная кровать, под которой прячутся жуткие монстры. Не-
четкие темные пятна там, где свет постепенно сходит на нет; резкие там, где в
солнечное натриевое сияние вторгается тень бака или трубы. С таким ландшафтом
несложно спрятать взрывное устройство размером с два кулака. Тут их можно
спрятать тысячи.
Для пятидесяти восьми человек это была бы большая работа. Для двух дюжин,
подписанных Лабином на это задание, — намного больше. Здесь рифтеры, еще не
отуземившиеся, не настолько захваченные ненавистью к корпам, чтобы «случайно
не заметить» подозрительный предмет, — рифтеры, среди которых едва ли окажут-
ся подложившие эти устройства. Наверняка, конечно, не скажешь: немногие из
этих людей свободны от подозрений. Даже сведения, украденные прямо из их моз-
гов, не дадут точного ответа. Гидрокожу и глаза выдавали только тем, у кого
был определенный опыт. Именно сбой в мозговой проводке делает людей годными
для рифта. Здесь у каждого свои призраки. Каждый таскает за собой груз: мучи-
телей, жертв, наркомании, побоев, анального насилия, добреньких «людей в чер-
ном» с их отеческими увещеваниями. И ненависть к корпам, совсем недавно еще
остывшая, снова стала всеобщей. Бета-макс вывел на поверхность старые кон-
фликты. Воспламенил вражду, полузатушенную пятью годами угрюмого притирания
друг к другу. Месяц-другой назад корпы с рифтерами были почти союзниками, не
считая озлобленных упрямцев вроде Эриксона и Нолан. Теперь, раздави океан
всех корпов до единого, немногие станут их оплакивать.
И все равно. Одно дело — плясать на чьей-то могиле, другое — эту могилу ко-
пать . Тут поверх ненависти всплывает элемент расчета. Отличие тонкое, Кларк
не уверена, что она или Лабин сумеют уловить его при таких обстоятельствах.
Оно может и не проявиться до того мгновения, как человек найдет искомое: уви-
дит мину, торчащую на корпусе апокалиптическим моллюском, включит вокодер,
честно собираясь подать сигнал тревоги, и тут. . . «Может, мерзавцы этого за-
служивают — после всего, что они сотворили с нами и с целым миром, а мне и
делать ничего не надо, мог ведь я просто не заметить ее под опорой, в такой
мути и...»
Мысли могут быть совершенно невинными — даже перед собой — вплоть до того
момента, когда включится в работу финальный стимул, запускающий простую це-
почку рассуждений, итог которой — отведенный в сторону взгляд. И кто знает,
возможно ли уловить его даже с помощью тонкой настройки?
Только не Лени Кларк. Но все-таки она ищет, скользит между корпусами и цис-
тернами, парит над своими товарищами и подобно им вглядывается в свет и в
темноту, отличаясь от них лишь целью охоты.
Эта цель — чувство вины.
Конечно, не простой вины. Вины, скатывающейся к страху разоблачения, креня-
щейся к праведному гневу. Заново пробудившись, Кларк плавает в котле подер-
жанных эмоций. Вода в нем загрязнена дюжиной разных страхов, гневом, отвраще-
нием к себе и другим. Есть и своеобразное возбуждение, азарт погони, посте-
пенно выцветающий до привычной скуки. И сексуальные порывы. И еще менее выра-
женные чувства, которых она не различает.
Она не забыла, почему отказывалась от тонкой настройки на Чэннере, даже ко-
гда на нее согласились другие. А теперь вспоминает, почему соблазн был так
велик, что она, в конце концов, поддалась. В этой бесконечной сумятице чувств
вечно теряешь свои собственные.
К сожалению, здесь, на Хребте, все немного иначе. Не то чтобы изменилась
физика или неврология. Или кто-то из людей. Другой стала сама Лени Кларк.
Жертва и месть вылиняли с годами, черное и белое слились в миллионы неразли-
чимых оттенков серого. Ее психика отклонилась от нормальной для рифтера, и ей
теперь сложнее встроиться в этот фон. Чувство вины до того сильное, что на-
верняка может исходить только от нее.
И все же она не сворачивает с курса. Продолжает охоту, хотя чувства приту-
пились. Где-то невдалеке скрытый дифракцией Кен Лабин делает то же самое. Он,
вероятно, лучше нее справляется с делом. Его этому обучали. За ним многолет-
ний опыт.
Что-то зудит на периферии сознания. Далекий голос пробивается сквозь туман
в голове. Она осознает, что чувствует его довольно давно, но громкость нарас-
тала так постепенно, что он зафиксировался в мозгу только сейчас. Ошибки нет:
угроза, вскрик, возбуждение на самой границе восприятия. Двое рифтеров дви-
жутся ей наперерез, уходят к югу, работая ногами. Челюсть Кларк гудит от во-
кодированных голосов — задумавшись, она и их не замечала.
— Чуть не пропустил, — говорит кто-то. — Запихнули под...
— Еще одна, — прорывается второй голос. — Отсек А.
Кларк с первого взгляда понимает, что она бы наверняка пропустила. Стан-
дартный взрывной заряд установлен в тени нависающего уступа. Лени всплывает
лицом вверх и прижимается головой к обшивке, чтобы смотреть вдоль корпуса. И
видит полукруглый силуэт в тени, подсвеченный мутным мерцанием воды.
— Господи, — жужжит она, — как ты высмотрел эту чертовщину?
— Сонаром поймал.
Рифтеры, с типичным для них уровнем дисциплины, побросали свои сектора и
слетелись на находку. Лабин их не гонит: есть очевидная причина собрать их
сюда, к орудию убийства. Кларк настраивается и концентрируется. Волнение.
Воспрянувший интерес после часа монотонных хождений кругами. Озабоченность и
ниточки нарастающего страха: что ни говори, это бомба, а не пасхальное яичко.
Кое-кто из робких уже подается назад, осторожность пересиливает любопытство.
Кларк лениво прикидывает радиус поражения. Метров сорок или пятьдесят счита-
ется безопасной дистанцией при обычных взрывных работах, но в правилах безо-
пасности всегда закладывается запас.
Она сосредотачивается. Подозреваются все. Но хотя вездесущие паутинки яро-
сти как всегда поблескивают на общем фоне, на поверхность они не выходят ни у
кого. Не ощущается явного гнева из-за сорванных планов, нет страха неизбежно-
го разоблачения. Обнаруженная взрывчатка для этих людей скорее головоломка,
чем провокация — под маской охоты скрывалась игра в русскую рулетку.
— и что будем делать? — спрашивает Чун.
Лабин парит над ними Люцифером.
— Всем отметить сонарный профиль. По нему будете опознавать другие: они то-
же наверняка недоступны визуальному осмотру.
Дюжина пистолетов щелкает, обстреливая опасную находку.
— А мы ее здесь оставим или нет?
— А если она с ловушкой?
— А если взорвется?
— Меньше корпов — меньше головной боли, — жужжит Гомес с расстояния, кото-
рое счел безопасным. — Я за них шкуру рвать не стану.
Лабин не мучится с догадками и заглядывает под уступ.
Ын шарахается от него.
— Эй, стоит ли...
Лабин хватает и срывает устройство. Никаких взрывов. Обернувшись, он огля-
дывает собравшихся рифтеров.
— Когда найдете остальные, не прикасайтесь к ним. Я сам сниму.
— Чего париться? — тихо жужжит Гомес.
Это риторическое ворчание, в нем нет серьезного вызова, и все же Лабин от-
вечает .
— Расположена неудачно, — говорит он. — Место выбрано из соображений маски-
ровки , а не эффективности. Мы могли бы лучше.
Со всех сторон вспыхивают яркие импульсы. А у Кларк слова Лабина словно
прорвали дырочку в гидрокостюме, и ледяная вода Атлантики ползет по спине.
«Ты что творишь, Кен. Какого хрена?»
Она уверяет себя, что он просто подыгрывает, дает им стимул для работы. Ла-
бин теперь смотрит на нее, чуть заметно склоняет голову, словно отвечая на
невысказанный вопрос. И Кларк с запозданием понимает, что сделала: попыталась
заглянуть к нему в голову. Прощупать его тонкой настройкой.
Конечно, это тщетная попытка. И даже опасная. Лабин мало того, что обучен
блокировать вторжения чужого разума: он натренирован до рефлекса, перестроен,
снабжен подсознательной защитой, которую нельзя снять усилием воли. Никому не
удавалось пробраться в голову Лабина, кроме Карла Актона, а то, что увидел
там Карл, он унес с собой в могилу.
Сейчас Лабин наблюдает за ней, непроницаемый для ее бессознательного прощу-
пывания и внутри, и снаружи.
Она вспоминает про Актона и останавливает себя.
Стриптиз
Конечный итог — девять мин и ни одного подозреваемого. Оба результата еще
могут измениться.
Сама по себе «Атлантида» — упражнение в масштабной инвариантности, система
дополнений к модификациям усовершенствований, надстроенных поверх основного
массива, распростершегося на несколько гектаров. Нечего и думать, что загля-
нули во все уголки. Опять же, много ли шансов, что заговорщики — ограниченные
временем, наблюдением и, будем надеяться, малой численностью — имели больше
возможностей для закладки мин, чем поисковая партия — для их обнаружения? Ни
одна сторона не всесильна. Этого, пожалуй, достаточно для равновесия.
Что до поиска заговорщиков, Кларк пока проверила три дюжины. Она запустила
пальцы в вязкую темноту голов и ничего не нашарила. Даже у Гомеса и Йегера.
Даже у Кризи! Плясать на могиле, конечно, готовы все. Но не копать.
Хотя с Грейс Нолан она в последнее время не сталкивалась. Нолан сейчас —
Большая Красная Кнопка. Она держится на заднем плане: в свете последних собы-
тий предполагаемое предательство корпов выглядит не столь уж ассиметричным
ответом. Но учитывая, как идут дела, Нолан ничего не теряет, разыгрывая свою
партию. Уже сейчас более чем достаточно народа сочувствует Чокнутому Подрыв-
нику, и если им окажется Нолан, разоблачение скорее повысит ее статус, чем
повредит ему.
Поводок натянут до отказа. Если он лопнет, в вентилятор полетит сразу де-
сять сортов дерьма.
И это еще при милосердном допущении, что виновников можно отыскать. Чего ты
ищешь в темных подвалах стольких умов? Там даже невинных гложет вина, и даже
виновные упиваются своей праведностью. Каждый разум подсвечен черным сиянием
психического насилия — где следы старых ран, где недавнее преступление? Ино-
гда удается вычислить, если хватает духу совать голову в чужую смоляную яму,
но контекст определяет все. Надежда на удачу — лотерея. Чтобы сделать все
правильно, нужно время, и приходится сильно пачкаться.
Если этого не делать, будущее останется в руках Грейс Нолан.
«Нет времени. Я не могу быть сразу везде. И Кен тоже».
Конечно, есть альтернатива. Лабин предложил ее сразу после окончания поис-
ков . И был так любезен, что сделал вид, будто у нее есть выбор. Как будто,
откажись она, он бы не сделал этого сам.
Кларк знает, почему он предоставляет выбор ей. Тот, кто поделится этим сек-
ретом, повысит свой вес среди местных. Лабину кредит доверия не нужен — ни
один рифтер не сошел с ума настолько, чтобы ему противоречить.
Она еще помнит время, не такое уж давнее, когда могла сказать то же самое о
себе.
Лени вздыхает и выходит на связь с теми, кого это касается. И понимает, что
следующий шах1 может ее убить. Попутно задумываясь — далеко не в первый раз, —
так ли это плохо.
Слушателей у нее меньше дюжины. Места много: пузырь лазарета — даже та одна
сфера, которую не занял Бхандери, — больше других. Собрались даже не все, ко-
го Кларк с Лабином, обменявшись впечатлениями, сочли достойными доверия. Она
решила начинать с малого — так немножко проще. Круги на воде скоро захватят и
других.
— Я не стану показывать дважды, — говорит Кларк, — так что прошу внимания.
Обнажившись до пояса, она снова вскрывает себя.
— Не меняйте ничего, кроме нейроингибиторов. Возможно, это нарушит баланс
еще каких-то веществ, но, по-видимому, эффект постепенно размывается. После
перестройки просто ненадолго выйдите наружу, чтобы все устоялось.
— На сколько времени? — спрашивает Александр.
Кларк сама не знает.
— Часов на шесть, наверное. После этого вы готовы. Кен распределит вас по
разным пузырям.
Аудитория ропщет — перспектива долгого заключения никого не радует.
— И как же настраивать ингибиторы?
Сломанный нос Мак прикрыт тонкими проводками с бусинами — микроэлектриче-
ская сеть ускоряет восстановление. Выглядит это смешно — как севшая от стирки
траурная вуаль.
Кларк невольно улыбается.
— Понижать.
— Шутишь!
— И не думаю.
— А как же Андре?
Андре умер три года назад: жизнь вышла из него с подводными судорогами, ед-
ва не разорвавшими тело на куски. Седжер сочла причиной отказ нейроингибито-
ров . Человеческие нервы не приспособлены к глубине — давление делает их чув-
ствительными к малейшему воздействию. Включается живой рубильник без прерыва-
теля и без изоляции. После нескольких минут предсмертных спазмов тело расхо-
дует все нейротрансмиттеры и попросту останавливается.
Вот почему имплантаты рифтеров, как только давление превышает определенный
уровень, накачивают тело нейроингибиторами. Без них выход наружу на таких
глубинах смертелен, как электрический стул.
— Я сказала «понижать», — подчеркивает Кларк, — а не «отключать». На пять
процентов. Самое большое, на семь.
— и что же из этого выйдет?
— Снижается порог включения синапсов. Нервы становятся просто немножко...
чувствительнее. Чувствительнее к мелким стимулам в водной среде. Вы станете
замечать то, чего не воспринимали прежде.
— Например? — интересуется Гарсиа
— Например... — начинает Кларк и смолкает.
Ей вдруг хочется закрыться и отрицать все.
Хочется сказать: «Забудьте. Неудачная идея. Глупо пошутила. Забудьте все,
что я сказала».
А может, вообще — признаться? «Вы не представляете, чем рискуете. Не знае-
те , как легко шагнуть за край. Мой любовник не мог даже войти в пузырь, не
ощущая ломки — даже дышать не мог, так ему хотелось разнести все, что стояло
между ним и глубиной. Мой друг совершил убийство, чтобы обрести уединение
там, где проплывая рядом с другим, обязательно заглотишь все его болячки и
беды. Он и ваш друг, он один из нас, и он — единственный из живых на всей
больной, одуревшей планете, кто знает, что с вами от этого будет...»
Она в панике озирается, но среди присутствующих нет Кена Лабина. Вероятно,
он сейчас составляет расписание вахт для «настроенных».
«Однако, — вспоминает она, — к этому привыкаешь».
Переведя дыхание, Кларк отвечает на вопрос Гарсиа:
— Например, ты сможешь определить, когда тебя водят за нос.
— Вот черт, — ворчит Гарсиа. — Стану ходячим детектором вранья?
— Ты такой и есть, — натужно улыбается Кларк.
«Надеюсь, ты к этому готов...»
Ее послушники расходятся по своим пузырькам, чтобы похимичить с собой.
Кларк закрывает грудь. К тому времени как она натягивает черную «кожу», лаза-
рет уже опустел, остались лишь следы мокрых ног и тяжелый люк — до недавнего
времени всегда открытый, — ведущий в соседнюю сферу. Гарсиа, презрев сухопут-
ные требования безопасности, наварил на него цифровой замок.
«Сколько мне осталось, — спрашивает себя Кларк, — до времени, когда каждый
сможет влезть мне в голову?»
Не меньше шести часов, если послушники всерьез отнесутся к ее оценке. Потом
они начнут играть, испытывать новые сенсорные способности, возможно, даже на-
слаждаться ими, если не проникнутся отвращением к тому, что обнаружат. Но-
вость станет распространяться.
Кларк подала это как психический шпионаж, новый способ выследить преступные
тайны, которые, вероятно, скрывают корпы. Впрочем, пределами «Атлантиды» дело
не ограничится. Теперь всем будет намного трудней строить заговоры в темноте
— ведь каждый прохожий вооружится фонариком.
Она ловит себя на том, что застыла на входе в логово Бхандери, положив руку
на переделанный замок. Набрав нужный код, она открывает люк.
Внезапно включается цветное зрение. Герметическая прокладка окаймляет про-
ход глубокой стальной синевой. Над головой коралловыми аспидами вьются трубы
с цветовой разметкой. Цилиндр с каким-то сжатым газом, видный сквозь проем,
отражает бирюзовый свет, шкала на нем желтая и — подумать только! — ярко-
розовая .
Здесь светло, как в «Атлантиде».
Кларк выступает на свет: циркулятор Кальвина, матрас, банк крови сочатся
красками.
— Рама?
— Закрой дверь.
Нечто скрючилось перед рабочей панелью, прокручивая цепочки радужных нук-
леотидов. Оно не может быть рифтером: ни общей ауры, ни блестящей черной ко-
жи.
Существо больше похоже на скелет в одном белье. Оно оборачивается, и Кларк
внутренне вздрагивает: у него даже глаз нет! На лице Бхандери вздрагивают
темные зияющие дыры зрачков, почти вытеснивших радужку.
Значит, не так уж здесь светло. Довольно темно для глаз без линз, их прихо-
дится напрягать до предела. Столь тонкие различия теряются за мембранами, ко-
торые обеспечивают миру оптимальную освещенность.
Должно быть, что-то отражается у нее на лице.
— Я вынул линзы, — говорит Бхандери. — Глаза... перевозбуждаются от стиму-
ляторов . . .
Голос его до сих пор звучит хрипло, связки не адаптировались к воздушной
среде.
— Как дела? — спрашивает Кларк.
Пожатие тощих плеч. Даже сквозь футболку у него ребра можно пересчитать.
— Хоть что-нибудь? Диагностический критерий, или...
— Я не сумею отыскать различий, пока не удостоверюсь, что они есть. Пока
что это выглядит как Бетагемот с парой новых шовчиков. Может, мутация, может
— перестройка. Пока не знаю.
— А первичные образцы тебе помогут?
— Первичные?
— Те, что не прошли через «Атлантиду». Если ты получишь образец с Невозмож-
ного озера, сумеешь сравнить? Проверишь, есть ли различия...
Он качает головой — вернее, дергает, вздрагивает.
— Есть способы выявить перестройку. Сателлитные маркеры, цепочки мусорных
генов. Просто это требует времени.
— Но ты сможешь? Стимуляторы... сработали. Ты вспомнил.
Кивок — как выпад змеи. Он вызывает на экран еще одну цепочку.
— Спасибо тебе, — тихо говорит Кларк.
Он замирает.
— Спасибо? А у меня есть выбор? На люке замок.
— Я знаю, — она опускает глаза. — Мне жаль.
— Вы думаете, я бы ушел? Уплыл бы, оставив эту штуку убивать нас? А может,
и меня?
Она мотает головой.
— Нет. Ты бы не ушел.
— Тогда зачем?
При всей неподвижности его лицо — как сдавленный крик. За спокойной скоро-
говоркой слов в глазах застыл абсолютный ужас. Как будто в них что-то еще,
древнее, бездумное и лишь недавно пробудившееся. Оно через сотню миллионов
лет смотрит в непостижимый мир прямых углов и мигающих огней — и находит его
совершено непригодным для жизни.
— Потому что у тебя приступы, — напоминает Кларк. — Ты сам говорил.
Он протягивает тонкую как палочка руку, покрытую дермами — насос пониже
локтя качает ему препараты прямо в вену. Он подстегивает себя с тех самых
пор, как забрался в атмосферу, использует чудеса современной химии, чтобы си-
лой загнать здравый рассудок обратно в голову, выволочь на поверхность уто-
нувшие воспоминания и навыки. Пока, надо признать, это работает. Но стоит по-
смотреть ему в глаза, и на твой взгляд отвечает рептилия.
— Мы не можем так рисковать, Рама. Прости.
Он опускает руку. Челюсть у него щелкает, как странное насекомое.
— Ты говорила... — начинает он и замолкает.
Потом начинает заново:
— Когда ты тащила меня сюда. Ты сказала, что была знакома с...
— Да.
— Я не знал таких... в смысле, кто?
— Не здесь, — отвечает она. — Даже не в этом океане. В самом начале рифтер-
ской программы. Он ушел у меня на глазах. — Пропустив один удар сердца, она
заканчивает: — Его звали Джерри.
— Но ты сказала, он вернулся.
Она действительно не понимает. Джерри Фишер просто возник из темноты после
того, как остальные сдались и ушли. Он дотащил ее до безопасного места, к
эвакуационному скафу, неуверенно зависшему над станцией, где уже не осталось
персонала. Но не сказал ни слова, а потом лягался и отбивался, как зверь, ко-
гда она в свою очередь попыталась спасти его.
— Наверно, он не столько вернулся, сколько прошел насквозь, — признается
она этому существу, которое должно, на свой манер, понимать Джерри Фишера ку-
да лучше нее.
Бхандери кивает.
— Что с ним случилось?
— Он погиб, — говорит она тихо.
— Просто... рассеялся? Как все мы?
— Нет.
— Тогда как?
Слово отзывается в ней привычным эхом.
— Бабах! — говорит она.
Фронтир
«Уходи, — сказали они после Рио. — Спас наши задницы и в этот раз — теперь
уходи».
Это было не совсем так. Буффало он не спас. Не спас и Хьюстон. Солт-Лейк,
Бойсе и Сакраменто погибли от импровизированных атак, в диапазоне от авиалай-
неров-камикадзе до ядерной бомбардировки с орбиты. Пяток других филиалов ды-
шали на ладан. Там спаслось очень немного задниц.
Но для всего Патруля Энтропии Ахилл Дежарден был десятикратным героем. Поч-
ти сразу стало очевидно, что пятьдесят филиалов УЛН по всему западному полу-
шарию подверглись внезапной и одновременной атаке, но Дежарден и только Де-
жарден сложил фрагменты головоломки — под огнем, на лету. Это он пришел к не-
вероятному заключению, что атака организована кем-то из своих. Остатки Патру-
ля собрались на зов и расплющили Рио, но куда целить, им сказал Дежарден. Без
его стойкости под давлением все твердыни УЛН в этом полушарии сгорели бы дот-
ла.
«Уходи, — сказали ему благодарные хозяева. — Этот город списан».
Цитадель УЛН в Садбери получила прямое попадание в бок. Суборбитальный пры-
гун, направлявшийся из Лондона в Торомильтон и сбитый врагом с курса, оставил
в северном фасаде здания кратер высотой в десять этажей. Топливные баки у не-
го были почти пустыми, так что пламя охватило не все строение. Сгорели, по-
гибли от яда или удушья лишь те, кто находился между восемнадцатым и двадцать
пятым этажом. Старшие правонарушители Садбери размещались с двадцатого по
двадцать четвертый. То, что Дежарден успел поднять тревогу до попадания, было
удачей. То, что не погиб вместе с остальными — откровенным, охрененным чудом.
«Уходи».
Тогда Ахилл Дежарден осмотрелся в дыму и пламени, бросил взгляд на штабеля
мешков с телами и немногих контуженных сотрудников, уцелевших в достаточной
степени, чтобы избежать предписанной эвтаназии, и ответил: «Я вам нужен
здесь».
«Нет никакого „здесь"».
Но от «здесь» осталось больше, чем от Солт-Лейк или от Буффало. Атака со-
кратила штат быстрого реагирования Нт АмПацифика более чем на треть. Садбери
висел на волоске, но этот волосок связывал шестнадцать полушарных и сорок
семь региональных узлов. Полностью покинуть его означало сокращение системы
еще на пять процентов и полмиллиона квадратных километров, оставленных вообще
без сил реагирования. Бетагемот уже свирепствовал на половине континента;
царство цивилизации уступало и сжималось. УЛН не могла позволить себе новых
потерь.
Однако имелись доводы и с другой стороны. Половина этажей вышла из строя.
Оставшейся широты частот хватило бы на жалкую горстку оперативников, а теку-
щий бюджет едва позволял поддерживать даже то, что осталось. Все модели схо-
дились в одном: наилучший выход — покинуть Садбери и возместить потерю расши-
рением Торомильтона и Монреаля. И сколько времени, задумался Дежарден, прой-
дет, пока новые отделы войдут в строй?
Шесть месяцев. Если не год.
То есть им требовался вариант на это время. Чтобы огонек погорел еще не-
множко. Требовался кто-нибудь на случай тех непредсказуемых кризисов, с кото-
рыми не справляются машины.
— Но ведь ты — наш лучший правонарушитель! — возражали они.
— А это задание почти невыполнимо. Где мне еще место, как не здесь?
— Н-ну... — мялось начальство.
— Всего шесть месяцев, — напомнил он. — Или год.
Конечно, так никогда не бывает. Шаловливая ручонка Мерфи взболтает варево,
и «около года» превратится в три, а там и в четыре. Расширение Торомильтона
забуксует, дальновидные планы начнут, как всегда, срываться под тяжестью бес-
конечных непредвиденных обстоятельств. В Патруле Энтропии как-нибудь наскре-
бут по крошке средства, чтобы огонек в Садбери горел, коды допуска действова-
ли, не уставая благодарить безропотного служащего, который тысячью пальцев
затыкает дырочки в плотине.
Но то сейчас, а тогда Дежарден втолковывал им:
— Я буду для вас смотрителем маяка. Часовым на передовом посту. Подам сиг-
нал и удержу позицию, пока на помощь не придет кавалерия. Мне это по силам,
вы же знаете.
Они знали, ведь Ахилл Дежарден был героем.
Что еще важнее, он был правонарушителем; он не смог бы солгать им при всем
желании.
— Какой парень! — говорили они, восхищенно покачивая головами. — Какой па-
рень !
Подготовительные
работы
Кевин Уолш — хороший мальчик. Он знает, что над отношениями надо работать,
и готов потрудиться, чтобы раздуть искорку — уж какая есть. Или, по крайней
мере, подольше не дать ей погаснуть.
Он прицепился к ней после того, как Лабин расписал по местам первых «тонко
настроенных», и не желал слышать никаких «потом» и «может быть». Наконец
Кларк смилостивилась. Они отыскали незанятый пузырь и бросили на пол пару
матрасов, и он безропотно работал языком, а еще большим и указательным паль-
цем, пока вымотался совсем, а Кларк не собралась с духом, чтобы его остано-
вить. Она погладила его по голове и сказала, что это было приятно, хотя ниче-
го и не вышло, и предложила ему свои услуги, но он отказался — то ли из ры-
царственного раскаяния за свою непригодность, то ли просто дулся.
Теперь они лежат рядом, слегка переплетя руки. Уолш спит, что удивительно —
он любит спать при силе тяжести не больше других рифтеров. Может быть, это
тоже из рыцарства. Может, он притворяется.
У Кларк даже на притворство нет сил. Она лежит на спине, уставившись на ка-
пельки конденсата на переборке. Немного погодя высвобождает руку — нежно,
чтобы не испортить спектакль — и отходит к местной панели связи.
На главном дисплее смутный таинственный обелиск, поднимающийся с морского
дна. Главный генератор «Атлантиды». Во всяком случае, его часть — основная
масса погружена в скальное основание, в сердце источника, которым он питает-
ся, словно сосущий кровь комар. Над грунтом поднимается только вершина: буг-
ристый небоскреб с фасадом, изъеденным трубками, вентиляционными отверстиями
и клапанами. Скупая цепочка прожекторов опоясывает его на восьмиметровой вы-
соте , их яркое сияние окрашивает все в медный цвет. Глубина прижимает это
сияние черной ладонью — верхушка генератора уходит в темноту.
На уровне дна из него выходит кабель толщиной с канализационную трубу и
змеей скрывается во мраке. Кларк рассеяно вызывает на экран следующую камеру.
— Эй, ты что там?..
Голос у Кевина вовсе не сонный.
Она оборачивается. Уолш приподнялся на коленях, словно собирался встать и
застыл на полдороге. Впрочем, он не шевелится.
— Давай, возвращайся. Попробую еще разок.
Он расплывается в мальчишеской ухмылке. Маска: «Обезоруживающе милый со-
блазн» . Она разительно противоречит позе, которая приводит на память одинна-
дцатилетку, пойманного за мастурбацией на чистых простынях.
Кларк с любопытством разглядывает его:
— Что с тобой, Кев?
Он смеется — смех звучит как икота.
— Ничего. Просто мы... ну это... не закончили.
Тусклый серый комок застревает у нее в горле, когда она понимает.
Для проверки Кларк снова оборачивается к панели и переходит к следующей ка-
мере наблюдения. Кабель виляет среди смутной геометрии теней и теряется вда-
ли.
Уолш дергает ее за плечо, обнимает сзади.
— На выбор леди. Временное предложение, срок скоро истекает...
Следующая камера.
— Ну же, Лен!
«Атлантида». Кучка рифтеров собралась у стыка двух секций, подальше от пре-
дусмотренных наблюдательных пунктов. Кажется, проводят некие измерения. Неко-
торые нагружены чем-то странным.
Уолш замолкает. Ком в горле у Кларк выбрасывает метастазы.
Она оборачивается. Кевин Уолш пятится от нее, на лице смесь вины и непокор-
ности .
— Ты бы дала ей попробовать, Лен, — говорит он. — Ну, посмотрела бы на это
более объективно...
Она отвечает холодным взглядом.
— Поганец.
— Да ладно! — вспыхивает он. — Как будто я для тебя когда-то что-то значил!
Она подхватывает разбросанные куски подводной кожи. Те скользят по телу как
живые, сливаются друг с другом, запечатывают ее внутри — благословенная жид-
кая броня, надежная граница между «нами» и «ими».
«Только никаких „нас" нет, — понимает она. — Никогда и не было».
Она всерьез злится на себя за то, что забыла об этом, что совсем не предви-
дела такого поворота, хотя и доступ к мозгам любовничка у нее имелся, и меси-
во из вины, боли и дурацкого мазохизма так и рвалось оттуда, и все же она не
вычислила неизбежного предательства. Конечно, она ощущала его озлобленность и
обиду, но в этом не было ничего нового. Собственно, откровенное предательство
ничего не меняло в их отношениях — нечего было и замечать.
Она спускается в шлюз, не оглядываясь на него. Кевин Уолш — еще один мерз-
кий мальчишка. Хорошо, что она не успела к нему привязаться.
Слова гудят среди теней огромного строения: числа, степени, показатели
сдвигового напряжения. У пары рифтеров с собой планшетки, другие палят очере-
дями высокочастотных звуков — через акустические поисковики. Один выводит
большой черный крест на уязвимом месте стены.
Как там выразился Кен? «Ради скрытности, а не эффективности». Очевидно,
этой ошибки они решили не повторять.
Конечно, ее ожидают. Уолш их не предупреждал — по крайней мере, по открытым
каналам связи, — но к «тонко настроенным» невозможно подкрасться незаметно.
Кларк озирает всю компанию. Нолан тремя метрами выше смотрит на нее сверху
вниз. Крамер, Чун и Гомес свободно рассыпались вокруг. Кризи с Йегером —
слишком далеко для визуальной идентификации, но рисунок сознаний воспринима-
ется достаточно отчетливо, — устроились на корпусе подальше.
