




































































































































































































































Текст
экология НАШЕСТВИЙ жи вотн ых и РАСТЕНИЙ
Ч. ЭЛТОН экология НАШЕСТВИИ ЖИВОТНЫХ И РАСТЕНИЙ
THE ECOLOGY OF INVASIONS BY ANIMALS AND PLANTS by CHARLES S. ELTON
METHUEN AND GO LTD London 1958
Ч. ЭЛТОН
экология НАШЕСТВИЙ животных И РАСТЕНИЙ
Перевод с английского Ю. И. Лашкевича
Под редакцией и с предисловием проф. Н. П. Наумова
ИЗДАТЕЛЬСТВО
ИНОСТРАННОЙ ЛИТЕРАТУРЫ
АННОТАЦИЯ
Книга посвящена проблеме массового распространения и «вспышек размножения» чужеземных видов животных и растений и отчасти бактерий и вирусов после их случайного проникновения или преднамеренного завоза в страну или на отдельные участки местности, где их раньше не было. Подробно рассматриваются взаимоотношения пришлых чужеземных видов с местной фауной и флорой и в связи с этим вопросы структуры и динамики популяций отдельных видов, ценозов и природного комплекса в целом. Значительное место отводится освещению экономических и других последствий случайного завоза сельскохозяйственных и лесных вредителей. В заключительных главах автор пытается найти рациональные пути поддержания относительной стабильности популяций животных и растений, а также подавления их размножения.
Книга предназначена для зоологов и ботаников, в частности для работников эпидемиологических и противочумных станций и станций защиты растений.
Редакция биологической литературы
Чарльз С. Элтон, профессор Оксфордского университета, член Королевского общества (Академия наук Англии),—один из виднейших зоологов мира, своими работами оказавший большое влияние на развитие экологии животных. Его книги «Экология животных» («Animal Ecology», 1927, 1930), «Полевки, мыши и лемминги, очерки по динамике популяции» («Voles, Mice and Lemmings», 1942), «Экология животных и эволюция» («Animal Ecology and Evolution», 1930) и небольшая «Экология животных» («Ecology of Animals»), в 1934 г. переведенная на русский язык под редакцией проф. Д. Н. Кашкарова, так же как и многочисленные статьи в редактируемом Элтоном английском журнале Journal of Animal Ecology и других изданиях, свидетельствуют об очень широком диапазоне его интересов. Тут и вопросы внутривидовых, в частности территориальных, отношений, и роль заболеваний в жизни популяций, и вопросы биоценологии и географии животных, проблемы эволюции и многие другие. Все же основное внимание Элтон всегда уделял и уделяет сейчас самой важной и основной проблеме современной экологии животных — проблеме численности популяций и их динамики. Ч. Элтона с полным правом можно назвать пионером в разработке очень многих вопросов, связанных с этой проблемой, имеющей не только выдающийся теоретический интерес, но и огромное практическое значение.
Элтона в равной степени интересуют обе стороны проблемы. Он — организатор и руководитель Бюро популяций животных в Оксфорде (Bureau of Animal Population), представляющего собой один из наиболее крупных международных центров по изучению популяций животных и динамики их численности.
Последняя книга Элтона («The Ecology of Invasions by Animals and Plants», 1958), русский перевод которой предлагается советским читателям, посвящена исключительно интересному и важному вопросу о расселении и «нашествиях» живых существ. Это более или менее полная и хорошо документированная сводка данных об изменениях ареалов ряда видов животных и растений, о «нашествиях чужеземцев», во многих случаях причиняющих огромный ущерб хозяйству или здоровью людей.
Это в то же время живой рассказ о победах, к сожалению еще немногих, над «захватчиками», рассказ, полный упреков тем, кто по незнанию, а иногда и умышленно игнорировал целительные силы самой природы и тем самым усугублял бедствие. Книга Элтона — страстный призыв к изучению, охране и активному использованию этих сил с целью избежать в будущем еще больших неприятностей. В этом основной смысл и интерес единственной в своем роде сводки, длительно подготовлявшейся и опубликованной Ч. Элтоном в знаменательный для биологов 1958 год.
Зоологам и ботаникам хорошо известны случаи как быстрого увеличения численности и подчас очень широкого расселения отдельных видов животных и растений, так и, наоборот, сокращения их ареалов и исчезновения видов во многих местах прежнего обитания. Отдельные примеры того и другого рода разбросаны в специальной литературе и обычно приводятся в зоогеографических и фитогеографических сводках и руководствах. Они особенно многочисленны в литературе по борьбе с вредителями сельского хозяйства и переносчиками болезней. Во многих случаях появление или исчезновение животных и растений объясняется изменением природной обстановки. Однако все большее влияние на это оказывает деятельность человека, все более сокращающего расстояние между (разными точками земной поверхности и разрушающего преграды, бывшие ранее непреодолимыми.
Хотя в общих чертах явление расселения и нашествий животных и растений было известно, книга Ч. Элтона производит на читателя огромное впечатление. Это объясняется тем, что даже известные ранее отдельные факты, соединенные вместе рукой мастера, превратились в яркую картину, поражающую размахом явле-
нпя и темпами его развития и, несомненно, заставляющую глубоко задуматься над возможными последствиями этого явления. Автор вскрывает неосознанные, а иногда и сознательно допущенные ошибки, вызвавшие губительные нашествия вредителей, и горячо желает предотвратить их повторение. Хотя в книге Ч. Элтона нет упоминания о том, что она связана с юбилеем эволюционного учения Ч. Дарвина и идей А. Уоллеса, она вышла удивительно ко времени. Ч. Элтон и начинает изложение с тех изменений, которые за сравнительно небольшой промежуток времени произошли в так называемых уоллесовских биогеографических областях, основная фауна и флора которых сложилась еще в третичное время и отличалась высокой специфичностью (главы I и II).
В последующих главах (III—V) подробно описаны эти изменения видового состава фауны и флоры — значительное уменьшение степени специфичности, а иногда и эндемизма, свойственного разным областям.
В завершающих главах не только разобран механизм возникновения подъема численности, или «экологического взрыва», и сопровождающего его «нашествия» (главы VI и VII), но и показаны пути и способы, с помощью которых можно было бы предотвратить эти нашествия вредителей или по крайней мере резко ограничить их размах и наносимый ими ущерб. Основная идея книги заключается в том, что необходимо, разумно относясь к живой природе, использовать ее естественные силы для борьбы с разрушительными нашествиями вредных видов. По мнению автора, этим можно не только ограничить, но и предотвратить нашествия. «Необходимо тщательное изучение взаимоотношений между популяциями, а также тех типов и комбинаций местообитаний, которые лучше всего могут обеспечивать устойчивость и ослаблять взрывную силу вспышек размножения и нашествий чужеземных организмов» (стр. 143). С этой точки зрения Элтон смотрит и на охрану природы. «Охрана природы... может означать, с одной стоишь!, защиту диких видов от все возрастающего использования их человеком, а с другой — способ достижения наивысшей продуктивности используемой территории (стр. 203). На этом основана охрана природы и в СССР.
Возможность добиться относительной устойчивости популяций и высокой продуктивности территории обеспечивается, по Элтону, сохранением богатства и разнообразия занимающих ее естественных сообществ. «Я попробую привести некоторые доказательства того, что устойчивость относительно простых сообществ растений и животных нарушить легче, чем устойчивость более богатых биоценозов; другими словами, простые сообщества более склонны к нарушающим их целостность колебаниям численности популяций... и более уязвимы для нашествий». Поэтому в экономическом отношении сомнителен и даже опасен путь «упрощения ради эффективности» (стр. 191). С этими мыслями также трудно не согласиться. Сохранение богатой и сложной живой природы в районах с любой формой хозяйства не только желательно с эстетической точки зрения, не только важно для охраны здоровья, но и целесообразно с утилитарной сельскохозяйственной точки зрения.
Эти мысли далеки от пресловутого «невмешательства в природу», от принципа «laisser faire, laisser passer». Элтон во многих местах своей книги говорит о разумном подходе к природе, о необходимости и неизбежности ее изменения под влиянием человека, о необходимости сохранения не разнообразия биоценозов вообще, а сохранения видов, полезных и нужных нам в разных отношениях.
Следует остановиться и на некоторых других мыслях Ч. Элтона, во многом созвучных идеям наших экологов. Элтон — убежденный сторонник углубленных многолетних стационарных исследований всей сложной совокупности живого населения какой-либо территории. Опыт двенадцатилетнего комплексного изучения Ч. Элтоном и его коллегами небольшого холмистого участка Уитхэм-Вудс близ Оксфорда как бы перекликается с призывом К. Ф. Рулье «изучить три вершка прилежащего болота».
Среди многочисленных и разнообразных биологических механизмов колебаний численности животных и особенно ее крупных подъемов («взрывов») Ч. Элтон решающим считает биотические отношения, так как возможности размножения и расселения зависят прежде всего от наличия пищи, существования конкурентов, врагов, паразитов и других сожителей и их обилия, т. е. прежде
всего от соотношения разных звеньев в цепях питания; физические факторы, главным образом погода, служат лишь фоном, на котором развиваются эти взаимоотношения, фоном очень важным, но не определяющим характер биологических событий. Той же точки зрения придерживаются и многие советские экологи. Одновременно Ч. Элтон считает попытки математической интерпретации борьбы за существование со стороны А. Лотки и В. Вольтерры чрезмерным упрощением и далекой от действительности схематизацией, рассматривающей не явление в целом, а искусственно вырванный отдельный его элемент.
Ч. Элтон обращает большое внимание на опасность для других животных неумеренно широкого применения губительных инсектицидов и ядов. Слабая избирательность этих веществ приводит к гибели множества организмов, среди которых лишь немногие по-настоящему вредны; остальные же либо непосредственно полезны, либо играю^ важную роль в жизни почвы и подстилки. Их истребление может вызвать серьезные и неблагоприятные нарушения «почвенного метаболизма».
Множество других ценных и интересных выводов найдут читатели на страницах этой книги. В ней, разумеется, есть и спорные положения, по которым высказывались и иные суждения. Это касается многих частных вопросов охраны природы и некоторых других соображений Элтона. В каждой интересной и оригинальной книге это и неизбежно, и необходимо. Но ее фактическая основа великолепна; она представляет, конечно, основную ценность. Мы надеемся, что перевод книги Ч. Элтона привлечет внимание не только зоологов и ботаников, но и всех читателей, интересующихся биологией и ее важными современными проблемами, а также и природой в целом.
Проф. И. П. Наумов
Моей жене Е. Сковелл
ИЗ ПРЕДИСЛОВИЯ АВТОРА
В этой книге я пытался свести воедино три направления исследований. которые меня непосредственно занимали в течение последних 30 лет. Первое — это история фаун^, обычно расцениваемая как чисто академический предмет; между тем ряд серьезных неблагополучий, происходящих сейчас на земном шаре, обусловлен именно некоторыми событиями в истории фауны. Второе — это экология,, в частности структура и динамика численности популяций. Третье — охрана природы. Впервые я опубликовал некоторые свои мысли относительно роли нашествий в 1943 г., в обзорной статье «Изменение зоогеографических областей земного шара», нацечатанной в журнале военного времени, выходившем под названием «Польская наука и образование». Впоследствии мне приходилось много заниматься вопросами сохранения природы, так как я участвовал в создании и развитии Комитета охраны природы (Nature Conservancy). В марте 1957 г. по третьей программе Би-би-си было передано три моих выступления под названием «Устойчивость в природе и преграды» («Balance and Barrier»). Впоследствии эти выступления были опубликованы в журнале «Слушатель» (The Listener, 1957, v. 57, рр. 514—515, 556—557, 596— 597, 600). Эти выступления и послужили основой для написания данной книги.
Глава I
УЧАСТНИКИ НАШЕСТВИЙ
j^Jbi живем в мире, полном взрывов, и хотя время и место очередной вспышки нам не всегда известны, вероятно, мы могли бы найти способ предотвратить ее или по крайней мере уменьшить ее силу. Нам угрожают не только термоядерное оружие и атомная война, хотя сейчас эта угроза — одна из самых серьезных. Есть и другие, «экологические взрывы», и о них-то и пойдет речь в этой книге. «Экологическим взрывом» мы называем непомерное увеличение численности какого-либо живого организма — будь то вирус гриппа, чумная бактерия, гриб, вызывающий болезнь картофеля, зеленое растение вроде кактуса опунции или животное, подобное серой (каролинской) белке. Я намеренно употребляю слово «взрыв» (explosion), так как оно означает внезапное освобождение сил, проявление которых сдерживалось ранее другими силами (первоначально это слово означало взрыв насмешек, которыми публика осыпала актеров, когда те не могли больше сдерживать ее силой своей игры).
Экологические взрывы отличаются от некоторых других тем, что они не производят столь сильного шума и протекают не так быстро. Иными словами, они могут медленно развиваться и так же медленно затухать; но иногда их последствия оказываются весьма внушительными и вызывают разорение, гибель или вынужденную эмиграцию множества людей. В конце первой мировой войны на Западном фронте вспыхнула эпидемия гриппа, которая пере|росла в пандемию и охватила буквально весь мир, не пощадив даже эскимосов Лабрадора и Гренландии; считают, что она унесла 100 млн. человеческих жизней. До сих пор еще не совсем потухла великая пандемия бубонной чумы, которая началась в Китае в конце прошлого столетия, была занесена корабельными крысами в Индию,
Фиг. 1. Графства США, в которых была зарегистрирована чума человека [22].
фиг. 2. Распространение чумы грызунов в западных графствах США
[22].
Ф и г. 3. Суслик Citellus beecheyi — один из носителей чумы в Калифорнии; снимок сделан в свободном от чумы районе Сьерра-Невады (фото автора, 1938).
Южную Африку и на другие континенты и ныне тлеет там средн сотен видов диких грызунов, так же как и в главном очаге своего первоначального возникновения, в восточной Азии Ч В Китае она время от времени дает весьма крупные вспышки, проявляясь в легочной форме и напоминая «черную смерть» средневековой Европы. В 1911 г. в Маньчжурии от такой вспышки погибло около 60 тысяч человек. К счастью, эта форма болезни, передающаяся непосредственно от человека к человеку без промежуточного звена (блохи), редко встречалась на других охваченных пандемией континентах. Всюду, где чума проникла в естественные экологические сообщества, она способна давать более или менее сильные вспышки, хотя, к счастью для человеческого рода, цепь контактов, вызы-
1 Чума в Индии и Южной Африке существует издавна; в порты этих стран она могла заноситься в результате завоза больных чумой крыс, но в популяции местных крыс (в портах) она могла проникать и из рядом расположенных природных очагов и отсюда развозиться по разным странам. Чума диких грызунов (ее природные очаги), обнаруженная в горных странах, пустынях и степях Азии, Африки и Америки, видимо, существо-
Г ЛА В А
вающих подобную вспышку, не так легко привести в действие. Многие популяции песчанок, населяющих южноафриканские степи, являются постоянными носителями бактерий, и среди них часто вспыхивают естественные эпизоотии. От песчанок бактерии могут быть переданы блохами многососцовым мышам (Mastomys nata-lensls Smith). Этот вид, в отличие от песчанок, живет в соприкосновении с синантропными крысами; последние могут случайно заразиться, и в результате возникнут отдельные случаи бубонной чумы у человека [4]. Такие случаи в свою очередь могут послужить началом небольших местных эпидемий; но этого может и не быть. В США и Канаде такой же «подпольный» мир носителей чумы (представленный иными видами) установлен на огромных пространствах западных районов (фиг. 1—3), однако вспышки среди людей немногочисленны [22]. Таким образом, цепь контактов здесь еще слабее, чем в Южной Африке, хотя потенциальная опасность чумы существует. Нет сомнений, что в начале XX в. в Калифорнию завозились на судах и больные чумой люди, и зараженные крысы, однако возможно, что в это время в Северной Америке возбудитель чумы уже существовал. Проф. Мейер, который впервые начал систематическое экологическое изучение очагов чумы в Америке, пишет: «Единственный возможный вывод состоит в том, что вопрос о первоначальном источнике и времени появления области эндемической дикой чумы на североамериканском континенте требует дальнейшего исследования и критического анализа» [24].
Другой пример «взрыва» — происшедшее 100 лет назад распространение фитофторы из Европы в Ирландию, которое уничтожило урожай картофеля, вызвав голод. Большинству людей приходится столкнуться с каким-либо, пусть не столь значительным, нашествием чужеземного вида. Подобные взрывы сами по себе беззвучны, но часто вызывают большой шум в прессе; нередко можно увидеть набранные крупным шрифтом заголовки вроде таких. как «Эпидемия малярии в Бразилии», «Повреждение леса вала еще до проникновения чумного микроба в городские популяции крыс. Ее возникновение связано с появлением песчанок и сурков (третичный период).— Прим. ред.
в Кэннок-Чэйз» или «Кроличья болезнь в Кенте». Появление кроличьей болезни — миксоматоза — и ее последующее распространение было одним из крупнейших экологических взрывов, испытанных в нашем столетии Великобританией; разносторонние последствия этого взрыва будут ощущаться еще много лет.
Подобные события стали для нас более обычными, чем они были в прошлом столетии, не только благодаря газетным заголовкам и усовершенствованию средств связи и информации. Они и в самом деле происходят теперь гораздо чаще; в наше время они настолько обычны па всех материках и островах и даже в океанах, что нам необходимо понять, чем они бывают вызваны, и попытаться создать себе какие-то общие представления обо всех этих вещах в целом. Почему вирус, благополучно размножавшийся на бразильских диких кроликах, внезапно уничтожил значительную часть популяций кролика в Западной Европе? Почему колорадский картофельный жук беспокоит нас только сейчас, спустя более чем 300 лет после введения в культуру самого картофеля? Почему сосновая пяденица два года назад вдруг необычайно размножилась в посадках сосны в Стаффордшире и Мэришире? Ведь на континенте подобные вспышки размножения сосновой пяденицы происходили уже на протяжении более чем полутора столетий, да и в Англии она не принадлежит к только что завезенным видам.
Приведенные примеры показывают, что возможны два существенно различных типа вспышек размножения: одни возникают в результате успешного распространения вида, занесенного в страну извне, другие происходят в аборигенных или давно существующих популяциях. В этой книге речь будет идти главным образом о вспышках первого рода, связанных с нашествиями. Но для того, чтобы понять взаимодействие вновь появившихся форм с местной фауной и флорой, придется рассмотреть экологические исследования и идеи, касающиеся равновесия внутри отдельных сообществ и между различными сообществами, взятыми в целом. Другими словами, наша тема шире любых практических цроблем борьбы с вредителями, хотя многие приводимые нами примеры взяты из области прикладной экологии. Дело в том, что мы живем в такой период мировой истории, когда смешение многих тысяч различных
организмов из разных частей земного шара приводит к потрясающим сдвигам в природе. Мы наблюдаем огромные изменения в устойчивости естественных популяций земного шара. Борьба с вредителями, конечно, очень важна, ибо мы должны охранять необходимые для нашей жизни ресурсы и защищать себя от болезней и последствий экономических бедствий. Но следует попытаться охватить всю проблему в гораздо более широком плане. Наиболее крупные экологические взрывы способствовали изменению хода истории, и, как мы покажем дальше, нередко можно установить, что они вызывались нарушением той изоляции материков и островов, которая создалась в начале и середине третичного периода.
Для того чтобы конкретизировать проблему, мы приводим истории семи видов, занесенных из другой страны и бурно размножившихся.
Около 1929 г. несколько африканских комаров случайно достигли северо-восточной части Бразилии, завезенные, вероятно, из Дакара на высокоскоростном французском истребителе. Им удалось перебраться на землю и основать небольшую колонию в прибрежном болоте. Вначале на них не обратили особого внимания, хотя в близлежащем городке возникла довольно острая вспышка малярии, причем переболели почти все жители. В последующие несколько лет насекомые довольно спокойно распространялись вдоль прибрежной полосы, пока, наконец, примерно в 320 км от исходного пункта не вспыхнула эпидемия малярии, продолжавшаяся до 1938—1939 гг.; оказалось, что к тому времени комары проникли еще на 300 с лишним километров в глубь страны по долине реки Жагуариби (фиг. 4). Это была одна из тяжелейших эпидемий,, когда-либо происходивших в Бразилии: малярией болели сотни тысяч человек, около 12 тысяч, как полагают, умерло, и жизнь сельского населения была частично парализована. Биологические причины этого бедствия были до крайности просты: в стране всегда существовали малярийные комары, но не было таких, которые, подобно африканскому виду, регулярно залетали бы в жилища, а также могли бы столь успешно плодиться в прогреваемых солнцем открытых водоемах вне леса. К счастью, обе эти особенности делали воз-
Фиг. 4. Области распространения африканского малярийного комара Anopheles gambiae в Бразилии в 1938, 1939 и 1940 гг.; в дальнейшем он был полностью истреблен на территории Южной Америки [28].
можной борьбу, с комарами, и Рокфеллеровский фонд совместно с бразильским правительством провел поистине изумительную кампанию, в которой было занято более 3 тысяч человек. Была произведена сплошная обработка всех мест выплода и опрыскивание внутренних помещений домов. Это грандиозное мероприятие позволило менее чем за 3 года полностью истребить Anopheles gambiae на южноамериканском континенте [28].
Здесь можно видеть три главных элемента, характерных для такого рода ситуаций. Во-первых, исторический момент: данный вид комаров водился только в тропической Африке, а в Южную Америку был занесен человеком. Во-вторых, экологические особенности — биология размножения, а также места отдыха и питания, связанные с человеком. Разумеется, проведенная кампания не была бы успешной без интенсивных экологических исследований этих особенностей, легших в основу приемов эпидемиологического обследования и борьбы с инфекцией. В-третьих, катастрофические последствия интродукции нового вида. В связи с этим был введен карантинный осмотр самолетов, в результате чего на одном самолете обнаружили муху це-це {Glosslna palpalls) — африканского переносчика сонной болезни, в настоящее время не встречающегося нигде, кроме Африки [28].
Второй пример касается одной из болезней растений. В начале XX в. на востоке США появилась губительная болезнь, поражающая деревья каштана, которая вызывалась грибом Endothia parasitica (фиг. 5). Болезнь была завезена из Азии с саженцами из питомников. В 1913 г. гриб обнаружили на каштане в Азии, где он не причиняет своему естественному хозяину никакого вреда. Однако восточноамериканский вид каштана — Castanea dentata — оказался настолько восприимчивым, что в большей части области распространения гриба был почти нацело уничтожен (фиг. 6). На этом виде каштана паразитируют два местных вида Endothia, не причиняющие ему вреда. Они безвредны и для некоторых других деревьев, на которых они тоже встречаются (например, для дуба); один из этих видов заражает также деревья Castanea sativa в Европе [27]. Как показывает карта (фиг. 7), уже к 1911 г. болезнь, распространяясь переносимыми ветром спорами, охватила
Ф п г 5. Мицелий гриба Endoihia parasilica белые I бурые веерообразные разрастания, видные после снятия коры с больных американских каштанов [10].
Ф и г. 6. Американский каштан Castanea dentata, почти погибший в резуль-татезаражения грибом Endothia parasitica, завезенным из Азии [10]. Новые побеги, растущие от ствола, в свою очередь будут инфицированы и отомрут.
Ф и г. 7. Распространение азиатской болезни каштанов (возбудитель Endothia parasitica) в 10 штатах США, где она поражала американский каштан Castanea dentata [23].
Горизонтальной штриховкой отмечена область, где большая часть деревьев уже погибла; вертикальной штриховкой — область почти сплошной зараженности; черными кружками — изолированные очаги, многие пз которых ликвидированы.
по крайней мере десять штатов, и причиненные ею убытки составляли к тому времени не менее 25 млн. долларов [23]. В 1926 г. она все еще распространялась к югу, и к 1950 г. повсюду, кроме крайнего юга, большая часть каштанов погибла; то же самое происходит и на Тихоокеанском побережье. До сих пор единственной .ответной мерой была интродукция китайского каштана (С. mollis-sima), обладающего высокой, хотя и не полной устойчивостью к грибу: в процессе эволюции между этим видом и его паразитом создались такие же устойчивые взаимоотношения, какие выработались у американских деревьев и их собственных паразитов.
По этим же причинам крупные дикие животные Африки не страдают от присутствия в их крови трипаносом, убивающих таких интродуцированных в эту страну домашних животных, как корова п лошадь. В биологическом смысле трипаносомиаз африканских
Фиг. 8. Расширение области гнездования европейского скворца (Sturnus vulgaris) в США и Канаде с 1891 по 1926 гг. [3].
Кружки за пределами линии 1926 г.— главные пункты, где скворец был зарегистрирован в зимнее время и еще не закрепился.
животных представляет собой ситуацию, подобную той, которая, вероятно, создалась бы в случае интродукции американского каштана в Азию. Китайский каштан устойчив к грибу как в Азии, так и в Америке. Но уже в 1911 г. выяснилось, что европейские каштаны, растущие в Америке, восприимчивы к болезни [23]. В 1938 г.
болезнь 1 появилась в Италии, где она дала внезапную вспышку п теперь угрожает каштанам, выращиваемым в чистых насаждениях для получения урожая плодов; она проникла также в Испанию и рано или поздно, весьма вероятно, появится и в Англии [8]. К несчастью, китайский каштан не может хорошо расти в Италии^ и остается лишь надеяться, что когда-нибудь удастся вывести устойчивый сорт путем гибридизации.
Третий пример — европейский скворец (Sturnus vulgaris), распространившийся по США и Канаде за период в 60 лет (он обосновался еще на двух других континентах — в Южной Африке и Австралии, а также в Новой Зеландии). Естественный ареал этого вида скворцов охватывает Сибирь, а с севера на юг простирается от Норвегии и севера Европейской части СССР до Средиземного моря. Таким образом, можно ожидать, что эта форма способна адаптироваться к самым различным видам континентальных местообитаний и климата. Однако первые попытки ее интродукции в США не имели успеха. Затем в Центральный парк в Нью-Йорке завезли около 80 скворцов, и несколько пар в 1891 г. начали размножаться. После этого их численность и ареал неуклонно увеличивались, если не считать очень холодной зимы 1917—1918 гг., когда много птиц погибло. Однако до 1916 г. область обитания постоянных популяций еще не перешла за Аллеганы. Составленная Куком карта (фиг. 8), отражающая положение дел к 1926 г., показывает, что область размножения расширялась концентрически, причем далеко за пределами этой области постоянно регистрировались не размножавшиеся птицы; к 1926 г. граница области размножения перевалила за Аллеганы, но нигде не вышла на запад от линии, идущей от озера Мичиган приблизительно с севера на юг [3]. К 1954 г. процесс расселения почти закончился и скворцов можно-было встретить, по крайней мере во время перелетов, почти на всей территории США, хотя они еще не вполне закрепились в? отдельных частях западного побережья. Во время перелетов скворцы проникали в северную Мексику, а в 1953 г. одну птицу видели в Аляске [17]. Это был настоящий экологический взрыв, начавшийся
1 Занесенная в Европу из Америки.— Пр им. ред.
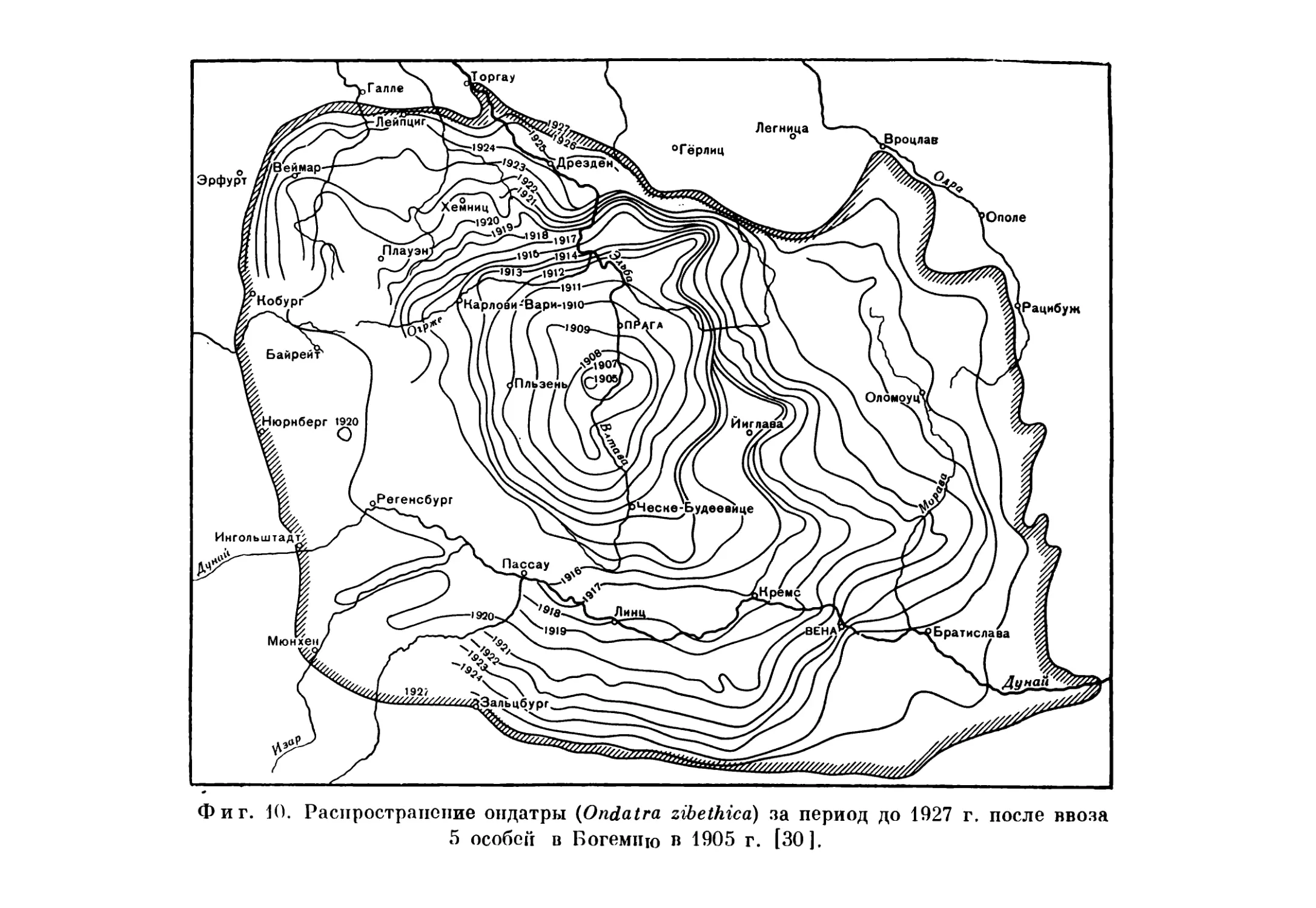
с размножения нескольких пар птиц в городском парке,— совершенно так же, как распространение североамериканской ондатры Ondatra zibethlca (фиг. 9) в Европе началось всего лишь с пяти особей, которых держал один землевладелец в Чехословакии в 1905 г. (фиг. 10). Ныне численность ондатры в Европе (это наш четвертый пример) достигает многих миллионов, и ареал этого
Фи г. 9. Ондатра {Ondatra zibethlca) в своем природном местообитании в болотах Монтезума, штат Нью-Норк.
вида увеличился в результате дополнительных интродукций с целью использования его меха: рассеявшиеся животные и их потомки основали новые очаги распространения (фиг. И). В Финляндии после 1922 г. произвели более 200 выпусков ондатры (в 1922 г.—непосредственно из Чехословакии), и ежегодная добыча ее теперь составляет от 100 до 240 тысяч штук [1]. В результате интродукции, произведенной в Советском Союзе, ондатра стала важным пушным зверем в бассейнах великих рек Сибири, на севере европейской части СССР и в Казахстане [18]. Говоря языком зоогеографов, чисто палеарктический вид (скворец) и чисто неарктический вид (ондатра) за полстолетия превратились в голарктические формы (фиг. 12).
Фиг. 10. Распространение ондатры (Ondatra zibethicd) за период до 1927 г. после ввоза 5 особей в Богемию в 1905 г. [30].
Фи г. И. Распространение ондатры (Ondatra zibethica) во Франции [6]. Крестиком указано место поимки одной особи.
У Ч А С Т II И К И Н А III ЕСТВИ И
Пятый пример — растение, изменившее часть ландшафта Англии — высокий и сильно растущий болотный злак Spartina town-sendil, который во многих местах заселил илистые пространства литорали у берегов Англии [14]. Это естественный гибрид английского вида S. maritima с американским видом S. alternifolia, завезенным на южное побережье Англии и закрепившимся там в начале XIX в. Этот устойчивый, хорошо размножающийся гибрид
Фи г. 12. Распространение ондатры (Ondatra zibethica) в Европе и Азии [5 ].
впервые обнаружили в Саутгемптоне в 1870 г. В течение первых 30 лет он распространялся не очень быстро, но в XX в. захватил огромные пространства па побережье Ламанша, не только в Англии, но и на севере Франции (фиг. 13). Этот гибрид культивировали в некоторых других районах Англии, а также завезли в Северную и Южную Америку, в Австралию и Новую Зеландию. Американская родительская форма была в значительной степени подавлена или вытеснена гибридом. Здесь перед нами любопытное следствие распространения вида растений человеком: от случайного скрещивания неарктической и палеарктической форм возник новый полиплоидный гибридный вид, ставший в результате дальнейшей сознательной интродукции человеком почти космополитом.
A
Фиг. 13. Глинистое засоленное болото с доминирующим растением Spar-tina townsendii', заметно разрастание зарослей Spartina за последние 50 лет (А и Б).
Причем это — в общем довольно полезное растение, так как оно закрепляет голый подвижный ил приливной зоны, на котором не могут расти никакие другие сосудистые растения, помогает формированию новых участков суши, где нередко на первых порах образует солончаковые пастбища. Влияние его на структуру побережья еще не вполне выяснено географами и экологами; Тэнсли, однако, отмечает, что «никакой другой вид солончаковых растений,
Ф в г. 14 Молодь морской миноги Petromyzon marinus, нападающая на гольца Salvelinus fontinalis в аквариуме [21].
по крайней мере в северо-западной Европе, не дает при осушении литорали сколько-нибудь сравнимого по быстроте и силе эффекта» [29].
Сходные по масштабам изменения, вызываемые нашествием видов из других областей, наблюдались в пресноводных озерах и реках.
Наш шестой пример касается морской миноги (Petготуzon marinus) в области Великих озер Северной Америки [7]. Это североатлантический вид, живущий преимущественно в море, но для
нереста заходящий в реки. Еще в прошлом он естественным образом обосновался в озере Онтарио, а также в некоторых малых озерах штата Нью-Йорк. Ниагарский водопад явился непреодолимым препятствием для проникновения морской миноги дальше, в Великие озера. В 1829 г. была закончена постройка Уэлландского судоходного канала, создавшего обходный путь в озеро Эри, но лишь спустя еще сто лет морскую миногу обнаружили в этом озере. Дальнейшее расселение происходило ночти со скоростью взрыва. В 1930 г. миноги достигли реки Септ-Клэр, а к 1937 г. прошли через нее в озера Гурон и Мичиган и начали входить для икрометания в реки, впадающие в эти озера. В 1946 г. они уже достигли озера Верхнее. Все это время они уничтожали гольца Salveltnus namaycush, имеющего огромное хозяйственное значение. Морская минога — это одновременно подвижный хищник и эктопаразит: она присасывается к рыбе (фиг. 14), выделяет в рану антикоагулянт и растворяющую ткани жидкость, выгрызает мясо своей жертвы и высасывает из нее соки до тех пор, пока она не погибнет; иногда жертва гибнет через несколько часов, а иногда — только через неделю. Уловы гольца всегда колебались в некоторых пределах; поэтому данные рыболовной статистики начиная с 1889 г. были тщательно проанализированы. Оказалось, что никогда еще уловы не падали столь стремительно: в течение 10-лет после того, как начали ощущаться последствия вторжения миноги, ежегодный улов гольца в американских водах озер Гурон и Мичиган упал с 3 900 000 до всего лишь 12 000 кг\ Не лучше обстояло дело и в канадских водах [12]. Это не было вызвано изменением интенсивности лова. Сильно пострадали и другие виды рыб, в том числе озерный сиг, налим и представители чукучановых (Catasto-midae); численность всех этих видов уменьшилась. Таким образом, постройка судоходного канала, предпринятая для того, чтобы создать возможность вывоза продукции Среднего Запада, нанесла огромный ущерб рыбному хозяйству Великих озер, сказавшийся спустя более чем сто лет. Однако в озере Эри миноги не размножались — отчасти потому, что там мало гольца, а также, вероятно, потому, что впадающие в это озеро реки мало пригодны для нереста [19].
Фиг. 15. Мохноногий краб (Eriocheir sinensis) [26].
А. Самец с поднятыми клешнями. Б и В. Планктонные личинки.
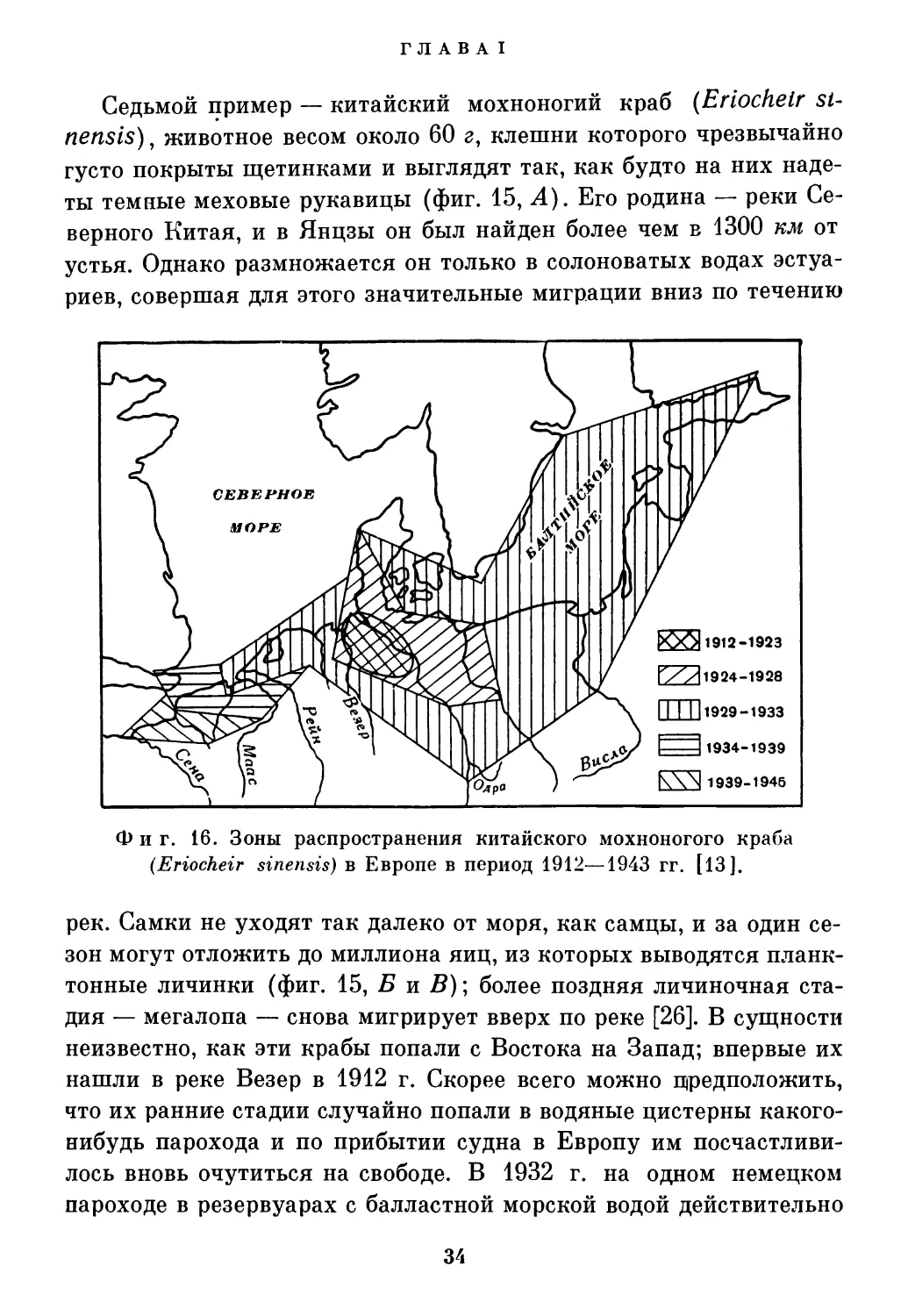
Седьмой пример — китайский мохноногий краб (Eriochelr sinensis) , животное весом около 60 г, клешни которого чрезвычайно густо покрыты щетинками и выглядят так, как будто на них надеты темные меховые рукавицы (фиг. 15, 4). Его родина — реки Северного Китая, и в Янцзы он был найден более чем в 1300 км от устья. Однако размножается он только в солоноватых водах эстуариев, совершая для этого значительные миграции вниз по течению
Фиг. 16. Зоны распространения китайского мохноногого краба (Eriocheir sinensis) в Европе в период 1912—1943 гг. [13].
рек. Самки не уходят так далеко от моря, как самцы, и за один сезон могут отложить до миллиона яиц, из которых выводятся планктонные личинки (фиг. 15, Б и В); более поздняя личиночная стадия — мегалопа — снова мигрирует вверх по реке [26]. В сущности неизвестно, как эти крабы попали с Востока на Запад; впервые их нашли в реке Везер в 1912 г. Скорее всего можно предположить, что их ранние стадии случайно попали в водяные цистерны какого-нибудь парохода и по прибытии судна в Европу им посчастливилось вновь очутиться на свободе. В 1932 г. на одном немецком пароходе в резервуарах с балластной морской водой действительно
были найдены два крупных экземпляра; полагают, что их захватили вместе с водой в гамбургской гавани, хотя обычно эти резервуары хорошо защищены сетками. За последние 45 лет эти крабы заселили другие европейские реки от района Балтики до Сены (фиг. 16). Крабы, проникшие в Эльбу, распространились до самой Праги, подобно саламандрам Карела Чапека. На британские острова этот краб еще не проник, но весьма вероятно, что когда-нибудь это произойдет, так как один живой экземпляр уже был пойман в Челси в 1935 г.
Этих семи примеров достаточно, чтобы показать, к чему приводит намеренная и случайная интродукция различных видов человеком, в особенности завоз их на другие континенты. Вторжению новых видов подверглись воды морей, эстуариев, рек и озер, побережья морей и эстуариев, леса тропической и умеренной зоны, сельские местности и города. В XVIII в. океанских судов водоизмещением свыше 300 т было мало. Теперь их тысячи. Правительственная карта, отражавшая местонахождение всех океанских судов Британской Империи на 7 марта 1936 г., показывает, что в этот день 1462 судна находилось в море и 852 — в портах; следует также учесть, что на эту карту не было нанесено положение судов каботажного плавания. Некоторое представление о том, какое это может иметь значение для распространения животных, дают результаты экологических наблюдений, сделанных видным исследователем тропических насекомых Майерсом во время плавания от Тринидада до Манилы на рангунском судне, перевозившем рис. Этот ученый развлекался тем, что составлял список всевозможных животных, обнаруженных им на борту,— от тараканов и рисовых долгоносиков до блох и комнатных животных [25]. Всего он насчитал 41 вид таких путешественников, главным образом насекомых. Когда он распаковывал свои чемоданы в гостинице в Маниле,, из них выползло несколько жуков. Это были жуки Trlbollum casta-neum — известные вредители запасов муки и зерна, один из видов, живших в рисе на судне.
