/




























































































Автор: Николаев В.А. Глезер З.И. Макарова И.В. Моисеева А.И.
Теги: растения биология флора растениеводство водоросли флора ссср
Год: 1988




Текст
ДИАТОМОВЫЕ
ВОДОРОСЛИ
СССР
11(1)
THE DIATOMS
OF THE
USSR
FOSSIL
AND RECENT
VOL.
II
FASC.
1
PYXIDICULACEAE,THALASSIOSIROPSIDACEAE,
TRICERATIACEAE, THALASSIOSIRACEAE
6
Leningrad
„NAUKA”
Leningrad Branch
1988
ДИАТОМОВЫЕ
ВОДОРОСЛИ
СССР
ИСКОПАЕМЫЕ
И СОВРЕМЕННЫЕ
том
II
вып.
1
PYXIDICULACEAE.THALASSIOSIROPSIDACEAE,
TRICERATIACEAE, THALASSIOSIRACEAE
В
Ленинград
„НАУКА”
Ленинградское отделение
1988
561.26
>мовые водоросли СССР (ископаемые и современные). Т. II, вып. 1. - Л.: Наука, 1988.-
1уске помещены вводные разделы, посвященные морфологии и терминологии диатомовых
ослей, предлагаемая система и принципы ее построения, описания 2 порядков, 4 семейств,
дов и 109 видов (древних и современных), обнаруженных на территории Советского Союза,
ключи, описания видов и внутривидовых таксонов (таксономия, морфологическая характе-
ра, экология, географическое и геологическое распространение, таблицы иллюстраций).
:к рассчитан на альгологов, геологов, гидробиологов, ихтиологов, преподавателей, аспирантов
;ентов. Библиогр. 217 назв. Ил. 60 + 7.
Редакционная коллегия:
3. И. ГЛЕЗЕР, И. В. МАКАРОВА (ответственный редактор),
А. И. МОИСЕЕВА, В. А. НИКОЛАЕВ
Edited by
S. I. GLESER, I. V. MAKAROVA (editor-in-chief),
A. I. MOISSEEVA, V. A. NIKOLAEV
Рецензенты:
M. M. ГОЛЛЕРБАХ, H. H. ДАВЫДОВА
>00000-752 . RR _ fV
(02)-88 6
12-026627-2 (т. II, вып. 1)
>2-026624-8
©Издательство „Наука”, 1988 г.
ПРЕДИСЛОВИЕ
Настоящим выпуском продолжается публикация руководства „Диатомовые,
водоросли СССР. Ископаемые и современные”, первый том которого издан
в 1974 г. и посвящен общим вопросам диатомологии и истории развития и форми-
рования флоры диатомовых водорослей от мела до современности в пределах
Советского Союза. Первоначально было задумано издать зто руководство в трех
томах. Но со времени опубликования первого тома прошло более 10 лет. За это время
у нас и за рубежом появилось много новых данных по морфологии панциря современ-.
ных и ископаемых диатомовых водорослей, полученных при помощи сканирующего
и трансмиссионного электронных микроскопов (СЭМ, ТЭМ), позволивших не только
выявить новые признаки структуры панциря и створок, но и переоценить таксономи-
ческую значимость ранее используемых признаков. Новые данные значительно?
дополнили наши представления о структуре панциря, внесли коррективы в систе-'
матику многих таксонов. Все это значительно увеличило предполагаемый объем „
издания. Учитывая это и прочие обстоятельства, редколлегия пришла к выводу
о целесообразности подготовки и публикации последующих материалов отдель-
ными выпусками.
Издаваемое руководство является результатом многолетнего изучения диатомой
вых водорослей большим коллективом авторов в пределах Советского Союза, а также
обобщения и анализа многочисленных зарубежных исследований последнего деся-
тилетия.
План издания выпусков и инструкций для авторов разработаны А. И. Прошкиной-
Лавренко, В. С. Шешуковой-Порецкой, А. П. Жузе, 3. И. Глезер, И. В. Макаровой,
А. И. Моисеевой. При подготовке инструкций был использован опыт работы авторов
руководства „Диатомовый анализ” (1949—1950). Существенными отличиями от
последнего являются цитирование при виде первоисточника, привлечение при морфо-
логических описаниях новейших данных электронной микроскопии, более полных
сведений о местонахождениях и использование по мере возможности оригиналь-
ных иллюстраций. Все выпуски включают более 40 семейств и более 200 родов.
Предложенная для настоящего издания классификация диатомовых водорослей
составлена коллективом авторов: 3. И. Глезер, Н. И. Караевой, И. В. Макаро-
вой, А. И. Моисеевой, В. А. Николаевым. Виды внутри родов расположены по
алфавиту.
Первый выпуск посвящен морфологии и терминологии панциря диатомовых
водорослей, их классификации и описанию видов с ключами из 4 семейств двух
порядков. Pyxidiculales и Thalassiosirales , обнаруженных в пределах Советского
Союза.
Издание выпусков осуществляется по-прежнему Научным советом по про-
блеме: „Биологические основы рационального использования, преобразования
и охраны растительного мира”, подготовлено и обеспечено Ботаническим инсти-
тутом (БИН) АН СССР, Всесоюзным геологическим институтом Министерства
геологии СССР (ВСЕГЕИ) и Ленинградским государственным университетом
(ЛГУ).
В составлении первого выпуска приняли участие следующие авторы: С. И. Генкал
(Ин-т биологии внутренних вод АН СССР, Борок), 3. И. Глезер (ВСЕГЕИ, Ленин-
град), Р. Н. Джиноридзе (ЛГУ), Н. И. Караева (Институт ботаники АН АзССР,
Баку), И. В. Макарова (БИН АН СССР), А. И. Моисеева (ВСЕГЕИ), В. А. Ни-
колаев (БИН АН СССР), Н. И. Стрельникова (ЛГУ), В. С. Шешукова-Порец-
кая (ЛГУ), Н. Б. Цой (Тихоокеан. океанол. ин-т ДВНЦ АН СССР, Влади-
восток) .
Электронные микрофотографии были выполнены при участии Н. В. Ченцо-
вой, Л. А. Карцевой, Е. В. Дрожжиной (БИН АН СССР), а рисунки художницей
И. Г. Гай.
Авторы будут признательны за все обнаруженные погрешности и критические
замечания.
И. В. Макарова
6
ОБЩАЯ ЧАСТЬ х
МОРФОЛОГИЯ ПАНЦИРЯ ДИАТОМОВЫХ ВОДОРОСЛЕЙ
Электронно-микроскопические исследования диатомовых водорослей, широко
развернувшиеся в последнее десятилетие, особенно в связи с введением в практику
диатомологов сканирующего электронного микроскопа (СЭМ), дали возможность
получить новые, ценные для систематики сведения о морфологии панциря, что
привело к изменению в значительной степени прежних представлений о его тонкой
структуре: позволило установить строение многих деталей панциря не только
с наружной, но и с внутренней поверхности, недостаточно хорошо различимых в свето-
вом микроскопе (СМ), а также выявить ряд новых, ранее неизвестных элементов
панциря. Важным моментом для совершенствования систематики и филогении диато-
мей было разделение сборного понятия „шипы” на несколько самостоятельных
морфологических структур, выполняющих различные функции. К ним относятся
двугубые выросты, выросты с опорами, замкнутые выросты и собственно шипы
и шипики. Принципиально новые сведения получены о поясковой зоне панциря,
труднодоступной при исследовании объекта в СМ. Много новой информации дало
электронно-микроскопическое изучение шовной системы пеннатных диатомей. Все это
требовало усовершенствования терминологии структурных элементов панциря
и введения новых терминов, что было сделано коллективом диатомологов разных
стран, объединенных в рабочую группу на 3-м и 4-м Международных симпозиумах
по современным и ископаемым диатомовым водорослям. Результаты работы
этих групп были опубликованы (Anonymous, 1975; Предложения..., 1977; Росс
и др., 1983).
В настоящем выпуске описывается морфология панциря диатомовых водорослей
с использованием современной международной терминологии (Росс и др., 1983),
но с некоторыми изменениями и дополнениями, основывающимися на данных авторов
этой главы. Морфологические структуры иллюстрированы оригинальными микро-
фотографиями.
Панцирь. Панцирь диатомовых водорослей представляет собой наружную
кремнеземную оболочку клетки и состоит из двух створок и пояска, составляющих
две половины панциря, которые наподобие коробки с крышкой надвигаются одна
на другую. Известны роды, у которых поясок отсутствует или образуется только
в период деления клетки, в таком случае обе створки панциря плотно смыкаются друг
с другом непосредственно краями загиба створок. Одна половина панциря (находящая
на другую), полученная от материнской клетки,называется э пи т е к о й, другая,
меньшая (заходящая под эпитеку), образованная после цитокинеза, — гипотекой.
Эпитека и гипотека состоят из двух частей: створки и пояскового ободка. Створка,
принадлежащая эпитеке, называется эпивальвой, створка, принадлежащая гипо-
теке, — гиповальвой.
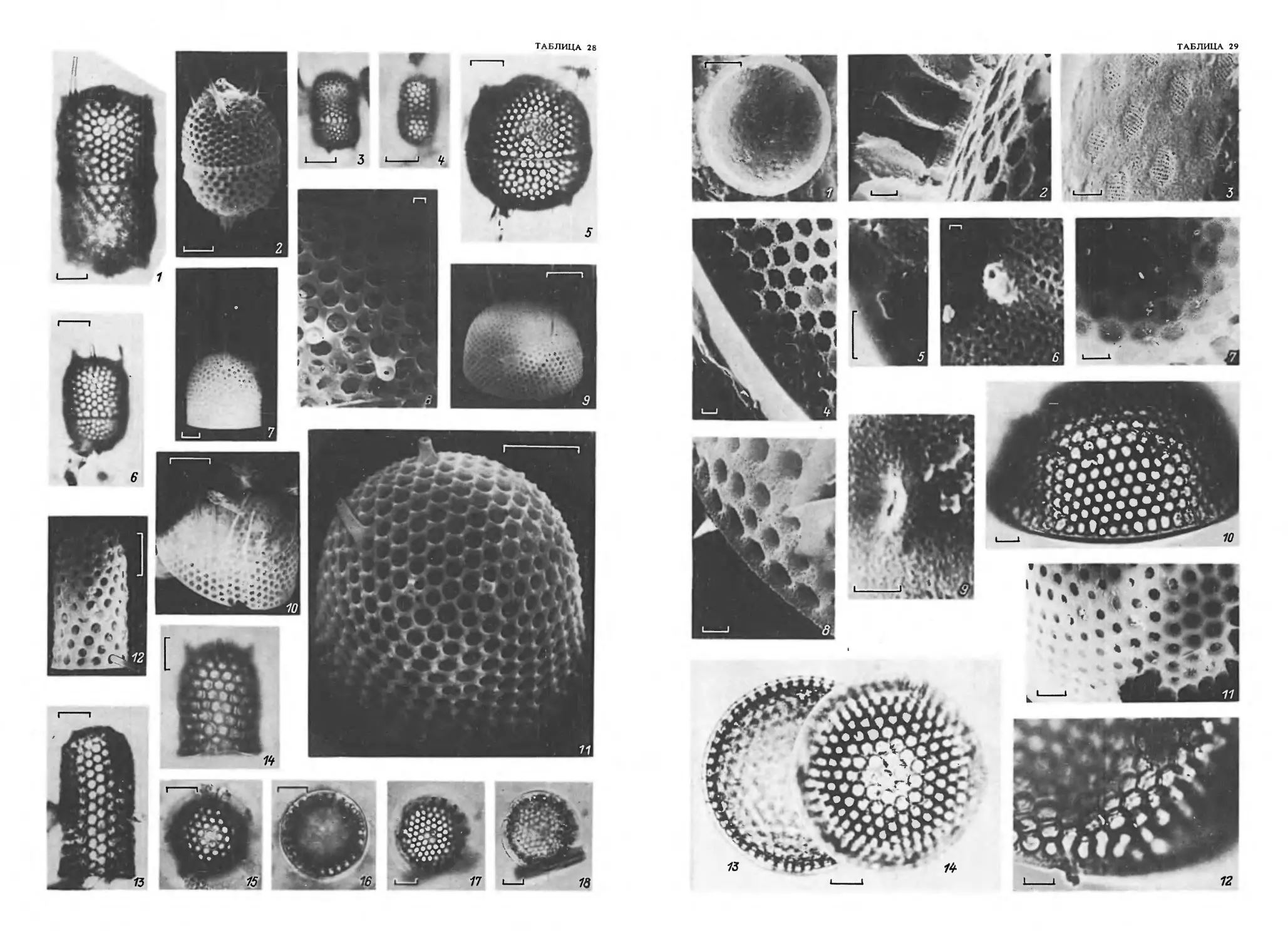
Панцирь имеет почти правильную геометрическую форму: шаровидную, линзо-
видную, дисковидную, цилиндрическую, призматическую, палочковидную, клино-
видную, седловидную и пр. (табл. 1, 1-6). У центрических диатомей с круглыми
створками множество осей, являющихся диаметром створки. У пеннатных диато-
мей различают три плоскости симметрии: продольную (апикальную), поперечную
7
(трансапикальную) и створковую (вальварную), которым соответствуют три основные
взаимно перпендикулярные оси симметрии: продольная, поперечная и центральная
(первальварная). Центральная ось перпендикулярна к поверхности обеих створок пан-
циря и проходит через центры. Исключение составляет асимметричный панцирь рода
Rhoicosphenia, который не имеет ни одной плоскости симметрии, так как его гетеро-
польный панцирь изогнут по всем трем плоскостям симметрии (Диатомовый анализ,
1949, кн. 1 : 13, 14, рис. 4, 5). Панцири бывают изопольными, гетеропольными и дорси-
вентральными. У гетеропольного панциря широкий конец называется головным,
узкий — базальным, а симметричен он по продольной и створковой осям (род
Gomphonema). Дорсивентральный панцирь изогнут по продольной оси, поэтому имеет
поперечную и створковую оси симметрии, и несимметричен по продольной оси
(роды Amphora, Cymbella). Выпуклая часть такого панциря и створки называется спин-
ной, а вогнутая или плоская — брюшнрй.
Створка — одна из двух противоположных дистальных пластинок панциря, кото-
рая может быть плоской, вогнутой, выпуклой, тангенциально-, радиально- или концентри-
чески-волнистой (табл. 2,1-8). Створки-бывают различных размеров и разнообразной
формы: круглые, эллиптические, ланцетные, ромбические, линейные, прямоугольные,
многоугольные, четырехугольные, треугольные, волнистые, овальные, гитаровидные, сиг-
мовидные, серповидные, ладьевидные, булавовидные, клиновидные и пр. (табл. 1, 7—16).
Центрические диатомеи характеризуются обычно актиноморфными створками, объеди-
няющими формы с круглыми, треугольными и многоугольными очертаниями и с ра-
диальной структурой, пеннатные — большей частью удлиненными створками разно-
образных очертаний с различным отношением длины к ширине створки и, как правило,
симметрично-перистой структурой (табл. 1,12, 14). Помимо того имеются комбиниро-
ванные варианты: продолговато-эллиптические, продолговато-, полу-, линейно-ланцет-
ные, удлиненно-эллиптические, удлиненно-яйцевидные, удлиненно-клиновидные и пр.
Концы створок также разнообразны: тупые, острые, остро, тупо, широко закруг-
ленные, тупые, клиновидные, остро- и тупоклювовидные, головчатые и т. д. (табл. 2,
12-19).
Стенка створок пронизана ареолами, порами и трубковидными полыми выроста-
ми, в расположении которых наблюдаются характерные особенности. Через ареолы
осуществляется обмен протопласта с окружающей средой. Помимо ареол на створках
имеются и другие перфорации: глазки, ложные узелки, ложные глазки, стигмы.
Лицевая часть ствгорки— часть створки, окруженная загибом и снаб-
женная различными структурными элементами, имеющими разное строение на наруж-
ной и внутренней ее поверхностях.
Загиб створки — краевая часть створки, отличающаяся наклоном или
отогнутостью, а иногда и структурой. Загиб бывает низким или высоким, иногда
образует большую часть боковой поверхности панциря (род Aulacosira — табл. 2, 9—11).
Поля. Гиалиновые поля — участки базального кремнеземного слоя, не пер-
форированного ареолами. Они имеют различные форму, величину и расположение,
наиболее часты следующие их варианты.
Осевое поле проходит по апикальной оси створки, изредка сдвинуто экс-
центрически (виды рода Achnanthes). У большинства навикулоидных диатомей на нем
расположен шов (табл. 3,1, 5), а на бесшовных створках в родах Cocconeis, Achnanthes
и др. осевое поле часто утолщено (табл. 3, 2). Известны редкие случаи, когда это поле
имеет специализированную структуру (Navicula schirschowii Sechkina и др.) . Осевое поле
присуще всем пеннатным видам, за редким исключением (табл. 3,4).
Среднее поле — расширение осевого поля на его середине. У диатомей с утол-
щенным центральным узелком средним полем называют неутолщенный гиалиновый учас-
ток вокруг центрального узелка. В случае если среднее поле вытянуто поперек до краев
створки, оно называется фасцией (Achnanthes brevipes Ag. и др. — табл. 3,3). Среднее
поле иногда может быть односторонним за счет не развития штрихов на одной половине
створки или только слабо асимметричным (Navicula cincta (Ehr.) Ralfs, N. eidrigiana
8
Carter и др.). В последнем случае Картер (Carter, 1979) предлагает ту сторону
створки, на которой центральное поле более расширено, называть „с т а ф ф”,
а противоположную — „д и с т а ф” (табл. 3,5). Данные о размере и форме сред-
него, а иногда и осевого поля используются как диагностические признаки на видо-
вом уровне.
Конечные поля — расширения осевого поля на концах створки, у шовных
пеннатных диатомей на них расположены конечные узелки (табл. 3, б).
Центральное поле — более или менее четко выраженная централь-
ная гиалиновая часть створки центрических диатомей (некоторые виды родов
Coscinodiscus, Actinocyclus, Actinoptychus и др.). У них нередки также расположенные
радиально поля — гиалиновые лучи.
Боковые поля — участки, прерывающие поперечные штрихи и представляю-
щие апикально направленные отроги среднего поля (табл. 3,7, 8, 10). Ряды обычно
коротких штрихов, симметрично располагающихся по обе стороны вдоль шва между
боковыми и осевым полем, Росс (Ross, 1972) называет продольными лентами. По
имеющимся немногочисленным электронно-микроскопическим данным, боковые поля
обычно перфорированы порами (табл. 3,77). Помимо пор эти поля могут иметь
и другую структуру (Lyrella hennedyi (W. Sm.) Kar., L. sandriana (Grun.) Kar. и др.),
характер которой неясен, так как пока она не изучена электронно-микроскопически.
Боковые поля являются характерной особенностью рода Lyrella, встречаются они и у еди-
ничных видов других родов: Mastogloia, Achnanthes, Navicula. У видов рода Amphora они
развиваются иногда лишь на спинной стороне и часто не соединены со средним полем
(табл. 3, 10). Структура, размеры и форма боковых полей — важный диагностический
признак на видовом уровне.
Подковообразное поле — гиалиновый участок, сдвинутый к краю
створки, при этом на внутренней поверхности створки ближний к шву край поля
утолщен вплоть до образования кремнеземной капотообразной структуры, торчащей
над поверхностью створки. Известно с достоверностью у Achnanthes rostrata 0strup
(табл. 3, 72, 13). В светооптических исследованиях с трудом отличается от углубления
на этом же участке створки, именуемом синусом (Moss, Carter, 1982), которое уста-
новлено для A. lanceolata (ВгёЪ.) Grun.
Конопеум — вытянутый бесструктурный кремнеземный лоскут, расположен-
ный поверх кремнеземного слоя и обычно прикрепленный к нему только одним своим
краем вдоль осевого поля. Край, противоположный прикрепленному, обычно не дости-
гает загиба, свободен, между ним и базальным слоем различается щель, расширяю-
щаяся на концах. Структура стенки створки под конопеумом отличается от структуры
остальной части ее (табл. 3, 9). Конопеум известен у некоторых видов рода
Lyrella, у которых он прикрывает боковые поля, а также отмечен для Navicula bacillum
Ehr., Nitzshia spathulata Breb., по-видимому развит у Navicula americana Ehr., N. pelagica Cl.
(Poulin, Cardinal, 1982).
Элементы створки. Выпуклость — приподнятая часть створки, не выдаю-
щаяся латерально за ее край (роды Biddulphia, Hemiaulus, Odontella, Briggera и др.). Вы-
пуклость может иметь ту же структуру, что и створка (табл. 9,72), или особую струк-
туру. Частный, случай — рог — длинная узкая выпуклость, которая оканчивается пря-
мой или изогнутой вершиной (род Hemiaulus и др. — табл. 9,13).
Щетинка — полый, обычно очень длинный вырост, выступающий далеко за пре-
делы края створки и имеющий иную структуру, нежели створки (табл. 4, 7-6). У ко-
нечных клеток колонии они называются конечными щетинками. Расстояние между
створками смежных клеток в колонии называется окном, или апертурой (роды
Chaetoceros, Eucampia — табл. 4,2, 7).
Складка — внутренняя складка стенки створки (Hemiaulus capitatus Grev.).
Специализированная складка у видов рода Aulacosira называется бороздой.
Шип- сплошное кремнеземное образование, выступающее обычно из наружной
поверхности створки. Они бывают различной величины, но обычно остроконечные,
9
иногда с гранями у основания, расположенные кольцом по краю или по всей поверхно-
сти створки (табл. 5,7-5).
Соединительный шип — шип, который, взаимно сцепляясь с шипом
соседнего панциря, объединяет панцири в цепочки. Они бывают различных размеров
и форм: остроконечные, тупые, скошенные, расширенные, головчатые, выемчатые,
разветвленные, лапчатые и пр. Эти образования присущи как центрическим, так и пен-
натным диатомеям (табл. 5,6—18).
Ш и п и к — маленький шип, часто многочисленные образования, покрывающие
поверхность створки (табл. 4,13,14).
Гранула — маленький округлый кремнеземный выступ на поверхности створки
(табл. 4,75).
Ребро — вытянутое в радиальном, апикальном или трансапикальном направле-
нии утолщение базального кремнеземного слоя створки. Ребра разнообразны по
строению и расположению. У пеннатных диатомей очень широко распространены
поперечные ребра (табл. 6, 7, 2), редко продольные (табл. 6,5), у центрических —
радиальные. Специализированными ребрами являются следующие их варианты.
Псевдосепта — ребро или кремнеземная складка створки, вдающаяся
в полость панциря в виде короткой перегородки. Псевдосепта, отходящая от краев
загиба створки, расположена в створковой плоскости (некоторые виды родов
Gomphonema, Stauroneis, Navicula и в роде Rhoicosphenia — табл. 6,8, 9). У центрических
диатомей (роды Biddulphia, Anaulus, Hemiaulus), а также в роде Rhopalodia псевдосепты
расположены на внутренней поверхности створки перпендикулярно к ней и часто
трактовались как „внутренние ребра” (табл. 6,10).
Осевые ребра — элементы, между которыми заключена щель шва (табл. 6,
5-5). Они развиваются в большей или меньшей степени у всех шовных диатомей.
Как правило, осевые ребра расположены на внутренней, редко на наружной стороне
створки. Еще реже осевые ребра развиваются на обеих поверхностях, что особенно
характерно для группы Undulatae рода Mastogloia и для рода Brachysira.
Кольцевидная диафрагма — кремнеземная перегородка, высту-
пающая внутрь створки у края загиба параллельно плоскости створки (род
Aulacosira).
Краевой гребень — ребро, расположенное обычно на границе поверхности
лицевой части створки и загиба, повторяющее ее очертания и имеющее различное
строение и структуру. Так, в роде Lithodesmium гребень перфорирован мелкими отвер-
стиями, тогда как у Ditylum brightwellii (West) Grun. он сплошной у молодых клеток,
а у зрелых свободный край его рассечен на петли (табл. 6, 6). Структуру, ранее
рассматриваемую Россом и др. (1983) как воротничок (табл. 6, 7), мы считаем воз-
можным трактовать также как краевой гребень.
Гребешок — высокое тонкое ребро, бахромчатое или зубчатое по свободному
краю, расположенное на наружной поверхности створки (Qdontotropis cristata Grun.).
Ушко (отариум) — каждое из пары коротких мембрановидных ребер, распо-
ложенных друг против друга у основания двугубого выроста или вблизи него
(род Rhizosolenia).
Поясок. Поясок (цинктура) — часть панциря, расположенная между эпи-
и гиповальвой, он состоит из двух поясковых ободков: э п и- и гипоцингулюма.
Каждый из них одним краем соединен с соответствующей створкой, образуя эпи-
и гипотеку.
Поясковый ободок (цингулюм) — часть пояска, обычно состоящая
из двух и более (до 30) единичных элементов — ободков, ближний к створке заходит
краями под ее загиб, следующий ободок — под край первого и т. д. Таким образом,
следует различать термины „поясок”, „поясковый ободок” и „ободок”, означающие
целое и его части. Имеющиеся в литературе указания о существовании видов, имеющих
поясковые ободки, состоящие лишь из одного ободка, повергались сомнению (Stosch,
1975). Форма и структура ободков,'их число в цингулюме, частота (величина для
Рис. 1. Схема строения пояскового ободка.
1 - створка; 2-5 - элементы пояскового ободка: 2 - вальвокопула, 3 - вставочный ободок
(копула) ,4, 5 - соединительные ободки (плевры).
каждого вида постоянная) отличаются большим разнообразием и являются важными
диагностическими признаками (рис. 1). Ободки в пределах одного пояскового ободка
могут быть одинаковыми (табл. 7, 1-3), но очень часто отличаются по форме или
.структуре (табл. 7,4—7), менее важно их отличие по ширине.
В соответствии с особенностями строения ободки в пределах одного пояскового
ободка различаются терминологически. В настоящей работе изменен объем понятий
„вальвокопула”, „поясковый ободок”, „вставочные и соединительные ободки” сравни-
тельно с прежними светооптическими представлениями и уточнена их трактовка
(Росси др., 1983).
Вальвокопула — вставочный ободок, прилегающий непосредственно
к створке, если он по строению или структуре отличается от следующего за ним.
В противном случае этот ободок терминологически не выделяется. Вальвокопула
отмечается в панцире у видов родов Cocconeis, Grammatophcга, Berkeleya rutilans
(Trentep.) Grun. (табл. 7, 4—11) и мн. др. На вальвокопуле у отдельных видов могут
развиваться фимбрии или септы (см. ниже). Если все последующие за вальвокопулой
ободки (или все ободки панциря, когда нет оснований выделять вальвокопулу) неоди-
наковы по строению, то среди них различают вставочные и соединительные ободки.
Вставочными ободками (копулами) именуют элементы (один или
несколько), сходные по строению и структуре и расположенные проксимально
к створке. Соединительным (плеврой) называется следующий за вставоч-
ными, отличный от них ободок, расположенный дистально к створке. У некоторых
видов таких ободков два или больше, но, так как они одинаковы морфологически,
нет смысла разделять их терминологически.
11
Если все ободки в панцире по структуре одинаковы, то их следует трактовать
как соединительные (или плевры), хотя иногда они несколько отличаются по ширине.
Одинаковые ободки развиваются у Berkeleya scopulorum (Breb. et Kiitz.) Сох, у некото-
рых видов Fragilaria, многих видов рода Navicula, у большинства, а возможно, и у всех
представителей секции Halamphora Cl. рода Amphora (табл. 7,2, 5) и др.
Поясок может состоять из цельных или двучленистых ободков, реже он состоит
из сегментов. Цельные ободки чаще всего бывают открытыми, редко замкнуты-
ми. Последние, имеющие вид кольца, известны у Epithemia, Hantzschia amphioxys (Ehr.)
Grun., у некоторых популяций Н. marina (Donk.) Grun., у представителей рода Cocconeis,
у Biddulphiopsis titiana (Grun.) Stosch et Simonsen (Mann, 1982; Stosch, Simonsen, 1984),
наконец замкнуты лишь отдельные ободки в пояске Rhabdonema arcuatum (Lyngb.) Kiitz.
(Pocock, Cox, 1982). Открытые цельные ободки имеют вид кремнеземных
лент (табл. 7, 12, 17), длина их немного меньше периметра створки, и меэвду концами
их, обычно суживающимися, остается щель. Открытые концы таких ободков у центри-
ческих диатомей с цилиндрическим панцирем сдвинуты относительно друг друга,
а у пеннатных всегда располагаются на концах панциря (табл. 7, 13), альтер-
нативно у соседних ободков. Открытые цельные ободки отмечены в родах Anaulus,
Melosira, у видов секции Halamphora Cl. рода Amphora (табл. 7, 12), в роде
Navicula.
Двучленистыми называются ободки, состоящие из двух частей, одна
из которых по длине превосходит половину периметра створки. Они отмечаются
нечасто, таковы ободки многих видов рода Cymbella (Krammer, 1981) или отдельные
ободки в панцире Rhabdonema arcuatum (Lyngb.) Kiitz. (Pocock, Cox, 1982), Coscinodiscus
granii Gough. Двучленистость ободков трудноразличима в СМ, возможно, что они встре-
чаются значительно чаще. Отмечены немногочисленные случаи, когда ободки обра-
зованы почти равными по длине фрагментами, это полукольцевидные ободки
(Rhizosolenia).
Поясковый ободок, образованный многочисленными сегментами — кремнезем-
ными пластинками, обычно более или менее сходными в пределах одного панциря
по величине и форме, называется сегментированным. Ни один из сегментов
не превосходит по длине половину периметра створки. Сегменты имеют форму корот-
ких лент, ромбов, чешуек, они черепитчато налагаются друг на друга (роды
Lithodesmium, Rhizosolenia, Ditylum - табл. 7, 14-16). Принадлежность отдельных сег-
ментов к вальвокопуле или плевре установить очень трудно.
Во всех случаях, когда ободки не являются цельным кольцом, зазоры, остаю-
щиеся между концами ободка или краями сегментов, всегда прикрываются снизу
нижележащим ободком, глубоко подстилающим предыдущий (табл. 16, 77), или же
лигулой — наиболее широкой частью следующего ободка, имеющей форму выступа
(табл. 7,77—79). В интактном панцире лигула всегда направлена к створке. Встречаются
виды, у которых ободки кроме лигулы имеют еще и антилигулы - противоположный
выступ, направленный от створки. У пеннатных диатомей лигула всегда расположена
в пояске близ одного из концов панциря и поэтому в интактном панцире особенно трудно-
различима даже в ЭМ. Ободки с лигулой встречены у большого числа диатомей, близких
или далеких в родственном отношении (роды Fragilaria, Rhoicosphenia, Coscinodiscus,
Thalassiosira, Pinnularia и др.), антилигула наряду с лигулой наблюдается у некоторых
видов родов Rhizosolenia, Thalassiosira, а у Cyclotella striata (Kutz.) Grun. отмечается
только антилигула (Helmcke et al., 1974).
Структура элементов пояска чаще всего отличается от структуры лицевой части
створки. Обычно она состоит из пор, реже ареол, расположенных рядами. У единичных
видов один или все ободки имеют неперфорированную складку, проходящую вдоль
одного края ободка, которая накладывается поверх ареолированной их части, прикры-
вая ее на всем протяжении или частично. Обычно элементы поясковых ободков
у большинства диатомовых водорослей однослойные, но в роде Undatella (Paddock, Sims,
1980) они полые, трубчатые.
12
Фимбрии — короткие выросты вальвокопулы в направлении створковой
плоскости (Holmes et al., 1982) (табл. 7, 8, 9). Они расположены более или менее регу-
лярно по всей окружности или лишь вдоль средней трети ободка. Фимбрии наиболее
обычны в роде Cocconeis на вальвокопуле у бесшовной теки, а у С. scutellum Ehr. разви-
ваются и на вальвокопуле у шовной теки, известны они и у Navicula mutica Kutz. (Phipps,
Rosowsky, 1983).
У некоторых диатомей вставочные ободки несут дополнительные образования —
септы. Септа — это однослойная тонкая кремнеземная пластинка различной формы,
выступающая в створковой плоскости из вставочного ободка (табл. 7, 10, 11}. У цент-
рических диатомей септы редки, известны пока только в роде Thalassiosira* они имеют
вид неширокой диафрагмы, окаймляющей ободки по одному или обоим краям
(Syvertsen, 1977; Fryxell et al., 1981). У пеннатных они широко распространены в различ-
ных родах, всегда имеют отверстие (Rhabdonema arcuatum (Lyngb.) Kiitz., Grammato-
phora — табл. 7,11} или же располагаются полярно (роды Licmophora, Striatella, некото-
рые виды Rhabdonema и др. — табл. 7, 20}. Более сложной формы септы отмечаются
в роде Epithemia, у видов Climacosphenia (Round, 1982). Полярные септы в гетерополь-
ных панцирях располагаются на широком конце ободков (табл. 7, 20}, в изопольных
же на обоих концах (роды Striatella, Tabellaria, некоторые виды Rhabdonema и др.).
Число септ в панцире, их ширина, расположение и форма варьируют у разных
видов и родов и являются диагностическим признаком на видовом и родовом
уровнях.
Ободок особой формы, несущий два или более партектумов, называется
партектальным кольцом и характерен только для рода Mastogloia (табл. 16,
12, 13}. Партектумы представляют собой отсеки, которые с помощью партек-
тальных каналов сообщаются с внешней средой. Партектумы в каждом кольце могут
быть равных или разных размеров. Наружные отверстия партектальных каналов
хорошо видны в ЭМ на поясковой стороне панциря в виде квадратных, овальных или
треугольных отверстий (табл. 16, 13}. Поверхность партектумов пороидная (табл. 16,
72). Они осуществляют какую-то еще не выясненную функцию, возможно выде-
лительную.
Поясок, важный в таксономическом отношении элемент панциря, еще недоста-
точно изучен; терминология составляющих его частей разработана неудовлетвори-
тельно и неполно и нуждается в ревизии на основании дальнейшего электронно-микро-
скопического изучения. Особенности строения пояска имеют большое значение, так как
именно за счет него объем жесткого кремнеземного панциря может увеличиваться
в период жизни клетки. Происходит это как вследствие увеличения числа слагающих
его элементов, так и в результате расхождения элементов, ранее глубоко накладываю-
щихся друг на друга, уменьшения области их наложения, вплоть до соединения встык.
Перфорации створки. Пора — сквозное, открытое с обоих концов отверстие
в базальном кремнеземном слое. Поры разделяются по форме на поровые каналы,
которые иногда в литературе называются межштриховыми (Stosch, Simonsen, 1984)
или простыми (Li, Volcani, 1985) порами, а по местоположению — на крибральные поры
и пореллы глазка.
Поровый канал - поры в толстом базальном кремнеземном слое, открытые
на наружной и внутренней поверхностях округлым отверстием (роды Corona, Lepidodis-
cus, Rhizosolenia, Actinoptychus, Gyrodiscus и др. — табл. 8, 7). Они располагаются на
створке беспорядочно (табл. 8, 2), образуют радиальные ряды (табл. 8, 3} или ряды,
сгруппированные в пучки (табл. 8, 4}. Часто поровые каналы являются дополнитель-
ными перфорациями между пороидными или пекулярными ареолами (роды
Trigonium, Eucampia, Isthmia и др. - табл. 8,5). В СМ поровые каналы видны в виде мел-
ких точек.
Крибральные поры — отверстия тонкого слоя кремнезема на крибруме
(см. ниже).
Пореллы глазка — отверстия в пластинке глазка (см. ниже).
13
Ареола — регулярно повторяющаяся перфорация базального кремнеземного
слоя, обычно замкнутая велумом, рикой или тем и другим. По своему строению ареолы
разделяются на пороидные и локулярные.
Пороидная ареола, или п о р о и д — проход цилиндрической или
слабо конусовидной формы, пронизывающий базальный кремнеземный слой и закры-
тый на наружной или внутренней поверхности велумом различного типа. Велум обычно
расположен на суженной части прохода. Его диаметр меньше диаметра прохода, кото-
рый никогда не расширяется в толще базального кремнеземного слоя, или равен
ему (роды Biddulphia, Briggera, Odontella, Sheshukovia, Trinacria и др. — табл. 8,6). Отвер-
стие, противоположное велуму, называется фораменом. Проход пороидной ареолы
гладкий или с волой (см. ниже) .
В о л а — радиальные, различной формы выросты боковых стенок ареолы (табл. 8, 7).
У некоторых видов к воле прикрепляется крибрум (роды Stictodiscus, Epithemia, Isthmia
и др. — табл. 9, 7) или тонкая мембрана без видимых перфораций (род Arachnoidiscus).
Вола может быть от слабо развитой, в виде небольших бугорков на стенках ареол,
до сильно развитых выростов с разветвленными несрастающимися концами.
У центрических диатомовых водорослей ареолы располагаются на створке беспо-
рядочно, в радиальных рядах, иногда собранных в пучки, а у пеннатных обычно в парал-
лельных поперечных рядах. Иногда пороидные ареолы находятся только на отдельных
участках створки, например на ее загибе, в то время как остальная часть створки
имеет локулярные ареолы.
Локулярная ареола, или л о к у л а - камера в толще базального крем-
неземного слоя, закрытого на наружной или внутренней поверхности велумом различ-
ного типа. Диаметр камеры всегда больше диаметра форамена (роды Pyxidicula, Thalas-
siosira, Hemidiscus, Asteromphalus, Coscinodiscus и др. - табл. 8, &). В отличие от пороид-
ной ареолы локула всегда расширена в толще створки. Локулярные ареолы различа-
ются по форме<>по положению велума и его строения.1
Форма локул определяется строением их стенок и хорошо видна на поперечных
сломах створок. Правильные полигональные локулы (роды Pyxidicula, Triceratium, Cos-
cinodiscus, Thalassiosira и др.) имеют вид пчелиных сот, образованы тонкими вертикаль-
ными стенками одинаковой длины (табл. 8,9). Неправильная многоугольная форма
локул (род Melosird) зависит от различной длины наружных перегородок (табл. 8,10).
Цилиндрическая форма локул, суженная к обеим поверхностям, характерна для родов
Podosira, Hyalodiscus и др. (табл. 8, 77). Локулы, расширенные различным образом
в толще створки (роды Cyclotella, Aulacodiscus и др.), образованы толстыми стенками
ареол (табл. 8,12). Иногда локулы закрыты на одной или обеих поверхностях неперфо-
рированным слоем кремнезема (роды Hyalodiscus, Actinocyclus и др. — табл. 8, 13),
и тогда камеры являются полузамкнутыми или замкнутыми в толще кремнезем-
ного слоя.
Велум — тонкий перфорированный слой кремнезема, закрывающий ареолу на
наружной (роды Stellarima, Actinocyclus, Coscinodiscus и др. — табл. 9, 7) или внутрен-
ней (ceM.Pyxidiculaceae, Thalassiosiraceae и некоторые другие - табл. 9,2) поверхности.
Перфорации велума обычно не менее 30 нм в поперечнике и часто не округлые.
Имеются следующие типы велума.
Крибрум — сетчатый велум или перфорированный регулярно расположен-
ными крибральными порами. Различают сплошной и прерывистый крибрум. Сплошной
1 В литературе (Ross, Sims, 1972; Предложения..., 1977; Росси др., 1983) использовался тер-
мин „псевдолокула”, под которой понимали камеру, образованную расширением дистальных частей
анастомозирующих или сетчато переплетенных ребер на наружной или внутренней поверхности
створки. Последующие исследования (Николаев, 19846) показали, что псевдолокула соответствует
локулярной ареоле с велумом, расположенным на внутренней поверхности створки. Анастомози-
рующие ребра являются дополнительными элементами структуры, которые отделяют ареолы, или
небольшие их группы, или группы поровых каналов друг от друга (табл. 9, 9, 10). Поверхность
между анастомозирующими ребрами обычно вогнутая.
14
крибрум не прерван в зоне межареолярных перегородок (виды родов Pyxidicula, Trice-
ratium, Pontodiscus и др.). Крибральные пэры располагаются радиальными рядами
(табл. 9. 5) или беспорядочно (табл. 9, 4). Прерывистый крибрум имеет округлую
или полигональную форму (роды Thalassiosira, Pyxidicula, Trigonium, Coscinodiscus и др.),
а между крибрумами соседних ареол находится гладкая бесструктурная зона базаль-
ного слоя кремнезема (табл. 9,2). Крибральные поры располагаются прямолинейными
или концентрическими рядами. У некоторых видов родов Triceratium, Coscinodiscus
и др. они закрыты дополнительной мембраной — крибеллой (табл. 9,5).
Рота— велум, состоящий из перекладины, пересекающей ареолу, или из несколь-
ких радиальных перекладин (роды Podosira, Trigonium и др.), в обоих случаях с расши-
рением в центре или без него (табл. 9,6).
Р и к а — очень тонкий перфорированный слой кремнезема, замыкающий ареолу;
перфорации обычно округлые и не более 15 нм в диам., часто перекрестно расположен-
ные. Рика встречается обычно у пеннатных диатомей (Росс и др., 1983).
Альвеола — удлиненная камера или одна из серии камер, проходящих от осе-
вого поля или центральной части створки к ее краю и открывающаяся внутрь панциря
большим отверстием, а с наружной поверхности закрытая тонким перфорированным
слоем (роды Cyclotella и Pinnularid). Наружный слой обычно перфорирован поровыми
каналами. Большое отверстие альвеолы образовано расширением дистальных частей
ребер на внутренней поверхности створки (табл. 9,8).
Ложный глазок — небольшой участок створки со сгруппированными арео-
лами, размер которых обычно меньше размера ареол на основной части створки (виды
родов Terpsinoe, Trigonium и др. - табл. 16, 8).
Глазок — пластинка кремнезема обычно с утолщенным бесструктур-
ным краем, пронизанная плотно расположенными порами — пореллами (роды
Triceratium, Auliscus, Odontella, Zygoceros и др. — табл. 16, 7). Глазки различаются по их
положению на створке, строению пластинки и характеру бесструктурного края. Глазок
может находиться на хорошо развитой (роды Odontella, Triceratium и др. — табл. 16, 7)
или слабо развитой (род Auliscus - табл. 16, 2) выпуклости. Пластинка глазка обычно
округлой формы, плоская, вогнутая или выпуклая. У центрических диатомей пореллы
образуют радиальные ряды или концентрические круги (роды Odontella, Auliscus и др. —
табл. 16,5) или расположены беспорядочно Zygoceros и др. — табл. 16, 4). Иногда
на поверхности пластинки имеются дополнительные параллельные ребра (табл. 16,5).
У пеннатных диатомей (роды Synedra, Fragilaria и др.) пореллы расположены чаще
в прямых продольных рядах (табл. 16, 6), а иногда разделены продольным бесструк-
турным ребром (род Entopyla), являющимся продолжением осевого поля (табл. 16, 7).
Бесструктурный край глазка гладкий, иногда состоит из нескольких концентрических
ребер (табл. 16,5).
Ложный узелок — краевое или прикраевое образование, обычно одно на
створку. Иногда он ареолирован (Actinocyclus ingens Rattray — табл. 16, 9), иногда
с крышечкой, иногда блестящий (Pontodiscus miocenicus (Crasske) Moiss. et Sheshuk. —
табл. 9,4; Actinocyclus actinochilus (Ehr.) Simonsen - табл. 16,10).
Стигма — расположенная на среднем поле особая перфорация базального слоя,
отличающаяся от ареолы отсутствием велума или рики и незамкнутая на наружной
поверхности створки (табл. 17, 7, 2). Вследствие очень слабой изученности этим терми-
ном пока обозначаются элементы довольно различного строения. Так, у Navicula mutica
Kutz. (табл. 17,5, 4) стигма на внутренней поверхности створки прикрыта кремнезем-
ной тонкой пластинкой (как бы крышечкой) и очень напоминает по строению двугубый
макровырост у Coscinodiscus granii. У представителей рода Cymbella (подрод Cymbella)
стигма имеет неравномерно бахромчатые края. Пока недостаточно изучено строение
стигмы в роде Proschkinia, где внутреннее отверстие замкнуто сферическими кремне-
земными образованиями (табл. 17, 6), а иногда может быть и без них (Караева, 1978).
Одиночная стигма очень характерна для некоторых видов секции Punctatae Cl. рода
Navicula, встречается также и у других представителей этого рода, а также в родах
15
Gomphonema, Neidium, Caloneis, Climaconeis и т. д. В роде Cymbella стигмы обычно раз-
виваются на брюшной стороне створки в числе от одной до нескольких (табл. 17,2, 5).
Стигма встречается только у навикулоидных диатомей. Наличие стигмы является важ-
ным диагностическим признаком при определении вида. Стигму следует отличать от
стигмой да.
Стигмоид в отличие от стигмы, будучи частью штриха, по внутреннему строе-
нию полностью повторяет ареолу, на наружной же поверхности отверстие его не замк-
нуто, как и у стигмы (Krammer, 1982). Этот элемент встречается в роде Cymbella, пред-
ставляя проксимальный конец центрального штриха на спинной стороне.
Буллула — пузырьковидная пустота в базальном кремнеземном слое, располо-
женная среди полностью изолированных ареол (род Aulacodiscus, Cyclotella и др. —
табл. 9, 77).
Гипокауст — непрерывающееся пространство в базальном кремнеземном
слое между полностью изолированными ареолами с тонкими стенками.
Продольные каналы — трубкообразные полости в стенке створки,
простирающиеся на большей части ее длины, иногда прерванные в центре. Границы кана-
лов при светооптических исследованиях трактовались как „продольные линии”. Про-
дольные каналы расположены у края створки (виды родов Dyctioneis и Neidium) или
вдоль шва {Scoliotropis и Diploneis - табл. 10, 7, 2) (Sims, Paddock, 1979). У видов
рода Diploneis стенка канала на внутренней поверхности створки вздута и иногда от-
крывается наружу и внутрь панциря порами (табл. 10,3,4), каналы сообщаются также
проходами с прилегающими ареолами. У отдельных видов рода Neidium, например
N. iridis (Ehr.) Cl., каналов несколько и они также соединяются проходами друг с дру-
гом. Канал, близкий к осевому полю, имеет больший диаметр, чем остальные.
Штрихи. Штрих — это ряд ареол или альвеол (реже 2—3 ряда ареол) или же оди-
ночная альвеола, если она не является частью ряда. Соответственно этому штрихи назы-
вают 1-3- или многорядными (табл. 10, 5-7). У единичных видов штрихи в полном
смысле слова отсутствуют, а вдоль края' створки проходит лишь один ряд ареол
{Thalassionema nitzschioides Grun. — табл. 10, 8). Участки базального кремнеземного
слоя между штрихами называются интерштрихами. Если интерштрихи утолщены на
внутренней поверхности створки, то они являются ребрами (табл. 10, 9). Очень часто
штрихи просто углублены в базальный кремнеземный слой створки {Navicula linckei
Hust.,7V. yarrensis Grun., виды Pinnularia — табл. 10, 10), в таких случаях интерштрихи
между ними следует отличать от ребер, что с уверенностью можно сделать лишь с по-
мощью СЭМ. Деление штрихов на гладкие, линеолированные, точечные, основанное на
светооптических исследованиях, не очень надежно и при использовании ЭМ не всегда
подтверждается. Поэтому при описании ультраструктуры указывают обычно формулу
ареол, из которых состоят штрихи. Расположение и направление штрихов на створке
у каждого вида постоянно, так же как и частота их. У центрических диатомей с круглы-
ми створками штрихи обычно располагаются радиально. Большинство пеннатных диа-
томей характеризуется штрихами, расположенными по обе стороны от осевого поля
симметрично. Лишь у единичных родов {Eunotia, Actinella и др.) штрихи несиммет-
ричны по отношению к осевому полю.
Штрихи пеннатных диатомей различаются по направлению их на створке.
Параллельные штрихи располагаются перпендикулярно апикальной оси створки
или шву {Navicula plicata Donk., N. normalis Hust.) .Радиально-симметричные
направлены от края створки к центру {N. hasta Pant., N. plasty stoma Ehr., N. lanceolata
(Ag.)Kiitz. и др. - табл. 10, 5-7). Как правило, направление штрихов на протяжении
длины створки чаще всего неодинаково. У многих диатомовых штрихи могут быть
параллельными или радиальными только на середине створки, у концов же меняют
направление (табл. 10, 6, 9). Конвергентными бывают штрихи обычно на
концах створки, они направлены от края створки к ее концу {N. tripunctata (Mill.)
Bory, N. gottlandica Grun., N. costulata Grun. и др.). Значительно реже
ареолы на створках образуют поперечные и диагональные взаимопересекающиеся
16
ряды (табл. 10,77), что характерно для рода Pleurosigma, некоторых Navi-
cula и др.
Указанная систематизация направления штрихов обычно разрабатывается для
навикулоидных диатомей, у которых шов расположен посередине створки апикально.
Менее систематизировано направление штрихов у каналовидных диатомей, возможно
в связи с тем, что у них этот признак не имеет такой большой таксономической ценно-
сти, как у навикулоидных видов.
Дефект Вуа - нарушение в упорядоченном расположении штрихов, разви-
вающееся только по одну сторону от осевого поля и на равном расстоянии от концов
створки (табл. 10, 6, 9). В панцире у большинства видов или целиком родов это нару-
шение проявляется чаще всего по одну сторону от апикальной плоскости, реже по раз-
ные стороны (Mann, 1983). Дефект Вуа возникает в процессе морфогенеза в результате
неодновременного отложения кремнезема. Именно в области дефекта завершается
отложение кремнезема вокруг шовной щели (Chiappino, Volcani, 1977). Это нарушение
характерно для многих видов рода Navicula (особенно для секции Lineolatae), встреча-
ется также на створках представителей родов Rhoicosphenia, Cymbella, Mastogloia и др.,
у которых иногда развивается лишь на одном из концов и с одной стороны (табл. 10,72).
Выросты. Двугубый вырост (лабиатный) — специализированная
трубка, пронизывающая стенку створки насквозь (табл. 11, 1—3). Как установлено
электронно-микроскопически, морфология его сильно варьирует. Внутренняя часть
выроста у всех диатомей короткая, не превышает 2 мкм, разнообразна по строению.
Она имеет вид прямой или изогнутой щели, чаще окруженной двумя кремнеземными
утолщениями — „губами” (роды Odontella, Detonula, Asteromphalus, Actinoptychus, Aula-
codiscus, многие пеннатные - табл. 11,7), или вид короткой воронкообразной сплющен-
ной трубки (роды Roperia, Actinocyclus, некоторые Thalassiosira, Coscinodiscus —
табл. 11,4; 12, 8, 9, 7 7, 18). Наружная часть выроста в виде трубки, более или менее
выдающейся над поверхностью створки (иногда до 25 мкм), наблюдается в основном
у центрических диатомей — у большинства представителей родов Thalassiosira, Trinacria,
в родах Odontella, Lithodesmium, Ditylum (табл. 11,5, 6).
У многих центрических видов на наружной поверхности створки трубка не разви-
вается и вырост заканчивается лишь округлым или щелевидным отверстием с утолще-
нием по краю или без него, что наблюдается у видов родов Melosira, Coscinodiscus, Stella-
rima, Bacteriastrum (табл. 11, 7, 8, 13), а также у большинства пеннатных видов. Обычно
выросты у каждого вида одинаковы по форме и размерам. Но в группе видов рода
Coscinodiscus (С wailesii Gran et Angst, C. jonesianus (Grev.) Ostf., C. concinnus W. Sm.,
C. granii, C. asteromphalus Ehr.) на створке выделяются крупными размерами и особым
строением два макровыроста ( в СМ — „отростки”) (табл. 11, 9-72). В роде
Pyxidiculaua наружной поверхности створки трубки имеют различную длину или совсем
не развиты (Николаев, 1984). Отметим также, что в этом роде отдельные выросты
могут располагаться в центре ареолы, пронизывая велум (табл. 1 1,14-16) (Макарова,
Моисеева, 1986). У представителей сем. Lithodesmiaceae (Stosch, 1977; Джафарова, 1984)
известен двойной двугубый вырост, который отличается строением лишь
внутренней части выроста, представляющий собой бугорок, открывающийся в полость
панциря двумя щелями (табл. 12, 7).
Двугубые выросты имеются на створке у центрических и пеннатных диатомей,
как современных, так и ископаемых. У центрических они не найдены у единичных
представителей многих родов и у некоторых родов в целом (Coretron, Pyxilla и др.).
У пеннатных этот элемент известен лишь у бесшовных и у большинства представителей
с примитивным швом (роды Eunotia, Peronia, Actinella). Не найдены они пока, по свиде-
тельству Симонсена (Simonsen, 1979) и других авторов, в родах Tetracyclus, Plagiogram-
та, Sceptroneis, Trachysphenia, Dimerogramma, Climacosphenia
Число и расположение двугубых выростов на створке у центрических диатомей
чрезвычайно разнообразны. Они могут быть многочисленными, одиночными, парными
(табл. 12,2-9).
2 Заказ 518
17
В настоящее время выделяется три типа локализации двугубых выростов на
створке: 1) неопределенно расположенное большое число, 2) краевое кольцо (иногда
их два), 3) центральная группа. По В. А. Николаеву (1984), эти типы дают возможность
характеризовать крупные таксоны. Отметим, что между тремя типами имеются и пере-
ходные варианты, возникшие за счет редукции части выростов.
Многочисленные выросты, расположенные беспорядочно, в основном наблюда-
ются у древних видов, реже у современных. Иногда выросты упорядочены лишь
частично, чаще всего по краю створки, где образуют кольцо (Coscinodiscus asteromphalus
Ehr., С. perforates Ehr., Druridgea compressa (West) Donk., некоторые виды Podosira, Hyalo-
discus и др.). В других случаях выросты локализованы только в виде краевого кольца
и один в центре (Azpeitia nodulifera (A. S.) Fryx. et Sims, A. elegantula (Grev.) Sims).
Краевым кольцом располагаются двугубые выросты у многих диатомовых из
родов Actinocyclus, Brightwellia, Par alia, Pontodiscus и др. (табл. 12,5). Нечасто на створ-
ках находится один (многие виды родов Thalassiosira, Dactyliosolen, Detonula, Porosira,
Cymatotheca) или два прикраевых выроста. В последнем случае у видов с круглыми
створками они могут быть краевыми противостоящими (Eupodiscus oculatus Qxqn., Tha-
lassiosira elsayedii Fryx., T. symmetrica Fryx. et Hasle, Planctoniella sol (Wall.) Schutt
(табл. 12,4) (Fryxell, 1975; Simonsen, 1979, и др.). Ко второму типу расположения относит-
ся и вариант, наблюдаемый у центрических видов, имеющих ланцетную и 3-4-угольную
форму створки (некоторые виды из родов Pseudo trie era tium, Trigonium, Trinacria), у ко-
торых выросты тяготеют к полюсам. Именно эта тенденция нашла развитие у пеннатных
диатомей.
Центральная группа выростов отмечена у видов родов Eunotogramma, Stictodiscus,
у некоторых видов Hemiaulus,Stellarima и др. (табл. 12, 6, 7). Производным является
вариант, когда в центре развивается лишь один вырост, как это отмечается для некото-
рых видов родов Hemiaulus, Cerataulina, Anaulus и др.
Наконец, у нескольких родов имеется единичный вырост, расположенный на
различном расстоянии от края створок (Eucampia, некоторые Thalassiosira и т. д. —
табл. 12,£).
У многих диатомей расположение краевых двугубых выростов может быть опре-
деленным образом связано с другими элементами створки. Так, у Eupodiscus oculatus
Grev. два противоположных выроста всегда расположены по концам диаметра, перпен-
дикулярно линии, соединяющей глазки. У большинства видов Thalassiosira двугубый вы-
рост располагается в кольце выростов с опорами (табл. 12,9). В роде Actinocyclus и у не-
которых Coscinodiscus (С. curvatulus Grun., С. crenulatus Grun.) двугубые выросты часто
располагаются на конце самых длинных рядов ареол в пучках. В родах Asteromphalus,
Asterolampra, Discodiscus и др. выросты расположены на дистальных концах камер.
Направление щели двугубого выроста обычно характеризуется определенным
постоянством. У диатомей с круглыми створками преобладает радиальное направление
(многие Thalassiosira, Porosira и др.), реже тангентальное (Actinocyclus, некоторые
Thalassiosira, Roperia tesellata (Roper) Grun., Azpeitia nodulifera и др. — табл. 12, 8, 9).
У колониальных видов двугубый вырост может развиваться лишь в определенных
клетках цепочки. Так, у Chaetoceros didymus Ehr., Bacteriastrum furcatum Shadbolt
(Fryxell, 1978) он отмечен на конечных створках колонии и отсутствует на всехинтер-
калярных. Лишь на одной створке панциря развивается двугубый вырост у видов
родов Odontotropis, Pseudostictodiscus, Arachnoidiscus.
Двугубые выросты у пеннатных диатомей чаще всего немногочисленны. Один
вырост на створке характерен для родов Meridion, Tabellaria, Hustedtiella, некоторых
видов Licmophora, Diatoma и др. (табл. 12,10, 77). Два выроста встречены у некоторых
представителей родов Synedra, Diatoma, Grammatophora (табл. 12,12-14), Actinella и др.
Только в сем. Protoraphidaceae отмечены две группы выростов (Hasle, 1974).
В большинстве случаев двугубые выросты или их группы у пеннатных локализу-
ются близ окончания осевого поля, а щель их ориентирована под углом к нему. В родах
Tabellaria и Hustedtiella вырост расположен эксцентрически. Если на створке имеются
два двугубых выроста, то они, как установил Симонсен (Simonsen, 1979), распо-
лагаются на противоположных сторонах осевого поля диагонально-симметрично
(Raphoneis, Delphineis, Grammatophora — табл. 12,14) или по одну сторону осевого поля,
т. е. зеркально-симметрично (Gephyria, Asterionella, Podocystis, Striatella, Synedra -
табл. 12, 12, 13). Число и характер симметрии двугубых выростов у пеннатных диато-
мей (Simonsen, 1979) постоянны на родовом уровне, но в литературе приводятся
и отдельные противоречащие данные (Jurily, Jercovic, 1973; Караева, Джафарова, 1982).
Наблюдается изменение числа выростов и ориентации их щели под влиянием условий
культуры и солености среды (Grammatophora marina (Lyngb.) Kiitz., Coscinodiscus
granii — табл. 12,15).
Периплектон — вырост, у которого наружная часть вверху разделена на две
ветви, охватывающие ствол подобного выроста смежной створки (Rutilaria radiata Grove
et Sturt).
Кольцевидный (многопоровый) вырост — трубка, проходящая
сквозь кремнеземный слой; на наружной поверхности створки она короткая, слегка
конусовидной формы, на внутренней расширенная на свободном конце, с многочислен-
ными порами, пронизывающими ее основание. Кольцевидный вырост расположен
в центре или близ центра створки (Thalassiosiropsis wittiana (Pant.) Hasle — табл. 12, 16).
Вырост с опорами, или подпертый вырост — трубка, открытая
с обоих концов и проходящая через стенку створки (табл. 13, 9); наружный конец
трубки сильно или слегка выступает над наружной поверхностью створки или заканчи-
вается круглым, иногда очень маленьким отверстием, не выступающим над поверхно-
стью створки, а внутренний слегка или сильно выдается за пределы внутренней поверх-
ности створки и окружен 2—5 сопутствующими порами (поры сателлиты), разделен-
ными поддерживающими опорами. У некоторых видов рода Thalassiosira (Т. angulata
(Greg.) Hasle, Т. aestivalis Gran) в наружную часть трубки входит другая трубочка, мень-
шая по диаметру (табл. 13,5). У многих видов Thalassiosira, Detonula, Lauderia и особенно
Sceletonema наружные трубки выростов с опорами в виде вытянутого цилиндра или
желоба значительной длины, до 5 мкм, иногда расширенного на верхушке, или окру-
женного воронкой (Thalassiosira nordenskioeldii Cl., Thalassiosira sp.), или вздутого у осно-
вания (Т. bulbosa Syvertsen — табл. 13,7, 2, 4-7). У части родов (Porosira, Bacterosira,
Planctoniella, Stephanodiscus) и некоторых видов Thalassiosira трубка с наружной и внут-
ренней поверхностей створки небольшой длины (табл. 13, 3) или наружные трубки
отсутствуют (Т. confusa Makar., Т. ignota Makar., Т. oliveriana (О. Meara) Makar, et Niko-
laev, T marujamica Sheshuk. emend. Makar, и др., а также представители родов Cyclostepha-
nos, Cyclotella — табл. 13, 8\ 14, 77).
