/





























































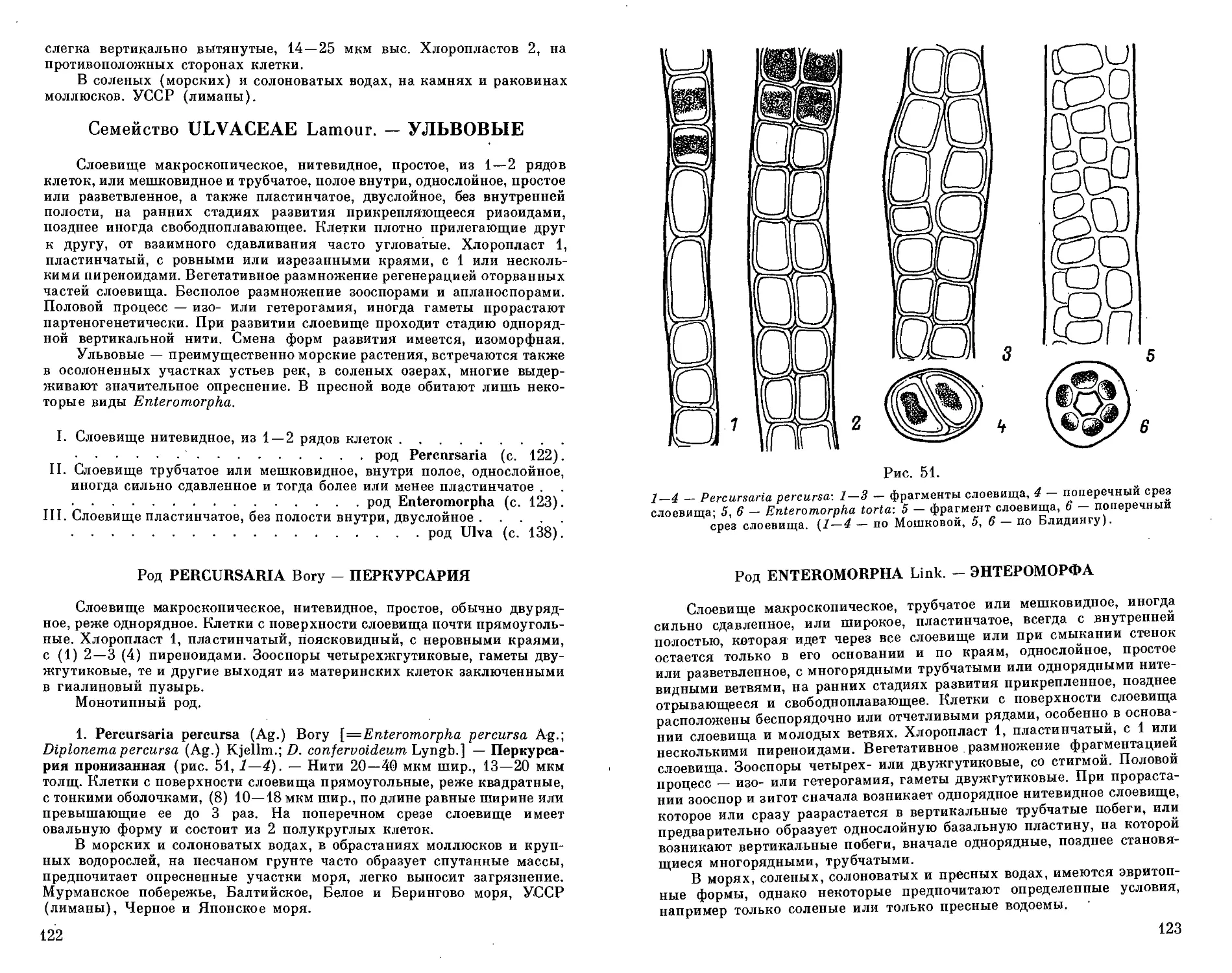










































































































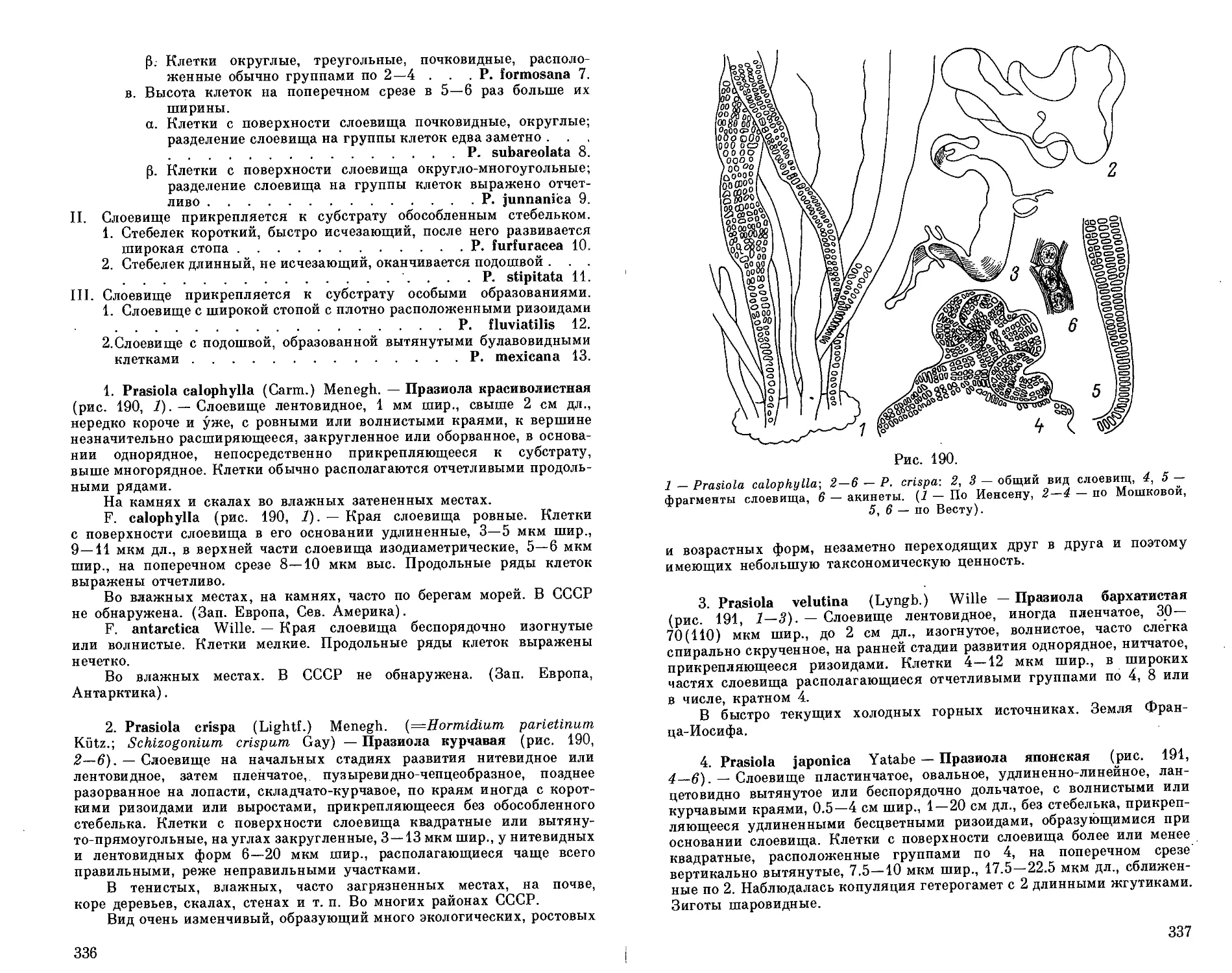












Автор: Голлербах М.М. Мошкова Н.А.
Теги: систематика растений растения биология растениеводство водоросли
Год: 1986
Похожие




Текст
ОПРЕДЕЛИТЕЛЬ
ПРЕСНОВОДНЫХ
ВОДОРОСЛЕЙ
СССР
ю
Зеленые
водоросли
АКАДЕМИЯ НАУК СССР
БОТАНИЧЕСКИЙ ИНСТИТУТ ИМ. В. Л. КОМАРОВА
ОПРЕДЕЛИТЕЛЬ
ПРЕСНОВОДНЫХ
ВОДОРОСЛЕЙ СССР
В ЧЕТЫРНАДЦАТИ ВЫПУСКАХ
Ответственн ый редактор
заслуженный деятель науки РСФСР
проф. М. М. ГОЛЛЕРБАХ
1
L—J
ЛЕНИНГРАД
ИЗДАТЕЛЬСТВО «НАУКА»
ЛЕНИНГРАДСКОЕ ОТДЕЛЕНИЕ
1986
ОПРЕДЕЛИТЕЛЬ
ПРЕСНОВОДНЫХ
ВОДОРОСЛЕЙ СССР
ВЫПУСК 10 (1)
Н. А. МОШКОВА, М. М. ГОЛЛЕРБАХ
ЗЕЛЕНЫЕ ВОДОРОСЛИ
КЛАСС УЛОТРИКСОВЫЕ (1)
ПОРЯДОК УЛОТРИКСОВЫЕ
CHLOROPHYTA: ULOTRICHOPHYCEAE,
ULOTRICHALES
Редактор К. Л. ВИНОГРАДОВА
И
ЛЕНИНГРАД
ИЗДАТЕЛЬСТВО «НАУКА»
ЛЕНИНГРАДСКОЕ ОТДЕЛЕНИЕ
1986
УДК 582.26/27 (0.83.71)
Определитель пресноводных водорослей СССР. Вып. 10(1). Мош-
кова Н. А., Г о л л е р б а х М. М. Зеленые водоросли. Класс улотрик-
совые (1). — Л.: Наука, 1986. — 360 с.
Очередной выпуск серии «Определитель пресноводных водорослей
СССР» охватывает первую часть обширной группы, составляющей
самостоятельный класс в отделе зеленых водорослей. Здесь представлены
все водоросли, входящие по современным классификациям в порядок
улотриксовых, которые являются в большинстве микроскопическими
многоклеточными или, реже, одноклеточными формами. В книге
сначала сообщаются краткие общие сведения, преимущественно по
морфологии этих водорослей, необходимые при пользовании Опреде-
лителем, затем даются таблицы для определения и диагнозы таксонов
всех рангов — от отдела до видов, разновидностей и форм. Для видовых
и внутривидовых таксонов приводятся краткие данные о распростра-
нении в пределах СССР. Включены также некоторые виды, которые
до сих пор в СССР еще не найдены, но могут быть обнаружены или
представляют большой таксономический интерес, для них указывается
район распространения вне СССР. Всего в Определителе описано
392 вида. Ил. — 195.
Рецензенты:
Ю. Е. ПЕТРОВ, Т. Ф. КОЗЫРЕНКО
О
2004000000-508
042 (02)-86
256-86-7
© Издательство «Наука», 1986 г.
ОТ РЕДАКЦИИ
Настоящий выпуск Определителя продолжает собой серию
пособий для научного распознавания водорослей, населяющих
наши континентальные водоемы и наземные субстраты, издава-
емую Ботаническим институтом им. В. Л. Комарова АН СССР
с 1951 г. В ней принимают участие альгологи не только института,
но и других учреждений Советского Союза. По первоначальному
замыслу, утвержденному в свое время редакционной коллегией,
весь Определитель был рассчитан на 14 выпусков, из которых
за истекшее время удалось опубликовать 11. Таким образом,
данный выпуск является двенадцатым по счету, но десятым
по нумерации общего плана издания. Он посвящен повсеместно
распространенным водорослям, выделяемым по принятой в дан-
ном Определителе системе в класс улотриксовых, но не всем
известным таксонам этой группы, а только первой части их
в объеме порядка собственно улотриксовых. Остальные 2 порядка
этого класса — микроспоровые и эдогониевые — будут опублико-
ваны отдельной книгой как вторая часть десятого выпуска.
В основу данного выпуска была положена соответствующая
часть книги киевского альголога Н. А. Мошковой, изданной
в серии украинских определителей пресноводных водорослей
(Н. А. Мошкова. Визначник пршноводних водоростей Укра1н-
ськоГ РСР. VI. Улотриксов! i кладофоров! водоростг Кшв, 1979).
Для нашего издания материалы этой книги были значительно
дополнены Н. А. Мошковой. Работа над русским текстом была
выполнена ею по договору о содружестве между Ботаническим
институтом им. В. Л. Комарова АН СССР (Ленинград), изда-
ющим' всю серию «Определитель пресноводных водорослей
СССР», и Институтом ботаники им. Н. Г. Холодного АН УССР
(Киев), взявшим на себя подготовку к печати рукописи и предо-
ставившим иллюстративный материал. Некоторые дополнения
и общая переработка текста и иллюстраций в связи с целями
и задачами общесоюзной серии определителей, существенно
отличающейся по ряду показателей от украинской серии, выпол-
нены М. М. Голлербахом.
Улотриксовые водоросли в том объеме, как они представлены
в данной книге, — группа обширная, по ряду разделов трудная
для таксономической идентификации изучаемого материала.
По современным классификациям сюда отнесены теперь некото-
5
рые роды и виды, ранее входившие в класс хлорококковых,
но характеризующиеся вегетативным клеточным делением,
нередко приводящим к образованию временных комплексов
клеток или коротких, легко распадающихся нитей. При определе-
нии таких водорослей нередко возникают затруднения даже
в отнесении их к одному из указанных классов. Кроме того,
к настоящему времени среди улотриксовых описано большое
количество монотипных родов, часто трудно различаемых, что
невольно заставляет думать о наличии здесь не самостоятельных
родовых таксонов, а лишь стадий развития ранее описанных
родов. Поэтому необходимо соблюдать большую осторожность
при определении и тщательно сопоставлять описания близких
форм. Критические замечания, которые могут при этом возник-
нуть, следует направлять в адрес редакции: 197022, Ленинград,
П-22, ул. проф. Попова, 2. Ботанический институт им. В. Л. Кома-
рова АН СССР, лаборатория альгологии.
Для общей ориентировки приводим перечень всех выпусков
серии (звездочкой отмечены опубликованные выпуски).
* Вып. 1. Общая часть (1951).
* Вып. 2. Синезеленые водоросли — Cyanophyta (1953).
* Вып. 3. Золотистые водоросли — Chrysophyta (1954).
* Вып. 4. Диатомовые водоросли — Bacillariophyta (1951).
* Вып. 5. Желтозеленые водоросли — Xanthophyta (1962).
* Вып. 6. Пирофитовые водоросли — Pyrrophyta (1954).
* Вып. 7. Эвгленовые водоросли — Englenophyta (1955).
* Вып. 8. Зеленые водоросли: Вольвоксовые — Chlorophyta: Volvoco-
phyceae (1959).
Вып. 9. Зеленые водоросли: Хлорококковые — Chlorophyta: Chioro-
coccophyceae.
* Вып. 10. Зеленые водоросли: Улотриксовые — Chlorophyta: Ulotriho-
phyceae (1), Ulotrichales. (Опубликована только первая
часть выпуска — 1986).
* Вып. 11. Зеленые водоросли: Десмидиевые — Chlorophyta: Desmidia-
les (2). (Опубликована только вторая часть выпуска —
1982).
Вып. 12. Зеленые водоросли: Мезотениевые, Гонатозиговые, Зигнемо-
вые — Chlorophyta: Mesotaeniales, Gonatozygales, Zygne-
matales.
* Вып. 13. Зеленые водоросли: Сифонокладовые, Сифоновые — Chloro-
phyta: Siphonocladophyceae, Siphonophyceae. Красные водо-
росли — Rhodophyta. Бурые водоросли — Phaeophyta
(1980).
* Вып. 14. Харовые водоросли — Charophyta (1983).
6
I. ОБЩ АН ЧАСТЬ
1. ОБЩАЯ ХАРАКТЕРИСТИКА
УЛОТРИКСОВЫХ ВОДОРОСЛЕЙ
Основные признаки улотриксовых водорослей. К водорослям
порядка Ulotrichales относятся многоклеточные организмы,
обычно прикрепленные, изредка пассивно плавающие, нит-
чатого, пластинчатого, трубчатого или разнонитчатого (гетеро-
трихального) строения, но встречаются также и одноклеточные
формы или группы из 2—4 и более клеток как производные
от нитей. На талломах нередко образуются щетинки или волоски,
иногда луковицеобразно вздутые у основания. Рост талломов
верхушечный (апикальный) или вставочный (интеркалярный).
Нити часто покрыты слоем слизи различной толщины, гомоген-
ной или фиброзной структуры. С выделением талломами боль-
шого количества слизи связано вторичное возникновение шаро-
видных, полушаровидных или лопастных форм.
Клетки талломов одноядерные, преимущественно со сплош-
ной оболочкой, тонкой или толстой, в последнем случае нередко
слоистой; оболочка из двух равных половинок встречается только
у рода Radlofilum. Цитоплазма прилегает к клеточным стенкам,
в середине клетки центральная вакуоля с клеточным соком.
Хлоропласт зеленого цвета разных оттенков, обычно один,
пристенный, у одних видов в форме широкого или узкого
цилиндра (кольца), цельного или продырявленного, у других —
в форме полуцилиндра или слегка согнутой пластинки или
диска, реже центральный, звездчатый. Пиреноиды имеются, один
или несколько, реже отсутствуют. Продукт ассимиляции — крах-
мал или маслоподобное вещество. Питание преимущественно
автотрофное, однако некоторые представители наряду с фото-
синтезом могут использовать и готовые органические вещества.
Вегетативное размножение распадением талломов на фраг-
менты или отдельные клетки с утолщенной оболочкой и большим
количеством запасных питательных веществ (акинеты). Бес-
полое размножение дву- или четырехжгутиковыми зооспо-
рами, реже апланоспорами или гипноспорами. Половой процесс
изогамный, гетерогамный и оогамный. У некоторых видов
7
установлено изоморфное или гетероморфное чередование форм
развития. Пальмеллевидное состояние наблюдается довольно
часто.
Улотриксовые водоросли — преимущественно микроскопи-
ческие формы, в то же время некоторые виды родов Stigeoclonium,
Drapamaldia, особенно Draparnaldiella, Enteromorpha и Ulva,
обладают крупными талломами (слоевищами), достигающими
в длину нередко нескольких дециметров.
Происхождение и родственные связи. Вопрос о происхожде-
нии улотриксовых водорослей решается по-разному. Существует
точка зрения, что возникновение нитчатого таллома зеленых
водорослей было связано с непосредственным переходом древних,
примитивных жгутиковых или коккоидных форм к прикреплен-
ным условиям существования: у прикрепленной клетки посте-
пенно возникла и закрепилась способность к неограниченному
росту ее верхушечной части с последующей сегментацией
протопласта и образованием поперечных перегородок. Однако
возможен и другой путь — возникновение порядка Ulotrichales
из хлоросарциновых водорослей, которые, будучи тесно связан-
ными с хлорококковыми, отличаются вегетативным делением
клеток, сопровождающимся образованием клеточных пакетов.
Процесс эволюции в группе улотриксовых шел в направлении
от простейшей формы многоклеточного таллома — нити к слож-
ной разнонитчатой структуре. В пределах нитчатого типа строе-
ния водорослей различают одно- и многорядную нить, последняя
возникает в результате определенного усложнения однорядной
нити. Производными многорядной нити являются, с одной
стороны, одно- и двуслойная пластинчатая форма тела, а с дру-
гой — трубчато-пузыревидная. Разнонитчатая форма строения
таллома обусловлена совмещением двух форм вегетативного
размножения клетки — деления и почкования, а также возник-
новением функционального расчленения таллома в пределах
однорядной нитчатой структуры. Однослойная пластинчатая
форма тела могла возникнуть также в результате редукции
прямостоячей части разнонитчатого таллома и усиленного разви-
тия ползучей по субстрату горизонтальной части или в результате
срастания обильно разветвленных в плоскости однорядных нитей.
Наконец, в пределах улотриксовых водорослей наблюдается как
временное состояние и пальмеллоидная структура, характе-
ризующаяся образованием большого количества слизи, объеди-
няющей неподвижные клетки, непосредственно между собой
не связанные.
В пределах класса Ulotrichophyceae порядок Ulotrichales
занимает центральное место и является прогрессивной линией
развития, давшей начало через разнонитчатую структуру всем
остальным 'чисто-зеленым растениям. Его родственные связи
с двумя другими порядками этого класса неясны. Наиболее
близок к нему несомненно порядок Microsporales, характеризую-
щийся более всего Н-образным строением клеточных оболочек,
8
так как некоторое подобие такого строения наблюдается и среди
типичных улотриксовых (род Binuclearia), но по форме хлоро-
пласта они резко различны. Третий порядок класса — Oedo-
goniales своеобразен во всех отношениях: по вегетативным
и особенно генеративным признакам, не имеющим аналогов
в порядке Ulotrichales. Поэтому наиболее обоснованным, по-види-
мому, следует считать предположение, что порядки Ulotrichales,
Microsporales и Oedogoniales являются тремя самостоятельными
ветвями, развившимися независимо друг от друга от какого-то
общего улотриксового предка.
, Распространение. Улотриксовые водоросли приспособились
к разнообразным условиям существования. Особенно широко
распространены они в пресных проточных и стоячих водах,
нередки также в солоноватых, щелочных, известковых и кислых
водах. Улотриксовые преимущественно прикрепленные формы,
подавляющее большинство — эпифиты. Их можно встретить
на листьях и стеблях водных цветковых растений (вегетирующих
и отмерших), на крупноклеточных нитчатых водорослях,
обработанной и необработанной древесине, а также на камнях,
раковинах моллюсков и т. п. Поселяясь на растительных и живот-
ных организмах, улотриксовые используют их лишь как место
для прикрепления. Некоторые улотриксовые являются эндофи-
тами, но, проникая внутрь других организмов, они сохраняют
самостоятельное автотрофное питание (виды родов Endoclo-
nium, Entocladia, Coleochaete и др.). Своеобразную экологическую
группу представляют собой эндолитофильные улотриксовые,
связанные с известковым субстратом. С одной стороны, это
сверлящие водоросли. В пресных водах СССР найдены Gomon-
tia codiolifera, внедряющаяся в известковые отложения, G. hol-
denii, живущая на раковинах перловницы (Unio), G. perforans,
сверлящая раковины живых беззубок (Anadonta). Внедряясь
в известковый субстрат, эти водоросли делают его рыхлым, легко
поддающимся воздействию физических и химических факторов.
Более многочисленной является группа туфообразующих водо-
рослей, переводящих растворенные в воде соли кальция в не-
растворимые и отлагающих их на поверхности своих талломов,
благодаря чему на субстрате возникают мелкие твердые известко-
вые полушаровидные подушечки. Располагаясь близко одна около
другой, подушечки вскоре сливаются в известковые корочки
с бугристой или зернистой поверхностью (виды родов Fridea,
Chlorotylium, некоторые виды рода Gongrosira и т. п.).
В подавляющем большинстве улотриксовые развиваются
в теплый период года, некоторые вегетируют лишь при низкой
температуре. Криофильные формы (виды родов Koliella, Raphido-
пета) обитают на поверхности слежавшегося снега или на неко-
торой глубине в его толще, а также на поверхности льда.
Незначительная часть улотриксовых приспособилась к вне-
водным условиям существования на влажных кирпичных и дере-
вянных стенах, оконных стеклах теплиц, коре хвойных и широко-
9
лиственных деревьев, полусгнившей древесине, влажных уча-
стках скал, камнях, железобетонных блоках, а также на влажной
почве и непосредственно в почве.
Практическое значение. Практическое значение улотрик-
совых водорослей многообразно и еще не полностью выяснено.
Наравне со многими другими группами водорослей они играют
определенную роль в процессах естественного самоочищения
вод, усиливая окисление и окончательную минерализацию орга-
нических веществ. Некоторые виды весьма чутко реагируют
на гидрохимические изменения среды и используются как
биологические индикаторы для быстрого определения степени
загрязненности воды.1 Четко выраженными а-мезосапробами
являются Stigeoclonium tenue, Enteromorpha intestinalis, к 0-ме-
зосапробам относятся Chlorhormidium subtile, Microthamnion
kuetzingianum, Chaetophora elegans, к олигосапробам — Ulothrlx
zonata, Draparnaldia glomerata, D. plumosa, Coleochaete pulvinata.
Некоторые из улотриксовых являются постоянными компонен-
тами обрастаний судов и различных гидротехнических соору-
жений, погруженных в воду. Развиваясь в большом количестве
на протяжении всего летнего периода, они способствуют кор-
розии металлических и бетонированных частей и гниению
деревянных строений. В почвах улотриксовые участвуют в про-
цессах накопления органических веществ, влияя тем самым
на повышение их плодородия.
Методы сбора и изучения. Сбор и изучение улотриксовых
водорослей проводятся теми же методами, которые применяются
и к водорослям других систематических групп.1 2 Поскольку
улотриксовые являются преимущественно прикрепленными фор-
мами, основное внимание при сборе материала необходимо
обращать на обрастания различных субстратов, погруженных
в воду (стебли и листья водных цветковых растений, сваи мостов,
бревна, ветви, камни, раковины моллюсков и т. д.). Налет с суб-
стратов можно соскабливать ножом или специальным скребком,
но при таком способе сбора материала теряется много интерес-
ных организмов, а главное нарушается картина взаимного раз-
мещения компонентов обрастания. Лучше всего собирать
водоросли вместе с субстратом.
Соскобленный налет, кусочки субстратов с водорослями,
сплетения нитчаток и т. п. необходимо поместить в стеклянные
баночки (желательно из тёмного стекла) с заранее налитым
в них 4%-ным раствором формальдегида. Этим исключается
1 Унифицированные методы исследования качества воды. Ч. III.
Методы биологического анализа воды. Приложение 2. Атлас сапробных
организмов. М., 1977. 176 с.
2 Голлербах М. М., Полянский В. И. Пресноводные водоросли
и их изучение. — В кн.: Определитель пресноводных водорослей СССР.
М., 1951, вып. 1 (Общая часть). 199 с.
-in
возможность заноса в исследуемый материал взвешенных в воде
организмов, нежелательных при обработке обрастаний. В склян-
ках оказываются лишь прикрепленные к субстрату и заведомо
осажденные среди них водоросли.
Обрастания раковин моллюсков необходимо фиксировать
вместе с самими раковинами, предварительно удалив моллюска,
так как для консервации его тела требуется более высокая
концентрация формальдегида (около 10 %), губительно дей-
ствующая на водоросли.
Аэрофитные водоросли, обитающие на коре деревьев,
листьях, хвое, полусгнивших пнях, обработанной и необработан-
ной древесине, а также на скалах, влажных камнях и т. ш,
следует собирать вместе с субстратом не только в специально
приготовленные для них бумажные пакеты, но и в баночки
с 4%-ным раствором формальдегида. Сбор и изучение напочвен-
ных и почвенных водорослей проводится по общепринятой
методике.3
Поскольку улотриксовые водоросли в большинстве своем
мелкие формы, то присутствие или отсутствие их в собранном
материале можно установить лишь с помощью прямого микро-
скопического анализа. В тех случаях, когда материал не фикси-
руется на месте сбора, предварительный просмотр проб необхо-
димо проводить в день сбора, чтобы ориентироваться в качествен-
ном составе организмов до наступления существенных измене-
ний — образования и выхода гамет, зооспор, перехода многих
форм в пальмеллевидное состояние и т. д. Материал вначале
необходимо просматривать под стереомикроскопом для общей
ориентировки и выборки наиболее удачных мест, а потом под
обычным световым микроскопом, используя большие увели-
чения.
При определении улотриксовых водорослей весьма часто
возникает необходимость в окрашивании материала. Для выявле-
ния и изучения слизи, покрывающей талломы многих водорослей,
слизистых влагалищ и обверток нитей или отдельных клеток
применяются слабые растворы анилиновых красок: метилено-
вого синего, генцианвиолета, анилинового синего, сафранина,
карболового фуксина, а также разведенная тушь. Пектиновые
вещества интенсивно окрашиваются метиленовым синим и саф-
ранином.
Для выявления целлюлозы в составе оболочек клеток приме-
няют хлор-цинк-иод (раствор Люголя). Ядра наиболее эффек-
тивно окрашивать нейтральным красным (нейтральрот), ген-
цианвиолетом, гематоксилином Бёмера. Пиреноиды окраши-
ваются в красный цвет 1%-ным раствором кислого фуксина либо
0.1%-ным спиртовым или крепким водным раствором эозина.
3 Голлербах М. М., Штина Э. А. Почвенные водоросли. Л., 1969.
228 с.
И
При изучении анатомического строения талломов крупных,
сложно организованных водорослей необходимо делать срезы
лезвием бритвы или на микротоме по общепринятой методике.4
Декальцинацию талломов улотриксовых водорослей лучше
проводить молочной кислотой. Для просветления материала
можно применять 5%-ный раствор едкого кали.
2. МОРФОЛОГИЯ УЛОТРИКСОВЫХ ВОДОРОСЛЕЙ
Строение клетки. Клетки улотриксовых разнообразны по
форме и величине. У подавляющего большинства они цилиндри-
ческие.
По соотношению длины и ширины их условно можно разде-
лить на 3 группы: изодиаметрические — длина приблизительно
равна ширине, в оптическом разрезе клетки имеют форму
квадрата; удлиненные или удлиненно-цилиндрические — длина
в несколько раз больше ширины, в оптическом разрезе клетки
имеют вид прямоугольника, вытянутого по продольной оси
таллома; укороченные или короткоцилиндрические — длина
в несколько раз меньше ширины, в оптическом разрезе клетки
имеют вид прямоугольника, вытянутого по поперечной оси
таллома. Цилиндрические клетки, вздутые в средней части,
называются бочонковидными. Довольно часто встречаются также
клетки шаровидные, яйцевидные, грушевидные, эллипсоидные,
веретеновидные, иголковидные, прямые или изогнутые, на концах
тупые или заостренные. Шаровидные клетки чаще всего можно
обнаружить у растений, перешедших в пальмеллевидное состо-
яние.
Оболочка клеток твердая, в основном цельная, глад-
кая, тонко- или толстостенная, обычно бесцветная, реже окра-
шенная соединениями железа в бурый цвет. Она состоит из двух
слоев: внутреннего — тонкого, целлюлозного и наружного —
более толстого, с примесью пектиновых веществ, гомогенного
или слоистого, с продольным или косым расположением слоев,
часто легко ослизняющегося. Двустворчатая оболочка с равными
створками, но не имеющими формы буквы Н, свойственна лишь
видам рода Radiofilum. Все клетки в нитях одеты снаружи еще
своеобразной обверткой (влагалищем) из пектиновых веществ,
хорошо заметной лишь у поперечных перегородок, обычно тонкой,
реже довольно толстой, гомогенной или отчетливо слоистой,
нередко легко ослизняющейся.
Расположение клеток в нитях может быть одно-
или многорядным. При однорядном расположении у одних видов
4 Наумов Н. А., Козлов В. Е. Основы ботанической микротехники.
М., 1954. 312 с.; Паушева 3. П. Практикум по цитологии растений.
М„ 1970. 255 с.
12
клетки чаще всего последовательно соприкасаются друг с другом,
у других сближены попарно. При многорядном расположении
клетки обычно теряют свою правильную геометрическую форму
от взаимного сдавливания и становятся многогранными (много-
угольными) .
Размеры клеток колеблются от 1—2 до 500 мкм
и более. Наиболее мелкие клетки встречаются у представителей
рода Stichococcus, наиболее крупные — у видов родов Draparnal-
dia и Drapamaldiella.
Цитоплазма обычного строения, располагается постен-
ным слоем, середина клетки занята вакуолями клеточного
сока. Величина вакуолей, число их и расположение внутри
цитоплазмы зависят главным образом от строения и характера
расположения хлоропластов.
Ядро преимущественно одно, по нескольку ядер встре-
чается лишь в старых клетках некоторых видов трентеполий
и в клетках Ulvella. Большей частью оно шаровидное или слегка
эллипсоидное, обычно незначительных размеров, малозаметное.
У одних видов ядро находится в пристенном слое цитоплазмы,
у других размещается в особом слизистом тяже, проходя-
щем посередине клетки или смещенном к одной из ее сторон.
Хлоропласты по форме кольцевидные, дисковидные,
пластинчатые и тогда прямые или изогнутые в форме полуцилин-
дра, реже лентовидно вытянутые и слегка спирально изогнутые,
с ровными или лопастными краями, иногда глубоко рассеченными
на дольки, цельные или продырявленные, нередко настолько
сильно, что принимают вид сеточки, выстилающей продольные
стенки клеток. Чашевидный хлоропласт встречается редко,
преимущественно у репродуктивных клеток. Обычно хлоропласт
один, иногда их несколько. По местоположению в клетках они
бывают периферическими, или постенными, и осевыми, или
центральными. Постенные хлоропласты большей частью тон-
кие, располагаются в периферическом слое цитоплазмы непосред-
ственно под оболочкой или на некотором расстоянии от нее.
Осевые хлоропласты обычно массивные, звездчатые, занимают
в клетке центральное положение. Хлоропласты улотриксовых
водорослей содержат хлорофилл а и Ъ и каротиноиды (каротин,
ксантофилл).
Внутри хлоропластов или на их поверхности располагаются
особые белковые тельца — пиреноиды. Различают пиреноиды
скорлупчатые, окруженные покровом из ассимилятов, и голые,
лишенные такого покрова. Улотриксовым водорослям свой-
ственны преимущественно скорлупчатые пиреноиды.
Продуктом ас си милиции является главным обра-
зом крахмал, который откладывается вокруг пиреноидов или
непосредственно в строме хлоропласта. У некоторых водорослей
(Microthamnion, Trentepohlia) образуется маслоподобное веще-
ство, бесцветное иди окрашенное.
Нитчатые формы. Нить является наиболее простой формой
13
многоклеточного таллома улотриксовых водорослей. При этом
у форм, живущих в свободном состоянии как в водной, так
и в воздушной среде, отсутствует морфологически уловимое
различие между вершиной и основанием нити {Binuclearia,
Geminella, Gloeotila, Radiofilum, Chlorhormidium и др.) и каждая
клетка в равной мере способна к делению надвое в одном
поперечном направлении. Рост таких водорослей происходит
вставочно (интеркалярно), а верхушечный (апикальный) рост
отсутствует. У прикрепленных форм (виды родов Ulothrix,
Uronema и др.) базальная и верхушечная клетки отличаются
от остальных клеток таллома. Базальные клетки утеряли способ-
ность к дальнейшему делению и обычно имеют форму довольно
вытянутого конуса, суженной частью обращенного к субстрату.
На нижнем конце этих клеток могут возникать пектозные или
целлюлозные образования (дермоиды) в виде полушаровидных
подушечек или дисковидных стоп с ровными или изрезанными
краями, служащие для более прочного прикрепления водо-
рослей к субстрату. Иногда базальная клетка вытягивается
в ризоид.
Верхушечные клетки большей частью полушаровидные,
тупые или разнообразно заостренные, прямые, реже клювовидно
изогнутые, иногда с утолщенной оболочкой на самой верхушке.
У прикрепленных нитевидных талломов верхушечный рост
совмещается со вставочным.
Среди улотриксовых встречаются дву- и многорядные
нитевидные формы. Они образуются в результате продольного
или косого вставочного деления клеток первоначально одноряд-
ных нитей. Двурядные нити можно наблюдать у видов рода
Cylindrocapsa. Однако многорядность здесь еще окончательно
не установилась и возникает лишь при определенных условиях
существования. Отчетливо многорядность выражена у взрослых
талломов Schizomeris leibletnii. Деление клеток здесь происходит
не в двух, а в трех направлениях, благодаря чему микро-
скопическая однорядная нить молодого растения постепенно
превращается в многорядный макроскопический нитевидный
таллом (слоевище) взрослого организма, состоящий из мелких
полигональных клеток.
Пластинчатые формы. Пластинчатые талломы улотриксовых
водорослей преимущественно однослойные, реже двуслойные,
изредка многослойные. Они образуются из одноряднонитчатых
форм в результате продольного деления их клеток (Ulva, Рга-
siola) или вследствие срастания простых или ветвящихся в одной
плоскости нитей (Protoderma, Epibolium, Coleochaete). В послед-
нем случае они часто имеют вид паренхимной ткани. Очертания
талломов разнообразны, они могут быть правильно дисковидными
или, чаще, неправильными. Двуслойные пластинчатые талломы
вследствие расхождения слоев иногда принимают вид мешка,
пузыря (Monostroma) или трубки, простой либо разветвленной
(Enterombrpha).
14
Разнонитчатые, или гетеротрихальные, формы. Разнонитча-
тая структура талломов является дальнейшим морфологическим
усложнением нитчатого тела водорослей и характерна для боль-
шого числа родов улотриксовых. Она обусловлена совмещением
двух форм вегетативного размножения клетки — деления и поч-
кования. ° Это совмещение привело не только к образованию
разветвленных в плоскости талломов, но и к возникновению
у них восходящих нитей, вначале простых, позднее разнообразно
разветвленных. При типичном гетеротрихальном строении тал-
лома должны быть хорошо выражены как горизонтальная, так
и вертикальная части (Stigeoclonium). В тех случаях, когда
одна из двух частей таллома недоразвивается или исчезает
вовсе, возникают различные модификации гетеротрихальной
структуры.
При более или менее полном исчезновении вертикальных
нитей таллом приобретает вид разнообразных однослойных
пластинок (Protoderma, Epibolium) или дисков (Coleochaete),
при недоразвитии или выпадении горизонтальной части интен-
сивно развитыми оказываются вертикальные нити (Draparnaldta,
Draparnaldiella, Cloniofora). Разветвленные формы обычно
кустиковидные, при этом у одних представителей нет резкого
различия между главными нитями (стволами) и боковыми вет-
вями (Stigeoclonium, Ireksokonia, Microthamnion), у других
эти различия выражены ярко (Draparnaldta, Draparnaldiella,
Draparnaldiopsis, Myxonemopsis). В последнем случае главные
нити, или стволы, состоят из одного ряда крупных цилиндри-
ческих или слегка бочонковидных клеток, в которых хлоропласт
занимает незначительную часть их длины, а боковые ветви
образованы мелкими клетками с хлоропластом, почти полностью
выстилающим клеточные стенки.
Для некоторых разветвленных водорослей характерно образо-
вание большого количества слизи, объединяющей нити в полу-
шаровидные или лопастные колонии (Chaetophora). Слизь обра-
зуется чаще всего в результате видоизменения пектиновых слоев
оболочки клеток или обверток нитей, бывает вязкой, эластичной,
крящевидной, гомогенной или слоистой (фиброзной), иногда
настолько нежной и так мало отличающейся от воды, что без
окраски ее заметить трудно.
Разветвленным формам улотриксовых свойствен в основном
боковой тип ветвления, протекающий преимущественно в форме
моноподиального ветвления. В этом случае главная нить (ствол)
не прекращает роста в длину и образует боковые ветви ниже
5 В первом случае происходит деление клетки надвое в одном
направлении, поперечном к продольной оси таллома; во втором клетка
образует вырост, который позже отделяется поперечной перегородкой
от образовавшей его клетки, но отпочковавшаяся клетка не теряет
связи с материнским организмом.
15
своей точки роста, обычно в восходящей (акропетальной)
последовательности. При типичном моноподии боковые ветви раз-
виты слабее главной нити, однако у ряда улотриксовых водорос-
лей вследствие быстрого дорастания дочерних клеток до размеров
материнских происходит резкое сдвигание в сторону боковой
ветвью главной нити, в результате чего возникает впечатление
дихотомии.
Значительно реже в пределах улотриксовых наблюдается
симподиальное ветвление, обычно также возникающее из моно-
подия. Главная нить при этом ветвлении сдвигается вбок, а ее
место занимает развившаяся боковая ветвь, растущая по направ-
лению главной нити. В дальнейшем эта ветвь в свою очередь
сдвигается вбок, ее заменяет новая боковая ветвь и т. д. Таким
образом, основная ось растения, по виду похожая на моноподий,
состоит из последовательного соединения осей разных порядков.
Довольно часто наблюдается также смешанное ветвление.
Расположение боковых ветвей на нитях водорослей может
быть очередным, противостоящим и мутовчатым. При очередном
размещении ветви бывают одиночными, если от любой клетки
нити отходит только по одной ветви, и парными, если от клетки
отходит с одной какой-либо стороны по две рядом возникшие
ветви. Если отходящие две ветви располагаются против друг
друга, ветви являются противостоящими, или супротивными.
При мутовчатом размещении от клетки нити отходят по 3—4
и более ветвей, расположенных по кругу. Ветви образуются
не на всех клетках нити. Первые 2—4 клетки, лежащие непосред-
ственно под верхушечной, лишены ветвей. Нередко в нитях
наблюдается более или менее правильное чередование клеток,
образующих ветви и лишенных их.
Верхушечные клетки главных нитей и многих веточек
некоторых разнонитчатых водорослей (сем. Chaetophoraceae)
часто оканчиваются бесцветными одноклеточными и заострен-
ными или многоклеточными и тупыми волосками, длина клеток
которых возрастает по мере удаления их от основания к вер-
шине, а ширина меняется незначительно. Содержимое клеток
волосков сильно редуцируется, особенно в самых верхушечных
клетках.
Одноклеточные волоски, нередко луковицеобразно вздутые
у основания, не отделяющиеся или отделяющиеся поперечными
перегородками от материнских клеток, образуются большей
частью на ползучих по субстрату нитях, а также однослойных
пластинчатых талломах. Возникают они вначале как бугорчатые
образования на оболочках клеток. По мере роста внешний слой
оболочки клеток на вершине бугорка разрывается, волосок же
образуется из внутреннего слоя оболочки, способного к дальней-
шему растяжению. В него из клетки переходит плазма без ядра.
У ряда водорослей (Coleochaete, Chaetosphaeridium) внешний,
прорванный слой оболочки образует вокруг волоска своеобразное
гомогенное или отчетливо слоистое влагалище.
16
Размножение. Разнообразные формы размножения улотрик-
совых водорослей могут быть сведены к трем типам: вегетатив-
ному, бесполому и половому.
Вегетативное размножение, в основе которого
лежит неограниченная способность водорослей к возобновлению
ими утраченных частей таллома или к развитию из отдельных
частей тела вновь растения, весьма широко распространено
среди улотриксовых. Простейшим способом является фрагмента-
ция — распадение тела водорослей на отдельные многоклеточные
или малоклеточные участки, которые затем снова разрастаются
до размеров взрослой особи. Фрагментация наиболее часто
встречается у форм с нитчатой (подпорядок Ulotrichineae) или
пластинчатой (подпорядок Ulvineae) структурой таллома и про-
исходит обычно без предшествующего этому процессу видимого
изменения в состоянии отдельных клеток. У некоторых водорос-
лей (Stichococcus, Koliella и др.) распадение нитей на отдельные
клетки является основным способом размножения.
Нередко, вне прямой зависимости от размножения, может
иметь место образование особых покоящихся стадий — акинет,
приспособленных к перенесению неблагоприятных условий.
В таких случаях отдельные клетки или все клетки некоторых
особей с нитчатой или разнонитчатой структурой таллома при-
нимают более или менее округлую форму, одеваются толстыми
оболочками (утолщение стенок клеток носит вторичный харак-
тер), в протопластах их откладывается большое количество
запасных питательных веществ (крахмал, масло). При благо-
приятных условиях эти клетки могут снова продолжить нормаль-
ное развитие без распадения нити. Такие акинеты получили на-
звание покоящихся акинет. В тех случаях, когда происходит
распадение нити на отдельные клетки и, следовательно, процесс
связан с размножением, они называются акинетами размноже-
ния. Подобные акинеты весьма часто встречаются у представите-
лей родов Ulothrix, Binudearia, Gloeotila, Chlorhormidium,
Cylindrocapsa, а также многих хетофоровых водорослей. После
периода покоя они непосредственно прорастают в новое растение
или сначала образуют зооспоры или апланоспоры.
Бесполое размножение осуществляется большей
частью подвижными четырех- или двужгутиковыми зооспорами,
реже неподвижными апланоспорами и гипноспорами. Клетки,
в которых образуются споры (спорангии), в большинстве случаев
ничем не отличаются от остальных вегетативных клеток таллома,
реже они имеют иную форму и более крупные размеры (Gongro-
sira, Trentepohlia и др.). У трентеполиевых водорослей спорангии
сидят на особых субспорангиальных клетках, прямых или крюч-
ковидно изогнутых. Зооспоры шаровидные, эллипсоидные или
грушевидные, одноядерные, лишенные обособленной оболочки,
с 2—4 жгутиками на переднем, более заостренном конце и хлоро-
пластом в расширенном заднем конце. Обычно они имеют
пульсирующие вакуоли и стигму. Зооспоры образуются одиночно
2 Зеленые водоросли. Улотриксовые
17
или, чаще, в числе нескольких из внутреннего содержимого
материнской клетки, выходят наружу через образующееся
в оболочке круглое или щелевидное отверстие, реже вследствие
ее общего ослизнения. В момент выхождения из материнской
клетки зооспоры иногда окружены тонким слизистым пузырем,
в скором времени расплывающимся (Ulothrix, Aphanochaete
и др.).
У многих улотриксовых водорослей вместо зооспор или
наряду с ними образуются неподвижные споры — апланоспоры.
Они также возникают из протопласта клетки в числе одной
или нескольких, но не вырабатывают жгутиков, а, приняв
шаровидную форму, одеваются собственной оболочкой, в образо-
вании которой оболочка материнской клетки не участвует.
Этим они существенно отличаются от акинет. Апланоспоры
освобождаются вследствие разрыва или ослизнения оболочек
материнских клеток, прорастают после некоторого периода покоя.
Апланоспоры с очень толстыми оболочками называются гипно-
спорами (Ulothrix). Они обычно принимают на себя функцию
покоящейся стадии.
Половое размножение осуществляется посред-
ством гамет, возникающих в неизмененных, слегка измененных
или значительно преобразованных клетках — гаметангиях. Под-
вижные гаметы обычного монадного строения, двужгутиковые.
У улотриксовых известны все три формы полового процесса —
изогамия, гетерогамия и оогамия. При изогамии, если обе копули-
рующие гаметы развиваются на одной особи, водоросли назы-
ваются гомоталличными, если копуляция возможна только между
гаметами, образовавшимися на разных особях. — гетероталлич-
ными. При этом гаметы морфологически совершенно подобны
друг другу и различия между ними являются чисто физиологиче-
скими (разного полового знака). Зигота одевается толстой оболоч-
кой, нередко со скульптурными украшениями, содержит большое
количество запасных питательных веществ и прорастает
сразу или после некоторого периода покоя. При прорастании
содержимое зиготы у большинства представителей делится
на 4 части, которые в виде зооспор выходят из оболочки зиготы
и прорастают в новые особи. Значительно реже гаметы разви-
ваются в новый организм без слияния, сами по себе, без обра-
зования зиготы. Такое размножение называется партеногенезом,
а споры, образующиеся из отдельных гамет, — партено-
спорами.
При гетерогамии обе гаметы также подвижны, но разли-
чаются между собой по величине и иногда по форме. Более
крупные гаметы (макрогаметы), часто менее подвижные, принято
считать женскими, меньшие по величине (микрогаметы) и более
подвижные — мужскими. У форм с гетерогамным половым про-
цессом наблюдается разная степень дифференциации мужских
и женских гамет. В одних случаях различия эти невелики,
и тогда говорят просто о гетерогамии, в других весьма значи-
18
тельны. Если женская гамета неподвижна и напоминает больше
яйцеклетку, то подвижная мужская становится сперматозоидом,
а половой процесс получает название оогамии. Гаметангии, в кото-
рых возникают яйцеклетки, называются оогониями, от вегета-
тивных клеток они отличаются как по форме, так и по величине.
Гаметангии, в которых образуются сперматозоиды, называются
антеридиями. Зигота, получившаяся в результате оплодотворения
яйцеклетки сперматозоидом, одевается толстой оболочкой и носит
название ооспоры.
У некоторых водорослей с гетерогамным половым процессом
наблюдается своеобразный переход от гетерогамии к оогамии.
Так, у видов рода Aphanochaete в оогонии развивается подвижная
четырехжгутиковая макрогамета. Выйдя из оогония, она некото-
рое время двигается, затем теряет жгутики, превращаясь в непод-
вижную яйцеклетку, которая и оплодотворяется сперматозоидом.
У видов рода Chaetonema неподвижная яйцеклетка при созрева-
нии выскальзывает наружу из оогония и остается лежать рядом
в виде темного неподвижного тела. Оплодотворение совершается
также вне оогония.
При типичной оогамии женские гаметы (яйцеклетки)
крупные, неподвижные и развиваются чаще всего по одной
в оогонии, мужские гаметы (сперматозоиды) мелкие, подвижные,
образуются в антеридии в большом количестве. Оогонии и анте-
ридии могут развиваться на одной особи, в этом случае водоросли
однодомные; если они развиваются на разных особях — двудом-
ные. Настоящая оогамия в порядке Ulotrichales встречается
в семействах Cylindrocapsaceae и Coleochaetaceae. У видов рода
Cylindrocapsa оогонии представляют собой увеличившиеся в раз-
мерах вегетативные клетки шаровидной или яйцевидной формы,
окруженные несколькими отстающими друг от друга слизистыми
оболочками; сперматозоиды образуются здесь в клетках той же
нити; при созревании в оболочке оогония возникает отверстие,
через которое проникают сперматозоиды. Однако наиболее ярко
выражена оогамия у видов рода Coleochaete. Здесь оплодотворен-
ная яйцеклетка одевается толстой бурой оболочкой, а соседние
с ней клетки дают короткие веточки, которые обрастают ооспору,
оплетая ее однослойной корой.
Смена форм развития. У некоторых улотриксовых водорослей
одна и та же особь может давать начало как зооспорам, так
и гаметам. В других случаях зооспоры и гаметы образуются
н’а разных особях, т. е. жизненный цикл водорослей включает
в себя как половую (гаметофит), так и бесполую (спорофит)
форму развития. Спорофит обычно диплоидный, т. е. имеет
в клетках удвоенный набор хромосом, гаметофит гаплоидный,
т. е. имеет одинарный набор хромосом. Это наблюдается в тех
случаях, когда редукционное деление (мейозис) происходит
при образовании спор (спорическая редукция) и часть жизнен-
ного цикла водоросли от зиготы до образования спор проходит
в диплофазе, а часть от споры до образования гамет в гаплофазе.
2*
19
Такой диплогаплофазный цикл развития свойствен видам родов
Ulva и Enteromorpha.
В пределах улотриксовых водорослей широко распространена
также зиготическая редукция, когда мейозис происходит при
прорастании зиготы. Диплоидной в этом случае оказывается
только зигота, весь остальной жизненный цикл протекает
в гаплофазе. Такой гаплофазный цикл развития характерен
для многих водорослей, принадлежащих к подпорядкам Ulotri-
chineae и Chaetophorineae.
Значительно реже встречается гаметическая редукция, когда
мейозис происходит при образовании гамет. В этом случае
гаплоидными являются только гаметы, весь остальной цикл
диплоидный. Такой цикл развития получил название дипло-
фазного.
В морфологическом отношении смена форм развития сво-
дится лишь к двум типам: изоморфной, когда обе формы —
бесполая (спорофит) и половая (гаметофит) морфологически
вполне подобны друг другу, и гетероморфной, когда они мор-
фологически различны. Изоморфная смена форм развития
известна у родов Ulva, Enteromorpha, гетероморфная — у рода
Monostroma. Гаметофит у видов этого рода многоклеточный,
пластинчатый, крупный, а спорофит одноклеточный, мелкий.
Смена форм развития, проявляясь обычно с большой правиль-
ностью, может резко нарушаться в зависимости от условий среды.
3. СИСТЕМА УЛОТРИКСОВЫХ ВОДОРОСЛЕЙ
До сего времени не существует единого мнения об объеме
порядка Ulotrichales и его месте в системе нитчатых зеленых
водорослей. Одними исследователями улотриксовые /рассматри-
ваются в широком объеме, и все же в ранге порядка, другими
в том же объеме, но в ранге класса. Наконец, многие..исследова-
тели принимают порядок Ulotrichales в узком объеме, как один
из порядков класса Ulotrichophyceae, но и в этом слу'чае неодно-
значно.
Расценивая таксономический ранг улотриксовых водорослей
как порядок, одни исследователи (Bohlin, 1901; Oltmanns, 1904;
Heering, 1914, и др.) включают в него лишь одноядерные, нитча-
тые, простые или разветвленные, пластинчатые и трубчато-
пластинчатые формы с изогамным или оогамным половым
процессом, клетки которых имеют цельную оболочку и содержат
пристенный или центральный хлоропласт. Представители дру-
гого направления (Smith, 1933, 1938, 1955; Fritsch, 1935; Iyengar,
1951; Randhawa, 1962, и др.) причисляют к порядку Ulotrichales
не только одноядерные, простые нитчатые или пластинчатые,
но и многоядерные простые или разветвленные нитчатые формы.
Некоторые (Prescott, 1951; Round, 1966) принимают порядок
Ulotrichales в очень узком объеме, относя к нему только
20
одноядерные, простые, однорядные или многорядные нитчатые
формы с изогамным половым процессом, клетки которых
обладают лишь цельной оболочкой. По мнению других (Мейер,
1961; Bourrelly, 1966, и др.), сюда надо включать нитчатые формы
не только с изогамным, но и оогамным половым процессом,
клетки которых имеют как цельную оболочку, так и состоящую
из двух Н-образных половинок.
В ранге класса улотриксовые водоросли впервые были рас-
смотрены Пашером (Pascher, 1931). В его представлении класс
Ulotrichineae включал 3 подкласса: Ulotrichlnae, Microsporinae,
Oedogoniinae. Подкласс Ulotrichlnae объединял 2 порядка: Ulo-
trichales и Chaetophorales. К порядку Ulotrichales относились
4 семейства: Ulotrichaceae, Cylindrocapsaceae, Ulvaceae, Blasto-
sporaceae, т. e. одноядерные, простые нитчатые, а также
пластинчатые, трубчато-пластинчатые формы с изогамным
и оогамным половым процессом, клетки которых, обладая
цельной оболочкой, содержали пристенный или центральный
хлоропласт. Выделение водорослей с разнонитчатой структурой
талломов в самостоятельный порядок Chaetophorales оправдыва-
лось, по мнению Пашера, их гетеротрихальным строением,
которому придавалось особое филогенетическое значение.
Взглядов Пашера придерживались в своих классификацион-
ных построениях Фрич (Fritsch, 1935), Н. Н. Воронихин
и Е. В. Шляпина (1949), Иенгар (Iyengar, 1951), Фотт (Fott,
1956, 1959, 1967), Буррелли (Bourrelly, 1966) и др., внося в то же
время существенные изменения и дополнения.
В соответствии с классификацией, принятой для данной
серии определителей, порядок Ulotrichales рассматривается здесь
как один из порядков класса Ulotrichophyceae (остальные два —
Microsporales и Oedogoniales). В таких рамках он представляет
собой, по нашему мнению, весьма четко очерченную системати-
ческую группу, которая по своим морфологическим особенностям
хорошо отличается от остальных систематических групп. Поря-
док Ulotrichales объединяет как нитчатые простые и разветвлен-
ные, так и пластинчатые и трубчатые структуры, которые могут
быть правильно поняты только как производные нити.
Основными признаками, определяющими объем порядка,
являются: преобладание многоклеточных талломов; одноядер-
ность нормальных вегетативных клеток; их деление с участием
материнской оболочки (истинное деление); как правило, один
пристенный кольцевидный, реже пластинчатый хлоропласт
(исключение составляют празиоловые водоросли, обладающие
центральным звездчатым хлоропластом); цельность оболочек
(в тех случаях, когда они построены из отдельных частей, то
в оптическом разрезе не имеют формы буквы Н); присутствие
у подвижных репродуктивных клеток 2—4 жгутиков.
Наиболее важными видовыми и внутривидовыми при-
знаками являются: форма и ширина вегетативных клеток; отно-
шение у них ширины к длине; строение оболочек вегетативных
21
клеток и зигот; положение, количество и форма хлоропластов;
наличие и характер ризоидных выростов, волосков, волосовидных
образований. Ни один из вышеуказанных признаков, рассматри-
ваемый изолированно, не может иметь абсолютного значения,
а потому любая таксономическая единица, в том числе и вид,
характеризуется определенным комплексом морфологических
признаков с учетом некоторых биологических особенностей.
Ниже приводится принятая нами система порядка. Характе-
ристики таксономических единиц помещены в соответствующих
разделах «Специальной части».
Порядок Ulotrichales
Подпорядок Ulotrichineae
Сем. Ulotrichaceae
Сем. Schizomeridaceae
Сем. Cylindrocapsaceae
Подпорядок Ulvineae
Сем. Monostromataceae
Сем. Ulvaceae
Подпорядок Chaetophorineae
Сем. Chaetophoraceae
Сем. Microthamniaceae
Сем. Trentepohliaceae
Сем. Aphanochaetaceae
Сем. Coleochaetaceae
Сем. Chaetosphaeridiaceae
Подпорядок Prasiolineae
Сем. Prasiolaceae
4. ОСНОВНЫЕ СИСТЕМАТИЧЕСКИЕ СВОДКИ
Арнольди В. М. Введение в изучение низших организмов.
Изд. 3-е. М„ 1925. 355 с.
Виноградова К. Л. Класс улотриксовые (Ulotrich.opb.y-
сеае). — В кн.: Жизнь растений. М., 1977, т. 3, с. 281 — 305.
Воронихин Н. Н., Шляпина Е. В. Водоросли. — В кн.:
Жизнь пресных вод СССР. М.; Л., 1949, т. 2, с. 357—477.
Мошкова Н. О. Ulotrichales. — В кн.: Визначник npicHO-
водних водоростей УкраТнськоТ РСР. КиТв, 1979, вып. 6.
498 с.
BourrellyP. Les algues d’eau douce. Initiation a la systema-
tique. T. I. Les algues vertes. Paris, 1966. 456 p.
F о tt B. Algenkunde. Jena, 1971. 581 S.
Fritsch F. E. The structure and reproduction of the algae.
Cambridge, 1956, vol. 1. 791 p.
Hazen T. E. The Ulotrichaceae and Chaetophoraceae of the
United States. — Mem. Torrey Bot. Club, 1902, vol. 11, N 2.
121 p.
22
lie ering W. Chlorophyceae III. — In: Pascher’s Siisswasser-
Flora Deutschlands, Osterreichs und der Schweiz. Jena, 1914,
H. 6. 250 S.
I s 1 a m N. A revision of the genus Stigeoclonium. — Beih. Nova
Hedwigia, 1963, H. 10. 164 S.
P r i n t z H. Chaetophorales. — In: Engler A., Prantl K. Die natiir-
lichen Pflanzenfamilien. Bd 3. Chlorophyceae. Leipzig, 1927.
411 S.
Printz H. Vorarbeiten zu einer Monographic der Trentepohlia-
ceen. — Nytt Mag. Naturv., 1939, Bd 80. 210 S.
Printz H. Die Chaetophoralen der Binnengewasser. — Hydro-
biologia, 1964, vol. 24, N 1—3, S. 1—9.
Ramanathan K. R. Ulotrichales. — In: I. C. A. R. Mono-
graphs on Algae. New Delhi, 1964. 188 p.
Smith G. M. The fresh-water algae of the United States. New
York, 1950. 719 p.
Starmach K. Chlorophyta III — Zielenice nitkowate. — In:
Flora sbodkowodna Polski. Warszawa; Krakow, 1972, t. 10.
750 s.
23
II. СПЕЦИАЛЬНАЯ ЧАСТЬ
Отдел CHLOROPHYTA-
ЗЕЛЕНЫЕВОДОРОСЛИ1
Класс ULOTRICHOPHYCEAE -
УЛОТРИКСОВЫЕ
Формы многоклеточные, реже упрощенные до немногих клеток
или даже одноклеточные, прикрепленные или свободноживущие. Таллом
микро- или макроскопический, в виде простых или разветвленных
нитей, у части форм с резким различием между основным стволом
и боковыми ветвями, нередко окруженный слизью, иногда в виде пласти-
нок или полых трубочек, редко в виде пакетиков клеток или отдельных
клеток. Клетки одноядерные. Размножение клеток путем типично веге-
тативного деления с участием материнской оболочки. Бесполое раз-
множение зооспорами и апланоспорами. Половой процесс изо-, гетеро-
и оогамный, у многих форм отсутствует.
I. Клеточная оболочка цельная.
1. Формы многообразные — от нитевидных до одноклеточных; при
делении клеток на оболочке никаких колпачков не возникает;
половой процесс — изо-, гетеро- и оогамия.
.......................пор. Ulotrichales (с. 19).
2. Формы только нитевидные; при делении клеток на верхнем конце
каждой из них на оболочке возникают характерные колпачки,
отмечающие количество произошедших делений; половой про-
цесс — только оогамия.......пор. Oedogoniales.1 2
II. Клеточная оболочка из 2 частей, имеющих в продольном разрезе
форму буквы Н и соединяющихся краями посередине каждой клетки;3
половой процесс отсутствует..пор. Microsporales.2
1 Характеристику отдела и деление его на классы см. в вып. 8.
2 Порядкам Oedogoniales и Microsporales будет посвящен вып. 10(2).
3 При установлении этого признака следует иметь в виду, что
он бывает не всегда ясно выражен и, кроме того, свойствен также
нитчатым формам из отдела Xanthophyta, которые, однако, легко узна-
ются по дисковидным хлоропластам.