Развитие средств транспорта в течение последних ста лет непрестанно поддерживало и усиливало эту «бомбардировку» всех
стран чужеземными видами, перевезенными случайно или намеренно по морю, по воздуху или по суше из мест, которые ранее были разобщены. Конечно, не всем растениям и животным, разносимым по земному шару, удается закрепиться в тех местах, куда они попадают; и не все закрепившиеся виды вредны для человека, хотя они неизбежно как-то изменяют соотношения между местными видами. Но этот всемирный цроцесс, усиливающийся с каждым годом, ведет к постепенной ломке того распределения видов, которое существовало всего сто лет назад.
Для того чтобы понять все значение происходящего, необходимо обратить свой взор на много миллионов лет назад. Внимание к тому, что в различных частях мира существуют обширные фаунистические области, в основном соответствующие континентам, впервые привлек Альфред Уоллес. Эти области известны как «области Уоллеса», хотя в общих чертах размещение их наметил еще орнитолог Склэтер. Однако Уоллес проделал огромную работу по сбору и классификации данных об этих областях. Он предполагал, что эти пространства оставались разобщенными на протяжении столь долгих периодов времени, что в их пределах сохранялись или возникали вновь многие специфические группы животных. В детских книжках о животных это бывает выражено просто и ясно: тигр живет в Индии, кенгуру — в Австралии, а бегемот — в Африке. Иногда в них сообщают также, что болотный бобр, или нутрия, живет в Южной Америке (фиг. 17). В книге, рассчитанной на более подготовленного читателя, могло быть сказано, что этот крупный водяной грызун, вероятно, впервые появился во внутренней части Южной Америки (теперь мы знаем это наверное). Однако теперь, при переизданиях, следовало бы добавить примечание о том, что нутрия отлично существует также в штатах Орегон, Калифорния и Нью-Мексико [11], а также в Луизиане (где недавно за один год было поймано 374 тысячи штук), на юго-востоке СССР [32], во Франции [6] и в речных поймах графства Норфолк в восточной Англии (фиг. 18). В мехе нутрий, населяющих эти места, живет паразит — вошь Putrifquenia coypus из семейства Gyropidae, который тоже происходит из Южной Америки [9]. Эти вши обладают усиками, напоминающими по форме раздвижной
Фиг. 17. Семья нутрий Myocaslor coypus па своей родине в Южной Америке |2|.
Фиг. 18. Местообитание южноамериканской нутрии (Myocastor coypus) па канале средн равнины в восточной Англии (фото автора, 1957).
Укрытие и пищу дает пышная болотная растительность, в данном слу чае главным образом манник (Glyceria maxima').
гаечный ключ, благодаря которым они, вероятно, и смогли удержаться на зверьках на всем их пути из Южной Америки.
Но в очень давние времена, скажем 100 млн. лет назад, в меловом периоде, фауна всего земного шара была гораздо более космополитической и не была в такой степени разделена океанами, пустынями и горами, как теперь. Если бы в те времена, когда в теплых мелких морях формировались меловые отложения Мальборо или Дувра, печатались детские книжки, то в них можно было бы прочитать: «Огромные динозавры водятся во всем мире, кроме Новой Зеландии; пе попадайтесь им на пути». В них было бы также указано, что водяные чудовища живут не только в каком-нибудь одном узком морском заливе. С точки зрения зоогеографии это была бы довольно скучная книга, но картинки и описания нравов животных были бы очень интересны. В те времена не было бы большой нужды в зоопарках: достаточно было бы просто выйти из дому, чтобы, соблюдая известные предосторожности, можно было тут же, на месте, наблюдать за динозаврами и регистрировать на карточках число отложенных ими яиц. Вместе с тем эти динозавры имеют большое значение для серьезного обсуждения истории Земли: поскольку животное величиной с грузовик не могло попасть с одного материка на другой иначе как по суше, приходится предполагать, что материки были связаны друг с другом — если не все время, то хотя бы достаточно часто (в масштабах геологического времени).
Этот ранний период более или менее космополитической наземной и пресноводной фауны и флоры продолжался примерно в три раза дольше, чем промежуток времени от мелового периода до наших дней. Лишь после мелового периода сформировались области Уоллеса, так как моря, а позже и такие участки суши, как Гималаи и пустыни Центральной Азии, создали для очень многих видов непреодолимые преграды. Фактически на земном шаре была уже не одна, а пять или шесть больших фаун, помимо бесчисленных более мелких, сформировавшихся на изолированных островах (например, на Гавайских островах, Новой Зеландии или Новой Каледонии) и в таких огромных озерах, как Байкал или Танганьика. Нарушение такого распределения фаун началось задолго до появления человека. Значительное смешение фаун происходило в
последние миллионы лет до ледникового периода и после него: двумя важными факторами этого были: поднятие из океана Панамского перешейка и неоднократно возникавшее в различные времена соединение материков в районе нынешнего Берингова пролива. Однако человек искусственно форсирует весь этот процесс, и за это ему приходится испытывать на себе его многосторонние последствия.
Вот уже тридцать лет я читаю сообщения об этой огромной волне нашествий; во многих публикациях чувствуется, что их авторы сами были очевидцами описываемых событий, и можно ощутить актуальность и масштабы всей этой проблемы — то, чего не может передать сухой и сжатый текст учебников. Мы не имеем права ошибаться: на наших глазах происходит одно из величайших исторических потрясений в мировой фауне и флоре. Мы могли бы повторить слова конандойлевского профессора Челленджера, произнесенные им на плато «затерянного мира»: «На нашу долю выпало счастье присутствовать при одной из тех битв, которые определяют дальнейший ход истории, решают судьбы мира». Но как решится теперь эта судьба? Не обречен ли наш мир на гибель? Таковы вопросы, на которые должны попытаться ответить экологи.
Глава II
ЗООГЕОГРАФИЧЕСКИЕ ОБЛАСТИ УОЛЛЕСА
Г\дин из главных тезисов этой книги состоит в следующем: если ” мы хотим предвидеть, что может произойти с экологическим равновесием на Земле, мы должны проникнуть взором не только в будущее, но и в прошлое. Если бы за последние 100 млн. лет флора и фауна земного шара развивались таким образом, что каждый организм имел бы возможность распространиться во все части мира, где условия для него оказались бы пригодными, так что каждому типу экологической обстановки соответствовал бы только один вид, то возможности будущего изменения флоры и фауны под влиянием деятельности человека были бы иными. Эти возможности были бы гораздо меньше, хотя все же оставались бы значительными, так как человек не только передвигает виды, как фигуры на шахматной доске, но и изменяет их места обитания. В описанном только что мпре, где не существовало бы препятствий для расселения, обитали бы главным образом паптропические виды, виды умеренных широт (сейчас мы имеем преимущественно «панарктические» виды) или формы с биполярным распространением; континентальные виды заселяли бы все острова, пресноводные — свободно проникали бы во все изолированные водоемы, а ареалы морских животных поясами охватывали бы весь мир как в северном, так и в южном полушарии. Кролик, возможно, давно уже жил бы в Австралии, нутрия — в восточной Англии, китайский краб — в Эльбе, а гигантская улитка — па Марианских островах, разумеется, если бы эти виды вообще появились в этих местах и закрепились там в борьбе с конкурирующими формами.
Очень многие виды способны распространиться по всему миру — либо потому, что экологические преграды, сдерживающие расселение других форм, не действуют на них, либо потому, что
они обладают исключительно эффективными средствами распространения, что по существу одно и то же. Calanus finmarchicus — наиболее многочисленный представитель веслоногих рачков в планктоне морей северной Атлантики — может попасть в Индийский океан в районе Мадагаскара, так как холодные северные течения, достигнув тропических морей, уходят на глубипу и продолжают свой путь под более теплыми поверхностными водами; таким способом этот рачок действительно пересек экватор, или, точнее, прошел под ним. Многие птицы мигрируют чуть ли не через весь мир: полярная крачка способна перелетать из Арктики в Антарктику, золотистая ржанка — из Северной Америки в Южную и обратно, ласточка — из Европы в Южную Африку; некоторые водоплавающие птицы совершают перелеты между Австралией и Японией. Ареал микроорганизмов, споры или высохшие тела которых переносятся ветром или птицами (застревают в перьях), тоже нередко обнимает весь земной шар. Таковы многие простейшие, коловратки и ветвистоусые рачки, не говоря уже о семенах многих растений. Кроме того, существует много подвижных форм, постепенно заселивших весь земной шар, несмотря на океаны, горы и пустыни. Одним из наиболее известных примеров служит сова-сипуха.
Однако огромное множество других растений и животных никогда не получало возможности расселиться по всему миру. Сущность областей Уоллеса в том и состоит, что в различных частях земного шара такие виды оказались изолированными на достаточно долгий срок и подверглись благодаря этому глубокому изменению, что наложило определенную печать на местные флоры и фауны. К этому добавились два других процесса, еще более усложнивших современную картину распространения организмов. Во-первых, ареалы некоторых групп, прежде очень широко распространецных, сократились и теперь ограничиваются одним материком, островом пли озером. Во-вторых, после длительных периодов изоляции в третичный период, когда образовались области Уоллеса, произошло некоторое смешение фаун еще до того, как на сцену выступил человек, сильно ускоривший этот процесс. Примером первого процесса может служить секвойя, которая была распространена от
восточных до западных берегов Северной Америки и в Евразии, теперь же растет только в Калифорнии и Орегоне. Пример второго процесса — распространение тапиров: как показывают ископаемые находки, тапиры возникли в некогда изолированной Северной Америке, но распространились в Центральную и Южную Америку и в юго-восточную Азию, где они живут и поныне, хотя на своей первоначальной родине исчезли. Итак, три причины усложнения общей картины таковы: нарушение древнего космополитического распространения организмов, появление специфических региональных групп и частичное проникновение элементов этих региональных групп в другие области земного шара. Если бы такая диффузия доходила до полного перемешивания фаун, то мы не могли бы сейчас обнаружить области Уоллеса или, если бы их и можно было заметить, они представляли бы собой нечто совершенно иное.
В этой книге мы не намереваемся критиковать основные положения зоогеографии — мы лишь попытаемся коснуться некоторых из многих воззрений, в которых существо дела наполовину скрыто от рядового исследователя глубокими наносами неточно установленных «фактов», сомнительных теорий и сведений, которые еще никогда не были использованы для решения зоогеографических проблем. Обозревая этот огромный неудобоваримый материал, мы почти завидуем самому Уоллесу, который по крайней мере мог лежать в своем лагере, пораженный лихорадкой, и размышлять об основной концепции борьбы за существование и естественного отбора, которую до него полностью разработал только один человек! Мы считаем нужным привести здесь скромную оценку, которую сам Уоллес дал своим областям, или «Regions», как он называл их: «Наша цель — отразить, с наибольшим приближением к истине, основные особенности распространения современных животных, но не тех, которые жили в какой-либо один пли все прошлые геологические периоды. Если когда-нибудь мы получили бы достаточно данных по географии и биологии Земли в прошлые эпохи, мы действительно смогли бы приближенно установить, каковы были зоогеографические области в плиоцене, миоцене или эоцене; но любая попытка представить все эти области в связи с
областями нашей собственной эпохи неизбежно приведет к путанице» [57].
Прежде чем описывать области Уоллеса, необходимо коснуться распределения организмов на Земле до мелового периода. Почти все геологи согласны в том, что на протяжении всей истории с палеозойских времен до наших дней, несмотря на множество изменений в деталях береговой линии, всегда существовали участки суши, соответствующие современным материкам. Значительные части этих масс суши вообще никогда не были покрыты морем. Жизнь, существовавшая в морях, начала завоевывать сушу и пресные воды, возможно, еще в силуре и уже наверное в девоне, т. е. (с довольно большой возможной ошибкой в оценках) около 315 млн. лет назад или раньше. В последующие 230 млн. лет, т. е. до конца мелового периода, не было такой эпохи, когда, судя по ископаемым, существовало бы хоть что-нибудь, похожее на области Уоллеса. Часто появлялись региональные различия, особенно между фаунами северного и южного полушарий; происходили климатические изменения, вроде Пермской ледниковой эпохи, были сухие и влажные периоды, изменялась площадь, занятая сушей. Но совершенно очевидно, что каждая значительная новая группа организмов в конце концов распространялась по всему миру, заселяя все или почти все области, соответствующие современным материкам, и некоторые участки, превратившиеся ныне в острова (например, Мадагаскар). Это время можно назвать периодом космополитического распространения. Когда мы подходим к мезозойской эре, особенно к ее середине и концу (юрскому и меловому периодам), появляются убедительные данные в пользу того, что климат на Земле был и более однотипным и более теплым, чем теперь. Приходится допустить, что произошли какие-то очень большие изменения, чтобы объяснить появление мощных лесов в Гренландии и на Шпицбергене и многочисленных аммонитов и других морских животных в арктическом океане [33].
В меловом периоде или немного позднее, в зависимости от того, о какой части земного шара идет речь, имела место значительная трансгрессия моря, прервавшая сухопутные связи во многих частях земного шара — в районах Панамского перешейка и Берип-
гова пролива, около Австралии и в других местах. Время этих величайших событий, превративших материки в архипелаг, имело особо важное значение для истории. Множество групп растений и животных к тому времени прошло уже значительную часть эволюционного пути к своему нынешнему состоянию. К концу мелового периода существовали многие современные роды деревьев, нанример дубы, тоноли, березы, смоковница, магнолии, лавры, сосны, ели и кедры. Возникли ночти все современные семейства, очень многие роды и даже некоторые виды насекомых. Но млекопитающие, нтицы и пресноводные рыбы находились, если можно так выразиться, еще на пороге гигантского эволюционного скачка, когда материки разделились и каждый из них нолучил свою часть первичных форм млекопитающих и нтиц, а некоторые — и пресноводных рыб, развитие которых носле разобщения областей ношло совершенно самостоятельными путями.
Уоллес разделил мир на шесть областей, именуя их но названиям материков или используя греческие и латинские корни. В этом ему помогли его необычайно широкие познания в ряде наиболее важных вопросов зоогеографии, приобретенные в результате собственных исследований на Амазонке и Рио-Негро в 1848— 1852 гг. и на Малайском архипелаге в 1854—1862 гг. Своп первые мысли о географическом распространении животных он записал, находясь еще па Востоке; они были опубликованы в 1860 г. Он вернулся к ним позже, когда собрал 125 660 экземпляров животных и установил то, что впоследствии получило название «линии Уоллеса». Его шесть областей можно обрисовать очень коротко, хотя описание их особенностей заняло два тома общим объемом в 1110 страниц и вряд ли может быть резюмировано здесь. Неотр(эпическая область охватывает Южную и Центральную Америку до линии, проходящей через Мексику, и Вест-Индию (фиг. 19). Северная Америка, ее арктические острова и Гренландия образуют Неарктическую область (фиг. 20). Палеарктическая область обнимает Европу и часть Азии, простираясь от Британских островов до Японии (фиг. 21). В Азии Уоллес выделил Восточную область (фиг. 22), в которую входят Индийский полуостров, юго-восточная Азия с южным Китаем и Тайванем, а также Филиппинские
Ф и г. 19. Фауна лесов верхней Амазонки.
Слева направо: головач (Cephalopterus); два кракса (Стах); два летящих тукана (Pteroglossus); трубач (Psophia); два голпбрп-эльфа (Lophornis); вдали виден ягуар.
Ф иг. 20. Млекопитающие североамериканских прерий.
Вдали — стадо бизонов; ближе — два вилорога (Antilocapra); слева — гофер (Geomys), который вообще ведет почти всецело подземный образ жпзпп; справа — три луговые собачки (Су поту з).
Фи г. 21. Млекопитающие средней части палеарктической области.
Вдали-- волки; ближе — три сайги; слева — слепыш (Spalax)— грызун, ведущий подземный образ жизни; справа - выхухоль (Myogale), водяное насекомоядное длиной 30—40 см.
Фиг. 22. Фауна лесов Малайского полуострова.
Вверху — три белоруких гиббона (Hylobales lar); слева — два рогоклюва (Corydon); справа — два Edolius; ниже — аргус в брачном наряде перед самкой; внизу — птица-носорог (Uuceros).
острова и все острова Малайского архипелага к западу от Целебеса и Ломбока. Северную границу Восточной области образуют Гималаи и великие горные цепи, расположенные дальше к востоку; разобщенность областей усиливают лежащие к северу от гор пустыни. Эфиопская область (фиг. 23) охватывает Африку к югу от Сахары и Мадагаскар. К Австралийской области (фиг. 24) относятся острова к востоку от Борнео, Явы и Бали, Новая Гвинея и Австралия, а также Новая Зеландия и острова Тпхого океана. Плодотворность такого разделения на области полностью подтвердилась в последующие сто лет, несмотря на бесконечные споры о деталях их границ, подразделений, фаунистического состава и истории. Сам Уоллес полагал, что Новая Зеландия представляет своего рода «аномалию». Я добавил бы, что и огромное пространство на востоке Тихого океана за пределами Австралийской континентальной дуги, с его млечным путем островов, населенных в большинстве случаев небольшим числом видов (таковы даже Гавайские острова), заслуживает самостоятельного рассмотрения. Так что фактически существует семь «царств жизни».
История некоторых из этих областей вполне выяснена (обе Америки и Европа), в отношении других (Австралия и Африка) мы встречаемся с относительной скудостью ископаемых данных; для Азии картина весьма неполна, и история ее основывается главным образом на косвенных данных. Нам удобно будет начать с Неотропической области. Когда «Бигль» вышел из Баия-Бланкп летом 1833 г., Чарлз Дарвин остался на берегу, чтобы совершить путешествие в Буэнос-Айрес по суше [36]. На этом пути он и обнаружил скопление крупных костей ископаемых животных, среди которых были кости четырех видов огромных вымерших неполнозубых из группы Gravigrada, двух других неполнозубых, лошади, токсодонта, а также остатки, которые он определил как зубы животного из группы, известной ныне под названием Litoptefna. Позже, в январе следующего года, он нашел половину скелета «величиной не меньше верблюда», погребенную в красноцветных плейстоценовых глинах щебнистой равнины в пампасах Патагонии. Это была Macrauchenla — последний представитель Litop-terna, который, по замечанию Скотта, «был, как мы можем себе
Ф и г. 23. Фауна западной части экваториальной Африки.
Слева внизу— выдровая землеройка (Potamogale); вдали две гориллы; слева на ветке сидит птл да Т uracus; над рекой угпт вдовушка (Vidia); на берегу лежит кпстеухан свинья (Potamochocrus); над ней на дереве— лемур потто (Perodi('ticus).
представить, вероятно, одним из самых причудливых членов этой группы чудовищ» [51]. Litopterna (так же как и Toxodonta) — одна из нескольких странных групп копытных, никогда не распространявшихся за пределы Неотропической области, возникших и вымерших там во время третичной изоляции. Геологи располагают прямыми данными о существовании морского пролива между Северной и Южной Америкой с середины эоцена до середины плиоцена— в течение периода огромной длительности (фиг. 25). Напомним читателю последовательность эпох третичного периода — эоцен, олигоцен, миоцен, плиоцен — и четвертичного периода — плейстоцен (ледниковая эпоха) и голоцен (послеледниковая эпоха). Предки плацентарных млекопитающих, от которых произошли Litopterna и другие эндемичные группы Южной Америки, по-видимому, попали на этот материк в течение сравнительно недолгой эпохи, называемой палеоценом (между концом мелового периода п эоценом), а также в начале эоцена. Сумчатые тоже обитали в Южной Америке до образования пролива, и позже среди них возникли многочисленные формы, часть которых сохранилась кое-где до сих пор в виде мелких сухопутных и водных животных. Существовали и сумчатые хищники, причем один из них поразительно напоминал саблезубого тигра. Происходила бурная эволюция необычайных форм неполнозубых, из которых ныне сохранились муравьеды, ленивцы и броненосцы; однако в давно прошедшие времена существовали также гигантские броненосцы и гигантские ленивцы самого странного облика. В Южной Америке жили и до сих пор живут многие своеобразные группы грызунов; Симпсон предполагает, что исходные формы грызунов и южноамериканских обезьян перебрались через пролив, «перепрыгивая» с острова на остров [52]. В самом деле, хорошо известно, что тропические реки выносят в море целые «плоты» из растительного материала, на которых иногда находятся животные. Так, в 1883 г. на древесных стволах, плывших близ Явы, видели мартышку [39]; после изменения силы экваториального противотечения на побережье Перу на обломках корабля приплыла змея Bothrops atrox [45]; Даммерман приводит и другие подобные примеры [35]. Но какого-либо представителя Toxodonta или Litopterna или же шиншиллу величиной
Фиг. 24. Фауна равнин Нового Южного Уэльса.
Слева направо: дв; »епгуру (Macropus); пара лирохвостов (Мепита); два эму (Dromnpws); дпа хохлатых Осуркаря; белоног (Podargus); сумчатая белка (Petaurus).
Фиг. 25. Разрыв связи через Панамский перешеек в третичном периоде, по заключениям геологов [44].
Ни один из проливов не существовал в течение всего третичного периода; их существование было разделено во времени промежутком, продолжавшимся с середины эоцена до середины плиоцена.
с носорога не переправишь через пролив на небольшом плотике из частей растений.
Таким образом, солидные геологические и палеонтологические данные, а также особенности современной фауны доказывают существование в прошлом длительной изоляции. Если бы лучше сохранилась ископаемая орнитофауна, мы, несомненно, смогли бы проследить сходную эволюцию многих семейств птиц, эндемичных теперь для Неотропической области,— например американских страусов, паламедей, гуахаро, туканов, представителей семейства Coerebidae, древолазов, ленивок и других. Изоляция наложила глубокий отпечаток на состав всей флоры и фауны. Среди третичных ископаемых Аргентины и Чили нет ни одного рода североамериканских растений, за исключением гинкго. Большая часть северных родов и видов до сих пор не встречается в Чили, хотя, будучи завезены человеком, они успешно растут там [34]. Но эта изоляция сохранялась только до середины плиоцена, когда восстановление Панамского перешейка позволило фаунам севера и юга перемешаться. Многие животные, называемые теперь южноамериканскими, в действительности возникли в других местах, а в Южную Америку проникли с севера лишь после этого воссоединения.
В группу таких плиоценовых пришельцев входили тапиры, ламы, пеккари, олени, лисицы, собаки, выдры, медведи, еноты и скунсы; некоторые из них сохранились доныне. На юг проникла также весьма интересная группа, включавшая мастодонтов, лошадей, американских антилоп, полевок и настоящего саблезубого тигра, но эти животные вымерли и только человек снова ввез лошадей. Абсолютно достоверно, что часть плиоценовых пришельцев и аборигенов Неотропической и Неарктической областей, смешавшись друг с другом после длительной изоляции этих фаун, подверглись массовому уничтожению. Обязательно ли смешение видов, эволюи-ровавших отдельно и не приспособленных друг к другу, должно сопровождаться такими катастрофическими последствиями, к каким привело проникновение морской миноги в Великие озера или распространение азиатского гриба-паразита каштанов в Америке? Хотя масштабы времени совершенно различны (в одном случае —
миллионы лет, а в другом — десятилетия), в обоих случаях мог действовать один и тот же принцип.
Еще более удивительные палеонтологические данные накопились со времен Уоллеса относительно Северной Америки; здесь происходило сходное в некоторых отношениях развитие специфических фаун [51]. Налицо несомненные признаки длительной изоляции, которая, однако, сочеталась со столь же несомненными вселениями — вероятно, из Восточной Азии через область Берингова пролива. С одной стороны, некоторые семейства млекопитающих возникли и развивались только в Северной Америке и не найдены за ее пределами. Таковы гоферы — многочисленные в наше время грызуны, ведущие подземный образ жизни, а также давно вымершие свиноподобные ореодонты; здесь же возникли верблюды и тапиры, но теперь они встречаются только в других странах, куда онп сумели попасть в плиоцене или плейстоцене. С другой стороны, пришельцы из Палеарктической области, например олени и представители отряда хоботных, пришли сюда в миоцене. Однако жирафы и страусы из Азии никогда не проникали в Америку. Это объясняется, по-видимому, тем, что сухопутный мост через Берингово море, по словам де Шардена, «никогда не был достаточно широким и удобным. Преодолеть его удалось лишь тем, кому благоприятствовал случай, а также подвижным, легко приспособляющимся животным или при наличии сильного биологического давления» [37]. Симпсон на основании обзора ископаемых находок заключил, что «во все времена после раннего эоцена сходство между фаунами Евразии и Северной Америки было значительно меньшим, чем в настоящее время. Это сходство было особенно мало со среднего до позднего эоцена и с позднего олигоцена до раннего миоцена. Уж во всяком случае к этим периодам понятие «Голарктическая область неприложимо; оно вряд ли применимо и к другим эпохам, кроме раннего эоцена и современности» [52].
Некоторым из неотропических животных тоже удалось проникнуть в Северную Америку по вновь образовавшемуся перешейку. Пглошерст (Erethlzori), успешно заселявший леса до Канады и Аляски включительно, принадлежит к одному из 14 семейств грызунов, возникших в Южной Америке, где и поныне живут другие
лесные виды дикобразов. Один вид водосвинок — крупных водных грызунов, принадлежащих к одному из таких же эндемичных семейств,— в ледниковый период проник в южные части Северной Америки и заселил их, но впоследствии вымер, как и огромные Gravigrada и глиптодонты (гигантские броненосцы), распространившиеся далеко на север из первоначального центра их эволюции на юге. В 1941 г. была найдена кость стопы одного из крупных ленивцев в том, что Шток назвал «мерзлым навозом» Аляски. Ее извлекли из ледниковых отложений, в которых здесь же были найдены кости сайги, бизона и мамонта (роды Палеарктической области), а также других животных, возникших в пределах Северной Америки (например, одного крупного верблюда) [53].
В кратком очерке можно лишь указать на то, что история областей Уоллеса в действительности вырисовывалась постепенно, подобно фотографическому изображению в медленно действующем проявителе; теперь уже доказано, что в течение части третичного периода, когда суша представляла собой архипелаг разобщенных континентов, существовали огромные отдельные «природные запо^ ведпики». Человек продолжает обмен видами, происходивший еще около 15 млн. лет назад, когда некоторые материки вновь соединились, и ускоряет его. Но эффективность наиболее значительных физических и экологических преград убедительно подтверждается тем, что эти «царства или области жизни» до сих пор сохраняют глубокий отпечаток независимой эволюции, нередко происходившей в экологических системах сходного типа (лес, пустыня, степь, озеро, река и др.). Вряд ли можно объяснить чистой случайностью то, что стирание первоначальных различий в результате двусторонних миграций наиболее заметно в относительно простых экологических системах арктической тундры, фауна и флора которой во многом носят циркумполярный характер; что второе место по сходству занимают бореальные леса Канады, Аляски, Камчатки, Сибири и северной части Европы [54]; и, наконец, что в Северной Америке и Евразии это сходство уменьшается по направлению к югу и сильнее всего черты прошлого сохранены в тропических лесах Центральной и Южной Америки, Африки и Восточной области — в зоне, наиболее богатой видами.
На Палеарктическую, Восточную и Эфиопскую области чрезвычайно сильное влияние оказали три обстоятельства: существование моря Тетис, горообразовательные процессы третичного периода и периоды оледенения. Море Тетис простиралось в виде пояса от Атлантики до Индо-Пацифики. Это было очень древнее море, более мелкое, чем главные океаны,— своего рода континентальное море, отделявшее Евразию от Африки и Индии, но служившее широким путем сообщения для тепловодного морского населения двух океанов. Ныне остатками его являются Средиземное и Черное моря, изолированные Каспийское и Аральское моря, Красное море и Индо-Пацифическая область. На геологических картах мира его часто довольно произвольно изображают как постоянную преграду между северными и южными материками, существовавшую на всем огромном протяжении палеозоя и мезозоя, т. е. по крайней мере до конца мелового периода. Но былое всемирное распространение столь многих наземных и пресноводных животных и наземных растений не позволяет поверить, что море Тетис никогда не пересекалось сухопутными «мостами». Временами они, вероятно, существовали.
В третичном периоде эта геосинклиналь частично поднялась, образовав столь же колоссальные горные цепи, протянувшиеся от Пиренеев до Гималаев и дальше, в юго-восточную Азию и Китай. К миоцену горообразовательный процесс зашел уже довольно далеко п создал преграду на северной границе Восточной области. Так как эти горные цепи занимают большую полосу примерно под одинаковыми широтами (пока не сворачивают к югу у восточного конца), они образуют довольно четкую границу между тропической и умеренной зонами. Таким образом, Восточная область — это область сравнительно поздняя, образование которой обусловлено появлением преграды для распространения видов к сев.еру и ее однотипным тропическим характером. Если бы не возникли горы п лежащие за ними пустыни, то эта область была бы теперь всего лишь тропическим поясом Палеарктической области и уже не сохранила бы характера столь богатого живого музея специфических форм прошлого. Во времена плиоцена, а возможно, и в последующий период ближайшие крупные острова Малайского архипелага
соединялись с материком; они отделились сравнительно недавно, значительно позднее, чем Мадагаскар или Новая Зеландия, но раньше, чем Великобритания.
Поднятие дна моря Тетис перерезало также основной канал между восточным и западным океанами в районе Среднего Востока, что в свою очередь означало появление широкой сухопутной связи между Палеарктической и Эфиопской областями. То, что это действительно имело место, подтверждается, помимо естественных логических следствий поднятия горных цепей, еще двумя группами данных. Во-первых, сходство между средиземноморской частью моря Тетис и восточными морями начинает резко уменьшаться в миоцене. До конца олигоцена во всех этих морях существовала весьма богатая тропическая фауна с коралловыми рифами, моллюском Nautilus и мечехвостами, которые теперь отсутствуют в западной Атлантике, a Nautilus — вообще во всей Атлантике [38]. Изменение ископаемых убедительно доказывает, что в миоцене море Тетис было разделено.
Во-вторых, мы находим ископаемые остатки наземных животных, перешедших в течение третичного периода из Африки в Палеарктическую область или в обратном направлении. Полагают, что даманы и слоны возникли на африканском континенте — хотя в раннетретичное время северная Африка находилась к югу от моря Тетис и поэтому фауна ее была «эфиопской», тогда как теперь она отделена от Эфиопской области Уоллеса пустынями и фауна ее носит палеарктический характер. Ископаемый материал по Африке пока еще слишком скуден, чтобы позволить нам датировать эти переселения (например, у нас нет плиоценовых остатков) [40]. Но за последнее десятилетие собраны весьма богатые коллекции миоценовых ископаемых, в том числе млекопитающих, насекомых и растений, из восточной Африки, особенно с островов озера Виктория; когда эти материалы будут полностью опубликованы, заполнится большой пробел в наших знанпях [41]. В раннетретичные времена в Африке, по-видимому, развивалась своеобразная фауна, хотя масштабы этой эволюции, вероятно, не были столь велики, как в Северной и Южной Америке. Мадагаскар представляет собой музей некоторых из этих своеобразных форм,
так как он был отрезан от материка или откололся от него еще до великих нашествий, происходивших в миоцене и плиоцене. Среди вселившихся в Африку форм находилась, по-видимому, большая часть тех крупных животных (например, антилоп), которых мы обычно считаем типично африканскими. Известно, что бегемот и жирафа появились в Африке в нижнем плейстоцене и почти несомненно пришли из Азии [40].
Плейстоценовый ледниковый период (или скорее ряд ледниковых периодов) оказал огромное влияние на распространение животных в Европе. По мере наступления льдов жизнь, конечно, уничтожалась (сохраняясь лишь на выступающих, подобно нунатакам, горах); различные зоны жизни отодвигались к югу, навстречу одновременному продвижению ледников с горных систем (которые, как уже было сказано, сами возникли лишь в третичном периоде), п в то время как на севере и в горах умеренной зоны изменялось количество выпадавшего снега и положение снеговой линии, в Африке происходили параллельные изменения влажности климата — дождливые эпохи чередовались с длительными засушливыми периодами. Еще одно косвенное следствие оледенений состояло в потере океанами огромного количества воды. В результате образовалось множество сухопутных связей, соединявших острова друг с другом и с материками; в тропиках это повлияло на рост коралловых рифов. Особенно тяжелую катастрофу испытала Европа: ее флора и фауна до сих пор гораздо беднее флоры и фауны Восточной Азии, где бедствие носило более или менее локальный характер. Каждый год наши сады обогащаются, черпая из обильного источника китайской растительности, сохранившейся со времен плиоцена. Не будь ледникового периода, мы имели бы, вероятно, чудесные смешанные леса с дикими магнолиями и лаврами и с эпифитными орхидеями, подобные тем, какие около 110 лет назад описал Гукер, путешествовавший по Сиккиму. В Китае растет около 500 видов деревьев [42]!
Хотя Австралия — очень древний материк, данные о том, что в конце мелового периода в ней обитали некоторые крупные динозавры, а также другие факты доказывают, что до того времени она находилась в контакте с остальным миром. К моменту открытия
ее европейцами там не было никаких плацентарных млекопитающих, кроме летучих мышей, грызунов и собаки динго, и наблюдался расцвет эволюции сумчатых — группы млекопитающих, вымершей во всех других местах, за исключением Южной Америки (где многообразие их значительно уменьшилось по сравнению с прошлым) и Северной Америки, где сохранился лишь один опоссум. Другая, еще более примитивная группа — яйцекладущие однопроходные (утконос и ехидна) — тоже имеет своим последним прибежищем Австралию и Новую Гвинею. Эти факты, по-видимому, доказывают, что Австралийская область порвала связь с Азией уже к третичному периоду или в самом его начале. Карта суши и морского дна района Малайского архипелага на первый взгляд не позволяет объяснить, каким образом такая изоляция могла сохраняться в течение примерно 80 млн. лет. Однако летом 1856 г. Уоллес проплыл от острова Бали до лежащего к востоку от него острова Ломбок, и вот что он писал: «Холмы покрыты густыми низкими зарослями бамбука, Buclda spinosa и других кустарников; равнины украшены сотнями величавых пальм, а во многих местах — пышной кустарниковой растительностью. Птицы многочисленны и необычны; здесь я впервые увидал много австралийских форм, совершенно отсутствующих на островах, лежащих западнее. В изобилии встречаются маленькие белые какаду, и их пронзительные крики, заметная белая окраска и изящные желтые гребешки делают их очень яркой чертой ландшафта. Это самая западная точка на земном шаре, где можно встретить представителей данного семейства. Некоторых мелких медососов из рода Ptilotis и необычного большенога Megapodius gouldil вы тоже впервые увп-дите лишь здесь, если вы держите путь на восток» [56].
Это внезапное изменение фауны Уоллес подробно описал в 1860 г., а позднее Т. Гексли назвал эту границу «линией Уоллеса». К северу она проходит между Борнео и Целебесом, по глубокому Макассарскому проливу — очень древнему геологическому образованию, служившему, по-видимому, препятствием для распространения пресноводных рыб на восток. Американский зоолог Рейвен [48], который провел несколько лет на Борнео и Целебесе, собирая животных, впоследствии нанес на карту все находки млекопитаю
щих па всем Малайском архипелаге п в прилежащих районах. Его карты показывают, как далеко распространилась каждая группа из своей первоначальной области, находившейся в Азии или в Австралия, и как убывает число видов по направлению к месту встречи двух фаун в центральной зоне, образуемой Целебесом, Малыми Зондскими и Молуккскими островами. Из Азии расселялись
Ф и г. 26. Распространение одного австралазпйского семейства птиц — какаду -- к востоку от линии Уоллеса [49 J.
Крестиком помечено место единственной находки к западу от этой линии.
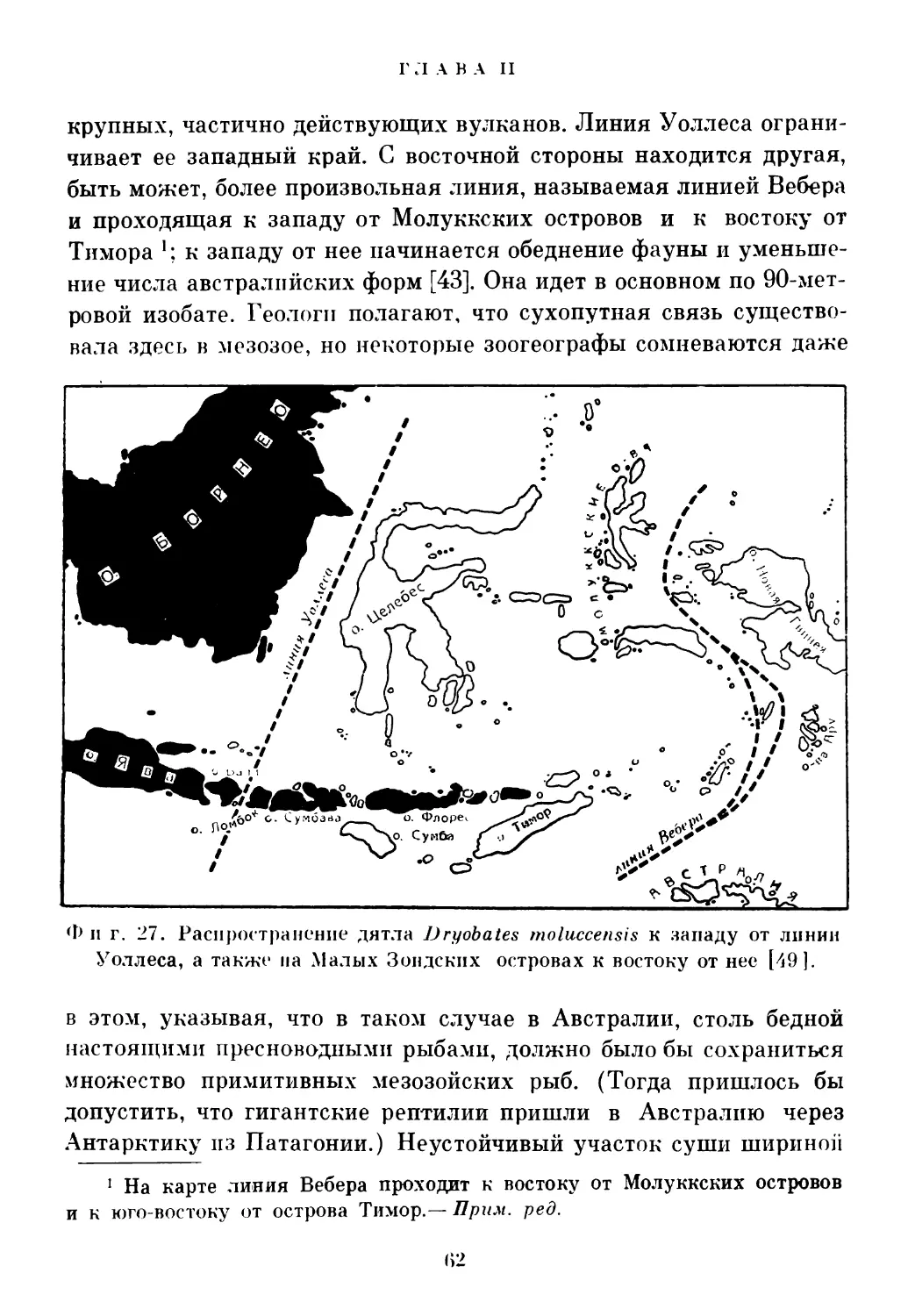
такие группы, как хищные, насекомоядные, грызуны семейства беличьих, слоны, такие копытные, как носорог и тапир, и приматы; пз Австралии — различные группы сумчатых, составлявших часть более богатой фауны материка. Только летучие мыши, как можно было ожидать, распространены повсюду. Три карты, составленные Реншем (фиг. 26—28), иллюстрируют такое распространение птиц и других групп животных. Как известно, центральная зона имела очень бурную геологическую историю; там и сейчас много
крупных, частично действующих вулканов. Линия Уоллеса ограничивает ее западный край. С восточной стороны находится другая, быть может, более произвольная линия, называемая линией Вебера и проходящая к западу от Молуккских островов и к востоку от Тимора к западу от нее начинается обеднение фауны и уменьшение числа австралийских форм [43]. Она идет в основном по 90-метровой изобате. Геологи полагают, что сухопутная связь существовала здесь в мезозое, но некоторые зоогеографы сомневаются даже
Ф и г. 27. Распространение дятла Dryobates moluccensis к западу от линии Уоллеса, а также па Малых Зондских островах к востоку от нес [49].
в этом, указывая, что в таком случае в Австралии, столь бедной настоящими пресноводными рыбами, должно было бы сохраниться множество примитивных мезозойских рыб. (Тогда пришлось бы допустить, что гигантские рептилии пришли в Австралию через Антарктику из Патагонии.) Неустойчивый участок суши шириной
1 На карте линия Вебера проходит к востоку от Молуккских островов и к юго-востоку от острова Тимор.— Прим. ред.
1000—1300 км, время от времени погружавшийся под уровень моря и впоследствии отрезавший Австралию от Азии, послужил преградой, обусловившей образование еще одной уоллесовской зоогеографической области. В начале третичного периода Борнео и Ява, вероятно, еще не поднялись над морем, так что эта зона была в то время шире. Но можно было бы предположить (насколько мне известно, этого еще никто не делал), что эффективность этой преграды определялась еще одной причиной — экологическими пертурбациями, возникавшими при происходивших время от времени
Фиг. 28. Распространение австралазнйских родов млекопитающих, птиц, рептилий, амфибий, бабочек и наземных улиток в районе Малайского архипелага и в восточной части Новой Гвинеи [49}.
Проценты показывают относительное число таких родов на различных островах.
вторжениях животных с той и с другой стороны на острова с относительно ненасыщенными сообществами.
Для того чтобы придать законченность определению Австралийской области, нужно еще отметить, что удивительный ряд островов к востоку от Австралии, включающий Новую Зеландию, Новую*
Каледонию, Новые Гебриды, Соломоновы острова и острова Фиджи, составляет часть того, что Зюсс назвал «осколками затонувшего материка». Дальше к востоку все острова — чисто океанического происхождения: они образовались из вулканов или выраставших на них коралловых рифов. Эти океанические острова никогда не были соединены с сушей, и поэтому их фауна и флора бедны и состоят из потомства случайных пришельцев, чрезвычайно редко попадающих на тот или иной остров. Мы могли бы назвать этот район Тихоокеанской областью. И хотя здесь никогда не было материка, вряд ли можно оставить без рассмотрения зоогеографию островов океана, площадь которого больше площади всех материков и островов мира, взятых вместе! С точки зрения экологии современные нашествия животных и растений на эти острова принадлежат к самым интересным, хотя и плачевным событиям нашей эпохи (см. гл. IV).
Распространение пресноводных рыб замечательнейшим образом подтверждает (если в этом еще есть необходимость) сделанные выводы о времени и ходе развития областей Уоллеса. Настоящие пресноводные рыбы, т. е. виды, не выходящие в эстуарии пли в море, вероятно, могут преодолевать морские преграды только в результате каких-нибудь редких случайностей. Их медленное расселение в очень большой степени зависит от изменений водных систем — смещения водоразделов, соединения устьев рек при поднятиях суши, а иногда происходит путем «перепрыгивания» через водоразделы рек и озер при случайном переносе икры ветром и птицами или в результате деятельности человека [47]. В конце мелового периода началась быстрая эволюция обширной группы современных рыб, так называемых карпообразных, или костнопузырных (Ostariophysi), к которой относится большинство пресноводных рыб земного шара и около четвертой части всех видов рыб вообще; эта эволюция продолжалась и в рапнем эоцене. Известны п две группы более древних пресноводных рыб; двоякодышащие и Osteoglossidae, отдельные представители которых до сих пор живут в разных частях света (двоякодышащие — в Австралии, Африке и Южной Америке, Osteoglossidae — в северной Австралии, на Борнео и Суматре, на полуострове Малакка, в верховьях Нила, в
западной Африке и Южной Америке). Двоякодышащие существовали еще в палеозое, и австралийская форма в течение всего мезозоя была распространена по всему свету. Ранние формы Osteoglos-sidae встречаются в эоцене Северной Америки и в меловых и эоценовых отложениях Англии [50]. Но 30 семейств карпообразных практически совершенно отсутствуют в Австралазии. Майерс [46] пишет: «Макассарский пролив образует одну из самых поразительных зоогеографических границ между пресноводными ихтиофаунами мира. К востоку от него лежит Борнео с его 17 семействами и 300 или более видами первично пресноводных рыб. А в каких-нибудь 140 км к западу находится Целебес, на котором такие рыбы представлены лишь двумя видами, вероятно завезенными человеком».