Внутренняя часть выростов с опорами характеризуется различным строением
и размерами. Виды, не обладающие трубкой, выступающей на наружной поверхности,
обычно имеют трубку на внутренней поверхности створки значительной длины
(Thalassiosira confusa, Т. ignota, Т. latimarginata Makar., Т frenquelli KozL, T. oestrupii
(Ostf.) Hasle и др. — табл. 13,77, 72). У других Thalassiosira внутренняя часть трубки
мало выступает над поверхностью (Т. constricta Gaarder, Т. lineata Jouse, Т. bramaputrae
(Ehr.) Hakansson et Locker, T. nordenskioeldii и др. — табл. 13, 10). Известны виды, у ко-
торых выросты с опорами очень маленькие (Т. leptopus (Grun.) Fryx. et Hasle, T. nativa
Sheshuk., T. orientalis Sheshuk. emend. Makar., виды Cyclotella)^ незаметны в CM. Виды
и роды характеризуются различным числом сопутствующих пор. Виды рода
Thalassiosira имеют от 3 до 5, чаще 4 сопутствующие поры, Porosira — 3, Cyclotella - 2—3
(табл. 13,76-27). У некоторых видов Thalassiosira, имеющих длинную внутреннюю
трубку (Т. frenquelli, Т. poroseriata (Ramsfjell) Hasle, Г. latimarginata), опоры, разделяю-
щие сопутствующие поры, продолжаются на трубку выроста, в результате чего она
приобретает трехгранную форму (табл. 13, 72).
Выросты с опорами - многочисленная структура на створке, за исключением рода
Minidiscus, Thalassiosira conferta Hasle, T. profunda (Hendey) Hasle, имеющих всего несколь-
ко выростов. Располагаются они обычно кольцом на границе лицевой части створки
19
с загибом (сем. Thalassiosiraceae) или на загибе (сем. Stephanodiscaceae и некоторые виды
рода Thalassiosira — табл. 14, 7—5). У некоторых видов известно два краевых кольца
(Т. oliveriana, Т. antarctica Comber) или помимо краевого кольца на створке имеется еще
одно (Г. diporocyclus Hasle) или несколько колец отступя к центру створки (Т. subtilis
(Ostf.)Gran — табл. 14, 6, 8, 9). Иногда выросты с опорами располагаются по всей
поверхности створки (Т. eccentrica (Ehr.) Cl., Т. symmetrica Fryx. et Hasle, T. gravida CL,
T. rotula Meunier, T. mendiolana Hasle et Heimdal, роды Porosira, Lauderia — табл. 14,
12-17). При этом если ареолы расположены в тангенциальных рядах, то выросты
с опорами находятся между ними, а если в радиальных рядах, то выросты занимают
место одной из ареол (Thalassiosira lentiginosa (Jan.) Fryx.). Иногда они расположены
в краевой зоне в шахматном порядке (Т. baltica (Grun.) Ostf., Т. gravida, Т. mendiolana -
табл. 14, 7, 10). Помимо краевого кольца выросты с опорами присутствуют в центре
створки или близ него. У одних таксонов в центре один вырост с опорами (табл. 15,
1—3), у других их более одного, и они могут образовывать один или два ряда, или груп-
пу (виды родов Thalassiosira, Cyclotella — табл. 15, 4-12), или центральное замкнутое
кольцо (Thalassiosira plicata Schrader, Т. weissflogii (Grun.) Fryx. et Hasle — табл. 15,13-15),
или несомкнутое кольцо (Т. anguste-lineata (A. S.) Fryx. et Hasle — табл. 15,16), или дугу
(некоторые виды Cyclotella), иногда многочисленные центральные выросты с опорами
образуют пучок (Thalassiosira rotula, Т. gravida, Т. hyalina (Grun.) Gran, род Bacterosira —
табл. 15,77,18).
Краевые и центральные выросты с опорами могут быть одинаковыми или разными
по строению. Те и другие имеют или длинную наружную трубку (Thalassiosira norden-
skioeldii, Т. pacifica Gran et Angst, T. aestivalis, T. allenii Takano, T nidulus (Temp, et Brun)
Jouse, panDetonula — табл. 13, 7, 2, 4, 5), или трубку, не выступающую над наружной
поверхностью створки, а простирающуюся внутрь (Thalassiosira confusa, Т. ignota, Т. oest-
rupii, Т. poroseriata, Т. plicata, Т. oliveriana — табл. 13, 77, 12, 15). У многих видов крае-
вые выросты с опорами имеют наружную часть трубки значительной длины, а централь-
ные выросты заканчиваются только отверстием на наружной поверхности створки
(Т. coronifera (Pr.-Lavr.) Pr.-Lavr., Т. angulata, Т. baltica, Т. decipiens (Grun.) J0rg. и др. —
табл. 14, 7). Некоторые представители (роды Sceletonema, Porosira) не имеют централь-
ных выростов с опорами. Выросты с опорами свойственны только порядку Thalassio-
sirales, включающему 3 семейства: Thalassiosiraceae (9 родов), Stephanodiscaceae
(5 родов) и Ectodyctionaceae (1 род).
Замкнутый вырост — полая трубка, сильно выступающая над наружной
поверхностью створки, имеющая отверстие на свободном конце и замкнутая на другом
(т. е. трубка не пронизывает стенку створки насквозь). Этот тип выростов свойствен
ограниченной группе диатомовых водорослей: некоторым видам рода Thalassiosira
(Т. australis М. Per., Т. punctigera (Castr.) Hasle, Т antarctica, Т. marujamica, Т. lundiana
Fryx. и др. — табл. 4,8-12), а также Lauderia annulata Cl. Располагаются они или на краю
створки (T.marujamica) или между двумя кольцами выростов с опорами (Т. antarctica)
или образуют редкое кольцо внутрь от кольца выростов с опорами (Т. punctigera, Т. tu-
bifera Fryx., Т. bramaputrae NM.bramaputrae - табл. 4, 77, 12). Направлены они обычно
вертикально и служат для соединения соседних клеток в колонию.
Шов. Куникулюс — структура, состоящая снаружи из простой поры на цент-
ральном поле между проксимальными концами ветвей шва и соединяющаяся с удли-
ненным возвышенным участком, пронизанным щелью, которая заканчивает внутрен-
нюю щель на центральном поле (Navicula delognei V. Н.) (Росс и др., 1983).
Шов— элемент структуры, развивающийся на одной или обеих створках, разно-
образного, иногда довольно сложного строения. Все многообразие строения шва можно
свести к двум основным типам: щелевидному и каналовидному.
Щелевидный шов представляет собой удлиненную щель, проходящую
через стенку (табл. 3, 7). Щель может быть прямой и пересекать створку перпендику-
лярно или под каким-либо углом (Rouxia californica М. Perag., Achnanthes pusilia Grun.)
(Krammer, 1981) или складчатой, тогда она коленчато изогнута поперечно (Mastogloia,
20
Stauroneis, Navicula, Cymbella). В складчатой щели различаются внутренняя и наружная
части, направление которых, а следовательно, и проекция часто не совпадают. У видов,
имеющих складчатую щель, по обе стороны от нее стенка створки в поперечном сечении
несимметрична: с одной стороны она имеет вид паза, с другой — входящего в него
клина. Обычно осевые ребра, между которыми заключена щель шва, параллельны,
но иногда они могут быть наклонены друг к другу и образуют канал, более или менее
закрытый на всем протяжении створки (род Proschkinia) (Караева, 1978). В роде
Amphora одно или значительно реже оба наружных осевых ребра образуют уступ, чаще
всего отгибающийся под прямым углом и называемый осевой складкой
(табл. 18, 7) (Караева, 1975).
Хорошо развитый шов характеризуется наличием центрального узелка между его
двумя ветвями, а на их концах — конечных узелков. Центральный узелок —
утолщение базального кремнеземного слоя между двумя ветвями шва, к узелку при-
мыкают две центральные поры, представляющие собой расширение щелей
ветвей шва (табл. 18, 2, 5). поверх центрального узелка или в нем расположены
не пронизывающие насквозь створку продолжения щелей шва — центральные
щели. Особо крупный узелок, на внутренней поверхности ограниченный с двух сторон
скобковидными кремнеземными утолщениями, имеет место у видов Pleurosigma, Gyro-
sigma (табл. 18,5) и, по-видимому, в роде Plagiotropis.
Ставрос — центральный узелок, который расширен трансапикально и достигает
краев створки или приближен к ним. У исследованных в СЭМ видов, имеющих ставрос,
среднее поле расширено в виде фасции. Ставрос является характерной особен-
ностью рода Stauroneis, изредка наблюдается он также у других щелешовных
(Amphora staurophora (Castr.) Cl., Л. ostrearia Breb. — табл. 18,4, 5). Встречающийся очень
редко высокий и узкий ставрос, иногда с отрогами, называется тигиллятным,
а боковые отроги его тигиллумом (Capartogramma crucicula (Grun.) Ross.) (Росс
и др., 1983). Псевдоставрос (Hendey, 1964) отличается от ставроса тем, что является
утолщением поперечных ребер на внутренней поверхности створки и не сопровождается
фасцией на наружной (круг форм Haslea crucigera (W. Sm.) Simons.).
Конечные узелки и конечные щели по строению аналогичны
центральным. Особенно длинные конечные щели развиваются у некоторых
Pleurosigma, Diploneis, у большого числа видов рода Mastogloia они переходят и на загиб
створки (табл. 16, 72). У многих диатомей шов на внутренней поверхности заканчи-
вается хоботком (хеликтоглоссой) — губообразно вытянутым образова-
нием (роды Pleurosigma, Gyrosigma, Caloneis amphisbaeena Cl., многие виды родов
Navicula, Cymbella) или кремнеземным утолщением (табл. 18, 6, 7).
У группы видов, имеющих слабо развитый шов, он отличается короткими
щелями, заметно (роды Peronia, Krasskiella, Rouxia), а иногда и очень сильно удаленны-
ми друг от друга, причем в последнем случае отсутствует центральный узелок
(сем. Eunotiaceae).
Щелевидный шов характерен для большого числа пеннатных диатомей, чаще всего
он расположен посередине створки апикально, значительно реже (Eunotiaceae) сдвинут
к краю ее. Наличие шва, тип его строения служит важным признаком для различения
таксонов высокого ранга.
Каналовидный шов представляет собой трубку, проходящую в стенке
створки и более или менее выступающую на ее внутренней поверхности. На наружной
поверхности этой трубки расположена щель, с помощью которой она сообщается
с внешней средой (табл. 18, 8, 9). На внутренней поверхности створки щель чаще всего
сопровождается слабо развитыми ребрами и иногда имеет узелки, что можно видеть
через отверстия в стенке канала (табл. 18, 10, 77). Эти отверстия, с помощью которых
каналовидный шов соединяется с полостью панциря, называются интерфибулами
(в СМ — килевые точки). Кремнеземные образования между интерфибулами, по суще-
ству участки стенки трубки, называются фибулами ив случаях частичной редукции
этой стенки иногда имеют вид мостиков (табл. 18,77).
21
Каналовидный шов располагается посередине створки апикально (Bacillaria, Gom-
photeca, очень редко в роде Nitzschia), или в различной степени сдвинут от средней
линии, или, наконец, иногда расположен вокруг створки по спирали (Cylindrotheca). Чаще
всего каналовидный шов расположен на киле или крыле. Килем называется участок
створки, на котором стенка ее резко изогнута под углом (табл. 18, 12). У каналошов-
ных диатомей, как правило, киль расположен на границе перехода лицевой части створ-
ки в загиб, этот загнутый под углом участок створки бывает более или менее оттянут,
и по вершине этого киля проходит каналовидный шов. Наличие киля с каналовидным
швом в нем является важной диагностической особенностью родов пеннатных диато-
мей, особенно част он у диатомовых водорослей, относящихся к сем. Nitzschiaceae и от-
дельным видам сем. SurireUaceae. Как установлено с помощью СЭМ, киль, по-видимому,
имеют и отдельные виды, относимые к роду Entomoneis Ehr. (Paddock, Sims, 1981). Пока
известен только один род (Simonsenia), который имеет канал, расположенный на киле,
но не имеет фибул. У диатомей с сильно оттянутым килем следует отличать фибулы
как участки стенки каналовидного шва от иных образований, не имеющих отношения
к системе шва. К ним Пэддок и Симс (Paddock, Sims, 1981) относят „вторичные”,
или „килевые” фибулы, расположенные в киле, но вне канала, а также
„случайные” фибулы. Последние более крупные, расположены менее пра-
вильно, имеют различную форму и не столь регулярны. Их происхождение у форм
с латерально сжатой створкой объясняется возникновением анастомозов. Согласно
Россу и др. (1983), киль называется крылом, если в нем чередуются окна (участки
слияния лицевой части створки и загиба) и каналы крыла (проходы между полостью
панциря и каналом шва). Крыло со швом окружает створку (табл. 17, 7-1Г, 18, 13),
при этом шов имеет лишь два узелка, одинаковых или немного отличающихся,
причем в последнем случае невозможно различить центральный, как это можно
видеть на табл. 17, 12, 13. Строение киля и крыла еще не вполне хорошо иссле-
довано в ЭМ.
ОСНОВНЫЕ ПРИНЦИПЫ КЛАССИФИКАЦИИ
ДИАТОМОВЫХ ВОДОРОСЛЕЙ
Система диатомовых водорослей разрабатывается исследователями разных стран
около 150 лет. Подробный обзор по этому вопросу сделан И. В. Макаровой (1986).
В данном очерке мы кратко остановимся лишь на отдельных принципах, положенных
в основу новейших классификаций диатомовых водорослей, и на характеристике
системы, предлагаемой в настоящем издании редколлегией.
Классификация Шютта (Schutt, 1896), являющаяся наиболее полным обобщением
работ по систематике диатомовых водорослей XIX в., широко использовалась диатомо-
логами более 80 лет. Дополнения и изменения, внесенные в нее различными исследова-
телями (Oestrup, 1910; Hustedt, 1927—1930; Karsten, 1928;Hendey, 1937,1964; Диатомо-
вый анализ, 1949—1950; Топачевский, 1962; Основы..., 1963; Шешукова-Порецкая,
1967, и др.), не затрагивали ее основ.
В настоящее время благодаря изучению панциря диатомей в электронном микро-
скопе выяснилось, что ряд признаков, используемых Шюттом для выделения родов,
семейств, порядков, нередко носит формальный характер и не отражает родства
таксонов.
В последние годы предприняты попытки разработки новой филогенетической
классификации для центрических диатомовых водорослей (Ross, Sims, 1973; Simonsen,
1979; Глезер, 1981; Николаев, 1984а). Хотя многие таксоны Шютта вошли в новые
классификации, но их характеристика и объем часто существенно иные. Значи-
тельные расхождения имеются и между системами указанных выше авторов,
поскольку они основываются на разной оценке таксономической значимости
признаков.
22
Росс и Симс (Ross, Sims, 1973) для обоснования своей системы впервые
использовали тонкое строение ареол, к важным таксономическим признакам, так же
как и позднее Симонсен, отнесли наличие и характер расположения двугубых выростов
и выростов с опорами, присутствие глазков и ложных глазков.
Симонсен (Simonsen, 1979) строит свою классификацию (наиболее полную из всех
предложенных) с учетом признаков, которые он считает у диатомей примитивными
(образование покоящихся спор, сильное развитие створок по первальварной оси, коло-
ниальный образ жизни, разное строение створок в одном панцире, один центральный
или почти центральный вырост на створке). Однако другие исследователи (Ross, Sims,
1973; Беклемишев, Семина, 1982) не подтверждают примитивного характера этих
признаков.
3. И. Глезер (1981, 1986а) предложена классификация центрических диатомей,
во многом принципиально отличающаяся от систем Симонсена, Росса и Симс. Ею
(Глезер, 1975а, 19756, 1978, 1979, 1981, 1983, 1986а, 19866) рассмотрено таксо-
номическое значение целого ряда признаков в свете исторического развития диатомей.
Установлено, что каждому таксону свойственно сочетание признаков, унаследованных
от предков, и признаков, специфических для данного таксона (мозаичность признаков
по Майру (1971, 1974) и Раупу и Стэнли (1974) или эволюционная разновозрастность
признаков по А. Л. Тахтаджяну (1954)). Больше унаследованных признаков сохраня-
ется у наиболее древних представителей диатомей. Наличие общих, широко распростра-
ненных признаков (например, строение ареол и характер расположения их на створке)
обычно свидетельствует о родстве таксонов высокого ранга, у подчиненных же,
более мелких таксонов эти признаки имеют свое специфическое выражение. Сходство
признаков часто обусловлено не единством происхождения, а параллелизмом в разви-
тии или конвергенцией. Отдельные признаки (форма панциря и створок, выпуклости
на створках, глазки и ложные глазки, соединительные шипы и др.) могли неоднократно
возникать в процессе эволюции в различных ветвях филогенетического древа. При
выделении таксонов высокого ранга они имеют меньшее значение, чем широко
распространенные признаки.
В. А. Николаев (1984а) в основу своей классификации центрических диатомей
положил иерархию признаков, выявленных им при детальном изучении панцирей совре-
менных и вымерших диатомей при помощи СЭМ. Особенно большое внимание он
уделяет таксономическому значению перфораций панциря, к которым относит ареолы,
двугубые выросты, выросты с опорами и др. Принципиально новой является высокая
оценка такого признака, как положение велума у ареол. Этот признак наряду с формой
и полярностью панциря используется им для выделения порядков. К ведущим призна-
кам для выделения семейств относятся тип ареол, положение двугубых выростов
на створке и отчасти форма панциря. Для установления родов важными признаками
являются форма створок, положение и число двугубых выростов, строение ареол
и расположение их на створке.
В предлагаемой в настоящем издании классификации диатомовых водорослей
использованы новейшие достижения советских и зарубежных исследователей по морфо-
логии, биологии, экологии, биогеографии и закономерностям эволюционного развития
диатомовых водорослей. Сравнительно-морфологическому анализу подвергалась сово-
купность признаков, характерных для того или иного таксона. Для установления гене-
тических связей таксонов выявлялись причины их сходства: родство, конвергенция,
параллелизм развития. Учитывалось, что отдельные приспособительные признаки могли
неоднократно возникать в процессе эволюции в различных филумах, один и тот же
признак у таксонов разного ранга может иметь неодинаковое таксономическое значе-
ние. Для выявления более примитивных и эволюционно продвинутых таксонов прини-
малось во внимание явление эволюционной разновозрастности признаков у современ-
ных и древних их представителей.
При выделении порядков в первую очередь учитывались типы ареол (пекулярные,
пороидные, с велумом на наружной или внутренней поверхности створки), наличие
23
или отсутствие выростов с опорами, число и локализация двугубых выростов. Для
обоснования семейств принималось во внимание главным образом наличие специали-
зированных признаков, связанных с одиночным или колониальным, планктонным
или бентосным образом жизни (камеры, выпуклости створки, глазки, ложные
глазки, соединительные шипы, краевой гребень, периплектон, рельеф створки,
форма панциря и створок). К ведущим родовым признакам отнесены форма пан-
циря и створок, особенности расположения на них различных элементов и их мор-
фология.
Диатомовые водоросли рассматриваются нами в качестве самостоятельного отдела
Bacillariophyta, который делится на 2 класса: Centrophyceae и Pennatophyceae, как это
было предложено А. П. Жузе и В. С. Шешуковой-Порецкой (Основы..., 1963). Эти
классы отличаются помимо биологических и морфологических признаков особенностя-
ми эволюционного развития (Глезер, 1986а). Класс Centrophyceae развивался преиму-
щественно в конце мезозоя—кайнозое, а расцвет класса Pennatophyceae в основном
приходится на поздний кайнозой.
Класс Centrophyceae в зависимости от типа ареол (пороидные, локулярные,
с внутренним или наружным велумом), характера их расположения, числа и положения
двугубых выростов, наличия выростов с опорами, специальных приспособлений для
образования колоний или прикрепления клеток к субстрату, формы панциря и створок
предлагается делить на 7 порядков.
Порядок Pyxidiculales объединяет 3 семейства диатомей с пороидными и популяр-
ными ареолами, имеющими велум с внутренней и форамен с наружной стороны
створки. Наиболее простыми шаровидными, эллипсоидными, цилиндрическими панци-
рями с двугубыми выростами, расположенными по краю и всей поверхности створки,
характеризуется древнее семейство Pyxidiculaceae. Оно включает одиночные и коло-
ниальные планктонные формы, у последних соединение клеток в колонии осуществля-
ется при помощи наружных трубок двугубых выростов. Первые представители этого
семейства известны с поздней юры. Сем. Thalassiosiropsidaceae включает один планктон-
ный род, существовавший в раннем мелу—позднем палеоцене и характеризующийся
своеобразным кольцевидным выростом в центре створки и одним двугубым выростом.
Сравнительно молодое монотипное семейство Triceratiaceae отличается более сложной
призматической формой панциря с треугольными и полигональными створками и нали-
чием особых элементов панциря — выпуклостями с глазками на вершинах, участвую-
щими в соединении клеток в колонии и прикреплении их к субстрату. Двугубые
выросты находятся по краю створки и на ее углах. Первые представители этого
семейства известны с середины эоцена.
Обширный порядок Thalassiosirales, также с ареолами, имеющими велум на внут-
ренней поверхности створки, четко обособлен от порядка Pyxidiculales благодаря
наличию выростов с опорами. Функция их пока окончательно не выяснена, но широкое
распространение этого элемента структуры свидетельствует о его важном значении
в жизни клеток. Порядок включает 3 семейства. Сем. Thalassiosiraceae, более древнее,
объединяет преимущественно морские роды, их находки отмечаются главным образом
с неогена. Исключение представляют род Thalassiosira, известный с эоцена, и род
Skeletonema, единичные виды которого указываются для палеоцена. К сем. Stephanodis-
сасеае относятся пресноводные, относительно молодые роды, первые представители
этого семейства описаны с олигоцена. Сем. Ectodyctionaceae, не включенное в данный
раздел, будет охарактеризовано в следующем выпуске.
Другие порядки класса Centrophyceae характеризуются ареолами с велумом на
наружной и фораменом на внутренней поверхности.
В порядок Melosirales включены условно 3 семейства: Pseudopodosiraceae, Melosira-
сеае и Aulacosiraceae, их генетические связи неясны и требуют изучения. К общим
признакам относится цилиндрическая форма панциря преимущественно с круглыми
створками, сильно развитыми по первальварной оси, что вообще характерно для
колониальных форм. Для каждого из семейств характерны специфические особенности
24
расположения элементов структуры на лицевой части и загибе створки и своеобразные
приспособления к соединению клеток в колонии, строение и расположение перфораций
также особые у каждого из семейств.
Наиболее древним является сем. Pseudopodosiraceae, достоверно известное с конца
раннего—начала позднего мела. Его самые древние представители отличались уже высо-
кой специализацией и несомненно прошли длительный путь эволюции. Особенностью
родов этого семейства является дифференциация поверхности створки на „ассимили-
рующую” (загиб створки, на котором, как правило, расположены ареолы и имеется
кольцо двугубых выростов) и „соединительную” (лицевая часть створки с соедини-
тельными шипами, часто сложной конфигурации: с радиальными ребрами, бугорками).
Если ареолы и имеются на лицевой части створки, то обычно только в ее краевой части.
В образовании клеток в колонии участвуют кроме краевых шипов радиальные ребра,
благодаря чему образуются очень прочные, довольно плотные колонии — цепочки.
Высокая степень специализации в этом семействе подчеркивается также различным
строением панцирей в зависимости от положения клеток в колонии, что было установ-
лено у древнего вида Paralia sulcata Heib. (Crawford, 1979). Семейство объединяет пре-
имущественно морские виды.
В сем. Melosiraceae также имеется дифференциация в структуре и функции лицевой
части и загиба створки. Двугубые выросты, продуцирующие слизь подушечек, нахо-
дятся на уплощенной средней части створки, окруженной венцом соединительных
шипов или воротничком, также участвующих в образовании колоний. Остальная
поверхность створок покрыта более или менее одинаковыми ареолами. Это семейство
сравнительно молодое, находки его первых представителей относятся, вероятно,
к позднему олигоцену; виды рода Melosira преимущественно солоноватоводно-морские,
единичные пресноводные.
В отличие от двух других для сем. Aulacosiraceae характерно более или менее оди-
наковое расположение ареол как на лицевой части створки, так и на загибе створок.
Соединение клеток в цепочки осуществляется при помощи краевых шипов; двугубые
выросты образуют кольцо по краю загиба створки. Специфическим признаком является
кольцевидная диафрагма. Виды исключительно пресноводные, впервые отмечаются
начиная с олиго цена.
Порядок Coscinodiscales отличается от других порядков Centrophyceae наличием
у большинства представителей сплошной сети л окулярных ареол с велумом на наруж-
ной и фораменом на внутренней поверхности и краевым кольцом двугубых выростов.
Редко имеется один или несколько выростов в центре створки. Порядок объединяет
преимущественно свободноживущие одиночные формы с относительно низкоцилиндри-
ческими панцирями с круглыми створками. Деление на семейства основано на различ-
ных признаках, приспособительное значение которых не всегда ясно.
Сем. Hyalodiscaceae в отличие от других семейств характеризуется дифференциа-
цией структуры в центральной части створки, связанной с приспособлением к коло-
ниальному или прикрепленному образу жизни.
Монотипное семейство Stellarimaceae включает один из . наиболее древних мор-
ских планктонных родов, для которого характерно центральное расположение одного
или нескольких двугубых выростов своеобразного строения (Николаев, 1983). Оно
тесно примыкает к сем. Coscinodiscaceae, у которого двугубые выросты имеют
различное положение на створке: образуют кольцо по краю створки, или много-
численные выросты расположены по всей поверхности створки, а нередко
кроме обычных развиваются макровыросты. Структура, как правило, более
или менее одинаковая на всей поверхности створки, только у родов
Brightwellia и Craspedodiscus намечается небольшая дифференциация в строении и форме
ареол. Большинство представителей сем. Coscinodiscaceae морские, планктонные, их рас-
цвет приходится на палеогеновый и неогеновый периоды.
Сем. Trigoniumaceae включает единственный род Trigonium, который в противопо-
ложность большинству представителей порядка Coscinodiscales имеет призматический
25
панцирь, ложные глазки на вершинах выпуклостей и двугубые выросты в центре
створки. Виды этого рода бентические, колониальные, известны с эоцена.
Монотипное семейство Azpeitiaceae включает один морской род, виды которого
существуют с эоцена до настоящего времени. Для рода особенно характерно кроме
краевого кольца крупных двугубых выростов наличие одного слегка эксцентрично
расположенного центрального двугубого выроста.
Близко в морфологическом отношении к сем. Coscinodiscaceae стоит сем. Hemidis-
сасеае, отличительной особенностью которого является наличие ложного узелка и коль-
ца краевых двугубых выростов особого строения. Оно объединяет морские планктон-
ные и пресноводные роды, находки его первых представителей относятся к олигоцену.
Своеобразное семейство Asterolampraceae отличается развитием особых камер
в стенке створки, открывающихся внутрь панциря узкой щелью, а наружу — крупным
овальным отверстием. Двугубые выросты образуют по краю створки кольцо, часто
располагаясь у основания этих отверстий. Как правило, поверхность камер обычно
бесструктурная, типичные же для порядка Coscinodiscales локулярные ареолы образуют
сплошную сеть между камерами (Gombos, 1980). Отмечается внезапное появление
почти всех представителей этого семейства в среднем эоцене, в современном океаниче-
ском планктоне обитают виды только 2 родов: Asterolampra и Asteromphalus.
Характерной чертой сем. Eupodiscaceae является наличие глазков и кольца двугу-
бых выростов, расположенных по краю створки между глазками. Первые представи-
тели семейства известны с палеоцена.
В порядок Coscinodiscales включено и сем. Heliopeltaceae, характеризующееся часто
сложной радиальной и концентрической волнистостью створок и крупными краевыми
двугубыми выростами. Однако положение в системе этого семейства не совсем ясно
из-за недостаточной изученности тонкого строения ареол. Семейство включает
древние роды, известные с позднего мела и дожившие до настоящего времени.
Порядок Biddulphiales объединяет преимущественно колониальные формы, общей
чертой которых являются пороидные ареолы, немногочисленные двугубые выросты,
располагающиеся обычно в центре, реже у края створки, соединение клеток в колонии
осуществляется благодаря выпуклостям стенки створок, имеющих на своих вершинах
особые элементы в виде ложных глазков, глазков, соединительных шипов. Панцири
от дисковидных, чаще всего с эллиптическими створками, до призматических с тре-
угольными и полигональными створками. В зависимости от характера приспособления
к колониальному образу жизни или к прикреплению клеток к субстрату выделяется
8 семейств, и одно семейство (Stictodiscaceae) включает одиночные формы главным об-
разом с круглыми створками.
Сем. Biddulphiaceae объединяет роды, виды которых ведут колониальный образ
жизни. В качестве приспособлений для соединения клеток в колонии у них на полюсах
или углах створок имеются выпуклости с ложными глазками или глазками на верши-
нах, двугубые выросты обычно расположены в центре створки.
В близкое к нему семейство Hemiaulaceae включены древние роды, имеющие на
вершинах полярных выпуклостей (выростов — рогов) кроме глазков и ложных глаз-
ков еще и соединительные шипы, единичные двугубые выросты обычно в центре
или у края створки (Ross, Sims, 1985).
У вымерших эоценовых видов рода Kitt onia, выделенного в самостоятельное моно-
типное семейство Kittoniaceae, соединение клеток в колонии происходит при помощи
своеобразных трубковидных выростов. Они расположены не на полюсах створок,
как в других семействах порядка Biddulphiales, а отходят от средней части створок, их
боковая поверхность бесструктурная, на вершинах же, так же как у некоторых предста-
вителей Hemiaulaceae, имеются глазки и венец соединительных шипов.
Сем. Isthmiaceae характеризуется асимметричным строением створки благодаря
наличию выпуклости на одном из ее полюсов. Клетки соединяются в кустистые рыхлые
колонии при помощи слизистых подушечек на выпуклостях соседних клеток. Много-
26
численные двугубые выросты находятся у одного из полюсов створки. Виды рода
Isthmia известны с конца эоцена.
Для сем. Chaetoceraceae характерно наличие особых образований - щетинок со свое-
образной структурой, способствующих соединению клеток в рыхлые колонии — цепочки.
Более сложное соединение клеток в колонии наблюдается у остальных семейств
порядка Biddulphiales. В сем. Cymatosiraceae соединение клеток в колонии происходит
при помощи краевых шипов и глазков на вершинах полярных выпуклостей ство-
рок или особых соединительных волосков; в центральной части створки имеется один
двугубый или трубковидные выросты (Hasle et al., 1983). У представителей сем. Rutila-
riaceae оно осуществляется двояким образом — при помощи периплектона и краевых
зубчиков (Hasle, 1975). В образовании колоний у представителей сем. Lithodesmiaceae
принимает участие краевой гребень сложного строения; для этого семейства типичен
также один центральный двойной двугубый вырост с хорошо развитой наружной
трубкой.
Большая часть представителей порядка Biddulphiales древние, господствовавшие
во флорах позднего мела—палеогена; исключением является сем. Lithodesmiaceae, виды
которого неизвестны в ископаемом состоянии; сравнительно молодые роды составляют
и сем. Chaetoceraceae, начало развития которого приходится, по-видимому, на олигоцен.
Порядок Auliscales с одним семейством Auliscaceae включает преимущественно
морские вымершие роды, только виды рода Auliscus дожили до настоящего времени.
Характерными особенностями порядка являются цилиндрические панцири с круглыми
створками, наличие глазков, расположение 1—2 двугубых выростов по краю створки
между соседними глазками. Ареолы ичень мелкие, обычно в коротких радиальных
штрихах. В. А. Николаев обнаружил у одного из видов рода Auliscus, что перфорации
представляют собой поровые каналы.
Порядок Rhizosoleniales с одним семейством Rhizosoleniaceae объединяет роды,
виды которых имеют ярко выраженные приспособления к планктонному образу жизни
и образованию колоний. Удлинение панцирей по первальварной оси происходит за счет
многочисленных ободков, соединение клеток в колонии у рода Rhizosolenia осуществля-
ется, по данным Хасле (Hasle, 1975), при помощи отростка — наружной части двугубого
выроста, при этом отросток одной клетки входит в соответствующее углубление сосед-
ней клетки. У видов сем. Rhizosoleniaceae установлены локулярные ареолы с велумом
на наружной и фораменом на внутренней поверхности, редко поровые каналы. В это
семейство объединены род Rhizosolenia, известный с раннего палеоцена и доживший до
настоящего времени, род Riedelia, представители которого найдены в среднем эоцене,
и 2 современных морских рода.
В порядок Rhizosoleniales условно включено монотипное семейство Рух iliac еае.
Виды рода Pyxilla — типичные представители эоценовой флоры. По характеру соедине-
ния клеток в колонии они несколько напоминают некоторые виды Rhizosolenia, но име-
ют и ряд отличительных особенностей: створки у них палочковидные, сильно удлинен-
ные по первальварной оси, отсутствует отросток, в соединении клеток в колонии
принимают участие соединительные шипы, ареолы пороидные. Образования, открытые
Н. И. Стрельниковой и В. А. Николаевым, сходные с ободками пояска видов рода
Rhizosolenia, возможно, являются остатками пояска материнской клетки, которые
образуются у некоторых диатомей во время деления.
Пеннатные диатомовые водоросли рассматриваются как прогрессивно развиваю-
щаяся до настоящего времени группа, два его порядка представляют основные этапы
развития класса. Генеральная линия развития пеннатных шла по пути приспособления
их к подвижному образу жизни, возникновению шва различного типа строения.
Класс Pennatophyceae по наличию или отсутствию шва традиционно делится
на 2 порядка: Araphales и Raphales. Порядок Araphales может рассматриваться как про-
межуточный между классами Centrophyceae и Pennatophyceae, поскольку он совмещает
в себе черты обоих классов. Форма панциря и створок, билатеральная симметрия в рас-
положении штрихов, осевое поле у него характерные для пеннатных, но наряду с этим,
27
как и у центрических, многие виды имеют двугубые выросты, глазки, ложные глазки
и шипы, которые соединяют клетки в столь распространенные у бесшовных колонии.
В порядке Araphales, у представителей которого в отличие от остальных пеннатных
на осевом поле нет шва, выделено 5 семейств, основными критериями их разграниче-
ния являются симметрия створок, наличие или отсутствие таких признаков, как
изогнутость панциря, различия в строении створок одного панциря, развитие грубых
ребер на внутренней поверхности створок, развитие септ на соединительных ободках,
а также число двугубых выростов на полюсах. Основное ядро порядка составляют
3 близких семейства: Fragilariaceae, Diatomaceae и Tabellariaceae. В литературе отмечают-
ся разногласия относительно необходимости разделения этих семейств и положения
отдельных родов в них (Hendey, 1964; Simonsen, 1979).
Большое семейство Fragilariaceae характеризуется отсутствием грубых ребер на
внутренней поверхности створок и септ на соединительных ободках. Роды в семействе
различаются по симметрии створок, форме створки и осевого поля, развитию среднего
поля и т. д. У большинства исследованных в ЭМ родов отмечается наличие глазков или
ложных глазков, двугубые выросты не найдены лишь в единичных родах, у большин-
ства же их (1)2. В этом семействе особняком стоят 2 рода: Thalassionema и Thalassiothrix,
которые, по-видимому, заслуживают выделения в самостоятельное семейство по особо
грубой роте и отсутствию глазков, последнее Симонсен (Simonsen, 1979) считает
результатом редукции.
В сем. Diatomaceae включены лишь роды, отличающиеся наличием на внутренней
поверхности створки грубых ребер, по строению приближающихся к псевдосептам
и чередующихся с несколькими рядами штрихов, двугубых выростов у них 1(2),
поясок состоит из многочисленных соединительных ободков без септ. В пределах
семейства 2 рода, Diatoma и Meridion, очень близки и отличаются лишь гетеропольностью
створок у последнего. Род Plagiogramma стоит особняком в семействе, отличаясь отсут-
ствием двугубых выростов и наличием фасции, ограниченной на внутренней поверхно-
сти створки двумя псевдосептами.
Сем. Tabellariaceae характеризуется развитием септ на ободках. Роды в пределах
семейства отличаются по симметрии панциря, числу септ в панцире, альтернативности
их расположения. Двугубые выросты у большинства родов расположены апикально —
по одному на каждом конце, редко один субцентральный. Род Tetracyclus по наличию
грубых поперечных ребер сближается с сем. Diatomaceae. По одному субцентральному
выросту роды Tabellaria и Hustedtiella, по Симонсену (Simonsen, 1979), напоминают
Huttoniella из сем. Eupodiscaceae, что может указывать на полифилетическое развитие
бесшовных диатомей из последнего семейства.
Небольшие семейства Entopylaceae и Protoraphidaceae, возможно, представляют со-
бой боковые ветви. Первое, пока почти не исследованное в ЭМ, резко выделяется
в этом порядке изогнутостью панциря по трансапикальной оси и различиями в струк-
туре двух его створок. В сем. Protoraphidaceae, отличающемся осевым полем, S-образ-
ным у концов, двумя апикальными группами двугубых выростов, род Protoraphis стоит
наиболее близко по морфологии к указанному выше ядру порядка.
В порядке Raphales, объединяющем диатомеи, имеющие шов, выделено 11 семейств.
Критериями деления порядка являются такие признаки, как тип строения шва, развитие
его на одной или обеих створках, особенности симметрии панциря и створки, наконец,
наличие или отсутствие двугубых выростов, в целом для порядка нехарактерных.
Как известно, почти все виды Raphales подвижны, редкие случаи неподвижности, по-
видимому, вторичного происхождения. Колониальный образ жизни у Raphales довольно
редок.
Центральным в порядке является большое семейство Naviculaceae, включающее
и наиболее древние роды с щелевидным швом. Морфология представителей этого
семейства, характеризующихся в подавляющем их большинстве зеркальной симметрией
(случаи ее нарушения, по-видимому, вторичны) и щелевидным швом, расположенным
апикально, очень пестра, отличается большим разнообразием формы и структуры
28
створок, но при этом главным является постоянство симметричности панциря
и створок по апикальной и трансапикальной осям. Форма и детали строения щеле-
видного шва, центрального узелка, гиалиновых полей, положение шва, рельеф створки
и другие признаки имеют большое значение для разграничения родов семейства. Хотя
не все представители семейства исследованы электронно-микроскопически, но уже
сейчас ясны коренные различия в ультраструктуре отдельных родов. Здесь встречаются
роды, имеющие ареолы пороидные или пекулярные, реже альвеолы. В последние годы
на основании электронно-микроскопического исследования был пересмотрен объем
некоторых родов, и прежде всего наиболее обширного рода Navicula, из которого выде-
лены новые Haslea, Lyrella и восстановлены старые роды Climaconeis, Brachysira и др.
В семействе можно выделить группы близких по морфологии родов.
I группа объединяет роды Navicula, Lyrella, Anoeomoneis, Brachysira, Trachyneis.
Роды в этой группе отличаются по наличию и отсутствию боковых полей, конопеума,
продольных ребер и других элементов. Несмотря на выделение из рода Navicula ряда
мелких родов, все же он до сих пор еще остается гетерогенным. Об этом свидетель-
ствует, в частности, наличие в этом роде отдельных видов с альвеолярным строением
створки (Schoeman, Archibald, 1978), возможно являющихся переходными к III группе
по этому признаку.
Во II группе роды Gyrosigma, Pleurosigma, Donkinia объединяет общность сигмоидно
изогнутых шва и створки, строение центрального узелка, а у достаточно изученных
с помощью ЭМ первых двух родов еще и локулярное строение ареол. У некоторых
представителей этих родов осевые ребра изогнуты на протяжении части створки друг
к другу, образуя канал над щелью шва. Этот признак, а также неразвитое осевое поле
несколько сближают Pleurosigma и Gyrosigma с родом Proschkinia,^ строение центрально-
го узелка и ареол — с родом Haslea, ввиду чего последние два рода можно рассматривать
как связующие I и II группы.
Ill группа объединяет роды, имеющие альвеолы: Pinnularia, Dimidiata, Caloneis,
Oestrupia, резко отличающиеся в этом семействе от остальных и, возможно, заслуживаю-
щие выделения в том или ином объеме в самостоятельное семейство.
IV группа характеризуется наличием продольных каналов, расположенных вдоль
шва — роды Diploneis, Scoliotropis или вдоль края створки — роды Dictyoneis, Neidium.
V группа объединяет роды со слабо развитым швом, обычно имеющим короткие
ветви, дистальные концы которых удалены от концов створки, а проксимальные —
часто друг от друга. Это роды Rouxia, Amphipleura, Fnistulia, Berkeleya. У всех родов
в V группе ареолы замкнуты рикой, а шов завершается развитым хоботком. Эта группа,
возможно, близка к I, судя по сходству в строении шва ее представителей и рода
Brachysira.
Не все роды достаточно легко вписываются в указанные группы. Так, несколько
особняком в семействе стоят роды Mastogloia, близкий по структуре створки к 1 группе,
но характеризующийся специализированной вальвокопулой в виде партектального
кольца, и Stauroneis, отличающийся кроме ставроса также нередким развитием псевдо-
септы на концах створки и т. д. О многих родах до сих пор нет достаточных ультра-
структурных данных. Так как мы пока не можем с достаточной уверенностью судить
о гомологичности отдельных структур друг другу, по-видимому, следует воздержаться
от утвердительных заключений о степени родства групп между собой.
Сем. Achnanthaceae прошло регрессивный путь развития. Вторичность характерной
для него одношовности панциря подтверждена данными ряда авторов (Mann, 1984;
Boyle et al., 1984), наблюдавшими у видов Achnanthes и Cocconeis на начальных этапах
процесса морфогенеза заложение шовной щели на обеих створках с последующим
заполнением щели на одной из створок кремнеземом. Роды семейства отличаются
по характеру симметрии панциря и створок, изогнутости створки, развитию шва, нали-
чию грубых ребер.
Небольшое семейство Eunotiaceae характеризуется слабо развитым щелевидным
швом и в отличие от остальных шовных диатомей развитием двугубых выростов
29
(Hasle, 1973c) (исключение — род Semi or bis). Установлено, что Eunotiaceae следует рас-
сматривать как слепую эволюционную ветвь, независимую от Naviculaceae (Simonsen,
1979).
Два семейства: CymbeUaceae и Gomphonemataceae — отличаются от Naviculaceae ха-
рактером симметрии створок и панциря, строением шва и ультраструктурой. Имеются
виды, могущие рассматриваться как связующее звено между родами Cymbella и Navicula.
Таковы Cymbella pusilia (по ультраструктуре, наличию дополнительного кремне-
земного ребра, сопровождающего шов, и другим признакам близкий к секции
Lineolatae рода Navicula), С. angustata и С. amphicephala со слабо выраженной дорсивент-
ральностью створки (близкие по структуре группе видов из секции Punctatae того же
рода). Два рода, входящие в сем. CymbeUaceae, характеризующиеся симметричностью
створки лишь по поперечной оси, дифференцируются по различиям в строении ареол,
положению велума и ребер, наличию осевой складки в роде Amphora и по хорошо раз-
витым конечным щелям, наличию порового поля, стигм и стигмоидов у Cymbella. С дру-
гой стороны, С. sinuata рассматривается как близкий к роду Gomphonema (Krammer,
1982), возможно, он является переходным между семействами CymbeUaceae и Gompho-
nemataceae. Сем. Gomphonemataceae отличается клиновидным панцирем, асимметричной
по поперечной оси створкой. Роды в пределах семейства дифференцируются по наличию
или отсутствию продольных ребер, пересекающих штрихи, шипов, расположенных на
широком конце створки, и др.
Небольшой род Rhoicosphenia, у представителей которого створки одного панциря
отличаются по строению шва, обоснованно выделен в самостоятельное семейство
Rhoicospheniaceae Манном, считающим, что процесс морфогенеза створки с короткими
шовными веточками принципиально отличается от этого же процесса бесшовных ство-
рок (Mann, 1984). Манн рассматривает виды рода Rhoicosphenia как отличающиеся от
Gomphonema не только по деталям ауксоспорообразования, но и по строению терми-
нального поля пор и мембраны, замыкающей ареолы. Действительно, у Rhoicosphenia
ареолы замкнуты рикой, тогда как у Gomphonema велум имеет одно подковообразное
отверстие.
Сем. Entomoneidaceae отличается сильно развитым килем с щелевидным швом на
его вершине. Два рода его различаются по наличию и отсутствию соединительных обод-
ков и формой киля. У некоторых представителей семейства в киле обнаружены вторич-
ные фибулы. Факт обнаружения в роде видов с каналовидным швом пока не нашел
отражения в системе ввиду необходимости дополнительных исследований.
Роды с типичным каналовидным швом относятся к 4 семействам, различающимся
симметрией створки, развитием киля или его отсутствием, позицией шва и т. д. При
этом 2 семейства: Epithemiaceae и Auriculaceae — имеют основной общий признак —
дорсивентральное строение панциря, различаясь между собой позицией шва, наличием
или отсутствием киля, особенностями строения поясковых элементов. Следует
согласиться с Симонсеном (Simonsen, 1979) в том, что Epithemiaceae — слепая эволю-
ционная ветвь, развившаяся из Naviculaceae. Предполагается, что каналовидный шов
развился в этом семействе конвергентно со швом Nitzschiaceae. Представители
Rhopalodiaceae в отличие от Epithemiaceae обладают килем с проходящим в нем швом,
расположенным на границе спинной стороны створки и загиба. Роды в Auriculaceae диф-
ференцируются по развитию псевдосепт на створках, сочленяющихся с модифициро-
ванными соединительными ободками, или их отсутствию, характеру мембран, замыкаю-
щих ареолы. В сем. Nitzschiaceae центральным является наиболее широко распростра-
ненный и трудноопределяемый его род Nitzschia, представители которого ввиду отсут-
ствия резких морфологических отличий, крайней вариабельности видов и большого
числа переходных форм очень трудно дифференцируются. Успешная попытка упорядо-
чить видовые границы в этом роде может быть осуществлена только на основе элект-
ронно-микроскопического исследования (Lange-Bertalot, Simonsen, 1978). Наличие
у Simonsenia крыла с каналовидным швом (но без фибул) рассматривается в настоящее
время как результат конвергенции с крылом Surirellaceae (Simonsen, 1979).
30
В сем. Surirellaceae каналовидный шов расположен в круговом киле и имеет
лишь два слаборазвитых узелка. Мы считаем, что современные электронно-микро-
скопические данные полностью подтверждают теорию К. Мережковского о проис-
хождении Surirellaceae из Лмпсм/а-подобных предков. Следует отметить, что осе-
вое поле у Surirellaceae негомологично осевому полю навикулоидных диатомей.
КЛАССИФИКАЦИЯ ДИАТОМОВЫХ ВОДОРОСЛЕЙ,
ПРИНЯТАЯ В НАСТОЯЩЕМ ИЗДАНИИ
Отдел В ACILLARIOPH YT А
Класс CENTROPHYCEAE
1. Порядок PYXIDICULALES
1. Сем. PYXIDICULACEAE Nikolaev
Роды: Pyxidicula Ehr.
Costopyxis Gies.
Endictya Ehr.
2. Сем. THALASSIOSIROPSIDACEAE
Nikolaev
Род: Thalassiosiropsis Hasle
3. Сем. TRICERATIACEAE Schutt
Род Triceratium Ehr.
Род неясного систематического
положения в порядке Pyxidiculales:
Gladius Schulz
2. Порядок TH AL ASSIOSI RALES
1. CeM.THALASSIOSIRACEAE Lebour
emend. Hasle
Роды: Thalassiosira Cl.
Sceletonema Grev.
Cymatotheca Hendey
Planctoniella Schutt
Porosira J0rg.
Bacterosira Gran
Detonula Schutt
Lauderia Cl..
Minidiscus Hasle
2. Сем. STEPHANODISCACEAE
Makar.
Роды: Stephanodiscus Ehr.
Cyclostephanos Round
Cyclotella Kutz.
Concentrodiscus Churs.,
Moiss. et Suchova
Stephanocostis Genkal et
Kuzmina
3. Сем. ECTODYCTIONACEAE Churs.
et Tschem.
Род Ectodyction Churs. et Tschem.
3. Порядок MELOSIRALES
1. Сем. PSEUDOPODOSIRACEAE
(Sheshuk.) Gies.
Роды: Paralia Heib.
Pseudopodosira Jouse emend.
Vekschina
Bipalla Gies.
Distephanosira Gies.
2. Сем. MELOSIRACEAE Kutz.
Род. Melosira Ag.
3. Сем. AULACOSIRACEAE Moiss.
Роды: Aulacosira Thw.
Alveolophora Moiss. et
Nevretd.
Роды неясного систематического
положения в порядке Melosirales:
Trochosira Kitt.
Gleseria Lupikina et Dolmatova
4. Порядок COSCINODISCALES
1. Сем. HYALODISCACEAE Gies.
Роды: Hyalodiscus ,Ehr.
Podosira Ehr.
2. Сем. STELLARIMACEAE Nikolaev
nom. nov.
Род Stellarima Hasle
3. Сем. COSCINODISCACEAE Kutz.
Роды: Coscinodiscus Ehr.
Craspedodiscus Ehr.
Brightwellia Ralfs
Pseudotriceratium Grun.
Cosmiodiscus Grev.
Porodiscus Grev.
Ethmodiscus Castr.
4. Сем. TRIGONIUMACEAE Gies.
Род Trigonium Cl.
5. Сем. AZPEITIACEAE Gies, et Makar.
Род Azpeitia M. Perag.
6. Сем. HEMIDISC ACE AE Hendey
emend. Simonsen
Роды: Actinocyclus Ehr.
Hemidiscus Wallich
Pontodiscus Temnisk. et She-
shuk.
Roperia Grun.
31
7. Сем. ASTEROLAMPRACEAE Smith
emend. Gombos
Роды: Discodiscus Gombos
Bergonia Temp.
Rylandsia Grev. et Ralfs
Asterolampra Ehr.
Asteromphalus Ehr.
8. Сем. EUPODISCACEAE Kiitz.
Voi&v. Eupodiscus Bail.
Zygoceros Ehr.
9. Сем. HELIOPELT ACE AE Smith
emend. Ross
Роды: Aulacodiscus Ehr.
Actinopty chus Ehr.
Lepidodiscus Witt
Род неясного систематического по-
ложения в порядке Coscinodiscales'.
Undatodiscus Lupikina
Порядок BIDDULPHIALES
1. Сем. STICTODISCACEAE Schutt
Vojybv. Stictodiscus Grev.
Pseudostictodiscus Grun.
Arachnoidiscus Deane ex
Pritch.
Rocella Hanna
2. Сем. BIDDULPHIACEAE Kutz.
Роды:Biddulphia Gray
Sheshukovia Gies.
Lisitzinia Jouse
Anaulus Ehr.
Eunotogramma Weisse
Euodia Bail.
Peponia Grev.
Pseudocerataulus Pant
Striatocellina Gies.
Terpsinoe Ehr.
Odontella Ehr.
Eucampia Ehr.
Amphitetras Ehr.
Cerataulus Ehr.
Pleurosira (Menegh.) Trev.
3. Сем. HEM1AULACEAE Jouse, Kiss
et Poretzky
Роды: Hemiaulus Ehr.
Trinacria Grev.
Monobrachia Schrader
Arcus Oest.
Keratophora Pant.
Briggera Ross et Sims
Strelnikovia Ross et Sims
Goniothecium Ehr
Dicladiopsis De Toni
Climacodiutn Ehr.
Ceratauhna Perag.
Attheya West
4. Сем. KITTONIACEAE Gies.
Род Kittonia Grove et Sturt
5. Сем. ISTHMIACEAE Schutt
Род Isthmia Ag.
6. Сем. CHAETOCERACEAE Smith
Роды: Chaetoceros Ehr.
Bacteriastrum Schadb.
Corethron Castr.
7. Сем. CYMATOSIRACEAE Hasle,
Stosch et Syvertsen
Роды: Cymatosira Grun.
Praecymatosira Strel.
Campylosira Grun.
8. Сем. RUTILARIACEAE Pant.
Роды: Rutilaria Grev.
Kisseleviella Sheshuk.
9. Сем. LITHODESMIACEAE
H. et M. Perag.
Роды: Lithodesmium Ehr.
Bellerochea V. H.
Ditylum Bail.
Род неясного систематического по-
ложения в порядке Biddulphiales:
Hydrosera Wallich
6. Порядок AULISCALES
1. Сем. AULISCACEAE Hendey
Роды:/ItzZfsctzs Ehr
Rattrayella De Toni
Glyphodiscus Grev.
Craspedoporus Grev.
Corona Lef. et Chen.
Plicatodiscus Gies.
Pseudocerataulus Pant.
Pseudoauliscus Leud.-Forti
7. Порядок RHIZOSOLENIALES
1. Com. RHIZOSOLENIACEAE Petit
Роды: Rhizosolenia (Ehr.) Brightw.
Riedelia Jouse et Sheshuk.
Dactyliosolen Castr.
Guinardia Perag.
2. Сем. PYX1LLACEAE Schutt
Род Pyxilla Grev.
Род неясного систематического по-
ложения в порядке Rhizosoleniales:
Leptocylindrus Cl.
Роды неясного систематического
положения в классе Centrophyceae:
Monopsia Gr. et St.
Pomphodiscus Bork. et Mean.
Truania Pant.
Odontotropis Grun.
Споры
Acanthodiscus Pant.
Actinodictyon Pant.
Chasea Hanna
Chaeloniodiscus Pant.
Cladogramma Ehr.
Dicladia Ehr.
Epith elion Pant.
Gyrodiscus Witt
Haynaldia Pant.
Hyalopyxis Makar.
Kentrodiscus Pant.
Liradiscus Grev.
Mastogonia Ehr.
Muelleriella V. H.
Pantocsekia Grun.
Periptera Ehr.
Ploiaria Pant.
Poretzkia Jouse
Pseudopyxilla Forti
Pt его theca (Grun.) Forti
Pyrgodiscus Kitt.
Stephanogonia Ehr.
Xanthiopyxis Ehr.
Класс PENNATOPHYCEAE
1. Порядок A RAPHALES
1. CeM.FRAGILARIACEAE (Kiitz.) D.T.
Роды: Clavicula Pant.
Grunowiella V. H.
Rhaphoneis Ehr.
Delphineis Andrews
Sceptroneis Ehr.
Detonia Freng.
Fragilaria Lyngb.
Toxarium Bail.
Synedra Ehr.
Ardissonia De Notaris
Thalassionema Grun.
Thalassiothrix (Cl.) Grun.
Opephora Petit
Asterionella Hass.
Centronella Voigt
Ikebea Komura
Hannaea Patr. (= Ceratoneis
Grun.)
Dimerogramma Ralfs
Podocystis Bail.
Glyphodesmis Grev.
Falcula Voight
Amphicampa Ehr.
2. Сем. DIATOMACEAE Dumortier
?qw>v. Diatoma Bory
Meridion Ag.
Plagiogramma Grev.
3. Сем. TABELLARIACEAE Schutt
Роды: Tabellaria Ehr.
Tetracyclus Ralfs
Rhabdonema Kiitz.
Grammatophora Ehr.
Licmophora Ag.
Striatella Ag.
Cyclophora Castr.
Climacosphenia1 Ehr.
4. Сем. ENTOPYLACEAE Grun.
Род Entopyla Ehr.
5. Сем. PROTORAPH1DIACEAE
Simonsen
Род Protoraphis Simonsen
2. Порядок RAPHALES
1. Сем. NAVICULACEAE Kiitz.
Роды: Lyrella Kutz.
Navicula Bory
Anomoeoneis Pfitz.
Brachysira Kiitz.
Trachyneis Cl.
Stauroneis Ehr.
Haslea Simonsen
Gyrosigma Hass.
Pleurosigma W. Sm.
Pinnularia Ehr.
Dimiditata Hajos
Caloneis Cl.
Oestrupia Heiden
Diploneis Ehr.
Scoliotropis Cl.
Neidium Pfitz.
Rouxia Brun et Herib
Amphipleura Kiitz.
Frustulia Rabh.
Berkeleya Grev.
Brebissonia Grun.
Mastogloia Thw.
Progonoia Schrader
Diatomella Grev.
Scoliopleura Grun.
Stenoneis Cl.
Proschkinia Kar.
2. Сем. ACHNANTHACEAE Kutz.
Роды: Cocconeis Ehr.
Achnanthes Bory
Eucocconeis CI.
Anorthoneis Grun.
Campyloneis Grun.
3 Заказ 518
33
3. Сем. EUNOTIACEAE Kutz. Роды: Eunotia Ehr. Peronia Breb. et Arn. ActineUa F. M. Lewis 4. Сем. RHOICOSPHENIACEAE Mann Род Rhoicosphenia Grun. 5. Сем. CYMBELLACEAE (Kutz.) Grun. Роды: Cymbella Ag. Amphora Ehr. 6. Сем. GOMPHONEMATACEAE (Kutz.) Grun. Роды: Gomphonema Ehr. Gomphocymbella 0. Mull. Gomphopleura Reich. Gomphoneis Cl. Didymosphenia M. Schmidt 7. Сем. ENTOMONEIDACEAE Reim. Роды: Entomoneis Ehr. Plagiotropis Pfitz. Pseudoamphiprora Cl. 8. Сем. EPITHEMIACEAE Grun. Роды: Epithemia Breb. Denticulopsis Simonsen Denticila Kutz. 9. Сем. RHOPALODIACEAE Topatsch. Роды: Rhopalodia 0. Mull. Auricula Castr. 10. Сем. NITZSCHIACEAE Grun. Рощл: Nitz schia Hass. Hantzschia Grun. Bacillaria Gmelin Cylindrotheca Rabh. Gomphonitzschia Grun. Mediaria Sheshuk. Pseudoeunotia Grun. Simonsenia Lange-Bertalot Fragilariopsis Hust. И. Сем. SURIRELLACEAE Kutz. Роды: Surirella Turp. Cymatopleura W. Sm. Campylodiscus Ehr. Stenopterobia Br6b. Род неясного систематического по- ложения в классе Pennatophyceae-. Pseudonitzschia Perag.
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Отдел В ACILLARIOPH YTA
Клетки одиночные или соединены в разнообразные колонии. Протопласт клетки
заключен в кремнеземный панцирь, состоящий из двух половинок (створок), налегаю-
щих одна на другую. Между створками часто возникают вставочные и соединительные
ободки (поясок), увеличивающие объем панциря. Стенки панциря снабжены перфора-
циями: порами, пороидными или л окулярными ареолами, альвеолами, глазками,
ложными глазками, ложными узелками, а также различного рода выростами:
двугубыми, замкнутыми, выростами с опорами, шипами, рогами, щетинками и другими
элементами структуры: ребрами, камерами, септами и пр. Расположение структурных
элементов панциря, их качественная и количественная характеристики имеют большое
таксономическое значение и служат основными критериями при их систематике.
Размножение осуществляется путем деления клетки на две дочерние и половым
путем с образованием ауксоспор и покоящихся спор.
Обитают в морях, солоноватых и пресных водоемах во всех географических
зонах, в планктоне, бентосе и обрастаниях. Вне водных биотопов встречаются на влаж-
ных скалах, коре деревьев, на сырой земле, в почве, на снегу и льду.
I. Панцирь актиноморфного строения. Створки круглые, эллиптические, 3—6-уголь-
ные. Структура створок из пор и ареол, расположенных радиально, реже танген-
циально. Створки снабжены различными выростами, шипами, щетинками, рогами.
Шов отсутствует. Виды преимущественно планктонные, морские и пресноводные ....
...........................................класс Centrophyceae (с. 35).
II. Панцирь зигоморфного строения. Створки вытянутые по продольной оси, от широ-
коэллиптических до узколинейных. Структура створок из пор, ареол, штрихов, ребер,
расположенных перисто. Створки менее структурированы различными элементами.
Имеется шов или бесструктурное осевое поле. Виды преимущественно бентосные,
подвижные и прикрепленные, морские и пресноводные..класс Pennatophyceae.
Класс CENTROPHYCEAE
Клетки одиночные или образуют различного типа колонии. Панцирь шаровидный,
эллипсоидный, цилиндрический, линзовидный, дисковидный, призматический, со
вставочными и соединительными ободками кольцевидной или чешуевидной формы.
Хлоропласты в виде мелких многочисленных зерен или более или менее крупных
пластинок, реже одной пластинки Створки круглые, эллиптические, 3—6-угольные.
Лицевая часть створки плоская, выпуклая, вогнутая, реже концентрически-
или радиально-волнистая, снабженная различными структурными элементами. Струк-
тура створок из пор, пороидных или локулярных ареол, расположенных в радиаль-
ных рядах, иногда собранных в пучки, тангенциальных рядах, реже в концент-
рических кругах, редко без определенного порядка. Велум ареол с наружной
или внутренней поверхности створок. Загиб высокий или низкий, мелко ареоли-
рованный.