24
Порядок ULOTRICHALES-
УЛОТРИКСОВЫЕ
Водоросли нитчатые, разнонитчатые, пластинчатые или мешко-
видные, реже одноклеточные, преимущественно прикрепленные, реже
живущие свободно, микроскопические или различимые невооруженным
глазом, нередко макроскопические. Окрашены обычно в зеленый цвет,
немногие виды оранжево-красные, у некоторых оранжево-красный цвет
наблюдается лишь на известных стадиях развития. Одноклеточные
формы обычно образуют короткие нити из небольшого числа клеток
или плотные скопления. Нитчатые талломы простые или разветвленные,
одно-, реже многорядные, часто покрытые слизью разной толщины
и структуры, при определенных условиях легко распадающиеся на
малоклеточные фрагменты и даже отдельные клетки. Разнонитчатые
водоросли вследствие усиленного или ослабленного развития стелющейся
или вертикальной частей таллома разнообразны по форме, имеют вид
кустиков или полушаровидных и лопастных образований или пластин.
Однослойные пластинчатые талломы неопределенных очертаний или
правильно дисковидные с радиально расходящимися рядами клеток.
Двуслойные пластинчатые талломы обычно плоские, листовидно рас-
ширенные или лентовидные, или вследствие расхождения слоев в сере-
дине с полостью и тогда пузыревидные, мешковидные или трубчатые
с однослойными стенками, простые или разветвленные. На поверхности
многих клеток образуются волоски (хеты) или щетинки. Клетки обычно
одноядерные. Хлоропласт пристенный, пластинчатый или дисковидный,
ноясковидный или кольцевидный, цельный или продырявленный, с пи-
реноидами или без них; центральный звездчатый хлоропласт встречается
только у празиоловых. Продукт ассимиляций — крахмал или масло-
подобное вещество. Вегетативное размножение разрывом талломов на
фрагменты или акинетами. Бесполое размножение зооспорами и аплано-
спорами. Половой процесс изогамный, гетерогамный, оогамный. Некото-
рые виды характеризуются правильной сменой половой и бесполой
форм развития. Известны пальмеллевидное состояние и некоторые другие
стадии покоя.
Улотриксовые водоросли широко распространены в природе. Пре-
обладающее большинство обитает в бентосе пресных водоемов, значи-
тельно меньшее число встречается в солоноватых водоемах и морях,
а также в почве и на наземных субстратах, часть — типичные эпифиты,
встречаются и эндофиты.
1. Организмы типично многоклеточные, нитчатые или пластинчатые,
не распадающиеся на отдельные клетки.
1. Талломы нитчатые.
А. Нити не разветвленные, без волосков и щетинок.
а. Нити однорядные, реже многорядные...............
..........подпорядок Ulotrichineae (частично) (с. 26).
б. Нити двурядные, реже однорядные.................
.....................подпорядок Ulvineae (частично) (с. 112).
Б. Нити разветвленные, с одноклеточными или многоклеточными
волосками или щетинками, реже без них..................
..........подпорядок Chaetophorineae (частично) (с. 142).
2. Талломы пластинчатые.
А. Талломы микроскопические, однослойные, правильно диско-
видные или разнообразных очертаний, часто с волосками
25
или щетинками........................................
..........подпорядок Chaetophorineae (частично) (с. 142).
Б. Талломы макроскопические, одно- или двуслойные, волоски
или щетинки отсутствуют.
а. Хлоропласт пристенный, пластинчатый; талломы пузыре-
видные, или трубчатые с однослойными стенками и по-
лостью в середине, или листовидно расширенные, дву-
слойные, без полости в середине ...................
..........подпорядок Ulvineae (частично) (с. 112)
б.- Хлоропласт центральный, звездчатый; талломы пластин-
чатые, лентовидные, однослойные, реже нитевидные .
.....................подпорядок Prasiolineae (с. 333)
II. Организмы одноклеточные или образующие короткие нити, легко
распадающиеся на отдельные клетки............................
.................подпорядок Ulotriehineae (частично) (с. 26)
Подпорядок ULOTRICHINEAE - УЛОТРИКСОВЫЕ
Таллом нитевидный, реже одноклеточный. Нити простые, одно-,
реже многорядные, за немногими исключениями микроскопические,
преимущественно прикрепленные, реже живущие свободно; одноклеточ-
ные формы слагаются в короткие, легко распадающиеся нити, иногда
объединенные общей слизью. Клетки одноядерные, с тонкими или
утолщенными оболочками; базальные и верхушечные клетки у при-
крепленных нитей обычно отличаются по форме от остальных клеток.
Хлоропласт пристенный, пластинчатый или дисковидный, большей
частью в виде полуцилиндра, или поясковидный до отчетливо кольце-
видного, располагающийся только в средней части клеток или почти
полностью выстилающий ее продольные стенки, цельный или продыряв-
ленный, с 1 или несколькими пиреноидами, реже без них. Вегетативное
размножение распадением нитей или акинетами. Бесполое размножение
зооспорами, апланоспорами и гипноспорами. Половой процесс пре-
имущественно изогамный, реже гетерогамный или оогамный.
Большей частью это пресноводные организмы, обычно встречаю-
щиеся в обрастаниях на водных цветковых растениях, крупных нитчатых
водорослях, древесном и каменистом субстратах, реже свободно лежащие
на дне водоемов или плавающие *на поверхности или в толще воды.
Некоторые из них обитают на коре деревьев, влажных скалах, стенах
и других наземных субстратах, а также в почве и на ее поверхности.
Водоросли всех семейств, принадлежащие к подпорядку улотрик-
совых, на начальных фазах развития не имеют типичного для них
строения и поэтому очень сходны между собой. При определении
это следует иметь в виду и внимательно просматривать значительную
часть материала, отыскивая кроме ювенильных взрослые формы. При
определении решающее значение имеют форма, строение .и положение
хлоропласта, форма базальных и верхушечных клеток, а также способ
прикрепления водорослей.
I. Нити всегда однорядные, прикрепленные или пассивно плавающие;
имеются также одноклеточные свободноживущие формы . . . .
.......................................сем. Ulotrichaceae (с. 27).
II. Нити во взрослом состоянии многорядные, на ранних стадиях одно-
рядные, прикрепленные, но легко отрывающиеся от субстрата и
пассивно плавающие; одноклеточные формы отсутствуют.
26
1. Нити большей частью двурядные, прикрепляющиеся к субстрату
целлюлозной подошвой; клетки шаровидные или эллипсоидные,
иногда короткоцилиндрические, с толстыми слоистыми оболоч-
ками, часто попарно сближенные; верхушечная клетка округлая
.................................сем. Cylindroeapsaeeae (с. 107).
2. Нити многорядные, прикрепляющиеся к субстрату конусовидной,
нередко очень вытянутой базальной клеткой, реже ризоидными
выростами; клетки цилиндрические, с тонкими оболочками, плотно
соединенные между собой; верхушечная клетка заостренная .
...............................сем. Sehizomeridaeeae (с. 110).
Семейство ULOTRICHACEAE Kiitz. —
УЛОТРИКСОВЫЕ
Талломы многоклеточные, в виде простых нитей, реже одноклеточ-
ные, неподвижные. Нити обычно однорядные, длинные или короткие,
прямые или разнообразно изогнутые, одинаковой ширины на всем
протяжении, часто легко распадающиеся на фрагменты или отдельные
клетки, прикрепленные или свободноживущие, иногда окруженные
довольно толстым слизистым влагалищем гомогенной или фиброзной
структуры, реже влагалище трубчатое, тонкостенное, без наружного
слизистого покрова. Клетки одноядерные, короткие или длинные, циь
линдрические, эллипсоидные, шаровидные, бочонковидные, веретено-
видные, иголковидные, прямые или изогнутые, на концах тупые или
заостренные, иногда несколько неправильные, со сплошными (не дву-
створчатыми) 4 целлюлозными, довольно толстыми, нередко слоистыми,
легко ослизняющимися оболочками. Хлоропласт пристенный, кольцевид-
ный, поясковидный или дисковидный, цельный или продырявленный,
с 1 или несколькими пиреноидами, иногда без пиреноидов, с ровными
или изрезанными краями, частично или почти полностью выстилающий
продольные стенки клеток. Все клетки нитей, за исключением базальной,
образующей целлюлозный дермоид или ризоидный вырост, способны
к делению надвое. Вегетативное размножение распадением нитей на
малоклеточные отрезки, а также акинетами. Бесполое размножение
дву- или четырехжгутиковыми зооспорами, апланоспорами и гипно-
спорами. Половой процесс преобладающе изогамный,. гаметы двужгути-
ковые, гетерогамия указана лишь для некоторых видов рода Chlorhor-
midium, оогамия обнаружена только у Koliella longiseta. Известно паль-
меллевидное состояние.
Преимущественно пресноводные водоросли, встречающиеся в об-
растаниях различных субстратов или оторванными в планктоне, а также
в поверхностной пленке ила. Аэрофитные представители обитают на коре
деревьев, влажных кирпичных и деревянных стенах, оконных стеклах
и т. п., некоторые виды часто встречаются на поверхности почвы
и в почве.
I. Нити всегда многоклеточные, не распадающиеся на отдельные клетки.
4 Исключение составляет род Radiofilum, представители которого
обладают клеточными оболочками, состоящими из. двух простых поло-
винок, не складывающихся в Н-образные фигуры.
27
1. Поперечные перегородки клеток одинаковой толщины.
А. Нити прикрепленные; верхушечная и базальная клетки нитей
по форме отличаются от остальных клеток.
а. Нити прикрепляются непосредственно базальной клеткой;
верхушечная клетка преимущественно округленная .
.........................род Ulothrix (частично) (с. 3f).
б. Нити прикрепляются дермоидом; верхушечная клетка
к вершине суженная.
а. Дермоид в виде полушаровидной подушечки или круп-
ного диска, с ровными или лопастными краями; верху-
шечная клетка вытянутая, явственно заостренная . .
..............................род Uronema (с. 43).
Р- Дермоид в виде небольшой плоской стопы на конце
ножки (стебелька); верхушечная клетка не вытянутая,
слегка конусовидная . . . род Hormidiella (с. 48).
Б. Нити свободноживущие, реже прикрепленные; все клетки
нитей по форме одинаковые.
а. Оболочка клеток обычного строения, без пектиновых колец
или муфт у поперечных перегородок,
а. Нити без слизистого влагалища.
* Нити прямые или слегка изогнутые, но не спи-
рально.
+ Хлоропласт с пиреноидом.
X Хлоропласт пластинчатый, дисковидный;
нити иногда коленчато согнутые .
род Chlorhormidium (частично) (с. 49).
XX Хлоропласт поясковидный или кольце-
видный; нити без коленчатых изгибов
. . . род Ulothrix (частично) (с. 31).
+ + Хлоропласт без пиреноидов.
X Клетки преимущественно длинные, с тон-
кими оболочками, нити всегда однорядные;
в стоячих и проточных водах..............
. . . род Gloeotila (частично) (с. 58).
XX Клетки короткие, с толстыми оболочками,
нити одно- и двурядные; на коре деревьев,
влажной почве ..............................
. . . род Hormidiopsis (частично) (с. 63).
** Нити спирально изогнутые.
+ Витки спирали сжатые, клетки короткоцилинд-
рические или короткоэллипсоидные . . . .
.род Chlorospira (с. 65).
+ + Витки спирали растянутые, клетки удлиненно-
цилиндрические или удлиненно-эллипсоидные
.......................род Gloeotila (частично) (с. 58).
р. Нити со слизистым влагалищем.5
* Влагалище гомогенное.
+ Влагалище хорошо заметное без окраски.
X Хлоропласт с пиреноидом.
О Хлоропласт пластинчатый, дисковидный.
5 В некоторых случаях наличие влагалища бывает столь неяв-
ственным, что для его установления требуется применение окраски
или туши.
28
Л Клетки только эллипсоидные, рас-
положенные во влагалище попе-
речно, разобщенные или сближен-
ные в группы по 2—4 ....
род Hormidiopsis (частично) (с. 63).
дд Клетки эллипсоидные или цилинд-
рические, расположенные во влага-
лище продольно.
§ Нити обычного строения, с плот-
но соединенными клетками .
..................род Chlor-
hormidium (частично) (с. 49).
§§ Нити необычного строения, с
разобщенными клетками — рав-
номерно поодиночке или по-
парно ......................род
Geminella (частично) (с. 65).
ААЛ Клетки шлемовидно-шаровидные,
разобщенные или соединенные друг
с другом слизистым тяжом .
род Radiofilum (частично) (с. 72).
00 Хлоропласт поясковидный или кольце-
видный .................................
. . . род Ulothrix (частично) (с. 31).
XX Хлоропласт без пиреноидов, лентовидный,
слегка спирально изогнутый.................
. . . . род Gloeotila (частично) (с. 58).
+ + Влагалище плохо заметное и выявляется только
при окраске или в туши.
X Хлоропласт поясковидный или кольцевид-
ный, с 1 или несколькими пиреноидами
. . . род Ulothrix (частично) (с. 31).
XX Хлоропласт пластинчатый, дисковидный,
без пиреноидов ............................
. . . род Gloeotila (частично) (с. 58).
** Влагалище фиброзное.
+ Клетки цилиндрические, бочонковидные или
вытянуто-эллипсоидные, со сплошной оболоч-
кой, плотно соединенные друг с другом.
X Клетки соприкасаются друг с другом всей
поверхностью поперечных перегородок;
хлоропласт поясковидный или кольцевид-
ный, широкий, с пиреноидом ....
. . . род Ulothrix (частично) (с. 31).
XX Клетки соприкасаются друг с другом не-
значительной частью поперечных пере-
городок; хлоропласт лентовидный, узкий,
слегка спирально изогнутый, без пире-
ноидов ....................................
. . род Gloeotila (частично) (с. 58).
+ + Клетки шаровидные или эллипсоидные, с дву-
створчатой оболочкой, разобщенные или со-
единенные друг с другом только слизистыми
тяжами.........................................
. . . . род Radiofilum (частично) (с. 72).
*** Влагалище трубчатое, тонкостенное, нежное;
клетки удлиненно-цилиндрические, с округлыми
29
концами, разобщенные..........................
.................................род Geminellopsis (с. 76).
б. Оболочка клеток с пектиновыми кольцами или муфтами
у поперечных перегородок . . род Ulotriehopsis (с. 79).
2. Поперечные перегородки клеток разной толщины . . . .; .
...................................род Binuclearia (с. 77).
II. Нити всегда малоклеточные, легко распадающиеся на фрагменты
или отдельные клетки, реже отдельные клетки преобладают.6
1. Нити и клетки лишены слизи (или слизь тонким слоем покрывает
оболочку).
А. Клетки цилиндрические, без кольцевых утолщений на обоих
концах,
а. Свободные концы нитей не вытянутые, округлые,
а. Клетки прямые.
* Хлоропласт с пиреноидом.
+ Хлоропласт 1.
X Хлоропласт поясковидный; водные формы
род Gloeotillopsis (с. 80).
XX Хлоропласт дисковидный; аэрофитные
формы..................
род Chlorhormidium (частично) (с. 49).
+ + Хлоропластов несколько.......................
.............род Heterotrichopsis (с. 81).
* * Хлоропласт без пиреноидов....................
. . . род Stichococcus (частично) (с. 81).
0. Клетки изогнутые.
* Клетки дуговидно изогнутые, их оболочка покрыта
тонким слоем слизи . . . род Chodatia (с. 84).
* * Клетки слегка полукругло или S-образно изо-
гнутые, их оболочка лишена слизи.................
. . . род Stichococcus (частично) (с. 81).
б. Свободные концы нитей вытянутые, заостренные или при-
тупленные .......................род Raphidonema (с. 87).
Б. Клетки цилиндрические, с кольцевидными утолщениями на
обоих концах и коническими выростами на поперечных стен-
ках ...................................род Catena (с. 89).
В. Клетки эллипсоидные или шаровидные.......................
род Stichococcus (частично) (с. 81).
Г. Клетки веретеновидные, иголковидные, прямые или изогну-
тые, щетинковидно заостренные на одном или обоих концах
...........род Koliella (с. 90).
2. Нити и клетки заключены в слизь.
А. Преобладает нитчатая стадия...............................
...........................род Geminella (частично) (с. 65).
Б. Преобладает одноклеточная стадия с образованием слизистых
колоний.
а. Клетки цилиндрические, с округлыми концами; слизистые
колонии преимущественно округлые.........................
....................................род Fottea (с. 101).
б. Клетки веретеновидные или цилиндрические, заостренные
на обоих концах; слизистые колонии округлые, эллипти-
6 В группу одноклеточных форм входит еще род Marvania Hind.,
но его строение настолько своеобразно, что относить его к улотриксовым,
на наш взгляд, преждевременно.
30
ческие, веретеновидные или в виде цилиндрических тяжей
.............................род Elakatothrix (с. 101).
Род ULOTHRIX Kutz. - УЛОТРИКС
Нити однорядные, простые, иногда окруженные слизистым гомо-
генным или фиброзным влагалищем, в некоторых случаях значительно
превосходящим ширину клеток, прикрепляющиеся к субстрату непо-
средственно базальной клеткой или свободноживущие. Клетки цилиндри-
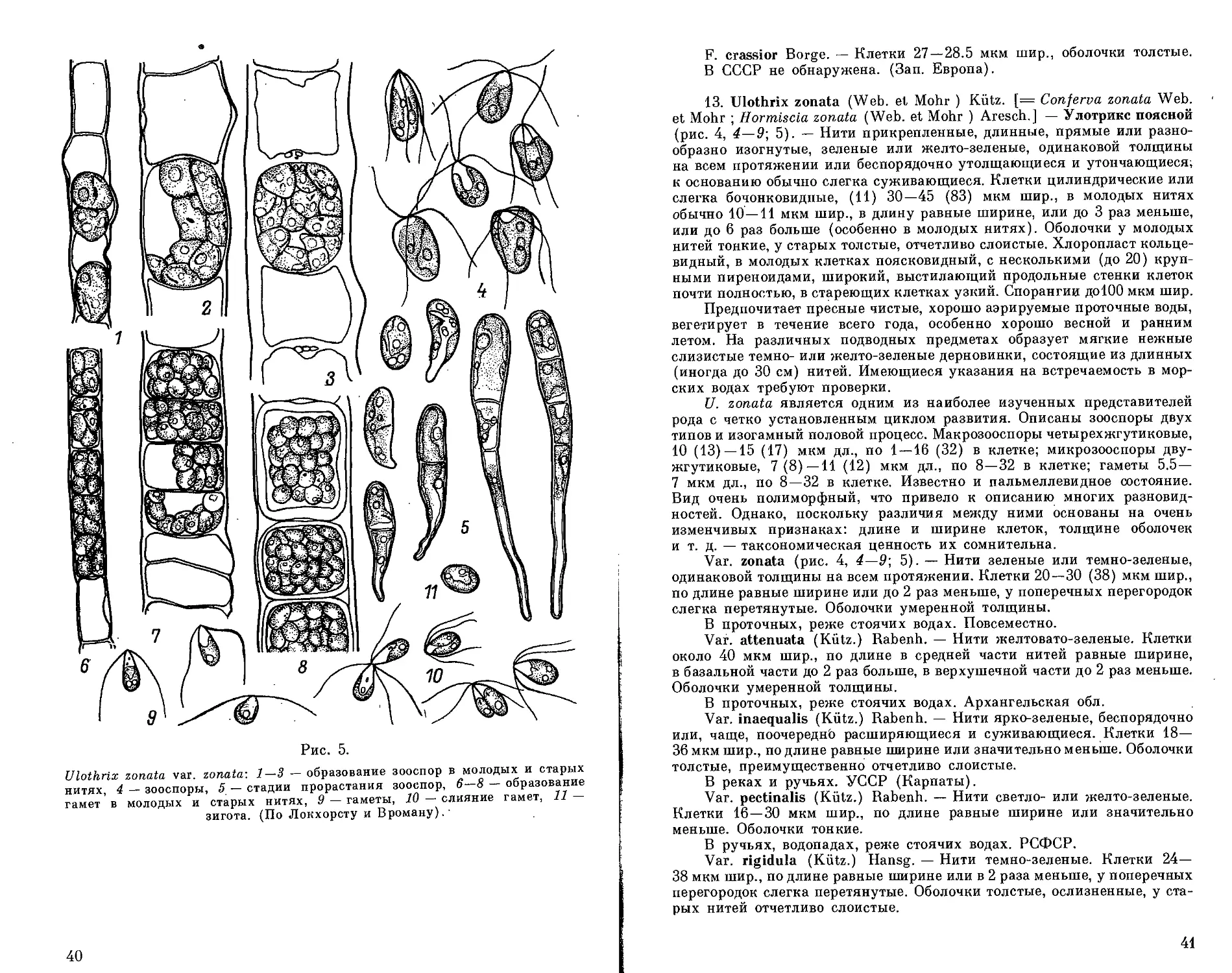
ческие, короткие или длинные, реже бочонковидные, верхушечная
обычно широкоокруглая, иногда удлиненная и слегка заостренная,
базальная удлиненная, к основанию суженная. Оболочки бесцветные,
разной толщины. Хлоропласт поясковидный или кольцевидный, с 1 или
несколькими пиреноидами, выстилающий продольные стенки клеток
почти полностью или только частично. Зооспоры четырехжгутиковые,
иногда двужгутиковые, со стигмой. Известно образование апланоспор,
гипноспор и акинет. Половой процесс изогамный. Гаметы двужгути-
ковые, грушевидные, со стигмой, развивающиеся на тех же нитях, что
и зооспоры. Зигота теряет жгутики, округляется, покрывается толстой
оболочкой и прорастает после периода покоя. Наблюдались случаи
партеногенетического развития гамет, иногда гаметы, находясь в мате-
ринской клетке, превращаются в партеноспоры. При .неблагоприятных
условиях может происходить переход талломов в пальмеллевидное
состояние — клетки нитей округляются, теряют связь друг с другом,
но продолжают делиться в разных направлениях, в результате чего
возникает бесформенное скопление клеток, погруженных в слизь.
Представители рода обитают преимущественно в континентальных
проточных, реже стоячих водах, некоторые в морях, а также в почве
и на ее поверхности. Много эвритопных форм. Род содержит ряд недо-
статочно изученных, трудно идентифицируемых видов. В затруднитель-
ных случаях необходимо тщательное сличение определяемого материала
с несколькими видовыми описаниями. В почвенных культурах мелко-
клеточные виды могут быть смешаны с видами родов Chlorhormidium
и Stichococcus, имеющими дисковидные хлоропласты, поэтому точное
установление формы хлоропластов является обязательным.
I. В пресных, реже минерализованных водах, некоторые виды также
и в почве.
1. Нити не более 10 мкм шир.
А. Нити без слизистого влагалища.7
а. Хлоропласт с 1 пиреноидом.
а. Клетки 4—6 мкм шир., по длине равные ширине или
до 5 раз больше......................U. subtilissima 1.
р. Клетки 5—10 мкм шир., по длине равные ^ширине
или только до 2.5 раза больше.
* Оболочки клеток часто слегка ослизняющиеся.
Хлоропласт широкий, выстилающий продольные
стенки клеток почти полностью................
............................U. tenerrima 2.
* * Оболочки клеток не ослизняющиеся.
7 Слизистое влагалище не следует смешивать.с. легким ослизнением
оболочек клеток, свойственным ряду видов как постоянный признак
или при определенных условиях.
31
+ Клетки 8—10 мкм шир. Хлоропласт узкий,
выстилающий продольные стенки клеток
только частично . . . . U. idiospora 3.
+ + Клетки 5—8.4 мкм шир. Хлоропласт ши-
рокий, выстилающий продольные стенки
клеток почти полностью........................
........................U. variabilis 4.
б. Хлоропласт с 2—4 (6) пиреноидами . . . U. quaternaria 5.
Б. Нити окружены слизистым влагалищем.
а. Влагалище гомогенное; клетки 2—4 мкм шир., по длине
до 6 раз больше.........................U. limnetica 6.
б. Влагалище фиброзное; клетки (5)8 — 10 мкм шир., по
длине только до 2 раз больше................U. mucosa 7.
2. Нити от 10 мкм шир. и более (только изредка меньше).
А. Оболочки клеток тонкие, не слоистые.
а. Оболочки легко ослизняющиеся; клетки обычно укорочен-
ные, длина их меньше ширины, реже равна ей...............
........................................U. osciHarina 8.
б. Оболочки не ослизняющиеся; клетки обычно вытянутые,
длина их больше ширины или равна ей, реже они укоро-
ченные.
а. Клетки узкого диапазона размерности, только И —
12 мкм шир...........................U. pamirica 9.
р. Клетки широкого диапазона размерности, (6.5) 10 —
22 (34) мкм шир......................U. tenuissima 10.
Б. Оболочки клеток толстые, слоистые (только в молодом состоя-
нии иногда тонкие).
а. Клетки 9—14 (19) мкм шир............U. moniliformis И.
б. Клетки 12—22 (28.5) мкм шир............U. aequalis 12.
в. Клетки (И) 30—45 (83) мкм шир. . . . U. zonata 13.
II. В солоноватых и морских водах (один вид изредка и в пресноводных
водоемах).
1. Хлоропласт с 1 пиреноидом; нити до 22 мкм шир.
А. Клетки (5) 10 — 15 (21.8) мкм шир. . . . U. implexa 14.
Б. Клетки 8 — 22 мкм шир...................U. pseudoflacca 15.
2. Хлоропласт с 1—3 (4) пиреноидами; нити 10—25 (30) мкм шир.
...................................................U. Пасса 16.
1. Ulothrix subtilissima Rabenh. (= U. subtilis Kiitz. f. subtilissima
Rabenh.) — Улотрикс тончайший (рис. 1,1,2). — Нити длинные, тонкие,
желто-зеленые. Клетки цилиндрические, 4—6 мкм шир., по длине равные
ширине или до 2—3 (5) раз больше, у поперечных перегородок не пере-
тянутые или слегка перетянутые. Оболочки тонкие. Хлоропласт с 1 пире-
ноидом, выстилающий продольные стенки клеток более чем наполовину.
В пресных и минерализованных стоячих и проточных водах, в об-
растаниях различных субстратов, нередко в планктоне (особенно в ослиз-
ненном состоянии), обнаружен также в почвах. Повсеместно.
По размерам клеток близок к U. variabilis, но отличается более
длинными клетками. В водных культурах, засеянных почвой, развива-
ется на дне сосудов.
2. Ulothrix tenerrima Kutz. [= U. subtilis Kiitz. var. tenerrima
Kirchn.; V. tenuis Kiitz.; Conferva tenerrima Kiitz.; Hormiscia subtilis
(Kiitz.) De-Toni var. tenerrima De-Toni] — Улотрикс нежнейший (рис. 1,
3—9). — Нити длинные, часто до 10 см, в водных лабораторных культу-
32
Рис. 1.
1,2 — Ulothrix subtilissima', 3—9 — U. tenerrima', 3—6 — f. tenerrima (3, 4 —
вегетативные нити, 5, 6 — нити с макрозооспорами), 7—9 — f. dentatospora
(7 — вегетативная нить, 8, 9 — апланоспоры); 10,11 — U. idiospora, нити с аки-
нетами. (1, 3 — по Геерингу, 2 — по Прескотту, 4 — по Газену, 5 — по Гс, 6, 10,
11 — по Весту, 7—9 — по Голлербаху).
3 Зеленые водоросли. Улотриксовые
33
pax часто короткие, прямые или изогнутые, светло-зеленые. Клетки
цилиндрические, 6—10 мкм шир., по длине равные ширине, несколько
меньше или до 1.5 раза больше, у поперечных перегородок слегка
перетянутые. Оболочки тонкие, часто слегка ослизняющиеся, иногда
значительно, до 4 мкм толщ., и тогда поверхность нитей становится
неровной. Хлоропласт широкий, слабо изрезанный по краям, с 1 пире-
ноидом, выстилающий продольные стенки клеток почти полностью. Аки-
неты шаровидные или эллипсоидные, до 12.6 мкм шир., до 16.8 мкм дл.,
с толстой, гладкой или крупнозубчатой оболочкой.
В стоячих и проточных водах, а также в почве.
F. tenerrima (рис. 1, 3—6). — Апланоспоры шаровидные, с бесцвет-
ной гладкой оболочкой.
В пресных и минерализованных проточных и особенно стоячих
водах, на различных субстратах, одиночно среди других водорослей,
нередко в почве. Европ. ч. СССР, Кавказ, Полярн. и Сев. Урал, Ср. Азия,
Зап. и Вост. Сибирь, Дальн. Восток, нередко.
Размеры апланоспор не описаны, но врад ли они больше тех,
которые установлены для f. dentatospora.
F. dentatospora Hollerb. (рис. 1, 7—9). — Апланоспоры эллипсоид-
ные, на концах нитей почти шаровидные, 10.5 — 12.6 мкм шир., 12.6—
16.8 мкм дл., одиночные или по 2—3 в ряд, с1 бесцветной крупнозубчатой
оболочкой.
В песчаных почвах. Ленинградская обл.
От типовой формы отличается только крупными зубчиками на
оболочке апланоспор.
3. Ulothrix idiospora G. S. West — Улотрикс особоспоровый (рис. 1,
10, Л). —Нити длинные, слегка изогнутые. Клетки цилиндрические,
8—10 мкм шир., по длине равные ширине, несколько меньше или
до 2.5 раза больше, у поперечных перегородок не перетянутые. Оболочки
тонкие. Хлоропласт узкий, явственно изрезанный по краям, с 1 пире-
ноидом, выстилающий продольные стенки клеток лишь частично. Аки-
неты одиночные или по нескольку в ряд (до 20), эллипсоидные, бочонко-
видные или цилиндрические, 10—13 мкм шир., 18—36 мкм дл., с толстой
мелкоямчатой оболочкой.
В прудах и мелких проточных лужах, среди других нитчатых
зеленых водорослей. В СССР не обнаружен. (Австралия).
Малоизученный вид, отличающийся своеобразием строения акинет.
4. Ulothrix variabilis Kiitz. [= U. subiilis Kutz. var. variabilis Kirchn.;
Hormiscia subttlis (Kiitz.) De-Toni var. variabilis (Kiitz.) Kirchn.] —
Улотрикс изменчивый (рис. 2,1,2). — Нити короткие, обычно до 500 мкм,
реже до 2 мм дл., прямые или изогнутые. Клетки коротко- или удлиненно-
цилиндрические, 5—7 (8.4) мкм шир., по длине равные ширине, вдвое
меньше или до 2 (3) раз больше, у поперечных перегородок не пере-
тянутые или чуть заметно перетянутые. Оболочки очень тонкие. Хлоро-
пласт с 1 нередко крупным пиреноидом, выстилающий продольные
стенки клеток обычно почти полностью.
В стоячих и проточных водах, а также в различных почвах. Аркти-
ческие о-ва, Европ. ч. СССР, Кавказ, Полярн. и Сев. Урал, Ср. Азия,
Зап. и Вост. Сибирь.
Водоросль хорошо развивается в водных культурах, засеянных
почвой, образуя рыхлые сплетения из длинных нитей или затягивая
поверхность питательного раствора плотной матовой, слегка складчатой
пленкой. Может пышно развиваться в культурах с источником азота
34
Рис. 2.
1, 2 — Ulothrix variabilis', 3,4 — U. quaternaria f. quaternaria: 3 — образование
и выход зооспор, 4 — зооспоры; 5, 6 — V. limnetica: 5 — var. llmnetica, 6 —
var. minor. (7, 5 — по Газену, 2 — по Прескотту, 3, 4 — по Плейфайру, 6 —
по Тейлиигу).
в виде аммонийной соли, но при резком подкислении среды (до pH ЗА)
происходит сильная деформация клеток. В старых нитях клетки обычно
с толстой оболочкой, перетянут!ые у поперечных перегородок и часто
заполненные крупными зернами крахмала. Вид близок к U. subtilisstma,
но отличается более короткими клетками и особенно короткими нитями.
По размерам клеток близок также к U. tenerrima.
5. Ulothrix quaternaria Playf. — Улотрикс четверной (рис. 2, 3,4). —
Нити длинные, темно-зеленые. Клетки цилиндрические, 4—9 (12) мкм
шир., по длине равные ширине или до 2—4 (6) раз больше. Хлоропласт
кольцевидный, с 2—4 (6) пиреноидами, выстилающий продольные
стенки клеток полностью.
В пресных водах. На поверхности субстрата образует шелковистые
пленки.
Вид мало изучен. По ширине клеток приближается к U. tenerrima,
но отличается большей их длиной и особенно большим числом пиреноидов
в хлоропластах.
F. quaternaria (рис. 2, 3, 4). — Клетки 7—9 мкм шир., по длине
равные ширине или до 2 (4) раз больше.
В СССР не обнаружена. (Австралия).
3»
35
F. angustissima Playf. — Клетки 4 мкм шир., по длине в 2.5—6 раз
больше.
В СССР не обнаружена. (Австралия).
6. Ulothrix limnetica Lemm. — Улотрикс озерный (рис. 2, 5, 6). —
Нити длинные, прямые или слегка искривленные, окруженные толстым
бесцветным гомогенным слизистым влагалищем, вместе с которым могут
достигать 27 мкм толщ. Клетки цилиндрические, 2—4 мкм шир., по длине
равные ширине или до 6 раз больше.
В стоячих, реже проточных водах.
От остальных видов рода отличается мелкими размерами клеток
и характером влагалища.
Var. limnetica (рис. 2, 5). — Клетки 4 мкм шир., по длине равные
ширине или до 6 раз больше.
В различных водоемах, в планктоне, обрастаниях и среди других
свободноживущих нитчатых водорослей. Местами в европ. ч. СССР,
единично в Ср. Азии и на Дальн. Востоке.
Var. minor Lemm. (рис. 2, 6). — Клетки 2 мкм шир., по длине
в 3—6 раз больше.
В стоячих водоемах. Московская обл.
7. Ulothrix mucosa Thur. — Улотрикс слизистый (рис. 3, 1, 2). —
Нити длинные, прямые или слегка изогнутые, окруженные очень толстым
слизистым влагалищем фиброзной структуры, с волокнами, направлен-
ными поперечно; в местах поперечных клеточных перегородок волокна
отсутствуют. Клетки цилиндрические, (5) 8 — 10 мкм шир., по длине
до 2 раз больше, у поперечных перегородок не перетянутые, или бочонко-
видные и тогда заметно перетянутые. Оболочки довольно толстые, бес-
цветные, отчетливо двуконтурные. Хлоропласт широкий, с 1 пиреноидом,
выстилающий продольные стенки клеток почти полностью.
В проточных и стоячих водах. Единично в Архангельской обл.,
ЛатвССР, ЛитССР, редко в УССР, УзССР.
Редкий малоизученный вид, легко отличается от видов рода харак-
терным слизистым влагалищем фиброзной структуры.
8. Ulothrix oscillarina Kutz. — Улотрикс колеблющийся (рис. 3,
3, 4). — Нити длинные, ярко-зеленые. Клетки цилиндрические, большей
частью короткие, 10—14 мкм шир., по длине равные '/t, '/2 или> реже,
всей ширине клетки, у поперечных перегородок-обычно не перетянутые.
Оболочки тонкие, легко ослизняющиеся. Хлоропласт широкий, с 1 пире-
ноидом, выстилающий продольные стенки клеток более чем наполовину.
В стоячих и слабо проточных водах, а также в местах, орошаемых
капельной влагой (водопады, фонтаны), где образует ярко-зеленые
слизистые мягкие дерновинки. МолдССР, УССР, Кавказ, УзССР, Дальн.
Восток, единично.
9. Ulothrix pamirica (Wille) Printz — Улотрикс памирский (рис. 3,
5—7). —Нити длинные, прямые или слегка разнообразно изогнутые.
Рис. 3.
1,2 — Ulothrix mucosa; 3, 4 — U. oscillarina; 5—7 — U. pamirica: 5 — нить
с незрелыми апланоспорами, 6 — нить co зрелыми апланоспорами, 7 — нить
с овальными акинетами; 8, 9 — U. tenuissima; 10—14 — U. moniliformis. (2,
10—13 — по Скуе, 2 — по Ольтманнсу, 3 — по Весту, 4 — по Мошковой, 5—7 —
по Вилле, 8, 9 — по Газену, 14 — по Геерингу).
36
37
Клетки цилиндрические, 11 — 12 мкм шир., по длине равные ширине или
до 4 раз больше, у поперечных перегородок не перетянутые. Оболочки
тонкие. Акинеты яйцевидные или цилиндрические, с равномерно утол-
щенной оболочкой. Апланоспоры шаровидные или яйцевидные, по 1—3
в каждой клетке, на ранних стадиях развития содержат крахмал, при
дозревании приобретают кирпично-красный цвет и капли масла.
В пресных водах. Памир.
Редкая, недостаточно изученная водоросль.
10. Ulothrix tenuissima Kiitz. (= (J. tenuis Kiitz.; Hormiscia tenuis
De-Toni) — Улотрикс тончайший (рис. 3, 8,9). — Нити длинные, прямые
или слегка искривленные, темно-зеленые. Клетки цилиндрические,
(6.5) 10—22 (34) мкм шир., по длине равные ширине (особенно в моло-
дых нитях), или до 2 раз меньше, или до 2 раз больше, у поперечных
перегородок не перетянутые; встречаются нити, состоящие как из корот-
ких, так и из длинных клеток, постепенно сменяющих друг друга.
Оболочки тонкие. Хлоропласт широкий, с (1.) 2—4 крупными пире-
ноидами, выстилающий продольные стенки клеток почти полностью.
Зооспоры возникают во вздутых, отчетливо перетянутых клетках.
В пресных минерализованных стоячих и особенно проточных водах,
а также на влажной почве. Европ. ч. СССР, Кавказ, Полярн. и Сев. Урал,
КазССР, УзССР, Зап. Сибирь, Дальн. Восток, нередко.
Вид изучен недостаточно. Близок к V. aequalis, от которого хорошо
отличается тонкими, неслоистыми оболочками, а также цветом хлоро-
пласта. Очень напоминает молодые нити U. zonata, однако нити U. tenuis-
sima темнее, клетки с более тонкой оболочкой и значительно короче.
Эти отличительные признаки весьма сомнительны, поскольку могут легко
изменяться в зависимости от экологических условий. Значительно боль-
шего внимания заслуживает количество пиреноидов; у U. zonata обычно
больше 4 пиреноидов, а у U. tenuissima по 2—4.
И. Ulothrix moniliformis Kiitz. — Улотрикс четковидный (рис. 3,
10—14). — Нити разной длины, светло-зеленые. Клетки бочонковидные,
9 — 14 (19) мкм шир., по длине равные ширине или несколько меньше,
реже до 2 раз больше, у поперечных перегородок отчетливо перетянутые.
Оболочки толстые, бесцветные, у крупноклеточных нитей часто слоистые
и ослизняющиеся. Хлоропласт широкий, выстилающий продольные
стенки клеток почти полностью, однако он нередко сжимается и в виде
округлой или эллипсоидной желто-зеленой массы смещается к какой-
либо одной продольной стенке.
В пресных водоемах разного типа.
Редкий малоизученный вид. По размерам и способности к ослиз-
нению оболочек клеток близок к U. oscillarina, но отличается формой
клеток и толстыми слоистыми оболочками.
Var. moniliformis (рис. 3, 10—14). — Клетки 11 — 14 (19) мкм шир.
В проточных и стоячих водах, чаще всего в болотах, рассеянно
среди других водорослей. ЭССР, ЛатвССР, УССР, Ср. Азия.
Var. braunii (Kiitz.) Rabenh. — Клетки 9—12 мкм шир.
В реках с быстрым течением. УССР.
12. Ulothrix aequalis Kiitz. (= U. cateniformis Kiitz.) — Улотрикс
ровный (рис. 4, 1—3). — Нити длинные, желтовато-зеленые. Клетки
цилиндрические, 12—22 (28.5) мкм шир., по длине равные ширине или
до 2 раз больше, реже значительно меньше, у поперечных перегородок
не перетянутые или слегка перетянутые, при ослизнении нитей и
38
Рис. 4.
1—3 — Ulothrix aequalis f. aequalis: 1, 2 — вегетативные нити, 3 — акинеты;
4—9 — U. sonata var. zonata: 4 — кольцевидный хлоропласт с пиреноидом,
5, 6 — фрагменты взрослой нити, 7 — фрагмент старой нити, 8 — верхушка
молодой нити, 9 — базальная часть нити. (7,3 — по Геерингу, 2 — по Прескотту,
4 — по Рич, 5, 9 — по Гааену, 6—8 — по Локхорсту и Вроману).
образовании акинет бочонковидные. Оболочки довольно толстые, часто
слоистые. Хлоропласт широкий, с (1) 2 пиреноидами или без них,
выстилающий продольные стенки клеток почти полностью.
В пресных проточных и стоячих водоемах разного типа.
F. aequalis (рис. 4, 1—3). — Клетки 12—22 мкм шир., оболочки
довольно толстые.
В мелководных участках водоемов может развиваться в массе,
чаще встречается рассеянно среди других водорослей. Во многих местах
СССР, нередко.
39
Рис. 5.
Ulothrix zonata var. zonata: 1—3 — образование зооспор в молодых и старых
нитях, 4 — зооспоры, 5 — стадии прорастания зооспор, 6—8 — образование
гамет в молодых и старых нитях, 9 — гаметы, 10 слияние гамет, 11
зигота. (По Локхорсту и В роману).
40
F. crassior Borge. — Клетки 27 — 28.5 мкм шир., оболочки толстые.
В СССР не обнаружена. (Зап. Европа).
13. Ulothrix zonata (Web. el Mohr ) Kutz. [= Conferva zonata Web.
et Mohr ; Hormiscia zonata (Web. et Mohr ) Aresch.] — Улотрикс поясной
(рис. 4, 4—9\ 5). — Нити прикрепленные, длинные, прямые или разно-
образно изогнутые, зеленые или желто-зеленые, одинаковой толщины
на всем протяжении или беспорядочно утолщающиеся и утончающиеся;
к основанию обычно слегка суживающиеся. Клетки цилиндрические или
слегка бочонковидпые, (И) 30—45 (83) мкм шир., в молодых нитях
обычно 10—11 мкм шир., в длину равные ширине, или до 3 раз меньше,
или до 6 раз больше (особенно в молодых нитях). Оболочки у молодых
нитей тонкие, у старых толстые, отчетливо слоистые. Хлоропласт кольце-
видный, в молодых клетках поясковидный, с несколькими (до 20) круп-
ными пиреноидами, широкий, выстилающий продольные стенки клеток
почти полностью, в стареющих клетках узкий. Спорангии доЮО мкм шир.
Предпочитает пресные чистые, хорошо аэрируемые проточные воды,
вегетирует в течение всего года, особенно хорошо весной и ранним
летом. На различных подводных предметах образует мягкие нежные
слизистые темно- или желто-зеленые дерновинки, состоящие из длинных
(иногда до 30 см) нитей. Имеющиеся указания на встречаемость в мор-
ских водах требуют проверки.
U. zonata является одним из наиболее изученных представителей
рода с четко установленным циклом развития. Описаны зооспоры двух
типов и изогамный половой процесс. Макрозооспоры четырехжгутиковые,
10 (13) —15 (17) мкм дл., по 1 — 16 (32) в клетке; микрозооспоры дву-
жгутиковые, 7 (8) —11 (12) мкм дл., по 8—32 в клетке; гаметы 5.5 —
7 мкм дл., по 8—32 в клетке. Известно и пальмеллевидное состояние.
Вид очень полиморфный, что привело к описанию многих разновид-
ностей. Однако, поскольку различия между ними основаны на очень
изменчивых признаках: длине и ширине клеток, толщине оболочек
и т. д. — таксономическая ценность их сомнительна.
Var. zonata (рис. 4, 4—9; 5). — Нити зеленые или темно-зеленые,
одинаковой толщины на всем протяжении. Клетки 20—30 (38) мкм шир.,
по длине равные ширине или до 2 раз меньше, у поперечных перегородок
слегка перетянутые. Оболочки умеренной толщины.
В проточных, реже стоячих водах. Повсеместно.
Var. attenuate (Kiitz.) Rabenh. — Нити желтовато-зеленые. Клетки
около 40 мкм шир., по длине в средней части нитей равные ширине,
в базальной части до 2 раз больше, в верхушечной части до 2 раз меньше.
Оболочки умеренной толщины.
В проточных, реже стоячих водах. Архангельская обл.
Var. inaequalis (Kutz.) Rabenh. — Нити ярко-зеленые, беспорядочно
или, чаще, поочередно расширяющиеся и суживающиеся. Клетки 18—
36 мкм шир., по длине равные ширине или значительно меньше. Оболочки
толстые, преимущественно отчетливо слоистые.
В реках и ручьях. УССР (Карпаты).
Var. pectinalis (Kiitz.) Rabenh. — Нити светло- или желто-зеленые.
Клетки 16—30 мкм шир., по длине равные ширине или значительно
меньше. Оболочки тонкие.
В ручьях, водопадах, реже стоячих водах. РСФСР.
Var. rigidula (Kiitz.) Hansg. — Нити темно-зеленые. Клетки 24—
38 мкм шир., по длине равные ширине или в 2 раза меньше, у поперечных
перегородок слегка перетянутые. Оболочки толстые, ослизненные, у ста-
рых нитей отчетливо слоистые.
41
Рис. 6.
7, 2 — Vlothrlx implexa: 3, 4 — U. pseudojlacctr, 5—8 — U. flacca: 5, 6 —
вегетативные нити, 7 — образование гамет, 8— гамета. (7, 2 — по Газену,
3 —8 — по Каргеру).
42
В горных ручьях и быстро текущих реках. УССР (Карпаты).
14. Ulothrix implexa (Kutz.) Kiitz. (= U. submarina Kiitz.; U. sub-
flacca Wille; Hormidium implexum Kiitz.; Hormtsda implexa Rabenh.) —
Улотрикс перепутанный (рис. 6, 1, 2). — Нити прикрепленные или
свободноплавающие, короткие, до 3 см, разнообразно изогнутые, бледно-
желто-зеленые, в ватообразных массах или собранные в мягкие пучки.
Клетки цилиндрические, иногда слегка вздутые, (5) 10 — 15 (21.8) мкм
шир., по длине равные ширине, или до 2 раз меньше, или до 4 раз больше.
Оболочки тонкие. Хлоропласт поясковидный, с 1 крупным пиреноидом,
выстилающий продольные стенки клеток полностью или частично.
На поверхности нитей часто откладываются в больших количествах соли
кальция.
В морских, солоноватых, изредка и пресных водах, на твердых
грунтах, водных цветковых растениях, крупных водорослях или среди
них. Во многих районах СССР, довольно часто.
Отличается от U. pseudoflacca тонкими нитями и более длинными
клетками.
15. Ulothrix pseudoflacca Wille — Улотрикс ложноповислый (рис. 6,
3, 4). — Нити прикрепленные, обычно длинные, зеленые или светло-
зеленые. Клетки цилиндрические, 8—22 мкм шир., по длине равные
ширине или до 4 раз меньше. Оболочки тонкие. Хлоропласт с 1 пире-
ноидом, выстилающий продольные стенки клеток почти полностью.
При подсыхании возникают акинеты, в которые нревращается все содер-
жимое клетки, при этом внутренние слои оболочки клеток утолщаются,
а наружные растрескиваются. Прикрепляется к субстрату удлиненной
базальной клеткой, закругленной внизу.
В соленых водоемах, а также в морях, на камнях и растениях.
Вегетирует на протяжении всего года. УССР (лиманы Черного моря),
Каспийское, Охотское (Сахалин, Курильские о-ва) и Японское моря.
16. Ulothrix Пасса (Dillw.) Thur. — Улотрикс повислый (рис. 6,
5—8). — Нити прикрепленные, обычно длинные, светло- или темно-
зеленые. Клетки цилиндрические, 10—25 (30) мкм шир., по длине равные
ширине или до 4 раз меньше, у поперечных перегородок не перетянутые
или слегка перетянутые, при образовании гамет вздувающиеся, 50—
60 (80) мкм шир. Оболочки тонкие. Хлоропласт широкий, с 1—3 (4) пи-
реноидами, выстилающий продольные стенки клеток почти полностью.
Иногда от нескольких нижних клеток нити отходят ризоиды.
В соленых водах, а также в морях, на скалистых, илисто-песчано-
галечных грунтах, на водорослях, раковинах моллюсков. Выдерживает
опреснение и загрязнение. УССР (Славянские соленые озера, лиманы
Черного моря), Каспийское, Аральское, Берингово, Охотское (Сахалин,
Курильские о-ва) и Японское моря.
Род URONEMA Lagerh. - УРОНЕМА
Нити однорядные, простые, иногда в верхушечной части крючко-
видно согнутые, без слизистого влагалища, прикрепляющиеся к суб-
страту с помощью особого пектинового образования — дермоида,
выделяемого базальной клеткой. Клетки цилиндрические или слегка
вздутые, короткие или удлиненные, верхушечная вытянутая, заострен-
ная или, реже, округлая, прямая или изогнутая, иногда вздутая, базаль-
43
ная обычно удлиненная, к основанию большей частью отчетливо
суженная. Дермоид в форме лолушаровидной подушечки или диско-
видной стопы, с ровными или лопастными краями. Оболочки тонкие,
бесцветные. Хлоропласт пластинчатый, лентовидный или кольцевидный,
часто сильно изрезанный, с 1—2 (5) пиреноидами, выстилающий
продольные стенки клеток почти полностью. Зооспоры четырехжгути-
ковые, со стигмой, по 1—2 в любой клетке слоевища, включая
верхушечную и базальную; они освобождаются через округлое отвер-
стие в оболочке материнской клетки, возникающее к моменту их созре-
вания. Апланоспоры шаровидные или эллипсоидные, зеленые, с бес-
цветной оболочкой. Известно также образование акинет. Половое раз-
множение неизвестно.
Виды рода обитают преимущественно в пресных и минерализо-
ванных стоячих и проточных водах на растительном субстрате.
Род наиболее близок к роду Ulothrix, отличается наличием дермоида,
с помощью которого нити прикрепляются к субстрату, а также присут-
ствием в базальных клетках хорошо развитого хлоропласта и способ-
ностью базальных клеток к образованию зооспор.
I. Длина клеток равна ширине, меньше или до 4(7) раз больше;
верхушечная клетка не вздутая (иногда вздутая).
1. Дермоид полушаровидный, с ровными краями.
А. Нити одинаковой ширины, на всем протяжении ровные.
а. Нити в верхушечной части прямые или слегка изогну-
тые, иногда крючковидно согнутые; дермоид преимуще-
ственно бесцветный...................U. confervicola 1.
б. Нити в верхушечной части всегда слегка серповидно со-
гнутые; дермоид преимущественно бурый.................
...................................U. intermedium 2.
Б. Нити не ровные...........................U. gigas 4.
2. Дермоид дисковидный, с лопастными краями . . . U. terrestre 3.
II. Длина клеток превосходит ширину в 4—13 раз; верхушечная клетка
обычно вздутая..................................U. elongatum 5.
1. Uronema confervicola Lagerh. — Уронема конфервная (рис. 7, 7—
17). — Нити до 1.5 мм дл., прямые или слегка искривленные,
в верхушечной части иногда крючковидно изогнутые. Клетки цилин-
дрические, 3.5—9 мкм шир., по длине равные ширине, меньше или
до 2—4 (7) раз больше, у поперечных перегородок не перетянутые,
реже слегка перетянутые, верхушечная (12) 20—35 мкм дл., прямая,
к вершине заостренная, клювовидная, иногда заметно вздутая, базальная
(10) 18 — 32 мкм дл., к основанию .заметно или лишь слегка суженная.
Дермоид полушаровидный, с ровными краями, бесцветный. Хлоропласт
с 1 — 2 пиреноидами. Зооспоры 6—7 мкм шир., 9 — 10 мкм дл., по 1
в клетке.
В пресных и минерализованных стоячих и проточных водах, на
растительных субстратах.
Var. confervicola (рис. 7,1—4). — Нити до 1.5 мм дл., в верхушечной
части прямые или слегка искривленные. Клетки 3.5—9 мкм шир.,
по длине в 2—3 раза больше, верхушечная 20—35 мкм дл., прямая,
заостренная.
На водных цветковых растениях и крупноклеточных нитях зеленых
водорослей. В разных районах СССР, нечасто.
Var. uncinatum Popova (рис. 7, 5—11). — Нити до 0.5 мм дл., в вер-
хушечной части крючковидно изогнутые. Клетки (3.5) 4—6 мкм шир.,
44
Рис. 1.
1—17 — Uronema confervicola: l—4—v&r. confervicola (1, 2 — вегетативные
нити, 3 — образование зооспор, 4 — зооспора), 5—11 — var. uncinatum (5, 6 —
вегетативные нити, 7—10 — верхушки нитей, 11 — основание нити), 12—17 —
var._africanum (12, 13 — молодые нити, 14,— взрослая нить, 15, 16 — верхушки
нитей, 17' — основание нити); 18—20 — U. intermedium: IS — вегетативная
нить, 19 — верхушка нити, 20 — основание нити. (1—4 — по Лагерхейму, 5—
11 — по Поповой, 12—17 — по Шредеру, 18—20 — по Буррелли).
45
по длине равные ширине или до 4 (5) раз больше, верхушечная 8 — 9 мкм
шир., вытянутая, иногда заметно вздутая или клювовидно согнутая,
с небольшим утолщением оболочки на свободном конце.
В минерализованных озерах, на крупноклеточных нитчатых водо-
рослях. Зап. и Вост. Сибирь.
Var. africanum (Borge) Printz (=U. africanum Borge) (рис. 7, 12—
17). — Нити короткие, приблизительно из 12 клеток, слегка искривлен-
ные. Клетки 4.5—6 мкм шир., по длине до 3—6 (7) раз больше,
верхушечная слегка клювовидно согнутая, реже прямая, к вершине
заостренная, по длине в 3.5—4 раза больше ширины.
В пресных водоемах. В СССР не обнаружена. (Вост. Африка).
2. Uronema intermedium Bourr. — Уронема промежуточная (рис. 7,
18—20). — Нити прямые, в верхушечной части постоянно слегка серпо-
видно согнутые. Клетки цилиндрические, 5.8—8 мкм шир., 7—25 мкм
дл., верхушечная на конце заостренная, клювовидная, базальная 10—
15 (25) мкм дл., к основанию отчетливо суженная. Дермоид полушаро-
видный, с ровными краями, преимущественно бурый. Хлоропласт с 2 пи-
реноидами.
В пресных стоячих водах, на нитях эдогониума и кладофоры.
УССР.
По размерам клеток близка к U. confervicola и U. terrestre, отли-
чается от первой короткими малоклеточными нитями и цветом дермо-
ида, от второй — формой дермоида.
3. Uronema terrestre Mitra — Уронема наземная (рис. 8, 1, 2). —
Нити до 6 (30) см дл. Клетки цилиндрические или слегка вздутые,
3—5 мкм шир., 22—34 (60) мкм дл., верхушечная прямая, иногда
изогнутая, шиповидно заостренная, базальная суженная. Дермоид диско-
видный, с лопастными краями и буроватой концентрической зоной.
Хлоропласт с четким пиреноидом. Зооспоры 5—8 мкм шир., 6—15 мкм дл.
Апланоспоры шаровидные.
На очень влажной почве рисовых полей, водных цветковых расте-
ниях и крупноклеточных нитчатых водорослях. В СССР не обнаружена.
(Индия).
4. Uronema gigas Vischer — Уронема гигантская (рис. 8, 3—7).—
Нити до 5 см дл., обычно искривленные, неровные. Клетки цилиндри-
ческие, иногда слегка вздутые, 8—11 мкм шир., 8—25 мкм дл.,
верхушечная прямая, на конце заостренная, базальная к основанию
отчетливо суженная. Дермоид полушаровидный, с ровными краями.
Хлоропласт с 1 — 5 пиреноидами, выстилающий продольные стенки
клеток почти полностью.
В стоячих и проточных водах, в обрастаниях растений. В СССР
не обнаружена. (Зап. Европа).
5. Uronema elongatum Hodg. — Уронема вытянутая (рис. 8, 8—
12). — Нити прямые, реже слегка искривленные, до 5 мм дл. Клетки
цилиндрические, иногда слегка вздутые, 4.4—9.6 мкм шир., 22—80 мкм
дл., верхушечная заостренная, более или менее искривленная или,
реже, прямая, к концу обычно слегка вздутая, базальная 60—90 мкм дл.,
к основанию суженная. Дермоид дисковидный, цельнокрайный. Хлоро-
пласт с 1 — 2 (3) пиреноидами, выстилающий продольные стенки клеток
на 'Д—3Л их длины.
46
Рис. 8.
1,2 — Uronema terrestre-. 1 — общий вид нити, 2 — образование и выход
зооспор; 3—7 — U. gigas: 3 — общий вид нити, 4 — верхушка нити, 5 — средняя
часть нити, 6 — основание нити, 7 — образование зооспор; 8—12 — U. elonga-
tum: 8 — верхушка нити, 9 — апикальный конец верхушечной клетки, 10 —
основание нити, 11 — образование и выход зооспор, 12 — зооспора. (7, 2 —
по Митра, 3—7 — по Вишеру, 8—12 — по Готгетсу).
47
В стоячих и проточных водах, на крупноклеточных нитях зеленых
водорослей и водных цветковых растениях. В СССР не обнаружена.
(Зап. Европа, Сев. Америка).
Отличается от всех видов рода более всего формой верхушечной
клетки нитей, а также очень длинными клетками их, в которых хлоро-
пласт обычно занимает лишь незначительную часть лумена.
Род HORMIDIELLA Iyengar et Kanthamma —
ГОРМИДИЕЛЛА
Нити однорядные, простые, прикрепляющиеся к субстрату с по-
мощью дермоида, имеющего вид небольшой плоской стопы на конце
ножки (стебелька). Клетки бочонковидные, явственно перетянутые у по-
перечных перегородок, верхушечная и базальная клетки слегка конусо-
видные. Оболочки тонкие. Хлоропласт пристенный, поясковидный,
с пиреноидом, выстилающий продольные стенки клеток почти пол-
ностью. Зооспоры двужгутиковые, слегка дорсивентральные, без стигмы;
они образуются по 1 в обычно более мелких клетках, возникающих
из основных в результате их быстрого деления, и освобождаются через
округлое отверстие в боковой стенке каждой клетки. Апланоспоры
известны. Половое размножение неизвестно.
Рис. 9.
Hormidiella parvula: 1—3 — взрослая нить, образование и выход зооспор,
4 — зооспоры, 5—10 — стадии прорастания зооспоры в новую нить. (По Иенгару
и Кантаме).
48
1. Hormidiella parvula Iyengar et Kanthamma — Гормидиелла кро-
шечная (рис. 9). — Нити прямые или слегка изогнутые, обычно мало-
клеточные, иногда до 250 мкм дл. Клетки 8—9 мкм шир., 3.2—
8 мкм дл. Зооспоры 5—5.5 мкм шир., 6.7 — 7 мкм дл.
В почве. В СССР не обнаружена. (Индия: Мадрас).
Водоросль весьма своеобразна по строению, но еще мало изучена.
Обнаружена в лабораторной культуре, засеянной почвой, среди других
почвенных водорослей.
Род CHLORHORMIDIUM Fott -
ХЛОРГОРМИДИУМ
[=Hormidium Kiitz. emend. Klebs (non Hormidium Lindley);
Klebsormidium Silva, Mattox et Blackwell]
Нити однорядные, простые, лишь иногда с зачаточным ветвле-
нием в виде коротких боковых выростов клеток, не отделяющихся
поперечной перегородкой и возникающих главным образом в местах
коленообразных перегибов нитей, реже на их концах, свободноживу-
щие, у одного вида со слизистым влагалищем. Клетки цилиндриче-
ские, реже бочонковидные, на обоих концах нитей одинаковые, на конце
округленные. Оболочка тонкая до толстой, иногда отчетливо двуслойная,
часто липкая, но никогда не ослизняющаяся. Хлоропласт пристенный,
пластинчатый, обычно эллиптический или округлый, прилегающий
в форме полуцилиндра к боковой стенке клетки, выстилая ее вдоль
частично или полностью. Пиреноид 1, обычно удлиненный, не всегда
явственный. Продукт ассимиляции — крахмал, откладывающийся вокруг
пиреноида и в строме хлоропласта. Имеются 2 крупные, полярно
размещенные вакуоли, в которых часто находятся зернышки вещества
неизвестного химического состава, иногда заполняющие всю полость
клетки. Зооспоры двужгутиковые, асимметрично-грушевидные, с вы-
пуклой спинной и плоской брюшной стороной, без стигмы; они
образуются по 1 в более коротких клетках, возникающих из основных
в результате их быстрого деления, и освобождаются через разрыв
или отверстие в оболочке материнских клеток. Апланоспоры по 1
в клетке, способные прорастать без периода покоя. Акинеты эллипсо-
идные, с утолщенной оболочкой. Половой процесс гетерогамный (пока
известен только для С. jlaccidum). Гаметы двуя<гутиковые, грушевидные,
двух размеров (макро- и микрогаметы), без стигмы, по 1 в клетке.
Наблюдались случаи партеногенетического развития гамет.
В пресных стоячих и проточных водах, а также на камнях, коре
деревьев, влажной почве и в толще почвенного слоя.
Название Chlorhormidium введено Фоттом (В. Fott) в 1960 г. вместо
названия Hormidium — рода, описанного Кютцингом (F. Т. Kiitzing)
в 1843 г. и затем обработанного Клебсом (G.* Klebs) в 1896 г., так
как оно оказалось гомонимом названия одного из родов семейства
орхидных (Hormidium Lindley), описанного в 1840 г. Однако, как дока-
зывают в публикации 1972 г. американские альгологи (Р. С. Silva,
К. R. Mattox, W. Н. Blackwell), название Фотта неправомерно, так
как лектотипом рода был выбран Н. velutinum, который оказался
схизогониевой водорослью, а сейчас род Hormidium общепринят в пони-
мании Клебса, т. е. как типично улотриксовая водоросль. Для этого
рода они предложили название Klebsormidium, что влечет за собой
установление новых комбинаций для всех видов, входящих в этот род,
4 Зеленые водоросли. Улотриксовые
49
ими же это было сделано только для 5 видов. Мы считаем формаль-
ное переименование этих видов в настоящем Определителе еще прежде-
временным, так как этот род в целом явно нуждается в монографи-
ческой обработке вместе с близкими родами Ulothrix и Stichococcus
в свете новых данных об их тонком строении, получаемых с помощью
электронного микроскопа. Основное отличие рода Chlorhormidium от
Ulothrix — это небольшой, преимущественно дисковидный хлоропласт,
всегда с 1 пиреноидом, а род Stichococcus, близкий к роду Chlorhor-
midium по форме и размерам хлоропласта, отличается отсутствием
пиреноида.