Одна обширная группа карпообразных — сомовидные (Siluroi-dea) —содержит несколько австралийских видов, по-видимому перешедших в пресные воды вторично, после промежуточной эволюции в море. В Неотропической области ныне имеется 9 эндемичных семейств Siluroidea, в Неарктической — одно семейство, не считая одного вида в Китае; эндемичные семейства есть также в Эфиопской и Восточной областях. Пресноводные рыбы очень медленно расселяются через новые связующие участки суши; их можно было бы назвать «живыми ископаемыми» в том смысле, что их современное распространение часто на один или два геологических периода отстает от распространения более подвижных млекопитающих.
Глава III
НАШЕСТВИЯ НА МАТЕРИКИ
Т£ огда мы размышляем о вселениях растений, животных и их ми-крое конических паразитов на новые для них материки и острова, мы воспринимаем их как некие пертурбации, влекущие за собой неожиданные последствия, которые в свою очередь приводят к усложнению экологических систем, и без того достаточно сложных для понимания, не говоря уже об их контролировании и о нагромождении новых трудностей для человека. Эти трудности особенно возросли за последние 150 лет, и нам приходилось бороться с ними при помощи ряда довольно поспешных временных мер паллиативного характера, лишь иногда подкрепленных основательным биологическим изучением популяций или хотя бы систематическим наблюдением за ходом событий. Действительно легко почувствовать то, о чем писал Эдвард Гиббон в конце своей книги «The Decline and Fall of the Roman Empire»: «Историк может похвалиться важностью и разнообразием описываемых им событий; но, сознавая несовершенства своего труда, он нередко должен в этом винить неполноту самих источников» !. Мы, однако, не имеем в виду критиковать биологов, которым приходилось со скудными средствами решать нескончаемый ряд непредвиденных и не терпящих отлагательства проблем; к тому же у нас есть несколько превосходных, тщательно скомпилированных историй экологических «нашествий» — особенно следует отметить работу различных ведомств Сельскохозяйственного департамента США, которые впервые внесли известную методичность в изучение этих вопросов. В настоящей главе мы рассмотрим лишь несколько примеров, вы-
1 Э. Гиббон, «История упадка и разрушения Римской империи», перевод В. Н. Неведомского, ч. VII, изд. К. Т. Солдатенкова, Москва, 1886.
бранных не столько ввиду их практического значения для хозяйства, медицины или ветеринарии, сколько главным образом потому, что они хорошо иллюстрируют идеи этой книги, или просто потому, что по ним имеются хорошие карты, отражающие процесс заселения материков чуждыми видами.
Никто по существу не знает, сколько видов распространилось за пределы своих естественных ареалов, но таких видов, вероятно, сотни тысяч, причем тысячи из них оказали заметное влияние на жизнь человека — вызвали гибель людей или повысили стоимость жизни. Если говорить о достаточно далеком будущем, то биологический мир станет в конце концов не сложнее, а проще и беднее. Вместо шести континентальных областей с их второстепенными компонентами — фаунами горных вершин, островов и пресных водоемов,— разделенных преградами, препятствующими расселению организмов, образуется единый мир, в котором сохранившиеся дикие формы будут распространены до границ, определяемых только их наследственными свойствами; механические же преграды будут преодолены. Если бы мы построили шесть огромных резервуаров, наполнили их водой и при помощи узких трубок, снабженных кранами, соединили каждый из них со всеми другими; если бы мы добавили в эти резервуары различные смеси ста тысяч разнообразных химических веществ в растворе, а затем ежедневно открывали каждый из кранов на одну минуту, то вещества начали бы постепенно диффундировать из одного резервуара в другой. Если бы трубы были узкими и имели тысячи километров в длину, то процесс шел бы очень медленно. Могло бы потребоваться чрезвычайно много времени, чтобы вся система пришла к окончательному равновесию, причем к этому времени множество исходных веществ в результате химических реакций перестало бы существовать, уступив свое место новым соединениям или веществам, пришедшим из других резервуаров. Эти резервуары — материки, а трубы — транспортные средства, обеспечивающие торговые связи; но человеку не удалось полностью закрыть краны, несмотря на то, что во многих случаях он хотел бы сделать это. И хотя есть закон сохранения материи, закона сохранения видов не существует.
Одна из главных причин расселения и акклиматизации видов заключалась в том, что человек распространял по всему миру растения, особенно те, которые он намеренно ввозил для развития сельского и лесного хозяйства или для украшения садов. Фэрчайлд, бывший руководителем ведомства по интродукции растений в США, в своей книге о путешествии по тропикам между прочим упоминает, что работа этой организации «привела к интродукции почти 200 000 различных видов и разновидностей растений из всех частей света» [78]. Это весьма солидное дополнение к местным флорам! И как торговцы следуют за завоевателями, так животные следовали за растениями. Например, летом 1916 г. в одном питомнике в Нью-Джерси было найдено около дюжины странных майских жуков; их определили как Popillia japonica и назвали «японскими жуками». Из этого очага жуки начали быстро распространяться [102]. В первый год они населяли участок площадью меньше 0,5 га. В последующие семь лет их ареал занимал соответственно 8, 18, 120, 257, 675, 1833 и 6105 кв. км, причем последняя цифра относится к 1923 г. (фиг. 29). Дальнейшее распространение японского жука вплоть до 1941 г., когда он населял свыше 50 000 кв. км, показывает карта на фиг. 30. Эти жуки были завезены, вероятно, в 1911 г. с партией ирисов или азалий из Японии. В Японии они редко причиняют вред, но в Америке их численность вначале была чудовищной [66]. Уже в 1919 г. один человек мог собрать до 20 000 жуков в день; сообщают, что в одном фруктовом саду, где было 156 не очень старых персиковых деревьев, за два часа собрали 208 галлонов 1 жуков, а на следующий день число их на деревьях, казалось, не уменьшилось! Популяция жуков, способных питаться на более чем 250 видах деревьев и других растений (в их числе — более десяти важных культур, от сои и клевера до яблони и персика), причем они нередко уничтожают всю листву, поистине представляет собой нечто страшное [64]. Столь же серьезным оказалось нашествие другого восточного насекомого, завезенного из Японии с посадочным материалом в начале 20-х годов нашего века,— червеца Pseudaonidia duplex. Он размножился в Луизиане, Техасе и
1 Галлон — мера емкости, равная 3,8 л.— Прим. ред.
Алабаме. Этот вид может жить почти на 200 видах растений, средн которых наиболее важны цитрусовые [72]. Однако в Японии он не является серьезным вредителем; то же самое можно сказать об азиатском жуке Autoserica castanea, который живет в Китае и
Фиг. 29. Границы последовательного распространения японского жука (Popillia japonica) в 1916—1923 гг. из пункта его первоначального появления в Нью-Джерси [102].
Японии, но в 1921 г. попал в Нью-Джерси и впоследствии стал вредителем садов в окрестностях Нью-Йорка [86]. В этом районе в дерновом слое иногда совместно встречаются личинки Autoserica cas-ranea, Popillia japonica и еще одного жука, завезенного сюда в 20-х годах,— Anomala orientalis [58].
ГЛАВА Ш
Новые виды проникают в ту или иную страну из всех частей света. Едва ли не половина встречающихся в США важнейших вредителей (около 180 видов) занесена из других стран [100]. Больше
Ф и г. 30. Расселение жука Popillia japoniea в США в период 1916—1941 гг. [105].
Впоследствии он распространился еще дальше — в Северную Каролину, Западную Виргинию и Огайо, а также на север; отдельные «островки» появились и за пределами этих районов.
всего «нашествий» в Северную Америку происходило из Европы, что и следовало ожидать из-за существующих между ними давних и интенсивных транспортных связей. К самым ранним пришельцам
относятся гессенская муха (Mayetiola destructor) — вредитель пшеницы — и Яблоновая плодожорка (Carpocapsa pomonella), наносящая вред главным образом насаждениям яблони; за ними последовала длинная цепь других иммигрантов [90]. Одни, как, например, долгоносик Hylastinus obscuras, появившийся около 1878 г., заселили в основном всю страну (так же, как и скворец, а до него домовый воробей) [98]. Другие, например кукурузный мотылек (Pyrausta nubilalis), начавший распространяться в 1917 г., двигались довольно медленно, но неуклонно и, вероятно, до сих пор расширяют свои ареалы [59, 106]. Некоторые виды, завезенные сравнительно недавно, возможно, приобретут гораздо большее значение в будущем; таковы, например, европейский майский хрущ (Melo-lontha vulgaris, в Америке называемый Amphimallon majalls), который достиг штата Нью-Йорк в 1942 г. [85], и картофельная нематода Heterodera rostochlensis, обнаруженная на Лонг-Айленде примерно в зто же время, но проникшая туда, вероятно, десятью годами раньше [65]. Эти два вида вредят растениям как в Европе, так и в Америке: личинки хруща живут среди корней, взрослые жуки уничтожают листву деревьев, а нематода повреждает картофель. В настоящее время нематодой заражено лишь около 320 га площади, занятой картофелем, но искоренить ее пока не удается.
Обращаясь к другим материкам, пославшим своих представителей в Северную Америку, можно назвать долгоносика Listroderes obliquus и аргентинского муравья Iridomyrmex humllis, прибывших из Южной Америки. Долгоносик появился в США в 1922 г. п заселил три южных штата [87]; он проник также в Австралию и Южную Африку. Муравья впервые обнаружили близ Нью-Орлеана (штат Луизиана) в 1891 г., но, вероятно, он был завезен туда несколько раньше — возможно, на судах, доставлявших кофе из Бразилии. Этот необычайно агрессивный муравей имеет своей родиной Южную Америку и был первоначально описан по экземплярам’из Аргентины, а позже найден в Бразилии, Уругвае и других местах. Ньюэлл и Барбер [94] замечают: «о том, что Аргентина — родина этого муравья, свидетельствует также то обстоятельство, что в этой стране он, по-видимому, обычно не приносит существенного вреда». Спустя менее 50 лет после его появления в районе Нью-Орлеана
аргентинский муравей заселил значительную часть южных штатов и проник также (к 1905 г.) в Калифорнию, где широко распространился (фиг. 31). Повсюду он неимоверно размножался и массами вторгался в дома и сады (в том числе и в фруктовые), поедая пищевые продукты или — вне помещений — других насекомых,
Фиг. 31. Распространение аргентинского муравья Iridomyrmex humilis в США [103].
Точками обозначены очаги распространения, обнаруженные в результате обследования; крестами — местности в Калифорнии, из которых поступили сообщения о наличии муравья; квадратиком — изолированный очаг муравьев.
а также питаясь выделениями червецов и тлей на различных деревьях; к этим деревьям и обратно муравьи бегают по определенным «тропинкам», точно так же, как у нас в Англии лесные муравьи ходят, например, к соснам и березам. Аргентинские муравьи забираются также в ульи за медом. Характерно, что эти свирепые и многочисленные муравьи полностью изгоняют местных муравьев. Смит пишет, что он «часто наблюдал сражения между местными и аргентинскими муравьями... Так как аргентинские муравьи по мере своего продвижения уничтожают фактически всех местных
муравьев, то определить границы захваченной ими области сравнительно легко... Как только аргентинские муравьи начинают исчезать, местные муравьи вновь появляются на территории и спустя несколько лет становятся столь же многочисленными, как и прежде» [103]. Так же, быть может, волки, лисицы и ягуары продвигались во времена плиоцена в Южную Америку, вытесняя местных плотоядных сумчатых семейства Borhyaenidae. Аргентинский муравей, в сущности, не так уж быстро распространяется в природе, поскольку спаривание у него происходит почти только в гнезде, а за год он мог бы проползти не больше нескольких сот футов. Столь быстрое расселение его в США произошло, по-видимому, с помощью торгового транспорта, особенно железнодорожного.
Стремительному вторжению аргентинского муравья подверглись и другие страны. В Южной Америке и Австралии наблюдалось такое же исчезновение местных муравьев. На юге и западе Австралии аргентинский муравей появился к 1939—1941 гг., а в 1951 г. он захватил еще один плацдарм, в Новом Южном Уэльсе. В 1955—1956 гг. заселенные муравьем площади составляли в Западной Австралии свыше 100 кв, км, в штате Виктория — 25 кв, км и в Новом Южном Уэльсе 8,5 кв. км, В Америке нашествие было несколько ослаблено с помощью отравленных приманок, но в Западной Австралии муравьи не трогали приманку; однако здесь с большим успехом применяли контактные инсектициды, например ДДТ и диэлдрин. После опрыскивания домов высокими концентрациями диэлдрина инсектицид продолжал вызывать гибель муравьев в течение четырех с половиной лет; кампания с целью окончательного истребления муравьев все еще в разгаре. В очищенных районах (опрыскивание губит не только муравьев, но попутно и всех мух, комаров, тараканов и блох и, возможно, многие другие виды) вытесненные местные виды муравьев быстро появляются вновь и занимают местность, как только территория обеззараживается [84].
Лучшим примером вида, завезенного из Австралии, служит червец Icerya purchasi, который появился в Калифорнии около 1868 г. и начал угрожать всей будущности цитрусовых насаждений штата.
Этот червец хорошо известен среди сельскохозяйственных энтомологов тем, что его удалось за два года решительно ограничить с помощью многочисленного потомства, полученного всего от 139 особей божьей коровки Novlus (Vedalla) cardinalis — естественного врага этого червеца в Австралии. Австралия была источником яда, но она же дала и противоядие, и это чудо экологического исцеления впоследствии было совершено во всех других странах, в которые проник червец leery a purchasl и где он начал вредить (например, в Европе, Сирии, Египте, Южной Африке, Японии, Новой Зеландии, Южной Америке и на Гавайских островах). Об открытии такого метода борьбы (идея принадлежала Райли, а полевую работу провел Кёбеле) Ховард замечает: «Вероятно, столь поразительного успеха никогда больше не удастся достигнуть в США», и добавляет, что этот успех породил слишком много оптимизма относительно легкости, с которой можно использовать интродукцию в|рагов и паразитов для борьбы с завезенными вредителями [90]. Например, тщательно продуманная интродукция паразитов и естественных врагов из Японии для борьбы с жуком Popil-На japonlca не дала такого повсеместного и быстрого результата, какой был достигнут в борьбе с червецом.
Перемещение вредителей культурных растений через Атлантический океан не было односторонним. Примерно 90 лет назад во Франции размножилась американская тля филлоксера (Phylloxera vitlfolii), до этого мирно существовавшая на диких виноградных лозах в США, к востоку от Скалистых гор. Попав в страну через Бордо, а может быть, также и через другие порты, филлоксера очень быстро проникла в винодельческие районы Европы и в Алжир; она переносится с виноградными лозами и часто распространяется с ними из одного района в другой [90]. Филлоксера не менее 18 раз появлялась и на Британских островах [110]. Образуемые ею корневые галлы вызывали гибель лозы в Европе. После того как упадок виноделия достиг крайней степени и 1 200 000 га французских виноградников были уничтожены, одному французу пришла в голову мысль прививать европейские сорта винограда на американские сорта, устойчивые к корневой фазе развития филлоксеры, и экономическая опасность миновала. Подобное же нашествие про
изошло в Австралии в 1875 г. и в Калифорнии в 90-х годах прошлого века.
Из США пришел и другой знаменитый «интервент» — колорадский жук (Leptinotarsa decemllneatd), в природных условиях живущий в восточной части района Скалистых гор, от Колорадо до
Фиг. 32. Естественное распространение колорадского жука (Leptinotarsa decemlineata) на западе Северной Америки до перехода его на культурный картофель [104].
Встречавшийся первоначально в Нью-Мексико и Аризоне, этот жук в конце XVIII в. начал медленно расселяться к северу по прериям вдоль восточной части Скалистых гор (показано крестиками и буквой d). На западе распространение ограничивали Скалистые горы и засушливая долина реки Колорадо, на востоке — область, в которой нет растений рода Solanum. Буквами i,j и т показано распространение трех других видов Leptinotarsa.
Мексики, и питающийся главным образом на дикорастущем паслене Solanum rostratum (фиг. 32). Он может также кормиться на других представителях семейства пасленовых и на некоторых иных растениях [104]. Когда энтомологи впервые нашли этого жука в Колорадо, там еще не выращивали культурный картофель (Solanum tuberosum), и жук начал распространяться только после того, как его будущее новое кормовое растение — картофель — продвину-
лось на запад и достигло естественного ареала жука. После этого, т. е. начиная с 1859 г. и в течение примерно 20 лет, колорадский жук распространялся на восток и к 1874 г. достиг берегов Атлантического океана (фиг. 33). Некоторое время думали, что в области расселения этого жука невозможно будет выращивать картофель.
Фи г. 33. Нашествие колорадского жука (Leptinotarsa decemlineata) на культурный картофель в Северной Америке (показано вертикальной штриховкой) [104].
На севере (косая штриховка) — зона серьезного повреждения картофеля; ее окружает зона значительного, но не очень сильного повреждения (клетка). Распространению жука на запад отчасти препятствовали высокие горы (горизонтальная штриховка) и пустыня (П);------------------------крайняя граница расселения жука.
Первый заокеанский плацдарм был захвачен жуком в 1876 г., когда он проник в Германию, но его там уничтожили. Последующие небольшие вторжения, в том числе в районе Тильбюри в 1901 г., тоже не имели успеха; только в 1920 г. популяциям, проникшим (подобно филлоксере) из-за океана в Бордо, удалось размножиться, и к 1935 г. жук распространился по всей Франции, а позже и в других странах (фиг. 34). Время от времени он попадал и в Англию (в 1947 г. было 264 сообщения о размножении жука), но до
сих пор с ним удавалось справляться. В 1955 г., когда в Англии и Уэльсе под картофелем было занято около 242 000 га земли, там было обнаружено всего лишь 53 колорадских жука; по-видимому, все жуки были уничтожены, так как вспышек внутри страны не было. В Шотландии на ее 70 000 га нашли одного жука, а в Ирландии на 47 000 га — ни одного [75].
Итак, в XVI—XVII вв. неарктический вид Solatium ввезли в Европу; там из него вывели новые формы, эти формы вновь увезли в Северную Америку, где они достигли ее западных районов, что вызвало распространение жука на восток, приведшее в конце концов к заселению им Европы. Но растительноядный жук до сих пор еще не достиг равновесия со своим кормовым растением.
Последним из крупных североамериканских «интервентов» была гусеница Hyphantrla сипеа, попавшая в Венгрию во время второй мировой войны, в 1940 г. Через несколько лет она быстро распространилась по Австрии, Чехословакии, Румынии и Югославии [76]. Эта гусеница способна совершенно уничтожать листву многих деревьев и других растений, что она делает и в Европе. Она не наводняет леса, а останавливается у опушек — по крайней мере в настоящее время. Одна из своеобразных ее особенностей состоит в том, что в Европе она предпочитает тутовые деревья, которые почти не трогает в Америке.
Описывая нашествия чужеземных форм на нескольких примерах, пусть даже самых выдающихся, трудно дать полное представление о бурном натиске видов, вырывающихся или уже вырвавшихся из пределов родного материка на просторы земного шара. Поистине мы могли бы повторить слова Уолта Уитмена из его поэмы с весьма подходящим названием «As consequent, etc.»:
Текут по полям Огайо и по его лесам,
Текут вниз по каньонам Колорадо от источников вечных снегов, Текут, исчезая, в Орегоне и там, на юге, в Техасе,
Текут на севере, пробивая себе путь к Эри, Ниагаре, Оттаве, Текут в заливы Атлантики, в великий соленый простор.
Во всех тех случаях, когда нам известна история вторжения, оно начиналось с появления очень небольшого ядра популяции, рост которой со временем становился подобным бурному потоку, а затем
Фиг. 34. Распространение колорадского жука (Leptinotarsa decemlineata) в Европе в 1956 г.
Штриховкой показана область современного распространения;|белыми оставлены области, где жука нет или отмечаются лишь немногие изолированные «вспышки»;’покрыты точками области, для которых нет сведений.
нередко и наводнению. Когда популяция достигает таких размеров, ее распространению редко может помешать что-либо, кроме естественных преград в окружающей среде. Последовательное расселение вредителей сельскохозяйственных культур по земному шару иллюстрируют шесть карт (фиг. 35—40) из серии, составленной Государственным институтом энтомологии [68]. (Аналогичную серию карт для микозов сельскохозяйственных культур и деревьев опубликовал Государственный институт микологии.) Эти карты показывают ряд различных этапов распространения видов. Конечно, многие виды еще не вышли за пределы своего материка, даже если они широко расселились на нем вместе с распространением культурных растений. Жук Poplllla japonica (фиг. 35) и пилильщик Gllpinia hercyniae (фиг. 36) — палеарктические виды, проникшие в восточную часть Северной Америки. Первый из этих видов — восточноазиатский, второй — европейский. Колорадский жук, как уже говорилось, служит примером экспансии в противоположном направлении (фиг. 37). «Люцерновая блоха» Sminthurus virldis (это не блоха, а ногохвостка) достигла юга Австралии и Дальнего Востока, но не проникла в Северную Америку (фиг. 38). Бабочка репница (Pieris гараё) — тоже палеарктический вид, который в середине прошлого века распространился по всей умеренной зоне Северной Америки, а также прочно обосновался на Бермудских и Гавайских островах, в Австралии и Новой Зеландии (фиг. 39). Австралийский червец leery a purchasi проник, по-видимому, почти во все страны, в которых он может существовать (фиг. 40). Мы видим, что, хотя таких же конечных результатов достигнет, возможно, и расселение других видов, этот процесс еще очень далек от своего завершения, особенно ввиду того, что человек теперь стойко охраняет границы зоогеографических областей с помощью массовых систем карантина и истребительных мероприятий. И все же, несмотря на карантин (по крайней мере в США), одному весьма серьезному вредителю хлопчатника удалось в течение примерно 25 лет проникнуть на четыре новых для него материка. Это розовый червь (Pectlnophora gossypielld) — небольшая коричневая бабочка, получившая свое видовое название по последней фазе своей гусеницы, имеющей розовую окраску; первоначально она жила,
Фиг. 35. Распространение жука Popillia japonica (на 1952 г.).
Фиг. 36. Распространение елового пилильщика Gilpinia hercyniae (на 1953 г.).
вероятно, в Индии, а может быть, и во всей юго-восточной Азии, на дикорастущем и культурном хлопчатнике. Она, возможно, была и в центральной Африке, прежде чем началось ее расселение по всему миру; но полагают, что ее сначала завезли в Египет с импортированным хлопчатником или его семенами из Индии в 1906 г.; из Египта, где она привлекла к себе внимание в 1911 г., она распространилась в восточную и западную Африку, а также в Китай и на различные острова, в частности Филиппинские и Гавайские.
Ф иг. 37. Распространение колорадского жука (Leplinotarsi decemlineata) (на 1951 г.).
В Мексику она попала в 1911 г., а в Техас (с мексиканскими семенами хлопчатника) — к 1917 г. Позже она проникла в Бразилию и Австралию [91].
Можно заметить, что вселениям чаще всего подвергаются земледельческие области или территории, существенно измененные в результате деятельности человека. Но некоторые виды (таких пока меньшинство) проникают дальше, в природные водоемы и леса— в биоценозы, которые все еще сохраняют свое богатство и разнообразие даже после воздействия таких связанных с человеком факторов, как пожары, смена растительности после рубок леса, регулирование стока вод или прокладка осушительных каналов. Из этих видов я уже описал североамериканскую ондатру, южноаме-
Ф п г. 38. Распространение ногохвостки Sminthurus riridis («люцерновой блохи»), происходящей из Европы (па 1956 г.).
Фиг. 39. Распространение бабочки репницы (Pieris гарае), происходящей из Европы (па 1952 г.).
риканскую нутрию и китайского мохноногого краба и упомянул о каролинской белке. Мы можем дополнить этот список примерами из небольшого контингента видов, завезенных из Европы и случайно проникших в лесные области Северной Америки [71]. Прежде всего это насекомые, которые, подобно уже описанному японскому жуку, нападают не столько на естественные леса, сколько на сады, зеленые изгороди и другие искусственные насаждения. К группе бабочек, в разное время появившихся и размножившихся в лесах
Ф и г. 40. Распространение червеца Icerya purchase, происходящего из Австралии (па 1955 г.).
восточных штатов, относятся непарный шелкопряд (1869 г.), златогузка Nygmia phaeorrhoea (1897 г.) и завезенная совсем недавно (около 1949 г.) зимняя пяденица (Operophtera brumata). Широколиственные деревья тоже приобрели новых иноземных вредителей: таковы два вида пилильщиков, минирующих листья березы,— Phyllotoma nemorata и Fenusa pusilia (первый обнаружен в 1905 г., время появления второго не вполне ясно); ивовая волнянка (Stilpnotia salicis) — вредитель тополей и ив (с 1920 г.); жук Plagiodera versicolora — довольно обычный вид, скелетирующий листья подстригаемых ив по берегам английских рек, а в Америке [89] наносящий такие же повреждения как местным, так и иноземным видам ив (с 1911 г.); короед Scolytus multistriatus (с 1909 г.)
и листоед Gallerucella xanthomeleana (примерное 1840г.), хотя и не проникающий в глубину лесов; и, наконец, червец Cryptococcus fagi, появившийся к 1890 г. [81]. Хвойные деревья также подверглись колоссальным нашествиям вредителей, до сих пор продолжающих распространяться: появились еловый пилильщик Gilpinia (Dipriori) herciniae (по крайней мере с 1922 г.); три вида соснового пилильщика — Diprion simile (в 1914 г.), Neodiprion sertifer (в 1925 г.) и Diprion frutetorum (время вселения не вполне установлено); тля Adelges (Chermes) piciae-, лиственничная моль Coleophora larlcella (с 1909 г.); моль Rhyacionia buoliana, проникшая в США к 1914 г., а в Канаду — в 1925 г. [69].
Обычно считали несомненным, что лиственничный пилильщик (Pristophora erichsonii) — страшнейший вредитель леса в Северной Америке — тоже пришел из Европы. Бесспорно, он имеет теперь голарктический ареал, так же как и лиственница, представленная четырьмя видами — североамериканским, сибирским \ японским и европейским. Но, по заключению Коппеля и Лейюса [70], фактические данные «указывают только на то, что лиственничный пилильщик уже некоторое время существовал в Северной Америке. Его происхождение нельзя определить на основании имеющихся сведений». Возможно, что он, подобно гессенской мухе,— очень ранний пришелец.
Лесные энтомологи Канады и США составили ряд превосходных сводок данных и карт, характеризующих нашествия вредителей; четыре такие карты мы приводим здесь (см. также фиг. 69). Зимняя пяденица появилась настолько недавно, что едва успела попасть в учебники [79, 82]. Она завладела лишь небольшим плацдармом в Новой Шотландии (фиг. 41). Червец Cryptococcus fagi имеет центры распространения в различных местах. Его медленное наступление прямым фронтом, начавшееся от Галифакса в Новой Шотландии, куда он попал до 1890 г. (фиг. 42), отличалось удивительной регулярностью. Это насекомое способствует естественному
1 В Сибири имеются два вида лиственницы — сибирская (Larix sibiri-са), растущая к востоку до среднего течения Нижней Тунгуски, и даурская (L. daurica), занимающая верховья Нижней Тунгуски и всю территорию Якутии и Дальнего Востока.— Прим. ред.
Фиг. 41. Начало одного вторжения на новый континент— распространение в Канаде зимней пяденицы (Operophtera brumata), завезенной из Европы около 1949 г.
заражению древесины бука пагубным паразитом — грибом Nectrla, вызывающим рак деревьев, В Новой Шотландии в 1948 г. более 80% буков были поражены |раком; не лучше обстояло дело и на острове Принца Эдуарда. Этого червеца нередко можно видеть в трещинах коры некоторых буков на Британских островах, но только в Дании и на востоке Северной Америки экологические условия способствуют развитию губительной болезни.
t Болезнь вязов тоже вызывается совместным действием насекомых и одного гриба — Cerastomella ulmi. Она повредила и погубила многие деревья в Европе; место ее происхождения не известно. С 1927 г. она распространилась также в Англии, где получила название «голландской болезни вязов»; вначале от нее гибло устрашающее число живых изгородей из вязов, но теперь она перешла в хроническую форму и дает в отдельные годы лишь местные вспышки в разных частях страны [95]. Такие погибшие деревья часто можно видеть (фиг. 43). В Англии гриб распространяют и частично вносят в ткани деревьев два вида короедов: Scolytus multlstrlatus и S. destructor (= scolytus)-, это связано с тем, цто взрослые жуки кормятся в свежей коре молодых веточек, часто в местах разветвлений, и могут жить на совершенно здоровых деревьях, тогда как ходы, служащие для размножения, встречаются обычно под корой менее здоровых, отмирающих или срубленных деревьев [83]. Один из этих короедов, S. multlstrlatus, еще в начале нашего столетия был завезен в США, вероятно, с неокоренным лесом, прибывавшим в морские порты и доставлявшимся, быть может, также и в глубь страны [67]. Впервые жука нашли в Массачусетсе в 1909 г., и с тех пор он проник во многие части Новой Англии; в 1938 г. его распространение выглядело так, как будто оно началось из двух главных центров — одного в районе Нью-Йорка и другого, почти смежного, в южной части Нью-Гэмпшира. В этих участках и за их пределами живет местный ильмовый короед (Hylurgoptnus ruflpes), тоже способный переносить гриб, хотя, может быть, и не столь эффективно, как европейский вид. Недавно было обнаружено весьма широкое распространение третьего короеда, Scolytus sulcatus, который не был известен в течение многих лет и был обнаружен лиш^ после того, как успел
Фиг. 42. Медленное распространение европейского червеца Cryptococcus fagi в Канаде (за период около 60 лет) [81].
К 1926 г. Новая Шотландия была сплошь заселена червецом. Остров Принца Эдуарда оказался сплошь заселенным к 1929 г., аостров Кейп-Б ретон— к 1926 г.
широко распространиться [96]. Этот вид живет в основном на яблонях, но переходит и на вязы; его возможная роль в распространении заболевания изучена еще мало. Сама болезнь (т. е. сочетание вяза, короедов и гриба) распространена в зоне, гораздо
Ф и г. 43. Отмирающий вяз и группа здоровых вязов в Оксфордшире (фото автора, 1957).
Слева от больного дерева видна сохранившаяся часть другого вяза, отмершая вершина которого отвалилась раньше. Болезнь вызывается грибом Cerastomella ulmi, который переносят короеды.
более узкой, чем ареал короеда (фиг. 44). Полагают, что гриб проник в страну с зараженной вязовой древесиной, употреблявшейся для изготовления фанеры; он был впервые обнаружен в Огайо в 1930 г. За период с 1934 по 1940 г. основная область, охваченная заболеванием, возросла примерно с 6250 почти до 97 500 кв. км, не считая отдельных «островков» за ее пределами. Эта основная область в 1940 г. включала половину штата Нью-Джерси, юго-восточный угол штата Нью-Йорк и часть восточного Коннектикута. В экологическом отношении это крупное нашествие интересно тем, что насекомое-переносчик, по-видимому,
появилось на континенте и распространилось раньше гриба — подобно тому, как переносчик желтой лихорадки, комар Aedes aegypti, имеет гораздо более широкий ареал, чем область современного распространения вируса, который еще не достиг Азии. Гибель вязов, вызванная самой болезнью, а также
Фиг. 44 .'^Проникновение короеда Scolytus multistriatus из Европы в Север-jyro Америку и более позднее, более ограниченное распространение переносимого им гриба Cerastomella ulmi, вызывающего болезнь вязов (данные на 1937 г.) [67].
Штриховкой показана основная область распространения голландской болезни вязов; белыми кружками — изолированные очаги болезни; черными кружками — находки Scolytus multistriatus.
в результате применения мер по борьбе с нею, была огромной. Бруэр [62] сообщает, что с 1933 по 1940 г. в ходе санитарных прочисток было вырублено 4250 тысяч вязов, однако, по его мнению, «нельзя утверждать, что уже происходит искоренение голландской болезни вязов в основной области ее распространения», хотя в ряде крайних пунктов ее удалось подавить. Для того чтобы на основании этих цифр не создалось ложного впечатления о полном уничтожении вяза в данном районе, следует добавить, что среднее
число зараженных экземпляров среди сохранившихся деревьев в этой области составляло всего лишь 1 на 8000; это дает также представление о роли вяза в защитных полосах и лесах в Америке.
Переходим к вредителям хвойных лесов. В 1941 г. Браун опубликовал список в 101 вид насекомых, питающихся хвоей ели в Канаде [63]. Это почти все местные формы, и некоторые из них,
Ф п г. 45. Уничтожение белой ели (Picea glauca.) европейским еловым пилильщиком {Gilpinia hercyniae) в верховьях реки Каскаппдия, Нью-Брансуик (Канада), октябрь 1932 г.
Мертвые и отмирающие ели кажутся серыми; темные деревья — здоровые бальзамические пихты (Abies balsamea).
например листовертка Archips fumiferana, причиняют серьезный вред. В 1930 г. на полуострове Гаспе (южный Квебек) был обнаружен европейский вид сосновых пилильщиков, уничтожавший там хвою белой и черной ели. Известно, что он появился в Северной Америке в 20-х годах, а может быть, и раньше [74]. Ко времени обследования вспышки в районе Гаспе погибли почти две трети деревьев белой ели (фиг. 45) и четверть деревьев черной ели на площади 7500 кв. км\ повреждения распространились, кроме того, на
другие части Квебека, приморские провинции Канады и Новую Англию до штата Нью-Йорк. К концу 1937 г. на полуострове Гаспе высокая численность вредителя наблюдалась на площади почти в 25 000 кв. км, и Балч указывает, что к тому времени этот пилильщик был самым многочисленным из питающихся на ели насекомых
Ф и г. 46. Распространение европейского елового пилильщика (Gilpinia hercyniae) в Канаде и США в 1938 г. [74].
в северо-восточной части Северной Америки (фиг. 46 и 47). Ввиду различий в биологии пилильщиков в Европе и Америке были проведены тщательные исследования [60], показавшие, что в Европе существуют два вида — Gilpinia polytomiun и G. hercyniae — и что именно последний завезен в Америку; но в более ранних сообщениях часто употребляется название G. polytomum. Разные виды ели составляют больше одной пятой части товарного древостоя лесов Канады, и этот пилильщик питается только на них. Перед нами
чужеземный вид/ занявший доминирующее положение в естественном лесном сообществе, хотя «естественные» леса Канады настолько сильно изменились в результате длительной истории лесоразработок и лесных пожаров, что это слово следует понимать главным образом в том смысле, что всякое лесное сообщество отличается более сложной экологией, чем любая полевая культура, и обладает поэтому многими чертами девственной растительности. По Доудену «самая удивительная особенность вспышки размножения елового
фиг. 47. Подробная карта распространения и численности европейского елового пилильщика в Канаде в 1942 г. [80].
Произошло резкое снижение численности по сравнению с необычайно высоким уровнем, достигнутым несколькими годами раньше.
пилильщика в Канаде и США состояла в том, что на него почти совершенно не нападали паразиты, хотя различные животные, например землеройки, мыши и белки, возможно, уничтожали до 50% зимующих коконов. Напротив, в Европе, где это насекомое известно уже больше столетия и почти не причиняет вреда, оно имеет ряд весьма важных паразитов». В 1932 г. была начата интродукция в Америку его европейских паразитов, и в подходящее время
было выпущено в природу более 20 видов. Размах этих операций был гигантским: выпустили около 7 млн. особей хальциды Micro-plectron fuscipennls, паразитирующей на коконах пилильщика. Впоследствии были разведены и выпущены еще сотни миллионов особей, но результат операции пока еще трудно оценить [275].
В североамериканских лесах, так же как и в садах, один за другим начинают появляться и вредители азиатского происхождения. Я упомяну лишь короеда Xylosandrus germanus из Японии и Китая, который проник к 1932 г. в восточную часть США, а двадцатью годами позже — в Германию. Он прогрызает свои ходы в таких деревьях, как ольха, бук и дуб, причем его распространение, в сущности, только началось [107, 108].
В запасах зерна и иных продуктов и изделий появляются все новые обитатели с других материков, однако многие чужеземные насекомые и клещи распространились очень давно, так что время их появления не было зарегистрировано или же не был точно определен их вид, а поэтому место их происхождения установить нелегко. Многие из них стали почти космополитами, а так как они живут в помещениях, то среди них есть и субтропические виды, выживающие и в умеренных широтах. Однако среди обитателей складов в Англии есть и недавние пришельцы. К ним относятся австралийский «ковровый» жук Anthrenocerus australis — один из вредите* лей текстиля и домашних вещей в Австралии и Новой Зеландии [61], который появился у нас (правда, главным образом в морских портах) с 1933 г., и бабочка Aphomla gularis из Восточной области, ныне распространившаяся в Европе и Северной Америке, хотя тоже главным образом в прибрежных районах (фиг. 48), где ее гусеницы пожирают на складах такие продукты, как миндаль, грецкий орех, арахис и чернослив (фиг. 49); пока еще нет сведений о проникновении ее на южные материки [101].
Почти все насекомые-иммигранты, которых мы рассмотрели, были завезены случайно; часто это происходило несмотря на строгие карантинные меры. Но большую часть млекопитающих (кроме крыс и мышей), птиц, лягушек, жаб и рыб сначала ввозили намеренно, хотя многие из них впоследствии стали чрезвычайно вредными или приобрели дурную славу. Только длинный обзор позво
лил бы проследить историю всех этих интродукций; пожалуй, то, что было сказано об ондатре, дает довольно верное представление о типичном ходе событий. В одной новой монографии прослежена (в значительной мере до наших дней) подобная история целых 200 видов млекопитающих! Но большинство намеренных завозов животных не имело успеха или не давало настоящей
Фиг. 48. Распространение Aphomia gularis, расселившейся из восточной области по северному полушарию, но, вероятно, еще не проникшей в южное полушарие [101].
вспышки численности [73]. Наиболее способными к росту популяции в различных странах, куда они были ввезены, оказались, пожалуй, следующие млекопитающие: из Палеарктической области — кролик, заяц-русак, лисица, лань, благородный олень и олень-сика; из Неарктической — каролинская белка и ондатра; из Неотропической — нутрия; из Восточной области — мангуста и олень-аксис. Австралия, по-видимому, внесла существенный вклад только в фауну Новой Зеландии, а Новая Зеландия представляет собой самый необычный из особых случаев: это страна, в которой нет никаких аборигенных млекопитающих, кроме летучих мышей !. Происходящее там ныне столкновение некоторых
1 И одного вида грызунов (маорийской крысы), видимо завезенного туда туземцами. — Пр им. ред.
Фиг. 49. Aphomia gularis (сем. Galleriidae)— амбарный вредитель, в последние годы завезенный с Востока в Европу и Северную Америку [101].
А. Взрослая форма (вид сверху). Б. Взрослая форма (вид сбоку). В. Гусеницы, поедающие миндаль на складе.
ной Америки. Теперь она, видимо, является наиболее широко распространенной из пресноводных рыб мира [92].
Третий стимул привел совсем недавно к возникновению новой тенденции в расселении рыб. Ныне тысячи людей содержат у себя дома, и даже в магазинах и ресторанах, аквариумы с крошечными яркоокрашенными тропическими рыбками. Сотни тропических форм рыб становятся предметом растущей торговли, и некоторые из них, как отметил Майерс, случайно попадают в природные водоемы и заселяют их [93]. Согласно устным сообщениям, китайская рыбка Macropodus opercularis и мексиканские виды Platypoecllus maculatus и Xiphophorus hellerii размножаются в районе Эверглейдс, в штате Флорида.
Глава IV
СУДЬБА ОТДАЛЕННЫХ ОСТРОВОВ
Т£огда в марте 1774 г. капитан Кук бросил якорь близ острова
Пасхи, он записал в своем дневнике [122]: «Природа чрезвычайно скупо одарила своими милостями это место». И это была чистая правда, ибо живой природе лишь с большим трудом удалось проникнуть на остров. Ближайший материк — Южная Америка — находится в 3650 км и даже ближайший покрытый растительностью остров Тихого океана (остров Дюси) удален на 1600 км. Этот осколок вулканических горных пород (из которых были высечены знаменитые статуи, фиг. 51) с его холмами п поросшими травой низинами примерно в три раза меньше острова Уайт, площадь которого составляет лишь около’ 9% общей площади Маркизских островов — одной из отдаленнейших групп гористых островов Океании; площадь этих последних в свою очередь равна примерно 8% общей площади Гавайского архипелага. Гавайская группа — самая крупная, разнообразная и богатая жизнью среди настоящих океанических островов центральной части Тихого океана, хотя площадь ее достигает всего 16 000 кв. км, тогда как площадь Африки равна почти 30 млн. кв. км.
Растения и животные не только сумели попасть на эти отдаленные архипелаги и острова без помощи человека, но местами даже образовали роскошную тропическую растительность, а иногда, хотя и не часто, здесь появлялись своеобразные эндемичные группы растений и животных. Остров Кракатау, на котором при разрушительном извержении 1883 г. вся жизнь была полностью уничтожена дождем горячего вулканического пепла, образовавшего на некоторых склонах горы слои толщиной более 30 м, был вновь заселен растениями и животными с ближайшей суши, и спустя 50 лет на нем рос густой тропический лес с эпифитами и множеством видов
Фиг. 51. «Монументы острова Пасхи» — гигантские статуи, сооруженные ранними обитателями острова, полинезийцами.
На переднем плане слева, по-впдпмому, местное дерево Sophora toromiro, ныне почти исчезнувшее.
Фиг. 52. Покрытые травянистой растительностью наружные склоны древнего кратера Рано Рараку на восточной стороне острова Пасхи; видны огромные статуи, воздвигнутые ранними обитателями острова.
животных. К 1933 г. на острове обитало по меньшей мере 720 видов насекомых, 30 видов постоянно живущих птиц и несколько видов рептилий и млекопитающих, хотя не было лягушек и жаб. Но этим видам достаточно было перебраться различными способами с соседних тропических островов — Явы и Суматры, отделенных всего лишь 40 км моря [123].
В то время, когда на острове Пасхи впервые собирали коллекции, там было чрезвычайно мало местных растений и животных
Фиг. 53. Эндемичные животные острова Пасхи.
До сих пор на этом острове было найдено лишь 5 эндемичных видов наземных и пресноводных животных, в том числе: 1 — наземная улитка Melampus pascus [135]; 2—4 — три формы Pacificella variabilis — наземной улитки, которая была описана как эндемичный вид, но впоследствии была сведена в один вид с Tornatellinops impressa с островов Фиджи (ни один из этих трех видов не превышает по размерам 5 мм) [146]; 5 — долгоносик Pentar thron paschale [ИЗ].
