/






































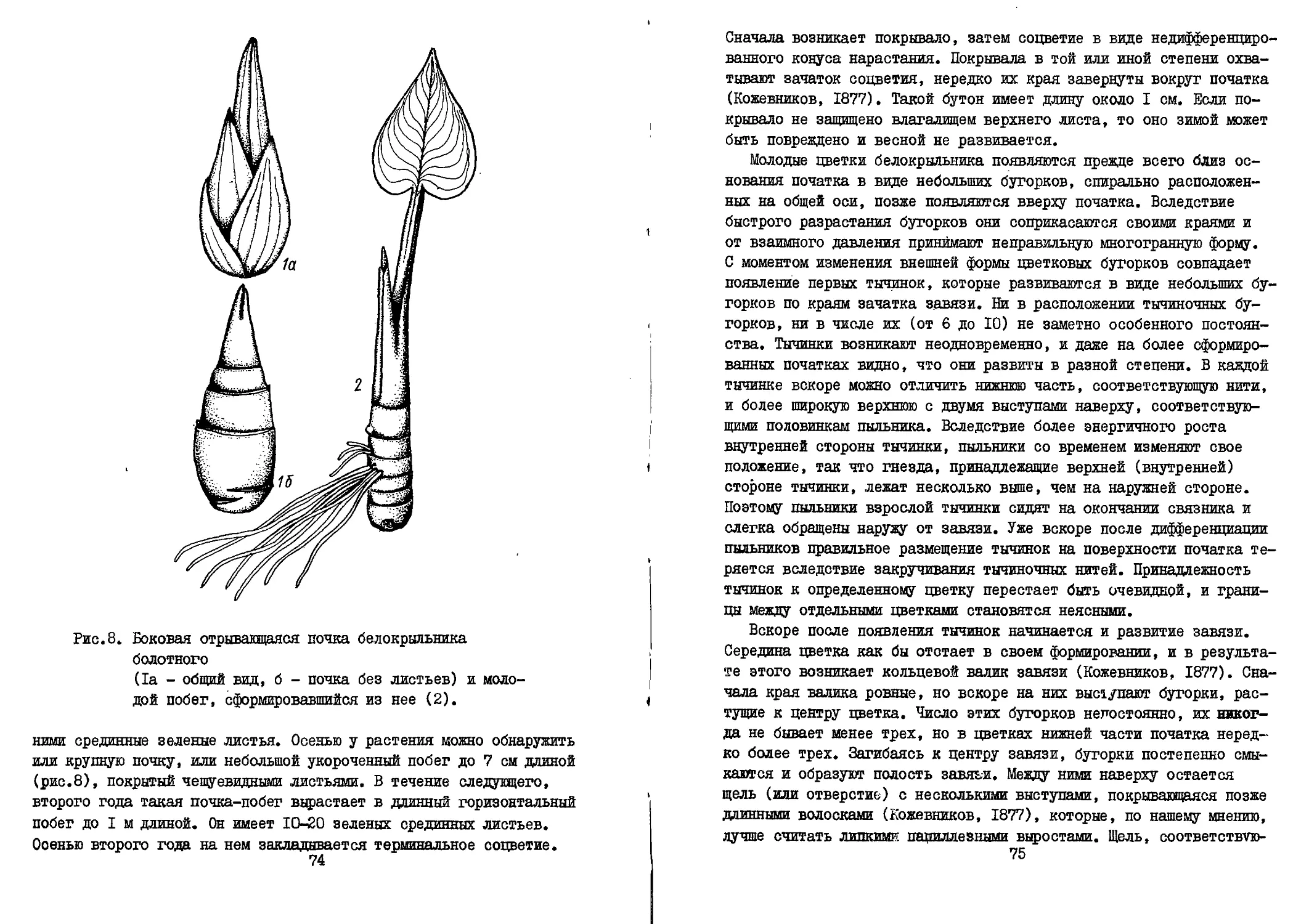





















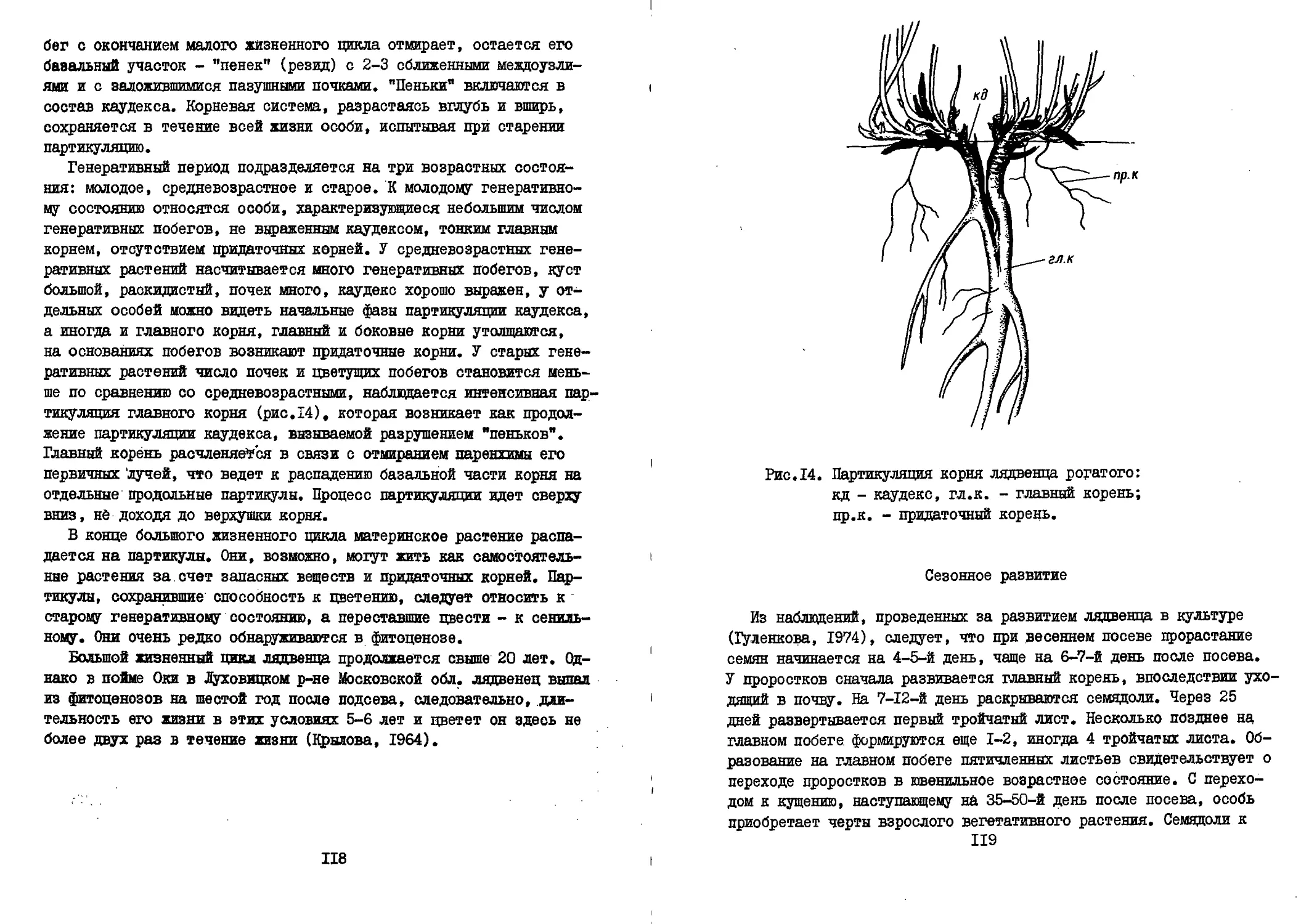











































































Текст
БИОЛОГИЧЕСКАЯ
ФЛОРА
Московской области
БИОЛОГИЧЕСКАЯ
ФЛОРА
МОСКОВСКОЙ ОБЛАСТИ
Под редакцией
профессора Т.А.Р аботнова
Выпуск седьмой
Издательство Московского университета
1983
УДК Б8179 (47I73II)
Биологическая флора Московской области, вып.7.
Под род. Т.А.Работнова. - М.: Изд-во Моск, ун-та, 1983. - 263с.
Сборник продолжает серию публикаций о сосудистых растениях
Подмосковья. В нем обобщены итоги экспериментальных работ, ориги-
нальных наблюдений и литературных данных о систематике растений,
их географическом распространении, морфологии, внутривидовом раз-
нообразии, экологии, фитоценологии, хозяйственном значении и т.д.
В выпуске содержатся материалы о многих хозяйственно важных ви-
дах: лесных деревьях (липа), кормовых травах (лядвенец, тимо-
феевка), ягодниках.
Для широкого круга специалистов-биологов, работников сельского
и лесного хозяйства.
Печатается по постановлению
Редакционно-издательского совета
Московского университета
Рецензенты:
проф. Т.С.Серебрякова; доц. И.М.Культиасов
Редакторы выпуска
И.А.Рубанов и М.Г.Вахрамеева
BIOLOGICAL PLOBA OP THE MOSCOW HEGIOH
Chief Editor
prof. T.A.Babotnov
Part III
Edited by
I.A.Gubanov and M.G.Vakhrameeva
(o') Издательство Московского
университета, 1983 г.
И.И.Науялис, В.Р.Филин
ЩИТОВНИК ГЛУБОКОЙ
Номенклатура, систематическое положение,
внутривидовая систематика
Drycpterie filix-mas (L.) Schott, 1834, Gen. fil.:t.9; ФО-
МИН, 1934, Фл. СССР 1:36; Назаров, 1940, в Маевок., Фл., изд.7:
43; Пояркова, 1954, там же, изд.8:52; Муравьева, 1964, там же,
изд.9:56; Heywood, 1964, Fl. Eur. 1:21; Ворошилов, 1966, Опр.
раст. Моск. обл.:47; А.Бобров, 1974, Фл. евр. части СССР 1:81. -
Pclypodium filix-mas L., 1753, Sp. pl. 2:1090. - Polystichum
filix-тав Roth, 1800, Tent. Fl. Germ. 3:82; Кауфм., 1866, MOCK,
фл., изд. 1:622, 1889, изд.2:652; Цингер, 1886, Сб. свед.:504. -
Aspidium filix-mas Sw. ар. Schrad., 1800, Journ. Bot. 2:38;
Шмальг., 1897, Фл. 2:694; Сырейщ., 1906, Илл. фл. 1:42; он же,
1927, Опр. раст. Моск. губ.:39. - Щитовник мужской.
В отечественных "Флорах" и определителях род включали и не-
редко включают сейчас в сем. Polypodiaceae в.1., но большинство
птеридологов в настоящее время относят его к сем. Aspidiaceae.
Во "Флоре европейской части СССР" (т.1, 1974) род отнесен к сем.
Dryopterldaceae.
Латинское название рода представляет латинскую транслитерацию
греческого слова, происходящего от drys - дуб и pterie - папо-
ротник, следовательно, Drycpterie - папоротник, растущий под ду-
бом. Русское название рода представляет собой перевод латинского
названия Aspidium, происходящего от греческого aspidion - не-
большой щит, или щиток; название Aspidium было дано по форме
покрывальца соруса. Видовой эпитет происходит от латинского fi-
lix - папоротник и тав - мужчина, следовательно filix-тав -
"мужской папоротник", более крупный, чем женский (Каден, Терен-
тьева, 1975).
И.Мэнтон ( Manton, 1950 ) впервые обнаружила, что западноев-
ропейские растения, определяемые обычно как D.filix-mas (L.)
Schott, в действительности относятся к нескольким видам, обра-
зующим комплекс и способным в ряде случаев скрещиваться между со-
бой: двум видам, размножающимся половым путем, - D.filix-mas (L.)
3
llohott e.etr. (4n) И D.abbreviata (Lam. et DC.) Newm. (2П)1 И
апогамно размножающемуся D.borreri Newm.2 * 4 (2n, 3n). Полагают,
что в Англии D.filix-mas в.str. представлен тремя разновидностя-
ми, D.abbreviata - скорее всего тремя подвидами, a D.borreri в
действительности состоит из четырех апогамно размножающихся видов
( Corley, 1967 ). Из распространенных в Европе видов к комплексу
D.filix-mas относится также D.caucasica Fraser-Jenkine et Cor-
ley (D.filix-mas var.athyriformis Fomin ); к нему же принадлежит,
по-видимому» и распространенный на Пиренейском п-ове D.resendea-
na Rez.Pinto, который раньше определяли как D.borreri или D.fi-
lix-mas f.vallongensis (Rezendo Pinto, 1968-1969 ).
Объем комплекса D.filix-mas в других частях света еще недоста-
точно выяснен, но бесспорно, что он довольно обширен. Например, в
Гималаях в.него входит по меньшей мере 10 видов, объединенных в
три группы ( Mehra, Loyal, 1965; Loyal, 1966 ). В Московской обл.
D.filix-mas (L.) Schott - единственный представитель одноименно-
го комплекса.
В литературе приведено большое число разновидностей и форм D.
filix-mas (L.) Schott в.1j (Флора СССР, т.1, 1934). Некоторые из
этих таксонов теперь сведены в синонимы других видов комплекса D.
filix-mas. Выяснение правильного названия и статуса остальных
разновидностей и форм в свете современных данных о комплексе - де-
ло весьма сложное.
D.filix-mas - аллотетраплоид, один из родителей которого не-
сомненно D.oreades, а другим, по всей вероятности, является D.
caucasica, хотя и не исключено, что какой-то представитель комп-
лекса D.villarii (Manton, 1950; Manton, Walker, 1954; Widen,
Britton, 1971; Fraser-Jenkins, Corley, 1972; Widen, 1972; Widen
a.a., 1973).
В Европе известно несколько гибридов щитовника мужского с дру-
гими видами^: D. х brathaica Fraser-Jenkins et Reichst (D.filix-
mas x D.carthusiana), D. x digenea Dornin (D.filix-mas x D.oreop-
I Название незаконное, его следует заменить на D.oreades Fomin
(Fraser-Jenkins, Jermy, 1976).
p
Законное название вида - D.pseudomas (Wollaston) Holub et Pou-
r zar (Holub, 1967).
° Некоторые исследователи за гибрид D.filix-mas х D.carthusiana
принимали Aspidium remotum A.Br., но хроматографическое изуче-
ние аутентичного образца ( widen a.a., 1976 ) показало, что на-
звание D,remote следует относить к триплоидному апогамному так-
сону, возникшему, но-видимому, при скрещивании D.pseudomas х D.
assimilis (Beni,-Eschelmuller, 1973).
4
teris)'1, D. x euxinensis Fraser-Jenkins et Corley (D.filix-mas
x D.oaucasica), D. x mantoniae Eraser-Jenkins et Corley (D.filix-
mas x D.oreades)2 *, D. x subaustriaca Rothm. (D.filix-mas x D.aus-
triaca)2, D. x tavellii Rothm. (D.filix-тав x D.pseudomas)4. В
Северной Америке известен гибрид D.filix-mas х D.marginalis (Win-
slow, 1910; Wagner, Chen, 1965 ). Гибриды щитовника мужского с
другими видами из Московской обл. не описаны.
Географическое распространение
Нередко полагают, что щитовник мужской - "почти космолитичес-
кий лесной неморально-бореальный вид" (Дедов и др., 1974, с.37),
но на самом деле точные границы ареала вида неизвестны и на тех
картах, где показано распространение D.filix-mas (L.) Schott в.
str. (Hulten, 1962; Meusel u.a., 1965; Гричук, Моносзон, 1971),
приведен ареал одноименного комплекса. Более точные сведения о
распространении собственно щитовника мужского приведены лишь для
Западной Европы ( Atlas florae Europaeae, 1972 ) и СССР (Атлас
ареалов ..., 1976).
Ареал щитовника мужского охватывает почти всю Западную Европу
и значительную часть Восточной Европн. В европейской части СССР,
где вид отмечен во всех флористических районах (Флора европейской
части СССР, т.1, 1974), северная граница сплошного ареала, охваты-
вающего главным образом лесные области, протягивается от границы
с Финляндией на Карельском перешейке на восток вдоль 61-й паралле-
ли северной широты до Северного Урала, восточная граница проходит
по восточному склону Урала до верховьев р.Урал, а южная граница
почти совпадает с южной границей лесостепной зоны. Кроме того,
обособленные участки ареала имеются на Кольском п-ове, Северном
Кавказе, в Закавказье, Крыму, на юге Западной Сибири и Краснояр-
ского края, в Казахстане и горах Средней Азии. Изолированные мес-
1 В правильном определении природы растения, описанного Доминым в
качестве гибрида, можно усомниться, так как у D.oreopteris
(=Oreopteris limbosperma) 2n = 34 (Manton, 1950).
2 Гибрид получен также искусственно ( Manton, 1950).
2 Если D.austriaca = D.dilatata, TO D. x subaustriaca = D.subal-
pinum.
4 Гибрид представлен в природе двумя циторасами с 2n = 164 и
205, эти расы получены и экспериментально ( Dopp, 1955).
х 5
I -1964
тонахождения отмечены во многих районах, самое восточное из них -
в Саянах (Флора Центральной Сибири, 1979). Не исключено, что неко-
торые указания на местонахождения щитовника мужского в Средней
Азии относятся к другому виду комплекса. Например, исследованные
Л.С.Рябковой (1973, 1974) споры растений из Таджикистана много
мельче спор щитовника мужского из Европы и приближаются по своим
размерам к спорам диплоидных членов комплекса.
Распространен щитовник мужской и в Северной Америке, поэтому
определять ареал этого вида как дизъюнктивный европейско-западно-
азиатский (Атлас ареалов..., 1976) можно, лишь рассматривая рас-
пространение вида в Евразии. Исходя из того, что споры растений с
запада Северной Америки меньше похожи на споры образцов щитовника
мужского из Западной Европы, чем споры растений с востока Север-
ной Америки, некоторые исследователи предположили, что североаме-
риканские растения из восточных и западных районов, относимые к
щитовнику мужскому, представляют собой разные таксоны (Crane,
1956; Wherry, 1961 ). Была даже выдвинута гипотеза, согласно кото-
рой восточный щитовник, в отличие от европейского, является ауто-
тетраплоидным дериватом D.abbreviata (Wid&i, Britton, 1971 ). Но
более тщательный морфологический и хроматографический анализ вос-
точно-североамериканских растений указывает на то, что они пред-
ставляют соббй аллотетраплоидный таксон, длительная изоляция кото-
рого привела к небольшим химическим и морфологическим отличиям
его от европейского D.filix-mas (Britton, Jenny, 1974).
В СССР щитовник мужской приурочен в основном к средним широтам
и едва заходит в пределы Арктики. Это мезофильное лесное растение
обитает главным образом в хвойных (еловых, елово-пихтовых), широ-
колиственных (дубовых, буковых, грабово-буковых), смешанных и мел-
колиственных лесах. Растение предпочитает районы с выраженным оке-
аническим или умеренно континентальным климатом, поэтому в облас-
тях, где западный перенос осадков ослабевает, оно встречается ред-
ко. Изолированные местонахождения щитовника мужского могут иметь
реликтовый характер. Например, на юго-востоке европейской части,
в северном Прикаспии (в районах южных степей и полупустынь) щитов-
ник мужской - реликт, скорее всего рисс-вюрмского межледниковья
(Иванов, 1972). Указания на встречаемость щитовника мужского очень
высоко в горах Средней Азии, возможно, относятся к другие видам
комплекса. В некоторых случаях современные местонахождения щитов-
ника мужского в горах в тех районах, где лесов сейчас нет, могут
свидетельствовать о наличии лесов в этих пунктах в историческое
6
время (Коннов, 1971).
В Московской обл. щитовник мужской встречается во всех бота-
нико-географических районах.
Морфологическое описание
Многолетний корневищный гемикриптофит. Корневище мощное, 20-
35 см, редко до I м ( Karpowi.cz, 1972 ) длиной и до 5-10 см диа-
метром, у молодых растений вертикальное, позже полегающее, косое
до дуговидного, редко почти горизонтальное, при отклонении от
вертикали слегка уплощенное, обычно простое, плотно усаженное
расположенными по спирали светло-бурыми, с возрастом чернеющими
основаниями черешков отмерших вай. Молодые участки корневища по-
крыты также бурыми чещуями. На основании черешка на его спинке
или чуть сбоку от нее развивается почка, которая может дать нача-
ло новому растению. Стебель на поперечном сечении многоугольный,
по своей структуре радиально-симметричный, с диктиостелой из 8-
13 пучков. В стебле и основаниях черешков есть схизогенные полос-
ти с железистыми волосками (Де Бари, 1877).
Корни 20-50 (100) ом длиной, волокнистые, придаточные, отхо-
дят по 1-5 от оснований черешков, расположенных на нижней сторо-
не корневища. Стела диархная, поэтому боковые корни располагают-
ся обычно двурядно. Корни первого порядка до 1,5-2 мм диаметром.
Поверхность корня, утерявшая корневые волоски, темно-бурая до
почти черной.
Ваи 15-140 см длиной и 5-40 см шириной, собранные по З-П на
верхушке стебля в виде воронки, травянистые, как правило летне-
зеленые. Черешок до 5 мм толщиной, в 3-5 раз короче пластинки,
покрытый чещуями и волосками, более густыми к основанию черешка.
На поперечном сечении черешок почти четырехугольный, с неглубо-
кой адаксиальной выемкой и закругленными абаксиальными углами, с
5-8 пучками близ своего основания, в средней части черешка число
пучков может увеличиваться до 9 ( Luerseen, 1889 ); адаксиальная
пара пучков крупнее остальных. Чешуи на черешке до 3,5 мм длиной,
треугольно-ланцетные, продолговатые или линейные, с редкими мел-
кими зубцами по краю. Молодые чешуи соломенно-желтые, с возрас-
том они буреют, иногда имеют темную полосу близ основания.
Пластинка ваи сверху невогнутая, темно-зеленая, снизу более
светлая, на большем своем протяжении дважды перистая, в очерта-
7
нии от овально-ланцетной до продолговато-ланцетной, с усеченным
основанием и заостренной верхушкой, с наибольшей шириной в своей
средней части или чуть выше середины (самые длинные перья 4-14-е
снизу). Главный рахис густо покрыт (особенно снизу) соломенно-
желтыми чешуями и светлыми, почти белыми волосками, рахисы перьев
опушены в меньшей степени. Адаксиальный желобок главного рахиса
открыт к желобкам рахисов перьев. Перья очередные, отходят почти
под прямым углом от главного рахиса по 15-35 пар с каждой сторо-
ны, плоские, 1,5-3 см шириной, сидячие или на очень коротких чере-
шочках, в очертании от продолговато-ланцетных до линейных, усечен-
ные в основании и постепенно заостренные к верхушке. Срединные на-
иболее широкие перья ваи с 15-30 перышками с каждой стороны свое-
го рахиса.
Перышки от округлых до продолговато-ланцетных, с усеченным ос-
нованием, часто с почти параллельными краями и закругленной, реже
слегка заостренной верхушкой, цельные, с городчато-зубчатыми кра-
ями или с немногими лопастями или даже долями, в свою очередь,
зубчатыми по краю; зубцы острые, но без оттянутого остроконечия,
направленные вверх. Два базальных перышка основного пера по длине
почти равны между собой или же базископичёские, особенно у расте-
ний влажных и тенистых мест, длиннее акроскопических. Базальные
перышки у нижних перьев анадромные, у срединных - почти супротив-
ные, у верхних - катадромные. Сидячие на коротких черешочках два
базальных перышка крупнее и четко отделены от остальных сомкнутых
и со слившимися основаниями перышек того же пера. Базальные лопас-
ти или сегменты базальных перышек обычно образуют ушки. Пластинка
перышка сверху голая, снизу покрыта редкими волосками и узкими че-
шуями, приуроченными к жилкам. Перышки гипостомные, устьица 48-70
мкм длиной ( Corley, 1967; Fraser-Jenkins, Corley, 1972 ). Жилко-
вание перышек перистое, открытое; боковые жилки нерезкие, один,
реже два-три раза вильчато ветвятся и заканчиваются близ края лис-
та в булавовидных гидатодах.
Сорусы смешанного типа ( Bower, 1923 ), округлые, слегка зиго-
морфные, сближенные, сидят в нижней части перышка близ середины
акроскопических веточек вильчато разветвленных боковых жилок. На
даждом перышке до 12 сорусов. Плацента в очертании сверху полулун-
ная. Покрывальце в очертании почковидное, 1-2 мм диаметром, при-
креплено в месте выемки по желобчатой складке, плоское, реже слег-
ка выпуклое, голое, цельнокрайнее, реже с редкими различимыми в
микроскоп зубчиками. У молодого соруса покрывальце почти белое, с
8
краями, не подогнутыми под спорангии; при созревании соруса по-
крывальце буреет, ссыхается (но не опадает) и не полностью при-
крывает ’'сорус. ‘
Спорангии полипоидного типа, ножки их с 1-2 железистыми волос-
ками, опадаквдими при созревании спорангия. Споры коричневатые,
билатерально-симметричные, однолучевые, по форме напоминающие се-
мя фасоли, от 36-46 до 50-72 мкм длиной (вместе с периспорием).
Периспорий с бородавчатыми и гребневидными выступами-складками,
образующими замкнутые или незамкнутые петли; длина щели равна при-
мерно половине длины большей экваториальной оси (Давлиниадзе,
1965; Гричук, Моносзон, 1971; Кутафьева, 1972; Praseг-Jenkins,
Corley, 1972; Karpowicz, 1972 ). Хромосомное число 2п=1б4.
Обоеполые заростки сердцевидные, с глубокой апикальной выемкой,
с ризоидами, антеридиями и архегониями на нижней стороне и с одно-
клеточными железистыми волосками по краю заростка и на его нижней
стороне.
Онтогенез
Сорусы щитовника мужского созревают во второй половине лета, и
большая часть спор оседает на почву во время массового раскрыва-
ния спорангиев. Прорастание спор и развитие заростков в естествен-
ных условиях наблюдать трудно, но данные, полученные при изучении
этих процессов в лабораторных условиях, имеют не меньшее, а даже
большее значение для понимания физиологических особенностей на-
чальных этапов онтогенеза гаметофита. Химический состав спор при-
водит А.Р.Кизель ( Kiesel, 1925 ). Большое значение для понимания
биологии вида имеют сведения о том, что в качестве запасного пита-
тельного вещества споры щитовника мужского содержат жиры (Cour-
bet, Metche, 1971), что они не содержат хлоропластов ( Miller,
1968), а периспорий их не пропускает лучи с длиной волны меньше
570 нм (Orth, 1937). Споры щитовника мужского содержат мало во-
ды - около 4% (Courbet, Metche, 1971), поэтому устойчивы к воз-
действию низких температур. Известно, что они сохраняли жизнеспо-
собность после пребывания в течение нескольких часов в жидком ге-
лии (Becquerel, 1930). Ненабухшие споры щитовника, по-видимому,
без ущерба для себя переносят низкие зимние температуры и другие
экстремальные условия.
Сведения о способности спор щитовника мужского сохранять жизне-
9
способность при хранении противоречивы. После года хранения про-
растало до 94% спор (Mohr,"1956). По одним данным (Dopp, 1927),
спустя 2,5 года после сбора споры сохраняли жизнеспособность пол-
ностью, но по другим источникам (Laage, 1907), спустя 1,5-2 г
после сбора споры не прорастают.
Время выхода спор из состояния покоя регулируется разными фак-
торами внешней среды, в первую очередь оветом. Споры щитовника
Мужского не прорастают в темноте. Многочисленными опытами доказа-
но, что красный свет стимулирует прорастание спор большинства па-
поротников, в том числе и щитовника, а синий и дальний красный
свет тормозят этот процесс (Miller, 1968). Модель, Объясняющая
регуляцию прорастания спор светом, включает: I - абсорбирующий
красный и дальний красный свет фитохром; 2 - пигмент, поглощающий
синий свет и лучи, близкие к ультрафиолетовой области спектра;
3 - неизвестный фактор X (Furuya, 1978). Ингибирование процесса
прорастания мало зависит от температуры, но выход спор из состоя-
ния покоя в темноте прямо зависит от нее.
Известно, что споры одного растения различаются между собой по
степени реакции на тот или иной фактор, способствующий прораста-
нию (Furuya, 1978). Наблюдения в природе показывают, что у щитов-
ника мужского, по-видимому, лишь небольшая часть спор прорастает
осенью (Schneller, 1975), а основная масса их начинает прорас-
тать весной по мере изменения спектрального состава света и повы-
шения ночных температур.
В лабораторных условиях споры щитовника мужского начинают про-
растать на 4-41-й день после посева (Karpowicz, 1927, I960; Du
Buy, Neurnbergk, 1938; Карнишина, 1975; Zirner, 1976). Такая
разница в сроках прорастания зависит не только от условий опыта,
но объясняется также и физиологической гетерогенностью опор, в
том числе и спор одного растения, что увеличивает шансы на выжива-
ние данной популяции заростков. Споры прорастают лучше в условиях
длинного дня, чем короткого. Они могут прорастать на среде с ин-
тервалом pH от 3 до 9,5, но слабокислая среда (pH 5,2-5,4) опти-
мальна для прорастания (Mohr, 1956). Хотя споры и могут прорас-
тать при очень высокой интенсивности света (Orth, 1937), зарост-
ки вряд ли способны вынести сильную инсоляцию.
Тип прорастания спор щитовника описывается как центростреми-
тельный (Мотове, 1942), Сипплярпнй (Atkinson, Stokey, 1964) ИЛИ
аспидиоидный (Nishida, 1965). Но если учитывать направление рос-
та пророотковой нити не только по отношению к первичному ризоиду,
10
но и к полярной оои споры, то правильнее характеризовать прораста-
ние спор щитовника как полярное и относить его к типу Vittaria
(Kayar, Kaur, 1968, 1971).
Как и прорастание спор, ранние этапы морфогенеза заростков па-
поротников контролируются фотореактивной системой (Miller, 1968).
Слабый дневной или красный свет обусловливает формирование про-
ростковой нити (протонемы), состоящей из немногих удлиненных кле-
ток, а интенсивный белый или интенсивный синий свет способствует
переориентации клеточных делений в апикальной клетке протонемы и
переходу от однонаправленного роста к двунаправленному (формирова-
ние пластинки). Нить растет по направлению к источнику красного
света, который воспринимается фитохромом, локализованным в плазма-
лемме апикальной клетки (Furuya, 1978). в лабораторных условиях
длину нити можно ограничить тремя клетками, но в природе у щитов-
ника обычно вырастает более длинная нить.
При переходе к двунаправленному росту верхушечная клетка нити
делится перпендикулярно продольной оси, образуя терминальный воло-
сок и вздутую субтерминальную клетку, которая впоследствии обычно
не делится. Третья от конца клетка делится продольно, а ее дерива-
ты продольно и поперечно, вследствие чего образуется неравнобокая
пластиночка, на одной из сторон которой вычленяется меристемати-
ческая клетка. Далее ПО типу Adiantum (Nayar, Kaur, 1969, 1971)
возникает сердцевидный заросток с верхушечной меристемой, располо-
женной в выемке, со срединным ребром (подушкой), с ризоидами и по-
ловыми органами (антеридиями и архегониями) на нижней стороне и с.
одноклеточными железистыми волосками по краю и на поверхности.
Наряду с такими обоеполыми сердцевидными заростками у щитовни-
ка мужского могут развиваться и другие типы заростков: сердцевид-
ные обоеполые, сердцевидные женские, языковидные мужские (Karpo-
wicz, 1927; Bittmannova-Nogova, 1962). По-видимому, У него можно
обнаружить те же типы заростков, что и у D.ludovicianas I) серд-
цевидные женские (на очень старых женских заростках могут появ-
ляться веточки с антеридиями), 2) сердцевидные протерацдрические
обоеполые, 3) сердцевидные синхронно обоеполые, 4) америстемати-
ческие нитевидные и лопатчатые мужские, 5) сердцевидные мужские
(Couaena, Horner, 1970). Заростки разного типа отличаются друг
от друга скоростью роста и временем созревания, что зависит от
многих факторов, в том числе и от плотности посева. Гетерогенный
состав популяций заростков одного растения увеличивает вероят-
ность межзаросткового скрещивания, межзаросткового спаривания и
II
уменьшает возможность внутризаросткового оплодотворения (Kiekow-
ski, 1969), что в конечном счете способствует гетерозиготности
зародышей.
В лабораторных условиях первые антеридии появляются на зарост-
ках на 59-65-й день после посева спор, первые архегонии - на 65-
121-й день, зародыш - на IOO-IIO-й день (Karpowicz, 1927, I960;
Бриеде, 1972). У щитовника мужского отмечено образование и апо-
гамных спорофитов (Manton, Walker, 1954; Fries, 1970а,b).
Первая вал пророотка двулопастная или двураздельная с двуло-
пастными долями, вильчатой жилкой и одноклеточными волосками по
краю. По мере появления новых вай у проростков постепенно уста-
навливается перистая структура ваи, самыми широкими перьями явля-
ются базальные.
Ваи проростков сидят по спирали (1/3 или 1+2), стебель облада-
ет гаплостелой, от стебля отходят 14 придаточных корней (Hofmeis-
ter, 1862). Над пятой или шестой ваей стебель сильно утолщается,
система филлотаксиса усложняется (3+5), а пучки образуют диктио-
стелу. На второй год жизни растение становится более мощным, ваи
сидят по спирали 5/13 (5+8). Эта же спираль сохраняется у взрос-
лого растения или она переходит в 8/21 (8+13). Корни теперь обра-
зуются только на основании черешков. Корневище с возрастом начи-
нает полегать и покрывается частицами опада так, что основная ее
часть у зрелого растения располагается в подстилке и в верхнем
слое почвы. Придаточные корни в связи с этим начинают расти толь-
ко с одной стороны корневища.
Не исключено, что у щитовника мужского, как и у щитовника
австрийского (Cutter, Voeller, 1959), помимо простой правой или
левой спирали по основному ряду Фибоначчи изредка могут встречать-
ся и другие типы листорасположения: спираль по дополнительному ря-
ду, парная спираль, трехмерные ложные мутовки. Высокий индекс фил-
лотаксиса у корневища щитовника мужского обусловлен большим раз-
растанием субапикальной области в радиальном направлении, чем по
вертикали (Wardlaw, 1963). Если рост субапикальной зоны по верти-
кали превышает ее рост по радиусу, то возникает столон. Такие сто-
лоны, столь характерные для Matteuccia struthiopteris, изредка
встречаются и у щитовника австрийского (Wardlaw, 1963), отмечены
они нами и у щитовника мужского.
Как указано выше, у взрослого растения на абаксиальной сторо-
не оснований черешков развиваются почки, стела которых связана с
одной из меристем корневища или теряется в коре корневища (рис.1).
12
2см
Рис.I. Схема последовательного развития побегов
щитовника мужского:
I - почка в основании черешка;
2-9 - развивающиеся из нее разновозрастные
побеги.
Наши наблюдения в Московской обл. показывают, что побеги;, возни-
кающие из почек, расположенных у поверхности почвы, как правило;
погибают; побеги же, возникающие из почек-, расположенных на доста-
точной глубине (5-6 см), в первые 3-4 года (а иногда и до 10 лет)
развиваются в почве и образуют стебель с катафиллами. Когда ось
таких побегов достигает 2-5 см в длину и 1-1,5 см в диаметре, у
них появляются придаточные корни (по одному на черешок возникаю-
щих катафиллов), а затем над поверхностью почвы разворачивается
первая зеленая вая типичной для взрослого растения формы, но мень-
ших размеров - с пластинкой 10-20 см длиной и 3-8 см шириной.,- Та-
кие особи клона с 1-3 ваями можно условно назвать имматурными.
Примерно через 4-6 лет "имматурные" особи становятся взрослыми ве-
гетативными. При этом резко увеличивается диаметр корневища (до 3-
4 см), увеличивается число вай и их размеры (до 30 см в длину и
до 15 см в ширину), возрастает число придаточных корней (до 2-3
13
жа каждый из появляющихся черешков). На пластинках вай появляются
первые сорусы (5-10 на особь). Видимо, вскоре начинается разруше-
ние базальной части корневища.
Наблюдения в культуре показывают, что возникшее из зиготы рас-
тение может приступить к спороношению уже на шестой год (Manton,
Walker, 1954). У фертильных особей каждый год разворачивается до
10 вай длиной до 100-120 см и шириной до 25-30 см. Диаметр корне-
вища достигает 5-8 см в апикальной части. В начале генеративного
периода растение приступает к вегетативному размножению, на череш-
ках в апикальной части корневища начинают прорастать адаксиальные
почки. Наши наблюдения показывают, что почки прораотают не на каж-
дом основании черешка,а в среднем на десятом-двенадцатом, что со-
гласуется с данными В.Гофмейстера (Hofmeieter, 1862).
Рост щитовника мужского, вероятно, детерминирован. При старе-
нии организма, по-видимому, происходит уменьшение объема апекса,
что ведет к уменьшению диаметра корневища. Появляющиеся корни те-
ряют способность ветвиться. Ваи уменьшаются в размерах, вероятно,
происходит и возврат к системе филлотаксиса, свойственной молодо-
му растению.
Максимальный возраот особей, рассчитанный по среднему числу
листьев, появляющихся каждый год, с учетом диаметра корневища, до-
стигает в Московской обл. в 90-летнем ельнике волосисто-осоковом з
50 лет. Длина корневищ таких старых особей достигает 30-35 см. По-
скольку в литературе сообщалось о корневищах до I м дайной, можно
думать, что максимальный возраст растения, обитающего в условиях
ценотического и экологического оптимума, достигает 100-120.лет.
4
i
Сезонное развитие
Ранней весной перезимовавшая верхушечная почка мощных спороно- ;
сящих особей содержит многочисленные зачатки вай, самые молодые
из которых развиваются в ваю на третий год после своего заложения
(Hofmeieter, 1862). Таким образом, в почке содержится три набора
зачатков вай. У мощного растения около 10 самых внешних густо по-
крытых буроватыми чешуями улиткообразных зачатков вай будут разво-
рачиваться в текущем году. Эти зачатки отличаются от более моло-
дых большей величиной и наличием хорошо выраженных зачатков перь- 4
ев и перышек. Самые внешние из зачатков текущего года уже содер-
жат зачатки сорусов, поэтому выделение фенофаз появления споранги-
14
ев или появления сорусов следует принимать с оговорками.
В Московской обл. зачатки вай внешнего набора начинают активно
разворачиваться в начале мая. Поскольку максимум ауксина выраба-
тывается в самых молодых перышках (White, 1971), то повреждение
морозом молодых вай ведет к нарушению нормального хода морфогене-
за - возникают уродливые ваи с более широкими пластинками и более
рассеченными перьями. В случае полного повреждения зачатков вай
текущего года весной разворачиваются зачатки следующего (второго)
набора, но образующиеся из них ваи, как правило, несут небольшое
число сорусов, так как заложение сорусов на примордиях второго на-
бора зависит от активности фотосинтеза и (или) фотопериодических
реакций в уже развернувшихся ваях предыдущего набора (Wardlaw,
Sharma, 1963). При срезании развернувшихся вай в июне у растения,
как показали наши наблюдения в Московской обл., через 1,5 мео.раз-
ворачиваются новые ваи. При этом число новых вай в среднем в два
раза меньше по сравнению с числом срезанных, а общая масса их в 9
раз меньше по сравнению с ваями ненарушенного растения. Сорусы на
новых ваях до сентября не появились. Повторное удаление вай, по-
видимому, приведет к истощению растения и к его гибели.
При Ю°С за 14 недель у ювенильного растения развиваются в
среднем 2 листа, увеличение температуры до 20°С приводит к увели-
чению числа листьев до 8 (Zimmer, 1976). В течение первого года
жизни у растения образуется 2-3 листа ювенильного типа с наиболее
широкой частью листовой пластинки в нижней части вай. Полностью
пластинка ваи в Московской обл. разворачивается в конце мая - на-
чале июня, после чего отмечается наиболее активный рост корней,
продолжающийся до поздней осени.
Мейоз в большинстве спорангиев протекает во второй половине
июня, споры созревают к концу июля. Наличие июньского минимума.и
осеннего максимума содержания крахмала в корневище (Блукет, 1964)
становится понятным в свете сезонного ритма-развития ваи. Так как
сорус содержит разновозрастные спорангии, то процесс высеивания
спор растянут во времени, хотя основная масса их выбрасывается из
спорангиев, видимо, в течение одной-двух недель.
При наступлении морозов ваи полегают на землю. На них в это
время находится значительное число спор, так как некоторые самые
внутренние спорангии соруса часто вовое не раскрываются, а в дру-
гих уже вскрывшихся спорангиях часть спор осталась. Кроме того,
много высыпавшихся спор прилипает к пластинке ваи. Ваи к зиме бу-
реют и отмирают, но ифюторые (в Московской обл., как правило,
15
J стерильные), особенно в условиях мягкой зимы, уходят под снег и
jj. перезимовывают в зеленом состоянии, отмирая весной при таянии сне-
2 га. Отмершие пластинки разлагаются очень быстро, но верхушка че-
решка сохраняется еще два-три года вокруг почки, защищая ее.
TI
Wi
jj Способы размножения и распространения
J В ельнике волосистоосоковом в Московской обл. на зрелом 20-30-
J летнем растении, несущем 5-8 листьев, по нашим подсчетам, может
J образоваться 16063+5805 сорусов. Одна зрелая особь может продуци-
п ровать до 10 млн. спор (Погорелова, Работнов, 1978). Основная мас-
J са выброшенных из спорангиев спор разносится воздушными потоками,
но значительная часть спор остается/на ваях и попадает на почву
J лишь после отмирания вай (см. выше). У вай, собранных в Москов-
ской обл. в сентябре, иногда около трети спорангиев в некоторых
j сорусах не раскрыты. Хотя единичные споры папоротников, в том чис-
i ле и щитовника, могут разноситься ветром на несколько сотен кило-
метров (Заклинская, 1951), все же большая часть выброшенных спор
оседает близ материнского растения. Дальность разноса спор и ско-
л рость их переноса зависят от скорости ветра, интенсивности пере-
мешивания атмосферы, рельефа местности и характера растительного
' покрова (Hirst, 1959). Естественно, что при одинаковых погодных
< условиях споры особей, обитающих на открытом месте, будут разно-
Л ситься дальше, чем споры особей, обитающих под пологом густого ле-
] са. В широколиственном лесу "Тульские засеки” под кронами куртины
из пяти особей щитовника мужского число спор превышает 2,5 тыс.
на I см^, на расстоянии 1,5 м от края кроны число спор падает до
950 на I сь^, а с удалением от кроны на 4 м это число снижается
до 50 и даже 25 (Заклинская, 1951). Сходные результаты получены и
другими исследователями (Schueller, 1975).
Оседающие из воздуха споры могут разноситься дальше потоками
воды или насекомыми. Поскольку споры щитовника мужского не прорас-
тают в темноте, то значительная часть их гибнет, будучи погребена
в течение осенне-весеннего периода под лесным опадом или занесена
в почву дождевыми и талами водами. Прорастанию спор, осевших под
кроной материнского растения, у некоторых папоротников препятству-
ют также ингибиторы, выделяемые корнями и листьями спорофита (Co-
urbet, 1957; Davidonle, Ruddat, 1973; Davidonis, 1976). ВОЗМОЖНО,
такие ингибиторы присущи также и щитовнику мужскому, под кронами
16
особей которого заростков мы не находили.
По наблюдениям в Московской обл. наиболее пригодны для разви-
тия заростков и первых этапов развития спорофита пионерные обна-
женные субстраты, способные длительное время удерживать влагу:
вывалы, разрыхленная гумусированная почва в лесу, гниющая древе-
сина и т.п. При благоприятных погодных условиях на таких субстра-
тах в год оплодотворения появляется до 18 тыс. на I м^ молодых
спорофитов (Науялис, 1979). Дальнейшая судьба этих спорофитов за-
висит от воздействия большого числа биотических и абиотических
факторов. Можно лишь отметить, что в Московской обл. молодые рас-
тения 2-4-летнего возраста и истинно имматурные особи встречаются
крайне редко. Таким образом, появление щитовника на новом месте,
т.е. расширение ареала вида определяется условиями, благоприятны-
ми не только для развития гаметофита (который один только и может
заноситься в новые места), но и для развития спорофита.
Появившись на новом месте, щитовник может далее захватывать
территорию, размножаясь вегетативно и образуя обширные клоны.
Очень редко у него наблюдается вильчатое ветвление корневища (Hof-,
meister, 1862), обычно же новые особи возникают из почек, закла-
дывающихся на черешках фертильных растений. Такой способ размноже-
ния весьма эффективен. Например, в ельнике волосистоосоковом Ма-
линского лесничества Московской обл. в елово-щитовниковой микро-
группировке особи с почками достигают 76% общего числа растений
(максимальное число почек на особь 23), а количество особей с уже
развитыми дочерними растениями составляет 18% общего числа расте-
ний (максимальное число дочерних особей на одно взрослое растение
II). В елово-щитовниково-волосистоосоковой микрогруппировке в том
же ельнике соответствующие данные следующие: 41% (14) и 18% (14),
в елово-березово-щитовниково-лютиковой микрогруппировке - 65% (21)
и 35% (9). В еловой с мелким подростом микрогруппировке, где щи-
товник представлен молодыми растениями, особи, имеющие почки, со-
ставляют 8% общего числа папоротников, число почек при этом не
больше одной на корневище, дочерние особи отсутствуют вовсе.
Поскольку дочерние особи имеют более молодой возрастной уро-
вень по сравнению с материнской, вегетативное размножение играет,
по-видимому, главную роль в самовозобновлении популяций щитовника
мужского в Московской обл. в условиях, не соответствующих в насто-
ящий момент фитоценотическому и экологическому оптимуму вида.
2-1964
17
Экология
Встречаемость щитовника мужского на обширной территории в раз-
ных формациях указывает в целом на довольно широкий диапазон эко-
логической приспособляемости как гаметофита, так и спорофита.
Большое число разносимых спор и малые размеры заростков обуслов-
ливают в конечном счете возможность "подбора" видом благоприятных
микрониш для роста гаметофита и дая оплодотворения - затененных,
влажных, более или менее обнаженных субстратов. В хвойно-широко-
лиственном лесу в горах Швейцарии заростки распределены по место-
обитаниям следующим образом; на твердой лесной почве - 15,3%, на
листьях и хвое - 0,1%, на камнях - 2,4%, на свежей древесине -
0,3%, на гниющей древесине - 81,9% (Schneller, 1975).
Случайно найденные у щитовника мужского апогамные спорофиты
характеризуются карликовым ростом и более быстрым развитием по
сравнению со спорофитами, возникшими половым путем (Manton, Wal-
ker, 1964). Эти различия указывают на слабое генетическое укло-
нение апогамных растений от нормы и одновременно позволяют пред-
полагать, что в тех пунктах ареала, где условия для полового вос-
произведения неблагоприятны, апогамные расы могут встречаться ча-
ще, а быстро заканчивающие свое развитие гаметофиты этих рас мо-
гут расти и в более засупживых, более солнечных местообитаниях по
сравнению с местообитаниями гаметофитов половых рас.
Заростки могут, видимо, успешно перезимовывать. Об этом кос-
венно свидетельствуют результаты наших наблюдений за молодыми
растениями на искусственно созданных площадках с перекопанной
почвой в ельнике беспокровном в Подмосковье. В конце сентября на
площадке в I м2 в среднем мы насчитали 450 проростков и ювениль-
ных растений папоротников, а в июне следующего года было уже 666
таких растений, т.е. число их увеличилось на 48%. При этом у поя-
вившихся с весны спорофитов при основании сохранилось зеленое сло-
евище гаметофита.
Гаметофиты чувствительны к загрязнению воздуха - они прекраща-
ют рост через 2 ч после помещения в атмосферу с хлором в концен-
трации 10', а спустя еще 2 ч погибают (Bittmannova, 1964). Хотя
гаметофиты и могут расти на субстрате с pH 3 (Mohr, 1956), кис-
лая реакция среды, вероятно, подавляет активность сперматозоидов.
Поэтому "кислые" дожди могут в конечном счете оказать неблагопри-
18
ятное воздействие на состав популяций щитовника, уменьшая количе-
ство генетических рекомбинаций.
Спорофиты щитовника шуйского лучше всего чувствуют себя на по-
стоянно и умеренно влажных хорошо аэрируемых рыхлых почвах широко-
лиственных и смешанных лесов о развитой подстилкой. Например, в
крупнопапоротниковой микрогруппировке в "окне” в лесу Pague syl-
vatica - Asperula odorata - Oxalie acetoeella на Карпатах на 10
мы насчитали 41 растение в среднем с 6 ваями, средняя длина ко-
торых 75,3+1,4 см и ширина 23,8+0,5 см. В елово-щитовниково-воло-
систоосоковой микрогруппировке ельника волосистоосокового в Мос-
ковской обл. на 10 м2 встречается до 9 растений папоротника в сред-
нем с 5 ваями, длина которых 68,3+1,4 см и ширина 26,7+0,4 см. В
крупнопапоротниковой микрогруппировке в "окне" ельника-кисличника
в Подмосковье число особей щитовника на 10 м2 не превышает 5, дли-
на их вай достигает 61,6+1,6 см, ширина - 19,8+0,4 см.
Щитовник мужской - гомойогидрический вид, не выдерживающий вы-
сыхания (Карреп, 1965). Его относят к акцессивным видам, положи-
тельно реагирующим на осушение (Аболинь, 1978), избыток влаги ве-<
дет к вымоканию корневищ. На сухих почвах у щитовника мужского
прорастает больше придаточных почек, чем в обычных, умеренно влаж-
ных местах (Hofmeieter, 1862), и этот переход к усиленному веге-
тативному размножению свидетельствует об отклонении условий суще-
ствования в неблагоприятную для растения сторону.
Щитовник мужской относят обычно к теневым растениям (Ellenberg,
1974). Когда широколиственные породы в лесу еще не оделись лист-
вой, разворачивающиеся молодые ваи хорошо защищены от сильной ин- .
соляции и иссушения чещуями и волосками, густо покрывающими';абак-
сиальную сторону рахисов и зачатков перьев. Темпы развёртывания
вай, по-видимому, хорошо сопряжены со скоростью облиствения леса.
К тому времени, когда пластинка ваи развернется полностью.и будет
обращена к свету адаксиальной стороной, лес уже оденется листвой. '
Интенсивность дыхания у особей щитовника, обитающих в лесу, летом
снижается, нетто-ассимиляция достигает положительных величин при
оптимальной для фотосинтеза травянистых растений лиственных лесов
температуре в 15-18° (Daxer, 1934). Клетки эпидермиса пластинки
ваи содержат хлоропласты, а жидки далеко отстоят одна от другой,
как в листьях мезофильных двудольных. Устьица и гидатоды приуроче-
ны к нижней стороне пластинки.
Щитовник может расти и на открытых местах - во влажном климате
в горах и на вырубках, но в последнем случае он быстро выпадает,
19
не выдерживая конкуренции со светолюбивыми травами и кустарниками
(например, малиной). У растений открытых мест ваи развиваются
быстрее, но имеют меньшие размеры и большее число устьиц (Tabor-
ska, Bednarova, 1973).
В Московской обл. в ельнике волосистоосоковом щитовник мужской
наилучшего развития достигает в местах, где освещенность в июле
составляет 3-5% от освещенности открытого места, но встречается и
в местах, где освещенность достигает лишь 1-2% открытого места.
При увеличении освещенности (при образовании "окон" в древостое)
щитовник мужской вытесняется кочедыжником женским. В очень тенис-
тых местах численность вида падает. Например, в благоприятном по
здафическим и водным показателям дубо-липняке снытево-волосисто-
осоковом на 10 м^ приходится в среднем всего 3 растения щитовни-
ка, несущих по 3-4 ваи размером 42x16 см, что объясняется хорошо
развитым подлеском и вторым ярусом древостоя.
Напочвенные поздневесенние заморозки повреждают молодые ваи щи-
товника, а повторение их может привести к истощению растения. В
морозильных камерах щитовник выдерживает понижение температуры до
-20°, полностью отмирает при -27° (Карреп, 1965), поэтому в мес-
тах с незначительным снежным покровом и слабо развитой подстилкой
корневище вымерзает. Редкие находки щитовника мужского в Арктике
и высоко в'горах приурочены к местам, хорошо прогреваемым летом и
укрытым мощным снеговым покровом зимой.
Оптимальны для щитовника богатые дерново-подзолистые, бурые и
серые лесные почвы, но сн может произрастать и на подзолах, пес-
ках, скелетных почвах, на выходах известняков и на сложенных из
известняка стенках (Быков, I960; Цутафьева, 1972; wiiimot, 1979).
К приводимым в литературе показателям реакции почвы: рН=5,5-6,5
(Быков, I960), 6,1-6,4 (Kallio е.а., 1969), 4,5-6,4 (Vaicys е.
а., 1979) - следует подходить с учетом того, что корни щитовника
располагаются в основном в горизонте Aj, а авторы обычно не сооб-
щают, pH какого горизонта они определяли. Щитовник мужской являет-
ся индикатором удовлетворительной аэрации почв (Работнов, 1950).
Фитоценология
Заростки щитовника мужского в борьбе за свет и влагу с мхами и
травами напочвенного покрова леса мало конкурентоспособны, поэто-
му встречаются главным образом на обнаженных субстратах. В стадии
20
зрелого спорофита щитовник - чаще всего субдоминант третьего яру-
са ("средних трав" Б.А.Быкова, I960) - удерживает за собой однаж-
ды захваченную территорию в течение многих десятилетий. Характе-
ризуясь достаточной конкурентоспособностью, вид принимает учас-
тие в сложении различных микрогруппировок широколиственных и сме-'
шанных лесов. Многочисленные ассоциации, в которые входит вид,
объединяются иногда в несколько групп (или циклов), при этом ука-
зывается, что везде слой щитовника мужского входит в состав глав-
нейших формаций, что подчеркивает древний характер всего ряда ас-
социаций (Быков, I960). В лесах Подмосковья вид наиболее характе-
рен для коренных ассоциаций еловых и елово-широколиственных ле-
сов Picea abies - Carex pilosa, Quercus robur + Picea abies -
Carex pilosa, Picea abiee - Oxalis acetosella.
Устойчивое положение вида в ассоциациях связано с его средо-
образующей способностью, которая во многом определяется крупными
размерами растения. Воронковидная до 2 м в диаметре крона эффек-
тивно перехватывает свет и осадки, направляя воду к верхушке кор-
невища. Длительно (до 30 лет) функционирующая корневая система
выходит за пределы кронн на расстояние 2-2,5 радиусов кроны
(рис.2). По нашим подсчетам, у растения примерно 30-летнего воз-
раста с семью ваями и кроной высотой до 90 см, откопанного в ель-
нике волосистоосоковом, в приповерхностном слое почвы (0-5 см)
сосредоточено до 23% корней, основная масса корней (до 58%) при-
ходится на горизонт 5-10 см, остальные корни идут глубже, причем
некоторые из них достигают глубины 40 см. В подстилку корни не
заходят. Общая масса корней в абсолютно сухом состоянии 49,3 г,
масса корневища - 84,5 г. Общая поверхность корней достигает
10,4 м^, а их длина - 10033 м. Если учесть, что проективное по-
крытие вида в его зарослях обычно колеблется от 80 до 100% (Бы-
ков, I960), то следует признать, что слой щитовника мужского мо-
жет быть сомкнутым не только надземно, но и подземно. При связы-
вании кроны в плотный пучок на поверхность почвы поступает в
2,8 раза больше света и в 1,8 раза больше осадков.
Хорошо приспособленный к жизни в широколиственных и смешан-
ных лесах в условиях слабой освещенности летом (в буковых лесах
вид удерживается при полноте древостоя до 0,85 -Долуханов, I960)
щитовник мужской при достаточной численности создает еще более
суровый световой режим для более низкорослых сопряженных с ним
травянистых растений, выступая в качестве виолента в своем яру-
се. С ЩИТОВНИКОМ мужским сопряжены Carex pilosa, Asarum europa-
2X-I964 21
Рис.2. Корневые системы щитовника мужского на площадке
0,25 м2 в слое почвы 0-6 см в ельнике волосисто-
осоковом в Московской обл.
• eum, Galeobdolon luteum, Oxalis acetosella, Fragaria vesca, Aego-
podium podagraria и др., корневая система которых располагается
преимущественно в приповерхностном (0-5 см) слое почвы. В лесах
со значительной примесью мелколиственных пород на плохо аэрируе-
мых почвах с щитовником мужским часто сопряжен кочедыжник женский.
Этот более светолюбивый и имеющий поверхностную корневую систему
вид вытесняет щитовник в более тенистые места.
Средообразуюцая роль вида во многом зависит от возрастного со-
става популяций, который не остается неизменным в разных микро-
22
группировках одного типа леса (табл.1). С увеличением возраста
группы отмечается падение надземной биомассы цветковых трав под
Таблица I
Состав возрастных групп парцеллярных ценопопуляций
щитовника мужского в ельнике волосистоосоковом
(Московская обл.) в % от общего числа особей
1 Возраст- Микпогоуппиповки 1
1 ная груп- 1 па (воз- . раст в ! годах) (ГДо 10 лет березово- .елово-щи- елово-березо- еловая с для типа
елово-щи- товнико- товниково— волосисто- во-щитовнико- во-лютиковая мелким подрос- леса в 1 целом 1
вая 4,3 осоковая 12,8 3,0 том 61,5 14,9 J
J П-20 40,0 50,7 28,8 38,5 43,0 ।
1 21-30 45,7 21,6 53,0 — 30,0 1
I 31-40 5,7 13,5 13,6 — 10,2 ।
' 41-50 2,9 1,4 1,6 - 1,5 !
} 51-60 1,4 - - - 0,4 j
1 Общее чис- 1 ло иссле- 71 148 66 37 402 !
1 дованных । особей
пологом особей соответствуищей возрастной группы. Например, в ело-,
во-щитовниково-волосистоосоковой микрогруппировке эта биомасса
(абсолютно сухая) равнялась под пологом 10-^летних особей
80,2+7,1 г/ьг, II-20-летних - 55,0+5,5, 21-30-летних - 35,7+3;4,
31-40-летних - 30,4+4,5, 41-50-летних - 19,1+2,8; контроль -„
96,3+5,5 г/м^. Щитовник мужской подавляет семенное возобновление
трав и древесных пород. Его отрицательное воздействие на пророст-
ки древесных растений склонны связывать с выделением бластоколи-
нов, в частности кумаринов (Быков, I960).
В Талыше отмечено произрастание щитовника мужского в качестве
эпифита на стволах Carpinus caucasicus и Pterocarya pterocarpa?
(Аскеров, Бобров, 1972). ?
Консортивные связи
По наблюдениям И.А.Селиванова (1976), у некоторых особей щитов-
ника мужского имеется микориза везикулярно-арбускулярного типа. .
23
По—видимому» микориза развивается лишь на почвах, бедных некото-
рыми элементами. Ее наличие положительно сказывается на росте
спорофита при недостатке в почве фосфора (cooper, 1977).
Некоторые ногохвостки поедают споры щитовника (Schneller,
1975). Можно предполагать, что и почвенные клещи используют в пи-
щу споры щитовника, так как они успешно питаются пыльцевыми зер-
нами цветковых и спорами грибов (Chmielewski, 1978; Петрова и
др., 1979). На щитовнике мужском живут некоторые тли, например
Amphorophora ampullata (Определитель насекомых..., 1964; Schnel-
ler, 1975), а в черешках вай - дичинки Biasticotoma filiceti
(Ермоленко, 1972). Не исключено, что щитовник мужской, как и дру-
гие изученные виды рода, содержит экдизоны, наличием которых можно
объяснить слабую поедаемость папоротников насекомыми.
На щитовнике мужском может паразитировать Cuscuta europaea L.
(Gams, 1938 ). Отмечены случаи поражения спорофитов грибами, как
сумчатыми - Taphrina vestergrenii (Giesenh.)•Sacc. et Sydow,
так И ржавчинными - Milesia kriegeriana (Magn.) Pauli (Sorauer,
1928). Во влажной атмосфере наблюдается поражение вай грибом Сог-
ticium anceps (из базидйбМицетов), что приводит к значительному
сокращению фотосинтезирующей поверхности растения (Gregor, 1938).
Болезни листьев вызывают также сумчатые грибы Mycosphaerelia ро-
lypodii (Rah.) Jaap И M.filicum (Deem) Starb., базидиомицет,
Herpobasidium filicinum (Rostr.) Lind., его мицелий найден И В
корневище щитовника (Sorauer, 1932; Gregpr, 1938). Вирусные за-
болевания приводят к уменьшению размеров вай, их побледнению и
преждевременному увяданию (Blattny, Pilous, 1953).
Биологическая продуктивность
Наземная абсолютно сухая биомасса в крупнопапоротниковой микро-
группировке в "окне" липо-ельника зеленомощно-волосистоосокового
Малинского лесничества Московской обл. составляет 5,0 г/м2; в
крупнопапоротниковой микрогруппировке в "окне" липо-ельника зелен-
чуко-волосистоосокового - 45,2 г/м2, в дубово-снытьевой микрогруп-
пировке - 0,3 г/м2 (Бязров и др., 1971).
По нашим подсчетам, масса вай в абсолютно сухом состоянии у мо-
дельной 30-летней оссби щитовника мужского в ельнике волосистоосо-
ковом в Московской обл. 33,9 г. На I м3 почвы под той же особью
приходится 444 г корней (абсолютно сухая масса). Масса корневищ
24
папоротника очень варьирует в зависимости от плотности популяции
вида. В елово-щитовниково-волосистоосоковой микрогруппировке того
же ельника волосистоосокового при плотности популяции 9 особей на
10 м2 масса корневищ в абсолютно сухом состоянии 210 кг/га.
Хозяйственное значение
Щитовник мужской - одно из древнейших лекарственных растений.
Сейчас в медицине и ветеринарии употребляется экстракт из корне-
вища и препарат из него - филиксан - в качестве сильного средст-
ва против ленточных глистов. Действующим веществом являются про-
изводные флороглюцина (Атлас ареалов..., 1976; Губанов и др.,
1976). Состав производных флороглюцина в настоящее время успешно
используется и в целях систематики и диагностики ( Widen, 1969;
и др.).
Корневище, содержащее до 8$ дубильных веществ, использовали
для дубления кожи и окраски ее в желтый цвет (Полезные растения
СССР, 1951). К рассеянным в разных источниках данным химических
анализов щитовника мужского следует подходить с осторожностью,
учитывая сезонную и географическую изменчивость биохимического
состава и возможность анализа других видов комплекса, произраста-
ющих в тех же районах (например, на Кавказе).
Щитовник мужской давно используется в качестве объекта науч-
ных исследований по морфогенезу растений. Ценится он и как деко-
ративное растение (Киселев, 1964; Стеценко, 1979). Из-за декора-
тивных качеств вай вид уже истреблен в некоторых местах Молдавии
(Гейдеман, Киртока, 1975). Растение ядовито для скота (Гроздов,
1963).
25
И.И.Науялис, В.Р.Филин
КОЧЕДЫЖНИК ЖЕНСКИЙ
Номенклатура, систематическое положение,
внутривидовая систематика
Athyrium filix-femina (L.) Roth, 1800, Tent. Fl. Germ. 3:65;
Сырейщ., 1906, Илл. фл. 1:38; он же, 1927, Опр. раст. Моск, губ.:
38; Фомин, 1934, Фл, СССР 1:53; Назаров, 1940, в Маевск., Фл.
изд. 7:44; Пояркова, 1954, там же, изд. 8:56; Муравьева, 1964,
также, изд. 9:59; Jenny, 1964, Fl. Eur. 1:18; Ворошилов, 1966,
Опр. раст. Моск. обл.:48; А.Бобров, 1974, Фл. евр. части СССР I:
75. - Polypodium filix-femina L. 1753, Sp. pl» 2:1090. - Aspleni-
um filix-femina Bernh. , 1806, Schrad. Joura. 1,2:2б; Кауфм.,
1866, Моск. фл., изд. 1:624, 1899, изд. 2:655; Цингер, 1886, Сб.
свед.:506; Шмальг., 1897, Фл. 2:688; Петунн., 1901, Крит, обзор
3:147. - Кочедыжник женский.
Границы и объем рода еще недостаточно выяснены (Kato, 1977),
к нему причисляют около 180 видов (Willis, 1973). Род относили
раньше к Bolypodiaceae или К Aspleniaceae, в настоящее время
большинство птеридологов включают его в Athyriaceae. Некоторые
исследователи предлагают выделять подсемейство Athyrioideae в
пределах Dryopteridaceae (Науаг, 1970; Яауаг, Kaur, 1971). Коче-
дыжник женский и некоторые близкие виды объединяют в комплекс или
группу Athyrium filix-femina (Bir, 1972; Kato, 1977).
Латинское название рода неясной этимологии (Каден, Терентьева,
1975), русское - от слова кочедыг (шило для плетения лаптей), ве-
роятно, тюркское по происхождению (Меркулова, 1967). Видовой эпи-
тет происходит от латинского filix - папоротник и femina - жен-
щина, т.е. "женский папоротник", меньший по размеру, чем мужской
(Каден, Терентьева, 1975).
В пределах A. filix-femina, обладающего обширным циркумполяр-
ным ареалом и характеризующегося большой изменчивостью, уже дав-
но выделяют много форм и разновидностей ( Luerssen, 1889; Gold-
schmidt, 1906), отмечаемых и в СССР. Лишь для азиатской части
СССР А.В.Фомин (1930) указывал II разновидностей, таксономичес-
кий ранг некоторых из них был впоследствии повышен (на наш
26
взгляд, без достаточного основания).
Гибридов в пределах рода Athyrium мало (В1т, 1972). Извес-
тен гибрид A. filix-femina х A.distentifolium (Флора европейской
части СССР, т.1, 1974). Предположение, что одна из разновиднос-
тей кочедыжника женского представляет собой результат скрещива-
ния вида с Dryopteris dilatata (Сипдивинский, 1974), вряд ли
можно принимать всерьез.
Географическое распространение
Голарктический неморально-бореальный вид (Hulten, 1962; Meu-
sel е.а., 1965). Приведенную в работе В.П.Гричука и М.Х.Монос-
зон (1971) карту ареала нужно уточнить, так как вид найден в
Гренландии (Feilberg, 1975). Распространение вида связано с лес-
ной зоной умеренных областей Европы, Сибири, Дальнего Востока,
Северной Америки и с горными лесами Средней и Юкной Европы, Севе-
ро-Западной Африки, Малой Азии, Кавказа, Северного Ирана, Север-
ного Тянь-Шаня, Гималаев, севера Китая и Японии, Северной Амери-
ки. Вид заходит также в пределы Арктики, а в горах - выше верх-
ней границы лесного пояса. В Московской обл. встречается во всех
ботанико-географических районах.
Морфологическое описание
Многолетний корневищный гемикриптофит-хамефит. Корневище у мо-
лодого растения полегающее, почти горизонтальное, у зрелого почти
вертикальное, вильчато-ветвящееся, толщиной до 8 см. Радиально-
симметричная диктиостелическая ось корневища до 1,5 см в диаметре,-
многоугольная на поперечном сечении, плотно покрыта расположенны-
ми по спирали веретеновидными черными основаниями черешков, а
близ верхушки также и бурыми чешуями до 3 см в длину. Корни перво-
го порядка отходят, как правило, по одному близ суженного основа-
ния черешка по медиане его абаксиальной стороны. Они черные, во-
локнистые, диархные, до 2,5 мм в диаметре и 10-40 (60) см в длину,
несут довольно многочисленные боковые корни, которые, в свою оче-
редь, могут ветвиться один-два раза. Ветвление корневищ приводит
к образованию мощных кочек, насчитывапцих до 40 вершин. В Москов-
ской обл. кочки достигают 15-20 см в высоту и 30-50 см в диаметре,
27
имеют 8-15 вершин и муфту 2-4 см толщиной из густо переплетенных
корней.
Ваи 20-150 см в длину и 6-50 см в ширину, собраны по 3-13 на
верхушке корневища в виде воронки, летнезеленые. Черешок в 2-3
(4) раза короче пластинки. Основание черешка расширенное, сплю-
щенное в трансверсальной плоскости и дуговидно загнутое, черное,
с двумя белыми полосами-пневматофорами на боковых ребрах, на по-
перечном срезе веретеновидное с более сильновыпуклой абаксиаль-
ной стороной. Остальная часть черешка, толщиной до 4 мм, на попе-
речном срезе округло-прямоугольная с закругленными абаксиальными
углами, с ровной в нижней части и желобчатой выше адаксиальной
стороной. Окраска этой части черешка варьирует от бледно-желтова-
то-зеленой до фиолетово-красной (красный цвет при сушке теряет-
ся). В некоторых популяциях преобладают особи с красноватой ок-
раской черешка (дрейф генов), которая, как показали опыты
( Schneller, 1979), доминирует над зеленой. Черешок'покрыт чещуя-
ми до 5 мм длиной, очертания чещуй варьируют от линейно-ланцетных
до овальных, а цвет (в пределах популяции) - от светло-коричнево-
го до черноватого (у особи цвет чещуй более или менее постоянен).
Плотность расположения чешуй и их размеры возрастают к прокси-
мальному концу черешка, дистальная же часть его может быть почти
совсем без чешуй. В черешок входит один листовой след, который,
отделяя корневой след, вскоре разделяется на два пучка, соединяю-
щихся в один в дистальной части черешка.
Пластинка ваи обычно светло-зеленая, нежная, реже (у растений
освещенных местообитаний) более темная и жесткая, в основании су-
женная и усеченная, верхушка ее заостренная, очертание пластинки
варьирует от продолговато-яйцевидной до ланцетовидной, с наиболь-
шей шириной чуть выше середины. Большая часть пластинки, как пра-
вило, дважды перистая. Главный рахис и стержни следующих порядков
с открытыми один к другому адаксиальными желобками, с рассеянными
короткими и узкими ланцетно-линейными или линейными чещуями и с
нежелезистыми одноклеточными булавовидными волосками с гранистой
головкой. Волоски обычно располагаются на адаксиальной стороне ра-
хисов в дистальной части ваи, реже они покрывают обе поверхности
и заходят на черешок. Обычно их немного (следует учесть, что при
подсыхании часто опадают), но иногда они образуют на пластинке
как бы мучнистый налет. Главный рахис желтовато-зеленый или крас-
новатый, реже черноватый. Изменчивость вай по величине, цвету и
форме чещуй, плотности опушения в пределах одного небольшого гео-
28
графического района не меньше различий между растениями из дале-
ко отстоящих один от другого пунктов, что указывает на генетичес-
кую гетерогенность популяций (Schneller, 1979).
Перья до 4 см в ширину, в очертании продолговато-ланцетные до
линейных, заостренные, очередные (базальные почти супротивные),
расставленные, сидячие или с очень короткими (1-3 мм) черевичка-
ми, отходят по 15-40 с каждой стороны главного рахиса почти под
прямым углом к нему,при этом пластинка пера располагается в од-
ной плоскости с рахисом или заметно наискось к нему так, что ба-
зископический край пера выдвинут адаксиально. Перышки перьев,
расположенных в средней части ваи, почти сомкнутые, очередные
(базальные почти супротивные). Акроскопическое базальное перышко
крупнее базископического, а акроскопическая часть пластинки каж-
дого перышка крупнее базископической его части. Перышки в очерта-
нии от продолговатых до ланцетных, острые или заостренные, с кли-
новидным основанием, перисто-лопастные или перисто-раздельные,
реже в основании почти рассеченные. Лопасти и доли (или сегменты)
их от продолговато-яйцевидных до продолговато-ланцетных, с 2-3
прямыми или изогнутыми зубцами. Жилкование перышек перистое, от-
крытое, боковые жилки простые или одинажды-дважды вильчатые.
Сорусы смешанного типа, до I мм в ширину, подковообразные,
крючковидные, косополосовидные, реже округлые или почти почковид-
ные. Зрелые сорусы желтоватые. Плаценты асплениоидного типа, си-
дят вдоль акроскопической стороны боковых жилок перышка, иногда
пересекая их своей изогнутой дистальной частью. Как правило, на
акроскопической части лопасти или доли перышка образуется одна
плацента, так что каждое перышко несет до 30 сорусов, располагаю-
щихся двумя продольными рядами. Покрывальце прикреплено одним
краем вдоль жилки и открыто на стороне соруса, обращенной к сред-
ней жилке перышка. Свободный край покрывальца выемчато-зубчатый
или бахромчатый. Спорангии полиподиоидного типа.
Споры светло-коричневые, билатерально-симметричные, однолуче-
вые, формой напоминают семя фасоли. Периспорий плотно прилегает к
экзоспорию и почти не заметен, зернистый или мелкоморщинистый.
Длина спор с периспорием от 23 до 55 мкм (Erdtman е.а., 1961 ;
Гричук, Моносзон, 1971; Кутафьева, 1972; Karpowicz, 1972; Bel-
ling, Heusser, 1974; Schneller, 1979), при этом размах индивиду-
альной изменчивости очень велик - от 26,5 до 50,6 мкм (Schneller,
1979).
Хромосомное ЧИСЛО 2п =80 (х = 10?) (Fabbri, 1963; Schneller,
29
1979), У var.pecWn.ata Cl. 2n = 82 (Roy, Pandey, 19бЗ), У расте-
ний иг Эльзаса соматическое число оказалось равным 50 (Kempf,
1967).
Обоеполые заростки симметричные, сердцевидные, до 3,5 мм в ;ши-
рину, с хорошо развитой подушкой и широкими крыльями, без волос-
ков (если не считать ризоиды).
Онтогенез
Споры не содержат хлоропластов (Miller, 1968), в качестве за-
пасного питательного вещества включают капли лира (Courbet, Met-
che, 1971). В темноте споры обычно не прорастают, в запечатанной
пробирке при комнатной температуре они сохраняют-жизнеспособность
в течение трех лет (Schneller, 1979). При комнатной температуре
и дневном свете они прорастают на 4-33-й день после посева (Каг-
powicz. 1927; Schneller, 1979). Прорастают споры по типу Vitta-
ria, а заростки развиваются по типу Aspidium, реже по типу Adi-
antum (Nayar, Kaur, 1971).
‘Онтогенез заростков находится под контролем системы, включаю-
щей антеридиоген. В многоспоровой культуре отмечено два типа онто-
генеза заростков: I) стерильные заростки становятся женскими, а
затем обоеполыми; 2) стерильные заростки становятся мужскими, а
затем последовательно обоеполыми, женскими и вновь обоеполыми.
Заростки разного пола различаются размерами, морфологией, ско-
ростью роста И развития (Bittmannova-Kogova, I960, 19б2; Schnel-
ler, 1979). У A.fiiix-femina преобладает межзаростковое скрещи-
вание, но у некоторых разновидностей отмечена апогамия, сопряжен-
ная в цикле развития с апоспорией (Farmer, Digby, 1907).
