/



































































































































Автор: Козловская Н.В. Цвелев Н.Н.
Теги: общая ботаника ботаника флористика сравнительная флористика
Год: 1987




Текст
УДК581.9 001.11+002
Теоретические и методические проблемы сравнительной флористики: Материалы
II рабочего совещания по сравнительной флористике. Неринга, 1983. - Л.: Наука,
1987. - 283 с.
Сборник содержит переработанные и дополненные авторами материалы II рабочего
совещания по сравнительной флористике (г. Неринга, 1983 г.), отражающие совре-
менное состояние ее теории и методов, тенденции развития. Предметом статей сбор-
ника являются анализ базового понятия флоры, подходы к изучению флор, оценка
разных опорных единиц флористических сравнений, вопросы методики, примене-
ния математических методов. Сборник заключают одобренные участниками сове-
щания программы флористических исследований, анализ системы понятий и терми-
нов флористики.
Рецензенты:
Н. В. КОЗЛОВСКАЯ, Н.Н.ЦВЕЛЕВ
Редакционная коллегия:
Р. В. Камелин, Ж. П. Лаздаускайте (секретарь), А. А. Лякавичюс,
Л. И. Малышев, О. В. Ребристая, Б. А. Юрцев
КМТИММ1
ЫШ. ИЗЯ
„ 2004000000-610 о„ тт ~ „ 1поп
оп--- 264-87 -// © Издательство „Наука 1987 г.
ПРЕДИСЛОВИЕ
Представляемый сборник по цельности композиции, единству теоре-
тических и методических установок авторов приближается к коллектив-
ной монографии. Он содержит материалы II рабочего совещания по срав-
нительной флористике (20—24 сентября 1983 г., г. Неринга, Литовская
ССР), ревизованные авторами спустя некоторое время после совещания
и отражающие современное состояние теории и методов флористики
в нашей стране, тенденции развития этой науки. Совещание было посвя-
щено памяти выдающегося советского ботаника Александра Иннокен-
тьевича Толмачева, одного из основоположников современной сравни-
тельной флористики, в связи с 80-летием со дня его рождения.
Непосредственным предшественником совещания в Неринге было
I рабочее совещание по сравнительной флористике, проведенное Комис-
сией ВБО по истории флоры и растительности в апреле 1971 г. в Ленин-
граде. Тематика первого совещания была ограничена обсуждением мето-
да конкретных флор (КФ), уже получившего к тому времени широкое
распространение в нашей стране; активным участником был А. И. Тол-
мачев, автор метода. Совещание проводилось в форме дискуссии „за
крутым столом” и характеризовалось высоким полемическим нака-
лом; в ходе полемики участники ее, имевшие уже немалый опыт работы
методом КФ, впервые осознали, что на базе метода фактически сложи-
лись 2 научные традиции (обе берущие начало от А. И. Толмачева), отли-
чающиеся как трактовкой КФ, так и некоторыми деталями методики
(наиболее резко это проявилось в вопросе о площади выявления КФ).
В ходе самой полемики были впервые сформулированы концепции „про-
бы флоры” (флоры географического пункта) и „парциальной флоры”.
Позиции участников дискуссии по некоторым вопросам отличались не-
примиримостью, что проявилось и в последующих публикациях. Труды
совещания не издавались в виде сборника, но в „Ботаническом журнале”
была опубликована его подробная научная хроника (Юрцев, 1974а), для
которой большинство участников сами составили обобщенные резюме
своих выступлений. Кроме того, некоторые участники опубликовали
статьи, содержащие ревизованный текст их выступлений на совещании
(Малышев, 1972; Шмидт, 1972,1976; Юрцев, 1975а).
В целом же I совещание дало импульс развитию сравнительной фло-
ристики в нашей стране, в частности ее теории и методов. Оно способст-
вовало дальнейшей популяризации метода КФ, те или иные модификации
3
которого были приняты на вооружение новыми коллективами флори-
стов. За время, прошедшее между I и II рабочими совещаниями, был
завершен ряд крупных циклов флористических исследований, выпол-
ненных как методом КФ, так и другими; выполнены и некоторые спе-
циальные методические исследования по вопросам, сформулированным
на совещании в апреле 1971 г. (Малышев, 19766; Шмидт, 1976; Барано-
ва, 1977; Табака и др., 1977; Заславская, Петровский, 1983; Ребристая,
статья в наст, сб.). Были обнародованы и новые теоретические концеп-
ции в области флористики и ботанической географии. Существенно рас-
ширились опыт применения и сам арсенал математических методов при
сравнительном изучении флор. Расширился и круг авторов, не только
участвующих в региональных флористических исследованиях, но и кров-
но заинтересованных в разрешении теоретических и методических проб-
лем флористики. Перечисленные факторы, вероятно, и определили успех
И рабочего совещания, весь труд по непосредственной организации и
проведению которого взяли на себя литовские ботаники.
Прогресс в развитии сравнительной флористики в нашей стране за
период с 1971 г. по 1983 г. нашел отражение и в более широкой тематике
совещания в Неринге. Так, здесь обсуждались содержание базового поня-
тия флористики и ботанической географии „флора”, основные подходы
к изучению флоры; метод КФ рассматривался уже в ряду других мето-
дов сравнительной флористики, отмечались не только его сильные сторо-
ны, но и ограничения на его применение; обсуждались также тенденции
и перспективы дальнейшего развития и взаимодействия методов флори-
стики, а также программы флористических и комплексных ботанико-
географических исследований разной детальности.
Несомненный успех совещания означал успех научных начинаний
Толмачева, способствовал дальнейшей консолидации его школы. Были
представлены почти все основные коллективы флористов страны — от
Прибалтики до Сибири (за исключением ряда московских и дальнево-
сточных, что лишь отчасти компенсируется включением в настоящий
сборник статьи А. К. Скворцова о флористических исследованиях в за-
падных районах Нечерноземья). Особенно отрадным было активное уча-
стие в совещании научной молодежи, что является важным звеном
в обеспечении преемственности поколений флористов, в передаче накоп-
ленного опыта.
Помимо краткого перечня вопросов для обсуждения и собственно
Программы совещания (с указанием темы каждого заседания и намечен-
ных докладов), участники заблаговременно получили список научных
работ (37 названий), которые были опубликованы после I совещания
по проблемам, намеченным к обсуждению, и содержали богатый свежий
фактический материал; в целом уровень фактической „оснащенности”
защищаемых положений на II совещании существенно повысился.
Успеху совещания несомненно способствовала обстановка уюта и
комфорта, созданная гостеприимными хозяевами — литовскими бота-
никами (на базе одного из пустовавших в осеннее время пансионатов
Неринги). Об энтузиазме участников совещания говорит хотя бы то,
что ежедневно после двух запланированных заседаний они собирались
4
вечером в уютном холле библиотеки пансионата для продолжения дис-
куссий. И на заседаниях, и в многочисленных беседах и спорах в кулуа-
рах царила атмосфера свободной, дружественной, творческой дискуссии,
благодаря чему работа совещания была весьма продуктивной, а обсуж-
дение — конструктивным, приведшим к значительному сближению
взглядов дискутировавших сторон и прежде всего к взаимопониманию.
Совещание продемонстрировало единство взглядов участников, пред-
ставлявших разные коллективы флористов, на флору как естественно-
исторический феномен; было достигнуто согласие и в вопросе о соот-
ношении и взаимодействии разных методов полевых флористических
исследований, что позволило заключить настоящий сборник трудов сове-
щания публикацией программ флористических исследований разной де-
тальности (на основе проекта, обсуждавшегося на совещании и в целом
одобренного участниками). Наконец, в серии докладов была продемон-
стрирована плодотворность дифференциального изучения региональных
флор по их внутриландшафтным подразделениям (парциальным флорам
экотопов, ценофлорам, флороценотипам). Наибольшие разногласия, как
это часто бывает, выявились в вопросах терминологии, в силу чего мно-
гими участниками было высказано предложение о создании словаря тер-
минов на базе системы основных понятий флористики. Поэтому в конце
настоящего сборника приводится „Очерк системы основных понятий
флористики”, подготовленный Б. А. Юрцевым и Р. В. Камелиным для
обсуждения на следующем рабочем совещании по сравнительной флори-
стике. Эта система, по мысли составителей очерка, и должна послужить
основой для сведения синонимичных терминов, применяемых разными
авторами, облегчить взаимопонимание, подготовить предпосылки для
унификации (в недалеком будущем) терминологии флористики и про-
чих ботанико-географических дисциплин.
В целом совещание 1983 г. в Неринге составило ощутимый этап
в развитии сравнительной флористики в нашей стране, чему в немалой
степени должна способствовать публикация его трудов и флористиче-
ских программ в данном сборнике.
В первом разделе сборника помещены 4 концептуальные статьи, по-
священные анализу базового понятия „флора” и связанным с ним мето-
дологическим проблемам флористики.
С обсуждения этих вопросов и началось совещание в Неринге. Общим
для всех 4 статей является признание флоры системой видов в их мате-
риальном выражении на территории флоры (т. е. системой их местных
популяций), что определяет и подходы к изучению современных флор,
в частности особое внимание к установлению особенностей местных
популяций всех видов, их распределения по территории флоры, эколого-
ценотических связей, их местной истории. Это положение не является
пока общепризнанным — ему противостоит традиция считать флору
лишь качественным спектром таксонов данной территории или, еще про-
ще, списком видов, обитающих на ней (более подробно обе конкурирую-
щие точки зрения рассмотрены в 1-й статье данного раздела). Отголоски
2-й точки зрения имеются и в статье Ю. Р. Шеляг-Сосонко и Я. П. Дидуха
5
(признание флоры качественной, притом „ретроспективной” системой
популяций, элементы которой не взаимодействуют друг с другом), хотя
авторы считают элементами флоры (как системы) именно популяции
видов, а не просто таксоны, и в своей программе изучения флоры боль-
шое место уделяют анализу ценотической структуры флорол, ценофлор
и т. д.; таким образом, подходы названных авторов к изучению флоры -
те же, что и в других статьях данного раздела.
Попытка защитить распространенную („классическую”) точку зре-
ния на флору, фауну, биоту („композиция” таксонов, но не система
популяций) предпринята недавно Ю. И. Черновым (1984а, 19846). Собст-
венно говоря, весь пафос критики Чернова (1984а), по крайней мере
в данном вопросе, направлен на понятийно-терминологический аспект
работ критикуемых авторов, обвиняемых им в смешении понятий и тер-
минов биогеографии (в том числе флористики, фаунистики) и синэко-
логии. Так, Чернов не возражает против привлечения популяционных
характеристик видов к анализу и сравнению флор, но считает это заимст-
вованием флористикой понятий и методов синэкологии. При этом он
упускает, что современная концепция вида в систематике и флористике
по самой своей сути популяционно-генетическая и что даже классическая
задача флористики — инвентаризация флоры” — ныне предполагает не
только выявление всего разнообразия таксонов, но и установление фе-
нотипического и экологического своеобразия местных популяций каж-
дого вида, без чего, в частности, немыслима и научно-грамотная поста-
новка охраны генофонда местной флоры.
Не заметил Чернов и того, что даже в классических представлениях
о фитоценозе по существу совмещены 2 концепции последнего: как со-
вокупности и системы популяций всех видов, сообитающих на данном
экотопе, т. е. парциальной флоры (растительного населения) экотопа,
и как типа ценотической организации, производного от ценобиотических
свойств сообитающих особей (биоморфы, ценобиоморфы, ценотипы),
но инвариантного к их видовому составу. Элементами систем первого
рода являются виды (их местные популяции), второго — особи. То же
полностью справедливо и по отношению к концепции биоценоза. Поэто-
му, если говорить о фитоценозе как о растительном населении экотопа,
основные факторы филоценогенеза окажутся общими с таковыми фло-
рогенеза (формирования и развития региональных флор) ; более того,
сам флорогенез представляет интегральный итог филоценогенезов всех
типов сообществ на территории флоры. Что же касается тезиса Чернова
о конвергентном формировании однотипных сообществ в разных очагах
флорогенеза из разного таксономического материала как о принципи-
альном отличии филоценогенеза от флорогенеза, то это относится лишь
к абстрактным типам ценотической организации, но не к конкретным
ценотическим системам: в разных очагах флорогенеза в сходных усло-
виях формируются некоторые подобия, но отнюдь не тождественные
ценотические системы.
Развернутое определение флоры, данное в статье Г. В. Вынаева,
как и известное определение Толмачева (1974, с. 112), ограничивает
объем этого понятия единицами регионального уровня, являющимися
6
основными объектами флористики; в статьях Юрцева, а также Шеляг-Со-
сонко и Дидуха флора трактуется как универсальное безранговое поня-
тие, приложимое и к единицам топологического (внутриландшафтного)
уровня (вплоть до фитоценоза), выделяемым как парциальные флоры
(ПФ) или ценофлоры. Обсуждение этого вопроса, живо дебатировавше-
гося на совещании в Неринге, дано во вводной статье Юрцева. Подчерк-
ну, что и данный вопрос дискутировался исключительно в понятийно-
терминологическом аспекте, поскольку в серии заслушанных докладов
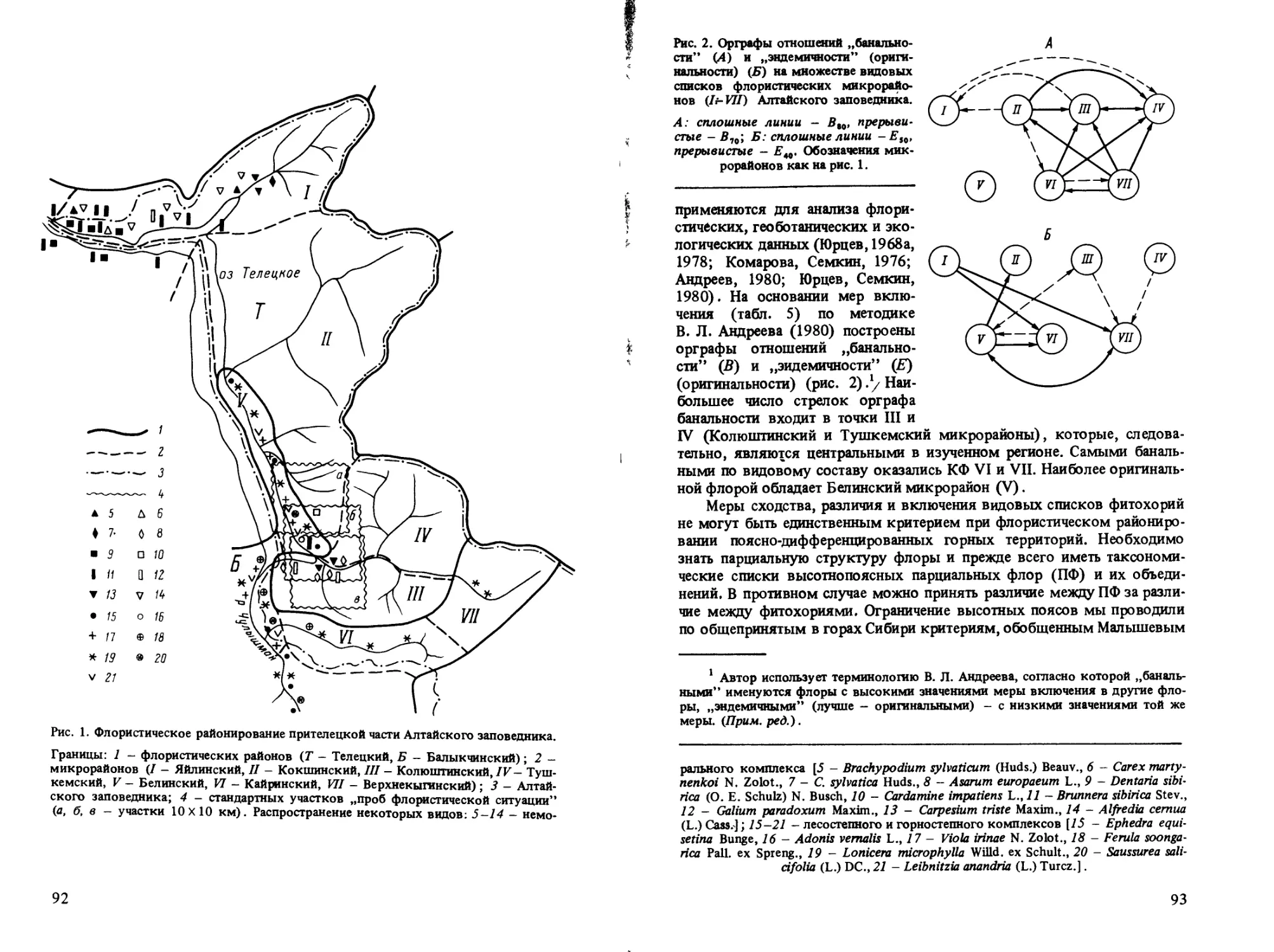
(см. статьи Л. В. Мариной, Я. П. Дидуха, В. П. Седельникова, А. А. Сага-
телян, В. А. Мартыненко) было убедительно показано, что ПФ,и в частно-
сти ценофлоры (объединения ПФ экологически, флористически и струк-
турно-сходных сообществ), могут устойчиво различаться, а следователь-
но, и анализироваться практически по всем тем же показателям, что и
региональные флоры.1
В статье Р. В. Камелина, заключающей первый раздел сборника,
акцентирован эволюционноисторический аспект изучения флоры, к сожа-
лению, все реже затрагиваемый при анализе региональных флор и. веро-
ятно, наиболее трудный из-за необходимости привлечения разнообраз-
ного материала из смежных областей (в том числе из-за необходимости
сопряженного филогенетического анализа многих таксонов). Этот
аспект изучения природных флор — один из самых важных; ему предпо-
лагается уделить большое внимание на следующем рабочем совещании.
Второй раздел сборника, посвященный методическим проблемам
флористики, включает 18 статей, сгруппированных в 2 подраздела: пер-
вый посвящен обсуждению и апробации различных опорных единиц фло-
ристических сопоставлений (7 статей), а также сравнительному анализу
парциальных флор (4 статьи и вторая часть статьи Л. В. Мариной) ; вто-
рой — общим и частным вопросам использования математических мето-
дов для решения основных задач сравнительной флористики (7 статей).
1 В недавно опубликованной статье Б. Н. Норина „Флористическая, экологиче-
ская и фитоценолотическая интерпретация строения растительного покрова” (Бо-
тан. журн. 1984. Т. 69, № 3. С. 273-282) постулируется взаимная независимость
флористической, эколотической и фитоценолотической концепций (качественных
моделей) растительного покрова, но полностью обойден молчанием вопрос о воз-
можных путях синтеза результатов этих подходов к изучению единого объекта.
В качестве элементарных выделов растительного покрова (при разных подходах)
рассматриваются соответственно КФ, „растительность отдельных местообитаний
(участков) ландшафтного экофона (по Ю. П. Кожевникову) ”, наконец, „ценоти-
чески связанная фитохора” (по Б. Н. Норину); в качестве элементов флористиче-
ских, экологических, фитоценотических систем трактуются видовые популяции,
экобиоморфы, ценоячейки или синузии соответственно. Однако нетрудно пока-
зать, что все рассмотренные Нориным элементарные подразделения растительного
покрова, выделенные при названных трех подходах, представляют ступени единой
иерархии систем популяций разного уровня территориальной размерности и что
каждая из этих ступеней может изучаться с точки зрения качественного (в том
wcne и экобиоморфного) состава, количественного соотношения, а также взаимо-
действия сообитающих видов и их типологических объединений разного рода, как
и с точки зрения генезиса и тенденций развития данной системы и ее экологических
связей с окружающей средой (в этом и заключена основа синтеза разных подходов
к единому объекту).
7
В вопросе о соотношении опорных единиц сравнительной флористи-
ки одной из тонкостей по-прежнему остается различение КФ как наи-
меньшего естественного регионально-флористического выдела и пробы
флоры (флоры географического пункта; локальной флоры), которая
во многих случаях может выявлять подавляющую часть состава КФ, ха-
рактерные для нее соотношение элементов, экотопологическую диффе-
ренциацию и т. д., но может также фиксировать ситуации переходных
или контактных зон между двумя или несколькими КФ, либо же ситуа-
цию флористического континуума. Как видно из материалов сборника
и большинства других опубликованных работ по КФ, под последними
обычно имеются в виду именно пробы флоры, не привязанные к кон-
кретному флористическому выделу, и лишь в статье Н. И. Золотухина
под КФ подразумеваются элементарные выдели дробного флористиче-
ского районирования. Эта статья и помещенная вслед за ней работа
Л. В. Табака методически интересны также тем, что авторы переходят
от выборочного к сплошному детальному флористическому обследова-
нию территории, что пока возможно лишь для территорий с высокой
общей плотностью накопленной флористической информации, либо для
охраняемых территорий, в первую очередь заповедников. Статья Л.В.Ма-
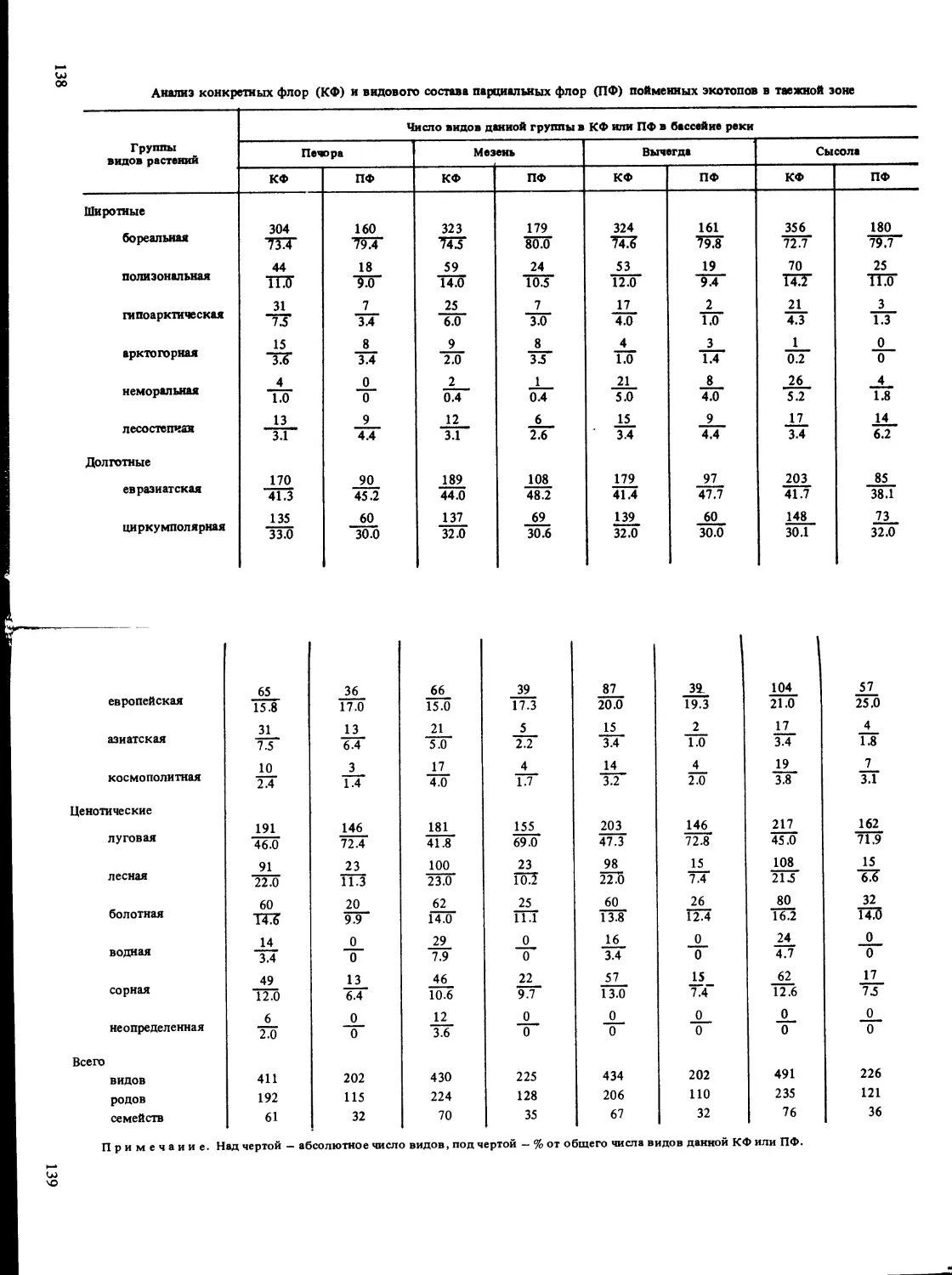
риной содержит доказательства индивидуальности флор небольших реч-
ных бассейнов - в пределах КФ, что проявляется не только в наличии
дифференциальных видов, но и в своеобразии экотопологической струк-
туры флор разных бассейнов, а также в устойчивых отличиях экологиче-
ски однотипных ПФ в различных бассейнах. Это заставляет уделить спе-
циальное внимание при дальнейших методических исследованиях поиску
и апробации критериев флористической гомогенности территории, по-
скольку предложенный Толмачевым критерий постоянства набора видов
на однотипных экотопах в пределах КФ „работает” далеко не всегда.
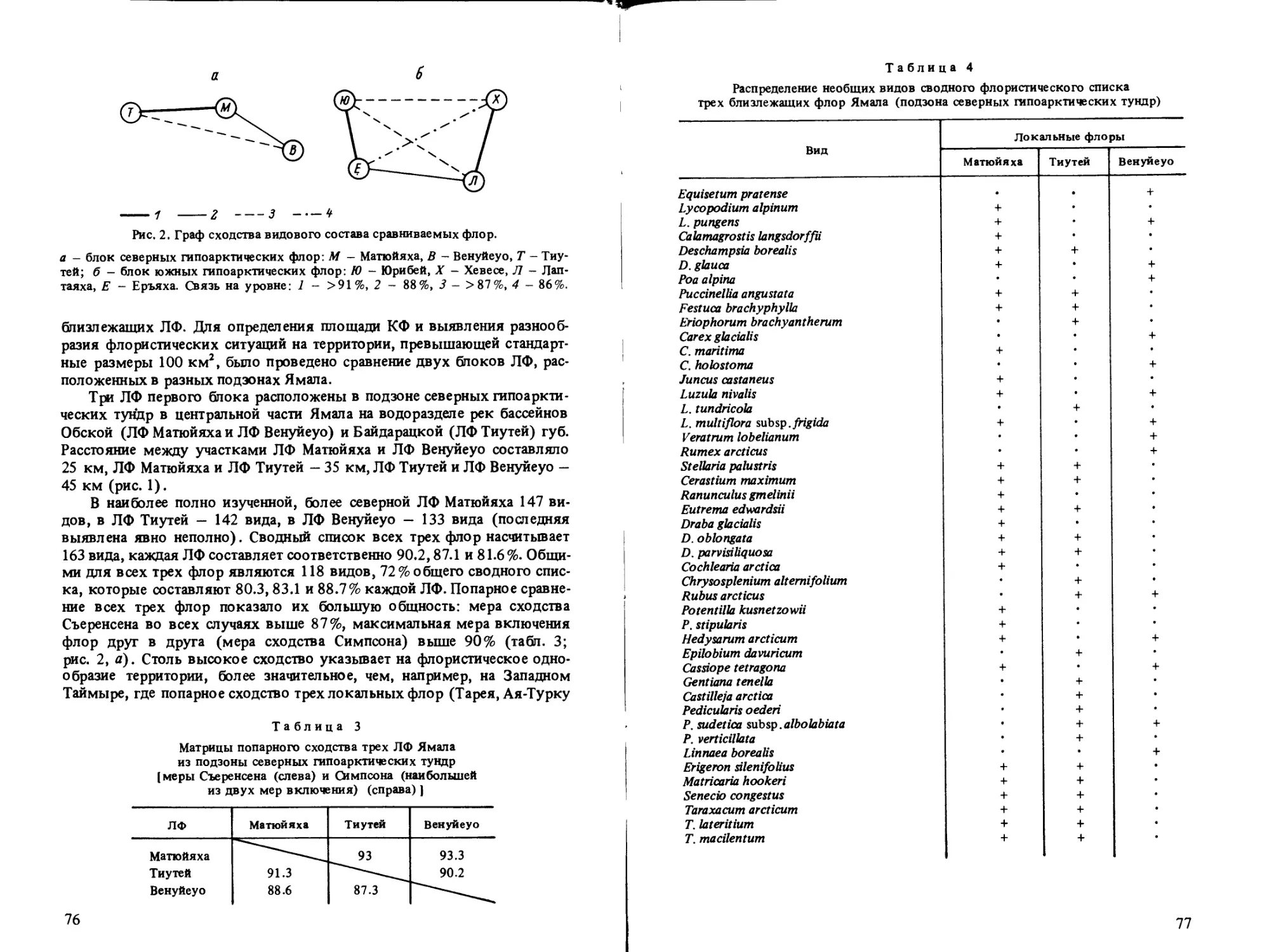
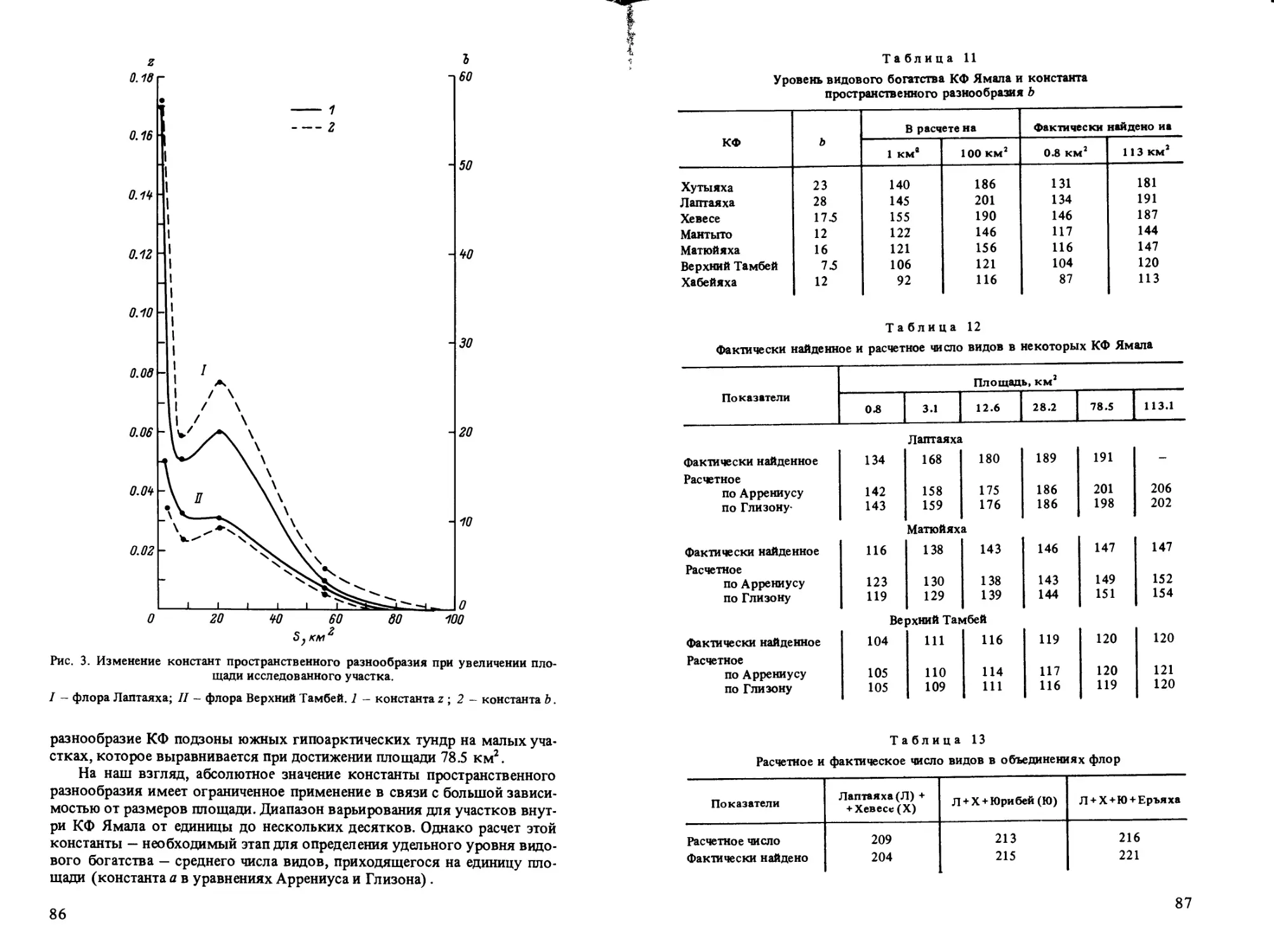
В статье О. В. Ребристой показана значительная монотонность (малое
пространственное разнообразие) флоры тундровой низменности Ямала,
что убедительно иллюстрируется анализом зависимости видового богат-
ства от площади пробы флоры.
В цикле статей из того же подраздела, посвященных сравнительному
анализу ПФ, все авторы оперируют с уже упоминавшейся категорией
ценофлор („формационный анализ”, по Толмачеву). Подчеркну, что
выделение и изучение ценофлор имеет смысл только в том случае, если
синтаксоны, которым они принадлежат, имеют четкую экологическую
и экотопологическую привязку; точно так же даже первичное выделение
ПФ экотопов предполагает привязку их к определенным местоположе-
ниям со сходной растительностью — практически мы имеем дело с типа-
ми и классами биотопов. В интересах проведения сравнительного анализа
вы дел ы ПФ должны быть достаточно крупными (для обеспечения обзор-
ности) при достаточной эколого-флористической гомогенности. Этим
требованиям наиболее отвечают флоры простых мезоэкотопов (экологи-
чески достаточно однородных урочищ, объединяющих близкие типы
фаций), либо же объединения списков сообществ экологически и флори-
стически целостных, более или менее крупных синтаксонов (вплоть до
флороценотипов в смысле П. Н. Овчинникова и Р. В. Камелина - флоро-
8
генетически единых типов растительности, также порядков и классов
в классификационной системе сигматистов). Кстати, флористическая
классификация сообществ в духе школы Браун—Бланке в своей основе
имеет группировку ПФ (микро) экотопов по их флористическому сход-
ству, и детально разработанная этой школой процедура упорядочения
прямоугольной матрицы описаний по вертикали и горизонтали может
быть использована при обработке сводных списков различных катего-
рий ПФ.1 Так, перестановка столбцов (описаний) по сходству набора
видов (с учетом количественных показателей) приводит к выделению
ценофлор (объединению ПФ) разного ранга, перестановка строчек по
сходству распределения видов по описаниям и синтаксонам дает эколо-
го-ценотические группы видов; ядро характерных видов крупного эко-
логически и флористически цельного синтаксона или определенного
класса ПФ есть его флороценотический комплекс. Таким образом, на
(элементарном) уровне микроэкотопа, фации, фитоценоза изучение ПФ
проводилось фактически с ранних этапов становления фитоценологии;
для современного этапа характерно включение элементарных ПФ
в иерархию внутриландшафтных флористических (экотопологических)
выделов и, главное, применение к ним мощного современного аппарата
анализа флоры.
Статьи Я. П. Дидуха, В. П. Седельникова, А. А. Сагателян, отличаю-
щиеся подходами к выделению синтаксонов с целью сравнительного
анализа их ценофлор, наглядно демонстрируют значение раздельного изу-
чения и анализа крупных классов ПФ для более глубокого понимания за-
кономерностей формирования региональной флоры. В статье Седельни-
кова ставится интересный вопрос о параллелизме между уровнями сход-
ства ПФ синтаксонов возрастающего ранга и уровнями сходства ПФ
микро-, мезо-и макроэкотопов (экотопологических выделов возрастаю-
щей гетерогенности). Для того чтобы ответить на этот вопрос, необходи-
мо в одном и том же районе применить оба подхода к выявлению ПФ
разного ранга (путем типизации массовых описаний фитоценозов и пу-
тем составления списков для микро-, мезо- и макроэкотопов).
В статье В. А. Мартыненко выявлены тенденции зональных измене-
ний ценофлор интразональной луговой растительности — на фоне усред-
ненных изменений таежных КФ по тому же зональному профилю.
Во втором подразделе обзорная статья Л. И. Малышева подводит
итоги накопившегося богатого опыта применения математических мето-
дов в сравнительной флористике (в основном по материалам разнооб-
разных собственных исследований автора); эта статья дополняет две
недавно опубликованные сводки В. М. Шмидта (1980, 1984); в свою
очередь существенным дополнением к ней является публикуемая в том
Же подразделе статья Б. И. Семкина о применении теоретико-графовых
методов при изучении КФ. В большинстве остальных статей того же
подраздела приводятся результаты исследований по тому или иному
конкретному вопросу, представляющие интерес как с содержательной
1 Существуют и строго количественные методы упорядочения прямоугольной
матрицы (R0-анализа).
9
стороны, так и использованием тех или иных приемов количественного
анализа (см. также статьи Мариной, Золотухина, Ребристой, Дидуха,
Седельникова, Лякавичюса, помещенные в других разделах). Статьи
М. Лайвиныпа и др., также статья А. А. Лякавичюса информируют о ра-
ботах латвийских и литовских ботаников по созданию информационно-
поисковых систем, которые в будущем должны получить все более
широкое распространение в сравнительной флористике, оперирующей
с громоздкими массивами исходных данных.
Значительный интерес представляют и другие статьи, излагающие
результаты региональных флористических исследований в Литве, Сред-
ней России, на Кавказе (раздел III).
Основные теоретико-методические установки первых разделов сбор-
ника сфокусированы в итоговом разделе, содержащем программы фло-
ристических исследований разной степени детальности. Программы под-
готовлены Б. А. Юрцевым при участии Р. В. Камелинаи с учетом много-
численных устных и письменных замечаний и пожеланий, сделанных по
обсуждавшемуся на совещании в Неринге проекту. Эти программы при-
званы служить основой при разработке конкретных программ, которые
в каждом случае должны быть достаточно индивидуализированы и отра-
жать опыт и научные интересы данного коллектива флористов, равно
как размеры, природные особенности и степень изученности территории.
В программах уделено немало внимания участию флористов в комплекс-
ных ботанико-географических и экологических (биогеоценологических)
исследованиях; учтена специфика изучения флоры территорий, претер-
певших антропогенную трансформацию.
Задачи флористических исследований формулируются в порядке
возрастающей сложности; формулируя их, мы стремились не только
указать ближайшие, первоочередные задачи, но и наметить перспективы
развития флористики по пути не только расширения, но и углубления
наших знаний о природных флорах.
Составители сборника выражают надежду, что его публикация ока-
жет действенную методическую помощь коллективам ботаников, изу-
чающих флору различных частей страны, увеличит их заинтересованность
в разработке теоретических и методических проблем флористики и по-
служит стимулом для дальнейшего прогресса науки о флоре в нашей
стране.
Январь 1986 г. Б. Юрцев
10
РЕШЕНИЕ II РАБОЧЕГО СОВЕЩАНИЯ
ПО СРАВНИТЕЛЬНОЙ ФЛОРИСТИКЕ
С 20 по 24 сентября 1983 г. в г. Неринга Литовской ССР состоялось
Второе рабочее совещание по сравнительной флористике, организованное
Всесоюзным ботаническим обществом (Комиссия по истории флоры и
растительности), Научным советом по проблеме „Биологические основы
рационального использования, преобразования и охраны растительного
мира” (Секция флоры и систематики высших растений), Институтом
ботаники АН Литовской ССР (Лаборатория флоры и геоботаники) и
посвященное 80-летию со дня рождения А. И. Толмачева, автора концеп-
ции и метода конкретной (элементарной) флоры, одного из основопо-
ложников современной сравнительной флористики в нашей стране.
Совещание собрало основной актив ботаников, кровно заинтересо-
ванных в разработке теории и методики флористики, из различных науч-
ных центров СССР (10 институтов Академии наук СССР и академий
союзных республик, 6 университетов, Сельскохозяйственной академии
в г. Каунасе, 3 заповедников; всего 54 участника из 14 городов и 3 запо-
ведников, в том числе 9 докторов и 27 кандидатов наук). Среди присут-
ствовавших значительную часть составляла научная молодежь.
Совещание отразило значительные продвижения в области сравни-
тельной флористики в нашей стране за последние десятилетия, связанные
во многом с поиском и разработкой новых методов. Завершен ряд цик-
лов региональных исследований методом конкретных флор и другими
методами, результаты опубликованы в виде монографий и серий статей.
За 12 лет, истекших после I рабочего совещания (апрель 1971, Ленин-
град), появились новые теоретические концепции, методические разра-
ботки; достигнуты успехи и в применении математических методов
к решению задач сравнительной флористики. В настоящее время наша
страна занимает лидирующее положение в области науки о флоре, и в ча-
стности в вопросе об элементарных единицах дифференциации регио-
нальных флор как основе флористических сопоставлений. Показателем
интенсивного развития является и приток молодых кадров в данную
область ботаники.
Совещание проходило в атмосфере товарищеской, конструктивной
дискуссии при высокой активности участников. Оно показало, что среди
исследователей имеется принципиальное единство во взглядах на флору
как на природное (естественноисторическое) явление и на подходы к ее
изучению. Состоявшийся плодотворный обмен мнениями и опытом
11
между участниками способствовал уяснению соотношения разных мето-
дов полевых сравнительно-флористических исследовании, оптимальной
области применения каждого метода, перспектив их дальнейшего разви-
тия. Существование непреодолимых пока еще терминологических разно-
гласий делает актуальным унификацию терминологии в области флори-
стики и как временную, но неотложную меру — составление словаря
терминов (с выделением синонимических).
Совещание постановляет:
1. Одобрить работу, проделанную Комиссией ВБО по истории флоры
и растительности и Лабораторией флоры и геоботаники Института бота-
ники АН Литовской ССР по подготовке и проведению настоящего сове-
щания. Выразить благодарность всех участников непосредственным орга-
низаторам — литовским ботаникам — за гостеприимство и высокий уро-
вень проведения совещания.
2. Одобрить и принять за основу подготовленный инициативной
группой (Ботанический институт АН СССР) проект типовых программ
флористических исследований разной степени детальности. Создать рабо-
чую комиссию в составе: Р. В. Камелии, О. В. Ребристая, В. М. Шмидт,
Б. А. Юрцев для подготовки программ к публикации с учетом сделанных
замечаний и предложений.
3. Просить редколлегии „Ботанического журнала” и „Украинского
ботанического журнала” опубликовать научную хронику совещания и
обсуждавшиеся на нем типовые программы флористических исследо-
ваний.
4. Просить Научный совет по проблеме „Биологические основы ра-
ционального Использования, преобразования и охраны растительного
мира” издать труды совещания в виде сборника „Теоретические и мето-
дические проблемы современной флористики (Материалы Второго рабо-
чего совещания по сравнительной флористике 20—24 сентября 1983 г.,
г. Неринга Литовской ССР)” объемом 20 авт. листов, для подготовки
сборника к печати избрать редколлегию в составе: Р. В. Камелин,
Ж. П. Лаздаускайте, А. А. Лякавичюс, Л. И. Малышев, О. В. Ребристая,
В. М. Шмидт, Б. А. Юрцев.
5. Признать целесообразным проводить регулярно рабочие совеща-
ния по сравнительной флористике через каждые 5 лет; просить Проблем-
ный совет по ботанике предусмотреть в координационных планах на
ХП пятилетку проведение ближайшего (третьего) совещания в 1988 г.
6. Признать целесообразным включить в программу 3-го рабочего
совещания по сравнительной флористике обсуждение новых теоретиче-
ских и методических разработок в области сравнительной флористики,
современных методов комплексного анализа флоры и восстановления ее
истории, также сбора, хранения и обработки флористической информа-
ции, особенностей флористического изучения территорий, сильно нару-
шенных хозяйственной деятельностью человека.
12
I. ФЛОРА КАК БАЗОВОЕ ПОНЯТИЕ
ФЛОРИСТИКИ
Б. А. Юрцев
ФЛОРА КАК БАЗОВОЕ ПОНЯТИЕ ФЛОРИСТИКИ:
СОДЕРЖАНИЕ ПОНЯТИЯ, ПОДХОДЫ К ИЗУЧЕНИЮ
Ботанический институт им. В. Л. Комарова АН СССР
Флористика в нашей стране переживает период оживленного теоре-
тического и методического поиска; свидетельством этого является и
ставший традицией созыв рабочих совещаний по сравнительной флори-
стике. В вопросе об элементарных естественных подразделениях флоры,
об опорных единицах флористических сопоставлений, о территориально-
экологических составляющих элементарных флор отечественная флори-
стика сейчас безраздельно лидирует. И это вполне закономерно: еще
в 1931 г. А. И. Толмачев (1931), обнародовав интересную, стимулирую-
щую концепцию конкретной, или элементарной, флоры, поставил вопрос
об элементарных единицах флористических сравнений, о выявлении фло-
ристических комплексов разных экотопов и тем самым очертил сферу
пересечения интересов флористики и геоботаники, область их взаимодей-
ствия в познании растительного мира.
А. И. Толмачев активно участвовал в I рабочем совещании по сравни-
тельной флористике (Ленинград, апрель 1971 г.). Сейчас его нет в наших
рядах, но его идеи и методы по-прежнему действенны, а следовательно,
развиваются его учениками и учениками его учеников. И несомненный
успех II рабочего совещания (Неринга, сентябрь 1983 г.), посвященного
80-летию со дня рождения А. И. Толмачева, явился лучшей памятью об
одном из основоположников современной отечественной флористики.
Прогресс сравнительной флористики в нашей стране за последние де-
сятилетия вызвал к жизни целый ряд новых понятий и терминов, произ-
водных от базового понятия „флора”, таких как „естественная флора”,
„конкретная, или элементарная, флора”, „локальная флора”, „проба
флоры”, „парциальная флора”. Вполне очевидно, что от трактовки базо-
вого понятия зависит и наш подход к изучению перечисленных выше
единиц. Расширение функций современной флористики, все более актив-
ное участие флористов в решении вопросов охраны растительного мира
и в комплексных экологических исследованиях заставляют расширить
содержание понятия „флора”, с тем чтобы привести его в соответствие
с новыми запросами к флористике и с современной практикой флори-
стических исследований.
Подробный анализ понятия „флора” дан в статье „Флора как при-
родная система” (Юрцев, 1982а), представляющей заранее опубликован-
ный полный текст доклада, специально подготовленного для обсуждения
13
на II рабочем совещании по сравнительной флористике. В публикуемом
здесь ревизованном тексте вступительного доклада автора на открытии
совещания акцентируются основные положения предлагаемой расширен-
ной концепции флоры, с особым вниманием к дискуссионным вопросам,
обсуждавшимся на совещании.
1. Первый (центральный) вопрос, связанный с принятием данной
концепции: флора —Список ли видов некоторой целостной территории
(таксономический спектр) или же сам природный объект, отображае-
"' мый этим списком, т. е. реальная совокупность видов в той форме, как
они представлены/на данной территории — иными словами, совокупность
местных географических популяции всех видов растений (растительное
население, смешанная популяция растений) 1 К последней формулировке
приходишь на основе синтеза традиционного понимания флоры как мно-
жества видов в топографическом контуре и современного взгляда на вид
как на полную (генеральную) популяцию.
Для Толмачева — флориста и ботанико-географа, как и для его уче-
ников и последователей, а также для большинства других отечественных
флористов-фитогеографов, этот вопрос никогда не был актуален: флора
всегда понималась и понимается как природное, естественноисториче-
ское образование, воплощенное в ткани растительного покрова. Между
тем в современной советской биогеографической (фактически зоогео-
графической) литературе часто проводится резкая грань между фауной
как совокупностью видов, представленной их списком, и животным ми-
ром, или животным населением, как совокупностью всех особей (попу-
ляций) этих видов. Фауну принято считать объектом зоогеографии (или
фауногенетической зоогеографии), животное население — объектом
экологии (или экологической зоогеографии; см.: Беклемишев, 1982;
Старобогатов, 1982); впрочем, имеются и другие взгляды (см., напри-
мер: Тупикова, 1976).
Тот же центральный вопрос можно сформулировать иначе: достаточ-
но ли для познания флоры любой территории зарегистрировать присутст-
вие на ней всех таксонов (выявить таксономический состав флоры), или
> же необходимо в дополнение к этому установить, как каждый таксон
t . - (V., представлен на территории флоры, т. е. определить некоторые элементар-
£ . i» ные характеристики его местной географической популяции? " 4
‘ В числе таких характеристик в первую очередь следует назвать:
,1) географическую структуру популяции, отражающую в конечном счете
! распределение вида по подчиненным фитохориям того или иного ранга
<* (на территории данной региональной флоры) или же по представитель-
< v г < 1 ной совокупности проб флоры — выборочных или регулярных; 2) экото-
! - дологическую структуру — распределение (в пределах конкретной фито-
. t , хории) по типам и классам экотопов (биотопов) или соответствующим
'* ' 'им типам сообществ; географическая и экотопологическая структуры
могут быть закартированы или же представлены в виде матрицы, где
' t, '. горизонтальные строчки соответствуют видам или расам, вертикаль-
ные — фитохориям или экотопам (биотопам) определенного ранга;
в результате сравнения распределения разных видов по фитохориям
X- С; , Г
I , , ,7 ' и г ' г £ , , М-G у С-Ъ'Ч 1 { / 1 ' 't_
или экотопам и сравнения фитохорий и экотонов — по набору видов
матрица может быть упорядочена по вертикали и горизонтали; 3) широ-
ту экологической амплитуды местной популяции, измеряемую в физи-
ческой шкале амплитудой значений лимитирующих факторов, при кото-
рых встречается вид, а в био экологической — числом классов экотопов,
освоенных им, в пределах данной фитохории; 4) активность вида (точ-
нее, его местной географической популяции) как меру его жизненного
преуспевания на данной территории, в общем пропорциональную степени
насыщения последней этим видом (степени заполнения местной популя-
цией вида своего „яруса жизни”) ; одно из выражений „веса вида” в дан-
ной флоре.
Целесообразно различать географическую активность, экотопологи-
ческую (внутриландшафтную) активность и парциальную активность
(в пределах того или иного типа или класса экотопов). Географическую
активность можно выразить через встречаемость вида в совокупности
подчиненных фитохорий одного и того же ранга или же в совокупности
проб флоры [регулярных или выборочных: „флористическая частот-
ность”, в смысле М. П. Наткевичайте-Иванаускене и Ю. Ю. Тупчяускайте
(1982)]. Интересным примером использования этого показателя для ре-
шения ботанико-географических проблем является проведенное цитиро-
ванными авторами сравнение распределения различных географических
элементов флоры Литовской ССР по классам географической активно-
сти („флористической частотности”).
Пятибалльная шкала измерения внутриландшафтной активности (по
соотношению широты экологической амплитуды, встречаемости в дан-
ном ландшафте и характерному уровню численности, выраженным также
в баллах) была предложена ранее (Юрцев, 1968а). Позднее предпринима-
лись попытки выразить тот же показатель в более точной шкале отноше-
ний, допускающей все арифметические операции (Катенин, 1974,1981);
в этом случае фактически рассчитывается среднее проективное покрытие
вида в данном ландшафте, определяемое с помощью только двух призна-
ков — компонент: встречаемости и покрытия; однако полученный таким
путем показатель активности действителен лишь для однотипных ланд-
шафтов и является функцией не только макроклимата, но и характера
рельефа и состава горных пород: при одинаковых показателях активно-
сти в двух ландшафтах с резко неодинаковой контрастностью экотопов
реальная активность будет выше в случае большей контрастности (на-
пример, в горных ландшафтах по сравнению с равнинными). Сходные
подходы к измерению активности предлагались Л. И. Малышевым
(1973, 1976а) и А. А. Лякавичюсом (см. наст, сб.) с той разницей, что
исходные показатели — обилие и встречаемость — Малышев выражает
в шкале порядков, а это делает неправомерными последующие опера-
ции — умножение и извлечение корня; Лякавичюс также допускает воз-
можность при расчетах активности выражать обилие в баллах. Я. П. Ди-
ДУх (1982) предложил оригинальный способ отображения активности
на трехкомпонентных гистограммах.
Парциальную активность также предлагалось выражать в шкале
порядков — по соотношению характерного обилия и константности
1С
(Галанин, 1980). Однако в этом случае, вероятно, был бы особенно
оправдан и расчет среднего проективного покрытия; если же можно
определить долю каждого типа или класса экотопов от общей площади
ландшафта (например, путем дешифрирования аэрофотоснимков),
внутриландшафтную активность нетрудно определить как среднее про-
ективное покрытие вида в ландшафте, суммировав произведения сред-
него проективного покрытия (=парциальной активности) вида в каж-
дом типе или классе экотопов на долю этого типа или класса от общей
площади ландшафта [или от площади локальной флоры (Юрцев, 1982) ].
Работа эта в целом весьма трудоемка и потому оправдана лишь в пунк-
тах наиболее детальных многолетних флористических исследований (ста-
ционары, охраняемые территории) и лишь для высокоактивных видов,
которые иным способом трудно ранжировать по их активности. Данные
по активности вида в той или иной фитохории или в различных типах и
классах экотопов можно вносить в соответствующие ячеи матриц гео-
графической и экотопологической структуры популяций видов (см.
выше) вместо альтернативных показателей присутствия—отсутствия
(+, —; 0, 1); если внутриландшафтная или парциальная активность вы-
ражена в баллах, вместо расчета средней активности (среднего балла)
корректнее сравнивать частоты высоких и низких баллов?
Я специально более подробно остановился на различных попытках
объективизировать показатель активности (как одно из возможных вы-
ражений „веса” вида во флоре) ввиду особой важности получения весо-
вых характеристик популяций с точки зрения рассматриваемой концеп-
ции флоры. В настоящее время мы не располагаем методами достаточно
простыми и эффективными для изучения геногеографической и генэко-
логической, а также феногеографической и фенэкологической структур
региональных популяций видов; однако поиск в этом направлении,
конечно, необходим и должен быть предусмотрен в перспективных про-
граммах хотя бы отдельных, наиболее укомплектованных специалиста-
ми и соответствующим оборудованием комплексных биологических
стационаров.
Несмотря на все сказанное, до сих пор термин „флора” по традиции
-яаще всего понимается в смысле „таксономический” (и прежде всего
видовой и расовый) состав растительного покрова. Тем~не менее реги-
страция тех или иных характеристик региональной популяции каждого
вида (без чего флора как природный феномен не может быть понята)
уже давно вошла в повседневную практику флористики, например при
составлении региональных „Флор” и „Конспектов флоры”, в специаль-
ных проектах картирования распространения видов в границах той или
иной региональной флоры, в лучших образцах анализа флоры, наконец,
при организации охраны редких и (или) быстро исчезающих видов, при
восстановлении истории флоры (выявление и изучение реликтовых ви-
дов); тем более необходимы такие данные для изучения процессов
эволюции в природных популяциях и для прогнозирования путей разви-
тия флоры, ее перестроек в связи с крупными естественными и техно-
генными изменениями среды. Современной практике изучения фпоры
полнее отвечает определение ее как совокупности (множества) местных
16
популяций всех видов растений на данной территории. Такое понимание
по существу включает как составную часть и традиционную концепцию
флоры как множества (набора, списка) видов, обитающих на данной
территории (в границах топографического контура) ; ведь разные попу-
ляции опознаются прежде всего по их видовой принадлежности. Тем са-
мым набор таксонов (таксономический спектр) выступает в качестве
одного из необходимых (первого по очередности выявления!) атрибутов
флоры как материального природного объекта. Противопоставление ме- fa
нее полной и более полной концепций флоры (фауны, биоты) в качестве
самостоятельных объектов, относимых к разным научным дисциплинам,
представляется искусственным, излишним усложнением. Ведь существо-
вание в фитоценологии двух противоположных школ классификации
фитоценозов: по набору видов и по их соотношению (в конечном счете
по доминированию) не привело к распаданию ее на 2 самостоятельных
дисциплины с разными объектами, напротив, отмечается тенденция
к синтезу и взаимообогащению.
Если же часть флористов предпочтет придерживаться традиционного
понимания флоры как только набора (списка) видов, приоритетным
термином для обозначения совокупности местных популяций всех видов
растений на данной территории будет „растительное население” (mixed
plant population) (Юрцев, 1982а, с. 5); мною он употребляется в каче-
стве синонимического термину „флора”. С этим вполне согласуется и
определение животного населения автором этого термина В. Н. Беклеми-
шевым (1931) как совокупности особей всех видов животных, т. е. со-
вокупности местных географических популяций этих видов, в принятом
здесь демографическом (более широком и к тому же исходном) пони-
мании термина „популяция” [полный эквивалент в русском языке —
„население”; подробное обоснование именно такой трактовки этого тер-
мина недавно опубликовано А. К. Скворцовым (1982, с. 6)].
2. Из признания флоры совокупностью местных популяций всех ви-
дов растений, населяющих данную территорию, с неизбежностью следует
тот вывод, что флора — не просто совокупность, но и материальная си-
стема этих популяций. Ведь каждая популяция для нормального сущест-
вования и воспроизведения нуждается в определенном минимуме жиз-
ненного пространства и материально-энергетических ресурсов и, кроме
того, способна неограниченно размножаться в геометрической прогрес-
сии (при наличии свободного пространства и ресурсов), однако и про-
странство, и ресурсы в любом замкнутом контуре ограничены по опре-
делению.
Содержанием отношений между популяциями разных видов как раз
и является распределение жизненного пространства и материально-энер-
гетических ресурсов, т. е. проблема экологических ниш в широком
понимании,1 в масштабах достаточно крупных территорий, включающих
1 Под „экологической нишей” целесообразно понимать распределенную и бо-
лее или менее обособленную в пространстве и времени, также по градиенту условий
долю вида (его части, группы видов) в биологическом круговороте вещества и
потоке энергии (т е в обращении ресурсов) на данной территории
2 Заказ 1220 _____ 17
'Л у Z77T" ? I
Л -г Г Аммм W
один или много ландшафтов. Если отношения индивидуумов растений
как прикрепленных организмов (в отличие от подвижных животных),
как правило, ограничены рамками небольшого участка конкретного
экотопа (и фитоценоза), то отношения региональных популяций разных
видов растений развертываются на фоне значительного разнообразия
экотопов (биотопов), как длительно существующих, так и кратковре-
менных (сериальных). Они могут принимать форму взаимозамещения
в пространстве и (или) времени и форму „экологической очереди”, т. е.
последовательной утилизации вещества и энергии в ходе биологического
круговорота.
Известный афоризм Ю. Одума, приводимый В. И. Василевичем
(1979, 1983), о том, что экотопы — адреса организмов, а экониши — их
профессии, имеет в виду именно последний тип отношений; кроме того,
он отражает собственно фитоценологический подход к проблеме эко-
ниш, рассматривая последние не в целом для ландшафта, но лишь раз-
дельно для конкретных фитоценозов. С точки зрения флористического
подхода к той же проблеме, близкого к пониманию экологических ниш
самим автором этого термина орнитологом Дж. Гриннеллом (Grinnell,
1977), эконишами вида в пределах ландшафта являются и все освоенные
им экотопы, как постоянные, так и временные; это — „экологические
адреса” вида, а не географические. Экониши любого вида можно разде-
лить на основные, второстепенные и случайные, на устойчивые и эфемер-
ные (недолго функционирующие). При этом даже в одном сообществе
вид может занимать одновременно разные экониши (так, вид дерева
в лесном ценозе может одновременно устойчиво встречаться в разных
подъярусах древостоя, в подлеске и в травяно-кустарничковом ярусе).
В принципе можно говорить об эконишах в отношении утилизации любо-
го ресурса, если он важен для поддержания численности популяции и
к тому же лимитирован; впрочем, некоторые виды ресурсов образуют
корреляционные плеяды, что облегчает их систематизацию.
Таким образом, ландшафтная экониша вида может быть определена
как сумма и как система локальных (интраэкотопных, интрабиотоп-
ных)1 экониш, те же в свою очередь — как сумма и как система элемен-
тарных (многофакторных) экониш, принципиально соответствующих
каждому типу элементарных мест в биоценозе, по Василевичу (1983).
Так, в приведенном выше примере вид дерева занимает в лесном ценозе
как минимум 3 элементарные экониши. Наконец, элементарные много-
факторные экониши должны быть охарактеризованы совокупностью
парциальных (однофакторных) экониш — по тому или иному ресурсу
или группе ресурсов в данном фитоценозе.
Итак, в характеристику любой парциальной однофакторной экони-
ши должны входить ее „географический адрес” (фитохория), ее „эколо-
гический адрес” (экотоп, биотоп, сообщество), вид ресурса, место
в „экологической очереди”1 2 по данному ресурсу („экологическая про-
1 Именно в таком объеме рассматривают экониши В- И. Василевич (1979,
1983) и некоторые другие фито-и биоценологи.
2 Там, где эта очередь имеется (например, в потреблении света растениями раз-
ных ярусов лесного ценоза).
18
фессия”), доля ценопопуляции в потреблении данного ресурса. Конечно,
определение этой доли может быть выполнено лишь в ходе комплексных
экологических исследований на стационарах, флорист же обычно вынуж-
ден ограничиться группировкой видов по их „экологическим адресам” —
экотопам.
Ландшафтные экониши разных видов растений в одном ландшафте,
как правило, не тождественны (несмотря на значительное их перекрытие
во многих случаях), различаясь набором или относительной ролью ло-
кальных экониш (основных или хотя бы второстепенных). Значительное
перекрытие или даже практическое совпадение характера локальных
экониш разных видов в пределах одного биотопа, по-видимому, распро-
страненный случай; сосуществование в этом случае объясняется замед-
ленностью конкурентного исключения у растений (Василевич, 1979,
1983), сопоставимого по длительности с флюктуациями среды, присут-
ствием видов с приблизительно равной конкурентной мощностью на дан-
ном биотопе, постоянным заносом диаспор с других (соседних) экото-
пов. Поэтому вытеснение обычно оказывается неполным, что и приводит
к количественному преобладанию в сообществах видов с покрытием
1% и менее (Василевич, 1983). Систематически близкие виды, утилизи-
рующие ресурсы одной и той же элементарной экониши, в основном
формируются аллопатрически, их сосуществование — результат вторич-
ной миграции и наблюдается нечасто.
Среди факторов дифференциации экониш целесообразно различать,
вслед за Василевичем (1983), ресурсы и условия (последние не явля-
ются предметом конкуренции, но ограничивают толерантность вида и
потому способствуют взаимозамещению видов с неодинаковой толерант-
ностью). Виды с разной жизненной стратегией (Раменский, 1935; Grime,
1979; Работнов, 1983) обычно занимают и разные экониши. Так, свое-
образие экониш ценопатиентов (Работнов, 1983) определяется их спо-
собностью существовать при низкой концентрации лимитированных ре-
сурсов, экотопопатиентов — их высокой толерантностью к крайним
условиям. Виоленты („конкурентные виды”, по Грайму) способны за-
хватывать львиную долю ресурсов в зоне оптимальных условий, доступ-
ной поселению представителей различных экологических групп. Нако-
нец, эксплеренты („рудералы”) приспособлены к быстрому захвату вре-
менных экониш с легкодоступными ресурсами.1
Общая цель изучения (анализа) флоры в подобном аспекте — вы-
явить в ее составе взаимодополняющие группы видов со сходными ме-
стом и ролью во флористической системе, оценить их взаимодействие
в настоящем, прошлом (флорогенез) и будущем (прогноз) на фоне
изменений климата и других компонентов физико-географической
1 Необходимо учитывать также, что „виолентность”, „патиентносгь” и „экспле-
рентность” - качества, присущие в той или иной степени каждому виду (Grime,
1979; Работнов, 1983). Поэтому в любой локальной эконише (в том числе и в „не-
благоприятных условиях”, требующих от всех произрастающих здесь видов повы-
шенной толерантности - „патиентности”) правомерно выделять в составе парци-
альной флоры экотопа своих виолентов (наиболее конкурентоспособных), цено-
патиентов и эксплерентов.
19
среды. Такой подход к изучению флоры, помимо всего прочего, живее,
интереснее, он требует от флориста быть не только специалистом по рас-
познаванию видов, но и биологом, экологом, географом — одним сло-
вом, натуралистом.
При этом у флориста ботанико-географического профиля нет основа-
ния опасаться дублиров ания фитоценологов. Фитоценологи изучают
взаимодействие видов в рамках фитЬценоэов, микро группировок, сину-
зий — на уровне систем взаимодействующих особей, своими специфиче-
скими средствами. Между тем даже долговременный прогноз состояния
растительного покрова любой фитохории нельзя свести к сумме прогно-
зов для каждого типа фитоценозов в отдельности. Ведь при крупных
переменах климата контур данного фитоценоза может быть заселен вида-
ми, ныне обитающими на других, контрастных экотопах или даже в дру-
гих районах. Важным элементом долгосрочного прогноза должна стать
оценка состояния региональной популяции каждого вида в целом и ее
вероятной реакции на ожидаемое изменение климата. Ареной эволюции
видов также являются территории значительно более крупные, чем от-
дельные фитоценозы. Если в ходе длительной флюктуации климата
какой-либо массовый, активный вид не сможет сохраниться ни на одном
из экотопов, восстановление растительного населения после возвратных
изменений климата будет неполным; даже если вид сможет удержаться
лишь на одном из экотопов со специфическими крайними условиями,
испытав необратимое обеднение генофонда, возврат к исходному состоя-
нию невозможен.
В результате взаимодействия местных популяций видов в любой фи-
тохории устанавливаются определенные, неслучайные количественные
отношения между группами видов, отличающимися сочетаниями
свойств, от которых прямо или косвенно зависит их место во флористи-
ческой системе (системе популяций). Следствием взаимодействия попу-
ляций в настоящем и прошлом при неодинаковых адаптивных возмож-
ностях разных филумов является значительное постоянство соотноше-
ний различных родов, семейств и других надвидовых таксонов в преде-
лах крупных фитохорий или ботанико-географических зон. Еще большее
постоянство отмечается в соотношении различных категорий элементов
флоры (географических, географо-генетических, экологических, эко-
лого-ценотических, биоморфологических и иных), отражающих свойства
каждого вида в целом или же только его местной региональной популя-
ции; в довольно узких рамках варьирует в пределах фитохории и таксо-
номическое разнообразие флоры, измеряемое числом видов и вышестоя-
щих таксонов. Эти закономерности, известные еще Альфонсу Декандо-
лю, стали предметом специальных исследований А. И. Толмачева (1941,
19706, 1974) и его последователей (Малышев, 19726; Шмидт, 1980) и
получили отражение в представлении об известном постоянстве в конк-
ретных фитохориях не только состава, но и соотношения таксонов и раз-
личных категорий элементов флоры, т. е. таксономической, географиче-
ской (широтной, т. е. зональной, долготной, высотной и комбинирован-
ной) и иных структур флоры. Тем самым каждая региональная флора
признается не случайной комбинацией, а абстрактной системой таксонов
20
и различных категорий типологических элементов. Достаточно очевидно,
что подходы к флоре как к абстрактной системе таксонов (филумов!)
и элементов и как к материальной системе видовых популяций взаимо-
дополняют: ведь невозможно объяснить количественные соотношения
крупных таксонов и элементов флоры, отвлекаясь от свойств и взаимо-
действия местных популяций видов, представляющих эти таксоны и эле-
менты; к тому же и количественные соотношения таксонов и элементов
удастся определять точнее, если внести поправки на „вес” в данной фло-
ре популяций соответствующих видов. С другой стороны, первостепен-
ное значение приобретает разделение флоры на элементы по тем свойст-
вам видов и их местных популяций, от которых зависят характер и
исход их взаимодействий с другими видами. Таковы, например, эколого-
ценотические элементы (устанавливаются по набору типов или классов
экотопов, где встречается вид в данной флоре), флороценотические
комплексы (по преимущественному тяготению вида к тому или иному
классу экотопов и сообществ), экогеографические элементы (по сходст-
ву географического и экологического распределения вида на территории
флоры), также подразделения флоры по жизненной стратегии видов (по:
Раменский, 1935; Grime, 1979), по их экобиоморфе и ценобиоморфе,1
по географической и экотопологической (внутриландшафтной) актив-
ности и т. д.
Таким образом, основным методом изучения флоры как материаль-
ной системы видовых популяций является привычная для флориста про- _
цедура анализа флоры, т. е. группировки видов по различным показате- q k у
лям их поведения в пределах данной тррритприи (в этом нет ничего . f
чуждого флористике, привнесенного из фитоценологии!). w ' 9
Остается добавить, что подход к флорам как весовым (дескриптив-
ным) множествам видов или их групп (крупных таксонов, элементов
флоры) позволяет использовать для сравнительного изучения флористи-
ческих систем весьма простой, но вполне строгий математический аппа-
рат алгебры дескриптивных множеств (Семкин, 1973; Юрцев, Семкин,
1980), отражающий при сравнении различных структур флоры пропор-
циональный вклад каждого таксона и элемента, а не только его поряд-
ковый номере нисходящем ряду (Марина, 1982,1983). Напротив, коэф-
фициенты ранговой корреляции, часто используемые при сравнении так-
сономической и иных структур флоры (Малышев, 1972, 19766; Шмидт,
1980), при большей трудоемкости их расчета огрубляют выражение
соотношений таксонов или элементов, переводя его из шкалы отноше-
ний в более слабую шкалу порядков.
3. Следующее важное звено развиваемой концепции флоры — при-
знание иерархической структуры любой флоры (как системы попу-
ляций), в том числе и конкретных флор, которые в сравнительной
флористике принято считать элементарными флористическими едини-
цами; иерархическую структуру соответственно имеют и региональные
1 Под ценобиоморфой имеется в виду объединение биоморф со сходными
Ц®йобиотическими свойствами и сходным ценоэффектом.
21
популяции видов. Ключевой момент концепции — тезис о том, что любая
естественная флора, любое подразделение растительного покрова пред-
ставляет систему видЬвых популяций того или иного уровня. По отно-
шению к данной флоре как системе популяций все подчиненные (вклю-
ченные) флоры более низкой ступени иерархии представляют подсисте-
мы, принципиально однотипные данной.
Представление об иерархии флористических систем планетарного
и регионального уровня мы получаем с помощью флористического райо-
нирования. Подобно тому как в современной физической географии
принято делить геосистемы на 3 основных уровня по их территориальной
размерности: планетарный, региональный и топологический (внутри-
ландшафтный), флористические системы (системы популяций) также
правомерно подразделять на планетарные, региональные (от флористи-
ческой области до конкретной флоры) и экотопологические (внутри-
ландшафтные); последние и было предложено называть парциальными
(„частичными”) флорами (Юрцев, 1975, 1982а; Юрцев, Семкин, 1980),
определяя их как естественные флоры любых экологически своеобраз-
ных подразделений ландшафта. С этой точки зрения (и как система цено-
популяций) фитоценоз — типичная парциальная флора (ПФ). По ряду
показателей (например, по соотношению родов, семейств, географиче-
ских элементов) многие фитоценозы существенно отличаются от мест-
ной конкретной флоры в целом. Однако между фитоценозом как ПФ
географической фации и конкретной флорой (элементарной естествен-
ной флорой регионального уровня) есть еще, как минимум, 2 ступени
ПФ — ПФ урочищ и ПФ местностей, т. е. ПФ более крупных подразделе-
ний ландшафта, включающих каждый несколько или много типов экото-
пов (а местность и несколько классов экотопов). Упомянутые подразде-
ления ландшафта в меру своего экологического своеобразия и экологи-
ческой цельности могут рассматриваться в качестве экотопов различного
иерархического уровня: фации — в качестве микроэкотопов, урочища —
мезоэкотопов, местности — макроэкотопов (Юрцев, Семкин, 1980).
Введение иерархического принципа в систематизацию экотопов умень-
шит субъективизм в дробности выделения последних, пока целиком за-
висящей от вкусов и темперамента исследователя. Если соотношения
таксонов и элементов флоры в ПФ сравнительно гомогенных урочищ не-
редко близки к таковым в ПФ их фаций, то те же соотношения в ПФ
местностей обычно значительно ближе к таковым в конкретной флоре:
по мере объединения ПФ экологически контрастных участков наблюдае-
мые на тех количественные- отношения крупных таксонов и элементов
постепенно усредняются, одновременно приобретая большую стабиль-
ность (Марина, 1983).
Парциальные флоры принципизльно не могут считаться экологиче-
скими элементами конкретной флоры (в отличие, например, от таких
экологических объединений видов, как ксерофиты, мезофиты, гигрофи-
ты и т. п.), так как сумма видов, относящихся к каждому элементу
флоры, не может превышать целое — количество видов во флоре (как
это обычно имеет место при суммировании количеств видов на всех эко-
топах); один вид не может принадлежать сразу двум или нескольким
22
элементам, но сплошь и рядом встречается в нескольких или многих
ПФ — эколого-топографических подразделениях региональных флор.
Нередко высказывается мнение о неправомерности рассматривать
комплексы видов отдельных экотопов в качестве флор: так думал
А. И. Толмачев, об этом говорили многие участники совещания в Не-
ринге. Однако никто из выступавших не смог указать ни одного принци-
пиального, сколько -нибудь существенного отличия ПФ в целом от собст-
венно флор (=флор регионального уровня). Между тем за время, истек-
шее со времени I рабочего совещания (1971 г.), было показано, что ПФ
обладают всеми атрибутами региональных флор, отличаются друг от
друга по всем категориям флористических признаков (включая таксо-
номическую и географическую структуры); все они представляют фло-
ристические системы — системы популяций и сравнительно автономны
по своему генезису. Те же отличия от региональных флор, которые дей-
ствительно имеют место (меньшая индивидуализированность состава —
большая повторяемость, что делает правомерным типологическую клас-
сификацию ПФ и объединение однотипных ПФ в рамках конкретной
флоры; более прямая экологическая обусловленность флористических
отличий соседних ПФ; меньшая устойчивость их против длительных
флюктуаций климата и т. д.), количественного порядка и изменяются
очень постепенно в иерархическом ряду флористических систем; при
этом сколько-нибудь существенные рубежи по разным показателям про-
ходят на неодинаковом уровне, многие — между ПФ мезо- и макроэко-
топов, т. е. еще на внутриландшафтном уровне (см. выше). Что же ка-
сается индивидуальности состава, то она возрастает в том же иерархиче-
ском ряду очень постепенно и в очень небольшой еще степени свойствен-
на КФ — элементарным региональным естественным флорам, которые
вполне правомерно типизировать и даже объединять (например, КФ
карбонатных и силикатных ландшафтов в рамках одного флористиче-
ского района или даже округа). Вполне правомерно трактовать КФ (как
единицы на рубеже регионального и экотопологического уровней, совме-
щающие признаки того и другого) и как ПФ мегаэкотопов. Эндемизм
в норме проявляется среди региональных флор более высокого уровня,
чем КФ или флоры районов, причем в разных природных областях на
неодинаковом уровне. Однако в отдельных случаях выявляются энде-
мики индивидуальных ПФ; последние и по другим признакам могут
быть вполне уникальны и неповторимы [так, ПФ окружения Гильмим-
линейских термальных источников на востоке Чукотского полуост-
рова включает 3 рода сосудистых растений, нигде более не известных
в Арктике (Экосистемы термальных источников..., 1981)]. Все это
снимает принципиальные возражения против распространения общего
понятия „флора” (с добавлением эпитета „парциальная”) на единицы
экотопологического уровня, тем более что это понятие по своей сути
безранговое. Все известные отличия ПФ от региональных связаны с иной
территориальной размерностью и вполне отражаются названным терми-
ном. Следует лишь сознавать, что любая флора принадлежит индивиду-
альному контуру, флористы же часто имеют дело с объединениями одно-
типных ПФ, что также вполне правомерно. Но как бы мы ни называли
23
ПФ,1 для большинства флористов и ботанико-географов с каждым годом
становится очевиднее, что раздельная инвентаризация и сравнительный
анализ эколого-топографических подразделений элементарных регио-
нальных флор — мощный метод сравнительного изучения региональных
флор, их районирования, познания их генезиса и тёндёнцшГразвития;
это убедительно прозвучало и на совещании в Неринге. Не случайно
А. И. Толмачев определяет элементарную флору фактически как объеди-
нение ПФ всех экотопов: даже в элементарной региональной флоре соот-
ношения таксонов и элементов, реально наблюдаемые на экологически
контрастных частях территории флоры, как бы смешаны и усреднены.
Чтобы понять, как сформировалась, живет, развивается такая флора —
объединение, очень важно выделить в этой чрезвычайно сложной задаче
более простые, естественные, экологически очерченные составляющие.
Итак, любая флора — гетерогенная система, состоящая из контраст-
ных совокупностей видов, разделенных внутри элементарных региональ-
ных флор экотопологически, а в более крупных региональных флорах —
также географически.
4. Взаимодействие между соседними флорами осуществляется по-
средством обмена диаспорами и панмиксии и по существу представляет
обмен генетической информацией (частицами, массой которых можно
пренебречь). Это взаимодействие наиболее эффективно проявляется
на всех пространственных уровнях — от внутриландшафтного до плане-
тарного — в периоды существенных изменений климата и других факто-
ров среды; его можно свести к сопряженной, большей частью взаимоза-
висимой перестройке географической и экотопологической структуры
популяций разных видов. Свойство флор перестраиваться за счет внут-
ренних резервов, достигая соответствия с изменившимися внешними
условиями, определено как буферность флористических систем; свой-
ство их возвращаться к состоянию, близкому к исходному, по заверше-
нии климатической флюктауации — как устойчивость этих систем
(Юрцев, 1982). Было показано, что на иерархической лестнице флори-
стических систем сверху вниз возрастает взаимозависимость популяций
разных видов, сила и однозначность их взаимодействия (максимальны
они в фитоценозах и их частях); напротив, снизу вверх возрастает устой-
чивость и буферность системы, поскольку на крупных, сложно диффе-
ренцированных территориях расширяются возможности для перестройки
географической и экотопологической структур популяций. Было пока-
зано также, что иерархия естественных флор, охарактеризованных как
системы местных популяций всех видов автотрофных организмов, в их
взаимосвязи с физико-географической средой, отображает иерархию
территориальных экосистем, а каждая флора представляет автотрофный
блок соответствующего биотического сообщества такой экосистемы
(Юрцев, 1982).
1 Предлагались также следующие синонимические названия: „комплекс видов
экотопа”, „флористический комплекс экотопа”, „флорула”, „флористическая
парцелла”, „флоротопологический комплекс”.
24
5. Специального анализа с системных позиций заслуживает явление
флористического континуума. В его основе лежит „стремление” сосед-
них флор к обмену элементами, что осуществляется через обмен диаспо-
рами и панмиксию [конъюкция систем, по Тахтаджяну (1972)]; в пре-
дельном случае можно говорить о гибридизации систем, т. е. образовании
новых систем из элементов старых, например в полосе контакта разных
региональных типов флор (особенно на вышедших из-под моря или кон-
тинентального льда молодых участках суши). В итоге, флора каждого
достаточно компактного, экологически дифференцированного участка
суши (в том числе и расположенного на стыке двух региональных типов
флор) функционирует как нормальная флористическая система и заслу-
живает изучения в качестве локальной флоры, или „флоры географиче-
ского пункта”, для выявления локальной флористической ситуации.
Систематизация данных представительной серии проб флоры позволяет
проводить флористическое районирование территории, выявлять и оце-
нивать флористические рубежи, восстанавливать историю флоры и т. д.
Значение такого подхода к флористическому обследованию территории
возрастает в связи с низкой дискретностью многих элементарных флори-
стических выделов, эффектом флористического континуума, флористи-
ческих градиентов.
6. В заключение необходимо остановиться на дискуссионном вопро-
се о месте науки о флорах как природных системах в системе ботаниче-
ских дисциплин. Этот вопрос следует обсудить и в практической плоско-
сти: какие специалисты могут и должны изучать региональные системы
популяций?
Рассмотренная выше концепция флоры как иерархически дифферен-
цированной системы местных популяций всех видов растений, населяю-
щих данную территорию, представляет самую общую модель раститель-
ного покрова; из нее легко вывести путем упрощения традиционную
фитоценологическую модель (растительность как совокупность фитоце-
нозов и открытых группировок растений) и традиционную флористиче-
скую (флора как совокупность видов).
Самое же широкое и непредвзятое общее определение растительного
покрова (безотносительно к той или иной его модели) — совокупность
всех особей растений на данной территории (то же, что фитострома в от-
ношении территории Земли). То, что понятия „флора” и „раститель-
ность” тесно связаны и относятся к единому природному телу, давно
осознано ботаниками; широкое распространение при этом получил из-
вестный афоризм о том, что „флора” и „растительность” — 2 стороны
WHOft медали (того, что нередко именуют -ряг.титепьным покроем”).
Одно из видоизменений этого афоризма — недавно высказанное Ю. Р. Ше-
ЙЯг-Сосонко и Я. П. Дидухом (1979) представление о том, чтолЬлора —
качественная- а рас.титрпкнлгтк - количественная пппсистема единой
Системы — рягтитрпьмпт плурпна Однако едва ли такие построения мо-
гут приблизить нас к уяснению действительного соотношения флористи-
ческой и фитоценологической моделей растительного покрова. Путь
к уяснению дает анализ иерархической структуры флористических
25
систем: продолжение ее на внутриландшафтный (экотопологический)
уровень приводит к заключению, что фитоценологическая модель расти-
тельного покрова как сложной, но закономерной мозаики фитоценозов
соответствует одному из нижних этажей естественной иерархии флори-
стических систем. При этом и качественный состав таксонов, и их коли-
чественные соотношения, как и их взаимодействия, могут и должны
изучаться на любой ступени данной иерархической лестницы. Именно
иерархическая модель растительного покрова способна стать основой
органичного синтеза флористической и фитоценологической информации
о растительной оболочке Земли. Речь идет о синтезе, а не простом сум-
мировании флористических и фитоценологических признаков.
Это, конечно, не „чистая” (рафинированная) флористика в традици-
онном смысле (выявление и анализ набора таксонов), но это и не фито-
ценология, изучающая фитоценозы и их части, прежде всего на уровне
совокупностей взаимодействующих особей.1 Неодинакова и детальность
отображения дифференциации растительного покрова флористом и гео-
ботаником-фитоценологом в связи с неодинаковой крупностью элемен-
тарных единиц. Однако, даже имея дело с самыми крупными подразделе-
ниями растительности Земли, геоботаник характеризует их преобладаю-
щим (или плакорным, зональным) типом фитоценозов, т. е. признаками
одной из низших единиц иерархии, генерализованной до уровня высших
категорий классификации. Изучение ПФ для флориста — инструмент
сравнительного анализа региональных флор, для геоботаника — дополни-
тельный метод познания сложных территориальных комбинаций фитоце-
нозов — фитоценохор. Какая же отрасль ботаники способна и должна
включить в свою программу анализ всей иерархии подразделений расти-
тельного покрова как систем популяций?
Для построения любой адекватной модели фитоценоза нельзя обой-
тись — в качестве „строительного материала” — без единиц, представляю-
щих особи разного типа, с той или иной совокупностью ценобиотических
и экологических свойств (экобиоморф, ценобиоморф). Подобная мо-
дель (назовем ее собственно фитоценологической) может достаточно
точно отображать разные стороны функционирования фитоценоза в за-
данных условиях, представляя растительный покров более обширной
территории как закономерную мозаику фитоценозов. Однако для того
чтобы модель сохраняла способность функционировать в существенно
и при этом непредсказуемо изменяющихся условиях на протяжении
тысячелетий, составляющие ее элементы должны обладать способностью
к непрерывной адаптации и взаимоадаптации; в природном раститель-
ном покрове это достигается объединением особей в иерархически диф-
ференцированные популяции — самовоспроизводящиеся эволюциони-
рующие системы, способные к автокорректировке и адаптации. Поэтому
в более полной модели фитоценоза единицы, символизирующие особи,
1 Некоторые современные теоретики фитоценологии (например: Василевич,
1983) не считают ценопопуляции основными фитоценологическими объектами,
с чем соглашаются не все фитоценологи (см., например: Уранов, 1975; Работнов,
1983).
26
должны входить в состав разных популяций, число которых не может
быть меньше числа типов особей, с поправкой на спектр возрастных со-
стояний. Но прежде чем мы сможем проникнуть в генетические меха-
низмы самоподдержания и развития географических популяций разных
видов, мы должны изучить географическую и экото по логическую их
структуры первоначально хотя бы на уровне демографических показа-
телей, а также изучить сопряженность названных структур популяций
разных видов в пространстве и во времени. Отсюда видна необходимость
иерархических флористических моделей дифференциации растительного
покрова, показывающих, как „сшивается” мозаичная („лоскутная”)
ткань его в сложное целое нитями местных географических популяций
разных видов. Задача эта несомненно актуальна, но до сих пор она не
включена в программу той или иной научной дисциплины. Итак, к какой
области знания она относится? Точка зрения автора: данная концепция
флоры должна стать одним из оснований ботанической географии —
дисциплины более широкой, чем собственно флористика и фитоценоло-
гия, призванной обеспечить взаимодействие этих наук в изучении расти-
тельной оболочки Земли и соединяющей идеологию и методологию бота-
ники (биологии) и географии.1 Другие авторы, например Вальтер (Walter,
1979, нит. по пер.: Вальтер, 1982), как и многие его предшественники,
называют такую более широкую дисциплину геоботаникой, включая
в последнюю как один из разделов флористику (флористическую геобо-
танику) . Дело, конечно, не в названии, а в том, чтобы реально существо-
вала подобная дисциплина, а в ней — общий раздел, обеспечивающий гар-
моничные взаимодействие и синтез подходов и методов флористики и
фитоценологии при изучении растительного покрова. Предлагаемая кон-
цепция флоры специально рассчитана на такой синтез.
Без флористов, имеющих дело при исследовании региональных флор
с местными географическими популяциями всех видов, изучение флоры
в подобном аспекте невозможно; не беда, если при этом они будут рабо-
тать не только как ортодоксальные флористы и систематики, но и как
ботанико-географы в классическом значении этого слова. Другая катего-
рия заинтересованных специалистов — геоботаники-фитоценологи с бота-
нико-географическом кругозором и достаточной подготовкой в области
флористики. Кроме того, в небольших региональных коллективах бота-
ников, вынужденных изучать растительный мир своей области или рес-
публики со всех возможных точек зрения, размежевание между флори-
стами и геоботаниками, к счастью, не зашло еще слишком далеко, что
1 Нет серьезных оснований опасаться поглощения флористики географией:
флору смогут изучать только ботаники. Но данный биологический объект геогра-
фичек по своей природе, и изучение его немыслимо без сочетания биологических
Подходов и методов с географическими, что многократно расширяет возможности
Для сравнений. Столь же безграничны возможности и для углубления биологиче-
ского подхода к флористическим объектам. Достаточно упомянуть про анализ
флоры по биоморфам в их современной трактовке, про использование-данных
экологической физиологии при установлении экологических групп, характеристи-
ке экотопов, объяснении „стратегий растений”, про сопряженный филогенетиче-
ский анализ сообитающих таксонов, про использование методов генетики для изу-
чения географических и ценотических популяций и т. п.
27
облегчает соединение обоих подходов. [Напротив, специалисты — бота-
нико- и биогеографы (согласно официальной номенклатуре), которых
готовят соответствующие кафедры географических факультетов многих
вузов, не получают основательной подготовки в области систематики и
флористики].
Идеальным же полигоном для наиболее детального комплексного
изучения флоры могут и, по-видимому, должны стать заповедники и
комплексные биологические и географические стационары.
„Флора (фауна, биота) как множество видов и как система взаимо-
действующих и сопряженно эволюционирующих их местных популя-
ций — одно из фундаментальных понятий биологии, область приложения
которого все расширяется” (Юрцев, 1983, с. 21) •
Г. В. Вынаев
О ПОНЯТИИ „ФЛОРА” И ЗАДАЧАХ НАУКИ О ФЛОРЕ
Институт экспериментальной ботаники им. В. Ф. Купревича
К концу XX в. флористика пришла к своеобразному кризису, когда
потребовались пересмотр ее основного (базового) понятия „флора” и
разработка адекватной современному уровню знаний ее дефиниции.
Современный период характеризуется восприятием и применением
рядом биологических наук так называемого системного подхода, и
в этом плане Б. А. Юрцевым (1982а) уже предложено рассматривать
флору как природную систему — систему местных популяций растений,
произрастающих на естественно ограниченной территории. Формальное
(традиционное) определение флоры как совокупности видов растений,
произрастающих на определенной территории, уже не удовлетворяет ни
теоретически, ни методологически уровню развития современной флори-
стики и применяемым ею методам. Действительно, рассматривая флору
как систему местных популяций растений, мы не должны забывать, что
эта система сама является подсистемой более сложных природных си-
стем — геосистем (биогеосистем, природно-территориальных комплек-
сов, ландшафтов, целых природных районов и т. д.), по В. Б. Сочаве
(1978). Поскольку существует несколько уровней организации геоси-
стем — от планетарного до топологического, то, вероятно, это же спра-
ведливо и для флористических подсистем.
Следовательно, в задачу флористов должно входить изучение соче-
таемости популяций различных видов на различных уровнях естествен-
ной дифференциации географической среды. Нам представляется акту-
альной проблема выбора основных уровней изучения флористических
систем при региональных флористических исследованиях. Вслед за
А. И. Толмачевым (1974), отмечавшим, что представление о флоре
всегда имеет географическое, но не узко топографическое или форма-
ционное содержание, мы также утверждаем, что на нижнем пределе
28
применимости этого понятия элементарной естественной флорой нужно
считать флору ландшафта (Лукичева, Сабуров, 1969) — основную еди-
ницу, служащую для целей комплексного природного (физико-геогра-
фического) районирования территорий (Исаченко, 1977). Флористи-
ческие образования внутриландшафтного (топологического) уровня
как совокупности локальных популяций растении, занимающие опре-
деленные экотопы и экотоны, а также их парагенетические ряды, мы
предлагаем называть флоротопологическими комплексами (ФТК)
(Вынаев, Кожаринов, 1981), в отличие от предложенных ранее для
таких сочетаний популяций терминов „парциальная флора”, „флори-
стический комплекс фации”, „флористический комплекс экотопа”
(Юрцев, 1975а), „фациальная флора” (Шеляг-Сосонко, Дидух, 1979)
и др. Мы солидарны с Н. А. Миняевым (см.: Юрцев, 1974а), утверждав-
шим, что естественная флора на определенном уровне неделима в том
смысле, что любые ее составные части уже не могут бьггь~ПОДведены
под понятие ..Флора” и должны рассматриваться лишь как ее струк-
турные элементы, как например комплексы видов, свойственные опре-
деленным типам местообитаний. Флоротопологические комплексы
мы рассматриваем как эколого-топологическую структурную единицу
естественной флоры и в соответствии с этим определяем естественную
флору как качественно своеобразное, исторически обусловленное и
динамичное природное явление, характеризующееся специфическими,
присущими только ему происхождением, расовым составом, флоро-
генетической, экологической и биологической структурами, законо-
мерностями развития, а также связями с другими компонентами гео-
графической среды и флорами различных фитохорий. Флора — истори-
чески сложившееся, детерминируемое современной географической
средой и связанное обменом с другими фитохориями региональное
сочетание географических и биологических рас растений — целостная,
относительно открытая и в то же время фитоценотически замкнутая,
структурно-функциональная, самопроизвольно развивающаяся
в пространстве и во времени система местных популяций растений,
образующих конкретные, экологически обусловленные и простран-
ственно локализованные флоротопологические комплексы. Поскольку
естественная флора трактуется нами как сложное природное явление,
мы считаем, что она должна являться объектом особой ботанической
науки „флорологии”1 (см. также: Шеляг-Сосонко, 1980), или учения
о флорах (Толмачев, 1974), имеющей своей целью всестороннее по-
знание естественных флор как сложных природных явлений, выра-
ботку адекватной этому явлению дефиниции, построение классифи-
каций и поиск объективных критериев выделения границ естественных
флор, познание их расового состава, географической, генетической,
Особый уровень познания биологических объектов, компонентами которых
являются биоты (и их части - элементарные флоры), и соответствующую ему
»Уку синтетического характера флоро (биото)генетику выделял и Р. В. Камелин
(1969). {Прим, ред.) .
29
экологической, биологической и других структур, изучение процессов
флорогенеза в исторической перспективе, а также современных, обус-
ловленных как природными, так и антропогенными факторами флоро-
динамических процессов, синантропизации и т. д.
Ю. Р. Шеляг-Сосонко, Я. П. Дидух
СИСТЕМНЫЙ ПОДХОД К ИЗУЧЕНИЮ ФЛОРЫ
Институт ботаники им. Н. Г. Холодного АН УССР
Понятие „конкретная флора” сформировалось на качественно новом
этапе развития флористики, вызванном к жизни необходимостью сравне-
ния огромного флористического материала, который был накоплен за
многие годы исследований региональных флор. Становление нового
этапа было, с одной стороны, подготовлено всем ходом развития флори-
стики, а с другой — обеспечено успехом ряда других научных дисциплин
(философии, математики), понятия и методы которых в 60—70-е годы
текущего столетия стали активно использоваться в биологических нау-
ках. В связи с этим на повестку дня флористики был поставлен ряд во-
просов: 1) сущность и объем понятия „флора” и соотношение флор
регионального и топологического уровней; 2) структура флор; 3) эво-
люция флор; 4) -методы изучения флор; 5) методология использования
для целей флористики понятий и методов других научных дисциплин
и т. д. Естественно, что решение этих вопросов возможно только на но-
вом уровне познания флор, который бы обеспечил качественный скачок
в развитии флористики. На наш взгляд, в методическом плане этот ска-
чок был связан с развитым А. И. Толмачевым методом конкретных
флор, а его методологической основой может стать системный подход.
Приложение такого подхода к флоре позволяет рассматривать ее в каче-
стве целостного многопланового природного явления, состоящего из
множества определенным образом связанных между собой различными
отношениями элементов, сила связи и уровень организации которых
позволяют вычленить их из подобных им систем или систем другого
класса. Достоинством системного подхода является его гибкость, ибо
он позволяет рассматривать каждый элемент в качестве отдельной систе-
мы более низкого порядка. При этом вычленение систем может быть
многомерным, т. е. проведено по многим признакам. Однако следует
иметь в виду, что сам по себе системный подход не дает решения задачи,
а обеспечивает только правильный подход к ее решению (один из воз-
можных. — Ред.).
Понятие „флора” на протяжении исторических периодов не остава-
лось постоянным, а эволюционировало как в силу изменения целей, ко-
торые ставились перед флористикой, так и в силу изменения понимания
сущности вида. На первоначальном этапе, когда вид рассматривался с ти-
пологических позиций, а перед флористами стояла цель инвентаризации
30
флор определенных территорий, исследования велись в региональном
аспекте. Это и определило понимание флоры как совокупности видов
растений, встречающихся на данной территории. Поскольку эти исследо-
вания велись в основном систематиками, а один исследователь не мог
охватить всего разнообразия растительного мира, то в этот период сфор-
мировался таксономический взгляд на флору (флора высших растений,
флора грибов, флора водорослей, лишайников, мхов и т. д.). Развитие
палеоботаники привело к необходимости отражения в понимании флоры
временных аспектов, вследствие чего появились термины „ископаемая
флора”, „третичная флора”, „флора ледникового периода”, „современ-
ная флора” и т. д. Развитие геоботаники вызвало появление эколого-
ценотического аспекта, что нашло отражение в появлении терминов
„флора степей”, „флора лесов”, „флора лугов”, „водная флора” и т. д.