по сравнению с Кракатау, площадь которого в 8 с лишним раз меньше. Шведская экспедиция Скоттсберга, на короткое время посетившая остров Пасхи в 1917 г., опубликовала ряд очень интересных сообщений об этом острове, так же как и о Хуан Фернандесе — острове Робинзона Крузо [144—146]. Помимо удаленности этого острова, необходимо учитывать еще два обстоятельства. Во-первых, он сильно изменился в результате деятельности человека, особенно выпаса овец и коров и усиленного уничтожения деревьев
(фиг. 52). По-видимому, первоначально ландшафт представлял собой лесостепные саванны с травянистым покровом. Поэтому некоторые из местных растений и животных, возможно, исчезли еще до того, как их могли собрать биологи. Единственная древесная порода — Sophora toromiro — сейчас почти уничтожена. Во-вторых, не было собрано сколько-нибудь полных коллекций насекомых и других мелких животных, так как даже шведские исследователи находились там всего две недели. Тем не менее на острове, несомненно, неу местных дождевых червей (есть лишь один интродуцированный вид) и нет сухопутных птиц и других позвоночных, кроме нескольких завезенных человеком видов. Флора включает 31 вид цветковых растений (не считая культурных растений — пизанга, батата, сахарного тростника и других), 15 видов папоротников, из которых 4 эндемичны, и 14 видов мхов, в том числе 9 эндемичных.
Таким образом, число видов, которые могли бы принадлежать к аборигенной флоре острова, не достигает даже 50 — беднейшая флора! Невероятно мало и число видов животных, которых можно счесть аборигенами. До сих пор известно лишь 5 эндемичных форм: один вид сетчатокрылых [139], один вид мух [125], один долгоносик [ИЗ], один водяной жук и одна наземная улитка (фиг. 53) [135]. Водяной жук Bidessus skottsbergi был найден среди водорослей в кратерном озере Рано-Као, где обнаружены также эндемичные виды водяных мхов [157]. Никаких других пресноводных животных пока не найдено, хотя мир микроорганизмов окажется, вероятно, достаточно богатым, так как они легко переносятся ветром в самые отдаленные страны. Об остальных наземных животных либо известно, что они завезены из других мест, либо можно предполагать это, поскольку все они — космополитические виды, распространяемые человеком; к ним относятся примерно 44 вида насекомых, пауков и других беспозвоночных (в том числе одна стрекоза, возможно, залетевшая на остров самостоятельно), две интродуцированные ящерицы, два вида птиц, завезенных из Чили, и крысы. Таким образом, число сохранившихся местных видов в 10 раз меньше числа интродуцированных форм, причем разница в численности особей еще значительнее.
Существует много тысяч мелких островов, которые, подобно острову Пасхи, были настолько удалены от центров активной эволюции на материках, что могли приобрести лишь горсть случайных иммигрантов, прежде чем прибытие на эти острова человека упро* стило и ускорило процесс расселения видов. На небольшом острове Пальмира в экваториальной зоне Тихого океана было лишь 14 видов местных растений. Энтомофауна атолла Мидуэй, лежащего у западного конца цепи Гавайских островов, тоже была крайне скудной: жуков там было лишь 6 видов, а мух — всего 9 [128].
Прежде чем перейти к более катастрофическим вторжениям иноземных видов на океанические острова, полезно рассмотреть, что происходит на трех очень отдаленных островах южной части Атлантического океана — на группе островов Тристан-да-Кунья. Весьма полное биологическое обследование этих островов произвела Норвежская научная экспедиция, происходившая в 1937— 1938 гг. под руководством Христоферсена [121]. Имеются также более ранние данные, особенно в отношении растений и птиц. Эти три острова (Тристан, Найтингел и Инаксессибл) еще больше удалены от ближайшего континента, чем остров Пасхи,— они находятся в 4650 км от Южной Африки, более чем в 5000 км от Бразилии и в 7200 км от мыса Горн. Сам Тристан представляет собой верхнюю часть вулкана, поднимающегося более чем на 3600 м над морским дном и примерно наполовину выступающего над поверхностью океана. На его вершине находится древний кратер с озером внутри. Склоны покрыты довольно богатой растительностью, включающей лишь одно небольшое дерево, но целых три вида древовидных папоротников и множество грубых злаков и сухих кустарников. На прибрежной полосе суши длиной около 5 км есть небольшое селение.
На этих островах цроизрастает около 50 местных и 70 иноземных видов цветковых растений, а также почти 300 видов папоротников, мхов и печеночников [120]. Животная жизнь, если не считать морских птиц, очень бедна. Есть пять видов птиц, часть которых образовала разновидности на разных островах: один представитель семейства мухоловок, внешне похожий на дрозда, два вьюрка, нелетающий коростель и лысуха (коростель на острове Инак-
сессибл, лысуха, сейчас почти вымершая, на острове Тристан [129]. Несколько примеров позволят получить представление о фауне насекомых. Из 12 зарегистрированных бабочек в сборах экспедиции имеются только 8; лишь 5 из 12, видимо, аборигены, остальные завезены человеком [150]. Из 4 найденных видов тлей по крайней мере 2 прибыли сюда с человеком [141]. С особой тщательностью были исследованы жуки. Из 20 видов, относящихся, как полагают, к аборигенной фауне, только два являются хищниками, а остальные — растительноядные формы, многие из которых принадлежат к семейству долгоносиков, широко распространенных и на других океанических островах (все 15 долгоносиков острова Тристан — нелетающие эндемичные формы). Кроме этих местных жуков, есть еще 6—7 видов, относительно которых известно (или можно предполагать), что они завезены человеком [116].
Следует учитывать, что транспортная связь между островами Тристан-да-Кунья и внешним миром чрезвычайно слаба. И тем не менее в составе их фауны скоро будет столько же иноземных форм, сколько в ней было местных видов. В 1882 г. близ островов произошло кораблекрушение, и несколько серых крыс перебралось с судна на берег. Местный настор настаивал на уничтожении этих крыс, но им дали проникнуть на остров, и теперь они размножились во многих частях Тристана, поедают картофель (основная сельскохозяйственная культура на этих островах) и нападают на гнездящихся птиц в более диких участках горной области [114]. Они, как предполагают, истребили и лысуху на острове Тристан, возможно, при содействии одичавших кошек. К счастью, на два других острова крысы пока еще не проникли.
До сих пор не отмечалось крупных вспышек размножения завезенных извне насекомых, и главным врагом картофеля является местная бабочка, совсем не имеющая паразитов. Но одна из завезенных тлей, Myzus persicae, в других странах известна как переносчик двух из наиболее опасных вирусов картофеля, а одна из бабочек — вредитель крестоцветных. Следует отметить, что некото-рые завезенные виды встречаются пока лишь в узкой полосе, заселенной человеком, где имеются пастбища, сады и картофельные поля, однако другие, например крысы, проникли и в природ
ные местообитания. Завезенный жук Quedius tnesomelinus (семейство Staphylinidae) и европейская многоножка Cylindroiulus lates-triatus (занесенная человеком также в Северную Америку, Южную Африку и на Азорские острова) заселили очень широкую область папоротниковых зарослей с деревьями, болота, прибрежную приливную полосу и другие места. В то же время другая европейская многоножка, Blantulus guttulatus, была найдена на обрабатываемой земле [130].
Итак, перед нами океанический остров, где человек отвоевал у природы небольшой участок, оставив основную часть в первозданном состоянии. Его влияние на относительно дикие части острова сводится к добыче древесины и использованию морских птиц, но, кроме того, выражается также в распространении по острову завезенных на судах иноземных видов. Чтобы проследить тот же процесс на более обширной группе островов, обратимся к Гавайскому архипелагу.
Нет надобности описывать Гавайские острова — удаленные от материка, гористые, вулканические, с тропическим климатом и богатой растительностью, сохранившие до наших времен одну из самых своеобразных среди известных нам островных фло(р и фаун. Теперь почти никто не отрицает, что это настоящие океанические острова, такие же, как Тристан-да-Кунья и остров Пасхи, и что населяющие их эндемичные виды произошли от редких иммигрантов, которым пришлось проделать несколько тысяч километров пути по океану. До Америки и Австралии больше 6500 км; до ближайших материковых островов — Японии — 5000 км; до островов Фиджи, находящихся где-то у края древнего, погрузившегося в океан остатка Австралазийского материка,— около 4500 км. До того как лодки полинезийцев примерно в XII в. пашей эры или раньше достигли Гавайских островов, острова эти были, вероятно, покрыты роскошными лесами, исключая засушливые места и лавовые потоки недавнего происхождения. С тех пор леса постепенно отступали от побережья и теперь занимают лишь четвертую часть своей первоначальной территории, сохранившись главным образом в горах. В этом повинны и лесные пожары, и одичавшие коровы овцы, козы и лошади, и расчистка земли под посевы. Весьма зна
чительная часть видов все же сохранилась, и если добавить к ним формы, отмеченные первыми исследователями, получится довольно внушительный список, хотя многие виды, возможно, вымерли еще до прихода европейцев, и высказывалось даже предположение, что чуть ли ие третья часть первоначальной фауны насекомых исчезла до того, как ее успели зарегистрировать. Из 1729 видов цветковых растений более 90% не встречаются нигде, кроме этих островов.
Фиг. 54. Раковппы’гавапскпх наземных улиток, поеденные интродуцированными крысами [147 ].
3—7 — Achatinella; 9 и 10 — Amastra.
Циммерман [158], в книге которого («Насекомые Гавайских островов») я нашел много критически рассмотренных данных, полагает, что примерно из 6000 найденных видов насекомых 3722 эндемичны, тогда как остальные встречаются и в других местах, а здесь появились в результате естественного пли искусственного заноса.
На Гавайских островах обитают два крупных семейства наземных улиток: семейство Achatinellidae с 215 видами эндемично для Гавайских островов, семейство Amastridae с 294. видами — почти эндемично (фиг, 54), Представители Achatinellidae живут только на деревьях; раковины их отличаются пестрым рисунком п яркой
окраской. Среди Amastridae наблюдается эволюция различных экологических форм, как древесных, так п ползающих по земле.
Из 77 эндемичных форм птиц, обнаруженных на всем Гавайском архипелаге, около 43 видов и подвидов принадлежит к Drepaniidae — семейству, внутри которого на Гавайских островах выделилось свыше десяти экологических групп — медососы, охотники за древесными насекомыми, насекомоеды, семеноеды, орехо-еды, плодоеды и т. п.; на материке подобные различия развились бы между отдельными отрядами, а не родами птиц. Неудивительно, что капитан Кинг был озадачен, увидев одну из таких птиц. В 1779 г. он писал [131]: «Желтоголовая птица, которую по строению клюва мы сочли за попугая, тоже очень обычна. Она, однако, отнюдь не принадлежит к этой группе, но весьма похожа на желтого клеста, Loxia flavicans Линнея». Это была Psittacirostra pslt-tacea (семейство Drepaniidae) (фиг. 55). Систематики пытались подсчитать, от какого числа предков могли произойти все эти гавайские группы организмов, т. е. сколько исходных иммигрантов попало различными путями на острова. Для цветковых растений их оказалось около 272 видов; для насекомых — 233—254; для Achatinellidae — только один; для наземных улиток — 22—24 [158]; для птиц — приблизительно 14, причем Drepaniidae произошли только от одной из этих форм [133].
Какова была дальнейшая судьба этой удивительной флоры и фауны? Прежде всего перечень видов необычайно увеличился за счет завоза, как преднамеренного, так и случайцого. Составление полного списка насекомых еще не закончено, но примерно из 1100 видов, вошедших в первые пять томов «Насекомых Гавайских островов», 420 считают пришельцами. Это лишь грубая оценка, так как остается рассмотреть еще девять отрядов насекомых, в том числе такие чрезвычайно важные группы, как бабочки, жуки, мухи и перепончатокрылые. В 1953 г. опубликован список 49 «экономически важных» насекомых, обосновавшихся на островах после 1939 г. [137]. Происхождение этих видов весьма различно: они прибыли из Калифорнии, Мексики, с островов Самоа, Филиппинских островов, Фиджи, Гуама, Сайпана, с Новой Гвинеи, а происхождение некоторых из них просто неизвестно. Среди этих иммигрантов
Ф и г. 55. «Оу» (Psittacirostra psittacea) — представитель сем. Drepaniidae, возникшего на Гавайских островах [155 [.
был аргентинский муравей, завезенный (вероятно, из Калифорнии) в 1940 г. Когда Уилер [153] в 1934 г. составлял список муравьев Гавайских островов, он упомянул, что этот вид был перехвачен при карантинном обследовании и к тому времени его еще не было на островах. Уилер сообщил также, что на острове Оаху муравей-листорез Pheidole megacephala в то время вытеснял ранее завезенного муравья Solenopsis rufa. Циммерман в 1948 г. писал: «Прожорливый Pheidole tnegacephala один учинил небывалое массовое уничтожение насекомых. В пределах ареала этого губителя местной энтомофауны можно найти лишь немного эндемичных видов насекомых. Он обитает почти повсюду — от морского побережья до болотистых лесов. Ниже 600 м ныне можно встретить лишь несколько видов местных насекомых». Было известно, что этот муравей-листорез ранее проник на Кана|рские острова и остров Мадейру, а затем там появился аргентинский муравей, вытеснивший не только Pheidole, но практически всех местных муравьев в поясе ниже 900 м [153]. Быть может, то же самое произойдет теперь на Гавайских островах.
При появлении нового вредителя обычно делают попытки бороться с ним путем интродукции его врагов. Сообщение о таких насекомых, завезенных на Гавайские острова в период с 1953 по 1955 г., дает весьма яркое представление об этом процессе [152]. Для борьбы с кустарником Lantana был ввезен мексиканский жук-усач, буравящий стебли, а также центральноамериканский жук-листоед и калифорнийская бабочка из семейства Phalaenidae-, личинки этих видов поедают листья Lantana, С той же целью были интродуцированы некоторые другие насекомые, в том числе муха, поедающая семена. Из Мексики ввезли муху, личинки которой питаются соцветиями Eupatorlum glandulosum — растения, родственного нашему посконнику коноплевому (Е. cannabinum) и сделавшегося на Гавайских островах, а также на Филиппинах и в других местах сорняком; из Бразилии — совку, способную поедать листья другого вредного в местных условиях растения — кустарника Heteromelon arbutlfolia. Интродуцированы четыре вида мексиканских жуков-навозников, личинки которых могли бы помочь в борьбе с личинками некоторых мух; два вида ос из семейства Scoliidae
с острова Гуам для уничтожения различных пластинчатоусых жуков; два паразита из Аризоны — для повторного их испытания (спустя 10 лет после первой безуспешной попытки их интродукции) в борьбе с бабочкой, повреждающей цветки мескитового де рева; мексиканская божья коровка — для уничтожения тлей на плантациях сахарного тростника; и, наконец, две хищные улитки (одна с Марианских островов, а другая — из Флориды) как возможные враги гигантских африканских улиток. Это настоящий рынок для обмена видами, беспорядочное смешение форм с различных материков и островов мира, быстрое разрушение зоогеографических областей и островных фаун, описанных Уоллесом!
Большинство растительноядных насекомых было завезено вслед за интродукцией растений (полевых и садовых культур, плодовых и лесных деревьев). Обычно проходит несколько лет, прежде чем насекомое сможет догнать свое кормовое растение, как это недавно произошло при интродукции Leucaena glauca [143]. Первоначальной родиной этого крупного кустарника из семейства бобовых была, вероятно, Южная Америка, но его завезли и в другие части земного шара, в том числе на Гавайские острова (в 1888 г.). Здесь он ценится как кормовая культура, богатая белком; семена его собирали и высевали, а позже стали использовать для местного производства украшений. Он стал быстро развивающейся добавочной культурой на Гавайских островах, но в 1954 г. обнаружили, что в его семенах обитает маленький жучок, принадлежащий к семейству Anthribidae и происходящий из Индо-Китая и Филиппин, который на острове Оаху иногда губит весь урожай семян. Бороться с ним пока не удается.
Не следует думать, что местная фауна просто отступает вместе с лесом, сохраняясь при этом полностью, хотя и на меньшей территории. Конечно, довольно значительная час^ъ леса и некоторые другие стации на возвышенных участках сохраняются, так как их невозможно использовать для земледелия. Однако бродячие коровы и другие одичавшие животные и здесь наносят им большой ущерб. И черные крысы, сильное влияние которых — часто повторяющийся мотив в современной истории островов, тоже проникли в леса. Местные бабочки сильно уменьшились в числе, а некоторые из них
вымерли. Циммерман объясняет это проникновением в леса паразитических перепончатокрылых — ихневмонид, завезенных для борьбы с вредителями полей, и указывает, в частности, на три вида: Caslnaria infesta, Cremastus flavoorbitalis и Hyposoter exi-guae, имеющие очень широкий круг хозяев. Ведь насекомые Гавайских островов в естественных условиях не подвергались нападению паразитов и других врагов и поэтому не были приспособлены к защите от них, как континентальные виды. Это снижение численности местных бабочек, возможно, было причиной того, что численность некоторых видов Odynerus (род ос-охотниц, представленный на Гавайских островах многими видами) также сократилась; дело в том, что эти осы откладывают яйца на гусениц, которые служат запасом пищи для их личинок.
Среди многочисленных иммигрантов, появившихся на Гавайских островах, ни один не проделал столь длительного и упорного путешествия через всю Индо-Пацифическую область, как гигантская улитка A chatiпа fulica. К роду Achatlna, в основном распространенному в Эфиопской области, относится более 65 видов, живущих главным образом в тропических лесах. К этому роду относятся крупнейшие из современных наземных улиток, самая крупная из которых, Achatina achatina, до сих пор живет только в западной Африке, где служит излюбленным лакомством местного населения. A. fulica несколько мельче (фиг. 56). Это восточноафриканский вид, который, возможно, уже давно был завезен на Мадагаскар и начал распространяться на близлежащие острова Маврикий (к 1800 г.) и Реюньон (к 1821 г.), а затем на Сейшельские (к 1840 г.) и Коморские (к 1860 г.) острова. Несколько улиток было выпущено в Калькутте в 1847 г., и Бекверт [115], изучавший расселение этого вида и разработавший систематику всей группы, писал, что «вначале распространение улитки в южной Азии было очень медленным». Она прибыла на Цейлон к 1900 г., появилась на полуострове Малакка к 1922 г. (а возможно, на 12 лет раньше), на Борнео — к 1928 г., в Сиаме — в 1937—1938 гг. и в Гонконге — в 1941 г. Через Индонезию она прошла в 20-х и 30-х годах XX в. К 1925 г. она очутилась в Японии, хотя и не особенно процветала там, в 1938 г. достигла островов Палау, затем перешла на Мариан
ские острова, вскоре став серьезным сельскохозяйственным вредителем на многих островах Микронезии. Настоящим бичом эта улитка сделалась на острове Гуам, куда она была завезена в 1946 г. с Сайпана (Марианские острова). К 1948 г. она захватила три плацдарма на Новой Гвинее и поблизости от этого острова. Тем временем (около 1947 г.) несколько улиток попало в Калифорнию. Считают, однако, маловероятным, что она сможет прочно обосноваться
Фиг. 56. Гигантские африканские улитки [111].
Вверху крупная Achatina achatina, %о сих пор встречающаяся только в западной Африке; внизу—A. fulica, распространив-, шаяся благодаря человеку из восточной Африки по всей области Индийского и Тихого океанов. Гавайские острова — крайний восточный пункт, в котором она прочно закрепилась.
в США, так как тамошний климат неблагоприятен для тропических улиток. Это грандиозное расселение отчасти происходило в результате случайной перевозки с продуктами и растительными материалами, но объясняется также и ценностью улитки в качестве пищевого продукта. Величина этих улиток, их прожорливость и численность придают их нашествиям характер необычайного события. Некоторые виды, появляясь в массовом количестве, представ
ляют серьезную опасность для автотранспорта — вплоть до того, что заставляют буксовать колеса автомобилей! В вытянутом состоянии улитка может достигать 23 см в длину, не считая раковины.
Гигантская улитка проникла на Гавайские острова в 1936 г., и, несмотря на большие усилия, затрачиваемые на борьбу с нею, она теперь населяет остров Оаху, где повреждает сельскохозяйственные культуры; полагают, что это нашествие началось всего с двух особей, завезенных с Тайваня. Сейчас в широких масштабах ведется испытание эффективности естественных врагов Achatina — плотоядных улиток — для борьбы с нею; этих плотоядных улиток завозят не только с первоначальной родины вредителя — из восточной Африки, но также с одного из Марианских островов и из Флориды [154]. Тем временем многие из красивых местных улиток, принадлежащих к семейству Amastridae, стали редко встречаться в низинах, а некоторые виды вымерли. Одной из причин этого было, по-видимому, уничтожение их завезенными крысами [147]; местная полинезийская крыса с существенно иным типом питания не могла играть решающей роли в вымирании этой группы.
Среди птиц тоже наблюдается рост числа иноземных видов и уменьшение числа местных видов. В 1940 г., когда Брайен [117] подвел итог составу фауны Гавайских островов, вселившиеся формы составляли уже около одной трети всех птиц; на этих островах пробовали акклиматизировать 94 вида птиц, причем в отношении 53 видов эта попытка увенчалась успехом. Эти птицы происходят из самых различных стран: фазан Phasianus colchlcus — из Европы; фазан Ph. versicolor — из Японии; перепел Lophortyx California — из Калифорнии; перепел Coturnlx coturnix — из Японии; голубь Streptopella chinensis — из восточной Азии; и, наконец, голубь Geopella striata — с Малайского архипелага — вот лишь небольшой перечень охотничьих птиц, которые теперь прочно обосновались на Гавайских островах [142]. Оба фазана не только широко распространились, но в некоторых местах дали помеси (фиг. 57). Банкивская курица (Gallus gallus), вероятно, была завезена из Малайи еще полинезийскими мореплавателями, так как ко времени открытия капитаном Куком Гавайских островов эта птица уже обитала на них. Обыкновенный голубь (Columba llvia) является здесь
не только городской птицей, как в США, а обитает в диком состоянии на скалах. Индийский розовый скворец (Acridotheres tristis), завезенный из Индии в 1865 г., хорошо известен экологам благодаря роли, которую он играл в первоначальном распространении семян Lantana.
Согласно Манро [134], который всю жизнь наблюдал гавайских птиц и вместе с Перкинсом и другими ранними натуралистами
Ф и г. 57. Распространение фазанов, интродуцированных на Гавайские острова в 1947 г. [142].
Серым показано распространение Phasianus colchicus; точками— Ph. versicolor; черным — главным образом гибриды (остров Ниихау не обследован).
исследовал фауну и собирал коллекции в конце XIX в., несколько интродуцированных птиц более или менее глубоко проникли в леса. «Пекинский соловей» Leiothrix lutea (китайский вид, завезенный в 1918—1920 гг.) живет теперь на большинстве главных
островов; между тем известно, что в Японии у этого вида обнаружена птичья малярия. Примерно с 1900 г. живет на островах китайский дрозд Trochalopterum сапогит — обитатель кустарников, проникший в лес глубже всех других видов; правда, по поступившим в 1944 г. сообщениям, численность его местами уменьшилась. С 1890 г. появилась японская синица Paras varius, которая вполне освоилась в лесу. Было высказано предположение, хотя и не подкрепленное прямыми данными, что подобные виды переносят болезни птиц из нижней зоны в верхние и, возможно, причиняют вред птицам семейства Drepaniidae. Численность почти всех этих замечательных птиц уменьшилась, даже там, где сохранились их излюбленные места обитания. Лишь около трети видов и островных подвидов имеют достаточные шансы на выживание в будущем. Оказалось, что некоторые формы, считавшиеся вымершими, все еще существуют, но в свое время не были обнаружены, так как поиски их связаны с большими трудностями. Так, медососа Pal-meria dolei не видели с 1907 г., и только в 1950 г. его удалось сфотографировать телеобъективом на высоких горах острова Мауи. Птицу Pseudonestor xanthophrys, которую не видели в течение 50 лет, в 1950 г. наблюдали на том же острове на высоте 1900 м.
«Попугай» капитана Кинга, или «оу» (Psittacirostra psittacea), всегда встречался в изобилии на главных островах, но Перкинс в 1903 г. указал, что эта птица, широко распространенная на других островах, почти исчезла на Оаху. Это, по его мнению, могло быть вызвано конкуренцией со стороны черных крыс Rattus rattus, распространившихся в лесах острова: «Теперь на обширных пространствах трудно найти хотя бы один красный плод «йене», который не был бы в той или иной степени объеден или попорчен крысами; таким образом, возможно, что крысы были косвенной причиной вымирания «оу» [138]. Действительно, клюв этой птицы особецно хорошо приспособлен для выклевывания плодов «иеие» (Freycine-tia ornotti) — лианы с яркими цветками, оплетающей лесные деревья; а основное местообитание «оу» на всех островах находилось в той зоне, где встречалась эта лиана. Однако число этих птиц уменьшилось и на других островах, где они могли бы питаться другими плодами, а также гусеницами (которыми они кормят
своих птенцов); известно, что они поедают и плоды некоторых интродуцированных растений, например гуайявы, успешно распространяющейся на Гавайских островах. Известно также, что домашние птицы принесли с собой болезни, из которых по крайней мере одна была отмечена у птиц семейства Drepaniidae. На острова проникла также птичья малярия, которая, возможно, распространилась там при помощи комаров [112]. Комары, по-видимому, представлены только чужеземными видами — первоначально на островах их не было. Брайен [118] в своей «Естественной истории Гавайских островов» пишет, что комар, кусающий ночью (Culex fati-gans), как полагали, был завезен из Мексики еще в 1826 г. Есть также несколько видов Stegomyia, кусающих днем [118]. Все это указывает на возможность сложного взаимодействия различных факторов, которые никто еще не смог как следует проанализировать. Каково, например, влияние недостатка гусениц местных бабочек на птенцов лесных птиц? «Оу» все еще существует, хотя в 1944 г. полагали, что этот вид близок к вымиранию. В 1950— 1951 гг. несколько птиц видели в одном горном лесном заповеднике и в Национальном парке острова Гавайи, на высоте в несколько тысяч футов. Судьба отдаленных островов в нашем столетии может быть смягчена, если часть видов избегнет уничтожения. Особенно это относится к птице «оу», о которой Перкинс писал: «Иногда она поет во время полета, и нередко, когда птицы летят небольшой стайкой, они поют все вместе (это бывает и в других случаях), и притом очень красиво» [138].
Из других отдаленных островов с Гавайским архипелагом может соперничать только Новая Зеландия. Этот остров выглядит как часть материка, хотя он никогда, вероятно, не имел прямой связи ни с каким материком и был изолирован на протяжении периода огромной длительности. Таким образом, по условиям жизни этот остров отчасти сходен с материком, но его история типична для океанического острова. Ни одно другое место земного шара не подвергалось в течение столь долгого времени такому упорному наплыву агрессивных пришельцев, особенно млекопитающих; вначале эти пришельцы успешно заселяли остров, но во многих случаях они оказывали губительное влияние на свои собственные
местообитания. Первоначально на Новой Зеландии не было никаких местных млекопитающих, кроме летучих мышей. В 1950 г. Водзицкий [156] имел возможность написать монографию по экологии 29 «животных, представлявших проблему», среди которых было 11 копытных: 4 из Японии, 4 из Северной Америки и 3 из Европы (включая Англию); к ним следует добавить одичавший домашний скот, в том числе и свиней. Перечень успешно закрепившихся на Новой Зеландии млекопитающих и птиц, составленный Водзицким, приведен на фиг. 58. В период между 1851 и 1910 гг. на Новой Зеландии не раз выпускали благородного оленя (Cervus elaphus), и он быстро размножился на обоих островах. Заселяемые им из разных центров отдельные участки сейчас в пределах каждого острова слились между собой, но наибольшая заселенность до сих пор имеется на Южном острове (фиг. 59 и 60). Благородный олень уже оказал глубокое влияние на местные леса, особенно в сравнительно сухих местах; однако к наиболее сильной эрозии почвы приводит повреждение лесов в относительно влажных районах. Весьма вероятно, что на многих водоразделах олени при содействии домашнего скота нарушили прежние соотношения, способствуя катастрофической эрозии почвы, последствия которой ощущаются не только в горах, но и в тех частях нижних долин, куда смывается избыточное количество наносов сверху.
Прослеживая историю вторжения насекомых на Новую Зеландию, пришлось бы повторить то, что уже говорилось о других странах. Считая только формы, занявшие видное положение, в период с 1929 по 1939 гг. появилось 14, а к 1949 г. еще 9 иноземных видов [124]. Среди последних была обычная для Европы оса Vespula ger-manica, устраивающая гнезда в земле [136]. В 1945 г. в одном пункте Северного острова пчеловоды заметили каких-то странных ос, летающих около ульев. С тех пор эти осы быстро распространялись и увеличивали свою численность, чему, вероятно, способствовало отсутствие зимних холодов. В 1945 г. было уничтожено 7 осиных гнезд, на следующий год — 140, а к 1948 г.— более 3000, но и это не остановило расселения. Когда объявили вознаграждение за самок, собранных в период покоя, один школьник менее чем за неделю принес 2400 штук!
Опоссум
Кенгуру валлаби
Австралийский кури ный гусь Черный лебедь Австралийский пере* пел
Птица Platycercus
Белый какаду Попугай
Белоспинная сорока Черноспинная соро-
Серна
Домовый сыч
Полинезийская собака Маорийская крыса
3 и
ллингтон
о
41
КРАЙСТЧЕРЧ
ДАНИДИВ
НОВАЯ ЗЕЛАНДИЯ
Еж Горностай Хорек Ласка Европейская собака Кошка Черная крыса. Серая крыса Мышь
Кролик Заяц Парковый скот Дикий гусь Каменный баран Благородный олень Лань Кабан
Тар Олень аксис Олень аамбар Олень сика Каменная куропатка Китайская горлица Индийский розовый скворец Павлин
Вапити Виргинский олень Мазам OdocoiHeus henr tonus Американский лось
Е Р А
11.3%
Канадская казарка Калифорнийская куропатка Виргинский перепел
Кряква Фазан
Луговой жаворонок
Певчий дрозд Черный дрозд Завирушка лесная Грач
Скворец
Домовый воробей Зяблик Коноплянка Щегол Зеленушка Обыкновенная овсянка
Л
Ф и г. 58. Интродуцированные птицы и млекопитающие, акклиматизировавшиеся на Новой Зеландии, с указанием места их происхождения (процент форм, происходящих из различных стран, показан зачерненным сектором соответствующего круга) |156].
Фиг. 59. Распространение интродуцированного благородного оленя (Cervus elaphus) на Северном острове Новой Зеландии в 1947 г. [156].
Буковые леса (Nolhofagus) обведены сплошной линией.
Фиг. 60. Распространение интродуцированного благородного оленя (Cervus elaphus) на Южном острове Новой Зеландии в 1947 г. [156].
Буковые леса (Nothofagus) обведены сплошной линией.
Несмотря на мероприятия, проводившиеся и проводимые на отдаленных островах в течение последнего времени, судьба этих островов довольно печальна. Для перестройки их флоры и фауны в сбалансированную устойчивую систему видов потребуется очень длительное время. Пока никто еще не пытался представить себе, каким будет конечный результат. Какова полноценная экологическая система в таких местах, как Гуам, Кауаи или остров Пасхи? Сколько видов могут сосуществовать в одном месте? Как достигается равновесие между ними? Можем ли мы сочетать простые сообщества сельскохозяйственных культур с естественной сложностью природы, особенно при наличии почти неисчерпаемого резервуара видов на материках, создающих возможность новых вторжений, нарушающих сложившуюся картину? Все эти вопросы не за горами, хотя большинство экологов до сих пор относилось к ним без всякого энтузиазма; если экологи и замечали их, то их брала оторопь и они отворачивались, предпочитая трудную и утомительную работу по изучению биологии отдельных видов, ныне живущих или вымерших.
В конце этой главы мне хотелось бы оглянуться на более приятный и уравновешенный экологический мир, существовавший до того, как европейцы вторглись в эту далекую галактику островных сообществ. В те времена на множестве тихоокеанских островов жили в огромном количестве мелкие крысы; они происходили от одной малайской формы и были, очевидно, завезены полинезийцами при их великих переселениях на восток и на юг несколько сот лет назад [149]. Rattus exulans (так же, как и близкородственные формы вроде R. hawaliensls, см. фиг. 61) — небольшая крыса, гораздо более кроткого нрава и менее агрессивная, чем крупные «корабельные» черные и обычные серые крысы; установлено, например, что на Новой Зеландии местная крыса не причиняет вреда островным птичьим заповедникам. На многих островах Ти1ого океана местная крыса была либо истреблена кошками, либо вытеснена более крупными видами крыс. Долгое время думали, что она вымерла и на Гавайских островах, пока не было установлено, что ее смешивали с молодыми особями серой формы черных крыс и что она сохранилась на островах наряду с двумя чужеземными
видами крыс [148]. Из книг миссионеров мы узнали, что эта мелкая крыса занимала видное место в жизни людей. На острове Раротонга, в центральной части Тихого океана, ее высоко ценили как объект спортивной охоты. «В те дни — до того, как была завезена кошка,— крысы были весьма многочисленны. Охота на крыс была серьезным занятием взрослых мужчин, а мясо этих животных считалось деликатесом» [127]. На островах Тонга в самом начале
Фиг. 61. Гавайская крыса Rattus hawaiiensis [147 ].
Мелине крысы из этой группы видов были завезены первоначально из Малайи на тихоокеанские острова полинезийскими мореплавателями. Эта крыса до сих пор живет на некоторых островах, в том числе на Гавайских и на Новой Зеландии, но во многих местах вымерла, отчасти в результате вытеснения крысами, завезенными европейцами.
XIX в. король и придворные часто отправлялись стрелять крыс на лесных тропах, используя огромные луки и стрелы длиной около 2 м. «Выигравшей считается та партия охотников, которая первой убила десять крыс. Если крыс много, то обычно проводят три или четыре игры» [132]. Существовали тщательно разработанные правила, вроде наших правил футбола или охоты, и одна удивитель-
нал льгота — убив птицу, можно было зачесть ее за крысу! Даже во второй половине XIX в. на острове Мангаиа еще сохранилась поговорка «сладок, как крыса» [126]. Фон Гохштеттер [151] писал в 1867 г. о маленькой новозеландской крысе: «Этих туземных крыс было так мало уже ко времени прибытия первых европейцев, что местный вождь, заметив крупных европейских крыс на борту одного из кораблей, умолял капитана пустить этих крыс на берег и тем самым создать возможность разведения новой, более крупной дичи». Капитан Кук [122], посетивший в 1774 г. остров Пасхи, сообщает: «У них есть также крысы, которых они, по-видимому, едят; ибо я видел человека, державшего в руке несколько мертвых крыс; он, казалось, не желал с ними расставаться и дал мне понять, что они предназначаются в пищу». Можно ли сейчас найти отдаленный остров, где людям не хотелось бы расстаться с новыми крысами, появившимися там за последние 180 лет? Пожалуй, уместно было бы вспомнить рассказ Бакстона и Гопкинса [119] о появлении более 100 лет назад человеческих блох на одном тихоокеанском островке: «Мирные жители острова Аитутаки, заметив, что эти крошечные создания постоянно проявляют беспокойство и любопытство, а временами даже назойливы, пришли к логичному заключению, что это души умерших белых людей». Мы можем надеяться, что беспокойная любознательность в научном исследовании позволит выяснить, как можно восстановить нарушенное равновесие и сохранить его в дальнейшем.
Глава V
ИЗМЕНЕНИЯ, ПРОИСХОДЯЩИЕ В МОРЕ
Пусть мудрость Океана мне смешна, Но я скорблю, утратив большее, чем царства: Дни мира и дремотной тишины;
Тогда не знали мы, что ведь победа — птица, И упустить ее возможно, и поймать!
Китс, Гиперион
Т> отличие от суши и пресных вод море кажется почти нетрону-той стихией. Однако в результате деятельности человека за последние сто лет и здесь начались крупные изменения в распространении видов. Эту деятельность можно разбить на три типа: во-первых, постройка новых каналов; во-вторых, случайный перенос организмов на судах; и, в-третьих, намеренная их интродукция. Панамский канал, в формальном смысле вновь отрезавший Неарктическую область от Неотропической, вряд ли явился серьезным препятствием для наземных форм или существенным путем для переселения морских организмов из одного океана в другой. В 1935 и 1937 гг. Гильдебранд [179] обследовал фауну шлюзов и отдельных участков этого канала и обнаружил, что очень многие рыбы и некоторые другие животные уже прошли часть пути с того и с другого конца. В самом деле, нет никаких физических преград, которые могли бы помешать этому; Гильдебранд приводит фотографию, где изображены люди, поднимающие несколько очень крупных рыб со дна осушенной шлюзовой камеры. Реальным барьером служат 65 км пресной воды, в особенности обширное озеро Гатун. Из рыб туда смогли проникнуть, как и следовало ожидать, только те, которые могут жить в солоноватой и даже пресной воде — различные бычки и некоторые другие рыбы. Единственный
вид, о котором известно, что он прошел через весь канал,— это атлантический тарпун (Tarpon atlanticus)-, 4 экземпляра его были найдены в самом нижнем шлюзе со стороны Тихого океана при осушении этого шлюза в 1937 г. Тарпунов находили также у тихоокеанского конца канала уже на уровне моря, но не вылавливали (в 1939 г.) в самом Панамском заливе. По-видимому, они весьма обычны в озере Гатун.
Совсем иначе обстоит дело с Суэцким каналом, хотя и здесь есть ряд серьезных преград для прохождения морских форм. По характеру эти препятствия противоположны существующим в Панамском канале. Суэцкий канал имеет около 160 км в длину, и в середине его находится Большое Горькое озеро протяжением около 23 км. Это озеро имеет очень высокую соленость вследствие растворения осадков каменной соли, отложившихся в более ранний период. Однако, согласно Фоксу [176], руководившему Кембриджской экспедицией в район Суэцкого канала в 1924 г., соленость воды уменьшилась в результате примешивания воды из океана и продолжала снижаться. Как мы уже говорили в гл. II, огромный участок моря Тетис, соединявший область Средиземного моря с Индийским океаном, к миоцену отделился, после чего начали появляться большие различия в ископаемых фаунах к востоку и к западу от этой области. Индийский океан сохранил свою богатейшую фауну. Средиземное море значительно обеднело. Это, без сомнения, объясняется главным образом тем, что оно стало частью огромного солоноватого Сарматского, а позже Понтийского моря, занимавших значительную часть Центральной Европы, район Черного моря и Арало-Каспийскую область. Детали истории Суэцкого залива сложны и еще до конца не выяснены [186]. Известно, однако, что в течение какого-то времени он соединялся с Красным морем, так как в его среднеплиоценовых отложениях найдены морские ежи и другие ископаемые, свойственные этому морю. Кажется несомненным, что после этого он был отрезан от восточных морей на протяжении всего четвертичного периода или большой его части. В новейшие времена фауны Средиземного и Красного морей были совершенно различны — они имели и до сих пор имеют сравнительно мало
общих видов. Другой канал (из Красного моря), который египетские фараоны построили несколько тысяч лет назад, не мог служить путем для переселения морских видов, так как имел длинный пресноводный участок и не пропускал суда непосредственно в Средиземное море [176].
С тех пор как был открыт Суэцкий канал (т. е. с 1869 г.), многим животным удалось перейтп из Красного моря в Суэцкий залив и распространиться, иногда довольно широко, по Средиземному морю [197]. Обмен видами главным образом в этом направлении объясняется направлением течений, а также приливных волн, которые на протяжении большей части года идут по каналу со стороны Красного моря. Так. например, только два из 16 крабов, собранных экспедицией на канале, были средиземноморскими [164]. Многие из них, вероятно, смогли пройти канал благодаря движению судов, облегчивших им пересечение Горького озера, однако имела место, по-видимому, и непосредственная миграция. Распространение краба Neptunus pelagtcus — вида, свойственного Красному морю,— было прослежено работниками Компании Суэцкого канала, вылавливавшими его на пищу [164]. В 1889—1893 гг. впервые наблюдалось повышение его численности в канале, к 1898 г. он достиг Порт-Саида и 4 года спустя был там обычным животным. К 1930 г. он стал обычен и в Палестине. Ныне он служит одним из основных продуктов питания египтян, его ловят в районах Порт-Саида, Александрии и Хайфы, и оп распространился по крайней мере до Кипра [174]. Сходным образом шло успешное расселение другого краба — Myrax fug ах. Красноморские виды — краб Neptunus sanguinolentus и рак The-nus orientalls — обнаружены в 1896 г. в гавани Риека на Адриатическом море. Морская жемчужница Pinctada vulgaris распространилась из Красного моря до самого Туниса [176]. Таким образом, в последние 90 лет мы были свидетелями вновь начавшегося обогащения фауны моря Тетис. Однако мне кажется весьма вероятным, что Большое Горькое озеро, соленость которого более чем вдвое превышает соленость морей, помешает прохождению многих растений и животных через канал или надолго задержит их распространение.
Случайный перенос организмов судами, в цистернах для водяного балласта или просто на поверхности корпуса, был и остается мощным, постоянно действующим фактором в расселении морских растений и животных по всему миру; по-видимому, таким путем был, например, завезен в Европу китайский мохноногий краб. В 1946 г. в пробах планктона из южной части Северного моря впервые были обнаружены личиночные формы креветки Processa aequimana, и число их в период 1946—1948 гг. непрерывно возрастало; взрослые животные здесь еще не найдены. Известно, что эта креветка водится в Красном море; ее личинки находили в Суэцком канале, а взрослую форму — в районе Неаполя.
Днища судов быстро обрастают сидячими морскими водорослями и животными, среди которых могут находить убежище и пищу более подвижные формы; в этом своеобразном местообитании были обнаружены целые сообщества [178]. Капитан Слонам [195] рассказывал, что, когда он один пересекал Атлантический океан на паруснике «Спрей», сопровождавшие его рыбы и дельфины повернули в сторону и последовали за крупным парусным судном с сильно обросшим днищем; «рыбы всегда следуют за обросшим судном», добавляет он [195]. Такое обрастание создает условия для переноса животных на огромные расстояния, причем многие современные суда не составляют исключения, несмотря на широкое применение химических средств, противодействующих обрастанию^ Достоверно известно, что улитка-туфелька (Crepidula), о которой мы будем говорить ниже, поселяется на днищах судов, долго стоящих в порту, и может быть завезена в другие места при перемене мест стоянки этих судов [161]. Подобным же образом объясняют появление индо-пацифи-ческой диатомовой водоросли Biddulphia sinensis в Северном мрре примерно в 1903 г. Данные о ее последующем распространении и поистине астрономическом размножении в этом море сведены у Харди [177], который приводит также хорошие изображения этой четырехклеточной микроскопической водоросли, плавающей у поверхности воды. Ее распространение интересно не только как пример захвата нового ареала, но и потому, что она стала одним
из доминирующих видов в фитопланктоне этой части Северного моря, а кроме того, проникла в Ирландское море и в скандинавские воды.
Водоросли прибрежной зоны тоже завозятся из одного океана в другой. Существует небольшая, мало заметная красная водоросль Asparagopsls armata, называемая также Falkenbergta rufo-lanosa (фиг. 62), которая обитает в нижней части литорали
Фиг. 62. Falkenbergia rufolanosa — австралийская водоросль, недавно появившаяся в Европе и Северной Америке [175].
и в изобилии встречается вдоль южного побережья Австралии; ее можно найти также на Тасмании и на Новой Зеландии. Всесторонние исследования И. Фельдмана и Г. Фельдман [175] показали, что «виды», известные под этими двумя названиями, представляют собой чередующиеся фазы жизненного цикла одной и той же водоросли и что Falkenbergta — это тетраспоровая фаза. Сильным подкреплением этого вывода послужило происшедшее недавно одновременное проникновение обеих форм в Средиземное
море п Западную Европу, куда водоросль, несомненно, была завезена судами (фиг. 63). Asparagopsls впервые обнаружили на крайнем юге атлантического побережья Франции в 1923 г.; в том же году Falkenbergla была найдена у Шербура, a Aspara-gopsis в Алжире. На фиг. 63 показаны места последующих
Фиг. 63. Одновременное распространение двух фаз жизненного цикла австралийской водоросли [175].