I. Нити без слизистого влагалища; клетки от 4(5) мкм шир. и более.
1. Нити более или менее прямые, коленчато не согнутые.
А. Оболочки клеток тонкие.
а. На влажных наземных субстратах .... С. flaccidum 1.
б. В стоячих и проточных водах ..... С. subtile 4.
Б. Оболочки клеток толстые.
а. Клетки 8—20 мкм шир., оболочки очень толстые . . . .
.......................................С. mucosum 7.
б. Клетки (5.6) 7 — 11 мкм шир., оболочки умеренно
толстые..................................С. dissectum 8.
2. Нити часто коленчато согнутые.
А. Нити не образуют боковых выростов ... С. fluitans 2.
Б. Нити часто образуют боковые выросты ... С. rivulare 3.
II. Нити без слизистого влагалища; клетки не более 5 мкм шир.
1. Оболочки тонкие; длина клеток обычно равна ширине или
несколько больше ее......................С. pseudostichococcus 6.
2. Оболочки толстые; длина клеток обычно значительно больше
ширины...............................С. tribonematoideum 5.
III. Нити в толстом слизистом влагалище; клетки 9.5 мкм шир .
...............................................С. korschikovii 9.
1. Chlorhormidium flaccidum (Kiitz.) Fott [=Ulothrix flaccida Kiitz.;
Hormiscia flaccida (Kiitz.) Lagerh.; Hormococcus flaccidus (Kiitz.-) Chod.;
Stichococcus flaccidus (Kiitz.) Gay; Hormidium flaccidum (Kiitz.) A. Br.
s. str.; H. nitens (Kiitz.) Rabenh.; Klebsormidium flaccidum (Kiitz.)
Silva, Mattox et Blackwell] — Хлоргормидиум повислый (рис. 10, И). —
Нити длинные, реже короткие, прямые или разнообразно изогнутые.
Клетки цилиндрические, у поперечных перегородок не перетянутые или
только слегка перетянутые, реже бочонковидные, (4.5) 5—14 мкм шир.,
по длине равные ширине, меньше ее или до 3 раз больше, обычно
с тонкими оболочками. Хлоропласт большей частью крупный, дисковид-
ный, с ровными или волнисто изрезанными краями и довольно
крупным пиреноидом, выстилающий продольные стенки клеток напо-
ловину, несколько больше или меньше.
Один из наиболее распространенных видов рода, встречающийся
преимущественно в аэрофитных условиях и в почве, реже в воде.
Он весьма изменчив по строению нитей и размерам клеток, что дало
повод различать в нем несколько разновидностей и форм, связанных,
однако, между собой переходами. В частности, для типовой разно-
видности Геерингом (W. Heering) в 1914 г. был выделен ряд форм,
которые представляют собой скорее всего лишь отклонения, вызванные
условиями существования. Таксономическая ценность их сомнительна.
Var. flaccidum (=Hormidium flaccidum A. Br. s. str.) (рис. 10). —
Нити длинные, реже короткие. Клетки цилиндрические, у поперечных
50
Рис. 10.
Chlorhormidium flaccidum var. flaccidum: 1 — типичная нить, 2 — поперечный
разрез клетки,' 3, 4 ~ нити с клетками разного размера, 5 — фрагментация
нити, 6 — зооспоры, 7 — образование акинет, 8—10 — образование апланоспор,
// — апланоспоры, 12—15 — деление апланоспор, 16 — макрогаметы, 17 —
микрогаметы, 18 — слияние гамет, 19—21 — прорастание зиготы. (1, 2, 5, 6 —
по Фарукви, 3, 4, 7—21 — по Вилле).
4*
51
перегородок не перетянутые, слегка перетянутые или бочонковидные,
5—14 мкм шир.
На влажных наземных субстратах, в стоячих, реже проточных
водах и в почвах. Повсеместно.
В лабораторных культурах в водных питательных растворах растет
в погруженном состоянии в толще раствора и на дне.
F. flaccidum (= f. typica Heering.) — Нити длинные. Клетки ци-
линдрические, у поперечных перегородок не перетянутые, 5—9 мкм шир.,
по длине равные ширине или до 3 раз больше.
Повсеместно.
F. tumidum Heering (=Stichococcus flaccidus Hazen). — Нити ко-
роткие. Клетки обычно слегка вздутые, бочонковидные, у поперечных
перегородок заметно перетянутые, 6—9.5 мкм шир., по длине равные /4
ширины или до 2 раз больше, с довольно толстыми оболочками. Хлоро-
пласт массивный, с крупным пиреноидом.
На влажных скалах и коре деревьев; встречена также на Бал-
тийском побережье. В СССР не обнаружена. (Зап. Европа).
F. montanum Heering [= Hormiscia flaccida (Kiitz.) Lagerh. var.
montana Hansg.]. — Нити длинные. Клетки цилиндрические, у попереч-
ных перегородок не перетянутые, 8—14 мкм шир., по длине равные
ширине, реже в 2 раза меньше или до 2 раз больше. Хлоропласт
довольно крупный, желтовато-зеленый. Оболочки клеток липкие, но не
ослизненные, благодаря приклеивающимся частицам детрита шерохова-
тые.
На влажных субстратах. ЛатвССР.
F. aquaticum Heering — Нити длинные. Клетки цилиндрические,
у поперечных перегородок не перетянутые, 10—14 мкм шир., по длине
обычно до 2 раз больше, тонкостенные. Хлоропласт светло-зеленый,
пиреноид слабо заметен.
В стоячих водах. В СССР не обнаружена. (Зап. Европа).
Var. lubricum Chod. (= Hormidium lubricum Chod.) (рис. 11, 1—
3). — Нити длинные. Клетки цилиндрические, у поперечных перегородок
слегка перетянутые, 5—6 мкм шир., (8) 12—15 (25) мкм дл., с тонкими
оболочками. Хлоропласт дисковидный, с волнисто изрезанными краями,
выстилающий продольные стенки клеток наполовину. Пиреноид не всегда
отчетлив.
В почвах. В СССР не обнаружена. (Зап. Европа).
Водоросль весьма своеобразна, но пока изучена только в культурах,
засеянных почвой. При культивировании на агар-глюкозе образуются
дисковидные колонии, которые через некоторое время становятся
скользкими, а клетки сплошь заполняются зернами крахмала. В лабора-
торных культурах на минеральных питательных растворах развивается
в виде шелковисто блестящей поверхностной пленки, чем сближается
с var. nitens.
Var. nitens Menegh. emend. Klebs [=Ulothrix nitens Menegh.;
Hormiscia flaccida (Kiitz.) Lagerh. var. nitens (Menegh.) Hansg.; Hormi-
dium nitens (Menegh.) emend. Klebs] (рис. 11, 4—7). — Нити длинные,
большей частью разнообразно изогнутые. Клетки цилиндрические, у по-
перечных перегородок не перетянутые или слегка перетянутые, (4.5)
5—7 мкм шир., по длине равные ширине или до 3 раз больше,
с тонкими оболочками. Хлоропласт округлый или эллиптический, высти-
лающий продольные стенки клеток на 2/з- Пиреноид почти всегда
хорошо заметен.
На влажных наземных субстратах и почвах. В разных районах
СССР, довольно часто.
52
Рис. И.
Chlorhormidium flaccidum: 1—3 — var. lubricum, 4—7 — var. nitens, 8,9 —
var. crassum. (1—6, 8, 9 — по Шода, 7 — по Фарукви).'
В лабораторных культурах в водных питательных растворах обра-
зует шелковисто блестящие пленки, плавающие по поверхности раствора
и заходнщие на стенки сосуда. В молодых культурах нити длинные,
разнообразно петлевидно изогнутые, расположенные параллельно друг
другу, в стареющих легко распадающиеся на мелкоклеточные фраг-
менты и даже отдельные эллипсоидные клетки, густо заполненные
блестнщими зернами запасного питального вещества.
F. nitens. — Клетки 5—7 мкм шир., 6—18 мкм дл.
Европ. ч. СССР, Ср. Азия, Зап. Сибирь.
F. tenuis Boye-Pet. — Клетки 4.5 мкм шир., 5—15 мкм дл.
Известна только из культур, засеянных почвой. В СССР не обнару-
жена. (Зап. Европа).
Var. crassum Chod. (= Homidium, crassum Chod.) (рис. 11, 8—9). —
Нити длинные, прямые или слегка изогнутые. Клетки 6.5—7.5(8) мкм
тир., 15—20 мкм дл. Хлоропласт с четким пиреноидом.
Известна только из культур, засеянных почвой. В СССР не обна-
ружена. (Зап. Европа).
2. Chlorhormidium fluitans (Gay) Starmach [=Stichococcus fluitans
Gay; Homidium fluitans (Gay) Heering] — Хлоргормидиум плавучий
(рис. 12, 1—4). — Нити недлинные, часто коленчато изогнутые, легко
распадающиеся на фрагменты или отдельные клетки. Клетки цилиндри-
53
54
ческие или слегка бочонковидные, у поперечных перегородок обычно
несколько перетянутые, 6.5—9 мкм шир., по длине равные ширине или
до 3 раз больше. Образование зооспор наблюдается редко.
В местах, орошаемых капельной влагой, в виде желтовато-зеленых
налетов. В СССР не обнаружен. (Зап. Европа).
Вид изучен слабо. По размерам клеток близок к С. rivulare., но
отличается местообитанием.
3. Chlorhormidium rivulare (Kutz.) Starmach [= Ulothrix rivularis
Kutz.; Hormiscia rivularis De-Toni; Stichococcus rivularis (Kiitz.) Hazen;
Hormidium rivulare Kiitz.] —Хлоргормидиум ручейный (рис. 12, 5—
8). — Нити длинные, часто коленчато согнутые; в местах сгибов с их
наружной стороны нередко образуются небольшие, отходящие под неко-
торым углом тупые выросты, напоминающие начальные стадии образо-
вания боковых ветвей. Клетки цилиндрические, у поперечных перего-
родок не перетянутые или едва заметно перетянутые, 4—11(13.5) мкм
шир., по длине равные ширине или до 3 раз больше, с довольно
толстыми оболочками. Хлоропласт круглый или эллиптический, четкий,
с крупным, 3.5 X 4.3 мкм, эллиптическим, часто неявственным пиренои-
дом, выстилающий продольные стенки клеток частично, даже полностью.
В быстро текущих водах — ручьях, источниках в виде светло-зеле-
ных пучков, прикрепляющихсн к камннм; обнаружен также в водных
культурах, засеннных почвой из поверхностных слоев. Ленинград-
ская обл., ЛатвССР, УССР, Ставропольский край, Кавказ, УзССР,
ТаджССР, Приморский край.
Типичный обитатель быстро текущих вод, для почв, по-видимому,
нехарактерен, но все же довольно часто встречается в водных культурах,
засеянных почвой, в виде отдельных нитей, растущих в смеси с другими
водорослями. Образование парных коленчато согнутых клеток наблюда-
ется постоянно. Фрагментация нитей этому виду несвойственна.
4. Chlorhormidium subtile (Kiitz.) Starmach [=Ulothrix subtilis
Kiitz.; Hormiscia subtilis (Kiitz.) De-Toni; Stichococcus subtilis Kleriker:
Hormidium subtile Heering] — Хлоргормидиум тонкий (рис. 12, 9—12). —
Нити длинные, прямые или слегка изогнутые. Клетки цилиндриче-
ские, у поперечных перегородок обычно не перетянутые, иногда слегка
бочонковидные, 4.5—7 (11) мкм шир., по длине равные ширине или
до 3 раз больше, с тонкими нежными оболочками. Хлоропласт эллип-
тический или округлый, с отчетливым мелким пиреноидом. Акинеты
шаровидные или эллипсоидные, 7—9 мкм шир.
В быстро текущих водах — водопадах, фонтанах и т. п. в виде
зеленых слизистых хлопьев, в стоячих водах нередко образует канато-
видно переплетенные массы нитей, во влажных местах, например в тепли-
цах на цветочных поддонах и т. п., растет в виде курчавых пучков.
Олиго-р-мезосапроб. В разных районах СССР, довольно часто.
Рис. 12.
1—4 — Сhlorhormidium fluitans: 1—3 — фрагменты нитей, 4 — акинеты; 5—8 —
С. rivulare', 9—12 — С. subtile: 9 — общий вид нити, 10i — отдельная клетка,
11 — акинеты, 12 — вырост на нити; 13—15 — С. tribonematoideum: 13, 14 —
общий вид нитей, 15 — фрагмент нити с пустыми зооспорангиями; 16—18 —
С. pseudostichococcus: 16 — общий вид нити, 17 — фрагмент нити, 18 — аки-
неты. (1—4, 18 — по Ге, 5—8 — по Газену, 9 — по Фричу, 10—12, 16, 17 —
по Клерку, 13—15— по Скуе).
55
Многочисленные указания о нахождении в СССР водорослей под
названием Ulothrix subtilis Kiitz. и Hormiscia subtilis (Kiitz.) De-Toni
весьма неопределенны, поскольку могут относиться как к Chlorhormidium
subtile, так и к одному из мелкоклеточных видов улотриксов —
Ulothrix subtilissima Rabenh., U. limnetica Lemm., U. mucosa Thnr.,
U. variabilis Kiitz., U. tenerrima Kiitz.
5. Chlorhormidium tribonematoideum (Skuja) Starmach — Хлор-
гормидиум трибонемовидный (рис. 12,13—15). — Нити длинные, прямые
или слегка изогнутые. Клетки цилиндрические, иногда бочонковидные,
у поперечных перегородок более или менее перетянутые, 2.5—5 мкм
шир., по длине до 6 раз больше, с относительно толстыми гладкими,
у молодых растений бесцветными, у зрелых — светло-бурыми обо-
лочками. Хлоропласт неопределенных очертаний, с более или менее изре-
занными краями, обычно с 1 пиреноидом, выстилающий продольные
стенки клеток наполовину, реже полностью. Зооспорангии по форме
не отличаются от обычных вегетативных клеток, 4.7 мкм шир., 7 —
10 (17) мкм дл., отверстие для выхода зооспор 2 мкм в диам. Акинеты
эллипсоидные, с утолщенными оболочками, 4.5—5.5 мкм шир., 6 — 20 мкм
дл.
В планктоне стоячих водоемов. В СССР не обнаружен. (Швеция).
Редкий, слабо изученный вид. Принадлежность к роду Chlorhormi-
dium довольно сомнительна. Хорошо отличается от остальных видов
рода оболочками клеток, окрашенными в бурый цвет окислами железа,
сильно вытянутыми клетками и характерными зооспорангиями, весьма
сильно перетянутыми у поперечных перегородок. Образование зооспор
не прослежено.
6. Chlorhormidium pseudostichococcus (Heering) Fott (= Hormidium
pseudostichococcus Heering) — Хлоргормидиум ложностихококковый
(рис. 12, 16 18). — Нити большей частью короткие, легко распадаю-
щиеся на отдельные клетки. Клетки обычно цилиндрические, у попе-
речных перегородок явственно перетянутые, при распадении нитей
эллипсоидные, 2.5—3 мкм шир., по длине до 3(4.5) раз больше,
с тонкими оболочками. Хлоропласт бледно-желтовато-зеленый, с 1 пире-
ноидом.
На влажных кирпичных стенах и древесине образует зеленые налеты,
состоящие из коротких нитей или отдельных клеток, в воде обычно
встречаются длинные нити; обнаружен также в почвах. Европ. ч. СССР,
местами.
7. Chlorhormidium mucosum (Boye-Pet.) Starmach (—Hormidium
mucosum Boye-Pet.) — Хлоргормидиум слизистый (рис. 13, 1, 2).—
Нити длинные, прямые или слегка изогнутые, с ровной или беспоря-
дочно бородавчатой поверхностью. Клетки обычно цилиндрические, 8—
20 мкм шир., по длине равные ширине или укороченные, с довольно
толстой двуслойной оболочкой; внутренний слой оболочки целлюлозный,
сильно разбухающий под действием хлор-цинк-иода или едкого кали.
На сырой почве образует зеленые войлоковидные налеты, которые
при высыхании становятся бледно-зелеными. В СССР не обнаружен.
(Зап. Европа).
8. Chlorhormidium dissectum (Gay) Fareoqui [—Stichococcus dis-
sectus Gay; Hormidium dissectum (Gay) Chod.]. — Хлоргормидиум рас-
сеченный (рис. 13, 3—9). — Нити короткие (до 10 клеток), разнообразно
56
Рис. 13.
1,2 — Chlorhormidium mucosum: 1 — клетки с содержимым, 2 — фрагмент
нити с толстой оболочкой; 3—9 — С. dissectum: 3—5 — фрагменты нитей,
6 — нить, состоящая из клеток разного размера, 7 — отдельная клетка, 8 —
образование акинет, 9 — акинеты; 10, 11 — С. korschlkovil: 10 — нить с эпифи-
том Stipitochrysis monorhiza в слизистом влагалища нити, 11 — фрагмент
нити (слизь не изображена). (1, 2 — по Бойе-Петерсену, 3, 6,7 — по Фарукви,
4, 5, 8, 9 — по Ге, 10, 11 — по Коршикову).
изогнутые, легко распадающиеся на фрагменты или отдельные клетки.
Клетки обычно цилиндрические, у поперечных перегородок нередко
довольно сильно перетянутые, и тогда нити могут становиться четко-
видными, при распадении нитей эллипсоидные до почти шаровидных,
(5.6) 7—11.2 мкм шир., 5 — 23 мкм дл., с довольно толстыми оболочками.
Хлоропласт дисковидный, с лопастными, иногда слегка завернутыми
краями, выстилающий продольные стенки клеток наполовину или
на 3/i. Пиреноид крупный, отчетливый.
На влажных твердых субстратах образует темно- или ярко-зеленые
налеты, встречается также в различных почвах, наиболее характерен
для целинных дерново-подзолистых и торфяно-болотных почв. Ленин-
градская и Кировская обл., ЛатвССР, ЛитССР, БССР, УзССР, Зап.
Сибирь.
Самостоятельность этого вида до сих пор ставится под сомнение,
поскольку по многим признакам он очень близок к С. flaccidum.
Отличается клетками, обладающими более толстыми оболочками и за-
кругленными на концах нитей, а также очень легким распадением нитей
на отдельные клетки. Водоросль хорошо развивается в водных культурах,
57
засеянных почвой, образуя на дне сосудов рыхлые хлопья. В молодых
культурах клетки короткие, в стареющих удлиненные, заполненные
зернами запасного питательного вещества (акинеты?).
9. Chlorhormidium korschikovii Hollerb. (см. с. 343) (—Hormidium
mucosum Korsch, non H. mucosum Boye-Pet.) — Хлоргормидиум Kop-
шикова (рис. 13, 10, 11). — Нити длинные, прямые или слегка
изогнутые, окруженные толстым слизистым влагалищем. Клетки ци-
линдрические, у поперечных клеточных перегородок не перетянутые,
9.5 мкм шир., 13—15.5 мкм дл., с тонкими нежными оболочками.
Хлоропласт тонкий, нежный, почти эллиптический, с ровными краями
и утолщенной средней частью, содержащей пиреноид, выстилающий
продольные стенки клеток почти полностью в длину и до половины
по окружности; перед делением клеток он делается выемчатым в сере-
дине. По концам клеток 2 очень крупные вакуоли, между ними отчетливо
выраженное ядрб, прижатое к клеточной стенке. Наружное слизистое
влагалище очень толстое, до 35 мкм, по обе стороны нити. Размноже-
ние не прослежено.
В болотистых водоемах и озерах. Карельская АССР (Ковда), Горь-
ковская обл.
По наличию очень толстого слизистого влагалища этот интересный
вид резко отличается от остальных представителей рода. К сожалению,
он долго оставался незамеченным, так как был описан А. А. Корти-
ковым в 1941 г. в подстрочном примечании как Hormidium mucosum
в статье, посвященной флагеллатам, будучи субстратом для поселения
Stipitochrysls monorhiza. Он нами переименован как гомоним. Не исклю-
чено, что он все же ближе к роду Ulothrix.
Род GLOEOTILA Kutz. - ГЛЕОТИЛА
Нити однорядные, простые, короткие или длинные, нередко окру-
женные слизистым влагалищем, большей частью незаметным без окраши-
вания или туши. Клетки цилиндрические или эллипсоидные, реже шаро-
видные, у поперечных перегородок не перетянутые или слегка перетяну-
тые. Оболочки тонкие, нежные. Хлоропласт без пиреноида, обычно
пластинчатый, дисковидный, прилегающий к одной из сторон продольных
стенок клеток в форме полуцилиндра или, реже, узкой, спирально
изогнутой ленты. Акинеты в форме отдельных округлившихся клеток.
Зооспоры двужгутиковые, по 1 в клетке. Половое размножение не-
известно.
В пресных стоячих и проточных водах, в планктоне и обрастаниях,
одиночно или в скоплениях.
I. Нити спирально изогнутые.
1. Клетки 1 мкм шир., у поперечных перегородок не перетя-
нутые..............................................G. spiroides 1.
2. Клетки 1.5—2.6 мкм шир., у поперечных перегородок перетя-
нутые ...........................................G. spiralis 2.
II. Нити прямые или разнообразно слабо изогнутые.
1. Клетки 1—4 мкм шир., слизистое влагалище имеется, гомогенное.
А. Слизистое влагалище большей частью заметно без окраши-
вания.
а. Нити короткие, клетки по длине в 2—4 раза больше
ширины; хлоропласт пластинчатый, дисковидный ....
................................... . . G. curta 5.
58
б. Нити длинные, клетки по длине в 10—15 раз больше
ширины; хлоропласт лентовидный, слабо спирально изог-
нутый ........................................G. turfosa 6.
Б. Слизистое влагалище без окрашивания не заметно. Клетки
по длине в 3—25 раз больше ширины . . . . G. pelagica 7.
2. Клетки 3—5 мкм шир., слизистое влагалище имеется, фиброзной
структуры, или отсутствует.
А. Слизистое влагалище отсутствует.
а. Клетки по длине до 12 раз больше ширины, у поперечных
перегородок не перетянутые . . . . G. scopulina 3.
б. Клетки по длине до 3 раз больше ширины, у поперечных
перегородок слегка перетянутые . . . . G. protogenlta 4.
Б. Слизистое влагалище имеется, широкое, фиброзной струк-
ту ры .......................................G. pulchra 8.
3. Клетки 4.5—10 мкм шир., слизистое влагалище имеется, гомо-
генное, или отсутствует.
А. Клетки шаровидные или эллипсоидные; слизистое влагалище
имеется, довольно широкое, гомогенное . . . G. mucosa 10.
Б. Клетки цилиндрические; слизистое влагалище имеется, но без
окраски не заметно, или отсутствует.
а. Слизистое влагалище имеется.
а. Клетки 4.5—6.5 мкм шир., по длине равные ширине
или немного больше.........................G. pallida И.
р. Клетки 5—10 мкм шир., по длине равные ширине или
до 3 раз больше............................G. caldaria 12.
б. Слизистое влагалище отсутствует; клетки 6.8—7.9 мкм
шир., по длине в 2—4 раза больше ширины, в средней
части слегка суженные...................G. subconstricta 9.
1. Gloeotila spiroides (G. S. West) Printz (= Ulothrix spiroides
G. S. West) — Глеотила спиралевидная (рис. 14, 1). — Нити спирально
изогнутые, с растянутыми витками; глубина спирали 17—23 мкм, рас-
стояние между витками 23—35 мкм. Клетки цилиндрические, 1 мкм
шир., 4.5—8.5 мкм дл., у поперечных перегородок не перетянутые.
Хлоропласт пластинчатый, выстилающий продольные стенки клеток
весьма значительно.
В планктоне. В СССР не обнаружена. (Англия).
По внешнему облику вид очень сходен с G. spiralis, хорошо отли-
чается более тонкими нитями, правильными удлиненно-цилиндриче-
скими клетками без перетяжек у поперечных перегородок.
2. Gloeotila spiralis Chod. (= G. contorta Chod.) — Глеотила спи-
ральная (рис. 14, 2). — Нити спирально изогнутые, с растянутыми
витками, иногда окруженные едва заметным гомогенным слизистым
влагалищем. Клетки цилиндрические, 1.5—2.6 мкм шир., по длине
до 5 раз больше, у поперечных перегородок отчетливо перетянутые,
с тонкими оболочками, окрашивающимися от хлор-цинк-иода в светло-
фиолетовый цвет. Хлоропласт пластинчатый, выстилающий продольные
стенки клеток лишь частично.
В стоячих и проточных водах, в обрастаниях среди других водо-
рослей и в планктоне. Во многих районах СССР.
От весьма близкого G. spiroides отличается несколько большей
шириной клеток и отчетливыми перетяжками у поперечных перего-
родок.
59
Рис. 14.
1 — Gloeotila spiroides; 2 — G. spiralis; 3 — G. scopulina; 4—6 — G. protogenita'.
4 — вегетативная нить, 5 — нить с акинетами, 6 — образование зооспор. (1 —
по Весту, 2 — по Шода, 3 — по Газену, 4 — 6 — по Борзи).
3. Gloeotila scopulina (Hazen) Heering (^Stichococcus scopulinus Ha-
zen) — Глеотила скалистая (рис. 14, 3). — Нити длинные, прямые или
очень слабо изогнутые. Клетки цилиндрические, 3—3.5 мкм шир.,
по длине до 12 раз больше, у поперечных перегородок не перетянутые.
Хлоропласт лентовидный, узкий, с довольно отчетливо изрезанными
краями. Зооспоры часто.
В стоячих и проточных водах, реже в почвенных культурах,
на влажных скалах образует зеленые скользкие налеты. Во многих
районах СССР.
Вид хорошо отличается узким лентовидным хлоропластом с изре-
занными краями.
'60
4. Gloeotila protogenita Kiitz. — Глеотила протогенитная (рис. 14,
4—6). — Нити длинные, прямые или слегка изогнутые. Клетки цилинд-
рические, 3.5—5 мкм шир., по длине обычно до 3 раз больше, у попе-
речных перегородок слегка перетянутые. Хлоропласт в виде пояска,
широкий, с ровными краями. Акинеты бочонковидные. Зооспоры
часто, эллипсоидные, 2.5—3 мкм шир., 3—4 мкм дл. Половое размножение
неизвестно.
В стоячих и проточных водах, на влажной почве, иногда на крышах
домов. Европ. ч. СССР, КазССР, Зап. Сибирь.
От весьма близкой G. scopulina отличается более короткими клет-
ками, слегка перетянутыми у поперечных перегородок, и более широким
хлоропластом с ровными четкими краями.
5. Gloeotila curta Skuja — Глеотила укороченная (рис. 15, 1). —
Нити короткие, прямые или слегка изогнутые, обычно окруженные
узким, 5—5.8 мкм, гомогенным слизистым влагалищем. Клетки ци-
линдрические, 2—2.8 мкм шир., по длине в 2—4 раза больше, у попе-
речных перегородок отчетливо перетянутые. Хлоропласт пластинчатый,
выстилающий продольные стенки клеток обычно почти полностью.
В планктоне. В СССР не обнаружена. (Зап. Европа).
6. Gloeotila turfosa Skuja — Глеотила торфяная (рис. 15, 2).—
Нити обычно длинные, прямые или слегка изогнутые, окруженные
гомогенным слизистым влагалищем, 18—20 мкм шир. Клетки цилиндри-
ческие, сильно вытянутые, 1.8—4 мкм шир., 11—20 (57) мкм дл., у по-
перечных перегородок слегка перетянутые. Хлоропласт лентовидный,
узкий, спирально изогнутый на 1—2 оборота. Зооспоры не наблюдались.
В планктоне озер, чаще в болотах. В СССР не обнаружена.
(Швеция).
Хорошо отличается от других видов рода очень сильно вытяну-
тыми клетками и лентовидным, спирально изогнутым хлоропластом.
7. Gloeotila pelagica (Nyg.) Skuja (= Ulothrix pelagica Nyg.) —
Глеотила пелагическая (рис. 15, 3). — Нити длинные, обычно прямые,
изредка слегка спирально изогнутые, без заметного слизистого влага-
лища, обнаруживаемого только при окрашивании или в туши. Клетки
цилиндрические, 1 — 3 мкм шир., 6—28 мкм дл., у поперечных перего-
родок слегка перетянутые. Хлоропласт пластинчатый, более или менее
поясковидный, широкий, выстилающий продольные стенки клеток при-
мерно на 3/4. В протоплазме встречаются мелкие зернышки крахмала.
В планктоне озер. В СССР не обнаружена. (Зап. Европа).
Отличается от G. turfosa широким, более или менее поясковидным
хлоропластом.
8. Gloeotila pulchra Skuja — Глеотила красивая (рис. 15, 4). —
Нити длинные, окруженные очень широким, около 100 мкм, слизистым,
радиально исчерченным фиброзным влагалищем; у поперечных перего-
родок исчерченность отсутствует. Клетки цилиндрические, 3.5—4 мкм
шир., 20—40 мкм дл., прямые или слегка изогнутые, на концах закруг-
ленные. Хлоропласт лентовидный, тонкий, обычно слегка спирально
изогнутый. В маленьких вакуолях, расположенных у полюсов клеток,
встречаются мелкие зернышки.
В планктоне озер. В СССР не обнаружена. (Швеция).
От остальных видов рода хорошо отличается широким, радиально
исчерченным слизистым влагалищем.
61
Рис. 15.
1 — Gloeotila curta; 2 — G. turfosa', 3 — G. pelaglca; 4 — G. pulchra', 5 — G. sub-
constricta; 6 — G. mucosa; 7 — G. pallida; 8 — G. caldaria. (1—4 — по Скуе,
5 — по Весту, 6—8 — по Кютцингу).
9. Gloeotila subconstricta (G. S. West) Printz (— Ulothrix subcon-
stricta G. S. West) — Глеотила слабоперетянутая (рис. 15, 5). —
Нити длинные, большей частью прямые, без слизистого влагалища.
Клетки цилиндрические, 6.8—7.9 мкм шир., по длине в 2—4 раза
больше, в средней части слегка суженные, к концам несколько
расширенные, у поперечных перегородок незначительно перетянутые.
Хлоропласт пластинчатый, узкий, со слегка изрезанными краями,
выстилающий продольные стенки клеток примерно на ‘/з в средней части.
В стоячих водах, чаще всего в прудах. В СССР не обнаружена.
(Зап. Европа, Сев. Америка).
От остальных видов рода отличается формой клеток и более круп-
ными их размерами.
10. Gloeotila mucosa Kiitz. — Глеотила слизистая (рис. 15, 6). —
Нити длинные, четковидные, окруженные гомогенным слизистым вла-
галищем. Клетки почти шаровидные или удлиненно-эллипсоидные, 4.5—
7.5 мкм шир., по длине равные ширине или несколько больше. Хлоро-
пласт широкий, поясковидный, выстилающий продольные стенки клеток
почти полностью.
В стоячих водах, в планктоне, изредка в почве. БССР, КазССР.
62
11. Gloeotila pallida Kiitz. — Глеотила бледная (рис. 15, 7). —
Нити слизистые, светло-зеленые. Клетки цилиндрические, 4.5—6.5 мкм
тир., по длине равные ширине или несколько больше.
В стоячих водах, в планктоне. УССР.
Сомнительный малоизученный вид. По размерам клеток весьма
сходен с предыдущим видом, хорошо отличается отсутствием
слизистого влагалища и формой клеток.
12. Gloeotila caldaria Kiitz. (= Ulothrix caldaria Collins) — Глеотила
тепличная (рис. 15, 8). — Нити длинные, прямые или слегка изогнутые,
нежные, слизистые, блестящие, светло- или темно-зеленые. Клетки
цилиндрические, 5—10 мкм шир., по длине равные ширине или, чаще,
до 3 раз больше, у поперечных перегородок не перетянутые или
слегка перетянутые. Хлоропласт широкий, дисковидный, выстилающий
продольные стенки клеток почти полностью.
В теплицах и теплых источниках. В СССР не обнаружена.
(Зап. Европа).
Вид очень близкий к G. protogenita, хорошо отличается размерами
и местообитанием.
Род HORMIDIOPSIS Heering - ГОРМИДИОПСИС
Нити короткие, однорядные, иногда местами двурядные. Клетки
эллипсоидные, квадратные или прямоугольные, иногда шаровидные или
неправильной формы. Хлоропласт пристенный, пластинчатый, с пире-
ноидом или без него. Распадение нитей сопровождается одновремен-
ным ослизнением их. Апланоспоры известны.
Типичный аэрофит.
Род сомнительный, еще малоизученный, с трудом идентифицируе-
мый в массе зеленых водорослей, поселяющихся на коре деревьев
или почве. Возможно, представляет собой лишь стадии развития
других нитчаток.
I. Клетки квадратные до прямоугольных, с округленными углами;
пиреноид отсутствует.............................Н, crenulatum 1.
II. Клетки только эллипсоидные; пиреноид имеется.................
...........................................Н. ellipsoideum 2.
1. Hormidiopsis crenulatum (Kiitz.) Heering (=Ulothrix crenulata
Kutz.; Hormidium crenulatum Kutz. sec. Brand) — Гормидиопсис мелко-
городчатый (рис. 16, 1 — 4). — Нити волнистые, довольно прочные, одно-
рядные, местами двурядные. Клетки квадратные до прямоугольных,
с округленными углами, иногда шаровидные или неправильной формы,
9—15 мкм шир., 5—30 мкм дл., у поперечных перегородок слегка
перетянутые. Хлоропласт массивный, с ровными или лопастными
краями, без пиреноида, выстилающий продольные стенки клеток
полностью. Продукт ассимиляции — масло. Оболочки двуслойные, вну-
тренний слой толстый, внешний тонкий, при высыхании нитей еле
заметный, во влажных условиях сильно разбухающий, и тогда нити
оказываются заключенными в толстый слой слизи. Ослизнение
внешнего слоя оболочки часто сопровождается усиленной фрагментацией
нитей, благодаря чему образуется беспорядочное скопление клеток,
окруженных слизью. Разрыву нитей способствует отмирание в них
некоторых клеток, принимающих при этом менисковидную форму.
Апланоспоры шаровидные, 8—9 мкм в диам., по 8 и более в любой
63
Рис. 16.
1—4 — Hormidiopsis crenulatum: 1—3 — разновозрастные нити, 4 — образова-
ние апланоспор; 5—8 — Н. ellipsoideum. (1—4 — по Бранду, 5—8 — по Прес-
котту) .
клетке нити; они освобождаются вследствие разрыва и ослизнения
оболочек материнских клеток и прорастают без периода покоя, при этом
могут делиться сразу на 4 клетки.
На коре деревьев и влажной земле образуют тонкие налеты, встреча-
ются также на игольчатых листьях пихты белой (Abies alba). ЛатвССР,
Кавказ.
2. Hormidiopsis ellipsoideum Prescott — Гормидиопсис эллипсоид-
ный (рис. 16, 5—8). — Нити слизистые, преимущественно очень
короткие. Клетки эллипсоидные, расположенные перпендикулярно к про-
дольной оси нити, поэтому ширина их больше длины, 8 мкм шир.,
5.5 мкм дл., соединенные в группы по 4, каждая из которых окружена
широким бесцветным гомогенным слизистым влагалищем д'о 14.8 мкм
шир. Хлоропласт с загнутыми, но несмыкаютцимися краями, с пире-
ноидом, выстилающий продольные стенки клеток только частично.
Оболочки тонкие. Бесполое и половое размножение не наблюдалось.
В торфяных болотах, среди других водорослей. В СССР не обна-
ружен. (США).
64
Род CHLOROSPIRA Korsch. - ХЛОРОСПИРА
Нити однорядные, простые, спирально изогнутые, без слизистого
влагалища, свободноживущие. Клетки цилиндрические или эллипсоид-
ные, с широко закругленными концами. Хлоропласт пластинчатый,
выстилающий продольные стенки клеток в форме полушара, без
пиреноида. Продукт ассимиляции — крахмал или масло. Зооспоры
и гаметы неизвестны.
Типичный фитопланктер. Виды Chlorospira на первый взгляд можно
спутать с некоторыми видами Gloeotila со спирально извитыми
нитями, отличаются более короткими нитями.
I. Клетки 2—2.3 мкм шир., 4.7 —6.5 мкм дл........С. irregularis 1.
II. Клетки 5 мкм шир., 10—11 мкм дл....................С. crassa 2;
1. Chlorospira irregularis Korsch. — Хлороспира неправильная
(рис. 17, 7). — Нити большей частью короткие, беспорядочно
спирально изогнутые, с плотно сдвинутыми витками, часто спутанные
в клубок. Клетки короткоцилиндрические или эллипсоидные, иногда
слегка изогнутые, 2—2.3 мкм шир., 4.7—6.5 мкм дл., обычно по 3 на каж-
дый виток спирали. Хлоропласт широкий, с вырезом посередине,
выстилающий продольные стенки клеток почти полностью.
В стоячих водах, в планктоне. УССР (окр. Харькова).
2. Chlorospira crassa Korsch. — Хлороспира толстая (рис. 17, 2). —
Нити короткие, правильно спирально изогнутые, с широко раздвинутыми
витками, глубина спирали около 20 мкм, расстояние между витками
около 17 мкм. Клетки цилиндрические, с тупо закругленными концами,
5 мкм шир., 10—11 мкм дл., по 4 на каждый виток спирали. Хлоропласт
широкий, выстилающий продольные стенки клеток полностью, чуть
не доходя до их концов.
В проточных водах, в планктоне. РСФСР (Горьковская обл.,
р. Ока).
Род GEMINELLA Тигр. - ГЕМИНЕЛЛА
(= Hormospora Nag.)
Нити однорядные, простые, короткие или длинные, окруженные
толстым слизистым влагалищем, гомогенным или радиально исчер-
ченным, обычно хорошо видимым без окрашивания, свободноживущие.
Рис. 17.
1 — Chlorospira irregularis', 2 — С. crassa. (По Коршикову).
5 Зеленые водоросли. Улотриксовые
65
Клетки эллипсоидные, реже цилиндрические, равно удаленные друг
от.друга, попарно сближенные или последовательно соединенные друг
с другом преимущественно частью поперечных перегородок. Хлоропласт
’ поясковидный, с ровными или слегка изрезанными краями, иногда диско-
видный, с пиреноидом или, реже, без него. Дочерние клетки
после деления некоторое время лежат сближённо, что особенно отчет-
ливо заметно у форм с рассеянным размещением клеток. При неблаго-
приятных условиях иногда наблюдается деление клеток в трех направ-
лениях, в результате чего внутри слизистого влагалища возникает
беспорядочное скопление клеток, которые после освобождения прорас-
тают в новые нити. Зооспоры с точностью неизвестны. Акинеты
с утолщенными бурыми оболочками (известны только у G. interrupta).
Преимущественно в пресных стоячих и проточных водах, нередко
в болотах, канавах, на влажной земле, иногда в морских водах.
I. В воде.
1. Оболочки клеток ровные.
А. Клетки в слизистых влагалищах отчетливо разобщенные.
а. Клетки равноудаленные друг от друга....................
........................................G. ordinata 1.
б. Клетки попарно сближенные. —
а. Клетки цилиндрические, 3 мкм шир....................
................................G. subtilissima 2.
Рр. Клетки эллипсоидные,-4.7—8 мкм шир................
...................................G. interrupta 3.
Б..Клетки в слизистых влагалищах чаще всего соприкасающиеся
друг с другом.
а. Влагалища до 20 мкм шир., гомогенные.
а. Клетки 2.5—3 мкм шир................. G. smithii 4.
р. Клетки (2) 4—8 (10) мкм шир. . . . . . . G. minor 5.
б. Влагалища (20) 30—50 мкм шир., гомогенные, реже фиб-
розные.
а. Клетки 4—7 мкм шир..................G. planctonica 8.
р. Клетки 12—20 мкм шир..................G. mutabilis 6.
в. Влагалища свыше 100 мкм шир., гомогенные, клетки
4.5—5 мкм шир..........................G. monospora 7.
2. Оболочки клеток волнистые или с вмятинами....................
............................................G. crenulatocollis 9.
II. На влажной почве; клетки 5.5 мкм шир., 4.4—8.5 мкм дл..........
................................................G . terricola 10.
1. Geminella ordinata (W. et G. S. West) Heering (= Hormospora
ordinata W. et G. S. West) — Геминелла правильная (рис. 18, 1). —
Нити прямые или слегка изогнутые, с гомогенным слизистым влага-
лищем. Клетки эллипсоидные, 5.4—5.9 мкм шир., равноудаленные друг
от друга. Хлрропласт небольшой, без пиреноида.
В стоячих водах. УССР (Крымская обл., в предгорных районах).
2. Geminella subtilissima (Lagerh.) Printz (= Hormospora subti-
lissimd Lagerh.) — Геминелла тончайшая (рис. 18, 2). — Нити изогнутые,
реже прямые, с гомогенным слизистым влагалищем. Клетки цилиндри-
ческие, на концах закругленные, 3 мкм шир., 9 — 12 мкм дл., в молодых
нитях сближенные попарно, в старых сильно удаленные друг от друга.
Хлоропласты в числе 1—2, дисковидные, без пиреноида.
В планктоне прудов. В СССР не обнаружена. (Зап. Европа).
66
Рис. 18.
1 — Geminella ordinata', 2 — G. subtilissima', 3—6 — G. Interrupta: 3 — общий
вид нити, 4, 5 — образование акинет, 6 — зрелые акинеты; 7 — G. smithii.
(.1 — по Весту, 2—6 — по Лагергейму, 7 — по Буррелли).
3. Geminella interrupta (Тигр.) Lagerh. — Геминелла прерывистая
(рис. 18, 3—6). — Нити более или менее прямые, большей частью
длинные, с гомогенным слизистым влагалищем 10—18 мкм шир., ровным
или местами с легкими перетяжками. Клетки эллипсоидное, 4.7—
6 (8) мкм шир., 5.5—15 мкм дл., непосредственно после деления
сближенйые попарно, впоследствии удаленные друг от друга. Хлоропласт
довольно крупный, без пиреноида. Акинеты эллипсоидные, 9—10 мкм
шир., до 15 мкм дл., с довольно толстой шероховатой бурой оболочкой.
В пресных стоячих и проточных водах, в обрастаниях между другими
водорослями, иногда в планктоне. Местами в разных районах СССР.
Слабо изученный вид, по внешнему облику чрезвычайно сходный
со стадиями развития Geminella planctonica (Bolochonz.) Tiwary et
Pandey.
5»
67
4. Geminella smithii Bourr. — Геминелла Смита (рис. 18, 7). —
Нити короткие, из 4—8 клеток, разнообразно изогнутые, с гомогенным
слизистым влагалищем 15 — 20 мкм шир., свободноживущие. Клетки
цилиндрические, 2.5—3 мкм шир., 5—8 мкм дл., на концах закруг-
ленные, соприкасающиеся друг с другом. Хлоропласт небольшой, с пире-
ноидом.
В планктоне прудов. В СССР не обнаружена. (Франция).
5. Geminella minor (Nag.) Heering (= Hormospora minor Nag.;
H. mutabilis Nag. var. minor Hansg.) — Геминелла меньшая (рис. 19,
7). — Нити прямые или слегка изогнутые, с гомогенным слизистым
влагалищем 8—16 (18) мкм шир. Клетки цилиндрические, (2) 4—
8 (10) мкм шир., 3—14 (25) мкм дл., у поперечных перегородок
заметно перетянутые, обычно соприкасающиеся друг с другом. Хлоро-
пласт более или менее поясковидный, с пиреноидом.
В стоячих, реже проточных водах. ЭССР, ЛатвССР, УССР, Красно-
дарский край, КазССР, Ср. Азия, Зап. Сибирь, Тувинская АССР.
6. Geminella mutabilis (Nag.) Wille (= Hormospora mutabilis
Nag.) — Геминелла изменчивая (рис. 19, 2, 3). — Нити более или менее
прямые, с гомогенным слизистым влагалищем до 46 мкм шир. Клетки
эллипсоидные или цилиндрические, закругленные на концах, 12»
20 мкм шир., по длине равные ширине или до 2 раз больше, сопри-
касающиеся друг с другом или, реже, равномерно и незначительно
разобщенные.
В болотах и канавах, среди других водорослей, мхов, водных цвет-
ковых растений. ЛатвССР, ЛитССР, Чукотский авт. окр., Тувинская
АССР.
7. Geminella monospora J. W. G. Lund — Геминелла односпоровая
(рис. 19, 4, 5). — Нити короткие, из 4 (12) клеток, изогнутые, с гомоген-
ным слизистым влагалищем более 100 мкм шир. Клетки цилиндри-
ческие, 4.5—5 мкм шир., 14—40 мкм дл., на концах закругленные,
слегка разобщенные друг от друга. Хлоропласт поясковидный, без
пиреноида. Апланоспоры шаровидные, 5—7 мкм в диам., по 1 в'клетке,
освобождающиеся через отверстие в оболочке материнской клетки.
В планктоне озер. В СССР не обнаружена. (Англия).
8. Geminella planctonica (Bolochonz.) Tiwary et Pandey (=Gloeo-
tila planctonica Bolochonz.). —Геминелла планктонная (рис. 20).—
Нити длинные или короткие, прямые или слегка изогнутые, со слизистым
влагалищем 20—50 мкм шир., гомогенным или, реже, радиально
исчерченным, ровным или с перетяжками, свободноживущие. Клетки
цилиндрические или слегка вздутые, (4) 6 — 7 мкм шир., по длине равные
ширине или в 1.5—3 раза больше, обычно соприкасающиеся друг
с другом полностью или группами по нескольку или разобщенные
поодиночке. Хлоропласт поясковидный, с ровными или слегка
изрезанными краями, без пиреноида, выстилающий продольные стенки
клеток наполовину. Апланоспоры известны.
Типичный фитопланктер.
Var. planctonica (рис. 20, 1—3). — Слизистое влагалище 30—35 мкм
шир., гомогенное. Клетки 6—7 мкм шир., 6—21 мкм дл., полностью
соприкасающиеся друг с другом.
В планктоне стоячих водоемов. Кольский п-ов, Карельская АССР,
Ладожское озеро, окр. Ленинграда.
68
Рис. 19.
1 — Geminella minor’, 2, 3 — G. mutabilis’, 4—5 — G. monospora: 4 — участок
нити в слизистом влагалище, 5 — одиночные клетки. (7 — по Буррелли, 2 —
по Весту, 3 — по Прескотту, 4, 5 — по Ланду).
Var. indicaTiwary et Pandey (рис. 20, 4—9). — Слизистое влагалище
20 — 50 мкм шир., гомогенное или радиально исчерченное, иногда только
на отдельных участках, ровное или с перетяжками. Клетки 4—7 мкм
шир., 5—12 мкм дл., соприкасающиеся друг с другом, иногда разобщен-
ные.
В планктоне рисовых чеков. В СССР не обнаружена. (Индия).
От типовой формы отличается более толстым слизистым влагалищем,
довольно часто радиально исчерченным на всем протяжении или
только на отдельных его участках.
69
70
9. Geminella crenulatocollis Prescott — Геминелла мелкогородчато-
волнистая (рис. 21, 1, 2). — Слизистое влагалище гомогенное, широкое.
Клетки эллиптические или квадратные, с волнистыми продольными
стенками, всегда попарно сближенные. Хлоропласт в виде неправильной,
иногда волнистой пластинки, выстилающий продольные стенки клеток
частично или, реже, полностью.
В торфяных болотах. В СССР не обнаружена. (США: Висконсин).
По форме и расположению клеток несколько напоминает G. mutabi-
lis, отличается неровными оболочками клеток и формой хлоропласта.
10. Geminella terricola Boye-Pet. — Геминелла наземная (рис. 21,
3). — Нити короткие, разнообразно изогнутые, с гомогенным слизистым
влагалищем 9.5—10 мкм шир. Клетки эллипсоидные или яйцевидно-
шаровидные, 5.5 мкм шир., 4.4—8.5 мкм дл., соприкасающиеся друг
с другом довольно плотно. Хлоропласт нередко со слегка изрезанными
краями, с пиреноидом, выстилающий продольные стенки клеток не пол-
ностью. Однорядность в размещении клеток иногда нарушается.
На влажной почве. БССР.
Рис. 21.
7, 2 — Geminella crenulatocollis} 3 — G. terricola. (1, 2 — по. Прескотту, 3 —
по Бойе-Петерсену).
Рис. 20.
Geminella planctonlca- 1—3 — var. planctonlca (7 — общий вид нити, 2, 3 —
образование апланоспор), 4—9 — var. indica (4 — общий вид нити, 5—8 —
фрагментация клеток в нитях, 9 — нить с радиальной штриховатостью слизи-
стого влагалища). (1—3 — по Болохонцеву, 4—9 — по Тивари и Пандей).
71
Род RADIOFILUM Schmidle - РАДИОФЦЛУМ
Нити однорядные, иногда многорядные, обычно простые, реже беспо-
рядочно разветвленные, с анастомозирующими ветвями, короткие
и хрупкие или длинные и гибкие, окруженные толстым слизистым
влагалищем гомогенной или фиброзной структуры, свободноживущие
или прикрепленные. Клетки линзовидно-шаровидные, эллипсоидные или
эллипсоидно-веретеновидные, почти чечевицеобразные, расположенные
наибольшим диаметром перпендикулярно к продольной оси нити,
разобщенные или соединенные друг с другом слизистыми тяжами.
Оболочка клеток состоит из двух половинок, при делении каждая
из дочерних клеток получает только одну из половинок оболочки
материнской клетки, а вторую образует наново. Хлоропласт пластин-
чатый, иногда колоколообразный, с 1—2 пиреноидами. Специализиро-
ванные клетки размножения достоверно не установлены.
Виды Radiofilum обитают главным образом в пресных стоячих
и проточных водах, в обрастаниях разных субстратов и в планктоне.
Своеобразие в строении оболочек и делении резко выделяет этот род
среди других представителей сем. Ulotrichaceae.
I. Клетки соединены при помощи прочного слизистого тяжа.8
1. Слизистое влагалище отчетливо двуслойное....................
...........................................R. conjunctivum 1.
2. Слизистое влагалище гомогенное...............R. paradoxum 2.
II. Клетки соединены только с помощью общего слизистого влагалища,
тяж между ними отсутствует.
1. Нити длинные, клетки расположены последовательно одна
за другой.
А. Слизистое влагалище только немного шире клеток; оболочки
клеток без экваториальной утолщенной кромки.................
..........................................R. flavescens 3.
Б. Слизистое влагалище намного шире клеток; оболочки клеток
с экваториальной утолщенной кромкой.........................
.......................................R. mesomorphum 4.
2. Нити короткие и благодаря способности клеток делиться в двух
направлениях часто становятся неправильными . ..................
. . . . .................................R. irregulare 5.
1. Radiofilum conjunctivum Schmidle — Радиофилум соединитель-
ный (рис. 22, 1, 2).— Нити короткие, однорядные, простые, очень
редко разветвленные, легко распадающиеся на фрагменты. Слизистое
влагалище толстое, около 25 мкм шир., состоящее из двух слоев:
внешнего более или менее гомогенного и внутреннего фиброзной
структуры с поперечным расположением фибрилл. Клетки округло-
линзовидные, почти шаровидные, 6 мкм шир., 4 мкм дл., обычно
раздвинутые и связанные между собой толстым слизистым тяжом.
После деления новые клетки некоторое время остаются сближенными,
слизистый тяж между ними возникает постепенно.
В стоячих водах — озерах, прудах, в обрастаниях различных суб-
стратов или оторванными в планктоне. ЛатвССР, РСФСР (Москов-
ская обл.), УССР.
8 Для достоверного установления наличия тяжей и строения слизи
нередко бывает необходимо применять окрашивание объекта.
72
Рис. 22.
1, 2 — Radiofilum conjunctivum: 1 — нить при малом увеличении, 2 — то же
при большом увеличении (после окраски); 3—7 — Я. paradoxum: 3, 4 — нити
разного строения, 5 — делящиеся клетки, 6 — формирование слизистого тяжа
при делении клетки, 7 — нить в слизистом влагалище. (1 — по Болину, 2 —
по Бруннталеру, 3—7 — по Шода).
73
74
Вид известен пока из немногих мест, но несомненно распространен
значительно шире. Хорошо отличается от других видов рода отчетливо
двуслойным слизистым влагалищем.
2. Radiofilum paradoxum (Chod. et Topali) Printz [= Interfilum
paradoxum Chod. et Topali; Geminella paradoxa (Chod. et Topali)
Printz] — Радиофилум парадоксальный (рис. 22, 3—7). — Нити одно-
рядные, простые или разветвленные. Слизистое влагалище гомогенное,
мягкое, без четкого контура, значительно шире клеток. Клетки почти
шаровидные, иногда слегка угловатые, 5—7 мкм шир., далеко раздви-
нутые и связанные между собой довольно толстым слизистым тяжом.
В стоячих водах, в слизи колоний Ophrydium versatile. В СССР
не обнаружен?* (Зап. Европа).
3. Radiofilum flavescens G. S. West — Радиофилум желтоватый
(рис. 23, 1, 2). — Нити однорядные, простые, длинные, прямые или
слегка изогнутые. Слизистое влагалище немного шире клеток, фиброз-
ной структуры. Клетки разобщенные, широкоэллипсоидные, 7.5—
10.5 мкм шир. Хлоропласт желтовато-зеленый.
В планктоне озер. ЛатвССР, РСФСР (Валдайское озеро).
4. Radiofilum mesomorphum Skuja — Радиофилум среднеформный
(рис. 23, 3, 4). — Нити однорядные, простые, длинные. Слизистое вла-
галище 33—40 мкм шир., с легкой радиальной исчерченностью или
почти гомогенное, без четкого контура. Клетки округло-линзовидные
до почти шаровидных, 13—17 мкм шир., 5—14 мкм дл., разобщенные,
с четко выраженной утолщенной экваториальной кромкой оболочки.
В планктоне озер. В СССР не обнаружен. (Зап. Европа).
Близок к R. conjunctivum, отличается отсутствием четкой дифферен-
циации слизистого влагалища на два слоя и отсутствием слизистого
тяжа, связывающего клетки. Некоторые клетки могут слегка выдви-
гаться в сторону, что иногда приводит к возникновению ложного
ветвления.
5. Radiofilum irregulare (Wille) Brunnth. (= Hormospora irregula-
ris Wille) — Радиофилум неправильный (рис. 23, 5—8). — Нити корот-
кие, обычно одно-, реже многорядные, простые или нередко беспоря-
дочно разветвленные, с анастомозами между ветвями. Клетки эллипсо-
идно-веретеновидные, (7)12 — 15 мкм шир., 4—6 мкм дл., способные
делиться в двух направлениях. Слизистое влагалище четко очерченное,
до 30 мкм шир., бесцветное, фиброзной структуры.
В стоячих водах — озерах, прудах, в обрастаниях и в планктоне.
ЭССР, ЛатвССР, УССР.
Один из наиболее изменчивых видов по форме и размерам слое-
вища.
Рис. 23.
1, 2 — Radiofilum flavescens-, 3, 4 — R. mesomorphum; 5—8 — R. irregulare:
5 — нить при малом увеличении, 6—8 — фрагменты нити при большем увеличе-
нии. (7, 2 — по Весту, 3, 4 — по Скуе, 5, 6 — по Вилле, 7, 8 — по Мёбиусу).
75
Род GEMINELLOPSIS Korsch. - ГЕМИНЕЛЛОПСИС
Нити однорядные, простые, короткие, с тонкостенным нежным
влагалищем, в котором клетки расположены на более или менее равных
расстояниях или парами. Клетки удлиненно-цилиндрические, округлые
на концах, с очень тонкой, плотно прилегающей к влагалищу
оболочкой. Хлоропласт пластинчатый, без пиреноида, прилегающий
к одной из продольных стенок клетки в виде полуцилиндра. Рост
нитей интеркалярный, с последующим расхождением поделившихся
клеток и удлинением влагалища. Зооспоры и гаметы неизвестны. Покоя-
щееся состояние — цисты.
В пресных или слабо минерализованных водоемах и стоячих водах.
Близок, с одной стороны, к Geminella, с другой — к Binuclearia,
отличается от первой отсутствием в хлоропласте пиреноида и толстого
Рис. 24.
1—4 — Gemtnellopsls fragtlis-. 1, 2 — общий вид нитей, 3 — образование цисты,
4 — отдельная клетка при большом увеличении; 5—9 — Binuclearia lauterbornii.
(1—4 — по Коршикову, 5 — по Скуе, 6, 7 — по Шмидле, 8, 9 — по Когану).
76
слизистого влагалища, от второй — отсутствием толстых слоистых по-
перечных перегородок между клетками. В плотной тонкостенной
трубке клетки отделены друг от друга пустыми участками. Концы
влагалища могут быть открытыми, если первая клетка находится на не-
котором расстоянии от конца, или же закрытыми, если первая клетка
лежит в самом конце влагалища.
1. Geminellopsis fragilis Korsch. — Геминеллопсис ломкий (рис. 24,
1—4). — Нити 100—400 мкм дл., из 8—32 (64) клеток, прямые или
почти прямые. Клетки цилиндрические, на концах округлые, 2.5
3 мкм шир., 13—14 мкм дл., расположенные одна от другой на расстоя-
нии 6_7 мкм. Хлоропласт с выемкой посередине, выстилающий продоль-
ные стенки клеток полностью. Цисты образуются из выходящих наружу
протопластов вегетативных клеток, шаровидные, с гладкой оболочкой.
В планктоне рек и прудов. РСФСР (Горьковская обл.: р. Ока),
УССР (Харьковская и Донецкая обл.).
Род BINUCLEARIA Wittr. - БИНУКЛЕАРИЯ
Нити однорядные, простые, с разной толщиной перегородок между
клетками, на ранних стадиях развития прикрепленные с помощью
полушаровидного целлюлозного дермоида, выделяемого базальной
клеткой, позднее свободноживущие. Клетки одноядерные, цилиндри-
ческие или эллипсоидные, расположенные равномерно или попарно
сближенные. Хлоропласт поясковидный, иногда кольцевидный, с полу-
круглыми вырезами, обращенными к полюсам клетки, вследствие
чего на концах ее образуются округлые пространства, заполненные
зернами или каплями жироподобного вещества. Пиреноид 1, располо-
женный в одной из боковых лопастей хлоропласта, не всегда заметный.
Продукт ассимиляции — маслоподобное вещество, встречаются и зерна
крахмала. Рост нитей интеркалярный, только что образовавшиеся при
делении клеток поперечные стенки тонкие, затем они постепенно утол-
щаются и нередко далеко отодвигают клетки друг от друга. Иногда
наблюдается образование вокруг старых нитей своеобразных студенистых
поясков. Акинеты известны. Зооспоры четырехжгутиковые, со стигмой,
по-видимому, по 4 в клетке; они освобождаются через отверстие
в боковых стенках. Апланоспоры шаровидные. Половое размножение
неизвестно.
Преимущественно пресноводные, реже солоноватоводные водоросли,
встречающиеся в стоячих и проточных водоемах, на почве и в поверх-
ностных слоях почвы. Строение протопласта изучено еще недостаточно.
Важнейшими таксономическими признаками являются ширина клеток,
соотношение между их длиной и шириной, характер расположения
клеток, а также толщина поперечных перегородок.
I. Клетки 2—4 мкм шир............................В. lauterbornii 1.
II. Клетки 5—36 мкм шир.
1. Клетки 5—13 мкм шир., обычно попарно сближенные............
.............................................В. tectorum 2.
2. Клетки 34 — 36 мкм шир., обычно равномерно удаленные друг
от друга........................................В. zaisanica 3.
1. Binuclearia lauterbornii (Schmidle) Pr.-Lavr. (= Planctonema
lauterbornii Schmidle; Binuclearia eriensis Tiff.; B. tenuis Kogan) —
Бинуклеария Лаутерборна (рис. 24, 5—9). — Нити прямые, реже слегка
77
Рис. 25.
1—6 — Binuclearia tectorum: 1 — вегетативная нить, 2 — образование акинет,
3 — прорастание акинет, 4, 5 — нити с олиготрофных водоемов, 6 — нить
с эвтрофных водоемов; 7—10 — Ulothrichopsis cylindrica: 7—9 — фрагменты
нитей с хлоропластами разных очертаний, 10 — нить с апланоспорами. (1 —
по Виттроку, 2, 3 — по Вилле, 4, 5 — по Скуе, 6 — по Мошковой, 7—10 —
по Вихманну).
изогнутые, большей частью из равномерно удаленных друг от друга
клеток, попарно сближенных только непосредственно после деления.
Клетки цилиндрические, на концах закругленные, или эллипсоидно-
цилиндрические, 2—4 мкм шир., по длине в 2—5 раз больше. Оболочки
тонкие, неслоистые, неослизняющиеся. Протопласт плотно прилегает
к оболочкам клеток. Акинеты неизвестны.
В планктоне рек, крупных озер, реже в континентальных солоно-
ватых и опресненных морских водах. Европ. ч. СССР, Таганрогский
залив Азовского моря, сев. Каспий, ТуркмССР, оз. Байкал.
78
Вид распространен, очевидно, значительно шире, чем указывается
в литературе. Близок к В. tectorum, отличается не только размерами,
но и местообитанием.
2. Binuclearia tectorum (Kiitz.) Beger (= Gloeotila tectorum Kiitz.;
Binuclearia tatrana Wittr.) — Бинуклеария покрытая (рис. 25, 1—6). —
Нити прямые или слегка изогнутые, большей частью из попарно сближен-
ных клеток, отделенных друг от друга разными по толщине поперечными
перегородками. Клетки эллипсоидные или эллипсоидно-цилиндрические,
5—13 мкм шир., по длине вдвое меньше или до 8 раз больше, у поперечных
перегородок более или менее перетянутые, с протопластом, неплотно
прилегающим к оболочкам. Пространство у поперечных перегородок
обычно заполнено слизью. Утолщенные поперечные перегородки у старых
растений могут выделять вокруг нитей студенистое вещество в форме
поясков, иногда сливающихся в сплошную слизистую муфту, из которой
выступают лишь конечные клетки нитей. Акинеты образуются путем
утолщения внутреннего слоя оболочек, длительное время остаются свя-
занными в нить и распадаются в результате кольцевидных разрывов
внешнего слоя оболочки клеток.