35
Обитают в основном в морях и океанах, реже в пресных водоемах, обычно
в планктоне Многие роды и виды вымершие.
I. Клетки одиночные или образуют рыхлые нитевидные колонии. Панцирь шаровидный,
дисковидный или призматический. Двугубые выросты многочисленные, образуют
краевое кольцо, центральную группу или разбросаны по всей створке, реже одиноч-
ные, расположенные эксцентрично, иногда они редуцированы, реже имеются замкну-
тые и кольцевидные выросты. Выросты с опорами отсутствуют ...............................
.........................................................порядок Pyxidiculales (с.36).
II. Клетки образуют плотные нитевидные или гибкие цепочковидные колонии. Панцирь
цилиндрический. Двугубые выросты обычно одиночные, расположены близ края
створки, реже отступя от края или ближе к центру, редко их несколько. Имеются
замкнутые выросты и шипы. По краю створки или на загибе кольцо из выростов
с опорами, присутствующих также в центре створки, редко по всей поверхности
створки...................................................порядок Thalassiosirales (с. 56).
Порядок PYXIDICULALES Nikolaev
Клетки одиночные или соединены в колонии. Хлоропласты многочисленные,
мелкие, в виде зерен. Панцирь шаровидный, дисковидный, цилиндрический или призма-
тический. Створки круглые, эллиптические или полигональные, плоские, выпуклые,
иногда волнистые. Ареолы локулярные, реже пороидные, с велумом, расположенным
на внутренней поверхности, и фораменом на наружной. Имеются двугубые выросты,
кольцевидный вырост и шипики, редко, двугубые выросты редуцированы. У поли-
гональных створок на выпуклостях имеются глазки. Известны покоящиеся споры
и ауксоспоры.
Виды морские, современные и вымершие.
Типовое семейство: Pyxidiculaceae Nikolaev.
I. Створки круглые.
1. Створки плоские, с кольцевидным выростом в центре, редко с одним двугубым
выростом близ края............................. сем. Thalassiosiropsidaceae (с. 52).
2. Створки выпуклые, двугубые выросты с наружными трубками, реже наружные
трубки отсутствуют..................................сем. Pyxidiculaceae (с. 36).
II. Створки эллиптические или полигональные .............сем. Triceratiaceae (с. 52).
Сем. PYXIDICULACEAE Nikolaev
Клетки одиночные или соединены в колонии. Панцирь сферический, дисковидный
или цилиндрический. Поясок отсутствует. Створки круглые, разнообразно выпуклые,
реже почти плоские. Ареолы слагают сплошную сеть, реже разомкнутые. Многочислен-
ные двугубые выросты расположены беспорядочно по всей створке или образуют
одно краевое кольцо, реже несколько колец. Одиночные двугубые выросты формируют
центральную группу. Наружные трубки двугубых выростов служат для соединения
клеток в колонию, реже они отсутствуют. Известны покоящиеся споры.
Виды морские, преимущественно вымершие, реже современные.
Тип: Pyxidicula Ehr. emend. Streln. et Nikolaev. .
I. Створки колпачковидные, выпуклые, с ребрами и иногда отогнутым краем. Ареолы
иногда разделены анастомозирующими ребрами..............род Costopyxis (с. 48).
36
II. Створки полусферические, чашевидные, линзовидные, выпуклые или почти плоские,
без радиальных ребер. Ареолы образуют сплошную сеть.
1. Створки полусферические, чашевидные, линзовидные, выпуклые, загиб низкий.
Двугубые выросты по краю створки, беспорядочно по всей створке или несколько
в центре.................................................род Pyxidicula (с. 37).
2. Створки плоские, загиб высокий. Двугубые выросты по краю неотчетливые
............................................................род Endictya (с. 51).
Род PYXIDICULA Ehr. (Ehrenberg, 1838b : 165) emend. Streln. et Nikolaev
Stephanopyxis Ehr. (Ehrenberg, 1844 : 264). — Creswellia Grev. (Greville in Gregory,
1857 : 64).
Клетки одиночные или образуют колонии в виде цепочек. Панцирь шаровидный,
эллипсоидный, линзовидный или короткоцилиндрический. Створки полусферические,
чашевидные, в виде усеченного конуса, равновыпуклые или выпуклые в разной
степени. Структура из пороидных или л окулярных ареол с внутренним сплошным или
прерывистым крибрумом. Двугубые выросты многочисленные, на наружной поверх-
ности створки в виде трубок различной длины (в СМ — шипы), или в виде округлого
отверстия, или снаружи закрытые (в СМ — небольшие шипики или бугорки), распо-
ложены по всей поверхности створки или образуют одно, реже несколько колец. На
внутренней поверхности створки двугубые выросты открываются щелью с утолщен-
ным валиковидным краем или округлым отверстием без утолщения по краю. Покоя-
щиеся споры образуются эндогенно или полуэндогенно.
Виды морские, преимущественно вымершие.
Известны с мела доныне. Есть указание (Rotpletz, 1896; Векшина, 1960)
на находки в юрских отложениях.
Тип: Р. cruciata Ehr.
П р и м е ч. В диагнозах некоторых видов не указан тип ареол вследствие их
неизученное™.
I. Двугубые выросты с длинными трубками на наружной поверхности створок (в СМ —
шипы).
1. Двугубые выросты на створке расположены беспорядочно, редко образуют коль-
цо близ края створки.
А. По краю створки ареолы удлиненной формы.
а. Створки в виде усеченного конуса. Ареолы очень крупные, меньше 1
вЮмкм.......................................................17. Р.megapora.
б. Створки выпуклые до полусферических. Ареолы мельче, 1—2 в 10 мкм.
Край широкий, гиалиновый.
а. Створки слабо выпуклые. Несколько крупных двугубых выростов распо-
ложено беспорядочно . . .’............ . . 2.... 16. Р. maiginata.
0. Створки выпуклые до полусферических. Крупные двугубые выросты
образуют на середине радиуса или близ края створки кольцо..............
..................................................... 12. P.grunowii.
Б. Ареолы одинаковые по всей створке или слегка удлиненные к краю.
а. Двугубые выросты на наружной поверхности створки с длинными заострен-
ными трубками..................................................11.Р. ferox.
а. Ареолы недоразвитые, край широкий, гиалиновый..........8. Р. defecta.
/3. Ареолы отчетливые, по краю створки вытянутой формы . . 15. Р. lavrenkoi.
б. Двугубые выросты на наружной поверхности створки с короткими труб-
ками ....................................................26. Р. spinosissima.
37
2. Двугубые выросты расположены группой или образуют кольцо
А. Двугубые выросты расположены на вершине створки группой...............
........................................................19. Р. nipponica.
Б. Двугубые выросты расположены кольцом по краю или на середине радиуса
створки.
а. Кольцо двугубых выростов расположено по краю створки.
а. Острые шипики по углам ареол отсутствуют.
+ На загибе 2 ряда почти прямоугольных ареол, двугубые выросты
короткие, расположены плотно.
— Ареолы в центре неправильной формы или разрежены . . 6.Р. crenata.
— Ареолы в центре правильной шестиугольной формы, плотно сомк-
нутые ............................................3. Р. biseriata.
++ На загибе 2 ряда прямоугольных ареол отсутствуют, двугубые выросты
длинные, расположены плотно или редко.
— Двугубые выросты прямые, расположены плотным кольцом по
самому краю створки................................29. Р. uralensis.
— Двугубые выросты расположены редко по краю створки.
О Гиалиновый край узкий .........................2. Р. barbadensis.
ОО Гиалиновый край широкий...........................27. Р. superba.
Р. Острые шипики по углам ареол имеются.
+ Створки выпуклые, край узкий, со структурой..........23. Р. schenkii.
++ Створки слабо выпуклые, край широкий, гиалиновый ................
................................................ 14. Р. joynsonii
б. Кольцо двугубых выростов расположено ближе к середине радиуса створки.
а. Трубки двугубых выростов расширены или разветвлены на свободном
конце.
+ Двугубые выросты редкие, расширены на конце. Ареол 2—3 в 10 мкм
.................................................... 22. Р. punctata.
++ Двугубые выросты многочисленные, разветвленные на конце. Ареол
5—6 в 10 мкм.............................................5. Р. corona.
р. Трубки выростов не разветвленные на конце.
+ Ареолы одинакового размера по всей створке или уменьшаются
к краю.
— Панцирь линзовидный или сферический.
О Створки с широким гиалиновым краем.
X Бугорки по краям ареол образуют кольцевые линии..........
.............................................10. Р. edita.
XX Бугорки не образуют кольцевых линий............21. Р. polaris.
ОО Створки с узким гиалиновым краем.
X Створки линзовидные или полусферические.......l.P.arctica.
XX Створки трапециевидные или полусферические................
......................................... 30. Р. weissflogii.
— Панцирь цилиндрический, реже сферический.
О Острые шипики по углам ареол имеются.
X Ареолы одинакового размера по всей створке . . 25. Р. spinosa.
XX Ареолы к краю створки уменьшаются.............24. Р. similis.
ОО Острые шипики по углам ареол отсутствуют.........28. Р. turris.
++ Ареолы сильно уменьшаются на загибе..............20. Р. palmeriana.
И. Двугубые выросты с короткими или слабо развитыми трубками на наружной поверх-
ности створок, или трубки отсутствуют.
1. Поверхность створки бугристая.............................4. Р. charkoviana.
2. Поверхность створки ровная.
А. Панцирь цилиндрический....................................13. Р. inermis.
Б. Панцирь линзовидный или сферический.
38
а. Ареолы неправильной формы, рядов не образуют.............9. Р. dissona.
б. Ареолы правильной округло-шестиугольной формы, расположены в рядах.
а. Ареолы одинакового размера по всей створке ...........7. Р. cruciata.
р. Ареолы уменьшаются к краю.
+ Ареолы в изогнутых тангенциальных рядах ............18. Р. moelleri.
++ Ареолы в прямых тангенциальных, вторично косо пересекающихся
рядах.
- Гиалиновый край широкий, 3—5 мкм.............31. Р. weyprechtii.
— Гиалиновый край узкий, 1—2 мкм..................32. Р. zabelinae.
1. Р. arctica (Grun.) Streln. et Nikolaev (Стрельникова, Николаев, 1986 : 952).-
Stephanopyxis turris (Grev. et Arnott) Ralfs var. arctica Grun. (Grunow, 1884 : 91, tab. V,
20-22). — 5. barbadensis Grev. sensu Strelnikova (Стрельникова, 1974 : 58, табл. Ill,
27,22; VI, 1-8).
Панцирь сферический или линзовидный. Створки более или менее выпуклые, от
часового стекла до полусферы, 40—70 мкм в диам., с узким гиалиновым краем. Ареолы
локулярные, с прерывистым крибрумом, в прямых или слегка изогнутых тангенциаль-
ных рядах, в центре 2—3 ареолы в 10 мкм, у некоторых створок ареолы уменьшаются
к краю, до 3—4, иногда 5—6 в 10 мкм Кольцо редких двугубых выростов с длинными
трубками на наружной поверхности (в СМ — шипы) расположено ближе к середине
радиуса створки, второе кольцо по краю створки без наружных трубок. Двугубые
выросты открываются на внутренней поверхности округлым или щелевидным отвер-
стием с утолщенными краями. (Табл. 19,1-15*, 20,1-5).
Морской вид.
Поздний мел — вост, склон Урала. Палеоген — Зап. Сибирь. Поздний миоцен —
вост. Камчатка. Четверт. период — Карелия. Ныне — Норвежское море. Вне СССР.
Эоцен — Дания, Швеция, Норвежское море.
2. Р. barbadensis (Grev.) Streln. et Nikolaev (Стрельникова, Николаев, 1986 : 952). —
Creswellia barbadensis Grev. (Greville, 1865 : 3, tab. 1, 11). — Stephanopyxis barbadensis
(Grev.) Grun. (Grunow, 1884 : 91).
Панцирь эллипсоидный. Створки равномерно выпуклые, 20—40 мкм в диам.,
с узким отогнутым гиалиновым краем. Ареолы в прямых, слегка изогнутых танген-
циальных рядах, 5—6 в 10 мкм. Двугубые выросты с наружными трубками (в СМ —
острые шипы) по краю створки. (Табл. 20,13-14).
Морской вымерший вид.
Поздний эоцен—ранний олигоцен — Норвежское море, Малые Антильские о-ва
(о-в Барбадос), Новая Зеландия (у г. Оамару), Тасманово море.
3. Р. biseriata (Streln.) Streln. et Nikolaev (Стрельникова, Николаев, 1986 : 952).—
Stephanopyxis biseriata Streln. (Стрельникова, 1974 : 69, табл. VII, 8a, 6, 9).
Панцирь эллипсоидный до почти сферического. Створки выпуклые, 50—70 мкм
в диам., с вертикальным загибом 5—7 мкм выс. Ареолы правильной шестиугольной
формы, в прямых или слегка изогнутых тангенциальных рядах, образуют плотную
сомкнутую сеть, 2 ареолы в 10 мкм в центре, 4—5 по краю. На загибе 2 ряда округло-
четырехугольных ареол. На границе диска створки с загибом кольцо шипиков (дву-
губых выростов?), 4—6 в 10 мкм. Край отогнутый, гиалиновый. (Табл. 20, 6, 7).
Морской вымерший вид.
Поздний мел — вост, склон Урала.
4. Р. charkoviana (Jouse) Streln. et Nikolaev (Стрельникова, Николаев, 1986 : 952). —
Stephanopyxis charkovianus Jouse (Жузе, 1949 : 42, табл. 8, 6а-в).
Панцирь сферический. Створки выпуклые, с бугристой поверхностью, 2—3 мкм
шир., 50—65 мкм в диам., 50—55 мкм выс., с широким гиалиновым краем. Ареолы
39
локулярные, со сплошным крибрумом, его поры расположены радиально, 5—6 ареол
в 10 мкм. Двугубые выросты без развитой трубки на наружной поверхности створки,
по всей створке в средней части бугорков, с внутренней поверхности створки откры-
ваются округлым отверстием. (Табл. 21,5-8).
Морской вымерший вид.
Эоцен — Курская, Воронежская, Волгоградская обл., УССР.
5. Р. corona (Ehr.) Streln. et Nikolaev (Стрельникова, Николаев, 1986 : 952). —
Systephania corona Ehr. (Ehrenberg, 1844 : 272; 1854, tab. 33, XV/22, XVIII/16). -
Stephanopyxis corona (Ehr.) Grun. (Grunow, 1884 : 90).
Панцирь от линзовидного до низкоцилиндрического или сферического, с неодина-
ковыми створками. Створки в разной степени выпуклые, 17—95 мкм в диам. Ареолы
в прямых тангенциальных рядах/5—6 в 10 мкм, у края створки ареолы разрежены
и немного мельче. Венец многочисленных шипов (двугубых выростов?), разветвленных
на концах, на границе лицевой части створки и загиба, 4—5 шипов в 10 мкм. Выросты
соединяются концами и тонкой гиалиновой пленкой. Иногда имеется второе кольцо
выростов ближе к краю створки, или выросты на створке расположены беспорядочно.
(Табл. 20 Л-12).
Морской вымерший вид.
Эоцен — Курская, Воронежская, Волгоградская обл., УССР. Неоген — Камчатка.
Вне СССР. Олигоцен — Норвежское море. Неоген — Венгрия, США (штаты Калифорния
и Мериленд), Мексика, Бермудские о-ва.
6. Р. crenata (Sheshuk.) Streln. et Nikolaev (Стрельникова, Николаев, 1986 : 952). -
Stephanopyxis crenata Sheshuk. (Шешукова-Порецкая, Глезер, 1964 I 81, табл. I, 6-8).
Створки слабо выпуклые, 52—60 мкм в диам., с вогнутым центром и узким,
около 3 мкм, гиалиновым краем. Загиб невысокий, 10—12 мкм. Ареолы со сплошным
крибрумом. На середине створки ареолы крупные, 2—3 в 10 мкм, неправильной формы,
образуют сомкнутую сеть и расположены тангенциальными рядами или разрежены;
к периферии ареолы уменьшаются, до 3—4.5 в 10 мкм, образуют концентрические ряды.
По углам краевых ареол имеются бугорки. На границе лицевой части створки и загиба
(между вторым и третьим рядами ареол от края загиба) кольцо двугубых выростов, 2—3
в 10 мкм, с грубыми невысокими трубками, окруженными гиалиновым кантом,
имеющих вид трезубца, с внутренней стороны выросты открываются округлыми
отверстиями с утолщенными краями. Второе кольцо более редких двугубых выростов
без видимых наружных трубок близ края створки. (Табл. 21,7-4).
Морской вымерший вид.
Эоцен — Воронежская, Харьковская обл., УССР, КазССР.
7. Р. cruciata Ehr. (Ehrenberg, 1843 : 422, tab. Ill, 7/6). — Dietyopyxis cruciata Ehr.
(Ehrenberg, 1844 : 267). — Coscinodiscus cruciatus (Ehr.) Kiitz. (Kiitzing, 1844 : 132, tab. 28,
10). —Pyxidicula cruciata Ehr. (Hustedt, 1930 : 301).
Створки выпуклые, почти полусферические, 19—50 мкм в диам. Ареолы локу-
лярные, с прерывистым крибрумом, одинакового размера по всей створке, 2.5—3
в 10 мкм, в прямых тангенциальных и вторичных, косо пересекающихся под углом
60° рядах (такое расположение ареол создает в СМ оптическую фигуру шести-
лучевого креста). Двугубые выросты без наружных трубок, расположены беспоря-
дочно на створке, на внутренней поверхности створки в виде округлого отверстия.
(Табл. 21,9—15).
Морской вымерший вид.
Эоцен — Воронежская, Харьковская, Ростовская обл., КазССР, Зап. Сибирь.
Неоген — Камчатка. Вне СССР. Эоцен — Атлантический океан (Бермудские о-ва,
Фолклендское плато), Тихий океан (Тасманово море), Новая Зеландия (у г. Оамару).
Неоген — Чехословакия, Румыния.
40
8. P. defecta (Streln.) Streln. et Nikolaev (Стрельникова, Николаев, 1986 : 952). —
Stephanopyxis defectus Streln. (Джиноридзе, Жузе, Стрельникова, 1979 : 35, рис. 24—27).
Створки слабо выпуклые, 30—45 мкм в диам., с широкими отогнутыми гиалино-
выми краями. Ареолы пороидные или недоразвитые, 2—3 в 10 мкм, расположены
без особого порядка. Межареольные перегородки приподняты по углам в виде ши-
пиков. У некоторых форм сохраняемся очертание ареолы, крибрум незаметен.
Несколько двугубых выростов, расположенных без особого порядка. (Табл. 19, 16;
20, 75,26).
Морской вымерший вид.
Вне СССР. Эоцен — Норвежское море.
9. Р. dissona (Schulz) Streln. et Nikolaev (Стрельникова, Николаев, 1986 : 952).
Створки сильно выпуклые, полусферические, 25—100 мкм в диам. Ареолы локу-
лярные, с фораменом неправильной, преимущественно округлой формы, сплошным
крибрумом, с радиально расположенными порами. По краям ареол небольшие бугорки.
Двугубые выросты расположены кольцом ближе к центру створки, без хорошо разви-
той трубки на наружной поверхности створки, с внутренней поверхности створки
в виде округлого или слабо сплющенного отверстия. Край гиалиновый, 2—3 мкм шир.
Морской вымерший вид.
Поздний мел — Пензенская обл., вост, склон Урала. Вне СССР. Сенон — Польша
(Гданьская бухта).
F. dissona. — Coscinodiscus dissonus Schulz (1935 : 387, tab. 1,5).
Створки 25—100 мкм в диам. Ареолы крупные, 1.5 в 10 мкм, плотно сомкнутые,
расположены без особого порядка. (Табл. 22,1—8).
F. dissoniformis (Streln.) Streln. et Nikolaev (Стрельникова, Николаев, 1986 : 952). —
Coscinodiscus dissoniformis Streln. (Стрельникова, 1965 : 34, табл. 4,3,4; 5,1,2).
Створки 30—70 мкм в диам. Ареолы мелкие, 3—5 в 10 мкм, расположены беспо-
рядочно в центре и с тенденцией к концентрическим рядам у края. (Табл. 22, 9—12).
Встречается вместе с типовой формой.
10. Р. edita (Jouse) Streln. et Nikolaev (Стрельникова, Николаев, 1986 : 952). —
Stephanopyxis edita Jouse (Жузе, 1955 : 85, табл. 1,7).
Створки выпуклые до полусферических, 20—51 мкм в диам., с гиалиновым
краем. Ареолы переходного типа от пороида к локуле, 2—4 в 10 мкм, с очень толстыми
стенками, что в СМ создает впечатление разреженных ареол. Крибрум прерывистый.
На наружной поверхности по углам ареол крупные бугорки, образующие на створке
ясные кольцевые линии. Редкие двугубые выросты с наружными трубками образуют
кольцо близ середины радиуса створки. Закрытые двугубые выросты разбросаны
на внутренней поверхности створки, иногда наблюдаются на крибруме. (Табл. 23,1—6).
Морской вымерший вид.
Эоцен — Зап. Сибирь, КазССР. Вне СССР. Норвежское море, южн. часть Атлантиче-
ского океана (Фолклендское плато).
И. Р. ferox (Grev.) Streln. et Nikolaev (Стрельникова, Николаев, 1986 : 952). -
Cresswellia ferox Qxqn. (Greville, 1859 : 166, tab. VIII, 75,16).
Створки более или менее выпуклые до полусферических, 12—75 мкм в диам.,
толстостенные, с широким отогнутым гиалиновым краем. Ареолы локулярные, 2—5
в 10 мкм, расположены плотно сомкнутой сетью. Многочисленные двугубые выросты
с длинными острыми трехгранными трубками на наружной поверхности створки
расположены без особого порядка, на внутренней поверхности они открываются
округлым отверстием. (Табл. 23, 7-9).
Морской вымерший вид.
Поздний мел — вост, склон Урала. Палеоцен — Средн. Поволжье, вост, склон
Урала. Неоген — Дальний восток. Вне СССР. Неоген — Калифорния.
41
12. P. grunowii (Gr. et St.) Streln. et Nikolaev (Стрельникова, Николаев, 1985 :952). —
Stephanopyxis grunowii Gr. et St. (Grove, Sturt, 1888, in Schmidt, 1874-1959,
tab. 130,7-4).
Створки более или менее выпуклые до полусферических, 50—150 мкм в диам.,
с отогнутым гиалиновым краем. Ареолы в намечающихся тангенциальных рядах,
1.5—2 в 10 мкм, по краю они более крупные, вытянутые. Межареольные перегородки
с мелкими шипиками и бугорками. Кольцо грубых двугубых выростов на середине
радиуса створки или ближе к краю. (Табл. 25,5-5).
Морской вымерший вид.
Вне СССР. Поздний эоцен — южн. часть Атлантического океана (Фолклендское
плато), южн. часть Тихого океана (Тасманово море), Новая Зеландия (у г. Оамару).
Ранний олигоцен — Атлантический океан.
П р и м е ч. Вид, близкий к Р. marginata (Grun.) Streln. et Nikolaev.
13. P. inermis (Jouse) Streln. et Nikolaev (Стрельникова, Николаев, 1986 : 952). —
Stephanopyxis inermis Jouse (Жузе, 1961 : 60, табл. 1,2).
Панцирь цилиндрический, 19—27 мкм в диам. Створки 18—37 мкм выс., колпачко-
видные, одна из них выше другой. Ареолы локулярные, разреженные, 3—5.5 в 10 мкм.
На загибе ареолы в более или менее прямых рядах, перпендикулярных или наклонных
по отношению к центральной оси створки. Двугубые выросты отсутствуют.
(Табл. 24,1-4).
Морской вымерший вид.
Неоген — Камчатка, Сахалин, о-в Итуруп.
П р и м е ч. Вид, близкий к Р. turris f. cylindrus по форме створки, отличается раз-
реженным расположением ареол. Видимо, спора.
14. P.joynsonii (A. S.) Streln. et Nikolaev (Стрельникова, Николаев, 1986 : 952). —
Stephanopyxis joynsonii A. S. (Schmidt, Atlas, 1888, tab. 123, 9).
Створки равномерно слабо выпуклые, 100—111 мкм в диам., с широким,
10—12 мкм, гиалиновым краем. Ареолы в прямых тангенциальных рядах, 3—4
в 10 мкм, с небольшими шипиками по углам. По краю створки кольцо редких длинных
двугубых выростов, имеющих на наружной поверхности створки вид заостренных тру-
бок, на внутренней открывающихся щелевидным отверстием. (Табл. 24,5—9).
Морской вымерший вид.
Эоцен — Воронежская, Харьковская, Ростовская обл. Вне СССР. Эоцен —
о-в Барбадос. Поздний эоцен — южн. часть Атлантического океана (Фолклендское
плато) , южн. часть Тихого океана (Тасманово море), Новая Зеландия.
15. Р. lavrenkoi (Jouse) Streln. et Nikolaev (Стрельникова, Николаев, 1986 : 952). —
Stephanopyxis lavrenkoi Jouse (Жузе, 19496 : 40, табл. 10,9а-б).
Створки выпуклые, 13—45 мкм в диам., с отогнутым краем 4—7 мкм шир.
Ареолы пороидные, 3-5 в 10 мкм, группируются в слегка косые ряды. По краю
створки ареолы вытянутой формы. Углы ареол с небольшими шипиками или бугорка-
ми. Двугубые выросты с наружной поверхности в виде трехгранной трубочки, с внут-
ренней с округлыми отверстиями. (Табл. 23,10-12).
Морской вымерший вид.
Поздний мел — вост, склон Урала. Палеоцен — Средн. Поволжье, вост, склон Урала.
16. Р. marginata (Grun.) Streln. et Nikolaev (Стрельникова, Николаев, 1986 : 952). —
Stephanopyxis marginata Grun. (Grunow, 1884 : 90, tab. V, 77).
Створки слабо выпуклые, в виде часового стекла, 28—83 мкм в диам., с широким
гиалиновым краем. Ареолы 1—2 в 10 мкм, по краю вытянутой формы и более крупные.
Межареольные перегородки с шипиками. Несколько длинных двугубых выростов
расположено на створке без особого порядка. (Табл. 25, 7, 2).
42
Морской вымерший вид.
Эоцен — Пензенская обл., КазССР, Зап. Сибирь. Поздний олигоцен — п-ов Мангы-
шлак, Командорские о-ва. Вне СССР. Поздний эоцен—ранний олиго цен — юго-вост,
часть Индийского океана. Поздний олигоцен — Норвежское море.
17. Р. megapora (Grun.) Streln. et Nikolaev (Стрельникова, Николаев, 1986 : 952). -
Stephanopyxis megapora Grun. (Grunow, 1884 : 89, tab. V, 24a, b).
Панцирь почти сферический. Створки выпуклые, в виде усеченного конуса,
28—80 мкм в диам. Ареолы очень крупные, меньше 1 ареолы в 10 мкм, иногда сильно
вытянутые по краю. Межареольные перегородки с мелкими шипиками. Несколько
крупных трубок двугубых выростов расположено на вершине створки. (Табл. 24,
10—13\ рис. 2,7, 2).
Морской вымерший вид.
Палеоцен — Средн. Поволжье, вост, склон Урала. Эоцен - УССР, Зап. Сибирь.
Вне СССР. Эоцен — Норвежское море.
18. Р. moelleri (A. S.) Streln. et Nikolaev (Стрельникова, Николаев, 1986 : 952). —
Coscinodiscus moelleri A. S. (Schmidt, 1878, tab. 59,77; Rattray, 1890 : 19). — C. moelleri
var. macroporus Grun. (Grunow, 1884 : 84, tab. IV, 25).
Панцирь почти сферический. Створки сильно выпуклые, 50—180 мкм в диам.
Ареолы локулярные, с прерывистым крибрумом, в слегка изогнутых тангенциальных
рядах, на середине 2, у края 3—4 в 10 мкм, краевые отличаются почти четырехугольной
формой, центральная ареола обычно более крупная. Двугубые выросты без наружной
трубки расположены в беспорядке по всей створке, на внутренней поверхности створки
открываются короткой щелью с утолщенными краями. (Табл. 25, 6, 7\ 29, 13, 14).
Морской вымерший вид.
Эоцен — Калининградская обл., Зап. Сибирь, КазССР. Вне СССР. Палеоцен—эоцен —
Дания.
П р и м е ч. Вид, очень близкий к Р. weyprechtii Grun.
19. Р. nipponica (Gran et Yendo) Streln. et Nikolaev (Стрельникова, Николаев, 1986 :
952). — Stephanopyxis nipponica Gran et Yendo (1914 : 27, fig. 16).
Клетки соединяются в короткие колонии посредством наружных трубок двугу-
бых выростов. Панцирь эллипсоидный, 24—36 мкм в диам. Створки до 55 мкм выс.,
высота больше диаметра. Структура из ареол, 4—5 в 10 мкм, несколько более крупных
на вершине. Двугубые выросты с длинной наружной трубкой, на вершине створки,
второе кольцо двугубых выростов, открывающихся маленьким отверстием, — по
самому краю створки. Выросты расположены на крибруме, открываются в локулу.
С внутренней стороны двугубые выросты заканчиваются округлыми отверстиями.
(Табл.27,7-77).
Неритический арктобореальный вид.
Преимущественно в дальневосточных морях, а также в Тихом океане у Алеутских
о-вов и у берегов Сев. Америки.
20. Р. palmeriana (Grev.) Streln. et Nikolaev (Стрельникова, Николаев, 1986 : 952). -
Cresswellia palmeriana Grev. (Greville, 1865 : 137, tab. I, 9). - Stephanopyxis palmeriana
(Grev.) Grun. (Grunow, 1884 : 90).
Клетки соединяются в длинные цепочки. Панцирь низкоцилиндрический.
Створки с более или менее вертикальным загибом, 27—150 мкм в диам. Ареолы
локулярные, в центре диска крупные, 1.5-2 в 10 мкм, к загибу мельче, расположены
в намечающихся изогнутых тангенциальных рядах. На загибе ареолы расположены
плотно сомкнутой сетью, в верхней части загиба 3—4 ареолы, по краю до 5—6 в 10 мкм.
На границе лицевой части створки с загибом, кольцо из 10-22 двугубых выростов, на внут-
ренней поверхности створки открывающихся щелью с утолщенным краем. Покоящиеся
43
6
споры сдвоенные, крупные, толстостенные, с мелкими шипиками по краям ареол.
Первичная створка с длинными шипами, с помощью которых смежные споры соеди-
няются в пары, вторичная с острыми длинными шипами, расположенными беспорядочно
и не служащими для соединения с другими створками. (Табл. 25,5, 9; рис. 2,5-5).
Планктонный неритический тепловодный тропический вид. В ископаемом состоя-
нии неизвестен.
Ныне — Японское море. Вне СССР. Ныне — Атлантический и Тихий океаны.
21. Р. polaris (Grun.) Streln. et Nikolaev (Стрельникова, Николаев, 1986 : 952). —
Stephanopyxis turris var. polaris Grun. (Grunow, 1884 : 89, tab. V, 79, 25, 25). —5. grunowii
sensu Jouse (Жузе, 1955 : 85, табл. 2,1-4).
Панцирь сферический или эллипсоидный. Створки 35—115 мкм в диам., обычно
с широким, 1.7—5 мкм, отогнутом краем. Ареолы в неотчетливых тангенциальных
рядах, 1—2 в 10 мкм. По углам ареол имеются бугорки или шипики. На створке
несколько крупных двугубых выростов. (Табл. 26, 75,14).
Морской вымерший вид.
Палеоцен - Средн. Поволжье, вост склон Урала. Эоцен — Зап. Сибирь. Неоген —
Камчатка. Четверг, период — Вост. Сибирь.
22. Р. punctata (Jouse) Streln. et Nikolaev (Стрельникова, Николаев, 1986 : 952). -
Stephanopyxis punctata Jouse' (Жузе, 1955 : 87, табл. 5,10a, 6).
Панцирь более или менее эллипсоидный, с пояска в очертании шестиугольный.
Створки низкие, с плоской, вогнутой или слегка выпуклой вершиной, 25—65 мкм
в диам., около 15 мкм выс., с отогнутым краем. Ареолы округло-многоугольные,
расположены в намечающихся, не всегда правильных тангенциальных рядах или без
особого порядка, 2—3 в 10 мкм, иногда сильно разреженные на середине створки.
Отогнутый край с одним рядом слегка вытянутых ареол, 3—4 в 10 мкм. В месте
перегиба створки в загиб кольцо редких грубых шипов, расширенных на свобод-
ном конце в виде трезубца (видимо, двугубые выросты), 1-1.5 шипа в 10 мкм.
(Табл. 26, 7-5).
Морской вымерший вид.
Эоцен — УССР, КазССР, Зап. Сибирь, Сев .-Вост. СССР.
23. Р. schenkii (Kanaya) Streln. et Nikolaev (Стрельникова, Николаев, 1986 : 952). —
Stephanopyxis schenkii Kanaya (1950 : 67, tab. 2,2-4). —S. neogena Sheshuk. (Шешукова-
Порецкая, 1959, табл. 1,5a-г; 1961, табл. 3,4а, б).
Панцирь эллипсоидный, толстостенный. Створки выпуклые, 30—120 мкм в диам.,
12—35 мкм выс., с плоским отогнутым краем 3—5 мкм шир. Ареолы пороидные,
с острыми шипиками по углам, на диске, 0.75—3 в 10 мкм, в прямых тангенциальных
рядах, перекрещивающихся в трех направлениях. На загибе ареолы расположены
беспорядочно или более или менее прямыми поперечными рядами. Размеры ареол
на загибе одинаковые или постепенно уменьшаются к краю. В верхней части загиба
2—3 ряда ареол с более длинными грубыми прямыми острыми шипами (двугубыми
выростами?). По краю имеются мелкие шипики против границ ареол и иногда один
ряд удлиненных ареол или мелких точек по периферии. (Табл. 26,10-12).
Морской вымерший вид.
Неоген — Камчатка, Сахалин. Вне СССР. Неоген — Япония.
Рис. 2.
1, 2- Pyxidicula megapora (Grun.) Streln. et Nikolaev: 1 - вид co створки, 2 - целый панцирь. Эоцен,
Зап. Сибирь (ориг. Н. И. Стрельниковой) ; 5-5 - Р. palmeriana-(Giev.) Streln. et Nikolaev: 5 - вид co
створки, 4 — вегетативная клетка сразу после деления, 5 — покоящаяся спора (Cupp, 1943);
6, 7 - Р. turris (Grev. et Arnott) Streln. et Nikolaev f. turris’. 6 - покоящаяся спора, 7- вегетативная
клетка в момент деления (Cupp, 1943). (Шкала: 10 мкм).
45
24. P. similis (Sheshuk.) Streln. et Nikolaev (Стрельникова, Николаев, 1986 : 952). —
Stephanopyxis similis Sheshuk. (Шешукова-Порецкая, 1967 : 135, табл. 12,5a, 6).
Створки сильно выпуклые до полушаровидных, 40—110 мкм в диам., 17—
45 мкм выс., обычно со слегка отогнутым, очень узким, 1 мкм шир., гиалиновым
краем. Ареолы популярные, с прерывистым крибрумом, в центре 1.5—2.5, у края
2—4 в 10 мкм, в прямых тангенциальных рядах. По углам ареол короткие шипики.
На середине радиуса кольцо грубых двугубых выростов, 2.5—3 выроста в 10 мкм,
с наружной трубкой, загнутой кверху, с внутренней стороны створки двугубые
выросты открываются округлым отверстием. (Табл. 26,4—9).
Морской вымерший вид.
Эоцен — Чукотка (р. Анадырь). Неоген — Сахалин.
25. Р. spinosa (Jouse) Streln. et Nikolaev (Стрельникова, Николаев, 1986 : 952). —
Stephanopyxis spinosa Jouse (Жузе, 1955 : 87, табл. 1,8).
Створки цилиндрические или полусферические, толстостенные, 25—50 мкм в диам.,
27—40 мкм выс. Ареолы пекулярные, с острыми шипиками по углам, 2—3 ареолы
в 10 мкм. Двугубые выросты с длинными наружными трубками, расположены ближе
к вершине створки. (Табл. 23,13,14).
Морской вымерший вид. Скорее всего, спора.
Эоцен — Зап. Сибирь, Чукотка (р. Анадырь).
26. Р. spinosissima (Grun.) Streln. et Nikolaev (Стрельникова, Николаев, 1986 :952). —
Stephanopyxis spinosissima Grun. (Grunow, 1884 : 90,91).
Створки равномерно выпуклые, 35—60 мкм в диам. Ареолы 5—7 в 10 мкм,
образуют плотно сомкнутую сеть. Многочисленные двугубые выросты с наружными
трубками беспорядочно разбросаны по створке. (Табл. 24,14-15).
Морской вымерший вид.
Ранний олигоцен—поздний миоцен — п-ов Мангышлак. Вне СССР. Ранний олиго-
цен — Атлантический океан (субтропическая зона). Поздний олигоцен - Норвежское
море. Поздний миоцен — Калифорния.
27. Р . superba (Grun.) Streln. et Nikolaev (Стрельникова, Николаев, 1986 : 953). —
Creswellia superba Grev. (Greville, 1861 : 68, tab. 8,5—5). — Stephanopyxis superba Grun.
(Grunow, 1884: 91).
Створки выпуклые, 24—75 мкм в диам., полусферические, с широким гиалино-
вым краем. Ареолы в параллельных и косо пересекающихся тангенциальных рядах,
3—3.5 в 10 мкм, по краю ареолы крупнее, вытянутой формы, 2—3 в 10 мкм. Один ряд
грубых шипов расположен около края створки. (Табл. 28,15-18).
Морской вымерший вид.
Эоцен — европ. часть СССР. Вне СССР. Поздний эоцен—ранний олигоцен -
о-в Барбадос, южн. часть Тихого океана (Тасманово море), Новая Зеландия (у г. Оамару).
Поздний олигоцен—ранний миоцен — Норвежское море.
28. Р. turns (Grev. et Arnott) Streln. et Nikolaev (Стрельникова, Николаев,
1986 : 953).
Панцирь цилиндрический. Створки выпуклые, от формы часового стекла до
высокого цилиндра. Ареолы пекулярные, расположены плотно сомкнутой сетью, на
вершине в прямых или изогнутых тангенциальных, на загибе в косо пересекающихся
рядах. Двугубые выросты расположены кольцом на границе выпуклого диска и загиба.
Покоящиеся споры эллипсоидной или почти сферической формы, с толстыми стенками,
с грубыми наружными трубками двугубых выростов на вершине.
Вид полиморфный.
Планктонный неритический бореально-тропический вид. Редко в северных
и дальневосточных морях. Нередко в пелагиали более теплых морей.
46
Поздний мел—ныне.
F. turris. — Creswellia turris Grev. et Arnott (Gregory, 1857 : 536, tab. 14, 109). —
Stephanopyxis turris (Grev. et Arnott) Ralfs (Pritchard, 1861 : 826, tab. 5. 74). — S. turris
var. intermedia Grun. (Grunow, 1884 : 87, tab. V, 15, 16). — S. appendiculata Ehr. (Ehrenberg,
1854, tab. 18,4).
Панцирь низкоцилиндрический. Створки 7—60 мкм в диам., 4—50 мкм выс.
Диаметр равен высоте или немного превышает ее. Ареол 3—8 в 10 мкм. (Табл. 28,
1-10\ рис. 2,6, 7).
Планктонная неритическая умеренно тепловодная бореально-тропическая форма.
Поздний мел — Пензенская обл., вост, склон Урала. Палеоцен — Средн. Поволжье,
вост, склон Урала. Эоцен — европ. часть СССР, Зап. Сибирь, КазССР, Дальний Восток
(Чукотка). Поздний олигоцен — южн. Мангышлак. Неоген — юг европ. части СССР,
Дальний Восток. Четверг, период — север европ. части СССР, Зап. Сибирь, Дальний
Восток. Ныне — в пелагиали теплых морей, редко в сев. и дальневосточных морях.
Вне СССР. Поздний мел (сенон) — Польша (Гданьская бухта); Маастрихт — Кали-
форния, южн. часть Тихого океана (плато Кэмпбелл). Палеоцен — южн. часть Атланти-
ческого океана (Фолклендское плато), южн. часть Тихого океана (Тасманово море).
Эоцен — Норвежское море, Дания, Швеция, Калифорния, о-в Барбадос, Куба, юг
Атлантического океана (Фолклендское плато), южн. часть Тихого океана (Тасманово
море). Олигоцен — юг Атлантического (Фолклендское плато), Индийского и Тихого
океанов. Неоген - Япония. Ныне — Атлантический, Тихий океаны.
F. cylindrus (Grun.) Streln. et Nikolaev (Стрельникова, Николаев, 1986 : 953). —
Stephanopyxis turris var. cylindrus Grun. (Grunow, 1884 : 87, tab. V, 7—9).
Панцирь узкоцилиндрический, co створками 12—35 мкм в диам., 18—60 мкм выс.
Высота створки превышает диаметр, поэтому створки ложатся преимущественно
с загиба. (Табл. 28,11—14).
Морская, по-видимому вымершая форма.
Поздний мел — вост, склон Урала. Палеоцен — Поволжье, вост, склон Урала.
Эоцен — европ. часть СССР, Зап. Сибирь, КазССР, Дальний Восток. Поздний олигоцен —
Мангышлак. Неоген — юг европ. части СССР, Дальний Восток. Четверт. период — север
европ. части СССР, Зап. и Вост. Сибирь, Японское море (поверхностный слой донных
осадков). Вне СССР. Эоцен — Дания, Швеция. Неоген — Калифорния, о-ва Прибылова,
Берингово море.
29. Р. uralensis (Streln.) Streln. et Nikolaev (Стрельникова, Николаев, 1986 : 953). —
Stephanopyxis uralensis Streln. (Стрельникова, 1971 : 42, табл. 1,67).
Панцирь линзовидный. Створки слабо выпуклые, 20—59 мкм в диам. Ареолы
популярные, в прямых тангенциальных рядах, 3 в 10 мкм, одинакового размера
по всей створке, образуют плотно сомкнутую сеть. Крибрум прерывистый. Двугубые
выросты, 3 в 10 мкм, с заостренными наружными трубками, расположены плотным
кольцом по самому краю створки, на внутренней поверхности они открываются
округлым отверстием. (Табл. 22,13-15).
Морской вымерший вид.
Поздний мел (кампан) — вост, склон Урала.
30. Р. weissflogii (A. S.) Streln. et Nikolaev (Стрельникова, Николаев, 1986 : 953). —
Stephanopyxis weissflogii A. S. (Schmidt, Atlas, 1888, tab. 123, 2). — S. grunowii Gr. et St.
sensu Jouse (Жузе, 1949 : 39, табл. 8,4).
Панцирь очень выпуклый, сферический. Створки 56—150 мкм в диам., сбоку
в виде трапеции с более или менее выпуклой вершиной, с косо опадающим загибом
и узким гиалиновым краем. Высота загиба 35—40 мкм. Ареолы 1.5—2 в 10 мкм. Кольцо
крупных двугубых выростов расположено в месте перехода лицевой части створки
в загиб. (Табл. 29,10-12).
Морской вымерший вид.
47
Эоцен — Воронежская, Харьковская, Ростовская и Волгоградская обл. Вне СССР.
Эоцен — Норвежское море, Атлантический океан (плато Сан-Пауло, Фолклендское
плато). Поздний эоцен—ранний олигоцен — южн. часть Тихого океана (Тасманово
море), Новая Зеландия (у г. Оамару).
31. Р. weyprechtii Grun. (Grunow, 1884 : 92, tab. V, 5).
Створки равномерно выпуклые, 60—75 мкм в диам., с широким гиалиновым
краем. Ареолы локулярные, на загибе пороидные, с прерывистым крибрумом, на
середине створки 2-3 ареолы, к краю мельче, 4-5 в 10 мкм, расположены в танген-
циальных и вторично косо пересекающихся рядах. Двугубые выросты без наружной
трубки, расположены кольцом по краю створки, а отдельные выросты — беспорядочно
по всей створке, на внутренней поверхности они открываются щелью с утолщенными
валиковидными краями. (Табл. 29,7-9).
Морской вымерший вид.
Поздний мел — вост, склон Урала. Палеоцен — Средн. Поволжье. Эоцен —
Свердловская и Челябинская обл. Плиоцен — Дальний Восток.
32. Р. zabelinae (Jouse) Makar, et Moiss. (Макарова, Моисеева, 1986 : 244-245,
табл. I, 7-75; 11,7-75).— Thalassiosira zabelinae J ouse (Жузе, 1961 : 66, табл. II, 7-7).—
Pyxidicula weyprechtii Grun. sensu Gapon. (Гапонов, 1927, табл. I, 75, 76). — Thalassiosira
usatschevii Jouse (Жузе, 1961 : 64, табл. 1,10).
Панцирь линзовидный, с неодинаково выпуклыми створками: одна равномерно
или конусовидно выпуклая, другая менее выпуклая или уплощенная. Створки круглые,
30—120 мкм в диам., 15—40 мкм выс. Край загиба отогнутый, гиалиновый, 1—2 мкм
шир. Ареолы локулярные, в тангенциальных рядах, на утолщенных створках, переходя-
щих иногда в косо пересекающиеся ряды; на середине створки 4—5 ареол, близ края
5—6.5 в 10 мкм. Крибрум прерывистый, поры на крибруме расположены в прямых
рядах, 7—8 пор в 1 мкм. На вершине более выпуклых створок иногда имеются
1—3 двугубых выроста с наружной трубкой. По краю загиба створки кольцо очень
маленьких двугубых выростов, 1 в 10 мкм, заканчивающихся на наружной поверхности
створки отверстием, а на внутренней — короткой щелью с небольшим утолщением.
В средней части створки несколько редких двугубых выростов, расположенных на
крибруме ареолы, открывающихся в локулу ареолы. Наружная трубка двугубых вы-
ростов выступает из ареолы, а на внутренней поверхности двугубый вырост открывается
узкой щелью с валиковидным утолщением, окруженным порами крибрума. (Табл. 30).
Морской вымерший вид.
Поздний миоцен—плиоцен — сев.-вост, побережье Чукотского моря; Кам-
чатка, о-в Карагинский (тюшевская серия), Сев. Сахалин (маямрафская свита),
Южн. Сахалин (маруямская свита). Плиоцен — о-в Итуруп. Донные осадки дальне-
восточных морей и сев.-зап. части Тихого океана. Вне СССР. Поздний миоцен—плиоцен —
Япония.
Род COSTOPYXIS Gies. (Глезер, 1984 : 291)
Stephanopyxis Ehr. (Ehrenberg, 1844 : 294), р. р.
Панцирь сферический или эллипсоидный. Створки более или менее выпуклые,
колпачковидные, с кольцевой бороздой или шейкой, отделяющей хорошо выраженный
бесструктурный край. Структура из мелких пороидных ареол, разделенных сетью
наружных анастомозирующих ребер, редко ребра отсутствуют. Двугубые выросты на
наружной поверхности створок с длинной трубкой (в СМ — шипы), расположены
на вершине или на середине радиуса створки. С внутренней стороны двугубые выросты
открываются щелью. Видимо, это споры.
48
Виды морские, вымершие.
Известны в мелу и палеогене.
Тип: С. costata (Gies.) Gies.
I. Ребра в параллельных вертикальных рядах, горизонтальные ребра не параллельные
или отсутствуют.
1. Ребра сплошные.
А. На створках имеются вертикальные и горизонтальные ребра.
а. На вершине несколько двугубых выростов.....................1. С. antiqua.
б. На вершине 1 (2) двугубых выроста ..........................7. С. trochlea.
Б. На створках имеются только вертикальные ребра.
а. Двугубые выросты имеются, расположены кольцом по краю диска.............
................................................'......... 4. С. mirabilis.
б. Двугубые выросты отсутствуют............................... 3. С. costata.
2. Ребра прерывистые................................................6. С. schulzii.
II. Ребра в параллельных горизонтальных рядах, вертикальные ряды не параллельные.
1. Система ребер простая ...........................................5. С. reticulata.
2. Система ребер сложная............................................2. С. broschii.
1. С. antiqua (Jouse) Gies. (Глезер, 1984 : 291). — Stephanopyxis antiqua Jouse (Жузе,
1951a : 46, табл. I, За, 36; Стрельникова, 1971 : 56, табл. 3,18-20).
Панцирь сферический. Створки колпачковидные, 42—50 мкм в диам.,
28—40 мкм выс. Ареолы пороидные, 6—7 в 10 мкм, в радиальных рядах. Сеть верти-
кальных и горизонтальных ребер образует почти прямоугольные ячеи, 3 в 10 мкм.
На вершине створки ребра менее грубые, часто едва заметные, образуют ячеи неправиль-
ной формы, иногда ребра отсутствуют. В верхней части загиба, ближе к вершине, одно
горизонтальное ребро выступает в виде неширокого канта. Двугубых выростов 2—3,
расположены группой на вершине створки, с внутренней стороны они открываются
щелью с валиковидными утолщенными краями. Гиалиновый край по периферии густо
заштрихован, 11—12 штрихов в 10 мкм. (Табл. 31,1—3, 6).
Морской вымерший вид.
Поздний мел — Пензенская обл. (р. Мокша), вост, склон Урала.
2. С. broschii (Grun.) Streln. et Nikolaev comb. nov. — Stephanopyxis broschii Grun.
(Grunow, 1884 : 90, tab. 5,26-30).
Панцирь сферический. Створки 11—34 мкм в диам., одинаковой или разной
высоты, 8—35 мкм, со слегка отогнутым краем до 3 мкм шир. Ареолы пороидные,
27—30 в 10 мкм, в косо пересекающихся рядах. На створке сложная система анастомо-
зирующих ребер. Грубые ребра на лицевой части створки образуют ячеи неправильной
многоугольной формы, на загибе ребра в отчетливых горизонтальных и вертикальных
рядах образуют прямоугольные ячеи, 3—5 в 10 мкм. Тонкие ребра разделяют ячеи
сетки грубых ребер на более мелкие ячейки, что придает в СМ ячеям сетки грубых
ребер пятнистый вид. Углы полигональных ячей приподняты в виде бугорков или шипи-
ков. На середине радиуса или ближе к вершине створки кольцо крупных шипов
(в СЭМ — наружные трубки двугубых выростов). (Табл. 31,4, 5, 7, 8).
Морской вымерший вид.
Поздний мел — вост, склон Урала. Палеоцен — Средн. Поволжье, вост, склон
Урала. Эоцен — Калининградская обл., Пензенская обл., УССР, КазССР, УзССР,
Зап. Сибирь. Вне СССР. Эоцен — Швеция.
3. С. costata (Gies.) Gies. (Глезер, 1984:291). - Stephanopyxis costatus Gies. (Глезер,
1967 : 22, фиг. 1-3).
Панцирь сферический. Створки сильно выпуклые, 22 мкм в диам., 25—27 мкм выс.
с ясно выраженной шейкой. Структура из параллельных вертикальных ребер, 4 в 10 мкм,
4 Заказ 518
49
Рис. 3.
1-4- Costopyxis mirabilis (Forti et Schulz) Gies.: 1,2 —
створки с загиба, 3 ~ вид co створки, 4 - целый
панцирь. Ранний мел, альб - ФРГ, у г. Ганновера
(Forti, Schulz, 1932). (Шкала: 10 мкм).
не доходящих до вершины створки, и плот-
ных параллельных рядов мелких пороидов,
28 в 10 мкм, между ними на вершине
створки среди пороидов многочисленные
мелкие шипики. Грубые шипы отсутству-
ют. Край загиба створки бесструктурный.
(Табл. 31,9).
Морской вымерший вид.
Поздний эоцен - КазССР.
4. С. mirabilis (Forti et Schulz) Gies. (Глезер, 1984 : 291). — Stephanopyxis mirabilis
Forti et Schulz (1932 : 246, fig. 6).
Створки колпачковидные, выпуклые, 30-77 мкм в диам., 30—44 мкм выс.,
с более или менее глубокой шейкой. Структура из ребер (на некоторых створках
ребра отсутствуют), между ними расположены радиальные ряды мелких пор, 18—20
в 10 мкм. Кольцо грубых шипов (возможно, наружных трубок двугубых выростов)
расположено по краю створки. Ребра отчетливо видны и на шейке. (Рис. 3).
Морской вымерший вид.
Вне СССР. Ранний мел (альб) — ФРГ (у г. Ганновера).
5. С. reticulata (Long, Fuge et Smith) Gies. (Глезер, 1984 : 292). - Stephanopyxis refz-
culata Long, Fuge et Smith (1946 : 111, tab. 19, 9).
Панцирь сферический. Створки круглые, выпуклые, 20-60 мкм в диам., вершина
створки выпуклая, плоская или слегка вогнутая. Ареолы пороидные, 18 в 10 мкм,
в радиальных рядах. Система ребер простая: на лицевой части створки ребра образуют
сеть с ячеями неправильной формы, на загибе расположены четкими горизонтальными
рядами и менее четкими вертикальными, образуя сеть с прямоугольными ячеями. В мес-
те перехода лицевой части створки в загиб расположен венец двугубых выростов с гру-
быми короткими наружными трубками, 3 в 10 мкм. Край гиалиновый. (Табл. 31,10-14).
Морской вымерший вид.
Поздний мел — Пензенская обл.,вост. склон Урала. Вне СССР. Поздний мел —
Калифорния.
6. С. schulzii (Stein.) Gies. (Глезер, 1984 : 291).
Панцирь сферический. Створки колпачковидные, с кольцевым углублением
(шейкой) и отогнутым гиалиновым краем. Структура из прроидных ареол и шипова-
тых ребер.
Морской вымерший вид, возможно споры.
F. schulzii. - Stephanopyxis schulzii Stein, in Schulz (Schulz, 1935 : 396, tab. 2,
7, 2). — S. schulzii f. schulzii Streln. (Стрельникова, 1974 : 55, табл. IV, 1-3, 8).
Створки 12-80 мкм в диам., 11—50 мкм выс., с низкой шейкой и узким
гиалиновым краем. Шиповатые ребра в параллельных вертикальных и менее правиль-
ных горизонтальных рядах образуют сеть. Ареолы расположены сдвоенными рядами
в ячеях сети, 5—7 ареол в 10 мкм. Двугубые выросты на вершине створки, на внутрен-
ней поверхности створки открываются щелью. (Табл. 31,15-19).
Поздний мел — Пензенская обл., Зап. Сибирь, Гыданский п-ов. Вне СССР. Сенон —
Польша (Гданьская бухта).
50
F. omata (Schulz) Streln. comb. nov. — Stephanopyxis omata Schulz (1935 : 397,
tab. 2,4).
Створки от колпачковидных до почти плоских, 12-60 мкм в диам. Структура
из шиповатых коротких ребер, расположенных беспорядочными вертикальными ряда-
ми. Ареолы пороидные, 5—7 в 10 мкм, в намечающихся радиальных рядах. Край широ-
кий, гиалиновый или с ребрами, отогнутый наподобие полей шляпы или вертикальный.
Наружные трубки двугубых выростов расположены на вершине створки, на внутренней
поверхности створки двугубые выросты открываются щелью. (Табл. 32,1-4, 9-16).
Встречается вместе с типовой формой.
F. nuda Streln. comb. nov. — Stephanopyxis schulzii f. nuda Streln. (Стрельникова,
1974 : 55, табл. IV, 10-12', V,7).
Створки в виде сильно выпуклого колпачка с шейкой и широким отогнутым
гиалиновым краем, 9—100 мкм в диам. Структура из беспорядочно расположен-
ных пороидных ареол, 5—8 в 10 мкм. Двугубые выросты и ребра отсутствуют.
(Табл. 32,5-8).
Встречается вместе с типовой формой.
7. С. trochlea (Hanna) Streln. comb. nov. — Trochosira trochlea Hanna (1927 : 123,
tab. 21,8, 9).
Панцирь эллипсоидальный. Створки колпачковидные, 15—30 мкм в диам.
Ареолы пороидные, 20 в 10 мкм, в радиальных рядах. Ребра на загибе в вертикальных
параллельных рядах и более или менее правильных горизонтальных рядах. К вершине
створки ребра образуют ячеи неправильной формы, 6—7 ячей в 10 мкм. В верхней
части загиба, ближе к вершине, одно горизонтальное ребро выступает в виде неширо-
кого канта. На вершине створки 1 (2) наружные трубки двугубых выростов (в СМ —
шипов), окаймленных гиалиновыми гребнями. (Табл. 32,17,18).
Морской вымерший вид.
Эоцен - Курило-Камчатский желоб. Вне СССР. Средний эоцен — Норвежское
море, южн. часть Атлантического океана, Калифорния, юго-зап. часть Тихого океана.
Род ENDICTYA Ehr. (Ehrenberg, 1845 : 76)
Клетки образуют короткие плотные цепочки, соединенные створками. Хлоро-
пласты в виде крупных пластинок. Панцирь в виде барабана. Поясок образуется перед
делением клетки. Створки круглые, почти плоские, с высоким загибом. Структура
створок из пекулярных ареол с велумом на внутренней поверхности.
Виды морские, литоральные и глубоководные.
Плейстоцен—ныне.
Тип: Е. oceanica Ehr.
1. Е. oceanica Ehr. (Ehrenberg, 1845 : 76).
Створки 80-205 мкм в диам. Ареолы в тангенциальных рядах, на середине
створки 2.3—2.7, у края 3—3.5 ареолы в 10 мкм. Форамены круглые, велум сплошной.
На границе с загибом кольцо двугубых выростов, 3—4 в 10 мкм. Загиб створки высо-
кий, со структурой из ареол, расположенных в горизонтальных и вторично косых
рядах, 3.5-5 ареол в 10 мкм. (Табл. 34, 7-10).
Морской неритический и литоральный, преимущественно бореальный вид.
Поздний эоцен—олигоцен — р. Анадырь. Поздний эоцен — Зап. Сибирь. Миоцен —
Камчатка (тюшевская серия). Поздний миоцен - юг европ. части СССР. Плейстоцен —
Сев.-Зап. СССР. Донные осадки Берингова моря. Ныне — Черное, Японское моря.
Вне СССР. Неоген — Венгрия. Ранний миоцен Испания, Франция. Миоцен — Венгрия,
Индия, Никобарские о-ва. Плиоцен — Калифорния, Перу. Плейстоцен — Швеция, Голлан-
дия. Донные осадки экваториальной зоны Индийского океана. Ныне — северные моря
у берегов Англии, Голландии, Дании, Средиземное море.
51
Сем. THALASSIOSIROPSIDACEAE Nikolaev
Клетки одиночные. Панцирь дисковидный. Створки круглые, слабо выпуклые
или со складкой в середине. Ареолы популярные, в прямых тангенциальных ря-
дах. Один кольцевидный (многоопорный) вырост расположен в самом центре
створки или несколько эксцентрично. Двугубый вырост иногда имеется близ края
створки.
Один морской вымерший род.
Поздний мел, палеоцен, ранний эоцен.
Тип: Thalassiosiropsis Hasle.
Род THALASSIOSIROPSIS Hasle (Hasle, Syvertsen, 1985 : 89)
Панцирь дисковидный. Створки круглые. Ареолы пекулярные, с фораменом на
наружной поверхности створки и крибрумом на внутренней, расположены в прямых
тангенциальных рядах. Ряды крибральных пор на вёлуме в радиальных и взаимо-
перёсекающихся рядах. Один кольцевидный вырост находится в центре или около
центра створки.
Тип: Т. wittiana (Pant.) Hasle.
1. Т. wittiana (Pant.) Hasle (Hasle, Syvertsen, 1985 : 89, tab. 1—5). — Coscinodiscus
lineatus sensu Weisse (1854, tab. 1,2a, b) . — C. lineatus Lfossilis Jouse (Жузе, 1948 : 350).
Створки слабо выпуклые, с небольшим углублением в центре или слабо волни-
стые, 30—220 мкм в диам. У крупных створок ареолы крупные, 3-5 в 10 мкм,
у мелких 6—7 в 10 мкм. Кольцевидный вырост на наружной поверхности створки
имеет короткую конусовидную трубку, которая на внутренней поверхности воронко-
видно расширена и перфорирована у основания многочисленными отверстиями. Вокруг
выроста 2—7 ареол, лишенных крибрума. Загиб створки с одним рядом ареол
и рядом мелких пор. (Табл. 33).
Морской вымерший вид.