Эти две фазы были описаны как две самостоятельные формы Asparagopsis armatci (белые кружки) в юго-западной Европе и Falkenbergia rufolanosa (крестики) в северной Африке; черные кружки с крестиками— места, где были найдены одновременно обе формы.
находок до 1934 г. В 1941 г. водоросль была найдена па западе Ирландии, в 1950 г. она вполне закрепилась на полуострове Корнуолл, в 1951 г.— на островах Силли [180], а к 1954 г. уже имелась колония на острове Мэн [199]. Известен другой вид Aspara-gopsls, распространенный в тропических морях всего мира, но возможно, что это его естественный ареал.
Усоногий рак Eltninius modestus живет в приливной зоне скалистых берегов Новой Зеландии и Австралии. В 1945 г. его заметили на юго-восточном побережье Англии [171]. Вероятно, он появился там по крайней мере за несколько лет до этого, поскольку обследование, проведенное в 1947 г., показало, что он широко распространен от Норфолка до Дорсета и живет также в одном пункте Южного Уэльса. Этот усоногий рак, несомненно, способен путешествовать на подводных частях судов, так как он встречается на них в большом числе и давно уже был найден на корпусе судна, курсировавшего между Голландией и Англией. Ныне он заселил большую часть северного побережья Франции и живет также у берегов Бельгии и Голландии [162]. Единичная особь на скале была найдена в 1954 г. у острова Камбрей в Клайде при обширных полевых исследованиях усоногих этого района [170]. В 1949 г. этот вид впервые обнаружили в Южной Африке, в Кейптауне [193]. Это доминирующий вид, способный заселить побережье, несмотря на конкуренцию других видов усоногих; однако совершенно вытеснить их он способен лишь в определенных зонах. Е. modestus живет преимущественно в нижней части литорали и еще ниже, процветая в защищенных и мутных водах и вступая таким образом в конкуренцию также и с устрицами [167]. «ELminius является доминирующим литоральным организмом в эстуариях Колна, Блэкуотера, Крауча и Темзы» [171]. Из других усоногих столь же дальние путешествия на обшивке судов совершили Balanus eburneus. попавший из восточной части Северной Америки в Средиземное море, а оттуда на Британские острова, и В, tmprovlsus, завезенный из северного полушария в Австралию [161].
Но самым важным фактором, участвующим в расселении морских животных по различным частям света, является, вероятно,
устричное хозяйство — очень древний распространенный во всем мире промысел, теперь постепенно превращающийся и в прикладную отрасль науки. Устричный промысел связан со значительно большими изменениями природных местообитаний, чем любой вид рыбного хозяйства, и в этом отношении напоминает использование суши для земледелия или скотоводства, тогда как почти все чисто морские рыбные промыслы до сих пор остаются на уровне охотничьего хозяйства — они основаны на знании биологии объекта и на ограничении его лова, но не на тщательно продуманном изменении местообитания 1. Два приема устричного хозяйства оказали глубокое влияние на распространение видов. Один из них состоит в том, что свободноплавающим молодым устрицам дают оседать на искусственные субстраты («коллекторы»)— раковины, черепицы, бамбук и т. п. [191]; в дальнейшем эти предметы помещают на дно в местах, богатых планктоном, для «откорма» устриц. Другой сводится к ввозу устриц из других стран, после чего их таким же образом откармливают перед продажей. В Англии может жить и размножаться только местная устрица Ostrea edulis. Однако в прошлом многократно привозили на судах португальских устриц О. angulata и восточноамериканских устриц О. vlrginica, хотя размножающиеся популяции этих видов в наших водах не возникают. Интересный случай ненамеренной перевозки устриц на новое место произошел около 1870 г., когда в Бискайском заливе близ Аркашона затонуло судно с грузом португальских устриц [191]. Образовавшаяся здесь новая устричная колония стала одним из мест регулярной добычи устриц. Таким образом, эти моллюски — нечто вроде сидячих овец, которых можно перемещать с одного морского «пастбища» на другое.
Массовое перемещение устриц, проводимое без особых предосторожностей, неизбежно вело к распространению и других видов. Первым из имеющих значение видов была улитка-туфелька
1 Это справедливо лишь отчасти, так как все большее место в охотничьем п-рыбном хозяйстве занимают разведение, подкормка, защита промысловых животных и другие биотехнические меры.— Пр им. ред.
(Crepidula fornicata), завезенная с восточного побережья Северной Америки на тихоокеанское побережье п в Западную Европу [200]. Мы не знаем точно, когда она проникла в Англию, но известно, что она впервые была замечена близ Брайтлпнгси в Эссексе около 1890 г. [167]. С тех пор она распространилась вдоль всего побережья Англии до Шотландии на востоке и до Корнуолла на западе [166]. В 1953 г. несколько экземпляров было в первый раз найдено в Милфорд-Хейвене, на юге Уэльса [169]. Этот моллюск, особи которого сидят друг на друге, располагаясь целыми ярусами, несколько сходен по своим потребностям с устрицей, так как он питается, отцеживая планктон. Поэтому он серьезный конкурент устриц в отношении места на субстрате, особенно ввиду того, что он предпочитает такие же илистые берега. Я еще раз упомяну об этом виде в гл. VL
Выступил на сцену, хотя гораздо позднее, и один серьезный враг устриц. Во всем мире устричники подвергаются нападению некоторых улиток; в Англии встречаются два их вида: первый Purpura (или Nucella) laplllus, которую часто можно видеть в местах скопления мидий; она же истребляет усоногих раков; второй — Ocenebra erlnacea. В 1928 г. был обнаружен американский моллюск «устричное сверло» (Urosalplnx cinerea) (фиг. 64), который позже проник на устричные банки в Эссексе и Кенте, но не распространился дальше (фиг. 65). Он не имеет свободноплавающей стадии, и расселение его осуществляет главным образом человек. Теперь мы знаем, что он проник в нашу страну, вероятно, в конце прошлого века [165]. Его следует считать вполне процветающим иммигрантом, питающимся молодыми устрицами и другими животными и успешно размножающимся: плотность его популяций достигает примерно 5 особей на 1 кв. м. Популяции устриц в Англии за последние десятилетия пережили тяжелые бедствия, и появление нового врага, способного уничтожить половину годичного прироста устричной банки, крайне нежелательно. Устрицы плохо переносят очень холодные зимы п понесли большие потери в 1928—1929, 1939—1940 и 1946—1947 гг. Численность Ocenebra тоже уменьшилась, а в результате последней неблагоприятной зимы этот вид почти вымер в Эссексе и Кенте,
хотя и сохранился на южном побережье. Urosalpinx, мепее чувствительный к холоду, не пострадал и, таким образом, занял доминирующее положение в сообществе [167], В 1955 г, Ocenebra только еще начинала вновь появляться в прежних местах [167], Urosalpinx проник и на тихоокеанское побережье США [184].
Это перемещение устриц и связанных с ними организмов приводит к последствиям, о которых в нашем кратком очерке можно
Ф и г. 64. Интродуцированный американский моллюск «устричное сверло» (Urosalpinx cinerea) на съедобной устрице (Ostrea edulis) [167].
сказать лишь несколько слов. В 1949 г. несколько партий Ostrea edulis было завезено на американское побережье в штате Мэн; устрицы начали размножаться, и есть надежда на появление постоянных популяций [184]. Японскую устрицу Ostrea gigas в 1905 г. впервые завезли на побережье штата Вашингтон, а значительно позднее — в Британскую Колумбию, Орегон и Калифорнию; создалась возможность широкой эксплуатации «тихоокеанских устриц» [183]. Однако молодь до сих пор выращивают в Японии и перевозят через океан в Америку, так как размножение
Фиг. 65. Известные области распространения (зачернены) американского «устричного сверла» (Urosalpinx cinerea) на английских устричных банках [1651.
этого вида в новом местообитании происходит лишь спорадически. С этой молодью, как это всегда бывает, были завезены и другие впды, в том числе японский двустворчатый моллюск Paphia phi-lippinarutn, который по крайней мере съедобен, и японская улитка Trltonalia japonica, нападающая как на привозных, так и на местных устриц. В 1947—1948 тг. японских устриц завезли в Австралию; те из них, которые были выпущены на побережье Тасмании, благополучно обосновались там и даже размножились, хотя пока еще неизвестно, насколько постоянными окажутся их популяции [198]. Подобную историю можно было бы повторять бесконечно; например, на Гавайские острова (как будто тропические моря еще недостаточно богаты!)- завозили устриц Ostrea virginica и О. gigas, которые начали неплохо приживаться, а также австралийскую О. cucullata, которая погибла [173]. Если бы была создана мощная корпорация, которая занялась бы расселением по всему миру специально подобранных организмов, живущих в разных частях света у нижней границы приливной зоны, то и она не смогла бы сделать больше, если учесть, что процесс этот идет полным ходом всего лишь сто лет, а может быть, и меньше того.
Немало пришлось попутешествовать и другим съедобным двустворчатым моллюскам. На тихоокеанском побережье США живет теперь восточноамериканский вид Му a arenaria (естественный ареал которого включает также Европу); он был завезен туда еще к 1874 г., вероятно, случайно вместе с устрицами [196]. На Гавайских островах появились два восточноазиатских вида — Paphia philippinarum и Cytherea meretrlx [173]. Но эти переселения незначительны в сравнении с интенсивными перевозками устриц во все концы света. Последним примером искусственного завоза, не имеющего, однако, никакого хозяйственного значения, служит небольшой (не более 2,5 см) краб Rhithropanopeus harrisi (семейство Xanthidae). Примерно в 1938 г. он был завезен с восточного побережья Северной Америки, вероятно с устрицами, в Калифорнию. Здесь он живет в довольно мутных водах эстуариев, но только в тех местах, где происходящее время от времени опреснение воды губит местного краба Hetnigrapsus oregonensls, очень
сходного с ним по своей биологии. Rhithropanopeus harrlsl любит селиться среди известковых трубок червя Mercierella enlgmatica (семейство Serpulidae) [182]. В 1953 г. этого краба обнаружили в копенгагенской гавани, где он жил вместе с тем же самым червем Л4. enlgmatica, тоже завезенным сюда! Он проник и в другие части Европы, в том числе в Черное море [201, 202].
Можно не запомнить все эти даты, названия видов и места, но трудно не заметить возникновения упорной тенденции к взаимному обмену между береговыми фаунами материков, а иногда и между планктоном различных морей. Процесс этот только еще начинается, однако некоторые виды уже заняли выдающееся положение в новых сообществах, к которым они присоединились: Biddulphia — в фитопланктоне, Elminius — в биоценозе литорали, устрицы — на различных низменных илистых берегах; и там же — их основные враги, вроде Urosalpinx, и конкуренты, вроде Crept-dula; следует вспомнить и болотный злак Spartina townsendii.
Ряд поразительных экологических взрывов произошел в Каспийском море. Этот сильно изменившийся остаток моря Тетис претерпел много превращений, прежде чем достиг современного экологического состояния; однако он все еще сохраняет огромное богатство жизни. Это крупнейшее солоноватоводное озеро в мире — длина его достигает 1300 км; его соленость вдвое 1 меньше солености океана, химический состав воды тоже несколько иной; на больших глубинах, как и в Черном море, живут только микроорганизмы; в северной замерзающей части моря обитает каспийский подвид тюленей. В прибрежных районах весьма богат бентос и развито рыболовство. Аральское море — озеро с еще меньшей соленостью воды — тоже представляет собой остаток моря 2. В пустынях этой области до сих пор есть колодцы, содер
1 Почти в 2,5 раза.— Прим. ред.
2 Советские исследователи рассматривают Аральское море как молодое (среднечетвертичное) пресноводное озеро, осолоненное вследствие малого стока речных вод (см. Герасимов и Марков, Четвертичная геология, М., 1939). Фауна Арала молодая и представлена преимущественно выходцами из Каспия (Л. С. Берг, Аральское море, Изв. Турк. отд. геогр. о-ва, вып. 5, 1908).— Прим. ред.
жащие морских фораминифер. Хотя Черное море — соленоводное, в его лиманах живут многие из солоноватоводных видов, живших в Каспийском море, а еще раньше — в огромном Понтийском море, объединявшем во времена плиоцена все моря этого района. В 1934 г. советские гидробиологи впервые предложили произвести интродукцию животных Азовского и Черного морей в Каспийское и Аральское моря в интересах рыбного хозяйства [203]. Эта идея возникла на основе двух необычных событий, полной истории которых мы, к сожалению, не знаем. Когда-то, не очень давно, двустворчатый моллюск Mytllaster lineatus из Черного моря и креветка Leander adspersus из Азовского моря случайно попали в Каспийское море и размножились там в колоссальных количествах. Оба эти вида живут также в Средиземном море. Интродуцировали и различных рыб, из которых кефаль (Mugil), как сообщают, успешно акклиматизировалась. Но когда в Аральское море вселили один вид осетров, он принес с собой паразитического червя Nitzschla sturionis, который причинил серьезный ущерб другому осетру, живущему в этом море.
В 1937 г. проводилось изучение эколого-физиологических особенностей солоноватоводного многощетинкового червя Nereis succinea из Черного и Азовского морей [190], а примерно в 1940 г. его с поразительным успехом интродуцировали в Каспийское море [160]. К 1952 г. завершена программа разностороннего экологического изучения этого вида, который к тому времени был уже одним из доминирующих представителей бентоса. Подобно злаку Spartina в Англии, он нашел такую область илистого дна, в которой не было других доминирующих видов. К 1946 г. популяции Nereis расселились до предела пригодных для него местообитаний в водах с низкой соленостью, и можно было заявить, что «на долю Nereis падает от 74 до 7s всей калорийной массы бентоса северной части Каспия в июне». Червь стал важной дополнительной пищей для нагула двух видов осетровых рыб; утверждают, что это не нарушило соотношений других донных животных. Черви живут в поверхностном слое органического материала на илистом или песчаном дне; они скрываются в этом материале и питаются им. Если сказать, что самка Nereis succinea отклады
вает 80—100 тысяч яиц, то это позволит получить представление о том, насколько быстро червь может заселить любую среду, дающую простор для его экспансии [190].
Остается закончить наш очерк несколькими примерами успешной интродукции проходных и даже чисто морских рыб. В северной части Тихого океана, от Японии до западных берегов Америки, обитает группа тихоокеанских лососей Onchorhynchus. Здесь сосредоточен один из крупнейших лососевых промыслов мира. Лососи живут в море, но для размножения поднимаются вверх по рекам. Этих лососей всего пять видов: чавыча (Опсhorny псhus tschawytscha), нерка, или красная (О. nerka), кижуч (О. kisutch), горбуша (О. gorbuscha) и, наконец, кета (О. keta). Нас интересуют первые четыре вида. За период с 1872 по 1930 г. Бюро рыболовства США с благожелательными намерениями передало свыше 100 млн. икринок тихоокеанских лососей в другие страны, рассчитывая на акклиматизацию этих рыб в новых местах. Дело было поставлено очень хорошо, и в отличие от многих других подобных мероприятий результаты тщательно документировались [172]. Опыты проводили многие страны, хотя Норвегия отказалась от них. Так как эти северные рыбы могут жить лишь в ограниченном диапазоне температур воды, интродукция была успешной только в северной и южной умеренных зонах п не удалась в таких местах, как Гавайские острова; в некоторых других местах, например в Аргентине, акклиматизации, вероятно, помешало слишком высокое содержание в реках взвешенного ила. Регулярные миграции лососей из моря в реки установились в Чили (кижуч или нерка), Новой Зеландии (чавыча), штате Мэн (горбуша), провинциях Нью-Брансуик и Онтарио в Канаде (чавыча) ; некоторые популяции полностью перешли к жизни в реках или озерах, например нерка на Новой Зеландии, чавыча на Тасмании и ряд популяций на востоке Северной Америки. На Новой Зеландии с 1905 г. существуют регулярно размножающиеся стада чавычи, выведшейся из икры, отложенной в 1901 г.; эти стада используют для нереста многие реки восточного побережья (фиг. 66) и обитают в прилегающих участках моря, где половозрелые лососи проводят большую часть своей жизни [159]. В ре
зультате этого грандиозного эксперимента род рыб, ранее встречавшийся только в северной части Тихого океана, проник в другие океаны земного шара — в зоны, где летние температуры морской
Фиг. 66. Распределение популяций интродуцированной чавычи (Onchorhynchus tschawytscha) на Новой Зеландии [159].
Сплошными линиями показаны реки с хорошо закрепившимися стадами чавычи»: прерывистыми линиями — реки с небольшим числом особей; пунктиром — реки, где чавыча отсутствует.
воды не поднимаю гея выше 15—20°. После многих безуспешных попыток, предпринятых за последние 90 лет, на Новой Зеландии удалось также создать размножающуюся популяцию семги (Sal-mo salar), но она прижилась только в одной речной системе [159].
В период с. 1871 по 1880 г. больше миллиона штук молоди шэда (Alosa sapidissitna) из восточной части Северной Америки было выпущено в реку Сакраменто (Калифорния) и почти миллион — в реку Колумбия в 1885—1886 гг. К 1879 г. эта рыба начала там ловиться в количествах, достаточных для продажи, а в последние годы средний ежегодный улов достигал нескольких миллионов фунтов [185, 196]. Шэд встречается теперь от северной границы Мексики до Аляски и острова Врангеля, хотя область его промыслового лова не так широка. Нив [189] пишет: «Быть может, лучшим свидетельством в пользу того, что шэд ведет себя подобно местной рыбе, являются недавние жалобы на истощение его запасов в реке Колумбия, сопровождаемые требованием изучить надлежащим образом состояние его стада».
Последний пример такого массового размножения рыб касается морского окуня (Roccus saxatllis). Это весьма крупная рыба: официальный «чемпион» из штата Каролина весил свыше 55 кг и достигал почти 2 м длины — мечта рыболова! Обычный крупный экземпляр весит около 4,5 кг, но, по-видимому, нередки экземпляры и по 9—12 кг [187]. Будучи морской рыбой, он все же заходит для нереста в менее соленые воды эстуариев. Его естественный ареал охватывает атлантическое побережье Северной Америки — от Флориды до залива св. Лаврентия. В 1879 г. он был впервые завезен в Калифорнию, а в 1882 г. там была выпущена вторая и последняя партия (435 особей). Популяция росла очень быстро и распространялась в другие участки тихоокеанского побережья [194]. Хотя морской окунь особенно ценится как объект спортивного рыболовства, в 1926 г., помимо добычи рыболовов-любителей, было выловлено около 500 000 кг этой рыбы. Но с 1935 г. в Калифорнии ловить ее разрешалось только любителям. «Ежегодные уловы в этом штате с 1942 г. были более илп менее одинаковы и составляли около 1 500 000 штук. Подсчитали, что каждый год на поездки для лова морского окуня тратится около 10 млн. долларов и что этот вид доставляет около 2 млн. человеко-часов развлечения в год» [189]. Мир, который начинает оценивать свои развлечения в человеко-часах, вероятно, не слишком озабочен разрушением зоогеографических областей Уоллеса;
но интересно проследить за результатами проводимых в Калифорнии исследований по биологии морского окуня: не окажутся ли его не особенно четко выраженные миграции сходными с наблюдаемыми в Атлантике? Не скажется ли на других промысловых рыбах тот факт, что он питается в значительной мере анчоусами и креветками [181]? Сколько еще важных хищных рыб можно было бы переселить подобным же образом — с успехом и без дурных последствий? Нив [189] говорит: «В некоторых отношениях эти успехи так же ясно демонстрируют наше невежество в вопросах динамики популяций, как и неудачи, которыми часто сопровождались наши попытки интродуцировать виды в новую для них среду». Теперь, когда мы закончили обзор нашествий чужеземных видов на материки, острова и в моря, естественно перейти к рассмотрению отношений между популяциями.
Глава VI
ВЗАИМООТНОШЕНИЯ МЕЖДУ ПОПУЛЯЦИЯМИ
"О гл. I этой книги я описал некоторые организмы, успешно обо-сновавшиеся в новых для них областях суши или моря, подобно тому как военный корреспондент мог бы написать серию сообщений, повествующих о скрытом просачивании подвижных отрядов, о неожиданных нападениях, о последовательных волнах подкреплений, прибывающих в том случае, если передовым отрядам пе удалось захватить плацдарм, об атаках и контратаках и, наконец, о широком наступлении войск и захвате территории, с которой их уже вряд ли можно будет вновь вытеснить. Читатель мог видеть, что в результате прежней изоляции материков и до некоторой степени океанов на Земле возникло столько видов растений и животных, что она вряд ли смогла бы обеспечить им всем существование, если бы эти виды вновь перемешались; образовались почти безграничные резервы видов, двигающихся в атаку на другие части света,— резервы, которых хватит еще на тысячи лет. Создается примерно такое же впечатление, как при чтении фантастического романа Уэллса «Пища богов», в котором автор писал об этой «пище»: «Из Англии она вместе со славой и тенью своего могущества весьма скоро перебралась на континент, а пройдя через Европу, появилась и в Америке, и в Японии, и в Австралии, всюду производя одинаковый эффект. Проникала она в жизнь медленно, окольными путями, и перескакивала с места на место, несмотря ни на какие противодействия. «Крупное» как бы возмутилось против «мелкого». Несмотря на предрассудки, на законы и регламентацию, несмотря на упрямый консерватизм, лежащий в основе всякого общественного строя, «пища богов» тихими, но верными шагами завоевала себе положение». Как
хотелось бы, чтобы Уэллс мог описать распад областей Уоллеса в 42-м веке!
Когда мы описывали нашествия животных и растений, речь шла прежде всего о видах, экспансия которых имела успех. Но неизмеримо большее число нашествий вообще терпит неудачу — иногда сразу же, иногда по прошествии даже многих лет (как это было с полевым жаворонком в Америке). Пришельцы встречают сопротивление. Это-то сопротивление, оказываемое либо непосредственно человеком, либо природой, либо человеком, мобилизующим себе в помощь природу, мы и должны сейчас рассмотреть: что оно собой представляет, как его понять, каким образом его можно, если потребуется, регулировать и в случае надобности усилить. В конце этой книги я намерен подвести читателя к мысли о том, что мы стоим перед жизненной необходимостью не только найти новые средства и методы подавления численности того или иного растения или животного, но и по-новому осмыслить и перестроить значительную часть мирового ландшафта, с которым человек так бесцеремонно обращался и который он уже так сильно изменил; в то же время мы должны сохранить возможно большую часть настоящей дикой природы, содержащей богатые естественные биоценозы. Другими словами, необходимо тщательное изучение взаимоотношений между популяциями, а также тех типов и комбинаций местообитаний, которые лучше всего могут обеспечивать поддержание устойчивости и ослаблять взрывную силу вспышек размножения и нашествий чужеземных организмов.
Чтобы изучить это сопротивление и обусловливающие его силы, необходимо заглянуть по другую сторону поля битвы. Можно пытаться дать отпор пришельцам на трех различных этапах. Можно напасть на них еще до того, как они проникли в страну или во время их попыток прорваться через охрану — это карантин. Можно ликвидировать их первые небольшие плацдармы — это истребление. Иногда можно уничтожить и более крупную популяцию, как это произошло в борьбе с африканскими малярийными комарами в Бразилии (см. гл. I), но это бывает очень редко. Обычно же, если нашествие получило дальнейшее разви
тие, остается только удерживать численность особей в известных границах, т. е. осуществлять контроль.
Хотя во многих странах существуют системы карантина с целью предупреждения (или попытки предупреждения) проникновения чужеземных видов, представляющих опасность, очень часто тот или иной вид вовсе не кажется нам нежелательным. Мы не стремимся исключить новые породы лесных деревьев; п жаль было бы не допускать в страну крупных бабочек-голубянок, вврзенных из Голландии в восточную Англию после того, как английская популяция их вымерла. Но даже лесное дерево может нести с собой определенные опасности. При акклиматизации эвкалипта в Калифорнии с ним не было занесено ни одно пз питающихся на нем австралийских насекомых, так как в Калифорнии эти деревья выращивали из семян. Однако в Южную Африку и Новую Зеландию с молодыми саженцами эвкалипта завезли несколько видов таких насекомых, и они сделались там вредителями [226]. Ныне в США запрещен ввоз вишневых деревьев с четырех других материков ввиду опасности завезти с ними вирусы [224]. Было подсчитано, что семена пшеницы могут нести около 55 различных бактериальных и грибных болезней, часть которых поражает не только пшеницу [217].
В том, как легко можно завезти в страну чужеземное насекомое, я убедился, когда перед самой войной привез домой в Англию несколько крупных желудей из Америки, из штата Висконсин. Я хотел просто держать их на своем письменном столе в качестве сувениров. Однако через несколько дней я увидел, что из этих желудей вылезли личинки хруща. Конечно, я сейчас же бросил все желуди и личинки в кипящую воду, и с ними было покончено. Между тем когда таможенный чиновник спрашивал, нет ли среди моих вещей чего-либо, что следует предъявить, мне, конечно, не могло прийти в голову сказать о желудях; более того, я не уверен, что чиновник заинтересовался бы этими желудями, если бы даже я и сказал о них.
Но не всегда такие вещи кончаются благополучно. Девяносто лет назад Л. Трувело, французский астроном, работавший в Гарвардской обсерватории и занимавшийся также изучением раз
личных шелкопрядов, привез к себе домой, в штат Массачусетс, некоторое количество грены европейского непарного шелкопряда (Lymantrla dispar) [210]. Несколько яиц или гусениц случайно ускользнули и положили начало одному из крупнейших нашествий гусениц в Новой Англии (фиг. 67). Гусеницы уничтожали листву деревьев в лесах, садах и плодовых насаждениях, и, несмотря на огромную практическую и исследовательскую работу, в том числе интродукцию многих паразитов и некоторых врагов, этот вредитель до сих пор представляет хроническую проблему. К 1944 г. непарный шелкопряд в основном занял всю область, в которой он может быть многочисленным (фиг. 68 и 69). К числу кормовых растений, предпочитаемых гусеницами этого шелкопряда, относятся ольха, яблоня, липа, американский клен, два вида березы, боярышник, все дубы, все виды тополей и ив, не считая многих других, тоже часто повреждаемых видов [204]. За нашествием непарного шелкопряда последовала намеренная интродукция многочисленных паразитов и нескольких врагов этого вида пз Европы — немалое дополнение к фауне Северной Америки.
Приятель одного моего друга, только что вернувшийся из Египта, был порядком удивлен, когда из пуговиц его рубашки начали вылупляться мелкие жучки. Оказалось, что пуговицы были сделаны из скорлупы плодов одной пальмы и личинки продолжали жить в материале, по-видимому пройдя невредимыми через все технологические процессы — подобно Чарли Чаплину в фильме «Новые времена».
Таким образом, хотя теперь вряд ли кто-нибудь вновь прибудет в Новую Зеландию в сопровождении живого благородного оленя, нам приходится допускать, что проникновение животных, растений и их паразитов (а также паразитов человека) в новые для них страны будет продолжаться в ближайшую тысячу лет и, вероятно, в последующие тысячелетия. Каждый год будет приносить нам что-нибудь новое в этой области. Иными словами, взаимоотношения между видами будут непрерывно изменяться в каждой стране. Карантин и массовые истребительные мероприятия позволяют лишь выиграть время, и хотя они полезны и необходимы, они все же чрезвычайно дорого обходятся. Ведь так мало
Фиг. 67 Самки непарного шелкопряда (Limantria dispar), откладывающие яйца на ствол дуба в Массачусетсе в 1895 г. [210].
нужно особей, чтобы заложить основу популяции, и так много труда, чтобы впоследствии ее уничтожить!
Было около двух десятков очень крупных нашествий, которые имели с точки зрения человека удовлетворительный исход (здесь я, конечно, не говорю об интродукции культурных растений, домашних животных и врагов и вредителей). Таково было проникновение средиземноморской мухи-пестрокрылки Ceratitls capitata
Ф и г. 68. Распространение непарного шелкопряда (Lymantria dispar) — палеарктического вида, случайно завезенного в восточную часть Северной Америки в конце XIX в.
во Флориду в 1929 г., африканского малярийного комара в Бразилию в 1938—1939 гг. (см. гл. I), колорадского жука на Британские острова и вируса ящура в США. Историю борьбы с ящуром и победы над ним в Калифорнии живо описал Поль де Крюи в своей книге «Охотники за микробами»; она подтверждается официальными документами. За период до 1940 г. в США произошли и были ликвидированы 10 случаев заноса этого вируса, крупнейшим из которых была эпизоотия 1914 г., распространившаяся на чикагские скотопригонные дворы [213]; в борьбе с ящуром пришлось уничтожить 3556 партий крупного рогатого скота. Различные вспышки имели самое разное происхождение; иногда инфекцию заносили из Азии, например в 1884 г. из Японии,
иногда из Южнрй Америки, в других случаях из Европы. В 1924 г. в Калифорнии началась эпизоотия не вполне ясного происхождения, которая была подавлена с помощью исключительно же-сюких мер — было забито около 110 000 голов крупного рогатого
Фиг. 69. Распространение непарного шелкопряда (Lymantria dispar) на востоке США [204].
За последние 30 лет ареал его расширился очень незначительно.
скота и других сельскохозяйственных животных; кроме того, болезнь распространилась на диких оленей в Национальном лесном парке Стэнислоу, где за всю кампанию было отстреляно около 22 000 оленей (примерно у 10% животных были видимые признаки болезни). Общая стоимость всех этих мер по искоренению инфекции в Калифорнии составила около 7 млн. долларов [215].
На Британских островах удалось уничтожить ондатру (Ondatra zibethlca), пока она еще не успела по-настоящему закрепиться в реках и прудах (фиг. 70). Благодаря принятым мерам через несколько лет у нас не было ни одной ондатры: правда, общая численность этого вида в Англии никогда не превышала нескольких тысяч [231]. Эти ондатры сбежали со звероводческих ферм Англии, Шотландии и Ирландии [230]. В Шотландии при истреблении ондатры погибло множество особей других видов, попадавших в ловушки для ондатр, установленные у края воды. Так, в системах рек Форт и Эрн в Шотландии, кроме 945 ондатр, было убито 5783 местных животных, в том числе 2305 водяных крыс и 2178 шотландских белых куропаток [220].
Хотя знание экологии имеет чрезвычайно важное значение для проведения карантинных и истребительных мероприятий, все же эти мероприятия в конечном счете сводятся к искусственным методам, направленным на прямое умерщвление организмов или изменение местообитаний; подобные методы часто бывают эффективны и даже остроумны, но с точки зрения эколога не слишком тонки. Длительная регуляция численности — вещь совершенно иного рода. Она состоит в снижении численности вида до уровня, исключающего его доминирование в сообществе, и в ослаблении или даже полном подавлении значительных колебаний численности и вспышек размножения. Для осуществления этой регуляции используются не только искусственные приемы, но и силы самой природы. Нередко одни эти силы ограничивают распространение вида и в конце концов обусловливают то или иное стойкое равновесие. Так, например, трудно было бы утверждать, что распространение канадской элодеи (Elodea canadensis), захватившее в прошлом веке всю Великобританию, было поставлено под контроль человеком. Это зеленое водяное растение с мягкими стеблями было впервые замечено в пруду на шотландской границе в 1842 г. и вскоре появилось в одном месте в Лестершире, будучи случайно завезено с американским лесом. После этого элодея с необычайной быстротой заселила реки, каналы, озера, канавы и пруды по всей стране; она распространялась вниз по течению рек, вдоль каналов, ее переносили птицы, а в отдельных случаях
Ф и г. 70. Проникновение ондатры (Ondatra zibethica) в графство Шропшир (Англия) по данным до 1933 г. включительно [230].
Спустя год или два ондатра была полностью истреблена в местах ее выпуска в Англии, Шотландии и Ирландии.
(например, в Кембридже) ее специально интродуцировали в качестве научной диковинки. Согласно Дрюсу, наибольшее изобилие элодеи наблюдалось примерно в 80-х годах прошлого века, а после этого наступил значительный и повсеместный регресс, и ее никогда больше не рассматривали как настоящее бедствие [208]. Однако в эпоху своего расцвета она образовала такие густые заросли в реке Трент, что рыбаки не могли орудовать своими сетями; в районе Кембриджа она заполнила реку Кэм, мешала грести и настолько затрудняла движение барж на конной тяге, что приходилось впрягать добавочных лошадей; один или два купальщика запутались в ней и утонули; она препятствовала оттоку дренажной воды в Фенах 1 [221]. В наши дни элодея, конечно, не могла бы вызвать тревогу среди шотландских рыболовов [218] или сделать непроходимыми отдельные участки Темзы [208], как об этом сообщали в то время. Ее и теперь легко найти во многих водоемах, где она постоянно растет, но в умеренных количествах. Причины сокращения ее численности совершенно неизвестны. Возможно, что они имеют генетическую природу или обусловлены истощением запасов какого-то редкого элемента, необходимого для питания. В некоторых местах борьбе с элодеей помогали поедавшие ее коровы и водяные птицы [218]. Одно совершенно несомненно: человек непосредственно не регулировал численность этого растения.
Какого рода экологическая среда ожидает вид, вторгающийся в новое местообитание? Если он проникает в склад, полный пищевых продуктов, он уже найдет там небольшую компанию других животных; если попадет на земли, занятые сельскохозяйственными культурами, то это будет несколько более разнообразное сообщество; если эти земли окружены сетью придорожных полос, живых изгородей и лесных участков, он встретит гораздо более богатый комплекс растений и животных, а в более или менее естественном лесу найдет исключительно сложный мир живых организмов. Большинство людей просто не знает, какое богатство видов и какое огромное число отдельных животных сосуществуют
1 Болотистая местность в Кембриджшире и Линкольншире.—Лрилс. ред.
бок о бок в одном месте. Отчасти это связано с тем, что животные в значительной степени скрываются в укрытиях — эволюционное следствие напряженной борьбы между хищниками или паразитами и их жертвами, борьбы, загоняющей животных в укрытие, которое служит также физической защитой от других факторов. Частично это обусловлено и тем, что большинство животных — довольно мелкие существа, как, например, насекомые, пауки и клещи, илп (в воде) насекомые, ракообразные, коловратки и простейшие. Мир микроскопических грибов разнообразнее, чем мир крупных зеленых растений, хотя эти грибы, конечно, не образуют тех макроскопических растительных структур, которые создают живописный фон для большей части сообществ. В воде микроскопические водоросли также более разнообразны и многочисленны, нежели цветковые растения.
Больше 12 лет я изучал небольшой холмистый участок земли близ Оксфорда и регистрировал факты, из которых очень многие были сообщены мне другими экологами университета, изучавшими там экологические или иные проблемы [209]. Уитхэм-Вудс занимает меньше 5 кв. км. Это типичный уголок центральных графств Англии: леса и поля, ручьи и болота, несколько участков, заросших травой Роа flabellata, на вершине холма и Темза, омывающая этот холм с двух сторон. Мы уже знаем, что в этом обычном (и поэтому очень красивом) сельском уголке Англии, который пока не слишком испорчен прогрессом лесного и сельского хозяйства XX в., обитает примерно 2500 видов животных, причем, по всей вероятности, в будущем мы здесь обнаружим еще столько же, если не больше видов. Число особей на всей этой площади, несомненно, исчисляется миллиардами: мы до некоторой степени уже знаем об этом из подсчетов популяций, проведенных здесь экологами.
Естественно, большинство животных заселяет не все местообитания, а лишь одно или несколько из них; но и в этом случае животным приходится внедряться в чрезвычайно сложное сообщество различных популяций, они ищут места для размножения и обнаруживают, что эти места заняты, ищут пищу, которой уже питаются другие виды, и убежища, в которых скрываются
другие животные, ломятся в эти убежища, врываются в них — и часто их отбрасывают назад. Кроме того, фауна каждого местообитания частично совпадает с фауной соседнего местообитания. Экологическая система имеет отдельные «центры деятельности» — таковы, например, почва и кроны деревьев, болото и ручей, упавший ствол и птичье гнездо,— по они всегда в каком-то отношении связаны между собой, и эти связи могут влиять на отношения между популяциями. Таким образом, пришелец тем или иным способом пробивает себе дорогу внутрь сложной системы. Самое короткое описание такой ситуации (и удобное к тому же, если вспомнить, что нам приходится описывать не столько известное, сколько неизвестное) состоит в утверждении, что пришелец встречает экологическое сопротивление. Что же представляет собой это сопротивление? И почему некоторые виды внезапно преодолевают его?
Сопротивление новым пришельцам можно наблюдать в установившихся типах растительных сообществ; в самом деле, конкуренция составляет одну из центральных проблем экологии растений — конкуренция из-за света, пространства и химических компонентов почвы; правда, благодаря причинам, не особенно хорошо вскрытым даже современной фитоэкологией, можно найти полсотни и больше различных видов растений, постоянно растущих вместе в растительном сообществе одного типа. Однако большая часть чужеземных растений, в том числе, конечно, и сельскохозяйственные культуры, растут у нас в местообитаниях, резко упрощенных человеком: на пахотных землях, на пустырях и свалках, вдоль железнодорожных линий, стен и т. п. «Кроме полевых сорняков, лишь немногие ввезенные виды действительно настолько укоренились, что сделались частью британской флоры» [223] — иначе говоря, они в большинстве своем не пронцкли в насыщенные растительные сообщества Британских островов. Теперь известно, что некоторые из наших сорняков, в изобилии растущих или росших раньше на открытом грунте полей или придорожных полос, были широко распространены в стране еще в начале послеледникового периода [212]. Когда они впервые колонизировали Британские острова, перед ними был открытый грунт тундры, где конкурен
ция между растениями не так сильна, как в полностью развитом луговом или лесном сообществе. Позже их численность значительно сократилась. Такова же история по крайней мере одного вида животных — улитки Succinea oblonga. Эта улитка была широко распространена во внутренних частях Англии и найдена в позднеледниковых отложениях близ Кембриджа, к северу от Лондона и в ряде других мест [216, 227]. Теперь же она встречается лишь на нескольких приморских песчаных дюнах вроде Бронтон-Барроус в Девоншире. Подорожник Plantago maritima тоже имел довольно сходную историю и ныне растет около моря, а также на некоторых горах. Лишенная запаха ромашка Matricaria inodora была позднеледниковым растением, а теперь растет как сорняк на полях и пустырях. Один из ее близких подвидов — обычное приморское растение [212].
Глухая крапива (Lamium album) — одно из тех растений, которые так и не смогли проникнуть в естественные фитоценозы Британских островов. Это губоцветное похоже на жгучую крапиву, но лишено жгучих волосков, хотя его листья имеют острый вкус. Его белые цветки на протяжении значительной части года посещаются шмелями и ранней весной служат одним из важных источников их пищи. Глухая крапива в изобилии растет на обочинах дорог, по краям полей, на пустырях и т. п.— всегда вне наших природных сообществ; Тэнсли едва упоминает о ней в своей «Растительности Британских островов». На своей родине, на Кавказе,— это лесная форма, отлично растущая в богатом сообществе других видов. Тем не менее в Англии, где она, несомненно, существует сотни лет, она так и не стала лесным растением. Но предположим, что она стала бы таким растением. Вытеснила бы она при этом какой-нибудь местный вид или была бы просто добавлением к списку местной флоры? Этот вопрос, настоятельно требующий ответа, вновь и вновь возникает в нашем уме и является, пожалуй, той единственной важнейшей проблемой, которая лежит в основе всех описанных в этой книге фактов.
Вселившиеся животные часто действительно вытесняют местных или уменьшают их численность. В простейшей форме это можно наблюдать в море, где многие животные, отцеживающие
пз морской воды планктон или органический детрит, конкурируют из-за места на побережье или на дне моря. Примером могут служить устричные банки, о которых говорилось в гл. V; но здесь пришелец вступает в контакт не только с устрицами. Об этом можно судить по данным экологического обследования двух устричных банок, проведенного Мистакидисом [219] в эстуариях Эссекса (такого рода полное обследование сообщества, более 50 лет терпевшего экологические невзгоды, произведено впервые). Ми-стакидис изучил два участка, один площадью 6 кв. м, другой — 13 кв. м, и нашел, что число видов животных достигало на них соответственно 92 и ИЗ, хотя большинство видов было представлено лишь редко рассеянными единичными особями или колониями. Но 24 вида попадались на обоих участках более чем в половине мест сбора проб. Таким образом, это было сообщество устричной банки — система, для обозначения которой немецкий эколог Мёбиус много лет назад впервые ввел описательный термин биоценоз,— и в такое сообщество входил в качестве конкурента моллюск Crepidula (улитка-туфелька). На этих устрични-ках Эссекса Crepidula была доминирующим животным, популяция которого достигла уже поразительной плотности — 446 особей на 1 кв. м на первом участке и 179 — на втором (фиг. 71), что соответствует живому весу 10 и 4 т на 1 акр. Особи Crepidula живут ярусами, друг на друге, и в данных популяцих число таких ярусов достигало 2—8, а в исключительных случаях — даже 18. Забавно, что люди и эти моллюски на восточном побережье Северной Америки выработали один и тот же принцип небоскребов для полного использования ценной территории. Для Crepidula это означает получение большего количества пищи и высокие темпы размножения. Здесь нужно пояснить, что в Эссексе были обследованы заброшенные устричные банки, которыми начти не занимались по меньшей мере 8 лет и популяции которых слабо пополнялись молодью. Но, как можно видеть, заброшенный уст-ричник весьма богат другими видами, хотя многие из них в свою очередь, несомненно, пострадают от дальнейшего распространения Crepidula. В хорошо поставленных устричных хозяйствах улиток-туфелек вылавливают и дробят машинами, производя, в сущности,
«прополку». Территория быстро вернется к состоянию, удачно характеризуемому как «ил и улитки-туфельки», если не осуществлять отлов и удаление Crepidula, и снова достигнет климакса через 10—15 лет. На хорошо обслуживаемых устричных банках в Эссексе донное сообщество можно описать как сообщество Ostrea с Elminius в качестве эпифауны, так как этот усоногий
Ф и г. 71. Карта плотности популяции устрицы Ostrea edulis (А) и ее случайно завезенного конкурента — американского моллюска Crepidula (Б) на устричной банке площадью 13 га в реке Роч в Эссексе [219].
Эта банка была запущена, и на ней встречалась лишь молодь устрицы. Цифры указывают число устриц на 1 кв. .и; средняя плотность популяции Crepidula — 179 особей на 1 кв. м.
рак теперь встречается повсеместно как на устрицах, так и на «коллекторах», и популяции его, по-видимому, еще не достигли максимальной плотности [219].
Конкуренция из-за пространства обычна и среди более подвижных животных. Хотя европейский скворец, очевидно, нашел для себя в США более или менее новую пищевую нишу и благодаря этому избежал слишком активной конкуренции с местными птицами из-за пищи, он, однако, изгнал некоторых птиц из мало
численных мест гнездования в городах. В результате птица Sialia stalls и дятел Colaptes auratus встречаются реже в тех местах, где города заселены скворцом, хотя нет оснований опасаться за пх популяции в целом [17]. Очень интересные соотношения наблюдались в ходе расселения двух видов стеблевых пилильщиков в США [229]. Хлебный пилильщик (Cephas pygmaeus) проник
Фиг. 72. Расселение в США черного хлебного пилильщика (Cephus tabidus), завезенного из Европы к 1889 г. [229].