Проростки и ювенильные растения кочедыжника женского имеют
листья, сходные по очертанию с листьями соответствующих стадий
щитовника (рис.З), но отличаются от них отсутствием железистых
волосков. Наблюдения в природе показывают, что в первый год у юве-
нильного растения в Московской обл. развивается 2-3 (4) ваи дли-
ной не более 5 см, корни длиной 2-3 см, обычно не ветвятся. На
2-й год число вай возрастает до 7-8, реже до 12, длина их не пре-
вышает 10 см, появляющиеся корни начинают ветвиться.
- На 2-3-й год растение вступает в имматурную фазу, в начале ко-
торой у него ия год образуется обычно небольшое (2-5) число
вай длиной до 15 см, но зато активно растет корневище (за три го-
30
Рис.З. Проростки и ювенильные растения кочедыжника
женского.
да длина его достигает I см). Ваи обычно стерильные, но при ис-
кусственном улучшении условий даже двухлетнее растение может об-
разовать ваи с сорусами, правда, сорусы содержат абортивные спо-
ры. 10-20-летние особи образуют ежегодно 4-5 (12) вай длиной в
среднем около 50 см (максимально до 82 см) и шириной около 20 см.
31
Корневище у особей этой возрастной группы может начать ветвиться.
Около 30$ особей этой группы образуют спороносные ваи (спороноше-
ние начинается у некоторых особей с II лет). По-видимому, особи
этой группы можно считать виргинильными.
С 20-30 лет особи становятся взрослыми. Показатель филлотакси-
са достигает' 5/13, ваи достигают максимальных для данного место-
обитания размеров, а корневище начинает активно ветвиться. С уве-
личением возраста в основном происходит увеличение мощности кор-
невища и числа вай у особи в силу увеличения числа вершин корне-
вища. В кочке примерно 70-летних особей начинается деградация от-
дельных вершин, очевидно, растение вступает в это время в сениль-
ную фазу.
Сезонное развитие
Поздней осенью в почке содержится, по-видимощу, три набора за-
чатков вай, самые внешние из которых начинают активно разверты-
ваться следующей весной (в Подмосковье в начале мая). Скорость
развертывания зависит от погодных условий. Так, в одной из микро-
группировок кочедыжника в Московской обл. большинство вай развер-
нулось в 1975 г. к 18-20 мая, в 1976 - к 14-20 июня, в 1978 - к
8-10 июня, в 1979 - к 2-4 июня.
Массовое спороношение в Московской обл. отмечено в конце
июля - начале августа. При наступлении первых заморозков ваи по-
гибают и полегают, образуя, вокруг кочек слой ветоши, хорошо защи-
щающий кочку от зимних морозов. Проведенное нами удаление ветоши
способствовало повреждению почек морозами (следующей весной обра-
зовались уродливые ваи) и даже полной гибели некоторых растений.
Способы размножения и распространения
При размножении спорами кочедыжник женский ведет себя как ти-
пичный эксплерент. По нашим подсчетам, одна 20-30-детняя особь в
ельнике крупнопапоротниковом в Подмосковье может продуцировать
около I млрд, спор (14-15 тыс. сорусов на ваю или 260-280 тыс. на
особь). По подсчетам И.Шнеллера (Schneller, 1979), одна особь с
пятью фертильными ваями продуцирует около 75 млн. спор. Основная
масса спор рассеивается примерно за 20 дней, но даже до апреля
32
следующего года на вае может сохраниться несколько тысяч спор,
жизнеспособность которых в условиях США (штат Айова) превышает
50% (Farrar, Gooch, 1975; Farrar, 1976). В Западной Европе при
посеве на среду прорастает 50-95% высыпаемых осенью спор; споры,
собранные с вай весной, также оказались живыми (Schneller, 1979).
В природе из высыпавшихся осенью спор, видимо, лишь единичные
прорастают осенью, большая часть пропадает в лесной подстилке, но
какая-то часть попадает в благоприятные условия и прорастает сле-
дующей весной, рпоры, попавшие в почву, сохраняют жизнеспособ-
ность весьма длительный срок, во всяком случае до момента высыпа-
ния новых спор (Schneller, 1979), но большая часть спор в почве,
очевидно, погибает. Тем не менее, учитывая высокую популяционную
плотность кочедыжника женского, в местах его произрастания на по-
верхность, подходящих для развития заростков субстратов, выпадает
большое число спор.
Споры, проросшие первыми, как правило, дают начало женским за-
росткам, которые, выделяя физиологически активные вещества (анте-
ридиоген), способствуют возникновению в популяции мужских зарост-
ков. После прекращения выделения антеридиогена женскими заростка-
ми мужские заростки через некоторое время станут обоеполыми (точ-
нее, физиологически женскими) и таким образом в среде вновь поя-
вится антеридиоген, способствующий формированию мужских заростков
из недавно проросших спор. Более того, как показали опыты и на-
блюдения в природе (Schneller, 1979), выделения зрелых заростков
вызывают прорастание спор, находящихся в почве. При этом образует-
ся почти бесцветная протонема, на кончике которой, когда он про-
бьется на поверхность почвы, появляется антеридий с жизнеспособ-
ными сперматозоидами.
Хотя количество женских и мужских заростков в микропопуляции
зависит от плотности посева, запаса спор в почве, условий освещен-
ности и минерального питания и водоснабжения, в целом.большая спо-
ровая продуктивность, растянутость процессов созревания и высыпа-
ния спор, способность спор длительное время сохранять жизнеспособ-
ность и, наконец, наличие системы, включающей антеридиоген, ведут
к тому, что в микропопуляциях заростков, появляющихся на новом
месте, образуется большое число мужских заростков, а потому увели-
чиваются шансы на межзаростковое скрещивание и падают шансы на
внутризаростковое оплодотворение. В старых же установившихся мик-
ропоцуляциях количество мужских и женских заростков будет пример-
но одинаковым, но следует учесть, что при гомозиготности - при
3-1964 33
внутризаростковом и межзаростковом оплодотворен^ (siekowskl,
1969) - генетический груз у кочедыжника женского очень велик и
проявляется он не только на стадии зиготы и ранних этапах онтоге-
неза, но и значительно позже (Schneller, 1979). Преобладание ге-
терозиготных особей в популяциях объясняет большую гетерогенность
этих популяций.
Поселившись на новом месте, особи кочедыжника женского могут
несколько десятков лет успешно удерживать территорию, чему способ-
ствуют вильчатое ветвление корневища и образование мощных кочек.
Наступающую в сенильном периоде партикуляцию этих кочек вряд ли
следует считать приспособлением к вегетативному размножению осо-
бей. В естественных условиях у кочедыжника не отмечено образова-
ния почек вегетативного размножения, но у вай, поставленных в во-
ду и помещенных во влажную атмосферу, такие почки могут появить-
ся (Hofmeister, 1862).
Экология
Экология гаметофитов кочедыжника женского сходна в основных
чертах с экологией гаметофитов мужского (Schneller, 1975, 1979;
Науялис, 1979), но заростки и проростки кочедыжника успешнее раз-
виваются и чаще встречаются не на гниющих древесных остатках, а
на обнаженной почве (вывала, кротовины, лесные тропинки, дороги).
Содержание в атмосфере хлора в концентрации 1СГ^ подавляет рост
заростков и при длительном воздействии ведет к их отмиранию (Bed-
narova-Kogova, 1968),
Кочедыжник женский - гигромезофит, он является индикатором от-
носительно достаточного обеспечения водой (Eilenberg, 1974) и
хорошо чувствует себя при сочетании большого количества осадков с
относительно малой испаряемостью (Долуханов, I960). В листопадных
лесах растение увядает при 3 атм водного потенциала, в то время
как большинство других лесных трав при 5 атм (Eilenberg, 1939).
Высокие требования к влагообеспечению объясняют приуроченность и
пышное развитие кочедыжника в понижениях рельефа, по берегам пото-
ков, окраинам водоемов и болот и т.п., в лесной зоне.
Освещенность до 5% от освещенности открытого места вполне удов-
летворяет взрослое растение кочедыжника (Eilenberg, 1974), новее
же спорофиты этого папоротника более светолюбивы, чем спорофиты
щитовника мужского. Кочедыжник раньше выпадает из сообществ с
34
сомкнутым древостоем и положительно реагирует на увеличение осве-
щенности в лесных сообществах, вытесняя щитовник мужской в "ок-
нах" древостоя. О светолюбии кочедыжника отчасти свидетельствуют
и результаты подсчета числа вай на I $ поверхности почвы в раз-
ных микрогруппировках в Московской обл.: в елово-лещиново-коче-
днжниковой микрогруппировке их 51,6+7,3, в'елово-березово-коче-
дыжниковой - 57,1+6,2, в кочедыжниковой в-"окне" - 80,7+8,3. В
"окнах" резко возрастает и число особей с сильно разветвленными
корневищами, например, в сосново-лещиново-кочедыжниковой микро-
группировке в Подмосковье из 116 особей 45 имели корневище с од-
ной вершиной, 27 - с двумя и 12 - с четырьмя, а в кочедыжниковой
в "окне" сосняка иэ 46 особей 30 имели корневище с тремя вершина-
ми, по 13 особей - с двумя, четырьмя и пятью вершинами.
На вырубках кочедыжник выпадает, не выдерживая конкуренции с
травами. Хотя при благоприятных для него фитоценотических услови-
ях кочедыжник может расти и на открытом месте, все же он лучше
чувствует себя в слегка затененных влажных местах. Точка компен-
сации у лесных особей лежит между 300 и 500 лк (Schulae, 1972).
Наиболее крупные кочки кочедыжника были встречены нами в оврагах,
выше верхней границы леса, в Карпатах - из 29 особей 15 имели
корневища с пятью вершинами, 2 - с пятью - десятью, 12 - с один-
надцатью и больше вершинами (2 из них имели больше 30 вершин). В
лесах рост кочек, вероятно, сдерживается в первую очередь конку-
ренцией корней древесных растений.
Температурный режим Центральной Европы, видимо, благоприятен
для вида, так как в этой области он индифферентно относится к
температурному режиму (Eilenberg, 1974). Летние и осенние замо-
розки до -3° ведут к обмерзанию вай. Корневища, собранные в сен-
тябре, выдерживают промораживание в камерах до -8°, а собранные с
декабря по март - до -16° (карреп, 1964). Согласно Л.Каппену,
этот вид лучше других папоротников Европы переносит низкие темпе-
ратуры, но мы думаем, что этому в первую очередь способствует ук-
рытие кочки полегающими осенью ваями. В суровых температурных ус-
ловиях Приполярного Урала у кочедыжника не развиваются споры (Иго-
шина, 1966).
По одним данным кочедыжник женский индифферентно относится к
реакции почвы (Eilenberg, 1974), но Б.А.Быков (I960) указывает,
что вид обитает на влажных лесных торфяно-перегнойных почвах с
рН=4-6, часто с признаками оглеения; в то же время вид встречен и
на выходах известняков (Brooks, Roth, 1977). Замечания, сделан-
35
иые нами относительно связи щитовника мужского с почвами разной
кислотности (см. выше), справедливы и по отношению к кочедыжнику.
А.А.Ниценко (1969) относит вид к нитрофильной теневой свите. В Ле-
нинградской обл. самые мощные заросли вида встречены на слабо дре-
нированных песчаных, супесчаных или суглинистых почвах с близким
уровнем грунтовых вод и со слоем торфа мощностью до 50 см (Полян-
ская и др., 1935). На Кавказе вид приурочен к темно-бурым выщело-
ченным почвам (Осорин, 1951). Вместе со страусником обыкновенным
он образует заросли вдоль ручьев на иловатых, иногда торфяно-бо-
лотных почвах (Порфирьев, 1975). В Московской обл. растения луч-
ше всего чувствуют себя на слабодерновых-среднеподзолистых и
среднедерновых-слабоподзолистых почвах в кочедыжниковых ельниках,
которые занимают места, переходные от слегка пониженных равнин к
низинным болотам или небольшие повышения среди черноолыпатниковых
трясин (Юркевич и др., 1971).
Фитоценология
Лишь в экстремальных условиях - в субальпийском поясе Урала
(Игошина, 1952, 1966) или на молодых субстратах и в "окнах" дре-
востоя кочедыжник женский может образовывать самостоятельные
группировки, обычно же вид является субдоминантой третьего яруса
в широколиственных, смешанных и мелколиственных лесах (Быков,
I960). В Московской обл. наилучшего развития кочедыжник достига-
ет в таком демутационно-сукцессионном типе леса, как ельник круп-
нопапоротниковый. В коренных ассоциациях еловых и широколиствен-
ных лесов и в производных осинниках и березняках он устойчивый,
но не доминантный вид. На 10 м2 в ельнике крупнопапоротниковом
мы насчитали 64+7 особей, в сосняке крупнопапоротниковом - 46+5,
в ельнике-кисличнике - 3, в ельнике волосистоосоковом - 2, в ель-
нике-зеленомошнике одна особь приходится более чем на 10 м2.
Популяции кочедыжника женского состоят из большого числа осо-
бей. Так, в Западной Европе на 10 м2 приходится 14-57 особей,
среднее расстояние между которыми 1,4-3 м, изолированные растения
встречаются очень редко (Schneller, 1979).
При поселении вида на новом месте максимальная гибель особей,
по нашим наблюдениям в Подмосковье, происходит на рубеже 1-2 го-
да, а затем у стерильных особей корневище отмирает чаще всего в
возрасте 11-20 лет, а у фертильных - в возрасте 21-30 лет.
36
Кочедыжник активно влияет на состав и структуру сообщества,
особенно его нижних подъярусов. Вид может быть отнесен к виолен-
там с эксплерентными свойствами в молодом состоянии. Виолентность
основана на воронковидном расположении крупных вай и мощно разви-
той корневой системе, перехватывавших свет, воду и питательные
вещества (рис.4). Кочедыжник перехватывает до 80$ осадков и около
Рис.4. Корневые системы кочедыжника женского на
площадке 0,25 м^ в слое почвы 0-1 см в
ельнике крупнопапоротниковом в Московской
обл.
90% света от поступающих на верхнюю границу зарослей папоротника.
В итоге освещенность под зарослями кочедыжника не превышает 1% ст
освещенности открытого места (Науялис, 1976). Заросли кочедыжника
3Х-1964 37
снижают и температуру воздуха приземного слоя на 2-3° (Карреп,
1964).
Формирование кочедыжником кочек также увеличивает его виолент-
ные свойства, так как занимакщие обширную площадь кочки механичес-
ки исключают возможность поселения многих видов. В елово-лещиново-
кочедыжниковой микрогруппировке ельника крупнопапоротникового в
Подмосковье площадь Основания кочки варьирует от 900 до 3500 см2.
Основная масса (до 90%) корней у зрелого растения сосредоточена
в подстилке и приповерхностном горизонте (0-5 см) почвы (Науялис,
1976), где они заполняют до 5% объема почвы. Неудивительно, что
под кустом папоротника образуется "мертвая" (без травянистых рас-
тений) зона, радиус которой достигает иногда 70-90 см. Граница та-
кой зоны четко совпадает с границей распространения (по горизонта-
ли) корневой системы папоротника. При поднятии и связывании вай
кочедыжника в плотный пучок количество света и осадков, достигаю-
щих поверхности почвы, увеличивается в несколько раз, в результа-
те чего, как показали наши подсчеты, за июнь - август число стеб-
лей кислицы под "кронами" папоротника возросло почти на 50% по
сравнению с началом опыта' (в Контроле число стеблей кислицы даже
уменьшилось на 1%). ''
Группировки кочедыжника женского бедны флористически (Науялис,
1977). Семенное возобновление трав древесных пород в зарослях это-
го папоротника подавлено (Кац, 1936; Ссорин, 1951; Соколов, 1952;
Орлов, 1953; Дылис, 1968; Карпов, 1969; Науялис, 1977). В Москов-
ской Обл. С ЭТИМ ВИДОМ сопряжены Ajuga reptans, Crepis paludosa,
Dryopteris carthusiana, Pilipendula ulmaria, Geum rivals, Ranun-
culus repens, R.cassubicus и др. виды. Кочки кочедыжника часто
заселяют кислица И мхи: Atrichum undulatum, Climacium dendroides,
Hylocomium splendens, Cirriphyllum piliferum, виды Eurhynchium,
Braohythecium, Mnium И некоторые другие.
Консортивные связи
Споры кочедыжника женского могут служить пищей для Collembola,
Myriopoda, Dermaptera и Triciioniscidae (Schneller, 1979). У спо-
рофита обнаружена везикулярно-арбускулярная микориза (Селиванов,
1976). Галлы на ваях и уродства вай вызывают Synchytrium phegopte-
richis И S.athyrii (Gregor, 1938), а Также муха Chortophila
signata (Lawalree-Colloris, 1961). Ваи поражают аскомицет Val-
38
densia heterodoxa, несовершенные грибы Cercosporella filicia-fe-
minae и Leptoetroma sp. (Gregor, 1938). Верхушки МОЛОДЫХ вай
охотно обкусывают лоси. Это приводит к истощению и в конце концов
к гибели особей папоротника, о чем_свидетельствуют результаты на-
ших постоянных наблюдений над 30 особями кочедыжника в одном из
"окон" древостоя в Малинском лесничестве Московской обл.:
1 1 Год на- । бладе- । ния 1 Число особей с отмершими верхуш- ками корневищ Число вай на всех особях до скусы- вания лосями (в скобках процент уродливых вай) Число вай, появив-1 шихся после по- . вреждения особей лосями (в скобках процент уродливых 1 вай) ।
' 1975 0 672 (1,0) 104 (24,0) 1
1 1976 0 649 (6,5) 83 (31,4) ।
1 1977 I 549 (7,6) 82 (38,8) 1
1 1978 3 496 (9,4) 46 (48,4) 1
' 1979 1 9 401 (9,7) 34 (62,1) 1
Кочки кочедыжника могут служить местом устройства гнезд птица-
ми (нами обнаружено гнездо славки-черноголовки).
Биологическая продуктивность
Фитомасса вай, корневищ и корней, откопанных до глубины 40-
50 см в Подмосковье в кочедыжниковой микрогруппировке в "окне" со-
ставляет соответственно 166, 4021 и 1547 г/лг (в абсолютно сухом
состоянии), в дубово-липово-кочедыжниковой - 26, 721 и 319 г/л?,
в сосново-лещиново-кочедыжниковой - 16, 687 и 306 г/м^. Масса од-
ной ваи длиной 120 см в сыром состоянии может достигать 35 г. У
модельного куста с 18 ваями и кроной высотой до 90 см общая длина
корней достигала 31552 м, а их масса 1073 г (в абсолютно сухом со-
стоянии) .
Хозяйственное значение
Споры кочедыжника женского употребляются в экспериментальной
биологии (Dyer, 1979). Корневища содержат активные глистогонные
соединения (Патарая, 1968), их используют для фальсификации корне-
39
вкщ мужского папоротника (Мау, 1978). В народной медицине ими
лечили ревматизм (Роллов, 1908). Корневища пригодна для дубления
кои (Патарая, 1968), в них содержится да 12% таннидов (Lietuvos
tsr flora, 1959). В Приморье молодке разворачивающиеся ваи
("улитки") папоротника используют в пищу (Лучник, 1938). Извест-
но много разновидностей этого вида, обладающих высокой декоратив-
ностью и устойчивостью в условиях города (Роллов, 1908; Foerster,
1957; Стеценко, 1979). Подземные части кочеднжника - хороший суб-
страт для выращивания орхидей. Есть указания, что ваи поедает
крупный рогатый скот (Патарая, 1968).
40
А.Р.Матвеев
РОД ТИМОФЕЕВКА
Род Тимофеевка рыеит ь. (сем. Роасеае - злаки) содержит
около 20 видов, распространенных во внетропических странах обоих
полушарий. Латинское название рода происходит от древнегреческого
phieoa - злак (Флора СССР, т.2, 1934). В СССР встречается 9 ви-
дов. В Московской обл. растут 2 вида.
Ключ для определения московских видов
I. Веточки колосовидных метелок, сросшиеся с осью метелки. Ме-
телки при сгибании сохраняют цилиндрическую или эллипсоидальную
форму. P.pratense.
2. Веточки колосовидных метелок, не сросшиеся с осью. Метелки
при сгибании становятся лопастными. P.phleoides.
I. ТИМОФЕЕВКА ЛУГОВАЯ
Номенклатура^ и географическое распространение
Phleum pretense L. 1753, Sp. pl. 59s Кауфм., 1866, Моск. фл.,
изд. 1:543; 1889, иэд. 2:574; Дингер, 1886, Сб. свед.:491; Шмальг.,
1897, Фл. 2:600; Петувн., 1901, Крит, обзор 3:105; Сырейщ., 1906,
Илл. фл. 1:111; он же, 1927, Опр. раст. Моск. губ.:60; Овчинников,
1934, Фа. СССР 2:132; Рожевиц, 1940, в Маевок., Фл., изд. 7:104;
Цвелев» 1954, там же, изд. 8:840, изд. 9:801; Скворцов, 1966, Опр.
раст. Моск. обл.:74; Цвелев, 1974, Фл. евр. части СССР 1:246; он
же, 1976, Злаки СССР:366, Humphries, 1980, Fl. Eur. 5:240. - Ти-
мофеевка луговая.
Помимо типового подвида - P.pratense subsp. pratense выделяют
P.pratense subsp. nodosum (JJ.) Arcang. (P.nodosum L.), однако
1 Для всех охарактеризованных в данном выпуске видов (за исключе-
нием щитовника мужского, кочедыжника женского и белокрыльника
болотного) номенклатуру составил (или проверил, уточнил и допол-
нил) В.Н.Тихомиров.
41
статистически достоверную разницу между указанными подвидами йо
диагностическим признакам не удалось установить (Ходунов, 1932).
В начале ХУШ столетия Тимофей Гансон ввел это растение в культу-
ру, отсюда и пошло русское название "Тимофеева трава", или "тимо-
феевка" .
Ряд авторов (Meusel, 1943; Meusel е.а., 1965) характеризуют
ареал тимофеевки луговой как бореальыо-субмеридиональный, Е.Хуль-
тен (Huiten, 1950) считает его циркумполярным. В естественных
условиях вид распространен на большей части территории Европы,
Азии и Америки. Он встречается во всех районах европейской части
СССР в лесной и лесостепной зонах, в горных районах Кавказа,
Средней Азии, Казахстана, западной и южной части Восточной Сиби-
ри и Дальнего Востока. В Московской обл. растет во всех районах.
Морфологическое описание
Тимофеевка луговая относится к гемикриптофитам, подгруппе про-
тогемикриптофитов. Это рыхлокустовый розеткообразующий поликарпи-
ческий симподиально нарастающий многолетник с монокарпическими
моноциклическими и озимыми (редко дициклическими) экстравагиналь-
ннми полурозеточными побегами (Evans, 1927; Смелов, 1947, 1966;
Langer, 1955; Серебрякова, 1956, 1971а).
Корневая система мочковатая. У проростков она состоит из коле-
оризы, т.е. метаморфизированного главного корня и придаточных
корней (семенного и стеблевых). Корневые волоски на колеоризе и
корнях сходны по строению и происхождению. Отмечается полиморфизм
стеблевых придаточных корней, выделяют маловетвящиеся толстые
(диаметр в среднем 0,82) и тонкие сильноветвящиеся (диаметр в
среднем 0,46 мм). Первые закладываются в верхнем ярусе, вторые -
в нижнем ярусе зоны кущения. Семенной придаточный корень функцио-
нирует весь вегетационный сезон и отмирает через 3-4 мес в следу-
ющий сезон вегетации. Большая часть корней живут 1-2 года, мень-
шая часть (только толстые) отмирают на 3-4-й год. Глубина проник-
новения корней до 90 см и более (Evans, 1927; Штреккер, 1931;
Koennekamp, 1934; Lincola, Turikka, 1936; Roberts, Hunt, 1936;
Смелов, 1947, 1966; Лебедев, Углов, 1961; Мельник, 1969; Матвеев,
1972а; Матвеев, Шуман, 1974).
Надземная часть стебля развивается по моноциклическрму и ози-
мому типу, реже дициклическому. Базальная зона побега состоит из
42
диагестропнсго участка, переходящего на изгибе в "дугу укорочен-
ных междоузлий" (Смирнов, 1958), и гаплокорма (Evans, 1927),
т.е. части стебля, состоящей из 1-2, реже 3 удлиненных утолщен-
ных междоузлий. Основания побегов как главной оси, так и боковых
имеют сбратноконическую форму. В суженной части основания побега
в так называемой "шейке корневища" первые (по происхождению) 1-2
укороченных междоузлия не несут придаточных корней. Каждый побег
имеет чещуевидные влагалищные листья, предлист, 5-7 ассимилирую-
щих листьев у вегетативных розеточных побегов, 7-24 листа у полу-
розеточного побега, 7-10 листьев у генеративного полурозеточного
побега.
Стебель полый, толщиной от I до 3,5 мм. Паренхимные ткани гап-
локорма заполняются в течение сезона вегетации запасными вещест-
вами в виде флеина, тритицина, гемицеллюлозы, моносахаров (Vol-
kart, Kirchner, 1913; Смелов, 1947, 1966; Осецкий, 1969; Colby
е.а., 1973). После использования запасных веществ боковыми побе-
гами, развивающимися весной следующего сезона, гаплокорм отмира-
ет (июнь - июль). Длительность жизни гаплокорма около I года (2
сезона вегетации), зона кущения не отмирает сразу после плодоно-
шения, а живет 3-4 года (Смелов, 1947, 1966; Михайловская, 1965;
Матвеев, 1972а).
По направлению и расположению листьев на стебле Н.П.Голубев
(1931) выделяет 3 типа: с побегами, имеющими прямостоячие листья,
с горизонтальными листьями, с поникшими листьями. М.С.Щенкова
(1961) различает 5 типов кустов: I) с преимущественно лежащими
стеблями, 2) с приподнимающимися в верхней части стеблями, 3) со
стеблями, слегка приподнимающимися в нижней части и направленны-
ми прямо в верхней части (вазообразная форма куста), 4) со стеб-
лями косо вверх направленными (куст слегка развалистый), 5) с
прямо вверх направленными отеблями (куст прямой сжатой формы).
Е.Н.Синская (1948) указывает на существование 2 климатических
экотипов: северный характеризуется простратной формой роста и
меньшей высотой, южный имеет прямостоячую или форму роста с более
быстрым отрастанием. В пределах экотипической популяции она выде-
ляет 3 экоэлемента: прямостоячую, простратную и промежуточную
формы.
В образовании боковых побегов из почек зоны кущения характерна
мезотония (Смелов, 1947, 1966; Серебрякова, 1967, 1971а). Отмеча-
ется способность побегов к надземному кущению (точнее ветвлению)
при повышенной влажности почвы, воздуха, внесении удобрений, в
43
уоловжях короткого дня (Evans, 1927; Сметанникова, 1950; Тарков-
ский, Артамонова, 1953; йейсле, 1956; lanfer, Byle, 1958; Коря-
кина, 1964). Побега тимофеевки преимущественно зкстравагииальнне,
однако в условиях пойми при отложении иаилка свыше 4-5 см наблю-
дали образование интравагииальннх побегов, мевдоувлия ми чаете |
удлиняются, образуя гипоподии. Проявляется 2 ^периода кущения: ве-
сенний и летне-осенний, или фаза нрегеиеративного и постгенера- I
тивного кущения (Серебрякова, 1967, 1971а). Можно выделить весен-
нее и летне-осеннее прегеиеративное кущение 1-го года жизни (при
развитии из семени), весеннее прегеиеративное кущение 2-го и 3-го
года кивни (при развитии из семени); постгеиеративное летне-осен-
нее кущение 1-го, 2-го (при развитии побега из семени) и 3-го
(при развитии побега из почки) года жизни побега; постгенератив-
ное весеннее кущение 2-го, 3-го (при развитии из семени) и 4-го
(при развитии из почки) года кивни. Куст может иметь от 2-3 до
250 побегов.
Листья тимофеевки луговой в почкосложении свернуты направо
(Volcart, Kirchner, 1913). Ассимилирующие листья взрослого рас-
тения. имеют ширину 3-10 мм л длину около 15 см, они гладкие, го-
лые, верхняя сторона серо-зеленая, матовая, нижняя потемнее, лос- I
нящаяся, но не блестящая. На верхней стороне посредине проходит j
пологая складочка, на просвет имеет вид ярко-белой широкой жилки.
Листовая пластинка с четкими неглубокими бороздками, разделенны-
ми низкими плоскими ребрами. Листья без ушков, светло-зеленые, в
основании с длинными обращенными назад щетинками, у вегетативных
- побегов проходит узкая (ухе высоты шипиков) кайма (Голубинцева, *
Лебедев, 1959). Наличие шипиков обусловливает шероховатость верх-
ней части края листа. Влагалища листа вверх шероховатые, не взду-
тые, открытые. Анастомозы на влагалищах пе видан. Влагалища ниж-
них листьев красновато-бурые, волокнистые, верхних - зеленые.
Площадь ассимилирухщей листовой поверхности полурозеточного веге- i
тативиого побега изменяется по параболической кривой и корреля- I
тивно связана с поглощающей поверхностью корней и условиями внеш- *
ней среда (Мельник, 1969), причем внесение азота увеличивает дли-
ну и ширину листьев, но не увеличивает их число (Смелов и др.,
1951). Понижение освещения и короткий день увеличивают ассимили-
рухщую поверхность листьев побегов I-го и 2-го порядков ж умень-
шают у побегов 3-го порядка. Язычок большой, 2-3 мм длины, белый,
тупой, у верхних листьев островатый.
Соцветие - колосовидная цилиндрическая метелка (султан) обнч-
44
но одинаковой толщины (до I см) по всей длине, 3-15 (25) см длины.
Веточки соцветия почти по всей длине срослись с главной осью й
при сгибании султана лопастей не видно. Размеры соцветий зависят
от условий освещения - при длинном дне развиваются короткие сул-
таны (Ржанова, 1957; Корякина, 1964). Колоски маленькие, 3-5 мм
длины, продолговато-клиновидные. Колосковые чешуи продолговатые,
пленчатые, с твердым раснитчатым килем, который продолжается в
ость. Цветковые чешуи в 2-3 раза короче колосковых, на верхушке
плоско обрезанные и слегка зазубренные, без остей, пленчатые, с
продольными жилками, нижняя (наружная) шире верхней, 1,6 мм дли-
ны, с пятью жилками, верхняя (внутренняя) с двумя жилками.
Тычинок 3, они с тонкими нитями, которые быстро удлиняются не-
задолго до раскрытия цветка (Godron, 1873). Пыльники фиолетовые,
четырехгранные, с очень узким связником. Пальцевые зерна разме-
ром 31,3-36,5 мкм, сферические или эллипсоидальные.
Перекрестноопылякщееся растение с низкой автофертильностью
(Синская, Шебалина, 1936). При уединенном произрастании завязыва-
ние семян отсутствует или крайне недостаточно.'Цветение двухразо-
вое в течение суток: утреннее (с 2 ч 30 мин до 9 ч) и вечернее
(с 15 до 20 ч). Время цветения зависит от погодных условий (God-
ron, 1873; Кернер, 1902; Jones, Novell, 1946; Пономарев, Труба-
чева, 1962; Пономарев, 1964). Внесение азотных удобрений и опти-
мальная влажность увеличивают продолжительность цветения на 7-15
дней (Мельник, 1969). Раскрывание цветков происходит в базипе-
тальном направлении (Godron, 1873; Пономарев, 1964). Отмечено
более раннее появление рылец-протогиния (Пономарев, 1964), наблю-
дается также и одновременное выдвижение тычинок и рылец (Troll,
1931), т.е. в каждой популяции имеются как гомогамные, так и пре-
тогиничные растения. Растение обильно продуцирует пыльцу. Срок
прорастания пыльцы в зависимости от температуры, влажности возду-
ха и других факторов от I до 4 и более часов (Овеснов, 1966).
Пестик способен принимать пыльцу один день. В некоторых естест-
венных и искусственных условиях (влажное лето, короткий день и
др.) наблюдается израстание колосков и пролификация (Evans, 1927;
Киршин, 1963).
Пленчатая зерновка тимофеевки луговой безостая, яйцевидной
фермы, к вершине заостренная, у основания притупленная, в попе-
речном сечении почти круглая, матовая, серебристо-серая или свет-
ло-серая, длиной 1,5-2,25 мм, диаметром 0,75-1 мм. Масса I тыс.
пленчатых зерновок около 600 мг, в I кг до 1,6 млн. зерновок (Доб-
45
рохотсп, 1961). Величина и вес семян неодинаковые в различных
частях султана: в верхней части семена обычно крупнее, чем в
нижней (Корякина, 1964).
Число хромосом X = 5,7. Указывается различный диплоидный на-
бор хромосом - 14, 21, 28, 35, 36, 40, 42, 48, 56, 63, 70, 84
(Хромосомные числа..., 1969).
Онтогенез
Семена тимофеевки относятся к группе с коротким и слабым поко-
ем и требуют для послеуборочного дозревания 1-1,5 мес (Овеснов,
1965). При хранении в лабораториях оии способны сохранять всхо-
жесть 10 и более (до 16) лет (Корякина, 1930; Соколова, 1934;
Овеснов, 1965, 1966). Наличие цветковых чещуй на семенах способ-
ствует сохранению всхожести более длительный срок (Коenod, 1965).
В естественных условиях прорастание семян обычно происходит
поздним летом или ранней осенью в год образования семян, при за-
сушливой осени проростки появляются весной. Оптимальная темпера-
тура прорастания от 20 до 30°, причем отмечено различное отноше-
ние семян к постоянной и переменной температуре, что, вероятно,
частично связано с неодинаковым возрастом семян. Только что уб-
ранные семена прорастали лучше при средней постоянной температу-
ре, через 2-3 мес до 3 лет - лучше при переменной (от 15 до 30°),
после 3 лет хранения семена к температуре безразличны (Овеснов,
1965). Яровизация с-емян ускоряет прохождение основных фаз разви-
тия (Белоножко, Чеботарева, 1951). Противоречивые данные имеются
по влиянию света на прорастание. Это можно объяснить тем, что
свежие семена при прорастании положительно реагируют на свет, а
через несколько месяцев индифферентно (Овеснов, 1965). На прорас-
тание могут влиять и аллелопатические выделения (Никольская, Со-
колов, 1966).
У тимофеевки наблюдается проторизония (Татаринова, 1953; Овес-
нов, 1965; Добрынин, 1969). При прорастании семян выделяют фазы:
I) водопоглощения, 2) колеоризы, 3) развития придаточного семен-
ного корня, 4) развития колеоптиля и 5) первого зеленого листа
(Овеснов, 1965; Строна, 1966). Колеориза корневидная, клиновид-
ная. Емкость зародышевой почки тимофеевки равна 2 листовым обра-
зованиям (Овеснов, 1965; Мельник, 1969). Размер колеоптиля сильно
варьирует и зависит от глубины посева (Evans, 1927), достигает
46
длины 0,8 см (Мельник, 1969).
Проростки представляют собой однопобеговые растения с 2-3
листьями, сохранившейся зерновкой, колеоптилем (в естественных
условиях длиной 0,5-0,7 см) и слабо развитой корневой системой,
состоящей из одного зародышевого, семенного и стеблевых придаточ-
ных корней. Глубина проникновения корней 2-4 см. Высота пророст-
ка в среднем 4,3 см. При выходе колеоптиля на поверхность емкость
верхушечной почки равняется 5 листовым образованиям (Лебедев,
1968). Состояние проростков может длиться 1-1,5 мес, реже до 4
мес.
Ювенильное состояние тимофеевки начинается с начала самостоя-
тельного питания растения и продолжается до начала кущения (имма-
турного состояния) или же до перехода в генеративное состояние.
Ювенильные растения в отличие от проростков питаются самостоя-
тельно и развивают более мощную систему придаточных корней. Лис-
товые пластинки длиннее и шире, чем у проростков. Ювенильные розе-
точные растения представляют собой однопобеговые розетки высотой
до 17 см с 4-5 листьями, глубина проникновения корневой системы
10-15 см. Это состояние может продолжаться от 1,5 мес в случае
перехода к кущению до II мес и более в случае перезимовки в виде
розетки. Ювенильные полурозеточные растения - однопобеговые рас-
тения с 2-3 вытянутыми междоузлиями и 5-6 листьями. К осени у
этих побегов образуется гаплокорм, это состояние может продол-
жаться от 2-2,5 мес (при переходе в генеративное состояние) до 4-
5 мес (при гибели соломенной части удлиненного побега от замороз-
ков).
Имматурные растения в отличие от ювенильных представляют собой
первичные простые кусты, состоящие из побегов 1-го и 2-го поряд-
ков, или сложные кусты, состоящие из побегов 1-го, 2-го, 3-го,
4-го и т.д. порядков. Имматурное состояние продолжается или до
отмирания удлиненных междоузлий вегетативных побегов 1-го (иног-
да 2-го) порядков, или до перехода в генеративное состояние. Имма-
турные розеточные растения представляют систему розеток 2-го и бо-
лее порядков и составляют соответственно простой и сложный куст.
Это возрастное состояние продолжается от 1,5 (в случае образова-
ния удлиненного побега) до II мес (в случае перезимовывания) и не
является обязательным в жизненном цикле растения. Имматурные полу-
розеточные растения имеют простой или сложный куст из одного или
нескольких полурозеточных побегов 1-го и 2-го порядков, имеющих,
в свою очередь, один или несколько розеточных побегов (2-4-го по-
47
рядков). Корневая система состоит из зародышевого корня, проника-
ющего до 20 ом, и 10-15 стеблевых придаточных корней (Лебедев,
1966), Это возрастное состояние продолжается от 2-2,5 (при пере-
ходе в генеративное состояние) до 4 мес (при гибели удлиненных
побегов от заморозков), оно также необязательно в жизненном цик-
ле тимофеевки.
Взрослые виргинильные растения - это вегетативные растения се-
менного происхождения, которые имеют один или несколько гаплокор-
мов, оставшихся после отмирания стеблевой части удлиненных побе-
гов предыдущего сезона вегетации. Они представляют собой систему
из двух-трех парциальных кустов. Образование виргинильных расте-
ний начинается с отмирания соломины удлиненных побегов имматур-
ных растений и продолжается до перехода особи в генеративное со-
стояние. Это возрастное состояние не является обязательным в жиз-
ненном цикле растения. В виргинильнсм состоянии особи могут нахо-
диться 1-2 вегетационных сезона.
К группе молодых генеративных растений отнесены генеративные
особи семенного происхоадения, которые еще не распались на вто-
ричные системы парциальных кустов. Это состояние начинается с мо-
мента перехода в генеративное состояние и продолжается до образо-
вания клона. Группу молодых генеративных растений можно подразде-
лить на три подгруппы: а) однопобеговые растения, б) однокусто-
вые генеративные, представляющие собой первичный простой или слож-
ный куст, в) многокустовые генеративные растения, являющиеся си-
стемой из 2-3 и более простых или сложных парциальных кустов. Ге-
неративные побеги таких растений не достигают максимальной высо-
ты. Соцветие небольших размеров, у однопобеговых растений в есте-
ственных условиях в красноовсяничнсй формации в Калужской сбл.
насчитывалось в соцветии 100-300 колосков. Однокустовые особи в
тех условиях продуцируют 200-500 семян, многокустовые - 400-800
(редко больше), семян. В среднем, многокустовые особи имеют 235
придаточных корней.
К группе средневозрастных генеративных растений относятся рас-
тения вегетативного происхождения, образовавшиеся при клонирова-
нии первичных систем парциальных кустов. Средневозрастные генера-
тивные состояния начинаются с момента формирования клона и продол-
жаются до образования вторичных систем из 5-6 (не менее 3) прос-
тых (реже в системе попадаются отдельные сложные) парциальных кус-
тов. Генеративные побеги достигают максимальной высоты (до 120 см),
развивают соцветия дс 25 см. Каждая особь образует 17-20 тыс. и
48
больше семян. Среднее по мощности растение в естественных услови-
ях образует 300-400 и более корней на одну особь.
К группе старых генеративных растений относятся старые особи,
представляющие собой вторичную систему, состоящую из сочетания
одиночных побегов и простых парциальных кустов, которые по мере
старения образуют систему одиночных побегов. Старыми генератив-
ными растениями мы считаем особи с момента образования вторичных
систем из 2-5 простых парциальных кустов до полной утраты генера-
тивной функции, т.е. до перехода в сенильное состояние. У этих
растений неоднократно наблюдалось отсутствие возобновления оди-
ночных побегов. Генеративные побеги с небольшими соцветиями 0,5-
6 см длины. Каждая оссбь способна дать около 2-4 тыс. семян. Ста-
рые генеративные растения в пойме р.Угры в Калужской обл. при
краткосрочном заливании имели в среднем 165 корней на одну особь.
Генеративные временно нецветущие растения - это взрослые веге-
тативные растения, которые, достигнув половой зрелости, в теку-
щем году не зацвели. Они по морфологическим признакам соответст-
вуют растениям, находящимся в генеративном состоянии.
Постгенеративный период характеризуется тем, что растения не
цветут и не плодоносят вследствие возрастной утраты.этой способ-
ности. Постгенеративный период начинается с момента прекращения
образования генеративных побегов и продолжается до отмирания по-
следнего побега особи. Эти растения могут представлять собой:
I) систему, состоящую обычно из сочетания простых парциальных
кустов и одиночных побегов; 2) систему, состоящую из 2-6 одиноч-
ных побегов, у которых только осенью из пазушных почек зоны куще-
ния образуется одна редко две розетки (такие особи обычно преоб-
ладают среди субсенильных растений); 3) отдельные простые парци-
альные кусты, соединенные с небольшим числом гаплокормов отмер-
ших побегов. У сенильных растений часто отсутствует возобновле-
ние побегов. Внешне побеги растений очень похожи на побеги имма-
турных и ювенильных растений. Удлиненные побеги субсенильных рас-
тений имеют 2-3 вытянутых междоузлия. Листья узкие, ювенильного
типа, с шириной пластинки 2-3 мм. В одном фитоценозе у субсениль-
ных растений в среднем было 67 корней на одну особь. Сенильные
растения состоят из системы двух отдельных розеток или одиночных
розеток, связанных с зонами кущения отмерших побегов.
Старого генеративного, субсенильного и сенильного состояний
тимофеевка достигает в средней полосе в 7-10-летнем возрасте, а
за Полярным кругом в посеве (при отсутствии семенного возобнсв-
49
4-1964
двнжя) старые’генеративные и особи постгенеративного периода
держались в травостое более 17 лет, т.е. прохождение онтогенеза
удинялооь. У большей части особей главный побег обязательно за-
цветает и по этому признаку относится ко второй категории вари-
антов болыюго жизненного цикла не очень долговечных злаков с
относительно коротким виргинильным Периодом. У некоторых особей
побег 1-го порядка не зацветает и прохождение большого жизненно-
го цикла является производным от второй категории вариантом,
обычным при ухудшении условий жизни (Серебрякова, 1971а,б; Мат-
веев, 1972а, 1975)..
В большом жизненном цикле особи тимофеевки проходят несколь-
ко этапов морфогенеза: первичный одиночный побег, первичный куст,
первичная система парциальных кустов, вторичная система парциаль-
ных кустов, система одиночных побегов (иногда отдельный парциаль-
ный куст), вторичный одиночный симподиально нарастающий парциаль-
ный побег (Матвеев, 1972а, 1975; Смирнова, 1976). У тимофеевки
выявлена поливариантность онтогенеза, которая проявляется в раз-
ной последовательности процессов морфогенеза (это связано с усло-
виями произрастания и биотипическим разнообразием - Годунов,
1972), выпадении этапов морфогенеза, разных темпах развития осо-
бей, т.е. в неодинаковой скорости морфогенетических изменений за
один и тот же отрезок времени (Матвеев, 1972а, 1975).
Сезонное развитие
По ритму побегообразования куст тимофеевки представляет ообой
симподиально нарастающую систему озимых 2-го и 3-го года зацвета-
ния и моноциклических монокарпических побегов (Evans, 1927; Го-
лубев, 1931; Смелов, 1947, 1966; Серебрякова, 1956, 1967, 1971).
В почках возобновления насчитывается 2-3 зачатка, состоящих из
предлиста и 1-2 почечных чешуй. К августу почки имеют 7-8 листо-
вых зачатков, I предлист и 5-6 почечных чешуй. К началу августа
формируется розеточный побег. После перезимовки в начале мая мо-
гут формироваться побеги прегенеративного весеннего кущения. Фор-
мирование соцветия начинается в конце мая. Предельная емкость
зрелой боковой закрытой почки от 6 до 8, а открытой верхушечной
от 4-6 до 8-10 (Evans, Grover, 1940; Серебрякова, 1971). Нерав-
номерность развитости почек в зоне кущения (с наиболее развитыми
в средней части - мезотония) сохранилась и при выращивании почек
50
отдельно от побегов (Смелов, 1947, 1966).
В полном онтогенезе побега тимофеевки луговой можно выделить
фазы: I) почки, в которой предлагается различать два этапа ее
формирования, - незрелой и зрелой почки; 2) развертывания низо-
вых листьев (растущей почки); 3) неветвящегося розеточного побе-
га; 4) первичного,или прегенеративного, кущения; 5) формирования
соцветия и цветоносного стебля (для генеративных побегов) или ве-
гетативной соломины (для полурозеточных вегетативных побегов);
6) цветения и плодоношения; 7) постгенеративного кущения и вто-
ричной деятельности корневища и зоны кущения (Смелов, 1947, 1966;
Серебрякова, 1971; Матвеев, Шуман, 1974). Седьмую фазу, в свою
очередь, можно подразделить на 3 самостоятельные: I) собственно
постгенеративного летне-осеннего кущения, она начинается с отми-
рания соломины материнского побега, заканчивается с наступлением
зимы; 2) постгенеративного кущения следующего сезона вегетации,
в эту фазу 17$ почек тимофеевки луговой способны давать побеги
(Смелов, 1966), в летний период подобные почки уже погибают;
3) деятельность зоны кущения, Связанная лишь с запасанием и про-
ведением питательных веществ из гаплокормов и других органов за-
паса в ассимилирующие боковые побеги и осуществлением всесторон-
ней физиологической связи между побегами поколений (Смелов, Тата-
ринова, 1958; Татаринов, 1958, 1961; Смелов, 1966).
Длительность прохождения отдельных фаз различна. Третью фазу
побег проходит в вегетационных опытах из семени до начала куще-
ния 25 дней (Мельник, 1969), в естественных условиях - от 1,5 до
6 мес. Четвертая фаза у моноциклического .побега до формирования
генеративных органов длится от 0,5 до 4 мес, у озимого побега -
8-9 мес. Пятую фазу однопобеговое растение проходит от 1,5-2 мес
до 3 лет, шестую - 10-20 дней, седьмую - около 2 лет (постгенера-
тивное летне-осеннее кущение - около 3 мес, кущение следующего
сезона вегетяции - около 2 мес, вторичная деятельность - от 15
до 23 мес. Таким образом, полный онтогенез побега завероается за
3-4,5 г.
Ритм сезонного развития складывается из определенных этапов
дифференциации и роста вегетативных и генеративных органов, про-
ходящих в определенной последовательности и с различной скорос-
тью, и состоит из 12 этапов органогенеза. У побегов тимофеевки
луговой выделены следующие этапы (Ржанова, 1957; Куперман, Ржа-
нова, 1963): I) недифференцированного конуса нарастания (фаза
всхода); 2) дифференциации конуса нарастания (фаза розетки), у
51
моноцикличооких яровых побегов весеннего кущения он сравнительно
короткий, а у озимых побегов летне-осеннего кущения продолжается
я течение осенне-зимнего времени (фаза кущения); 3) дифференциа-
ции осей соцветия, который начинается у тимофеевки при разверты-
вании 3-5-го листа (фаза выхода в трубку); 4) закладки колосков
(та же фаза); 5) формирования цветков в колосьях (фаза стеблева-
ния); 6) формирования пыльников и пестика (та же фаза); 7) фор-
мирования половых клеток (та же фаза); 8) выколашивания, или га-
метогенез (фаза колошения); 9) цветения и оплодотворения, или зи-
готогенез (фаза цветения); 10) формирования зерновки (фаза нали-
ва); II) накопления запасных веществ в эндосперме (фаза молочной
спелости); 12) созревания семян (фаза созревания).
У тимофеевки ввделяют 5 этапов формирования и роста листа:
возникновение бугорка на конусе нарастания, формирование валика,
образование пленчатого примордия, превращение пленчатого примор-
дия в зеленый колпачковый листочек, развертывание колпачкового
листа в зеленый лист (Лебедев и др., 1966). Продолжительность жиз-
ни каждого листа от 25 до 60 дней (Evans, 1927; Смелов, 1966;
Мельник, 1969), причем в нижнем ярусе главного побега (I-3-й
лист) длительность жизни 20 дней, в среднем (4-7-й лист) - 28
дней и в верхнем ярусе (8-10-й лист) - 44 дня (Мельник, 1969).
Число междоузлий сильно варьирует и может достигать у удлиненных
побегов 28, у генеративных более стабильно - 5-8 (Evans, 1927;
Смелов, 1947, 1966; Татаринова, 1953; Серебрякова, 1971). Тимофе-
евка луговая не обладает органическим периодом покоя. Выше отме-
чалась двухвслновая ритмика цветения.
Увеличение массы корней происходит в два этапа: до конца фазы
кущения и после колошения до фазы полного цветения, причем у
взрослых растений в начальный период формирования побега рост
корней значительно отстает от роста надземных органов (Коеппё-
kamp, 1934; Смелов, 1947, 1966; Сметанникова, 1950; Смелов и
др., 1951; Клапп, 1961; Лебедев, 1966 и др.). На сезонную динами-
ку изменения количества корней оказывают влияние температура и
влажность почвы ( Darrow, 1939; Смелов, 1947, 1966; Мельник,
1969). '
52
Способы размножения и распространения
Основной способ размножения тимофеевки - семенной. На восхо-
дящей части кривой большого жизненного цикла благодаря глубокому
омоложению отделившиеся части (вырезанные зоны кущения и отдель-
ные почки разного ярусного положения) могут развиваться в нор-
мальные генеративные особи (Смелов, 1947, 1966; Волкова, 1961).
С 3-4-го года способность к кущению и образованию почек ослабева-
ет и у таких особей происходит вегетативный распад. Однако внесе-
ние высоких доз полных минеральных удобрений на естественных пой-
менных лугах при искусственном орошении приводило часть особей
старых генеративных растений к глубокому омоложению и переходу
их в молодое генеративное состояние (опыты Мещерского филиала
ВНИИГиМ в Рязанской обл.). По характеру дезинтеграции особи тимо-
феевка относится к моноцентрическощу типу биоморф с полной отно-
сительно поздней неспециализированной дезинтеграцией, не образу-
ет специализированных для вегетативного размножения органов.
Семенная продуктивность особи изменяется в онтогенезе по одно-
вершинной кривой с максимумом в естественных условиях у средне-
возрастных растений (Матвеев, 1972, 1975).
Экология и фитоценология
Тимофеевка луговая - мезофит, способный произрастать в услови-
ях от лугостепного до сыролугового увлажнения (47-84-я ступени
шкалы увлажнения Л.Г.Раменского). Преимущественно встречается на
свежих местообитаниях, на крайне сухих, на чрезмерно влажных не
растет (Eilenberg, 1952; Hundt, 1964). Может переносить и засуш-
ливые периоды (Балаева, 1968; Добрынин, 1969; и др.). В отноше-
нии изменчивости водного режима нетребовательна - встречается от
высокообеспеченного водного питания до сильно переменного увлаж-
нения (3-12-я ступени шкалы Л.Г.Раменского). Способна переносить
весеннее затопление до 15-30 дней (Терехов, 1955). К подтоплению
устойчива, но для полей орошения пригодна условно (Клапп, 1961).
Встречается при отложении наилка от 0,1 до 2 см. По нашим наблю-
дениям, в пойме р.Угры отрастает при наилке 5-9 и даже 12 см.
Тимофеевка растет преимущественно на глинистых, суглинистых,
торфянистых почвах, реже на легких почвах и песках. Встречается
на 1^>чвах 6-20-й ступеней шкалы богатства почв Л.Г.Раменского.
4 -1964 53
На внесение удобрений реагирует положительно (Голубев, 1931; Сме-
лов, 1966; Работнов, 1974; и др.), однако внесение значительных
доз азота уменьшает зимостойкость тимофеевки, особенно в траво-
смеси (Работнов, 1974). К кислотности почв довольно безразлична,
произрастает при pH от 5,0 до 8,3, однако предпочитает pH = 5,5-
5,9 (Hund-t, 1964; Клапп, 1965). Тимофеевка - индикатор довольно
высокого плодородия, хорошей аэрации, рвЕВДИи почвы, близкой к
нейтральной (Матвеева, 1971).
Тимофеевка светолюбива, переносит.лишь незначительное затене-
ние (Eiienberg, 1952) и в зависимости от влажности занимает от
2-й до 4-й ступени светолюбия (Кашкарова, 1972). В молодом состо-
янии (от проростков до имматурного) чувствительна к затенению
(Щенкова, 1961). Изучение фотопериодической реакции показало,
что. пребывание тимофеевки в условиях короткого дня (12-13 ч) за-
держивает в год посева рост и развитие, но способствует усилению
кущения и облиственности, сказывается на углеводном обмене, со-
держании сушы углеводов, хлорофилла, каротина и т.п. (Смелов,
1947, 1966; Ржанова, 1951; и др.).
Тимофеевка луговая участвует в сложении и иногда доминирует в
сообществах материковых и горных лугов. Может быть доминантой в
искусственных сообществах (особенно с клевером), однако продук-
тивность, жизненное состояние, численность особей и значимость
этого вида в травостое зависят от системь'. использования, состава
травосмеси, метеорологических условий и т.д. (Работнов, 1974; и
др.). Исследования многих авторов показали, что добавление к ти-
мофеевке других злаков в посеве повышает общую продуктивность
травостоя, но не всегда повышает долю тимофеевки в травосмеси.
Она обычно более конкурентоспособна, чем овсяница луговая, но ме-
нее, чем райграсе многолетний и ежа сборная (Frame е.а., 1971;
и др.).
Тимофеевка положительно влияет на биологическую активность
почв. Установлено, что при ускоренном залужении с посевом тимофе-
евки (4-й год жизни) на гриве окской поймы биологическая актив-
ность почвы уменьшалась с переходом от верхних горизонтов к ниж-
ним в соответствии с уменьшением содержания корней и корневых ос-
татков. В межгривном понижении биологическая продуктивность в
слое от 0 до 60 см была выше (39,1$), чем на гриве (11,7%), и ве-
сеннее затопление в течение одного месяца повысило биологическую
продуктивность почвы в 20 раз (Куркин, Бобнева, 1978).
Взаимоотношения тимофеевки луговой с некоторыми растениями бы-
54
ли изучены на естественном участке полидоминантного кострово-ов-
сянично-разнотравного луга среднего уровня в центральной пойме
р.Угры в Калужской обл. Были отмечены все пять типов сопряженнос-
тей. Наиболее часто встречается двузначная сопряженность тимофе-
евки луговой (действующий вид), т.е. с возрастанием численности
тимофеевки численность подчиненного вида растет, затем падает
(отмечена у овсяницы луговой, горошка мышиного, люцерны желтой,
будры плющевидной, вербейника монетчатого). Отрицательная сопря-
женность отмечена у овсяницы красной, тысячелистника обыкновенно-
го, мятлика лугового (с увеличением численности тимофеевки их чис-
ленность падала). Положительная сопряженность с тимофеевкой отме-
чена у лисохвоста лугового. Сложную сопряженность (с увеличением
численности тимофеевки численность подчиненного вида сначала
уменьшается, затем возрастает) наблюдали у свербиги восточной,
ежи сборной, костра безостого, клевера ползучего, одуванчика
обыкновенного. У погремка отмечена безразличная сопряженность.
В качестве подчиненного вида тимофеевка луговая имеет также
различную сопряженность: двузначную - с овсяницей луговой, крово-
хлебкой лекарственной, лисохвостом луговым, мятликом луговш;
сложную - со свербигой восточной, ежой сборной, костром безостым,
тысячелистником обыкновенным, горошком мышиным, будрой шшцевид-
ной; положительную - с клевером луговым; отрицательную - с оду-
ванчиком обыкновенным и люцерной желтой. Следует отметить, что в
разные годы складывается разный характер взаимоотношений между
одними и теми же парами видов при сходных экологических и ценоти-
ческих условиях. В целом тимофеевка не оказывает заметного отри-
цательного воздействия на другие травы (Василевич, 1969).
Консортивнне связи
Всходы и молодые растения могут повреждать большая злаковая
тля, кукурузная тля, хлебный клопик, странствующий клопик, поло-
сатая цикадка, личинки стеблевых блошек, полосатая хлебная блош-
ка, личинки проволочников (Моляков, 1916; Evans, 1927). Листья,
стебли, зона кущения и корни тимофеевки повреждаются различными
группами гетеротрофов. Наземные части повреждают ячменная тля,
злаковая тля, кукурузная и большая злаковая тля. Корни и зону ку-
щения повреждают корневая тля, тимофеевичный долгоносик, личинки
щелкунов, личинки жука-красуна, вредная долгоножка, тли (Моляков,
55
.1'916; Ячевский, 1926; Зорина, 1955; Машек, 1967, 1977). Генера-
тивные органы повреждают хлебный клещик, тимофеевичный трипс,
жук-красун, полооатая хлебная блошка, северная ржаная совка,
обыкновенная зерновая совка, шведская муха, тимофеечные мухи, а
по нашим наблюдениям в Подмосковье, - личинки пилильщика и клопы.
Эпибионтными и эндобионтными консортами надземных и подземных
органов ЯВЛЯЮТСЯ также эубактерии (Xanthomonas translucens
(I.J.R.) Dowson, Pseudomonas sp.), из микобактерий - молочнокис-
лые бактерии (Lactobacterium, Streptococcus и др.) (Федоров, Не-
помилуев, 1954; Шилова, 1955). Среди грибных консортов тимофеев-
ки известны микоризные грибы из родов Fusarium, Pythium и пара-
зитные: Claviceps purpurea Tul., Epichloe typhina Tul., Phylla-
chora graminis Fuck., Sclerotinia borealis, Urocystis agropyri
(Preuss) Schroet., Puccinia phlei-pratensis (Erikss. et Henn.),
Mastigosporium kitzenbergense Schlos. (МОЛЯКОВ, 1916; Дидусь,
1947; Крюгер, 1959, 1961а,б; Селиванов и др., 1961, 1962; Году-
нов, 1971).
На пойменных и суходольных лугах Московской обл. к тимофеевке
луговой могут присасываться Odontites serotina (Lam.) Rehb.,
Euphrasia tenuis (Brenn.) Wettst., Melampyrum arvense L., Pedi-
cularis kaufmannii Pinzer., Rhinanthus major subsp. vernalis
Zing. К раннему выпасу неустойчива, но встречается в ряду паст-
бищной дигрессии от очень слабого выпаса до полусбоя (Eilenberg,
1952; Раменский и др., 1956).
Биологическая продуктивность
Тимофеевка луговая - высокопродуктивный вид, который высевают
как один из основных компонентов травосмеси в полевых и лугопаст-
бищных севооборотах. В посевах тимофеевка может существовать и
давать относительно высокую урожайность в течение 10 и более лет
(Клинген, 1911; Смелов, 1966). Биологическая продуктивность в
пойме р.Яхромы (Московская обл.) на осушенном торфянике составля-
ла на 7-м году жизни 40 ц/га, а в травосмеси с другими видами ко-
лебалась от I до 43 ц/га, причем самый высокий урожай был полу-
чен в первом укосе на 2-й год пользования (Путятин, 1939). Абсо-
лютный урожай первого укоса сильно колеблется - от 8-9 до 60-75
ц/га. По обобщенным данным ВНИИ кормов урожай тимофеевки (посев)
при сенокосном использовании на суходольных лугах составляет 35-
56
45 ц/га сена, на низинных - 40-50 ц/га и на осушенных болотах -
55-75 ц/га. В травосмесях с клевером или сложных травосмесях уро-
жайность повышается до 3-го, реже 4-го года жизни.
Урожай тимофеевки в чистом посеве (первый укос третьего года
пользования) в Подмосковье (Качалкино) состоял из 1718 побегов
на I кг сена (Смелов, 1966), причем масса I генеративного побега
равнялась 0,76 г, полурозеточных вегетативных - 0,32 и розеточ-
ных - 0,17 г. Олиственность (в весовых процентах) генеративных
побегов 29%, полурозеточных вегетативных - 56%. Иная структура
урожая характерна для тимофеевки в смеси с овсяницей луговой и
ежой сборной - 3500 побегов/кг сена, масса генеративного побега -
0,53 г, полурозеточного вегетативного - 0,15 г. Олиственность ге-
неративных побегов была выше - 38%, полурозеточных вегетативных -
56%.
Хозяйственное значение
Тимофеевка луговая - прекрасное кормовое растение, имеющее вы-
сокую питательную ценность. Она широко используется в луговом и
полевом травосеянии, для создания долголетних культурных пастбищ
и сенокосов (Минина, 1949; Эрингис, 1964; Работнов, 1974). Тимо-
феевка может быть использована для закрепления склонов (Черкасо-
ва, 1957; Воронцова и др., 1976). В условиях культуры урожай ко-
леблется в довольно значительных пределах (от 10 до 60 ц/га се-
на), максимальный урожай обычно наблюдается на второй год (Ларин,
1956). Отмечено вредное влияние пыльцы тимофеевки на человека
(Вульф, Малеева, 1969).
2. ТИМОФЕЕВКА СТЕПНАЯ
Номенклатура и географическое распространение
Phleum phleoides (Ь.) Karst., 1881, Deutschl. Р1.:374; ОВЧИН-
НИКОВ, 1934, Фл. СССР 2:131; Рожевиц, 1940, в Наевск., Фл. изд.
7:104; Цвелев, 1954, там же, 8:840, 1964, изд. 9:801; Скворцов,
1966, Опр. раст. Моск. обл.:74; Цвелев, 1974, Фл. евр. части
СССР 1:244; он же, 1976, Злаки СССР:364; Humphries, 1980, Fl.
Bur.5:240. - Phalaris phleoides L. 1753, Sp. pl.:55. - Phleum bo-
57
elimer.t Wlbel, 1799» Prim. Pl. Werth.:125; Кауфм. 1866, Моск.
фЛ>» И8Д. 1:550, 1889, изд. 2:574; Цингер, 1886, Сб. свел.:491;
Шмальг., 1889, Фл. 2:601; Петунн., 1901, Крит, обзор 3:105; Сы-
рейщ., 1906, Илл. фл. 1:112; он же, 1927, Опр. раст. Моск, губ.:
60. - Тимофеевка степная.
По мнению ряда авторов (Meueel, 1943; Meusel а.а., 1965),
тимофеевка степная относится к евразиатско-бореально-средиземно-
морско-(суб)горно-континентальному флористическому элементу. По
классификации Л.М.Носовой (1966, 1973), это европейско-среднеази-
атский вид, сформировавшийся в конце третичного и начале четвер-
тичного периода. Тяготение ареала вида к Средиземноморью предпо-
лагает древнесредиземноморские корни вида и его происхождение от
третичных мезофильных элементов (Носова, 1966, 1973). Распростра-
нена по всей Евразии. В Московской обл. встречается в южных и
юго-восточных районах, реже, чем предыдущий вид.
Морфологическое описание
Тимофеевка степная - многолетний поликарпический рыхлокусто-
вый злак с короткими гипогеогенными корневищами и экстравагиналь-
ной рыхлой дерновиной. Растение с симподиально нарастающими мо-
но-, ди- и трициклическими, реже тетра- и пентациклическими (Мат-
веев, 1965) полурозеточными генеративными, скрытогенеративными
(редко) и розеточными вегетативными побегами. Отмечают розеточ-
ные побеги с неполным циклом развития (Борисова, I960). Корневая
система мочковатая, у проростков состоит из колеоризы, семенного
и стеблевых придаточных корней. Стеблевые придаточные корни поли-
морфны: тонкие ксрни закладываются в нижнем ярусе зоны кущения,
распространены в поверхностных горизонтах до 50 см; толстые про-
никают глубже - до 1,5-2 м (Зозулин, 1959; Матвеев, 1975). Колео-
риза живет до 1,5 мес, толстые корни - свыше 4 лет.
Соломина прямая или приподнимающаяся, 1,5-2 мм толщины, до
85 см длины, с 2-3 узлами, пурпурного иди лиловатого цвета, кни-
зу. ст середины гладкая, под соцветиями более или менее шершавая.
Внешне морфологическое строение побега сходно со строением тимо-
феевки луговой. Префдоральные и флоралыше зоны существуют у ди-
циклического побега 3-4, у трициклического - 4-5 лет, полный он-
тогенез .завершается через 5-8 лет после возникновения.
Лиотья без опушения, серо-зеленые, длиннозаостренные, плоские,
58
частично вдоль сложенные, без пологой складочки посередине, по
краям остропиловидно-зазубренные, кайма края листа широкая, шире
высоты шипиков (Флорова, Раменский, 1937). Лист в почкосложении
свернут, 2-3 (5) мм ширины. Нижние листья до 12 см длины, верх-
ние намного короче и в 2-4 раза короче стебля. Язычок тупой,
2-3 мм. Влагалища листьев гладкие, округлые со спинки.
Соцветие - колосовидная метелка (точнее ложный колос метельча-
того происхождения), беловато-зеленое, зеленое, иногда пурпурное,
узкое, длинноцилиндрическое, иногда к обоим концам слегка сужен-
ное, 13-15 см длины, 5-8 мм в диаметре. Веточки лопастей не срос-
шиеся, иногда немного отклонены. Колоски продолговатые или лан-
цетные, маленькие, 2,5-3 (до 5) мм ширины, одноцветковые, при со-
зревании разрывающиеся над цветковой чешуей, с остроугольной вы-
резкой между остевидными заострениями. Колосковые чешуи пленча-
тые, острошершавые, киль их изогнутый, твердый, с зазубринками
до 0,5 мм длины, переходящий в остевидное острошершавое заостре-
ние. Цветковые чещуи на спинке без ресничек, пленчатые, с малоза-
метными жилками, на I/4-I/3 короче колосковых чешуй, яйцевидные,
туповато-заостренные. Верхняя цветковая чещуя с килем, острой
или остистой усеченной верхушкой, с 5 жилками. Нижняя цветковая
чещуя короче верхней, I,7-2,3 мм длины, без ости, более широкая
и узкая, охватывает верхнюю, короткопушистая, с 2 жилками. Тычи-
нок 3, нити их волосовидные, длиннее колосковых чещуй; пыльники
продолговатые, раздвоенные, 0,6-1,3 мм длины. Завязь полуокруг-
лая, волосовидная, столбиков 2, рыльца перьевидные.
Пленчатая зерновка тонкоморщинистая, голая, матовая, серебрис-
то-серая или соломенно-серая, длиной 1-1,75 мм и диаметром
0,5 мм. Стерженек тонкий, круглый, до I/5-I/6 длины зерновки. Пя-
точка прямоусеченная, едва заметная, есть остевидное заострение.
Масса I тыс. пленчатых зерновок около 0,3 г. В I кг до 3,3 млн.
пленчатых зерновок (Доброхотов, 1961).
Число хромосом х = 5,7. Указывается различный диплоидный на-
бор 2х = 14, 28, I4+I-4B, 28+I-2B, I4+I-IB: 21 (Хромосомные чис-
ла..., 1969).
Онтогенез
Семена тимофеевки степной встречаются в почве редко (Голубева,
1962, 1966), несмотря на довольно высокую семенную продуктивность
59
UMAM. Семена способны прорастать в год образования, без периода
ПОКОЯ (Соколова, 1934; Овеснов, 1965). Прораотание может происхо-
дить в широком диапазоне условий. Свет увеличивал всхожесть в
1,6-3 раза при малоблагоприятной температуре (Панферов, 1939) и
угнетал прорастание старых оемян (Овеснов, 1965). Стратификация
(1-4°) повышает энергию прорастания, но снижает всхожесть (Овес-
нов, 1965). Семена способны всходить даже в фазе молочной спелос-
ти. Всхожесть сохраняется 7-10 лет. Прорастание обычно наблюдает-
ся поздним летом и в начале осени. Всходы могут появиться на 3-
10-й день после попадания семян в почву (Каменецкая, 1952).
Проростки представляют собой одиночные розеточные побеги, со-
храняющие зерновку и колеоризу, с колеоптилем, 2-3 листьями, од-
ним семенным и 2-4 стеблевыми придаточными корнями. Длительность
существования семенного корня неизвестна, стеблевых - свыше 5-6
лет. Пластинка первого листа до 30 мм длины и 2 мм ширины, второ-
го - длиннее, опушена редкими и длинными волосками (Покровская,
I960). Состояние проростков длится 1-9 мес и более (Матвеев,
1975).
Ювенильные растения в отличие от проростков питаются самосто-
ятельно и развивают более мощную систему придаточных корней: тон-
ких сильноветвящихся и толстых слабоветвящихся. Это состояние на-
чинается с отмирания зерновки и продолжается от 2 мес до 2,5-5
лет, до начала весеннего (реже осеннего) прегенеративного куще-
ния 1-го или 2-го года развития или до перехода в генеративное
состояние на 2-3-й (реже 4-5-й) год жизни. Ювенильные растения
первого года представляют собой однопобеговые розетки с 3-4 зеле-
ными листьянш, корневая система состоит из одного семенного и 4-6
стеблевых придаточных корней. Ювенильные растения 2-го года и
следующих, лет жизни - розеточный побег с 2-4 зелеными и 6-10 сухи-
ми листьями, семенного корня нет, стеблевые имеют боковые корни
пяти и более порядков.
Имматурные растения в отличие от ювенильных образуют первич-
ные простые или сложные кусты. Длина и ширина листьев не достига-
ет размеров взрослого растения. Диаметр дерновинки 1-6 см. Имма-
турное состояние продолжается до отмирания розеточных побегов
1-го и 2-го порядков или до перехода в генеративное состояние.
Имматурные первой подгруппы представляют собой дуст из розеточ-
ных побегов - главного и боковых 2-го порядка прегенеративного
летне-осеннего 2-го сезона или весеннего 2-3-го сезонов вегета-
ции. Имматурные второй подгруппы - это подобный куст, но с побе-
60
гами как I-2-го, так и 3-4-го и т.д. порядков прегенеративного
весеннего и летне-осеннего кущений 3-4-го и т.д. лет жизни. В
имматурном состоянии особь может находиться от 10 мес до 2-4 лет.
Взрослые виргинидьные растения семенного происхождения пред-
ставляют собой систему из 2-5 и более парциальных кустов и имеют
небольщую плешину отмерших побегов. Это состояние начинается с
момента отмирания надземной части побегов и заканчивается перехо-
дом особи в генеративное состояние, продолжается от I до 5 (воз-
можно и более) лет.
Молодые генеративные растения - это особи семенного происхож-
дения, которые могут существовать в виде: одиночных генеративных
побегов (такие особи развиваются из ювенильных растений на 2-3-й,
реже 5-й год и после цветения приступают к постгенеративному лет-
не-осеннему кущению 2-3-го и т.д. года жизни); однокустовых гене-
ративных растений, представляющих собой простой или чаще сложный
куст; многокустовых генеративных растений, представляющих систе-
му из 2-4 и белее сложных или реже простых парциальных кустов,
соединенных зонами кущений отдоерищ^адземных част-й генерат з-
ных или чаще вегетативных пр'бегОв (обычно в системе число живых
побегов преобладает над- числом отмерших). Особь имеет от I до
6-8 генеративных побегов и обра~-учт 200-500 и более семян. Сред-
няя высота генеративных побегов и размер соцветий еще не достига-
ют максимальной величины. Предельная глубина проникновения корне-
вой системы достигает НО см (Голубев, 1962). В молодом генера-
тивном состоянии растения могут находиться 2-6 лет.
Средневозрастные генеративные растения представляют собой вто-
ричные системы парциальных кустов, образовавшиеся при клонирова-
нии. Средневозрастное генеративное состояние начинается с момен-
та образования клона и продолжается до образования вторичннх си-
стем из 8-10 сложных, реже простых парциальных кустов. По нашим
данным, в Калужской обл. образовавшийся клон имеет 10-20 и более
генеративных растений, средняя семенная продуктивность достигает
максимума 2-5 тыс. семян на клон. Системы парциальных кустов со-
ставляют дерновину, по форме напоминающую круг (не всегда цель-
ный) , в центре которого имеется довольно большая беспобеговая
''плешина'', состоящая из остатков базальных частей отмерших побе-
гов, т.е. представляют собой молодой клон. Корни достигают глуби-
ны 170 см и более. В зтом состоянии растения находятся 4-5 и бо-
лее лет.
Старые генеративные растения представляют отдельные вторичные
61
ожотомы сложных и простых парциальных кустов, отдельные простые
кусты или одиночные генеративные побеги. Системы парциальных кус-
тов составляют отдельные, четко отделенные друг от друга сегмен-
ты (парциали) круга, нарастающие, центробежно. (В некоторых случа-
ях форму круга в клоне установить невозможно.) В этом состоянии
растения находятся до полной потери ими генеративной функции и
могут существовать 10-30 лет и более. Генеративные побеги по мощ-
ности и внешним признакам очень похожи на побеги молодых генера-
тивных особей. Отделившаяся партикула способна дать свыше 2 тыс.
семян, при большей степени распада (вплоть до одиночных побегов)
семенная продуктивность уменьшается до 240 семян на особь и мень-
ше.
У генеративных особей возможны перерывы в цветении и это по-
зволяет выделить подгруппу временно не цветущих растений, находя-
щихся в соответствущем генеративному молодом, средневозрастном
и старом состояниях.
Постгенеративный период начинается с момента прекращения гене-
ративной функции и продолжается до отмирания последнего побега
особи. У растений постгенеративного периода часто отсутствует
возобновление побегов и выражено много полусгнивших и отмирапцих
зон кущения отмерших побегов. Внешне побеги очень похожи на побе-
ги. имматурных и ювенильных растений. Субсенильные растения пред-
ставляют собой небольшие системы простых розеточных, реже слож-
ных парциальных кустов или системы одиночных йобегов (обычно п+2,
п+з и более порядка), нарастающих односторонне, соединенные зо-
нами кущения отмерших побегов. Побеги внешне очень напоминают по-
беги имматурных оообей. В атом состоянии растение может находить-
ся 5-6 лет и более. Сенильные растения представляют системы оди-
ночных розеточных побегов (редко в системе только 2 побега, иног-
да даже’одиночные побеги), связанные с эонами кущений побегов,
отмерших в предыдущие сезоны вегетации. Розеточные побеги непол-
ного цикла некоторых субсенильных и сенильных растений могут раз-
виваться как тетра- и пентациклические. Подобное удлинение полно-
го онтогенеза побега позволяет особям дольше сохранять за видом
территорию. Это состояние обычно продолжается 3-5 лет.
Общая длительность жизни тимофеевки степной около 30 лет и бо-
лее (Матвеев, 1955). У некоторых растений главный побег- (1-го по-
рядка) обязательно становится генеративным и по этому признаку
они относятся ко 2-й категории вариантов большого жизненного цик-
ла относительно долголетних злаков. По нашим данным, такой вари-
62
ант прохождения большого жизненного цикла на остепненных поймен-
ных лугах р.Угры в Калужской обл. составляет 56% молодых генера-
тивных растений. Ход онтогенеза других особей (около 44%) соот-
ветствует особому варианту 2-й категории злаков, у которых побег
1-го порядка никогда не становится генеративным. По длительности
жизни и особенностям развития тимофеевка степная занимает проме-
жуточное положение между I-м и 2-м вариантом 2-й категории зла-
ков (Серебрякова, 1967, 1970, 1971а,б) с ярко выраженными преге-
неративным (зацветают на 2-6 год жизни) и постгенеративным пери-
одами.