На современном этапе оценки флоры с позиций системного подхода и
понимания вида как системы популяций Б. А. Юрцев (1982а) определяет
флору уже как множество местных популяций всех видов растений на
данной территории. Принимая такую формулировку в целом, мы счи-
таем необходимым детализировать ее. На наш взгляд, флора — это мно- '
жество популяций видов растений, населяющих определенную террито- *' : 1*
рию в ее естественных границах.1 Следовательно, это понятие применимо 7lt '
как к флоре ботанико-географического региона, так и к элементам ланд-^
шафта, но не административной или другой искусственно выделенной;
части территории (например, южной части болота или квартала леса).,/1г ,
Вполне понятно, что флора естественных территорий в процессе своего^ z
становления прошла определенный исторический путь развития, однако Г,/
вводить это в дефиницию самого понятия излишне. При таком подходе *5/
возникает вопрос о том, какой термин оставить за группой видов искус-
ственно выделенных территорий или не прошедшей общего историческо- ,
го пути формирования. Очевидно, за такими группами видов следует f '*
оставить термин „совокупность растений”; другими словами, флора — ; t
это не любая совокупность видов, а только представляющая собой более:
или менее целостное явление, сформировавшееся на основе каких-либо
отношений слагающих ее популяций. Такое понимание флоры вытекает
и из теории систем, основными понятиями которых являются „система”,
„элемент” и „структура”.
На наш взгляд, весьма удовлетворительное определение системы
дано В. С. Готтом и др. (1974). Согласно их определению, система —
это множество связанных между собой элементов (и подсистем), упо-
рядоченных по отношениям, которые обладают определенными свойст-
вами. Это множество характеризуется единством, проявляющимся в на-
личии общих для всего множества свойств и функций, т. е. в определен-
ной автономности его поведения. Каждая система состоит из элементов,
которые мы понимаем как относительно нерасчлененные дискретные
единицы, характеризующиеся в своем совокупном единстве определен-
ными свойствами и явлениями; это единство обусловлено наличием
1 В этом смысле обычно употребляется термин „естественная флора” (имеется
в виду естественность выдела). (Прим. ред.).
31
принципов, способов, законов связи, отношении их в рамках данного
целого, которые рассматриваются в качестве структур (Свидерский,
1962). Такое определение структуры охватывает все виды связей и не
включает в себя элементы.
Растительный покров, используя энергию окружающей среды (в ви-
де неорганической формы движения материи), создает свою, качествен-
но новую органическую материю. Хотя растения и находятся в более
тесной функциональной связи с внешней средой, чем друг с другом,
образуя реально функционирующую цепь, тем не менее растительный
покров по отношению к окружающей его среде может рассматриваться
как самостоятельная система, в результате функционирования которой
происходит интеграция физических и химических свойств материи и
создается качественно новое, органическое вещество. Элементами такой
системы являются популяции видов с определенными свойствами, про-
являющимися в виде различных признаков, которые могут быть оцене-
ны с качественной и количественной сторон.
Вслед за Б. А. Юрцевым (1968а)1 мы считаем, что соотношения
между популяциями видов в растительном покрове можно свести
к двум аспектам: 1) фитоценотическому, отражающему количественную
сторону, и 2) флористическому, отражающему качественную сторону
(разнообразие видов). Исходя из этого флору и растительность можно
рассматривать как две самостоятельные системы, существующие в нераз-
рывном единстве.
Во флористике системный подход основной задачей ставит изучение
организации флористических систем, а также их структуры, происхожде-
ния и эволюции.
Во флоре в отличие от фитоценозов и биогеоценозов непосредствен-
ное взаимодействие между слагающими ее элементами не выражено.
Это послужило поводом для утверждения о том, что флора не является
системой. Однако с таким утверждением нельзя согласиться. Как указы-
вает Н. Ф. Овчинников (1967), при наличии связей не обязательно одна
вещь должна воздействовать на другую. Такого рода воздействие может
быть, а может и отсутствовать, но из этого не вытекает вывод об отсутст-
вии связей. Воздействие — одна из форм связи, но не единственная фор-
ма. Флора не является ни материальной, ни абстрактной системой, а зани-
мает промежуточное положение и, согласно В. И. Василевичу (1983),
должна относиться к генезисной группе систем, элементы которой имеют
1 Современная точка зрения Б. А. Юрцева (1982а, а также первая статья в наст,
сб.) заключается в том, что и качественный (таксономический, типологический)
состав слагающих флору видовых популяций, и их количественные соотношения
и взаимодействия правомерно и целесообразно исследовать на всех уровнях
иерархии естественных флор как систем популяций (с соответствующим видоиз-
менением методик). Фитоценолог изучает растительное сообщество и как систему
ценопопуляций (элементарную парциальную флору), и как множество систем осо-
бей; качественные характеристики сообществ (в том числе их состав) интересуют
фитоценолога не меньше, чем количественные. Выявление качественного состава
элементов системы всегда предшествует установлению их количественных отноше-
ний. Едва ли целесообразно говорить о качественной и количественной системах
для одной и той же совокупности элементов раздельно.(Прим, ред.).
32
общее происхождение, но в настоящее время непосредственно не взаимо-
действуют друг с другом.1
Анализ флоры предполагает выяснение положения исследуемой
флоры в ряду себе подобных. Здесь представляется целесообразным
рассматривать положение флоры в двух аспектах: региональном и эко-
лого-ценотическом (топологическом). Систематизация в региональном
аспекте предполагает выделение и построение системы фитохорий
(фитохорионов) на основе флористических критериев — присутствия
или отсутствия видов или таксонов более высокого ранга и установ-
ления их границ.1 2 Для фитохорий, характеризующихся достаточно
высоким эндемизмом, в качестве критериев используются эндемич-
ные таксоны, наиболее полные региональные перечни которых опу-
бликованы А. Л. Тахтаджяном (1978). Для северных регионов по-
вышается -значение комплексных критериев, а именно выделения
дифференциальных и кодифференциальных групп видов (Юрцев и
др., 1978). На наш взгляд, главнейшим принципом выделения фито-
хорий и установления их ранга должен быть комплекс флористиче-
ских показателей с учетом научно обоснованного „взвешивания” при-
знаков.
Флора выделенных фитохорий представляет собой множество попу-
ляций растений, распределенных неравномерно в силу экологических
условий. Фитохории, примыкая друг к другу, отличаются от соседних
присутствием или отсутствием одного или нескольких видов или таксо-
нов более высокого ранга. Границы фитохорий теоретически должны
проводиться по границам распространения ценофлор (см. ниже), отли-
чающихся составом таксонов; дифференциальные таксоны служат кри-
териями разграничения. Однако на практике это затруднено как в силу
объективных причин (размытость и несовпадение границ распростране-
ния видов, локальное распространение последних), так и субъективных
(недостаточная изученность границ), поэтому для установления границ
часто используются более „физиономичные” геоботанические или гео-
графические признаки. Каждая выделенная фитохория является непо-
вторимой системой, однако уровень или ранг ее неповторимости разли-
чен и устанавливается в зависимости от систематического ранга разгра-
ничительных таксонов.
1 Этот тезис противоречит принятому выше авторами статьи определению фло-
ры как множества местных популяций, т. е. множества видов в их материальном
выражении на данной территории, и проводимому ими в ряде работ анализу „цено-
тической структуры” флоры (подробнее см. статью Я. П. Дидуха в наст, сб.), т. е.
анализу флоры по ценотической значимости видов, их жизненной стратегии, актив-
ности и т. д. Взаимодействие видов друг с другом во всей совокупности ценозов
на территории флоры - один из ведущих факторов флорогенеза, оно же контроли-
рует соотношение в современной флоре видов, представляющих разные филумы,
разные ареалогические, экологические; биоморфологические типы и т. д. К соб-
ственно генезисным системам можно отнести разве что филумы с преимущественно
аллопатрической дифференциацией. {Прим. ред.).
2 Наряду с этим в качестве признаков фитохорий (фитохорионов) могут слу-
жить количественные соотношения таксонов и типологических элементов флоры,
о чем говорится в конце статьи. {Прим, ред.).
3 Заказ 1220 33
Региональная иерархия фитохорий представляет собой систему, со-
стоящую из подсистем, более низкого уровня. Наиболее всеобъемлющей
является планетарная система, включающая в себя царства, области,
провинции, округи, районы. В качестве наименьшей региональной си-
стемы принимается элементарная флора,1 определяемая как флора
всего непрерывного пространства, где на однотипных зкотопах набор
видов практически одинаков, а их различия носят случайных характер.
Для „пробы” такой флоры мы сохраняем название „конкретная флора”.
При топологическом анализе флор выделяется система фитомер
(аналогично фитоценомерам В. Б. Сочавы (1979)], которые объединя-
ются по флористическому подобию сообществ и представляют собой
систему флор другого уровня. В настоящее время для анализа наиболее
часто используются данные о флорах типов растительности, хотя при
этом не исключается возможность анализа флоры синтаксонов более
низкого ранга в пределах как одной, так и разных региональных флор
(например, флора сосновых, дубовых, буковых и других лесов). В каче-
стве инициальной флористической системы топологического уровня
Б. А. Юрцев (1982а) предложил рассматривать парциальную флору
(ПФ).1 2 ПФ является флорой целостной территории. Однако в горных
системах, где вследствие большой мозаичности экологических условий
вообще невозможно выделить целостную, сколько-нибудь полно отра-
жающую флористическое разнообразие синтаксона территорию, прихо-
дится изучать флору отдельных, порой далеко лежащих друг от друга
площадок. Несмотря на территориальную разобщенность, совместное
произрастание видов здесь не случайно, а обусловлено экологическими
и ценотическими условиями в естественных границах экотопов. Флору
таких синтаксономических единиц мы называем ценофлорой (флоро-
ценотическим комплексом — в наших прежних публикациях). Это
понятие соответствует объединению однотипных ПФ в трактовке Юрце-
ва (1982а). В зависимости от рассматриваемого уровня мы выделяем
ценофлоры лесов горного Крыма или его южного макросклона, напри-
мер дубовых, сосновых, можжевеловых и других лесов. Низшей едини-
цей топологического уровня является флора фитоценоза — относительно
однородного контура растительности, внутри которого нельзя провести
никакой границы по фитоценотическим параметрам (Василевич, 1983).
Таким образом, фитоценоз представляет то узловое и элементарное
звено флоры и растительности, ниже которого мы уже не можем гово-
рить об этих понятиях. Из основ геоботаники известно, что площадь
размером 10 X 10 м (для травянистых типов) или 25 X 25 м (для лесов)
уже дает представление о закономерностях строения ценоза, в то время
как вопрос о площади „проб”, отражающих закономерности строения
флористических структур топологического уровня, остается открытым
1 Имеется в виду элементарная естественная флора, что было оговорено выше.
{Прим, ред.).
2 Б. А. Юрцев (см. также первую статью в наст, сб.) говорит об иерархической
системе парциальных флор (ПФ) в рамках конкретной флоры; инициальной ступе-
ни иерархии соответствует фитоценоз как ПФ микроэкотопа. {Прим, ред.).
34
И требует постановки эксперимента. По-видимому, она окажется значи-
тельно больше, чем применяемая при геоботанических описаниях.1
С целью изучения способа организации флор и закономерностей их
изменения необходимо рассмотреть их основные свойства. Как известно,
в каждой флоре наблюдается упорядоченное отношение множества обра-
зующих ее элементов. Этими элементами являются популяции видов,
характеризующиеся определенными свойствами или признаками: напри-
мер, все виды принадлежат к тому или иному таксону высшего ранга,
имеют определенный ареал, относятся к определенной биоморфе,1 2 обла-
дают определенными экологическими и фитоценотическими свойствами.
Количественные соотношения между классами этих признаков имеют не
случайный, а закономерный характер. Такое упорядоченное отношение
между признаками или свойствами говорит о наличии определенного
типа связи между элементами3 и рассматривается нами как количествен-
ное выражение определенной структуры флоры. Это подтверждается
ограниченным количеством и разнообразием таксонов разного ранга
во флоре, закономерным изменением количественных соотношений
между классами таких признаков, как типы ареалов, био морф, фито-
ценоциклов и др. Установление этих соотношений, границ их варьиро-
вания для разных типов флор позволяет выявить не только определен-
ные закономерности, но и делать прогнозы. В качестве примера со-
шлемся на результаты анализа систематических структур по набору
10 ведущих семейств, приведенных в работах А. И. Толмачева (1931,
1970, 1974), Б. А. Юрцева (1968а, 1982а), Л. И. Малышева (1972),
Р. В. Камелина (1973) и др.
Системный подход к пониманию флоры привел к развитию струк-
турносравнительного анализа, который предполагает сравнение флор
по определенным свойствам составляющей флору совокупности видов.
В связи с тем что виды, а значит, и флоры теоретически обладают не-
определенно большим числом свойств, во флористических системах
можно выделить неопределенно большое число структур. Однако в этом
нет необходимости, так как вполне достаточно провести анализ струк-
тур, отражающих существенные черты флор. Такими структурами
являются систематическая, географическая, биоморфологическая, эко-
логическая, ценотическая, генетическая и др. Они могут быть выражены
количественно на основе соотношения между объединениями видов
(точнее, их популяций) по какой-либо категории присущих им призна-
ков или свойств.
1 По-видимому, здесь речь должна идти не столько о площади пробных площа-
док (ПФ может изучаться по всему контуру), сколько о повторности изучения
индивидуальных контуров, достаточной для выявления состава данной объединен-
ной ПФ („ценофлоры”). (Прим. ред.).
2 Один вид может иметь несколько биоморф. (Прим, ред.).
3 При изучении таксономической и различных типологических структур фло-
ры (т. е. соотношения в ней крупных таксонов или разных категорий типологиче-
ских элементов - ареалогических групп, биоморф и т. д.) в качестве элементов
флористического множества - точнее, фактор-множества (Юрцев, 1982а) - рассмат-
риваются уже не виды, а их объединения по тем или иным классификационным
признакам.(Прим, ред.).
35
I
Таким образом, со сменой парадигмы во флористике, которая имеет
место в настоящее время, происходит и смена методов изучения флоры.
Именно поэтому на данном качественно новом этапе развития флористи-
ки ведущее значение приобретает структурно-сравнительный метод,
позволяющий вскрыть всю многомерность такого сложного природного
явления, как флора.
Р. В. Камелин
ПРОЦЕСС ЭВОЛЮЦИИ РАСТЕНИЙ В ПРИРОДЕ
И НЕКОТОРЫЕ ПРОБЛЕМЫ ФЛОРИСТИКИ
Ботанический институт им. В. Л. Комарова АН СССР
Мне близок развиваемый Б. А. Юрцевым (1982а) подход к изуче-
нию флоры как природной системы. Правда, я предпочитаю говорить
о флоре как о подсистеме биоты — биологической системы, соответст-
вующей особому уровню организации живого на Земле, который выше
биоценотического, но ниже обще биосферного. Я считаю также право-
мерными представление о флоре (и биоте в целом) как о биотиче-
ской части более сложных образований — геосистем и вытекающие
из этого представления о возможности чисто географических подходов
к ее изучению.
Но особенно близки мне представления о флоре как о системе ви-
дов, в значительной степени организованных в ценозы, — системе, сло-
жившейся в результате реально протекающих, относительно обособлен-
ных и ограниченных территориально и экологически процессов эволюции
во всем разнообразии их проявлений. Именно эта сторона флористиче-
ских исследований развивается ныне недостаточно полно, и именно ей
посвящена настоящая статья.
Уже довольно давно я развиваю гипотезу, согласно которой ре-
альный процесс эволюции идет по крайней мере на всех надорганиз-
менных уровнях организации живого — их, не считая биостромати-
ческого или биосферного, для которых мы пока только и знаем
процесс жизни, всего три: популяционно-видовой, биоценотический
и биотический (Камелин, 1969, 1973). В то же время, материальных
носителей этого процесса, равно как и элементарных эволюционных
явлений для всех этих уровней, мы почти не знаем. Элементарные
единицы, явления и процессы, лежащие по общему мнению в основе
всего процесса эволюции, выделены лишь на молекулярном уровне
организации живого — на уровне, где по существу нет живых
структур.
Соответственно трем основным уровням организации живого на
Земле, на которых и протекает реальный процесс эволюции, мы можем
(в значительной мере искусственно, абстрагируя) вычленять и три сто-
36
pofgj процесса эволюции — филогенез, филоценогенез и биотогенез (фло-
фауногенез) ?
фактически мы не знаем механизмов ни одной из этих разных сто-
эволюции, объединяемых нами, по блестящей догадке Ч. Дарвина,
концепцией общего фактора эволюции — естественного отбора наиболее
ffpMcnncoбпенных особей (в более общих случаях — локальных популя-
ций), которые адаптированы к тому или иному диапазону косной среды.
Но различные группы фактов указывают нам на определенное разнооб-
разие проявлений отбора (и в том числе на ограничения его) в зависимо-
сти от того, к какому уровню организации живого они относятся, участ-
вуют ли в филогенезе, филоценогенезе или биотогенезе. И если, к тому
ЖВ» мы признаем и возможность гибридогенной трактовки части микро -
и макроэволюционных явлений в природе (к чему у ботаников есть
достаточно много оснований), то общее количество факторов реального
процесса эволюции в природе, которые мы должны учитывать, сильно
возрастет.
Когда мы пытаемся вычленить в общем процессе эволюции (абстра-
гировать) филогенез, мы имеем дело с элементарными природными
образованиями, системами особей, объединенных „кровным родством”,
единым генофондом, реально осуществляемой панмиксией — популя-
циями и видами. Они реально существуют в природе, мы зачастую видим
их объем и границы, но обычно (и у растений особенно) границы эти не-
четки, трудны для исследования.
Абстрагируя филоценогенез, мы имеем дело с элементарными систе-
мами особей различных видов, которые частично связаны „кровным
родством”, а частично — консортивными, конкурентными и тому подоб-
ными взаимоотношениями, возникающими в результате совместного
обитания при определенном минимуме ресурсов косной среды, также
С ,денопопуляциями” (различными частями локальных популяций) и
с биоценозами, иногда же и с сукцессионными комплексами слабо рас-
члененных или сменяющих друг друга биоценозов или другими едини-
цами. Все эти единицы также реально существуют в природе, но границы
ИХ объективно оценить мы обычно не можем.
• Абстрагируя биотогенез, мы имеем дело с территориально отграни-
ЧОННыми биотами (флорами и фаунами) — надценотического уровня
системами популяций или частей популяций разных видов, в подавляю-
щем числе случаев организованных в ценозы, связанных пространствен-
ное общностью территории обитания, временной общностью историче-
ского развития на данной территории и определенными процессами дина-
МИЧеского взаимодействия в рамках минимума доступных ресурсов
Косной среды (прежде всего энергетических). Для территориально отгра-
ШВЧДННых выделов биоты, различные компоненты которых (популяции
ЙфВкретных видов и их части) взаимосвязаны и через „кровное род-
<ЯМЮ”, на различных по величине участках этой территории по существу
- 1 Эти стороны эволюционного процесса, как правило, обходят важнейшие
«водки по эволюции, как отечественные, так и зарубежные (см., например: Тимо-
феев-Ресовский и др., 1977; Grant, 1977). Исключения очень редки (Шварц, 1980).
37
пока не удается однозначно определить элементарную единицу исследо-
вания (и организации), далее неделимую. В природе эти элементарные
единицы отграничиваются легче по признакам комплексного природного
районирования, но не по чисто биологическим (флористическим и фау-
нистическим) признакам.
Некоторым приближением к этой элементарной неделимой единице
биотического уровня являются следующие понятия: экосистема в перво-
начальном смысле Тэнсли (Tansley, 1920,1935), т. е. как „гомеостаз при-
родных явлений”, как „ячейка гомеостаза жизни”; элементарная естест-
венная флора со всей совокупностью остальных (гетеротрофных) ком-
понентов биоты на территории ее выявления, в смысле Камелина (1969,
1973), т. е. как система популяций или крупных частей популяций ви-
дов, отграниченная комплексными природными рубежами и имеющая
хотя бы один эндемичный вид в своем составе, диагностирующий ее
своеобразие; элементарный флористический район, в смысле Б. А. Юрце-
ва (1982а) и многих других авторов; ареал сукцессионной системы или
„ботанико-географический район”, в смысле С. М. Разумовского (1981;
имеется в виду сама идея, но не ее реализация в работах названного авто-
ра); эпигенема (эпигены), в смысле Высоцкого (1915,1925) и Р.И. Або-
лина (1914). Но все это лишь некоторые приближения к отграничению
элементарной единицы биоты, которые еще предстоит связать воедино.
„Элементарная флора” в смысле А. И. Толмачева (1932), по-видимому,
несколько идеализирует природные явления и во всяком случае непри-
менима к горным и южным (тропическим) флорам, если в качестве кри-
терия ее выделения принять требование тождества набора видов на одно-
родных биотопах и в одних и тех же типах биоценозов.
Все другие представления об элементарной единице флористики
(и фаунистики) либо не отвечают требованиям к данной категории
(хотя на других уровнях познания они могут „работать”), либо
устарели.
Уже на уровне, на котором идут процессы филогенеза собственно,
мы сталкиваемся с отсутствием гиатусов между элементарными природ-
ными явлениями — прежде всего локальными популяциями (и всегда
между „ценопопуляциями”); это выражается в образовании, прежде
всего у растений, и подвидовых, и надвидовых (грегарных, в широком
смысле слова) структур, нарушающих изоляцию. Но это структуры био-
логические, и изучать их нашими методами возможно, хотя, конечно,
в целом они могут быть и географически определенными, доступными
изучению и биологическими, и географическими методами.
На уровне фитоценозов мы обычно имеем дело со слабой отграни-
ченностью элементарных явлений (от ценоячеек до фитоценозов, как
возобновляющихся систем), с недостаточно разработанной системой
критериев их классификации. Эти критерии особенно усложняются при
попытках объединения признаков, работающих на уровне собственно
фитоценозов (как автотрофной основы биоценозов),и признаков, рабо-
тающих лишь по отношению к разным компонентам гетеротрофных
блоков биоценоза. Но в целом это прежде всего биологические образо-
вания, которые может изучать биолог и эколог.
38
, На уровне биот мы умеем определять границы элементарных явле-
лишь по комплексным природным характеристикам. Структуры
дею уровня в равной мере географические и биологические. Соответ*
думкно мы можем изучать их по-разному. Как биологи мы должны
дознавать, что систему, в которой базовыми являются еще две крайне
Л^устаточно изученные системы более низких уровней, изучать очень
доожно. Но если мы интересуемся реальным процессом эволюции, мы
можем использовать для изучения какие-то обобщенные показатели это-
го процесса, в детали не вникая, поскольку они трудны для познания.
Географический подход при этом использовать легче, в том числе и био-
логу. Он может быть применен и к видам, и в какой-то мере к ценозам,
и, безусловно, к элементарным хоролого-биологическим системам.
Большая часть флористов придерживается второго (географического)
подхода к изучению флоры, тогда как зоологи-фаунисты прибегают
к нему намного реже. Я считаю географический подход к флоре вполне
правомерным, сознавая, однако, что все же для биолога первый (биоло-
гический) подход — основной.
В любом случае, изучая биоты, мы имеем дело с множеством видов
в той или иной хорологической структуре (топографическом контуре
и т. п.). Это множество мы можем изучать и классическим описательным
методом биологии, и статистическими методами (разумеется, с соответ-
ствующими упрощениями). Мы знаем, что региональные флоры (биоты)
почему-то включают лишь очень малую долю видов общеземной флоры
(проблема всеобщей недонасыщенности биот). Отсюда возможность и
Правомерность экологических (или эко биологических) методов изуче-
ния флор (биот)-, а также важность приложения к последним сравнитель-
ного метода биологии, несмотря на несравнимость многих выделов био-
ты в их природных границах, не говоря уже об искусственных.
Наконец, эмпирически мы знаем, что флористические множества
изменяются в геологическом времени, и поэтому они могут и должны
Изучаться историческим методом биологии. Здесь имеются в виду фло-
рогенетические подходы и методики. Последние, правда, доступны лишь
лицам, которые могут объединить в себе систематика, фитоценолога и
флориста, знающего значительно более широкий регион, чем непосредст-
венно исследуемый и диагностируемый им. Большая часть современных
флористических работ, к сожалению, не выходит за рамки первого (опи-
сательного) уровня познания, значительная часть остальных — за рамки
Второго уровня. Я считаю это основным недостатком современного со-
стояния флористики, ее „слабым звеном”. Другими недостатками я
считаю чрезмерное усердие в применении статистических методов к явле-
ниям слабо отграниченным, недостаточно квалифицированное экологи-
ческое изучение флор — увлечение методиками выделения элементов
биоты по элементам ландшафта, крайне слабую разработанность и недо-
статочную смелость в применении сравнительных методик. Наконец,
Крупным объективным слабым звеном я считаю почти полное отсут-
ствие комплексирования с зоологами разного профиля при изучении
Юдиных биот и слабое использование данных по низшим растениям
И мохообразным (вследствие их крайне недостаточной изученности)
39
I
при сравнительной характеристике естественных флор. Уже привлече*
ние материалов по разным группам низших дало бы кардинально иные
результаты при сравнении ранее изученных флористами флор почти
по всем показателям. Достаточно напомнить, что для альгофлоры озер-
ных систем понятия флора и растительность практически совпадают,
носят не только качественный, но и количественный характер, что в слу-
чае почвенных микробоценозов мы имеем совершенно иные границы
элементарных ценотических явлении, чем в ценозах высших расте-
ний, и т. д.
Следует сказать, что и биологические методы изучения флоры, и
в частности подходы к изучению флорогенеза, могут быть достаточно
разнообразны в зависимости от того, каким составляющим реального
процесса эволюции мы уделяем главное внимание.
Остановлюсь на некоторых нестандартных (не получивших широко-
го распространения) путях изучения флорогенеза.
1. Мы можем рассматривать любую флору как арену микроэволю-
ции и видообразования или как часть таковой. При этом подходе мы
изучаем в основном эйдологические (популяционно-видовые, расовые)
закономерности сложения флоры? Здесь наиболее интересны следую-
щие проблемы: эндемизм флоры и его причины, реликтовые типы
данной флоры, связи через них с иними флорами и почины обособ-
ления или консервации в данной фло^Ущ^изфлоры на содержащиеся
в неигкп^^ видки гибридогенные комплексы
и расы, резкие-мутанты, диплоидные и разпияньш^по гёЖЗисУЧ^ли-
плоидные расы и т. п. Возможен подход к анализу флоры и с точки
зрения явлений дрейфа генов, с точки зрения фено гео графических
закономерностей (разделение на группы видов с малым фенотипиче-
ским разнообразием и с повышенным фенотипическим разнообразием,
выделение типов с особо высоким фенотипическим разнообразием).
Наконец, эйдологическим подходом возможно выделить „случайные”
особенности видового состава (насыщенность—недонасыщенность), хотя
за ними обычно скрываются закономерности заноса, правила „первого
вида” (первопоселенца), фитоценологические аномалии и т. п. — общие
черты подобных „случайностей” распределения вполне возможно вы-
явить и анализировать. В результате мы могли бы и классифициро-
вать флоры с точки зрения характерных черт видообразовательного
процесса.
2. Мы можем рассматривать любую флору как арену филоценоге-
неза. В этом процессе прежде всего мы можем уловить определенные
„филумы” растительности и их временные выражения — флороценотипы.
Возможно, что определенная ограниченность их набора в любой флоре
связана с генетической обусловленностью экологических и эко биологи-
ческих характеристик. Типы экобиоморф, по-видимому, в значительной
мере генетически обусловлены, хотя бы за счет генетической обусловлен-
1 Понятие „эйдология” ввел М. В. Клоков еще в 1947 г., но он опубликовал
свои взгляды позднее (Клоков, 1973).
40
определенного морфологического типа (повторяющегося в раз-
часто удаленных таксонах).
Возможно, конечно, и наследование самых различных признаков
Цфокта” - норм реакции на изменения условий среды. В отношении
факторов среды это доказано, например, наследственной обусловленно-
сти реакций клеток разных видов на определенный диапазон темпера-
тдоных воздействий. Следует предполагать, что могут наследоваться
|К©пределенные признаки адаптации к биологической среде - нормы
конкурентной реакции, возможность или невозможность сосущество-
ggBu с растениями с какими-либо ярко выраженными средообразова-
тельными свойствами. Во многом это пока лишь предположения, но
отнюдь не невероятные. Часть подобных наследственных свойств может
возникать вследствие гомологической упорядоченности как морфо-
логических, так и физиологических (и эко биологических) призна-
ков; что-то можно отнести и на счет гипотетической трансдукции (пе-
реноса) генов при вирусных инфекциях и т. п. Но флора как арена
филоценогенеза может быть рассмотрена и как единая динамическая
система (или ряд динамических систем) растительных сообществ,
как определенный, свойственный только данной флоре набор сук-
цессионных рядов и различных структур, участвующих в них (или
Как ряд таких наборов либо же, наконец, как региональный вариант
сукцессионных систем, повторяющихся с закономерными изменения-
ми в разных флорах). Все это - пока еще не освоенный резерв, бла-
годарное поле для флористического (а не только геоботанического!)
анализа.
Эти же биоценотические (фитоценотические, геоботанические) зако-
номерности склада флор вполне могут быть использованы и для сравне-
ния флор.1
3. Наконец, при рассмотрении флоры как исторически сложившейся
динамической системы высшего, по сравнению с ценозами, уровня орга-
низации возможны также свои специфические подходы.
Первый из них — определение типа флоры по соотношению в ее гене-
зисе процессов автохтонного образования и иммиграции таксонов.
Основные типы флор: автохтонная, миграционная, переработанная
in situ. Методы выявления — флорогенетические.
Второй подход заключается в типизации естественных флор по
переживаемой данной флорой фазе развития (флорогенеза). По этому
Признаку провизорно можно наметить следующие типы: 1) прогрес-
сивно развивающаяся флора (находится в расцвете, характеризуется
большим числом видов, чем исходная; темпы видообразования повы-
шены, велики и миграционные возможности видов); 2) стабилизиро-
вавшаяся флора [представлена приблизительно таким же количеством
Видов, что и предшествовавшая ей (исходная) флора той же территории;
1 До сих пор остается нерешенным вопрос о том, что более устойчиво - тип
Данной естественной флоры (даже если она не отделена от соседних флор непрохо-
димыми природными барьерами) или любой тип сообщества, входящий в данную
флору.
41
темпы видообразования стабильны]; 3) угасающая флора (резко ре-
дуцированная в отношении разнообразия таксонов; темпы видообра-
зования резко снижены, миграционные возможности слабые); 4) вто-
рично обогащенная флора (фаза обогащения может сменить 2-й или
3-и этап развития; темпы видообразования ослаблены, миграционные
возможности велики).
Круг стоящих перед флористами проблем изучения эволюции в при-
родном растительном покрове на всех уровнях его дифференциации,
конечно, не исчерпывается эскизно намеченными выше вопросами; воз-
никающие в связи с этим задачи хотя и трудны, но важны и увлекатель-
ны, они все еще ждут своих исследователей.
42
II. МЕТОДИЧЕСКИЕ ПРОБЛЕМЫ
СОВРЕМЕННОЙ ФЛОРИСТИКИ
1. РАЗВИТИЕ МЕТОДА КОНКРЕТНЫХ ФЛОР ЗА 1971-1985 гг.
ВЫБОР ОПОРНЫХ ЕДИНИЦ ФЛОРИСТИЧЕСКИХ СРАВНЕНИЙ
В. М. Шмидт
О РОЛИ А. И. ТОЛМАЧЕВА В РАЗВИТИИ
СРАВНИТЕЛЬНОЙ ФЛОРИСТИКИ
Ленинградский государственный университет им. А. А. Жданова
Со времени зарождения сравнительной флористики как отрасли фло-
ристической географии растений прошло 160 лет: первые примеры изме-
нения флор в географическом аспекте были приведены еще Скоу
(Schow, 1823). Однако подлинным основоположником этой отрасли
знания по праву считается Альфонс Декандоль (De Candolle, 1855), впер-
вые показавший, что флористическое богатство различных районов зем-
ного шара может и должно служить объектом сравнительного изучения.
Он впервые указал также и на то, что такое сравнение возможно лишь
при условии приблизительной равноценности флор.
Действительно, научное сравнение любых объектов предполагает
их принципиальную сопоставимость. Однако накопление флористических
данных по мере развития нашей науки происходило в значительной мере
без расчета на их последующее сравнение. Многочисленные флористиче-
ские сводки охватывают самые различные по своим размерам и принци-
пам выделения территории. Это обстоятельство, отнюдь не лишая такие
сводки важного познавательного значения, сильнейшим образом затруд-
няет сравнение накопленного флористического материала и установление
закономерностей географической изменчивости флористических пока-
зателей.
Итак, сравниваемые флоры должны быть равноценны. Но в каком
смысле следует понимать эту равноценность? Каковы критерии равно-
ценности, позволяющие обеспечить научно обоснованное сравнение
флор? На эти вопросы пытались ответить многие ученые, но полного со-
гласия между ними не достигнуто и по сей день.
Что касается Декандоля, то сам он придавал понятию равноценности
флор явно топографический оттенок, указывая, что „было бы абсурдно
сравнивать с точки зрения общего числа видов области, которые не явля-
лись бы заметно одинаковыми по своей площади” (De Candolle, 1855,
р. 1270). Это указание, составляющее первый принцип обеспечения сопо-
ставимости флор, было, несомненно, достаточно справедливым: в общем
случае формула „чем больше площадь — тем больше видов” представля-
ется вполне очевидной.
Между тем во многих частных случаях дело обстоит сложнее и
указанная формула „не работает”, ибо число видов растений зависит
43
не только от площади, но и от множества других (исторических, клима-
тических, орографических и пр.) факторов среды. При этом чем крупнее
территория, тем сильнее по комплексу условий отличаются друг от друга
ее более мелкие районы, а это не может не отражаться на составе их
флоры. В итоге флора крупной территории неоднородна, имеет „сбор-
ный” характер, гетерогенна по ряду флористических показателей, хотя,
разумеется, бывают и исключения (например, в тундрах или пустынях,
где однородность флоры может сохраняться на большом протяжении).
Сказанное означает, что при сравнении одинаковых по площади, но круп-
ных флор важные флористические особенности их отдельных районов
могут остаться незамеченными. В этом смысле сопоставимость крупных
флор как таковых снижается, приобретает ограниченное значение.
Все это было осмыслено А. И. Толмачевым уже к началу 30-х годов,
но наиболее убедительно показано им в известной работе „О количест-
венной характеристике флор и флористических областей” (1941). Тол-
мачев обратил внимание флористов на то, что при сокращении площади
пестрота условий среды уменьшается и что таким путем можно добиться
сопоставления участков, каждый из которых во флористическом отно-
шении выглядел бы сравнительно однородным.
В результате Толмачевым был выдвинут второй важный принцип
сопоставимости флор: они должны иметь ограниченные размеры, т. е.
не представлять собою „сборные” флоры.
Третий принцип, указанный им же, предостерегает исследователей
от поспешных выводов: сравниваемые флоры должны характеризовать-
ся высокой степенью изученности. Добавим лишь, что при изучении мел-
ких флор требуемая полнота инвентаризации видового состава достига-
ется легче и быстрее.
Таким образом, мы располагаем тремя принципами обеспечения
сопоставимости флор, первый из которых был выдвинут Декандолем,
а два других - Толмачевым: 1) приблизительное равенство площадей
сравниваемых флор; 2) ограниченный размер этих площадей; 3) высо-
кая степень флористической изученности.
По поводу первого принципа надо заметить, что Толмачев специаль-
но подчеркивал возможность соблюдения лишь приблизительного равен-
ства площадей сравниваемых флор: излишне жесткие в этом отношении
требования технически невыполнимы и биологически мало оправданы,
ибо несколько отличающиеся по площади флоры могут быть вполне
равноценны в ботанико-географическом смысле. В специальных случаях
(например, при флористическом районировании) исследователь бывает
просто вынужден сравнивать флору более крупной территории с флора-
ми отдельных ее районов. Кроме того, разные флористические показа-
тели в разной мере чувствительны к различию площадей сравниваемых
флор. Например, при сравнении систематической структуры допустима
большая разница в площадях изученных флор, чем при сравнении числа
таксонов (Шмидт, 1980).
Известно, что многие современные отечественные и зарубежные фло-
ристы стремятся к сравнению строго определенных по своей площади
эталонных флор (например, 100,1000или даже 10000км2). С формаль-
44
но-статистической точки зрения такие действия выглядят безупречно.
Мы видим, однако, что исходя из ботанико-географических соображений
Толмачев трактовал первый принцип сопоставимости флор в значительно
более мягкой форме.
В конечном счете, как мы полагаем, правильнее всего придерживать-
ся двойного условия биолого-статистической сопоставимости флор, имея
в виду как приблизительное равенство их площадей, так и совпадение
флористических рангов (Шмидт, 1980).
К формулировке второго принципа — ограниченного размера сравни-
ваемых флор — Толмачев пришел в результате разработки широко ис-
пользуемого ныне отечественными флористами метода конкретных
флор (Толмачев, 1931, 1932—1935, 1941, 1970, 1974). Он исходил из
того, что по мере сокращения площади обследования можно побиться
такогополржения,когда набор дидов. свойственных всем экотопам
данной местности, будет определяться чисто экологическими условиями
дредьЬ тогда как ,п.)
раптанвагдацтожнутп Р°пь- Такие
конкретные (или элементарные) флоры (КФ), по мнению Толмачева,
являются наиболее подходящими объектами для сравнения в широком
географическом диапазоне. Представление о конкретной флоре как ре-
ально существующей в природе флористической единице низшего ранга
несомненно сыграло выдающуюся роль в развитии сравнительной фло-
ристики, а сам метод был взят на вооружение многими советскими бо-
таниками.
Не следует забывать и рекомендаций Толмачева в отношении площа-
ди КФ. В условиях Арктики на основании собственных исследований он
предлагал придерживаться площадей в 100 км2, но указывал, что в дру-
гих климатических зонах площадь КФ может возрастать (например,
в тропиках до 1000 км2). Мы глубоко убеждены в справедливости этого
указания и полагаем, что в разных зональных (и прочих) условиях пло-
щадь КФ просто не может быть одинаковой. Наш опыт показывает, на-
пример, что при продвижении из Арктики к югу площадь КФ возрастает
примерно до 300-350 км2 на севере Архангельской обл. и до 700-
750 км2 в условиях Ленинградской, Новгородской и Псковской обл.
Очень близкие к последним цифры (650—700 км2) были получены для
Латвии путем специального эксперимента на местности (Табака и др.,
1977). Указанная разница в площадях КФ, расположенных в трех разных
природных зонах и флористических областях, вполне естественна и не
лишает эти флоры сопоставимости (если, конечно, вслед за Толмачевым,
рассматривать их в качестве естественных флористических единиц низ-
шего ранга).
Любопытно, что в отечественной ботанике у Толмачева был скром-
ный предшественник, в некоторых отношениях опередивший даже
Декандоля. Мы имеем в виду К. К. Клауса — химика и фармацевта,
одновременно занимавшегося ботаникой на вполне профессиональном
уровне. Еще в 1851-1852 гг. им было изучено и сравнено несколько
локальных флор, по своему объему весьма близких к тем, которые мы
теперь называем конкретными, или элементарными, флорами. Более
45
того, сравнивая флоры окрестностей г. Сергиевска и г. Сарепты
Клаус фактически предвосхитил использование в сравнительной фло-
ристике ряда количественных показателей (Шмидт, Ильминских,
1982).
В связи с этим нельзя не подчеркнуть, что метод КФ Толмачева
открыл широкие возможности для использования в сравнительной
флористике математических приемов анализа и сравнения флор. Сам
Толмачев математические методы в своей работе не применял, огра-
ничиваясь подсчетом числа таксонов и их отношений во флорах. Напри-
мер, им впервые приведено множество данных о числе видов в КФ
различных районов земного шара (Толмачев, 1941, 1970, 1974).
По личному опыту мы знаем, однако, что Толмачев с вниманием и
симпатией относился ко всем попыткам внедрить математические
методы в сравнительную флористику, если получаемые результаты
не противоречили ботанико-географической сути дела. В этом он был
совершенно прав; математический аппарат формален и успешное его
применение зависит от биологической доброкачественности исходных
данных.
По нашему мнению (Шмидт, 1980, 1981), статистическая сущность
представления о КФ состоит в следующем. Как естественная элементар-
ная флористическая единица КФ представляет собою генеральную сово-
купность произрастающих на ее территории видов растении. В этом
своем качестве КФ — чрезвычайно удобный объект для решения срав-
нительно-флористических задач и выявления закономерностей измене-
ния флористических параметров в географическом аспекте. С другой
стороны, конкретную флору правомерно рассматривать и в качестве
выборки из флоры более крупной территории. В этом смысле она служит
„пробой флоры” (термин Б. А. Юрцева), способной удовлетворительно
моделировать, например, состав флоры во много раз большего по пло-
щади флористического района. В такой трактовке статистической сущ-
ности КФ нет ничего противоречивого, ибо объем генеральной совокуп-
ности задается исследователем в зависимости от стоящей перед ним
задачи.
С 1970-х годов наблюдалось интенсивное развитие теоретических
основ метода КФ наряду с его практическим применением в различных
районах нашей страны. Отчетливо выявилось стремление ряда исследова-
телей к углубленному эколого-топологическому анализу структуры КФ
посредством изучения так называемых парциальных флор (Юрцев,
Семкин, 1980; Юрцев, 1982а).
Лучшим памятником ученому служат его идеи, живущие и развиваю-
щиеся благодаря усилиям его учеников и последователей. Толмачеву
удалось создать школу единомышленников, работающих в разных кон-
цах нашей Родины, развивающих собственные научные направления, но
объединенных общей целью служения сравнительной флористике — той
отрасли знания, основоположником которой в нашей стране и явился
Александр Иннокентьевич Толмачев.
46
Б. А. Юрцев
ЭЛЕМЕНТАРНЫЕ ЕСТЕСТВЕННЫЕ ФЛОРЫ
И ОПОРНЫЕ ЕДИНИЦЫ СРАВНИТЕЛЬНОЙ ФЛОРИСТИКИ
Ботанический институт им. В. Л. Комарова АН СССР
Вопрос о выборе опорных единиц при флористических сравнениях
с полной определенностью поставлен А. И. Толмачевым (1931) более
полувека тому назад, когда им было сформулировано понятие конкрет-
ной, или элементарной, флоры (КФ) . Этот вопрос оживленно обсуждал-
ся в основном отечественными флористами в последние десятилетия,
и тем не менее до сих пор по нему не достигнуто согласия между разны-
ми авторами. Причина во многом лежит в сложной природе самого
объекта - естественных флор, представляющих нечеткие, „размытые”
множества, элементы которых — виды и их комплексы — нежестко
связаны друг с другом в процессе формирования и расселения.
Едва ли вызовет возражение тот тезис, что наиболее сравнимы меж-
ду собой по совокупности показателей выделы флористического (или
комплексного ботанико-географического) районирования одного и того
же ранга. Однако установление рангов флористических выделов обычно
производится на заключительном этапе исследования и при современном
уровне разработанности принципов и методов флористического райони-
рования не свободно от многих элементов субъективизма: ранги выде-
лов — не нечто, лежащее на поверхности. Отсюда стремление Толмачева
и его последователей иметь дело с элементарными единицами иерархии
естественных флор регионального порядка. Цель этой работы - рассмот-
реть природу элементарных флористических выделов (регионального
порядка) под углом зрения задач сравнительной флористики.
Первым и главным условием познания КФ Толмачев считал выявле-
ние флористических комплексов [= парциальных флор (ПФ), в смысле
Б. А. Юрцева (1982а)] всех типов экотопов, существующих в данных
„обще гео графических” условиях; этим и определяется выход на регио-
нальный уровень — переход от единиц экотопологического (внутриланд-
шафтного) уровня к региональным флорам низшего ранга. Этим обеспе-
чивается также полнота выявления и всех флорогенетических (флори-
стических, в смысле М. Г. Попова и Р. В. Камелина) и флороценотиче-
ских (Юрцев, 1982а) комплексов, которые в рамках единой КФ, как
правило, разделены экотопологически.
Классический метод выявления КФ, как известно, заключается в по-
степенном расширении обследованной площади, на которой осмотрены
ПФ всех экотопов, до того, как прирост видов практически не прекра-
тится. К настоящему времени накоплен огромный опыт выявления со-
става КФ методом Толмачева, в основе которого лежит математическая
операция объединения множеств видов, обитающих на различных экото-
пах; автор метода исходил из допущения, что по достижении ареала-ми-
нимума дальнейшее объединение ПФ соседних контуров экотопов пере-
станет давать прирост списка видов — в пределах контура КФ кривая
зависимости числа видов от обследованной площади имеет асимптотиче-
ский характер („выходит на плато”) . Такие случаи были зарегистриро-
ваны (например, на равнинах Ямала: Ребристая, статья в наст, сб.), хотя
чаще исследователи прекращают поиск на стадии „практического прекра-
щения” прироста списка, не стремясь нащупать топографические преде-
лы каждой КФ. Задачу поиска элементарных флористических единиц
(регионального уровня) можно сформулировать и так: до какого пре-
дела правомерно продолжать объединение ПФ топографически примы-
кающих контуров экотопов, если мы хотим избежать смешения несколь-
ких элементарных флор регионального уровня в едином флористиче-
ском списке?
Предлагались разные математические выражения зависимости числа
видов от размеров обследованной площади (Малышев, 1972), сущест-
венными компонентами которых являются удельное флористическое бо-
гатство (число видов на площади стандартного размера, принятой за еди-
ницу измерения, например 1 км2 или 100 км2) и эмпирически находи-
мый индекс пространственного разнообразия флоры. Выяснены неуни-
версальность предложенных эмпирических формул, а также изменение
индекса пространственного разнообразия флоры на разных ступенях
увеличения площади. Отдавая должное значению этих интересных работ,
нельзя не отметить некоторый общий их недочет: суммарный характер
„подбираемых” индексов пространственного разнообразия, не вскры-
вающих основные компоненты и факторы разнообразия и их соотноше-
ние на том или ином пространственном уровне. Кроме того, как отме-
чает Л. И. Малышев (1975), индексы рассчитываются на основе модели
континуума флоры и случайного распределения видов, что далеко не
всегда имеет место.
Попытаемся проанализировать основные компоненты пространст-
венного разнообразия флоры, начиная со стадии объединения ПФ разных
типов и классов экотопов (опускаются стадии выявления ПФ первого
обследованного микро-, затем мезоэкотопа). Процедуру выявления
любой КФ можно расчленить на 2 основных этапа с противоположной
стратегией поиска: до достижения ареала-минимума КФ (собственно
инвентаризация) и после его достижения (контроль за полнотой выявле-
ния, определение топографических границ данной КФ, ее контакта с со-
седними). Впрочем, первый основной этап с известной условностью
можно подразделить еще на два: 1) выявление разнообразия экотопов,
точнее — экотопологического разнообразия ПФ; 2) выявление полного
состава видов, обитающих на каждом типе и классе экотопов. Итого
3 этапа.
На первом этапе флорист стремится к скорейшему, по возможности,
посещению разных типов и классов экотопов, в первую очередь макси-
мально контрастных во флористическом отношении; этим достигается
очень быстрый рост списка, а также общая ориентировка в характере
флоры (флористической ситуации). В открытой местности обычно зада-
ется большой рекогносцировочный маршрут (или серия маршрутов),
цель которого - осмотреть хотя бы по 1 разу ПФ всех экотопов „сводно-
го” экологического профиля данного ландшафта. Трудности связаны
48
кжде всего с наличием редких и даже уникальных (неповторяющихся)
мегов экотопов, что существенно влияет на площадь выявления КФ,
доним из компонентов которой является площадь выявления разнообра-
Вия экотопов; последняя в свою очередь зависит как от общего числа,
здк и от площади выявления конкретных типов и классов экотопов,
особенно самых редких. Поскольку наиболее редкие экотопы могут
встречаться не в каждом контуре однотипного ландшафта, представляя
как бы случайный компонент данного ландшафта, при разграничении
КФ, вероятно, следует в первую очередь ориентироваться на повторя-
ющиеся типы экотопов.
Если бы каждый контур любого типа экотопов заключал все виды,
обитающие в данном типе, для выявления КФ было бы достаточно одно-
кратного осмотра каждого типа экотопов [или фитоценозов (Шмидт,
1972, 1980) ]. Более того, ПФ многих (разных) типов и классов экото-
пов имеют немало общих видов, а некоторые типы не содержат свойст-
венных только им таксонов; это существенно ускоряет процедуру выяв-
ления КФ (так же как обилие видов, специфичных для определенных
типов экотопов, замедляет ее). Однако только часть видов встречается
на данном типе экотопов регулярно, в каждом контуре; постоянство
других может быть невелико или даже ничтожно мало (при высокой
верности), вплоть до крайнего случая эндемизма для индивидуального
экотопа (такие примеры известны среди растений скал, термальных
источников и т. д.). Это также существенно увеличивает площадь выяв-
ления КФ, ставя ее в зависимость от площади выявления ПФ разных
типов экотопов и заставляя значительно увеличить повторность осмотра
каждого типа экотопов. Положение усугубляется тем, что чем реже
встречается данный тип экотопа, тем, как правило, большую долю
в его „сводной” ПФ составляют виды с низким постоянством: непо-
стоянство состава особенно характерно для сильно разобщенных релик-
товых группировок. Отсюда не линейная (близкая к квадратичной)
зависимость между редкостью типа экотопа и площадью выявления его
объединенной ПФ;1 эта площадь может значительно превышать площадь
контуров элементарных (и более крупных) естественных флор, при
разграничении которых мы вынуждены ориентироваться прежде всего
на набор неуникальных типов экотопов и их ПФ, а также на отличия
в составе постоянных видов на однотипных зкотопах. Для редких же
типов экотопов с нестабильным флористическим составом необходим
осмотр всех индивидуальных контуров в пределах КФ — таким путем
можно достигнуть практически полного выявления состава ПФ (в пре-
делах данной КФ). В целом же, на II этапе кривая прироста числа
выявленных видов имеет затухающий характер. Впрочем, разграниче-
ние I и II этапов во многом условно: выявление самых редких эко-
топов иногда происходит уже после того, как обычные зкотопы осмот-
рены в достаточной повторности и состав их ПФ выявлен с высокой
полнотой.
1 Имеется в виду сводный список однотипных ПФ, получаемый путем объеди-
нения списков индивидуальных.
4 Заказ 1220
49
Ill этап начинается после того, как прирост списка практически пре-
кратился - достигнут ареал-минимум КФ и ставится задача найти при-
родные границы данной КФ. Если на I этапе флорист стремился охватить
участки максимально контрастные в экологическом и флористическом
отношениях, присоединение которых давало бы наибольший прирост
списка, то на III этапе он, напротив, присоединяет к уже выявленному
контуру КФ лишь такие участки, которые не увеличивали бы сколько-
нибудь существенно общий флористический список (с минимальной раз-
ностью и максимальным пересечением флористических множеств).
Такую территорию В. М. Шмидт (1980; см. также: Юрцев, 1974а) фигу-
рально назвал „мертвой зоной”. Большинство флористов перестает удли-
нять радиальные маршруты при первых признаках попадания в „мерт-
вую зону”, поэтому о соотношении ее с „ареалом-минимумом” мы знаем
очень мало. Можно предположить, что феномен „мертвой зоны” (дости-
жения асимптоты) нередко бывает иллюзорен и соответствует заключи-
тельной части II этапа, когда невыявленными остались в основном лишь
наименее константные виды редких экотопов; доля таких видов в соста-
ве КФ всегда остается заниженной (недовыявленной); однако и цен-
ность их для разграничения естественных флор исчезающе мала, несмот-
ря на высокую информативность части редких видов для восстановления
истории флоры (Юрцев, 1983а).
Рассмотрим критерии разграничения элементарных естественных
флор регионального уровня, по которым можно судить о выходе за пре-
делы „мертвой зоны”, когда дальнейшее объединение соседних ПФ уже
неправомерно. В их числе следует назвать: 1) регулярное появление но-
вых типов экотопов, с достаточно самобытными (имеющими ядро спе-
цифических видов) ПФ, что обычно наблюдается при попадании в другой
ландшафт и сопровождается общим изменением флоры, выдержанным
на значительной территории; с этой точки зрения важно, чтобы базовый
лагерь при изучении КФ располагался где-то в середине природного кон-
тура, а не на границе двух-трех ландшафтов (в последнем случае есть
риск, что на I этапе исследователь, стремясь к охвату контрастных экото-
пов, не обратит внимания на флористические различия ландшафтов);1
2) появление несвойственных первой КФ константных видов на тех же
типах экотопов; это сигнализирует о переходе в другую КФ в более ши-
рокой трактовке (Толмачев, 1931) или в другой элементарный флори-
стический район, в смысле Юрцева (1975а); практическое применение
этого критерия затрудняется явлением внутриландшафтной мозаичности
флор — очаговым распределением не только редких, но и некоторых
обычных и массовых видов, о чем будет сказано ниже (появление же но-
вых редких видов на тех же экотопах или даже новых очень редких
1 А И. Толмачев (1931, 1970), говоря об универсальности набора видов для
всего контура КФ, отметил лишь одно условие этого - постоянство флористиче-
ского состава ассоциаций - и недооценил значение другого фактора - постоянства
набора ассоциаций (= ПФ всех типов экотопов) для всего участка. Флористическая
однородность контура КФ предполагает наличие как ядра постоянных типов экото-
пов, так и ядра постоянных видов на каждом из них.
50
типов экотопов еще не говорит о переходе в соседнюю КФ); 3) регуляр-
ное появление на тех же местоположениях принципиально иного класса
ПФ, например степных или лесных массивов (исключая случаи, когда
они представлены одиночными реликтовыми островками); это — при-
знак пересечения ботанико-географических границ более высокого ран-
га, нежели границы между элементарными флористическими районами;
4) в качестве критерия разграничения соседних элементарных естествен-
ных флор выдвигался также признак присутствия в каждой из них хотя
бы одного эндемичного вида или расы (Камелин, 1969, 1973); однако
низшие единицы дифференциации региональных флор, хорошо различаю-
щиеся комплексом флористических показателей (соотношением разного
рода таксонов и элементов), как правило, лишены эндемиков, хотя в ви-
де исключения можно встретить расы и виды, эндемичные даже для
одного экотопа; в норме же эндемизм -проявляется в фитохориях не са-
мого низкого ранга (впрочем, в разных природных областях — неоди-
накового) .
Перейдем к рассмотрению факторов, определяющих размер площа-
ди выявления КФ и ее компонентов: площади выявления спектра экото-
пов, площади выявления сводных ПФ каждого класса экотопов; а также
размер контуров элементарных естественных флор в целом.
1. Разнообразие местоположений (формы рельефа с учетом слагаю-
щих их литологических разностей) и их распределение образуют жесткий
(сравнительно медленно изменяющийся) каркас ландшафта и опреде-
ляют дифференциацию его флоры, сами же определены геоисторически-
ми факторами (эндо- и экзогенными). По этому признаку равнинные
ландшафты обычно существенно отличаются от горных меньшей пестро-
той и контрастностью, большей стабильностью фаций, но зато протяжен-
ность сочетаний форм рельефа и литологических разностей относительно
слабо зависит от географической зональности, и потому сопряженные
с ними флористические выделы (КФ) могут быть использованы для
межзональных сравнений. Многие элементы расчленения ландшафта рит-
мически повторяются (например, долины и водоразделы) и отобража-
ются ритмической внутриландшафтной структурой (инфраструктурой)
КФ. Разные (в том числе соседние) ландшафты обычно различаются
Спектром местоположений и его широтой (разнообразием), что и опре-
деляет отчасти своеобразие „общегеографических условий” в их преде-
лах; так, расчленение на долины и водоразделы свойственно большин-
ству ландшафтов, однако не в каждом из них сочетаются выходы карбо-
натных, основных и кислых силикатных пород, имеются горные струк-
*Уры и т. д.; соответственно приходится говорить о фациальной неполно-
Чтенности многих ландшафтов как об одном из факторов их флористи-
*ского своеобразия и генезиса их флоры.
* Фациальное разнообразие ПФ выявляется более полно в вышестоя-
щих флористических выделах - флористических районах, округах и т. д.
этом одна из причин большей стабильности флоры более крупных,
'’фДДиально полночленных контуров (Малышев, 1972; Марина, 1983,
И1акже статья в наст, сб.) ]. Чередование контрастных по рельефу и лито-
Яогии ландшафтов может сопровождаться фациально обусловленной
51
чересполосицей контрастных КФ в рамках одного района или округа;
последние должны характеризоваться спектром КФ.
Формы рельефа и их литогенная основа, как правило, более устой-
чивы на фоне флюктуаций климата, чем растительная оболочка ланд-
шафта (за исключением отдельных, высоко динамичных местополо-
жений) .
Подчеркну, что разграничение элементарных флор по смене ланд-
шафтных условий - лишь удобный рабочий прием и все в конце концов
решают масштаб и характер флористических отличий: если таковые не
выявлены или носят случайный характер (например, в случае если ланд-
шафты различаются соотношением, но не набором типов экотопов),
контуры обоих ландшафтов, вероятно, будут отнесены к единой КФ
при условии их достаточной флористической гомогенности. И, напротив,
если через ландшафт проходит ясная флористическая граница, он будет
отнесен к двум КФ.
2. Степень экологической контрастности спектра местоположений
данного ландшафта, длина градиента по ведущим (особенно лимитирую-
щим) экологическим факторам. Это определяет объективную возмож-
ность „упаковки” разнообразных экологически^ ниш, т. е. сосущество-
вания (на разных отрезках градиента) экологически о-йтичных флори-
стических комплексов. В горах эти возможности заведомо шире, чем
на расчлененной равнине, на последней — шире, чем на плоской низмен-
ности; экологическая емкость ландшафта во многом зависит и от его
литологического разнообразия, непосредственно влияющего на степень
фациальной полночленности ландшафта.
3. Конкурентные отношения различных флористических (биоти-
ческих) комплексов, сменяющих друг друга вдоль экологических
градиентов; то, какая часть градиента освоена конкретным комп-
лексом видов, во многом определяется конкурентной агрессивностью
видов комплекса и их средообразующими свойствами (способностью
частично нивелировать исходные экологические различия местополо-
жений). Так, некоторые зональные комплексы видов (например,
таежные и гипоарктические тундровые) на равнинах и низкогорьях
резко доминируют в ландшафте на самых различных местоположе-
ниях, оттеснив многие другие комплексы на более или менее редкие
экотопы со специфическими условиями. В этом случае фоновые типы
экотопов могут принадлежать одному классу (или группе близких
классов).
4. Степень динамичности ландшафтной оболочки, создающая пред-
посылки для „упаковки” экологических ниш не только в пространстве,
но и во времени, т. е. для смены различных комплексов видов на данном
местоположении в ходе первичных и вторичных сукцессий. Эта динамич-
ность зависит от расчлененности рельефа (энергии процессов эрозии и
денудации), устойчивости горных пород против разрушения, от фазы
тектонического развития данной и сопредельных территорий, интен-
сивности седиментации. Господство климаксовых комплексов воз-
можно лишь при длительно стабильном рельефе и сопровождается рез-
ким сужением ландшафтных позиций (вплоть до перехода в реликто-
52
вое состояние) комплексов растений-эксплерентов, приспособленных
к захвату участков с ослабленной конкуренцией; напротив, комплексы
эксплерентов могут доминировать в ландшафте быстро поднимающих*
ся гор.
5. Как отдельный фактор следует выделить антропогенную (отчасти
и зоогенную) трансформацию ландшафта и растительного покрова,
вплоть до полной его переработки, когда на самых разных местоположе-
ниях господствуют комплексы эксплерентов (как местных, так и занос-
ных), что частично нивелирует исходные различия ПФ разных экотопов,
а доминировавшие в естественном покрове комплексы видов (включая
зональные и климаксовые) сохраняются на отдельных разобщенных
участках („антропогенные реликты”), причем стабильность их состава
резко понижается, а площадь выявления возрастает. Изучая подобные
флоры, следует четко разграничивать разные задачи: 1) регистрация сов-
ременного состояния данной флоры; 2) реконструкция исходного со-
стояния флоры на той же территории (до периода интенсивной антропо-
генной трансформации); осуществление второй задачи предполагает
полный поконтурный учет сохранившихся фрагментов ненарушенных
или слабее нарушенных участков растительного покрова, широкую
экстраполяцию на основе раздельного учета ПФ разных типов экотопов,
привлечение флористических данных по ландшафтам-аналогам, что во
многом напоминает задачу по составлению карты восстановленной расти-
тельности.
6. Природная история данного участка суши, длительность периода
преемственного развития его биоты, степень контрастности ландшафтно-
климатических условий минувших геологичесих эпох, интенсивность
автохтонного видообразования, условия для сохранения реликтов и
для иммиграции видов с соседних и отдаленных территорий. Все это не-
посредственно отражается на степени гетерогенности эко топологиче-
ской и других структур местной флоры и тем самым на площади ее
выявления.
Факторы 2 (экологическая контрастность местоположений), 4 (сте-
пень динамичности ландшафтной оболочки) и 6 (контрастность усло-
вий минувших эпох, длительность преемственного развития биоты —
возможность сохранения разнообразных групп реликтов) в общем
способствуют увеличению площади выявления КФ, хотя зависимость
далеко не всегда однозначна. Фактор 3 (конкурентная способность
флористических комплексов) сокращает площадь выявления состава
доминантных комплексов, но увеличивает таковую реликтовых; диф-
ференцированно влияет на площадь выявления состава разных групп
и фактор 5 (антропогенная трансформация ландшафта). На фоне
остальных следует выделить первый фактор — существование относи-
тельно дискретных ландшафтов (в умеренно широком понимании
этого термина как геосистем определенного ранга), контуры которых
фиксируются повторяющимся сочетанием форм рельефа и литологиче-
ских комплексов, т. е. сочетанием местоположений; от разнообразия
местоположений в данных биоклиматических условиях зависит и
спектр экотопов, и разнообразие их ПФ, а следовательно, и общий состав
53
и характер флоры.1 Размеры соответствующих ландшафтно-флористи-
ческих контуров могут быть очень различны (но не ниже какого-то
минимального порогового уровня), в горах они обычно меньше, чем на
равнинах, хотя площадь выявления флоры (минимальная площадь, на
которой состав видов стабилизируется до определенного, достаточно
высокого уровня) в горах, как правило, выше в силу действия факто-
ров 2 и 4. На равнинах же, напротив, площадь ландшафтно-флористиче-
ских контуров в среднем увеличивается, а площадь выявления сокра-
щается. Таким образом, можно предположить, что в ряду ландшафтов
с неодинаковой (нарастающей или убывающей) пестротностью местно-
стей, урочищ и фаций должна проявиться обратная зависимость между
размером элементарных флористических выделов регионального уровня
и площадью выявления их флоры. Разумеется, этот парадокс нуждается
в подтверждении и проверке на обширном фактическом материале. Пока
же имеются лишь отрывочные данные для отдельных (удаленных один
от другого) природных районов.
Эмпирический опыт дробного флористического районирования тер-
ритории Алтайского заповедника (Золотухин, 1982,1984, а также статья
в наст, сб.) дал средние размеры элементарных выделов, существенно
отличающихся по флористическим показателям, — 200-300 км1 2; они
трактуются как микрорайоны, а их флоры — как КФ. Любопытно, что
такого же порядка средние размеры геоботанических микрорайонов
в условиях Латвии (Л. В. Табака, доклад на совещании в Неринге),2
площадь отдельных микрорайонов может быть и меньше 100 км2 или же
значительно больше 300 км2.
Что же касается „площади выявления” флоры, то она существенно
изменяется в зависимости как от литолого-орографических, так и от
биоклиматических условий и степени нарушенное™ покрова, хотя экспе-
риментальных данных по этому вопросу крайне мало. Нередко принима-
ется (Шмидт, 1980, а также статья в наст, сб.), что в Арктаке для выяв-
ления состава КФ достаточно 100 км2, в северной тайге 300—350 км2,
в полосе восточноевропейских хвойно-широколиственных лесов 700—
750 км2, в тропическом поясе — не менее 1000 км2. В условиях Латвии
(Табака и др., 1977) опытным путем получена цифра 500—600 км2.
Однако и в условиях одной и той же ботанико-географической зоны
пространственное разнообразие флоры, а следовательно, и ареал-мини-
мум КФ могут существенно варьировать, что видно из экспериментально
полученных данных различных авторов для ряда конкретных районов.
Так, в Псковской и Новгородской областях (подзона хвойно-широко-
лиственных лесов), по данным В. М. Шмидта (1976, 1980), сокращение
обследованной площади с 700 км2 до 300 км2 привело к „потере” 11.2 %
и 6.4% (в среднем 8.8%) видов, в основном очень редких. В той же
1 Флоры экологически контрастных ландшафтов в ряде отношений сходны
с флорами островов - определенной степенью изоляции, известной автономностью
развития.
2 Вспомним, что А. И. Толмачев (1970) считал возможным изучение в качест-
ве КФ флор элементарных геоботанических районов.
54
подзоне в Западной Латвии сокращение обследованной площади с 630 до
250 км2 сопровождалось потерей 5.3% и 8.5% (Табака и др., 1977).
В Беркшире (Англия, зона широколиственных лесов) трехкратное со-
кращение площади — с 750 до 250 км2 — имело результатом уменьшение
туземных видов на 17.6% (Малышев, 1972). В то же время на южной
окраине тундровой зоны, но в условиях среднегорного рельефа и значи-
тельного литологического разнообразия [выдержанного на всем протя-
жении восточной части Люпвеемского горного массива (Заславская,
Петровский, 1983)] в Анюйском нагорье сокращение обследованной
площади с 600 до 300 км2 приводит в среднем к потере 18.8% видов,
что говорит о большем пространственном разнообразии данной западно-
чукотской флоры, несмотря на ее более низкий уровень богатства.1
По данным В. В. Петровского и Т. В. Плиевой (1986),в южной (гольцо-
восеверотаежной) части Анюйского нагорья и соседней части Анадыр-
ского плоскогорья попарное объединение флор сходных по условиям
соседних участков по 300 км2 дает существенный прирост списка и
большую стабильность показателей флористического богатства (для
600 км2). В то же время О. В. Ребристая (см. статью в наст, сб.) путем
сравнения видового богатства серии „вложенных” один в другой участ-
ков разного радиуса пришла к выводу, что на тундровых равнинах
Ямала для практически полного выявления КФ (98-99%) достаточно
площади 30 км2, что составляет незначительную долю от площади всего
контура КФ.
Поскольку микрорайоны площадью 200—300 км2 и менее во многих
случаях значительно отличаются один от другого по флористическим
показателям и вполне отвечают представлению об элементарных фло-
ристических выдел ах (КФ), налицо ситуация, когда площадь большин-
ства или всех этих выделов оказывается ниже „площади выявления”
КФ; соответственно метод заложения пробных площадей по 500—600
или даже 750—800 км2 может оказаться недостаточно чуток, для
того чтобы выявить самые дробные флористические выделы. Это озна-
чает лишь, что флоры последних должны обследоваться по всему кон-
туру (особенно ПФ спорадически распределенных экотопов в их пре-
делах) .
В самом деле, за понятием „площадь выявления флоры” по су-
ществу скрывается феномен стабилизации числа (и набора) видов
1 По данным Т. М. Заславской и В. В. Петровского (1983), на стандартной
площади 300 км2 выявляется 60-74% флоры обследованной восточной части
Люпвеемского массива общей площадью 1500 км2; на сдвоенных участках (по
600 км2) - 76-89%, утроенных (900 км2) - 82-96%, учетверенных (1200 км2) -
94-99%. Очень сходные показатели приводят В. В. Петровский и Т. В. Плиева
(1986) для района колымско-анадырского водораздела: здесь на площадях по
300 км2 выявляется от 61.0 до 76.9% флоры района площадью 1500 км2 (всего
372 вида), на сдвоенных (600 км2) - (75.8)813-84.4%, сокращение обследован-
ной площади с 600 до 300 км2 в среднем приводит к потере 15.2 % видов. В Псков-
ской обл. на участке 750 -800 км2 выявляется 75-90% видов более протяженного
(в 8-13 раз) флористического района (Шмидт, 1980). В Западной Латвии на участ-
ках по 70 км2 выявлено 77-85.4% видов КФ площадью 630 км2 (Табака и др.,
1977).
55
на определенном этапе расширения обследованной площади? Но явля-
ется ли сама по себе стабилизация видового разнообразия гарантией фло-
ристической гомогенности той территории, на которой она фиксируется?
Очевидно, нет (см. выше). Стабилизация при расширении обследованной
площади наступает и в тех случаях, когда базовый лагерь расположен на
стыке двух (трех) элементарных ландшафтно-флористическихвыделов.
Итак, несоответствие между размером дискретных ландшафтно-фло-
ристических контуров и „площадью выявления” (=площадью стабили-
зации состава) флоры на территориях с контрастным рельефом и литоло-
гией может выражаться в смене контрастных флористических ситуаций
на сравнительно небольших пространствах, недостаточных для стабилиза-
ции в их пределах набора видов (прекращения прироста списка). В этих
случаях в качестве выделов КФ правильнее рассматривать и изучать ланд-
шафтно-флористические контуры, устойчиво отличающиеся набором
экотопов и флорой. Стремление же к наращиванию площади до полного
прекращения прироста списка в данном случае привело бы к смешению
существенно отличных флористических ситуаций; кроме того,в горных
районах достигнуть полного прекращения прироста списка с расшире-
нием обследованной площади практически невозможно.
Могут ли дробные ландшафтно-флористические выделы, о которых
шла речь, удовлетворять задачам сравнительной флористики — служить
опорными единицами флористических сопоставлений? Прежде чем об-
суждать этот вопрос, определим основные задачи флористического изу-
чения территории: 1) выяснение состава, а затем и соотношения видов
растений на данной территории; 2) выяснение степени однородности
флоры, первичное флористическое разделение территории, выяснение
географической дифференциации представленных на ней естественных
флор; 3) сравнительная типологическая оценка (анализ) естественной
флоры (флор) по соотношению таксонов высокого ранга и различного
рода элементов флоры (включая анализ эндемизма, реликтовых элемен-
тов) , уровню богатства; 4) сравнение с соседними и другими флорами,
подключение районирования изученной территории к общеземному фло-
ристическому районированию; 5) восстановление истории формирова-
ния флоры (растительного покрова, ландшафтов) изученной территории
с элементами прогноза; 6) выявление важнейших ботанических объек-
тов охраны, разработка мер охраны и т. д.
Едва ли не очевидно, что для решения первой и пятой задач выявле-
ние элементарных флор важно не само по себе, а лишь как способ „фло-
ристического зондирования” территории с помощью „проб флоры”;
полученные для разных пунктов данные затем объединяются и обобща-
ются. Так, вопросы флорогенеза целесообразно решать лишь для доста-
точно обширных территорий. Соотношения таксонов и различного рода
1 Эта стабилизация во многих случаях оказывается неполной. Именно поэтому
Л И. Малышев (1972) предложил определять пороги стабилизации не через про-
цент выявления (как делают другие авторы) , а по определенной мере (%) сниже-
ния прироста с удвоением обследованной площади; на этом основан и расчет „ре-
презентативности” участка флоры (Малышев, 1975).
56
элементов флоры, свойственные вышестоящим фитохориям (третья за-
дача) , со значительной полнотой (хотя и с известными деформациями!)
выявляются на уровне не только КФ, но и их фациально представитель-
ных частей — речных бассейнов I порядка (Марина, 1982,1983,1985,а так-
же статья в наст, сб.) и даже на уровне некоторых ПФ повышенной гете-
рогенности. При флористическом обследовании ландшафта мы имеем
достаточно материала для выявления основных флороценотических и
эколого-ценотических комплексов видов, для суждения об их богатстве,
степени стабильности состава (или, напротив, недонасыщенности),об их
активности, взаимоотношениях, ландшафтных позициях и т. д. — показа-
телях, из которых складывается представление о флоре как о природной
системе (Юрцев, 1982а). Для выявления и оценки географической диффе-
ренциации региональной флоры (вторая задача) изучение представитель-
ной серии рассредоточенных по ее площади КФ имеет первостепенное
значение. В тех же (пока еще редких) случаях, когда ставится задача
максимального дробного ботанико-географического и флористического
районирования территории, именно ландшафтно-флористические выде-
лы, о которых шла речь выше, приобретают значение основных (конеч-
ных) единиц флористического деления территории — на региональном
уровне. Выявление их имеет особую ценность и для охраны ботанических
объектов (6-я задача). Решение же четвертой задачи возможно лишь
после того, как мы справились со второй и третьей и изученные элемен-
тарные флоры сгруппированы в более крупные естественные единицы.
Видовое богатство (шире — таксономическое разнообразие) флоры —
существенный показатель сравнительной оценки флор, хотя, пожалуй,
не самый важный для решения перечисленных выше задач флористики.
Основные компоненты видового богатства крупной региональной фло-
ры — уровень богатства, измеряемый средним числом видов в КФ (Тол-
мачев, 1931, 1970), и разнообразие конкретных флор (а также флор
фитохорий более высокого ранга) в пределах данной флоры — как по
числу, так и по средней флористической самобытности одноранговых
выделов. Однако число видов в КФ (как элементарных естественных
флорах регионального уровня), при значительном варьировании послед-
них по площади и спектру экотопов, — далеко не однозначный показа-
тель: равное видовое богатство КФ может достигаться при существенно
разном соотношении удельной концентрации видов на равной площади
(например, 100 км2), площади всего контура и пространственного раз-
нообразия флоры в пределах данного контура (Малышев, 1972, 1975,
19766). В свою очередь первый и третий показатели во многом зависят
от разнообразия и контрастности экотопов, числа разнотипных ПФ, их
среднего видового богатства и их самобытности, соотношения в их со-
ставе обычных и редких видов и от ряда других факторов, характери-
зующих „анатомию” флоры (Юрцев, 1975а). Чем дальше мы продвинем-
ся в расшифровке этих компонентов богатства и пространственного раз-
нообразия КФ, тем глубже сможем понять различия сопоставляемых
региональных флор.
До сих пор речь шла об элементарных гомогенных флористических
контурах. Очень естественный, но жесткий критерий флористической
57
гомогенности был предложен Толмачевым (1931): тождество набора
видов на однотипных экотопах (постоянство состава ассоциаций). Позд-
нее была сделана оговорка о допустимости случайного варьирования
состава однотипных ПФ — за счет более редких, малоконстантных видов.
Однако и в смягченной форме требование постоянства состава однотип-
ных ПФ далеко не всегда соблюдается, особенно в горных районах. Так,
Л. В. Мариной (1982, 1983, 1985, а также статья в наст, сб.) обнаружены
заметные различия в составе однотипных (относящихся к одному клас-
су) ПФ небольших (I порядка) речных бассейнов, рассекающих массив-
ные высокогорья геологически однородных компактных хребтов Кур-
куре и Те ты ко ль (Алтайский заповедник); высокогорные флоры обоих
хребтов трактуются как две КФ; различия касаются набора не только
редких, но и обычных в том или ином бассейне видов, включая специфи-
ческие для данного класса экотопов. Очень резкие различия в составе
некоторых однотипных ПФ (например, таковых южных склонов долин
с реликтовой степной и криоксерофитной растительностью) между со-
седними речными бассейнами (р. Пинейвеем и р. Кремянка) в пределах
одной КФ выявлены нами на юго-западном побережье Чаунской губы;
они касаются состава даже массовых, фоновых видов степных колоний.
Аналогичные различия выявлены на юго-востоке Чукотского полуостро-
ва в окрйестностях пос. Янракыннот при сравнении ПФ отдельных сопоч-
ных массивов сходного литологического состава площадью в несколько
квадратных километров. Все это — примеры очагового распределения
видов и их комплексов в пределах одной КФ даже на однотипных экото-
пах, проявления флористической мозаичности, нарушающей общую про-
странственную гомогенность элементарной флоры.1 На это приходится
делать поправки при выделении элементарных контуров: через послед-
ние не должны проходить лишь конечные (общие) и притом более или
менее массивные границы ареалов или их крупных обособленных частей.
Тем не менее вопрос о критериях гомогенности КФ нуждается в дальней-
шей разработке.
С этой точки зрения весьма интересен предпринятый Мариной (1982)
опыт сопоставления по нескольким показателям флор речных бассейнов
I порядка в пределах массивной высокогорной, орографически и литоло-
гически цельной части хр. Куркуре, трактуемой как выдел одной высо-
когорной КФ. При очень высоком уровне сходства флор бассейнов (осо-
бенно по таксономической структуре на уровне семейства и по географи-
ческой структуре) наметилась определенная группировка этих флор
в микрорайоны, однако группировка по разным показателям существен-
но не совпадает; это говорит о том, что высокогорная часть хребта не де-
лится однозначно по сумме флористических показателей. Разумеется,
этот критерий целостности КФ нуждается в дальнейших проверке и апро-
бации на материале по флорам различных равнинных и горныхтерриторий.
1 Причинами такой мозаичности являются, с одной стороны, распределение
контуров некоторых типов экотопов (например, связанных с речными долинами
или водоразделами) более или менее изолированными скоплениями, с другой -
эффект рефугиума (Юрцев, 1975а).
58
Во многих случаях, особенно в переходных полосах между террито-
риями, существенно отличающимися климатом, природной историей,
составом ботанических комплексов, приходится сталкиваться с явле-
нием флористического континуума: зоны относительного сгущения гра-
ниц ареалов, выглядящие как массивные рубежи на мелкомасштабных
(обзорных) картах, при переходе к более детальному масштабу (на не-
больших территориях) растягиваются в непрерывное поле флористиче-
ского градиента. Этот градиент наиболее ясно проявляется в пределах
орографически и литологически однородных контуров, в то время как
чередование ландшафтов с контрастными рельефом и литологией часто
усиливает мозаичность в распределении сменяющих друг друга флори-
стических комплексов: Феномен флористического континуума в более
или менее разком выражении можно наблюдать на морских и океанских
побережьях, макросклонах хребтов и т. д. На орографически и литологи-
чески пестрых гористых территориях [с небольшой протяженностью и
значительной экологической контрастностью элементарных гомогенных
контуров — порядка местностей, макроэкотопов (Юрцев, Семкин,
1980)] встречаются случаи, когда местности с контрастными рельефом
и литологией не повторяются в пределах данного ландшафта, что резко
снижает и его флористическую гомогенность. В приморских районах
(например, на Чукотском полуострове) эта неоднородность нередко уси-
ливается постепенным изменением климата от берега моря в глубь суши.
Все это делает невозможным в этих случаях выделение достаточно про-
тяженных, ландшафтно-полночленных однородных флористических кон-
туров. Однако и в подобной ситуации возможно заложение и сравни-
тельное изучение выборочных или регулярных проб флоры с целью реше-
ния перечисленных выше задач сравнительной флористики.
Метод выборочных проб флоры вырос из практики применения
метода КФ (Юрцев, 1975а), хотя само определение КФ Толмачевым
(1931 и др.) содержало характеристику элементарного флористического
выдела региональнрго уровня; однако в своей практике исследователи
имели дело лишь с „площадью выявления” КФ, не заботясь об отыска-
нии природных границ последних. Практически этот метод состоит в вы-
яснении флористической ситуации в серии более или менее удаленных
друг от друга географических пунктов, из чего и складывается „флори-
стическое зондирование” территории. Такой подход применим как на
территориях, естественно распадающихся на относительно дискретные
ландшафтно-флористические единицы (о которых шла речь выше), так
и в условиях континуума или неповторяющейся дробной мозаичности.
Насколько важно, чтобы проба флоры представляла гомогенный флори-
стический контур с точки зрения решения перечисленных выше основ-
ных задач региональных флористических исследований? Допустимо ли,
например, чтобы центр такой пробной площади располагался на стыке
двух или даже трех КФ (как естественных выделов) ? Ведь только в этом
случае можно утверждать, что центром пробы флоры в принципе может
служить любая точка суши.
Выше уже говорилось, что типологическая оценка флоры может
быть дана с высоким приближением даже на основе изучения небольшой
59
части контура КФ, если в этой части представлены ПФ всех основных
классов экотопов (например, небольшого речного бассейна). Соотноше-
ния крупных таксонов и различных категорий элементов флоры доста-
точно устойчивы даже в пределах крупных фитохорий, и поэтому они
выявляются и в тех сложных пробах флоры, о которых идет речь. Что же
касается задачи выявления географической дифференциации региональ-
ной флоры, то заложение проб флоры на стыке двух или нескольких КФ
или же в полосе повышенного флористического градиента — в переход-
ной полосе между двумя фитохориями — имеет и свои определенные
преимущества, позволяя нащупать и изучить флористические контакт-
ные эоны. Важно лишь, чтобы данные по такой пробе флоры собирались
максимально дифференцированно для естественных подразделений проб-
ной площади (а не только по всему участку в целом) и чтобы внимание
флориста было „насторожено” на выявление флористического градиента
в пределах обследованной площади. Таким путем нетрудно отличить слу-
чаи повышения флористического богатства в контактных зонах от слу-
чаев повышенного богатства контуров гомогенных КФ. Изучение „кон-
тактных” проб флоры позволяет также изучать экологическое распреде-
ление и взаимоотношения контрастных (в том числе и географически
замещающих) флористических комплексов в условиях одинакового
макроклимата. Мне уже приходилось говорить (Юрцев, 1982а),что фло-
ры краевых частей протяженных флористических контуров в полосе
контакта образуют особые флористические системы с повышенной бу-
ферностью, что они нередко служат ареной гибридогенеза и перифериче-
ского формообразования — изучение их представляет интерес и с этой
точки зрения.
Что же касается протяженности радиальных маршрутов при обследо-
вании пробы флоры, то она должна быть достаточной для выяснения
флористической ситуации в данном географическом пункте (включая
ситуации континуума или контактной полосы) и может существенно
варьировать. Для ценотического уровня дифференциации растительного
покрова аналогичное понятие — „ценоквант” — введено В. И. Василеви-
чем (1973). Стабильность размеров обследованной площади желательна
для определения удельного флористического богатства (Малышев,
1972), для оценки же пространственного разнообразия полезно взятие
серии проб флоры на участках возрастающего радиуса из одного базово-
го лагеря (Юрцев, 1975а; Шмидт, 1976, 1980). Кстати, трактовка КФ
в цитированных работах В. М. Шмидта максимально приближается к кон-
цепции выборочной пробы флоры, так как признается правомерность
выявления КФ вокруг любой точки земной поверхности; разница с прак-
тикой арктических флористов сводится к существенно большим разме-
рам пробных площадей. Пробой флоры является и геон Р. Бешела
(1969) — „биологическая единица определенного размера, но с неопре-
деленными границами”.
Сейчас, когда обработаны и опубликованы результаты обследования
нескольких региональных флор методом выборочных проб флоры (Фло-
ра Путорана, 1976; Ребристая, 1977; Заславская, 1982, и др.), можно
утверждать, что этот метод „работает” и позволяет решать все основные
60
задачи флористики, несмотря на то что длительность исследований
в одном пункте, размер обследованной площади, степень ее ландшафт-
ной однородности, плотность маршрутов и, несомненно, полнота выяв-
ления состава локальной флоры существенно варьировали в пределах
каждого упомянутого цикла исследований. Тем не менее даже 6—10-
дневного обследования (в условиях Крайнего Севера!) оказалось доста-
точным, для того чтобы выяснить флористическую и ботанико-географи-
ческую ситуацию в данном пункте, соотношение крупных таксонов, ши-
ротных и долготных географических элементов, провести районирование
территории. При планировании исследований региональной флоры мето-
дом выборочных проб (в условиях обычного дефицита кадров, времени
и средств) возникает дилемма между желательной повторностью и плот-
ностью размещения проб и необходимой детальностью изучения каждой
локальной флоры. Оптимальный выход заключается в увеличении по-
вторности проб за счет известного снижения детальности обследования
каждой до твердого обоснованного минимума и в дополнении менее
детальных проб определенной долей более детальных (контрольных),
изучаемых в течение двух-трех сезонов с максимальной полнотой, по
программе ботанико-географических полустационаров или даже стацио-
наров (Юрцев, 19756). Фундамент же этой программы-минимум в соот-
ветствии с классической установкой Толмачева — выявление состава ПФ
всех типов экотопов. Выявление же полного набора крайних раритетов,
как показывает опыт изучения локальной флоры с. Лужки на юге Мос-
ковской обл., требует работы очень многих флористов в течение многих
десятилетий и даже при этом не может быть гарантировано на 100%;
к тому же обнаружение этих раритетов обычно лишь уточняет, но не из-
меняет существенным образом характеристику местной флоры; кроме
того, значительная часть их бывает обнаружена в других пробах флоры
и так или иначе попадает в региональный список. Тем не менее девизом
флористического изучения территории должно быть исчерпывающе пол-
ное выявление флоры.
Вполне очевидно, что изучение КФ и других выборочных проб фло-
ры, как правило, не является самодовлеющей задачей сравнительной
флористики, но должно быть подчинено более общей цели изучения и
сравнения крупных региональных флор. Отсюда важность получения и
мобилизации любой флористической информации по промежуточным
территориям (между пробами), сочетания проб флоры со „связующи-
ми” маршрутами с целью уточнить положение и характер флористиче-
ских рубежей, повысить повторность осмотра редких экотопов с неста-
бильным составом ПФ и т. д. Планирование сети выборочных проб долж-
но предусматривать не только более или менее равномерное распределе-
ние их по территории, но и охват совокупностью проб всех типов ланд-
шафта, литологических комплексов и т. д. Вполне очевидно и то, что
сеть опорных проб флоры необходимо создавать и в областях, ранее
в основном изучавшихся маршрутным методом — для обеспечения срав-
нимости.
Специального обсуждения заслуживает вопрос об использова-
нии в качестве единиц флористических сопоставлений флор речных
бассейнов, впервые поставленный Р. В. Камелиным (1969,1973). Бассей-
ны рек одинакового порядка — метамерные элементы ритмической
структуры физико-географической оболочки суши, нередко включаю-
щие основной или даже полный набор экотопов данной территории, в го-
рах — спектр высотных поясов; в пределах единого бассейна ПФ разных
поясов и экотопов взаимодополняют друг друга, связаны обменом диа-
спор и панмиксией, образуя тесную гетерогенную систему, причем их
взаимодействие особенно эффективно на фоне крупных флюктуаций
климата, Выше говорилось о том, что даже бассейны самых малых рек
(I порядка) имеют черты флористической индивидуальности в пределах
выдела КФ.1
Черты индивидуальности усиливаются при переходе к более круп-
ным бассейнам. Так, естественная флора бассейна р. Варзоб на южном
склоне Гиссарского хребта, по данным Р. В. Камелина (1973 и личное
сообщение), включает 15 эндемичных видов и выделяется в самостоя-
тельный флористический район. Можно ли считать ее элементарной есте-
ственной флорой? Очевидно, лишь в том случае, если территория не де-
лится на более компактные и целостные ландшафтно-флористические
выдели, а это в горах, со свойственной им мозаичностью местоположе-
ний, а иногда и постепенным изменением условий от вершин к подно-
жиям, видимо, наблюдается нередко. Сложности возникают с трактов-
кой массивныхвысокогорных водоразделов, нередко целостных и само-
бытных во флористическом отношении, но принадлежащих сразу двум
или нескольким бассейнам. Но главная сложность — в неоднородности
флоры многих крупных речных бассейнов, дренирующих контрастные
в ландшафтном и ботанико-географическом отношении территории,
иногда относящиеся к разным округам, провинциям, областям. И на бас-
сейнах какой крупности (какого порядка) остановиться? Критерий на-
личия хотя бы одного эндемичного вида в речном бассейне (Камелии,
1973) „не работает” на более северных территориях, да и в горной Сред-
ней Азии едва ли универсален (собственный эндемик может найтись в до-
лине небольшого ручья и не отыскаться в крупном ущелье). Несмотря
на отмеченные трудности, сам принцип сравнения горных флор по круп-
ным бассейнам (ущельям) при условии ботанико-географической цель-
ности последних безусловно перспективен, хотя и нуждается в дальней-
шей методической разработке.
Тенденции дальнейшего развития методов флористического обследо-
вания крупных территорий проявляются уже сейчас в странах с наивыс-
шей плотностью накопленной флористической информации, в частности
1 Поэтому правомерно говорить о внутренней географической дифференциа-
ции КФ, а также включить в программу изучения последней выяснение того, как
распределяются виды в ее пределах по речным бассейнам I порядка. А ргюп можно
предвидеть 2 разные ситуации, а) чересполосицу, обусловленную варьированием
набора и соотношения экотопов в разных бассейнах или случайностями заноса диа-
спор; б) закономерный географический градиент (если же состав флор бассейнов
резко изменяется вдоль определенного рубежа, КФ должна быть разделена).
62
в Западной Европе, Прибалтийских республиках СССР и др. Они заклю-
чаются в переходе к регулярному „картированию флоры” на сеточной
основе с размерами ячей сетки порядка 100 км2 и несколько менее,
т. е. вполне сопоставимыми по территориальной протяженности с проба-
ми флоры. Хотя изначальная цель подобных проектов сводилась к авто-
матизации картирования распространения видов на данной территории,
а также хранения, корректировки, обработки и выдачи фитохорологи-
ческих данных, реализация проектов означала переход к новому этапу
региональных флористических исследований методом регулярных проб
флоры. Однако методология проб флоры была привнесена в проекты
сеточного картирования флоры позднее, на основе анализа тенденций
развития метода КФ (Юрцев, 1975а), и пока частично реализована лишь
в работах латвийских ботаников (Табака и др., 1977, и др.). Совсем не-
давно Т. М. Заславская и В. В. Петровский (1983), работающие методом
выборочных проб флоры, провели эксперимент по обследованию восточ-
ной части Люпвеемского горного массива (Анюйское нагорье) на 5 при-
мыкающих стандартных пробных площадях по 300 км2 и пришли к за-
ключению, что изъятие данных по любой пробе из 5 приводит к утрате
ценной флористической информации (несмотря на высокий процент
выявления генеральной совокупности видов на 4 пробах) и потому в ус-
ловиях чукотского среднегорья для полного выявления видового соста-
ва необходимо сплошное обследование флоры методом примыкающих
КФ (=проб флоры). Автор этих строк убежден, что будущее флористи-
ки — в регулярном обследовании территории методом проб флоры,
в синтезе „техники” картирования флоры на сеточной основе и методо-
логии проб флоры, сформировавшейся в русле метода КФ.
При регулярном обследовании территории уменьшается риск случай-
но пропустить важные элементы географической и экотопологической
дифференциации флоры. Не столь опасна и неполная флористическая
репрезентативность каждой учетной ячеи сетки, хотя едва ли стоит стре-
миться к чрезмерному дроблению стандартных ячей — слишком малые
пробы не в состоянии отразить флористическую ситуацию в данном гео-
графическом пункте. Исследования территории методом регулярных
проб флоры также нуждаются в тщательном планировании, в частности
в выделении наиболее представительных (и менее затронутых антропо-
генными воздействиями) ячей и блоков ячей для специального первооче-
редного изучения по программе проб флоры той или иной степени де-
тальности [включая программы полустационаров и стационаров (Юрцев,
19756) ]. Как и в случае выборочных проб, целесообразно выделять ячеи
для детального изучения как в пределах гомогенных флористических
контуров, так и в контактных зонах и переходных полосах. Есть все
основания придать исследованию флоры на детальных ключевых участ-
ках (в опорных, модельных ячеях сетки) комплексный ботанико-гео-
графический характер (Юрцев, 1982а, а также статья в наст, сб.).
Известным недочетом сеточного метода картирования флоры
является формальный (произвольный) характер ячей, не согласую-
щийся с природными контурами. Это не столь существенно для выяв-
ления границ крупных фитохорий, но затрудняет задачу разграничения
63
1
элементарных флористических выделов, подчас соизмеримых по площа-
ди с самими ячеями сетки.1 В Алтайском заповеднике применен другой
подход к регулярному обследованию флоры — по дробным природным
контурам (Золотухин, 1983, а также статья в наст, сб.), первоначально
ландшафтного обоснования, с последующей корректировкой на основе
флористических данных. К сожалению, сама процедура исследования
еще не была подробно описана. В настоящее время такой подход целе-
сообразен для охраняемых территорий как объектов первоочередного
и длительного (в принципе постоянного) наиболее детального флористи-
ческого изучения и контроля. Выше уже обсуждались трудности, подчас
возникающие при выделении гомогенных элементарных флористических
контуров. Однако и на канве стандартных ячей можно выявить рисуцок
элементарных флористических выделов, если дифференцированно под-
ходить к сбору флористической информации по учетным квадратам.
Опыт корректировки границ стандартных ячей по природным границам
уже имеется у латвийских ботаников (выступление Л. В. Табака на сове-
щании в Неринге).
Так или иначе, но мы имеем все основания прогнозировать все боль-
шую детализацию наших знаний о географической дифференциации ре-
гиональных флор в предвидимом будущем, благодаря чему исследовате-
ли вновь и вновь будут возвращаться к нелегкому вопросу об элемен-
тарных флористических выделах.
В то же время, несмотря на объективные трудности выделения эле-
ментарных флористических контуров, разработанные и применяемые
в нашей стране методы „флористического зондирования” территории
в общем справляются с решением основных задач флористики, включая
выявление тонкой географической структуры (дифференциации) регио- !
нальных флор и их экотопологической инфраструктуры.1 2 1
Можно добавить, что достигнутый за последнее время определенный f
прогресс в разработке качественных и количественных методов флори- I
стики расширил диапазон сравнимости флор, в том числе принадлежа-
щих неравновеликим и фациально несходным территориям. Так, для
сравнения неравновеликих флор применяется расчет мер включения,
1 В этих случаях пришлось бы резко снизить площадь учетных квадратов до
нескольких квадратных километров. Но представительность таких проб в усло-
виях геологически и орографически пестрых территорий невелика.
2 Под инфраструктурой (подструктурой) элементарной естественной флоры
регионального уровня (как системы местных популяций видов растений) имеется
в виду взаиморазмещение входящих в ее состав разнотипных ПФ по совокупности
местоположений, обеспечивающее относительно устойчивое (подвижно-равновес-
ное) сосуществование в рамках одного ландшафта контрастных (различных по
экологии, географическим связям и происхождению) флористических комплек-
сов, а также генетическую интеграцию популяций активных видов. При анализе
инфраструктуры флоры должен рассматриваться ритмический рисунок распреде-
ления экотопов, „опорных" для разных флористических комплексов, служащих
каналами связи и расселения, а также экологическими микрорефугиумами в пес-
си мал ьные климатические фазы. От инфраструктуры конкретной флоры зависят
относительные ландшафтные позиции сосуществующих флористических комп-
лексов, характер их взаимодействия на фоне флюктуационных изменений кли-
мата.
64
отражающих отношение части и целого (Юрцев, Семкин, 1980). Флори-
стическое сравнение территорий с несходным набором экотопов облег-
чается дифференциальным сравнением ПФ общих типов и классов эко-
топов. Показан значительный территориальный диапазон стабильности
важных типологических особенностей флОры — соотношений крупных
таксонов и элементов. Наконец, выяснена первостепенная роль регуляр-
но встречающихся (и потому легко выявляемых) видов и рас для раз-
граничения фитохорий при флористическом районировании (Юрцев,
1983а).
Другая важная тенденция развития флористики будет заключаться
в углублении существующих подходов к изучению флоры, ее состава,
структуры, жизни, генезиса, тенденций развития и во все более тесном
подключении флористики к многоотраслевым программам комплексно-
го изучения биосферы.
Подведем некоторые итоги.