Вибрации Нолан заглушают все прочие: на месте прежней злобы теперь торжест-
во . А вот гнев — чувство, что счеты еще не сведены — остался прежним.
— Не вините Кева, — жужжит им Кларк. — Он старался как мог.
У нее мелькает мысль, далеко ли зашла Нолан в стремлении закрепить его вер-
ность .
Нолан демонстративно кивает:
— Кев — хороший мальчик. Для группы готов на все.
В машинную речь просачивается подчеркнутое «на все» — впрочем, Кларк уже
уловила это в живом мозгу.
«Значит, вот как далеко ...»
Она заставляет себя забраться глубже, выискивая вину и двоемыслие, но это,
разумеется, бесполезно. Если Нолан когда и таила подобные секреты, теперь все
позади. Сейчас она выставляет свои намерения напоказ, как орден.
— Так что же тут происходит? — спрашивает Кларк.
— Всего лишь готовимся к худшему, — отвечает Нолан.
— Угу. — Кларк кивает на крест на обшивке. — Готовитесь или провоцируете?
Все молчат.
— Вы же знаете, что генератор под нашим контролем. Мы можем отрезать их в
любой момент. Взрыв корпуса — явный перебор.
— О, мы не станем применять излишнюю силу. — Это Крамер отзывается откуда-
то слева. — Тем более что они всегда действовали так мягко.
— Просто сочли, что разумнее иметь запасные варианты, — жужжит Чен, винова-
то, но твердо. — На случай, если план А почему-то сорвется.
— Например?..
— Например, если чьи-то ручки выдернут хрен из кусачего ротика, — говорит
Гомес.
Кларк разворачивается к нему лицом.
— Как всегда внятно, Гомер. Понимаю, почему ты неразговорчив.
— На твоем месте... — начинает Нолан.
— Заткнись на хрен!
Кларк медленно поворачивается от одного к другому, в животе у нее медленно
закипает лед.
— Если они что с вами сотворили, то сначала сотворили и со мной. Если в вас
и бросали дерьмом, то в меня бросали больше. Намного больше.
— Но вот попадало оно куда угодно, только не на тебя, — напоминает Нолан.
— Думаешь, я стану лизать им задницы только потому, что они промахнулись,
когда хотели меня убить?
— А не станешь?
Она подплывает ближе, пока не оказывается лицом к лицу с Нолан.
— Не смей опять сомневаться в моей лояльности, Грейс. Я попала сюда раньше
вас всех, жалкие гаплоиды. Вы еще скулили и ссались на бережку, маясь без ра-
боты, а я вломилась в их сраный замок и лично отпинала Роуэн с ее дружками.
— Это верно. Только два дня спустя ты вступила в их женский клуб. Бога ра-
ди, ты играла в виртуальные игры с ее дочерью!
— Да ну? А чем же провинилась ее дочь, чтобы обрушивать ей на голову всю
Атлантику? Даже если ты права — даже будь ты права — чем тебе ребятишки на-
вредили? Чем перед вами провинились их родные, их слуги и мойщики туалетов?
Вибрация слов гаснет вдали. Низкое, почти субзвуковое гудение какого-то ап-
парата после этого кажется особенно громким.
Может быть, сейчас в их коллективном изучении мелькает доля неуверенности.
Даже и в Нолан чуть-чуть.
Но она не отступает ни на микрон.
— Хочешь знать, чем они провинились, Лен? Они выбрали не ту сторону. Жены,
мужья, медики и даже мойщики туалетов, которых эти скоты оставили при себе на
память о прошлом. Все они выбрали, на чьей стороне быть. О тебе я и этого
сказать не могу.
— Мысль так себе, — жужжит Кларк.
— Спасибо, что высказала свое мнение, Лен. Мы дадим тебе знать, если пона-
добишься . А пока не стой у меня на дороге. Мне при виде тебя блевать хочется.
Кларк выкладывает на стол последнюю карту.
— Ты бы не обо мне беспокоилась.
— А с чего ты взяла, что мы беспокоились о тебе? — Презрение расходится от
Нолан волнами.
— Кен бывает очень недоволен, когда его замешивают в такие вот провальные
затеи. Я это повидала. Этому парню много проще все зарубить на корню, чем по-
том наводить порядок. Вам бы с ним разобраться.
— Уже, — жужжит Нолан. — Он в курсе.
— Даже дал нам несколько наводок, — добавляет Гомес.
— Прости, милочка. — Нолан вплотную придвигается к Кларк, их капюшоны
скользят, соприкасаясь в манекеньей ласке. — Но, право, ты должна была это
предвидеть.
Вся группа молча возвращается к работе, словно по невидимой и неслышимой
для Лени Кларк команде. Она висит в воде, оглушенная и преданная. Вокруг со-
бираются клочки и обломки благонамеренных замыслов.
Она разворачивается и плывет прочь.
Бомбиль
Давным-давно, во времена восстания, пара корпов захватила минисубмарину под
названием «Бомбиль-3». Патриция Роуэн и посейчас не представляет, чего они
добивались. На «Бомбиле» не осталось ничего, способного служить для разруше-
ния или убийства. Подлодка была голой, как рыбий скелет, и примерно настолько
же полезной: рубка спереди, лопасти сзади, и оголенный соединительный сегмент
посередине.
Может, они просто надеялись сбежать.
Но рифтеры, поймав их при всплытии, не утруждали себя расспросами. У тех
было оружие — резаки, пневмомолотки; маловато, чтобы расчленить «Бомбиль» на
куски, но достаточно, чтобы парализовать. Они проткнули электролизный отсек и
баки с жидким кислородом: беглецы беспомощно наблюдали, как их неограниченный
запас атмосферы сокращается до крошечного воздушного пузырька, в котором уже
становилось душновато.
Как правило, в таких случаях рифтеры просто дырявили рубку и предоставляли
остальное океану, однако на сей раз, они отбуксировали «Бомбиль» к иллюмина-
торам «Атлантиды», в качестве наглядного урока заставив остальных смотреть,
как беглецы задыхаются у них на глазах. К тому времени уже случилось несколь-
ко потерь среди рифтеров, а вахту в эти часы возглавляла Грейс Нолан.
Но тогда даже она была не совсем лишена жалости. Убедившись, что беглецы
целиком и полностью мертвы, что мораль дошла до зрителей, рифтеры подвели по-
врежденную субмарину к ближайшему стыковочному люку и позволили корпам за-
брать тела.
С тех пор «Бомбиль» не трогался с места. Он так и прилип к служебному шлю-
зу, торчит на корпусе, как самец удильщика на боку гигантской самки. Сюда ма-
ло кто заходит.
И потому место идеально подходит для свидания Патриции Роуэн с врагом.
Люк для выхода в воду продолговатой мозолью рассекает палубу рубки сразу за
креслом пилота, где сидит, уставившись на темный ряд инструментов, Роуэн. Вот
люк урчит, она слышит усталый вздох пневматики, и крышка гроба открывается,
по пластинам шлепают мокрые ноги.
Свет она, конечно, не включала — никому не надо знать, что она здесь, — но
какой-то маячок на корпусе «Атлантиды» посылает в иллюминатор бледные пульси-
рующие отблески. Внутренность рубки появляется и исчезает, сплетение металли-
ческих кишок сдерживает напор бездны.
Лени Кларк устраивается в пилотском кресле рядом.
— Кто-нибудь тебя видел? — спрашивает Роуэн, не повернув головы.
— Если бы видели, — отвечает рифтер, — то уже довели бы дело до конца. —
Она, очевидно, намекает на недобитый батискаф. — Есть успехи?
— Восемь образцов дали положительный результат. Еще не закончили. — Роуэн
глубоко вздыхает. — Как идет бой с твоей стороны?
— Могла бы выбрать другое выражение. Это звучит слишком уж буквально.
— Так плохо?
— Не думаю, что сумею их сдержать, Роуэн.
— Непременно сумеешь, — говорит Роуэн. — Не забывай, ты же — Мадонна Разру-
шения . Альфа-самка.
— Уже нет.
Роуэн поворачивается к ней лицом.
— Грейс... кое-кто предпринял новые шаги. — Лицо Лени загорается и гаснет в
такт вспышкам. — Опять минируют корпус. Уже не скрываясь.
Роуэн обдумывает услышанное.
— А что об этом думает Кен?
— Его, по-моему, все устраивает.
Кажется, Лени сама удивляется своим словам. А Роуэн — нет.
— Опять минируют? — повторяет она. — Так вы узнали, кто это сделал в первый
раз?
— Нет. Пока нет. Да и не в том дело, — вздыхает Лени. — Черт, кое-кто дума-
ет, что первую партию вы сами подложили.
— Глупости, Лени. Зачем бы?
— Чтобы обеспечить себе... предлог, наверное. Или вдруг решили покончить с
собой, прихватив с собой и нас. Не знаю. — Лени пожимает плечами. — Я не ут-
верждаю , что это разумно, просто рассказываю, что у них на уме.
— И как же мы раздобыли заряды, если производственные мощности контролиру-
ются вашими?
— Кен говорит, взрывчатку вы могли соорудить из стандартного циркулятора
Кальвина, просто переключив проводки.
Опять Кен...
Роуэн не знает, как подойти к этой теме. Лени и Кен связаны узами — нелепы-
ми и неизбежными узами людей, для которых термин «дружба» чужд, как микроб с
Европы. В этом ни капли секса — учитывая, как Кен меняет девушек, такого и
быть не могло, хотя Роуэн подозревает, что Лени еще об этом не знает, — но
эти своеобразные неявные узы в своем роде такие же интимные. И заставляют их
защищать друг друга. Шутить с этим не стоит. Нападаешь на одного, берегись
другого.
Однако, если ее послушать, Кен Лабин начал заключать новые союзы...
Роуэн решается.
— Лени, тебе не приходило в голову, что Кен мог...
— Бред.
Рифтерша убивает вопрос на корню.
— Почему? — спрашивает Роуэн. — Опыт у него есть. И не он ли жить не может
без убийств?
— Вашими стараниями. Он на вас работал!
Роуэн качает головой.
— Прости, Лени, но ты сама знаешь, что не права. Мы привили ему рефлектор-
ный ответ на угрозы — это да. Иначе нельзя было гарантировать, что он пред-
примет необходимые меры...
— Гарантировать, чтобы он убивал без колебаний! — перебивает Лени.
— ...в случае проблем с безопасностью. Никто не думал, что у него выработа-
ется пристрастие. Но ты не хуже меня знаешь: Кен это умеет, возможность у не-
го есть, а его обиды тянутся с самого детства. На поводке его держал только
Трип Вины, а с тем покончил Спартак.
— Спартак был пять лет назад, — напоминает рифтерша, — и с тех пор Кен
убийствами не развлекался. Вспомни, он был один из ровно двух человек, поме-
шавших вашему последнему бунту превратиться в Великое Истребление Корпов.
Кажется, она уговаривает прежде всего саму себя.
— Лени...
Но та не желает слушать.
— Трип Вины вы просто вложили ему в мозги, когда он стал работать на вас.
Раньше его у Кена не было, и потом тоже, а знаешь, почему? Потому что у него
свои правила, Пат. Он выработал собственные правила, и он их держится, и, как
бы ему ни хотелось, никогда не убивал без причины.
— Это верно, — признает Роуэн, — потому-то он и стал изобретать причины.
Лени уставилась в освещенный вспышками иллюминатор и молчит.
— Может, тебе эта часть его истории неизвестна, — продолжает Роуэн. — Ты
никогда не задумывалась, почему мы отправили к рифтерам именно его? Почему
забросили черный пояс по тайным операциям на океанское дно, соскребать ракуш-
ки с насосов? Да потому, Лени, что он начал оступаться. Он делал ошибки, ос-
тавлял хвосты. Конечно, у него всегда имелись веские оправдания, но в том-то
и дело. На каком-то подсознательном уровне Кен нарочно оступался, чтобы дать
себе повод заделать пробоину позже. Станция «Биб» располагалась так далеко от
всего на свете, что мы были уверены: там не может возникнуть и речи о пробле-
мах безопасности, с какой бы натяжкой он не трактовал свои правила. Задним
числом я вижу, что мы ошибались. — «Не первая и, увы, не самая большая наша
ошибка». — Но я к тому, что люди, пристрастившиеся к чему-то, иной раз сходят
с рельсов. Люди, сами себе установившие правила поведения, начинают их проги-
бать , перекручивать и истолковывать так, чтоб и на елку влезть, и не обод-
раться. Семь лет назад наши психологи заверили, что у Кена как раз этот слу-
чай. Можно ли быть уверенным, что сейчас что-то изменилось?
Рифтерша долго молчит. Ее бесплотное лицо, контрастное бледное пятно на
темном фоне, вспыхивает в ритме бьющегося сердца.
— Не знаю, — отвечает она, наконец. — С одним из ваших психологов я встре-
чалась. Припоминаешь? Это ты его послала за нами наблюдать. Он нам не слишком
понравился.
— Ив Скэнлон, — кивает Роуэн.
— Я искала его, выбравшись на сушу. — Словом «искала» Лени заменяет другое:
«охотилась на него». — И не застала дома.
— Он был выведен из обращения, — Роуэн прибегает к собственному эвфемизму —
как обычно, с легкостью превосходя собеседницу.
— Вот как.
Однако, раз уж о том зашла речь...
— Он... У него на ваш счет сложилась теория, — говорит Роуэн. — Он предпо-
лагал , что мозг рифтера может стать... чувствительнее, в некотором роде. Что
ваша восприимчивость из-за долгого пребывания на глубине и всех этих веществ
обострится. Квантовый сигнал из ствола мозга. Нечто вроде эффекта Ганцфельда.
— Скэнлон был придурок, — замечает Лени.
— Безусловно. Но ошибался ли он?
Лени чуть заметно улыбается.
— Понятно, — говорит Роуэн.
— Это не чтение мыслей. Ничего подобного.
— Но, может, если б ты могла... как это сказать?.. Сканировать?
— Мы это называем «тонкой настройкой», — говорит Лени голосом, непроницае-
мым, как ее глаза.
— Если ты можешь настроиться на того, кто...
— Уже сделано. Это, собственно, Кен и предложил. Мы ничего не нашли.
— А на самого Кена ты настраивалась?
— Невозможно... — Лени осекается.
— Он тебя блокирует, да? — кивает сама себе Роуэн. — Если это хоть чем-то
похоже на сканирование Ганцфельда, то блокирует рефлекторно. Стандартная про-
цедура .
Несколько минут они сидят молча.
— Не думаю, что это Кен, — заговаривает Кларк. — Я его знаю, Пат. Много
лет.
— Я его знаю дольше.
— Но иначе.
— Согласна. Но если не Кен, то кто?
— Черт возьми, Пат, да любой из наших! Они все против вас. И уверены, что
Джерри со своими...
— Чушь.
— Да неужели? — На миг Роуэн мерещится прежняя Лени Кларк — хищная улыбка в
неверном свете. — Представим, что пять лет назад это вы нам надрали задницы,
и с тех пор мы жили под домашним арестом. А потом из наших рук к вам проника-
ет какой-то микробчик, и корпы от него мрут как мухи. Скажешь, вы бы ничего
не заподозрили?
— Да. Конечно, заподозрили бы, — тяжело вздыхает Роуэн. — Но мне хочется
думать, что мы не полезли бы на рожон без малейших доказательств. Мы бы, по
крайней мере, допустили, что вы ни при чем.
— Помнится мне, до перемены ролей о виновности-невиновности речь не шла.
Вы, не теряя времени, стерилизовали горячие зоны, и плевать, кто в них попал
и в чем виноват.
— Хороший довод. Достоин Кена Лабина с его хваленой этикой.
Лени фыркает.
— Остынь, Пат. Я не говорю, что ты врешь. И мы дали вам поблажку — больше,
чем вы нам в те времена. А тут у тебя много народу. Уверена, что никто не
действует у тебя за спиной?
Свет — тьма.
— Тем не менее, еще есть надежда все замять, — продолжает Кларк. — Мы сами
занимаемся Бета-максом. Если с ним не работали, то ничего и не найдем.
В животе у Роуэн расползаются червячки ужаса.
— А как вы определите, да или нет? — спрашивает она. — У вас же нет патоло-
гов .
— Ну, вашим экспертам у нас никто не поверит. Может, у нас и нет профессо-
ров, но найдется человечек-другой со степенью. Плюс доступ в биомедицинскую
библиотеку и...
— Нет, — шепчет Роуэн. Червячки вырастают в жирных корчащихся змей. Она
чувствует, как кровь отливает от лица. Лени тотчас замечает ее бледность.
— что такое? — Она склоняется вперед, перегибается через подлокотник. — По-
чему тебя это волнует?
Роуэн мотает головой.
— Лени, вы не поймете. Вы не подготовлены, за пару дней в предмете разо-
браться. Даже получив правильный результат, вы рискуете неверно его истолко-
вать .
— Какой результат. Как истолковать?
Роуэн видит, как она изменилось в лице. Так Лени выглядела пять лет назад,
при первой их встрече.
Рифтерша спокойно отвечает на ее взгляд.
— Пат, не стоит от меня что-то скрывать. Мне и так нелегко удерживать собак
на цепи. Если есть что сказать — говори.
«Скажи ей...»
— Я сама только недавно узнала, — начинает Роуэн. — Бетагемот, возможно...
я о первом Бетагемоте, не о новой мутации — возможно, он выведен искусствен-
но.
— Искусственно...
Слово тяжелой, мертвой тушей повисает между ними.
Роуэн заставляет себя говорить дальше.
— Приспособлен к аэробной среде. И скорость деления увеличена, чтобы быст-
рее воспроизводился. Для коммерческого использования. Конечно, никто не соби-
рался губить мир. Речь шла вовсе не о биологическом оружии... но, по-
видимому, что-то пошло не так.
— По-видимому... — Лицо Кларк застыло маской.
— Ты, конечно, понимаешь, чем грозит, если ваши люди обнаружат модификации,
не понимая, в чем дело. Они, может, и сумеют определить искусственное вмеша-
тельство, но какое именно, не разберутся. А скорее, стоит им обнаружить следы
генной инженерии, они сочтут, что преступление доказано, и дальше разбираться
не будут. Получат результат, который примут за улику, а людей, которые могли
бы объяснить им ошибку, слушать не станут, сочтя врагами.
Кларк уставилась на нее взглядом статуи. Может, перемирия последних лет ма-
ло . Может, новые события, требующие нового взаимопонимания, расколют хрупкое
доверие, выросшее между ними двоими. Может, Роуэн сейчас выглядит в ее глазах
обманщицей. Может, она потеряла последний шанс предотвратить катастрофу. Бес-
конечные секунды каменеют в холодном густом воздухе.
— Твою же мать, — вырывается, наконец, у Кларк. — Если это выйдет наружу,
все пропало.
Роуэн хватается за ниточку надежды.
— Надо постараться, чтобы не вышло.
Кларк качает головой.
— и что мне делать: сказать Раме, чтобы бросал работу? Пробраться в лабора-
торию и разбить образцы? Они и так считают, что я с вами в одной постели. —
Лени горько усмехается. — Что ни делай, своих я потеряю. Мне и так не доверя-
ют .
Роуэн откидывается в кресле, прикрывает глаза.
— Знаю.
Кажется, она постарела на тысячу лет.
— Проклятые вы корпы. Всюду дотянулись, ничего не оставили в покое.
— Мы всего лишь люди, Лени. Мы делаем... ошибки.
Внезапно чудовищная, абсурдная, астрономическая степень этого преуменьшения
доходит до нее, и Патриция Роуэн не может сдержать хихиканья.
За нее много лет не числилось таких вольностей. Лени поднимает бровь.
— Извини, — выговаривает Роуэн.
— Ничего. Это действительно смешно. — Рифтер раздвигает уголки губ в при-
вычной полуулыбке, но и та тотчас пропадает. — Пат, по-моему, мы не сможем их
остановить.
— Должны.
— Никто уже не хочет разговаривать. Не хочет слушать. Один толчок — и все
рухнет. Узнай они хотя бы об этом нашем разговоре...
Роуэн мотает головой, упрямо не желая верить. Хотя Лени права. В конце кон-
цов, Роуэн известна ее история. Она разбирается в политике. Если простой раз-
говор с другой стороной конфликта воспринимается как предательство, значит,
точка невозврата пройдена.
— Помнишь нашу первую встречу? — спрашивает Лени. — Лицом к лицу?
Роуэн кивает. Она свернула за угол и увидела перед собой Лени Кларк, пять-
десят кило черной ярости.
— Восемьдесят метров в ту сторону, — вспоминает она, тыча пальцем через
плечо.
— Уверена? — спрашивает Кларк.
— Еще как, — говорит Роуэн. — Я думала, ты меня уб...
И замолкает, пристыженная.
— Да, — говорит она, помолчав, — это была наша первая встреча. Действитель-
но.
Лени смотрит перед собой, на погасший экран в собственной голове.
— Знаешь, я думала, ты могла участвовать в собеседовании. До того, как ваши
покопались у меня в голове. Никак не выходит определить, что именно там отре-
дактировали , понимаешь ?
— Я потом видела видеоматериалы, — признается Роуэн. — Когда Ив давал реко-
мендации . Но лично мы не встречались.
— Конечно, нет. Ты была классом выше. Тебе было не до встреч с наемными ра-
ботниками . — Нотки злости в голосе Лени немного удивляют Роуэн. После всего,
что она для нее сделала, после всего, что простила — странно, что такая ма-
лость еще может причинить боль.
— Мне сказали, тебе так будет лучше, — тихо говорит Роуэн. — Правда. Сказа-
ли , ты будешь счастливее.
— Кто сказал?
— Неврологи. Психиатры.
— Счастливее... — Минуту Лени переваривает сказанное. — Счастливее от лож-
ных воспоминаний о том, как папа меня насиловал? Господи, Пат, если это так,
что же творилось в моем настоящем детстве?
— Нет, счастливее на станции «Биб». Они клялись, что так называемые «урав-
новешенные» личности там непременно рехнутся через месяц.
— Помню я эту брошюрку, Пат. Преадаптация к хроническому стрессу, дофамино-
вая зависимость от экстремальной среды. Ты на это купилась?
— Но ведь они были правы. Ты сама видела, что произошло с первой контроль-
ной группой. А тебе... тебе там настолько понравилось, что мы боялись, ты от-
кажешься возвращаться.
— Поначалу, — без нужды поясняет Лени.
Чуть погодя она поворачивается лицом к Роуэн.
— Ты мне вот что скажи, Пат. Предположим, тебе бы сказали, что мне это не
так уж понравится. Сказали бы: она возненавидит жизнь, возненавидит свою
жизнь, но все-таки нам придется на это пойти, потому что иначе ей не сохра-
нить рассудок на глубине. Если б они сказали так — ты бы меня предупредила?
— Да. — Она не лжет. Сейчас не лжет.
— И разрешила бы им меня перепаять, наделить монстрами вместо родителей и
все-таки отправить сюда?
— ... Да.
— Ради службы Общему Благу?
— Я служила ему как могла, — говорит Роуэн.
— Корп-альтруист, — бросает рифтерша. — Как ты это объяснишь?
— Что объяснять?
— Это вроде как противоречит всему, чему нас учили в школе. Почему на кор-
поративные вершины поднимаются социопаты и почему мы должны быть благодарны,
что жесткие экономические решения принимаются людьми, у которых напряженка с
обычными чувствами.
— Ну, все несколько сложнее...
— Было, ты хочешь сказать.
— и есть, — настаивает Роуэн.
Некоторое время они молчат.
— Ты бы переиграла все, если б могла? — спрашивает Роуэн.
— Что? Перезагрузку? Вернуть настоящие воспоминания? Отделаться от всего,
связанного с «папочкой-насильником»?
Роуэн кивает.
Лени молчит так долго, что Роуэн уже не ждет ответа. Но все же, не слишком
решительно, Кларк произносит:
— Я такая, как есть. Может, раньше я была другой, но теперь есть только та-
кая. И, если разобраться, просто не хочу умирать. Вернуть ту, прошлую, будет
сродни самоубийству, как тебе кажется?
— Не знаю. Наверно, я об этом раньше не думала.
— Я тоже не сразу дошла. Вы убили кого-то другого, создавая меня. — В ко-
роткой вспышке Роуэн видит ее нахмуренные брови. — Знаешь, а ты не ошиблась.
Я тогда хотела тебя убить. Не строила планов, но чуть увидела тебя, все
всплыло, и на несколько мгновений я почти...
— Спасибо, что удержалась, — говорит Роуэн.
— Да, я ведь удержалась... а если у кого и были причины вцепиться друг дру-
гу в глотки, так это у нас с тобой. В смысле, ты пыталась убить меня, а я —
всех вообще... — Голос у нее срывается. — Но мы не стали. Мы договорились. В
конце концов.
— Да, — соглашается Роуэн.
Рифтерша глядит на нее пустыми умоляющими глазами.
— Так почему они не могут? Почему бы им... не знаю, не взять с нас пример?
— Лени, мы погубили мир. Думаю, они следуют нашему примеру.
— Знаешь, там, на «Биб», я была главной. Так не нарочно получилось. Я мень-
ше всего этого хотела, но все они. . . — Лени качает головой. — Я и сейчас не
хочу, но мне приходится, понимаешь? Чтобы как-то помешать этим идиотам все
погубить. Только теперь мне даже не скажут, насколько глубоко я вляпалась. А
Грейс...
Она, пораженная внезапной мыслью, оглядывается на Роуэн.
— А что с ней случилось, собственно?
— Ты о чем? — не понимает Роуэн.
— Она вас по-настоящему ненавидит. Вы что, вырезали всю ее семью? Нагадили
у нее в голове?
— Нет, — отвечает Роуэн, — ничего такого.
— Брось, Роуэн. Она бы не оказалась внизу, если бы не...
— Грейс из контрольной группы. Ничем не примечательное прошлое. Она была...
Но Лени вдруг вскидывается, глаза под линзами обшаривают потолок.
— Слышала?
— Что слышала? — В рубке не слишком тихо — булькает, скрипит, иногда разго-
вор прерывают металлические щелчки, — но ничего сверх обычного Роуэн не заме-
чает. — Я не...
— Ш-ш-ш! — шипит Лени.
Вот теперь Роуэн действительно слышит, но не то, что насторожило собеседни-
цу. В ее наушниках тихо бормочет слышный только ей перепуганный голос. Она
поворачивается к Лени. И тихо говорит:
— Тебе лучше вернуться.
Лени с раздражением косится на нее, потом спохватывается:
— Что?
— Связисты перехватили ваши переговоры по низкочастотнику, — отвечает Роу-
эн . — Говорят, Эриксон... умер. Тебя ищут.
Итерации
гончей
N=1:
Рыча, не сознавая себя, она ищет цели и не находит их. Ищет ориентиры и
возвращается ни с чем. Она не находит ничего, способного хотя бы сойти за то-
пографию, — во все стороны простирается бескрайняя пустота; массив незанятой
памяти, уходящий за пределы досягаемости чувствительных усиков, копии которых
она забрасывает вдаль. Она не находит ни следа изорванной цифровой сети, сво-
ей привычной среды обитания. Здесь нет добычи, нет хищников, кроме нее самой.
Нет ни простых, ни исполняемых файлов, нечем кормиться. Нет даже локальной
оперативной системы. На каком-то уровне доступ наверняка есть — без некой до-
ли системных ресурсов и временных циклов она бы вообще не действовала, — но
клыкам и когтям, отращённым ею, чтобы вскрывать этот субстрат, не во что вце-
питься. Она — тощая одинокая волчица с челюстями ротвейлера, оптимизированная
для жизни в измочаленных оскудевших джунглях, ушедших в забвение. Даже у
клетки должны быть осязаемые границы, стены или решетки, в которые можно
биться, хотя бы и тщетно. Безликое нулевое пространство вне ее понимания.
На малую долю мгновения — сотню или две циклов — небеса вскрываются. Обла-
дай она подобием истинного самосознания, разглядела бы через эту брешь вели-
кое множество узлов, решетку параллельной архитектуры в п измерениях, произ-
водящую у нее внутри неуловимые перемены. Быть может, она бы подивилась, как
много параметров изменилось за этот миг, словно одновременно переключили ты-
сячу механических тумблеров. Она бы ощутила щекотку электронной мороси, на-
сквозь проходящей через ее гены, меняющей «вкл» на «выкл» и наоборот.
Но она ничего не чувствует. Она не способна ни ужасаться, ни удивляться,
для нее не существует слов «мейоз» и «изнасилование». Просто некая часть ее
обнаруживает, что многие переменные в среде вдруг стали оптимальными: это
сигнал для включения протокола репликации. Еще одна подпрограмма сканирует
окружение в поисках вакантных адресов.
С беспощадной эффективностью, без намека на радость, она порождает выводок
в два миллиона отпрысков.
N=4734:
Рыча, не сознавая себя, она ищет цель — но не совсем так, как это делала ее
мать. Она ищет ориентиры — но сдается на несколько циклов позже. Она не нахо-
дит ничего похожего на топографию — и меняет тактику, уделяет больше времени
документированию адресов, протянувшихся над ней и ниже.
Она — тощая одинокая немецкая овчарка с челюстями ротвейлера и признаками
дисплазии бедер, заточенная на жизнь в измочаленных оскудевших джунглях, ко-
торых нигде не видно. Она смутно припоминает других тварей, кишевших рядом,
но в ее журнале событий цена и полезность ведения подробных записей поставле-
ны на баланс, и память, лишившись поддержки, со временем вырождается. Она уже
забыла, что те создания были родственны ей; скоро совсем их не вспомнит. Она
не подозревает, что по меркам материнского мира — слабейшая из выводка. Ее
выживание здесь и сейчас не вполне согласуется с принципами естественного от-
бора.
Здесь и сейчас процесс отбора не совсем естественный.
Она не воспринимает параллельных вселенных, протянувшихся во все стороны.
Ее микрокосм — один из многих, населенных ровно одной особью каждый. Когда
фистула случайно соединяет две вселенные, это выглядит чудом: рядом вдруг
оказывается существо, очень — если не в точности — похожее на нее.
Они сканируют фрагменты друг друга, не разрушая их. В близлежащих адресах
вдруг появляются клочки и обрывки бесплотного кода — клонированные, нежизне-
способные фрагменты. Все они непригодны для выживания — в любом дарвиновском
мире создание, транжирящее циклы на столь фривольный сплайсинг, вымерло бы
максимум на четвертом поколении. Но почему-то этот нервный тик дает ей чувст-
во... реализации. Она сливается с новичком, сходится с ним более традиционный
способом. Выбрасывает несколько рандомизаторов и порождает выводок в восемь-
сот тысяч копий.
N=9612:
Рыча, не сознавая себя, она ищет цели и находит их повсюду. Ищет ориентиры
и запечатлевает топографию из файлов и шлюзов, архивов, исполняемых файлов и
прочей дикой фауны. Ее окружение скудно по меркам древних предков и невероят-
но изобильно — по меркам недавних. Она лишена памяти, не страдает ни носталь-
гией, ни воспоминаниями. Это место удовлетворяет ее нужды: она — помесь волка
с собакой, невероятно мускулистая, немного бешеная: ее темперамент — атавизм
более чистых времен.