сюда из Европы к 1887 г. и в дальнейшем распространился в некоторых из северо-восточных штатов. Другой европейский вид — черный хлебный пилильщик (С, tabidus) — в 1889 г. был найден в Нью-Джерси, после этого неуклонно продвигался на запад и па юг и к 1940 г. достиг пределов, показанных на фиг. 72. Во время своего распространения второй вид заселил часть лежащей к северу территории, уже занятой первым, тогда как в восточной Пенсильвании С. pygmaeus постепенно вытеснял нового пришельца; на другой карте (фиг. 73) показан результат — два отдельных
ареала, лишь слегка перекрывающихся на довольно узкой полосе. Недавние наблюдения показывают, что в районах, где живут оба вида, у С. pygmaeus взрослые особи выводятся примерно на неделю раньше, чем у С. tabidus. Если учесть эти наблюдения, а также то обстоятельство, что в стебле пшеницы часто
Ф и г. 73. Взаимоисключающее распространение двух завезенных из Европы пилильщиков— Cephus pygmaeus и С. tabidus— на востоке США [229].
С. tabidus сначала проник дальше на север, в область, уже занятую С. pygmaeus. Узость зоны перекрывания ареалов, возможно, обусловлена конкуренцией (в частности.конкуренцией между личинками за пространство в стеблях пшеницы).
находят несколько яиц пилильщиков, хотя зрелости достигает лишь одна личинка, можно предположить, что личинки С. pygmaeus уничтожают яйца и вылупляющихся из них личинок независимо от видовой принадлежности. Этим можно было бы объяснить уменьшение численности С. tabidus в районах, где ветре-
чается С. pygtnaeus. Именно такого рода результат дали некоторые из классических экспериментов со смешиванием лабораторных популяций двух различных насекомых, личинки которых живут в зернах пшеницы.
Я привел примеры, в которых конкуренция возникает из-за жизненного пространства, мест питания или мест размножения, потому что эти виды конкуренции легче всего понять. Но весьма вероятно, что конкуренция обычно представляет собой вещь гораздо более сложную. Говоря о «конкуренции», нужно четко различать значения этого слова. В обычном смысле оно означает прямую борьбу между особями двух видов, которую экологи обыкновенно называют «столкновением». Но возможно множество косвенных влияний, либо осуществляющихся через посредство других видов, например паразитов или врагов, либо зависящих иг сравнительной приспособленности двух видов животных или от целой цепи причин и следствий, которую иногда очень трудно проследить. Эти влияния, так же как и столкновения (если они •называют решающее воздействие на размножение или выживание), могут вести к вытеснению одного вида или части его популяции другим видом — явлению, о внутренних причинах которого мы обычно почти ничего не знаем. Главная трудность для понимания этих концепций состоит в том, что вытеснение может происходить без прямых столкновений — как в случаях, указанных выше, или в таких случаях, когда понижение численности одного вида чисто случайно совпадает с повышением численности другого вида; кроме того, столкновение может не сопровождаться вытеснением, как, например, в тех случаях, когда особи двух видов птиц сталкиваются из-за места гнездования, хотя кругом достаточно других мест, которые может занять потерпевшая поражение сторона. В тех случаях, когда совершенно ясно (как, например, в случае с аргентинским муравьем), что жестокие сражения постоянно приводили к неукоснительному и катастрофическому вытеснению всех других видов (в том числе других муравьев), перед нами вполне типичный пример вытеснения в результате столкновения. В случае с белой глухой крапивой мы ясно видим неспособность растения проникнуть в высокоорганизованный
насыщенный фитоценоз, но не знаем причин этого. В случае вымирания английских устриц мы видим действие целого комплекса обстоятельств, среди которых, как показывает прямое наблюдение, очень важную роль играет конкуренция за пространство с американским моллюском и австралийским усоногим раком. Но нужно помнить, что здесь действуют и многие другие факторы, например появление нового врага устриц — моллюска Urosalpinx, неодинаковое влияние холодных зим, а также иные причины смертности, о которых мы практически ничего не знаем (как, скажем, в случае гибели устриц в 1920 г.) [191]. Примером нашего незнания характера конкуренции может служить также история двух видов белки в Англии. Завезенная из Америки каролинская белка (Sciurus carolinensis) вытеснила обыкновенную белку (Sciurus vulgaris) в центральных графствах и в части южной Англии [225]. Местами это происходило очень быстро; но нередко для завершения этого процесса, какова бы ни была его природа, требовалось до 15 лет. Мы до сих пор не имеем представления о том, что, собственно, происходит,— мы только констатируем изменение; поэтому естественно проявлять осторожность в прогнозах; тем не менее существует огромная область, где обыкновенная белка так и не появилась вновь, а именно в тех местах, где впервые распространилась каролинская белка и где она является теперь постоянным обитателем. Вне этой области до сих пор есть еще много популяций обыкновенной белки, хотя численность их обнаруживала колебания, часто очень сильные.
Иногда иноземным видам удавалось проникать в сообщество, не вызывая заметных пертурбаций или вымирания близких видов местной фауны. За последние 25 лет в реках и каналах Англии спокойно расселялись различные пресноводные рачки. Один из них, Eucrangonyx gracilis, происходит из Северной Америки. В Оксфорде я заметил, что его переносили из одного пруда в другой с декоративными водяными растениями; вероятно, таким же образом его завезли из Америки. Этот рачок имеет довольно тонкое тело голубоватого цвета и обычно двигается прямо, тогда как наш обычный Gammarus pulex толще, имеет коричневую окраску и часто лежит или плавает на боку, особенно во время спаривания
самца с самкой. Eucrangonyx gracilis широко распространился в Англии и Уэльсе [228]. Другой североамериканский рачок — Gammarus fasciatus — в Англии прижился лишь в отдельных местностях и только в соленой воде, в Ирландии же он поселяется п в пресных водах. Еще два вида происходят из района Каспийского моря. Corophium curvispirvum первоначально обитал в соленых и солоноватных водах Каспийского и Черного морей, а в последние десятилетия проник в реки Европы. В 1935 г. самой западной точкой его распространения был район Тьюксбери (Глостершир), где он жил в трубках из ила в реке Эвой [207]. Второй вид — Orchestia bottae, весьма подвижный рачок, способный прыгать. Он был найден в Темзе [205], в одной из рек в графстве Норфолк [222] и в Йоркшире [211]. Таковы четыре новых представителя британской фауны ракообразных, три из которых способны поселяться в пресных водах. Нам пока еще трудно судить о том, какова будет их дальнейшая судьба и в какой мере их присутствие отразится на нашей собственной жизни. Но кажется почти несомненным, что конкуренция среди подобных ракообразных возможна [214]. Наш Gamtnarus pulex отсутствует в Ирландии и на некоторых мелких островах близ западного побережья Великобритании. В этих местах экологическую нишу G. pulex в пресных водах занимает другой вид — G. duebenl, который почти на всей территории самого острова Великобритания живет только в солоноватых водах, например в эстуариях. На мысе Лизард (Корнуолл) G. duebeni населяет пресные воды, a G. pulex, по-видимому, отсутствует. Оба вида живут бок о бок только в одном месте — на острове Мэн, где соотношения между ними, возможно, находятся еще в процессе становления. Таким образом, когда где-либо появляются и распространяются новые виды, то даже если их распространение не носит характера «взрыва», они могут предвещать изменения в соотношениях популяций. Распутывание любого подобного клубка связей — интересная, но часто очень трудная задача.
Глава VII
ЗАМЕНА ПРЕЖНИХ ЦЕПЕЙ ПИТАНИЯ НОВЫМИ
природе живые организмы связаны между собой в очень сложные системы, известные под названием цепей питания. Такие цепи состоят обычно из нескольких звеньев, проходя от растения через различные виды животных; число звеньев часто достигает пяти, но редко бывает большим. Альфред Лотка назвал эти цепи видов объединенными преобразователями энергии, так как каждый вид использует на поддержание жизни, движение и рост часть энергии, первоначально уловленной растениями из солнечного света, и передает ее следующему виду в этом круговороте. «Тела организмов, относящихся ко всем этим видам, вместе с определенными неорганическими структурами, составляют один огромный всемирный преобразователь. Полезно приучиться представлять себе этот преобразователь как одно громадное целое, одну великую империю» [257]. Каждый вид превращает энергию органических веществ в теплоту и частично, при жизни или после смерти особей, используется в пищу другими видами. Даже животных, составляющих последнее звено цепи питания, после их смерти пожирают другие организмы. Растение, пока оно живет, сохраняет большую часть своих тканей, хотя довольно значительная часть их переходит в цепи питания животного мира. Но, видимо, гораздо больше растительного материала переходит туда после гибели растения, а также с опадающими листьями; в меньшей степени это относится к останкам животных. Если бы дело обстояло иначе, мы не видели бы сплошной массы зеленой растительности, жизненная база всех биоценозов была бы ненадежной и существование целых сообществ оказалось бы весьма непрочным, чего никак нельзя сказать о большинстве природных биоценозов. Растения и животные при жизни могут терять некото
рые части своего тела (например, листья, пыльца, слинявшие покровы насекомых), выделять секреты (например, нектар и выделения тлей) или экскреты (они особенно обильны у крупных жвачных, но имеют такое же значение, хотя и менее заметны, у других животных). Все это служит началом новых, побочных связей в экологической системе. Система всех этих цепей, этих объединенных преобразователей энергии необъятна и сложна, но мы еще очень далеки от ее понимания и представляем ее себе лишь в самых общих чертах. Нам необходимо знать, какую роль играют цепи питания в регуляции величины и плотности популяции, так как в наше время они непрерывно изменяются и нарушаются, а прежние виды замещаются новыми.
Первым, кто наглядно изобразил картину цепей питания, был Питер Брейгель Старший. Он обладал точным глазом художника и в своих красочных, динамичных полотнах, на которых богато представлены и пейзаж и жанровые сцены, он подчас достигал почти устрашающего проникновения в сущность природы и в природу человека. В 1556 г. он создал поразительный рисунок, с которого была сделана гравюра (фиг. 74). На рисунке, в частности, изображен некто, кто мог бы быть, возможно, одним из первых экологов, производящий анализ питания (очевидно, при помощи огромного хлебного ножа), а в правом верхнем углу — поспешно удаляющийся ранний представитель прикладной биологии, частично превратившийся в рыбу в результате питания рыбой, которая тоже ела рыбу (это, вообще говоря, верно, хотя лишь в строго определенном смысле). В целом эта картинка говорит о пищевых отношениях в природе больше, чем можно было бы передать с помощью любых многословных логических рассуждений. Основой для рисунков Брейгеля, по-видимому, послужили пословицы, бывшие в употреблении во времена средневековья. В «Перикле» Шекспира два рыбака говорят между собой о крупной рыбе, поедающей более мелкую. Как я упоминал в своей другой книге, у китайцев есть меткие «экологические» пословицы на подобные темы [245]. Уортингтон установил, что баньоро живущие у озера
1 Народность из группы банту.— Прим. ред.
Альберта, очень искусно ловят рыбу, используя одну за другой ряд наживок, составляющих цень питания [273].
Однако на культурных землях, будь то пастбища, нашнн пли сады, человек всегда стремился укоротить цени нптания, уменьшить их число и заменить старые цепи новыми. Нам нужны растения, которые не шли бы в пищу никаким другим травоядным
Фиг. 74. «Крупная рыба поедает мелкую» (гравюра с’рисунка Питера Брейгеля Старшего, 1556; по [249]).
животным, кроме нас самих. Нам нужны травоядные животные, мясо которых нам не приходилось бы делить с другими плотоядными. Только наши основные продукты питания морского происхождения — это до спх пор конечные звенья почти полных естественных пищевых цепей: камбала ест двустворчатых моллюсков, питающихся детритом; сельдь поедает планктон, представляющий сложное сообщество многих видов; кит питается ракообразными из семейства Euphauslidae, использующими в пищу более мелкий планктон. Эти три примера кажутся па первый
взгляд очень простыми, а между тем эти три цепи обеспечивают пищей и другими материалами множество людей.
Мы должны расчищать джунгли, вспахивать целину или вести устрпчное хозяйство; подавлять, уничтожать или изгонять всех конкурентов; укорачивать цепи питания и получать больше богатых энергией веществ; улучшать домашних и полуодомашненных животных. Мы не всегда укорачиваем пищевые цепи до предела, становясь вегетарианцами, потому что наше слабое пищеварение, наши вкусы и содержание в пище ряда важных химических веществ находятся в некотором противоречии с наиболее экономичным способом получения одних лишь калорий. Я наблюдал в роще из молодых березок в Нью-Форесте лесных муравьев, «пасших» на ветках небольшие стада тлей. Муравьи уничтожили на деревьях абсолютно всех других насекомых. Это были их пастбища, и около каждого стада тлей стояли «часовые», в то время как другие муравьи приходили и «выдаивали» из тлей их сладкие выделения. Я обломал нёсколько небольших веточек, так что сок из них начал вытекать наружу, и очень скоро некоторые из муравьев по-кпнулп свои стада и начали собирать сок у его первоисточника. Если бы нам позволяло наше пищеварение, мы, несомненно, ча-сто ели бы траву и обходились без овец и коров.
Некоторые из глубочайших изменений в цепях питания возникли вследствие интродукции и распространения домашних животных, питающихся подножным кормом. Еще сто лет назад по травянистым равнинам Северной Америки (фиг. 75) бродили огромные стада бизонов. Буффало Билл1 умер только в 1917 г. Бизон был главным травоядным животным в центральной части континента, но в течение сравнительно немногих лот был полностью вытеснен крупным рогатым скотом и овцами; уничтожению его способствовало и развитие других отраслей сельского хозяйства. Изменились также структура и состав растительности прерий. Но один вид птиц — воловья птица (Molothrus ater), которая обычно сопровождала бизонов, по-видимому, для ловли
1 Прозвище одного охотника, известного своими «рекордами» в уничтожении бизонов.— Прим. ред.
Ф и г. 75. Старые пылевые ванны’американского бизона на возвышенных равнинах Канзаса в 1899 г. (последние бизоны были убиты в Канзасе в 1879 г.) [248].
Фиг. 76. Огромные штабеля черепов и других костей бизонов (числом около 25 000) у обочины Саскачеванской железной дороги [252].
насекомых, вспугиваемых ими при движении, и часто садилась на этих животных, переменила объект своей привязанности. Эта птица, которую видели на равнинах первые поселенцы, в наши дни стала «коровьей птицей» пастбищ [247]. Старая фотография (фиг. 76), изображающая тысячи бизоньих черепов, сложенных штабелями, дает живое представление о размерах этого экологического сдвига. Вытеснение бизона домашним скотом происходило косвенным путем благодаря тому, что человек сменил источник животной пищи, разводя с этой целью домашних животных. Процесс совершался в грандиозных масштабах — миллионы бизонов вытеснялись миллионами голов крупного рогатого скота и овец 261]. Могло бы показаться, что, создав этот новый комплекс це-лей питания, совсем просто поддерживать их в устойчивом состоянии. Они коротки, и их легко регулировать. Однако именно ?та устойчивость во многих частях мира нарушалась в результате чрезмерного или неправильного использования пастбищ, а эрозия почвы нередко завершала процесс, который может привести к самой короткой пищевой цепи — цепи с нулевым числом звеньев, г. е. тому, что математики любят называть «предельным случаем». Как показывают любопытные фотографии (фиг. 77, А и Б), подобное положение может сложиться, например, в Канаде, даже в местах, благоприятных для сельского хозяйства. И если людям не удается управлять столь простой цепью связанных между собой популяций, то неудивительно, что более сложные цепи создают еще больше забот и затруднений.
Леопольд и Дарлинг [256] проследили очень интересную историю северного оленя в Аляске. Этих животных завезли в 1891— 1902 гг. из Лапландии, чтобы расширить жизненные ресурсы эскимосов, и они распространились там, достигнув численности свыше полумиллиона голов. В настоящее время численность оленей сократилась в 20 раз (фиг. 78). Причина, по-видимому, проста: оленям позволили уничтожить запасы лишайников, необходимые для их прокорма в зимнее время; лишайники растут очень медленно, для их полного возобновления требуется не менее 25 лет, а экология их весьма сложна, так как большую роль в ней играют и такие факторы, как, например, пожары [260].
A
Ф п г. 77. Полная эрозия верхнего слоя почвы в одном сельскохозяйственном районе па юге провинции Онтарио, частично обусловленная нерегулируемым выпасом скота после вырубки существовавшего здесь леса (фото автора, 1938).
А. Пни вепмутовоп сосны Pinus strobus) стоят, приподнятые на обнаженных корнях. Б. Видны новые посадки (слева) п реколонизация голых песков растительностью, которые должны привести н восстановлению почвенного покрова.
Использование живых организмов для накопления энергии и получения специальных веществ, необходимых человеку, до сих пор остается центральной отраслью производства во всем мире, так как мы еще не можем получать синтетические пищевые вещества сколько-нибудь приемлемой стоимости. Ученым удалось
Фиг. 78. Распространение северного оленя в Аляске [256].
Олень был завезен в Аляску в конце XIX в.; максимальная численность его превысила 500 тысяч голов (1930—1938 гг.), после чего катастрофически упала, главным образом вследствие истощения запасов его зимней пищи — лишайников.
уже найти замену некоторым дарам природы, например растительным красителям, шелку, тягловой силе лошадей и почтовым голубям. Кое-какие новые вещества и машины в значительной мере вытеснили использование живых организмов, хотя механического коня приходится кормить нефтью. Запасы нефти и угля в земной коре позволяют нам изготовлять или приводить в действие большую часть таких заменителей, и пока эти запасы не
исчерпаны, нам гораздо проще обходиться без лишних хлопот с живыми организмами. До сих пор еще существует во всем мире производство таннина, необходимого для обработки кожи. Для этого используют природный таннин, находящийся в достаточной концентрации в определенных растениях (травах, кустарниках и деревьях), которые растут на всех материках и служат источником для поставок таннина на мировой рынок. Однако дубильные вещества для некоторых целей можно теперь получать синтетическим путем. Уже одно это изменит цепи питания на всех пяти материках, где собирают или возделывают растения, содержащие таннин. Тем не менее все еще остается важнейшей проблемой сохранение, регуляция и, где это необходимо, изменение системы цепей питания без ломки взаимоотношений между популяциями. Именно последняя проблема еще не получила никакого решения, хотя она становится с каждым годом все более острой вследствие расселения видов на новые территории.
В «каменный век» экологии животных, т. е. примерно 25 лет назад, казалось разумным предполагать, что каждая природная цепь питания сама по себе позволяет объяснить регуляцию численности популяций. Допустим, что растение А служит пищей для травоядного животного Б, Б — добычей для хищника В, В — добычей для Г, а Г — добычей для Д. Каждый «ярус» потребителей ограничивает численность особей на нижележащем ярусе, а каждый нижележащий ярус как источник пищи ограничивает рост популяций яруса, расположенного над ним. На <го, что эти рассуждения не замыкаются в круг, независимо друг от друга указали в тот период два математика — Лотка [258] и Вольтерра [267], уравнения и предположения которых произвели глубокое впечатление на их современников. Будучи математиками, они не пытались рассмотреть всю цепь питания с усложнением отношений в пяти звеньях. Они взяли только два ее звена — хищника и его добычу. Дальнейшие их рассуждения показывали, что в действительности два таких вида должны поочередно регулировать численность друг друга с вытекающими отсюда колебаниями численности. Эта теория казалась весьма правдоподобной, поскольку она появилась в то время, когда уже было замечено
существование таких колебаний в природе, хотя это, строго говоря, не было достаточным доказательством. Но теперь, спустя 30 лет, мы имеем больше фактов для ее проверки, и не остается сомнений в том, что теории, объясняющие регуляцию численности организмов с помощью отношений в цепях питания, чрезмерно упрощают дело и часто даже неверны для определенных видов. Здесь действуют и другие факторы, в том числе случайные «катастрофы»; существуют — и даже, пожалуй, встречаются чаще, чем мы раньше думали,— механизмы регуляции, действующие через посредство отношений внутри популяции данного вида.
Тем не менее потенциальная сила пищевых связей, несомненно, действует во многих случаях биологической борьбы с вредителями. Когда численность популяции чрезмерно возрастает, ее удается иногда снизить путем интродукции врага (фиг. 79 и 80), паразита (фиг. 81) или — в случае борьбы с сорняками — растительноядного животного. Такого рода специфические мероприятия практикуются теперь очень широко, и во все страны мира, где сколько-нибудь развито земледелие или лесное хозяйство, непрерывно завозят живых помощников в борьбе с вредителями. О некоторых случаях мы уже упоминали. В борьбе с червецом Icerya purchasi и с кактусом опунцией такие операции оказались поразительно успешными, с европейским сосновым пилильщиком — дали весьма обнадеживающие результаты, с жуком Poplllia japonica и непарным шелкопрядом — были не вполне удачны. На примере Новой Зеландии и Гавайских островов мы убедились также в том, что чужеземные виды с этой целью можно завозить из любой части света, если есть хоть малейшая надежда, что они будут полезны. Все это изменяет распределение и группировку видов на земном шаре.
Многие из ввозимых врагов и паразитов вредителей не приживаются вовсе. Многие другие закрепляются в новой для них стране, но не всегда подавляют размножение вредителя. В *гех случаях, когда ввоз таких видов, по-видимому, дал положительные результаты, мы все равно не можем быть твердо уверены, что они сделали это самостоятельно, без содействия каких-то неизвестных нам факторов или событий. Но тем не менее несо-
Ф п г. 79. Червец Icerya purchasi, подвергающийся нападению божьей коровки Novius (Vedalia) cardinalis, на цитрусовом дереве в Калифорнии (как вредитель, так п его враг происходят из Австралии).
Ф и г. 80. Нападение улитки Gonaxias на крупную африканскую наземную улитку Achatina julica* [232]. Gonaxias интродуцировали на Марианские острова, а оттуда на Гавайские острова с целью использовать се в борьбе с Achatina.
Фиг. 81. Силы, регулирующие численность в природе, используемые в биологической борьбе с вредителями [238 ].
Тахина Centeter cinerea стремительно опустилась на 1—2 секунды на спаривающихся жуков Popillia japonica и отложила на грудь самки два яйца, из которых разовьются паразитические личинки. Эту муху интродуцировали из Японии в США с целью использовать ее в борьбе с жуком.
мнение, что биологический метод борьбы иногда давал очень значительный эффект даже при катастрофическом положении. В таких случаях биолог не укорачивает, а удлиняет цепи питания, но по возможности старается добавить к ним не более одного нового звена. Например, исключительный успех борьбы с опунцией в Австралии, главным образом с помощью южноамериканской бабочки Cactoblastts cactorum, возможно, не был бы достигнут, если бы вместе с этой бабочкой был завезен какой-нибудь из ее паразитов или если бы паразиты местных австралийских насекомых могли уничтожать больше гусениц (они уничтожают менее 25% их) [242}. Но есть и такие чужеземные сорняки (например, крестовники на Новой Зеландии или зверобой в Австралии), бороться с которыми при помощи ввезенных насекомых не удавалось, хотя сами насекомые успешно акклиматизировались.
Иногда вселившееся растение может постепенно подавляться в результате последующей случайной интродукции животных. Возможно, что это начинает сейчас происходить с Rhododendron ponticum — крупным кустарником с пурпурными цветками, происходящим из средиземноморской области. Уже к 1763 г. он был завезен на Британские острова и теперь прочно закрепился как дикорастущий вид на песчаных и кислых почвах и даже в качестве подлеска в лесах. Например, в дубовых лесах Килларни он на большой площади вытесняет падуб [268]. Лесоводы считают этот кустарник серьезным сорняком, мешающим лесовозобновлению [237]. Его редко едят кролики, чем, как и у бузины и боярышника, отчасти обусловлено его преимущество в конкуренции с другими кустарниками. Один специалист по пчеловодству писал: «Там, где нужна свободно растущая, не подстригаемая живая изгородь и где достаточно места, вряд ли какое-либо растение может соперничать с обыкновенным рододендроном (/?. ponticum)» [253]. Автор имеет здесь в виду использование рододендрона для защиты от ветра. Мед, собираемый с этого вида, иногда ядовит, что еще в древности отметил Ксенофонт во время прохождения его армии через Малую Азию. Однако в Англии едва ли были случаи отравления — вероятно, потому, что пчелам трудно
добывать нектар из цветков /?. ponticum, тогда как шмели, имеющие длинный хоботок, посещают их регулярно.
Что происходит с кустарником, завезенным извне около 200 лет назад и обладающим такой способностью к внедрению? На его рост влиял ряд местных насекомых, но это не имело сколько-нибудь серьезного значения [271]. В лесах на рододендрон могут нападать два вида долгоносиков — Otiorhynchus singulars и О, sulcatus (известные вредители плодово-ягодных
Фиг. 82. Естественный ареал рода Rhododendron, основная масса видов которого встречается в юго-восточной Азии [271 ].
кустарников) — и некоторые бабочки семейства Tortricidae, к которому относится ряд видов, сильно повреждающих листву дубов. Гусеницы некоторых из этих бабочек питаются листьями рододендрона, а одна скручивает их и окукливается в них, после того как закончит свое развитие на дубе. На рододендроне находили и некоторых других местных бабочек и жуков, но в незначительном количестве. Кроме того, на Британских островах на этом растении обосновались четыре иноземных вида насекомых [270, 271]. Естественный ареал всего рода Rhododendron (с которым более или менее совпадает ареал рода Azalea) охватывает Северную Америку, отдельные районы Европы и юго-восточную Азию
вместе с Новой Гвинеей, а один вид встречается на севере Австралии (фиг. 82). Таким образом, есть очень большой запас возможных иммигрантов-насекомых, если учесть, что для них известно более 900 видов кормовых растений. Моль Gracilaria azaleella, обитавшая в Японии, теперь проникла в Северную Америку и Европу и зарегистрирована в Англии. Алейродида Dialeurodes chittendenl, происходящая, вероятно, из области Гималаев (хотя это не установлено достоверно), распространилась в Северной Америке и Канаде, а в 30-х годах этого столетия была завезена из Америки в Англию, где расселялась с юга и юго-востока. Медвяная роса, выделяемая ее личинками, служит питательной средой для роста черной плесени на листьях. Третий иммигрант — Stephanitls rhododendri, клоп из семейства Tingi-dae, впервые обнаруженный в Англии в 1901 г.; он распространился очень широко (фиг. 83) и стал нападать на многие виды рододендронов, в том числе /?. pontlcum, хотя последний не относится к предпочитаемым видам. Этот клоп прокалывает хоботком нижнюю сторону листа, нарушая структуру тканей и вызывая появление крапчатости, известной под названием «ржавчины» [272]. Он происходит из Северной Америки и, прежде чем попасть на Британские острова, проник в Европу. Четвертый, самый важный вид — это Graphocephala coccinea, крупная цикадка из семейства Jassidae с блестящим, окрашенным в красный и зеленый цвета телом, тоже североамериканского происхождения; она достигла Англии к 1933 г. и распространилась в южных и юго-восточных графствах [233]. Эта цикадка сосет сок главным образом с верхней стороны листа, обычно через жилки, и редко питается на цветочных почках. Но она откладывает яйца в желобок, пропиливаемый самкой в нижней чешуйке этих почек, и весной здесь выводятся личинки. С распространением Graphocephala в Англии совпало появление так называемой «болезни бутонов», тоже, по-видимому, завезенной из Америки [243], которая начинается осенью и ведет к полной гибели цветочной почки; ее возбудитель — гриб Pycnosteanus azaleae, Считают чрезвычайно вероятным, что заражение происходит при откладке яиц цикадкой, хотя прямого доказательства этого еще нет [236].
Фиг. 83. Распространение в Англии клопа Slephanilis rhododendri (сем. Tingidae), случайно завезенного из Северной Америки через Европу [272].
Этот нлоп питается на листьях Rhododendron ponticum и других видов.
Во всех этих событиях, начинавшихся случайным образом и развивавшихся в разное время и с разной скоростью, мы можем с несомненностью усмотреть образование вокруг Rhododendron ponticum естественного сообщества растительноядных насекомых, которые снижают жизнеспособность растения, поедая листья или высасывая из них соки, а одно из них, кроме того, переносит микоз. Эти насекомые происходят из Азии и Америки, но к нпм присоединяются и несколько британских видов. Может быть, одна из причин столь энергичного внедрения R. ponticum в фитоценозы Англии именно в том п заключается, что он не захватил с собой всю свою цепь питания? И как скоро насекомые приобретут паразитов и врагов или начнут конкурировать между собой? Может быть, это уже происходит? Сравните этот зарождающийся биоценоз на листьях рододендрона с тем, что нашел Тпль-ден [266] на Baccharis pilularis — кустарнике песчаных дюн Калифорнии и Орегона. В результате двух лет систематического наблюдения он обнаружил 257 видов членистоногих, в том числе 221 насекомое; 65 из них были паразитами, и это еще не полный список. Там было 53 растительноядных вида (грызущие листья, минирующие их, пробуравливающие стебель, сосущие сок листьев, питающиеся корнями, галлообразователи), 23 вида хищных членистоногих, 55 видов первичных паразитов, 9 вторичных паразитов и даже один третичный паразит. R. ponticum в Англии еще только начинает приобретать свою фауну, и вполне возможно, что он окажется не в состоянии преодолеть растущее сопротивление и погибнет.
R. ponticum захватил на Британских островах определенную экологическую позицию в конкуренции с другими кустарниками, а аргентинский муравей — на большинстве материков в конкуренции с другими видами муравьев. Но иногда пришелец находит экологическую нишу, вовсе не занятую сколько-нибудь сходным организмом. Это произошло в некоторых горных лесах Кипра [269]. Корабельные, или черные, крысы (Rattus rattus), вероятно, попали на остров сотни лет тому назад — предполагают, что в Западную Европу они вселились в раннем средневековье. На Кипре они живут не только вблизи человека, но и далеко от него,
в лесах и зарослях кустарников. Там, где леса образованы алеппской сосной (Pinus halepensis), крысы едят ее шишки — питание, совершенно необычное для этого вида, хотя оно обычно для белок. Но па Кипре вовсе нет белок. Первоначальной родиной черной крысы были тропические леса Азии, и во многих районах мира она живег не на деревьях, а в кровлях туземных построек; даже в Англии она обитает в более высоких частях домов, чем серая крыса. В процессе всестороннего «обследования» Кипра она нашла себе свободную нишу. Клест (Loxia curvirostra) на Кипре тоже использует шишки, ио другого вида сосны — Pinus nigra. На многих тихоокеанских островах черная крыса живет также на пальмах и питается молодыми кокосовыми орехами.
Иногда биологический метод борьбы состоит в попытке разорвать прочную цепь симбиоза между муравьем и тлей или червецом. В Калифорнии был произведен эксперимент с аргентинским муравьем на цитрусовых плантациях, где он обитает в больших количествах [241]. В этих садах на деревьях жила лимонная щитовка (Aonidiella aurantii) — азиатский вид. При помощи специфических инсектицидов на некоторых деревьях удавалось уничтожить муравьев, не причиняя вреда щитовкам. В результате на деревьях с муравьями оказалось в среднем в 5, а в сезон максимальной численности в 150 раз больше щитовок, чем на деревьях без муравьев. Это было вызвано тем, что аргентинские муравьи уничтожали многих (хотя и не всех) естественных врагов и паразитов червеца. Такого же рода ситуация возникла при борьбе с вызываемым вирусом вздутием побегов какао — одним из бичей культуры какао на Золотом Береге. В распространении этой болезни участвуют два местных вида насекомых: Pseudococcus njalensis — наиболее часто встречающийся на деревьях какао червец — передает вирус при высасывании сока, а муравьи рода Crematogaster защищают червеца [251]. Под защитой муравьев червецы становятся многочисленными; важное значение имеет то обстоятельство, что сахаристые выделения червецов служат питательной средой для бактерий и плесневых грибов и что червецы гибнут, если эти выделения пе удаляются муравьями. Сами муравьи живут в ходах, прогрызаемых другими насеко
мыми в отмерших ветвях деревьев какао. Другим примером сложности взаимоотношений, которые могут влиять на распространение и поддержание болезни, служит американская болезнь каштанов (см. гл. I). Замечено, что один из наиболее обычных путей проникновения гриба внутрь дерева — это ход, проделанный каким-либо насекомым, живущим в коре или древесине. «Во многих
Ф и г. 84. Усач Tetropium gabrieli, личинки которого повреждают лиственницу преимущественно в искусственных насаждениях (за пределами естественных ареалов как паразита, так и хозяина) [250].
частях страны, пораженных этой болезнью, весьма прямые данные указывают на то, что насекомые, прогрызающие ходы в коре, особенно златка Agrilus bilineatus, имеют непосредственное отношение к болезни не менее чем в 90% всех ее случаев» [259].
Таким образом, сложность естественных взаимоотношений популяций достаточно очевидна каждому, кто обратит на нее внимание. Даже тогда, когда в другую страну перенесена вся цепь питания «растение — животное», возможны сдвиги, так как системе, которая сама по себе несложна, приходится действовать в новой среде. Европейская лиственница (Larix decidua) растет
в Швейцарии в природных условиях, и на ней живет местный жук-усач Tetropium gabrleli (фиг. 84). В пределах естественного ареала случаев повреждения деревьев жуками не зарегистрировано. Но вне этой области, особенно в Германии, а иногда и
Фиг. 85. Распространение усача Tetropium gabrieli и его кормового растения — лиственницы [250].
Заштрихованные области I—IV — естественный ареал европейской лиственницы (Larix decidua); V — ареал сибирской лиственницы; белые кружки — места, где были найдены жуки, но не были обнаружены повреждения деревьев; черные кружки — места вспышек массового размножения жука (все они находятся вне естественного ареала жука и дерева).
в Англии насаждения лиственницы часто повреждаются там, где появился жук (фиг. 85). Следовательно, что-то изменяет взаимоотношения дерева с насекомым в новых местообитаниях. Гориус [250] нашел, что у деревьев обнаруживаются признаки физиоло
гического изменения до того, как в них проникнут жуки; поэтому возможно, что ослабление деревьев вызывается каким-то несоответствием среды — климата или почвы, и ослабленные деревья становятся более уязвимыми для насекомого.
Примерно лет 80 назад, когда стали серьезно разрабатывать идею биологической борьбы с вредителями, одновременно начало входить в моду применение различных ядов для борьбы с микозами растений и насекомыми — вредителями сельского хозяйства. Насколько мне известно, никто еще не нашел эффективного антагониста для микроскопических грибов, паразитирующих на растениях, хотя недавно открыли, что ржавчинный гриб, растущий на пшенице и других злаках, имеет паразита — бактерию, которая в свою очередь может содержать бактериофаг, т. е. мы имеем цепь из паразита и гиперпаразита [254]. Лучшее средство против микозов и других паразитарных болезней растений заключается в том, чтобы найти или вывести устойчивую линию или же дать ей возможность самой возникнуть под более общим воздействием естественного отбора. Один микоз спаржи, не носящий в Европе эпидемического характера, был занесен в США, где он «охватил всю страну и фактически полностью уничтожил выращивание спаржи. Постепенно, однако, болезнь становилась менее губительной, и к настоящему времени культура спаржи восстановилась, а болезнь стала второстепенной инфекцией» [264]. К такому счастливому исходу привели, по-видимому, генетические изменения в популяциях гриба, спаржи или обоих этих видов, а не опрыскивание химикатами. Хотя генетика такого отбора обычно сложна, «практическая селекция обыкновенно не ждет результатов глубокого анализа отношений между растением-хозяином и грибом» [262].
То же самое можно было бы сказать о чрезвычайно интенсивном применении инсектицидов, практикуемом сейчас во всех странах, где занимаются земледелием. Если раньше с этой целью использовали лишь несколько хорошо испытанных химических веществ, то теперь с каждым годом увеличивается число новых, особенно (в последние 15 лет) порошкообразных и жидких препаратов синтетических хлорированных углеводородов (в том
числе ДДТ) и органических фосфатов (в том числе паратиона). Вместе с тем очень сильно расширилось применение опрыскивания веществами, ядовитыми для различных растений (в том числе фитогормонами, например 2,4-Д). Обладая средствами для уничтожения паразитических грибов, растительноядных насекомых и сорняков, биолог-практик стремится обойти все досадные затруднения, связанные со сложным взаимодействием естественных популяций,— по существу, просто ликвидировать природные пищевые цепи, оставив лишь культурные растения, которые придавали бы ландшафту упорядоченный и утилитарный характер. Кроме того, он способен уничтожить переносчиков болезней растений, например тлей и червецов, передающих и распространяющих фитопатогенные вирусы, и кровососущих насекомых — переносчиков заболеваний человека и домашних животных. В исключительных случаях опыливание и опрыскивание действительно полностью уничтожает довольно значительную популяцию насекомых, как это было, например, при эпидемии малярии в Бразилии (см. гл. I).
Существуют — как я полагаю, к счастью для будущего мировой флоры и фауны и для мира, в котором будут жить наши потомки,— два явления, противодействующие полному успеху этой химической войны. Первое из них — развитие устойчивых линий, а второе — компенсаторные реакции в экологических сообществах. Популяции некоторых видов насекомых, в течение ряда лет систематически подвергавшиеся действию ядов, стали менее чувствительными к определенным химическим веществам, а иногда и к целым группам родственных веществ [234, 235]. Это, в частности, относится к таким «звездам» среди вредителей, как яблонная плодожорка (Carpocapsa pomonella), червецы Saissetia oleae и Aonidiella aurantii, вредящие цитрусовым, хлопковая тля (Aphis gossypii) и калифорнийская щитовка (Aspidiotus рег-niciosus), повреждающая плодовые деревья. Кроме того, устойчивость появилась у комнатной мухи (Musca domestica)— во многих странах, у африканского клеща Boophilus decoloratus, у короеда Scolytus multistriatus — в Америке, и у некоторых малярийных комаров. Цитрусовая щитовка (Aonidiella aurantii) —
главный вредитель цитрусовых деревьев в южной Калифорнии — стала настолько устойчива к газации посадок парами синильной кислоты, что деревья часто вновь заселяются ею в течение первого же года. В 1941 г. было доказано, что устойчивость зависит от единичного гена, сцепленного с полом. В последующие годы для борьбы с червецом применяли масляное опрыскивание ветвей.
В последнее время главным средством уничтожения личинок малярийных комаров служили инсектициды, нередко распыляемые с самолетов. Но в северной Нигерии возникла линия Anopheles gambiae, устойчивая к диэлдрину, причем эта устойчивость наследуется по законам Менделя [240]. У другого вида малярийных комаров — A. sacharovi — развилась устойчивость к ДДТ в Греции и Ливане. С 1945 г. засоленные болота на востоке Флориды опрыскивали ДДТ. «До обработки ДДТ в области засоленных болот было столько комаров, что они препятствовали полному ее освоению. Освобождение от комаров после обработки рассматривалось как один из ярких примеров эффективности современных методов борьбы с насекомыми. Однако недавно сообщили, что у двух самых плодовитых комаров — Aedes solicitans (Wlkr.) и A. taeniorhynchus Wied.— вырабатывается устойчивость к ДДТ» [235]. Правда, эти два вида не переносят малярию. Примечателен случай с клещом Boophilus decoloratus-. в его популяциях в одной части южноафриканского побережья сначала развилась высокая устойчивость к жидкостям, содержащим мышьяк; тогда с большим успехом стали применять гамма-гексахлорциклогексан (гам-мексан); однако сейчас начинает появляться устойчивость п к этому препарату.
По-видимому, каждая популяция насекомых является генетически смешанной в отношении различных признаков, обычно свойственных виду, и у некоторых видов эти признаки, например строение кутикулы или какие-то свойства ферментов, влияют на способность выдерживать действие ядов. В результате интенсивного отбора, обусловленного высокой смертностью от отравления, сохраняются наиболее устойчивые линии; возможно, что возникают и новые мутации, как это происходит у бактерий, приобретающих устойчивость к антибиотикам. Такой же процесс, несо
мненно, происходит и в популяциях паразитических грибов, подвергающихся воздействию химических агентов. Устойчивость к ядохимикатам появилась пока лишь у немногих видов членистоногих, да и то лишь к одному или нескольким инсектицидам — правда, часто к очень важным. И все же Бейбере и Прэтт [235] в 1951 г. писали: «В настоящее время, однако, можно думать, что любое позитивное утверждение относительно устойчивости окажется ошибочным или требующим видоизменения. Мы присутствуем при самом начале процесса, который по силе своей, возможно, станет подобен лавине».
Второй недостаток инсектицидов и фунгицидов состоит в том, что они никогда не действуют только на тот вид, с которым ведется борьба. Они влияют на численность его конкурентов, паразитов, врагов и других видов, вызывая изменения, которые могут нарушить соотношения популяций во всем сообществе (несколько больше об этом будет сказано в гл. VIII). Есть и другие цепи последствий. «Ежегодное массовое применение ДДТ, технического гексахлорциклогексана и, вероятно, других стойких хлорированных углеводородов создает определенную опасность того, что за сравнительно небольшое число лет снизится плодородие почв на обрабатываемых этими веществами участках» [246]. Опыты, проведенные в штате Мэриленд, доказали, что ДДТ, гексахлорциклогексан, хлордан и токсафен (все это хлорированные углеводороды) угнетают рост культур, выращиваемых из семян, например бобов, пшеницы и ячменя. Кроме того, у некоторых сортов растений снижается урожай семян [239]. Поэтому такие последствия использования инсектицидов приходится сопоставлять с прямой выгодой от их применения, т. е. снижением ущерба, наносимого насекомыми. Это, однако, совсем новое и чрезвычайно сложное направление исследований; еще мало ‘Сделано для выяснения всех путей возможного влияния интересующих нас факторов [265]. Мы упомянули об этом только для того, чтобы показать на примере, что при помощи ядов с избирательным действием вряд ли можно контролировать даже довольно упрощенную экологическую систему, не вызывая целого ряда непредвиденных следствий; эти следствия нужно изучить и
предусмотреть, прежде чем вводить меры по регуляции численности популяции. Были проведены эксперименты с целью выяснить, как действуют на почвенных клещей и насекомых дозы ДДТ и других ядов, применяемых в борьбе с вредителями культурных растений [263]. Опыты ясно показали всю тонкость соотношений между популяциями мелких членистоногих, населяющих культурную почву. Гаммексан (гамма-изомер гексахлор циклогексана) снижал численность ногохвосток (Collembola) и клещей; однако после одновременной обработки участков почвы также и ДДТ число клещей уменьшалось, тогда как чпсло ногохвосток вскоре увеличивалось. «Лабораторные испытания показали, что ДДТ, даже в наивысших концентрациях, совершенно не действует на Collembola, но определенно токсичен для всех исследованных Mesostigmata. Эти Mesostigmata — хищные клещи, активно преследующие ногохвосток, так что их устранение, вероятно, и позволило ногохвосткам увеличить свою численность. Зная, что численность этих насекомых во всякой обычной почве огромна и что они питаются главным образом микроскопическими грибами, мы не можем сомневаться в том, что остатки ядохимикатов способны изменить матаболическую активность почвенного биоценоза и тем самым повлиять па урожайность культур. Это связано также с изменениями бактериальной флоры.
Об этом удивительном дожде смерти, которым поливают столь значительную часть земной поверхности, мы вкратце рассказали здесь для того, чтобы подготовить читателя к развитым в двух последних главах нашей книги идеям о том, что возможны иные методы охраны богатств органического мира, способные дать более стойкие результаты. Ни один реально мыслящий человек не может отказаться от биологических или химических методов борьбы с вредителями, однако оба метода можно существенно видоизменить и приспособить к реальному положению вещей на экологической сцене с ее весьма тонко организованной системой связанных между собой популяций. Быть может, когда-нибудь мы будем расценивать нынешнее массовое уничтожение вредителей и несистематический выпуск врагов-хищников или паразитов так же, как мы расцениваем сейчас ошибки века индустриализа
ции, эксцессы колониальной эксплуатации или хищническую вырубку естественных лесов. Леопольд [255] писал: «Один принципиальный дефект системы охраны природы, всецело основанной на экономической выгоде, связан с тем, что большинство членов наземных биоценозов не имеет никакого экономического значения. Примером служат дикорастущие цветы и дикие певчие птицы. Сомнительно, чтобы из 22 000 видов высших растений и животных местной флоры и фауны Висконсина больше 5% можно было продавать, использовать в качестве корма для животных, пищи для человека или каким-либо иным образом. Однако все эти организмы являются членами биотического сообщества, и если (как я полагаю) стабильность сообщества зависит от его целостности, они имеют право на дальнейшее существование». Прежде чем рассмотреть практические стороны этого обстоятельства, нам предстоит исследовать причины и мотивы, побуждающие человека охранять природу.