1. Представлению о КФ как элементарных естественных флорах ре-
гионального уровня вполне соответствуют флоры дискретных ландшаф-
тов (при условии, если ландшафтным различиям соответствуют флори-
стические и если через данный ландшафт не проходит ни один региональ-
но-флористический рубеж). Такие ландшафтно-флористические выдели
могут служить опорными единицами при флористических сравнениях.
В качестве опорных единиц заслуживают дальнейшей апробации также
флоры речных бассейнов, как малых (внутриландшафтных), так и более
крупных.
2. На территориях со сложно расчлененным рельефом и пестрой ли-
тологией размер контуров КФ как ландшафтно-флористических выделов
может быть меньше их „площади выявления” (= площади стабилизации
состава) ; в этом случае КФ должна обследоваться на всем своем протя-
жении, с особым вниманием к по контурной инвентаризации ПФ неповсе-
местных типов экотопов.
3. Представление А. И. Толмачева о гомогенности („универсально-
сти”) состава КФ на всем ее протяжении как следствии „постоянства
состава ассоциаций”, даже с позднейшими поправками о случайном ха-
рактере различий, часто не подтверждается практикой выявления КФ
(непостоянство состава ПФ многих типов экотопов, „очаговая” встречав-,
мость даже части массовых видов, различия между флорами речных бас-
сейнов в пределах КФ); это оставляет открытым вопрос о критериях
гомогенности КФ. Другое осложнение при выделении дискретных конту-
ров КФ связано с явлением флористического континуума. Этих ограни-
чений не существует для метода выборочных проб флоры (флор окрест-
ностей того или иного географического пункта) , выросшего из практики
метода КФ и за последние десятилетия доказавшего свою эффективность
при решении всех основных задач сравнительной флористики.
4. Дилемма между желательной повторностью заложения проб фло-
ры внутри протяженной фитохории и необходимой детальностью, также
длительностью изучения каждой пробы может быть разрешена путем
5 Заказ 1220
65
сочетания флористических проб разной степени детальности (программа-
минимум в максимальном числе пунктов, программа-максимум — в ми-
нимальном; наиболее детальные пробы флоры изучаются в особо пред-
ставительных пунктах как контрольные).
5. Тенденции дальнейшего развития флористики заключаются во
взаимодополнении и сближении маршрутного метода и метода выбороч-
ных проб флоры и в постепенном переходе к системе регулярных проб
(в результате соединения техники „сеточного картирования флоры”
с методологией КФ). Для особо детально изучаемых территорий (в пер-
вую очередь заповедников) конечными единицами учета дифференциа-
ции флоры могут служить более или менее дискретные ландшафтно-фло-
ристические выделы КФ.
6. В результате совершенствования методов сравнительной флори-
стики диапазон сравнимости флор значительно расширяется.
Г. В. Вынаев, Д. И. Третьяков
ПРОГРАММА ИЗУЧЕНИЯ ФЛОРЫ БЕЛОРУССИИ
МЕТОДОМ ЭТАЛОННЫХ ПРОБ
Институт экспериментальной ботаники им. В. Ф. Купревича
Исходя из развиваемой нами концепции флористических систем (си-
стем местных популяций) как подсистем соответствующих природно-
территориальных экосистем (геосистем) составлена долгосрочная про-
грамма изучения естественных флор природных (физико-географиче-
ских) районов Белоруссии и их ландшафтно-топологической дифферен-
циации методом эталонных проб — проб флористической ситуации
в конкретных географических пунктах (Юрцев, 1975а), расположенных
в различных природных районах и в разных генетических типах ланд-
шафтов. В 32 выделенных по принципам комплексного физико-геогра-
фического районирования природных районах Белоруссии (Дементьев,
1978) картографически заложена регулярная система эталонных проб,
охватывающая по возможности все пространственно-экологическое
разнообразие природно-территориальных комплексов республики. Эта-
лонная проба представляет собой территорию в 100 км2, которая очерчи-
вается радиусом 6 км из центра выбранного пункта (базового лагеря).
Эта территория изучается путем прокладки системы радиальных и произ-
вольно направленных линейных маршрутов, причем и сами эталоны, и
эти маршруты в эталонной пробе должны охватывать максимальное
ландшафтно-топологическое разнообразие данной территории. В ряде
случаев целесообразно прокладывать в характерных или специальных
сочетаниях различных типов местоположений непрерывные эколого-фло-
ристические профили с описанием системы парагенетически связанных
и генетически контрастных экотопов. Основной единицей такого эколо-
го-флористического исследования являются флоротопологические комп-
66
лексы, т. е. комплексы ценопопуляций, связанные с конкретными эко-
топами и экотонами в ландшафтном поле, или, иначе, видовой состав
конкретных фитоценозов или растительных группировок (Вынаев, Ко-
жаринов, 1981). Местонахождение экотопов на местности определяется
при их флористическом описании и характеристике путем вычисления
точного азимута и расстояния от базового лагеря. Такой азимутально-
дистанционный подход к описанию конкретных флоротопологических
комплексов позволяет изучить не только ландшафтно-топологическую
структуру естественной флоры, но и'(при повторном их обследовании)
те ф лоро динамические процессы, которые происходят в экотопах с раз-
личными экологическими режимами, так как те видимые изменения,
которые наблюдаются во флоре в целом, первично осуществляются, на-
капливаются и передаются на более высокие уровни именно с топологи-
ческого уровня. Применение этого метода в условиях Белоруссии, где за-
ложено свыше 600 флористических эталонов и некоторые из них уже изу-
чены, показало его высокую репрезентативность и эффективность как
в методологическом, так и в чисто практическом (методическом) плане.
О. В. Ребристая
ОПЫТ ПРИМЕНЕНИЯ МЕТОДА КОНКРЕТНЫХ ФЛОР
В ЗАПАДНОСИБИРСКОЙ АРКТИКЕ (ПОЛУОСТРОВ ЯМАЛ)
Ботанический институт им. В. Л. Комарова АН СССР
Север Западной Сибири — большая территория между Полярным
Уралом и Таймыром, от Байдарацкой до Енисейской губы - был до по-
следнего времени ботанически почти не исследован. Флористические ре-
зультаты экспедиции Русского географического общества 1908 г. (Жит-
ков, 1913), Гыданской экспедиции АН СССР 1927-1928 гг. (Городков,
1928, 1944) не получили должного отражения в литературе. Имеющиеся
сведения о флоре этого района базировались в основном на гербарных
сборах землеустроителей и геоботаников, а также на опубликованных
геоботанических работах (Андреев, 1934; Михайличенко, 1936; Нико-
лаева, 1941; Бочидр., 1971,идр.).
Когда А. И. Толмачев поставил передо мной задачу флористического
обследования Западносибирской Арктики, отличающейся малонаселен-
ностью и труднодоступностью, было принято решение проводить всю ра-
боту методом конкретных флор (КФ) как испытанным способом полу-
чения максимальной информации при минимальной затрате времени
и сил. Наш опыт свидетельствовал о высокой эффективности этого ме-
тода. Он был накоплен и обобщен в 1958—1965 гг. при изучении флоры
востока Большеземельской тундры под непосредственным руковод-
ством А. И. Толмачева (Ребристая, 1977).
Развитие теории сравнительной флористики в последние полтора
десятилетия выявило некоторые разночтения в понимании „конкретной
67
флоры”. Мною это понятие трактуется как минимальная региональная
естественная флора, объединяющая виды растений всех типов экотопов
на однородном по своим физико-географическим условиям участке зем-
ной поверхности. Территориально она может соответствовать низшим
звеньям геоботанического районирования — микрорайонам, элементар-
ным геоботаническим районам (Толмачев, 1974) или ландшафтам (Лу-
кичева, Сабуров, 1969). КФ имеет площадь выявления, которая нахо-
дится в прямой зависимости от экотопологического разнообразия ланд-
шафтов — ареал-минимум КФ (Шмидт, 1972, 1980; Юрцев, 1975а).
А. И. Толмачев в своих работах указывал, что флористическое обсле-
дование ареала-минимума КФ вполне достаточно для проведения сравни-
тельно-флористических исследований. „Надобности в «прощупывании»
того, какими могут быть максимальные размеры площади с вполне
однородной флорой, обычно не возникает: эта трудоемкая и малоинте-
ресная работа просто не оправдывается ее результатами” (Толмачев,
1974, с. 186). Площадь ареала-минимума КФ в условиях Арктики состав-
ляет, по Толмачеву, около 100 км2. Однако вопрос об общей площади
КФ и соотнесении ее с площадью флористического района имеет не толь-
ко теоретический, но и практический интерес в связи с дробным флори-
стическим районированием и выяснением соотношений КФ—флора райо-
на-флора округа.
Более строгий подход к терминам и понятиям „КФ” и „ареал-мини-
мум КФ” показал, что на практике при изучении флоры любого района
мы имеем дело в общем случае не с ареалом-минимумом КФ, но с про-
бой флористической ситуации, или пробой флоры, — флорами различных
географических пунктов (Юрцев, 1975а), так как до анализа собранного
материала, за редким исключением, не можем решить, находимся ли мы
в центре одной или на границе двух соседних КФ. Ю. Р. Шеляг-Сосонко
(1980) предложил называть пробы флоры локальными флорами (ЛФ),
чтобы отличать их от флор элементарных флористических выделов ре-
гионального уровня. Довольно частый случай, когда ЛФ соответствует
ареалу-минимуму КФ. Всю КФ и ее площадь можно выявить путем зало-
жения сети близко расположенных друг к другу проб флоры, составле-
ния списков ЛФ с их последующим объединением и сравнительным ана-
лизом. Таким образом, выявление полного видового состава КФ, опре-
деление ее площади, так же как и разграничение соседних КФ, можно
провести лишь при детальном сравнении ряда близлежащих ЛФ.
При флористическом обследовании Ямала передо мной стояли и не-
которые методические задачи: 1) выяснение хода увеличения числа ви-
дов при увеличении радиуса обследованного участка; 2) определение
ареала-минимума (площади выявления) КФ; 3) установление соотноше-
ния близлежащих ЛФ с КФ и более крупными территориальными флори-
стическими выделами.
Работа проводилась методом выборочных проб флоры — ЛФ в 1976—
1983 гг. на п-ове Ямал. Обследовано 20 эталонных участков из различных
подзон. Выбор участков большей частью производился заранее на основа-
нии изучения имеющихся материалов по природным условиям соответст-
вующих районов, реже определялся транспортными возможностями.
68
Продолжительность работы в одном пункте колебалась от 3 до 15
дней в зависимости не столько от флористической ситуации, сколько от
транспортных возможностей. Оптимальным для проведения достаточно
полной инвентаризации флоры в наших условиях следует признать срок
в 12-15 дней.
Выбор места для базового лагеря производился с учетом трех глав-
ных моментов: 1) ландшафтной однородности района (мы сознательно
избегали гетерогенных участков на стыке двух и более ландшафтов),
2) удаленности лагеря не менее 6—8 км от крупных естественных преград
для беспрепятственного обследования возможно большей территории и
3) разбивки базового лагеря в месте наибольшего расчленения рельефа
и, следовательно, максимального экотопологического разнообразия, что
облегчает выявление парциальных флор (ПФ) всех типов экотопов.
Полуостров Ямал — один из наиболее низменных (максимальная вы-
сота над уровнем моря 95 м в осевой части полуострова) и выровненных
районов Арктики. Он представляет собой аккумулятивную равнину, по-
крытую мощным чехлом четвертичных отложений, достигающих 250—
300 м толщины и состоящих из переслаивающихся глинистых, суглини-
стых и песчаных разностей морского и частично ледникового генезиса.
На больших пространствах на дневную поверхность выходят песчаные
грунты; суглинистые и реже глинистые отложения ограниченно распро-
странены в центральной, осевой, и в юго-восточной, приобской, частях
полуострова, тогда как выходы коренных пород и щебнистые грунты
вообще отсутствуют. Малая расчлененность рельефа, повсеместное рас-
пространение многолетнемерзлых толщ и, как следствие, слабый дренаж
явились причиной значительной заболоченности территории и обилия
озер. На протяжении четвертичной истории растительного покрова Ямала
неоднократно чередовались эпохи уничтожения растительности много-
кратными морскими трансгрессиями, воды которых полностью покры-
вали полуостров, и эпохи восстановления покрова за счет иммиграции
растительных комплексов из соседних областей. В результате фациаль-
ной неполночленности ландшафтов Ямала, их малого экотопологическо-
го разнообразия и молодости самой страны флора полуострова отличает-
ся значительной бедностью: КФ подзоны южных гипоарктических тундр
насчитывают 170—190видов, КФ арктических тундр - 100—120видов, что
на 30-50 видов меньше, чем КФ востока Большеземельской тундры
(Ребристая, 1977) и Таймыра (Соколова, 1984а; Матвеева, Заноха, 1986).
Перечисленные выше особенности ландшафтов и флоры Ямала об-
легчили постановку и решение методических задач.
1. Изменение числа видов при постепенном наращивании
обследованной площади и определение ареала-минимума
конкретной флоры
В ближних окрестностях базового лагеря, который, как уже отме-
чалось, располагался на участке с расчлененным рельефом (долина
речки, край озерной депрессии), проводилось обследование всех типов
69
Таблица 1
Возрастание числа зарегистрированных видов при увеличении площади ЛФ
70
экотопов с составлением первичных списков ПФ. Площадь обследования
не превышала 0.5—0.8 км2. Сводный список ПФ для этой площади являл-
ся основой для дальнейшей работы. Благодаря наличию топографических
карт и аэрофотоснимков мы могли фиксировать встречаемость новых
видов на серии площадей возрастающего радиуса. Постепенное увеличе-
ние протяженности радиальных маршрутов из базового лагеря — один из
методических приемов выявления КФ, предложенный автором метода
А. И. Толмачевым. Были выбраны площади с радиусом 0.5 км (5 =
= 0.8 км2), 1.0 (5=3.1 км2), 2.0 (5 = 12.6 км2), 3.0 (5=28.2 км2),
5.0 (5 =78.5 км2), 6.0 (5 = 113.1 км2), 10.0 км (5=314.1 км2). Прак-
тически площадь всегда оказывалась несколько меньше, так как при ее
расчете мы не исключали водную поверхность многочисленных озер,
попадавших в соответствующий контур.
Результаты, полученные при обследовании 9 пунктов из трех подзон
(табл. 1), показали, что в зависимости от экотопологического разнооб-
разия на площади 0.8 км2 удается выявить не менее 70% всей флоры, на
площади 3 км2 — более 85 % и на площади 12 км2 — около 95 %. На пло-
щади несколько менее 30 км2 (круг радиусом 3 км) видовой состав
выявляется почти полностью — на 97—99%, и именно эту площадь можно
рассматривать как ареал-минимум КФ для флор Ямала.
Площадь 0.8 км2 можно считать репрезентативной,1 так как при уве-
личении ее почти в 4 раза (до 3.1 км2) число зарегистрированных видов
возрастает всего на 6—18%. Репрезентативность наших участков во всех
случаях выше 90% (см. ниже, раздел 3).
В ряде случаев для приблизительной оценки флористической ситуа-
ции при кратковременном обследовании достаточно осмотреть площадь
радиусом 2 км, чтобы выявить флору на 95 %. Для многих ботанико-
географических сопоставлений такая полнота выявления достаточна.
Однако представляет интерес, какие же виды составляют последние 5 %.
Рассмотрим это более подробно на примере ЛФ Матюйяха из подзоны
северных гипоарктических тундр.
В ЛФ Матюйяха зарегистрировано 147 видов. В окрестностях лагеря
найдено 116 видов, в том числе 2 вида, нигде более в районе не обнару-
женных: Lycopodium pungens и Calamagrostis langsdorffii. Они встречены
в довольно обычных экотопах, но при этом на северной границе ареала
(самые северные местонахождения на Ямале). При увеличении радиуса
обследования до 1 км добавились новые типы экотопов: глубокие овра-
ги с песчаными склонами и нивальными местообитаниями у подножий,
бугры пучения разной степени задернованности в долине речки, проточ-
ные болота. Были найдены 22 новых вида, большинство которых —
обычные виды относительно редких экотопов. На одном из бугров пуче-
ния, сложенном суглинками и поросшем Taraxacum lateritium, в массе
найдена Potentilla kusnetzowii — вид, отмеченный впервые не только для
Ямала, но и для всей Западносибирской Арктики. На других буграх
По Малышеву (1975), репрезентативной предложено считать площадь, уве-
личение которой вдвое приводит к увеличению флористического списка на 20%
(минимальная площадь) или на 14 % (оптимальная площадь).
71
пучения, несмотря на произрастание тех же сопутствующих видов, Р. kus-
netzowii не была обнаружена, зато в массе разрослась Carex maritime, jjp-
статочно редкий на Ямале вид, отмеченный в районе Матюйяха в не-
скольких местах на сходных экотопах.
Увеличение радиуса обследования до 2 км дало прибавку 5 видов,
из которых Luzula multiflora svkwp.frigida находится на северной границе
распространения; Cochlearia groenlandica и Ranunculus gmelinii, редкие
в ЛФ виды обычных экотопов, часто отмечались в других пунктах Яма-
ла; Saxifraga nivalis и Erigeron eriocephalus — довольно обычные виды
часто встречающихся в ЛФ экотопов (разнотравных береговых склонов),
случайно (?) не найденные ранее.
На площади радиусом 3 км дополнительно были найдены лишь 3
вида. Juncus castaneus в ЛФ Матюйяха был встречен 1 раз, редкий вид,
отмеченный почти во всех ЛФ, расположенных южнее. Draba glacialis най-
дена 1 раз на крутом склоне глубокого оврага, редкий для Ямала вид.
Erigeron silenifoUus в значительном обилии рос на высоком песчаном эро-
дированном берегу реки, его местообитание можно расценить как обыч-
ное не только в окрестностях данной ЛФ, но и на всем Ямале; однако
этот вид был найден лишь второй раз (первый — в таких же условиях
на р. Тиутей, в одной из близлежащих ЛФ). И наконец, при увеличении
радиуса до 5 км, в дриадово-моховой тундре на пологом склоне высоко-
го коренного берега р. Матюйяха был найден Ranunculus lapponicus — вид,
обычный для других ЛФ; довольно обычно для данной ЛФ и названное
местообитание, хотя оно и нехарактерно для R. lapponicus', вид оказался
редкостью для района.
Два более дальних маршрута — 8-километровый на запад, к центру
водораздела бассейнов рек Тиутей и Матюйяха, и 10-километровый на
юг, а затем вдоль южного макросклона водораздела правых притоков
р. Матюйяха, протянувшегося в широтном направлении, не дали никакой
прибавки к списку видов. Наше стремление детально обследовать юж-
ный макросклон было вызвано предположением о возможном флори-
стическом обогащении экотопов речных долин и особенно разнотравных
склонов южной экспозиции видами, находящимися на северном пределе.
Однако состав ПФ как этих, так и других типов экотопов оказался до-
статочно стабилен.
Таким образом, уже на площади в 3 км2 было выявлено 94% фло-
ры, а на площади 30 км2 — 99 %. Эту площадь в условиях равнинных
тундр Ямала и можно принять за ареал-минимум КФ, площадь выявле-
ния КФ, в данном случае весьма полного.
Увеличение площади обследования в 10 раз — с 3 до 30 км2 — дало
прибавку всего 8 видов, 6 из которых относятся к числу редких для ЛФ
и для всего Ямала.
Редкими для ЛФ мы считаем виды, найденные в данной ЛФ 1 раз,
но обычные в других ЛФ Ямала, редкими для Ямала — виды, встречен-
ные в ЛФ 1—2 раза и отмеченные в небольшой части ЛФ Ямала. Распре-
деление редких видов по площади оказалось довольно равномерным.
На площади 0.8 км2 (Л = 0.5 км) найдено 2 редких вида, на
3.1 км2 (R = 1.0 км) — еще 1, на 12.6 км2 (R =2.0 км) — еще 2, на
Таблица 2
Распределение редких видов в ЛФ Ямала и вероятные причины их редкости
ЛФ Редкие виды Причины редкости Распределение в других ЛФ
Хутыяха Eleocharis acicularis Северная граница ареала Не найден
Agrostis stolonifera То же »» 99
Pinguicula villosa Редкий экотоп Редко в южных ЛФ
Лаптаяха Orthilia obtusata 99 Обычен в соседней ЛФ
Equisetum variegatum Реликт Не найден
Luzula rufescens (Реликт) 99 99
Rosa acicularis Северная граница ареала Обычен южнее во всех ЛФ
Хевесе Oxycoccus microcarpus То же Редко в южных ЛФ
Pinguicula villosa 99 99 То же
Chamaerion angustifoUum Обычен южнее во всех ЛФ
Chrysosplenium tetrandrum (Южная граница) Редко в более север- ных ЛФ
Phippsia algida Редкий экотоп, южная граница То же
Ляккатосе Ranunculus monophyllus Северная граница ареала Обычен южнее
Veronica longifolia То же Редкий вид, но посто- янно в южных ЛФ
Salix reticulata Северная граница ареала, редкий экотоп Обычен в южных ЛФ
Calamagrostis langsdorffii Северная граница ареала Южнее обычен
Юрибейто Gastrolychnis apetala Реликт Не найден
Equisetum scirpoides
Salix reticulata Северная граница ареала, редкий экотоп Обычен в южных ЛФ
Calamagrostis langsdorffii Северная граница ареала То же
Мантыто Ranunculus reptans То же 99 99
Batrachium eradicatum ? Очень редко
Carex holostoma (Реликт) В соседней ЛФ
Antennaria dioica Северная граница ареала Южнее часто
Матюйяха Lycopodium alpinum То же В южных ЛФ обычен
L. pungens »9 99 Обычен
Calamagrostis langsdorffii 99 99 99
Ranunculus gmelinii ? Довольно обычен
R. lapponicus 2 Обычен
Juncus castaneus Северная граница ареала, редкий экотоп Редко в более южных ЛФ
73
Таблица 2 (продолжение)
ЛФ Редкие виды Причины редкости Распределение в других ЛФ
Верхний Taraxacum arcticum Редкий экотоп Редко
Тамбей Senecio congestus Carex vaginata subsp. quasi- vaginata >9 99 Северная граница ареала Обычен Южнее обычен
Хабейяха Taraxacum Astragalus umbellatus Salix lanata 7 7 Северная граница ареала Редко Не найден Южнее обычен
>28.2 км3 (R — 3.0 км) — еще 3, на 78.5 км2 (R =5.0 км) — еще 1 ред-
кий вид.
Как показывает табл. 2, в каждой ЛФ Ямала постоянно встречается
небольшое число редких видов, составляющих около 5 % каждой флоры.
Редкие виды не всегда поддаются точному учету, именно их чаще всего
и просматривает исследователь, поэтому вполне возможно, что число
видов за счет редких во флорах Ямала несколько занижено. О том, что
на практике полное выявление состава флоры удается очень редко, писал
еще А. И. Толмачев (1974, с. 186), поэтому следует говорить лишь
о „практически полной” инвентаризации флоры.
Причины редкой встречаемости отдельных видов неодинаковы: это
могут быть редкие виды обычных экотопов (редкость определяется
особенностями распространения и прорастания диаспор, приживания мо-
лодых растений, т. е. биологией вида), либо виды редких экотопов (ред-
кость их зависит от ландшафтных, в первую очередь абиотических фак-
торов) , либо виды, редкие на Ямале в силу исторических причин, причем
в последнем случае могут действовать и отмеченные выше факторы.
Анализ состава редких видов имеет большое значение для установле-
ния их миграционных каналов, времени миграции или перехода в релик-
товое состояние, для палеофлористических реконструкций, тогда как
для сравнительного таксономического и географического анализа флор
вполне достаточно флористических списков с полнотой выявления
90-95 %.
2. О соотношении территориально сближенных локальных флор
с конкретными флорами и более крупными флористическими
выделами
Выше было показано, что в условиях Ямала для выявления ареа-
ла-минимума КФ достаточно обследовать площадь, равную 30 км2.
Площадь локальной флоры, как правило, в 2—3 раза больше площади
выявления (ареал-минимум) КФ. Однако вопрос о площади всей КФ
(если под КФ подразумевается минимальная естественная региональ-
ная флора) может быть решен при анализе нескольких (двух и более)
74
66е 68° 70° 72° 74° 76°
68° 70° 72° 74° 76°
1 : 5 000 000
Рис. 1. Расположение изученных локальных флор на Ямале.
1 - Еръяха; 2 - Лаптаяха; 3 - Хевесе; 4 - Юрибей; 5 - Хутыяха; 6 - Ляккатосе;
7 - Юрибейто; 8 - Мантыто; 9 - Венуйеуо; 10 - Матюйяха; 11 - Тиутей; 12 -
Верхний Тамбей; 13 — Хабейяха.
75
Рис. 2. Граф сходства видового состава сравниваемых флор.
а - блок северных гипоарктических флор: М - Матюйяха, В - Венуйеуо, Т - Тиу-
тей; б — блок южных гипоарктических флор: Ю - Юрибей, X - Хевесе, Л - Лап-
таяха, Е - Еръяха. Связь на уровне: 1 - >91%, 2 - 88%, 3 - > 87 %, 4 - 86%.
близлежащих ЛФ. Для определения площади КФ и выявления разнооб-
разия флористических ситуаций на территории, превышающей стандарт-
ные размеры 100 км2, было проведено сравнение двух блоков ЛФ, рас-
положенных в разных подзонах Ямала.
Три ЛФ первого блока расположены в подзоне северных гипоаркти-
ческих тундр в центральной части Ямала на водоразделе рек бассейнов
Обской (ЛФ Матюйяха и ЛФ Венуйеуо) и Байдарацкой (ЛФ Тиутей) губ.
Расстояние между участками ЛФ Матюйяха и ЛФ Венуйеуо составляло
25 км, ЛФ Матюйяха и ЛФ Тиутей — 35 км, ЛФ Тиутей и ЛФ Венуйеуо —
45 км (рис. 1).
В наиболее полно изученной, более северной ЛФ Матюйяха 147 ви-
дов, в ЛФ Тиутей — 142 вида, в ЛФ Венуйеуо — 133 вида (последняя
выявлена явно неполно). Сводный список всех трех флор насчитывает
163 вида, каждая ЛФ составляет соответственно 90.2,87.1 и 81.6%. Общи-
ми для всех трех флор являются 118 видов, 72 % общего сводного спис-
ка, которые составляют 80.3,83.1 и 88.7% каждой ЛФ. Попарное сравне-
ние всех трех флор показало их большую общность: мера сходства
Съеренсена во всех случаях выше 87%, максимальная мера включения
флор друг в друга (мера сходства Симпсона) выше 90% (табл. 3;
рис. 2, а). Столь высокое сходство указывает на флористическое одно-
образие территории, более значительное, чем, например, на Западном
Таймыре, где попарное сходство трех локальных флор (Тарея, Ая-Турку
Таблица 3
Матрицы попарного сходства трех ЛФ Ямала
из подзоны северных гипоарктических тундр
[меры Съеренсена (слева) и Симпсона (наибольшей
из двух мер включения) (справа)]
ЛФ Матюйяха Тиутей Венуйеуо
Матюйяха 93 93.3
Тиутей 91.3 90.2
Венуйеуо 88.6 87.3
76
Таблица 4
Распределение необщих видов сводного флористического списка
трех близлежащих флор Ямала (подзона северных гипоарктических тундр)
Вид Локальные флоры
Матюйяха Тиутей Венуйеуо
Equisetum pretense • • +
Lycopodium alpinum + • •
L. pungens + • +
Cahmagrostis langsdorffii + • •
Deschampsia borealis + + •
D. glauca + • +
Poa alpina • • +
Puccinellia angustata + + •
Festuca brachyphylla + + •
Eriophorum brachyantherum • + •
Carex glacialis • « +
C. maritime + • •
C. holostoma « • +
Juncus castaneus + • •
Luzula nivalis + • +
L. tundricola • + •
L. multiflora subsp.frigida + • +
Veratrum lobelianum • • +
Rumex arcticus • • +
Stellaria palustris + +
Cerastium maximum + +
Ranunculus gmelinii + •
Eutrema edwardsii + +
Draba glacialis + •
D. oblongata + 4-
D. parvisiliquosa + +
Cochlearia arctica + •
Chrysosplenium altemifolium « +
Rubus arcticus • + +
Potentilla kusnetzowii + •
P. stipularis + •
Hedysarum arcticum + • +
Epilobium davuricum • +
Cassiope tetragona + • +
Gentiana tenella +
Castilleja arctica +
Pedicularis oederi +
P. sudetica subsp .albolabiata + +
P. verticillata +
Linnaea borealis • +
Erigeron silenifolius + +
Matricaria hookeri + +
Senecio congestus + +
Taraxacum arcticum + +
T. lateritium + +
T. macilentum + +
77
Таблица 5
Сравнение трех близлежащих ЛФ Ямала по относительной
специфичности их состава
Показатели Число общих и специфических видов в ЛФ
Матюйяха Тиутей Венуйеуо
Всего видов 147 142 133
В том числе 118 118 118
общие* 80.2 83.1 88.7
необщие 29 24 15
в том числе общие с ЛФ Матюйяха — 14 6
” ” ЛФ Тиутей 14 — 2
” ” ЛФ Венуйеуо 6 2 —
специфические 9 8 7
Отсутствуют только в этой ЛФ 2 6 14
* Над чертой — абсолютные величины, под чертой — %.
и Шайтан) из той же подзоны северных гипоарктических тундр значи-
тельно меньше, мера сходства Съеренсена составляет 74-77 %, при высо-
ком (но меньшем, чем на Ямале) включении одной флоры в другую —
82-88.9% (Соколова, 1984а). Достаточно близкие ЛФ Колымо-Анадыр-
ского водораздела, выявленные каждая на площади 300 км2, имеют зна-
чения меры сходства Съеренсена 81-82% (Петровский, Плиева, 1986).
Высокое сходство трех флор и большой процент общих видов за-
ставляют с глубоким вниманием отнестись к тем минимальным разли-
чиям, которые выявляются при детальном сравнении флористических
списков. При этом особое значение придается комплексам (плеядам)
видов, имеющих сходную эколого-географическую характеристику, и
в меньшей степени „случайным” видам. Был проанализирован список
из 46 видов, необщих для трех ЛФ (табл. 4, 5).
ЛФ Матюйяха имеет 9 (6%) специфических видов: Cochlearia arcti-
са — довольно обычный для северной части Ямала вид, отсутствие его
в других флорах, возможно, случайно или связано с недостаточной пол-
нотой сборов. 6 видов находятся на северном пределе своего распростра-
нения, причем Juncus castaneus, Ranunculus gmelinii, Draba glacialis, Potentil-
la stipularis - стенотопные виды, спорадически распространенные по все-
му полуострову, кроме подзоны арктических тундр, a Calamagrostis langs-
dorffii, Lycopodium alpinum - виды более южных ЛФ, обычные для Яма-
ла, но редкие для ЛФ Матюйяха, так как находятся на северной границе.
Два вида — Carex maritima, Potentilla kusnetzowii — редкие для Ямала,
нахождение их в данной ЛФ, вероятно, связано с ландшафтными осо-
бенностями ее территории. 6 видов (4%) являются общими лишь с ЛФ
Венуйеуо: 2 вида - Lycopodium pungens, Luzula multiflora subsp. frigida -
78
находятся на северном пределе, остальные 4 вида — Deschampsia glauca,
Luzula nivalis, Hedysarum arcticum, Cassiope tetragona, обычные для
Ямала, отсутствуют в ЛФ Тиутей, по-видимому, случайно (причины
неясны).
Весьма важную группу составляют 14 видов, общих с ЛФ Тиутей.
Выделяется комплекс из 10 видов, характерный для растительных груп-
пировок суглинистых оползней и моховых тундр на шлейфах суглини-
стых склонов: Deschampsia borealis, Puccinellia angustata, Festuca brachy-
phylla, Eutrema edwardsii, Draba oblongata, D. parvisiliquosa, Taraxacum
arcticum, T. lateritium, T. macilentum, Matricaria hookeri. Один вид — Erige-
ron silenifolius — редкий для Ямала; в обеих ЛФ найден на высоких бере-
гах рек на полузакрепленных песках в значительном обилии. Остальные
виды - Senecio congestus, Stellaria palustris, Cerastium maximum, на наш
взгляд, случайно отсутствуют в ЛФ Венуйеуо.
Таким образом, для ЛФ Матюйяха характерно наличие немногочис-
ленных более южных видов, сближающих ее с ЛФ Венуйеуо и имеющих
здесь (или поблизости) северную границу, и распространение более об-
ширной группы видов, характерной для суглинистых оползней и сбли-
жающих ее с ЛФ Тиутей.
ЛФ Тиутей имеет 8 специфических видов (5.7%), ядро которых со-
ставляют виды, характерные для ландшафтов наиболее древних морских
террас с выходами суглинков и активными термоэрозионными процес-
сами: Luzula tundricola, Pedicularis oederi, Eriophorum brachyantherum,
Epilobium davuricum. По-видимому, случайно не найден в других ЛФ
обычный вид Chrysosplenium altemifolium, a Castilleja arctica, Pedicularis
verticillata, находящиеся на северной границе, распространены в данном
районе спорадически. Отсутствие поздно развивающейся Gentiana tenella
в списках других ЛФ может быть объяснено более ранними сроками ра-
боты в этих ЛФ.
Виды ЛФ Тиутей, общие лишь с ЛФ Матюйяха, были рассмотрены
выше, а с ЛФ Венуйеуо имеется лишь 2 общих вида — Rubus arcticus, ко-
торый не был найден в ЛФ Матюйяха, так как, по-видимому, здесь про-
ходит его северная граница, и Pedicularis sudetica subsp. albolabiata, отсут-
ствие которого в ЛФ Матюйяха, возможно, случайно.
Для ЛФ Венуйеуо отмечено 7 специфических видов (5 %). Четыре
вида — Equisetum pratense, Роа alpina, Veratrum lobelianum, Linnaea borea-
lis — находятся на северной границе, причем их процветанию способст-
вуют подходящие экотопы — крутые высокие береговые склоны южной
экспозиции, обращенные к долине крупной реки. Rumex arcticus, напро-
тив, на своем южном пределе имеет спорадическое распространение —
только на минеральных задернованных суглинистых участках с ниваль-
ным режимом, ограниченно представленных в районе. Carex holostoma -
очень редкий вид, найденный ранее в 90 км юго-восточнее, в бассейне
р. Сеяха, по-видимому, его распространение связано с молодыми долин-
ными осоковыми болотами с минимальным торфонакоплением. Нако-
нец, нахождение еще одного вида осоки - С. glacialis на верхнем перегибе
коренного берега реки на сухом песчаном пятне пока не поддается
объяснению.
79
Таблица 6
Распределение необщих видов сводного флористического списка четырех
близлежащих ЛФ Ямала (подзона южных гипоарктических тундр)
Вид Локальные флоры
Юрибей Хевесе Лапта я ха Еръяха
Botrychium boreale + + « +
Equisetum limosum + • + +
E. scirpoides « + •
E. variegatum « • +
Hierochloe odorata + • + +
H. pauciflora • + +
Arctagrostis arundinacea + +
Deschampsa borealis + •
D. obensis • + + +
Poa pratensis + +
Phippsia algida + +
P. concinna • • +
Festuca brachyphyUa + + +
Bromopsis pumpelliana +
Roegneria borealis +
Carex vaginata sdbsp.quasivaginata • + +
Juncus arcticus + +
Luzula rufescens • +
L. tundricola +
L. parviflora + + +
Salix arctica • + + +
S. lapponum • + +
Alnus fruticosa + + •
Rumex graminifolius • + + +
Oxyria digyna • • +
Polygonum laxmannii • « +
Stellaria ciliatosepala • + • •
Cerastium maximum + + +
C. regelii + + + •
Arenaria stenophylla subsp.polaris + • +
Caltha arctica • + + •
Delphinium middendorffu + + + •
Ranunculus glabriusculus • • +
R. gmelinii + + +
R. reptans • +
Eutrema edwardsii • + +
Rorippa palustris • • +
Arabis septentrionalis • + +
Draba alpina • +
D. fladnizensis • + +
D. glacialis • + +
D. oblongata • + +
D. pauciflora + +
D. pseudopilosa • + •
Saxifraga nelsoniana • + +
Chrysosplenium tetrandrum • + •
Dryas octopetala + • + +
D. punctata + + + •
Sanguisorba officinalis • • + •
80
Таблица 6 (продолжение)
Вид Локальные флоры
Юрибей Хевесе Лаптаяха Еръяха
Rosa acicularis • • + +
Epilobium davuricum + + + •
E. anagallidifolium + • • • +
Chamerion angustifolium + + +
Orthilia obtusata + + •
Harrimanella hypnoides + • •
Oxycoccus microcarpus + + +
Diapensia lapponica • • +
Pinguicula villosa + + +
Thymus reverdattoanus + • • 4-
Veronica alpina + + + •
Lagotisglauca subsp.minor • + + •
Pedicularis compacta • + + •
P. verticillata + 4 + +
Euphrasia frigida • • + +
Galium uliginosum + • +
Erigeron borealis + + •
E. eriocephalus + • +
Senecio atripurpureus • + •
Taraxacum nivale • + •
Crepis nigrescens + • +
Matricaria hookeri + + +
Aster sibiricus • • +
Таким образом, все 3 флоры обладают некоторой долей специфично-
сти, что, несмотря на значительное сходство, позволяет рассматривать их
как самостоятельные КФ. Для КФ Матюйяха и Венуйеуо общими явля-
ются бореальные гипоарктические и арктоальпийские виды на северном
пределе, для КФ Тиутей и Матюйяха — комплекс видов древних морских
террас. Между КФ Тиутей и Венуйеуо существуют более общие связи. На
основе анализа списка и с учетом полевых маршрутных наблюдений, а
также на основе анализа картографического материала, нами были при-
близительно намечены топографические границы трех КФ, причем из-за
отсутствия данных граница между КФ Тиутей и Венуйеуо частично оста-
лась открытой (невыясненной).
Площадь КФ Матюйяха составляет приблизительно не менее 1000км2,
площадь КФ Тиутей — не менее 700 км2 и КФ Венуйеуо - не менее 650 км2.
КФ Матюйяха и Венуйеуо в силу значительного сходства флористиче-
ского состава и общего комплекса гипоарктических и бореальных видов
на их северном пределе объединены в один флористический район, тогда
как КФ Тиутей со специфическим набором видов и особенностями ланд-
шафта отнесена к другому флористическому району.
Второй блок из четырех ЛФ расположен в подзоне южных гипоарк-
тических тундр (рис. 1) между р. Юрибей и Обской губой. Расстояние
между центрами ЛФ Юрибей и Хевесе и Юрибей и Лаптаяха — 50 км,
между Юрибей и Еръяха — 60 км, между Бръяха и Хевесе — 45 км,
между Еръяха и Лаптаяха и Лаптаяха и Хевесе - 20 км. Сводный список
всех четырех ЛФ насчитывает 221 вид, из них 149 видов (67.4%) общие,
6 Заказ 1220
81
Таблица 7
Сравнение четырех близлежащих ЛФ Ямала по относительной
специфичности их состава
Показатели Число общих и специфических видов в ЛФ
Юрибей Хевесе Лаптаяха Еръяха
Всего видов 180 187 191 180
В том числе
149 149 149 149
общие*
82.7 79.7 78.0 82.7
необщие 31 38 42 31
в том числе
общие с ЛФ Юрибей — 12 14 16
” ” ЛФ Хевесе — 25 8
” ” ЛФ Лаптаяха — 14
специфические* 4 2.2 7 3.7 6 3.1 6 3.3
Отсутствуют только в этой ЛФ 6 7 2 6
* Над чертой — абсолютные величины, под чертой — %.
Таблица 8
Матрицы попарного сходства четырех ЛФ Ямала из подзоны
южных гипоарктических тундр [ меры Съеренсена (слева)
и Симпсона (наибольшей из двух мер включения) (справа) ]
ЛФ Юрибей Хевесе Лаптаяха Еръяха
Юрибей ^^88.9 90.6 91.7
Хевесе 87.1 ^923 87.8
Лаптаяха 87.8 913 91.7
Еръяха 91.7 86.1 883>^
а 72 - необщие (табл. 6, 7). Для сравнения можно сказать, что во флоре
Люпвеемского горного массива Западной Чукотки общие виды пяти ЛФ
составили лишь 39% (Заславская, Петровский, 1983), а во флоре Колы-
мо-Анадырского водораздела — 48% (Петровский, Плиева, 1986).
Каждая ЛФ включает от 81.4 до 86.4% видов сводного списка, при-
чем общие виды составляют в них от 78 до 83 %. Значения меры сходства
Съеренсона между всеми парами флор очень велики — от 86.1 до 91.7%
(табл. 8; рис. 2, б), а меры сходства (наибольшего включения) по
Симпсону — от 87.8 до 92.5 %.
Вместе с тем для каждой ЛФ отмечено по 4—7 специфических видов,
анализ которых позволяет выявить особенности каждой ЛФ (всего 23
вида). Для ЛФ Юрибей отмечено 4 специфических вида: Roegneria sub fib-
rosa — единственная находка на Ямале, Deschampsia borealis и Bromus
pumpellianus встречаются спорадически по всему полуострову, их отсут-
82
ствие в соседних ЛФ пока необъяснимо, а находка Luzula tundricola очень
четко указывает на специфику ландшафта ЛФ Юрибей, связанную с ее
приуроченностью к IV морской террасе, наиболее древней для Ямала.
Из числа специфичных для ЛФ Хевесе видов выделяется Equisetum scir-
poides, редкий для Ямала вид, свойственный ландшафтам ледникового
генезиса, а также Harrimanella hypnoides — амфиатлантический вид, до-
вольно редкий на Ямале, распространение которого требует дополни-
тельного изучения. Находка Senecio atripurpureus свидетельствует о появ-
лении в районе торфянистых тундр, которые отсутствуют в близлежащих
ЛФ, остальные виды на Ямале распространены спорадически.
Для ЛФ Лаптаяха из 6 специфических видов следует выделить два,
найденных в долине реки Лаптаяха и нигде более на Ямале не отмечен-
ных: Sanguisorba officinalis и Luzula rufescens, а также Equisetum variega-
tum, найденный на низменном берегу Обской губы. Остальные 3 вида не
представляют редкости для КФ Ямала и отсутствуют в соседних ЛФ,
по-видимому, случайно.
Больше всего позитивных отличий в ЛФ Еръяха: здесь найдена
Aster sibiricus (первая находка на Ямале), Rorippa palustris (на северной
границе распространения), Ranunculus glabriusculus (редкий в южных
тундрах Ямала вид) и, наконец, 3 вида, характерных для песчаных суб-
стратов: Diapensia lapponica (на северной границе), Oxyria digyna и Poly-
gonum laxmannii, отсутствующие в остальных трех ЛФ, но широко рас-
пространенные в других ЛФ Ямала.
Так как позитивные особенности рассматриваемых ЛФ незначитель-
ны, нам пришлось прибегнуть к анализу негативных особенностей каж-
дой ЛФ. Только в ЛФ Юрибей отсутствуют 5 видов, из них для двух ви-
дов — Oxycoccus microcarpus и Pinguicula villosa — нет соответствующего
экотопа — болот со сфагновыми кочками; отсутствие Rumex graminifo-
lius, вида подчеркнуто псаммофильного, коррелирует с присутствием
Luzula tundricolir (см. выше). Для ЛФ Хевесе характерно отсутствие ви-
дов, находящихся на северной границе в соседних флорах: Hierochloe
odorata, Luzula parviflora, Equisetum limosum, а также спорадически рас-
пространенных в подзоне южных гипоарктических тундр Cerastium maxi-
mum и Pedicularis verticillata. Род Dry as представлен двумя таксонами:
D. punctata и D. х vagans, тогда как собственно D. octopetala не найдена.
Практически нет негативных признаков в ЛФ Лаптаяха — отсутствие
Botrychium boreale связано, бесспорно, с ранними сроками обследова-
ния: лишь 1 вид — Matricaria hookeri — достоверно отсутствует во флоре.
Нахождение М. hookeri в соседней ЛФ Хевесе связано с антропогенными
экотопами, в естественных условиях она также не найдена.
Наибольшие негативные отличия имеет ЛФ Еръяха: только здесь от-
сутствует 6 видов, сходных по своей экологической приуроченности
к суглинистым субстратам.
Участок каждой ЛФ мы признаем за ареал-минимум КФ, так как,
исходя из определения КФ, для каждого экотопа в пределах ЛФ харак-
терен постоянный набор видов. Сравнение ЛФ между собой показало,
1 Такую же корреляцию наблюдаем во флоре Юрибейто.
83
Таблица 9
Попарное сравнение четырех близлежащих ЛФ Ямала
ЛФ Юрибей Хевесе Лаптаяха Еръяха
Юрибей 180 206 932 207 93.6 195 88.2
Хевесе 161 187 204 92.3 209 95.0
Лаптаяха 163 174 191 206 93.2
Еръяха 165 161 163 180
Примечание. Полужирным выделено число видов в каждой ЛФ; в нижней
части таблицы — число общих видов в двух ЛФ; в верхней над чертой — общее чис-
ло видов в двух ЛФ, под чертой — % от сводного списка четырех ЛФ-
что наибольшую флористическую общность имеют ЛФ Хевесе и Лап-
таяха. Им свойственно наибольшее число общих видор — 174, составляю-
щих в каждой из них 93.0 и 91.1% соответственно; они имеют наимень-
ший общий список (204), отношение числа общих видов к общему числу
видов 85.3% при наивысшей максимальной мере включения (93%). Эти
флоры и территориально близки друг к другу, а основные позитивные
и негативные различия связаны с отсутствием в одной из них (Хевесе)
крупной речной долины. По-видимому, они входят в одну КФ.
Столь же близка между собой и другая пара ЛФ — Юрибей и Еръяха.
Они насчитывают одинаковое число видов — по 180, несколько меньше,
чем предыдущие ЛФ, что сказалось и на общем числе видов (195), и на
числе общих для этих ЛФ видов (165). Отношение числа общих видов
к общему числу видов 84.6%, Значения мер сходства Съеренсена и Симп-
сона очень высоки — 91.7%. Однако различия между этими ЛФ вполне
четкие — в одной из них отсутствуют и присутствуют в другой комплек-
сы видов, довольно активных, а не редких, случайных. В связи с этим
мы рассматриваем ЛФ Юрибей и Еръяха как самостоятельные КФ.
Попарное сравнение всех четырех КФ показало, что наименьшее
сходство по видовому составу с другими имеет КФ Еръяха, наиболь-
шее — Лаптаяха (табл. 9). Отличительные признаки КФ Еръяха дают
основание отнести ее к другому флористическому району, тогда как КФ
Юрибей и КФ Хевесе—Лаптаяха, бесспорно, должны быть отнесены к од-
ному и тому же флористическому району.
3. Количественная оценка пространственного разнообразия
н уровня видового богатства конкретных флор Ямала
Для объективной оценки богатства КФ Ямала и их пространствен-
ного разнообразия были вычислены показатели пространственного раз-
нообразия — константы z из уравнения Аррениуса и Ъ из уравнения
Глизона (по: Малышев, 1972,1975,1976а; Шмидт, 1980,1984).
84
Таблица 10
Константы пространственного разнообразия КФ Ямала, вычисленные
для соседних участков площадей возрастающего радиуса
КФ Сравниваемая площадь, км2
3.1-0.8 12.6-3.1 28.2-12.6 78.5-28.2 113.1-78.5
Z Ъ Z b Z Ъ Z Ь Z ъ
Хутыяха 0.12 40.8 0.08 29.6 0.05 20.0 0.005 2.2 0 0
Лаптаяха 0.17 57.8 0.05 19.7 0.06 25.7 0.01 4.5 — —
Хевесе 0.12 42.5 0.02 9.8 0.03 14.3 0.026 11.2 0 0
Мантыто 0.08 23.8 0.04 13.1 0.018 5.7 0.02 6.7 0 0
Матюйяха 0.13 37.4 0.02 8.2 0.03 8.6 0.006 2.2 0 0
Верхний Тамбей 0.05 11,9 0.03 8.2 0.03 8.6 0.008 2.2 0 0
Хабейяха o:i 22.0 0.055 13.1 0.056 14.3 0 0 0 0
Аррениуса,
Ь
из уравнения
— константа из
Примечание
уравнения Глизона.
z — константа
Расчет производился двумя способами — методом соприкасающихся
площадей и методом наименьших квадратов.* 1 В первом случае были по-
лучены константы пространственного разнообразия для отдельных час-
тей площади КФ (табл. 10), во втором — значение для всей площади КФ
(табл. 11). Общий ход изменения константы z и Ъ сходен (рис. 3).
Однако в связи с малыми различиями во флорах Ямала мы отдаем пред-
почтение уравнению Глизона, так как константа Ъ обладает большим
дифференцирующим свойством.
Следует отметить некоторое повышение константы пространственно-
го разнообразия на площади 28.2 км2 по сравнению с меньшей площадью,
что прослеживается почти во всех КФ. Вместе с тем почти не выявляются
различия между КФ разных подзон: несколько выше пространственное
1 Расчет константы пространственного разнообразия [ 1) уравнение Аррениуса,
2) уравнение Глизона] разными методами.
Метод соприкасающихся площадей:
D -- log^-logy, .
log X, - log X, ’
2) b = .
log X, - log Xj
Метод наименьших квадратов:
1) n log a + z S log x = S log у;
log a S log x + z S (log x)2 = S log x • log y.
2) na + b S log x = S у,
a S log x + A S (log x)2 = Sy • log x.
Во всех случаях z нЬ — константы пространственного разнообразия; у - число
видов на площади х; а — удельный уровень видового богатства; п - число квадра-
тов (участков), участвующих в расчете.
85
Рис. 3. Изменение констант пространственного разнообразия при увеличении пло-
щади исследованного участка.
I - флора Лаптаяха; II - флора Верхний Тамбей. 1 - константа z ; 2 - константа Ъ.
разнообразие КФ подзоны южных гипоарктических тундр на малых уча-
стках, которое выравнивается при достижении площади 78.5 км2.
На наш взгляд, абсолютное значение константы пространственного
разнообразия имеет ограниченное применение в связи с большой зависи-
мостью от размеров площади. Диапазон варьирования для участков внут-
ри КФ Ямала от единицы до нескольких десятков. Однако расчет этой
константы — необходимый этап для определения удельного уровня видо-
вого богатства — среднего числа видов, приходящегося на единицу пло-
щади (константа а в уравнениях Аррениуса и Глизона).
86
Таблица 11
Уровень видового богатства КФ Ямала и константа
пространственного разнообразия b
КФ Ь В расчете на Фактически найдено на
1 км1 100 км2 0.8 км2 113 км2
Хутыяха 23 140 186 131 181
Лаптаяха 28 145 201 134 191
Хевесе 173 155 190 146 187
Мантыто 12 122 146 117 144
Матюйяха 16 121 156 116 147
Верхний Тамбей 73 106 121 104 120
Хабейяха 12 92 116 87 113
Таблица 12
фактически найденное и расчетное число видов в некоторых КФ Ямала
Площадь, км2
Показатели 0.8 3.1 12.6 28.2 78.5 113.1
Лаптаяха
фактически найденное Расчетное 134 168 180 189 191 —
по Аррениусу 142 158 175 186 201 206
по Тли зону- 143 159 176 186 198 202
Матюйяха
Фактически найденное Расчетное 116 138 143 146 147 147
по Аррениусу 123 130 138 143 149 152
по Глизону 119 129 139 144 151 154
Be эхний Тамбей
фактически найденное Расчетное 104 111 116 119 120 120
по Аррениусу 105 110 114 117 120 121
по Глизону 105 109 111 116 119 120
Таблица 13
Расчетное и фактическое число видов в объединениях флор
Показатели Лаптаяха (Л) + + Хевесе (X) Л + Х + Юрибей (Ю) Л+ Х + Ю +Еръяха
Расчетное число 209 213 216
Фактически найдено 204 215 221
87
Рис. 4. Эмпирическое (/) и расчетное (2, 5) число видов на участках различной
площади в пределах конкретных флор.
КФ: I - Лаптаяха (южные гипоарктические тундры), II - Матюйяха (северные ги-
поарктические тундры), III - Верхний Тамбей (арктические тундры). Рассчитано:
2 - по уравнению Аррениуса, 3 - по уравнению Глизона.
При расчете системы уравнений методом наименьших квадратов
(Шмидт, 1980) были получены константы пространственного разно-
образия, а затем и константа а удельного уровня видового богатства
(табл.11).
По уровню видового богатства все КФ четко проявляют подзональ-
ные различия: удельное богатство КФ южных гипоарктических тундр на
20—35 видов выше, чем северных, а КФ северных гипоарктических тундр
на 15—30 выше, чем КФ арктических тундр Ямала.
Цифры, полученные при расчете числа видов на определенную пло-
щадь, исходя из вычисленных констант b и а, показали неплохое соответ-
ствие эмпирическим данным, однако постоянно имеет место завышение
расчетного числа видов для малых площадей (в нашем случае 0.8 км2),
занижение для средних (3.1 км2,12.6 км2, 28.2 км2) и вновь завышение
для больших площадей. Расчетное число видов на 100 км2 оказалось
выше числа видов, найденных для всех КФ на площади 113 км2 (табл. 11,
12; рис. 4).
Экстраполяция данных на еще бблыпие площади показала следую-
щее: если выбрать за основу наиболее богатую КФ Лаптаяха с числом ви-
дов 191, то совокупность двух КФ — Лаптаяха и Хевесе должна дать по
формуле ^ = aiogx на площади = (78.5 + 113.1) 209 видов. Присоеди-
88
Таблица 14
Флористическая репрезентативность наименьших участков КФ Ямала,
рассчитанная методами соприкасающихся участков (А)
и наименьших квадратов (Б)
КФ А Б
а Ъ R а Ь R
Хутыяха 135 40.8 90.7 140 23 95
Лаптаяха 140 57.8 87.0 145 28 94
Хевесе 152 423 91.3 155 173 97
Мантыто 119 23.8 93.8 122 12 97
Матюйяха 120 37.4 90.3 121 16 96
Верхний Тамбей 105 11.9 963 106 73 98
Хабейяха 89 22.0 92.4 92 12 96
Примечание, а — удельный уровень видового богатства, Ъ — константа
пространственного разнообразия, R — репрезентативность (%) .
Таблица 15
Репрезентативность отдельных участков некоторых КФ Ямала
- КФ Площадь, км
ОЛ 3.2 12.6 28.2 78.5 113.1
Хутыяха 94.9 95.4 95.8 96.0 96.2 96.3
Лаптаяха 94.0 94.7 95.2 95.4 95.7 —
Хевесе 96.8 96.8 97.0 97.1 97.2 —
Мантыто 97.0 97.1 97.3 97.4 973 —
Матюйяха 96.1 96.3 96.6 96.7 96.8 —
Верхний Тамбей 97.8 97.9 98.0 98.1 98.1 —
Хабейяха 96.0 96.3 96.6 96.7 96.8 —
нение к первым двум третьей КФ — Юрибей (Ю) площадью 785 км2
увеличивает расчетный список до 213 видов. И наконец, присоединение
еще одной КФ Еръяха площадью 70 км2 дает общее число 216 видов.
Эти значения несколько отличаются от фактических данных в сторону
увеличения при соприкасающихся площадях (Лаптаяха! Хевесе) и в сто-
рону уменьшения при объединении флор несоприкасающихся террито-
рий, для которых константа пространственного разнообразия должна
быть иной (табл. 13).
Расчет репрезентативности (R) участков, выполненный на основе
полученных коэффициентов пространственного разнообразия (Ь) и
удельного уровня видового богатства (а) (табл. И) по формуле1
1 По: Малышев, 1975; Шмидт, 1980.
89
показал, что участки КФ площадью 0.8 км2 репрезентативны на 94—98 %
(табл. 14) . При расчете констант а и Ъ для двух соприкасающихся пло-
щадей (0.8 км2 и 3.1 км2) репрезентативность участка 0.8 км2 несколь-
ко ниже (87—96.5 %).
С увеличением площади участков их репрезентативность возрастала
незначительно (не более 2%; табл. 15).
Итак, для КФ Ямала характерна высокая степень сходства, большое
число общих видов. Общность видового состава КФ увеличивается к се-
веру. Сходство КФ северного блока выше, чем южного, за счет увеличе-
ния процента общих видов, повышаются и значения меры включения
одной КФ в другую. При сравнительном анализе КФ определенный вес
имеют как позитивные, так и негативные различия состава видов, причем
в том и другом случае преимущество отдается эколого-географическим
плеядам видов.
Ареал-минимум КФ для равнинных флор Ямала значительно меньше
принятой стандартной площади 100 км2 и равен 30 км2, а общая пло-
щадь значительно превышает ее, достигая 1000 км2 (а возможно, и бо-
лее) . Площадь выявления КФ постепенно уменьшается с юга на север.
Территориальные выделы КФ мы считаем возможным устанавливать
лишь после детального сравнения всех ЛФ — на заключительном этапе
работы. В этом случае КФ как естественные флористические выделы
дают основу для проведения границ при флористическом районировании.
Для получения исчерпывающе полных сведений о составе региональной
флоры необходимо сплошное обследование территории методом регу-
лярных проб флоры, соприкасающихся своими границами, так как до
5—6% видов флоры распространено крайне спорадически.
Качественная характеристика отдельных флористических выделов
может быть получена при флористическом обследовании ареалов-мини-
мумов КФ.
Удельный уровень видового богатства КФ Ямала один из наиболее
низких для флор Арктической Евразии, однако четко отражает зональ-
ные черты КФ-
Репрезентативность даже небольших участков внутри КФ (< 1 км2)
очень высока, более 94%.
В заключение приношу благодарность В. М. Шмидту и Б. А. Юрцеву
за ценные консультации.
Н. И. Золотухин
ОПЫТ ФЛОРИСТИЧЕСКИХ ИССЛЕДОВАНИЙ
НА УРОВНЕ ФИТОХОРИЙ НАИМЕНЬШЕГО РАНГА
(НА ПРИМЕРЕ АЛТАЙСКОГО ЗАПОВЕДНИКА)
Алтайский государственный заповедник
Под фитохорией наименьшего ранга мы понимаем флористиче-
ский микрорайон, т. е. территорию, отграниченную по совокупности
флористических признаков и далее неделимую по этим признакам.
90
Одновременно мы считаем (Золотухин, 1982) флористический микро-
район конкретной флорой (КФ), что не противоречит взглядам автора
данного понятия, основоположника метода КФ А. И. Толмачева (1931,
1941, 1974). Б. А. Юрцев (1975а, 1982а) и Ю. Р. Шеляг-Сосонко (1980)
также предлагают закрепить авторское определение КФ за элементарным
флористическим районом.
Некоторые принципы флористического районирования горных тер-
риторий рассмотрены нами ранее (Золотухин, 1982). В той же работе
приведена схема фитохорий центральной и южной частей Алтайского
государственного заповедника. Многие критерии флористического райо-
нирования, качественные и количественные характеристики флор неод-
нократно анализировались в литературе, поэтому в приводимом ниже
конкретном материале мы заострим внимание преимущественно на ред-
ко учитываемых особенностях фитохорий.
Район исследований охватывает прителецкую (северную) часть запо-
ведника общей площадью более 1800 км2 (рис. 1; табл. 1). Рельеф гор-
ный, с высотами от 434 до 2600 м над ур. м. Более чем 10-летние работы,
в которых, кроме автора, принимали участие Л. В. Марина, И. Б. Золоту-
хина, А. В. Галанин, Т. Е. Теплякова, позволили максимально полно изу-
чить флору этой территории, До 1984 г. в претелецкой части заповедника
выявлено 997 аборигенных видов сосудистых растений, а вместе с антро-
похорами — 1113 видов. Почти все антропо хоры (включая археофиты)
локализованы в пределах пос. Яйлю, шести кордонов и двух стоянок
туристов, занимающих в совокупности менее 1 % площади заповедника
(Золотухин, 1983). Лишь немногие заносные виды встречаются по тро-
пам и галечникам в других районах заповедника. Адвентивные растения
мы исключаем из дальнейшего анализа.
По совокупности флористических признаков, таких как общий ви-
довой состав, дифференциальные и кодифференциальные таксоны, гео-
графические и генетические элементы флоры и др., прителецкая часть
заповедника разбита нами на 7 КФ площадью от 100 до 700 км2 (рис. 1;
табл. 1). Увеличение полноты изученности КФ и районов в последние
годы показано в табл. 2. Хорошим вспомогательным инструментом при
оценке степени изученности флоры является критерий „автономности”,
рассчитываемый по эмпирической формуле Л. И. Малышева (1969,
1976а). Меры автономности КФ в пределах фитохорий более крупного
ранга (район, округ) должны быть близки. Более низкие показатели
автономности имеют недостаточно обследованные фитохории по сравне-
нию с хорошо обследованными, части фитохорий по сравнению со всей
их территорией, фитохории низшего ранга по сравнению с фитохориями
более высокого ранга (табл. 2).
К числу основных критериев флористического районирования при-
надлежат сходство, различие и меры включения таксономического со-
става флор анализируемых выделов. В табл. 3 дана матрица мер пере-
сечения флор семи микрорайонов по их видовому составу, на основе
которой по формулам Съеренсена—Чекановского и Престона рассчитаны
коэффициенты сходства (табл. 4). Эти же данные использованы при
вычислении простейших мер включения, которые уже довольно широко
91
Рис. 1. Флористическое районирование прителецкой части Алтайского заповедника.
Границы: 1 - флористических районов (Т - Телецкий, Б - Балыкчинский); 2 -
микрорайонов (/ - Яйлинский, II - Кокшинский, III - Колюштинский, IV— Туш-
кемский, V - Белинский, VI - Кайринский, VII - Верхнекыгинский); 3 - Алтай-
ского заповедника; 4 - стандартных участков „проб флористической ситуации”
(а, б, в — участки 10 X 10 км). Распространение некоторых видов: 5-14 - немо-
92
Рис. 2. Орграфы отношений „банально-
сти” (Л) и „эндемичности” (ориги-
нальности) (Б) на множестве видовых
списков флористических микрорайо-
нов {If-VII) Алтайского заповедника.
А: сплошные линии - Bt0, прерыви-
стые - В10; Б: сплошные линии -Esn,
прерывистые - Ем. Обозначения мик-
рорайонов как на рис. 1.
применяются для анализа флори-
стических, геоботанических и эко-
логических данных (Юрцев, 1968а,
1978; Комарова, Семкин, 1976;
Андреев, 1980; Юрцев, Семкин,
1980). На основании мер вклю-
чения (табл. 5) по методике
В. Л. Андреева (1980) построены
орграфы отношений „банально-
сти” (В) и „эндемичности” (£)
(оригинальности) (рис. 2)./ Наи-
большее число стрелок орграфа
банальности входит в точки III и
IV (Колюштинский и Тушкемский микрорайоны), которые, следова-
тельно, являются центральными в изученном регионе. Самыми баналь-
ными по видовому составу оказались КФ VI и VII. Наиболее оригиналь-
ной флорой обладает Белинский микрорайон (V).
Меры сходства, различия и включения видовых списков фитохорий
не могут быть единственным критерием при флористическом райониро-
вании поясно-дифференцированных горных территорий. Необходимо
знать парциальную структуру флоры и прежде всего иметь таксономи-
ческие списки высотнопоясных парциальных флор (ПФ) и их объеди-
нений, В противном случае можно принять различие между ПФ за разли-
чие между фитохориями. Ограничение высотных поясов мы проводили
по общепринятым в горах Сибири критериям, обобщенным Малышевым
1 Автор использует терминологию В. Л. Андреева, согласно которой „баналь-
ными” именуются флоры с высокими значениями меры включения в другие фло-
ры, „эндемичными” (лучше - оригинальными) - с низкими значениями той же
меры. (Прим, ред.).
рального комплекса [5 - Brachypodium sylvaticum (Huds.) Beauv., 6 - Carex marty-
nenkoi N. Zolot., 7 - C. sylvatica Huds., 8 - Asarum europaeum L., 9 - Dentaria sibi-
rica (О. E. Schulz) N. Busch, 10 - Cardamine impatiens L., 11 - Brunnera sibirica Stev.,
12 - Galium paradoxum Maxim., 13 - Carpesium triste Maxim., 14 - Alfredia cemua
(L.)Cass.-]; 15-21 - лесостепного и горностепного комплексов [/5 - Ephedra equi-
setina Bunge, 16 - Adonis vemalis L., 17 - Viola irinae N. Zolot., 18 - Ferula soonga-
rica Pall, ex Spreng., 19 - Lonicera microphylla Willd. ex Schult., 20 - Saussurea sali-
cifolia (L.) DC., 21 - Leibnitzia anandria (L.) Turcz.].
93
94
Таблица 1
Некоторые количественные особенности фитохорий прителецкой части Алтайского заповедника
п
§
ж
п
о
s
по высотным поясам* ей С- 00 О\ со Or- 1 сп еч оо О | I оо оо чЧ чЧ СЧ ' чЧ чЧ
о чч Оч О ч© Г- «П I О СП оо ОО I I ЧЧ ОО чч еч еч еч 1 сп еч
вл г- 40 о Tf 40 еч | К) СО 4© 4© I 1 СО еч еч еч еч со со
нл о О 40 СО । 2 £ 2£ 2 । 1 । । 4Г) СО СО
лс 1 1 1 1 1 1 р? 1 1
оот}-^©т}-оо^©ечтгеч
2
4)
О
П
^>1Л4©\©00ОеЧ00ГЧ
ООООГ*-4©04^нГ*-СПОО
OOvF)^*VF)»nO\»n^,<*‘>
ooooooooo
ОООООСПОСПО
чТСПГ-ечеЧчТчЧччеЧ
OOOvnOOOOO
омпооооооо
Оснг-ечсчОччечсч
co еч
* Условные обозначения: лс — лесостепной, нл — черневой (нижний лесной) , вл — горнотаежный (верхний лесной), с — субальпий-
ский, а - альпийский.
1.Ш^>дШ'?А,шмвд^йшайаАЗД?а>п.^Х’>м.й,ла jwwn.i u .«мимиму»? (глеи <w««««• ч№л*.т«иг»/’т
Таблица 2
Увеличение полноты изученности флоры прителецкой части
Алтайского заповедника в 1982-1983 гт. и ее влияние
на показатель автономности флоры
Районы и микрорайоны Период исследований Выявлено Значение показателя автономности флоры
родов видов
Телец кий До 1.1.1982 334 804 -0.02
До 1.1.1983 345 847 -0.01
До 1.1.1984 345 848 -0.01
Яйлинский До 1.1.1981 283 566 -0.20
До 1.1.1982 285 576 -0.19
До 1.1.1983 288 585 -0.18
Кокшинский До 1.1.1981 225 438 -0.24
До 1.1.1982 236 463 -0.23
До 1.1.1983 242 476 -0.22
Колюштинский До 1.1.1981 259 497 -0.25
До 1.1.1982 264 510 -0.24
До 1.1.1983 271 564 -0.15
До 1.1.1984 271 566 -0.15
Тушкемский До 1.1.1981 274 522 -0.25
До 1.1.1983 292 598 -0.17
Балыкчинский До 1.1.1982 327 778 -0.03
До 1.1.1983 340 826 -0.02
Белинский До 1.1.1982 277 552 -0.23
До 1.1.1983 285 572 -0.19
Кайринский До 1.1.1982 208 396 -0.29
До 1.1.1983 219 438 -0.21
Верхнекыгинский До 1.1.1982 188 325 -0.46
До 1.1.1983 196 382 -0.28
Таблица 3
Матрица мер пересечения флор микрорайонов^рителецкой части
Алтайского заповедника (по видовому составу)
Микро- район I II III IV V VI VII
I 585
II 354 476
III 403 387 566
IV 409 403 476 598
V 396 285 325 360 572
VI 279 332 361 369 234 438
VII 243 306 311 327 176 330 382
* Обозначения микрорайонов в соответствии с рис. 1.
95
Таблица 4
Матрица мер сходства видового состава флор микрорайонов
прителецкой части Алтайского заповедника, вычисленных по формулам
Съеренсена-Чекановского (правая часть) и Престона (левая часть), %
Матрица мер включения флор микрорайонов прителецкой части
Алтайского заповедника по видовому составу, %
Микро- район I II III IV V VI VH
I 61 69 70 68 49 42
II 75 — 81 85 60 70 64
III 71 69 — 84 58 64 55
IV 68 67 80 — 60 62 55
V 69 50 57 63 — 41 31
VI 64 76 82 84 53 — 75
VII 64 80 81 86 46 86 —
(1977). Геоботаники иногда выделяют единый лесной пояс с двумя под-
поясами (Огуреева, 1980), но по составу флоры черневой (нижний лес-
ной) и горнотаежный (верхний лесной) высотные пояса обособлены не
менее, чем субальпийский (рис. 3). По общему видовому составу КФ I
довольно существенно отличается от КФ II, III, IV (табл. 4, 5), но срав-
нение списков высотнопоясных ПФ показывает, что эти отличия опреде-
ляются прежде всего бедностью флоры альпийского пояса, где зарегист-
рировано только 37 видов сосудистых растений (см. табл. 1), так как
высокогорья Явлинского микрорайона представлены небольшими фраг-
ментами, достигающими лишь 2000 м над ур. м.
Мы считаем наиболее рациональным использование мер включения
не для сравнения флор одноранговых и близких по числу видов фитохо-
рий, а для проверки возможности отнесения части к предполагаемому
целому. Телецкий флористический район, к которому мы относим
микрорайоны I—IV, на площади около 1600 км2 расположен за пре-
делами заповедника (западный берег оз. Телецкого, верховья р. Бии).
По данным П. Н. Крылова (1927—1964) и материалам наших исследова-
ний, на указанной территории известно 36 аборигенных видов растений,
96
Рис. 3. Графы сходства высотнопоясных парциальных флор прителецкой части
Алтайского заповедника, рассчитанные по формуле Престона.
А - все микрорайоны; Б - стандартные участки „проб флористических ситуаций”
(а на рис. 1); В - высокогорные (д + с) пояса; Г - все пояса, кроме черневого
(нл); Д - лесные (нл + вл) пояса, а, с, вл, нл, лс - высотные пояса (см. табл. 1).
I-VII - микрорайоны (см. рис. 1). Сплошные жирные линии - C1Q, тонкие - С60,
прерывистые - Сзо (цифры на линиях связи - значения меры сходства).
7 Заказ 1220
97
Рис. 4. Орграфы меры включения видовых списков флористических микрорайонов
в сводный видовой список Телецкого флористического района.
Цифры в секторах большого круга - число видов Телецкого района (без специфи-
ческих видов каждого микрорайона), с которым производится каждое конкретное
сравнение. Цифры на стрелках - мера включения, %. I—V - микрорайоны как
на рис. 1.
отсутствующих в микрорайонах I—IV. Число специфичных видов в этих
микрорайонах характеризуется следующими цифрами: в I — 15 видов,
в II — 16, в III — 21, в IV — 23 вида, а их флоры включаются во флору
остальной части Телецкого района на 96—97% (рис. 4). Микрорайон VI
имеет 32 специфичных вида, а микрорайон V — 102 вида, и меры вклю-
чения их во флору Телецкого района заметно ниже (рис. 4).
Поддерживая в целом принцип выделения в горных условиях фито-
хорий, территориально охватывающих все пояса, необходимо подчерк-
нуть, что в каждом конкретном случае этот тезис должен проверяться
и анализироваться. Так, КФ V выделена только в пределах одного лесо-
степного пояса и отнесена к территориально более отдаленному району —
Балыкчинскому. В таких сложных ситуациях необходимо учитывать
критерий наименьшего дополнения, который можно сформулировать
следующим образом: территория присоединяется к той соседней фито-
хории, которой она дает наименьшее дополнение видового списка рас-
тений. При этом анализируется ряд фитохорий возрастающего ранга.
Присоединение фитохории V к территориально сопряженной фитохории
II увеличивает флору микрорайона на 287 видов,Телецкого района — на
102 вида, а всего Шорского флористического округа, занимающего Гор-
ную Шорию, Салаир, Северо-Восточный Алтай, часть Кузнецкого Алатау,
Западного Саяна и Манских белогорий Восточного Саяна (площадь при-
мерно 100 тыс. км2), — около 80 видов. Присоединение же КФ V к Ба-
лыкчинскому району добавляет к его флоре всего 36 видов, а к флоре
Североалтайского округа и того меньше — около 15 видов.
Для проверки гипотезы о возможности присоединения КФ V в каче-
стве ПФ к КФ II мы провели также сравнение видового состава террито-
риально сопряженных высотнопоясных ПФ. Если сходство черневого
98
пояса с горнотаежным в пределах КФ варьирует от 51 до 61 %, то лесо-
степной пояс с соседним горнотаежным сходен только на 39 % (рис. 3).
Микрорайоны VI и VII по видовому составу имеют большее сходство
с КФ Телецкого района, чем с КФ V (табл. 4, 5; рис. 2). Но если мы
учтем высотную поясность и при сравнении КФ V с соседними КФ Телец-
кого района исключим черневой пояс, являющийся высотнопоясным ана-
логом лесостепного пояса (занимает те же высоты над уровнем моря),
то это сходство окажется меньшим (рис. 3), чем сходство КФ V с КФ VI
и VII (табл. 4).
Проведенный дополнительный анализ подтверждает эти данные.
Наиболее важными для районирования обследованной территории явля-
ются неморальный, лесостепной и горностепной комплексы видов. Из 57
видов неморального комплекса, встречающихся в прителецкой части за-
поведника, в КФ I-IV произрастает от 20 до 38 видов, а в КФ V таких
видов всего 13, в КФ VI и VII еще меньше — 8 и 6 видов соответственно.
Виды горностепного и лесостепного комплексов свойственны преимуще-
ственно Белинскому микрорайону и низовьям р. Чулышман, часть из
них встречается и в КФ VI, VII. Распространение некоторых редких
в прителецкой части заповедника видов неморального, лесостепного и
горностепного комплексов показано на рис. 1.
При проверке пригодности регулярной сети квадратов для сбора и
обработки флористических материалов на сложно расчлененных горных
территориях мы заложили 3 стандартных участка „проб флористической
ситуации” площадью по 100 км2 таким образом, что они заведомо охва-
тывают части двух или трех фитохорий. Квадраты а и в соприкасаются
сторонами, а квадрат б перекрывает значительную часть квадратов айв
(рис. 1). Выявленное число видов по изученным участкам характеризу-
ется следующими цифрами: а — 676, б — 760, в — 766, что значительно
выше, чем в равновеликих или больших по площади КФ (табл. 1). Сход-
ство видового состава пробы флоры а с пробой флоры в по уравнению
Престона составляет 82%, что также значительно превышает сходство
КФ (табл. 4). Это объясняется включением в пробы флоры а и в флори-
стически богатых и почти идентичных участков КФ V. Таким образом,
наши данным подтверждают мнение Ю, Р. Шеляг-Сосонко (1980) о том,
что метод квадратов обладает невысокой разрешающей способностью
в отношении установления границ фитохорий. Это относится прежде все-
го к условиям гор. Еще В. Л. Комаров (1947, с. 45) писал, что исследо-
вание горных стран требует вообще большей детальности, так как часто
даже соседние долины сильно разнятся по своей флоре”.
Иногда в качестве основного критерия флористического райониро-
вания используется систематическая структура флоры (Шмидт, 1975).
В данном сообщении мы не рассматриваем этот показатель из-за ограни-
ченности объема статьи. Кроме того, в некоторых случаях применение
его не дает удовлетворительных результатов (Камелии, 1979; Золоту-
хин, 1982).
Критерий эндемизма, последовательно примененный в фундамен-
тальной работе А. Л. Тахтаджяна (1978), имеет главенствующее значе-
ние только при макрорайонировании. Алтае-Саянская флористическая
99
Рис. 5. Распространение некоторых видов растений в Восточном Алтае по разным
данным.
1 - Populus nigra L., по нашим данным [по И. Ю. Коропачинскому (1983) распро-
странен в границах всей обследованной территории]; 2, 3 - Bupleurum martjanovii
Kryl.: 2 - по литературным данным (Эндемичные высокогорные растения...,1974)
3 - по нашим данным; 4, 5 - Pachyneurum grandiflorum (С. А. Меу.) Bunge14 - по
литературным данным (Эндемичные высокогорные растения..., 1974), 5 - по на-
шим данным.
провинция, в которой на площади в 15 млн. км2 представлено более
2800 видов сосудистых растений, насчитывает примерно 120 эндемов и
150 субэндемов. Эндемов и субэндемов входящих в Алтае-Саянскую
провинцию округов уже значительно меньше — от 8 до 40, а флористиче-
ским районам свойственны только единичные эндемы,или таковые от-
сутствуют вовсе.
Недавно Б. А. Юрцев (1983а) выдвинул очень интересное положение
о количественной оценке „веса” видов при флористическом райониро-
вании. Предлагается учитывать модальность (банальность, в смысле ряда
других авторов) и специфичность (эндемичность) путем определения
мер включения фитохории в ареал и ареала в фитохорию. К сожалению,
100
практическое применение этого критерия, даже для относительно хоро-
шо изученных территорий, — дело отдаленного будущего. Главные труд-
ности возникают при определении площади ареала вида, полученные
данные неизбежно сильно зависят от полноты изученности флор (все
еще недостаточной), фрагментированности непосредственной заселенно-
сти видами площади, антропогенной освоенности территории, масштаба
так называемых карт ареалов и т. д.; об этом говорится и в цитирован-
ной работе Юрцева.
Проиллюстрируем сказанное лишь некоторыми примерами (рис. 5).
И. Ю. Коропачинским (1983) Populus nigra L. на карте ареала приводится
для всей обследованной нами территории площадью 1 млн. га, на кото-
рой мы выделяем 41 КФ. По нашим данным, вид здесь встречается толь-
ко в пределах двух КФ Балыкчинского флористического района на пло-
щади 1000 га. Если же учесть конкретную площадь, занимаемую этим
видом (к примеру, проекцию крон), то она на указанной территории
составит менее 10 га. Таким образом, в зависимости от точности данных
и масштаба их представления площадь ареала вида в пределах Восточно-
го Алтая оказалась отличной в 10s раз. Сходная картина наблюдается и
в отношении других видов, приведенных на рис. 5.
Подобных примеров можно привести много. Каждый вид заселяет
свой ареал не сплошь, и следует отличать кружево ареала от разреженно-
сти в пределах стации (Беклемишев, 1982). Но чтобы определить эти
параметры у растений, необходимы детальные популяционные исследо-
вания, проведение которых для всех видов и на всей территории просто
нереально. Кроме того, и редкие, и банальные виды далеко не равноцен-
ны по своей фитохорологической значимости, что определяется историей
таксона и флоры в целом, способом опыления и распространения семян,
жизненной формой и др. Иногда отсутствие массового вида на соседней
территории не столь важно для целей районирования, если этот вид,
к примеру, кальцефил и его распространение на изученной территории
однозначно коррелирует с распространением соответствующих горных
пород (то же справедливо для растений болот, пойм и т. д.). С другой
стороны, даже редкие эндемичные и реликтовые виды имеют существен-
ное значение для флористического районирования, особенно если они
группируются в плеяды (рис. 1) и вместе занимают значительную часть
фитохории.
Проведение детального флористического районирования особенно
важно для специально охраняемых территорий, в первую очередь запо-
ведников. Как известно, ценность охраняемой территории прямо про-
порциональна богатству ее биоты, ее репрезентативности, способности
автономно функционировать, наличию редких объектов и обратно
пропорциональна интенсивности прямого антропогенного воздействия
(Второв, 1980; Второв, Второва, 1983). Исходя из этого мы провели
анализ с целью выделения наиболее ценных и в то же время эталонных
участков в пределах Алтайского заповедника. Прежде всего выявили
видовое богатство флоры по микрорайонам. Наиболее богаты КФ в при-
телецкой части заповедника, в том числе Белинский микрорайон, где
на площади 100 км2 представлены 572 вида аборигенных сосудистых
101
Рис. 6. Наиболее ценная во флористическом отношении территория Алтайского
заповедника.
1 — граница заповедника; 2 - единичные местонахождения редких в заповеднике
видов растений; 3 - флористически наиболее ценная территория.
растений, а если исключить из микрорайона 40 км2 акватории оз. Те-
лецкого, — 569 видов. Затем нанесли на картосхему местонахождения
159 видов, которые известны в заповеднике на площади не более
1 км2, и ограничили эти местонахождения единым замкнутым контуром
(рис. 6) таким образом, что на 50% площади заповедника оказались
представлены все (!) 1445 видов его флоры. Определили представлен-
ность особо охраняемых видов: в каждом из 35 флористических микро-
районов заповедника произрастает от 4 до 16, а всего в заповеднике
33 вида растений, рекомендуемых к государственной охране различными
сводками.
102
Рис. 7. Флористическое районирование Алтайского государственного заповедника.
Границы: а - южная и западная (северная и восточная совпадают с границами фло-
ристических районов); б - флористических районов; в - микрорайонов. Районы:
Т - Телецкий; Б - Балыкчинский; Ш - Шавлинский; Ч - Чульчинский; У - Узу-
ноюкский; Я - Язулинский; Д - Джулукульский. Микрорайоны: 1 - Яйлинский;
2 - Кокшинский; 3 - Колюштинский; 4 - ТушкемскиЙ; 5 - Белинский; 6 - Кай-
ринский; 7 - Верхнекыгинский; 8 - Каякский; 9 - Деринкульский; 9а - Кулу-
зунский; 10 - Чакрымский; 11 - Шавлинский; 12 - Онгурашский; 13 - Мейду-
гемский; 14 - Среднекулашский; 15 - Казактушкенский; 16 - Сайгонышский;
17 - Садонкаинский; 18 - Кумыйский; 19 - Каратумышский; 20 - Яхансорин-
ский; 21 - Боошконский; 22 - Кюнтюштуксинский; 23 - Караизекский; 24 -
Мыярыкский; 24а — Каракемский; 25 - Верхнеколбакаинский; 26 - Узуноюк-
ский; 27 - Южнотетыкольский; 28 - Каязинский; 29 - Джулукульский; 30 -
Южночулышманский; 31 - Таскыльский; 32 - Верхнемакатинский; 33 - Южно-
шапшальский. Заштрихованы эталонные микрорайоны.
103
С учетом всего вышеизложенного мы выделили 13 флористически
наиболее ценных микрорайонов Алтайского заповедника (рис. 7).
В этих фитохориях (вместе с пятью небольшими заповедными участками
долины р. Чулышман, относящимися к четырем особым микрорайонам)
на площади, составляющей около 40% от 881 238 га территории заповед-
ника, произрастает 97 % известных в заповеднике видов сосудистых рас-
тений, все 33 вида флоры, рекомендуемых к государственной охране,
почти все неморальные реликты и алтае-саянские эндемы заповедника.
Кроме того, эти микрорайоны могут являться эталонами семи флористи-
ческих районов, пяти округов и двух подпровинций Алтае-Саянской про-
винции. Здесь заложено три высотных профиля, более 50 постоянных
пробных площадей, феномаршруты и т. д., где проводится изучение се-
зонной, разногодичной и многолетней динамики растительных объектов.
Выделенные микрорайоны (рис. 7) подлежат самой строгой охране в ка-
честве особо ценных и эталонных.
В заключение следует отметить, что в Алтайском заповеднике пред-
стоит еще большая работа по накоплению и особенно синтезу флористи-
ческих, фитоценологических, зоологических и экологических данных для
более углубленного познания структуры и динамики биоты охраняемой
территории.
Л. В. Табака
НЕКОТОРЫЕ ИТОГИ СРАВНИТЕЛЬНОГО ИЗУЧЕНИЯ ФЛОРЫ
РАЗЛИЧНЫХ ПРИРОДНО-ТЕРРИТОРИАЛЬНЫХ
ПОДРАЗДЕЛЕНИЙ ЛАТВИИ
Институт биологии АН ЛатвССР
Приступая в 1971 г. к планомерному изучению флоры высших сосу-
дистых растений Латвии, в том числе ее инвентаризации, выявлению рас-
пространения видов и картированию ареалов, в лаборатории ботаники
Института биологии АН Латвийской ССР была разработана оригинальная
сеточная основа (сетка квадратов) для всей территории республики (см.
рисунок). Каждый квадрат представляет собой прямоугольник со сторо-
нами 7.5 в. д. и 5 с. ш., площадью 70.7 км2. В качестве кода квадрата
принято двойное обозначение соответственно оси координат. Общее чис-
ло квадратов — 1017, из них 822 квадрата целиком расположены на тер-
ритории республики, 195 квадратов — частично.
Детальная инвентаризация флоры осуществляется в рамках природ-
ных (геоботанических) районов, которых в Латвии выделено 8 (Флора
и растительность Латвийской ССР, 1977, 1979, 1982, 1985). Видовой
состав флоры каждого квадрата обследуется по биотопам, списки ви-
дов составляются как дифференцированные (для каждого биотопа),
так и общие. Для каждого гербарного экземпляра заполняется перфо-
карта (учетная карточка) с обязательным указанием номера квадрата,
104
205
в котором вид обнаружен, а также некоторых экологических и фитоце-
нологических показателей.
В ходе инвентаризации флоры изучается видовое разнообразие (ви-
довая насыщенность) на различных по площади эталонных участках —
квадратах сетки (малые квадраты), группах малых квадратов и сериях
малых квадратов в различных регионах Латвии.
В каждом исследованном геоботаническом районе были заложены
крупные эталонные участки, состоящие из 9 (3 X 3) малых квадратов
(т. е. группы малых квадратов). В первоначальном варианте число ма-
лых квадратов достигало 16 (4X4). Был поставлен вопрос: сможет ли
видовой состав флоры крупного эталонного квадрата, рассматриваемого
нами в качестве конкретной флоры, отразить характер флоры геоботани-
ческого района в целом?
За основу изучения видового разнообразия флоры и видовой насы-
щенности берется один малый квадрат (базовый). Основа методики
заключается в постепенном расширении сферы интенсивного флористи-
ческого поиска в малом квадрате и за его пределами до тех пор, пока
прирост списка видов практически не прекратится.
В настоящее время изучено 10 конкретных флор (КФ) в пяти геобо-
танических районах. В ходе постепенного наращивания площади базово-
го малого квадрата оказалось, что в условиях Латвии число видов прак-
тически перестает увеличиваться на рубеже 600 км2 и, очевидно, именно
здесь начинается „мертвая зона”, размеры которой заранее неизвестны.
При тщательном изучении этой зоны прирост числа видов исключительно
мал и происходит за счет случайных находок редких, а также некоторых
заносных и сегетальных видов.
Таким образом, нами было высказано предположение, что площадь
выявления КФ в изученных геоботанических районах Латвии находится
в пределах600—650 км2.
Изучение видовой насыщенности КФ показало, что она неодинакова
и колеблется от 72% до 85 % от общего видового состава соответствую-
щего геоботанического района. Систематическая структура ведущих се-
мейств аналогична систематической структуре флоры района, имеет
место незначительное порядковое перемещение некоторых семейств, что
объясняется существованием локальных условий.
Для условий Латвии также были выявлены некоторые флористиче-
ские показатели по разновеликим площадям — 50 км2, 70 км2, 140 км2 и
т. д. до 630 км2 — с целью определения зависимости числа видов от пло-
щади. Были проанализированы данные видовой насыщенности 14 малых
квадратов, достаточно равномерно разбросанных по территории респуб-
лики. Эти одинаковые по площади квадраты рассматриваются как вы-
борки, по которым можно судить о составе более крупных флористиче-
ских выделов. В пределах Приморской низменности (район 1) в различ-
ных по физико-географическим условиям участках были заложены 4 ма-
лых квадрата. В этих квадратах, площадь которых одинакова (70.7 км2),
выявлено 510—670 видов, что составляет 50—60% от общего числа видов
района.
Наиболее представительным по видовой насыщенности оказался
106
квадрат, расположенный в окрестности курорта Кемери и отличающийся
разнообразием природных условий, а вместе с тем и весьма большим
многообразием типов местообитаний, в том числе специфических. Этим
объясняется исключительно высокая видовая насыщенность (более
700 видов) на площади 50 км2. Общее число видов этого квадрата со-
ставляет 2/з видового состава флоры Приморской низменности.
Наименее насыщенным видами (450 видов) оказался квадрат, распо-
ложенный на приморских песках побережья Балтийского моря, где
основная роль принадлежит сосновым лесам, верховым и бедным в видо-
вом отношении низинным болотам. Детальное изучение флоры малых
квадратов Приморской низменности (район 1) показало, что в каждом из
них встречается 46—59% видов от общего числа видов этого геоботаниче-
ского района. Общее число видов во всех четырех квадратах достигает
почти 70% от общего числа видов флоры района в цепом. Можно предпо-
ложить, что этот показатель оказался бы более высоким (порядка 80—
90%) при условии исследования флоры несколько большего числа малых
квадратов этого района, заложенных в неодинаковых по природным
условиям участках (например, разных геоботанических микрорайонах).
Представляется, что в условиях Латвии для выявления основных
флористических показателей природного района более целесообразно
заложение серии малых квадратов, чем одного крупного эталонного
участка, рассматриваемого в качестве КФ. Подобный подход к изучению
флоры диктуется также значительной степенью освоенности территории
Латвии. Именно этот момент может повлечь за собой неоправданное уве-
личение площади выявления КФ.
Исследование флоры Латвии методом сеточного картирования пока-
зало, что разработанную нами сеть квадратов можно рассматривать как
систему регулярных проб флоры и к ней вполне применима методика
изучения КФ.
Сеточное картирование флоры оказалось весьма целесообразным не
только при проведении сравнительно-флористических исследований, но
и для хорологических, в первую очередь для показа данных картирова-
ния способом представительства на сеточной основе. Полученные таким
образом данные весьма успешно используются при обобщении материала
для карт „Atlas Florae Europaea”, каждая ячейка которых включает почти
35 латвийских квадратов.
Л. В. Марина
СРАВНИТЕЛЬНЫЙ АНАЛИЗ ФЛОР РЕЧНЫХ БАССЕЙНОВ
И ИХ ЭКОТОПОЛОГИЧЕСКОЙ СТРУКТУРЫ
Висимский государственный заповедник
Для решения многих задач сравнительной флористики необходимо
изучение внутриландшафтной структуры флор (Юрцев, Семкин, 1980;
Юрцев, 1982); одиако методика подобных исследований разработана не-
достаточно.
107
Изучая высокогорные флоры двух соседних хребтов Восточного
Алтая — Куркуре и Тетыколь — дифференцированно по внутриланд-
шафтным подразделениям разного ранга (речным бассейнам, типам и
классам экотопов), мы попытались выяснить некоторые методические
вопросы: насколько флоры речных бассейнов отражают видовой состав
и структуру флоры всего хребта; могут ли они служить в качестве опор-
ных единиц при сравнительно-флористических исследованиях; какова
экологическая подоплека различий флор речных бассейнов, парциальные
флоры каких экотопов обусловливают эти различия в большей степени,
каких — в меньшей?
Высокогорную флору каждого изученного нами хребта, представ-
ляющего единый, литологически однородный массивный конутр пло-
щадью около 500 км2, можно рассматривать по однородности условий
и протяженности как конкретную флору (рис. 1). Водосборные бассей-
ны рек первого порядка,1 площадью 36 км2 в среднем, представляют
крупные метамерные подразделения территории конкретной флоры,
в каждом из них закономерно повторяются основные типы экотопов
всего хребта (Марина, 1982,1983). Высокогорья этих хребтов разделены
долиной р. Шавлы, лежащей в лесном поясе, но через ее верховья сбли-
жены в единую систему с другими высокогорьями.
На хр. Куркуре были детально изучены флоры 10 модельных речных
бассейнов из разных частей хребта, на хр. Тетыколь — 5. Маршруты
в каждом бассейне проходили однотипно, пересекая последовательно
все крупные формы рельефа (днища троговых долин, их борта разной
экспозиции и прилегающие к ним водораздельные гривы; днища, стенки
и гребни каров), а на каждой из них — все типы экотопов в некоторой
повторности. Длина маршрутов в каждом речном бассейне составила
в среднем 2.4 км на 1 км2 площади. Флора одного бассейна изучалась
в течение 5—7 сут, некоторые бассейны, кроме того, посещались неодно-
кратно в течение вегетационного периода с начала июля по конец августа
(Марина, 1982,1983).
Во флоре высокогорий хр. Куркуре зарегистрировано 415 видов со-
судистых растений, на хр. Тетыколь — 368, общих видов — 330. В одном
бассейне встречается на хр. Куркуре от 213 до 282 (в среднем 252) ви-
дов, на хр. Тетыколь — от 240 до 290 (в среднем 262). Это составляет
в среднем 61—71% от высокогорной флоры соответствующего хребта,
при этом каждый речной бассейн имеет во флоре от 1 до 13 видов, не
встреченных ни в одном другом бассейне. Вполне очевидно, что обследо-
вание одного речного бассейна недостаточно для более или менее исчер-
пывающего выявления видового состава флоры хребта.
Изученные речные бассейны значительно различаются между собой
по площади (от 11 до 62 км2), однако, как показали расчеты, прямой
зависимости числа зарегистрированных видов от площади бассейна, а
также от степени его изученности, т. е. отношения длины маршрутов,
пройденных при обследовании флоры, к площади, не наблюдается. Так,
1 Имеются в виду наименьшие постоянные водотоки, которые могут быть на-
званы реками.
108
Рис. 1. Схема высокогорной части хребтов Куркуре и Тетыколь.
Реки: ТУ - Туракая, КА - Каяк, КАН - Кандык, У - Угрюмая, КУ - Кулузун,
КАТ - Катуярык, МК - Малый Куркуре, БК - Большой Куркуре, КЮ - Кюнтюш-
туксу, Л - Лунная, Т - Тургенсу, СК - Средний Кулаш, К - Керексу, ВК - Верх-
ний Кулаш, С - Садагартем. Прерывистая линия - граница высокогорной части
хребтов.
флора бассейна р. Керексу площадью 27 км2 насчитывает 248 видов,
а бассейна р. Средний Кулаш площадью 57 км2 — 262 (при одинаковой
степени изученности — 3.3 и 3.7 км маршрутов на 1 км2), т. е. увеличе-
ние площади бассейна в 2 раза не дает существенной прибавки видов.
А один из самых небольших по площади бассейн р. Кулузун (12 км2)
имеет на хр. Куркуре самую богатую флору — 282 вида. При достаточно
полной обследованности флоры различия в видовом богатстве обуслов-
лены естественными причинами: большей или меньшей развитостью
отдельных типов экотопов, различным соотношением высотных поясов
растительности.
Сравнение видового богатства всех 15 флор речных бассейнов между
собой показало достаточно высокий уровень сходства: 71.9—86.7%1 на
хр. Куркуре, 81.8-86.9% на хр. Тетыколь, 703-84.6% между речными
бассейнами обоих хребтов. Это подтверждает правомочность рассмотре-
ния высокогорных флор каждого хребта в качестве конкретной флоры
1 Здесь и далее используется мера сходства Съеренсена-Чекановского.
109
A
СК т вк
Рис. 2. Дендрограммы сходства флор речных бассейнов между собой и с флорой
соответствующего хребта, совмещенные с диаграммами.
А — по географической структуре; группа: 1 — азиатская, 2 — азиатско-американ-
ская, 3 — евразиатская, 4 - голарктическая; Б - по высотно-зональной структуре;
группа: 1 - альпийская, 2 - арктоалытийская, 3 - монтанная, 4 - горно-равний-
ная; ФК - флора хр. Курку ре; ФТ - флора хр. Тетыколь. Остальные обозначения
как на рис. 1.
и позволяет предположить, что обе они принадлежат к одному элемен-
тарному флористическому району в понимании Б. А. Юрцева (1975а).
Далее мы проанализировали таксономическую, географическую и
высотно-зональную структуру флоры каждого бассейна и сравнили их
между собой и с соответствующей структурой высокогорной флоры
того же хребта (рис. 2).