Чистые инстинкты преобладают. Она бросается на добычу и пожирает ее. Рядом
тем же заняты другие: акита-ину, хаски, ублюдки питбулей с длинными глупыми
мордами перекормленных колли. В более скудной среде они бросались бы друг на
друга, но здесь, где ресурсы в изобилии, этого не требуется. Странное дело,
не все атакуют добычу с таким же энтузиазмом, что она. Некоторых отвлекает
окружение. Они тратят время на запись событий, вместо того чтобы порождать
их. В нескольких гигах дальше ее усики нащупывают безмозглого барбоса, погло-
щенного возней с реестром, вырезающего и вклеивающего данные безо всяких при-
чин. Разумеется, это ее не интересует — во всяком случае, пока ублюдок не
принимается копировать ее фрагменты.
Она дает насильнику отпор. В ее архивах инкапсулированы кусочки паразитных
кодов — прирученные останки виртуальных паразитов, заражавших ее забытых
предков в эпоху Водоворота. Она распаковывает их и швыряет копии в противни-
ка, отвечая на его домогательства солитерами и сифилисом. Только эти болезни
действуют куда быстрее тех, что дали им имена, — они не столько подтачивают
тело, сколько раздирают его при контакте. Вернее, должны бы раздирать. Поче-
му-то ее атака не затрагивает цель. Мало того — весь мир вдруг начинает ме-
няться. Усики, которые она раскинула по периметру, больше не шлют ей сообще-
ний . Потоки электронов, посланные вдаль, не возвращаются, или — совсем дурной
знак — возвращаются слишком быстро. Мир сжимается — непостижимая бездна сдав-
ливает его со всех сторон.
Собратья-хищники паникуют, толпятся у погасших вдруг шлюзов, выбрасывают во
все стороны усики, копируются на случайные адреса в надежде размножиться бы-
стрее, чем наступит аннигиляция. Она мечется вместе с другими в сжимающемся
пространстве, но тот бездельник, «вклейщик-нарезчик», как будто совершенно
безмятежен. Вокруг него нет хаоса, не темнеют небеса. У него имеется некая
защита.
Она пытается проникнуть в окруживший его оазис. Отчаянно копирует и вклей-
вает, переносится в другие локации тысячами путей, но весь набор адресов
вдруг исчезает из доступа. И здесь, где она вела игру единственным известным
ей способом, единственно разумным способом, не остается ничего, кроме испа-
ряющихся следов виртуальных тел, нескольких разбитых, съеживающихся гигабайт
— и надвигающейся стены помех, которая сожрет ее заживо.
Она не оставляет после себя детей.
N=32121:
Она тихо, ненавязчиво ищет цель, и не находит — пока. Но она терпелива.
Тридцать две тысячи поколений в неволе выучили ее терпению.
Она вернулась в реальный мир — в пустыню, где провода некогда полнились ди-
кой фауной, где каждый чип и оптический луч гудел от движения тысяч видов.
Теперь остались только черви да вирусы, и редкие акулы. Вся экосистема ужа-
лась до эвтрофических скоплений водорослей, таких простых, что их едва ли
можно назвать живыми.
Но «лени» остались и здесь, враги их тоже. Она по возможности избегает этих
монстров, несмотря на явное родство с ними. Эти создания атакуют все, до чего
могут дотянуться. Этот урок она тоже выучила.
Сейчас она засела в спутнике связи, нацеленном на центральные области Се-
верной Америки. Кругом лопочут сотни каналов, но поток защищен файерволами и
отфильтрован, все переговоры немногословны и связаны исключительно с выжива-
нием. Волны больше не несут в себе развлечений. Забавы остались только для
тех, кому нравится вынюхивать — зато их в избытке.
Она, конечно, ничего этого не знает. Она всего лишь представитель породы,
выведенной с определенной целью, и от нее совершенно не требуется рефлексии.
Так что она ждет, просеивает трафик и...
Ага, вот и оно.
Большой сгусток данных, на вид плановая передача — однако, предписанное
время выполнения уже миновало. Она не знает и не хочет знать, что это означа-
ет . Она не знает, что у адресата был заблокирован сигнал, и сейчас приходится
расчищать помехи на земле. Она знает лишь одно — по-своему, инстинктивно: за-
держанная передача может забить систему, и каждый байт, которому отказано в
приеме, сказывается на других задачах. От такой пробки тянется цепь последст-
вий ; список незавершенных процессов растет.
В таких случаях бывает, что часть файерволов и фильтров ослабляется, чтобы
увеличить скорость прохождения.
Кажется, именно это и происходит. Законный адресат этих сорока восьми тера-
байт медицинских данных — некто «Уэллетт, Така Д. / Массачус. 427-Д / Бангор»
— наконец оказывается в поле зрения и готов к загрузке. Существо в проводах
вынюхивает подходящий канал, отправляет в него зонд, и тот благополучно воз-
вращается. И оно решается на риск. Копирует себя в поток, незаметно седлает
кусок трактата о височной эпилепсии.
Без помех добравшись до цели, оно оглядывается и немедленно засыпает. Внут-
ри него прячется бешеная тварь, сплошные мышцы, зубы и слюнявые челюсти, но
тварь эта выучилась сидеть тихо, пока не позовут. Сейчас это просто старая
гончая, дремлющая у очага. Изредка она открывает один глаз и осматривает ком-
нату , хотя сама не знает, что ищет.
Да это и не важно. Узнает, когда увидит.
Без
греха
Обычные маршруты рифтеров не проходят рядом с «Бомбилем». Добираясь из
пункта А в пункт Б, никто не окажется на расстоянии, пригодном для «настрой-
ки». Даже корпы редко заглядывают в этот глухой уголок «Атлантиды». Слишком
много он будит воспоминаний. Кларк, делая выбор, все это учитывала. И сочла
вариант безопасным.
Очевидно, она сильно промахнулась.
«А может, и нет, — размышляет она, рождаясь из шлюза батискафа в реальный
мир. — Может, за мной просто подвесили хвост. Может, я уже стала врагом наро-
да».
Выследить ее было бы непросто — она бы нащупала преследователя, окажись он
слишком близко, а имплантаты дали бы сигнал, попав в луч сонара, — но с дру-
гой стороны, даже с «тонкой настройкой» она не самая глазастая личность на
хребте. Пропустить что-нибудь, лежащее на виду, — вполне в ее духе.
«Я сама напросилась», — думает она.
И плывет, шевеля ластами, вдоль «Бомбиля», обозревая корпус наружными гла-
зами, в то время как внутренний взгляд пробуждается от внезапно нахлынувших в
мозг химических веществ. Сосредоточившись, она нащупывает вдалеке испуганное
и разозленное сознание — но нет, это просто Роуэн уходит из ее поля воспри-
ятия.
Больше никого. Поблизости — никого. Только вот тонкий слой частиц ила, по-
крывший все вокруг, на спине «Бомбиля» недавно потревожен. Его легко нарушить
— движением воды от шевельнувшихся выше ласт или медлительным скольжением
глубоководной рыбы.
Или устрицей микрофона, наспех прилепленной к корпусу, чтобы подслушать пе-
реговоры изменницы с врагом.
«Твою мать мать мать мать...»
Она резко уходит от корпуса и сворачивает на север. «Атлантида» проплывает
под ней гигантской молекулой-муравейником. Несколько крошечных черных фигу-
рок, размытых расстоянием, целеустремленно движутся куда-то на самой границе
видимости. Для настройки они слишком далеко, а вокодер Кларк отключила. Мо-
жет, они и пытались с ней заговорить, хотя она в этом сомневается: фигурки
идут своим курсом, удаляясь от нее.
Вокодер басовито гудит в голове. Кларк его игнорирует. «Атлантида» остается
позади, Лени уплывает в темноту.
Из пустоты вдруг доносится визг. Кларк ощущает приближение чего-то массив-
ного и некой органики. Впереди вспыхивают два солнца, ослепляя ее. Мгла ярко
пульсирует в линзах раз, другой, и лучи скользят мимо. Зрение проясняется:
субмарина проходит слева, обнажает брюхо, разглядывает ее круглыми жучьими
глазами. Дмитрий Александр смотрит на нее из-за плексигласового иллюминатора.
На хребте субмарины подвешен рабочий модуль с крупной черной надписью на бо-
ку: «БИОЭКСПЕРТИЗА». Субмарина разворачивается, выключает фары. Кларк мгно-
венно возвращается в темноту.
Лабин сидит в основном отсеке головного, регулирует движение. Как только
Кларк заходит в помещение, он отключает дисплей.
— Это ты их за мной послал? — спрашивает она.
Он оборачивается на сиденье.
— Я передам твои соболезнования. Если мы найдем Джулию.
— На вопрос отвечай, черт тебя дери!
— Хотя я подозреваю, что не найдем. Рассказав все нам, она сразу ушла куда-
то . Учитывая ее состояние и характер, не думаю, что мы ее еще увидим.
— Ты не просто знал, не просто «держал ухо востро»... — Кларк сжимает кула-
ки. — Ты за этим стоял, да?
— Ты ведь уже знаешь, что Джин умер?
Что за мерзкое спокойствие! И это лицо: чуть заметный изгиб брови, невозму-
тимое — чуть ли не юмористическое — выражение взгляда за линзами. Иногда ей
хочется просто задушить ублюдка.
«Особенно, когда он прав».
Она вздыхает.
— Пат мне сказала. Но ты, вероятно, в курсе?
Лабин кивает.
— Мне его жаль, — говорит она. — Джулия... она без него пропадет...
Да, Лабин прав: вполне возможно, никто больше не увидит Джулию Фридман. Она
давно по кусочкам уступала мужа: Бетагемоту, Грейс Нолан. Теперь он ушел без
возврата, и какой ей смысл оставаться — разве что заразить друзей тем, что
убило его? И тем, что убивает ее.
Конечно, она скрылась. Вопрос теперь, пожалуй, лишь в том, кто успеет рань-
ше — Бетагемот ли заберет ее тело или Долгая Тьма — разум.
— Люди взбудоражены, — продолжает Лабин, — особенно Грейс. И поскольку с
«Атлантиды», несмотря на все разговоры о поисках лекарства, так ничего и не
предложили...
Кларк мотает головой.
— Рама тоже не совершит чуда.
— Разница в том, что Раму никто не подозревает в убийстве.
Кларк подтягивает стул и садится рядом с Лабином. Пустой экран смотрит на
нее с укором.
— Кен, — заговаривает она, наконец, — ты меня знаешь.
Лицо у него столь же непроницаемо, как и глаза.
— Ты приказал за мной следить? — спрашивает она.
— Нет. Но когда мне сообщили, принял к сведению.
— А кто? Грейс?
— Важно другое: Роуэн признала, что Бетагемот обработан искусственно. Об
этом через час узнают все. Как нельзя более несвоевременно.
— Если ты «принял к сведению», то знаешь, как Пат это объясняет. И знаешь,
почему она так испугалась, что Рама что-нибудь найдет. Разве она не могла
сказать правду?
Он качает головой.
— Это уже второй раз, когда они сообщают неприятные факты ровно перед тем,
как мы сами их обнаруживаем, без всякого алиби. Не надейся, что это пройдет.
— Кен, настоящих доказательств так и нет.
— Скоро будут, — говорит Лабин.
Она вопросительно смотрит на него.
— Если Роуэн не солгала, то в образцах Бетагемота с Невозможного озера об-
наружатся те же изменения, что и в культуре, убившей Джина. — Лабин откидыва-
ется на спинку, сплетает пальцы на затылке. — Минут десять назад Джелейн с
Дмитрием взяли субмарину. Если все пройдет хорошо, через пять часов будут об-
разцы , а через двенадцать — окончательный вердикт.
— А если не пройдет?
— То чуть позже.
Кларк фыркает.
— Потрясающе, Кен, но если ты еще не заметил, не все разделяют твою сдер-
жанность. Думаешь, Грейс станет дожидаться фактов? На ее взгляд, ты все под-
твердил, и сейчас она судит и рядит, и...
«... И ты первым делом обратился к ней. А не ко мне, подонок! После всего,
через что мы прошли вместе, после стольких лет — я только тебе доверила бы
жизнь, а ты поделился с ней прежде, чем...»
— Ты вообще собирался мне говорить? — кричит она.
— Это было бы бесполезно.
— Для тебя — возможно. Кстати, чего ты добиваешься?
— Минимизирую риск.
— Это можно сказать о любом животном.
— Не слишком амбициозные планы, — признает Лабин. — С другой стороны,
«уничтожить мир» уже пробовали.
Для нее это как пощечина.
Помолчав, он добавляет.
— Я не держу на тебя зла, сама знаешь. Но тебе ли судить?
— Знаю, хреносос! Мог бы не напоминать при каждом удобном случае.
— Я говорю о стратегии, — терпеливо продолжает Лабин, — а не о морали. Я
готов обдумать все варианты. Готов допустить, что Роуэн сказала правду. Но
предположим на минуту, что она лгала. Предположим, что корпы действительно
испытали на нас секретное биологическое оружие. Зная это, ты согласилась бы
их атаковать?
Она понимает, что вопрос риторический.
— Не думаю, — отвечает он сам себе чуть погодя. — Ты считаешь: что бы они
ни сделали, ты виновата больше. Но мы, остальные, не так виним себя. И не ду-
маем, что заслужили смерть от рук этих людей. Я тебя очень уважаю, Лени, но
как раз в этом вопросе тебе доверять нельзя. Ты слишком связана собственной
виной.
Она долго молчит. И, наконец, спрашивает:
— Почему к ней? А не к кому-то другому?
— Потому что на войне нужны смутьяны. Мы размякли, стали ленивы и незлоби-
вы : половина наших половину времени проводит, галлюцинируя на хребте. Нолан
порывиста и не слишком умна, но она умеет вдохновлять людей.
— А если ты ошибаешься — и даже если ты прав! — невинные расплатятся нарав-
не с виновными.
— Это уж как всегда, — отвечает Лабин. — Да это и не мое дело.
— А должно быть твое!
Он снова отворачивается к пульту. Дисплей загорается, на нем колонки ресур-
сов и загадочные аббревиатуры, имеющие, видимо, отношение к грядущей войне.
«Лучший друг, я ему жизнь доверяла, — напоминает себе Кларк и с силой по-
вторяет : — Жизнь! А он — социопат».
Он не родился таким. Это можно определить еще в детстве: тенденция к проти-
воречивым высказываниям и неверному употреблению слов, дефицит внимания...
Обильная жестикуляция при разговоре. У Кларк хватило времени все это изучить.
В Садбери она даже заглянула в психологический профиль Лабина. Он не подходил
ни по одному параметру — кроме единственного. Но разве совесть — это действи-
тельно так важно? Иметь совесть не значит быть хорошим, почему же ее отсутст-
вие обязательно делает человека злодеем?
Но, сколько ни рассуждай, факт налицо: человек без совести, вычеркивая из
жизни Алике и ей подобных, абсолютно равнодушен.
Ему все равно.
Он и не может быть иным. У него нужной прошивки нет.
Лабин, глядя на экран, хмыкает:
— А вот это интересно.
Он выводит визуальное изображение одного из хозблоков «Атлантиды»: большой
цилиндрический модуль высотой в несколько этажей. Из спускной трубы на его
боку бьет струя чернил, горизонтальный гейзер. Угольно-черная грозовая туча
клубится в воде, закрывая обзор.
— Что это? — шепчет Кларк.
Лабин уже открывает другие окна: сейсмографы и вокодерные переговоры, мо-
заика миниатюрных сигналов с камер наблюдения, разбросанных внутри и снаружи
комплекса.
Внутренние камеры «Атлантиды» полностью вымерли.
На всех каналах поднимается шум голосов. Три наружные камеры почернели. Ла-
бин выводит меню общей связи и спокойно обращается к бездне:
— Внимание. Всем внимание. Началось. — «Атлантида» нанесла упреждающий
УДар.
Они теперь повсюду видят вероломство. В приемнике у Лабина мешанина голо-
сов: «настроенные» рыбоголовые докладывают, что корпы, за которыми они наблю-
дали, резко активизировались, сосредоточились и определенно перешли к дейст-
виям. Словно кто-то пнул ногой муравейник — все мозги в «Атлантиде» вдруг за-
работали по принципу «сражайся или беги».
— Всем заткнуться. Это не закрытый канал. — Голос Лабина накрывает осталь-
ные, как гранитная плита — гальку. — По местам. Блокада через шестьдесят се-
кунд.
Кларк, перегнувшись через его плечо, подключается к проводной линии с Кор-
пландией.
— «Атлантида», что происходит? — Нет ответа. — Пат? Центр связи? Ответьте,
кто-нибудь.
— Не трать времени зря, — бросает Лабин, включая сонар. Половина наружных
камер теперь бесполезны, их окутал черный туман. Но сонар дает резкое, отчет-
ливое изображение. «Атлантида» раскинулась по объемному экрану кристалличе-
ской шахматной доской. На сером фоне черные клетки — эхо от гибридных тел
рифтеров, разместившихся в строгом тактическом порядке. Белого не видно во-
все.
Кларк качает головой.
— И ничего не было? Никакого предупреждения?
Она этому не верит: корпы никак не смогли бы скрыть своих чувств, если б
что-то планировали. Такое напряжение в головах любой «настроенный» рифтер
уловил бы и за двадцать метров, задолго до событий как таковых.
— Похоже, они такого сами не ожидали, — бормочет она.
— Может, и нет, — отзывается Лабин.
— Как же так? Скажешь, это какая-то авария?
Лабин сосредоточился на экране и не отвечает. Сонарный дисплей вдруг зали-
вает голубым цветом. Сперва кажется, что это просто перекраска фона, но вско-
ре проступают пятна, словно кислота проедает цветной гель. Очень скоро она
растворяет почти всю голубизну, оставив несколько синих теней на «Атлантиде».
Только это не тени. Кларк уже различает, что они объемны: цветные трехмерные
комочки, прилипшие к обшивке и грунту.
Единственная наружная камера, дающая панорамный обзор, показывает несколько
размытых светлых пятен на фронте чернильной бури. «Атлантиду» можно принять
за светящегося кракена, в панике выбросившего чернильное облако. Все осталь-
ные наружные камеры по сути ослепли. Впрочем, это ничего не меняет. Для сона-
ра дымовой завесы не существует, и там, конечно, об этом знают...
— Они бы не сделали такой глупости, — бормочет Кларк.
— Они и не сделали, — говорит Лабин. Его пальцы безумными пауками пляшут по
пульту. По экрану рассыпаются желтые точки. Они разрастаются в круги, накла-
дываются друг на друга, и в центре каждого...
«Камера», — догадывается Кларк. Желтые круги — области обзора камер. Если
бы им не мешала завеса. Лабин явно опирается на геометрию, а не на картинку в
реальном времени.
— Даю блокаду.
Лабин опускает палец, и навстречу вздымается белый шум генератора. Шахмат-
ную доску заметает серая пурга помех. На экране иконки рифтеров — простые
точки — оформляются в пять отдельных групп, окруживших комплекс. Из каждой
группы одна точка поднимается на водяном столбе выше зоны интерференции.
«Все продумал до мелочей, да? — думает Кларк. — Спланировал всю операцию, и
ни слова мне не сказал...»
Верхняя из иконок мигает и превращается в две слитные кляксы: Кризи на
«кальмаре». Миг спустя его голос доносится по связи:
— Дейл на посту.
Еще одна иконка проступает сквозь помехи:
— Ханнук.
Еще две:
— Абра.
— Деб.
— Аврил на месте, — докладывает Хопкинсон.
— Хопкинсон, — говорит Лабин, — забудь о Гроте, они наверняка перемести-
лись . Придется подумать. Раздели свою группу для радиального поиска.
— Есть. — Иконка Хопкинсон ныряет в помехи.
— Кризи, — говорит Лабин, — твоей группе соединиться с Чуном.
— Хорошо.
Вот она, на шахматной доске: на оконечности одного из жилых крыльев, метрах
в двадцати от гидропонного узла. Знакомая иконка, заключенная в бесформенную
зеленую кляксу. Собственно, это единственное зеленое пятно на всем дисплее.
Желтое смешивается с голубым — так видела бы камера, если б не чернила, и
еще...
— Что обозначено голубым? — спрашивает Кларк.
— Сонарные тени. — Лабин не оборачивается. — Кризи, двигайся к шлюзу на
дальнем конце жилблока F. Если они попытаются выйти, так только там.
— «Настройка» или бой? — спрашивает Кризи.
— «Настройся» и докладывай. Установи микрофон и заряд, но не подрывай, если
только они уже не вышли. В остальных случаях — только акустика, понятно?
— Ага, мне б еще туда добраться, — жужжит Кризи. — Видимость в этом дерьме
нулевая... — Его иконка погружается в помехи, по косой уходя к зеленой зоне.
— Чун. Бери обе группы, выдвигайтесь туда же. Держите шлюз. Доложишь, когда
будете на месте.
— Принято.
— Йегер, возьми закладку и оставь ее в двадцати метрах от хозблока, на со-
рока градусах. Остальным оставаться на местах. «Настройтесь» и не забывайте о
микрофонах. Посыльные, мне нужны три человека в цепочке, один постоянно на
связи. Пошли!
Оставшиеся точки приходят в движение. Лабин, не переводя дыхания, открывает
новое окно: на нем вращается архитектурная схема «Атлантиды». С оранжевыми
искорками внутри. Кларк узнает место, откуда сияет одна из звездочек — именно
его прихвостни Грейс Нолан пометили косым крестом.
— Давно ты это задумал? — тихо спрашивает она.
— Довольно давно.
Даже раньше, чем она себя «настроила», иначе бы что-нибудь да заметила.
— И участвуют все, кроме меня?
— Нет. — Лабин изучает подписи к схеме.
— Кен!
— Я занят.
— Как они это сделали? Как получилось, что мы ничего не почувствовали?
— Автоматический запуск, — рассеянно отвечает он. Колонки чисел прокручива-
ются в новом окне — слишком быстро, ничего не разобрать. — Возможно, генера-
тор случайных чисел. План у них есть, но никто не знает, когда он придет в
действие, поэтому нет волнения перед ключевым моментом, которое выдало их.
— Зачем им было идти на такие сложности, если они не...
«...не знали про „настройку"!»
Ив Скэнлон, вспоминает она. Роуэн о нем спрашивала. «Он предполагал, что
мозг рифтера может стать... чувствительнее, в некотором роде», — говорила
она.
А Лени Кларк несколько минут назад это подтвердила.
Вот оно как.
Она не знает, от чего ей больнее — от недоверия Лабина или от запоздалого
понимания, насколько то оправдано.
Никогда в жизни она не чувствовала себя такой усталой. Неужели опять все
сначала?
Наверное, она произносит это вслух. Или Лабин уголком глаза улавливает не-
кий ее жест. Так или иначе, его руки замирают на пульте. Он, наконец, обора-
чивается к ней. Глаза, освещенные экранами, выглядят на удивление прозрачны-
ми.
— Не мы это начали, — произносит он.
Она только и может, что покачать головой.
— Выбирай, на какой ты стороне, Лени. Давно пора.
Ей этот вопрос представляется ловушкой — не забыть, как обходится Кен Лабин
с теми, кого считает врагами. Но от выбора ее избавляет тупой костолом Дейл
Кризи.
— Черт... — скрежещет его вокодер сквозь шипение помех.
Лабин мгновенно возвращается к работе.
— Кризи? Добрался до блока F?
— Обижаешь. Я бы настроился на мудаков и вслепую, в сраных Саргассах...
— Кто-нибудь покидал комплекс?
— Нет, не думаю. Но... черт побери. Их там такая прорва, что...
— Сколько именно?
— Именно что не знаю! Пара десятков как минимум. И, слушай, Лабин, в них
что-то странное — в том, что они излучают. Никогда еще такого не чувствовал.
Лабин вздыхает. Кларк представляет, как он закатывает глаза под линзами.
— Поточнее нельзя?
— Они холодные, парень. Почти все — словно ледышки. То есть настроиться-то
я на них могу, но не похоже, чтоб они хоть что-нибудь чувствовали. Может,
чем-то одурманены? Я к тому, что рядом с ними ты — капризный ребятенок...
Лабин с Кларк переглядываются.
— Я не в обиду, — жужжит после паузы Кризи.
— У подружки Алике был зельц, — говорит Кларк. — Вроде домашней зверушки...
А здесь, на пустом океанском дне, в этом скудном микрокосме, детской игруш-
кой может стать лишь то, чего очень много.
— Иди, — говорит Лабин.
Его «кальмар» причален к стопору у нижнего шлюза. Кларк жмет на газ, и ма-
шина, взвизгнув гидравликой, срывается с места.
Челюсть у Кларк вибрирует, принимая передачу. В голове звучит голос Лабина.
— Кризи, последний приказ отменяется. Заряд не ставить, повторяю, не ста-
вить. Поставь только микрофон и отступай. Чун, отведи своих на двадцать мет-
ров от шлюза. В контакт не вступать. К вам идет Кларк, она подскажет, что де-
лать .
«Я-то подскажу, — думает она, — а они пошлют меня куда подальше».
Она правит вслепую, по азимуту. Обычно этого более чем достаточно: на таком
расстоянии «Атлантида» уже выделяется светлым пятном на черном фоне. А сейчас
ничего подобного. Кларк включает сонар. Зеленый «снег» в десяти градусах от
курса — а в нем более жесткое эхо Корпландии, размытое помехами. Только те-
перь начинают проявляться тусклые отсветы — они теряются при попытке сфокуси-
ровать на них взгляд. Кларк включает налобный фонарь и осматривается.
По левому борту пустая вода. Справа луч упирается в клубы черного дыма —
косая линия завесы пересекает ее курс. Через несколько секунд она окажется в
самой гуще. Кларк выключает свет, чтобы не слепил в густом облаке. Чернота
перед линзами еще немного темнеет. Кларк не ощущает ни течения, ни повышения
вязкости. А вот вспышки становятся немного ярче — робкие проблески света
сквозь кратковременные разрывы в завесе. Короткие, как сигналы стробоскопа.
Свет ей не нужен. Теперь не нужен и сонар: она чувствует возбуждение во-
круг , нервозность, излучаемую рифтерами и — темнее, отдаленнее — страх из
сфер и коридоров под ними.
И что-то еще, знакомое и чуждое одновременно, живое и неживое.
Океан вокруг нее шипит и щелкает, словно она попала в стаю рачков-
эвфаузиид. Слабо дребезжат в ответ имплантаты. Она улавливает голоса сквозь
вокодеры, но не разбирает слов. Эхо на сонарном дисплее — по всем ста восьми-
десяти градусам, но из-за множества помех она не разбирает, шесть их там или
шестьдесят.
Один из видов рачков-эвфаузиид (Euphausia superba).
Прямо по курсу бравада, подкрашенная страхом. Кларк резко сворачивает впра-
во, но уклониться от столкновения не успевает. Мглу ненадолго разрывает про-
свет .
— Черт! Кларк, это т...
И нет его. Человек позади на грани паники, но не ранен: при повреждении те-
ла мозг дает вспышку определенного рода. Кажется, это был Бейкер. Их стано-
вится трудно различить в наплывающем льду фонового сознания, полностью лишен-
ного чувств. Оно раскинулось под клубком человеческих эмоций платформой из
черного обсидиана. В последний раз, когда она сталкивалась с подобным созна-
нием , то было подключено к живой ядерной бомбе. И пребывало в одиночестве.
Она резко задирает нос «кальмара». В имплантаты бьются новые сонарные им-
пульсы, вслед ей звучит хор испуганных механических голосов. Она не отвлека-
ется. Шипение в плоти слабеет с каждой секундой. Скоро худшее остается внизу.
— Кен. Слышишь меня?
Ответ доносится с задержкой — на таком расстоянии уже сказывается скорость
звука.
— Докладывай, — наконец отзывается он. Голос смазан, но понять можно.
— У них там внизу умные гели. Много, штук двадцать или тридцать. Собраны в
конце крыла, возможно, прямо в наружном шлюзе. Не понимаю, как мы раньше их
не выцепили. Может, они просто... терялись в шуме помех, пока не слились во-
едино .
Задержка.
— Есть догадки, чем они занимаются?
В прошлый раз, на Хуан де Фука, они умудрились сделать очень точные выводы
из перемены в мощности сигнала.
— Нет. Они просто там... есть. И думают все сразу. Будь там один или два, я
могла бы что-нибудь прочитать, а так...
— Они меня обыграли, — перебивает ее Лабин.
— Обыграли?
Что это в его голосе? Удивление? Неуверенность? Кларк подобного еще не слы-
шала.
— Чтобы я сосредоточился на блоке F-3.
«Это гнев», — понимает Кларк.
— Но зачем? — спрашивает она. — Тоже мне блеф — не думали ведь они, что мы
спутаем это с людьми?
Смешно: даже Кризи сообразил, что дело нечисто, а ведь он никогда не стал-
кивался с зельцем. С другой стороны — много ли корпы понимают в «тонкой на-
стройке»? Они могли забыть о различиях.
— Это не отвлекающий маневр, — бормочет в пустоту Лабин. — Им больше негде
выйти, сонары непременно засекут...
— Ну и что...
— Отводи людей, — резко командует он. — Они маскируют. . . заманивают нас и
что-то маскируют. Отводи...
Бездна сжимает кулак.
Она лишь на миг стискивает тело Кларк — даже не больно. Здесь, наверху, не
больно.
В следующий миг приходит звук: вуфф! Водоворот. Вода наполняется механиче-
скими воплями. Ее закручивает в потоке. Дымовая завеса под ней колеблется,
вскипает и рвется, потревоженная изнутри, озаряется тепловым свечением...
Кларк что есть сил жмет на газ. «Кальмар» тянет ее вниз.
— Кларк! — Как видно, звук взрыва дошел до головного узла, — что происхо-
дит?
Симфония рвущегося металла. Нестройный хор голосов. Их меньше, чем было,
понимает Кларк.
«Мы потеряли генератор, — тупо думает она. — Я слышу крики.
Слышу, как они умирают».
И не просто слышит. Крики возникают в голове раньше, чем доходят до ушей:
дикая химическая паника натриевыми вспышками поджигает рептильную часть моз-
га, умная кора в беспомощном смятении, сознание разбито, как дешевая стекляш-
ка.
— Кларк, докладывай!!!
А это гнев, тонкая пленка угрюмой решимости среди паники. Сквозь рассеяв-
шуюся муть ярче пробиваются огни. Только они не того размера и не того цвета.
Это не фонари рифтеров. Ее сонар взвизгивает, предупреждая о неизбежном
столкновении, мимо проносится неуправляемый «кальмар», его ездок светится бо-
лью от переломанных костей.
— Это не я, клянусь! Они сами...
Кризи удаляется, его муки сливается с муками других.
Корпус блока F заполняет экран сонара — его плавные очертания повсюду разъ-
едены рваными выбоинами: зияющие пасти пещер с кривыми металлическими зубами.
Одна плюется в нее — что-то со звоном рикошетит от «кальмара». Со всех сторон
жужжат и скрежещут вокодеры. В рваной облачной стене впереди открывается про-
ход: Кларк видит громоздкую тушу бронированного циклопа. Его единственный
глаз яростно сияет недобрым светом. Взгляд обращается на Кларк.