Глава VIII
ПРИЧИНЫ, ПОБУЖДАЮЩИЕ ОХРАНЯТЬ ПРИРОДУ
Т£ ак-то я посетил одну очень хорошую школу, директор которой •Ч, стремился заинтересовать своих учеников работой в большом огороде. Огород был отличный, и видно было, что детям очень нравилось это занятие. Я спросил учителя, имеет ли он время рассказывать им что-нибудь и о животных, на что он ответил: «О да, я учу их, что животные являются вредителями!» Это вполне понятная точка зрения практического человека, ставящего перед собой сравнительно узкие задачи; совершенно иной взгляд выражен в словах Роберта Броунинга:
Я сын Земли!
Не троньте ж лик ее...
И тем не менее один великий литературный критик сказал, что гений Броунинга имеет здоровые, цепкие корни в реальной жизни. На Британских островах существует более 25000 различных видов аборигенных наземных и пресноводных животных, а во всем мире число их, вероятно, превышает миллион. Характер предстоящего нам длительного сосуществования с этими животными в очень большой степени зависит от нашего отношения к живой природе и к природе вообще. Жизнь человечества я сравниваю с очень долгим путешествием по железной дороге в компании со всеми этими другими пассажирами, и мне кажется, что есть три фундаментальных вопроса, до сих пор ожидающих своего решения. Первый из них, который обычно не считают основным, является, в сущности, этической проблемой. В мире есть несколько миллионов человек, которые полагают, что животные имеют право на существование и на то, чтобы в их жизнь не вмешивались, или что люди по крайней мере не должны пре
следовать животных и истреблять целые виды. Некоторые люди не отступят от такой точки зрения даже тогда, когда она грозит серьезной опасностью им самим. Усилия по борьбе с чумными крысами на некоторых товарных складах Индии иногда оставались тщетными потому, что работники складов выставляли крысам воду для питья. Практическому человеку Запада подобные идеи покажутся безрассудными или сентиментальными. Но можно ли просто отмахнуться от них, назвав их пустой сентиментальностью, если такой большой ученый и своего рода пророк, как д-р Швейцер [287], говорит: «Огромным недостатком всех этических систем до сих пор было то, что в них рассматриваются лишь взаимоотношения человека с человеком»?
Второй вопрос можно назвать эстетическим или интеллектуальным. Мы можем сказать, что природа — все виды дикой живой природы с окружающей ее средой — интересна, а также красива п способна возбуждать эмоции. Это источник вдохновения для поэтов и художников, источник материалов и удовольствия для натуралистов и ученых и, наконец, место отдыха и развлечений. Во всем этом интересы человека решительно выступают на первое место.
Третий вопрос — практический: земля, культурные растения, лес, вода, морские промыслы, болезни и пр. Этот третий вопрос так угрожающе нависает над всем миром, что чуть ли не заслоняет собой два других. Причина очень проста: «червячок в сердце розы» — это проблема народонаселения. Человеческий род до сих пор размножался подобно водяным крысам или гигантским улиткам, и слишком многие из нас оказались в местах, не слишком подходящих для жизни. Один из непререкаемых фактов нашего столетия состоит в том, что человек увеличивает не только свою численность, но и свои потребности. Энергичнее, чем когда-йибо в истории мира, он стремится проникнуть в местообитания, которые раньше не использовались или использовались слабо. И каждый его шаг по этому пути влечет за собой новые экологические пертурбации, в том числе такие, которые проистекают от новых нашествий животных, растений и их паразитов.
Итак, есть три точки зрения. Вы можете считать, что поразительно разнообразная жизнь на земном шаре развилась не для того только, чтобы ее использовать, истощать ее резервы и, быть может, в значительной части уничтожать ее. Вы можете держаться мнения, что всякая живая природа настолько интересна и красива, что ее следует сохранить,, в особенности для того, чтобы ею могло наслаждаться потомство. Такая точка зрения довольно распространена в экономически развитых странах, но встречает гораздо меньше сочувствия там, где даже простое поддержание человеческой жизни составляет проблему. Но где бы вы ни жили, с практическими проблемами приходится считаться в первую очередь. Для того чтобы жить и добывать себе хлеб насущный, люди должны выращивать растения, им нужна земля и хорошие сельскохозяйственные культуры. Нельзя рассчитывать на то, что охрана природы для удовольствия, поучения или обеспечения высших прав животным когда-либо получит приоритет перед интересами выживания самого человека. Да этого и не должно быть.
Но что, если конфликт между этими интересами совсем не так велик, как это кажется на первый взгляд? Предположим, что можно было бы сохранить разнообразие природы, основываясь на всех трех мотивах: потому, что это справедливое соотношение между человеком и другими живыми созданиями, потому, что это позволяет сделать жизнь более интересной, и потому, что это способствует экологической устойчивости — экологическому сопротивлению нашествиям чужеродных видов и «взрывам» в естественных популяциях. Это была бы четвертая точка зрения — попытка согласовать позиции, бывшие до сих пор противоположными. Если только отказаться от мысли, что человеку предназначено быть всепобеждающей и обеспложивающей силой на Земле, то можно прийти к какому-то соглашению. Это означало бы поиски некоего мудрого принципа сосуществования между человеком и природой, даже если речь пойдет об изменившемся человеке и видоизмененной природе. Это я и имею в виду под «охраной природы». Когда-нибудь, в следующем тысячелетии, эколог, возможно, будет в состоянии объявить, что эта проблема
снята с повестки дня. Сейчас мы можем только сообщить о текущем положении дел и дать обнадеживающий прогноз, т. е. сказать примерно то, что инженер-атомник мог сказать десять лет назад об атомных электростанциях. Но этот экологический прогноз, для того чтобы он оправдался, требует уменьшения прямого вмешательства человека в жизнь природы, а не усиления его; необходимо переложить на природу часть той работы, которую сейчас с таким неистовым рвением пытаются делать инженеры и химики. И это вполне трезвый прогноз, а не сентиментальные мечты.
Сейчас я попробую привести некоторые доказательства того, что устойчивость сравнительно простых сообществ растений л животных нарушить легче, чем устойчивость более богатых биоценозов; другими словами, простые сообщества более склонны к нарушающим их целостность колебаниям численности популяций (особенно животных) и более уязвимы для вторжения посторонних организмов. Ведь если удастся показать, что это недалеко от истины, то придется признать, что в нашей современной манере обращения с культурными землями есть нечто весьма опасное и что опасность станет еще больше, если мы и дальше пойдем по пути «упрощения ради эффективности». Следует помнить, что краткий обзор данных, в сущности, позволяет только предварительно обосновать ту или иную точку зрения, но не доказать ее справедливость. Но мы выдвигаем отнюдь не надуманную идею; даже если бы она и казалась надуманной, нам все-таки следовало бы проверить ее путем научного исследования, ибо вся эта проблема в высшей степени важна для будущности каждого вида, обитающего на Земле.
Прежде всего у нас есть результаты математического анализа динамики численности популяций. Один из путей анализа упрощенных «моделей» популяций привел главным образом к тому, что выявилась тонкость ожидаемой устойчивости отношений внутри популяции и между популяциями. Самое крупное теоретическое (а оно и есть только теоретическое) открытие состояло в том, что популяции из одного вида жертвы и его врага-хищника или из одного вида хозяина и его паразита обнаруживали бы
заметные колебания численности, даже при отсутствии стимулов извне, связанных, например, с капризами климата. Эти простые модели почти лишены внутренней стабильности, но их флуктуации не обязательно приводят к вымиранию видов. В переводе на обычный язык это значит, что в сообществе животных, состоящем только из двух таких видов, численность популяции никогда не была бы постоянной — возникали бы периодические «пики» численности то одного, то другого вида.
Эти выводы, основанные на алгебре и особенно на дифференциальном и интегральном исчислении, были в известной степени подкреплены лабораторными экспериментами с мелкими животными, главным образом с простейшими, клещами, дафниями и жуками, которых содержали в тщательно стандартизированных условиях. Особенно соответствуют теоретическим расчетам опыты с простейшими и клещами, так как в этих опытах популяция одного вида животного-жертвы содержится вместе с одним видом животного-хищника, хотя в действительности опыты приходилось повторять и проводить в разнообразных вариантах. И здесь результаты ясно показывают одно: очень трудно поддерживать равновесие между небольшими популяциями в этой простой смеси, так как не только происходят колебания численности обоих видов, но нередко один или оба вида вымирают. Эти эксперименты почти не получили дальнейшего развития; в сущности, до сих пор имеется лишь одна значительная серия таких опытов, проведенная советским экологом Г. Ф. Гаузе в 30-х годах нашего века [279, 280]. Быть может, техническая трудность постановки опытов и поддерживания смешанных популяций была причиной того, что эта работа не была повторена. Но она имеет, по моему мнению, фундаментальное значение.
Гаузе ввел в эксперимент еще один экологический принцип: он использовал убежище [279, 281]. Предоставляя жертве убежище, он смог изменять ее шансы на выживание. Но и в этом случае созданные им системы никогда не были стабильными — либо «хищник» вымирал, так как не мог добыть себе пищу, либо эффективность убежища была такова, что лишь отдаляла окончательное истребление жертвы. Но в природе второй результат
мог бы иметь большое значение, так как позволил бы местным иммигрантам из другой части системы прибыть и помочь поддержанию первоначальной популяции жертвы. Гаузе действительно пробовал вводить несколько дополнительных особей извне, как бы из природного резерва, и таким способом ему удавалось поддерживать смесь популяций, хотя это не устраняло больших колебаний их численности. Именно так обстоит дело, когда весной плодовый сад химически стерилизуют, а летом он вновь заселяется организмами из окружающих живых изгородей и лесов (см. гл. IX).
Третья группа данных состоит в том, что природные местообитания на небольших островах, по-видимому, более доступны для нашествия иноземных видов, чем на материках. В особенности это относится к океаническим островам, на которых сравнительно немного аборигенных видов (см. гл. IV). Даже на биологически хорошо заселенном острове Великобритании сейчас примерно вдвое больше видов диких оленей и грызунов, чем было зо время завоевания его норманнами; кроме того, мы знаем, что Британские острова отделились от материка около 7000 лет назад и только после этого приобрели полный комплект видов, рассеявшихся на север и запад по окончании последнего оледенения. Эти добавочные виды были завезены человеком, и некоторые из них, расселяясь по стране, давали значительные вспышки размножения, а отдельные лесные популяции оленей даже теперь, возможно, накапливают силы для будущих крупных «извержений».
Четвертый довод состоит в том, что нашествия и вспышки чаще всего происходят на обрабатываемых землях и в искус-, ственных насаждениях, т. е. в местообитаниях и сообществах, очень сильно упрощенных человеком. Это упрощение зависело главным образом от трех моментов: от того, что человек обычно способствует проникновению чужеземных культурных растений, уже лишенных полной фауны связанных с ними форм; от того, что он выращивает их в условиях частичной или полной монокультуры; и от того, что он старается уничтожить все другие виды, которые считает вредными, причем попутно уничтожает
или угнетает множество видов, судьба которых ему просто безразлична. Кроме того, генетический отбор культурных растений, а также, как упоминалось в предыдущей главе, некоторых вредителей, на свой лад нарушает биологическое адаптивное равновесие. Иноземные виды, как мы видели на некоторых примерах, приведенных в нашей книге, проникают иногда и в природные местообитания. Это произошло с опунцией в Австралии, с европейским еловым пилильщиком в Канаде и с морским окунем в ‘районе Калифорнии. В двух первых случаях ситуация, по-видимому, выправляется в результате продуманной интродукции врагов или паразитов, отчего сообщество становится чуть-чуть сложнее, а не проще: морской окунь не вызвал никаких зарегистрированных сдвигов в сообществе, в которое он столь стремительно проложил себе путь. В общем можно было бы ожидать, что либо численность иммигрантов с необычным «экологическим потенциалом», проникающих в более или менее богатые природные биоценозы, можно будет регулировать экологическими методами, либо они найдут для себя место, не вызвав значительных нарушений соотношения других популяций. Однако это отнюдь не всеобщее правило, как показывает история каролинской белки в лесах Англии и американского моллюска Crepidula на устричных банках. В этих случаях мы, по-видимому, присутствуем при «решающих битвах истории». Тем не менее поразительно, что в собственно лесной части экологической системы Уитхэм-Вудс (см. гл. VI) богатые естественные сообщества приняли в свою среду лишь трех или четырех важных иммигрантов, в том числе каролинскую белку и европейский вид клена — явор (Acer pseudo-platanus). Первая появилась в районе Оксфорда в XX в., а второй проник в Англию несколько сот лет назад. Главным врагом клена является каролинская белка, которая губит деревья, обгрызая кору с ветвей, и это единственный серьезный фактор, причиняющий им вред. Эта система из двух расселяющихся видов еще не достигла устойчивости.
Пятая группа данных касается, тропиков. С этими данными я впервые столкнулся несколько лет назад, когда однажды целый час излагал идеи о вспышках размножения насекомых трем
лесным инспекторам, приехавшим из-за границы. Наконец один пз них вежливо заметил, что этот вопрос, в сущности, их не интересует, поскольку в их лесах не бывает вспышек размножения насекомых! Оказалось, что он прибыл из Британской Гвианы, второй инспектор — из Британского Гондураса, а третий — из тропической зоны Индии. В настоящее время голландский специалист по лесной экологии Вут полагает, что это — общее правило, отражающее экологическую стабильность влажных тропических лесов [289]. Такие леса очень богаты видами. По представлениям Вута, в них всегда достаточно врагов и паразитов, готовых накинуться на любой вид, численность которого начинает необычно увеличиваться, и «осадить» его, образовав сложную систему тормозов и буферов. Конечно, это только теория, и, по всей вероятности, будут найдены и другие причины. Но экологическая устойчивость влажного тропического леса, по-видимому, представляет собой вполне установленный факт. Оди, руководящий исследованиями по медицинской экологии, проводимыми англичанами в Малайе, установил, почему клещевой тиф сделался в этой стране опасной болезнью. В Малайе используемое для покрытия построек растение лаланг (Imperata cylindrica), полевая крыса, или «тикус-сава» (Rattus argentlventer), и один из переносчиков клещевого тифа (Trombicula akamushi) являются доминирующими видами на открытых территориях. Все это — не аборигены Малайи, и всюду, где тот или иной из этих видов встречается на участках, лишенных леса, он образует более плотные и мощные популяции, чем то же растение или животное в лесу. Резервуаром инфекции служит, вероятно, лес, но в лесу она практически теряется в сложном сообществе; необходимо было упрощение биоценоза и концентрирование популяций, происходящее после вырубки леса, чтобы эта инфекция активировалась и сделалась наконец опасной для человека [274].
Шестая группа данных — это сведения, полученные в результате недавних исследований по борьбе с вредителями плодовых садов. Такие сады особенно подходящи для изучения влияния экологического многообразия, так как они занимают промежуточное положение между естественными лесами и полевыми посе-
вами — они проще по структуре, чем леса, но сложнее, чем посевы, и более постоянны. Они гораздо больше подвергаются различным искусственным воздействиям, чем леса, и страдают от нашествий множества животных, грибов и других вредных организмов. Большинство этих вредных организмов теперь проникло во все страны, где развито плодоводство, так что вся проблема представляет огромный интерес для каждого, кто хочет получать хорошие урожаи. Здесь я рассматриваю ее главным образом как пример вмешательства человека в относительно простую экологическую систему.
Наиболее полное исследование было проведено в Новой Шотландии группой канадских энтомологов, которые пытались выяснить причины появления нескольких следующих друг за другом волн вредителей на яблонях [284, 286]. Некоторые из таких организмов наносят особенно большой вред, причем все это — виды, первоначально завезенные из других стран: гриб Venturia inae-quails, вызывающий паршу яблок; гусеница яблонной плодожорки (Carpocapsa pomonella), червец Lepidosaphes ulmi, и, наконец, европейский паутинный клещик Metatetranychus ulmi* Примерно с 1930 г. начался ряд необъяснимых вспышек размножения вредителей. Затем было сделано необычайное открытие: эти вспышки почти несомненно вызывались побочным действием фунгицидной жидкости на врагов и паразитов вредителей [282]. После изменения химического состава одной из таких жидкостей, применяемой против парши яблок, было отмечено пеимоверное размножение червецов на коре и ветвях яблонь. Оказалось, что прежняя жидкость для опрыскивания убивала червецов, одного из их врагов и одного из паразитов, новая же не вредила червецам, но по-прежнему уничтожала их врага и паразита, еще раз заставляя нас вспомнить поговорку: «игра не стоит свеч».
Выявились и другие интересные последствия опрыскивания. В последние годы во всем мире применяли ДДТ в качестве инсектицида в плодовых садах, в частности для уничтожения яблонной плодожорки. Оказалось, однако, что он убивает врагов клеща Metatetranychus ulmi, не уничтожая самого клеща [283]. В результате этот клещ размножился в плодовых садах всего
мира. Коллиер нашел, что в садах Эссекса клещ М. ulml служит добычей по меньшей мере для 45 видов насекомых и клещей [276, 277]. Как у нас, так и в Канаде в запущенных плодовых садах мы находим очень малочисленные популяции Л1. ulmi п хороший комплекс его естественных врагов, так как все его враги питаются не только одним этим вредителем, а имеют определенный круг жертв. Никто, конечно, не думает, что яблоневые сады можно было бы без всякого риска оставить в покое и прекратить всякое опрыскивание, чтобы в них установилось естественное равновесие между видами. Но совершенно очевидно, что искусственное поддержание стабильности — проблема очень тонкая; как заметил один ведущий канадский специалист по экологии плодовых садов, «мы движемся от кризиса к кризису и только обмениваем одну проблему на другую» [285].
Приведенные выше шесть групп данных можно подытожить следующим образом. Математические теории относительно свойств цепей питания и упрощенные лабораторные эксперименты подготавливают наш ум к представлению о нестабильности даже очень простых систем популяций. В таких системах мы можем ожидать сильных колебаний численности, а часто и вымирания видов. Если структура местообитания дополнительно усложнена введением укрытий, возможно некоторое ослабление этой неустойчивости, хотя полный успех в эксперименте достигается очень редко. Океанические острова и монокультуры сельскохозяйственных растений представляют собой простые экологические системы, весьма подверженные вторжениям организмов извне (из других стран или из других местообитаний в той же стране) и последующим частым вспышкам размножения иммигрантов. Однако во влажных тропических лесах эти явления в поразительной степени ослаблены. В плодовом саду, который долго не.обрабатывали инсектицидом, устанавливаются устойчивые экологические отношения между сотней или большим числом обитающих в нем видов животных, хотя такой сад не удовлетворяет требованиям в отношении качества и количества плодов. Вспышки размножения вредителей в плодовых садах частично бывали обусловлены проникновением новых видов извне, частично — многочис
ленными случайностями и изменениями взаимодействия видов, которые имеют место в любом сообществе животных, а нередко также вследствие избирательного действия ядохимикатов, нарушающего взаимоотношения между вредителями, их естественными врагами и паразитами. Все эти шесть групп различных данных, по-видимому, говорят об одном и том же, хотя каждая требует более детального анализа, для которого не хватило бы места в нашей книге. Мы аргументируем наш тезис потому, что если он правилен, то мы сможем улучшать наше биологическое хозяйство, улучшая взаиморасположение местообитаний и создавая более или менее сложные сообщества растений и животных (см. гл. IX). А мы должны пытаться сделать это из трех побуждений, уже перечисленных выше: чтобы создать убежища для диких видов, чтобы сделать окружающую нас среду интересной и удовлетворяющей наши духовные запросы и чтобы способствовать устойчивости популяций и создать разнообразные биоценозы, в которых популяции испытывали бы сложные компенсаторные воздействия со стороны всех сочленов сообщества.
Наши аргументы отнюдь не направлены на поддержку идеи полного невмешательства в экологические системы мира. Разрушение областей Уоллеса, стремительный рост народонаселения и достижения техники сделали любую такую идею неприемлемой. Нужно управлять будущим органического мира, но это управление не должно быть чем-то вроде игры в шахматы — оно должно скорее напоминать управление кораблем. Нам необходимо научиться более мудро использовать огромные потенциальные силы размножения популяций растений и животных, научиться предоставлять некоторым из этих сил достаточную свободу, чтобы они могли взаимодействовать между собой, научиться конструировать такую окружающую среду (например, определенные типы укрытий), которая поддерживала бы постоянную устойчивость в том или ином сообществе. Я думаю, что в области борьбы с вредителями уже начинается такая смена идей. Можно привести два примера. Как упоминалось в гл. VII, самым злостным вредителем цитрусовых насаждений в южной Калифорнии является цитрусовая щитовка (Aonidiella aurantii), с которой приходится
бороться путем регулярных опрыскиваний деревьев различными инсектицидами. Пытались применять биологические методы борьбы, но они оказались не особенно эффективными. Отделение биологической борьбы Цитрусовой опытной станции Калифорнийского университета начало в 1948 г. интересный эксперимент [278]. Из 17 лимонных и апельсинных рощ, не подвергавшихся опрыскиванию, 3 не опрыскивались в течение 1—2 лет, а 5 других — в течение 8 лет и более. После того как было установлено, что принадлежность деревьев к разным сортам или наличие у них болезней не оказывает влияния на результаты, внимание было сосредоточено на численности врагов и паразитов вредителей в каждой роще. Опыты с защитой щитовок на целой ветви дерева, на которой у них не было ни врагов, ни паразитов, доказали, что последние играют очень большую роль в подавлении численности вредителя. На опрыскиваемых деревьях быстро вновь разводилось множество щитовок, а на деревьях, которые не опрыскивали, их появлялось гораздо меньше. Главным антагонистом щитовки был его азиатский паразит — хальцида Aphytis chry-somphall, к тому времени уже широко распространенная в Калифорнии. Хотя процент хозяев, содержащих личинки паразита, был не особенно высок, оказалось, что взрослая форма паразита просто питается щитовками, обусловливая их значительную дополнительную гибель. Есть также три вида хищных насекомых, которые становятся многочисленными, когда возрастает численность щитовок, но эти виды сами по себе не могут регулировать популяцию щитовки, по крайней мере в условиях данной местности. В отчете об этих исследованиях говорится, что «ежегодная обработка инсектицидами для борьбы со щитовкой, вероятно, исключает возможность установления удовлетворительного естественного равновесия».
Стоит только усвоить представление о том, что сложность видового состава — свойство сообщества, подлежащее изучению и использованию в целях защиты растений, как перед нами открывается почти безграничное поле деятельности. Та же группа калифорнийских исследователей попробовала использовать еще одну идею, которая на первый взгляд кажется весьма неожиданной
[288]. Червец Saissetla oleae, также происходящий из Азии, тоже является серьезным вредителем цитрусовых садов. Он особенно многочислен во внутренних частях штата, потому что в связи с более континентальным климатом там ежегодно наблюдаются резкие циклические изменения численности червецов и их обычного паразита — хальциды Metaphycus helvolus. В прибрежных областях сезон длиннее и генерации сильнее перекрываются во времени, что сглаживает изменения численности, вызываемые паразитом. В первом случае происходят резкие колебания и при низкой численности хозяина численность паразита настолько падает, что уже не может быстро возрасти снова. Предполагали, однако, что если сохранять один вид паслена, Solanum douglasli, часто растущий под цитрусовыми деревьями, то червецы развивались бы раньше, так как на этом растении они растут быстрее; таким образом, сезон существования хозяев удлинился бы. Раньше на Solanum douglasil смотрели как на вредный сорняк именно по той причине, что он тоже служит кормовым растением для червеца. Эксперименты были прерваны из-за того, что в конце июля большинство растений паслена было уничтожено чересчур усердными работниками сада, не знавшими об этих экспериментах. Это не были полноценные опыты с тщательной регистрацией времени наступления тех или иных событий, а также искусственным заражением червецом паслена и цитрусовых деревьев. Достоинство этих экспериментов, практическая польза которых не была еще подтверждена, состояло в том, что они строились на принципах, на основе которых действительно работает природа, а не тех, что используются инженером или химиком.
От небольшой группы из четырех видов — лимонного дерева, паслена, червеца и паразитирующей на нем хальциды — очень далеко до почти непостижимого и неисчерпаемого богатства форм влажного тропического леса или даже до нескольких тысяч видов, населяющих Уитхэм-Вудс в Беркшире. Выяснить, до какого уровня нужно довести сложность, чтобы получить мало-мальски' устойчивое сообщество,— это задача будущих исследований, и притом неотложная. В основе всей этой проблемы лежит также вопрос о скорости приобретения видом генетических приспосо
бительных изменений. Появление устойчивых линий насекомых и клещей после сравнительно немногих лет применения ядохимикатов доказывает, что процесс этот может идти очень быстро. О том же свидетельствует все расширяющаяся работа по селекции сортов растений и пород животных, устойчивых к определенным болезням или насекомым-вредителям. Увидим ли мы подобное же взаимоприспособление между вредителями и их врагами или паразитами? Это вполне возможно и еще раз указывает на необходимость давать простор для воздействия всех сил сообщества по различным экологическим каналам. Даже если отдельные взаимоотношения изменятся вследствие генетических изменений, остальные могут сохраняться в течение длительного времени. В связи с этой идеей встает вопрос о том, как можно выявить все способы поддержания или создания достаточно богатых растительных и животных сообществ в условиях нашего меняющегося ландшафта, т. е. как найти пути сохранения экологического многообразия.
Глава IX
СОХРАНЕНИЕ МНОГООБРАЗИЯ ПРИРОДЫ
«И сосуд, который горшечник сделал из глины, развалился в руке его; и он снова сделал из него другой сосуд, какой горшечнику вздумалось сделать».
Книга пророка Иеремии, XVIII, 4.
Т) сюду, кроме большей части океана, дикая природа отступает или меняет свой облик. Еще через тысячу лет почти вся поверхность суши и значительная часть пресных вод будут изменены человеком или во всяком случае их фито- и зооценозы будут глубоко отличаться от тех, которые населяли их всего несколько сот лет назад. Мы должны привыкнуть к мысли, что даже .там, где растительность по внешнему виду сохранит как будто бы свой первоначальный характер (как это часто бывает в лесу, восстановившемся после пожара или смены культур), популяции растений и животных изменят свой состав, свою сложную структуру и взаимоотношения (что касается животных, то это будет почти незаметно для случайного взгляда). И все время в эти сообщества будут продолжать вторгаться виды из других частей мира. До сих пор основная тяжесть этих нашествий падала на сообщества, значительно измененные и упрощенные человеком. Однако некоторые организмы проникают и в более устойчивые и зрелые биоценозы океана и естественного леса. На эксплуатируемых землях всего мира мы наблюдаем уменьшение богатства и разнообразия видов: монокультуры, подвергаемые опрыскиванию по строгому плану, пастбища с чистыми популяциями трав, чистые посадки деревьев, замещение многоярусных зрелых лиственных лесов быстро растущими хвойными с их относительно простой структурой и бедным населением, расчистка пустырей,
опрыскивание обочин дорог фитогормонами и насаждение экзотических видов, многие из которых могут быть буквально стерильны в отношении животной жизни — по крайней мере вначале. Мы могли бы охарактеризовать этот поток событий словами Ис&ии: «Горе вам, прибавляющие дом к дому, присоединяющие поле к полю, так что другим не остается места, как будто вы одни поселены на земле» (Книга пророка Исаии, V, 8).
Но человек не может жить одними огорчениями. Есть множество способов создать в странах, в которых мы живем, лучшие условия для сохранения дикой жизни, более интересную живую природу и более надежную базу для фермера, лесовода и рыбака. Если дикая природа отступает, мы должны научиться передавать часть ее стойкости и богатства ландшафтам тех земель, с которых мы снимаем наши урожаи. «Охрана природы» — понятие многостороннее, ибо оно может означать, с одной стороны, защиту диких видов от все возрастающего использования их человеком, а с другой — способ достижения наивысшей продуктивности используемой территории. Говоря об охране природы, следует ясно указывать, какого рода «охрана» имеется в виду. Я думаю, что если доводы, развитые в этой книге, основательны, то под охраной природы нужно было бы понимать сохранение или придание ландшафту максимального экологического многообразия — во всем мире, на каждом материке или острове и, насколько это возможно, в каждой местности. И если будет обеспечено существование местных видов, то я не вижу причин, почему бы реконструкция сообществ, направленная на то, чтобы сделать их богатыми, интересными и устойчивыми, не могла бы включать интродукцию тщательно отобранных экзотических форм, тем более что многие из них все равно рано или поздно появятся и займут какую-нибудь нишу.
Эта идея, которая может показаться довольно туманной, станет более понятной, если пояснить ее примером. Значительная часть ландшафта Англии, весьма окультуренного, до сих пор изрезана замечательной сетью живых изгородей и придорожных полос. Эти длинные извилистые ленты, тянущиеся вдоль дорог, тропинок и по краям полей, представляют собой последнее дей
ствительно обширное прибежище природы, которое мы имеем в Англии, если не считать диких вересковых пустошей в северных горах и окружающих нас морей. Нам необходимо большое число более мелких заповедников для специальных целей — для того чтобы способствовать сохранению того или иного животного, растения или местообитания. Но наши придорожные живые изгороди и пустоши занимают совершенно особое место, так как, имея протяженность около 300000 км, они расположены среди наших обрабатываемых земель, а также части городских земель. Полосы еще большей общей протяженности окаймляют наши отдельные поля. На живые изгороди большинство людей смотрит как на нечто само собой разумеющееся. Во многих частях страны они составляют неотъемлемую часть пейзажа (фиг. 86 и 87). К лучшему, что когда-либо написал Ричард Джеффрис [296], относится его простое описание живой изгороди, ведущей к меловым холмам. Карел Чапек писал: «Я ходил по дорогам, окаймленным живыми изгородями, настоящими живыми изгородями, которые делают Англию настоящей Англией, ибо они ограничивают, но не стесняют» [290]. До сих пор можно найти множество хороших, регулярно подстригаемых живых изгородей, большей частью из боярышника, дополняемого другим кустарником и некоторым числом деревьев; но многие из таких изгородей начинают исчезать под влиянием того, что я назвал бы «сельским хозяйством в мечтах инженера». Один очень крупный фабрикант машин недавно писал в одном из наших сельскохозяйственных журналов: «Что касается конструктивных предложений, позвольте мне прежде всего подчеркнуть, что нам нужно повыдергать некоторые из наших живых изгородей». Можно не сомневаться, что уж он-то не думал о сохранении экологического многообразия.
Несколько лет назад луговым полосам вдоль наших сельских дорог угрожала массовая обработка гербицидами, которые губят многие из наших цветов, оставляя одну траву. Эту кампанию могли начать, не посоветовавшись с обыкновенными людьми, не спросив, безразлично ли им, что у нас не будет синей луговой герани или других красивых цветов около дорог, пе выяснив,
Фиг. 86. Дорожка в Оксфордшире в мае во время цветения купыря (Anthriscus sylvestris) (фото автора, 1957).
Живая изгородь состоит из вяза, орешника п клена, а также лазящих стеблей ломоноса и двух падубов, оставленных там, где они росли. Летом такие места полны цветов п насекомых.
Ф и г. 87. Обочина дороги в Гэмпшире в конце июня, в самом начале сенокоса (фото автора, 1954).
Несмотря на ветры, обдувающие этот меловой холм (что можно видеть по форме кроны тисса), вдоль дороги протянулась полоса высокого смешанного луга с борщевником (Негас-leum sphondylium)tna цветках которого скопляется много насекомых для сбора пыльцы и нектара.
возможны ли неожиданные побочные эффекты вроде тех, которые наблюдаются в плодовых садах после опрыскивания фунгицидом или инсектицидом (см. гл. VIII). Одна из фирм, производящих опрыскивание, включила в список сорняков, подлежащих уничтожению, шиповник. К счастью, Ведомство охраны природы, в обязанности которого входит внимательно следить за всем, что касается экологии страны, смогло отсрочить это мероприятие, воззвав к здравому смыслу правительственных ведомств, сельскохозяйственных энтомологов, коммерческих фирм, производящих опрыскивание, и инженеров-дорожников графств. Кампания была частично отложена, и тем временем велись кое-какие ботанические исследования, которые содействовали пересмотру программы. В настоящее время, по-видимому, достигнут более или менее разумный компромисс: на определенной полосе растения опрыскивают или, чаще, скашивают (сейчас нередко машиной), оставляя за этой полосой «естественный сад» по краю поля.
Живые изгороди и обочины дорог поистине необычайно разнообразны по структуре и составу их сообществ. Каждый придорожный участок чем-нибудь отличается от другого. Но почти все они экологически богаты и обычно устойчивы; иногда эта устойчивость временно нарушается лишь в местах ремонта дороги, что только увеличивает разнообразие наблюдаемых экологических сукцессий. Разве не стоило бы подумать над тем, что это одно из самых драгоценных наших сокровищ. В Англии я не вижу никакой другой экологической системы, у которой были бы столь очевидны все качества, необходимые для сохранения многообразия. Это прибежище для дикой жизни: здесь и гнездовья мелких птиц, и обильные популяции насекомых — не только живущих на листьях и ветвях, но и посещающих цветы в начале лета. Очень многие из встречающихся здесь видов обитают также на опушках лесов или на краях лесных прогалин — в местообитаниях, которые при современном ведении лесного хозяйства все в большей степени лишаются обитателей. Затем, это источник огромного удовольствия для путешественника — цветущая живая изгородь из боярышника с сопутствующими ему кустарниками, такими, как кизил, шиповник и бузина, цветы на обочинах дороги
и посещающие их насекомые, вроде бабочек из семейства белянок, размножающихся на кустах крушины. Я знаком с одной датской семьей, которая приезжала в Англию, особенно на время летнего отдыха, чтобы видеть и фотографировать на обочинах наших дорог дикие цветы, исчезнувшие во многих частях Дании в результате интенсивного использования земли и применения гербицидов. Для натуралиста и эколога существует, помимо этих удовольствий, еще очарование стаций, в которых можно найти чуть ли не половину наших британских видов растений и животных. Возможно, что после леса вполне развитый злаково-разнотравный луг является одним из богатейших сообществ нашей страны.
Можно привести и целый ряд экономических соображений в пользу сохранения живых изгородей, а вместе с ними и придорожных полос, помимо, использования первых как заграждений, а вторых — как места для пешеходов и «зон безопасности» для экипажей. Прежнее представление о том, что обочины дорог представляют собой резервуар сорняков, быстро утрачивает свое значение, так как сельскохозяйственные культуры подвергаются все более и более систематической обработке гербицидами. В нашей стране экология живых изгородей изучалась очень мало, и единственная общая карта распространения в Англии различных типов лесных полос, окружающих поля, составлена немецким географом [295]. Мы обычно видим в живой изгороди лишь укрытие, и далеко не всем известно, что в Шлезвиг-Гольштейне эти изгороди на 20% повышают урожаи зерна и настолько уменьшают вызываемое ветром испарение воды с поверхности полей, что это равносильно увеличению количества осадков на одну треть [297]. Живые изгороди служат там также источником лесоматериалов, который по продуктивности сравнивают с лесными насаждениями. И у нас в Англии пятую часть нашей собственной древесины дают деревья лесных полос, главным образом дуб, вяз и ясень (фиг. 88) [222]. Эти деревья обеспечивают также тень домашним животным и людям. В какой степени лесные полосы служат местом обитания врагов и паразитов вредных насекомых и клещей, должны выяснить будущие исследования. Но одна
Ф и г. 88. Деревья живых изгородей дают пятую часть древесины, выращиваемой в Англии (фото автора, 1957).
Эти дубы на первый взгляд растут на земляном валике, покрытом травой, но в действительности здесь есть также остатки хорошей живой изгороди из кустарников, растущих на известковых почвах, в том числе кизила, орешника, бересклета, шиповника, тёрна и калины (Viburnum). Эта дорога в Гэмпшире сфотографирована в начале июня. Справа заросли жгучей крапивы (Urtica dioica) — растения, обладающего исключительно богатой собственной фауной насекомых. Вдали цветущий боршевнпк (Heracteum sphondytium). В середине на переднем плане немного глухой крапивы (Lamium album).
недавняя работа показывает, что это не только теоретическое предположение [291]. Сравнение численности пауков в опрыскиваемых и неопрыскиваемых плодовых садах Кента и Эссекса показало, что опрыскивание почти нацело уничтожает пауков, нормально живущих в необрабатываемых садах, но затем начинается заселение садов пауками из окружающих лесов и живых изгородей. Поэтому в опрыскпваемых садах было найдено только 20 видов пауков, а в неопрыскиваемых — 41. Практическая сторона этого заключается в том, что некоторые пауки уничтожают клещей, вредящих плодовым деревьям, а число пауков в опрыскиваемых садах было примерно в 25 раз меньше, чем в неопрыскиваемых, и, если бы не проникновение пауков извне, они там вовсе бы исчезли. Действительное значение этого для борьбы с вредителями плодовых деревьев еще не установлено.
Живые изгороди — сравнительно недавнее добавление к британскому ландшафту, появившееся главным образом в результате огораживания полей и развития дорог за последние 2— 3 века. Однако то обстоятельство, что им свойственны богатые и устойчивые биоценозы, содержащие значительную часть флоры и фауны луга, лесной опушки, а иногда и самого леса, доказывает, что можно с успехом создать такого рода хорошо заселенные местообитания в виде лент, вьющихся среди более интенсивно используемых полей и лесов. Они образуют нечто вроде соединительной ткани, связывающей между собой отдельные элементы ландшафта. К ним следовало бы добавить полосы отчуждения железных дорог; хотя в них нет свободного доступа они служат хорошим убежищем для животных и растений и услаждают взор пассажира. Они отличаются от придорожных полос тем, что в значительной части представляют отдельные фазы восстановления живого покрова после гарей. Следует упомянуть еще одну крупную сеть диких местообитаний — внутренние водные пути страны. В Англии около 2500 км каналов, пе считая примерно такой же протяженности судоходных рек; еще гораздо большее протяжение имеют речки, ручьи и канавы; все эти водоемы увеличивают многообразие и сложность экологии страны.
Только что приведенные примеры наводят на мысль о необходимости планового создания лучшего, более разнообразного ландшафта, обеспечивающего всестороннюю охрану природы. Ни в одной стране это не пытались делать с такой энергией и таким размахом, как в США 1, где побудительной причиной служило предохранение почвы от эрозии, а также горячее желание сохранить местообитания для дичи. Общая картина этой работы и связанных с ней экологических идей лучше всего дана в книге Эдуарда Грэема «Принципы использования земли» [293]. В недавно появившемся кратком очерке [294] тот же автор отмечает необычайные успехи в усложнении ландшафта, достигнутые в некоторых частях США за последнее десятилетие, и в то же время указывает, что во избежание риска при осуществлении подобных планов нам еще очень многого недостает в области экологических знаний. Он говорит: «Необходимо дополнительное изучение экологических взаимоотношений на площадях, предоставляемых исключительно природе, и на землях, используемых иным образом, например для земледелия, скотоводства или лесного хозяйства». Значительная часть работы Службы Охраны Почвы в США касается восстановления того, что потеряно, или создания совершенно новых типов взаиморасположения местообитаний (фиг. 89 и 90), в то время как в Англии мы, возможно, все еще имеем шансы сохранить наш собственный замечательный ландшафт, прежде чем он утратит свое экологическое многообразие. Этот тип ландшафта был создан отдельными людьми, главным образом сельскими жите^®р, которые действовали инстинктивно и устраивали себе место' для жилья, а не только для возделывания растений. Его еще можно видеть вполне сохранившимся, например в Херефордшире (фиг. 91). Жизненно важно, чтобы отныне каждый, кто собирается как-то изменить, отрезать, осушить, подвергнуть опрыскиванию или засадить любую полоску или уголок
1 С автором нельзя согласиться. Масштабы плановой реконструкции ландшафта, особенно степного и пустынного, с помощью лесоразведения в нашей стране давно превзошли усилия западных стран. Да и сама идея мелиорации климата с помощью разведения лесов родилась в России и связана с именами В. В. Докучаева и А. И. Воейкова.— Прим. ред.
Ф и г. 89. Террасированные склоны и живые изгороди, устроенные для борьбы с эрозией почвы на одной ферме в штате Индиана (вид с самолета).
Живые изгороди состоят из Rosa mulliflora. Видны также несколько прудов; за последние 12 лет в США было устроено около 2 млн. таких прудов, и они тоже увеличивают разнообразие животного п растительного мира.
Фиг. 90. Пятилетние живые изгороди из цветущих растений Rosa multiflora на одной ферме в штате Мэриленд.
На востоке США эту интродуцированную азиатскую розу очень широко использовали для устройства живых изгородей. Она образует эффективное заграждение от скота, дает приют диким млекопитающим, птицам п насекомым и красива во время цветения и плодоношения.
Фиг. 91. Богатый, сложный и устойчивый в экологическом отношении ландшафт (фото автора, 1953).
Поля, живые изгороди и леса, главным образом из дуба и ясеня с орешником (вид с вершины холма Мпдсаммер-Хилл, рай-он.Малверн-Хиллс в Херефордшире). Для этого ландшафта характерно обилие диких как растительных, так и животных форм; многие виды являются местными формами, интересны и красивы; сельское и лесное хозяйство ведется хорошо, леса и культуры — в здоровом состоянии. Сохраним ли мы все это для потомства — не только своего, но и потомства всех видов животных и растений?
земли, задал себе три вопроса: какие животные и растения существуют здесь, что красивое или интересное может быть утрачено и насколько предполагаемое изменение может увеличить все возрастающую неустойчивость биоценозов? Итак: убежище, красота и интерес и, наконец, устойчивость. Эта концепция, возможно, позволит нам внести в измененный ландшафт некоторые из экологических особенностей дикой природы. Разве не хорошо было бы иметь возможность сказать, как сказал Джон Мьюир [298], шотландец, сделавшийся великим американским проповедником охраны природы: «Здравомыслящим и свободным людям вряд ли покажется необходимым пересекать материк в поисках естественных красот природы, какой бы легкой ни была дорога, ибо они найдут их в изобилии на месте, где бы они ни находились» [298]. Сможем ли мы через полвека рассуждать так, как мог рассуждать Мьюир полвека назад!
ЛИТЕРА ТУРА
Глава I
1. Artimo A., Finland a profitable muskrat land. Preliminary report., Suom. Riista, 4, 7—61 (1949).
2. Cabrera A., Y e p e s J., Historia natural ediar. Mammiferos Sud-Americanos (vida, costumbres у descripcion), Buenos Aires, 1940.
3. Cooke M. T., The spread of the European starling in North America (to 1928), Circ. U. S. Dep. Agric., 40, 1—9 (1928).
4. D a v i s D. H. S., Plague in Africa from 1935 to 1949. A survey of wild rodents in African territories, Bull. World Hlth Org., 9, 665—700 (1953).
5. D e V о s A., M a n v i 1 1 e R. H., Van G e 1 d e r R. G., Introduced mammals and their influence on native biota, Zoologica, N. Y., 41, 163— 194 (1956).
6. D о r s t J., Giban J., Les mammiferes acclimates en France depuis un siecle. Terre et la Vie, 101, 217—229 (1954).
7. East B., Is the lake trout doomed? Nat. Hist., N. Y., 58, 424—428 (1949).
8. Forestry Commission. Chestnut blight caused by the fungus Endothia parasitica, Bookl. For. Comm., 3, 1—6 (1950).
9. Freeman R. B., Pitrufquenia coypus Marelli (Mallophaga, Gyropi-dae): an ectoparasite on Myocastor coypus Mol., Ent. Mon. Mag., 82, 226— 227 (1946).