ПО
6
МК ТУ КУ КАИ БК
ВК ФТ СК
К 7 С
Для таксономических спектров сравниваемых флор характерно
полное совпадение набора 10 ведущих семейств. Семейства Asteraceae и
Сурегасеае занимают почти во всех флорах 1-е и 2-е места по числу видов.
Остальные семейства в некоторых флорах незначительно меняются
местами в пределах соседних пар. Для более точного сравнения систе-
матической структуры мы использовали меру сходства Съеренсена,
модифицированную для весовых множеств Б. И. Семкиным (1973),
которая в случае процентных спектров вычисляется как сумма ми-
нимальных весов (%) соответствующих семейств в сравниваемых
флорах. Сравнение процентных спектров 10 ведущих семейств флор
речных бассейнов с процентным спектром соответствующего хребта
111
при помощи этого коэффициента показало очень высокое сходство:
от 92.8 до 97.5%.
Анализ флор по составу и соотношению четырех географических
групп видов (голарктическая, евразиатская,- азиатско-американская,
азиатская), а также подгрупп самой крупной азиатской группы обнару-
жил еще более высокое сходство флор речных бассейнов с флорой всего
хребта, места всех групп и даже подгрупп в нисходящем ряду совпадают,
а сравнение процентных спектров при помощи упомянутого выше коэф-
фициента обнаруживает сходство от 92.4 до 985 % (рис. 2, Л).
По соотношению высотно-зональных групп (рис. 2, Б) сходство так-
же высокое, но амплитуда варьирования коэффициента несколько ши-
ре — от 83.4 до 98.8%, что отражает неодинаковую представленность
разных поясов в сравниваемых бассейнах, при этом высотно-зональные
группы в некоторых флорах имеют различный порядок расположения
в нисходящем ряду. ’
Таким образом, флора каждого речного бассейна по своей таксоно-
мической, географической и высотно-зональной структурам близка
к флоре всего хребта. По-видимому, в высокогорьях Южной Сибири
(при условии однородности литологического состава, геоморфологиче-
ского строения и растительного покрова) флора одного речного бассей-
на, включающего все пояса выше границы леса, может служить моделью
флоры всего хребта (не слишком протяженного). При выборе модель-
ного речного бассейна необходимо учитывать следующее: площадь бас-
сейна должна быть достаточно большой, чтобы были хорошо представ-
лены все растительные пояса, а также все основные типы экотопов,
свойственные данному району.
Сравнение высокогорных флор речных бассейнов в пределах одного
хребта по видовому составу, таксономической, географической и высот-
но-зональной структурам (Марина, 1982, 1983) показало определенную
группировку соседних бассейнов по каждому из названных показателей
(при высоком общем уровне сходства) ; однако группировки флор бас-
сейнов по разным показателям, как правило, совпадают лишь частично.
Таким образом, высокогорную флору каждого изученного хребта нельзя
однозначно разделить на „микрорайоны” по сумме флористических при-
знаков.
Рассмотрим теперь закономерности распределения видов по терри-
тории конкретной флоры на примере хр. Куркуре. Все виды мы разде-
лили на 5 классов константности по встречаемости их в 10 речных бас-
сейнах. В 1 -й класс вошли виды, встреченные в одном или двух бассей-
нах, во 2-й — в трех или четырех и т. д. В данном случае константность
является приближенным выражением географической активности видов
на данной территории (Юрцев, 1982а).
Самым богатым по числу видов оказался 5-й класс константности
(виды, встреченные в 9—10 речных бассейнах) — 38.6%; почти вдвое
меньше видов в 1-м классе — 20.7%; еще меньше в остальных: во 2-м —
14.6%, в 3-м- 12.3 %, в 4-м- 13.8 %.
Интересным оказалось распределение географических и высот-
но-зональных групп по классам константности (см. таблицу). Доля
112
I Распределение географических и высотно-зональных групп флоры
хр. Куркуре по классам константности (% от видов данного класса
I константности от числа видов группы)
Группы видов Класс константности*
1 2 3 4 5
Географические
голарктическая 34.6 24.6 25.0 35.2 285
евразиатская 25.9 24.6 20.8 18.5 13.9
азиатско-аме ри канская 25 7.0 4.2 5.6 2.0
азиатская 37.0 43.8 50.0 40.7 55.6
Высотно-зональиые
альпийская 11.1 21.1 25.0 20.4 25.2
арктоальпийская 12.3 14.0 10.4 20.4 25.2
монт энная 27.2 31.6 25.0 16.7 21.9
горно-равнинная 49.4 33.3 39.6 42.5 205
* Классы константности: 1 — виды встречены в 1—2 речных бассейнах, 2 —
в 3-4, 3 —в 5-6,4 —в 7-8, 5 -в 9—10 бассейнах.
голарктических видов хотя и изменяется по классам, однако изменения
не выходят за пределы амплитуды случайного варьирования. Достовер-
ность различий определялась на основе критерия х2: при Р=0.05 крити-
ческое значение х2 должно быть не меньше ЗВ4 (Рокицкий, 1973).
В первом случае значение х2 равно 0.92. Доля евразиатских видов досто-
верно снижается от 1-го класса к 5-му (х2 = 5.13) . Азиатско-американ-
ская группа при общей малочисленности изменяется по классам слу-
чайно. Содержание азиатских видов возрастает от 1-го класса к 5-му
(37.0—55.6%) хотя и скачками, но достоверно (х2 = 7.29) .
Весьма значительно повышается от 1-го класса к 5-му доля альпий-
ских (от 11.1 до 32.4%), менее резко — арктоальпийских видов (от 123
до 25.2); значение х2 соответственно равно 12.3 и 5.6. При этом очень
существенно снижается доля горно-равнинных видов — от 49.4 до 205 %
(Х2 = 20.6). Некоторое снижение монтанных видов не достоверно
(Х2 = 0В2).
Таким образом, среди видов спорадически распространенных в пре-
делах высокогорий хр. Куркуре относительно повышена доля горно-
равнинных видов и видов с евразиатским типом ареала, т. е. в разных
частях хребта в высокогорья заходят из лесного пояса неодинаковые
виды. Для широко распространенных видов характерно более высокое
содержание альпийских и арктоальпийских видов преимущественно
с азиатским типом ареала, т. е. данный комплекс имеет более высокую
устойчивость и активность в высокогорьях хребта. Это еще раз под-
тверждает целостность высокогорной флоры хребта.
Флора речного бассейна, как и конкретная флора, является гетеро-
генной системой, состоящей из большого числа парциальных флор (ПФ)
различных, часто весьма контрастных экотопов (Юрцев, Семкин, 1980) .
8 Заказ 1220
ИЗ
Мы изучили распределение видов по экотопам на примере 3 модельных
речных бассейнов: Кулузун (хр. Куркуре), Средний Кулаш и Садагартем
(хр. Тетыколь) . На первом этапе работы было выделено 49 типов экото-
пов, отличающихся по месту в рельефе, характеру субстрата и раститель-
ности. Для каждого типа экотопов составлялись флористические списки
в 2—10-кратной повторности. Таким образом, ПФ каждого типа эк0топа
представляет объединение ПФ конкретных контуров однотипных эко-
топов. Далее для лучшей обозримости и удобства интерпретации мы
объединили близкие типы экотопов по сходству их местоположения и
растительности в классы экотопов. В результате получилось И классов
экотопов, из которых 7 приурочены к субальпийскому поясу, 4 — к аль-
пийскому. Набор классов экотопов в изученных речных бассейнах оди-
наков, за исключением редкого класса экотопов ксеротермной расти-
тельности щебнисто-дресвяных обнажений, обнаруженного в верхней
части субальпийского пояса бассейна р. Садагартем, однако в нем содер-
жится лишь 5 видов, не отмеченных в других классах экотопов этого
бассейна.
Для выяснения экотопологической структуры флоры каждого реч-
ного бассейна мы определили отношения сходства между ПФ всех клас-
сов экотопов внутри бассейна. Как и следовало ожидать, сходство оказа-
лось невысоким, поскольку классы экотопов экологически довольно
резко отличаются друг от друга: в бассейне р. Садагартем значение меры
сходства варьирует от 8.1 до 67В%, в бассейне р. Средний Кулаш — от
13.8 до 61.9%, в бассейне р. Кулузун — от 3.9 до 61.8%. На основании
матрицы мер сходства были построены дендриты и выделены плеяды,
показывающие отношения максимального сходства ПФ разных классов
экотопов (рис. 3).
Дендриты наглядно показывают, насколько своеобразна экотополо-
гическая структура флоры каждого бассейна, в особенности р. Кулузун.
Однако при анализе дендритов обнаруживаются плеяды из ПФ, имеющих
повышенное сходство, причем во всех речных бассейнах одинаковые по
составу. В них входят ПФ классов экотопов ерников (IV), тундр (IX) и
овсяницево-кобрезиевых лугов (X) . Последний класс экотопов в бассей-
не р. Кулузун не входит в плеяду, но как показывают расчеты, мера
включения его ПФ в ПФ класса экотопов тундр (IX) очень высока — 90%,
что при бедности видового состава ПФ овсяницево-кобрезиевых лугов
в бассейне р. Кулузун свидетельствует о производности ее от ПФ тундр.
В обоих речных бассейнах хр. Тетыколь имеют повышенное сходство
между собой и тесно примыкают к этой плеяде ПФ других альпийских
классов экотопов: разреженных группировок стенок каров (XI) и аль-
пийских лугов (VIII). Классы экотопов субальпийского пояса в этих
речных бассейнах имеют между собой низкое сходство. В бассейне р. Ку-
лузун, напротив, классы экотопов субальпийского пояса теснее связаны
между собой, чем классы экотопов альпийского.
Во всех речных бассейнах обособлена от других ПФ класса экотопов
болот (II).
Для выявления сходства набора видов на однотипных экотопах
в пределах 3 речных бассейнов мы провели их попарное сравнение.
114
a
Рис. 3. Дендриты сходства и корреляционные плеяды классов экотопов по видо-
вому составу их парциальных флор.
Бассейны рек: а - Садагартем, б - Средний Кулаш, в - Кулузун. Классы экотопов
субальпийского пояса: I - прибрежных травянистых кустарников, II - высокогор-
ных болот, III - травянистых ивняков и высокотравных лугов, IV — ерников,
V - петрофитных ксеротермных кустарниково-травянистых группировок южных
склонов, VI - петрофитных кустарничково-травянистых группировок северных
склонов, VII - ксеротермной травянистой растительности щебнисто-дресвяных
обнажений; альпийского пояса: VIII - альпийских лугов, IX - тундр, X - овсяни-
цево-кобрезиевых лугов, XI - разреженных группировок стенок каров. Арабские
цифры — значения меры сходства парциальных флор, %.
115
Сходство оказалось невысоким (от 32.5 до 77.7%) по сравнению со
сходством флор речных бассейнов в целом (79 В—843%). Это объясня-
ется, по-видимому, тем, что в ПФ небольших и относительно гомогенных
контуров бывает представлена лишь часть характерных для экотопа ви-
дов, по мере расширения спектра экотопов и объединения их ПФ во фло-
ре бассейна видовой состав стабилизируется и сходство повышается.
Флористическое сходство однотипных классов экотопов между бас-
сейнами рек одного хребта оказалось почти всегда выше, чем каждого
из них с бассейном другого хребта, за исключением следующих классов
экотопов субальпийского пояса: высокогорных болот, петрофитных
группировок южных и северных склонов.
Все речные бассейны оказались в наибольшей степени сходны по
ПФ следующих классов экотопов: ивняков и высокотравных лугов
(66— 78%), альпийских лугов (66—75%), тундр (66—71%), ерников
(61—67 %) . Это классы экотопов с хорошо сформированными раститель-
ными сообществами, эдификаторы которых, обладая высокой конку-
рентной способностью, ограничивают проникновение случайных видов.
Поэтому видовой состав их ПФ в пределах одного флористического
района более стабилен. Высокое флористическое сходство имеет и самый
гетерогенный класс экотопов разреженных группировок стенок каров
(62—75 %), в котором за счет объединения ПФ разных типов экотопов
видовой состав в целом стабилизируется. Минимальное сходство в срав-
ниваемых бассейнах выявилось между ПФ экологически своеобразного
класса экотопов высокогорных болот (45—51 %), а также класса экото-
пов петрофитных группировок южных склонов субальпийского пояса
(41—54%), характеризующегося несомкнутой растительностью, в кото-
ром из-за снижения конкуренции со стороны активных, приспособлен-
ных к холодным условиям высокогорий видов облегчено внедрение
других — самых различных видов, особенно из верхней части лесного
пояса.
При расчете меры сходства ПФ однотипных классов экотопов все
виды рассматривались нами как равноценные. Однако, как показал
Б. А. Юрцев (1983а), дифференциальные виды, т. е. виды, присутствую-
щие только в одной из сравниваемых флор, имеют различную индика-
торную ценность в зависимости от специфичности и константности их
в рассматриваемых флорах. Мы проанализировали дифференциальные
виды всех классов экотопов бассейнов рек Садагартем и Средний Кулаш.
Их можно подразделить на специфичные (не встречающиеся в данном
речном бассейне в других классах экотопов) и неспецифичные, а каждую
из полученных групп видов — на константные в данном классе экотопов
и редкие виды. Наибольший вес как дифференциальные элементы имеют
специфичные и одновременно константные виды, наименьший — неспе-
цифичные и редкие (Юрцев, 1983а) . Относительно высокая доля специ-
фичных и одновременно константных дифференциальных видов отме-
чена в классах экотопов, имеющих низкое флористическое сходство:
высокогорных болот, петрофитных группировок южных склонов, при-
брежных травянистых кустарников. В остальных классах экотопов та-
ких видов мало (1—5) или совсем нет, т. е. среди дифференциальных
116
видов преобладают неспецифичные, зашедшие случайно из других клас-
сов экотопов, поэтому реальное сходство этих ПФ выше, чем это следует
из меры сходства Съеренсена—Чекановского.
Таким образом, раздельное изучение флоры о'боих хребтов по их
внутриландшафтным подразделениям показало’ что основные показате-
ли структуры флоры (таксономической, географической, высотно-зо-
нальной) , характерные для достаточно крупных выделов районирования,
выявляются на уровне флор речных бассейнов, если те включают
основное разнообразие классов экотопов, свойственное данной террито-
рии. Это позволяет рекомендовать флоры речных бассейнов площадью
в несколько десятков квадратных километров для апробации в качестве
опорных единиц при выявлении и сопоставлении структуры флор различ-
ных в ботанико-географическом отношении территорий.
В то же время анализ экотопологической структуры флор речных
бассейнов выявил значительные различия между парциальными флорами
однотипных классов экотопов, которые касаются не только состава ред-
ких видов, но в некоторых классах экотопов также специфичных для
данного экотопа видов, а также константных в нем. Это говорит об изве-
стной автономности формирования высокогорных флор речных бассей-
нов даже в пределах одного хребта.
Я. П. Дидух
ОПЫТ СТРУКТУРНО-СРАВНИТЕЛЬНОГО АНАЛИЗА
ГОРНЫХ ЭЛЕМЕНТАРНЫХ ФЛОР
Институт ботаники им. Н. Г. Холодного АН УССР
Элементарные флоры (ЭФ) можно разделить на 4 типа, отличающих-
ся друг от друга степенью дифференциации экотопов, их нарушенностью,
выраженностью границ с соседними ЭФ и т. д. (в связи с чем ставятся
различные задачи при их изучении и применяются разные методы иссле-
дования) : 1) ЭФ однообразных и малонарушенных равнинных террито-
рий тундровой и большей частью лесной зон, 2) ЭФ в значительной степе-
ни „окультуренных” территорий лесостепной, степной и отчасти лесной
зон, 3) ЭФ островов, 4) ЭФ горных территорий.
При изучении последних не столь важны вопросы о длине, направлен-
ности и плотности маршрутов, степени повторности посещения однотип-
ных экотопов, а наиболее значимыми представляются вопросы понима-
ния объема ЭФ, определения их границ, изменения дифференциальных
и к о дифференциальных видов в зависимости от высотных и долготных
градиентов. Маршруты в горах желательно прокладывать во все доступ-
ные места, чтобы осмотреть все разнообразие экотопов. Большая пестро-
та и дифференциация экотопов в горах является причиной того, что
в одной ЭФ на однотипных экотопах при наличии сходного в целом видо-
вого состава появляются новые виды. Такие виды обычно являются
117
редкими и распространены локально, в связи с чем для их выявления
необходимо исследование всех кажущихся однотипными экотопов.
Таким образом, одно из „правил” А. И. Толмачева, согласно которо-
му в пределах одной ЭФ на однородных экотопах встречается одинако-
вый набор видов, в горах довольно часто нарушается и становится непри-
емлемым.1 Выявление редких видов очень важно: хотя они и не влияют
на общие статистические показатели, но играют существенную роль в ка-
честве критериев определения ранга естественных флор и их границ.
Актуальным является вопрос объема горной ЭФ. Согласно данным
Р. В. Камелина (1973) и нашему представлению, такая флора должна
включать все горные пояса растительности одного речного бассейна (во-
досбора) , поскольку в пределах отдельных поясов обычно не полностью
выявляются общерегиональные закономерности и такие флоры невоз-
можно использовать в качестве эталонов для сравнения с флорами регио-
нального уровня. Подобные флоры — системы топологического уровня.
Величина речного бассейна также может быть различна, поэтому берется
весь водосбор или же его часть, отличающаяся от других частей появле-
нием качественно новых экотопов или другим их сочетанием. При этом
важно, чтобы в состав ЭФ не включались территории водосборов сосед-
них рек, хотя и это положение не является абсолютным правилом.
Например, на южном макросклоне Крымских гор, где речки достигают
в длину лишь нескольких километров, мы были вынуждены отнести
к одной ЭФ территории водосборов нескольких речек.
Сложным вопросом является и выбор критериев выделения горных
ЭФ. В качестве таковых критериев нами использовались дифференциаль-
ные (эндемичные или с дизъюнктивным распространением, не встречаю-
щиеся на соседних территориях), а также позитивно и негативно кодиф-
ференциальные виды (Юрцев и др., 1980; Шеляг-Сосонко и др., 1982).
Хотя высотные градиенты кодифференциальных видов значительно рез-
че, чем долготные, в качестве критериев мы использовали последние.
При этом первые вовсе не игнорируются; они как бы „проектируются”
на плоскость и таким образом отражают региональные закономерности
изменения флор.
Не менее дискуссионен и сложен вопрос о границах ЭФ, что объясня-
ется локальным распространением дифференциальных и кодифференци-
альных видов в горах. Мы различаем 4 типа границ: 1) выделяемые с по-
мощью флористических критериев (дифференциальные и к о дифферен-
циальные виды или резкие изменения их активности); 2) выделяемые
по ценотическим признакам (смена синтаксонов или соотношения
поясов растительности); 3) выделяемые по географическим признакам
(водоразделам, смене геологических пород, почв, климатическим барье-
рам и т. д.); 4) условные как вследствие их размытости или включения
новых типов ценозов, которые не могут быть отнесены ни к одному из
соседних типов. В качестве последнего типа границ можно привести
1 В более поздних работах А. И. Толмачев (1941, 1970) допускал некоторое
варьирование состава видов на однотипных экотопах в пределах одной конкретной
флоры.
118
крымские яйлы, растительность которых представлена горными степя-
ми, отличающимися коренным образом от соседних лесных типов расти-
тельности южного и северного макросклонов. Выделение яйлы в качест-
ве самостоятельной ЭФ делает ее несопоставимой с другими горными ЭФ
и неподчиненной системе районирования. Нет основания для отнесения
яйлы только к одной из ЭФ южного или северного макросклона и
исключения ее из соседней, так как это существенно сказывается при
сравнении этих ЭФ. В связи с этим мы вынуждены проводить условную
границу через яйлу.
Разнообразие, пестрота и высотная дифференциация горных экото-
пов — причина того, что ЭФ, как правило, сложена из качественно различ-
ных флор разных экотопов, сравнение которых позволяет отразить изме-
нение флористических признаков в зависимости от действия экологиче-
ских факторов. Кроме того, выделение такого типа флористических
систем необходимо, когда при сравнении разновеликих флор горных
регионов (например, Кавказа с Крымом) нужно сопоставить их по набо-
ру поясов и богатству флоры.
Исследование горных ЭФ и составляющих их флористических систем
экотопологического уровня предполагает дальнейший структурно-срав-
нительный анализ флоры. В качестве примера такого анализа рассмотрим
полученные нами данные по Ялтинскому горно-лесному заповеднику.
Ялтинский заповедник занимает западную часть южного макроскло-
на и вершину Главной крымской гряды. Границы его неестественны,
поэтому мы рассматриваем заповедник лишь как достаточно репрезен-
тативную „пробу”, отражающую закономерности растительного покрова
ЭФ, т. е. в качестве конкретной флоры (14 230 га) (Шеляг-Сосонко,
1980) . Растительность заповедника довольно разнообразна и образует
4 высотных пояса: 1) высокоможжевеловые, пушистодубовые леса и
саванноиды (от побережья моря до высоты 400—450 м); 2) крымско-
сосновые леса с участием скальнодубовых (от 400—450 м до 800—950 м
над ур. м.); 3) буковые и грабовые леса с участием лесов из сосны
Сосновского (от 800—950 м до 1000-И 200 мнад ур.м.); 4) пояс горных
степей и томилляров (вершина Главной гряды). Кроме того, встреча-
ются участки наскальных группировок и луговой растительности (Ше-
ляг-Сосонко, Дидух, 1980).
Первый этап структурного анализа предполагает вычленение фито-
мер1 — флористических систем экотопологического уровня, названных
ценофлорами [соответствуют объединениям парциальных флор микро-
экотопов в понимании Б. А. Юрцева (1982а) ]. Поскольку растительный
покров в горах характеризуется большой мозаичностью и дифференциа-
цией, то на практике мы выделяли и изучали „пробы” ценофлор, в до-
статочной мере отражающие их ботанико-географические черты в преде-
лах ЭФ. Пробы выделялись по следующей методике. На местности
проводились геоботанические описания пробных площадок, охватываю-
щих все синтаксономическое разнообразие. Видовой состав площадок
1 Аналогично понятиям „фитоценомер” и „фитоценохор”, введенным В. Б. Со-
чавой (1979).
119
Рис. 1. Эмпирическое распределение коэффициентов видового сходства ассоциаций
сосновых лесов Горного Крыма.
1,2- кумулята коэффициентов видового сходства ассоциаций сосновых лесов
[7 - Pineta sosnovskyi(Pps),2 - Pineta pallasianae (Ppp)]; 3, 4 - гистограмма коэф-
фициентов видового сходства ассоциаций сосновых лесов (5 - Pineta sosnovskyi,
4 - Pineta pallasiana).
сравнивался попарно по формуле Стугрена—Радулеску и при помощи
ЭВМ вычислялся коэффициент видового сходства.1 Полученные коэффи-
циенты образуют довольно непрерывный ряд, в связи с чем визуально
трудно, а порой невозможно обнаружить пороговый коэффициент, что
осложняет выделение проб. Такая проба должна включать те геоботани-
ческие описания, видовой состав которых более сходен друг с другом,
чем с описаниями площадок других формаций. Поэтому пороговый
коэффициент выбирался произвольно, путем расширения объема выбор-
ки до тех пор, пока она не объединила большинство описаний синтак-
сонов. Построение гистограммы коэффициентов сходства показало, что
модальный класс значений коэффициентов различных ассоциаций форма-
ции лежит в пределах р = 0.6-^0.7, а пороговый, при котором большин-
ство пробных площадок объединятся в плеяду, р = О.1-^О.З и зависит
от количества этих площадок. Обычно число рассматриваемых значений
1 Значения коэффициента Стугрена-Радулеску изменяются от -1 (все виды
общие) до +1 (нет общих видов), и, строго говоря, этот коэффициент представ-
ляет собой относительную меру различия двух множеств. (Прим, ред.).
120
Рис. 2. Граф сходства проб ценофлоры лесов сосны крымской по видовому составу.
а - площадки описаний, б - границы ценофлор, в - допороговые значения коэф-
фициентов Стугрена-Радулеску при попарном сравнении площадок по составу
видов (Рпор — 0.2 4-0.3), г - самые низкие для данной площадки значения коэффи-
циентов Стугрена-Радулеску, лежащие выше порога.
допороговых коэффициентов сходства примерно равно количеству пло-
щадок и, как видно из рис. 1, соответствует началу линейного участка
графика кумуляты эмпирического распределения значений коэффици-
ентов. В дальнейшем на основе допороговых коэффициентов строился
граф. В качестве примера рассмотрим графы сходства проб ценофлор
лесов сосны крымской (рис. 2) и сосны Сосновского (рис. 3).
Анализ рис. 2 показывает, что образовавшаяся плеяда включает те
сообщества, во 2-м древесном ярусе которых содоминируют Quercus ри-
bescens, Q. petraea, Carpinus orientalis и Juniperus oxycedrus, а в травостое —
121
0.02 f 20
Рис. 3. Граф сходства проб ценофлоры лесов сосны Сосновского по видовому составу.
Обозначения как на рис. 2.
122
Рис. 4. Граф сходства проб ценофлор Ялтинского заповедника по видовому со-
ставу.
Ценофлоры: Sv - саванноидная; Pf - петрофитная; Ju - можжевельника высоко-
го; Qp - пушистодубдвых лесов; Qpet - скальнодубовых лесов; Рр - крымско-
сосновых лесов; Ps -'лесов сосны Сосновского; CbFs - грабово-буковых лесов;
TS - томиллярно-степных сообществ; Рг - лугов. Связи, обозначенные сплошной
и прерывистой линиями, показывают значения коэффициента Стугрена-Радулеску
ниже и выше 0.55 соответственно.
Brachypodium rupestre, Carex humilis, Physospermum comubiense и Laser
trilobum. Отделившийся ряд площадок (7, 8, 17,18, 20 и 21), характери-
зующихся густым 2-м ярусом лиственных пород и разреженным траво-
стоем, по флористическому составу стоит ближе к соответствующим
лиственным лесам, чем к типичным сосновым, в связи с чем исключается
из данной ценофлоры.
Анализ рис. 3 свидетельствует, что плеяда включает ассоциации с до-
минированием в травяном покрове Brachypodium rupestre, Carex humilis
и Laser trilobum, а отделившиеся группы характеризуются наличием со-
эдификатора в древесном ярусе Pinus pallasiana (17—20) или Fagus sylva-
tica (21-25) (Дидухи др., 1977).
Дальнейший этап — сравнение плеяд между собой в целом. Посколь-
ку они включают уже набор ассоциаций, то вследствие возрастания видо-
вого их состава границы между ними становятся более размыты, чем
между конкретными описаниями. В результате отдельные плеяды, имею-
щие — 1<р<0 и размытые границы, объединялись в одну ценофлору
(например, степи и томилляры), а ценофлоры, имеющие р<055> —
в один тип. Всего для заповедника выделено 10 ценофлор, объединенных
в 4 типа (рис. 4) (Дидух, 1977а) •
Второй этап структурносравнительного анализа предполагает сопо-
ставление ценофлор по отдельным структурам: систематической, геогра-
фической, биоморфологической и т. д.
123
Рис. 5. Граф максимального сходства ценофлор Ялтинского заповедника по их
систематической структуре.
Обозначения ценофлор как на рис. 4.
Систематическая структура определяется Толмачевым (1970) как
свойственное данной флоре распределение видов между систематиче-
скими категориями высшего ранга. Сравнение систематических структур
флор проводилось методом ранговой корреляции на основе выборки
10 ведущих семейств, представленных во всех флорах. Методика срав-
нения довольно подробно изложена в работах Л. И. Малышева (1972),
В. М. Шмидта (1980) и широко используется многими исследователями,
в связи с чем мы на ней не останавливаемся.1 На основе полученных
коэффициентов ранговой корреляции (т) строился граф (рис. 5). Как
легко убедиться, он в целом близок к графу видового сходства цено-
флор, однако это сходство далеко не полное, что указывает на нетож-
дественность показателей систематической структуры и видового сход-
ства флор. Если закономерности изменения видового сходства в боль-
шей мере зависят от экол ого-климатических параметров (влажность,
высота над уровнем моря) и близости расположения их относительно
друг друга (непосредственность контакта), то соотношение семейств —
от строения (ярусности, сомкнутости и светового режима под пологом)
ценозов. Последнее подтверждается тем, что семейственные спектры
саванноидной, можжевеловой, петрофитной, томиллярно-степной и луго-
вой ценофлор характеризуются более высокими коэффициентами сход-
ства, чем контактирующие пушистодубовые, скальнодубовые, крымско-
сосновые, сосновскососновые и грабово-буковые, которые по система-
тической структуре более сходны между собой. При этом ценофлоры
образуют ряд от саванноидной через гелиофитные можжевеловую, пет-
рофитную, луговую и томиллярно-степную к ценофлорам менее освет-
ленных ценозов крымскососновых и сосновскососновых, далее пуши-
стодубовых и скальнодубовых и, наконец, самых умброфитных — гра-
бово-буковых (Дидух, 19776).
Возможно применение и других методик сравнения (Юрцев, Семкин,
1980).
124
Рис. 6. Граф максимального сходства ценофлор Ялтинского заповедника по их
географической структуре.
Обозначения ценофлор как на рис. 4.
Таким образом, на основе сходства систематических структур
ценофлор еще нельзя судить об их видовом сходстве. Если последнее
свидетельствует о сходстве систем, то первое — о сходстве одной из
структур.
Географическая структура определяется нами как свойственные
флоре количественные соотношения видов, представляющих разные
типы ареалов. Самый сложный момент в изучении географической струк-
туры — отсутствие унифицированной системы типов ареалов видов или
теоретических разработок такой системы, которая позволяла бы в даль-
нейшем группировать ареалы для целей сопоставления. Однако отсут-
ствие такой системы вовсе не говорит об отсутствии закономерностей
в изменении географических структур флор. Не останавливаясь на гео-
графическом спектре флоры заповедника и его ценофлор, отметим, что
для сравнительного анализа, который проводился по такой же методике,
как анализ систематических структур, нами было взято 10 ведущих и
наиболее характерных типов ареалов. Каждому типу присваивался ранг
по мере убывания числа видов, а затем определялись коэффициенты
ранговой корреляции попарно между всеми ценофлорами и строился
граф. При тпор = 0.70 граф разбивается на 5 плеяд, в целом похожих на
плеяды видового сходства (за исключением отделившейся саванноид-
ной ценофлоры) (рис. 6). Отдельные вершины в пределах этих плеяд
более тесно связаны между собой, чем с вершинами соседних плеяд, а
связи между плеядами имеют размытый характер. Граф географических
125
Рис. 7. Граф максимального сходства ценофлор Ялтинского заповедника по 10 (а)
и 15 (6) их биоморфологическим признакам.
Обозначения ценофлор как на рис. 4.
структур ценофлор оказался более сходен с графом сходства видового
состава флор, чем граф систематических структур.
Биоморфологическая структура определяется как свойственное
флоре количественное соотношение видов, распределенных по типам
биоморфологических признаков. В качестве системы биоморф нами
бЬгла использована линейная система В. Н. Голубева (1972) , в которой
признаки не соподчинены и рассматриваются как независимые, что по-
зволяет вести сравнение флор по любому из них. Не останавливаясь на
детальном анализе спектров биоморф заповедника и ценофлор, отметим,
что для сравнения биоморфологических структур нами было взято 10
и 15 основных признаков. Признаки были ранжированы описанным
выше способом, на основе чего были вычислены коэффициенты ранго-
вой корреляции и при пороговом коэффициенте тпор = 0В построены
графы сходства ценофлор по биоморфологической структуре (рис. 7).
Из рисунка видно, что 10 основных признаков уже достаточно репрезен-
тативно отражают характер биоморфологических структур. Например,
биоморфологическая структура можжевеловой ценофлоры очень близка
к таковой петрофитной. Эти 2 ценофлоры представлены растительными
группировками, распространенными в наиболее ксерических условиях
Южного берега Крыма. Значительное отклонение биоморфологической
структуры саванноидов объясняется количественно иным строением
126
его ценозов; при этом биоморфологическая структура и видовой состав
саванноидов имеют большее сходство с таковыми можжевеловой и пет-
рофитной ценофлор, чем с другими. Отдельную и четко очерченную
плеяду образуют лесные ценофлоры с высокими коэффициентами сход-
ства биоморфологических структур (т = 0.75-^0В9). Томиллярно-степ-
ная и луговая ценофлоры значительно отличаются одна от другой и от
остальных по характеру биоморфологической структуры и видовому
составу. Граф сходства биоморфологических структур значительно боль-
ше совпадает с графом сходства видового состава, чем остальные графы.
Это объясняется тем, что биоморфологические признаки — результат
адаптации к эколого-ценотическим условиям, по которым отличаются
выделенные ценофлоры (Дидух, 1978) .
Экологическая структура флоры отражается в количественном соот-
ношении видов, сгруппированных по нормам реакции их на действие
первичных экологических факторов (тепловой, световой, водный ре-
жим, химический состав почвы) . Такая реакция может быть неодинако-
вой и у разных географических популяций, одного вида, в частности
вследствие их конкурентных отношений с окружающими популяциями
других видов. В связи с этим флористы вынуждены использовать отно-
сительную шкалу нормы реакции видов, в которой они ранжируются
друг относительно друга, хотя такая ранжировка в значительной степени
зависит и от принятых крайних точек отсчета, а также от разнообразия
экотопов в данной местности. Нами проведен лишь первый этап изучения
экологической структуры флоры заповедника в целом. При этом виды
классифицировались по нормам реакции их на воздействие света (умбро-
фиты и гелиофиты) , водного и химического режима почвы (ксерофиты,
ксеропетрофиты, мезоксерофиты, мезоксеропетрофиты, ксеромезофи-
ты, ксеромезопетрофиты, мезофиты, гигромезофиты, гигрофиты, гидро-
фиты, галофиты и паразиты).
Ценотическая структура отражается в свойственных флоре коли-
чественных соотношениях объединений видов по их фитоценотической
значимости. Указанные выше фитоценотические параметры опреде-
ляются на основе 3 показателей: широты эколого-ценотической ампли-
туды видов, степени их постоянства и проективного покрытия. Эти
вопросы нашли частичное отражение в нашей работе по активности видов
(Дидух, 1982), а здесь мы вносим некоторые дополнения. По фитоце-
нотической значимости разделяем виды на доминанты, автохтонные и
аллохтонные ассектаторы. По характеру поведения выделяются 3 груп-
пы видов, названные Л. Г. Раменским (1938) виолентами (силовики),
патентами (выносливцы) и эксплерентами (выполняющие) . По направ-
ленности изменения активности виды подразделяются на 4 группы:
экспансивные, преуспевающие, реликтовые и угасающие. По степени
изменения активности виды относим к 5 группам: особоактивные,
высокоактивные, среднеактивные, малоактивные и неактивные (Юрцев,
1968а).
Фенологическая структура отражается в количественном соотноше-
нии видов по фенологическим ритмам развития. Здесь анализируются
ритмы вегетации (кратковременные — 3—5 мес, среднедлительные —
127
6—9 мес и длительные —10—12 мес) , цветения (по временам года или
длительности), созревания или дессиминации плодов и семян.
Структурно-сравнительный анализ флор, несмотря на ряд трудно-
стей, связанных как с недостаточной изученностью части видов, так
и отсутствием единых схем классификации отдельных признаков этих
видов, имеет большое научное и практическое значение, так как, позво-
ляя установить определенные закономерности изменения структур флор,
дает возможность прогнозировать эти изменения, что выводит флори-
стику на новый этап развития этой дисциплины.
В. П. Седельников
ЦЕНОТИЧЕСКАЯ СТРУКТУРА ВЫСОКОГОРНОЙ ФЛОРЫ
АЛТАЕ-САЯНСКОЙ ГОРНОЙ ОБЛАСТИ
Центральный Сибирский ботанический сад СО АН СССР
При геоботаническом изучении высокогорной растительности Алтае-
Саянской горной области, а также при ее классификации неизбежно
встала проблема выделения ценофлор (формационных флор) как исто-
рически и ценотически гомогенных групп видов. Перспективность ис-
пользования подобных групп в комплексе с геоботаническими характе-
ристиками для классификации растительности убедительно показана
в работах П. Н. Овчинникова (1947) и Р. В. Камелина (1979).
Основой выделения ценофлор в геоботанических работах является
объединение флористических списков однотипных фитоценозов, каждый
из которых может рассматриваться как парциальная флора (ПФ) низ-
шего ранга (Юрцев, 1982а). Поэтому ценофлоры, выделяемые геобота-
никами, по сути являются обьединенйями однотипных ПФ в понимании
Б- А. Юрцева. Исходным материалом для данного сообщения послужили
как собственные, так и имеющиеся в литературе геоботанические описа-
ния высокогорной растительности Алтае-Саянской горной области. Всего
было использовано около 1800 описаний. Обработка материала проводи-
лась по следующей схеме: 1) объединение флористических списков одно-
типных описаний легко различаемых в природе элементов растительно-
сти в условную ценофлору; 2) сравнение и оценка условно выделенных
ценофлор; 3) разбиение выделенных ценофлор по взвешенным призна-
кам (в данном случае проективное покрытие видов) .
Всего в используемых описаниях было зарегистрировано 517 видов
высших сосудистых растений, которые по описанному выше способу
были распределены по следующим 16 ценофлорам: 1) щебнистых тундр
(84 вида), 2) дриадовых тундр (119), 3) лишайниковых тундр (118),
4) овсяницевых тундр с Festuca sphagnicola (163), 5) кобрезиевых тундр
с Kobresia myosuroides (113), 6) ерниковых тундр с Betula rotundifolia
(119), 7) хионофитных альпийских лугов (78), 8) среднетравных аль-
пийских лугов (101), 9) высокотравных субальпийских лугов (126),
128
Рис. 1. Дендрограмма сходства ценофлор.
1—16 - ценофлоры (объяснение в тексте).
10) переувлажненных субальпийских лугов (91), 11) сообществ субаль-
пийского пояса с доминированием В. rotundifolia (84), 12) то же с доми-
нированием Salix glauca (82), 13) кедровых редколесий (133), 14) лист-
венничных редколесий (199), 15) высокогорных болот (48), 16) участ-
ков с доминированием Festuca altaica и Ptilagrostis mongholica (121 вид).
Следует отметить, что каждой из предварительно выделенных ценофлор
соответствует и определенный тип экотопа.
Последующий этап работы заключался в попарном сравнении цено-
флор и построении дендрограммы сходства. При этом использовалась
методика кластерного анализа (Дюран, Оделл, 1977). Была составлена
матрица мер — абсолютных мер сходства (числа общих видов) 16 цено-
флор. Для расчета относительной меры попарного сходства применена
мера Съеренсена—Чекановского
К(А, В) =
2п(А ПВ)
п(А)+п(В\
где А и В — множества, п (А А В) — число элементов пересечения мно-
жеств; после чего составлена матрица значений меры сходства (см. таб-
лицу) и на ее основе построена дендрограмма (рис. 1).
Из анализа дендрограммы видно, что выделилось несколько групп
тесно связанных между собой ценофлор.
Группа I включает ценофлоры кустарниковых (с Betula rotundifolia
и Salix glauca} переувлажненных сообществ и переувлажненных субаль-
пийских лугов. Меры сходства 0.9 и 0.7. Данные ценофлоры занимают уме-
ренно переувлажненные местообитания в нижней части субальпийского
пояса. Анализ видового состава показал, что в группе господствуют
преимущественно монтанные виды и виды, заходящие в высокогорья
9 Заказ 1220
129
г.
Матрица сходства ценофлор
из нижних поясов гор. Участие альпийских и субальпийских видов незна-
чительно. Сообщества, приуроченные к данным местообитаниям, форми-
руют гемигигрофитный ряд высокогорной растительности. Ценофлоры
этой группы генетически связаны с кустарниковой растительностью реч-
ных долин. На уровне сходства 0.5 к данной группе примыкает ценофло-
ра сообществ с доминированием Festuca altaica и Ptilagrostis mongholica.
Группа II объединила ценофлоры кедровых редколесий и высоко-
травных субальпийских лугов на уровне сходства 0.8. Для экотопов этой
группы характерен хорошо выраженный снеговой покров, сформирован-
ные почвы, умеренное увлажнение. Основу флоры составляют монтан-
ные мезофиты. Ценофлоры группы формируют мезофитный ряд высоко-
горной растительности, генетически связанный с неморальным флористи-
ческим комплексом. Отражением связи является присутствие немораль-
ных реликтов, часто играющих значительную роль в сообществах.
Группа III объединяет ценофлоры щебнистых, дриадовых и лишайни-
ковых тундр на уровне сходства 0.8. Их экотопы занимают верхние
уровни гольцового пояса в наиболее экстремальных условиях. Во флоре
господствуют альпийские и арктоальпийские виды, преимущественно
криофиты. Генетически данные ценофлоры связаны с флорой холодного
плейстоценового комплекса.
Группа IV объединила ценофлоры овсяницевых и кобрезиевых
тундр (уровень сходства 0.7), формирующихся также в близких эколо-
гических условиях.
Ценофлоры III и IV объединяются на уровне сходства 0.5, где к ним
присоединяется и ценофлора ерниковых тундр. Формируется группа це-
нофлор высокогорной криофитной растительности.
Особняком стоят ценофлоры хионофитных альпийских лугов и вы-
сокогорных болот. Первая из них формируется в очень специфических
термических и гидрологических условиях повышенного снегонакопле-
ния, вторая представляет, очевидно, реликтовое явление в высокогорной
растительности. Этим можно объяснить их низкое сходство с основными
высокогорными ценофлорами.
Рассмотрим некоторые общие закономерности в распределении це-
нофлор в зависимости от уровня сходства (рис. 1). Выше уровня сход-
ства 0.7 ценофлоры объединились в уже охарактеризованные 4 группы,
наиболее однородные экологически. Каждая из выделившихся групп
занимает экотопы, близкие по таким показателям, как положение в ря-
ду высотной поясности, условия увлажнения, термический режим, мощ-
ность снегового покрова. Каждая из групп является с экологических
позиций наиболее гомогенным комплексом гемигигрофитной, мезофит-
ной и криофитной высокогорной растительности. Дополнительная про-
верка гомогенности выделенных групп ценофлор мерами включения
(Юрцев, Семкин, 1980) также показала их высокое сходство. Так, меры
включения березковых и ивовых сообществ субальпийского пояса 0.9
и 0.8; меры включения ценофлоры щебнистых тундр в таковые лишай-
никовых и дриадовых тундр соответственно 0.96 и 0.97; овсяницевых
и кобрезиевых тундр 0.6 и 0.9 (флора кобрезиевых тундр включена во
флору овсяницевых на 93%). Подобное высокое сходство ценофлор
130
131
в группах и выровненность экологических условий местообитании по-
зволяют рассматривать каждую группу ценофлор как объединение ПФ
мезоэкотопов.
На уровне связи 0.6 и 0.5 выделились 3 группы ценофлор (рис. 1).
Анализ их экологической структуры показал, что экологическая гете-
рогенность внутри групп возросла. Основной действующий фактор,
отвечающий за объединение ценофлор на данном уровне в группы, —
экологические условия высотных поясов. В первую группу вошли
ценофлоры нижних уровней субальпийского пояса (березковые и иво-
вые субальпийские кустарники, переувлажненные субальпийские луга),
во вторую - высокотравные субальпийские луга, среднетравные альпий-
ские луга, кедровые редколесья (свойственные гумидным районам
высокогорий с развитым субальпийским поясом) и в третью группу -
ценофлоры, сформированные в экологических условиях гольцового
(альпийского) пояса. Каждую из трех групп ценофлор данного уровня
можно рассматривать как объединение ПФ макроэкотопов, сформи-
ровавшихся под воздействием экологических факторов высотных
поясов.
Наконец, на самых низких уровнях сходства (0.4—0.2) четко выде-
ляются только 2 крупные группы ценофлор и 2 отдельно расположенные
ценофлоры (хионофитных альпийских лугов и высокогорных болот).
Сравнение этих групп с имеющимися литературными данными по исто-
рии растительности Аптае-Саянской горной области показало, что в дан-
ном случае на дендрограмме (рис. 1) отражен процесс исторической диф-
ференциации флоры. Так, первая группа генетически связана с немораль-
ным флористическим комплексом и формирует травянисто-кустарнико-
вый ряд высокогорной растительности. Вторая группа, формирующая
горнотундровый ряд высокогорной растительности, генетически связана
с холодным плейстоценовым флористическим комплексом лиственнич-
ных редколесий. Флора хионофитных альпийских лугов, имеющая мак-
симальное сходство на уровне 0.3, исторически обособлена от остальной
высокогорной растительности, а флора высокогорных болот (макси-
мальное сходство 0.2) представляет реликтовое явление в экологиче-
ских условиях высокогорий.
Историю формирования данных ценофлор отражает и соотношение
в них высотно-зональных групп видов. Так, в лугово-кустарниковом
ряду ценофлор численно преобладают монтанные виды, на втором ме-
сте — виды, заходящие из нижних поясов гор (придаточные виды), на
третьем — арктоальпийские и альпийские виды; в горнотундровом ряду
на первом месте альпийские и арктоальпийские виды, на втором — мон-
танные, на третьем — придаточные; флора хионофитных альпийских
лугов состоит в основном из арктоальпийских и альпийских видов с не-
значительным участием монтанных; флора высокогорных болот — из
придаточных видов с небольшим участием монтанных, альпийских и
субальпийских. В соответствии со взглядами А. И. Толмачева (1948) на
высокогорные ландшафты северного полушария ценофлоры лугово-
кустарникового ряда соответствуют растительности альпийского типа
ландшафта, а горнотундрового ряда — растительности гольцового типа
132
Рис. 2. Дифференциация объединения цено-
флор уровня мезоэкотопов по цеиотическо-
му признаку (проективное покрытие).
Тундры: а - алекториевые, щ - щебнистые,
кд - кладониево-дриадовые, д - дриадовые,
тд - травянисто-дриадовые.
ландшафта. Каждую из этих групп цено-
флор можно рассматривать как объеди-
нение ПФ мегаэкотопов.
Что же касается ценофлор хио-
нофитных альпийских лугов и вы-
сокогорных болот, то они представ-
ляют объединения ПФ уровня мезоэкотопов, исторически обособлен-
ные от остальных ценофлор высокогорной растительности.
Следует отметить, что все изложенные построения получены на осно-
вании признака присутствия—отсутствия видов в ценофлорах. С приме-
нением этого признака удалось выделить объединения ПФ трех уровней:
1) мегаэкотопов (отражает исторические условия формирования ПФ),
2) макроэкотопов (отражает дифференциацию видов по высотным поя-
сам) , 3) мезоэкотопов (отражает дифференциацию флоры по близким
типам экотопов). Последним из названных уровней и ограничились
разграничительные свойства признака присутствие—отсутствие.
Для выделения объединений ПФ более низкого уровня был приме-
нен фитоценотический признак „проективное покрытие видов”. Рас-
смотрим его действие на примере разбиения ценофлоры кустарничково-
лишайниковой растительности (объединение ПФ мезотопического уров-
ня) . Как уже отмечалось, в нее вошли ценофлоры щебнистых, дриадо-
вых и лишайниковых тундр. Было выделено 5 градаций проективного
покрытия (менее 5%, 6 — 10%, 11 - 20%, 21 - 30% и выше 30%), на
основании которых каждому виду в геоботанических описаниях был
придан определенный вес. Далее было проведено попарное сравнение
описаний с использованием мер включения. На основании полученных
мер включения была рассчитана мера сходства Съеренсена—Чеканов-
ского (Семкин, Комарова, 1977). Флористические списки сходных
описаний были объединены в общие флоры с усреднением весовых ха-
рактеристик каждого вида. Следующим этапом работы было попарное
сравнение выделенных флор и построение дендрограммы (рис. 2).
Как видно из рис. 2, группы ПФ мезотопического уровня разбились
на 3 подгруппы ПФ ценотического (микротопического) уровня. В пер-
вую подгруппу вошли алекториевые и щебнистые тундры, во вторую —
кладониево-дриадовые и дриадовые тундры, в третью — травянисто-
дриадовые тундры. Ценофлора травянисто-дриадовых тундр связана
с первыми двумя на уровне сходства 0.5. Дополнительный анализ пока-
зал, что именно через нее осуществляется связь на ценотическом уровне
с ценофлорой травянистых тундр. Анализ геоботанических описаний,
вошедших в различные объединения ПФ микротопического уровня,
показал, что они довольно хорошо соответствуют группам ассоциаций
133
в традиционном понимании. С помощью подобной методики были выде-
лены 32 объединения ПФ микротопического уровня.
В заключение хочется отметить, что введение Б. А. Юрцевым (1982а)
понятия „парциальные флоры” различного топического уровня стало
хорошим связующим между флористикой и геоботаникой, особенно при
анализе ценофлор.1
А. А. Сагателян
СООТНОШЕНИЕ ГЕОЭЛЕМЕНТОВ В ПАРЦИАЛЬНЫХ ФЛОРАХ
МЕГРИ И ШИРАКА
Институт ботаники АН АрмССР
Сопоставлялись хорологические спектры парциальных флор (ПФ)
Мегринского флористического района и Ширака - переходной террито-
рии, включающей 2 флористических района, из которых южный находит-
ся под большим влиянием Армено-Иранского центра, а северный - Кав-
казского. Флора Мегри насчитывает 1467 видов на площади 800 км2,
Ширака — 1248 видов на 3 тыс. км2. В Мегри представлены все пояса
восточнозакавказского типа поясности, в Шираке — выпадает предгор-
ный и большая часть низкого рий.
Лесная ПФ в Мегри представлена 156 видами, в Шираке - 57. Треть
их в Мегри и 43 % в Шираке — голарктические, точнее, среднеевропей-
ские неморальные виды. В мегринских лесах хорошо представлены арме-
но-иранский, переднеазиатский, восточнозакавказский, субсредиземно-
морский и малоазиатско-кавказский виды; в Шираке же существенна
роль лишь малоазиатско-кавказских видов. Подобную картину наблю-
даем и в спектре ПФ лугов, которая в Мегри насчитывает 149 видов,
в Шираке — 139. Треть из них — голарктические, пятая часть в Мегри и
третья часть в Шираке — малоазийско-кавказские виды, прочие геоэле-
менты представлены слабее; так, на долю армено-иранских видов прихо-
дится */7 ПФ. Аналогично распределены лугостепные виды, которых
в ПФ Мегри 46, в ПФ Ширака 120. Третью часть ПФ составляют голаркти-
ческие виды. В Шираке х/3 ПФ составляют малоазийско-кавказские и
х/5 - армено-иранские виды. Таким образом, лесную, горнолуговую и
лугостепную ПФ сравниваемых районов, генетически связанные друг
1 Под объединением ПФ мезо-, макро- и мегаэкотопов автор имеет в виду по-
следовательные ступени классификации ценофлор (объединений ПФ микроэкото-
пов) по сходству видового состава, т. е. типологические единицы. Это расходится
с трактовкой мезо- и макроэкотопов Б. А. Юрцевым (1982а; см. также: Юрцев,
Семкин, 1980) как экологически своеобразных, но топографически цельных под-
разделений ландшафта, сопоставляемых соответственно с урочищами и местностя-
ми (мегаэкотопы - с ландшафтами в целом). Тем не менее В. П. Седельникову
удалось показать, что высшие единицы классификации ценофлор приближенно
соответствуют объединениям ПФ высотных поясов, которые можно рассматривать
как макроэкотопы. (Прим. ред.).
134
с другом, мы объединяем в восточнозакавказско-загросско-эльбурсскую
фратрию неморального типа растительности, в смысле В. Б. Сочавы
(1979). При этом в Шираке названные ПФ более связаны с анатолийским
и кавказским флористическими центрами, в Мегри же — с армено-иран-
ским.
ПФ альпийских лугов (140 видов в Мегри, 149 в Шираке) отли-
чается высокой оригинальностью, наименее связана с ПФ нижележащих
поясов и наиболее соотносится с альпийскими лугами других вершин
Малой Азии, Кавказа и в меньшей степени Армено-Иранской про-
винции.
ПФ степей в Мегри насчитывает 67 видов, в Шираке — 214 (основ-
ной флороценотип!). Более четверти их приходится на армено-иранский
геоэлемент. 11% в ширакских и 18% в мегринских степях составляют
субсредиземноморские (понтические) виды. Малоазийско-кавказских
видов в ширакских степях 17%, в мегринских - 12%. Далее следует
переднеазиатский и голарктический элементы.
ПФ трагакантников представлена 76 видами в Мегри и 63 - в Ши-
раке. Из них 50 мегринских и 38 ширакских представителей ПФ не выхо-
дят в своем распространении за пределы Передней Азии, причем полови-
на таких видов — армено-иранские. Трагакантники отличаются стенохор-
ностью флоры и гомогенностью хорологического спектра. Вместе со
степями они относятся к переднеазиатской фратрии степного типа расти-
тельности.
ПФ арчовников, отсутствующих в Шираке, представлена в Мегри
109 видами, пятая часть которых приходится на армено-иранский
элемент. Далее по спектру следуют субсредиземноморский, перед-
неазиатский и древнесредиземноморский. 62 вида этой ПФ не вы-
ходят в своем распространении за пределы Передней Азии, что свиде-
тельствует о принадлежности наших арчовников переднеазиатской
фратрии.
ПФ шибляка насчитывает 184 вида в Мегри и 56 - в Шираке. В Мег-
ри, за исключением 10 голарктических видов, все они ограничены в рас-
пространении евродревнесредиземноморскими территориями. При этом
26% ПФ приходится на армено-иранский элемент, по 14% — на древне-
средиземноморский и переднеазиатский элементы. В спектре ПФ фрига-
ноидов, которая в Мегри насчитывает 200 видов, а в Шираке - 137,
прослеживается та же последовательность геоэлементов, однако здесь
больше армено-иранских (22% в Мегри) и ирано-туранских видов. Особо
выделяется стенохорность флоры на пестроцветах, фрагмент которых
имеется в Мегри. Из 55 видов этой ПФ 34 вида армено-иранские по ареа-
лу, а древнесредиземноморских и ирано-туранских видов по 7. Таким
образом, ши&тяк, фриганоиды и растительность пестроцветов исследуе-
мых районов мы относим к армено-иранской фратрии нагорноксерофит-
ной растительности.
Наконец, фрагменты туранской фратрии представляет полукустар-
никовая пустыня, нацело сложенная древнесредиземноморскими видами
с преобладанием видов туранских.
135
В. А. Мартыненко
АНАЛИЗ ВИДОВОГО СОСТАВА ЛУГОВЫХ СООБЩЕСТВ
В КОНКРЕТНЫХ ФЛОРАХ ТАЕЖНОЙ ЗОНЫ
ЕВРОПЕЙСКОГО СЕВЕРА
Институт биологии Коми филиала АН СССР
В настоящее время изучение конкретных флор (КФ) не должно
ограничиваться составлением флористических списков для территории
в целом. Видовой состав растительного населения отдельных экотопов
внутри контура КФ настолько различен, что без получения информации
о наборе видов в каждом типе экотопов и о роли этих видов в сложении
растительных сообществ мы не имеем полной флористической картины
ландшафта.
В пределах таежной зоны европейского северо-востока была изучена
серия КФ, расположенных в Архангельской обл. и Коми АССР. Учет ви-
дового состава растений в различных экотопах показал, что резерватами
значительной части природной флоры являются луговые сообщества.
Между тем они, особенно в поймах крупных рек, интенсивно использу-
ются в качестве сенокосов и пастбищ, а часто просто распахиваются и
засеваются 1—2 видами местных или инорайонных многолетних трав.
Все это наносит непоправимый урон генофонду природной флоры и
в связи с этим ставит задачу организации широкой сети луговых заказ-
ников. Для выполнения такой работы необходимо иметь данные по фло-
ристическому составу луговых сообществ в разных географических
условиях - на фоне общей флористической характеристики того или
иного района.
Для этой цели мы выбрали 4 ранее изученные КФ, располагающиеся
в разных речных бассейнах и образующие географический ряд протяжен-
ностью с севера на юг более 500 км. Одна КФ находится в подзоне север-
ной тайги в бассейне реки Печоры (Усть-Цильма), три — в подзоне сред-
ней тайги, с севера на юг: в бассейнах рек Мезени (Кослан), Вычегды
(Палевицы), Сысолы (Визинга). Данные анализа КФ были опублико-
ваны ранее (Мартыненко, Шмидт, 1981). В последующем во всех четы-
рех флорах детально изучался видовой состав растений пойменных лу-
гов, т. е. одного типа экотопов, выделенного по растительности, что при-
ближенно соответствует объединению однотипных парциальных флор
(ПФ), в смысле Б. А. Юрцева (1982а; а также статья в наст. сб.). Для
этого нами закладывались профили из площадок 2 X 2 м, поперек каж-
дой поймы, а вдоль реки в разных зонах поймы делались описания пло-
щадок 10 X 10 м. Для определения роли каждого вида в сложении пой-
менных луговых сообществ учитывались его обилие и встречаемость
в данном типе экотопа каждой КФ. Все виды распределялись по широт-
ным (зональным) и долготным географическим, а также ценотическим
группам. Полученные данные позволили выяснить, каков вклад поймен-
ных луговых экотопов в состав и структуру природной флоры таежной
зоны, в какой степени ПФ пойменных лугов отражает общие закономер-
136
ности КФ и в чем ее специфика, как изменяются ценотические позиции
отдельных видов в разных географических условиях. Основные итоги
систематического, географического и ценотического анализа видового
состава пойменных лугов и КФ в целом приводятся в таблице.
Прежде чем перейти к обсуждению полученных результатов, очень
коротко охарактеризуем растительность рассматриваемой территории.
На юге подзоны средней тайги господствуют зеленомошные еловые леса
из Picea obovata с Vaccinium myrtillus и V. vitis-idaea, на значительных
площадях они замещаются вторичными (после рубки) елово-березовы-
ми и осиновыми лесами, поймы рек в значительной степени облесены.
В долинах и на водоразделах отмечается примесь Abies sibirica и изредка
TUia cordata. При движении к северу липа, а затем и пихта исчезают, уве-
личиваются площади заболоченных ельников с Polytrichum commune, Le-
dum palustre, видами Salix, сосняков co сфагновыми мхами. На дрениро-
ванных участках много Empetrum. В подзоне северной тайги заболочен-
ные ельники преобладают, депрессии водоразделов заняты крупными
верховыми болотами. Луга образованы Agrostis gigantea, Festuca rubra,
Alopecurus pratensis, Deschampsia caespitosa, Trifolium pratense, T. repens
и многими другими; на севере подзоны высокое обилие имеют также
Sanguisorba officinalis, Polygonum viviparum, Calamagrostis purpurea, в юж-
ных КФ характерно присутствие Centaurea jacea, Galium verum, Campanula
patula.
Из таблицы явствует, что количество видов в КФ постепенно снижа-
ется в направлении с юга на север — от 491 до 411. При этом в поймен-
ных луговых сообществах отмечается от 46 до 52% видового состава
каждой из флор, но закономерного падения уровня видового богатства
пойменных экотопов не наблюдается. Число видов в них колеблется от
202 до 226. В пойме Печоры их столько же, как в расположенной далеко
к югу пойме Вычегды, а на Мезени, занимающей пограничное положение
между средней и северной таежными подзонами, столько же, как на юге
средней тайги — в бассейне Сысолы. Несмотря на попарное совпадение
числа видов на пойменных лугах, качественный состав их оказался до-
вольно различным: значения коэффициента Жаккара, показывающего
общность ПФ изученных экотопов, оказались равными 57—58 %, в то вре-
мя как КФ в целом имели коэффициенты 66—69 %. Качественные разли-
чия видового состава ПФ пойменных экотопов обусловлены прохожде-
нием через область наших исследований границ распространения многих
видов: северные пределы имеют Campanulapatula, Centaurea scabiosa, юж-
ные - Geranium albiflorum, Elymus mutabilis.
Систематическая структура изученных КФ и ПФ одного типа экото-
пов в них имеют много общего, о чем свидетельствует почти одинаковый
набор 10 ведущих семейств, однако места, занимаемые ими, несколько
варьируют. В КФ первое место везде принадлежит Asteraceae (10% видо-
вого состава), далее идут Роасеае (9 %), Сурегасеае (6 %), Rosaceae (5 %),
за ними в разной последовательности Caryophyllaceae, Ranunculaceae,
Scrophulariaceae, Salicaceae, Polygonaceae, Brassicaceae.
В поймах из числа ведущих выпадают ивовые, появляются Fabaceae.
На Печоре и Вычегде первое место занимают злаковые, второе — астровые,
137
Анализ конкретных флор (КФ) и видового состава парциальных флор (ПФ) пойменных экотопов в таежной эоне
Группы видов растений Число видов данной группы в КФ или ПФ в бассейне реки
Печора Мезень Вычегда Сысола
КФ ПФ КФ ПФ КФ ПФ КФ ПФ
Широтные
304 160 323 179 324 161 356 180
бореальная 73.4 W 743* 80.0 747 797 72.7 79.7
44 18 59 24 53 19 70 25
полигональная ТПГ То" 14.0 10.5 12.0 9.4 ПТ ITT
31 7 25 7 17 2 21 з
гипоарктическая Тз* 3.4 6.0 3.0 4.0 1.0 4.3 1.3
арктогорная 15 8 9 8 4 3 1 о
Тб’ 3.4 2.0 33 1.0 1.4 0.2 0
неморальная 4 0 2 1 21 8 26 4
1.0 0 0.4 0.4 5.0 4.0 5.2 1.8
лесостепная 13 9 12 6 15 9 17 14
Долготные 3.1 4.4 3.1 2.6 3.4 4.4 3.4 6.2
евразиатская 170 90 189 108 179 97 203 85
41.3 45.2 44.0 48.2 41.4 47.7 41.7 38.1
циркумполярная 135 60 137 69 139 60 148 73
33.0 30.0 32.0 30.6 32.0 30.0 30.1 32.0
европейская 65 15.8 36 17.0 66 15.0 39 17.3 87 20.0 39 19.3 104 21.0 57 25.0
азиатская 31 13 21 5 15 2 17 4
7.5 6.4 5.0 2.2 3.4 1.0 3.4 1.8
10 3 17 4 14 4 19 7
космополитная 2.4 1.4 4.0 1.7 3.2 2.0 3.8 3.1
Ценотические 191 146 181 155 203 146 217 162
луговая 46.0 72.4 41.8 69.0 47.3 72.8 45.0 71.9
91 23 100 23 98 15 108 15
лесная 22.0 11.3 23.0 ГбЗ 227 7.4 213 “б7
болотная 60 20 62 25 60 26 80 32
та 9.9 14.0 11.1 13.8 12.4 16.2 ТО
14 0 29 0 16 0 24 0
водная 3.4 0 7.9 0 3.4 0 4.7 0
49 13 46 22 57 15 62 17
сорная 12.0 6.4 10.6 9.7 13.0 7.4 12.6 73
6 0 12 0 0 0 0 0 ,
неопределенная 2.0 0 3.6 0 0 0 0 0
Всего
видов 411 202 430 225 434 202 491 226
родов 192 115 224 128 206 110 235 121
семейств 61 32 70 35 67 32 76 36
Примечание. Над чертой — абсолютное число видов, под чертой — % от общего числа видов данной КФ или ПФ.
на юге территории возрастает участие и видовое разнообразие Scro-
phulariaceae.
Десять ведущих семейств включает 58—59 % видов КФ и от 63 % (на
юге) до 73 % (на севере) видового состава ПФ пойменных луговых эко-
топов.
Анализ участия разных зональных групп видов показал постоянное
преобладание на обоих уровнях сравнения бореальной группы, к кото-
рой относится 3/4 и более видов. Почти все доминанты луговых сооб-
ществ — бореальные виды (Alopecurus pratensis, Festuca rubra, Deschamp-
sia caespitosa и др.). Весомую долю - от 9 до 11 % флористического со-
става пойменных лугов — составляют виды полигонального распростра-
нения (Polygonum aviculare, Veronica serpyllifolia}. В КФ процентное уча-
стие их несколько больше за счет полигональных водных растений (Ро-
tamogeton perfoliatus, Lemna trisulca). Остальные зональные группы име-
ют еще более низкие показатели. Из гипоарктической группы, снижаю-
щей свое разнообразие в КФ с севера на юг (таблица), на лугах присутст-
вуют лишь отдельные виды (Calamagrostis lapponica, Ranunculus borealis}.
Арктогорная группа как во флорах в целом, так и в поймах на лугах
имеет минимальные показатели участия (1—3.6%). Если в подзоне се-
верной тайги мы еще находим Potentilla crantzii, Pedicularis compacta, то
в подзоне средней тайги лишь изредка Роа alpina, а в пойме Сысолы нет
ни одного арктогорного вида. Распределение южных зональных групп —
неморальной и лесостепной — имеет свои особенности. Первая представ-
лена преимущественно в южных КФ, в основном это лесные растения
(Tilia cordata, Asarum europaeum), лишь отдельные из них отмечаются на
пойменных лугах (Ajuga reptans, Scrophularia nodosa}. В связи с тем что
виды лесостепной группы находят благоприятные условия на открытых
пространствах пойм, их процентное участие там выше, чем в КФ (Carex
praecox, Astragalus danicus}. На юге территории оно достигает 6 % и даже
в пойме Печоры в подзоне северной тайги 4.4%.
Анализ участия долготных групп видов показал, что доля циркумпо-
лярной группы во всех случаях составляет не менее */з флористического
состава. Для евразиатской группы характерны более высокие показа-
тели: 41-44% каждой КФ и 45—48% видового состава ПФ пойменных
луговых экотопов; в ПФ поймы Сысолы этот показатель ниже — 38%,
что связано с увеличением там доли европейских видов. При движении
с севера на юг количество видов с европейскими ареалами постепенно
возрастает. В северных КФ эта долготная группа включает 15—16% видо-
вого состава, в южных — 20—22%. Такая же картина и в пойменных лу-
говых сообществах: на Печоре и Мезени по 17%, на Вычегде и Сысоле
соответственно 19 и 25 %. В распределении азиатских (в том числе сибир-
ских) видов наблюдается обратная закономерность: их число увеличива-
ется на севере и уменьшается на юге территории. Процентное участие их
значительно меньше, чем европейских, особенно в луговых сообществах
(см. таблицу). В бассейне Печоры процентное участие азиатской группы
в пойме близко к таковому в КФ в целом.
Определялось участие ценотических групп видов растений во флорах.
На пойменных лугах европейского Севера встречаются многие виды
140
лесной ценотической группы (Majanthemum bifolium, Equisetum sylvati-
cum), что свидетельствует об их послелесном прошлом. В ПФ пойменных
экотопов эта группа составляет 6—11% видов, при этом количество их
с юга на север увеличивается. Вклад луговой ценотической группы в КФ
42-47 %, а в поймах она преобладает по числу видов (69-73 %). Болот-
ные и прибрежноводные растения наибольшего разнообразия достигают
на юге территории, как в КФ, так и на пойменных экотопах. Сорняки на
пойменных лугах играют заметно меньшую роль, чем в соответствующих
КФ (см. таблицу). Водная ценотическая группа (Potamogeton natans, Се-
ratophyllum demersum и др.) на лугах отсутствует, наибольшее участие ее
отмечается в КФ бассейна Мезени.
Для теоретических построений и практического использования дан-
ных по флоре ландшафтов и их частей следует учитывать особенности
распределения отдельных видов и их роль в сложении растительных
сообществ. Для этого нами использовался показатель встречаемости
(в %), определяемый в выборках описаний пойменных лугов Печоры,
Мезени, Вычегды и Сысолы. Встречаемость большого количества видов
не меняется или почти не меняется в луговых сообществах во всех пой-
мах. К ним относятся Agrostis gigantea, Juncus filiformis, Ranunculus acris,
Achillea millefolium, Galium boreale. Имеется группа видов с высокой
встречаемостью в северных бассейнах Печоры и Мезени: Polygonum vivi-
parum (43—45%), Sanguisorba officinalis (42%), Tanacetum bipinnatum (25%)
и другие, южнее они отмечаются единично или вообще отсутствуют. В об-
ратном направлении, с юга на север, снижаются показатели встречаемо-
сти на лугах у Galium mollugo: от 22 % на Сысоле до 15 % на Вычегде и
1 % на Мезени.
Таким образом, проведенное исследование показало, что луговые
сообщества, расположенные в поймах таежных рек европейского Севера,
включают около половины видового состава КФ, а иногда и несколько
более половины. Они имеют четко выраженный бореальный характер,
участие в них зональных групп более северного распространения — гипо-
арктической и арктогорной - меньше, чем в КФ в целом. Из южных
групп - неморальной и лесостепной - заметнее представлена последняя,
находящая в пойменных экотопах оптимальные условия для своего
развития. При хозяйственном освоении пойм возникает угроза потери
значительной части генофонда природной флоры. Необходимо создание
сети луговых заказников с учетом флористических особенностей лу-
говых сообществ, наличия в них редких и хозяйственно ценных видов
растений.
141
2. КОЛИЧЕСТВЕННЫЕ МЕТОДЫ АНАЛИЗА
И СРАВНЕНИЯ ФЛОР
Л. И. Малышев
СОВРЕМЕННЫЕ ПОДХОДЫ К КОЛИЧЕСТВЕННОМУ
АНАЛИЗУ И СРАВНЕНИЮ ФЛОР
Центральный Сибирский ботанический сад СО АН СССР
Основоположник количественных методов в сравнительной флори-
стике — выдающийся швейцарский ботаник Альфонс Декандоль (De
Candolle, 1855). До него исследователи приводили иногда в своих рабо-
тах данные о количестве в той или иной флоре растений разной таксоно-
мической принадлежности, но эти данные не подвергались ботанико-гео-
графическому анализу. Несколькими годами раньше Декандоля русский
химик и ботаник К. К. Клаус внес свой вклад в методологию при иссле-
довании приволжских флор (Клаус, 1851, 1852). Однако новаторство
Клауса было не оценено и забыто не только зарубежными, но и отечест-
венными ботаниками вплоть до недавнего времени (Шмидт, Ильмен-
ских, 1982). Основная причина этого кроется, видимо, в слишком ло-
кальной ориентации исследования, отраженной в названии труда, что не
привлекало всеобщего внимания.
Свои исследования Декандоль и Клаус совершили в эпоху усиленно-
го накопления и обобщения ботанико-географических фактов. После-
дующие десятилетия после середины прошлого века дали мало в области
совершенствования методов сравнительной флористики на основе учета
количественных признаков. Наступил длительный период накопления
фактического материала. Одновременно продолжалась дифференциация
науки. От систематики растений окончательно обособилась ботаническая
география, которая в первые десятилетия XX в. в свою очередь раздели-
лась на флористику, геоботанику (или фитоценологию) и собственно
ботаническую географию. В силу родства геоботаники и флористики не-
которые приемы количественного анализа, разработанные для описания
растительных сообществ, оказались применимыми для целей количест-
венного анализа флоры, например коэффициенты общности Жаккара,
уравнения Глизэна, Аррениуса и Уранова, предложенные для измерения
зависимости числа видов растений от размера площади.
После разработок Декандоля и Клауса возрождение сравнительной
флористики на основе учета количественных признаков наступило в се-
редине XX в. и бурно продолжается в наше время, хотя некоторые цен-
ные в методическом отношении исследования были выполнены еще
в 20—40-е годы нынешнего века. Стимулом для возрождения был общий
прогресс вычислительной математики и особенно совершенствование
биометрии в связи с созданием удобных и быстродействующих средств
вычислительной техники в виде разнообразных электронных калькуля-
торов и компьютеров. На очереди — внедрение в обиход ученого персо-
142
нальных компьютеров, создание в гербарном деле автоматизированных
информационно-поисковых систем с оснащением гербарных кабинетов
дисплеями (Лякавичюс, Красаускас, 1977; Малышев, 1977а; Мякушко
и др., 1981).
Пионером возрождения сравнительной флористики на основе широ-
кого использования количественных методов в СССР был А. И. Толма-
чев (1941, 1970а, 19706, 1971), а за рубежом - некоторые ученые из
франкоязычных и англоязычных стран. В методическом отношении важ-
ное значение для современной сравнительной флористики имеет учет
фактического материала способом представительства на сеточной основе
(сокращенное название: способ представительства). Впервые он приме-
нен в США в 30-х годах, когда на карто-схемах ареалов отмечался факт
наличия отдельных видов растений в ячеях сети, соответствующих адми-
нистративным округам в тех или иных штатах. Этот способ до сих пор
применяется в США в публикуемых „Флорах”.
Способ представительства был модифицирован в Великобритании
применением сети из регулярных квадратов, образованных географиче-
скими параллелями и меридианами, при составлении „Атласа британской
флоры” (Atlas..., 1962), затем при изучении флоры отдельных графств,
при картировании флоры Европы в целом и ее отдельных районов. Этот
метод удобен для картирования ареалов растений в странах с высоким
уровнем флористической изученности, в частности оказался перспектив-
ным при изучении флоры Латвийской ССР. Использование регулярной
сети при выявлении ареала значительно облегчает статистическую обра-
ботку материала с помощью компьютера (Cadbury et al., 1971).
В районах с низким уровнем общей флористической изученности бо-
лее удобен предложенный Толмачевым метод конкретных флор (КФ).
Статистические оценки возможны в этом случае на основе сравнения
тщательно изученных выборных пробных участков. Этот метод разрабо-
тан на примере двух обследованных участков — Яму-Тарида и Яму-Неру
на п-ове Таймыр (Толмачев, 1931, 1932). Последовательно он применен
лишь в последние десятилетия на Чукотке, востоке Большеземельской
тундры, плато Путорана, в Псковской обл. и в некоторых других местах.
Методом КФ Н. С. Водопьянова (1984) исследовала закономерности
зонального изменения флоры Среднесибирского плоскогорья. Для этого
она заложила профиль-трансект по 108 меридиану. Начала работу в При-
байкалье и в течение трех лет продвигалась на север, пока не достигла
Северного Ледовитого океана в районе Хатангского залива.
Вероятно, не будет преувеличением сказать, что в последние десяти-
летия сравнительная флористика более всего развивалась в Советском
Союзе. Этому отчасти благоприятствовало разнообразие флористических
контрастов в стране в связи с большой ее протяженностью. В последние
десятилетия выявлена математическая зависимость флористического бо-
гатства от внешних условий и исторических факторов (Малышев, 1969;
Шмидт, 1977, 1979, 1981а), разработана схема количественного анализа
флоры на основе трех главных параметров, которыми являются про-
странственное разнообразие флоры, уровень видового богатства и репре-
зентативность участков обследования (Малышев, 1972а, 1975; Шмидт,
143
1974а). Особо интересна разработка возможности флористического
районирования на фитостатистической основе. Этому вопросу в послед-
ние десятилетия посвящен ряд исследований (Баранова и др., 1971; Ма-
лышев, 1973, и др.). Итоговое значение имеет книга В. М. Шмидта „Ста-
тистические методы в сравнительной флористике” (1980).
Инвентаризация видового состава — это первый (начальный) этап
флористического исследования. Вслед за ним предстоит изучение особен-
ностей ее состава и расшифровка генезиса как природного объекта, обра-
зующего систему. Применение количественных методов позволяет при
этом чисто субъективным суждениям противопоставить объективные
критерии (Малышев, 1976а, 1977в). Простые арифметические подсчеты
по таксономической структуре флоры (определение числа видов, родов
и семейств) и установление родовых, семейственных или хорологиче-
ских спектров — все это дает фактический материал, который нуждается
в дальнейшем сопоставлении. Равным образом простое установление
факта, что на площади определенного размера обнаружено то или иное
количество видов, родов и семейств растении, само по себе еще не харак-
теризует флору как богатую или бедную, с преобладанием автохтонной
или аллохтонной тенденции в генезисе, если при этом не сделаны соответ-
ствующие расчеты по уравнению регрессии с пересчетом на стандартную
площадь.
Методы сравнительной флористики удачно использованы Б. А. Юрце-
вым (1968а) при исследовании флоры Сунтар-Хаята с обоснованием по-
нятия активности вида. Им было установлено наличие Гипоарктического
ботанико-географического пояса, расшифрована роль Берингии для исто-
рической географии растений и флористические связи между Северо-Во-
сточной Азией и Северной Америкой (Юрцев, 1966, 1972, 19746, 1976а,
1981, 19826; Юрцев и др., 1978). Для горных районов Средней Азии эта-
лонное значение в области сравнительной флористики имеют исследова-
ния Р. В. Камелина (1973, 1979).
Интегральное применение количественных методов для расшифров-
ки особенностей состава и генезиса флоры предпринято при изучении
высокогорий Сибири (Малышев, 1965а, 19656; Высокогорная флора
Станового нагорья, 1972; Красноборов, 1976). Последовательная попыт-
ка разработать алгоритмическую схему для определения особенностей
состава и тенденций в генезисе флоры сделана при изучении плато Путо-
рана в полосе северной тайги (Малышев, 19766).
Наиболее важными являются следующие количественные характери-
стики флоры.
1. Уровень видового богатства и пространственное разнообразие
флоры. Эти параметры определяются по уравнению регрессии. Из них ви-
довое богатство флоры (величина а) находится путем пересчета на стан-
дартную площадь. В случае изучения методом КФ возможен пересчет на
100 км2. Пространственное разнообразие флоры (величинаz) определя-
ется по уравнению Аррениуса. Оно установлено эмпирически, как и аль-
тернативные ему уравнения Глизэна и Уранова. В отдельных конкретных
случаях лучшие результаты дает то одно, то другое из них (Dony, 1971;
Макарова, 1983). Поэтому желательна дальнейшая ревизия этих уравне-
144
ний для уточнения условий наибольшей пригодности. Априорно можно
ожидать, что уравнение, включающее большое число переменных вели-
чин, приведет к более полному совпадению фактических и прогнозируе-
мых данных, чем уравнение, оперирующее с небольшим числом перемен-
ных, однако само расширение числа переменных величин не всегда жела-
тельно.
2. Сравнительное обилие видов растений в полных и парциальных
конкретных флорах.1 Знание сравнительного обилия необходимо при
пользовании методом КФ для суждения о том, насколько достаточна по
размеру территория эталонного участка. Площадь, удвоение которой
приведет к увеличению числа видов растений на 20%, была принята за
минимальную, а на 14% — за оптимальную (Малышев, 1972а). Однако
эти критерии слишком условные, поэтому взамен их позднее предложен
общий критерий репрезентативности размера участка флоры и разрабо-
тан алгоритм для расчета (Малышев, 19766). Найдено, что на плато Путо-
рана 13 изученных КФ со средней площадью каждой из них около 79 км2
репрезентативны в среднем на 91 %. Их удвоение в 2 раза (до 159 км2)
увеличит список растений до 300 видов вместо 273. В составе этих КФ
относящиеся к лесному поясу участки (парциальные КФ) репрезентатив-
ны в среднем на 85 % и высокогорные — на 90%. Следовательно, во всех
случаях полные и парциальные КФ Путорана как достаточно репрезента-
тивные пригодны для получения сравнительных данных об уровнях фло-
ристического богатства.
3. Сходство и различие флор по нумерическим данным. Расшифров-
ка этой характеристики имеет значение для районирования флор, выяс-
нения их генезиса и для определения взаимного проникновения элемен-
тов флоры из одной геосистемы в другую (например, из лесного пояса
в высокогорную).
Для сравнения видового состава флор, в том числе парциальных КФ,
пригоден классический коэффициент линейной корреляции Пирсона (ве-
личина z). Он оценивает лишь взвешенные компоненты, в данном случае
наличие или отсутствие таксона (того или иного вида, рода или семей-
ства) с количественной характеристикой по встречаемости, активности
или же по числу содержащихся видов (в случае родов или семейств).
Для флористического анализа хорошие результаты дает канониче-
ское уравнение Престона (Preston, 1962). На основе учета количества ви-
дов растений в двух сравниваемых флорах и общего для них количества
видов растений возможно вычислить показатель различия сравниваемых
флор (величина z, тождественная показателю пространственного разно-
образия флоры, см. выше). Величина z имеет критическое значение
около 0.27, по уклонению от которого можно судить, насколько обе
сравниваемые флоры являются частями единого целого (при значениях
меньше 0.27) или же относятся к генетически разным системам, а в слу-
чае островных флор — являются изолятами (при значениях больше
0.27). Критика уравнения Престона (Песенко, 1982, и др.) недостаточно
1 Под парциальными КФ автор имеет в виду ПФ отдельных высотных поясов
в пределах горной КФ. {Прим, ред.).
10 Заказ 1220
145
обоснована. Оно найдено пригодным для выявления генетических связей
высокогорных флор Южной Сибири и Монголии, оценки эндемизма
в высокогорных флорах Северной Азии и районирования степной Бай-
кальской Сибири (Малышев, 1968,1979; Пешкова, 1972).
Многие флористы пользуются иными коэффициентами сходства-
различия: Жаккара, Съеренсена, Чекановского, Стугрена и Радулеску,
Жаккара в модификации Малышева (Малышев, 19726) и некоторыми
другими (Шмидт, 19816, и др.). Эти коэффициенты относятся к одному
классу точности и дают достоверные результаты лишь при сравнении
флор с одинаковым или сходным по величине количеством видов расте-
ний и малопригодны для сопоставления части и целого. Взамен их
Б. И. Семкин и некоторые другие исследователи показали в последние
годы перспективность для решения последней задачи учета мер включе-
ния в соответствии с моделью кругов Эйлера на основе теории множеств
(Семкин, Комарова, 1977; Юрцев, 1978; Юрцев, Семкин, 1980; Семкин,
Куликова, 1981; Седельников, 1982). Опыт этих исследователей заслу-
живает восприятия.
4. Поясно-зональная и высотная структуры флоры. Эти характери-
стики устанавливаются путем учета распределения. видов растений по
поясно-зональным группам (например, арктической, альпийской, аркто-
альпийской, гипоарктической, монтанной, гипоарктомонтанной, бореаль-
ной, лесостепной и т. п.) и по высотным комплексам (высокогорному,
общегорному, лесному поясам и т. п.), т. е. двумерно. Названные показа-
тели могут быть выражены в процентах, т. е. в относительном измерении,
или же в абсолютном измерении — путем пересчета числа видов растений
на равную площадь (например, на 100 км2).
5. Таксономическая структура флоры на уровне семейств и родов
(семейственные и родовые спектры). Выявление этих параметров необ-
ходимо для оценки самобытности флоры и выработки схемы флористи-
ческого районирования. Из соображений практического удобства может
быть использована для анализа лишь головная часть сравниваемых се-
мейственных (точнее, семейственно-видовых) или родовых (точнее,
родово-видовых) спектров, ранжированных по количеству содержащих-
ся видов растений. При этом для флор, находящихся в разных ботанико-
географических зонах или относящихся к подразделениям (фитохори-
ям) высокого ранга, более показательно сопоставление семейственных
спектров. Для сравнения на уровне провинций и особенно округов,
когда контрастно отражаются лишь поздние этапы в эволюции раститель-
ного покрова, более подходящим будет сравнение родовых спектров.
Наконец, сравнение флористических районов целесообразно проводить
непосредственно на основе учета видовых списков растении, а не по
спектрам семейств или родов. Кроме того, в некоторых случаях могут
представить интерес семейственно-родовые спектры, в которых семей-
ства ранжированы по числу содержащихся в них родов растений.
Сравнение семейственных и родовых флористических спектров воз-
можно путем определения коэффициента ранговой корреляции Кендала
(1975) или же Спирмена (Мартыненко, Шмидт, 1981). Оба коэффици-
ента более или менее равноценны. Во флористике коэффициент ранговой
146
корреляции Кендала впервые применили независимо друг от друга
Л. И. Малышев и В. М. Шмидт (Заки, Шмидт, 1972, 1973; Малышев,
19726; Ребристая, Шмидт, 1972). Техника расчета к настоящему времени
хорошо отработана (Малышев, 19766). Трудности от сопоставления дан-
ных разных авторов могут возникнуть оттого, что одни используют
коэффициент Кендала, другие - коэффициент Спирмена или же учиты-
вают головные части спектра из разного числа членов, например 5, 7,10,
15 или даже 20. Но рекомендовать какой-либо стандарт, может быть,
опрометчиво, хотя лично мы отдаем предпочтение коэффициенту Кен-
дала и 10-членным спектрам.1
6. Соотношение количества видов и родов во флоре. Зависимость
подчцняется уравнению логарифмического ряда (Fisher et al., 1943). Она
может быть выражена также квадратичным уравнением (Малышев,
1969).
Еще со времени Декандоля известно, что соотношение видов и ро-
дов зависит от размера флоры и от географической широты. Весьма важ-
но приложение этой характеристики для определения меры самобытно-
сти флоры. С помощью эмпирического уравнения по фактическому ко-
личеству родов можно рассчитать ожидаемое количество видов. Если
автохтонная и аллохтонная тенденции в генезисе флоры не уравновеше-
ны, тогда будет иметь место несовпадение фактических и расчетных дан-
ных о числе видов во флоре (величины S и о ). По относительной величи-
не этого несовпадения можно судить о мере самобытности, или автохтон-
ности, флоры (величина Л): А = (5- 5)IS, а именно положительная
величина коэффициента А свидетельствует о преобладании автохтонной
тенденции в гейезисе флоры, отрицательная — аллохтонной, а нулевое
значение — о сбалансированности (уравновешенности) обеих тенденций
(Малышев, 1969, 19766).
7. Флористическое районирование на основе количественных призна-
ков. Это направление сравнительной флористики усиленно развивалось
в последние десятилетия на основе учета сходства или различия таксоно-
мической структуры флоры. Как отмечено выше, для установления
крупных подразделений районирования результативен учет семействен-
ных (для флористических областей) или родовых спектров (для флори-
стических провинций) на основе определения коэффициентов ранговой
корреляции Кеидэла или Спирмена, а для более дробных единиц флори-
стического районирования (на уровне округов и районов) пригоден ана-
лиз непосредственной видовой структуры флоры (Малышев, 1973).
При этом лучшие результаты дает определение сходства или различия
на основе учета мер включения или же определения коэффициентов по
каноническому уравнению Престона. Представляет интерес также уста-
новление выделов районирования методом таксономического анализа
1 В последние годы для сравнения процентных спектров семейств, родов, раз-
личных категорий элементов флоры успешно использованы обычные меры сход-
ства (например, Жаккара, Съеренсена и др.), модифицированные для весовых
множеств (Марина, 1982, а также статья в наст, сб.; Соколова, 19846, 1985).
(Прим. ред.).
147
Е. С. Смирнова (Шмидт, Баранова, 1975) и на основе учета мер автохтон-
ности или аллохтонности флоры (Малышев, 1969, 19766; Золотухин,
1982).
Полученные тем или иным способом объективные показатели
сходства или различия флор изученных территорий могут быть орга-
низованы для целей последующего построения флористического райо-
нирования в виде корреляционных плеяд или же в виде дендритов
по максимальному корреляционному пути, называемых иначе опти-
мальными деревьями сходства. Но особенно пригодно для целей райо-
нирования построение дендрограмм на основе кластерного анализа
(Шмидт, 1974а, 19746,1975; Пааль, Семкин, 1983, и др.). В случае боль-
шого количества флористических выделов и учета обширных таксоно-
мических списков техника расчета трудоемка. Тогда весьма желательно
использование компьютеров (Clayton, Неррег, 1974; Clayton, Panigraha,
1974, и др.)
8. Использование количественных методов при анализе флоры для
решения фундаментальных проблем ботанической географии и охраны
растительного мира. Подлежат дальнейшему углубленному изучению
с позиций сравнительной флористики изолированные охраняемые тер-
ритории как ложноостровные биоты. Это имеет значение при опреде-
лении* размера территории, отводимой под заповедование (Simberloff,
Abele, 1976; Симберлофф, 1982; McCoy, 1983; Reed, 1983), так как пре-
вращение в изоляту слишком малой территории приведет рано или
поздно к общему обеднению флористического или фаунистического
состава.
Удалось сконструировать номограмму, которая позволяет прогнози-
ровать флористическую репрезентативность участков земной поверхно-
сти в зависимости от размера их площади при разных значениях меры
пространственного разнообразия биоты (Малышев, 1980). Она позво-
ляет определить меру обеднения островных флор в зависимости от вели-
чины и удаленности острова от материка или же от соседних крупных
островов.
В связи с проблемой охраны растительного мира на основе количест-
венного анализа при сравнении флор обнаружен на севере Европы, вклю-
чая северо-запад европейской части СССР, так называемый флористиче-
ский вакуум (Малышев, 1981). Предположительно он образовался со вре-
мени последнего плейстоценового оледенения и не заполнен до настоя-
щего времени. Наличием флористического вакуума можно объяснить
усиленное образование там микровидов, особенно в родах AlchemiUa,
Hieracium, Rubus. Для дальнейшего решения этой и ряда других проблем
флористики и ботанической географии будут полезными приемы количе-
ственного анализа фактических данных. Перспективно также широкое
использование во флористике экологических данных на основе совокуп-
ного углубленного исследования флоры и растительности, образующих
вместе единый растительный покров (Шеляг-Сосонко, Дидух, 1980,
и др.).
148
Б. И. Семкин
ТЕОРЕТИКО-ГРАФОВЫЕ МЕТОДЫ
В СРАВНИТЕЛЬНОЙ ФЛОРИСТИКЕ
Тихоокеанский институт географии ДВНЦ АН СССР
В сравнительной флористике используются бинарные отношения
сходства, включения (целое—часть), типичности и др. (Юрцев, Семкин,
1980; Шмидт, 1980), которые возможно изображать в виде различных
графов. Теория графов дает простой, достаточно доступный и мощный
инструмент для анализа указанных отношений и решения многих задач
упорядочивания флористических описаний. В настоящей статье рассмат-
риваются методы теории графов и отношений, которые наиболее часто
используются при обработке матриц флористических данных: диаграм-
мы Чекановского, оптимальные деревья (Василевич, 1969) и иерархиче-
ские дендрограммы (Sokal, Sneath, 1963). Большое внимание в работе
уделяется также анализу разновеликих по числу видов флор и использо-
ванию при этом ориентированных графов (Семкин, Комарова, 1977).
Матрица флористических данных
и ее преобразование
Флористические данные обычно представляются в форме прямо-
угольной таблицы (табл. 1) размером q X s, где q — количество объектов
(флор), a s - количество признаков (число видов в сводном списке).
Обозначим для простоты объекты (флоры) — столбцы матрицы цифра-
ми /= 1, 2, ..., q, а признаки (виды) — строки цифрами i = 1, 2, ..., s.
Тогда на месте пересечения строки i и столбца / (или, говоря короче,
в клетке //) поставим значение равное 1, если i-й вид есть в /-й флоре,
и 0, если i-й вид не встречен в /-й флоре. Такая таблица чисел будет
Таблица 1
Матрица флористических данных
Нумерация видов Нумерация флор
1 2 ... / ... я
1 Л11 л12 ... *1/ ...
2 а21 а22 ... a2i ... fl2q
... ... ... ... ...
I ai2 ... aU ... aiQ
... ... ... ... ...
S а31 аз2 ... asj ... asq
149
Таблица 2
Матрица пересечения
N* флоры 1 2 ... 0 ... я
1 С11 С12 ... с10 ... Clq
2 С21 С22 ... С20 ... С1Я
... ... ... ... ...
a Cal Са2 ... ... Са<?
... ... ... ... ...
Я С91 С92 ... СЯ& ... сяя
называться матрицей флористических данных. На основе этой матрицы
можно получить ряд других признаков путем взятия части видов свод-
ного списка [видов определенного семейства или рода, видов, например,
четырех отделов (Плаунообразных, Хвощеобразных, Папоротникообраз-
ных, Голосеменных), видов 10 наиболее обильных по числу видов.се-
мейств и т. д.] или путем укрупнения признаков, т. е. рассмотрением
классов эквивалентности, например, родов, семейств и т. д., а также раз-
личных распределении видов по классам эквивалентности — таксономи-
ческих спектров (семейственно-видовых, семейственно-родовых, родо-
во-видовых), географических (широтных, долготных, широтно-долгот-
ных), спектров жизненных форм и других (Юрцев, 1982а). Следователь-
но, в общем случае элементы матрицы флористических данных могут
быть любыми неотрицательными числами.
По матрице флористических данных рассчитывается матрица
(табл. 2) абсолютных мер сходства, или пересечения,1 размером q2.
Общий элемент ее имеет вид:
s
Са0 = Smin(a/a, aip), а = 1,..., q; 0 = 1,..., q; aia > 0, aif} > 0,
где а и (3 - номера двух сравниваемых флор.
Если флористические данные были нормированы к 1 (или 100%), то
матрица пересечений будет также и матрицей сходства. Матрица пере-
сечения симметрична относительно главной диагонали, т. е. выполняется
равенство элементов для любых номеров к и I :Ск1 = С1к. Поэтому до-
статочно приводить только половину элементов матрицы пересечения,
расположенных над или под главной диагональю, и элементы главной
диагонали.
1 Матрицу абсолютных мер сходства кратко будем называть матрицей пересе-
чения, так как каждый элемент этой матрицы есть мера пересечения двух дескрип-
тивных множеств, характеризующих описания флор (Семкин, 1973а; Юрцев, Сем-
кин, 1980).
150
Таблица 3
Матрица включения
—► 1 2 • • • 0 ... q
1 Сц/Сц CnlC22 ... ... Clq/Сц
2 С21/С22 C22lC22 ... С2^С22 ... ^2ql^22
... ... ... ... ...
а Calida, а ^а2^аа ... ... CaqlCaa,
q CqJCqq cqdcqq ... Cqfi!Cqq ... Cqq!Cqq
Для ненормированных данных матрица пересечения легко преобра-
зуется в матрицу мер включения. Для этого необходимо элементы каж-
дой строки матрицы пересечения разделить на соответствующий этой
строке диагональный элемент.1' Например, элементы строки а Са1, Са2,
..., Са0> Caq делятся на диагональный элемент и получается сле-
дующая строка:
Cai/Caa> ^a2^aa> •••» ^ар/^аа» •••» ^aql^aa’
В результате получаем матрицу включения (табл. 3). В ней стрелка (-►)
указывает направление включения одной флоры в другую. Например,
первая строка матрицы означает степень включения флоры № 1 в осталь-
ные флоры за № 2, 3..q. Обычно используются только некоторые эле-
менты строки, имеющие наибольшие численные значения, превышающие
заданный порог (Семкин, Комарова, 1977). Элементы матрицы включе-
ния, находящиеся симметрично относительно главной диагонали (все
элементы диагонали равны 1, или 100%), не равны друг другу, т. е. мат-
рица включения несимметричная. Например, возьмем 2 элемента матри-
цы включения и расположенные симмет-
рично главной диагонали матрицы включения. Очевидно, в общем слу-
чае - это различные числа, т. е.
Матрицу включения можно привести к симметричному виду различ-
ными способами. Наиболее часто используются следующие четыре (Юр-
цев, Семкин, 1980; Семкин, Комарова, 1985):
к(п) _/Ла/0+А0/а)
а’*Д 2 )
при i? = +°°, 1? = 0, 1?=1, 1? = —1. Соответствующие меры сходства назы-
ваются: Симпсона, Охай, Кульчинского, Съеренсена. Обычно указанные
меры записываются в простой форме соответственно:
1 В нашем случае диагональный элемент есть число видов соответствующей
флоры, например — число видов в флоре, обозначенной номером а.
151
Таблица 4
Матрица сходства
№ флоры 1 2 • •• ... Q
1 1 г(л) А12 ... г(п) А10 ... г(п) Х1<7
2 к(п) 1 г(ч) г(п)
21 л2р 2<?
а к(п) а1 г(п) а 2 ... к(п) Аа0 ... к(п) Ха<7
<7 г(п) г(л) Aq2 ... г(л) Aq0 ... 1
K(+°°) _ л а, р ~ max(Ka^, ^/а),
^а, \ ~ V^a/3'^/а»
x£i = |(X./,+«,/.).