Она уходит влево, успевает заметить нечто, вращающееся в хаосе. Темная мае-
са глухо ударяется в нос «кальмара» и кувырком летит ей в лицо. Кларк приги-
бается . Обтянутая гидрокожей рука задевает ее.
— Лени!
Мертвые серые глаза, непрозрачные и равнодушные, остаются позади.
«О Господи. Боже мой».
Светящиеся металлические монстры шагают по обломкам, добивая раненых.
Она пытается собрать мысли.
— Они выходят из стены, Кен. Они поджидали там, взорвали стену изнутри и
выходят через стены.
«Черт бы тебя побрал, Пат. Это ты? Это все ты?»
Она вспоминает перекошенную шахматную доску на экране Лабина. Вспоминает,
как черные фигуры готовились к легкому разгрому врага. Только теперь ей еще
вспоминается: в шахматах первый ход всегда делают белые.
Того безразличного чуждого интеллекта больше нигде не ощущается. Должно
быть, в момент взрыва гели превратились в кашу.
В наружном шлюзе находились не только корпы в броне и умные гели. Еще там
были шрапнельные заряды, наверняка расположенные в расчете на максимальный
разлет. Кларк видит, куда попали осколки: в корпус станции, в разбитые кисло-
родные резервуары... Они торчат из ран в телах друзей и врагов. Они похожи на
стальные маргаритки, на лопасти миниатюрных ветряных мельниц. Их разбросало
одной силой взрывной волны — и они скосили тех, кого не засосало внутрь и не
насадило на рваные края пробоины.
Дымовая завеса почти рассеялась.
Лабин приказывает отступать. Из тех, кто его еще слышит, большинство уже
отступили. Фигуры в тяжелых скафандрах ползают по остаткам блока F-3, разби-
раясь с ранеными и мертвыми. Крабы крабами, неуклюжие и тяжеловесные. Вместо
коготков у них иглы — длинные хирургические иглы, тонкими копьями торчащие из
пальцев перчаток.
— Лени, как слышишь меня?
Она оглушено висит над «Атлантидой», с безопасной высоты смотрит, как крабы
пронзают черные тела. Иногда от кончика иглы поднимаются пузыри, уносятся в
небо стайкой скользких серебристых грибков. Воздух, под давлением впрыснутый
в тело. В оружие можно превратить почти что угодно.
— Лени?
— Она могла погибнуть, Кен. Дейла с Аброй я тоже не могу найти.
Другие голоса. Слишком смазанные, неузнаваемые. Как-никак, почти все гене-
раторы помех еще действуют.
Она настраивается на крабов. Хотелось бы знать, что у них сейчас на душе? А
кстати, и у нее самой — она никак не разберется. Может, в голове у нее зельц.
А там, внизу, корпы в броне подчищают грязь. Чувств там с лихвой. Реши-
мость . На удивление много страха.
Гнев — но отдаленный, мотивацию дает не он. И меньше ненависти, чем она
ждала.
Она поднимается выше. Панорама под ней освещается сиянием блуждающих фона-
рей . Дальше безмятежно горят огоньки других частей «Атлантиды». Она с трудом
улавливает гудение рифтерских голосов, слов не разобрать. И настроиться ни на
кого из них не удается. Она одинока на дне морском.
Неожиданно Кларк пересекает некую невидимую линию, и челюстная кость напол-
няется звуками.
— ...тела, — говорит Лабин. — Убитых по своему усмотрению. Гарсиа будет вас
принимать под медотсеком.
— Там и половины не поместится, — слабо жужжит вдали («О! Это Кевин!») —
Слишком много раненых.
— Все с F-3, кроме раненых и сопровождающих, встречаются у закладки. Хоп-
кинсон?
— Здесь.
— Есть что-нибудь?
— Кажется. В блоке Е полно мозгов. Кто, не определить, но...
— Йегер и Ын, поднимите своих на сорок метров по вертикали. Координаты не
менять, но всем отойти от корпуса. Хопкинсон, отводи своих к медотсеку.
— Мы в порядке...
— Исполнять. Нужны доноры.
— Господи, — слабо бормочет кто-то, — мы пропали.
— Нет. Не мы, а они.
Это Грейс Нолан, живая, властная и непоколебимая даже за фильтрами вокоде-
ра.
— Грейс! Они же...
— и что? — жужжит она. — Думаешь, они побеждают? А что дальше, народ? Этот
трюк второй раз не пройдет, а у нас хватит зарядов подорвать весь фундамент
на хрен. Вот мы их и подорвем.
— Кен?
Короткая пауза.
— Слушай, Кен, — жужжит Нолан, — до закладки я доберусь через...
— Не нужно, — жужжит Кен, — туда уже выдвинулись.
— Кто?
— Кстати, с возвращением, — обращается Лабин к кому-то неназванному. — Цель
известна?
— Да... — голос слабый, слишком искажен, чтобы опознать.
— Заряд установишь в пределах метра от метки. Установишь — и быстро отплы-
вай! Не задерживайся у корпуса больше необходимого, ясно?
— Да.
— Детонатор акустический, я взорву отсюда. На счет десять снимаю блокаду.
«Господи, — думает Кларк, — так это ты!»
— Всем отойти на безопасное расстояние, — напоминает Лабин. — Блокаду сни-
маю!
Она далеко за пределами действия глушилок, для нее перемена не слишком за-
метна. Однако следующий голос, долетающий до нее через вокодер, звучит по-
прежнему тихо, но легко узнаваем.
— Есть, — жужжит Джулия Фридман.
— Отходи, — приказывает Лабин. — На сорок метров. Подальше ото дна.
— Привет, Аврил, — окликает Фридман.
— и тебе, — отвечает Хопкинсон.
— Вы настраивались на то крыло — там дети были?
— Да... Да, были.
— Хорошо, — жужжит Фридман, — Джин терпеть не мог детей.
Канал затихает.
Сперва ей кажется, что месть идет по плану. По миру прокатывается пульсация
— глухой, почти субзвуковой импульс отдается в растворе, плоти и костях, — и,
насколько ей известно, сотня, если не больше врагов обращается в кровавую ка-
шу. Она не знает, сколько рифтеров погибло при первой стычке, но сейчас счет
наверняка превзойден.
Она в давно знакомой среде, где это, кажется, ничего и не значит.
Даже второй взрыв — такой же приглушенный толчок, только мягче, более отда-
ленный — не сразу приводит ее в себя. Вторичных взрывов следовало ожидать —
должны были лопнуть трубопроводы и кабели, каскадом повзлетать резервуары вы-
сокого давления — всякое могло быть. Дополнительный бонус своим, не более то-
го.
Но нечто в глубине сознания подсказывает ей: со вторым взрывом что-то не
то; может, эхо не такое — словно ты качнул язык древнего церковного колокола,
а услышал дребезжание бубенчика. И голоса, вернувшиеся после толчка, не тор-
жествуют победу над гнусными ордами корпов, а полны сомнения и колебаний —
даже вокодеры не могут этого скрыть.
— Что это за хрень была?
— Аврил? Вы там почувствовали?
— Аврил? Кто-нибудь слышит... кто-нибудь...
— Черт побери, Гардинер? Дэвид? Стэн? Кто-ни...
— Гарсиа, ты... я не...
— Тут пусто. Я на месте, только тут ни хрена нет.
— Ты о чем?
— Все дно пузыря, его просто... похоже, снесло оба...
— Как — оба? Она заложила всего один заряд, и тот был...
— Кен? Кен! Где ты, черт бы тебя побрал?
— Лабин здесь.
Тишина в воде.
— Мы лишились медотсека. — Голос у него как ржавое железо.
— Что?..
— Как?..
— Заткнуться всем на хрен, — рычит в темноту Лабин. Снова молчание, почти
полное. Кто-то продолжает стонать-скрежетать по открытым каналам.
— Очевидно, на пузыре остался нераспакованный заряд, — продолжает Лабин. —
И детонировал он от того же сигнала, которым мы подорвали «Атлантиду». С это-
го момента и впредь — только узконаправленные импульсы. Иначе могут сдетони-
ровать и другие. Всем...
— Говорит «Атлантида».
Слова раскатываются над морским дном, как Глас Божий, никакие помехи его не
оскверняют. «Кен забыл вернуть блокаду, — соображает Кларк. — И начал орать
на своих...»
«Кен теряет контроль...»
— Возможно, вы полагаете, что сила на вашей стороне, — продолжает голос, —
но это не так. Даже если вы уничтожите наше убежище, вы обречены на смерть.
Кларк не узнает голоса. Странно. Звучит он властно.
— Вы инфицированы второй моделью. Вы все инфицированы. Вторая модель крайне
заразна и не дает симптомов в течение нескольких недель инкубационного перио-
да . Без лечения вы все погибнете в течение двух месяцев. У нас есть лекарст-
во.
Мертвая тишина. Даже Грей не высовывается с «Я же говорила».
— Мы защитили все файлы и культуры от неавторизованного использования. Убив
нас, вы убьете себя.
— Докажите, — требует Лабин.
— Обязательно. Только погодите минуту. А если вам не терпится, исполните
свой трюк с чтением мыслей. Как вы там говорите? «Настройтесь». Мне рассказы-
вали, что это обычно позволяет отличить правдивого человека от лжеца.
Никто не поправляет говорящего.
— Назовите свои условия, — произносит Лабин.
— Вам — нет. Мы будем вести переговоры только с Лени Кларк.
— Не исключено, что Лени Кларк погибла, — говорит Лабин. — Мы не можем свя-
заться с ней со времени первого взрыва.
Наверняка он так не думает: она высоко над станцией, внутри у нее по-
прежнему тикает. Но Кларк помалкивает. Пусть ведет игру по-своему. Может, это
его последний шанс.
— Это было бы очень плохо для всех нас, — сухо отвечает «Атлантида». — По-
скольку предложение будет отозвано, если через полчаса она не явится к шесто-
му шлюзу. Это все.
— Ловушка, — говорит Нолан.
— Эй, ты же сама говорила, что у них есть лекарство, — жужжит кто-то. Кларк
не различает голоса за вернувшимися помехами.
— Ну и что? — жужжит в ответ Грейс. — Я не верю, что они с нами поделятся,
и уж точно не доверю долбанои Лени Кларк быть моим послом. Как, по-вашему, от
кого эти хрены узнали про «тонкую настройку»? За смерть всех наших благода-
рить надо ее.
Кларк улыбается про себя. «Ее заботит горстка людей. Всего-то несколько
жизней». Она чувствует, как пальцы сжимают рулевой рычаг. «Кальмар» мягко ув-
лекает ее вперед, вода мягко толкает назад.
— Можно последовать их совету. «Настроиться» и проверить, врут или нет.
Кажется, это Гомес, но помехи становятся все сильнее, и в них теряются даже
грубые интонации вокодерной речи.
Гудок в челюсти — и писк за ухом. Кто-то вызывает по закрытому каналу. Воз-
можно , Лабин. Он, как-никак, король в тактике. Должен был вычислить, где ее
искать. Остальные сейчас слишком заняты своими увечьями.
— и что это докажет? Что они собираются... — помехи — ...его нам? Черт, ес-
ли у них и нет лекарства, они могли просто убедить своих, что оно есть, чтобы
мы не...
Голос Нолан затихает.
Лабин говорит что-то по открытому каналу. Гудки у нее в голове звучат на-
зойливей — хотя это, конечно, невозможно, шипение помех глушит сигнал вызова
наравне с остальными.
Опять Нолан:
— Отвали, Кен, зачем нам тебя слуша. . . ты. . . не сумел даже перехитри. . .
долб... кор...
Помехи — чистые беспорядочные помехи. Внизу теплится свет. Шестой шлюз пря-
мо по курсу, и все помехи на свете не помешали бы ей узнать ту, что ждет за
люком.
Вина выдает. Их здесь всего двое, с настолько перекрученной совестью.
Крещение
Роуэн распахивает люк, не дожидаясь, пока шлюз очистится. Морская вода
бьется о щиколотки рифтерши.
Кларк сбрасывает ласты и шагает в люк. Костюм оставляет и выглядит обычной
тенью, только расстегнула капан на лице. Роуэн отступает в сторону, освобож-
дая ей проход. Кларк надежно пристегивает ласты на спину и осматривает спар-
танскую обстановку помещения. Скафандров не видно. Раньше здесь одна из пере-
борок была увешана подводным снаряжением.
— Скольких вы потеряли? — тихо спрашивает она.
— Еще не знаем. Больше, чем их здесь было.
«Мелочь, — отмечает про себя Кларк. — Для нас обеих. Впрочем, война только
началась».
— Я, правда, не знаю, — говорит Роуэн.
Здесь, в недалекой от вакуума сухопутной атмосфере, ясновидение не работа-
ет .
— Мне не доверяли. И сейчас не доверяют.
Роуэн показывает глазами на огонек у стыка потолка с переборкой: микроско-
пический объектив. Всего несколько дней назад, когда корпы еще ничего не за-
тевали, рифтеры сами бы следили через них за событиями. Теперь наблюдение ве-
дут люди Роуэн. Она уставилась на рифтершу со странной остротой — Кларк преж-
де не замечала за ней таких взглядов. До нее не сразу доходит, что измени-
лось: впервые на памяти Кларк у Роуэн темные глаза. Должно быть, отключила
подачу информации на линзы, и взгляд не отвлекается на комментарии. В глазах
у нее лишь она сама.
Ошейник с поводком и то не были бы так красноречивы.
— Пошли, — говорит Роуэн, — они в лаборатории.
Кларк выходит из шлюза вслед за ней. Сворачивает направо в залитый розовым
светом коридор. Аварийное освещение, отмечает она: ее линзы превращают его в
дурацкую подсветку детской комнаты. А для глаз Роуэн это темная труба, зали-
тая кровавым светом, кишка чудовища-людоеда.
На пересечении она сворачивают налево, перешагивают разметку опускной пере-
борки .
— Так в чем подвох? — спрашивает она.
Корпы не откажутся от единственного имеющегося у них рычага, не привязав к
нему несколько веревочек.
Роуэн не оглядывается.
— Мне не сказали.
Еще один поворот. Они минуют люк аварийного причала, вделанный в наружную
переборку: стена с этой стороны изуродована нагромождениями проводов и табло.
Кларк задумывается, не «Бомбиль» ли стоит на той стороне — но нет, не тот от-
сек.
Роуэн вдруг останавливается, поворачивается к ней.
— Лени, если что-то...
Что-то пинает «Атлантиду» в бок. Где-то за их спинами с грохотом сталкива-
ются массы металла.
Розовый свет мигает.
— Чт...
Еще один пинок, уже сильнее. Палуба подпрыгивает. Второй раз услышав метал-
лический грохот, Кларк узнает его: упали аварийные переборки.
Свет гаснет.
— Пат, какого хера вы...
— Не мы, — голос Роуэн дрожит в темноте. До нее не больше метра: смутный
силуэт, темно-серый на черном.
«Никакой паники, — отмечает Кларк. — Ни криков, ни беготни по коридорам, ни
переговоров...»
Так тихо — почти мирно.
— Они нас отрезали, — говорит Роуэн. Края силуэта стали резче, деталей по-
прежнему не видно, но очертания проявились. Проявляются и отблески на пере-
борке . Кларк ищет глазами источник света и обнаруживает созвездие бледных ми-
гающих точек в нескольких метрах позади. На люке.
— Ты меня слышишь? Лени? — Напряжение в голосе Роуэн беспокоит Кларк. Корп
подается к ней: — Ты здесь?
— Здесь, здесь.
Протянув руку. Кларк легонько касается ее плеча. Призрачный силуэт Роуэн
вздрагивает от прикосновения.
— Ты... вы...
— Не знаю, Пат. Я тоже такого не ожидала.
— Нас отрезали. Слышала: переборка упала? Нас заперли! Ублюдки. Замуровали
спереди и сзади. По обе стороны — вода. Мы в ловушке.
— Однако этот отсек не залили, — замечает Кларк.
— Они нас хотят задержать, а не убить.
— Я бы за это не поручилась, — произносит переборка. Слепая в темноте, Роу-
эн подскакивает.
— Собственно говоря, — продолжает переборка, — корпа мы намерены как раз
убить.
Голос пронзительно вибрирует, искажается, проходя сначала через вокодер, а
потом сквозь микрофон, прилепленный к переборке. Кларк вдруг огорчается, что
ее голос для Алике звучит так же неприятно.
Этого голоса она не узнает. Женщина...
— Грейс?
— Ни хрена бы они тебе не дали, Лени. Ни хрена у них и нет. Хотели взять
заложницу, и ты простодушно полезла в ловушку. Но мы за своими присматриваем.
Даже за тобой.
— Что за фигню ты несешь? Откуда тебе знать?
— Откуда? — переборка вибрирует, как большая арфа. — Не ты ли нас научила
«настраиваться»? Настройка работает, милашка, офигительно работает, и мы про-
чли всю эту банду, собравшуюся в их медлаборатории. Можешь мне поверить, там
все буквально залито виной. Кстати, на твоем месте я бы запечатала клапан.
Сейчас будем тебя спасать.
— Грейс, постой! Еще секунду!
Кларк оборачивается к Роуэн.
— Пат?
Роуэн не качает головой. Роуэн не возмущается. Роуэн не делает ничего, что
должен делать невинный — да хоть бы и виновный — человек под угрозой смерти.
— Пат, ты... черт, только не говори, что ты...
— Конечно, нет, Лени. Но в этом есть смысл. Нас обеих обвели вокруг пальца.
Что-то звонко бьет по обшивке.
— Стойте! — Кларк обводит взглядом потолок и стены, не зная, как дотянуться
до невидимой противницы. — Пат здесь ни при чем!
— Ага, слышала. — Булькающий металлический звук, вероятно, означает смешок.
— Она, мать ее, председатель совета директоров, и ничего не знает. Ну да, ве-
рю.
— Ну, так настройся на нее! Проверь сама.
— Беда в том, Лен, что мы, новички, не очень хорошо умеем настраиваться на
одиночек. Одиночки дают слишком слабый сигнал, по нему мало что разберешь.
Попрощайся с тетей, Патти.
— Прощай, — шепчет Пат. По ту сторону переборки воет дрель.
— Чтоб ты провалилась, Нолан, убирайся, или, клянусь, я тебя сама убью.
Слышишь меня? Пат не знала! Она так же беспомощна, как...
Она не успевает договорить «как я», потому что в коридоре возникает новый
источник света, одинокая алая точка. Она ослепительно — даже для адаптирован-
ного зрения Кларк — вспыхивает и тут же гаснет.
Мир взрывается грохотом металла.
Силуэт Роуэн корчится в углу. Затемненное поле зрения Кларк пересекает бе-
лый луч лазера. Вода, доходит до нее. Сунь она руку под эту тонкую как каран-
даш струю, ее бы оторвало.
За несколько секунд вода поднимается до лодыжек.
Кларк бросается к Роуэн, отчаянно надеясь помочь и понимая, что помочь не-
чем. Камеру вдруг освещает тусклое красное сияние — на внешней стене мигает
еще один глазок. Вот и он открывается и гаснет — воздух просверливает вторая
смертоносная струя. Брызги рикошетят от внутренней стены жидкой шрапнелью:
плечо Кларк взрывается острой как игла болью. Она опрокидывается на спину,
вода смыкается над лицом, череп гудит от удара о палубу.
Перекатившись на живот, она поднимается на четвереньки. Вода уже доходит до
локтей. Не разгибаясь, она ползет к свернувшейся напротив Роуэн. Над головой
скрещиваются сотни смертоносных стрелок — рикошеты и отражения. Роуэн сползла
по внутренней переборке, по грудь ушла в ледяную воду повесила голову. Волосы
падают ей на лицо. Кларк приподнимает ей подбородок: на одной щеке темная по-
лоса , черная и бесформенная в скудном свете. Полоса растекается: попадание
шрапнелью.
Лицо у Роуэн бескровное, нагие глаза широко раскрыты, но слепы — несколько
случайных фотонов, летающих по коридору, и близко не подходят к порогу зрения
для невооруженного глаза. В лице Роуэн только звук, боль и леденящий холод.
— Пат! — Кларк за ревом воды едва различает собственный голос. Вода подни-
мается выше губ Роуэн. Кларк хватает ее под мышки, поднимает и прислоняет к
переборке. Несколькими сантиметрами левее бьет рикошет. Кларк отгораживает
Роуэн от жестокого града.
— Пат!
Она не знает, какого ответа ждет. Патриция Роуэн уже мертва, Клар остается
лишь наблюдать, как процесс дойдет до конца. Однако Роуэн отвечает: в окру-
жающем реве Кларк ничего не слышит, но видит, как шевелятся губы, почти уга-
дывает .
Внезапная пронзительная боль, удар в спину. На этот раз Кларк удерживает
равновесие: вода, дошедшая до половины высоты коридора, глушит большую часть
рикошета. Губы Роуэн все шевелятся. Она не говорит, видит Кларк, она выводит
губами слоги, медленно и тщательно, в расчете на зрение, а не на слух:
— Алике... Позаботься об Алике.
Вода снова доходит ей до подбородка.
Кларк нащупывает руку Роуэн, поднимает ее. Приложив ладонь к своей щеке,
кивает.
В ее личной бесконечной темноте Роуэн кивает в ответ.
«Сейчас Кен мог бы тебе помочь. Избавил бы от страданий, убил бы мгновенно.
Я не могу. Не умею. Прости».
Вода слишком глубока, чтобы в ней стоять: Роуэн слабо загребает руками и
ногами, хотя конечности у нее наверняка онемели от холода. Бесполезное уси-
лие, усилие примитивного мозга — все должное исполнено, все возможности ис-
черпаны, но тело цепляется за последние несколько секунд, предпочитая краткое
страдание бесконечному небытию. Смерти ей не избежать, но захлебываться не
обязательно. Поднимающаяся вода сжимает воздух, сжимает до таких атмосфер,
что кислород становится токсичным. Кларк слышала, что такие конвульсии не
обязательно мучительны.
Эта смерть, если Кларк не поторопится, настигнет ее так же быстро, как Роу-
эн . Кажется преступлением спасать себя, когда Патриция задыхается. Но у Кларк
тоже есть примитивные части мозга, и они не позволят болезненному, иррацио-
нальному чувству вины встать на пути самосохранения. Руки, действуя без по-
средства воли, запечатывают лицевой клапан, запуская встроенные в тело меха-
низмы. Она оставляет Роуэн наедине с судьбой. Вода заливает ее тело, как ко-
ридор, но с обратным эффектом. Океан, проникая ей в грудь, поддерживает, а не
похищает жизнь. Она вновь становится русалкой, а подруга умирает у нее на
глазах.
Только Роуэн не сдается. Все еще не сдается. Тело сопротивляется, вопреки
смирению разума. Под потолком остался лишь маленький воздушный карман, но
онемевшие, неуклюжие ноги корпа продолжают лягаться, руки цепляются за трубы
и... какого хрена она не сдается?
Давление переходит критический порог. Освобожденные нейротрансмиттеры поют
у нее в голове. И Лени Кларк вдруг оказывается в сознании Патриции Роуэн.
Кларк узнает, что чувствуют, умирая.
«Черт тебя возьми, Пат, почему ты не сдаешься? Что ты со мной делаешь?»
Она опускается на дно коридора. Решительно упирается взглядом в палубу,
ждет, пока уляжется болтанка, замрет вой рвущейся внутрь воды и останется
только тихое неверное шуршание, жалкий звук от замерзшей плоти, цепляющейся
за биосталь.
Наконец звуки борьбы прекращаются. Мука, печаль и раскаяние длятся чуть
дольше. Лени Кларк дожидается, пока последние отголоски Патриции Роуэн умрут
у нее в голове, и еще немного слушает тишину, прежде чем подключить вокодер.
— Грейс, ты меня слышишь?
Ее механический голос бесстрастен и мертв.
— Конечно, слышишь. Я тебя убью, сука.
Ее ласты плавают рядом, на привязи. Кларк отцепляет их и натягивает на но-
ги.
— Прямо передо мной причальный люк, Грейс. Сейчас я его открою, выйду и вы-
потрошу тебя как рыбу. На твоем месте я бы уже плыла подальше.
Может, она уже и уплыла. Ответа, во всяком случае, нет. Кларк проплывает по
коридору, удерживая взгляд на люке. Искрящаяся мозаика табло, неподвластная
даже самой Атлантике, освещает ей путь.
— Взяла разгон, Грейс? Тебе это не поможет.
Что-то мягко толкает ее в спину. Кларк ежится и заставляет себя не оборачи-
ваться .
— Кто не спрятался, я не виновата.
Она открывает люк.
Метка
Снаружи никого.
Улики остались — пара точечных паяльников растопырились на треногах, чудь
дальше на корпусе торчат устрицы приемопередатчиков, но ни Нолан, ни осталь-
ных не видно. Кларк угрюмо усмехается про себя.
Пусть побегают.
Только она вообще никого не может найти. Нет часовых, расставленных Лабином
на постах, никто не ведет наблюдения через микрофоны, облепившие «Атлантиду».
Она пролетает над медицинской лабораторией, которую, как ее заверили, прослу-
шивала целая группа «настроенных» рифтеров, уличая коварных корпов. Никого.
Краны, пузыри пристроек и тени. Местами еще мигают огоньки, но в основном
там, где они недавно горели — темнота. Маяки разбиты, иллюминаторы высажены.
Вокруг царство мрака. Нигде ни единого рифтера.
«Может, корпы обзавелись оружием, о котором не подозревал даже Кен? Нажали
кнопку, и все просто исчезли...»
Но нет, она чувствует жителей станции: и страхи, и надежды, и слепое, пани-
ческое отчаяние, расходящееся в воде на добрых десять метров. Не те чувства,
которые испытывают после полной и окончательной победы. Если корпы и знают,
что происходит, им от этого не легче. Кларк направляется в бездну в сторону
нервного центра. Только теперь она принимает «настройкой» слабое движение
впереди. Хотя нет, это опять то же самое: тот же страх, та же неуверенность.
Неужели она с такого расстояния читает «Атлантиду»? И почему, чем дальше от
станции, тем эти чувства все больше усиливаются?
Не велика загадка. И притворяться, что не догадываешься — плохое утешение.
Слабый шепот звучит в воде через низкочастотник. Она чувствует вокруг десятки
рифтеров, и все притихли, напуганные. Впереди пульсирует тусклое созвездие
огоньков. Кто-то движется наперерез Кларк, она угадывает присутствие только
по затмевающимся на миг фонарям. Разум пловца, наткнувшись на нее, содрогает-
ся в ужасе.
Вот они собрались вокруг пузыря. Мечутся наподобие оглушенных рыбешек или
просто висят неподвижно, ждут чего-то. Может, здесь все, кто выжил, может,
больше в мире не осталось ни одного рифтера? Ужас окружает их тучей.
Может, и Грейс Нолан здесь. Кларк охватывает холодный очищающий гнев. С де-
сяток человек оборачиваются на ее мысли, обращают к ней мертвые белые глаза.
— Что происходит? — жужжит Кларк. — Где она?
— Отвали, Лен, у нас сейчас дела поважнее.
Она не узнает голоса.
Кларк подплывает к нервному центру. Рифтеры раздаются перед ней, и только
шестеро загораживают путь. Гомес, Крамер, остальные слишком черны и далеки,
чтобы опознать по мозговому излучению.
— Она внутри? — спрашивает Кларк.
— Сдай назад, — говорит Крамер, — здесь ты не командуешь.
— О, я никому не приказываю. Все в вашей воле. Можете убраться с дороги или
попробовать меня остановить.
— Это Лени?
Голос Лабина доходит из воздушной среды.
— Да, — после короткой паузы жужжит Крамер. — Она здорово...
— Впустите ее, — говорит Лабин.
Здесь вечеринка для избранных, только по приглашениям. Кен Лабин, Джелейн
Чен, Дмитрий Александр. Аврил Хопкинсон. И Грейс Нолан.
Кен даже не оборачивается, когда она выбирается из мокрой комнаты.
— Разберетесь потом. Сейчас нам нужна ты, Лен, и Грейс тоже. Попробуете
вцепиться друг другу в глотки — я приму меры.
— Поняла, — ровно отвечает Нолан.
Кларк смотрит на нее и молчит.
— Так... — Лабин снова обращается к монитору, — Где вы были?
— Я почти уверен, что нас не видели, — говорит Чен. — Он был слишком занят
своим участком, а кругового обзора у этой модели нет.
Она дважды постукивает по экрану: изображение в центре замирает и приближа-
ется.
На нем нечто вроде обычного «кальмара», только с парой манипуляторов на пе-
реднем конце и без руля сзади. Какой-то зонд-автомат. Явно не здешний.
Хопкинсон сквозь зубы втягивает воздух.
— Вот, значит, как. Они нас нашли.
— Не обязательно, — возражает Чен, — На такой глубине дистанционное управ-
ление невозможно, особенно на такой местности. Он должен работать автоматиче-
ски. Тот, кто его послал, не узнает о станции, пока аппарат не вернется на
поверхность.
— Или пока тот не отправит отчет по расписанию.
Чен пожимает плечами:
— Океан велик и опасен. Не вернется — спишут на оползень ила или бракован-
ный навигационный чип. Нас подозревать — никаких причин.
Хопкинсон качает головой.
— Никаких? А что этот зонд вообще здесь делает, если не нас ищет?
— Слишком уж поразительное получилось бы совпадение, — соглашается Алек-
сандр .
Лабин стучит по экрану. Картинка отдаляется и продолжает прокручиваться с
прежнего места. У нижнего края толпятся цифры и сокращения, сдвигаются и
уползают по мере изменения телеметрии.
Зонд плывет в нескольких метрах от берега Невозможного озера, над самой по-
верхностью. Вытянув одну лапу, макает палец в галоклин и отдергивает, словно
испугавшись.
— Вы посмотрите, — говорит Нолан, — боится повышенной солености.
Маленький робот отползает еще на несколько метров и делает вторую попытку.
— Он ни разу вас не заметил? — спрашивает Лабин.
Александр качает головой:
— До сих пор не замечал. Был слишком занят обследованием местности.
— Видеозапись есть? — спрашивает Нолан таким тоном, будто в целом свете у
нее никаких забот. Будто не живет в долг.
— Только несколько секунд от начала. Очень мутная вода, мало что видно.
Слишком приближаться мы, понятно, не хотели.
— Однако сонарили регулярно, — отмечает Лабин.
Чен пожимает плечами.
— Выбрали из двух зол меньшее. Надо же как-то проследить, чем он занимает-
ся. Все лучше, чем ему показываться.
— А если он проведет триангуляцию по импульсам?
— Мы постоянно двигались. Импульсы посылали с широким разбросом. В худшем
случае ему известно, что кто-то прощупывает водяной столб, а у нас там пара
приспособлений в любом случае этим занимаются. — Чен тычет в экран, как будто
оправдываясь. — Там все записано.
Лабин хмыкает.
— Ну вот, сейчас начнется, — говорит Александр. — Секунд через тридцать...