В большом жизненном цикле особи тимофеевки проходят несколько
сменяющих друг друга этапов морфогенеза: первичный одиночный по-
бег, первичный куст, первичная система парциальных кустов, вто-
ричная система парциальных кустов, система одиночных побегов
(или парциальный куст), вторичный одиночный симподиально нарас-
тающий парциальный побег. У тимофеевки степной большой жизненный
цикл может осуществляться по различным вариантам, что проявляет-
ся в разной последовательности процессов морфогенеза, выпадении
отдельных этапов морфогенеза и некоторых возрастных состояний
(особенно в прегенеративный и постгенеративный периоды) и разных
темпах развития особей, т.е. разной скорости перехода иэ одного
возрастного состояния в другое эа один и тот же промежуток време-
ни (Матвеев, 1975).
Сезонное развитие
Всходы тимофеевки отепной появляются чаще всего осенью - в
сентябре (Каменецкая, 1949, 1952), в отдельные годы, по нашим на-
блюдениям, и весной. Имея в основном ди- и трициклические побеги
(как с полным, так и с неполным циклом развития), в ювенильном и
сенильном состояниях побеги могут развиваться как тетра- и пента-
циклические. Заложение соцветий происходит осенью: сильно вытяги-
вается конус нарастания (до 0,6 мм) и закладываются валики буду-
щих крокщих чещуй. Перезимовывают с одним зеленым листом в лет-
них розетках и с зелеными осенними розетками (Борисова, I960).
Зимующие почки находятся в почве. Молодые побеги могут зимовать
с 12 зелеными листьями (Прозоровский, 1940а).
Тимофеевку считают малоустойчивым к скашиванию видом, произ-
растающим преимущественно на одноукооннх, реже двуукосных лугах.
63
Homo скашивания она отрастает незначительно и пригодной для ис-
польвования отавы не образует (Борисова-Туленкова, 1959). При от-
сутствии скашивания число особей увеличивается (Прозоровский,
19406).
Тимофеевка относится к группе полуэфемероидных растений, так
как она в период засухи сокращает до минимума число нижних
листьев и у нее отмирают генеративные побеги с листьями. По рит-
му листообразования относится к группе поликарпических многолет-
ников с двумя генерациями листьев. Такая ритмичность связана с
развитием весенних и летне-осенних розеточных побегов. Средний
срок отмирания перезимовавших листьев в лесостепной зоне 28 мая.
По реакции на'снятие воздействия низких осенне-зимних температур
относится к группе, не имеющей органического периода покоя (Голу-
бев, 1962, 1965).
Зацветание происходит базипетально. За 1-2 дня до цветения со-
цветие становится более раскидистым за счет отклонения цветоно-
жек от главной оси. Тимофеевка относится к группе коротко цвету-
щих растений - цветение отдельного цветка продолжается 3-5 ч
(Оловянникова, 1949). Преобладает перекрестное опыление (Понома-
рев, 1964). По ритму цветения тимофеевка степная находится между
группами раннелетних (30Д1) и среднелетних (31/УП) растений,
причем в культуре все фазы цветения наступают раньше. При небла-
гоприятных условиях (климатических, ценотических и пр.) растения
могут перейти во временно нецветущее состояние (Матвеев, 1975) и
в состояние летнего покоя (Куркин, 1971). Изучение ценопопуляций
тимофеевки показало, что относительное участие временно нецвету-
щих растений соответствующего генеративного состояния с возрастом
повышается: среди молодых генеративных их оказалось 5,3%, среди
средневозрастных - 12,2%, а среди старых - 42,9%.
Способы размножения и распространения
Размножается исключительно семенами. Зерновки переносятся вет-
ром на очень небольшие расстояния (Сорные растения СССР, 1934;
Elienberg, 1952). Вегетативно в естественных условиях не размно-
жается и лишь со средневозрастного генеративного состояния обра-
зуется клон, при этом чаще всего наблюдается вегетативный распад
особей, не сопровождающийся глубоким омоложением.
Семенная продуктивность тимофеевки изменяется по одновершин-
64
ной кривой. Пр набладениям в Калужской обл. среднее число семян
на одну молодую генеративную особь составляло 668, средневозраст-
ную - 2608,8, старую - 1441,4; общая семенная продуктивность по-
пуляции около 2 тыс. семян на I м?. Семенное размножение расте-
ний происходит обычно не ежегодно и зависит от погодных условий,
фитоценотической обстановки и т.д.
Экология, фитоценология и консортивные связи
Тимофеевка степная - ксеромезофит, встречается от сухостепно-
го до влажнолутового увлажнения (37-70-я ступени шкалы увлажне-
ния Л.Г.Раменского), от переменно-обеспеченного до сильно пере-
менного увлажнения (7-12-я ступени шкалы Л.Г.Раменского). В силь-
ную засуху переходит в состояние вторичного покоя. Встречается
только на почвах 7-17 ступеней шкалы богатства почв Л.Г.Раменско-
го, на нейтральных или щелочных (Eiienberg, 1952). Индикатор до-
вольно высокого плодородия, хорошей аэрации, реакции почвы, близ-
кой к нейтральной, постоянной или временной сухости почв (Матве-
ева, 1971). Светолюбивое растение, выносит только очень слабую
затененность (Eiienberg, 1952; Кашкарова, 1972). Тимофеевка теп-
лолюбива, к холоду чувствительна.
Тимофеевка степная может быть доминантой (чаще кодоминантом)
или принимать лишь небольшое участие в создании ценозов. В подзо-
не северных степей В.Н.Голубев (1962, 1965) считает ее компонен-
том фитоценоза 2-й категории, т.е. субдоминантой.
Изучение ценопопуляций тимофеевки степной проводилось в раз-
личных условиях: от смешанных лесов до типчаково-ковыльных сте-
пей. Большинство спектров возрастных состояний (кроме типчаково-
ковыльных степей и специфических условий некосимой степи в Цент-
рально-Черноземном заповеднике) были полночленными с высокой чис-
ленностью как старых, так и отдельных групп молодых растений, с
двувершинной кривой, отражающей неравномерность возобновления.
Одна группа спектров с хорошо выраженными флюктуациями в левой
части спектра, вторую группу ’составляют ценопопуляции, в кото-
рых ежегодные флюктуации выражены слабо (Матвеев, 1975; Ценопопу-
ляции растений, 1976).
Тимофеевка степная - микосимбиотроф (Селиванов и др., 1966;
Селиванов, Утемова, 1968). Часто поражается фитогельминтами
(Кирьянова, Кралль, 1969; Кралль, Кралль, 1971). Выдерживает в
5-1964 65
малом обилии умеренное влияние выпаса, а иногда даже сильное (Ра-
менский и др., 1956), к вытаптыванию чувствительна.
66
А.П.Ефремов, Ю.Е.Алексеев
БЕЛОКРЫЛЬНИК БОЛОТНЫЙ
Номенклатура и географическое распространение
Calla palustris L., 1753, Sp. pl.:968; Кауфм., 1866, Моск,
фл., изд. 1:593, 1889, изд. 2:622; Цингер, 1886, Об. свед.:406;
Шмальг., 1897, Фл. 2:525; Сырейщ., 1906, Илл. фл. 1:217; он же,
1927, Опр. раст. Моск. губ.:84; Кузенева, 1935, Фл. СССР 3:482;
Назаров, 1940, в Маевок., Фл., изд. 7:207; Бобров, 1954, там а»,
изд. 8:702, 1964, изд. 9:689; Тихомиров, 1966, Опр. раст. Моск,
обл.:107; Иконников, 1979, Фл. евр. части СССР, 4:316; Prime,
1980, Pl. Eur. 5:269. - Белокрыльник болотный (семейство Ага-
сеае - Аронниковые).
Голарктический бореальный вид, широко распространенный в Ев-
разии и Северной Америке. В СССР северная граница ареала идет от
центральной части Карелии на Архангельск, Верхнюю Золотницу Усть-
Почу, Мезенскую и Печерскую Пижму, доходит на север до р.Ижмы в
Коми АССР. В Западной Сибири она спускается к югу до 63°30 с.ш.
на р.Енисее у с.Аменское. В Якутии идет по р.Вилюй и поднимается
по р.Лене до Жиганска, направляясь на восток по р.Алдан. Южная
граница ареала белокрыльника в СССР проходит фрагментарно через
Львовскую, Ворошиловоградскую, Киевскую, Харьковскую, Воронеж-
скую (Старый Оскол), Тамбовскую, Пензенскую, Оренбургскую (Бузу-
лукский бор) области. Далее она идет через Северный Казахстан
(оз.Камышлово), Алтайский край (Дулундинские боры), юг Краснояр-
ского края, Прибайкалье, Приамурье и Приморье к оз.Ханка. Бело-
крыльник встречается на Камчатке, о.Кунашир (на других Куриль-
ских о-вах отсутствует), о.Сахалин (Крылов, 1929; Флора СССР,
т.З, 1935; Попов, 1957; Ворошилов, 1966; Флора Северо-Востока...,
т.2, 1976; Флора Центральной Сибири, 1979; Флора европейской час-
ти СССР, т.4, 1979). В Московской обл. распространен во всех рай-
онах, но на правобережье Оки, видимо, очень редок.
Ископаемые остатки белокрыльника известны лишь из отложений
рисс-вюрмской межледниковой эпохи: у г.Галич Костромской обл.,
близ д.Пышки Смоленской обл. и на р.Неман к северу от г.Гродно
(Арискина, 1950).
67
Морфологическое описание
Многолетнее травянистое поликарпическое растение с горизон-
тальным белым корневищем, диаметром 0,8-2 см, симподиально ветвя-
щимся (рис.5). Если корневище стелется по поверхности субстрата,
Рис.5. Общий вид белокрыльника болотного в начале
цветения.
оно имеет темно-зеленую окраову^. Его междоузлия неравные, от 0,4
до 6 см длины. На узлах после отмирания листьев остаются кольце-
образные рубцы. На живых участках корневищ от узлов отходят мочки
многочисленных придаточных корней, длина которых различна в раз-
ных условиях, часто достигает 60 см. Корни белые, неветвящиеся в
воде или с тонкими боковыми корешками в торфянистой почве, имеют
Оригинальные материалы собраны в окрестностях оз.Высоковское
. (ивановская обл., Фурмановский р-н).
68
на всем протяжении одинаковую толщину (1,5-2 мм) и тупой кончик.
Корневые волоски и микориза у них не отмечены (raetsavainto,
1931). На большинстве узлов находятся пазушные почки возобновле-
ния, по одной на каждом узле. Живое корневище со стеблем достига-
ет I м длины, а на сплавинах - до 1,4 м (Арискина, 1950). Граница
между корневищем и стеблем нечеткая, так как растение часто сте-
лется по поверхности субстрата, а корни развиваются и на молодых
частях побегов.
Листья очередные, направленные вертикально вверх, в числе ТО-
20 на главном побеге. Черешок длинный, вогнут на адаксиальной
стороне, внизу расширен во влагалище (в почке оно свернуто), рав-
ное половине длины черешка. Влагалище кожистое, плотно охватывает
стебель, переходящее в язычок (лигулу). Язычки молодых листьев
имеют форму конуса и покрывают передний конец поднимающегося по-
бега с молодыми листьями и пазушными почками. Листовая пластинка
сердцевидная, тонкая, длиной 8-16 см и шириной 7-14 см, дутонерв-
ная, блестящая, сверху темно-зеленая, снизу немного светлее. Обе
ее стороны имеют приблизительно одинаковое количество округлых
устьиц. Молодые пластинки в почке закручены, причем последующий
лист закручен в противоположном направлении по отношению к ниже-
расположенному листу.
Белокрыльник имеет терминальное соцветие (рис.6), очень редко
наряду с ним возникает боковое соцветие (Hock, 1911). Цветонос-
ный побег прямостоячий, цилиндрический, до 30 см высотой. Под по-
чатком расположен кроющий лист (покрывало), имеющий влагалище и
яйцевидную пластинку 4-6 см длиной, наверху суженную в линейный
кончик, сверху белую, снизу зеленую; влагалище без язычка или он
очень маленький, реже влагалище длинное, сросшееся с цветоносом.
Основание пластинки покрывала имеет два ушка, одно ниже другого.
Покрывало охватывает соцветие до его распускания, образуя бутон.
Обычно покрывало I, но изредка встречаются на соцветии 2 (такой
случай отмечен и нами), 3 или 4 покрывала, причем расположены они
очередно, а их размеры уменьшаются снизу вверх (Hock, 1911).
Початок удлиненный, цилиндрический, зрелый до 6 см длины, не-
много короче покрывала. Он состоит из многочисленных, плотно при-
легающих друг к другу, расположенных по спирали, цветков. Цветки
обоеполые, без околоцветника, обычно имеют 6 (иногда до 10) тычи-
нок и короткояйцевидную завязь с 6-12 удлиненными семязачатками
(рис.6). Завязь образована 3 или более (до 6) плодолистиками,
число последних зависит от положения цветка в початке и, как пра-
Рис.6. Генеративные органы белокрыльника болотного:
I - соцветие; 2 - цветки; 3 - тычинка; 4 - за-
вязь (а - продольный разрез); 5 - семяпочка;
6 - семя (а - продольный разрез).
70
вило, увеличивается сверху вниз, менее 3 плодолистиков в завязи
не обнаружено (Кожевников, 1887). Рыльце сидячее, густо покрытое
прозрачными папилл езннми липкими выростами. На вершине початка
имеются только мужские цветки, состоящие из 6 тычинок в двух кру-
гах (Hock, 1911). Тычиночные нити плоские, 1,5-2 мм дайной,
0,5 мм шириной, вдвое длиннее пыльников. Пыльники белые, широко-
эллиптические, двухкамерные (см. рис.6). Существует мнение, что
цветки белокрыльника однопалые (Крылов, 1929; Некрасова, 1949;
Попов, 1957), т.е. женские состоят из одного пестика, окруженно-
го однотычиночннми мужскими цветками. Это мнение ошибочно. Оно
возникло, вероятно, потому, что в сформировавшихся початках труд-
но провести границу между отдельными цветками. По наблюдениям
Д.А.Кожевникова (1877), цветки белокрыльника обоеполые, но грани-
ца между ними хорошо видна только на ранних стадиях развития.
Плод - сочная ярко-красная ягода до 8 мм в диаметре, с 3-12
семенами. Спелые ягоды заполнены прозрачной, студенистой, упругой
массой, расположенной преимущественно над семенами (см. рис.6).
Толщина стенок завязи, из которой образуется внешняя мясистая
часть ягоды, неодинакова в разных местах. Наиболее тонка боковая
стенка ягоды в месте ее прикрепления к оси початка.
Зрелые семена коричневые, овальные, размером 3x2 мм, с парал-
лельными продольными неглубокими бороздками (см. рис.6). На хала-
зальном конце семени между"бороздками расположены правильные ря-
ды бугорков..Есть указания, что семена растения фиолетовые (Некра-
сова, 1949). В действительности такой цвет имеют семена, не осво-
бодившиеся от мякоти ягоды, тогда как после освобождения от нее
они становятся коричневыми. Одновременно поверхность бугорков се-
мян разрушается и на месте последних образуются ямки, дно которых
состоит из губчатой ткани. Возможно, ямки обеспечивают впитывание
воды прорастающим семенем (Dudley, 1937). По семяпочке к халазе
тянется семенной рубчик, который образует гребень приблизительно
0,4 мм шириной. В халазальной части семени рубчик расширяется в
бугорок диаметром около I мм. Семенная оболочка толстая, прочная,
содержит много воздушных полостей, благодаря чему семена бело-
крыльника хорошо плавают. Поверхность семян покрыта восковым на-
летом и поэтому они совершенно не смачиваются водой. Семя содер-
жит обильный эндосперм, в котором заключен крупный зародыш, почти
равный длине семени и дифференцированный на трубковидную семядолю,
зачаток зародышевого корешка и зародышевую почечку.
71
Указывают следующие диплоидные числа хромосом белокрыльника:
?.п - 36 (Dudley, 1937), 2п = 72 (Weislo, 1970).
Онтогенез
При прорастании семян сначала вытягивается вперед самая нижняя
часть семядоли, охватывающая зародышевую почку, верхняя часть се-
мядоли остается в семенной оболочке и поглощает питательные веще-
ства эндосперма (рис.7). После этого нижняя часть семядоли начина-
ет изгибаться вниз и от нее отрастает зародышевый корешок. В ниж-
ней части семядоли, в месте отхождения от нее зародышевого корня,
находится кольцевой валик ("воротничок"), опоясывакщий семядолю.
Затем из отверстия в семенной оболочке выступает верхняя часть
семядоли, из которой, прорывая ее, выходит первый лист. Он ма-
ленький и устроен проще, чем последующие листья (см. рис.7), сла-
бо дифференцирован, но имеет среднюю жилку (Dudley, 1937). Вто-
рой лист не имеет средней жилки, его листовая пластинка переходит
во влагалище, не образуя черешка. У третьего листа появляются при-
знаки типичного срединного листа: средняя жилка, короткий черешок
и более широкая листовая пластинка (см. рис.7). Четвертый лист
развивается', как взрослый лист. По нашим наблюдениям и опытам
М.Дадлея (Dudley, 1937), ни один из проростков не доживал до ста-
дии пяти и более листьев, поэтому их развитие неизвестно. По дан-
ным М.Дадлея (Dudley, 1937), примерно в -половине изученных им
случаев первичный корень не развивался совсем, но проросток полу-
чал питание через придаточный корень, возникший из центрального
вдлиндра непосредственно под семядольными лопастями немного выше
"воротничка".
Кроме семенного возобновления белокрыльник имеет вегетативное
размножение при помощи корневищ или отламывающихся почек возобнов-
ления (рис.8). Первоначальное заложение почек происходит в пазу-
хах чешуевидных листьев крупной материнской почки (почка в почке
или система годичного побега в почке), из которой на будущий год
сформируется надземный побег. Следовательно, в нормально развитой
почке осенью можно обнаружить зачатки маленьких боковых почек, ко-
торые на следующий год сами превратятся в нормальные крупные поч-
ки, а еще через год в надземные побеги.
Молодой побег, образующийся из почки возобновления, имеет дву-
килевой предлист, за которым следуют 2-5 бесцветных листьев, а за
72
1
2
3
Рис.7. Прорастание семени и формирование проростка белокрыльника
болотного: 1-2 - появление нижних частей семядолей; 3 - по-
явление верхних частей семядолей и главного корня; 4-6 -
появление первого листа; 7 - появление второго листа;
8 - появление третьего листа.
Рис.8» Боковая отрывающаяся почка белокрыльника
болотного
(1а - общий вид, б - почка без листьев) и моло-
дой побег, сформировавшийся из нее (2).
ними срединные зеленые листья. Осенью у растения можно обнаружить
или крупную почку, или небольшой укороченный побег до 7 см длиной
(рис.8), покрытый чешуевидными листьями. В течение следующего,
второго года такая почка-побег вырастает в длинный горизонтальный
побег до I м длиной. Он имеет 10-20 зеленых срединных листьев.
Осенью второго года на нем закладывается терминальное соцветие.
74
Сначала возникает покрывало, затем соцветие в виде недифференциро-
ванного конуса нарастания. Покрывала в той или иной степени охва-
тывают зачаток соцветия, нередко их края завернуты вокруг початка
(Кожевников, 1877). Такой бутон имеет длину около I см. Если по-
крывало не защищено влагалищем верхнего листа, то оно зимой может
быть повреждено и весной не развивается.
Молодые цветки белокрыльника появляются прежде всего близ ос-
нования початка в виде небольших бугорков, спирально расположен-
ных на общей оси, позже появляются вверху початка. Вследствие
быстрого разрастания бугорков они соприкасаются своими краями и
от взаимного давления принимают неправильную многогранную форму.
С моментом изменения внешней формы цветковых бугорков совпадает
появление первых тычинок, которые развиваются в виде небольших бу-
горков по краям зачатка завязи. Ни в расположении тычиночных бу-
горков, ни в числе их (от 6 до 10) не заметно особенного постоян-
ства. Тычинки возникают неодновременно, и даже на более сформиро-
ванных початках видно, что они развиты в разной степени. В каждой
тычинке вскоре можно отличить нижнюю часть, соответствующую нити,
и более широкую верхнюю с двумя выступами наверху, соответствую-
щими половинкам пыльника. Вследствие более энергичного роста
внутренней стороны тычинки, пыльники со временем изменяют свое
положение, так что гнезда, принадлежащие верхней (внутренней)
стороне тычинки, лежат несколько выше, чем на наружней стороне.
Поэтому пыльники взрослой тычинки сидят на окончании связника и
слегка обращены наружу от завязи. Уже вскоре после дифференциации
пыльников правильное размещение тычинок на поверхности початка те-
ряется вследствие закручивания тычиночных нитей. Принадлежность
тычинок к определенному цветку перестает быть очевидной, и грани-
цы между отдельными цветками становятся неясными.
Вскоре после появления тычинок начинается и развитие завязи.
Середина цветка как бы отстает в своем формировании, и в результа-
те этого возникает кольцевой валик завязи (Кожевников, 1877). Сна-
чала края валика ровные, но вскоре на них выступают бугорки, рас-
тущие к центру цветка. Число этих бугорков непостоянно, их никог-
да не бывает менее трех, но в цветках нижней части початка неред-
ко более трех. Загибаясь к центру завязи, бугорки постепенно смы-
каются и образуют полость завяьи. Между ними наверху остается
щель (или отверстие) с несколькими выступами, покрывающаяся позже
длинными волосками (Кожевников, 1877), которые, по нашему мнению,
лучше считать липкими пациллезными выростами. Щель, соответствую-
75
Ulftrt рыльцу, ведет в канал, токе усаженный слизевыделякщими волос-
ками и служащий для прохождения пыльцевых трубок внутрь завязи.
Одновременно с изменениями формы завязи внутри нее развивают-
ся плаценты и семяпочки. На продольном разрезе завязи в период,
когда края ее не сомкнулись, видно, что дно ее становится выпук-
лым. По краям конуса появляются бугорки - зачатки семяпочек. Сна-
чала они растут вверх, но вскоре загибаются к стенкам завязи, по-
том к ее дну. У семяпочки развивается сперва внутренний, затем
наружный интегументы, и она занимает обратное положение, свойст-
венное ей во взрослом состоянии. Число семяпочек колеблется от 4
до 12. Они сидят на плацентах, расположенных в виде кольца. Осно-
вания плацент срастаются, имея наверху небольшие щели. Выше пла-
центы свободны, на поперечном разрезе имеют трапециевидную или
треугольную форму, на внешней их стороне находится вырезка, дохо-
дящая почти до места прикрепления семяпочки (Кожевников, 1877).
На каком-то этапе развития молодого початка все его цветки
очень сходны, имеют тычинки и завязь. Но вскоре наступают разли-
чия, которые в общих чертах состоят в том, что в нижних цветках
початка завязи достигают наибольшего развития, тогда как в верх-
них они развиты слабее, а на самой вершине початка совершенно ат-
рофируются. Этим и объясняется наличие одних тычинок на вершине
зачатка. В самых верхних цветках завязь очень рано останавливает-
ся в своем развитии на стадии кольцевого валика и впоследствии
закрывается со всех сторон разрастающимися тычинками. Число тычи-
нок в цветках зависит от положения последних в початке. Нижние
цветки имеют не более 6 тычинок, а верхние - до 10. Сказанное по-
казывает, что у белокрыльника при наличии обоеполых цветков суще-
ствует тенденция к разделению полов (Кожевников, 1877). К концу
вегетационного периода цветки полностью формируются, но ваттветяет
растение весной следующего года.
Рядом с соцветием и одновременно с ним закладывается боковая
почка возобновления с двукилевнм предлистом в своем основании.
Почка сильно сдвинута к вершине побега и поэтому ее можно принять
за терминальную. Из нее на следующий год разовьется олиственный
побег 2-го порядка, который продолжит нарастание в длину главного
побега и осенью закончит свое развитие по уже описанной схеме. Пе-
резимовавшая прошлогодняя часть побега в течение лета постепенно
отмирает, и на зищу остается уже новый побег, выросший за лето из
почки возобновления.
76
Сезонное развитие
Белокрыльник болотный относится к труппе растений, в почках
возобновления которых побеги будущего года сформированы полностью,
включая соцветие. Весной происходит лишь рост побега, заложенного
в почке предшествующим летом. В Московской обл. рост побегов рас-
тения начинается в середине апреля, реже в начале мая, что опре-
деляется погодными условиями (Серебряков, Галицкая, 1951). Во вто-
рой половине мая белокрыльник зацветает, до цветения на побеге
формируется 1-2 срединных листа. Цветение продолжается до середи-
ны июня.
Растение характеризуется протогинией (Hock, 1911; Dudley,
1937). Пыльники начинают раскрываться в початке только тогда, ког-
да почти все завязи уже опылены пыльцой других особей. На первой
стадии рыльца покрыты папиллезными прозрачными выростами и готовы
к восприятию пыльцы. Тычиночные нити на этой стадии короткие и
пальники находятся ниже рылец. После того как большинство рылец в
соцветии опылено (при этом их папиллезные выросты подсыхают и
рыльца становятся темными), происходит удлинение тычиночных нитей
на 3-4 мм, пыльники оказываются выше рылец и затем раскрываются.
По данным Ф.Хека (Hock, 1911), нити до этого находятся в скручен-
ном состоянии, а перед раскрытием пыльников распрямляются. По на-
шим наблюдениям, удлинение тычиночных нитей происходит только в
результате их быстрого роста. У отдельного цветка раскрывание
пыльников происходит разновременно, а у початка в целом - акропе-
тально. Протогиния препятствует самоопылению цветков. В соцветиях,
закрытых нами марлевыми колпаками на стадии бутона, не развилось
ни одного плода. Следовательно, ни самоопыления, ни постоянной
гейтеногамии у белокрыльника нет. Раскрывание пыльников в почат-
ках может происходить и без определенной последовательности, иног-
да одновременно в верхних и нижних его цветках (Hock, 1911), но
развитие пылец осуществляется в початке строго акропетально. В
Зврхней части початка, а также в той части, которая обращена к по-
крывалу, может происходить гейтоногамия с помощью насекомых, осу-
ществляющих перекрестное опыление (пыльца растения склеена в комоч-
ки и совсем нелетуча).
В середине июля в верхушечных почках корневищ белокрыльника за-
кладываются новые соцветия, у которых к концу вегетащюнного пери-
ода полностью сформированы цветки, включая тычинки и пестики (Се-
77
рвбрлков, Галицкая, 1951). В июле заканчивается заложение новых
почек возобновления, но их размеры увеличиваются до осени. Форми-
рование почек возобновления у белокрыльника начинается за два го-
да до юс развития в побеги, т.е. в пазухах зачатков зеленых
листьев почек будущего года.
В середине августа появляются первые зеленые плоды и начинает-
ся отмирание листьев. К концу сентября листья и корневища прошло-
го года полностью отмирают. На зиму живым остается корневище, вы-
росшее в уходящем году и несущее боковые и верхушечную почки воз-
обновления.
Способы размножения и распространения
Белокрыльнику свойственно семенное и вегетативное размножение.
Осенью цветонос подгнивает и падает вместе с початком на влажную
почву или в воду. Плоды от разбухания студенистой прозрачной мас-
сы разрываются в месте прикрепления к оси початка, початок быстро
распадается, семена легко высвобождаются из ягод. В одном початке
содержится 350-400 семян. В основном семена распространяются во-
дой, они хорошо плавают благодаря наличию большого количества воз-
духоносных полостей в оболочке. Проростки растения встречаются
часто, но в небольшом числе и почти исключительно в местах, где
на поверхности имеется слой воды (Солоневич, 1956). Здесь они пла-
вают и укореняются в отмерших остатках растений.
Образование густых куртин белокрыльника площадью в несколько
квадратных метров осуществляется исключительно за счет вегетатив-
ного разрастания и размножения. Ежегодно на годичном приросте кор-
невища возникает 1-5 боковых ветвей. При отмирании главного корне-
вища боковые ветви теряют с ним связь и продолжают далее существо-
вать самостоятельно, нарастая в длину и образуя новые боковые по-
беги. Кроме того, вегетативное размножение происходит за счет тех
почек возобновления, которые не выросли в боковые побеги, а отло-
мились от основного корневища на ранней стадии своего развития
(рис.8). Отламывание почек происходит легко, так как основания по-
чек тонкие и хрупкие. Почки могут распространяться как внутри то-
го же водоема, так и путем переноса птицами в другие места, прили-
пая вместе с грязью к ногам птиц. У почек образуются придаточнца
корни и в благоприятных условиях они формируют новые растения.
78
Экология и фитоценология
Белокрыльник приурочен к сильно обводненным местообитаниям с
относительно богатым минеральным питанием: эутрофным и мезотроф-
ным болотам, берегам стариц, озер, ручьев, сплавинам, канавам.
Наиболее интенсивно вегетативное размножение происходит на при-
брежных участках озер, на мелководьях с хорошим грунтовым или на-
мывным питанием. Условно средняя степень развития особей вида от-
мечена на молодой сильно увлажненной сплавине, а наименьшая - на
торфянистой, мощной сплавине.
Белокрыльник - типичный гигрофит (Hock, 1911). В заостренном
кончике его листа, на верхней и нижних сторонах размещаются много-
численные гидатоды, через которые вода выделяется в виде капель.
Весьма теневыносливое растение, хорошо развивается под пологом
ольхи черной (Солоневич, 1956). На озерных сплавинах и островках
белокрыльник образует обширные заросли в совершенно безлесных со-
обществах. Здесь молодые листья растения обычно свернуты в трубку
и расположены вертикально. Свертывание пластинок и их вертикаль-
ное расположение можно расценивать как приспособление против силь-
ного испарения воды и перегрева лучами солнца.
По экологической классификации растений (Eiienberg, 1974) бе-
локрыльник - полутеневой-полусветовой вид (6-я ступень шкалы),
растущий на сырых, плохо аэрируемых почвах (3-я ступень шкалы
влажности), со слабокислой, нейтральной, в некоторых случаях кис-
лой или основной реакцией (6-я ступень по отношению к pH почвы),
бедных или промежуточных по содержанию азота (4-я степень шкалы
обеспеченности азотом). По Э.Котилайнену (Kotiiainen, цит. по Со-
лоневич, 1956), белокрыльник - слабоацедофильный вид, для него
благоприятны местообитания со значениями pH = 4,1-6,0. Согласно
данным А.А.Немчинова (1934), на болотах Ленинградской обл. бело-
крыльник обитает на субстратах с pH = 5,25-5,85.
В почве корневище белокрыльника всегда расположено горизонталь-
но, но на различной глубине в разных экологических условиях. В
водной среде оно размещается (плавает) по поверхности, как и в
почвах с очень высоким уровнем грунтовых вод. В местообитаниях,
где уровень грунтовых вод летом снижается на 10-20 см, корневище
расположено приблизительно на такой же глубине. Редки случаи, ког-
да корневище растения лежит в рыхлом слое сфагнового ковра несколь-
ко выше уровня воды.
79
Многочисленные корни сосредоточены у белокрыльника на переднем
конце корневища, где они отходят от его узлов вниз или наклонно в
глубь торфа. В воде корни плавают в поверхностном слое. Длина кор-
кой зависит от экологических условий: в водоемах они наиболее
длинные - до 70-85 см (Солоневич, 1956). На сфагновых болотах со
олаборазложившиися поверхностным слоем торфа белокрыльник имеет
более короткие корни - до 20-25 см. В плотном и сильнее разложив-
шемся торфе корни растения становятся извилистыми и еще более ко-
роткими - 10-15 см. Следовательно, чем плотнее субстрат, на кото-
ром растет растете, тем короче его корни (Metsavainio, 1931).
Несмотря на значительную массу белекрыльника в некоторых болот-
ных фитоценозах (например, в черноольшанниках), его роль как тор-
фообразователя ничтожна вследствие быстрого разрушения его отмер-
ших частей (Солоневич, 1956).
Белокрыльник - компонент гигрофильных и болотных фитоценозов.
В эутрофных местообитаниях, например в черноольшанниках, вероят-
но, оптимальные условия для растения: он образует здесь густые и
больше заросли, побеги достигают 35-40 см высоты и имеют листья
с крупными пластинками. Также успешно вид развивается на сплави-
нах, образуя чистые заросли. Наиболее крупны его особи по краям
сплавины. Гораздо хуже развивается белокрыльник в травяно-сфагно-
вых сообществах. Здесь уменьшается его численность, он не образу-
ет чистых зарослей. Побеги размещены рассеянно, их высота лишь
10-15 см, листовые пластинки некрупные. Признаки сильного угнете-
ния выражены у белокрыльника особенно сильно в местообитаниях с
мощным сфагновым покровом. В этих случаях из мха торчит только
полупогребенная листовая пластинка желтоватого цвета небольших
размеров (Солоневич, 1956).
Консортивные связи
Белая окраска верхней стороны покрывала и слабый неприятный
запах цветков, лишенных нектара (Kugler, 1970), способствуют при-
влечению к цветущим початкам насекомых, любящих гниль. Среди них
особенно характерны двукрылые: Drosophilla graminum Fall., Hyd-
rella griseola Fall. И ВИДЫ ИЗ родов Chironomus И Tachydromia,
а также мелкие жуки Cassida nobilis L., Aphthona coerulea Payk.,
Hypera polygon! L., Sitona sp., Melygetes sp. (Knuth, 1899). Жу-
ки из рода Oxythyrea осуществляют опыление цветков белокрыльника
(Hock, 191т). По нашим наблюдениям, на цветках растения чаще
всего встречались мелкие жуки из рода Melygetes (сем. Nitiduii-
dae), которые питаются пыльцой и опыляют цветки. Они значитель-
но преобладали по числу особей над другими видами насекомых, по-
сещавших цветки, и поэтому их можно считать основными опылителями
в районе исследования. Кроме того, на цветках зарегистрированы жу-
ки Anisostica novemdecimpunctata L. (Coccinellidae) и мухи Helo-
philus lineatus F. (Syrphidae), которые также могут опылять
цветки. Опылителями могут быть трипсы Thysanoptera (Hook, 1911)
и улитки Helix hortensis l>., которые, ползая по початкам, пачка-
ются в пыльце (Knuth, 1899).
Белокрыльник повреждается некоторыми чещуекрылыми. Гусеницы
Arctia cajja L. И Pergesa elpenor L. повреждают ТОЛЬКО ЛИСТЬЯ,
а гусениця листовертки Clepsos spectrana Tr. (C.costana F.) по-
вреждают листья и початки. Первые два вида встречаются редко и не
приносят существенного вреда растению. Гусеницы листовертки встре-
чаются гораздо чаще и в годы своего массового размножения могут
сильно повреждать початки.
Биологическая продуктивность и хозяйственное значение
Для определения биологической продуктивности белокрыльника бы-
ли сделаны нр уровне водной поверхности три укоса надземной фито-
массы, два - на оз.Спасском, один - на оз.Высоковском (Ивановская
обл., Фурмановский р-он) (табл.2).
Таблица 2.
Продуктивность надземных (надводных) побегов белокрыльника
болотного в различных местообитаниях
। Место укоса и на- ! звание ассоциации I ' Время укоса Проектив- ное по- крытие , % Высота побе, гов, см Фитомасса, г j
сырая сухая ]
1 Оз.Спасское, вехо- I во-белокрыльнико- । вая 21/У1 60 25-30 800 92 |
1 Оз.Спасское, ооо- । ково-белокрыльни- । ковая 14/УП 80 30 1550 180 ।
। Оз.Высоковское, бе- । локрыльниковая 31/УП 100 30-35 2100 250 1
6-1964 81
Вое части растения в сыром виде ядовиты - содержат острое жгу-
чее вещество. Ткани его содержат кристаллы щавелевокислого каль-
ция (Hook, 1911). В корневище много крахмала (Флора СССР, т.З,
1936), а так как высушенное или сваренное оно становится безвред-
ным, тс в старину крестьяне северных губерний России примешивали
его к ржаной муке. Отсюда и произошло название растения - "хлеб-
ница" (Анненков, 1878). Но некоторым указаниям (Некрасова, 1949),
белокрыльник употребляется на корм свиньям, тогда как ряд. других
авторов считают, что скот его не поедает из-за неприятного запаха
(Hock, 1911). Существует мнение, что по той же причине семена не
поедают и водоплавающие птицы (Hock, 1911). По нашим наблюдени-
ям, живые органы растения не имеют неприятного запаха - он харак-
терен только для отмерших разлагающихся частей.
82
М.Г.Вахрамеева, С.В.Никитина, Л.В.Денисова
РОД РЯБЧИК
Род рябчик (Fritillaria L.) относится к семейству лилейных
(Liliaceae). Многолетние луковичные травянистые растения с поник-
шими цветками. Околоцветник простой венчиковидный 6-членный с нек-
тарниками. Тычинок 6, с продолговатыми пыльниками. Столбик I,
рыльце 3-раздельное. Плод - коробочка. Семена плоские, в 2 ряда в
гнезде, светло-коричневые или золотисто-желтые, В роде около 80
видов, в СССР - 26, в Московской обл. - 2. Латинское название ро-
да происходит от fritilius - шахматная доска.
Ключ для определения московских видов
I. Листья мутовчатые, супротивные или очередные, линейно-лан-
цетные, плоские, отклонение прямостоячие, верхние линейные, с тон-
кой усиковидной закрученной верхушкой. Плод 6-гранный с крылатыми
углами. F.ruthenica.
2. Листья очередные, широколинейные, желобчатые, отклоненные
или вниз загнутые. Плод 3-гранный, с тупыми углами. F.meleagris.
РЯБЧИК РУССКИЙ
Номенклатура и географическое распространение
Fritillaria ruthenica Wikstrom, 1821, Kungl. Sv. Vet.-Acad.
Handl.:353; Кауфм., 1866, Моск. фл., изд. 1:495, 1899, изд. 2:
512; Дингер, 1886, Сб. свел.:431; Шмальг., 1897, Фл. 2:507; Пе-
тунн., 1901, Крит, обзор, 3:68; Сырейщ., 1906, Илл. фл. 1:239; он
же, 1927, Опр. раст. Моск. губ.:90; Лозина-Лозинская, 1935, Фл.
СССР 4:306; Назаров, 1940, в Маевок., Фл., изд. 7:227; Победимова,
1954, там же, изд. 8, 1954:675, 1964, изд. 9:553; Скворцов, 1966,
Опр. раст. Моск. обл.:115; Артюшенко, 1979, Фл. евр. части СССР,
4:237; Hix, 1980, Fl. Bur. 5:33, - Рябчик русский.
Евразиатский вид, ареал занимает значительную территорию в ев-
ропейской и азиатской частях СССР. Западная его граница проходит
по Украине: Киевская обл. - Кировоградская обл. - Криворожская
83
одд., далее на юг она идет через Запорожскую, Ростовскую, Волго-
градскую, Саратовскую, Уральскую, Актюбинскую, Кустанайскую и Ка-
рагандинскую области. Северная граница ареала немного не доходит
до верховьев Дона, затем идет через Тамбовскую обл., Татарскую
АССР, Куйбышевскую обл., Башкирскую АССР, Оренбургскую, Кустанай-
скую области. Самое южное местонахождение вида в Ставропольском
крае, в окрестностях г.Невинномнсска (Флора СССР, т.4, 1935;
Гроссгейм, 1940; Поляков, 1941); самое северное - в Серпуховском
р-не Московской обл. Указание Н.Н.Кауфмана (1886) о произрастании
рябчика русского на правобережье Оки между реками Беспутйй и Скни-
гой А.К.Скворцов (1969) счиигет относящимся к искусственно переса-
женным особям с левого берега р.Оки. В последние годы на правобе-
режье Оки вид не обнаружен (Алексеев, Губанов, 1980).
Морфологическое описание
Рябчик русский - геофильный эфемероид (Ротов, 1972а). Луковица
до I см в поперечнике, сплюснутая, состоит из донца, одной запаса-
ющей мясистой обратнояйцевидной чешуи и двух пленчатых чещуй.
Пленчатые чешуи расширены у основания и сужены к вершине, края их
смыкаются, они с двух сторон охватывают лист с красновато-коричне-
вым черешком и буро-зеленой свернутой листовой пластинкой. В осно-
вании листа находится замещающая луковица, состоящая из одной мя-
систой чешуи и зачатка другой чешуи.
Стебель круглый, 20-65 см высоты, прямой, начиная со второй
трети олиственный. Листья в числе II-I9, супротивные или очеред-
ные, слегка стеблеобъемлющие, линейные, острые, 6-9 см длины, 3-5
мм ширины; верхние супротивные, нитевидные, более короткие, сбли-
женные, с тонкой усиковидной закрученной верхушкой.
Цветки в пазухах верхних листьев. Цветоносы изогнутые, значи-
тельно короче околоцветников. Околоцветник колокольчатый, темно-
красный с темным неясным шахматным рисунком, внутри желтый, с зе-
леноватой полоской, расширяющейся книзу посреди доли. Наружные до-
ли околоцветника узкоовальные, до 3 см длины, до 6 мм ширины;
внутренние - обратнояйцевидные, с оттянутой тупой верхушкой, до
3,5 см длины и до 1,5 ом ширины в самой широкой части. Нектарники
округло-овальные, резко выдающиеся наружу в виде бугорков. Тычин-
ки короче околоцветника более чем в два раза. Пестик до середины
разделен на 3 рыльца, превышает длину тычинок. Коробочка до 2 см
длины и 1,8 см ширины, сверху притупленная, внизу слегка суженая.
Масса I тыс. семян 1,8 г.
Диплоидное число хромосом - 18 (Хромосомные числа..., 1969).
Онтогенез, сезонное развитие, способы размножения
Онтогенез рябчика русского изучен слабо. По наблюдениям
В.М.Двораковской (1969, 1973а,б), семена имеют недоразвитый заро-
дыш, в связи с чем они отличаются замедленным прорастанием. В
культуре для их успешного развития необходимы сначала теплый, а
потом холодный периоды. Во время теплого периода (оптимум 10-12°)
происходит доразвитие зародыша, поэтому при ранних сроках посева
(июль - август) наблюдается и лучшая всхожесть. При позднем сроке
посева (конец августа - сентябрь) зародыш в семенах часто не успе-
вает развиться в связи с низкими ночными температурами, что явля-
ется одной из причин низкой всхожести. В первый год семядоля с се-
менной кожурой на конце выносится на поверхность, выполняя функ-
цию листа. На второй год появляется один зеленый лист, на третий -
функционируют два листа. Вначале формируется укороченный вегета-
тивный побег с розеточными листьями, при переходе в генеративное
состояние из терминальной почки розеточного побега образуется уд-
линенный цветоносный побег с зелеными листьями (Голубев, 1965).
Генеративный период наступает на 4-5-й год жизни (Ротов, 1973а).
По наблюдениям в Приокско-Террасном заповеднике (Московская
обл.), мы выделили следующие возрастные группы рябчика русского.
Ювенильные - особи с одним небольшим листом длиной от 2,4 до 7,5
(в среднем 5,2+0,7) см и шириной 0,3-0,9 (в среднем 0,5+0,!) см.
Имматурные - особи также с одним, но более крупным листом длиной
6-11,4 (в среднем 8,1+1,6) см и шириной 0,7-1,6 ( в среднем
1,0+0,2) см. Взрослые виргинильные особи имеют большее число
листьев. Среди них выделяются две подгруппы. В первую отнесены
особи, имеющие 2-4 листа, расположенных в основном мутовчато, но
встречается и очередное листорасположение (из 40 особей этой груп-
пы 3 имели очередные листья); высота растений - 12-27 (в среднем
17,3+4,5). Другая подгруппа - особи с 5-10 листьями, расположен-
ными также мутовчато, но встречается и смешанное мутовчато-очеред-
ное листорасположение. Генеративные особи имеют 7-18 листьев,
3-10 верхних нитевидных листа, 1-3 цветка (в единичных случаях до
7 цветков); высота растений от 18 до 85 см.
6х-!964 85
По ритму развития рябчик русский - средневесеннее растение с
одной генерацией листьев и побегов (Голубев, 1965). Гост листьев
начинается во второй декаде апреля и продолжается до конца второй
декады мая. К середине апреля, когда надземная часть рябчика толь-
ко начинает отрастать, у луковицы появляются придаточные корни.
Рост и развитие большинства листьев происходит :при сравнительно
невысокой средней температуре (9-15°). К концу первой декады мая
надземные органы большинства особей рябчика русского уже заканчи-
вают свой рост (Двораковская, 1973а).
За 20-летний срок (с 1946 по 1966 г.) наблюдений в Приокско-
Террасном заповеднике наиболее раннее начало цветения рябчика
русского отмечено 21 ДУ 1956 г., наиболее позднее - 13/У I960 г.,
средняя многолетняя дата приходится на 5Д (Каляев, 1971). Таким
образом, амплитуда колебаний между наиболее ранним и наиболее
поздним сроком зацветания составила 22 дня. Средняя многолетняя
дата массового цветения приходится на 17 мая, отцветания -
22 мая. Плодоношение в конце мая - начале июня.
Продолжительность вегетации рябчика составляет 75-80 дней (Ро-
тов, 1976). В зависимости от сроков и времени оттаивания почвы
колебания между сроками начала вегетации в отдельные годы могут
достигать 20-22 дня. Отмирание надземных органов начинается в на-
чале июня, одновременно отмирает и корневая система. В период
летнего покоя сохраняется только луковица, состоящая из мясистой
чешуи и зачатка пленчатых чещуй. В первую декаду сентября у луко-
вицы появляются корни, и перед уходом под снег луковица имеет за-
чаток стебля или листа. По данным В.Н.Голубева (1965), рост
листьев и стеблей происходит еще под снегом. Развитие этих орга-
нов в зимний период представляется важным этапом, обусловливаю-
щим раннее цветение растения.
Рябчик русский размножается как семенным, так и вегетативным
путем. При вегетативном размножении происходит либо заложение в
пазухах запасающих луковичных чешуй двух почек возобновления, каж-
дая из которых дает начало самостоятельной дочерней луковице, ли-
бо идет образование нескольких придаточных выводковых почек (Ро-
тов, 1973а).
Наблюдения над цветением и плодоношением рябчика в Приокско-
Террасном заповеднике показали, что наибольшее число плодоносящих
особей приходится на опушку сосново-дубового леса (97,4% от цвету-
щих особей), наименьшее - на типчаково-разнотравную степь со степ-
ной вишней (88,11%). В среднем в коробочке содержится 59,7 семян.
86
По нашим подсчетам, на I м остепненного луга приходится 1200 се-
мян рябчика русского.
Экология
Рябчик русский - ксеромезофит, встречается обычно в относи-
тельно увлажненных местообитаниях: по склонам и .днищам балок и
оврагов, в микропонижениях. В основном он использует атмосферную
влагу, практически не получая капиллярно-поднимапцуюся почвенную
влагу. Это связано с тем, что его луковица с придаточными корнями
находится в самом верхнем горизонте (Голубев, 1965). Раннее пре-
кращение вегетяции и переход растения в состояние покоя являются
хорошим приспособлением к перенесению засухи. Перенос растения в
мезофильные условия способствует увеличению его общей продуктив-
ности. В степях под Курском средняя высота рябчика 36 см, сред-
нее число цветков у одной особи - 1,08, а в посадках в Москве
(Главный ботанический сад АН СССР) соответственно 53 и 3,4 (Ро-
тов, 1974). В естественных местонахождениях в Московской обл.,
по нашим наблвдениям, средняя высота рябчика 48,9 см, среднее чис-
ло цветков у особи - 1,4.
Требователен к почвам. В Приокско-Террасном заповеднике рябчик
произрастает на богатых гумусом (до 10%) аллювиальных наносах пой-
мы Оки (Смирнов, 1958). В лесостепной и степной зонах чаще встре-
чается на выщелоченных черноземах, избегает более бедные серые
лесные почвы. Нередко растет на меловых и известковых субстратах.
К низким температурам и резким колебаниям дневных температур
ранней весной рябчик русский устойчив, он развивается в сравни-
тельно холодный отрезок вегетационного периода. Последние замороз-
ки почвы в Московской обл., по средним многолетним данным, прихо-
дятся на 16 мая, т.е. на период цветения рябчика. В малоснежные
зимы могут повреждаться луковицы, причем в такие зимы не наблюда-
ется характерного для рябчика Подснежного развития листьев и побе-
гов (Голубев, 1965).
Обычно рябчик встречается на хорошо прогреваемых, но затенен-
ных участках: опушках, под отдельно стоящими деревьями, среди за-
рослей степных кустарников. И.К.Пачоский (1915) относил рябчик
русский даже к теневым растениям. Умеренное затенение оказывает
благоприятное воздействие на рост и развитие рябчика.
Наши исследования (табл.З) позволили выделить два экоэлемента -
87
лооной и степной, что находится в соответствии с данными Р.А.Го-
това (1974, 1976) о существовании светового (степного) и тенево-
го (лесного) экоэлементов рябчика русского. В лесных сообществах
рябчик заметно выше, чем в степных (остепненных лугах), у него
большее число зеленых листьев, и обычно большее число цветков и
размеры их крупнее.
Таблица 3.
Морфологические особенности рябчика русского в различных
фитоценозах Приокско-Террасного заповедника
. Морфологические признаки Типчаково- разнотрав- ный луг Ковыльно- разнотрав- ный луг Липняк сны- тево-пер- ловниковый Молодой осинник перловни- ковый
1 Высота, см 32,4+7,4 37,8+6,4 53,9+1,8 41,6+6,8
Число нижних 1 листьев 9,7+1,7 9,2+1,6 9,3+1,4 9,7+1,5
Число верхних 1 листьев 5,6+1,6 4,8+1,7 5,1+2,1 5,4+1,9
Число цветков I,6+0,7 1,5+0,7 I,5+0,4 1,6+0,8
। Длина околоцветни- । ка, см 2,2+0,3 2,3+0,3 2,3+0,3 2,5+0,3
Фитоценология
Наиболее широко распространен рябчик русский в лесостепи, до-
вольно часто встречается в степи, реже в подзоне широколиственных
лесов (Поляков, 1941; Крашенинников, Кучеровская-Рожанец, 1941).
В лесостепи и северной степи рябчик русский отмечен как на водо-
раздельных плато, так и по склонам логов (чаще киной экспозиции).
В степной зоне наиболее часто рябчик растет не на открытых степ-
ных участках, а на лесных полянах, опушках дубрав и березняков
вместе с луговыми и лесолуговыми видами. В подзоне южных степей
рябчик русский встречается редко и не обильно, чаще в лесах и по
склонам балок. На территории Казахстана он тяготеет к долинам рек,
склонам балок и оврагов.
В Московской обл. рябчик русский приурочен к ее южной остепнен-
ной части. Наибольший интерес представляют два местонахождения
рябчика русского - лужковская степная колония, расположенная в юж-
ной части Приокско-Террасного заповедника, и никифоровская - к
88
востоку от первой. Они являются наиболее северными и приурочены к
особым локальным местообитаниям, к так называемым "долам" - участ-
кам поймы с богатой колонией окской флоры, обогащенной степными
•видами. Популяции рябчика русского находятся в наиболее удаленной
от Оки части поймы, непосредственно примыкающей к боровой окраи-
не. Рябчик русский является здесь пойменным растением, обитающим
на богатых гумусом (до 10%) аллювиальных наносах. Он очень типи-
чен для зарослей кустарников из вишни степной, для разреженных
лесных участков из дуба и березы, для разнотравно-типчаково-ко-
выльных сообществ. Численность его в данных местообитаниях дости-
гает максимальных величин, отмечается прекрасное развитие и обык-
новенно (кроме очень сухих лет) обильное плодоношение (Смирнов,
1958).
В табл.4 приведены численность и возрастной состав популяций
рябчика в различных фитоценозах Приокско-Террасного заповедника
(по данным обследования 1976 г.).
Наибольшая численность особей всех возрастных групп в 1976 г.
отмечена в типчаково-разнотравном фитоценозе с вишней степной,
наименьшая - на типчаково-разнотравном остепненном лугу. Здесь,
как и в ассоциации на опушке сосново-дубового леса, .в популяции
рябчика преобладают взрослые виргинильные и генеративные особи.
Наилучшее возобновление рябчика отмечено в лесных ассоциациях. В
целом можно сказать, что в Приокско-Террасном заповеднике рябчик
русский находится в хорошем состоянии и хорошо возобновляется.
Хозяйственное значение и охрана
Рябчик русский - красиво цветущее весеннее растение - пред-
ставляет большой интерес для интродукции. Наблюдения Р.А.Ротова
(1974) показали, что в культуре у рябчика увеличиваются размеры
вегетативных органов и число цветков. Его можно рекомендовать для
посадки в садах, парках., на открытых или слабозатененных участках
с хорошо удобренными почвами. Введение его в культуру будет спо-
собствовать и охране вида.
Численность и ареал рябчика в последние годы значительно сокра-
тились главным образом за счет изменения или разрушения мест его
обитания. Сбор рябчика как декоративного растения вызывает сокра-
щение численности его вида. В связи с этим он внесен в "Красную
книгу СССР" (1978) как исчезающий вид. Он охраняется в ряде запо-
89
Таблица 4.
Средняя численность рябчика русского (на I м2) в различных
фитоценозах Приокско-Террасного заповедника
1 Название 1 фитоценоза ' Возоастное состояние Общее число
ювениль- ное имматур- ное взрослое вирги- нильное генера- тивное
Глипняк снытево- . перловниковнй 10,0 (42,4)s 3,6 (15,2) 6,8 (28,8) 3,2 (13,6) . 23,6
Молодой осинник 1 перловниковнй 8,0 (29,0) 4,4 (15,0) 5,2 (18,0) 10,4 (33,0) 28,0
1 Опушка сосново-ду- 1 бового леса с гсс- । подством перловни- ка в травянистом 1 ярусе 0,8 (6,9) 0,4 (3,4) 3,2 (27,6) 7,2 (62,1) 11,6
1 Ковндьно-разно- । травный остепнен- . ный луг 2,4 (7,6) 4,0 (12,6) 12,8 (40,6) 12,4 (39,2) 31,6
1 Типчаково-разно- 1 травный остепнен- I ный луг с чемери- 1 Чей 4,8 (17,6) 4,8 (17,6) 12,4 (45,6) 5,2 (19,2) 27,2
1 Типчаково-разно- 1 травный остепнен- 1 ный луг с вишней । степной II,2 (17,6) 13,6 (21,4) 21,6 (34,0) 17,2 (27,0) 63,6
* В скобках указан процент особей соответствующего возрастного
состояния от общей численности рябчика в ценопопуляции.
ведников и заказников лесостепной и степной зон СССР. В Москов-
ской обл. следует уделить особое внимание охране этого редкого и
декоративного растения в связи с тем, что это наиболее северная
изолированная часть ареала.
90
2. РЯБЧИК ШАХМАТНЫЙ
Номенклатура, внутривидовая систематика
и географическое распространение
Fritillaria meleagris L., 1753, Sp. pl.:3O4; Шмальг., 1897,
Фл. 2:508; Лозина-Лозинская, 1935, Фл. СССР 4:306; Победимова,
1954, в Маевок., Фл. изд. 8:675; 1964, изд. 9:663; Артюшенко,
1979, Фл. евр. части СССР 4:237; Rix, 1980, Fl. Eur. 5:31. - Ряб-
чик шахматный.
Отмечают большую изменчивость формы и окраски цветка рябчика
шахматного. Выделена разновидность var.brachypoda Back, с ворон-
ковидным околоцветником, широко отстоящими листочиами околоцвет-
ника с зелеными пятнами, очень длинными пыльниками (Hegi, 1939).
Широко распространена форма с белым околоцветником. А.Бушман
(Buschmann, 1951) дает характеристику формы "albino", имеющей
околоцветник от белого до зеленого цвета с более мелкими размера-
ми, особи этой формы чаще имеют не один, а 2-3 цветка.
Среднеевропейский вид, его ареал охватывает значительную часть
Европы (Loew, Kirchner, 1934; Hegi, 1939; Buschmann, 1951; Clap-
ham e.a., 1952). В Советском Союзе рябчик шахматный указывается
для Волжско-Донского, Нижне-Донского, Нижне-Волжского, Заволжско-
го районов (Флора СССР, т.4, 1935). Е.Г.Победимова во "Флоре"
П.Ф.Маевского (1964) указывает на произрастание рябчика шахматно-
го в Калужской, Орловской, Липецкой, Курской областях, считая его
распространение в Куйбышевской, Саратовской, Волгоградской облас-
тях сомнительным. Есть еще несколько островных местонахождений
рябчика шахматного в Латвийской ССР и Ленинградской обл. (Ротов,
19736).
В Московской обл. рябчик шахматный обнаружен недавно (Алек-
сандров, 1971; Ротов, 19736; Курлович, Алексеев, 1975). Самая
крупная популяция найдена в Серпуховском р-не на границе с Калуж-
ской обл.
Морфологическое описание
Рябчик шахматный имеет много общего с рябчиком русским. Луко-
91
лица шаровидная, немного сплюснутая, одета бурой пленчатой оболоч-
кой, до 7-15 мм в диаметре, имеет две мясистые чешуи, располагает-
ся на глубине 5-7 см. По поводу структуры луковицы рябчика сущест-
вует разногласие: И.Г.Серебряков (1952) и Р.А.Ротов (1976) счита-
ют , что она .характеризуется моноподиальной структурой только в
ювенильном возрасте, все остальное время она симподиальна; З.П.Бо-
чанцева (1963) полагает, что луковицы рябчика имеют всегда монопо-
диальное строение. По нашему мнению, первая точка зрения бодее
правомерна.
Высота стебля 12-26,6 (33) см. Листья в числе 2-6 (чаще 4), в
отличие от предыдущего вида все очередные, почти равно отстоящие
друг от друга, расположены в верхней части стебля, широколинейные,
желобчатые, на конце туповатые, отклоненные или вниз загнутые,
при основании полустеблеобьемлющие 6(8)-10,5 (13) см длиной, 3-7
(10) мм шириной.
Цветки обычно одиночные (реже 2-3), колокольчатые, на цветонож-
ке, которая короче верхнего листа. Доли околоцветника продолгова- .
то-эллиптические, кверху слегка суженные, тупые, все почти одина-
ковой величины - 3-4 см в длину, 1-1,5 см в ширину, с четким шах-
матным рисунком темно-пурпурового цвета, чередующимся с беловатым.
Тычинки снизу более бледные, почти в 2 раза короче долей около-
цветника, с линейными желтоватыми пыльниками на беловатых, в 1,5- .
2 раза более коротких, книзу постепенно расширяющихся нитях. Пес-
тик немного длиннее тычинок, до 1/3 рассечен на 3 рыльца (Loew,
Kirchner, 1934; Флора СССР, т.4, 1935). Имеются нектарники (Tur-
rill, 1950). Коробочка в отличие от предыдущего вида 3-гранная с
тупыми гранями, содержит многочисленные семена. Семена мелкие,
длина их 4,3-5,4 мм, ширина 3-3,8 мм, масса I тыс. семян 2,2 г
(Ротов, 19736, 1974, 1976).
Диплоидный набор хромосом 24 (Clapham е.а., 1952; Beck, 1953;
Garcke, 1972).
Онтогенез и сезонное развитие
Семена прорастают обычно осенью. Ювенильные особи имеют один
ассимилирующий лист с длинным, расположенным в почве черешком, пе-
реходящим в ланцетную по форме надземную пластинку, --4-I4 см дли-
ной, 0,4-1 см шириной (Ротов, 19736, 1974), луковица - 4-9 мм в
диаметре. По нашим наблюдениям, в Серпуховском р-не на лесных и
92
луговых участках имматурные особи имеют 1-3 листа, нижний лист
длиной 6-16 см, шириной 8,5-18 см. Взрослые виргинильные особи
высотой 10-22 см, с 3-7 листьями, длина нижних листьев 8-18 си,
ширина 0,5-1,2 см. Преобладают особи с 4-6 листьями (до 93$). Ди-
аметр луковиц 8-15 см. Генеративные особи высотой 15-37 ом, с
4-5 зелеными Листьями, дайна нижних листьев 7-18 см, ширина 0,6-
1,4 см. Все встреченные нами особи имели только один цветок, дли-
на его 2-4,5 см, диаметр 2-3,8 см, однако есть указания на разви-
тие двух цветочных побегов (Loew, Kirchner, 1934).
Рябчик шахматный начинает вегетировать в конце марта, рост зе-
леных листьев опережает цветение. Р.А.Ротов (19726, 1976) относит
этот вид к луковичным геоэфемероидам. Цветение начинается в конце
апреля - первых числах мая и продолжается от 5 дней до двух не-
дель. На относительно сухих солнечных местах рябчик зацветает на
3-5 дней раньше. По наблюдениям Ю.В.Александрова (1971), в Мос-
ковской обл. рябчик шахматный цвел в 1965 г. (после холодной вес-
ны) в I-2-й декаде мая, в 1968 г. зацвел 18 апреля, в 1969 г. -
около 25 мая. Р.А.Ротов (1976) в 1973 г. там же наблюдал начало
вегетации 2 апреля, начало цветения 28 апреля, окончание цвете-
ния 7 мая, окончание вегетации 12 июня. Нами в этом же месте в
1977 г. массовое цветение наблюдалось 5-6 мая, в 1978 г. - 7-8
мая. Задержка в цветении отдельных особей отмечается, если их лу-
ковица расположена слишком глубоко (Loew, Kirchner, 1934). В
июле надземная часть отмирает, в почве остается лишь замещающая
луковица. Июль и август луковицы отдыхают, корней у них нет.
Осенью из основания молодой луковицы образуются придаточные корни.
Первые корни - всасывающие с многочисленными корневыми волосками.
По наблюдениям в Западной Европе в марте после перезимовки образу-
ются контрактильные корни (Loew, Kirchner, 1934).
Размножается рябчик шахматный как семенным, так и вегетативным
путем с помощью дочерних луковиц, которые закладываются в пазухах
чещуй. Перекрестноопыляемое растение с протогиничными цветками,
опыляется шмелями. У культивируемых растений изредка отмечают про-
тандрию (Loew, Kirchner, 1934). Как правило, рябчики цветут каж-
дый год, особенно при условии достаточного освещения (Beck, 1953).
Семян образуется много, благодаря наличию воздушных мешков они хо-
рошо переносятся ветром и водой (Loew, Kirchner, 1934).
93
Экология
Рябчик шахматный растет только в условиях значительного увлаж-
нения, часто даже на затопляемых участках в долинах рек, на забо-
лоченных лугах (Loew, Kirchner, 1934; Hegi, 1£39; Garcke, 1972) -
8-я ступень шкалы увлажнения (Eilenberg, 1974). Только в таких
условиях он интенсивно цветет и плодоносит (Ротов, 19726). В ус-
ловиях степной зоны (Центрально-Черноземный заповедник, Курская
обл.) он, как правило, растет в нижней трети или у основания скло-
нов северной экспозиции, т.е. в местах с повышенным почвенным ув-
лажнением (Ротов, 19726).
На изменение освещенности реагирует очень чутко, предпочитая
достаточно освещенные места (8-я стуяень шкалы Элленберга). К поч-
вам требователен (Garcke, 1972), растет на серых лесных, черно-
земных и других богатых почвах. Предпочитает почвы нейтральной
реакции (7-я ступень шкалы Элленберга), в Румынии распространен
при pH 6-7 (Bujorean, Grigore, 1965). Растет обычно на равнине,
поднимается в горы не более.чем на 800 м над уровнем моря (Loew,
Kirchner, 1934; Hegi, 1939; Bujorean, Grigore, 1965). К НИЗКИМ
температурам и резким колебаниям температуры, по-видимому, мало
чувствителен, так как его цветение приходится на то время, когда
часто Высокие дневные температуры сопровождаются ночными замороз-
ками на почве.
Р.А.Ротов (19736) ввделил 3 экоэлемента рябчика шахматного
(табл.5): теневой, открытых, освещенных полян и мезогигрофидьный
(растения, произрастающие вблизи понижений рельефа, заполненных
водой и заросших кустарниками).
Таблица 5.
Морфологические признаки вегетативных и генеративных органов
рябчика шахматного (Ротов, 19736)
Is '' _______1 Экоэлемент__________________I
I Морфологический признак 1 теневой открытых полян гигромезо- фильный
1 Высотаj см 26,8 16,0 28,0
1 Число листьев 3-5 3-4 4-5
। Длина листьев, см 10,5 7,7 15,0
1 Ширина листьев, см 8,4 7,8 12,0
1 Длина долей околоцветни- 1_КЙ>_Я£ , 30,0 28,0 35,0
94
Теневой и мезогигрофидьный экоэлемент превосходит экоэлементы
открытых полян по всем основным показателям, что свидетельствует
о требовательности рябчика к условиям почвенного увлажнения и по-
ложительной реакции на умеренное затенение, имеющее место в лесу
до полного распускания листьев. Р.А.Ротов (1976) считает, что
экологический оптимум рябчика шахматного находится в лесных фито-
ценозах, а не в луговых и степных. На открытых солнечных местах
число листьев обычно меньше, стебли искривлены, иногда околоцвет-
ник недоразвит (Ротов, 1973).
Нами были проведены измерения рябчика шахматного по возрастным
группам в осиново-березовом купальницево-разнотравном лесу у Прот-
вино и на поляне среди этого леса (табл.6).
Таблица 6.
Размеры вегетативных особей рябчика шахматного
в лесу (I) и на поляне (П)
1 1 ! т Морфологические показатели — — — — — — — — — — — — р — — — — — — Возрастные группы особей 1
ювенильные имматурные взрослые 1 виргинильные!
I" 1 Длина нижнего листа, см 6,83+0,17 10,78+0,29 13г2*0,24 S
1 I Ширина нижнего листа, см 0,44+0,14 0,73+0,02 0,78+0,02 1
1 Высота растения, см . 12,6+0,31 13,45+0,38 16,95+0,28 (
1 1 Длина нижнего листа, см 7,03+0,18 8,92+0,18 1 10,89^0,28 ।
1 1 П Ширина нижнего лис- та, см 0,58+0,03 0,82+0,02 0,7+0,03 J
1 1 Высота растения, см 10,82+0,35 12,8+0,22 13,98+0,42 [
Как видно из табл.6, растения в лесу по всем изученным показа-
телям крупнее, чем на поляне, исключение составляют лишь размеры
листьев у ювенильных растений, которые несколько больше на поляне.
Таким образом, наши данные также подтверждают мнение Р.А.Ротова о
том, что экологический оптимум рябчика шахматного находится в лес-
ных фитоценозах, а не на открытых солнечных участках.
Фитоценология
Рябчик шахматный раот'ет на лугах, в степях (чаще по днищам
95
отолных оврагов), в разреженных лесах, на лесных полянах, опуш-
ках (HormuBaki, 1931; Hegi, 1939; Beck, 1953; Александров, 1971).
Цонопопуляциж рябчика в Серпуховском р-не Московской обл. находит-
ся в разреженном березово-осиновом лесу (сомкнутость крон 0,3-0,4)
с примесью сосны и липы. Подлесок развит слабо (бересклет бородав-
чатый, шиповник, крушина). Травяной покров развит хорошо, но к мо-
менту цветения рябчика большая часть видов только начинает вегети-
ровать. Небольшая ценопопуляция, но с высокой плотностью (около
80 особей на I м^) нами была встречена на участке почти чистого
липового леса. Довольно крупная ценопоцуляция нами встречена так-
же в заболоченном березняке с мощным слоем торфа. Причем на участ-
ках с нарушенным травяным покровом рябчик рос особенно обильно. С
увеличением примеси сосны рябчик исчезает, что, вероятно, в пер-
вую очередь связано с большой сухостью почвы под сосняками в этом
районе. Оптимальными условиями, по мнению Р.А.Ротова, следует счи-
тать условия разреженного светлого леса на сырых почвах.
Ценопопуляции на юге Московской обл. (Серпуховский р-н) много-
численные, с высокой плотностью особей на I м~. Р.А.Ротов (1974)
указывает 40 (в среднем 28) ювенильных особей, 36-96 (в среднем
63) взрослых виргинильных и 1-5 (в среднем 2) генеративных особи.
Всего на I м^ им было встречено от 50 до 140 особей. Нами был
проведен подсчет численности рябчика шахматного на трех участках:
в разреженном (сомкнутость 0,3-0,4) осиново-березовом купальнице-
во-черноосоково-разнотравном лесу у Протвино (I), на поляне среди
этого леса (П) и в осиново-березовом снытево-злаково-разнотравном
лесу в Приокско-Террасном заповеднике (Ш); данные сведены в табл.7.
Из трех изученных ценопопуляций (цифры средние по 100 площад-
кам) самая молодая - в лесу Приокско-Террасного заповедника, где
наибольший процент ювенильных особей. Две другие ценопопуляции
сходны. Численность генеративных особей на всех трех участках до-
вольно мала (особенно на Ш, где растут лишь единичные генератив-
ные особи). Однако, судя по значительной численности вида в изу-
ченных условиях (29-55 особей в среднем на I м^), семенной продук-
тивности даже единичных генеративных экземпляров достаточно для
поддержания популяций рябчика в этих условиях.
Хозяйственное использование и охрана
Рябчик шахматный культивируют с древних времен (Hegi, 1939;
Beck, 1953). Хороший ранний медонос (Hegi, 1939). Луковицы
96
Таблица 7.
Численность и возрастной состав ценопопуляций рябчика
шахматного в различных фитоценозах
1______Группы возрастных состояний
Показатели Число особей на I м^ Участок I П Ш ювениль- ные II 5 18 имматур- ные 20 10 10 взрослые виргинжль- ные 24 13 I генера- 1 тивные 1 I 1 I J единично ।
I 19 40 43 I 1
Возрастной со- П 16 36 44 4
став, % Ш 64 340 2 — 1
съедобны, их употребляли в пищу римляне, поэтому скопления рябчи-
ка часто находят поблизости от древних римских лагерей (Beck,
1953). Рябчик усиленно истребляют на букеты. В связи с этим во
многих странах ставится вопрос о необходимости его охраны (Bujo-
rean, Grigore, 1965; Bradshaw, 1977). Состояние популяции рябчи-
ка в Серпуховском р-не вызывает большие опасения, так как там
срываются практически все цветущие особи, с другой стороны, идет
прямое уничтожение из-за строительных работ.
7-1964
97
В.А.Романова
ЛУННИК ОЖИВАЮЩИЙ
Номенклатура и географическое распространение
Lunaria rediviva L., 1753, Sp. pl.:653; Кауфм., 1866, Моск,
фл., изд. 1:60, 1889, изд. 2:51; Дингер, 1886, Об. свед.:69;
Шмальг., 1895, Фл. 1:84; Петунн., 1896, Крит, обзор 1:71; Сырейщ.,
1907, Илл. фл. 2:199; он же, 1927, Опр. раст. Моск. губ.:146;
Буш, 1939, Фл. СССР 8:335; Иконников-Галицкий, 1940, в Маевск.,
Фл., изд. 7:398; Васильченко, 1954, там же, изд. 8:349, 1964,
изд. 9:340; Ball, 1964, Fl. Bur. 1:296; Ворошилов, 1966, Опр.
раст. Моск. обл. :181; Котов, 1979, Фл. евр. части СССР 4:76. -
Лунник оживающий (сем. Cruciferae или Brassicaceae - кресто-
цветные ).
Род Lunaria L. насчитывает 3 вида, из них на территории СССР
встречается только один. Родовое название происходит от латинско-
го luna - луна, что указывает на форму плода. Видовое название,
видимо, связано с тем, что блестящая перепончатая перегородка
плодов сверкает на солнце до поздней осени.
Европейский вид, третичный реликт (Флора СССР, т.8, 1939). На
север доходит до Балтийского моря, на запад - до Португалии, на
юг - до Италии и Югославии. Как заносное встречается в Северной
Америке (Hegi, 1928'; Meusel а.а., 1965).
На территории СССР ареал лунника занимает почти всю европейскую
часть. Северная граница его проходит по северным районам Эстонии,
через пос.Гостилицы Ломоносовского р-на и д.Баляшково, Крестецко-
го р-на, Ленинградской обл., поворачивает к юго-восточной части
Новгородской обл. (г.Демянск), через Калининскую и Горьковскую
обл. (где он указан только в урочище Слуде на правом берегу Оки),
далее поворачивает на г.Чебоксары (по правому берегу р.Кумашки) и
идет до г.Куйбышева. Отсюда граница поворачивает на запад до юж-
ной части Тульской обл., потом идет на юг до городов Сумы и Харь-
кова, далее по югу Украины и Молдавии, где лунник отмечен близ
с.Теленешты (Раевский, 1884; Коркинский, 1892; Meusel u.a., 1965;
Чопик, 1978; Бибикова и др., 1978). Нами в гербариях БИН АН СССР
и МГУ обнаружены сборы лунника, характеризующие самую северную
(пос.Гостилицы, сбор 1959 г.) и самую южную (с.Теленешты, сбор
98
1948 г.) точки, расположенные за пределами известной из литерату-
ры границы ареала.
Для лунника характерно рассеянное распространение в пределах
ареала. На территории СССР он принадлежит к числу весьма редких
видов, известны лишь отдельные его местонахождения, обычно распо-
ложенные на значительном расстоянии друг от друга. В Московской
обл. наиболее крупные популяции лунника находятся в Ступинском
р-не в долине р. Не сочни и в Загорском р-не близ с.Взгляднево. В
небольшом количестве лунник встречается еще в 4 пунктах: в Руз-
ском р-не около д.Мосеевка, в Солнечногорском р-не близ с.Камен-
ка, в Истринском р-не около Ново-Иерусалимского монастыря и в
Дмитровском р-не в долине р.Яхрома.
Морфологическое описание
Лунник оживающий - многолетнее травянистое поликарпическое ко-
роткокорневищное растение, гемикриптофит. Корневая система прида-
точная. Корни размещены относительно равномерно на протяжении
каждого годичного прироста корневища. Диаметр придаточных корней
1,5-2 мм, средняя длина 15-20 см, расположены они в пределах гу-
мусового горизонта. Молодые придаточные корни светлые, почти бе-
лые, затем опробковевают и становятся светло-бурыми или бурыми,
старые разрушающиеся - темно-бурые. Корневище бурое, в молодом
состоянии светло-бурое. Оно обычно находится в поверхностном слое
почвы на глубине 5-10 см. Средняя длина корневища у взрослой ге-
неративной особи (имеющей 2-3 побега) - от 10 до 15 см.
Возобновление побегов симподиальное. К осени надземная часть
побега отмирает, а сохраняется лишь плагиотропная подземная часть,
представленная участками монокарпических побегов предыдущих лет,
которые начинают уже со второго года функционировать как органы
отложения пластических веществ. На базальной части ортотропного
побега, расположенной у поверхности почвы, находятся 1-2 почки
возобновления, из которых на следующий год разовьются моноцикли-
ческие побеги. Основание многолетнего ортотропного побега имеет
несколько укороченных междоузлий, которые не отмирают к концу ве-
гетационного периода и превращаются в восходящее или горизонталь-
ное корневище. На корневище закладываются почки. Основная масса
почек становится спящими. Лишь 1-2 верхние боковые оказываются
почками возобновления, из них на будущий год разовьются новые по-
99
беги. Кбрневшце лунника может ветвиться - в базальной части орто-
тропного побега закладываются две боковые почки возобновления, ко-
торые на следующий год дают начало двум моноциклическим ортотроп-
ным побегам.