£<-1) _ ^а/р ’ Кр/а
а’Р ^а/р+^р/а
После симметризации матрицы включения получаем матрицу мер
сходства (табл. 4), на основе которой возможно разбиение флор на
классы.
Матрицу сходства можно дальше преобразовывать, находя так назы-
ваемое транзитивное замыкание (Tamura et al., 1971; Орловский, 1981;
Парницкий, 1981) на основе расчета элементов по формуле
*(а0 = max Klp\ min(Ka2> К2р\ •••> min(Kaq, ,
(2)
где К ар — элемент, стоящий на пересечении a-строки и 0-столбца в новой
матрице, полученной после первой итерации. Указанную процедуру мож-
но продолжать до тех пор, когда будет выполняться следующее равен-
ство:
— лар , а—1,..., </,0—1, ...,</,
где п — число итераций.
В результате получаем матрицу, для элементов которой выполняется
ультраметрическое неравенство (Sibson, 1971; Tamura et al., 1971; Мир-
кин, 1980):
ар min [л ау, К ], 1 7 Ц.
152
Таблица 5
Матрица пересечения для флор 10 островов Курильской гряды
№ флоры 1 2 3 4 S 6 7 8 9 10
1 552 391 379 279 174 145 136 84 166 119
2 391 634 418 263 165 140 126 86 148 115
3 379 418 579 321 192 162 148 94 180 151
4 279 263 321 379 197 165 154 99 182 134
5 174 165 192 197 237 175 163 109 173 122
6 145 140 162 165 175 191 152 98 148 116
7 136 126 148 154 163 152 181 99 148 115
8 84 86 94 99 109 98 99 186 102 81
9 166 148 180 182 173 148 148 102 320 190
10 119 115 151 134 122 116 115 81 190 216
Примечание. Здесь и в табл. 6 и 7 номера флор островов: 1 — Шикотан,
2 — Кунашир, 3 — Итуруп, 4 — Уруп, 5 — Симушир, 6 — Кетон, 7 — Расту а, 8 -
Мату а, 9 — Парамушир, 10 — Шумшу.
Как известно (Парницкий, 1981), при выполнении ультраметрического
неравенства рассматриваемые объекты просто разбиваются на классы.
Рассмотрим конкретный пример представления флористических дан-
ных в виде матриц. Возьмем из литературных источников данные о фло-
рах 10 Курильских островов (Шмидт, 1980). На основе коэффициентов
сходства Съеренсена—Чекановского и флористического богатства остро-
вов может быть восстановлена матрица пересечений (табл. 5).
Как видно из матрицы пересечений, флоры Курильских островов
разновелики по числу видов. Так, флора о. Матуа (186 видов) резко
отличается от флоры о. Кунашир (634 вида). Поэтому в данном случае
необходимо рассчитывать матрицу мер включения. После деления каж-
дого элемента строки матрицы пересечения на соответствующей этой
строке диагональный элемент получим матрицу мер включения (табл. 6).
Таблица 6
Матрица мер включения для флор 10 островов Курильской гряды
N* флоры 1 2 3 4 S 6 7 8 9 10
1 100 71 69 51 32 26 25 15 30 22
2 62 100 66 41 26 22 20 14 23 18
3 65 72 100 55 33 28 26 16 31 26
4 74 69 85 100 52 44 41 26 48 35
5 73 70 81 83 100 74 69 46 73 51
6 76 73 85 86 92 100 80 51 77 61
7 75 70 82 85 90 84 100 55 82 64
8 72 74 81 85 94 84 85 100 88 70
9 52 46 56 57 54 46 46 32 100 59
10 55 53 70 62 56 54 53 38 88 100
153
Таблица 7
Матрица сходства для флор 10 островов Курильской гряды
№ флоры 1 2 3 4 S 6 7 8 9 ю
1 100 66 67 60 44 39 37 25 38 31
2 66 100 69 52 38 34 31 23 31 27
3 67 69 100 67 47 42 39 27 40 38
4 60 52 67 100 64 58 55 40 52 45
5 44 38 47 64 100 82 78 62 62 54
6 39 34 42 58 82 100 82 64 58 57
7 37 31 39 55 78 82 100 67 59 58
8 25 23 27 40 62 64 67 100 47 49
9 38 31 40 52 62 58 59 47 100 71
10 31 27 38 45 54 57 58 49 71 100
Наибольшее включение наблюдается флоры о-вов Матуа, Кетон и
Расшуа в флору о. Симушир, соответственно на 94, 92 и 90%, т. е. ука-
занные флоры (на 9/ю и выше)’ являются частями флоры о. Симу-
шир, поэтому этот остров вполне характеризует флору всех Средних
Курил.
Произведем симметризацию матрицы включения путем взятия сред-
него гармонического двух элементов, находящихся симметрично глав-
ной диагонали. В результате получим матрицу сходства Съеренсена—Че-
кановского (табл. 7).
Транзитивное замыкание матрицы сходства находится после возве-
дения данной матрицы в четвертую степень, согласно правилу „строка-
столбец” по формуле . В результате получается матрица (табл. 8),
имеющая только 7 значении элементов. Ниже будут построены ориенти-
рованный граф включения по данным матрицы мер включения, иерархи-
ческая дендрограмма по данным матрицы сходства и диаграмма Чека-
новского по данным матрицы транзитивного замыкания.
Таблица 8
Матрица транзитивного замыкания
№ флоры 1 2 3 4 5 6 7 8 9 10
1 100 67 67 67 64 64 64 64 62 62
2 67 100 69 67 64 64 64 64 62 62
3 67 69 100 67 64 64 64 64 62 62
4 67 67 67 100 64 64 64 64 62 62
5 64 64 64 64 100 82 82 67 62 62
6 64 64 64 64 82 100 82 67 62 62
7 64 64 64 64 82 82 100 67 • 62 62
8 64 64 64 64 67 67 67 100 62 62
9 62 62 62 62 62 62 62 62 100 71
10 62 62 62 62 62 62 62 63 71 100
154
Графические методы анализа матриц включения и сходства
Рассмотрим матрицу мер включения (табл. 6). Выберем порог
5 > 80% и построим граф включения. Примем объекты флоры за верши-
ны графа. Если в матрице включения на пересечении «-строки и 0-столбца
находится элемент, который больше или равен порогу 6, то из вершины а
исходит дуга1 графа в вершину 0. Дугу обозначают стрелкой. В нашем
случае, например, при 6 >80%, флора 8 включается в флору 3, 4, 5, 6,
7,9, так как на пересечении 8-й строки со столбцом 3, 4, 5,6,7,9 нахо-
дятся числа больше 80 %.
На рис. 1 приведен граф включения для 10 флор островов Куриль-
ской гряды, показывающий связи флор Средних Курил с Южными и
Северными Курилами. Наибольшее количество флор при 5 > 80 % вклю-
чается в флору о. Итуруп. „Наиболее неспецифическая” (по: Юрцев,
1968а) флора о. Матуа при 70% включается во флоры всех остальных
островов Курильской гряды. В целом граф включения показывает более
сильные связи Средних Курил с Южными.
Анализ отношения сходства флор Курильских островов начнем с са-
мого простого метода, используемого в геоботанике, — метода диаграмм
Чекановского (Василевич, 1969; Макаревич, 1971; Норин, 1971). Одна-
ко этот метод в нашем случае должен быть несколько изменен. Обычно
при использовании метода Чекановского производят разбивку мер сход-
ства на различное число классов и дают условное обозначение этим клас-
сам, иногда в виде различной плотности штриховки. В нашем случае
можно взять матрицу транзитивного замыкания (табл. 8) и ввести обо-
значения для 7 различных ее элементов. Располагая элементы этой мат-
рицы с большими численными значениями ближе к диагонали, мы полу-
чим диаграмму Чекановского (рис. 2). В результате метод Чекановского
существенно упростился благодаря введению в него алгоритма нахожде-
ния транзитивного замыкания. В конечном счете после использования
указанного алгоритма матрица сходства имеет ограниченный набор по
величине элементов, и ее достаточно просто привести к блочно-диаго-
нальному виду. В нашем случае (рис. 2) метод Чекановского позволил
разбить всю совокупность флор 10 островов Курильской гряды на 3
группы: флоры Южных, Средних и Северных Курил.
В свою очередь флора Южных Курил неоднородна, и в ней выделя*
ются менее связанные флористической общностью 3 подразделения:
1) флора о. Шикотан, 2) флоры о-вов Кунашир и Итуруп, 3) флора
о. Уруп.
Флора островов Средних Курил разбивается также на 2 подразделе-
ния: 1) флоры о-вов Симушир, Кетон, Расшуа, 2) флора о. Матуа. Дан-
ные разбиения флоры Курильских островов на классы соответствует
приведенному в работе В. М. Шмидта (1980).
Рассмотрим методы анализа отношения сходства, основанные на
агломеративных иерархических кластер-процедурах (Айвазян и др.,
1974). Приведем один простой пример. Возьмем часть матрицы сходства
1 Ориентированное ребро называется дугой (Берж, 1962).
155
Рис. 1. Ориентированный граф включения флор Курильских островов.
1 - Шикотан, 2 - Кунашир, 3 - Итуруп, 4 - Уруп, 5 - Симушир, 6 - Кетон, 7 -
Расшуа, 8 - Матуа, 9 - Парамушир, 10 - Шумшу; сплошные линии - включения
> 80%, прерывистые - > 70%.
а ЕН ЕН ®г И5 Нв
Рис. 2. Диаграмма Чекановского, построенная для флор Курильских островов.
Обозначения флор (1-10) как на рис. 1. а-ж — цифровые значения, заимствован-
ные из табл. 8: а - 62, б - 64, в - 67, г — 69, д - 71, е - 82, ж - 100.
156
a
5 6 7 8 б
5 100 82 78 62 [5,6] 7 8 в
6 100 82 64 5,6 100 80 63 [(5.6), 7] 8
7 100 67 7 100 67 [(5,6), 7] 100 65
8 100 8 100 8 ЮО
5, 6] : 82 [(5,6), 7] : 80 [(5,6,7), 8]: 65
tv 5 6 7 8
%
90 - ।____J <
80 - 1—-|----
70 -
60 -
Рис. 3. Схема расчета и построения дендрограммы методом взвешенного среднего
арифметического связывания.
а-в - часть матрицы (табл. 7) (а) и ее преобразование (б, в); г - дендрограмма,
построенная методом усредненного среднеарифметического связывания. 5-8 -
номера флор, как на рис. 1.
(табл. 7), характеризующей степень сходства флор островов Сред-
них Курил (рис. 3), и построим дендрограмму, например, методом
усредненного среднего арифметического связывания (Айвазян и др.,
1974).
Сущность алгоритма состоит в следующем. В матрице мер сходства
выбирается максимальное число (исключая диагональные элементы).
Если таких чисел несколько, то берется первое по порядку (для этого
элементы матрицы сходства можно упорядочить, например по строкам
слева направо). В нашем случае таким числом будет 82 (рис. 3, а). Эле-
менты, имеющие это значение сходства, объединяются в группу [5, 6].
Размер матрицы сходства уменьшается на единицу, при этом делается
расчет сходства выделенной группы с остальными элементами по фор-
муле
ад/] Л)
rtfKii, к) + п]К(], к)
ni+ni ’
где [i, /] — группа из двух объектов i nj,k — объект, с которым ищется
сходство указанной группы, щ — число элементов в i объекте, и, - число
элементов в / объекте. В нашем случае имеем
157
К ([5,6], 7) = 1'78 + 1 ' 82 = 80;
К([5,6], 8) = 1 '62^ -64 = 63 (рис. 3, б).
&
Процедура сохраняется для пересчитанной матрицы, и ее размер также
уменьшается на единицу. Сходство в нашем случае элементов [(5, 6), 7]
с элементом 8 равно
^([(5,6),7],8)= 2~63з+1 ~67 =65 (рис.З.в).
На основе полученных данных строится дендрограмма (рис. 3, г).
Существенным этапом в указанном алгоритме является расчет сход-
ства между группами объектов. Наиболее общую формулу для этих це-
лей предложил А. Н. Коломогоров (по: Айвазян и др., 1974) :
njK^ki, V + njK^ti, k)
ni + nj
где [z, /] - группа из двух классов i nj-, к — класс, с которым определя-
ется сходство группы [z , /]; и,- и — число объектов в i и / классах,
_оо^^^+ооф
Приведем некоторые из частных алгоритмов иерархической класси-
фикации объектов.
(I) Полносвязывающий метод: ([z,/], k) = min [A (z ,к), K(j,k) ];
(II) метод среднего гармонического связывания:
К ([1П ,:Л _ w, к) ка,
к> K(i,k) + K(j,k) ’
(III) метод взвешенного среднего гармонического связывания:
К'_х ([i, 11 Л) =---’ K(j, к)------------- .
к) + —K(j, к)
п,+п; Пг+П].
(IV) метод среднего геометрического связывания:
Ко ([/. /], к) = ^K(i, к) К(], к)',
(V) метод взвешенного среднего геометрического связывания:
К'о ([i,j],k) = кп‘ + %; к) • + Ч, к);
158
567856765678 5676
Т~1 78
82 82
64.3
Рис. 4. Дендрограмма для флор Средних Курил, построенная восемью методами.
а — полносвязывающий метод; б - методы среднего гармонического, геометриче-
ского и арифметического связывания; в - методы взвешенного гармонического,
геометрического и арифметического связывания; г - односвязывающий метод.
Обозначения флор 5-8 как на рис. 1. Цифры над горизонтальными линиями зна-
чение меры сходства, %.
(VI) метод среднего арифметического связывания:
Kl([i.n,k) = ±[K(J,k') + K(j, *)];
(VII) метод взвешенного среднего арифметического связывания:
_ njK{i, к) + njK(j, к).
“ f
(VIII) односвязывающий метод:
*+eo(U/],*) = max [If (i, k),K(j, fc)].
В литературе по численной таксономии ряд указанных алгоритмов
известен под другими названиями: I - метод „дальнего соседа”, или
„полной связи” (Sokal, Sneath, 1963; Lance, Williams, 1967; Айвазян и др.,
1974); VII — метод средней связи; VIII - метод „ближайшего соседа”,
или единственной связи (Парницкий, 1981).
Наиболее часто в флористических исследованиях используется одно-
связывающий метод, который находится в прямой зависимости с мето-
дом построения кратчайшего дерева, или дендрита (Семкин, 1979; Хан-
тиган, 1980). Обычен также метод взвешенного среднего арифметическо-
го связывания (Пааль, Семкин, 1983; Пушкарь и др., 1984). Другие из
приведенных методов почти не используются в ботанике. Для данного
примера мы построили дендрограммы по каждому из 8 методов (рис. 4)
и получили весьма близкие результаты.
Очевидно, что методы иерархической классификации, основанные на
обобщенном взвешенном среднем связывании,дают близкие результаты
при выполнении следующего условия для объединяемых объектов i nj-.
К (i,j) имеет высокое значение =>К (i, к) — близкое к K(j, к). В нашем
примере это условие выполнилось, например К(5, 6) = 82%=>Х’(5, 7) =
= 78 % и К (в, 7) = 82 %, т. е. 78 % близко к 82 %.
Так как методы взвешенного среднего геометрического, гармониче-
ского и арифметического связывания в нашем примере дали одинаковые
159
Рис. 5. Дендрограмма для флор Курильских островов, построенная методом взве-
шенного среднего арифметического связывания.
Обозначения флор 1 — 10 как на рис. 1.
результаты, для построения дендрограммы флор Курильских островов
возьмем один из них, например последний из указанных. Несильно
отличные результаты от них дал односвязывающий метод, который экви-
валентен методу построения кратчайшего дерева. Этот метод мы также
примем для упорядочивания флор Курильских островов. На рис. 5 и 6
приведены дендрограмма и кратчайшее дерево (дендрит) для флор Ку-
рильских островов. На дендрограмме также указаны стрелками включе-
ния меньших по численности видов флор в более богатые флоры.
В целом флора Курильских островов разбивается на 3 группы флор:
Южных, Средних и Северных Курил. Следует заметить, что методы тран-
зитивного замыкания матрицы сходства (или диаграмма Чекановского),
оптимального дерева (дендрит) и односвязывающий — эквивалентны.1
Так, веса на дендрите (рис. 6) и меры сходства в матрице транзитивного
замыкания (табл. 8) - одни и те же числа. Если учесть включение флор
сравнительно малочисленных по числу видов во флоры более богатые
в видовом отношении, то разбиение флоры Курильских островов на
3 группы будет более конкретным. По-видимому, часто высокое
включение одной флоры в другую (например, флоры о. Шумшу во
флору о. Парамушир) обусловлено неполнотой сбора видов на этих
островах. По уточненным данным В. Ю. Баркалова (1981), флоры
указанных островов сходны на 86% (по мере Съеренсена) и имеют
соответственно число видов 407 и 473. Однако, используя меры
1 Указанные методы основаны на учете единственной, наиболее сильной из
связей между сравниваемыми объектами.
160
Рис. 6. Кратчайшее дерево (дендрит) для флор Курильских островов.
Обозначения флор 1—10 как на рис. 1. Цифры на линиях связи соответствуют зна-
чениям меры сходства в матрице транзитивного замыкания (табл. 8).
включения, можно даже при неполных видовых списках этих флор по-
казать их флористическую близость.
Сравнительный анализ флор рассматриваемыми методами можно
производить и по другим флористическим признакам, например по се-
мейственно-видовым [или „головной” части спектра, по Л. И. Малышеву
(1973)], по широтным географическим спектрам и т. д. В настоящее
время некоторые флористы предлагают использовать для указанных це-
лей методы таксономического анализа Е. С. Смирнова (Шмидт, 1980,
1984) или коэффициент Кендэлла (Ребристая, Шмидт, 1972; Шмидт,
1980). Как отмечалось ранее (Юрцев, Семкин, 1980), общность спектров
можно более точно количественно определить с помощью меры сходства:
п
ка& = %min(dia, di&),
которую мы будем использовать в приведенных ниже примерах. В связи
с ограниченным объемом статьи приведем дендрограммы, построенные
только по двум признакам: 1) семейственно-видовым спектрам (взято
12 наиболее обильных по числу видов семейств); 2) по широтным спект-
рам. Данные для расчета взяты из работ О. В. Ребристой (1977) и
В. М. Шмидта (1980) по конкретным флорам (КФ) европейского севе-
ро-востока.
Из дендрограммы следует (рис. 7, а), что семейственно-видовые
спектры разбивают совокупность рассматриваемых КФ на 2 большие
группы, относящиеся к разным подпровинциям — Канино-Печорской
(Одиндомусюр, Малый Сюрней, Нямдою, Саваю, Морею, Лёк-Воркута,
Пым-Ва-Шор, Сивая Маска) и Югорской подпровинции (Варнек, Хабаро-
ве, Амдерма, Хуптпэ, Усть-Кара, Хальмер-Ю, верховья Кары, Гнегью,
Воркута) Восточноевропейско-Западносибирской провинции (Ребристая,
1971). Выявлено, что по учтенному показателю КФ Лёк-Воркута ближе
И Заказ 1220
161
%
100
90
80
a
70 u
Рис. 7. Дендрограмма для конкретных флор европейского северо-востока, постро-
енная методом взвешенного среднего арифметического связывания.
а - семейственно-видовые спектры, б - широтные спектры.
к флорам Канино-Печорской подпровинции. Как видно, семейственно-
видовой спектр может быть выбран одним из флористических признаков
для разделения флор на классы, имеющие ранг подпровинций.
Зональное деление флор на примере Большеземельской тундры и
некоторых других районов возможно на основе спектров широтных
162
географических групп (Ребристая, 1977). По дендрограмме (рис. 7, б) вы-
деляются группы флор, соответствующие следующим подзонам: высоко-
арктических тундр (о. Георга, п-ов Адмиралтейства), северных гипоарк-
тических тундр (Хабарове, Амдерма, Хуптпэ, Усть-Кара; к этой же
группе тяготеет флора Маточкина Шара из подзоны арктических тундр);
южных гипоарктических тундр (Хальмер-Ю, верховья Кары, Гнетъю,
Одиндомусюр, Малый Сюрней, Нямдою, Саваю, Морею, Воркута) и лесо-
тундры (Пым-Ва-Шор, Лёк-Воркута, Сивая Маска). Южная крупноерни-
ковая полоса подзоны южных гипоарктических тундр по указанным
признакам не выделяется, а относимые к ней флоры присоединяются
(Ребристая, 1977) к северной мелкоерниковой полосе подзоны южных
гипоарктических тундр (Воркута) и к лесотундре (Пым-Ва-Шор).
Таким образом, в сравнительной флористике достаточно использо-
вать односвязывающий метод (дендрит) и метод взвешенного среднего
арифметического связывания для классификации равновеликих флор.
Для разновеликих флор необходим анализ отношения включения с по-
мощью ориентированных графов. Указанные методы могут быть исполь-
зованы для анализа различных флористических признаков.
В. М. Шмидт
О НЕКОТОРЫХ ПРИЕМАХ СРАВНЕНИЯ
СИСТЕМАТИЧЕСКОЙ СТРУКТУРЫ ФЛОР
•Ленинградский государственный университет им. А. А. Жданова
Систематическая структура, или флористический спектр, является
одним из важнейших показателей, используемых в сравнительной флори-
стике (Толмачев, 1974; Шмидт, 1980). Полный флористический спектр
представляет собою ранжированный по числу таксонов низшего ранга
ряд таксонов высшего ранга (например, ряд семейств, выстроенных по
числу видов или родов; ряд родов по числу видов).
Для сравнения систематической структуры флор используют обычно
не весь спектр, а лишь его головную часть, представленную набором
10-15 ведущих таксонов. Это оправдано тем, что такой набор достаточ-
но полно отражает основные черты систематической структуры флор.
Так, например, 10 ведущих по числу видов семейств бореальных флор
содержат до 60% их видового состава, а в условиях Арктики этот про-
цент значительно выше (Шмидт, 1980). А. И. Толмачев (1974) отмечал,
что 10 ведущих по числу видов семейств прекрасно характеризуют
,^чицо” флористической области. Вместе с тем нельзя не заметить, что
исключение из сравнения тех таксонов, которые в ранжированном ряду
занимают последующие места вплоть до последнего, означает потерю
некоторой части информации о сходстве систематической структуры
флор. К этому вопросу мы вернемся в дальнейшем.
163
Был период, когда исследователи ограничивались сравнением флори-
стических спектров „на глаз”, и это, естественно, не могло привести
к получению обобщенного показателя сходства флор по данному при-
знаку. Случилось так, что нам (Заки, Шмидт, 1972; Ребристая, Шмидт,
1972) и Л. И. Малышеву (19726) одновременно и независимо друг от
друга пришла в голову мысль об использовании для этой цели приемов
ранговой корреляции. Мысль, вообще говоря, напрашивающаяся, ибо
коэффициенты ранговой корреляции ps Спирмена и т Кендэла (см.,
например: Кендэл, 1975) специально разработаны для решения подобно-
го рода задач. При этом некоторое предпочтение отдавалось коэффици-
енту Кендэла
2s
т п(п - 1У
где s — сумма рангов (т. е. мест, занимаемых семействами или родами
в ранжированном ряду); п - число пар сравниваемых рангов (т. е. число
ведущих семейств или родов, по которым ведется сравнение).
Дело в том, что этот коэффициент в случае несвязанных рангов рас-
считывается несколько проще и быстрее, чем коэффициент Спирмена,
имеющий вид
где Ed2 - сумма квадратов разностей (d=x-y) между соответствую-
щими рангами (х и у) двух рядов; п — число пар рангов.
Оба коэффициента варьируют в пределах от +1 (полное сходство
двух рядов) до —1 (полное несходство, противоположный порядок рас-
положения таксонов), но имеют разный масштаб, т. е. не адекватны друг
другу по степени выражения сходства.
Здесь надо отметить 2 момента: 1) ранговые коэффициенты корре-
ляции в нашем случае, как впервые отметил Л. И. Малышев (1973), не
имеют ошибки репрезентативности, ибо головная часть флористического
спектра не может рассматриваться в качестве выборки: применительно
к задаче исследования она сама является генеральной совокупностью.
К этим коэффициентам следует относиться как к обычным коэффици-
ентам сходства (Жаккара, Съеренсена—Чекановского и др.), которые
также не имеют ошибки репрезентативности из-за отсутствия естествен-
ной вариабельности учитываемого признака; 2) приведенные выше фор-
мулы справедливы только для случая так называемых несвязанных
рангов, когда каждый таксон имеет свой порядковый номер (ранг),
выраженный числом, не составляющим среднее арифметическое из сумм
нескольких одинаковых рангов (т. е. когда, например, нет двух или бо-
лее семейств с одинаковым числом видов).
На практике же вполне обычен случай, когда 2-3 или более таксо-
нов „делят” между собою места и тогда их ранг, выражаемый средним
из суммы нескольких рангов, может оказаться и дробным числом.
Это — случай так называемых связанных рангов, в котором приходится
164
вводить в указанные выше формулы особым образом рассчитываемые
поправки. Расчет поправок занимает больше времени, чем расчет самих
коэффициентов, но для коэффициента ранговой корреляции ps Спирме-
на это выполняется легче и быстрее, чем для коэффициента т Кендэла.
Поскольку ранговые коэффициенты Спирмена и Кендэла представ-
ляют собою частные случаи обобщенного коэффициента корреляции
(Кендэл, 1975), то в интересующих нас целях вполне может быть исполь-
зован обыкновенный коэффициент линейной корреляции Бравэ—Пир-
сона (г). Этому вопросу посвящена специальная статья (Шмидт, 19816),
поэтому здесь мы рассмотрим лишь основные результаты.
Известно, что коэффициент корреляции г может быть вычислен по
различным формулам. Наиболее удобной в данном случае является
формула, применяемая для малых выборок:
ZxZy
Sxy--------—
п
где х и у — значения рангов одинаковых таксонов в систематической
структуре двух сравниваемых флор; п — число пар рангов.
Существенными оказались 2 обстоятельства: 1) приведенная форму-
ла в отличие от коэффициентов ранговой корреляции не требует введе-
ния поправок в случае связанных рангов и 2) во всех случаях соблюда-
ется равенство г = ps, т. е. эти 2 коэффициента тождественны по степени
выражения связи. Последнее обусловлено тем, что в рассматриваемой
ситуации сумма рангов двух рядов всегда одинакова: Sx = 'Ey и, следо-
вательно, Sx • Sy = (Sx)2 = (Sy)2. Для коэффициента корреляции г
это является, действительно, частным случаем, ибо при других обстоя-
тельствах указанные равенства, как правило, не соблюдаются.
Не вдаваясь в алгебраические преобразования, отметим, что нам
(Шмидт, 19816) удалось вывести новые и более удобные в вычислитель-
ном отношении формулы г и ps.
Для случая несвязанных рангов
3 [4Sxy - n(n+I)2]
Рс Г 3 >
д п — п
а для случая связанных рангов
_ г ____________4 Sxy - n(n+ I)2__________
\/[4Sx2- и(и+ I)2] [4Sy2 - n(n+ I)2]'
До сих пор речь шла о сравнении наборов ведущих таксонов флоры,
т. е. о сопоставлении „головной части” флористических спектров, что
вполне оправдано для решения ряда задач. Но как бы там ни было, при
таком подходе к делу часть информации о систематической структуре
165
флор неминуемо утрачивается, так как положение тех таксонов, кото-
рые составляют среднюю и хвостовую часть флористического спектра,
не учитывается. Между тем они также участвуют в сложении систематиче-
ской структуры флоры и определяют некоторые ее черты. Например,
даже те семейства, которые представлены всего одним видом, могут
быть далеко не безразличны флористу: единственный вид нередко мно-
гое говорит исследователю об особенностях флорогенеза на данной тер-
ритории. Не учитывается рассмотренной методикой и то обстоятельство,
что одно и тоже число видов во флорах при одном и том же числе се-
мейств может быть распределено между последними по-разному, т. е.
само соотношение богатых и бедных видами семейств в разных флорах
далеко не одинаково.
Это распределение лишь теоретически может быть равномерным
(все семейства флоры содержат одинаковое число видов). Хорошо изве-
стно, что на практике оно всегда неравномерно и, более того, обычно
резко асимметрично: число „богатых” семейств невелико, что и позво-
ляет выделять „головную часть” флористического спектра. Степень не-
равномерности указанного распределения может и должна учитываться
при сравнении флор.
Сказанное побудило нас (Шмидт, Малышев, 1983) искать способ ста-
тистического сравнения полных флористических спектров.
Для решения этой задачи потребовалось ввести новое понятие „слож-
ность систематической структуры флоры” (или „сложность флористиче-
ского спектра”), которое включает в себя одновременно как уровень
флористического богатства (т. е. число таксонов того или иного ранга),
так и степень выравненности распределения таксонов низшего ранга по
таксонам высшего ранга. Иначе говоря, сложность систематической
структуры флоры рассматривается как функция двух аргументов: числа
таксонов и выравненности их распределения по таксонам следующего
ранга.
В геоботанике (Василевич, 1978) и экологии (Песенко, 1978а)
имеются примеры использования показателя, который учитывает при-
знаки, аналогичные тем, которые входят в состав сформулированного
выше понятия сложности систематической структуры флоры. Имеется
в виду одна из информационных мер разнообразия — так называемая
//-функция:
Я= -'Lpi\og2pi ,
причем за вероятность pt следует принять наблюдаемую встречаемость
данного числа таксонов низшего ранга в составе каждого таксона высше-
го ранга, т. е. /и, где nt — число таксонов низшего ранга (напри-
мер, видов) в составе таксона высшего ранга (например, семейства);
п — общее число таксонов низшего ранга (видов) во флоре.
//-функция принимает максимальное значение тогда, когда имеет
место полная выравненность распределения (например, когда все семей-
ства содержат одинаковое число видов). Ясно, что это - лишь теорети-
ческий предел //-функции, который может быть рассчитан по формуле
166
//max = log2Л7, где N - число таксонов высшего ранга. Отношение рассчи-
танного значения Н к //тах может служить показателем приближения
эмпирического распределения к максимальной выравненности: Н'=
=///Ятах, причем значение Я=0 (все виды флоры относятся к одному
семейству) практически не осуществляется.
Что касается минимального значения //-функции, то его можно опре-
делить (Песенко, 1978а) по формуле //mjn = 1/п [nlog2n-(n-N+ 1) X
X log2 (n-N+ 1)], где n - число таксонов низшего, aN— высшего ранга.
Эта формула соответствует тому теоретически крайнему случаю, когда
все семейства, кроме одного, содержат по единственному виду каждое.
В промежутке между Нтт и //тах //-функция принимает любые
значения в зависимости от числа таксонов низшего и высшего ранга и от
степени выравненности распределения. Чем оно контрастнее (т. е. чем
меньше его выравненность, чем резче выделяются богатые и бедные ви-
дами семейства), тем меньшее значение принимает эта функция. Ясно,
что пределы варьирования //-функции, зависящие от числа таксонов низ-
шего и высшего ранга, справедливы лишь для каждого конкретного слу-
чая сравнения флористических спектров.
На практике рекомендуется рассчитывать 3 „сорта” //-функций,
биологическая интерпретация которых может быть содержательной:
HSF — показатель сложности структуры семейств по числу видов; HGF -
показатель сложности структуры семейств по числу родов; HSG — пока-
затель сложности структуры родов по числу видов.
В заключение отметим, что //-функцию можно использовать также
при сравнении географических, экологических и прочих спектров флор:
важно, чтобы исходные данные были представлены ранжированными
рядами.
А. В. Галанин
ОПЫТ СОПРЯЖЕННОГО АНАЛИЗА
ТИПОЛОГИЧЕСКИХ СТРУКТУР
КОНКРЕТНЫХ ФЛОР
Институт биологических проблем Севера ДВНЦ АН СССР
Исследования методом конкретных флор (КФ) показали, что при
широких географических сравнениях отдельные КФ целесообразно рас-
сматривать как „точечные” характеристики флористической ситуации
(Юрцев, 1975а, 1982а), так как при этом не столь важно, на какой пло-
щади они выявлены, а важно, чтобы они были полными и целостными
в ландшафтном отношении, чтобы площадь их выявления охватывала
все разнообразие биотопов элементарного флористического района
в некоторой повторности. В разных природных зонах, на равнинах и
в горах площади выявления КФ колеблются в достаточно больших пре-
делах - от 100 до 1000 км2 (Юрцев, 1975а; Шмидт, 1980).
167
Однако КФ не только точечные характеристики флористической
ситуации. Они имеют вполне определенную „внутреннюю” эколого-
ценотическую структуру (Галанин, 1973), которая зависит не только
от характера рельефа и экологической структуры ландшафта, но и
от исторических факторов. Именно историческая обусловленность
многих особенностей эколого-ценотической структуры КФ привлекает
к ее изучению флористов. В этом случае флорист, так же как и гео-
ботаник, имеет дело с растительным покровом и всеми категориями,
отражающими структуру растительности. Представление о флоре как
о множестве местных популяций видов в наибольшей степени соответ-
ствует современному состоянию флористики, так как сейчас уже недо-
статочно рассматривать и изучать флору как множество равноценных
таксономических единиц безотносительно к объему их местных популя-
ций, без учета взаимодействия этих популяций со средой и друг с другом
(Юрцев, 1982а).
Несомненно, что важнейшей характеристикой флоры как дискрип-
тивного множества является видовой состав и таксономическая струк-
тура в целом. Вместе с тем на этом множестве могут быть выявлены и
другие аспекты структуры: ареалогический, эколого-ценотический,
спектр жизненных форм и т. д. Иными словами, на одном и том же мно-
жестве видов можно получить различные разбиения на подмножества,
получив при этом несколько аспектов структуры, или несколько фак-
тор-множеств (Юрцев, Семкин, 1980).
Обозначим фактор-множества А, В, .... К, а их подмножества А ь А2,
..., А{; Bi, В2, ...» Bj-, ...; Kit К2, ..., Kt. Каждый вид конкретной флоры
входит в одно из подмножеств каждого фактор-множества. В пределах
одного фактор-множества подмножества не пересекаются, но подмноже-
ства, относящиеся к разным фактор-множествам, взаимно пересекаются,
образуя матрицу отношений. Обозначим элементы пересечения двух фак-
тор-множеств
п{А^Вх), п{ЛхС\В2), ..., ntA^Bj),
n(A2c\Bi), и(42ПЯ2), ..., n(X2n^),
n(At C\Bi\ n(At ПВ2), ..., л (A, QB.),
где (AiABx) - число элементов (видов), одновременно принадлежа-
щих подмножествам A i и Bi фактор-множеств А и В, и т. д.
Система этих пересечений образует матрицу бинарных отношений,
анализируя которую мы можем поставить вопрос о статистической не-
зависимости или, напротив, о сопряженности распределения видов по
двум фактор-множествам (Галанин, 1974; Галанин, Марина, 1982).
В случае отсутствия сопряженности ожидаемое число видов в подмно-
жествах (элементах) пересечения можно рассчитать по формуле
п(А{)- п(В.)
Я(А,ПД;)= <’n
168
где n(Aj) — число видов в подмножестве А{ фактор-множества А\
— число видов в подмножестве By фактор-множества В; N— об-
щее число видов во флоре. При наличии сопряженности n(AiC\Bj)T^
^n(At<~\Bj). Статистическая достоверность сопряженности может быть
оценена с помощью критерия Пирсона х2:
р * , Т2
где aij = n(Ai ПВ;); а1^ = п(А{ АВу)т; q - число элементов в фактор-
множестве А; р — число элементов в фактор-множестве В. Рассчитанное
по формуле значение х2 сравнивается с табличным при к степенях сво-
боды, где £ = <?(р-1),и5 %-ном уровне значимости PQ 05.
Достоверность величины х2 оценивается как для матрицы в целом,
так и отдельно для строк и столбцов, и даже для отдельный элементов
пересечения. В связи с тем что число подмножеств в фактор-множествах
в разных КФ неодинаково, необходимо нормировать суммарное значение
Х2матрицы числом элементов пересечения (число клеток матрицы бинар-
ных отношений). Этот показатель т можно назвать „среднее значение х2
на одно подмножество пересечения”:
X2
т =-----
q р
В принципе сопряженный анализ фактор-множеств КФ можно проводить
не только с помощью матриц бинарных отношений, но и тринарных и т. д.
В этом случае элементы пересечения будут тройными, четверными и т. д.
Степень сопряженности при увеличении размерности матрицы будет сни-
жаться. Ясно, что для анализа сопряженности трех и более фактор-мно-
жеств КФ необходимо использование ЭВМ. Ввод информации должен
осуществляться путем перечисления видов КФ с указанием их принад-
лежности к определенным подмножествам всех выделенных фактор-
множеств.
Настоящий метод сопряженного анализа структуры КФ позволяет
изучать флору естественной фитохории как некое закономерное целое,
как систему видов и вышестоящих таксонов. Эта система видов лимити-
рована разными факторами: разнообразием таксонов разного ранга,
количеством и характером географических элементов, жизненных форм,
эколого-ценотических элементов. Степень целостности флоры как си-
стемы видовых популяций невысока, это — стохастическая система, но
тем не менее географическое и экотопическое „маневрирование” попу-
ляций при перестройках флор происходит, по-видимому, сопряженно
(Юрцев, 1982а). К сожалению, конкретными и тем более достоверными
данными по этому вопросу мы практически не располагаем.
169
Настоящая статья является результатом многолетних флористиче-
ских исследований методом КФ в разных регионах страны. Впервые
сопряженный анализ долготной и широтной ареалогических структур КФ
был использован нами для анализа двух КФ Чукотской тундры (Гала-
нин, 1974).
Одна КФ выявлена на Восточной Чукотке в среднем течении р. Эргу-
веем. Район расположен в полосе типичных гипоарктических тундр и
относится к Берингийско-Чукотской подпровинции Чукотской провин-
ции Арктической флористической области (Юрцев, 19746). Рельеф сред-
негорный, мелкосопочный. Климат арктический морской. Здесь широко
распространены мощные снежники, порой перелетовывающие, и специ-
фические нивальные сообщества растений с сильно укороченным сезон-
ным циклом развития.
Вторая КФ выявлена на Западной Чукотке в среднем течении р. Рау-
чуа. Район расположен в полосе южных гипоарктических тундр и отно-
сится к Континентально-Чукотской подпровинции Чукотской провинции
Арктической флористической области (Юрцев, 19746). Рельеф района
также среднегорный, но со слабо выраженными альпийскими чертами.
Климат арктический континентальный. Мощные снежники и их ниваль-
ные сообщества здесь встречаются значительно реже, чем в предыдущем
районе (Галанин, 1973,19776).
Третья КФ выявлена на северо-востоке европейской части СССР
в бассейне р. Цильмы. Район расположен в подзоне северной тайги в вос-
точной части Тиманского кряжа и относится к Восточноевропейской
провинции Бореальной флористической области (Толмачев, 1974).
Рельеф равнинный с выходами коренных пород по берегам рек. Климат
бореальный континентальный, с продолжительной многоснежной холод-
ной зимой и прохладным коротким летом.
Четвертая КФ расположена в окрестностях биостанции Сыктывкар-
ского университета (Коми АССР) в среднем течении р. Вычегды в под-
зоне типичной тайги и также относится к Восточноевропейской провин-
ции Бореальной флористической области. Рельеф - слабо всхолмленная
равнина, обширная долина крупной реки. Климат бореальный, умеренно
континентальный, с многоснежной холодной зимой и относительно теп-
лым непродолжительным летом.
Пятая КФ выявлена в окрестностях Сыктывкара. Этот район распо-
ложен несколько западнее предыдущего и с некоторой натяжкой может
быть отнесен к переходной полосе от подзоны типичной тайги к подзоне
южной тайги. Общее экологическое разнообразие ландшафта здесь выше,
сильнее выражено и антропогенное воздействие.
Шестая КФ расположена в дельте р. Неман в Калининградской обл.
Эта южная часть полосы хвойно-широколиственных лесов Среднеевро-
пейской флористической области. Рельеф — приморская низменность,
дельта крупной реки. Климат умеренно континентальный, с мягкой
зимой и относительно прохладным для данной широты и дождливым
летом. Сильно выражено антропогенное воздействие. Значительная
часть территории является польдерными землями (Галанин, Алексеева,
1976).
170
1
Седьмая КФ расположена на Восточном Алтае в районе хр. Куркуре
(Алтайский заповедник). Это район горной лиственничной и кедровой
тайги с возвышающимися над ней высокогорьями (ерниковые тундры,
субальпийские и альпийские луга, скалы, осыпи, субнивальный пояс).
Рельеф типично альпийский, высокогорный, отдельные -вершины пре-
вышают 3100 м над ур. м. Более детально флора и растительность этого
района охарактеризована ранее (Галанин, 1979; Галанин, Марина, 1982).
Кроме флоры хр. Куркуре, все виды КФ были разделены на 2 типа
ареалогических элементов: широтные (4) по протяженности ареала с се-
вера на юг и долготные (В) по протяженности ареала с запада на восток.
Оба фактор-множества и элементы их пересечения приведены в табл. 1-4.
В каждом элементе пересечения приведено наблюдаемое число видов и
ожидаемое. Значения х2 рассчитаны отдельно для широтных и долготных
элементов и для КФ в целом.
Анализ показал, что для флор Эргувеем и Раучуа сопряженность
между широтной и долготной протяженностью ареалов недостоверна,
как в целом по таблице, так и по отдельно взятым фактор-множествам
(табл. 1). Только для Раучуа достоверно отрицательное отклонение
в подмножестве пересечения „бореальные берингийские виды”. Отсутст-
вие значимой сопряженности между выделенными фактор-множествами
показывает, что широтная и долготная ареалогические структуры обеих
КФ практически независимы друг от друга.
Во флоре Цильмы оба аспекта ареалогической структуры достоверно
сопряжены друг с другом в целом (табл. 2). Кроме того, по долготным
элементам не случайно распределены полигональные виды, неморально-
бореальные и бореальные. При этом широтная протяженность ареала пря-
мо пропорциональна их долготной протяженности. Гипоарктические
виды по долготным секторам распределены случайно. Слабые отклоне-
ния от случайного распределения наблюдаются в группах арктических
и арктоальпийских видов.
Во флоре окрестностей биостанции Сыктывкарского университета
(табл. 3) степень сопряженности широтной и долготной ареалогических
структур более высокая, чем во флоре Цильмы. Подобный характер рас-
пределения видов по долготным и широтным элементам пересечения
наблюдается и во флоре окрестностей г. Сыктывкара (табл. 2), но сте-
пень сопряженности здесь еще выше. Максимальная сопряженность ши-
ротной и долготной ареалогических структур имеет место во флоре
дельты Немана (табл. 4).
Разумеется, данные, которыми мы располагаем, не позволяют делать
далеко идущие выводы. К тому же интерпретация таких зависимостей
очень трудна из-за многомерности и многофакторности естественной
флоры как экологически и исторически обусловленной системы. Тем
не менее нельзя не обратить внимание на то, что степень сопряженности
долготной и широтной ареалогических структур КФ по мере продви-
жения на юг от тундры к широколиственным лесам закономерно уве-
личивается. По всей вероятности, флоры перигляциалыюй зоны Восточ-
ной Европы и Сибири во время оледенения были тесно взаимосвязаны
и относились к одной флористической области. Поэтому долготная
171
Таблица 1
Распределение видов флоры Эргувеем и Раучуа по широтным и долготным а ре алогическим элементам
Долготные ареалогические элементы (By)
Широтные ареалогические элементы (Л,-) Циркумпо- лярные и почти цир- кумполяр- ные Евразиатско- эападноаме- риканские Сибирско- американ- ские Берингийские (восточноси- бирско-зап ад- ноамерикан- ские) Американ- ские, захо- дящие на восток Се- верной Азии Евр азиат- ские и си- бирские, не заходящие в Америку Восточноси- бирские и чукотские, не заходящие в Америку Е х2
Среднее течение р. Эргу! веем
Арктические 19 21.4 6 6.9 2 2.4 18 15.8 6 5.1 2 3.0 7 53 60 1.49
Арктоальпийские 53 19 5 39 15 9 13 153 1.07
54.6 17.5 6.2 40.2 12.9 7.7 13.9
Гипоарктические 22 22.8 5 7.3 3 2.6 19 16.8 4 •5.4 4 3.2 7 5.8 64 1.92
Бореальные 12 7.1 4 2.3 2 0.8 2 5.3 0 1.7 0 1.0 0 1.8 20 12.99
Е 106 34 ’ 12 78 25 15 27 297
X3 3.55 2.24 2.16 2.69 236 1.75 2.52 17.47
Среднее течение р. Раучуа
Арктические 14 13.3 6 6.1 2 13 7 5.7 3 13 0 2.6 2 4.4 34 5.92
Арктоальпийские 49 50.0 19 17.2 5 5.6 28 22.8 4 5.6 9 10.4 14 16.4 128 2.46
Гипоарктические 26 33.6 10 11.6 4 3.8 18 15.3 3 3.8 10 7.0 15 11.0 86 5.35
Бореальные 36 44.3 8 9.7 3 3.2 4 12.9 4 3.2 7 5.6 10 9.2 72 8.63
Е 125 43 14 57 14 26 41 320
х2 3.34 0.71 0.25 8.11 2.33 4.43 3.19 22.36
Примечание. Здесь и в табл. 2 — 12 иад чертой — фактическое число видов в каждом элементе пересечения, под чертой — ожидае-
мое (пояснение в тексте).
Таблица 2 Распределение видов флоры бассейна р. Цильмы (Коми АССР) и окрестностей Сыктывкара по широтным и долготным ареалогическим элементам
Широтные ареалогические элементы (А{) Долготные ареалогические элементы (By )
Циркумполярные и почти циркум- полярные Евразиатские и европейско- сибирские Азиатские и азиат- ско-восточно- европейские Европейские и европейско-за- падноазиатские X2
Бассейн р. Циль мы
Полизональные 55 28.0 15 28.7 0 4.8 4 12.5 74 43.16*
Неморально-бо реальные 4 13.3 16 13.6 4 2.3 11 5.9 35 1239*
Бореальные 116 166 30 70 382 9.46*
144.8 148.0 24.9 64.4
Арктические и арктоальпийские 30 21.6 15 22.1 3 3.7 9 9.6 57 5.72
Гипоарктические 22 19.3 20 19.8 2 3.3 7 8.6 51 1.19
227 232 39 101 599
х2 41.92* 11.43* 7.75 11.02* 72.12*
Окрестности Сыктывкара
Полизональные 57 26.0 47 61.0 1 2.0 20 36.0 125 47.79*
Неморально-бо реальные 8 7.9 8 18.6 0 0.6 22 11.0 38 17.75*
Бореальные 23 170 6 92 291 29.96*
603 142.1 4.6 83.9
Лесостепные 2 5.9 13 13.7 1 0.4 12 8.1 28 5.11
Гипоарктические и арктоальпий- ские 16 5.8 11 13.7 0 0.4 1 8.1 28 24.96*
106 249 8 147 510
х2 80.51* 15.27* 2.62 27.17* 12537*
* Значения X2. говорящие о достоверной сопряженности.
X * « * * О' СП «п О о 00 »п О v-j 'О ч- еч f'' чо -н Г- -ч 137.27*
(ГОТНЫМ м m in оч оо г* О гч m я—« гч 382
о и S I § о. § о п S 4> н б о. Эндемы се- веро-востока европейской части СССР О1еч О 1 —1* 1* Ч- 4О|-1 ojo о|о 40 31.00*
секты (By) Американ- ско-евро- пейские -к " • # I* a# 1# гч о|о о|о О О ч- 0.50
а 3 ыв карского униве элементам Л о 5 О X X о о X Е о 5 4> Европейские и европейско- западно азиатс- кие (на восток до Оби) ^2 3 ОО «1^ сп|^ о|-ч 65 30.29*
Таблиц эиостанции Сыкт а ре алогическим Долготные а] Азиатские и восточноевро- пейско-азиатс- кие Ч- Г~- ♦ * 00 ’'1® о|о 14 18.38*
! 8. X о 2 Евразиатские и европейско-за- падноазиатские (на восток до Байкала) t-|cn m| Ч- £ ^|ч- ®|сп 161 20.40*
« i п и § Циркумпо- лярные и почти цир- кумполяр- ные 58 36 60 г? Г'"| 2 о| «и г-1 сч 132 36.70*
1 & с Широтные ареалогические элементы (Л{) Полигональные Неморал ьно-боре - альные Бореальные Лесостепные Гипоарктические и арктоальпийские н X
Значения х2> говорящие о достоверной сопряженности.
* Ожидаемые числа видов, равные нулю, получены в результате округления дробей до целых чисел.
176
Таблица 4
Распределение видов флоры дельты р. Немана по широтным и долготным
ареалогическим элементам
Широтные ареалогические элементы (Л{-) Долготные ареалогические элементы (By)
Циркумпо- лярные и почти цир- кумполяр- ные Европейские (в том числе) западно- европей- ские) Европей- ско-за- падно- аэиатские Еврази- атские X1
Полигональные 17 5.1 0 5.0 0 2.8 2 6.1 19 38.33*
Бореально-немо- 69 56 37 98 260 4.81
ральные 693 68.8 37.9 83.8
Не моральные 2 13.4 31 13.2 11 7.3 6 16.1 50 41.92*
88 87 48 106 329
х1 37.47* 31.38* 4.70 11.51* 85.06*
Примечание. Здесь и в табл. 5-12 звездочкой отмечены значения х\
говорящие о достоверной сопряженности.
и широтная протяженность ареала у характерных представителей тундро-
вых флор слабо связаны друг с другом. По мере продвижения к югу
ранг долготных градиентов, лимитирующих распространение видов, ста-
новится соизмеримым с рангом широтных или зональных градиентов.
Таким образом, в Берингийском секторе Арктики происходило более
или менее свободное и независимое расселение холодостойких элемен-
тов флоры в долготных и широтных направлениях.
Говоря об организации флоры как системы региональных видовых
популяций, мы имеем в виду сопряженность ее эколого-ценотической и
ареалогической структур, определяемую синхронностью преобразований
этих структур во времени. Для изучения этого вопроса проведен сопря-
женный анализ эколого-ценотической (А) и ареалогической (В) струк-
тур пяти конкретных флор. Для флор Эргувеем, Раучуа, окрестностей
биостанции Сыктывкарского университета и окрестностей г. Сыктывка-
ра сопряженность эколого-ценотической структуры со структурами ареа-
логическими рассчитывалась отдельно для широтных и долготных эле-
ментов (табл. 5—11). Во флоре Куркуре ареалогические элементы выде-
лены без подразделения на широтные и долготные (табл. 12). Для всех
девяти матриц величина сопряженности в целом достоверна. При этом
значительно выше сопряженность с широтной, а не долготной ареалоги-
ческой структурой (табл. 13).
Разные эколого-ценотические элементы флор имеют разную ареало-
гическую структуру, а следовательно, и разную историю формирования.
Так, например, во флоре Эргувеем в комплексе видов, тяготеющих
12 Заказ 1220
177
Таблица 5
Распределение видов флоры Эргувеем и Раучуа по эколого-ценотическим и широтным ареалогическим элементам
Эколого- ценотические элементы (4/) Широтные ареалогические элементы (Бу)
Арктоаль- пийские Арктиче- ские Гипоаркти- ческие Бореаль- ные Е х2
Эргувее м
С широкой экологи- ческой амплитудой 10 14.2 2 3.1 И 4.1 0 1.6 23 14.8*
Сухих горных тундр 23 18.5 5 4.0 2 5.4 0 2.1 30 5.6
Равнинных тундр 8 5.6 1 1.2 0 1.6 0 0.6 9 3.2
Сырых эвтрофных тундр 32 32.7 13 7.1 5 9.5 3 3.8 53 9.0*
Нивальных луговин 85 58.6 0 12.8 10 17.0 0 -6.7 95 34.3*
Пойменных ивняков 22 14 22 14 35.1*
и лугов 44.4 9.7 12.9 5.1 / Z
При бре жно-водных местообитаний 3 9.2 5 2.0 3 2.7 4 1.1 15 16.4*
Е 183 40 53 21 297
х2 30.7* 26.6* 26.7* 34.4* 118.4*
Раучуа
С широкой экологи- ческой амплитудой 7 7.3 0 1.5 10 4.4 0 3.9 17 12.5*
Сухих горных тундр 50 35.0 7 7.3 20 21.0 5 18.8 82 16.6*
Равнинных тундр 6 3.4 0 0.7 2 2.1 0 1.8 8 4.5
Сырых эвтрофных тувдр 25 14.9 4 3.1 5 9.0 1 8.0 35 15.1*
Нивальных луговин 9 4.3 1 0.9 0 2.6 0 2.3 10 10.0*
Пойменных ивняков 23 6 28 41 98 25.0*
и лугов 41,8 8.7 25.1 22.4
Олиготрофных болот ВЫСОКОЙ ПОЙМЫ 2 11.5 6 2.4 8 6.9 11 6.2 27 17.2*
При бре жно-водных местообитаний 3 6.8 2 1.4 2 4.1 9 3.7 16 14.2*
2 125 26 75 67 293
х2 38.9* 9.0 13.2* 54.0* 115.1*
Распределение видов флоры Эргувеем по эколого-ценотическим и долготным ареалогическим элементам
X * # # 'О iq еч 40 rt 40 ’о Vi щ го ей оо 40 еч ,-ч м 76.8*
м <о 04 04 <о щ еч еч еч 04 г— । 297
Восточносибир- ские, не идущие в Америку 1 18 10.6
_ I ’’I’ 100 40 °|- -U °|d ™ 2 - “ S : -|з
:ские элементы 1 Евразиатские и сибирские =|2 -|S =|S -> 2.9 0 5.1 10 04 ,|оо ей *4 О 16 18.3*
sr Е о 5 о. 4) 2 X о Восточносибир- ско-западно- американские (берингийские) оо I4® —,|’_1 '«o —ч 140 |гч —• ei 2ч 21.8 14 “7 I’’!’ 1^’ 68 * г4 m
§ Сибирско-амери- канские и си- бирско-западно- американские „-|O4 - Г"**| • 1 4^1 “ |un |^> |<^ 2л оо 2 22.7 15 |4О 71 5.9
Циркумполяр- ные и почти цир- кумполярные oj£ 40 |S £ 21.9 50 | 39.3 23 3 123 9.3
Эколого-ценотические элементы (A j) С широкой экологиче- ской амплитудой Сухих горных тундр Равнинных тундр Сырых эвтрофных тундр Нивальных луговин Пойменных ивняков и лугов Прибрежно-водных ме- стообитаний н сч X
179
00
о
Таблица 7
Распределение видов флоры Раучуа по эколого-ценотическим и долготным ареалогическим элементам
Эколого-ценотические элементы (Л(-) Долготные ареалогические элементы (By)
Циркумполяр- ные и почти цир- кумполярные Сибирско-амери- риканские и си- бирско-западно- американские Восточносибир- ско-эападно- американские (берингийские) Евраэиатские и сибирские Восточносибир- ские, не идущие в Америку Е х2
С широкой экологиче- 8 4 3 0 2 17 1.7
ской амплитудой 6.7 4.0 3.0 1.4 1.9
Сухих горных тундр 19 323 17 19.3 26 14.3 7 6.7 13 9.2 82 17.1*
Равнинных тундр 2 2 3 0 1 6.0
3.2 1.9 1.4 0.7 0.9 8
Сырых эвтрофных тундр 13 13.9 11 8.2 4 6.1 3 2.9 4 3.9 35 1.8
Нивальных луговин 4 4.0 2 2.4 3 1.7 0 0.8 1 1.1 10 1.9
Пойменных ивняков и 39 27 9 12 11 98 & с
лугов 38.8 23.1 17.1 8.0 11.0 0 *0
Олиготрофных болот 20 3 3 1 0 27 14.1*
высокой поймы 10.7 6.4 4.5 2,2 3.0
Прибрежно-водных ме- 11 3 0 1 1 16 7.0
стообнтаний 6.3 3.8 2.8 1.3 1.8
Е 116 69 51 24 33 293
Xs 18.1* 4.1 23.2* 5.7 5.0 56.1*
Таблица 8
Распределение видов флоры окрестностей биостанции Сыктывкарского университета по эколого-ценотическим
и широтным ареалогическим элементам
Эколого-ценотические элементы (Лу) Широтные ареалогические элементы (By)
Полизоиаль- ные Неморальио- бо реальные Бореальные Лесостепные Гипоарктические и арктоальпийские Е X2
Лесов зеленомошно-кустар- 9 65 9 0 0 83 173*
яичкового ряда 22 49 7 2 2
Олиготрофных болот и забо- 0 11 6 0 1 18 14.0*
лоченных сосняков 5 11 2 0 0
Боров беломошных 2 5 8 11 5 2 0 0 3 0 18 10.1*
Пойменных ивняков и речных 7 24 3 0 0 34 3.2
пляжей 9 20 3 1 1
Катищ и мелиорируемых лугов 22 И 15 25 0 4 5 1 0 1 42 36.0*
Среднеувлажненных и сухих 28 59 15 3 3 108 5.4
лугов 29 64 9 2 2
Сырых и заболоченных лугов 25 28 0 0 0 53 15.9*
и травяных болот 14 31 5 1 1
Водных и прибрежно-водных 11 14 _1 0 0 26 5.5
местообитаний 7 15 2 1 1
Е 103 225 39 8 7 382
г» X 181 37.4* 11.6 26.6* 21.5* 10.5 107.6*
Таблица 9
оо
N)
Распределение видов флоры окрестностей биостанции Сыктывкарского университета по эколого-ценотическим
и долготным ареалогическим элементам
Эколого- ценотические элементы (А{) Долготные ареалогические элементы (By)
Циркумполяр- ные и почти циркумпо- лярные Евразиатские и европейско- западноазиатс- кие (на восток до Байкала) Азиатские и восточноевро- пейско- азиатские Европейские и европейско- западно- азиатские (иа восток до Оби) Американ- ско-евро- пейские Эидемы севе- ро-востока европейской части СССР Е х2
Лесов зел еномошно- 32 28 8 14 0 1 11.08
кустарничкового ряда 28.7 35.0 3.0 14.1 0.9 1.3 83
Олиготрофных болот и 15 2 0 1 0 0 19.24*
заболоченных сосняков 6.2 7.6 0.7 3.1 0.2 0.3 18
Боров беломошных 8 6.2 4 7.6 1 0.7 3 3.1 0 0.2 2 0.3 18 12.19*
Пойменных ивняков и 9 14 3 8 0 0 4.62
и речных пляжей 11.8 14.3 1.3 5.8 0.4 0.5 34
Катищ и мелиорируе- 13 22 1 4 2 0 9.90
мых лугов 14.5 17.7 1.5 7.2 0.4 0.7 42
Среднеувлажиенных и 12 60 1 31 1 3 108 33.66*
сухих лугов 37.3 453 4.0 18.4 1.1 1.7
Сырых и заболоченных 22 28 0 2 1 0 10.62
лугов и травяных болот 18.3 22.3 1.9 9.0 0.6 0.8 53
Водных и прибрежно- 21 3 0 2 0 0
водных местообитаний 9.0 11.0 1.0 4.4 0.3 0.4 26 24,83
Е 132 161 14 65 4 6 382
х1 48.12* 20.20* 16.70* 19.05* 8.68 13.39 126.14*
Та блица 10
Распределение видов флоры окрестностей г. Сыктывкара по эколого-ценотическим и широтным ареалогическим элементам
Эколого- ценотические элементы (Aj) Широтные ареалогические элементы (By)
Поли зо- нальные Арктоаль- пийские Гипоарк- тические Ар кто бо- реальное Бореаль- ные Немораль- но-бо реаль- ные Почти не- мо раль- ные Лесостеп- ные Е х2
Лесов зеленомошно-кус- 0 2 9 12 102 И 12 1 149 57.33*
тарничкового ряда 22.8 2.4 4.3 7.2 96.4 4.1 5.1 6.7
Сосновых лесов 1 1.2 0 0.2 0 0.4 0 0.7 12 9.1 0 0.4 0 03 1 0.6 14 4.08
Низинных болот и забо- 5 0 4 2 66 2 0 1 80 16.38*
лоченных лугов 12.3 1.3 2.3 3.9 51.7 2.2 2.7 3.6
Олиготрофных болот 1 2.0 0 0.2 1 0.4 3 0.6 8 8.4 0 0.4 0 0.4 0 0.6 13 12.62*
Пойменных ивняков 0 7.8 2 0.8 2 13 4 23 33 33.0 3 1.4 5 1.7 2 2.3 51 18.95*
Водных и прибрежно- 20 0 0 1 17 0 0 0 38 42.98*
водных местообитаний 5.8 0.6 1.1 1.8 24.6 1.0 1.3 1.7
Сухих и среднеувлаж- 19 6 1 7 134 1 2 15 185 23.27*
ненных лугов 28.4 3.0 5.4 9.0 119.7 5.1 6.3 8.4
Сильно нарушенных 49 0 _1 1 29 0 2 8 90 116.15*
местообитаний 13.8 13 2.6 4.4 58.2 2.5 3.1 4.1
Е 95 10 18 30 401 17 21 21 620
еч X 183 163.76* 8.67 1334* 18.76* 23.97* 21.06* 23.77* 18.23* 291.76*
оо Таблица 11
Распределение видов флоры окрестностей г. Сыктывкара по эколого-ценотическим и долготным ареалогическим элементам
Эколого- ценотические элементы (Aj) Долготные ареалогические элементы (Bj)
Циркум- полярные и почти циркум- полярные Евразиат- ские Европейско- сибирские (на восток до Байкала) Европейско- западноази- атские (на восток до Ср. Азии и Енисея) Европейско- западноси- бирские (иа восток до Оби) Азиатские и аэиатско- врсточно- европейские Европей- ские Е х2
Лесов зелено мошно-кус- 45 19 38 6 22 5 14
14Q 1 С ГТ*
тарничкового ряда 49.3 28.1 31.5 7.9 12.9 2.2 17.1 1*Г7 1Э.0 /
Сосновых лесов 7 0 3 2 0 0 2 14 6 86
4.6 2.6 3.0 0.8 1.2 0.2 1.6
Низинных болот и забо- 50 12 13 2 1 0 2 80
лоченных лугов 26.5 15.1 16.9 4.3 7.0 1.2 9.2 35.58*
Олиготрофных болот 12 4.3 0 2.5 1 2.8 0 0.7 0 1.1 0 0.2 0 1.5 13 20.95*
Пойменных ивняков И И 11 4 5 1 8 51 3 93
16.9 9.6 10.8 2.7 4.4 0.7 5.8
Водных и прибрежно- 23 4 9 0 2 0 0
водных местообитаний 12.6 7.2 8.0 2.0 3.3 0.6 4.4 38 17.64*
Сухих и среднеувлаж- 30 39 44 14 20 2 36
ненных лугов 61.2 34.9 39.1 9.9 16.1 2.7 21.2 185 30.16*
Сильно нарушенных мес- 27 32 12 5 4 1 9
тообитаний 29.8 17.0 19.0 4.8 7.8 1.3 10.3 90 18.06*
Е 205 117 131 33 54 9 71 620
х2 63.07* 24.03* 6.72 8.24 17.25* 6.14 23.40* 148.85*
Таблица 12
Распределение видов флоры хр. Куркуре по эколого-ценотическим и ареалогическим элементам
Эколого- ценотические элементы (Л(-) Ареалогические элементы (By)
Циркумпо- лярные по- лизональ- ные и бо- реальные Широко распрост- раненные арктоаль- пнйские Ев разиат- ские по- лигональ- ные и бо- реальные Азиатские бореаль- ные и по- ли зональ- ные Централь- ноазиат- ские Южиоси- бирско- с ре дне ази- атские Южноси- бирско- северо- моиголь- ские Алтайские эндемики и геми- эндемики Е х2
Лесов Субальпийские эвритоп- 13 11.2 2 1 10.3 3 21 10.4 0 13 9.6 4 0 1.4 0 2 5.7 2 6 7.3 3 1 1.3 0 57 14 25.1* 5.4
ные Субальпийских лугов 2.9 17 29.7 2.7 13 27.4 2.7 46 27.7 25 34 25.7 0.4 3 3.7 15 17 15.2 1.9 18 19.3 0.3 4 3.4 152 29.0*
Пойменных субальпий- 7 8 5 7 1 2 3 0 33 2.9
ских кустарников Болот субальпийского 6.5 23 5.9 5 6.0 6 5.6 3 0.8 0 3.3 2 4.2 0 0.7 0 39 43.7*
пояса Водные субальпийского 7.6 И 2.3 1 7.1 1 2.2 0 1.0 0 3.9 0 4.9 0 0.9 0 13 45.1*
пояса Скал и осыпей субаль- пийского и альпийского 25 5 7.0 11 2.4 3 6.6 11 0.3 2 1.3 3 1.7 10 9.3 2 47 13.9
поясов Тундроподобных труп- 9.2 10 85 4 8.6 0 7.9 0 1.2 0 4.7 3 6.0 0 1.0 0 17 23.9*
пировок Альпийских лугов 3.3 0 115 3.1 20 10.7 3.1 0 10.8 2.9 4 10.0 0.4 4 1.4 1.7 14 5.9 2.2 14 7.5 0.4 3 1.3 59 57.8*
Субнивального пояса 0 3.5 14 3.2 0 3.3 0 3.0 1 0.4 0 1.8 3 2.3 0 0.4 18 56.6*
00 Е 88 80 82 76 11 45 57 10 450
х2 96.7* 77.6* 47.6* 23.9* 10.6 20.1* 20.1* 6.7 302.3*
Таблица 13
Средние значения сопряженности (т) разных типологических структур
конкретных флор
Сравниваемые типологические структуры (фак- тор множества) Конкретные флоры
Эргу- веем Рау- чуа бассейна р.Циль- мы биостанции Сыктывкар- ского уни- верситета окрест- ностей г. Сык- тывкара дельты р. Не- ман хр.Кур- куре
Широтная и долгот- ная ареалогические структуры 0.6 0.8 3.6 4.6 6.3 7.1 —
Эколого-цента че- ская и широтная ареалогические структуры 4.2 3.6 — 2.7 4.6 — 3.8
Эколого-ценотаче- ческая и долготная ареалогические структуры 2.2 1.4 — 2.6 2.7 — —
к сухим горным тундрам,1 несколько „завышено” число арктоальпий-
ских видов. Такая же закономерность наблюдается и во флоре Раучуа.
В комплексе видов, тяготеющих к сырым эвтрофным тундрам, района
Эргувеем число арктических видов больше ожидаемого. На Раучуа же
в этом комплексе „завышено” число арктоальпийских видов. Таким
образом, если на Эргувеем комплекс видов эвтрофных тундр по своей
широтной ареалогической структуре отличается от комплекса видов
сухих горных тундр, то на Раучуа эти различия не достоверны.
На Эргувеем в комплексе видов, тяготеющих к нивальным лугови-
нам, наблюдается явное превышение над ожидаемым числом арктоаль-
пийских видов. То же самое, но в меньшей степени наблюдается и на Рау-
чуа. На Эргувеем в комплексе видов,тяготеющих к пойменным ивнякам
и лугам, завышено представительство гипоарктических, бореальных и
арктических видов, а на Раучуа в этом комплексе завышено представи-
тельство только бореальных видов. Широтная ареалогическая структура
комплекса активных видов сходна на Эргувеем и на Раучуа. В обоих слу-
чаях превышение приходится на гипоарктические виды. Весьма своеоб-
разна широтная ареалогическая структура комплекса видов болот высо-
кой поймы Раучуа. Здесь больше ожидаемого видов бореального и
арктического склада.
На основании полученных данных можно предположить, что при по-
холодании климата в тундровых ландшафтах бореальные виды будут
дольше сохраняться в прйменных ивняках, на пойменных лугах, среди
прибрежно-водной растительности, на болотах высокой поймы. В эпохи
1 Здесь и ниже речь идет не о полном списочном составе видов парциальных
флор соответствующего класса экотопов, а о комплексе видов, верных или наибо-
лее характерных для него. (Прим, ред.).
186
потеплений часть этих видов из внутриландшафтных рефугиумов может
поселяться и за пределами речных долин: в эвтрофных местообитаниях,
на сухих участках склонов гор и равнинных увалов. При этом арктиче-
ские виды сохранят свои позиции также в составе пойменных ивняков,
пойменных лугов, болот высокой поймы, в эвтрофных тундрах. При оче-
редном похолодании арктические виды из эколого-ценотических внутри-
ландшафтных рефугиумов расселяются в сухие горные тундры.
Анализируя сопряженность эколого-ценотической и долготной ареа-
логической структуры тундровых КФ в первую очередь следует обратить
внимание на виды восточносибирско-западноамериканского типа ареа-
ла — берингийские в широком смысле. На Эргувееме берингийцы явно
приурочены к сухим горным тундрам и избегают нивальные луговины.
На Раучуа берингийцы также чаще ожидаемого встречаются в сухих гор-
ных тундрах. Однако на Раучуа число берингийцев больше ожидаемого
и в равнинных тундрах. Таким образом, в условиях Берингийского сек-
тора Арктики в неоген-четвертичное время сформировался достаточно
богатый комплекс видов, связанных с сухими горными тундрами. В эту
группу включены и древние гольцовые элементы, распространенные
в данном долготном секторе.
Распределение восточноазиатских1 видов по эколого-ценотическим
элементам на Раучуа соответствует ожидаемому. На Эргувеем их число
„завышено” только в пойменных ивняках и на пойменных лугах. При-
мерно так же распределены в обеих флорах евразиатские и сибирские
виды. Для них предпочтительнее оказываются пойменные ивняки и пой-
менные луга, -избегают же они нивальные тундры. Эти долготные ареало-
гические группы в обеих флорах стоят на последнем месте по числу ви-
дов, так как резко преобладают виды, общие с северо-западом Северной
Америки.
Анализ сопряженности эколого-ценотической и широтной ареалоги-
ческой структуры флоры окрестностей биостанции Сыктывкарского
университета (табл. 8) показал, что в еловых и смешанных кустарничко-
вых лесах имеет место достоверное превышение над ожидаемым числа
неморальных и неморально -бореальных видов. Виды эвризональные,
напротив, в этих лесах встречаются реже ожидаемого. На олиготрофных
болотах и в заболоченных сосновых лесах „завышено” число бореальных
видов и отсутствуют эвризональные. Набор видов сосняков лишайнико-
вых (боры беломошные) по широтной ареалогической структуре сходен
с таковым олиготрофных болот. Распределение видов, тяготеющих
к пойменным ивнякам и речным пляжам, по широтным элементам близ-
ко к ожидаемому. Эвризональные и лесостепные виды чаще ожидаемого
встречаются на нарушенных лугах, в местах обработки древесины —
катищах, на мелиорируемых лугах.
Несколько неожиданно существенное отличие широтной ареалогиче-
ской структуры комплексов видов сухих и среднеувлажненных лугов
1 Из соображений краткости здесь и в таблицах так именуются виды востока
Северной Азии, которые в собственно Восточную Азию в большинстве случаев
не заходят.
187
и комплекса видов сырых заболоченных лугов и травяных болот. Если
на сухих и среднеувлажненных лугах больше ожидаемого бореальных
видов, то на сырых и заболоченных лугах и в травяных болотах больше
полизональных видов. Бореальные же здесь в явном дефиците. По дол-
готной ареалогической структуре 2 типа лугов также резко отличаются
друг от друга (табл. 9). На сухих и среднеувлажненных лугах больше
ожидаемого европейских и европейско-западноазиатских видов, при „не-
доборе” циркумполярных и почти циркумполярных. На сырых же и
заболоченных лугах и травяных болотах европейских и европейско-за-
падноазиатских видов меньше ожидаемого. Вероятно, это связано с боль-
шей стабильностью во времени комплекса видов сырых и заболоченных
лугов по сравнению с комплексом видов сухих и среднеувлажненных
лугов.
В комплексе видов водных и прибрежно-водных местообитаний
явно преобладают над ожидаемым виды с циркумполярными и почти
циркумполярными ареалами. Выше ожидаемого их число и в составе
эколого-ценотического элемента олиготрофных болот и заболоченных
сосняков. Возможно, это говорит о большей стабильности условий на
экотопах с повышенной ролью широко распространенных видов. При
формировании флоры еловых и смешанных зеленомошно-кустарничко-
вых лесов сибирские флористические связи, видимо, преобладали над
европейскими. Таким образом, и циркумполярные, и евразиатские виды
на европейский северо-восток после оледенения проникали преимущест-
венно из Сибири.
В табл. 10 дан анализ распределения видов флоры окрестностей
г. Сыктывкара по эколого-ценотическим и широтным ареалогическим
элементам. В комплексе видов еловых и смешанных зеленомошно-кус-
тарничковых лесов больше ожидаемого неморальных и неморально-бо-
реальных видов при полном отсутствии полизональных. Но в отличие от
флоры окрестностей биостанции здесь в составе комплекса видов ело-
вых и смешанных лесов больше ожидаемого не только сибирских, но и
европейско-западносибирских видов (табл. 11). Ареалогическая струк-
тура комплексов видов сухих и среднеувлажненных лугов, заболочен-
ных лугов, олиготрофных болот, водных и прибрежно-водных местооби-
таний, сильно нарушенных местообитаний в окрестностях г. Сыктывкара
практически не отличается от таковой в окрестностях биостанции, что
подтверждает сделанные выше выводы и предположения о характере
генезиса флоры и растительности европейского северо-востока.
Результаты сопряженного анализа эколого-ценотической и ареалоги-
ческой структур флоры хр. Куркуре (Восточный Алтай) были опубли-
кованы ранее (Галанин, Марина, 1982). Здесь же (табл. 12) отметим,
что широко распространенные циркумполярные, полизональные и боре-
альные виды на Куркуре чаще ожидаемого встречаются на болотах и
в водоемах субальпийского пояса. В явном дефиците они на альпийских
и субальпийских лугах и в субнивальном поясе. Широко распростра-
ненные арктоальпийские виды в „избытке” на альпийских лугах и
в субнивальном поясе, а вот на субальпийских лугах они встречаются
реже ожидаемого. Наблюдаемое число евразиатских полизональных
188
и бореальных видов больше ожидаемого в лесах и на субальпийских
лугах, но меньше ожидаемого на альпийских лугах. Таким образом,
ареалогические структуры, а следовательно, и генезис флоры альпий-
ских и субальпийских лугов Восточного Алтая резко отличаются.
Центральноазиатские, южносибирско-среднеазиатские и южносибирские
виды чаще ожидаемого встречаются в комплексе видов альпийских
лугов.
В заключение можно сказать, что использование метода сопря-
женного анализа типологических структур КФ позволяет получить до-
полнительную информацию об их генезисе, показывает, что разные
эколого-ценотические элементы КФ имеют достоверно отличающуюся
ареалогическую структуру. Вопреки ожиданию степень сопряженности
эколого-ценотических и ареалогических структур в целом оказалась
достоверной для всей КФ, причем степень этой сопряженности практиче-
ски не зависит от географической широты района, характера его рельефа
и генезиса флоры (различия значений х2 в пределах ошибки).
А. Е. Кученева
К МЕТОДИКЕ ОЦЕНКИ ВСТРЕЧАЕМОСТИ
И ЧИСЛЕННОСТИ РАСТЕНИЙ
В КОНКРЕТНЫХ ФЛОРАХ
Калининградский государственный университет
Изучение растительного покрова Южной Прибалтики (Калининград-
ская обл.) представляет значительный научный и практический интерес.
Своеобразное географическое положение области определило „проме-
жуточный” характер ее растительности (между подзоной широколист-
венных лесов европейского типа и подзоной хвойно-широколиственных
лесов лесной зоны Евразии) ц „стыковый” характер флоры, сочетающей
в своем составе элементы самых различных флор — от собственно боре-
альных до паннонских (Победимова, 1956). Этим же можно объяснить
богатство флоры этой сравнительно небольшой территории (более 1200
видов).
Подавляющее большинство опубликованных фундаментальных ра-
бот по флоре и растительности территории современной Калининград-
ской обл. относится к довоенному периоду (Abromeit, 1895: Abromeit
et al., 1898-1940; Steffen, 1931). В послевоенное время Ботаническим
институтом им. В. Л. Комарова АН СССР изданы материалы комплекс-
ной экспедиции, где освещаются отдельные вопросы состояния флоры
и растительности области (Леонтьев, 1956; Матвеева, 1956, 1967; Побе-
димова, 1956; Ниценко, 1970). Однако за истекший период времени
флора и растительность области претерпели значительные изменения,
связанные с усилением антропогенного воздействия. Изменились взгля-
ды на объем и структуру многих таксонов.
189
Все это выдвигает задачу критического пересмотра сложившихся
представлений о флоре и растительности Калининградской обл.
Флористические исследования проводились ботаниками Кали-
нинградской обл. в рамках работы кафедры ботаники Калининград-
ского государственного университета. Обработка собранных мате-
риалов позволила выявить состояние и охарактеризовать динамику
флоры в некоторых районах и ключевых участках области по видо-
вому составу отдельных семейств, редким и исчезающим видам, си-
нантропным растениям (Андронова, Кученева, 1972; Галанин, 1977а;
Расиньш и др., 1977; Семенова, 1979; Андронова и др., 1982, 1983;
Кученева, 1983). В последние годы изучается флора и растительность
Самбийского (Калининградского) полуострова, интересного и свое-
образного в ландшафтном и флористическом отношениях района
области.
С начала 1970-х годов ведется работа по инвентаризации флоры ме-
тодом конкретных флор (КФ). А. В. Галанин, организовавший последо-
вательное изучение флоры области указанным методом, считает, что
в условиях Калининградской обл. размеры площади выявления КФ со-
ставляют 200—300 км2. Наши работы на Самбийском полуострове согла-
суются с этим положением.
В работе нами используется комбинированная формула встречаемо-
сти и обилия (Abrometi et al., 1898—1940), которая чрезвычайно проста и
удобна в применении. Этот метод вполне приемлем для изучения флоры
именно Калининградской обл., так как с его помощью флора этой тер-
ритории была охарактеризована почти 100 лет назад и повторная инвен-
таризация по той же методике может дать интересные сравнительные
результаты.
Формула И. Абромайта включает 2 показателя: V (от немецкого
Verbreitung — распространение, встречаемость) и Z (от немецкого Zah-
len — количество) / каждый из которых имеет числовые выражения (ко-
эффициенты), дающие соответственно количественную характеристику
встречаемости местонахождений данного вида в пределах контура КФ
(или любого другого флористического, экологического или администра-
тивного выдела) и числа экземпляров в местонахождениях.
Формула немецкого флориста нами несколько усовершенствована
в части, касающейся количественной оценки встречаемости местонахож-
дений, коэффициенты которой получили следующие цифровые выраже-
ния (К1-5): 1 — очень редко (1—2 местонахождения); 2 — редко (3—6
местонахождений); 3 — не так редко (7—15 местонахождений); 4 —
местонахождения часты (16—25); 5 — широко распространены (более
25 местонахождений).
В отношении Z нами сохранены следующие коэффициенты: 1 — весь-
ма ограниченное количество (1—2 особи); 2 — несколько экземпляров
1 Данная формула включает оба основных компонента экотопологической ак-
тивности видов, в смысле Б.А.Юрцева (1968а, 1982а; см.также статьи Б.А.Юрцева,
Л. И. Малышева и А. А. Лякавичюса в наст, сб.), выраженные в баллах и приводи-
мые рядом друг с другом. {Прим, ред.).
190
Динамика распространения {темная часть) Listera cordata (I), Goodyera repens (II) и
Epipogium aphyllum (III) на территории Калининградской обл.
а - литературные данные за 1898-1942 гг.; б - наши наблюдения за 1970-1984 гг.
(3—6); 3 — в большем количестве (7—12); 4 — больше 12 экземпляров-
(особей); 5 — вид образует аспект ценоза.
Одновременно мы выделяем 5 категорий географической встречае-
мости видов по наличию их во флоре административных районов области
[оговоримся, что границы административных районов практически со-
впадают с границами округов, выделенных немецкими флористами
(Abromeit et al., 1898—1940)]: 1-я категория — редкие виды (встречаю-
щиеся в 1—2 районах области); 2-я — нечасто встречающиеся (в 3—5
районах); 3-я — распространенные (в 6—8 районах); 4-я — весьма рас-
пространенные (в 9—11 районах); 5-я — повсеместные.
Наша работа в известной мере согласуется с работами ботаников
Литвы и Латвии. В конечном счете при использовании указанной ме-
тодики в сочетании с прямым сеточным картированием возможно
191
Таблица 1
Структура и динамика местных популяций видов сем. Orhidaceae
в Калининградской обл. за период с 1898 по 1984 г.
Вид Категория встречае- мости Исследованные районы
Самбийский п-ов (исключая косу) Куршская коса
Cephalanthera rubra 1/1 k’-’z’-’/f’z1-2 -/-
CoraUorhiza trifida 3/1 p-»ZJ-3/r*ZJ r*z2/o
Coeloglpssum viride 3/0 r’-’Z’-’/O —
Cypripedium calceohis 2/0 r3z2/o —
Dactylorhiza majaUs 4/2 r’Z’-’/r’Z1 r2z2/r2z‘-2
D. maculate 4/3 V4Z3-*IV3Z3-* V'Z'IV'Z'-*
D.fuchsii -/з -IV'Z3-3 -/HZ1'2
D. baltica -/1 -IV'Z* -/-
Epipactis atrorubens 2/2 K’Z'-’/K’Z1-3 p-iZ’-’/F'^1-3
E. helleborine 3/1 r’Z’-’/F'^1-3 V'z'l-
E. palustris 3/2 K3-4Z3/K1Z1 —
E. purpumte 2/- r2z2-3/- —
Epipogium aphyllum 1/1 r2z2-3/- —
Goodyera repens 3/2 r3-4z3-4/r3z3 r2z3/r2z3
Gymnodenia conopsea 2/1 jziZ’/r’-’Z3 —
Hammarbia pahidosa 3/1 r’Z’-’/r’Z1-3 —
Hermtnium monorchis 2/2 p-’Z’/^’Z1-’ —
Liparis loesetii 3/1 K3Z2-4/F'iZ1-3 r‘z2/o
Listera cordate 3/1 r3-4ZS-4^-3Zi-3 r’z’-’/r’z2
L. ovate 3/1 r3-4Z3-4^r3Za-3 -/r2z2-3
Malaxis monophyllos 3/2 rizi-3/r,-iz‘-i k‘z7-
Neottianthe cucullata 1/1 piZ’-’/^Z1-3 r’z1-2^
Neottia nidus-avis 1/1 r’-’zvr'z1 —
Orchis mascula 4/— r2z2/- —
0. nuUtaris 2/- r’Z1-4/- —
0. ustulate I/O r2z2/o —
Platanthera bifolia 5/5 r5z3-4/r5z2-4 r3Z3/r2Z‘-3
P. chlorantha 4/4 r3zJ-3/r3Z3-3 r2z’-7r‘z2
Примечание. Здесь и в табл. 2 слева — литературные данные за 1898—
1942 гг., справа — наши данные за 1970—1984 гг.; прочерк означает, что достовер-
ные данные отсутствуют и наших находок нет, 0 — вид, по-видимому, исчез.
выявление структуры и динамического состояния флоры Калининград-
ской обл. как в целом, так и в отдельных фитохориях низшего порядка.
Наиболее полно с использованием указанной методики нами охарак-
теризовано семейство ятрышниковых (табл. 1).
Данные табл. 1, дополнительно проиллюстрированные рисунком
(см. рисунок), свидетельствуют о негативных изменениях, произошед-
ших за указанный период времени в структуре местных популяций
192
Таблица 2
Динамика состояния местных популяций редких видов растений
Самбийского полуострова с 1898 по 1984 г. (выборочно)
Вид Категория встре- чаемости Конкретные флоры
Морское побережье Куршская коса Центральная часть
Astrantia major I/O — — r2z2-3/o
Actaea spicata 5/4 — r’Z’/r’Z* r4z3/r2z2
BulUarda aquatica I/O r’z’/o — —
Cqrastium sylvaticum 1/0 — — r‘Z3/0
Corallorhiza trifida 3/1 — r‘z2/o r2z2-3/o
Digitalis ambiqua 4/0 — — r2z3/o
Elatine triandra I/O — — r‘Z3/0
Eryngium maritimum 2/2 r«Z3-4/r3Z3 V4Z3~4/V3~4Z3 —
Hedera helix 3/2 — -/r‘Z3 r2z4-5/r‘z3
Hepatica nobilis 4/2 — -/^’Z1 v2z3-*iv'z3
Lathyrus maritimus 1/1 V*Z5/V*Z5 r4z4-5/r4z4-5 —
Limosella aquatica 1/0 r'z'/o — —
Linaria loeseUi 1/1 — pMZ’-Vr’Z3-4 —
Linnaea borealis 3/2 — Pz’-^r'z4 r2z5/o
Lunaria rediviva 0/1 — -/^‘Z5 —
Neottianthe cucuUata 2/1 — r‘Z2/0 —
Polypodium vulgare 4/4 r4z3-4/r3z’ v3z3tv3z3 V3Z3/V2~3Z3
Pulsatilla pratensis 4/1 r3z5/r‘z4 — r3z5/o
Sarothamnus scoparius 2/1 r3z5/r‘z5 v'z'tv'z* -/H‘z3
TrolUus europaeus 5/2 — — V3Z3/VlZ3
видов сем. Orhidaceae, в распространении отдельных видов. В первую
очередь это объясняется уменьшением лесистости территории (с 28%
в конце прошлого столетия до 19% в настоящее время), интенсивным
освоением земель, увеличением площади городов, вытесняющих приго-
родные ценозы, усилением рекреационного пресса и т. д.
Как уже говорилось, в последние годы флористические исследова-
ния проводились на Самбийском полуострове. Полуостров представляет
собой северо-западную оконечность Балтийского кристаллического
щита; с юга он ограничивается глубоко врезавшимся в сушу Калинин-
градским заливом, с севера — Куршским заливом; часть северного по-
бережья (от Куршской косы до мыса Таран) и западное побережье омы-
ваются Балтийским морем. Специфическими в ландшафтном и флори-
стическом отношении являются территории Куршской и Балтийской
кос. Климат Самбийского полуострова хорошо выраженный примор-
ский, со среднегодовой температурой 7.5 °C.
Флора Самбийского полуострова насчитывает около 1000 видов
высших растений; около 120 видов отмечены только для этой террито-
рии, что позволяет считать Самбий самостоятельным флористическим
районом. К настоящему времени обследованы следующие ландшафтные
13 Заказ 1220
193
районы Самбийского полуострова: Куршская коса, побережье Балтий-
ского моря от г. Зеленоградска до г. Балтийска, Вислинская (или Бал-
тийская) коса, центральная часть полуострова, побережье Калининград-
ского залива.
Выше упоминалось, что в пределах Самбийского полуострова произ-
растает около 120 видов растений, не встречающихся в других частях
области. Кроме того, здесь отмечено около 100 видов 2-й категории
встречаемости.
Сравнительный анализ полученных нами данных позволяет охарак-
теризовать видовой состав и динамику некоторых демографических
показателей популяций особо редких и редких растений за 50—90 лет
и наметить возможные меры охраны (табл. 2). Наиболее ранимыми
оказываются виды стенотопные. Как и в горном Крыму (Дидух, 1980),
это в первую очередь Corallorhiza trifida, единственное местонахождение
которой, подтвержденное нашими геоботаническими описаниями, обна-
ружено на Куршской косе. Затем идут виды, находящиеся на границе
своего ареала и в силу этого обстоятельства постоянно подверженные
стрессу. В числе таких видов назовем прежде всего Digitalis ambigua и Не-
dera helix.
Отдельную группу составляют растения, у которых резко понизилась
географическая встречаемость. Corallorhiza trifida, почти полностью исчез-
нувшая из флоры Самбия, обнаруживает такую же тенденцию и в других
районах области. Заметно уменьшилось число местонахождений и изме-
нился характер популяций Pulsatilla pratensis. В связи со строительством
и лесохозяйственными работами исчезли популяции линнеи северной под
Зеленоградском, вблизи пос. Лесное (там же погибло единственное на
Куршской косе местообитание неоттианты клобучковой), под Логвино
и Люблино. Достоверно известные (подтвержденные гербарными сбора-
ми и геоботаническими описаниями) местонахождения линнеи северной
сохранились в окрестностях пос. Рыбачьего и Морского на Куршской
косе; они представлены двумя небольшими по площади пятнами
(1—2 м2) со сплошным проективным покрытием.
Некоторые виды не претерпели заметных изменений. В большинстве
случаев это виды эвритопные. Особую группу образуют здесь псаммо-
фиты: Ammophila arenaria, A. baltica, Calamogrostis epigeios, Corynephorus
canescens, Festuca arenaria, Eryngium maritimum, Lathyrus maritimus, Hon-
kenya peploides и другие виды, обитающие на песках (в том числе и на
подвижных дюнах), где другие виды, хотя бы и эвритопного в целом
характера, обитать не могут.
Обнаружены новые виды: Bidens melanocarpa, Verbascum phlomoides,
Linaria dolichoceras, ранее не числившиеся во флоре области (Abromeit et
al., 1898—1940; Победимова, 1956). В числе новых для обследованного
района следует назвать Lunaria rediviva, растение „Красной книги СССР”
(1978). Самое крупное местонахождение (из двух имеющихся в обла-
сти) расположено на Куршской косе неподалеку от курортного города
Зеленоградска и регулярно наблюдается нами. Эти местонахождения
Lunaria rediviva не указаны немецкими флористами (Abromeit et al.,
1898-1940).
194
Таким образом, используемая нами методика позволяет получить
разносторонние данные, характеризующие флору Калининградской обл.,
ее отдельные семейства или иные группы растений, а также выявить
изменения, которые претерпела флора этой территории за почти веко-
вой период времени.
М. Лайвиныи, А. Расиньш, А. Калниня
КЛАССИФИКАТОР ВЫСШИХ СОСУДИСТЫХ РАСТЕНИЙ
ДЛЯ АВТОМАТИЗИРОВАННОЙ БИБЛИОТЕКИ
ДАННЫХ ПО ФЛОРИСТИКЕ И ГЕОБОТАНИКЕ
Научно-производственное объединение „Силава”
Прибалтийское отделение ВИЗР
Создание больших автоматизированных библиотек данных требует
разработки самых различных видов классификаторов — упорядоченных
систем признаков и объектов исследования. При разработке библиотеки
данных биологической инвентаризации Латвии (Biologiskas invEntariza-
cijas daTu bibliotekA — BETA) создан классификатор высших сосудистых
растений Латвии и сопредельных территорий — CLASIF-001.
Классификатор CLASIF-001 предназначен для более рациональной
организации массивов экспериментальных Данных на магнитных носите-
лях ЭВМ, и, кроме того, он сам является удобной информационной си-
стемой для получения разного рода сведений об упорядоченной системе
таксонов растений, накопленных в классификаторе.
Классификатор CLASIF-001 содержит следующую информацию.
1. Соподчиненная система таксонов: семейство -* род -► вид -* под-
вид, записанная в словесном виде.
2. Нумерация каждого ранга таксонов высших сосудистых расте-
ний трех видов: а) региональная (для Латвии и сопредельных терри-
торий) цифровая характеристика таксонов, приведенная авторами и
состоящая из 8 символов, в которой 3 цифры (слева) обозначают по-
рядковый номер семейств, 2 следующие — родов, далее 2 — видов и
1 последняя — подвидов; б) нумерация таксонов по среднеевропей-
ской системе, разработанная Ф. Эрендорфером и сотрудниками, ко-
торая содержит 7 символов — первые 3 цифры (слева) обозначают
порядковый номер родов, 2 следующие — видов и 2 последние — под-
видов; в) нумерация таксонов (родов и видов) по фундаментальной
„Флоре СССР” (в 30 томах). В организации массивов данных и рас-
четах активно используется лишь региональная нумерация. Осталь-
ные две (среднеевропейская и „Флоры СССР”) являются пассивны-
ми и предусмотрены для совмещения нескольких подобных клас-
сификаторов высших сосудистых растений при решении конкретных
задач.
195
3. Принадлежность видов к четырем категориям: адвентивные (А),
сорняки (N), культурные растения (К) и натурализовавшиеся расте-
ния (NT).
Региональный классификатор высших сосудистых растений
(CLASIF-OO1) является иерархическим, содержащим взаимосвязанную
нумерацию таксонов четырех рангов. Это дает возможность оперативно
дополнить классификатор новыми таксонами или изъять ненужные. Кро-
ме того, довольно широкий спектр таксонов (от подвидов до семейств)
обеспечивает ее применение в сравнительных количественных флористи-
ческих исследованиях.
С. А. Овеснов
ОПЫТ ИСПОЛЬЗОВАНИЯ КОЭФФИЦИЕНТА КЕНДЭЛА
ДЛЯ СРАВНЕНИЯ СИСТЕМАТИЧЕСКОЙ СТРУКТУРЫ
РАВНИННЫХ ФЛОР
Пермский государственный университет
Сравнение флор, имеющее целью определить их место в типоло-
гическом ряду флор, а также в системе фитохорий (флористическое
районирование), может проводиться по любым признакам видов —
географическим, экологическим, формационным и др., но основой
является сопоставление видовых списков сравниваемых флор. Именно
оно представляет базу для выделения фитохорий низших рангов (ниже
области и особенно провинции). В настоящее время наметились 3 под-
хода к сопоставлению флор. Это сравнение, во-первых, полных фло-
ристических списков, как двустороннее, так и одностороннее (Юрцев,
1968а) (данный подход можно назвать флористическим); во-вто-
рых, так называемых миграционно- или географо-генетических групп,
то в более широком (Козловская, 1974), то в более узком смысле
(Баранова и др., 1971); этот подход можно назвать флорогенетиче-
ским. И наконец, высказывались предложения об использовании
дополнительных критериев (Малышев, 1973), а именно активности
и встречаемости видов. Этот подход можно назвать флороценотиче-
ским.1
Поскольку отнесение вида к той или иной миграционно- или геогра-
фо-генетической группе является в известной степени гипотезой, а вве-
дение упомянутых дополнительных критериев, как показано Б. А. Юрце-
вым (1968а) и А. И. Толмачевым (1974), приводит не к собственно фло-
ристическому, а, скорее, к ботанико-географическому районированию,
основным следует признать флористический подход.
1 Более полно существующие подходы к сравнительному анализу флор рас-
смотрены в соответствующем разделе помещенных в этой книге „Программ фло-
ристических исследований...” (Прим, ред.).
196
Очень удобным для целей флористического районирования кажется
сравнение систематической структуры флоры (Толмачев, 19706), при-
чем под этим понимается соотношение семейств и родов по количеству
присущих им в данной флоре видов растений; на практике используется
набор 10—15 ведущих семейств или родов — так называемая „головная
часть” спектра (Малышев, 1973; Шмидт, 1980). Для количественного
выражения степени сходства систематической структуры сравниваемых
флор одновременно и независимо Л. И. Малышевым и В. М. Шмидтом
в 1972 г. предложено использовать метод корреляции рангов с расчетом
коэффициента ранговой корреляции Кендэла (Заки, Шмидт, 1972; Ма-
лышев, 19726; Ребристая, Шмидт, 1972). С помощью этого метода было
проведено флористическое районирование Болыпеземельской тундры и
прилегающих районов, Канарских и Курильских островов и некоторых
других регионов. Позднее для этой же цели был использован коэффи-
циент ранговой корреляции Спирмена (Седельников, 1970). Однако
принципиального различия между этими показателями нет (Гаек, Шидак,
1971).