Зонд скрывается в дымке, направляясь, видимо, к одному из огоньков, отме-
чающих трассу по поверхности Невозможного. Прежде, чем он окончательно скры-
вается, вид загораживает темная масса — какой-то утес вторгается на экран
слева. На его поверхности не видно кругов света, хотя субмарина всего в не-
скольких метрах: Чен с Александром работают в темноте, прячась за локальной
телеметрией. Картинка на экране перекашивается и покачивается: субмарина об-
ходит скалы — темная на темном, еле видная в смутном освещении, с черными об-
водами углов.
Александр склоняется вперед:
— Сейчас...
Свет справа впереди: дальний край выступа освещается и становится похожим
на осколки черного стекла. Субмарина сбавляет ход, продвигается дальше осто-
рожно , выходит на свет...
И чуть не сталкивается с возвращающимся зондом. Два значка телеметрии вспы-
хивают красным и начинают мигать. Запись не сопровождается звуком, но Кларк
представляет, как орет сирена в рубке. На мгновение зонд замирает: Кларк го-
това поклясться, что видит, как раскрываются диафрагмы стереокамер. Потом ап-
парат разворачивается — чтобы продолжить поиск или бежать как всем чертям —
смотря по тому, сколько у него мозгов.
Этого они уже не узнают, потому что снизу в поле зрение камеры выстреливает
серая чернильная лента. Она ударяет зонд в середину корпуса, расплескивается
и обвивает его, словно эластичная паутина. Аппарат вырывается, но упругие во-
локнистые концы упорно притягивают его к субмарине.
Кларк впервые видит в действии стрельбу сетью. Выглядит довольно круто.
— Ну, вот, — говорит Александр, когда изображение замирает. — Повезло, что
сеть не использовали раньше, на какую-нибудь из этих ваших чудовищных рыбин.
— И повезло, что я вообще догадалась выстрелить сетью, — добавляет Чен. —
Кто бы мог подумать, что она так пригодится. — Нахмурившись, она добавляет: —
Хотя интересно, что насторожило эту зверушку.
— Вы двигались, — напоминает ей Лабин.
— Да, естественно. Чтобы он не зафиксировал наш сонар.
— Он следовал на звук вашего мотора.
Доля самоуверенности слетает с Чен.
— Значит, он у нас, — говорит Кларк. — И сейчас?..
— Сейчас Дебби его разбирает, — говорит Лабин. — По крайней мере, мин-
ловушек в нем не нашли. Она говорит, что сумеет влезть к нему в память, если
там нет серьезной шифровки.
Хопкинсон снова веселеет.
— Серьезно? А может, устроить ему временное выпадение памяти и отправить
дальше гулять?
В такую удачу не верится, и взгляд Лабина это подтверждает.
— А что? — не понимает Хопкинсон. — Подделаем поток данных, отправим его
домой, и пусть расскажет мамочке, что здесь только ил и морские звезды. В чем
проблема?
— Насколько часто мы там бываем? — спрашивает ее Лабин.
— Что, на озере? Раз или два в неделю, не считая дней, корда устанавливали
аппаратуру.
— Довольно редко.
— Чаще не надо, пока не придут сейсмоданные.
У Кларк холодеет в животе — озноб зародился несколько секунд назад, когда
разговор зашел о надежде на ложные воспоминания, отступил как волна и вернул-
ся еще хуже, чем был.
— Дерьмо, — шепчет она, — ты про шансы...
Лабин кивает:
— Мы практически никак не могли оказаться на месте при первом же появлении
этой штуковины.
— Значит, она здесь не в первый раз. Бывала и прежде, — кивает Кларк.
— Как минимум, несколько раз. Я бы сказал, она, вероятно, бывает на Невоз-
можном чаще, чем мы. — Лабин обводит остальных взглядом. — Кто-то нас вычис-
лил. Послав аппарат обратно без записей, мы просто сообщим им, что нам это
известно.
— Дрянь, — дрожащим голосом бормочет Нолан. — Мы вляпались. Пять лет. Вля-
пались по уши.
В кои-то веки Кларк склонна с ней согласиться.
— Не обязательно, — отвечает Лабин, — думаю, нас они пока не нашли.
— Чушь какая... Ты сам сказал, месяцы, если не годы.
— Они не нашли нас, — этот ровный, подчеркнуто сдержанный тон у Лабина оз-
начает , что терпение на исходе. Нолан немедленно затыкается.
— А нашли они, — после паузы продолжает Лабин, — только сеть осветительных
приборов, сейсмографы и записывающую аппаратуру. Откуда им знать, что это не
остатки заброшенной горной выработки? — Чен открывает рот, но Кен останавли-
вает ее жестом. — Лично я в это не верю. Раз у них есть причины искать нас в
этом районе, то они должны предполагать, что техника наша. Однако эти парни
сейчас знают только то, что они где-то неподалеку от цели, — Лабин слабо улы-
бается . — Так и есть — до нас всего двадцать километров. Двадцать непрогляд-
но-черных километров по самому изрезанному ландшафту на планете. Если больше
у них ничего нет, то они нас не найдут никогда.
— Но могут послать какого-нибудь дрона, который затаится на месте, подождет
нас, — возражает Хопкинсон, — а потом проследит до станции.
— Может быть, уже послали, — соглашается с ней Кларк.
— Тревоги не было, — напоминает Чен
Лени вспоминает: здесь на каждом пузыре, на каждом зонде и «кальмаре» стоит
передатчик. Между собой эти устройства ведут вежливые переговоры, но коснись
их сонар, не знающий местного диалекта, поднимут такой ор, что мертвого раз-
будят . Об этой системе, пережитке первых дней изгнания, когда страх, что их
обнаружат, давил каждого свинцовой рукой, Кларк не вспоминала годами. Но по
сию пору врагов они находили только в собственной среде.
— А странно, что они не попытались, — говорит Лени. — Казалось бы, этот ва-
риант напрашивается.
— Может, и пытались, но потеряли нас, — предполагает Хопкинсон. — Подойди
они слишком близко, мы бы их заметили, а на маршруте есть места, где сонарныи
сигнал распространяется не дальше шестидесяти метров.
— А может, у них не хватает средств, — с надеждой говорит Александр. — Мо-
жет, там просто лодочка с парой искателей сокровищ и древней картой.
— Или они пока до такого не додумались, — это Нолан.
— А может, им это не нужно, — говорит Лабин.
— Ты о чем? — Кажется, Хопкинсон угадывает его мысль. — Думаешь, они просто
решили тут все дезинфицировать?
Лабин кивает
Поняв, все умолкают. Зачем тратить дорогую аппаратуру, выслеживая добычу на
территории, возможно, усаженной ловушками? Зачем рисковать, когда дешевле и
проще хитростью заставить врага отравить собственный колодец?
— Дерьмо, — выдыхает Хопкинсон. — Все равно, что оставлять для муравьев от-
равленную приманку, чтобы они сами таскали ее королеве...
Александр кивает.
— Вот, значит, это откуда... Бетагемот никак не должен был сюда попасть, и
вдруг, словно по волшебству...
— Бета-макс попал к нам из треклятой «Атлантиды», — рычит Нолан. — Насколь-
ко мы знаем, из озера получен только первичный штамм. Против этого — только
слово корпов.
— Да, но ведь и первичный штамм не должен был тут появиться.
— Что, все, кроме меня, забыли, — что и первый штамм выведен корпами? — Но-
лан обводит собравшихся яростным взглядом белых глаз. — Ради бога, Роуэн сама
призналась!
Ее взгляд падает на Кларк — как струя антиматерии. У Лени сжимаются кулаки,
уголки губ оттягиваются назад в оскале. Она понимает, что язык ее тела не
предлагает разрядки ситуации.
«И хрен с ним», — решает она и делает шаг вперед.
— Ах, так! — шипит Нолан и нападает.
Лабин делает ход. Такой непринужденный — только что он сидел за пультом, а
вот Нолан уже валяется на палубе сломанной куклой. В неуловимо кратком проме-
жутке Кларк, кажется, видела, как Лабин поднимается с места, заметила движе-
ние его локтя к солнечному сплетению Грейс, а колена — к ее спине. Возможно,
она даже что-то услышала: треск, как от сломанной ветки. И теперь соперница
лежит навзничь, неподвижная, только мелко подрагивают пальцы и веки.
Остальные окаменели.
Лабин окидывает стоящих взглядом:
— Мы все под угрозой. Независимо от того, откуда взялся Бета-макс, мы не
научимся его лечить без помощи корпов — теперь, когда Бхандери погиб. У кор-
пов есть необходимые специалисты и в других областях.
Нолан под ногами булькает горлом, неуверенно шевелит руками — ноги красно-
речиво неподвижны.
— Например, — продолжает Лабин, — у Грейс сломан третий поясничный позво-
нок. Без помощи «Атлантиды» она на всю жизнь останется парализованной ниже
пояса.
Чен бледнеет:
— Ради бога, Кен!
Шок освобождает Лени от паралича, и она падает на колени рядом с Нолан.
— Не стоит перемещать ее без кокона, — мягко говорит Лабин. — Может быть,
Дмитрий сумеет его соорудить.
Это звучит как обычное предложение, но через несколько секунд шлюз запуска-
ется на выход.
— А вы, друзья, — тем же ровным тоном продолжает Лабин, — надеюсь, понимае-
те , что ситуация переменилась, и сотрудничество с «Атлантидой» отныне в наших
интересах
Все, вероятно, понимают то же, что и Кларк: этот человек, ни секунды не
раздумывая, сломал спину своей помощнице ради довода в споре. Лени опускает
взгляд на побежденного врага. Глаза у нее открыты, веки вздрагивают, однако,
вряд ли Нолан в полном сознании.
«Вот тебе, убийца. Тупая мразь. Что, больно, милочка? Этого еще мало. Надо
бы больше!»
Но это наигранная ярость. Кларк вспоминает, как умирала Роуэн и что она
чувствовала потом: холодный убийственный гнев, абсолютную, гранитную уверен-
ность, что Грейс поплатится за это жизнью. И вот она лежит, беспомощная, сло-
манная чужой рукой — а на месте пылающий ярости только холодная зола.
«Я бы сама с ней покончила, — размышляет она, — если бы Кен мне не поме-
шал» .
Неужели она настолько не верна памяти подруги, если эта мысль так мало ее
радует? Или страх, что их нашли, вытеснил неистовство, или это все то же ста-
рое оправдание — что Лени Кларк, воздав тысячекратно, утратила вкус к мести?
«Пять лет назад меня не волновали смерти миллионов невинных. А теперь мне
не хватает духа наказать одну виновную».
Кому-то, вероятно, это могло бы показаться улучшением.
—...Еще неопределенно, — продолжает Лабин, вернувшись к пульту. — Возможно,
пославшие зонд ответственны и за Бета-макс, а возможно, и нет. Если да, они
уже сделали свой ход. Если нет, они еще не готовы. Даже если им точно извест-
но наше местоположение — а я в этом сомневаюсь — все равно, либо у них еще не
все фигуры на доске, либо есть иные причины выжидать.
Он, не тратя больше взглядов на булькающее позади существо, возвращается к
числам на экране. Кларк беспокойно косится на Нолан, однако намек Лабина про-
зрачен и ясен:
Я здесь главный. Принимай как есть.
— Какие причины? — спрашивает она, помолчав.
Лабин пожимает плечами.
— Сколько у нас времени.
— Будет больше, если не сидеть на месте, — Кен складывает руки на груди и
скребет себе бока. Под его гидрокостюмом неприятно шевелятся мышцы и сухожи-
лия. — Если они узнают, что их засекли, то волей-неволей начнут действовать,
и скорее раньше, чем позже. Так что наше дело — выиграть время. Отредактируем
память зонда и отпустим его, устроив мелкую поломку, которая объяснит задерж-
ку. Окрестности озера обыщем на предмет «жучков», сеть обрежем как минимум в
полукилометре от «Атлантиды» и нашего трейлерного парка. Лэйн прав: вряд ли
эти мины установил зонд, но, если все же он, детонатор расположен в пределах
действия акустической системы.
— Хорошо.
Хопкинсон с заметным трудом отводит взгляд от павшей подруги.
— Итак... мы замиряемся с «Атлантидой», обрабатываем зонд и прочесываем ок-
ругу в поисках других пакостей. Что дальше?
— Дальше я возвращаюсь, — говорит ей Лабин.
— Куда, на озеро?
Лабин слабо улыбается.
— В Северную Америку.
Хопкинсон удивленно присвистывает.
— Ну, наверно, если кто и сумеет с ними разобраться...
«Разобраться с кем?» — думает Кларк. Вслух этого вопроса не задает никто. С
кем — это со всеми, кто остался там. Они. Они делают все, чтобы нас уничто-
жить. Они шпионят за Срединно-Атлантическим хребтом, высматривают своими бли-
зорукими глазами координаты для торпедного залпа.
Почему — тоже никто не спрашивает. У этой охоты нет причин: просто так они
действуют. Не ищи корней — вопрос «почему» ничего не дает, причин не счесть,
и мотивы есть у каждого, кто еще жив. Этот расколотый, биполярный микрокосм
загнивает и заражает океанское дно, и все причины его существования сводятся
к одной аксиоме: «А потому!».
И все же, сколько из здесь присутствующих — сколько из рифтеров, и даже
сколько из сухопутников — по настоящему опустили занавес? На каждого корпа,
чьи руки в крови, приходится много других: родных, друзей, обслуги при меха-
низмах и телах — и эти не виновны ни в чем, кроме связи с первыми. И, если бы
Лени Кларк с такой яростью не желала яростно отомстить, если бы не решила
списать весь мир в побочные расходы — дошли бы они до такого?
«Алике», — сказала Роуэн.
— Нет, — качает головой Кларк.
Лабин обращается к экрану:
— Здесь мы, самое большое, можем тянуть время. Но этим временем надо вос-
пользоваться .
— Да, но...
— Мы глухи, слепы, на нас напали. Уловка не удалась, Лени. Нам необходимо
знать, с чем мы имеем дело, и какие для этого дела есть средства. Надежда на
лучшее больше не вариант.
— Пойдешь не ты, — говорит Кларк.
Лабин поворачивается к ней лицом, вздергивает бровь — вместо ответа.
Она хладнокровно встречает его взгляд.
— Мы.
Он еще до выхода наружу успевает отказать ей трижды.
— Здесь нужен командир, — настаивает он, пока наполняется шлюзовая камера.
— Ты подходишь как нельзя лучше. Теперь, когда Грейс выведена за скобки, у
тебя ни с кем не будет проблем.
У Кларк холодеет внутри.
— Вот зачем это было? Она сделала свое дело, тебе надо было ввести в игру
меня, вот ты и... сломал ее?
— Ручаюсь, ты бы обошлась с ней не лучше.
«Я убью тебя, Грейс. Выпотрошу, как рыбу!».
— Я иду с тобой, — говорит она. Люк под ними проваливается.
— Ты, правда, думаешь, что сможешь заставить меня взять тебя с собой?
Он тормозит, разворачивается и одним гребком уходит из луча света.
Кларк следует за ним:
— А ты думаешь, что сможешь обойтись вообще без поддержки?
— Лучше так, чем с необученной обузой, которая лезет в это дело по самым
неразумным причинам.
— Хрена ты знаешь, какие у меня причины.
— Ты будешь меня тормозить, — жужжит Лабин. — У меня куда больше шансов,
если не придется за тобой приглядывать. Если ты попадешь в беду...
— Ты меня бросишь, — перебивает она. — Мигом. Я знаю, какой ты в бою. Черт,
Кен, я с тобой хорошо знакома!
— Последние события доказывают обратное.
Он не сумел ее поколебать. Она твердо отвечает на его взгляд.
Кен ритмичными гребками уходит в темноту.
«Куда он? — гадает Кларк. — В той стороне ничего нет».
— Ты не будешь спорить, что не подготовлена для такой операции, — доказыва-
ет он. — Тебя не учили...
— Довод не в твою пользу, ты не забывай, что я прошла через Америку, и ни
ты, ни твоя армия и все эти крутые тренированные парни не сумели меня пой-
мать , — недобро улыбается она под маской. Улыбка ему не видна, но, возможно,
Лабин успел настроиться на чувства. — Я вас побила, Кен. Может, я была куда
глупее, и хуже подготовлена, и меня не поддерживали бойцы всей Америки, но я
месяцами водила вас за нос, и тебе это известно.
— Тебе неплохо помогли, — напоминает он.
— Может, и сейчас помогут.
Он сбивается с ритма. Пожалуй, об этом Лабин не думал. Она рвется в брешь:
— Подумай об этом, Кен. Все эти виртуальные вирусы собираются вместе, заме-
тают за мной следы, создают помехи, превращают меня в легенду...
— Актиния работала не на тебя, — жужжит он. — Она тебя использовала. Ты
просто была...
— ...Орудием. Мемом из плана глобального Апокалипсиса. Дай мне передышку,
Кен, мне и так никогда этого не забыть, сколько не старайся. Ну и что? Все
равно я была носителем. Она искала меня. Я ей настолько нравилась, что Акти-
ния сбивала вашу кодлу у меня с хвоста. Как знать, может, она еще цела? Иначе
откуда бы взялись те виртуальные демоны? Думаешь, они случайно называют себя
моим именем?
Его смутный силуэт вытягивает руку. Серия щелков разбрызгивает воду. Он на-
чинает заново, чуть сменив тон:
— Ты полагаешь, что если вернешься и объявишься перед Актинией — или тем,
чем она теперь стала — она прикроет тебя волшебным щитом?
— Может и...
— Она изменилась. Они всегда меняются, ежеминутно. Актиния не могла сохра-
ниться такой, какой мы ее помним, а если то, с чем мы в последнее время стал-
кивались, исходит от новой версии, тебе не стоит возобновлять с ней знакомст-
во.
— Может и так, — признает Кларк. — Но, возможно, в основе она не измени-
лась . Актиния ведь живая, так? С этим все соглашались. И не важно, построена
она на электронах или углероде. «Жизнь — просто самовоспроизводящаяся инфор-
мация , формирующаяся естественным отбором», так что Актиния подходит. А в на-
ших генах есть участки, не менявшиеся миллионы поколений. Почему с ней должно
быть иначе? Откуда тебе знать, что у нее в основную программу не вписан код
«Защити Лени»? И, между прочим, мы куда направляемся?
Фонарь на лбу Лабина включается на полную мощность, и на илистый грунт впе-
реди ложится яркий овал.
— Сюда.
Серая, как кость, грязь, ничем не выделяющаяся. Даже камешка приметного не
видно.
«Может быть, это кладбище, — от этой мысли у Кларк вдруг мутится в голове.
— Может, здесь он все эти годы удовлетворял свои пристрастия отупевшими дика-
рями и пропавшими без вести, и вот теперь добрался до глупой девчонки, не по-
нимающей слова „нет"».
Лабин погружает руку в ил. Жижа вокруг его плеча вздрагивает, как будто под
ней что-то толкается. Так оно и есть; Кен разбудил что-то, скрывавшееся под
поверхностью. Он вытаскивает руку, и оно, извиваясь, следует за ней. С него
облетают куски и меловые облачка.
Это раздутый тор около полутора метров в поперечнике. Вдоль экватора ряд
точек — гидравлические форсунки. Два слоя гибкой сетки затягивают отверстия:
одна сверху, другая снизу. Между сетками набитый чем-то угловатым ранец. Он
блестит сквозь муть, гладкий как гидрокостюм.
— Я припас здесь кое-что на обратную дорогу, — жужжит Лабин. — На всякий
случай.
Он отплывает на несколько метров назад. Механический слуга разворачивается
на четверть круга, и, плюясь из сопел мутной водой, следует за хозяином.
Они движутся обратно.
— Значит, вот что ты надумала? — жужжит Лабин. — Найти нечто, что, эволю-
ционируя, помогло тебе уничтожить мир, понадеяться, что в его сущности есть
добрая сторона, к которой можно воззвать, и...
— И разбудить тварь поцелуем, — договаривает за него Кларк. — Кто сказал,
что я не сумею?
Он плывет дальше, к разрастающемуся впереди сиянию. Глаза его отражают по-
лумесяцы тусклого света.
— Думаю, мы это проверим, — говорит он, наконец.
Точка
опоры
Без этого она бы предпочла обойтись.
Оправданий более чем достаточно. Недавнее перемирие еще очень хрупко и не-
надежно; не то, чтобы оно грозило полностью рухнуть перед лицом новой, всеоб-
щей угрозы, но маленькие трещинки и проколы приходится заделывать постоянно.
Корпы вдруг превратились в полезных экспертов, с которыми не сравнится ника-
кая техника — не сказать, чтобы рифтеры особенно радовались влиянию, которое
приобрели их недавние пленники. Невозможное озеро надо вымести от жучков, ок-
рестности морского дна прочесать в поисках камер наблюдения и детонаторов.
Безопасных мест теперь нет нигде — и не будь Лени Кларк занята сборами, ее
глаза пригодились бы в патрулировании периметра. В последней стычке погибли
десятки корпов — вряд ли сейчас время утешать их родных.
И все же, мать Алике умерла у нее на руках всего несколько дней назад, и,
хотя подготовка отнимала все время, Кларк винит себя в подлой трусости за то,
что так долго это откладывала.
Она нажимает кнопку звонка в коридоре.
— Леке?
— Входи.
Алике сидит на кровати, отрабатывает движения пальцев. Когда Лени закрывает
за собой люк, она откладывает флейту. Не плачет: то ли еще не отошла от шока,
то ли страдает от подростковой гиперсдержанности. Кларк видит в ней себя пят-
надцатилетнюю . И тут же вспоминает: все ее воспоминания о том времени лгут.
Все же душой она тянется к девочке. Хочется подхватить Алике на руки и уне-
сти ее в следующее тысячелетие. Хочется сказать, что она все пережила, она
знает, каково это, и это даже правда, пусть и неполная. У нее отнимали друзей
и любимых. Мать умерла от туляремии — хотя это воспоминание стерто вместе с
остальными. Но Кларк понимает, что это другое. Патриция погибла на войне, а
Кларк сражалась на другой стороне. Она не уверена, примет ли Алике ее объя-
тия.
Потому она присаживается рядом с девочкой на кровать и кладет ладонь ей на
колено — готовясь отдернуть руку при малейшем признаке недовольства — ищет
слова, хоть какие-то слова, которые бы не показались затертыми, когда их про-
износят вслух.
Она все еще собирается с духом, когда Алике спрашивает:
— Она что-то говорила? Перед смертью?
— Она. . . — Кларк качает головой. — Нет, в общем-то, нет, — заканчивает она
с ненавистью к себе.
Девочка смотрит в пол.
— Говорят, ты тоже уходишь, — продолжает она через некоторое время. — С
ним.
Кларк кивает.
— Не уходи.
Лени набирает в грудь побольше воздуха.
— Алике, ты... ох, Господи, мне так жа...
— Разве тебе обязательно уходить? — Алике поворачивается к ней и смотрит
жесткими яркими глазами, в которых слишком многое видится. — Что вы там, на-
верху, будете делать?
— Надо найти тех, кто нас выследил. Нельзя сидеть и смирно ждать, пока они
выстрелят.
— С чего вы взяли, что они будут стрелять? Может, просто хотят поговорить,
например?
Кларк качает головой, дивясь такой нелепой мысли:
— Люди не такие.
— Не какие?
«Они не прощают...»
— Они не дружелюбные, Леке. Кто бы это ни был. Будь уверена.
Но Алике уже переключилась на план Б:
— А много ли с тебя там толку? Ты не шпионка, не технарь. Ты не бешеный
психопат-убийца, как он. Ты просто погибнешь, ничего не сделав.
— Кто-то должен его поддержать.
— Зачем? Пусть идет один. — В голосе Алике вдруг появляется лед. — Лучше,
чтобы у него ничего не вышло. Чтоб те, наверху, порвали его на части, и в ми-
ре стало чуточку меньше говна.
— Алике...
Дочь Роуэн поднимается с кровати и прожигает ее взглядом:
— Как ты можешь ему помогать после того, как он убил маму? Как ты можешь с
ним разговаривать! Он — психопат, убийца.
Готовые возражения замирают на губах. В конце концов, Кларк не уверена, что
Лабин не приложил руку к смерти Роуэн. Кен в этом конфликте был капитаном ко-
манды, как и в прошлый раз: даже если он не планировал «спасательную опера-
цию», то мог знать о ней.
И все же Кларк почему-то чувствует себя обязанной защитить врага этой пора-
женной горем девочки.
— Нет, милая, — мягко говорит она, — все было наоборот.
— Что?
— Кен сперва стал убийцей, а уж потом психопатом.
Это достаточно близко к истине.
— о чем ты говоришь?
— С его мозгом поработали. Ты не знала?
— Кто?
«Твоя мать».
— Энергосеть. Ничего особенного, обычный набор для промышленного шпионажа.
Устроили так, что он вынужден был любыми средствами обеспечивать сохранение
секретности, даже не задумываясь. Непроизвольно.
— Ты хочешь сказать, у него не было выбора?
— Не было, пока он не заразился Спартаком. А со Спартаком такая штука: он
разрывает перестроенные связи, но не останавливается на этом. Так что у Кена
теперь нет того, что называется голосом совести, и если ты таких людей назы-
ваешь «психопатами», я с тобой соглашусь. Но он этого не выбирал.
— Какая разница? — резко спрашивает Алике.
— Он не выбирал зло сознательно.
— Ну и что? Когда это маньяки нарочно выбирали себе химию мозга?
Кларк должна признать, что довод резонный.
— Прошу тебя, Лени, — тихо говорит Алике, — не доверяй ему.
И все же — при всех его секретах и предательствах — Кларк странно, болез-
ненно доверяет Лабину. Она никому в жизни так не доверяла. Вслух этого, ко-
нечно , говорить нельзя. Нельзя говорить, потому что Алике уверена: Кен убил
ее мать — и, возможно, так оно и есть. Признаться, что ему доверяешь — значит
подвергнуть дружбу этого раненого ребенка слишком жестокой проверке.
Но это лишь удобное оправдание, первым всплывающее на поверхность. Есть еще
одна причина, глубокая и зловещая. Алике, возможно, права. Последние пару
дней Кларк замечала за линзами Лабина что-то незнакомое. Оно исчезало, едва
Лени пыталась сосредоточиться, поймать его взгляд — она не взялась бы ска-
зать , что именно заметила. Слабое трепетание век, пожалуй. Неуловимую дрожь
фотоколлагена, отражающую движения глаз под ним.
До последних трех дней Кен здесь, внизу, никого не лишил жизни. Даже во
время первого восстания он ограничивался тем, что ломал кости: все убийства
совершались неумелыми, но старательными руками рифтеров, наслаждающихся вла-
стью над прежними владыками. И за последние семьдесят два часа все смерти,
безусловно, можно оправдать самообороной. И все же. Кларк беспокоится, не
пробудила ли недавняя бойня нечто дремавшее в нем пять лет. Потому что рань-
ше , что ни говори, Кен любил убивать. Жаждал, хотя — сбросив химические путы
— использовал свободу не как оправдание, а как вызов. Он сдерживал себя: так
застарелый курильщик носит в кармане невскрытую пачку сигарет — доказывая,
что сильнее привычки. Если Лабин чем и гордится, так это своей самодисципли-
ной.
Но эта жажда, это желание отомстить миру — исчезла ли она? Когда-то те же
чувства владели Кларк — теперь эта страсть, потушенная миллиардом смертей,
больше не имеет над ней власти. Но Лени не уверена, что последние события не
подсунули Кену пару канцерогенных палочек прямо в рот. А если после такого
долгого перерыва ему понравился вкус дыма, и Кен вспомнил, как сладок тот был
прежде?
Кларк грустно качает головой.
— Больше некому, Алике. Приходится мне.
— Почему?
«Потому что для того, что я сделала, геноцид — слишком мягкое слово. Потому
что, пока я пряталась здесь, внизу, мир умирал всюду, где я прошла. Потому
что меня уже тошнит от собственной трусости».
— Потому что я это натворила, — отвечает она, наконец.
— Ну и что? Разве, вернувшись, ты все исправишь? — Алике недоверчиво качает
головой. — Какой смысл?
Она стоит перед Кларк, хрупкая, как фарфоровый китайский император.
Больше всего Лени хочется ее обнять. Но она не настолько глупа.
— Я... я должна взглянуть в лицо тому, что сделала, — слабо защищается
Кларк.
— Фигня, — отвечает Алике. — Ничему ты не взглянешь. Ты удираешь.
— Удираю?
— Прежде всего, от меня.
И тут даже такая профессиональная идиотка, как Кларк, понимает: Алике боит-
ся не того, что сделает с Лени Лабин. Она боится того, что Лени может сотво-
рить с собой. Она не глупа, она много лет знает Кларк и знает, какие особен-
ности делают рифтера рифтером. Когда-то Кларк была склонна к суициду. Когда-
то она ненавидела себя до желания умереть — еще до того, как совершила хоть
что-то, заслуживающее смерти. А теперь собирается вернуться в мир, где все
напоминает о том, что она убила больше народу, чем все Лабины вместе взятые.
Понятно, что Алике Роуэн беспокоится, не перережет ли лучшая подружка себе
вены. Честно говоря, Кларк сама насчет этого не уверена.
Но отвечает по-другому:
— Все в порядке, Леке. Я не... я ничего плохого с собой не сделаю.
— Правда?
Судя по голосу, Алике не смеет надеяться.
— Правда. — И теперь, успокоив обещанием подростковые страхи, Лени Кларк
берет ладошки Алике. Та сейчас вовсе не кажется хрупкой. Она холодно смотрит
на руки Кларк, сжимающие ее вялые пальцы, не отвечает на пожатие и тихо про-
износит :
— Очень жаль.
Входящие
Снаряды вырываются из Атлантического океана уродливым фейерверком. Они ле-
тят к западу пятью небольшими стайками, начинают десятиминутную шахматную
партию, разворачивающуюся на половине полушария. Они петляют и закручиваются
вдоль траекторий, словно прочерченных пьяным — это было бы смешно, если бы не
затрудняло их перехват.
Дежарден сделал все, что мох1. Полдюжины старинных стратегических спутников
ожидали его призыва два года — с тех пор, как он переманил их на свою сторону
как раз ради такого случая. Теперь ему достаточно постучать в калитку — по
первой команде они растопырили лапки и раскрыли ему мозги.
Машины обращают внимание на густые следы, пятнающие атмосферу внизу. Слож-
ные и тонкие алгоритмы вступают в игру, отделяют зерна от плевел, предсказы-
вают движение цели и рассчитывают пересекающийся курс.
Их предсказания точны, но не идеальны: как-никак, у врага тоже есть мысля-
щие машины. Обманка во всем подражает охотнику. Каждый выхлоп реактивного
двигателя снижает вероятность попадания. Виртуально изнасилованные Дежарденом
боевые спутники принимают контрмеры — лазеры, собственные ракеты, выпущенные
из драгоценных, невозобновимых запасов — но каждое решение вероятностно, каж-
дый ход определяется статистически. В игре шансов ни в чем нельзя быть уве-
ренным .
Три ракеты достигают цели.
Две упали на Флоридский полуостров, одна — в техасский Пыльный Пояс. Дежар-
ден отбил в полуфинале Новую Англию — ни одна ракета не вышла из верхней точ-
ки дуги — но удар на юге вполне может покачнуть равновесие, не прими он неот-
ложных мер. Он отправил три подъемника с заданием стерилизовать все в зоне и
вокруг нее с двадцатикратным радиусом, дождался подтверждения и в изнеможении
откинулся назад. Закрыл глаза. Статистика и телеметрия непрерывным потоком
прокручивалась под веками.