Стебли имеют высоту 30-100 ом, они прямые, ребристые, шерша-
вые от разбросанных волосков, в соцветии ветвятся. Листорасполо-
жение нижних листьев супротивное, верхних очередное. Черешок лис-
та с желобком, коротковолосистый. Листья на верхней стороне тем-
но-зеленые, на нижней - блестящие, голубовато-зеленые, усеяны с
обеих сторон короткими белесыми волосками, основная масса кото-
рых сконцентрирована вдоль жилок. Самые нижние листья, расположен-
ные супротивно, у всех растений (за исключением проростков) округ-
ло-яйцевидные с заостренной верхушкой и пильчатым краем (см.
рис.10). Они обычно в 1,5-2 раза мельче, чем следующая за ними
пара листьев. Срединные листья ддинночерешковые, сердцевидные, с
зубчато-пильчатым краем; верхушечные - короткочерешковые, яйцевид-
но-заостренные (см. рис.II).
Соцветие метельчатое, состоит из верхушечной и боковых кистей.
В каждой кисти от 3 до 18 цветков. На оси соцветия у нижних цвет-
ков могут формироваться кроющие короткочерешковые листья яйцевид-
но-продолговатой формы (см. рис.11-12). Цветки правильные, с двой-
ным околоцветником, 4-членные, крупные, 15-17 мм в диаметре, ду-
шистые. Чашелистики при основании мешковидные, .длиной 5-7 мм. Ле-
пестки лиловые, крупные - до 20 йи длины, с длинным (примерно в
1/3 лепестка) ноготком. Тычинок шесть, нити их свободные, простые
иди с зубцом. При основании коротких тычинок по одной большой коль-
цевидной, снаружи дву-, изнутри трехлопастной медовой железке, ко-
торая иногда разделена на внешнюю и внутреннюю части. Завязь на
ножке, столбик длинный, рыльце двулопастное.
Плод - стручочек, очень большой, 4-5 см длили, эллиптический,
С обоих КОНЦОВ острый, СО стороны СПИНОК сплюснутый, сидящий на
плодоножке (гинофоре) до 35 мм длины (рис. 12). Створки плода плос-
кие, с сетчатым рисунком. Перегородки оо слоем волокон, железисто-
блестящие. При созревании створки плода раскрываются снизу вверх,
отделяются от перегородки. При падении они захватывают с собой се-
мена, которые таким образом могут переноситься ветром на неболь-
шие расстояния. В каждом стручочке от 2 до 8 (чаще 3-4) семян.
100
Онтогенез, сезонное развитие и способы размножения
Опавшие осенью семена прорастают весной следующего года. Про-
растание происходит по надземному типу (рис.9) в конце апреля -
Рис.9. Проростки (1-3) и ювенильная особь (4) лунника
оживающего.
начале мая. Первым появляется корешок. Затем одновременно с рос-
том корешка происходит интенсивное вытягивание гипокотиля. Семя-
доли освобождаются от семенной кожуры и начинают зеленеть. Они
эллиптической формы, цельнокрайние, первоначально имеют длину 10-
7^-1964 101
12 ом и ширину 6-7 мм, средняя жилка едва заметна. Через месяц
они увеличиваются в размерах примерно в 3 раза. От средней жилки
семядолей с каждой стороны отходят по 2-3 боковых жилки. Уже в
начале мая главный корень проростка может образовывать боковые
корни. Первые листья формируются в середине мая. Листья супротив-
ные, черешковые, яйцевидно-заостренные с пидовидно-зубчатнм кра-
ем. К моменту их появления гипокотиль имеет длину 15-20 мм и диа-
метр 1,5-2 мм, эпикотиль небольшой - 3-5 мм. Средние размеры лис-
тьев проростков 2,5 на 3,5 см, а у наиболее крупных экземпляров
листья могут достигать размеров 5,5 на 4,5 см. Длина черешка 4-5
(7) см.
Семядоли засыхают к середине - концу июля. Проростки лунника
превращаются в ювенильные растения (рис.9). Их листья достигают
8 см в длину и 7 см в ширину. Нарастая моноподиально, ювенильные
растения к концу вегетационного периода формируют эпикотиль дли-
ной до 3 см. В начале августа иногда начинает развиваться вторая
пара листьев, но полностью развиться не успевает и отмирает в
связи с наступлением осени. Однако в культуре к октябрю у пророст-
ков формируются 2-3 пары листьев. В ювенильном состоянии растения
находятся два года. К концу первого вегетационного периода орто-
тропный надземный побег отмирает. Сохраняется лишь его нижняя
часть с 1-2 боковыми почками возобновления. Из них на следующий
год разовьются I или 2 новых побега. У ювенильного растения кор-
невище еже не развито. Корневая система представлена главным кор-
нем, который может достигать в длину 15’ см, в'толщину 2-3 см. Юве-
нильные растения второго года жизни имеют две пары супротивно рас-
положенных листьев.'Уже на второй год жизни становится заметна
контрактильная деятельность длинных упругих придаточных корней,
отходящих от базальной части ортотропного побега, а позже во.
взрослом виргииильном и генеративном состоящих - от плагиотроп-
ной части монокрапичеоких побегов предыдущих лет. Эти корни неред-
ко в верхней части имеют зеленую окраску, длина их 15-20 ом. При
сокращении корней апикальная часть корневища втягивается в почву
(на 2-3 мм в год)вследствие этого формируется коленообразный
изгиб корневища.
Особи третьего года жизни можно относить к группе взрослых
виргинильных (рис.10). Их высота примерно 15-25 см. Побег у трех-
летней особи несет три пары супротивно расположенных листьев.
Средние размеры первой пары 4,5-5 на 4,5-5 см, второй пары - 10
на 8 см. Третья пара листьев если начинает развиваться, то обыч-
102
Рис.10. Надземная часть взрослой виргинильной особи
лунника оживающего.
но не успевает полностью сформироваться к концу вегетационного
периода. В таком состоянии лунник, как правило, может находиться
до 7 лет, но в благоприятных условиях до 4 лет (например, в Туль-
ских засеках), а в неблагоприятных - до 12 лет. Особи в возрасте
от 4 до 7 лет практически не отличаются по признакам надземных
побегов друг от друга. Побеги имеют высоту 40-60 см, лишь у от-
дельных 7-8-летних особей достигают высоты 80 см. Листья у веге-
103
тативных растений расположены только супротивно. Но мере продви-
жения вверх по стеблю листья становятся более вытянутыми. Так,
если у первой пары сердцевидных листьев длина превышает их шири-
ну на 1,5-2 см, то у верхушечных листьев длина нередко превышает
ширину в 1,5-2 раза, т.е. они становятся сердевидно-длиннозаост-
ренными., В конце ию;я первая пара листьев опадает. К седьмому го-
ду жизни длина корневища 3-5 см, толщина 5 мм. Главный корень
уходит вглубь на 15 см. От плагиотропного корневища почти верти-
кально вниз отходят длинные (до 25 см) упругие белые придаточные
корни.
В генеративное состояние основная масса особей лунника перехо-
дит на седьмом году жизни, однако в благоприятных условиях уже в
6 лет растения приобретают облик зрелой генеративной особи. Мож-
но выделить подгруппу молодых (7-10 лет) и подгруппу зрелых
(старше 10 лет) генеративных особей. Молодые генеративные расте-
ния имеют более крупные размеры, чем вегетативные - их высота 70-
80 (реже 100) см. Для особей этой подгруппы (рис.II) характерно
наличие .1-2 ортотропных побегов, на которых формируются, кроме
2-3 пар' сердцевидных листьев, 1-2 верхушечных листа яйцевидно-за-
остренной формы, имеющих Очередное расположение, и образование
одной терминальной кисти с 4-9 (до 15) цветками, причем в пазу-
хах яйцевидно-заостренных листьев могут развиваться и отдельные
цветки.' Корневище молодых генеративных особей продолжает увеличи-
ваться в длину и толщину, достигая 13 см длины и I см толщины.
Прирост корневища за год составляет в среднем I (реже до 3) см,
длина придаточных корней до 35 см.
Зрелые генеративные особи (рис.12) имеют высоту в среднем 90-
100 (до 150) см. Для них характерно образование 2-4 ортотропных
побегов, на каждом из которых закладываются 8-12 листьев, причем
2-3 нижние пары расположены супротивно, а верхние листья очеред-
ные (у особей старше 16 лет нередко только одна нижняя пара листь-
ев расположена супротивно). На верхушке ортотропного побега фор-
мируются 2-3 яйцевидно-заостренных листа, ниже находящиеся листья
могут.иметь форму, переходную от сердцевидной к яйцевидно-заост-
ренной. Максимальный размер листьев 22x20 см. У зрелых генератив-
ных особей сложное метельчатое соцветие, состоящее из верхушечной
и боковых кистей. Как правило, на одном побеге образуется 20-50
цветков,, но иногда их значительно больше - до НО. Корневище
взрослых' генеративных особей может достигать в длину 20-33 см, в
толщину 1,5-2 см, как правило, интенсивно ветвится. В 16 лет в ба-
104 .
Рис.II. Верхняя часть молодой генеративной особи
лунника оживающего.
зальной части корневищ становятся заметны признаки распада.
Возраст, в котором особи переходят в сенильное состояние, уста-
новить не удалось. Мы обнаружили всего несколько сенильных особей
в возрасте от 22 до 24 лет.
В апреле, как только стает снег, молодые побеги лунника появля-
ются над поверхностью почвы. Развитие проходит довольно быстро,
так как в почках возобновления побеги будущего года бывают пол-
ностью сформированы еще осенью. К концу мая растения достигают вы-
105
Рио.12. Верхняя часть зрелой генеративной особи
лунника оживающего.
соты 80-100 см и зацветают. Цветение продолжается до середины
июля, изредка наблюдается повторное цветение поврежденных скотом
побегов в августе. К середине июня у многих растений уже завязы-
ваются плоды, которые созревают в конце августа - начале сентяб-
ря и осыпаются в конце сентября - начале октября. С середины
июня начинает опадать нижняя лара листьев, к середине октября по-
беги стоят обезлиственные. Почки возобновления в зто время дости-
106
тают размеров 2-3 см.
Лунник размножается в основном семенным путем, однако не ис-
ключена возможность распространения участками корневища при его
механическом повреждении. Семена из-за большой массы не могут пе-
реноситься ветром на большие расстояния и опадают поблизости от
материнского растения.
Семенная продуктивность молодых генеративных особей невелика.
Как правило, только 25-30$ цветков дают плоды. У зрелых генера-
тивных особей завязывание происходит у 30-50$ цветков. При этом
число цветков и плодов, образующихся на одной особи, изменяются
в больших пределах. Так, в лунниково-черноольховой ассоциации на
р.Песочне у 114 зрелых генеративных особей, изученных нами, чис-
ло цветков колебалось от 30 до 350, а плодов - от 10 до 98. Сле-
довательно, одна особь может дать от 30 до 380 оемян, но не все
завязавшиеся семена созревают. Например, в лунниково-вязовой ас-
социации на источнике Гремячем в августе мы насчитали 50-80 семян
на I м2, а в октябре число опавших семян там же составляло всего
от 20 до 42, т.е. только 30-50$ семян достигли зрелости. Часть
семян, видимо, поедается личинками длинноусой моли.
Экология и фитоценология
Лунник - мезофит. В его местообитаниях, по Е.Ландольту (Lan-
dolt, 1976), увлажнение варьирует от средней сухости до влажно-
го (3~я ступень). По шкале Г.Элленберга (Ellenberg, 1974) лунник
занимает 6-ю ступень увлажнения, произрастая на почвах между све-
жими и влажными.
Предпочитает тенистые леса, но его можно встретить и на совер-
шенно открытых местах. Г.Элленберг располагает лунник между тене-
выми и полутеневыми растениями (от 5 до 100$ полной освещеннос-
ти - 4-я ступень шкалы освещенности). Е.Ландольт указывает, что
этот вид растет преимущественно в затененных местах при относи-
тельной освещенности, равной 10$ (2-я ступень шкалы). В Москов-
ской обл. он чаще входит в состав фитоценозов со степенью сомкну-
тости крон 0,6-0,9, а также произрастает на открытых местах в пой-
мах рек. К температуре требователен, не выносит поздних замороз-
ков и крайне низких температур. На территории с континентальным
климатом встречается только на участках с благоприятными условия-
ми увлажнения, освещенности и почвенными.
107
Занимает участки с почвами, богатыми доступным азотом (4-я
ступень шкалы богатства почв Ландольта), со средним содержанием
гумуса (3-я ступень шкалы Ландольта), даже считается индикатором
значительного количества азота (4-я ступень шкалы Элленберга).
Предпочитает песчаные хорошо аэрируемые почвы, встречается на
почвах разного механического состава: тяжелосуглинистых, супесча-
ных, щебнистых (Moor, 1975), иногда на склонах, усеянных базаль-
товыми камнями (Eilenberg, 1974). Служит индикатором почв, бога-
тых основаниями (4-я ступень шкалы Ландольта). Кислотность почв,
на которых растет лунник, колеблется от 5,5 до 8 (Hegi, 1928).
Г.Элленберг указывает на приуроченность лунника к нейтральным
почвам (7-я ступень шкалы). В условиях Московской обл. лунник
растет на дерново-слабоподзолистой и светло- серой почвах с pH
от 5,1 до 7,9 (чаще 6-7,9), отдельные особи встречены на почвах
с pH 4,3.
В Западной Европе лунник чаще встречается в горах, чем на рав-
нине, поднимаясь до 1400 м над ур.моря (Hegi, 1928). На террито-
рии СССР лунник встречается как в горах, так и на равнине. Но на
равнине, в том числе и в Московской обл., предпочитает крутые .
склоны оврагов или берега рек.
Лесной вид, в горах он обычно растет в буковых лесах, но
встречается также в смешанных, хвойно-широколиственных, а иногда
и чисто сосновых лесах (Hegi, 1928). Есть указание на доминиро-
вание лунника в травяном покрове кленовых фитоценозов (Moor,
1975). Из травянистых растений его верными спутниками в горных
европейских лесах являются Mycelis muralis, Impatiens nolitange-
re, Merourialis perennis, Aruncus silvestre (Eberle, I960).
На востоке ареала в пределах СССР лунник растет в елово-широ-
колиственных и широколиственных лесах смешанного состава (дуб,
липа, клен, вяз, ясень). В Калининской обл. лунник входит в сос-
тав елово-широколиственного леса (Невский, 1947), а в Московской
обл. лунник в основном встречается в лесах с сомкнутостью крон
0,6-0,9, состоящих из вязов (гладкого и шершавого), клена, чере-
мухи, а также в ольшаниках и осинниках. Из травянистых растений
луннику часто сопутствуют такие неморальные виды, как копытень
европейский, пролесник, зеленчук желтый, медуница неясная, сныть,
колокольчик широколистный. В травяном покрове лесов для лунника
характерно образование почти чистых зарослей, в которых участие
других видов ничтожно. В качестве содоминантов изредка могут вы-
ступать крапива, страусник, сныть.
108
В поймах рек лунник встречается небольшими группами в ольшани-
ках, в зарослях кустарников вместе с другими травянистыми спутни-
ками широколиственных пород: зеленчуком желтым, колокольчиком ши-
роколистным, таволгой вязолистной. В поймах встречаются также не-
большие участки чистых зарослей лунника площадью в несколько м^,
обычно среди крапивы.
В спектре возрастных состояний большинства ценопопуляций на
территории Московской обл. отмечены два пика численности: пер-
вый - проростки, второй - зрелые генеративные особи. Встречены
различные типы ценопопуляций: зрелые, стареющие, регрессивные,
переходные от стареющих нормальных к регрессивным. Определен ба-
зовый спектр ценопопуляций лунника (на материале Московской обл.),
в доверительную зону которого вошли все изученные ценопопуляций:
взрослые виргинильные - 9#, молодые генеративные - 5%, зрелые ге-
неративные - 38%, сенильные - 8%, Нами установлено, что нормаль-
ные ценопопуляций формируются в условиях низкой освещенности
(сомкнутость крон 0,8-0,9), а стареющие нормальные и регрессив-
ные - цри высокой освещенности (сомкнутость крон 0,4-0,6). Это
объясняется тем, что при большом освещении корневище лунника уси-
ленно ветвится - одна особь начинает формировать до 20-30 побегов
(а в иных условиях 4-6), плотность ценопопуляций резко возрастает
и создаются условия, неблагоприятные для развития проростков, так
как проективное покрытие лунника в таких фитоценозах составляет
100# (в фитоценозах с меньшей освещенностью цроективное покрытие
лунника не превышает 70-90#).
Консортивные связи, биологическая продуктивность и
хозяйственное значение
Лунник - перекрестноопыляемое растение, автогамия наблюдается
очень редко (Hegi, 1928), опыляется шмелями и. бабочками (Eberle,
I960). Нами замечено посещение цветков пчелами, которых привлекал
нектар, находящийся в медовых железках. Нектаром лунника питается
и длинноусая моль (Adella rufimitrella). Гусеницы зтой моли пита-
ются семенами лунника (Eberle, 1960), возможно, что низкая уро-
жайность семян лунника в некоторых популяциях связана с поврежде-
нием их этой молью.
Нами определена надземная фитомасса особей лунника в Ступин-
ском р-не Московской обл. Воздушно-сухая масса проростков и юве-
109
пильных оообей О,I-O,5 г, взрослых виргинильных - от I до 2,6
(3,8) г, генеративного побега молодого растения - 2,3-4,5
(6,9) г, генеративного побега зрелого растения - 5-13 г. Изуче-
ние продуктивности ценопопуляций лунника в различных местообита-
ниях Загорского и Ступинского р-нов Московской обл. показало, что
максимальная надземная биомасса накапливается в пойменных лунни-
ково-шершавовязовых, лунниково-черноольховых и лунниково-серооль-
ховых фитоценозах - от ИЗ до 184 г сухого вещества на I м2. Ос-
новная часть биомассы (70-94$) ценопопуляций формируется зрелыми
генеративными особями. Эти фитоценозы характеризуются средними
условиями освещенности и богатыми почвами, часто формирующимися
на выходах известняков. Менее благоприятные условия складываются
для лунника на склонах оврагов. Здесь его надземная биомасса в
лунниково-гладковязовых, лунниково-сероольховых и лунниково-чере-
муховых фитоценозах колеблется от 66 до 96 г/м2. Для этих фитоце-
нозов характерна низкая освещенность и меньшее богатство почв.
Лунник - высокодекоративное растение. Он внесен в "Красную
книгу СССР" (1978). Сокращение его численности связано с вырубкой
лесов, а также со сбором его как декоративного растения. Наиболее
крупные и жизнеспособные популяции лунника на территории Москов-
ской обл. встречаются на двух ограниченных по площади участках:
в долине р.Песочни и на источнике Гремячем около д.Взгляднево.
ПО
Н.П.Писковацкова, И. С. Михайловская'1'
ЛЯДВЕНЕЦ РОГАТЫЙ
Номенклатура, систематическое положение
и географическое распространение2
Lotus cornioulatus L., 1753, Sp. pl.:775; Кауфм., 1866, Моск,
фл., изд.1:131, 1889, изд.2:129; Цингер, 1886, Сб. свед.:135;
Шмальг., 1895, фл. 1:248; Сырейщ., 1907, Илл. фл. 2:301; он же,
1927, Опр. раст. Моск. губ.:173; Федченко, 1940, в Маевск., Фл.,
изд. 7:466; Куприянова, 1945, Фл. СССР, 11:291; Борисова, 1954, в
Маевск., Фл., изд. 8:153, 1964, изд. 9:155; Тихомиров, 1966, Опр.
раст. Моск. обл.:2I4; Ball et Chrtkova-Zertova, 1968, Fl. Bur. 2:
174. - Лядвенец рогатый (семейство бобовых - Fabaceae, триба Lo-
teae Benth.).
Вид очень изменчив. По характеру опушения, форме листочков,
строению чашечки, длине ее зубцов, размерам цветка, форме паруса
(флага), характеру роста побегов и ряду других признаков в преде-
лах вида издавна выделяли несколько разновидностей. В последние
десятилетия в нашей стране особенно благодаря усилиям М.В.Клоко-
ва (1953, 1’961) и Н.А.Миняева (1957; Миняев, Улле, 1977) получи-
ло распространение мнение о целесообразности выделения из соста-
ва L.cornioulatus L. в.1. довольно многочисленных "мелких" видов.
Если разделять эту точку зрения, то в Московской обл. нужно при-
знавать по крайней мере три вида: L.cornioulatus L. в.str., L.ar-
vensis Pers., L.zhegulensis Klok. Однако реальность существова-
ния этих "видов" требует подтверждения. Ясно, что лядвенец измен-
чив, но характер этой изменчивости требует специальных исследова-
ний, прежде всего генетических и цитологических. А пока заключе-
ния систематиков, довольствующихся для выделения серии видов толь-
ко набором морфологических признаков в разных комбинациях, хотя и
на определенном географическом фоне, не представляются достаточно
убедительными. Кроме того, распространение и экологическая приуро-
ченность рас лядвенца у нас, в условиях Центральной России, не яс-
ны, поэтому в настоящей работе принята широкая трактовка вида.
И.С.Михайловская написала разделы о корнях, каудексе и большом
жизненном цикле.
2 Автор раздела В.Н.Тихомиров.
III
Lotua coraicuiatus L. s.l. - широко распространенный еврази-
атокий бореально-субмеридиональный вид. Ареал его простирается
от Атлантического побережья Европы до Центральной Азии и от суб-
арктических районов до Малой Азии, Крыма, Кавказа и Индии. Расте-
ние интродуцировано в Америку и Исландию, занесено в Австралию
(Флора СССР-, Т.П, 1945; Flora Europaea, vol.2, 1968). Приуро-
чен к лесной полосе, лесостепи и степи. В пределах Московской обл.
распространен повсеместно, но тяготеет к хорошо дренированным поч-
вам более легкого механического состава, поэтому малочислен или
может отсутствовать на больших площадях в заболоченных районах.
Морфологическое описание
Многолетнее поликарпическое травянистое длинностержнекорневое
многоглавое растение с ортотропными и анизотропными моноцикличес-
кими побегами, развивающимися по яровому или озимое типу и возоб-
новляющимися базисимподиально (Голубев, 1962; Денисова, 1903; Бу-
ленкова, 1974). Корневая система стержневая. Главный корень дости-
гает глубины 1,5-2 м в зависимости от степени увлажненности суб-
страта. Диаметр корня взрослого растения 1-2 см. Ветвление сравни-
тельно слабое, скелетных боковых корней образуется немного. Ба
утолщающемся главном корне возникают в большом количестве прида-
точные ризогенные сосущие корни, располагающиеся вдоль по основ-
ным ризостикам корня. Поперечный срез главного корня, обладающего
вторичным строением, дает возможность по признакам его анатомичес-
кого строения (годичным кольцам древесины, слоям рогового луба и
сериям радиальных лучей) определить возраст растения (Михайлов-
ская, I960, 1968).
Многолетние растения лядвенца, как и многие другие травянистые
стержнекорневые бобовые (Ильина, 1964), обладают каудексом. Он
представляет собой базальную побеговую часть куста, образованную
системой симпбдиально связанных между собой оснований боковых от-
мерших побегов прошлых лет - "пеньков". Эти "пеньки" несут на се-
бе многочисленные почки, которые обусловливают ежегодное возобнов-
ление новых побегов растения. Основание каудакса, включающее в се-
бя гипокотильную и эпикотильную части растения, тесно связано с
базальной частью главного корня, который, обладая втягивающей спо-
собностью, углубляет и прижимает к почве основания побегов с поч-
ками (Михайловская, 1967, 1972). Наличие камбия вызывает значи-
112
тельное утолщение и разрастание каудекса в диаметре.
У взрослых особей различаются генеративные и вегетативные удли-
ненные побеги. В пределах одной особи наблюдается разнообразие
структуры побегов, связанное с длительностью пребывания его в по-
чечной фазе, со временем прорастания из почки (Гуленкова, 1974),
а также с разной степенью ветвления скелетного побега и образова-
ния побегов обогащения. Одни побеги в основании имеют чешуйчатые
или недоразвитые листья с редуцированными пластинками (листья ни»
зовой формации), сидящие на сближенных узлах; у других листья ни-
зовой формации отсутствуют, гипоподий сильно вытянут (Туленкова,
1974). Большинство побегов возобновлении сильно ветвится с .обра-
зованием побегов обогащения, разветвленных до 3-го, 4-го, иногда
5-го порядков. На некоторых побеги обогащения отсутствуют вовсе
или боковые побеги представлены цветоносами.
В основании побегов возобновления закладываются пазушные поч-
ки. Этому виду свойственно образование сериальных почек и их вет-
вление. Сериальные почки имеются в пазухах всех низовых листьев,
начиная от предлиста, при этом в пазухах самых нижних листьев их
бывает по 3-4, а в пазухах выше расположенных по побегу их закла-
дывается по 2-3. Наибольшего развития достигают крайние почки,
слабее сформированы средние (Денисова, 1963).
По характеру роста побеги возобновления могут быть ортотропны-
ми и приподнимающимися, чем обусловлена конфигурация куста ляд-
венца рогатого. Наиболее часто встречается развалистая и полураз-
валистая форма куста, оформляющаяся ко второму году жизни расте-
ния (Зазоев, 1937). Н.А.ОДухина (1956), изучавшая лядвенец в куль-
туре (Ленинградская обл.), отмечает разные формы куста: прямосто-
ячую, почти прямостоячую, рыхлую, приподнимающуюся или прижатую к
земле. По мнению автора, форма куста может служить систематичес-
ким признаком разновидностей. М.А.Т'уленкова (1974), наблюдая за
развитием лядвенца рогатого в посевах на территории Московской
обл., обратила внимание на преобладание ортотропных побегов у осо-
бей первого года жизни и наличие плагиотропных побегов с припод-
нимающейся верхушкой, обусловливающих раскидистую форму куста в
будущем.
Стебли тонкие, нежные, голые или заметно опушенные, достигают
длины в начале цветения 30-60 см, в период плодоношения 75 см и
насчитывают соответственно 15-18 или 18-22 междоузлий (Зазоев,
1937). Высота стебля дикорастущих форм равна 10-40 см, в культу-
ре - 60-80 см (Дмитриева и др., 1974). Строение узла стебля трех-
8-1964 113
лакуиное, трехпучковое. Латеральные пучки листового следа снача-
ла идут по коре междоузлия и затем входят в центральный цилиндр.
На отебле в свяви с этим заметны два боковых ребра.
Одни авторы считают, что листья лядвенца рогатого тройчатые с
прилистниками, сходными с листочками (Кауфман, 1866; Шмальгаузен,
1895; Зазоев, 1937; Ларин и др», 1951; Еленевский, 1969; и др.).
Другие рассматривают лист как сложный непарноперистый пятилисточ-
ковый, у которого нижняя пара листочков находится у основания
листа и имеет вид прилистников (Пайпер, 1930; Флора СССР, т.П,
1945; Гроссгейм, 1952; Голубинцева, Лебедев, 1’954; и др.). Листья
сидячие. Верхние листочки обратнояйцевидные или ланцетные, Id-
15 мм длины, 6-10 мм ширины; нижние - косояйцевидные,-7-ТП мм дли-
ны, 4-6 мм ширины. Каждая долька сложного листа имеет свой чере-
шок.
Соцветие лядвенца рогатого характеризуется как головка (Кауф-
ман, 1866; Маевский, 1940; Flora Europaea, vol.2, 1968), зонтик
(Шмальгаузен, 1895; Сырейщиков, 1927; Флора СССР, т.П, 1945),
зонтиковидное соцветие (Пайпер, 1930; Маевский, 1954, 1964), зон-
тиковидная головка (Зазоев, 1957), зонтиковидно-головчатое соцве-
тие (Флора Северо-Востока, 1976). В.Тролль (Troll, 1964), рас-
сматривая соцветие как чаоть генеративного побега, входящего в
общую систему осей особи, считает соцветие представителей семей-
ства бобовых двойной кистью. Исходя из концепции В.Тролля, в со-
цветии лядвенца рогатого выделяются структурные элементы - парци-
альные, частные, или элементарные соцветия, представленные пазуш-
ными зонтиками.
Наблюдения показывают, что степень разветвленности цветоносной
части генеративного побега неодинакова (рис.13). В отдельных слу-
чаях боковые побеги обогащения (паракладии, по Троллю), не образу-
ются, а от оси скелетного побега (условно главной оси) отходят
длинные цветоносы, заканчивающиеся зонтиками. У этих побегов гене-
ративная часть соответствует главному (собственно) соцветию, пред-
ставленному непрерывно нарастающей скелетной осью с 2-4 элементар-
ными соцветиями - зонтиками. У большинства побегов в генеративной
части, помимо главного (собственно соцветия), формируются генера-
тивные побеги обогащения (паракладии). Наиболее развитые из них
насчитывают 7-9 узлов и ветвятся до 3-го, 4-го, реже 5-го поряд-
ков, менее развитые включают 2-5 узлов и ветвятся до 3-го порядка.
Оси последних порядков заканчиваются элементарными соцветиями.
Благодаря образованию паракладий формируется сборное соцветие.
114
Рис.13. Два типа соцветий лядвенца рогатого (схема):
гл.с. - главное (собственно) соцветие;
эл.с. - элементарное соцветие (зонтик);
пк - паракладии; в.п. - вегетативный побег.
Таким образом, соцветие лядвенца рогатого отличается от описанных
В.Троллем (Troll, 1964) соцветий других бобовых и представлено
открытой сложной метелкой, или сборным метельчатым соцветием с
парциальными (элементарными) соцветиями - зонтиками.
Цветки мотылькового типа, свойственного бобовым, сидят на ко-
ротких (1-2,5 мм) цветоножках. Чашечка сростнолистная, 5-6 мм дай-
ны, зубцы чашечки почти равные, при основании треугольные, кверху
шиловидные, до распускания цветков сходящиеся. Венчик 10-18 мм
дайны, свободный, опадаиций, желтый, иногда с оранжевым флагом.
Флаг 10-13 мм дайны, 6-10 мм ширины с широкоокруглым отгибом,
быстро переходящий в клиновидный ноготок. Крылья около 10 мм дай-
ны, о обратнояйцевиднрй, на верхушке усеченной пластинкой, в четы-
115
ро раза длиннее ноготка. Лодочка по длине почти равна крыльям,
вогнута под прямым углом и вдруг заострена в носик. Тычинок 10,
9 ороолиоь, одна свободная с изогнутой тычиночной нитью. У пред-
ставителей семейства бобовых сначала тычинки закладываются в два
круга, но при дифференциации соединяются и срастаются (Имс, 1964).
У лядвенца рогатого 5 внутренних тычинок короткие, 5 наружных -
длинные, на свободном конце утолщенные в виде булавы (Knuth,
1895; Кернер, 1902). Для бобовых характерны одиночные микроспоры,
мономерный гинецей, кампилотропные семязачатки, короткий фунику-
луо (Тахтаджян, 1966). Эмбриогенез и микроспорогенез Lotus corni-
oulatus L. описаны М.М.Чубирко (1964, 1967).
Плод - двустворчатый, линейный, прямой или согнутый дугой, од-
ногнездный или с поперечными перегородками боб 15-25 мм длины, 2-
3 ш ширины, с более или менее изогнутым носиком. В каждом бобе
10-30, иногда 50 семян. Семена темно-коричневые или бурые, иног-
да темно-мраморно-пятнистые, гладкие, блестящие, шаровидные или
слегка сплюснутые, мелкие - 1,25-1,75 мм в диаметре. Семенной руб-
чик почти незаметный. Масса I тыс. семян в среднем 1,2-2 г (Зазо-
ев, 1937; Дмитриева и др., 1974).
Число хромосом 12, 24, 26 (Хромосомные числа..., 1969).
Онтогенез
Одни авторы считают, что всхожесть семян лядвенца рогатого мо-
жет сохраняться 18-26 лет (Африкян, 1957, ссылаясь на нянина
Н.В.Егеровой) и даже 28-43 года (Мещеряков, 1957). По другим дан-
нш, при благоприятных условиях семена сохраняют всхожесть лишь в
течение 3-4 лет (Люшинский, Прижуков, 1969). Из-за твердой кожуры
прорастание семян может задерживаться до второго года после посе-
ва (Мухина, 1956). Скарификация повышает всхожесть семян до 70-
8$ (Ненароков, Раменокая, 1951). В культуре появление первых про-
ростков отмечалось на 4-5-й день после посева. Период прорастания
растянутый, новые растения появлялись ж 20-25-й день после посе-
ва (Туленкова, 1974). Явление растянутости прорастания семян име-
ет место и в естественной обстановке. На лугах проростки могут по-
являться весной, но в массе - только летом.
Прорастание надземное, семвдми мясистые. На главном побеге
вслед за семядолями развертываются 2-3 тройчатых листа и несколь-
ко листьев пятерного тина, разделенных даиннжи междоузлиями. В
•116
пазухах семядолей и тройчатых листьев закладываются почки возоб-
новления. С первых дней у молодых особей быстро растет главный
корень, длина его у растений, проживших один месяц, достигает 15-
20 см СГуленкова, 1974).
Через 35-50 дней после посева у лядвенца рогатого образуется
куст, состоящий из 5-6 побегов (Чуенкова, 1954; Туленкова, 1974).
При кущении сначала формируются побеги возобновления из почек,
заложенных в семядольных узлах. Несколько позднее, а иногда и па-
раллельно с ними развертываются листья побегов обогащения главной
оси. К этому времени семядоли подсыхают, в большинстве случаев от-
падают. Главный корень этих растений углубляется в почву до 60-
70 см, боковые корни ветвятся до 3-го порядка. На главном и боко-
вых корнях уже имеются клубеньки.
Таким образом, в первый год жизни особь лядвенца проходит не-
сколько этапов, соответствующих трем возрастным состояниям по
классификации Т.А.Работнова (1950, 1964). Проростки - однопобего-
вые растения с семядолями и тройчатыми листьями, ювенильные осо-
би - однопобеговые растения с тройчатыми и пятерными листьями,
взрослые виргинильные - двух- или многопобеговые растения (Гулен-
кова, 1974). Так же, как и в культуре, в травостое суходольного
луга (Истринский р-н Московской обл.) к концу первого года жизни
особь лядвенца рогатого переходит от однопобегового состояния к
кущению, образует 2-3 боковых побега и развивает глубоко уходящий
в почву главный корень с немногочисленными боковыми корнями (Ми-
хайловская, 1967). При изучении онтогенеза лядвенца рогатого в
условиях пойменных лугов Северной Двины были встречены растения,
у которых еще до момента кущения главная ось разветвилась с обра-
зованием 2-3 побегов обогащения. Такие растения могут быть отнесе-
ны к имматурнощу возрастному состоянию.
Как в культуре, так и в естественных местообитаниях лядвенец
рогатый обычно зацветает на второй год жизни, единичные экземпля-
ры могут перейти к цветению в первый год (Дуленкова, 1974), в при-
родных сомкнутых ценозах не ранее, чем на второй, а часто на пя-
тый год. Начало цветения свидетельствует о переходе особи в гене-
ративный период. У генеративных особей ежегодно симподиально от-
растает серия генеративных и вегетативных побегов возобновления,
начало которым дают пазушные боковые почки. Вое побеги отличаются
однотипностью развития. Большинство побегов с момента раскрытия
почки завершают свое развитие в течение одного вегетационного се-
зона, т.е. являются моноцикличесними, иногда озимыми. Каждый по-
8X-I964 Н7
бег о окончанием малого жизненного цикла отмирает, остается его
базальный участок - "пенек" (резцд) с 2-3 сближенными междоузли-
ями и с наложившимися пазушными почками. "Пеньки" включаются в
состав каудекса. Корневая система, разрастаясь вглубь и вширь,
сохраняется в течение всей жизни особи, испытывая при старении
партикуляцию.
Генеративный период подразделяется на три возрастных состоя-
ния: молодое, средневозрастное и старое. К молодому генеративно-
му состоянию относятся особи, характеризующиеся небольшим числом
генеративных побегов, не выраженным каудексом, тонким главным
корнем, отсутствием придаточных корней. У средневозрастных гене-
ративных растений насчитывается много генеративных побегов, куст
большой, раскидистый, почек много, каудекс хорошо выражен, у от-
дельных особей можно видеть начальные фазы партикуляции каудекса,
а иногда и главного корня, главный и боковые корни утолщаются,
на основаниях побегов возникают придаточные корни. У старых гене-
ративных растений число почек и цветущих побегов становится мень-
ше по сравнению со средневозрастными, наблюдается интенсивная пар-
тикуляция главного корня (рис.14), которая возникает как продол-
жение партикуляции каудекса, вызываемой разрушением "пеньков".
Главный корень расчленяется в связи с отмиранием паренхимы его
первичных 'лучей, что ведет к распадению базальной части корня на
отдельные продольные партикулы. Процесс партикуляции идет сверху
вниз, не доходя до верхушки корня.
В конце большого жизненного цикла материнское растение распа-
дается на партикулы. Они, возможно, могут жить как самостоятель-
ные растения за счет запасных веществ и при,дат очных корней. Пар-
тикулы, сохранившие способность к цветению, следует относить к
старому генеративному состоянию, а переставшие цвести - к сениль-
ному. Они очень редко обнаруживаются в фитоценозе.
Большой жизненный цикл лядвенца продолжается свыше 20 лет. Од-
нако в пойме Оки в Луховицком р-не Московской обл. лядвенец выпал
из фитоценозов на шестой год после подсева, следовательно, дли-
тельность его жизни в этих условиях 5-6 лет и цветет он здесь не
более двух раз в течение жизни (Крылова, 1964).
118
Рис.14. Партикуляции корня лядвенца рогатого:
кд - каудекс, гл.к. - главный корень;
пр.к. - придаточный корень.
Сезонное развитие
Из наблюдений, проведенных за развитием лядвенца в культуре
(Пуленкова, 1974), следует, что при весеннем посеве прорастание
семян начинается на 4-5-й день, чаще на 6-7-й день после посева.
У проростков сначала развивается главный корень, впоследствии ухо-
дящий в почцу. На 7-12-й день раскрываются семядоли. Через 25
дней развертывается первый тройчатый лист. Несколько позднее на
главном побеге формируются еще 1-2, иногда 4 тройчатых листа. Об-
разование на главном побеге пятичленных листьев свидетельствует о
переходе проростков в ювенильное возрастное состояние. С перехо-
дом к кущению, наступающему на 35-50-й день после посева, особь
приобретает черты взрослого вегетативного растения. Семядоли к
119
атому времени подсыхают и отпадают, семядольный узел втягивается
в почву благодаря контрактильным свойствам главного корня. Таким
обрааом, в культуре, а в равной степени и в естественной обста-
новке, лядвенцу рогатому свойственны быстрые темпы развития. Пе-
реход от состояния проростка до взрослого вегетативного возраст-
ного состояния происходит в течение одного сезона.
В одной популяции особи отличаются разным темпом своего разви-
тия. Так, генеративного состояния могут достичь растения уже на
первом году жизни. Часто особи одного возрастного состояния зна-
чительно отличаются друг от друга по своему календарному возрас-
ту. Например, в средневозрастном генеративном состоянии могут
быть и 5-, и 10-летние растения, а хронологический возраст старых
генеративных особей оказывается равным 10 и 20 годам (Михайлов-
ская, 1967),
В генеративном состоянии растения проводят большую часть жиз-
ни. Длительное цветение, видимо, связано с постоянной сменой и
обновлением побегов, ежегодно возникающих из разных сериальных
почек каудекоа. Омолаживающему действию растений может также спо-
собствовать и постоянное нарастание в толщину главного корня (за
счет работы камбия), связанное о накоплением новых питательных
веществ. Все это позволяет говорить о том, что ритм развития ляд-
венца рогатого отличается большой вариабельностью и пластичностью
и делает его весьма устойчивым в местах обитания.
Культурные и дикорастущие формы лядвенца рогатого отрастают и
зацветают раньше других бобовых. В средней полосе европейской
части СССР массовое цветение его приурочено к третьей декаде июня
(Дмитриева и др., 1974), цветущие особи можно встретить в. августе
и даже в сентябре. По ритму цветения этот вид отнесен к рання-,
среднепозднему циклу (Голубев, 1965). Растянутость периода цвете-
ния вызвана различиями во времени заложения элементарных соцветий
на скелетной оси и боковых побегах обогащения, а также неодновре-
менным развертыванием почек и развитием из них побегов. Зачатки
нижних зонтиков главного соцветия заметны в почке, т.е. формиру-
ются в почечную фазу побега. Развитие других элементарных соцве-
тий происходит во внепочечную фазу, в период роста скелетной оси
и силлептических побегов обогащения.
Г.М.Дениоова (I960), проведя наблюдения на пойменных лугах Се-
верной Двины, относит лядвенец к группе растений, у которых гене-
ративные и вегетативные побеги длительно нарастают верхушками,
причем продолжительность роста тех и других побегов почти одина-
120
коза. В.Н.Голубев (1965), изучив лядвенец рогатый в луговой сте-
пи, характеризует его побеги как длительно вегетирующие, образую-
щие две генерации листьев. Вторая генерация листьев возникает за
счет дополнительного верхушечного нарастания и образования побе-
гов обогащения. Отмечен, третий способ возникновения листьев в ре-
зультате летнего развертывания почек и развития из них побегов
возобновления.
М.А.Гуленкова (1974) различает три типа побегов по времени их
отрастания и развития: летние (июль - август), летне-осенние (ав-
густ - сентябрь) и осенние (вторая половина сентября - октябрь).
Летние побеги - генеративные и вегетативные, сохраняют зеленые
листья до ооени и достигают длины 18-40 см; летне-осенние - веге-
тативные, 6-II см длины с 10-12 зелеными листьями; осенние побеги
недоразвиты, вытягиваются от 0,2 до II ом, несут всего 2-8 зеле-
ных листьев или представлены едва тронувшимися в рост почками и
развиваются по озимому типу.
Детальные наблюдения за заложением почек и развитием из них
побегов были проведены Г.М.Денисовой (1963) на лугах Северной Дви-
ны и Подмосковья. Почки возобновления различаются временем зало-
жения и длительностью внутрипочечного развития. Почки, располо-
женные в пазухах предлистьев скелетного побега, закладываются в
июле - августе в качестве внучатых и трижды зимуют в состоянии
почки. Почки, лежащие в пазухах листьев, выше расположенных по
побегу, отличаются меньшим (два года и менее) периодом внутрипо-
чечного развития. Емкость зрелой закрытой почки равна 10-12 лис-
товым зачаткам, из них 4-5 - зачатки низовых листьев, а 5-7 - за-
чатки стеблевых листьев, т.е. вегетативная сферы в почке заложе-
на не полностью. В зрелой генеративной почке в августе - сентяб-
ре формируются зачатки соцветий в паузах 6-9-го листовых зачатков,
в сентябре цветки заметны в виде бугорков (Денисова, 1963), В вер-
хушечной почке зеленых побегов, развивавшихся осенью, насчитывает-
ся 6-7 листовых зачатков, а в почках, только что тронувшихся в
рост (почках-побегах), число их равно 8 (Гуленкова, 1974). Таким
образом, степень сформированнооти побегов будущего года у лядвен-
ца рогатого неодинакова. Они находятся в состоянии почек, почек-
побегов, либо о ооени успевают развить базальные участки с низовы-
ми и зелеными листьями и сохраняются под снегом зелеными (озимые
побеги).
По способу перезимовки в условиях луговой степи этот вид отне-
сен к летнезеленым. Нижние листья на побеге подсыхают в июле, а
121
усыхание всего побега происходит в октябре (Голубев, 1965).
М.Ф.Заэоев (1937), ссылаясь на данные Рюбеля, указывает, что на
высокогорных лугах лядвенец зимует в зелейом состоянии.
Большинство почек возобновления у лядвенца рогатого оказывает-
ся в почве за счет втягивания корней каудекоа с базальными участ-
ками побегов. Часть почек может располагаться на поверхности поч-
вы (Гуленкова, 1974). По способу перезимовки почек этот вид отне-
сен к головчатым гемикриптофитам и рассматривается как молодое
отчленение рода Lotus (Gluch, 1969). Каудекс ежегодно обогащает-
ся новыми почками до включения в него "пеньков” от отмерших веге-
тативных и генеративных побегов. Втягивание и углубление его в
почну способствуют образованию придаточных корней на базальных
частях побегов. Корневая система в связи о этим приобретает чер-
ты смешанной.
Способы размножения и распространения
Семенное размножение . О.Хагерун (цит. по
Шамурину, 1956), изучая опыление лядвенца рогатого на Фарерских
о-вах, обнаружил у него автогамию - опыление происходит в бутоне
незадолго до распускания цветка, пыльца попадает большей частью
прямо на рыльце. Основной же способ опыления у этого растения -
энтомофилия (Knuth, 1895). О приспособленности вида к энтомофи-
лии свидетельствуют "мотыльковое” устройство цветка и наличие в
нем нектара (Кернер, 1902).
Нектар в цветках* бобовых выделяется нектарными железками, рас-
положенными в основании короткой цветочной трубки, выстилая углуб-
ление на дне ее или образуя небольшое выпуклое кольцо. Н.Н.Карта-
шова (1965) обнаружила однотипность корреляции между секрецией
нектара и фазами развития цветка у представителей зтого семейства:
нектарники начинают функционировать при раскрывании цветка, наи-
большее количество нектара выделяется в фазе, когда пыльники от-
пилили и пыльца осыпалась, в конце развития цветка нектароввделе-
ние сокращается.
Подобно другим бобовым цветки лядвенца протераидричны. В буто-
нах величиной 3-4 мм тычинки полностью сформированы. Перед распус-
канием цветка размеры бутонов увеличиваются до 10-13 мц, к этому
моменту вскрываются пыльцевые мешки. В раскрывшемся цветке пыльни-
ки продолжают пылить, впоследствии подсыхают, отгиб паруса рас-
122
прямляется и постепенно боковые его стороны завертываются назад.
В последней фазе морфогенеза цветка наблюдается падение тургора
венчика, парус опускается вниз и желтая окраска лепеотков меняет-
ся на оранжевую. В дальнейшем парус, весла и в последнюю очередь
лодочка опадают. Продолжительность жизни цветка равна 3-5 дням.
А.А.Щибря и А.И.Мартьянова (I960) провели специальные наблюде-
ния за способами опыления и плодоношением лядвенца рогатого в пой-
ме Оки близ Дединова, Луховицкого р-на, Московской обл. Авторы
установили, что длительность жизни цветков зависит от опыления: у
цветков, изолированных от посещения насекомыми, этот период длит-
ся 5-6 дней; у цветков, свободно посещаемых пчелами, лепестки вен-
чика опадают на третий день. Отмечена многократность (до 20 раз)
посещения пчелами одних и тех же цветков, наибольшее количеотво
плодов и семян образуется при свободном посещении цветков пчелами,
под изолятами плоды почти не завязываются. При свободном посеще-
нии цветков пчелами плодоношение составляет В0% (Зазоев, 1937). В
опытах А.Тоума и Д.Джонсона (loom, Jonson, 1945) при самоопыле-
нии образовалось 3,5% бобов, при перекрестном - 71,6%. Исследова-
ния.К.Роланда (Boland, 1949) показали, что при самоопылении в по-
левых условиях образовалось 2f> бобов от числа опыленных цветков и
ни одного плода не завязалось в теплице, перекрестное опыление в
полевых условиях дало 13% бобов, а в теплице '-' 23%. Обнаружено,
что формы атташе и hireutus имеют высокую самостерильность, а
filicaulus и utiginosuB практически самофертильны.
В литературе приводятся сведения о числе стеблей на одном рас-
тении лядвенца рогатого, однако нельзя понять, стебель какого по-
бега (возобновления или обогащения) берется за счетную единицу.
Так, по данным М.Ф.Зазоева (1937), число стеблей на растении варь-
ирует от 30 до 750, в среднем 100. В первый год на растении обра-
зуется 16-150 стеблей, во второй - 25-388 (Африкян, 1957). Эти
цифры, видимо, отражают число генеративных и вегетативных побегов,
приходящихся на одну особь. Принимая за очетщю единицу побег..
возобновления, мн выяснили, что наибольшее число генеративных пр-: < у
бегов образуется у средневозрастных генеративных бсобей (23^2,2)i’
минимально у молодых (6,6+0,6), у старых этот показатель остается <?
высоким (17,2+0,5), уступая несколько средневозрастным особям. Та- .,
кая же закономерность прослеживается и в числе цветков на одной?
особи, определенном в первую половину цветения (Уранов, Писковац-
кова, 1974). В культуре на одном растении может образоваться от
13 да 177 элементарных соцветий, а в естественных местообитаниях
123
встречаются особи о 400 соцветиями (Мухина, 1956). Как ухе упоми- '
иалооь выше, каждое элементарное соцветие состоит из 2-8 цветков.
По наблюдениям Н.М.Мухиной (1956), наиболее обильное завязывание
плодов отмечено в соцветиях ранних сроков цветения, часть образо-
вавшихся плодов на растении не дозревает.
Семенное размножение является основным и практически единст-
венным способом размножения лядвенца рогатого. По способу диоое-
минации бобовые отнеоены к автомеханюхсрам (Левина, 1967). В стен-
ках плода лядвенца рогатого дифференцируются разные слои тканей,
среди которых выделяется твердый слой клеток, косо тянущихся от
одного края створки к другому. Каждая из этих клеток при высыха-
нии окручивается, что приводит к закручиванию всего твердого слоя
и стенок плода. Створки плода скручиваются в 2-3 оборота, остают-
ся прикрепленными к цветоножке. Семена при этом могут разбрасы-
ваться на значительное расстояние (Кернер, 1902).
Вегетативное размножение. Н.А.Мухи-
на (1956) набладала образование корневых отпрысков на боковых
корнях лядвенца рогатого. Такие растения чаще встречались в куль-
туре, единично, в естественных местообитаниях. Образование корне-
вых отпрысков происходило в тех случаях, когда корни выходили на
поверхность почвы. Мы встретили только две особи лядвенца, на бо-
ковых корнях которых при обнажении появлялись корневые отпрыски.
Однако нам не удалось найти отделившиеся от материнского растения
корневые отпрыски и пронаблюдать их дальнейшее развитие, поэтому
нет основания говорить о вегетативном размножении. Появление кор-
невых отпрысков на боковых корнях и возможность вегетативного раз-
множения корневыми черенками следует понимать, очевидно, как тен-
денцию к корнеотпрысковости.
Редкое отделение партикул от материнской особи в конце большо-
го жизненного цикла свидетельствует о старческом распаде особи и
не приводит к вегетативному размножению.
Экология, фитоценология и консортивные связи
Лядвенец рогатый к воде мало требователен, обычно встречается
при увлажнении от сухо- до влажнолугового (59-72-я ступени шкалы
Л.Г.Раменского, 4-я ступень шкалы Элленберга, 2-я ступень шкалы
Ландолъта). Может выдерживать длительное затопление (35-50 дней),
но избыточное увлажнение действует на него отрицательно (Ларин и
124
др., 1951).
К свету довольно требователен - как правило, встречается при
относительной, освещенности более 10%, Ландольт и Элленберг отно-
сят его у полусветовым растениям. Отличается зимостойкостью и вет-
ровыносливостью (Ларин и др., 1951; Ненароков, Раменская, 1951).
Не требователен к богатству почв питательными веществами,
встречается на почвах от довольно бедных до богатых (13-21-я сту-
пени шкалы Л.Г.Раменокого, 2-я ступень шкалы Элленберга). Растет
на разных типах почв - от черноземных до подзолистых, с различным
механическим составом (от песчаных до глинистых). Выносит как со-
лонцеватсоть, так и сильную выщелоченность (Ларин и др., 1951;
Раменский и др., 1956). Предпочитает почвы от нейтральных до ще-
лочных, с 'рН=5,4-8,0 (7-я ступень шкалы Эллёнберга, 4-я ступень
шкалы Ландольта), однако способен мириться с.избыточной кислот-
ностью, встречаясь на почвах с рН=4,0-4,2 (Родионов, 1952).
Лядвенец рогатый - обычный компонент фитоценозов пойменных и
суходольных лугов, встречается на полянах, склонах, выгонах, в
светлых борах, вдоль дорог. В луговых сообществах он редко высту-
пает доминантой, чаще играет второстепенную роль (Ларин и др.,
1951). На почвах со слабой кислотностью, но при хорошем воздушно-
водном режиме образует рогатолядвенцовую формацию мелко-низкобобо-
вого луга (Матвеева, I960). В пойме Северной Двины близ Холмо-
горски мы описали нормальные ценопопуляции лядвенца, различавшие-
ся по соотношению численности молодых и старых особей (Писковац-
кова, 1976). На пастбищных угодьях прирусловой и центральной пой-
мы господство принадлежит молодым особям (молодые нормальные цено-
популяции). На сенокосном участке отмечено преобладание старых
растений (старая нормальная ценопопуляция).
Лядвенец рогатый - симбиобактерофаг (Родионов, 1952). Его ста-
рые побеги иногда поражаются ржавчинным грибом, а корень поврежда-
ют личинки жуков, живущие в его внутренних тканях, и полевые мыши.
На семена нападает лядвенецевая зерновка Bnmchus lotipogy (Зазс-
ев, 1937). В бутоны откладывает яйца комарик Diplosis loti, вызы-
вая образование галлов. Венчики при этом не раскрываются, чашечка
оказывается вздутой, иногда мясистой, цветки внешне напоинают
почку или маленькую луковицу (Кернер, 1902). Опыление цветков про-
изводится насекомыми, собиращими пыльцу и нектар. В составе фау-
ны опылителей из перепончатокрылых - медоносные пчелы (44,0%), ре-
же шмели (9,5%), посещаются цветки также представителями отряда
двукрылых (45,9%) (Смарагдова, 1969).
125
Биологическая продуктивность
и хозяйственное значение
Урожайность сена в чистых посевах лядвенца рогатого на Москов-
ской опытной станции составила в среднем 20 ц/га (Родионов, 1936).
На торфяной болотной почве он дал в сумме за три укоса 67 ц/га
сена (Куприянов, 1939). Урожай сена колеблется по годам иопользо-
вания: в первый год он составил 42,5 ц/га, во второй - 53,9 и в
третий - 38,3 ц/га (Родионов, 1952). На Краснодарской опытной
станции получен урожай сена от 21 до 102 ц/га, средний годовой
(за 5 лет) - 76,4 ц/га (Зазоев, I937). На Сочинской сельскохозяй-
ственной опытной станции урожай сена лядвенца составил в суше за
4 укоса 131,1 ц/га (Куприянов, 1939). Этот вид дает высокую про-
дуктивность в смеси с другими культурами (Родионов, 1936; Морозо-
ва, 1955). Урожай семян в разных экологических условиях от 4 до
12 ц/га (Африкян, 1965).
Установлено, что I га чистого посева лядвенца в нечерноземной
полосе может дать 28-30 кг сахара в нектаре (Копелькиевский,
1954; Пономарева, 1954). Содержание сахара в нектаре одного цвет-
ка в первый год жизни растения составляет 0,183 мг, на второй
возрастает до 0,713 мг (Смарагдова, 1969). Исследования, прове-
денные на холмогорских заливных лугах низовий Северной Двины, по-
зволили установить, что нектаропродуктивность цветков лядвенца в
теплые солнечные дни составляет 0,7-0,9 мг. Количество нектара
зависит от численности генеративных особей, возрастности ценопо-
пуляций и в связи о этим варьирует от 67,7 до 263,3 мг на 0,25 №
(Уранов, Писковацкова, 1974).
Лядвенец ценится как хоромее кормовое сенокосное и пастбищное
растение. Лучшие сорта содержат 18% сырого протеина, 12-14% чис-
того белка, 7,0-7,5% золы и 25-30% клетчатки в пересчете на сухое
вещество (Родионов, 1936). Кормовое значение лядвенца несколько
снижается из-за наличия в цветках и листьях горького цианогенно-
го глюкозида, выделяющего синильную кислоту (Африкян, 1957). Мак-
симальное количество синильной кислетм накапливается во время мас-
сового цветения, однако количество ее не опасно для скота. Содер-
жание ее падает в конце цветения и сокращается в сене (Мухина,
1956).
Лядвенец рогатый - ценный медонос (Копелькиевский, 1954; Поно-
марева, 1954; Африкян, 1965). Он испытывался как сидеральное
126
удобрение и признан прекрасным предшественником табака, овоа и
картофеля (Зазоев, 1937; Родионов, 1952). Рекомендуется ддя посе-
вов в травостоях полевых и кормовых севооборотов нечерноземно*
полосы (Симонов, 1951), представляет интерес для возделывания на
песчаных, супесчаных и вновь осваиваемых землях (Родионов, 1936).
Он хорошо переносит сенокошение и стравливание (Ларин и др.,
1951).
127
Л.П.Рыоин
ЛИПА. СЕРДЦЕВИДНАЯ
Номенклатура и внутривидовая систематика
Tilia cordata Mill. 1768, Gard. Diet. ed. 8, № I; ШмалЬГ.,
1895, Фл. 1:179; Сырейщ., 1907, Илл. фл. 2:354; он же, 1927, Опр.
раст. Моок. губ.:184; Назаров, 1940, в Маевок. Фл., изд. 7:506;
Малеев, 1949, Фл. СССР 15:20; Пояркова, 1954, в Маевск., Фл.,
изд. 8:296, 1964, изд. 9:289; Васильев, 1958, Деревья и кустарни-
ки СССР 4:722; Ворошилов, 1966, Опр. раст. Моок. обл.:230; Bro-
via*, 1968, Fl. Bur. 2:248. - T.parvifolia Ehrh. 1790, Beitr.
Katurk. 5:159; Кауфм., 1866, Моск. фл., изд. 1:102, 1889, изд.
2:99; Цингер, 1886, Сб. свед.:114; Петунн., 1896, Крит, обзор
1:103. - Липа сердцевидная, или мелколистная (семейство липовых -
Tiliaceae).
По мнению В.Н.Сукачева (1934), липа сердцевидная в природе ма-
ло варьирует, но в культуре известны ее многие аберрации, имеющие
садоводственное значение. И.Н.Васильев (1958) называет следующие
формы: I) f.ovalifolia (Spach.) Ig. Vassil. отличается более или
менее продолговатыми листьями, у основания косо усеченными; 2) f.
pendula (Beissn., Schnelle et Zbl.) Ig. Vassil. с плакучими
листьями; 3) f.aureavariegata (C.K.Schneid.) V.Engl. с золотисто-
пестрыми листьями весьма разнообразной формы; 4) f.vulgaris (Нау-
ne) Ig. Vassil. Иначе представлял внутривидовое разнообразие ли-
пы Вагнер (Wagner, 1933; цит. по Васильеву, 1958) - он называл
32 разновидности и 26 видов гибридного происхождения.
Географическое распространение
Евро-сибирский вид. В Западной Европе отсутствует лишь на запа-
де и юге Пиренейского п-ова, а также на Аппенинском и Балканском
п-вах (исключая их северные районы), а на севере Европы - в север-
ных частях Шотландии и Скандинавии (Rubner, 1960). на территории
СССР граница ареала липы проходит через северную оконечность Онеж-
ского озера, пересекает р.Онегу в ее среднем течении, р.Северную
Двину у устья р.Ваги; за северным изгибом р.Камы она поворачивает
128
к югу в уходит за Урал к р.Иртышу, а затеи к р.Таре. Липа, встре-
чающаяся "островами" в западных предгорьях Кузнецкого Алатау, в
горных районах южной части Средней Сибири, выделяется в особый
вид Tilia Bibirica Bayer1. КЖная граница ареала проходит через
Кишинев, направляется в почти широтном направлении к Волгограду,
затем уходит по р.Волге на Саратов и Куйбышев, а оттуда поворачи-
вает к Оренбургу. Липа встречается также в Крыму и на Кавказе,
преимущественно в верхнем лесном поясе.
На территории Московской обл. встречается довольно часто на до-
статочно плодородных влажных и дренированных почвах; по этой при-
чине редка в Мещерской низменности, где эдафический фон создают
бедные пески и где очень велико заболачивание. Всего липняками в
Московской обл. занято около 7 тыс. га, что составляет 0,5% лесо-
покрытой площади (Вакуров, Надеждин, 1968). Как правило, ныне су-
ще ствукщие липовые древостои - производные от вырубленных сложных
ельников и сосняков. На значительно большей площади липа встреча-
ется в подмосковных лесах в качестве сопутствующей или подчинен-
ной породы; в хвойных лесах она обычно образует второй ярус дре-
востоев, а в менее благоприятных условиях входит в состав подлес-
ка и не достигает высоты древесного полога.
М.И.Нейштадт (1957), рассматривая историю ареала липы сердце-
видной и основываясь на данных пыльцевых спектров, считает, что в
днепровско-валдайское межледниковье липа на территории Русской
равнины была распространена примерно в тех же границах, что и в
настоящее время, но ее участие в лесах было более значительным.
В конце межледниковья в связи с ухудшением климатических условий
липа стала исчезать. Во время валдайского оледенения она сохрани-
лась лишь в отдельных убежищах. 0 распространении липы в древнем
голоцене (9800-12000 лет назад) данных очень мало. В раннем голо-
цене (7700-9800 лет назад) ареал липы вновь стал расширяться, при-
ближаясь к водоразделу бассейна Северного Ледовитого океана и Чер-
номорско-Каспийского баосейна. В среднем голоцене (2500-7700 лет
назад) липа достигла границ современного ареала, а может быть про-
двинулась несколько севернее, одновременно проникнув за Урал. В
Т - “
Морфологически липа сердцевидная и липа сибирская очень близки,
многие авторы считают их за один вид. Однако в работах послед-
них лет это разделение проводится. Во "Флоре СССР" (т.15, 1949)
разделительными признаками приняты форма листовой пластинки и
окраска нижней стороны листьев. И.В.Васильев (1958) добавляет к
этому характер опушения завязи, а Ю.П.Хлонов (1965) - форму и
направленность лопастей рылец.
9-1964 129
иовднем голоцене северная граница ареала липы несколько отступила
к игу, а количество липы в лесах заметно уменьшилось.
Ю.Р.Шеляг-Сосонко (1968) к территории, где липа занимает устой-
чиво фитоценотические позиции, относит только Предуралье. В.Б.Со-
чава и А.М.Семенова-Тяншанская (1956) считают, что роль доминанта
или субдоминанта лива : играет в приволжских и приуральских широко-
лиственных лесах. Иную точку зрения высказывает С.Ф.Курнаев
(1968). Он называет леса из липы сердцевидной "главнейшей форма-
цией" коренных широколиственных лесов Русской равнины. В зоне ши-
роколиственных лаооВ они занимали пдакорные условия и, следова-
тельно, должны рассматриваться как зональные. Современное располо-
жение ооновных массивов липовых лесов совершенно не отвечает их
доагрикультурноцу распространению, причиной этому послужило интен-
сивнейшее истребление липы. По мнению того же автора, липа имела
безраздельное господство на плакорах северной части зоны широко-
лиственных лесов, тогда как в южной части преобладал дуб: липовые
леса связаны с достаточно влажным и умеренно теплым летом, в то
время как дубовые - с более жарким и сухим. В области распростра-
нения липовых лесов среднегодовое количество осадков составляет
540^600 мм,, наибольшая часть их выпадает в июле - августе; в об-
ласти оптимального развития дуба в эти месяцы количество выпадаю-
щих осадков, напротив, резко снижается. Таким образом, в северной
части зоны широколиственных лесов коренные липняки замещают ельни-
ки о липой, являющиеся зональными в подзоне смешанных лесов, в
свою очередь, уступая южнее место дубовым лесам.
В доагрикультурннй период липовые леса по зональным экотопам
сплошной полосой тянулись вплоть до р.Волги, а далее, вследствие
возрастающей континентальности, климата, распадались на отдельные
массивы, приуроченные к наиболее возвышенным водоразделам, полу-
чавшим большее количество осадков. На западном склоне Южного Ура-
ла, где общая влажность увеличивалась, липовые леса вновь форми-
ровали сплошную полосу, а за Уралом, где значительно возрастала
континентальность климата, встречались только отдельными "острова-
ми". В Западной Европе полосу липовых лесов продолжают, по мнению
С.Ф.Вурнаева (1968, 1980), буковые леса.
130
Морфологическое описание
Дерево первой величины, достигающее в высоту 30-32 и при диа-
метре ствола до I м; старые (300-400-^летние) липы, растущие в пар-
ках или на открытых местах, могут иметь в поперечнике до 2 м. В
лесной среде стволы липы отличаются прямизной, хорошей очищенноо-
тью от сучьев. Кора темная, продольно-бороздчатая, с хорошо выра-
женной коркой.
У липы мощная крона, создающая сильное затенение. Верхние вет-
ви обычно направлены кверху, средние имеют горизонтальную направ-
ленность, нижние свисают, что особенно заметно у деревьев, расту-
щих на опушках. Окраска побегов отчасти зависит от освещения.
Вследствие большой протяженности кроны ее отдельные части (уров-
ни) функционируют в существенно неодинаковых условиях фитосреды -
разнятся освещенность, влажность, температурный режим. Эта верти-
кальная дифференциация кронового пространства позволяет говорить
о трех основных биогеоценотичеоких горизонтах (Днлис, Носова,
1977): верхний горизонт наибольшего развития листьев и молодых по-
бегов, а также цветков и плодов, преобладают листья оветового ти-
па (плотные и кожистые); масса ветвей ненамного превышает массу
листьев и молодых побегов; средний горизонт с заметным уменьшени-
ем развития ассимилирующей поверхности, меньшей интенсивностью
цветения и плодоношения; скелетные части кроны по массе в значи-
тельной степени превосходят фотосинтезирующие органы; нижний гори-
зонт слабого развития ассимилирующих органов, листья типично тене-
вой структуры, линейный прирост молодых побегов очень мал, и по
массе, и по поверхности скелетная часть кроны немного превышает
фотосинтезирующую часть.
Нарастание побегов оимподиальное; верхушечная точка роста еже-
годно теряется, начинают развиваться побеги из боковых почек. Ве-
гетативные и цветоносные побеги существенно не различаются и в
равной мере выполняют ассимиляционные функции (Серебряков, 1952).
Листорасположение очередное. По наблюдениям В.Р.Васильева и
И.С.Антоновой (1979), на одной и той же ветви в одинаковом коли-
честве имеются побеги с правой и левой спиралью. Лиотья простые,
с сердцевидно-выемчатым основанием, по краям зубчатые, с вытяну-
той заостренной верхушкой, длина и ширина листовых пластинок 4-
8 см. Листовые почки яйцевидной формы, тупые, голые, коричневые,
около 6 мм длины и 4 мм ширины. Формирование листовой пластинки
131
липы обстоятельно описано И.Г.Серебряковым (1947). Сверху листья
томно-зеленые, голые; снизу сизо-зеленые, о отчетливыми базальны-
ми жилками, в углах которых пучки рыжеватых волосков. Уже при вы-
ходе лиота из почки начинают проявляться различия между световыми
и теневыми листьями, достигая максимума к окончанию их роста -
лиотья о теневой структурой менее кожисты, со слабее развитой па-
лисадной тканью.
;Цветочные почки располагаются вдоль материнского побега. Со-
цветия-полузонтики из 5-9 цветков прикреплены к общей цветоножке
со светло-зеленым (желтоватым) удлиненным прицветником. Венчик из
5 желтоватых голых лепестков, чашечка из 5 светло-зеленых опушен-
ных чашелистиков, у их оснований пучки волосков и нектарники. Ты-
чинок 20-30, отчасти они срастаются в пучки. Пестик о верхней 5-
гнездной завязью, густо покрытой волосками, о голым столбиком и
5-зубчатым рыльцем. Орешковидные обратнояйцевидные плоды имеют по
1-2 семени. Масса I тыс. плодов 31-33 г (Гроздов, 1952). Прицвет-
ный лист остается при соплодиях и своей парусностью способствует
разносу семян ветром.
Липа обладает мощной корневой системой,' обеспечивающей деревь-
ям ветроустойчивость. По наблюдениям В.В.Смирнова и В.Г.Семеновой
(1970), в липняке разнотравно-волосистоосоковом на супесчано-су-
глинистых почвах в Серебряноборском лесничестве Московской обл.
корни липы уходят на глубину более 2 м. При этом корневые плапып
располагаются в верхних 40 ом, корни толщиной более 2 ом проника-
ют до глубины 60 см, 1-2 см - до 80 ом, 0,5-1*ом - до I м. Что же
касается корней с диаметром менее 0,5 см, то максимум их (более
75%) приходится на верхние 40 ом, тогда как глубже I м располага-
ется не более 5%. Сходное явление мы обнаружили в сосняке с липой
чернично-разнотравном на песчано-супесчаной почве в том же лесни-
честве, несмотря на различие в почвенных условиях - 75-85% корней
липы диаметром менее 0,5 см находится в верхних 50 см почвы. На
следующий полуметровый сдой приходится 10-15% их общего количест-
ва, а на толщу 100-150 ом - 5-10%.
Морфологию корневых систем липы изучали С.Н.Карандина (1950),
К. М. Смирнова (1952), Е.М.Самойлова (1965), В.Е.Наумова (1972) и
др. Как правило,, обращается внимание не только на мощность корне-
вой системы, но и на ее пластичность - способность реагировать на
специфику почвенных условий, на характер корневых систем сопутст-
вующих пород, что позволяет липе с наибольшей полнотой использо-
вать почвенный объем.
132
По данным А.А.Чистяковой (1978а), у деревьев семенного проис-
хождения корневая система в процессе онтогенеза из стержнекорне-
вой становится киотекорневой; у деревьев вегетативного происхож-
дения корневая система только кистекорневого типа. А.А.Чистякова
(1978а) выделяет три экологические и морфологические формы корне-
вой системы липы: поверхностная, поверхностно-якорная и поверх-
ностно-стержнево-якорная. Некоторые авторы относят липу к корне-
отпрысковым растениям, ио, по мнению А.А.Чистяковой (19786), эта
точка зрения ошибочна - на корнях липы почек нет и возникают они
только на органах побегового происхождения. Из проснувшихся почек
формируются эпигеогенные и гипогеогенные корневища, древеснеющие
уже в первый год своего существования. Глубина гипогеогенных кор-
невищ 5-15 см, скорость роста - 1,5-3,5 см/г. На них развиваются
чешуевидные листья, в пазухах которых находятся почки. Образова-
ние гипогеогенных корневищ протекает тем интенсивнее, чем меньше
.плотность субстрата.
Согласно наблюдениям ряда авторов (Ну, 1960а; и др.), весной,
во время распускания листьев, корни растут медленно, причем вна-
чале появляются всасывающие, а позже ростовые корни. В периоды
бутонизации и цветения липы корни растут быстрее. И.Н.Рахтеенко
(I960) одним из периодсв активного роста корней липы считает
июль - август.
А.А.Чистякова (1979) считает, что одноствольное прямостоящее
дерево - это не единственный и, возможно, не основной вариант жиз-
ненной формы липы. Есть и другие варианты, существование которых
обусловлено способностью липы к вегетативному разрастанию и раз-
множению. Значительно чаще липа растет куртинами, имеющими доволь-
но сложную структуру и состоящими из парциальных образований раз-
ного возраста, соединенных друг с другом одревесневшими корневи-
щами - ксилоризомами. Особой модификацией жизненной формы липы яв-
ляется кустовидная форма, появлякщаяся как результат угнетения.
Онтогенез
Схема онтогенеза липы сердцевидной и отдельные возрастные ста-
дии изучены А.А.Чистяковой (1978а, 1979), предложившей различать
в большом жизненном цикле этого вида семенной ряд и два варианта
вегетативного ряда.
Первой стадией семенного ряда являются всходы. Прорастание на-
133
9Х-1964
ХОЖПОЯ * зависимости от степени зрелости семян, а она, в свою
очзрздь, связана с прохождением стратификации (Любченко, 1959,
I960). Повой семян в значительной степени обусловлен состоянием
зародыша (Любченко, 1963). В лесу всхода появляются с мая по июль.
Вначале проростки несут только почти округлые, пальчато-пятило-
пастные семядоли диаметром 1,5-2 ом, они сохраняются до осени.
Позже формируется первый удлиненно-яйцевидный трехлопастный, с го-
родчатыми краями.
Ранние всходы к осени формируют до 5-7 (чаще 1-3) листьев,
поздние - имеют только семядольные листья. В пазухах листьев и се-
мядолей закладываются почки, размеры которых увеличиваются к вер-
шине побега (Грохольская, 1950). Высота всходов-однолеток 4-7 см.
У 50-70% всходов хорошо заметен стержневой корень, у остальных
развивается стержне-киотекорневая система. Главный корень вначале
растет вертикально вниз, но на глубине 4-6 см резко изменяет на-
правление; к концу сезона вегетации окончание корня, а иногда и
большая часть его отмирает. От главного корня отходят боковые дли-
ной 0,5-1,5 ом. На базальной полегающей части стебля появляются
придаточные корни, растущие горизонтально.
Ювенильные растения - одноосные сеянцы. Продолжительность этой
стадии - 5-7 лет (в неблагоприятных условиях несколько дольше).
Листочки удлиненно-яйцевидной формы, теневой структуры. У подавля-
ющего числа растений в этом возрасте корневая система стержне-кис-
текорневого типа, у остальных - кистекорневая; последняя формиру-
ется только в том случае, если главный корень отмирает полностью
или сохраняет лишь свою базальную часть. Гипокотиль, который у
всходов располагается над поверхностью почвы, у ювенильных расте-
ний постепенно втягивается в почву. Высота особей, относящихся к
этой возрастной стадии, 3-30 см, диаметр стволика 1-5 мм.
Имматурные растения отличаются появлением боковых осей, в ре-
зультате чего начинает формироваться крона. При благоприятных ус-
ловиях она приобретает удлиненно-пирамидальную форму, а в случае
угнетения - зонтиковидную, поскольку главная ось не выделяется, а
окончания боковых осей оказываются примерно на одном уровне. Верх-
ние листья постепенно перестраивают свою структуру от теневой к
световой. Преобладает корневая система кистекорневбго типа, и
только у 10-20% особей сохраняется стержне-кистекорневая система.
Четко обособляются якорные корни. К концу этой стадии (она может
длиться до 40-летнего возраста) высота молодых липок составляет
0,7-2,5 м, диаметр ствола - 1-3 см.
134
Взрослые виргинильные растения представляют собой узкокронные
деревца, заметно (по сравнению с имматурными растениями) убыстря-
ющими свой рост. Верхние листья в кроне имеют световую структуру,
нижние и внутрикроновые - теневую. Высота таких особей 4-6 м,
диаметр стволика до 6 ом, диаметр кроны 1,5-2,5 м. Стержне-киоте-
корневая система на этой стадии встречается крайне редко. Главный
корень, если он сохранился, становится якорным. Придаточных ске-
летных корней 5-10, из них 1-2 корня направлены вертикально вниз,
остальные - горизонтально.
Молодые генеративные особи цветут, хотя еще и не очень обильно,
в основном в средней части кроны, начинающейся на высоте 3,5-8 м
и занимающей около половины длины ствола; диаметр кроны 2-4,5 м.