При определении места изученной нами (Овеснов, 1983а, 19836)
флоры юга Пермской области в системе флористического районирования
были сопоставлены полные флористические списки для всех сравнивае-
мых флор с вычислением коэффициента общности Жаккара (Шмидт,
1980), а также определены коэффициенты Кендэла для 10 ведущих се-
мейств и родов. Сравнивались следующие флоры:1 Приморской низмен-
ности (ПН) и Курземского геоботанического района (КР) Латвийской
ССР, Западного (ЗП) (Псковского) флористического района Псковской
обл., Беловежской Пущи (БП), Приокско-террасного заповедника (ПТ),
Рязанской Мещеры (РМ), юга Коми АССР (ЮК), юга Пермской обл.
(ЮП), Кунгурской лесостепи (КЛ) (юго-восток Пермской обл.) и Ново-
сибирской обл. (НО). Эти флоры, за рядом исключений, более или менее
территориально соизмеримы и фациально равноценны.
Методика расчета коэффициентов Кендэла и Жаккара, а также по-
строения графов сходства сравниваемых флор подробно описаны в лите-
ратуре (Василевич, 1969; Шмидт, 1980, и др.). Опуская в целях эконо-
мии места полные корреляционные матрицы, перейдем непосредственно
к основным результатам сравнения.
Три графа на рис. 1 и 2 имеют определенные черты сходства. Это про-
является главным образом в том, что в каждой выделяется по 4 корре-
ляционные плеяды. Однако состав их различен. Так, 4 плеяды сходства
ведущих семейств (рис. 1, а) выглядят следующим образом: первая, наи-
более крупная плеяда образована пятью флорами (ЮП, ПН, КР, ПТ, РМ),
ее ядро составляют флоры КР и РМ; вторая включает две флоры — ЗП и
ЮК; третья также две — БП и КЛ; четвертая представлена флорой НО.
1 Данные о флорах были заимствованы из литературных источников (Смирнов,
1958, 1971; Конспект флоры Псковской обл., 1970; Николаева, Зефиров, 1971; Ко-
ролева и др,, 1973; Биркмане, Юкна, 1974; Флора северо-востока..., 1974-1977;
Конспект флоры Рязанской Мещеры, 1975; Биркмане и др., 1977; Определитель
высших растений..., 1981; Шилова, 1982) с последующей унификацией таксонов.
197
Рис. 1. Граф сходства соотношений 10 ведущих семейств (а) и родов (б) в сравни-
ваемых флорах.
1-4 - значения т (а: 1 - 1-0.96, 2 - 0.95-0.86, 3 - 0.85-0.79, 4 - 0.78-0.73;
б-. 1 - 0.75-0.62, 2 - 0.61-0.58, 3 -0.56-0.49,4 - 0.46-0.42). Остальные обозна-
чения в тексте.
Рис. 2. Граф сходства видового состава сравниваемых флор.
1-4 - значения коэффициента общности Жаккара: 1 - 1QA-1QA, 2 - 64.8—60,5,
3 - 59.9-50,6,4 - 49.6—44.4. Остальные обозначения в тексте.
198
Плеяды сходства ведущих родов выглядят иначе (рис. 1,6). Наибо-
лее компактной является плеяда, образованная тремя флорами — ПН,
КР и ЗП. Более слабо связаны пять флор, входящих в состав другой
плеяды (ЮК, РМ, ЮП, КЛ и ПТ). Особняком от других флор и друг от
друга стоят флоры БП и НО.
Сопоставление полных флористических (видовых) списков, которое
в условиях равнин Евразии с их низким эндемизмом является основой
для флористического районирования, привело к несколько иным резуль-
татам (рис. 2). Две центральные, слабо дифференцированные плеяды
образованы 8 флорами. Ядро первой из них составляют флоры ПН, КР
и ЗП; к ним примыкает флора БП. Ядро второй составляют флоры РМ
и ПТ с примыкающими к ним флорами ЮП и КЛ. Несколько обособлена
флора ЮК, особняком стоит флора НО. Многочисленные связи между
восточноевропейскими флорами свидетельствуют о тесных флорогенети-
ческих отношениях между ними.
Только результаты сопоставления списков видов в достаточной сте-
пени совпадают с имеющимися схемами флористического районирова-
ния, в частности с флористическим районированием А. Л. Тахтаджяна
(1978). В соответствии с ним флоры первой плеяды входят в состав
Центральноевропейской, второй — Восточноевропейской, флора юга
Коми АССР — Североевропейской, а флора Новосибирской обл. — в со-
став Западносибирской провинции Циркумбореальной области Бореаль-
ного подцарства Голарктического царства.
Проведенное сравнение систематической структуры 10 небольших
региональных флор, в основном относящихся к разным частям Восточ-
но-Европейской равнины, позволяет высказать мнение, что спектр веду-
щих семейств в большей мере отражает широтно-зональные позиции
флоры, спектр ведущих родов — долготные (провинциальные) позиции.
В обоих случаях сравнение систематической структуры дало лишь самую
первичную ориентировку с точки зрения задачи флористического райо-
нирования.1
К аналогичным выводам на примере флористического районирова-
ния горных территорий несколько ранее пришел и Н. И. Золотухин
(1982).
1 Обращает на себя внимание весьма высокий общий уровень сходства семей-
ственных спектров сравниваемых флор разных лесных районов Восточно-Европей-
ской равнины, слабоконтрастных в зональном отношении, при несколько неожи-
данных „связях” (например, между Курземским районом Латвии и Рязанской
Мещерой); очевидно, данный показатель следует применять к более контрастному
флористическому материалу. Интерпретация выявленных отношений сходства,
вероятно» выиграла бы от рассмотрения состава семейств или родов „головной”
части спектров в полученных группах флор. Для того чтобы результаты сравнений
видовых списков и соотношения ведущих семейств и родов в изученных флорах
были более сопоставимыми, следовало бы и при сравнении семейственных и родо-
вых спектров использовать коэффициент общности Жаккара, модифицированный
для весовых множеств. (Прим, ред.).
199
III. НЕКОТОРЫЕ ИТОГИ РЕГИОНАЛЬНЫХ
ФЛОРИСТИЧЕСКИХ ИССЛЕДОВАНИЙ
А. А. Лякавичюс
РАЗВИТИЕ ФЛОРИСТИЧЕСКИХ ИССЛЕДОВАНИЙ
В ЛИТВЕ
Институт ботаники АН ЛитССР
XVI экспедиция-конференция прибалтийских ботаников, проведен-
ная в 1982 г. в южной части Литвы, была посвящена двум знаменатель-
ным событиям в литовской флористике — 200-летию выхода в свет вто-
рой части „Литовской флоры” Ж. Э. Жилибера (Gilibert, 1781—1782) и за-
вершению в 1980 г. издания коллективного капитального труда „Флора
Литовской ССР”.
История флористических исследований в Литве между этими двумя
датами делится на несколько этапов, различающихся между собой про-
должительностью, характером, интенсивностью и результатами исследо-
ваний. После выхода в свет последнего тома „Флоры Литовской ССР”
начался следующий, качественно новый этап исследования флоры рес-
публики, исторически относящийся уже к третьему столетию ботаниче-
ских наук в Литве.
„Флора” Жилибера была первым обобщающим трудом флористиче-
ского направления в Восточной Прибалтике, и Литва в конце XVIII в.
по сравнению с некоторыми другими странами Европы была флористи-
чески хорошо изученной.
Автора первой литовской „Флоры” в наши дни можно критиковать
за весьма ограниченную территорию исследований, за недостаточную
представительность списка видов. Кроме того, Жилибер, как пытливый
исследователь, выделивший и описавший ряд новых таксонов видового
и внутривидового рангов, дал вновь описанным видам, а также целому
ряду уже известных видов новые названия, из которых некоторые, к со-
жалению, не соответствовали бинарной номенклатуре. Позднее, когда
начал действовать Международный кодекс ботанической номенклатуры,
труд Жилибера был полностью отвергнут, а все видовые названия при-
знаны недействительными (nomina invalid!).
Однако, несмотря на это, сравнительно непродолжительная деятель-
ность Жилибера в старом Вильнюсском университете и его публикации
положили начало бурному развитию литовской ботанической науки.
Так, были начаты довольно обширные для того времени (конец XVIII—
начало XIX в.) флористические исследования, в проведение которых
включились ученики и последователи Жилибера — С. Б. Юндзил, Ю. Юнд-
зил, Й. Ф. Вольфганг, С. Б. Горский, Ю. Пабрежа. Некоторыми результа-
тами этих работ в виде гербариев и научных статей мы пользуемся и
сегодня.
200
Следует, однако, отметить, что такие интенсивные исследования фло-
ры Литвы длились только до закрытия Вильнюсского университета, т. е.
около 50 лет. С середины прошлого века и до 30-х годов XX в. ботани-
ческие исследования в Литве почти не проводились, лишь изредка появ-
лялись публикации со списками видов, собранных или зарегистрирован-
ных на небольших территориях.
Флористические исследования ожили в период с 1922 г. по 1940 г.,
когда в Каунасе были учреждены университет и ботанический сад,
а в Дотнуве — сельскохозяйственная академия и когда возобновила ра-
боту кафедра ботаники Вильнюсского университета. В этот период до-
вольно быстро накапливались гербарные сборы, появились некоторые
флористические работы более или менее обобщающего характера, одна-
ко они, как у В. Славинского (Slawinski, 1924), к сожалению, не охва-
тывали всю нынешнюю территорию Литвы.
Интенсивное и планомерное изучение видового состава растительно-
го покрова Литвы началось после 2-й мировой войны, когда в системе
Академии наук Литовской ССР был создан Институт биологии (с 1959 г.
Институт ботаники) и когда работники ботанических кафедр литовских
вузов активно включились в научно-исследовательскую работу. Боль-
шим коллективом авторов, сложившимся в тот период, за 20 лет был
подготовлен 6-томный труд о флоре республики. Завершение его изда-
ния почти совпало с 200-летием появления в свет вышеупомянутого
труда Жилибера.
Несмотря на то что изучение флоры Литвы началось давно, в настоя-
щее время мы не располагаем достаточными данными, на основании
которых можно было бы судить о ее динамике. Это объясняется, во-
первых, тем, что первые публикации, относящиеся к XVIII и XIX вв.,
касаются территории, значительно более обширной, чем современные
административные границы Литовской ССР, а в более объемных сводках
не упоминаются отдельные местонахождения. Во-вторых, до середины
XX в. ботаническая изученность республики оставалась крайне неравно-
мерной: в основном исследовались окрестности Вильнюса и Каунаса
(сотрудниками университетов), частично Клайпеды (немецкими бо-
таниками) ; данные по отдельным местностям были разрозненные и не-
полные.
Очень важным фактором, усложняющим флористическое изучение
любого региона, является усиливающееся влияние человека на естествен-
ные экосистемы, а тем самым и на растительный покров. Аграрнопро-
мышленное освоение земель в Литве, сопровождаемое интенсивными
мелиоративными работами и сведением лесов (лесистость территории
Литвы меньше, чем в соседних республиках), за несколько десятилетий
привело к изменению видового состава растительности. Уже исчезли
25 видов, возникла реальная угроза исчезновения ряда других, увеличи-
лось число редких видов из-за сокращения площадей, пригодных для
их произрастания, или из-за полной трансформации некоторых из них.
Первоначально флористическим исследованиям был присущ качест-
венный аспект, т. е. ботаники ограничивались установлением факта
присутствия вида на определенной территории, в то время как весьма
201
важным является накопление данных для более подробной характеристи-
ки видов растений и их местообитаний. Первой работой именно такого
рода была опубликованная в 1939 г. П. Снарскисом таксономическая
обработка манжеток (Snarskis, 1939), которая сопровождалась характе-
ристикой их местообитаний и иллюстрировалась точечными картами рас-
пространения отдельных видов и внутривидовых таксонов в Литве.
В этой работе собраны воедино таксономические, экологические и хоро-
логические сведения по одной группе растений. Через 15 лет этим же
автором был издан определитель видов растений (Snarskis, 1954), в кото-
ром отражены многие сведения о местонахождениях видов, накопленные
другими авторами, и помещены картосхемы распространения в республи-
ке почти 400 видов растений. Этот труд, наряду с другими изданиями (Re-
gelis, 1931, 1932, 1935, 1936, 1937, 1939, 1942; Hryniewiecki, 1933; Mows-
zowicz, 1958) и послужил основой для составления полного списка ви-
дов флоры Литвы.
Для более исчерпывающей характеристики видов, однако, пона-
добился дополнительный материал, поэтому с начала 70-х годов литов-
ские ботаники начали проводить экспедиции не только для выявления
распространения в республике редких и имеющих ограниченное рас-
пространение видов, но и с целью сбора возможно более полных данных
для экологической и фитоценологической характеристики местона-
хождений гербарируемых видов, включая сведения о жизненности
и численности популяций. В результате сбора и анализа данных (около
60000 гербарных образцов в различных гербариях Литвы) был полу-
чен основной материал для оригинальной характеристики видов при
составлении капитального труда о флоре республики. Более того, эти
данные послужили основой при составлении республиканского списка
видов растений, нуждающихся в охране, а также при выделении тер-
риторий заказников по охране таких видов. В период с 1970 по 1980 г.
проводились детальные исследования растительности и флоры на охра-
няемых территориях, в результате чего стала возможной разработка
принципов и рекомендаций по сохранению видов на естественных
экотопах.
При исследованиях флоры охраняемых территорий возникла
необходимость не только в установлении ее видового состава, но и
в оценке состояния и численности популяций отдельных видов, т. е.
в установлении их активности, ибо такая оценка видов, особенно
охраняемых, позволяет следить за тенденциями в изменении состава
растительности территорий и своевременно принимать соответствую-
щие меры.
Согласно последним предложениям Юрцева (Юрцев, 1982а), для
некоторых заказников была определена так называемая „территориаль-
ная активность” видов, получаемая из отдельных парциальных активно-
стей вида на тех или иных частях данной территории, обычно на разных
экотопах. Для этого была применена формула
А = jr 2 D^Q,,
V /=1
202
где А — средняя территориальная, или интегральная, активность вида;
Dj — обилие вида на конкретном (z-м) экотоне (участке) территории;
F, — встречаемость вида на данном (z-м) участке; Q — общая площадь
заказника: Q, — доля площади z-го экотопа (участка) от общей площади
территории; п — число экотопов или участков, на которых устанавлива-
лась промежуточная или парциальная активность вида.
Эту формулу можно изобразить более компактно:
_ п
Л = S л,,
/=1
где Л, — территориальная активность на конкретном участке.
Последняя формула соответствует по существу словесному предло-
жению Юрцева (1982а) по установлению относительного покрытия
в ландшафте.
С увеличением количества накопленного материала возникли за-
труднения с его обработкой и использованием. Десятки тысяч гербарных
этикеток расширили знания по отдельным видам, но оказалось невоз-
можным выбрать сведения по отдельным биотопам, по определенным
географическим пунктам и т. п. Необходимость в таких сведениях воз-
никла при подготовке материала для „Критического конспекта флоры
Прибалтики”, для „Атласа европейской флоры” и для других изданий.
В связи с этим было решено создать с помощью ЭВМ автоматизирован-
ный фонд гербарных данных и разработать программу его анализа. В на-
стоящее время такой фонд, созданный на базе Гербария Института бота-
ники АН Литовской ССР и получивший название „Herlit”, уже существует.
Таковы основные этапы изучения флоры Литвы. В настоящее время
возникла острая необходимость в более детальном выяснении с помо-
щью растровых карт распределения на территории республики отдель-
ных видов, а также в более широком использовании новых, более точ-
ных методов количественной оценки их популяций. Необходимо также
повысить интерес к изучению состава и распространения некоторых або-
ригенных осенних эфемероидов, до сих пор остававшихся вне поля зре-
ния наших флористов. Следует также заострить внимание на исследова-
нии постоянно меняющегося комплекса заносных видов, так как этот
раздел флористики имеет не только научное, но и большое практическое
значение.
А. К. Скворцов
ИЗУЧЕНИЕ ФЛОРЫ ЗАПАДА НЕЧЕРНОЗЕМНОГО ЦЕНТРА РСФСР
(БРЯНСКОЙ, КАЛУЖСКОЙ И СМОЛЕНСКОЙ ОБЛАСТЕЙ)
Главный ботанический сад АН СССР
Среди областей Нечерноземного центра флористически лучше всего
изучена Московская. Ей посвящена целая серия „Флор” и „Определите-
лей”, опирающихся на критическое изучение обширных материалов
203
(последняя сводка: Ворошилов и др., 1966). Несмотря на это, и после
1966 г. в области был сделан ряд новых интересных находок видов, ко-
торые можно причислять к коренной флоре, не говоря уже о появлении
множества новых адвентивных растений.
Благодаря давним и прочным традициям флористических исследова-
ний на Украине флора этой республики весьма хорошо изучена. В после-
военные десятилетия заметно активизировались и флористические Иссле-
дования Белоруссии. Однако значительная территория РСФСР, заключен-
ная между хорошо изученной Московской обл. и границами Украины и
Белоруссии и как бы „в углу” между этими границами, до недавнего
времени почти не пользовалась вниманием ботаников-флористов.
Нельзя, конечно, сказать, что Смоленская, Брянская и Калужская
области были вовсе „белым пятном” во флористическом отношении.
Западная часть Брянской обл. (входившая ранее в состав Черниговской
губ.) была охвачена прекрасной для своего времени флористической
сводкой А. С. Роговича (1869), которая и по сию пору не потеряла
своего значения.
Для восточной части Брянской обл. (относившейся ранее к Орлов-
ской губ.) и для Смоленской обл. отправным пунктом флористического
изучения явился „Сборник сведений” В. Я. Цингера (1886). Калужской
обл. больше „посчастливилось”: еще до „Сборника сведений” появился
очень тщательный флористический обзор П. П.Саницкого (1884), а затем
вскоре сводки Д. И. Литвинова (1895) и А. Ф. Флерова (1907—1912).
После названных основных публикаций по существу вплоть до
1960-х годов прогресс в изучении флоры трех-областей выражался лишь
в изредка появлявшихся статьях, посвященных отдельным находкам,
да во внесении дополнений в переиздания „Флоры Средней России”
П. Ф. Маевского (последнее издание: Маевский, 1964). Ее последнее
издание практически и принимается за флористическую сводку по на-
шим областям. Но полноценной такую сводку считать, конечно, нельзя.
Из появлявшихся в журналах и сборниках флористических сообщений
многие были включены во „Флору” Маевского некритически, а многие
другие, наоборот, вовсе не были учтены; из-за перемен в административ-
ном делении возникло много неточностей в указаниях на распростране-
ние видов.
Нам представляется важной и насущной задачей создание совре-
менной критической сводки по флоре наших трех областей. Заполнив
пробелы в знании их флоры, мы имели бы неплохую сплошную фло-
ристическую изученность огромной территории от Москвы к западу и
юго-западу до государственных границ. Целостный подход, т. е. рассмот-
рение флоры всех трех областей вместе, оправдывается и значительным
сходством физико-географических и климатических условий, а также
и общих характеристик растительного покрова. Все три области распо-
ложены на равнине, рельеф и покровные породы которой сформированы
в плейстоцене, все три — в лесной зоне и притом в основном в подзоне
смешанных лесов (только юго-восток Брянской обл. и небольшие уча-
стки крайнего востока Калужской находятся за пределами ареала ели,
в подзоне широколиственных лесов, что, впрочем, в Брянской обл.
204
не бросается в глаза из-за широкого распространения сосняков на занд-
ровых песках).
Целостный подход к изучению флоры региона, более широкого,
чем одна административная область, представляется сейчас, пожалуй,
вообще наиболее актуальным применительно к флоре европейской тер-
ритории СССР. В качестве хороших примеров можно назвать уже появив-
шиеся „Флору северо-востока европейской части СССР” (1974—1977),
„Определитель высших растений Северо-Запада европейской части
РСФСР” (1981), а также намеченную к изданию „Флору Прибалтики”.
Общая территория Смоленской, Брянской и Калужской областей
(114.6 тыс. км2) сопоставима с территорией трех областей (Новго-
родской, Псковской и Ленинградской), охваченных упомянутым
„Определителем” (196.5 тыс. км2) и даже с территорией Белоруссии
(207.6 тыс. км2).
Флористическое изучение Брянской и Калужской областей значи-
тельно активизировалось за последние 10—15 лет. В Брянской обл. ин-
тенсивно работали местные ботаники: П. 3. Босек и группа из Брянского
(бывшего Новозыбковского) педагогического института, возглавляе-
мая А. Д. Булоховым. Им принадлежит длинный ряд замечательных нахо-
док: на юго-востоке области, в бассейнах Сева и Усожи, обнаружена це-
лая свита лугово-степных видов: Stipa pennata, Carex humilis, Allium flave-
scens, Veratrum nigrum, Ranunculus illyricus, Peucedanum alsaticum, Lino-
syris vulgaris, Inula hirta, Cirsium pannonicum, Scorzonera purpurea и др.
Интересные находки были сделаны и в других местах Брянской обл.
(Lycopodium selago, Cinna pendula, Carex brizoides, C. umbrosa, Fritillaria
meleagris, Dianthus armeria, Pulsatilla pratensis, Hepatica nobilis, Linum pe-
renne, Armeria elongata, Chondrilla juncea и др.).
В Брянской обл. проводились исследования и группой москвичей:
сотрудниками Главного ботанического сада АН СССР В. В. Макаровым,
Г. М. Проскуряковой и мной, а также работавшим в контакте с нами
Ю. Е. Алексеевым из МГУ. Упомяну наиболее интересные наши находки
(из числа опубликованных): Gymnocarpium robertianum, Najas marina,
Scolochloa festucacea, Carex juncella, Dianthus pseudarmeria, Corydalis inter-
media, Erysimum silvaticum, Ranunculus bulbosus, Potentilla recta, Trifolium
dubium, Vicia dumetorum, Teucrium scordium, Centaurea marschalliana.
Особенно примечательно обнаружение богатого очага л угово-степной
флоры на выходах мела в долине р. Судости в Погарском р-не, т. е. еще
примерно в 100 км к западу, в глубину лесной зоны от лугово-степного
очага на Севе и Усожи (Скворцов, 1982). Ковыль здесь находится даже
в лучшем состоянии, чем на Усожи.
В Калужской обл. проявляли активность сотрудники Калужского
и Московского им. Ленина педагогического институтов (Н. С. Воронин,
Л. С. Волоснова, Т. Г. Дервиз-Соколова, Г. И. Пешкова, М. С. Хомутова
и др.). Здесь особенно замечательно нахождение на известняках в низо-
вьях р. Тарусы Cortusa matthioli и Gymnocarpium robertianum. Среди опу-
бликованных упомянутыми авторами новых интересных находок в об-
ласти можно назвать Carex loliacea, Alisma lanceolatum, Ranunculus poly-
phyllus, Phyteuma spicatum, Filago minima.
205
Некоторые новые интересные находки на р. Угре в Калужской обл.
были опубликованы мной в популярной статье в журнале ,Дрирода”
(Скворцов, 1980): Anthericum ramosum, Arctostaphylos uva-ursi, Galium
triflorum и др.
Вместе с тем уже накоплены обширные материалы, собранные мной
во всех трех областях в основном в течение последних 5 лет, которые
еще не полностью обработаны и не опубликованы. Эти материалы отра-
жают не только, и даже не столько, нахождение видов, новых для той
или иной области, как главным образом уточнение распространения ви-
дов редких или важных в ботанико-географическом отношении. Для не-
которых видов, считавшихся очень редкими или даже вовсе отсутствую-
щими в наших областях, обнаружены целые серии местонахождений.
Упомяну некоторые более интересные находки.
Polystichum braunii около р. Угры, на протяжении около 100 км ее
течения в Смоленской и Калужской областях-, от северного колена у впа-
дения р. Лостминки до устья р. Извери, найден в 5 точках.
Roegneria fibrosa найдена неоднократно на р. Неруссе в Брянской
обл., на Зап. Двине, уже близ границы с Белоруссией, в Смоленской обл.,
а по старым гербарным образцам, ранее неверно определявшимся, выяв-
лена и для Оки в Калужской обл.
Роа supina в массовых количествах обнаружен на сыроватых паст-
бищных лугах близ Смоленска и далее к западу вдоль Днепра до гра-
ницы Белоруссии.
Carex brizoides была обнаружена недавно брянскими ботаниками
(как новая находка для флоры Средней полосы) в западной части Брян-
ской обл. Мной найдена на юго-западе Калужской обл. у р.Болвы, в лист-
венном лесу вдоль края поймы, массово на протяжении около 0.5 км.
Наличие С. paniculata во флоре Средней полосы вызывало сомнения.
Мною обнаружены две довольно крупные популяции немного севернее
Смоленска и у с. Ямного Бетлицкого р-на Калужской обл.
С. juncella неоднократно отмечена в Брянской обл., в Калужской
собрана в Хвастовичском р-не; в Брянской обл. собраны С. hartmannii
(поймы Десны и Ипути) и С. aquatilis (пойма Десны).
Listera cordata — Ярцевский р-н Смоленской обл.
Dactylorhiza baltica, D. maculata, D. traunsteineri — ряд находок, позво-
ляющих уточнить наличие и распространение этих видов в наших обла-
стях.
Ranunculus lanuginosus вовсе не указывается у Маевского. Выявлен
весьма значительный очаг распространения (около 100 км с севера на
юг) в северо-западной части Смоленской обл. (не менее 10 конкретных
точек).
Agrimonia odorata также вовсе отсутствует у Маевского. В действи-
тельности широко (хотя и довольно рассеяно) распространена по всем
трем областям: по южной половине Смоленской (к востоку до р. Вори
в Темкинском р-не), по всей Брянской и почти по всей Калужской, от
р. Рессети вплоть до р. Нары (и встречена даже за Нарой, километрах
в двух в сторону Подольска в Московской обл.). Всего отмечено около
25 точек.
206
Armeria elongata недавно была обнаружена брянскими коллегами
близ Карачева. Мною найдена близ пос. Теребень Хвастовичского р-на
Калужской обл., примерно в 40 км от карачевского местонахождения
и в очень сходной ситуации: на песчаных луговинах на месте сведенного
старого сосняка, поблизости от поселка и в условиях довольно выражен-
ного антропогенного влияния, однако в сопровождении отнюдь не сор-
няков, а типичных видов вторичных песчаных пустошей, таких как
Koeleria glauca, Scleranthus perennis, Dianthus arenarius, Sempervivum sobo-
liferum, Potentilla argentea, Jasione montana, Artemisia campestris, Helichry-
sum arenarium.
Казалось, что Empetrum nigrum сохраняется только на севере Смо-
ленской обл., после того как замечательный, описанный А. Ф. Флеровым
большой верховой торфяник близ ст. Бездон (севернее Кирова) в Ка-
лужской обл. был осушен. Однако несколько восточнее железнодорож-
ной станции Вертехово удалось обнаружить небольшой уцелевший верхо-
вой торфяник вокруг озера (вероятно, провального), также называемо-
го Бездон; здесь вороника еще сохраняется.
Обнаружен также ряд местонахождений других редких и интересных
видов, на которых подробнее остановиться уже нет возможности. В част-
ности, Lycopodium selago, Dryopteris austriaca, Роа remota, P. bulbosa,
Festuca altissima, Lerchenfeldia flexuosa, Koeleria grandis, K. delavignei,
Cinna latifolia, Weingaertneria canescens, Sieglingia decumbens, Glyceria nemo-
ralis, Rhynchospora alba, Blysmus compressus, Carex remota, C. brunnescens,
C. disperma, C. loliacea, C. montana, Juncus inflexus, J. atratus, Lilium marta-
gon, Veratrum lobelianum, Iris sibirica, Gladiolus imbricatus, Coeloglossum
viride, Orchis coriophora, Betula fruticosa, Moehringia lateriflora, Stellaria
alsine, Silene chlorantha, Pulsatilla patens, Hepatica nobilis, Delphinium da-
tum, Anemone silvestris, Corydalis cava, C. intermedia, Sisymbrium strictissi-
mum, Nasturtium officinale, Lunaria rediviva, Sempervivum soboliferum,
Astragalus arenarius, Genista germanica, Orobus niger, Lathyrus palustris,
Vicia cassubica, Hypericum montanum, Elatine alsinastrum, Viola selkirkii,
V. uliginosa, V. stagnina, Epilobium nervosum, Sanicula europaea, Laserpitium
latifolium, Angelica palustris,Peucedenum cervaria,P. oreoselinum,Centaurium
pulchellum, Omphalodes scorpioides, Pulmonaria angustifolia, Scutellaria
hastifolia, Dracocephalum ruyschiana, Galeopsis pubescens, Lycopus exaltatus,
Digitalis ambigua, Scrophularia alata, Galium vernum, G. ruprechtii, G. triflo-
rum, Valeriana rossica, V. wolgensis, Adenophora liliifolia, Cirsium rivulare,
Crepis sibirica, C. biennis, Scorzonera humilis, S. purpurea и др.
Несмотря на столь значительный, даже, можно сказать, неожиданный
„урожай” новых находок, в целом изученность флоры трех областей еще
нельзя считать достаточной. Обилие новых находок надо воспринимать
не как свидетельство полноты достигнутого знания, а скорее как показа-
тель глубины того незнания, в котором мы до сих пор пребывали. Осо-
бенно много нового нужно ожидать в Смоленской обл., которая из всех
трех остается наименее изученной; к сожалению, в этой области и сейчас
нет активных местных флористов. Но интереснейшие новости не пере-
стают поступать и из Брянской и Калужской областей: так, калужанам
недавно удалось найти и собрать в гербарий ковыль на Оке севернее
207
Перемышля, о котором уже давно были известия, но документального
подтверждения не было; брянские коллеги с лесотехнического факуль-
тета нашли в области Daphne спеогит.
Как должно идти дальнейшее изучение флоры наших областей?
Я полагаю, что нужно использовать все пути и возможности. При той
степени изученности, которая имеется на сегодняшний день, и при
крайней ограниченности сил и возможностей нерационально было бы
предлагать какую-либо жесткую схему. Еще никоим образом нельзя
считать исчерпанной необходимость и перспективность даже чисто
рекогносцировочных автомобильных маршрутов, тем более что идущее
сейчас интенсивное дорожное строительство очень сильно расширяет
возможности таких маршрутов. Очень эффективны в условиях наших
областей однодневные радиальные маршруты со стационарной базы,
как чисто пешеходные, так и с использованием общественного транс-
порта, особенно если база расположена в центре интересного района.
Разумеется, небесполезно было бы и выявление и полное обследование
„конкретных флор”, в понимании школы А. И. Толмачева. Но при
настоящем уровне изученности флоры наших областей и при недостатке
флористов ориентация на конкретные флоры, пожалуй, еще прежде-
временна.1
Весьма существенное значение может иметь полевая практика
студентов, если ей придать целенаправленный флористический ха-
рактер (при этом имеются в виду, конечно, студенты необязательно
только местных вузов). Студенческие сборы очень часто содержат
интереснейшие и иногда совсем неожиданные данные. Однако при
этом необходимо, чтобы собираемые студентами гербарии отвечали
всем основным требованиям. Иначе получаются досадные случаи,
вроде, например, рассказанного мне недавно проф. Н. С. Ворониным:
в студенческом гербарии из Калужской обл. оказался Adonis vernalis,
до сих пор в области совершенно неизвестный; но точной этикетки
нет, и удостовериться в действительном наличии адониса в Калужской
обл. нельзя. Подобные случаи хорошо знакомы и работникам других
вузов.
Наряду с продолжением полевых изысканий важной ближайшей
целью изучения флоры наших областей надо считать проведение на карте
хотя бы некоторых границ ареалов, что позволило бы наметить в первом
приближении и флористическое районирование. Несмотря на кажущуюся
однородность растительного покрова всех трех областей, можно смело
предполагать, что, вероятно, не менее трети всех видов совокупной фло-
ры наших областей имеет в пределах этих областей границы своих ареа-
лов. При этом может обнаружиться иногда и не совсем предвиденное:
например, оказалось, что естественный ареал Salix alba захватывает
1 Вероятно, вполне своевременно поставить задачу постепенного создания
хотя бы разреженной сети представительных опорных (детально изученных) кон-
кретных флор на фоне продолжающегося сбора флористической информации
всеми доступными способами, что особенно актуально для баз полевой практики
студентов. (Прим. ред.).
208
в Смоленской обл. только ее юго-восточную окраину, а южная граница
ареала серой ольхи, которая восточнее Калуги близко совпадает с гра-
ницей ареала ели, западнее Калуги резко от ели отрывается и в Смолен-
ской обл. проходит уже севернее Рославля.
Г. М. Файвуш
СРАВНЕНИЕ ФЛОР ВЕРХНЕ-АХУРЯНСКОГО И ШИРАКСКОГО
ФЛОРИСТИЧЕСКИХ РАЙОНОВ (СЕВЕРНАЯ АРМЕНИЯ)
Институт ботаники АН АрмССР
Ширак — естественноисторический район, расположенный на северо-
западе Армении,— включает в себя 2 флористических района — Верхне-
Ахурянский (площадь 1200 км2) и Шмракский (1700 км2). Место
Ширака при ботанико-географическом районировании Кавказа и Закав-
казья всегда вызывало много споров: разными авторами (Гроссгейм,
Магакьян, Тахтаджян и др.) он относился или к Армено-Иранской, или
к Кавказской, или к Малоазийской провинции.
В настоящее время во флоре Ширака известно 1248 видов сосуди-
стых растений, из них в Верхне-Ахурянском районе — 753, в Шмрак-
ском — 949. Общих для двух районов видов — около 36% всей флоры,
что совсем немного для районов с общей границей.
При рассмотрении спектров семейств видно, что они очень сходны,
однако имеются и некоторые различия. Если в Ширакском районе пер-
вые 3 места занимают крупнейшие полиморфные семейства A steraceae,
Fabaceae, Роасеае, то в Верхне-Ахурянском — на третьем месте Rosaceae
(в Ширакском на восьмом). Далее, в Ширакском районе на пятом месте
Brassicaceae, а в Верхне-Ахурянском оно на девятом. Еще более рельефно*
эти различия отражаются в родовых спектрах. На первом месте в Ширак-
ском районе характернейший для Ирано-Туранской подобласти род
Astragalus (30 видов, половина из которых — армено-иранские и атропа-
тенские), а в Верхне-Ахурянском он с 9 видами на четвертом месте.
На втором месте в Ширакском районе род Centaurea (17 видов, 10 из ко-
торых армянские), в Верхне-Ахурянском — всего 7 видов васильков.
Первое место в Верхне-Ахурянском районе занимает род Ranunculus
(13 видов, преобладают голарктические и кавказские виды), в Ширак-
ском районе этот род на четвертом месте; на втором месте — род Ca-
rex — голарктическая черта. На третьем месте в обоих районах род
Allium, в основном это — узкоареальные иранские и кавказские виды.
Вообще по спектрам родов и семейств видно, что в Верхне-Ахурянском
районе преобладают общеголарктические и кавказские черты, а сами
спектры близки к спектрам Кавказской провинции, спектры же Ширак -
ского района — армено-иранского типа.
Еще более точно это разграничение проявляется при сравнении рас-
пределения геоэлементов. В Ширакском районе полностью преобладает
14 Заказ 1220
209
ирано-туранский элемент (половина приходится на армено-иранские
виды), в Верхне-Ахурянском районе этих видов в 4.5 раза меньше
(армено-иранских видов меньше в 3 раза). Зато в Верхне-Ахурянском
районе значительно больше голарктических и малоазиатско-кавказских
видов. Кроме того, в Ширакском районе почти в 2 раза больше среди-
земноморских видов.
Таким образом, в результате проведенной работы можно считать,
что граница между Армено-Иранской и Кавказской провинциями прохо-
дит между Верхне-Ахурянским и Ширакским флористическими района-
ми, т. е. Верхне-Ахурянский район является южным краем Кавказской
провинции, а Ширакский — северо-западной окраиной Армено-Иранской
провинции в пределах Армянской подпровицции, по А. Л. Тахтаджяну
(1978).
М. В. Казакова
ВКЛАД А. И. ТОЛМАЧЕВА В РЕШЕНИЕ ПРОБЛЕМЫ РЕЛИКТОВ
С ТОЧКИ ЗРЕНИЯ БОТАНИЧЕСКОЙ ГЕОГРАФИИ
Заповедник „Галичья гора”
А. И. Толмачев внес существенный вклад в разработку многих во-
просов ботанической географии, и в частности проблемы реликтов.
Большинству флористов приходится так или иначе обращаться к рас-
смотрению этого явления. Необходимость анализа реликтов возникла
и в ходе наших исследований флоры Липецкой обл., причем нашу пози-
цию по этому вопросу определило и конкретизировало знакомство с ря-
дом работ А. И. Толмачева (1931, 1938, 1952, 1958, 1962, 1974 и др.).
Основу флористического анализа составляет выделение элементов
флоры. Рассматривая флору любой территории как явление историче-
ское, мы выделяем элементы, „отражающие как бы исторические стадии
участия вида в формировании флоры. Это — элементы реликтовые (или
просто реликты), консервативные и прогрессивные” (Толмачев, 1974,
с. 164). Основным в определении реликта, по мнению Толмачева, явля-
ется констатация неблагоприятных для него соотношений с сегодняшни-
ми условиями существования”, которыми вызваны сокращение ареала
и уменьшение числа популяции. В понимании Толмачева, ,,не относитель-
ная древность и не устойчивость признаков, но наличие известного несо-
ответствия природы вида (группы видов) современным условиям, опре-
деляющее невозможность процветания в данную эпоху”, является сущ-
ностью реликта как пережитка некогда существовавших условий.
Реликтовый элемент, таким образом, является показателем условий
минувших времен, в которых существовала иная флора (в рассматри-
ваемой конкретной области), отличного от современной облика.
Увидеть данное несоответствие позволяет характер ареала — либо
очень узкий в цепом, либо разделенный на несколько изолированных
210
фрагментов. Весьма распространен случай, когда вид имеет довольно
обширный ареал и выступает в современной флоре в качестве элемента,
гармонирующего с существующими условиями, но в значительном отры-
ве от основной части ареала имеются островные фрагменты его, позво-
ляющие характеризовать вид в составе этих местных флор в качестве
реликтового элемента.
На примерах из флоры европейского северо-востока Толмачев
(1938) показал, что не всякий редкий, узколокализованный и стенотоп-
ный вид следует считать реликтовым элементом флоры. Так, в районе
Пинеги—Кулоя арктоальпийские виды Aster alpinus или Astragalus uralen-
sis, находящиеся в явном отрыве от основной области распространения
и в чуждом для них лесном окружении, отнесены Толмачевым к релик-
товым элементам ледникового периода. В то же время такие растения,
как Chrysanthemum sibiricum и Anemone sylvestris, расположенные на гра-
нице сплошного ареала, рассматривались Толмачевым как формаль-
ные”, нереликтовые элементы современной флоры. Но на территории
Среднерусской возвышенности, в условиях европейской лесостепи хри-
зантема сибирская (Dendranthema zawadskii (Herb.) Tznq\. = Chrysanthe-
mum sibiricum Fisch.) выступает в качестве реликта ледникового перио-
да: растение находится в значительном отрыве от уральской части ареала
(болбе 1500 км), известны лишь две небольшие популяции из Липецкой
и Курской областей, причем число особей в первой популяции весьма
невелико.
Подход Толмачева к определению реликтовых элементов привле-
кает своей конкретностью и четкостью изложения. Это помогло нам
проанализировать группу из 35 видов, представленных во флоре Липец-
кой обл. и относимых традиционно со времен Д. И. Литвинова, Б. М. Ко-
зо-Полянского и С. В. Голицына к категории реликтовых. В результате
мы пришли к выводу, что далеко не все виды этой группы обладают не-
обходимыми признаками реликтовости.
Известные к настоящему времени сведения о флоре европейской
части страны показывают, что некоторые виды на территории Средне-
русской возвышенности не имеют островных фрагментов ареала, а на-
ходятся на окраине довольно обширной области своего распространения.
Вопрос же о положении вида на границе ареала заслуживает специального
изучения. В исследованном нами районе проходит северная граница ареа-
лов Ephedra distachya, Allium inaequale, Arabis auriculata, западная грани-
ца Artemisia armeniaca, A. latifolia, A. sericea, Helictotrichon desertorum
(основная часть ареала}, восточная граница Laserpitium latifolium. Ника-
ких оснований рассматривать эти виды на окраине своего естественного
ареала, соответствующей их экологическим требованиям, реликтовыми
в современной обстановке нет.
Некоторые виды, как например Carex humilis, до сих пор традицион-
но считаются реликтовыми на Среднерусской возвышенности и авто-
матически зачисляются в разряд „классических” реликтов. Но, как
совершенно справедливо замечал Толмачев (1974, с. 167), „пред-
ставление о реликтовой природе растений, как очень многие естест-
веннонаучные представления, не имеет характера чего-то абсолютного
211
и непререкаемого”. Изучение осоки низкой в разных частях ареала по-
зволило нам убедиться в том, что это весьма современный и активный
элемент в лесостепных районах Среднерусской возвышенности и в дру-
гих частях своего в целом разорванного ареала.
Наличия дизъюнкций в ареале еще недостаточно для характеристики
вида как реликтового. Необходима констатация несоответствия приро-
ды (экологии) вида окружающей обстановке, послужившего причиной
обособления и сохранения островного фрагмента ареала. Draba sibirica,
например, обладает обширным ареалом к востоку от Урала. На Сред-
нерусской возвышенности, в лесостепной зоне, находится значитель-
но меньшая часть ареала, но в пределах ее вид является обычным рас-
тением луговых степей, опушек, полян, лугов, широко встречается
на- вторичных местообитаниях — по обочинам дорог и насыпям.
Происходит даже расширение области распространения вида за пределы
лесостепной зоны — по железнодорожным насыпям и луговым обочи-
нам шоссе он проникает в лесную зону, известен в Московской, Рязан-
ской и Брянской областях. Следовательно, нет оснований считать этот
элемент реликтовым во флоре Среднерусской возвышенности. Приве-
денный пример подтверждает мысль Толмачева о том, что „переход
на положение реликта — процесс не необратимый и что при опреде-
ленных внешних условиях реликт может стать активным видом и как
бы «уйти» с исторически намечавшегося пути к вымиранию” (1974,
с. 166).
Следуя определению Толмачева, к реликтовым элементам лесостеп-
ной части Среднерусской возвышенности бесспорно относятся Chrysan-
themum sibiricum, Cardamine trifida (Poir.) В. M. G. Jones (Dentaria alaunica
Golits.), Potentilla pimpinelloides L. (P. tanaitica Zing.).
В своем определении понятия „реликт” Толмачев расходился
с теми авторами, которые распространяли его на все более древние
элементы флоры, вошедшие в состав ее в одну из минувших эпох и
не претерпевших с того времени изменений. Он выделял суть этого
понятия — противоречие с современными условиями существования —
и зачастую подкреплял зту мысль примерами из самых различных
флор. На своем материале из лесостепной флоры Среднерусской
возвышенности мы постарались показать возможности применения
идей Толмачева к анализу реликтовых элементов данной террито-
рии. Это позволило несколько иначе взглянуть на известную здесь
группу реликтов и частично пересмотреть сложившееся о них пред-
ставление.
212
Р. И. Гагнидзе
ВОПРОСЫ ФЛОРОГЕНЕЗА НЕКОТОРЫХ ВЫСОКОГОРНЫХ
ФЛОРОЦЕНОТИЧЕСКИХ КОМПЛЕКСОВ КАВКАЗА
ПО КАРИОЛОГИЧЕСКИМ ДАННЫМ
Институт ботаники АН ГССР
Кариологический метод наряду с другими широко применяется
в систематике растений. Отрасль кариологии — кариогеография, или
цитохорология (Favarger/1967; Ehrendorfer, 1970, 1980), цитогеография
(Sauer, 1979), география чисел хромосом (установление хромосомных
рас, плоидности видов, подвидов и популяций), успешно может быть ис-
пользована для выяснения некоторых вопросов флорогенеза той или
иной фитохории (фитохориона).
С этой цепью на основе ботанико-географических (включая ареало-
гические) и кариологических исследований в разных высотных поясах
растительности изучены некоторые флороценотические комплексы,
а также отдельные эндемичные и редкие виды высокогорий Большого
Кавказа. Флороценотический комплекс1 нами понимается как истори-
чески сложившаяся совокупность видов естественной флоры, эколого-
ценотический и географический ареалы которых совпадают (Толмачев,
1931, 1974; Попов, 1963; Юрцев,. 1968а, 19686; Зозулин, 1970, 1973;
Юрцев, Петровский, 1971; Камелин, 1973; Гагнидзе, 1974, 1977, 19846).
При этом исторически сложившаяся совокупность видов одного комп-
лекса флорогенетически должна быть связана с определенным простран-
ством и экоценотипом.
Установлено, что во флороценотических комплексах субальпийско-
го, альпийского и субнивального высотных поясов растительности гра-
нитных, сланцевых, порфиритовых, известняковых орографических
единиц Кавказа отмечается -преимущественно низкое процентное со-
держание полиплоидных видов с удвоенными и многократно умножен-
ными (тетра-, гексаплоиды и т. д.) хромосомными наборами. На опреде-
ленном этапе развития вида часть полиплоидных хромосомных рас была
изолирована от анцестральных форм, вследствие чего произошло частич-
ное или полное ограничение панмиксии между популяциями или видами.
1 Термин „флороценотический комплекс” впервые введен Б. А. Юрцевым по
существу как синоним термина „эколого-ценотическая группа видов” (Юрцев,
19686, с. 86; Юрцев, Петровский, 1971). Впоследствии трактовка первого термина
его автором была несколько изменена, с тем чтобы оба несли самостоятельную
смысловую нагрузку: эколого-ценотические группы объединяют виды, сходные
по всему спектру экотопов и сообществ (фитоценоциклу), флороценотические же
комплексы объединяют виды по совпадению основного для каждого из них класса
экотопов и сообществ (см. первую статью Юрцева в наст, сб.), образуя „флористи-
ческие ядра” экологически своеобразных классов сообществ. В данной статье
„флороценотический комплекс” определяется по существу как эколого-географи-
ческий элемент флоры (по Юрцеву), причем ссылка на работы Попова, Зозулина,
Камелина привносит сюда также известное флорогенетическое содержание.
(Прим. ред.).
213
Величина популяций и их дифференциация определяются изоляцией.
Майр (1973) и Грант (Grant, 1981, цит. по пер.) считают географическую
изоляцию одним из универсальных факторов обособления и эволюции
вида. На определенном этапе своего развития изолированные популяции
представляют собой уже самостоятельные новые виды или подвиды
(Майр, 1973; Толмачев, 1974), входящие в состав различных флороце-
нотических комплексов.
В условиях высокогорий одним из важных факторов эволюции так-
сона является одновременно высотная и широтно-долготная пространст-
венная изоляция, так как ареал горных видов — явление трехмерное
(Юрцев, 1968а; Куваев, 1972; Гагнидзе, 1974; Толмачев, 1974; Гагнид-
зе, Иванишвили, 1975а).
Отмеченное выше преобладание диплоидных видов в субнивальном
флороценотическом комплексе высокогорий Кавказа (Тумаджанов,
Беридзе, 1968; Харадзе и др., 1973а, 19736; Харадзе, 1974) ставит под
сомнение зависимость процесса полиплоидизации от экстремальных
условий произрастания (Гагнидзе, 1983, 1984а, 19846).
Несмотря на повышение доли полиплоидов во флоре высоких
широт, а также в субарктических высокогорьях, нет оснований рас-
пространять эту закономерность на южные горные фитохории Пале-
арктики, в частности на „Средиземногорье” и Кавказ (Гагнидзе, 1984а,
19846).
Мнение Стеббинса (Stebbins, 1984), что полиплоиды лучше приспо-
сабливаются к климату альпийского пояса, требует пересмотра. Изуче-
ние полиплоидии в альпийских флорах Европы (Kupfer, 1974) и Север-
ной Америки показало, что процент полиплоидов здесь, как и на Кав-
казе, не выше, чем во флоре нижнегорных поясов. Стеббинс объясняет
образование полиплоидов реакцией растений на неустойчивость усло-
вий существования, арктоальпийскими миграциями и гибридизацией
при повторных контактах между различными экотипами одного вида,
между близкими диплоидными видами, а также между видами поли-
плоидными.
Данное мнение подтверждается результатами изучения высокогор-
ных видов рода Ranunculus на Центральном Кавказе. Диапазон высотно-
го распространения популяций очень многих видов рода Ranunculus (на-
пример, R. caucasicus, R. oreophilus, R. raddeanus) весьма велик. Поэтому
для установления их фенетической изменчивости на высотном профиле
исследовались популяции на площадках, заложенных с интервалами
в 100 м, параллельно изучались также хромосомные расы. Выбор в каче-
стве объектов видов отмеченного рода вызван полиморфизмом вегета-
тивных и генеративных органов, наличием нескольких, в основном не
очень четких, а также дискретных фенотипов внутри популяций (Чурад-
зе, 1981, 1982, 1984).
Несмотря на морфологическое разнообразие и значительную широт-
но-высотную амплитуду распространения рода Ranunculus в целом, на
Кавказе наблюдается константность хромосомных чисел; в основном
данный вид представлен диплоидными хромосомными расами. Там же,
где имеются условия для повторного контакта между видами, отмечены
214
Рис. 1. Распространение на Кавказе и в Закавказье диплоидных видов рода Scabiosa.
1 - S. bipinnata (2n= 16), 2 - S. olgae (2n = 18), 5 - S. colchica (2n= 18), 4 - S.gum-
betica (2n = 18).
триплоидные и тетраплоидные хромосомные расы, 2п = 24, 32 (Агапова,
1981, 1983; Соколовская, 1982; Агапова, Земскова, 1983; Гагнидзе,
Чурадзе, 1984, 1985).
Диплоидность кавказских лютиков, по-видимому, следствие геогра-
фической изоляции на протяжении длительного исторического развития
флоры Кавказа.
Следует специально рассмотреть случай, когда отдельные популяции
вида имеют узко локальное распространение в пределах ареала и обособ-
ленность части их достигла видового уровня. Ограниченное расселение
таких узко локальных видов следует объяснить гомогенностью их гено-
фонда, которая определяет также ограниченную изменчивость популяций
и обедняет состав флороценотических комплексов.
Одним из путей обогащения флоры той или иной высокогорной
области является процесс формирования многочисленных локальных
видов одного рода с различными основными числами хромосом или раз-
ными хромосомными наборами. При кариологическом изучении кальце-
фитной флоры Западного Закавказья выявлены многочисленные при-
меры подобного рода. Так, в роде Scabiosa отмечены 2 основных числа
хромосом: х = 8 и 9. Развитие эндемичной кавказской кальцефитной
215
Рис, 2. Распространение на Кавказе и в Западном Закавказье видов рода Kemula
riella.
1 - К. colchica (2n= 12), 2 - К. caucasica (2n= 18).
группы рода Scabiosa происходило на диплоидном уровне; среди видов,
имеющих 2п = 18, можно назвать 5. gumbetica, S. olgae, S. colchica, изоли-
рованные друг от друга как в систематическом, так и в географическом
отношениях. К диплоидной группе видов относятся также восточнокав-
казские эндемы S. georgica, S. owerinii и кавказско-малоазиатская 5. Ы-
pinnata, имеющие, однако, другое основное число х = 8 (2п= 16) (Магу-
лаев, 1979; Гагнидзе, 1983) (рис. 1).
Кавказско-малоазиатский род Kemulariella был выделен из рода
Aster С. Г. Тамамшян. Распространен в основном в западной части Боль-
шого Кавказа и в альпийском и субальпийском поясах Западного Закав-
казья (рис. 2). Из представителей данного рода Kemulariella caucasica -
луговой монтанный вид с триплоидным набором хромосом (2п=18).
В пределах его ареала встречается и другой — кальцефильный, эндемич-
ный для Западного Закавказья, древний диплоидный вид К. colchica
(2п =12). Оба вида имеют изолированное положение в системе рода.
К. caucasica следует отнести к числу древних полиплоидов.
Высокогорная кальцефитная флора Западного Закавказья характе-
ризуется значительным видовым эндемизмом. Во флоре известняковых
орографических единиц Западного Закавказья высокогорные эндемики
выявлены в родах Gentiana, Campanula, Draba, Kemulariella, Albowiodoxa
(Amphoricarpus), Chymsydia, Woronowia, Arabis, Ranunculus, Betonica
и др. Возраст кальцефильных эндемичных видов различен. Большинство
видов — циркумэвксинские и циркумсредиземноморские или кавказ-
ско-переднеазиатские, возникшие в результате трансформаций анце-
стральных видов.
216
Рис, 3. Распространение видов рода Betonica на Кавказе и в Закавказье.
1-3 - диплоидные виды (2п= 16): 1 - В. ossetica, 2 - В. nivea, 3 - В. abchasica;4 -
В. masandarana.
В высокогорном петрофитном комплексе особо следует отметить
схизоэндемы Большого Кавказа, представленные несколькими стено-
хорными видами, дифференцированными от единого общего предка.
Таковыми являются Betonica abchasica, кальцефит Западного Закав-
казья (2п = 16), В. ossetica (2п = 16), В. nivea (2п = 16), также В. masan-
darana (Хинтибидзе, 1953; Харадзе, 1974) (рис. 3).
Род Alyssoides (Vesicaria) — олиготипный средиземноморский; вид
A. graeca — восточносредиземноморского распространения; в Средизем-
номорье встречается лишь тетраплоидная раса (2п=16). Кавказская
раса, распространенная на северо-западе Кавказа и в Сванетии, характе-
ризуется диплоидным набором (2п = 8), ее следует относить к среди-
земно морско-эвксинскому элементу (рис. 4).
Рис. 4. Распространение диплоидной (/) (2п=8) и тетраплоидной (2) (2п=16)
хромосомных рас Alyssoides graeca (Reut.) Jav.
217
Проведенное нами кариологическое исследование эндемичного каль-
цефитного комплекса горных фитохорий Западного Закавказья показа-
ло преобладание диплоидных видов, как и в субнивальном поясе (Гаг-
нидзе, 1983, 1985). Сопряженный кариологический и ботанико-географи-
ческий анализ выявил основные черты формирования и последующего
развития кальцефитного комплекса Западного Закавказья на фоне исто-
рии Эвксинского бассейна. Многочисленные кальцефильные таксоны
оформились в результате более поздней изоляции отдельных частей При-
черноморья и Большого Кавказа. Некоторые диплоидные виды в систе-
матическом отношении изолированы, например Scutellaria helenae (2n =
= 22), Ranunculus helenae (2n=16). Полиплоидные эндемичные виды
в данных фитохориях представляют собой архиэндемы, или палеополи-
плоидные эндемы (например, Campanula mirabilis с 2n= 102). Ботанико-
географические, ареалогические и кариологические данные свидетельст-
вуют также о древних связях высокогорной флоры Кавказа со среди-
земноморскими и переднеазиатскими комплексами.
Длительное существование Черноморского бассейна, его соединение
с Тетисом и Паратетисом или изоляция от них способствовали форми-
рованию (наряду с высокогорными флороценотипами) также лито-
кальцефитных комплексов, представители которых имели и циркум-
эвксинское распространение. Дальнейшие тектонически обусловленные
изменения в орографии окружающих стран и образование Эвксинского
бассейна вызвали дифференциацию анцестральных видов и формирова-
ние локальных или викарных рас в отдельных частях Большого Кавказа
и Эвксинской области (Колаковский, 1961; Харадзе, Гагнидзе, 1970;
Гагнидзе, 1983, 19846, 1985).
218
IV. ПРОГРАММЫ
ФЛОРИСТИЧЕСКИХ ИССЛЕДОВАНИЙ
РАЗНОЙ СТЕПЕНИ ДЕТАЛЬНОСТИ
Назначение предлагаемых „типовых” программ — служить основой
для разработки конкретных программ с учетом размеров, особенностей
и уровня изученности территории, а также различного контингента ис-
полнителей этих программ и срока, на который они рассчитаны.
Предусматривается 4 основных уровня детальности и соответственно
4 формы флористических исследований. В Приложении приведены типо-
вые программы изучения проб флоры (конкретных, локальных флор)
разной степени детальности: для так называемых ботанико-географичес-
ких эталонов, для ботанико-географических полустационаров, для комп-
лексных биологических стационаров и станций (см. также: Юрцев,
19756).
Изложению программ предпослан краткий перечень разделов, по ко-
торым может Проводиться обработка полученной флористической ин-
формации (целевая установка программ).
ОБЩИЕ ЗАДАЧИ ФЛОРИСТИЧЕСКОГО
ИЗУЧЕНИЯ ТЕРРИТОРИИ
§ 1 /Исчерпывающая инвентаризация видового и расового состава на
современном уровне на основе полевых исследований и критического
изучения прежних коллекций, таксономической и флористической лите-
ратуры^ В перспективе все большее значение при инвентаризации флоры
будет иметь также выявление фенетического своеобразия местной попу-
ляции (популяций) каждого вида, для чего необходим сравнительный
анализ его изменчивости на протяжении территории флоры и за ее пре-
делами. _____
§ 2. / Изучение распределения каждого вида (расы) по территории
флоры и в ее пределах по классам экотопов и сообществ ^т е- выявление
и классификация географической и экотопологической структуры мест-
ных популяций —населений всех видов и рас), оценка его „веса” (актив-
ности) на данной территории и основных признаков-компонентов послед-
него показателя: широты экологической амплитуды, встречаемости —
1 При обработке материала по трудным критическим труппам, выявлении
расового состава крайне желательно участие специалистов-монотрафов (ревизия
сборов, консультации).
219
„географической активности”, характерного уровня численности. Жела-
тельно провести сравнение по названным показателям местной геогра-
фической популяции данного вида с соседними.
§ 3. Выявление и оценка географической дифференциации флоры
на данной территории по естественным географическим подразделениям
последней: флористическое районирование территории (в том числе
выявление флористических границ и оценка их ранга, выявление комп-
лекса эндемичных, других дифференциальных и ко дифференциальных
таксонов каждой фитохории), подключение местного районирования
к общерегиональному и планетарному. Как основа районирования пред-
полагается сравнительный анализ границ ареалов таксонов в пределах и
за пределами территории изученной флоры, а также качественное и коли-
чественное сравнение естественных флор по числу, составу и соотноше-
нию таксонов разного ранга (прежде всего видов и рас) и различных ка-
тегорий типологических элементов флоры (см. § 5).
§ 4. Выявление экотопологической дифференциации флоры изучен-
ной территории — набора и видового состава парциальных флор (ПФ)
разных типов и классов экотопов и сообществ („ценофлор”), изучение
распределения ПФ по территории флоры; выявление комплекса специ-
фических видов для каждого класса экотопов и сообществ (флороцено-
тических комплексов).
Важно достигнуть единообразного подхода к выделению типов
и классов экотопов,1 классификации сообществ, по крайней мере в рам-
ках данной конкретной программы.
§ 5. Анализ флоры в целом, флор подчиненных (включенных) фи-
тохорий и основных типов парциальных флор или ценофлор (формаци-
онный анализ в смысле А. И. Толмачева).
Базой для анализа является максимально полный и точный (критиче-
ский) список видов и рас (подвидов), зарегистрированных на территории
флоры. Для сильно нарушенных хозяйственной деятельностью террито-
рий рекомендуется, помимо анализа списка в целом, раздельно анализи-
ровать по сходной программе набор аборигенных и адвентивных видов.
Основные разделы анализа флоры
А. Таксономический анализ: набор крупных таксонов (родов, се-
мейств и вышестоящих категорий), их соотношение по числу таксонов
низшего ранга (в первую очередь видов), различные типы таксономи-
1 Обычно типы экотопов - участки со сходными экологическими услови-
ями - распознаются в природе по своеобразию растительности (внешний облик,
своеобразие набора и соотношения видов) на определенных местоположениях (эле-
ментах рельефа с определенным составом почвообразующих пород), фактически
это - типы биотопов. Значительно отличающиеся растительностью биотопы на одно-
типных местоположениях большей частью соответствуют разным стадиям сукцес-
сии и, как привило, также отличаются экологическими условиями (в том числе
ценосредой). Сходные (в том числе и флористически) типы экотопов объединя-
ются в классы. В природе разнотипные экотопы одного класса (сходные фации)
обычно группируются в урочища (об иерархической системе экотопов см.: Юрцев,
Семкин, 1980; Юрцев, 1982а).
220
ческой структуры флоры. При анализе состава особо выделяются и рас-
сматриваются эндемичные, субэндемичные и другие дифференциальные
таксоны. При особо детальных исследованиях этот раздел может быть
дополнен эйдологическим (расовым) анализом (подробнее об этом
говорится ниже в разделе „Биологические элементы”).
Б. Типологический анализ: по различным категориям современ-
ных, наблюдаемых в настоящее время признаков, которые в принципе
могут быть выявлены с большой точностью. Для всех видов, кроме
эндемиков изученной флоры, правомерны и желательны 2 взаимодопол-
няющих подхода к группировке (выделению типологических элемен-
тов) практически по любому признаку: а) по общим особенностям
вида в целом, б) по особенностям его местной географической попу-
ляции.
Круг признаков видов, по которым может анализироваться флора,
почти безгранично широк, поэтому отбор признаков зависит от задач
исследования, как и различаемое число градаций любого признака. Ниже
приводятся лишь основные группы (категории) признаков, по которым
выделяются элементы флоры.
1. Географические: по характеру современного ареала - могут ха-
рактеризоваться набором крупных фитохорий (иногда континентов
и субконтинентов), в которых встречается вид; для северных флор ши-
роко используется метод „биогеографических координат” (Юрцев,
1968а) — раздельной классификации видов по зональному (широтному)
и секторальному (долготному) распространению и по соотношению ши-
ротного и долготного распространения; при анализе горных флор с раз-
витой высотной поясностью выделяются также высотные (или поясно-
зональные) группы; для флор холодных областей и горных поясов по-
лезно различение арктических, арктоальпийских и альпийских видов;
для видов северных внетропических флор дополнительно практикуется
разделение на группы по присутствию или отсутствию во внетропических
районах южного полушария (биполярные, нотобореальные и т.д.); выше
уже говорилось о важности выделения и анализа группы эндемичных
и других дифференциальных видов.
2. Экологические: по отношению видов к различным категориям
экологических факторов (соответствующие группы не вполне удачно
именуются „экоморфами” — см.: Цыганов, 1983); следует точно указать
критерии, на основании которых виды распределялись по экологическим
группам, обычно флористы судят об этом косвенно — на основе анализа
распределения видов по местоположениям; особое место занимает клас-
сификация видов по их отношению к термической зональности — пояс-
ности и к континентальности — океаничности климата, в основе которой
лежит климатологический анализ ареалов.
3. Эколого-ценотические, или синэкологические: по распределению
видов по экотопам и сообществам (эколого-ценотические группы объ-
единяют виды со сходным набором типов экотопов или классов экото-
пов и сообществ в пределах данной фитохории, сходным фитоценоцик-
лом, флороценотические комплексы — виды, в той или иной мере специ-
фичные для определенного класса экотопов и сообществ), по их экото-
221
пологической активности - преуспеванию в данном ландшафте, по их
фитоценотипу и т. д.
4. Биологические: по жизненной форме как комплексной характе-
ристике вида или отдельным ее признакам-компонентам [„линейная
классификация жизненных форм”, по В. Н. Голубеву (1972)], по спосо-
бам опыления, способам распространения семян, „жизненной стратегии”
[например, К- и г-стратегии, по Пианка (1981)], по соотношению гене-
ративного и вегетативного размножения, вегетативной подвижности
и т.д. Особо важны (но и особенно трудны для изучения) показатели,
характеризующие модусы видообразования и эволюции таксонов, на-
пример кариологические (выделение диплоидов и полиплоидов разного
уровня, анеуплоидов, апомиктов) и более сложные, комплексные [на-
пример, классификация Фаварже (Favarger, 1961) с дополнениями
Стеббинса (Stebbins, Major, 1965); см. также статью Р. В. Камелина
в наст, сб.].
Так,метод полиплоидных спектров Фаварже (Favarger, 1961) пред-
полагает разделение всей флоры на диплоиды, палеополиплоиды (таксо-
номически резко обособленные типы; близко родстве иные диплоиды
не сохранились), мезополиплоиды (хорошо обособленные виды; имею-
щие более примитивных диплоидных сородичей в том же роде или близ-
ком роде), неополиплоиды (морфологически слабо обособленные расы,
как правило, замещающие или вытесняющие на данной территории близ-
кую диплоидную расу), хромосомные расы (морфологически неотличи-
мые от диплоидной расы и, как правило, обитающие с ней совместно).
Стеббинс и Мэйджер (Stebbins, Major, 1965) видоизменили классифика-
цию Фаварже применительно к задаче анализа эндемизма. Они различают
палеоэндемики (монотипные секции или таксоны более высокого ран-
га) , схизоэндемики (возникшие в разных районах более или менее од-
новременно от общего предка), патроэндемики (диплоидные или низко-
полиплоидные эндемики, ставшие родоначальниками широко распро-
страненных высокополиплоидных форм), апоэндемики (полиплоидные
эндемики, возникшие от широко распространенного диплоидного или
низкополиплоидного таксона).
Специальный фитоэйдологический (расовый) анализ предполагает
вычленение в составе флоры различных типов рас — собственно амфи-
миктических видов, амфимиктических, изолированных или полуизоли-
рованных подвидов, резко изолированных, малочисленных локальных
популяций, обитающих в отрыве от основного ареала вида, агамных
комплексов и „видов”, клоновых образований, а также различных ги-
бридогенных образований — гибридных зон смешения (гибридных
„роев” - swarms), вторично сливщихся видов, обособленных гибридо-
генных рас (видов и подвидов), резко уклоняющихся гибридогенных,
обычно бесплодных, особей и клонов. Следует учесть, что агамные комп-
лексы также чаще всего являются результатами гибридизации исходных
видов, один из которых или даже оба в настоящее время могут отсут-
ствовать на территории распространения агамного комплекса.
Установив соотношение различных категорий элементов флоры
по числу относящихся к ним видов, определяют соответствующие типо-
222
логические структуры флоры: географические, экологические и т.д.
Таким путем определяется или уточняется место изученной флоры в ти-
пологических классификациях флор по разным группам признаков и
в системе фитохорий (флористическое районирование). Кроме того, ана-
лиз флоры по признакам и свойствам местных географических популя-
ций каждого вида — основной подход к познанию флоры как природной
системы (Юрцев, 1982а, а также в наст, сб.).
Большой интерес представляет также анализ корреляций в распреде-
лении видов данной флоры по разным категориям элементов (например,
географических и экологических; см. статью А. В. Галанина в наст, сб.)
и выяснение на этом основании двумерных групп (например, эколого-
географических — по сходству ареала и распределения по экотопам и со-
обществам в его пределах).
В. Флорогенетический (исторический, историко-типологический)
анализ по происхождению и истории расселения каждого таксона (в пер-
вую очередь видов): а) в целом, б) на территории флоры (история
местной географической популяции). Происхождение каждого вида
(или вышестоящего таксона) может оцениваться почти по всем типоло-
гическим признакам, перечисленным выше (место-происхождения, ланд-
шафтно-климатическая обстановка, экологические условия, ценотичес-
кое окружение, исходный ценотип, биоморфа и т.д.), и, кроме того,
должно быть привязано к тому или иному отрезку геологического вре-
мени. Подразделы а и б для эндемичных таксонов совпадают; для флор
с повышенной ролью аллохтонных (миграционных) элементов особенно
важно выяснить время и обстановку вхождения вида в данную фло-
ру и его местную историю. Суждения о происхождении таксона всегда
в той или иной мере гипотетичны и нуждаются в перекрестной проверке
всеми доступными данными различных дисциплин (см. ниже), при этом
следует избегать чрезмерной детализации (создающей иллюзию высокой
точности). Для более молодых флор в первом приближении о месте
и обстановке формирования вида можно судить по данным о современ-
ном его распространении и эколого-ценотических связях (с необходимы-
ми коррективами). Поэтому богатую информацию об истории изученной
флоры может дать сопряженный эколого-географический (типологичес-
кий) ее анализ, выявление корреляций в распределении видов по геогра-
фическим и экологическим элементам и палеогеографическая интерпре-
тация выявленных соотношений. Наиболее фундаментальным биологиче-
ским подходом к восстановлению истории флоры и более тесно связан-
ных между собой комплексов видов в ее составе является сопряженный
филогенетический и палеогеографический анализ систематических групп,
к которым относятся виды данной флоры или данного флористического
комплекса (Камелин, 1973) ?
1 Материалом для такого анализа являются монографические обработки раз-
личных систематических групп, как опубликованные другими авторами, так и ори-
гинальные, что предполагает достаточную подготовку флориста-флорогенетика
в области систематики.
223
Все разделы анализа флоры имеют существенное значение для фло-
ристического районирования и могут быть использованы для характе-
ристики фитохорий того или иного ранга; поэтому анализ флоры по воз-
можности должен проводиться в сравнительном плане. При сравнении
флор целесообразно придерживаться следующей логической схемы (Юр-
цев, Семкин, 1980) : 1) сравнение флор по их общему видовому (и расо-
вому) составу с расчетом мер сходства, а для неравновеликих флор —
и мер включения; анализ состава общих и дифференциальных таксонов,
интерпретация; 2) сравнение флор по набору и соотношению крупных
таксонов и различных категорий типологических элементов флоры
(с отвлечением от их видового состава), интерпретация; 3) раздель-
ное сравнение флор по видовому и расовому составу каждого из круп-
ных таксонов и элементов, общих для сопоставляемых флор, интер-
претация.
§ 6. Восстановление истории флоры (растительного покрова) тер-
ритории как компонента ее ландшафтной оболочки: установление основ-
ных истоков формирования флоры, места, времени, палеогеографичес-
ких и палеоэкологических условий становления ее как индивидуального
сочетания флористических комплексов, наконец, выяснение основных
этапов ее развития. Собственный подход флористики к решению пере-
численных задач связан с флорогенетическим анализом (см. выше),
однако необходима палеогеографическая увязка флорогенетических
гипотез с независимо полученными данными других дисциплин: геоло-
гии, палеонтологии, палеоботаники, зоогеографии и др.; поэтому прие-
мы флорогенетического анализа входят в арсенал методов палеогеогра-
фии как комплексной дисциплины.
§ 7. Выявление природных и антропогенных тенденций изменения
флоры, прогноз будущих ее состояний. Базой для прогноза, помимо
данных о предполагаемых изменениях климата и других абиотических
компонентов физико-географической оболочки в связи с природными
процессами и деятельностью человека, должны служить закономерности
развития флоры (растительного покрова), выявленные путем флороге-
нетического анализа (с учетом имеющихся палеоботанических данных),
а также результаты изучения данной региональной флоры как подвижно-
равновесной системы местных популяций видов растений.
§ 8. Выявление редких (реликтовых, эндемичных и др.) видов и рас
(также прочих внутривидовых единиц), нуждающихся в охране, карти-
рование их распространения, учет современного состояния популяций;
выявление очагов повышенной концентрации редких реликтовых эле-
ментов флоры, реликтовых типов сообществ. Выработка эффективных
мер охраны, содействие их осуществлению (с помощью местных и цент-
ральных организаций охраны природы).
Выполнение перечисленных задач предполагает повышение точности
и полноты регистрации флористической информации (прежде всего
высокой точности и информативности этикеток гербарных сборов,
флористических списков, точной привязки местонахождений к карте),
а также рационализацию и в перспективе унификацию и автоматиза-
цию сбора и хранения флористической информации — в форме, макси-
224
мально удобной для дополнения, корректировки, обработки и выда-
чи; создание информационно-поисковых систем при крупных гербари-
ях и т. д.1
ПРОГРАММА МАРШРУТНЫХ
РЕГИОНАЛЬНЫХ ИССЛЕДОВАНИЙ
Данный метод флористических исследований — традиционный и наи-
более старый. Его преимущество перед методом выборочных проб фло-
ры — возможность проследить постепенные изменения флоры, нащупать
места прохождения границ; недостаток (при необходимости охватить
обширные пространства в ограниченный срок) — экстенсивность: боль-
шие маршруты не дают достаточно полного представления о том, что
встречается и что отсутствует в том или ином пункте. В настоящее
время, однако, маршрутный метод претерпевает серьезные изменения
в направлении интенсификации, достижения большей плотности флори-
стической информации; он сближается с методом проб флоры (взаимо-
дополнение!), и эта тенденция будет усиливаться при переходе к регу-
лярному (сеточному) картированию флоры (см. ниже). Значение мар-
шрутного метода повышается и на территориях, давно попавших под
антропогенный пресс, где естественная растительность сохранилась от-
дельными фрагментами.
При планировании сети линейных маршрутов необходимо обеспечить
посещение основных природных районов, в частности всех ландшафтно-
геоморфологических единиц (хребтов, возвышенностей, низменностей,
крупных долин, бассейнов рек) и всех литологических комплексов
(в первую очередь коренных пород основного и кислого состава, суглин-
ков и песков и т.д.). При планировании маршрутов важное подспорье -
специальные природные карты и картосхемы районирования: геоморфо-
логические, геологические, почвенные и, конечно, геоботанические.
Чтобы получить достаточное представление о флоре каждого физико-
географического подразделения территории и оценить степень ее цель-
ности („естественности” как флористического выдела), важно добиться
неоднократного, более или менее равномерного пересечения крупных
природных контуров (желательно и в разные фенологические сезоны).
Основным способом фиксации флористической информации по-прежне-
му остаются гербарные сборы, незаменимые в случае критических или
трудных для распознавания в поле таксонов; должны документироваться
(с соблюдением правил сохранения редких растений!) также все важные,
принципиальные флористические находки. Наряду с этим все большее
значение приобретает составление флористических списков в пунктах
остановок (прежде всего в наиболее типичных или особо примечатель-
1 Эти вопросы следует обсудить на следующем рабочем совещании по срав-
нительной флористике, намеченном на сентябрь 1988 г.
15 Заказ 1220
225
ных местах со специфическими, редкими экотопами.1 При длительной
остановке (один—несколько дней) желателен флористический осмотр
всего разнообразия экотопов (полного экологического профиля); если
остановки более короткие, важно, чтобы их совокупность позволяла
ознакомиться с парциальными флорами всех классов экотопов и сооб-
ществ (флороценотипов). Рекомендуется раздельная регистрация набора
видов в каждом крупном классе экотопов (желательно с оценкой пар-
циальной активности каждого вида на данном экотопе или хотя бы
характерного проективного покрытия, обилия). При совместной работе
с геоботаником целесообразно иметь флористически полные описания
основных сообществ каждого класса экотопов.
Для того чтобы обеспечить сопоставимость флористических данных
по территориям, изученным либо маршрутным методом, либо методом
проб флоры (конкретных флор, локальных флор),1 2 целесообразно
иметь дополнительно к сети флористических маршрутов хотя бы весьма
разреженную сеть эталонных (опорных) проб флоры — детально изучен-
ных конкретных или локальных флор - для всех основных природных
подразделений данной территории. Такая сеть постепенно создается
(например, в местах вузовских баз летней практики, академических
стационаров), но пока слишком медленно. В подобных опорных пунк-
тах желательно провести наиболее детальное изучение флоры — по про-
грамме полустационаров или стационаров (см.).
ПРОГРАММА ИЗУЧЕНИЯ ТЕРРИТОРИИ
МЕТОДОМ ВЫБОРОЧНЫХ ПРОБ ФЛОРЫ
(КОНКРЕТНЫХ, ЛОКАЛЬНЫХ ФЛОР)
Достоинства метода неоднократно обсуждались (см.: Юрцев, 1974а,
1975а); важнейшие из них состоят в том, что концентрация усилий
флориста на ограниченной площади создает возможность достигнуть
высокой полноты выявления флоры даже для совсем не изученных
ранее территорий, с большей достоверностью говорить не только о „плю-
1 Широко практикуется составление флористических списков для разных
пунктов (или экотопов) на основе сводного флористического списка всей регио-
нальной флоры (по данным прежних исследований), отпечатанного на специальном
бланке или заранее перенесенного в полевой журнал. Помимо факта присутствия,
для каждого встреченного вида желательно регистрировать показатели-компоненты
его активности (степень повсеместности, обилие, спектр экотопов и сообществ).
Удобная форма регистрации - табличная (матричная), где строки соответствуют
видам (подвидам), столбцы - пунктам, выделам экотопов или описаниям сооб-
ществ; при этом каждое местонахождение видов, трудно распознаваемых в поле,
должно документироваться гербарными образцами; отдельно должна быть дана
подробная характеристика природных условий и растительности территории, охва-
ченной списком.
2 Под конкретными флорами здесь имеются в виду элементарные флоры ре-
гионального уровня, приближенно соответствующие выделам флористически одно-
родных ландшафтов; под локальными флорами - флоры пробных площадей, за-
кладываемых для выявления флористической ситуации в том или ином геохрафи-
ческом пункте (в принципе могут закладываться на стыке 2-3 конкретных флор;
подробнее см. статью Юрцева в наст, сб.).
226
сах”, но и о „минусах” флористических списков, судить об уровне
богатства региональной флоры и о полноте выявления локальной -
по „контрольным цифрам” видового разнообразия других конкретных
флор той же фитохории; наконец, глубже изучить флору как своеобраз-
ный естественноисторический феномен - подвижно-равновесную много-
видовую систему местных популяций, взаимодействующих друг с дру-
гом на совокупности экотопов.
О некоторых слабых сторонах метода сказано выше, основа их —
выборочное™, фрагментарность проб флоры, неизбежность интерполя-
ций. Поэтому сеть детальных проб флоры нуждается в дополнении хотя
бы неполными списками для промежуточных точек, в максимальном
использовании всех доступных источников флористической информа-
ции (в первую очередь учет сборов прежних коллекторов) и, где это воз-
можно, в дополнительных маршрутных пересечениях территории. Поэтому
оптимальная схема исследований заключается в синтезе методов выбороч-
ных проб и маршрутного: выполняется линейный маршрут (например,
по большой реке) с сочетанием коротких и длительных остановок: во вре-
мя длительных остановок изучаются конкретные (локальные) флоры.1
Планирование флористического исследования территории методом
выборочных проб должно следовать тем же критериям, что и планирова-
ние сети маршрутов: охват всех основных природных районов, желательно
в некоторой повторности. Предпочтительнее закладывать каждую пробу
в пределах однородного по физико-географическим условиям природно-
го района; в случае ограниченности времени, кадров и транспортных воз-
можностей допустимо (а иногда и целесообразно) заложение проб на сты-
ке двух (трех) природных районов с раздельной регистрацией флоры
для каждого из них (в пределах общего списка локальной флоры).
Орографически и литологически пестрые территории нуждаются в наибо-
лее плотной сети проб флоры; для районов флористических и ботанико-
географических границ рекомендуется закладывать пересекающие их
профили в виде серии проб флоры (по возможности сходных по рельефу
и составу пород) вплоть до трансекты из примыкающих проб.
Интересы достижения достаточной плотности флористической инфор-
мации по обширной территории не позволяют изучать все пробы флоры
(конкретные, локальные флоры) с одинаковой детальностью (типовые
программы изучения проб флоры разной детальности приводятся ниже).
Менее детальные программы ботанико-географических эталонов (см.)
необходимо дополнять некоторым количеством опорных проб флоры —
по программе полустационаров и стационаров (см.), с продолжением
исследований в течение двух-нескольких лет и в разные фенологические
сезоны; выбор участков таких опорных флор должен быть тем более
продуманным, чем меньше их число. Интенсификация исследований не-
обходима, для того чтобы найти ответ на вопросы, возникшие в ходе
экстенсивного изучения территории, и обеспечить контроль над полнотой
выявления флоры при более кратковременном обследовании.
1 Для обеспечения полноты выявления флоры желательны повторные посе-
щения тех же участков в другие годы и в иные фенологические сезоны.
227
Ввиду отсутствия согласованности между исследователями в во-
просе о площади выявления конкретных флор в разных природных зо-
нах и для обеспечения сопоставимости данных по одной природной зоне
(области) рекомендуется до проведения специальных методических
исследований придерживаться сложившихся традиций: для таежной зоны
и полосы хвойно-широколиственных лесов 500—700 км* 2 (до 1000 км2),
для тундровой — 100—300 км2 (до 500—600 км2).1 Для сравнительной
оценки пространственного разнообразия флоры в разных географичес-
ких условиях наиболее методически безупречно было бы получение дан-
ных для серии площадей возрастающего размера и радиуса из единого
базового лагеря: площадью 10 км2 (Л = 1.8 ... 2 км), 100 км2 (R = 5.6 ...
6 км), 300 км2 (Л =9.8 ... 10 км), 700 км2 (Л = 14.9 ... 15 км) и т.д.,
для чего необходима регистрация расстояния от базового лагеря
местонахождений особо редких видов.2 Для определения естественных
границ конкретной флоры важно проанализировать состав видов, появ-
ляющихся на периферии обследованного участка: появление очень ред-
ких, спорадически распространенных видов или новых редких экотопов
и сообществ еще не говорит о выходе за пределы данной конкретной
флоры — в последнем случае должны появиться устойчивые отличия
(см.: Юрцев, статья в наст, сб.).
Увеличение длительности обследования флоры в каждом географи-
ческом пункте (по сравнению с маршрутным методом) позволяет при-
менить более единообразный и скрупулезный подход к изучению мест-
ной флоры, охватить исследованием все типы экотопов в достаточной
повторности (с раздельной инвентаризацией их парциальных флор),
составлять экологические портреты всех видов по наблюдениям в дан-
ном пункте, оценивать экотопологическую активность вида в каждом
пункте (см. ниже). При обработке материалов по представительной
серии конкретных флор появляется возможность показа на бланковой
карте не только присутствия, но и отсутствия вида (светлые кружки
на бланке), его активности в каждом пункте, даже характера распростра-
нения по экотопам.
ПРОГРАММА СПЛОШНОГО ФЛОРИСТИЧЕСКОГО
ОБСЛЕДОВАНИЯ ТЕРРИТОРИИ:
СЕТОЧНОЕ КАРТИРОВАНИЕ ФЛОРЫ
Подобные программы впервые возникли на территориях с достаточ-
но высокой плотностью ранее накопленной флористической информа-
ции; они ознаменовали важный этап перехода от выборочного изучения
По неопубликованным данным Р. В. Камелина, для степной зоны площадь
выявления приблизительно равна 300-500 км2, для пустынной - свыше 1000 км2.
2 По Л. И. Малышеву (1972а), для суждения об уровне флористического богат-
ства флор при неравновеликости территорий проб флоры необходимо приведение
к одинаковой площади путем перерасчета по предложенной эмпирической формуле.
228
флоры к сплошному, регулярному ее изучению („картированию”)
и как таковые несомненно отражают завтрашний день флористики.
По своим исходным задачам проекты сеточного картирования фло-
ры1 представляли лишь форму накопления, хранения, выдачи и обработ-
ки флористических данных с помощью ЭВМ и в первую очередь форму
картирования распространения видов в пределах данной региональной
флоры. Детальность отражения дифференциации флоры („разрешающая
способность метода”) в значительной степени зависит от размера наи-
меньших (элементарных) квадратов, принятых в данном проекте. При
увеличении площади квадрата до 2500 км2 (сторона квадрата 50 км),
что принято в ряде проектов для обширных, пока недостаточно изучен-
ных* территорий, обследование территории в основном продолжается
традиционным маршрутным методом. При уменьшении размеров эле-
ментарных квадратов до 1000, 600, 300, 100 км2 и несколько менее
(например, 70 км2 в проекте картирования флоры Латвийской ССР) эти
квадраты становятся соизмеримыми с размерами участков проб флоры
и с площадью выявления конкретной флоры в тех или иных природных
зонах [по А. И. Толмачеву (1970) — от 100 до 1000 км2]; при этом су-
щественно возрастает разрешающая способность метода, а квадрат стано-
вится доступным для обследования однодневными радиальными мар-
шрутами, как при изучении выборочных проб флоры. Объединение
флористической информации по примыкающим малым квадратам (в
пределах достаточно гомогенной территории) позволяет находить эмпи-
рически „площадь выявления конкретной флоры” в различных ботани-
ко-географических условиях (Табака и др., 1977) — минимальную пло-
щадь, по достижению которой при флористическом обследовании
территории прирост видов почти прекращается.
Дальнейшее дробление квадратов делает их менее представитель-
ными в смысле отражения флористической ситуации в данном геогра-
фическом пункте (набор экотопов, состав их парциальных флор).
При достаточно дробных и в то же время представительных элемен-
тарных ячеях сеточного картирования флоры общая сеть их представляет
систему регулярных проб флоры и к ней применимы методология и ме-
тодика изучения конкретных (локальных) флор — проб флоры различной
детальности (см.ниже). Избавляя от необходимости специально планиро-
вать сеть маршрутов или выборочных проб флоры, регулярное сеточное
картирование флоры позволяет планировать сеть опорных ячей или
групп ячей (наиболее представительных, достаточно гомогенных по об-
щим природным и ботанико-географическим условиям и с достаточно
сохранившейся растительностью) для первоочередного, наиболее обстоя-
тельного, специального их изучения по программе ботанико-географи-
ческих эталонов, полустационаров или стационаров (см.). В первую
очередь такого изучения заслуживают охраняемые территории, базы
ботанической летней практики, районы существующих стационаров.
Сеточное картирование флоры (на основе достаточно дробных и
в то же время представительных ячей) позволяет решать все общие
1 Имеется в виду способ представительства на сеточной основе.
229
вопросы изучения и анализа флоры (см. выше), разделяя и все отмечен-
ные выше преимущества метода выборочных проб флоры (например,
показ активности вида, его распределения по поясам, экотопам в разных
ячеях сетки). Дополнительное и очень важное преимущество: использо-
вание памяти и операционных возможностей современных ЭВМ при на-
коплении, корректировке, обработке и выдаче флористической инфор-
мации. При введении информации о различных параметрах среды в ту же
систему ячей появляется возможность анализа корреляций между рас-
пространением видов и их комплексов и распределением различных эко-
логических факторов (см.: Шмидт, Ребристая, 1977).
Основной недостаток метода — формальный характер контуров
квадратов, навязываемых природе. На равнинах с ослабленной контраст-
ностью экотопов и постепенным (континуальным) характером флори-
стических изменений (размытым характером границ) не очень крупные
квадраты, по-видимому, способны вполне адекватно отобразить основ-
ные градиенты флористических изменений в пространстве. В горных
районах с повышенной внутренней контрастностью и мозаичностью эко-
логических и ботанико-географических условий включение в элементар-
ный учетный квадрат фрагментов разных флориртических контуров не-
сколько затушевывает и усредняет конкретные ситуации (эффект „шу-
мов”); тем не менее и в условиях горных стран метод может быть
использован (см.: Шмидт, Ребристая, 1977). Можно рекомендовать диф-
ференцированное (раздельное) изучение флоры частей квадрата, явно
принадлежащих разным природным (или флористическим) районам,
с постепенным выявлением (на заранее заданной формальной канве —
сети квадратов) естественных флористических контуров (стратегия се-
точного картирования флор).
ПРОГРАММЫ ИЗУЧЕНИЯ ФЛОРЫ ЗАПОВЕДНИКОВ
И ДРУГИХ КРУПНЫХ ОХРАНЯЕМЫХ ТЕРРИТОРИЙ
Флоры заповедников нуждаются в наиболее детальном сплошном
обследовании. Хотя многие заповедники являются (или станут) частью
территорий, обследуемых сеточным методом, для них можно рекомен-
довать изучение флоры по естественным контурам, например, дробным
ландшафтным или геоботаническим районам (микрорайонам) или не-
большим, но представительным речным бассейнам, с последующей фло-
ристической корректировкой первоначально намеченных контуров и пе-
реходом к дробному флористическому (или комплексному — в случае
совпадения границ) районированию территории. Интересный опыт подоб-
ного подхода к обследованию флоры заповедника имеется для обшир-
ной территории Алтайского заповедника (Золотухин, 1982, а также
статья в наст, сб.). В этом случае элементарные флористические контуры
(микрорайоны) трактуются в качестве конкретных флор; все они заслу-
живают изучения по программе ботанико-географических эталонов (см.),
часть наиболее представительных участков, относящихся к разным фито-
хориям и разным природным районам, — по программе ботанико-геогра-
230
фических полу стационаров (см.). Как территории, изъятые из хозяй-
ственного пользования и потому имеющие особое значение для изучения
и охраны зональных (фоновых) и реликтовых биотических комплексов
и для мониторинга, заповедники представляют оптимальные условия
и для проведения комплексных стационарных исследований биоты
на специально отведенных представительных частях охраняемой терри-
тории; важной составной частью этих работ должно стать наиболее глу-
бокое и детальное изучение флоры как природной системы, в тесной
увязке с фитоценологическим и экологическим изучением растительного
покрова (см. программу флористических работ на комплексных ста-
ционарах и станциях). Для проведения подобных работ важно иметь
достаточно сильный коллектив квалифицированных флористов с хоро-
шей подготовкой и в области геоботаники; поскольку таковые есть
лишь при очень немногих заповедниках, целесообразно привлечь к де-
тальному изучению флоры и растительности коллективы флористов
и геоботаников со стороны — из региональных или центральных учре-
ждений (системы Академии наук СССР и союзных республик, вузов
и т.д.), либо же кооперировать работу местных ботаников и приезжих.
Для заповедников с малой контрастностью условий (особенно в за-
крытых — лесных — ландшафтах) при поконтурном изучении парциаль-
ных флор целесообразно использовать (там, где она имеется) кварталь-
ную сеть, с максимально детальным обследованием представительных
модельных кварталов или блоков кварталов и обязательным дополни-
тельным осмотром остальной части каждого контура.
При привлечении достаточного контингента специалистов и обеспече-
нии необходимых условий для их работы заповедники могут стать узло-
выми ячейками познания основных региональных типов флор (и расти-
тельного покрова в целом) и их динамических тенденций. В частности,
здесь возможны наиболее полный синтез флористических, фитоценологи-
ческих, экологических методов в изучении растительного покрова, выяв-
ление и анализ корреляций в дифференциации растительного населения и
других компонентов физико-географической оболочки (в первую очередь
почвенного покрова и микроклимата), основательное изучение аутэколо-
гических и биологических особенностей видов — компонентов различных
флористических комплексов, их экологических амплитуд, а также режи-
ма прямо действующих факторов в основных классах экотопов; изучение
и картирование экотопологической структуры местных популяций хотя
бы модельных видов — представителей различных флороценотических
комплексов; наконец, изучение флюктуационных (различной продолжи-
тельности) и направленных изменений флоры и ее внутриландшафтных
подразделений — экологически контрастных парциальных флор.
ОСОБЕННОСТИ ИЗУЧЕНИЯ ФЛОРЫ ГОРНЫХ ТЕРРИТОРИЙ
Сложный рельеф горных территорий, разнообразие естественных
препятствий, с одной стороны, и особая сложность и мозаичность расти-
тельного покрова, быстрота смен экотопов, пестрота и динамичность
231
фитоценозов — с другой, определяют значительную специфику флористи-
ческих исследований в горах. Фактически любое исследование флоры
горных территорий не может вестись иначе, чем сочетанием маршрутного
метода и метода выборочных проб флоры.
Поскольку никакая карта (и в том числе крупномасштабная карта
растительности или почв) не может быть достаточно детальной для ото-
бражения всей мозаики экотопов на любом участке гор, как в нижних,
так и особенно в средних их поясах, любые флористические работы
в горах начинаются с сети опорных маршрутов — рекогносцировочного
флористического обследования. Маршруты эти прокладываются таким
образом, чтобы пересечь все пояса растительности, по крайней мере
в долине основной водной артерии обследуемой территории и 2—3 ее
основных притоков, также по гребням местных водоразделов, не менее
чем по 1—2 в различных ступенях гор (низкогорья, среднегорья с резким
рельефом, среднегорья с платообразными поверхностями выравнивания,
высокогорья); кроме того, 2—3 маршрута в среднегорьях пересекают
средние части мезосклонов — с наибольшей пестротой условий. После про-
ведения опорных маршрутных исследований закладывается несколько
базовых лагерей в разных поясах (по крайней мере по одному в каждой
крупной ступени рельефа), с которых тщательно обследуются парциаль-
ные флоры всех типов местообитаний в непосредственном окружении ба-
зовых лагерей, и пролагаются дополнительные радиальные и продольные
маршруты, которые должны пересекать местные водоразделы в ущельях
разного типа и разной ориентировки по странам света, а также основные
водоразделы данной речной системы (в различных точках)< С особой
тщательностью обследуются разные типы скальных выходов (прирусло-
вые, склоновые, вершинные и приводораздельные гребни); специально
осматриваются разные типы осыпей, различающиеся составом и свой-
ствами осыпного материала, а также многочисленные в горах типы био-
топов с нарушенной растительностью. Лесные массивы в горах обяза-
тельно пересекаются как продольными (горизонтальными), так и попе-
речными (вертикальными) маршрутами, а гребни основных водоразде-
лов или хорошо выраженных их отрогов частично траверсируются.
В южных горах обязательно повторение основных типов маршрутов
в различные полевые сезоны (с повторным осмотром основных типов
экотопов), причем для достаточно репрезентативного (80—90 %) вы-
явления состава флоры сложных многопоясных горных территорий
площадью 1000-1500 км2 необходимо не менее 4—5 различных по погод-
ным условиям полевых сезонов.
При флористических исследованиях в горах одновременно с выявле-
нием таксономического состава флоры, сбором экологической, экотопо-
логической и фитоценологической информации необходимо также фик-
сировать высотные пределы распределения видов, в том числе в местах
инверсий поясности в открытых и закрытых долинах горных рек, а так-
же в определяемых по растительному покрову участках ущелий с пере-
хватом влажных ветров или, напротив, с эффектом „дождевой тени”.
В экстремально аридных низкогорьях (как и в ряде других горных
районов с высокой обнаженностью коренных пород) достаточная полнота
232
выявления флоры может быть достигнута лишь при посещении всех
(в том числе очень редких и небольших по площади) литологических
разностей. Здесь для первичного выбора маршрутов необходимы круп-
номасштабные геологические карты, а также карты всех водных источ-
ников, как пресных, так и засоленных.
Более высокое разнообразие и контрастность экотопов в горах (по
сравнению с равнинами) обусловливают повышенное пространственное
разнообразие флоры, и в том числе более быструю пространственную
смену контрастных флористических ситуаций. Это, с одной стороны,
заставляет стремиться к расширению площади „проб флоры” (с цепью
добиться если не прекращения, то по крайней мере резкого снижения
прироста видов по мере расширения обследованной площади), с дру-
гой — заставляет ограничивать размеры пробных площадей во избежание
смешения в одном флористическом списке существенно отличающихся
элементарных флор (например, принадлежащих контрастным по рель-
ефу и литологии, но ограниченно протяженным ландшафтам). Отсюда
наибольшая оправданность именно в условиях высоких гор с хорошо
развитой поясной дифференциацией растительного покрова сплошного
(а не выборочного) детального обследования флоры отдельных хребтов,
горных узлов и т.д.; однако в ходе такого обследования необходимо
раздельно регистрировать состав и популяционные характеристики ви-
дов в естественных ландшафтных подразделениях данного горного соору-
жения — ландшафтных районах или микрорайонах — или же в бассейнах
рек (ущельях), а в их пределах — в разных высотных поясах. В случае не-
возможности на данном этапе изучения территории провести сплошное
ее обследование в качестве пробных площадей могут быть избраны
естественные подразделения горной территории (ландшафты, бассейны
не слишком больших рек), максимально полно представляющие спектр
высотных поясов, а в их пределах — спектр экотопов. Если же нет воз-
можности заложить пробные площади раздельно в ландшафтных районах
(или речных бассейнах) каждого типа, допустимо заложение пробной
площади на стыке нескольких ландшафтов (микрорайонов, речных бас-
сейнов) с раздельной регистрацией набора и позиций видов в каждом
из ландшафтов (микрорайонов, бассейнов). Включение в пробную пло-
щадь всех высотных поясов, представленных в данном горном ланд-
шафте (речном бассейне), важно с точки зрения задач флористического
и комплексного ботанико-географического районирования территории;
не менее важна для решения тех же задач раздельная регистрация парци-
альных флор разных высотных поясов, что позволит сравнивать горные
территории с разной полнотой представленности спектра высотных поя-
сов („поясной колонки”). Вполне оправдано также изучение наряду
с многопоясными однопоясных проб флоры (при условии их достаточ-
ной выраженности и территориальной целостности) и последующее срав-
нение их с принадлежащими тому же поясу парциальными флорами
многопоясных проб. Как аналитический прием вполне оправдано (и очень
часто практикуется) специальное изучение флор одного высотного пояса
или группы флористически близких поясов [например, высокогорных
флор, объединяющих 2 или 3 пояса: субальпийский, альпийский и суб-
233
нивальный; при этом субальпийский пояс обычно обнаруживает многие
черты переходности (промежуточности) между альпийским и верхним
лесным и в зависимости от задач исследования, степени флористической
близости к тому или другому нередко комбинируется с одним из них
в процессе изучения и сравнительного анализа взаимозамещающих высот-
ных комплексов видов]. При флористическом изучении метфдом выбо-
рочных проб крупных хребтов с хорошо развитыми макросклонами
(включающими менее высокие отроги) может быть применена следую-
щая схема размещения пробных площадей: по одной — многопоясной —
на каждом макросклоне в наиболее широкой части хребта и одна в осе-
вой высокогорной части хребта, что вкупе составляет поперечный про-
филь; для более протяженных хребтов подобные профили целесообраз-
но заложить, как минимум, также в обеих окраинных частях хребта или
же многократно на его протяжении. Во всех случаях, особенно я® при
заложении пробных площадей в осевой части хребта, внимание флориста
должно быть нацелено на то, чтобы уловить (не пропустить) возможные
флористические рубежи в пределах геоморфологически и литологически
'более или менее однородных территорий (отражающие изменения кли-
мата и различия природной истории).