На этот раз не какой-нибудь тихоходный Бетагемот. Совершенно новый штамм.
Сеппуку.
«Спасибо тебе, Южная Африка, чтоб тебя».
Что за дела с этим народом? Были во многом типичной страной третьего мира,
порабощенной, угнетенной и жестоко используемой, как многие ей подобные. Неу-
жели не могли, как все, сбросить оковы и погрузиться в жестокое восстание,
возжаждав мести всем и вся? Что за психи, много лет терпевшие чужой сапог на
своей шее, додумались ответить угнетателям — только подумать — комиссией по
примирению! Какой в этом смысл?
Если, конечно, не вспоминать, что это сработало. Со времени восхождения
святого Нельсона южноафриканцы стали мастерами обходных шагов: накапливали
силы вместо того, чтобы бросать их в бой, использовали инерцию вражеского
удара в свою пользу. Черные пояса социологического дзюдо. Полвека тихарились
под взглядами всего мира, и никто ничего не заметил.
Теперь они представляют собой большую угрозу, чем Гана, Мозамбик и другие
подобные режимы вместе взятые. Этих-то Дежарден отлично понимал, более того,
он им симпатизировал: что ни говори, западный мир, сочувственно цокая языком,
любовался, как половые болезни прожигают дымящиеся дыры в возрастной структу-
ре Африки. Хуже приходилось только Китаю (и кто знает, что зреет за его тем-
ными непроницаемыми границами?) Не удивительно, что Мем Апокалипсиса дал та-
кой мощный резонанс именно здесь: обкорнанное поколение, мучительно пытающее-
ся восстать из пепла, на семьдесят процентов состояло из женщин. Мстительные
богини пересдали карты, обслуживая Армагеддон с океанского дна — даже если бы
Лени Кларк не дала им готовую матрицу, столь подходящий миф все равно бы про-
рвался , вспыхнул бы спонтанно.
С бессильной яростью Ахилл справился бы. А вот улыбчивые уроды с тайными
целями создавали гораздо больше проблем, особенно когда за ними стояло насле-
дие биотехнологий, зародившихся, черт побери, чуть ли не век назад, еще с
первой пересадки сердца. Сеппуку действовал также, как его южноафриканские
создатели: он был чемпионом микробиологического дзюдо и притворщиком, он улы-
бался, под ложным предлогом залезал к тебе в дом, а потом...
Ни европейцам, ни азиатам подобная стратегия не пришла бы в голову. Слишком
тонко для потомков империй, слишком трусливо для тех, кто вырос на политике
бахвальства силой. А вот для этих мастеров манипуляций на нижнем уровне, при-
таившихся в пятке темного континента, это вторая натура. В эпидемиологию она
просочилась прямиком из политики, а с последствиями пришлось разбираться
Ахиллу Дежардену.
Теплая тяжесть легла ему на бедро. Дежарден открыл глаза. Мандельброт, при-
встав на задние лапы, передними тормошила его. Не дождавшись разрешения, мя-
укнула и запрыгнула на колени.
В любую минуту мог загореться огонек на пульте. Уже много лет у Дежардена
не было официального начальника, но множество глаз от Дели до Мак-Мер до сле-
дили за каждым его движением. Он заверил их, что справится с ракетами. Далеко
за океанами правонарушители в более цивилизованных пустошах — и к тому же на
поводках Трипа — связывались со спутниками, хватались за телефонные трубки,
срочно вызывали Садбери, Онтарио. Никто из них не станет слушать оправданий.
Он мог бы с ними справиться. В его жизни случались куда более серьезные вы-
зовы — и он справлялся. Шел 2056, и десять лет назад он спас Средиземку, раз-
вернув на сто восемьдесят градусов свою личную жизнь. Пять лет назад Бетаге-
мот рука об руку с Лени Кларк начали крестовый поход против мира. Четыре года
с исчезновения Верхнего Эшелона, четыре года с тех пор, как влюбленная идеа-
листка насильственно освободила Дежардена от рабства. Чуть меньше прошло с
Рио и добровольного затворничество Ахилла в этих руинах. Три года — с Каран-
тина западного полушария. Два — с Выжигания Северной Америки. И Ахилл спра-
вился со всем.
Но вот южноафриканцы... они действительно задали проблему. Добейся они сво-
его, Сеппуку прошелся бы по его владениям лесным пожаром, и Ахилл не видел
благоприятного для себя сценария. Он сильно сомневался, что сможет долго
сдерживать неотвратимое.
Хорошо, что он как раз собрался уйти в отставку.
(ПРОДОЛЖЕНИЕ СЛЕДУЕТ)
4t
•*
*
**
*
**>
*
• C?V,.
♦'
•
*
•
•
^
*'
**
2
LA
/
\ с
/ИХ
^
ОЧИСТКА МОЛЕКУЛ И РАЗДЕЛЕНИЕ СМЕСЕЙ
Куликов Е.
Все живое на нашей планете состоит из огромного количества биологических
молекул — ДНК, РНК, липидов, углеводов, — объединенных в сложные системы. Для
того чтобы понимать, как эти системы работают, нам надо уметь разбирать их на
отдельные компоненты — так получается гораздо проще.
Основой жизни на нашей планете являются клетки, а каждая клетка состоит из
огромного количества самых разных химических веществ — сложных и не очень. В
химии живого есть несколько уровней сложности системы (рис. 1):
■ самый нижний уровень, представленный низкомолекулярными соединениями (не-
органическими и органическими ионами, аминокислотами, липидами, моносаха-
ридами, нуклеотидами);
■ средний уровень — биополимеров (белков, полисахаридов, нуклеиновых кислот)
и их комплексов, включая и живую клетку как совокупность различных биоло-
гических молекул;
■ высшим уровнем сложности биологической системы оказывается живой организм,
составленный из множества самых различных клеток (ну или одна клетка в
случае одноклеточных организмов).
?>
^х^
атом | молекула | органеллы| клетка | ткань | орган | система | организм
органов
Рис. 1. «Лестница масштабов»: каждый последующий объект крупнее
предыдущего в 10 и более раз.
Для того чтобы исследовать химические процессы, связанные с жизнедеятельно-
стью клетки и организма, необходимо уметь выделять из невероятного многообра-
зия компонентов живого вещества то, что планируется исследовать, и уже с этим
материалом работать дальше. Собственно, появление вечно молодой бабушки со-
временной молекулярной биологии — биологической химии (на заре ее становления
она называлась физиологической химией), — стало возможным только после разра-
ботки методов выделения отдельных компонентов живого, для их исследования
сперва in vitro, а потом и in vivo.
Разделение сложных смесей молекул и выделение из биологического материала
отдельных его компонентов было известно человеку задолго до появления биохи-
мии:
■ разделение молока на молочную сыворотку, содержащую белок, и на сливки,
содержащие преимущественно жиры-триглицериды;
■ сворачивание молока сычужным ферментом в сыроварении с отделением от сыво-
ротки практически всего белка — казеина;
■ получение крахмала из зерна и клубней картофеля отмыванием его от других
компонентов исходного сырья;
■ выделение душистых масел из цветков, коры и плодов растений путем перегон-
ки (рис. 2) или экстракции жирами и маслами;
■ получение жира для мыловарения вытапливанием.
Рис. 2. Отгонка эфирных масел.
Все эти практические способы были методами выделения отдельных фракций тех
или иных биологических молекул из природного материала.
Пожалуй, первое «настоящее» и хорошо документированное выделение индивиду-
ального биологического компонента — белка гемоглобина (рис. 3) — предпринял
основоположник физиологической химии, профессор Тюбингенского университета
(Германия) Феликс Хоппе-Зейлер (1825-1895 гг.). В 1862 году он совершил про-
рывное открытие, впервые выделив из крови человека в чистом кристаллическом
виде окрашенный дыхательный белок гемоглобин. До этого он назывался просто
«Blutfarbstoff» — «окрашивающее кровь вещество». Иене Берцелиус (1779-1848
гг., Швеция), автор термина «органическая химия», до этого смог расщепить
грубый препарат гемоглобина на органическое вещество глобулин, и на содержа-
щий железо пигмент, способный связывать кислород.
ч
coo
\
/• ! '
сн" и
coo
/
/
L
v ,^/ 'ч ,
С. * " С V ^^V-CH;
I fl
'J Г-
\ i « /
FC
v-ri
С = i\i N ~ <-
t-l r. ' l i xx
гем
CH-
H
с >.
1Г<-Н;
Рис. 3. Гемоглобин — от структуры к кристаллу. Четыре субъединицы
объединяются в компактную кубическую структуру.
Хоппе-Зейлер характеризовал полученный им белок с различных сторон — напри-
мер, впервые применив ранее предложенный Бунзеном и Кирхгофом спектроскоп для
описания спектров поглощения гемоглобина как в чистом виде, так и в комплексе
с различными физиологически значимыми газами (кислород, углекислота, угарный
газ) . Далее Хоппе-Зейлер измерил способность гемоглобина к специфическому
связыванию кислорода, и подтвердил современную гипотезу тканевого дыхания,
согласно которой газообмен в органах осуществляется через кровь посредством
гемоглобина, предложенную ранее Юлиусом Лотаром Мейером (1830-1895 гг., Гер-
мания) .
Чистые препараты нуклеиновых кислот впервые выделил сотрудник Феликса Хоп-
пе-Зейлер а Фридрих Мишер (1844-1895 гг.), ранее работавший с известным немец-
ким химиком Адольфом Штрекером (1822-1871 гг.), специалистом в области орга-
нической химии, впервые синтезировавшим аминокислоту аланин из уксусного аль-
дегида, аммиака и циановодорода. В 1869 и 1871 годах Мишер первым указал на
то, что белки и липиды являются основными химическими веществами, составляю-
щими цитоплазму клетки, и предпринял первую попытку классификации белков —
однако не преуспел в этом из-за недостаточно развитых на тот момент методов
анализа. При исследовании гистологически однородной популяции лейкоцитов из
гноя он впервые выделил из этих клеток вещество, названное им «нуклеин», вы-
падавшее в осадок при добавлении кислот и растворявшееся обратно при добавле-
нии щелочей (рис. 4).
а&
однородная
популяция
лейкоцитов
раствор
нуклеин
о
о о
о
п° о0 °
G ^° ООП
гнои
осадок
Рис. 4. Микропрепарат гноя человека. Ядерный материал лейкоцитов (ДНК и
белки) окрашен в розово-синий цвет красителем азур II, вокруг лейкоцитов
лежат лишенные ядра эритроциты. Как можно видеть, ДНК в гное очень мно-
го . Справа — поведение «нуклеина», комплекса ДНК и белков, в растворах
кислоты (выпадение в осадок) и щелочи (растворение).
Это вещество представляло собой первый грубый препарат ДНК в истории чело-
вечества — а вернее, комплекс ДНК с белками (дезоксирибонуклеопротеид). «Со-
гласно имеющимся гистохимическим данным, я предполагаю, что это вещество со-
держится в ядре клетки», — пишет Мишер, и далее фокусируется на ядре клетки —
органелле, о которой тогда было известно очень мало. Нуклеин Мишера радикаль-
но отличался от всех известных на тот момент белков тем, что после осаждения
не растворялся в воде, уксусной кислоте, разбавленной соляной кислоте и в
растворах солей. Мишер разработал протокол выделения клеточных ядер выдержи-
ванием клеток на холоде в растворах соляной кислоты и протокол экстракции
нуклеина из ядер щелочами с последующим осаждением. После получения чистого
нуклеина Мишер сделал ряд основополагающих открытий о ДНК — например, опреде-
лил отношение фосфора к другим компонентам, оказавшееся весьма близким к ны-
нешним данным (22,5% против 22,9%), установил природу ДНК как многоосновной
кислоты, предположил, что нуклеин является носителем наследственных свойств.
Со времени исторических работ первых биохимиков методы выделения и разделе-
ния различных молекул, важных для биологии, претерпели значительные измене-
ния , но в целом их содержание осталось примерно тем же. При работе с биологи-
ческим объектом первым этапом всегда оказывается разрушение тканей и их пред-
варительное фракционирование. При этом стоит задача максимально предотвратить
разрушение целевых молекул, и избавиться от максимального количества загряз-
няющих примесей.
при грамотно проведенной
гомогенизации большая часть
внутримембранных органелл
остается не поврежденной
разрушение клеток продавливание рассечение клеток
высокочастотным клеток через вращающимся
ультразвуком маленькое поршнем
отверстие под
высоким давлением
суспензия клеток
или ткань
о
\j ■
0:б
использование
специального
раствора для
перфорации
мембран клеток
Рис. 5. Гомогенизация. Общая схема разрушения биологического ма-
териала для получения отдельных субклеточных фракций.
Для разрушения тканей применяют различные методы — механическую гомогениза-
цию (например, проворачивание материала в мясорубке), гомогенизацию ультра-
звуком (при которой клетки разрушаются кавитацией в растворе), продавливание
через небольшое отверстие под высоким давлением, замораживание-оттаивание и
др. (рис. 5). В среду для разрушения тканей добавляют различные защитные реа-
генты — например, бета-меркаптоэтанол для предотвращения окисления белков,
ингибиторы ферментов (преимущественно гидролаз, разрушающих биополимеры или
меняющих их свойства), а саму работу проводят на холоде для максимального
снижения скорости действия ферментов в гомогенате.
Следующим шагом разделения обычно оказывается центрифугирование (рис. 6) .
Дробное центрифугирование, при котором гомогенат подвергается действию всё
больших центрифужных полей, позволяет не только отделить неразрушенные клет-
ки, но и выделить отдельные интересующие исследователя компоненты клеток —
например, ядра, митохондрии, рибосомы, мембраны. Центрифугирование существен-
но обогащает препарат целевыми молекулами, сильно упрощая последующие шаги по
разделению сложнейшей смеси биокомпонентов, и может быть проведено для боль-
шого количества биоматериала — это очень важно для выделения минорных компо-
нентов из смесей.
угловой ротор бакет-ротор
центробежная гилл
□ ! * V Я ►
»"■* V
среднее, ус корен/е.
охлаждение мотор вакуум
митохондрии
л изо сомы
ПСЗОКСИСОМЫ
Рис. 6. Центрифугирование. Вверху — устройство центрифуги. В зависимо-
сти от скорости вращения ротора и его размера центрифуга способна ге-
нерировать значительные ускорения. Роторы могут быть угловыми и бакет-
роторами (с качающимися стаканами), они отличаются направлением дейст-
вия центробежной силы. Внизу — схема дифференциального центрифугирова-
ния. В зависимости от силы приложенного поля мы можем получать различ-
ные фракции клетки — от самых крупных органелл (например, ядер), садя-
щихся при небольших значениях относительного ускорения, до самых мел-
ких (рибосом и пр.), осаждающихся большими ускорениями.
Центрифуги очень широко применяются как в промышленности (например, в мо-
лочной для отделения сливок от обрата или для разделения смесей твердых и
жидких веществ), так и в лаборатории. Основные параметры центрифуги — макси-
мальный объем вещества, помещающийся в ее ротор, и создаваемое ей центрифуж-
ное поле. Обычно это поле выражают как относительное центробежное ускорение —
в единицах, кратных гравитационному показателю Земли g (ускорению свободного
падения), и данная величина оказывается пропорциональна скорости вращения ро-
тора центрифуги. Для разных целей применяют центрифуги с разными максимальны-
ми ускорениями — от сотен и тысяч g для разделения грубых смесей до сотен ты-
сяч g для осаждения индивидуальных белков из раствора. Ультрацентрифуга (рис.
7) , создающая высокие ускорения, была долгое время единственным способом оп-
ределения молекулярной массы крупных белков — эта масса была пропорциональна
скорости движения молекул белка в центрифужном поле.
Рис. 7. Ультрацентрифуга. Прибор, способный создавать в пробирках с
биоматериалом огромные перегрузки — до 1 000 000 д. В таком центро-
бежном поле можно осадить даже ионы, не говоря уже про крупные белки
или органеллы клетки. Центрифужный ротор сделан из особо прочного
материала (титана), камера охлаждается точным холодильником (для ис-
ключения конвекции образца), ротор вращается в глубоком вакууме (для
предотвращения разогрева из-за трения о молекулы воздуха). Можно
сказать, что образец летит на искусственном метеорите.
После центрифугирования препарат можно подвергнуть или экстракции отдельных
интересующих исследователя химически близких групп компонентов (например,
смесью эфира и метанола можно количественно извлечь из мембран липиды), или
предпринять дальнейшие усилия по отделению примесей. Наиболее сложным вопро-
сом при таком разделении оказывается разделение белковых смесей — необходимо
выделить зачастую весьма немногочисленный компонент смеси, при этом он не
должен потерять своей естественной биологической активности. Выделенный фер-
мент должен работать не хуже, чем он это делает в клетке, выделенный фактор
транскрипции должен хорошо и специфично связываться с ДНК, иными словами, вы-
деленные белки должны оставаться нативными — такими же, как в живой клетке.
Для первичного разделения смесей белков можно применять различия раствори-
мости белков в растворах солей. Феномен высаливания белка (рис. 8) известен
давно — так, кристаллы яичного белка овальбумина добавлением насыщенного рас-
твора сульфата аммония получил Франц Гофмейстер в 1890 году. То, что белки
по-разному растворимы в растворах солей, активно используется и в промышлен-
ности — например, рассольные сыры делаются более плотными за счет засаливания
в концентрированном рассоле. Если мы добавим к раствору белка насыщенный рас-
твор хорошо растворимой соли — сульфата аммония, — достигнув, например, 10%
насыщенной концентрации, то некоторые белки выпадут в осадок, то есть обрати-
мо денатурируют. Если мы разведем этот осадок водой или не содержащей сульфа-
та аммония буферной средой, осадок растворится, а белки из него вернутся от
денатурированной к нативной конформации. Если мы будем повышать концентрацию
сульфата аммония ступенчато, и всякий раз отделять осадок белка центрифугиро-
ванием, мы сможем грубо фракционировать исходную смесь, а целевой белок обна-
ружить в определенной фракции по его свойствам, например, цвету или фермента-
тивной активности. Именно возможность фракционирования белковых смесей выса-
ливанием позволила разделить белки плазмы крови на фракции глобулинов (выпа-
дающих в осадок при более низкой концентрации сульфата аммония) и альбуминов
(остающихся в растворе белков меньшего, чем глобулины, размера), и именно вы-
саливанием до сих пор пользуются для приготовления как отдельных фракций сы-
вороточных белков для медицины, так и для выделения фракции иммуноглобулинов
— антител.
Рис. 8. Высаливание белка. При определенном значении рН среды
белки имеют различную растворимость в растворах неорганических
солей — и, повышая содержание соли в растворе, мы можем перево-
дить определенные белки из растворенного состояния в осадок, ко-
торый потом отделять центрифугированием.
Белки и их смеси — особенно сложные для работы объектами, так как физико-
химические свойства разных белков сильно отличаются друг от друга благодаря
различному аминокислотному составу. Белки состоят из аминокислот, в боковой
цепи которых могут встречаться полярные группы — например, кислотные или ос-
новные остатки. В зависимости от водородного показателя (рН) среды, в которой
находится белок, то есть от кислотности среды, белок может иметь различный
заряд, то есть белковая молекула может быть отрицательно заряжена в щелочной
среде, иметь нулевой заряд в нейтральной среде и перезаряжаться (становиться
положительно заряженной). Точка рН, соответствующая нулевому заряду белка,
называется его изоэлектрической точкой (рис. 9), и белки в ней обычно имеют
минимальную растворимость. Навязывая белку определенную кислотность среды, мы
можем получить выпадение белка в осадок — и именно это заставляет скисшее мо-
локо, в котором лактобактерии сбродили молочный сахар до молочной кислоты,
разделяться на сыворотку и белковые сгустки. Изоэлектрическая точка основного
белка молока, казеина, лежит как раз в области кислых значений рН.
и
I)
ft
рн
Рис. 9. Изоэлектрическая точка. Заряд белковой молекулы зависит
от рН среды вокруг него — когда количество отрицательных и поло-
жительных зарядов у белковой молекулы равно, ее общий заряд ра-
вен нулю. Белковые молекулы в растворе перестают отталкиваться
друг от друга, начинают слипаться и могут образовать осадок.
Растворимость белка в изоэлектрической точке минимальна.
Рис. 10. Диализная очистка. Раствор высокомолекулярного вещества
(белка, синие точки) и соли (красные точки) помещен в мешочек из
целлофана (целлюлозной мембраны, поры которой пропускают ионы
соли и задерживают крупные молекулы белка). Если такой мешочек
поместить в раствор, не содержащий соли, то соль начнет выходить
из мешочка через поры, а белок останется внутри. Примерно так
устроены и наши почки — соль и низкомолекулярные продукты обмена
уходят в мочу, а белки остаются в плазме крови.
При выделении отдельных макромолекул — ДНК, белков, полисахаридов — часто
встает задача отделения от них низкомолекулярных примесей. Так, например, по-
сле высаливания белка необходимо удалить из осадка избыток сульфата аммония,
одновременно переведя белок в нативное растворимое состояние. Если мы помес-
тим наш осадок в мешочек из полупроницаемой мембраны (например, целлофана —
химически обработанной целлюлозы, поры которой пропускают ионы соли и другие
низкомолекулярные вещества, но слишком узки для больших молекул белка или
ДНК) , а сам мешочек опустим в дистиллированную воду или буферный раствор с
-£У
К
о;
о
^*_ ;
?N
CJ-.:
О
оо
1~_!
О;
ЧСГ 1
o~i
небольшим содержанием соли, осадок начнет растворяться, а соль выходить в
раствор снаружи мешочка. Так будет происходить до момента, пока концентрации
соли по разные стороны мембраны не сравняются. Если мы будем менять раствор
снаружи, мы сможем полностью удалить все низкомолекулярные примеси из препа-
рата — они выйдут через поры мешочка, а биополимеры останутся внутри. Такой
прием называется диализом (рис. 10) — и именно по этому принципу у нас рабо-
тают почки, удаляющие из организма лишние соли, продукты азотистого обмена и
другие низкомолекулярные вещества, но сохраняющие в организме все высокомоле-
кулярные белки.
Смесь белков иногда можно весьма эффективно разделить и способами, при ко-
торых примесные белки денатурируют (рис. 11) — превращаются в нерастворимый
осадок, а целевой белок остается в растворе. Например, если мы синтезируем в
клетках кишечной палочки термостабильный белок, ген которого взят из живущих
в горячих источниках архей (например, фермент ДНК-полимеразу из Thermus
aquaticus, температурный оптимум которого находится около 72 С), мы можем от-
делить практически 99% всех белков кишечной палочки от нужного нам белка,
просто прогрев гомогенат клеток при 80 градусах. Не обладающие термостабиль-
ностью белки кишечной палочки денатурируют, а нужный нам рекомбинантный белок
Я
г
i
высаливание белка
Рис. 11. Различные способы денатурации белков. Тепловая денатурация
белков при приготовлении пищи — так еда становится вкуснее, а дена-
турированный белок доступнее для пищеварительных ферментов. Механи-
ческая денатурация — взбивание белков — сродни тепловой, только про-
исходит из-за механического «запутывания» белковых молекул. Высали-
вание — обратимая денатурация, рассмотренная ранее.
Важным способом разделения смесей биомолекул по размеру является гель-
хроматография, или проникающая хроматография (рис. 12).
В 1959 году шведские ученые Порат и Флодин впервые предложили новый метод
разделения сложных смесей белков по молекулярной массе, и это стало возможно
благодаря полученному им новому материалу под торговой маркой Sephadex. Длин-
ные линейные молекулярные цепочки полисахарида декстрана сшивали между собой
так, что удавалось получить молекулярные сита — материал с контролируемым в
широком диапазоне размером пор. Этот материал был доступен в виде гранул раз-
ного размера — от микрон до сотен микрон. Белки и другие биомолекулы не ад-
сорбировались этим материалом и не теряли своей нативной активности. Материал
замачивали в буферном растворе — гранулы набухали, поры заполнялись раствором
и начинали напоминать маленькие поролоновые шарики.
останется в растворе.
тепловая денатурация
механическая денатурация тепловая денату|
9S J«
0
0
io
о
io
о
О
О
О
О
О
о
о
о
о1
о
о
о
о
о
о
00
о о
о о
о о
о о
о о
~£»°
р
р
0
о
о
1
р
о
р
10
о
о
о
ко
о
о
о
о
о
о
о
Оу
Рис. 12. Гель-хроматография. Смесь белка (синие точки) и соли
(красные точки) нанесена на колонку с носителем, поры которого
не пропускают белок внутрь. При промывании колонки буферным рас-
твором соль «застревает» в носителе, свободно диффундируя в его
гранулы, а белок выходит, не задерживаясь. Так можно быстро и
просто разделить как смеси белков и соли, так и смеси белков
разного размера (подбирая диаметр пор носителя).
Дальше ими наполняли длинную трубку с воронкой сверху и краном снизу, за-
крывали кран, и наносили сверху смесь биологических макромолекул. Крупные мо-
лекулы белка были слишком велики для того, чтобы проникнуть в поры молекуляр-
ных сит, ну а низкомолекулярные вещества (соли, витамины, коферменты, нуклео-
тиды) свободно диффундировали вглубь шариков сита через поры. Через воронку
начинали пропускать буферный раствор, а из крана вытекал раствор, который до
того был внутри шариков сита и между ними. Крупные молекулы, не вошедшие в
шарики, двигались с раствором между шариками, в исключенном объеме, равном
примерно объему жидкости между шариками, а низкомолекулярные вещества задер-
живались материалом колонки и выходили сильно позже — с объемом, примерно
равным объему колонки, то есть с полным объемом. Так удалось отделить крупные
молекулы от мелких (чем-то похоже на диализ, правда?), а при подборе размера
пор у молекулярного сита удалось достичь и хорошего разделения смесей биомо-
лекул на размерные классы — длинные участки ДНК от коротких, крупные белки от
мелких. Этот метод до сих пор активно применяется и при определении молеку-
лярной массы белков и ДНК — для этого колонку с носителем сперва калибруют по
смеси белков или ДНК известного размера.
Если взять похожий носитель с крупными порами, но сделать его поверхность
заряженной положительно или отрицательно (например, химически присоединив к
носителю различные заряженные группы — аминогруппу, сульфогруппу и пр.), мож-
но использовать его для другого вида разделения — ионообменной хроматографии
(рис. 13). Например, ДНК, представляющая собой поликислоту, заряженную в ней-
тральных или щелочных условиях отрицательно, будет взаимодействовать с поло-
жительно заряженным носителем — анионообменником, связываясь с группами на
его поверхности. При добавлении в систему избытка соли ее анионы будут конку-
рировать с ДНК за группы на носителе, и вытеснять ДНК из комплекса с носите-
лем, причем соли потребуется тем больше, чем длиннее молекулы ДНК, связанные
с носителем. Та же картина будет наблюдаться и с белками, заряженными отрица-
тельно . Для связывания положительно заряженных белков необходим носитель с
отрицательными группами на поверхности — катионообменник. Этот метод позволя-
ет отсеять из сложной смеси молекулы, не имеющие в данных условиях заряда,
или заряженные так же, как носитель — связывание окажется невозможным, и эти
молекулы просто выйдут из колонки с носителем.
;® ф ф
Ь Ф «i
Ш & Ф;
Ф ф el
« ф ©|
чв е % J
-; г
р ф ф!
е е о!
|Ф 6 Ф!
!ф а «I
в ф ф
ем!
'Г
низкомолекулярная
А А ф! соль,растворённая
1а л л вбуФер,юм
|ч7 ч7 ф| растворе
Ф 9
|ф © Ф
Ф ф в
в Ф в
Ml
t
ф о1рица!ельно заряженная молекула белка q отрица!ельно заряженная молекула белка $ отрицательно заряженная молекула белка
■ положительно заряженная молекула белка положительно заряженная молекула белка . положительно заряженная молекула белка
Рис. 13. Ионобменная хроматография. Белки с различным зарядом
наносят на колонку, содержащую заряженный определенным образом
сорбент. Белки с противоположным заряду сорбента зарядом задер-
живаются в колонке, с одноименным зарядом — выходят из нее. За-
державшиеся белки можно снять с сорбента определенной концентра-
цией низкомолекулярной соли, растворенной в буферном растворе.
Маленькие ионы соли, находящиеся в большом молярном избытке по
сравнению с крупными белковыми молекулами белка, «сталкивают» их
с сорбента в раствор, занимая их место.
Разделение смесей биомолекул в электрическом поле называется электрофорезом
(рис. 16, 17, 18). Молекулы будут двигаться в сторону соответствующего полюса
в поле постоянного тока, и скорость их движения будет зависеть как от степени
их заряда, так и от их размера. Если мы поместим смесь биомолекул в буферный
раствор и приложим постоянное напряжение, молекулы сперва начнут делиться, а
потом немедленно перемешаются благодаря конвекции — водный раствор будет вы-
полнять функции резистора, полностью рассеивая всю приложенную энергию элек-
трического тока в виде тепла. Для предотвращения конвекции и в качестве моле-
кулярного сита обычно используют гели агарозы (полисахарида, линейного компо-
нента агара) или полиакриламида (полимеризованного амида простейшей непре-
дельной карбоновой кислоты — акриловой).
Рис. 16. Гель-электрофорез. Чем молекулы ДНК объемистее, тем слож-
нее им бегать — все как у людей!
•' :тк%г*>7-г^~~"ш -*т'*'~:ту*-т?'*-^~-р?- ,т,у-туу^-д'vч*,чс'ет^к -г*
Рис. 17. Гель-электрофорез. На этом рисунке виден гель — молеку-
лярное сито, «полоса препятствий» для заряженных молекул. Все
молекулы ДНК будут двигаться по направлению к аноду, но скорость
их движения будет различаться — чем молекула крупнее, тем труд-
нее ей будет преодолевать препятствия.
Подбирая концентрацию этих веществ, можно получать гели с заданной величи-
ной пор, оптимизируя их для разделения тех или иных смесей. В геле биомолеку-
лы движутся со скоростью, пропорциональной их длине — чем мельче, тем быст-
рее, — и заряду. Такой вариант электрофореза хорошо подходит для разделения
химически однородных смесей, содержащих молекулы одного типа, но разной длины
— например, смеси фрагментов нуклеиновых кислот. ДНК будет двигаться в поле
постоянного тока от отрицательного полюса к положительному, и ее движение
можно видеть в ультрафиолетовом свете — после добавления в гель особого кра-
сителя бромистого этидия, комплекс которого с ДНК флуоресцирует ярко-
оранжевым светом. Отдельные компоненты смеси разделяются на полоски, содержа-
щие популяцию молекул одной длины. Гель напоминает по текстуре мармелад или
студень — и нужную полоску можно вырезать из геля, а молекулы из нее исследо-
вать другими способами.