Нижняя часть ствола имеет корку с глубокими трещинами. Прироста
еще относительно велики, но уже постепенно начинают снижаться. К
концу этой возраотной стадии (к 60-80 годам) высота молодых де-
ревьев составляет 6-17 м при диаметре ствола 5-25 ом. Корневая
система исключительно кистекорневого типа (от главного корня со-
храняется только короткая утолщенная базальная часть). Глубина
проникновения корней 1-1,5 м.
Средневозрастные генеративные особи имеют более раскидистые
кроны - 3-9 м в диаметре. Нижние 5-15 м ствола уже очищены от
сучьев, кора с отчетливо выраженными трещинами. Форма лиотьев ста-
новится широкояйцевидной (вместо округлояйцевидной). Обильное цве-
тение и плодоношение в средней и верхней частях кроны становятся
довольно обычными. Увеличивается радиус корневой системы, но тип
ее и глубина проникновения корней остаются прежними. Эта стадия
завершается к 80-100 годам.
Старые генеративные особи имеют широкопирамидальную крону, уже
почти полностью лишившуюся первичных скелетных осей; их заместили
ветви, возникшие из спящих почек (вторичная крона). Верхние боко-
вые ветки находятся на одном уровне с главной осью, почти прекра-
тившей рост. Цветение и плодоношение продолжаются, несмотря на то,
что многие деревья поражены сердцевидной гнилью, однако с возрас-
том увеличивается число пустозернистых плодов. Высота старых гене-
ративных особей (возраста до 150-300 лет) может достигать 30 м,
диаметр кроны нередко превышает 10 м, глубина корневой система до-
стигает 2,5 м. Корневая система сохраняет кистекорневой тип.
Под пологом леса значительную часть составляют особи вегетатив-
ного происхождения. Они имеют опецифичеокий онтогенез и несколько
иные возрастные стадии, однако биоморфологичеокие особенности воз-
135
рвотных групп особей вегетативного происхождения близки соответст-
вующим группам семенных особей (Чистякова, 1978а, 1979). Семенным
ювенильным оообям соответствуют ювенилоподобные образования веге-
тативного происхождения. Они также имеют удлиненно-яйцевидные
трехлопастные листья, но отличаются заметно большей высотой (до
I м), величиной годичного прироста, размерами листьев, а также ха-
рактером корневой системы. Возрастные пределы этой группы - 1-6
лет.
В следующих возрастных группах (имматурной, взрослой, вирги-
нильной, молодой, средневозрастной и старой генеративной) сходст-
ва между особями семенного и вегетативного происхождения больше:
у них близкие размеры надземных органов, аналогичная форма кроны,
сходные темпы роста. Однако формирующиеся парциальные образования
качественно неодинаковы. Особи, возникающие из почек непосредст-
венно на материнском стволе (пневая поросль), растут быстрее, чем
особи, появившиеся из почек на корневищах (ксилоризомах), но они
раньше стареют, в большинстве своем загнивая уже в среднем возрас-
те. В 45-90 лет такое дерево переходит в старое генеративное со-
стояние, а затем выпадает. Что же касается ксилбризомных особей,
то они менее подвержены быстрому старению, причина этого - удален-
ность от материнского растения и определенная автономность разви-
тия (Чистякова, 1979). По своему габитусу они не уступают особям
семенного происхождения.
Куртины липы со временем могут распадаться на части. Это может
произойти в результате разрушения материнской особи, под влиянием
абиотических факторов среды. В результате возникает клон, состоя-
щий из нескольких партикул разного возрастного состояния, причем
ими могут быть и отдельные особи, и сложные ксилоризомные индиви-
ды. Часть партикул обнаруживает признаки омоложения. Этот процесс
постоянного омоложения популяций липы имеет особенно большое зна-
чение в северных частях ее ареала, где почти полностью отсутству-
ет семенное возобновление этой порода, поскольку только благодаря
ему липа обеспечивает стабильное существование. Однако явления
омоложения наблюдаются и в центральной, и в южной частях ареала
липы (Чистякова, 1979). Преобладание омоложения над старением оп-
ределяет ее фитоценотическую устойчивость.
М. С.Двораковокий и М.Д. Алтухов (1963) предельным возрастом для
липы считают 700 лет.
136
Сезонное развитие
На территории Подмосковья набухание листовых почек у липы при-
ходится на третью декаду апреля (Елагин, 1976). Распускание листь-
ев и рост листовой пластинки продолжаются до начала июня. Позднее
распускание листьев предохраняет лицу от повреждения весенними за-
морозками.
Цветение липы начинается в конце июня - начале ивм. Цветочные 'J
почки закладываются в апреле - начале мая того же годе, когда про-
исходит цветение (Морохин, 1926; Устинова, 1952; Артяменко, Соко-
лов, 1955). По многолетним средним данным липа зацветает в г.Вое-
кресенске (юг Московской обл.) 29 июня, в Приокско-Терраонои за-
поведнике 6 июля, в г.Пущино 7 июля; в Наро-Фоминском р-не 9 июля
(Сезонная жизнь природы Русской равнины, 1979). Расхождение в да-
тах наступления этой фенологической фазы определяется не только
различным географическим положением названных пунктов, но и не-
одинаковой продолжительностью наблюдений (от 12 до 22 лет). В раз-
ные годы цветение отмечается в сроки, нередко довольно далеко от-
стоящие друг от друга: в г.Воокресенске - от 16 июня (1967) до
9 июля (1965), в Приокско-Террасном заповеднике - от 24 июня
(1948) до 28 июля (I960), в г.Пущино - от 26 июня (1967) до 18
июля (1965), в Наро-Фоминском районе - от I июля (1966) до 25
июля (1965). В период 1924-1938 гг. средним сроком зацветания ли-
пы в Подмосковье было 6-10 июля (Шиголев, Шиманюк, 1949). Для
сравнения заметим, что в этот же период на линии Кишинев - Ростов
липа зацвела 15 июня, а на линии Ленинград - Вологда - Киров -
Пермь - 15 июля. По данным В.И.Долгошова (1957), за 70 лет наблю-
дений в Подмосковье вероятность зацветания липы во второй декаде
июня составила 6%, в третьей декаде того же месяца - 19, в первой
декаде июля - 36, во второй декаде июля - 33 и в третьей декаде -
7%. И.С.Молукадо (1950) сообщает, что на Киевщине различают по
времени цветения две формы липы: рано- и поздноцветущую (разница
в сроках составляет 15-20 дней).
Цветение продолжается 10-15 дней, после чего завязавшиеся пло-
ды начинают созревать. Этот процесс завершается к концу октября,
но плоды остаются на ветвях и рассеиваются главным образом только
весной следующего года.
Осеннее расцвечивание листьев начинается в Подмосковье во вто-
рой половине августа (Елагин, 1976). В сентябре большая часть
137
листвы опадает, но листопад продолжается и в октябре. Общая дли-
тельность процесса -регятяции липы в центральной части Русской рав-
нины составляет около шести месяцев.
Способы размножения и распространения
Липа размножается и распространяется как. оеменннм, так и веге-
тативным путем. Успех оплодотворения в большой степени зависит от
интенсивности цветения, численности наоекомых-опылителей, погод-
ных условий: неблагоприятна, о одной стороны, дождливая и прохлад-
ная погода, с другой - слишком жаркая и сухая. Урожай семян зави-
сит от полноты и возраста древостоев. Например, в Среднем По-
волжье средний урожай в спелых липняках составляет 25 кг/га, в
приспевающих - 15 кг/га, в средневозрастных - 10 кг/га. В высоко-
урожайные годы выход оемян достигает 100 кг/га, но такие обильные,
урожаи повторяются редко (Напалков, 1952). Семенному размножению
препятствуют мышевидные грызуны, поедающие или повреждающие зна-
чительную часть опавших плодов.
Большая часть всходов появляется в первой половине вегетацион-
ного периода, непроросшие семена быстро теряют всхожесть. Многие
всходы вскоре погибают, особенно в условиях недостаточной освещен-
ности. Как правило, нормально развитие всходы обнаруживаются поч-
ти исключительно в "окнах", где световой режим значительно благо-
приятнее. Большой ущерб всходам липы наносят грибные заболевания
(Гусева, 1965). В конечном итоге семенное размножение липы чаще
всего очень затруднено и существенного значения для сохранения це-
нопопуляции не имеет. ‘
Значительно успешнее липа размножается вегетативно - за счет
развития почек как на надземных, так и подземных побегах, но не
корнях; как уже отмечалось, вопреки мнению некоторых авторов, ли-
па не является корнеотпрнсковнм растением (Литвинов, 1957; Хлонов,
1965; Чистякова, 19786). Из почек, находящихся на надземных орга-
нах, формируются эпигеогенные корневища. Именно таково, например,
происхождение пневой поросли, которую липа способна давать до глу-
бокой старости (Хлонов, 1965; Чистякова, 19786). Эпигеогенное кор-
невище может сформироваться не из спящей, а из обычной пазушной
почки в случае плагиотропного роста побега, если он соприкасается
с поверхностью почвы, а тем более погружен во влажную подотидку.
При таком положении очень быстро появляются придаточные корни.
138
Спящие почки на подземных побегах могут дать начало гипогеоген-
ным корневищам. В результате появления на корневищах обоих типов
ортотропных побегов возникает куртина - система плагиотропных и
ортотропных побегов, до известного времени не теряющих связи о ма-
теринской особью и друг с другом. Многие куртины имеют сложную
структуру, представляя собой коротко- и длинноксилоризомные инди-
виды. Куртины со временем распадаются, образуя клоны - совокупнос-
ти партикул разного возрастного состояния. Степень вегетативной
подвижности липы увеличивается с юга на север - вое более распро-
страненным вариантом жизненной формы становится факультативный
стланик. Однако эта закономерность нарушается в интразональных
экотонах, где условия существования становятся "нетипичными" (Чис-
тякова, 19786). Высокая побегопроизводительнооть липы сохраняется
на протяжении практически всей жизни особи (Рябчинский, 1964; и
др.).
Экология
Липа предпочитает свежие и влажные, хорошо аэрируемые почвы,
не испытывающие периодического иссушения, однако летние засухи пе-
реносит вполне удовлетворительно благодаря мощной и глубокой кор-
невой системе. По данным Ю.П.Бялловича (1957), может выдерживать
русловое затопление в течение 10-20 дней, на длительно заливаемых
участках пойм не встречается. На затопление паводковыми водами
иногда реагирует образованием в нижней части ствола придаточных
корней (Шиманюк, 1964). Не растет на заболоченных почвах.
Г.Ф.Морозов (1949) писал, что в нашем континентальном климате
липа как бы замещает бук. Эту точку зрения полностью разделяет
С.Ф.Дурнаев (1968, 1980).
Липа - очень теневыносливая порода, но не тенелюбивая даже на
ранних возрастных стадиях - недостаток света отрицательно сказыва-
ется на возобновительном процессе. С возрастов потребность в све-
те увеличивается. Большинство авторов (Сукачев, 1938; Гроздов,
1952; Шиманюк, 1967; и др.) считает липу холодостойкой породой,
редко повреждающейся морозами. Однако М.Е.Ткаченко (1950) замеча-
ет, что в молодости на открытые местах липа нередко страдает от
заморозков. Ю.П.Хлонов (1965) относит липу к холодостойким поро-
дам, но считает ее достаточно требовательной к температурным усло-
виям в период вегетации.
139
Липа требовательна к плодородию почвы, но встречается как на
богатых суглинках, так и на более бедных песках, при наличии в
них оупосчано-оутлинистых прослоек. В лесной зоне является пока-
зателем лучших почв (Сукачев, 1938); другие породы в этих место-
обитаниях формируют древостои наивысшей продуктивности (Ткаченко,
1952). Не выносит засоления.
Липа принадлежит к числу относительно газоустойчивнх пород, че-
щу отчасти способствует характер жилкование листовой пластинки -
места газовых ожогов постепенно выкрашиваются, но продырявленные
листья остаются живыми (Кулагин, 1974). Особенности фотосинтеза
чистых и запыленных листьев липы исследованы М.Ф.Ершовнм (1957),
который пришел к выводу, что промышленное загрязнение листовой по-
верхности - один из факторов, подавляющих этот процесс вплоть до
полного прекращения фотосинтеза в жаркие дни. Причина состоит в
изменении температурного режима листьев - темная окраска осаждаю-
щейся пыли способствует увеличению поглощения тепла и 'может вы-
звать их перегрев с последующими структурными изменениями в хлоро-
пластах. Характерно смещение максимума интенсивности фотосинтеза
запыленных листьев на утренние часы, в это время затемненная по-
верхность листьев приближает их температуру к величинам, оптималь-
ной для ассимиляционной деятельности.
Пластичность липы, с одной стороны, и ее эстетические достоин-
ства - с другой, позволяют широко использовать липу при озелене-
нии городов и других населенных пунктов (Грохольская, 1950; и
др.). В уличных посадках у липы уменьшаются размеры листовых плас-
тинок, снижается интенсивность фотосинтеза и дыхания, замедляется
рост. Но тем не менее липа активно адаптируется к городской среде,
что, в частности, выражается в усилении энергетического обмена
(Молоток и др., 1970). Корневая система липы приспосабливается да-
же к асфальтированному покрытию (Ду, 19606, 1961, 1962), однако
высокое наоыщение корнями ограниченного объема почвы (посадочной
ямы) сказывается отрицательно. По данным Н.В.Лобанова (1953), у
лип, растущих на улицах городов, корни лишена микоризы.
Фитоценология
Липа принадлежит к числу древесных пород, способных активно вы-
полнять здификаторнне функции. Влияние на среду в первую очередь
осуществляется посредством перехвата значительного количества све-
140
та. Наши наблюдения в липняке разнотравно-волосистоосоковом в Се-
ребряноборском лесничестве Московской обл. показали, что если в
конце апреля, когда чешуи на листовых почках липы только начинают
раздвигаться, относительная освещенность на уровне травяного по-
крова составляет в среднем 48$, то к середине последней декады
мая она снижается до 22$, а еще спустя несколько дней - до 10$.
В июне - августе освещенность под пологом липового леса составля-
ет 3-6$ от освещенности открытого места. Это соотношение сохраня-
лось и в безоблачные, и в пасмурные дни, хотя иным становился ка-
чественный состав проникающей радиации (Рысин, 1964). Большое вли-
яние на световой режим ценоза липа оказывает и в тех случаях, ког-
да она входит в качестве примеси в состав первого яруса или форми-
рует второй ярус.
Как показали исследования Ю.Л.Цельникер (1969), и световые, и
теневые листья липы относительно хорошо пропускают радиацию с дли-
нами волн порядка 750 нм (ближние инфракрасные лучи) и почти не
пропускают радиацию с длинами волн порядка 410 нм (синие лучи).
Поэтому под густым пологом липы (как, впрочем, и других листвен-
ных пород) радиация бедна синими лучами, их участие возрастает с
увеличением ажурности полога. Оптические свойства листьев липы в
течение вегетационного периода заметно меняются. Если в конце мая
пропускание фотосинтетически активной радиации листом липы состав-
ляло 16$, то в июле этот показатель составлял только 9$, увеличи-
ваясь к началу сентября до 35$. Ю.Л.Цельникер (1969) связывает
зти изменения с сезонной динамикой содержания пигментов в листьях.
Плотный полог липы служит основной деятельной поверхностью при
формировании температурного режима и режима влажности воздуха,
оказывая соответствующее влияние и на температурный режим почвы.
По сравнению с безлесными участками суточные колебания температур
и влажности фитосреды имеют значительно меньшие амплитуды.
Давно известно, что липа увеличивает плодородие почвы. Ряд ра-
бот по этому вопросу появился в последние годы. Например,
Р.М.Алексахин (1961), исследуя изменение свойств почв осинников
при смене их древостоями с липой, отметил повышение содержания
азота, подвижных соединений фосфора, увеличение насыщенности осно-
ваниями, в то же время снижение кислотности почв. Л.С.Травникова
(1959), изучавшая влияние поселения липы под пологом сосняка дуб-
някового на свойства почвы, установила их значительное улучшение,
о чем косвенным образом свидетельствует характер травяного покро-
ва, в котором суборевый комплекс видов сменился неморальным.
141
Почвоулучшапцее влияние липы осуществляется через опад. Сред-
нее содержание азота в лиотьях липы (в % на абсолютно сухую мас-
су) составляет 2,81,кальция - 1,48, магния - 0;29, калия - 1,72,
фосфора - 0,22 (Ремезов, Погребняк, 1965). Разумеется, в разных
типах леса, формирующихся на различных почвах, зти показатели в
той или иной степени разнятся, но то, что липа переводит из глу-
боких слоев почвы в верхние горизонты значительное количество
элементов питания, является непреложным фактом. Например, по рас-
четам И.И.Судницына (1964), выполненным в разнотравно-волосисто-
осоковом липняке Серебряноборского лесничества, в год наблвдений
в почву с опадом поступило 51,2 кг/га СаО, 3,6 кг/га MgO,
18,6 кг/га К2О, 1,2 кг/га Па2о, 7,7 кг/га Р2О5. Общее количе-
ство опада составило свыше 2 т/га сухой массы. По данным В.Г.Се-
меновой (1975), годичный опад липы имеет следующий фракционный
состав (по массе): листья 67,1$, ветви 20,6$, семена 2,4$, при-
цветники 4,4$, кора 0,2$, почечные чешуи 2,0$, смесь 1,9$. На по-
верхность почвы опад поступает в течение всего года, конечно, пик
приходится на сентябрь - октябрь (время листопада).
В наибольшем количестве липа берет из .почвы азот и кальций,
меньше потребляется калия, магния, фосфора, оеры и т.д. В очень
небольшом количестве поглощаются железо, марганец, натрий (Реме-
зов и др.', 1959). С возрастом потребление азота и зольных элемен-
тов питания возрастает. По расчетам К.М.Смирновой (1952), в лип-
няке волосистоосоково-снытевом в Мордовоком заповеднике потребле-
ние за год азота 13-летним деревом липы составило 5,7 г, 25-лет-
ним - 19,7, 40-летним - 43 , 74-летним - 140 г. Потребление каль-
ция составило соответственно 7,6/ 25,7, 56,8 и 184,6 г, магния -
0,9, 3,5, 7,9 и 27,9 г, калия - 2,9, 11,6, 24,9 и 73,3 г, фосфо-
ра - 0,6, 2,2, 4,9 и 17,1 г, азота - 0,7, 3,0, 6,5 и 14,9 г. В
74-летнем липняке накапливается азота 1052 кг/га, кальция 1289,
магния 181, калия 647, фосфора 78 кг/га (Ремезов, Погребняк,
1965).
Опад липы очень быстро разлагается (обычно уже к середине сле-
дующего года) вследствие энергичной деятельности почвенной мезо-
фауны и микроорганизмов (Утенкова, 1959; Судницын, 1964; и др.).
Происходит почти полная минерализация подстилки, что существенно
ускоряет круговорот веществ в лесу. В сообществах, где липа вхо-
дит в соотав древостоя только как примесь, даже небольшое количе-
ство ее опада (5$ от массы образца) заметно ускоряет разложение
всей подстилки (Самойлова, 1961).
142
К.М. Смирнова (1952) исследовала круговорот элементов питания в
оипняках Мордовского заповедника. В Московской обл. аналогичное
исследование провели В.Г.Семенова и В.В.Смирнов (1973). Авторы
сделали попытку сбалансировать потребление и расход азота и золь-
ных элементов в одном из наиболее распространенных типов липовых
лесов - липняке разнотравно-волосистоосоковом. Установлено, что
древостой удерживает от 20 до 40% веществ, извлеченных из почвы
(в большей мере удерживается железо - 61%). В наибольших количест-
вах возвращаются в почву кальций, азот, калий - элементы, играю-
щие особенно важную роль в жизни растений. Надземная часть древо-
стоя удерживает гораздо больше веществ, чем подземная часть, в си-
лу значительно большей своей фитомассы. Есть основания говорить о
специфической ритмике содержания элементов питания в почвах липо-
вый лесов (Ремезов и др., 1959).
Почвоулучшакщая роль липы столь велика, что при ее поселении
под пологом других пород (сосна, ель) довольно быстро и притом оп-
ределенным образом начинают меняться состав и структура травяного
покрова. Если же липа становится господствующей породой, то ниж-
ние ярусы растительности видоизменяются практически полностью. На
территории Серебряноборского лесничества есть два участка липово-
го леса: один - производный от сосняка с липой - расположен на
древней пеочаной террасе р.Москвы, другой - производный от ельни-
ка с липой - находится на водораздельном поднятии с двучленными
супесчано-суглинистыми почвами, подстилаемыми мореной. Несмотря
на различия в происхождении и в лесорастительных условиях оба лип-
няка имеют очень сходную по составу и структуре растительность:
почти чистый липовый древостой, отсутствие подлеска, хорошо разви-
тый травяной покров с преобладанием осоки волосистой и неморально-
го разнотравья, практически полное отсутствие мохового Покрова.
Причина этого сходства в мощном эдификаторном воздействии липы на
фитосреду, следствием чего является заметное выравнивание условий
в подиологовом пространстве, В обоих случаях полностью отсутству-
ет возобновление хвойных пород, что свидетельствует о большой ус-
тойчивости этих сообществ и позволяет отнести их к категории "ус-
тойчиво производных". Именно такой характер имеет большинство .гип-
няков Подмосковья. Травяной покров, формирующийся под лиловыми
древостоями, в свою очередь, увеличивает интенсивность биологичес-
кого круговорота веществ.
0 влиянии липы на микрофлору почвы дают представление данные,
полученные С.В.Егоровой (1979) в четырех типах леса восточного
143
Подмосковья, один из которых был условно коренной (ельник с липой
кколичко-зеленчуковый), а три других производные (в числе послед-
них липняк зеленчуково-волосистоосоковый). Почва одна и та же -
дерново-ореднеподзолистая супеочано-суглинистая на карбонатной мо-
рене. Установлено, что в липняке заметно выше микробиологическая
активность почвы, в результате чего минерализация растительных ос-
татков идет здесь более быстро. Среда изучавшихся четырех типов
леса липняк по биогенности минеральных горизонтов почвы можно по-
ставить на первое место. Если подстилка ельника бедна микрофлорой
вследствие ее повышенной кислотности и значительного содержания
терпенов, смол и ряда других соединений, то в липняке микрофлора
почвы значительно богаче, причем микроорганизмы проникают на боль-
шую глубину. В подстилке липняка (в отличие от ельника) намного
выше содержание нитрифицирующих, денитрифицирующих, азотфиксирую-
щих бактерий, что свидетельствует об интенсификации кругооборота
азота. Что же касается почвы (ее верхних горизонтов), то там чаще
встречаются желтопигментные бактерии, микобактерии, неспорообразу-
ющие бактерии, актиномицеты. Несколько обильнее, чем в ельнике,
развиваются целлюлозоразрушающие бактерии и менее обильны грибы.
На этих же участках леса Т.И.Алексахина (1979) исследовала поч-
венные водоросли. Число клеток водорослей в почве липняка было за-
метно выше, чем в почве ельника, разнообразнее был и видовой со-
став. Так, например, в липняке значительно больше участие желто-
зеленых, сине-зеленых и особенно диатомовых водорослей. Все зто
признаки увеличивающегося почвенного плодородия. На участие липы
в составе древостоев определенным образом реагирует и почвенная
мезофауна (Перель, 1964).
Если влияние липы на световой режим, на фитоклимат подпологово-
гб пространства и на лесорастительные свойства почвы является бес-
спорным, то пока только предположительно можно судить о степени и
направленности биохимического воздействия этой породы на другие
виды растений. Опыты в этом направлении проделаны Г.Г.Барабанец-
ким (1973а,б), который считает возможным положительное влияние ко-
линов корней липы на другие древесные породы, в частности ясень.
Однако неубедителен вывод автора о том, что корневые выделения ли-
пы в некоторой степени обусловливают вытеснение корней других дре-
весных пород в верхние горизонты почвы. Сомнительно и другое пред-
положение Г.Г.Барабанецкого, что в чистых липовых насаждениях мо-
жет иметь место угнетение липы собственными выделениями в резуль-
тате их повышенной концентрации. Вряд ли кому доводилось видеть
144
вырождение чистого липняка семенного происхождения. Напротив, ли-
па принадлежит к числу пород, не только прочно удерживающих свои
позиции, но и усиливающих их со временем.
Л.А.Крот (1972) сделал вывод о положительном влиянии корневых
выделений липы на саженцы дуба. Им установлено, что интенсивность
фотосинтеза у этих саженцев возрастает на 42%. В свою очередь,
корневые выделения дуба примерно в такой же степени усиливают ин-
тенсивность фотосинтеза саженцев липы. М.В.Колесниченко и Ю.М.Спа-
хов (1970) исследовали биохимическое влияние липы на интенсив-
ность поглощения дубом фосфора.
Липа успешно сосуществует со многими породами: елью, сосной,
дубом, осиной, березой, а в ряде случаев и активно замещает эти
породы. Обследуя сложные боры на территории Московской и сопре-
дельных областей (Владимирской, Рязанской, Калужской), мы постоян-
но констатировали полное отсутствие возобновления сосны в сосня-
ках с липой, в то время как липа возобновлялась там достаточно
удовлетворительно для сохранения ценопопуляций. Главную причину
мы видим в крайне неблагоприятном для всходов сооны световом ре-
жиме - сильном затенении, которое в значительной степени обуслов-
лено наличием в составе древостоя липы. Очевидно, сосновые части
древостоев современных сосняков с липой сформировались в услови-
ях, когда липа отсутствовала. Под пологом сосны она поселилась
только спустя несколько десятков лет, о чем свидетельствует ана-
лиз возрастной структуры древостоев - как правило, липа на 40-50
лет моложе сосны. Такие древостои могли формироваться на вырубках
и залежах. Можно предположить, что нередко поселению сосны благо-
приятствовало временное отсутствие липы.
Бкце в недавнем прошлом липа усиленно истреблялась на различные
хозяйственные нужды. Нам уже доводилось (Рысин, 1968) цитировать
статью, опубликованную в 1835 г. в "Лесном журнале", автор кото-
рой подсчитал, что в те времена в России только на лапти уничто-
жалось около 3 млрд, молодых липок ежегодно. Помимо лаптей из ли-
пы изготавливали огромное количество (многие миллионы) кулей и ро-
гож, посуду, различные поделки и т.д. Более 200 лет назад один из
первых русских лесоводов А.Т.Болотов (1766) предлагал разводить
липу искусственно, чтобы хоть частично возместить и уменьшить ее
вырубку в лесу, а автор другой статьи того же времени советовал
вместо лубяных (липовых) коробьев делать осиновые, а лапти поста-
раться заменить кожаной обувью. Только в последние десятилетия
спрос на липу резко упал, и ее количество в наших лесах отало
10-1964 145
быстро увеличиваться.
Если при совместном произрастании липа несомненно вытесняет
оосну, делая невозможным ее возобновление (Рысин, 1969), то ее
взаимоотношения с елью носят иной характер. Ель - теневыносливая
порода, ж влияние липы на ее всходы и подрост не столько пагубно,
как на соону. Поэтому елово-липовые леса вполне устойчивы, а со-
вместное произрастание в них пород с различными фитоценотическими
и экологическими особенностями предопределяет формирование четко
выраженной парцеллярной структуры (Бязров и др., 1971; Культиа-
оов, Минина, 1971; Алейникова и др., 1979). В липняках, сформиро-
вавшихся на месте сложных ельников в результате хозяйственной де-
ятельности человека, еловый подрост появляется, хотя обычно в
очень небольшом количестве. Липа, конечно, затрудняет его рост,
однако он вполне жизнеспособен и с годами увеличивает прирост (Ры-
син, 1979). Тем не менее процесо восстановления ели очень затянут,
и подавляющее большинство липняков являются длительно и устойчиво
производными. В.П.Крайнев (1939), анализируя причины малочислен-
ности всходов ели при участии липы в составе древостоев, одним из
главных факторов считает наличие мощного липового опада, препят-
ствующего прорастанию семян и нормальному развитию всходов.
В свою очередь, еловые древостои оказывают угнетапцее влияние
на подрост липы. Оно состоит как в сильном затенении, так и в кор-
невой конкуренции за влагу и элементы питания. В.И.Абражко (1968,
1969) показал, что 7-10-летние сеянцы липы под пологом ельников
отчетливо реагируют на конкурентные отношения в сфере корневых
систем снижением скорости роста и изменением структуры надземных
и подземных органов. Эта форма угнетения подроста липы в еще боль-
шей степени была выражена в березняках.
М.И.Гордиенко (1972, 1973) изучал взаимодействие липы, ясеня и
дуба в искусственных и естественных насаждениях на территории Ук-
раины. Им установлено, что липа не конкурирует с дубом за влагу и
питательные вещества. Дуб при совместном произрастании с липой
развивает более мощную корневую систему в верхних горизонтах поч-
вы. Со своей стороны липа ускоряет разложение опада и способству-
ет повышению плодородия почвы, тем самым благоприятно влияя на
дуб. Этому выводу не противоречат наши наблюдения в дубово-липо-
вых леоах Подмосковья. Как замечает С.Ф.Курнаев (1955), в дубовом
лесу подрост липы появляется, если есть семенники липы, но его ко-
личество и характер развития в значительной степени определяются
структурой ценоза и мощностью подстилки. Успешному сосуществова-
146
нию дуба и липы, по мнению И.Н.Рахтеенко (I960), благоприятствует
то обстоятельство, что максимумы и минимумы интенсивности роста
корней у дуба и липы приходятся на разные сроки: период макси-
мального роста активных корней дуба есть в то же время период ми-
нимального роста корней липы.
В березняках с полнотой не более 0,6, где достаточно благопри-
ятен световой режим, липа растет "успешно", но в высокополнотных
березняках большая часть липового подроота испытывает угнетение
вследствие затенения. Аналогичная ситуация складывается и в осин-
никах, только после распада осинового древостоя липа получает не-
обходимые условия для нормального развития. Со своей стороны,
сформировав сомкнутый полог, липа уже не допускает развития ооины.
Нельзя не признать, что взаимоотношения липы с другими порода-
ми изучены далеко не достаточно. Многие заключения основаны на
частных наблюдениях и нередко противоречат друг другу. Так, на-
пример, довольно единодушно принимается вывод, что между липой и
дубом нет напряженных конкурентных отношений в сфере корневых си-
стем. Однако, если М.И.Гордиенко (1972, 1973) указывает, что зна-
чительная часть корней дуба развивается в верхней чаоти почвы, по
данным других авторов (Карандина, I960; Солдатов, 1955; Самойлова,
1965), проводивших наблюдения в дубово-липовых лесах, основная
масса корней липы по сравнению с корнями дуба располагается бли-
же к поверхности почвы.
Описания основных типов липовых лесов, встречающихся на терри-
тории Московской обл., опубликованы Н.А.Коноваловым (1929),
С.А.Никитиным (1961), С.Ф.Курйаевым (1968, 1980), Л.П.Рысиным
(1974, 1979). Наиболее распространенными являются липняки волосис-
тоосоковый, снытево-волосистоосоковый, снытевый, гравилатовнй, та-
волговый, широкотравный. Под влиянием запаса и высоких рекреацион-
ных нагрузок постепенно увеличивается площадь липняков, где харак-
терный для них покров меняет свой состав и структуру - в нем начи-
нают доминировать леоо^лутовые злаки и разнотравье.
Консортивные связи1
Липа - энтомофильное растение. Процесс опыления требует непре-
менного участия насекомых, среди которых главную роль играет пче-
I Соавтор раздела - Г.В.Линдеман.
147
ла, собирающая липовый нектар во время цветения.
Липа - микотроф. Микориза вначале эктотрофного типа (Казакова,
1978; Лобанов, Трошина, 1978), позднее эктоэндотрофного (Лобанов,
1953). Возрастные состояния микоризы отличаются не только анато-
мическим строением, но и цветом: "желтые" микоризы - наиболее мо-
лодые и активные, "черные" - уже закончили свое развитие (Казако-
ва, 1978).
Вредителем плодов является клоп-солдатик, он может повреждать
до 98% урожая (Положенцев и др., 1952). Как уже отмечалось, опав-
шие плоды в большом количестве поедаются мелкими млекопитающими:
лесной и желтогорлой мышами, полевками и землеройками. По наблю-
дениям Л.В.Заблоцкой (1957а), в Приокско-Террасном заповеднике
широкое распространение лесной мыши связано с участием липы в со-
ставе древостоев. Что же касается желтогорлой мыши, то ее встре-
чаемость в сосновых лесах заповедника также оказалась пропорцио-
нальной количеству плодоносящих лип. Плоды липы зачастую служат
основным кормом этого вида, после неурожаев численность желтогор-
лой мыши быстро снижается (Заблоцкая, 19576).
За счет листьев липы существует значительное число насекомых.
Среди них в примерно равном соотношении представлены и специали-
зированные монофаги, обитающие только на липе, и полифаги, потреб-
ляющие листья различных широколиственных и мелколиственных пород.
Первые, как правило, сравнительно малочисленны и не наносят де-
ревьям сколько-нибудь значительных повреждений, среди вторых есть
виды, способные давать вспышки массового размножения, при которых
может получать сильные повреждения и липа. В их числе лунка сереб-
ристая, непарный шелкопряд, березовая и зимняя пяденица и другие
виды. Могут быть многочисленными несколько видов галлообразующих
клещей из рода Eryophyes. В целом липа слабо и редко страдает от
листогрызущих членистоногих и почти не нуждается в специальных
защитных мероприятиях.
До достижения возраста спелости липа относительно устойчива к
грибным заболеваниям. Однако к I00-II0 годам в липняках Белорус-
сии поражено гнилями до 65% деревьев (Чичин, 1956, 1958), а в Баш-
кирии - до 100%. Древесину ствола у старых деревьев разрушает пре-
имущественно обыкновенный трутовик Pomes fomentarius (Fr.)
Kicks.; он вызывает внутреннюю гниль, нередко ведущую к излому
ствола на разной высоте. На сухобочинах обычен Schizophyllum com-
mune Ft. Корни ослабленных и отмирающих деревьев часто поражает
опенок Armillariella mellea (Fr.) Karst. Плесневение плодов вызы-
148
вает гриб Oedocephalum glomeruloeum Chev. (Черемисинов и др.,
1970).
На ослабленных, отмирающих и свежеотмерших стволах и ветвях ли-
пы поселяются многие виды насекомых-ксилофагов; представлены все
основные группы, кроме рогохвостов. Чаще это многоядные виды: ко-
роеды Xyleborus diepar Р., Trypodendron aignatum Р., златка Di-
cerca alni Piech., Дровосек Meaoaa myope Dalm. И др.; ЛИШЬ коро-
ед Emoporus tiliae Panz. и златка Lampra rutilans Р. живут
только на липе. По-видимому, роль этих видов в жизни липы невели-
ка, случаев их массового размножения неизвестно.
Своеобразный характер имеет орнитофауна липовых лесов. По на-
блюдениям Т.И.Корнеевой (1979), в нескольких типах липняков вос-
точного Подмосковья население птиц в липовых сообществах сходно и
отчетливо отличается от состава орнитофауны хвойных лесов. Кроме
зяблика в .липняках доминируют большая синица и мухоловка-пеструш-
ка, что обусловлено обилием дупел в старых липах. Довольно обыч-
ны другие обитатели широколиственных лесов - клинтух и сойка.
Вместе с тем гнездятся виды, характерные для хвойных лесов: буро-
головая гаичка, московка, королек, зеленая пеночка, пеночка-тень-
ковка. В целом преобладают дуплогнездники, в то время как птиц,
гнездящихся в кронах деревьев и в подлеске, относительно немного.
Биологическая продуктивность
Как у всякой древесной породы, фитомасса, продуцируемая липой,
определяется условиями местообитания и возрастом деревьев. Липня-
ки приурочены к плодородным почвам и поэтому отличаются хорошим
ростом. В Московской обл. липа обычно растет по П классу боните-
та, запас древостоев в возрасте 80-90 лет при полноте 0,8-0,9 мо-
жет приближаться к 400 м3/га (Курнаев, 1968).
В.В.Смирнов и В.Г.Семенова (1968, 1970) провели детальный ана-
лиз фитомассы липы в 77-летнем липняке разнотравно-волосистоосоко-
вом в Серебряноборском лесничестве. Общий запас абсолютно сухой
фитомассы составил 141 т/га: масса стволов 122 т/га (86,4%),
толстых ветвей 11,4 т/га (8,1%), тонких ветвей 4,9 т/га (3,5%),
листьев 2,5 т/га (1,8%), плодов с прицветниками 0,2 т/га (0,2%).
Подземная фитомасса на этом же участке леса составила 50,8 т/га,
т.е. немногим более 25% от суммарной массы. Авторами установлены
тесные прямолинейные зависимости меаду массой разных надземных
ЮХ-1964 149
частей дерева, что позволило вывести ряд уравнений и рассчитать
коэффициенты корреляции. Ряд уравнений для .липы предложен П.А. Со-
коловым (1975, 1978).
Н.В.Дылис и Л.М.Носова (1977) опубликовали характеристики фито-
массы средних модельных деревьев липы разного возраста из несколь-
ких типов леса. Например, в липняке волосистоосоковом в Малинков-
оком лесничестве Московской обл. анализ 78-летнего дерева липы по-
казал, что на ствол приходится 83,5% всей надземной массы, на вет-
ви - 12,6, на листья - 1,3, на плоды с прицветниками - 0,4%. Как
видно, эти данные очень близки результатам, полученным В.В.Смирно-
вым и В.Г.Семеновой (1970). С возрастом меняются не Только общие
запасы фитомассы, но и вертикальное распределение массы и поверх-
ности отдельных фракций, а также соотношение фотосинтезирующих и
скелетных частей (Носова, 1970). П.А.Соколов (1975) отмечает, что
с увеличением высоты липового древостоя в кроне уменьшается доля
участия тонких олиственных побегов. Накопление фитомассы идет от-
носительно медленно в молодом древостое, но значительно убыстря-
ется после прохождения стадии жердняка (Ремезов и др., 1959).
По мнению В.Г.Семеновой (1975), положение листьев в кроне мало
отражается на содержании в них зольных элементов и азота, хотя и
прослеживается незначительное увеличение содержания ряда элемен-
тов в листьях нижней части кроны по сравнению с ее верхней частью.
Однако Л.Б.Корецкая (1970), проанализировав особенности вертикаль-
ного распределения зольных элементов у деревьев липы, находит оп-
ределенную связь его с внутриярусной стратификацией кроны, объяс-
няя эту зависимость различными свойствами биогеоценотических гори-
зонтов. Зальный состав органов липы меняется и во времени (Бурков,
1966, 1967, 1968). К концу периода вегетации содержание зальных
элементов уменьшается в листьях, но увеличивается в побегах и осо-
бенно в корнях. Содержание азота в побегах к фазе одревеснения по-
степенно уменьшается, а затем вновь начинает увеличиваться. Срав-
нительно мало изменяется содержание фосфора. Сезонную изменчи-
вость содержания азота и ряда зольных' элементов нужно учитывать
при проведении листовой диагностики.
Хозяйственное значение
Как уже отмечалось, в нашей стране липа издавна считается од-
ной из наиболее ценных древесных пород. Различную домашнюю утварь,
150
обувь, строительный материал, лыко для кулей и рогож - все давала
липа. Не случайно в ряде губерний она ценилась дороже сосны, и по-
этому некоторые лесохозяева отказались от бессистемной рубки липы
и приняли тот или иной оборот рубки липовой поросли (чаще семи-
или вооьмилетний). Неоднократные призывы к сохранению липы в ле-
сах Центральной России успеха не имели.
С липой в огромной степени связано пчеловодство, и этому вопро-
су посвящена обширная литература (Монахов, 1969; Соколов, 1972;
Мурахтанов, 1977; и др.). Липа - самый продуктивный медонос, даю-
щий в год меда до I т/га. Цветки липы широко используют в медици-
не и легкой промышленности. Плоды содержат целебное липовое масло.
Листья и почки богаты витаминами и могут быть использованы на
корм скоту. Сохраняется, хотя и не в таком размере, как прежде,
потребность в липовой коре. Древесина липы используется для изго-
товления фанеры, чертежных досок, музыкальных инструментов, в
строительстве, в токарном деле и т.д.
Липа - традиционная порода наших парков. В недавно опубликован-
ной сводке "Древесные растения парков Подмосковья" (Александрова
и др., 1979) приводятся сведения о породном составе 222 парков -
в 210 из них есть липа. Она же широко используется для посадок в
городских скверах, садах, на бульварах и улицах.
Очень велико лесоводственное значение липы как породы, улучша-
ющей качество древостоя и повышающей плодородие почвы. Так, напри-
мер, о пользе совместного произрастания сосны и липы свидетельст-
вует многолетний опыт создания лесных культур в Лесной опытной да-
че ТСХА (Тимофеев, 1965, 1966а, 19666, 1975; Кротова, 1964; Югай,
1979). Липа способствует лучшему очищению стволов сосны от сучьев,
создает благоприятный фитоклимат под пологом леса, способствует
активизации деятельности почвенной мезофауны, прежде всего дожде-
вых червей, обогащает верхние слои почвы элементами питания. Слож-
ные древостои всегда успешнее противостоят засухам, пожарам, моро-
зам, нападениям вредителей. Однако при определенных условиях (в
случае необходимости содействия естественному возобновлению сосны
или при создании подпалоговых культур этой породы) липа начинает
играть отрицательную роль вследствие создаваемого ею сильного за-
тенения, и поэтому приходится идти на ее временную частичную вы-
рубку.
М.И.Гордиенко (1979) считает, что в культурах дуба и ясеня
обыкновенного в условиях равнинной части Украины липа является оп-
тимальной подгонной породой, поскольку она заметно улучшает рост
151
этих пород: к тому же ее присутствие в составе древостоев повыша-
ет их хозяйственную ценность. На целесообразность введения липы
в состав хвойных лесов Средней Европы указывают западноевропей-
ские лесоводы. Главную ценность этой породы они видят в ее отно-
сительно невысокой требовательности к почве, большой теневыносли-
вости и способности увеличивать почвенное плодородие. Вместе с
тем определенную ценность имеет и липовая древесина. Культуры ли-
пы оказались выгоднее других мягколиственных пород и в отношении
затрат, поскольку они нуждаются только в эпизодическом уходе.
Липовые леса - наше национальное достояние. В Московской обл.
уже в течение ряда лет предпринимаются усилия с целью качествен-
ного улучшения существующих .липняков и по переводу в липовые дре-
востои с помощью комплекса лесохозяйственных мероприятий малоцен-
ных осинников и березняков, в составе которых есть липа. Сейчас
на территории Русской равнины и Урала липняки занимают около
2 млн. га. По мнению С.Ф.Курнаева (1980), за счет реконструкции
мелколиственных древостоев зту площадь можно увеличить вдвое не
более чем за 25 лет. Огромное значение имеет организация рацио-
нального комплексного лесопользования (Мурахтанов, 1972).
152
Т.В.Багдасарова, М.Г.Вахрамеева, С.В.Никитина, Л.В.Денисова
РОД ГРУШАНКА
Род грушанка (Pyrola L.) относится к семейству грушанковых
(Pyrolaceae). Латинское название рода происходит от Pyrus - гру-
ша за сходство листьев этого растения с листьями груши. Такая же
этимология и русского названия. Pyrola - циркумполярный бореаль-
ный род, распространенный от Арктики до Гималаев и Мексики, Он со-
держит около 20 видов, из которых в СССР произрастает 7, в Москов-
ской обл. - 4 вида.
Грушанки - многолетние растения, розеточные гемикриптофиты,
жестколистно-вечнозеленые или факультативно-летнезеленые, с хоро-
шо развитым длинным корневищем, расположенным неглубоко, от кото-
рого отходят многочисленные придаточные корни. Розетки из 3-5 ко-
жистых листьев, живущих 2-4 года. Цветонос с 1-2 чешуевидными
листьями, цветки собраны в кисть. Чашечка 5-листная, остается при
плодах; венчик из 5 свободных лепестков, опадакщий; тычинок 20;
вскрываются двумя отверстиями; пыльца в тетрадах; завязь верхняя
5-гнездная. Плод - многосеменная коробочка, раскрывается пятью
створками. Семена очень мелкие, желтоватые или светло-коричневые,
с не очень плотной наружной и внутренней оболочками (Grevillius,
Kirchner, 1923; Флора СССР, т.18, 1952; Ворошилов и др., 1966).
По мнению А.И.Толмачева (1951), предки грушанковых вошли в со-
став древних лесов таежного типа как вечнозеленые автотрофные рас-
тения, а в условиях тайги у них выработался смешанный автотрофно-
сапротрофный тип питания. Наиболее разнообразные формы грушанок
встречаются в горах Восточной Азии и на западе Северной Америки.
Ключ для определения московских видов'1
I. Столбик прямой, очень короткий (до 2 мм длины), не выступающий
из венчика, с 5-лопастным рыльцем без утолщенного кольца под
ним; пыльники до I мм длины; венчик шаровидный, почти не приот-
крытый; листья удлиненные, в очертании овальные,.реже почти
ОКруТЛЫе..............................................P-m-inn-n,
- Столбик более длинный, 3-6 мм длины, выступающий из венчика,
I Составил В.Н.Тихомиров.
153
отклоненный вниз или почти прямой, но в последнем случае с
утолщенным кольцом под неясно 5-лопастным рыльцем; пыльники
1,5-3 мм длины, венчик раскрытый; листья широкоовальные или
округлые, иногда ширина листовой пластинки превышает ее дли-
ну .................................... *................2
2. Венчик зеленовато-желтый; цветоносный стебель и черешки при-
корневых листьев обычно красноватые; прикорневые листья обрат-
нояйцевидные или округлые, нередко ширина их превышает длину;
крощие листья цветков короче цветоножек, линейно-ланцетные;
стебель (по крайней мере в нижней части) острогранистый или
ребристый ....................................... P.chlorantha.
- Венчик белый или слегка розоватый; цветоносный стебель и череш-
ки прикорневых листьев обычно (не всегда!) без красноватого от-
тенка; прикорневые листья округлые или широкоовальные; кроящие
листья цветков примерно равны цветоножкам или несколько длин-
нее их.........................................................3
3. Столбик резко отклоненный вниз; все тычинки отогнуты кверху;
доли чашечки до 3 мм длины, треуготгйго-шиловидные или яйцевид-
но-ланцетные; цветок тпироко раскрытый ..... P.rotundifolia.
- Столбик почти прямой; тычинки не отклоненные кверху, сходящие-
ся; доли чашечки 1,5-2 мм длиной, треугольные; цветок полурас-
крытый,' шаровиднО-колокольчатый ....................... p.media.
I. ГРУШАНКА КРУГЛОЛИСТНАЯ
Номенклатура и географическое распространение
Pyrola rotundifolia L., 1753» Sp. pl.:396; Кауфм., 1866, Моск,
фл., изд. 1:314, 1889, изд. 2:323; Цингер, 1886, Сб. свед.:289;
Шмальг., 1897, Фл. 2:187; Сырейщ., 1910, Илл. фл, 3:6; он же,
1927, Опр. раст. Моск. губ.:202; Федченко, 1940, в Маевск., Фл.,
изд. 7:560; Буш, 1952, Фл. СССР 18:10; Бобров, 1954, в Маевск.,
Фл., изд. 8:413; Скворцов, 1964, там же, изд. 9:399; он же, 1966,
Опр. раст. Моск. обл.:256; Krisa, 1972, Pl. Eur. 3:4. - Грушанка
круглолистная.
Отмечена большая склонность данного вида к гибридизации с дру-
гими видами рода, а иногда и с другими родами сем. Pyrolaceae
(Knaben, Engelakj^n, 1968).
Грушанка круглолистная имеет обширный голарктический ареал, ко-
154
торий занимает всю территорию Европы, большую часть Азии и Север-
ной Америки. В Европе она произрастает на севере до Исландии,
Шотландии, Северной Скандинавии (в Норвегии до 70°7* сев.широты),
на юге - до Северной Испании, Северной Италии, средней части Бал-
канских гор. На территории СССР встречается в европейской части,
Сибири, Казахстане и горах Средней Азии (Grevillius, Kirchner,
1923; Hegi, 1927; Флора СССР, т.18, 1952).
Северная граница ареала вида в Советском Союзе проходит по по-
бережью Северного Ледовитого океана, но от устья Оби спускается
несколько к югу по р.Полуй, а по р.Енисею вновь поднимается на
север до побережья океана. В бассейне р.Лены она сворачивает на
юго-восток и проходит по бассейнам рек Алдан и Амта. Восточная
.граница проходит по бассейнам Амги и Учура, спускаясь к оз.Байкал
и далее на запад, почти совпадая с государственной границей СССР,
затем через Северный Казахстан, по южным частям Челябинской и
Оренбургской областей, через Саратовскую, Воронежскую, Харьков-
скую, Днепропетровскую, Кировоградскую, Хмельницкую и Черновицкую
области. Западная граница вида совпадает с государственной грани-
цей СССР. Грушанка круглолистная растет также на большей части
Кавказа, в горах Киргизии и Узбекистана. Более подробно ее ареал
показан в работе В.Г.Тебса (1964). В Московской обл. встречается
повоеместно.
Морфологическое описание
Основная масса корней грушанки круглолистной находится на глу-
бине 8-12 ом. В условиях средней полосы корневище сохраняется до
8 лет, ежегодный его прирост достигает в среднем 18 см. Придаточ-
ные корни не ветвятся, корневые волоски расположены только на са-
мом их конце (Тебе, 1964). Листья округлые, 3-3 см длиной, темно-
зеленые.
Цветоноо длиной до 16 см, кисть длинная, разносторонняя, 8-14-
цветковая. Цветки отклоненные, реже поникающие. Цветоножки почти
равны цветкам; кроющие листья перепончатые ланцетные, равны или
немного длиннее цветоножек. Чашелистики яйцевидно-ланцетной фор-
мы, длиной 3,5-5,5 мм, концы их отогнуты книзу. Венчик широко рас-
крытый, душистый, в поперечнике 15-20 мм, состоит из округло-яйце-
видных, тупых, вогнутых, белых лепестков длиной 6,5-10 мм, шири-
ной 4-6 мм, в 1,5-2 (Зр раза длиннее долей чашечки. Тычинки с.
155
оранжевыми пыльниками, пригнутые к верхней стороне венчика, пыль-
цевые верна диаметром 37 мк (Сладаов, 1954). Завязь с длинным
(7,5-10 мм) отолбиком, при основании отогнутым вниз, столбик по
длине равен или несколько больше лепестков. Коробочка длиной 4,5-
5 мм, шириной 7-8 мм.
Диплоидный набор хромосом - 46 (Knaben, Engelskjtfn, 1968).
Онтогенез
Семена грушанки круглолистной пылевидные, масса одного семени
0,0007 мг (Grevillius, Kirchner, 1923). Всхожесть их довольно вы-
сока. В естественных местообитаниях при осеннем посеве семена про-
растают через 8 мес, всхожесть от 38 до 72%; через II мес в усло-
виях ботанического сада всхожесть до 92% (Luck, 1940). Для про-
растания требуются сначала пониженные температуры в течение 3 мес,
а затем температура должна находиться в пределах от 8 до 20° (оп-
тимальная 15°).
Точка роста стебля в зародыше семени развита значительно сла-
бее точки роста корня. Поэтому при прорастании семени клетки кор-
невого полюса начинают усиленно делиться, образуя зачаток главно-
го корня, а затем и главный корень растения. Стеблевой же полюс
остаетоя без изменений и бнотро замирает. В последующем на базаль-
ной части формирующегося корня появляется эндогенно одна или не-
сколько придаточных почек, вырастающих в надземные побеги. Таким
образом, развитие стеблевого полюса, или точки роста, прекращает-
ся еще* в прозмбриональном состоянии, а побеги развиваются как при-
даточные образования. О таких растениях с редуцированными главны-
ми побегами иногда говорят, как о "свободно живущих корнях" (Се-
ребряков, 1952).
При прорастании семян грушанок происходит заражение проростка
(прокаулома) микоризным грибом (Luck, 1940, 1941; Lihnell, 1942;
Harley, 1959). В асептических условиях (при отсутствии гриба)
прорастает в лучшем случае не более 10-20% семян (Lihneii, 1942).
фи этом образуется проросток такой же морфологической структуры,
как и при наличии микоризы, который способен расти и ветвиться в
течение 3,5 лет, достигая 30 см в длину, но затем все же погибает
(Harley, 1959). Развитие прокаулома в природных условиях длится
2 года (Luck, 1941).
Последующие этапы онтогенеза грушанки круглолистной не изуче-
156
ны. Это связано с тем, что особи семенного происхождения в приро-
де встречаются крайне редко. Однако развитие парциальных особей,
возникших путем вегетативного размножения, изучено достаточно
подробно.
Ветвление подземных корневищ грушанки круглолистной по А.П.Хох-
рякову (1961) и Е.Г.Чернову (1959) симподиальное, по И.Г.Серебря-
кову (1952) - моноподиальное. Мы считаем, что в начале развития
подземный побег ветвится моноподиально, но затем переходит к сим-
подиальному ветвлению. Подземные побеги возникают в числе 3-5 в
пазухах чешуйчатых листьев корневищ предыдущего порядка ветвле-
ния близ их вершины лишь тогда, когда вершина отомрет или когда
эти корневища от горизонтального роста перейдут к вертикальному,
чтобы дать начало новой парциальной особи (Хохряков, 1961). Как
правило, чем ближе к верхушке материнского побега возникают под-
земные дочерние побеги, тем быстрее они заканчивают свой рост и
тем они короче. В течение года может образоваться до трех генера-
ций таких побегов. Однако изредка моноподиальное нарастание кор-
невища может длиться не один год, а два-три, причем никаких боко-
вых побегов не образуется. Все корневища одного порядка, т.е.
возникшие из одного материнского корневища появляются, как прави-
ло, одновременно. Изменение направления роста корневища может
происходить в любое время вегетационного периода, с ранней весны
до поздней осени. Зима не влияет на дифференциацию подземных кор-
невищ, но их окончания, находящиеся близко к поверхности почвы,
замерзают и отмирают. По выходе подземного корневища на поверх-
ность междоузлия его заметно сокращаются и образуется первая ро-
зетка листьев. Затем в течение 2-3 лет происходит моноподиальный
ортотропный рост, причем каждый год у вершины годичного прироста
образуется новая розетка листьев (Хохряков, 1961).
Из почек, расположенных в пазухах листьев первой розетки, час-
то уже на следующий год развиваются новые добавочные ортотропные
побеги, в дальнейшем образующие последовательные мутовки в таком
же числе, что и главный-стебель. Через 2-3 (редко 4) года на вер-
шине годичного побега закладывается терминальное соцветие и на
следующий год особь зацветает. Вопреки мнению многих авторов (He-
gi, 1927; Серебряков, 1952; Каверзнева, 1955), побег после цве-
тения и плодоношения зачастую не отмирает. Одновременно с заложе-
нием терминальной цветочной почки или на следующий год после цве-
тения в пазухах верхних розеток яякля девают с я- вегетативные почки
(Хохряков, 1961). По нашим наблюдениям, в ельниках Подмосковья у
157
грушанки круглолистной обычно в год цветения закладываются боко-
вые побеги. Почки крупные (0,5-0,7 ом), удлиненно-конические,
почкосложение объемлющее (Василевская, Шилова, I960). Наружные
первые чещуй 3-5 мм, по зеленой окраске и консистенции напомина-
ют лист. Четвертая и пятая чешуи отличаются от первых более раз-
витой проводящей системой. Пятая и шестая чещуй достигают 7 мм
длины и имеют более развитую сеть жилок 2-го и 3-го порядков. Они
еще больше, чем наружные, напоминают лист..
Такая специфика строения почечных чещуй грушанки объясняется
тем, что в них, в отличие от зеленых листьев, быстрее проходит
дифференциация всех тканей и быстрее начинаются процессы старения.
Об этом можно оудить по следующему. Осенью в почках у смежных лис-
товых зачатков, один из которых является чешуей, а другой на сле-
дующий год разовьется в зеленый лист, сильно выражена различная
степень дифференциации тканей. В чешуе имеется вполне развитая
проводящая система. С такой степенью развития всех тканей чещуя
отмирает. В это время в зачатке листа ткани еще находятся в состо-
янии деления, жилки не сформированы. Укороченный онтогенез чещуй
влечет редукцию тканей чешуй по сравнению с листом. Чещуй почек
грушанки можно назвать листоподобными, так как они еще очень на-
поминают лист, но уже защищают внутренние органы почки (Василев-
ская, Шилова, I960).
Уже на ранних стадиях развития верхушечной почки в сентябре -
октябре в пазухе пятой-шестой чешуй часто имеются боковые почки
длиной до I мм, с несколькими листовыми зачатками. Развитие боко-
вых почек, по-видимому, становится возможным лишь благодаря тому,
что они закладываются на очень ранних стадиях развития чешуй. Поч-
ки, находящиеся в пазухе чешуй, резко отличаются от верхушечной
почки небольшими размерами конуса нарастания, мелкоклеточностью и
сильно растянутым первым междоузлием. Формирование зачатков внут-
ри боковых почек происходит медленно. Побеги из этих почек разви-
ваются лишь на четвертый год, после того как на третий год отцве-
тает главный побег.
Сформировавшаяся цветочная почка состоит из чещуй и цветков с
прицветниками. Чещуй близки по своему строению к зеленым листьям
и имеют хорошо развитую проводящую систему. Междоузлия между че-
щуями и листьями одногодичного вегетативного побега сильно укоро-
чены. Пазувиае почки возникают часто и закладываются кии в пазу-
хе чещуй, так ж ц пазухе листьев. Ветвление надземных побегов у
грушанки круглолистной наблюдается крайне редко. Продолжитель-
158
ность развития побегов в семействе грушанковых тесно связана с
продолжительностью жизни их многолетних листьев (Шилова, 1959,
I960).
Однолетние побеги имеют высоту 4-8 см, с I или 2 листьями дли-
ной 1,7-2,! см, шириной 1,7 см, средняя площадь листа 3,6 см^.
Двулетние побеги с 3-4 листьями длиной 3,4-3,9 см, шириной 3,1-
3,9 см, средняя площадь листа 12 см^. Трехлетние вегетативные по-
беги имеют высоту 10-13 см, с 5-8 листьями длиной 4,5 см, шириной
4,3-4,5 см, средняя площадь листа 15-22 см^. Длина генеративных
побегов 22-30 см.
Сезонное развитие и способы размножения
Грушанка круглолистная относится к группе растений, у которых
в почках возобновления к концу лета побег будущего года сформиро-
ван полностью (Серебряков, 1952). По нашим наблюдениям, в ельни-
ке-зеленомошнике с августа по сентябрь постепенно развиваются и
растут цветочные почки, в которых происходит дифференциация эле-
ментов соцветия. К концу сентября заметна вполне сформировавшаяся
кисть маленьких бутонов. Цветки пребывают в бутонах II-I2 мес. За-
цветает грушанка круглолистная в Московской обл. 15-25 июня, от-
цветает 20-30 июля, плоды созревают к 15 августа. В холодное дожд-
ливое лето 1980 г. созревание плодов затянулось до середины сен-
тября.
Одновременно с терминальным соцветием в пазухах листьев розет-
ки закладываются 2-4-6 листовых почек, покрытых 4-6 светло-зелены-
ми пленчатыми чешуйками (Хохряков, 1961).
Рост начинается после таяния снега о конца апреля - начала мая.
Трогаются в роот цветочные и листовые почки. Парциальные особи,
на которых образовались цветочные почки, в данном году новых
листьев не образуют. В Подмосковье в начале мая начинают развер-
тываться молодые листья, из почек образуется иольшое число новых
побегов. В фазу плодоношения (конец июля - начало августа) листья
прекращают рост, происходит закладывание листовых и цветочных по-
чек будущего года. В конце октября растение переходит в состояние
относительного зимнего покоя.
Наибольшее влияние на фенотип вида летом имеет температура воз-
духа (Тебе, 1964). Набухание и пробуждение генеративной почки от
зимнего покоя и развитие соцветия начинаются при быстром повыше-
159
иии температурной кривой до 18° и более. Созревание коробочек и
разбрасывание семян происходят при более низкой температуре - от
7 до 10°. Влажность воздуха и осадки влияют на сроки наступления
фенологических фаз.
Грушанка круглолистная - энтомофил, с большой долей автогамии
в опылении (Терехин, 1962). Семенная продуктивность по сравнению
с другими грушанками не очень высока. По нашим данным, в услови-
ях Подмосковья у грушанки круглолистной 50 цветоносов имели от 6
до 14 зрелых коробочек. В одной коробочке в среднем 135 семян (от
НО до 200).
Преобладает вегетативное размножение, оно идет за счет ветвле-
ния корневищ.
Экология и фитоценология
Грушанка круглолистная - мезофит, встречается в условиях от
оухо- до влажнолугового увлажнения (60-74-я ступени шкалы Л.Г.Ра-
менского , 3-я ступень шкалы Ландольта).
Растет при самой различной освещенности - от глубокой тени до
открытых мест (buck, 1940), чаще встречается в затененных местах
с относительной освещенностью не более 10%. Г.Элленберг располага-
ет этот вид между теневыми и полутеневыми (4-я ступень шкалы).
Э.Ландольт относит к теневым (2-я ступень шкалы). В условиях Мос-
ковской обл. растет, как правило, при относительной освещенности
4-24%. Мы отметили тесную связь интенсивности освещения с длиной
цветоносного побега, диаметром цветка и площадью листовой пластин-
ки (табл.8). С уменьшением освещенности длина цветоносного побега
увеличивается - в ельнике-эеленомошнике с минимальной относитель-
ной освещенностью (7%) длина цветоносного побега достигла 26 см.
Минимальному значению освещенности соответствуют наименьший диа-
метр цветка (около I см), и минимальное число цветков в соцветии
(8). Площадь листовых пластинок увеличивается с уменьшением осве-
щенности. Произрастает на разных почвах, как правило, бедных мине-
ральным азотом (2—я ступень шкалы Ландольта), индикатор бедных
азотом почв. Требовательна к содержанию гумуса (5-я ступень шкалы
Ландольта), предпочитает хорошо.аэрируемые почвы (4-я ступень шка-
лы Ландольта). Предпочитает слабокислые почвы, но может расти на
нейтральных и слабощелочных (3-я ступень шкалы Ландольта). В усло-
виях Московской обл. раотет на почвах с pH (водным) гумусового го-
160
Таблица 8
Зависимость морфологических признаков грушанки
круглолистной от освещенности
1 । Тип леса 1 Относи- тельная осве- щен- ность , $ Длина цвето- носно- го по- бега, см Число цвет- ков в соцве- тии Диаметр цветка, мм Площадь ли-| стовсй пла-( СТИНКИ, С1^|
। Ельник-зеленомошник 4-7 26 8 10 14,4-19,2 ।
1 Ельник с примесью 1 липы разнотравно- । волосистоосоковой 10-12 23 10 . 17 13,6-18,6 ।
1 Сосняк кислично- । чернично-разнотрав- 15-20 22-25 9-13 14 14,1-20,3 {
1 । Сосняк с елью разно- . травный 18-20 22 10-15 15 9,8-21,3 1
। Березняк орляково- . разнотравный 22-24 23-24 9-12 19 11,7-13,7 1
ризонта от 4,4 до 6,1 (pH солевое тех же горизонтов 3,8-5,4). Со-
держание Са и Mg в этих горизонтах от 0,96 до 3,45 мгэкв/100 г
почвы. Влажность подстилки 45-60$, влажность верхнего горизонта
15-30$.
Растет в различных условиях рельефа, как на равнинах, так и в
горах, поднимаясь до 1800 м (Andrea, 1914). Произрастает обычно
в тенистых хвойных лесах, но встречается и в смешанных, преимуще-
ственно мелколиственно-хвойных, значительно реже в широколиствен-
ных лесах (Andres, 1914). На территории Московской обл. встреча-
ется в различных ассоциациях сосновых и еловых лесов большими пят-
нами, приуроченными к взрослым елям. По нашим наблвдениям, она ха-
рактерна для ельников-зеленомошников, ельников-марьянниково-зеле-
номошных, ельников-сосново-ожиково-пальчатоосоковых, ельников'-бе-
резово-вейниково-разнотравных, ельников-разнотравных, ельников-ор-
ляково-костянично-разнотравных, сосняков елово-березово-ожиково-
зеленомошных, сосняков елово-вейниково-чернично-зеленомошных, бе-
резняков, образовавшихся на месте вырубленных ельников, и других
лесов,
В ельнике-зеленомошнике на 100 мы насчитали 757 парциальных
особей грушанки круглолистной, из них 755 вегетативных и 2 генера-
тивные. В ельнике разнотравном - I4II парциальных особей, из кото-
11-1964 161
pwx 1340 вегетативных и 71 генеративная; в сосняке елово-разно-
травном - 553 парциальных особей, из которых 35 вегетативных и
18 генеративных. Наиболее высокая жизненность грушанки круглолист-
ной отмечена в ельнике разнотравно-вслосистоосоковом на дерново-
сильноподзолистых суглинистых почвах (Одинцовский р-сн Московской
обл.).
Консортивные связи
Грушанка круглолистная, как и все представители рода, - мико-
троф. Из ее корней изолировано 5 видов базидиальных грибов (Luck,
1940; Harley, 1959). Гифы проникают обычно неглубоко в эпидер-
мальные и субэпидермальные клетки, которые иногда могут быть це-
ликом заполнены мицелием. Более сильно микориза грушанок развива-
ется на бедных почвах, где рост корней ослаблен (Harley, 1959).
В жизненном цикле грушанок микориза играет очень существенную
роль. В.Кэмп (Camp, 1940) считает, что род Pyrola находится на
границе между автотрофами и паразитами. И.Беленовский (Velenov-
sky, 1907) отмечает, что первое время проросток (прокаулом ) ве-
дет подземный образ жизни, питаясь исключительно за счет микориз-
ного гриба,. Э.С.Терехин (1962) предполагает, что продукты фотосин-
теза нужны грушанкам лишь для прохождения генеративного цикла, а
все остальное время они могут питаться за счет мицелия грибов.
Э.С.Терехин считает, что основным фактором, определяющим расселе-
ние грушанок, является не режим влажности и освещенности, а их
взаимоотношения с микоризным грибом.
Цветки грушанки круглолистной энтомогамны, но посещаются насе-
комыми редко. Есть данные об опылении грушанки шмелями (ВотЪив
scrimshiranus К., В.hypnorum Ь. , B.terrestris L., В.agrorum F.),
мухами Platycheiris clypeatus Mg, и жуками Trichopticus hirsutu-
lus Zett. (Grevillius, Kirchner, 1923). Предполагают, что претки
грушанки опыляются насекомыми в первые фазы цветения, а в послед-
ние фазы происходит самоопыление, что связано с направлением рос-
та столбика в разные фазы цветения (Hegi, 1927). Грушанка круг-
лолистная (а также малая и средняя) имеет специальные приспособ-
ления для автогамии - пыльники имеют апикальные поры, ориентиро-
ванные таким образом, что через них в поникших цветках пыльца вы-
сыпается на рыльце (Пономарев, Верещагина, 1973). Предполагается,
что в среднетаежных темнохвойных лесах цветки грушанок не посеща-
162
ются насекомыми. В опытах с изоляцией цветков от насекомых уста-
новлено, что тем не менее завязывание плодов было почти полным: у
грушанки круглолистной 98%, средней - 91, малой - 100% (Понома-
рев, Верещагина, 1973).
Стебли, листья и плоды грушанок поражаются различными грибами:
Melampsora pyrolae Sohrol., Actinonema pyrolae Allesch., Septo-
ria pyrolae Ell. et M. , S.schelliana Thum., Phyllosticta pyrolae
Ell. et Ev. (Ростовцев, 1908).
Биологическая продуктивность
и хозяйственное значение
В зависимости от различных типов леса и экологических условий
урожайность надземных частей грушанки круглолистной колеблется
от 300 до 1260 г/м^ сырой массы, подземных частей - от 310 до
620 г/иг сырой масоы (табл.9).
Таблица 9
Фитомасса (г/м^) грушанки круглолистной в различных лесных
фитоценозах Одинцовского р-на Московской обл.
Число Сыра? фитомасса Воздушно-сухая фито- । масса i
Типы леса особей надзем- под- общая надзем- под- общая
I м^ ная зем- фито- ная зем- фито-
часть ная масса часть ная масса
часть часть
Ельник-зелено- мошник 6,8 640,8 312,8 953,6 296,0 252,8 548,8
Ельник с при- месью липы разнотравно- волосистоосо- ковый 11,6 1258,4 616,0 1874,4 480,8 278,4 759,2
Сосняк чернич- но-зеленомош- ный 6,4 680,8 389,6 1070,4 301,6 206,4 508,0
Сосняк елово- разнотравный 7,5 808,0 459,2 1267,2 366,4 296,6 663,0
Березняк орля- ково-разно- травный 5,0 300,6 337,6 683,0 111,3 134,2 245,5
Грушанка круглолистная применялась в народной медицине при
кровотечениях, болях в желудке, гнойных заболеваниях почек, рев-
163
мативме, лихорадке. Основное ее свойство, как и у всех грушанок,
кровоостанавливающее (Тебе, 1964). Есть указания на возможность
культивировать грушанку круглолистную в садах на почве, приближа-
ющейся по составу к лесной (Kleiner, 1946).
2. ГРУШАНКА МАЛАЯ
Номенклатура и географическое распространение
Pyrola minor L., 1753, Sp. pl.s396; Кауфм. , 1866, Моск. фл.,
изд. 1:315, 1889, изд. 2:324; Цингер, 1866, Об. свел.:290;
Шмальг., 1897, Фл. 2:187; Сырейщ., 1910, Илл. фл. 3:8; он же,
1927, Опр. раот. Моск, губ.:202; Федченко, 1940, в Маевок., Фл.,
изд. 7:560; Буш, 1952, Фл. СССР 18:13; Бобров, 1954, в Маевск.,
Фл., изд. 8:412; Скворцов, 1964, там же, изд. 9:398; он же, 1966,
Опр. раст. Моск. обл.:256; Krisa, 1972, Pl. Eur. 3:3, - Грушан-
ка малая.
Грушанка малая встречается в Европе, Азии и Северной Америке.
В Европе растет от Британских островов (кроме Шотландии), Голлан-
дии, Дании, Скандинавии (в Норвегии доходит до 71°Ю’с.ш.), Но-
вой Земли до Испании, Италии и Балканского п-ова. Она произраста-
ет по всей Азии, включая Малую, Среднюю и Восточную до Сахалина
и средней Японии. В Северной Америке вид идет от Гренландии на
севере до штата Орегон на юге.
В СССР в лесной зоне европейской части растет повсеместно (к
югу реже), в азиатской части - во всех районах Западной и Восточ-
ной Сибири (по Енисею доходит до 71°с.ш.), на Дальнем Востоке в
Казахстане и Средней Азии (Тянь-Шань) (Grevillius, Kirchner,
1923; "Флора СССР", T.I8, 1952; Flora Europaea, vol.3, 1972).
Морфологическое описание
Грушанка малая, как и предыдущий вид, имеет неглубокую корне-
вую систему, располагающуюся преимущественно на глубине до ТО-
15 см. Корневище имеет вид толстых белых нитей до 3-5 мм в диа-
метре,. от которых отходят тонкие неветвящиеся корни. Конец корней
светлее, корневые волоски находятся только на самом их конце.
По морфологическому строению она.сходна с круглолистной. Побе-
164
ги у нее развиваются по типу полициклических. Надземные стебли
12-30 см высоты, тонкоребристые, иногда несколько скрученные.
Листья собраны при основании надземных стеблей, реже более или
менее раздвинуты, яйцевидные, широко эллиптические, реже округ-
лые или округло-овальные, по краю едва заметно тупо городчато-
пильчатые, жестковатые. Пластинки их 2,5-6 см длины, 1-5 см шири-
ны, немного длиннее или почти равны черешку, иногда короче его.
Выше на стебле имеются 1-2 узких линейных заостренных буроватых
листа. Листья грушанки малой, как правило, живут 2 года (редко
до 4 лет).
Цветки поникающие, собраны по 7-20 в довольно густую кисть 2-
8,5 см длины, цветоножки 3-6 мм длины. Прицветники ланцетно-ли-
нейные перепончатые. Доли чашечки широкотреутольные или широкояй-
цевидные, заостренные или туповатые, при основании прикрывающие
друг друга краями, прижатые к венчику; длина их не более ширины
при основании. Венчик белый или розоватый, шаровидный, почти за-
крытый, 6-8 мм в поперечнике (вполне расправленный, не более
13 мм в диаметре); лепестки около 6 мм длиной, 3-3,5 мм ширины,
вогнутые. Тычинки прямостоячие, окружающие столбик. Столбик I-
2 мм длины, короче завязи и не выдается из венчика, рыльце широ-
кое (1,5-2 мм в диаметре). Коробочка 5,5 мм длины, 3,5-8 мм шири-
ны. Семена пылевидные, масса I семени 0,0007 мг.
Диплоидный набор хромосом - 46 (Flora Europaea, vol.3, 1972).
Онтогенез, сезонное развитие
и способы размножения
Семена грушанки малой сходны по своему строению с семенами гру-
шанки круглолистной. Развитие проростка (прокаулома) идет по тому
же типу. Корневище и надземные побеги развиваются так же, как и у
грушанки круглолистной. Особи семенного происхождения в природе
встречаются крайне редко, вследствие этого проследить развитие
особей на начальных этапах онтогенеза мы не имели возможности.
Нами были получены некоторые данные по характеристике разно-
возрастных побегов грушанки малой в Одинцовском р-не Московской
обл. Однолетние^ побеги имеют розетку из 2-5 листьев со средней
I
Возраст побегов определяли по остаткам от розеток листьев.
1^-1964
165
длиной черешка 1,55 си, длина листовой пластинки 2,37 си, ширина
2,06 ом. Двулетние побеги имеют две розетки с 5-9 листьями (в
обеих розетках) оо средней длиной черешка прошлогодних листьев
2,16 ом, длина листовой пластинки 3,39, ширина 2,73 см. Побеги
3-4- редко 5-летние имеют хорошо сохранившиеся розетки только за
два последних года, общее число листьев у них от 2 до 9, причем
листья текущего года оказались крупнее, чем соответствующие
листья побегов более молодых: длина черешка 1,96 см, длина листо-
вой пластинки 3,09 см, ширина 2,44 см.
Зацветает побег чаще всего на 3-4-й год (реже на 2-й или 5-й).
7С% всех обследованных генеративных побегов сохранили розетки
двух последних лет, 30% побегов - розетки трех лет. Общее число
листьев на генеративном побеге 2-8. Листья текущего года крупнее,
чем предыдущих лет, а также крупнее листьев вегетативных побегов:
длина черешка 2,71 см, длина листовой пластинки 4,08 см, ширина
2,96 см. Почти все обследованные нами побеги, отцветшие в преды-
дущий год, дали 1-2 новых вегетативных побега, образовавшихся из
пазушных почек в основном верхней розетки листьев.
Среди генеративных выделяются побеги с цветущей центральной
осью и особи более старые с отцветшей центральной осью и цветущи-
ми боковыми осями 2-го и 3-го порядков. Развертывание боковых по-
бегов, чабто довольно многочисленных, происходит до и после цве-
тения из пазушных почек листьев разных лет генерации, но чаще
всего из листьев верхней розетки. Высота цветочной стрелки от 9
до 25, в среднем 18,2 ом. Число цветков на особь от 3 до 21 (в
среднем по 100 обследованным оообям 11,7), число плодов также от
3 до 21 (в среднем 9,9 на особь).