Кампан—Маастрихт — вост, склон Полярного и Приполярного Урала (леплинская
свита). Ранний палеоцен — Средн. Поволжье (нижнесызранская подсвита), вост, склон
Урала (марсятская свита). Поздний палеоцен — Средн. Поволжье (камышинская
свита), вост, склон Урала (серовская свита). Вне СССР. Кампан-маастрихт — Тихий
океан (плато Кэмпбелл). Маастрихт — Калифорния (формация Морино).
Сем. TRICERATIACEAE Schutt
Формы колониальные. Панцирь призматический, с поясковыми ободками,
3—многоугольными, редко эллиптическими створками, с выпуклостями на углах,
заканчивающихся глазками. Структура створок, выпуклостей и поясковых ободков
неодинакова. На лицевой части створок пекулярные шестиугольные ареолы
с внутренним велумом, сплошным или прерывистым, и наружным фораменом, в более
или менее прямых рядах, параллельных краям створки или расположенных под
небольшим углом к ним. Иногда по краю створки ряд двугубых выростов с хорошо
развитой наружной частью в виде трубок или двугубые выросты по одному в углах
створки.
Виды морские, вымершие и современные.
Тип: Triceratium Ehr.
П р и м е ч. Семейство монотипное, роды, включенные в это семейство (подсемей-
ство) в системе диатомей Шютта—Хустедта, относятся к другим семействам (Глезер,
1986в).
52
Род. TRICERATIUM Ehr. (Ehrenberg, (1839) 1841 : 159)
Biddulphia Gray sensu Van Heurck (1895 : 466, p. p.).
Описание рода совпадает с описанием семейства.
Тип: Т. favus Ehr., современный.
П р и м е ч. В отличие от классификации Хустедта (Hustedt, 1927—1966), в кото-
рой в род Triceratium включены многочисленные виды, характеризующиеся большим
разнообразием признаков, в данном руководстве этот род принят в узком объеме
в соответствии с первоначальным пониманием его Эренбергом, затем Хэнди (Hendey,
1964). Эволюция взглядов на критерии выделения рода освещена в работе 3. И. Глезер
(1975а).
I. У основания выпуклостей имеется грубый шип (двугубый вырост) ................
. ..........................................................2. Т. unguiculatum.
II. У основания выпуклостей грубый шип (двугубый вырост) отсутствует ... 1. т. favus.
1. Т. favus Ehr. (Ehrenberg, (1839) 1841 : 159).
Створки 3 (4)-угольные. Внутренняя стенка шестиугольных ареол пронизана
коническими порами. Наружное отверстие каждой поры прикрыто выпуклым колпач-
ком, состоящим из нескольких фасеток. На наружной поверхности каждой фасетки
по 1—2 коротких тупых выроста и в основании полулунная щель 15—20 нм шир. От
наружных концов вертикальных стенок в углах ареол отходят раздвоенные или
сильно разветвленные шипы 5—9 мкм дл. По краям створок проходит канал (в СМ —
крыло), внутрь створок открывающийся порами, такими же, как на внутренней стенке
шестиугольных ареол, на наружной тонкой стенке — серия вертикальных щелей
и беспорядочно расположенных мелких круглых отверстий. Внутри канала — верти-
кальные опоры, делящие его на камеры. По краю створки ряд из 5—20 шпателевидно
расширенных на внешнем конце наружных трубок двугубых выростов, открывающихся
эллиптическим отверстием, а на внутренней стороне — щелью, окаймленной с двух
сторон утолщениями, реже щели нет. Поясковых ободков 3, разной структуры
(Miller, Collier, 1978). Самый широкий ободок является соединительным, его края
загибаются внутрь, внешняя кромка края 1—1.5 мкм шир., с нежными штрихами,
5—6 в 1 мкм, остальная часть — 3 мкм шир., бесструктурная. На поясковом ободке
шаровидные камеры (в СМ — точки, 15—16 в 10 мкм), 400—500 нм в диам., с боко-
выми стенками 10 нм толщ., они открываются наружу каналом 100 нм в диам.,
а внутрь — каналом 140 нм в диам. Два других поясковых ободка являются своеобраз-
ными образованиями, расположены последовательно над первым ободком, прикрывая
частично загиб створки. Второй ободок, 4—5 мкм шир., примыкает к первому
и закрывает область краевого внутреннего канала створки, отверстия в его стенке
90—100 нм в диам., 10 отверстий в 10 мкм. Третий ободок расположен выше внутрен-
него краевого канала, прикрепляясь при помощи нежно заштрихованного, завернутого
внутрь края, аналогичного краю первого пояскового ободка. Ширина его неодинаковая,
в середине 6—7 мкм, к углам створок, где расположены выросты, 1.5-3 мкм. Вдоль
наружного края ободка 2 ряда круглых отверстий, на остальной поверхности верти-
кальные пластинки, 22—26 в 10 мкм, а между ними вертикальные щели 50—80 нм шир.
(Табл. 34, 7, 2).
Четверт. период — Сев .-Зап. СССР. Ныне — Японское море. Вне СССР. Миоцен —
Япония. Ныне — все моря Европы. Данные о находках створок этого вида в эоценовых
осадках Атлантического океана требуют уточнения.
П р и м е ч. Росс и Симс (Ross, Sims, 1973) , впервые изучившие Т. favus в СЭМ,
считали, что ареолы у этого вида псевдолокулярного типа. Более детальные исследова-
ния ультра структуры створок вида, проведенные Миллером и Колли (Miller, Collier,
1978), показали, что стенка створки у Т. favus состоит из камер, дно которых образует
53
мембрана сложного строения. Наружу эти камеры открываются крупным круглым
отверстием — фораменом. Согласно принятой в руководстве терминологии, ареолы
у Т. favus пекулярного типа с внутренним велумом и наружным фораменом; велум
типа крибрум, его поры прикрыты крибеллой, которая у данного вида имеет сложное
строение в виде фасеток.
2. Т. ungiuculatum Grev. (Greville, 1864 : 85, tab. 11,9).
Створки четырехугольные, с прямыми или более или менее сильно вогнутыми
сторонами, 20—100 мкм дл. Выпуклости невысокие, конусообразные, с глазками
на вершине. Ареолы в прямых рядах, расположенных почти по диагонали створки,
2—3 ряда в 10 мкм. Крибрум сплошной, его поры в прямых радиальных рядах,
2—3 поры в 1 мкм. Двугубые выросты в углах створки, их наружная трубка (в СМ —
грубый шип) расположена у основания выпуклости, с ее внутренней стороны, внутрен-
няя часть выроста в виде сплющенной трубки со щелью. (Табл. 34,3-5).
Поздний эоцен — Воронежская обл. (касьяновская свита). Вне СССР. Средний—
поздний эоцен — Атлантический океан (о-в Барбадос, Фолклендское плато).
Вид, недостаточно изученный
Т. pulvinar A. S. (Schmidt, 1888, in Schmidt, Atlas, 1874-1959, tab. 126, 8; 168, 7).
Створки четырехугольные, с оттянутыми и заостренными углами, равномерно
слабо выпуклые, с прямыми или довольно сильно вогнутыми сторонами, ПО—
140 мкм дл. Выпуклости небольшие, конусообразные, с тупо срезанной вершиной,
заканчивающейся глазком. Шестиугольные ареолы в середине створки крупные,
2.5 в 10 мкм, у краев мельче, около 4 в 10 мкм, в прямых рядах, расположенных под
некоторым углом к краям створки и пересекающихся по ее диагонали. На внешних
стенках ареол, в их углах, мелкие шипики. Край гиалиновый, около 2 мкм шир.
Поздний эоцен — УССР (обуховская свита). Вне СССР. Поздний эоцен — Новая
Зеландия, Тасманово море. Ранний олигоцен — Тасманово море, Тихий океан
(приантарктическая часть).
П р и м е ч. Описание этого вида в литературе отсутствует. На современных микро-
фотографиях (Hajos, 1976 : 828, tab. 12, 8, 9; 13, 3, 4; 14,1-8; 21, 1, 12) видно его
большое сходство с Т. unguiculatum (Gombos, 1976 : 598, tab. 33, 1, 3; 34, 7-6). Основ-
ное отличие данного вида от предыдущего состоит в отсутствии грубого шипа (наруж-
ной трубки двугубого выроста), однако постоянство этого признака требует уточнения.
Род неясного систематического положения в порядке
Род GLADIUS Schulz (1935 : 391)
Панцирь эллипсоидный или высокоцилиндрический, палочковйдный или в виде
скалки с оттянутыми вершинами. Поясок отсутствует. Створки с расширенным
основанием (базальная часть) и оттянутой в виде выроста вершиной (апикальная
часть). Вершина расширена в разной степени, форма расширения округлая, груше-
видная, усеченная. Структура из пороидных или локулярных ареол с внутренним
крибрумом или шипиков. На конце оттянутой вершины расположен вырост в виде
трубки (в СМ — шипик), окруженный крибральными порами в радиальных рядах,
реже поры расположены беспорядочно или отсутствуют.
Морской вымерший род. характерный для мелового периода.
Тип: G. speciosus Schulz.
1. G. speciosus Schulz (1935 : 391, tab. 1,6-8).
Панцирь эллипсоидный или цилиндрический, палочковидный или в виде скал-
ки, 50—600 мкм дл. Створки от почти полусферических до палочковидных или
54
кеглевидных. Расширенное основание 20—100 мкм в диам., оттянутая вершина
4—60 мкм в диам. Структура базальной части створки из локулярных ареол с широким
фораменом, 2.5—5 ареол в 10 мкм, структура апикальной части варьирует от локулы
до пороида или шипиков, на основе чего выделяются внутривидовые таксоны. На конце
оттянутой вершины расположен вырост в виде трубки (в СМ — шипик), окружен-
ный крибральными порами.
Морфологически очень изменчивый вид. Возможно, это споры.
F. aculeolatus Streln. (Стрельникова, 1974 : 105, табл. 51,5-70). - Pyxilla speciosa
(Schulz) Streln. f. aculeolata Streln. (Стрельникова, 1966 : 35, табл. IV, 3, 4, 11).
Панцирь палочковидный или в виде скалки. Структура базальной части створки
из локулярных ареол, апикальной — из пороидных ареол с шипиками по углам, или
ареолы отсутствуют, а на их месте остаются только шипики. На вершине расположен
довольно длинный вырост в виде трубки (в СМ — шипик). (Табл. 35,1-10).
Морская вымершая форма.
Поздний мел (кампан) — вост, склон Урала.
П р и м е ч. Отличается от типовой формы структурой апикальной части из
пороидных ареол и шипиков.
F. ellipsoideus Streln. (Стрельникова, 1974 : 105, табл. 49, 7, 72, 73, 76, 77, 79). —
Pyxilla speciosa (Schulz)Streln. f. ellipsoidea Streln. (Стрельникова, 1966 :35, табл. Ill, 1-4).
Панцирь эллипсоидно-цилиндрический с закругленной или приостренной верши-
ной. Соотношение высоты створки и диаметра от 1 : 1 до 1 : 3. Ареолы локулярные,
одинаковые по всей створке. (Табл. 35,18-20).
Морская вымершая форма.
Поздний мел (кампан) — вост, склон Урала.
П р и м е ч. От типовой формы отличается эллипсоидно-цилиндрическим панцирем
без оттянутой в виде выроста вершины.
F.poratus Streln. (Стрельникова, 1974 : 106, табл. 49,22; 51,77, 72). - Pyxilla spe-
ciosa (Schulz) Streln. f. porata Streln. (Стрельникова, 1966 : 35, табл. IV, 5, 6, 72).
Панцирь высокоцилиндрический, в виде скалки с очень длинными и тонкими,
оттянутыми в виде выростов вершинами. Структура расширенной базальной части
створки из локулярных ареол, апикальной — из беспорядочно расположенных пор.
На середине вершины — вырост в виде трубки (в СМ — шип) 5—10 мкм дл. (Табл. 35,
11-17).
Морская вымершая форма.
Поздний мел (кампан) — вост, склон Урала.
П р и м е ч. Отличается от типовой формы структурой апикальной части из
редких пор.
F. speciosus Schulz (1935:391, tab. 1, 6-8) emend. Streln. (Стрельникова, 1974 : 104,
табл. 49,2-77, 74, 75, 73,20,27; 50,2-79; 51,7,4). - Gladius jouseanus Haj6s (1975 :
932, tab. 11, 7-72). - G. maximus Hajos (1975 : 933, tab. 11. 13). — G. pacificus Hajos
et Stradner (Hajos, 1975 : 933, tab. 11,7, 2; 26,2). — Tubularia pistillaris var.grossepunctata
Long, Fuge et Smith (1945 : 115, tab. 18, 72).
Панцирь цилиндрический, палочковидный или в виде скалки, с оттянутой
в виде выроста апикальной частью, вершина которой расширена в разной степени:
округлая, грушевидная, усеченная. Структура базальной и апикальной частей из
локулярных ареол. Маленький трубковидный вырост (в СМ — шипик) окружен
крибральными порами в радиальных рядах или расположенных беспорядочно.
(Табл. 36).
Морская вымершая форма.
Поздний мел (кампан, Маастрихт) — Пензенская обл., вост, склон Урала, Гыдан-
ский п-ов, Корякский хребет (р. Апука). Вне СССР. Поздний мел (сенон) -- Польша
(Гданьская бухта); Маастрихт — Калифорния, южн. часть Тихого океана (плато
Кэмпбелл).
Вид, недостаточно изученный.
55
G. antiquus Forti et Schulz (1932 : 242, tab. 3,6; texttab.3; Диат. анал. кн. 2, 1949 :
199, табл. 97,5а-в).
Эпитека цилиндрическая, 200—360 мкм дл., вершина ее головчатая, 30—62 мкм
в диам., суженная шейка 14—60 мкм в диам. На вершине головки 2 острых, более или
менее длинных шипа. Ареолы в 3 взаимно перекрещивающихся рядах, 4-6 в 10 мкм.
Мел (альб) - Свердловская обл. Вне СССР. Ранний мел (альб) — ФРГ (у г. Ган-
новера) .
Порядок TH ALASSIOSIR ALES Gies, et Makar.
Клетки образуют плотные нитевидные или гибкие колонии, в которые они
соединены непосредственно краевыми шипиками, краевыми выростами с опорами
или тяжами, выходящими из центральных выростов с опорами, редко клетки оди-
ночные. Панцири цилиндрические, со вставочными и соединительными ободками или
без них. Створки круглые, редко эллипсоидные, плоские, слегка выпуклые, вогнутые,
тангентально- или концентрически-волнистые. Ареолы л окулярные или пороидные,
с фораменами на наружной поверхности створки и крибрумом на внутренней (редко
наоборот), расположенные в тангенциальных или радиальных рядах или пучках. Иногда
имеются гиалиновые радиальные линии в виде ребер, заканчивающиеся шипом на заги-
бе, а в периферической зоне створки — альвеолы. По краю створки или на загибе обычно
одно кольцо (реже их более одного) выростов с 2-5 опорами; центральные выросты
с опорами от одного до многих. Двугубый вырост один, реже несколько, в краевой
зоне, иногда отступя от края ближе к центру.
Виды морские, пресноводные, современные и вымершие.
Типовое семейство: Thalassiosiraceae Lebour emend.Hasle,
I. Форамен на наружной поверхности створки, крибрум на внутренней.
1. Створки плоские, иногда выпуклые или вогнутые, очень редко с тангентальной
волнистостью. Ареолы в тангенциальных или радиальных рядах, иногда образую-
щих пучки. Выросты с опорами обычно хорошо заметные, с 3—5 сопутствующими
порами. Виды преимущественно морские..............сем. Thalassiosiraceae (с. 56).
2. Створки плоские, с тангентальной или радиальной волнистостью. Ареолы в ра-
диальных рядах. На створке имеются гиалиновые радиальные линии или ребра
или в периферической зоне альвеолы. Выросты с опорами малозаметные,
с 2—3 сопутствующими порами. Виды преимущественно пресноводные.............
....................................................сем. Stephanodiscaceae.
II. Форамен на внутренней поверхности створки, крибрум на наружной............
.......................................................сем. Ectodictyonaceae.
Сем. THALASSIOSIRACEAE Lebour emend. Hasle.
Клетки в колониях, образованных путем соединения трубок выростов с опорами
или при помощи хитиновых тяжей, выходящих из трубок. Панцирь от низко- до
высоко цилиндрического, иногда с многочисленными вставочными и соединительными
ободками, вследствие чего первальварная ось значительно превышает диаметр клетки.
Хлоропласты в виде зерен или округлых пластинок. Створки круглые, плоские,
выпуклые или вогнутые, очень редко с тангентальной волнистостью. Ареолы распо-
ложены в радиальных рядах, иногда группирующихся в пучки, или в тангенциальных
рядах. В центре створки один, несколько или много выростов с опорами, образующих
центральный пучок, иногда одна крупная ареола или центральный вырост, или вырост
находится отступя от центра. По краю створки выросты с опорами образуют 1,2 или
более колец, иногда они расположены по всей поверхности створки. Наружная часть
трубок равна по длине внутренней, больше или отсутствует. Краевые выросты
с опорами с 3—5 сопутствующими порами. Двугубый вырост часто крупнее
выростов с опорами, с длинной наружной трубкой, реже без нее, редко очень маленький,
56
незаметный; обычно расположен в краевом кольце выростов с опорами, реже отступя
от края или в средней части створки. Его внутренняя щель ориентирована радиально,
под углом 'или тангентально. Иногда имеются замкнутые выросты, расположенные
по краю створки. В некоторых родах известны споры.
Виды морские, солоноватоводные, редко пресноводные, вымершие и современ-
ные, известные с миоцена, за исключением нескольких видов, найденных в эоцене,
космополит.
Тип: Thalassiosira Cl.
I. Клетки одиночные, редко парные или собраны в группы.
1. Клетки только одиночные. Панцирь низкоцилиндрический, более 20 мкм в диам.
Выросты с опорами образуют краевое кольцо. Двугубый вырост у края створки
или на загибе.
А. Створки эллиптические, редко почти круглые, тангентально-волнистые по
продольной оси. Ареолы в радиальных, иногда в неотчетливых продольных
рядах Выросты с опорами крупные. Двугубый вырост один на загибе............
...................................................род Cymatotheca (с. 84).
Б. Створки круглые, плоские или слегка выпуклые. Ареолы в тангенциальных
рядах. Выросты с опорами маленькие. Двугубых выростов 2 у края створки,
расположены друг против друга по диаметру створки..........................
...................................................род Planktoniella (с. 85).
2. Клетки одиночные или парные, иногда собраны в группы. Панцирь высоко цилинд-
рический, менее 10 мкм в диам. Выросты с опорами вне краевого кольца. Дву-
губый вырост близ центра створки ........................род Minidiscus (с. 90).
11 Клетки обычно в плотных нитевидных или гибких колониях или погружены в сли-
зистую массу.
1. Клетки соединены в гибкую цепочку одним или несколькими хитиновыми тяжами
или погружены в слизистую массу.
А. Панцирь всегда с многочисленными вставочными и соединительными ободками.
Ареолы в радиальных рядах. Выросты с опорами по всей поверхности створки,
кроме центральной части. Двугубый вырост на наружной поверхности створки
без трубки, его отверстие крупное, радиально вытянутое . . род Porosira (с. 86).
Б. Панцирь с меньшим числом вставочных и соединительных ободков. Ареолы
в тангенциальных и радиальных рядах, иногда собранных в пучки. Выросты
с опорами образуют краевое кольцо, в центре створки один, несколько или
пучок выростов. Двугубый вырост на наружной поверхности с трубкой, реже
без нее,его отверстие круглое ........................род Thalassiosira (с.58).
2. Клетки соединены в прочные нитевидные цепочки непосредственно выростами
с опорами или замкнутыми выростами.
А. Клетки соединены в цепочки центральными выростами с опорами и краевыми
замкнутыми выростами. Панцирь высокоцилиндрический, тонкий, с многочис-
ленными вставочными и соединительными ободками .... род Lauderia (с. 88).
Б. Клетки соединены в цепочки краевыми или центральными выростами с опо-
рами. Панцирь низкоцилиндрический или дисковидный, довольно толстый,
с несколькими вставочными и соединительными ободками.
а. В центре створки пучок из выростов с опорами, при помощи которых клет-
ки соединяются в цепочки. Краевые выросты с опорами с короткой наруж-
ной трубкой.........................................род Bacterosira (с. 87).
б. В центре створки один вырост с опорами или он отсутствует, клетки соеди-
няются в цепочки краевыми выростами с опорами. Краевые выросты с опо-
рами с длинной наружной трубкой.
а. Панцирь цилиндрический. Краевые выросты с опорами в виде коротких
трубок, в центре створки один вырост. Цепочки короткие ...............
............................................... род Detonula (с. 87).
57
(3. Панцирь линзовидный или высокоцилиндрический. Краевые выросты
с опорами щелевидно-желобковидные или трубковидные, в центре створки
вырост отсутствует. Цепочки обычно длинные . . . род Sceletonema (с. 82).
Род THALASSIOSIRA Cl. (Cleve, 1873 : 6)
Клетки образуют гибкие цепочки,в которых они соединены между собой одним
или несколькими хитиновыми тяжами, выходящими через центральные выросты
с опорами. Иногда клетки погружены в слизистую массу или капсулы, редко клетки
одиночные. Хлоропласты пластинчатые, прилегающие к створкам или пояску. Панцирь
в виде высокого или низкого цилиндра, редко дисковидный, с пояска четырех- или
восьмиугольный. Поясковая зона теки обычно состоит из ареолированной вальво-
копулы, вставочного ободка с одним рядом пор и нескольких, обычно бесструктурных
соединительных ободков (СЭМ). Ободки воротничковидной формы, часто с лигулой,
иногда имеются септы. Створки круглые, плоские, слегка или очень выпуклые или
вогнутые посередине, ареолированные, иногда с радиальными ребрами. Ареолы
локулярные, в прямолинейных или изогнутых тангенциальных рядах либо радиаль-
ных, иногда собранных в пучки, в которых радиальные ряды ареол параллельны
среднему ряду. Краевые выросты с опорами образуют обычно одно кольцо, редко
больше, иногда расположены по всей поверхности створки. В центре створки один,
несколько или много выростов с опорами. Они имеют выступающую наружную трубку
или простирающуюся внутрь клетки, окруженную 2—5 сопутствующими порами. Дву-
губый вырост обычно один, редко два или более, находится чаще всего в прикраевой
зоне, иногда отступя от края или ближе к центру створки (у одного вида в центре
створки), он обычно с длинной трубковидной наружной частью, реже его крупное
наружное отверстие не выступает над поверхностью створки. Для некоторых видов
известны замкнутые выросты и шипы в прикраевой зоне. Загиб створки низкий
или высокий, со структурой из мелких ареол, часто край загиба заканчивается корот-
кими ребрышками. Известны ауксоспоры и споры. Некоторые ископаемые виды
сохранились только в виде спор.
Виды планктонные, морские, солоноватоводные, редко пресноводные, совре-
менные и вымершие.
Тип: Т. nordenskioeldii CL
Род разделен на 4 секции.
I. Структура створок в СМ обычно отчетливая, ареолы расположены в определенном
порядке.
1. Ареолы в тангенциальных или прямолинейных рядах, радиальные ряды выра-
жены неотчетливо................................1. Секция Tangentales (с. 58).
2. Ареолы в отчетливых радиальных рядах.
А. Радиальные ряды ареол образуют пучки, иногда у края заметны ареолы в тан-
генциальных рядах.............................2. Секция Fasciculigera (с. 69).
Б. Радиальные ряды ареол не образуют пучков, только иногда прослеживаются
неотчетливые пучки или заметны вторичные концентрические круги ...........
...................................................3. Секция Thalassiosira (с. 73).
II. Структура створок в СМ почти невидимая, расположение ареол неясное.........
..............................................4. Секция Inconspicuae (с. 81).
1. Секция TANGENTALES Makar. (Макарова, 1971 : 70)
Створки плоские или слегка вогнутые. Ареолы в прямых или изогнутых танген-
циальных рядах или расположены линейными рядами, иногда намечаются отдельные
радиальные ряды. По краю створки или на загибе одно кольцо крупных или мелких
выростов с опорами, в центре створки обычно один вырост с опорами, редко их более
58
одного, иногда они имеются по всей поверхности створки. Краевые выросты с опорами
обычно с более длинной наружной частью, центральные выросты часто не выступают
над наружной поверхностью створки. Двугубый вырост один, редко два или более,
расположен обычно на границе с загибом створки. В прикраевой зоне створки иногда
имеются шипы или замкнутые выросты
Тип: Т. eccentrica (Ehr.) CL
I. Краевые выросты сопорами малозаметные в СМ, находятся на загибе створки.
1. По краю створки кольцо отчетливых шипов или замкнутых выростов
А. Створки крупные, до 110 мкм в диам., в центре створки одна крупная ареола,
окруженная 6—8 более мелкими ареолами. Рядом с центральной ареолой один
очень маленький вырост с опорами .............................9. Т. eccentrica.
Б. Створки не более 50мкм в диам., крупная ареола в центре створки отсутствует,
а В центре створки несколько или группа маленьких выростов с опорами.
Ареолы расположены в тангенциальных, иногда с тенденцией к образованию
пучков, или в линейных рядах.
а. В центре створки одна маленькая ареола окружена кольцевидным же-
лобком, в котором расположены 5—6 выростов с опорами ..................
................................................... 20. Т. nodulolineata
0. В центре створки маленькая ареола, окруженная желобком, отсутствует.
+ Центральных выростов с опорами 3-5, расположены в центре створки
...........................................................12. Т. incerta.
++ Центральных выростов с опорами много, расположены свободно
......................................................... 18. Т. marujamica.
б. В центре створки вырост с опорами отсутствует. Ареолы расположены
в линейных рядах.............................................. 15. Т. leptopus.
2. По краю створки кольцо шипов или заметных выростов отсутствует .............
.....................................................................4. Т. antiqua.
II. Краевые выросты с опорами хорошо заметные в СМ, находятся на границе с загибом
створки.
1. Створки с одним кольцом крупных и отчетливых краевых выростов с опорами,
имеющих трубки на наружной поверхности створки значительной длины.
А. Краевые выросты с опорами расположены равномерно.
а. В центре створки один вырост с опорами.
а. Центральный вырост с опорами хорошо заметный в СМ, крупный, с разви-
той трубкой на наружной поверхности створки.............22. Т. pacifica.
0. Центральный вырост с опорами малозаметный в СМ, на наружной поверх-
ности створки его трубка заканчивается отверстием.
+ Краевые выросты с опорами на наружной поверхности створки имеют
вид конических трубок, 4—6 в 10 мкм. Двугубый вырост расположен
посередине между двумя выростами с опорами.
— Ареолы на створке почти одной величины, на загибе створки обра-
зуют косо пересекающиеся ряды . ... ?^.............6. Т. coronifera.
— Ареолы близ центра створки более крупные, уменьшающиеся
к краю, на загибе образуют вертикальные ряды .... 7. Т. decipiens.
++ Краевые выросты с опорами на наружной поверхности створки имеют
вид цилиндрических трубочек, вставленных коаксильно одна в другую,
3 выроста в 10 мкм. Двугубый вырост расположен ближе к одному
из выростов с опорами.................................. 2. Т. angulata.
б. Близ центра створки незамкнутое кольцо выростов с опорами..............
....................................................3. Т.anguste-lineata.
Б. Краевые выросты с опорами расположены неравномерно.
а. Имеются выросты с опорами на лицевой части створки. Ареолы мелкие,
14-16 в 10 мкм............................................17. Т. makarovae.
59
б. Выросты с опорами на лицевой части створки отсутствуют. Ареолы крупные,
6—9 в 10 мкм..............................................10. Т. heteromera.
2. Створки с одним кольцом не крупных краевых выростов с опорами (иногда они
незаметные), имеющих трубки на наружной и внутренней поверхностях створки
незначительной длины.
А. Краевые выросты с опорами довольно отчетливые в СМ, с трубкой, выступаю-
щей на наружной поверхности створки.
а. Центральный вырост с опорами один.
а. Центральный вырост с опорами малозаметный в СМ или отсутствует,
двугубый вырост отчетливый.
+ Двугубый вырост один, находится в одном кольце с выростами с опо-
рами, расположенными на границе с загибом створки ... 25. Т. tenera.
++ Двугубых выростов 3................................13. Т. intermedia.
0. Центральный вырост с опорами отсутствует, двугубый в СМ незаметен
..........................................................8. Т. delicatissima.
б. Центральных выростов с опорами более одного.
а. Ареол на середине створки 7—11, у края 12—18 в 10 мкм. Клетки соеди-
нены в цепочку несколькими тяжами..........................1. Т. aculeata.
ft. Ареол на середине створки 12—16, у края 18—20 в 10 мкм. Клетки соеди-
нены в цепочку одним тяжом ................................23.-Т. parva.
Б Краевые выросты с опорами не отчетливые в СМ, с трубкой, выступающей
внутрь клетки.
а. Внутренняя часть выростов с опорами с заметной трубкой.
а. В центре створки один вырост с опорами. Двугубый вырост близ центра
створки................................................ 21. Т. oestrupii.
р. В центре створки выростов с опорами более одного, они образуют 1 или 2
прямые или изогнутые линии. Двугубый вырост в одном кольце выростов
с опорами или отступя внутрь.
+ Загиб створки высокий, центральные выросты с опорами образуют
одну изогнутую линию...............................14. Т. latimarginata.
++ Загиб створки низкий, центральные выросты с опорами образуют
обычно 2 линии.
— Ареолы в неясных тангенциальных рядах, с тенденцией к образова-
нию пучков, 8-11 ареол в 10 мкм. Центральные выросты с опорами
образуют 1—2 линии....................................5.Т. confusa.
— Ареолы в тангенциальных, изогнутых к центру рядах, на середине
створки 6—9 в 10 мкм. Центральные выросты с опорами образуют
всегда 2 линии.........................................11. Т. ignota.
б Внутренняя часть выростов с опорами с малозаметной трубкой.
а. Створки плоские. Ареолы 5—6-угольные, довольно крупные, на середине
створки до 12 ареол в 10 мкм.
+ Створки до 20 мкм в диам. Ареолы резко уменьшающиеся к краю,
на середине 10—12, у края 16—20 в 10 мкм................. 19. Т. nativa.
++ Створки более крупные, 18—28 мкм в диам. Ареолы одинаковой
величины, в прикраевой зоне удлиненные, 6—8 в 10 мкм..................
..................................................24. Т. singularis.
р. Створки почти плоские. Ареолы круглые, на середине створки до 18
в 10 мкм............................................... 16. Т. levanderi.
1. Т.aculeata Pr.-Lavr. (Прошкина-Лавренко, 1956 : 59,рис. а—к).
Клетки одиночные или в коротких цепочках, соединенные 3—5 короткими
тяжами. Панцирь в виде барабана,9—11 мкм выс., с 2 вставочными ободками. Створки
слегка выпуклые, 8—38.5 мкм в диам. Ареолы в тангенциальных рядах с тенденцией
к радиальным, на середине створки 7— 11, у края 12—18 ареол в 10 мкм. В центре створки
60
несколько маленьких выростов с опорами. По краю створки кольцо выростов
с опорами, 3—5 в 10 мкм. Наружная часть выростов превышает внутреннюю. Дву-
губый вырост в одном кольце с выростами с опорами. Загиб створки невысокий,
с мелкими ареолами в косых, взаимопересекающихся рядах. (Табл. 37,1-4).
Солоноватоводный неритический низкобореальный вид.
Плиоцен (акчагыл) — г. Уральск. Ныне — Азовское, Каспийское моря, возможно
азово-каспийский эндемик.
2. Т. angulata (Greg.) Hasle (1978 : 93—101, fig. 4, 41, 70—99). — Orthosira angulata
Greg. (Gregory, 1857 : 498, tab. 10, 43). — Coscinodiscus decipiens Grun. (Grunow in Van
Heurk, 1880—1885, tab. 91,70). — Thalassiosira decipiens (Grun.) Jtfrg. sensu Hust. (Hustedt,
1927-1930: 322, fig. 158).
Клетки соединены длинным тяжом в рыхлые цепочки от 4 до 18 клеток. Панцирь
в виде цилиндра,8—28 мкм выс., с несколькими вставочными и соединительными обод-
ками. Створки почти плоские, 17—40 мкм в диам. Ареолы в тангенциальных изогнутых
рядах, в центре створки 8—12, у края 12—15 в 10 мкм. Форамены почти круглые,
крибрум неправильных очертаний. Центральный вырост с опорами обычно рядом
с центральной, более крупной ареолой. Краевые выросты с опорами крупные, на гра-
нице с загибом створки, 3 выроста в 10 мкм. На наружной поверхности трубка больше,
чем на внутренней, 1.6—1.7 мкм дл., и состоит из 2 трубочек различного диаметра,
вставленных одна в другую. Двугубый вырост в виде слегка изогнутого цилиндра
2.6—2.7 мкм дл., в одном кольце с выростами с опорами, ближе к одному из них, его
внутренняя часть в виде сплющенной трубки, щель ориентирована радиально. Загиб
створки довольно высокий, с более мелкими ареолами, 16—20 в 10 мкм. По краю
загиба короткие ребрышки, 17—18 в 10 мкм. (Табл. 37,5—9).
Морской неритический и океанический планктонный широкобореальный вид.
Плейстоцен — Ленинградская, Архангельская обл. Голоцен — Онежский и Коль-
ский п-ова. Ныне — Белое, Баренцево, Карское, Лаптевых, Чукотское, Берингово,
Охотское и Японское моря, бухта Патрокл, Амурский лиман, Чукотка (р. Яма).
Вне СССР. Средний миоцен — Япония. Плиоцен — Калифорния. Четверт. период —
Фенноскандия. Ныне — от Норвежского моря до берегов Португалии, Атлантический
океан: у вост берегов Сев. Америки, заливы Мэн, Наррагансетт, у берегов Греции,
залив Сароникос, Тихий океан: у берегов Сев. Америки, Калифорнийский залив,
Пьюджет Саунд.
3. Т. anguste-lineata (A. S.) Fryx. et Hasle (Fryxell, Hasle, 1977 : 73-75, fig. 22-34). -
Coscinodiscus anguste-lineatus A. S. (Schmidt, 1878, tab. 52, 34). — C. polychordus Gran
(1897 : 30, tab. 2, 33', tab. 3, 56). — Coscinosira polychorda Gran (1900 : 115). —
Thalassiosira polychorda (Gran) J0rg. (Jorgensen, 1899 : 15). — T. omata Pr.-Lavr.
(Прошкина-Лавренко, 1959 : 76, рис. 1—8).
Клетки соединены 4—9 довольно длинными тяжамй в цепочки. Панцирь в виде
барабана с 2 вставочными ободками. Створки почти плоские, 14—60 мкм в диам.
Ареолы в тангенциальных изогнутых или прямых рядах, переходящих иногда в прямо-
линейные или образующих пучки, в которых ряды ареол параллельны более длинному
среднему ряду, 8—12 в 10 мкм. Незамкнутое кольцо выростов с опорами близ центра
створки состоит из отдельных групп выростов; в каждой группе 2—5 выростов. Их
наружная часть в виде маленького отверстия, внутренняя — короткой трубки. Краевые
выросты с опорами расположены равномерно на границе с загибом створки, 3—4 вы-
роста в 10 мкм, имеют форму цилиндра с подставками близ наружной поверхности
створки. Двугубый вырост крупный, на наружной поверхности створки в виде цилинд-
ра, на внутренней — уплощенной трубки, в одном кольце с краевыми выростами с опо-
рами, его внутренняя щель ориентирована радиально. Загиб створки низкий, с мелкими
ареолами, расположенными в трех взаимопересекающихся рядах, 24 ареолы в 10 мкм.
Край загиба створки заканчивается четкими ребрышками, 14—16 в 10 мкм. (Табл. 38).
61
Морской и солоноватоводный неритический и океанический холодноводный
и тепловодный широкобореальный и нотальный вид.
Поздний миоцен (мэотис) — Крым. Плиоцен — сев .-зап. часть Тихого океана. Позд-
ний плейстоцен — Чукотка. Голоцен — Белое море (послеледниковые отл.). Ныне -
Белое, Баренцево, Черное, Чукотское, Охотское, Берингово, Японское моря, Куриль-
ский пролив, бухта Патрокл. Вне СССР. Миоцен—плиоцен - Японское море. Ныне -
Норвежское, Северное моря, проливы Скагеррак, Каттегат, Ла-Манш, сев. Атлантика,
заливы юго-вост, и сев. Канады, Гудзонов залив и канадская Арктика, Тихий океан:
сев. часть и вдоль берегов Японии, а также у берегов южн. Калифорнии и Чили.
4. Т. antiqua (Grun.) A. Cl. (Cleve-Euler, 1941 : 173, fig. 4). - Coscinodiscus antiquus
Grun. (Grunow, 1884 : 84, tab. 4, 24). — C. antiquus var. lapponicus A. Cl. (Cleve-Euler,
1951 : 72, 119, b-d). — C. antiquus var. lapponica f. inermis A. Cl. (Cleve-Euler, 1941 : 174,
fig. 5, 77).
Панцирь в виде барабана. Створки почти плоские, 17—64 мкм в диам. Ареолы
с круглыми или вытянутыми фораменами, в тангенциальных, вогнутых к центру
рядах, на середине створки 4—6, близ края 6—11 ареол в 10 мкм. В центре створки
одна крупная ареола, или ареолы мелкие, свободно расположенные. Центральный
вырост с опорами очень маленький, заканчивается отверстием на наружной поверхно-
сти. Краевые выросты с опорами небольшие, многочисленные, расположенные на загибе
створки (в СМ почти незаметные). Двугубый вырост со слегка удлиненной наружной
частью, расположен на границе с загибом. Загиб створки очень низкий, заканчивается
очень короткими ребрышками. (Табл. 37,10-13) .
Морской неритический и океанический вымерший вид.
Поздний миоцен—ранний плиоцен — Сев. Сахалин (маямрафская свита), Южн. Са-
халин (маруямская свита). Миоцен — Камчатка (тюшевская серия). Плиоцен —
о-в Итуруп. Плейстоцен — Сев.-Зап. СССР. Вне СССР. Ранний эоцен — Дания. Эоцен —
Швеция. Поздний миоцен — Калифорния (формация Монтерей). Миоцен—плиоцен —
Япония. Поздний плиоцен — о-в Хонсю.
5. Т. confusa Makar. (Макарова, 1975 : 149, рис., 7, 2).
Клетки образуют цепочки из 4—6 клеток, соединенных несколькими тонкими
тяжами. Панцирь в виде барабана, 16.5—20 мкм выс. Створки почти плоские, 21.6—
24.5 мкм в диам. Ареолы в неясных тангенциальных рядах, с тенденцией к образо-
ванию пучков, 8—11 ареол в 10 мкм в центре створки и 9—12 у края. В центре створки
5—7 отчетливых выростов с опорами, расположенных в 1—2 слегка изогнутые линии.
Краевые выросты с опорами на границе с загибом створки расположены равномерно,
2 в 10 мкм. Все выросты с опорами с внутренней стороны створки имеют трубку с 3 опо-
рами и более длинную у краевых выростов, с наружной — отверстие. Двугубый вырост
в одном кольце с выростами с опорами или немного отступя внутрь, занимает место
одного из них или ближе к одному из выростов, на внутренней поверхности створки
с короткой трубкой, переходящей в сильно расширенную уплощенную часть, с длинной
щелью, ориентированной тангенциально или под углом к радиусу створки, с наружной
поверхности он заканчивается отверстием. Загиб створки низкий, с вертикальными
рядами ареол, 12—14 в 10 мкм. (Табл. 39,1—8).
Морской высоко бореальный вид.
Поздний плейстоцен — Чукотка (бухта Провидения). Ныне — Баренцево море
(Дальние Зеленцы). Вне СССР. Ныне - сев .-вост, часть Тихого океана: Алеутский
желоб, Аляскинский залив, к юго-востоку от Курильских о-вов.
6. Т. coronifera (Pr.-Lavr.) Pr.-Lavr. (Прошкина-Лавренко, 1960а :48). - Т. coronata
Рг.-1^г.(Прошкина-Лавренко, 1955а : 45, табл. I, 6-10).
Клетки одиночные или соединены тяжом в короткие цепочки. Панцирь в виде
барабана. Створки слегка вогнутые, 11—48 мкм в диам., с неровной бугристой лицевой
62
частью. Ареолы в тангенциальных рядах с переходом в радиальные, на середине створки
8—10 ареол, у края 15—17 в 10 мкм. В центре створки один вырост с опорами (мало-
заметный в СМ), на наружной поверхности створки имеет очень маленькое отверстие,
а на внутренней — небольшую трубку. Краевые выросты с опорами в виде длинных
конических трубочек, скошенных вверху, расположены в верхней части загиба створки
очень равномерно, по 4—5 в 10 мкм. Двугубый вырост около 2 мкм дл., в одном
кольце с выростами с опорами. Загиб створки высокий, с отчетливыми косо пересекаю-
щимися рядами ареол, 16—17 в 10 мкм. (Табл. 37,14-19).
Солоноватоводный неритический планктонный низкобореальный вид, возможно
эндемик.
Поздний миоцен (поздний сармат) — Таманский п-ов. Плиоцен (акчагыл) —
г. Уральск. Ныне — Черное, Азовское, Каспийское моря.
7. Т. decipiens (Grun.) J<£rg. (Grunow, 1878 : 125). - Coscinodiscus eccentricus Ehr.
var.? decipiens Grun. (Grunow in Schneider, 1878 : 128, tab. 4,18), non Thalassiosira deci-
piens Grun. (Grunow in Jorgensen, 1905 : 96, fig. 3).
Клетки образуют колонии двух типов: соединены одним центральным тяжом
или непосредственно выростами створок. Панцирь от линзовидного до цилиндриче-
ского, со вставочными и соединительными ободками. Створки от выпуклых до вогну-
тых, 9—40 мкм в диам. Ареолы в тангенциальных, дуговидно изогнутых рядах с отдель-
ными радиальными рядами, близ центра створки более крупные, к краю уменьшаю-
щиеся, 8—12 в 10 мкм. Форамены в СМ шестиугольной формы, крибрум неправильных
очертаний. Центральный вырост с опорами близ центральной ареолы, на наружной
поверхности имеет очень маленькое отверстие, а на внутренней — небольшую трубку,
окруженную 4 сопутствующими порами. На загибе створки кольцо краевых выростов
с опорами, 4—6 в 10 мкм, с трубкой на наружной поверхности створки значительной
длины. Двугубый вырост крупный, расположен между 2 краевыми выростами с опора-
ми, с наружной поверхности створки его трубка по размерам значительно больше,
чем у выростов с опорами, его внутренняя щель ориентирована радиально. Загиб
створки невысокий, плавно изогнутый, с ареолами в вертикальных рядах, 10—15
в 10 мкм. Известны ауксоспоры. (Табл. 39, 9).
Солоноватоводный эвригалинный широко бореальный вид, обитающий как эпи-
фит, реже в планктоне.
Ныне — Каспийское и Баренцево моря. Вне СССР. Ныне — рр. Темза, Эльба,
в осадках рек и эстуариев вдоль побережья Северного моря, водоемы Сев. Америки.
8. Т. delicatissima Pr.-Lavr. (Прошкина-Лавренко, 19606 : 58, табл.11,21).
Панцирь низко цилиндрический. Створки плоские, 3.5—14 мкм в диам. Ареолы
мелкие, в неправильных тангенциальных рядах с тенденцией к образованию полукон-
центрических рядов, в центре створки 24—28 ареол, у края 30 в 10 мкм. Краевые
выросты с опорами очень маленькие. Двугубый вырост в СМ незаметен. (Табл. 39, 77).
Солоноватоводный вымерший вид.
Поздний миоцен (мэотис) — Таманский и Керченский п-ова.
9. Т. eccentrica (Ehr.) Cl. (Cleve, 1903—1904 : 216). — Coscinodiscus eccentricus Ehr.
(Ehrenberg, 1839 : 146). — C. eccentricus var. fasciculatus Hust. (Hustedt, 1927—1930 : 390,
fig. 202).
Клетки соединены тонким тяжом в непрочные цепочки. Панцирь низкоцилиндри-
ческий, с 2 вальвокопулами, со структурой из мелких ареол, расположенных вертикаль-
ными рядами, 4 ряда в 1 мкм, 2 вставочными ободками, имеющими один ряд пор,
и несколькими соединительными ободками с лигулами. Створки плоские, 18—110 мкм
в диам. Ареолы в тангенциальных, изогнутых к центру створки рядах, иногда с тенден-
цией к образованию пучков, на середине створки 5—7 ареол, у края 8—12 в 10 мкм.
В центре створки одна более крупная ареола, окруженная 6—8, обычно 7 ареолами,
63
не отличающимися по размерам от других ареол. Рядом с центральной ареолой малень-
кое отверстие центрального выроста с опорами. Краевые выросты с опорами в виде
коротких узких трубочек, расположены в 1—2 ряда на загибе створки, 6—7 в 10 мкм,
в СМ они незаметны. Имеются также выросты с опорами по всей створке, их отверстия
на наружной поверхности створки располагаются между рядами ареол. Двугубый
вырост с длинной трубкой на наружной поверхности створки, в кольце крупных,
неравномерно расположенных шипов, 3—4 шипа в 10 мкм, его внутренняя щель ориен-
тирована радиально. Загиб створки невысокий, с мелкими ареолами в прямых или
неправильно косых рядах, 18—20 ареол в 10 мкм. Известны ауксоспоры, часто наблю-
даемые в культурах. (Табл. 40,1—9).
Морской эвригалинный и эвритермный неритический планктонный широко-
бореальный тропическо-нотальный вид.
Палеоцен — Поволжье, вост, склон Урала. Майкоп — Краснодарский край. Нео-
ген — зап. и вост, побережье Камчатки, Сев. и Южн. Сахалин, Чукотка. Миоцен -
Камчатка (тюшевская серия). Поздний миоцен — Сев. Сахалин (маямрафская свита);
подний сармат — УССР; мэотис — Таманский п-ов. Плиоцен — о-в Итуруп, Сев. Сахалин.
Плейстоцен — Сев.-Зап. СССР (межледниковые отл.), р. Вага и Сев. Двина (бореальные,
межледниковые отл.), Сев. Двина (позднеледниковые отл.), Сев.-Вост. СССР, Берин-
гово и Охотское моря, Курило-Камчатская впадина, сев.-зап. часть Тихого океана.
Голоцен — Сев. Двина, Онежский п-ов (послеледниковые отл.), Охотское море. Ныне —
Балтийское, Белое моря, Сев. Ледовитый океан, Баренцево, Берингово, Охотское,
Японское моря, сев.-зап. часть Тихого океана, Черное, Азовское и Аральское моря,
Днестровский и Днепровско-Бугский лиманы и в устьях приазовских рек. Вне СССР,
Поздний эоцен—ранний олигоцен — Калифорния. Неоген — Япония, Никобарские о-ва.
Миоцен — Венгрия, Япония, Малайский архипелаг. Средний миоцен — вост, побережье
Сев. Америки. Средний миоцен—поздний плиоцен — о-в Ява. Поздний миоцен —
Югославия. Плиоцен — Калифорния. Плиоцен—плейстоцен — вост, побережье Сев. Аме-
рики. Четверт. период — Фенноскандия. Плейстоцен — экваториальная зона Тихого,
Атлантического и Индийского океанов. Ныне — Северное и Ирландское моря, Англий-
ский канал, Адриатическое, Мраморное моря, Босфор, устья европейских рек, Тихий
океан у берегов Сев. Америки, Чили и сев. Австралии, Атлантический и Индийский
океаны, Австралия (эстуарий р. Свен).
10. Т. heteromera Makar. (Макарова, 1975а : 151, 152, рис., 9).
Створки слегка вогнутые, 25.2—28.8 мкм в диам., грубо ареолированные. Ареолы
в тангенциальных и слегка радиальных рядах, в центре створки 6, у края 9 ареол
в 10 мкм. В центре створки 5 маленьких выростов с опорами, собраны в виде звезды.
По краю створки кольцо из коротких выростов с опорами, расположенных очень
неравномерно, 2—3 в 10 мкм. Двугубый вырост довольно заметный, в одном кольце
с краевыми выростами с опорами. Загиб створки низкий, с мелкими ареолами, обра-
зующими прямые вертикальные ряды, 10—12 в 10 мкм. (Табл. 39,10).
Морской планктонный высокобореальный вид.
Ныне — Баренцево море (Дальние Зеленцы).
11. Т. ignota Makar. (Макарова, 1975 : 150,151, рис., 5-8).
Клетки соединены толстым тяжом в цепочки:из 4—11 клеток. Панцирь в виде
невысокого, 16.5—20 мкм, цилиндра, с 2 вставочными ободками. Створки плоские,
19.8—39 мкм в диам. Ареолы в тангенциальных, изогнутых к центру рядах, в центре
створки 6—9 ареол, близ края 8—10 в 10 мкм. В центре створки 5—12 выростов
с опорами, образующих 2 прямые или изогнутые линии, занимающие место 1—2 цент-
ральных ареол, реже расположение их беспорядочное. Краевые выросты с опорами
расположены равномерно, 2—3 в 10 мкм. У всех выростов с опорами трубочки до
1.3 мкм дл., с 3 опорами на внутренней поверхности створки, на наружной по-
верхности выросты заканчиваются отверстием. Двугубый вырост с короткой трубкой
64
на внутренней поверхности створки и отверстием на наружной, расположен в одном
кольце с краевыми выростами с опорами, обычно занимая место одного из них или
слегка отступя внутрь от этого кольца, его внутренняя щель ориентирована танген-
циально. Загиб створки невысокий, с мелкими ареолами, 11 — 14 в 10 мкм. (Табл. 39,
72-75).
Морской высокобореальный вид.
Ныне — Баренцево (Дальние Зеленцы), Берингово моря, сев.-зап. часть Тихого
океана.
12. Т. incerta Makar. (Макарова, 1961 : 50, табл. I, 9-76).
Клетки соединены тонким тяжом в короткие цепочки из 2—4 клеток. Панцирь
цилиндрический, 11—18 мкм выс. Створки почти плоские, 11.4—33 мкм в диам., обычно
15—17 мкм. Ареолы в тангенциальных и радиальных рядах, иногда с тенденцией
к образованию пучков, в центре створки 10—16 ареол, у края 16—20 в 10 мкм. Фора-
мены круглые или неправильно округлые, крибрум с 20 крибральными порами
в 1 мкм. В центре створки свободно расположены 3—5 выростов с опорами. На наруж-
ной поверхности створки они имеют отверстие, на внутренней — короткую трубочку.
Краевые выросты с опорами очень маленькие, 3—4 в 10 мкм, образуют кольцо на
загибе створки. На границе с загибом створки кольцо крупных шипов до 1 мкм дл.,
расположенных неравномерно, 5—8 в 10 мкм. Двугубый вырост в одном кольце
с шипами, снаружи имеет трубку около 2 мкм дл., на внутренней поверхности
створки — короткую и сильно сплющенную трубку с длинной щелью, ориентированной
радиально. Загиб створки невысокий, с мелкими ареолами 16—20 в 10 мкм, располо-
женными в косых и волнистых рядах, по краю загиба очень короткие ребрышки,
12 в 10 мкм. (Табл. 41,7-72).
Солоноватоводный эвригалинный и эвритермный широкобореальный вид.
Ныне — Азовское, Каспийское, Аральское моря, в водохранилищах Волги
(Куйбышевском, Саратовском, Волгоградском), ее нижнем течении, в р. Каме, Перт-
озере (Карелия), оз. Белом (Вологодская обл.). Вне СССР. Ныне — эстуарии рр. Темзы
и Большой Уз, оз. Эри, реки Калифорнии, Миссисипи, Нил.
13. Т. intermedia Makar. (Макарова, 19886 : 59, табл. XXIX, 7, 8).
Клетки соединены в короткие цепочки. Панцирь в виде низкого барабана, со
вставочными ободками. Створки слегка выпуклые посередине, 18—25 мкм в диам.
Ареолы в тангенциальных рядах, у края створки с тенденцией к радиальным, на сере-
дине створки 10—11 ареол, у края 12—14 в 10 мкм. В центре створки один вырост
с опорами. Краевые выросты с опорами хорошо заметные, 4—5 выростов в 10 мкм,
образуют одно кольцо на границе с загибом створки. Двугубых выростов 3, в одном
кольце с выростами с опорами, два из них приближены друг к другу, третий отдален.
Загиб створки невысокий. (Табл. 40,10).
Солоноватоводный низкобореальный вид.
Ныне — Каспийское море (Южный Каспий).
П р и м е ч. В работе А. И. Прошкиной-Лавренко и И. В. Макаровой (1968, табл. II,
14) приведен под названием Т. aculeata.
14. Т. latimarginata Makar. (Макарова, 1975 : 150, рис., 5, 4). - Т. trifulta Fryx.
(Fryxell, Hasle, 1979 : 16-19, fig. 1-24).
Панцирь в виде барабана, со вставочными и соединительными ободками. Вальво-
копула с кольцом крупных пор и многочисленными мелкими нерегулярными рядами
пор, с двойной септой. Вставочный ободок с лигулой, одним кольцом пор и одной сеп-
той. Соединительные ободки бесструктурные. Створки почти плоские, 21.6—53 мкм
в диам. Ареолы в тангенциальных, вогнутых к центру рядах, с тенденцией к радиаль-
ным, иногда в почти линейных рядах, в центре створки 6—7 ареол в 10 мкм, близ края
8—10. В центре створки 3—5 выростов с опорами, образующих одну более или менее
5 Заказ 518 £5
прямую или изогнутую линию. Краевые выросты с опорами расположены равномерно,
2.5—3 в 10 мкм. Выросты с 3 поддерживающими опорами на внутренней поверхности
створки, с наружной заканчиваются отверстием. Двугубый вырост вне краевого кольца
выростов с опорами, обычно отступя на 2—3 ареолы внутрь, на наружной поверхности
створки заканчивается отверстием, его внутренняя щель ориентирована тангенциально.
Загиб створки 1.8—2 мкм выс., с мелкими, слегка удлиненными ареолами, 9—14
в 10 мкм, образующими волнистые ряды. (Табл. 40,11-17).
Морской высокобореальный и высоконотальный вид, возможно биполярный.
Поздний плейстоцен — Чукотка. Голоцен — Охотское и Берингово моря, Коман-
дорские о-ва. Ныне — Баренцево (Дальние Зеленцы), Берингово моря.Вне СССР.
Ныне — сев.-зап. часть Тихого океана, Алеутские о-ва, антарктическая зона Индийского
и Тихого океанов.
15. Т. leptopus (Grun.) Hasle et Fryx. (Hasle, Fryxell, 1977 : 20—22, tab. 1—4). —
Coscinodiscus leptopus Grun. (Grunow in Van Heurck, 1880—1885 : 131, fig. 5, 6). —
C. lineatus Ehr. (Ehrenberg, 1838 : 129).
Панцирь низкоцилиндрический. Створки плоские, 30—50 мкм в диам. Ареолы
в строгих линейных рядах, пересекающихся в трех направлениях, одинаковой
величины по всей створке, 4.5—6 в 10 мкм. Центральный вырост с опорами
отсутствует. Краевые выросты с опорами в виде небольших трубочек, расположены
на загибе створки зигзагообразно и неравномерно, от 1 до 5 в 10 мкм, с внутренней
поверхности створки они более заметны и окружены 5 сопутствующими порами. Дву-
губый вырост с крупной наружной трубкой у края створки, его внутренняя щель
ориентирована радиально. На загибе створки кольцо отчетливых, нерегулярно
расположенных замкнутых выростов, 2—5 в 10 мкм. Загиб створки невысокий,
с мелкими ареолами, расположенными изогнутыми горизонтальными рядами, 12—20
рядов в 10 мкм. (Табл. 42,12).
Морской океанический неритический вид, широко распространенный в морях
и океанах бореальной, тропической и нотальной зон.
Миоцен — Камчатка (тюшевская серия). Плиоцен — Камчатка (Усть-Камчат-
ский р-н), Южн. Сахалин (маруямская свита). Плейстоцен — Сев .-Зап. СССР. Голоцен —
Охотское море. Ныне — Белое море, Чешская губа, Карское, Берингово, Охотское,
Японское моря, Тихий океан у берегов Камчатки, Амурский лиман. Вне СССР.
Эоцен — Швеция, Дания. Миоцен — Испания, Франция, Югославия, Венгрия, Индия, Япо-
ния, Калифорния, Мексика, Сев. Америка. Миоцен—плиоцен — о-в Ява. Плейстоцен —
Польша, Фенноскандия. Донные осадки экваториальной зоны Индийского, Атлантиче-
ского и Тихого океанов. Ныне — во всех европейских морях, у атлантических
и тихоокеанских берегов Сев. Америки, Калифорнии, у берегов Чили, Индий-
ский океан.
16. Т. levanderi Van Goor (1924 : 322, fig. 11).
Клетки образуют непрочные цепочки. Створки почти плоские, 8—16 мкм в диам.
Ареолы круглые, в тангенциальных рядах, на середине створки 14—18 ареол, у края
23—24 в 10 мкм. Выросты с опорами не описаны.
Солоноватоводный среднебореальный вид.
Плейстоцен (межледниковые отл.) — р. Косма, р. Пеза (правый приток р. Мезень).
Ныне - Рижский залив, Белое и Охотское моря, у берегов Южн. Сахалина. Вне СССР.
Ныне - Финский залив, устье Эльбы, лагуны Южн. Африки.
П р и м е ч. Систематическое положение этого вида не совсем ясно, он нуждается
в дополнительном и более тщательном изучении.
17. Т. makarovae Mukchina (Jouse, Mukchina, 1978 : 914, tab. 6,14).
Створки плоские, 46—60 мкм в диам. Ареолы мелкие, 14—16 в 10 мкм, в линей-
ных и тангенциальных рядах. По краю створки кольцо нерегулярно расположенных
66
выростов с опорами, 8—9 в 10 мкм, которые имеются и на лицевой части створки.
Двугубый вырост в одном кольце с краевыми выростами с опорами.
Солоноватоводно-пресноводный вид.
Поздний плиоцен — Черное море.
18. Т. marujamica Sheshuk. emend. Makar. (Шешукова-Порецкая, 1959, табл. I, 7).
Створки слегка выпуклые, 12—35 мкм в диам. Ареолы в тангенциальных, изогну-
тых к центру рядах, иногда с тенденцией к радиальности, близ центра створки
8—10 ареол, у края 10—12 в 10 мкм. В центре створки многочисленные свободно распо-
ложенные выросты с опорами. Краевые выросты с опорами на загибе створки (невиди-
мые в СМ), все они открываются на наружной поверхности очень маленьким отверсти-
ем, а на внутренней представлены маленькой короткой трубкой. На границе с загибом
створки кольцо крупных, неравномерно расположенных замкнутых выростов, 1—2
в 10 мкм. Двугубый вырост в одном кольце с замкнутыми выростами, с длинной
наружной трубкой и сильно сплющенной внутренней частью, его щель ориентирована
радиально. Загиб створки низкий, с широким краем, заканчивающимся ребрышками,
10-14 в 10 мкм. (Табл. 41,13-21}.
Морской вымерший вид.
Миоцен — Камчатка (тюшевская серия). Поздний миоцен — Сев. Сахалин, зап. по-
бережье (маямрафская свита). Поздний миоцен—ранний плиоцен — Южн. Сахалин
(маруямская свита).
19. Т. nativa Sheshuk. (Шешукова-Порецкая, 1964 : 75, табл. 1,4, 5).
Створки плоские, 7—20 мкм в диам. Ареолы 5—6-угольные, со слегка волнистыми
тангенциальными прямыми или слегка вогнутыми рядами, на середине створки 10—12
ареол в 10 мкм, у края 16—20. Центральные выросты с опорами неотчетливые, их,
по-видимому, один или несколько. Краевые выросты с опорами редко расположенные,
на наружную поверхность створки открываются маленьким отверстием (в СМ неза-
метны) . Двугубый вырост неизвестен. Загиб створки невысокий, с вертикальными
рядами ареол, 20—24 ряда в 10 мкм. (Табл. 42,1-4}.
Морской вымерший вид.
Поздний миоцен — Сев. Сахалин, п-ов Шмидта и сев. побережье (маямрафская
свита). Поздний миоцен—плиоцен — Южн. Сахалин (маруямская свита). Вне СССР.
Неоген — Япония. Поздний миоцен — Калифорния, сев.-вост. Япония.
20. Т. nodulolineata (Hendey) Hasle et Fryx. (Hasle, Fryxell, 1977 : 35, 36,
fig. 86—93). — Coscinodiscus nodulolineatus Hendey (1957 : 39, tab. 5,4,5).
Клетки соединены в короткие цепочки. Панцирь низкоцилиндрический. Створки
плоские, 15—30 мкм в диам. Ареолы в прямых, почти линейных или тангенциальных
рядах, 5—6 в 10 мкм. Форамены круглые, часто с нитевидными радиальными выроста-
ми, крибрум округлой или угловатой формы, до 20 крибральных пор в 1 мкм.