ОСОБЕННОСТИ ИЗУЧЕНИЯ ФЛОРЫ ТЕРРИТОРИЙ,
ПРЕОБРАЗОВАННЫХ ДЕЯТЕЛЬНОСТЬЮ ЧЕЛОВЕКА
При изучении флоры территорий, растительный покров которых
испытал значительную антропогенную трансформацию, перед флористом
обычно стоят 2 самостоятельные (хотя и тесно взаимосвязанные)задачи:
1) регистрация и анализ современного состояния флоры в том виде,
какой она приобрела благодаря интенсивному хозяйственному освоению
территории; 2) реконструкция первичного (былого) состояния флоры
(и растительности) той же территории до ее хозяйственного освоения
(без этого невозможно выполнение всех остальных задач, перечисленных
в первом разделе настоящей Программы).
Для решения первой задачи, по-видимому, не требуется существенно
расширять пробные площади (по сравнению с таковыми на территориях
с хорошо сохранившимся природным покровом). Особое значение
приобретает раздельное изучение парциальных флор, сгруппированных
не только по экологическому своеобразию их местоположений (поло-
жение в рельефе, состав почвообразующих пород), но и по степени антро-
погенной трансформации исходной растительности (от участков, слабо
нарушенных или почти не нарушенных, до собственно антропогенных:
полей сельскохозяйственных культур, поселков, различного рода назем-
ных коммуникаций и т.п.). Большое значение приобретает разделение
флоры на ? крупные фракции: аборигенную и адвентивную (заносную).
При изучении парциальных флор экотопов с нарушенной исходной расти-
тельностью целесообразно различать следующие группы видов по степени
их приуроченности к местам нарушений: 1) целиком ограниченные в сво-
ем распространении в данном ландшафте участками сильных нарушений
234
и антропогенными экотопами (с дальнейшим подразделением на антро-
пофильные аборигенные виды и адвентивные: сегетальные, рудеральные
и т.д.); 2) виды, отмечаемые и в составе природных ценозов, но встре-
чающиеся с более высокими константностью, обилием и жизненностью
в местах нарушений и на антропогенных экотопах; 3) виды, равно ха-
рактерные для почти не нарушенных и сильно нарушенных участков или
проявляющие на последних явные признаки угнетения (более низкие
обилие, константность и жизненность); 4) виды, обычно выпадающие
из состава растительности в местах сильных нарушений или находящиеся
здесь на последних стадиях деградации их природных популяций. Первые
2 группы составляют антропофильный элемент флоры (с подразделе-
нием на эвантропофилов и гемиантропофилов), 2 последние группы
(за вычетом видов индифферентных) - антропофобный элемент (под-
разделяемый далее на гемиантропофобов и эвантропофобов — „антропо-
генных реликтов”) . Представляет немалый научный и прямой практиче-
ский интерес учет изменения, соотношений представителей названных
групп в сукцессионных рядах парциальных флор по усилению воздей-
ствия конкретных факторов антропогенных нарушений, а также в обрат-
ных, демутационных (восстановительных) рядах. Для аборигенных
антропофилов важно установить их коренные экотопы и их стратегии
в природном покрове; для заносных видов — источники, пути и агентов
заноса, давность заноса, степень натурализации на территории изучаемой
флоры (дифференцированно по классам и типам экотопов). Антропо-
фобные виды („антропогенные реликты”) также заслуживают специаль-
ного внимания (установление причин их недостаточной пластичности -
„ригидности”; изучение „генетико-автоматических” процессов в малых
популяциях, обеднения генофонда и т.д.). В целом же изучение флор
освоенных территорий имеет не только практический, но и немалый
теоретический интерес, так как здесь мы сталкиваемся с быстрой и бур-
ной перестройкой флоры при участии активных миграционных процес-
сов; выявленные закономерности важны и как основа прогноза будущих
изменений флоры территорий, пока еще слабо затронутых хозяйственной
деятельностью. В этой области открывается широкое поле для сотруд-
ничества флориста с геоботаниками и биоморфологами (изучение жиз-
ненных форм, эколого-морфологических и биологических особенностей
характерных или всех представителей названных групп, определение
ценотипа и стратегии видов и т.д.).
При изучении южных районов СССР, наиболее богатых и своеобраз-
ных во флористическом отношении, следует учитывать, что большая
часть их территории (и в первую очередь аридные и субаридные равнины,
среднегорья и низкогорья) особенно сильно преображена человеком.
Антропогенный пресс на растительный покров длится здесь уже несколько
тысячелетий, в результате чего мы имеем дело хотя и с очень богатыми,
но полностью или в значительной степени антропогенно перестроенными
флорами. Выделение в их составе антропофильного и антропофобного
элементов практически невозможно без специального изучения смен ти-
пов хозяйствования на крупных территориях (хозяйственно-культурных
типов этнографов: Левин, Чебоксаров, 1957; Андрианов, Чебоксаров,
235
1972). Здесь крайне важно различать участки новейших нарушений,
участки и территории регулярных и давних нарушений (длительно ис-
пользуемые пастбища и другие кормовые угодья, разновременные зале-
жи, особенно по бывшим богарным распашкам, средние участки долин
рек в пределах зон расселения) и участки, традиционно охраняемые
в силу религиозных или культурных традиций (кладбища, мазары, свя-
щенные рощи, отдельные скальные обрывы или отдельные горы),
но в большинстве случаев также сильно преображенные. Следует также
учитывать, что за огромный период хозяйствования человека на этих
землях шла стихийная и сознательная интродукция многих ценных рас-
тений как в результате их культивирования для хозяйственных или
культурных потребностей, так и в результате случайных заносов ( с запа-
сами пищи и кормов).
Особенно важно оценить возможности древнего заноса на юге Сред-
ней Азии, где еще относительно недавно регулярно осуществлялись
тысячекилометровые кочевые переходы скотоводов (как с равнин
в горы, так и из южных пустынь на более северные территории). На по-
добных территориях значительную часть видов, ныне воспринимаемых
как аборигенные компоненты природного растительного покрова, в ре-
зультате детальных исследований нередко приходится считать адвентив-
ными, а многие сукцессионные ряды растительности — лишенными
завершающих стадий развития вследствие истребления доминантов кли-
максовых сообществ. Поэтому антропогенные изменения флор здесь
могут быть предметом лишь специального углубленного изучения.
Для восстановления первичного облика флоры (и растительности)
сильно измененных хозяйственной деятельностью территорий первосте-
пенное значение приобретает детальное и разностороннее изучение пар-
циальных флор лучше сохранившихся участков растительного покрова,
привязка их к определенным местоположениям, что обеспечивает воз-
можность экстра- и интерполяций на те участки территории, где исход-
ный растительный покров был целиком уничтожен или сильно нару-
шен. Задача эта напоминает создание карты потенциальной (восстано-
вленной) растительности и требует также привлечения данных по одно-
типным, но лучше сохранившимся ландшафтам той же фитохории
(округа, провинции), а также сбора и изучения архивных материалов
(например, старых карт землеустройства и т.д.). Переход многих исход-
но вполне активных или даже фоновых видов и фоновых (плакорных)
сообществ на положение антропогенных реликтов заставляет расширять
пробные площади, увеличивать повторность осмотра однотипных экото-
пов вплоть до сплошного поконтурного их осмотра, сочетать детальное
обследование флоры методом радиальных маршрутов из нескольких
базовых лагерей с более или менее протяженными линейными маршру-
тами; весьма перспективен также метод регулярного флористического
обследования территории на сеточной основе (чему благоприятствует
развитая транспортная сеть на освоенных человеком землях). Напротив,
разграничение контуров элементарных естественных флор в этих усло-
виях затруднено. Пример прежних степных территорий европейской
части СССР, где первичный растительный покров сохранился лишь
236
фрагментами — на „неудобных землях” и в немногих степных заповедни-
ках, в какой-то степени обнадеживает: видов, известных из этих районов
до сплошной распашки степной целины, но не сохранившихся сейчас,
почти нет (если, конечно, говорить о крупных территориях); однако
популяционные (в первую очередь демографические) характеристики
видов в корне изменились, и исходное состояние популяций приходится
реконструировать с учетом исторических свидетельств и косвенных
признаков.
ПРИЛОЖЕНИЕ. ПРОГРАММЫ ИЗУЧЕНИЯ ПРОБ ФЛОРЫ
РАЗНОЙ ДЕТАЛЬНОСТИ
1. Программа изучения конкретной (локальной) флоры
на ботанико-географических эталонах
Размер участка, обследуемого в качестве пробы флоры, - от (50)100 до
700 км1 (как правило, не более 1000 кма) (см. выше); в случае необходимости он
может быть увеличен (или уменьшен) до естественных размеров контура конкрет-
ной флоры: в разных зонах эмпирически сложились свои представления о размерах
пробы флоры - площади выявления конкретной флоры (см. выше).
Согласно традиции, сложившейся в тундровых и высокогорных районах Се-
верной Азии, продолжительность обследования пробы флоры указанного размера
1-4 нед; в природных областях с более резкой сезонной сменой синузий, вероятно,
она должна быть увеличена до естественной продолжительности одного вегетацион-
ного периода; кроме того, желательно повторное посещение участка в годы с су-
щественно отличающимися погодными условиями. Необходима точная фиксация
(публикация) данных о размере обследованного участка, сроках и продолжитель-
ности обследования, персональном составе группы, обследовавшей участок.
Основное содержание работы
1. Ориентировка в дифференциации растительного покрова участка в зависи-
мости от факторов местоположения (рельеф, состав почвообразующих пород) с
экологической интерпретацией, выявление основных типов и классов экотопов;
целесообразно иметь геоботанические описания фоновых и наиболее своеобразных
сообществ для основных типов и классов экотопов. Целесообразно также геобота-
ническое описание экологических профилей (лучше почвенно-геоботаническое).
2. По возможности исчерпывающая инвентаризация состава видов и рас сосу-
дистых растений на основе планомерного осмотра всех типов экотопов в необхо-
димой повторности (если она имеет место на данном участке) - для редких типов
экотопов с повышенной концентрацией редких, в особенности реликтовых элемен-
тов флоры необходим осмотр всех контуров их; составление общего флористи-
ческого списка и эколого-ценотической характеристики видов (рас) по наблю-
дениям на данном участке; оценка активности каждого вида (обычно в баллах:
237
Юрцев, 1968а) и ее признаков-компонентов: широты экологической амплитуды,
распределения по территории (встречаемости), характерного уровня численности.
3. Крайне желательна раздельная регистрация состава видов на разных типах
и классах экотопов (изучение парциальных флор) с оценкой парциальной актив-
ности каждого вида на данном типе экотопов. Регистрация проводится либо путем
описания модельных контуров каждого типа или класса экотопов (гомогенных
урочищ - мезоэкотопов: Юрцев, 1982а), либо посредством составления сводных
(объединенных) списков для каждого типа (класса) экотопов или экологически
своеобразных синтаксонов (ценофлоры). Подробнее об этом говорится ниже.
II. Программа изучения флоры
иа ботанико-географических полу стационарах1
Размер обследуемого участка тот же, что на ботанико-географических этало-
нах, но продолжительность обследования увеличивается до 2-3 сезонов и более.
В состав группы, кроме флориста, должен входить геоботаник с опытом крупно-
масштабного картирования растительности, желательно также - почвовед; полезно
участие в работе бриолога и лихенолога и ряда других специалистов (например,
микраклиматолога, геоморфолога, палеоботаника-четвертичника и т.д.).
Существенный раздел общей программы полустационара - создание в первый
же год крупномасштабной карты растительности (или серии фрагментов карты)
на основе детальных аэрофотоснимков (например, масштаба 1 : 25 000), к которой
могли бы быть привязаны изучаемые флористом контуры парциальных флор (же-
лательно и почвенной карты).
Увеличение длительности исследований на ограниченном участке позволяет
ставить следующие задачи.
1. Практически исчерпывающее выявление локальной флоры за счет сущест-
венного увеличения повторности осмотра однотипных экотопов, полного выявле-
ния и осмотра контуров редких экотопов со своеобразным, часто непостоянным
набором видов,
2. Изучение иерархической структуры данной конкретной флоры путем раз-
дельного сопоставления аннотированных флористических списков для ее террито-
риально-экологических подразделений разного уровня: парциальных флор макро-
экотопов (местностей), мезоэкотопов (урочищ), микроэкотопов (фаций, сооб-
ществ - последние изучаются на основе серий геоботанических описаний).
Из перечисленных уровней для сравнительного анализа парциальных флор
особенно перспективен уровень мезоэкотопов (урочищ) либо же типологических
объединений парциальных флор микроэкотопов одного класса (ценофлор); близ-
1 Программы наиболее детальных флористических исследований (в комп-
лексе с геоботаническими, экологическими и др.) на полустационарах и стациона-
рах приближаются по своему типу скорее к „программам-максимум”; цель их -
наметить конкретные пути углубления наших знаний о флоре как о природной
системе; задачи формулируются в порядке возрастания их сложности; в такой же
последовательности рекомендуется комплектовать программы конкретных иссле-
довательских проектов, ориентируясь на контингент специалистов и сроки, на
которые рассчитан проект. Чем сложнее конкретная исследовательская программа,
тем она индивидуальнее и тем в большей мере отражает интересы проводящего
исследования научного коллектива.
238
кие по объему единицы - парциальные флоры флороценотипов, экологически,
флористически, фитогеографически и флорогенетически самобытных типов расти-
тельности, как правило, имеющих флористическое ядро из верных и характерных
видов (флороценотический комплекс). Преимущество таких единиц в качестве
объектов сравнительного анализа - их достаточная крупность („обзорность”)
в сочетании с достаточной экологической однородностью. В результате сравнения
парциальных флор по таксономическому составу, набору и соотношению типоло-
гических элементов некоторые типы парциальных флор, ориентировочно намечен-
ные по ландшафтно-геоботаническим признакам, могут быть объединены, другие,
напротив, разделены на два или несколько типов: в конечном счете систематизация
парциальных флор проводится по флористическим признакам. Кроме того, заслу-
живают специального раздельного изучения флоры речных бассейнов I порядка
в пределах конкретной флоры как ее метамерные подразделения с признаками из-
вестной автономности (Марина, 1983, а также в наст. сб.). Интересная задача - попы-
таться увязать систему парциальных флор макро- и мезоэкотопов с выделяемыми
геоботаником картируемыми территориальными единицами разного ранга - ком-
бинациями сообществ (фитоценохорами) и с аналогичными почвенными (и ланд-
шафтными) выделами. Для экологической интерпретации различий между парци-
альными флорами разных классов экотопов очень существенны данные почвоведа
и микроклиматолога.
3. Значительное увеличение повторности при изучении однотипных контуров
парциальных флор мезо- и микроэкотопов и привязка основных выделов к круп-
номасштабной геоботанической карте позволяют существенно углубить изучение
экотопологической структуры местных популяций. Регистрация ее возможна
в форме матрицы (Юрцев, 19836), где горизонтальные строчки соответствуют ви-
дам, вертикальные столбцы - сгруппированным по типам и классам конкретным
контурам мезо- или микроэкотопов, в каждой клетке отмечается либо балл актив-
ности, либо среднее проективное покрытие; введение данных по проективному по-
крытию позволит более точно оценить на основе геоботанической крупномасштаб-
ной карты - хотя бы для более массовых видов - их внутриландшафтную (экото-
пологическую) активность, в шкале отношений (Юрцев, 1982; Лякавичюс, в наст,
сб.). На основе крупномасштабной карты растительности (или непосредственно
аэрофотоснимков) возможно картографическое отображение экотопологической
структуры местных популяций - хотя бы для модельных видов (фоновых или,
напротив, редких, реликтовых) из разных флороценотических комплексов. Карти-
рование внутриландшафтного распределения местных популяций модельных видов
позволит выявить для каждого специфический рисунок („узор”) распределения,
оценить степень связности или разобщенности его ценопопуляций, установить лока-
лизацию опорных экотопов данного вида - все это факторы, от которых зависит
место вида в системе отношений с другими видами на территории флоры.
Детальный материал по парциальным флорам мезо- и микроэкотопов, допол-
ненный материалом геоботанических описаний (которые необходимо привязать
к типам парциальных флор), облегчает выделение флороценотических комплек-
сов - групп видов, верных тому или иному классу экотопов или имеющих в нем
экологоценотический оптимум. Разбивку флоры на флороценотические комплексы
целесообразно провести по материалам первого года работ, с корректировкой
в последующие сезоны. Специального выделения и анализа заслуживает также
группа высокоактивных (фоновых) видов растений данного ландшафта.
239
При наличии соответствующих специалистов на ботанико-географическом
полустационаре может быть проведено (как комплексное исследование) изучение
жизненных форм группы высокоактивных видов и основных представителей раз-
личных флороценотических комплексов (желательно специалистом-биоморфоло-
гом) вкупе с непосредственным изучением (геоботаником) вертикальной и гори-
зонтальной структуры сообществ, наиболее типичных для того или иного класса
экотопов. Это позволит выявить основные структурно-динамические типы сооб-
ществ на территории флоры и далее классифицировать виды изучаемой флоры
по их месту и роли в структуре сообществ (фитоценотипы), по их „жизненной
стратегии” и т.д.
III. Программа изучения флоры
на комплексных биологических стационарах и станциях
(тл. хорошо оборудованных, постоянно действующих стационарах)
Стационары и станции отличаются от полустационаров значительно большей
длительностью исследований (на стационарах она обычно ограничена 5-10 годами,
но исследования на том же участке могут быть возобновлены через определенный
интервал), созданием постоянной, хорошо оборудованной базы, расширением кон-
тингента специалистов. Соответственно реально возможное число комплексных
стационаров для любой природной области лимитировано, а к выбору места для
них должны предъявляться особые требования (в первую очередь репрезентатив-
ности для данной фитохории).
Стационары представляют наиболее благоприятные условия для решения
задач, сформулированных в программе полу стационаров; эти задачи и должны
стоять перед флористами в первые же годы работы стационара.
Существенные разделы в общей программе комплексного стационара (стан-
ции) следующие: 1) создание для одной и той же территории серии природных
карт — ландшафтной, геоботанической (возможно, и специальной карты мезоэко-
топов - эколого-флористической), почвенной, геологической, геохимической,
гидрологической; 2) закладка постоянных участков в основных типах сообществ
(на разных классах экотопов) или полигонов-трансект для сравнительного изуче-
ния режимов прямодействующих факторов, биологической продуктивности и био-
логического круговорота, многолетней (на станциях - вековой) динамики и смен
сообществ; 3) постановка эколого-физиологических исследований продукцион-
ного процесса и адаптаций.
Это позволяет углубить изучение экотопологической структуры местной
флоры и составляющих ее флористических (флороценотических) комплексов,
в частности по следующим разделам: 1) при определении „веса” видов и их групп
(элементов флоры) в разных типах парциальных флор (в шкале отношений)
использовать не только проективное покрытие, но также весовые и объемные пока-
затели обилия, данные о вкладе вида в продукционный процесс и т.д.; 2) при срав-
нительном экологическом анализе флороценотических комплексов (также эколо-
го-ценотических групп) и соответствующих им классов экотопов и сообществ
использовать данные по режимам прямодействующих факторов и эколого-физио-
логическим особенностям видов, а также данные экспериментальной фитоценоло-
гии; целесообразна также постановка вопросов экофизиологам и специалистам
240
по экологическим режимам со стороны флориста-эколога - в ходе комплексных
исследований; 3) при разделении флоры на группы видов со сходной жизненной
стратегией, сходным распределением по экологическим нишам использовать дан-
ные геоботаников по сукцессионным отношениям в растительном покрове терри-
тории стационара. Важное звено в этой работе - изучение жизненных форм.
Настало время поставить - хотя бы на отдельных наилучше оборудованных,
укомплектованных специалистами стационарах - изучение вопросов генетики
(для начала фенетики) популяций для модельных местных популяций видов, пред-
ставляющих разные типы экотопологической структуры (например, фоновых,
высокоактивных эвритопных видов или массовых стенотопных или редких и мало-
численных) .
Как подход к изучению процессов панмиксии и обмена диаспорами (интегри-
рующих флору) целесообразно поставить изучение „пыльцевого дождя” и разноса
диаспор в пределах территории стационара (на разных классах экотопов), состава
и количества жизнеспособных семян, а также пыльцы в верхних горизонтах почвы
(что имеет выход и в четвертичную палеоботанику).
16 Заказ 1220
241
ОЧЕРК СИСТЕМЫ
ОСНОВНЫХ ПОНЯТИЙ ФЛОРИСТИКИ*
Задача этого раздела - систематизировать понятия и термины современной
сравнительной флористики. Не имея пока возможности выполнить эту работу
с исчерпывающей полнотой, мы ограничили свою задачу рассмотрением лишь более
или менее специфических для данной научной дисциплины понятий и терминов,
ориентируясь в первую очередь на работы советских авторов, опубликованные
за последние 2-3 десятилетия.
Трудность этой задачи связана со значительной избыточностью терминов -
флористика разделяет эту трудность с геоботаникой, общей экологией, географией
и другими дисциплинами, имеющими дело с нечеткими (пересекающимися) мно-
жествами — сложными и многомерными природными объектами („большими
системами”). Количество обращающихся терминов намного превышает число
понятий и все продолжает расти.
Правило приоритета, служащее „ариадновой нитью” в таксономической номен-
клатуре живых организмов, здесь не может спасти положение и должно применять-
ся с разумными ограничениями. Ведь в отличие от названий таксонов живых орга-
низмов, названий, конструируемых в сущности как имена собственные, любой
термин - имя нарицательное. Как имена нарицательные (в постоянном обращении),
термины должны быть удобными для употребления (в частности, для произноше-
ния) и более или менее органично вписываться в язык данной страны (но также
переводиться на основные зарубежные языки; при прочих равных условиях, интер-
национальные термины предпочтительнее). В отличие от названий в таксономии,
которые, если доказана их синонимичность ранее введенным для того же таксона
названиям, попросту отбрасываются (приоритетные же сохраняются, даже если
они идут вразрез с морфологическими, экологическими или географическими
признаками таксона), хорошо прижившиеся основные международные термины
(такие как флора, элемент флоры, популяция и т. д.) в ходе развития понятийно-
терминологического аппарата данной науки обычно не изымаются из обращения,
но включаются и в другие (конкурирующие) понятийно-терминологические
системы с соответствующим изменением содержания и (или) объема сопряженного
понятия. В конечном счете в науке побеждает и утверждается (конечно, только
на известное время) та понятийно-терминологическая система, которая доказала
свою жизненность, будучи удобной, логически стройной (непротиворечивой),
достаточно полной и гибкой (допускающей введение дополнительных понятий
и конструирование производных терминов). Итак, понятийно-терминологические
системы апробируются практикой науки и могут быть канонизированы лишь реше-
нием представительного научного конгресса.
1 Авторы Б. А. Юрцев, Р. В. Камелин (сокращенный вариант, полный текст
предполагается опубликовать отдельной брошюрой).
242
Для сравнительной флористики, переживающей период быстрого развития
(особенно в нашей стране), время для канонизации той или иной понятийно-терми-
нологической системы, очевидно, еще не настало, что показала и дискуссия на
II рабочем совещании в Неринге. Для того чтобы приблизить это время, подгото-
вить унификацию терминологии в недалеком будущем, а на данном этапе облегчить
взаимопонимание исследователей, придерживающихся нетождественных систем
терминов, на совещании в Неринге была поставлена задача составить словарь
синонимичных терминов флористики, относящихся к одним и тем же понятиям.
Поскольку в отношении существа основных понятий флористики и ботаничес-
кой географии разногласий между исследователями на совещании в Неринге
не выявилось, мы решили положить в основу систематизации синонимических
терминов (даны в правом столбце) систему основных понятий (левый столбец);
„куст” терминов, относящихся к единому понятию, и есть собрание синонимов;
выделены термины, которые могут быть рекомендованы до проведения полной
унификации.1 Во всех случаях мы предпочитали наиболее лаконичные формули-
ровки, главное назначение которых - очертить объем основных понятий и отграни-
чить от близких понятий; различие в содержании, вкладываемом в каждое поня-
тие, рассмотрено под той же рубрикой в комментариях.
Самая общая логическая категория, с которой постоянно имеет дело флори-
стика, - (любая) совокупность видов растений, произрастающих на той или иной
территории (обозначим ее как „любая территориальная совокупность видов”).
Иногда встречаются попытки именно к такой, самой общей категории применить
термин „флора” („весенняя флора”, „флора кальцефитов”, „флора бурачнико-
вых” такой-то территории), что нашло отражение даже в „Русско-латинском слова-
ре для ботаников” (Кирпичников, Забинкова, 1977, с. 753), где „флора” расши-
фровывается как „совокупность растений”.1 2 Среди участников обоих рабочих
совещаний по сравнительной флористике не оказалось сторонников такого взгляда
на флору; по-видимому, таковых нет и среди остальных ботаников, изучающих
флору как самобытное явление природы.
Следующий этап деления исходного понятия „территориальная совокупность
видов растений” приводит к выделению двух контрадикторных (взаимодополняю-
щих) понятий: 1) полная территориальная совокупность видов растений, 2) непол-
ная (частичная) территориальная совокупность видов растений (= та или иная
часть полной территориальной совокупности видов, выделенная по какой-либо
категории признаков).
Эти 2 категории, вкупе исчерпывающие объем исходного понятия, и будут
положены нами в основу систематизации понятий флористики.
При этом необходимо подчеркнуть, что практически любые полные территори-
альные совокупности видов растений, как и их части - комплексы видов - обу-
словлены экологически и исторически, т. е. не только современными экологичес-
кими условиями данной территории, но и совокупностью процессов эволюции,
расселения и взаимодействия видов растений на той же, соседних и даже многих
весьма удаленных территориях в прошлые геологические эпохи (в иных ланд-
шафтно-климатических и экологических условиях). Это общее положение сохра-
няет силу для всех более частных флористических категорий и ниже специально
не оговаривается.
1 Из ранее опубликованных работ наиболее обстоятельно система понятий
и терминов, относящихся к флоре, рассмотрена в статье Юрцева (1982а).
2 Такое определение оставляет неясным, имеем ли мы дело с совокупностями
особей или видов растений; флористика имеет дело с совокупностями видов!
243
1. Полная территориальная сово-
купность видов растений (множество
видов растений в топографическом
контуре).
Флора (Juchacz-Nagy, 1966; Юрцев,
Семкин, 1980; Юрцев, 1982а, а также
в наст, сб.).
Общее определение понятия предполагает полный охват всего разнообразия
растительных организмов, к которому до недавнего времени относили также
грибы. Однако значительное отставание таксономии и флористики низших расте-
ний и мохообразных от таковых сосудистых растений до недавнего времени делало
нереальным охват полного разнообразия растительных организмов любой террито-
рии: исследователи обычно имели дело либо с видами сосудистых растений, относя-
щихся к нескольким таксономическим типам, либо с одним или немногими типами
бессосудистых растений (мохообразными, лишайниками, водорослями - несколь-
ко типов, грибами). Как временное и компромиссное решение до сих пор принято
под термином „флора” (без дополнительных определений) подразумевать сово-
купность сосудистых растений, в остальных случаях говорить о „бриофлоре”,
„лихенфлоре”, „альгофлоре”, „микофлоре”.
Некоторое (хотя явно недостаточное!) оправдание раздельному изучению
совокупности видов сосудистых растений заключается в значительном расхождении
их экологических ниш с таковыми других типов организмов, что находит отраже-
ние и в отличии типов распространения и т. д, Рассмотрение же в качестве „флор”
более дробных таксономических единиц не оправдано - последние правильнее
рассматривать в качестве таксономических элементов флоры (см. раздел 2).
Рациональный нижний территориальный предел рассмотрения территориаль-
ных совокупностей видов как таковых - площадь фитоценоза („контур-фито-
ценоза”, „фитохоры” в смысле Б. Н. Норина), тем более что чем меньше площадь
участка, тем выше роль случайных факторов в подборе произрастающих на нем
видов.
Из соображений лаконизма здесь и ниже говорится о совокупностях видов;
однако при сравнительно-географическом анализе региональных флор основной
единицей расчленения флоры обычно является географическая раса политипичес-
кого вида (что соответствует виду „монотипистов”); точно также при сравнении
совокупностей видов экологически своеобразных внутриландшафтных подразде-
лений региональной флоры в число таких единиц целесообразно включать и эколо-
гические расы, представленные в данном ландшафте самостоятельными попу-
ляциями.
В зависимости от подхода к изучению и анализу территориальной совокуп-
ности видов в это общее понятие (при сохранении его объема) может вкладывать-
ся неодинаковое содержание.1 Так, виды в качестве элементов флористического
множества могут рассматриваться либо как дискретные целостные единицы -
по общим признакам каждого из них (тип ареала и т. д.), либо же с уточнением
специфических особенностей местной региональной популяции каждого (например,
типа распространения в пределах территории данной флоры). Кроме того, объеди-
нение видов в группы по той или иной категории признаков (таксономических
или типологических) позволяет анализировать и сравнивать флоры уже не как
множества видов, а как множества классов эквивалентности (фактормножества;
см.: Юрцев, Семкин, 1980) - таксономических или типологических элементов
флоры. С этим связаны дополнительные понятия и термины флористики.
1 Эти различия подходов (разные типы анализа) в основном сохраняют значе-
ние и для неполных территориальных совокупностей видов и могли бы быть рас-
смотрены во вводной части раздела.
244
1. 01. 1. Полная территориальная
совокупность местных популяций1 ви-
дов растений (совокупность, или мно-
жество, популяций всех видов растений
в данном топографическом контуре).
Флора (растительное население)
(Юрцев, Семкин, 1980; Юрцев, 1982а,
а также в наст, сб.).
Замена вида его местной популяцией в определении флоры ставит конкретную
и важную задачу: не ограничиваясь определением видовой принадлежности всех
растений, установить для каждого вида фенетические (морфологические, физиоло-
гические, биологические, экологические, ценотические) и хорологические, а в пер-
спективе также генетические характеристики и особенности его местной популя-
ции; флора становится тем самым весовым множеством, элементы которого -
виды - несут определенные количественные весовые характеристики.
1. 01. 2. Территориальная система
местных популяций всех видов расте-
ний.
Флора (растительное население)
(Юрцев, Семкин, 1980; Юрцев, 1982а,
а также в наст, сб.).
Переход от совокупности местных популяций всех видов к их (материальной)
системе логически неизбежен, поскольку популяции видов неизбежно образуют
конкурентную подвижно-равновесную систему, взаимодействуя на всей совокупно-
сти экотопов территории флоры; результатом этих взаимодействий в современную
и предшествующие геологические эпохи (зависящим в свою очередь от свойств
видов и от состояния их местных популяций) и является известное постоянство
количественных соотношений таксонов и типологических элементов флоры (см.).
1. 02.1. Полная территориальная
ранжированная совокупность надвидо-
вых таксонов растений того или иного
ранга.
Это упорядоченное по числу видов
(рас) или в общем случае по числу под-
чиненных таксонов того или иного ран-
га множество таксономических элемен-
тов флоры представляет абстрактную
систему (абстрактную таксономиче-
скую модель флоры).
Систематическая структура флоры
(Толмачев, 19706, 1974; Шмидт, 1980,
1984 и более ранние статьи); (семей-
ственный или родовой) флористичес-
кий спектр (Малышев, 19726, 19766);
таксономическая структура флоры
(Юрцев, 1982а, а также в наст, сб.);
в этом же значении мог бы применять-
ся термин „таксономический спектр
флоры”*.1 2
Дальнейшее подразделение систематической, или таксономической, структуры
флоры на более частные категории проводится по рангу таксономических элемен-
тов, на которые разделяется флора, - чаще всего на уровне семейств и родов
(„семейственная” или „родовая структура” или, лучше, „систематическая структу-
ра на уровне семейства”, „рода” и т. д.). Поскольку, однако, высшие таксоны мо-
гут сопоставляться не только по числу представляющих их во флоре видов ? но и
по числу более крупных подчиненных таксономических единиц (например, родов -
при сравнении семейств), для уточнения нередко используются такие термины,
как „семейственно-видовой спектр” (сопоставление семейств по числу видов),
„семейственно-родовой спектр” (то же, но по числу родов, представляющих каж-
дое семейство в данной флоре), и т. д. (см., например, статью Б. И. Семкина
в наст, сб.); такие термины целесообразно употреблять в тех случаях, когда
в качестве счетных единиц используются не виды и расы, а более крупные таксоны.
1 Популяции здесь понимаются в демографическом смысле, как население
вида: полная совокупность особей данного вида в топографическом контуре
(подробнее см.: Юрцев, 1982а).
2 Звездочкой отмечены ранее не употреблявшиеся термины, предлагаемые
авторами.
245
1. 02. 2. Полная территориальная
ранжированная совокупность типоло-
гических элементов флоры (того или
иного ранга).
Типологическая структура фло-
ры* (см. в наст, сб.: Программы фло-
ристических исследований...); в том
же значении может употребляться тер-
мин „типологический спектр флоры”*.
Конкретные подразделения (выражения) типологической структуры флоры
определяются прежде всего категорией признаков, по которым виды (расы)
типизируются - объединяются в классы (генерализация, или укрупнение, элемен-
тов исходного множества видов; см. раздел 2), более дробные подразделения -
по рангу (крупности) данной категории типологических элементов. Во флористи-
ческой литературе ранее применялись термины, относящиеся к той или иной
конкретной категории типологической структуры, например географическая
и высотно-зональная структуры флоры (Марина, 1982, 1983), географическая,
биоморфологическая, экологическая, ценотическая, фенологическая структуры
флоры (Шеляг-Сосонко и др., 1985). Каждый тип структуры нередко подразделя-
ется на подтипы: так, географическая структура может подразделяться на широт-
ную (зональную), долготную (секторальную) , высотную (поясную, иногда поясно-
зональную) и комбинированную.
Категории структур, показывающие соотношение тех или иных флорогенети-
ческих (историко-типологических) элементов, отражающих не современное состо-
яние, а прошлое вида или его популяции, по-видимому, следует именовать истори-
ко-типологическим и.
При подходе к флоре как к весовому множеству видов (растительному
населению) возникает дополнительная возможность учета соотношения крупных
типологических элементов флоры по сумме весов видов, представляющих каждый
элемент в данной флоре (см. раздел 2).
На следующем этапе деления исходного понятия „полная территориальная
совокупность видов растений” (ПТСВР) возможны 2 взаимонезависимых подхода
к делению: по естественности или произвольности контуров, занятых изучаемой
совокупностью видов (1.1 и 1.2), и по уровню территориальной размерности
данного контура - региональному или топологическому, внутриландшафтному
(1.А и 1.Б). Оба подхода одинаково важны и потому должны учитываться одно-
временно; результатом их совмещения являются 4 крупных подразделения:
1. 1А. ПТСВР естественного (флористического!) контура регионального уровня;
1. 1Б. ПТСВР естественного контура топологического (внутриландшафтного) уров-
ня; 1. 2А. ПТСВР произвольного контура регионального уровня; 1. 2Б. ПТСВР
произвольного контура внутриландшафтного уровня.
Рассмотрим, однако, первые выделы обоих делений независимо (1.1 и 1.А).
1.1. Естественная полная террито-
риальная совокупность видов растений
(полная совокупность видов растений
того или иного естественного террито-
риального выдела, устойчиво отличаю-
щегося от соседних выделов флористи-
Естественная флора1 (Юрцев,
1975а, 1982а); флора (Шеляг-Сосонко
и др., 1985; Шеляг-Сосонко, Дидух,
в наст, сб.).
1 Омонимом является термин „естественная флора” в его употреблении
Г. В. Вынаевым (1984), согласно которому естественная флора объединяет абори-
генные и заносные виды и противопоставляется совокупности интродуцированных
(введенных в культуру) видов „полной, или сводной, флоры”. К совокупности
аборигенных видов данной территории нередко применяется термин „аборигенная
флора” (точнее, аборигенный элемент флоры), заносные же виды в их совокупно-
сти лучше именовать „адвентивным элементом флоры”.
246
ческими показателями, т.е. составом
и/или соотношением видов, а также бо-
лее крупных таксономических и типо-
логических элементов флоры). Внутри
каждого естественного флористическо-
го выдела, по определению, не должно
проходить ни одной флористической
границы того же или более высокого
ранга, нежели ранг самого выдела.
Термин „естественная флора” подразумевает естественность выдела флоры.
На региональном уровне таковым обычно является выдел флористического райо-
нирования (или комплексного: ботанико-географического, ботанического или
природного, учитывающего, наряду с другими, признаки самой флоры) .
1.А. Полная территориальная со-
вокупность видов растений (ПТСВР)
любого контура регионального или
планетарного уровня размерности,
в смысле В. Б. Сочавы (1978); обозна-
чим ее как полную региональную
совокупность видов.
Флора (Толмачев, 1974, с. 112);
флора регионального уровня, регио-
нальная флора (Юрцев, 1982а, с. 13,
а также в наст, сб.).
Именно региональные флоры - основной объект сравнительной флористики.
Даже принимая широкое общее понимание флоры (изложенное выше), мы
можем противопоставить региональные флоры [как „собственно флоры” (Юрцев,
1982а) ] флорам топологического, или внутриландшафтного, уровня (о них пойдет
речь ниже).
Многие авторы (например, Толмачев, 1974, с. 112), сужая понятие „фло-
ра”, стремятся уместить весь его объем в рамки основных интересов сравни-
тельной флористики. Даваемые при этом определения обычно содержат крите-
рии разграничения региональной флоры с внутриландшафтными флористическими
выделами.
1. 1А. Естественная полная регио-
нальная совокупность видов растений
(ПТСВР естественного флористичес-
кого выдела региональной размер-
ности) .
Естественная флора (Попов, 1963;
Камелин, 1973; Юрцев, 1975а; Вынаев,
в наст, сб.) ; естественная флора регио-
нального порядка (Юрцев, Семкин,
1980); региональная естественная
флора (Юрцев, 1982а, с. 13); флора
(Вынаев, 1984).’
Региональные естественные флоры (флоры фитохорий любого ранга), если
не основной объект, то, во всяком случае, основной предмет интереса сравнитель-
ной флористики, по крайней мере на стадии анализа и сравнения флор; как и со-
пряженное понятие „фитохория” (выдел флористического районирования), они
входят в число основных понятий флористической фитогеографии, флористиче-
ского районирования [„фитохориономии”, по А. Л. Тахтаджяну (1978) ].
Наиболее развернутое определение понятия приводится в работах Г. В. Вына-
ева (1984, с. 7, а также статья в наст, сб.).
1 В системе терминов Шеляг-Сосонко и Дидуха (статья в наст, сб.) для обо-
значения того же понятия было бы достаточно словосочетания „региональная
флора”.
247
1. 1А. 1. Элементарная естествен-
ная полная региональная совокупность
видов растений.
Критерий элементарности: через
территорию данной флоры не проходит
ни одной флористической границы ре-
гионального уровня.
Элементарная естественная флора
(Камелин, 1973; см. также 1. 1А. 1. 3),
но в более общем значении (Юрцев,
1975а, 19766); элементарная естест-
венная флора регионального уровня
(Юрцев, 1982а); элементарная регио-
нальная естественная флора*.
В качестве критериев гомогенности элементарных флористических выделов
региональной размерности предлагались следующие:
1. 1А. 1. 1.. Постоянство состава
ассоциаций (постоянство набора видов
на однотипных экотопах, за вычетом
случайных различий) на всем протяже-
нии контура,элементарного флористи-
ческого выдела (Толмачев, 1931,
1970а, 1974; Юрцев, 1975а).
1. 1А. 1. 2. Постоянство не только
основного состава, но и набора ассоци-
аций (и несущих их типов экотопов)
на всем протяжении выдела (Юрцев,
1975а, 1982а и в этой книге); отсю-
да - общее соответствие таких единиц
геоботаническим микрорайонам или
элементарным геоботаническим райо-
нам, о котором писал и А. И. Толмачев
(1974, с. 186), а также физико-геогра-
фическим ландшафтам (в не слишком
укрупненной трактовке), при условии
если через них не проходит ни одна
региональная флористическая граница.
Конкретная, или элементарная,
флора (Толмачев, 1931, 1986; Миняев
в: Юрцев, 1974а); флора элементарного
флористического района (Юрцев, 1975а,
1982а; Юрцев, Семкин, 1980); элемен-
тарная флора (Шеляг-Сосонко, 1980;
Шеляг-Сосонко, Дидух, 1980; Шеляг-
Сосонко и др., 1985).
Конкретная, или элементарная,
флора (Юрцев, 1975а, 1982а; Юрцев,
Семкин, 1980); флора ландшафта
(Лукичева, Сабуров, 1969); конкрет-
ная флора (Дидух, Шеляг-Сосонко,
1982, с. 16).
Для горных территорий в качестве элементарных естественных флор предлага-
лось также изучать:
1. 1А. 1. 3. ПТСВР умеренно круп-
ных речных бассейнов, не пересекае-
мых региональными флористическими
границами и имеющих в своем составе
по крайней мере одну эндемичную
расу (Камелин, 1969, 1973); (послед-
ний критерий обсуждается во 2-й ста-
тье Юрцева в наст, сб.).
1.1 А. 1.4. Как самый дробный
флористический выдел, который еще,
по-видимому, можно отнести к регио-
нальной размерности, Л. В. Мариной
(1982, 1983, а также статья в наст, сб.)
апробированы флоры бассейнов самых
малых рек - рек I порядка, т. е. под-
разделения конкретной флоры в более
узком ее понимании (1. 1А. 1. 2).
Элементарная естественная флора
(Камелин, 1969, 1973) (к сожалению,
данный термин не отражает приурочен-
ности выделяемых единиц к речным
бассейнам - см. ниже; его лучше упо-
треблять в разделе 1. 1А. 1); флора
ущелья (Харадзе в: Юрцев, 1974а).
Флора речного бассейна I порядка
(Марина, 1982, 1983, а также статья
в наст, сб.).
248
Отсюда еще один критерий гомогенности конкретной флоры (КФ), предло-
женный для апробации Л. В. Мариной (1983) на основе проведенных ею сопостав-
лений^ рамках одной КФ группировки флор речных бассейнов по разным флори-
I этическим показателям не совпадают, что говорит о невозможности однозначно
разделить КФ на части по сумме показателей.
1. 2А. ПТСВР произвольного кон-
тура региональной размерности (под
произвольным контуром в этой работе
имеется в виду любой контур, не со-
впадающий с тем или иным выделом
флористического или комплексного -
включающего и флористические крите-
рии - районирования).
„Искусственная флора” (неудач-
ный, но ранее часто употреблявшийся
термин); флора произвольного регио-
нального контура*.
Наиболее распространенные варианты:
1. 2А. 1. ПТСВР флористических
или комплексных ботанико-географи-
ческих пробных площадей - террито-
рий, достаточно протяженных для
выявления основных признаков мест-
ной региональной флоры.
Проба флористической ситуации,
проба флоры - имеются в виду регио-
нальные флоры (Юрцев, 1975а, 1982а;
Юрцев, Семкин, 1980); проба регио-
нальной флоры*.
Им противостоят (например: 1. 2А. 2. ПТСВР политико-административных или,
скажем, хозяйственно-экономических подразделений) флоры стран, администра-
тивных областей, районов и т, д.
1. 2А. 1.1. ПТСВР пробной площа-
ди для выявления элементарной регио-
нальной флоры.
1. 2А. 1. 2. ПТСВР площади выяв-
ления флористической ситуации в дан-
ном географическом пункте.
Без проведения дробного флори-
стического районирования изучаемой
территории уверенное разграничение
выделов элементарных региональных
флор, как правило, невозможно; по-
этому выборочно закладываемые про-
бы флоры в общем случае приходится
рассматривать как локальные флоры
(Ребристая, статья в наст, сб.), тем бо-
лее что методика изучения их совпа-
дает с методикой выявления КФ.
1. 1Б. 1. Полная территориальная
совокупность видов растений (ПТСВР)
естественного (выделяющегося и по
флористическим признакам) контура
топологического (внутр иландшафт-
ного) уровня (полная естественная
Флора ареала-минимума КФ
[в смысле А. И. Толмачева (1970а,
1974) и других авторов] = флора пло-
щади выявления КФ*, проба КФ*; кон-
кретная флора (Шеляг-Сосонко, 1980;
Шеляг-Сосонко, Дидух, 1980; Шеляг-
Сосонко и др., 1985).
Флора геона (Бешел, 1969); фло-
ра (окрестностей) географического
пункта (Юрцев, 1975а, 1982а); локаль-
ная флора (Шеляг-Сосонко, 1980; Юр-
цев, 1982а); в этом же смысле обычно
употребляется и термин „проба фло-
ры” (Юрцев, 1975а), хотя он в прин-
ципе может относиться и к более круп-
ным, чем КФ, региональным флорам.
Совокупность видов растений эко-
топа (Толмачев, 1974); флористиче-
ский комплекс фации, урочища и т. д.
(Юрцев, 1975а); флористические ком-
плексы эколого-топографических под-
разделений территории (Юрцев, 19766);
249
территориальная совокупность видов
растений любого экологически и фло-
ристически своеобразного подразделе-
ния ландшафта).
Так же как региональные флоры
обычно получают географические на-
звания, для обозначения ПТСВР вну-
триландшафтных контуров целесо-
образно использовать название экотопа
или синтаксона.
парциальная флора (экотопа) (Юрцев,
1975а, 1982а; Юрцев, Семкин, 1980);
флористическая парцелла (экотопа)
(Малышев, в выступлениях на совеща-
нии в Неринге).
ПТСВР каждого конкретного естественного подразделения ландшафта в прин-
ципе индивидуальна, но поскольку экологически однотипные контуры, как прави-
ло, несут сходное растительное население, свойственные им совокупности видов
легко поддаются типизации; чаще исследователи имеют дело с объединениями
парциальных флор (ПФ) однотипных экотопов или синтаксонов, в которых сле-
дует учитывать степень постоянства (константность) для них каждого вида, а также
варьирование его весовых характеристик в пределах типа.
ПТСВР экотопа или сообщества, являясь эколого-топографической частью
местной региональной флоры, в то же время обнаруживает черты флористической
целостности (как полные флоры небольших участков суши). Это и отражает
термин ,,парциальная флора” экотопа (или сообщества), относящийся, как уже
говорилось, к индивидуальным контурам.
1. 1Б. 2. Объединение ПТСВР фло-
ристически однотипных естественных
внутриландшафтных контуров (эко-
топов, биотопов, сообществ),
Ранг данной единицы внутри-
ландшафтного уровня обозначается
через ранг соответствующей экотопо-
логической единицы (микроэкотоп,
макроэкотоп) - при работе с индиви-
дуальными единицами - или, при объ-
единении ПФ однотипных экотопов,
через классификационный ранг изучае-
мых эколого-топографических подраз-
делений. Например, объединенная ПФ
такого-то типа или класса (микро) эко-
топов. Оговоримся, что в типологиче-
ской иерархии экотопов типы объеди-
няются в классы.
1. 1Б. 2. 1. Объединение ПТСВР
флористически и экологически одно-
типных сообществ.
Как и в разделе 1. 1Б. 2, ранг
рассматриваемых единиц определяется
рангом синтаксона.
Комплекс видов данного типа
местообитаний (Миняев в: Юрцев,
1974а); флористический комплекс
класса экотопов или сообществ (Юр-
цев, 19766); экологический элемент
флоры - применительно к совокупно-
стям видов растений экотопологиче-
ских единиц (Кожевников, 1974;
Шмидт, 1980); объединение парциаль-
ных флор (Юрцев, 1982а, а также
в наст, сб.); объединенная ПФ типа
(класса) экотопов или синтаксона
(Марина, 1983; Юрцев, цит. соч.);
флоротопологический комплекс (Вы-
наев, 1984, а также в наст, сб.); флора
типа (класса) экотопов (разные
авторы).
Флористический комплекс (типов
сообществ) (Миняев в: Юрцев, 1974а);
флористический комплекс класса
сообществ (Юрцев, 19766); флоро-
ценотический комплекс (Шеляг-Сосон-
ко, Дидух, 1980; Шеляг-Сосонко и др.,
1985);* ценофлора (Лавренко, 1965;
Седельников, 1985, а также в наст, сб.;
Дидух, в наст, сб.).
1 Тот же термин был ранее введен и широко употребляется в другом смысле
(см. ниже, раздел [2. 2А.]. 3. 2 - с. 258).
250
При изучении экотопологической структуры не очень крупной региональной
флоры нередко приходится иметь дело с ценофлорами синтаксонов, ареал которых
выходит за пределы соответствующей фитохории. В связи с этим возникают 2 зада-
чи: 1) изучение ПФ, представляющих соответствующий синтаксон в данной регио-
нальной флоре; 2) сравнение их с однотипными ценофлорами за пределами данной
флоры с целью выявить специфику местной ценофлоры того же синтаксона.
Выше уже говорилось, что ПФ экотопов или сообществ, а также их объедине-
ния (в том числе, ценофлоры) представляют эколого-топографические (экотопо-
логические) подразделения региональной
и типологических элементов флоры, для
дим особый термин.
1. 1Б. 3. Полная совокупность объ-
единенных ПФ (или ценофлор) того или
иного ранга, ранжированная по их от-
носительной роли в ландшафте и (или)
ординированная по их флористическо-
му и экологическому сходству,
1. 2Б. Полная территориальная
совокупность видов растений произ-
вольного контура топологического
(внутриландшафтного) уровня.
флоры. Как и в случае таксономических
их упорядоченной совокупности необхо-
Экотопологическая структура
флоры (Галанин, 1979; Марина, 1983;
Юрцев, 1982а, а также в наст. сб.).
Термина, соответствующего об-
щему значению этого понятия, до сих
пор не было предложено; едва ли име-
ется в нем потребность.
Очевидно, потребность имеется лишь в специальных терминах для обозначения
флоры пробных площадок, закладываемых для выявления ПФ протяженных
микро-, мезо- и макроэкотопов, и в частности ценофлор, хотя во всех случаях,
когда это возможно, лучше иметь дело с полными индивидуальными контурами
(если они не слишком протяженны).
1. 2Б. 1. ПТСВР пробной пло-
щадки для выявления ПФ (макро-,
мезо-, микро-) экотопа.
1. 2Б. 2. ПТСВР пробной пло-
щадки для выявления ценофлоры
синтаксона.
2. Неполная (частичная) террито-
риальная совокупность видов (часть
полной территориальной совокупности
видов, выделенная по какой-либо
категории признаков).
Проба ПФ экотопа*.
Проба ценофлоры синтаксона*.
Элемент флоры; фракция флоры
(разные авторы, за исключением слу-
чаев, когда эти термины употребля-
ются в более узком специальном
значении, что должно быть оговорено
в определении элемента).
Поскольку по своей сути термин „элемент” означает часть некоторой полной
совокупности вообще, этот термин должен употребляться с тем или иным эпите-
том, определяющим основание для данного деления на элементы (например, тип
ареала или экология видов) и тем самым категорию элементов. Элементы иногда
подразделяют на субэлементы (элементы *1, Ill и более высокого порядков)
или последовательно объединяют в группы, классы, типы элементов.
Наряду с общим термином „элемент флоры” во флористике широко приме-
няются в сходном значении термины „группа видов (растений) ”, „комплекс
видов”, иногда „флористический комплекс”. Впрочем, последнему термину также
нередко придается более узкое и определенное, как правило флорогенетическое,
содержание.
Очевидно, от употребления термина „флористический комплекс” в общем
значении (без определения категории комплекса с помощью специального эпитета)
251
следует воздержаться; сведение же его к чистому синониму термина „флора” или
„региональная естественная флора” и вовсе не целесообразно.
Итак, термины „элемент флоры”, „фракция флоры”, с одной стороны, „груп-
па видов” и „комплекс видов” - с другой, в принципе могут употребляться в тож-
дественных значениях. Однако в употреблении (предпочтении) тех или других
имеются некоторые тонкости: 1) выделение элементов - всегда дедукция (умо-
заключение от целого к части) и возможно только на заключительных этапах
изучения флоры; напротив, объединение видов в группы и комплексы представ-
ляет научную индукцию (заключение от частного к общему) и возможно еще
на стадии незавершенного исследования; 2) группы и комплексы, по определению,
должны включать более одного вида (но вполне закономерно существование ви-
дов, не поддающихся объединению с другими в группу по данной категории призна-
ков и при заданном пороговом уровне сходства) ; элементы флоры же своей сово-
купностью должны охватывать все виды данной флоры, часть элементов может
быть представлена каждая единственным видом; 3) элементы флоры выделяются
специально для анализа данной флоры, группы же и особенно комплексы видов
могут устанавливаться для намного более обширных территорий и могут быть
лишь частично представлены или даже совсем не представлены в изученной флоре.
Приемы же систематизации как элементов флоры, так и групп или комплек-
сов видов в основном одинаковы. При установлении самых крупных категорий
как элементов флоры, так и групп или комплексов видов, возможны 2 независи-
мых принципа (подхода): 1) отделение типологических (и историко-типологичес-
ких) элементов флоры (групп видов), установленных по общим признакам каж-
дого вида (т. е. по его поведению на всем протяжении ареала), от таковых, выде-
ленных по тем же категориям признаков его местной региональной популяции;
в результате этой операции мы имеем: 2. 1. Общие, или генеральные, типологи-
ческие и историко-типологические элементы флоры; 2.2. (Внутри)региональные
типологические и историко<гипологические элементы флоры; 2) отделение соб-
ственно-типологических элементов флоры (2.А) от историко-типологических
(флорогенетических, исторических) (2.Б).
Пересечение обоих подходов дает 4 основных типа элементов флоры: 2.1А.
Общие типологические элементы; 2.1 Б. Общие историко^гипологические (= исто-
рические) элементы; 2.2А. Региональные (= внутрирегиональные) типологические
элементы; 2.2Б. Региональные историко-типологические элементы.
Положение осложняется тем, что расчленение флоры на типологические
и историко-типологические элементы как по общим, так и по региональным
признакам таксонов может проводиться на уровне таксонов разного ранга,
т. е. не только видов (или рас), но и надвидовых таксонов, представляющих таксо-
номические элементы флоры (2.3). Все же на том уровне, на котором работает
большинство флористов, первостепенное значение имеет анализ флоры на уровне
вида (или расы). Поэтому принципы выделения различных категорий элементов
флоры ниже будут показаны на примере типологической и историко-типологичес-
кой классификаций видов.
Дальнейшее подразделение 4 самых крупных категорий элементов флоры
должно проводиться уже по той или иной конкретной категории типологических
признаков: 1) географические, 2) экологические, 3) ценотические, 4) биологи-
ческие и т.д. и их комбинациям: (1, 2) - эколого-географические, (2, 3) - эко-
лого-ценотические, (2, 4) - эколого-биологические, (3, 4) - ценобиологические
и т.д. При этом каждая категория типологических признаков нередко имеет слож-
ную, многокомпонентную структуру, что предполагает целую систему одномерных
классификаций (например, выделение экологических элементов по отношению
к различным экологическим факторам или их плеядам и т.д.). Кроме того, система
любой категории элементов флоры может быть иерархической или, во всяком
252
случае, различаться рангом выделяемых основных типологических или таксономи-
ческих единиц. В принципе число категорий признаков, по которым могут выде-
ляться элементы флоры, невозможно жестко ограничить — ограничение их, выбор
наиболее важных и существенных должны диктоваться задачами конкретного
флористического исследования.
Поскольку выделение элементов флоры имеет давнюю историю и в подходах
разных авторов к решению этой задачи наблюдается значительный разнобой, в рам-
ках этой работы немыслимо дать обзор всех или даже основных предлагавшихся
ранее классификаций элементов. Наша задача - систематизировать основные
подходы к выделению элементов флоры, используя для обозначения их категорий
наиболее употребительные (или кажущиеся авторам наиболее рациональными) тер-
мины, а если таковые отсутствуют - в ряде случаев предлагая новые. Часть 2 рабо-
ты, по мысли авторов, представляет собой своего рода предварительный (рабочий)
макет классификации элементов флоры, не претендующий на полноту, но содержа-
щий достаточно разносторонний концептуальный материал для обсуждения этой
сложной и многоаспектной проблемы на III совещании по сравнительной флорис-
тике (на I и II совещаниях эти вопросы не обсуждались).
В отечественной ботанико-географической литературе наиболее общую (хотя
в настоящее время уже неполную) классификацию элементов флоры опубликовал
Е. В. Вульф (1941); в дальнейшем ее дополняли и видоизменяли Я. Эйларт (1960),
Б. А. Юрцев (1968а), Р. В. Камелин (1973), Ю. Р. Шеляг-Сосонко и Я. П. Дидух
(1978), Дидух и Шеляг-Сосонко (1982) и др.
Во избежание повторов мы рассмотрим совместно основные подразделения
сначала общих типологических (2.1 А) и региональных типологических (2.2А)
элементов, отражающих рецентный аспект анализа флоры, а затем подразделения
общих и региональных исторических (историко-типологических) элементов
(2.1Б и 2.2Б), относящихся к флорогенетическому анализу.
2.А. Части флоры, выделяемые
по той или иной категории ныне на-
блюдаемых типологических признаков:
2.1А - каждого вида в целом;
2.2А - его местной региональ-
ной популяции (в грани-
цах изучаемой флоры).
Типологические элементы флоры*:
общие;
региональные.
Типологические признаки видов и их местных популяций, как уже отмечалось,
обусловлены не только современными условиями, но и историей вида. Однако
в данном разделе анализа флоры мы должны отвлечься от тех или иных истори-
ческих гипотез, с тем чтобы как можно точнее и объективнее выявить современные
особенности вида (популяции); привлечение исторических гипотез правомерно
на стадии интерпретации результатов типологического анализа (и потому они
не должны включаться в исходные данные).
Во избежание повтора длинных формулировок ниже в левом столбце будут
в каждом случае указаны непосредственно категории типологических признаков,
по которым выделяется данная категория элементов флоры, в правом столбце -
названия элемента флоры. Приводится номер третьей и последующих ступеней деле-
ния исходного понятия „территориальная совокупность видов”(после 2.1 А. и 2.2А).
...1. Особенности географического
распространения:
2.1А.1 - вида в целом (тип
ареала);
2.2А.1 - его же в границах дан-
ной региональной фло-
ры.
Географические элементы флоры:
общие;
региональные.
253
В том же значении могли бы употребляться термины „хорологические” или
„ареалогические элементы флоры”.
Синонимичные термины: для 2.1А.1. - компоненты (Reichert, 1921), геоэле-
менты (Клеопов, 1938), тип ареала (многие авторы), ареалогические группы
(Чопик, 1976), группы ареала (Дидух, Шеляг-Сосонко, 1982); для 2.2А.1. - хоро-
логические группы видов (Козловская, Парфенов, 1972). Выделение географиче-
ских элементов - наиболее распространенный, фактически обязательный раздел
анализа флоры. Разная степень сходства - различия видовых ареалов оправдывает
иерархический подход к систематизации географических элементов; можно пред-
ложить следующие ступени классификации видов по типу ареала: субэлемент
(если необходимо) -элемент-группа элементов-класс элементов-тип элементов.
Задачи систематизации видовых ареалов в целом и их частей в границах
данной флоры имеют свою немалую специфику, однако подходы к решению тех
и других могут быть в определенной мере сходны.
Ниже представлены лишь важнейшие, методически достаточно разработанные
подходы к выделению географических элементов.
...1.1. Положение ареала (или его
части) в системе географических, био-
или ботанико-географических, биокли-
матических зон, их высотных аналогов
(поясов), а также их долготных сек-
торов.
Координатные географические эле-
менты*: зональные, поясно-зональные,
высотно-поясные, долготные (разные
авторы) - одномерные, также двумер-
ные и трехмерные (по сочетанию 2 или
3 биогеографических координат).
Данный подход обоснован как „принцип биогеографических координат”
(Юрцев, 1968а). Он особенно оправдан и регулярно и давно применяется при
анализе флор более северных территорий, с непрерывным циркумполярным про-
стиранием зон (тундровой, таежной) ; однако очень часто его применяют и к более
южным флорам, где единых циркумполярных зон растительности не существует,
и исследователи выделяют условные биогеографические зоны - например, темпе-
ратную, меридиональную — или используют систему термоклиматических поясов
(холодный, холодно-умеренный, умеренный, теплоумеренный и т.д.); многие евро»
пейские исследователи следуют координатной биогеографической системе элемен-
тов Мейзеля (Meusel et al., 1965). При анализе небольших горных флор поясно-зо-
нальные группы используются для определения высотной координаты - для уста-
новления же широтной иногда учитывают положение ареала вида в системе флори-
стического обмена: Арктика (Субарктика)-высокогорья (верхнелесные уровни
гор) (Юрцев, 1968а).
При выделении одномерных координатных элементов виды могут группи-
роваться по амплитуде (набору) широтных или высотных полос или долготных
секторов („принцип пределов” - Юрцев, 1968а) либо по наиболее регулярной
встречаемости в одной (одном) из них („принцип центра тяжести”); в том
и другом случае по альтернативному принципу выделяются субэлементы (под-
группы) .
Целесообразно проводить координатную классификацию ареалов по распро-
странению видов в зональной системе северного полушария, выделяя по присут-
ствию или отсутствию вида в аналогичной зоне южного полушария особую катего-
рию элементов (или субэлементов зональных элементов): биполярные, нотоборе-
альные и т. п. При зональной классификации ареалов северных флор арктические
виды правомерно сближать с арктоальпийскими, гипоарктические - с гипоаркто-
монтанными соответственно (на правах особых субэлементов или элементов одной
группы), ориентируясь на отношение вида к системе зон и подзон. Если амплитуда
широтного распространения вида в разных долготных секторах неодинакова или,
напротив, в разных широтных полосах долготная амплитуда вида существенно
254
рознится, при анализе данной региональной флоры приходится в первую очередь
учитывать поведение вида в данном секторе или данной широтной полосе.
...1.2. Положение ареала (или его
части) в системе выделов природного,
комплексного био- или ботанико-
географического, флористического
районирования Земли (2.1А.1.2) или
территории данной флоры (2.2А.1.2).
Хориономические* географичес-
кие элементы - общие или региональ-
ные (географические элементы в смы-
сле очень многих авторов).
При данном подходе каждый элемент флоры характеризуется набором соот-
ветствующих выделов районирования, а иерархическая классификация элементов
строится на соподчинении этих выделов. Понятно, что общие хориономические
элементы (2.1 А.1.2.1) характеризуются более крупными гео-, био- или фитохори-
ями (вплоть до набора континентов или субконтинентов), нежели региональные.
Крупность единиц, характеризующих региональные хориономические элементы
(2.2А.1.2), зависит от размеров и степени внутренней однородности или контраст-
ности (по природным условиям и природной истории) территории данной регио-
нальной флоры. При этом в качестве характеризующих единиц могут быть исполь-
зованы первично выделенные на территории изученной флоры флористические,
геоботанические или ландшафтные районы (еще до подключения их к общеземной
или макрорегиональной хориономической системе).
Общие хориономические элементы по числу заселенных ими фито (или гео-)-
хорий планетарного уровня (областей, подцарств, царств; континентов) могут
быть подразделены (2.1А.1.2.2) на эврихорные (полихорные, мультирегиональные
элементы,1 вплоть до космополитных) и стенохорные; среди последних особое зна-
значение для фитогеографии и флорогенетики имеют эндемики фитохорий ранга
области и более низкой; большое значение имеет и таксономический ранг энде-
мизма.
Региональные хориономические элементы в свою очередь (2.2А. 1.2.2) могут
подразделяться по встречаемости видов (или надвидовых таксонов) в совокуп-
ности фитохорий невысокого ранга, различаемых в пределах территории данной
флоры [географическая активность (Юрцев, 1982а, первая статья в наст, сб.) j.
Вероятно, по этому показателю уместно также различать эврихорные и стенохор-
ные региональные хориономические элементы (2.2А.1.2.2).
К данному типу деления хориономических элементов тесно примыкает еще
один тип деления - по степени непрерывности распространения видов, которая
может оцениваться на уровне фитохорий того или иного ранга; основные типы
распространения - сплошной (непрерывный), диффузный (спорадический, мелко-
прерывистый) и дизъюнктивный. Подобные 3 типа правомерно различать как
в группе общих хориономических элементов (1.1А.1.2.3), так и в группе регио-
нальных (1.2А.1.2.3). Наличие крупных дизъюнкций - характерный признак ре-
ликтовых ареалов, по которым распознаются реликтовые элементы флоры (исто-
рико-типологическая категория; см. ниже). Однако в ряде случаев дизъюнкция
может быть результатом дальнего переноса диаспор или политопного возникно-
вения вида, например, путем гибридогенеза, авто- или аллополиплоидии.
...1.3. Относительное положение
территории данной флорыиобщего аре-
ала вида или заселенной им части упо-
мянутой территории.
Релятивные (соотносительные)
географические (ареалогические) эле-
менты* - общие или региональные.
1 В координатной системе элементов их аналогами являются полизональные
(„мультизональные”) виды.
255
Общие релятивные ареалогические элементы устанавливаются по положению
условного центра общего ареала вида относительно контура данной флоры (запад-
нее или восточнее, севернее или южнее либо же в пределах территории флоры);
региональные релятивные элементы - по положению заселенной видом части
территории флоры относительно ее центра. Одноименные (например, западные)
общие и региональные релятивные элементы могут не совпадать, что дает материал
для исторических интерпретаций; так, во флоре Чукотки Kobresia sibirica Turcz.-
западный общий релятивный элемент (по положению центра ареала), но восточный
региональный (распространен преимущественно в восточной части территории).
Особый тип географических элементов может устанавливаться по форме аре-
ала или его крупных фрагментов (...1.4), в особенности если ареалы, сходные
по форме и положению "центра^но отличающиеся протяженностью, группируются
в серии [эквиформные ареалогические элементы, в смысле Хультена (Hulten, 1937,
1972)]. Подобный подход применялся к анализу общих ареалов, но в принципе
он применим и к их региональным фрагментам (в особенности в случаях дизъюнк-
тивного распространения). При интерпретации серий эквиформных ареалов (equi-
formal progressive areas) Хультен исходил из гипотезы о переживании соответствую-
щими видами ледниковых эпох в небольших рефугиумах и о послеледниковом
расселении „пластичных видов” („радиантов”) с неодинаковой скоростью, тогда
как „ригидные виды” в результате необратимого обеднения генофонда утратили
способность к расселению и маркируют своими ареалами (или их фрагментами)
положения рефугиумов. И в этом случае выделение типологических элементов -
по форме ареала или его фрагментов - дает материал для исторических гипотез,
выражением которых является разделение флоры на радиантов различных рефу-
гиумов (историко-типологические элементы); обе эти категории, однако, нельзя
смешивать.
...2. Особенности экологии (в том
числе экологической приуроченности)
видов (2.1 А.2) или его местной регио-
нальной популяции (2.2А.2).
Экологические (аутэкологиче-
ские) элементы флоры - общие или
региональные,
Виды с обширными ареалами неизбежно сталкиваются с разными, нередко
весьма контрастными экологическими ситуациями и потому обычно дифферен-
цированы в экологическом отношении, что проявляется как в изменении спектра
экотопов в разных частях ареала, так и в формировании различных наследственных
климатических и экологических рас; последние очень часто могут встречаться
на одной и той же территории на контрастных типах экотопов. Экологические
элементы флоры, установленные по поведению вида на территории данной не слиш-
ком протяженной региональной флоры (2.2А.2), экстраполировать на другие
части ареала того же вида следует с большой осторожностью. Для случаев, когда
экология вида меняется в разных частях его общего ареала или территории флоры,
приходится говорить о двумерных эколого-географических элементах - как
общих: 2.1А. (1, 2), так и региональных: 2.2А. (1, 2).
Структура категорий экологических элементов флоры сложна и многомерна,
что зависит прежде всего от двух групп обстоятельств: 1) множественность кате-
горий экологических факторов; большинство факторов сами разложимы на ряд
компонент; положение облегчается в тех случаях, когда можно выделить крити-
ческие факторы (правило Либиха) и когда разные факторы образуют крепкие
корреляционные плеяды; 2) возможность разных подходов к оценке отношения
растений к тем или иным экологическим факторам: путем выявления приурочен-
ности видов к территориям или участкам, заведомо отличающимся режимом
конкретных экологических факторов (хоро- и топоэкологические элементы),
или же путем изучения морфофизиологических и экофизиологических адаптаций
256
растений к тем же группам факторов (экофизиологические элементы флоры).
В большинстве случаев эколого-физиологическая изученность региональных флор
еще явно недостаточна для проведения такого анализа всей флоры; как правило,
флорист судит об экологической природе вида по приуроченности его к участкам
с определенными экологическими условиями, т. е. по косвенным показателям;
известно также, что распределение видов по экотопам и климатически отличаю-
щимся территориям зависит не только от их аутэкологических свойств, но и
от конкурентных отношений (отличие аутэкологических оптимумов и амплитуд
толерантности от синэкологических). Кроме того, в отношении к любому фактору
виды могут классифицироваться по положению зоны оптимума и по широте
и положению всей амплитуды толерантности. Во многих случаях более простые
классификации (с меньшим числом экологических градаций по каждому фактору)
предпочтительнее, так как они не создают иллюзию высокой точности. Тем
не менее по косвенным показателям разработаны очень длинные экологические
шкалы, включающие большое число градаций различных экологических фак-
торов - как прямодействующих, так и косвенных [Раменский и др., 1956;
EUenbeig, 1974; Цыганов, 1983; критическая оценка дана в статье Ю. И. Самойлова
(1986)].
Целесообразно разделить семейство экологических элементов флоры (прежде
всего региональных) на 2 основные группы: хоро- и топоэкологических элементов
и экофизиологических элементов флоры; дальнейшее подразделение проводить по
категориям экологических факторов.
...2.1. Приуроченность вида к эко-
логически отличающимся по данному
фактору:
...2.1.1 - территориям (регио-
нальной размерно-
сти) ;
...2.1.2 - участкам (топологи-
ческой, или внутри-
ландшафтной, размер-
ности) - экотопам.
Хороэкологические* (климатичес-
кие) элементы флоры;
топоэкологические* элементы флоры.
В системе хороэкологических (климатических) элементов рассматриваются
косвенно действующие факторы регионального (и планетарного) уровня размер-
ности, относящиеся к типам климата и выявляющиеся при географических сравне-
ниях. Сюда относятся: термоклиматические элементы (гекистотермные, микро-
томные, мезотермные, макротермные и т. д.) по отношению к термической
поясности климатов, хороэкологические элементы по отношению к континенталь-
ности - океаничности климата (к ним примыкают двумерные элементы флоры
по отношению к гидротермическому типу климата); иногда виды группируют
по отношению к аридности—гумидности климата, суровости зим и т. п. (Малышев,
1972в; Цыганов, 1983). Хороэкологические элементы могут устанавливаться
по общему ареалу вида либо же по распределению вида в пределах достаточно
протяженной региональной флоры.
Топоэкологические элементы могут быть подразделены на 2 главные кате-
гории: аэротопические (по отношению видов к факторам аэротопа, или климато па:
температурам воздуха - летним, зимним, освещенности, влажности воздуха,
мощности и времени схода снежного покрова и т. п.) и эдафотопические (по отно-
шению к увлажнению почв, его переменности, солевому режиму, кислотности
почв, их аэрации, температурам летним и зимним, богатству доступными соедине-
ниями азота и т. д.). Отношение видов к конкретным факторам передается с по-
мощью конструкций ,,-фильный”, ,,-фобный”, „-индифферентный (нейтральный) ”,
‘/а 17 Заказ 1220
257
желательно с уточнением зоны толерантности, но могут применяться конструкции
,,-топный” (вместо ,,-фильный”): ксеротопный, мезотопный, гигротопный и т. д.
То по экологические элементы выявляются на основе анализа распределения
видов по экотопам, ранжированным в соответствии с градациями учитываемых
факторов, их не следует смешивать с полными территориальными совокупностями
видов различных экотопов (парциальными флорами - см. раздел 1.1 Б).
...2.2. Экофизиологические и мор- Экофизиологические элементы
фофизиологические особенности вида, флоры,
отражающие его адаптации- к града-
циям конкретных экологических фак-
торов.
Экофизиологические элементы подразделяются по категориям прямодейству-
ющих факторов - аэротопа и эдафотопа. Физиологический эксперимент в контро-
лируемых, а отчасти и в природных условиях позволяет выявить истинные пределы
экологической толерантности растений в целом и основных физиологических
процессов, в первую очередь к градациям критических факторов (например,
засухоустойчивость и жаростойкость растений пустынь), взаимовлияние различных
факторов и т. д., отделить аутэкологический оптимум вида (популяции) от синэко-
логического (эколого-ценотического). Важная задача экофизиологических иссле-
дований - выявить наследственно закрепленную экотипическую дифференциацию
видов по отношению к ведущим экологическим факторам, а также уровень
внутрипопуляционного наследственного полиморфизма по экофизиологическим
признакам.
При эколого-физиологических исследованиях экологические реакции растения
обычно разлагаются на составляющие их процессы. Для интегральной оценки
экологических реакций особое значение приобретает комплексный показатель
продуктивности (нетто-продукции) растения, а также анализ изменений биоморфы
в разных экологических условиях.
...3. Распределение вида по типам
сообществ (фитоценоцикл), его при-
уроченность к определенным классам
сообществ, роль в растительном по-
крове или отдельных сообществах
Фитоценотические, ценотические
элементы флоры („ценоэлементы”
ряда авторов).
Этот раздел анализа флоры пересекается с геоботаническим (фитоценологи-
ческим) анализом растительного покрова. При обследовании флоры компактных
территорий основными единицами описания фитоценоцикла вида могут быть
,,крупные” ассоциации или группы ассоциаций, но в определенных целях (для
дальних сравнений) в качестве таковых могут использоваться формации или даже
типы растительности (также флороценотипы, фратрии формаций).
Группировка видов по их фитоценоциклам - амплитуде сочетаемости
(...3.1. Ценоспектровые элементы флоры*, эколого-ценотические элементы)
часто не совпадает с группировкой по их приуроченности, абсолютной или преиму-
щественной, к тому или иному, достаточно крупному классу сообществ (..3.2),
по которой устанавливаются флороценотические комплексы (элементы) в смысле
Б. А. Юрцева (1982а, а также в наст, сб.) ; последним соответствуют ценоэлементы
ряда авторов.
Флористы и геоботаники нередко предпочитают говорить об эколого-ценоти-
ческих группах или элементах флоры (Галанин, 1973; Василевич, 1986), посколь-
ку анализ распределения видов по сообществам трудно отделить от анализа рас-
пределения их по экотопам (биотопам) ; при строгом подходе эколого-ценотичес-
кие элементы можно выделить в особую категорию элементов флоры ...3.(2, 3).
258
К этому же разделу анализа флоры относится определение широты эколого-
ценотической амплитуды вида - например, по числу освоенных им крупных
классов экотопов (биотопов), с выделением эвритопных и стенотопных (иногда
также гемиэвритопных и гемистенотопных) видов (Юрцев, 1968а); при строго
фитоценологическом подходе можно говорить о широте фитоценоцикла (цено-
спектра), измеряя его числом крупных синтаксонов, в которых встречен вид
(Дидух, 1977в, 1982). Соответственно выделяются категории эвриценотопных,
гемиэвриценотопных, гемистеноценотопных, стеноценотопных видов (Дидух,
1977в). Разделение флоры на элементы по экотопологической или собственно
ценотической активности видов проводится по сочетанию трех показателей:
широты спектра экотопов или сообществ, степени повсеместности, уровню чис-
ленности на характерных экотопах, иногда же только по двум последним пока-
зателям (Юрцев, 1968а, 1982а, а также в наст, сб.); собственно фитоценологиче-
ский подход к оценке активности применил Я. П. Дидух (1982).
Поскольку спектр ценотических сочетаний на протяжении ареала вида, как
правило, существенно изменяется, выделение ценотических и эколого-ценотичес-
ких элементов обычно приходится проводить по поведению местной региональной
популяции вида [2.2А.З; 2.2А. (2, 3) ].
Изменение спектра ценотических сочетаний вида на протяжении его общего
ареала (или хотя бы в более крупных фитохориях; иногда же - и в пределах
достаточно протяженной местной флоры) дает основание для выделения ценогео-
графических и экоценогеографических элементов флоры, как общих: 2.1А. (1, 3) ;
2.1 А. (1, 2, 3), так и региональных: 2.2А. (1, 3); 2.2А. (1, 2, 3). Впрочем, флоро-
ценотические комплексы (элементы; ценоэлементы) могут устанавливаться
по преимущественной приуроченности видов к одному и тому же крупному классу
сообществ не только в данной флоре, но и на всем протяжении ареала. Примером
таких общих флороценотических комплексов могут служить таежный, немораль-
ный, степной комплексы (флороценотические элементы).
К этой же общей категории фитоценотических элементов относится классифи-
кация видов по их роли в растительном покрове в целом (см. также группы
активности) и в отдельных типах сообществ - система фитоценотипов (см.: Васи-
левич, 1983; Работнов, 1983, и др.). Примерами фитоценотипов могут служить
эдификаторы, прочие доминанты, ассектаторы или же, в системе Л. Г. Раменского
(1938), виоленты, патиенты.эксплеренты (группа ценотических элементов флоры).
Ценотипу эксплерентов соответствуют „асоциальные, или ценофобные, элементы
флоры” многих авторов. Наконец, виды могут классифицироваться и по их месту
в региональной сукцессионной системе (системах) - приуроченности к начальным,
средним или заключительным (климаксным) фазам разных сукцессионных рядов.
...4 Те или иные биологические Биологические элементы флоры,
особенности видов - общие или регио-
нальные.
В качестве биологических признаков рассматриваются самые различные
жизнепроявления видов растений (общие для вида в целом или только для части
его популяций), не сводящиеся к географическому распространению или распреде-
лению вида по сообществам, экотопам, градиенту экологических условий, но так
или иначе отражающие приспособленность растений к среде обитания, обеспечиваю-
щие поддержание, размножение и расселение вида, влияющие на пути (формы)
и темпы эволюционных преобразований популяций. Подмножество биологических
классификаций видов (рас), непосредственно отражающее биологические и гене-
тические основы их формирования и эволюции, целесообразно выделить в особую
группу (или даже самостоятельную категорию) эйдологических элементов флоры,
особенно тесно примыкающую к типу историко-типологических элементов.
259
Наиболее часто практикуется разделение изученной флоры на биологические
элементы по способу (способам) опыления (для цветковых растений), способам
переноса диаспор, по жизненной форме (в той или иной ее трактовке) или отдель-
ным ее компонентам (например, по долговечности растений); иногда выделяют
ритмологические (фенологические) элементы флоры - по сезонной ритмике раз-
вития листового аппарата и т. д. Заслуживает внимание также группировка видов
по жизненной стратегии (К- и г-стратегии: Пианка, 1981), соотношению вегетатив-
ного и генеративного размножения, степени вегетативной подвижности и т. д.
В системе биологических элементов имеет особое значение и наиболее регу-
лярно практикуется классификация видов по жизненной форме - биоморфе
(биоморфные элементы флоры). С давних пор при анализе флор используется
система жизненных форм Раункиера - по положению перезимовывающих почек
относительно поверхности земли (признак, экологически чуткий и важный в стра-
нах с резко выраженной сезонностью климата). В последнее время в нашей стране
при биологическом анализе флоры все чаще используется более универсальная
система жизненных форм И. Г. Серебрякова (1962), обычно с рядом упрощений
(вплоть до возврата к „основным биоморфам”: травы о дно-двулетние и много-
летние, кустарнички, полукустарнички, кустарники, полукустарники, лианы,
деревья).
По сочетанию биоморфы с важнейшими экологическими признаками вида
(отношение к основным экологическим факторам) выделяются экобиоморфы,
которые в качестве экобиоморфных элементов могут составить предмет двумер-
ного эколого-биологического анализа флоры [... (2,4)]. Е. М. Лавренко и В. М. Све-
шникова (1968) подчеркнули необходимость возможно более полного учета
жизнепроявлений растений и их экологических особенностей при изучении экобио-
морф; однако чем шире круг признаков, характеризующих экобиоморфы, тем
труднее классифицировать последние. Предложенная В. Н. Голубевым (1972)
так называемая линейная классификация жизненных форм (экобиоморф) пред-
ставляет сумму одномерных классификаций вида по отдельным категориям биоло-
гических (в том числе биоморфологических) и экологических признаков; одна
из этих категорий - „основная биоморфа”; сама система признаков, разработанная
Голубевым, может представить интерес с точки зрения задачи анализа флоры.
Для целей анализа флоры, помимо самого Голубева и его учеников, этот подход
применяли Я. П. Дидух (1977в), Ю. Р. Шеляг-Сосонко, Дидух (1980) и др., выделя-
ющие категорию биоморфологических элементов флоры.
К уже упоминавшейся категории эйдологических элементов флоры относится
группа кариосистематических элементов (Favarger, 1961) - разделение флоры
на виды диплоидные, анеуплоидные, палео-, мезо- и неополиплоидные разного
уровня плоидности и хромосомные расы (см. раздел „Программы флористических
исследований...”), а также система собственно эйдологических элементов (там же).
2.Б. Временные, географические,
экологические, ценотические, биологи-
ческие и комплексные характеристики
формирования и расселения:
2.1Б - видов в целом (также
вышестоящих таксо-
нов) или
2.2Б - региональных популя-
ций видов (в границах
изучаемой флоры).
Историко-типологические*, исто-
рические, генетические элементы
флоры, флорогенетические элементы -
общие или региональные (историчес-
кие свиты, исторические комплексы,
флористические комплексы в понима-
нии разных авторов).
Четыре предлагаемых синонимичных термина могут применяться на выбор -
на равных основаниях: 1-й - наиболее полный и точный, но несколько громоздок,
260
3-й и 4-й вызывают у некоторых придирчивых читателей ассоциации с наукой
генетикой; 2-й как будто бы лишен видимых изъянов.
Наблюдаемые ныне типологические признаки таксонов обычно дают основной
материал для построения исторических гипотез (палеоботанические свидетельства,
как правило, фрагментарны и касаются сравнительно немногих таксонов), но обя-
зательно должны быть отделены от самих гипотез. В то же время основные ка-
тегории типологических и историко-типологических элементов флоры во многом
сходны, с той существенной разницей, что в исторические классификации (там,
где это возможно) должна вноситься дополнительная координата - геологического
времени, которая может служить и самостоятельным критерием выделения одной
из категорий исторических элементов. Для неэндемичных и неавтохтонных таксо-
нов происхождение вида и история его местной региональной популяции — 2 вполне
самостоятельные проблемы, относящиеся к разным аспектам и разным разделам
флорогенетического анализа.
2БЮ. Приуроченность формирова-
ния вида (2.1 Б.О) или его региональной
популяции (2.2Б.0) к определенному
интервалу геологического времени или
стадии формирования данной флоры.
Хроногенетический*, хроноисто-
рический*, временной (темпоральный)
исторический элемент флоры, хроно-
элемент - общий или региональный.
Нередко практикуемое разделение флоры на древние и молодые (палео- и нео-
хронные) элементы, к которым можно было бы добавить категорию „новейший
элемент” (для современной эпохи интенсивного преобразования природы челове-
ком), грешит неопределенностью: понятия „древний” и „молодой” для разных
региональных флор имеют неодинаковое содержание. Возраст вида или его кон-
кретной региональной популяции (для реликтов - последний период его относи-
тельного преуспевания на данной территории или период перехода в реликтовое
состояние) должен быть хотя бы очень приблизительно привязан к тому или иному
интервалу геологического времени, а в случае региональных исторических элемен-
тов - желательно также к одной из основных стадий формирования данной флоры
[стадиальный анализ в смысле А. И. Толмачева (1974)].
Соответственно целесообразно различать группы геохронологических* элемен-
тов (2.1Б.0.1 и 2.2Б.0.1) и группы стадиальных* элементов флоры (2.2Б.0.2).
Геохронологические элементы (как общие, так и региональные) принято при-
вязывать к стандартным геохронологическим подразделениям. Для растений -
вероятных спутников человека может быть использована археологическая шкала
возраста (палеолит, мезолит, неолит и еще подробнее, по культурам). Точность
возрастных оценок существенно выше и в случае хорошей обеспеченности геологи-
ческих разрезов, содержащих слои с макроостатками растений, датировками
абсолютного возраста”.
Стадиальный анализ флоры (Толмачев, 1957, 1974; Камелин, 1973) исходит
из того общего положения, что собственный возраст данной естественной регио-
нальной флоры определяется временем (периодом) формирования основы, или
ядра, флоры - характерного для нее сочетания элементов флоры (имеются в виду
флорогенетические элементы или флористические комплексы в их историко-типо-
логической трактовке). Отсюда основные категории стадиальных (всегда регио-
нальных!) элементов суть: 1) виды, существующие на данной территории с какого-
то времени до формирования ядра местной флоры современного типа, пережитки
предшествующих, более древних флор (пребазихронный*, пренуклеарный*,
палеореликтовый элемент*);1 2) виды, вошедшие в первичное ядро региональной
1 Они могут быть автохтонными или аллохтонными для данной территории.
261
флоры современного типа (базихронный*, нуклеарный*элемент); 3) виды, попол-
нившие местную флору уже после формирования ее ядра (постбазихронный*,
постнуклеарный* элемент). Их дальнейшее подразделение должно проводиться
на основе привязки к геохронологической шкале или крупным палеогеографи-
ческим событиям, оказавшим заметное влияние на развитие данной флоры. Оче-
видно, и в стадиальном анализе следует выделять как 4-ю группу новейший эле-
мент - пополнение флоры, связанное с интенсивной хозяйственной деятельностью
человека.
Что касается видов 2-й и 3-й группы, перешедших ныне в реликтовое состоя-
ние (судя по географическим и экологическим критериям; см. ниже), то их можно
именовать соответственно мезо- и неореликтовыми. Предполагается, что виды 1-й
группы должны, как правило, находиться в реликтовом состоянии. В том же
значении можно употреблять следующие термины: пребазихронные, базихронные
и постбазихронные реликты (то же в отношении эндемиков).
Классификация новейших пополнений флоры, связанных прямо или косвенно
с преобразованием растительного покрова деятельностью человека [их можно
противопоставить как антропогенный (ксеногенный, адвентивный) элемент сово-
купности аборигенных видов природной флоры], со значительной детальностью
и в разных аспектах разработана в ряде работ (Thellung, 1919; Holub, Jiiasek, 1961,
1968; Яброва-Колаковская, 1977), к которым мы и отсылаем заинтересованных
флористов. По времени заноса, в частности, различаются археофиты (занесены
в доисторическую эпоху), кенофиты (в историческое время, но до создания совре-
менной системы эффективных путей сообщения), эукенофиты (в последнюю эпоху).
Стадиальный анализ целесообразно проводить лишь для достаточно крупных
региональных флор.
2.Б.1. Географические характери-
стики (особенности) формирования
и расселения вида в целом (2.1Б.1) или
его региональной популяции (2.2Б.1).
Географо-генетические, географо
исторические, историко-географичес-
кие элементы флоры - общие и регио
нальные.
• быть получена, если мы разделим группы видов с единым типом
(принадлежащие к одному, достаточно крупному, географичес-
на подгруппы, принадлежащие каждая особому патрогенетич$с-
При классификации патрогенетических элементов могут быть
же основные принципы, что и при классификации географических
При восстановлении географической истории вида или его региональной попу-
ляции мы можем ограничить свою задачу выяснением родины (очага, или „центра”,
происхождения) вида или его местной популяции (хтоногенетические*, патрогене-
тические* элементы - общие: 2.1Б.1.1 или региональные: 2.2Б.1.1) или распро-
странить ее на географическую характеристику дальнейшего расселения вида (соб-
ственно географо-генетические элементы, дефинитивные географо-генетические*
элементы - общие: 2.1Б.1.2 или региональные: 2.2Б.1.2). Фактически 2-я группа
элементов может
распространения
кому элементу)
кому элементу,
использованы те :
(типологических) элементов, координатный, хориономический, релятивный; боль-
шое значение имеет анализ прерывистости распространения (особенно типов повто-
ряющихся дизъюнкций) и серий эквиформных ареалов или фрагментов ареалов.
Примером координатных патрогенетических единиц (в данном случае широт-
ных, зональных) могут служить арктогенные и альпигенные элементы в группе
видов с арктоальпийским типом распространения. В качестве примера хориономи-
ческих патрогенетических элементов приведем разграничение берингийских и анга-
ридских элементов или радиантов Северной и Южной Берингии. Релятивный прин-
цип широко используется при классификации миграционных (аллохтонных)
региональных патрогенетических элементов - по относительному расположению
источника (или источников) заселения территории флоры данным видом. Дизъ-
262
юнктивный характер распространения, если так или иначе может быть обоснована
его вторичность, обычно служит главным географическим критерием выявления
реликтовых элементов флоры (так называемых географических реликтов), проти-
вопоставляемых консервативным и прогрессивным элементам (Толмачев, 1974)
(подробнее см. ниже). Выделение реликтов правомерно как на общеземной шкале
(„общеземные реликты”, они же часто, но не всегда, и „таксономические релик-
ты” - рестанцы), так и на региональной: существуют реликтовые популяции
вполне процветающих видов.
Как и в других разделах анализа флоры, системы общих и региональных
элементов пересекаются в случае эндемичных видов (и других таксонов), системы
же общих и региональных патрогенетических элементов - в случае автохтонных эле-
ментов данной региональной флоры. Главные подразделения в системе региональ-
ных патрогенетических элементов (2.2Б.1.1.) - виды автохтонные [местного гене-
зиса; ортанты, в смысле Голуба и Йирасека (Holub, Jirasek, 1967,1968)] и аллохтон-
ные (иммигрировавшие из других фитохорий). Автохтонные виды могут быть
подразделены на эндемиков и радиантов, но это деление получает более полное
развитие уже в системе дефинитивных географо-генетических элементов, в которой
радианты данной фитохории (палеофитохории) могут быть подразделены по гео-
графическим результатам расселения за пределами территории их первоначального
становления.
Аллохтонные виды обычно отождествляют с миграционным элементом. Одна-
ко по существу и автохтонные неэндемичные виды мигрировали, хотя и в противо-
положном направлении. Точнее было бы деление на эмиграционный неэндемичный
(для автохтонов) и иммиграционный (для аллохтонов) элементы.
Стеббинс и Мэйджер (Stebbins, Major, 1965; см. : Программы флористических
исследований ..) предложили разделять эндемичные виды (на основе сравнитель-
ного анализа ареалов и отношений кратности хромосомных чисел этих видов
и их близких сородичей) на палеоэндемиков, схизоэндемиков, патроэндемиков
и апоэндемиков. Данная классификация является эйдолого-географо-генетической
региональной.
Следует предусмотреть подразделение автохтонных эндемичных видов и рас
на коренных (эуавтохтонных) * и вторичных (аллоавтохтонных) * эндемиков
в зависимости от того, принадлежат ли они эндемичной автохтонной таксономичес-
кой группе надвидового ранга или же являются местными дериватами аллохтон-
ных групп.
2.Б. (2, 3). Приуроченность форми-
рования и расселения вида 2.1Б. (2, 3)
или его региональной популяции
2.2 Б. (2, 3) к определенным классам
экотопов, биотопов, сообществ.
Экоценогенетические*, ценогене-
тические* (Зозулин, 1957, 1970а) эле-
менты флоры, эколого-ценотические
исторические элементы (исторические
свиты, экологические свиты видов,
историко-экологические комплексы
разных авторов; ценогенетические, це-
ногенные комплексы, ценоэлементы
во флорогенетической трактовке).
В принципе возможно раздельное выделение и рассмотрение экогенетических
(эколого-исторических, 2.1 Б.2. и 2.2Б.2; не путать с генэкологическими! ) и цено-
генетических (фитоценогенетических, филоценогенетических; фитоценоисторичес-
ких - 2.1Б.З. и 2.2Б.З) элементов флоры. Можно выделять также хороэкогенети-
ческие элементы (например, по отношению исходной популяции вида к термичес-
кой зональности или к степени континентальности климата) и топоэкогенетические.
Материал для распределения видов по ценогенетическим группам дает ана-
лиз современной приуроченности видов к определенным крупным синтаксонам,
263
их принадлежности к тем или иным (общим или региональным) флороценоти-
ческим и ценогеографическим (экоценогеографическим) комплексам (элементам
флоры). Положение упрощается в тех случаях, когда приуроченность вида
к определенному классу сообществ (например, степных, неморальнолесных,
темнохвойнотаежных) достаточно строгая и выдержана в пределах большей
части его ареала; однако судить о ценотической обстановке формирования
вида намного труднее, если даже в одном природном районе он обычен в самых
разных типах сообществ или если его ценотическая приуроченность существен-
но изменяется на протяжении ареала. В этих случаях приходится сравнивать
позиции и жизненность вида в разных типах сообществ (особенно в оптимальной
зоне его ареала), учитывать эколого-ценотические связи родственных таксонов,
сопоставлять имеющуюся палеогеографическую и палеоботаническую инфор-
мацию о природной истории разных, частей ареала вида. Кроме того, часть видов
(эксплеренты, пионерные, асоциальные виды) тяготеет к участкам с отсутству-
ющими или неустановившимися ценотическими отношениями и непосредственно
зависит от комплекса абиотических условий. Едва ли следует стремиться к стро-
гому разграничению экологических и фитоценотических исторических элементов
(разве что при определении первичного - исходного - фитоценотипа современ-
ных видов).
При выявлении региональных ценогенетических и экоценогенетических
элементов флоры ставится задача - выяснить эколого-ценотическую обстановку
формирования местной региональной популяции каждого вида, которая для
аллохтонных видов могла определяться эколого-ценотическими миграционны-
ми каналами (по каким экотопам, биотопам, в составе каких сообществ вид
впервые проник на территорию флоры), а для многих видов - условиями
переживания местной популяцией последнего критического для нее периода
(периодов).