В мелких озерах, лужах и болотах, в обрастаниях растений, на
влажных каменных стенах, облицовывающих источники, изредка
в планктоне и в поверхностных слоях почвы, чаще всего ранней весной
и осенью. Европ. ч. СССР, местами.
Сообщения о нахождении В. tectorum в планктоне Волги и Байкала ,
(Прошкина-Лавренко, 1966), а также сведения о массовом развитии
этой водоросли в планктоне Днепра и особенно Днепро-Бугского лимана
(Приймаченко, 1953) нам кажутся сомнительными. Местообитаниями
В. tectorum являются чаще всего дистрофные водоемы.
3. Binuclearia zaisanica Skvortz. — Бинуклеария зайсанская. — Нити
прямые. Клетки цилиндрические, 34—36 мкм шир., по длине в 5—8 раз
больше, с толстыми поперечными перегородками.
В планктоне оз. Зайсан (Восточно-Казахстанская обл.).
Вид сомнительный, очень своеобразный, хорошо отличающийся
от других видов рода очень крупными размерами клеток.
Род ULOTHRICHOPSIS Wichmann - УЛОТРИХОПСИС
Нити однорядные, простые. Клетки цилиндрические, не перетяну-
тые или слегка перетянутые у поперечных перегородок. Хлоропласт
поясковидный, с 1 — 2 заворотами и четким пиреноидом. Рост нитей
интеркалярный, при делении клеток происходит разрыв внешнего, пекти-
нового слоя оболочки, располагающегося затем вокруг нити в виде
своеобразного кольца у поперечных стенок клеток. Продукт ассимиля-
ции — крахмал и волютин (?). Апланоспоры шаровидные, по 1—4
в клетке. Зооспоры и гаметы неизвестны.
1. Ulothrichopsis cylindrica Wichmann — Улотрихопсис цилиндри-
ческий (рис. 25, 7—10). — Клетки 7.5—8.5 мкм шир., по длине равные
ей, меньше или в 2.5 раза больше.
В солоноватых водах. В СССР не обнаружен. (Зап. Европа).
Вид сомнительный, слабо изученный, скорее всего — стадия раз-
вития других нитчаток.
79
Род GLOEOTILOPSIS Iyengar et Philipose -
ГЛЕОТИЛОПСИС
Нити однорядные, простые, короткие, без слизистого влагалища,
легко распадающиеся на малоклеточные фрагменты, свободноживущие.
Клетки цилиндрические, с широко закругленными концами. Хлоропласт
поясковидный, с отчетливым пиреноидом, выстилающий продольные
стенки клеток полностью и примерно на 3/4 по периметру. Зооспоры
двужгутиковые, яйцевидные, с палочковидной стигмой, по 1—8 (16)
в клетке; они освобождаются через разрыв оболочки. Апланоспоры
эллипсоидные, толстостенные. Половое размножение неизвестно.
1. Gloeotilopsis planctonica Iyengar et Philipose — Глеотилопсис
планктонный (рис. 26, 1—3). — Клетки 3—5 (7.5) мкм шир., 8—
24 (55) мкм дл.
В проточных лужах. В СССР не обнаружен. (Индия).
Рис. 26.
1—3 — Gloeotilopsis planctonica-. 1 — фрагмент нити, 2 — зооспора, 3 — аплано-
споры; 4 — Heterothrichopsls vlridis, вегетативные нити и образование аплано-
спор. (1—3 — по Иенгару и Филипозу, 4 — по Иенгару и Кантаме).
80
Род HETEROTHRICHOPSIS Iyengar et Kanthamma -
ГЕТЕРОТРИХОПСИС
(= Ulothrichopsis Iyengar et Kanthamma, non Wichmann)
Нити однорядные, простые, очень короткие, не более 4 клеток.
Клетки цилиндрические, на концах закругленные, с (1) 2—4 (8) хлоро-
пластами. Хлоропласты с отчетливым пиреноидом, выстилающие про-
дольные стенки клеток полностью и примерно на 3/ь по периметру.
Апланоспоры эллипсоидные до шаровидных. Зооспоры и гаметы не-
известны.
Род описан крайне недостаточно, только по материалу из культур.
1. Heterothrichopsis viridis Iyengar et Kanthamma — Гетеротрихоп-
сис зеленый (рис. 26, 4). — Нити 6.2—7.9 мкм шир., 15—33 мкм дл.
Из культуры водорослей на агаре. В СССР не обнаружен. (Индия).
Систематическое положение вида крайне неопределенное. Отличие
от Gloeotilopsis по числу хлоропластов легко объяснить условиями
культуры, как деление их при образовании апланоспор.
Род STICHOCOCCUS Nag. - СТИХОКОККУС
Нити однорядные, простые, короткие, легко распадающиеся на
одиночные или соединенные попарно клетки. Клетки цилиндрические,
прямые или слегка изогнутые, эллипсоидные, на свободных концах
обычно широко закругленные, реже почти шаровидные. Оболочки клеток
преимущественно тонкие, но способные ослизняться или образовывать
слизистый покров на поверхности нитей или клеток. Хлоропласт пластин-
чатый, нередко правильно дисковидный, с цельными или слегка из-
резанными краями, без пиреноида, прилегающий в виде полуцилиндра
к одной из продольных стенок клетки или, реже, к одному из ее полюсов.
В клетках часто образуются 2 полярно расположенные вакуоли. Раз-
множение только распадением нитей на отдельные клетки и поперечным
делением клеток надвое.
Преимущественно аэрофитные водоросли, встречающиеся также
в проточных и стоячих водах, главным образом в пленке поверхностного
натяжения. Виды рода нередко идентифицируются с трудом и требуют
критического пересмотра. Разными авторами они классифицируются
по-разному.
I. Клетки правильной формы, без вздутий.
1. Клетки по длине равны ширине или слегка превышают ее.
А. Клетки 0.5—0.7 мкм шир., 1 —1.5 мкм дл. . . . S. atomus 1.
Б. Клетки 1—2.5 мкм шир.
а. Клетки 1 — 1.5 мкм шир., 1.5—4 мкм дл...............
.....................................S. minutissimus 2.
б. Клетки 1.9—2.5 мкм шир., 6—8 мкм дл.................
..................................S. epiplanctonicus 3.
В. Клетки 2—4.5 мкм шир.
а. Клетки преимущественно шаровидные.
а. Клетки 2—4.5 мкм шир., 3.2—8 мкм дл.; в почве . . .
.........................................S. chodatii 4.
р. Клетки 3.5—4 мкм шир., 4—6 (10) мкм дл.; на извест-
няках ...............................S. chlorelloides 5.
6 Зеленые водоросли. Улотриксовые 81
б. Клетки преимущественно эллипсоидные, 2.5—3 мкм шир.,
4—6 мкм дл.; на скалах......................S. minutus 6.
в. Клетки преимущественно короткоцилиндрические, 2.6—
3 мкм шир., 2.8—4 мкм дл.; в воде и на влажной почве
..........................................S. lacustris 7.
2. Клетки по длине значительно превышают ширину, до 10 раз
и более.
А. Клетки до 3 мкм шир.
а. Свободные концы клеток закругленные.
а. Клетки 1.2—2.7 (3) мкм шир..............S. minor 8.
р. Клетки 2—3 мкм шир......................S. dubius 9.
б. Свободные концы клеток обычно срезанные, клетки 1.8—
2 мкм шир........................S. membranaefaciens 10.
Б. Клетки от 3 мкм шир. и более.
а. Клетки большей частью в рыхлых цепочках, 3—6 мкм шир.,
4.2— 12 мкм дл........................S. bacillaris И.
б. Клетки большей частью одиночные.
а. Клетки (2.5) 3.5—4 мкм шир., 8—12 мкм дл...........
.....................................S. palescens 12.
р. Клетки 5 мкм шир., 7—25 мкм дл. . . . S. fragilis 13.
3. Клетки по длине в 10—20 раз превышают ширину.
А. Клетки 1.8—2.7 мкм шир....................S. exiguus 14.
Б. Клетки 3—4 мкм шир........................S. mirabilis 15.
II. Клетки неправильно цилиндрические, со вздутиями в разных участ-
ках, иногда согнутые под углом, (3) 4.7—7.3 мкм шир.................
...............................................S . variabilis 16.
1. Stichococcus atomus Skuja — Стихококкус крохотный (рис. 27,
1). — Клетки одиночные или попарно связанные, 0.5—0.7 мкм шир.,
1—1.5 мкм дл., короткоцилиндрические, яйцевидные, на концах закруг-
ленные или слегка заостренные.
В озерах, в наннопланктоне. В СССР не обнаружен. (Зап. Европа).
Вид изучен недостаточно и, вероятно, легко просматривается.
2. Stichococcus minutissimus Skuja — Стихококкус наименьший
(рис. 27, 2).— Клетки одиночные, после деления некоторое время
попарно связанные, 1 — 1.5 мкм шир., 1.5—4 мкм дл., эллипсоидные,
короткоцилиндрические, на концах слегка закругленные или несколько
заостренные. Хлоропласт заполняет около половины просвета клетки.
В озерах, в наннопланктоне. В СССР не обнаружен. (Зап. Европа).
Отличия в размерах от S. atomus настолько незначительны, что,
скорее всего, это один и тот же вид.
3. Stichococcus epiplancticus Skuja — Стихококкус. эпипланктонный
(рис. 27, 3). — Клетки одиночные или попарно связанные, прикрепляю-
щиеся с помощью слизи к колониям Chrysosphaerella, 1.9—2.5 мкм шир.,
6—8 мкм дл., эллипсоидные или цилиндрические, на концах обрубленные
или слегка закругленные.
В озерах, преимущественно весной. В СССР не обнаружен. (Зап.
Европа).
От остальных видов рода отличается эпифитным способом су-
ществования.
4. Stichococcus chodatii (Bial.) Heering [=Diplosphaera chodatii
Bial. pr. p.; Stichococcus diplosphaera (Bial.) Chod.] — Стихококкус Шода
82
Рис. 27.
I — Stichococcus atomus', 2 — S. minutissimus; 3 — S. epiplancticus; 4 — S. cho-
datii', 5 — S. chlorelloides; 6 — S. minutus; 7 — S. lacustris; 8 — S. minor; 9 —
S. dubius; 10 — S. membranaefaciens. (1—3 — по Скуе, 4—6, 9 — по Гринтческо
и Петерфи, 7, 9, 10 — по Шода).
(рис. 27, 4). — Клетки одиночные, после деления некоторое время
попарно связанные, реже в нитях по 3—.5, 2—4.5 мкм шир., 3.2—
8 мкм дл., шаровидные, реже эллипсоидные, иногда расширенные на
одном и суженные до почти заостренных на другом конце. Хлоропласт
бледно-зеленый, дисковидный, с ровными краями, заполняющий просвет
клетки обычно частично, реже полностью.
В поверхностных слоях разных почв. Ленинградская обл., Зап.
Сибирь, местами.
Близок к <S. chlorelloides.
6*
83
5. Stichococcus chlorelloides Grintz. et Peterfi — Стихококкус хлорел-
ловидный (рис. 27, 5). — Клетки одиночные, реже попарно связанные,
3.5—4 мкм шир., 4—6 (10) мкм дл., преимущественно шаровидные или
эллипсоидные, реже широкоцилиндрические. Хлоропласт желто-зеленый,
дисковидный, заполняющий только часть клетки.
В расщелинах известняков. В СССР не обнаружен. (Румыния).
6. Stichococcus minutus Grintz. et Peterfi — Стихококкус маленький
(рис. 27, 6). — Клетки одиночные или в нитях по 3—5, 2.5—3 мкм шир.,
4—6 мкм дл., эллипсоидные, реже широкоцилиндрические. Хлоропласт
бледно-зеленый, дисковидный, расположенный большей частью у одного
из полюсов клетки.
На скалах. В СССР не обнаружен. (Румыния).
7. Stichococcus lacustris Chod. — Стихококкус озерный (рис. 27, 7). —
Клетки одиночные, после деления некоторое время попарно связанные,
2.6—3 мкм шир., 2.8—4 мкм дл., короткоцилиндрические, реже почти
шаровидные или несколько эллипсоидные. Хлоропласт дисковидный,
с ровными краями, заполняющий просвет клетки частично, реже пол-
ностью.
В воде и на влажной почве. В СССР не обнаружен. (Зап. Европа).
8. Stichococcus minor Nag. s. str. — Стихококкус меньший
(рис. 27, S). — Клетки одиночные, реже в рыхлых нитях по 2—4,
1.2—2.7 (3) мкм шир., 2.8—14.6 мкм дл., цилиндрические, с закруглен-
ными концами, иногда слегка изогнутые. Хлоропласт ярко-зеленый,
в виде изогнутой пристенной пластинки с ровными краями, занимающий
обычно 1 /з—'/s клетки. Почти во всех клетках имеются мелкие зернистые
вкрапления, которые с возрастом становятся все более четкими. У полю-
сов некоторых, наиболее крупных клеток хорошо заметны прозрачные
капли жира.
В поверхностных слоях разных почв, на влажных каменных и
деревянных стенах, коре деревьев и т. п. Во многих местах СССР.
9. Stichococcus dubius Chod. — Стихококкус сомнительный (рис. 27,
9). — Клетки одиночные или в нитях по 2—3, 2—3 мкм шир., 6—
10 мкм дл., цилиндрические, прямые, на концах закругленные или
слегка обрубленные. Хлоропласт дисковидный, с ровными краями.
Известен из культур. В СССР не обнаружен. (Зап. Европа).
Сомнительный малоизученный вид. По размерам близок к S. minor.
10. Stichococcus membranaefaciens Chod. — Стихококкус пленчато-
видный (рис. 27, 10). — Клетки одиночные или по 2 вместе, 1.8—
2 мкм шир., 5.5—8.6 (9) мкм дл., цилиндрические, прямые, иногда
слегка изогнутые, на концах более или менее прямо срезанные, реже
слегка закругленные, иногда эллипсоидные, с небольшим количеством
капелек жира. Хлоропласт дисковидный, с довольно изрезанными
краями.
Известен из культур. В СССР не обнаружен. (Зап. Европа).
Сомнительный вид. 11
11. Stichococcus baeillaris Nag. s. str. — Стихококкус палочковидный
(рис. 28,1). — Клетки одиночные или соединенные по 2—4 (10) в разно-
образно изогнутые, легко распадающиеся нити, 3—6 мкм шир., 4.2—
12 мкм дл., цилиндрические, прямые, на концах слегка закругленные
84
Рис. 28.
1 — Stichococcus bacillaris; 2 — S. pallescens; 3 — S. fragilis; 4 — S. exiguus;
5 — S. mirabilis; 6 — S. variabilis; 7 — Chodatia tetrallantoidea. {1—3 — no
Шода, 4, 5 — по Гринтческо н Петерфи, 6 — по Весту, 7 — по Коль).
85
или более или менее прямо срезанные. Хлоропласт дисковидный, с ров-
ными или слегка изрезанными краями, занимающий большую часть
просвета клетки.
В почве, на влажной почве, кирпичных, деревянных стенах, полу-
сгнившей древесине, реже в воде. Повсеместно.
12. Stichococcus pallescens Chod. — Стихококкус бледноватый
(рис. 28, 2). — Клетки одиночные, (2.5) 3.5—4 мкм шир., 8—12 мкм дл.,
цилиндрические, с закругленными или слегка срезанными концами,
прямые, иногда несколько изогнутые, реже эллипсоидные.
Известен из культур. В СССР не обнаружен. (Зап. Европа).
13. Stichococcus fragilis Gay — Стихоккокус ломкий (рис. 28, 3'). —
Клетки одиночные, реже в коротких, легко распадающихся нитях,
5 мкм шир., 7—25 мкм дл., цилиндрические, прямые, на концах более
или менее закругленные. Хлоропласт дисковидный, с волнистыми
краями, согнутый в форме полуцилиндра.
На оконных стеклах теплиц в виде нежного, слегка слизистого
зеленоватого налета. Кольский п-ов, ЛитССР.
14. Stichococcus exiguus Gem. — Стихококкус мизерный (рис. 28,
4). — Клетки одиночные, реже соединенные попарно, 1.8—2.7 мкм шир.,
(8) 15—20 (30) мкм дл., цилиндрические, прямые или слегка изогнутые,
на концах закругленные. Хлоропласт бледно-зеленый, в виде продольной
пластинки с небольшой выемкой посередине. В старых клетках на обоих
концах иногда заметны крупные вакуоли.
В поверхностных слоях разных почв, а также на плодовых телах
трутовиков. РСФСР (Ленинградская обл.), УССР.
15. Stichococcus mirabilis Lagerh. — Стихококкус удивительный
(рис. 28, 5). — Клетки одиночные, реже соединенные попарно, 3—
4 мкм шир., 8— 30 (50) мкм дл., цилиндрические, прямые или слегка
дуговидно изогнутые, на концах закругленные. Хлоропласт узкий, с из-
резанными краями. В стареющих клетках в большом количестве капельки
жира.
В стоячих водах, на пнях, в почвах, а также на оконных стеклах
теплиц. РСФСР (Кировская обл.), УССР.
16. Stichococcus variabilis W. et G. S. West — Стихококкус измен-
чивый (рис. 28, 6). — Клетки обычно одиночные, иногда по 2 и более
в коротких нитях, (3) 4.7—7.3 мкм шир., 6.3 —16.2 (32) мкм дл., не-
правильной формы, преимущественно цилиндрические, на концах за-
кругленные, с вздутиями в разных местах, реже согнутые под углом.
Хлоропласт с небольшими выемками по краям, заполняющий почти
весь просвет клетки.
На влажных камнях и в почвах. РСФСР (Ленинградская обл.),
БССР, УзССР.
Род CHODATIA Ко! - ШОДАТИЯ
Нити однорядные, простые, короткие, легко распадающиеся на
одиночные или соединенные попарно клетки. Клетки цилиндрические,
дуговидно изогнутые, на концах закругленные, реже обрубленные.
Оболочки клеток тонкие, бесцветные, покрытые нежным бесцветным
слоем слизи. Хлоропласт пластинчатый, желто-зеленый, с ровными
86
или слегка волнистыми краями, без пиреноида. На концах клеток
бывают заметны вакуоли. Продукт ассимиляции — масло. Размножение
только распадением нитей на отдельные клетки и поперечным делением
клеток надвое.
Криофильная водоросль.
1. Chodatia tetrallantoidea Koi — Шодатия четырехколбасовидная
(рис. 28, 7). — Клетки 1.5 мкм шир., 6—7 мкм дл.
На поверхности плотного снега. В СССР не обнаружена. (Зап.
Европа: Альпы).
Род RAPHIDONEMA Lagerh. emend. Hind. —
РАФИДОНЕМА
Нити однорядные, простые, короткие, из (2) 4—16 (32) клеток,
к обоим концам суженные, вытянуто-веретеновидные, прямые или слегка
изогнутые, без слизистого влагалища. Клетки цилиндрические, у по-
перечных перегородок не перетянутые или бочонковидные, перетянутые,
нередко до .почти шаровидных, на свободных концах нитей обычно
вытянутые, заостренные и нередко отогнутые в сторону, иногда на одном
конце тупые. Оболочки клеток тонкие, бесцветные. Хлоропласт пластин-
чатый, дисковидный или лентовидный, без пиреноида. В протоплазме
заметны капельки жира. Размножение только распадением нитей на
малоклеточные фрагменты; в месте разрыва нити клетки вначале округ-
ляются, затем удлиняются и заостряются.
Криофильные водоросли, обитающие преимущественно в высоко-
горных и арктических районах. К роду Raphidonema, следуя взглядам
Гиндака (Hindak, 1963), мы относим лишь те виды, которые в есте-
ственных условиях встречаются в виде коротких нитей. Остальные
виды этого рода перешли в род Koliella.
I. Конечные клетки нитей коротко заостренные йли тупо закругленные
...............................................R. brevirostre 1.
II. Конечные клетки нитей удлиненно заостренные.
1. Клетки 1 — 2.5 мкм шир.......................R. sabaudum 2.
2. Клетки (1) 3—4.5 (6) мкм шир....................R. nivale 3.
1. Raphidonema brevirostre Scherff. — Рафидонема короткокончико-
вая (рис. 29, 1, 2). — Нити из 4—8 (32) клеток, до 120 мкм дл., прямые,
реже слегка изогнутые. Клетки цилиндрические, 1—4 мкм шир., 0.8—
3 мкм дл., у поперечных перегородок не перетянутые или заметно
перетянутые. Конечные клетки чаще всего на одном конце тупые
или коротко заостренные, на другом удлиненно заостренные.
На снежных полях.
Var. brevirostre (рис. 29, 7). — Клетки 3—4 мкм шир., 1 — 3 мкм дл.
В горных районах. В СССР не обнаружена. (Зап. Европа: Татры,
Альпы).
Var. canadense Koi (рис. 29, 2). — Клетки 1 — 1.5 мкм шир., 0.8—
1.5 мкм дл.
В СССР не обнаружена. (Канада).
2. Raphidonema sabaudum Koi — Рафидонема сабаудская (рис. 29,
3, 4). — Нити из 8—16 (32) клеток, прямые или слегка S-образно
изогнутые. Клетки цилиндрические, 1—2.5 мкм шир., 0.6—2 мкм дл.,
87
88
заметно перетянутые у поперечных перегородок, конечные клетки обычно
одинаково удлиненные и заостренные.
На поверхности снега, чаще всего на фирновых полях.
Var. sabaudum (рис. 29, 3). —Клетки в средней части нитей 2 —
2.5 мкм шир., около 2 мкм дл., конечные клетки 1.5 мкм шир., 5.8 мкм дл.,
равномерно суженные и заостренные.
В высокогорных районах. В СССР не обнаружена. (Франция,
Монблан).
Var. minus Koi (рис. 29, 4). —Клетки в средней части нитей
1 —1.5 мкм шир., 0.6—1.2 мкм дл., конечные клетки резко суженные
и вытянутые в тонкие, шиповидно заостренные отростки, часто слегка
изогнутые в разные стороны.
В СССР не обнаружена. (Канада).
3. Raphidonema nivale Lagerh. — Рафидонема снеговая (рис. 29,
5, 6). — Нити из 2—4(7 и более) клеток, разнообразно изогнутые.
Клетки цилиндрические, (1) 3 — 6 мкм шир., конечные клетки постепенно
удлиненные и заостренные, нередко слегка изогнутые, или резко сужен-
ные и шиповидно вытянутые.
На поверхности снега и льда.
Var. nivale [= R. nivale (Lagerh.) Chod. sensu Printz] (рис. 29, 5). —
Нити из 3—8 клеток. Клетки 3—6 мкм шир., по длине в 2—4 раза
больше, конечные клетки равномерно суженные и заостренные.
На фирновых полях. Арктические о-ва, Кавказ, Амурская обл.
(верховья р. Зеи).
Var. taylori Koi (рис. 29, 6). — Нити из 2—4 клеток. Клетки
1 — 1.5 мкм шир., 2—7 мкм дл., конечные клетки резко суженные и
шиповидно вытянутые.
На поверхности снега. В СССР не обнаружена. (Канада).
Род CATENA Chod. - КАТЕНА
(non Catena Maslov, alg. fossil.)
Нити однорядные, простые, короткие, из 2—4 клеток, чаще клетки
одиночные. Клетки цилиндрические, с кольцевидными утолщениями
на обоих концах и коническими выростами в центре поперечных стенок.
Хлоропласт пластинчатый, ярко-зеленый, без пиреноида. Размножение
неизвестно.
Существенно отличается от других родов своеобразными кольце-
видными утолщениями и коническими выростами на поперечных стенках.
Систематическое положение водоросли неясно, некоторыми исследовате-
лями относится к хлорококковым водорослям.
1. Catena viridis Chod. — Катена зеленая (рис. 29, 7). —Клетки
3—4 мкм шир., 4—6 мкм дл., соединяющиеся друг с другом коническими
выростами.
Рис. 29.
1,2 — Raphidonema brevirostre: 1 — var. brevirostre, 2 — var. canadense; 3,4 —
R. sabaudum: 3 — var. sabaudum, 4 — var. minus; 5, 6 — R. nivale: 5 — var.
nivale, 6 — var. taylori; 7 — Catena viridis. (1 — по Шерффелю, 2—4, 6 —
по Коль, 5 — по Лагерхейму, 7 — по Шода).
89
В пресных водах, в планктоне. В СССР не обнаружена. (Зап.
Европа).
Редкий вид, своеобразием облика резко отличающийся от всех
остальных представителей семейства. Изучен еще крайне недостаточно.
Род KOLIELLA Hind. - КОЛИЕЛЛА
(= Raphidonema Lagerh. р. р.; Ankistrodesmus Corda р. р.;
Raphidium Kiitz. р. р.)
Нити временные, возникающие в процессе размножения, не более
чем из 8 клеток, обычно клетки одиночные или, реже, соединенные
попарно. Клетки веретеновидные, или веретеновидно-цилиндрические,
прямые или изогнутые, часто дуговидно, иногда S-образно или спирально,
на концах заостренные, шиловидно или волосовидно вытянутые, реже
закругленные. Оболочки тонкие, бесцветные. Слизи вокруг клеток нет.
Хлоропласт 1, пластинчатый, более или менее дисковидный, часто
согнутый в виде полуцилиндра, или лентовидный, иногда спирально
изогнутый в один или несколько оборотов, с ровными или слегка
изрезанными краями, без пиреноида (только у одного вида описан
пиреноид), никогда не заполняющий концов клетки; перед началом
деления клетки наблюдаются 2 и больше хлоропластов. Вегетативное
размножение поперечным делением клеток надвое, дочерние клетки
вначале обычно с одним заостренным и другим закругленным концом.
У некоторых видов известны двужгутиковые зооспоры и половой процесс
в форме оогамии.
Большинство видов — криофилы, обитающие в высокогорных и
арктических районах на поверхности плотного снега и льда, часто на
фирновых полях, некоторые виды встречаются также в пресных проточ-
ных и стоячих водах в планктоне и обрастаниях, реже на влажных
скалах и почве, иногда среди мхов. В пределах СССР виды рода изучены
крайне недостаточно.
I. Клетки прямые или очень слабо изогнутые.
1. Клетки относительно широкие, веретеновидные, иногда заострен-
ные только на одном конце.
А. Клетки длинные, 3—5 мкм шир., до 80 мкм дл.................
..............................................К. nivalis 1.
Б. Клетки умеренно длинные, 2—4 мкм шир., не более 50 мкм дл.
а. Клетки 10—18 (28) мкм дл.....................К. Helvetica 2.
б. Клетки 26—50 мкм дл.
а. Клетки 2.5—3 мкм шир., 26—36 мкм дл., на концах
оттянутые в тонкие длинные шиповидные отростки
...........................................К. bernina 3.
р. Клетки 3—4 мкм шир., 30—50 мкм дл., на концах
постепенно суженные и заостренные ... К. viretii 4.
у. Клетки 2.8—3.2 мкм шир., 35—45 мкм дл., на концах
довольно резко суженные и слабо заостренные............
...................................... К. chodatii 5.
2. Клетки относительно широкие, цилиндрические, заостренные на
обоих концах.
А. Клетки 1.9—2.4 мкм шир., 8.5—18 мкм дл.......К. stagnalis 6.
Б. Клетки 1.5—2.5 мкм шир., 15—34 мкм дл......................
........................................К. planctonica 7.
90
3. Клетки тонкие, игловидные или вытянуто-веретеновидные, очень
длинные.
А. Хлоропласт пластинчатый или лентовидный, прямой или
слегка спиральный.
а. Концы клеток вытянутые, заостренные, но не переходящие
в волосок..............................К. spiculiformis 8.
б. Концы клеток волосовидно вытянутые.
а. Клетки игловидные, очень узкие.
* Концы клеток волнисто изогнутые или отогнутые
..................................К. longiseta 9.
* * Концы клеток прямые, иногда раздвоенные . . .
• .................................К. norvegica 10.
3. Клетки вытянуто-веретеновидные, в средней части до-
вольно широкие.............................К. crassa И.
В. Хлоропласт лентовидный, отчетливо спиральный . . . .
...................................................К. spirotaenia 12.
II. Клетки отчетливо и разнообразно изогнутые.
1. Хлоропласт без пиреноида, клетки разной длины.
А. Клетки веретеновидные, иногда червеобразные.
а. Клетки короткие, до 17 мкм дл.
а. Клетки заостренные на обоих концах.
* На снегу; клетки сильно заостренные ....
...............................К. transit vanica 13.
* * В планктоне; клетки слабо заостренные..........
...............................К. sigmoidea 14.
Р. Клетки заостренные преимущественно только на одном
конце...................К. sempervirens (частично) 15.
б. Клетки длинные, до 50 мкм дл.
а. Клетки только дугообразно изогнутые, 1 — 1.5 мкм шир.,
21—40 мкм дл...............................К. alpina 16.
р. Клетки разнообразно, но слабо изогнутые, 1—3 мкм шир.
12—40 мкм дл............................К. tatrae 17.
у. Клетки сильно спирально изогнутые, 1.5—2 мкм шир.,
(4) 20—35 (50) мкм дл.................К. spirulinoides 181.
Б. Клетки цилиндрические, суженные на концах.
а. Клетки на концах суженные в острие . .................
.......................................К. closterioides 19.
б. Клетки на концах слабо суженные и закругленные.
а. Клетки 4—17 мкм дл...................................
. ...................К. sempervirens (частично) 15.
р. Клетки 14—38 мкм дл.................К. corcontica 20.
2. Хлоропласт с пиреноидом, клетки короткие . . .................
............................................К............................................ pyrenoidifera 21.
1. Koliella nivalis (Chod.) Hind. [= Raphidium nival'e Chod.; Ari-
kistrodesmus nivalis (Chod.) Brunnth.; Raphidonema cryophilum R. Chod.
et F. Chod.] — Колиелла снеговая (рис. 30;. 1). — Клетки веретено-
видные, в средней части более или менее цилиндрические, 3—5 мкм шир.,
10—80 мкм дл., прямые или слегка изогнутые, на концах обычно резко
суженные и шиповидно заостренные. Хлоропласт пластинчатый.
На поверхности плотного снега в горных районах. В СССР не
обнаружена. (Швейцария: зап. Альпы).
2. Koliella helvetica (Koi) Hind. (= Raphidonema helveticum Koi) —
Колиелла швейцарская (рис. 30, 2). — Клетки коротковеретеновидные,
91
Рис. 30.
1 — Koliella nivalis', 2 — К. Helvetica', 3 — К. bernina; 4 — К. viretii; 5 — К. cho-
datii. (1, 4 — по Шода, 2, 3, 5 — по Коль).
92
2—3 мкм шир., 10—18 (28) мкм дл., обычно прямые, реже слегка
серповидно изогнутые, на концах резко суженные и оттянутые в тонкие
длинные шиповидные отростки. Хлоропласт пластинчатый, со слегка
изрезанными краями.
На поверхности плотного снега в высокогорных районах. В СССР
не обнаружена. (Швейцария: Бернские Альпы).
3. Koliella bernina (Koi) Hind. (= Raphidonema berninum Koi) —
Колиелла бернская (рис. 30, 3). — Клетки веретеновидные, в средней
части более или менее цилиндрические, 2.5—3 мкм шир., 26—36 мкм дл.,
прямые или слегка серповидно изогнутые, на концах резко суженные
и оттянутые в тонкие длинные шиповидные отростки. Хлоропласт
пластинчатый, с ровными или слегка волнистыми краями.
На поверхности плотного снега в высокогорных районах. В СССР
не обнаружена. (Швейцария: Бернские Альпы).
4. Koliella viretii (Chod.) Hind. [= Raphidonema viretii Chod.;
R. viretii R. Chod. et F. Chod.; Ankistrodesmus viretii (Chod.) Brunnth.] —
Колиелла Вирета (рис. 30, 4). — Клетки веретеновидные, 3—4 мкм шир.,
30—50 мкм дл., прямые или слегка серповидно или S-образно изогнутые,
на концах постепенно суженные и заостренные. Дочерние клетки, воз-
никшие в результате поперечного деления материнской клетки надвое,
имеют вид разнообразно вытянутых треугольников; в тех случаях,
когда они остаются длительное время соединенными друг с другом,
возникают звездчатые колонии. Хлоропласт пластинчатый.
На фирновых полях в горных районах, реже в пресных водах.
ЛатвССР.
Местонахождение в ЛатвССР (под названием Ankistrodesmus viretii)
сомнительно.
5. Koliella chodatii (Koi) Hind. (= Raphidonema chodatii Koi) —
Колиелла Шода (рис. 30, 5). — Клетки веретеновидные, в средней части
более или менее цилиндрические, 2.8—3.2 мкм шир., 35—45 мкм дл.,
прямые или разнообразно изогнутые, на концах довольно резко сужен-
ные и слабо заостренные. Изредка встречаются уродливые формы
с 3 вытянутыми концами. Хлоропласт пластинчатый.
На поверхности плотного снега в горных районах. В СССР не обна-
ружена. (Франция, Швейцария: зап. Альпы).
6. Koliella stagnalis Hind.—Колиелла прудовая (рис. 31, 1). —
Клетки на большем протяжении цилиндрические, 1.9—2.4 мкм шир.,
8.5—18 мкм дл., прямые, на обоих концах резко суженные и обычно
коротко заостренные. Хлоропласт пластинчатый, с ровными краями.
В стоячих водах, в весеннем планктоне. В СССР не обнаружена.
(Чехословакия).
7. Koliella planctonica Hind. — Колиелла планктонная (рис. 31,
2). — Клетки цилиндрические, 1.5—2 (2.5) мкм шир., (15) 21 — 26
(34) мкм дл., прямые, на обоих концах резко суженные, коротко шипо-
видно заостренные или слегка вытянутые. Хлоропласт пластинчатый,
с ровными краями.
В стоячих водах, в весеннем и осеннем планктоне. В СССР не обнару-
жена. (Чехословакия).
93
Рис. 31.
1 — Koliella stagnalis; 2 — К. planctontca. (По Гиндаку).
8. Koliella spiculiformis (Vischer) Hind. (= Raphidonema spiculi-
forme Vischer) — Колиелла пиковидная (рис. 32, 1, 2). — Клетки игло-
видные или вытянуто-веретеновидные, 0.8—2 (2.5) мкм шир., 40—
80 мкм дл., большей частью прямые; на концах постепенно суженные,
вытянутые и сильно заостренные. Хлоропласт лентовидный или пластин-
чатый, прямой или слегка спиральный.
В мелких водохранилищах, в планктоне. В СССР не обнаружена.
(Швейцария, Чехословакия).
94
Рис. 32.
1,2 — Koliella spiculiformis: 1 — клетки из естественных местонахождений,
2 — клетки из культур; 3—10 — К. longlseta: 3 — взрослые клетки, 4 — раз-
множение делением, 5 — четырехклеточная стадия, 6 — клетки после деления,
7 — выход зооспоры, 8 — молодая клетка, образованная зооспорой, 9 — крупная
клетка (яйцеклетка), оплодотворяющаяся сперматозоидом, 10 — ооспора; 11 —
К. norvegica; 12 — К. crassa; 13 — К. spirotaenia. (7, 2, 11, 12 — по Гиндаку,
3, 6 — по Вишеру, 4, 5, 7—10 — по Коршикову, 13 — по Весту).
9. Koliella longiseta (Vischer) Hind. (= Raphidonema longlseta
Vischer) — Колиелла длиннощетинковая (рис. 32, 3—10). — Клетки
игловидные, 1.2—3 (5) мкм шнр., (50) 100—140 мкм дл., прямые, реже
слегка спирально изогнутые, на концах постепенно суженные, заострен-
ные, волосовидно вытянутые, при делении обычно соединенные по
95
2—4 (8). Концы клеток ровные, волнистые или отогнутые в сторону.
Хлоропласт узкий, пластинчатый или лентовидный и тогда более или
менее спиральный. Зооспоры двужгутиковые, по 1 в клетке. Половой
процесс оогамный, сперматозоид похож на зооспору, яйцеклетка не
отличается от вегетативной клетки, ее содержимое после (или во время?)
оплодотворения выходит через боковое отверстие в оболочке клетки
и прикрепляется снаружи к ее поверхности. Зигота 10 мкм в диам.,
с шероховатой оболочкой.
В стоячих и проточных водах, часто в болотах, в планктоне и
в обрастаниях среди других водорослей. УССР, РСФСР (Иркутская обл.:
Братское водохранилище).
По общему облику клеток очень напоминает некоторых предста-
вителей рода Ankistrodesmus (Chlorococcales), хорошо отличается спо-
собом размножения и хлоропластом, занимающим лишь среднюю часть
клетки.
10. Koliella norvegica Hind. — Колиелла норвежская (рис. 32, 11). —
Клетки игловидные, очень узкие и длинные, 0.8—1.5 мкм шир., 42—
96 мкм дл., прямые или слегка дугообразно изогнутые, в средней части,
содержащей хлоропласт, 10—20 мкм дл., слегка веретеновидные до
цилиндрических, к концам резко суживающиеся. Концы клеток щетинко-
образные, 17—40 мкм дл., одиночные или иногда раздвоенные, гибкие,
прямые или слабо изогнутые. Хлоропласт пластинчатый, без пиреноида.
В планктоне олиготрофных горных озер. В СССР не обнаружена.
(Сев. Норвегия).
11. Koliella crassa Hind. — Колиелла толстая (рис. 32,12). — Клетки
вытянуто-веретеновидные, 2.5—5 мкм шир., 36 — 102 мкм дл., в средней
части довольно широкие, к концам довольно резко суживающиеся.
Концы клеток щетинкообразные, до 40 мкм дл., прямые или чуть
отогнутые. Хлоропласт пластинчатый, корытовидный, выстилающий
стенки у молодых клеток только частично, у старых полностью, без
пиреноида.
В планктоне удобряемых рыбоводных прудов. В СССР не обнару-
жена. (Словакия: Братислава).
12. Koliella spirotaenia (G. S. West) Hind. [= Ankistrodesmus spiro-
taenia G. S. West; Raphidonema spirotaenia (G. S. West) Korsch.] —
Колиелла спиральноленточная (рис. 32, 13). — Клетки игловидные,
(1.8) 2—3 мкм шир., (135) 156—255 (340) мкм дл., прямые или слегка
изогнутые, на концах постепенно суженные, заостренные, волосовидно
вытянутые. Хлоропласт лентовидный, спирально завернутый, с 2—
5 оборотами спирали.
В стоячих и проточных водах, в планктоне. УССР, РСФСР (Иркут-
ская обл.: Братское водохранилище).
13. Koliella transilvanica (Koi) Hind. (= Raphidonema transil-
vanica Koi) — Колиелла трансильванская (рис. 33, 1). — Клетки вере-
теновидные, 2—2.5 мкм шир., (2) 15 мкм дл., дугообразно или серповидно
изогнутые, на концах постепенно суженные и сильно заостренные,
иногда слегка вытянутые, при интенсивном делении соединенные по 2,
4, 8. Хлоропласт пластинчатый, перед делением клеток их может
быть 2—8.
На поверхности плотного снега в высокогорных районах. В СССР
не обнаружена. (Румыния: южн. Карпаты).
96
Рис. 33.
1 — Koliella transilvanica-, 2 — К. sigmoidea; 3, 4 — К. sempervirens, клетки
из разных культур. (1 — по Коль, 2—4 — по Гиндаку).
Занимает промежуточное положение между родами Koliella и
Raphidonema, однако многоклеточное состояние неустойчивое, наблюда-
ется лишь при интенсивном делении.
14. Koliella sigmoidea Hind. — Колиелла сигмовидная (рис. 33, 2). —
Клетки веретеновидные, 1 — 1.4 мкм шир., 4—10 мкм дл., слабо изогнутые
до сигмовидных, постепенно суживающиеся от середины к концам,
после деления часто соединенные концами по 2. Концы клеток конусо-
видно заостренные, на вершине притупленные. Хлоропласт пластин-
чатый, без пиреноида.
В планктоне небольших эвтрофированных лагун. В СССР не об-
наружена. (Куба).
15. Koliella sempervirens (Chod.) Hind. (== Raphidonema semper-
virens Chod.) — Колиелла вечнозеленая (рис. 33, 3, 4). — Клетки
веретеновидные, часто на одном конце широко закругленные, на другом
постепенно суженные и заостренные, реже до цилиндрических, на обоих
концах слабо суженные и закругленные, 2—4 мкм шир., 4(10) —
15 (17) мкм дл., разнообразно изогнутые, соединенные в 2—4-клеточные,
легко распадающиеся нити. Хлоропласт пластинчатый, перед делением
клеток их может быть 2—4.
В стоячих и слабо проточных водах, в планктоне и в обрастаниях,
а также на влажной почве. ЭССР, ЛатвССР, УССР, Якутская АССР
(бассейн среднего течения р. Лены).
Вид изучен главным образом в лабораторных культурах и в этих
условиях обнаруживает сильную изменчивость формы клеток.
7 Зеленые водоросли. Улотриксовые 97
16. Koliella alpina (Koi) Hind. (= Raphidonema alpina Koi) —
Колиелла альпийская (рис. 34, 1). — Клетки веретеновидные, 1 —
1.5 мкм шир., 21—40 мкм дл., дугообразно изогнутые, к концам посте-
пенно суженные и заостренные. Хлоропласт пластинчатый.
На поверхности плотного снега в горных районах. В СССР не
обнаружена. (Зап. Европа:. Альпы).
17. Koliella tatrae (Koi) Hind. (= Ankistrodesmus tatrae Koi) —
Колиелла татрская (рис. 34, 2—4). — Клетки веретеновидные, 1 —
3 мкм шир., 12—40 мкм дл., разнообразно изогнутые, к концам посте-
пенно суженные, заостренные и нередко слегка вытянутые, при делении
часто соединенные по 2—4. Хлоропласт пластинчатый, с ровными или
изрезанными краями, обычно 1, реже их 2 или несколько. Известны
акинеты.
На поверхности плотного снега и фирновых полей в высокогорных
районах, а также в пресных стоячих и проточных водах.
Var. tatrae (рис. 34, 2). — Клетки 1.5—2 мкм шир., 20—35 мкм дл.,
серповидно, спирально или S-образно изогнутые.
На поверхности плотного снега. В СССР не обнаружена. (Швейца-
рия: зап. Альпы, Чехословакия, Польша: Татры).
Var. fogarasensis (Koi) Hind. (= Raphidonema tatrae var. fogarasen-
sis Koi) (рис. 34, 3). — Клетки 1—2 мкм шир., 20—40 мкм дл., серпо-
видно, дуговидно или слегка S-образно изогнутые, на концах постепенно
суженные, заостренные и иногда слегка вытянутые. Хлоропласт
пластинчатый, с ровными краями. '
На поверхности плотного снега. В СССР не обнаружена. (Румыния:
южн. Карпаты).
Var. bratislavensis Hind. (рис. 34, 4). — Клетки (2) 2.5—3 мкм шир.,
(12) 23 (25) мкм дл., разнообразно, чаще спирально изогнутые, к концам
постепенно суженные, вытянутые и заостренные. Известны акинеты.
В пресных водах. В СССР не обнаружена. (Чехословакия: пруд
в окр. Братиславы).
18. Koliella spirulinoides Hind. — Колиелла спнрулиновидная
(рис. 34, 5). — Клетки червеобразные, 1.5—2 мкм шир., (4) 20—
35 (50) мкм дл., сигмовидные или спирально изогнутые до улитковидно
скрученных, с 1 — 6 изгибами на клетку, в средней части цилиндрические,
к концам суживающиеся, после деления часто соединенные концами
по 2 (в лабораторной культуре до 10 в ряд). Концы клеток конусовидно
заостренные, на вершине притупленные, реже несуживающиеся, закруг-
ленные. Хлоропласт пластинчатый, без пиреноида.
В речном планктоне. В СССР не обнаружена. (Куба).
19. Koliella closterioides (Kuff.) Hind. (= Raphidonema closterloides
Kuff.) — Колиелла клостериевидная (рис. 35, 7). —Клетки в средней
части более или менее цилиндрические, 2 мкм шир., 15 — 17 мкм дл.,
на концах постепенно суженные, заостренные и изогнутые в одну
сторону. Хлоропласт пластинчатый, с ровными краями.
Среди мхов. В СССР не обнаружена. (Южная Америка).
20. Koliella corcontica Hind. — Колиелла корконтская (рис. 35, 2). —
Клетки цилиндрические, 2.4 —2.8 мкм шир., 14—38 мкм дл., дуговидно
изогнутые, иногда почти прямые, на концах еле заметно суженные
и закругленные. Хлоропласт пластинчатый, согнутый в виде полу-
цилиндра.
98
Рис. 34.
1 - Koliella alpina; 2-4 - К. tatrae: 2 - var. tatrae, 3- yar. fogarasensis,
4 — var. bratislavensis; 5 — K. spirulinoides. (1 — по Коль, 2—5 по Гиндаку).
99
7*
Рис. 35.
1 — Koliella closterioides-, 2 — К. corcontica; 3 — К. pyrenoldifera. (1 — по Куф-
ферату, 2 — по Гиндаку, 3 — по Коршикову).
На влажных скалах. В СССР не обнаружена. (Чехословакия).
21. Koliella pyrenoidifera (Korsch.) Hind. (= Raphidonema pyrenoidi-
fera Korsch.) — Колиелла пиреноидоносная (рис. 35, 3). — Клетки
веретеновидные, короткие, 2.2—2.8 мкм шир., 7—13 мкм дл., в средней
части прямые, на концах резко суженные и хвостовидно оттянутые
в заостренные и отогнутые в одну сторону отростки. Хлоропласт пластин-
чатый, с 1 пиреноидом в расширенной средней части.
В стоячих и слабо проточных водах, а также в смешанных культурах
водорослей. УССР, Вост. Сибирь.
Формой клеток вид несколько напоминает К. sempervirens, отлича-
ется присутствием в хлоропласте пиреноида. В связи со слабой изучен-
ностью рода пока нет оснований для выделения К. pyrenoldifera
в отдельный род, несмотря на то что остальные виды рода пиреноида
не имеют.
100
Род FOTTEA Hind. - ФОТТЕЯ
Нити не образуются. Клетки цилиндрические, с широко закруглен-
ными концами, одноядерные, беспорядочно располагающиеся по не-
скольку или одиночно в гомогенной слизи, образуя микроскопические
колонии разнообразной формы. Оболочки клеток тонкие. Хлоропласт
пристенный, корытовидный, без пиреноида. Размножение поперечным
делением клеток надвое с последующим дорастанием до первоначаль-
ного размера. Зооспоры и половой процесс неизвестны.
Способ размножения сближает род Fottea с родами Koliella, Chodatia
и Elakatothrix, хотя по ряду других признаков его следовало бы относить
к Chloro со ccales.
1. Fottea cylindrica Hind. — Фоттея цилиндрическая (рис. 36). —
Колонии преимущественно в виде округлых или овальных масс гомо-
генной слизи, 8—12 мкм толщ., с беспорядочно расположенными по
2—12 и более клетками, реже клетки одиночные. Клетки 8—9 мкм шир.,
21—32 мкм дл., прямые, реже несколько изогнутые. Хлоропласт зеленый
или темно-зеленый, пластинчатый, с завернутыми краями, выстилающий
Рис. 36.
Fottea cylindrica-1 — отдельные клетки с поверхности, 2 — клетка в поперечном
разрезе, 3—5 — слизистые колонии разного облика. (По Гиндаку).
101
продольные стенки клеток почти полностью. Ядро шаровидное, в центре
клетки. В протоплазме заметны капельки жира.
В почве. В СССР не обнаружена. (Чехословакия: Татры, на высоте
около 2100 над ур. м.).
Род ELAKATOTHRIX Wille - ЭЛАКАТОТРИКС
Нити не образуются. Клетки веретеновидные, цилиндрические,
эллипсоидные, прямые или слегка изогнутые, иногда асимметричные,
на концах суженные, заостренные или тупоконические, покрытые
толстым слоем слизи, одиночные или соединенные в колонии различной
формы. В колониальной слизи клетки обычно сближены попарно, нередко
расположены на одной линии или смещены в стороны, часто лежат
рядом или беспорядочно. Слизистые покровы клеток разных поколений
в большинстве случаев сливаются в общую бесструктурную слизистую
массу, реже остаются индивидуализированными в виде системы вставоч-
ных дочерних слизистых пузырей. Хлоропласт пристенный, пластин-
чатый или лентовидный и спирально изогнутый, с пиреноидом или
без него, не доходящий до концов клетки. Размножение поперечным
делением клеток надвое, вновь образовавшиеся концы дочерних клеток
вначале широко закругленные. Зооспоры и половой процесс неизвестны.
В пресных стоячих и проточных водах, а также на влажной почве
и в почве.
Формой клеток представители рода напоминают виды родов Ankistro-
desmus, Fusola и Quadrigula из порядка Chlorococcales, однако раз-
множение последних происходит с помощью автоспор и у них никогда
не наблюдается поперечного деления клеток надвое, что составляет
характерную черту улотриксовых. Род наиболее близок к Koliella, Fottea
и Chodatia.
I. Колониальная слизь с внутренними пузырями из слизистых оболочек
дочерних колоний разных поколений . . Е. gloeocystiformis 1.
II. Колониальная слизь гомогенная.
1. Колонии округлые, с большим количеством беспорядочно располо-
женных клеток.....................................Е. subacuta 2.
2. Колонии эллипсоидные или веретеновидные, с малым количеством
клеток.
А. Колонии большей частью правильно эллипсоидные.
а. Клетки цилиндрические, на концах резко суженные и
коротко заостренные........................Е. acuta 3.
б. Клетки веретеновидные, прямые или слегка изогнутые,
часто асимметричные, на концах слабо суженные и закруг-
ленные . . . ..............................Е. biplex 4.
Б. Колонии веретеновидные.
а. Колониальная слизь жидкая, контуры колоний заметны
только в туши; хлоропласт пластинчатый до лентовидного
Е. genevensis 5.
б. Колониальная слизь более плотная, контуры колоний отчет-
ливые; хлоропласт лентовидный, спирально извитой.
а. Клетки на концах постепенно суженные; в сфагновых
болотах, среди мхов......................Е. parvula 6.
р. Клетки на концах резко суженные, коротко заостренные;
в пресных водоемах в планктоне . . . Е. spirochroma 7.
102
Рис. 37.
Elakatothrix gloeocystiformis. (1 — по Гиндаку, 2 — по Коршикову).
3. Колонии в виде цилиндрического тяжа или неопределенной
формы.
А. Хлоропласт с пиреноидом...................Е. gelatinosa 8.
Б. Хлоропласт без пиреноида..............Е. pseudogelatinosa 9.
1. Elakatothrix gloeocystiformis Korsch. — Элакатотрикс глеоцисто-
видный (рис. 37). — Колонии округлые, до 60 мкм в диам., с большим
числом беспорядочно расположенных клеток, колониальная слизь с си-
стемой вставочных дочерних пузырей, снабженных на поверхности
тонким кожистым слоем. Клетки веретеновидные, 3.5—5.5 мкм шир.,
10—21 мкм дл., в средней части более или менее цилиндрические,
на концах суженные, закругленные или слегка заостренные. Хлоропласт
пластинчатый, с пиреноидом.
В стоячих водах, чаще всего в прудах, в планктоне. ЭССР, БССР,
УССР.
2. Elakatothrix subacuta Korsch. — Элакатотрикс островатый
(рис. 38, 1—3). — Колонии округлые, с гомогенной слизью, 30—70 мкм
в диам., нередко еще крупнее, с 2 —32 беспорядочно расположенными,
часто попарно сближенными клетками. Клетки веретеновидные, 4 —
4.5 мкм шир., 15—16 мкм дл., на концах суженные и закругленные.
Хлоропласт пластинчатый, с пиреноидом.
В стоячих и проточных водах, в планктоне. РСФСР (Горьков-
ская обл., Зап. Сибирь), УССР, УзССР.
103
Рис. 38.
1 — 3 — Elakatothrix subacuta; 4, 5 — E. acuta', 6 — E. biplex. (2 — 5 — по Корши-
кову, 6 — по Нигаарду).
3. Elakatothrix acuta Pasch. — Элакатотрнкс острый (рис. 38, 4, 5). —
Колонии удлиненные, эллипсоидные, но могут быть и иных очертаний,
30—120 мкм дл., с гомогенной слизью и малым количеством клеток,
расположенных более или менее в одном направлении. Клетки веретено-
видно-цилиндрические, 3—4 мкм шир., 10—21 мкм дл., на концах резко
суженные и коротко заостренные. Хлоропласт пластинчатый, с пире-
ноидом.
В реках и стоячих водоемах, в планктоне. РСФСР (Ленинград-
ская обл., Зап. Сибирь), УССР (Харьковская обл.), ГрузССР.
4. Elakatothrix biplex (Nyg.) Hind. (— E. gelatinosa Wille f. biplex
Nyg.) — Элакатотрнкс двусплетенный (рис. 38, 6). — Колонии эллип-
соидные, с гомогенной слизью и малым количеством клеток, нередко
клетки одиночные. Клетки веретеновидные, 2.5—5.5 мкм шир., 7—27 мкм
дл., прямые или слегка изогнутые и асимметричные, на концах слабо
суженные и закругленные. Хлоропласт пластинчатый, с пиреноидом.
В протоплазме встречаются капельки масла.
В озерах, в планктоне. В. СССР не обнаружен. (Зап. Европа).
5. Elakatothrix genevensis (Reverd.) Hind. (= Ankistrodesmus gene-
vensis Reverd.; Elakatothrix lacustris Korsch.) — Элакатотрнкс женевский
(рис. 39, 1). —Колонии веретеновидные, с гомогенной слизью с неяс-
ными внешними контурами, заметными лишь в растворе туши, из
2 (4—6) клеток; многоклеточные колонии не образуются вследствие
легкого разрушения колониальной слизи. Клетки веретеновидные,
3—4.5 мкм шир., 21—38 (55) мкм дл., прямые, на концах постепенно
суженные и заостренные. Хлоропласт пластинчатый до лентовидного,
с пиреноидом.
104
' В стоячих и проточных водах, а также в болотах, в планктоне
и в обрастаниях различных субстратов. Повсеместно.
6. Elakatothrix parvula (Arch.) Hind. (= Sptrotaenia parvula
Arch.) — Элакатотрнкс крохотный (рис. 39, 2). — Колонии веретено-
видные, 17—20 мкм шир., 60—115 мкм дл., с гомогенной слизью и малым
количеством клеток, чаще всего двуклеточные. Клетки веретеновидные,
(3.1) 3.5—4.5 (6.5) мкм шир., 17—35 (40) мкм дл., прямые или слегка
изогнутые, на концах постепенно суженные и довольно заостренные.
Хлоропласт лентовидный, спирально изогнутый до 2 оборотов, с пире-
ноидом. В протопласте встречаются капельки масла.
В сфагновых болотах, между дерновинками мха. В СССР не обна-
ружен. (Чехословакия).
Вид малоизученный, очень близкий к Е. genevensis.
7. Elakatothrix spirochroma (Reverd.) Hind. (= Raphldium spiro-
chroma Reverd.; Ankistrodesmus spirochroma Reverd.) — Элакатотрнкс
спиральноокрашенный (рис. 39, 3). — Колонии веретеновидные, с гомо-
генной слизью и малым количеством клеток, чаще всего двуклеточные.
Клетки веретеновидные, 3—3.5 мкм шир., 21—59 мкм дл., на концах
резко суженные и вытянутые в короткий, довольно тупой гиалиновый
кончик. Хлоропласт лентовидный, спирально изогнутый до 2 оборотов,
с пиреноидом. В протоплазме много вакуолей разной величины.
В стоячих и проточных водах, в планктоне. В СССР не обнаружен.
(Швейцария, Чехословакия).
Вид малоизученный, очень близкий к Е. parvula.
8. Elakatothrix gelatinosa Wille — Элакатотрнкс слизистый (рис. 40,
7). — Колонии в виде цилиндрического длинного тяжа из гомогенной
Рис. 39.
1 — Elakatothrix genevensis-, 2 — Е. parvula-, 3 — Е. spirochroma. (По Ревер-
Дину).
105
Рис. 40.
1 — Elakatothrix gelatinosa-, 2, 3 — E. pseudogelatinosa. (1, 3 — по Коршикову,
2 — по Гиндаку).
слизи с продольно расположенными обычно в один ряд и попарно
сближенными клетками. Клетки веретеновидные, 3—6 мкм .шир., 11 —
22 мкм дл., к обоим концам равномерно суженные и заостренные.
Хлоропласт пластинчатый, с пиреноидом. В протоплазме часто встреча-
ются капельки масла.
В стоячих и проточных водах, в планктоне. В различных районах
СССР, нередко.
9. Elakatothrix pseudogelatinosa Korsch. — Элакатотрикс ложно-
слизистый (рис. 40, 2, 3). — Колонии в виде длинного цилиндрического
тяжа из гомогенной слизи, около 14 мкм шир., с продольно расположен-
ными в один ряд и попарно сближенными клетками; при интенсивном
делении клеток их количество в колонии может сильно возрастать,
однорядность расположения и контуры колонии меннются. Клетки
веретеновидные, 3 мкм шир., 14—28 мкм дл., заостренные на одном
или обоих концах. Хлоропласт пластинчатый, без пиреноида.
В стоячих и проточных водах, в планктоне. Север Европ. ч. СССР.
106
Семейство CYLINDROCAPSACEAE Wille -
ЦИЛИНДРОКАПСОВЫЕ
Водоросли многоклеточные, в виде простых нитей. Нити одно-,
дву- и трехрядные, прикрепляющиеся к субстрату широкой короткой
целлюлозной подошвой или отрывающиеся и свободноплавающие, окру-
женные толстым слизистым слоистым влагалищем. Клетки одноядерные,
преимущественно округлых очертаний, с толстыми слоистыми оболоч-
ками; верхушечная клетка округлая. Хлоропласт центральный (осевой),
массивный, звездчатый или неправильно продырявленный, с пиреноидом.
Вегетативное размножение фрагментацией слоевища и акинетами. Бес-
полое размножение зооспорами, реже апланоспорами. Половой процесс
оогамный. Ооспора коровой тканью не оплетается и прорастает после
периода покоя.
В пресных стоячих и проточных водах.
Род CYLINDROCAPSA Reinsch - ЦИЛИНДРОКАПСА
Нити на ранней стадии развития однорядные, позднее, в результате
продольных или косых делений клеток, могут стать неправильно много-
рядными, лентовидными и даже неравномерно пластинчатыми, при-
крепленные, но легко отрывающиеся от субстрата. Клетки цилиндри-
ческие, шаровидные, эллипсоидные, яйцевидные или почти конусовид-
ные, реже короткоцилиндрические, часто сближенные попарно, с тол-
стыми слоистыми оболочками. При вегетативном размножении нити
распадаются на акинеты или группы клеток. Зооспоры двужгутиковые,
шаровидные или эллипсоидные, со стигмой и 2 пульсирующими вакуо-
лями, расположенными в переднем конце; они возникают по 1 или
по 2—4 в каждой клетке слоевища, за исключением базальной. Аплано-
споры эллипсоидно-яйцевидные, иногда со стигмой; они освобождаются
вследствие ослизнения оболочек материнских клеток. Растения одно-
домные. Половой процесс оогамный. Оогонии шаровидные или яйце-
видные, с 1 яйцеклеткой, окруженные несколькими отстающими друг
от друга слизистыми оболочками; они образуются из вегетативйтлх клеток
путем их разрастания, одиночные или собранные рядами. Яйцеклетки
шаровидные, зеленые, с многочисленными зернами крахмала, часто
имеющие сбоку светлое пятно. Перед оплодотворением оболочки оогонйя
на одной его стороне несколько выпячиваются и на вершине выпуклины
вследствие ослизнения образуется отверстие, через которое проникают
сперматозоиды. Антеридии маленькие, шаровидные или слегка сплюс-
нутые в дорсивентральном направлении, со слоистыми оболочками и
светло-оранжевым содержимым; они также возникают из вегетативных
клеток в результате быстро следующих друг за другом делений и
располагаются рядами или соединены попарно в небольшие группы,
окруженные общим слоистым влагалищем. В каждом антеридии образу-
ется по 2 сперматозоида, которые выходят наружу через боковое отвер-
стие, будучи заключенными в слизистом пузыре, быстро расплывающемся
в воде. Сперматозоиды веретеновидные, на заднем конце округленные
или слегка вытянутые и заостренные, светло-оранжевые, с 2 жгутиками
и 2 пульсирующими вакуолями в переднем, бесцветном конце. Ооспоры
с толстой студенистой, быстро отвердевающей оболочкой и маслянистым
красноватым содержимым. Нередки случаи партеногенетического раз-
вития яйцеклеток, когда они, одевшись студенистой оболочкой, делятся
на 2—4 дочерние клетки, каждая из которых вырастает в новую нить.
107
Рис. 41.
1—5 — Cylindrocapsa conferta'. 1 — вегетативная нить, 2, 3 — молодые нити
с подошвой, прикрепляющейся к субстрату, 4 — апланоспоры в молодой нити,
5 — образование оогониев; 6—10 — С. involuta: 6 — нить с оогонием и антери-
диями, 7 — оогоний с яйцеклеткой и сперматозоидами, 8 — освобождение
сперматозоидов, 9 — сперматозоид отдельно, 10 — два сперматозоида в пузыре.
(1, 5 — по Прескотту, 2—4 — по Принтцу, 6—10 — по Ценковскому).
Известно пальмеллевидное состояние, во время которого вместо нитей
возникают бесформенные скопления клеток, окруженные большим
количеством слизи.
В пресных стоячих и проточных водах.
Недостаточно изученный род. Кроме приведенных здесь видов для
водоемов Зап. Европы указаны еще С. атоепа Wolle и С. nuda Reinsch,
крайне сомнительные, изученные очень слабо.