В центре створки одна небольшая ареола, вокруг которой расположены 5—6 очень
маленьких центральных выростов с опорами, имеющих небольшие отверстия на наруж-
ной поверхности створки и короткую трубочку, окруженную 3 сопутствующими
порами, — на внутренней. Краевые выросты с опорами очень маленькие, расположены
равномерно на загибе створки, 4 в 10 мкм. На границе с загибом створки кольцо
довольно крупных шипов, 2—3 в 10 мкм. Двугубый вырост крупный, в одном кольце
с шипами, его щель ориентирована почти радиально. Загиб створки невысокий, с мел-
кими ареолами, расположенными в косо пересекающихся рядах, 12—14 в 10 мкм.
(Табл. 42,5—11}.
Солоноватоводный и морской вид.
Ныне - Азовское море. Вне СССР. Ныне — Атлантический океан у берегов
Зап. Африки и Уругвая.
67
21. T. oestrupii (Ostf.) Hasle (1972 : 544). — Coscinosira oestrupii Ostf. (Ostenfeld,
1900 : 52). — Thalassiosira antiqua (Grun.) A. Cl. var. septata Pr.-Lavr. (Прошкина-Лавренко,
1955:48,табл.1,12-14; 11,7, 2). — T. oestrupiiplana Jouse (Жузе, 1968: 15,табл. 1,8,9).
Клетки соединены 4 довольно длинными тяжами в короткие цепочки. Панцирь
в виде высокого или низкого цилиндра с закругленными краями. Вальвокопула доволь-
но широкая, со структурой из мелких ареол в прямых вертикальных рядах, 18—21 ряд
в 10 мкм, вставочные ободки узкие, с одним рядом пор, соединительные ободки бес-
структурные. Имеются септы. Створки почти плоские, 15—35 мкм в диам. Ареолы
в тангенциальных рядах, с круглым или шестиугольным фораменом, в центре 7—
9 ареол, у края 9—11 в 10 мкм, крибрум из мелких крибральных пор, расположенных
в прямых или слегка искривленных рядах, пересекающихся в трех направлениях.
В центре створки обычно одна более крупная ареола, рядом с которой расположен
центральный вырост с опорами. Краевые выросты с опорами имеют длинную трубку
с 3 опорами на внутренней поверхности створки, а на наружной заканчиваются малень-
ким отверстием, 2—4 или 6—8 в 10 мкм. Двугубый вырост близ центра створки на рас-
стоянии 1—5 ареол от центрального выроста с опорами. На наружной поверхности
створки он заканчивается маленьким отверстием, его внутренняя щель ориентирована
тангенциально или под углом к радиусу створки. (Табл. 42,13—17).
Морской эвритермный океанический и неритический, широко распространенный
от Арктики до Антарктики вид, космополит.
Поздний миоцен—ранний плиоцен — сед.-зап. часть Тихого океана. Плейстоцен —
Черное море. Голоцен — Охотское море. Ныне — Баренцево, Японское, Черное, Азов-
ское моря. Вне СССР. Неоген — сев .-вост. Япония. Ранний миоцен—поздний
плиоцен — сев. часть -Тихого океана, США (штат Орегон). Миоцен—плиоцен —
сев.-вост. часть Тихого океана, Калифорния, Италия. Плиоцен — Индийский океан.
Плейстоцен — субтропическая и тропическая зоны Тихого и Индийского океанов, сев.-
вост. Япония. Ныне — Северное и Норвежское моря, сев. и южн. Атлантика, Мексикан-
ский залив, Карибское море, море Уэдделла, Индийский океан, Тихий океан (субаркти-
ческая, бореальная и нотальная зоны) .
22. Т. pacifica Gran et Angst (1931 : 437, fig. 12). — T. pulchella Takano (1963 : 5,
fig. l,a, b; tab.1,77, a, b).
Клетки соединены тонким тяжом в гибкие цепочки. Панцирь в виде низкого,
8—12 мкм, цилиндра со скошенными углами, со вставочными и соединительными обод-
ками. Вальвокопула с мелкими круглыми порами, около 40 в 10 мкм, вставочный
ободок с одним рядом пор, 5—6 в 1 мкм, и примыкающей к нему узкой септой. Створ-
ки плоские или слегка вогнутые в центре, 10—50.5 мкм в диам. Ареолы в линейных,
тангенциальных, слегка изогнутых к центру рядах, иногда с тенденцией к радиальным,
в центре 10—16 ареол, а близ края 12—18 ареол в 10 мкм. Форамены от круглых до
угловатых, на крибруме поры занимают центральную часть, 16—18 в 1 мкм. В центре
створки рядом с наиболее крупной ареолой один вырост с опорами, на наружной по-
верхности створки он имеет заметную трубку, на внутренней — небольшую трубку
с 4—5 сопутствующими порами. Краевые выросты с опорами почти цилиндрические,
0.8—1.1 мкм дл., 0.2—0.4 мкм в диам., равномерно расположенные, 6 в 10 мкм. Двугу-
бый вырост в одном кольце с выростами с опорами, отличается от них снаружи большей
величиной, 1.2—1.4 мкм дл., и формой — в виде расширенной трубки, его внутренняя
щель ориентирована радиально. Загиб створки довольно высокий, с отчетливыми ради-
альными ребрышками, 12—14 в 10 мкм. (Табл. 43,1-13}.
Морской неритический широкобореальный и нотальный, в основном бореально-
пацифический вид.
Плейстоцен — Охотское море. Голоцен — Белое и Охотское моря. Ныне — Чукот-
ское, Берингово, Охотское, Японское моря. Вне СССР. Ныне — Тихий океан: Аляскин-
ский залив, Алеутский желоб, залив Пьюджет Саунд, у берегов Чили, Атлантический
океан, у берегов Сев. Америки.
68
23. T. parva Pr.-Lavr. (Прошкина-Лавренко, 1955a : 45, табл. 1,1— 5).
Клетки одиночные или в коротких рыхлых цепочках, соединены одним тяжом.
Панцирь в виде барабана с 2 вставочными ободками. Створки почти плоские, 12.5—
37 мкм в диам. Ареолы в тангенциальных рядах с тенденцией к образованию радиаль-
ных рядов и неотчетливых пучков, на середине створки 12—16, у края 18—20 ареол
в 10 мкм. В центре створки 5—6 очень маленьких выростов с опорами, с наружной
поверхности створки заканчивающихся только маленьким отверстием. Краевые вы-
росты с опорами на границе с загибом створки, их наружная часть в виде длинных
и узких трубочек, 5—6 выростов в 10 мкм. Двугубый вырост крупный, на внутренней
стороне кольца из выростов с опорами. Загиб створки низкий, с мелкими ареолами
в вертикальных рядах, 30—32 ряда в 10 мкм. (Табл. 43,14-17).
Солоноватоводный неритический низкобореальный вид.
Поздний миоцен (поздний сармат) — Таманский п-ов. Ныне — Черное, Азовское,
Каспийское моря, возможно эндемик южных морей СССР.
24. Т. singularis Sheshuk. (Шешукова-Порецкая, 1967 : 145, табл. 14, £).
Створки плоские, 18—28 мкм в диам. Ареолы шестиугольные, одинаковой вели-
чины, в тангенциальных прямых или слегка искривленных, реже вогнутых рядах, 7—
11 ареол в 10 мкм. Центральные выросты с опорами неотчетливые в СМ, возможно 3—
8 в центре створки. Прикраевая зона узкая, круто опадающая, с удлиненными арео-
лами, 6—8 в 10 мкм, более отчетливо заметными в наклонном положении, создающими
впечатление о наличии коротких тупых выростов по краю створки. Двугубый вырост
не описан. Загиб створки низкий, отогнутый. (Табл. 43,7S).
Морской вымерший вид.
Поздний миоцен — Сев. Сахалин (маямрафская свита). Поздний миоцен—плио-
цен — Южн. Сахалин (маруямская свита).
При меч. Вид недостаточно изученный, применение СЭМ позволит уточнить
морфологию, определить более точно его систематическое положение.
25. Т. tenera Pr.-Lavr. (Прошкина-Лавренко, 196la : 33, табл. 1,1—4’, II, 5—7).
Клетки соединены по 2 одним коротким тяжом. Панцирь цилиндрический,
6.8—11 мкм выс., поясок с мелко перфорированной вальвокопулой и 5—6 соединитель-
ными ободками, видимыми в СЭМ. Створки плоские, 6.5—15 мкм в диам. Ареолы
в тангенциальных, иногда почти линейных рядах, в центре 12—13 ареол, у края до 15
в 10 мкм. По данными ЭМ, центральная ареола намного больше остальных, внутри нее
находится очень маленький центральный вырост с опорами. На наружной и внутренней
поверхностях створки он имеет короткую трубку, окруженную 4—5 сопутствующими
порами. Краевые выросты с опорами очень маленькие, 4—6 в 10 мкм, на границе с заги-
бом створки, закрыты, вероятно, кремнеземной пластинкой, имеющей форму клина,
прикрепляющейся к каждому выросту, что придает краю волнистый вид. Двугубый
вырост небольшой, в одном кольце с выростами с опорами, расположен ближе
к одному из них. Загиб створки низкий, с ребрышками, между ними 2 ряда очень мел-
ких ареол, 23—26 в 10 мкм. (Табл. 43,79, 20).
Солоноватоводный, возможно морской неритический широкобореальный вид.
Поздний миоцен (мэотис) — Крым. Ныне — Черное море. Вне СССР. Ныне —
Атлантический океан, у берегов Норвегии, Английский пролив, реки Англии, Мексикан-
ский залив, Тихий океан: заливы Пьюджет Саунд, Микава (Япония), у берегов Кали-
форнии и Австралии.
2. Секция Fasciculigera Makar.(Макарова, 1971 : 70)
Створки плоские, выпуклые, редко со складкой в центре. Ареолы в радиальных
рядах, образующих отчетливые пучки, в которых ряды ареол параллельны среднему
ряду. На створке обычно одно кольцо краевых выростов с опорами, реже их 2 и более,
69
иногда выросты по краю расположены в шахматном порядке. В центре створки один
вырост с опорами или их несколько, с более длинной трубкой на внутренней или на-
ружной поверхности. Двугубый вырост обычно один, расположен в краевом кольце
выростов с опорами, иногда отступя от края или ближе к центру створки. Иногда име-
ются замкнутые выросты.
Тип: Т. baltica (Grun.) Ostf.
I. На створке одно краевое кольцо выростов с опорами.
1. В прикраевой зоне помимо выростов с опорами кольцо из 3—9 редких замкнутых
выростов.........................................................32. Т. punctigera.
2. Замкнутые выросты отсутствуют.
А. Двугубый вырост расположен в одном кольце с краевыми выростами
с опорами.
а. Ареолы очень мелкие, до 26 в 10 мкм.......................31. Т. ordinaria.
б. Ареолы более крупные. <
а. Пучки ареол отчетливые.
+ На середине створки 10—14 ареол, по краю 6—7 выростов с опорами
в 10 мкм.............................................29. Т. kryophila.
++На середине створки 18 ареол, по краю 3—5 выростов с опорами
в 10 мкм..............................................26. Т. aestivalis.
Р. Пучки ареол не отчетливые, на середине створки 16 ареол в 10 мкм.....
......................................................30. Т. maeotica.
Б. Двугубый вырост расположен близ центра створки..............28. Т. bioculata.
II. На створке несколько колец выростов с опорами.
1. Выросты с опорами расположены в шахматном порядке по краю створки, среди
них 2—4 двугубых выроста..........................................27. Т. baltica.
2. Выросты с опорами образуют 3 и более колец на створке, двугубый вырост один,
его местоположение различное.......................................33. Т. subtilis.
26. Т. aestivalis Gran (Gran a. Angst, 1931 : 436, fig. 10). - T. concaviuscula Makar. (Ма-
карова, 1978 : 222,223, табл. I, 7-77).
Клетки с пояска восьмиугольные, соединены тонким тяжом.Створки плоские с не-
большим углублением в центре, 20—32 мкм в диам. Ареолы собраны в отчетливые
пучки, на середине створки 18 ареол в 10 мкм, образуют плотную сеть, в центре створки
ареолы немного крупнее. Рядом с центральной ареолой центральный вырост с опорами
с 4—5 сопутствующими порами. Краевые выросты с опорами довольно крупные, равно-
мерно расположенные, 3—5 в 10 мкм, с 4 сопутствующими порами. На наружной по-
верхности их трубки более длинные, чем на внутренней. Двугубый вырост в одном
кольце с выростами с опорами, ближе к одному из них с более крупной наружной
трубкой, его внутренняя щель ориентирована радиально и окружена утолщенными кра-
ями. Загиб створки низкий, с ареолами в прямых и косо пересекающихся рядах, 20—
22 ареолы в 10 мкм. (Табл. 44).
Морской бореальный и нотальный вид.
Ныне — Черное море. Вне СССР. Ныне — Тихий океан: заливы Пьюджет Саунд,
Калифорнийский, у берегов Чили и Перу, Атлантический океан, шельф юго-зап.
Африки.
27. Т. baltica (Grun.) Ostf. (Ostenfeld, 1901 : 290). — Coscinodiscus polyacanthus
Grun. var. baltica Grun. (Grunow in Cleve a. Grunow, 1880 : 112). — Thalassiosira subsalina
Pr.-Lavr. (Прошкина-Лавренко, 1955a : 10—47, табл. 1,77).
Клетки образуют цепочки из 2—8 клеток, соединенных тонким тяжом. Панцирь
в виде барабана с несколькими встадочными и соединительными кольцевидными обод-
ками. Створки круглые, плоские, 16.5—60 мкм в диам. Ареолы в довольно отчетливых
пучках, в центре створки 12—24 ареолы, у края 24—26 в 10 мкм. Форамены круглые,
70
крибрум с 24 порами в 1 мкм. В центре створки от 2 до 8 свободно расположенных
выростов с опорами; с наружной поверхности створки они открываются небольшими
отверстиями, а с внутренней имеют слегка выступающие трубки, окруженные 3 или
4 сопутствующими; порами. Краевые выросты с опорами довольно крупные, располо-
жены в шахматном порядке, их наружная часть более длинная, до 1 мкм, чем внутрен-
няя. Двугубых выростов 2—4, расположены по краю створки и имеют с наружной по-
верхности створки длинную, до 1.6—1.8 мкм, трубку, а с внутренней — более корот-
кую уплощенную трубку с утолщенными краями, их щель ориентирована радиально.
Загиб створки довольно высокий, с мелкими ареолами в радиальных и вторично
косых рядах, 20—28 рядов в 10 мкм. Споры с равномерно выпуклыми створками, их
структура из мелких ареол, образующих пучки. По краю створки кольцо выростов
с опорами. (Табл. 45).
Солоноватоводный планктонный неритический, часто обитающий в опресненных
участках морей широкобореальный вид.
Поздний миоцен (поздний сармат) — Таманский п-ов. Плиоцен (акчагыл) —
р. Урал. Плейстоцен — Ленинградская, Архангельская обл., рр. Мга, Вытегра, Ладожское
озеро, Карелия, Черное море. Голоцен — Архангельская обл,, Белое море. Ныне —
Балтийское море, Финский, Нарвский, Рижский заливы, Невская и Лужская губы,
Белое море, Чешская губа, рр. Сев. Двина, Печора, Ома, Пеше, Карское, Черное, Азов-
ское, Каспийское моря, Днепровско-Бугский лиманы. Вне СССР. Четверт. период —
Швеция, Финляндия. Ныне — Балтийское море, Ботнический залив, проливы Каттегат
и Скагеррак, у берегов Дании, Швеции, Финляндии, южн. часть Северного моря, сев.
Атлантика.
28. Т. bioculata (Grun.) Ostf. (Ostenfeld, 1903 : 564, fig. 120, 121). — Coscinodiscus
bioculatus Grun. (Grunow, 1884 : 107, tab. 3,30).
Клетки одиночные или соединенные тяжом в рыхлые цепочки. Панцирь цилиндри-
ческий, 6—10 мкм выс., с несколькими вставочными и соединительными ободками.
Створки плоские, 30—54 мкм в диам. Ареолы в неясных пучках, 18—20 ареол в 10 мкм.
В центре створки ареолы слегка разрежены за t счет вклинивающихся рядов.
Близ центра створки один вырост с опорами. Краевые выросты с опорами
у загиба створки, 2—3 в 10 мкм, на внутренней поверхности трубка более длин-
ная, чем на наружной. Двугубый вырост близ центра створки, на наружной поверх-
ности створки открывается отверстием, на внутренней имеет уплощенную трубку
со щелью, ориентированной почти радиально. Споры эллипсоидные, с равно-
мерно выпуклыми и слегка уплощенными створками. Структура из ареол, образую-
щих пучки. (Табл. 46,7).
Морской неритический арктобореальный и нотальный вид.
Ныне — Белое, Баренцево, Карское моря, пролив Маточкин Шар, оз. Дерюгина (Но-
вая Земля), Берингово, Охотское и Японское моря, у Южн. Сахалина, в районе Куриль-
ских о-вов. Вне СССР. Ныне — Гренландское, Норвежское, Северное моря, сев. Атлан-
тика: заливы Св. Лаврентия, Шалер, Фанди, у берегов сев.-зап. Африки, Тихий океан:
у берегов Аляски и Канады, зал. Пьюджет Саунд, у берегов южн. Австралии и Новой
Зеландии.
29. Т. kryophila (Grun.) J0rg. (Jorgensen, 1905 : 96, tab. 6, b). — Coscinodiscus kry-
ophilus Grun. (Grunow, 1884 : 81, tab. 3,21).
Клетки соединены тяжом в короткие цепочки. Панцирь низкоцилиндрический.
Створки плоские, до 41 мкм в диам. Ареолы в отчетливых пучках, на середине створки
10—14, у края 14—18 ареол в 10 мкм, в центре створки ареолы разрежены. Централь-
ный вырост с опорами один. Краевые выросты с опорами равномерно расположенные,
6—7 в 10 мкм. Двугубый вырост в краевом кольце выростов с опорами, выделяется
более крупными размерами. Загиб створки низкий. (Табл. 46,2).
71
Морской неритический и пелагический арктобореальный вид.
Миоцен — Камчатка, Сев. Сахалин. Плиоцен — Камчатка, Южн. Сахалин. Плейсто-
цен — Сев.-Зап. СССР (морские межледниковые отл.). Голоцен — Сев.-Зап. СССР,
Кольский п-ов (реликтовые озера). Четверт. период — Охотское, Берингово моря,
Курило-Камчатская впадина и сев.-зап. часть Тихого океана. Ныне — Белое, Баренцево
моря, Сев. Ледовитый океан, Охотское, Японское моря и у вост, берегов Южн. Саха-
лина. Вне СССР. Неоген — Япония. Четверт. период — Финляндия. Ныне — проливы Ска-
геррак и Каттегат.
30. Т. maeotica Pr.-Lavr. (Прошкина-Лавренко, 1960 : 57, табл. 2,22—25).
Панцирь с 2 вставочными ободками. Створки почти плоские, 11—45 мкм в диам.
Ареолы образуют неярко выраженные многочисленные, до 26, пучки, близ центра
створки 16 ареол, у края 18—20 в 10 мкм. В центре створки несколько беспорядочно
расположенных ареол, среди которых находится, вероятно, один центральный вырост
с опорами. Краевые выросты с опорами небольшие, многочисленные, равномерно рас-
положенные, 7—8 в 10 мкм, образуют одно кольцо на границе с загибом створки.
Двугубый вырост довольно крупный, в одном кольце с краевыми выростами с опо-
рами. Загиб створки невысокий, с мелкими, взаимопересекающимися рядами ареол,
20—22 в 10 мкм. (Табл. 46,3).
Солоноватоводный неритический вымерший вид, возможно эндемик.
Поздний миоцен (мэотис) — Крым, Таманский и Керченский п-ова.
31. Т. ordinaria Makar. (Макарова, 1977а : 537, табл. 1,1—7; 2,1-6).
Клетки соединены тонким тяжом в короткие цепочки. Панцирь в виде низкого,
3.5—5.5 мкм, барабана, с 2 вставочными ободками. Створки плоские, 15—30 мкм
в диам. Ареолы одинаковой величины, в отчетливых пучках, образуют плотную сеть,
26 ареол в 10 мкм. В центре створки 1 (2) выроста с опорами. По краю створки, на гра-
нице с ее загибом одно кольцо краевых выростов с опорами, расположенных очень
равномерно, 5 в 10 мкм. Их наружная трубка длиннее внутренней, окруженной 4 по-
рами. Двугубый вырост в одном кольце с краевыми выростами с опорами, обычно
приближен к одному из них и отличается более крупной трубкой (около 1 мкм дл.)
на наружной поверхности; на внутренней поверхности трубка короткая, щель ориенти-
рована радиально. Загиб створки низкий, с мелкими, слегка удлиненными ареолами,
расположенными в вертикальных рядах, 35—38 в 10 мкм. (Табл. 47,1-7).
Солоноватоводный планктонный низко бореальный вид.
Ныне — Черное море.
32. Т. punctigera (Castr.) Hasle (1983 : 602, fig. 1-45). — Ethmodiscus punctiger Castr.
(Castracane, 1886 : 167, tab. 3, 1). - Coscinodiscus punctiger (Castr.) H. Perag. (Peragallo
in Tempere et Peragallo, 1889—1895 : 57, N 107, 108). - Ethmodiscus japonicus Castr.
(Castracane, 1886: 168, tab. 22, 2). - Coscinodiscus verecundus Mann(1907 :259, tab. 50,
1). - C. angstii Gran (Gran in Gran et Angst, 1931 : 443, fig. 19). — Thalassiosira japonica
Kiss. (Киселев, 1935 : 99, рис. 17—19). — T. angstii (Gran) Makar. (Макарова,1970:
13,14).
Клетки соединены тонким тяжом в колонии. Панцирь высоко цилиндрический.
Вальвокопула нежно ареолированная, вставочный ободок с рядом удлиненных пор и
лигулой, соединительный ободок бесструктурный, с лигулой и антилигулой. Створки
равномерно выпуклые, 50—130 мкм в диам. Ареолы в крупных пучках, 13—16 ареол
в 10 мкм. В центре створки ареолы часто разрежены, среди них центральный вырост
с опорами, открывающийся наружу отверстием. Краевые выросты с опорами на границе
с загибом створки, небольшие, довольно часто и равномерно расположенные, 5—6
в 10 мкм, в СЭМ они имеют форму рюмки: с перетяжкой у основания и суженным
дистальным концом. Внутрь от кольца выростов с опорами кольцо из 3—9 очень круп-
ных, часто изогнутых, неравномерно расположенных замкнутых выростов, иногда
72
они отсутствуют. Двугубый вырост крупный, в одном кольце с замкнутыми выростами,
его трубка на наружной поверхности во много раз превышает короткую сплющенную
трубку на внутренней поверхности, щель ориентирована радиально. Загиб створки
низкий, с ребрышками, 10—12 в 10 мкм. (Табл. 46,4—10}.
Морской неритический планктонный широко бореальный вид.
Ныне — Чукотское, Берингово, Охотское, Японское моря, зал. Петра Великого,
бухта Патрокл, Амурский лиман, осадки сев.-зап. части Тихого океана. Вне СССР.
Ныне - Карибское море (?), Тихий океан: зал. Пьюджет Саунд, Токийский залив.
33. Т. subtilis (Ostf.) Gran (1900 : 117). —Podosira subtilis Ostf. (Ostenfeld in Knudsen
a. Ostenfeld, 1899 : 55).
Клетки погружены в слизистую массу. Панцирь высокоцилиндрический, с пояска
с закругленными краями. Створки очень выпуклые, 16—35 мкм в диам. Ареолы в мно-
гочисленных пучках, около 30 ареол в 10 мкм. Форамены шестиугольные (в СЭМ),
крибрум из 6—8 регулярно расположенных пор. В центре створки несколько ареол,
обычно окруженных окремненным кольцом, рядом с которым находится центральный
вырост с опорами. На створке 3—4 кольца выростов с опорами, имеющих очень корот-
кие трубочки на внутренней поверхности створки и почти не выступающих с наружной
поверхности. Двугубый вырост небольшой, в краевом кольце выростов с опорами или
ближе к центру, с внутренней поверхности створки в виде сплющенной трубки
со щелью, ориентированной радиально, с наружной открывается узким отверстием
с утолщенным краем. (Табл. 47,8—13}.
Океанический планктонный бореальный тропический и низконотальный вид.
Ныне — Японское море. Вне СССР. Ныне — Северное море, пролив Ла-Манш, сев.
Атлантика, Карибское море, Мексиканский залив, Тихий океан: сев.-зап. часть,
Аляскинский залив, у берегов Калифорнии, Перу и Чили, Тасманово море, Индийский
океан у берегов Индии и Африки.
3. Секция Thalassiosira (Макарова, 1971 : 70)
Створки обычно плоские, редко выпуклые или волнистые. Ареолы в радиальных
рядах, иногда с тенденцией к образованию неотчетливых пучков или с выраженными
вторичными концентрическими рядами. По краю створки одно кольцо редко или часто
расположенных выростов с опорами, иногда они образуют более широкое кольцо, где
выросты расположены в шахматном порядке, у некоторых видов выросты имеются
по всей створке. В центре створки или близ центра один вырост сопорами или их много,
они образуют пучок или кольцо близ центра створки. Выросты с опорами обычно имеют
более длинную трубку на наружной поверхности створки. Двугубый вырост один, чаще
на границе с загибом створки, редко между центром и краем створки.
Т и п: Т. nordenskioeldii Cl.
I. Створки более 10 мкм в диам., с отчетливой структурой, видимой в СМ.
1. Панцирь с пояска восьмиугольный, краевые выросты с опорами крупные........
......................................................43. Т. nordenskioeldii.
2. Панцирь с пояска четырехугольный, краевые выросты с опорами короткие.
А. В центре створки выросты с опорами образуют пучок.
а. По краю створки одно кольцо из выростов с опорами, на лицевой части
створки выросты отсутствуют................. ...............40. Т. hyalina.
б. По краю створки более одного кольца из выростов с опорами.
а. Выросты с опорами по краю створки образуют 2 кольца, иногда между
ними кольцо из замкнутых выростов, на лицевой части створки выросты
с опорами отсутствуют..................................34. Т. antarctica.
/3. Выросты с опорами по краю створки образуют более 2 колец, расположены
в шахматном порядке, на лицевой части створки выросты с опорами
имеются.
73
+ Структура створки из отчетливых ареол, выросты с опорами
на лицевой части створки немногочисленные.........38. Т. gravida.
++Структура створки из радиальных ребер, выросты с опорами
на лицевой части створки многочисленные.............48. Т. rotula.
Б. В центре створки выросты с опорами не образуют пучка.
а. Центральные выросты с опорами одиночные, или их несколько, и они
разбросаны между ареолами; краевые выросты с опорами в СМ не
отчетливые.
а. Ареолы округлые, 10—12 в 10 мкм.....................42. Т. manifesta.
р. Ареолы вытянутые, 4—9 в 10 мкм......................44. Т. orientals,
б. Центральные выросты с опорами собраны в группы или образуют кольцо,
краевые выросты с опорами в СМ отчетливые.
а. Створки почти плоские, выросты с опорами близ центра створки образуют
неправильное кольцо.....................................49. Т. weissflogii.
р. Створки с тангентальной волнистостью, выросты с опорами близ центра
створки образуют группы из 1—5 выростов или второе кольцо на
1/2 радиуса створки.................................35. Т. bramaputrae.
II. Створки обычно менее 10 мкм в диам., с трудноразличимой в СМ структурой.
1. Двугубый вырост расположен в краевом кольце выростов с опорами.
А. Двугубый вырост меньше выростов с опорами.
а. Клетки с пояска восьмиугольные, структура створок из мелких ареол....
......................................................37. Т. conferta.
б. Клетки с пояска четырехугольные, структура створок из радиальных ребер
или ареол неправильных очертаний.
а. Структура створок из ареол в радиальных рядах, с тенденцией к танген-
циальным у края створки....................................41. Т. mala.
Р. Структура створок из радиальных ребер, дихотомически разветвленных
по направлению к краю створки.
+ Краевые выросты с опорами бульбообразно расширены у наружной
поверхности створки.................................36. Т. bulbosa.
++Краевые выросты с опорами не расширены у поверхности створки.
— Створки 2.2—7.5 мкм в диам. Краевые выросты с опорами располо-
жены редко. Двугубый вырост приближен к одному из краевых выро-
стов с опорами.....................................47. Т. pseudonana.
—Створки 8—17 мкм в диам. Краевые выросты с опорами расположены
часто. Двугубый вырост занимает -место одного из выростов
с опорами............................................39. Т. guillardii.
Б. Двугубый вырост крупнее выростов с опорами, заметный.......45. Т. parvula.
2. Двугубый вырост расположен близ центра створки............46. Т. proschkinae.
34. Т. antarctica Comber (1896 : 491, fig. 1—6). — Т. fallax Meunier (1910 : 268, tab.
30,7-4).
Клетки соединены в цепочку тонким и гибким тяжом, сильно разветвленным
близ поверхности створки. Панцирь в виде барабана, с широким пояском. Вальвоко-
пула довольно широкая, со структурой из пор, расположенных вертикальными рядами,
36 в 10 мкм, вставочный ободок с одним кольцом пор, 48 в 10 мкм, соединительных
ободков 3, бесструктурных. Створки плоские, 23—40 мкм в диам. Ареолы образуют
сомкнутую сеть, 20—22 в 10 мкм, радиальные ряды у края створки иногда дихотомиче-
ски разветвленные, что создает впечатление пучков. Форамены 4—6-угольные или
неправильной формы. В центре створки пучок из выростов с опорами. По краю створки
2 кольца из выростов с опорами, в наружном кольце 9—12 в 10 мкм, во внутреннем —
6—8 в 10 мкм. У некоторых экземпляров между этими кольцами выростов кольцо
из замкнутых выростов, 3—6 в 10 мкм. Двугубый вырост крупный, расположен между
двумя кольцами из выростов с опорами или в одном кольце с замкнутыми выростами.
74
Его наружная часть в виде рупора, внутренняя сильно сплющенная, с длинной узкой
щелью, ориентированной радиально. Загиб створки низкий, с мелкими неправильными
ареолами и гиалиновым краем. Споры эндогенные, полуэндогенные и экзогенные,
сильно окремнены, со структурой, имеющей значительные отличия от структуры веге-
тативных клеток. Первичная створка сильно выпуклая, с многочисленными выростами
с опорами, расположенными по всей ее лицевой части (Syvertsen, 1979). Ареолы в ради-
альных рядах, 10—14 в 10 мкм. По краю створки 1 или 2 кольца выростов сопорами.
Вторичная створка более сильно окремнена и грубо ареолирована, 7—10 ареол в 10 мкм,
менее выпуклая, чем первичная. Двугубый вырост в одном кольце с выростами с опо-
рами. (Табл. 48,7—5).
Морской неритический биполярный вид.
Средний и поздний плейстоцен — Чукотка. Голоцен — Охотское море. Ныне —
Берингово, Охотское моря, у берегов Чукотки, Камчатки и Курильских о-вов. Вне
СССР. Ныне — Атлантический океан: у берегов вост. Гренландии, у сев. и зап. берегов
Скандинавии, в фиордах Тронхейма и Осло, у берегов Шпицбергена, в море Уэдделла,
у Южн. Шетландских и Оркнейских о-вов; Тихий океан: сев.-зап. часть, у берегов
Аляски, в море Росса.
35. Т. bramaputrae (Ehr.) Hakansson et Locker (1981 : 123, tab. 6, 34—37; 7, 38—40;
12,69—75).
Клетки одиночные. Створки круглые, с тангентальной волнистостью. Ареолы тесно
расположенные, в радиальных рядах. Форамены небольшие, находящиеся в углублении
поверхности створки, крибрум с густо расположенными порами, иногда в радиальных
рядах. Близ края створки вклиниваются короткие ряды ареол. Краевые выросты
с опорами отчетливые, часто и равномерно расположены по краю лицевой части створ-
ки, с наружной длинной трубкой. Внутрь от краевого кольца кольцо редких замкнутых
выростов, или на */2 радиуса створки второе кольцо выростов с опорами, или по обе
стороны от ее центра 2 группы выростов с опорами, в одной группе 1—2 выроста, во вто-
рой 4—5, с очень короткой внутренней трубкой и с отверстием на наружной поверхности
створки. Загиб створки очень низкий, с короткими радиальными ребрышками,
14 в 10 мкм.
Пресноводно-солоноватоводный эвригалинный, арктобореальный и нотальный
вид, часто встречающийся в прибрежном морском планктоне, солоноватых и пресных
континентальных водоемах.
Плиоцен — ныне.
Var. bramaputrae. — Stephanodiscus bramaputrae Ehr. (Ehrenberg, 1854, tab. 35A,
9—10). — Coscinodiscus lacustris Grun. (Grunow in Cleve et Grunow, 1880 : 114). — C. lacu-
stris var. genuinus A. Cl. (Cleve-Euler, 1942 : 249, fig. 13). — Thalassiosira lacustris (Grun.)
Hasle (Hasle, Fryxell, 1977 : 40).
Створки тангентально-волнистые, 12.5—52 мкм в диам. Ареолы образуют доволь-
но густую сеть, 10—14 в 10 мкм, с внутренней поверхности створки отчетливые радиаль-
ные гиалиновые полосы, 10—16 в 10 мкм, у края створки дихотомически разветвлен-
ные, между ними радиальные ряды крибральных пор. Краевые выросты с опорами раз-
делены одной ареолой, 6—8 выростов в 10 мкм. Внутрь от краевого кольца кольцо
более крупных, редко расположенных замкнутых выростов, 2—4 в 10 мкм. Иногда
выросты с опорами по всей лицевой части створки. (Табл. 49,1—4).
Плиоцен — Закавказье, Дальний Восток, о-в Итуруп. Плейстоцен — Ленинградская
обл. (межледниковые отл.), Карелия (позднеледниковые отл.). Голоцен — Онежское
озеро (послеледниковые отл.), Балтийское море. Ныне — Финский залив, Белое,
Карское, Печорское, Чукотское, Берингово, Черное, Азовское, Каспийское, Аральское
моря, оз. Ильмень, Балхаш, рр. Даугава, Днепр, Урал, Амур, Амударья, водоемы
Туркмении. Вне СССР. Плейстоцен — Финляндия. Ныне — в прибрежных водах евро-
пейских морей, в солоноватых и пресных водоемах бореальной зоны, водоемы Индии,
Тибета, Японии, эстуарий р. Свен (Австралия).
75
Var. septentrionalis (Grun.) Makar. (Макарова, 19886 : 76; Макарова, 1983 : 24).-
Coscinodiscus lacustris var. septentrionalis Grun. (Grunow, 1884 : 85, tab. D, 28, 33).
Створки выпуклые, с тангентальной волнистостью, 15—40 мкм в диам. Ареолы
крупнее, чем у var. bramaputrae, 5—9 в 10 мкм. Краевые выросты с опорами разделены
между собой 2—3 ареолами, 3—4 выроста в 10 мкм. На */г радиуса створки по обе сто-
роны от центра по 1—2 или более выростов с опорами. (Табл. 49,5—76).
Плейстоцен — Сев.-Зап. СССР, Белое, Берингово и Охотское моря. Голоцен —
Архангельская обл. Ныне — обычно вместе с var. bramaputrae. Вне СССР. Балтика (лито-
риновые отл.). Плейстоцен — Финляндия. Ныне — вместе с var. bramaputrae.
Nax. proximus(Makar.)Makar. (Макарова, 19886 :77; Макарова, 1983 : 24). — Coscino-
discus proximus Makar. (Макарова, 1961 : 52, табл. II, 1-4).
Створки слегка выпуклые, с отчетливой тангентальной волнистостью, крупные,
81.2—92.4 мкм в диам. Ареолы мелкие, 8—10 в 10 мкм. Краевые выросты с опорами
отчетливые, очень равномерно расположенные, 6—8 в 10 мкм, разделенные одной
ареолой. На х/2 или 7з радиуса створки второе кольцо неравномерно расположенных
выростов с опорами, 2—3 в 10 мкм. (Табл. 50, 7—6).
Ныне — Каспийское море, оз. Балхаш.
36. Т. bulbosa Syvertsen (Syvertsen, Hasle, 1984 : 168-171, fig. 1—32).
Клетки одиночные или собраны в короткие цепочки. Панцирь низкоцилиндриче-
ский, со вставочными и соединительными ободками. Створки плоские, слабо окремнен-
ные, 6—10 мкм в диам. Ребра в радиальных рядах, на середине радиуса створки дихо-
томически разветвляются, 38—44 в 10 мкм. Крибрум с нерегулярно расположенными
крибральными порами. В центре створки, рядом с крупной ареолой вырост с опорами
с 3 сопутствующими порами. В верхней части загиба створки кольцо выростов с опора-
ми, с бульбообразным расширением у наружной поверхности створки, внутренняя
трубка узкая, с 2 сопутствующими порами, 5—6 в 10 мкм. Двугубый вырост малень-
кий, расположен между двумя выростами с опорами, его внутренняя щель ориентиро-
вана радиально. Загиб створки низкий, с вертикальными рядами ареол. Покоящиеся
споры с ареолами, имеющими маленькие форамены. (Табл. 50, 7, 8).
Морской арктобореальный вид.
Ныне — Берингово, Охотское моря. Вне СССР. Ныне — арктические моря.
37. Т. conferta Hasle (Hasle, Fryxell, 1977 : 239-244, fig. 1-23).
Клетки с пояска восьмиугольные. Панцирь высокоцилиндрический, с 1—2 вста-
вочными и многочисленными соединительными ободками. Вальвокопула с мелкими
ареолами, чередующимися с ребрами. Створки слегка выпуклые, с вогнутым центром,
12.5 мкм в диам. Ареолы в радиальных рядах, 18 в 10 мкм. Форамены многоугольные.
В центре створки несколько мелких ареол и один центральный вырост с опорами с 2—
4 сопутствующими порами. По краю лицевой части створки на границе с загибом с про-
тивоположных сторон по 2 краевых выроста с опорами, два из них, более крупных,
сближены, и между ними очень маленький двугубый вырост, два других на значитель-
ном расстоянии друг от друга. Выросты с опорами с длинной трубкой на наружной
поверхности, на внутренней окруженной 4—5 сопутствующими порами. Загиб створки
высокий, с мелкими ребрышками, 23—24 в 10 мкм. (Табл. 50, 9,10).
Морской бореально-тропический и нотальный вид.
Ныне — тундровые озера на п-ове Таймыр. Вне СССР. Ныне — Осло-фьорд, Вест-
фьорд, Троннхеймс-фьорд, сев. Атлантика, у берегов сев.-зап. Африки, Тихий океан:
у берегов Калифорнии, Чили.
П р и м е ч. В диагнозе этого вида (Hasle, Fryxell, 1977) указывается, что количе-
ство краевых выростов с опорами (от 3 до 14) зависит от диаметра створки.
38. Т. gravida Cl. (Cleve, 1896 : 12, tab. 2, 14-16). — T. tchemiae Manguin (1960 :
244, tab. 3,28—31).
76
Клетки с закругленными углами, соединены в цепочки толстым тяжом, который
у поверхности створки разделен на несколько тонких нитей. Панцирь цилиндрический,
11—30 мкм выс. Вальвокопула широкая, с нерегулярно расположенными в вертикаль-
ных и косых рядах порами, 30—36 в 10 мкм, вставочный ободок с одним кольцом пор,
34 в 10 мкм, соединительных ободков 3, бесструктурных, с широкой лигулой. Створки
слегка выпуклые с вогнутостью в центре, 14—65 мкм в диам. Ареолы в радиальных
рядах, образуют сомкнутую сеть, у крупных створок с тенденцией к образованию
неотчетливых пучков, 16—18 ареол в 10 мкм. Форамены круглые или неправильно
округлой формы, с сомкнутыми или одиночными нитевидными радиальными выроста-
ми. В центре створки несколько или много выростов с опорами, образующих пучок.
Краевые выросты с опорами довольно крупные, до 1 мкм дл., образуют несколько
колец по загибу и краю створки, располагаясь в шахматном порядке, 3—5 в 10 мкм.
По всей лицевой части створки также имеются выросты с опорами, расположенные
неправильными концентрическими кругами. Наружный конец трубки выроста сопора-
ми часто зубчатый. Двугубый вырост на границе с загибом створки, его наружная
часть в виде довольно высокого (до 1.6 мкм) цилиндра с расширенным наружным
концом, внутренняя короткая, крупная, сильно сплющенная, с длинной щелью,
ориентированной под небольшим углом к радиусу створки. Загиб створки невысокий,
с ареолами, продолжающимися со створки, и с выростами с опорами, расположенными
в шахматном порядке. Споры имеют неодинаково выпуклые створки, первичная створ-
ка очень выпуклая, вторичная менее выпуклая, с уплощенной серединой. Структура
из крупных ареол, расположенных в неотчетливых пучках и тангенциальных, вогнутых
к центру или радиальных и вторично косых рядах у края. По краю створки коль-
цо выростов с опорами, в центре створки группа выростов с опорами. (Табл. 51,
1—12).
Морской океанический и неритический биполярный вид.
Поздний миоцен—поздний плиоцен (?) — Дальний Восток. Плейстоцен — Ладож-
ское озеро, Карелия, Архангельская обл., сев. Сибирь, Чукотка, сев.-зап. часть Тихого
океана. Голоцен — Охотское море. Ныне — сев. и дальневосточные моря, Амурский
лиман. Вне СССР. Ныне — Сев. Ледовитый океан, у берегов сев. европейских морей
и проливов, сев. Атлантика: у берегов вост. Америки, заливы Св. Лаврентия, Шалёр,
Фанди, у Лабрадора, Ньюфаундленда и вост. Гренландии, вокруг Исландии,
Тихий океан у берегов Калифорнии; в южном полушарии: у берегов Южн.
Африки, в морях Антарктиды и антарктических секторах Тихого и Атлантиче-
ского океанов.
39. Т. guillardii Hasle (1978 : 274-278, fig. 28-50).
Клетки с пояска четырехугольные, одиночные. Панцирь низкоцилиндрический,
тонкий. Створки плоские, 8—17 мкм в диам. Кремнеземный базальный слой с радиаль-
ными, отчасти нерегулярными рядами пор. Наружная поверхность створки со слегка
выступающими кремнеземными ребрами различной ширины. В центральной части
створки ребра радиальные, к краю дихотомически разветвленные или образующие
нерегулярные круги, 3—4 ребра в 1 мкм. На 7з ~ lh радиуса 1—3 выроста с опорами
с 2—4 сопутствующими порами, или выросты отсутствуют. Краевые выросты с опорами
образуют одно кольцо, 8—10 в 10 мкм, их наружная трубка длиннее внутренней,
с 4 сопутствующими порами. Двугубый вырост в одном кольце с выростами с опорами
занимает место одного из них, его внутренняя щель прямоугольная, ориентирована
радиально. Загиб створки с мелкими или прямыми ребрами, 5—8 в 1 мкм. Иногда
на границе между лицевой частью створки и загибом одно кольцо гранул. (Табл. 52,
1-4).
Солоноватоводный вид, обитающий в морских и солоноватых прибрежных водах.
Ныне — Саратовское водохранилище, низовье Волги, рр. Ока, Сура, озера Севан,
Пертозеро (Карелия). Вне СССР. Ныне - Финский и Токийский заливы, реки Англии,
в притоках р. Везер (ФРГ) , рр. Тиса, Дунай (Венгрия), США (реки сев. Огайо) , оз. Эри.
77
40. T. hyalina (Grun.) Gran (1897 : 4, tab. 1,17, 18). — Coscinodiscus hyalinus Grun.
(Grunow in Cleve et Grunow, 1880 : 113, tab. 7,128).
Клетки образуют цепочки из 4—8 клеток, соединенных тонким тяжом. Панцирь
низкоцилиндрический, с 2 вставочными ободками. Створки почти плоские, слабо
вогнутые в центре, 12—42 мкм в диам. Ареолы мелкие, близ центра створки 18—24,
у края 16—20 в 10 мкм. В центре створки пучок из выростов с опорами. Краевые
выросты с опорами образуют одно кольцо, 8—10 в 10 мкм, на наружной поверхности
трубка 1—1.2 мкм дл. Двугубый вырост в одном кольце с краевыми выростами с опо-
рами, его наружная часть в виде слегка расширенной кверху трубки 1.3—1.5 мкм дл.,
внутренняя щель ориентирована радиально. Радиальный ряд ареол, направленный от
центра створки к двугубому выросту, близ его основания прерван и заканчивается
узкой гиалиновой полосой. Загиб створки низкий, с 2—3 горизонтальными рядами
ареол. Край загиба гиалиновый. (Табл. 48, 6-12).
Морской и солоноватоводный планктонный пелагический и неритический аркто-
бореальный вид.
Плиоцен — Камчатка. Плейстоцен — сев.-зап. часть Тихого океана, Берингово,
Охотское и Японское моря. Поздний плейстоцен — Чукотка. Голоцен — Охотское
море. Ныне — сев. и дальневосточные моря (от Анадырского залива до сев. части Япон-
ского моря). Вне СССР. Неоген — о-в Хоккайдо. Ранний плиоцен—поздний плейсто-
цен — сев.-вост. Япония. Ныне — Балтийское, Норвежское, Гренландское моря, Баф-
финов и Гудзонов заливы, у Лабрадора, Девисов пролив, у сев.-вост. берегов США.
41. Т. mala Takano (1965 : 1, fig. 1, а-т, tab. 1,1-8).
Клетки заключены в слизистую массу. Панцирь дисковидный. Вальвокопула
с 2—3 рядами пор, 10 в 1 мкм. Створки почти плоские, 4—10 мкм в диам. Ареолы
в центре створки более крупные, в радиальных рядах, ближе к краю в тангенциальных,
20—24 ареолы в 10 мкм. Форамены имеют 4—6-угольные очертания, крибрум с много-
численными порами. Близ центра створки один вырост с опорами с 3—4 сопутствующи-
ми порами. Краевых выростов с опорами 7—8 в 10 мкм, с короткой внутренней труб-
кой, 4 сопутствующими порами и наружным отверстием, окруженным гиалиновым
кольцом. Двугубый вырост между двумя выростами с опорами, на наружной поверх-
ности заканчивается округлым отверстием, на внутренней он в виде очень маленькой
трубки со щелью, ориентированной радиально. Загиб створки невысокий, с одним ря-
дом ареол, край его гиалиновый. (Табл. 50, 11—13).
Морской неритический бореальный и нотальный вид.
Ныне — Охотское море, Чукотка (Ямская губа и р. Яма, приливно-отливная зона).
Вне СССР. Ныне — Тихий океан: Аляскинский залив, у берегов Калифорнии, Японии
и сев. Австралии, Атлантический океан: Мексиканский залив, у берегов Сев. Америки,
южн. Бразилии и Дании, Индийский океан, у берегов Африки и Индии.
42. Т. manifesto Sheshuk. emend.Makar. (Шешукова-Порецкая, 1961 : 54).
Створки слегка выпуклые, 18—30 мкм в диам. Ареолы в слегка изогнутых ради-
альных рядах, иногда дихотомически разветвленных, близ центра створки образуют
концентрические ряды или расположены беспорядочно, 10—12 в 10 мкм, к краю створ-
ки ареолы уменьшаются в размерах, 22 и более в 10 мкм. Форамены маленькие, круг-
лые, неодинаковой величины, крибрум образует искривленные радиальные ряды, 8—
9 крибральных пор в 1 мкм. В центре створки один или несколько выростов с 2 сопут-
ствующими порами. Краевые выросты с более крупными отверстиями, чем форамены,
образуют несколько колец, в СМ плохо различимы, с внутренней поверхности с 2—
4 сопутствующими порами, иногда недоразвитыми. Двугубый вырост находится от-
ступя от края створки, его щель ориентирована радиально. Споры слегка выпуклые,
с вогнутым центром, гиалиновые, 13—20 мкм в диам. В центре створки 3—23 выроста
с опорами с 2 сопутствующими порами, на наружной поверхности заканчиваются отвер-
стием. Краевые выросты с опорами образуют на загибе створки одно или несколько
7R
колец, на наружной поверхности заканчиваются отверстием или небольшой трубкой,
с внутренней трубка очень короткая, с 4 сопутствующими порами. Двугубый вырост
не связан с выростами с опорами, на наружной поверхности заканчивается отверстием,
на внутренней — валиковидным утолщением, его щель ориентирована радиально. Загиб
низкий с загнутым краем. (Табл. 52,5—12).
Морской неритический вымерший вид.
Миоцен — Камчатка (тюшевская серия), Сев. Сахалин, зап. побережье Сахалина
(маямрафская свита). Поздний миоцен—ранний плиоцен — Южн. Сахалин (маруямская
свита). Вне СССР. Неоген — Япония. Поздний миоцен — о-ва Хоккайдо, Хонсю.
43. Т. nordenskioeldii Cl. (Cleve, 1873 : 7, tab. 1,1).
Клетки образуют короткие или длинные гибкие цепочки, соединенные тонким
тяжом. Панцирь с пояска восьмиугольный, 9—15 мкм выс., со вставочными и соедини-
тельными ободками. Вальвокопула из мелких ареол, расположенных вертикальными
рядами, около 30 рядов в 10 мкм. Створки выпуклые с углублением в центре, 11 —
45 мкм в диам. Ареолы довольно мелкие, в центре створки 18—20 ареол, иногда более
в 10 мкм. В СЭМ форамены округлой или слегка вытянутой формы, крибрум с 20 по-
рами в 1 мкм. В центре створки один вырост с опорами, окруженный несколькими
более крупными ареолами. Отступя от края на */з диаметра створки кольцо крупных,
равномерно расположенных краевых выростов с опорами, 2—3 в 10 мкм. На наружной
поверхности их трубка 0.2—0.4 мкм в диам., 1.8—3 мкм дл., слегка расширена у основа-
ния, с окаймляющей воронкой наверху до 1.2 мкм в диам., реже без нее. Двугубый
вырост в одном кольце с выростами с опорами, почти не отличается по размерам от них,
с наружной трубкой без воронки, на внутренней поверхности с короткой трубкой
и щелью, ориентированной радиально. На загибе створки ареолы в вертикальных рядах,
20—24 ареол в 10 мкм. Известны споры, по форме и структуре сходные с вегетативной
клеткой. (Табл. 53,7-77).
Морской пелагический и неритический арктобореальный вид.
Неоген — зап. и вост, побережье Камчатки, Сев. и Южн. Сахалин. Плиоцен — Кам-
чатка, сев.-зап. часть Тихого океана. Плейстоцен — Сев.-Зап. СССР, сев.-зап. часть Тихого
океана. Поздний плейстоцен — Чукотка. Голоцен — Берингово и Охотское моря. Ныне —
все сев. и дальневосточные моря и заливы. Вне СССР. Неоген — Япония. Ныне — Сев.
Ледовитый океан, сев. моря и проливы, сев. Атлантика: заливы юго-вост. Канады,
Наррагансетт, сев. и вост, части Тихого океана: зал. Пьюджет Саунд и у берегов Кали-
форнии.
44. К orientalis Sheshuk. emend. Makar. (Шешукова-Порецкая, 1959, табл. 1,9а, б) .
Створки почти плоские, 20—44 мкм в диам. Ареолы в радиальных и вторично
концентрических рядах, вытянутой формы, 4—9 в 10 мкм, к краю уменьшаются в раз-
мерах. В центре створки 2—3 (редко многочисленные) выроста с опорами, заканчиваю-
щиеся на наружной поверхности маленькими отверстиями. Краевые выросты с опорами
с короткой трубкой, 5 в 10 мкм, в СМ незаметны. Двугубый вырост с длин-
ной наружной трубкой, с внутренней поверхности створки с валиковидным утолщением
и щелью, ориентированной радиально. Загиб створки низкий, с ребровидными утолще-
ниями^—13 в 10 мкм. (Табл. 52,73-77).
Морской вымерший вид.
Поздний миоцен—плиоцен — Сев. Сахалин (маямрафская свита). Поздний мио-
цен-плиоцен — Южн. Сахалин (маруямская свита). Плиоцен — Камчатка. Вне СССР.
Неоген — Япония.
45. Т. parvula Makar. (Макарова, 19776 : 907,908, табл. 1,7-5).
Панцирь в виде низкого барабана. Створки плоские, 7—10.5 мкм в диам. Ареолы
(по данным ТЭМ) образуют сомкнутую сеть, их форма от округло-угловатой до слегка
вытянутой, в радиальных рядах в центре створки 24—26 ареол, у края 28 в 10 мкм.
79
Близ края створки ареолы в более или менее тангенциальных рядах. Крибрум с 20
порами в 1 мкм. В центре створки один вырост с опорами, окруженный крупными
и мелкими ареолами. Краевые выросты с опорами равномерно расположены на границе
с загибом створки, 8 в 10 мкм, с 4 сопутствующими порами. Трубка выростов на наруж-
ной поверхности створки более длинная, до 0.4 мкм. Двугубый вырост крупнее кра-
евых выростов с опорами, в одном кольце с ними. Загиб створки низкий, с мелкими,
вертикально расположенными ареолами, 40 в 10 мкм. (Табл. 57,10—14).
Солоноватоводный бореальный вид.
Ныне — Охотское и Азовское моря.
46. Т. proschkinae Makar. (Макарова, Генкал, Кузьмин, 1979 : 922, табл. 1,7-7).
Клетки одиночные. Панцирь цилиндрический, тонкокремнеземный, со вста-
вочными ободками. Створки круглые, плоские или слегка выпуклые. Ареолы полиго-
нальные, вытянутые, иногда створки с радиальными ребрами без поперечных перегоро-
док между ними. На ребрах или между ареолами иногда шипики. В центре створки
ареолы круглые, \ края вытянутой формы. Крибрум с многочисленными крибраль-
ными порами. Центральный вырост с опорами с 2 сопутствующими порами Краевые
выросты с опорами, редкие, 4—7 на створке, реже многочисленные, с 4 сопутствующими
порами. Двугубый вырост очень маленький, расположен близ центра створки, отделен
от центрального выроста с опорами одной ареолой, его щель ориентирована радиально.
Загиб створки низкий, с вытянутыми ареолами, 5—6 в 1 мкм.
Солоноватоводный и пресноводный бореальный вид.
Var. proschkinae.
Створки 3—11.5 мкм в диам. Ареолы полигональные, в центре створки 2.5—
3 в 1 мкм, у края 3—5. Крибрум с 21—23 крибральными порами в 1 мкм. Краевые
выросты с опорами от 4 до многочисленных. (Табл. 54,1-8).
Ныне — Азовское и Каспийское моря, оз. Аджикабул (АзССР). Вне СССР. Ныне —
реки Англии.
Var. spinulata (Takano) Makar. (Макарова, 19886:81, табл. LI, 27, 22). — Т. spinulata
Takano (1981 : 33, 34, fig. 1,B, 14-25).
Створки 3.3—4 мкм в диам. Структура из полигональных ареол илй радиальных
ребер без поперечных перегородок меаду ними, 4—5 в 1 мкм, с отчетливыми шипиками
на ребрах. (Табл. 54, 9,10).
Ныне — Охотское море, зал. Шелихова. Вне СССР. Ныне — Япония: реки, озера,
заливы префектур Оита, Фукуока, Шизуока, Аихи.
47. Т. pseudonana Hasle et Heimdal (1970 : 565, fig. 27—38).
Клетки очень малых размеров, иногда заключенные в слизь. Панцирь низкоцилин-
дрический, очень тонкий. Створки плоские, 2.2—7.5 мкм в диам. Структура створок из
радиальных ребер, 5—7 в 1 мкм, у края створки ребра дихотомически разветвляются;
иногда между ребрами поперечные перегородки, образующие ареолы. Базальный крем-
неземный слой створки с неправильно расположенными порами или в радиальных
рядах. На */2 или */з радиуса створки 1—2 центральных выроста с опорами, с 2—3 со-
путствующими порами, или выросты отсутствуют. Краевые выросты с опорами от 5
на створке до 5—12 в 10 мкм, на наружной поверхности трубка немного длиннее,
чем на внутренней, окруженной 3 сопутствующими порами. Двугубый вырост очень
маленький, расположен в краевом кольце выростов с опорами, обычно приближен
к одному из них, имеет округлую щель, ориентированную радиально. Загиб створки
относительно высокий, со структурой как на лицевой части створки. (Табл. 54,11-22).
Солоноватоводный бореально-тропический вид.
Ныне — Охотское, Черное, Каспийское, Азовское моря, водоемы Эстонии, Волж-
ское, Пестовское водохранилища (Московская обл.), рр. Яма (Чукотка), Енисей,
Селенга, озера Байкал, Севан, Соленый Ясхан (Туркмения). Вне СССР. Ныне — при-
брежные воды европейских морей, р. Везер (ФРГ), реки Англии, Венгрии, водоемы
Японии, у берегов сев. Австралии.
80
48. T. rotula Meunier (1910 : 264, tab. 29, 67—70).
Клетки соединены толстым тяжом, образованным несколькими тонкими нитями,
в прямые или слегка изогнутые цепочки. Панцирь от низко- до высокоцилиндрического,
13.6—38.4, обычно 17 мкм выс., иногда его высота превышает ширину в 3 раза, с пояска
прямоугольный, со слегка закругленными углами, с 2 вставочными ободками и септами.
Структура створок из отчетливых радиальных ребер, 20—24 в 10 мкм, несколько раз
дихотомически разветвленных на протяжении радиуса створки от центра к периферии.
У края створки между ребрами с заметными поперечными перегородками, иногда
наблюдаемыми и у центра створки, 26—28 ареол в 10 мкм в центре створки, 22—26
у края. Крибрум с порами в радиальных рядах, 12—15 пор в 1 мкм. В центре створки
плотный пучок из 9—24 выростов с опорами с 4—5 сопутствующими порами. По всей
лицевой части створки многочисленные выросты с опорами, образующие в прикраевой
зоне кольцо из 3 смещенных рядов, 8—12 выростов в 10 мкм, разделенных 1—3 ареола-
ми. Двугубый вырост крупный, в кольце краевых выростов с опорами. Его наружная
часть в виде рупора, внутренняя сплющенная, с длинной щелью, ориентированной
радиально. (Табл. 55, 7-7; 56,7-72).
Морской бореально-тропический низконотальный вид.
Голоцен — Белое море. Ныне — Баренцево, Охотское, Японское моря, у Куриль-
ских о-вов. Черное море. Вне СССР. Ныне — Атлантический океан: у берегов Зап. Евро-
пы, Карибское, Саргассово моря, залив Наррагансетт, Канарское течение, Тихий океан:
у берегов сев. Аляски, Калифорнийский залив, у сев. Австралии, у берегов Новой
Зеландии, пролив Кука, Индийский океан у берегов Африки.
49. Т. weissflogii (Grun.) Fryx. (Fryxell et Hasle, 1979 : 68—72, fig. 1—15). — Micro-
podiscus weissflogii Grun. (Grunow in Van Heurck, 1880—1885 : 231). — Thalassiosira fluviati-
lis Hust. (Hustedt, 1926 : 565, fig. 1—4). — T. fluviatilis f. mangrovii Manguin (Manguin in
Bourrelly et Manguin, 1952 : 35, tab. I, 6a—b). — T. hustedtii Poretzky et Anissim. (Порецкий,
Анисимова, 1933 : 47, рис. 9,2). — T. hustedtii var. vana Makar, et Pr.-Lavr. (Макарова,
Прошкина-Лавренко, 1964 : 39, рис. 5).
Клетки образуют короткие цепочки. Панцирь в виде барабана со вставочными
ободками. Вальвокопула с порами, образующими косые ряды, вставочный ободок
со значительной бесструктурной частью, 4 соединительных ободка имеют одинаковую
структуру, но различную ширину, последний, 5-й, соединительный ободок совсем узкий.
Створки почти плоские, 13—25 мкм в диам. Структура створок из радиальных ребер,
32—38 в 10 мкм, или с мелкой сеткой в виде перекрестного жилкования. Близ центра
створки неправильное кольцо или группа из нескольких выростов с опорами. Краевые
выросты с опорами расположены равномерно, 12—14 в 10 мкм. Двугубый вырост
крупный, в краевом кольце выростов с опорами, посередине между двумя соседними
выростами, его внутренняя щель ориентирована радиально. Загиб створки довольно
высокий, со слабо развитой сеткой ареол. (Табл. 57, 7—9).
Пресноводно-солоноватоводный широкобореальный и нотальный вид.
Ныне — Каспийское море, Мингечаурское и Иваньковское водохранилища,
соленые озера Старой Руссы, реки Украины. Вне СССР. Ныне — Балтийское море, южн.