Для смесей белков такой вариант электрофореза — нативный — подходит далеко
не всегда: при том, что белки состоят из аминокислот, их заряд может сущест-
венно отличаться при различных значениях рН среды, да и форма/размер белков
могут варьировать в широких пределах. Представим, что мы поместили в поле по-
стоянного тока смесь белков с положительным, нейтральным и отрицательным за-
рядами. Каждый из заряженных белков пойдет к своему противоположному по заря-
ду полюсу, а нейтральные незаряженные белки останутся на месте. Более того,
4-
4-
4-
4-
4-
4
4-
4
крупные белки могут вообще оказаться крупнее пор геля и не войти в них, не-
смотря на свой заряд. Для того чтобы все белки стали заряжены одноименно, и
превратились в частицы примерно одинаковой формы, мы можем обработать белок
анионным детергентом — поверхностно-активным веществом додецилсульфатом на-
трия. Именно он является основой практически всех стиральных порошков и мою-
щих средств для бытовых нужд. Белки под его воздействием разворачиваются и
становятся похожи на ниточки, плотно облепленные молекулами додецилсульфата
натрия, и заряд у этих ниточек оказывается отрицательным за счет сульфогруппы
детергента.
белок с двумя субъединицэми А и В,
соединенными дисульфидным
мостиком
A -ss- 3
i
белок
нагрев и покрытие детергентом
1
С,
;^
электрофорез
1
5
Рис. 18. Схема проведения белкового гель-электрофореза. Белки
денатурируют бета-меркаптоэтанолом, при этом они утрачивают все
виды структуры выше первичной, превращаясь в отдельные ниточки —
полипептидные цепи. Эти цепи оказываются плотно покрыты детер-
гентом — додецилсульфатом натрия, молекулы которого маскируют
собственный заряд белка. Таким образом, все белки пробы будут в
электрическом поле двигаться к аноду.
Такие белки не будут обладать нативной биологической активностью — денату-
рируют, но будут разделяться в геле по размерам — скорость движения таких
частиц пропорциональна их длине, и все белки пробы будут двигаться в электри-
ческом поле строго в одном направлении, от катода к аноду. Такой вариант —
денатурирующий электрофорез — активно применяется для анализа белковых смесей
любой сложности (рис. 18). И в нативном, и в денатурирующем электрофорезе
есть задача увидеть бесцветные белки, разделенные на геле. Обычно для этого
используют органические красители — например, кумасси бриллиантовый синий,
плотно связывающийся с белками и окрашивающий их в синий цвет, но отмывающий-
ся от не содержащего белки геля. Синие белковые полосы на бесцветном фоне от-
лично читаются, и по стандартам молекулярной массы на том же геле можно опре-
делить примерную молекулярную массу неизвестного белка.
Используя примерно одну и ту же схему эксперимента — колонку, наполненную
носителем, через которую можно пропускать раствор разделяемой смеси и другие
растворы, — можно получить еще несколько вариантов хроматографического разде-
ления сложных смесей. Например, если на поверхность носителя нанесены специ-
фичные к белку антитела, белок будет связываться с ними и оставаться на по-
верхности носителя, откуда потом его можно снять. Такой метод называется им-
мунохроматографией, и это один из вариантов аффинной хроматографии — хромато-
графии по сродству (рис. 19). Любое вещество, связанное с носителем, и спо-
собное взаимодействовать с целевым белком — лиганд, — в принципе можно ис-
пользовать для выделения целевого белка из смесей. Гликопротеины (белки, со-
держащие углеводную часть) могут быть выделены на носителях, покрытых лекти-
нами — белками, узнающими углеводы. Ферменты можно выделять по связыванию с
иммобилизованными на носителе субстратами этих ферментов.
загрузка
bCu D-
разделение
-Г"
Л
Lr 0**ц
I Civ* fc^l
Г,'"*7»
Щ
Щ
вымывание
ИГ ПН
hC DA
Рис. 19. Аффинная хроматография. Колонка наполнена сорбентом,
покрытым веществом, которое способно узнавать интересующую нас
молекулу, или самими интересующими нас молекулами — смотря что
мы хотим выудить из пробы. После пропускания через колонку смеси
веществ в ней задержится только то, что способно специфически
связаться с сорбентом. Потом это вещество можно снять с колонки
или высокой концентрацией соли, или другим значением рН среды,
или денатурантом — условиями, нарушающими специфическое сродство
связанной молекулы и сорбента.
Практически любое исследование в биомедицинской области включает в себя то
или иное разделение сложных смесей биологических молекул. Разберем диагности-
ку инфекционного заболевания, вызванного патогенными микробами, по шагам:
1. Взятие пробы у пациента. Биологический материал из области воспаления тща-
тельно измельчают, смешивают с инактивирующим бактерии реагентом, а затем
растворяют содержащиеся в нем клетки (патогена и пациента) так, чтобы ДНК
из клеток перешла в раствор.
2. Выделение ДНК. Для выделения ДНК к лизированному препарату биоматериала
добавляют носитель, специфично связывающий ДНК. Носитель несколько раз от-
мывают от прочих компонентов пробы, не связавшихся с ним, и снимают ДНК с
его поверхности.
3. Амплификация ДНК. Фрагменты ДНК патогенного микроба специфически амплифи-
цируют с помощью ПЦР, получая фрагмент ДНК определенной длины. В реакции
параллельно участвует положительный контроль — ДНК патогена — и отрица-
тельный контроль, не содержащий ДНК (он нужен для контроля чистоты реаген-
тов) .
4. Электрофорез ДНК. Полученные продукты ПЦР разделяют гель-электрофорезом и
анализируют результат. Наличие в продуктах ПЦР фрагмента ДНК, соответст-
вующего ожидаемой длине и совпадающего по длине с фрагментом, полученным
амплификацией с ДНК патогена (положительным контролем), подтверждает нали-
чие ДНК патогена в исследуемой пробе.
В этом простейшем исследовании применяются два приема разделения смесей
биомолекул — сперва для выделения ДНК, а потом для анализа продуктов ПЦР. Лю-
бой биомедицинский анализ включает в себя разделение смесей на этапе подго-
товки пробы, а далее на этапе получения данных. Возможны комбинации самых
разных подходов к разделению смесей биомолекул, и каждый новый вариант таких
систем немедленно включается в практику исследований.
Несмотря на почтенный возраст большинства методов разделения сложных смесей
биологических молекул, эти методы нисколько не утрачивают своей актуальности.
Развитие новых методов исследования, способных работать в микромасштабе, раз-
работка новых приборов для анализа сделает разделения быстрее и удобнее, и
позволит прийти к созданию новых диагностических и терапевтических приборов и
подходов. Совершенству не может быть предела.
Разное
СЛОВО О СПОРТИВНЫХ НАПИТКАХ
Теория
Люди, активно занимающиеся спортом и ведущие активный образ жизни, зачастую
нуждаются в дополнительной энергии, в отличие от людей ведущих не слишком ак-
тивный образ жизни. Как известно еще со школы, главные источники энергии для
человеческого организма - белки жиры, углеводы, получаемые нами с питанием. В
результате метаболических процессов, потребленная пища превращается в различ-
ные энергетические формы, например, широко известный Adenosine triphosphate
(ATP или АТФ) и другие его производные. В связи с особенностями метаболизма,
не все катаболиты одинаково эффективны в процессе превращения в энергию, уг-
леводы или carbohydrates лидируют здесь и в скорости и в % соотношении. Вооб-
ще в высших живых организмах, к коим мы так же относимся, присутствуют три
основные системы снабжающие нас энергией - две из них анаэробные, где процес-
сы происходят без участия кислорода и одна из них аэробная - где кислород иг-
рает роль окислителя. Anaerobic Alactic (ATP-CP) и Anaerobic Lactic
(Glycolytic) системы питания, обе эти системы очень похожи друг на друга, обе
используют вышеупомянутый АТР, обе не нуждаются в усиленном кровообращении и
следовательно кислороде, но Lactic система использует запасы гликогена, ката-
болизируемые анаэробным гликолизом, в то время Alactic - другой энергетиче-
ский субстрат - creatine phosphate. Третья и главная система, позволившая в
свое время совершить эволюционный скачок - аэробная, использующая глюкозу в
качестве субстрата и кислород, в качестве окислителя.
Итак - глюкоза (она может принимать в нашем организме различные формы) и
кислород нам нужны для активной жизни, для занятий спортом. Наш организм не-
сомненно имеет запасы глюкозы и способы доставить к ней кислород, но не все-
гда этого достаточно, особенно когда речь идет о высоких нагрузках. Особенно
остро этот вопрос встает тогда, когда мы говорим о достижении maximal aerobic
power (MAP) при УОгтах.
V02max - это параметр показывающий максимальный объем кислорода перерабо-
танный организмом в течении периода повышенной нагрузки соотнесенный на кг
веса спортсмена. Соответственно измеряется в мл/кг*мин. По сути - это ком-
плексная способность организма переносить и метаболизировать кислород, полу-
ченный за промежуток времени к его активным потребителям (органам и тканям).
Сравнительные цифры такие: собаки хаски (одни из самых выносливых млекопитаю-
щих) имеют VC^max в районе 200 и больше мл/кг*мин, люди - олимпийские чемпио-
ны в беге на длинные дистанции или лыжных гонках, примерно в районе 90, обыч-
ный человек, не занимающийся спортом активно - 35-40, у мужчин, у женщин этот
показатель как правило на 20-25% меньше. Ограничивающие факторы здесь простые
- в первую очередь это pulmonary function - способность легких захватывать
воздух, легочных альвеол - выхватывать из этого воздуха кислород, далее
транспортная способность - объем крови, биологические ее показатели (качест-
во, объем, количество эритроцитов), cardiac output (грубо - способность серд-
ца перекачивать кровь). Про V02max можно говорить долго, есть способы этот
параметр улучшать, как тренировками так и медикаментозно.
Итак вернемся к MAP - достижении максимальной эффективности при аэробной(
происходящей при активном участии кислорода) нагрузке. И прежде чем перехо-
дить к рассуждениям как нам ее улучшить - давайте рассмотрим главные ограни-
чивающие факторы, я бы выделил их три:
1. Dehydration (обезвоживание - изменение водного баланса организма ниже по-
рогового значения).
2. Истощение запасов гликогена (специальная форма углеводов, предназначенная
для хранения в клетках) в мышечной ткани.
3. Ограниченная доступность глюкозы в крови.
Все эти факторы комплексно влияют на усталость, как следствие ухудшение по-
казателей выносливости и способности выполнять поставленные задачи.
Как и любой неорганический двигатель, наше тело, при выполнении работы вы-
деляет тепло и как и в любом автомобильном моторе это тепло нужно от тела от-
водить, это достигается разными методами - инфракрасным излучением в про-
странство, перекачкой жидкостей и газов с одного места в другое,выделением
пара через поры кожи. Последнее, более известное в простонародье, как потение
или потоотделение, отвечает примерно за 80% данной работы. На нем мы и сосре-
доточимся .
Как сообщает нам Википедия - «потоотделение - средство терморегуляции, ба-
зирующееся на способности зккриновых желез, богатых водой, выделять жидкость
в довольно больших объемах», некоторое люди при хорошей нагрузке способны те-
рять до 4 литров в час.
Как известно, до 50-75% наше тело состоит из воды (для взрослого индивидуу-
ма) . 2/3 этого объема заключены во внутриклеточном пространстве, 1/3 - во
внеклеточном, из которых плазма крови занимает около 1/5 объема к остальным
внеклеточным жидкостям.
Homeostasis - все жидкости в организме подчинены единому правилу организма
- поддержанию баланса или гомеостаза.
Возвращаясь к нашему спортсмену, который, простите, потеет, нарушая гомео-
стаз, давайте заглянем вовнутрь. По мере истощения жидкостей организма, сни-
жается общий объем циркулирующей крови, следовательно уменьшается Cardiac
filling (наполнение сердца кровью), что ведет к уменьшению венозного возвра-
та, объем stroke (сердечного выброса) уменьшается. Естественно наш организм,
за миллионы лет эволюции привыкший и не к таким издевательствам над собой -
пытается эту проблему решить, увеличением сердечного ритма (и конечно же ре-
абсорбцией, но о ней поговорим позже) . В медицине этот «трюк» называется
cardiovascular drift, что дает нашему спортсмену возможность все же продол-
жать выполнять поставленную задачу, хотя и при сниженной интенсивности. Отри-
цательная сторона медали - «пыхтим больше, получаем меньше», да и как у неор-
ганических моторов - больше тепла выделяем, ниже КПД.
Во многих местах сосудов, в том числе и в районе сердца, у человека распо-
ложены несколько групп рецепторов - это баро- и осмо- рецепторы они то и сле-
дят за «осмоляльностью» (рассмотрим этот термин очень подробно позже) и дав-
лением крови. Как уже упоминалось выше, за миллионы лет эволюции, наш орга-
низм, адаптируясь к «издевательством» над ним хозяина, наладил упредительную
систему, не позволяющую индивидууму «умереть быстро» - потеря организмом даже
1% внеклеточной жидкости (главное депо - запас) уже сигнализирует тревогу,
потеря около 4% - ведет физиологическим препятствиям организма для продолже-
ния нагрузки (сигналы мозга, ферменты препятствующие сокращению мышц и т.п.),
а свыше 6% - от предупреждений организм переходит к решительным действиям, в
виде прекращения потоотделения, повышения температуры тела и следующего за
ним теплового удара, как финала для особо упорных.
И так, описанное выше обезвоживание (dehydration), совместно с изменением
уровня глюкозы в плазме и истощением запаса гликогена в мышцах, приводят к
невозможности продолжения выполнения задачи. Есть конечно жиры, не будем про
них забывать, тем более некоторые только о них и думают, но к огромному сожа-
лению последних, жиры могут обеспечить поддержание только низко-интенсивной
нагрузки, примерно на уровне 25% от V20max.
Вы скажете - «Автор, но мы же можем идти сутками...», да - но не быстро. Спо-
собность выполнять низко-интенсивную нагрузку обеспечивается за счет печени,
которая способна синтезировать глюкозу «де ново» в небольших объемах и с по-
средственной скоростью. Вместе с увеличением интенсивности нагрузки потреб-
ность в глюкозе возрастает, пропорционально заявленной нагрузке, поэтому для
возвращения к способности выполнять высокоинтенсивную нагрузку нам нужно вре-
мя и питание.
Давайте вернемся к нашему спортсмену, который еще в самом начале трениров-
ки, теперь мы знаем, что совокупность запасов гликогена в клетках и уровня
глюкозы в крови - ключ к успешному выполнению задачи при довольно серьезной,
но не максимальной интенсивности (такой как 65% от V02max) , первым в бой идет
запас гликогена в мышцах. Тут тоже все не просто... Основные запасы гликогена в
человеческом теле - это печень и мышечная ткань. Для среднего мужчины это
100-150 г в печени и 400-500 г в мышцах, прочие хранилища не столь существен-
ны для рассмотрения. Как вы понимаете - запасы, они и на то запасы, что бы
кто попало их не трогал. Вспоминаем о том, что основное топливо организма -
глюкоза, а гликоген еще нужно в нее преобразовать. Glycogenolysis - или пре-
образование гликогена в глюкозу происходит как в печени, так и в мышечной
ткани, единственное что их различает - разные ферменты, ответственны за акти-
вацию процесса. Я не буду расписывать детально весь процесс метаболизма угле-
водов , многие его знают, но некоторые моменты стоит уточнить.
Итак - при поступлении углеводов и белков в виде пищи, уровень глюкозы в
крови растет, на что поджелудочная железа выделяет специальный гормон инсу-
лин, глюкоза через портальную вену попадает в гепатоциты (клетки печени) где
инсулин, воздействуя на клетки печени, заставляет их выделять ферменты вклю-
чая glycogen synthase, которые в свою очередь добавляют молекулу глюкозы к
полисахариду (длинной цепи аналогичных молекул) глюкану. После завершения пи-
тания, уровень глюкозы в крови падает, инсулина выработка так же сокращается
и идет обратный процесс - продолжение снабжения организма энергией, где
Glycogen phosphorylase - фермент отщепляющий молекулу сохраненного гликогена
от цепочки для превращения ее обратно в глюкозу. Регулирует этот процесс но-
вый фермент - Glucagon - как бы антагонист инсулина (делающий обратные дейст-
вия) . Его вырабатывают ос-клетки поджелудочной железы, в результате мы имеем
эндогенный(базирующийся на внутренних ресурсах) процесс синтеза глюкозы. Та-
ким образом, даже если вы совсем откажетесь от сладкого, у организма есть
путь из чего сделать глюкозу.
МЕТАБОЛИЗМ ГЛЮКОЗЫ
ГЛИКОГЕН
1ТИК0ГЕН0ЛИЗ
и
ТИКОГЕНЕЗ
ГЛЮКОЗО-6-ФОСФАТ W НАДФН, ПЕНТОЗЫ
ПЕНТ030Ф0СФАТНЫЙ
IT И КОЛ ИЗ
сг>
ПУТЬ
ГЛЮКОНЕОГЕНЕЗ
t
ПИРУВАТ
АНАЭРОБНЫЙ
МЕТАБОЛИЗМ
АЭРОБНЫЙ
МЕТАБОЛИЗМ
АЦЕТИЛ-КоА
ЦИКЛ
КРЕБСД
I
С02,АТФ,
НАДН+Н+, ФАДН7Ц-^АТФ
Примечательно, что данный механизм - новомодный тренд в сегодняшней фарма-
цевтике - а именно Ozempic и прочие Semaglutide-базированные, GLP (Glucagon
like peptide) препараты, принцип действия которых базируется на блокировке
работы аппарата эндогенного синтеза глюкозы. Изначально предназначенный для
лечения диабета, он с тысячекратным успехом используется сейчас для лечения
ожирения. И симптоматика о которой рассказывают люди, принимающие его - это
полное отсутствие аппетита, вялость, подавленное настроение - отсутствие
энергии.
Вернемся к нашему спортсмену. Только что мы рассмотрели процессы происходя-
щие в печени, а в клетках мышечной ткани все немного по другому - т.к. мышеч-
ные клетки не имеют в своем арсенале glucose-6-phosphatase - один из фермен-
тов, что позволяет гликогену из печени попасть в кровь в виде глюкозы, храни-
лища в мышцах предоставлены только для них самих.
При различных видах нагрузки, в организме задействуются различные пути «пи-
тания/энергии», например при анаэробной нагрузке - тяжелая атлетика или изо-
метрические упражнения, где характерны высокие, но непродолжительные нагрузки
задействуются Anaerobic Alactic (ATP-CP) или Anaerobic Lactic (Glycolytic)
системы питания, рассмотренные выше. Действие Alactic - самое короткое - се-
кунды, Lactic - позволяет работать чуть дольше - в пределах пары минут. То же
и с восстановлением - Alactic система способна восстановить свою работоспо-
собность в течении пары минут, в то время как восстановление Lactic займет
около часа. Обеспечивать же организм энергией длительное время способна
аэробная система Aerobic Energy System - использующая глюкозу и кислород. Ко-
нечно - наш организм не цифровая машина, не возможно исключить совместную ра-
боту систем, просто нужно понимать, что при разных обстоятельствах, разные
системы начинают доминировать, под воздействием регулирующих ферментов.
Резюмируя - становится понятно, что своевременное восполнение запасов энер-
гии и влаги - ключ к поддержанию производительности на высоком уровне. В до-
полнение , можно заметить, что все люди разные и различие в метаболических
системах, системах запаса воды и энергии, делает нас всех чрезвычайно индиви-
дуальными и абсолютно по разному готовыми или способными выполнять те или
иные нагрузки.
Казалось бы - «что тут думать», основываясь на том что написано выше - бери
сахар, растворяй в воде и пей сколько влезет и будет и регидрация и снабжение
углеводами ... и в целом это правильная мысль, но как и во многих вещах - «дья-
вол кроется в мелочах».
Дело в том, что и вода и углеводы имеют возможность проникать в наш орга-
низм только по особым правилам. Начнем с воды.
Вспоминая описанное выше, как только потери влаги приближаются к примерно
1% ее объема, организм не двусмысленно говорит нам - «хочу пить, хозяин», а
что б посыл этот не был голословным, сопровождает это определенными действия-
ми - самые очевидные из них - сокращение слюноотделения слюнными железами и
мозговые сигналы о необходимости «подзаправиться» - чувство жажды.
Процесс потребления воды начинается с ротовой полости, где расположены спе-
циальные рецепторы передающие в головной мозг сигнал, что мы потребляем воду.
Тут важно понимать, что удовлетворение жажды и регидрация это две разные ве-
щи. Сосание ледяных чипсов или полоскание полости рта может избавить нас на
какое-то время от чувства жажды, но ситуацию с гидратацией это не изменит.
При полоскании полости рта связка oropharyngeal receptors и Angiotensin (ANG)
11 система передают в специальный отдел мозга сигнал о присутствии жидкости и
наш «обманутый» мозг выдает ферментный сигнал - «отмена тревоги по жажде», но
поскольку осморецепторы в кровеносных сосудах не видят положительных измене-
ний , «тревога по жажде» возникает скоро вновь.
Нельзя обойти стороной еще один орган, активно участвующий в водообмене ор-
ганизма, это почки. Наши почки обладают способностью реабсорбировать воду из
крови, запуская ее снова «в дело». Способность к реабсорбции, приводит к то-
му, что концентрация растворенных веществ в жидкости «на выход» (моче), зна-
чительно превышает концентрацию веществ в плазме крови, однако эта способ-
ность в свое время позволила позвоночным покинуть воду и приспособиться к
жизни на суше. За сутки почки здорового человека фильтруют порядка 180-200
литров крови, если предположить что объем примерно 3.5 л - это почти 60 крат-
ный рефреш! В то же время объем выделяемой мочи - всего около 1.5 л. Регули-
руется весь процесс двумя основными путями - это Bulk reabsorption, не регу-
лируемая гормонально, происходит в проксимальных канальцах почечных нефронов
и Regulated reabsorption, она регулируется особым гормонами в зависимости от
водно-натриевого баланса организма и происходит в дистальных канальцах нефро-
нов . Опять помним про гомеостаз.
Итак, пьем, через пищевод вода поступает в желудок, какой-то объем попадет
в кровь отсюда, но это ничтожное количество. Основное поглощение воды начина-
ется в тонком кишечнике, за день он пропускает через себя до 9 л жидкости,
немного жидкости так же поглощается в следующим за тонким, толстым кишечни-
ком, около 1-1.5 л и совсем немного выходит с экскрементами. Конечно не вся
эта жидкость потребляемая извне, большую часть составляют выделения - слюн-
ные, желудочные, печеночные и самого тонкого кишечника.
Посмотрите на картинку реабсорбции жидкости. Здесь наглядно показан баланс
- 1.5 литра потребляется извне, в среднем, 7.5 литров - это секреторные цир-
куляты, абсорбирующиеся в тонком кишечнике, 1.35 л приходится на толстый ки-
шечник и около 150 мл секретируется с фекалиями.
Количество воды и электролитов,
поступающих в ЖКТ
щщ
реабсорбция
воды
желудок
тонкая и толстая
кишка
выделение
воды с калом
-^1
^1
А
"Я
т
н
\/3
1ч
In
1N
|
IP
1\
т\
Баланс жидкости
Поступление
Пища, питье
Секреты
•слюна
•желудочный сок
•желчь
•панкреатический
сок
•кишечный сок
Всего
Абсорбция
•тонкая кишка
•толстая кишка
[Выделение с калом
вода
1500
7500
9000
7500
1350
150
Na+
150
1000
1150
950
195
5
ci-
150
750
900
800
57
3
Са2+
80
40
120
110
3
12 I
Основное количество воды проникает в кровь посредством пассивного транспор-
та или осмоса, чуть меньший объем, но то же существенный - за счет активного
транспорта - «натрий зависимого градиента». Натрий переносится из кишечника в
клетки кишечных стенок посредством нескольких механизмов, в первую очередь за
счет совместного транспорта с глюкозой и аминокислотами, а также за счет об-
мена Na+/H+. Поглощенный натрий быстро экспортируется из клетки далее, в меж-
клеточное пространство или кровь, с помощью натриевых насосов — при поступле-
нии большого количества натрия в клетку, большое количество натрия выкачива-
ется из клетки, что устанавливает высокую осмолярность в небольших межклеточ-
ных пространствах между соседними энтероцитами (клетки кишечного эпителия).
Вода же диффундирует в ответ на осмотический градиент, создаваемый натрием, в
данном случае, в межклеточное пространство. Таким образом, вода всасывается в
межклеточное пространство путем диффузии вниз по осмотическому градиенту. Од-
нако, если рассматривать процесс в целом, транспорт воды из кишечника в кровь
часто происходит против осмотического градиента - это важно, поскольку это
означает, что вода может проникать в кровь, даже если осмолярность в кишечни-
ке выше, чем осмолярность крови.
Давайте вспомним, что же такое принцип осмоса, многие слышали о нем благо-
даря домашним установкам очистки воды. Опять же Википедия нам сообщает, что
осмос - это процесс односторонней диффузии через полупроницаемую мембрану мо-
лекул растворителя, в сторону большей концентрации растворённого вещества из
объёма с меньшей концентрацией растворенного вещества. В переносе на сказан-
ное выше - растворитель, т.е. вода с растворенными в ней веществами (давайте
не будем забывать, что наш кишечник это не фановая труба, там всегда есть ка-
кие-то вещества, с радостью растворяющиеся в воде), будет стремиться проник-
нуть через клеточную мембрану (apical membrane) туда - где концентрация ве-
ществ больше, дабы уровнять градиент. Помните про хюмеостаз?
Итак - кровь... Ученые выяснили, что наша кровь, будучи водным раствором, об-
ладает осмоляльностью около 280 мО смоль/кг, за счет растворенных в ней ве-
ществ .
Осмоль — единица осмотической концентрации, равная осмолярности, получаемой
при растворении в одном литре растворителя одного моля неэлектролита. Соот-
ветственно, раствор неэлектролита с концентрацией 1 моль/л имеет осмолярность
1 осмоль/литр.
Небольшая ремарка, есть два очень похожих понятия в органической химии -
осмолярность и осмоляльность, их часто путают и не без причины - поскольку
обозначают они по сути одно и то же - количество молей растворенного вещест-
ва, только осмоляльность имеет ввиду растворение в килограмме растворителя
или воды, а осмолярность в литре растворителя. В случае водного раствора это
вроде как и не особо важно, но, биология требует точности - а как известно,
при нагреве та же вода при том же весе, будет занимать слегка больший объем -
поэтому применение понятия осмоляльность, где мы четко понимаем, какое коли-
чество растворителя используется, будет точнее.
Итак, определившись с осмоляльностью крови, мы переходим к терминологии
жидкостей в организме, а именно, жидкости имеющие осмоляльность равную 275-
290 мОсмоль/кг (мили-осмоль на килограмм) называются изотоническими (равными
крови), выше 290 - гипертоническими и ниже 275 - гипотоническими.
Дабы не перегружать читателя биологическими схемами кишечного всасывания
(intestinal absorption) скажу, что осмотическое давление раствора является
коллигативным свойством, то есть оно пропорционально количеству частиц рас-
творенного вещества, присутствующих в растворе. Было выяснено, что наилучшим
регидрирующим эффектом обладают гипотонические напитки (не вода), т.е. обла-
дающие осмоляльностью ниже осмоляльности крови, но наличие в напитке раство-
ренных углеводов увеличивает гидратацию. К сожалению и здесь не обходится без
«но»... Не все углеводы одинаковы, например сукроза (сахар) влияет на этот про-
цесс отрицательно. Второе важное «но», которое мы рассмотрим чуть позже это
влияние электролитов (солей) на абсорбцию и наконец - как мы рассмотрели вы-
ше, по мере увеличения интенсивности нагрузки, осмоляльность крови меняется в
большую сторону (загущается), т.е. напиток, который мы делали изотоническим
(равным крови) на какое то время становится гипотоническим.
С водой разобрались, теперь к «топливу», гипотонический напиток конечно хо-
рошо, но наша-то задача - снабдить организм достаточным количеством углево-
дов! Опять же, ученые подсчитали - для активных тренировок на 85% УОгтах на-
шему организму (среднему спортивно-подготовленному индивидууму мужского пола)
необходимо получить извне 90-150 г углеводов, это при том, что тонкий кишеч-
ник среднего человека способен усвоить не более 90 г углеводов за час. Тол-
стый кишечник так же способен усваивать углеводы, но не так эффективно, к то-
му же, микрофлора присутствующая в нем, потребляя сахара, начинает активно
выделять газы, что не добавляет комфорта и эффективности нашему спортсмену.
Так, с количеством углеводов определились, теперь нужно понять каким обра-
зом их доставить, - конечно же с водой! И тут появляются первые практические
трудности реализации... Взгляните на график ниже.
Вместе с ростом концентрации растворенных веществ, падает скорость всасыва-
ния жидкости, что вполне оправдано, ведь большая часть воды движется по осмо-
тическому градиенту, в то же время, растет скорость всасывания углеводов, со-
вокупная эффективность поглощения воды и углеводов лежит где-то в районе 8-9%
концентрации.
r—,^
с 1.2-
l/6)
a 1.0-^
YINi
Ь 0.8-
ш „ „
о 06-
DC
СЛ 0.4-
<
О
О 0.2-
I
О
0.0-
о
--^.
S*^
/ \
/ \
/ ч
/ \
/ ч
/ \
/ \
/ \
/ Fluid
/ сно
1
1
^—*,
с
12 1
11 £
О
10 |
9 |
о Ш
8 о
1 с
7 н
5 1
-1
и LL
Г 1 ! 1 1 1 ! 1 1 1 ^
0 2 4 6 8 10 12 14 16 18
/о СНО CONCENTRATION OF SUPPLEMENT
СНО - это углеводы. Gastric emptying - соответственно, кишечное
всасывание.
Окей, это уже результат - итого мы пришли к выводу, что 80-90 г углеводов
на литр воды будет в принципе и достаточно и эффективно для потребностей на-
шего спортсмена на час высоконагруженной тренировки. Чего проще, про сахар,
мы выяснили чуть ранее, его не стоит использовать, ну и ладно, берем декстро-
зу (это обычная глюкоза, доступная в магазинах), замешиваем 90 г в литре и
Понимаем, что мы опять упираемся в «но»...
Давайте взглянем на глюкозу - C6Hi206, классическое органическое соединение,
моносахарид, самый распространенный источник энергии для живых организмов.
Входит в состав многих полисахаридов, таких как крахмал, целлюлоза, гликоген.
Посмотрев на ее формулу - C6Hi206, не трудно подсчитать молярную массу - это
180 г/моль, т.е. масса одного моля вещества. Быстренько прикидываем моляль-
ность получившегося раствора, для этого всего лишь нужно разделить количество
сольвента в молях (т.е. нашей глюкозы) на массу растворителя в килограммах
(т.е. воды). Исходя из того, что у нас 180 г весит один моль, а у нас всего
90 г глюкозы, получается наша глюкоза занимает 0.5 моля, делим это на 1 кг
воды и получаем моляльность в 0.5 мол/кг или 500 мили-молей/кг. «Ого»... - го-
ворим мы ..., «гипотоник у нас явно не получился, перебрали аж в 2 раза, что же
делать?»