10. G r a v a 11 G. F., M a r s h a 1 1 R. P., Chestnut blight in the Southern Appalachians, Dep. Circ. U. S. Dep. Agric., 370, I—II (1926).
11. Harris V. T., The nutria as a wild fur mammal in Louisiana. Trans. 21st N. Amer. Wildl. Conf., 474—486, 1956.
12. H i 1 e R., E s c h m e у e r P. H., L u n g e r G. F., Decline of the lake trout fishery in Lake Michigan. Fish. Bull., U. S. Fish and Wildlife Service, 52 (No 60), 77—95 (1951).
13. Hoestlandt Н., Le crabe chinois (Eriocheir sinensis Mil. Ed.) en Europe et principalement en France, Ann. Epiphyt. N. S., 11, 223—233 (1945).
14. H u b b a r d С. E., Grasses. Harmondsworth, Middlesex, 1954.
15. J о h n s о n С. E., The muskrat in New York: its natural history and economics. Roosevelt Wild Life Bull., 24, 193—320 (1925).
16. Johnson Samuel, A journey to the western islands of Scotland, London, 1775.
17. Kalmbach E. R., Pigeon, sparrow and starling control. Pest Control, 22 (5), 9—10, 12, 31—32, 54 (1954).
18. Бобринский H. А., Кузнецов Б. А., Кузякин А. П., Определитель млекопитающих СССР, VIII, Отряд грызуны, М., 1944
19. Langlois Т.Н., The western end of Lake Erie and its ecology. Ann Arbor, 1954.
20. Laurie E. M. 0., The coypu (Myocastor coypus) in Great Britain, J. Anim. EcoL, 15, 22—34 (1946).
21. Lennon R. E., Feeding mechanism of the sea lamprey and its effect on host fishes. Fish. Bull., U. S. Fish and Wildlife Service, 56 (No 98), 247—293 (1954).
22. Link V. B., A history of plague in the United States of America, Publ. |Hlth Monogr., Wash., 26, 1—120 (1955).
23. M e t с a 1 f e H., С о 1 1 i n s J. F., The control of the chestnut bark disease, Farmers’ Bull. U. S. Dep. Agric., 467, 1—24 (1911).
24. M e у e r K. F., The known and the unknown in plague, Amer. J. Trop. Med., 22, 9—36 (1942).
25. Myers J.G., The arthropod fauna of a rice-ship, trading from Burma to the West Indies, J. Anim. EcoL, 3, 146—149 (1934).
26. Peters N., Panning A., Die chinesische Wollandkrabbe (Eriocheir sinensis H. Milne-Edwards) in Deutschland, Zool. Anz., 104 (Suppl.), 1—180.
27. S h e a r C. L., S t e v e n s N. E., T i 1 1 e r R. J., Endothia parasitica and related species, Bull. U.S. Dep. Agric., 380, 1—82 (1917).
28. S о p e r F. L., Wilson D. B., Anopheles gambiae in Brazil 1930 to 1940, New York, 1943.
29. T a n s 1 e у A. G., The British Islands and their vegetation, Cambridge, 1939.
30. UlbrichJ., Die Bisamratte: Lebensweise, Gang ihrer Ausbreitung in Europa, wirschaftliche Bedeutung und Bekampfung, Dresden, 1930.
31. U 1 m A., The Chinese chestnut makes good, Amer. Forests, 54, 491, 518, 520 and 522.
32. Верещагин H. К., Акклиматизация нутрии (Myocastor coypus Mol.) в западной Грузии, Tp. ин-та зоол. АН Груз. ССР, 4, 3—42 (1941).
Глава II
33. А г к е 1 1 W. J., Jurassic geology of the world. Edinburgh juid Londom 1956.
34. Campbell D. H., Relations of the temperate floras of North and South America, Proc. Calif. Acad. Sci., 25, 139—146 (1944).
35. D a m m e r m a n K. W., The fauna of Krakatau, Amsterdam, 1948.
36. D arw in C., Journal of researches into the natural history and geology of the countries visited during the voyage round the world of H. M. S., «Beagle», London, 1845.
37. De Chardin P. T., The movements of the fauna between Asia and North America since the Lower Cretaceous, Proc. 6th Pacif. Sci. Congr. 1939, 3, 647—648 (1940).
38. E к m a n S., Zoogeography of the sea, London, 1953.
39. H i с к s о n J. S., A naturalist in North Celebes, London, p. 190, 1889.
40. H о p w о о d A. T., In M a t t i n g 1 e у P. F. Distribution of animals and plants in Africa, Nature, 171, 639—640 (1953).
41. L e а к e у L. S. В., С 1 а г к W. E. Le Gros, British-Kenya Miocene Expeditions, Nature, 175, 234 (1955)
42. L e e Shu n-C h i n g, Forest botany of China, Changhai, 1935.
43. M а у r E., Wallace’s Line in the light of recent zoogeographic studies, Quart. Rev. Biol., 19, 1—14 (1944).
44. M а у r E., History of the North American bird fauna, Wilson Bull., 58, 3—41 (1946).
45. M u r p h у R. C., Oceanic and climatic phenomena along the west coast of South America during 1925, Geogr. Rev., 16, 26—54 (1926).
46. Myers G. S., Paleogeographical significance of fresh-water fish distribution in the Pacific, Proc. 7th Pacif. Sci. Congr., 1949, 4, 38—48 (1953).
47. N о r m a n J. R., A history of fishes, London, 1931.
48. Raven H. C., Wallaces Line and the distribution of Indo-Austra-lian mammals, Bull. Amer. Mus. Nat. Hist., 68, 177—293 (1935).
49. Rensch B., Die Geschichte des Sundabogens, eine tiergeographische Untersuchung, Berlin, 1936.
50. Schaeffer B., The evidence of the fresh-water fishes, Bull. Amer. Mus. Nat. Hist., 99, 227—234 (1953).
51. S с о t t W. B., A history of land mammals in the Western Hemisphere, New York, 1937.
52. S i m p s о n G. C., Life of the past. An introduction to palaeontology, New Haven, 1953.
53. S t о с к C., A ground sloth in Alaska, Science, 95, 552—553 (1942).
54. Van Dyke E. C., The origin and distribution of the coleopterous insect fauna of North America, Proc. 6th Pacif. Sci. Congr. 1939, 4, 255— 268 (1940).
55ч W а 1 1 а с е A. R., On the zoological geography of the Malay Archipelago, J. Linn. Soc. (Zool.), 4, 172—184 (1860).
56. Wallace A. R., The Malay Archipelago: the land of the orangutan and the bird of paradise. A narrative of travel with studies of man and nature, London, 1869.
57. Wallace A. R., The geographical distribution of animals, London, 2 vols, 1876.
Глава III
58. Adams J. A., The Oriental beetle as a turf pest associated with the Japanese beetle in New York, J. Econ. Ent., 42, 366—371 (1949).
59. В а к e r W. A., V a n с e A. M., Status of the European corn borer in 1937, J. Econ. Ent., 31, 348—353 (1938).
60. Balch R. E., The outbreak of the European spruce sawfly in Canada and some important features of its economics, J. Econ. Ent., 32, 412—418 (1939).
61. Bezant, E. T., Further records of the Australian carpet beetle, Anth-renocerus australis" (Hope) (Col., Dermestidae) in Britain, Ent. Mon. Mag., 92, 401 (1956).
62. В r e w e r E. G., The fight for the elms. Amer. Forests, 47, 22—25 (1941).
63. Brown A. W. A., Foliage insects of spruce in Canada, Tech. Bull. Dep. Agric. Can., 31, 1—29 (1941).
64. Butcher D., Your shade trees and the Japanese beetle, Amer. Forests, 47, 396—397 (1941).
65. C h i t w о о d B. G., The golden nematode of potatoes, Circ. U. S. Dep. Agric., 875, 1—48 (1951).
66. Clausen С. P., King J.L., Teranishi C., The parasites of Popillia japonica in Japan and Chosen (Korea) and their introduction into the United States, Dep. Bull. U. S. Dep. Agric., 1419, 1—55 (1927).
67. Collins C. W., Two elm scolytids in relation to areas infected with the Dutch elm disease fungus, J. Econ. Ent., 31, 192—195 (1938).
68. Commonwealth institute of entomology. Distribution maps of insect pests, London.
69. Coppell H. C., Arthur A. P., Notes on introduced parasites’ of the European pine shoot moth, Rhyacionia buoliana (Schiff.) (Lepidoptera: Tortricidae), in Ontario, 84th Rep. Ent. Soc. Ont., 55—58, 1953.
70. С о p p e 1 1 H. C., L e i u s K., History of the larch sawfly, with notes on origin and biology. Canad. Ent., 87, 103—111 (1955).'
71. С г a i g h e a d F. C. et. al., Insect enemies of Eastern forests. Misc. Publ. U.-S. Dep. Agric., 657, 1—679 (1949).
72. Сг essm an A. W., Plank H. К.» The camphor scale, Circ. U. S. Dep. Agric., 365, 1—19 (1935).
73. D e V о s A., M a n v i 1 1 e R. H., Van Geld er G., Introduced mammals and their influence on native biota, Zoologica, N. Y., 41, 163— 194 (1956).
74. Dowden P. B., Present status of the European spruce sawfly, Diprion polytomum (Htg.), in the United States, J. Econ. Ent., 32, 619—624 (1939).
75. European and Mediterranean plant protection organization, Leptinotarsa decemlineata Say, Colorado beetle, Europe — 1955, Paris. 1956.
76. ’European and mediterranean plant protection organization, Hyphantria cunea Drury. Report of the Fourth International Conference on fall web-worm, Paris, 1957.
77. European plant protection organization. Colorado beetle in Europe in 1952, Paris, 1953.
78. F a i r c h i 1 d D., Garden islands of the Great East, collecting from the Philippines and Netherlands India in the Junk «Cheng Ho», New York, 1945.
79. Forest insect and disease survey, Dept, of Agriculture, Canada, Annual Rep. 1955, 9 and 21 (1956).
80. Forest insect survey, Dept, of Agriculture, Canada, Annual Rep., 1942, 3—6 (1943).
81. Forest insect survey, Dept, of Agriculture, Canada, Annual Rep., 1948, 11 and 26 (1949).
82. Forest insect survey. Dept, of Agriculture, Canada, Annual Rep., 1950,9 (1951).
83. Forestry commission. Elm disease. Leaf!. For. Comm., London, 19, 1—8 (1938).
84. F о r t e P. N., The eradication of pests: some observations on the Argentine ant campaign in Western Australia, J. Dep. Agric. W. Aust., 5, Ser. 3, (5), 1—8 (1956).
85. Gambrell F. L., Mendall S. C., Smith E. H., A destructive European insect new to the United States, J. Econ. Ent., 35, 289 (1942).
86. Hallock H. C., Life history and control of the Asiatic garden beetle, Circ. U. S. Dep. Agric., 246, 1—20 (1936).
87. High M. M., The vegetable weevil, Circ. U. S. Dep. Agric., 530, 1—25 (1939).
88. Hodson W. E. H., Colorado beetle. N. A. A. S. Quart. Rev., I, 51—52 (1948).
89. Hood С. E., Life history and control of the imported willow leaf beetle, Circ. U. S. Dep. Agric., 572, 1—9 (1940).
90. Howard L.O., A history of applied entomology (somewhat anecdotal), Smithson. Misc. Coll., 84, 1—564 (1930).
91. Hunter W.D,., The pink bollworm. . . Dep. Bull. U. S. Dep. Agric., 1397, 1—30 (1926).
92. К г u m h о 1 z L. A., Reproduction in the western mosquitofish, Gam-busia affinis affinis (Baird and Girard), and its use in mosquito control. Ecol. Monogr., 18, 1—43 (1948).
93. M у e r s G. S., An American cyprinodont fish, Jordanella jloridae, reported from Borneo, with notes on the possible widespread introduction of foreign aquarium fishes, Copeia, 267—268 (1940).
94. Newell W., Barber T. C., The Argentine ant. Bull. U. S. Bur. Ent., 122, 1—98 (1913).
95. Peace T. R., Tree diseases in Great Britain, 1950—1951. A general review. Rep. For. Res., London for... 1951, 94—98 (1952).
96. Pechuman L. L., A preliminary study of the biology of Scolytus sulcatus LeC, J. Econ. Ent., 31, 537—543 (1938).
97. Phillips J. C., Wild birds introduced or transplanted in North America, Tech. Bull. U. S. Dep. Agric., 61, 1—63 (1928).
98. Rockwood L. P., The clover root borer. Dep. Bull. U. S. Dep. Agric., 1426, 1—48 (1926).
99. Scheffer T. H., Cottman C., The crested myna, or Chinese starling, in the Pacific Northwest, Tech. Bull. U. S. Dep. Agric., 467, 1—26 (1935).
100. Smith H. S., On some phases of preventive entomology, Sci. Mon., N. Y., 29, 177—184 (1929).
101. Smith K. G., The occurrence and distribution of Aphomia gularis (Zell.) (Lep., Galleriidae), a pest of stored products, Bull. Ent. Res., 47, 655—667 (1956).
102. Smith L. В., H a d 1 e у С. H., The Japanese beetle, Dep. Circ. U. S. Dep. Agric., 363, 1—66 (1926).
103. Smith M. R., Distribution of the Argentine ant in the United States and suggestions for its control and eradication, Circ. U. S. Dep. Agric., 387, 1—39 (1936).
104. Trouv elot B., Le dolyphore de la pomme de terre (Leptinotarsa der cemlineata Say) en Amerique du Nord, Ann. fipiphyt. N. S., I, 278—336 (1936).
105. United states bureau of entomology and plant quarantine, Japanese beetle. Insect Pest Surv. Bull. U. S., 21, 801—802 (1941).
106. Vance A. M., Studies on the prevalence of the European corn borer in the East North Central States, Circ. U.S. Dep. Agric., 649, 1—23 (1942).
107. Wichmann H.E., Im europaischen Grossraum eingeschleppte Bor-kenkafer, Z. Angew. Ent., 37, 92—109 (1955).
108. Wichmann H. E., Einschleppungsgeschichte und Verbreitung des Xylosandrus germanus Blandf. in Westdeutschland (nebst einem Anhang. Xyleborus adumbratus BJandf.), Z. Angew. Ent., 40, 82—99 (1957).
109, W illiamsC. В.» Beware of this beetle! Zoo., 11, 56—57 (1937).
110. Wilson G; F ox., Phylloxera on vines, a new British record, Proc. R. Ent. Soc. London, 10, 25—28 (1935).
Глава ГУ,
111. Abbot R. T., March of the giant African snail, Nat. Hist., N. Y., 80, 68—71 (1949).
112. A m a d о n D., The Hawaiian Honeycreepers (Aves, Drepaniidae), Bull. Amer. Mus. Nat. Hist., 95, 155—262 (1950).
113. Aurillius C., Coleoptera-Curculionidae von Juan Fernandez und der Oster-Insel. In The natural history of Juan Fernandez and Easter Island (Ed. C. Skottsberg). Uppsala, 3, 461—477 (1926).
114. В arrow К. M., Three years in Tristan da Cunha, London, 910.
115. Bequaert J.C., Studies in the Achatinae, a group of African land snails, Bull. Mus. Comp. Zool., 105, 1—216 (1950).
116. BrinckP., Coleoptera of Tristan da Cunha. Results of the Norwegian Sci. Exped. to Tristan da Cunha 1937—1938, No 17, 1—121 (1948).
117. Bryan E. H., A summary of the Hawaiian birds, Proc. 6th Pacif. Sci. Congr., 1939, 4, 185—189 (1940).
118. Bryan W. A., Natural history of Hawaii, Honolulu, 1915.
119. Buxton P. A., Hopkins G. H. E., Researches in Polynesia and Melanesia ... Parts I—IV. (Relating principally to medical entomology.), Mem. London, Sch. Hyg. Trop. Med., I, 1—260 (1927).
120 .*C hristophersen E., Problems of plant geography in Tristan da Cunha. Norsk Geogr. Tidsskr.. 7, 106—112 (And personal communication.), 1939.
121. Christophersen E., Tristan da Cunha: the lonely isle, London, 1940.
122. Cook1!., The three famous .voyages of Captain J ames Coor round the world. . .London and New York, 1890.
123. Dammerman K. W., The fauna of, Krakatau 1883—1933, Amsterdam, 1948.
124. Dumbleton L. J., Entomological aspects of insect quarantine in New Zealand. Proc. 7 th Pacif. Sci. Congr., 1949, 4, 331—334 (1953).
125. Enderlein G., Die Dipterenfauna der Juan-Fernandez-Inseln und der Oster-Insel. In The Natural History of Juan Fernandez and Easter Island (Ed. C. Skottsberg). Uppsala, 3, 634—680 (1938).
126. Gill W. W., Life in the Southern Isles, London, 1876.
127. Gill W. W., Historical sketches of savage life in Polynesia, Wellington, 1880.
128. Gulick A., Biological peculiarities of oceanic islands, Quart. Rev. Biol., 7, 405—427 (1932).
129. Hagen Y., Birds of Tristan da Cunha. Resultb of the Norwegian Sci Exped. to Tristan da Cunha 1937—1938, No 20, I—248 (1952).
130. J e e к e 1 C. A. W., Diplopoda Results of the Norwegian Sci. Exped. to Tristan da Cunha 1937—1938, No 32, 5—9 (1954).
131. К i n g J., см. Кук [122], стр. 1010.
132. Martin J., An account of the natives of the Tonga Islands in the South Pacific Ocean . . . compiled from the extensive communications of Mr. William Marine, London, 2 vols, I, Ch. 9, 1818.
133. M а у r E., The zoogeographic position of the Hawaiian Islands, Condor, 45, 45—48 (1943).
134. M u n r c G. C., Birds of Hawaii, Honolulu, 1944.
135. (Ldhnei N. H., Mollusca from Juan Fernandez and Easter jlsland. In The natural history of Juan Fernandez and Easter Island (Ed. C. Skotts-berg), Uppsala, 3, 219—254 (1926).
136. Paterson C. R., The establishment and spread in New Zealand of the wasp Vespa germanica, Proc. 7th Pacif. Sci. Congr., 1949, 4, 358—362 (1953).
137. Pemberton С. E., Economic entomology in Hawaii, Proc. 7th Pacif. Sci. Congr., 1949, 4, 91—94 (1953).
138. Perkins R.C. L., Fauna Hawaiiensis or the Zoology of the Sandwich (Hawaiian) Islands. Vol. I, Part. IV. Vertebrata, 363—466, Cambridge, 1903.
139. Petersen P. Esben, More Neuroptera from Juan Fernandez and Easter Island. In The natural history of Juan Fernandez and Easter Island (Ed. C. Skottsberg), Uppsala, 3, 309—313 (1924).
140. Richards L. P., Baldwin P. H., Recent records of some Hawaiian honeycreepers, Condor, 55, 221—222 (1953).
141. Ris Lambers D.H., Aphididae of Tristan da Cunha, Results of the Norwegian Sci/Exped. to Tristan da Cunha'1937—1938, No 34, 1—5 (1955).
142. Schwartz C. W., Schwartz E.R., A reconnaissance of the game birds in Hawaii, Hilo, Hawaii, 1949.
143. Sherman M., Tamashiro M., Biology and control of Araecerus levipennis Jordan (Coleoptera: Anthribidae), Proc Hawaii. Ent. Soc., 16, 138—148 (1956).
144. Skottsberg C., Notes on a visit to Easter Island. In The natural history of Juan Fernandez and Easter Island (Ed. C. Skottsberg), Uppsala, I, 1—20 (1920).
145. Skottsberg C., The vegetation of Easter Island. In The natural history of Juan Fernandez and Easter Island (Ed. C. Skottsberg), Uppsala, 2, 487—502 (1927).
146. Skottsberg C., Derivation of the flora and fauna of Juan Fernandez and Easter Island. In The natural history of Juan Fernandez and Easter Island (Ed. C. Skottsberg), Uppsala, I, 193—438 (1956).
147. Stokes J..F. G., Notes on the Hawaiian rat, Occ. Pap. Bishop Mus.. 3 (4), 11—21 (1917).
148. S v i h 1 a A., The Hawaiian rat, Murrelet, 17, 3—14 (1936).
149. T a t e G. H. H., Rodents of the genera Rattus and Mus from the Pacific Islands, Bull. Amer. Mus. Nat. Hist., 68, 145—178 (1935).
150. Viette P. E.L., Lepidoptera. Results of the Norwegian Sci. Exped. to Tristan da Cunha 1937—1938, No 23, 1—19 (1952).
151. Von Hochsterter F., New Zealand, its physical geography, geology and natural history, Stuttgart, 1867.
152, . Weber P. W., Recent introductions for biological control in Hawaii, I. Proc. Hawaii. Ent. Soc., 16, 162—164 (1956).
153. Wheeler W. M., Revised list of Hawaiian ants. Occ. Pap. Bishop Mus., 10 (21), 1—21 (1934).
154. Williams F. X., Some natural enemies of snails of the genus Acha-tina in East Africa. Proc. 7th Pacif. Sci. Congr., 1949, 4, 277—278 (1953).
155. Wilson S. В., E v a n s A. H., Aves Hawaiiensis. the birds of the Sandwich Islands, London, 1890—1899.
156. Wodzicki K. A., Introduced mammals of New Zealand, an ecological and economic survey, Bull. D. S. I. R., N. Z., 98, 1—255 (1950).
157. Zimmermann A., Coleoptera-Dytiscidae von Juan Fernandez und der Oster-Insel. In The natural history of Juan Fernandez and Easter Island (Ed. C. Skottsberg), Uppsala, 3, 298—304 (1924).
158. Zimm er m a n E.C., Insects of Hawaii. I. Introduction, Honolulu, 1948.
Г лава V
159. Allen К. R., The geography of New Zealand’s freshwater fish. N. Z. Sci. Rew., 14 (3), 3—9 (1956).
160. Anon (Introduction of Nereis into the Caspian Sea.) The Times, 7 November, 1940.
161. BishopM.W. H., Distribution of barnacles by ships, Nature, 167, 531 (1951).
162. Bishop M. W. H., Crisp D.J., The Australasian barnacle, Elmi-nius modestus, in France, Nature, 179, 482—483 (1957).
163. Calhoun A. J., Annual migrations of California striped bass. Calif. Fish Game, 38, 391—403 (1952).
164. Caiman W.T., Zoological results of the Cambridge Expedition to the Suez Canal, 1924. XIII. Report on the Crustacea Decapoda (Brachyura). (With Appendix by H. M u n г о Fox.) Trans. Zool. Soc. London, 22, 211— 219 (1927).
165. Cole H. A., The American whelk tingle, Urosalpinx cinerea (Say), on British oyster beds, J. Mar. Biol. Ass. U. K., 25, 477—508 (1942).
166. Cole H. A., The American slipper limpet (Crepidula fornicata L.) on Cornish oyster beds. Fish. Invest., London, Ser. 2, 17 (7), 1—13 (1952).
167. Cole Н. A., Benthos and the shellfish of commerce. In Sea fisheries: theii* investigation in the United Kingdom (Ed. M. Graham), London, 139—206 (1956).
168. Cole H. A., Oyster cultivation in Britain. A manual of current practice, London, 1956.
169. Cole H. A., Baird R. H., The American slipper limpet (Crepidula jornicata) in Milford Haven, Nature, 172, 687 (1953).
170. Connell J. H., Elminius modestus Darwin, a northward extension of range, Nature, 175, 954 (1955).
171. Crisp D. J., Chipperfield P. N. J., Occurrence of Elminius modestus (Darwin) in British waters, Nature, 161, 64 (1948).
172. Davidson F. A., Hutchinson S. J., The geographic distribution and environmental limitations of the Pacific salmon (genus On-chorhynchusY Bull. U. S. Bur. Fish., 48 (No 26), 667—692 (1938).
173. Edmondson С. H., Wilson I.H., The shellfish resources of Hawaii, Proc. 6th Pacif. Sci. Congr., 1939, 3, 241—243 (1940).
174. Ekman S., Zoogeography of the sea, London, 1953.
175. Feldman J., Feldman G., Recherches sur les Bonnemaisonia-cees et leur alternance de generations, Ann. Sci. Nat. Bot.’JSer. II, 3, 75— 175 (1942).
176. FoxH.Munro, Zoological results of the Cambridge Expedition to the Suez Canal, 1924. I. General part. Trans. Zool. Soc. London, 22, 1—64 (1926).
177. Hardy A. C., The open sea. Its natural history: the world of plankton, London, 1956.
178. Hentschel E., Der Bewuchs an Seeschiffen. Int. Rev. Hydrobiol., II, 238—264 (1923).
179. Hildebrands. F., The Panama Canal as a passageway for fishes, with lists and remarks on the fishes and invertebrates observed, Zoologia, N. Y., 24, 15—45 (1939).
180. Horridge G. A., Occurrence of Asparagopsis armata Harv. on the Scilly Isles, Nature, 167, 732—733 (1951).
181. Johnson W. C., Calhoun A. J., Food habits of California striped bass, Calif. Fish Game, 38, 531—534 (1952).
182. J о n e s L. L., An introduction of an Atlantic crab into San Francisco Bay. Proc. 6th Pacif. Sci. Congr., 1939, 3, 485—486 (1940).
183. Kincaid T., The acclimatization of the Pacific oyster (Ostrea lape-rousii Schrenck-0strea gigas Thunberg) upon the west coast of North America, Proc. 7th Pacif. Sci. Congr., 1949, 4, 508—512 (1953).
184. Loosanoff V.L., The European oyster in American waters, Science, 121, 119—121 (1955).
185. Mansueti R., Kolb H., A historical rewiew of the shad fisheries of North America, Publ. Chesapeake Biol. Lab., 97, 1—293 (1953).
186. Marshall ,N. В.. The «Manihine» Expedition to the Gulf of Aqata 1948—1949. IX. Fishes. Bull. Brit. Mus. (Nat. Hist.) Zool. Ser., I, 221-252 (1952).
187. Merriman D., Studies on the striped bass (Roccus saxatilis) of the Atlantic coast. Fish. Bull., U.S. Fish and Wildlife Service, 50 (No 35), 1—77 (1941).
188. Mistakidis M. N., Hancock D. A., Reappearance of Ocenebra erinacea (L.) off the east coast of England, Nature, 175, 734 (1955).
189. Neave F., Introduction of anadromous fishes on the Pacific coast. Canad. Fish Culturist, No 16, 25—26 (1954).
190. Сборник работ об акклиматизации Nereis succinea в Каспийском море (под ред. В. Н. Никитина), Изд. Моск. О-ва Исп. Природы, М., 1952.
191. Orton J. Н., Oyster biology and oyster-culture, being the Buckland Lectures for 1935, London, 1937.
192. Rees С. B., Cattley J. G., Processa aequimana Paulson in the North Sea, Nature, 164, 367 (1949).
193. Sandison E. E., Appearance of Elminius modestus in South Africa Nature, 165, 79—80 (1950).
194. Scofield N. В., В г у a n t H. C., The striped bass in California, Calif. Fish. Game, 12, 55—74 (1926).
195. Slocum J., Sailing alone around the world. London (Originally, publ. 1900).
196. S m i t h jjH. M., A review of the history and results of the attempts to acclimatize fish and other water animals in the Pacific states, Bull. U. S. Fish. Comm., 15, 379—472 (1895).
197. Steinitz W., Die Wanderung indopazifischer ’Arten ins Mittelmecr 'seit Beginn der Quartarperiode. Int. Rev. Hydrobiol., 22, 1—90 (1929).
198. Thompson J. M., The acclimatization and growth of the Pacific oyster (Gryphaea gigas) in Australia. Aust. J. Mar. Freshw. Res., 3, 64—73 (1952).
199. Walker M. I., Burrows E. M., Lodges. M., Occurrence of Falkeribergia rujolanosa in the Isle of Man, Nature, 174, 315 (1954).
200. W a 1 n e P. R., The biology and distribution of the slipper limpet Crepidula fornicata in Essex rivers with notes on the distribution of the larger epi-benthic invertebrates. Fish. Invest., London, Ser. 2, 20 (6), 1—50 (1956).
201. Wolff T., The estamerikanske krabber fundet i Danmark. Flora og Fauna, 60, 19—34 (1954).
202. W о 1 f f T., Occurrence of two East American species of crabs in European waters, Nature, 174, 188—189 (1954).
203. Зенкевич Л. А., Успехи изучения морской фауны СССР за двадцать лет, Зоол. журн., 16, 830—870, 1937.
Глава VI
204. Brown R. С., S h е а 1 s R. A., The present outlook on the gypsy moth problem, J. For., 42, 393—407 (1944).
205. Cain A. J., Cushing D. H., Second occurrence and persistence of the amphipod Orchestia bottae M. Edwards in Britain, Nature, 161, 483 (1948).
206. Cole H. A., The American slipper limpet (Crepidula fornicata L.) on Cornish oyster beds. Fish. Invest., London, Ser. 2, 17 (7), 1—13 (1952).
207. Crawford G. I., A review of the amphipod genus Corophium, with notes on the British species, J. Mar. Biol. Ass. U. K., 21, 589—630 (1937).
208. Druce G. C., The flora of Berkshire, Oxford, 1897.
209. Elton C. S., M i 1 1 e r R. S., The ecological survey of animal communities. with a practical system of classifying habitats by structural characters, J. Ecol., 42, 460—496 (1954).
210. Forbush E. H., F e r n a 1 d С. H., The gypsy moth. Porthetria dispar (Linn.), Boston, 1896.
211. Fryer G.; Distribution of British freshwater Amphipoda, Nature, 168, 435 (1951).
212. Godwin H., The history of the British flora: a factual basis for phytogeography, Cambridge, 1956.
213. Hull T. G., Diseases transmitted from animals to man. Springfield and Baltimore, 1941.
214. H у n e s H. B. N., British freshwater shrimps, New Biology, 21, 25—42, Harmondsworth, Middlesex, 1956.
215. К e a n e C., The epizootic of foot and mouth disease, Spec. Publ. Calif. Dep. Agric., 65, 1—54 (1926).
216. К i n g W. B. R., О а к 1 e у К. P., The Pleistocene succession in the lower parts of the Thames Valley, Proc. Prehist. Soc., N. S., 2, 52—76 (1936).
217. M с c u b b i n W. A., The plant quarantine problem, Ann. Cryptog. Phytopath., Copenhagen, 11, 1—255 (1954).
218. Maxwell H., Waterfowl and the American pond-weed (Elodea canadensis) Scot. Nat., 81—83 (1915).
219. Mistakidis M. N., Quantitative studies of the bottom fauna of Essex oyster grounds. Fish. Invest., London, Ser. 2, 17, (6), 1—47 (1951).
220. M u n г о T., Note on musk-rats and other animals killed since the inception of the campaign against musk-rats in October 1932, Scot. Nat., 11—16 (1935).
221. P г a 11 A., (Nineteenth century.) The flowering plants and ferns of Great Britain, London, 5.
222. Reid D. M., Occurrence of the amphipod Orchestia bottae and other organisms in Britain, Nature, 161, 609 (1948).
Л Й Т Е Р А Т У t> A
223. Ridley Н. N., The dispersal of plants throughout the world. Ashford, Kent, 1930.
224. Sherman R. W., New trends in plant quarantine, J. Econ. Ent., 49, 881—883 (1956).
225. Shorten M., Squirrels, London, 1954.
226. SmithH.S. etal., The efficiency and economic effects of plant quarantines in California, Bull. Calif. Agric. Exp. Sta., 553, 1—276 (1933).
227. SparksB. W., The non-marine Mollusca of the Interglacial deposits at t Bobbitshole, Ipswish. Phil. Trans. Ser., B., 241, 1—44 (1957).
228. SpoonerG.M., Distribution of British freshwater Amphipoda, Nature, 167, 530 (1951).
229. Udine E. J., The black grain stem sawfly and the European wheat stem sawfly in the United States, Circ. U. S. Dep. Agric., 607, 1—9 (1941).
230. Warwick T., The distribution of the muskrat (Fiber zibethicus) in the British Isles, J. Anim. Ecol., 3, 250—267 (1934).
231. WarwickT., A contribution to the ecology of the musk-rat (Ondatra zibethica) in the British Isles, Proc. Zool. Soc. London, Ser. A, 110, 165— 201 (1941).
Глава VII
232. Abbott R. T., Operation snailfolk, Nat. Hist., N. Y., 60, 280—285 (1951).
233. Allen A. A., Graphocephala coccinea Forst. (Hem., Tettigoniellidae) in Kent, etc., Ent. Mon. Mag., 89, 71 (1953).
234. Babers F. H., Development of insect resistance to insecticides. U. S. Bur. Ent. Plant Quarant. E-776, 1—31 (1949).
235. В a b e r s F. H., Pratt J. J., Development of insect resistance to insecticides. II. A critical review of the literature up to 1951. U. S. Bur. Ent. Plant Quarant, E-818, 1—45 (1951).
236. Baillie A. F. H., J e p s о n W. F., Bud blast disease of the Rhododendron, J. R. Hort. Soc., 76, 355—365 (1951).
237. Brown J. M. B., Rhododendron ponticum in British woodlands, Rep. For. Res., London . . . 1953, 42—43 (1954).
238. Clausen С. P., К i n g J. L., T e r a n i s h i C., The parasites of Po-pillia japonica in Japan and Chosen (Korea) and their introduction into the United States, Dep. Bull. U. S. Dep. Agric., 1429, 1—55 (1927).
239. Cullinan F. P., Some new insecticides—their effect on plants and soils, J. Econ. Ent., 42, 387—391 (1949).
240. Davidson G., Insecticide resistance in Anopheles ganibiae Giles: a case of simple Mendelian inheritance, Nature, 178, 863—864 (1956).
Литература
241. De Bach Р., Fleschner С. A., Dietrick E.J., A biological check method for evaluating the effectiveness of entomophagous insects, J. Econ. Ent., 44, 763—766 (1951).
242. D о d d A. P., The biological campaign against prickly-pear.Commonwealth Prickly Pear Board, Brisbane, 1940.
243. Dodge B. 0., R i с к e 11 H. W., Diseases and pests of ornamental plants, Lancaster, Pennsylvania, 1943.
244. Duffy E. A. J., A monograph of the immature stages of British and imported timber beetles, London, 1953.
245. E 1 t о n C., Animal ecology, London, 1927 (Элтон Ч., Экология животных, Биомедгиз, М.—Я., 1934).
246. F о s t е г А. С., Some plant responses to certain insecticides in the soil, Circ. U. S. Dep. Agric., 862, 1—41 (1951).
247. Friedmann H., The cowbirds. A study in the biology of social parasitism, Springfield and Baltimore, 1929.
248. G a r r e t s о n M. S., The American bison, New York, 1938.
249. Gluck G., Pieter Brueghel le Vieux, Paris, 1939.
250. G о r i u s U., Untersuchungen iiber den Larchenbock, Tetropium Gabrieli Weise mit besonderer Beriicksichtigung seines Massenwechsels, Z. Angew. Ent., 38, 157—205 (1955).
251. Hanna A. D., JudenkoE., Heatherington W., The control of Crematogaster ants as a means of controlling the mealybugs transmitting the swollen-shoot virus disease of cacao in the Gold Coast, Bull. Ent. Res., 47, 219—226 (1956).
252. Hewitt C. G., The conservation of the wild life of Canada, New York, 1921.
253. H о w e s F. N., Plants and beekeeping, London, 1945.
254. К 1 e m e n t Z., Kiraly Z., Hyperparasitic chain of a fungus, a bacterium and its phage on wheat, Nature, 179, 157—158 (1957).
255. L e о p о 1 d A., A sand county almanac, and sketches here and there, New York, 1949.
256. Leopold A. S., Darling F. F., Wildlife in Alaska, an ecological reconnaissance, New York, 1953.
257. L о t к a A. J., Elements of physical biology, Baltimore, 1925.
258. L о t к a A. J., Fluctuations in the abundance of species considered mathematically, With comment by V. Volterra, Nature, 119, 12—13 (1-927).
259. Metcalfe H., Collins J. F., The control of the chestnut bark disease, Farmers’ Bull. U. S. Dep. Agric., 467, 1—24 (1911).
260. P a 1 m e r L. J., R о u s e С. H., Study of the Alaska tundra with reference to its reactions to reindeer and other grazing, Res. Rep. U. S. Fis’i and Wildlife Service, 10, 1—48 (1945).
261. Roe F. G., The North American buffalo: a critical study of the species in jts wild state, Toronto, 1951.
262. Sansome -F.W., Breeding disease-resistant plants, Nature, 145, 690— 693 (1940).
263. S h e a 1 s J. G., Soil population studies. I. The effects of cultivation and treatment with insecticides, Bull. Ent. Res., 47, 803—822 (1956).
264. S m i t h H. S., et al., The efficacy and economic effects of plant quarantines in California, Bull. Calif. Agric. Exp. Sta., 553, 1—276 (1933).
265. Thomas E. J. D., The residual effects of crop-protection chemicals in the soil. In Plant Protection Conference 1956. Proceedings of the Second International Conference at Fenhurst Research Station, England, London, . 215—222 (1957).
266. Tilden J. W., The insect associates of Baccharis pilularis De Candolle, Microentomology, 16, 149—188 (1951).
267. Volterra V., Legons sur la th^orie mathematique de la lutte pour la vie, Paris, 1931.
268. Warburg E. F., A changing flora as shown in the status of our trees and shrubs. In The changing flora of Britain (Ed. J. E. Lousley), Arbroath, 171—180 (1953).
269. WatsonJ.S., The rat problem in Cyprus, Colonial Res. Publ., 9, 1—66 (1951).
270. W i 1 s о n G. Fox., Insect pests of the genus Rhododendron, Proc. 7th Int. Congr. Ent., 1938, 4, 2296—2323 (1939).
271. Wilson G. Fox., Insect pests of rhododendrons: their distribution in Britain, Proc. R. Ent. Soc. London, Ser. A, 14, 1—5 (1939).
272. Wilson G. Fox., Pests of flowers and shrubs, Bull. Minist. Agric., London, 97, 1—105 (1950).
273. WorthingtonS., Worthington E. B., Inland waters of Africa, London, 1933.
Глава VIII
274. A u d у J. R., Ecological aspects of introduced pests and diseases, Med. J. Malaya, 11, 21—32 (1956).
275. В ir d F. T., E Igee D.E.,A virus disease and introduced parasites as factors controlling the European spruce sawfly, Diprion hercyniae (Htg.), in central New Brunswick, Canad. Ent., 89, 371—378 (1957).
276. Collyer E., Biology of some predatory insects and mites associated with the fruit tree red spider mite (Metatetranychus ulmi (Koch) in southeastern England. II. Some Important predators of the mite, J. Hort. Sci., 28, 85—97 (1953).
277. CollyerE., Ibid. III. Further predators of the mite, J. Hort. Sci., 28, 98-113 (1953).
278. DeBach P., FleschnerC. A., Dietrick E.J., Natural control of the California red scale in untreated orchards in southern California, Proc. 7th Pacif. Sci. Congr., 1949, 4, 236—248 (1949).
279. Гаузе Г. Ф., The struggle for existence, Baltimore, 1934.
280. Гаузе Г. Ф., Verification experimentales de la theorie mathematique de la lutte pour la vie. Actualites Sci. Industr., No. 277, 1—63 (1935).
281. Гаузе Г. Ф., Смарагдова Н. П., В и т т A. A., Further studies of interaction between predators and prey, J. Anim. Ecol., 5, 1—18 (1936).
282. Lord F. T., The influence of spray programmes on the fauna of apple orchards in Nova Scotia: II. Oystershell scaleLepidosaphes ulmi (L.), Canada, Etit., 79, 196—209 (1947).
283. Lord F. T., Ibid. IX. Studies on means of altering predator populations, Canad. Ent., 88, 129—137 (1956).
284. Pickett A. D.,A critique on insect chemical control methods. Canad. Ent., 81, 67—76 (1949).
285. Pickett A. D., Controlling orchard insects, Agric. Inst. Rev., 8, 52— 53 (1953).
286. Pickett A. D., P a 11 e r s о n N. A., The influence of spray programmes on the fauna of apple orchards in Nova Scotia. IV. Review. Canad. Ent., 85, 472—478 (1953).
287. Schweitzer A., My life and thought: an autobiography (Transl. С. T. Campion.), London, 188, 1956.
288. Smith H. S., D eBach P., Artificial infestation of plants with pest insects as an aid in biological control, Proc. 7th Pacif. Sci. Congr., 1949, 4, 255-259 (1953).
289. V ou t e A. D., Regulation of the density of the insect-populations in virgin-forests and cultivated woods, Arch. Neerl. Zool., 7, 435—470 (1946).
290. С a p e к К., Letters from England, London, 1925 (Чапек К., Письма из Англии, Сочинения, т. 2, Гослитиздат, М., 1959).
291. С h a n t D. A., Predacious spiders in orchards in south-eastern England, J. Hort. Sci., 31, 35—46 (1956).
292. Forestry commission. Report of the Committee on Hedgerow and Farm Timber, 1955, London (1955).
293. Graham E.H., Natural principles of land use, New York, etc., 1944.
294. Graham E. H., Nature protection as a part of land development, Proc. 6th Meeting Int. Union Conserv. Nature and Nat. Res., 194—201 (1957).
295. Hartke W., Die Heckenlandschaft. Der geographische Charakter eines Landeskulturproblems, Erdkunde, 5, 132—152 (1951).
296. J effries R., Wild life in a southern country, London, Ch. 3, 1879.
297. Marquardt G., Die Schlcswig-Holsteinische Knicklandschaft, Schr. Geogr. Inst. Univ. Kiel., 13 (3), 1—90 (1950).
298. M u i r J., Our National Parks, Boston and New York, 1909.
ОГЛАВЛЕНИЕ
Предисловие к русскому изданию ...................... 5
Из предисловия автора........................* . . 11
Глава I Участники нашествий................................ 13
Глава II Зоогеографические области Уоллеса.................. 40
Глава III Нашествия на материки............................ 66
Глава IV Судьба отдаленных островов......................... 99
Глава V Изменения, происходящие в море ................ . 124
Глава VI Взаимоотношения между популяциями................. 142
Глава VII
Замена прежних цепей питания новыми . ..... 162
Глава VIII Причины, побуждающие охранять природу .... 188
Глава IX Сохранение многообразия природы................... 202
.Дцтература .............., . , ,................. 214
ч. ЭЛТОН Экология нашествий животных и растений
Редактор Н. О. Фомина Художник А. В. Шипов Художественный редактор
В. И. Шаповалов Технический редактор А. Г. Резоухова
Сдано в производство 11/IV 1960 г. Подписано к печати 31/V 1960 г.
Бумага 60 X 92J/ie = 7,3 бум. л.
14,5 печ. л. Уч.-изд. л. 11,9 Изд. № 4/5151. Цена 9 р. 85 к.
С 1/1-1961 г. Цена 99 к. Зак. 418 ИЗДАТЕЛЬСТВО
ИНОСТРАННОЙ ЛИТЕРАТУРЫ Москва, 1-й Рижский пер., 2
Первая Образцовая типография имени А. А. Жданова
Московского городского совнархоза Москва, Ж-54, Валовая, 28
ИЗДАТЕЛЬСТВО
ИНОСТРАННОЙ ЛИТЕРАТУРЫ
ИМЕЮТСЯ В ПРОДАМСЕ СЛЕДУЮЩИЕ КНИГИ ПО БИОЛОГИИ
1. К. Вилли „Биология"
2. А. Кэйн „Вид и его эволюция"
3. Д. Лэк „Численность животных и ее регуляция в природе"
4. Д. Лэк „Дарвиновы вьюрки"
5. Э. Майр, Э. Л инс л и, Р. Юз ингер „Методы и принципы зоологической систематики"
ЗАКАЗЫ НА КНИГИ СЛЕДУЕТ ВЫСЫЛАТЬ ПО АДРЕСУ:
Москва, В-313, Ленинский пр., 86 Магазин № 72 Москниготорга