побережье Англии, Осло-фьорд, Мексиканский залив и залив Микава (Япония),
рр. Темза, Большой Уз (Англия), Везер, Верра, Вюмме (ФРГ), Тиса, Дунай
(Венгрия), реки на юге Чили, водоемы США, Португалии и Сингапура, эстуарий р. Свен
(Австралия).
4. Секция INCONSPICUAE Makar. (Макарова, 1971 : 71)
Створки плоские или вогнутые посередине. Структура створок очень нежная,
видимая только в ЭМ. В центре створки один вырост с опорами, по краю кольцо
выростов с опорами.
Тип: Т. condensata Cl.
*/2 6 Заказ 518
81
I. Панцирь обычно высокоцилиндрический, с многочисленными вставочными ободка-
ми. Створки плоские ............................................50. Т. condensata.
II. Панцирь низкоцилиндрический, вставочные ободки отсутствуют. Створки волнистые
...................................................................51. Т. frigida.
50. Т. condensata Cl. (Cleve, 1900 : 22, tab. 8,12,13).
Клетки соединены коротким тяжом в цепочки. Панцирь от низко- до высоко-
цилиндрического, с многочисленными вставочными ободками. Створки плоские со
слегка вогнутой серединой, 25—30 мкм в диам., с невидимой в СМ структурой. В центре
один вырост с опорами (слизевая пора). По краю створки кольцо равномерно рас-
положенных выростов с опорами.
Морской океанический и неритический планктонный широкобореальный вид.
Ныне — Японское море, у берегов Южн. Сахалина. Вне СССР. Ныне — сев. Атлан-
тика, Северное и Ирландское моря, у норвежских и шведских берегов, Английский
канал, прол. Скагеррак, Тихий океан: зал. Пьюджет Саунд и у берегов Новой Зеландии.
П р и м е ч. До сих пор этот вид никем не исследовался в ЭМ и подробные данные
о его морфологии отсутствуют, что затрудняет определение его систематического
положения.
51. Т. frigida A. Cl. (A. Cleve, 1942 : 270, fig. 44).
Клетки мелкие, соединенные тонкой нитью в многоклеточные цепочки. Расстояние
между клетками немного больше диаметра клетки. Панцирь низкоцилиндрический,
с пояска с закругленными углами. Створки 15—25 мкм в диам., с неправильной волни-
стостью и с тонкой структурой, почти невидимой в СМ.
Морской океанический среднебореальный вид.
Предпоследнее ледниковье — Охотское море. Вне СССР. Ныне — Скагеррак
и Каттегат.
Сомнительный вид
52. Т. satumii Lohmann (1908 : 243, tab. 7,3).
Клетки низкоцилиндрические, с узким пояском. Створки с заметной выпуклостью посере-
дине, 20—30 мкм в диам. Структура неизвестна.
Морской планктонный вид.
Ныне — Японское море, у берегов Южн. Сахалина. Вне СССР. Ныне — Балтийское море.
Примем. Этот вид с ограниченным распространением и недостаточно изученной морфоло-
гией. Изображение, данное у Ломанна и у Г. И. Гайла (1950, табл. XII, 10), неудовлетворительное
и.свидетельствует, скорее, о принадлежности этого вида к другому роду, возможно Cyclotella. '
Род SCELETONEMA Grev. (Greville, 1865 : 43)
Клетки соединены обычно в длинные прямые цепочковидные колонии при
помощи выростов с опорами. Хлоропластов 1—2, редко больше, в виде пластинок
или дисков. Панцирь линзовидный или высокоцилиндрический, иногда с многочислен-
ными соединительными ободками, тонко структурированными, с лигулой. Створки
круглые, плоские или равномерно выпуклые. Ареолы пекулярные (?), в радиальных
рядах. На границе лицевой части створки и загиба кольцо выростов с опорами трубко-
видной или щелевидно-желобковидной формы, окруженных 3 сопутствующими пора-
ми. Двугубый вырост один, находится в кольце выростов с опорами, у конечных
клеток ближе к центру. Его наружная часть в виде короткой или длинной трубки.
Загиб створки довольно высокий, ареолированный, иногда с тонкими ребрами. Из-
вестны ауксоспоры.
Виды планктонцые, морские, солоноватоводные, реже пресноводные, современ-
ные и вымершие.
Т и п: 5. barbadense Grev.
82
£8
' (р-^ > ’эид) ’дилвс
иияоамя я iimoxadaii ‘чаквдиляи онявпц ‘hmJohio KBd}j 'иший я wmw 6'6~ S HMdoflij
*cl~£ ‘яолаиТГ ai/ия я iqiOBiniodoirx ’аня himhi g*g ‘иияаэы^Ннииий qdirtiHEjj
•(£ and
‘9£ : t^96I ‘OMHadflB£f-BHHMmod]j ‘вя<х!вявдо) чв^вдо ja *JABq--jj шпаэшридХэ •§ •£
BdBm олошмэе xKdow хэая оя — ашчц '^ЭЭЭ дИ8
’сЮЭЛ ИЯ9(1 эхчхвяоноиоэ ‘Bdohaij ‘(яо-и иияачиох) bwoliXi -dd dBMirah ‘(ний
-чин}! я-о) эончкилоэд Bdaco ‘.Kdow aoMaqiiBdy ‘эояаиинавх ‘аояаяояу ‘aoHdaj/aoMaHOiiif
‘аояаюхо ‘ояолшкЬд ‘эояаюяХ^ ‘aoxadB^ ‘oflaHHadBg ‘aonag ‘эомаиингвд — эшчц
’КИЯ (1ИЯОИОИ1ЭОЯ) шчннэн
-BdioodiDBd OModnin ‘Hi4HiAidoiHdflC и шчннишиибде шчнт/ояохвяонояоа иолас!одо
•(£-1 > ’and I ‘g$ ^9В1) 'HOMgXdi ионниШ/ э — BdiHan випд яохэяя хшньаноя Л
‘MOMgXdi HOHxeZdBH noMiodox э HWBdono о floioodi4H хмнИияоядокэж-онТ/ияэпэГп аПчпом
woflQBdM я ioodi4H шчдХлХя^ ’говий я himhi ‘эмилЛшчя аанаи! ини ээяод HMdoflij
1 HOiaBLfiiodoLfx 'Э1ЧЯ ‘ололээы^ШппгиП oir олонтГияоения io qdiiTiHBjj
•(9-f ‘8-qBJ‘Z.Z,: 9981
‘эшлэдэ) лэ1д dwisod шкоту - (gj : g/,g ] ‘элэо) 1Э -( лэлд) шгце^оэ s I
\unaaBjpuijXa •§ '£...................яомэий эйия я ‘этчннэюинолонгл] iqiaBiniodoLfx II
ihns|Bsqns ’S ’t7................................ 19H dagad H^doflia эдилвя вц -g
•souiBjod •§ £........................................Bdgad iDidoflia эдилвс вц -у
•ai4HtfHHOMgXdi HHiBdouo a maodiqg •£
‘mnjBjsoo •§ * i................. эннКияомдояэж-онИияэпэГп HWBdono a wiaodiqg • {
•яонихавш! ai/ия я { HoiaBiriiodoirx *1
•(1Л1Я1Л101 :внвящ) *(t?96I ‘OHHadeBif-BHHxmodg ‘ввоЗвявдо) adow эояэципэв>1’BMdosio - p ‘иинонол
«новь - g > riE^Bjv la-JAEq-’ij шпээплрщуСэ у - 9~pc. (8961 ‘ваоЗвяедо ‘OMHodeB£f-EHiixmodjj) adow
эояэиипэвх ’ииноЕоя ияхэня вэаитпвнэи - p ‘ииноноя - f 7 :*1Э (*ЛЭ1Э) wnimsoo тиэио^ээ^ — £—I
'P -эиа
Рис. 5.
1-4 - Sceletonema subsalsum (A. Cl.) Bethge : 1-3 - часть
колонии, 4 - створка. Каспийское море (Прошкина-
Лавренко, Макарова, 1968). (Шкала: 10 мкм).
Солоноватоводный тепловодный бореаль-
ный вид.
Ныне — Каспийское море.
3. S. potamos (Weber) Hasle (1976 : 74, fig. 1-
29). — Microsiphona potamos Weber (1970 : 151,
fig. 2, 3). — Stephanodiscus subsalsus (A. Cl.) Host.
(Hustedt, 1928: 372, fig. 195).
Панцирь цилиндрический, 2—2.5 мкм выс.,
с многочисленными соединительными ободками
с лигулой, покрытыми продольными и попереч-
ными ребрами. Хлоропластов 1—2(4). Створки
плоские или слегка выпуклые, 2.6—5.7 мкм
в диам., с многочисленными мелкими гра-
нулами. Двугубый вырост с длинной наруж-
ной трубкой. На загибе створки ребра.
(Табл. 58,6, 7).
Пресноводный тепловодный бореальный вид.
Ныне — рр. Днепр, Кама, Ангара, водохра-
нилища Пестовское, Чебоксарское. Вне СССР.
Ныне — оз. Эри, рр. Вюмме (ФРГ), Литл-Майами
(США).
4. S. subsalsum (A. Cl.) Bethge (1928 : 343,
tab. II, 7-77). - Melosira subsalsa A. Cl. (Cleve-
Euler, 1912 : 509, fig. 1). — Stephanodiscus sub-
tilis (Van Goor) A. Cl. (Cleve-Euler, 1951 : 54,
fig. 72a—b).
Панцирь цилиндрический, 2—18 мкм выс.,
с многочисленными соединительными ободками
с лигулой, покрытыми продольными и поперечными ребрами. Хлоропластов 1—2.
Створки плоские или слегка выпуклые, 2.6—10.5 мкм в диам. Двугубый вырост обычно
с длинной наружной трубкой. (Табл. 58,8-10\ рис. 5).
Солоноватоводный, пресноводный эвригалинный тепловодный бореальный вид.
Ныне — Балтийское, Черное, Азовское, Каспийское моря, оз. Байкал, р. Волга.
Вне СССР. Ныне — Балтийское море в районе Стокгольма и севернее Ростока, Финский,
Ботнический заливы, р. Вюмме (ФРГ), зал. Шалер, оз. Эри.
Род CYMATOTHECA Hendey (1958 : 4)
Клетки одиночные. Панцирь с пояска линейно-прямоугольный. Створки эллипти-
ческие, редко почти круглые, согнутые по продольной оси. Ареолы локулярные, с велу-
мом на внутренней поверхности створки. Форма, величина и расположение ареол двух
половин створки, по обе стороны от продольной оси, различны. Загиб створки с верти-
кальными рядами мелких ареол. Обычно имеются кольцо выростов с опорами и один
двугубый вырост на загибе створки.
Морской род, встречающийся на литорали преимущественно тропических морей.
Тип: С. weissflogii Hendey.
84
l.C. weissflogii (Grun.) Hendey (1958 : 4, tab. 5, 9). — Euodia weissflogii Grun.
(Grunow, 1881, in Van Heurck, 1880-1885, tab. 126, 13). — Coscinodiscus temperei Brun
(Brun et Tempere, 1889 : 33, tab. 8, 2). — Hemidiscus weissflogii (Grun.) Hust. (Hustedt,
1955 : 11, tab. 1,5-5).
Створки эллиптические, реже почти круглые, 27—100 мкм дл., 23—46 мкм шир.,
ясно тангентально-волнистые по продольной оси, одна половина створки возвышается
над другой. Ареолы уменьшаются от продольной оси створки к ее краю; они часто
различны на половинах створки, расположены радиальными рядами, 4—6 в 10 мкм,
иногда неотчетливо выраженными вторичными неправильными продольными рядами.
Между радиальными рядами вклиниваются более короткие ряды ареол. Выросты
с опорами расположены редко, по всей лицевой части створки. На загибе створки
9—11 ареол в 10 мкм, 3—4 выроста с опорами в 10 мкм, на-наружной поверхности
трубки имеют значительную длину (иногда различимы в СМ). Край загиба узкий, гиа-
линовый. (Табл. 58,11-15).
Морской бентосный вид, преимущественно в тропических морях.
Средний миоцен — Камчатка, Кроноцкий р-н, р. Тюшевка (тюшевская серия,
оленинская свита). Поздний миоцен—ранний плиоцен — Сев. Сахалин, п-ов Шмидта
(маямрафская свита), Южн. Сахалин (курасийская и маруямская свиты). Плиоцен
вост. Камчатка, Усть-Камчатский р-н. Ранний плиоцен — зап. побережье Камчатки. Сред-
ний плиоцен (?) — сев.-зап. побережье Камчатки, Рекинникская губа (этолонская
свита?). Голоцен (?) — сев.-зап. часть Тихого океана и залива Петра Великого Япон-
ского моря (возможно, переотложения из неогеновых отложений). Вне СССР.
Неоген — Япония (Сендай и бухта Иокогама). Миоцен — Япония. Ранний миоцен
(аквитан—бурдигал) — Испания (Севилья). Средний миоцен—плиоцен — Япония.
Ныне — тропические моря у берегов Африки, Азии и Южн. Америки, у атлантических
берегов США (Сев. Каролина).
П р и м е ч. Большая часть ископаемых створок с Дальнего Востока крупнее
современных из тропических морей и Атлантики.
Находка в плейстоцене Калининградской обл. (Черемисинова, 1970) требует
подтверждения.
Род PLANKTONIELLA Schutt (1893 : 20)
Клетки одиночные. Хлоропласты многочисленные, в виде мелких округлых
пластиночек, прилегающих к створкам. Панцирь дисковидный, окружен широкой
гиалиновой каймой („парашютом”), разделенной радиальными перегородками на
клиновидные участки. Створки плоские или слабо выпуклые. Ареолы полигональные,
в-тангенциально изогнутых рядах. По краю створки кольцо выростов с опорами. Загиб
низкий, с кольцом коротких ребрышек.
Виды морские, планктонные, современные.
Т и п: Р. sol (Wallich) Schutt.
1. Р. sol (Wallich) Schutt (1893 : '20, fig. 8). — Coscinodiscus sol Wall. (Wallich, 1860 :
38, tab. 2, 7, 2).
Панцирь дисковидный. Створки 19—98 мкм в диам., с „парашютом” 50—165 мкм.
Ареолы в тангенциальных, изогнутых к центру рядах, в центре 5—8, у края 8—12 ареол
в 10 мкм. В центре створки одна более крупная ареола, окруженная 7 ареолами,
и один очень маленький вырост с опорами. По краю створки кольцо маленьких
выростов с опорами, среди которых 2 двугубых выроста, расположенных по диаметру
друг против друга. (Табл. 34,6).
Стеногалинный тепловодный панталассный, широко распространенный в тропиче-
ских и субтропических водах вид.
Плейстоцен — вост. Камчатка (ольховская свита). Голоцен — Онежский п-ов.
Ныне — у южн. берегов Приморья и Сахалина. Вне СССР Средний-поздний плейстоцен —
85
тропическая зона Индийского и Тихого океанов. Ныне — Норвежское море, у берегов
Англии, Средиземное, Красное моря, в тропических и субтропических водах Атланти-
ческого, Индийского и Тихого океанов, у берегов Сев. и Южн. Америки, Австралии
и Африки.
Род POROSIRA J0rg. (Jorgensen, 1905, tab. 6,7)
Клетки соединены в рыхлые цепочки несколькими тяжами, выходящими из
выростов с опорами. Хлоропласты многочисленные, пластинчатые, прилегающие
к створке. Панцирь сферический или цилиндрический, со вставочными и соединитель-
ными ободками (до 4 ободков, прилегающих к створке), перфорированными мелкими
порами в искривленных рядах. Створки сильно или равномерно выпуклые. Централь-
ное поле окружено окремненным кольцом. Ареолы в радиальных рядах, образуют
сплошную сеть. Выросты с опорами разбросаны по всей поверхности створки, кроме
центральной части, более многочисленные у края створки, с 2—4 сопутствующими
порами, на наружной поверхности створки заканчиваются отверстием или короткой
трубкой. Двугубый вырост крупный на внутренней поверхности створки, на наружной
заканчивается короткой трубкой или широкой щелью.
Виды морские, распространенные в холодных водах, вымершие и современные.
Т и п: Р. glacialis (Grun.) J0rg.
I. Панцирь цилиндрический, створки равномерно выпуклые...............1. Р. glacialis.
II. Панцирь сферический, створки сильно выпуклые ................... 2. Р. punctata.
1. Р. glacialis (Grun.) J0rg. (J0rgensen, 1905, tab. 6, 7). — Podosira hormoides var.
glacialis Grun. (Grunow, 1884 : 108, tab. E, 32). — P. glacialis Cl. (Cleve, 1896 : 12, tab. 2,
17—20). — Lauderiaglacialis Gran (1900 : 11, tab. 9,10-14).
Панцирь цилиндрический. Створки с плоской серединой, слабо окремнены,
20—70 мкм в диам. Ареолы в извилистых рядах, 20—26 ареол в 10 мкм. Форамены
маленькие, округлые. Выросты с опорами на наружной и внутренней поверхностях
створки в виде коротких трубок, с 2—3 сопутствующими порами. Двугубый вырост
на наружной поверхности створки в виде простой или окаймленной щели, ориентиро-
ван по радиусу створки. Споры линзовидные. (Табл. 60,1—6).
Морской стеногалинный неритический биполярный вид.
Поздний миоцен — Сев. Сахалин (маямрафская свита). Плейстоцен — Сев.-Зап.
СССР, Коми АССР, север Зап. Сибири, Берингово, Чукотское, Охотское моря. Голо-
цен — Балтийское, Белое, Баренцево, Карское, Берингово, Охотское, Японское моря,
тихоокеанское побережье Камчатки. Ныне — дальневосточные моря СССР. Вне СССР.
Плиоцен—плейстоцен — Норвежское море, Япония, сев. часть Тихого и Атлантического
океанов. Ныне — европейские моря, Сев. Ледовитый океан, арктобореальная область
Тихого и Атлантического океанов, Индийский океан.
2. Р. punctata (Jouse) Makar. (Макарова, 1988а : табл. 1,1-16). — Thalassiosira punc-
tata Jouse (Жузе, 1961 : 64, табл. I, 78\ III, 3).
Споры. Панцирь сферический. Створки с выпуклой серединой, сильно окремнены,
26—47 мкм в диам. Ареолы в более или менее прямых радиальных рядах, 10—12 ареол
в 10 мкм. Форамены меньше наружных отверстий выростов с опорами, крибрум из
радиальных рядов крибральных пор, 6 в 1 мкм. Выросты с опорами образуют неотчет-
ливые концентрические ряды, на внутренней поверхности с 3—4 сопутствующими
порами. Двугубый вырост на наружной поверхности в виде отверстия, на внутрен-
ней крупный, с длинной щелью, ориентированной радиально, окруженной широким
валиковидным утолщением, от выроста к центру идет короткая гиалиновая полоса.
(Табл. 59).
Морской вымерший вид.
86
Миоцен — Камчатка (тюшевская серия). Поздний миоцен — Сев. Сахалин (маям-
рафская свита). Поздний миоцен—ранний плиоцен — Южн. Сахалин (маруямская
свита). Вне СССР. Поздний миоцен—плейстоцен — сев.-зап. часть Тихого океана,
Япония.
Род BACTEROSIRA Gran (1900 : 14)
Клетки образуют плотные цепочки, соединенные центральными выростами с опо-
рами. Хлоропласты мелкие, в виде зерен, прилегающие к поясковой стороне. Панцирь
цилиндрический, с 2 воротничковидными ободками. Створки слегка вогнутые, в центре
со структурой из радиальных, дихотомически ветвящихся гиалиновых линий и ареол
между ними в радиальных рядах. На границе с загибом створки кольцо выростов
с опорами и пучок выростов с опорами в центральной, вогнутой части створки, окру-
женной кремневой зоной. Центральные выросты с опорами с 3—4 сопутствую-
щими порами, краевые — с 4 сопутствующими порами. Двугубый вырост в одном
кольце с выростами с опорами, на наружной поверхности створки заканчивается
щелью с утолщенными краями, ориентированной радиально. Известны покоящиеся
споры.
Известен 1 вид, распространенный преимущественно в арктических морях.
Т и п: В. fragilis Gran.
1. В. fragilis Gran (1900 : 14). — Coscinodiscus bathyomphalus Cl. (Cleve, 1883 : 489,
tab. 38,81).
Створки 16—25 мкм в диам. Ареолы постепенно уменьшаются от центра к краю
створки, в центре около 20 ареол в 10 мкм, у края около 30. Краевых выростов с опо-
рами 5-6 в 10 мкм. Споры с сильно выпуклой центральной частью створки и широкими
пологими краями. Ареолы в радиальных рядах, 10—14 в 10 мкм, с очень маленькими
фораменами. По краю лицевой части створки кольцо редких выростов с опорами,
3 в 10 мкм. Загиб створки с мелкими ареолами, 30—32 в 10 мкм, в искривленных
рядах. (Табл. 60, 7-12).
Морской стеногалинный неритический арктобореальный вид.
Миоцен—плиоцен — сев.-зап. часть Тихого океана. Плиоцен — Камчатка, о-ва Иту-
руп, Сахалин. Плейстоцен — Сев.-Зап. СССР, север Зап. Сибири, побережье Карского
и Чукотского морей. Голоцен — Балтийское, Белое, Баренцево, Карское, Берингово,
Охотское, Японское моря. Ныне — Белое, Баренцево, Карское, Чукотское, Берингово,
Охотское, Японское моря, район Курильских о-вов. Вне СССР. Миоцен — Япония,
север Тихого океана. Плиоцен—плейстоцен — Норвежское море. Ныне — север Тихого
и Атлантического океанов, Ледовитый океан.
П р и м е ч. В ископаемом состоянии встречаются в основном споры.
Род DETONULA Sch'utt (1896 : 83)
Клетки соединены в плотные прямые короткие нитевидные цепочки краевыми
выростами с опорами. Хлоропласты — округлые или лопастные пластинки или одна
крупная пластина. Панцирь цилиндрический, с многочисленными воротничковидными
ареолированными вставочными и соединительными ободками. Створки плоские или
вогнутые. Ареолы в радиальных рядах. На границе с загибом створки кольцо выростов
с опорами, имеющих длинную трубку на наружной поверхности створки и корот-
кую — на внутренней, с 4 сопутствующими порами. В центре створки 1—2 вырос-
та с 3—4 опорами. Двугубый вырост в одном кольце с краевыми выростами
с опорами. Загиб створки с ребровидными утолщениями. Известны покоящиеся
споры.
Виды морские, неритические.
Т и п: D. confervacea (Cl.) Gran.
. 87
I. Створки плоские, 3—7.5 мкм в диам., по всему краю не более 15 выростов с опорами
...............................................................2. D. subtilissima.
II. Створки слегка вогнутые, 6—15 мкм в диам., по краю 10—12 выростов с опорами
в 10 мкм . .,.................................................. 1. D. confervacea.
1. D. confervacea (CL) Gran (1900 : 113). — Lauderia confervacea Cl. (Cleve, 1896,
tab. 2, 21). - Detonula cystifera Gran (1900 : 113, tab. 9, 15-20). - Melosira albicans
Sheshuk. (Шешукова-Поредкая, 1967 : 124, табл. 10,2a, б\ 11, la, б).
Панцирь 15—25 мкм выс. Хлоропласты многочисленные. Створки слегка вогну-
тые, 6—15 мкм в диам. Ареолы в радиальных и вторично концентрических рядах,
30—40 ареол в 10 мкм. Форамены неправильной формы, близ загиба и на загибе створки
вытянутые. Наружные трубки краевых выростов с опорами со скошенными или загну-
тыми концами, 10—12 выростов в 10 мкм. Двугубый вырост снаружи имеет вид прямой
трубки, на внутренней поверхности в виде узкой щели, окруженной 2 утолщенными
губами. Загиб створки довольно высокий. Покоящиеся споры отличаются от вегетатив-
ных клеток более грубым панцирем. Загиб створки у спор более пологий, чем у вегета-
тивной клетки, с бесструктурным краем. Выросты с опорами и двугубый вырост распо-
ложены так же, как на вегетативной клетке. (Табл. 60,14, 15\ рис. 6,1—3).
Морской стеногалинный неритический арктобореальный вид.
Миоцен—плиоцен — Дальний Восток (покоящиеся споры). Плейстоцен — север
европ. части СССР (покоящиеся споры). Ныне — сев. и дальневосточные моря СССР,
тихоокеанское побережье Камчатки. Вне СССР. Ныне — Северное, Норвежское моря,
сев. Атлантика.
2. D. subtilissima Pr.-Lavr. (Прошкина-Лавренко, 1959 : 78—79, рис. 1—5; 1963 :
98-99, рис. 1,2).
Панцирь 4—14 мкм выс. Хлоропласт один. Створки плоские, с невидимой в СМ
структурой, 3—7.5 мкм в диам. По краю створки кольцо из 6—15 выростов с опорами,
число которых зависит от диаметра створки. Загиб створки низкий. Споры не наблюда-
лись. (Рис. 6,4-6).
Морской неритический вид.
Ныне — Азовское море.
П р и м е ч. Вид изучен недостаточно.
Род LAUDERIA Cl. (Cleve, 1873 : 8, tab. I, 7)
Клетки в длинных, довольно прочных цепочках, в которых они соединены крае-
выми замкнутыми выростами и центральными выростами с опорами (?). Клетки
расположены на расстоянии друг от друга или плотно примыкают друг к другу. Хлоро-
пласты — мелкие многочисленные пластинки. Панцирь высокоцилиндрический, тонкий,
с многочисленными соединительными ободками. Створки круглые, слегка выпуклые,
центральное поле окружено кремневым кольцом. Ареолы в радиальных рядах, непра-
вильных очертаний у края створки. На границе лицевой части створки и загиба кольцо
замкнутых выростов с очень длинной трубкой, на загибе створки выросты с опорами,
расположенные в шахматном порядке, с длинной трубкой на наружной поверхности
створки и короткой — на внутренней. Выросты с опорами иногда имеются близ
центра створки или редкие по всей створке. Двугубый вырост один, на границе
с загибом, с длинной трубкой на наружной поверхности, короткий, широкий — на
внутренней.
Виды морские, неритические.
Т и п: L. annulata Cl.
1. L. annulata Cl. (Cleve, 1873 : 8, tab. I, 7). — L. borealis Gran (1900:110, tab. 9,5-9).
88
Рис. 6.
1-3 - Detonula confervacea (Cl.) Gran: 1 - часть колонии, 2 - клетки co спорами, 3 - створка
(Hustedt, 1929); 4-6 - D. subtilissima Pr.-Lavr.: 4 - створка, 5 - панцирь, 6 - цепочка. Азовское море
(Прошкина-Лавренко, 1963). (Шкала: 10мкм).
Рис. 7.
1, 2 - Lauderia annulata Cl.: 1 — часть колонии, 2 - створка (Hustedt, 1929). (Шкала: 10 мкм).
Створки 20—70 мкм в диам. Ареолы в радиальных рядах, 30 ареол в 10 мкм,
разделенных дихотомически ветвящимися гиалиновыми линиями, более 30 в 10 мкм.
Выросты с опорами с длинной трубкой на наружной поверхности и короткой — на внут-
ренней. Замкнутые выросты по краю лицевой части створки (Syvertsen, Hasle, 1982).
(Табл. 60,75; рис. 7,7, 2).
Морской стеногалинный неритический, бореально-тропический вид.
Ныне — сев. моря СССР. Вне СССР. Ныне — атлантическое побережье Зап. Европы
и Африки, сев. Атлантика у берегов Америки, Тихий океан у берегов Канады, Калифор-
нии, Чили, Индийский океан.
7 Заказ 518
89
Род MINIDISCUS Hasle (1973 : 29)
Клетки очень малых размеров, распознаваемы обычно в ТЭМ, одиночные, иногда
парные или собраны группами. Хлоропластов 1—2, пристенных. Панцирь высоко-
цилиндрический, со скошенными углами. Створки круглые, плоские или выпуклые.
Ареолы локулярные (?), с фораменом многоугольной формы, образуют сомкнутую
сеть, в радиальных или тангенциальных рядах. Выросты с опорами вне краевого кольца,
2—7 на створке, с более длинной трубкой на наружной поверхности створки, с 2—3
сопутствующими порами на внутренней. Двугубый вырост один, близ центра створки,
едва заметный на наружной поверхности створки или с большой трубкой. Край створки
гиалиновый или с короткими ребрами.
Виды планктонные, вероятно морские и солоноватоводные.
Т и п: М. trioculatus (Taylor) Hasle.
I. Створки плоские, край гиалиновый, выросты с опорами с 2 сопутствующими порами
................................................................2. М. trioculatus.
IL Створки плоские или выпуклые, край с короткими ребрами, выросты с опорами
с 3 сопутствующими порами....................................... 1. М. comicus.
1. М. comicus Takano (1981 : 32, fig. 1, А, 2-13).
Створки плоские или слегка выпуклые, 2—3.4 мкм в диам. Ареолы в радиальных
рядах, 5—6 в 1 мкм. На створке 3 и более выростов с опорами, расположенных ближе
к краю створки, с 3 сопутствующими порами. Двугубый вырост с заметной высту-
пающей трубкой на наружной поверхности створки. Край створки с радиальными ребра-
ми, 6 в 1 мкм. (Табл. 60,16—18).
Солоноватоводный планктонный вид.
Ныне — Азовское море. Вне СССР. Ныне — у берегов Японии и в р. Сумида.
2. М. trioculatus (Taylor) Hasle (1973 : 29, fig. 101408). — Coscinodiscus trioculatus
Taylor (1967 : 437, tab. 5,43).
Створки плоские, 5.7 мкм в диам. Ареолы в тангенциальных рядах, в центре
створки 3 ареолы, у края 4 в 1 мкм. На створке 3 выроста с опорами, один близ центра
створки, два других ближе к краю, с 2 сопутствующими порами. Двугубый вырост
очень маленький, не выступающий над наружной поверхностью створки, расположен
через одну ареолу от центрального выроста с опорами. Край створки гиалиновый.
(Табл. 60,19).
Морской планктонный, широко распространенный вид.
Ныне — Охотское море. Вне СССР. Ныне — Норвежское море, вдоль атлантиче-
ского прибрежья Европы и Африки, у Кейптауна, Мексиканский залив, у о-ва Кейп-
Бретон, Тихий океан у Ванкувера, берегов Австралии, Индийский океан.
90
ЛИТЕРАТУРА
Беклемишев К. В., СеминаГ. И. Органоиды створки и некоторые черты эволюции диатомей
{Bacillariophyta) Ц Журн. общ. биол. 1982. Т. 43, № 4. С. 497-507.
Векшина В. Н. Диатомовые водоросли верхнеюрских отложений Западно-Сибирской низменно-
сти// Тр. СНИИГГИМС. 1960. Вып. 8. С. 160-162.
Векшина В. Н. Новый род и новые виды диатомовых из меловых и палеогеновых отложений
Западно-Сибирской низменности // Тр. СНИИГГИМС. 1961. Вып. 15. С. 89-97.
Г а й л Г. И. Определитель фитопланктона Японского моря // Изв. Тихоокеан. н.-и. ин-та рыб. хоз-ва
и океаногр. 1950. Т. 33. С. 5-177.
Генкал С. И., Макарова И. В. О роли подпертых выростов в жизнедеятельности представите-
лей рода Thalassiosira Cl. {Bacillariophyta) // Новости систематики низших растений. Л., 1984.
Т. 20. С. 9-12.
Генкал С. И., Макарова И. В. Новые для планктона Каспийского и Азовского морей виды
диатомовых водорослей // Новости систематики низших растений. Л., 1985. Т. 22. С. 35-37.
Генкал С. И., Макарова И. В. Новые данные о морфологии панциря Cyclotella caspia Grun.
{Bacillariophyta) 11 Ботан журн. 1986. T. 71, № 3. С. 371-374.
Глезер 3. И. О находке диатомовых, силикофлагеллят и эбриидей на западном склоне Муго-
джар // Новости систематики низших растений. Л., 1967. Т. 4. С. 21-25.
Глезер 3. И. Диатомовые водоросли палеогена // Диатомовые водоросли СССР. Ископаемые
и современные. Л.: Наука, 1974. Т. 1. С. 109-146.
Глезер 3. И. К ревизии рода Triceratium Ehr. sensu Hustedt, 1930 // Ботан. журн. 1975а. Т. 60, № 9.
С. 1304-1310.
(Глезер 3. И.) G1 е s е г S. I. Principles of systematics and some features of Cretaceous and Paleogene
Bacillariophyta. Evolution: Тез. докл. XII Междунар. ботан. конгр. Л., 19756. С. 38.
Глезер 3. И. Некоторые принципы систематики диатомовых водорослей {Bacillariophyta) на при-
мере изучения рода Biddulphia Gray sensu Hustedt, 1930 // Морская микропалеонтология. М.,
1978. С. 67-72.
Глезер 3. И. Эволюция и систематика порядка Biddulphiales {Bacillariophyta) 11 Палеонтол. журн.
1979. № 1. С. 110-120.
Глезер 3. И. К разработке новой классификации диатомовых водорослей // Систематика, эволю-
ция, экология водорослей и их значение в практике геологических исследований: Тез. докл.
II Всесоюз. палеоальгол. совещ. Киев, 1981. С. 108-110.
Глезер 3. И. Таксономическая значимость признаков у диатомовых водорослей в свете разра-
ботки новой классификации Bacillariophyta // Ботан. журн. 1983. Т. 68, № 8. С. 993-1001.
Глезер 3. И. К разработке новой классификации диатомовых водорослей // Актуальные вопросы
современной палеоальгологии. Киев, 1986а. С. 65-69.
Глезер 3. И. Филогения диатомовых водорослей порядка Coscinodiscales в связи с разработкой
новой классификации // Ежегодн. Всесоюз. палеонтол. о-ва. Л., 19866. Т. 29. С. 180-187.
Глезер 3. И. О положении рода Triceratium s. str. {Bacillariophyta) в системе диатомовых водорос-
лей // Ботан. журн. 1986в. Т. 71, № 11. С. 1543-1545.
Глезер 3. И., Макарова И. В. Новый порядок и семейство диатомовых водорослей {Bacillario-
phyta) Ц Ботан. журн. 1986. Т. 71, № 5. С. 673-676.
Джафарова С.К. К морфологии Ditylum brightwellii {Bacillariophyta) 11 Ботан. журн. 1984. T. 69,
№ 5. С. 671-673.
ДжиноридзеР. Н., Жузе А. П., Стрельникова Н. И. Описание диатомовых водорослей //
История микропланктона Норвежского моря. Л., 1979. С. 32-71.
Диатомовый анализ. Л., 1949-1950. Кн. 1-3 (1949. Кн. 1. 273 с.; 1949. Кн. 2. 283 с. 1950. Кн. 3.
398 с.).
Диатомовые водоросли СССР. Ископаемые и современные. Л.: Наука, 1974. Т. 1. 400 с.
Жузе А.П. Дотретичные диатомовые водоросли // Ботан. журн. 1948. Т. 33, № 3. С. 345-356.
Жузе А. П. Диатомовые третичных отложений // Диатомовый анализ. Л., 1949а. Кн. I С. 114-153.
Ж у з е А. П. Диатомовые мезозойских отложений//Диатомовый анализ. Л., 19496. Кн. 1. С. 109-114.
91
Жузе А. П. Диатомовые и кремневые жгутиковые водоросли верхнемелового возраста из Север-
ного Урала // Ботан. материалы Отд. спор. раст. БИН АН СССР. М.; Л., 1951а. Т. 7. С. 42-65.
Жузе А. П. Диатомовые палеоценового возраста Северного Урала // Ботан. материалы Отд. спор,
раст. БИН АН СССР. М.; 19516. Т. 7. С. 24-42.
Жузе А. П. Новые виды диатомовых палеогенового возраста // Ботан. материалы Отд. спор. раст.
БИН АН СССР. М.; Л., 1955. Т. 10. С. 81-103.
Жузе А. П. Морские диатомовые миоценового и плиоценового возраста Дальнего Востока //
Ботан. материалы Отд. спор. раст. БИН АН СССР. М.; Л., 1961. Т. 14. С. 59-70.
Жузе А. П. Стратиграфические и палеогеографические исследования в северо-западной части Ти-
хого океана. М.: Изд-во АН СССР, 1962. 260 с.
Жузе А. П. Новые виды диатомовых водорослей в донных осадках Тихого океана и Охотского
моря // Новости систематики низших растений. Л., 1968. Т. 5. С. 12-21.
(Жузе А. П., Мухина В. В.) Jouse А. Р., М u k h i n а V. V. Diatom units and the paleogeography
of the Black Sea in the Late Cenozoik (DSDP, Leg 42 B) // Init. Repts. DSDP. Washington, 1978.
Vol. 42, pt 2. P. 903-950.
Исправленная терминология кремнеземных компонентов клетки диатомовых водорослей //
Науч. докл. высш, школы. Биол. науки. 1983. № 5. С. 65-76.
Караева Н. И. Диатомовые водоросли бентоса Каспийского моря: Автореф. дис. ... докт. биол.
наук. Л., 1975. 43 с.
Караева Н. И. Новый подпорядок дйатомовых водорослей // Ботан. журн. 1978. Т. 63, № 12.
• С. 1747.
Караева Н. И. Исследование диатомей Каспийского моря в сканирующем электронном микро-
скопе. II. Роды Epithemia, Rhopalodia // Ботан. журн. 1981. Т. 66, № 4. С. 515-525.
(Караева Н. И.) Karajeva N.I. Some species of the genus Amphora Ehr. in the Caspian Sea //
Arch. Protistenk. 1987. Vol. 133. P. 93-109.
Караева H. И., Джафарова С. К. Двугубые выросты диатомей, их строение и локализация //
Баку, 1982. 23 с. Деп. в ВИНИТИ 1982 г., № 2100-82 деп.
Караева Н. И., Джафарова С. К. К морфологии Coscinodiscus granii (Bacillario phyto) 11 Ботан.
журн. 1985. T. 70, № 8. С. 1078-1081.
Караева Н. И., Магеррамова Н. Р., Рзаева С. Г. Морфология панциря диатомей рода
Amphora (Bacillariophyta) по данным электронной микроскопии // Ботан. журн. 1984. Т 69,
№4. С. 492-498.
Козлова О. Г.,Стрельникова Н. И. Диатомеи в планктоне, толше воды и в донных осадках
северо-восточной части Тихого океана // Микропалеонтология океанов и морей. М.: Наука,
1974. С. 63-75.
Лосева Э. И., Шешу кова- ПорецкаяВ. С. Об идентичности Melosira albicans и спор Detonu-
la confervacea (Bacillariophyta) 11 Ботан. журн. 1985. Т. 70, № 7. С. 945-947.
Майр Э. Принципы зоологической систематики. М., 1971. 454 с.
Майр Э. Популяции, виды и эволюция. М., 1974. 460 с.
Макарова И. В. Новые виды диатомовых водорослей сем. Coscinodiscaceae из Северного Кас-
пия // Ботан. материалы Отд. спор. раст. БИН АН СССР. М.; Л.. 1961. Т. 14. С. 49-52.
Макарова И. В. О возрасте и морфологии некоторых каспийско-черноморских видов рода
Thalassiosira CL // Ботан. журн. 1962. Т. 47, № 7. С. 1015-1017.
Макарова И. В. О морфологии и таксономии рода Thalassiosira Cl. // Новости систематики низ-
ших растений. Л., 1971. Т. 9. С. 67-73.
Макарова И. В. О морфологии Thalassiosira rotula Meunier // Новости систематики низших расте-
ний. Л., 1974. Т. 11. С. 96-99.
Макарова И. В. Новые виды рода Thalassiosira CL из Баренцева моря // Новости систематики
низших растений. Л., 1975. Т. 12. С. 149-152.
Макарова И. В. Новые данные о морфологии нескольких видов рода Thalassiosira Cl. из Тихого
океана, изученных с помощью трансмиссионного электронного микроскопа // Ботан. журн.
1976а. Т. 61, № 5. С. 605-610.
Макарова И. В. Морфологическая изменчивость панциря некоторых видов рода Thalassiosira Cl.
и ее таксономическое значение // Ботан. журн. 19766. Т. 61, № 11. С. 1589-1594.
Макарова И. В. Новые материалы к познанию рода Thalassiosira Cl. (Thalassiosiraceae) из Черного
моря // Ботан. журн. 1977а. Т. 62, № 4. С. 537-538.
Макарова И. В. Новые данные о роде Thalassiosira Cl. (Bacillariophyta) из Азовского моря //
Ботан. журн. 19776. Т. 62, № 6. С. 907-909.
Макарова И. В. Новые данные к изучению видов рода Thalassiosira Cl. (Bacillariophyta} из Атлан-
тического океана // Ботан. журн. 1978. Т. 63, № 2. С. 222-225.
Макарова И. В. О таксономии и морфологии Thalassiosira angustedineata (Bacillariophyta) // Ботан.
журн. 1981. Т. 66, №10. С. 1473-1475.
Макарова И. В. О номенклатуре некоторых видов рода Thalassiosira Cl. (Bacillariophyta) // Ново-
сти систематики низших растений. Л., 1983. Т. 20. С. 23-26.
Макарова И. В. Классификация диатомовых водорослей на современном этапе и проблема
построения их филогенетической системы // Ботан. журн. 1986. Т. 71, № 6. С. 713-722.
92
Макарова И. В. Новые комбинации таксонов из позднемиоценовых-раннеплиоценовых диато-
мовых флор Сахалина // Ботан. журн. 1988а. Т. 73, № 0. С. 1183-1186.
Макарова И. В. Диатомовые водоросли морей СССР. Род Thalassiosira Cl. // Л.: Наука, 19886.
117 с., 52 табл.
Макарова И. В., ГенкалС. И., Кузьмин Г. В. Виды рода Thalassiosira Cl., (Bacillariophyta),
найденные в континентальных водоемах СССР // Ботан. журн. 1979. Т. 64, № 7. С. 921-927.
Макарова И. В., Гусляков Н. Е. О морфологии створок некоторых видов рода Licmophora
Ag. (Bacillariophyta) 11 Ботан. журн. 1984. T. 69, № 6. С. 870-872.
Макарова И. В., Моисеева А. И. Новый вид рода Pyxidicula Ehi. (Bacillariophyta) 11 Ботан.
журн. 1986. T. 71, № 2. С. 244-245.
Макарова И. В., Николаев В. А. Заметки о роде Schimperiella Karsten (Bacillariophyta) 11
Ботан. журн. 1984. T. 69, № 1. С. 81-90.
Макарова И. В., Прошкина-Лавренко А. И. Новые диатомовые водоросли в планктоне
Каспийского моря // Новости систематики низших растений. Л., 1964. Т. 1. С. 34-43.
Макарова И. В., Семина Г. И. Новые данные о морфологии и распространении Thalassiosira
hyalina (Bacillariophyta) // Ботан. журн. 1982. Т. 67, № 1. С. 109-111.
Макарова И. В., Семина Г. И., Зернова В. В. Новые данные о морфологии и распростра-
нении двух видов Thalassiosira Cl. (Bacillariophyta) // Ботан. журн. 1981. Т. 66, № 7. С. 1032—
1035.
Николаев В. А. О роде Symbolophora Ehr. {Bacillariophyta) // Ботан. журн. 1983. Т. 68, № 8.
С. 1123-1128.
Николаев В. А. К построению системы центрических диатомовых водорослей (Bacillariophyta) //
Ботан. журн. 1984а. Т. 69, № 11. С. 1468-1474.
Н и к о л а е в В. А. О значении строения ареол для таксономии диатомовых водорослей (Bacillario-
phyta) и Ботан. журн. 19846. Т. 69, № 8. С. 1040-1046.
Основы палеонтологии. Водоросли. М., 1963. С. 55-151.
Порецкий В. С., Анисимова Н. В. Материалы к экологии диатомовых старорусских соле-
ных водоемов // Исследов. озер СССР. Л., 1933. Вып. 2. С. 31-60.
Предложения для стандартизации терминологии и диагнозов по диатомовым водорослям //
Ботан. журн. 1977. Т. 62, № 2. С. 192-213.
Прошкина-Лавренко А. И. Реликтовые диатомовые в планктоне Черного моря // Ботан.
материалы Отд. спор. раст. БИН АН СССР. М.; Л., 1955а. Т. 10. С. 45-54.
Прошкина-Лавренко А. И. Диатомовые водоросли планктона Черного моря. М.; Л.: Изд-во
АН СССР, 19556. 222 с.
Прошкина-Лавренко А. И. Новый вид талассиозиры в планктоне Азовского моря // Ботан.
материалы Отд. спор. раст. БИН АН СССР. М.; Л., 1956. Т. 11. С. 57-59.
Прошкина-Лавренко А. И. Новый вид рода Thalassiosira Cleve из Черного моря // Ботан.
материалы Отд. спор. раст. БИН АН СССР. М.; Л., 1959. Т. 12. С. 76-78.
Прошкина-Лавренко А. И. Новые и интересные диатомовые водоросли из мэотических
отложений Причерноморья // Ботан. материалы Отд. спор. раст. БИН АН СССР. М.; Л., 1960.
Т. 13. С. 50-66.
Прошкина-Лавренко А. И. Новые диатомовые водоросли из Черного и Азовского морей //
Ботан. материалы Отд. спор. раст. БИН АН СССР. М.; Л., 1961. Т. 14. С. 33-39.
Прошкина-Лавренко А. И. Диатомовые водоросли планктона Азовского моря. М.; Л.:
Изд-во АН СССР, 1963. 184 с.
Прошкина-Лавренко А. И., Макарова И. В. Водоросли планктона Каспийского моря.
Л., 1968. 293 с.
Рауп Д., Стенли С. Основы палеонтологии. М., 1974. 390 с.
Росс Р., Кокс А., Караева Н. И., М а н н Д. Г., П а д д о к Т. Б. Б., Симонсен Р.,
Симс П. А. Исправленная терминология кремнеземных компонентов клетки диатомовых
водорослей // Науч. докл. высш, школы. Биол. науки. 1983. № 5. С. 66-76.
Семина Г. И., Макарова И. В. О морфологии и распространении двух биполярных видов рода
Thalassiosira Cl. (Bacillariophyta) 11 Ботан. журн. 1983. T. 68, № 5. С. 611-618.
Стрельникова Н. И. Новые и редкие виды позднемеловых диатомовых водорослей восточного
склона Полярного Урала // Новости систематики низших растений. Л., 1965. Т. 2. С. 29-37.
Стрельникова Н. И. Ревизия позднемеловых представителей родов Gladius Schulz и Pyxilla
Grev. (Bacillariophyta} // Новости систематики низших растений. Л., 1966. Т. 3. С. 23-36.
Стрельникова Н. И. Новые виды диатомовых водорослей из позднемеловых отложений
восточного склона Полярного и Приполярного Урала // Новости систематики низших расте-
ний. Л., 1971. Т. 8. С. 41-51.
Стрельникова Н. И. Диатомеи позднего мела. М., 1974. 202 с.
Стрельникова Н. И., Николаев В. А. К ревизии родов Stephanopyxis и Pyxidicula (Bacilla-
riophyta) Ц Ботан. журн. 1986. Т. 71, № 7. С. 950-953.
Т а х т а д ж я н А. Л. Вопросы эволюционной морфологии растений. Л., 1954. 214 с.
Топачевский А. В. Вопросы цитологии, морфологии, биологии и филогении водорослей //
Киев, 1962. 235 с.
93
Черемисинова Е. А. О диатомовой флоре миндельрисских межледниковых отложений юго-
восточной части Ленинградской области // Вопросы четвертичной геологии. Рига, 1970. Т. 5.
С. 49-63.
Шешукова-Порецкая В. С. К ископаемой диатомовой флоре Южного Сахалина (морской
неоген) // Вести. ЛГУ. 1959. № 15. Сер. биол. Вып. 3. С. 36-55.
Шешукова-Порецкая В. С. Диатомовые водоросли некоторых свит морского неогена
Камчатки // Вести. ЛГУ. 1961. № 15. Сер. биол. Вып. 3. С. 46-59.
Шешукова-Порецкая В. С. Неогеновые морские диатомовые водоросли Сахалина и Кам-
чатки. Л., 1967. 432 с.
Шешукова-Порецкая В. С., Глезер 3. И. Новые виды морских палеогеновых диатомовых
водорослей УССР // Новости систематики низших растений. М.; Л., 1964. Т. 1. С. 78-92.
Anonymous. Proposals for a standartization of diatom terminology and diagnoses // Beih. Nova Hedwi-
gia. 1975. H. 53. S. 323-354.
В e t hg e H. Uber die Kieselalge Sceletonema subsalsum 11 Ber. Dtsch. bot. Ges. 1928. Bd 40. S. 340-347.
Bourrelly P., M a ng u in E. Algues d’eau douce la Guadeloupe et dependances. Bacillariophyceae //
Cent. Nat. Rec. Sci. Soc. Paris. 1952. Vol. 98. 281 p.
Boyle J. A., Pickett-Heaps J. D., Czarnecki D. B. Valve morphogenesis in the pennate diatom
Achnanthes coarciata 11 J. Phycol. 1984. Vol. 20, N 4. P. 563-573.
Brun J., TempereJ. Diatomes fossilies du Japon // Mem. Soc. phys. hist, natur. Genbve. 1889. Vol. 30,
N9. P. 1-75.
Carter J. R. On the identity of Navicula cincta Ehrenberg // Bacillaria. 1979. Vol. 2. P. 73-83.
Castracane F. Report on the scientific results of the vojage of H. M. S. Challenger during the years
1873-1876 // Botany. 1886. Vol. 2.178 p.
Chiappino M. L., VolcaniB. E. Studies on the biochemistry and fine structure of silicia shell forma-
tion in diatoms. VII. Sequential cell wall development in the pennate Navicula pelliculosa // Proto-
plasma. 1977. Vol. 93, N 2-3. P. 205-221.
С 1 e ve P. T. On diatoms from the Arctic Sea // Bih. Kgl. Sven, vetenskapsakad. handl. 1873. Bd 1. S. 1-28.
Cleve P. T. Diatoms from the West Indian Archipelago // Bih. Kgl. sven. vetenskapsakad. handl. 1878.
Bd58.S. 1-22.
Cleve P. T. Diatoms collected during the expedition of the Vega // Nordenskiold’s Vega-Exp. Vetiskrb.,
Jactt. Stockholm. 1883. Bd 3. S. 455-517.
Cleve P. T. Diatoms from Baffin Bay and Davis Strait, collected by M. E. Nilsson // Bih. Kgl. sven. veten-
skapsakad. handl. 1896. Bd 22, Afd. 3. S. 1-22.
Cl e v e P. T. Plancton table for the North Sea // Bull. Cons. Explor. Mer. 1903-1904. 216 p.
Cleve P. T., G r u n о w A. Beitrage zur Kenntniss der arctischen Diatomeen. // Bih. Kgl. Sven, vetenskaps-
akad. handl. 1880. Bd 17.121 S.
Cleve-Euler A. Das Bacillariaceenplankton in Gewassern bei Stockholm. III. Uber Gemeinden des
schwach salzigen Wassers und eine neue Charakterart derselben // Arch. Hydrobiol. 1912. Bd 7.
S. 500-514.
Cleve-Euler A. Alttertiare Diatomeen und Silicoflagellaten im Inneren Schwedens, Gefunden und
Gezeichnet Von Ake Berg, begehrieben von Astrid Cleve-Euler // Palaeontographica A. 1941. Bd 92.
S. 165-212.
Cleve-Euler A. Die Diatomeen von Schweden und Finnland. Teil 1 -5 // Bih. Kgl. Sven, vetenskaps-
akad. handl. 1951. Teil 1, Bd 2, N 1. S. 1-162; 1953. Teil 2, Bd 4, N 1. S. 1-158; 1953. Teil 3,
Bd 4, N 5. S. 1-255; 1955. Teil 4, Bd 5, N 4. S. 1 -232; 1952. Teil 5, Bd 3, N 3. S. 1 -153.
Cox E.J. Symmetry and valve structure in naviculoid diatoms // Beih. Nova Hedvigia. 1979. H. 64.
S. 193-206.
Crawford R. Taxonomy and frustular structure of the marine centric diatom Paralia sulcata // J. Phycol.
1979. Vol. 15, N 2. P. 200-210.
Crawford R. M. The diatom genus Aulacosira Thwaites: its structure and taxonomy // Phycologia.
1981. Vol. 20, N 2. P. 174-192.
Cupp E. Marine plancton diatoms of the west coast diatoms of North America // Bull. Scripps Inst.
Oceanogr. Univ. California. 1943. Vol. 5, N l.P. 1-238
Dawson P. A. Observations on diatoms species transferred from Gomphonema C. A. Agardh to Gompho-
neis Cleve // Brit, phycol. J. 1974. Vol. 9. P. 75-82.
Ehrenberg C. G. Die Infusionsihierchen als vollkommende Organismen // Ein Blick in das tiefere
organische Leben der Natur. Leipzig, 1838. 548 S.
Ehrenberg C. G. Uber nach jetzt zahlreich lebende Tierarten der Kreidebildung und den Organismus
der Polythalamien // Abh. Kgl. Akad. Wiss. Berlin. 1839 (1841). S. 81 -174.
Ehrenberg C. G. Verbereitung und Einflus des microskopischen Lebens in Siid- und Nord-America //
Abh Kgl. Akad. Wiss. Berlin. 1843. S. 291-445.
Ehrenberg C. G. Untersuchungen uber die kleinsten Lebensformen in Quellenlande des Euphraats
und Araxes, so wieiiber eine an neuen Formen sehrreiche marine Tripelbildung von den Bermuda //
Inseln. Verhandl. Kgl. Akad. Berlin. 1844 (1845). S. 253-275.
Ehrenberg C. G. Microgeologie. Das Erden und Felsen schaffende Wirken des unsichtbar kleinen selb-
standigen Lebens auf der Erde. Leipzig, 1854. 374 S.
94
Forti A., Schulz P. Erste Mitteilung iiber Diatomeen aus dem hannoverschen Gault. // Beih. Bot.
Centralbl. 1932. Bd 50, Abt. 2. S. 241-246.
Fryxell G. A. Three new species of Thalassiosira with observations on the occluded process, a newly
observed structure of diatom valves // Beih. Nova Hedvigia. 1975. H. 53. S. 57-75.
Fryxell G. A. Chain-forming diatoms: three species of Chactoceraceae // J. Phycol. 1978. Vol. 14.
P. 62-71.
Fryxell G. A-, Doucette G.J., H u b b a r d G. F. The genus Thalassiosira: the bipolar T. antarctica
Comber // Bot. mar. 1981. Vol. 24, N 6. P. 321-335.
Fry xe 11 G., Ha sle G. Coscinodiscineae: some consistent patterns in diatom morphology // Beih. Nova
Hedwigia. 1974. H. 45. S. 69-84.
Fryxell G. A., Hasle G. R. The genus Thalassiosira: T trifulta sp. nova and other species with trico-
lumnar supports on strutted processes // Beih. Nova Hedwigia. 1979. H. 64. S. 13-32.
G о m b о s M. Paleogene and neogene diatoms from the Falklend plateau and Malvinas outer Basin: Leg 36.
Deep sea drilling project // Init. Repts DSDP. Washington, 1977. Vol. 36. P. 575-685.
G о m Ъ о s A. M. The early history of the Diatom familyAsterolampraceae // Bacillaria. 1980. Vol. 3. P. 227-272.
Gran H. Bemerkungen iiber einige Planktondiatomeen // Nytl mag. naturviden. 1900. Bd 38, N 2.
S. 103-128.
Gregory W. On new forms of marine diatomaceae found in the firth of Clyde and in Loch Fyne // Trans.
Roy. Soc. Edinburg. 1857. Vol. 21. P. 473-542.
Greville R. Descriprion of Diatomaceae observed in Californian Guono // Quart. Fourn. Microscop.
Sci. London. 1859. Vol. 7. P. 155-167.
Greville R. K. Descriprion of new and rare diatoms // Trans. Mier. Soc. London. N. S. 1864. Vol. 12.
P. 81-86; 1865. Vol. 13. P. 43-75. 1 866. Vol. 14. P. 77-86. .
G r u n о w A. Die Diatomeen von Franz Jozefs-Land // Denkschr. Osterr. Akad. Math.-naturwiss. KI. 1884.
Bd 48. S. 53-112.
H a j 6 s M. Late Cretaceous Archaeomonadaceae, Diatomaceae and Silicoflagellatae from the south Pacific
ocean. Deep sea drilling project, Leg 29. Site 275 // Init. Repts DSDP. Washington, 1975. Vol. 29.
P. 913-1009.
Hajos M. Upper Eocene and Lower Oligocene Diatomaceae, Archaeomonadaceae and Silicoflagellate
in Southestern Pacific sediments, DSDP Leg 29 // Init. Repts DSDP. Washington, 1976. Vol. 35.
P. 817-883.
Hanna G.D. The lowest known tertiary Diatoms in California // J. Paleontol. 1927. Vol. 1, N 2.
P. 103-127.
Hasle G. R. The inclusion of Coscinosira Gran (JBacillariophvceae) in Thalassiosira Cl. // Taxon. 1972.
Vol. 21(4). P. 543-544.
Hasle G. R. Thalassiosiraceae a new diatom family // Norw. J. Bot. 1973a. Vol. 20. N 1. P. 67-70.
Hasle G. R. Some marine plankton genera of the diatom family Thalassiosiraceae 11 Beih. Nova Hedwigia.
1973b.H.45.S. 1-68.
Hasle G. R. The mucilage pore of pennate diatoms // Beih. Nova Hedwigia. 1973c. H. 45. S. 167-186.
H a s 1 e G. R. Some living marine species of the diatom family of Rhizosoleniaceae 11 Beih. Nova Hedwigia.
1975. H. 53. S. 99-140.
Hasle G. R. Brackish water and freshwater species of the diatom genus Skeletonema. II. Skeletonema
potamos comb, nov.// J. Phycol.1976. Vol. 12, N 1. P. 73-82.
Hasle G. R. Thalassiosira punctigera (Castr.) Comn. nov., a widely distributed marine planktonic diatom //
Nord. J. Bot. 1983. Vol. 3.P. 593-608.
Hasle G. R., Fryxell G. A. The genus Thalassiosira: some species a linear areola array // Beih. Nova
Hedwigia. 1977. H. 54. S. 15-66.
Hasle G. R., S t о s c h H. A.. Syvertsen E. E. Cvmatosiraceae, a new diatom family // Bacillaria.
1983. Vol. 6. P. 9-156.
Hasle G. R., Syvertsen E. E. Thalassiosiropsis, a new diatom genus from the fossil records // Micro-
paleontology. 1985. Vol. 31, N 1. P. 82-91.
H e 1 m с к e T. G., Krieger W., G с r 1 о f f T. Diatomeenschalen im electronenmicroskopischen Bild.
Berlin, 1974. Bd 8.
Hendey N.I. The plankton diatoms of the southern seas // „Discovery” Repts. 1937. Vol. 16.
P. 151-364.
Hendey N.I. Marine diatoms from some West African ports // J. Roy. Microsc. Soc. Ser. 3. 1957. Vol. 77,
pt 1-2. P. 28-85.
H e n d e у N. I. An introductory account of the smaller algae of British coastal waters. Pt. 5. Bacillariophyceae
(Diatoms). London, 1964. 317 p.
Hendey N. I., С r a w f о r d R. M. Notes on the occurence, distribution and fine structure of Druridgea
compressa (West) Donkin // Beih. Nova Hedwigia. 1977. H. 54. S. 1-14.
Hoagland K. D., Roso wsky J. R. Valve and band morphology of some freshwater diatoms: I. Fra-
gile ria capucina var. mesolepta // J. Phycol. 1978. Vol. 14, N 4. P. 479-495.
Hoban M. A. Biddilphioid diatoms. II. The morphology and systematics of the pscudocellate species.
Biddulphia biddulphiana (Smith) Boyer, B. alternans (Bailey) Van Heurck and Trigonium arcticum
(Brightwell) Cleve // Bot. mar. 1983. Vol. 26. P. 271-284.
95
Holmes R. W., Crawford R. M., Round F. F. Variability in the structure of the genus Cocconeis
Ehr. (Bacillariophyta) with special reference to the cingulum // Phycologia. 1982. Vol. 21, N 3.
P. 370-381.
Hustedt F. Die Bacillariaceen // Vegetation des Luncer Seengebietes (Nieder-Osterreich) // Intern. Rev.
Hydrobiol. Leipzig, 1922. Bd 10. S. 40-112, 233-270.
Hustedt F. Die Kieselalgen. 1-3 // Rabenhorst’s Kryptogamen-Flora Deutschlands, Osterreichs u.d.
Schweiz. Leipzig, 1927-1930. Teil 1. 925 S.; 1931-1959. Teil 2. 845 S.; 1961-1966. Icil 3.