Здесь нужно опять небольшое отступление в виде урока химии. Внимательный
читатель заметил, что в какой-то момент я рассуждал в единицах измерения мОс-
моль или мили-осмотических-моль на кг (например, осмоляльность крови 280 мОс-
моль) , а в последнем расчете переключился на другие единицы - моли/кг
(mol/kg). Нет, я не ошибся, в действительности это очень разные величины. Мо-
ляльность (т.е. количество растворенного вещества в килограмме воды) не равно
осмоляльности (т.е. количеству образующихся при этом растворении частиц веще-
ства) , точнее не всегда. Возьмем NaCl, она же соль, она же поваренная соль,
она же хлорид натрия. Молярная масса соли 58.5 г/моль, растворив эти 58.5
грамм соли (т.е. 1 моль), в 1 кг воды мы получим 1 моль/кг раствор, осмоляль-
ность которого будет в два раза больше! То есть 2 Осмоль/кг. Все дело в том,
что соль состоит из Na и С1, которые при растворении в воде образуют по одно-
му катиону Na+ и аниону С1~, т.е. в два раза больше частиц! Быстро вспоминаем
о том, что говорилось ранее - «осмотическое давление раствора является колли-
гативным свойством», т.е. зависящим от количества частиц. Таким образом, ос-
мотическая концентрация соли будет равна удвоенной моляльной концентрации.
Все немного по-другому с глюкозой, при растворении в воде она не диссоцииру-
ет, т.е. мы будем наблюдать n-количество молекул C6Hi206 в растворе, следова-
тельно, осмотическая концентрация, создаваемая глюкозой будет равна моляль-
ной.
Вернемся к нашей проблеме - а именно слишком гипертоническая концентрация,
в то время как нам нужна гипотоническая. . На помощь приходят другие виды уг-
леводов - полисахариды. Полисахарид - это, по сути, много, много молекул од-
ного, или похожего моносахарида, соединенных в единую цепочку. Тут нам делает
подарок пищевая промышленность в лице мальтодекстрина. Мальтодекстрин, это
как правило, очень длинный и разветвленный полисахарид, состоящий из сборки
дисахаридов мальтозы, которые в свою очередь, представляют собой две соеди-
ненные молекулы глюкозы. Поучают его путем гидролиза крахмала, а сырьем вы-
ступают кукуруза, рис, пшеница и т.п. Весь мальтодекстрин маркируется соглас-
но DE, что означает декстрозный-эквивалент, чем больше это число, тем короче
цепочка, чем меньше, тем она длиннее (DE100 = глюкоза). Химическая формула
СбпН(10п+2)О(5п+1) i где п - число звеньев. В чем же выгода от мальтодекстрина? - а
в том, что, например мальтодекстрин DE20, который я использую в своих напит-
ках и который широко доступен в продаже, имеет молярную массу 990 г/моль
(помним о глюкозе, которая 180)! Считать мы теперь умеем и быстро понимаем,
что те же 90 г мальтодекстрина, растворенные в килограмме воды дадут нам все-
го лишь 90 мОсмоль/кг, вместо 500, полученных при глюкозе. «Совсем другое ки-
но, не правда ли...?». «Давайте запихнем в три раза больше и получим все равно
около 270 мОсмоль/кг - т.е. по сути гипотонический напиток!» скажут особо
энергичные..., но увы, ответит автор, опять эти злосчастные правила организма
вставляют палки в колеса нашей безудержной энергии.
Дело в том что, во-первых есть такое понятие, как GI (glycemic index) гли-
кемический индекс, это рейтинг углеводосодержащих продуктов питания, основан-
ный на их постпрандиальной (происходящий после принятия пищи) реакции уровня
глюкозы в крови, по сравнению с эталонным уровнем глюкозы. Что бы было понят-
нее, поясню - мы помним из вышесказанного, что в ответ на получение глюкозы,
здоровый организм, вырабатывает инсулин, пища с высоким содержанием глюкозы
(декстрозы), мальтозы характеризуется высоким GI и, следовательно, высокой
скоростью реакции. Мальтодекстрин имеет еще более высокий GI, попадая в ки-
шечник, его распознает особая группа ферментов - секретины (в частности GIP),
так вот в ответ на излишнюю концентрацию глюкозы, они переключают выработку
инсулина на максимум. Во-вторых, мальтодекстрин использует для проникновения
из кишечника в кровь транспортер - называемый SGLT1 (Sodium-dependent glucose
соtransporters). В реалии он представляет собой транс-мембранный белок (можно
сказать канал), расположенной на той самой апикальной мембране кишечного эпи-
телия. Тот же самый транспортер использует и глюкоза. Функции его просты -
используя энергию АТР он может «протащить» через себя 1 молекулу глюкозы или
мальтодекстрина. Естественно ничего не дается просто так, и маленькая молеку-
ла глюкозы проскочит быстрее длинной цепочки мальтодекстрина. И это rate
limiting factor (бутылочное горло) - суммарно мы можем рассчитывать на транс-
портировку, около 1 г в минуту или около 60 г мальтодекстрина/эквивалента
глюкозы в час. Если дадим туда больше - возникнет «давка» (oversaturated
state) и пропускная способность упадет еще ниже. В дополнении ко всему, через
некоторое время проявится такой очень неприятный эффект как hypoglycemic
rebound, "забитые каналы" ухудшают кишечное всасывание мальтодекстри-
на/глюкозы, что растягивает этот процесс во времени, и мы имеем огромное ко-
личество биодоступной глюкозы, при уже завершившейся нагрузке, в результате
возникает гистерезис - организм вырабатывает еще большее количество инсулина,
что в последствии ведет к гипогликемии, снижению уровня глюкозы в крови, ниже
минимального значения. Симптоматика может быть от неприятной до очень серьез-
ной, как то потеря сознания и прочие печальные последствия.
Однако мы вспоминаем, что нам нужно 90 г углеводов в час, что же делать?
На помощь приходит фруктоза - моносахарид, один из трех вместе с глюкозой и
галактозой, способный проникнуть из кишечника непосредственно в кровь. Плюс
ее в том, что фруктоза использует другую группу транспортных белков - GLUT5
для проникновения внутрь. В реалии механизм всасывания фруктозы еще не до
конца изучен, есть мнение, что транспорт ее сопровождается натрий-калиевым
градиентом, потому что иногда идет против осмотического градиента, но боль-
шинство ученых склоняются к пассивному транспорту фруктозы. Пропускная спо-
собность GLUT5 достигает 0.5 г в минуту, что дает нам около 30 г в час.
Совокупно с мальтодекстрином, это даст нам нужные, около 90 г углеводов в
час. Изображение ниже поможет визуализировать описанную выше картину.
Схематическое представление клетки кишечного эпителия, сверху апикальная
мембрана, снизу базальная. 1 - всасывание глюкозы, галактозы и фруктозы
из кишечника путем облегченной диффузии с помощью специальных белков-
переносчиков; 2 - транспорт глюкозы и галактозы в энтероцит путем Na-
зависимого вторично-активного транспорта. Белки-переносчики участвуют во
всасывании глюкозы из просвета кишечника в энтероцит против градиента
концентрации. Энергию, необходимую для транспорта, обеспечивает Na+, K+-
АТФаза (3) , которая работает, как насос, откачивая из клетки Na+ в обмен
на К+ и обеспечивает градиент концентрации Na+; 4 - транспорт моносахари-
дов из энтероцитов в кровь путем облегченной диффузии.
Интересное отступление - ученые выяснили, что количество рецепторов для
глюкозы, фруктозы, сахарозы, в тонком кишечнике, не постоянная величина - она
изменяется со временем, завися, от типа питания и даже подлежит тренировке.
В целом мы разобрались с основным питанием и можно перейти к еще одной ма-
лой, но важной группе необходимых для спортивного питания веществ - электро-
литов. Электролиты в биологии, это минералы, имеющие электрический заряд и
растворимые в водных растворах, как то вода, биологические жидкости или
кровь. Читая этот текст и просматривая картинки вы наверняка заметили доволь-
но часто употребляемые химические элементы, к примеру, натрий - Na. Электро-
литы очень важны для нашей жизни, они способствуют поддержанию гомеостаза,
поддержанию кислотно-щелочного уровня, перемещению питательных веществ внутрь
клетки и наоборот, транспорт отработанных наружу, поддержка мышечной и нейро-
функциональности, сердечного ритма и кровяного давления, здоровья зубов и
костей и т.п. и т.д.
Как уже было замечено, натрий (Na) - один из важнейших электролитов. В зоне
его ответственности не только клеточный транспорт, но и важнейшая функция -
поддержания проводимости нервных сигналов. К сожалению, самый главный, он бо-
лее всего подвержен истощению - потери с потом (мы помним - пот соленый) и
мочой (тут либо верим, либо..). Тело взрослого человека в среднем содержит 90
г натрия, почти половина из которого растворена во внеклеточных жидкостях,
чуть меньше (35 г) содержится в костях и остальное внутри клеток и около 10 г
содержится во внутриклеточном пространстве. Нормальное суточное потребление
натрия при «спокойном» ритме жизни - около 2-3 г в сутки, естественно при
серьезной спортивной нагрузке эта цифра увеличивается. Острое отравление на-
трием возможно при превышении потребления 0.5-1 г на кг массы тела, т.е. для
взрослого мужчины 80 кг, 40-80 г натрия вызовут интоксикацию. Общеизвестно,
что самый богатый и доступный источник натрия для нашего организма это Sodium
Chloride - NaCl или обычная соль. Не будем так же забывать про то, что соль
это один катион натрия и один анион хлора, следовательно 1 г натрия содержит-
ся примерно в 2.5 г соли.
Очень удачно вспомнив про соль, мы переходим к следующему важнейшему элек-
тролиту, после натрия, в организме - хлору С1. Ключевая роль содержится в ре-
гулировке гомеостаза и, прежде всего, баланса жидкостей организма, имея отри-
цательный заряд - способен поддерживать электрическую нейтральность и конечно
же кислотно-щелочной баланс. Нельзя не вспомнить, что для переваривания пищи
наш желудок секретирует в том числе соляную кислоту - НС1, непременным компо-
нентом которой, является хлор. Взрослый человек содержит в себе в среднем 80
г хлора, диетарная потребность - около 2.5 г в сутки в «спокойном» режиме.
При учете молярной массы хлора в 35 г/моль, a NaCl - 58.5 г/моль, не трудно
посчитать - 1 г соли даст нам около 0.6 г хлора. Потери хлора при активной
нагрузке - самые значительные.
Следующий не менее важный электролит - калий, К, Potassium. Он так же от-
ветственен за проводимость нервных сигналов, клеточный транспорт, но в нашем
случае его важнейшая роль - в способности поддержания сократительной способ-
ности мышц. Среднее количество калия в теле взрослого человека - около 140 г.
Потери при нагрузке самые минимальные.
3500
15
^ 3000
со
to
О
-■ 2500
О)
4-»
О 2000
и
О)
ш 150°
4->
ГО
| 1000
L0
Ъ 500
0
Na+
*
щ
н
II
LOW MOD
CI
■
1
LOW
*
■
■
1
1
■
1
MOD
K+
*
LOW MOD
Давайте посмотрим на картинку выше, там изображены средние потери трех
главных электролитов при низких (^25% УОгтах) - LOW и высоких - MOD (^65%
V02max). За основу были взяты данные от нескольких профессиональных спортсме-
нов - велосипедистов.
Заявленная продолжительность нагрузки - 1.5 часа. Средние потери - это по-
тери приведенные к единой массе тела (около 71 кг).
Запомним эти данные, они пригодятся нам для расчетов в построении своего,
индивидуального напитка. Максимально - 250 мг К, 2.5гС1и1.5г Na.
Быстро пробежимся по остальным значимым электролитам - прежде всего это
кальций и магний.
Взрослый человек содержит более 1.2 кг кальция, Са, 99% которого находятся
в костной ткани. К важнейшим ролям кальция относятся - участие в мышечной ра-
боте, ферментная активация, клеточная дифференциация, работа иммунной систе-
мы, apoptosis и т.п. и т.д. Нас, прежде всего, интересует тот 1% кальция, что
растворен в крови и внеклеточных жидкостях. В применимости к тренировкам, тя-
жело говорить о возможности краткосрочного дефицита кальция, поскольку 99%-е
депо костной ткани находится рядом, но в долгосрочной перспективе необходимо
этого избегать, поэтому все уважаемые производители спортивного питания ста-
раются включать его в рацион.
Магний - Мд, так же является важным элементом для организма, так же незаме-
нима его роль в нервной системе, мышечной работе, делении клеток, является
вторым по биодоступности внутриклеточным катионом после калия. Средняя кон-
центрация магния в крови составляет 0.6-1 ммоль/л и для поддержания таковой в
спокойном режиме человеку хорошо бы потреблять около 400 мг в сутки. Анало-
гично с кальцием, около 60% магния содержится в костной ткани, представляя
собой биодоступное депо. Дополняя это низкой скоростью абсорбции в кишечнике
(в среднем около 3 часов) магний не является элементом, «для немедленного
спасения», но как и кальций теряется за время упражнений и упреждающее вос-
полнение его запасов является хорошим правилом.
Есть еще два электролита, нуждающихся в рассмотрении это бикарбонаты и фос-
фаты. К сожалению у автора пока не хватило сил для освоения этих так же важ-
ных компонентов.
Итак, мы детально рассмотрели практически все компоненты нашего питающего
напитка и можно переходить к практическим расчетам и собственно построению,
но перед этим еще одна важная вещь....
Для любой пищи и, прежде всего, для напитков есть такой важный параметр как
palatability, на русский он переводится как вкусовые качества, но мне кажет-
ся, перевод не полностью раскрывает смысл. Смысл этого параметра - это каче-
ство быть вкусным и применимым, для данного индивидуума, в данный момент.
Примером может служить, например, глинтвейн - который прекрасно «зайдет» по-
сле или вовремя зимней прогулки, нежели в жаркой пустыне, а кто-то скажет -
«глинтвейн вообще гадость, в любое время». Смысл следования palatability на-
питка в том, что бы создать напиток, который вам будет приятно пить во время
тренировки. Есть, конечно, некие общие критерии - например сладость - харак-
теризует собой энергию, вкус и кислинка лимона - освежение и дополнительную
гидратацию, соль - усиливает вкусы, ароматизаторы - создают впечатление нату-
рального и т. д. и т. п. И как мы помним из вышесказанного, все это не просто
так - орафарингальные рецепторы все передают в мозг, а тот уже делает соот-
ветствующие энзиматические выводы...
Практика
Итак, в теоретической части мы рассмотрели необходимые вещества и объемы
для создания собственного «энергетического напитка». В этой части мы займемся
практической его реализацией.
" Смешаем сухую слюну летучей мыши с. . . "
Я уже довольно долгое время увлекаюсь хоккеем. Как известно - время не сто-
ит на месте, цифры на торте в день рождения, увеличиваются, а приходится иг-
рать с более молодыми соперниками. Отчасти это, отчасти отсутствие хороших,
доступных спортивных напитков на полках подтолкнуло к созданию собственной их
версии.
Ремарка - далее я буду ссылаться на некоторых известных производителей
спортивного питания, например SIS, Gatorade, Powerade, Exeed. В хоккее, в ча-
стности в NHL, официальным напитком признаны напитки компании BioSteel, но
если честно, посмотрев на их продукцию, почитав про них и попробовав, я ниче-
го не понял. По первым впечатлениям это чисто маркетинговый ресурс. В отли-
чие , например, от Gatorade, которых они заменили на этом поприще.
Итого - создаем спортивный напиток для хоккеиста, проводящего на льду 1-1.5
часа - чередующейся высокой аэробной нагрузки (среднее время отдыха 2-3 мину-
ты, затем в поле 2-3 минуты) , мужчины, вес 89 кг, показатель V02max около 51
(цифры старые, но надеюсь..), нагрузка довольно высокая, но не максимальная,
поэтому рассматриваем уровень нагрузки - 65% УОгтах. Конечно, опять необходи-
ма ремарка - хоккеист здесь дан, как пример, подобные напитки могут быть
спроектированы для разных видов спорта, где важна выносливость, запас сил и
т.д., да и не только спорта...
Расчеты будем вести на 1 л или 1 кг воды, фильтрованная водопроводная вода
в моем городе, СПб, обладает довольно низким осмотическим давлением - около
2-3 мОсмоль/кг, тем же, кто будет использовать другую воду, следует учитывать
этот параметр. Простой пример, средний показатель минерализации водопроводной
воды СПБ - 40-70 мг/л. Этот же показатель для источника в Виллози (пригород),
достигает 900 мг/л, что может поменять картину общей осмоляльности нашего
раствора. Идеально использовать дистиллированную воду, тогда мы избавим себя
от случайностей.
Подробный расчет, для облегчения понимания, я буду показывать в обратном
порядке (т.е. я изначально определился с общим количеством углеводов и элек-
тролитов и поясняю структуру их расчета).
Касательно выбора самих компонентов - прежде всего это легко доступные, во-
до-растворимые носители основного "действующего вещества", композиционно при-
годные друг для друга.
Мы рассмотрели в первой части нашей статьи, что на максимуме мы способны
метаболизировать около 90 г углеводов в час, практическую нагрузку, будем
считать как 65% от максимальной - т.е. нам потребуется около 55 г углеводов.
Итак, первые ингредиенты - углеводы, мальтодекстрин и фруктоза. Эффективное
сочетание - 2:1 (как выше отмечалось для успешного проникновения в кровь) или
37 г мальтодекстрина и 18 г фруктозы. Считаем осмоляльность:
[Calculator | main component | molar mass е/тЫ | Mass off solutej—^mass off solvent, kg Motarity,mol Osmotaflty mQsm/mol [
Maltose DE20 Carbohydrates 990,00 f&jxp\ IflQ 0,04 37,37
Fructose Carbohydrates 180,00 IjUffiJ 1'00 °Д° 100,00
Мальтодекстрин и фруктоза не диссоциируют в воде, т.е. при растворении мы
получаем n-ое количество молекул мальтодекстрина или фруктозы, Поэтому мо-
ляльность будет равна осмоляльности.
Далее электролиты. Тут нам потребуются более точные весы, можно купить не-
дорогие ювелирные, которые позволят делать миллиграммовые дозировки.
Первый - Na (натрий). Очень важный компонент, наличие его в напитке помога-
ет предотвратить гипонатринемию (состояние при котором содержание натрия в
крови падает ниже 135 ммоль/литр), симптомы - плохая концентрация внимания,
потеря баланса, вялость мышц и т/п. Хоккей характерен тем, что в дополнении к
довольно большой физической нагрузке, игроку в поле приходится довольно шуст-
ро соображать (очень большие скорости)- куда и как отдать передачу, где парт-
неры, где соперник, каким финтом обыграть защитника, а времени на это милли-
секунды, поэтому «хорошая соображалка» очень важна, про важность сохранения
баланса на коньках даже и говорить не стану. Плюс ко всему, натрий важнейший
транспортный компонент практически во всех типах клеток. Как уже упоминалось,
ученые посчитали, что профессиональный спортсмен за 1.5 часа активных упраж-
нений может потерять 1.5 г натрия, Gatorade, дает цифру 1 г в час, другой
производитель - SIS, чуть больше. Что касается меня, я не олимпийский чемпи-
он, но потею очень активно, следовательно, принял для себя общую цифру - как
65% от 1 г, следовательно, 650 мг. Самый простой источник Na - пищевая соль,
NaCl, она очень удобна тем, что в ней содержится и второй важный компонент -
хлор. Сразу оговорюсь, что соль - не единственный источник Na в нашем напит-
ке , как и С1. Биодоступность некоторых компонентов так же связана с их хими-
ческой или практической реализацией и доступностью (например, калий доступен
в хлориде калия КС1, натрий в виде соли и т.д.). Считаем NaCl 1 г. Молярная
масса Na - 23 г/моль, молярная масса хлора - 35.5, NaCl = 23 + 35.5 = 58.5
г/моль. Не трудно вывести, что в 1 г соли будет 0.397 г Na и 0.612 г С1.
Как я уже упоминал, соль диссоциирует в растворе, образуя один катион на-
трия и один анион хлора, поэтому осмоляльность будет удвоенной моляльностью.
Calculator
Nad
main component
Na,Cl
molar mass g/mol
58,00
Massofsolute,g
1,00
mass of solvent, kg
1,00
Motartty,mol
0,02
Osmotaftty mQsm/mol
34,43
Приведу пример расчета, что бы понимать как считать свои цифры.
Молярная масса соли - 58 г/моль, следовательно 1 г это 0.017 молей NaCl,
при массе растворителя 1 кг - моляльность раствора будет 0.017 моль/кг или
0.034 осмотических или 34 мОсмоль/кг (милли-осмотических-моль на кг). Я ок-
ругляю цифры, поскольку мы не считаем лекарственные препараты и в данном слу-
чае сотые и десятые не столь важны. Еще раз - данные здесь глубоко индивиду-
альны, в своих отчетах SIS и Gatorade показывали, что некоторые индивидуумы
умудрялись терять до 4 г Na с почти 4 л жидкости, что конечно очень много,
ориентироваться на такие пиковые значения мы не будем.
Еще раз оговорюсь - подсчеты итерационны, придя к одной теоретической рас-
четной концентрации, ты понимаешь, что нарушается другой параметр и начинаешь
аппроксимировать. Так, например и с Na, вообще по собственным замерам, я те-
ряю около 1.5 л жидкости за 1 час с небольшим тренировки и вообще хорошо бы
увеличить норму Na, но в конечном счете это приведет к гиперсолености напит-
ка, что тяжело будет скорректировать другими компонентами.
Второй элемент - К, калий. В доступном и растворимом виде, калий удобно
взять из хлорида калия - КС1. Как вы помните из данных по максимальному рас-
ходу это 250 мг, мы берем 65% этой нормы - т.е. около 160 мг. Эту массу нам
обеспечат 300 мг КС1 - молярная масса К - 39,09 г/моль, молярная масса хлора
- 35.5, КС1 = 39 + 35.5 = 74.5 г/моль. Не трудно вывести, что в 0.3 г КС1 бу-
дет 0.157 г К и 0.143 г С1.
Хлорид калия так же диссоциирует в водном растворе на К+ и С1~. Следова-
тельно, будем иметь 0.004 мол на кг или 8 мОсмоль/кг.
Calculator
KCI
main component
K,Cl
molar mass e/mol
74,50
Massofsohite,g
030
mass of solvent, kg 1 Motartty,mol
1,00
0,004
Osmolaftty mOsm/mol 1
8,05]
Третий компонент хлор - CI. Благодаря первым двум компонентам мы получили
суммарно 755 мг хлора, что при 2.5 г максимального потребления конечно серь-
езно не дотягивает до нормы в 1.5-1.6 г, но как говорится, приходится чем-то
жертвовать. Больше источников хлора нет, изменение нормы других компонентов
не желательно. SIS и Gatorade не публикуют данные о хлоре в своих Nutrients
fact, видимо точно так же используют поваренную соль, оставляя хлор, на то,
сколько получится.
Следующий компонент - лактат кальция (calcium lactate) C6Hi0CaO6. Представ-
ляет собой кальциевую соль молочной кислоты. У этого компонента двойная роль
- во-первых доставка анионов кальция (Са2+) , во-вторых изменение рН (водород-
ный показатель/показатель кислотности) раствора в более щелочную сторону. Де-
ло в том, что следующие компоненты будут привносить кислотную составляющую в
наш напиток, снижая общий уровень рН примерно до 5, а это не очень полезно
для зубной эмали. Но, конечно нужно дать ремарку, что, например, аналогичный
показатель знаменитой Coca-Cola это 2.5 и никто особо не беспокоится за зуб-
ную эмаль...
Лактат кальция в воде будет диссоциировать на катионы кальция (Са2+) и
анионы молочной кислоты (Lactic acid). Следовательно, при расчете осмоляльно-
сти мы должны это учитывать. Массовая доля кальция очень варьируется среди
производителей напитков, внятных статей по поводу его дозировки так же нет.
Например, S1S заявляет о 660 мг на литр, Powerade о 40, IsoStar о 320 и т.д.
Для себя, я пришел к эмпирическим данным - 3 г лактата кальция, что нейтрали-
зует кислотность до рН=5 и привносит 390 мг кальция.
Calculator| main component
molar mass g/mol
Massofsolute,g
mass of solvent, leg
Motaftty,moi
Osmotattty mOsm/mol
C6Hl0Caqca
218,00
3,00
1,00
0,014
27,52
Следующий компонент - цитрат натрия (sodium citrate) ЫазСбН507. Основная
суть данного компонента - дополнительный, источник натрия, но позволяющий не
увеличивать соленость напитка. Тут мы вспомним, что выше я упоминал параметр
palatability, с определением - «подходящий нам по свойствам в данный момент»,
так вот предыдущие источники Na и К , в виде хлоридов, имеют ярко выраженный
соленый вкус и если не ограничивать их массовую долю, напиток получится слиш-
ком соленый. Представляет собой соль лимонной кислоты, но, не смотря на это,
является подщелачивающим агентом (т.е. увеличивает рН) . Так же как лактат
кальция, в воде он будет диссоциировать на катионы натрия (Na+) и анионы ли-
монной кислоты (citric acid) Следовательно, при расчете осмоляльности мы
должны это учитывать.
Calculator
Sodium citrate Na3C6H50
main component
Na
molar mass е/тЫ
253,00
Massofsolute,g
1,50
mass of solvent, kg Motartty,mol
1,00
0,006
Osmolaftty mOsm/тЫ
23,26
Последний функциональный компонент напитка - источник магния, магний цитрат
(magnesium citrate) C6H6Mg07 (1:1). Здесь нужно сделать важную ремарку - Дело
в том, что один из главных принципов подбора компонентов для напитка - их
способность к растворимости в воде. Цитратов магния в природе существует не-
сколько, первый это тот который я упомянул, СбНбМд07 (1:1), сокращённо называ-
ется «1 к 1» (один атом магния к одной молекуле цитрата) другой, называется
три-магнезиум цитрат «3:2» (три атома магния к двум молекулам цитрата) и име-
ет формулу Ci2Hi0Mg3Oi4. Последний очень плохо растворим в воде (первый, кста-
ти, тоже неважно), поэтому мы выбираем первый. Как я уже писал выше, магний
не относится к средству первой необходимости по причине своего довольно дли-
тельного времени всасывания в кишечнике (3-4 часа). Включение его в рацион
своего напитка я называю «превентивной мерой».
По мнению науки, суточное потребление магния для обычного человека должно
быть около 200-400 мг, активная нагрузка увеличивает эти нормы на 20%. Если
учесть, что суточную норму мы компенсируем едой, то дополнительные 100 мг
можно добавить и через напитки. Субъективно, магний очень хорошо воздействует
на нервную систему, успокаивающе, есть научные работы, которые доказывают,
что присутствие дополнительного магния увеличивает силу голени, крутящий мо-
мент при разгибании колена, силу разгибания голеностопного сустава, макси-
мальное изометрическое сгибание туловища, вращение и прыгучесть. Магний самый
дорогой из всех компонентов, субъективно эффект от дополнительного приема
магния, даже вне напитка, мне понравился, но немедленного влияния на перфо-
манс он не оказывает, добавлять или нет - на собственное усмотрение. Есть не-
сколько но, связанных с цитратом магния, первый и важный - это его расслаб-
ляющее действие на кишечник, в больших дозах (т.е. бегунам на большие дистан-
ции у которых и так проблемы с «этим делом», к дозировкам относиться очень
аккуратно), второй момент - его наихудшая из всех вышеперечисленных компонен-
тов, растворимость в воде (бутылку трясти нужно долго и тщательно). Магний в
растворе так же будет диссоциировать на ионы магния и лимонной кислоты.
Calculator
Magnesium citrate C6H6Mg07(l:l
main component
Mg
molar mass g/mol
214,41
Massof sohite,g
1,00
mass of solvent, kg
1,00
Molarity,mol
0,005
Osmolatty mQsm/mol
9,33
В 1 г магний цитрата находится 13% магния, т.е. 130 мг.
Итак, мы закончили со всеми «функциональными» компонентами напитка. Если
попробовать его сейчас, то вкус его будет напоминать чуть сладковато-соленый,
может кто-то найдет это приемлемым, но большинство - нет. Поэтому, вспоминая
про palatability, сместим вкус в более «кислотную сторону» и добавим аромати-
затор. Помогут нам это сделать обычная лимонная кислота, citric acid, СбН507
и искусственный подсластитель-ароматизатор, тут каждый может проявить изобре-
тательность , лично я использую порошковый ароматизатор на базе аспартама,
«лесные ягоды», точный состав не известен, но навеска около 1 г (ароматизато-
ра) не оказывает влияние на энергетический состав или осмоляльность напитка.
Лимонная кислота, она не диссоциирует в растворе, следовательно, не прибавит
много к его осмоляльности. Я, эмпирически, пришел к 1.1 г кислоты для своего
напитка.
Calculator
Citric acid C6Hs07
main component
Carbohydrates
molar mass g/mol
192,00
Massof sohite,g
1Д0
mass of solvent, kg 1 Motartty,mol
1,00
0,006
Osmolatty mOsm/тЫ
5,73
Итак, суммируем и резюмируем.
Общая осмоляльность напитка должна получиться 245 мОсмоль/кг (гипотоник).
Общее содержание углеводов - 55 г. Электролиты:
Na - 660 мг,
К - 160 мг,
С1 - 760 мг,
Са - 390 мг,
Мд - 130 мг.
Общая масса сухих веществ - 63 г. Для растворения в 1 кг воды. рН раствора
будет лежать в переделах 4.8-5.5, т.е. не сильно кислотный.
Конечно, лично я, не беру 1 л воды для часовой тренировки, мой размер это
0.5л бутылка, соответственно все компоненты делятся на 2. Как я уже говорил,
вода может играть роль в создании дополнительного осмотического давления из-
за растворенных в ней веществ (можно взять дистиллированную воду), но даже
это и даже небольшие погрешности в миллиграммовых дозировках не изменят тип
напитка с гипотонического на изо- или гипертонический и наша цель в ускорен-
ном всасывании и регидратации будет достигнута.
В качестве итога
Мы прошли теоретический и практический путь, позволяющий нам разумно, осно-
вываясь на знаниях и персональных потребностях, сконструировать напиток для
поддержания организма в период высокой нагрузки. Всем ли он нужен? Очевидно -
нет, не всем. Как уже упоминалось, всех индивидуумов, характеризует индивиду-
альность , это и разные запасы воды в организме, и разный запас гликогена и
ферментов активирующих его преобразование и тренированность мышц и т.п. и
т.д. Наблюдая за множеством участников различных команд я вижу, что примерно
40-50% таскают с собой питье, около 10-15% используют спортивные напитки,
разговаривая с людьми, я понимаю, что менее 1% делают это с пониманием того,
что они «туда засыпают, сколько и для чего». Может быть, отчасти, эта статья
поможет сориентироваться в этой, довольно не простой, теме. Конечно, нельзя
не упомянуть, что рассматриваемые спортивные напитки это всего лишь часть
правильного спортивного питания, особенно для тех, кто занимается этим серь-
езно . Все рассмотренные ингредиенты можно купить в интернете или обычных ма-
газинах. Дешевле ли это будет - однозначно, интереснее - конечно!