Вегетативный рост грушанки малой начинается сразу после тая-
ния снега - в конце апреля или начале мая. В начале мая разверты-
ваются молодые листья, образуется много новых парциальных особей.
Развитие розеточного побега завершается образованием терминальной
почки, в которой закладывается вегетативный побег будущего года.
Лишь после 3-4 лет верхушечная почка розеточного побега формирует-
ся как генеративная и в июне - июле в ней уже заложены верховые
листья с сидящими в их пазухе зачатками цветков. Зимой соцветия
надежно прикрыты чешуями, весной в них хорошо развиты нижние цвет-
ки.
Зацветает грушанка малая в условиях Подмосковья 20-25 июня, от-
цветает 15-20 июля. К концу августа созревают плоды. В конце ок-
тября растение переходит в состояние покоя.
166
Грушанка малая размножается как семенным, так и вегетативным
путем, вегетативное размножение преобладает, оеменная продуктив-
ность особей более высока, чем у предыдущего вида. По нашим дан-
ным, в условиях Подмосковья у грушанки малой 50 цветоносов имели
по 8-15 плодов. В одной коробочке в среднем содержится 2600 се-
мян.
Экология, фитоценология и консортивные связи
Грушанка малая раотет на почвах от среднесухих до влажных (3-я
ступень шкалы Ландольта). Г.Элленберг (Eiienberg, 1974) распола-
гает этот вид между полутеневыми и полусветовыми растениями (6-я
ступень шкалы), а Э.Ландольт относит к теневым (2-я ступень шка-
лы). В условиях Подмосковья, по нашим наблюдениям, грушанка малая
произрастает при относительной освещенности 7-27%. Отмечается
тесная связь интенсивности освещения с морфологическими признака-
ми растения. При малой освещенности (7-8%) в ельнике-зеленомошни-
ке длина цветоносного побега наибольшая (25-27 см), число цвет-
ков в соцветии наименьшее (8-10).
Произрастает на почвах разнообразных по механическому составу,
хорошо аэрируемых, бедных минеральным азотом (2-я ступень шкалы
Ландольта), требовательна к содержанию гуцуса в почве (4-я сту-
пень шкалы Ландольта). Предпочитает кислые почвы, являясь индика-
тором кислых почв (2-я ступень шкалы Ландольта). В условиях Мос-
ковской обл. встречается на почвах с pH водной вытяжки в гумусо-
вом горизонте от 4,1 до 5,5 (pH солевой вытяжки в тех же горизон-
тах 3,5-4,6). Содержание Са и Mg в этих горизонтах от 1,4 до
4,1 мгэкв/100 г почвы.
Растет как на равнине, так и в горах. Встречается в субальпий-
ских криволесьях и стланиках, поднимаясь в Западной Европе до вы-
соты 2530 м над ур.моря (Hegi, 1927). Обычно растет в хвойных и
смешанных лесах, реже в широколиственных, преимущественно дубо-
вых. В Подмосковье грушанка малая произрастает в различных ассо-
циациях сосновых и еловых лесов большими пятнами, приуроченными
обычно к взрослым елям. Она характерна для ельников-зеленомошни-
ков, ельников с примесью липы разнотравно-волооистоосоковых, ель-
ников разнотравных, ельников сосново-разнотравных, сосняков чер-
нично-разнотравных, сосняков елово-разнотравных, березняков, на
месте вырубленных еловых и елово-сосновых лесов и др.
167
Наибольшее число особей отмечено нами в ельниках с сосной раз-
нотравных на дерново-среднеподзолистой супесчаной почве, где об-
щее число парциальных особей на 100 м2 составляет 1333, тогда
как в ельнике-зеленомошнике оно равно 527. При этом в первом слу-
чае число ювенильных особей составляло 9,8%, вегетативных - 75,3,
генеративных - 14,9$; во втором - ювенильных - 6,7%, вегетатив-
ных - 75,8, генеративных - 17,5%.
Грушанка малая, как и грушанка круглолистная, - микотроф
(Luck, 1940; Lichnell, 1942; Harley, 1959). Цветки, как у преды-
дущего вида, чаще автогамны, о чем было сказано выше, однако из-
редка могут опыляться насекомыми. Известны примеры опыления гру-
шанки малой в Шотландии жуками и мухами, в Скандинавии - шмелями
и бабочками (Grevillius, Kirchner, 1923).
Биологическая продуктивность
В зависимости от различных типов леса и экологических условий
фитомасса (сырой вес) надземных частей грушанки малой колеблется
от 740 до 1050 г/м2, подземных частей - от 380 до 500 г/м2
(табл.10). Максимальная фитомасса как надземных, так и подземных
Таблица 10
Фитомасса (г/м2) грушанки малой в различных фитоценозах
Одинцовского р-на Московской обл.
— —1 1 1 Фитоценоз 1 ! Число побегов н а I - — — — —— — — - Сырая масоа Воздушно-сухая масса
надзем- ная подзем- ная надзем- ная подзем- ная
1 Ельник с примесью
• липы разнотравно- 1 волосистоосоковый 12,2 1036 500,8 516,0 172,0
* Ельник зеленомош- 1 ник 5,6 887,2 391,2 380,0 139,2
1 Елово-березовый с 1 разнотравьем 5,2 736,8 427,2 274,4 142,4
1 Сосново-елово-чер- 1 нично-разнотравный 6,0 896,0 386,3 318,4 124,0
1 Сосново-еловый раз- । потравный 1 13,4 1040,8 413,6 272,0 137,6
168
частей набладается в сложном ельнике о примесью липы и в сосново-
елово-разнотравном лесу, в остальных фитоценозах цифры ниже и до-
вольно сходны между собой.
3. ГРУШАНКА СРЕДНЯЯ
Номенклатура и географическое распространение
Pyrola media Swartz, 1804, Kungl. Sv. Vet.-Akad. Handl., nov.
ser. 25:257; Кауфм., 1866, Моск. фл., изд. 1:315, 1889, изд. 2:
324; Цингер, 1886, Сб. свед.:290; Шмальг., 1897, Фл. 2:187; Не-
ту нн., 1900, Крит, обзор 2:86; Сырейщ., 1910, Илл. фл. 3:7; он
же, 1928, Опр. раст. Моск. губ.:202; Федченко, 1940, в Маевск.,
Фл., изд. 7:560; Буш, 1952, Фл. СССР 18:12; Бобров, 1954, в Ма-
евск., Фл., изд. 8:412; Скворцов, 1964, там же, изд. 9:398; он
же, 1966, Опр. раст. Моск. обл.:257; Krisa, 1972, Fl. Eur. 3:3,-
Грушанка средняя.
Бореальный евросибирско-североазиатский вид, ареал которого
охватывает большую часть Европы и значительную часть Азии. Встре-
чается обычно изолированными участками и рассматривается как ре- '
ликт последнего оледенения (Grevillius, Kirchner, 1923). В Запад-
ной Европе чаще произрастает в атлантической, центральной и север-
ной частях. На север грушанка средняя идет довольно далеко - до
Исландии, Британских о-вов, Скандинавии (где доходит до 70°41’сев.
широты). На юге Европы грушанка средняя встречается значительно
реже, преимущественно в горах, в Италии доходит до Тосканы, на Пи-
ренейском Пг-ове полностью отсутствует, на Балканском п-ове редка.
В Азии растет от Малой Азии до Дальнего Востока.
В СССР распространена довольно широко, однако встречается зна-
чительно реже и в меньшей численности, чем другие виды этого рода.
Северная ее граница проходит несколько севернее линии Олонец - Во-
логда - Пермь. На юг грушанка средняя заходит далеко’, захватывает
лесостепные части среднеднепровского и средневолжокого районов.
Встречается в горах Крыма, Кавказа, Урала. В Западной Сибири рас-
тет в лесах по Оби, Иртышу, в Восточной Сибири - по Енисею, рас-
пространена на Алтае, Камчатке и Сахалине (Grevillius, Kirohner,
1923; Hegi, 1927; Hulten, 1950; Флора СССР, T.I8, 1952; Guziko- '
wa, 1971; Flora Europaea, voi.3, 1972). В Московской обл. встре-
чается значительно реже других видов рода (возможно, ее путают с
169
P.minor) в Северном, Западном, Каинско-Дмитровском, Восточном,
Пином и Приокском районах (Ворошилов и др., 1966).
Морфологическое описание,
онтогенез и сезонное развитие
Грушанка средняя по общему облику сходна с грушанкой кругло-
листной, а по строению цветков - с малой. Имеет тонкое, бурое,
сильно разветвленное корневище. Листья округлые или округло-
овальные, слабогородчатые, о маленькими сосочковидными зубчиками,
представляющими собой окончание жилок листа; черешки длиннее
пластинки.
Цветоносные побеги 15-30 см высотой, ребристые, с кистью из
5-15 цветков. Цветоножки отогнутые. Доли чашечки до 2 мм длиной,
прижаты к венчику (а позже - плоду) ,, в 3 раза короче лепестков,
яйцевидао-ланцетнне, заостренные иди треугольные, зеленовато-крас-
новатые, часто с отогнутым концом, не прикрывающие друг друга кра-
ями. Лепестки округлые, 6-8 мм длиной, 4,5-6 мм шириной, (5) 8-
9 мм длиной, белые или при основании розоватые, венчик полураскры-
тый, почти шаровидный.
Цветки содержат нектар в углублениях при основании пестика.
Пыльники 1,5-2 мм дайной. Столбик 1,3-6 мм дайной, в отличие от
грушанки малой, выдается из венчика. Под рыльцем столбик расширен
в кольцо, обычно более широкое, чем пятилопастное бугорчатое рыль-
це (Grevillius, Kirchner, 1923; Флора СССР, T.I8, 1952). Рыльце
клейкое, особенно в молодом цветке. Предполагают, что грушанка
средняя - перекрестноопыляемое растение, но насекомых-опылителей
на ней почти не видно, лишь в Финляндии наблюдали на цветках жу-
ков beptura maculioornis Des. и Grammoptera tabacicolor Des.
(Grevillius, Kirchner, 1923). Самоопыление возможно, как и у пре-
дыдущих видов (Пономарев, Верещагина, 1973), особенно у старых
цветков. Зацветает грушанка средняя в Подмосковье во второй поло-
вине июня - начале июля. Нередко цветет в течение всего июля (Сы-
рейщиков, 1910).
Плод 4-5 мм длиной, 6,5-8 мм шириной. По депням для Великобри-
тании, на соцветие приходится от 4 до 12 (в среднем 8,75+0,33)
плодов; в коробочке от 330 до 2742, чаще от 1182 до 1576 (в сред-
нем I379+I97) семян, т.е. один генеративный побег производит от
10 ДО 14 ТЫС. семян (Salisbury, 1942). Амфиплоид. 2п » 92. Пред-
170
полагают, что этот вид возник путем удвоения числа хромосом у
пёрвИЧНОГО гибрида P.minor х P.rotundifolia (Knaben, Engelskjjfn,
1968; Хромосомные числа цветковых растений, 1968; Guzikowa, 1971).
Онтогенез грушанки средней практически не Изучен. По нашим на-
блюдениям, в Хибинах грушанка может зацвести на 5-8-й год. В год
цветения образует только соцветия, листья не формируются, но есть
розетки листьев прошлых лет.
Экология и фитоценология
Грушанка средняя - мезофит, растет в условиях увлажнения от
влажно- до сыролугового (72-84-я ступени шкалы Л.Г.Раменского),
избегает как слишком сухих, так и слишком влажных почв (3-я сту-
пень увлажнения шкалы Ландольта). По отношению к свету нетребова-
тельна, чаще встречается на затененных участках (2-я ступень шка-
лы освещенности Ландольта), но считается более светолюбивой, чем
грушанка круглолистная (Grevillius, Kirchner, 1923).
Мезотроф, растет на почвах от бедных до средних по содержанию
питательных веществ (4-10~я ступени богатства почв по шкале
Л.Г.Раменского), бедных минеральным азотом (2-я ступень шкалы
Ландольта), но богатых гумусом (2-я ступень шкалы Ландольта). Гру-
шанка предпочитает хорошо аэрируемые почвы (4-я ступень шкалы Лан-
дольта), слабокислые, иногда нейтральные или слабощелочные -
pH = 4,5-7,5 (3-я ступень шкалы Ландольта). В хвойных лесах Поль-
ши растет на почвах с высоким содержанием гумуса и pH гумусового
горизонта 5,6-6,0 (Guzikowa, 1971).
В Западной Европе грушанка оредняя - преимущественно годный
вид, редко спускающийся ниже 400 м, а чаще поднимающийся до 2000-
3000 м над ур.моря (Andres, 1914; Grevillius, Kirchner, 1923; He-
gi, 1927). Иногда может расти по краям болот, ореди бурелома, на
лесных полянах (Hegi, 1927), на вересковых пустошах (Flora Euro-
paea, voi.3, 1972). В средней полосе европейской части СССР
встречается редко, преимущественно в хвойных лесах. В Московской
обл. растет также в основном в хвойных лесах (Ворошилов и др.,
1966).
171
4. ГРУШАНКА ЗЕЛЕНОЦВЕТНАЯ
Номенклатура и географическое распространение
Pyrola chlorantha Swartz, 1810, Kungl. Sv. Vet.-Akad. Handl.,
nov. set*, 31:190, tab. 5; Кауфм., 186.6, Моск. фл., ИЗД. 1:314,
1889, изд. 2:323; Цингер, 1886, Сб. овед.:290; Шмальг., 1897, Фл.
2:186; Петунн., 1900, Крит, обзор 2:86; Сырейщ., 1910, Илл. фл.
3:6; он же, 1927, Опр. раст. Моок. губ.:202; Федченко, 1940, в
Маевск., Фл., ИЗД. 7:560; Krisa, 1972; Fl. Eur. 3:3 - P.vires-
cens Schweigg. 1811, Fl. Erlang. Add.:154; Буш, 1952, Фл. СССР
18:9; Бобров, 1954, в Маевск., Фл., изд. 8:413; Скворцов, 1964,
там же, изд. 9:399; он же, 1966, Опр. раст. Моск. обл.:257. - Гру-
шанка зеленоцветная.
Вид мало изменчив. Однако Г.Хеги (Hegi, 1927) отмечает нали-
чие двух форм: f.composite H.Ands., имеющей цветонос с 3-5 цве-
тущими побегами, и f.dichotoma H.Ands. о цветоносом из двух вет-
вей, сросшихся до середины.
Имеет обширный палеарктический ареал, охватывающий почти всю
Европу (кроме самых южных частей Средиземноморья), значительную
часть Азии и Северную Америку (Grevillius, Kirchner, 1923; Hegi,
1927; Флора СССР, т.18, 1952; Stevanovic, 1972). В пределах СССР
ареал занимает почти всю европейскую часть до Прибалтшм и Каре-
лии на севере, до Киевской, Черниговской, Харьковской, Саратов-
ской, Оренбургской областей на юге. Растет также на Урале, Кавка-
зе, Карпатах, в Крыму. В азиатской части СССР грушанка зелено-
цветная распространена по всей Западной Сибири, в Восточной Сиби-
ри (по Енисею, в Ангаро-Саянском районе, Даурии) и на Камчатке
(Grevillius, Kirchner, 1923; Флора СССР, T.I8, 1952). В основном
ареал этого вида совпадает о ареалом Pinus syivestris ь. в Мос-
ковской обл. распространена практически повсеместно по хвойным
(преимущественно сосновым) лесам, где встречается рассеянно не-
большими куртинами.
Морфологическое описание
Грушанка зеленоцветная имеет округлые, очень плотные, кожистые
листья, сверху темно-зеленые, снизу светло-зеленые, о черешком
172
длиннее пластинки. По. нашим данным, для Московской обл. длина че-
решка 0,8-5,2 (в среднем 2,2) см, длина листовой пластинки 1-2,2
(в среднем 1,46) см. В отличие от других грушанок для этого вида
характерна редукция вегетативной массы - листья часто уменьшены
до размеров чещуи, но не утрачивают при этом деления на пластин-
ку и черешок. Цветоносный стебель острогранистый, темно-красный,
с очень мелкими оотроланцетными чешуевидными листочками. Длина
цветочной стрелки от 3,2 до 10 (в среднем 6,1) ом. Кисть с 3-7
желто-зелеными цветками.
Интересно отметить, что грушанка зеленоцветная, растущая в За-
онежье (Карелия), в сосняке-зеленомошнике, заметно крупнее, чем
в Подмосковье. Так, у вегетативных особей листья генерации теку-
щего года имели в среднем длину черешков 2,36 см, листовые плас-
тинки длиной 2,24 см, шириной 1,9-2 см; листья генерации предыду-
щего года соответственно 2,55, 2,65, 2,32 см. То же и в отношении
генеративных особей - длина цветочной стрелки в среднем 22,6 ом
(число цветков на побег - 7, число плодов - 3,3).
Характерная особенность морфологии вида - своеобразное строе-
ние почечных чешуй. Это сильно редуцированные листовые органы с
сокращенной до минимума поверхностью. Края почечных чешуй почти
не разрастаются, но в них всегда можно различить верхнюю более
широкую часть, соответствующую пластинке, и нижнюю, соответствую-
щую черешку, они почти не укрывают внутренние части почки (Васи-
левская, Шилова, I960).
ДИПЛОИДНЫЙ набор хромосом - 46 (Knaben, Engelskjrfn, 1968).
Онтогенез и сезонное развитие
У грушанки зеленоцветной в первый год развития побега, а иног-
да и в течение последующих двух лет его жизни зеленые листья мо-
гут не образовываться (роль ассимилирующих органов тогда выполня-
ют чешуи). Часто уже на третий год у нее формируется генеративный
побег специализированного типа (Шилова, I960). В Московской обл.
и Карелии мы выделяли побеги с 1-3 маленькими лиоточками, однолет-
ние, возможно двухлетние (если принять во внимание стадию с чешуе-
видными листьями). 3-4-летние побеги характеризуются одной, чаще
двумя розетками с 2-7 развитыми листьями.
Примерно пятая часть обследованных нами генеративных побегов
имела розетку одного года • Если считать, что побеги
173
грушанки эеленоцветной зацветают на третий год, то у генератив-
ных побегов с одной розеткой в первый год образовались чешуи, а
даурозеточные побеги ухе в первый год жизни имеют настоящие
лиотья. Листья, образованные в год перед цветением, по нашим дан-
ным, несколько крупнее, чем листья более ранней генерации (длина
черешка в среднем соответственно 2,33 и 1,44 см, длина листовой
пластинки 1,-93 и 1,42 ом, ширина - 1,84 и 1,3 ом). Примерно 16$
обследованных генеративных побегов имели стрелки с засохшими до
распускания цветками, на остальных 84$ побегов число нормальных
цветков колебалось от 3 до 7 (в среднем 5,1), число плодов от I
до 5 (в среднем 2,3).
По А.П.Хохрякову (1961), в пазухах диотьев грушанки зелено-
цветной не закладываются (в отличие от других видов рода) боковые
почки, и после цветения особь отмирает. Мы неоднократно встреча-
ли экземпляры с засохшими цветочными стрелками побегов 1-го, 2-го
и даже 3-то порядков. Обычно при этом пазушная почка одного из
листьев начинала развиваться в новый побег. Большей частью такие
почки закладываются в пазухах нижних листьев, а иногда и чещуй.
Таким образом, после цветения парциальная особь отмирает не всег-
да.
В основном число побегов в куртинах невелико. Куртина, включа-
ющая 82 побега на площади 1,5x1,5 м, встречена нами только однаж-
ды в сильно вытоптанном сосново-еловом зеленомошно-черничном лесу
на меоте бывшего кострища и не может считаться типичной для Под-
московья. Обычно куртина занимает площадь не более 0,2x0,25 м.
Почти во воех обследованных куртинах преобладают двухлетние вир-
гинильнне побеги, на них приходится от 36,4 до 89,4$ всех особей
популяции. Генеративных побегов мало - от 0 до 25$, по 1-2 (ред-
ко 3) в куртине. Лишь в одной куртине из 82 побегов II были гене-
ративными.
Генеративные почки закладываются ранней весной. Генеративный
побег начинает развиваться в год перед цветением и в августе под-
нимается над зелеными листьями на 0,9-0,13 см. Сформированные
цветки собраны в рыхлое соцветие с кроющими листочками. Под этой
недостаточной защитой цветки зимой частью замерзают (Grevillius,
Kirchner, 1923; Hegi, 1927).
Размножается вид преимущественно вегетативно с помощью длинных
корневищ. Но есть и семенное размножение. По нашим данным, вызре-
вает от I до 4 коробочек на особь. Семена высыпаются из коробочек
только в сухую погоду. Предполагается, что протогиничные цветки
174
грушанки зеленоцветной опыляются насекомыми, но конкретных указа-
ний на насекомых-опылителей нет.
Экология и фитоценология
Грушанка зеленоцветная малотребовательна к увлажнению, чаще
встречается на сухих почвах и является их индикатором (2-я сту-
пень шкалы Ландольта), по Л.Г.Раменскому, растет в условиях от
сухо- до влажнолугового увлажнения (59-74-я ступени шкалы), менее
требовательна к увлажнению, чем другие грушанки. Обычно растет в
полутени - поблизости от "окон", дорог, при относительной освещен-
ности более 10%, редко на полностью открытых местах (3-я ступень
шкалы Ландольта).
К богатству почв малотребовательна, часто встречается в олиго-
трофных условиях (3-5-я ступени шкалы Л.Г.Раменского), являясь,
по Э.Ландольту, даже индикатором бедных почв (2-я ступень шкалы).
Более требовательна, чем другие грушанки, к pH среды, никогда не
встречается на очень кислых почвах, предпочитает щелочные, бога-
тые известью почвы с pH 5,5-8. По Э.Ландольту, она - индикатор
почв, богатых основаниями (4-я ступень шкалы). Как и другие, гру-
шанки, предпочитает богатые гумусом, хорошо аэрируемые почвы (4-я
ступень шкалы Ландольта).
В условиях Московской обл. обычно встречается при относитель-
ной освещенности 28-65%, на сильноподзолистой супесчаной почве с
pH 3,8-4,0 (солевой вытяжки).
Растет как на равнине, так и в горах, поднимаясь до 2000 и бо-
лее м над уровнем моря (Hegi, 1927). Довольно строго приурочена
к хвойным лесам, особенно сосновым. По мнению Г.Хеги (Hegi, 1927),
ее следует отнести к характерным видам сосновых лесов. В лесах
преимущественно встречается среди зеленых мхов, реже среди травя-
нистых растений. Интересно указание (stevanovic, 1972) на появ-
ление небольших групп из 6-8 особей в искусственных посадках сос-
ны 50-60-летнего возраста в районе, где она раньше не встречалась
(редко до 40).
Хозяйственное значение и охрана
Листья грушанки зеленоцветной употреблялись в народной медици-
175
не для лечения ран. Возобновляется это растение слабо. Для сохра-
нения вида нужны специальные меры по охране. Так как он встреча-
ется довольно редко во многих частях своего ареала, то предлага-
ется к охране в Московской и ряде других областей страны, а так-
же охраняется в ряде западноевропейских стран.
176
Т.П.Баландина
ГОЛУБИКА ОБЫКНОВЕННАЯ
Номенклатура, внутривидовая систематика
и географическое распространение
Vaccinium uliginosum L., 1753, Sp. pl.:35O; Кауфм. , 1866,
Моск. фл., изд. 1:312, 1889, изд. 2:321; Цингер, 1886, Об. свед.:
287; Шмальг., 1897, Фл. 2:181; Сырейщ., 1910, Илл. фл. 3:15; он
же, 1927, Опр. раст. Моск. губ.:204; Федченко, 1940, в Маевск.,
Фл., изд. 7:564; Некрасова, 1952, Фл. СССР 18:96; Пояркова, 1954,
в Маевск., Фл., изд. 8:415, 1964, изд. 9:402; Скворцов, 1966,
Опр. раст. Моск. обл.:260; Popova, 1972, Fl. Eur. 3:13. - Голу-
бика обыкновенная (сем. Vicciniaceae - брусничные).
Для голубики характерна значительная изменчивость. Выделяют
несколько разновидностей: var.uliginosum (var.genuinum Herd.) -
ветвистый кустарничек до I м высотой; var.alpinum Е.Busch - не-
высокий полустелкщийся кустарничек с обратнояйцевидными или оваль-
ными листьями, более мелкими, чем у типовой разновидности, растет
в тундре и на гольцах (Розанова, 1934; Флора СССР, т.18, 1952);
var.vulcanorum (Кот.) Е.Busch (V.vulcanorum Кот) - прижатый К
почве сильно ветвящийся кустарничек, образующий довольно плотные
подушки, отмершие листья долго (1-2 года) остаются на ветвях,
описана с Камчатки, приводится для востока Чукотского п-ова (Юр-
цев и др., 1979). Б.А.Федченко (1923) описал белоягодную разновид-
ность голубики var.leucocarpum B.Fedtsch., собранную ИМ В окрест-
ностях г.Рыбинска на Прощинском болоте; она отмечена также в Горь-
ковской обл. (близ г.Балахна), к западу от г.Ломоносова Ленинград-
ской обл., на переходном болоте в Куярском лесхозе Марийской АССР
(Данилов и др., 1975). Выделена промежуточная (между белоплодной
и типичной) форма голубики с голубовато-синими плодами.
Бореальный циркумполярный вид, широко распространенный в аркти-
ческой и лесной зонах Северного полушария и в высокогорьях. Вне
Советского Союза голубика встречается во многих районах Азии, Се-
верной Америки и Гренландии. В европейской части СССР ареал голу-
бики охватывает всю тундровую зону, включая Новую Землю, а также
Карело-Лапландский, Двино-Печорский, Ладого-Ильменский, Верхне-
Волжский, Волжско-Камский, Верхне-Днепровский, Верхне-Днестров-
ский и Волжско-Донской флористические районы; в Азиатской части
12-1964 I77
СССР - вое районы Сибири и Дальнего Востока, а также восточное и
западное Закавказье (Флора СССР, т.18, 1952; Солоневич, 1956). В
Московской обл. этот вид распространен в Северном, Западном,
Клинско-Дмитровском и Восточном районах, а также на западе Приок-
ского района (Ворошилов и др., 1966).
Морфологическое описание
Голубика - симподиально ветвящийся кустарничек (Серебряков,
Чернышова, 1955) до I м высоты. Корневая система поверхностная
(корни не уходят глубже 20-25 см), представлена совокупностью
корней 1-го, 2-го и 3-го порядков, образующихся на погребенных
сфагновыми мхами надземных стволиках. Длина корней 1-го порядка
колеблется от 7-10 до 35-40 ом, толщина их не превышает 0,4-
0,5 мм, старые корни могут иметь толщину до 2 мм. Корни 2-го по-
рядка имеют длину от 3-5 до 9-II см, очень тонкие корни 3-го по-
рядка - О,5-1,5 см длиной (Серебряков, 1952, I960; Серебряков,
Чернышева, 1955; Солоневич, 1956). Придаточные корни могут вет-
виться до 4-5-го порядков.
Листья очередные, 6-38 мм длиной и 4-24 мм шириной на очень ко-
ротких (0,5-1,5 мм) черешках, обратнояйцевидные, обычно на конце
округлые, реже островатые, цельнокрайние, с несколько подвернуты-
ми книзу краями, голые, сверху светло-зеленые, снизу сизоватые с
выдавшимися жилками.
Цветки на коротких поникающих цветоножках, расположены по 1-3
на концах коротких прошлогодних веточек, обладают слабым приятным
запахом. Чашечка С 4-5 короткими округлыми зубцами 0,5-1 мм дли-
ной и 1-1,5 мм шириной. Венчик кувшинчатый, беловатый или слабо-
розовый, 3,5-5,5 мм длиной, с 4-5 отгибами. Тычинок 8-10 с голы-
ми, в основании слегка расширенными нитями, пыльники с длинными
придатками. Столбик иногда видается из венчика, завязь нижняя,
4-гнездная, с центральным семяносцем.
Плод - округлая ягода, 9-12 мм длиной, синеватая, с сизым нале-
том, внутри с зеленой мякотью. Форма плодов очень варьирует, встре-
чаются неграненые и граненые плоды. Из неграненых плодов отмечены
плоокие, плоско-округлые, округлые, округло-овальные, длинно-оваль-
ные, округло-грушевидные, грушевидные, широкоцилиндрические, ци-
линдрические суженные к верхушке (ifakai, 1919, цит. по Флоре
СССР, т.18, 1952); из граненых - плоокие четырехгранные, плооко-
178
округлые, четырехгранные, грушевидные (Синокая, Щенкова, 1928;
Розанова, 1934; Флора СССР, т.18, 1952; Нестерович и др., 1967).
Плоды содержат от 13 до 75 (в среднем 34) семян, по нашим подсче-
там в Подмосковье от 5 до 44 (в среднем 17) семян. Семена 0,5-
1,6 мм длиной, 0,3-0,8 мм шириной, полуовальные, или клиновидные,
мелкоямчатые или мелкобороздчатые, оранжевые (Нестерович и др.,
1967). Средняя маоса I ягоды 429+18 мг, масса I тыс. семян -
0,3 г (Шабарова, 1968).
Онтогенез и сезонное развитие
Семена голубики прорастают в июне - июле. Прорастание надзем-
ное. Всходы имеют ланцетно-овальные семядоли 3x1 мм, сверху свет-
ло-зеленые, снизу красные с неясной главной жилкой. Побуревшие
семядоли сохраняются на сеянцах до 5 лет. Первый лист размерам
2x1,5 мм, овально-округлый или обратно-яйцевидный, с неясными ро-
зоватыми разветвленными жилками, главная жилка листа извилистая.
Второй лист слегка заостренный, с розоватыми краями. Еивые листья
сохраняются на стебельке 3-4 года. Подоемядольное колено и стебе-
лек темно-красного цвета, постепенно оветлепцие книзу. Главный ко-
рень сильноветвистый (Авдошенко, 1948).
Развитие всходов голубики в природных условиях происходит край-
не медленно. В течение 3-5 лет они почти не увеличиваются в раз-
мерах и не сбрасывают на зиму листьев. Их высота в этом возрасте
большей частью равна 1,5-2,5 см. В дальнейшем весной из самой
верхней пазушной почки вырастает побег 2-го порядка, который как
бы продолжает рост главной оси. Затем происходит ежегодное отмира-
ние верхней части очередного годичного побега. Образуется главная
ось, которая состоит из отдельных годичных побегов поеледущих по-
рядков. Главная ооь у голубики живет обычно 13-15 лет, а в небла-
гоприятных условиях - до 4-5 лет. На 4-5-й год жизни особи незави-
симо от возраста главной оси из пазушных почек, находящихся в ос-
новании главной оси, вырастают побеги 2-го порядка, которые живут
13-15 лет, после чего, как и главная ось, начинают отмирать. Так
же закономерно через 4-5 лет вырастают побеги 3-го и 5-го поряд-
ков.
Цветение и плодоношение в природных условиях начинается на 15-
17-м году жизни всего растения (Чуйкова, 1959), однако есть указа-
ния (.Авдошенко, 1948), что цветение не наблюдается ранее 30-лет-
179
него возраота всего растения.
В условиях сфагновых болот у голубики, в отличие от черники и
брусники, отсутствуют специализированные подземные корневища,
возникающие из спящих почек. Ее ветви ежегодно нарастают сверху,
но одновременно нижние части ветвей погребаются непрерывно нарас-
тающим сфагновым покровом. Погребенные ветви вскоре укореняются,
причем зона образования придаточных корней ежегодно поднимается
все выше по стеблям по мере погружения в сфагновый покров. Такое
нарастание ветвей с одновременным погружением их нижних частей в
сфагновую толщу может продолжаться многие десятки лет. Погружен-
ные части довольно долго сохраняются живыми, но затем отмирают
(Серебряков, Чернышева, 1955).
Подобный способ нарастания не единственный для голубики. В ус-
ловиях минеральных почв (в горных тундрах Хибин, Полярного Урала,
на Таймыре) сна образует компактные кустарнички. Из спящих почек
иногда образуются столоновидные корневища (как у черники) с мно-
голетним (до 6-7 лет) подземным ростом, но более короткие. Их
верхушки на поверхности почвы дают начало новым парциальным кус-
там. Кроме системы главного корня, функционирующего многие десят-
ки лет, образуются придаточные стеблевые корни. Нарастание побе-
гов симподиальное (Серебряков, 1952; Серебряков, Чернышева, 1955).
Н.В.Куйкова (1959) отмечает стелющуюся форму роста голубики, ха-
рактерную для гор, обнаженных плато и горных тундр, поднимающую-
ся, например, в Хибинах, почти до границы распространения цветко-
вых растений.
Генеративные почки у голубики располагаются не только на кон-
цах укороченных побегов; цветочными могут быть 2-я, 3-я, реже 4-я
и 5-я боковые почки, считая от верхушки. Они отличаются от вегета-
тивных почек меньшей заостренностью и большими размерами. В почке
голубики различаются две наружных укороченных чещуй и две внутрен-
них. В генеративной почке закладываются I, реже 2-3 цветка.
В период окончания цветения (середина июня) в генеративных поч-
ках будущего года уже происходит формирование зачатков цветка. В
первую очередь закладывается околоцветник (сначала чашелистики,
затем лепестки). Вслед за околоцветником формируются зачатки ты-
чинок, причем первыми закладываются пыльники. В пестике сначала
закладывается рыльце, формирование столбика происходит позднее.
Вскоре после образования столбика (в июле) становятся хорошо за-
метными семяпочки. Пыльца формируется весной следующего года за
полмесяца до начала цветения. Наиболее интенсивное развитие зачат-
180
ков цветка происходит в начале их формирования, затем оно идет в
уйеренном темпе (Торопова, Черкасов, 1973). В генеративных поч-
ках, взятых нами в середине ноября на сфагновом болоте в Одинцов-
ском р-не Московской обл., оказались сформированными все элемен-
ты цветка: околоцветник, тычинки и пестик, в завязи видны семя-
почки.
Средняя дата начала цветения в Московской обл. - 26 мая, в Бе-
лоруссии - 27-28 мая, в Костромской обл. - 22-31 мая, в Якутии -
середина июня (Юркевич, Смоляк, 1963; Торопова, Черкасов, 1973;
Михалева, 1972). В Костромской обл. массовое цветение голубики
наблюдается 3-12 июня, начало созревания ягод - 21-30 июля, мас-
совое созревание ягод - 3-10 августа (Торопова, Черкасов, 1973).
Наши наблюдения показали, что в Московской обл, эти фенологичес-
кие фазы приходятся на те же сроки. Сроки наступления фенофаз мо-
гут сдвигаться в ту или иную сторону в зависимости от погодных
условий и местообитания. По данным В.М.Михалевой (1972), в Якутии
голубика развивается в сосняках в более сжатые сроки, чем в лист-
венничниках.
Развертывание листьев у голубики начинается в начале мая. Рост
вегетативных побегов наиболее интенсивно происходит в конце мая.
К началу - середине июня рост побегов заканчивается (Серебряков,
Галицкая, 1951; Солоневич, 1956). Длина одногодичных побегов, по
нашим данным, составляет в среднем 5,9 см на Луцинском осушенном
болоте и 5,3 см на неосушенном верховом болоте (Одинцовский р-н
Московской обл.). В начале сентября листья голубики начинают жел-
теть и постепенно опадать. В середине - конце сентября наступает
стадия осенне-зимнего покоя.
Способы размножения и распространения
Голубика - насекомоопыляемое растение. Цветки протерандричны.
Обычно отверстия пыльников раскрываются в то время, когда бутон
еще не раскрыт. Рыльце помещено значительно выше пыльников.
Вследствие такого расположения и неодновременности созревания ан-
дроцея и гинецея, самоопыление, как правило, отсутствует (Розано-
ва, 1934).
По данным А.А.Скрябиной (1970), от 30 до 70% бутонов и завязей
опадает по различным причинам: часть бутонов поедается гусеница-
ми, гибель цветков обусловлена как повреждением гусеницами, так и
I2X-I964 181
опадением недоразвитых или неоплодотворенннх цветков (вследствие
малочисленности насекомых или дождливой погоды). После созревания
ягоды долго не осыпаются, постепенно заоыхают; опадение таких
ягод возможно только при механическом встряхивании.
Несмотря на большую семенную продуктивность, размножение голу-
бики семенами очень затруднено. Прорастание' семян возможно только
при внсокой влажности, отсутствии прямого солнечного освещения,
нарушении мохового и травяного покрова. Всходы голубики можно
найти на поваленных деревьях, старых пнях (Аздошенко, 1948; Соло-
невич, 1956, 1970).
Всхожесть семян голубики, по данным С.И.Шабаровой (1968), по-
рядка 11-12$. В лабораторных условиях всхожесть свежеообранных
семян - 70$ (условия: выдерживание в оухом состоянии 30 дней, по-
сев в мокрый пеоок, выдерживание 8 дней в холодильнике при темпе-
ратуре -5° до -10° и 20 даей при 0°). Семена, хранившиеся I и да-
же 2 года, не теряют всхожести. Лучшая всхожесть отмечена при
предварительном 24-часовом замачивании. В естественных условиях
под пологом сосняка после предварительного замачивания всхожесть
семян голубики 28$, меньше, чем у черники в 2 раза (Koziracki,
Tagronski, 1978).
Характерное для голубики распределение в растительном покрове
(пятнами1 и небольшими куртинами) является следствием свойственно-
го ей вегетативного возобновления. В процессе непрерывного роста
в течение длительного периода голубика сильно разрастается, так
как у нее легко укореняются отдельные ветви, которые развиваются
затем как самостоятельные особи. При старении и заоыхании надзем-
ной части куста еро возобновление неоднократно осуществляется пу-
тем образования порослевых побегов из спящих почек, находящихся
в основнии куста и развивающихся в новый надземный куст (Солоне-
вич, 1956, 1970). Обильное образование порослевых побегов у голу-
бики наблщцалооь нами весной й летом 1979 г. в Подмосковье, что
связано, по-видимому, с тем, что крона этого кустарничка очень
пострадала от сильных морозов зимой этого года.
За счет порослевых побегов происходит также быстрое омолажива-
ние голубичников после пожаров. При благоприятных погодных усло-
виях после ранневесенних пожаров у порослевых побегов в тот же
год закладываются генеративные почки и на следующий год после по-
жара растение зацветает и дает плоды. После осенних пожаров голу-
бичник начинает давать урожай на третий год (Скрябина", 1970).
182
Экология и фитоценология
Голубика характеризуется весьма широкой экологической амплиту-
дой: она характерна для онрых и болотистых торфяников, растет на
верховых и полуосушенных болотах, но может встречаться и на сухих
местах, заходя в горы до 2500-3000 м над ур.моря (Розанова, 1934).
Отношение голубики к степени увлажнения болотных местообитаний
ярко выражается в характере распределения ее по болоту. На олиго-
трофных болотах этот вид приурочен к местообитаниям с умеренным
увлажнением..На средне- и сидьноувлажненных местообитаниях голу-
бика встречается редко и в незначительном количестве. На резковы-
пуклых болотах голубика сильно разрастается по относительно хоро-
шо дренированным краевым склонам (Солоневич, 1956). По шкале
Л.Г.Раменского (Раменский и др., 1956) наибольшей численности го-
лубика достигает при 80-89 ступенях увлажнения (от сыролутового
до болотно-лугового). Г.Элленберг (Eilenberg, 1974) отмечает ин-
дифферентное отношение голубики к условиям увлажнения. По нашим
наблюдениям,'в Одинцовском р-не Московской обл. голубика встреча-
ется на почвах с относительной влажностью от 78,2 до 90,5%.
Листья голубики отличаются высоким содержанием воды на протя-
жении всего вегетационного периода. Максимальное количество воды
в них отмечается в фазу цветения, к концу вегетации обводненность
имеет минимальное значение. Отношение количества воды к сырой мас-
се у голубики составляет 80% в начале вегетации и 60-68% в конце
вегетационного периода. Наибольшая величина осмотического давле-
ния в клетках листьев наблюдается в период цветения и образования
плодов (Климов и др., 1973).
v.uilginosum - вид светолюбивый, растет на открытых или слабо-
затененных местах (Розанова, 1934), значительно лучше плодоносит
на хорошо освещенных участках, чем на затененных (Солоневич,
1956). По шкале Г.Элленберга (Eilenberg, 1974) голубика занимает
ступень светолюбия, т.е. относится к группе растений между "полу-
теневыми" и "полусветовыми".
В отношении температурных условий этот вид более холодостойкий,
чем черника и брусника. Цветки голубики почти не страдают от час-
тых на болотах весенних заморозков (Солоневич, 1956; Михалева,
1972). Для Западной Европы Г.Элленберг (Eilenberg, 1974) указыва-
ет на безразличное отношение голубики к температурным условиям.
Голубика - олиготрофный вид, приспособленный к обитанию на бед-
183
ных минеральных почвах, а также на почвах, образованных торфом
слабой степени разложения (Солоневич, 1970). Наибольшей числен-
ности она достигает на особо бедных почвах, соответствующих 1-3
ступеням шкалы Л.Г.Раменского. По отношению к содержанию в почве
азота голубика занимает 2-ю ступень шкалы Элленберга (между
"очень бедными" и "бедными" почвами). Хороший эффект для повыше-
ния урожайности голубики дает внесение суперфосфата, а также ран-
невесеннее выжигание голубичников, при котором происходит обога-
щение почвы зольными элементами (Уход..., 1971).
Голубика обычно растет на почвах с очень кислой реакцией -
pH = 2,85-4,18 (Немчинов, 1934; Солоневич, 1956), I-я ступень
кислотности почв по шкале Элленберга. В Московской обл. голубика
растет на почвах с рН^ди = 4,3-4,35, рНС0Л = 3,2-3,3.
Голубика встречается в различных заболоченных хвойных лесах
(сосняках, лиственничниках, ельниках), являясь доминантой и суб-
доминантой кустарничкового яруса. В лесной и тундровой зонах на
верховых болотах, крупно- и мелкобугристых торфяниках голубика
играет заметную роль в растительном покрове наряду с Betula папа,
Empetrum nigrum, Ledum palustre, Rubus chamaemorus, Vaccinium vi-
tis-idaea и др. Вместе c Ledum palustre, Chamaedaphne calycula-
ta, Oxycoccus palustris голубика входит в состав кустарничкового
яруса облесенных олиготрофных болот Западной Сибири (как правило,
они облесены Pinus silvestris f.litvinowii, реже f.wilkomii)
(Цинзерлинг, 1938).
В горах и тундровой зоне голубика образует формацию Vaccinie-
ta uliginosi (Быков, 1965). В моховых тундрах v.uliginosum яв-
ляется характерным представителем разреженного травяно-кустарнич-
кового яруса (совместно С Carex rigiaa, Dryas punctata, Роа ero-
tica, Polygonum viviparum, Salix polaris, S.reticulata и Др.),
йлесте с такими кустарничками, как Arctous alpina, Betula папа,
Ledum decumbens, Vaccinium vitis-idaea, голубика - характерней-
ший элемент пятнистых тундр. В кочкарных тундрах голубика совмест-
но с рядом гипоарктических кустарничков (Betula папа, B.exilis,
Ledum decumbens, Vaccinium vitis-idaea) также играет определен-
ную роль в растительном покрове (Городков, 1939). Голубика вхо-
дит в состав растительных сообществ гольцов Северо-Восточной Азии
(вместе с Arctous alpina, Cassiope ericoides, Empetrum nigrum,
Ledum decumbens, Rhododendron aureum и др.).
На территории Московской обл. голубика встречается на верховых
болотах (иногда облесенных сосной) вместе с Chamaedaphne calycu-
184
lata и Ledum palustre.
Консортивные связи
Голубика - микотрофное растение (Солоневич, 1970). Корни ее
оплетены мицелием гриба Phoma radicis-vaccinii (Розанова, 1937).
Опыление цветков происходит с помощью пчел, муравьев и бабочек из
родов Licaena - голубянка и Aporia - боярышница.
Стебли, листья, цветки и плоды голубики поражаются различными
видами патогенных грибов. На листьях паразитируют Podosphaera
myrtillina (Pr.) Kunze et Schmidt, Antennaria arctica Kostr.,
Mycosphaerella vaccinii (Cooke) Schroet. (этот вид поражает и
почки), M.eriophila (Nieasl.) Lindau (поражает также цветки и
побеги), Thekopsora myrtilli (Schum.) Tranz., Exobasidium vacci-
nii (Puck.) Woron., Phyllosticta rhododendricola Brun., Lepto-
stroma herbarum (Pr.) Link, Stagonospora lambottiana Sacc., Co-
niothyrium phyllogenum Sacc. (встречается также на побегах голу-
бики) , Phlacosphaeria arctostaphyli Ell. et Ev. Стебли поражают
Lophodermium oxycocci (Pr.) Karst., Cytospora delicatula Shear,
Heteropatella lacera Puck., Oamarosporium antarcticum Speg.;
ПЛОДЫ И цветки - Monilinia irnula (Weinm.) Wh., Cladosporium oxy-
cocci Shear. (Ростовцев, 1908; Жуков, 1977).
На голубике обнаружены также различные видя насекомых. В Вос-
точной Сибири листья голубики сильно повреждаются тремя видами
пилильщиков: наиболее серьезный ущерб наносит Pristiophora mol-
lis — голубичный изменчивый пилильщик, меньше Amauronematus
schluteri - голубичный зеленый пилильщик и Pteronidea parvula -
голубичный волосатый пилильщик (Вержуцкий, 1970).
Биологическая продуктивность
и хозяйственное значение
Изучение продуктивности голубики проводилось нами в Одинцов-
ском р-не Московской обл. в двух типах местообитаний: на осушен-
ном верховом болоте (Pinus sylvestris - V.uliginosum + Ledum pa-
lustre - Sphagnum sp.) и на окраине верхового:болота (P.sylves-
tris - V.uliginosum + Chamaedaphne calyculata - Sphagnum sp.).
Полученные данные представлены в табл.II.
185
Таблица II
Фитомасса и урожайность плодов голубики в различных условиях
местообитания в Московской обл., r/i^
1 • Органы 1 растения 1 Стебли г 7 1 1 Осушенное верховое болото Неосушенноё верховое болото ।
оырая масса 310,2 сухая W масса 184,8 сырая масса 184,8 сухая ’ масса 1 110,2 ।
1 Листья 106,0 54,8 56,0 23,0 1
1 Годичный при- кроет 31,2 18,0 15,6 8,6 ।
1 Отмершая часть 7,6 7,4 2,8 2,2 *
। Плоды 24,1 - 6,5 — 1
Величина надземной фитомассы и урожайность плодов голубики вы-
ше на осушенном болоте (влажность почвы 78,2%), чем на верховом
неосушенном (влажность почвы 90,5%). Поскольку этот вид тяготеет
к местообитаниям с умеренным увлажнением, вероятно, различие в
продуктивности голубики на данных местообитаниях можно объяснить
различием влажности почв. Вроме того, на неосушенном болоте нами
отмечена высокая пораженность листьев и годичных побегов патоген-
ными грибами и насекомыми.
Наиболее высокие урожаи голубики (I00-II0 кг/га) бывают на
Дальнем Востоке, в Центральном, Волго-Вятском и Поволжском райо-
нах европейской части СССР. Хороший урожай повторяется 4-5 раз в
десятилетие (Уход..., 1971). Имеются данные, что урожайность го-
лубики может достигать 500, а в отдельных случаях до 1300 кг/га
(Губанов и др., 1976).
Плоды голубики употребляются в пищу. Они содержат до 8% саха-
ров, в том числе 0,94% сахарозы, 2,78% глюкозы, 3,77% фруктозы,
а также 0,97% яблочной кислоты, 0,29% дубильных веществ, 1,36%
клетчатки, 0,71% азотистых 'веществ, 0,22% золы, около 20 мг*% ви-
тамина С, до НО мг«% каротина (Розанова, 1934; 1Убанов и др.,
1976). Кроме того, в ягодах голубики содержатся многие важные мик-
роэлементы: 0,21% от массы абсолютно сухого вещества Mg, 0,23%
Са, 0,08% Fe, 0,03% Мп, 0,05% S1, 0,21% А1 (Богданова, Пира-
тов, 1978). Удельный вес сока голубики 1,045-1,050 г/см8, pH кле-
точного сока 1,1-1,8 (Баранова и др., 1975). Ягодоносная площадь
голубики в РСФСР - 444,4 тнс.га, возможный сбор ягод - 15,4 тнс.т
186
в год (Уход..., 1971). Голубика - хороший медонос. Листья пригод-
ны для дубления кож (Губанов и др., 1976).
В США 9 американских видов голубики культивируются. Выведено
большое число сортов (в настоящее время в США выращивают 25 сор-
тов высокой Голубики). Размножение на плантациях производят черен-
ками. Уход предусматривает рыхление междурядий, внесение удобре-
ний, орошение, обрезку ветвей. Плодоносить голубика начинает.на
третий год, полной продуктивности достигает на 6-10-й год
(30 ц/га), затем урожайность падает (Усовский, I960; Бурмистров,
1968; Гладкова, 1972, 1974).
187
Л.А.Дукова
РОД ПОДОРОЖНИК
Род Plantago - самый крупный в семействе Plantaginaceae, в
нем насчитывается 260 видов, произрастающих главным образом в
умеренных областях обоих полушарий, лишь немногие из них встреча-
ются в тропиках. В СССР около 30 видов этого рода.
Подорожники - многолетние или однолетние травы, имеющие розе-
точные или удлиненные вегетативные побеги. Листья цельные с дуго-
видным или почти параллельным жилкованием, с пластинкой, постепен-
но переходящей в черешок. Соцветия - простой колос, иногда голов-
чатые. Цветки с прицветниками и двойным околоцветником, обычно
обоеполые, анемофильные или частично энтомофильные. Чашечка 4-ло-
пастная или 4-раздельная. Венчик пленчатый с цилиндрической труб-
кой, 4-лопастной или 4-зубчатый. Андроцей образован 4 тычинками,
прикрепленными к трубке венчика и чередующимися с его лопастями,
нити тычинок длинные, выдающиеся из трубки венчика. Гинецей обра-
зован двумя плодолистиками, пестик с тонким терминальным столби-
ком и двулопастным рыльцем, завязь верхняя двугнездная. Плод -
перепончатая коробочка с одним-несколькими семенами в каждом
гнезде (Флора СССР, т.23, 1958; Тахтаджян, 1966).
В Московской обл. встречаются 5 видов этого рода.
Ключ для определения московских видов1
I. Стебель ветвистый, с несколькими короткими колосовидными
соцветиями на вершинах верхних ветвей, олиственный; листья супро-
тивные, линейные ............................... ... P.indica L.
(P.arenaria Waldst. et Kit., P.ramosa Aschers.).
- Все листья в прикорневой розетке; цветоносный стебель без-
листный, с единственным колосовидным соцветием.................2
2. Листья ланцетные, линейно-ланцетные или почти линейные; сте-
бель доверху резко бороздчатый; чашечка 3-дольная из-за срастания
двух из четырех долей; тычинки белые, в 4-5 раз длиннее венчи-
ка.... ......................................... P.lanoeolata Ь.
1 Составил В.Н.Тихомиров.
188
- Листья более широкие, иногда почти округлые; стебель по край-
ней мере в нижней части не бороздчатый; чашечка 4-дольная; тычин-
ки розовые, лиловые, бурые или почти белые ....................3
3. Листовая пластинка голая (основание и черешок могут быть
опушены); венчик и пыльники бурые или буроватые; тычиночные нити
не длиннее столбика .............................. P.major L. s.l.
- Листовая пластинка опушенная; венчик и пыльники розовые, ли-
ловатые или почти белые; тычиночные нити много длиннее столби-
ка .......................................................... 4
4. Листья широколанцетные, с длинными черешками, торчащие
вверх; стебли прямостоячие; доли венчика до 2 мм длины
. ........ P.stepposa Kuprian.
- Листья обычно более широкие, на открытых местах прижатые к
почве; нижние листья с очень короткими черешками; стебли обычно
в основании восходящие; доли венчика до 1,5 мм длины. .P.media L.
I. ПОДОРОЖНИК БОЛЬШОЙ
Номенклатура и географическое распространение
Plantago major L., 1753, Sp. р1,:112; Кауфм., 1866, Моск. фл.
изд. 1:408, 1889, изд. 2:423; Цингер, 1886, Об. свед.:360;
Шмальг., 1897, Фл. 2:350; Сырейщ., 1910, Илл. фл. 3:180; он же,
1927, Опр. раст. Моск. губ.:238; Горжова, 1940, в Маевск. Фл.,
Изд. 7:667, 1954, изд. 8:527, 1964, изд. 9:509; Григорьев, 1958,
Фл. СССР, 23:138; Скворцов, 1966, Опр. раст. Моск. обл.:299; aba-
ter, 1976, Pl. Eur. 4:39. - Подорожник большой.
Подорожник большой - евроазиатский вид, имеющий обширный ареал,
на других континентах встречается лишь как заносное растение. В
СССР северная граница его ареала проходит несколько южнее Мурман-
ска, достигает Архангельска и далее по 69° сев.широты идет на вос-
ток до Салехарда и Игарки, затем по рекам Нижняя Тунгуска и Вилюй,
пересекает низовья Алдана и доходит до Камчатки. Южнее этой грани-
цы подорожник большой на территории СССР встречается повсеместно,
за исключением Памира, Устюрта и песчаных пустынь Каракумов и Кы-
зыдкума (Флора СССР, т.23, 1958; Атлас ареалов... 1976). Обычное
растение в Московской обл. (Ворошилов и др., 1966).
189
Морфологическое описание
Многолетнее травянистое поликарпическое растение, короткокор-
невищное, кистекорневое. Корневая система образована придаточны-
ми корнями приблизительно одинаковой длины (до I м), располагаю-
щимися на небольшом вертикальном корневище. Молодые корни - бе-
лые, старые - темно-коричневые. Ветвление корней незначительно,
часто отмечают их контрактильность. Есть указания на способность
развития запасающей ткани в корнях (Серебряков, 1952;-Sagar,
1958; Sagar, Harper, 1964). Неспециализированное эпигеогенное
вертикальное корневище возникает вследствие отмирания листьев
главного розеточного побега и погружения его базальной части в
почву благодаря втягивающей деятельности придаточных корней. Сред-
няя длина корневища взрослой особи 3-4 см, толщина 2-3 см. Ветвя-
щееся корневище встречается крайне редко, так как при образовании
боковых розеточных побегов происходит его разрушение и отделение
партикул.
У подорожника большого ось 1-го порядка, образующая вегетатив-
ный розеточный побег, сохраняется долго и нарастает моноподиально.
Ассимилирующих листьев на розеточном побеге взрослого растения 5-
7, реже 9. Листья цельные, голые, иногда опушенные редкими корот-
кими волосками, резко суженные в черешок. Черешки по длине равны
пластинкам или длиннее их, реже короче. Листовые пластинки взрос-
лых листьев с 3-7 жилками, эллиптические, овальные.или яйцевидные,
с тупой верхушкой, округлым или клиновидным основанием; края цель-
ные, иногда городчато-волнистые, в нижней части слегка неправиль-
но зубчатые. Как правило, листья зеленые, но иногда с розоватым
или пурпурным оттенком. Общая длина их колеблется от 5 до 30 см.
Генеративные побеги подорожника большого называют цветочными
стрелками. Они формируются как оси 2-го, редко 3-го порядка, как
правило, лишены листьев и представляют собой гипоподий - первое
вытянутое междоузлие побега от материнской оси до предлиста. Дли-
на их 3-35 (60) см, они несут длинные, узкие, густые ботрические
соцветия - простые колосья (I) 10-15 (30) см длиной (ветвящиеся
соцветия встречаются как исключение). Прицветники треугольно-яйце-
видные, острые, килеватые, по краям широкопленчатые, длиной до
4 мм, шириной до 5 мм. Цветки актиноморфные, 2-4 мм в диаметре.
Чашелистики свободные почти до основания. Доли венчика яйцевидные
или яйцевидно-ланцетные, около I мм длиной, туповатые, желто-бе-
лые или буроватые. Пыльники вначале лиловые, затем белые, подвиж-
190
ные, прикреплены к спинной стороне, не превышают дайну столбиков.
Пестик с верхней завязью, тонким столбиком и двухлопастным перис-
тым рыльцем 0,3-0,6 см дайны. Цветки без запаха и нектара, расте-
ние ветроопыляемое и способное к самоопылению. Каждый цветок про-
тогиничён, цветки раскрываются акропетально, поэтому только рас-
крывшиеся первыми цветки могут обеспечить перекрестное опыление.
Коробочка ценокарпная, яйцевидно-коническая, яйцевидная или
шаровидная, почти вдвое длиннее чашечки (до 5 мм), с 5-30 (34) се-
менами. Обнаружена вивипария на затопленных речных отмелях. Семе-
на эллиптические, 1-5 мм длины и 0,8 мм ширины (спинная сторона
выгнута меньше, чем брюшная), темно-коричневые или коричневые со
светлым поясом вдоль спинной стороны, рубчик,светлый. Иногда на
светлой поверхности семенной кожуры образуются темные несимметрич-
ные выпуклости в виде косых складок (Korsmo, 1935). В созревшем
семени есть хорошо сформированный зародыш, занимающий 2/3 семени,
и эндосперм (Майсурян, Атабекова, 1978). Масса I тыс. семян 0,2-
0,3 Г (Korsmo, 1935; Salisbury, 1952).
Число хромосом 2п = 12 (Cuiigagh, 1934; Хромосомные числа...,
1969). Гибриды не выявлены и попытки получить их искусственным пу-
тем потерпели неудачу (Rahn, 1954, 1957).
Онтогенез^, сезонное развитие
и способы размножения
В почве семена подорожника большого могут сохраняться 40-60
лет (Milton, 1943). Экспериментально доказано, что после 40 лет
хранения семян в почве всхожесть их составляла 10%, после 20 лет -
83,5$ (Crocker, 1938), после 5 лет - 30% (Kjaer, 1940), после
1-3 лет - 89-100$ (Blom, 1979). При сухом хранении семена теряют
всхожесть в течение 5-10 лет. Оптимальными условиями для прораста-
ния свежесобранных семян являются свет и достаточно высокая темпе-
ратура: при 25° всхожесть на свету составляет 70%, в темноте - 8%,
а при 15° на свету всхожесть всего лишь 2% (Sagar, 1958). В при-
родных условиях семена, как правило, не прорастают до весны, а за-
I Для характеристики онтогенеза всех трех видов подорожника ис-
пользованы материалы курсовых работ студентов Московского гос.
педагогического ин-та им.В.И.Ленина Н.Грачевой, М.Пелевиной,
И.Пушковой.
191
тем проростки появляются спорадически в течение всего вегетацион-
ного сезона, причем более активно на уплотненных, лишенных расти-
тельности почвах (Marsdon-Jones, Turrill, 1938; Васильченко,
1965).
Прорастание надземного типа, развивающийся гипокотиль выносит
над почвой.2 семядоли 3,5-6 мм длины, на которых первое время со-
храняется перикарп. Семядоли зеленые, продолговато-эллиптические,
срастающиеся при основании, с одной жилкой. На третий день после
прорастания в комнатных условиях при 22-25° длина семядолей со-
ставляет 6-7,5 мм, ширина - 1,5 мм. До появления первого настоя-
щего листа в течение 5-6 дней происходит активный рост семядолей,
на 8-9-й день длина их достигает 12-13 мм, ширина - 2-2,5 мм; по-
том рост их замедляется, хотя длина может несколько возрастать до
15 мм.
Проростки (рис.15) - небольшие растения до 1-2 см высотой, фор-
мируют полициклический розеточный побег с 2 продолговато-эллипти-
ческими семядолями, срастакщимися при основании, и 1-3 короткоче-
решковыми овальными мелкими листьями (длина пластинки 0,4-0,5 см,
ширина 0,2-0,3 см) с одной жилкой. Хорошо выражены гипокотиль (3-
4 мм) и эпикотиль. Главный корень с 2-3 боковыми. В условиях эк-
сперимента на 35-44-й день после развертывания 3-4-го листа про-
исходит завядание и отмирание семядолей.
Ювенильные растения (см. рис.15) имеют главный розеточный по-
бег с 3-4 овально-эллиптическими листьями, у которых 1-3 жилки,
черешок равен по длине листовой пластинке, край пластинки цельный
или слегка зубчатый. Из гипокотиля формируется небольшое (2-3 мм)
вертикальное корневище, на нем возникают придаточные корни. Корне-
вая система образована системой главного корня и придаточными кор-
нями.
Имматурные растения (см. рис.15) формируют розеточный побег
1-го порядка с 4-5 овальными или овально-яйцевидными листьями, у
которых 3-5 жилок, черешки несколько длиннее пластинок. Корневище
утолщается, появляется большое число првдаточных молодых корней,
ветвящихся до 2-го порядка. В оптимальных условиях это возрастное
состояние может сразу перейти в генеративное.
Взрослое виргинильное растение (см. рис.15) выражено не всегда,
чаще в более сомкнутых ценозах иди при неблагоприятных условиях
существования. Главный розеточный побег с 6-7 овально-яйцевидными
листьями, у которых 5 жилок, черешки длиннее пластинок в 1,5 раза.
Корневище до I см толщины, большое число молодых придаточных кор-
192
Рис.15. Возрастные состояния подорожника большого:
р - проросток; j - ювенильное; im - иммэтур-
ное; v - взрослое виргинильное; gj - молодое
генеративное; g2 - средневозрастное генератив-
ное; gj - стареющее генеративное; ss - субсе-
нильное; в - сенильное.
ней, появляются старые корни.
Молодые генеративные растения (см. рис.15) по-прежнему сохра-
няют розеточный побег 1-го порядка с 6-8 овально-яйцевидными
листьями, у которых 5 жилок, черешки равны по длине пластинке или
несколько длиннее. В пазухах 6-7-го листа образуются 1-2 генера-
тивных монокарпических побега 2-го порядка - цветочные стрелки.
От более утолщенного корневища отходят многочисленные светлые мо-
лодые придаточные корни, старых корней мало, они ветвятся до 3-го
порядка. Сохраняются остатки отмерших листьев.
У средневозрастных генеративных растений (см. рис.15) продол-
жает функционировать розеточный побег 1-го порядка с 8-10 яйцевид-
13-1964 193
но-овальными листьями, у которых 5, реже 7 жилок, черешок в 1,3-
1,5 раза короче пластинки или равен ей. Генеративных побегов от I
до 4, они возникают как монокарпические побеги 2-го порядка. Кор-
невище более мощное, плотное, темное, неразрушаицееся, с листовы-
ми рубцами и остатками черешков. В корневой системе преобладают
многочисленные старые придаточные корни, молодых корней мало.
В старом генеративном состоянии (см. рис.15) сохраняется поли-
циклический розеточный побег 1-го порядка, 8-12 яйцевидных листь-
ев с 7 жилками, площадь листовой пластинки максимальная. Генера-
тивных побегов от I до 3-4. Резко возрастают размеры корневища,
число рубцов отмерших листьев и полуразрушенных черешков; корневи-
ще менее плотное, начинается его разрушение. Корневая система
представлена только старыми придаточными корнями.
Субсенильные растения (см. рис.15) имеют один розеточный поли-
циклический побег 1-го или 2-го порядка, число листьев сокращает-
ся до 6-8, форма листовой пластинки яйцевидно-овальная, жилок 7,
реже 5. Корневище максимальных размеров, мягкое, разрушающееся,
снабженное "муфтой" из отмерших черешков листьев. Генеративные по-
беги отсутствуют. Старые придаточные корни более короткие, в мень-
шем числе.
У сенильных особей (см. рис. 15) формируется один розеточный по-
бег с 3-5 овальными листьями с 5 жилками, которые по размерам и
форме похожи на листья имматурных растений. Корневище несколько
вытянутое до 1,5-2 см, косо направленное, темное, разрушенное;
придаточных корней мало, глубина их проникновения небольшая. Про-
должительность жизни в культуре составляет всего лишь 4 года, в
естественных ценозах - до 15-20 лет.
Вегетативные розеточные побеги подорожника большого развивают-
ся по типу полициклических. Конус нарастания побега 1-го порядка,
моноподиально нарастая в течение всей жизни особи, остается веге-
тативным. Ежегодно он развертывает 5-12 листьев. Рост побегов и
корней начинается рано весной и прекращается в естественных усло-
виях в районах умеренного климата в августе. Осенью на конусе на-
растания закладывается только часть зачатков листьев следующего
года. Таким образом, вегетативные розеточные побеги подорожника
большого являются побегами с неполным циклом (Серебряков, 1952).
Зимой в комнатных условиях растения, выращиваемые из семян, могут
развиться до виргинильного состояния, очень редко они зацветают,
но, как правило, погибают, не переходя к цветению.
Генеративные побеги моноциклические, возникают в пазухах 7-10-
194
го зеленого листа текущего года, поэтому цветение начинается толь-
ко в июне (в условиях Подмосковья 5-15Д1) и продолжается 3 мес.
Семена завязываются вскоре после оплодотворения, но не рассеива-
ются из коробочки до следующей весны. Это зимне-летнезеленое рас-
тение, гемикриптофит.
Размножается преимущественно семенным путем. Сведения о семен-
ной продуктивности очень противоречивы: по данным Г.П.Верзилина
(1953), одна особь производит от. 8 до 60 тыс. семян, другие авто-
ры называют 960 (Salisbury, 1916), до 2 тыс. (Мальцев, 1933) и
14 тыс. (Кернер, 1902) семян на одно растение. Семена покрыты
слизью, которая разбухает в воде, что приводит к разрыванию сте-
нок коробочки. Липкая слйзь способствует распространению семян жи-
вотными и человеком. Например, семена прилипают к перьям птиц и
переносятся на значительное расстояние (Thompson, 1952).
Вегетативное размножение в природе происходит, как.правило,
при механическом повреждении. Взрослые генеративные особи, имею-
щие несколько розеточных побегов, разделяются на несколько парти-
кул, образуя клон. Проводились опыты по искусственному клонирова-
нию (Marsdon-Jones, Turrill, 1938).
Экология, фитоценология и консортивные связи
Подорожник большой произрастает при увлажнении почвы от влаж-
ностепного до болотно-лугового (60-86-я ступень шкалы Л.Г.Рамен-
ского), максимальное его участие в травостое отмечено при влажно-
луговом увлажнении. Э.Ландольт считает его индикатором средних ус-
ловий увлажнения (3-я ступень шкалы Ландольта, 5-я ступень шкалы
Элленберга). В засушливых районах приурочен к поймам рек.
К свету требователен, более обилен в открытых ценозах, погиба-
ет при сильном затенении (Sagar, Harper, i960). Располагается
между полусветовыми и полностью световыми растениями (8-я ступень
шкалы освещенности Элленберга, 4-я ступень шкалы Ландольта). До-
статочно морозоустойчив, как правило, его почки и живые листья хо-
рошо сохраняются зимой в подстилке, иногда почки перезимовывают в
почве, главным образом на участках, лишенных растительности.
Подорожник большой встречается на самых разнообразных почвах,
но максимальная высота растений, наибольшее число генеративных по-
бегов и самые крупные листья отмечены на глинистых почвах (Mars-
don-Jones, Turrill, 1938; Sagar, Harper, 1964). К богатству
195
почв довольно требователен (IO-17-я ступень шкалы Л.Г.Раменского),
произрастает преимущественно на почвах, богатых доступными форма-
ми азота (4-я ступень шкалы Ландольта), однако Т.Элленберг счита-
ет его безразличным к содержанию азота в почве. Предпочитает сла-
бокислые почвы, никогда не растет на сильнокислых, изредка встре-
чается на нейтральных и слабощелочных (3-я ступень шкалы Ландоль-
та).
В горы поднимается до высоты 2000-2500 м над ур.моря.
Подорожник большой - обычное рудеральное растение, в основном
встречается вдоль дорог, на пустошах, залежах, насыпях, вдоль ме-
лиоративных каналов, в населенных рунктах. Широко распространен
на различных типах лугов, отведенных под пастбища, реже на лугах
сенокосного использования. Обычен в лесах, нарушенных в результа-
те вытаптывания травяного покрова (Salisbury, 1916). Вытаптыва-
ние обычно хорошо переносят как взрослые особи, так и проростки
(Blom, 1979), поэтому подорожник встречается при различной паст-
бищной дигрессии - от слабого выпаса до сбоя (3-9-я ступень шка-
лы Л.Г.Раменского). Иногда засоряет посевы многолетних трав, про-
пашные культуры, паровые поля, огороды, сады. Не образует сплош-
ных зарослей на больших площадях, хотя вдоль дорог иногда тянут-
ся длинные полосы этого вида.
Подорожник большой - микосимбиотроф.
На листьях встречаются различные пятна, вызванные грибами Ре-
ronospora alta Fuck, Ascochyta plantaginis Sacc. et Speg., Ramu-
laria plantaginea Sacc. et Berl., R.plantaginis Ell. et Mart.,
а также порошковидный налет конидиального спороношения, возникаю-
щий в результате развития Erysiphe communis (Wallr.) Link. Из-
вестны также вирусные заболевания подорожника большого, вызванные
различными вирусами табачной мозаики. Иногда на подорожнике боль-
шом паразитирует Cuscuta trifolii Bab. (Sagar, Harper, 1964).
Биологическая продуктивность
Урожайность свежих листьев в Белгородской обл. на дорогах и
выгонах колеблется от 34 до 132 г/м2, на лугах достигает 230 г/м2,
в смешанных лесах Кемеровской обл. от 4 до 600 г/м2. Подорожник
большой введен в культуру, плантации используются 3-5 лет, уборка
листьев производится 2 раза за сезон, урожай достигает 70-90 ц/га
(Губанов и др., 1976).
196
При отсутствии внутривидовой и межвидовой конкуренции в тепли-
це биомасса одного взрослого растения равна 14,2 г, при этом 20-
23$ приходится на репродуктивные органы; листовая поверхность со-
ставляет 1355 бм^. Внутривидовая конкуренция оказывает на подо-
рожник большой значительное отрицательное воздействие: при высо-
ком уровне численности биомасса особей этого вида падает в 2-3 ра-
за (Hawthorn, Cavers, 1976, 1978). Интенсивный выпас вызывает
уменьшение биомассы корней на 23-43$, одновременно с этим проис-
ходит увеличение биомассы побегов на 24-35% (Blom, 1979).
2. ПОДОРОЖНИК СРЕДНИЙ
Номенклатура и географическое распространение
Plantago media !>., 1753, Sp. pl.:113s Кауфм., 1866, Моск, фл.,
изд. 1:409, 1889, изд. 2:423; Цингер, 1886, Об. свед.:360;
Шмальг., 1897, Фл. 2:351; Сырейщ., 1910, Илл. фл. 3:181; он же,
1927, Опр. раст. Моск. губ.:238; Горшкова, 1940, в Маевск., Фл.,
изд. 7:667, 1954, изд. 8:528, 1964, изд. 9:510; Григорьев, 1958,
Фл. СССР 23:148; Скворцов, 1966, Опр. раст. Моск. обл.:299; aba-
ter, 1976, Pl. Bur. 4:41. - Подорожник средний.
Широко распространен в Европе, Западной Азии, изолированные
местообитания есть в Японии и Китае, отмечен в Северной Америке и
Новой Зеландии. В СССР встречается во всех районах европейской
части, Западной и Восточной Сибири, на Дальнем Востоке (Флора
СССР, т.23, 1958). В Московской обл. обычен во всех районах.
Морфологическое описание
Многолетнее травянистое поликарпическое короткокорневищное,
стержнекорневое растение. Система главного корня сохраняется на
протяжении всего онтогенеза, достигает значительной глубины (до
90 см) (Sagar, Harper, 19б4; Казанфарова, Гасанов, 1979). Ветвле-
ние главного корня, как правило, не очень интенсивно. В верхних
слоях почвы функционируют тонкие придаточные корни, отходящие от
небольшого ортотропного корневища. Есть указания на контрактиль-
ную деятельность боковых корней (Серебряков, 1952), что вызывает
углубление в почву базальной части розеточного побега и приводит
13х-1964 197
к образованию неспециализированного эпигеогенного корневища. Дли-
на его 2,5-3 см. На протяжении всей жизни растения функционирует
розеточный побег 1-го порядка, нарастающий моноподиально; возмож-
но образование боковых розеточных побегов 2-го порядка.
У взрослого растения ассимилирующих листьев 5-7, они цельные,
простые, цельнокрайние или неправильно зубчатые, опушенные (гуще
по жилкам), длина волосков 0,2-0,3 мм (Ворошилов и др., 1966),
жилкование дуговидное. Листовые пластинки эллиптические или яйце-
видные, реже яйцевидно-ланцетные, с острой верхушкой, обычно дли-
на их не более чем в 2-2,5 раза превышает ширину (длина 6-6,5 см,
ширина 3,35 см). Основание пластинки сужено в короткий широкий че-
решок (в 3-10 раз короче пластинки), иногда листья почти сидячие.
Общая длина листьев от 3 до 8,5-10 см.
Генеративные побеги подорожника среднего представляют собой
цветочные стрелки высотой 15-70 см, возникающие как боковые удли-
ненные побеги 2-го, реже 3-го порядка. Они покрыты прижатыми во-
лосками, формируют ботрическое соцветие - простой цилиндрический
колос 2-15 см длины. Прицветники яйцевидные или продолговато-яй-
цевидные, туповатые, по краям пленчатые, по длине равные или поч-
ти равные чашечке. Чашелистики эллиптические или яйцевидно-эллип-
тические, 1,5-2 мм длины, пленчатые, с хорошо выраженным килем.
Доли венчика беловатые или бело-розовые, ланцетные, тупые, диа-
метр венчика 4 см. Лилово-белые пыльники прикреплены к тычиночным
нитям спинной стороной, подвижны; нити красновато-фиолетовые, дли-
ной 0,2-1 см. Гинецей ценокарпный, завязь двураздельная, столбик
длинный, рыльце длинное перистое. Цветки протерогиничны, продуци-
руют огромное количество пыльцы - до 3,3 млн. пылинок в одном со-
цветии (Kugler, 1970), опыляются ветром, но нередко и насекомы-
ми. Отмечено удлинение рылец до 1-2 см при отсутствии перекрест-
ного опыления, на основании этого предполагается возможность са-
моопыления.
Плод - ценокарпная коробочка яйцевидной формы 2-4 мм длины с
(I) 3-4 (7) семенами. Семена овальные, часто неправильной формы,
со спинной стороны выпуклые, край часто завернут, рубчик светлый,
поверхность морщинистая коричневая или черно-коричневая со свет-
лой полосой на спинной стороне. Масса I тыс. семян 0,35-0,79 г
(Korsmo, 1935; Salisbury, 1952).
Число хромосом 2п = 24 и 12. Зарегистрированы гибриды с подо-
рожником ланцетолистным (Druce, Britton, 1928; Sagar, Harper,
1964). Получить гибриды с другими видами подорожников не удалось
198
даже в условии эксперимента.
Онтогенез, сезонное развитие и способы размножения
У подорожника среднего по материалам, собранным на суходольных
лугах Московской обл., нами выделены 10 возрастных состояний. Се-
мена требуют периода покоя, после которого прорастание наблюда-
лось в течение всего вегетационного периода. При сухом хранении
семена теряют всхожесть через 3 года, после 2 лет хранения всхо-
жесть составляет 20%. Свежесобранные семена прорастают с трудом
на свету. После 2-недельного воздействия температурой +5° семена
полностью прорастали на свету при +20-25°. Если семена 2 недели
хранились при 16°, то свет уже не играл роли для их полного про-
растания.