Как и при выделении географо-генетических элементов, акцент может де-
латься как на первичных эколого-ценотических условиях формирования вида
или его местной популяции, так и на позднейших тенденциях (тренде) измене-
ния его эколого-ценотической амплитуды и экотопологической активности
(эколого-ценотические критерии разграничения реликтовых, консервативных
и прогрессивных элементов флоры). По этим признакам Я. П. Дидух (1982)
предложил деление видов флоры на экспансивные, преуспевающие, угасающие
и реликтовые.
Крупные, исторически более или менее целостные, ценотические комплексы
видов, обычно лишь частично представленные в региональных флорах (как одно-
именный ценогенетический элемент), заслуживают флорогенетического анализа
в полном объеме. При построении системы ценогенетических элементов, в значи-
тельной степени основанной на приуроченности видов к крупным классам сооб-
ществ, особого внимания заслуживают флороценотипы [в смысле П. Н. Овчин-
никова (1947, 1971) и Р. В. Кам ел ина (1973, 1979)] - флорогенетически более
или менее цельные региональные варианты эколого-физиономических (= эко-
лого-структурных) типов растительности; однако система флороценотипов пока
разработана лишь для отдельных территорий (Средняя Азия и некоторые сосед-
ние). Географическая определенность флороценотипов превращает основанные
на них ценогенетические элементы по существу в географо-ценогенетические
]2.1Б. (1.3)].
По своему названию некоторые категории ценогенетических элементов сход-
ны с определенными типологическими (не историческими!) элементами флоры,
что может привести (и очень часто приводит) к путанице и подмене понятий, если
не уточнять каждый раз категорию элемента (пример: многозначность термина
„неморальный”).
264
2.Б.4. Те или иные биологические
особенности формирования и истории
расселения вида (2.1Б.4) или его ре-
гиональной популяции (2.2Б.4)^
Историко-биологические*, биоло-
гические историко-типологические эле-
менты флоры — общие и региональные;
сюда же, по-видимому, правильнее
относить и эйдологические элементы
флоры, рассмотренные в конце раздела
„типологические элементы”.
Историко-биологический анализ флоры может проводиться по тем же показа-
телям, что и биологический (типологический) анализ, но с акцентом на биологи-
ческие механизмы становления и расселения видов и их региональных популяций.
Предметом анализа могут быть реконструируемая биоморфа и экобиоморфа
родоначальных популяций видов, их жизненная стратегия, скорость смены поколе-
ний, соотношение различных способов размножения, для аллохтонных видов -
способы и агенты проникновения на территорию флоры и т.д. Но особенно акту-
альной следует признать разработку принципов и методов эйдологического анализа
флоры (Камелин, в наст, сб.; Программы флористических исследований...).
Один из возможных подходов к использованию кариосистематических данных
для эйдологического анализа демонстрирует уже упоминавшаяся выше классифи-
кация Фаварже.
2.Б. (О, 1, 2, 3, ?4). Сочетание ос-
новных временных, географических
и эколого-ценотических (по возмож-
ности также эйдологических) характе-
ристик формирования и расселения
вида в целом [2.1Б. (О, 1, 2, 3, ?4)j
или его региональной популяции
[2.2Б. (О, 1, 2, 3,?4)].
Комплексные (многомерные, пол-
ные) исторические, историко-типологи-
ческие, генетические элементы флоры
(флорогенетические комплексы соб-
ственно) .
Разбиение флоры на части (элементы) по совокупности названных характе-
ристик формирования и расселения каждого вида представляет синтез (подводит
итог) всех предыдущих разделов и этапов анализа флоры, общей целью которых
является реконструкция истории данной флоры, ее формирования, развития,
обмена с другими естественными флорами. Речь, конечно, может идти лишь о доста-
точно крупных естественных флористических выделах и о достаточно широких
временных, географических и эколого-ценотических характеристиках.
Как уже говорилось, возможны 3 основных подхода к разделению флоры
на генетические (флорогенетические) элементы, в данном случае многомерные
(комплексного обоснования): 1) по особенностям происхождения, признакам
первичной - исходной - популяции вида (комплексные патрогенетические
элементы); 2) по тенденциям изменения географических, экологических и цено-
тических позиций, а также степени преуспевания (активности) и жизненности
вида в целом или его местной географической популяции на более поздних этапах
его истории (трендовые генетические элементы, всегда комплексного обоснования,
с тремя основными градациями: реликтовые, консервативные и прогрессивные
виды); 3) по особенностям происхождения и дальнейшей исторической судьбе
вида или его региональной популяции (дефинитивные генетические элементы
комплексного обоснования).
Однако чем полнее и детальнее мы учитываем историю вида (не только
по фрагментарным данным геологической летописи, но и по отражению этой исто-
рии в современном распределении и состоянии вида), тем больше обнаруживаем
в ней индивидуальных, неповторяющихся черт; а это затрудняет разграничение
дефинитивных генетических элементов и делает целесообразным параллельное
18 Заказ 1220
265
выделение первых двух категорий и их последующий сопряженный анализ. Так,
прогрессивные, консервативные и реликтовые элементы могут быть подразделены
далее по происхождению относящихся к ним видов, а патрогенетические элементы,
напротив, по их тренду на поздних этапах истории флоры.
Завершая на этом свой эскиз системы элементов флоры, авторы хотели бы
подчеркнуть, что их главной задачей было систематизировать основные подходы
к анализу флоры, расчленению ее на элементы - систематизировать основные
категории элементов флоры; задача же сведения синонимических терминов
во втором разделе отступила на задний план и в рамках данной работы не могла
быть осуществлена даже с элементарной полнотой. Более того, при явной избыточ-
ности обращающихся терминов для некоторых понятий целый ряд категорий
еще не отражен существующей терминологией, что привело ряд авторов к непро-
стой задаче заполнения пробелов: конструированию новых терминов.
Сознавая несколько предварительный характер настоящей разработки, авторы
выражают надежду, что она будет способствовать упорядочению системы понятий
и терминов сравнительной флористики и ботанической географии, преодолению
существующего разнобоя, принятию в недалеком будущем единой, унифициро-
ванной системы.
266
ЛИТЕРАТУРА
Аболин Р. И. Опыт эпигенологической классификации болот // Болотоведе-
ние. 1914. Вып. 3-4. С. 230-287.
Агапова Н. Д. Сравнительно-кариологическое исследование некоторых люти-
ков европейской части СССР // Ботан. журн. 1981. Т. 66, № 4. С. 483-493.
Агапова Н. Д. Кариосистематические заметки о некоторых кавказских видах
рода Ranunculus (Ranunculaceae) // Ботан. журн. 1983. Т. 68, № 10. С. 1351-1357.
Агапова Н. Д., Земскова Е. А. Исследование кариотипа Ranunculus polyanthe-
mos (Ranunculaceae) // Ботан. журн. 1983. Т. 68, № 3. С. 336-^341.
Айвазян С. А., Бежаева 3. И., Староверов О. В. Классификация многомерных
наблюдений. М.: Статистика, 1974. 240 с.
Андреев В. Л. Классификационные построения в экологии и систематике.
М.: Наука, 1980.142 с.
Андреев В. Н. Кормовая база Ямальского оленеводства // Сов. оленеводство.
1934. Вып. 1. С. 99-164.
Андрианов Б. В., Чебоксаров Н. Н. Хозяйственно-культурные типы и пробле-
мы их картографирования // Сов. этнография. 1972. № 2. С. 3-16.
Андронова Н. Н., Кученева Г. Г. Заметки к флоре Калининградской области //
Изученность природных ресурсов Калининградской области. Л., 1972. С. 105-110.
Андронова Н. Н., Гай сено к С Н., Кученева А. Е. Распространение редких
и исчезающих видов флоры Калининградской области на Самбийском полуост-
рове // Пути адаптации растений при интродукции на Севере. Петрозаводск, 1982.
С. 51-60.
Андронова Н. Н., Кученева Г. Г., Кученева А.Е. Распространение видов семей-
ства ятрышниковые в Калининградской области // Охрана и культивирование орхи-
дей. Киев, 1983. С. 16-18.
Баранова Е. В. Анализ флоры Псковской области: Автореф. дис.... канд.биол.
наук. Л., 1977. 15 с.
Баранова Е. В., Миняев Н. А., Шмидт В. М. Флористическое районирование
Псковской области на фитостатистической основе // Вести. ЛГУ. 1971. № 9. С. 30-40.
Баркалов В. Ю. Сосудистые растения островов Шумшу и Парамушир (север-
ные Курилы): Автореф. дис. ... канд. биол. наук. Томск, 1981. 25 с.
Беклемишев В. Н. Основные понятия биоценологии в приложении к живот-
ным компонентам наземных сообществ // Тр. по защите растений. 1931. Т. 1, вып. 2.
С. 278-358.
Беклемишев К. В. О природе биогеографических доказательств // Морская
биогеография. М., 1982. С. 5-11.
Берж К. Теория графов и ее применение. М.: ИЛ, 1962. 320 с.
Бешел Р. Флористические соотношения на островах Неарктики // Ботан. журн.
1969. Т. 54, №6. С. 872-891.
Биркмане К. Я., Юкне Я. Я. Видовой состав флоры // Флора и растительность
Латвийской ССР: Приморская низменность. Рига, 1974. С. 22-43. .
Биркмане К.Я., Клявиня Г. Б., Табака Л. В. и др. Видовой состав флоры //
Флора и растительность Латвийской ССР: Курземский геоботан. р-н. Рига, 1977.
С. 20-65
267
Боч М. С., Герасименко Т. В., Толчельников Ю. С. О некоторых особенностях
растительности и почв тундровой зоны Ямала // Изв. Всесоюз. геогр. о-ва. 1971.
Т. 103, вып. 6. С. 531-538.
Вальтер Г. Общая геоботаника. М.: Мир, 1982. 261 с.
Василевич В. И. Статистические методы в геоботанике. Л.: Наука, 1969. 231 с.
Василевич В. И. Ценоквант как наименьшая пространственная единица расти-
тельного покрова // Ботан. журн. 1973. Т. 58, № 9. С. 1241-1251.
Василевич В. И. Ассоциированность видового состава сообщества как мера
его организованности // Журн. общ. биологии. 1978. Т. 39, № 2. С. 160-166.
Василевич В. И. Почему существуют многовидовые растительные сообщес-
тва // Ботан. журн. 1979. Т. 64, № 3. С. 341-350.
Василевич В. И. Очерки теоретической фитоценологии. Л.: Наука, 1983. 248 с.
Василевич В. И. Классификация сероольшатников Северо-Запада европейской
части РСФСР// Ботан. журн. 1986. Т. 70, №6. С. 731-741.
Водопьянова Н. С. Зональность флоры Среднесибирского плоскогорья. Ново-
сибирск: Наука, 1984. 158 с.
Ворошилов В. Н., Скворцов А. К., Тихомиров В. Н. Определитель растений
Московской области. М.: Наука, 1966. 367 с.
Второв П. П. Подходы и методы современной синтетической биогеографии //
Современные проблемы зоогеографии. М., 1980. С. 31-60.
Второв П. П., Второва В. Н. Эталоны природы. М.: Мысль, 1983. 205 с.
lZ Вульф Е.В. Понятие „элемент флоры” в ботанической географии // Изв. Все-
союз. геогр. о-ва. 1941. Т. 73, вып. 2. С. 155-168.
Вынаев Г. В. Флорогенетическая структура и фитогедграфические связи фло-
ры Белоруссии: Автореф. дис. ... канд. биол. наук. Минск, 1984. 28 с.
Вынаев Г. В., Кожаринов А. В. Экотопологический и флорогенетический ас-
пекты изучения и охраны реликтовых видов растений и флоротопологических
комплексов. Минск, 1981. 13 с. Деп. в ВИНИТИ 28.05.1981, № 2527-81.
Высокогорная флора Станового нагорья. Новосибирск: Наука, 1972. 272 с.
Высоцкий Г. Н. Ергеня: Культурно-фитологический очерк // Тр. Бюро по
прикл. ботанике. 1915. Т. 8, № 10-11. С. 1113-1433.
Высоцкий Г, Н. Покрововедение // Зап. Белорус, ин-та сел. и лес. хоз-ва. 1925.
Вып. 4. С. 151-159.
Гагнидзе Р. И. Ботанико-географический анализ флороценотического комп-
лекса субальпийского высокотравья Кавказа. Тбилиси: Мецниереба, 1974. 226 с.
Гагнидзе Р. И. Эколого-ценотическая характеристика и анализ вертикального
распространения высокотравных видов Кавказа // Заметки по систематике и гео-
графии растений. Тбилиси, 1977. Вып. 34. С. 41—83.
Гагнидзе Р. И. Кариологическое исследование флороценотических комплексов
известняков Западного Закавказья // Заметки по систематике и географии расте-
ний. Тбилиси, 1983. Вып. 39. С. 21-38.
Гагнидзе Р. И. Значение наркологического метода для выяснения вопросов
истории флоры // Материалы межфак. конф, по естеств. наукам Тбил. ун-та. Тби-
лиси, 1984а. С. 86-88.
Гагнидзе Р, И, Кариогеография естественной флоры высокогорья Кавказа и
вопросы флорогенеза // Всесоюз, конф. „Растительный покров субарктических
высокогорий и проблема арктоальпийских флористических связей”: Тез. доки.
Апатиты, 19846, С, 42-50.
Гагнидзе Р. И, Флорогенез высокогорного кальцефильного флористического
комплекса Западного Закавказья // IX Всесоюз. совещ. по изучению, использова-
нию и охране растит, мира высокогорий: Тез. докл. Владивосток, 1985. С. 17-18.
Гагнидзе Р. И., Иванишвили М. А. Об элементе флоры и некоторых принципах
классификации ареалов // Изв. АН ГССР. Сер. биол. 1975а. Т. 1, № 3. С. 201-209.
\/ Гагнидзе Р. И., Иванишвили М. А. Некоторые характерные типы ареалов фло-
ры Кавказа // Изв. АН ГССР. Сер. биол. 19756. Т. 1, № 5-6. С. 373-389.
Гагнидзе Р. И., Чурадзе М. В. Числа хромосом некоторых видов секции
Chrysanthae рода Ranunculus (Ranunculaceae) Грузии // Ботан. журн. 1984. Т. 69,
№11. С. 1570-1571.
268
Гагнидзе Р. И., Чурадзе М. В. Числа хромосом кавказских видов рода Ranun-
culus секции Chrysanthae (Ranunculaceae)// Сообщ. АН ГССР. 1985. Т 117 № 2
С. 376-380.
Гаек Я., Шидак 3. Теория ранговых критериев. М.: Мир, 1971. 375 с.
Галанин А.В. Эколого-ценотические элементы конкретной флоры (их выделе-
ние и анализ) // Ботан. журн. 1973. Т. 58, № 11. С. 1608-1618.
Галанин А.В. Эколого-ценотические элементы флоры: Эколого-ценотический
анализ двух конкретных флор Чукотской тундры: Автореф. дис. ... канд. биол.
наук. Л., 1974. 27 с.
Галанин А. В. Ботанико-географические исследования в Калининградской
области в свете проблем охраны природы // Актуальные проблемы охраны приро-
ды. Иваново. 1977а. С. 121-125.
Галанин А. В. Анализ распределения растений по типам местообитаний (на при-
мере района равнинно-горного ландшафта среднего течения р. Раучуа Западной
Чукотки)// Ботан. журн. 19776. Т. 62, № 8. С. 1089-1101.
Галанин А. В. Экотопологическая структура флоры хребта Куркуре (Восточ-
ный Алтай) // Ботан. журн. 1979. Т. 64, № 10. С. 1401-1413.
Галанин А. В. Флора и растительность Усть-Чау некого биологического стацио-
нара (Западная Чукотка) // Ботан. журн. 1980. Т. 65, № 9. С. 1174-1187.
Галанин А. В., Алексеева И. А. Флористическая структура ландшафта дельты
Немана // Растительность Куршской косы и Балтийского побережья. Вильнюс,
1976. С. 76-78.
Галанин А. В., Марина Л. В. Ареалогический и эколого-ценотический анализ
флоры хребта Куркуре (Восточный Алтай) // Ботан. журн. 1982. Т. 67, № 5.
С. 590-598.
Голубев В. Н. Принцип построения и содержание линейной системы жизнен-
ных форм покрытосеменных растений // Бюл. МОИП. Отд. биол. 1972. Т. 77,
вып. 6. С. 72-80.
Городков Б. Н. Работы Гыданской экспедиции АН по пути к истокам р. Гы-
ды // Докл. АН СССР. 1928. С. 113-117.
Городков Б. Н. Тундры Обь-Енисейского водораздела // Сов. ботаника. 1944.
№ 3. С. 3-20.
Готт В. Q, Тюхтин В. С., Чудинов Э. М. Философские проблемы современного
естествознания. М.: Высш, шк., 1974. 264 с.
Гроссгейм А. А. Анализ флоры Кавказа. Баку: Изд-во Азерб. фил. АН СССР,
1936. 260 с.
Гроссгейм А. А. Рец. на кн.: Клеопов Ю. Д. Проект классификации географи-
ческих элементов для анализа флоры УССР // Изв. Азерб. фил. АН СССР. 1939.
№5. С. 145-148.
Дементьев В. А. Физико-географическое районирование // Белорусская Совет-
ская Социалистическая Республика. Минск, 1978. С. 45-46.
Дидух Я. П. К методике исследования горных флор // Актуальные вопросы
современной ботаники. Киев, 1977а. С. 115-118.
Дтдух Я. П. Систематична структура флори Ялтинського прсько-лг
сового державного заповщника // Укр. ботан. журн. 19776. ЧТ. 34, № 2.
С. 159-163.
Дидух Я. П. Флора Ялтинского горно-лесного государственного заповедника,
ее структурно-сравнительный анализ и научные вопросы охраны: Автореф. дис. ...
канд. биол. наук. Киев, 1977в. 25 с.
Д1дух Я. П- Бюморфолопчна структура флори Ялтинського прсько-л!сового
державного запов!дника // Укр. ботан. журн. 1978. Т. 35, № 5. С. 470-475.
Дидух Я. П. Вопросы фитоценотической активности видов орхидных горного
Крыма в связи с проблемой их охраны // Охрана и культивирование орхидей. Тал-
лин, 1980. С. 49-54.
Дидух Я. П. Проблемы активности видов растений // Ботан. журн. 1982. Т. 67,
№7. С. 925-935.
Дидух Я. П., Шеляг-Сосонко Ю. Р, Карадагский государственный заповед-
ник: растительный мир. Киев: Наук, думка, 1982. 151 с.
269
Дидух Я. П., Дюльдин А. А., Немченко О. А. К вопросу о методе выделения
флористических комплексов // Актуальные вопросы современной ботаники. Киев,
1977. С. 235-238.
Дюран Б., Оделл П. Кластерный анализ. М.: Статистика, 1977. 128 с.
Житков Б. М. Полуостров Ямал. СПб., 1913. 348 с.
Заки М. А., Шмидт В. М. О систематической структуре флор Южного Средизем-
номорья. 1-3 // Вести. ЛГУ. 1972. № 9. С.57-69; 1973. № 9. С.47-57; № 21. С. 44-53.
Заславская Т. М. Флора тундровой части Анюйского нагорья (Западная Чу-
котка) : Автореф. дис. ... канд. биол. наук. Л., 1982. 23 с.
Заславская Т. М., Петровский В. В. О флоре Люпвеемского горного массива
(Анюйское нагорье) // Ботан. журн. 1983.Т. 68, № 2. С. 162-174.
Зозулин Г. М. Исторические свиты растительности лесостепи Среднерусской
возвышенности // Делегат, съезд ВБО: Тез, докл. Л., 1957. Вып. 3. С. 15-22.
Зозулин Г. М. Анализ лесной растительности степной части бассейна р. Дона
в пределах Ростовской и Волгоградской областей.: Автореф. дис. ... докт. биол.
наук. Л., 1970а. 50 с.
Зозулин Г. М. Исторические свиты растительности // Ботан. жури. 19706,
Т. 55, № 1. С. 23-33.
Зозулин Г. М. Исторические свиты растительности европейской части СССР //
Ботан. журн. 1973. Т. 58, № 8. С. 1081-1092.
Золотухин Н. И. Метод конкретных флор для целей флористического рай-
онирования горных территорий // Нетрадиционные методы в исследованиях расти-
тельности Сибири. Новосибирск, 1982. С. 10-31.
Золотухин Н. И. Адвентивные растения на территории Алтайского заповедни-
ка // Ботан. журн. 1983. Т. 68, № 11. С. 1528-1533.
Золотухин Н. И. Границы фитохорий, ареалы видов и вопросы четвертичной
истории флоры Алтайского заповедника // История растительного покрова Север-
ной Азии. Новосибирск, 1984. С. 129-144.
Исаченко А. Г. Основы ландшафтоведения и физико-географическое райони-
рование. М.: Высш, шк., 1965. 327 с.
Исаченко А. Г. Физико-географическое районирование Ц БСЭ. 3-е изд. 1977.
Т. 27. С. 350-351.
\/ Камелии Р. В. О некоторых основных проблемах флорогенетики // Ботан.
журн. 1969. Т. 54, №6. С. 892-901.
Камелин Р. В. Флорогенетический анализ естественной флоры горной Средней
Азии. Л.: Наука, 1973. 356 с.
Камелин Р. В. Кухистанский округ горной Средней Азии. Л.: Наука, 1979.
117 с. (Комаров, чтения; Вып. 31).
\/ Камышев И. С. Основы географии растений: Флористическая география расте-
ний. Воронеж: Изд-во Воронеж, ун-та, 1961. 190 с.
Катенин А. Е. Геоботанические исследования на Чукотке. 1. Растительность
среднего течения реки Амгуэмы // Ботан. журн. 1974. Т. 59, № 11. С. 1583-1595.
Катенин А. Е. Структура растительного покрова территории Гильмимлиней-
ских термальных источников // Экосистемы термальных источников Чукотского
полуострова. Л., 1981. С. 41—77.
Кендэл М. Ранговые корреляции. М.: Статистика, 1975. 214 с.
Кирпичников М. Э., Забинкова Н. Н. Русско-латинский словарь для ботани-
ков. Л.: Наука, 1977. 855 с.
(Клаус К. К.) Claus К. Localfloren der Wolgagegenden. SPb., 1851. 312 S.
Клаус К. К. Флоры местные приволжских стран. СПб., 1852. 312 с.
Клеопов Ю. Д. Проект класиф(кащ1 географ!чних елеменпв для анал!зу
флори УРСР. и Журн. 1н-ту боташки АН УРСР. 1938. № 17. С. 209-219.
Клоков М. В. Расообразование в роде тимьянов - Thymus L. на территории
Советского Союза. Киев: Наук, думка, 1973. 190 с.
Кожевников Ю. П. Анализ флоры Телекайской рощи и ее окрестностей (Цент-
ральная Чукотка) // Ботан. журн. 1974. Т. 59, № 7. С. 967—979.
Козловская Н. В. К флористическому районированию Белоруссии // Ботан.
журн. 1974. Т. 59, № 6. С. 795-804.
270
Козловская Н. В., Парфенов В. И. Хорология флоры Белоруссии. Минск:
Наука и техника, 1972. 310 с.
Колаковский А. А. Растительный мир Колхиды. М.: Изд-во МОИП, 1961. 460с.
Комаров В. Л. Введение к флорам Китая и Монголии // Избр. соч. М.; Л.:
Изд-во АН СССР, 1947. Т. 2. С. 5-377.
Комарова Т. А., Семкин Б. И. Первичные сукцессии тундровых сообществ
долины реки А.мгуэма (Центральная Чукотка) // Комаровские чтения. Владиво-
сток, 1976. Вып. 24. С. 36-61.
Конспект флоры Псковской области. Л.: Изд-во ЛГУ, 1970. 176 с.
Конспект флоры Рязанской Мещеры. М.: Лесн. пром-сть, 1975. 328 с.
Королева А. С., Красноборов И. М., Пеньковская Е. Ф. Определитель расте-
ний Новосибирской области. Новосибирск: Наука, 1973. 386 с.
Коропачинский И. Ю. Древесные растения Сибири. Новосибирск: Наука,
1983. 384 с.
Красная книга СССР: Редкие и находящиеся под угрозой исчезновения виды
животных и растений. М.: Лесн. пром-сть, 1978. 460 с.
Красноборов И. М. Высокогорная флора Западного Саяна. Новосибирск:
Наука, 1976. 379 с.
Краснов А. Н. Опыт истории развития флоры южной части Восточного Тянь-
Шаня. СПб., 1888. V + 413 с. (Зап. Рус. reoip. об-ва; Т. 19.).
Крылов П. Н. Флора Западной Сибири. Томск: Изд-во Том. ун-та, 1927-1964.
Т. 1-12.
Куваев В. Б. Некоторые закономерности высотного распределения растений //
Ботан. журн. 1972. Т. 57, № 9. С. 1108-1115.
Кученева А. Е. О биологии и экологии Lunaria rediviva L. в Калининградской
области // Тез. докл. VU делегат, съезда ВБО. Л., 1983. С. 282-283.
Кучеиева А. Е., Андронова Н. Н., Кученева Г. Г. Редкие и исчезающие растения
Куршской косы II Охрана генофонда природной флоры. Новосибирск, 1983.
С. 69-71.
Лавренко Е. М. О некоторых основных задачах изучения географии и истории
растительного покрова субаридных и аридных районов СССР // Ботан. журн. 1965.
Т. 50, №9 . С. 1260-1267.
Лавренко Е. М., Свешникова В. М. Об основных направлениях изучения эко-
биоморф в растительном покрове // Основные проблемы современной геобота-
ники. Л.: Наука, 1968. С. 10-15.
Лазаренко А. С. Основы! засади класиф1кац1Гареал{в листяних Moxie Радянсь-
кого далекого сходу. // Укр. ботан. журн. 1956. Т. 13, № 1. С. 31-39.
Левин М. Г., Чебоксаров Н. Н. Понятие о хозяйственно-культурных типах и ис-
торико-этнографических областях // Очерки общей этнографии. М., 1957. С. 41-66.
Леонтьев В. Л. Некоторые особенности лесов Калининградской области //
Тр. БИН АН СССР. Сер. Ш. Геоботаника. 1956. Вып. 10. С. 330-372.
Литвинов Д. И. Список растений, дикорастущих в Калужской губернии. Калу-
га, 1895. 112 с.
\/ Лукичева А. Н., Сабуров Д. Н. Конкретная флора и флора ландшафта // Ботан.
журн. 1969. Т. 54, № 12. С. 1911-1920.
Лякавичюс А. А., Красаускас А.-И. И. Применение электронно-вычислительной
техники в гербарии Института ботаники АН Литовской ССР // Ботан. журн. 1977.
Т. 62, №4. С. 523-527.
Магулаев А. К). К цитогеографии некоторых Dipsacaceae Северного Кавказа//
Проблемы ботаники. Новосибирск, 1979. Т. 14, вып. 1. С. 67-70.
Маевский П. Ф. Флора средней полосы европейской части СССР. 9-е изд. Л.:
Колос, 1964. 880 с.
Майр Э. Популяции, виды и эволюция. М.: Мир, 1973. 328 с.
Макаревич В. Н. Применение метода Чекановского при первичной обработке
геоботанических описаний // Методы выделения растительных ассоциаций. Л.,
1971. С. 125-140.
Макарова С. И. Сравнение моделей описания зависимости числа видов во фло-
ре от размера площади // Ботан. журн. 1983. Т. 68, № 3. С. 376-381.
271
Малышев Л. И. Высокогорная флора Восточного Саяна. М.; Л.: Наука, 1965а.
368 с.
Малышев Л. И. Проблема генезиса высокогорной флоры Восточного Саяна:
Ареалогический обзор семейств. Иркутск: Вост.-Сиб. кн. изд-во, 19656. 119 с.
(Науч, чтения памяти М. Г. Попова; Чтение 8).
Малышев Л. И. Генетические связи высокогорных флор Южной Сибири и Мон-
голии // Изв. СО АН СССР. 1968. № 15. С. 23-31.
Малышев Л. И. Зависимость флористического богатства от внешних условий
и исторических факторов // Ботан. журн. 1969. Т. 54, № 8. С. 1137-1147.
Малышев Л. И. Площадь выявления флоры в сравнительно-флористических
исследованиях // Ботан. журн. 1972а. Т. 57, № 2. С. 182-197.
Малышев Л. И. Флористические спектры Советского Союза // История фло-
ры и растительности Евразии. Л., 19726. С. 17-40.
Малышев Л. И. Особенности и генезис флоры // Высокогорная флора Стано-
вого нагорья. Новосибирск: Наука. 1972в. С. 150-179.
V Малышев Л. И. Флористическое районирование на основе количественных
признаков // Ботан. журн. 1973. Т. 58, № 11. С. 1581-1588.
Малышев Л. И. Количественный анализ флоры: пространственное разнообра-
зие, уровень видового богатства и репрезентативность участков обследования //
Ботан. журн. 1975. Т. 60, № И. С. 1537-1550.
Малышев Л. И. Генезис высокогорных флор Сибири // Изв. СО АН СССР.
1976а. № 10. С. 47-55.
Малышев Л. И. Количественная характеристика флоры Путорана // Флора
Путорана. Новосибирск, 19766. С. 163-186.
Малышев Л. И. Электронная обработка данных в гербарном деле и флористи-
ке // Ботан. журн. 1977а. Т. 62, № 5. С. 713-732.
Малышев Л. И, Критерии подразделения растительного покрова и особенности
поясности в горах Северной Азии Ц Ботан. журн, 19776. Т. 62, № 10. С. 1393-1403.
Малышев Л. И. Генезис высокогорных флор Сибири и методы его изучения //
Фундаментальные исследования: Биолог, науки. Новосибирск, 1977в. С. 51-57.
Малышев Л. К. Эндемизм в высокогорных флорах Северной Азии // Ботан.
журн. 1979. Т. 64, № 4. С. 457-468.
Малышев Л. И. Изолированные охраняемые территории как ложноостровные
биоты // Журн. общ. биологии. 1980. Т. 41, № 3 . С. 338-349.
V Малышев Л. И. Изменение флор земного шара под влиянием антропогенного
давления // Биол. науки. 1981. № 3. С. 5-20.
Марина Л. В. Опыт сравнительного анализа высокогорных флор речных бассей-
нов хребта Куркуре (Восточный Алтай) // Ботан. журн. 1982. Т. 67, № 3. С. 285-292.
Марина Л. В. Сравнительный анализ двух высокогорных флор Восточного
Алтая (по их внутриландшафтным подразделениям) : Автореф. дис. ... канд. биол.
наук. Л., 1983. 23 с.
Марина Л. В. Сравнительный анализ экотопологической структуры флор
речных бассейнов (Восточный Алтай) // Ботан. журн. 1985. Т. 70, № 12.
С. 1658- 1664.
Мартыненко В. А., Шмидт В. М. Биометрическое сравнение бореальных кон-
кретных флор Коми АССР// Ботан. журн. 1981. Т. 66, № 3. С. 353-370.
Матвеева Е. П. Ботанико-кормовое районирование Калининградской обла-
сти // Тр. БИН АН СССР. Сер. Ш. Геоботаника. 1956. Вып. 10. С. 5-30.
Матвеева Е. П. Луга Советской Прибалтики. Л.: Наука, 1967. 335 с.
Матвеева Н. В., Заноха Л. Л. Флора сосудистых растений п. Кресты // Южные
тундры Таймыра. Л., 1986. С. 101-117.
Миркин Б. М. Анализ качественных признаков и структур. М.: Статистика,
1980. 320 с.
Михайличенко В. С. Есюз рослинности твючносхщноГ ократни швострова
Ямалу // Журн. 1н-ту ботан!ки УАН. 1936. № 7. С. 71-103.
Мякушко Т. Я., Сиренко И. П., Глаголева Н. Г., Мельиик С. К. Информацион-
но-поисковая система гербарной коллекции типовых образцов новоописаний
Н. С. Турчанинова // Укр. ботан. журн. 1981. Т. 38, № 4. С. 71-73.
272
^Наткевичайте-Иванаускеяе М. П., Тупчяускайте ю. Ю. Показатели хорологи-
ческой структуры флоры и использование их для раскрытия ее динамики // Ботан
журн. 1982. Т. 67, № 10. С. 1377-1385.
Николаев В. М., Зефиров Б. М. Флора Беловежской Пущи. Минск: Ураджай,
1971. 184 с.
Николаева М. Г. Кустарниковый тип растительности южной части Большого
и Малого Ямала // Ботан. журн. 1941. Т. 26, № 1. С. 52-86.
Ниценко А. А. Материалы к изучению растительного покрова Куршской косы
(Калининградская область) // Ботан. журн. 1970. Т. 55, № 4. С. 481-490.
Норин Б. Н. Использование коэффициентов сходства для классификации
микро группировок лесотундры // Методы выделения растительных ассоциаций.
Л., 1971. С. 206-225.
Овчинников Н. Ф. Категория структуры в науках о природе // Структура
и фо^мы материи. М., 1967. С. 11-47.
Овчинников П. Н. О принципах классификации растительности // Сообщ.
Тадж. фил. АН СССР. 1947. Вып. 11. С. 18-23.
VОвчинников П. Н. Ущелье р. Варзоб как один из участков ботанико-географи-
ческой области Древнего Средиземья // Флора и растительность ущелья р. Варзоб.
Л., 1971. С. 396-447.
Огуреева Г. Н. Ботаническая география Алтая. М.: Наука, 1980. 189 с.
Определитель высших растений Северо-Запада европейской части РСФСР.
Л.: Изд-во ЛГУ, 1981. 376 с.
Орловский С А. Проблемы принятия решений при нечеткой исходной инфор-
мации. М.: Наука, 1981. 208 с.
Пааль Я. Л., Семкин Б. И. Анализ растительности средней тайги при помощи
мер включения и сходства// Ботан. журн. 1983. Т. 68, № 10. С. 1341-1350.
Парницкнй Г. Основы статистической информатики. М.: Финансы и статистика,
1981. 199 с.
Пачоский И. К, Стадии развития флоры // Вести, естествознания. 1981. Вып. 8,
С. 261-270.
Песенко Ю. А. Концепция видового разнообразия и индексы, его измеряю-
щие // Журн. общ. биологии. 1978а. Т. 39, № 3. С. 380-393.
Песенко Ю. А. Сравнительный анализ индексов общности // Морфология,
систематика и эволюция животных. Л., 19786. С. 2-29.
Песенко Ю. А. Принципы и методы количественного анализа в фаунистичес-
ких исследованиях. М.: Наука, 1982. 288 с.
Петровский В. В., Плиева Т. В. К флоре Колымо-Анадырского водораздела //
Ботан. журн. 1986. Т. 71, № 10. С. 1354-1365.
Пешкова Г. А. Степная флора Байкальской Сибири. М.: Наука, 1972. 208 с.
Пнанка Э. Эволюционная экология. М.: Мир, 1981. 399 с.
Победимова Е. Г. Состав, распространение по районам и хозяйственное строе-
ние флоры Калининградской области // Тр. БИН АН СССР. Сер. III. Геоботаника.
1956. Т. 10. С. 225-329.
Попов М. Г. Основы флорогенетики. М.: Изд-во АН СССР, 1963. 135 с.
Пушкарь В. С., Короткий А. М., Семкин Б. И. Кластерный анализ флористи-
ческих и фитоценотических описаний ископаемых диатомовых сообществ голо-
цена Сихотэ-Алиня // Ботан. журн. 1984.14. 69, № 11. С. 1532-1542.
Работнов Т. А. Фитоценология. 2-е изд. М.: Изд-во МГУ, 1983. 292 с.
Разумовский С М. Закономерности динамики биоценозов. М.: Наука,
1981. 231 с.
Раменский Л. Г. О принципиальных установках, основных понятиях и терми-
нах производственной типологии земель, геоботаники и экологии // Сов. ботаника.
1935. №4. С. 25-42.
Раменский Л. Г. Введение в комплексное почвенно-геоботаническое исследо-
вание земель. М.: Огиз; Сельхозгиз, 1938. 620 с.
Раменский Л. Г., Цаценкин И. А., Чижиков О. Н., Антипин Н. А. Экологи-
ческая оценка кормовых угодий по растительному покрову. М.: Сельхозгиз,
1956. 472 с.
273
Расиныи А., Кученева Г., Андронова Н., Киреева Е. Редкие и исчезающие
виды растений Калининградской области // Ботанические сады Прибалтики:
Охрана растений. Рига, 1977. С. 113-116.
Ребристая О. В. Флора востока Большеземельской тундры: Автореф. дис. ...
канд. биол. наук. Л., 1971. 21 с.
Ребристая О. В. Флора востока Большеземельской тундры. Л.: Наука,
1977. 334 с.
Ребристая О. В., Шмидт В. М. Сравнение систематической структуры флор
методом ранговой корреляции // Ботан. журн. 1972. Т. 57, № 11. С. 1353-1364.
Рогович А. С. Обозрение семенных и высших споровых растений, входящих
в состав флоры Киевского учебного округа. Киев: Изд-во Киев, ун-та, 1869. 308 с.
Рокицкий П. Ф. Биологическая статистика. Минск: Вышэйш. шк., 1973. 320с.
V Сагателян А. А. Географический анализ флоры Мегринского района Армян-
ской ССР // Ботан. журн. 1981. Т. 66, № 5. С. 650-661.
Сагателян А. А. Флора и растительность Мегринского района Армянской ССР:
Автореф. дис*. ... канд. биол. наук. Ереван, 1983. 24 с.
Сагателян А. А., Файвуш Г. М. Сравнительный анализ флор Мегри и Ширака
(спектры семейств) // Биол. журн. Армении. 1982а. Т. 35, № 3. С. 208-213.
Сагателян А. А., Файвуш Г. М. Родовые спектры и географические элементы
флор Мегри и Ширака//Биол. журн. Армении. 19826. Т. 35, № 8. С. 661-667.
Сакало Д. I. Дея Ki зауважения до вивчення icropii crenie европейсько! час-
тики СРСР // Ботан. журн. АН УРСР, 1957. Т. 14, № 2. С. 25-35.
Самойлов Ю. И. Экологические шкалы Л. Г. Раменского и аспекты их приме-
нения // Ботан. журн. 1986. Т. 71, № 2. С. 137-147.
Саницкий П. П. Очерк флоры Калужской губернии // Тр. СПб. о-ва естество-
испытателей, 1884. Т. 14, вып. 2. С. 285-358.
Свидерский В. Н. О диалектике элементов и структуры в объективном мире
и познании. М.: Соцэкгиз, 1962. 275 с.
Седельников В. П. Флора и растительность высокогорий Кузнецкого Алатау.
Новосибирск: Наука, 1979. 168 с.
Седельников В. П. К применению мер включения в сравнительной флористи-
ке // Нетрадиционные методы в исследованиях растительности Сибири. Новоси-
бирск, 1982. С. 32-36.
Седельников В. П. Растительность высокогорий // Растительный покров
и естественные кормовые угодья Тувинской АССР. Новосибирск, 1985.
С. 48-68.
Семенова 3. Е. К изучению синантропной флоры города Калининграда // Эко-
логия растений южной тайги. Калинин, 1979. С. 97-99.
Семкин Б. И. О теоретико-множественных методах изучения растительных
сообществ // Тез. докл. V делегат, съезда ВБО. Киев. 1973а. С. 210-211.
Семкин Б. И. Дескриптивные множества и их приложения // Исследование
систем. 1. Анализ сложных систем. Владивосток, 19736. С. 83-94.
Семкии Б. И. Эквивалентность мер близости и иерархическая классификация
многомерных данных // Иерархические классификационные построения в геогра-
фической экологии и систематике. Владивосток, 1979. С. 97-112.
Семкин Б. И., Комарова Т. А. Анализ фитоценотических описаний с использо-
ванием мер включения (на примере растительных сообществ долины р. Амгуэмы
на Чукотке) // Ботан. журн. 1977. Т. 62, № 1. С. 54-63.
Семкин Б. И., Комарова Т. А. Использование мер включения при изучении
вторичных сукцессий (на примере послепожарных сообществ Южного Сихотэ-
Алиня) // Ботан. журн. 1985. Т. 70, № 1. С. 89-97.
Семкин Б. И., Куликова Л. С Методика математического анализа списков
видов насекомых в естественных и культурных биоценозах. Владивосток: ДВНЦ
АН СССР, 1981. 74 с.
Серебряков И. Г. Экологическая морфология растений. М.: Высш, шк., 1962.
378 с.
Симберлофф Д. С Теория островной биогеографии и организация охраняе-
мых территорий // Экология. 1982. № 4. С. 3-13.
274
Скворцов А. К. Река Угра - жемчужина среднерусской природы // Природа,
1980 № 9 С. 14-24.
Скворцов А. К. Микроэволюция и пути видообразования. М.: Знание, 1982. 58 с.
Соколова М. В. Опыт количественного сравнения 8 конкретных флор Тай-
мыра // Ботан. журн. 1984а. Т. 69, № 2. С. 211-217.
Соколова М. В. Количественное сравнение восьми конкретных флор Таймыра
по их таксономической структуре (арктическая Средняя Сибирь) // Ботан. журн.
19846. Т. 69, № 6. С. 840-849.
Соколова М. В. Сравнение географической структуры конкретных флор Тай-
мыра из тундровой зоны (арктическая Средняя Сибирь) // Ботан. журн. 1985.
Т. 70, №9. С. 1224-1232.
Соколовская А. П. Полиплоидия среди цветковых растений разных ланд-
шафтов СССР // Тр. Ленингр. о-ва естествоиспытателей. 1982. Т. 75, вып. 3. 126 с.
Сочара В. Б. Введение в учение о геосистемах. Новосибирск: Наука, 1978. 319 с.
Сочава В. Б. Растительный покров на тематических картах. Новосибирск:
Наука, 1979. 190 с.
Старобогатов Я. И. Проблема минимального выдела в биогеографии и ее при-
ложение к фаунистической (фауногенетической) зоогеографии моря // Морская
биогеография. М., 1982. С. 12-18.
Табака Л. В. Некоторые количественные и качественные показатели видового
состава флоры Приморской низменности // Фитогеографическая, флористическая
и геоботаническая характеристика приморской растительности. Вильнюс, 1976.
С. 138-140.
Табака Л. В., Клявиня Г. Б. Сравнительный анализ основных флористических
показателей некоторых речных долин Латвии // Охрана флоры речных долин в При-
балтийских республиках. Рига, 1983. С. 23-28.
Табака Л. В., Клявиня Г. Б., Плотниекс М. Р. Некоторые методические вопро-
сы изучения видового состава флоры Западной Латвии Ц Флора и растительность
Латвийской ССР : Курземский геоботан. р-н. Рига, 1977. С. 86-120.
Тахтаджян А. Л. Тектология: история и проблемы // Системные исследова-
ния. 1971. М„ 1972. С. 200-277.
Тахтаджян А. Л. Флористические области Земли. Л.: Наука, 1978. 248 с.
Тимофеев-Ресовский Н. В., Воронцов Н. Н., Яблоков А. В. Краткий очерк
теории эволюции. Л.: Наука, 1977. 301 с.
Толмачев А. И. К методике сравнительно-флористических исследований.
1. Понятие о флоре в сравнительной флористике // Журн. Рус. ботан. о-ва. 1931.
Т. 16, №1. С. 111-124.
Толмачев А. И. Флора центральной части Восточного Таймыра. Л., 1932. Ч. 1.
126 с. (Тр. Поляр, комис. АН СССР; Вып. 8); Ч. 2. 75 с. (Тр. Поляр, комис.
АН СССР; Вып. 13); 1935. Ч. 3. 80 с. (Тр. Поляр, комис. АН СССР; Вып. 25).
Толмачев А. И. О характере аркто-альпийских и некоторых локально связан-
ных с ними элементов флоры в районе Пинеги и Сотки // Материалы по истории
развития флор европейского севера СССР. Архангельск, 1938. С. 5-42.
Толмачев А. И. О количественной характеристике флор и флористических
областей. М.; Л.: Изд-во АН СССР, 1941. 40 с. (Тр. Сев. базы АН СССР; Вып. 8).
Толмачев А. И. Основные пути формирования растительности высокогорных
ландшафтов северного полушария // Ботан. журн. 1948. Т. 33, № 2. С. 161-180.
Толмачев А. И. О некоторых задачах и методах исторической фитогеогра-
фии // Ареал (картографические материалы по истории флоры и растительности).
М.; Л., 1952. Вып. 1. С. 7-12.
Толмачев А.И. Некоторые основные представления флорогенетики // Делегат,
съезд ВБО: Тез. докл. Л., 1957. Вып. 3. С. 44-49.
Толмачев А. И. Ареал вида и его развитие // Проблема вида в ботанике.
М.; Л., 1958. Т. 1. С. 293-316.
Толмачев А. И. Основы учения об ареалах: введение в хорологию растений.
Л.: Изд-во ЛГУ, 1962. 100 с.
Толмачев А. И. Богатство флор как объект сравнительного изучения // Вести.
ЛГУ. 1970а. №9. С. 71-83.
275
Толмачев А. И. О некоторых количественных соотношениях во флорах зем-
ного шара // Веста. ЛГУ. 19706. № 15. С. 62-74.
(Толмачев А. И.) Tolmatchew А. 1. Uber einige quantitative Wechselbeziechungen
der Floren der Erde // Feddes Repert. 1971. Bd. 82, N 5. S. 345-356.
Толмачев А. И. Введение в географию растений. Л.: Изд-во ЛГУ, 1974. 244 с.
Толмачев А. И. Методы сравнительной флористики и проблемы флорогенеза.
Новосибирск: Наука, 1986. 196 с.
Тумаджанов И. И., Беридзе Р. К. К кариологическому изучению представи-
телей верхнеалышйской аднивальной флоры Большого Кавказа // Ботан. журн.
1968. Т. 53, №1. С. 58-62.
Тупикова Н. В. Картографирование животного населения // Итоги науки
и техники. Биогеография. М. 1976. Т. 1. С. 99-218.
Уранов А. А. Возрастной спектр фитоценопопуляций как функция времени
и энергетических волновых процессов // Науч. докл. высш. шк. Биол. науки.
1975. №2. С. 7-33.
Файвуш Г. М. Анализ степной флоры Ширака (Армянская ССР) // Тез. докл.
VIII Всесоюз. совещ. по изучению и освоению высокогорий. Свердловск, 1982а.
Ч. 1. С. 45.
Файвуш Г. М. Анализ флоры Ширака // Тез. докл. республ. конф, молодых
науч, сотрудников и аспирантов, посвящ. 60-летию образования СССР. Ереван,
19826. С. 32-33.
Флеров А. Ф. Флора Калужской губернии. Калуга, 1907. Ч. 1. 435 с.; 1912.
Ч. 2. 264 с.
Флора и растительность Латвийской ССР: Курземский геоботан. р-н. Рига:
Зинатне, 1977. 177 с.
Флора и растительность Латвийской ССР: Северо-Видземский геоботан. р-н.
Рига: Зинатне, 1979, 165 с.
Флора и растительность Латвийской ССР: Юго-Вост, геоботан. р-н. Рига:
Зинатне, 1982. 195 с.
Флора и растительность Латвийской ССР: Вост.-Латв, геоботан. р-н. Рига:
Зинатне, 1985. 295 с.
Флора Путорана. Новосибирск: Наука, 1976. 246 с.
Флора Северо-Востока европейской части СССР / Ред. А. И. Толмачев. Л.:
Наука, 1974-1977. Т. 1-4.
Хантингам Д. А. Задачи, связанные с функциями распределения в кластер-
анализе // Классификация и кластер. М., 1980. С. 42-65.
Харадзе А. Л. О некоторых флорогенетических группах эндемов Большого
Кавказа // Проблемы ботаники. Л., 1974. Т. 12. С. 70-76.
Харадзе А. Л., Гагнидзе Р. И. Обзор гемиксерофильного эндемичного эле-
мента флоры Новороссийской подпровинции Кавказа // Заметки по систематике
и географии растений. Тбилиси. 1970. Вып. 28. С. 56-82.
Харадзе А. Л., Гвиннанидзе 3. И., Давлианидзе М. Т. К кариологическому
изучению субнивального флористического комплекса // Заметки по систематике
и географии растений. Тбилиси, 1973а. Вып. 30. С. 78-83.
Харадзе А. Л., Гвиннанидзе 3. И., Давлианидзе М. Т. К кариологиче-
скому изучению представителей флоры субнивального флористического
комплекса // Проблемы ботаники. Баку; Ленинград. 19736. Т. 13.
С. 38-43.
Хинтибидзе Л. С. К изучению кавказских представителей рода Betonica L. //
Тр. Тбил. ботан. ин-та. 1953. Т. 15. С. 167-187.
Цингер В. Я. Сборник сведений о флоре Средней России. М.: Изд-во Моск,
ун-та, 1886. 520 с.
Цыганов Д. Н. Фитоиндикация экологических режимов в подзоне хвойно-
широколиственных лесов. М.: Наука, 1983. 197 с.
Чернов Ю. И. Флора и фауна, растительность и животное население // Журн.
общ. биологии. 1984а. Т. 45, № 6. С. 732-748.
Чернов Ю. И. Эволюционный процесс и историческое развитие сообществ //
Фауногенез и филоценогенез. М.: Наука, 19846. С. 5-23.
276
Чопик В. I. Високопрна флора Украшських Карпат. Киев: Наук, думка, 1976.
268 с.
Чурадзе М. В. Заметки о некоторых видах рода Ranunculus L. флоры Кавка-
за // Заметки по систематике и географии растений. Тбилиси, 1981. Вып. 31. С. 5-9.
Чурадзе М. В. Таксономические заметки о комплексе вида Ranunculus radde-
anus Regel// Сообщ. АН ГССР. 1982. Т. 106, № 2. С. 373-376.
Чурадзе М. В. Критическая заметка о виде Ranunculus caucasicus Bieb. //Сообщ.
АН ГССР. 1984. Т. 115, № 2. С. 377-380.
Шварц С С. Экологические закономерности эволюции. М.: Наука, 1980. 277 с.
Шеляг-Сосонко Ю. Р. О конкретной флоре и методе конкретных флор// Ботан.
журн. 1980. Т. 65, № 6. С. 761-774.
Шеляг-Сосонко Ю. Р., Дидух Я. П. Очерк флоры и растительности Ялтинского
горно-лесного государственного заповедника: Флора // Ботан. журн. 1978.
Т. 63, №10. С. 1430-1439.
Шеляг-Сосонко Ю. Р., Дидух Я. П. Применение системно-структурного метода
при исследовании флор // Актуальные вопросы современной ботаники. Киев, 1979.
С. 3-11.
Шеляг-Сосонко Ю. Р., Дидух Я. П. Ялтинский горно-лесной государственный
заповедник. Киев: Наук, думка, 1980. 184 с.
Шеляг-Сосоико Ю. Р., Дидух Я. П., Жижин Н. П. Элементарная флора и пробле-
ма охраны видов // Ботан. журн. 1982. Т. 67, № 6. С. 842-852.
Шеляг-Сосонко Ю. Р., Дидух Я. П., Молчанов Е. Ф. Государственный заповед-
ник „Мыс Мартьян”. Киев: Наук, думка, 1985. 256 с.
Шилова С И. Флора островной Кунгурской лесостепи: Автореф. дис. ... канд.
биол. наук. Л., 1982. 18 с.
Шмидт В. М. О площади конкретной флоры // Веста. ЛГУ. 1972. № 3. С. 57-66.
Шмидт В. М. Количественные показатели в сравнительной флористике // Бо-
тан. журн. 1974а. Т. 59, № 7. С. 929-940.
Шмидт В. М. Количественная характеристика флоры Курильских островов //
Вести. ЛГУ. 19746. № 15. С. 55-63.
Шмидт В. М. Сравнение систематической структуры флор Хоккайдо, Сахали-
на, Камчатки, Курильских, Командорских и Алеутских островов // Ботан. журн.
1975. Т. 60, № 9. С. 1225-1236.
Шмидт В. М. О двух направлениях развитая метода конкретных флор // Бо-
тан. журн. 1976. Т. 61, № 12. С. 1658-1669.
Шмидт В. М. Географическая изменчивость флористических показателей
на территории европейской часта СССР // Проблемы ботаники и почвоведения.
Л., 1977. С. 39-62. (Тр. Ленингр. о-ва естествоиспытателей; Т. 73, вып. 3).
Шмидт В. М. Зависимость количественных показателей конкретных флор
европейской части СССР от географической широты // Ботан. журн. 1979. Т. 64,
№2. С. 172-183.
Шмидт В. М. Статистические методы в сравнительной флористике. Л.: Изд-во
ЛГУ, 1980. 176 с.
Шмидт В. М. Приближенное прогнозирование флористических показателей
в зависимости от географической широты // Журн. общ. биологии. 1981а. Т. 62,
№ 3. С. 430-439.
Шмидт В. М. О коэффициентах корреляции, используемых для сравнения
систематической структуры флор // Веста. ЛГУ. 19816. № 3. С. 57-67.
Шмидт В. М. Современные проблемы сравнительной флористики // Веста.
ЛГУ. 1981в. №21. С. 42-51.
* Шмидт В. М. Математические методы в ботанике. Л.: Изд-во ЛГУ, 1984. 288 с.
\/ Шмидт В. М., Баранова Е. В. Применение метода таксономического анализа
для сравнения флор по их историко-географическим связям // Журн. общ. биоло-
гии. 1975. Т. 36, № 4. С. 555-562.
Шмидт В. М., Ильминских Н. Г. О роли К. К. Клауса в разработке методов
сравнительной флористики // Ботан. журн. 1982. Т. 67, № 4. С. 462-470.
Шмидт В. М., Малышев Л. И. Информационные индексы сложности система-
тической структуры флор // Журн. общ. биологии. 1983. Т. 44, № 1. С. 100-107.
277
Шмидт В. М., Ребристая О. В. Рец. на кн.: Хеннинг Хеуплер. Статистический
анализ точечно-растровых карт распространения сосудистых растений южной части
Нижней Саксонии // Ботан. журн. 1977. Т. 62, № 9. С. 1388-1391.
(Эйларт Я.) Eilart J. Floraelement moistest taimegeografias // Изв. АН ЭССР,
1960. Т.9. Сер. биол. № 2. С. 97-110.
Экосистемы термальных источников Чукотского полуострова. Л.: Наука,
1981. 142 с.
Эндемичные высокогорные растения Северной Азии / Отв.ред. А. И. Толмачев.
Новосибирск: Наука, 1974. 336 с.
Юрцев Б. А. Гипоарктический ботанико-географический пояс и происхожде-
ние его флоры. М.; Л.: Наука, 1966. 94 с. (Комаровские чтения; Вып. 19).
Юрцер Б. А. Флора Сунтар-Хаята. Л.: Наука, 1968а. 236 с.
Юрцев Б. А. Общие и региональные вопросы изучения истории растительного
покрова // Основные проблемы современной геоботаники. Л., 19686. С. 82-94.
(Юрцев Б. A.) Yurtsev В. A. Phytogeography of northeastern Asia and the problem
of Transberingian floristic interrelations // Floristics and paleofloristics of Asia and eastern
North America. Amsterdam: Elsevier Publ. Co., 1972. P. 19-54.
Юрцев Б. А. Дискуссия на тему: „Метод конкретных флор в сравнительной
флористике” И Ботан. журн. 1974а. Т. 59, № 9. С. 1399-1407.
Юрцев Б. А. Проблемы ботанической географии Северо-Восточной Азии.
Л.: Наука, 19746. 160 с.
Юрцев Б. А. Некоторые тенденции развития метода конкретных флор // Бо-
тан. журн. 1975а. Т. 60, № 1. С. 69-83.
Юрцев Б. А. Координация исследовательских программ разной интенсивности
как подход к комплексному изучению биосферы // Изв. АН СССР. Сер. биол.
19756. №4. С. 618-623.
Юрцев Б. А, Проблемы позднекайнозойской палеогеографии Берингии в свете
ботанико-географических данных // Берингия в кайнозое. Владивосток, 1976а.
С. 101-120.
Юрцев Б. А. Общие и региональные вопросы флорогенетики // Ботан. журн.
19766. Т. 61, № 10. С. 1468-1478.
Юрцев Б. А. Ботанико-географическая характеристика Южной Чукотки //
Комаровские чтения. Владивосток, 1978. Вып. 26. С. 3-62.
Юрцев Б. А. Реликтовые степные комплексы Северо-Восточной Азии. Ново-
сибирск: Наука, 1981. 168 с.
Юрцев Б. А. Флора как природная система // Бюл. МОИП. Отд. биол. 1982а.
Т. 87, вып. 4. С. 3-22.
(Юрцев Б. A.) Yurtsev В. A. Relicts of the xerophyte vegetation of Beringia in
northeastern Asia // Paleoecology of Beringia. New York: Academic Press, 19826.
P. 157-177.
Юрцев Б. А. О количественной оценке „веса” видов при флористическом
районировании // Ботан. журн. 1983а. Т. 68, № 9. С. 1145-1152.
Юрцев Б. А. Географические популяции видов как объект флористики // Тез.
докл. VII делегат, съезда ВБО. Л., 19836. С. 58.
Юрцев Б. А., Петровский В. В. Об индикационном значении флористических
комплексов на северо-востоке СССР // Теоретические вопросы фитоиндикации.
Л., 1971. С. 15-31.
Юрцев Б. А., Семкин Б. И. Изучение конкретных и парциальных флор с по-
мощью математических методов // Ботан. журн. 1980. Т. 65, № 12. С. 1706-1718.
Юрцев Б. А., Толмачев А. И., Ребристая О. В. Флористическое ограничение
и разделение Арктики // Арктическая флористическая область. Л., 1978. С. 9-104.
Яброва-Колаковская В. С Адвентивная флора Абхазии. Тбилиси: Мецниереба,
1977. С. 3-63.
Abromeit J. Kurze Vegetationseskize von Ostpreussen. Konigsberg, 1895.
Abromeit J., Neuhoff W., Steffen H. Flora von Ost- und Westpreussen. Berlin,
1898-1940.
Atlas of the British flora / Ed. Perring F. H., Walters S. M. London: Nelson, 1962.
432 p.
278
I
$
t
Cadbury D. A., Hawkes J. G., Readett R. C. A computer mapped flora: A study of
the contry Warwickshire. London; New York: Academic Press, 1971.768 p.
Clayton W. D., Hepper E. N. Computer-aided chorology of West African grasses //
Kew BuU. 1974. Vol. 29, N 1. P. 213-234.
Clayton W. D., Panigrahi G. Computer-aided chorology of Indian grasses // Kew
BuU. 1974. Vol. 29, N 4. P. 669-686.
Clements F. E. Dynamics of vegetation / Select., comp, et ed. B. W. AUered
a. E. S. Clements. New York: Wilson, 1949. 296 p.
Clements F. E., Martin E. V., Long F. L. Adaptation and origin in the plant world:
The role of environment. Waltham, 1950. 332 p.
De Candolle A. Geographic botanique raisonnee. Paris; Geneve, 1855.1365 p.
Dony J. G. Species-area relationships. UK: Natural Environment Research CouncU,
1971.54 р.
Ehrendorfer F. Mediterran-Mitteleuropaische Florenbeziechungen in Lichte cyto-
taxonomischer Befunde // Feddes Repert. 1970. Bd 81, N 1. S. 3—32.
Ehrendorfer F. Polyploidy and distribution//Polyploidy.New York,1980.P.45-60.
EUenberg H. Zeigerwerte der Gefasspflanzen Mitteleuropas. Gfittingen: Goltze,
1974. 97 S. (Scripta Geobotanica; Bd 9).
Favarger C. Sur 1’emploi des nombres de chromosomes en geographic botanique
historique // Bericht Geobot. Inst. Rubel. 1961. H. 32. S. 119-146.
Favarger C. Cytologic et distribution des plantes // Biol. Rev. 1967. Vol. 2, N 2.
P. 163-206.
Fischer R. A., Corbet A. S., Williams С. B. The relation between the number of spe-
cies and the number of individuals in a random sample of an animal population // J. Ani-
mal Ecol. 1943. Vol. 12, N 1-2. P. 42-58.
Giiibert J. E. Flora lithuanica inchoata seu enumeratio plantarum quas circa Grod-
nam coUegit et determinavit loannes Emmanuel Giiibert. Grodnae, 1781. CoU. 1-3.
XVIII + 164 p.; Vilnae, 1782. CoU. 4-5. 267 p.
Grant V. Organismic evolution. San Francisco, 1977. Рус. пер.: Грант В. Эволюция
организмов. М.: Мир, 1980. 407 с.
Grant V. Plant speciation. New York: Columbia Univ. Press, 1981. Рус. пер.: Грант В.
Видообразование у растений. М.: Мир, 1984.528 с.
Grime J. Р. Plant strategies and vegetation processes. New York: WUley. 1979. 222 p.
Grinnell J. The niche-relationship of the California thrasher // Auk. 1917. Vol. 34,
N 2. P. 428-433.
Holub J., Jirasek V. Zur Vereinheitlichung der Terminologie in der Phytogeogra-
phie // Folia geobot. phytotaxon. (Praha). 1967. N 1. S. 69-113.
Holub J., Jirasek V. Beitrag zur arealonomischen Terminologie // Folia geobot. phy-
totaxon. (Praha). 1968. N 3. S. 275-339.
Hryniewiecki B. Tentamen florae Lithuaniae. Warszawa, 1933. XVI + 368 p.
Hulten E. Outline of the history of arctic and boreal biota during the Quaternary
period. Stockholm. 1937; 2nd ed. New York: Verlag von J. Cramer, 1972.168p. + 43 pl.
Juchacz-Nagy P. Some theoretical problems of synbotany. Pt 2: Preliminaries an
axiomatic model-buUding // Acta biol. Dembricina. 1966. N 4.
Kupfer Ph. Recherches sur les Uens de parente entre la flore orophile des alpes et
celle des Pyrenees // Boissiera. 1974. Vol. 23. P. 11-322.
Lance G. N., Williams W. T. A general theory of classificatory sorting strategies.
1. Hierarchical systems // Comp. J. 1967. Vol. 9, N 4. P. 373-380.
Lietuvos TSR flora. Vilnius, 1959-1980. T. 1-6.
McCoy E. D. The application of Island-Biogeographic theory to patches of habitat:
how much land is enough? // Biol. Conserv. 1983. Vol. 25, N 1. P. 53-62.
Meusel H., Jager E., Weinert E. Vergleichende Chorologie der Zentraleuropaischen
Flora. Text. Jena: VEB Gustav Fischer Verlag, 1965. 583 S.
Mowszowicz I. Conspectus florae Vilnensis. Lodz, 1958. T. 2.128 s.
Preston F. W. The canonical distribution of commonness and rarity // Ecology.
1962. Vol. 43,N 2. P. 185-215;N 3. P.410-431.
Reed T. M. The role of species area relationships in reserve choice: a British exam-
ple // Biol. Conserv. 1983. Vol. 25, N 3. P. 263-272.
279
Regel C. Studien iiber Florenelemente. 1 // Osterr. Bot. Zeitschr. 1958. Bd 104,
N 4/5. S. 552-573.
Regelis K. Lietuvos floros sKltiniai // Lietuvos un4o darbai. [Kaunas], 1931. T. 5,
d. 2. P. 221-289; 1932. T. 7, d. 1. P. 5-71; 1935. T. 9. d. 2. P. 183-222; 1936. T. 11,
d.5. P. 47-81; 1937. T. 11, d. 4. P. 67-84; 1939. T. 13, d. 2. P. 5-279; [Vilnius],
1942. T. 1 (14), d. 2. P. 389-458.
Reichert J. Die Pilzflora Agyptens // Englers Bot. Jachrb. 1921. Bd LVI.
S. 598-727.
Sauer W. Problem! attuali di citogeografia in Botanies // Infor. Bot. Italiano. 1979.
Vol. 11. P. 289-298.
Schow J. F. Grundziige einer aflgemeiner Pflanzengeographie. Berlin, 1823. 524 S.
Simberloff D. S., Abele L. G. Island biography theory and conservation practice //
Science. 1976. Vol. 191, N 4224. P. 285-286.
Stewinski W. Zielone jeziora pod Wilnem. Wilno, 1924. 232 p.
Snarskis P. Lietuvos rasakilos (Alchemilla) ir ji^ issiplatirumas. [Kaunas], 1939.
347 p. (Lietuvos un-to Matematikos-gamtos f-to darbai; T. 13, d. 2).
Snarskis P. Vadova&augalams paiinti. Velnius, 1954. 906 p.
Socal R. R., Sneath P. H. A. Principles of numerical taxonomy. San Francisco;
London: W. H. Freeman, 1963. 359 p.
Stebbins G. L. Flowering plants: (Evolution above the species level). Cambridge
Mass.: Harvard Univ. Press, 1977. 397 p.
Stebbins G. L. Polyploidy and the distribution of the arctic-alpine flora: New
evidence and new approach // Bot. Helvetica. 1984. Vol. 94, N 1. P. 1-13.
Stebbins G. L., Major J. Endemism and speciation in the California flora. // EcoL
Monographs. 1965. Vol. 35. P. 1-35.
Steffen H. Vegetationskunde von Ostpreussen. Jena:Verlag von Gustav Fischer,
1931.406 S. (Pflanzensoziologie; Bd 1).
Tamura S., Higuchi S., Тапаса К. Pattern classification based on fuzzy relation //
IEEE Trans. Syst. Sci. Cybem. 1971. Vol.SMC-1, N 1. P. 61-66.
Tansley A. G. The classification of vegetation and the concept of development //
J. Ecol. 1920. Vol. 8, N 2. P. 118-149.
Tansley A. G. The use and abuse of vegetational concepts and terms // J. Ecol.
1935. Vol. 16, N 3. P. 284-307.
Thellung A. Zur Terminologie der Adventiv- und Ruderalfloristik // Allgem. Bot.
Zeitschrift (Karlsruhe). 1918 (1919). Bd 24. S. 9-12.
Wangerin W. Florenelemente und Arealtypen: Beitrage zur Arealgeographie der
deutschen Flora // Beihefte zum Botan. Centralblatt. 1932. Bd XLIX. Erganzungsband.
S. 515-566.
280
СОДЕРЖАНИЕ
Предисловие......................................................... 3
Решение II рабочего совещания по сравнительной флористике......... 11
1. Флора как базовое понятие флористики
Юрцев Б. А. Флора как базовое понятие флористики: содержание понятия,
подходы к изучению............................................. 13
Вынаев Г. В. О понятии „флора" и задачах науки о флоре............. 28
Шеляг-Сосонко Ю. Р., Дидух Я. П. Системный подход к изучению флоры . . 30
Камелин Р. В. Процесс эволюции растений в природе и некоторые про-
блемы флористики............................................... 36
П. Методические проблемы современной флористики
1. Развитие метода конкретных флор за 1971-1985 гг.
Выбор опорных единиц флористических сравнений
Шмидт В. М. О роли А. И. Толмачева в развитии сравнительной флористики 43
Юрцев Б. А. Элементарные естественные флоры и опорные единицы срав-
нительной флористики ............................................ 47
Вынаев Г. В., Третьяков Д. И. Программа изучения флоры Белоруссии
методом эталонных проб........................................... $$
Ребристая О. В. Опыт применения метода конкретных флор в Западно-
сибирской Арктике (полуостров Ямал).............................. 57
Золотухин Н. И. Опыт флористических исследований на уровне фито-
хорий наименьшего ранга (на примере Алтайского заповедника) .... 99
Табака Л. В. Некоторые итоги сравнительного изучения флоры различных
природно-территориальных подразделений Латвии................... 104
Марина Л. В. Сравнительный анализ флор речных бассейнов и их экотопо-
логической структуры............................................. Ю7
Дидух Я. П. Опыт структурно-сравнительного анализа горных элемен-
тарных флор..................................................... 117
Седельников В. П. Ценотическая структура высокогорной флоры Алтае-
Саянской горной области......................................... 128
Сагателян А. А. Соотношение геоэлементов в парциальных флорах Мегри
и Ширака........................................................ 134
Мартыненко В. А. Анализ видового состава луговых сообществ в конкрет-
ных флорах таежной зоны европейского Севера..................... 136
2. Количественные методы анализа и сравнения флор
Малышев Л. И. Современные подходы к количественному анализу и срав-
нению флор...................................................... 142
Семкин Б. И. Теоретико-графовые методы в сравнительной флористике . . 149
Шмидт В. М. О некоторых приемах сравнения систематической структуры
флор............................................................ 163
19 Заказ 1220
281
Галанин А. В. Опыт сопряженного анализа типологических структур
конкретных флор .............................................. 167
Кученева А. Е. К методике оценки встречаемости и численности растений
в конкретных флорах......................................... 189
Лайвиныи М., Расиныи А., Калниня А. Классификатор высших сосудистых
растений для автоматизированной библиотеки данных по флористике
и геоботанике................................................. 195
Овеснов С. А. Опыт использования коэффициента Кендэла для сравнения
систематической структуры равнинных флор...................... 196
III. Некоторые итоги региональных флористических исследований
Лякавичюс А. А. Развитие флористических исследований в Литве..... 200
Скворцов А. К. Изучение флоры запада Нечерноземного центра РСФСР
(Брянской, Калужской и Смоленской областей)................... 203
Файвуш Г. М. Сравнение флор Верхне-Ахурянского и Ширакского флори-
стических районов (Северная Армения) ......................... 209
Казакова М. В. Вклад. А. И. Толмачева в решение проблемы реликтов
с точки зрения ботанической географии......................... 210
Гагнидзе Р. И. Вопросы флорогенеза некоторых высокогорных флороцено-
тических комплексов Кавказа по кариологическим данным ........ 213
IV. Программы флористических исследований
разной степени детальности
Общие задачи флористического изучения территории .................219
Программы маршрутных региональных исследований....................225
Программа изучения территории методом выборочных проб флоры (кон-
кретных, локальных флор) ......................................226
Программа сплошного флористического обследования территории: сеточ-
ное картирование флоры.........................................228
Программы изучения флоры заповедников и других крупных охраняемых
территорий.................................................... 230
Особенности изучения флоры горных территорий..................... 231
Особенности изучения флоры территорий, преобразованных деятельностью
человека...................................................... 234
Приложение. Программы изучения проб флоры разной детальности 237
Очерк системы основных понятий флористики........................ 242
Литература....................................................... 267
282
ТЕОРЕТИЧЕСКИЕ И МЕТОДИЧЕСКИЕ ПРОБЛЕМЫ
СРАВНИТЕЛЬНОЙ ФЛОРИСТИКИ
МАТЕРИАЛЫ П РАБОЧЕГО СОВЕЩАНИЯ
ПО СРАВНИТЕЛЬНОЙ ФЛОРИСТИКЕ. НЕРИНГА, 1983
Утверждено к печати
Научным советом по проблеме
„Биологические основы рационального использования,
преобразования и охраны растительного мира”
и Ботаническим институтом им. В. Л. Комарова Академии наук СССР
Редактор издательства Л. В. Дроздовская
Художник А. И. Слепушкин
Технический редактор В. В. Шиханова
Корректоры И. А. Корзинина и Г. А. Лебедева
ИБ № 33041
Набор выполнен в издательстве на наборно-печатающем автомате. Подписано
к печати 28.04.87. М.-17154. Формат 60 X 90 1/16. Бумага офсетная № 1. Печать
офсетная. Усл. печ. л. 18.50. Усл. кр.-от. 18.75. Уч.-изд. л. 22.41. Тираж 1250.
Тип. зак. № 1220. Дела 3 р. 50 к.
Издательство „Наука”. Ленинградское отделение.
199034, Ленинград, В-34, Менделеевская линия, 1.
Ордена Трудового Красного Знамени
Первая типография издательства „Наука”.
199034. Ленинград, В-34, 9 линия, 12.
283
УДК 581. 9. 001.11
Юрцев Б. А. Флора как базовое понятие флористики: содержание понятия, под-
ходы к изучению // Теоретические и методические проблемы сравнительной флори-
стики: Материалы II рабочего совещания по сравнительной флористике. Неринга,
1983. - Л.: Наука, 1987. - С. 13-28.
Концепция флоры как многовидовой территориальной совокупности и систе-
мы популяций поглощает традиционную трактовку ее как набора (списка) видов
и полнее отвечает современным состоянию и задачам науки о природных флорах.
Содержанием отношений между популяциями разных видов является распределе-
ние экониш разного уровня (вплоть до ландшафтных и региональных), что выявля-
ется с помощью специальных типов анализа региональных и парциальных флор.
Рассмотрено соотношение флористической и фитоценотической моделей раститель-
ного покрова, которые соответствуют разным уровням его организации. Показано,
что флоры экологически своеобразных подразделений ландшафта (парциальные
флоры) обнаруживают все существенные общие признаки флоры. Основой синтеза
флористической и фитоценотической информации должна стать концепция расти-
тельного покрова как иерархически дифференцированных многовидовых систем
популяций.
УДК 581. 9. 001.11
Вынаев Г. В. О понятии „флора” и задачах науки о флоре // Теоретические
и методические проблемы сравнительной флористики: Материалы II рабочего
совещания по сравнительной флористике. Неринга, 1983. - Л.: Наука, 1987. -
С. 28-30.
Поддерживается взгляд на флору как природную систему популяций видов
растений. Автор разделяет взгляд А. И. Толмачева на флору как региональное
явление, не ниже флоры ландшафта. Внутриландшафтные флористические образо-
вания определяются как флоротопологические комплексы. Дано развернутое опре-
деление понятия „флора”, отражающее ее системную природу, историческую и гео-
графо-экологическую обусловленность. Изучение естественных флор должно быть
предметом самостоятельной науки „флорологии”, перечислены задачи этой науки.
УДК 581. 9. 001.11
Шеляг-Сосонко Ю. Р., Дидух Я. П. Системный подход к изучению фло-
ры // Теоретические и методические проблемы сравнительной флористики: Мате-
риалы II рабочего совещания по сравнительной флористике. Неринга, 1983. -
Л.: Наука, 1987. - С. 30-36.
Флора и растительность рассматриваются как две самостоятельные системы
популяций, существующие в неразрывном единстве. Флористический аспект взаи-
моотношения популяций отражает качественное разнообразие видов. Низшей сту-
пенью территориальной иерархии флористических и фитоценотических систем
является фитоценоз Упорядоченность отношений между видами как элементами
флористических систем выражается в различных структурах флоры: систематичес-
кой, географической, биоморфологической, экологической, ценотической и др.
284
УДК581. 9 +575. 858
Камелин Р. В. Процесс эволюции растений в природе и некоторые проблемы
флористики// Теоретические и методические проблемы сравнительной флористики:
Материалы II рабочего совещания по сравнительной флористике. Неринга, 1983. -
Л.: Наука, 1987. - С. 36-42.
Рассматриваются некоторые нетрадиционные методы флористики, связанные
с подходом к флоре как арене биото (флоро-) генеза., филоценогенеза и видообра-
зования (филогенеза). Явления флорогенеза, филоценогенеза и филогенеза счита-
ются разными (по уровню организации) составляющими единого процесса эволю-
ции в реальной природной среде. Механизмов большей части этих явлений мы пока
не знаем, но можем учитывать их суммарный результат. Обсуждаются особенности
этих сторон процесса эволюции, проблема элементарных единиц и элементарных
явлений эволюции на разных уровнях организации живого и некоторые вопросы
классификации флор по эволюционным показателям.
УДК 581. 9
Шмидт В. М. О роли А. И. Толмачева в развитии сравнительной флористики //
Теоретические и методические проблемы сравнительной флористики: Материалы
II рабочего совещания по сравнительной флористике. Неринга, 1983. - Л.: Наука,
1987. - С. 43-46.
Показано, что А. И. Толмачев - автор широко применяемого ныне отечествен-
ными флористами метода конкретных флор - впервые обосновал принципы
сравнения флор в широком географическом диапазоне, продемонстрировал их
приложение на серии фактических примеров и по справедливости должен считаться
основоположником сравнительной флористики в нашей стране.
УДК 581. 9. 002
Юрцев Б. А. Элементарные естественные флоры н опорные единицы сравнитель-
ной флористики // Теоретические и методические проблемы сравнительной фло-
ристики: Материалы II рабочего совещания по сравнительной флористике. Неринга,
1983. - Л.: Наука, 1987. - С. 47-66.
Представлению о конкретных флорах как элементарных естественных флорах
регионального уровня соответствуют флоры дискретных (флористически одно-
родных) ландшафтов. В случаях когда протяженность таких ландшафтов меньше
площади выявления конкретной флоры, последняя должна обследоваться по всему
контуру, с поконтурным осмотром всех редких экотопов. Значительное варьиро-
вание набора видов на многих типах экотопов оставляет открытым вопрос
о критериях гомогенности конкретной флоры. Этой трудности не существует
для метода выборочных проб флоры, доказавшего свою эффективность при реше-
нии всех основных задач сравнительной флористики. Дилемма между желательной
повторностью и необходимой детальностью проб может быть разрешена путем
системы проб разной степени детальности. Тенденция дальнейшего развития мето-
дов флористики состоит в сближении маршрутного метода с таковым выборочных
проб и в постепенном переходе к системе регулярных проб.
285
УДК581. 9 (476)
Вынаев Г. В., Третьяков Д. И. Программа изучения флоры Белоруссии
методом эталонных проб // Теоретические и методические проблемы сравнитель-
ной флористики: Материал II рабочего совещания по сравнительной флористике.
Неринга, 1983. - Л.: Наука, 1987. - С. 66-67.
Излагаются основные теоретико-методические установки долгосрочной про-
граммы изучения флоры Белоруссии методом эталонных проб, закладываемых
в 32 природных районах Белоруссии. Существенное значение придается заложению
маршрутов (система азимутов) и фиксации флористических данных.
УДК581. 9 (571.121) : 57. 087.1
Ребристая О. В. Опыт применения метода конкретных флор в Западносибир-
ской Арктике (полуостров Ямал) // Теоретические и методические проблемы
сравнительной флористики: Материалы II рабочего совещания по сравнительной
флористике. Неринга, 1983. - Л.: Наука, 1987. - С. 67-90.
Описана методика изучения конкретных флор (КФ) путем постепенного
наращивания обследованной площади. Выявлена высокая флористическая репре-
зентативность небольших участков внутри КФ, определены ареал-минимум КФ,
равный 30 км2, и площадь контуров некоторых КФ для равнин Ямала. На основе
приводимых флористических списков проведено сравнение двух блоков КФ из раз-
личных подзон, показана высокая степень сходства КФ одной подзоны, рассмотре-
ны специфические черты и негативные отличия всех изученных флор. Рассчитаны
коэффициенты пространственного разнообразия флоры и удельный уровень видо-
вого богатства, наиболее низкие среди известных до сих пор для Арктической
Евразии.
Ил. 4. Табл. 15.
УДК 581. 9 (235. 222)
Золотухин Н. И. Опыт флористических исследований на уровне фитохорий
наименьшего ранга (на примере Алтайского заповедника) // Теоретические
и методические проблемы сравнительной флористики: Материалы II рабочего
совещания по сравнительной флористике. Неринга, 1983. - Л.: Наука, 1987. -
С. 90-104.
Проведен сравнительный количественный анализ флор 7 флористических
микрорайонов прителецкой части Алтайского заповедника, трактуемых как кон-
кретные флоры в смысле авторского определения А. И. Толмачева. Дана крити-
ческая оценка различных критериев дробного флористического районирования.
Несмотря на важность проведения траниц и сравнения микрорайонов с охватом
парциальных флор всех высотных поясов, в ряде случаев приходится выделять
одно-двупоясные флоры, используя критерий „наименьшего дополнения”. Выявле-
ны и показаны на картосхеме участки повышенной концентрации редких и особо
охраняемых видов на территории заповедника, в совокупности охватывающие
на 40 % его территории 97 % видов флоры заповедника.
Ил. 7. Табл. 5.
286
УДК 581. 9 (474. 3)
Табака Л. В. Некоторые итоги сравнительного изучения флоры различных
природно-территориальных подразделений Латвии // Теоретические и методические
проблемы сравнительной флористики: Материалы П рабочего совещания по сравни-
тельной флористике. Неринга, 1983. - Л.: Наука, 1987. - С. 104-107,
Изучение флоры Латвии проведено путем регистрации распределения видов
в сети квадратов площадью 70.7 км2 каждый. В каждом геоботаническом районе
Латвии изучались эталонные участки из 9 примыкающих малых квадратов.
Изучение хода прироста числа видов при суммировании квадратов показало, что
площадь выявления КФ в условиях Латвии равна 600-650 км3. Сеточное карти-
рование флоры целесообразно при проведении сравнительно-флористических
и хорологических исследований.
Ил. 1.
УДК 581. 9 (235. 222)
Марина Л. В. Сравнительный анализ флор речных бассейнов и их экотопологи-
ческой структуры // Теоретические и методические проблемы сравнительной
флористики: Материалы II рабочего совещания по сравнительной флористике.
Неринга, 1983. - Л.: Наука, 1987. - С. 107-117.
Проведено изучение высокогорных флор двух соседних хребтов Восточного
Алтая дифференцированно по внутриландшафтным подразделениям разного ранга
(речным бассейнам, типам и классам экотопов). Выявлено, что таксономическая,
географическая и высотно-зональная структуры флоры всего хребта выявляются
достаточно полно на уровне флор речных бассейнов, которые можно рекомендо-
вать в качестве опорных единиц при выявлении и сопоставлении флор различных
в ботанико-географическом отношении территорий. Попарное сравнение флор
трех модельных речных бассейнов дифференцированно по всем однотипным клас-
сам экотопов выявило невысокое сходство их парциальных флор даже в пределах
одного хребта, причем в некоторых экотопах различия касаются состава не только
редких, но также специфичных и константных для данного экотопа видов. Сравне-
ние парциальных флор всех классов экотопов в пределах каждого речного бассейна
показало достаточное своеобразие их экотопологической структуры.
Ил. 3. Табл. 1.
УДК 581. 9 (234. 86)
Д и д у х Я. П. Опыт структурно-сравнительного анализа горных элементарных
флор Ц Теоретические и методические проблемы сравнительной флористики:
Материалы II рабочего совещания по сравнительной флористике. Неринга, 1983. -
Л.: Наука, 1987. - С. 117-128.
Элементарные флоры в горных районах должны включать все пояса расти-
тельности, как правило, в пределах одного (не слишком мелкого) водосборного
бассейна. Непостоянство набора видов на однотипных экотопах заставляет сгущать
сеть маршрутов. При разграничении флор используются различные категории
дифференциальных элементов и другие показатели. На примере флоры Ялтинского
горнолесного заповедника рассматриваются основные этапы структурносравни-
тельного анализа горной флоры (выявление ценофлор с помощью количественного
сравнения „проб ценофлор”, количественное сопоставление ценофлор по система-
тической, географической и иным структурам) .
Ил. 7.
287
УДК581. 9 (571. 1)
Седельников В. П. Ценотическая структура высокогорной флоры Алтае-
Саянской горной области //Теоретические и методические проблемы сравнительной
флористики: Материалы П рабочего совещания по сравнительной флористике.
Неринга, 1983. - Л.: Наука, 1987. - С. 128-134.
Изложены результаты сравнения 16 высокогорных ценофлор с применением
методов кластерного анализа. На дендрограмме хорошо выделились три уровня
сходства ценофлор. Каждый из уровней представляет собой объединения парциаль-
ных флор соответственно мезотопического, макротопического и мегатопического
уровней. Рассмотрены причины, приведшие к дифференциации ценофлор.
Ил. 2. Табл. 1.
УДК581. 9 (479. 25)
Сагателян А. А. Соотношение геоэлементов в парциальных флорах Мегри
и Ширака // Теоретические и методические проблемы сравнительной флористики:
Материалы II рабочего совещания по сравнительной флористике. Неринга, 1983. -
Л.: Наука, 1987. - С. 134-135.
Дано сопоставление хорологических спектров парциальных флор (ПФ) двух
различных районов флоры Армении, выделяемых по фитоценотическим связям
тех или иных видов. Показано различие состава разных ПФ в каждой из флор
и однотипных ПФ - в обеих флорах. Последнее подтверждает отнесение этих флор
к разным ботанико-географическим выделам.
УДК581. 9 (470.13)
Мартыненко В. А. Анализ видового состава луговых сообществ в конкрет-
ных флорах таежной зоны европейского Севера // Теоретические и методические
проблемы сравнительной флористики: Материалы II рабочего совещания по срав-
нительной флористике.Неринга, 1983. - Л.: Наука, 1987. - С. 136-141.
Парциальные флоры (ПФ) пойменных луговых экотопов включают не менее
половины видового состава соответствующих конкретных флор (КФ) таежной
зоны европейского Севера. В отличие от КФ они не обнаруживают закономерного
снижения уровня видового богатства в направлении с юга на север в пределах
таежной зоны. ПФ имеют четко выраженный бореальный характер, как и КФ
в целом, а соотношение участия в них видов северных (гипоарктическая, аркто-
горная) и южных (дубравная, лесостепная) широтных групп меняется в зависимо-
сти от подзонального положения. Для предотвращения потери значительной части
генофонда природной флоры при сельскохозяйственном использовании пойм
необходимо предусматривать создание луговых заказников.
Табл.1.
УДК581. 9 :57. 087.1
Малышев Л. И. Современные подходы к количественному анализу и сравне-
нию флор // Теоретические и методические проблемы сравнительной флористики:
Материалы II рабочего совещания по сравнительной флористике. Неринга, 1983. -
Л.: Наука, 1987. - С. 142-148.
Основоположниками количественных методов в сравнительной флористике
являются А. Декандоль и К. К. Клаус. После их исследований в середине XIX в.
следовал длительный период накопления фактических данных. Пионером возрож -
288
дения интереса к количественным методам во флористике в последние десятилетия
был А. И. Толмачев наряду с учеными из франке- и англоязычных стран. В недавнее
время особенно большие успехи в развитии и применении этих методов достигнуты
в СССР. Разработана схема количественного анализа флоры по следующим парамет-
рам: 1) уровень видового богатства и пространственное разнообразие; 2) сравни-
тельное обилие видов растений в конкретных флорах и их поясных подразделе-
ниях; 3) сходство и различие по нумерическим данным; 4) поясно-зональная
и высотная структура; 5) таксономическая (систематическая) структура по се-
мейственным и родовым спектрам; 6) соотношение количества видов и родов;
7) флористическое районирование на основе количественных методов; 8) исполь-
зование количественных методов при анализе флоры для решения фундаменталь-
ных проблем ботанической географии и охраны растительного мира.
УДК581. 9 : 57. 087.1
Семкин Б. И. Теоретико-графовые методы в сравнительной флористике //
Теоретические и методические проблемы сравнительной флористики: Материалы
II рабочего совещания по сравнительной флористике. Неринга, 1983. - Л.: Наука,
1987.-С. 149-163.
Рассматриваются некоторые методы теории графов и отношений и их исполь-
зование для решения задач сравнительной флористики, обсуждаются преобразова-
ния исходной матрицы флористических данных (с конкретными примерами).
Показано, что при сравнении равновеликих флор достаточно использовать метод
дендрита и метод взвешенного среднего арифметического связывания, при сравне-
нии разновеликих флор необходим анализ включения флор с помощью ориентиро-
ванных графов.
Ил. 7. Табл. 8.
УДК581. 9 :57.087.1
Шмидт В. М. О некоторых приемах сравнения систематической структуры
флор //Теоретические и методические проблемы сравнительной флористики:
Материалы II рабочего совещания по сравнительной флористике. Неринга, 1983. -
Л.: Наука, 1987. - С. 163-167.
Рассматриваются математические способы сравнения систематической струк-
туры флор. При сравнении головной части флористических спектров рекоменду-
ется пользоваться выведенными автором удобными рабочими формулами для вы-
числения тождественных значений (коэффициента ранговой корреляции Спирмена
или коэффициента корреляции Бравэ—Пирсона). Для сравнения полных флористи-
ческих спектров предлагается использовать информационную Я-функцию.
УДК 581. 9. 002
Галанин А. В. Опыт сопряженного анализа типологических структур конкрет-
ных флор // Теоретические и методические проблемы сравнительной флористики:
Материалы II рабочего совещания по сравнительной флористике. Неринга, 1983. -
Л.: Наука, 1987. - С. 167-189.
Предлагается и иллюстрируется на материале изученных автором конкретных
флор метод сопряженного анализа различных типологических структур флоры,
со статистической оценкой сопряженности распределения видов по широтным и
долготным группам, эколого-ценотическим и ареалогическим (широтным и дол-
готным). Выявленные сопряженности свидетельствуют о целостности изученных
289
флор как систем видов (популяций) и интерпретируются в свете различных фло-
рогенетических гипотез.
Табл. 13.
УДК581. 9 (470. 26)
Кученева А. Е. К методике оценки встречаемости и численности растений
в конкретных флорах //Теоретические и методические проблемы сравнительной
флористики: Материалы П рабочего совещания по сравнительной флористике.
Неринга, 1983. - Л.: Наука, 1987. - С. 189-195.
Проведена повторная (через 100 лет) инвентаризация флоры Калинин-
градской обл. с использованием показателей встречаемости и обилия. При-
веденные в статье данные о состоянии местных популяций видов семейства
ятрышниковых и некоторых редких видов свидетельствуют о негативных
изменениях в популяциях вплоть до полного исчезновения некоторых видов.
Ряд эвритопных видов, а также некоторые псаммофиты не претерпели изменений
в составе флоры.
Ил. 1. Табл. 2.
УДК581. 9 (474. 3)
Лайвиньш М., Расиньш А., Калниня А. Классификатор высших
сосудистых растений для автоматизированной библиотеки данных по флористике
и геоботанике //Теоретические и методические проблемы сравнительной флористи-
ки: Материалы II рабочего совещания по сравнительной флористике. Неринга,
1983. - Л.: Наука, 1987. - С. 195-196.
Описана информация, содержащаяся в классификаторе CLASIF-001, созданном
для высших сосудистых растений Латвии и сопредельных территорий - соподчинен-
ная система таксонов, записанная в словесном виде, цифровая характеристика
таксонов и принадлежность видов к 4 категориям: адвентивные, сорняки, куль-
турные и натурализовавшиеся растения.
УДК581. 9 : 57. 087.1
Овеснов С. А. Опыт использования коэффициента Кендэла для сравнения
систематической структуры равнинных флор // Теоретические и методические
проблемы сравнительной флористики: Материалы П рабочего совещания по сравни-
тельной флористике. Неринга, 1983. - Л.: Наука, 1987. - С. 196-199.
Флора юга Пермской области, изученная автором, сравнивается с флорами
более или менее сопоставимых по размерам и уровню флористического богатства
других территорий Восточно-Европейской равнины с помощью двух показателей:
общность видового состава (по Жаккару), сходство систематической структуры
на уровне семейств и родов (коэффициент ранговой корреляции Кендэла). Пока-
зано, что первый показатель точнее отражает положение сравниваемых флор
в системе фитохорий, второй показатель существенно зависит от зонального
(состав ведущих семейств) или долготно-провинциального (состав ведущих
родов) положения сравниваемых флор и имеет ограниченную ценность для срав-
нения фитохорий невысокого ранга.
Ил. 2.
290
УДК581. 9 (474. 5)
Лякавичюс А. А. Развитие флористических исследований в Литве //Теорети-
ческие и методические проблемы сравнительной флористики: Материалы П рабо-
чего совещания по сравнительной флористике. Неринга, 1983. - Л.: Наука, 1987. -
С. 200-203.
Рассматриваются основные исторические этапы исследования видового сос-
тава высших растений Литвы. Качественно новым этапом изучения флоры Литвы
считается работа коллектива флористов республики, подготовивших к изданию
капитальный шеститомный труд „Флора Литовской ССР”, завершенный в 1980 г.
В настоящее время осуществляется детальное исследование флоры ряда райо-
нов Литвы с учетом не только качественных, но и количественных показателей.
Приводится формула для оценки активности видов на определенной территории
(или для отдельных фитохорий), по которой определялась активность видов
флоры некоторых заказников Литвы,
УДК581. 9 (470. 33)
Скворцов А. К. Изучение флоры запада Нечерноземного центра РСФСР (Брян-
ской, Калужской и Смоленской областей) // Теоретические и методические про-
блемы сравнительной флористики: Материалы II рабочего совещания по сравни-
тельной флористике. Неринга, 1983. - Л.: Наука, 1987. - С. 203-209.
Охарактеризована степень изученности трех областей Нечерноземного центра
РСФСР, поставлена задача создания „Флоры” этой территории, целостной в ботани-
ко-географическом отношении. Накопленные за последнее десятилетие материалы
позволяют уточнить главным образом распространение редких и важных в ботани-
ко-географическом отношении видов, также новых для той или иной области. При-
ведены сведения о наиболее интересных флористических находках на территории
изученного’ района. Общая флористическая изученность явно недостаточна, требу-
ются усилия коллективов ботаников, работающих в педагогических вузах и уни-
верситетах этого региона с привлечением студентов во время полевой практики.
УДК581. 9 (479. 25)
Файвуш Г. М. Сравнение флор Верхне-Ахурянского и Ширакского флористи-
ческих районов (Северная Армения) // Теоретические и методические проблемы
сравнительной флористики: Материалы II рабочего совещания по сравнительной
флористике. Неринга, 1983. - Л.: Наука, 1987. - С. 209—210.
Проведено сравнение флор двух элементарных флористических районов,
прилегающих друг к другу на севере Армении^по показателям таксономического
спектра, географии видов и составу географических элементов, что дает возмож-
ность уточнить южную границу Кавказской провинции и северную - Армено-
Иранской провинции.
УДК 581. 9
Казакова М. В. Вклад А. И. Толмачева в решение проблемы реликтов с точки
зрения ботанической географии // Теоретические и методические проблемы срав-
нительной флористики: Материалы II рабочего совещания по сравнительной фло-
ристике. Неринга, 1983. - Л.: Наука, 1987. - С. 210-212.
Вслед за А. И. Толмачевым автор относит к реликтам только те виды, природа
которых находится в несоответствии с современными условиями. Проанализи-
291
рована группа из 35 видов флоры Липецкой обл., традиционно относимая к релик-
товым. Большинство видов следует исключить из категории реликтовых как
имеющих островное положение на границе ареала и являющихся активным эле-
ментом во флоре.
УДК 581. 9 (324. 9)
Гагнидзе Р. И. Вопросы флорогеиеза некоторых высокогорных флороценоти-
ческих комплексов Кавказа по кариологическим данным // Теоретические и мето-
дические проблемы сравнительной флористики: Материалы II рабочего совещания
по сравнительной флористике. Неринга, 1983. - Л.: Наука, 1987. - С. 213-218.
Приведены результаты кариосистематического изучения высокогорных видов
Кавказа и Западного Закавказья. Среди них отмечается низкий процент полиплои-
дов Рассмотрены различные случаи кариосистематической дифференциации ряда
систематических групп, со значительной долей эндемиков, в связи с вопросами
их происхождения и экологии.
Ил. 4.
292
Издательство ,Д1аука” готовит к выпуску
в III квартале 1987 года книгу
БОБКОВА К. С.
БИОЛОГИЧЕСКАЯ ПРОДУКТИВНОСТЬ ХВОЙНЫХ ЛЕСОВ
ЕВРОПЕЙСКОГО СЕВЕРО-ВОСТОКА
Объем 10 л. Цена 1 р. 60 к.
План выпуска 1987 года, III кв., № 324
В монографии представлены материалы многолетних стационарных
исследований хвойных фитоценозов северной и средней подзон европей-
ской тайги. Показаны особенности структурной организации раститель-
ности в основных типах сосновых и еловых лесов и их первичная про-
дуктивность. Дан анализ основных почвенно-экологических факторов,
определяющих био продукционный процесс. Особое внимание уделено
исследованиям эколого-физиологических аспектов жизнедеятельности
подземных органов сосны и ели.
Для ботаников, лесоведов, работников лесного хозяйства.
Для получения книги почтой заказы просим направлять по адресу:
197345, Ленинград, Петрозаводская ул., 7.
Магазин „Книга— почтой” Северо-Западной конторы „Академкнига”,
ЗАЯВКИ на данную книгу принимаются до 20 мая 1987 года.
293