I. Клетки 21—30 мкм шир.
1. Клетки поперечно-эллипсоидные или короткоцилиндрические, их
длина меньше ширины или равна ей................С. conferta 1.
2. Клетки преимущественно шаровидные или эллипсоидные, их длина
обычно превосходит ширину до 1.5 раза, реже равна ей или
меньше..........................................С. involuta 2.
II. Клетки (7) 9 — 16 (25) мкм шир...............С. geminella 3.
1. Cylindrocapsa conferta W. West — Цилиндрокапса скученная
(рис. 41, 1—5). — Нити сначала однорядные, затем неправильно много-
рядные, 24—30 мкм шир., из плотно расположенных, от взаимного
сдавливания разных по форме клеток. Клетки поперечно-эллипсоидные
или короткоцилиндрические, 21—27 мкм шир., 12—20 мкм дл., на ранних
стадиях шаровидные, полушаровидные или округло-конусовидные. Обо-
лочки клеток толстые, двуслойные. Апланоспоры яйцевидные, по 4
в материнской клетке.
В болотах, прудах, озерах, мелководных участках водохранилищ.
ЭССР, ЛатвССР, юг Европ. ч. СССР (УССР: Киевское водохранилище),
по-видимому и в других районах.
Редкий, очень интересный, но слабо изученный вид, отличающийся
108
от других видов рода более слоистой слизистой оболочкой нитей и более
узкой и короткой целлюлозной подошвой.
2. Cylindrocapsa involuta Reinsch — Цилиндрокапса окутанная
(рис. 41, 6—10). — Нити обычно однорядные, местами двурядные.
Клетки шаровидные, поперечно-эллипсоидные, иногда короткоцилиндри-
ческие, 22—30 мкм шир., по длине до 1.5 раза больше ширины, равны ей
или меньше ее. Оболочки клеток, как и общее влагалище нитей, часто
явственно слоистые. Оогонии 38—42 мкм шир. Ооспоры шаровидные
или слегка удлиненные, с гладкой студенистой бесцветной оболочкой,
24 мкм в диам. (незрелые). Сперматозоиды 15 мкм дл.
В болотах и озерах, между мхами и другими водными растениями,
обычно единично, иногда в массе. ЭССР, центр (Московская, Ярослав-
ская обл.) и юг Европ. ч. СССР.
Малоизученный вид. По всей вероятности, распространен значи-
тельно шире, чем считалось до сих пор.
3. Cylindrocapsa geminella Wolle — Цилиндрокапса парная
(рис. 42). — Нити однорядные, часто поделенные довольно глубокими
перетяжками на четковидные двуклеточные участки. Клетки эллипсоид-
ные, неправильно конусовидные или почти шаровидные, реже цилиндри-
ческие, нередко разной формы в одной и той же нити, (7) 9—16 (25) мкм
шир. Оболочки клеток толстые, слоистые, что особенно хорошо заметно
на концах двух сближенных клеток. Оогонии одиночные или по 2—4
в ряд, шаровидные, с бесцветной слоистой оболочкой. Ооспоры шаро-
видные, 18—50 мкм в диам., не заполняющие оогоний.
Рис. 42.
Cylindrocapsa geminella: 1, 2 — var. geminella, 3—5 — var.
по Прескотту, 3—5 — по Гансгиргу).
minor. (1, 2 —
109
В болотах, озерах и реках, в обрастаниях на водных цветковых
растениях, древесине, камнях, раковинах моллюсков, единичными
экземплярами и в виде светло- или темно-зеленых хлопьев, часто
в переплетении с другими нитчатками.
В биологическом и экологическом отношении вид изучен недоста-
точно. Установлены 2 разновидности, различающиеся главным образом
размерами ооспор.
Var. geminella (рис. 42, 7, 2). — Клетки почти шаровидные или
конусовидные, реже цилиндрические, 13.3—16 (20) мкм шир. Ооспоры
около 50 мкм в диам.,- оболочка слоистая, до 10—15 мкм толщ.
Северо-запад, центр и юг Европ. ч. СССР, ЭССР, ЛатвССР, КазССР,
вероятно и в других районах.
Var. minor Hansg. (рис. 42, 3—5).— Клетки эллипсоидные или
почти шаровидные, иногда вследствие сдавливания почти цилиндри-
ческие, 12—15 (25), в молодых нитях обычно 9—12 мкм шир. Ооспоры
18—23 мкм в диам. Сперматозоиды 2.5 мкм шир., 9—10 мкм дл.
КазССР (Акмолинская обл.).
Семейство SCHIZOMERIDACEAE Smith -
СХИЗОМЕРИЕВЫЕ
Водоросли многоклеточные, в виде простых нитей. Нити во взрослом
состоянии макроскопические, в основании однорядные, выше много-
рядные, прикрепляющиеся к субстрату конусовидной, нередко очень
вытянутой базальной клеткой, реже ризоидными выступами. Клетки
одноядерные, преимущественно цилиндрические, плотно соединенные,
с тонкими оболочками; верхушечная клетка заостренная. Хлоропласт
пристенный, продырявленный, с 1 или несколькими пиреноидами. Веге-
тативное размножение распадением нитей на фрагменты. Бесполое
размножение зооспорами. Половой процесс неизвестен.
В пресных водах.
Род SCHIZOMERIS Kutz. - СХИЗОМЕРИС
Нити темно- или желто-зеленые, прикрепленные с помощью базаль-
ной клетки с утолщенной оболочкой, особенно в нижней ее части.
Образование ризоидов наблюдается редко, клетки их тонкостенные, почти
полностью лишенные содержимого. У взрослых, многорядных нитей
наружные, продольные стенки клеток сильно утолщенные, внутренние,
продольные и поперечные, тонкие. Хлоропласт в виде пристенного
сетчато продырявленного кольца с 1 или несколькими пиреноидами,
реже встречается несколько хлоропластов, из которых каждый с 1 пире-
ноидом, в базальной клетке чаще всего в виде неправильно разветвленных
узких лент, анастомозирующих друг с другом. Продукт фотосинтеза —
крахмал, располагающийся вокруг пиреноидов и в строме хлоропласта.
Центральная часть клетки занята вакуолью с клеточным соком, в сре-
динном мостике цитоплазмы находится небольшое ядро с ядрышком,
реже оно расположено у внутренней поверхности хлоропласта, и тогда
от него к противоположной стороне клетки тянется тонкая плазмати-
ческая пленка. Зооспоры четырехжгутиковые, продолговатые, заострен-
ные спереди и закругленные сзади, со стигмой; они образуются по 1
в клетках верхней части нитей и освобождаются вследствие ослизнения
оболочек клеток.
НО
Рис. 43.
Schizomeris leibleinii: 1, 2 — общий вид нитей, 3 — клетки с сетчатым хлоро-
пластом, 4 — фрагмент старой нити, 5—7 — базальные участки нитей. (2—4 —
по Коршикову, 5—7 — по Скуе).
1. Schizomeris leibleinii Kiitz. — Схизомерис Лейблейна (рис. 43). —
Нити на ранней стадии развития однорядные, микроскопические, во
взрослом состоянии макроскопические, до 20 см дл., и тогда у основания
однорядные, 20—30 мкм шир., в средней части многорядные, до 150 мкм
шир., ровные на всем протяжении или с перетяжками. Клетки цилиндри-
ческие, 5.7—20 мкм шир., по длине равные ширине или несколько
меньше или больше; верхушечная клетка заостренная, особенно у про-
ростков. Хлоропласт в мелких клетках взрослых нитей выстилает не
только продольные, но частично и поперечные стенки, благодаря чему
структура его становится неясной, клетки кажутся равномерно зелеными,
пиреноиды плохо различимы.
В реках, озерах и прудах, в обрастаниях на водных цветковых
111
растениях, древесине, камнях, раковинах моллюсков. Во многих местах
СССР.
Описанные в литературе в качестве самостоятельных видов 5. ir-
regularis Fritsch et Rich, Uronema indicum Ghose, Pseudoschizomeris
exudata Deason et Bold существенно от Schizomeris leibleinii не от-
личаются.
Подпорядок ULVINEAE - УЛЬВОВЫЕ
Слоевище одно- или двурядно нитчатое, однослойно трубчатое или
мешковидное, одно- или двуслойно пластинчатое, обычно макроскопи-
ческое, простое или разветвленное. Клетки одноядерные, с тонкими или
утолщенными оболочками, плотно прилегающие друг к другу или рыхло
расположенные в межклеточном веществе. Хлоропласт преимущест-
венно 1, пристенный, пластинчатый, с 1 или несколькими пиреноидами.
Вегетативное размножение регенерацией оторванных частей слоевища,
а также отчленением от его краев группы клеток или одиночных клеток.
Бесполое размножение зооспорами и апланоспорами. Половой процесс
изогамный и гетерогамный. Имеется смена форм развития, изоморфная
или гетероморфная, в последнем случае слоевище спорофита представ-
лено обычно 1 клеткой.
Виды пресноводные и морские.9
I. Спорофит одноклеточный, микроскопический, гаметофит много-
клеточный, макроскопический; слоевище гаметофита трубчатое,
мешковидное или пластинчатое, однослойное; однорядных нитевид-
ных ветвей нет.............сем. Monostromataceae (с. 112).
II. Спорофит и гаметофит многоклеточные, макроскопические, сходного
строения; слоевище нитевидное, трубчатое или мешковидное, одно-
слойное или пластинчатое, двуслойное; однорядные нитевидные ветви
имеются............................сем. Ulvaceae (с. 121).
Семейство MONOSTROMATACEAE Kunieda -
МОНОСТРОМОВЫЕ
Слоевище гаметофита макроскопическое, трубчатое, мешковидное
или пластинчатое, всегда однослойное, прикрепляющееся к субстрату
подошвой, образованной переплетенными ризоидами, или базальным
диском, иногда с ризоидными выростами по краям. Клетки нередко
с сильно утолщенными оболочками, расположенные в межклеточном
веществе довольно рыхло или сближенно. Хлоропластов 1—2, с 1, реже
2—3 (5) отчетливо видимыми пиреноидами. Размножение бесполое и
половое. Спорофит и гаметофит морфологически различны: спорофит
одноклеточный, микроскопический, гаметофит многоклеточный, макро-
скопический. Смена форм развития имеется, гетероморфная, реже отсут-
ствует.
9 Из морских форм в наш Определитель включены только виды,
выносящие опреснение. Подробнее см.: Виноградова К. Л. Уль-
вовые водоросли (Chlorophyta) морей СССР. Л.; Наука, 1974. 165 с.
112
I. Слоевище трубчатое, простое или разветвленное; клетки многоуголь-
ные, расположенные плотно...................род Blidingia (с. ИЗ).
II. Слоевище мешковидное или разорванное до основания и тогда листо-
видное; клетки многоугольные, расположенные плотно, или округ-
лые, расположенные рыхло....................род Monostroma (с. 116).
Род BLIDINGIA Kylin - БЛИДИИГИЯ
Слоевище макроскопическое, трубчатое, простое или разветвленное,
прикрепляющееся к субстрату маленьким базальным диском. Клетки
с поверхности слоевища многоугольные, расположенные сближенно или
почти плотно продольными и поперечными рядами или беспорядочно.
Наружные и внутренние части оболочки клеток могут сильно утолщаться.
Хлоропласт звездчатый, с крупным пиреноидом в центре. Зооспоры
четырехжгутиковые, без стигмы. Проросток на стадии 3—4 клеток начи-
нает ветвиться в горизонтальной плоскости, в результате чего образуется
многоклеточный однослойный диск, который в центральной части посте-
пенно становится многослойным. Позднее верхний слой диска отслаи-
вается и разрастается вверх с образованием мешковидной структуры,
формирующейся в дальнейшем в трубчатое слоевище. Половое размно-
жение неизвестно.
Морские водоросли, выносящие большую степень опреснения.
Род Blidingia можно легко спутать с Enteromorpha, но в отличие от
последнего ветви у него никогда не бывают однорядными. Главные
ветви образуются обычно на ранних стадиях развития, как результат
выпячивания полости слоевища и усиленного деления клеток в опре-
деленном направлении. Микроскопические шиповидные веточки обычно
берут начало от нескольких инициальных клеток, реже от одной, но сна-
чала делящейся в горизонтальной плоскости.
I. Клетки в слоевище расположены беспорядочно, внутренние части их
оболочки утолщенные.
1. Внутренние части оболочки клеток утолщены незначительно (до
9 мкм).........................................В. minima 1.
2. Внутренние части оболочки клеток утолщены весьма значительно
(до 60 мкм)...................................В. chadefaudii 2.
II. Клетки в слоевище расположены продольными и отчасти попереч-
ными рядами, внутренние и наружные части их оболочки одинако-
вые, не утолщенные................................В. marginata 3.
1. Blidingia minima (Nag.) Kylin (=Enteromorpha minima Nag.) —
Блидингия наименьшая (рис. 44, 1— 3). — Слоевище до 20 см выс., до
6 мм шир., простое, реже скудно или обильно разветвленное, иногда
только с мелкими ветвями в основании. Клетки с поверхности слоевища
многоугольные или почти округлые, 4—10 мкм шир., 4—13 мкм дл.,
расположенные беспорядочно, лишь в основании слоевища и узких вет-
вях прослеживаются нечеткие продольные ряды. На поперечном срезе
слоевища клетки прямоугольные или эллипсоидные, слегка вертикально
вытянутые, 4—7 мкм шир., 6—11 мкм дл., наружная часть их оболочки
1.3—5.5 мкм толщ., внутренняя 1.3—9 мкм толщ., с заметной по мере
утолщения слоистостью; стенки слоевища в целом 10—20 мкм толщ.
В морских, соленых и опресненных водах, на камнях вблизи уреза
воды.
8 Зеленые водоросли. Улотриксовые 113
Рис. 44.
1—3 — Blidingia minima f. minima'. 1 — общий вид слоевищ, 2 — ъап клеток
с поверхности слоевища в средней части, 3 — то же в нижней части; 4—6 —
В. chadefaudii: 4 — общий вид слоевищ, 5 — вид клеток с поверхности слоевища
в верхней части, 6 — то же в нижней части. (По Виноградовой).
F. minima [—Enteromorpha minima f. glacialis Kjellm.; E. папа
(Sommerf.) Sjostedt; E. папа var. minima (Nag.) Sjostedt] (рис. 44, 1—
3). — Слоевище до 15 см выс., до 6 мм шир., простое или с редкими вет-
вями в основании, ровное или неправильно закрученное. Клетки с по-
верхности слоевища в средней его части 4—6 (7) мкм шир., 4—7 (9) мкм
дл., ближе к вершине 5—8 (10) мкм шир., (5) 7—9 (И) мкм дл.
В верхнем горизонте валунной и скалистой литорали как открытых,
так и защищенных мест, выносит небольшую степень опреснения. Мур-
манское побережье, Белое, Берингово, Охотское и Японское моря,
Курильские о-ва.
F. subsalsa (Kjellm.) Vinogr. [=Blidingia minima var. subsalsa
(Kjellm.) Scagel.; B. minima var. ramifera Blid.; Enteromorpha micrococca
Ahln. f. subsalsa Kjellm.; E. папа (Sommerf.) Sjostedt var. subsalsa
(Kjellm.) Sjostedt], — Слоевище до 20 см выс., 0.1 — 3 мм шир., обильно
разветвленное. Основная ось расширяется кверху или остается узкой,
нередко складчато скрученная. Ветви 1-го порядка длинные, узкие,
довольно густо покрытые короткими веточками 2-го порядка, часто
шиповидными и крючковидными. Клетки с поверхности слоевища в сред-
ней его части 5—10 мкм шир., 5—13 мкм дл.
В супралиторали и на литорали, вблизи ручейков и рек или в лагунах
с солоноватой водой, а также в водоемах, потерявших связь с морем,
образует спутанные массы; наиболее характерна для арктических райо-
нов. Новая Земля, Карское море (о-в Тыртов), Новосибирские о-ва
(о-в Котельный), Берингово и Охотское моря, Курильские о-ва.
114
От типовой формы отличается обильным ветвлением всего слоевища
и наличием ветвей 2-го порядка, а также более упорядоченным распо-
ложением клеток в слоевище вследствие его узости.
2. Blidingia chadefaudii (J. Feldm.) Blid. \= Enteromorpha chade-
jaudii J. Feldm.; Ulva micrococca (Kiitz.) Gobi] — Блидингия Шадефо
(рис. 44, 4—6). — Слоевище до 25 см выс., простое или с редкими ве-
точками в основании. Клетки с поверхности слоевища неправильно много-
угольные, часто округлые и тогда с хорошо различимыми межклетни-
ками, в средней части слоевища обычно 4—6 мкм шир., 5—7 мкм дл.,
ближе к вершине 5—8 (9) мкм шир., 8—10 (12) мкм дл., однако растения
могут быть как мелкоклеточными, так и крупноклеточными по всему
слоевищу. На поперечном срезе слоевища клетки вертикально вытянутые,
эллипсоидные, реже прямоугольные, часто с перетяжкой посередине,
4—7 мкм шир., 5—11 мкм выс., наружная часть их оболочки может
утолщаться до 8 мкм, внутренняя утолщается очень значительно, до
60 мкм, приобретая конусовидную или полусферическую форму и слои-
стую структуру; стенки слоевища в целом 15—80 мкм толщ.
В морских, солоноватых и опресненных водах, на камнях, дере-
вянных балках и т. п., в защищенных, но хорошо промываемых чистых
местах. Мурманское побережье, Белое, Охотское и Японское моря,
вост. Камчатка, о-в Сахалин, Курильские о-ва.
По внешнему облику близка к В. minima, отличается сильно утол-
щенными совнутри оболочками клеток, имеющими своеобразное слоистое
строение.
3. Blidingia marginate (J. Ag.) P. Dang. (= Enteromorpha marginata
J. Ag.; E. micrococca Kiitz.) — Блидингия окаймленная (рис. 45).—
Слоевище 2—5 см. выс., до 200 мкм шир., у основания цилиндрическое,
Рис. 45.
Blidingia marginata: 1,2 — общий вид слоевищ, 3 — вид клеток с поверхности
слоевища в верхей части, 4 — то же в средней части, 5 — поперечный срез
слоевища, 6, 7 — разные стадии образования ветвей. (По Мошковой и Юнгер).
8*
115
выше сдавленное, простое или слабо разветвленное, часто скрученное и
спутанное, с неровной поверхностью; от своеобразных расширений и
вздутий нередко отходят пучками многорядные микроскопические шипо-
видные или слегка удлиненные веточки. Клетки с поверхности слоевища
большей частью прямоугольные, реже многоугольные или неправильной
формы, 4.3—8.6 мкм шир., (3) 4—9.3 мкм дл., расположенные преиму-
щественно в узких, молодых частях слоевища отчетливыми продольными,
а иногда и поперечными рядами. На поперечном срезе слоевища клетки
почти квадратные или слегка вертикально вытянутые, 4—5 мкм шир.,
7—9.3 мкм выс., с равномерно не утолщенными оболочками; стенки
слоевища в целом 8.4 — 10 мкм толщ.
В морских водах, выносит небольшую степень опреснения.
От В. minima и В. chadefaudii хорошо отличается мелкими разме-
рами слоевища, не утолщенными совнутри и снаружи оболочками кле-
ток, отчетливым расположением клеток продольными, а нередко и попе-
речными рядами, особенно в молодых участках слоевища.
F. marginata (рис. 45). — Слоевище простое или очень слабо развет-
вленное, клетки 4.3—8.6 мкм шир., (3) 4.3—9.3 мкм дл.
В обрастаниях камней вблизи уреза воды или в местах, периодически
увлажняемых волнами или брызгами, где образует темно-зеленые густые
дерновинки. Черное (Аджарская АССР: Зеленый Мыс) и Каспийское
моря (залив Кара-Богаз-Гол).
F. ramulosa Vinogr. [=В. marginata subsp. subsalsa (Kjellm.) Blid.;
Enteromorpha micrococca var. subsalsa Kjellm.; E. radiata J. Ag.; E. ramu-
losa Borge]. — Слоевище разветвленное, веточки мелкие, клетки 6.5—
8.5 мкм в поперечнике.
В обрастаниях твердых субстратов вблизи уреза воды. В СССР не об-
наружена. (Зап. Европа).
От типовой формы отличается обильным ветвлением слоевища.
Род MONOSTROMA Thnr. - МОНОСТРОМА
Слоевище макроскопическое, на ранних стадиях развития пузыре-
видное, затем мешковидное, но легко разрывающееся и тогда листовидно
распростертое, вначале прикрепленное, позднее нередко отрывающееся
и свободноплавающее. Клетки с поверхности слоевища в его верхней
части округлые или от взаимного сдавливания многоугольные, в нижней
части удлиненные, булавовидные, нередко вытянутые в ризоиды, обычно
сближенные по 2—4 и более, но никогда не складывающиеся в ясные
квадратные или прямоугольные участки. Хлоропластов 1—2, чаще всего
с 1 центральным пиреноидом. Зооспоры четырехжгутиковые, по 4—16
в клетке спорофита. Нередко содержимое клетки спорофита может пре-
вращаться непосредственно в многоклеточный диск, многослойный в цен-
тральной части; слоевище формируется в результате отслаивания и раз-
растания верхнего слоя диска. Половой процесс изогамный или гетеро-
гамный. Гаметы двужгутиковые, помногу в любой клетке гаметофита, за
исключением клеток с ризоидными выростами. Правильная смена форм
развития может нарушаться, гаметы, теряя способность к копуляции, мо-
гут прорастать как в спорофит, так и в гаметофит.
В пресных водах и в морях. Систематика рода разработана слабо.
I. Клетки с 1 хлоропластом.
1. Слоевище до 10 мкм толщ.
А. Клетки с поверхности слоевища округлые или полукруглые,
расположенные рыхло..........................М. bullosum 1.
116
Б. Клетки с поверхности слоевища округленно-многоугольные,
расположенные плотно........................М. amorphum 2.
В. Клетки с поверхности слоевища квадратные, расположенные
плотно......................................М. fischeri 3.
2. Слоевище свыше 10 мкм толщ.
А. Клетки с поверхности слоевища округлые, собранные груп-
пами по 2 —4..............................М. oxyspermum 4.
Б. Клетки с поверхности слоевища многоугольные или округло-
многоугольные, обычно не собранные группами.
а. Клетки от четырех- до многоугольных, угловатые, сбли-
женные и расположенные беспорядочно....................
..................................М. membranaceum 5.
б. Клетки округло-многоугольные, в молодых частях слое-
вища собранные группами по 2—4, в старых расположен-
ные беспорядочно.........................М. wittrockii 6.
в. Клетки 4—6-угольные, округлые или вытянутые, сближен-
ные и расположенные обычно беспорядочно, реже про-
дольными рядами, иногда группами по 2—4................
...................................................М. grevillei 7.
II. Клетки с 2 хлоропластами, расположенными на противоположных
сторонах клетки...................................М. fuscum 8.
1. Monostroma bullosum (Roth) Wittr. — Монострома пузырчатая
(рис. 46, 1—5). — Слоевище темно-зеленое, вначале пузыревидное, затем
мешковидное, позже разорванное, пленчатое, около 10 мкм толщ.,
очень нежное, слизистое, прикрепленное к субстрату базальным диском.
Клетки с поверхности слоевища округлые или эллиптические, 3.5—
12 мкм шир., 11 — 12.5 мкм дл., непосредственно после деления полу-
круглые, иногда многоугольные, расположенные под значительным
слоем слизи сближенными по 2—4, на поперечном срезе слоевища
округлые. Хлоропласт 1.
В пресных проточных водах. ЭССР. Возможно, встречается и в дру-
гих местах, но легко просматривается.
2. Monostroma amorphum Collins — Монострома аморфная (рис. 46,
6,7). — Слоевище светло-зеленое, пластинчатое, маленькое, нежное,
слабо ослизненное, 6—8 мкм толщ. Клетки с поверхности слоевища
округленно-многоугольные, 8—10 мкм в диам., плотно расположенные,
на поперечном срезе слоевища эллиптические, горизонтально вытянутые.
Хлоропласт 1.
В проточных водах. В СССР не обнаружена. (Калифорния).
Вид своеобразный, но недостаточно изученный.
3. Monostroma fischeri Borge — Монострома Фишера (рис. 46,
8,9).— Слоевище зеленое или темно-зеленое, пластинчатое, нежное,
9—10 мкм толщ. Клетки с поверхности слоевища квадратные, 5—8 мкм
шир., непосредственно перед делением слегка удлиненные, расположен-
ные по 2—4, на поперечном срезе слоевища неправильно эллиптические,
2—4 мкм шир., горизонтально вытянутые. Хлоропласт 1.
Хорошо отличается от остальных видов рода квадратной формой
клеток.
Var. fischeri (рис. 46, 8, 9). — Клетки на поперечном срезе слоевища
3—4 мкм шир., расположенные довольно плотно.
В проточных водах. Земля Франца-Иосифа.
Var. minor Borge. — Клетки на поперечном срезе слоевища 2—3 мкм
шир., расположенные довольно рыхло.
117
Рис. 46.
1—5 — Monostroma bullosum; 1 — общий вид слоевища, 2 — вид клеток с по-
верхности слоевища, 3 — поперечный срез слоевища, 4 — зооспорангии и зоо-
споры, 5 — стадии прорастания зигоспоры; 6, 7 — М. amorphum: 6 — вид
клеток с поверхности слоевища, 7 — поперечный срез слоевища; 8, 9 — М. fi-
scheri: 8 — вид клеток с поверхности слоевища, 9 — поперечный срез слоевища.
(1 — по Кютцингу, 2—4 — по Блидингу, 5 — по Шода, 6, 7 — по Фричу,
8, 9 — по Борге).
В проточных водах. В СССР не обнаружена. (Зап. Европа).
4. Monostroma oxyspermum (Kutz.) Doty (=M. quaternarium
Desm.) — Монострома остросемянная (рис. 47,1—3). — Слоевище
светло-зеленое или зеленое, пленчатое, нежное, неопределенных очерта-
ний, до 40 см в поперечнике, с крупными лопастями, складками и от-
верстиями, по краям почти гладкое, 18—24 мкм толщ., вначале при-
крепленное коротким стебельком, отходящим от основания пластины,
позднее свободноплавающее. Клетки с поверхности слоевища округлые,
6 —11 мкм в диам., с толстыми оболочками, собранные группами по 2—4,
на поперечном срезе слоевища вертикально вытянутые, эллиптические
или полукруглые, 15—18 мкм выс., сближенные по 2. Хлоропласт 1.
В морях и пресных водах. Черноморское побережье Крыма.
5. Monostroma membranaceum W. et G. S. West — Монострома
мембранная (рис. 47, 4, 5). — Слоевище бледно-зеленое, в разросшемся
состоянии листовидно распростертое, до 9 см дл., к основанию суживаю-
щееся, слизистое, по краям часто изрезанное, 18 мкм толщ. Клетки
с поверхности слоевища многоугольные, 8—20 мкм в диам., сближенные,
расположенные без особого порядка, на поперечном срезе слоевища
эллиптические, вертикально вытянутые. Хлоропласт 1.
В пресных водах. В СССР не обнаружена. (Англия).
118
Рис. 47.
1—3 — Monostroma oxyspermum: 1 — общий вид слоевища, 2 — вид клеток
с поверхности слоевища, 3 — поперечный срез слоевища; 4, 5 — М. тетЬгапа-
сеит: 4 — общий вид слоевища, 5 — поперечный срез слоевища. (7 — по Кют-
циигу, 2, 3 — по Виттроку, 4, 5 — по Весту).
6. Monostroma wittrockii Born. — Монострома Виттрока (рис. 48). —
Слоевище светло-зеленое или зеленое, на ранних стадиях развития
пузыревидное, прикрепляющееся к субстрату маленькой ножкой,
образованной длинными переплетенными ризоидными выростами клеток,
расположенными в его основании, в разросшемся состоянии пластин-
чатое неправильных очертаний, до 6 (8) см в диам., складчатое, с ровными
или волнистыми краями, мягкое, 16—20 (29) мкм толщ., прикрепляю-
щееся ризоидами. Клетки с поверхности слоевища округло-многоуголь-
ные, 8—17 мкм в диам., располагающиеся в молодых частях более или
менее отчетливыми группами по 2—4, в старых без особого порядка,
на поперечном срезе слоевища эллиптические или почти квадратные
с закругленными углами, 8—17 мкм выс., по ширине такие же или
вдвое меньше. Хлоропласт 1.
119
Рис. 48.
Monostroma urittrockii: 1 — общий вид слоевища, 2 — вид клеток с поверхности
слоевища, 3 — поперечный срез слоевища. (7 — по Лаковитцу, 2, 3 — по Борне).
В морских, соленых и солоноватых водах, на камнях и раковинах
моллюсков. Каспийское море.
7. Monostroma grevillei (Thur.) Wittr. (=Enteromorpha grevillei
Thur.) — Монострома Гревилля (рис. 49). — Слоевище ярко-или бледно-
зеленое, до 100 см выс., нежное, вначале мешковидное, удлиненно-
булавовидное, нередко энтероморфообразное, затем разрывается и стано-
вится воронкообразным, неправильно лопастным или пластинчатым,
в нижней части 15—50 (65) мкм толщ., в верхней 12—40 мкм толщ.
Клетки с поверхности слоевища в ризоидной его зоне удлиненно-
эллиптические или веретеновидные, часто сильно вытянутые, 15—30 мкм
Рис. 49.
Monostroma grevillei-. 1,2 — общий вид слоевища, 3 — поперечный срез слое-
вища, 4 — вид клеток с поверхности слоевища в средней части, 5 — то же
в надризоидной части. (1—3 — но Блидингу, 4, 5 — по Виноградовой).
120
шир., 25—140 мкм дл., постепенно переходящие в ризоидные отростки,
в надризоидной зоне прямоугольные или неправильно четырехуголь-
ные, довольно удлиненные, расположенные продольными рядами или
беспорядочно, в остальной части 4—6-угольные, угловатые или округлые,
изодиаметрические или вытянутые, 5—20 мкм шир., 6—27 мкм дл.,
расположенные сближенйо или иногда по 2—4, на поперечном срезе
слоевища прямоугольные или овальные, 7 — 20 мкм шир., в высоту меньше
ширины, равны ей или до 2—3 раз больше. Оболочки клеток, обращен-
ные к поверхности слоевища, тонкие или утолщенные. Хлоропласт 1.
В морских водах, на камнях и водорослях, выносит опреснение
и загрязнение, но не выносит обсыхания. Преимущественно в северных
морях СССР, встречается и на Дальнем Востоке.
8. Monostroma fuscum (Post, et Rupr.) Wittr. [= Ulvaria obscura
(Kiitz’) Gayral] — Монострома темная (рис. 50). — Слоевище на ранних
стадиях развития трубчатое, позднее разрывается по длине и превраща-
ется в пластину, в основании которой сохраняется стебелек с подошвой;
пластина темно-буровато-зеленая, неопределенных очертаний, 10—20 см
в диам., почти гладкая, с неровными волнистыми или изрезанными
краями, с отверстиями, довольно грубая, 20—25 мкм толщ. Клетки с по-
верхности слоевища 4—6-угольные, 16—21 мкм шир., расположенные без
особого порядка, на поперечном срезе слоевища почти квадратные или
Рис. 50.
Monostroma- fuscum: 1 — общий вид слоевища, 2 — вид клеток с поверхности
слоевища в верхней части, 3 — то же в нижней части. (По Виноградовой).
121
слегка вертикально вытянутые, 14 — 25 мкм выс. Хлоропластов 2, на
противоположных сторонах клетки.
В соленых (морских) и солоноватых водах, на камнях и раковинах
моллюсков. УССР (лиманы).
Семейство ULVACEAE Lamour. — УЛЬВОВЫЕ
Слоевище макроскопическое, нитевидное, простое, из 1—2 рядов
клеток, или мешковидное и трубчатое, полое внутри, однослойное, простое
или разветвленное, а также пластинчатое, двуслойное, без внутренней
полости, на ранних стадиях развития прикрепляющееся ризоидами,
позднее иногда свободноплавающее. Клетки плотно прилегающие друг
к другу, от взаимного сдавливания часто угловатые. Хлоропласт 1,
пластинчатый, с ровными или изрезанными краями, с 1 или несколь-
кими пиреноидами. Вегетативное размножение регенерацией оторванных
частей слоевища. Бесполое размножение зооспорами и апланоспорами.
Половой процесс — изо- или гетерогамия, иногда гаметы прорастают
партеногенетически. При развитии слоевище проходит стадию одноряд-
ной вертикальной нити. Смена форм развития имеется, изоморфная.
Ульвовые — преимущественно морские растения, встречаются также
в осолоненных участках устьев рек, в соленых озерах, многие выдер-
живают значительное опреснение. В пресной воде обитают лишь неко-
торые виды Enteromorpha.
I. Слоевище нитевидное, из 1 — 2 рядов клеток.................
...................................род Percnrsaria (с. 122).
II. Слоевище трубчатое или мешковидное, внутри полое, однослойное,
иногда сильно сдавленное и тогда более или менее пластинчатое .
...................................род Enteromorpha (с. 123).
III. Слоевище пластинчатое, без полости внутри, двуслойное......
..........................................род Ulva (с. 138).
Род PERCURSARIA Воту - ПЕРКУРСАРИЯ
Слоевище макроскопическое, нитевидное, простое, обычно двуряд-
ное, реже однорядное. Клетки с поверхности слоевища почти прямоуголь-
ные. Хлоропласт 1, пластинчатый, поясковидный, с неровными краями,
с (1) 2—3 (4) пиреноидами. Зооспоры четырехжгутиковые, гаметы дву-
жгутиковые, те и другие выходят из материнских клеток заключенными
в гиалиновый пузырь.
Монотипный род.
1. Percursaria percursa (Ag.) Bory [=Enteromorpha percursa Ag.;
Diplonema percursa (Ag.) Kjellm.; D. confervoideum Lyngb.] — Перкурса-
рия пронизанная (рис. 51, 1—4). — Нити 20—40 мкм шир., 13—20 мкм
толщ. Клетки с поверхности слоевища прямоугольные, реже квадратные,
с тонкими оболочками, (8) 10—18 мкм шир., по длине равные ширине или
превышающие ее до 3 раз. На поперечном срезе слоевище имеет
овальную форму и состоит из 2 полукруглых клеток.
В морских и солоноватых водах, в обрастаниях моллюсков и круп-
ных водорослей, на песчаном грунте часто образует спутанные массы,
предпочитает опресненные участки моря, легко выносит загрязнение.
Мурманское побережье, Балтийское, Белое и Берингово моря, УССР
(лиманы), Черное и Японское моря.
122
Рис. 51.
1—4 — Percursaria percursa: 1—3 — фрагменты слоевища, 4 — поперечный срез
слоевища; 5,6 — Enteromorpha torta: 5 — фрагмент слоевища, 6 — поперечный
срез слоевища. (1—4 — по Мошковой, 5, 6 — по Блидингу).
Род ENTEROMORPHA Link. - ЭНТЕРОМОРФА
Слоевище макроскопическое, трубчатое или мешковидное, иногда
сильно сдавленное, или широкое, пластинчатое, всегда с внутренней
полостью, которая идет через все слоевище или при смыкании стенок
остается только в его основании и по краям, однослойное, простое
или разветвленное, с многорядными трубчатыми или однорядными ните-
видными ветвями, на ранних стадиях развития прикрепленное, позднее
отрывающееся и свободноплавающее. Клетки с поверхности слоевища
расположены беспорядочно или отчетливыми рядами, особенно в основа-
нии слоевища и молодых ветвях. Хлоропласт 1, пластинчатый, с 1 или
несколькими пиреноидами. Вегетативное размножение фрагментацией
слоевища. Зооспоры четырех- или двужгутиковые, со стигмой. Половой
процесс — изо- или гетерогамия, гаметы двужгутиковые. При прораста-
нии зооспор и зигот сначала возникает однорядное нитевидное слоевище,
которое или сразу разрастается в вертикальные трубчатые побеги, или
предварительно образует однослойную базальную пластину, на которой
возникают вертикальные побеги, вначале однорядные, позднее становя-
щиеся многорядными, трубчатыми.
В морях, соленых, солоноватых и пресных водах, имеются эвритоп-
ные формы, однако некоторые предпочитают определенные условия,
например только соленые или только пресные водоемы.
123
При определении видов важными диагностическими признаками
являются форма клеток и характер их расположения, особенно в верх-
ней части ризоидной зоны и сразу над ней.
I. Слоевище преимущественно не разветвленное, иногда с пролифи-
кациями.
1. Слоевище очень узкое, длинное, из 3—12 продольных, иногда
косо сбегающих рядов клеток.........................Е. torta 1.
2. Слоевище широкое, мешковидное или сдавленное, почти пластин-
чатое.
А. Клетки с поверхности слоевища расположены без особого
порядка, на отдельных участках иногда концентрическими ря-
дами; слоевище трубчатое или мешковидное.
а. Слоевище трубчатое, простое или слабо разветвленное, от
правильно цилиндрического или удлиненно-мешковидного
до широкопластинчатого с клиновидным основанием, легко
отрывающееся............................Е. intestinalis 2.
б. Слоевище мешковидное, пузыревидное, продолговато-яйце-
видное, вытянутое, с отверстием на вершине, простое, на
очень короткой ножке......................Е. maeotica 3.
Б. Клетки с поверхности слоевища расположены продольными
рядами (преимущественно в нижней части слоевища); слое-
вище плоское, пластинчатое или лентовидное.................
................................... . . . Е. linza 4.
II. Слоевище разветвленное или обильно снабженное пролификациями.
1. Клетки с поверхности слоевища расположены большей частью
продольными рядами, поперечные ряды имеются или отсутствуют.
А. Поперечные ряды клеток имеются, преимущественно в моло-
дых частях слоевища.
а. Клетки расположены четкими продольными рядами по
всему слоевищу.
а. Клетки квадратные или прямоугольные, вытянутые по
длине всего слоевища; хлоропласт с 1 пиреноидом .
..................................Е. ahlneriana 5.
р. Клетки квадратные, реже неправильно прямоугольные,
вытянутые по длине слоевища лишь в самой нижней
его части; хлоропласт с 2—6 пиреноидами...............
......................................Е. pilifera 6.
б. Клетки расположены продольными рядами преимуще-
ственно в основании слоевища.
а. Клетки в средней части слоевища 10—16 мкм шир.,
13—22 мкм дл., на поперечном срезе слоевища верти-
кально вытянутые........................Е. Hexuosa 7.
р. Клетки в средней части слоевища 8—12 мкм шир.,
10—15 мкм дл., на поперечном срезе слоевища почти
изодиаметрические.......................Е. jugoslavica 8.
Б. Поперечные ряды клеток отсутствуют. В основании слоевища
и в молодых ветвях клетки расположены четкими продоль-
ными рядами, в средней части правильность расположения
нарушается....................................Е. prolifera 9.
2. Клетки с поверхности слоевища на большей его части располо-
жены без особого порядка, и только местами встречаются про-
дольные, а иногда и поперечные ряды.
А. Слоевище от почти простого до обильно разветвленного на
всем протяжении, с ветвями 1—3-го порядков.
124
а. Ветви длинные или короткие, но не шиповидные.
а. Слоевище 2 — 10 мкм шир............Е. clathrata 10.
р. Слоевище до 278 мкм шир............Е. gracillima 11.
б. Ветви 2-го и 3-го порядков короткие, шиповидные ....
.......................................Е. ramulosa 12.
Б. Слоевище разветвлено лишь в нижней части, веточки преиму-
щественно 1-го порядка.....................Е. basiramosa 13.
1. Enteromorpha torta (Mert.) Reinb. (= Conferva torta Mert.) —
Энтероморфа скрученная (рис. 51, 5, 6). — Слоевище цилиндрическое,
нитевидное, простое, одинаковое по всей длине, 25— 50 мкм шир., с узкой
центральной полостью. Клетки с поверхности слоевища почти прямо-
угольные, 11 — 16 мкм шир., 12—28 мкм дл., расположенные в 3—12 про-
дольных рядах, иногда косо направленных, на поперечном срезе слоевища
до 20 мкм дл., с утолщенными наружными стенками. Хлоропласт
с 1 (2—-3) пиреноидами.
Образует спутанную массу вместе с другими водорослями. Бал-
тийское и Каспийское моря.
2. Enteromorpha intestinalis (L.) Link. (—Ulva Intestinalis L.) —
Энтероморфа кишечнообразная (рис. 52, 53). — Слоевище темно- или
светло-зеленое до белесого, трубчатое, правильно цилиндрическое, не-
редко удлиненно-мешковидное, с ровной, гладкой или вздуто-бугор-
чатой гофрированной поверхностью, обычно простое, иногда разветвлен-
ное или с редкими пролификациями, вначале прикрепленное, позднее
отрывающееся и свободноплавающее; когда стенки мешковидного слое-
вища спадаются, оно широкопластинчатое с клиновидным основанием,
иногда с перетяжками в нижней части, с ровными или извилистыми,
почти курчавыми краями. Клетки с поверхности слоевища неправиль-
ной формы, округло-многоугольные, 6—15 (20) мкм шир., (6) 7 —
20 (27) мкм дл., короткие или слегка вытянутые, особенно в надризоид-
ной части, расположенные беспорядочно, местами концентрическими ря-
дами, на поперечном срезе слоевища вертикально вытянутые, по длине
больше ширины в 1.2—4 раза, часто с сильно утолщенными, 4—30 мкм,
внутренними стенками. Хлоропласт цельный, плотный, с грубыми
краями, с 1 (2) пиреноидами, располагается обычно в верхнем углу
клетки в виде колпачка. Ризоидные клетки округлые, изодиаметрические
или вытянутые, крупнее и темнее остальных.
В пресных, солоноватых, соленых, морских, чистых и загрязненных,
проточных и стоячих водах, на мелководье, у уреза воды и значительно
глубже, в обрастаниях камней, раковин моллюсков и других твердых
субстратов, часто в очень больших количествах.
Сильно изменчивый вид. На основании разнообразного облика
слоевищ был разбит на ряд форм, часть которых соответствует ранее
описанным и считавшимся самостоятельными видам. Поскольку между
формами существуют переходы, точное определение их большей частью
затруднительно. Мы приводим только 3 формы.
F. intestinalis [=Е. intestinalis f. attenuata Ahln.; Ulva intestinalis
L.; Monostroma balticum (Aresch.) Wittr.] (рис. 52). — Слоевище на
ранних стадиях развития в виде булавовидных прикрепленных мешочков,
во взрослом состоянии в виде простой кишкообразной свободноплаваю-
щей трубки, при спадании стенок трубки плоское, 0.1—4 см шир.,
2—80 см, иногда до 2 м выс., с ровной, гладкой или бугорчатой,
неправильно складчатой поверхностью. Веточки и пролификации встре-
чаются редко. Ножка слоевища длинная и узкая, постепенно расширяю-
125
Рис. 52.
Enteromorpha intestinalis f. intestinalis: 1 — общий вид слоевища, 2 — вид
клеток с поверхности слоевища в верхней части, 3 — то же в надризоидной
части, 4 — поперечный срез слоевища в верхней части, 5 — то же в средней
части, 6 — то же в надризоидной части. (7 — по Кютцингу, 2—6 — по Мош-
ковой).
щаяся кверху, или короткая, плавно или резко переходящая в основную
часть слоевища. Клетки с поверхности слоевища округло-многоугольные,
8—15 (19) мкм шир., 12—20 (27) мкм дл., в ризоидной зоне часто
вытянутые в длину, на поперечном срезе слоевища 6—12 мкм шир.,
16—30 мкм дл.; ризоидные отростки хорошо видны, спускаются парал-
лельно стенке слоевища. Стенки слоевища в нижней его части 18—
48 мкм толщ., слизистый внутренний слой 3—19 мкм толщ., в верхней
части 15—30 мкм, слизистый слой 2.5—13 мкм.
126
Рис. 53.
Enteromorpha intestinalis: 1—6 — f. saprobia (7 — общий вид слоевищ, 2 —
вид клеток с поверхности слоевища в верхней части, 3 — то же в надризоидной
части, 4 — поперечный срез слоевища в верхней части, 5 — то же в средней
части, 6 — то же в надризоидной части), 7 — 11 — f. ramosa (7 — общий вид
слоевища, 8 — вид клеток с поверхности слоевища в средней части, 9 — то же
в надризоидной части, 10 — поперечный срез слоевища в средней части, 11 —
то же в надризоидной части). (По Мошковой).
В морских, солоноватых, сильно опресненных, пресных, стоячих
и проточных водах. Повсеместно.
Одна из наиболее распространенных водорослей.
F. saprobia Vinogr. (рис. 53, 1—6). — Слоевище простое, уплощен-
ное, 3—60 см выс., 0.5—6 см шир., с гладкими или волнистыми
краями. Клетки с поверхности слоевища беспорядочно расположенные,
7 — 19 мкм шир., 9—22 мкм дл., в ризоидной зоне округлые, почти
изодиаметрические, 7—13 мкм шир., 7—19 мкм дл., несколько выше
127
также округлые или слегка вытянутые в длину, 8—19 мкм шир., 12 —
22 мкм дл., в верхней части 8—14 мкм шир., 9—20 мкм дл., на поперечном
срезе слоевища (ближе к нижней его части) длинные и узкие, 14—45 мкм
шир., по длине в 2—4 раза больше, слегка суживающиеся на концах,
внутри ограниченные толстым слизистым слоем. Стенки слоевища
в нижней его части 30—90 (95) мкм толщ.
В солоноватых и морских водах, в бухтах, вблизи источников орга-
нического загрязнения, на мелководье. Черное и Японское моря.
От типовой формы отличается уплощенной формой слоевища.
F. ramosa Vinogr. (рис. 53, 7—Л). — Слоевище узкомешковидное,
обильно разветвленное, 2—15 см выс., гладкое или слегка скрученное;
ветви подобны главному стволу и сосредоточены преимущественно в ниж-
ней части слоевища, реже по всему слоевищу. Клетки с поверхности
слоевища округлой формы, в верхней части растения 8—14 мкм шир.,
11 — 25 мкм дл., в нижней части 8—13 мкм шир., 10—18 мкм дл., на попе-
речном срезе слоевища по длине в 1.5—3 раза больше ширины, внутрен-
Рис. 54.
Enteromorpha maeotica: 1—3 — общий вид слоевищ, 4 — вид клеток с поверх-
ности слоевища в средней части, 5 — то же в надризоидной части, 6 — попереч-
ный срез слоевища в средней части, 7 — то же в надризоидной части. (По
Мошковой).
128
Enteromorpha linza: 1,2 — общий вид слоевищ, 3 — вид клеток с поверхности
слоевища в верхней части, 4 — то же в средней части, 5 — то же в надризоидной
части, 6 — поперечный срез слоевища в нижней части. (7, 3—5 — по Вино-
градовой, 2, 6— по Блидингу).
ний слизистый слой 8—43 мкм толщ. Ризоидные клетки кончаются
довольно резко, их отростки уходят внутрь слоевища и с поверхности не
видны, непосредственно над ними клетки вытянуты. Стенки слоевища
в нижней его части 30—80 мкм.
Характерна для открытого морского побережья, в обрастаниях скал,
камней, реже Cystoseira ba.rba.ta, встречается от уреза воды до глубины
0.4—0.8 м, растет обычно пучками, заросли редкие, никогда не образуют
сплошного покрова. Черное море.
От типовой формы отличается наличием ветвления.
3. Enteromorpha maeotica Pr.-Lavr. (=£'. intestinalis f. elliptica Volk.;
E. elliptica Volk.; E. compressa var. maeotica Pr.-Lavr.) — Энтероморфа
меотическая (рис. 54). — Слоевище пузыревидное, яйцевидное, продолго-
вато-яйцевидное или вытянутое, 0.5—8 см шир., 2 — 10 (20) см выс.,
простое, на вершине с отверстием, на очень короткой ножке, слегка
расширяющейся в подошву; переход от ножки к основной части слоевища
9 Зеленые водоросли. Улотриксовые
129
может быть резким или постепенным; изредка встречаются пролифика-
ции, располагающиеся по всему слоевищу или только в нижней
его части. Клетки с поверхности слоевища преимущественно многоуголь-
ные, расположенные без особого порядка, 13—19 (23) мкм шир.,
12 — 27 (38) мкм дл., у молодых слоевищ 14—19 мкм шир., 18—27 мкм дл.,
угловатые, у более старых и крупных 13—17 мкм шир., 13—16 мкм дл.,
с округленными углами, на поперечном срезе слоевища прямоугольные,
12—18мкм шир., 17—27 мкм дл. Хлоропласте (2) 3—5 (7) пиреноидами,
нередко плохо различимыми.
В опресненных и соленых водах, в обрастаниях раковин моллюсков,
на небольшой глубине. Черное море (лиманы, лужи на побережье,
Сиваш), Азовское море (лиманы).
4. Enteromorpha linza (L.) J. Ag. (==E. linza f. lanceolata J. Ag. et
f. crispa J. Ag.; Ulva linza L.) — Энтероморфа линзовидная (рис. 55). —
Слоевище плоское, разнообразной формы, от почти нитчатой до ланцето-
видной, ширококлиновидной и ульвообразной, до 20 см шир., 5—50 см
выс., гладкое, с ровными или волнистыми краями, нередко с перетяжками
и перекрученное в нижней части, обычно простое, иногда у основания
с несколькими веточками, короткими и узкими, подобными главному
стволу; ножка отчетливо выраженная, переходящая в клиновидную или
сердцевидную пластину. Клетки с поверхности слоевища в ризоидной
зоне вытянутые в вертикальном направлении, несколько выше вытянутые
или изодиаметрические, 9—21 мкм шир., 11 — 31 мкм дл., по всей осталь-
ной части слоевища 4—5-угольные, угловатые, 8—19 мкм шир., 10.5 —
21 мкм дл., расположенные рядами, нарушаемыми на отдельных участ-
ках, на поперечном срезе слоевища прямоугольно-округлые, 13—38 мкм
дл., по ширине в 1.2—2 (3) раза меньше. Оболочки клеток внутри
слоевища утолщенные, 5—27 мкм толщ., что наблюдается не всегда.
Хлоропласт обычно с 1 (2), в клетках ножки с 2—3 пиреноидами.
В морях, соленых и солоноватых водах, преимущественно в за-
грязняемых участках, на камнях, скалах, на небольшой глубине.
Широко распространенный вид в морях Советского Союза.
По форме и характеру расположения клеток, строению хлоропласта
и числу пиреноидов Е. linza близка к Е. prolifera и с трудом отличима
от ее неразветвленной формы.
5. Enteromorpha ahlneriana Blid. ( = Е. compressa var. racemosa
Ahln.; E. procera var. denudata Ahln.) — Энтероморфа Альнера
(рис. 56). — Слоевище цилиндрическое, до 40 (50) см выс., с хорошо раз-
личимым, слегка расширяющимся или суживающимся к вершине глав-
ным стволом или без него, густо разветвленное, реже неразветвленное.
Ветви 1—3-го порядков к вершине обычно суживаются, веточки послед-
него порядка короткие, волосовидные, часто однорядные или из 1—4 ря-
дов клеток. Клетки с поверхности слоевища квадратные или прямоуголь-
ные, нередко пятиугольные, (10)12—15(22) мкм шир., (12)15 —
30 (33) мкм дл., угловатые, вытянутые по длине слоевища, в ризоидной
зоне булавовидные, расположенные отчетливыми продольными рядами,
в молодых частях продольными и поперечными рядами, на поперечном
срезе слоевища длина клеток в 1.1—1.7 раза превышает ширину. Обо-
лочки клеток неутолщенные. Хлоропласт с 1 пиреноидом. Стенки слое-
вища 20 — 30 мкм толщ.
В морских, солоноватых и довольно значительно опресненных чи-
стых водах, на мелководье, в обрастаниях камней и других твердых
субстратов, часто у самого уреза воды. Белое, Балтийское, Берингово
и Черное (Севастопольская и Новороссийская бухты) моря, о-в Сахалин.
130
Рис. 56.
Enteromorpha ahlneriana: 1 — общий вид слоевища в нижней части, 2 — вид
клеток с поверхности слоевища в верхней части, 3 — то же в средней части,
4 — то же в нижней части, 5 — то же в надризоидной части, б — поперечный
срез слоевища. (По Мошковой).
По строению слоевища и типу ветвления Е, ahlneriana сходна
с Е. prolijera, отличается преобладанием в слоевище прямоугольных,
вертикально вытянутых клеток.
6. Enteromorpha pilifera Kiitz. [=£. intestinalis var. capillaris
Kiitz.; E. flezuosa snbsp. pilifera (Kiitz.) Blid.J — Энтероморфа волоско-
носная (рис. 57). — Слоевище цилиндрическое, длинное, узкое,-0.5—
1 мм шир., обильно ветвящееся, образующее спутанные массы. Вет-
вление беспорядочное, имеются и однорядные короткие веточки или ве-
точки с однорядной вершиной. Клетки с поверхности слоевища четырех-
угольные, почти квадратные, или прямоугольные, реже неправильной
9*
131
Рис. 57.
Enteromorpha pillfera: 1 — общий вид слоевищ, 2 — вид клеток с поверхности
слоевища в концевой части, 3 — то же в узкой части, 4 — то же в широкой
части, 5—8 — поперечные срезы слоевища в разных частях. (По Мошковой).
формы, 8—16 мкм шир., 12—25 мкм дл., в широких участках слоевища
11—16 мкм шир., 16—25 мкм дл., в узких участках 8—11 мкм шир.,
12—16.5 мкм дл., вытянутые по длине слоевища только в самой нижней
его части, расположенные продольными рядами, а в узких частях слое-
вища и поперечными рядами. Хлоропласт с (2) 3—4 (5—6) пиренои-
дами.
В пресных или опресненных водах, на мелководье. Балтийское море
(устье р. Пярну), УССР (р. Кальмиус).
По строению слоевища близка к Е. flexuosa.
7. Enteromorpha flexuosa (Wolf.) J. Ag. (=E. flexuosa subsp. flexuosa
Blid.; E. intermedia Blid.; E. tubulosa Kiitz.; Conferva flexuosa Wulf.;
Ulva flexuosa Wulf.) — Энтероморфа извилистая (рис. 58). — Слоевище
трубчатое, разветвленное, реже простое, 1—7 мм шир., до 25 см выс.;
главный ствол расширяется кверху, ветвление редкое, ветви преимуще-
ственно 1-го порядка, часто сосредоточенные в нижней части слоевища,
реже встречаются ветви 2-го порядка и ветви, подобные главному
стволу, густо покрывающие последний на всем протяжении. Клетки
с поверхности слоевища в его основании прямоугольные, вытянутые
вертикально, 13—25 мкм шир., 17—40 мкм дл., расположенные отчет-
ливыми продольными рядами, выше по слоевищу они постепенно укора-
чиваются, 10—16 мкм шир., 13—22 мкм дл., сохраняя прямоугольную,
132
Рис. 58.
Enteromorpha flexuosa: 1 — общий вид слоевищ, 2 — вид клеток с поверхности
слоевища в верхней части, 3 — то же в средней части, 4 — то же в нижней
части, 5 — то же в надризоидной части, 6 — поперечные срезы слоевища.
(1 — по Блидингу, 2—6 — по Мошковой).
более или менее квадратную или многоугольную форму, в верхней части
слоевища и в молодых веточках часто квадратные или прямоугольные,
9—16 мкм шир., 10—24 мкм дл., расположенные продольными и попереч-
ными рядами, которые на отдельных участках могут нарушаться бла-
годаря появлению новых клеток, отчленяющихся косыми перегородками,
на поперечном срезе слоевища вертикально вытянутые, округленные,
13—21 мкм дл., по ширине в 1 — 1.6 раза меньше. Оболочки клеток
внутри слоевища в нижней его части могут слегка слоисто утол-
щаться. Хлоропласт с (1) 2—3 (5) пиреноидами, заполняющий, как
правило, почти всю видимую поверхность клетки. Стенки слоевища
16—27 мкм толщ.
В морских, солоноватых, а также пресных, чистых или загрязненных
водах, обычно недалеко от уреза воды, в обрастаниях камней и других
твердых субстратов.
Один из широко распространенных видов на территории СССР.
8. Enteromorpha jugoslavica Blid. — Энтероморфа югославская
(рис. 59). — Слоевище узкотрубчатое, 0.3—8 мм шир., 5—20 см выс.,
слабо разветвленное; главный ствол прослеживается по всему слоевищу,
к вершине несколько расширенный, ветви преимущественно 1-го по-
рядка, веточки 2-го порядка развиваются редко, волосовидные, могут
быть с однорядной вершиной. Клетки с поверхности слоевища почти
прямоугольные или неправильной формы, расположенные продольными,
а на отдельных участках и поперечными рядами, что нередко нару-
шается, в ризоидной зоне (ризоиды развиты незначительно и очень бы-
стро исчезают) 10—13 мкм шир., 16—30 мкм дл., вытянутые, по длине
больше ширины в 1.5—2.5 раза, в средней части слоевища 8—12 мкм шир.,
10—13 мкм дл., в верхней 8—11 мкм шир., 11 — 16 мкм дл., на поперечном
срезе слоевища почти изодиаметрические, слабо округленные, 9—11 мкм
133
Рис. 59.
Enteromorpha jugoslauica: 1 — общий вид слоевищ, 2 — участок характерного
ветвления, 3 — вид клеток с поверхности слоевища в верхней части, 4 — то же
в нижней части. (По Блидингу).
шир., 10—12 мкм дл. Хлоропласт с 1—5 пиреноидами. Стенки слоевища
13—17 мкм толщ.
В морских и соленых водах. Черное море (Севастопольская
бухта).
По размерам клеток и форме хлоропластов близка к Е. prolifera,
отличается большим числом пиреноидов (2—5).
9. Enteromorpha prolifera (О. F. Mull.) J. Ag. (—Ulva prolifera
0. F. Miill.) — Энтероморфа пролиферирующая (рис. 60). — Слоевище
трубчатое, очень часто уплощенное, с узкой, постепенно расширяющейся
кверху ножкой, 0.1—7 см шир., до 50 см выс., иногда значительно больше,
довольно обильно разветвленное или простое. Клетки с поверхности
слоевища 6—24 мкм шир., 8—33 мкм дл., в ризоидной зоне булавовидные
или прямоугольные,, расположенные рядами, несколько выше укорочен-
ные, но сохраняющие продольное расположение, в средних частях
главного ствола неправильно многоугольные, беспорядочно расположен-
ные, в верхних его частях и молодых ветвях почти квадратные, рас-
положенные продольными рядами, на поперечном срезе слоевища
вертикально вытянутые, 10—25 мкм дл. Оболочки клеток слегка утол-
щенные, особенно с внутренней стороны слоевища. Хлоропласт с 1 (2)
крупными пиреноидами.
В пресных, солоноватых и морских водах, выдерживает Широкий
диапазон солености, температуры и загрязнения.
F. prolifera [=£'. prolifera subsp. prolifera В lid.; E. compressa
p. prolifera (Ag.) Grev.; E. salina Kiitz.; Ulva compressa p. prolifera Ag.]
(рис. 60, 1—6). — Слоевище до 50 см выс., реже длиннее, довольно
сильно разветвленное, изредка простое. Ветвление неправильное, очеред-
ное или супротивное, на всем протяжении слоевища или только в нижней
его части. Клетки с поверхности слоевища почти прямоугольные, 6—
15 (24) мкм шир., 8 — 19 (33) мкм дл.
134
Enteromorpha prolifera'. 1—6 — f, prolifera {1 — общий вид слоевища, 2 —
вид клеток с поверхности слоевища в верхней части, 3 — то же в средней
части, 4 — то же в нижней части, 5 — поперечный срез слоевища в средней
части, 6 — то же в надризоидной части), 7—11 — f. simplex (7 — общий вид
слоевищ, 8 — вид клеток с поверхности слоевища в средней части, 9'— то же
в нижней части, 10 — поперечный срез слоевища в средней части, 11 — то же
в надризоидной части. (1—6 — по Мошковой, 7—11 — по Мошковой и Юнгер).
В бухтах, лагунах, у причалов, на набережных, пляжах и т. п.
Одна из широко распространенных форм водорослей в солоноватых,
соленых и морских водах СССР.
Р. simplex Vinogr. (рис. 60, 7—11). — Слоевище (0.3) 0.4—7 см шир.,
уплощенное, простое или с редкими пролификациями, постепенно расши-
ряющееся к вершине, иногда с перетяжками, с очень узкой, спирально
закрученной ножкой. Клетки с поверхности слоевища преимущественно
многоугольные, 6.5—14 мкм шир., (6.5) 10—21 мкм дл.
В морях, изолированных от моря водоемах, а также в устьях рек,
впадающих в море, на заиленных сваях, камнях, песке и гравии, часто
при значительном загрязнении и опреснении воды. Баренцево, Белое,
Берингово, Охотское и Черное (лиман в окр. Одессы, коса у Новорос-
сийска) моря, Командорские и Курильские о-ва.
10. Enteromorpha clathrata (Roth) Grev. (= Conferva clathrata
Roth) — Энтероиорфа решетчатая (рис. 61). — Слоевище цилиндриче-
ское, многократно или скудно разветвленное, 2—10 мм шир., с ветвями
1 —3-го порядков, нередко простое, 8—40 мм шир. Однорядные веточки
имеются, но редкие. Ризоидные клетки кончаются довольно резко на не-
большом расстоянии от подошвы, с отростками, ориентированными
135
Рис. 61.
Enteromorpha clathrata: 1 — общий вид слоевища, 2 — вид клеток с поверхности
слоевища в верхней части, 3 — то же в средней части, 4 — то же в нижней
части, 5 — поперечный срез слоевища. (7 — по Блидингу, 2—4 — по Вино-
градовой, 5 — по Зиновой).