Проростки (рис.16) представлены розеточным побегом с двумя уз-
копродолговатыми семядолями, на верхушке слегка выемчатыми или
двураздельными, каждая с одной жилкой. Первые ассимилирующие
листья серовато-зеленые из-за обильного опушения, продолговатые
с 1-3 жилками, широкими черешками, расширенными в основании. Ги-
покотиль 0,7-1,4 см, книзу суживающийся, четко выражен главный ко-
рень и 1-2 боковых корня.
Ювенильные растения (см. рис.16) представлены главным розеточ-
ным полициклическим побегом с 3-4 продолговатыми черешковыми опу-
шенными листьями, имеющими по 3 жилки. Семядоли не сохраняются.
Гипокотиль утолщается незначительно, на нем появляются придаточ-
ные корни, одновременно функционирует главный корень, ветвящийся
до 3-го порядка. Корневая система смешанного типа.
Имматурные растения (см. рис.16) имеют главный розеточный по-
бег, формирующий 4-5 продолговатых или овальных черешковых листа
с 3 жилками, сильно опушенных. Сохраняется главный корень, корне-
вая система смешанного типа. Корневище еще не сформировано.
У взрослых виргинильных растений (см. рис.16) главный розеточ-
ный побег образует 5-6 продолговато-эллиптических листьев с 5 жил-
ками, значительно опушенных, на коротких или длинных черешках в
зависимости от условий обитания. Боковые и придаточные контрак-
тильные корни втягивают гипокотиль и нижнее междоузлие надземной
части побега, таким образом формируется вертикальное корневище.
У молодых генеративных растений (см. рис.16) продолжает функ-
ционировать главный розеточный побег, на нем 5-7 эллиптических
199
Рис.16. Возрастные состояния подорожника среднего
(обозначения те же, что и на рис.15).
опушенных листьев с 5 жилками. Появляются генеративные побеги
(стрелки) как оси 2-го порядка в пазухе 3-5-го листа. Утолщается
корневище, формируются многочисленные придаточные корни, одновре-
менно сохраняется система главного корня.
У средневозрастных генеративных растений (см. рис.16) сохраня-
ется главный розеточный побег. Он развивает 5-6 опушенных овально-
продолговатых черешковых листьев с 7 (реже 5) жилками. Генератив-
ные побеги - стрелки возникают как оси 2-го порядка. Возможно фор-
мирование 1-2 боковых побегов. Корневище разрастается в толщину,
несет многочисленные старые придаточные корни, сохраняется систе-
ма главного корня. Появляются остатки отмерших листьев, прошлогод-
них генеративных побегов.
У старых генеративных растений (см. рис. 16) главный розеточный
побег погибает, функционируют 2-3 розеточных побега 2-го порядка,
образующих по 4-5 овальных листьев с 7 жилками. Формируются 2-3
200
генеративных побега - стрелки. Корневище мощное, темное, с много-
численными старыми придаточными корнями. Главный корень не сохра-
няется или мало отличается от 2-3 мощных боковых корней.
Субсенильные растения (см. рис.16) имеют 1-2 розеточных побега
возобновления, генеративннх побегов нет. Листья несколько меньших
размеров, с 5 жилками, продолговатые или ланцетные. Корневище
темное, мягкое, с отмершими участками. Придаточные корни малочис-
ленны.
у сенильных растений (см. рис.16) один розеточный побег возоб-
новления, листья на нем короткочерешковые, продолговатые, мелкие,
с 3-5 жилками. Корневище старое, темное, разрушенное, с единичными
придаточными корнями. Система главного корня не сохраняется.
Вегетативные полициклические побеги подорожника среднего актив-
но растут в апреле - мае и в августе - сентябре, образуя в тече-
ние сезона 5-9 листьев. Генеративные побеги закладываются в пазу-
хах зеленых листьев этого года. В течение всего онтогенеза верху-
шечная точка роста не переходит в генеративное состояние, т.е. по-
беги 1-го порядка являются побегами с неполным циклом.
Цветение с начала июня ДО первых морозов (Sagar, Harper, 1964),
в Московской обл. - в июне - августе (Ворошилов и др., 1964). Се-
мена созревают с июля до сентября. После созревания плодов гене-
ративные побеги отмирают, но могут сохраняться до следующей весны.
Зимует подорожник средний с зелеными листьями. Это зимне-летнезе-
леный гемикриптофит.
Вегетативное размножение может осуществляться благодаря отделе-
нию от главного розеточного побега искусственным или естественным
путем дочерних розеточных побегов. В условиях нарушенных ценозов
при интенсивной обработке почвы их может образоваться до 20 за се-
зон (Sagar, Harper, 19б4). На лугах партикуляция гораздо слабее
и выражена лишь у отарых растений. Опиоано восстановление особей
подорожника среднего из отрезанных в конце лета участков корневи-
ща (Gustafson, 1948).
Преобладает размножение семенами. Семенная продуктивность со-
ставляет 960 семян на одну особь (Salisbury, 1952). Коробочек в
одном колосе - от 52 до 130, в среднем 79 (Dowling, 1935). Семе-
на не покрыты слизью (в отличие от семян подорожников большого и
ланцетовидного), способ распространения их не ясен.
201
Экология, фитоценология и консортивные связи
Подорожник средний - индикатор средней сухости почв (2-я сту-
пень шкалы увлажнения Ландольта, 4-я ступень шкалы Элленберга),
характерен для почв с увлажнением.от лугово-степного до влажно-
лугового (49-74-я ступень шкалы Л.Г.Раменского).
К свету довольно требователен, Г.Элленберг его относит к полу-
световым растениям (7-я ступень шкалы освещенности). На открытых
местообитаниях встречаются приземистые распростертые растения, в
густом травостое произрастают высокие растения с длинночерешковы-
ми поднятыми вверх листьями, несколько напоминакщими листья подо-
рожника ланцетовидного. Устойчив как к низким, так и высоким тем-
пературам (Sagar, Harper, 1964), при этом сильно варьирует высо-
та его розеточного побега (от 5 до 80 см).
Максимальной численности достигает на довольно богатых и бога-
тых почвах (IO-19-я ступень шкалы Л.Г.Раменского), может расти на
почвах, довольно бедных, минеральным азотом (3-я ступень шкалы Эл-
ленберга, 2-я ступень шкалы Ландольта). Вид характерен для нейт-
ральных и щелочных почв, чаще с pH = 5,5-8,0 (4-я ступень шкалы
Ландольта, 8-я ступень шкалы Элленберга). В СССР наиболее распро-
странен на почвах с pH = 5,5-7,5 (Смелов, Работнов, 1929). По
данным некоторых ученых, в Западной Европе вид считается строгим
кальцефилом (Salisbury, 1952). Подорожник средний часто растет
на тяжелых глинистых почвах (Brenchley, 1920), практически отсут-
ствует на песчаных (Sagar, Harper, 1964).
Хорошо выдерживает вытаптывание, поэтому широко распространен
на пастбищах, по дорогам, бывает пионером на отмелях и обнажениях
известняка. Подорожник средний нередко закрепляет почву в дюнных
сообществах. Вид типичен для пойменных, суходольных, высокогорных
лугов, разреженных лесов, залежей. Изредка доминирует в сообщест-
вах.
Поражается различными грибами: Peronospora alta Fuck, Erysiphe
communis (Wallr. ). Link, Sclerotina trifoliorum Erikss., Cyrooeras
plantaginis (Sagar, Harper, 1964), вызывающими пушистый или по-
рошковидный налет и пятнистость. Микосимбиотроф.
202
3. ПОДОРОЖНИК ЛАНЦЕТНЫЙ
Номенклатура и географическое распространение
Plantago lanceolate L., 1753, Sp. pl.:113; Кауфм., 1866, Моск,
фл., изд. 1:409, 1889, изд. 2:423; Цингер, 1886, Об. свед.:36Г;
Шмальг., 1897, Фл. 2:351; Петунников, 1900, Крит, обзор 2:151; Сы-
рейщ., 1910, Илл. фл. 3:182; он же, 1927, Опр. раст. Моск, губ.:
238; Горшкова, 1940, в Маевск., Фл., изд. 7:667, 1954, изд. 8:526,
1964, изд. 9:509; Григорьев, 1958, Фл. СССР 23:156; Скворцов,
1966, Опр. раст. Моск. обл.:298; Chater, 1976, Fl. Eur. 4:42. -
Подорожник ланцетный.
Встречается в Евразии, Австралии, Новой Зеландии, Полинезии,
Северной Африке (Sagar, Harper, 1964). В Московской обл. обычен
во всех районах (Ворошилов и др., 1966).
Морфологическое описание
Многолетнее травянистое поликарпическое растение с хорошо раз-
витой системой главного корня и коротким вертикальным или косо
вверх направленным корневищем. Розеточные побеги формируют 5-9
ланцетных, зллиптическо-ланцетных или линейно-ланцетных, простых,
густо опушенных или почти голых листьев. Верхушка листовой плас-
тинки острая, край цельный (реже с немногими зубцами), основание
пластинки клиновидное, сужено в короткий или длинный черешок, жил-
кование дуговидное, жилок 3-5 (7).
Генеративные побеги безлистные (цветоносные стрелки), прямо-
стоячие или восходящие, явно продольно-бороздчатые, опушены при-
жатыми вверх волосками, формируются в пазухе верхних листьев розе-
точного побега 1-го порядка, моноциклические, монокарпические. Со-
цветие - густой, компактный, головчатый или щаиндрический прос-
той колос 3-7 (Ю) см длины. Прицветники яйцевидные или овальные,
заостренные, пленчатые, с хорошо выраженным килем, не достигающим
верхушки прицветника, темно-зеленые, иногда черные, покрытые точ-
ками, голые или негусто волосистые, равны чашечке или больше ее;
нижние прицветники больше верхних.
Чашелистики 2,5-3,5 мм длины, пленчатые с травянистым килем,
голые, реже по спинке волосистые; два передних чашелистика срас-
203
таится в широкую двухлопастную чащу. Венчик около 4 ни в диамет-
ре, коричневато-белый с овальными или продолговато-яйцевидными
долями с острыми выступающими срединными жилками. Тычинки белые,
фиолетовые, реже желтовато-зеленые, на длинных нитях, подвижные.
Пестик с перистым рыльцем и длинным столбиком, завязь с 2 семяза-
чатками. Плод - эллиптическая коробочка, около 3-6 мм длины, со-
держащая 1-2 семени (Флора СССР, т.23, 1958; Sagar, Harper, 1964).
Семена овальные или продолговато-эллиптические, 2,5 на 1,5 мм,
блестящие, желтовато-коричневые или темно-коричневые, рубчик тем-
ный, брюшная сторона сильновогнутая, спинная - выпуклая. Масса
I тыс. семян около 1,6 г (Когвшо, 1935; Salisbury, 1942), иног-
да дается больший диапазон этого показателя: 1,04-2,87 г (Sagar,
Harper, 1964).
Диплоидный набор хромосом 2п - 12 (Хромосомные числа...,
1969).
Онтогенез, сезонное развитие
и способы размножения
Прорастание семян спорадическое, достигает максимальных преде-
лов после,12 мес покоя. Более длительное сухое хранение семена
переносят плохо, однако известно сохранение ими всхожести до 9
лет (Мальцев, 1933). В почвах после 10 лет покоя прорастает до 8%
семян (Brenchley, 1918), иногда жизнеспособность погребенных се-
мян сохраняется 50-60 лет (Milton, 1943). Наиболее высокая всхо-
жесть (до 90$) отмечается при температуре 20°, при более низких
(15°), как и при более высоких температурах (25°) всхожесть со-
ставляет 36-50$. Влияние света на прорастание менее заметное, чем
у двух других видов подорожника (Sagar, Harper, 1964), семена по-
дорожника ланцетного могут прорастать в полной темноте (Blom,
1979).
Проростки (рис.17) небольшие (до 1-2 см высоты) растения, обра-
зующие главный полициклический розеточный побег с 2 узколинейными
тупыми семядолями, срастающимися своими основаниями, и 2-3 продол-
говато-линейными очередными листьями, постепенно суженными в кры-
латые- черешки. Край листовой пластинки неровнолистый, верхушка за-
острённая,-жидок 1-3, длина пластинки 2-3 см, ширина 0,2-0,4 см,
с обеих сторон редкие волоски. Гипокотиль обратноконический, чет-
ко выражен главный корень с боковыми корнями второго порядка.
204
Рис.17. Возрастные состояния подорожника ланцетного
обозначения те же, что и на рис.15).
Ювенильные растения (см. рис.17) имеют главный розеточный по-
бег с 3-4 узколанцетными листьями, суженными в длинные черешки.
Листовая пластинка с 3 жилками. Семядоли отсутствуют. Хорошо вы-
ражен главный корень и боковые 2-3-го порядка.
Имматурные растения (см. рис.17) образуют главный розеточный
205
побег с ддинночерешковыми узколанцетными листьями с 3 жилками
(ювенильные) и ланцетными листьями с 5 жилками (взрослые). Общее
число листьев 3-4. Появляется небольшое вертикальное корневище до
0,7 см длины и 0,2 см ширины. От него отходят тонкие молодые при-
даточные корни. Одновременно продолжают функционировать главный и
боковые корни.
Взрослые виргинильные растения (см. рис.17) сохраняют главный
розеточный побег с 4-5 ланцетными длинночерешковыми листьями. По
форме и размерам листья типичные для взрослого растения, с 5 жил-
ками. Корневище небольшое (до I см), хорошо развиты молодые цри-
даточные корни, главный корень часто отмирает.
Молодые генеративные растения (см. рис.17) формируют в пазухах
3-5-го листа главного розеточного побега 1-2 монокарпических гене-
ративных побега (стрелки). Розеточный побег 1-го порядка образует
5-6 ланцетных листьев с 5 жилками. Корневище увеличивается в дли-
ну и толщину, корневая система образована главным образом молоды-
ми придаточными корнями.
Средневозрастные генеративные (см. рис.17) растения еще сохра-
няют розеточный полициклический побег 1-го порядка, на нем обра-
зуются 8-10 длинночерешковых ланцетных листьев с 5-7 жилками, од-
новременно возникают 3-5 боковых розеточных побега, каждый с 4-5
листьями.. 5-6 генеративных монокарпических побегов (стрелки) фор-
мируются как оси 2-3-го порядка на главном и боковых - более силь-
ных розеточных побегах. Корневище прочное, темное, косо направлен-
ное; благодаря деятельности контрактильных корней втягиваются в
почву и нижние междоузлия боковых побегов, формирующие боковые от-
ветвления. Хорошо видны остатки отмерших черешков листьев и кор-
ней. Корневая система состоит из старых и молодых придаточных кор-
ней.
Старые генеративные растения (см. рис.17) характеризуются нача-
лом разрушения и партикуляцией старого корневища, приводящей к об-
разованию клона из нескольких особей вегетативного происхождения
(от 2-3 до 10-12). Каждый розеточный побег имеет 4-5 ланцетных
листьев с 5-7 жилками. Пластинка листа несколько уменьшается. Ге-
неративных побегов 2-3. Наблюдается большое число отмерших листь-
ев и придаточных корней. Корневая система менее мощная.
Субсенильные растения (см. рис.17) - это особи вегетативного
происхождения или сохранившие жизнеспособность розеточные побеги
возобновления семенной особи. Корневище темное, старое, полураз-
рушенное с большим числом отмерших листьев. Розеточный побег воз-
206
обновления формирует 4-5 ланцетных листьев с 5-7 жилками, несколь-
ко меныпих размеров. Генеративных побегов не образуется. Сокраща-
ется число живых придаточных корней.
Сенильные растения (см. рис.17) представлены одиночными розе-
точными побегами, образующимися на полуразрушенных участках ста-
рых корневищ. Листья 2-3, напоминают листья ювенильного типа,
пластинки их узколанцетные с 3-5 жилками. Сохраняются 3-4 живых
придаточных корня.
Минимальная продолжительность жизни в культуре - 2 года, в ес-
тественных фитоценозах длительность жизни достигает 12 и более
лет.
Для подорожника ланцетного характерно появление всходов как
осенью, так и весной следующего года. Розеточные побеги зимуют с
зелеными .листьями. Максимальный рост листьев весной и в начале ле-
та. Генеративные побеги закладываются в пазухах 3-5-го листа розе-
точного побега, развивающихся в данном году, поэтому появлению
цветоносных побегов предшествует развертывание 3-5 листьев на по-
беге 1-го порядка. Цветение начинается в условиях Московской обл.
5-15 июня (Серебряков, 1952), заканчивается в июле, раньше, чем у
двух предыдущих видов подорожника. Молодые растения цветут позже,
чем старые.
Растения протерогиничны, ветроопыляемы (хотя часто посещаются
насекомыми в поисках пыльцы, но они не трогают рыльца). Продуктив-
ность пыльцы определена за 24 ч в 42.4.I06 пылинок на I г из рас-
чета, что на I м^ встречается в среднем 47 генеративных побегов с
517 открытыми цветками и в каждом пыльнике в среднем 20,5 тыс. пы-
линок (Hyde, Williams, 1945). Типичен амфимиксис. Вивипария не
отмечается. Семена созревают через 2-3 недели после оплодотворе-
ния.
Размножается как семенным, так и вегетативным способом. В осно-
вании соцветия 37% двусеменных коробочек и 63% односемянных, в се-
редине и на верхушке их соответственно 52 и 4Р% (Dowling, 1935);
Семена активно не распространяются, поэтому на следующий год на
растении можно увидеть коробочки с сохранившимися семенами на за-
сохших генеративных побегах. Перегнивание оснований побегов, ска-
шивание, перегрызание способствуют рассеиванию семян; \;СёмёНнад<;йо-л':
жура выделяет особое клейкое вещество (Кернер, 19О2).;.Поёдёй;тене,^;'.
ративные побеги подорожника ланцетного, животные активно содей-;?:;
ствуют его распространению - примерно половина семян способна,к
прорастанию после прохождения через пищеварительный тракт. -
207
Вегетативное размножение происходит при партикуляции старых
особей, что приводит к образованию клонов. В то же время извест-
на способность образовывать придаточные почки на искусственно от-
даленных и горизонтально посаженных участках корней, из которых
развиваются новые особи (Sagar, Harper, 1964). В естественных
фитоценозах это, по-видимому, не играет существенной роли, хотя
при сильном выпасе и отсутствии конкурентов вегетативное размно-
жение заменяет семенное. В теплице при полном срезании надземного
побега у молодых растений можно наблвдать развитие новых побегов
из придаточных почек на корнях.
Экология, фитоценология и консортивные связи
Подорожник ланцетный отличается довольно широкой экологической
амплитудой. К увлажнению малотребователен - встречается в услови-
ях от лугово-степного до влажнолугового увлажнения (47-72-я сту-
пени шкалы Л.Г.Раменского), по Э.Ландольту (Landolt, 1977), яв-
ляется индикатором средней сухости почв (2-я ступень шкалы). Г.Эл-
ленберг (Eiienberg, 1974) считает вид безразличным к увлажнению
почвы.
К освещенности менее требователен, чем два предыдущих вида
(б-я ступень шкалы освещенности Элленберга, 3-я ступень шкалы
Ландольта). Г.Элленберг помещает его между полутеневыми и полу-
световыми растениями. Увеличение продолжительности освещения и
непрерывное освещение может вызывать более раннее цветение. К вы-
соким температурам устойчив, низкая температура оказывает незна-
чительное угнетающее действие, немного задерживает цветение (Davi-
es, 1938).
К богатству почв относительно требователен, встречается на поч-
вах от среднебогатых до богатых (8-16-я ступени шкалы Л.Г.Рамен-
ского, 3-я ступень шкалы Ландольта). Предпочитает почвы слабокис-
лые (3-я ступень шкалы Ландольта), но может расти и на субстратах,
богатых известью (Brierley, 1956), встречается также и на солон-
цеватых и даже сильно солончаковых почвах (Раменский и др., 1956).
Произрастает на разнообразных по механическому составу почвах:
песчаных, супесчаных, сутопптиптн*, глинистых.
При сильном выпасе на лугах численность подорожника ланцетного
может уменьшаться, так как происходит постоянное повреждение
листьев розеточных побегов и скусывание цветочных стрелок. Однако
208
в районах с более сухим климатом численность этого вида при пасть-
бе может и увеличиваться (stapledon, 1925; Baker, 1937)» К ска-
шиванию достаточно устойчив.
Встречается во многих сообществах: в травяном ярусе дубовых и
грабовых лесов, в дюнных сообществах, в верещатниках, в луговых
степях, на субальпийских и альпийских лугах, особенно часто на
различных типах материковых и пойменных лугов. Постоянный вид мно-
гих открытых сообществ на залежах, на пустырях, около дорог, по
берегам рек и оросительных каналов. Обычен как в фитоценозах рав-
нин, так и в высокогорных районах. При посеве на лугах (25 кг се-
мян на I га) подорожник ланцетный дает урожай от 152 до 918 кг/га
(Sagar, Harper, 1964). Интенсивный выпас вызывает уменьшение био-
массы его корней на 72$ (Blom, 1979).
Микосимбиотроф. Известны поражения грибами-паразитами: в виде
желтых пустул на листьях (Synchytrium aureum Schroet.), порошко-
видного налета (Erysiphe communis (Wallr.) Link), пятен на
листьях живых (Ramularia plantaginea Sacc. et Berl.) или отмира-
ющих Septoria plantaginis (Ces.) Sacc. Иногда распространен ви-
рус табачной мозаики.
Хозяйственное значение
Подорожник - старинное лекарственное средство, которое было из-
вестно в древнегреческой, римской, арабской медицине. Используют
чаще всего листья подорожника большого. Они содержат гликозиды,
дубильные вещества, каротин, 20,7-42,2 мг.% аскорбиновой кислоты,
витамин К, манит, сорбит, лимонную кислоту. Лечебный препарат из
листьев "Плантоглюцид", а также сок из свежих листьев применяют
для лечения гастритов и язвенной болезни (Атлас ареалов..., 1976;
Губанов и др., 1976).
Свежие листья подорожников большого и ланцетного и приготовляе-
мые из них экстракты, настои, сок оказывают хорошее ранозаживляю-
щее, кровоостанавливающее и бактерицидное действие. Семена подо-
рожников, содержащие много слизи и жирных масел, рекомендуются
как противовоспалительное средство при кишечных заболеваниях, в
том числе дизентерии. Экстракт листьев оказывает успокаивающее и
снотворное действие, понижает артериальное давление. Настой листь-
ев применяют как отхаркивающее средство (Атлас ареалов..., 1976).
16-1964
209
М.Г.Вйхрамеёва
ЖИМОЛОСТЬ ЛЕСНАЯ
Номенклатура и внутривидовая систематика
Loaicera xyloateun L., 1753, Sp. pl.:174; Кауфм., 1866, Моск,
фл., изд.1:223; 1889, изд. 2:225; Цингер, 1886, Об. свеД.:210; .
Шмальг., 1897, Фл. 2:4; Сырейщ., 1910, Илл. фл. 3:203; он хе,.
1927, Опр. раст. Моск. губ.:242; Федченко, 1940, в Маевск., Фл.,
изд. 7:677; Пояркова, 1958, Фл. СССР 23:559; она хе, 1954, в Ма-
евск., Фл., изд. 8:537; изд. 9:519, 1964; Ворошилов, 1966, в Опр.
раст. Моск. обл.:306; Browics, 1976, Fl. Eur. 4:47; Пояркова,
1979, Фл. евр. части СССР 3:20. - Жимолость лесная или обыкновен-
ная (сем. жимолостные - Caprifoliaceae).
Жимолость лесная довольно изменчива по окраске и форме плодов
и листьев. А.Редер (Behder, 1927) различал несколько разновиднос-
тей: таг.тоШв Bagel - с густо опушенными листьями с обеих сто-
рон; var.glabrescenB Zabel - с листьями эллиптическими и продол-
говатыми, голыми; var.lutea Loisel. - с желтыми плодами и опушен-
ными листьями. Г.Н.Зайцев и В.В.Шульгина (1962). эти таксоны пере-
водят в ранг форм, добавляя к ним еще две: f.bretipedunculata Ba-
gel - с цветоносами почти одной длины с черешком (5-7 мм) и f.
longipedunculata Zabel - с цветоносами длинными (около 25 мм).
С.Шнейдер (Schneider, 1911) выделял две разновидности: var.typi-
са Hegel с 5 фермами (f.elliptioa Beck, f.thuringica Zabel, f.
caapacta Zabel, f.aontinifolia Mey., f.lutea loi.-Deev.) и таг.
leiophylla Behder.- П.Кириллов (Kirillow, 1842) выделял три раз-
новидности жимолости лесной в зависимости от окраски плодов: таг.
leiocarpa - с белыми, var.xanthocarpa - с желтыми, var.aelano-
сагра - с черными пледами. Б.Анцалоне (Anaalone, 1963) указыва-
ет на широкое распространение формы с черными плодами в пределах
Италии.
Географическое распространение
Ареал занимает почти всю территорию Европы и незначительную
часть Азии - Западную Сибирь (Hegi, 1918; Флора СССР, т.23,
210
19.58; Зайцев, Шульгина, 1962; Anzalone, 1963; Flora Europaea,
vol.4, 1976). На север далее всего заходит в Финляндии - 63°45*
сев.широты (Корреп, 1898).
В пределах СССР широко распространена на значительной террито-
рии европейской части. Северная граница ареала проходит от Архан-
гельска на восток через Северную Двину и Вычегду (по Архангель-
ской, Вологодской областям и Коми АССР), затем по р.Волоснице,
притоку Камы, и на с.Корепино (через Кировскую и Пермскую облас-
ти). Южная граница ареала в основном совпадает с. границей между
лесом и степью и идет от северной Молдавии на Каменец-Подольский,
затем поворачивает к северу на Новгород-Волынский и Новгород-Се-
верский, Харьков, проходит по югу Курской и Воронежской областей,
через северную часть Ростовской обл., далее отклоняется к северо-
востоку на Петровок и Хвалынск (Саратовская обл.), переходит че-
рез Волгу, далее по северной части Общего Сырта к Бузулуку на Уфу
и Златоуст. На восток жимолость лесная идет через Урал до Алтая.
Жимолость лесная растет и на Кавказе (Корреп, 1898; Флора СССР,
т.23, 1958; Зайцев, 1962). В Московской обл. обычна во всех райо-
нах.
Морфологическое описание
Кустарник I - 2(3)м высотой с отстоящими или немного дуговид-
но вниз отогнутыми темно-серыми или буровато-серыми ветвями. Мо-
лодые побеги тонкие, зеленые или красноватые, голые или густо опу-
шенные. Почки веретеновидные, отстоящие от стебля, с беловатыми
или желтовато-серыми чешуйками, покрыты длинными прижатыми волос-
ками, длиной от 0,5 до I мм. На вершине почки имеются наподобие
кисточки длинные беловатые волоски. Как правило, особенно на мощ-
ных побегах, в пазухах листьев развивается не одна, а несколько
(2-4, реже 6) почек, расположенных в вертикальный ряд. Наиболее
мощной является самая нижняя почка, из нее обычно развивается
цветоносный побег (Серебряков, 1952). У жимолости лесной нет чет-
кого разделения на цветоносные и вегетативные побеги, так как цве-
тоносные побеги одновременно являются и ассимилиругацими и сущест-
венно не отличаются от вегетативных. И.Г.Серебряков (1952) отме-
чает, что по характеру своей латеральной симметрии побеги жимолос-
ти относятся к дорзовентральным. Поскольку на разных сторонах оси
формируются неодинаковые листья, то побеги аниэофильны и при этом
211
эпитрофны (максимальное развитие листьев и пазушных почек проис-
ходит на верхней поверхности стебля). Цветоносные побеги распола-
гаются по всей длине материнского побега. Соцветия в пределах по-
бега приурочены только к нижней его части, верхняя остается веге-
тативной.
Листья 3-7 см длиной, 2-5 см шириной, большей частью широко-
эллиптические или яйцевидно-эллиптические, нередко с немного за-
остренной верхушкой или даже вытянутой в небольшое остроконечие,
реже продолговато-эллиптические, более длинно заостренные, с ок-
руглым, иногда слегка выемчатым, реже клиновидным основанием,
сверху матовые темно- или серовато-зеленые, снизу более светлые
или слегка сероватые или сизоватые, довольно густо (редко рассе-
янно) опушенные мягкими, согнутыми волосками, реже с обеих сторон
мягковолосистые, по краю реснитчатые. Черешок 3-8 мм длиной, обыч-
но густо опушенный (Флора СССР, т.23, 1958).
Цветоносы железистые, 8-14 (18) мм длиной. Прицветники линей-
ные или узколинейные, длинно заостренные, обычно длиннее завязей,
железистые и волосистые. Прицветнички от'округло-яйцевидных до
округлых, короче завязей, обычно свободные, изредка сросшиеся у
самого основания, снаружи усажены железками и иногда волосками,
а по краю с более или менее густыми щетинковидными ресничками,
чередующимися с железками.
Цветки зигоморфные, попарно расположенные на пазушных цветоно-
сах. Х.Кюглер (Kugler, 1970) считает, что цветки жимолости собра-
ны в простые дихазии с недоразвитым верхушечным цветком. Отгиб ча-
шечки цветка 5-зубчатый, около I мм длины, надрезан на широкие по
краю железистые зубцы. Венчик трубчатый, кверху расширенный с поч-
ти 2-губым 5-раздельным отгибом, сначала белый или желтовато-бе-
лый, позднее желтоватый, снаружи волосистый, широкий или почти ци-
линдрический, у основания с крупным округлым мешковидным выростом.
Верхняя губа надрезана обычно на 1/4 своей длины на широкие почти
округлые тупые лопасти; нижняя губа продолговатая, вниз отогнутая.
Тычинок 4-5, они немного короче венчика, с нитями опушенными лишь
при основании и голыми пыльниками, около 2 мм длины. Столбик не-
много короче тычинок, до верхушки густо волосистый. Завязь 2-3-
гнездная, обычно округло-яйцевидная, густо усаженная железками
(Флора СССР, т.23; 1958).
Плод - шаровидная ягода, 7-9 мм в диаметре, попарно срастающа-
яся основаниями, с 1-6 мелкими овальными сплюснутыми с боков семе-
нами, темно-красная, редко желтая. Б.Анцалоне (Anzalone, 1963)
212
указывает для Италии форму жимолости с черными ягодами. Семена
красновато-фиолетовые с розоватым оттенком, 3-5 мм длины, 0,6-
0,8 мм толщины, изогнутые, с небольшим продольным валиком.
Диплоидный набор хромосом равен 18 (Garcke, 1972).
Онтогенез, сезонное развитие
и способы размножения
Семена жимолости мелкие, масса I тыс. семян от 3 до 6,4 мг
(Мисник, 1949; Зайцев, 1962; Нестерович и др., 1967; Качалов,
1970). Пустых семян до 10% (Мисник, 1949; Заборовский, 1962).
Всхожесть семян сохраняется недолго: через 6 мес после созрева-
ния - 93%, через I год - 68, через 3 года - 3, через 5 лет - 0%
(Зайцев, 1962), но по другим данным удовлетворительная всхожесть
сохраняется до 2 лет (Хохлов, Ковалева, 1950; Заборовский, 1962).
Семенная кожура задерживает прорастание семян. При весеннем посе-
ве желательна стратификация. В целях ускорения прорастания семена
выдерживают 2-3 мес во влажном песке при температуре 2-5° (Забо-
ровский, 1962). При удалении части семенной кожуры у места распо-
ложения корешка зародыша семена начинают прорастать на 10-е сут,
а "заканчивают" прорастание на 30-35-е сут (Заборовский, 1962).
Семена, посеянные весной без стратификации, запаздывают с прорас-
танием. Семена, посеянные в середине сентября, хорошо всходят на
следующую весну (Заборовский, 1962). В лабораторных условиях се-
мена жимолости прорастали через 46 дней после посева, первые на-
стоящие листья появились через 84 дня (Зайцев, J962).
Всходы жимолости лесной, по И.Т.Васильченко (I960), имеют широ-
коовальные семядоли 7 мм длины и 5 мм ширины, на верхушке закруг-
ленные и слегка выемчатые, почти сидячие с тремя продольными жил-
ками. Подсемядольная часть буровато-зеленая, около 20 мм длины,
0,8-1 мм ширины. Первые листья широкоовальные, почти округлые, на
очень коротких черешках, сверху зеленые, снизу более бледные, по
краю белореснитчатые. Верхняя сторона листьев несет обильные мел-
кие бородавочки и редкие волоски, нижняя сторона с рассеянными
волосками. Последующие листья постепенно становятся вытянутыми и
более сильно опушены с обеих сторон, черешки их также несколько
удлинены по сравнению с черешками первых листьев. Надсемядольное
междоузлие, как и все последующие междоузлия, покрыто прижатыми
курчавыми волосками.
213
1бЧ964
В первые годы жизни в условиях подмосковных лесов (смешанные
елово-березовые и елово-широколиственные леса) со степенью сомкну-
тости крон 0,6-0,8 (относительная освещенность 8-17%) жимолость
растет довольно медленно. В 3 года она имеет высоту 27-35 см, ди-
аметр ствола около 4 мм, в 5 лет высоту от 5. до 70 см, диаметр 6-
8 см. До 3-4 лет отмечается мононодиальное нарастание надземной
части, затем симподиальное. Корневая система в первые 2-3 года
жизни имеет стержневой характер с хорошо выраженным главным кор-
нем. Но позже в связи с тем, что для жимолости лесной характерно
полегание и укоренение ветвей, главную рожь начинают играть при-
даточные корни, основная масса которых расположена на глубине 4-
10 см. Зацветает жимолость к 10 годам. К этому возрасту она до-
стигает в высоту примерно I м, формируются новые скелетные оси.
Годичные приросты колеблются очень значительно - от 6 до 12
(20) см, однако в благоприятных условиях освещения (например, в
"окнах") может достигать величины 30-40 (45) см. По нашим наблю-
дениям, -взрослые экземпляры жимолости могут иметь до 2-2,5 м в вы-
соту и 4-8 осей. Границы особи определить трудно в связи с укоре-
нением ветвей, от которых на- значительном расстоянии отходят орто-
тропные побеги, связь их с материнской особью устанавливается
лишь раскапыванием.
Особенности морфогенеза побегов жимолости лесной были изуче-
ны на территории Главного Ботанического сада АН СССР М.Т.Мазурен-
ко (1972). Побег формирования жимолости не превышает 150-180 см
в длину и имеет не более 18-20 узлов. В узлах закладываются 3
(редко 6) сериальных почек. На второй год продолжается, как пра-
вило, моноподиальное нарастание побега. Если верхушечная почка от-
мирает, то из нижней почки серии развиваются акротоннои мезотон-
но побеги ветвления 1-го порядка. На третий год либо продолжается
моноподиальное нарастание побега, либо его верхушечная почка отми-
рает. Прирост третьего года по размерам и структуре походит на по-
беги ветвления первого года. Также моноподиально нарастают побеги
ветвления. Для них характерна изменчивость размеров. В зависимос-
ти от мощности побега они могут нести от I до 6 пар двуцветников
в разных узлах. Наиболее характерны побеги 10-15 см с 2-3 соцве-
тиями в нижней части и 4-5 узлами без цветков в верхней.
На третий год малого цикла иэ второй или верхней почки серии
на побеге формирования развиваются побеги дополнения длиной в
среднем 15 см и с 2-3 узлами с двуцветниками. Верхняя часть побе-
гов без двуцветников имеет 1-5 узлов. Листья в этих узлах крупнее,
214
чем в узлах с двуцветниками. В пазухах листьев закладываются по
2 (иногда 3) сериальные почки. На четвертый год симподиальное
ветвление иногда продолжается, но побеги ветвления 3-го порядка
укорочены до 1-3 см и в основном без двуцветников. Моноподиальное
нарастание иногда наблюдается, но на четвертый год побеги были
слабые и по строению напоминали симподиальные третьего года раз-
вития. Цикл развития на четвертый год, за редким исключением, за-
канчивается, и начинают развиваться побеги суммирования следующе-
го порядка.
По данным Б.В.Гроздова (1964), жимолость лесная растет быстро,
в первые 2 года достигая 50-60 см, к 5 годам - 1,2 м. Плодоноше-
ние наступает в 3-5-летнем возрасте, но, по всей вероятности, эти
данные относятся к культуре.
Побеги трогаются в рост и начинают развертывать листья в конце
апреля - начале мая (в Главном Ботаническом саду АН СССР 20 апре-
ля - Стогова, 1968). Средняя дата зацветания за 12 лет для Подмос-
ковья - 22 мая, ранняя - 10 мая, поздняя - I июня (Шиманюк, 1964).
Средняя продолжительность массового цветения в средней полосе -
8 дней, весь период цветения - 12 дней.
Созревание плодов происходит в августе, висят плоды до октября.
В тенистых местах плоды созревают на 1-2 недели позже, чем на ос-
вещенных. Нередко во второй половине августа наблюдаются вторич-
ное цветение и новый прирост жимолости, но плоды обычно не завя-
зываются (Зайцев, 1962). В Подмосковье листья начинают желтеть во
второй половине сентября, опадают в конце сентября - начале октяб-
ря. Процесс заложения и внутрипочечное развитие цветка жимолости
в основном сходен с калиной и другими жимолостями (Артюшенко,
1951).
Жимолость лесная размножается как семенным, так и вегетативным
способом. В лесу, видимо, чаще встречаются особи семенного проис-
хождения. Урожайность плодов значительно колеблется. В Одинцов-
ском р-не Московской обл. в ельнике кислично-зеленомошном на пер-
вой террасе р.Москвы в благоприятных условиях максимальная урожай-
ность составляет 676 тыс. плодов и средняя 375 тыс. плодов на
I га. В ельнике-зеленомошнике на водоразделе в том же районе в ус-
ловиях более бедной почвы и низкой освещенности урожайность всего
5-8 тыс. плодов на I га. В березняке с елью разнотравно-орляковом
там же урожайность плодов колебалась от 37 до 107 тыс. плодов на
I га. Масса 100 плодов в благоприятных условиях 35 (26-43) г, в
ельнике-кисличнике 22 г (18,7-27,7). Плоды распространяют птицы й
215
грызуны на значительное расстояние (Левина, 1957).
На тенистых участках жимолость размножается преимущественно
вегетативно, при этом размножение происходит обычно благодаря по-
леганию и укоренению ветвей, а не образованию корневищ (Хохряков,
Мазуренко, I960). В искусственных условиях жимолость разводится
как семенным путем (осенние посевы), так и зелеными черенками,
корневыми отпрысками и отводками. При рубке возобновляется по-
рослью (Хохлов, Ковалева, 1950; Гроздов, 1964).
Экология и фитоценология
Жимолость лесная - мезофит, растет в условиях среднего увлаж-
нения (Garcke, 1972) 60-77-я ступени увлажнения по шкале Л.Г.Ра-
менского, 3-я ступень по шкале Ландольта (Landolt, 1977), 5-я
ступень по шкале Элленберга (Eiienberg, 1974). Высокой засухоус-
тойчивостью не отличается, при затоплении гибнет (Зайцев, 1962).
Теневынослива. Г.Элленберг (1974) относит ее к полутеневым рас-
тениям (5-я ступень шкалы освещенности), чаще встречается при 10%
относительной освещенности. В лесах Московской обл. обычно встре-
чается в условиях относительной освещенности до 20%, но не менее
5%; плодоносит чаще в условиях относительной освещенности более
15%. Нередко растет на открытых местах (Черкезишвили, 1952).
К почвам не особенно требовательна, растет на подзолистых, дер-
ново-подзолистых, серых лесных почвах, однако предпочитает доволь-
но богатые почвы (6-13-я ступень шкалы Л.Г.Раменского) со средним
содержанием азота (3-я ступень шкалы Ландольта, 2-я ступень шкалы
Элленберга) и со средним содержанием гумуса (3-я ступень шкалы
Ландольта). Растет обычно на слабокислых и основных почвах (pH
водное 4,6-7,5), но переносит и очень кислые (Харитонович, 1975).
В условиях Подмосковья встречается на почвах с широким диапазоном
pH водного - от 3,95 до 6,05. В Западной Европе лесная жимолость
часто растет на почвах, богатых известью (Garcke, 1972).
Морозоустойчива, даже в самые суровые зимы не страдает от моро-
зов (Андронов, 1953), газоустойчива и дымоустойчива (Галактионов
и др., 1967; Харитонович, 1975). В горы поднимается до 2000 м (He-
gi, 1918).
Жимолость лесная - весьма распространенный вид в подлеске как
хвойных, так и смешанных и широколиственных лесов. На плодородной
почве нередко доминирует в подлеске. На европейской части СССР
216
растет в подлеске еловых лесов, в азиатской части встречается в
елово-пихтово-кедровых лесах, но, как правило, единичными экзем-
плярами. Широко распространена в широколиственных лесах средней
и восточной части Русской равнины, где является одним из наиболее
холодостойких кустарников. В Заволжье и Предуралье растет как в
чистых дубравах, так и в дубово-липовых, липовых и кленовых лесах.
В смешанных широколиственно-еловых лесах (с дубом, липой, ильмом)
жимолость - один из типичных представителей подлеска наряду с ле-
щиной и бересклетом, растет также по склонам оврагов, холмов, на
опушках. Иногда пышно разрастается на вырубках (Гроздов, 1964).
В Московской обл. жимолость встречается повсеместно в еловых,
сосновых, смешанных, широколиственных лесах. В благоприятных ус-
ловиях образует довольно густые заросли (до 2 тыс. экземпляров на
I га), обычно же ее численность не превышает 100-200 особей на
I га.
Консортивные связи
Жимолость лесная обычно опыляется пчелами. Ее плоды поедают
рябчик, славки, певчий дрозд, рыжая полевка, желтогорлая мышь
(Левина, 1957). Жимолость повреждают многочисленные беспозвоноч-
ные, преимущественно насекомые. Листья поедают 28 видов, в том
числе гусеницы различных пядениц, совок, пилильщиков, шпанская
мушка, долгоносик, несколько видов молей и пр. Многие из них име-
ют видовое название "жимолостный", что свидетельствует об их тес-
ной связи с представителями рода жимолость. Побеги и почки повреж-
дают 5 видов: жимолостная узкотелая златка, жимолостный галловый
пилильщик, жимолостная побеговая •веерница, виноградная листоверт-
ка, двенадцатипалая веерница обыкновенная. Цветки повреждают 3 ви-
да галлицы: жимолостная, бузиновая, черноногая. Плоды поедает гу-
сеница виноградной листовёртки (Цусев, Римский-Корсаков, 1951).
Г.Н.Зайцев (1962) отмечает на жимолости лесной в условиях Ленин-
града 36 видов насекомых и 19 видов паразитических грибов.
Представители рода жимолость, в том числе и лесная, повреждают-
ся различными грибами. Пятнистость листьев жимолости вызывают 8ер-
toria xylostei Sacc. et Wint., S.obscurata Th., Phylloaticta cap-
rifolii Sacc., P.vulgarie Desmaz., Ramularia lonicerae Vogl.,
Cercospora periclymeni Wint., мучнистую росу листьев - Micro-
sphgg^*a lonicerae (DC. ex St.-Am.) Wint., Phyllactinia guttata
16 -1964 217
(Wallr. ex fr.) Lev., засыхание ветвей - Diplodia deflectens
Kara., D.asooohytula Sacc., Camarosporium xylostei Sacc., Rhabdo-
spora xylostei Laub, et Fautr., R.lonicerae Sacc., Phoma crypt1-
ca Sacc., гниль ствола и побегов - Phellinus conehatus (Fr.)
Quel, f.lonicerae (Weinm.) Bond., f.lonicerinus (Bond.) Bond, et
Sing (Ванин и др., 1950). Указаны также следующие грибы, встре-
ченные на жимолости лесной: Leptothyrium periclymeni var.xylo-
stei Pas. И Kabatia mirabilis Bubal (Conners, 1959).
Хозяйственное значение
Лесная жимолость декоративна, разводится в садах и парках, от-
личается также хорошими почвозащитными свойствами и используется
при закреплении склонов оврагов и оползней, для живых изгородей.
Хорошо переносит стрижку (Дроздов, 1964; Шиманюк, 1967). Древеси-
на обладает большой прочностью, твердостью (латинское название
"xylosteum" от "кость-дерево"), употребляется на сапожные гвоз-
ди, ружейные шомпола, кнутовища и прочее. Плоды ядовиты, облада-
ют слабительным и рвотным действием. Эссенция из свежих зрелых
ягод применяется в гомеопатии. Медонос, охотно посещается пчела-
ми. Плодц W1 на изготовление розовой и фиолетовой краски (Хох-
лов, Ковалева, 1950; Зайцев, Шульгина, 1962).
218
ЛИТЕРАТУРА
А б о л и н ь А.А. Влияние антропогенных факторов на динами-
ку болот. - В кн.: Генезис и .динамика болот, выл.2. М., Изд-во
Моск, ун-та, 1978.
А б р а ж к о В.И. Водный режим подроста и механизмы конку-
ренции из-за почвенной влаги в биогеоценозах южной тайги. Авто-
реф. дис. на соиск. учен, степени канд. биол. наук. Л., 1968.
Авдошенко А.К. Семенное размножение брусничных. -
Докл. АН СССР, 1948, т.60, № 5.
Алейникова М.М., Порфирьев В.С., Ут-
ро б и н а Н.М. Парцеллярная структура елово-широколиственных
лесов востока европейской части СССР. М.: Наука, 1979.
Александров Ю.В. Новый вид для флоры Московской
области рябчик шахматный (Fritillaria meleagris). - Биол. науки,
1971, Я 6.
Александрова М.С., Лапин П.И., Пет-
рова И.П. и др. Древесные растения парков Подмосковья. М.:
Наука, 1979.
Алексахин Р.М. О влиянии липы на лесорастительные
свойства почвы. - Тр. Воронежского заповедника, 1961; вып.ХЗ.
Алексахина Т.Н. Группировка почвенных водорослей в
коренном и производном типах леса. - В кн.: Леса Восточного Под-
московья. М., Наука, 1979.
Алексеев Ю.Е., Губанов И.А. Флора окрестнос-
тей Пущина-на-Оке. М., Изд-во Моск, ун-та, 1980.
Андронов Н.М. Зимостойкость деревьев и кустарников в
Ленинграде. - Тр. Ботанического ин-та АН СССР* 1953, оер.6, вып.З.
Анненков Н.И. Ботанический словарь. Спб., 1878.
Арискина Н.П. К характеристике важнейших ценозообра-
зователей сплавин. - Учен. зап. Казанского ун-та, 1950, т.ПО,
выл.4.
Артюшенко З.Т. Развитие цветка и плода у жимолост-
ных. - Тр. Ботанического ин-та АН СССР, 1951, сер.7, вып.2.
Артюшенко З.Т., Соколов С.Я. Формирование
почек и развитие годичных побегов у некоторых древесных пород. -
Тр. Ботанического ин-та АН СССР, 1955, сер.6, вып.4.
Аскеров А.М., Бобров А.Е. Папоротники Талыша. -
Ботанический журн., 1972, т.57, № II.
219
Атлас ареалов и ресурсов лекарственных растений СССР. М.: ГУГК
СССР, 1976.
А ф р и к я н С.В. Лядвенцы Армянской ССР и их биолого-эколо-
гическая и кормовая характеристика. Автореф. дис. на соиск. учен,
степени канд. биол. наук. Ереван, 1957.
Барабанецкий Г.Г. О химической природе биологичес-
кой активности водорастворимых веществ листового опада ясеня и ли-
пы. - В кн.: Физиолого-биохимические основы взаимодействия расте-
ний в фитоценозах, вып.4. Киев, 1973а.
Барабанецкий Г.Г. О фитоценотической роли колиноз
корней липы мелколистной. - В кн.: Физиолого-биохимические основы
взаимодействия растений в фитоценозах, вып.4. Киев, 19736.
Баранова И.И., Смирнова Л.М., Ершо-
ва Г.Ф. Содержание химических веществ у ягод черники и голуби-
ки в различных местообитаниях. - В кн.: Ресурсы ягодных и лекарст-
венных растений и методы их изучения. Петрозаводск, 1975.
Бибикова Ю.А. и др. Охраняемые растения Налибокской
рощи. - Вести. Белорусского ун-JTa, 1978, сер.2, ft I.
Б л у к е т Н.А. Некоторые данные о наличии крахмала в орга-
нах папоротникообразных. - Докл. ТСХА, 1964, вып.92, ч.2.
Богданова Г.А., Муратов Ю.М. Брусника в ле-
сах Сибири. Новосибирск, 1978.
Б о л ‘о т о в А.Т. О рублении, поправлении и заведении ле-
сов. - Тр. Вольного экон, общества, 1766, ч.4, 5.
Борисова М.А. Побегообразование некоторых видов зла-
ков Стрелецкой степи Курской области. - Учен. зап. Моск. гор. пед.
ин-та, I960, т.57, вып.4.
Борисова-Гуленкова М.А. Способы отрастания
побегов после скашивания. - Учен. зап. Моск. гор. пед. ин-та,
1959, т.50.
Б р и е д е Г.Р. Результаты интродукции папоротников в Бота-
ническом саду ЛГУ им.П.Стучки за 1968-1970 гг. - Тр. Ботаническо-
го сада Латвийского ун-та, 1972, т.18.
Бурков В.Н. К методике.листовой диагностики азотного,
фосфорного и калийного питания липы мелколистной. - Агрохимия,
1966, ft 10.
Б у р к-0 в В.Н. Валовое содержание азота, фосфора и калия в
органах липы мелколистной. - Лесоведение, 1967, № 5.
Бурк о в В.Н. Валовое содержание питательных веществ в
листьях саженцев липы мелколистной как показатель их питания и
220
роста. - Агрохимия, 1968, » 4.
Бурмистров М.С. Выращивание голубики в США. - Садо-
водство, 1968, № 6.
Быков Б.А. Доминанты растительного покрова Советского Со-
юза, т.1, 2, 3. Алма-Ата, I960, 1962, 1965.
Б я з р о в Л.Г., Д ы л и с Н.В., Жукова В.М. и
др. Основные типы широколиственно-еловых лесов и их производных
Малинского лесничества Краснопахорского лесхоза Московской обл. -
В кн.: Биогеоценологические исследования в широколиственно-еловых
лесах. М.: Наука, 1971.
Бяллович Ю.П. Шкала устойчивости древесных и кустарни-
ковых пород к затоплению. - Ботанический журн., 1957, т.42, № 5.
Вакуров А.Д., Надеждин В.В. Сосновые леса
Подмосковья, их площади, динамика и существующие формы хозяйствен-
ного освоения. - В кн.: Сложные боры хвойно-широколиственных ле-
сов. М.: Наука, 1968.
Ванин С.И., Журавлев И.И., Соколов Д.В.
Определитель болезней древесных пород и кустарников, применяемых
для полезащитных насаждений. М.-Л.: Гослесбумиздат, 1950.
Василевич И. В. Исследование структуры лугового сооб-
щества с помощью межвидовых сопряженностей. - Проблемы ботаники,
1969, т.П.
Василевская В.К., Шилова Н.В. Особенности
строения листовых органов Pyrolaceae bindl. и их значение для по-
бегообразования. - Вести. Ленингр. ун-та, I960, № 3, сер. биол.,
вып.1.
Васильев И.В. Семейство липовых. - В кн.: Деревья и
кустарники. М.-Л.: Изд-во АН СССР, 1958.
Васильев Б.Р., Антонова И.С. Некоторые зако-
номерности морфологического строения годичного побега. - Вести.
Ленингр. ун-та, 1979, сер. биол., вып.2.
Васильченко И.Т. Всходы деревьев и кустарников. М.,
Л.: Изд-во АН СССР, I960.
Васильченко И.Т. Определитель всходов сорных расте-
ний. Л.: Колос, 1965.
Вержуцкий Б.Н. Пилильщики, вредящие голубике. - Защи-
та растений, 1970, № 3.
Верзилин Р.П. По следам Робинзона. М., Детгиз, 1953.
Волкова В.Г. Изучение способов клонирования тимофеевки
луговой и ежи сборной. - В кн.: Сборник трудов аспирантов и моло-
дых науч, сотрудников Всесоюз. ин-та растениеводства, вып.2. Л.,
1961.
Воронцова Л.И., Григорьева Н.М., Его-
рова В.Н. и др. Растения-задернитёли при освоении эродирован-
ных склонов. - В кн.: Тезисы докладов научно-производственного се-
минара по залужению склонов. Курск, 1976.
Ворошилов В.Н. Флора Советского Дальнего Востока.
М.: Наука, 1966.
Ворошилов В.Н., Скворцов А.К., Тихо-
миров ^,Н. Определитель растений Московской области. М.: На-
ука, 1966.
By А.В. Когда растут корни липы? - Цветоводство, 1960а, > 4.
By А.В. Залегание и рост корней у липы мелколистной в город-
ских условиях. - В кн.: Озеленение городов, вып.5. М., 19606.
В у А.В. Исследование особенностей роста корневой системы ли-
пы мелколистной в зависимости от условий произрастания в город-
ских насаждениях. Автореф. дис. на соиск. учен, степени канд. би-
ол. наук. М., 1961.
By А.В. Особенности роста корневой системы липы мелколист-
ной на асфальтированных улицах. - Науч. тр. Акад, коммун, хоз-ва,
1962, внп.19.
Вульф Е.В., Малеева О.Ф. Мировые ресурсы полез-
ных растений. Л.: Наука, 1969.
Галактионов И.И., В у А.В., Осин В.А. Де-
коративная дендрология. М.: Высшая школа, 1967.
Гладкова Л.И. Выращивание низкорослой голубики в США.-
Достижения науки и передовой опыт в сельском хозяйстве, 1972,
сер.1, й 9.
Гладкова Л.И. Выращивание высокорослой голубики в
США. - Достижения науки и передовой опыт в сельском хозяйстве,
1974, сер.1, > 9.
Годунов И.А. Местная дикорастущая тимофеевка луговая
как исходный материал для селекции на устойчивость к линейной
ржавчине. - Тр. по прикл. ботанике, генетике и селекции, 1971,
т.43, вып.З.
Годунов И.А. Характеристика состава популяций дикорас-
тущей люцерны серпообразной и тимофеевки луговой Северного Кавка-
за. - Науч. тр. Майкопской опыт, станции, вып.5. Краснодар, 1972.
Голубев В.Н. Основы биоморфологии травянистых растений
центральной лесостепи. - Тр. Центр.-Черноземы, заповедника, 1962,
222
вып.7.
Голубев В.Н. Эколого-биологические особенности травя-
нистых растений и растительных сообществ лесостепи. М.: Наука,
1965.
Голубев Н.П. Тимофеевка. М., Сельхозгиз, 1931.
Голубева И.В. Некоторые данные о запасе живых оемян в
почвах под лугово-степной растительностью. - Бюл. МОИН. Отд. биол.,
1962, т.67, вып.5.
Голубева И.В. Возрастной состав и динамика численнос-
ти популяций некоторых видов растений в разных условиях луговой
степи. Автореф. дис. на соиск. учен, степени канд. биол. наук. М.,
1966.
Голубинцева В.П., Лебедев П.В. Определи-
тель кормовых злаков и бобовых в нецветущем состоянии. Изд. 2-е.
М., Учпедгиз, 1959.
Гордиенко М.И. Взаимодействие ясеня обыкновенного и
липы мелколистной. - Науч. тр. Укр. сельскохоз. акад., 1972,
вып.64.
Гордиенко М.И. Взаимодействие дуба черешчатого и ли-
пы мелколистной. -Науч. тр. Укр. сельскохоз. акад., 1973, вып.94.
Гордиенко М.И. Лесоводственная оценка липы мелколист-
ной, клена остролистного и граба обыкновенного. - Лесоведение,
1979, № I.
Городков Б.Н. Растительность Арктики и горных тундр
СССР. - В кн.: Растительность СССР, т.1. М.-Л., Изд-во АН СССР,
1938.
Г р и ч у к В.П., Моносзон М.Х. Определитель одно-
лучевых спор папоротников из семейства Polypodiaoeae R.Br., про-
израстающих на территории СССР. М.: Наука, 1971.
Г р о з д о в Б.В. Дендрология. М.-Л.: Гослесбумиздат, 1952.
Г р о з д о в Б.В. Лесные травы, их индикаторное, кормовое и
лекарственное значение. М.: Гослесбумиздат, 1963.
Г р о з д о в Б.В. Декоративные кустарники. М«: Стройиздат,
1964.
Гросогейм А.А. Флора Кавказа, т.2, 5. Изд. 2-е. М.-
Л.: Изд-во АН СССР, 1940, 1952.
Грохольская В.А. Использование липы в полезащитных
насаждениях и озеленении поселков. М.-Л.: Гослесбумиздат, 1950.
Губанов И.А., Крылова И.Л., Тихоне-
в а В.Л. Дикорастущие полезные растения СССР. М.: Мысль, 1976.
223
Туленкова М.А. Побегообразование и формирование жиз-
ненной формы у лядвенца рогатого в предгенеративный период. - В
кн.: Возрастной состав популяций цветковых растений в связи с их
онтогенезом. М.: изд. МГПИ им.В.И.Ленина, 1974.
Гусев В.И., Римский-Корсаков М.Н. Оп-
ределитель повреждений лесных и декоративных деревьев и кустарни-
ков европейской части СССР. Л.: Гослесбумиздат, 1951.
Гусева А.Н. Роль грибных заболеваний в процессе возоб-
новления липы и сосны в сложных борах Подмосковья. - В кн.: Леса
Подмосковья. М.: Наука, 1965.
Давлиниадзе М.Ф. Морфология спор кавказских пред-
ставителей рода Dryopterie Adane. - Зап. по систем, и геогр.
растений АН ГрузССР, 1965, выл.24.
Данилов М.Д., Кудрявцева М.В., Мар-
тыненко В.В. Белоплодная голубика в Марийской АССР. -
Растительные ресурсы, 1975, т.П, внп.2.
Двораковская В.М. Ритм развития всходов и ювениль-
ных растений некоторых видов рода Fritiiiaria. - Биол. науки,
1973а, № 8.
Двораковская В.М. О прорастании семян Fritiiia-
ria lutea Mill. И Fritiiiaria ruthenica Wiketr. - Вол. ГБС AH
СССР, 19736, выл.89.
Двораковский M.C., Алтухов М.Д. Сравни-
тельная характеристика семенного возобновления мелколистной липы
(Tilia cordata Mill.) в различных экологических условиях. -
Вести. Моск, ун-та. Сер. биол. и почв., 1963, №5.
Де Бари Н. Сравнительная анатомия вегетативных органов
явнобрачных и папоротникообразных растений. Спб., 1877.
Дедов А.А., Лащенкова А.Н., Толма-
чев А.И. Сем. Polypodiaceae. - В кн.: Флора северо-востока
европейской части СССР, т.1. Л.: Наука, 1974.
Денисова Г.М. Побегообразование и ритм сезонного раз-
вития луговых растений низовий Северной Двины. - Учен. зап. Моск,
гор. пед. ин-та, I960, т.57, вып.4.
Денисова Г.М. К биологии побегообразования у некото-
рых бобовых растений. - Бюл. ГБС АН СССР, 1963, выл.51.
Д и д у с ь В.И. Заболевание тимофеевки склероцинием. - Се-
лекция и семеноводство, 1947, №2.
Дмитриева Е.И., Игловиков В.Т., Ко-
ню ш к о в Н.С., Раменская В.М. Растения сенокосов
224
и пастбищ. М.: Колос, 1974.
Доброхотов В.Щ Семена сорных растений. М.: Сель-
хозгиз, 1961.
Добрынин Г.М. Рост и формирование хлебных и кормовых
злаков. Л.: Колос, 1969.
Долгошов В.И. Некоторые особенности цветения липы
мелколиственной. - Пчеловодство, 1957, № 7.
Долуханов А.Г. Папоротниковые бучины Кавказа. - Тр.
МОИП. Отд. биол., I960, т.З.
Д ы л и с Н.В. Парцеллярная структура лесных биогеоценозов и
ее лесоводственное значение. - В кн.: Ботаника. Исследования,
вып.10. Минск, 1968.
Д ы л и с Н.В., Носова Л.М. Фитомасса лесных биогео-
ценозов Подмосковья. М.: Наука, 1977.
Егорова С.В. Микрофлора почв коренного и производных
типов леса. - В кн.: Леса Восточного Подмосковья. М.: Наука,
1979.
Елагин И.Н. Сезонное развитие сосновых лесов. Новоси-
бирск, 1976.
Еленевский А.Г. Определитель растений Павловской
слободы по вегетативным органам. М.: изд. МГПИ им.В.И.Ленина,
1969.
Ермоленко В.М. Рогохвости та пильщики. - В кн.: Фау-
на Украины, т.Ю, вып.2. Киев, 1972.
Ершов М.Ф. О фотосинтезе частых и запыленных листьев ли-
пы мелколистной и вяза мелколистного. - Докл. АН СССР, 1957,
т.112, № 6.
Жуйкова Н.В. О некоторых особенностях роста и развития
видов рода Vaccinium в условиях Хибинских гор. - Ботанический
журн., 1959, т.44, № 3.
Жуков А.М. Патогенные грибы на растениях сем. Vaccinia-
сеае в южной Сибири. - В кн.: Природные комплексы низших расте-
ний Западной Сибири. Новосибирск, 1977.
Заблоцкая Л.В. Материалы по экологии основных видов
мышевидных грызунов Приокско-Террасного заповедника, 1957а, т.1.
Заблоцкая Л.В. Растаскивание семян хвойных и липы
землеройками-бурозубками. - Тр. Приокско-Террасного заповедника,
19576, т.1.
Заборовский Е.П. Плоды и семена древесных и кустар-
никовых пород. М.: Гослесбумиздат, 1962.
225
Зазоев М.Ф. Лядвенец рогатый. - Тр. Краснодарской селек-
ционной станции, 1937, выл.2.
Зайцев Т.Н. Интродукция жимолости в Ленинграде. - Тр.
Ботанического ин-та АН СССР, 1962, сер.6, выл.8.
Зайцев Г.Н., Шульгина В.В. Род Lonicera Ь. -
В кн.: Деревья и кустарники СССР, т.6. М.-Л.: Изд-во АН СССР,
1962.
Заклинская Е.Д. Материалы к изучению состава совре-
менной растительности и ее спорово-пыльцевых спектров для целей
биостратиграфии четвертичных отложений. - Тр. Ин-та геол, наук АН
СССР, 1951, выл.127.
Зозулин Г.М. Поземные части основных видов травянистых
растений и ассоциаций плакоров среднерусской лесостепи в связи с
вопросами формирования растительного покрова. - Тр. Центр.-Черно-
земы. заповедника, вып.5., Курск, 1959.
Зорина Л.М. Вредители тимофеевки в Ленинградской обл. -
В кн.: Сб. работ Ин-та прикладной зоологии и фитологии, вып.З. Л.,
1955.
Иванов В.В. Находки папоротников на крайнем юго-востоке
европейской части СССР. - Биол. науки, 1972, № 6.
Игошина К.Н. Растительность субальп Среднего Урала. -
Тр. Ботанического ин-та АН СССР, 1952, сер.З, вып.8.
Игошина К.Н. Флора горных и равнинных тундр и редколе-
сий Урала. - В кн.: Растения севера Сибири и Дальнего Востока.
М.-Л.: Наука, 1966.
Ильина Е.А. Морфологическая природа и возрастные изме-
нения зоны кущения люцерны синегибридной. - Ботанический журн.,
1964, т.49, № 5.
И м с А.Д. Морфология цветковых растений. М.: Мир, 1964.
Каверзнева Ю.Г. О некоторых особенностях биологии и
морфогенеза подмосковных видов грушанковых. Автореф. дис. на со-
иск. учен, степени канд. биол. наук. М., 1955.
Каден Н.Н., Терентьева Н.Н. Этимологический
словарь латинских названий растений, встречающихся в окрестностях
агробиостанции .’ЛГУ "Чашниково". М.: Изд-во Моск, ун-та, 1975.
Казаифарова В.К., Гасанов А.М. Сравнитель-
ная морфологическая характеристика Plantago media и P.atepposa. -
Изв. АН АзССР, 1979, сер.биол., J6 I.
Казакова Г.И. О микоризе липы мелколистной. - В кн.:
Лесная геоботаника и биология древесных растений. Тула, 1978.
226
Каляев А.И. Фенологические наблюдения над древесными,
кустарниковыми, травянистыми растениями и шляпочными грибами. -
Тр. Приокско-Террасного заповедника, 1971, вып.5.
Каменецкая И.В. Влияние метеорологических условий
на семенное возобновление травянистых растений Стрелецкой степи.-
Вол. МОИП. Отд. биол., 1949, т.54, вып.4.
Каменецкая И.В. Семенное возобновление растительнос-
ти северной степи. Автореф. на соиск. учен, степени канд. биол.
наук. Л., 1952.
Карандина С.Н. Корневые системы древесных пород ши-
роколиственных лесов лесостепи. - Учен. зап. Ленингр. ун-та,
1950, вып.25.
Карнишина Л.М. Особенности развития и морфологии га-
метофитов и юных спорофитов представителей рода Dryopteris Adana,
s.l. - В кн.: Природные процессы и ресурсы Подмосковья. М.: Про-
свещение, 1975.
Карпов В.Г. Экспериментальная фитоценология темнохвой-
ной тайги. Л.: Наука, 1969.
Карташова Н.Н. Строение и функции нектарников цветка
двудольных растений. Томск, 1965.
Кац Н.Я. О фитоценозах Кавказского госзаповедника в свете
комбицативных свойств видов. - Землеведение, 1936, т.38, вып.З.
Качалов А.А. Деревья и кустарники. М.: Лесная промыш-
ленность, 1970.
Кашкарова В.П. Опыт оценки степени светолюбия видов
травянистого яруса с помощью методики Л.Г.Раменского. - Бюл. МОИП.
Отд. биол., 1972, т.77, вып.З.
Кернер фон Марилаун А. Жизнь растений,
т.1, 2. Спб., 1902.
Киршин И. К. О вегетативной пролификации у многолетних
злаков. - Ботанический журн., 1963, т.48, № I.
Кирьянова Е.С., Краль Э.Л. Паразитические не-
матоды растений и меры борьбы с ними, т.1. Л.: Наука, 1969.
Киселев Г.Е. Цветоводство. М.: Сельхозгиз, 1964.
Клапп Э. Сенокосы и пастбища. М.: Сельхозгиз, 1961.
Климов Л.А., Черкасов А.Ф., Горбино-
в а Л.И. Сравнительная характеристика водного режима некоторых
растений сем. Vacciniaceae в условиях Костромской обл. - Сб. на-
уч. работ Костромского пед. ин-та, 1973, вып.ЗО.
Клоков М.В. Заметка о некоторых критических видах ляд-
.227
венца. - Ботанические мат-лы Гербария БИН АН СССР, 1953, т.15.
Клоков М.В. Новые украинские мотыльниковые. - Ботани-
ческие мат-лы Гербария БИН АН СССР, 1961, т.21.
Клинген И.Н. Кормовые растения и польза от них, ч.2.
Изд. 2-е. Спб., I9II.
Кожевников Д.А. К истории развития цветка в семей-
стве Агасеае. - Бюл. МОИН, 1877, Т.52.
Колесниченко М.В., Спадов Ю.М. Влияние фи-
тоцидов вяза обыкновенного и липы мелколиственной на интенсивность
фосфорного питания дуба летнего. - Зап. Воронежского сельскохоз.
ин-та, 1970, т.45.
Коннов А.А. О нахождении папоротника Dryopteris filix-
maa (L.) Schott, на западном Памире. - Изв. АН ТадаССР, отд. би-
ол., 1971, № 3.
Коновалов Н.А. Типы леса подмосковных опытных лесни-
честв Щ10С. - Тр. по лесному опытному делу, 1929, вып.5.
Копелькиевский Г.В. Уточнить нормы и методы оп-
ределения нектаропродуктивности растений. - Пчеловодство, 1954,
№ II.
Корецкая Л. Б. Особенности вертикально-фракционного
распределения зольных элементов у липы мелколистной. - Лесоведе-
ние, 1970, '№ 5.
Коркинский С.И. Флора востока европейской России.
Томск, 1892.
Корнеева Т.М. Анализ населения птиц северо-восточной
части лесопаркового пояса Мооквы. - В кн.: Леса восточного Под-
московья. М.: Наука, 1979.
Корякина В.Ф. Влияние продолжительности хранения се-
мян луговых трав на их всхожесть. - В кн.: Обзор новой литературы,
вып.7. Вологда, 1930.
Корякина В.Ф. Особенности роста и развития многолет-
них кормовых растений. М.-Л.: Наука, 1964.
Крайнев В.П. Роль липы в ходе естественного возобнов-
ления ели. - Лесное хозяйство, 1939, № 9.
Кралль Э., Крал ль X. Семенные галлы тимофеевки
степной, вызванные нематодой. - В кн.: Фаунистические заметки,
т.1, № 4-5. Тарту, 1971.
Красная книга СССР. Редкие и находящиеся под угрозой исчезнове-
ния виды животных и растений. М.: Лесная промышленность, 1978.
Крашенинников И.М., Кучеровская-
228
Р ой а н е ц С.Е. Растительность Башкирской АССР. М.-Л.; Изд-
во АН СССР, 1941.
Крот Л.А. О взаимовлиянии дуба, липы и клена через их
корневые выделения. - В кн.: Фитоценология. исслед. в Белоруссии.
Минск, 1972.
Кротова Н.Г. Опыт выращивания смешанных насаждений
сосны с липой в Лесной опытной даче ТСХА. - Докл. ТСХА, 1964,
вып.99.
Крылов П.Н. Флора Западной Сибири, т.1-3. ''Томск, 1928-
1929.
Крылова Н.П. Закономерности семенного размножения и
устойчивости бобовых растений природных заливных лугов. Автореф.
дис. на соиск. учен, степени канд. биол. наук. М., 1964.
Крюгер Л.В. Микоризы травянистых растений естественных
фитоценозов центрального/Цредуралья. Автореф. дис. на соиск. учен,
степени канд. биол. н^укЛЛермь, 1959.
Крюгер Л. В. ''’Эндотрофная микориза травянистых растений
некоторых фитоценозов центрального Предуралья. - Биологический
журн., 1961а, т.46, № 5.
Крюгер Л.В. Динамика микоризообразования у злаков. -
Агробиология, I96I6, № 2.
Кулагин Ю.З. Древесные растения и промышленная среда.
М.: Наука, 1974.
Культиасов И.М., Минина О.И. О парцеллярном
строении ельника с липой и березой волосистоосокового в Подмос-
ковье. - Вести. Моск, ун-та. Себ. биол., почвов., 1971, № 6.
Куперман Ф.М., Р ж а н о в а Е.И. Биология разви-
тия растений. М.: Высшая школа, 1963.
Куприянов И.М. Внимание лядвенцу. - Сов. агрономия,
1939, й I0-II.
Куркин К.А. Летний и многолетний покой травянистых рас-
тений многолетников Барабинской лесостепи. - Ботанический журн.,
1971, т.56, й II.
Куркин К.А., Б о б н е в а А.П. Изменение биологичес-
кой активности почв пойменных лугов под воздействием различных при-
емов их окультуривания. - Экология, 1978, № I.
Курлович Л.Е., Алексеев Ю.Е. Новая находка
Fritiiiaria meleagria в Московской области. - Вести. Моск, ун-та.
Сер. биол. и почвов., 1975, № 2.
К у р н а е в С.Ф. Роль липы в лесах Московской области. - В
229
кн»: днт реконструкции малоценных лесов Московской области. М.:
Гослесбумиздат, 1955.
К у р н а е в С.Ф. Основные типы леса средней части Русской
равнины. М.: Наука, 1968.
К у р н а е в С.Ф. Теневые широколиственные леса Русской рав-
нины и Урала. М.: Наука, 1980.
Кутафьева Т.Н. Морфология спор плаунов, хвощей и па-
поротников Средней Сибири. - Ботанический журн., 1972, т.58, № 10.
Ларин И.В. Луговодство и пастбищное хозяйство. М.: Сель-
хозгиз, 1956.
Ларин И.В., Анабабян Ш.М., Работ-
нов Т.А. и др. Кормовые растения сенокосов и пастбищ СССР,
т.1-3. М.-Л.: Сельхозгиз, 1950, 1951, 1956.
Лебедев П.В. Морфогенез луговых злаков и условия внеш-
ней средн. Автореф. на соиск. учен, степени канд. биол..наук,
Пермь, 1966.
Лебедев П.В. > Особенности Проявления морфогенеза луго-
вых злаков в зависимости от факторов внешней среды. - Учен. зап.
Уральск, ун-та, № 73, сер. биол., вып.4. Свердловск, 1968.
Лебедев П.В., Мельник Н.С., Серая Г.П.
и др. Морфогенез побегов луговых злаков и условия внешней среды. -
Зап. Свердловского отд. ВБО, 1966, внп.4.
Лебедев П.В., Углов Н.П. Биология и агротехни-
ка лугопДстбищных трав. Свердловск, 1961.
Левина Р.Е. Способы распространения плодов и семян. М.:
Изд-во Моск, ун-та, 1957.
Л е й с л е Ф.Ф. Влияние условий прохождения световой стадии
на изменение морфологических признаков у некоторых растений. -
Труды Ботанического ин-та АН СССР, 1956, сер.4, вып.2.
Литвинов М.К. Естественное размножение липы в услови-
ях Брянского лесного массива. - В кн.: Сб. аспирантских статей
Брянского ЛГИ, 1957, S 2.
Лобанов Н.В. Микотрофность древесных растений. М.: Нау-
ка, 1953.
Лобанов Н.В., Трошина Т.В. Динамика отношения
поверхности активных корней к поверхности листьев у некоторых по-
род. - В кн.: Лесная геоботаника и биология древесных пород. Ту-
ла, 1978.
Лучник З.И. Кормовые растения южноуссурийской тайги. -
Труды горнотаежной станции, т.2. Владивосток, 1938.
• 230
Любч-енко В.М. Прорастание зародышей липы мелколистной
и бересклета бородавчатого в зависимости от степени зрелости се-
мян. - Лесной зурн., 1959, № 3.
Любченко В.М. Прорастание зародышей липы и берескле-
та в связи со стратификацией. - Ботанический журн., I960, т.45,,
» I.
Любченко В.М. Роль зародыша в состоянии покоя семян
Euonymue verrucosa и Tilia cordate. - В кн.: Ботаника. Исследо^
вания, вып.5. Мийск, 1963.
Люкшинский В.В., Прижуков Ф.Б. Семеновод-
ство луговых кормовых. М.: Колос, 1969.
Маевский П.Ф. Флора средней полосы европейской части
СССР. Изд. 7-9. М., Л.: Сельхозгиз, 1940, 1954, 1964.
Мазуренко М.Т. Некоторые особенности морфогенеза
трех видов жимолости. - Бюл. ГБС АН СССР, 1972, вып.83.
Майсурян Н.А., Атабекова А.И. Определи-
тель семян и плодов. М.: Колос, 1978.
Мальцев А.И. Сорные растения. М., Сельхозгиз, 1933.
Матвеев А.Л. Большой жизненный цикл тимофеевки луго-
вой. т- Бкм. МОИП. Отд. биол., 1972, т.77, № 3.