816 S.
Hustedt F. Marine littoral diatoms of Beaufort, North Carolina // Duke Univ. Mar. Stat. Bull. 1955.
N6.S.5-67.
Jorgensen E. Protist plancton of Northern Norwegian fjords // Bergens Mus. Skr. 1905. S. 47-145.
Jurily Q., Jerkovic L. Les organelles locomotrices des diatomees et leurs types potentiels // Arch.
Hydrobiol. 1973. Suppl.-Bd 41. S. 283-288.
К a n а у a T. Miocene diatom assemblages from the Onnagawa formation and their distribution in the
correlative formation in North-East Japan // Sci. Repts Tohoku Univ., Senday. Ser. 2.1959. Vol. 28.
P. 1-130.
К a r s t e n G. Bacillariophyta (Diatomeae) // Engler A., Prantl K. Die naturlichen Pflanzenfamilien. Leipzig,
1928. Bd 2, Abt. 2. S. 105-303.
Krammer K. Morphologic investigations of the valve and girdle of the diatom genus Cymbella Agardh //
Bacillaria. 1981. Vol. 4. P. 125-147.
Krammer K. Valve morphology in the genus Cymbella C. A. Agardh // Micromorphology of diatom
valves. Lehre, 1982. Vol. 11.289 p.
Kiit zing F. T. Die kieselschaligen Bacillarien oder Diatomeen. Nordhausen, 1844.152 S.
Lange-Bertalot H., Simonsen R. A taxonomic revision of the Nitzschia lanceolatae Grunow.
2. European and related extra-european freshwater and brackish water taxa // Bacillaria. 1978.
Vol. l.P. 11-113.
L i C. W., V о 1 c a n i В. E. Studies on the biochemistry and fine structure of silica, shell formation in dia-
toms. VIII. Morphogenesis of the cell wall in centric diatom Dytilum brightwellii // Protoplasma.
1985. Vol. 124, N l.P. 10-29.
Long J. A., F u g e D. P., Smith J. Diatoms of the Moreno Shale // J. Paleontol. 1946. Vol. 20, N 2.
P. 89-118.
Mann A. Report on the diatoms of the Albatross voyages in the Pacific Ocean 1888-1904 // Contribs.
U.S.nath. Herb. 1907. Vol. 10, N 5. P. 221-419.
Mann D. G. Structure, life history and systematics of Rhoicosphenia (Bacillariophyta). I, II // Phycolo-
gia. 1982. Vol. 18, N l.P. 162-176; 1984. Vol. 20,N4.P. 544-555.
Mann D. G. Symmetry and cell division in rapid diatoms // Ann. Bot. 1983. Vol. 52, N 4. P. 573-581.
Mann D.G. Structure, life history and systematic of Rhoicosphaenia (Bacillariophyta). V // J. PhycoL
1984. Vol. 20, N 4. P. 544-555.
Miller W. S., Collier A. Ultrastructure of the frustule of Triceratium favus (Bacillariophyceae) 11
J. PhycoL 1978. Vol. 14, N l.P. 36-62.
Moss M. О., С a r t e r J. R. The resurrection of Achnanthes rostrata 0strup. // Bacillaria. 1982. Vol. 5.
P. 157-164.
О e s t r u p E. Danske Diatomeer. Kjtfbenhavn, 1910. 323 p.
Okuno H. Marine diatoms // Helmcke O.-G., Kriger W. Diatomeenschalen im elektronenmikroskopischen
Bild. Berlin, 1970. Bd 7.
Ostenfeld С. H. Jagttgelser over Plankton-Diatomeer // Nytt mag. naturviden. 1900. Bd 3, N 4.
S. 287-302.
P a d d о с к T. В. В., Sims P. Q. Observations on the marine diatom genus Auricula and two new genera
Undatella and Proboscideae 11 Bacillaria. 1980. Vol. 3. P. 161-197.
P a d d о с к T. В. B., S i m s P. Q. A morphological study of keels of various rarhe-bearing diatoms // Bacil-
laria. 1981. Vol. 4. P. 177-223.
Phipps D. W., Rosowsky J.K. The morphology and integration of valves and bands in Navicula
mutica var. mutica (Bacillariophyceae) 11 J. PhycoL 1983. Vol. 19, N 3. P. 320-333.
Pocock K. L., Cox E. J. Frustule structure in the diatom Rhabdonema arcuatum (Lyngb.) Kutz. //
Nova Hedwigia. 1982. Bd 36. S. 621 -640.
Pritchard A. A history of Infusoria, including the Desmidiaceae and Diatomaceae British and Foreigh.
London, 1861. 968 p.
Poulin M., Cardinal A. Sea ice diatoms from Manitounuk Sound, southeastern Hudson Bay (Quebec,
Canada). I. Family Naviculaceae 11 Can. J. Bot. 1982. Vol. 60, N. 7. P. 1263-1278.
R a t t г a у J. A revision of the genus Coscinodiscus Ehrb., and of some allied genera. Edinburg, 1890. 244 p.
Rosowsky J.R. Valve and band morphology of some freshwater diatoms. II. Integration of valves
and bands in Navicula confervacea var. confervacea 11 J. PhycoL 1980. Vol. 16, N 1. P. 88-101.
Ross R The current state of diatom taxonomy at the species level, with special reference to some species
of Navicula sect. Lyratae 11 Beih. Nova Hedwigia. 1972. H. 39. S. 1-36.
Ross R., Sims P. The fine structure of the frustule in centric diatoms: a suggested terminology // Brit.
J. PhycoL 1972. Vol. 7. P. 139-163.
96
Ross R., Sims P. A. Observation on family and generia limits in the Centrales 11 Beih. Nova Hedwigia.
1973. H. 45. S. 97-132.
Ross R., Sims P. Some genera of the Biddulphiaceae (diatoms) with interlocking linking spines // Bull.
Brit. Mus. (Natur. Hist.) Bot. 1985. Vol. 13, N 3. P. 277-381.
Rothpletz A. Uber die Flych-Fucoiden und einige andere fossile Algen, sowie uber liassische Diatomeen
fuhrende Homschwamme // Z. Dtsch. Geol. Ges. 1896. Bd 48, H. 1. S. 854-914.
Round F. F. The diatom genus Climacosphenia Ehr. // Bot. mar. 1982. Vol. 25, N 11. P. 519-527.
Schmidt A. Atlas der Diatomaceenkunde. Aschersleben; Leipzig; Berlin, 1874-1959. Tafelen 1-420,
433-480.
Schoeman F. R., Archibald R.E. M. The diatom flora of Southern Africa. Pretoria, 1978.
Schulz P. Diatomeen aus senonen Schwammgesteinen der Danziger Bucht. Zugleich ein Beitrag zur Ent-
wicklungsgeschichte der Diatomeen // Bot. Arch. 1935. Bd 37. S. 383-413.
S c h ii 11 F. Das Pflanzenleben der Hochsee. Kiel; Leipzig, 1893. S. 20.
Schutt F. Bacillariales (JDiatomeae) 11 Engler A., Prantl K. Die natiirlichen Pflanzenfamilien. Leipzig,
1896. Bdl, Abt. lb. S. 31-150.
Simonsen R. Ideas for a more natural systems of the centric diatoms // Beih. Nova Hedwigia. 1972.
H. 39. S. 37-54.
Simonsen R. The diatom system, ideas on phylogeny // Bacillaria. 1979. Vol. 2. P. 9-71.
Sims P. A., P a d d о с к T. В. В. Observations and comment on some prominent morphological features
of Naviculoid genera // Beih. Nova Hedwigia. 1979. H. 64. S. 169-191.
Smith M. Q. The frustular and cytoplasmic fine structure of vegetative cells of Sceletonema costatum
(Greville) Cleve // Bacillaria. 1981. Vol. 4. P. 223-240.
S t о s c h H. A. An amended terminology of the diatom girdle// Beih. Nova Hedwigia. 1975. H. 53. S. 1 -28.
S t о s c h H. A. Observations on Bellerochea and Streptotheca, including descriptions of three new plancto-
nic diatom species // Nova Hedwigia. 1977. Bd 4. S. 113-166.
Stosch H. A., Simonsen R. Biddulphiopsis, a new genus of the Biddulphiaceae. 11 Bacillaria. 1984.
Vol. 7. P. 9-36.
Syvertsen E. E. Thalassiosira rotula and T. gravida: ecology and morphology // Beih. Nova Hedwigia.
1977. H. 54. S. 99-112.
Syvertsen E. E. Resting spore formation in clonal cultures of Thalassiosira antarctica Comber, T. nor-
denskioeldii Cleve and Detonula confervacea (Cleve) Gran // Beih. Nova Hedwigia. 1979. H. 64.
S. 41-63.
Syvertsen E. E., Hasle G. R. The marine planctonic diatom Lauderia annulate Cleve with particular
reference to the processes // Bacillaria. 1982. Vol. 5. P. 234-256.
Takano H. Notes on marine littoral diatoms from Japan. I // Bull. Tokai Reg. Fish. Res. Lab. 1963.
N36.P. 1-9.
Takano H. New and rare diatoms from Japanese marine waters. I, VI // Bull. Tokai Reg. Fish. Res. Lab.
1965. N 42. P. 1-10; 1981. N 105. P. 31-43.
Taylor F. J. R. Phytoplankton of the south western Indian Ocean // Nova Hedwigia. 1967. Bd 12.
P. 433-476.
Tempere J., Peragallo H. Diatoms collection. (Texte et tables de la collection des diatomees du
monde entier). Paris, 1889-1895. 305 p.
Van Heurck H. Synopsis des Diatomees de Belgique. Atlas. Texte. Anvers, 1880-1885. 235 p.
VanHeurckH. A treatise on the Diatomaceae I Transl. by W. E. Baxter. London, 1896. 559 p.
Wallich G. C. On siliceous organisms found in the digestive cavities of the Salpae // Trans. Mier. Soc.
London. N. S. 1960. Vol. 8. P. 36-55.
W e b e г С. I. A new freshwater centric diatom Microsiphona potamos gen»et sp. nov. // J. PhycoL 1970.
Vol. 6. P. 149-153.
Weisse J.E. Mikroscopische Analyse eines organischen Polierschifers aus dem Gonvernement Simbirsk //
Bull. Akad. Sci. 1854. N 2. P. 237-250.
ОБЪЯСНЕНИЯ К ТАБЛИЦАМ
Таблица 1
1-6 - форма панциря: 1 - линзовидная (Pyxidicula zabelinae (Jous6) Makar, et Moiss.),2- низко-
цилиндрическая (Thalassiosira eccentrica (Ehr.) Cl.), 3 — призматическая (Entogonia weissii Ehr.), 4 - вы-
сокоцилиндрическая (Aulacosira granulata (Ehr.) Simonsen var. curvata (Grun.) Simonsen), 5.- седловид-
ная (Campylodiscus clypeus Ehr. var. biocostatus QW. Sm.) Hust.), 6 — в виде сектора шара (Amphora aff.
acutiuscula Kiitz.); 7—16 - форма створок:. 7 - булавовидная (Licmophora sp.), 8 - эллиптическая
(Achnanthes sp.), 9 — клиновидная (Rhoicosphenia curvata (Kiitz.) Grun.), 10 - треугольная (Trinacria si-
mulacrum Gr. et St.), 11 — палочковидная (Bacillaria paradoxa Gmel.), 12 - ланцетная (Navicula sp.), 13 —
четырехугольная (Triceratium unguiculatum Grev.), 14 - широкоромбическая (Fragilaria sp.), 15 - яйце-
видная (Surirella ovalis Breb.), 16 - полулунная (Amphora pediculus Kiitz.). (СЭМ; шкала: 1—3, 5, 10,
11, 13 — 10 мкм, 4, 6—9, 12, 14—16 - 1 мкм; 7, 2, 4, 5, 8, 9,11,12,14,16— ориг. И. В. Макаровой;
3, 13 - ориг. В. А. Николаева, 6, 7 - Караева, 1987, 10 - ориг. 3. И. Глезер, 15 - ориг.
Н. Е. Гуслякова).
Таблица 2
1-8 - поверхность створок: 1 - плоская (Thalassiosira eccentrica (Ehr.) Cl.), 2- равномерно сла-
бо выпуклая (Cosmiodiscus insignis Jouse), 3 — сильно выпуклая (Pyxidicula nipponica (Gran et Jendo)
Streln. et Nikolaev), 4 — вогнутая (Cocconeis pediculus Ehr.), 5 — выпуклая с вогнутым центром (Hyalo-
pyxis concava (Sheshuk.)Makar.), 6 — концентрически-волнистая (Stephanodiscus astraea (Ehr.) Grun.),
7 - тангентально-волнистая (Thalassiosira bramaputrae (Grun.) Hakansson et Locker var. septentrionalis
(Grun.) Makar.), 8 - радиально-волнистая (Actinopty chus undulatus (Bail.) Ralfs)\ 9-11 - загиб
створки: 9 — очень высокий (Aulacosira granulata (Ehr.) Simonsen), 10 - высокий (Actinocyclus
ehrenbergii Ralfs), 11 — низкий (Thalassiosira eccentrica (Ehr.) CL); 12-19 — концы ство-
рок: 12 — острый (piitzschia tryblionella Hantzsch), 13- остро закругленный (Navicula directa W. Sm.),
14 — тупо закругленный (Achnanthes minutissima Kiitz.), 75 — тупой (Caloneis glabenima Ostr.), 16 -
клювовидный (Achnanthes clevei Grun. var. rostrata Hust.), 17— широко закругленный (Navicula oblon-
ga Kiitz.), 18- оттянутый (Amphora coffeaeformis Ag.), 19 - головчатый (Navicula intricate Kar.). (СЭМ;
шкала: 1—3, 7, 10, 11 — 10 мкм, 4-6, 8, 9,12-19- 1 мкм; 1-3, 5-11 - ориг. И. В. Макаровой, 4 -
ориг. Н. И. Караевой, 12-19 - ориг. Н. И. Ахметовой).
Таблица 3
7—2 - осевое поле (7 - Navicula halophila (Grun.) Cl., 2— Cocconeis placentula Ehr. var. lineata
(Ehr.) Cl.); 3 — фасция (Achnanthes brevipes Ag. var. intermedia (Kiitz.) Cl.); 4 - отсутствие осевого поля
(Proschkinia complanatoides (Hust.) Kar.); 5 — несимметричное среднее поле (Mastogloia lanceolata Thw.);
6 - конечное поле (Amphipleura scopulorum (Breb.) Cox f. belgica (V. H.) Cox); 7, 8,10 - боковые поля
(7 - Lyrella lyra (Ehr.) Kar. var. atlantica (A. S.) Kar., 8 - L. brachium (Hust.) Kar., 10 - Amphora sp.); 9 -
створка с разрушенным канопеумом, покрывавшим боковые поля (Lyrella pygmeae (Kiitz.) Makar, et
Kar.); 77 - ультраструктура бокового поля (Lyrella pygmeae (Kiitz.) Makar, et Kar.); 72,13 - подково-
образное поле, наружная и внутренняя поверхности (Achnanthes rostrata 0strup). (1-6, 8, 9, 12,13 -
СЭМ, 7 — CM, 10,11 - ТЭМ; шкала: 7, 5-7, 10- 10 мкм, 2-4, 8, 9, 11-13 - 1 мкм; 1-6, 8-11 -
ориг. Н. И. Караевой, 7 — Прошкина-Лавренко, 1963, 72,13— ориг. Ш. Дж. Мухтаровой).
Таблица 4
1—6— щетинки: 7 — внутренняя и наружная структура щетинок (Chaetoceros sp.), 2,4 — соеди-
нение щетинок двух смежных створок (2 - Ch. atlanticus Cl., 4 - Ch. decipiens Cl.), 3, 6 - структура
щетинок (Ch. decipiens Cl.), 5 — кольцо щетинок (Bacteriastrum hyalinum Lauder); 7 - окно (Eucampia
zoodiacus Ehi.y,8—12 — замкнутые выросты: 8—10 — выросты, расположенные на границе с загибом
98
створки (Thalassiosira marujamica Sheshuk. emend. Makar.), 11, 12 — выросты, расположенные на створ-
ке внутрь от краевого кольца выростов с опорами (11 — Т. punctigera (Castr.) Hasle, 12 — Т. bramaput-
rae (Grun.)Hakanssor et Locker var. bramaputrae^ 13,14 - шипики на поверхности створки (13 - Cam-
pylodiscus noricus Ehr. var. hibernicus (Ehr.) Grun., 14 — Thalassiosira bramaputrae var. septentrionalis
(Grun.) Makar.); 15 — гранулы (T anguste-lineata (A. Schmidt) Fryx. et Hasle). (СЭМ; шкала: 1, 2, 5, 7,
13 - 10 мкм, 3, 4, 6, 8—12, 14, 15-1 мкм; 1, 2, 4-12, 14, 15 - ориг. И. В. Макаровой, 3- ориг.
Т. Ю. Орловой, 13- ориг. Ш. Дж. Мухтаровой).
Таблица 5
1-5 — расположение и форма шипов: 1,2- кольцо остроконечных шипов по краю створки
(1 - Thalassiosira nodulolineata (Hendey) Hasle et Fryx., 2 - Cyclotella meneghiniana Kiitz.), 3 - шип глад-
кий, 4 — шип с гранями (3, 4 — Thalassiosira eccentrica (Ehr.) Cl.), 5 — длинные остроконечные шипы
отдельных клеток колонии (Aulacosira grayiulata (Ehr.) Simonsen f. granulata); 6—18 — различная форма
соединительных шипов: 6 - длинные остроконечные (Stephanodiscus hantzschii Grun.), 7 — короткие
средних клеток колонии (Aulacosira granulata f. granulata), 8 - скошенные (Stephanodiscus minutula
(Kiitz.) Round), 9 - загнутые (Fragilaria pinnata Ehr.), 10, 11 - расширенные на конце (10 - Aulaco-
sira italica (Ehr.) Simonsen, 11 — A. islandica (O. Mull.) Simonsen), 12,13— с выемкой (A. granulata f. cu-
rvata (Grun.) Simonsen), 14,15,17- разветвленные (14 - Aulacosira sp., 75 - Stephanodiscus binderanus
(Kiitz.) Krieger var. (?), 17 — S. binderanus var. binderanus), 16— головчатые (S. invisitatus Hohn et Heller-
man var. incognitus Kuzmin et Genkal), 18 - лапчатые (5. binderanus var. baicalensis Popovsk. et Genkal.).
(СЭМ; шкала: 1,2 — 10 мкм, 3—18 - 1 мкм; 1 - Макарова, 19776, 2, 5, 6, 8,10,11,14-18— ориг.
С. И. Генкала, 3, 4 — ориг. И. В. Макаровой, 7, 12, 13 — ориг. И. В. Макаровой, С. С. Бари-
новой, 9 — ориг. Н. И. Караевой).
Таблица 6
1,2— поперечные ребра (1 — Navicula sp., 2 - Diatoma hiemale (Lyngb.) Heib. var. mesodon (Ehr.)
Grun.); 3,4- осевые ребра (5 - Mastogloia pumila (Grun.) CL, 4 - Amphora securicula Perag.); 5 -
осевые и продольные ребра на внутренней поверхности створки (Brachysira aponina Kiitz.); 6, 7 -
краевой гребень (6 — Ditylum bright wellii (West) Grun., 7 - Melosira nummuloides (Dillw.) Ag.);7? -10 -
створки с псевдосептами (8 — Rhoicosphenia curvata (Kiitz.) Grun., 9 — Stauroneis smithii Grun., 10 -
Rhopalodia gibba (Ehr.) O. Mull. var. ventricosa (Ehr.) Grun.). (СЭМ; шкала: 1 мкм; 1, 3,5, 7,10- ориг.
H. И. Караевой, 2, 9 - ориг. Ш. Дж. Мухтаровой, 4 — Караева, 1987, 6 - ориг. С. К. Джафаровой,
8 — ориг. Н. И. Ахметовой).
Таблица 7
Поясок
1—3— панцири с одинаковыми ободками (7 - Proschkinia complanatula (Hust.) Kar., 2 — Berkeleya
scopulorum (Breb.) Cox f. belgica (V. H.) Cox, 3 — Amphora holsatica Hust.); 4—7— панцири с вальвоко-
пулами: 4 — панцирь, поясковый ободок которого развит только у эпитеки и состоит из вальвоко-
пулы, бесструктурного вставочного ободка и двух структурированных соединительных ободков
разной ширины (Grammatophora marina (Lyngb.) Kiitz.), 5 - панцирь с вальвокопулой, отличающейся
от остальных соединительных ободков наличием кремнеземной складки (Berkeleya rutilans (Tr^ntep.)
Grun.), 6, 7— то же, детали; 8, 9 - вальвокопула с фимбриями (Cocconeis scutellum Ehr. var. adjuncta
A. S.): 8- изолированная, 9 - соединенная co створкой; ГО, 11 - вальвокопула с септой (Grammato-
phora marina (Lyngb.) Kiitz.); 10 - изолированная, фрагмент, 77 - соединенная со створкой; 12,13 —
открытые ободки: 72- изолированный ободок (Amphora delicatissima Krasske),75- свободные концы
ободков в панцире (Navicula humerosa (Breb.) W. Sm.); 14—16 — сегментированные поясковые ободки
(Ditylum brightwellii (West) Grun.y 14 — фрагмент пояскового ободка, 15, 16 — изолированные сег-
менты; 77- соединительный ободок с лигулой (Thalassiosira nordenskioeldii CL); 18,19- фрагменты
соединительных ободков с лигулами (18 - Coscinodiscus granii Gough, 79 - Pinnularia borealis Ehr.);
20 — створка с соединительным ободком, имеющим септу на широком конце (Licmophora sp.). (1-
13, 17, 18, 20 - СЭМ, 14-16, 19 - ТЭМ; шкала: 1, 14-16- 10 мкм, 2-13, 17-20- 1 мкм; 7, 2,
4—11,13,19, 20- ориг. Н. И. Караевой, 3, 12 - Караева, 1987,14-16,18- ориг. С. К. Джафаровой,
7 7 - ориг. И. В. Макаровой).
Таблица 8
7 - поровые каналы, расположенные равномерно (Actinoptychus senariusEhi.y,2 - поровые кана-
лы, расположенные беспорядочно (Gyrodiscus vortex Witt..y,3 — поровые каналы в радиальных рядах (Co-
rona retinervis Sheshuk. et Gies.); 4 - поровые каналы, собранные в пучки,и двугубые выросты (стрелки)
(Lepidodiscus elegans Witt.); 5 - поровый канал (стрелка) среди пороидных ареол (Eucampia balaustium
Castr.); б- пороидная ареола с крибрумом на наружной поверхности створки (Trinacria coronata Witt.);.
99
7 — пороидная ареола с вол ой (стрелка) {Arachnoidiscus ehrenbergii Bailey); 8 — локулярная ареола
с фораменом на внутренней поверхности створки (стрелка) {Coscinodiscus bouvet Karsten); 9 - стенки
локулярных ареол одинаковой длины и прерывистый крибрум на внутренней поверхности створки
{Thalassiosiropsis wittiana (Pant.) Hasle); 10 - локулярные ареолы с многоугольной формой локул (стрел-
ка) {Melosira moniliformis (О. Miill.) Ag.); 11 - локулы цилиндрической формы, суженные к обеим по-
верхностям {Hyalodiscus zonulatus М. Perag.); 12 - локулы, расширенные в толще створки (Aulacodis-
cus sp.); 13 - локулярная ареола (стрелка) и закрытые камеры в толще створки [Actinocyclus ас-
tinochilus (Ehr.) Simonsen). (СЭМ; шкала: 1, 2, 5-13 - 1 мкм, 3, 4 - 10 мкм; ориг. В. А. Николаева).
Таблица 9
1 - прерывистый крибрум на наружной поверхности створки {Coscinodiscus bouvet Karsten);
2 — прерывистый крибрум на внутренней поверхности створки {Pyxidicula arctica (Grun.) Streln. et
Nikolaev); 3 - сплошной крибрум на внутренней поверхности створки {Р. dissonus (Schulz) Streln. et
Nikolaev); 4 - сплошной крибрум на наружной поверхности створки и ложный узелок (стрелка)
{Pontodiscus miocenicus (Krasske) Moiss. et Sheshuk.); 5 - крибеллы, закрывающие крибральные по-
ры {Coscinodiscus bouvet Karsten); 6 - рота [Trigonuim arcticum (Brightw.) Cl.); 7 - вола с прикреплен-
ным к ней крибрумом {Isthmia enervis Ehr.); 8 - альвеола {Cyclotella sp.); 9 - отдельные ареолы меж-
ду наружными анастомозирующими ребрами {Stictodiscus californicus A. Schmidt); 10 — группа поро-
вых каналов между наружными анастомозирующими ребрами {Glorioptychus callidus Hanna ); 11 -
буллула (стрелки) {Cyclotella sp.); 12 - полярные выпуклости с глазками и центральная выпуклость
{Briggera affixa Ross et Sims); 13 - рог {Hemiaulus sp.). (СЭМ; шкала: 1-11, 13 - 1 мкм, 12 - 10мкм;
ориг. В. А. Николаева).
Таблица 10
1-4- продольные каналы {Diploneis didyma (Ehr.) Cl.): 7-2 - поперечный слом, 3 - внутрен-
няя поверхность,поры только на концах каналов, 4 — наружная поверхность (продольные каналы
пронизаны порами на всем протяжении); 5-11 - штрихи: 5 - однорядные, радиально-симметричные
на середине створки [Navicula lanceolata (Ag.) Kutz.), 6 - двурядные, дефект Вуа на правой половине
створки {N. pupula Kutz.), 7 — многорядные {Achnanthes delicatula Kutz.), 8 — один ряд из ареол по
краю створки {Thalassionema nitzschioides Grun.), 9 - ребра, чередующиеся со штрихами, дефект Вуа
на правой половине створки {Navicula cryptocephala var. intermedia Grun.), 10 - штрихи, углубленные
в базальный кремнеземный слой {Pinnularia borealis Ehr.), 11 — ареолы во взаимопересекающихся
рядах {Pleurosigma elongatum W. Sm.); 72 — дефект Вуа {Mastogloia pusilia Grun.). {1—4, 9—11 — СЭМ,
5-8, 12 - ТЭМ; шкала: 1,4-10,12- 1 мкм, 2,3,11 - 10 мкм; Г-7,10-12- ориг. Н. И. Караевой,
8 — ориг. С. К. Джафаровой, 9 - ориг. И. В. Макаровой).
Таблица 11
Двугубые выросты
7 — вырост с длинной наружной трубкой [Odontella weissflogii (Janisch) Grun.); 2 — выросты,
проходящие через стенку ареолы, справа {Trigonium arcticum Brightw.); 3 - продольный слом через
вырост {Hemidiscus cuneiformis Wallich); 4 — выросты со сплющенной внутренней трубкой {Actinocy-
clus ehrenbergiiRa\is)’,5 - два выроста с длинными наружными трубками {lYinacria coronata Witt); 6 —
створка с краевым гребнем и центральным двойным двугубым выростом с длинной наружной труб-
кой [Ditylum brightwellii (West ) Grun.); 7,8 - наружная часть выростов в виде щелевидного отверстия;
7 — фрагмент загиба {Melosira varians Ag.), 8 - центральное кольцо [Stellarima stellaris (Roper) Hasle et
Sims); 9-12 - макровыросты: фрагменты внутренней поверхности створки с выростами различного
размера и формы - Coscinodiscus wailesii Gran et Angst, 10, 11 - C. jonesianus (Grev.) Ostf., 72 -
C. granii Gough); 13— наружная поверхность створки с отверстием макровыроста (С. granii)-, 14—16—
выросты, проходящие через ареолу [Pyxidicula zabelinae (Jouse) Makar, et Moiss.). (СЭМ; шкала: 1—4,
7, 8, 10—16 - 1 мкм, 5, 6, 9 — 10 мкм; 1-3, 5, 8, 9 - ориг. В. А. Николаева, 4, 7,10, 11,14-16 -
ориг. И. В. Макаровой, 6,12,13— ориг. С. К. Джафаровой).
Таблица 12
7 — двойной двугубый вырост с внутренней поверхности створки (pitylum brightwellii (West.)
Grun.); 2 — многочисленные выросты на створке {Podosira maxima (Kutz.) Grun.); 3— створка с крае-
вым кольцом выростов (Actinocyclus actinochilus (Ehr.) Simonsen); 4 - два краевых противостоящих
выроста в кольце выростов с опорами [Thalassiosira elsayedii Fryx.); 5 — два выроста, расположенных
на полюсах створки {Odontotropis carinata Grun.); 6, 7 - центральная группа выростов: 6 — немного-
численные [Stellarima microtrias (Ehr.) Hasle et Sims.), 7 - многочисленные {Trigonium formosum {Bri-
ghtw.) Cl.), 8 — вырост, расположенный близ края, со щелью, ориентированной тангентально
{Thalassiosira latimarginata Makar.); 9 - вырост со щелью, ориентированной радиально (Т. orientalis
Sheshuk. emend. Makar.); 10-14— полярные выросты у пеннатных диатомей: 10, 11 -набазальном
100
конце створки с наружной (10) и внутренней (11) поверхности (Licmophora ehrenbergii (Kutz.)
Grun.), 12, 13 — зеркально-симметричные выросты, внутренняя поверхность двух концов одной
створки (Synedra tabulate (Ag.) Kutz. var. acuminata Grun.), 14 — диагонально-симметричные выросты
(Grammatophora marina (Lyngb.) Kutz.); 15 - появление третьего, субцентрального выроста под
влиянием повышения солености на 5 °/00 (G- marina); 16 — кольцевидный вырост на внутренней
поверхности створки (Thalassiosiropsis wittiana (Pant.) Hasle); 17, 18 — двугубый вырост на внутрен-
ней поверхности створки (17 — Actinocyclus sp., 18 - A. variabilis (Makar.) Makar.). (1-13, 16-18 -
СЭМ, 14, 15 - ТЭМ; шкала: 1, 4, 8-18 — 1 мкм, 2, 3,5-7 — 10 мкм; 1,14, 15 - ориг. С. К. Джафа-
ровой; 2, 3, 5—7, 16 - ориг. В. А. Николаева, 4, 8, 9, 17, 18 — ориг. И. В. Макаровой, 10, 11 —
Макарова, Гусляков, 1984,12,13 - ориг. Н. И. Караевой).
Таблица 13
1—7— краевые выросты с опорами на наружной поверхности створки: 1, 2, 4 - с длинными
наружными трубками (1 — Thalassiosira nordenskioeldii Cl., 2 — Т. aestivalis Gran, 4 — Sceletonema
costatum (Grev.) Cl.), 3 — с короткими наружными трубками (Thalassiosira bramaputrae (Grun.)
Hakansson et Locker var. septentrionalis (Grun.) Makar.), 5 - с внутренней узкой трубкой (T. nidulus
(Temp, et Brun) Jouse), 6 — с воронкой на конце (T. nordenskioeldii Cl.), 7 — расширенные у основа-
ния (Т. bulbosa Syvertsen); 8 - центральные выросты с опорами без наружной трубки (Т. marujamica
Sheshuk. emend. Makar.); 9 — продольный разрез через вырост с опорами (Thalassiosira sp.); 10-15 —
форма и расположение выростов с опорами на внутренней поверхности створки: 10, 11 — краевое
кольцо выростов (10 - Т. nordenskioeldii Cl., 72 - Т. oestrupii (Ostf.) Hasle), 12 - выросты с 3 опора-
ми ( Т. latimarginata Makar.), 13,14 - центральные выросты с опорами (13 - Т. constricta Gaaider, 14 -
Т. bramaputrae var. septentrionalis (Grun.) Makar.), 75 - центральные и краевые выросты с опорами
(Cyclotella caspia Grun.); 16-21 - количество сопутствующих пор у выростов с опорами: 16 - с 2 по-
рами (Stephanodiscus invisitatus Hohn et Hellermann), 7 7-19 - с 3 порами (7 7, 18 - S. triporus Genkal
et Kuzmin, 19 — Thalassiosira oestrupii (Ostf.) Hasle), 20,21 - c 4 и 5 порами (20 - Thalassiosira sp.,
21 — T. marujamica Sheshuk. emend. Makar.). (1-6, 8-15, 18-21 - СЭМ, 7, 16, 17 - ТЭМ; шкала: 1,
2, 5—9, 11—15, 18-21 - 1 мкм, 3, 4, 10 - 10 мкм, 16, 17 - 0.1 мкм; 1-6, 8-11, 13, 14, 19-21 -
ориг. И. В. Макаровой, 7 — ориг. В. И. Михайлова, 12 — ориг. В. А. Николаева, 75 - Генкал, Мака-
рова, 1986,16-18 - ориг. С. И. Генкала).
Таблица 14
1—5— одно краевое кольцо из выростов с опорами на границе с загибом (7 — Thalassiosira alle-
nii Takano, 2 - T. aestivalis Gran, 5 - Coenobiodiscus muriformisLo&\\ch, Wight, Darley, 4 — Thalassiosira
bramaputrae (Grun.) Hakansson et Locker var. septentrionalis (Grun.) Makar., 5 - Stephanodiscus sp.); 6 -
2 кольца выростов с опорами на загибе (Thalassiosira oliveriana (О. Meara) Makar, et Nikolaev); 7, 10 -
шахматное расположение выростов на загибе (7 - Т. baltica (Grun.) Ostf., 10 - Thalassiosira sp.) ; 8 -
2 кольца выростов с опорами: одно на загибе, второе на лицевой части створки (Т. diporocyclus
Hasle); 9-3 кольца выростов с опорами (Т. subtilis (Ostf.) Gran ) ; 77 - кольцо выростов с опорами
на загибе, стрелки (Т. oestrupii (Ostf.) Hasle var. venrickae Fryx. et Hasle); 12 — немногочисленные вы-
росты на лицевой части створки (Т. lineata Jouse); 13-17 - многочисленные выросты на лицевой
части створки (13,14 - Т. mendiolana Hasle et Heimdal, 75 - Porosira pseudodenticulata (Hust.) Jouse,
16-P.glacialis^rg.,17 - T. eccentrica (Ehr.) Cl.). (7, 8, 9, 12, 15, 17 - ТЭМ,2-7,10,11,13,14,16 -
СЭМ; шкала: 7, 3, 5, 8-12, 14-17 - 1 мкм, 2, 4, 6, 7, 13 - 10 мкм; 7, 3, 5, 7, 8, 10-14, 16, 17 -
ориг. И. В. Макаровой, 2 — Макарова, 1978, 4, 15 — ориг. С. И. Генкала, 9 - Макарова, 19766, 6 -
Макарова, Николаев, 1984).
Таблица 15
1,2 - один центральный вырост с опорами (7 — Thalassiosira ordinaria Makar., 2 — Т. eccentrica
(Ehr.) Cl.); 3, 4 - один или два выроста с опорами, расположенных близ центра, и кольцо по краю
(Т. pseudonana Hasle, Heimdal);5—7 - несколько выростов с опорами, расположенных в средней
части створки (5 — Т. oliveriana (О. Meara) Makar, et Nikolaev, 6 — T. bramaputrae (Grun.) Hakansson et Lock-
er var. septentrionalis (Grun.) Makar., 7 — Cyclotella meneghiniana Kutz.) ; 8—11 — центральные выросты
с опорами расположены без особого порядка или образуют изогнутую линию (8, 9 - Т. baltica
(Grun.) Ostf., 10 - Т. ignota Makar., 77 - T. poroseriata (Ramsphjell) Hasle) ; 12 - центральные выросты
с опорами образуют две линии (Т. strelnikovae Makar.); 13 — выросты с опорами образуют кольцо
вокруг центральной малой ареолы (Т. nodulolineata (Hendey) Hasle et Fryx.); 14, 15 — выросты с опо-
рами образуют правильное замкнутое (14 - Т. plicata Schrader) или неправильное центральное коль-
цо (75 - Т. weissflogii (Grun.) Fryx. et Hasle) ; 16 — выросты с опорами образуют незамкнутое кольцо
(16 — Т. anguste-lineata (A.S.) Fryx. et Hasle); 17, 18 - выросты с опорами образуют пучок в центре
створки (17 — Т. rotula Meunier, 18 — Bacterosira fragilis Gran ). (7, 5—7, 9, 13, 14, 16, 17 - СЭМ, 2-4,
101
8, 10-12, 15, 18 — ТЭМ; шкала: 1-4, 6, 8—18 — 1 мкм, 5, 7 — 10 мкм; 1 - Макарова, 1977а,
2—4, 6, 9-11,17 - ориг. И. В. Макаровой, 5 - Макарова, Николаев, 1984, 7, 18 - ориг. С. И. Ген кала,
8, 13 - Макарова, 19776, 12 — Макарова, 1976а, 14 — Макарова, 1978, 15 - Макарова, 19766, 16 —
Макарова, 1981).
Таблица 16
1 — глазок {Odontella aurita Ag.);2, 3 — глазок на невысоких выпуклостях с радиальным рас-
положением порелл и концентрическими прерванными ребрами по краю {Auliscus sculptus (W. Sm.)
Ralfs); 4 — глазок с беспорядочно расположенными пореллами {Zygoceros weissflogii Pant.);5 — гла-
зок с параллельными ребрами {Eucampia balaustium Castr.); 6 — глазок с параллельными рядами
порелл {Fragilaria californica Grun. var. antarctica M. Perag.); 7 - глазок с продольным ребром {Entору -
la australis Ehr.), 8 — ложные глазки на полосах {Trigonium arcticum (Brightw.) Cl.) ; 9 — ареолирован-
ный ложный узелок (стрелка) {Actinocyclus ingens Rattray); 10 - блестящий ложный узелок
{A. actinochilus (Ehr.) Simonsen); 11 - нижний ободок, подстилающий открытую часть верхнего
ободка {Melosira nummuloides Ag.); 12 — открытый панцирь, к нижележащей створке прикреплено
партектальное кольцо {Mastogloia pusilia Grun.); 13 - наружные отверстия партектумов на пояске
{М, angulata Lewis). (СЭМ; шкала: 1, 3—7, 10, 12 — 1 мкм, 2, 8, 9, 11, 13 — 10 мкм; 1—10 - ориг.
В. А. Николаева, 11-13 - ориг. Н. И. Караевой).
Таблица 17
1,2 — стигма с наружной поверхности створки: 7—6 стигм {Cymbella Helvetica Kutz.), 2 — одна
стигма {Navicula mutica Kutz.);3-6 — стигма с внутренней поверхности створки: 3, 4, 6 — одна стиг-
ма {3, 4 - N. mutica, 6 - Proschkinia complanatula (Hust.) Kar.), 5-7 стигм {Cymbella Helvetica Kutz.);
7—13 - крыло: 7, 8 — деталь панциря, окруженного крылом, 9, 10 — поперечные сломы крыла
{Campylodiscus noricus Ehr. var. hibernicus (Ehr.) Grun.), 11 — крыло, окружающее створку {Surirella
biseriata Breb. var. bifrons (Ehr.) Hust.), 12,13 - узелки одной створки {Cymatopleura elliptica (Breb.)
Sm.). (СЭМ; шкала: 1-6, 9, 10, 12, 13 — 1 мкм, 7, 8,11 - 10 мкм; 1, 5, 7-13 - ориг. Ш. Дж. Мухта-
ровой, 2-4, 6 - ориг. Н. И. Караевой).
Таблица 18
1—11 — шов и его элементы: 1 - осевая складка шва {Amphora aff. A. acutiuscula Kiitz.), 2, 3 —
центральный узелок, внутренняя поверхность (2 - Mastogloia braunii Grun., 3 - Gyrosigma attenuatum
(Kiitz.) Rabh.),4, 5 -ставрос {Amphora ostrearia Breb.), 6 - конечный узелок, внутренняя поверх-
ность створки {Cymbella minuta Hilse), 7 - хоботок {Pinnularia sp.),£, 9 - каналовидный шов с на-
ружной поверхности {8 — Nitzschia vidovichii (Grun.) Perag., 9 — Epithemia turgida (Ehr.) Kiitz.), 10,11 -
каналовидный шов, внутренняя поверхность, фибулы(70 — Hantzschia amphioxys (Ehr.) Grun., 77 -
Nitzschia obtusa W. Sm. var.scalpelliformis Grun.)j 72, 13 - киль и крыло: 72 - киль, суженный посере-
дине {Nitzschia sp.), 13 — крыло, окружающее створку {Campylodiscus noricus Ehr. var. hibernicus
(Ehr.) Grun.). (СЭМ; шкала: 1-3, 5, 6-12 - 1 мкм, 4, 13 - 10 мкм; 1-3, 10-12 - ориг. Н. И. Карае-
вой, 4, 5 — ориг. Н. Е. Гуслякова, 6. 7, 13 - ориг. Ш. Дж. Мухтаровой, 8 — ориг. И. В. Макаровой,
9 — ориг. Н. И. Ахметовой).
Таблица 19
1-15 - Pyxidicula arctica (Grun.) Streln. et Nikolaev: 7, 3, 6, 9, 10 — загиб створки, 2, 72 - целый
панцирь, 4, 5, 7, 8 — вид со створки, 77, 14 — внутренняя поверхность створки, прерывистый криб-
рум, щелевидное отверстие двугубого выроста, 13 - наружные трубки двугубых выростов, 75 -
наружная поверхность створки, крибрум с порами в радиальных рядах. 7, 2, 72, 13, 15 - эоцен,
Норвежское море, 3, 9 — эоцен, Зап. Сибирь, 4-8, 10, 11, 14 - поздний мел, вост, склон Урала;
16 -Р, defecta (Streln.) Streln. et Nikolaev: вид co створки. Эоцен, Норвежское море {1-9 - СМ,
10-16 - СЭМ; шкала: 1—12,16 — 10 мкм, 13—15 — 1 мкм; ориг. Н. И. Стрельниковой).
Таблица 20
1-5 - Pyxidicula arctica (Grun.) Streln. et Nikolaev: 1-4 - внутренняя поверхность створки (7 -
2 кольца двугубых выростов, 2 — прерывистый крибрум и щелевидное отверстие двугубого вы-
роста, 3 — краевое кольцо двугубых выростов, 4 - крибрум), 5 — строение локулярной ареолы.
Поздний мел, вост, склон Урала; 6,7 — Р. biseriata (Streln.) Streln. et Nikolaev: 6 - загиб створки, 7 -
вид со створки. Поздний мел, вост, склон Урала; 8-12 — Р. corona (Ehr.) Streln. et Nikolaev: 8 — це-
лый панцирь, 9, 10 - соединение створок, 77, 72 — разветвленные на конце шипики (створка в раз-
ных фокусах). 8, 10—12 — Камчатка, р. Тхонма, 9 - ранний олиго цен, Тасманово море; 13, 14 -
Р, barbadensis (Grev.) Streln. et Nikolaev: 13 - вид co створки, 14 — целый панцирь. Ранний олигоцен,
102
Тасманово море; 15, 16 - Р. defecta (Streln.) Streln. et Nikolaev: 75 - панцирь с ареолами, 16 — пан-
цирь без ареол, место ареолы ограничено бугорками. Эоцен, Норвежское море. (7—5, 9, 14 — СЭМ,
6-8, 10—13, 15, 16 - СМ; шкала: 1, 4, 6—16 — 10 мкм, 2, 3, 5 - 1 мкм; 7—7, 9, 13—16 — ориг.
Н. И. Стрельниковой, 8,10—12 - ориг. О. С. Короткевич).
Т а б.л и ц а 21
1—4 — Pyxidicula crenata (Sheshuk.) Streln. et Nikolaev: 7 - внутренняя поверхность створки, 2
кольца двугубых выростов, 2 — наружная поверхность створки, кольцо наружных трубок двугубых
выростов, 3 - загиб створки, 4 — структура створки. 1,2 — эоцен, Воронежская обл., 3, 4 — эоцен,
левый берег Днепра; 5-8 - Р. charkoviana (Jous£) Streln. et Nikolaev: 5 - вид co створки, 6, 7 - внут-
ренняя поверхность створки, сплошной крибрум, внутренние отверстия двугубых выростов, 8 —
загиб створки. Эоцен, Воронежская обл.; 9—15 — Р. cruciata Ehr.: 9, 12 — наружная поверхность
створки, 77 — отверстия двугубых выростов, 72 — соединение створок, 10, 13,14 — внутренняя
поверхность створки: 10 - отверстия двугубых выростов, 13,14 — прерывистый крибрум, 75 — вид со
створки. 9, 11, 12 — эоцен, Атлантический океан, Фолклендское плато, 10, 13, 14 — эоцен, Атланти-
ческий океан, Бермудские о-ва, 75 — эоцен, Зап. Сибирь. (7, 2, 5—14 — СЭМ, 3, 4,15 - СМ; шкала:
7-5, 7—9, 13, 15 — 10 мкм, 6, 10-12, 14 - 1 мкм; 7, 2, 5—8 — ориг. Н. И. Стрельниковой, 3, 4 —
Шешукова-Порецкая, Глезер, 1964, 9—14 - ориг. Н. И. Стрельниковой и В. А. Николаева, 75 - Гле-
зер, 1974).
Таблица 22
1-8 - Pyxidicula dissona (Schulz) Streln. et Nikolaev f. dissona: 1,2— створка в разных фокусах,
3, 4, 6 — наружная поверхность створки, отверстия двугубых выростов, 5, 7, 8 - внутренняя
поверхность створки (5 — отверстия двугубых выростов, 7, 8 — кольцо двугубых выростов, сплош-
ной крибрум). Поздний мел, вост, склон Урала; 9-12 - Р. dissona (Schulz) Streln. et Nikolaev f. dis-
soniformis (Streln.) Streln. et Nikolaev: 9 - внутренняя поверхность створки, остатки сплошного
крибрума, 10, 11 — створка в разных фокусах. Поздний мел, вост, склон Урала; 13-15 — Р. uralensis
(Streln.) Streln. et Nikolaev: 13 — створка сбоку, 14 — вид со створки, 75 — внутренние отверстия
двугубых выростов. Поздний мел (кампан), вост, склон Урала. (7, 2, 10-12, 14 — СМ, 3—9, 13,
15 - СЭМ; шкала: 1-4, 7, 9, 10-14 — 10 мкм, 5, 6, 8, 15 — 1 мкм; 1—8 — ориг. Н. И. Стрельнико-
вой и В. А. Николаева, 9—15 - ориг. Н. И. Стрельниковой).
Таблица 23
1-6 - Pyxidicula edita (Jouse) Streln. et Nikolaev: 1, 3 — наружная поверхность створки (7 -
кольцо наружных трубок двугубых выростов, 3 — строение ареол, бугорки по углам ареол), 2 -
загиб створки; 4, 5 - внутренняя поверхность створки, прерывистый крибрум, 6 — вид со створки.
1-3 - эоцен, Норвежское море, 4, 5 - эоцен, р. Эмба, 6 - эоцен, Зап. Сибирь; 7—9 - Р. ferox (Grev.)
Streln. et Nikolaev: 7 - створка, 8, 9 — створка в разных фокусах. Палеоцен, Ульяновская обл.
(Сенгилей) ; 10-12 - Р. lavrenkoi (Jouse) Streln. et Nikolaev: 10, 12 - внутренняя поверхность створ-
ки, округлые отверстия двугубых выростов по краю прерывистого крибрума, 77 — наружная
поверхность створки. Палеоцен, вост, склон Урала; 13, 14 — Р. spinosa (Jous£) Streln. et Nikolaev:
13 - загиб створки, 14 - шипики по углам ареол. Эоцен, р. Эмба. (7, 3—5, 7, 10—14 — СЭМ, 2, 6, 8,
9 - СМ; шкала: 1, 2, 4, 6-9, 11-13 - 10 мкм, 3, 5, 10, 14-1 мкм; ориг. Н. И. Стрельниковой).
Таблица 24
1-4 - Pyxidicula inermis (Jouse) Streln. ei Nikolaev: 1, 3, 4 — целый панцирь, 2 — строение ареол.
Неоген, о-в Сахалин (маямрафская свита); 5-9 - Р. joynsonii (A. S.) Streln. et Nikolaev: 5, 6 - вид со
створки, наружная поверхность, 7-9 — внутренняя поверхность створки, щелевидные отверстия
двугубых выростов. Эоцен, Воронежская обл.; 10—13 — Р. megapora (Grun.) Streln. et Nikolaev:
10-11 — створки в разных фокусах, 72 - загиб створки. 10—12 — эоцен, Норвежское море, 13 -
эоцен, Зап. Сибирь; 14,15 - Р. spinosissima (Grun.) Streln. et Nikolaev: створки в разных фокусах.
Эоцен, Новая Зеландия (у г. Оамару). (7, 2, 4, 6-9 - СЭМ, 3, 5,10-15 — СМ; шкала: 7, 3—7, 9—15 -
10 мкм, 2, 8 - 1 мкм); 1, 2, 4 - ориг. И. В. Макаровой, 3 - ориг. В. С. Шешуковой-Порецкой,
5-9 — ориг. Н. И. Стрельниковой и Т. Г. Федорищенко, 10-15 - ориг. Н. И. Стрельниковой).
Таблица 25
1,2 — Pyxidicula marginata (Grun.) Streln. et Nikolaev: створки в разных фокусах. Эоцен, Зап.
Сибирь; 3-5 - Р. grunowii (Gr. et St.) Streln. et Nikolaev: 3, 4 - створки в разных фокусах, 5 —
створка. Эоцен, Тасманово море; 6, 7 - Р. moelleri (A. S.) Streln. et Nikolaev: 6 — часть створки
103
с внутренней поверхности и двугубые выросты, 7 - крибрум и двугубый вырост. Эоцен, р. Эмба;
8,9 - Р. palmeriana (Grev.) Streln. et Nikolaev: 8 — панцирь, 9 — популярные ареолы со сплошным
крибрумом на внутренней поверхности створки. 8 — Тихий океан, побережье Перу, 9 — Мексикан-
ский залив (7-5 - СМ, 6—9 — СЭМ; шкала: 1-6, 5 — 10 мкм, 7, 9 - 1 мкм; ориг. Н. И. Стрельни-
ковой) .
Таблица 26
1—3 - Pyxidicula punctata (Jouse) Streln. et Nikolaev: 1,2— створка в разных фокусах, 3 —
створка с вогнутым центром. 1,2 — эоцен, р. Анадырь, 3 — эоцен, Зап. Сибирь; 4—9 — Р. similis
(Sheshuk.) Streln. et Nikolaev: 4, 5 — створка в разных фокусах, 6 - целый панцирь, 7 - наружная
поверхность створки, 8, 9 — внутренняя поверхность створки, прерывистый крибрум, округлые
отверстия двугубых выростов. Эоцен, р. Анадырь; 10-12 - Р. schenkii (Kanaya) Streln. et Nikolaev:
10 - шипики на створке, И, 12 - вид со створки. Неоген, о-в Сахалин; 13, 14 - Р. polaris (Grun.)
Streln. et Nikolaev: створки в разных фокусах. Эоцен, Зап. Сибирь. (7-5, 10, 11, 13, 14 -
СМ, 6-9, 12 - СЭМ; шкала: 1-7, 9-14 - 10 мкм, 5-1 мкм; 1, 2, 4-9, 12 - ориг.
В. С. Шешуковой-Порецкой, 3, 13—14 — ориг. Н. И. Стрельниковой, 10, 11 — Шешукова-
Порецкая, 1967).
Таблица 27
7-77 — Pyxidicula nipponica (Gran et Yendo) Streln. et Nikolaev: 1-10 - вегетативные клетки (7,
5, 6, 8 - створка с загиба, 2—4 — вершина створки, наружные трубки двугубых выростов, 7 —
соединение двух створок, 9, 10 — краевое кольцо двугубых выростов: 9 — внутренние отверстия,
10 - наружные трубки, открывающиеся в локулы), 77 - створка споры. 1, 2, 5, 6 - сев.-зап. часть
Тихого океана, 3, 4 — Японское море. {1—4, 7—11 — СЭМ, 5, 6 - СМ; шкала: 7—5, 77—10 мкм,
9,10 - 1 мкм; 1,2 - ориг. И. Н. Сухановой, 3,4 - ориг. И. В. Макаровой, 5-11 - ориг. Н. И. Стрель-
никовой и В. А. Николаева).
Таблица 28
1-10 - Pyxidicula turris (Grev. et Arnott) Streln. et Nikolaev f. turris-. 1-6 - целый панцирь, 7, 9,
10 - загиб створки, 5 - наружные трубки двугубых выростов. 7 - эоцен, Норвежское море,
2, 7-10 — палеоцен, Ульяновская обл. (Сенгилей), 3, 4-6 - поздний мел, вост, склон Урала;
11-14 -Р. turris (Grev. et Arnott) Streln. et Nikolaev f. cylindrus {Gnm) Streln. et Nikolaev: различные створ-
ки. 77,12,14 — эоцен, Норвежское море, 13 -ранний палеоцен, вост, склон Урала; 15-18 - Р. superba
(Grun.) Streln. et Nikolaev: 15, 16 — створка в разных фокусах {16 - край створки), 17, 18 — створка
в разных фокусах: 17 - кольцо шипов, 75 - гиалиновый край. 75, 76 - эоцен, Тасманово море,
77, 75 - эоцен, Новая Зеландия (у г. Оамару). (7, 3-6, 13-18 - СМ, 2, 7-12 - СЭМ; шкала:
1—7, 9—18 — 10 мкм, 5—1 мкм; ориг. Н. И. Стрельниковой).
Таблица 29
1-9 - Pyxidicula weyprechtii Grun.: 1-7,9 - внутренняя поверхность створки (7 - створка, 2 -
локулярная ареола, 3 — прерывистый крибрум, 4 — краевое кольцо двугубых выростов, 5 - щель крае-
вого двугубого выроста, 6,7,9 - отверстия двугубых выростов), 5 - наружная поверхность створки.
7-5, 5 - эоцен, Казахстан, р. Эмба, 6, 7, 9 - поздний мел, вост, склон Урала; 10-12 - Р. weissflogii
(A. S.) Streln. et Nikolaev: 10 — загиб створки, 77 — внутренняя поверхность створки, 72 - край
створки. Эоцен, Воронежская обл.; 13,14 — Р. moelleri (A. S.) Streln. et Nikolaev: створка в разных
фокусах. Эоцен, Зап. Сибирь. (7-77 - СЭМ, 12-14 - СМ; шкала: 7, 10, 12-14 - 10 мкм,
2-5, 7-9, 77-1 мкм, 6 - 0.1 мкм; 1-9 - ориг. Н. И. Стрельниковой и В. А. Нико-
лаева; 10-12 - ориг. Н. И. Стрельниковой, Т. Г. Федоршценко; 13, 14 - ориг. Н. И. Стрель-
никовой) .
Таблица 30
1-15 — Pyxidicula zabelinae (Jouse) Makar, et Moiss.: 1,2- створки в различных фокусах, 3,4-
панцирь, 5, 6 — створки с наружной поверхности, 7, 5 — створки с внутренней поверхности,
9, 10 - двугубый вырост на наружной поверхности (трубки выростов обломаны), 11,12 - строение
ареолы, 13, 14 - двугубый вырост на внутренней поверхности створки, 75 - наружная трубка
двугубого выроста, проникшая сквозь ареолу. 1,2- неоген, Сев. Сахалин (маямрафская свита),
3-15 - поздний миоцен-плиоцен, Южн. Сахалин (маруямская свита). (7, 2 - СМ, 3-15 - СЭМ;
шкала: 7-5-10 мкм, 9-15 — 1 мкм; ориг. И. В. Макаровой).
104
Таблица 31
1—3, 6 — Costopyxisantique (Jouse) Gies.: 1,3 — загиб створки, 2 — структура вершины створ-
ки, 6 - внутренняя поверхность створки, двугубые выросты и сплошной велум. Поздний мел
(кампан), вост, склон Приполярного Урала; 4, 5,7,8 - С. broschii (Grun.) Streln. et Nikolaev: 4, 7 —
целый панцирь, 5 — загиб створки, 8 — внутренняя поверхность створки. Поздний мел (кампан),
вост, склон Урала; 9 - С. costata (Gies.) Gies.: панцирь. Поздний палеоцен-ранний эоцен, Поволжье;
10-14 — С. reticulata (Long, Fuge et Smith) Gies.: 10 — загиб створки, 11—14 — структура вершины
створки, 12 - наружная поверхность створки с сетью ребер, 13 - внутренняя поверхность створки.
Поздний мел (кампан), вост, склон Урала; 15-19- С. schulzii (Stein.) Gies, i. schulzii'. 15, 16 — струк-
тура вершины створки (разные фокусы), 17—19 — структура загиба створки (18, 19 — разные
фокусы). Поздний мел (кампан), вост, склон Урала.(7, 2, 4, 5, 9, 10, 14-19 — СМ, 3, 6-8, 11—13 —
СЭМ; шкала: 6, 8, 11 — 1 мкм, остальные - 10 мкм; 1-3, 6, 10—19 - ориг. Н. И. Стрельниковой,
4, 5,7,8 — ориг. Н. И. Стрельниковой и В. А. Николаева, 9 - ориг. 3. И. Глезер).
Таблица 32
1-4, 9-16 — Costopyxis schulzii (Stein.) Gies. f. ornata (Schulz) Streln.: 1, 2, 13 — структура вер-
шины створки (разные фокусы), 3, 4, 10 — структура загиба створки, 9 — целый панцирь, 77 —
соединение створок и остатки вегетативной клетки, 12, 14, 15 — внутренняя поверхность створки
(72 - щелевидное отверстие двугубого выроста, 14 — расположение двугубых выростов),
16 — строение пороидных ареол. Поздний мел (кампан), вост, склон Урала; 5-8 - С. schulzii (Stein.)
Gies. f.nuda Streln.: 5, 6 — структура вершины (разные фокусы), 7, 8 - створки с загиба. Поздний
мел (кампан), вост, склон Урала; 17,18 — С. trochlea (Hanna) Streln.: створки. Эоцен, Норвежское
море. (1-8, 17 - СМ, 9-16, 18 — СЭМ; шкала: 77, 72, 14, 16 — 1 мкм, остальные — 10 мкм; ориг.
Н. И. Стрельниковой).
\
Таблица 33
1-9 - Thalassiosiropsis wittiana (Pant.) Hasle: 1,2- створки с наружной поверхности и трубка
кольцевидного выроста (стрелка), 3 — створка с внутренней поверхности и кольцевидный вырост
(стрелка), 4, 5 - локулярные ареолы и крибрум, 6 - наружная трубка кольцевидного выроста
и форамены ареол, 7, 8 — кольцевидный вырост и крибрум ареол, 9 — основание отломанного коль-
цевидного выроста. 1—3, 6, 7, 9 — поздний мел, вост, склон Урала, 5, 8 - палеоцен, Ульяновская
обл. (СЭМ; шкала: 1—3 — 10 мкм, 4—9 - 1 мкм; ориг. В. А. Николаева).
Таблица 34
1,2 - Triceratium favus Ehr.: 7 — створка, 2 - панцирь. 7 — Северное море, 2 - Атлантический
океан, у берегов Гвинеи; 3-5 - Т. unguiculatum Grev.: 3 - общий вид створки, 4 - выпуклость
с глазком и наружная трубка двугубого выроста, 5 — двугубый вырост с внутренней стороны
створки. Эоцен, Фолклендское плато; 6 - Planktoniella sol (Wallich) Schutt: наружная поверхность
створки. Юго-вост, часть Тихого океана; 7—10 — Endictya oceanica Ehr.: 7 - створка, 8 — загиб створ-
ки, 9 — наружная поверхность створки, 10 — внутренняя поверхность створки. Черное море.
(7, 7-10 - СМ, 2-6 — СЭМ; шкала: 1—3, 6-10 - 10 мкм, 4, 5 - 1 мкм; 7 - Глезер, 1975а, 2 -
ориг. И. В. Макаровой и Л. В. Кузьменко, 3—5 — ориг. В. А. Николаева, 6 — ориг. Г. И. Семиной,
7-10 - Прошкина-Лавренко, 19556).
Таблица 35
7-70- Gladius speciosus Schulz f.aculeolatus Streln.: 1-9 - детали створки (7, 2, 4-9 - апи-
кальная часть со структурой из пороидов и шипиков, 3 - соединение створок расширенными базаль-
ными частями, структура из локулярных ареол), 10 - целый панцирь. Поздний мел (кампан);
11-17- G. speciosus Schulz f.poratus Streln.: 77 - створка, 72 - целый панцирь, 13-17 - детали
створки (13-16 - апикальная часть со структурой из пор, 77 - переходная часть к расширенному
основанию). Поздний мел (кампан) ; 18-20 - G. speciosus Schulz f. ellipsoideus Streln.: 18, 20 - раз-
ные формы створок, 79 - целый панцирь. Поздний мел (кампан). (7—5, 7, 10—20 — СМ, 8, 9 —
СЭМ; шкала: 1-7, 10—20 — 10 мкм, 8,9 — 1 мкм; ориг. Н. И. Стрельниковой).