в различных направлениях. Клетки с поверхности слоевища сразу над
ризоидными округло-многоугольные, несколько вытянутые в длину,
расположенные беспорядочно, крупные, 13—32 мкм шир., 18—60 мкм дл.,
выше по слоевищу 4—6-угольные и в широких ветвях неправильных
очертаний, расположенные беспорядочно, в узких ветвях почти квадрат-
ные или прямоугольные, 8—32 мкм шир., 12—56 мкм дл., вытянутые
вдоль и поперек слоевища, образующие продольные ряды, в самых моло-
дых частях расположенные продольными и поперечными рядами, на
поперечном срезе слоевища прямоугольные с округленными углами, по
длине больше ширины в 1.2—3 раза, в конечных веточках меньше
ширины. Хлоропласт с непостоянным числом пиреноидов, в осно-
вании слоевища их до 12, в средних частях 2—4 (5), в конечных
веточках 1—2.
В морях, изолированных от моря водоемах, а также в устьях рек,
впадающих в море, на мелководье, на камнях, раковинах моллюсков,
крупных водорослях, а также свободноплавающие. Баренцево, Белое,
Балтийское, Охотское, Черное, Азовское, Каспийское, Аральское, Япон-
ское моря.
11. Enteromorpha gracillima G. S. West — Энтероморфа грациозней-
шая (рис. 62). — Слоевище цилиндрическое, разнообразно изогнутое,
9—16 см дл., реже еще длиннее, очень тонкое, главный ствол до
278 мкм шир., обычно скудно разветвленное, с довольно гладкой или
слегка гофрированной поверхностью. Ветви 1-го порядка тонкие, 52—
110 мкм шир., длинные, изогнутые или короткие, на концах коротко
заостренные, ветви 2-го порядка 20—25 мкм шир. Клетки с поверхности
слоевища неправильно многоугольные, .9—23 мкм шир., расположенные
обычно более или менее беспорядочно, реже слабо выраженными
рядами.
В водах, богатых сероводородом, а также в солоноватых водах.
Малоизученный вид, весьма своеобразный по строению.
F. gracillima (рис. 62, 1, 2). — Главный ствол до 250 мкм шир.,
беспорядочно изогнутый, с довольно гладкой поверхностью. Ветви 1-го
порядка 64 — 110 мкм шир., длинные, изогнутые.
136
2
Рис. 62.
Enteromorpha gracillima'. 1, 2 — i. gracillima (7 — общий вид слоевища, 2 —
вид клеток с поверхности слоевища), 3, 4 — 1. breviramosa (3 — общий вид
слоевища, 4 — вид клеток с поверхности слоевища). (По Весту).
На скалах, по которым стекает вода, богатая сероводородом.
В СССР не обнаружена. (Южн. Африка).
F. breviramosa G. S. West (рис. 62,3,4). — Главный ствол до
278 мкм шир., волнообразно изогнутый, со слегка гофрированной по-
верхностью. Ветви 1-го порядка 52—91 мкм шир., короткие, обычно
двурядные, ветви 2-го порядка 20—25 мкм шир.
В солоноватых водах. В СССР не обнаружена. (Южн. Африка).
12. Enteromorpha ramulosa (Engl. Bot.) Hooker [=£'. crlnita (Roth)
J. Ag.] — Энтероморфа многоветочковая (рис. 63, 1). — Слоевище труб-
чатое, около 1 мм шир., 10—30 см дл., обильно разветвленное. От
главного ствола отходят очень длинные ветви 1-го порядка с большим
количеством коротких шиповидных, обычно прямых или несколько
согнутых, простых или повторно разветвленных ветвей 2-го порядка.
Клетки с поверхности слоевища округло-многоугольные, около 30 мкм
шир., около 40 мкм дл., в верхних участках слоевища 15 мкм шир.,
20 мкм дл., расположенные обычно довольно беспорядочно и лишь в моло-
дых ветвях продольными рядами. Хлоропласт с 2 — 10 пиреноидами.
В морях и солоноватых водах, в виде плотных спутанных дерновинок.
Черное, Азовское, Каспийское и Охотское моря.
13. Enteromorpha basiramosa Fritsch — Энтероморфа базальноветви-
137
Рис. 63.
1 — Enteromorpha ramulosa, общий вид части слоевища; 2 — Е. basiramosa,
общий вид слоевища. (7 — по Гарвею, 2 — по Весту).
стая (рис. 63, 2). — Слоевище трубчатое, до 2 см выс., обильно развет-
вленное в нижней части, выше только с одиночными короткими 1—2-ряд-
ными веточками; главный ствол до 500 мкм шир., значительно толще
веточек, слегка изогнутый, к концу суживающийся. Клетки с поверх-
ности слоевища квадратные или неправильно многоугольные, в расши-
ренных частях слоевища расположенные беспорядочно, в более узких
молодых частях продольными рядами, сбегающими несколько косо.
Хлоропласт с 1—2 пиреноидами.
В солоноватых и пресных водах. В СССР не обнаружена. (Южн.
Африка).
Род ULVA L. emend. Thur. — УЛЬВА
Слоевище макроскопическое, пластинчатое, двуслойное, лишенное
полости, простое или лопастное, с ровными или волнистыми краями,
часто с перфорациями, к основанию суженное в стебелек, прикрепляю-
щееся к субстрату базальным диском (подошвой). Клетки с поверхности
слоевища расположены беспорядочно, только иногда рядами, одноядер-
ные, ризоидные отростки клеток базального диска многоядерные. Хлоро-
пласт 1, пластинчатый, с 1 или несколькими пиреноидами. Вегетативное
размножение фрагментацией слоевища и прорастанием клеток базаль-
ного диска с образованием дополнительных побегов. Зооспоры четырех-
жгутиковые. Половой процесс — изо- или гетерогамия, гаметы двужгути-
ковые. Зигота прорастает сразу в новое многоклеточное слоевище,
проходя кратковременную нитчатую и трубчатую стадии, наблюдаю-
138
щиеся лишь в раннем онтогенезе. Смена форм развития имеется, изо-
морфная. Гаметофит двудомный.
Виды морские, встречаются также в соленых водоемах, потерявших
или теряющих связь с морем.
Виды рода Ulva морфологически разграничивать чрезвычайно
трудно. В настоящее время их принято различать по морям.
I. Обитает в Черном море.................................U. rigida 1.
II Обитает в северных морях европейской части СССР..................
....................................................U. lactuca 2.
III. Обитает в дальневосточных морях СССР............U. fenestrata 3.
1. Ulva rigida Ag. (= U. lactuca d. rigida Le Jolis) — Ульва жесткая
( рис. 64, 1—5). — Слоевище грубое, жесткое, темно-зеленое, от не-
скольких сантиметров до 1 м выс., вначале овально-округлое, позднее
широкоокруглое, неправильное, закругленное в форме розетки, простое
или лопастное, с ровными или волнистыми краями, часто с микроскопи-
ческими зубчиками по краю и с перфорациями. Клетки с поверхности
слоевища в его основании округлые, в остальной части 4—7-угольные,
10—22 (33) мкм шир., (И) 13 — 27 (30) мкм дл., на поперечном срезе
слоевища в его средней части овальные, по длине равные ширине или
в 2 раза больше, в нижней части вытянутые, по длине превышающие
ширину в 1.5—3 раза. Оболочки клеток совнутри могут утолщаться.
Хлоропласт с 1—3 пиреноидами. Пластина в целом в ризоидной части
слоевища 150—200 мкм толщ., в средней 55—95 мкм толщ.
Один из самых обычных видов в прибрежной части Черного
моря и в лиманах Причерноморья, достигающий массового развития
в загрязненных участках.
2. Ulva lactuca L. (=77. crassa Kjellm.) — Ульва латукообразная
(рис. 64, 6—10). — Слоевище довольно мягкое, ярко-зеленое или желто-
ватое, широколопастное, до 50 см выс., 30 см шир., цельное или
с отверстиями, нередко разорванное на лопасти, с волнистыми складча-
тыми краями. Стебелек мало заметен или отсутствует, подошва малень-
кая. Клетки с поверхности слоевища, включая его основание, 4—5-уголь-
ные, округлые или угловатые, неправильной формы, в нижней части
слоевища 11 — 24 мкм шир., 16 — 28 мкм дл., в верхней части 12—21 мкм
шир., 15—27 мкм дл., расположенные беспорядочно и только на отдель-
ных участках рядами, на поперечном срезе слоевища почти квадратные
или вытянутые в горизонтальном направлении, в нижней его части вы-
тянутые вертикально, в ризоидной части 32 мкм шир., 50 мкм дл.,
над ризоидной частью 16—20 мкм шир., 32 мкм дл., по длине равные ши-
рине или превышающие ее в 1.5—2 раза. Хлоропласт чашевидный,
с 1—3 пиреноидами, заполняющий видимую часть клетки на '/з- Пла-
стина в целом в основании слоевища 150 — 200 мкм толщ., в верхней части
50—80 мкм толщ.
В северных морях, в защищенных и полузащищенных местах. Мур-
манское побережье, Белое море.
3. Ulva fenestrata Р. et R. — Ульва продырявленная (рис. 65). —
Слоевище темно-зеленое или светло-желтое, от нескольких сантиметров
до 1 м выс., округлое, удлиненно-овальное или широкое, неправильное,
часто с перфорациями, с клиновидным или сердцевидным основанием,
наиболее отчетливо выраженным у молодых растений, начинающимся не-
посредственно от базального диска или снабженным короткой цилиндри-
139
Рис. 64.
1—5 — Viva rigida: 1 — общий вид слоевищ, 2 — вид клеток с поверхности
слоевища в верхней- части, 3 — то же в нижней части, 4 — поперечный срез
слоевища в верхней части, 5 — то же в нижней части; 6—10 — V. lactuca'.
6 — общий вид слоевищ, 7 — вид клеток с поверхности слоевища в верхней
части, 8 — то же в нижней части, 9 — поперечный срез слоевища в верхней
части, 20 — то же в нижней части. (1—6, 8—10 —по Виноградовой, 7 —
по Блидингу).
140
Рис. 65.
Viva fenestrata: 1 — общий вид слоевища, 2 — вид клеток с поверхности
слоевища. (По Виноградовой).
ческой ножкой. Клетки с поверхности слоевища 4—6-угольные, непра-
вильной формы, округлые или угловатые, 8—24 мкм шир., 8—32 мкм
дл., в его основании неправильно многоугольные, с округленными уг-
лами, расположенные беспорядочно; ризоидные клетки значительно
крупнее вегетативных, темно-зеленые, ризоиды отходят от внутренних
стенок клеток и с поверхности не видны; на поперечном срезе слоевища
клетки вертикально вытянутые, реже квадратные, 10—27 мкм шир.,
20—49 мкм дл., длина их к вершине слоевища уменьшается. Оболочки
клеток совнутри могут утолщаться, и тогда между рядами клеток обра-
зуется слизистый слой до 30 мкм толщ. Хлоропласт с 1—3 пиреноидами.
Пластина в целом в ризоидной части слоевища 100—200 мкм толщ.,
в нижней половине 65—125 мкм толщ., в верхней 40—110 мкм толщ.
Один из распространенных видов в прибрежных частях морей
Дальнего Востока, обитает в защищенных и открытых местах, в чистых и
загрязненных водах. Чукотское, Берингово моря, Командорские о-ва,
вост. Камчатка, Охотское море, о-в Сахалин, Курильские о-ва, Япон-
ское море.
Подпорядок GHAETOPHORINEAE —
ХЕТОФОРОВЫЕ
Слоевище нитчатое или распростерто-пластинчатое, микроскопиче-
ское или довольно крупное. Нити ветвящиеся, однорядные, часто сужен-
ные на концах, обычно прикрепленные, прямостоячие или ползучие,
полностью распростертые по субстрату или частично приподнимающиеся
над ним. На многих клетках прямостоячих и ползучих нитей и пластин
образуются одно- и многоклеточные волоски или щетинки, или они отсут-
ствуют. Клетки обычно одноядерные, с довольно толстой, гомогенной или
слоистой оболочкой, неослизняющейся или легко ослизняющейся. Хлоро-
141
пласт преимущественно 1, пристенный, кольцевидный, поясковидный
или дисковидный, цельный или продырявленный, нередко сетчатый,
с цельными или изрезанными краями, с пиреноидами или без них. Про-
дукт ассимиляции — крахмал, реже маслоподобное вещество, бесцветное
или окрашенное в желтый, оранжевый или красный цвет. Рост водорос-
лей верхушечный (апикальный) или вставочный (интеркалярный).
Вегетативное размножение акинетами или случайным разрывом слое-'
вища. Бесполое размножение зооспорами, апланоспорами, гипноспорами.
Половой процесс изо-, гетеро- или оогамный.
Хетофоровые водоросли представлены водными и аэрофитными фор-
мами, многие — типичные эпифиты, некоторые — эндофиты, обитающие
внутри слизистых обверток, в слоистых оболочках и внутри клеток во-
дорослей. Основными диагностическими признаками являются форма и
строение таллома, форма, строение и положение в клетке хлоропласта,
а также экологические особенности. Определению поддаются лишь
взрослые, полностью сформированные растения.
I. Слоевище в виде микро- или макроскопических разветвленных ку-
стиков.
1. Конечные клетки веточек заостренные или заканчиваются одно-
или многоклеточными волосками; хлоропласт с пиреноидом, про-
дукт ассимиляции — крахмал .....................................
.....................сем. Chaetophoraceae (частично) (с. 143).
2. Конечные клетки веточек закругленные, волоски отсутствуют;
хлоропласт без пиреноида, продукт ассимиляции — масло.
А. Талломы в виде разветвленных кустиков; оболочки клеток тон-
кие, не слоистые, масло бесцветное; в водоемах...............
...........................сем. Microthamniaceae (с. 300).
Б. Талломы в виде войлокообразных дерновинок, подушечек,
порошковатых налетов, реже однослойных пластинок; оболочки
клеток толстые, слоистые, масло желтое, оранжевое или крас-
ное; на коре деревьев, скалах, валунах.......................
.......................................сем. Trentepohliaceae (с. 275).
II. Слоевище в виде эластичных слизистых или твердых, инкрустирован-
ных известью, шаровидных или полушаровидных подушечек, лопаст-
ных образований, бугристых корочек, а также плотных дерновинок,
с одно- или многоклеточными волосками или без них..................
.............сем. Chaetophoraceae (частично) (с. 143).
III. Слоевище в виде однослойных дисков или разнообразно ветвящихся
по субстрату нитей, реже паренхимных пластинок неправильных
очертаний или рыхло расположенных в слизи клеток.
1. Клетки без волосков и щетинок................................
......................сем. Chaetophoraceae (частично) (с. 143).
2. Клетки с волосками.
А. Волоски одноклеточные, преимущественно вздутые у основа-
ния ..........................сем. Aphanochaetaceae (с. 303).
Б. Волоски многоклеточные, у основания не вздутые............
. . . . . . . сем. Chaetophoraceae (частично) (с. 143).
3. Клетки с щетинками.
А. Щетинки заключены в трубчатое влагалище, отогнутое в форме
воротничка....................сем. Coleochaetaceae (с. 316).
Б. Щетинки обычно без трубчатого влагалища, или оно слизистое,
цилиндрическое или конусовидное, гомогенное или слои-
стое ........................сем. Chaetosphaeridiaceae (с. 325).
142
Семейство CHAETOPHORACEAE (Harv.) De-Toni
et Levi - ХЕТОФОРОВЫЕ
Слоевище обычно состоит из двух частей: горизонтальной, распро-
стертой по субстрату, и вертикальной, прямостоячей, образованных вет-
вящимися или простыми йитями. Горизонтальная часть (подошва)
слагается из тесно расположенных, ветвящихся в плоскости нитей,
срастающихся боковыми сторонами в однослойную паренхимную пла-
стинку разнообразных очертаний, в которой местами даже трудно заме-
тить следы нитчатого строения; пластинка может образовываться также
в результате неправильного деления клеток или сдвигания их по отно-
шению друг к другу. Вертикальная часть представлена одной или не-
сколькими простыми или обильно ветвящимися нитями, многие веточки
которых оканчиваются прозрачными одно- или многоклеточными тупыми
или острыми волосками; волоски могут отходить и непосредственно от
клеток подошвы. Усиленное развитие вертикальной или горизонтальной
частей или полное отсутствие одной из них приводит к возникновению
разнообразных форм слоевища. Вертикальная часть может быть редуци-
рована до нескольких коротких ветвей или представлена только волосо-
видными образованиями, а также отсутствовать полностью; в последнем
случае слоевище часто приобретает форму диска или неправильной
пластинки, обычно однослойной, реже участками многослойной. При
недоразвитии или полном выпадении горизонтальной части хорошо
развитой, обильно разветвленной оказывается вертикальная часть слое-
вища, при этом, как правило, наблюдаются различия между главной
(основной) нитью (стволом) и боковыми ветвями. У таких слоевищ от
нижних клеток прямостоячих нитей отходят длинные разветвленные
ризоидные выросты, с помощью которых они прикрепляются к субстрату.
Хлоропласт преимущественно кольцевидный, реже в виде полуцилиндра,
цельный или продырявленный, нередко настолько сильно, что приобре-
тает вид сеточки, с 1 или несколькими пиреноидами, иногда без них.
Вегетативное размножение акинетами, реже распадением слоевища на
отдельные фрагменты. Бесполое размножение дву- или четырехжгутико-
выми зооспорами со стигмой, образующимися по 1 — 32 в любой клетке,
за исключением верхушечных, базальных и ризоидных; зооспорангии
имеют такой же вид, как и вегетативные клетки, только несколько более
вздутые, зооспоры освобождаются в результате ослизнения стенок зоо-
спорангия или через отверстие, возникающее в его оболочке. Известны
также апланоспоры и гипноспоры. Половой процесс изо- или гетерогам-
ный. Гаметангии похожи на вегетативные клетки или зооспорангии.
В пресных, солоноватых, соленых и морских водах, преимущест-
венно в обрастаниях разных органических и неорганических субстратов,
реже в слизистых обвертках, оболочках или внутри клеток водорослей.
I. Слоевище в виде разветвленно-нитевидных, вертикально направлен-
ных кустиков, обособленных или объединенных в дерновинки, полу-
шаровидные или шаровидные подушечки и корочки, реже в виде пла-
стинок из горизонтально распростертых нитей.
1. Нити к вершине утончающиеся и заканчивающиеся одно- или
многоклеточными волосками, реже закругленными клетками . . .
подсем. Chaetophoroideae (с. 144).
2. Нити без волосков, к вершине не утончающиеся и заканчиваю-
щиеся закругленными или коническими клетками...............
.............................подсем. Leptoslroideae (с. 218).
143
II. Слоевище в виде разветвленных ползучих нитей, обособленных или
смыкающихся в одно- или многослойные пссвдопаренхимные пла-
стинки .............................подсем. Ulvelloideae (с. 233).
Подсемейство CHAETOPHOROIDEAE Bourr.
Нити ветвящиеся, реже простые, прямостоячие, обычно с четко обо-
собленной главной (основной) нитью и короткими или длинными,
простыми или повторно разветвленными ветвями, реже ползучие, пол-
ностью распростертые по субстрату или частично приподнимающиеся над
ним, не сливающиеся или сливающиеся в паренхимные пластинки.
Конечные клетки нитей заостренные, реже закругленные или вытяги-
вающиеся в одно- или многоклеточный волосок.
I. Водоросли с хлоропластами, обычной зеленой окраски.
1. Хлоропласт 1, с пиреноидами.
Л. Слоевище в виде прямостоячих разветвленных кустиков,
а. Кустики объединены в дерновинки........................
..................род Stigeoclonium (частично) (с. 145).
б. Кустики объединены в слизистые колонии или обособ-
ленные.
а. Боковые ветви не отличаются от главных нитей.
* Прямостоячие нити заключены в обильную слизь,
формирующую слоевища шаровидной, полушаро-
видной или лопастной формы.....................
.......................род Chaetophora (с. 179).
* * Прямостоячие нити не заключены в слизь, слоевища
обособленные, кустиковидные.
+ Хлоропласт цельный или продырявленный,
с неровными краями, с 1 или несколькими пи-
реноидами; в пресных и солоноватых водах .
. род Stigeoclonium (частично) (с. 145).
+ + Хлоропласт сетевидный, со многими крупными
пиреноидами; эндем Байкала....................
...................род Ireksokonia (с. 186).
р. Боковые ветви отличаются от главных нитей.
* Боковые ветви простые или скудно разветвленные,
рассеянные...............род Cloniophora (с. 187).
* * Боковые ветви обильно разветвленные, преимуще-
ственно в мутовках.
+ Главные нити и длинные побеги состоят из оди-
наковых по размерам клеток; конечные клетки
веточек часто оканчиваются многоклеточным
волоском.
X Клетки главных нитей с тонкими оболочками.
О Вокруг главных нитей в большей или
меньшей степени развивается покров из
обильно разветвленных ризоидов; эндем
Байкала ..............................
. . . род Draparnaldiella (с. 190).
00 Вокруг главных нитей покров отсут-
ствует; повсеместно в проточных и стоя-
чих пресных водоемах...................
............род Draparnaidia (с. 196).
144
XX Клетки главных нитей с толстыми слои-
стыми оболочками ........................
. . . . род Myxonemopsis (с. 187).
+ 4- Главные нити и длинные побеги состоят из
чередующихся длинных и коротких клеток .
............род Draparnaldiopsis (с. 207).
Б. Слоевище в виде ползучих по субстрату обособленных ветвя-
щихся нитей с отходящими от них вверх простыми или
скудно разветвленными прямостоячими нитями или волосо-
видными ветвями.
а. От клеток нитей, распростертых по субстрату, отходят
вверх простые или разветвленные прямостоячие нити,
а. Прямостоячие нити преимущественно простые, верху-
шечные клетки конусовидные, волоски отсутствуют;
зооспоры двужгутиковые . . . род Iwanoffia (с. 210).
р. Прямостоячие нити разветвленные, верхушечные клетки
притупленные, заостренные или вытянутые в длинный
многоклеточный острый волосок; зооспоры четырехжгу-
тиковые.................... . . род Caespitella (с. 212).
б. От клеток нитей, распростертых по субстрату, отходят
вверх под прямым углом волосовидно заостренные ветви
..................................род Pseudochaete (с. 212).
В. Слоевище в виде паренхимной пластинки, распростертой
по субстрату и частично внутри субстрата, где переходит
в нити или в скопления клеток.
а. От поверхности пластинки отходят вверх короткие малокле-
точные простые или изредка разветвленные нити, реже
бесцветные волоски.....................................
..................род Stigeoclonium (частично) (с. 145).
б. От поверхности пластинки и по ее краям отходят короткие,
преимущественно простые нити, часто заканчивающиеся
волоском.....................рбд Endoclonium (с. 214).
2. Хлоропластов несколько, без пиреноидов....................
................................род Thamniolum (с. 216).
II. Водоросли без хлоропластов, бесцветные......................
.....................................род Saprochaete (с. 216).
Род STIGEOCLONIUM Kutz. - СТИГЕОКЛОНИУМ
Слоевище обычно состоит из горизонтальной части (подошвы),
образованной ползучими по субстрату и ветвящимися в плоскости ни-
тями, срастающимися боковыми сторонами в паренхимную пластинку
более или менее радиального строения, и вертикальной части — прямо-
стоячих разветвленных или простых нитей (главных нитей, или стволов),
отходящих от клеток подошвы; обе части окружены тонким слоем
слизи, заметным лишь при окрашивании. В большинстве случаев
преобладают прямостоячие нити, обычно обильно разветвленные,
создающие кустиковидную форму. Боковые ветви рассеянные, оче-
редные (одиночные или парные), реже супротивные, иногда сбли-
женные по нескольку пар или в мутовках, двум видам (5. fasciculare
и У. glomeratum) свойственны плотные кистевидно-пучковидные ветвле-
ния. Главные нити и боковые ветви у большинства видов существенно
не различаются, только у немногих (5. flagelliferum, S. атоепит. и др.)
главные нити (особенно в нижней части) выполняют лишь функцию
10 Зеленые водоросли. Улотриксовые
Л 45
носителей ветвей, так как их клетки утратили способность к образова-
нию зооспор. Ветви всех порядков оканчиваются одноклеточными или,
чаще, многоклеточными, острыми или тупыми волосками, у некоторых
видов волоски отсутствуют, их конечные клетки заостренные, слегка су-
женные или закругленные. В ряде случаев прямостоячие нити почти пол-
ностью редуцируются, значительного развития достигает подошва,
а волоски часто возникают из клеток ползучих нитей. Хлоропласт
пристенный, кольцевидный или поясковидный, с неровными краями,
цельный или сетчато продырявленный, с 1 или многими пиреноидами;
в веточках и на верхушках главных нитей он выстилает продольные
стенки клеток почти полностью, в средних участках главных нитей и осо-
бенно у основания занимает обычно только среднюю их часть в виде
узкого кольца. Акинеты образуются преимущественно в клетках по-
дошвы, их содержимое часто окрашено в оранжевый или красный цвет,
в клетках ветвей образуются еще покоящиеся акинеты, которые долгое
время остаются соединенными в цепочках -и прорастают при наступлении
благоприятных условий. Часто наблюдается пальмеллевидное состояние,
иногда связанное с начавшимся, но задержавшимся процессом обра-
зования зооспор. Зооспоры дву- и четырехжгутиковые, последние двух
сортов: макро- и микрозооспоры. Известны и апланоспоры. Половой про-
цесс изогамный, изучен недостаточно.
В пресных и солоноватых, преимущественно проточных водах,
главным образом в обрастаниях водных цветковых растений, разных
древесных субстратов, камней, раковин моллюсков и т. п.
Виды Stigeoclonium трудно различаются между собой вследствие
широкой морфологической изменчивости таксономических признаков.
Для определения видов наибольшее значение имеют форма и величина
клеток главных нитей, от которых отходят боковые ветви, однако ни один
из указываемых в диагнозах признаков нельзя использовать самостоя-
тельно, только в комплексе они позволяют правильно идентифицировать
виды.
I. Слоевище с хорошо развитыми прямостоячими нитями.
1. Клетки главных нитей по форме и величине не отличаются от
клеток ветвей.
А. Ветви 1-го порядка преимущественно очередные.
а. Ветви 2-го порядка только очередные; конечные клетки ту-
пые, шиловидно заостренные или вытянутые в заостренный
или тупой волосок.
а. Главные нити более или менее прямые; волоски имеются.
* Клетки главных нитей 6—20 мкм шир.
+ Главные нити скудно ветвящиеся, ветви 1-го
порядка немногочисленные.
X Ветви 2-го порядка длинные.................
......................S. subsecundum 1.
XX Ветви 2-го порядка короткие .......
............................S. thermale 2.
+ + Главные нити обильно ветвящиеся, ветви 1-го
порядка многочисленные.
X Ветви 2-го порядка длинные.................
............................S. setigerum 3.
XX Ветви 2-го порядка короткие..............
...........................S. aestivale 4.
* * Клетки главных нитей 25—47 мкм шир...........
.............................S. nudiusculum 5.
146
р. Главные нити разнообразно изогнутые; волоски отсут-
ствуют ............................S. biasolettianum 6.
б. Ветви 2-го порядка не только очередные, но и супротивные,
реже в мутовках; конечные клетки заостренные или вытя-
нутые в тонкий заостренный волосок . . . S. attenuatum 7.
Б. Ветви 1-го порядка не только очередные, но и супротивные,
реже в мутовках.
а. Главные нити и длинные ветви 1-го порядка с рассеянным
беспорядочным ветвлением.
а. Ветви 1-го порядка преимущественно очередные.
* Подошва отсутствует или развита очень слабо.
+ Ветви 1-го порядка в нижней части слоевища
удалены друг от друга; конечные клетки
заостренные......................S. elongatum 8.
+ + Ветви 1-го порядка в нижней части слоевища
сближены по 2—3 и направлены вверх; конеч-
ные клетки преимущественно вытянутые
в длинный многоклеточный волосок . . . .
................................S. longipilum 9.
** Подошва развита хорошо, в виде правильного диска
....................................S. nanum 10.
р. Ветви 1-го порядка очередные, одиночные или парные
и супротивные, реже в мутовках.
* Ветви 1-го порядка обычно короткие, рассеянные,
простые или очень скудно ветвящиеся; волоски
имеются, одноклеточные . . . S. subspinosum И.
* * Ветви 1-го порядка длинные, большей частью сбли-
женные попарно, обильно ветвящиеся.
+ Волоски имеются, многоклеточные, реже конеч-
ные клетки заостренные.
X Длина клеток главных нитей равна ширине,
меньше или несколько больше ее; конечные
клетки веточек шиловидно заостренные или
вытянутые в длинный многоклеточный воло-
сок ...................S. subuligerum 12.
XX Длина клеток главных нитей в 2—6 раз
больше их ширины; концы веточек раз-
личные.
О Конечные клетки веточек заостренные,
реже вытянутые в 1—3-клеточный воло-
сок ....................S. tenue 13.
00 Конечные клетки веточек большей частью
переходят в длинный многоклеточный во-
лосок ...................S. protensum 14.
XXX Длина клеток главных нитей в (4) 8—
10 (12) раз больше ширины; конечные
клетки большей частью вытянутые в длин-
ный многоклеточный волосок..................
.......................... S. segare 15.
-Г+ Волоски отсутствуют; конечные клетки веточек
туповато-конусовидные или заостренные, иног-
да сильно вытянутые.
X Ветви 1-го порядка обильно ветвящиеся,
ветви 2-го порядка весьма многочисленные
..............................S. lebelii 16.
10»
147
XX Ветви 1-го порядка не ветвящиеся или очень
скудно ветвящиеся, ветви 2-го порядка край-
не немногочисленные.
О На главных нитях и ветвях 1-го порядка
дополнительных новообразований нет . .
.......................S. stagnatile 17.
00 На главных нитях и ветвях 1-го порядка
образуются грозди вздутых эллипсоидных
клеток.................S. carolinianum 18.
б. Главные нити и длинные ветви 1-го порядка на вершине
с кистевидным ветвлением.
а. Кистевидные ветвления равномерные, плотные, прямые,
главная нить выражена' отчетливо на всем их протя-
жении; волоски имеются, многоклеточные.
* Длина клеток главных нитей равна ширине или до
2 раз больше....................S. fasciculare 19.
* * Длина клеток главных нитей в 2—7 раз больше
ширины............................S. glomeratum 20.
р. Кистевидные ветвления односторонние, рыхлые, дуго-
видно согнутые, главная нить не выражена на их протя-
жении; волоски отсутствуют, конечные клетки тупокону-
совидные или вытянуто заостренные . . . S. nelsonii 21.
2. Клетки главных нитей по форме и величине отличаются от клеток
ветвей.
А. Клетки главных нитей в нижней части растения цилиндри-
ческие или бочонковидные; ветви 1-го порядка очередные,
супротивные или в мутовках.
а. Клетки главных нитей бочонковидные, по длине равные
ширине или до 2 раз больше...............S. pusilium 22.
б. Клетки главных нитей цилиндрические, по длине равные
ширине или до 8 раз больше.
а. Ветви 2-го порядка изогнутые в форме завитка,
сближенные к верхушкам ветвей; конечные клетки
заостренные..........................S. helveticum 23.
р. Ветви 2-го порядка прямые, рассеянные; конечные
клетки коротко заостренные, реже вытянутые в длинный
волосок..............................S. lubricum 24.
Б. Клетки главных нитей в нижней части растения только ци-
линдрические; ветви 1-го порядка обычно супротивные,
а. Ветви 2-го порядка преимущественно очередные, оканчи-
вающиеся короткой заостренной клеткой; волоски обычно
отсутствуют................................S. amoenum 25.
б. Ветви 2-го порядка очередные или супротивные, сближен-
ные по нескольку вместе, оканчивающиеся шиловидно за-
остренной клеткой или длинным многоклеточным тупым
волоском........................ . S. flagelliferum 26.
II. Слоевище с редуцированными прямостоячими нитями.
1. Слоевище состоит из ползучих по субстрату нитей, образующих
подошву, и отходящих вверх простых или скудно разветвленных
коротких прямостоячих нитей; волоски отсутствуют...............
...........................................S. variabile 27.
2. Слоевище состоит из хорошо развитой подошвы и отходящих вверх
прямостоячих нитей, представленных только 1 или несколькими
клетками; волоски имеются.
А. Прямостоячие нити папилловидные, из 1 (2—3) коротких
148
клеток...........................S. moebiusianum 28.
Б. Прямостоячие нити из 1 — 5 удлиненных клеток.
а. Прямостоячие нити из 1—2 (5) клеток, обычно простые; во-
лоски в основании слегка вздутые.......................
....................................S. prostratum 29.
б. Прямостоячие нити из 3—5 и больше клеток, простые,
реже скудно ветвящиеся; волоски в основании не вздутые
........................................S. farctum 30.
1. Stigeoclonium subsecundum (Kiitz.) Kutz. [=Conferva subsecunda
Kutz.; Myxonema subsecundum (Kutz.) Hazen] — Стигеоклониум почти-
однобокий (рис. 66). — Слоевище кустиковидное. Кустики светло-,
желто- или темно-зеленые, образующие нежные слизистые пучковидные
дерновинки. Главные нити скудно ветвящиеся, прикрепляющиеся
к субстрату с помощью ризоидных выростов, на ранних стадиях развития
отходящие от коротких нитей подошвы. Ветви 1-го порядка очередные,
большей частью длинные, к концам постепенно суживающиеся, ближе
к вершине главных нитей короткие, нередко все направленные в одну
сторону; они отходят от обычных или слегка укороченных клеток.
Ветви 2-го порядка длинные. Клетки главных нитей цилиндрические,
6—20 мкм шир., по длине в 2—12 раз больше, у поперечных перегородок
не перетянутые, клетки боковых ветвей цилиндрические или слегка бо-
чонковидные, по длине равные ширине или до 2 (4) раз больше. Конечные
клетки тупые или вытянутые в заостренный 3—4-клеточный волосок.
Указывается для сернистых вод, однако довольно часто встречается
в холодных проточных водах, не содержащих сероводорода.
Var. subsecundum (рис. 66, 1—4). — Кустики преимущественно
темно-зеленые. Клетки главных нитей 10—20 мкм шир., по длине в 2—
12 раз больше. Конечные клетки боковых ветвей обычно вытянуты в за-
остренный волосок.
В реках, источниках. ЭССР, ЛатвССР, РСФСР (Ленинградская обл.),
АрмССР, Ср. Азия.
Var. tenuis Nordst. [=Stigeoclonium falklandicum var. longearticula-
tum Hazen; S. longearticulatum (Hansg.) Heering] (рис. 66,5).—
Кустики преимущественно светло-зеленые, скудно разветвленные, ветви
в их нижней части очень редкие, длинночленистые, в верхней более
многочисленные, состоящие из коротких клеток. Клетки главных нитей
6 — 10 (12) мкм шир., по длине в 4—9 (12) раз больше. Конечные
клетки веточек преимущественно тупые.
В водохранилищах. В СССР не обнаружена. (Чехословакия, Юго-
славия, Румыния).
2. Stigeoclonium thermale A. Br. [=Myxonema thermale (А. Вг.)
Hazen] — Стигеоклониум тепловодный (рис. 67,1, 2)Слоевище ку-
стиковидное. Кустики ярко- или светло-зеленые, слизистые, 2 —5 см выс.
Главные нити скудно ветвящиеся, отходящие от коротких нитей подошвы,
образованных более или менее шаровидными клетками, прикрепляющи-
мися к субстрату ризоидными выростами, которые отходят также и от
некоторых клеток при основании главных нитей. Ветви 1-го порядка
очередные, удаленные друг от друга, обычно длинные. Ветви 2-го по-
рядка обычно короткие, рассеянные, в верхушечной части кустиков
сближенные, преимущественно простые, реже с очередными веточками
3-го порядка. Клетки главных нитей цилиндрические, у поперечных
перегородок не перетянутые или слегка перетянутые, 7—12 мкм шир.,
по длине в нижней части слоевища равные ширине или до 2 раз больше,
149
Рис. 66.
Stigeoclonium subsecundum: 1—4 — var. subsecundum (1 — общий вид слоевища,
2 — одностороннее ветвление молодой нити, 3, 4 — фрагменты базальной части
слоевища), 5 — var. tenuis, фрагменты ветви. (По Исламу).
в средней части и в ветвях 2-го порядка до 3—5 раз больше. Концы глав-
ных нитей и боковых ветвей всех порядков оканчиваются длинным
многоклеточным тупым волоском, реже заостренной клеткой. При наступ-
лении неблагоприятных условий наблюдается переход в пальмеллевидное
состояние.
150
Рис. 67.
7, 2 — Stigeoclonium thermale: 1 — общий вид слоевища, 2 — фрагмент базаль-
ной части; 3, 4 — S. setigerum, фрагмент слоевища. (1, 3 — по Кютцингу,
2, 4 — по Исламу).
В стоячих и проточных водах, в том числе в теплых источниках
с температурой воды до -|-35 °C, на различных субстратах. УССР (Крым-
ская обл.).
3. Stigeoclonium setigerum Kiitz. — Стигеоклониум щетинконосный
151
(рис. 67,3, 4). — Слоевище кустиковидное. Кустики желтовато-зеленые,
нежные, образующие слизистые дерновинки до 1 см выс. Главные нити
обильно разветвленные, особенно в верхушечной части. Ветви 1-го по-
рядка очередные, удлиненные, оканчивающиеся шиловидно заостренной
клеткой или длинным тонким волоском. Ветви 2-го порядка очередные,
простые, тонкие, длинные. Клетки главных нитей и ветвей 1-го порядка
преимущественно цилиндрические, у поперечных перегородок не перетя-
нутые, 7 — 10 (12) мкм шир., по длине равные ширине или до 2—4 раз
больше, клетки ветвей 2-го порядка 3.5—6 мкм шир., по длине в 2—3 раза
больше.
В реках и ручьях, на водных цветковых растениях и древесном
субстрате. УССР, АрмССР, ТаджССР, Якутская АССР (среднее течение
р. Лены).
4. Stigeoclonium aestivale (Hazen) Collins [=Myxonema aestivate
Hazen; Stigeoclonium. autumnale Collins; S. longipilum var. lacustre
Chod.] — Стигеоклониум летний (рис. 68). — Слоевище кустиковидное.
Кустики светло-зеленые, образующие дерновинки 2—5 мм, реже до
1.5 см выс. Главные нити обильно и беспорядочно разветвленные, от-
ходящие от пальмеллевидного основания, образованного округлыми клет-
ками с ризоидными выростами. Ветви 1-го порядка длинные, очеред-
ные. Ветви 2-го порядка немногочисленные, очередные, короткие, пря-
мые, в верхушечной части слоевища обычно более сближенные. В базаль-
ной части кустиков от многих клеток главных нитей отходят длинные,
обычно разветвленные многоклеточные ризоидные выросты; они могут от-
ходить и от ветвей 1-го порядка, расположенных недалеко от основания
кустика. Клетки главных нитей цилиндрические, у поперечных перего-
родок не перетянутые, или бочонковидные, 7 —12 (15) мкм шир., по длине
в 2—6 раз больше, в верхушечной части обычно изодиаметрические.
Все ветви оканчиваются довольно длинным тонким острым волоском
или шиловидно заостренной клеткой.
В проточных водах — источниках, фонтанах, на стенках желобов,
водных цветковых растениях, раковинах моллюсков. В СССР не обнару-
жен. (Зап. Европа, Африка, Азия, Сев. и Южн. Америка).
5. Stigeoclonium nudiusculum Kiitz. — Стигеоклониум оголенный
(рис. 69,7,2). — Слоевище кустиковидное. Кустики светло-зеленые,
образующие слизистые, хлопьевидные дерновинки до 1.5 см выс., реже
длиннее. Главные нити скудно разветвленные, особенно в нижней части.
Ветви 1-го порядка длинные, очередные. Ветви 2-го порядка короткие,
очередные, рассеянные, реже сближенные. Клетки главных нитей ци-
линдрические или бочонковидные, толстостенные, 25—47 мкм шир., по
длине равные ширине или до 3 раз больше, клетки ветвей цилиндриче-
ские, чаще всего 9 мкм шир., по длине равные ширине или до 2 раз
больше. Конечные клетки в большинстве случаев переходят в длинный
многоклеточный волосок. Макрозооспоры 16—18 мкм дл., со стигмой,
расположенной несколько выше середины клетки. Микрозооспоры 11 —
14 мкм дл., на переднем конце вытянутые, со стигмой, расположенной
значительно ниже середины клетки.
Рис. 68.
Stigeoclonium. aestivale: 1 — 3 — базальные части слоевищ, 4 — фрагмент средней
части, 5 — верхняя часть. (7, 4, 5 — по Газену, 2, 3 — по Исламу).
152
153
В стоячих водах — прудах и канавах, в обрастаниях водных цвет-
ковых растений. УССР.
6. Stigeoclonium biasolettianum (Kiitz.) Kiitz. (—Draparnaldia biaso-
lettianum Kiitz.) — Стигеоклониум Биазолетта (рис. 69, 3). — Слоевище
кустйковидное. Кустики светло- или желто-зеленые, нежные, слизистые.
Главные нити вытянутые, беспорядочно изогнутые, с рассеянным,
местами односторонним ветвлением, по форме и размерам клеток не
отличающиеся от ветвей. Ветви 1-го порядка очередные, длинные, изогну-
тые, удаленные друг от друга. Ветви 2-го порядка часто односторонние.
Клетки главных нитей и боковых ветвей цилиндрические, у поперечных
перегородок не перетянутые, 12—15 мкм шир., по длине в 2—5 (8) раз
больше. Конечные клетки заостренные, волоски отсутствуют.
В проточных водах, в обрастаниях камней. В СССР не обнаружен.
(Сев. Европа).
154
7. Stigeoclonium attenuatum (Hazen) Collins (=Myxonema attenua-
tum Hazen) — Стигеоклониум оттянутый (рис. 70). — Слоевище ку-
стиковидное. Кустики темно-зеленые, слизистые, часто разрастающиеся
в пряди до 40 см дл. Главные нити в нижней части разветвлены обильно,
в верхней умеренно; они отходят от клеток подошвы, прикрепляю-
щихся к субстрату ризоидными выростами. Ветви 1-го порядка длинные,
очередные. Ветви 2-го порядка одиночные, реже парные, короткие,
шиповидные или плетевидные, очередные, супротивные или в мутовках.
Клетки главных нитей цилиндрические, у поперечных перегородок
не перетянутые, 5—7 мкм шир., по длине в 2—5 раз больше. Конечные
клетки заостренные или вытянутые в тонкий острый волосок.
В проточных водах, на разных субстратах, с ранней весны до позд-
ней осени. УССР (Донецкая обл., в роднике).
Некоторые авторы считают этот вид синонимом S. elongatum.
8. Stigeoclonium elongatum (Hass.) Kiitz. (=Draparnaldia elongata
Hass.; Stigeoclonium falclandicum Kiitz.) — Стигеоклониум вытянутый
(рис. 71, 1). — Слоевище кустиковидное. Кустики темно- или желто-зе-
леные, слизистые, до 4—5 см выс. Главные нити разветвлены неравно-
мерно, в верхней части обильнее, чем в нижней. Ветви 1-го порядка
преимущественно очередные, реже супротивные, в нижней части
слоевища длинные, удаленные друг от друга, в верхней короткие и сбли-
женные, разветвленные так же, как и главные нити. Ветви 2-го порядка
плетевидные. Клетки главных нитей в базальной части слоевища ци-
линдрические, у поперечных перегородок не перетянутые или слегка пе-
ретянутые, 6—12 мкм шир., по длине в 2—3 (4) раза больше. Конечные
клетки заостренные.
В проточных, реже стоячих водах, на водных цветковых растениях.
УССР, Абхазская АССР, АзербССР, Ср. Азия.
Малоизученный вид. От S. attenuatum отличается более крупными
клетками главных нитей, характером ветвления, отсутствием многокле-
точных волосков.
9. Stigeoclonium longipilum Kutz. — Стигеоклониум длиииоволоси-
стый (рис. 71, 2).— Слоевище кустиковидное. Кустики ярко-зеленые,
слизистые, образующие подушковидные дерновинки до 2 см выс. Главные
нити разветвлены наиболее обильно в нижней части, слагаясь в своеобраз-
ные пучки из лучеобразно направленных вверх ветвей. Ветви 1-го по-
рядка очередные, сближенные по 2—3, реже супротивные, длинные,
простые или слабо разветвленные. Ветви 2-го порядка немногочислен-
ные, местами пучковидно сближенные, очередные или супротивные,
с одиночными веточками 3-го порядка. Клетки главных нитей и ветвей
бочонковидные, у поперечных перегородок отчетливо перетянутые, в ниж-
ней части слоевища (4) 9 — 15 (19) мкм шир., по длине равные ширине
или до 2 (4) раз больше, у веточек последнего порядка 5—6 мкм шир.,
по длине в 1.5—2 раза больше; иногда наблюдается двурядное разме-
щение клеток, такие участки достигают 16 — 17 мкм шир. Конечные
клетки ветвей вытянутые в длинный многоклеточный волосок. Зооспоры
четырехжгутиковые, макрозооспоры 12—16 мкм дл., со стигмой, распо-
ложенной в середине клетки, при прорастании прикрепляющиеся
к субстрату косо, микрозооспоры 9—12 мкм дл., с продолговатой стигмой
в передней части клетки, после освобождения превращающиеся в аплано-
споры или иные стадии покоя; известны еще и другие двужгутиковые
зооспоры около 8 мкм дл., слабо суженные к переднему концу, со стигмой,
расположенной в центре клетки, но дальнейшее развитие их неизвестно.
155
Рис. 70.
Stigeoclonium attenuatum'. 1,2 — средняя часть слоевища, 3
(По Гаэену).
156
В стоячих и проточных водах, на различных субстратах.
На ранних стадиях развития этот вид бывает трудно отличить от
молодых талломов хетофоры. От остальных видов рода хорошо отличается
расположением на главных нитях пучковидно сближенных ветвей,
лучеобразно направленных вверх. Вид полиморфный.
Var. longipilum (рис. 71, 2). — Кустики до 2 см выс. Клетки главных
нитей 9—15 (19) мкм шир.
В реках, прудах, болотах, на водных цветковых растениях, крупных
нитчатых водорослях, раковинах моллюсков. РСФСР (Карельская АССР,
Московская обл., оз. Байкал), ЛатвССР, УССР, Кавказ.
Var. minus Hansg. — Кустики 3—5 мм выс. Клетки главных нитей
4—6 мкм шир.
В проточных водах. УССР (Карпаты).
157
10. Stigeoclonium nanum (Dillw.) Kiitz. [=Conferva папа Dillw.;
Myxonema nanum (Dillw.) Hazen; Stigeoclonium nanum f. subsimplex
Collins] — Стигеоклониум низкорослый (рис. 72). — Слоевище кустико-
видное. Кустики слагаются в дерновинки, состоящие из паренхимной
подошвы в виде правильного диска и довольно обильно разветвленных
прямостоячих главных нитей, отходящих почти от каждой клетки по-
дошвы. Ветви 1-го и 2-го порядков большей частью короткие, очередные,
изредка супротивные, к вершинам суженные. Клетки главных нитей ци-
линдрические или слегка бочонковидные, 6—8 мкм шир., подлине равные
ширине, меньше или до 2 раз больше ее. Конечные клетки тупые или
заостренные, волоски обычно отсутствуют. Размножение неизвестно.
Рис. 72.
Stigeoclonium nanum: 1—3 — фрагменты взрослого слоевища, 4 — молодое слое-
вище. (1—3 — по Исламу, 4 — по Фричу).
158
Рис. 73.
Stigeoclonium subspinosum: 1 — общий вид слоевища, 2 — базальная часть,
3 — фрагмент ветвления. (По Воронихину).
В стоячих и проточных водах, на разных субстратах. ЭССР, УССР.
Малоизученный, довольно сомнительный вид. Возможно, является
стадией развития какого-либо иного вида. На ранних стадиях развития
легко спутать со S. farctum.
11. Stigeoclonium subspinosum Kiitz. — Стигеоклониум колючкова-
тый (рис. 73). — Слоевище кустиковидное. Кустики бледно-зеленые, сли-
зистые, 0.5 —1.5 см выс. Главные нити скудно разветвленные. Ветви 1-го
порядка простые, короткие, шиповидные, очередные, одиночные, реже
парные, или супротивные. Клетки главных нитей цилиндрические,
6.4—10 мкм шир., по длине в 2—6 раз больше, наиболее длинные
в средней части главных нитей, в верхней — укороченные. Конечные
клетки шиловидно заостренные или вытянутые в одноклеточный острый
волосок.
В холодных проточных водах. УССР (горные водоемы Крыма и
Карпат).
От близкого S. stagnatile отличается скудным ветвлением, более
длинными клетками главных нитей, одноклеточными волосками и эко-
логией.
159
12. Stigeoclonium subuligerum Kiitz. [=5. protensum (Dillw.) Kiitz.
var. subuligerum (Kutz.) Rabenh.; Myxonema subuligerum (Kiitz.) Ha-
zen] — Стигеоклониум шилоносный (рис. 74). — Слоевище кустиковид-
ное. Кустики зеленые или светло-зеленые, пушистые, до 5 мм выс.,
иногда больше, прикрепляющиеся к субстрату подошвой, от которой под-
нимаются вверх многочисленные нити. Главные нити обильно разветвлен-
ные. Ветви 1-го порядка обычно длинные, супротивные, реже очередные,
рассеянные или сближенные по нескольку пар. Ветви 2-го порядка оче-
редные или супротивные, большей частью короткие, шиповидные, с ши-
роким основанием и суженным верхом. Встречаются веточки и 3-го по-
рядка. Клетки главных нитей цилиндрические, у поперечных перегородок
слегка перетянутые, реже сильно перетянутые, бочонковидные, 11 —
16 (18) мкм шир., по длине равные 'ширине или несколько меньше
или больше, у ветвей 6 — 10 мкм шир. Конечные клетки заостренные, мно-
гие вытянуты в относительно длинный бесцветный многоклеточный
волосок.
В проточных и стоячих водах, на водных цветковых растениях, дре-
весном субстрате, раковинах моллюсков и т. п. РСФСР (Ростовская
обл.), УССР, Ср. Азия.
13. Stigeoclonium tenue (Ag.) Kiitz. [= Draparnaldta tenuis Ag.; My-
xonema tenue (Ag.) Rabenh.; Stigeoclonium irregulare Kiitz.; S. tenue var.
irregulare (Kiitz.) Rabenh.; 5. (tenue) klebsii Heering; S. (tenue) pascheri
Heering; S. (tenue) westii Heering; S. stellare Kiitz.; S. gracile Skvortz.] —
Стигеоклониум тонкий (рис. 75). — Слоевище кустиковидное. Кустики
светло-зеленые, слизистые, образующие пучковидные дерновинки от не-
скольких миллиметров до 5 см выс., на ранних стадиях развития
прикрепляющиеся к субстрату нитями подошвы. Главные нити в верхней
части разветвлены обильнее, чем в нижней. Ветви 1-го порядка очередные
или супротивные, сближенные, но не более чем по 2 пары. Ветви 2-го по-
рядка многочисленные, очередные, реже супротивные, большей частью
короткие, прямые, шиповидные. Клетки главных нитей цилиндрические
или несколько вздутые в средней части, у поперечных перегородок
слегка перетянутые, 6—15 мкм шир., по длине в 3—5 (6) раз больше,
с тонкими бесцветными или утолщенными буроватыми оболочками,
у ветвей изодиаметрические, 5—6 мкм шир. Конечные клетки заострен-
ные, шиловидные, или вытянутые в тонкий острый, довольно короткий
1—3-клеточный волосок. Размножение исследовано недостаточно.
В стоячих и проточных водах, на различных субстратах, а-мезо-
сапроб. Одна из наиболее обычных, повсеместно распространенных
водорослей.
Var. tenue [=5. tenue var. genuinum Kirchn.) (рис. 75). —
Клетки главных нитей с тонкими бесцветными оболочками. Конечные
клетки шиловидно заостренные или оканчиваются тонким 1—3-клеточ-
ным волоском.
В реках, ручьях, озерах, прудах, болотах, лужах, на древесине, вод-
ных цветковых растениях, камнях, раковинах моллюсков. Повсеместно.
Var. uniforme (Ag.) Kiitz. (= Drapornaldia uniformis Ag.;
Stigeoclonium uniforme Rabenh.; 5. tenue var. gracile Kiitz.). — Клетки
главных нитей с утолщенными буроватыми оболочками, часто слегка
инкрустированными известью. Конечные клетки шиловидно заостренные,
волоски отсутствуют.
В проточных, реже стоячих водах, на различных субстратах.
РСФСР (Оренбургская обл., Башкирская АССР), УССР (Львовская и
Тернопольская обл.).
160
Рис. 74.
Stigeoclonium subuligerum. (По Газену).
11 Зеленые водоросли. Улотриксовые
161
Рис. 75.
Stigeoclonium. tenue. (По Газену).
162
14. Stigeoclonium protensum (Dilw.) Kiitz. — Стигеоклониум вытя-
нутый (рис. 76). — Слоевище кустиковидное. Кустики светло-зеленые,
нежные, образующие пушистые дерновинки, прикрепляющиеся к суб-
страту с помощью ризоидных выростов. Главные нити разветвлены
весьма обильно. Ветви 1-го порядка большей частью длинные, очеред-
ные, одиночные или, реже, парные, иногда супротивные, реже в мутов-
ках, беспорядочно разветвленные. Ветви 2-го порядка обычно короткие,
очередные, одиночные или, реже, парные, изредка супротивные. Встреча-
ются веточки и 3-го порядка. Клетки главных нитей цилиндрические
или слегка бочонковидные, 11.4—16 (23) мкм шир., по длине равные
ширине или до 2—4 (5) раз больше. Конечные клетки веточек большей
частью переходят в длинный, до 210 мкм и более, многоклеточный
тонкий острый волосок, реже заостренные.
В проточных, реже стоячих водах, на водных цветковых растениях.
УССР, местами.
15. Stigeoclonium segare Islam — Стигеоклониум сегаре (рис. 77). —
Слоевище кустиковидное. Главные нити вытянутые, беспорядочно
разветвленные. Ветви 1-го порядка очередные, супротивные или, реже,
в мутовках, рассеянные, местами сближенные. Ветви 2-го порядка
очередные, реже супротивные. Встречаются веточки и 3-го порядка,
очередные. Клетки главных нитей и ветвей цилиндрические, у попереч-
ных перегородок не перетянутые, в базальных частях главных нитей
и крупных ветвей сильно вытянутые, (8) 9.5—11 мкм шир., по длине
в (4) 8—10 (12) раз больше. Конечные клетки заостренные или вытяну-
тые в многоклеточный волосок.
В проточных водах, на периодически заливаемых полях, на цветко-
вых растениях. В СССР не обнаружен. (Новая Зеландия).
16. Stigeoclonium lebelii Islam — Стигеоклониум Лебеля (рис. 78). —
Слоевище кустиковидное. Кустики светло-зеленые, слизистые, до 2 см
выс. Главные нити разветвлены обильно и беспорядочно. Ветви 1-го по-
рядка длинные, обильно ветвящиеся, обычно супротивные или в му-
товках, реже очередные, местами сближенные по нескольку вместе.
Веточки 2-го порядка многочисленные, очередные или супротивные,
часто сближенные, реже в мутовках, обычно с короткими очередными
веточками 3-го порядка. Клетки главных нитей бочонковидные, у попе-
речных перегородок заметно перетянутые, 15—25 мкм шир., по длине до
2—5 раз больше, клетки ветвей цилиндрические, у поперечных перегоро-
док слегка перетянутые, короткие, 9—13 мкм шир., по длине равные
ширине или до 3 раз больше. Конечные клетки заостренные.
В пресных водах. В СССР не обнаружен. (Зап. Европа).
17. Stigeoclonium stagnatile (Hazen) Collins (=Myxonema stagnatile
Hazen) — Стигеоклониум прудовой (рис. 79, 1, 2). — Слоевище кустико-
видное. Кустики вначале прикрепленные, позднее свободноплавающие,
собранные в хлопьевидные дерновинки. Главные нити длинные, беспоря-
дочно разветвленные. Ветви 1-го порядка довольно короткие, часто слегка
изогнутые, очередные, парные, реже одиночные, или супротивные, из-
редка мутовчатые. Ветви 2-го порядка крайне немногочисленные, ко-
роткие, слегка искривленные. Клетки главных нитей цилиндрические,
7.5 — 11 мкм шир., по длине равные ширине или до 3 (6) раз больше,
клетки ветвей 7—9 мкм шир. Конечные клетки шиловидно заостренные.
В стоячих и слабо проточных водах, на водных цветковых растениях
и других субстратах, часто плавает в спутанных массах. Ср. Азия (в ри-
совых чеках).
11*
163
Малоизученный вид. Весьма близок к 6’. subspinosiim и S. protensum.
От первого отличается обильным ветвлением, более широкими клетками
главных нитей, многоклеточными волосками и экологией, от второго —
более узкими клетками главных нитей, довольно короткими шиповид-
ными веточками 1-го порядка и предпочтительным обитанием в стоя-
чих водах.
18. Stigeoclonium carolinianum Islam — Стигеоклониум каролинский
(рис. 79,3,4). — Слоевище кустиковидное. Кустики светло-зеленые, до
1 см выс., прикрепляющиеся к субстрату с помощью ризоидных выростов.
Главные нити разветвлены беспорядочно. Ветви 1-го порядка рассеян-
ные, очередные, иногда супротивные, изредка сближенные, длинные или
короткие, к вершинам заостренные. Ветви 2-го порядка крайне немного-
165
Рис. 79.
1, 2 — Stigeoclonium stagnatile: 1 — средняя часть слоевища, 2 — базальная
часть; 3, 4 — S. carolinianum. (По Исламу).
166
численны или отсутствуют. Клетки
главных нитей цилиндрические,
у поперечных перегородок не пере-
тянутые или слегка перетянутые,
6.6—13 мкм шир., 16—45 мкм дл.
Конечные клетки туповато конусо-
видные. На главных нитях и ветвях
1-го порядка образуются эллипсоид-
ные клетки 8—9.6 мкм шир., 22—
29 мкм дл., имеющие вид споран-
гиев; их значение не установлено.
В пресных водах. В СССР не об-
наружен. (США: Сев. Каролина).
19. Stigeoclonium fasciculare
Kutz. — Стигеоклониум пучковатый
(рис. 80).—Слоевище кустиковид-
пое. Кустики зеленые, слизистые,
в дерновинках до 5 см выс. Главные
нити и длинные ветви 1-го порядка
к вершинам кистевидно разветвлен-
ные; кистевидные ветвления равно-
мерные, плотные, прямые, с хорошо
прослеживаемой главной нитью.
Ветви 2-го порядка короткие, вос-
ходящие, иногда слегка изогнутые.
Клетки главных нитей 10—11 мкм
шир., по длине равные ширине или
до 2 раз больше. Конечные клетки
главных нитей, ветвей 1-го и некото-
рых ветвей 2-го порядков переходят
в длинный многоклеточный волосок
или заостренные.
Рис. 80.
Stigeoclonium fasciculare. (По
Кютцингу).
В стоячих и слабо проточных водах, на живых и отмерших водных
цветковых растениях, раковинах моллюсков. МолдССР, УССР, Ср. Азия,
Якутская АССР (среднее течение р. Лены).
Малоизученный вид. Близок к S. glomeratum, отличается более круп-
ными размерами слоевища и короткоцилиндрическими клетками глав-
ных нитей.
20. Stigeoclonium glomeratum (Hazen) Collins (= Myxonema glome-
ratum Hazen) —Стигеоклониум клубковидный (рис. 81).—Слоевище
кустиковидное. Кустики зеленые, нежные, до 1 см выс., прикрепляю-
щиеся к субстрату переплетенными нитями подошвы. Главные нити раз-
ветвлены неравномерно. Ветви 1-го порядка на большем протяжении
главных нитей очередные, реже супротивные, рассеянные, ближе к вер-
шине сближенные по нескольку вместе. Конечные ветвления главных
нитей и длинных ветвей 1-го порядка напоминают сложную кисть, равно-
мерную и плотную, с отчетливо выраженной средней осью с отходящими
от нее на всем протяжении ответвлениями, в которых можно наблюдать
ветви вплоть до 5—6-го порядков. От некоторых нижних клеток главных
нитей отходят к субстрату длинные ризоидные выросты. Клетки
главных нитей снизу сначала удлиненно-цилиндрические или несколько
вздутые, у поперечных перегородок иногда едва заметно перетянутые,
167
Рис. 81.
Stigeoclonium glomeratum: 1 — общий вид слоевища, 2 — фрагмент главной
нити с верхушечными разветвлениями. (По Мошковой).
168
11.4—14 мкм шир., по длине
в (2) 4—6 (7) раз больше, в
средней их части и в основании
длинных ветвей 1-го порядка
9—11 мкм шир., по длине в 2—
5 раз больше, у веточек цилинд-
рические, 6—8 мкм шир., по
длине равные ширине или до
2 раз больше. Конечные клетки
главных нитей и ветвей 1—3-го
порядков переходят в длинный
многоклеточный волосок или
слегка заостренные. Размноже-
ние не прослежено.
В стоячих и проточных во-
дах, на древесном субстрате.
УССР.
Близок к S. fasciculare, от-
личается меньшими размерами
слоевища и более удлиненными
цилиндрическими клетками
главных нитей.
Рис. 82.
Stigeoclonium nelsonii. (По Прескотту).
21. Stigeoclonium nelsonii Presrott — Стигеоклониум Нельсона
(рис. 82). — Слоевище кустиковидное. Главные нити длинные, неравно-
мерно скудно разветвленные. Ветки всех порядков очередные, рассеян-
ные. Верхушки главных нитей и длинных ветвей 1-го порядка заканчи-
ваются своеобразным, слегка дуговидно согнутым односторонним рых-
лым ветвлением кистевидно-гребенчатого типа, в котором встречаются
веточки до 4—5-го порядков. На протяжении ветвления обособленная
нить не просматривается, на вершине она резко теряется в разветвле-
ниях. Клетки главных нитей цилиндрические или слегка бочонковид-
ные, у поперечных перегородок не перетянутые, реже перетянутые,
12—20 мкм шир., по длине равные половине ширины или до 2—5 раз
больше. Конечные клетки веточек тупоконусовидные или вытянуто
заостренные.
В стоячих и проточных водах, на раковинах моллюсков. В СССР
не обнаружен. (США: штат Монтана).