Матвеев А.Р. Большой жизненный цикл, численность и
возрастной состав популяций тимофеевки луговой и тимофеевки степ-
ной. Автореф. дио. на соиск. учен, степени канд. биол. наук. М.,
1975.
Матвеев А.Р., Шуман Т.П. Изменчивость в. большой
и малом жизненных циклах анатомической структуры корней тимофеев-
ки луговой. - В кн.: Возрастной состав популяций цветковых расте-
ний в связи с их онтогенезом. М.: Изд-во Моск. гос. пед. ин-та
им.В.И.Ленина, 1974.
Матвеева В.П. Фитоценологическая классификация лугов
и травянистых болот советской Прибалтики. - Тр. Ботанического ин-
та АН СССР, I960, сер.З, вып.12.
Матвеева В.П. Фитоиндикаторы общего комплекса условий
существования, частных особенностей среды обитания и отдельных
почвенных горизонтов и теоретические вопросы фитоиндикации. Л.:
Наука, 1971.
Машек А.А. Изменение фауны вредителей тимофеевки разного
года пользования в чистом пооеве и в травосмесях. - Зап. Ленингр.
сельскохоз. ин-та, 1967, т.Ш.
Машек А.А. Вредители злаковых трав. Л.: Колос, 1977.
231
Мельник Н.С. Особенности органогенеза луговых злаков
ярового типа развития в связи с факторами внешней среды. Автореф.
дис. на соиск. учен, степени канд. биол. наук. Свердловск, 1969.
Меркулова В.А. Очерки по русской народной номенкла-
туре растений. М.: Наука, 1967.
Мин и.н а И.Т. Рациональная техника посева и нормы высева
лугопастбйщных травосмесей в различных природных условиях лесной
зоны. - Вопросы кормодобывания, вып.2. М.: Сельхозгиз, 1949.
Миняев Н.А. К систематике лядвенца северо-запада евро-
пейской части СССР. - Ботанические мат-лы Гербария БИН АН СССР,
1957, т.18.
Миняев Н.А., Улле З.Г. О видах лядвенца, встреча-
ющихся на северо-востоке европейской части СССР. - Новости систе-
матики высш, раст., 1977, т.14.
Мисник Г.Е. Производственная характеристика семян де-
ревьев и кустарников городских насаждений. М.: Изд-во Минкомунхо-
за РСФСР, 1949.
Михайловская И. С. Возрастные анатомические струк-
туры стержневых корней некоторых видов Leguminosae. - Ботаничес-
кий журн., I960, т.45, № 6.
Михайловская И. С. Изменение анатомической струк-
туры корней некоторых злаков в связи с их старением. - Бюл. МОИП.
Отд. биол., 1965, т.70, вып.З,
Михайловская И.С. О возрасте особей популяции Lo-
tus corniculatus L. - Ботанический журн., 1967, т.52, № 3.
Михайловская И.С. Разнообразие корней стержнекор-
невых бобовых и их,анатомическая структура. - Биол. науки, 1968,
№ 12.
Михайловская И.С. Возрастные анатомо-морфологи-
ческие изменения подземных органов желтой люцерны. - Бюл. МОИП.
Отд. биол., 1972, т.77, вып.1.
Михалева В.М. О биологии брусники и голубики в Якут-
ской АССР. - В кн.: Почвенные и ботанич. исслед. в Якутии. Якутск,
1972.
Молоток Г.П., В у А.В., Синюхин А. Адапта-
ция липы мелколистной в условиях Москвы. - Бюл. ГБС АН СССР,
1970, вып.76.
Молукало И.С. Две формы липы мелколистной. - Лесное
хозяйство, 1950, № II.
Моляков Л.И. Вологодская тимофеевка (сеянка). - В кн.:
232
Материалы по организации и культуре кормовой площади, вып.15. М.,
1916.
Монахов А. Липы - богатство России. - Пчеловодство,
1969, № 6.
Морозов Г.Ф. Учение о лесе. Изд. 7-е. М., Л.: Гослес-
бумиздат,. 1949.
Морозова З.В. Долголетние бобовые травы в луговых
травосмесях в нечерноземной полосе. Автореф. дис. на соиск. учен,
степени канд. сельскохоз. наук. М., 1955.
Морохин Л.И. Ход развития годичного бокового побега
липы (Tilia cordata Mill.) по наблюдениям 1924 и 1925 гг. - В
кн.: Сб. науч, статей Казанск. ин-та сельского хоз-ва и лесовод-
ства, вып.З и 4. Казань, 1926.
Мурахтанов Е.С. Основы организации комплексного хо-
зяйства в липняках Средней Волги. Л.: Изд-во Ленингр. ун-та,
1972.
Мурахтанов Е.С. Пчеловодство в липняках. М.: Лесная
промышленность, 1977.
Мухина Н.А. Изучение лядвенца рогатого различного гео-
графического происхождения в условиях Ленинградской обл. Автореф.
дис. на соиск. учен, степени канд. сельскохоз. наук. Л., 1956.
Напалков Н.В. Плодоношение древесно-кустарниковых по-
род и семенное хозяйство в лесах Среднего Поволжья. - Труды по
леей, хоз-ву Татарской ДОС, выл.10. Казань, 1952.
Наумова В.Е. Строение корневой системы липы в условиях
Средней Сибири. - Бюл. ГБС АН СССР, 1972, вып.85.
Науялис И. И. Вертикальная структура кустов Athyrium
filix-femina (L.) Roth. - Бюл. МОИП. Отд. биол., 1976, т.81, № 6.
Науялис И.И. 0 крупнопапоротниковых микрогруппировках
хвойно-широколиственных лесов Подмосковья. - Вестн. Моск, ун-та.
Сер. б,иол., 1977, № I.
Науялис И. И. Некоторые черты экологии проростков и юве-
нильных растений Athyrium filix-femina (L.) Roth и Dryopteris
filix-mas (L.) Schott в хвойно-широколиственных лесах Московской
области. - Бюл. МОИП. Отд. биол., 1979, т.84, вып.4.
Некрасова В.Л. Агасеае. - В кн.: Флора БССР, т.1.
М.: Сельхозгиз, 1949.
Немчинов А.А. Геоботаническая и агрохимическая харак-
теристика болот Ленинградской области. Л., 1934.
Нестерович Н.Д., Чекалинская Н.И.,
17-1964 233
Сироткин Ю.Д. Плоды и семена лиственных древесных расте-
ний. Минск, 1967.
Нейштадт М.И. История лесов и палеогеография СССР в
голоцене. М.: Изд-во АН СССР, 1957.
Ненароков М.И., Раменская В.М. Лядвенец
рогатый. - В кн.: Многолетние травы в лугопастбищных севооборо-
тах. М.: Сельхозгиз, 1951.
Никитин С.А. Типы лесов Серебряноборского опытного
лесничества. - Тр. Лаб. лесоведения АН СССР, 1961, т.2, вып.1.
Николь окая Л.Г., Соколов В.Н. О влиянии
корневых остатков некоторых многолетних трав на всхожесть семян
и рост тимофеевки луговой. - Тр. по прикл. ботанике, генетике и
селекции, 1966, т.38, № I.
Носова Л.М. Особенности вертикального распределения фи-
томассы липы разного возраста в лесных биогеоценозах. - Бюл. МОИП.
Отд. биол., 1970, т.75, вып.З.
Носова Л.М. Флоро-географический анализ северной степи
европейской части СССР. М.: Наука, 1973.
Овеснов А.М. Морфология и экология прорастания семян
дикорастущих злаков. Автореф. дис. на соиск. учен, степени докт.
биол. наук. Пермь, 1965.
Овеснов А.М. Покой семян дикорастущих злаков и пути
его устранения. - ’Учен. зап. Пермск. ун-та, 1966, т.130.
Оловянникова И.Н. Экология и ритм цветения расте-
ний Стрелецкой степи. - В кн.: Научно-метод. зап. Главн. упр. по
заповедникам, выл.12. М., 1949.
Определитель насекомых европейской части СССР, т.1. М., Л.:
Наука, 1964.
Орлов А.Я. Буковые леса Северо-Западного Кавказа. - В
кн.: Широколиственные леса Западного Кавказа. М., 1953.
О с е ц к и й С.И. Значение различных органов запаса для от-
растания многолетних злаков после дефолиации. - Бюл. МОИП. Отд.
биол., 1969, т.74, Jfc I.
П а й п ер Ч.В. Многолетние кормовые травы и их культура.
М.-Л.: Сельхозгиз, 1930.
Панферов В.П. Всхожесть семян дикорастущих трав и вли-
яние различных стимуляторов на их прорастание. - Труды Новочерк.
зоовет. ин-та, выл.5. Ростов н/Д, 1939.
П а т а р а я Л.М. Хозяйственное значение папоротников Аджа-
рии. - Учен. зап. Моск. обл. пед. ин-та, 1968, вып.181.
234
Пер е л ь Т.С. Комплексы почвенных беспозвоночных в неко-
торых типах леса Серебряноборского опытного лесничества. - В кн.:
Стационарные биогеоценотические исследования в южной подзоне тай-
ги. М.: Наука, 1964.
Петрова А.Д., Сизова Т.П., М и р ч и н к Т.Г.,
Покровская И.В. Изучение пищевой специализации клещей
Acotyledon obsolovii (Acariformes, Acarodae) в связи с их массо-
вым размножением в лабораторных культурах термитов. - Зоол. журн.,
1979, т.58, № 8.
Писковацкова Н.П. Онтогенез, возрастной состав
ценопопуляции и нектаропродуктивность колокольчика сборного и
лядвенца рогатого. Автореф. дис. на соиск. учен, степени канд.
биол. наук. М., 1976.
Погорелова Л.Д., Работнов Т.А. Опыт опре-
деления возраста особей Dryopteris filix-mas (L.) Schott. - Рас-
тит, ресурсы, 1978, т.14, вып.4.
Покровская В.М. Материалы по морфологии проростков
и всходов растений северных степей. - Труды Центр.-Черноземы, за-
поведника, I960, вып.6.
Полезные растения СССР, т.1. М., Л.: Изд-во АН СССР, 1951.
Положенцев П.А., Шемякин И.Я., Коро-
вина Н.И. 0 вредителях плодов ильмовых пород и липы. - На-
уч. зап. Воронежского лесохоз. ин-та, 1952, вып.4.
Поляков П.П. Растительность Джунгарского Алатау в рай-
оне рек Саркона и Баскана. - В кн.: Растительность Казахстана.
М., Л.: Изд-во АН СССР, 1941, т.2.
Полянская О.С., Корнилов М.Р., Трут-
нев А.Г. Почвенно-геоботанические исследования на территории
колхозов Крестецкого района Ленинградской области. - Труды Бота-
нического ин-та АН СССР, 1935, сер.З, вып.2.
Пономарев А.Н. Цветение и опыление злаков. - Учен,
зап. Пермск. ун-та, 1964, т.П4.
Пономарев А.Н., Верещагина В.А. Антэко-
логический очерк темнохвойного леса. - В кн.: Проблемы биогеоце-
нологии, геоботаники и ботанической географии. Л.: Наука, 1973.
Пономарев А.Н., Трубачева Т.П. Взрывча-
тое и порционное цветение злаков. - Докл. АН СССР, 1962, т.146,
£ 6.
Пономарева Е.Г. Лядвенец рогатый - ценная кормовая
культура и хороший медонос. - Пчеловодство, 1954, Л» 8.
235
Попов М.Г. Флора средней Сибири. М.-Л.: Изд-во АН СССР,
1957.
Порфирьев В. С. К синэкологии страусника обыкновенно-
го, индикатора почвенно-грунтовых вод. - В кн.: Водный обмен в
основных типах растительности СССР как элемент круговорота ве-
ществ и энергии. Новосибирск, 1975.
Проз оровский Н.А. Наблюдения над осенним и зим-
ним состоянием степных растений. - Труды Центр.-Черноземы. запо-
ведника, вып.1, М., 1940а.
Проз оровский Н.А. Изменение растительности Стре-
лецкой степи при отсутствии пастьбы скота и сенокошения. - Труды
Центр.-Черноземы, заповедника, вып.1. М., 19406.
Путятин М.Д. Многолетние кормовые травы на осушенных
низинных болотах. М.: Сельхозгиз, 1939.
Работнов Т.А. Жизненный цикл многолетних травянистых
растений в луговых ценозах. - Тр. Ботанического ин-та АН СССР,
1950, сер.З, выл.6.
Работнов Т.А. Определение возрастного состава популя-
ций видов в сообществе. - В кн.: Полевая геоботаника, т.З. М.-Л.:
Изд-во АН СССР, 1964.
Работнов Т.А. Луговедение. М.: Изд-во Моск, ун-та,
1974.
Раевский В. Растения Нижегородской губернии. Ниж. Нов-
город, 1884.
Раменский Л.Г., Цаценкин И.А., Чижи-
ков О.Н., Антипин Н.А. Экологическая оценка кормовых
угодий по растительному покрову. М.: Сельхозгиз, 1956.
Рахтеенко И.Н. Сезонный цикл роста активных корней
древесных пород. - Лесное хоз-во, I960, № 9.
Ремезов Н.П., Быкова Л.Н., Смирно-
в а К.М. Потребление и круговорот азота и зольных элементов в
лесах европейской части СССР. М.: Изд-во Моск, ун-та, 1959.
Ремезов Н.П., Погре бняк Н.С. Лесное почво-
ведение. М.-: Лесная промышленность, 1965.
Р ж а н о в а Е.И. Об изменчивости признаков тимофеевки луго-
вой в зависимости от световых условий. - Агробиология, 1951а, № 5.
Ржанова Е.И. Формирование генеративных органов у тимо-
феевки луговой и зависимости от продолжительности освещения и ка-
чества света. - Докл. АН СССР, I95I6, т.80, № 6.
? ж а н о в Е.И. Биологические основы культуры многолетних
236
злаков. М.: Изд-во Моск, ун-та, 1957.
Родионов В.А. Лядвенец рогатый - ценная многолетняя
бобовая трава. - Селекция и семеноводство, 1952, № 9.
Розанова М.А. Обзор литературы по родам Vaccinium L.
и Oxycoccus Adans. - Тр. по прикл. ботанике, генетике и селек-
ции, 1934, сер.8, № 2.
Роллов А.Х. Дикорастущие растения Кавказа, их распро-
странение, свойства и применение. Тифлис, 1908.
Ростовцев С. И. Пособие к определению паразйтных гри-
бов по растениям-хозяевам. М., 1908.
Ротов Р.А. К экологии рябчиков (Fritiiiaria L.) евро-
пейской части СССР. - Бюл. ГБС АН СССР, 1972а, вып.84.
Ротов Р.А. Некоторые итоги интродукции видов рода Fri-
tiiiaria s.l. - Бюл. ГБС АН СССР, 19726, вып.86.
Ротов Р.А. Перспективы интродукции отечественных, видов
подтрибы Fritiliariinae сем. лилейных. - В кн.: Успехи интродук-
ции растений. М.: Наука, 1973а.
Ротов Р.А. Рябчик шахматный (Fritiiiaria meleagris L.)
в Московской области. - Бюл. ГБС АН СССР, 19736, выл.90.
Ротов Р.А. Ценотические популяции видов Fritiiiaria ев-
ропейской части СССР как источники интродукции. - В кн.: Ботани-
ко-географические районы СССР. М.: Наука, 1974.
Ротов Р.А. Морфолого-биологические особенности лукович-
ных эфемероидов на примере рода Fritiiiaria L. - Тр. МОИП. Отд.
биол., 1976, т.42.
Р ы с и н Л.П. Световой режим в некоторых хвойных и листвен-
ных типах леса. - В кн.: Стационарные биогеоценотические исследо-
вания в южной подзоне тайги. М.: Наука, 1964.
Р ы с и н Л.П. Взаимоотношения сосны и широколиственных по-
род в лесах хвойно-широколиственной подзоны. - В кн.: Сложные бо-
ры хвойно-широколиственных лесов. М.: Наука, 1968.
Р ы с и н Л.П. Сложные боры Подмосковья. М.: Наука, 1969.
Р ы с и н Л.П. Типы леса Серебряноборского опытного лесниче-
ства. - В кн.: Природа Серебряноборского лесничества. М.: Наука,
1974.
Р ы с и н Л.П. Типы леса Восточного Подмосковья. - В кн.: Ле-
са Восточного Подмосковья.. М.: Наука, 1979.
Рябкова Л. С. Строение периспориев и экзоспория спор па-
поротников Таджикистана. - В кн.: Морфология пыльцы и спор совре-
менных растений. Л.: Наука, 1973.
17х-Х964 237
Рябкова Л.С. Морфология спор папоротников Таджикиста-
на. - В кн.: Растительность Таджикистана и ее освоение. Душанбе,
1974.
Рябчинский А.Е. Типы леса и естественного возобнов-
ления липы в БАССР. - Труда Башкирской ЛОС, 1964, выл.7.
Самойлова Е.М. Корневые системы липы в Воронежском
заповеднике. - Биол. науки, 1965, № 3.
Самойлова Е.М. Динамика разложения опада лиственных
пород. - Труды Воронежского заповедника, выл.13. Воронеж, 1961.
Селиванов И.А. Микосимбиотризм как форма консортив-
ных связей в растительном покрове Советского Союза. Автореф. дас.
на соиск. учен, степени докт. биол. наук. М., 1976.
Селиванов И.А., Андросимова Л.И.,
Любченко Т.М. Материалы и характеристика микориз субаль-
пийских растений Кавказа. - Учен. зап. Пермск. ун-та, 1962, т.22,
Ji 4.
Селиванов И.А., Верещагина В.А.,
Мельникова С.Л., Саламатова Н.Г. Материа-
лы к изучению микориз некоторых злаков, произрастаищих в степном
поясе. - Учен. зап. Пермск. ун-та, 1961, т.18, № 3.
Селиванов И.А., Калина Р.Г., Пашко-
ва А.П. и др. Материалы к инвентаризации микотрофных растений
склонов Эльбруса. - Учен. зап. Пермск. ун-та, 1966, т.130.
Селиванов И.А., Утемова Л.Д. Материалы ха-
рактеристики микориз злаков. - Учен. зап. Пермск. пед. ин-та,
1968, т.64.
Семенова В.Г. Влияние рубок главного пользования на
почвы и круговорот веществ в лесу. М.: Лесная промышленность,
1975.
Семенова В.Г., Смирнов Б.В. Круговорот азота
и зольных алементов в липняке волосистоосоковом. - В кн.: Лесовод-
ственные исследования в Серебряноборском опытном лесничестве. М.:
Наука, 1973.
Серебряков И.Г. 0 ритме сезонного развития растений
подмосковных лесов. - Вести. Моск, ун-та, 1947, > 5.
Серебряков И.Г. Морфология вегетативных органов
высших растений. М.: Сов. наука, 1952.
Серебряков И.Г. Типы развития побегов у травянистых
многолетнййов и факторы их формирования. - Учен. зап. Моек. гор.
пед. ин-та, 1959, т.100, вып.5.
238
Серебряков И.Г. О распространении корневых систем
некоторых арктических растений. - Учен. зап. Моск. гор. пед. ин-
та, I960, т.57, вып.4.
Серебряков И.Г., Галицкая Т.М. К биологии
сезонного развития болотннх растений Подмосковья в связи с усло-
виями их жизни и происхождением. - Учен. зап. Моск. гор. пед. ин-
та, 1951, т.19, вып.4.
Серебряков И.Г., Чернышева М.Б. О морфо-
генезе жизненной формы кустарничка у черники, брусники и некото-
рых болотных Ericaceae. - Бал. МОИН. Отд. биол., 1955, т.60,
вып.2.
Серебрякова Т.И. Побегообразование и ритм сезонно-
го развития растений заливных лугов Средней Оки. - Учен. зап.
Моск. гор. пед. ин-та, 1956, т.97, вып.З.
Серебрякова Т.И. Морфогенез вегетативных органов
и эволюция жизненных форм в семействе злаков. Автореф. дис. на
соиск. учен, степени докт. биол. наук. М., 1967.
Серебрякова Т.И. Морфогенез побегов и эволюция
жизненных форм злаков. М.: Наука, 1971а.
Серебрякова Т.И. Типы большого жизненного цикла и
структура наземных побегов у цветковых растений. ~.Бюд. МОИН.
Отд. биол., I97I6, вып.1.
Симонов С.Н. Результаты освоения новых видов дикорасту-
щих трав в культуре. - В кн.: Вопросы производства кормов, вып.З.
М.: Сельхозгиз, 1951.
С и н с к а я В.Н. Динамика вида. М.-Л.: Сельхозгиз, 1948.
С и н с к а я Е.Н., Шебалкина М.Л. Селекция кор-
мовых культур. М.: Сельхозгиз, 1936.
С и н с к а я Е.Н., Щ е н к о в а М.К. К вопросу о поли-
морфизме некоторых видов Vaccinium 1>. - Труды по прикл. ботани-
ке, генетике и селекции, 1928, т.18, вып.4.
Сипливинский В.Н. Заметки о байкальской флоре. -
Новости сист. высш, раст., т.11. Л.: Наука, 1974.
Скворцов А.К. О распространении элементов окской фло-
ры в южных районах Московской обл. и в соседних районах Тульской
и Калужской обл. - В кн.: Растительность и почвы Нечерноземного
Центра европейской части СССР. М.: Изд-во Моск, ун-та, 1969.
С л а д к о в А.Н. Морфологическое описание пыльцы грушанко-
вых, вертляницевых, вересковых, брусничных и ворониковых европей-
ской части Союза ССР. -Труда Ин-та географии АН СССР, 1954, т.61.
239
Скрябина А.А. Цветение и плодоношение Vaccinium uti-
ginosum. - Растит, ресурсы, 1970, т.6, вып.2.
Смарагдова Н.П. Клевер и пчелы в нечерноземной по-
лосе. М.: Изд-во Моск, ун-та, 1969.
Смелов С.П. Биологические основы луговодства. М.: Сель-
хозгиз, 1947.
Смелов С.П. Теоретические основы луговодства. М.: Ко-
лос, 1966.
Смелов С.П., Работнов Т.А. Материалы к изуче-
нию реакции луговых почв и распределению в связи с ней луговой
растительности. - Изв. Гос. лугового ин-та, 1928, > 1-3.
Смелов С.П., Татаринова Н.К. Связь между
побегами куста луговых злаковых и мульчирование. - Бюл. науч,-
техн. информ., 1958, № 5.
Сметанникова А.И. Сравнительное эколого-физиоло-
гическое изучение многолетних трав в чистом и смешанном посевах,-
Тр. Ботанического ин-та АН СССР, 1950, сер.4, вып.7, 8.
Смирнов В.В., Семенова В.Г. Вес надземных
частей деревьев в липняке волосистоосоковом. - Лесохоз. информа-
ция, 1968, вып.18.
Смирнов В.В., Семенова В.Г. Продуктивность
древостоев подзоны широколиственно-еловых лесов. - Растит, ресур-
сы, 1970,‘т.6, вып.2.
Смирнов П.А. Флора Приокско-Террасного гос. заповедни-
ка. - Тр. Приокско-Террасного заповедника, вып.2. М., 1958.
Смирнова К.М. Потребление и круговорот элементов пи-
тания в липняках Мордовского государственного заповедника. -
Вести. Моск, ун-та, 1952, № 6.
Соколов С.Я. Некоторые ценные древесные и кустарнико-
вые породы северной части Черноморского побережья Кавказа. - Тр.
Ботанического ин-та АН СССР, 1952, сер.5, вып.З.
Соколов П.А. Липняки Марийской АССР - кормовая база
пчеловодства. - В кн.: Сб. трудов Марийского политехи, ин-та,
1972, внп.З.
Соколов П.А. Учет фитомассы крон древостоев липы. -
Лесоведение, 1975, № 5.
С о к о лов П.А. Состояние и теоретические основы формиро-
вания липняков. Йошкар-Ола, 1978.
Соколова О.И. 0 всхожести семян дикорастущих кормовых
трав. - Соц. растениеводство, 1934, сер.А, й II.
240
Солдатов А.Г. Корневые системы древесных пород. Киев,
1955.
Солоневич Н.Г. Материалы к зколого-биологической ха-
рактеристике болотных трав и кустарников. - В кн.: Растительность
Крайнего Севера и ее освоение, вып.2. М.-Л.: Изд-во АН СССР,
1956.
Солоневич Н.Г. Экология и биология растений восточ-
ноевропейской лесотундры. - В кн.: Растительность Крайнего Севера
СССР и ее освоение, вып.10. Л.: Наука, 1970.
Сорные растения СССР, т.1. Л.: Изд-во АН СССР, 1934.
Ср я а в а В.Б., Семенова-Тяншан-
с к а я А.М. Широколиственные леса. - В кн.: Растительный по-
кров СССР, т.1. М.-Л.: Изд-во АН СССР, 1956.
С с о р и н В.А. Пихтовые леса Верхней Сванетии. - Ботани-
ческий журн., 1951, т.36, № 5. _
Стеценко Н.М. Интродукция папоротников в Ботаническом
саду Киевского государственного университета. - Бюл. ГБС АН СССР,
1979, вып.112.
Стогова Н.В. Сезонный ритм интродуцированных видов жи-
молости. - Бюл. ГБС АН СССР, 1968, выл.69.
Строна И.Г. Общее семеноведение полевых культур. М.:
Колос, 1966.
Судницын И.И. Динамика лесорастительных условий и
свойства почв в различных типах леса Серебряноборского опытного
лесничества. - В кн.: Стационарные биогеоценотические исследова-
ния в южной подзоне тайги. М.: Наука, 1964.
Сукачев В.Н. Дендрология с основами лесной геоботани-
ки. Изд. 2-е. Л., 1938.
Сырейщиков Д.П. Иллюстрированная флора Московской
губернии, чл.1-4. М., 1906, 1907, 1910, 1914.
Тарковский М.Н., Артамонова Е.И. Массо-
вое надземное кущение тимофеевки из верхнего узла. - Агробиология,
1953, № 5.
Татаринова Н.К. Формирование куста лугового злака
в первые годы его жизни. Автореф. дис. на соиск. учен, степени
канд. биол. наук. М., 1953.
Татаринова Н.К. "О взаимоотношениях побегов в кусте
лугового злака. - Физиология растений, 1958, т.5, вып.5.
Татаринова Н.К. О длительности жизни корней луговых
злаков. - Ботанический журн., 1961, т.46, # 7.
241
Тахтаджян А.Л. Система и филогения цветковых расте-
ний. Л.: Наука, 1966.
Тебе В. Г. Материалы по биологии и географическому распро-
странению грушанки круглолистной. - Тр. Ленингр. хим.-фарм. ин-
^а, 1964, выл.17.
Терехин Э.С. Эмбриология грушанок и вертляниц в связи
с их биологией и систематическим положением. Автореф. дис. на со-
иск. учен, степени докт. биол. наук. Л., 1962а.
Терехин Э.С. Влияние некоторых экологических факторов
на развитие эмбриональных структур Pyrolaceae-Monotropaceae. -
Ботанический журн., 19626, т.47, № 4.
Толмачев А.И. О некоторых архаических чертах расте-
ний тайги, их экологической и исторической обусловленности. - Тр.
Томск, ун-та, т.116. Томск, 1951.
Ткаченко М.Е. Общее лесоводство. Изд. 2-е. М., 1952.
Тимофеев В.П. Природа и насаждения Лесной опытной да-
чи ТСХА за 100 лет. М..: Лесная промышленность, 1965.
Тимофеев В.П. Роль липы в поднятии устойчивости и
продуктивности лесов. - Изв. ТСХА, 1966а, № I.
Тимофеев В.П. Влияние липы на устойчивость и продук-
тивность насаждений. - Лесное хозяйство, 19666, № 2.
Тимофе ев В.П. Взаимоотношения соснн и липы при сов-
местном'их произрастании в лесных культурах. - Иэв. ТСХА, 1975,
> I.
Торопова М.И., Черкасов А.Ф. Динамика раз-
вития генеративных почек у некоторых видов семейства Vacciniaceae
в условиях Костромской области. - Учен. зап. Костромского пед.
ин-та, 1973, вып.ЗО.
Травников а Л.С. Влияние поселения липы под пологом
сосняка дубнякового на свойства почвы. - Тр. Воронежского запо-
ведника, 1959, вып.8.
У р а н о в А.А., Писковацкова Н.П. Опыт при-
менения возрастного анализа ценопопуляций для прогноза нектаро-
продуктивности лугов. - Бюл. МОИП. Отд. биол., 1974, т.78, вып.6.
У совский В.Н. Голубика (вопросы культивирования). -
Природа, I960, S 12.
У стинова Е.И. Биология цветения лиственных древесных
пород. - Лесное хозяйство, 1952, № I.
У тенкова А.П. Результаты изучения разложения опада в
дубовом лесу. - Тр. Воронежского заповедника, 1959, вып.8.
242
Уход за дикорастущей голубикой. - Лесное хозяйство, J6.5, 1971.
Федоров М.В., Непрмилуев В.Ф. Основные
формы ризосферных бактерий тимофеевки (Phieun: pratense L.) и из-
менение количества их клеток в ризосфере по фазам развития и го-
дам жизни всего растения. - .Микробиология, 1954, т.23, выл.2.
Федченко Б.А. Белая голубика Vaccinium utiginosum
var.leucarpum nova var. - Бот. мат-ЛЫ Гербария ГБС, 1923, вып.4.
Флора европейской части СССР, т.1-4. Л.: Наука, 1974-1979.
Флора Северо-Востока европейской части СССР, т.1-4. Л.: Наука,
1974-1977.
Флора СССР, т.1, 2, 3, 4, 8, II, 15, 18, 23. М.-Л.; Изд-во АН
СССР, 1934, 1935, 1939, 1940, 1949, 1952.
Флора Центральной Сибири, т.1. Новосибирск, 1979.
Флорова В.М., Раменский Л.Г. Определитель
растений в нецветущем состоянии для средней части СССР. Изд. 2-е.
М.: Сельхозгиз, 1937.
Фомин А.В. Папоротникообразные - Filices. - В кн.: Фло-
ра Сибири и Дальнего Востока, вып.5. Л., 1930.
Харитонович Ф.Н. Биология и экология лиственных
кустарников. Минск, 1975.
X л о н о в Ю.П. Липы и липняки Западной Сибири. Новоси-
бирск, 1965.
Хохлов С.С., Ковалева А.С. Деревья и кустар-
ники Нижнего Поволжья. Саратов, 1950.
Хохряков А.Н. Некоторые особенности морфогенеза сред-
нерусских Pyrolaceae. - Ботанический журн., 1961, т.63, № 3.
Хохряков А.П., Мазуренко М.Т. Эволюция ти-
пов побегов у жимолостных. - Бюл. ГБС АН СССР, I960, выл.70.
Хромосомные числа цветковых растений. Л.: Наука, 1969.
Ц в е л е в Н.Н. Злаки СССР. Л.: Наука, 1976.
Цельникер Ю.Л. Радиационный режим под пологом леса.
М.: Наука, 1969.
Ценопопуляции растений. М.: Наука, 1976.
Цинзерлинг Ю.Д. Растительность болот. - В кн.: Рас-
тительность СССР, т.1. М., Л.: Изд-во АН СССР, 1938.
Черемисинов Н.А., Негруцкий С.Ф.,
Лешковцева И.И. Грибы и грибные болезни деревьев и кус-
тарников.. М.: Лесная промышленность, 1970.
Черкасова В.А. Улучшение сенокосов и пастбищ на скло-
нах балок в лесостепной эоне Украины. - В кн.: Улучшение природ-
243
ных кормовых угодий. М., 1957.
Черкезишвили Т.С. Представители рода Lonicera в
Грузии. - Вести. Тбил. бот. сада, 1952, вып.60.
Чернов Е.Г. Pyrolaceae. - В кн.: Флора Мурманской обл.,
1959, т.4.
Чистякова А.А. Большой жизненный цикл и фитоценоло-
гическая роль липы сердцелистной (Tilia cordata Bill,) в разных
частях ареала. - Автореф. дис. на соиск. учен, степени канд. би-
ол. наук. М., 1978а.
Чистякова А.А. О жизненной форме и вегетативном раз-
растании липы сердцевидной. - Бюл. МОИН. Отд. биол., 19786, т.83,
вып.2.
Чистякова А.А. Большой жизненный цикл Tilia cordata
Mill. -Бюл. МОИП. Отд. биол., 1979, т.84, вып.1.
Ч и ч и н М.И. Условия произрастания и продуктивность липня-
ков БССР. Автореф. дис. на соиск. учен, степени канд. биол. наук.
Минск, 1956.
Ч и ч и н М.И. Липа в Белоруссии. - В кн.: Труды БеЛНИИЛХ,
1958, выл.12.
Чопик В.Н. Редкие и исчезающие растения Украины. Киев,
1978.
Ч у б и р к о М.М. Эмбриогенез Lotus cornioulatus L. - Укр.
ботанический журн., 1964, т.21, №3.
Ч у б и р к О М.М. Микроспорогенез Lotus cornioulatus L. -
Бюл. МОИП. Отд. биол., 1967, т.72, вып.6.
Чуенкова О.С. Биологические особенности кормовой куль-
туры лядвенца рогатого. - Тр. Саратовского зооветеринарного ин-та,
1954, т.5.
Хабарова C.I. Про вар1абильнисть плодив та можливють
насДннэвого поновлення видйв з родини брусничник. - Укр. ботани-
ческий журн., 1968, т.25, № 3.
Ш а м у р и н В.Ф. Вопросы опыления у растений. - Ботаничес-
кий журн., 1956, т.41, № 9.
Шеляг-Сосонко Ю.Р. Популяционные фитоценотичес-
кие части ареалов дуба обыкновенного (Quereus гоbur L.) и липы
сердцелистной (Tilia cordata Mill.). - Укр. ботанический журн.,
1968, т.25, № 2.
Щиголев А.А., Ш и м а н ю к А.П. Сезонное развитие
природы. М.: Географгиз, 1949.
Шилова Е.И. Метод непосредственного наблюдения микро-
244
флоры ризосферы тимофеевки. - Вести. Ленингр. ун-та, 1955, № 4.
Шилова Н.В. Характеристики жизненных форм в семействе
Pyrolaceae Lindl. - В кн.: Рост растений, Львов, 1959.
Шилова Н.В. Побегообразование и особенности жизненных
форм в сем. Pyrolaceae Lindl. - Ботанический журн., I960, т.45,
№ 6.
Ш и м а н ю к А.П. Биология древесных и кустарниковых пород
СССР. Изд. 2-е. М.: Просвещение, 1964.
Ш и м а н ю к А.П. Дендрология. М.: Лесная промышленность,
1967.
Штреккер В. Луговые злаки. М.-Л.: Сельхозгиз, 1931.
Щенкова М.С. Дикорастущие многолетние кормовые травы
Коми АССР в естественных условиях и в культуре. М., Л.: Изд-во
АН СССР, 1961.
Щ и б р я А.А., Мартьянова А.И. Особенности опы-
ления лядвенца рогатого. - Агробиология, I960, № 5.
Э р и н г и с К.И. Долголетние культурные пастбища Литвы.
Вильнюс, 1964.
Югай А.Н. Динамика роста сосны и липы на мощнодерновом
среднеоподзоленном легком суглинке. - Докл. ТСХА, 1979, № 253.
К) р к е в и ч И.Д., Голод Д.С., Парфенов В.И.
Типы и ассоциации еловых лесов. Минск, 1971.
Ю р к е в и ч И.Д., Смоляк Л.П. Сезонное развитие
некоторых видов растений на болотах. - В кн.: Выращивание высоко-
продуктивных лесов. Минск, 1963.
Ю р ц е в Б.А. и др. Обзор географического распространения
сосудистых растений Чукотской тундры. - Бюл. МОИП. Отд. биол.,
1979, т.84, № 6.
Ячевский А.А. Карманный определитель грибов. I. Голо-
сумчатые. Л., 1926.
Andres Н. Piroleen-Studien. - Verhandl. Bot. Prov.
Brandenburg, 1914, 4 6.
Anzalone B.A. Ulteriori osservazioni sulla distribu-
zione in Italia di Lonicera xylosteum L. - Giom. Bot. Ital.,
1963, vol.70, N 5-6.
Atkinson L.R., S t о k e у A.G. Comparative mor-
phology of the gametophyte of homosporous ferns. - Phytomorpholo-
gy, 1964, vol.14, 4 1.
Atlas Florae Europaea, vol.1. Helsinki, 1972.
Baker' H.G. Alluvial meadows comparative study of grazed
245
and sown meadows. - Jouzn. Ecology, 1937, vol.25, И 43.
Beck С» yritillaries. London, 1953.
Becquerel P. La vie latente des spores de fougere
dans le vide aux basses temperatures de 1’helium liquide. -
Compt. Rend. Acad. Soi., 1930, vol.190, 3 19.
Bednarova-Hogova J. Opakovana pusobenl chlo-
ru na prothallia Athyrium filix-femina (L.) Both. - Acta Univ.
Palaek. Olomuc., 1968, vol.28.
Belling A.J., И e u в a e r C.J. Spore morphology
of the Polypodlaoeae of northeastern Sorth America. - Bui. Torrey
Bot. Club., 1974, vol.101, H 6.
Beni G. , Eschemuller A. Uber "Dryopteris
remote” und ihr Verkommen in Bayern. - Ber. Bayer. Bot. Ges.,
1976, vol.44.
Bittmanova J. Vliv chloru na vyvoj prothallii kap-
radin. - Acta Univ, palaek. olomuc., 1964, vol.16.
Bittmanova-Hogova J. Druhy prispevek к vy-
voj i gametofytu n^kterych naaich Polypodiacei. - Acta Univ. Pa-
lack. Olomuc., I960, vol.5.
Bittmannova-Kogova J. Vycoj gametofytu na-
eich kapradin celed? Polypodiaceae Dost. - Acta Univ. Palaek. Olo-
muc., 1962, vol.11.
Bla.ttny C., Pilous I>. Prispevek к poznani
ochuravXni, pohladanych za virony, nebo z viros podozrelych u me-
choree1% (Bryophyte) a kapradoroatu (Pteridophyta). - Xeskosl. bi-
ol., 1953, vol.2, H 2.
Blom C.We Trampling зоЛ compaction and Plantago. Rotter-
dam, 1979.
Bower P.O. The ferns, vol.1. Cambridge, 1923.
Brenchley W.E. Buried weed seeds. - J. Agric. Sci.,
1918, vol.9.
Brenchley W.E. Weeds of farm. London, 1920.
Brierley J.K. Some preliminary observations on the
ecology of pit heaps. - J. Bool., 1956, vol.44.
Britton В.И., J e r m у A.C. The spores of Dryop-
teris filix-mas and related taxa in Sorth America. - Can. J. Bot.,
1974, vol.52, Я 8.
Bradshaw A.D. Conservation problems in the future. -
Proc. Roy. Soo. 'London, 1977, vol.197, Я 1126.
Brooks Й.В., Both J.B. The discovery of Athyri-
• 246
urn filix-femina and other interesting pteridophytes in Southern
Kansas. - Amer. Fem. J., 1977, vol.67, Я 2.
Bujorean G., Grigore S. Fritiiiaria meleag-
ris L. si ocrotirea el. - Ocrotirea naturii, 1965, t.9, Я 1.
Buschmann A. Zur Kenntnis von Fritiiiaria meleagris
L. - Phyton, 1951, vol.3, fasc.3.
Camp W.H. Aphyllous forms in Pyrola. - Bui. Torrey Bot.
Club, 1940, vol.67, Я 6.
Chmielewski W. Plodnosci rozwoj niektorych gatun-
kow roztaczy ulowych na pylku. - Pszczel. zesz. nauk. Odd.
pszczel. Inst. Sad. Pulawach, 1978, Я 22.
Conners I.L. Species of Leptothyrium and Kabatia on
Lonicera. - Canad. J. Bot., 1959, vol.37, Я 3.
Cooper K.M. Endomycorrhizos affect growth of Dryopte-
ris filix-mas. - Trans. Brit. Hycol. Soc., 1977, vol.69, Я 1.
Corley H.V. Dryopteris filix-mas agg. in Britain. -
Proc. Bot. Soc. Brit. Isles, 1967, Я 7.
Courbet H. Action de quelques sucres sur la germinati-
on des spores et la croissance des prothalles d’Athyrium filix-
femina Roth en culture aseptique. - Compt. Rend. Acad. Sci.,
1957, vol.244, Я 1.
Courbet H., lletche M. Dosage de 1'azote mine-
ralisable et des acides amines libres des spores de quelques fou-
gerea. - Compt. Rend. Acad. Sci«, 1971, vol.272, Я 14.
С о u в e n s M.I., Horner H.T. Gametophyte ontoge-
ny and sex expression in Dryopteris ludoviciana. - Amer. Fern.J.,
1970, vol.60, H 1.
Crane F.H. Spore studies in Dryopteris. - Amer. Fem.
J., 1956, vol.46, Я 3.
Crane F.M. A key tb American Dryopteris species based
on characters of the perispore. - Amer. Fem. J., 1956, vol.46,
Я 4.
Crocker V, Live span of seeds. - Bot. Rev., 1938, H 4.
Cullgagh W. Chromosomes end chromosome morphology in
Plantaginaceae. - Genetics, 1934, Я 16.
Cutter E.G., V о e 1 1 e r B.R. Changes in leaf ar-
rangement in individual fem apices. - J. Linn. Soc. London,
1959, vol.56, Я 366.
Darrow R.A. Effects of soil temperature, pH and nitro-
gen nutrition on the development of Poa pratensis. - Bot. Gas.,
247
1939, Я Ю1.
Davidonis G.H. The occurence of thelypterin in
ferns. - Amer. Fem. J., 1976, vol.66, Я 3.
Davidonis G.H., R u d d a t M. Allelopathic com-
pounds thelypterin A and В in the fem Thelypteris normalis. -
Planta, 1973, vol.Ill, Я 1.
Davies W. Vegetation of grass verges and other excessi-
vely trodden habitats. - J. Ecol., 1938, Я 26.
D a x e r H. Uber die Assimilationsokologie der Waldboden-
flora. - Jb. Wise. Bot., 1934, vol.80.
D о p p W. Untersuchungen uber die Entwicklung von Prothali-
en einheimischer Polypodiaceen. - Pflanzenforschung, 1927, vol.8.
D о p p W. Experimentell erseugte Bastarde zwischen Dryopte-
ris filix-mas (L.) Schott und D.paleacea (Sw.) C.Chr. - Planta,
1955, vol.46, Я 1.
Dowling R.E. The ovary in the genus Plantago. - J.
Linn. Soc. (Bot.), 1935, vol.50.
Druce G.C., Britton G. Plantago lanceolata L.
var.sphaerostachya Rohl. - J. Bot., 1928, H 44.
Du Buy H.G., Heurnber.gk E.L. Growth, tro-
pisms and other movements. - In: Manual of Pteridology. Hague,
1938.
Dudley M.G. Morphological and cytological studies on
Galla palustris L. - J. Bot. Gaz., 1937, vol.98, H 3.
Dyer A.F. The experimental biology of fems. - Trans.
Bot, Soc. Edinb., 1979, vol.43, Я 2.
Eberle G. Silberblatt. - Hatur und Volk, i960, Bd 90,
Я 6.
Ellenberg H. Uber Zusammensetzung, Standort und
Stoffproduction bodenfeuchter Eichen- und Buchen-Mischwaldgesell-
schaften, Hordwestdeutschlands. - Mitt. Flor.-Soz. Arb. Rieder- .
sachsen, 1939, Heft 5.
Ellenberg H. Wiesen und Weiden und ihre standortli-
che Bewertung. - Landwirtsch. Pflanzensoziol., 1952, Bd 2.
Ellenberg H. Zeigerwerte der Gefasspglanzen Mittele-
uropas. - Scripts Geobotanica, 1974, vol.9.
Erdtman G,, Berglund В., P r a g 1 о w -
ski J. An introduction to a Scandinavien pollen flora. - Gra-
na Palynol., 1961, vol. 2, Я 3.
Evans M.W. The life history of timothy. - Bui. US depar-
248
tament of agriculture, 1927, К 1450.
Evans M.W., .Grover F. Developmental morphology
of the growing point of the shoot and the inflorescence in gras-
ses. - J. Agr. Res., 1940, vol.61, В 7.
Fab b ri F. Primo supplement©, alle Tavole chromoaomiche
delle Pteridophyta di Alberto Chiarugi. - Caryologia, 1963,
vol.16, В 2.
Farmer J.B., Digby I*. Studies on apospory and
apogamy in ferns. - Ann. Bot., 1907, vol.21.
Farrar D.R. Spcre retention and release froni overwinte-
ring fem fronds. - Amer. Fem J., 1976, vol.66, В 2.
Farrar D., Gooch R. Fem reproduction at Wood-
man Hollow, Central Iowa; preliminary observations and a conside-
ration of the feasibility of atuding fem reproductive biology
denature. - Proc. Iowa Acad. Sei., 1975, vol.82, В 2.
Feilberg J. Athyrium filix-femina in Greenland. -
Bot. Tidsskr., 1975, vol.70, В 2-3.
Flora Europaea, vol. 1-5. Cambridge, 1964, 1968, 1972, 1976,
1980.
Foerster K. Einzug der. Graserund Fame in die Garten.
Leipzig, 1957»
F r a me J., Harkless R.D., Hunt J.7.
The role of timothy (Phleum pretense L.) in high yieding conser-
vation leys. - In: 4th General Meeting. Lausanne, 1971.
Fraser-Jenkins C.R., Corley H.V. Dry-
opteris eaucasica - ancestral diploid in the male fem aggrega-
te. - Brit. Fem. Gaz., 1972, vol.10, В 5.
Fraser-J enkins C.R., J e r m у A.C. nomen-
clatural notes on Dryopteris Adans. - Taxon, 1976, vol.25, В 516.
Fries D. Induction de I’apogamie chez Dryopteris filix-
mas (L.) Schott cultive in vitro. - Bui. Soc. Bot. Belg., 1970a,
vol.103, В 2.
Fries D. Morphogenese des structures apogames induites
chez Dryopteris filix-mas (L.) Schott cultive in vitro. - Bui.
Soc. Bot. Belg., 1970b, vol.103, В 2.
F u r u у a M. Photocontrol of developmental processes in
fem gametophytes. - Bot. Mag. Tokyo, spec. Issue, 1978, В 1.
G'a in в H. Okologie der extratropischen Pteridophyten. - In:
Manual of Pteridology. Hague, 1938.
G a г с к e A. Illustrierte Flora Deutschland und angrenzen-
18-1964 249
de Gebiette. Berlin-Hamburg, 1972.
Gluck W. Wuchsformstudien an zentral europaischen Faba-
ceen. - Feddes Report., 1969, vol.79, И 6.
G о d г о n A. De la floraison des Graminees. - Mem. Soc.
Ration Cherbourg, 1873, eer.2, vol.7, R 11.
Goldschmidt M. Tabelle zur Bestimmung der in Mit-
teleuropa wild wachsenden Abarten und Fonnen von Athyrium filix-
femina Roth. - Hedwegia, 1906, Bd 65, H 2.
Gregor M.J. Associations with fungi and other lower
plants. - In: Manual of Pteridology. Hague, 1938.
Grevillius A., Kirchner 0. Pyrolaceae. -
In: Iiebensgeschichte der Bliitenpflanzen Mitteleuropas, 1923, Bd 4
Abt.1•
Griffits A. Polyploidy life of form and vegetative
reproduction. - Hereditas, 1948, N 34.
Gustafson A. Polyploidy life form and vegetative
reproduction. - Hereditas, 1948, R 34.
Guzikowa M. Wystepowanie gruszyczki posredniej (Pyro-
la media Sw.) w Polsce i jej rozmieszczenie ogolne. - Fragm. flo-
rist. et geobot., 1971, vol.17, R 4.
Harley J.Ii. Biology of Mycorrhiza. London, 1959.
Hawthorn W.R., Cavers P.B. Population dyna-
mics of perennial herbs Plantago major and P.rugellii Decne. - J.
Ecol., 1976, vol.64.
Hawthorn W.R., Cavers P.B. Resources alloca-
tion in young plant of two perennial species of Plantago. - Can.
Bot. J., 1978, vol.56, H 20.
Hegi G. Illustrierte Flora von Mitteieuropa, Bd 4, Abt.1;
Band 6, Abt.1. Munchen, 1918, 1927.
Hirst J.M. Spore liberation and dispersal. - In; Plant
pathology. Madison, 1959.
Hock F. Calla palustris L. - In: Lebensgeschichte der
Bliitenpflanzen Mitteleuropas, 1911, Bd 1, Abt.3.
Hofmeieter W. On the germination, development and
fructification of the higher Cryptogamia and on the fructificati-
on of the Coniferae. London, 1862.
Holub J. Remarks on the nomenclature of "Dryopteris bor-
reri Newman 1854". - Folia Geob. Phytotaxon, 1967, vol.2, H 3.
H u 1 t ё n E. Flora of Kamtchatka and the adjacent is-
lands. - Kongl. Sv. Vet. Akad. Handl, Tredje. Ser.3, 1930, R 8
250
(2).
H u 1 t ё a E. Atlas of the distribution of vascular plants
in North-West Europe. • Stockholm, 1950.
H u 1 t ё a E. The circumpolar plants. - Kungl. Sv. Vet.
Akad. Handl.' Ser.4, 1962, Bd 8, N 5.
Hundt R. Die Bergwiesen des Harzes, Thuringen Waides und
Erzgebirges. Jena, 1964.
Hyde H.A., Williams D.A. Studies in atmospe-
ric pollen. - New Phytol., 1945, N 45.
Jones M.D., Newell L.C. Pollination celles and
pollendispersal in relation to grass improvement. - Nebr. Agr.
Expz. Sta. Res. Bui., 1946, N 148.
Kallio P., Laine U., Makinen J. Vascu-
lar flora of Inari Lapland. - Turun yliopiston julk., ser.A,
1969, vol.11, N 42.
Happen L. Untersuchungen uber die Widerstandsfahigkeit
der Gametophyten einheimischer Polypodiaceen gegen uber Prost,
Hitze und Trockenheit. - Plora, 1964, Abt.A, N 156.
Karpowicz W. Studien uber die Entwicklung der Pro-
thallien und der ersten Sporophytblatter der einheimiscen Parn-
krauter. - Acad. Polon. Sci., ser.B, 1927, vol.27, N 1.
Karpowicz W. Stadium porownawcze nad krajowymi
przedstawkielami gatunkow rodzaju Dryopteris i Thelypteris. - Ac-
ta Soc. Bot. Polon., I960, vol.29, N 2.
Karpowicz W. Paprocie. 2 ed. Warzawa, 1972.
Kato M. Classification of Athyrium and allied genera of
Japan. - Bot. Mag. Tokyo, 1977, vol.90, N 1017.
Kempf C. Pougeres d’Alsace. - In: Inf. Ann. Caryosyst.
et Cytogenet., 1967, vol.1.
Kiese! A.P. Untersuchungen uber pflanzliche Portpflan-
zungszellen. - Zeitschr. Physiol. Chemie, 1925, N 149,
К 3 a e r A. Germination buried and dry stored seeds. -
Proc. Int. Seed. Test. Ass., 1940, N 12.
Kleiner P. Les Pyrolee, plantes de jardin? - Revue hor-
ticole. Nov. ser., 1946, t.30, N 5.
Klekowski E.I. Reproductive biology of the Pterido-
phyta. - Bot. J. Linn. Soc., 1969, vol.62, N 3.
Knaben G., Engelskj^n T. Studies in Pyro-
laceae, especially in the Pyrola rotundifolia complex. - Acta
Univ. Bergensis, Ser. math. rer. natur., 1968, N 4.
251
Knuth P.Handbuch der Bliitenbiclogie, Bd 1-2. Leipzig,
1895, 1899.
К о e n о d K.L. Germination of normal and hulled grass se-
ed stored under three conditions. - Crop Science, 1965, Я 5.
Корреп T. Geographische Verbreitung der Holzgewachse
des europaischen Russlands und des Kaukasus, Bd 1. Spt., 1898.
Korsmo E. Weed seeds. Oslo, 1935.
Koziracki L.A., Tagronski P.N. Dcswrade-
zenia nad generatywnym odnowieniem dziko rasnacych krzewinek z
radzajn Vaccinium (borowka) na Poiesin Zytomierskim. - Sylwan,
1978, vol.122, N 9.
Kugler H. Blutenokologie. Stuttgart, 1970.
L a a g e A. Bedingungen der Keimung von Fam- und Moosspc-
res. - Beih. Bot. Centralbl., 1907» vol.21, Я 1.
Landolt E. Okologieche Zeigerwerte zur Schweizer Flo-
ra. - Veroff. Geobot. Inst. Rubel, Zurich, 1977, Heft 64.
Langer R.H. Ear formation in tymcthy grass (Phleum pre-
tense) following vernalization and schort-day treatments. - Natu-
re, 1955, vol.176, N 4475.
L a n f e r R.H., Ryle J.A. Vegetative proliferati-
ons in herbage grasses. - J. Brit, grassl. Soc., 1958, vol.13,
Я 1.
L e p i к E. Verzeichnis der aus Estland gesammelten Pflan-
zengallen. - In: Mitt. Phytopathol. Versuchsstat. Univ. Tartu,
1943, vol.64.
Lietuvos TSR flora, t.1. Vilnius, 1959.
Lichnell D. Keimungsverauche mit Pyrolasamen. - Sym-
bolae Bot. Upsaliensis, 1942.
Lincola К., T u r r i к a A. Uber Wurzelsystem und
Wurzelaufrhatung der Wiesenpflanzen auf verschiedene Wiesenstan-
dorten. - Ann. Bot., 1936, N 6.
Loew E., Kirchner 0. Fritillaria. .- In: Leben-
sgeschichte der Bliitenpflanzen Mitteleuropas, 1934, Bd 1, Abt.3.
Loyal D.S. Apomixis in ferns with particular reference
to Dryopteris Adanson and some heterosporous members. - J. Indi-
an Bot. Soc., 1966, vol.45, N 1-2.
Luck R. Zur Biologie der heimischen Pirola-Arten. - Schr.
Phys.-ofon. Ges. Konigsl., 1940, vol.71.
Luerssen C, Die Farnpflanzen oder Gefassbundelkrypto-
gamen. - In: Rabenhorst's G.L. Kryptogamenarten von Deutschland,
252
Ostereich und der Schweiz, Bd 3. Leipzig, 1889»
M a m о s e S. Studies on the Gametophytes of ferns. - Taf.
J. Bot., 1942, vol.18.
M a n t о n X. Problems of cytology and evolution in the
Pteridophyta. Cambridge, 1950.
Hanton I., Walker 3. Induced apogamy in Dryop-
teris dilatata (Hoffm.) A.Gray and D.filix-mas (L.) Schlecht.
emend, and its significance for the interpretation of the two
species. - Ann. Bot., 1954, vol.18, Я 71.
Marsdon-Jones E.M., T u r r i 1 1 W.B. Plan-
tago major. - J. Ecol., 1938, vol.26<
May L.W. The economic uses and associated folklore of
ferns and fern allies. - Bot. Rev., 1978, vol.44, И 4.
M e h r a P.N., Loyal D.S. Cytological investigati-
ons in the Hymalayan Dryopteris Adanson. - Cytologia, 1965,
vol.18, Я 3.
Metsavainio K. Untersuchungen uber des Wurzelsys-
tem der Moorpflanzen. - Ann. Bot. Soc. Zool.-Bot. Penn. Vanamo,
1931, t.1, Я 1.
M e u s e 1 H. Vergleichende Arealkunde, Bd 1-2. Berlin,
1943.
Meusel H., Jager E., Weinert E. Ver-
gleichende Chorologie der Zentraleuropaischen Flora. Jena, 1965.
Miller J.H. Pern gametophytes aa experimental materi-
al. - Bot. Rev., 1968, vol.34.
M i 1 t о n W.E. The yeilds of ribwort plantain when sown
in pure plots and with grasses and clover species. - Weish. Ag-
ric., 1943, Я 17.
Mohr H. Die Beeinfluseung der Keimung von Famsporen
durch Licht und andere Faktoren. - Planta, 1956, N 46.
Я a у a r B.K. A phylogenetic classification of the homospo-
rous ferns. - Taxon, 1970, vol.19.
Я a у a r B.K., Kaur S. Spore germination in homospo-
rous ferns. - J. Palynol., 1968, vol.4, 11,
Я a у a r B.K., • Kaur S. Types of prothallial develop-
ment in homosporous fems. - Phytomorphology, 1969, vol.19, Я 2»
Я a у a r B.K., Kaur S. Gametophytes of homosporous
ferns. - Bot. Rev., 1971, vol.37, Я 3.
Я i s h i d a M. Types of spore germination in the ferns. -
J. Jap. Bot., 1965, vol.40.
I8x-I964 253
Orth R. Zur Keimungsphysiologie der Famsporen in vers-
ohiedenen Spektrallezirken. - Jarb. Wise. Bot., 1937, vol.84.
Rahn R. Experimental and cytological studies in Plantago
media. - Bot. Tiddekr., 1954, vol.31.
Rahn R. Chromosomes numbers in Plantago. - Bot. Tiddekr.,
1957, vol.53.
Rehder A. Manual of cultivated trees and shrubs. N.Y.,
1927.
Rezende Pinto M.C. Dryopteris resendeana. - Por-
tug. acta Biol., 1968-1969, Bd 9, N 3-4.
Roberts R.A., Hunt J.V. The effect of the stool
cutting oii the growth of root and stool of perennial ryegrass
(Lolium perenne) and thymothy (Phleum pretense) Welsh. - J. Ag-
ric., 1936, vol.12.
Roland M.K. Fertilization relationships in the genus
Lotus. - Agronomy J., 1949, vol.41, N 7.
R u b n e r K. Die Pflanzengeographischen des Waldbaues.
5 Auf. Neuman Verlag, I960.
Sagar C.R.,Plantago studies. - Bot. Proc. Soc. Brit. Is-
les, 1958, vol.3.
Sagar G.R., Harper J.L. Factors affecting the
gemination and early establishment of Plantagins. - In: Biology
of Weeds. Oxford, I960.
Sagar G.R., Harp er J.L. Plantago major L.,
P.media L. and P.lanceolata. - J. Ecology, 1964, vol.52, N 1.
Salisbury E. The oakhorn beam Woods of Herfordshire.-
J. Ecol., 1916, vol.4, N 83.
Salisbury E.J. The reproductive capacity of plants.
London, 1942.
Sal isbury E.J. Downs and dunes. London, 1952.
Schneider C.K. Illustriertes Handbuch der Laubholz-
kunde, Bd 2. Jena, 1911.
Schneller 1.1. Untersuchungen an einheimischen Far-
nen insbesondere der Dryopteris filix-mas Gruppe. - Ber. Schweiz.
Bot. Ges., 1975, vol.85, N 2.
Schneller 1.1. Biosystematic investigations on the
lady fern (Athyrium filix-femina). - Plant, syst. and evol.,
1979, vol.32, N 4.
Schulze F.D. Die Wirkung von Licht und Temperatur auf
den CO9-Geswechsel verachiedener Lebensfomen aus der Krauts-
S4
chicht eines montanen Buchenwaldea. - Oecologia, 1972, N 8, 3.
Staple don R.G. Permanentes grass farm groups, vol.3.
London, 1925.
Stevanovio V. Ekolosko-cenoloska analiza stanista
zelne kruscice (Pirola chlorantha Sw.) u Deliblatekoj peaceri. -
Glaenik .Inst. Bot., 1972.
Tabor sZk a B., Bednarova J. Priepevek к
stovnevaci anatomii pokozek kapralin. - Acta Univ, palaek. olomuc
Рас. rerum. natur., 1973, vol.43.
T h о m p s on G.M. The naturalization of animals and
plants in New Zealand. Cambridge, 1952.
Toom Y.A., Jonson I.J. Self- and crossfertility
relationships in Lotus comiculatus L. and Lotus tenuis Waidet.
et Kit. - J. Amer. Soc. Agron., 1945, vol.37, Я 12.
Troll. H.J. Untersuchungen uber Selbststerilitat Selbst-
fertilitat bei Grasam. - Zeitschr. fur Zuchtung, Reihe A, 1931,
vol.16.
Troll W. Die Infloreszenzen, Bd 1. Jena, 1964.
Turrill W.B. Character combinations and distribution
in the genus Fritiiiaria and allied genera. - Evolution, 1950,
vol.4, N 1.
Vaicys M., Raguotris A., Sleinys R
Misko dirvozemiu zinynas. Vilnius, 1979.
Wagner W.H., Chen K.L. Abortion of spores and
sporangia as a tool in the detection of Dryopteris hybrids. -
Amer. Fem. J., 1965, vol.55, N 1.
W a r d la w C.W. Apical organization and differentional
growth in ferns. - J. Linn. Sec. London (Bot.), 1963, vol.58,
M 373.
W a r d 1 a w C.W., Sharma D.N. Experimental and
analytical studies of pteridophytes. - Ann. Bot., 1963, vol.27,
N 105.
W e 1 s 1 о H« Karyological studies in Polish representati-
ves of spadiciflorae. - Acta Biol. Ser. bot., Cracov, 1970,
vol.13, H 2.
Wherry E.T. The fem guide. N.Y., 1961.
White R.A. Experimental and developmental studies of the
fem sporophyte. - Bot. Rev., 1971, vol.37, N 4.
W i d ё n C.J. Chromatographic investigations of European
and North American Dryopteris fems. - Farm. Aikak, 1972, vol.81.
255
W i d ё n C.J., Britton D.M. A chromatographic and
cytological study of Dryopteris filix-mas and relatives taxa in
North America. - Canad. J. Bot., 1971, vol.49, N 9.
W i d ё n C.-J., Fraser-Jenkins C.R.,
Lounasmaa M., E u w J.V., Reichstein T.
Bie Phloroglucide von Dryopteris caucasica (A.Br.) Fraser-Jen-
kins et Corley. - Helv. Chim. Acta, 1973, vol.56, N 3.
W i d ё n C.J., Lounasmaa M.G., J e r -
m у A.G., E r i w J., Reichstein T. Die Phlo-
rologlucide von zwei Famhybriden aus England und Scotland von
authentischen. - Helv. Chim. Acta, 1976, vol.59, fasc.5.
Willis I.C. A dictionary of the flowering plants and
ferns. 8 ed. Cambridge, 1973.
Willmot A. An ecological survey of the ferns of the
Barren Co. Clare, Eire. - Fern. Gaz., 1979, vol.12, N 1.
Winslow E.I.A new hybrid fern. - Amer. Fern. J.,
1910, vol.1.
Zimmer A.L. A dictionary of the flowering plants and
ferns. Cambridge, 1976.
256
Сводный указатель названий видов, охарактеризованных
в 1-7 выпусках "Биологической флоры Московской области"1
(первая цифра - № выпуска, через двоеточие - страницы)
Acer campestre L. 1:120-123
platanoides L. 1:106-119
Adonis vernalis L. 2:36-47
Adoxa moschatellina L. 5:147-155
Aegopodium podagraria L. 1:131-141
Agrostis canina L. 3:52-55
- "- gigantea Roth 3:43-47
- "- stolonifera L. 3:37-43
- "- stolonizans Bess. CM. A.stolonifera
- "- syreistschikowii P.Smirn. 3:55-61
- "- tenuis Sibth. 3:47-52
- "- vulgaris With. CM. A.tenuis
Allium uralnum L. 4:52-61
Anemone nemorosa L. 3:129-134
ranunculoides L. 3:120-129
- "- sylveetris L. 3:134-138
Asarum europaeum L. 1:41-51
Athyrium filix-femina (L.) Roth 7:26-40
Bromopsis inermis (Leys.) Holub CM. Bromus inermis
Bromus inermls Leys. 5:58-73
Bunias orientalis L. 1:52-68
Galla palustris L. 7:67-82
Campanula glomerate L. 5:156-168
Capsella bursa-pastoris (L«) Medik. 4:86-95
Carex acuta L. 6:120-127
- "- acutoformis Ehrh. 6:39-40
- "- appropinquate Schum. 6:143-145
- "- a qua til is Wahlenb.. 6:127-130
atherodes Spreng. 6:34-36
- "- brunnescens (Pers.) Poir. 6:186-188
canescene auct. CM. C.cinerea
- "- capillaris L. 6:111-113
caryophyllea Latour. 6:93-100
Составил И.А.ТУбанов.
257
ceepitosa L. 6:137-141
chordorrhiza Ehrh. 6:167-169
cinerea Poll. 6:183-186
contigua Hoppe 6:151-154
cuprina (Sand, ex Heuff.) Kendtv. CM. C.muricate
diandra Schrank 6:145-148
digitate L 6:85-90
dioica L. 6:177-180
disperma Dew. 6:192-194
disticha Hude. 6:157-160
diversicolor auct. cm. C.flacca
echinata Murr. 6:175-177
elongata L. 6:180-182
emasculate V.Krecz. CM. C.hartmanii
ericetorum Poll. 6:100-102
flacca Schreb. 6:62-66
flava L. 6:79-82
globularis 'L. 6:106-109
goodenoughii J.Gay CM. C.nigra
gracilis Curt. см. C.acuta
hartmanii Cajand. 6:118-120
helponastes Ehrh. 6:190-192
hirta I>. 6:30—34
hylaea V.Krecz. CM. C.cinerea
inflate duct. cm. C.rostrata
inumbrata V.Krecz. см. C.cespitose
irrigua (Wahlenb.) Smith ex Hoppe cm. C.magellanica
juncella (Fries) Th.Fries 6:141—143
lasiocarpa Ehrh. 6:41-44
leporina L. 6:172-175
limosa L. 6:1X3-116
loliacea L. 6:189-190
lumnitzeri Boiiy CM. C.contigua
magellanica Lam. 6:116-118
montana L. 6:103-106
muricata L. 6:155-157
nigra (L.) Beichard 6:130-133
obtusata Liljebl. 6:28-30
omskiane Meinsh. 6:134—137
orthostachys C.A.Mey. см. C.atherodes
258
- "- pallescens Ь. 6:85-85
panicea I>. 6г 7^—77
— "- pauciflora Lightf. 6:24-28
paupercula Michx. cm. C.magellanica
pediformis auct. CM. C.rhizine
— "- pilosa Scop. 6:66-74
— praecox Schreb. 6:160—166
- "- pseudocyperua L. 6:44-48
— remote I«. 6:169-172
— rhizina Blytt ex Llndbl. 6:90-93
- "- rhynchophysa C.A.Mey. 6:52-54
- "- riparia Curt. 6:36-58
- "- rostrata Stokes 6:48-52
— "- ruthenica V.Krecz. CM. C.caryopbyllea
- "- spicata Hude. см. C.contigua
- "- aupina Wahlenb. 6:109-111
— "- sylvatica Hude. 6:'58—62
- "- vaginate Tausch 6:78-79
- "- verna Chaix CM. C.caryopbyllea
- "- vesicaria L. 6:54-58
- "- vulpina 1.6:148-151
wiluica Meinsh. CM.' C.juncella
Centaurea scabiosa 1. 5:151-161
Cerasua fruticoaa Pall. 1:78-87
Convallaria ma jalia I>. 1:21—55
Corydalis bulboaa (1.) DC. 2:49-54
- "- cava (I.) Schweigg. et Koerte cm. C.buibcsa
- "- halleri (Willd.) Willd. CM. C.solida
- "- intermedia (1.) Merat 2:70-72
marschalliana Pera. 2:55-60
aolida (L.) Clairv. 2:60-70
Corylus avellana 1. 2:18-28
Cypripedixm calceolus L. 4:62-68
guttatum Sv. 4:68-70
Dactylia glomersta L. 3*76-89
Daphne mezereum L. 1:124—130
Deschaarpsia ceepitosa (L.) Beauv. 5:62-75
-"- flexuosa (L.) Heea 5:46^57
Dryopteris filix-mas (L.) Schott 7;3-25
Peatuca pratenais Hude. 3:105-118
259
Frangula alnua Mill» 4:158—144
Fraxinus excelsior L. 1:142-159
Fritillaria meleagris L. 7:91-97
ruthenioa Vikstr. 7:83-90
Galeobdolon lute urn Hude. 5:159-150
Glyceria aquatics (L.) Wahlenb. CM. G.maxima
-”— maxima (C.Hartm.) Holmb. 5:90-104
Goodyera repens '(L.) H.Br. 2:5-10
Helichrysum arenarium (L.) Moench 1:160-168
Hepatica nobills Mill. 4:7I-78
Heraoleum sibiricum L. 2:'I24-I36
Junous alpino-artioulatus Chaix ex Vill. CM. J.alpinus
-"- alpinus Vill. 4:44-47
-"- artioulatus L. 4:59-44
-"- attains Krock. 4:47-49
-"- bufonius L. 4:23-28
-"- compressus Jadq. 4:30-55
-"- conglomeratus L. 4:14-16
-"- effusus L. 4:6-14
-"- filiformis L. 4:18-23
fuscoater Schreb. CM. J.alpinus
-"- geniculatus Schrank cm. J.alpinus
—"— inflexus L. 4:16-18
-"- juzepczukii V.Krecz. et Gontsch. CM. J.ranarius
-"- lamprocarpus Ehrh. cm. J.articulatus
-"- leersii Mara. CM. J.conglomeratus
-"- macer S.F.Gray CM. J.tenuis
-"- nastanthus V.Krecz. et Gontsch. 4:28-29
-"- paniculatus Hoppe CM. J.inflexua
—!*- ranarius Song, et Perr. ex Billot 4:29-5°
-"- tenuis Willd. 4:35-39
Juniperus communis L. 3:28-35
Lactuoa tatarica (L.) C.'A.Mey. CM. Mulgedium tataricum
Latbyrus pratensis L. 4:127-157
-"- vernus (L.) Bernh. cm. Orobua vernus
Lerchenfeldia flexuosa (L.) Schur CM. Deschampaia flexuosa
Libanotis intermedia Hupr. CM. Seseli libanotis
-"— montana Crantz CM. Seseli libanotis
Lonicera xylosteum L. 7:210-218
Lotus oornioulatus L. 7:111-127
Lunaria rediviva L. 7:98-110
Luzula albida (Hoffm.) DC* CM. L.luzuloidea
-"- luzuloidea (Lam.) Dandy et Wilm. 5*83-86
multiflora (Eetz.) Lej. 5*86-91
nemoroaa (Poll.) E.Mey. CM. L.luzuloidea
palleacena Sw. 5*91-95
piloaa (L.) Willd. 5*76-8?
Mstrioaria chamomilla L. CM. M.recutita
recutita L. 4s198-205
Medicago falcata L. 4:96-^104
lupulina L. 4:107-110
romanica Prod. 4:105
aativa L. 4:105-10?
Mercurialia perennis L. 2:111-123
Mulgedium tataricum (L.) DC. 2:175-186
Nardua atricte L. Is 6-20
Onobrychia arenaria (Kit.) DC. I$88-97
Orobus vernus L. 1:98-105
Padua avium Mill. 'CM. Prunua padua
-n- racemoaa (Lam.) Gilib. CM. Prunua padua
Paria quadrifolia L. Is34—40
Phleum phleoidea (L.) Karat. 7:57-66
-"- pratenae L. 7:41-57
Picea abiea (L.) Karat. 3$4-27
-"- excelaa (Lam.) Link CM. P.abiea
Pinua aylveatria Lf 5*5-45
Plantago lanceolate L.' 7:203-209
-"- major L. 7:189-197
-"- media L. 7:197-202
Platanthara bifolia (L.) Eich. 2:11-17
Polygonum biatorta L. 2:29-55
Potentilla erecta (L.) Eaeuach. 1:67—77
Prunua frutiooaa. Pall. CM. Cerastia fruticoaa
-"- padua L. 2:81-88
Pulmonaria obacura Dumort. 4:179-190
Pulaatilla patena (L.) Mill. 4:79-85
Pyrola chlorantha Sw. 7:172-176’
media Sw. 7:169-171
minor L. 7:164-169
-"- rotundifolia L. 7:154-164
virescena auct. CM. P.chlorantha
261
Ilhamnus cathartics L. 4:145-151
frangula L. CM. Frangula alnus
Нишах confertus Willd. 5:105-124
Sambucus racemose L. 2:169-174
Seseli libanotia (L.) Koch 4:152-166
Sorbua aucuparia L. 2:75-80
Spergula arvanaia L. 5*125-131
linicola Boredu CM. S.srvensis
maxima Weihe CM. S.arveusis
vulgaris Boerm, см. S.arveusis
Thymus marschsllianus Willd. 2:138-151
pulegioides L. 2:151-159"
-"- serpyllum L. 2:159-168
Tilia cordata Mill. 7:128-152
Trifolium hybridum L. 2:102-110
prateuse L. 2:89-101
Tuaailago farfara B. 1:169-181
Vaccinium myrtillus L. 5:Ij2-I46
uliginosum L, ^177-187
vitis-idaea L. 4:167-178
Veratzrum lobelianum Bernh. 5:94-104
Viburnum opulus L. 4:191-197
Vioie cracca L. 4:114-126
Zarna inermis (Leys.) Lindm. cm. Bromua inermis
262
СОДЕРЖАНИЕ
Щитовник мужской. И.И.Науялис, В.Р.Филин................ g
Кочедыжник женский. И.И.Науялис, В.Р.Филин ............ 26
Род Тимофеевка. А.Р.Матвеев . .........................41
I. Тимофеевка луговая . . . . .....................41
2. Тимофеевка степная.............................. .57
Род Рябчик. М.Г.Вахрамеева, С.В.Никитина, Л.В.Денисова ,
I. Рябчик русский ........................... . . . . gg
2. Рябчик шахматный................................S3
Белокрыльник болотный. А.П.Ефремсв, Ю.Е.Алексеев .... 91
Лунник оживающий. В.А.Романова ........................ 98
Лядвенец рогатый. Н.И.Писковацкова, И.С.Михайловская . .
Липа сердцевидная. Л.П.Рысин...........*................Х28
Род Грушанка. Т.В.Багдасарова, М.Г.Вахрамеева, С.В.Никити-
на, Л.В.Денисова . . . ....................... .153
I. Грушанка круглолистная..........................
2. Грушанка малая..................................184
3. Грушанка средняя................................169
4. Грушанка зеленсцветная . ...................... J72
Голубика обыкновенная. Т.П.Баландина .................. . 177
Род Подорожник. Л.А.Жукова . . . .....................188
I. Подорожник большой...............................189
2. Подорожник средний.............................. . 197
3. Подорожник ланцетный............................203
Жимолость лесная. М.Г.Вахрамеева ....................... 210
Литература . . ......................................... 219
Сводный указатель названий видов, охарактеризованных в 1-7 .
выпусках "Биологической флоры Московской области” . 257
263
Биологическая флора
Московской области, вып.7.
Зав. редакцией Н.М.Глазкова
Редактор Н.М.Горелик
мл. редактор Т.А.Бритвина
Художественный редактор М.Ф.Евстафьева
Технический редактор Г.Д.Колоскова
Подписано к печати 30.09.82 Л-110169 Формат 60x90/16
Бумага офс. I Уел. печ. л. 16;5 Уч.-изй. л. 15,37
Тираж 400 экз. Заказ/до? Цена I р. 10 к. Заказная
Ордена "Знак Почета" издательство Московского, университета
103009, Москва, ул. Гердена. 5/7.
Типография ордена "Знак Псчета" изд-ва МТУ. Москва, Ленинские горн