Таблица 36
1-10 -Gladius speciosus Schulz f. speciosus1, 2 - целый панцирь, 3, 6, 7 - строение вершины
створки: трубковидный вырост, окруженный крибральными порами, 4 — апикальная часть, 5, 10 -
базальная часть, 8, 9 - строение локулярных ареол. Поздний мел (кампан), вост, склон Урала.
(7-5, 10 - СМ, 6-9 - СЭМ; шкала: 10 мкм; ориг. Н. И. Стрельниковой).
8 Заказ 518
105
Таблица 37
1-4 — Thalassiosira aculeata Pr.-Lavr.: различные створки. 1, 2 - Каспийское море, 3, 4 - Азов-
ское море; 5-9 - Т. angulata (Greg.) Hasle: 5-7 - различные створки, 8 - структура наружной по-
верхности створки, 9 — часть створки с двугубым выростом и 2 выростами с опорами. 5 - у берегов
Дании, эксиккат, 6, 7 — Чукотка (р. Яма), 8, 9 — Белое море; 10—13 — Т. antiqua (Grun.) А. Cl.:
10, 11 — различные створки, 12 - структура наружной поверхности створки, 13 - край створки
с 2 выростами с опорами. Миоцен, Сев. Сахалин (маямрафская свита); 14—19 — Т. coronifera
(Pr.-Lavr.) Pr.-Lavr.: 14-16 — различные створки, 17 — структура наружной поверхности створки,
18 - часть створки с двугубым выростом и 5 выростами с опорами, 19 - часть створки с отверстием
центрального выроста с опорами и кольцом краевых выростов с опорами. 14 — поздний сармат,
Таманский п-ов, 15 — Каспийское море. 16-19 — Азовское море. (7-5, 10, 11, 14-16 - СМ, 6,7 —
ТЭМ, 8, 9, 12, 13, 17—19 - СЭМ; шкала: 7-5, 10-12, 14-16 - 10 мкм, 6-9, 13, 17-19 - 1 мкм;
7, 2, 15 - Прошкина-Лавренко, Макарова, 1968, 3,4 — Прошкина-Лавренко, 1956, 5, 8-13, 17—19 -
ориг. И. В. Макаровой, 6, 7 - ориг. В. И. Михайлова, 14 — Макарова, 1962, 76 - Прошкина-
Лавренко, 1963).
Таблица 38
7-77 - Thalassiosira anguste-lineata (A. S.) Fryx. et Hasle: 1,2- различные створки, 3, 4 - струк-
тура наружной поверхности створки, 5 — структура внутренней поверхности створки, 6 - маленькие
отверстия центральных выростов с опорами на наружной поверхности створки, 7, 8 — центральные
выросты с опорами на внутренней поверхности створки, 9 — 2 краевых выроста с опорами, 10 -
структура загиба и створки с гранулами, двугубый вырост и 4 выроста с опорами на наружной
поверхности створки, 77 — форма выростов с опорами на наружной поверхности и форма двугубого
выроста с наружной и внутренней поверхностей. 7 - голоцен, Белое море, 2 - Черное море, 3, 6-9,
11 — Белое море, 4, 10 — Японское море, Амурский залив. (7, 2 - СМ, 3~11 - СЭМ; шкала: 1-5 —
10 мкм, 6-11 — 1 мкм; 7 - ориг. Р. Н. Джиноридзе, 2 - ориг. А. И. Прошкиной-Лавренко, 3—5, 7,
10 - ориг. И. В. Макаровой, 6, 8, 9,11 - Макарова, 1981).
Таблица 39
1-8 - Thalassiosira confusa Makar.: 1,2 — створка в различных фокусах, 3 - часть створки
и ее структура, 4 - створка с внутренней поверхности, 5 - створка с наружной поверхности, 6 -
центральные выросты с опорами, 7—2 краевых выроста с опорами с отломанными трубками,
8 - двугубый вырост. Сев.-вост. часть Тихого океана, Алеутский желоб; 9 - Т. decipiens (Grun.)
J0rg.: створка. Баренцево море; 10 — Т. heteromera Makar.: створка. Баренцево море; 77 - Т. delica-
tissima Pr.-Lavr.: створки. Мэотис, Таманский п-ов; 12—15 — Т. ignota Makar.: 12 - створка, 13 —
внутренняя поверхность створки, 14 — наружная поверхность створки, 75 - центральные и краевые
выросты с опорами и двугубый вырост на внутренней поверхности створки. 12 - Баренцево море,
13 — сев.-вост. часть Тихого океана, 14 — Берингово море, 75 - Тихий океан, к юго-востоку
от о-ва Шикотан. (7, 2, 9-12 - СМ, 3 — ТЭМ, 4-8, 13-15 — СЭМ; шкала: 1-4, 9-15 — 10 мкм,
5-8 — 1 мкм; 1,2- Макарова, 1981, 3-9, 13 - ориг. И. В. Макаровой, 10, 12 — Макарова, 1975,
77 - Прошкина-Лавренко, 1960, 14,15 - Макарова и др., 1981).
Таблица 40
1-9 - Thalassiosira eccentrica (Ehr.) Cl.: 7 - колония, 2 - панцирь, 3 - створка с наружной по-
верхности, 4 — створка, 5 — лицевая часть створки с выростами с опорами, 6, 7 — центральный вы-
рост с опорами на внутренней поверхности створки, 8 - двугубый вырост и 2 шипа с наружной по-
верхности створки, 9 - часть створки с внутренней поверхности, двугубый вырост. 7 - Тихий
океан, к ЮВ от Галапагосских о-вов, 2 - Японское море, Амурский залив, 3-6 - Черное море,
7 - Атлантический океан, шельф юго-зап. Африки, 8, 9 - Ионическое море; 10 - Т. intermedia Makar.:
створка. Каспийское море; 11—17- Т. latimarginata Makar.: 77 - створка, 12, 13 - наружная поверх-
ность створок, 14, 15 — внутренняя поверхность створки, 16 — край створки и загиб, 17 - краевой
вырост с опорами и двугубый. 11 — Баренцево море, 12, 15, 17 — поздний плейстоцен, Чукотка,
13, 14, 16 - голоцен, Охотское море. {1—3, 7-9, 12—17 — СЭМ, 4,10,11 - СМ, 5, 6 - ТЭМ; шкала:
1-4, 10-15 — 10 мкм, 5-9, 16, 17 - 1 мкм; 1-3, 5—9 — ориг. И. В. Макаровой, 4 — Прошкина-
Лавренко, 19556, 10 — Прошкина-Лавренко, Макарова, 1968, 77 - Макарова, 1975, 12-17 — ориг.
И. В. Макаровой и Т. Л. Невретдиновой).
Таблица 41
1-12— Thalassiosira incerta Makar.: 1-4 — различные створки, 5 - наружная поверхность створки,
6, 7 — внутренняя поверхность створки, 8, 9 - центр створки с 2 и 3 выростами с опорами,
10, 11 - маленькие отверстия центральных выростов с опорами на наружной поверхности, 12 —
106
двугубый вырост на внутренней поверхности створки. 1-3 - Каспийское море, 4 - Белое озеро,
5—12 — Азовское море; 13—21 — Т. marujamica Sheshuk. emend. Makar.: 13,14 — створки, 75 - створ-
ка с наружной поверхности, 16 — створка с внутренней поверхности, 17 — часть створки с наружной
поверхности с 2 замкнутыми выростами и 3 маленькими отверстиями краевых выростов с опорами
на загибе, 18 - часть створки с внутренней поверхности с центральными и краевыми выростами
с опорами, 19,20 — край створки с двугубым выростом, 27 — замкнутые выросты. Поздний миоцен,
Южн. Сахалин (маруямская свита). {1-3, 13, 14 - СМ, 4,8, 9 - ТЭМ, 5-7, 10-12, 15-21 - СЭМ;
шкала: 1—3, 5—7, 13, 14 — 10 мкм, 4, 8—12,15—21 — 1 мкм; 1-3- Прошкина-Лавренко, Макарова,
1968,4 — ориг. С. И. Генкала, 5—21 - ориг. И. В. Макаровой).
Таблица 42
1—4 — Thalassiosira native Sheshuk.: 1, 2 — створка в различных фокусах, 3 — наружная поверх-
ность створки, 4 — край створки и загиб с маленькими отверстиями краевых выростов с опорами.
Поздний миоцен, Южн. Сахалин (маруямская свита); 5—11 — Т. nodulolineata (Hendey) Hasle et Fryx.:
5 — створка с наружной поверхности, 6 — створка с внутренней поверхности, 7 — часть створки,
8-10 - центральные выросты с опорами (9-е наружной, 10 - с внутренней поверхности), 77 -
двугубый вырост с наружной поверхности створки и шипы. Азовское море; 12 - Т. leptopus (Grun.)
Hade et Fryx.: створка с наружной поверхности. Голоцен, Охотское море; 13-17 — Т oestrupii
(Ostf.) Hasle var. venrickae Fryx. et Hasle: 13 — створка, 14 — часть панциря с поясковым ободком,
75 — створка с наружной поверхности, 16 — створка с внутренней поверхности, 17 - двугубый
вырост. 13 — Черное море, 14—17 — Японское море, Амурский залив. (7, 2, 13 — СМ, 3—6, 9-12,‘
14—17 - СЭМ, 7, 8 - ТЭМ; шкала: 1, 2, 5, 6, 12, 13 - 10 мкм, 3, 4, 7-11,14-17- 1 мкм; 1-4,12,
14—17 — ориг. И. В. Макаровой, 5—11 - Макарова, 19776 (под названием Т. incerta Makar.), 13 -
Прошкина-Лавренко, 19556 (под названием Г. antiqua (Grun.) A. Cl. var. septate Pr.-Lavr.). *
Таблица 43
1-13 - Thalassiosira pacifica Gran et Angst: 1-4 - различные створки, 5-7 - створки с наруж-
ной поверхности, 8, 9, 11 - часть створок, структура, 10 - центральный вырост с опорами и криб-
рум, 72 - центральная часть створки, 13 - краевые выросты с опорами и двугубый вырост с наруж-
ной поверхности. 1—3, 6, 10, 11 - сев.-вост. часть Тихого океана, Алеутский желоб, 4,9 - Охотское
море, 5, 7—12, 13 — голоцен, Охотское море, 8 - голоцен, Белое море; 14—17 — Т. parva Pr.-Lavr.:
14, 15 — различные створки, 76 — створка с наружной поверхности, 17 — край створки. 14,15 — Чер-
ное море, 16,17 — Азовское море; 18 — Т. singularis Sheshuk.: створка. Миоцен, Сев. Сахалин,
п-ов Шмидта (маямрафская свита); 19, 20 — Т. tenera Pr.-Lavr.: различные створки. Черное море.
(7, 2, 14, 15, 18—20 - СМ, 3, 4, 8-10 - ТЭМ, 5-7, 11-13, 16, 17 - СЭМ; шкала: 1, 2, 4-7,14-16,
18—20 — 10 мкм, 3, 8—13, 17 — 1 мкм; 1—3, 10 — Макарова, 1976а, 4, 9 — ориг. В. И. Михайлова,
5-8, 11-13, 16, 17 - ориг. И. В. Макаровой, 14, 15 - Прошкина-Лавренко, 19556,18 - Шешукова-
Порецкая, 1967,19,20 — Прошкина-Лавренко, 1961.
Таблица 44
1-13 - Thalassiosira aestivalis Gran: 7 - створка, 2-4 - створки с наружной поверхности, 5 -
створка с внутренней поверхности, 6, 7 — край створки с краевыми выростами с опорами и дву-
губым выростом, 8 — двугубый вырост и 2 выроста с опорами на наружной поверхности створки,
9, 10 — двугубый вырост и краевые выросты с опорами на внутренней поверхности, 77 — централь-
ный вырост с опорами на внутренней поверхности, 72,13 — центральный вырост с опорами на наруж-
ной поверхности. 1, 3, 5, 13 - Атлантический океан, шельф юго-зап. Африки, 2, 4, 6-12 - Черное
море. (7 - СМ, 2-13 — СЭМ; шкала: 7—5 - 10 мкм, 6—13 — 1 мкм; 7, 5, 13 - Макарова, 1978,
2—4, 6—12 — ориг. И. В. Макаровой).
Таблица 45
1—13 - Thalassiosira baltica (Grun.) Ostf.: 1-5 - различные створки, 6 - наружная поверхность
створки, 7, 8 — центральные выросты с опорами на внутренней поверхности створки, 9 - край
створки с трубками краевых выростов с опорами и двугубого, 10 - двугубый вырост и краевые
выросты с опорами на внутренней поверхности створки, 77 - створка с 4 двугубыми выростами,
72, 13 — часть створки. 7 - Балтийское море, 2 - Азовское море, 3 - плейстоцен, Сев.-Зап. СССР,
4, 5 — поздний сармат, Таманский п-ов, 6-11 — Черное море, 72 —Азовское море, 13 - плейстоцен,
Сев.-Зап. СССР. (7-5 - СМ, 6, 8-11 - СЭМ, 7,12,13 - ТЭМ; шкала: 7-6-10 мкм, 7-13 - 1 мкм;
1, 3, 4, 12, 13 - ориг. И. В. Макаровой, 2 — Прошкина-Лавренко, 1963,5 - Макарова, 1962, 6—11 —
Макарова, 1977а).
107
Таблица 46
1 - Thalassiosira bioculata (Grun.) Ostf.: створка. Баренцево море; 2 - T. kryophila (Grun.)
створка. Сев.-вост. часть Тихого океана, Алеутский желоб; 3 - Т maeotica Pr.-Lavr.: створка.
Мэотис, Таманский п-ов; 4-10- Т punctigera (Castr.) Hasle: 4 - створка, 5 - створка с наружной
поверхности, 6 - край створки с соединительными ободками, 2 замкнутыми выростами и краевыми
выростами с опорами, 7 - структура загиба створки и форма краевых выростов с опорами на
наружной поверхности створки, 8 - краевые выросты с опорами на внутренней поверхности
створки, 9 — центральный вырост с опорами на внутренней поверхности створки, 10 - трубка дву-
губого выроста с наружной поверхности. 4^ - сев.-зап. часть Тихого океана, Алеутский желоб,
5—10 — Японское море, Амурский залив. {1-4 — СМ, 5-10 - СЭМ; шкала: 1-5 - 10 мкм, 6—10 -
1 мкм); 1, 4-10 - ориг. И. В. Макаровой, 2 — Козлова, Стрельникова, 1974, 3 - Прошкина-
Лавренко, 1960).
Таблица 47
1—7— Thalassiosira ordinaria Makar.: 1 — створка, 2 - часть створки и ее структура, 3 - двугу-
бый вырост, 4 - створка с наружной поверхности, 5 - панцирь, 6 — центральный вырост с опорами,
7 - створка с внутренней поверхности. Черное море; 8-13 — Т. subtilis (Ostf.) Gran: 8 - створка с на-
ружной поверхности, 9 - створка с внутренней поверхности, 10 - часть створки с 3 кольцами
выростов с опорами. 11-13 - структура створок с 4 кольцами выростов с опорами (77 - двугубый
вырост между вторым и третьим кольцом, 13 — двугубый вырост в краевом кольце выростов
с опорами). 8 — Атлантический океан, юго-вост. Африка, 9, 13 - Мозамбикский залив, 10 —
Индийский океан, экватор, 77 — Аравийское море, 12 — Тихий океан, Каролинские о-ва. (1—3,
10-13 - ТЭМ, 4-9 - СЭМ; шкала: 7, 4, 5,7,9 - 10 мкм, 2, 3, 6, 8, 10-13 - 1 мкм; 7, 3, 4, 7 -
Макарова, 1977а, 2, 5, 6, 8-10 - ориг. И. В. Макаровой, 11-13 - Макарова, 19766).
Таблица 48
1—5- Thalassiosira antarctica Comber: 7 — панцирь, 2 - створка с внутренней поверхности, 3 —
часть створки с наружной поверхности с краевыми выростами с опорами, 4 - центральные выросты
с опорами на наружной поверхности створки, 5 — спора. 1,2- сев.-зап. часть Тихого океана;' 3 —
Берингово море, 4 — Антарктика, 5 - голоцен, Охотское море; 6—12 — Т. hyalina (Grun.) Gran:
6 — структура створки, 7 — наружная поверхность створки, 8 - внутренняя поверхность створки,
9 — край створки, 10 - центральные выросты с опорами, 77, 72 - двугубый вырост (стрелка)
и краевые выросты с опорами. 6 - Чукотка (р. Яма), 7-10 — голоцен, Охотское море, 11, 12 -
Берингово море. (7-5, 7-11 - СЭМ, 6, 12 - ТЭМ; шкала: 7, 2 - 5 мкм, 3, 4, 9-12 - 1 мкм,
5—8 — 10 мкм; 1-4 - Семина, Макарова, 1983, 5, 7-10 - ориг. И. В. Макаровой и Т. Л. Невретди-
новой, 6 - ориг. В. И. Михайлова, 77 - Макарова, Семина, 1982, 72 - ориг. С. И. Генкала).
Таблица 49
1-4 - Thalassiosira bramaputrae (Ehr.) Hakansson et Locker var. bramaputrae-. 1 - наружная по-
верхность створки, 2 — часть створки с кольцом выростов с опорами и замкнутых выростов,
3 - край створки с 2 замкнутыми выростами, 4 — структура наружной поверхности створки. 1,3,4 -
Каспийское море, 2 - оз. Балхаш; 5-16 — Т. bramaputrae var. septentrionalis (Grun.) Makar.: 5 - створ-
ка, 6, 7, 9,16- наружная поверхность створок, 8 - структура створки, 10, 11 - внутренняя поверх-
ность створки, 72 - часть створки с краевыми выростами с опорами и группой их на створке, 13 -
3 краевых и I створочный вырост с опорами, 14 - двугубый вырост, 75 - край створки с наружной
поверхности с 3 выростами с опорами и двугубым выростом. 5 - Охотское море, 6, 7, 9-11, 13,14 -
осадки Белого моря, 8 - Пертозеро, 72 - низовье Волги, 75, 76 - плейстоцен, Сев.-Зап. СССР. (1-4,
6, 7, 9-11, 13-16 - СЭМ, 5 - СМ, 8,12 - ТЭМ; шкала: 7, 5-77. 76 - 10 мкм, 2-4, 12-15 - 1 мкм;
1-4, 15, 16 — ориг. И. В. Макаровой, 5 - ориг. В. И. Михайлова, 6-11, 13, 14 - ориг. С. И. Генкала
и И. В. Макаровой, 72 - ориг. С. И. Генкала).
Таблица 50
1-6 - Thalassiosira bramaputrae (Ehr.) Hakansson et Locker var. proximus (Makar.) Makar.: 7 - створ-
ка, 2,3 - часть створки с кольцом краевых выростов с опорами, 4 - внутренняя поверхность створки,
5-2 кольца выростов с опорами на внутренней поверхности створки, 6 - часть створки с внутрен-
ней поверхности с кольцом выростов с опорами и сплошным крибрумом. 1-4 - Каспийское море,
5, 6 - оз. Балхаш; 7, 8 - Т. bulbosa Syvertsen: 7 - створка вегетативной клетки, 8 — створка покоя-
щейся споры. Охотское море, Ямская губа; 9,10 - Т. conferta Hasle: 9 - створка, 10 - часть створ-
ки с центральным выростом и 2 краевыми выростами с опорами. П-ов Таймыр; 11—13 - Т. mala
108
Takano: 11 - створка, 12 - панцирь, 13 - край створки и поясок. 11 — Охотское море, Ямская
губа, 12, 13 - Атлантический океан, шельф Гвинеи. {1-3 - СМ, 4-6, 12, 13 - СЭМ, 7-11 - ТЭМ;
шкала. 1-4 - 10 мкм, 5-13 - 1 мкм; 1-3 — Макарова, 1961, 4, 12, 13 - ориг. И. В. Макаровой,
5, 6 - ориг. И. В. Макаровой и Н. И. Ахметовой, 7, 8,10,11 - ориг. В. И. Михайлова, 9 - Макарова
и др., 1979).
Таблица 51
1-12 - Thalassiosira gravida Cl.: 1,2- различные створки, 3 — центр створки, 4 - часть створ-
ки, структура, 5, 6 - наружная поверхность створки, 7 - центральные выросты с опорами с выходя-
щими из них тяжами, 8 — внутренняя поверхность створки, 9 - край створки с наружной поверх-
ности, трубка двугубого выроста и выросты с опорами на загибе и лицевой части створки, 10 —
структура части створки с внутренней поверхности, на загибе двугубый вырост, 11 - панцирь, 12 —
спора. 1-3 - Охотское море, 4 — сев.-вост. часть Тихого океана, Алеутский желоб, 5 - Берингово
море, 6, 9 - Японское море, Амурский залив, 7, 10, 11 - Антарктика, 8, 12 — голоцен, Охотское
море. {1 - СМ, 2-4 - ТЭМ, 5-12 - СЭМ; шкала: 1, 6, 8, 12 — 10 мкм,2-4, 7, 9, 10 - 1 мкм,
5, И - 5 мкм; 1—3 - ориг. В. И. Михайлова, 4, 6, 8, 9, 12 - ориг. И. В. Макаровой, 5, 7, 10 -
Семина, Макарова, 1983,11 - ориг. Г. И. Семиной).
Таблица 52
1 -4 - Thalassiosira guillardii Hasle: 1,2— различные створки, 3 - створка с наружной поверх-
ности, 4 - вырост с опорами и двугубый вырост. 1,3- Рыбинское водохранилище, 2,4 - Саратов-
ское водохранилище; 5—12 — Т. manifesto Sheshuk. emend. Makar.: 5 - створки, 6 — часть створки
с наружной поверхности, несколько колец из краевых выростов с опорами, 7 - часть створки с внут-
ренней поверхности и загиб створки с наружной поверхности, велум и краевые выросты с опорами,
8-12 - покоящиеся споры {8-10 — наружная поверхность створки: 8 - створка с одним краевым
кольцом и многочисленными центральными выростами с опорами, 9, 10 - створки с 2 кольцами,
расположенными в шахматном порядке, и 4 центральными выростами с опорами,отступя от края
отверстие двугубого выроста (стрелка); 11, 12 — створки с внутренней поверхности: краевые
и центральные выросты с опорами и двугубый вырост). Поздний миоцен, Сев. Сахалин (маямраф-
ская свита); 13-17 - Т. orientalis Sheshuk. emend. Makar.: 13, 14 - створка в различных фокусах,
75 — створка с внутренней поверхности, 16 - центральные выросты с опорами и велум, 17 —
краевые выросты с опорами и двугубый вырост. Поздний миоцен, Сев. Сахалин (маямрафская
свита). (7, 2, 4 - ТЭМ, 3, 6-12, 15-17 - СЭМ, 5,13,14 - СМ; шкала: 7, 2, 4, 6-12,16,17-1 мкм,
3, 5, 13-15 - 10 мкм; 1, 2, 4 - ориг. С. И. Генкала, 3 — Генкал, Макарова, 1984, 5-17 — ориг.
И. В. Макаровой).
Таблица 53
7-77 - Thalassiosira nordenskioeldii Cl.: 1-3, 5, 6 - различные створки, 4 - панцирь, 7 - крае-
вой вырост с опорами, 8 — центральный вырост с опорами, 9 — наружная поверхность створки,
10 — внутренняя поверхность створки и кольцо выростов с опорами, 77 — загиб створки с 2 вы-
ростами с опорами и двугубый вырост. 1, 4, 5, 7, 8, 10 - сев.-вост. часть Тихого океана, Алеутский
желоб, 2 - залив Посьета, 3 — Охотское море, 6 — Японское море, 9, 11 — залив Петра Великого.
(7, 2 - СМ, 3-8 - ТЭМ, 9-77 - СЭМ; шкала: 1, 2, 10 - 10 мкм, 3-9, 11 - 1 мкм; 7, 2, 4, 5, 7, 8,
10 - Макарова, 1976а, 3 - ориг. В. И. Михайлова, 6 - ориг. С. И. Генкала, 9,11 - ориг. И. В. Мака-
ровой) .
Таблица 54
1-8 - Thalassiosira proschkinae Makar, var. proschkinae'. 1-5 - различные створки, 6 - централь-
ный вырост с опорами и двугубый вырост, 7 — двугубый вырост, 8 — краевой вырост с опорами.
7 - Каспийское море, 2-4, 6 - Азовское море, 5, 7, 8 - оз. Аджикабул (АзССР) ; 9,10 — Т proschki-
nae var. spinulata (Takano) Makar.: различные створки. Охотское море; 11-22 - Т. pseudonana Hasle et
Heimdal: 77 - створка с наружной поверхности, 12 - створка с внутренней поверхности, 13-15 -
створки с одним центральным выростом с опорами, 16-20 - створки с 2 центральными выростами
с опорами (79. 20 - створки с шипиками), 27, 22 - створки без центрального выроста с опорами.
77, 12 — оз. Плещееве, 13, 15, 16 - Черное море, 14, 17-20 - Чукотка (р. Яма), 27 - оз. Севан,
22 - Саратовское водохранилище. {1-10, 13-22 - ТЭМ, 77, 72 - СЭМ; шкала: 1-6, 9—22 - 1 мкм,
7, 8 - 0.1 мкм; 1-4, 6 - ориг. С. И. Генкала и И. В. Макаровой, 5, 7, 8, 21, 22 — Макарова и др.,
1979, 9, 10, 14, 17-20 — ориг. В. И. Михайлова, 77, 72 - ориг. С. И. Генкала, 13, 15, 16 - ориг.
И. В. Макаровой).
109
Таблица 55
1-7 - Thalassiosira rotula Meunier: 1,2- часть створки и ее структура, 3 - центр створки с хи-
тановыми тяжами, 4—7 — центральные выросты с опорами у различных створок. 1, 6, 7 - Атланти-
ческий океан, Канарское течение, 2-5 — Ионическое море.(ТЭМ; шкала: 1 мкм; 1 — ориг. И. В. Ма-
каровой, 2—7 — Макарова, 1974).
Таблица 56
1—12— Thalassiosira rotula Meunier: 1,2- различные створки с наружной поверхности, 3 —
часть створки с наружной поверхности, 4 — створка с внутренней поверхности, 5 — центральные
выросты с опорами с наружной поверхности, 6,7 — соединительные ободки, 8 — вставочный ободок,
9 — вальвокопула и вставочный ободок, 10, 11 — двугубый вырост с наружной и внутренней поверх-
ностей створки, 12 - двугубый вырост с внутренней поверхности створки. 1-2, 4, 6-10, 12 -
Черное море, 3 — Японское море, Амурский залив, 5,11 — Атлантический океан, Канарское течение.
(СЭМ; шкала: 1, 2, 4,6,7 — 10 мкм, 3, 5, 8-12 - 1 мкм; ориг. И. В. Макаровой).
Таблица 57
1—9 — Thalassiosira weissflogii (Grun.) Fryx.: 1 — створка, 2—4 — центральные выросты с опора-
ми различных створок, 5 - край створки с двугубым выростом и кольцом краевых выростов
с опорами, 6—8 — структура наружной поверхности створки, 9 — двугубый вырост и выросты
с опорами на внутренней поверхности створки. Каспийское море; 10—14 — Т. parvula Makar.:
10, 12 — различные створки, 11 — часть створки с центральным и краевым выростами с опорами,
13 — центр створки, 14 — край створки. 10, 13, 14 — Азовское море, 11 — Чукотка (р. Яма),
12 — Охотское море, бухта Гертнера. (1—5,10—14 — ТЭМ, 6-9 - СЭМ; шкала: 1—14 — 1 мкм; 1, 3, 5, 8,
9 — Макарова, 19766, 2, 4, 6, 14 — ориг. И. В. Макаровой, 7 - ориг. Н. И. Караевой, 10, 13 —
Макарова, 19776,11,12 — ориг. В. И. Михайлова).
Таблица 58
1—5 — Sceletonema costatum (Grev.) Cl.: 1 — часть колонии с пояском, 2 - соединение выростов
с опорами двух створок различных панцирей, 3 - наружная поверхность створки с кольцом вы-
ростов, 4 — загиб створки и часть пояска, 5 — часть створки с внутренней поверхности, двугубый
вырост (стрелка). Японское море; 6, 7 — S. potamos (Weber) Hasle: 6 — створка, 7 — выросты с опо-
рами, загиб створки и вставочные ободки. 6 — оз. Хубсугул, 7 - р. Кама; 8—10 — S. subsalsum (А.С1.)
Bethge: 8 — структура загиба створки, 9 - структура поверхности створки, 10 - наружная поверх-
ность створки, выросты с опорами. Волга; 11—15 — Cymatotheca weissflogii (Grun.) Hendey: 11, 12 —
наружная поверхность створок, 13 - внутренняя поверхность створки, 14 - вырост с опорами
(стрелка) и крибрум ареол на внутренней поверхности, 15 - двугубый вырост на загибе внутренней
поверхности створки. Поздний миоцен,Сев.Сахалин, п-ов Шмидта (маямрафская свита). (7-5, 9,
12—15 - СЭМ, 6, 7, 8, 10 - ТЭМ, 77 - СМ; шкала: 7, 11-13 - 10 мкм, 2-10, 14, 15 - 1 мкм;
1-5 — ориг. И. В. Макаровой, 6—10 — ориг. С. И. Генкала, 77 — Шешукова-Поредкая, 1967,12—15 —
ориг. В. А. Николаева).
Таблица 59
1-10 - Porosira punctata (Jouse) Makar.: 1,2- створка в разных фокусах, 3 - створка с наруж-
ной поверхности, 4 — створка с внутренней поверхности, 5 - структура края и загиба створки,
6 — 2 крупных отверстия краевых выростов с опорами на наружной поверхности створки, 7 —
2 крупных отверстия створочных выростов с опорами, 8 - крибрум и выросты с опорами на внут-
ренней поверхности створки, 9 — двугубый вырост, 10 — край створки и слом через ареолы с внут-
ренней поверхности. Поздний миоцен, Сев. Сахалин (маямрафская свита). (7, 2 - СМ, 3-10 -
СЭМ; шкала: 1-4 — 10 мкм, 5-10 — 1 мкм; 1,2- ориг. И. В. Макаровой, 3—10 — Мака-
рова, 1988а).
Таблица 60
1-6- Porosira glacialis (Grun.) J0rg.: 7 — створка, 2 -форамены и отверстия двугубого выроста
на наружной поверхности створки, 3 — створка с внутренней поверхности, 4 — двугубый вырост
и велум с внутренней поверхности, 5 — выросты с опорами, 6 — слом через ареолы. 7 — голоцен,
ПО
Белое море, 2 - поздний плейстоцен, Чукотка (бухта. Провидения), 3, 4, 6 — голоцен. Охотское
море, 5 — поздний миоцен, Сев. Сахалин (маямрафская свита); 7-12 — Bacterosira fragilis Gran:
7 — структура створки, 8—12 — споры (5, 9 - наружная поверхность, 10 - внутренняя поверхность,
11 — наружные отверстия выростов с опорами, 12 — выросты с опорами с внутренней поверхности
створки). 7 - Чукотка (р. Яма), 8-12 - голоцен, Охотское море; 13 - Lauderia annulate CL: соеди-
нение двух панцирей. Индийский океан; 14,15 — Detonula confervacea (Cl.) Gran: спора {14 - створка
с наружной поверхности, 75 - створка с внутренней поверхности). 14 - голоцен. Охотское море,
75 - плейстоцен, север европ. части СССР; 16—18 - MinidiscuscowZcws Takano: 76, 77 - различные
створки, 18 - центральная часть створки с 3 выростами с опорами и двугубым выростом близ цент-
ра. Азовское море; 19 — М. trioculatus (Taylor) Hasle: створка. Охотское море. (7 — СМ, 2-6, 8-15 -
СЭМ, 7, 16-19 - ТЭМ; шкала: 1,3- 10 мкм, 2, 4-19 — 1 мкм; 7 - ориг. Р. Н. Джиноридзе,
2-6, 8-12, 14 - ориг. И. В. Макаровой и Т. Л. Невретдиновой, 7, 19 - ориг. В. И. Михайлова, 13 -
ориг. Т. Ю. Орловой, 75 — Лосева, Шешукова-Порецкая, 1985, 16-18 - ориг. С. И. Генкала
и И. В. Макаровой).
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
К СИСТЕМАТИЧЕСКОЙ ЧАСТИ*
А
aculeata Pr.-Lavr. (Thalassiosira) 60,65
aestivalis Gran (Thalassiosira) 70
albicans Sheshuk. (Melosira) 88
angstii Gran (Coscinodiscus) 72
angstii (Gran) Makar. (Thalassiosira) 72
angulata (Greg.) Hasle (Thalassiosira) 61
angulata Greg. (Orthosira) 61
anguste-lineata (A. S.) Fryx. et Hasle (Thalassio-
sira) 61
anguste-lineatus A. S. (Coscinodiscus) 61
annulata Cl. (Lauderia) 88
antarctica Comber. (Thalassiosira) 74
antiqua (Grun.) A. Cl. (Thalassiosira) 62
— var. septata Pr.-Lavr. 68
antiqua (Jous6) Gies. (Costopyxis) 49
antiqua (Jous6) (Stephanopyxis) 49
antiquus Grun. (Coscinodiscus) 62
— var. lapponica A. Cl. 62
- - f. inermis A. Cl. 62
- var. lapponicus A. Cl. 62
antiquus Forti et Schulz (Gladius) 56
appendiculate (Ehr.) (Stephanopyxis) 47
arctica (Grun.) Streln. et Nikolaev (Pyxidicula) 39
В
Bacillariophyta 35
Bacterosira Gran 87
baltica (Grun.) Ostf. (Thalassiosira) 70
barbadense Grev. (Sceletonema) 82
barbadensis Grev. (Creswellia) 39
barbadensis (Grev.) Grun. (Stephanopyxis) 39
barbadensis Grev. sensu Streln. (Stephanopyxis) 39
barbadensis (Grev.) Streln. et Nikolaev (Pyxidicula)
39
bathyomphalus Cl. (Coscinodiscus) 87
Biddulphia Gray sensu Van Heurck 53
bioculata (Grun.) Ostf. (Thalassiosira) 71
bioculatus Grun. (Coscinodiscus) 71
biseriata Streln. (Stephanopyxis) 39
biseriata (Streln.) Streln. et Nikolaev (Pyxidi-
cula) 39
borealis Gran (Lauderia) 88
bramaputrae (Ehr.) Hakansson et Locker (Thalas-
siosira) 7 5
- var. bramaputrae 75,76
— var. proximus (Makar.) Makar. 76
— var. septentrionalis (Grun.) Makar. 76
bramaputrae Ehr. (Stephanodiscus) 75
broschii Grun.(Stephanopyxis) 49
broschii (Grun.) Streln. et Nikolaev (Costo-
pyxis) 49
bulbosa Syvertsen (Thalassiosira) 76
C
Centrophyceae 35
charkoviana (Jouse) Streln. et Nikolaev (Pyxidi-
cula) 39
charcovianus (Jous6) (Stephanopyxis) 39
comicus Takano (Minidiscus) 90
concaviuscula Makar. (Thalassiosira) 70
condensata Cl. (Thalassiosira) 81, 82
conferta Hasle (Thalassiosira) 76
confervaceae (Cl.) Gran (Detonula) 87, 88
confervaceae Cl. (Lauderia) 88
confusa Makar. (Thalassiosira) 62
corona (Ehr.) Grun. (Stephanopyxis) 40
corona (Ehr.) Streln. et Nikolaev (Pyxidicula) 40
corona Ehr. (Systephania) 40
coronata Pr.-Lavr. (Thalassiosira) 62
coronifera (Pr.-Lavr.) Pr.-Lavr. (Thalassiosira) 62
costata (Gies.) Gies. (Costopyxis) 49
costata Grev. (Melosira) 83
costatum (Grev.) Cl. (Sceletonema) 83
costatus Gies. (Stephanopyxis) 49
Costopyxis Gies. 48
crenata Sheshuk. (Stephanopyxis) 40
crenata (Sheshuk.) Streln. et Nikolaev (Pyxidicula)
40
Creswellia Grev. 37
cruciate Ehr. (Dictiopyxis) 40
cruciata Ehr. (Pyxidicula) 37,40
cruciata Ehr. (Pyxidicula) 40
cruciatus (Ehr.) Kiitz. (Coscinodiscus) 40
Cyclotella Kiitz. 82
cylindraceum Pr.-Lavr. et Makar. (Sceletonema)
83
Cymatotheca Hendey 84
cystifera Gran (Detonula) 88
D
decipiens Grun. (Coscinodiscus) 61
decipiens (Grun.) Jdrg. (Thalassiosira) 63
decipiens (Grun.) J0ig. (Thalassiosira) 61
decipiens Grun. (Thalassiosira) 63
defecta (Streln.) Streln. et Nikolaev (Pyxidicula)
41
defectus Streln. (Stephanopyxis) 41
delicatissima Pr.-Lavr. (Thalassiosira) 63
Detonula Shutt 87
dissona (Shulz) Streln. et Nikolaev (Pyxidicula)
41
- - f. dissona 41
- - f.dissoniformis (Streln.) Streln. et Nikolaev
(Pyxidicula) 41
dissoniformis Streln. (Coscinodiscus) 41
dissonus Schulz (Coscinodiscus) 41
1 Курсивом выделены синонимы
112
Е
eccentrica (Ehr.) Cl. (Thalassiosira) 59,63
eccentricus Ehr. (Coscinodiscus) 63
- var. decipiens Grun. 63
- var. fasciculatus Hust. 63
edita Jouse (Stephanopyxis) 41
edita (Jouse) Streln. et Nikolaev (Pyxidicula) 41
Endictya Ehr. 51
F
fallax Meunier (Thalassiosira) 74
Fasciculigera Makar. 69
favus Ehr. (Triceratium) 53
ferox Grev. (Cresswellia) 41
ferox (Grev.) Streln. et Nikolaev (Pyxidicula) 41
fluviatilis Hust. (Thalassiosira) 81
- — f. mangrovii Manguin 81
fragilis Gran (Bacterosira) 87
frigida A. Cl. (Thalassiosira) 81
G
glacialis Cl. (Podosira) 86
glacialis Gran (Lauderia) 86
glacialis (Grun.) Jtfrg. (Porosira) 86
Gladius Schulz 54
gravida Cl. (Thalassiosira) 76
grunowii Gr. et St. (Stephanopyxis) 42
gruncwii Gr. et St; sensu Jousd (Stephanopyxis)
45,47
grunowii (Gr. et St.) Streln. et Nikolaev (Pyxidi-
cula) 42
guillardii Hasle (Thalassiosira) 77
H
heteromera Makar. (Thalassiosira) 64
hormoides (Mont.) Kiitz. (Podosira) 86
— var. glacialis Grun. 86
hustedtii Poretzky et Anissim. (Thalassiosira) 81
— var. vana Makar, et Pr.-Lavr. 81
hyalina (Grun.) Gran (Thalassiosira) 78
hyalinus Grun. (Coscinodiscus) 78
I
ignota Makar. (Thalassiosira) 64
incerta Makar. (Thalassiosira) 65
Inconspicuae Makar. 81
inermis Jouse (Stephanopyxis) 42
inermis (Jouse) Streln. et Nikolaev (Pyxidicula) 42
intermedia Makar. (Thalassiosira) 65
J
japonica Kiss. (Thalassiosira) 72
japonicus Castr. (Ethmodiscus>72
jouseanus Hajos (Gladius) 55
joynsonii A. S. (Stephanopyxis) 42
joynsonii (A. S.) Streln. et Nikolaev (Pyxidi-
cula) 42
К
kryophila (Grun.) J0rg. (Thalassiosira) 71
kryophilus Grun. (Coscinodiscus) 71
L
lacustris Grun. (Coscinodiscus) 75
— var. genuinus A. Cl. 75
- var. septentrionalis Grun. 76
lacustris (Grun.) Hasle (Thalassiosira) 75
- var. proximus (Makar.) Makar. 76
latimarginata Makar. (Thalassiosira) 65
Lauderia Cl. 88
lavrenkoi Jouse (Stephanopyxis) 42
lavrenkoi (Jous!) Streln. et Nikolaev (Pyxidicula)
42
leptopus Grun. (Coscinodiscus) 66
leptopus (Grun.) G. Fryx. et Hasle (Thalassiosira)
66
levanderi Van Goor (Thalassiosira) 66
lineatus Ehr. (Coscinodiscus) 66, 52
- — f. fossilis Jous£ 52
M
maeotica Pr.-Lavr. (Thalassiosira) 72
makarovae Mukchina (Thalassiosira) 66
mala Takano (Thalassiosira) 78
manifesta Sheshuk. emend. Makar. (Thalassiosira)
78
marginata Grun. (Stephanopyxis) 42
marginata (Grun.) Streln. et Nikolaev (Pyxidicula)
42
mamjamica Sheshuk. emend. Makar. (Thalassio-
sira) 67
maximus Hajos (Gladius) 55
megapora Grun. (Stephanopyxis) 43
megapore (Grun.) Streln. et Nikolaev (Pyxidicula)
43
Minidiscus Hasle 90
mirabilis (Forti et Schulz) Gies. (Costopyxis) 50
mirabilis Forti et Schulz (Stephanopyxis) 50
moelleri A. S. (Coscinodiscus) 43
— var. macroporus Grun. 43
moelleri (A. S.) Streln. et Nikolaev (Pyxidicula) 43
N
nativa Sheshuk. (Thalassiosira) 67
neogena Sheshuk. (Stephanopyxis) 45
nipponica Gran et Yendo (Stephanopyxis) 43
nipponica (Gran et Yendo) Streln. et Nikolaev
(Pyxidicula) 43
nodulolineata (Hendey) Hasle et G. Fryx. (Thalas-
siosira) 67
nodulolineatus Hendey (Coscinodiscus) 67
nordenskioeldii Cl. (Thalassiosira) 58,73, 79
О
oceanica Ehr. (Endictya) 51
oestrupii Ostf. (Coscinosira) 68
oestrupii (Ostf.) Hasle (Thalassiosira) 68
- var. plana Jousd 68
ordinaria Makar. (Thalassiosira) 72
orientalis Sheshuk. emend. Makar. (Thalassiosira)
79
ornata Pr.-Lavr. (Thalassiosira) 61
omata Schulz (Stephanopyxis) 51
113
р
pacifica Gran et Angst (Thalassiosira) 68
pacificus Hajos et Stradner (Gladius) 55
palmeriana Grev. (Cresswellia) 43
palmeriana (Grev.) Grun. (Stephanopyxis) 43
palmeriana (Grev.) Streln. et Nikolaev (Pyxidicula)
43
parva Pr.-Lavr. (Thalassiosira) 69
parvula Makar. (Thalassiosira) 79
pistillaris (Tubularia) 55
- var. grossepunctata Long, Fuge et Smith 55
Planktoniella Schtitt 85
polaris (Grun.) Streln. et Nikolaev (Pyxidicula) 45
poly acanthus Grun.(Coscinodiscus) 70
— var. baltica Grun. 7 0
poly chorda Gran (Coscinosira) 61
poly chorda (Gran ) J0rg. (Thalassiosira) 61
polychordus Gran (Coscinodiscus) 61
Porosira J0rg. 86
potamos (Weber) Hasle (Sceletonema) 84
potamos Weber (Microsiphona) 84
proschkinae Makar. (Thalassiosira) 80
— var. proschkinae 80
— var. spinulata (Takano) Makar. 80
proximus Makar. (Coscinodiscus) 76
pseudonana Hasle et Heimdal (Thalassiosira) 80
pulchella Takano (Thalassiosira) 68
pulvinar A. S. (Triceratium) 54
punctata (Jouse) Makar. (Porosira) 86
punctata Jous£ (Stephanopyxis) 45
punctata (Jouse) Streln. et Nikolaev (Pyxidicula)
45
punctata Jous6 (Thalassiosira) 86
punctigera (Castr. ) Hasle (Thalassiosira) 72
punctiger Castr. (Ethmodiscus) 72
punctiger (Castr.) H. Perag. (Coscinodiscus) 72
Pyxidicula Ehr. emend. Streln. et Nikolaev 37
Pyxidiculaceae Nikolaev 36
Pyxidiculales Nikolaev 36
— — f. schulzii 50
schulzii Stein. (Stephanopyxis) 50
— — f. nuda Streln. 51
— — f. schulzii Streln. 50
similis Sheshuk. (Stephanopyxis) 46
similis (Sheshuk.) Streln. et Nikolaev (Pyxidicula)
46
singularis Sheshuk. (Thalassiosira) 69
sol Wall. (Coscinodiscus) 85
sol (Wallich ) Schutt (Planctoniella) 85
speciosa (Schulz) Streln. (Pyxilla) 55
— — f.aculeolata Streln. 55
— — f. ellipsoidea Streln. 55
— — f. porata Streln. 55
speciosus Schulz (Gladius) 54
- - f. aculeolatus Streln. 55
— — f. ellipsoideus Streln. 55
- - f. poratus Streln. 55
- - f. speciosus 55
spinosa Jouse (Stephanopyxis) 46
spinosa (Jous6) Streln. et Nikolaev (Pyxidicula) 46
spinosissima Grun.(Stephanopyxis) 46
spinosissima (Grun.) Streln. et Nikolaev (Pyxidi-
cula) 46
spinulata Takano (Thalassiosira) 80
Stephanopyxis Ehr. 37,48
subsalina Pr.-Lavr. (Thalassiosira) 70
subsalsa A. Cl. (Melosira) 84
subsalsum (A. Cl.) Bethge (Sceletonema) 84
subsalsus (A. Cl.) Hust. (Stephanodiscus) 84
subtilis Ostf. (Podosira) 73
subtilis (Ostf.) Gran (Thalassiosira) 73
subtilis (Van Goor) A. Cl. (Stephanodiscus) 84
subtilissima Pr.-Lavr. (Detonula) 88
superba Grev. (Creswellia) 46
superba Grun. (Stephanopyxis) 46
superba (Grun.) Streln. et Nikolaev (Pyxidicula) 46
R
reticulata (Long, Fuge et Smith) Gies. (Costopyxis)
50
reticulata Long, Fuge et Smith (Stephanopyxis) 50
rotula Meunier (Thalassiosira) 81
S
satumii Lohmann (Thalassiosira) 82
Sceletonema Grev. 82
schenkii Kanaya (Stephanopyxis) 45
schenkii (Kanaya) Streln. et Nikolaev (Pyxidicula)
45
schulzii (Stein.) Gies. (Costopyxis) 50
— - f. nuda Streln. 51
- - f. ornata (Schulz) Streln. 51
Tangentales Makar. 58
tchemiae Manguin (Thalassiosira) 76
temperei Brun (Coscinodiscus) 85
tenera Pr.-Lavr. (Thalassiosira) 69
Thalassiosira 7 3
Thalassiosira Cl. 58
Thalassiosiraceae Lebour emend. Hasle 56
Thalassiosirales Gies, et Makar. 56
Thalassiosiropsidaceae Nikolaev 52
Thalassiosiropsis Hasle 52
Triceratiaceae Schutt 52
Triceratium Ehr. 52
trioculatus Taylor (Coscinodiscus) 90
trioculatus (Taylor) Hasle (Minidiscus) 90
trifulta Fryx. (Thalassiosira) 65
trochlea (Hanna) Streln. (Costopyxis) 51
trochlea Hanna (Trochosira) 51
turris Grev. et Arnott (Creswellia) 47
turris (Grev. et Arnott) Ralfs (Stephanopyxis) 47
114
- var. arctica Grun. 39
- var. cylindrus Grun. 47
— var. intermedia Grun. 47
— var. polaris Grun. 45
turris (Grev. et Arnott) Streln. et Nikolaev (Pyxidi-
cula) 46
- - f. cylindrus (Grun.) Streln. et Nikolaev
42,47
- f. turris 47
U
unguiculatum Grev. (Triceratium) 54
uralensis Streln. (Stephanopyxis) 47
uralensis (Streln.) Streln. et Nikolaev (Pyxidicula)
47
usatschevii Jous4 (Thalassiosira) 48
V
verecundus Mann (Coscinodiscus) 72
W
weissflogii A. S. (Stephanopyxis) 47
weissflogii (A. S.) Streln. et Nikolaev (Pyxidicula)
weissflogii Grun. (Euodia) 85
weissflogii (Grun.) G. Fryx. (Thalassiosira) 81
weissflogii (Grun.) Hendey (Cymatotheca) 84, 85
weissflogii (Grun.) Hust. (Hemidiscus) 85
weissflogii Grun. (Micropodiscus) 81
weyprechtii Grun. (Pyxidicula) 43,48
wittiana (Pant.) Hasle (Thalassfosiropsis) 52
Z
zabelinae (Jouse) Makar, et Moiss. (Pyxidicula) 48
zabelinae Jous6 (Thalassiosira) 48
СОДЕРЖАНИЕ
Предисловие...............................................................................
Общая часть
Морфология панциря диатомовых водорослей (Н. И. Караева, И. В. Макарова, В. А. Ни-
колаев) ...................................................................................... 7
Основные принципы классификации диатомовых водорослей (3. И. Глезер, Н. И. Караева) 22
Классификация диатомовых водорослей, принятая в настоящем издании (3. И. Глезер,
Н. И. Караева, И. В. Макарова, А. И. Моисеева, В. А. Николаев)............................... 31
Систематическая часть
Отдел Bacillariophyta (И. В. Макарова) ...................................................... 35
Класс Centrophyсеае {И. В. Макарова)....................................................... 35
Порядок Pyxidiculales (В. А. Николаев).................................................... 36
Сем. Pyxidiculaceae (Я А. Николаев) .................................................... 36
Роды: Pyxidicula Ehr., Costopyxis Gies. (Я. И. Стрельникова, В. А. Николаев)........ yj
Endictya Ehr. (И. В. Макарова) .................................................. 51
Сем. Thalassiosiropsidaceae (Я А. Ни колаев) ........................................... 52
Род Thalassiosiropsis Hasle (Я А. Николаев) ........................................... 52
Сем. Triceratiaceae (3. И, Глезер)...................................................... 52
Род Triceratium Ehr. (3. И. Глезер) ................................................... 53
Род неясного систематического положения в порядке: Gladius Schulz (Я. Я. Стрель-
никова) ............................................................................... 54
Порядок Thalassiosirales (3. И. Глезер, И. В. Макарова) .................................. 56
Сем. Thalassiosiraceae (Я. В. Макарова) ................................................ 56
Роды: Thalassiosira Cl. (Я. В. Макарова) .............................................. 58
Sceletonema Grev. (С. Я. Генкал, И. В. Макарова) ................................ 82
Cymatotheca Hendey (В. С. Шешукова-Порецкая, В. А. Николаев)..................... 84
Planktoniella Schutt (Я. Б. Цой, И. В. Макарова)................................. 85
Porosira Jtfig., Bacterosira Gran, Detonula Schutt, Lauderia Cl. (P. H. Джино pud зе,
И. В. Макарова).................................................................. 86
Minidiscus Hasle (Я. Я Макарова)................................................. 90
Литература................................................................................... 91
Объяснения к таблицам........................................................................ 98
Указатель латинских названий к систематической части........................................ 112
Таблицы 1-60 помешены между 88 и 89 стр.
Научное издание
ДИАТОМОВЫЕ ВОДОРОСЛИ СССР
ИСКОПАЕМЫЕ И СОВРЕМЕННЫЕ
Т о м II
В ы п. 1
Утверждено к печати
Ботаническим институтом им. В. Л. Комарова
Академии наук СССР
Редактор издательства Г. Т. Никифорова
Художник О. М. Разулевич
Технический редактор М. Э. Карлайтис
Корректоры Л. Я. Комм и Н. В. Романенкова
ИБ № 33284
Набор выполнен в издательстве на наб о рно-печатающем
автомате. Подписано к печати 09.11.88. М-38374. Формат
70 X 100 х/16- Бумага офсетная № 1. Печать офсетная.
Усл. печ. л. 9.75. + 5.2 (вкл.). Усл. кр.-от. 15.17. Уч.-изд. л.
17.79. Тираж 850. Тип. эак. № 518. Цена 2 р. 80 к.
Ордена Трудового Красного Знамени
издательство „Наука”. Ленинградское отделение.
199034, Ленинград, В-34, Менделеевская лин., 1.
Ордена Трудового Красного Знамени
Первая типография издательства „Наука”.
199034, Ленинград, В-34, 9 линия, 12.
/
КНИГИ ИЗДАТЕЛЬСТВА „НАУКА” МОЖНО ПРЕДВАРИТЕЛЬНО
ЗАКАЗАТЬ В МАГАЗИНАХ КОНТОРЫ „АКАДЕМКНИГА”, В МЕСТНЫХ
МАГАЗИНАХ КНИГОТОРГОВ ИЛИ ПОТРЕБИТЕЛЬСКОЙ КООПЕРАЦИИ.
Для получения книг почтой заказы просим направлять по адресу:
117192 Москва, Мичуринский пр., 12. Магазин „Книга—почтой” Централь-
ной конторы „Академкнига”.
197345 Ленинград, Петрозаводская ул., 7. Магазин „Книга—почтой”
Северо-Западной конторы „Академкнига”.
или в ближайший магазин „Академкнига”, имеющий
отдел „Книга-тючтой”:
480091 Алма-Ата, ул. Фурманова, 91/97 („Книга—почтой”);
370005 Баку, Коммунистическая ул., 51 („Книга—почтой”);
232600 Вильнюс, ул. Университете, 4;
690088 Владивосток, Океанский пр., 140 („Книга—почтой”);
320093 Днепропетровск, пр. Гагарина, 24 („Книга—почтой”);
734001 Душанбе, пр. Ленина, 95 („Книга—почтой”);
375002 Ереван, ул. Туманяна, 31;
644033 Иркутск, ул. Лермонтова, 289 („Книга—почтой”);
420043 Казань, ул. Достоевского, 53 („Книга—почтой”)
252030 Киев, ул. Ленина, 42;
252142 Киев, пр. Вернадского, 79;
252030 Киев, ул. Пирогова, 2;
252030 Киев, ул. Пирогова, 4 („Книга—почтой”);
277012 Кишинев, пр. Ленина, 148 („Книга—почтой”);
343900 Краматорск, Донецкой обл., ул. Марата, 1 („Книга—почтой”);
660049 Красноярск, пр. Мира, 84;
443002 Куйбышев, пр. Ленина, 2 („Книга—почтой”);
191104 Ленинград, Литейный пр., 57;
199034 Ленинград, Таможенный пер., 2;
194064 Ленинград, Тихорецкий пр., 4;
220012 Минск, Ленинский пр., 72 („Книга—почтой”);
103009 Москва, ул. Горького, 19а;
117312 Москва, ул. Вавилова, 55/7;
630076 Новосибирск, Красный пр., 51;
630090 Новосибирск, Морской пр., 22 („Книга—почтой”);
142284 Протвино,Московской обл., ул. Победы, 8;
142292 Пущино^Московской обл., МР „В”, 1;
620161 Свердловск, ул. Мамина-Сибиряка, 137 („Книга—почтой”);
700000 Ташкент, ул. Ю. Фучика, 1;
700029 Ташкент, ул. Ленина, 73;
700070 Ташкент, ул. Шота Руставели, 43;
700185 Ташкент, ул. Дружбы народов, 6 („Книга—почтой”);
634050 Томск, наб. реки Ушайки, 18;
634050 Томск, Академический пр., 5;
450059 Уфа, ул. Р. Зорге, 10 („Книга—почтой”);
450025 Уфа, Коммунистическая ул., 49;
720000 Фрунзе, бульв. Дзержинского, 42 („Книга—почтой”);
310078 Харьков, ул. Чернышевского, 87 („Книга—почтой”).
МАГАЗИН № 2 „КНИГА—ПОЧТОЙ” СЕВЕРО-ЗАПАДНОЙ КОНТОРЫ „АКАДЕМКНИГА”
ПРЕДЛАГАЕТ СЛЕДУЮЩИЕ КНИГИ, ВЫПУЩЕННЫЕ ИЗДАТЕЛЬСТВОМ „НАУКА”:
1. Артюшенко 3. Т., Федоров А. А. Атлас по описательной морфологии высших растений. Плод. -
Л., 1986. 6 р. 50 к.
2. Бондарцева М. А., Пармасто Э. X. Семейства гименохетовые, лахнокладиевые, кониофоровые,
щелелистниковые. (Определитель грибов СССР; Порядок афиллофоровые. Вып. 1). - Л., 1986.
1 р. 70 к.
3. Буданцев Л. Ю. История арктической флоры эпохи раннего кайнофита. - Л., 1983. 3 р. 40 к.
4. Бюллетень Главного ботанического сада. - М., 1985. Вып. 135.1 р. 20 к.
5. Владимир Николаевич Сукачев. Очерки, воспоминания современников. - Л., 1986. 2 р.
6. Гармаев Р. Б., Николаев С. М., Найдакова Ц. А. Зубчатка поздняя. - Новосибирск, 1982. 70 к.
7. Гецен М. В. Водоросли в экосистемах Крайнего Севера. - Л., 1985.1 р. 80 к.
8. Грумм-Гржимайло А. Л. В поисках растительных ресурсов мира. Некоторые научные итоги
путешествий академика Н. И. Вавилова. - Л., 1986.1 р. 20 к.
9. Декоративные растения для зеленого строительства. - Новосибирск. 1986. 65 к.
10. Козубов Г. М., Муратова Е. Н. Современные голосеменные: Морфолого-систематический обзор
и кариология. - Л., 1986. 2 р. 20 к.
11. Мазуренко М. Т. Вересковые кустарники Дальнего Востока: Структура и морфогенез. - М.,
1982. 2 р. 30 к.
12. Новости систематики высших растений. Т. 23. - Л., 1986. 3 р. 30 к.
13. Полевой В. В. Роль ауксина в системах регуляции у растений: Доложено на 44-ом ежегодном
Тимирязевском чтении 3.06.1983 г. - Л., 1986. 60 к.
14. Растительный покров высокогорий. - Л., 1986. 2 р. 90 к.
15. Растительный покров и естественные кормовые угодья Тувинской АССР. - Новосибирск, 1985.
3 р. 20 к.
16. Спрыгин И. И. Материалы к познанию растительности Среднего Поволжья. Научное наследство
Т. XI. - М., 1986. 9 р.
17. Степная растительность Сибири и некоторые черты ее экологии. - Новосибирск, 1982. 1 р. 50 к.
Заказы на книги просим направлять по адресу:
197345,Ленинград, Петрозаводская ул., 7
Магазин „Книга-почтой” Северо-Западной конторы „Академкнига”
ТАБЛИЦА 1
1ИГА”
:А”:
й. Плод. -
эфоровые,
-Л., 1986.
3 р. 40 к.
982. 70 к.
ные итоги
кий обзор
1ез. - М.,
жегодном
зек, 1985.
1следство.
1 р 50 к.
I. Заказ 518
ТАБЛИЦА 2
ТАБЛИЦА 3
•✓/л
ТАБЛИЦА 6
ТАБЛИЦА 7
ТАБЛИЦА 8
ТАБЛИЦА 9
ТАБЛИЦА 10
ТАБЛИЦА 11
ТАБЛИЦА 12
ТАБЛИЦА 13
ТАБЛИЦА 14
ТАБЛИЦА 15
ТАБЛИЦА 16
ТАБЛИЦА 17
II. Заказ 51 8
ТАБЛИЦА 18
ТАБЛИЦА 19
i г vtiHirsvi
or viiHirgvi
ТАБЛИЦА 22
ТАБЛИЦА 23
ТАБЛИЦА 24
ТАБЛИЦА 25
ТАБЛИЦА 26
ТАБЛИЦА 27
ТАБЛИЦА 28
ТАБЛИЦА 29
12
ТАБЛИЦА 30
ТАБЛИЦА 31
ТАБЛИЦА 32
15
ТАБЛИЦА 33
Ш.Заказ 518
ТАБЛИЦА 34
ТАБЛИЦА 35
ТАБЛИЦА 36
10
9
ТАБЛИЦА 37
ТАБЛИЦА 38
ТАБЛИЦА 39
ТАБЛИЦА 40
тилпил fi
ТАБЛИЦА 42
ТАБЛИЦА 43
ТАБЛИЦА 44
I
ТАБЛИЦА 45
ТАБЛИЦА 47
ГАБПИЦА 48
ТАБЛИЦА 49
I
IV Заказ 518
ТАБЛ ЩА 50
ТАБЛИЦА 51
ТАБЛИЦА S3
ТАБЛ!
ЦА 54
ТАБЛИЦА 55
ТАБЛИЦА 55 (продэлжение)
1й
12
ТАБЛИЦА 57 (продолжение)
11
I
ТАБЛИЦА 58
6
ТАБЛИЦА 58 (продолжение)
ТАБЛ4Ц^ 59
ТАБЛИЦА 60
9