22. Stigeoclonium pusilium (Lyngb.) Kiitz. (—Conferva pusilia
Lyngb.) — Стигеоклониум крохотный (рис. 83, 1—3). — Слоевище ку-
стиковидное. Кустики ярко-зеленые, в слизистых дерновинках до 1 см
выс. Главные нити обильно ветвящиеся. Ветви 1-го порядка многочислен-
ные, очередные, одиночные или изредка парные, реже супротивные,
отходящие от укороченных угловатых, слегка вздутых клеток. Ветви
2-го порядка короткие. Клетки главных нитей и ветвей 1-го порядка
преимущественно бочонковидные, у поперечных перегородок отчетливо
перетянутые, 15—20 (25) мкм шир., по длине равные ширине или до
2 раз больше, у ветвей 2-гопорядка 6 мкм шир., преимущественно изодиа-
метрические. Конечные клетки главных нитей и ветвей 1-го порядка
обычно вытянутые в длинный многоклеточный волосок, у ветвей 2-го по-
рядка конусовидно заостренные.
В стоячих водах — прудах, канавах, болотах. УССР.
Вид мало изучен, известен пока из немногих мест.
169
Рис. 83.
1-3 - Stigeoclonium. pusilium-, 4-7 - S. helveticum-, 8 - S. lubricum var.
yarians. (1 — 7 — по Исламу, 8 — по Газену).
170
23. Stigeoclonium helveticum Vischer — Стигеоклониум швейцар-
ский (рис. 83,4—7).—Слоевище кустиковидное. Кустики светло-
зеленые, в пучковидных дерновинках до 5 см выс. Главные нити беспоря-
дочно разветвленные или простые. Ветви 1-го порядка длинные, очеред-
ные, реже супротивные, большей частью сближенные по нескольку
вместе, отходящие обычно от укороченных клеток главных нитей. Ветви
2-го порядка очередные, разветвленные, характерно изогнутые к концам
в форме завитка, к вершинам ветвей 1-го порядка сближенные. Ветви
3-го порядка короткие, простые. Клетки главных нитей цилиндрические,
у поперечных перегородок обычно не перетянутые, 6 —12.5 мкм шир., по
длине равные ширине или до 8 раз больше. Конечные клетки заостренные.
В проточных, часто теплых водах. В СССР не обнаружен. (Швей-
цария, Сев. Америка).
24. Stigeoclonium lubricum (Dillw.) Kutz. [=Conferva lubrica Dillw.;
Stigeoclonium tenue Kiitz. var. lubricum Rabenh.; Myxonema lubricum
(Dillw.) Fries] — Стигеоклониум скользкий (рис. 83, 8; 84). — Слоевище
кустиковидное. Кустики темно-зеленые, блестящие, 0.5—30 см выс. Глав-
ные нити обильно разветвленные. Ветви 1-го порядка очередные, супро-
тивные или в мутовках, часто сближенные по нескольку пар; они отходят
от слегка вздутых угловатых клеток, более коротких, чем остальные
клетки главных нитей. Ветви 2-го порядка многочисленные, прямые,
очередные или супротивные, рассеянные, но в верхней части слоевища
обычно сближенные по нескольку вместе. Встречаются также ветви 3-го
и 4-го порядков. Клетки главных нитей в нижней части растения ко-
роткоцилиндрические, слегка вздутые, с утолщенными стенками, или
удлиненно-цилиндрические, тонкостенные, 11 — 19 мкм шир., по длине
в 2 (4—5) раз больше, у ветвей 1-го порядка 9.5—11 мкм шир.,
2-го порядка 6—7 мкм шир., 3-го порядка 5—6 мкм шир., по длине равные
ширине, несколько меньше или несколько больше ее. Конечные клетки
короткие, заостренные, реже переходящие в многоклеточный волосок.
В проточных и стоячих водах.
Var. lubricum (рис. 84). — Кустики до 30 см выс. Клетки главных
нитей в нижней части слоевища преимущественно короткоцилиндри-
ческие, слегка вздутые, с утолщенными стенками, 13—19 мкм шир., по
длине обычно до 2 (4) раз больше.
В проточных, реже стоячих водах, на различных субстратах. В раз-
ных районах СССР, местами.
Var. varians (Hazen) Collins (—Myxonema lubricum var. vartans
Hazen) (рис. 83, 8). — Кустики до 2 см выс. Клетки главных нитей
в нижней части слоевища тонкостенные, большей частью удлиненно-
цилиндрические, лишь изредка вздутые, 11 — 14 мкм шир., по длине
в 2—5 раз больше.
В проточных, реже стоячих водах, на камнях, древесном субстрате,
стеблях травянистых растений; часто вместе с типовой формой. ЛитССР,
МолдССР, УССР, Бурятская АССР.
25. Stigeoclonium amoenum Kiitz. [=5'. amoenum f. biforme Collins;
Myxonema amoenum (Kiitz.) Hazen] — Стигеоклониум прелестный
(рис. 85, 86). —Слоевище кустиковидное. Кустики светло- или желто-
зеленые, пушистые, обычно объединенные в слизистые хлопьевидные
дерновинки 0.5 —1.5 (15) см выс. Главные нити обильно разветвленные.
Ветви 1-го порядка преимущественно супротивные, сближенные по
2—3 пары, отходящие от укороченных угловатых клеток. Ветви 2-го по-
рядка большей частью короткие, супротивные или очередные, сбли-
171
Рис. 85.
Stigeoclonium атоепит var. атоепит. (По Газену).
Рис. 84.
Stigeoclonium lubricum var. lubricum, средняя и верхушечная части главной
нити слоевища. (По Газену).
173
172
женные по нескольку пар. Клетки главных нитей, особенно в нижней
части слоевища, тонкостенные, удлиненно-цилиндрические, у попереч-
ных перегородок не перетянутые или слегка перетянутые, только иногда
толстостенные, бочонковидные или более или менее короткоцилиндриче-
ские, у поперечных перегородок заметно перетянутые, 11.4—18 (38) мкм
шир., по длине в 1.5—8 раз больше, ближе к вершине по длине лишь
в 4 раза больше, у ветвей 1-го порядка при основании 6—11.4 мкм шир.,
по длине равные ширине или в 2—4 (8) раз больше, у ветвей 2-го по-
рядка 5—7 мкм шир., по длине равные ширине, несколько больше или
несколько меньше ее. Конечные клетки коротко- или вытянуто-конусо-
видные, заостренные или туповатые; волоски обычно отсутствуют. От
нижних клеток главных нитей нередко отходят ризоиды, из клеток кото-
рых могут возникать обычные вегетативные побеги.
В проточных и стоячих водах.
Var. amoenum (рис. 85). — Клетки главных нитей в нижней части
слоевища 11.4—18 мкм шир.
174
В реках, ручьях, реже прудах, на камнях, различных растительных
субстратах, нередко в массе. В разных районах СССР, довольно часто.
Var. insigne (Nag.) Islam (=5. insigne Nag.) (рис. 86). — Клетки
главных нитей в нижней части слоевища 28—38 мкм шир.
В стоячих и проточных водах, на растительном субстрате. ЛатвССР,
РСФСР (Московская и Калининская обл.).
26. Stigeoclonium flagelliferum Kiitz. — Стигеоклониум плетеносный
(рис. 87). — Слоевище кустиковидное. Кустики желто- или ярко-зеленые,
образующие дерновинки или пучки, скользкие на ощупь, до 2—5 (15) см
выс. Ветви 1-го порядка обильно разветвленные, обычно супротивные,
часто сближенные по 2—5 пар, реже рассеянные, отходящие от укорочен-
ных угловатых, слегка вздутых клеток главных нитей. Ветви 2-го порядка
такого же характера, но угловатые клетки, от которых они отходят,
здесь не всегда короче остальных клеток. Встречаются ветви и 3-го по-
рядка. Клетки главных нитей удлиненно-цилиндрические, в средней ча-
сти слоевища слегка вздутые, более или менее толстостенные, 14—
23 (25) мкм шир., по длине в 3—8 (12) раз больше, у ветвей 1-го порядка
9—12 (15.2) мкм шир., по длине до 4—6 (10) раз больше. Конечные
клетки шиловидно заостренные или переходят в длинный многоклеточ-
ный волосок. Размножение четырехжгутиковыми зооспорами и двужгути-
ковыми гаметами.
В проточных, реже стоячих водах, на разных субстратах. В разных
районах СССР, местами.
27. Stigeoclonium variabile (Nag.) Elenk. emend. — Стигеоклониум
изменчивый (рис. 88). — Слоевище состоит из ползучих, горизонтально
распростертых ветвящихся нитей, прижатых к субстрату и сросшихся
между собой боковыми сторонами в паренхимную пластинку разнообраз-
ных очертаний, и коротких прямостоячих нитей с редкими очередными
веточками. Клетки ползучих нитей неправильной формы, до 11 мкм шир.,
прямостоячих — цилиндрические, иногда в средней части слегка вздутые,
тонкостенные, 4.4—7.7 (9) мкм шир., по длине в 2—4 раза больше.
Конечные клетки вздуто-конические, тупые; волоски отсутствуют.
Макрозооспоры грушевидные или почти шаровидные, 6.6—7.6 мкм шир.,
8—8.6 мкм дл., со стигмой, микрозооспоры и гаметы не наблюдались.
В стоячих и слабо проточных водах, на разных субстратах, чаще
всего в колодцах и аквариумах, где образует тонкие ярко-зеленые
налеты или дерновинки. РСФСР (Ленинградская обл.). УССР.
28. Stigeoclonium moebiusianum (Collins) Printz (=Endoclonium
moebiusianum Collins) — Стигеоклониум Мёбиуса (рис. 89, 1—3). —
Слоевище состоит из хорошо развитой подошвы, образованной радиально
расходящимися, разветвленными в плоскости нитями, прижатыми к суб-
страту и сросшимися боковыми сторонами в паренхимный диск до
500 мкм в диам., и очень коротких, 1 (2—3)-клеточных, папилловидных
прямостоячих нитей. Клетки подошвы около 5 мкм шир., по длине
равные ширине или до 2 раз больше, клетки прямостоячих нитей
короткие, бочонковидные или булавовидные. Конечные клетки на вер-
шине закругленные, иногда вытянутые в тонкий бесцветный волосок.
Размножение не исследовано.
В различных водоемах, на водных цветковых растениях. В СССР не
обнаружен. (Зап. Европа, Америка).
175
Рис. 87.
Stigeoclonium flagelliferum. (По Мошковой).
176
Stigeoclonium variabile: 1 — общий вид слоевища в профиль, 2 — зооспоры,
3—6 — стадии прорастания зооспор. (По Еленкину).
29. Stigeocloninm prostratum Fritsch — Стигеоклониум стелющийся
(рис. 89, 4—8). — Водоросль образует дерновинки до 30 мкм выс.
Слоевище состоит из хорошо развитой подошвы, образованной ползучими,
ветвящимися в плоскости нитями, плотно прижатыми к субстрату, и
сросшимися боковыми сторонами в паренхимную пластинку разнообраз-
ных очертаний, и очень коротких, 1 — 2 (5)-клеточных, прямостоячих
нитей, возникающих почти от каждой клетки подошвы, обычно простых
и лишь изредка образующих 1—2-клеточные веточки. Клетки подошвы
цилиндрические, слегка вздутые на дорсальной стороне, 5—7 мкм шир.,
7 — 16 мкм дл., 5—12 мкм выс., клетки прямостоячих нитей цилиндри-
ческие, у поперечных перегородок не перетянутые, 5—8 мкм шир.,
по длине равные ширине или в 2—3 раза больше. Конечные клетки слегка
суженные, тупые. Волоски многочисленные, длинные, до 400 мкм, слегка
вздутые у основания, беспорядочно отходящие от клеток подошвы.
Размножение не исследовано.
В стоячих водах, преимущественно в озерах, реже в проточных водах,
на растительных субстратах, раковинах моллюсков. УССР.
По строению слоевища и размерам клеток близок к S. farctum,
возможно является лишь стадией развития одного из видов этого рода.
12 Зеленые водоросли. Улотриксовые
177
Рис. 89.
1—3 — Stigeoclonium moebiusianum: 1 — фрагмент слоевища сверху, 2, 3 —
то же в профиль; 4—8 — S. prostratum: 4 — фрагмент паренхимной пла-
стинки, 5 — стелющиеся разветвленные нити, 6, 7 — то же с прямостоячими
веточками, 8 — поперечный срез слоевища; 9—11 — S. farctum var. farctum:
9 — общий вид молодого слоевища сверху, 10 — фрагмент слоевища, 11 —
стелющаяся нить с прямостоячими веточками. (1—3 — по Мёбиусу, 4—8 —
по Фричу, 9, 11 — по Бертхольду, 10 — по Мошковой).
30. Stigeoclonium farctum Berth. (=S. huberi Heering; S. najadeanum
Skvortz.) — Стигеоклониум заполненный (рис. 89, 9—11). — Водоросль
образует подушков'идные дерновинки до 1.5 мм выс. Слоевище состоит из
ползучих, ветвящихся в плоскости нитей, плотно прижатых к субстрату
и обычно сросшихся боковыми сторонами в паренхимную, более или
менее дисковидную пластину, и коротких прямостоячих, простых или
скудно ветвящихся нитей, отходящих от клеток подошвы, иногда по 2—3
от одной клетки. Ветви сравнительно короткие, из 3—5 и больше клеток.
Между прямостоячими нитями находятся длинные многоклеточные
волоски, вырастающие из клеток подошвы. Клетки подошвы 6 — 12 мкм
шир., 8—12 мкм дл., слегка вздутые на дорсальной стороне, у прямостоя-
чих нитей цилиндрические, 4—8.6 мкм шир., 6—16 мкм дл., у поперечных
перегородок не перетянутые или слегка перетянутые, у волосков по
178
длине в 2—5 раз больше ширины. Конечные клетки слегка суженные,
тупые. Размножение не исследовано.
В стоячих и проточных водах.
Широко распространенный вид.
Var. farctum (рис. 89, 9—11). — Прямостоячие нити до 8.6 мкм шир.,
скудно разветвленные, боковые ветви одно- или двуклеточные.
В реках, прудах, озерах, болотах, на разных субстратах. Повсеместно.
Var. simplex Fritsch. — Прямостоячие нити простые, очень редко
с одной боковой веточкой.
В проточных водах. Полярный Урал.
Var. rivulare Butcher. — Прямостоячие нити тонкие, около 4 мкм
шир., волоски обычно отсутствуют.
В проточных водах. В СССР не обнаружена. (Англия).
Род CHAETOPHORA Schrank - ХЕТОФОРА
Слоевище слизистое, шаровидное, полушаровидное или лопастное,
содержащее нити, отходящие от подошвы. Слизь эластичная или
довольно плотная, почти хрящеватая. Нити прямостоячие, обильно вет-
вящиеся. Ветвление моноподиальное или симподиальное, однако вследст-
вие быстрого роста молодой боковой ветви и перемещения ее к вершине
материнской клетки нередко возникает впечатление дихотомии. Конеч-
ные ветвления разнообразные, у некоторых видов в форме плотных
кисточек. Клетки нитей цилиндрические или несколько вздутые, у по-
перечных перегородок не перетянутые или слегка перетянутые. Конечные
клетки заостренные или переходят в длинный бесцветный многоклеточ-
ный волосок, часто далеко выдающийся за пределы колониальной слизи.
Подошва развита слабо, состоит из рыхло расположенных шаровидных
клеток (пальмеллевидное состояние) или коротких нитей из сильно
вздутых клеток. В тех случаях, когда главные нити расходятся радиально
и одинаково быстро растут, образуются шаровидные или полушаровидные
(подушковидные) слоевища, при расположении главных нитей в одной
плоскости и неодинаково быстром их росте — неправильно лопастные
слоевища. В старых частях слоевища главные нити выполняют в основном
функцию носителей ветвящейся системы, способность к образованию
репродуктивных элементов ими утрачена, и часто можно обнаружить
длинные ризоидные выросты, отходящие от довольно высоко располо-
женных клеток и направленные к субстрату. Хлоропласт пристенный,
в нижних частях главных нитей в виде узкого кольца, в ответвлениях
заполняющий весь просвет клетки, с 1 или несколькими пиреноидами.
Бесполое размножение четырехжгутиковыми зооспорами, апланоспо-
рами, гипноспорами и акинетами. Последние, как и зооспоры, возникают
в клетках конечных ветвлений, реже в лежащих более глубоко. Половое
размножение двужгутиковыми гаметами.
В стоячих и проточных пресных, преимущественно чистых водах,
на водных цветковых растениях, древесном субстрате, раковинах мол-
люсков, камнях.
В качестве таксономических признаков используются форма колоний
и характер клеточных ветвлений. К указаниям об инкрустации слоевищ
известью следует относиться с большой осторожностью, так как этот
признак недостаточно постоянен.
I. Ветвление не равномерное, на вершине нитей ветви уплотненные.
1. Слоевище шаровидное или полушаровидное.
12*
179
А. Конечные ветвления нитей разнообразные по форме, но никогда
не имеют вида кисточек.........................С. elegans 1.
Б. Конечные ветвления нитей в виде плотных или рыхлых кисто-
чек.
а. Кисточки плотные.
а. Слоевище с бугорчатой поверхностью; главная нить па
конечных ветвях обычно продолжается в длинный много-
клеточный волосок....................С. tuberculosa 2.
р. Слоевище с гладкой поверхностью; волоски на конечных
ветвях отсутствуют...................С. punctiformis 3.
б. Кисточки рыхлые, веточки короткие, растопыренные . . .
...................................С. pisiformis 4.
2. Слоевище лопастное или кустиковидное .... С. incrassata 5.
II. Ветвление равномерное, на вершине нитей ветви не уплотненные.
1. Клетки главных нитей 18—20 мкм шир............С. flagellifera 6.
2. Клетки главных нитей 5 — 8 мкм шир.............С. attenuate 7.
1. Chaetophora elegans (Roth) Ag. (=Rivularia elegans Roth) —
Хетофора изящная (рис. 90). — Слоевище шаровидное или полушаро-
видное, с гладкой поверхностью, темно- или светло-зеленое. Главные
нити обильно ветвящиеся, поднимающиеся вверх и расходящиеся ра-
диально от подошвы. Ветви очередные, реже супротивные, в базальной
части нитей далеко удаленные друг от друга, в верхушечной части
сближенные. Конечные ветви разнообразные, но никогда не собранные
в виде плотных кисточек. Клетки подошвы шаровидные. Клетки главных
нитей цилиндрические или слегка бочонковидные, 6—15 мкм шир.,
по длине в 3—10 раз больше, у конечных ветвей цилиндрические,
5.7—11 мкм шир., по длине в 1.5—2 раза больше. Конечные клетки
конусовидно заостренные, реже вытянутые в волосок.
В стоячих и проточных водах, на водных цветковых растениях,
древесном субстрате, камнях, раковинах моллюсков. Слоевища часто
располагаются близко друг около друга и могут сливаться в большие
бесформенные массы, обволакивающие субстрат. Повсеместно.
2. Chaetophora tuberculosa (Roth) Ag. (=Rivularia tuberculosa
Roth) —Хетофора бугорчатая (рис. 91, 1, 2). —Слоевище на ранних
стадиях развития шаровидное, с гладкой поверхностью, во взрослом
состоянии в результате неравномерного роста главных нитей и их
ответвлений приобретает неправильную форму, с бугорчатой или бородав-
чатой поверхностью, обычно инкрустированное известью. В промежутках
между бугорками находятся рыхлые, скудно разветвленные нити, в бу-
горках их конечные обильные ветвления собраны группами, которые
при слабых увеличениях микроскопа видны на поверхности слизи как
зеленые точки. Главные нити прикрепляются к субстрату с помощью
ризоидных выростов, отходящих от многих их клеток, даже высоко
расположенных. Конечные ветвления имеют форму плотных кисточек,
в которых главная нить часто продолжается в длинный многоклеточный
волосок. Клетки нитей цилиндрические, у поперечных перегородок
иногда слегка перетянутые, 11.4—13 мкм шир., по длине до 2—3.5 раз
больше. Конечные клетки ветвей лишены волосков, слегка заостренные.
Известны акинеты и двужгутиковые зооспоры (гаметы?).
В проточных и стоячих водах, довольно часто в торфяных болотах,
на водных цветковых растениях, камнях. РСФСР (Кольский п-ов,
Ленинградская и Московская обл., Якутская АССР), УССР, Кавказ.
180
Рис. 90.
Chaetophora elegans: 1 — общий
вид слоевищ на субстрате (при
малом увеличении), 2 — иижняя
часть слоевища, 3 — верхушеч-
ная часть слоевища, 4, 5 —
форма ветвления, 6 — образова-
ние акинет. (I, 4—6 — по Мош-
ковой, 2, 3 — по Тиффани).
Рис. 91.
1, 2 — Chaetophora tuberculosa:
1 — общий вид слоевища (при
малом увеличении), 2 — конеч-
ное разветвление; 3 — С. puncti-
formis, конечное разветвление.
(1, 2 — по Мошковой, 3 — по
Мёбиусу).
181
Рис. 92.
Chaetophora pisiformis,
фрагмент слоевища,
(По Мошковой).
У некоторых исследователей проявляется тенденция к слиянию
С. tuberculosa с С. elegans, однако первая хорошо отличается от второй
наличием волоска у главной нити кисточек, бугорчатым слоевищем и
нередко значительным его обызвествлением.
3. Chaetophora punctiformis Kiitz. — Хетофора точкообразная
(рис. 91, 3). — Слоевище шаровидное, около 1 мм в диам., с гладкой
поверхностью. Главные нити около 140 мкм дл., разветвленные лишь
в нижней части, радиально расходящиеся от клеток подошвы. Конечные
ветвления в форме плотных кисточек, волоски отсутствуют. Клетки
главных нитей в их нижней части бочонковидные, выше цилиндрические,
у поперечных перегородок слегка перетянутые, 6—8 мкм шир., по длине
равные ширине или до 2 раз больше. Конечные клетки ветвей заострен-
ные, с редуцированным содержимым. Ризоидные выросты отсутствуют.
В разных водоемах. В СССР не обнаружена. (Зап. Европа, Америка).
Редкий и малоизученный вид.
4. Chaetophora pisiformis (Roth) Ag. (=Rivularia pisiformis Roth) —
Хетофора гороховидная (рис. 92). — Слоевище шаровидное или полуша-
ровидное, до 5 мм в диам., темно-, реже желто-зеленое, с гладкой
поверхностью. Главные нити прямостоячие, многочисленные, радиально
расходящиеся от подошвы, обильно разветвленные. Конечные ветвления
в виде рыхлых кисточек, состоящих из прямых, довольно коротких
ветвей. Клетки прямостоячих нитей 5.5—8 (10) мкм шир., по длине
до 3—6 раз больше, клетки ветвей 4—6 (7) мкм шир., по длине
равные ширине или до 1.5—2 раз больше. Конечные клетки конусо-
видно заостренные, реже переходящие в длинный многоклеточный
волосок.
В проточных, реже стоячих водах, на водных цветковых растениях;
встречается также в колодцах, на рисовых полях, в торфяных болотах.
ЛатвССР, РСФСР (преимущественно Европ. ч.), БССР, МолдССР, УССР.
182
5. Chaetophora incrassata (Hudson) Hazen [=Ulva incrassata Hudson;
Rlvularia cornu-dame Roth; R. endiviifolia Roth; Chaetophora indiviaefolia
(Roth) Ag.; C. cornu-dame (Roth) Ag.] — Хетофора утолщенная
(рис. 93). — Слоевище лопастное или разнообразно кустиковидно раз-
ветвленное, от нескольких миллиметров до нескольких сантиметров
высотой, нередко инкрустированное известью. Главные нити многочис-
ленные, прямостоячие, располагающиеся параллельно друг другу,
наружные из них односторонне ветвящиеся. Боковые ветви короткие,
с конечным ветвлением в форме рыхлых кисточек. Веточки кисточек
довольно короткие, слегка изогнутые, большей частью направленные
в одну сторону. Клетки главных нитей удлиненно-цилиндрические,
слабо вздутые в средней части, 8—16 мкм шир., по длине до 8 раз больше,
клетки конечных ветвлений цилиндрические, у поперечных перегородок
иногда слегка перетянутые, 6 — 11 мкм шир., по длине до 2 раз больше.
Конечные клетки заостренные или переходят в длинный многоклеточный
тупой волосок. Апланоспоры 8.9—11 мкм в диам.; они возникают
по 2—4 в клетке и могут длительное время лежать цепочками. Наблю-
далось образование акинет.
В стоячих и проточных водах, на водных цветковых растениях,
древесине, камнях.
Вид полиморфный. Описан ряд внутривидовых форм, в основном по
внешнему строению слоевища. Таксономическая ценность их вызывает
сомнения.
F. incrassata (=f. genuina) (рис. 93, 1—3). — Слоевище лопастное,
извилистое, с широкими долями, к основанию суженное.
Рис. 93.
С haetophora incrassata: 1—3 — f. incrassata (2 — общий вид слоевища, 2 —
главные нити с боковой ветвью, 3 — боковая ветвь отдельно), 4 — f. crystallo-
phora, общий вид слоевища, 5 — f. clavata, общий вид слоевища. (1, 5 —
по Мошковой, 2 — по Газеиу, 3 — по Смиту, 4 — по Кютцингу).
183
В реках, озерах, прудах, болотах. В ряде районов СССР, нередко.
F. polyelados Kutz. — Слоевище у основания вильчато разветвленное,
в верхней части расширенное, повторно перисторассеченное, по краям
с многочисленными короткими дольками.
В различных водоемах. В СССР не обнаружена. (Зап. Европа).-
F. crystallophora Kutz. (рис. 93, 4). — Слоевище разветвленное
в форме оленьих рогов со слабо повернутыми кончиками, инкрустиро-
ванное известью.
В реках, прудах, озерах. УССР.
F. incrustans Babenh. — Слоевище распростертое, извилисто-доль-
чатое, по краям с короткими дольками, слабо инкрустированное известью.
В реках. УССР.
F. crassa (Ag.) Kiitz. — Слоевище короткое, толстое, почти ровное,
с шиповидными кончиками.
В различных водоемах. В СССР не обнаружена. (Зап. Европа).
F. linearis Kiitz. — Слоевище более или менее плоское, нередко
узковильчато разветвленное.
В различных водоемах. В СССР не обнаружена. (Зап. Европа).
F. clavata (Horn.) Babenh. (=С. clavata Horn.) (рис. 93, 5).—
Слоевище с многочисленными, многократно повернутыми короткими
отстоящими и почти булавовидными дольками.
В прудах. УССР.
F. elongata Kiitz. — Слоевище кустиковидное, с узкими удлинен-
ными, простыми или слегка разветвленными дольками.
В ручье между торфяными карьерами. УССР.
6. Chaetophora flagellifera Kiitz. — Хетофора плетеносная (рис. 94,
1). — Слоевище сначала шаровидное, затем распростертое, мягкое. Глав-
ные нити длинные, почти параллельные, с удлиненными конечными
ветвями. Ветви плетевидные, нежные, почти параллельные. Клетки глав-
ных нитей в базальной части слоевища слегка вздутые, 18—20 мкм шир.,
по длине в 1.5—2.5 раза больше, выше заметно удлиненные. Конечные
клетки переходят в длинный многоклеточный заостренный волосок.
В пресных водоемах. ЛатвССР, ЛитССР, МолдССР.
Редкий малоизученный вид, по расположению нитей в колониальной
слизи и характеру конечных ветвей напоминает С. attenuata, однако
хорошо отличается короткими и широкими клетками главных нитей.
7. Chaetophora attenuate Hazen — Хетофора оттянутая (рис. 94, 2,
3). — Слоевище полушаровидное, зеленое или светло-зеленое, со слегка
бугристой поверхностью, до 2—5 мм в диам., с хорошо выраженными
подошвой и прямостоячими нитями. Подошва из угловато-шаровидных,
рыхло соединенных клеток, от которых отходят к субстрату многочислен-
ные ризоидные выросты. Главные нити прямые, почти параллельные
ДРУГ другу. Ветвление скудное, моноподиальное, но вследствие сдвигания
растущих ветвей к вершине материнской клетки и более или менее
параллельного расположения их по отношению друг к другу наблюдается
ложная дихотомия. Плотные кистевидные ветвления отсутствуют. Клетки
нитей цилиндрические, у поперечных перегородок иногда слегка пере-
тянутые, 5—8 мкм шир., у главных нитей по длине в 4 — 10 раз больше,
у ветвей по длине в 2—4 (6) раз больше. Конечные клетки заостренные
или переходят в длинный многоклеточный волосок. Инкрустация
известью наблюдается как в подошвенной, так и в периферической части
слоевища.
184
Рис. 94.
1 - Chaetophora flagellifera, ве₽ху“еч®® по Кютцингу, 2,3-
2 _ верхушечная часть слоевища,
185
Преимущественно в стоячих водах, на водных цветковых растениях.
УССР.
Хорошо отличается от других представителей рода отсутствием
плотных кистевидных конечных ветвлений. По-видимому, распростра-
нена значительно шире, но просматривалась из-за малых размеров.
Род IREKSOKONIA Meyer - ИРЕКСОКОНИЯ
Слоевище нитевидное. Нити ветвящиеся, прямостоячие, прикреп-
ляющиеся к субстрату ризоидами. Ветвление нитей моноподиальное,
ветви очередные, изредка супротивные. Ризоиды длинные, разветвлен-
ные. Клетки нитей обычно тонкостенные, цилиндрические, несколько
вздутые в средней части, у поперечных перегородок слегка перетяну-
тые, по длине равные ширине или до 1.5—3 раз больше, в нижней
части главных нитей толстостенные, их оболочки слоистые, бесцветные.
Конечные клетки ветвей притупленные или переходят в длинный тонкий
многоклеточный, преимущественно тупой волосок. Клетки ризоидов
толстостенные, цилиндрические или слегка вздутые. Хлоропласт сетчато
продырявленный, с многочисленными крупными пиреноидами, в клетках
главных нитей в виде нежной тонкой сеточки, полностью выстилающей
продольные стенки. Крахмал откладывается вокруг пиреноидов и
в строме хлоропласта. Бесполое размножение зооспорами. Зооспорангии
образуются из клеток боковых ветвей и верхушечных клеток главных
нитей. .Половое размножение не прослежено.
Этот род близок к роду Stigeoclonium, хорошо отличается размерами
слоевища, строением хлоропласта, мощным развитием ризоидной си-
стемы. Отчасти он близок и к роду Draparnaldia, а именно к D. simplex, но
отличается менее резко выраженной дифференциацией слоевища на ствол
и боковые ветви. Эндем Байкала.
Рис. 95.
1—3 — Ireksokonia formosa: 1 — фрагмент средней части слоевища, 2 — базаль-
ная часть, 3 — клетка главной нити; 4, 5 — Myzonemopsls crassimembranacea;
4 — боковая ветвь слоевища, 5 — клетка главной нити. (По Мейеру).
186
1. Ireksokonia formosa Meyer — Ирексокония красивая (рис. 95,
1—3). — Слоевище 10—12 см выс. Клетки главных нитей в базальной
части 120—135 мцм шир., в средней 50—80 мкм шир., на концах
14—17 мкм шир.; клетки волосков 9—13 мкм шир.; клетки ризоидов
18—45 мкм шир. Оболочки клеток в нижней части главных нитей
13—26 мкм толщ.
Оз. Байкал, на скалах, на глубине 2—5 м образует густые дерно-
винки.
По мнению К. И. Мейера, I. formosa можно рассматривать как
форму, связывающую род Stigeoclonium с байкальскими драпарналь-
диеллами и, возможно, исходную для них.
Род MYXONEMOPSIS Меуег - МИКСОНЕМОПСИС
Слоевище кустиковидное. Нити ветвящиеся, прямостоячие, с обособ-
ленной центральной или главной нитью и длинными, обильно разветвлен-
ными, очередными или супротивными боковыми ветвями. Клетки глав-
ных нитей цилиндрические или слегка бочонковидные, с толстой
слоистой оболочкой, клетки ветвей короткоцилиндрические, у попереч-
ных перегородок часто сильно перетянутые и: тогда Почти шаровидные,
тонкостенные. Конечные клетки заостренные или переходят в длинный
многоклеточный волосок. В базальной части слоевища от некоторых
клеток главных нитей отходят обильно разветвленные ризоидные вы-
росты, довольно часто возникающие и от основания низко расположенных
боковых ветвей. Хлоропласт сетчато продырявленный, с 2—4 крупными
пиреноидами. Размножение не прослежено.
Этот род близок к роду Ireksokonia, хорошо отличается длинными,
обильно разветвленными боковыми ветвями, напоминающими ветви
драпарнальдий. По мнению К. И. Мейера, занимает промежуточное
положение между родами Draparnaldia и Ireksokonia. Эндем оз. Байкал.
1. Myxonemopsis crassimembranacea Meyer — Миксонемопсис толсто-
стенный (рис. 95, 4, 5). —Кустики около 2 см выс., пуши-
стые. Клетки главных нитей и крупных боковых ветвей 90—150 мкм
шир., клетки ветвей в нижней части 70 мкм шир., в средней 30—40 мкм
шир., ближе к вершине 15 мкм шир.; клетки волосков 7.5—11 мкм шир.
Оболочки клеток бесцветные,, слоистые, 5.5—10 мкм толщ.
Оз. Байкал,, на скалах на глубине до 2 м.
Род CLONIOPHORA Tiff. - КЛОНИОФОРА
Слоевище кустиковидное. Нити ветвящиеся, прямостоячие, прикреп-
ляющиеся к субстрату ризоидами, с обособленной центральной осью —
главным стволом и многочисленными рассеянными длинными и корот-
кими боковыми побегами, повторно разветвленными. Повторные боковые
веточки короткие, часто искривленные, простые или разветвленные,
слегка оттопыренные или несколько прижатые, восходящие, очередные,-
одиночные, супротивные или в мутовках. Клетки главного ствола и
крупных боковых побегов цилиндрические, бочонковидные или слегка
булавовидные, у поперечных перегородок не перетянутые или заметно
перетянутые, клетки веточек цилиндрические или бочонковидные, из-
редка булавовидные, у поперечных перегородок не перетянутые или пере-
тянутые. Конечные клетки конусовидно суженные, тупые; волоски отсут-
187
1 — Cloniphora plumosa; 2—6 — C. spicata: 2—5 — верхние части слоевища,
6 — основание слоевища с ризоидами. (7 — по Исламу, 2—6 — по Тиффани).
ствуют. Хлоропласт поясковидный, по краям нередко изрезанный,
с многочисленными пиреноидами. Зооспорангии и гаметангии обра-
зуются из вздутых клеток боковых ветвей.
Род Cloniophora близок к родам Stigeoclonium и Drapamaldia и,
по всей видимости, является промежуточным между ними.
I. Клетки главных нитей цилиндрические, реже едва заметно булаво-
видные; боковые веточки многоклеточные, простые или повторно
разветвленные.
1. Клетки главных нитей 20—25 (32) мкм шир., по длине в 2 — 7 раз
больше..........................................С. plumosa 1.
2. Клетки главных нитей 30 — 40 мкм шир., по длине в 2 раза меньше
или до 3 раз больше...............................С. spicata 2.
188
II. Клетки главных нитей бочон-
ковидные, вздутые или слегка
булавовидные; боковые ве-
точки малоклеточные, про-
стые ... С. macrocladia 3.
1. Cloniophora ‘ plumosa
(Kiitz.) Bourr. (^Stigeoclonium
plumosum Kiitz.) — Клониофора
перистая (рис. 96, 1). — Кустики
2—5 см выс. Главные нити тон-
кие, обильно разветвленные пре-
имущественно в верхней части
слоевища. Боковые веточки мно-
гоклеточные, супротивные или,
чаще, в мутовках. Клетки глав-
ных нитей и длинных боковых
побегов цилиндрические, реже
слегка булавовидные, у попереч-
ных перегородок не перетянутые
или едва заметно перетянутые,
20—25 (32) мкм шир., по длине
в 2—7 раз больше. Конечные
клетки конусовидно суженные,
тупые, несколько длиннее осталь-
ных клеток.
В разных водоемах. В СССР
не обнаружена. (Зап. Европа,
Сев. Америка).
2. Cloniophora spicata (Schmi-
dle) Islam (=Stigeoclonium spica-
turn Schmidle; S. askenasii Schmid-
le; Cloniophora ivillei Tiff.; C. ca-
pitata Tiff.) — Клониофора коло-
систая (рис. 96, 2—6). — Ку-
стики 2—5 (10) см выс. Глав-
ные нити в нижней части обычно
Рис. 97.
Cloniophora macrocladia. (По Норд-
стедту).
неразветвленные или с очень
редкими короткими веточками и
обильными ризоидами, которые
могут отходить и от базальных клеток нижних длинных боковых побегов;
в верхней части слоевища главные нити и длинные боковые побеги
с многочисленными восходящими, более или менее прижатыми, простыми
или повторно разветвленными, обычно многоклеточными боковыми вет-
вями. Клетки главных нитей и длинных побегов цилиндрические или
бочонковидные, у поперечных перегородок не перетянутые или слегка
перетянутые, с довольно толстой оболочкой, в базальной части 16 мкм
шир., выше 30—40 мкм шир., по длине в 2 раза меньше или до 3 раз
больше, непосредственно после деления изодиаметрические, клетки боко-
вых веточек 10 — 12 мкм шир., с тонкими оболочками. Конечные клетки
конусовидно суженные, тупые.
В разных водоемах. В СССР не обнаружена. (Зап. Европа).
189
3. Cloniophora macroeladia (Nordst.) Bourr. [=Stigeoclonlum macro-
cladium (Nordst.) Schmidle] — Клониофоракрупноветвистая (рис. 97).—
Кустики 15—30 см выс. Главные нити довольно обильно разветвленные,
начиная с базальной части. Длинные боковые побеги очередные, супро-
тивные или, реже, в мутовках, почти такой же ширины, как и главные
нити. Боковые веточки малоклеточные, короткие, очередные. Клетки
главных нитей бочонковидные, у поперечных перегородок заметно пере-
тянутые, 25—50 (60) мкм шир., по длине в 1.5—2 (3) раза больше,
клетки веточек 6—10 (14) мкм шир., по длине равные ширине или до 2 раз
больше. Конечные клетки конусовидно суженные, тупые.
В разных водоемах. В СССР не обнаружена. (Зап. Европа, США).
Род DRAPARNALDIELLA Meyer et Scabitsch. —
ДРАПАРНАЛЬДИЕЛЛА
Слоевище кустиковидное. Нити обильно ветвящиеся, прямостоячие,
с отчетливо обособленной центральной осью — главным стволом, при-
крепляющимся к субстрату подошвой (у некоторых видов неясно выра-
женной), и многочисленными крупными, большей частью длинными
боковыми побегами, повторно разветвленными. Повторные боковые ветви
образуются как на главном стволе, так и на длинных боковых побегах,
короткие, одиночные, супротивные или по 3—4 в мутовках. У одних
видов в коротких боковых ветвях имеется четко выраженная средняя
ось, от которой почти по всей длине отходят простые или разветвленные
веточки, при этом часто она далеко выдается за пределы всей ветви;
у других видов в боковых ветвях четко выраженной средней оси нет,
она довольно быстро становится незаметной, переходя в разветвления.
Главный ствол состоит из ряда крупных цилиндрических или бочонко-
видных клеток, клетки длинных боковых побегов такие же, но клетки
веточек мелкие, обычно цилиндрические. Конечные клетки закругленные,
реже суженные, слегка заостренные или переходят в длинный многокле-
точный тупой волосок. Вокруг главного ствола и его ответвлений
развивается своеобразный рыхлый, так называемый гифовый покров,
состоящий из многочисленных, обильно разветвленных ризоидов; в верх-
них частях кустиков он тонкий или отсутствует, в нижних частях
толстый, мощный. Хлоропласт кольцевидный, в клетках ствола и круп-
ных побегов в виде тонкой сеточки, прилегающей к продольным стенкам,
с многочисленными пиреноидами, в клетках веточек неясно сетчатый,
с несколькими крупными пиреноидами. Размножение исключительно
половое, со строго выраженной гетероталличностью. Гаметы образуются
в многокамерных гаметангиях, возникающих из многих клеток средней
оси и крупных ветвей; к моменту образования гамет в этих клетках
происходят вначале поперечные, а затем продольные деления, и в каждой
маленькой клеточке возникшего многокамерного гаметангия образуется
по четырехжгутиковой широкогрушевидной гамете со стигмой и сетевид-
ным хлоропластом, освобождающейся через отверстие в боковой стенке
материнской клетки. Различия между гаметами разных полов незначи-
тельны, они только слабо отличаются по величине (слабая анизогамия).
Зооспоры не обнаружены.
Род Draparnaldiella близок к роду Drapamaldia, но, по мнению
К. И. Мейера и А. П. Скабичевского, филогенетически с ним не связан,
оба рода возникли независимо в разных эволюционных рядах, а их
сходство является результатом конвергенции. Важными диагностиче-
190
скими признаками для разграничения видов служат форма боковых
ветвей и наличие или отсутствие в них средней оси. Для определения
видов следует отбирать взрослые слоевища, вступившие в стадию
формирования гаметангиев, а в них — ветви средней части слоевища.
Эндем оз. Байкал.
I. Боковые ветви с четко выраженной средней осью.
1. Средняя ось в боковых ветвях выдается за их пределы.
А. Веточки в боковых ветвях отходят от средней оси почти на
всем ее протяжении.
а. Боковые ветви обильно разветвленные, в общих очертаниях
широкояйцевидные или полукруглые; веточки оттопырен-
ные ........................................D. pumila 1.
б. Боковые ветви скудно разветвленные, в общих очертаниях
ланцетовидные, вытянутые; веточки прижатые ....
D. simplex 2.
Б. Веточки в боковых ветвях отходят от средней оси только в ее
базальной части...............................D. arenaria 3.
2. Средняя ось в боковых ветвях за их пределы не выдается.
А. Боковые ветви перисторазветвленные, в общих очертаниях
широколанцетовидные; их средняя ось оканчивается волоском,
веточки отходят на всем ее протяжении......................
.....................................D. goroschankinii 4.
Б. Боковые ветви веерообразно разветвленные, в общих очерта-
ниях округлые; их средняя ось оканчивается закругленной
клеткой, веточки отходят преимущественно в ее базальной
части.........................................D. arnoldii 5.
II. Боковые ветви без средней оси, или она выражена слабо.
1. Конечные клетки веточек слегка суженные, закругленные или
переходят в многоклеточный волосок; боковые ветви отходят не от
каждой клетки ствола.
А. Клетки ствола в основании слоевища 400—450 мкм шир.,
в средней части 240— 360 мкм шир..............D. baicalensis 6.
Б. Клетки ствола в основании слоевища 160 — 300 мкм шир.,
в средней части 100—200 мкм шир.
а. Клетки ствола в его основании около 300 мкм шир.,
в средней части около 200 мкм шир...........D. vilosa 7.
б. Клетки ствола в его основании 160—180 мкм шир., в сред-
ней части 100 — 150 мкм шир.................D. pilosa 8.
2. Конечные клетки веточек широко закругленные, тупые, волоски
отсутствуют или встречаются редко; боковые ветви отходят от
каждой клетки ствола и плотно соприкасаются друг с другом .
...............................................D. lubrica 9.
1. Draparnaldiella pumila (Meyer) Meyer et Skabitsch. (=Draparnal-
dia pumila Meyer) — Драпарнальдиелла карликовая (рис. 98,1,2). — Ку-
стики слизистые, 7—10 см выс. Главный ствол обильно разветвленный,
прикрепляющийся к субстрату маленькой круглой подошвой. Боковые
ветви обильно разветвленные, рыхлые, в общих очертаниях широкояйце-
видные или полукруглые, супротивные или по 4 в мутовках, с четко
выраженной средней осью, далеко выдающейся за пределы ветвей в це-
лом. Веточки оттопыренные, изогнутые внутрь. Клетки главного ствола
цилиндрические или слегка бочонковидные, у поперечных перегородок
едва заметно перетянутые, с тонкой оболочкой, в основании слоевища до
191
Рис. 98.
1, 2 — Draparnaldiella pumila: 1 — фрагмент ствола с молодой ветвью, 2—
отдельная ветвь; 5, 4 — D. simplex; 5 — D. areaaria. (По Мейеру).
250 мкм шир., в средней части его 135—175 мкм шир., в верхушечной
45—50 мкм шир., по длине равные ширине, до 1.5 раза больше или,
реже, меньше ее (преимущественно в верхних участках слоевища),
клетки ветвей цилиндрические, 10—13.5 мкм шир. Конечные клетки
слегка суженные, закругленные или переходят в длинный многоклеточ-
ный волосок. Ризоидный покров хорошо развит, особенно в нижней части
кустика, благодаря чему главный ствол и длинные побеги достигают
2—3 мм толщ.
Оз. Байкал, на камнях.
2. Draparnaldiella simplex (Meyer) Meyer et Skabitsch. (=Draparnal-
dia simplex Meyer) — Драпарнальдиелла простая (рис. 98, 3, 4). — Кус-
тики бледно-зеленые, слегка слизистые, (10) 15—20 см выс. Главный
192
ствол обильно разветвленный, прикрепляющийся к субстрату с помощью
неясно выраженной подошвы. Длинные боковые побеги по 3—5 в мутов-
ках. Боковые ветви скудно разветвленные, вытянутые, с прижатыми
веточками, в общих очертаниях более или менее ланцетовидные, с четко
выраженной средней осью, выходящей за пределы ветвей в целом, реже
простые, одиночные, супротивные или в мутовках. Клетки главного
ствола цилиндрические или бочонковидные, у поперечных перегородок
слегка перетянутые, в нижней части слоевища 290—375 мкм шир.,
в средней 90—150 мкм шир., в верхушечной 28—37 мкм шир., по длине
меньше ширины (преимущественно в нижней части), равны ей (преиму-
щественно в средней части) или в 2—4 раза больше ее (преимущественно
в верхушечной части), клетки веточек цилиндрические, 10—35 мкм
шир., по длине большей частью равные ширине. Конечные клетки
заостренные или переходят в длинный многоклеточный волосок 6.7 —
10.3 мкм шир. Ризоидный покров выражен слабо и только в нижних
частях слоевища.
Оз. Байкал, на скалах, на глубине 2—10 м.
3. Draparnaldiella arenaria (Meyer) Meyer et Skabitsch. (=--Drapar-
naldia arenaria Meyer) — Драпарнальдиелла песчаная (рис. 98, 5). —
Кустики темно-зеленые, слизистые, 15—20 см выс. Главный ствол
обильно разветвленный, прикрепляющийся к субстрату хорошо выра-
женной подошвой. Боковые ветви скудно разветвленные, прижатые
к стволу, с длинной, четко выраженной средней осью, выходящей за
пределы ветвей в целом, от которой лишь у основания отходит несколько
простых или скудно разветвленных прижатых веточек. Клетки главного
ствола бочонковидные, у поперечных перегородок перетянутые, с тонкой
оболочкой, у основания слоевища 300— 340 мкм шир., в средней части
его 177.5—246 мкм шир., в верхушечной 46.2—70 мкм шир., по длине
равные ширине, несколько меньше (преимущественно в базальной
части) или до 3 раз больше ее, клетки веточек цилиндрические, 22 —
45 (58) мкм шир., по длине равные ширине или до 2 раз больше. Конеч-
ные клетки почти всегда переходят в длинный многоклеточный волосок
11.5—18 мкм шир. Ризоидный покров хорошо развит преимущественно
в нижней трети слоевища, в виде толстого рыхлого чехла, одевающего
главный ствол и основания длинных и коротких боковых побегов.
Оз. Байкал, на каменистом субстрате, находящемся на песчаных
участках дна, на глубине 5 м.
4. Draparnaldiella goroschankinii (Meyer) Meyer et Skabitsch. (=Dra-
parnaldia goroschankinii Meyer) — Драпарнальдиелла Горожанкина
(рис. 99, 7). —Кустики светло-зеленые, слизистые, 10—15 см выс.
Главный ствол обильно разветвленный, прикрепляющийся к субстрату
неясно выраженной подошвой. Боковые ветви одиночные, супротивные
или по 3—4 в мутовке, в общих очертаниях широколанцетовидные,
с четко выраженной средней осью, не выходящей за пределы ветвей
в целом, от которой на всем ее протяжении отходят простые или повторно
разветвленные веточки. Клетки главного ствола цилиндрические или
слегка брчонковидные, у поперечных перегородок перетянутые, с тонкой
оболочкой, у основания слоевища 300 — 350 мкм шир., по длине в 2—3 раза
меньше, в средней части его 200—280 мкм шир., по длине равные
ширине или до 2 раз больше, ближе к вершине 30—40 мкм шир., по
длине до 3 раз больше, клетки веточек цилиндрические, 12 — 17.7 мкм шир.
Конечные клетки слегка заостренные или переходят в длинный много-
клеточный волосок 5—8 мкм шир. Ризоидный покров выражен слабо,
13 Зеленые водоросли. Улотриксовые 193
Рис. 99.
1 — Draparnaldiella goroschankinii', 2 — D. arnoldii; 3—5 — D. baicalensis; 3 —
боковая ветвь, 4 — фрагмент ветви, 5 — клетка главного ствола. (По Мейеру).
развивается преимущественно в самых нижних частях слоевища.
Оз. Байкал, на камнях.
5. Draparnaldiella arnoldii (Meyer) Meyer et Skabitsch. (=Draparnal-
dia arnoldii Meyer) — Драпарнальдиелла Арнольди (рис. 99, 2). —
Кустики 10—15 см выс. Главный ствол обильно разветвленный, прикреп-
ляющийся к субстрату круглой подошвой. Боковые ветви супротивные
или по 3 в мутовке, веерообразно разветвленные, в общих очертаниях
округлые, с четко выраженной короткой средней осью, от которой
преимущественно от ее базальной части отходят растопыренные, простые
или повторно разветвленные веточки, по длине значительно превосходя-
щие среднюю ось. Клетки главного ствола цилиндрические, у поперечных
перегородок слегка перетянутые, с оболочкой 170—180 мкм шир., по
длине равные ширине или в 1.5 раза больше, клетки длинных боковых
побегов 90—100 мкм шир., клетки боковых ветвей бочонковидные,
преимущественно изодиаметрические, у поперечных перегородок заметно
перетянутые, 27—47 мкм шир. Конечные клетки слегка заостренные
или, чаще, переходят в длинный многоклеточный волосок, состоящий из
относительно коротких клеток, у средних веточных осей закругленные.
Ризоидный покров хорошо развит, преимущественно в нижней части
главного ствола и длинных боковых побегов.
Оз. Байкал, на камнях.
6. Draparnaldiella baicalensis (Meyer) Meyer et Skabitsch. (=Dra-
parnaldia baicalensis Meyer) — Драпарнальдиелла байкальская (рис. 99,
3—5). — Кустики темно-зеленые, слизистые, 15—25 (40) см выс. Глав-
ный ствол обильно разветвленный, прикрепляющийся к субстрату
круглой подошвой. Боковые ветви обильно разветвленные, довольно
плотные, в общих очертаниях полукруглые или округло-треугольные,
обычно по 3 в мутовке, реже супротивные или одиночные. Средняя
ось на протяжении боковой ветви скоро становится незаметной, теряясь
194
в разветвлениях. Веточки слегка дуговидно изогнутые внутрь. Клетки
главного ствола и длинных боковых побегов цилиндрические, у попереч-
ных перегородок слегка перетянутые, с довольно тонкой оболочкой,
у основания слоевища 400— 450 мкм шир., в средней части 240—360 мкм
шир., выше 110—150 мкм шир., у вершины около 75 мкм шир., по длине
равные ширине или в 1.5 раза больше, клетки ветвей цилиндрические
или слегка вздутые, 12—16.5 мкм шир. Конечные клетки слегка
заостренные или, чаще, переходят в длинный многоклеточный волосок
2.5—12.5 мкм шир. Ризоидный покров развит очень сильно, до 3 мм толщ.,
особенно в нижней части главного ствола, длинных и коротких боковых
побегов.
Оз. Байкал, на камнях.
7. Draparnaldiella villosa (Meyer) Meyer et Skabitsch. (=Dra-
parnaldta villosa Meyer) — Драпарнальдиелламохнатая (рис. 100,1,2).—
Кустики 6—8 см выс. Главный ствол обильно разветвленный, мохнатый.
Боковые ветви 1 — 2 мм дл., обильно разветвленные, компактные,
в общих очертаниях полукруглые или широкояйцевидные, обычно
по 3 в мутовке; они отходят почти от каждой клетки ствола и побегов и
тесно соприкасаются друг с другом, что придает слоевищам своеобразный
мохнатый вид. Средняя ось в полностью сформированных ветвях отсут-
ствует, она быстро теряется в разветвлениях, но в молодых ветвях
отчетливо просматривается на всем протяжении ветвей и даже выходит
за их пределы. Веточки слегка дуговидно изогнутые внутрь, повторно
разветвленные. Клетки главного ствола и длинных побегов цилиндриче-
ские, у поперечных перегородок слегка перетянутые, с толстыми оболоч-
ками, у основания слоевища около 300 мкм шир., в средней части
около 200 мкм шир., у вершины 47 мкм шир., по длине равные ширине
или несколько больше, клетки ветвей цилиндрические, 10—15 мкм шир.
Конечные клетки слегка суженные, закругленные, некоторые переходят
в длинный многоклеточный волосок. Ризоидный покров развит очень
сильно, образует вокруг главного ствола и длинных боковых побегов
толстый рыхлый чехол.
Оз. Байкал, на камнях.
1,2 — Draparnaldiella villosa: 1 — боковая ветвь, 2 — молодая веточка; 3— 5 —
D. pilosa: 3 — боковая ветвь, 4 — фрагмент молодой ветви, 5 — фрагмент глав-
ного ствола; 6 — D. tubrica. (По Мейеру).
13»
195
8. Draparnaldiella pilosa (Meyer) Meyer et Skabitsch. (=Draparnaldia
pilosa Meyer) — Драпарнальдиелла волосистая (рис. 100, 3—5). —
Кустики ярко-зеленые, слизистые, 10—20 см выс. Главный ствол
обильно разветвленный, прикрепляющийся к субстрату небольшим
диском. Длинные боковые побеги шнуровидные, к вершине посте-
пенно утончающиеся и становящиеся нитевидными. Боковые ветви
обильно разветвленные, довольно плотные, в общих очертаниях по-
лукруглые, по 3 в мутовке, почти на каждой клетке главного ствола и
длинных боковых побегов. Средняя ось отсутствует, быстро переходит
в разветвления. Веточки слегка дуговидно изогнутые внутрь, преиму-
щественно простые. Клетки главного ствола цилиндрические, у попереч-
ных перегородок слегка перетянутые, с довольно толстыми оболочками,
у основания слоевища 160—180 мкм шир., в средней части 100—150 мкм
шир., у вершины 40 — 55 мкм шир., по длине равные ширине, в 1.5—2 раза
больше или, реже, несколько меньше, клетки ветвей цилиндрические,
10.7—12.9 мкм шир. Конечные клетки суженные, закругленные, многие
переходят в длинный многоклеточный волосок. Ризоидпый покров развит
очень сильно, образует вокруг главного ствола, длинных и коротких
побегов толстый чехол, особенно мощный в нижних частях слоевища.
Оз. Байкал, на камнях.
9. Draparnaldiella lubrica (Meyer) Meyer et Skabitsch. (=Draparnal-
dia lubrica Meyer) — Драпарнальдиелла скользкая (рис. 100, в).—
Кустики темно-зеленые, слизистые, 12—20 см выс., окутанные плотным
слоем слизи, делающим их упругими и очень скользкими. Главный ствол
обильно разветвленный, прикрепляющийся к субстрату круглой подош-
вой. Боковые ветви короткие, обильно разветвленные, компактные, в об-
щих очертаниях полукруглые, округло-яйцевидные, обычно в мутовках;
они отходят почти от каждой клетки ствола и боковых побегов, тесно
смыкаются друг с другом и образуют вокруг них своеобразный сплошной
покров. Средняя ось в ветвях отсутствует, она быстро переходит в раз-
ветвления. Клетки главного ствола цилиндрические, у поперечных пере-
городок слегка перетянутые, с толстыми оболочками, 200—240 мкм шир.,
по длине равные ширине или до 2 раз больше, реже несколько меньше,
клетки длинных боковых побегов 120—150 мкм шир., по длине равные
ширине или до 1.5 раза больше, клетки средних осей в боковых ветвях
в базальной части 60—70 мкм шир., по длине равные ширине или
несколько больше, клетки веточек 9—10 мкм шир. Конечные клетки
слегка суженные, широко закругленные. Волоски встречаются редко.
Ризоидный покров развит очень сильно, длинные, обильно разветвленные
ризоиды густо оплетают главный ствол и все боковые побеги, неопле-
тенными остаются лишь самые верхушки ветвей па 3—4 мм.
Оз. Байкал, на камнях.
Род DRAPARNALDIA Богу - ДРАПАРНАЛЬДИЯ
Слоевище кустиковидное. Нити обильно ветвящиеся, прямостоячие,
с обособленной центральной осью — главным стволом, прикрепляю-
щимся к субстрату небольшой базальной клеткой и разветвленными
ризоидами, и многочисленными, длинными или короткими боковыми
побегами, повторно разветвленными. Повторные боковые ветви обра-
зуются как на главном стволе, так и на длинных боковых побегах,
короткие, одиночные, супротивные или по 3—4 в мутовках. У одних
видов в коротких боковых ветвях имеется четко выраженная средняя
196
ось, от которой почти по всей ее длине отходят простые или разветвленные
веточки, при этом часто она далеко выдается за пределы всей ветви;
у других видов в боковых ветвях четко выраженной средней оси нет,
она довольно быстро становится незаметной, переходя в разветвления.
Главный ствол состоит из одного ряда крупных цилиндрических или
слегка бочонковидных клеток, клетки длинных и коротких боковых
побегов такие же, но отличаются размерами, клетки веточек мелкие,
обычно цилиндрические. Конечные клетки заостренные или переходят
в длинный многоклеточный волосок. От нижних клеток не только глав-
ного ствола, но и многих боковых побегов также отходят ризоиды.
Хлоропласт кольцевидный, с довольно сильно изрезанными краями,
обычно продырявленный, часто настолько, что в крупных клетках глав-
ного ствола и боковых ветвей имеет вид тонкой нежной сеточки. Пире-
ноидов от 1 до нескольких. Макрозооспоры четырехжгутиковые, яйце-
видные или удлиненно-цилиндрические, с 2 пульсирующими вакуолями
и стигмой в переднем конце клетки; они возникают в боковых ветвях
по 1—4 в каждой клетке. Микрозооспоры яйцевидные, четырехжгутико-
вые, со стигмой в заднем конце клетки; потеряв жгутики, они переходят
в амебоидное состояние или сливаются друг с другом. Вместо зооспор
в клетках боковых ветвей нередко образуются апланоспоры с утолщен-
ными оболочками и желтовато-оранжевым или оранжево-красным
содержимым, известны также акинеты, прорастающие после периода
' покоя.
Преимущественно в холодных проточных, реже стоячих водах,
в обрастаниях камней, древесного субстрата, водных цветковых растений,
раковин моллюсков и т. п.
I. Хлоропласт в клетках ствола и длинных побегов в виде тонкой нежной
сеточки, почти полностью заполняющей продольные стенки клеток
D. platyzonata 1.
II. Хлоропласт в клетках ствола и длинных побегов в виде узкого или
широкого продырявленного кольца с изрезанными краями.
1. Боковые ветви с четко выраженной средней осью.
А. Средняя ось длинная, далеко выступающая за пределы боковых
ветвей; волоски многоклеточные, тупые или отсутствуют.
а. Боковые ветви в общих очертаниях широколанцетовидные
или заостренно-яйцевидные, веточки оттопыренные . . .
..........................................D. plumosa 2.
б. Боковые ветви в общих очертаниях полукруглые, веточки
оттопыренные........................D. champlainensis 3.
в. Боковые ветви в общих очертаниях узколанцетовидные,
вытянутые, веточки прижатые...............D. appressa 4.
Б. Средняя ось прослеживается на всем протяжении боковой
ветви, но не выступает за ее пределы; волоски одноклеточные,
острые или отсутствуют.
а. Боковые ветви восходящие, обильно разветвленные, в об-
щих очертаниях широкояйцевидные или широколанцето-
видные; волоски острые.......................D. acuta 5.
б. Боковые ветви отходят под прямым углом, в общих очерта-
ниях полукруглые, широкояйцевидные; волоски острые
или отсутствуют.
а. Боковые ветви скудно разветвленные, рыхлые, волоски
острые; клетки главного ствола 20—25 (40) мкм шир.
............................D. sparsifasciculata 6.
р. Боковые ветви обильно разветвленные, плотные, волоски
197
обычно отсутствуют; клетки главного ствола 150—
170 (200) мкм шир.......................D. ravenelii 7.
2. Боковые ветви без средней оси, или она выражена слабо.
А. Средняя ось прослеживается лишь в базальной части боковых
ветвей, где веточки постоянно отсутствуют . . . D. opposite 8.
Б. Средняя ось отсутствует, быстро теряясь в разветвлениях.
а. Боковые ветви восходящие, обильно разветвленные, в об-
щих очертаниях округлые, полукруглые или широкояйце-
видные; волоски, если присутствуют, тупые................
.......................................D. glomerate 9.
б. Боковые ветви отходят под прямым углом, скудно развет-
вленные; волоски, если присутствуют, острые..............
, .......................................D. judayi 10.
1. Draparnaldia platyzonata Hazen — Драпарнальдия широкопоясная
(рис. 101). — Кустики нежные, пушистые, 2—3 (7) см выс. Главный
ствол обильно разветвленный. Длинные боковые побеги и боковые ветви
отходят от главного ствола под прямым углом. Боковые ветви обильно,
большей частью односторонне разветвленные, в общих очертаниях
широко округло-треугольные или широкояйцевидные, одиночные, супро-
тивные или в мутовках, средняя ось на протяжении боковых ветвей
быстро теряется в разветвлениях. Клетки главного ствола цилиндри-
ческие или слегка бочонковидные, у поперечных перегородок заметно
перетянутые, 50—95 мкм шир., по длине равные ширине или в 2—3 раза
больше, клетки длинных побегов до 38 мкм шир., по длине равные
ширине, реже до 2 раз больше, клетки средней оси цилиндрические,
Рис. 101.
Draparnaldia platyzonata-. 1 — боковая ветвь, 2 — базальная часть слоевища
с ризоидными выростами, 3 — клетки главного ствола. (По Мошковой).
198
18 мкм шир., клетки веточек 6—11 мкм шир. Конечные клетки Веточек
заостренные или, чаще, переходят в тонкий длинный многоклеточный
тупой волосок. Хлоропласт в клетках ствола и длинных боковых побегов
в виде нежной широкоячеистой сеточки со многими пиреноидами,
выстилающей продольные стенки клеток почти полностью, в клетках
боковых ветвей неясно сетчатый. В нижних частях слоевища от клеток
ствола и крупных побегов отходят длинные многоклеточные разветвлен-
ные ризоидные выросты.
В проточных водах, особенно в ручьях, вытекающих из болот,
в озерах, на разных субстратах. УССР, Вост. Сибирь (Байкал).
199
2. Draparnaldia plumosa (Vauch.) Ag. [=Batrachospermum plumosum
Vauch.; Conferva mutabills Roth; Draparnaldia mutabills (Roth) Ce-
dercr.] — Драпарнальдия перистая (рис. 102, 103, 7). — Кустики светло-
или темно-зеленые, до 10 см выс. Главный ствол обильно разветвленный.
Боковые ветви обильно разветвленные, в общих очертаниях широколан-
цетовидные или заостренно-яйцевидные, с четко выраженной длинной
средней осью, далеко выступающей за пределы ветви. Веточки оттопы-
ренные, отходят по всей длине оси, оканчивается она длинным много-
клеточным тупым волоском 6—7.6 мкм шир. Клетки главного ствола
цилиндрические или слегка бочонковидные, у поперечных перегородок
не перетянутые или едва заметно перетянутые, 38—107 (114) мкм шир.,
по длине равные ширине или до 4 раз больше, с довольно толстыми,
заметно слоистыми оболочками, клетки длинных побегов такие же, 26.6—
68 мкм шир., по длине равные ширине или до 2 раз больше, клетки
веточек 5.7—10 мкм шир., по длине равные ширине или до 2.5 раза
больше. Конечные клетки заостренные или переходят в длинный много-
клеточный тупой волосок 3.8—5 мкм шир. Хлоропласт кольцевидный.
Макрозооспоры 8—10 мкм шир., 12—16 мкм дл., образуются по 1 в клет-
ках боковых ветвей.
В стоячих и проточных водах, на камнях, водных цветковых расте-
ниях, древесном субстрате, реже раковинах моллюсков.
Var. plumosa (рис. 102). — Боковые ветви плотные, в общих очерта-
ниях широколанцетовидные.
В реках, прудах, озерах, водохранилищах. Повсеместно. Вегетирует
с весны до осени.
Var. pulchella (Kiitz.) Rabenh. (=Draparnaldia pulchella Kiitz.) (рис.
103, 1). — Боковые ветви рыхлые, в общих очертаниях заостренно-яйце-
видные.
В стоячих и слабо проточных водах. УССР.
3. Draparnaldia champlainensis Cook — Драпарнальдия шамплейн-
екая (рис. 103, 2). — Кустики до 4—8 см выс. Главный ствол опутан
довольно густым покровом из длинных, обильно разветвленных ризоид-
Рис. 103.
1 — Draparnaldia plumosa var. pulchella; 2 — D. champlainensis. (1 — по Мош-
ковой, 2 — по Куку).
200
ных выростов, благодаря чему в базальной части достигает 375—800 мкм
толщ., в верхушечной 190 — 330 мкм толщ. Боковые ветви отходят
почти от каждой клетки ствола, располагаясь тесно сближенными
мутовками, в общих очертаниях полукруглые, средняя ось четко выра-
жена и далеко выдается за пределы ветви. Веточки оттопыренные.
Клетки главных стволов цилиндрические или слегка бочонковидные,
40—95 мкм шир., клетки веточек 5.2—10 мкм шир. Конечные клетки
заостренные или переходят в волосок 9—14.5 мкм шир., до 2 мм дл.
Хлоропласт кольцевидный.
В стоячих водах. В СССР не обнаружена. (США).
Редкий вид, обнаружен в оз. Шамплейн (штат Нью-Йорк) на
глубине около 25 м. Отличается мощным покровом из ризоидных
выростов, что характерно для рода Draparnaldiopsis, куда, по-видимому,
его и следует отнести.
4. Draparnaldia appressa Moschk. — Драпарнальдия прижатая
(рис. 104). — Кустики темно-зеленые, косматые, 5—7 (10) мм выс.
Главный ствол с длинными узкими боковыми побегами. Боковые ветви
длинные, скудно разветвленные, в общих очертаниях узколанцетовидные,
одиночные, супротивные, реже в мутовках, с четко выраженной средней
осью, выступающей за пределы ветви. Боковые веточки отходят почти
по всей длине средней оси, большей частью простые, прижатые, состоя-
щие обычно из 1—5 клеток, реже встречаются более длинные, слегка
оттопыренные веточки. Клетки ствола бочонковидные, у поперечных
перегородок заметно перетянутые, 26—60 мкм шир., по длине вдвое
меньше ширины или равны ей, клетки длинных боковых побегов бочонко-
видные, 20—32 мкм шир., по длине меньше ширины, равны ей или
несколько больше, клетки средней оси боковых ветвей у основания
9.5—11.4 мкм шир., ближе к вершине 7.6—9.5 мкм шир., клетки веточек
6—7.6 мкм шир., у поперечных перегородок слегка перетянутые.
Конечные клетки заостренные, у средней оси и некоторых веточек
боковых ветвей переходят в длинный 1 — 5-клеточный тупой волосок.
Хлоропласт кольцевидный. От нижних клеток главного ствола и базаль-
ных клеток низко расположенных крупных побегов отходят длинные,
обильно разветвленные ризоидные выросты.
В проточных водах, на древесном субстрате. УССР (Южн. Буг).
Вид близок к D. plumosa, хорошо отличается малыми размерами
слоевища и особенно формой боковых ветвей.
5. Draparnaldia acuta (Ag.) Kiitz. (=D. glomerata var. acuta Ag.;
D. confervoides Bory; D. spinosa Kiitz.) — Драпарнальдйя острая
(рис. 105). — Боковые ветви главного ствола и длинных боковых побегов
обильно разветвленные, в общих очертаниях округлые, широкояйцевид-
ные или широколанцетовидные, одиночные, супротивные или, реже,
в мутовках, обычно с ярко выраженной средней осью, не выступающей
за пределы ветви. Веточки отходят от средней оси почти по всей ее
длине, оттопыренные, нередко слегка изогнутые. В некоторых боковых
ветвях четко выраженной средней оси нет, она довольно быстро переходит
в разветвления. Клетки главного ствола бочонковидные или цилиндри-
ческие, у поперечных перегородок слегка перетянутые, 50 — 90 (110) мкм
шир,, по длине равные ширине или до 3 (4) раз больше, клетки длинных
боковых побегов несколько меньших размеров, клетки ветвей цилиндри-
ческие, 6—10 мкм шир. Конечные клетки заостренные или переходят
в легко отламывающийся острый одноклеточный, слегка изогнутый
волосок. Хлоропласт кольцевидный.
201
Рис. 104.
Draparnaldia appressa. (По Мошковой).
В проточных и стоячих водах, на водных цветковых растениях и
древесном субстрате. ЭССР, УССР, ГрузССР.
6. Draparnaldia sparsifasciculata Prescott — Драпарнальдия редко-
пучковатая (рис. 106, 7—3). — Боковые ветви главного ствола и длинных
боковых побегов одиночные или супротивные, редкие, рыхлые, скудно
асимметрично разветвленные, с довольно четко выраженной средней
202
осью, не выступающей за пределы ветви. Клетки главного ствола
бочонковидные, у поперечных перегородок заметно перетянутые, 20—
25 (40) мкм шир., по длине в 2—4 раза больше. Конечные клетки
веточек часто переходят в волосок. Хлоропласт кольцевидный.
В стоячих и проточных водах, на водных цветковых растениях.
В СССР не обнаружена. (Сев. Америка).
7. Draparnaldia ravenelii Wolle — Драпарнальдия Равенела (рис. 106,
4). — Боковые ветви главного ствола и длинных боковых побегов обильно
разветвленные, плотные, в общих очертаниях полукруглые или широко-
яйцевидные, супротивные или по 3 в мутовках, отходящие под прямым
углом, с четко выраженной средней осью, не выступающей за пределы
ветви. Клетки главного ствола цилиндрические или слегка бочонковид-
ные, 150—170 (200) мкм шир., по длине меньше ширины, равны ей или
несколько больше, клетки длинных побегов 60—100 мкм шир., по
длине вдвое меньше или до 2 раз больше, клетки веточек цилиндри-
ческие, 6—8 мкм шир. Конечные клетки заостренные, реже переходят
в волосок. Хлоропласт кольцевидный.
В стоячих и проточных водах. В СССР не обнаружена. (Зап. Европа,
Сев. Америка).
8. Draparnaldia opposite (Lyngb.) Ag. (=D. plumosa var. opposita
Lyngb.) — Драпарнальдия супротивная (рис. 106, 5). — Кустики до 5 —
Рис. 105.
Draparnaldia acuta. (По Мошковой).
203
Рис. 106.
1—3 — Draparnaldta sparsifasciculata: 1 — фрагмент главного ствола, 2 — от-
дельная клетка с ризоидным выростом, 3 — волосок; 4 — D. ravenelii; 5 —
D. opposite. (1 — 3 — по Прескотту, 4 — по Нигаарду, 5 — по Кютцингу).
8 см выс. Длинные побеги главного ствола одиночные, супротивные или
в мутовках. Боковые ветви компактные, в общих очертаниях широко-
ланцетовидные, короткие. Средняя ось в полностью сформиро-
ванных боковых ветвях прослеживается преимущественно в базальной
их части, где она лишена веточек. Клетки главного ствола цилиндри-
ческие, у поперечных перегородок не перетянутые или слегка перетя-
нутые. Конечные клетки заостренные или переходят в волосок. Хлоро-
пласт кольцевидный.
В проточных водах, в обрастаниях разных субстратов. В СССР
не обнаружена. (Зап. Европа, Сев. Америка).
9. Draparnaldia glomerate (Vauch.) Ag. — Драпарнальдия клубочко-
вая (рис. 107, 108,7). — Кустики до 8—10 см выс. Главный ствол
обильно разветвленный, прикрепляющийся к субстрату с помощью
базальной клетки и ризоидных выростов. Боковые ветви обильно, реже
скудно разветвленные, в общих очертаниях широкояйцевидные, округ-
лые, полукруглые, реже овальные, восходящие. Четко выраженной
средней оси в полностью сформированных ветвях нет, она довольно
быстро теряется в разветвлениях. Клетки главного ствола бочонковидные,
204
у поперечных перегородок заметно перетянутые, реже цилиндрические,
неперетяпутые, (20) 32—99 (125) мкм шир., по длине меньше ширины
или до 3 (4) раз больше, клетки длинных побегов обычно цилиндриче-
ские, 36 —40 мкм шир., по длине равные ширине или до 2 (3) раз
больше, клетки веточек цилиндрические, 6—9 мкм шир., по длине
в 1.2—2.5 раза больше. Конечные клетки заостренные или переходят
в длинный многоклеточный тупой волосок до 760—800 мкм дл. Хлоро-
пласт кольцевидный. Макрозооспоры 8—10 мкм шир., 12 — 18 мкм дл.
Апланоспоры с толстой, гладкой или бородавчатой оболочкой и оранжево-
красным содержимым.
В стоячих и особенно проточных холодных, хорошо аэрированных
водах, на разных субстратах. Массовая вегетация большей частью
приурочена к ранней весне, однако в водах с низкой температурой
обильное разрастание наблюдается нередко и летом.
Var. glomerate (=var. genuina Kirchn.) (рис. 107). — Боковые ветви
обильно разветвленные, рыхлые, в общих очертаниях широкояйцевидные
или округлые, расположенные близко друг к другу. Клетки главного
ствола бочонковидные, 32—125 мкм шир., по длине вдвое меньше или до
3 раз больше. Конечные клетки веточек часто переходят в длинный
многоклеточный тупой волосок.
В реках, ручьях, озерах. Повсеместно.
Var. remote Rabenh. |—£>. distans Kiitz.; D. glomerata var. distans
(Kutz.) Hansg.] (рис. 108, 7). — Боковые ветви обильно разветвленные,
плотные, в общих очертаниях полукруглые, далеко отстоящие друг от
друга. Конечные клетки веточек преимущественно слегка заостренные.
Рис. 107.
Draparnaldta glomerata var. glomerata: 1 — средняя часть главного ствола,
2 — нижняя часть с риэоидными выступами. (По Мошковой).
205
Рис. 108.
1 — Draparnaldia glomerata var. remota; 2, 3 — D. judayi. (1 — по Мошковой,
2. 3 — по Прескотту).
В стоячих водах, а также в реках, в местах с замедленным тече-
нием. РСФСР (окр. г. Белгорода), УССР (Днепр).
Var. groenlandica (Bachm.) Printz (==D. groenlandica Bachm.). —
Боковые ветви скудно разветвленные, рыхлые, в общих очертаниях
округлые или овальные. Клетки ствола цилиндрические, слегка вздутые
в средней части, 20—70 мкм шир., по длине до 4 раз больше.
В стоячих и проточных водах. В СССР не обнаружена. (Гренландия).
10. Draparnaldia judayi Prescott — Драпарнальдия Юдая (рис. 108,
2, 3). — Боковые ветви главного ствола и длинных боковых побегов
скудно разветвленные, супротивные или в мутовках, отходящие под
прямым углом. Клетки главного ствола и длинных побегов цилиндри-
ческие или бочонковидные, перетянутые у поперечных перегородок,
12 — 15.2 мкм шир. Конечные клетки заостренные или переходят в одно-
клеточный острый волосок, нередко слегка луковицеобразно вздутый
у основания. Хлоропласт кольцевидный.
В торфяных болотах, на водных цветковых растениях. В СССР не
обнаружена. (США).
206
Род DRAPARNALDIOPSIS Smith et Klyver —
ДРАПАРНАЛЬДИОПСИС
Слоевище кустиковидное. Нити ветвящиеся, прямостоячие, с обо-
собленной центральной осью — главным стволом, прикрепляющимся
к субстрату ризоидами, и обильно разветвленными боковыми ветвями.
Главный ствол состоит из правильно чередующихся длинных (межузло-
вых) и коротких (узловых) цилиндрических клеток. Боковые ветви
супротивные или до 5 в мутовках, реже одиночные, большей частью
обильно разветвленные; они образуются преимущественно на коротких
узловых клетках главного ствола и отходят под прямым углом. Базаль-
ная клетка ветвей клиновидная, дает начало 2—5 клеткам, от которых
в свою очередь отходят следующие ветви. Клетки веточек цилиндри-
ческие или бочонковидные, у поперечных перегородок перетянутые.
Конечные клетки конусовидно суженные, закругленные, или переходят
в бесцветный длинный многоклеточный тупой или в коротко заострен-
ный одноклеточный волосок. Хлоропласт пластинчатый, цельный или
продырявленный, у некоторых видов настолько сильно, что приобретает
вид тонкой широкоячеистой сеточки, затягивающей частично или
полностью продольные стенки клеток, по краям цельный или изрезанный.
Пиреноидов несколько, в клетках боковых ветвей обычно 1 — 2. Бесполое
размножение зооспорами, возникающими в обычных клетках боковых
ветвей.
В стоячих водах, на разных субстратах.
I. Главный ствол состоит из правильно чередующихся длинных и ко-
ротких клеток.
1. Боковые ветви супротивные.
А. Конечные клетки веточек обычно переходят в длинный много-
клеточный волосок............................D. alpinis 1.
Б. Конечные клетки веточек конусовидно суженные.........
...........................................D. indica 2.
2. Боковые ветви в мутовках.
А. Конечные клетки веточек обычно непосредственно переходят
в длинный волосок.........................D. intermedia 3.
Б. Конечные клетки веточек обычно непосредственно переходят
в короткий острый волосок.................D. salishensis 4.
II. Главный ствол состоит из беспорядочно расположенных длинных и
коротких клеток....................................D. simplex 5.
1. Draparnaldiopsis alpinis Smith et Klyver — Драпарнальдиопсис
высокогорный (рис. 109). — Главный ствол состоит из правильно чере-
дующихся длинных и коротких цилиндрических клеток. Боковые ветви
образуются только на коротких клетках, в общих очертаниях округло-
треугольные, супротивные. Длинные клетки ствола 18—26 мкм шир.,
18—33 мкм дл., короткие 18—27 мкм шир., 6—15 мкм дл., клетки
веточек слабо бочонковидные. Конечные клетки переходят в длинный
многоклеточный тупой волосок. Слоевище с волосками 700—1800 мкм
шир., без волосков 220—350 мкм шир. Ризоиды развиты слабо.
В стоячих водах. В СССР не обнаружен. (США: Калифорния).
2. Draparnaldiopsis indica Bharadwaja — Драпарнальдиопсис индий-
ский (рис. 110). — Главный ствол состоит из правильно чередующихся
длинных и коротких цилиндрических клеток. Боковые ветви супротив-
207
Рис. 109.
Draparnaldiopsls alpinis: 1 — фрагмент слоевища, 2,3 — стадии развития боко-
вой ветви, 4 — нижняя часть ствола с ризоидами. (По Смиту и Клайверу).
Рис. 110.
Draparnaldiopsls indtca: 1 — фрагмент главного ствола, 2 — боковая ветвь.
(По Барадвайя).
208
Рис. 111.
Draparnaldiopsls intermedia: 1 — фрагмент слоевища, 2 — узловая клетка с ве-
точками. (По Обуховой).
Рис. 112.
1 — Draparnaldiopsls salishensis: 2 — D. simplex. (1 — по Прескотту, 2 — по
Жао).
14 Зеленые водоросли. Улотриксовые
209
ные, образуются только на коротких клетках, в общих очертаниях
полукруглые, состоящие из базальной клетки и отхбдящих от нее
2—6 коротких, скудно разветвленных веточек. Длинные клетки ствола
20—60 мкм шир., 28—63 мкм дл., короткие 16—60 мкм шир., 8—33 мкм
дл.( клетки веточек 4.2—8.4 мкм шир. Конечные клетки конусовидно
суженные, волоски отсутствуют. Ризоиды развиты обильно, густо опуты-
вая нижнюю часть ствола.
В стоячих водоемах, на водных растениях. В СССР не обнаружен.
(Индия).
3. Draparnaldiopsis intermedia Obnch. — Драпарнальдиопсис проме-
жуточный (рис. 111). — Главный ствол состоит из правильно чередую-
щихся длинных и коротких цилиндрических клеток. Боковые ветви
образуются только на коротких клетках, преимущественно в мутовках до
5 в каждой, реже супротивные или одиночные, 33 — 35 мкм дл., их
базальная клетка клиновидная, от нее отходят 2—3 повторно разветвлен-
ные веточки. Длинные клетки ствола 26—32 мкм шир., 14—22.5 мкм дл.,
короткие 22 — 32 мкм шир., 6—8 мкм дл., клетки веточек 4.2—8.5 мкм шир.
Конечные клетки непосредственно переходят в длинный волосок,
В воде рисовых полей, среди других водорослей. КазССР.
4. Draparnaldiopsis salishensis Prescott — Драпарнальдиопсис сали-
схенский (рис. 112, 1). — Главный ствол состоит из правильно чере-
дующихся, почти изодиаметрических и более коротких цилиндрических
клеток. Боковые ветви образуются только на дисковидных клетках,
в мутовках по 3—4, скудно разветвленные. Длинные клетки ствола
20—25 мкм шир., 25 — 35 мкм дл., клетки веточек цилиндрические или
бочонковидные, у поперечных перегородок заметно перетянутые,
5—8 (10) мкм шир. Конечные клетки непосредственно переходят в
короткий острый волосок, луковицеобразно расширенный у основания,
реже заостренные.
В озерах. В СССР не обнаружен. (Сев. Америка).
5. Draparnaldiopsis simplex Jao — Драпарнальдиопсис простой
(рис. 112, 2). — Главный ствол состоит из длинных и коротких клеток,
расположенных без определенной последовательности. Боковые ветви
образуются не только на коротких, но и на длинных клетках ствола,
в общих очертаниях полукруглые, их базальная клетка несет 3—5 клеток,
в свою очередь ветвящихся далее. Клетки главного ствола 32—37 мкм
шир., 10—75 мкм дл., клетки веточек 5.4—7.2 мкм шир., 12—27 мкм дл.
Конечные клетки обычно конусовидно суженные, изредка продолжаются
в длинный многоклеточный волосок. Спорангии 8—10.8 мкм шир.
В прудах. В СССР не обнаружен. (Китай).
Род IWANOFFIA Pasch. - ИВАНОВИЯ
Слоевище нитевидное. Нити ползучие, ветвящиеся, прикрепляю-
щиеся к субстрату ризоидными выростами и отходящие от них, прямо-
стоячие, простые, изредка разветвленные, постепенно суживающиеся
к вершине. Клетки ползучих нитей обычно бочонковидные, прямостоя-
чих — цилиндрические, у поперечных перегородок не перетянутые или
слегка перетянутые. Конечные клетки конусовидные, тупые; волоски
отсутствуют. Хлоропласт пристенный, пластинчатый, с 1 или несколь-
кими пиреноидами. Размножение преимущественно двужгутиковыми
210
Рис. ИЗ.
1—6 — Iwanoffia terrestris: 1 — слоевище, возникшее из макрозооспоры, 2, 3 —
макрозооспоры, 4, 5 — микрозооспоры, 6 — образование микрозооспор в паль-
меллевидном слоевище; 7—12 — Caespitella pascheri: 7 — общий вид слоевища,
развившегося в культуре, 8, 9 — фрагмент слоевища, 10 — фрагменты нитей,
находящихся в стадии покоя, 11, 12 — зооспоры. {1—6 — по Иванову, 7—12 —
по Вишеру).
14*
211
зооспорами двух сортов. Макрозооспоры образуются по 1 в клетке,
имеют хлоропласт с 1 пиреноидом и стигму, расположенную в срединной
части клетки; при прорастании они образуют 1—3 выроста, развиваю-
щиеся в дальнейшем в ползучие и прямостоячие нити. Микрозооспоры
значительно уже макрозооспор, при их образовании материнская клетка
сначала делится на 2 клетки, в каждой из которых возникает по
1 двужгутиковой зооспоре с палочковидной стигмой в переднем конце;
при прорастании они образуют короткие нити, клетки которых в даль-
нейшем превращаются в акинеты. Известно пальмеллевидное состояние.
1. Iwanoffia terrestris (Iwanoff) Pasch. (=Stigeoclonium terrestre
Iwanoff) — Ивановия наземная (рис. ИЗ, 1—6). — Клетки 7 мкм шир.
Макрозооспоры 6.5—8 мкм шир., 8—И мкм дл., микрозооспоры 3—4 мкм
шир., 6—7 мкм дл.
На влажной земле. РСФСР (Ленинградская, Калининская обл.),
Ср. Азия.
Род CAESPITELLA Vischer - ЦЕСПИТЕЛЛА
Слоевище нитевидное. Нити ползучие, ветвящиеся, прикрепляю-
щиеся к субстрату ризоидными выростами, и отходящие от них прямо-
стоячие, разветвленные, к вершине постепенно суживающиеся. Клетки
ползучих нитей бочонковидные, у поперечных перегородок сильно пере-
тянутые, прямостоячих — цилиндрические, у поперечных перегородок
не перетянутые или слабо перетянутые. Конечные клетки конические,
притупленные или заостренные, яли вытягиваются в длинный острый
многоклеточный волосок. Хлоропласт пристенный, пластинчатый,
с 1 — 3 пиреноидами. Размножение преимущественно четырехжгутико-
выми грушевидными зооспорами со стигмой, образующимися по 2
в клетке. Известны также апланоспоры и гаметы.
1. Caespitella pascheri Vischer — Цеспителла Пашера (рис. ИЗ,
7—12). — Слоевища слагаются в дерновинки до 2 мм выс. Клетки
прямостоячих нитей 6—10 мкм шир., 12 — 20 мкм дл. Жгутики зооспор
в 1.5—2 раза длиннее их тела.
В стоячих водах. В СССР не обнаружена. (Швейцария).
Род PSEUDOCHAETE W. et G. S. West - ПСЕВДОХЕТЕ
Слоевище нитевидное. Нити ползучие, ветвящиеся, образованные
крупными клетками, и отходящие от них прямостоячие, волосовидные,
заостряющиеся, состоящие из 3—8 клеток. Клетки ползучих нитей тонко-
стенные, цилиндрические или бочонковидные, прямостоячих — цилинд-
рические, их длина возрастает по мере удаления от ползучих нитей.
Конечные клетки на верхушке заостренные или туповатые. Хлоропласт
поясковидный, продырявленный, нередко слегка изрезанный по краям,
с пиреноидом; в клетках ползучих нитей он прилегает к стороне,
обращенной к субстрату, в нижних клетках волосовидных нитей более
или менее полностью выстилает продольные стенки, в верхушечных
редуцируется и почти совершенно исчезает. У старых слоевищ конечные
клетки ветвей могут вздуваться в удлиненно-шаровидные спорангии,
содержимое которых последовательно распадается на 8—16 подвижных
спор.
В стоячих и слабо проточных водах, на разных субстратах.
212
Виды рода Pseudochaete легко спутать с видами рода Stigeoclonium,
у которых слабо развиты прямостоячие нити.
I. Длина клеток волосовидных нитей в 7—10 раз больше их ширины
...................................................Р. crassiseta 1.
II. Длина клеток волосовидных нитей в 12—18 раз больше их ширины
...................................................Р. gracilis 2.
1. Pseudochaete crassiseta (W. et G. S. West) Printz (=Herposteiron
crassisetum W. et G. S. West) — Псевдохете толстощетииковая (рис. 114,
1). — Клетки ползучих нитей 6.5—7.5 мкм шир., по длине в 1.5—2 раза
больше, волосовидных — 2.3—5 мкм шир., по длине в 7 — 10 раз больше,
В различных водоемах, в обрастаниях. В СССР не обнаружена.
(Цейлон, Центр. Азия).
2. Pseudochaete gracilis W. et G. S. West — Псевдохете изящная
(рис. 114, 2). — Клетки ползучих нитей 5.7—7.7 мкм шир., по длине
равные ширине или в 1.5 раза больше, волосовидных — 1.5—1.8 мкм
шир., по длине в 12 — 18 раз больше.
В водоемах с замедленным течением, на водных цветковых расте-
ниях, раковинах моллюсков. УССР, местами.
213
Род ENDOCLONIUM Szym. - ЭНДОКЛОНИУМ
Слоевище эпифитное или эндофитное, в форме пластинки, образо-
ванной нитями, или нитевидное. При эпифитном способе существования
нити ползучие, ветвящиеся, срастающиеся в паренхимные плоские,
распростертые по субстрату пластинки с отходящими от них прямостоя-
чими или только слегка приподнимающимися от краев разветвленными
нитями, которые довольно часто заканчиваются бесцветным волоском.
При эндофитном способе существования слоевище состоит из разветвлен-
ных нитей или скопления клеток, распространяющихся между клетками
в тканях водных цветковых растений (преимущественно ряски), иногда
выходящих наружу через устьица листьев и заканчивающихся волоском.
Клетки слоевища цилиндрические, слегка вздутые, или многоугольные
от взаимного сдавливания. Хлоропласт 1, с пиреноидом. Бесполое
размножение двужгутиковыми микрозооспорами и четырехжгутиковыми
макрозооспорами. Половое размножение двужгутиковыми гаметами,
которые, однако, могут прорастать и партеногенетически.
В стоячих и проточных водах.
I. Слоевище только эндофитное . . . . . . Е. ehroolepiforine 1.
II. Слоевище эпифитное, только частично эндофитное.
1. Эпифитное слоевище имеет вид паренхимного диска с короткими
простыми или скудно разветвленными прямостоячими или слегка
приподнимающимися от краев нитями . . . Е. polymorphum 2.
2. Эпифитное слоевище имеет вид дерновинок или подушечек,
инкрустированных известью.....................Е. pygmaeuni 3.
1. Endoelonium ehroolepiforine Szym. [—Stigeoclonium. chroolepi-
forme (Szym.) Heering] — Эндоклониум хроолепиформный. — Слоевище
эндофитное, нитевидное. Нити ветвящиеся. Клетки цилиндрические или
слегка вздутые. Макрозооспоры после выхода из материнской клетки
сразу прорастают в новые нити, разрастающиеся между клетками
в теле ряски. Микрозооспоры около 3 мкм шир., 5 мкм дл., при про-
растании образующие беспорядочные скопления клеток; из крупных
делящихся клеток возникают короткие ветвящиеся нити, заканчиваю-
щиеся одноклеточными волосками, торчащими наружу из тела растения-
хозяина.
Преимущественно в стоячих водах, эндофитно в теле ряски. РСФСР
(Калининская обл.).
Малоизученный вид, описанный поверхностно и без рисунка. Инте-
ресен своеобразием строения и образом жизни.
2. Endoelonium polymorphum Franke [=Stigeoclonium polymorphum
(Franke) Heering] — Эндоклониум полиморфный (рис. 115, 1—4)1 —
Слоевище эпифитно-эндофитное. Эндофитная часть состоит из беспоря-
дочного скопления клеток, поселяющихся в теле водного цветкового
растения — внутри его устьиц и под ними. Клетки в скоплениях шаро-
видные или многоугольные, около 27 мкм шир., с утолщенными оболоч-
ками. В этих клетках образуются двужгутиковые микрозооспоры 3.5 мкм
шир., около 7.5 мкм дл., со стигмой; они выходят наружу через устьица
растения-хозяина и прорастают на его поверхности. Эпифитная часть
слоевища имеет вид паренхимных пластинок, от поверхности и по краям
которых отходят вверх короткие, иногда скудно разветвленные нити.
Клетки пластинок преимущественно 4 мкм шир., 5 мкм дл., клетки
нитей цилиндрические, длина их равна ширине или несколько больше.
214
Рис. 115.
1—4 — Endoelonium polymorphum: 1 — эпифитное слоевище, выросшее в куль-
туре, 2 — эпифитное слоевище на ряске (вид сверху), 3 — образование зооспор,
4 — образование и копуляция гамет; 5 — Е. pygmaeum. (1—4 — по Франке,
5 — по Гансгиргу).
Конечные клетки тупоконические или вытянутые в многоклеточный
волосок. Макрозооспоры 10.8 мкм шир., около 13.5 мкм дл., со стигмой;
они образуются по 1 в клетках эпифитного слоевища и прорастают
в новые эпифитные слоевища без периода покоя. Там же образуются
и гаметы, по 2—4 и более в клетке, сливающиеся друг с другом или
развивающиеся партеногенетически. Зиготы превращаются в покоящиеся
споры, прорастающие в двужгутиковые микрозооспоры. Последние
прорастают в нити, которые проникают в устьица растения-хозяина и
образуют там эндофитную часть водоросли. Вследствие ограниченного
пространства клетки слоевища плотно соприкасаются друг с другом и
постепенно становятся многоугольными.
В стоячих и проточных водах, на ряске и других водных цветковых
растениях. ЛатвССР, РСФСР (Калининская обл.), БССР, УССР,
ТуркмССР.
Многие сообщения о нахождении этого вида кажутся сомнитель-
ными, поскольку не указываются ни условия, ни субстрат, ни форма,
в какой было встречено растение.
3. Endoelonium pygmaeuni Hansg. (=Stigeoclonium pygmaeum
Hansg.) — Эндоклониум карликовый (рис. 115, 5). — Слоевище эпи-
фитно-эндофитное. Эндофитная часть в форме нитей, развивающихся
в межклетниках паренхимы водных растений. Эпифитная часть в виде
тонкой светло-зеленой дерновинки или подушечки, инкрустированной
известью, с отходящими вверх нитями. Подошва состоит из многочислен-
ных, разветвленных в плоскости нитей, плотно прижатых к субстрату
и срастающихся между собой боковыми сторонами в диск обычно
неправильной формы. Прямостоячие нити до 200 мкм дл., из клеток
4—5 мкм шир., часто разветвленные уже у основания. Ветви короткие,
нередко слегка изогнутые, оканчивающиеся заостренной клеткой или
длинным многоклеточным волоском. Клетки эндофитной части слоевища
нередко превращаются в шаровидные или яйцевидные акинеты 6—15 мкм
шир., окрашенные в красный цвет, с довольно тонкой бесцветной
оболочкой..
215
В стоячих и проточных водах, на водных цветковых растениях и
других субстратах. БССР, УССР.
Малоизученный вид, детали строения которого во многом остаются
еще неясными.
Род THAMNIOLUM Woronich. - ТАМНИОЛУМ
Слоевище кустиковидное. Нити ветвящиеся, прямостоячие, с обособ-
ленной центральной осью — главным стволом, прикрепляющимся
к субстрату ризоидами, и более тонкими, чем главный ствол, восходя-
щими, очередными или часто односторонне размещенными длинными
боковыми побегами, довольно скудно разветвленными. Веточки очеред-
ные, вытянутые, скудно повторно разветвленные. Клетки главного
ствола, боковых побегов и веточек цилиндрические, у поперечных пере-
городок не перетянутые или слегка перетянутые, в главном стволе
нередко вздутые на концах и тогда отчетливо перетянутые, с тонкими
бесцветными оболочками. Конечные клетки заостренные. Ядро 1, округ-
лое или овальное, около 3 мкм в диам., с заметным ядрышком. Хлоро-
пласты многочисленные, пристенные, нежные, в виде пластинок,
довольно рыхло расположенных, без пиреноидов. Продукт ассимиля-
ции — масло (?). Размножение неизвестно.
1. Thamniolum elegans Woronich. — Тамниолум изящный (рис. 116,
1—5).— Дерновинки нежные, бледно-зеленые, до 1 см выс. Клетки
главного ствола 20—22 мкм шир., во вздутых местах 24—27.4 мкм шир.,
по длине в 5—8 (12) раз больше, клетки веточек 3—3.4 мкм шир., по
длине в 10—23 раза больше. Ризоиды отходят от базальных и вышераспо-
ложенных клеток.
В болотах, на древесном субстрате. Окр. Ленинграда.
Малоизученный своеобразный вид. Форма хлоропластов остается
неясной.
Род SAPROCHAETE Coker et Shanor - САПРОХЕТЕ
Слоевище кустиковидное. Нити бесцветные, ветвящиеся, прямостоя-
чие, с обособленной центральной осью — главным стволом, прикрепляю-
щимся к субстрату ризоидами, и многочисленными разветвленными
длинными и короткими боковыми побегами. Боковые веточки длинные,
повторно разветвленные, восходящие, очередные, супротивные или, реже,
в мутовках. Клетки главного ствола и боковых побегов цилиндрические
или слегка булавовидные, у поперечных перегородок не перетянутые
или отчетливо перетянутые, клетки веточек удлиненно-цилиндрические,
чаще булавовидные. Конечные клетки слегка заостренные или закруглен-
ные. Хлоропластов нет. В оболочках клеток не обнаружено ни целлюлозы,
ни хитина. Размножение фрагментацией слоевища. Бесполое и половое
размножение неизвестно.
1. Saprochaete saecharophila Coker et Shanor — Сапрохете сахаролю-
бивая (рис. 116, 6—9). — Слоевище в виде белых дерновинок до 5 см выс.
Клетки главных нитей около 33 мкм шир., до 175 мкм дл. Ядро крупное,
с ядрышком.
В проточных водах, содержащих стоки сахарных заводов, преиму-
щественно в зимние и весенние месяцы. В СССР не обнаружен. (США:
Сев. Каролина).
Малоизученный вид. Своеобразием облика резко отличается от всех
остальных представителей подсемейства.
216
Рис. 116.
1—5 — Thamniolum elegans: 1 — общий вид слоевища, 2 — клетки главного
ствола, 3 — боковая ветвь, 4, 5 — базальные части главного ствола с ризоидами;
6—9 — Saphochaete saccharophila: 6 — общий вид слоевища, 7 — клетки глав-
ного ствола и крупных побегов, 8 — боковая ветвь, 9 — базальная часть глав-
ного ствола с ризоидами. (1—5 — по Воронихину, 6—9 — по Кукеру и Шанору).
217
Подсемейство ULVELLOIDEAE Вопгг. —
УЛЬВЕЛЛОВЫЕ
Нити ветвящиеся, ползучие, разобщенные или тесно смыкающиеся
в псевдопаренхимные плоские однослойные или частично многослойные
микроскопические пластинки, за исключением рода Jaoa, характеризую-
щегося макроскопическим многослойным, более или менее полушаровид-
ным слоевищем. Волоски и щетинки отсутствуют.
Представители подсемейства в значительной мере являются организ-
мами солоноватых, соленых и морских вод. Включает формы, поселяю-
щиеся внутри растений и животных.
I. Слоевище микроскопическое, состоящее из нитей, разветвленных
в плоскости, или пластинчатое, одно- или многослойное, но без
полости внутри.
1. Нити разобщенные, редко анастомозирующие друг с другом или
срастающиеся частично или полностью в однослойную пластинку
с обособленными по краям простыми или разветвленными веточ-
ками, распростертыми по субстрату.
А. Эпифиты и эндофиты (только 3 вида на животных); хлоро-
пласт с 1 или несколькими пиреноидами, реже без пиренои-
дов .................................род Entocladia (с. 218).
Б. Эпизооиды и эндозооиды; хлоропласт постоянно с 1 пиренои-
дом .................................род Epieladia (с. 226).
2. Нити полностью сросшиеся в однослойную пластинку или правиль-
ный диск.
А. Пластинки неопределенных очертаний, с обособленными по
краям короткими веточками, распростертыми по субстрату
.....................................род Protoderma (с. 227).
Б. Пластинки дисковидные или линзовидные, с более или менее
ровными краями.
а. Пластинки в периферической части однослойные, в цен-
тральной многослойные; центральные клетки не выше
краевых.
а. Краевые клетки вытянутые . . . род Ulvella (с. 230).
р. Краевые клетки не вытянутые.......................
...........................род Pseudulvella (с. 231).
б. Пластинки однослойные; центральные клетки выше крае-
вых ..........................род Pringsheimiella (с. 231).
II. Слоевище макроскопическое, бляшковидное, слегка выпуклое или
почти полушаровидное, многослойное, с полостью внутри...........
............................................род Jaoa (с. 233).
Род ENTOCLADIA Reinke - ЭНТОКЛАДИЯ
Слоевище микроскопическое, нитевидное, эпифитное или эндофит-
ное, реже поселяющееся на неживом зоосубстрате. Нити беспорядочно
ветвящиеся или, реже, простые, ползучие. При эпифитном способе
существования нити разветвленные, полностью распростертые по суб-
страту, обособленные или соприкасающиеся друг с другом и тогда
образующие одно- или многослойные пластинки с отходящими во все
стороны ветвями. При эндофитном способе существования нити простые
или разветвленные, развивающиеся в толстой слоистой оболочке клетки
218
растения-хозяина, непосредственно под оболочкой или внутри на
некотором расстоянии от нее. Ядро 1. Хлоропласт 1, пристенный,
с 1 или несколькими пиреноидами, реже без них. Бесполое размножение
зооспорами и акинетами. Зооспоры четырехжгутиковые, яйцевидные,
со стигмой; они образуются по 1—8 в клетках вблизи центра слоевища.
Половое размножение двужгутиковыми яйцевидными изогаметами, воз-
никающими по 4—16 (32) в гаметангии. Любая клетка слоевища, за
исключением конечных, несколько увеличившись в размерах, может
превращаться в зооспорангий или гаметангий.
Виды рода Entocladia обитают в солоноватых, соленых, морских,
реже пресных водах.
I. Хлоропласт с 1 (2) пиреноидами.
1. Слоевище нитчатое, нити простые или разветвленные.
А. Нити не анастомозируют друг с другом; клетки цилиндриче-
ские, эллипсоидные или шаровидные.
а. Клетки цилиндрические, прямые, слегка искривленные
или неправильных очертаний, у поперечных перегородок
не перетянутые или отчетливо перетянутые.
а. Клетки 2—4 мкм шир., по длине-в 2 — 3 раза больше;
на нитях кладофоровых водорослей......................
....................................Е. gracilis 1.
р. Клетки 3—8 мкм шир., по длине до 6 раз больше; на
красных водорослях.......................Е. viridis 2.
б. Клетки шаровидные или широкоэллипсоидные..............
..................................Е. pithophorae 3.
в. Клетки удлиненно- или яйцевидно-эллипсоидные, реже
многоугольные.
а. Клетки тонкостенные, 2.4—4.3 мкм шир.; в слизи сине-
зеленых водорослей.................Е, cyclostoma 4.
р. Клетки толстостенные, 6—17 мкм шир.; на кладофоро-
вых водорослях.....................Е. cladophorae 5.
Б. Нити анастомозируют друг с другом; клетки неправильных
очертаний..................‘.................Е. polymorpha 6.
2. Слоевище пластинчатое полностью или только в центральной части.
А. Клетки цилиндрические или эллипсоидные, у поперечных
перегородок перетянутые.....................Е. perforans 7.
Б. Клетки цилиндрические или неправильно многоугольные,
у поперечных перегородок не перетянутые . . Е. basicladiae 8.
II. Хлоропласт со многими пиреноидами.
1. Слоевище нитчатое, нити простые или разветвленные.
А. Клетки цилиндрические,.эллипсоидные или шаровидные . . .
.............................................Е. inimane 9.
Б. Клетки удлиненно-цилиндрические.
а. Клетки 5—8 мкм шир., по длине в 4—8 раз больше . . . .
....................................Е. tenuis 10.
б. Клетки 7—10 мкм шир., по длине в 3—5 раз больше . . .
........................................Е. testarum И.
2. Слоевище пластинчатое, одно- или многослойное................
................................................Е. reineckei 12.
III. Хлоропласт без пиреноидов.................Е. spongophila 13.
1. Entocladia gracilis Hansg. [—Endoderma gracilis (Hansg.)
De-Toni] — Эитокладия тонкая (рис. 117, 1—3). — Нити обильно ветвя-
219
Рис. 117.
1—3 — Entocladia gracilis-. 1,2 — взрослые слоевища отдельно, 3 — слоевища
в разных стадиях развития на поверхности и внутри клетки кладофоры;
4 — Е. viridis, слоевище на клетке багрянки; 5 — Е. pithophorae, слоевище
на клетке питофоры. (1—3 — по Гансгиргу, 4 — по Рейнке, 5 — по Весту).
щиеся, ползучие, поселяющиеся на поверхности оболочек и внутри клеток
растения-хозяина. Клетки цилиндрические, 2—3.5 (4) мкм шир., по
длине в 2—3 раза больше, с тонкими оболочками, обычно бесцветными,
у некоторых эпифитных форм окрашенными окислами железа в ржаво-
желтый цвет. Конечные клетки суженные, широко закругленные.
Хлоропласт с 1 пиреноидом. Зооспоры 3 мкм шир., 5 — 6 мкм дл., по
1 в зооспорангии. Гаметангии 6—10 мкм в диам. Гаметы 1—2.5 мкм шир.,
4 мкм дл., по 4—32 в гаметангии. Зигота окружена толстой оболочкой,
прорастает после длительного периода покоя.
В пресных водоемах, эпифитно или эндофитно на нитях кладофоро-
вых водорослей. В СССР не обнаружена. (Зап. Европа).
2. Entocladia viridis Reinke [=Endoderma viridis (Reinke) La-
gerh.] — Энтокладия зеленая (рис. 117, 4). — Нити обильно и беспоря-
дочно ветвящиеся, поселяющиеся в оболочках растения-хозяина. Клетки
цилиндрические, часто неправильных очертании, прямые или искривлен-
ные, 3 — 6 (8) мкм шир., по длине равные ширине или до 6 раз больше.
Конечные клетки тупые или слегка заостренные. Хлоропласт с 1 пире-
ноидом. Зооспоры 5 мкм в диам.
В морских и соленых водах, в оболочках наружных клеток красных
водорослей. УССР (лиманы), Черноморское побережье Крыма и Кавказа,
Каспийское, Японское и Охотское моря, Сев. Ледовитый океан.
220
3. Entocladia pithophorae (G. S. West) Starmach (=Endoderma
pithophorae G. S. West) — Энтокладия питофоровая (рис. 117, 5). —
Нити короткие, скудно и беспорядочно ветвящиеся, поселяющиеся на
поверхности оболочек растения-хозяина. Веточки слегка спутанные,
к концам суженные. Клетки шаровидные или широкоэллипсоидные,
чдсто удлиненные, 25—38 мкм шир., с толстыми, до 3—3.8 мкм, оболоч-
ками. Конечные клетки 10.5—16 мкм шир. Хлоропласт с 1 пиреноидом.
Слоевище в целом нередко достигает 210 мкм в диам.
В пресных водоемах на нитях Pithophora cleveana. В СССР не обна-
ружена. (Сев. Америка).
4. Entocladia cyclostoma Waern — Энтокладия круглоустьичная
(рис. 118, 1—3). — Нити тонкие, простые или слабо ветвящиеся, внед-
ряющиеся в слизь растения-хозяина конечной клеткой, на больших
участках бесцветные, из 1 или нескольких клеток. Клетки удлиненно-
эллипсоидные, 2.4—4.3 мкм шир., тонкостенные, часто разобщенные.
Конечные клетки, погруженные в субстрат, вздутые, нередко слегка
Рис. 118.
1 — 3 — Entocladia cyclostom-a: 1,2 — взрослые нити в слизистом влагалище
ривулярии, 3 — молодой проросток; 4 — Е. cladophorae, нити в оболочке клетки
кладофоры (продольный разрез через часть клетки). (7—3 — по Верну, 4—
по Хорнби).
221
Рис. 119.
1 — Entocladia polymorpha, слоевище на клетке кладофоры; 2 — Е. perforans,
слоевище на листе зостеры. (7 — по Весту, 2 — по Губеру).
разветвленные. Хлоропласт со слабо изрезанными краями и 1 пиренои-
дом.
В морских и соленых водах, в слизистых влагалищах синезеленых
водорослей. В СССР не обнаружена. (Зап. Европа; побережье Балтий-
ского моря).
5. Entoeladia eladophorae (Hornb.) Starmach (=Endoderma clado-
phorae Hornb.) — Энтокладия кладофоровая (рис. 118, 4). — Нити
скудно и беспорядочно ветвящиеся, поселяющиеся в оболочках растения-
хозяина. Веточки к концам слегка суженные, чаще всего спутанные.
Клетки яйцевидно-эллипсоидные, позднее вследствие взаимного сдавли-
вания многоугольные, толстостенные, 6 — 17 мкм шир. Конечные клетки
3.5—4 мкм шир. Хлоропласт с 1 (2) пиреноидами. Слоевище в целом
нередко достигает 480 мкм в диам.
В соленых и морских, а также пресных водах, на нитях кладофоры
и ризоклониума. В разных районах Арктики.
6. Entocladia polymorpha (G. S. West) Starmach 1=Endo derma
polymorpha G. S. West) — Энтокладия многообразная (рис. 119, 1). —
Нити скудно и беспорядочно ветвящиеся, распростертые на поверхности
оболочек растения-хозяина, часто анастомозирующие друг с другом.
Клетки неправильных очертаний, обычно вытянутые, дорсивентрально
сплющенные, 6.1—9.5 мкм шир., 15—39 мкм дл., 3—7.5 мкм выс.,
с тонкими твердыми оболочками. Хлоропласт с 1, реже несколькими
пиреноидами. Слоевище в целом достигает 250 мкм в диам.
В пресных водоемах, на нитчатых, преимущественно кладофоровых
водорослях. В СССР не обнаружена. (Сев. Америка).
7. Entocladia perforans (Huber) Levr. (—Endoderma perforans
Hnber) — Энтокладия пронизывающая (рис. 119, 2). — Нити беспоря-
дочно ветвящиеся, радиально расходящиеся от морфологического центра
222
слоевища, проникающие в эпидермальные клетки и межклеточные поло-
сти растения-хозяина. Клетки цилиндрические, у поперечных перегоро-
док перетянутые, часто сильно вздутые и тогда шаровидные или эллип-
соидные, прямые или слегка изогнутые, 3—5 (14) мкм шир. Хлоропласт
с изрезанными краями и 1 пиреноидом. Зооспорангии с бесцветными
выростами, пронизывающими наружные оболочки клеток растений-
хозяев. Зооспоры четырехжгутиковые, со стигмой и хлоропластом с 1 пи-
реноидом, по 8 в зооспорангии.
В морях, на старых листьях Zostera. Черноморское побережье
Крыма.
8. Entocladia basicladiae (Jao) Starmach (=Endoderma basicladiae
Jao) — Энтокладия базикладиевая (рис. 120, 7). — Нити беспорядочно
ветвящиеся, на поверхности или, реже, внутри растения-хозяина, ме-
стами вытянутые, в центральной части слоевища тесно сросшиеся
в однослойную пластинку с отходящими во все стороны ползучими
нитями, слегка суженными к концам. Клетки короткоцилиндрические
или неправильно многоугольные, 7—12 мкм шир., 7—17 мкм дл.,
молодые удлиненно-цилиндрические или клиновидные, 4—9 мкм шир.,
10—31 мкм дл., вначале с тонкими, позднее утолщенными оболоч-
ками. Хлоропласт изрезанный по краям, с 1 — 2 пиреноидами.
В пресных водоемах, на кладофоровых водорослях, преимущественно
на нитях Basicladia. В СССР не обнаружена. (Китай).
9. Entocladia immane (Schmidle) Starmach (=Endoderma immane
Schmidle) — Энтокладия огромная (рис. 120, 2, 3). — Нити простые или
скудно ветвящиеся, распростертые на поверхности или внутри оболочек
растения-хозяина, короткие, обычно прямые, одиночные, реже образую-
щие небольшие скопления. Клетки в молодых нитях и на к.(ах старых
нитей цилиндрические, 16 мкм шир., 40—60 мкм дл., в средней части
нитей обычно шаровидные, 20 —40 мкм шир., окруженные толстой
слоистой оболочкой 3—4 мкм толщ. Конечные клетки нитей при рассма-
тривании сверху суженные, широко закругленные, при рассматривании
сбоку заостренные. Хлоропласт с несколькими пиреноидами. Старые
слоевища целиком состоят из вздутых шаровидно-эллипсоидных клеток.
В пресных водоемах, на Pithophora pachyderma. В СССР не обнару-
жена. (Индия).
10. Entocladia tenuis Kylin — Энтокладия тонкая (рис. 121, 7). —
Нити беспорядочно ветвящиеся, распростертые на поверхности субстрата.
Клетки цилиндрические, 5—8 мкм шир., по длине в 4—8 раз больше,
молодые 3—5 мкм шир., по длине в 8—20 раз больше. Хлоропласт со
многими пиреноидами. Зооспора при прорастании образует вначале
длинный трубчатый вырост, который в дальнейшем начинает ветвиться.
В морях, на пустых раковинах моллюсков. В СССР не обнаружена.
(Побережье Балтийского моря).
11. Entocladia testarum Kylin — Энтокладия раковинная (рис. 121,
2—6). — Нити беспорядочно ветвящиеся, распростертые на поверхности
субстрата. Клетки цилиндрические, часто односторонне вздутые и тогда
неопределенных очертаний, 7 — 10 мкм шир., по длине в 3—5 раз больше,
молодые 5—8 мкм шир., по длине в 4—8 раз больше. Хлоропласт
с многочисленными пиреноидами, нередко сетчато продырявленный.
Зооспоры 3.9—4.1 мкм шир., 7.5—8 мкм дл., образуются в любых
клетках слоевища.
223
Рис. 120.
1 — Entocladia basicladiae', 2, 3 — E. immane: 2 — слоевище с поверхности,
3 — слоевище внутри оболочек питофоры (в разрезе). (7 — по Жао, 2, 3 —
по Шмидле).
Рис. 121.
1 Entocladia tenuis-, 2 — 6 — Е. testarum'. 2 — слоевище с зооспорангиями,
3—6 — зооспора и стадии ее прорастания; 7, 8 — Е. reineckel, молодое и взрос-
лое слоевища на клетках питофоры. (1—6 — по Кюлину, 7, 8 — по Шмидле).
224
Рис. 122.
Entocladia spongophila, нити среди игл пресноводной губки. (По Веберу ван
Боссе).
В морях, на пустых раковинах моллюсков. В СССР не обнаружена.
(Побережье Балтийского моря).
12. Entocladia reineckei (Schmidle) Starmach (== Endoderma reineckei
Schmidle) — Энтокладия Рейнеке (рис. 121, 7, 8). — Нити обильно и
беспорядочно ветвящиеся, распростертые на поверхности или внутри
растения-хозяина; разрастаясь, они плотно соприкасаются друг с другом,
образуя одно- или многослойные плотные псевдопаренхимные слоевища.
Клетки неправильных очертаний, реже цилиндрические, часто слегка
вздутые, 16—36 мкм шир., по длине равные ширине или в 2—3 раза
больше. Хлоропласт с несколькими пиреноидами.
В соленых водах, на нитях Pithophora reineckei. В СССР не обна-
ружена. (О-ва Самоа).
13. Entocladia spongophila (Web. van Bos.) Starmach (=Endoderma
spongophila (Web. van Bos.) Printz; Cladophora spongophila Web. van
Bos.; Gongrosira spongophila (Web. van Bos.) Wille] — Энтокладия
губколюбивая (рис. 122). — Нити беспорядочно ветвящиеся, распростер-
тые по субстрату; тесно соприкасаясь друг с другом, они образуют
местами однослойные пластинки. Клетки цилиндрические, реже слегка
вздутые, 5.5—6.4 мкм шир., по длине равные ширине или в несколько
раз больше, иногда до 85 мкм дл. Хлоропласт без пиреноида. Спорангии
7—9 мкм шир., по длине равные ширине, вставочные или конечные,
нередко по 4—6 в ряд. В спорангиях образуется большое количество
зооспор (или гамет?), которые освобождаются через отверстие в стенке
клетки; дальнейшее их развитие не прослежено. Вегетативное раз-
множение распадением слоевища на одно- или малоклеточные фраг-
менты.
В пресных водоемах, на губках и Ephydatia fluviatilis. В СССР
не обнаружена. (Индонезия; о-в Суматра).
15 Зеленые водоросли, Улотриксовые 225
Рис. 123.
7 — Epicladia flustrae', 2 — E. pontlca', 3 — Protoderma uiride. (7 — по Кюлину,
2 — по Рохлиной, 3 — по Прескотту).
Род EPICLADIA Reinke - ЭПИКЛАДИЯ
Слоевище микроскопическое, нитевидно-пластинчатое, эпизооидное
и эндозооидное. Нити ветвящиеся, ползучие, полностью распростертые по
субстрату, тесно смыкающиеся в псевдопаренхимные однослойные
пластинки, с обособленными ползучими ветвями по краям. Клетки
цилиндрические, многоугольные или неправильных очертаний. Хлоро-
пласт 1, пристенный, пластинчатый, с 1 пиреноидом. Бесполое размно-
жение зооспорами, развивающимися помногу в клетке и выходящими
наружу через округлое отверстие в оболочке материнской клетки.
В морских, соленых и солоноватых водах.
I. Клетки псевдопаренхимной части слоевища 12—20 мкм шир.........
..............................................Е. flustrae 1.
II. Клетки псевдопаренхимной части слоевища 2.7—5.3 мкм шир. . . .
.................................................Е. pontica 2.
1. Epicladia flustrae Reinke — Эпикладия флюетровая (рис. 123,
7). — Клетки псевдопаренхимной части слоевища 12—20 мкм шир., по
длине равные ширине или несколько больше, обычно многоугольные.
Клетки нитей, обособленных по краям пластинки, цилиндрические,
22Г.
у поперечных перегородок слегка перетянутые, 5—10 мкм шир., по
длине несколько больше.
В оболочках мшанок Flnstra foliacea и Membranipora crustalenta,
обитающих преимущественно на нитях Cladophora. rupestris. Арктические
о-ва, побережье Новой Земли.
2. Epicladia pontica Roehl. — Эпикладия черноморская (рис. 123,
2). — Клетки псевдопаренхимной части слоевища с толстыми оболоч-
ками, 2.7—5.3 мкм шир., по длине равные ширине или вдвое больше.
Клетки нитей, обособленных по краям пластинки, цилиндрические,
5.4—10.8 мкм шир., по длине в 2—4 раза больше. Нити обычно 270—
300 мкм дл.
В морях и лиманах, на раковинах моллюсков, преимущественно
Pecten. УССР (лиманы), Черноморское побережье Крыма.
Род PROTODERMA Kiitz. - ПРОТОДЕРМА
Слоевище микроскопическое, нитевидно-пластинчатое. Нити корот-
кие, ветвящиеся, ползучие, полностью распростертые по субстрату,
часто радиально расходящиеся от центральной группы многоугольных
клеток, тесно смыкающихся в псевдопаренхимные однослойные пла-
стинки, по краям обособленные, расположенные группами более или
менее параллельно друг другу. Клетки цилиндрические, многоугольные
или неправильных очертаний. Конечные клетки веточек слегка заострен-
ные, реже несколько вздутые. Хлоропласт пристенный, пластинчатый,
с 1 иногда плохо заметным пиреноидом, реже без него. Бесполое
размножение двужгутиковыми яйцевидными или шаровидными зооспо-
рами с 2 пульсирующими вакуолями и стигмой в переднем конце,
возникающими по 4, 8 (16) в любых, чаще центральных клетках псевдо-
паренхимной части слоевища и освобождающимися вследствие ослизне-
ния оболочек клеток. Апланоспоры шаровидные или эллипсоидные.
Известны мелкие гаметы, которые копулируют. Зигота прорастает после
периода покоя. Нередко наблюдается пальмеллевидное состояние.
В стоячих и проточных пресных водах, в обрастаниях различных
субстратов.
Виды рода Protoderma, легко спутать с некоторыми видами Stigeoclo-
nium, слоевище которых на ранних стадиях развития имеет вид псевдо-
паренхимных пластинок.
I. Клетки в слоевищах, по крайней мере в молодых, расположены
правильными рядами, оболочки не ослизняющиеся.
1. Пластинчатая часть слоевища однослойная, хлоропласт с пиренои-
дом.
А. Клетки большей частью цилиндрические или неправильно
цилиндрические, 6—12 мкм шир., по длине до 2 раз больше
..............................................Р. viride 1.
Б. Клетки изодиаметрические, 6—8 мкм в диам...............
.......................................Р. daphnieola 2.
2. Пластинчатая часть слоевища в центре двуслойная, хлоропласт
без пиреноида...................................Р. beesleyi 3.
П. Клетки в слоевищах большей частью расположены беспорядочно,
оболочки ослизняющиеся.
1. Пластинчатая часть слоевища всегда однослойная; клетки 4—5 мкм
шир., пиреноид почти не заметен.................Р. frequens 4.
15*
227
2. Пластинчатая часть слоевища одно- или многослойная; клетки
5—12 мкм шир., пиреноид виден отчетливо . Р. eohaerens 5.
1. Protoderma viride Kiitz. — Протодерма зеленая (рис. 123, 3). —
Слоевище на ранних стадиях развития округлое, затем неопределенных
очертаний. Клетки в молодых слоевищах 3—6 мкм шир., по длине
в 2—3 раза больше, во взрослом состоянии в центральной части 6—12 мкм
шир., по длине равные ширине или до 2 раз больше. Конечные
клетки обособленных по краям коротких ползучих нитей слегка сужен-
ные, широко закругленные, тупые. Хлоропласт с пиреноидом. Зооспоры
3—3.5 мкм в диам. Апланоспоры 2—3 мкм в диам.
В реках, озерах, пойменных водоемах, на водных цветковых ра-
стениях, древесном субстрате, раковинах моллюсков, камнях. В ряде
мест СССР, нередко.
2. Protoderma daphnieola Fott — Протодерма дафниевая
(рис. 124). — Слоевище в виде псевдопаренхимной дисковидной пла-
стинки, образованной радиально расходящимися, разветвленными в пло-
скости нитями. Клетки изодиаметрические, 6—8 мкм в диам., у попе-
речных перегородок слегка перетянутые, в основании нитей непра-
вильно многоугольные. Конечные клетки обособленных по краям
нитей широко закругленные. Хлоропласт с пиреноидом. Зооспоры
вначале прорастают в короткие, малоклеточные нити, которые затем
начинают ветвиться.
В прудах, на дафниях. В СССР не обнаружена. (Зап. Европа).
3. Protodermabeesleyi (Fritsch) Printz (=Ulvella beesleyi Fritsch) —
Протодерма Биелея (рис. 125). — Слоевище более или менее правильно
дисковидное, однослойное, кроме средней части, где в результате по-
вторных делений возникает второй слой толстостенных округлых клеток,
превращающихся в дальнейшем в зооспорангии. Клетки цилиндри-
ческие, по длине в 2—4 раза больше ширины. Конечные клетки коротких,
распростертых по субстрату нитей широко закругленные. Хлоропласт
без пиреноида. Зооспоры по 4 в зооспорангии, грушевидные, двужгути-
ковые, со стигмой в заднем конце. Половое размножение не просле-
жено. Известно пальмеллевидное состояние с образованием шаровидно-
яйцевидных клеток с 4 (6) линзовидными хлоропластами; в каждой
клетке образуется по 4 зооспоры или снова по 4 новые клетки, осво-
Protoderma daphnieola, слоевища разного возраста. (По Фотту).
228
Рис. 125.
Protoderma beesleyi. 1 — взрослое слоевище с зооспорангиями, 2 — молодое
слоевище, 3 — зооспора,. 7 — пальмеллевидное состояние. (По Бислею).
Рис. 126.
1 — Protoderma jrequens',. 2, 3 — Р. eohaerens, фрагменты разных форм слое-
вища. (7 — по Батчеру, 2, 3 — по Фричу).
бождающиеся в результате ослизнения оболочки материнской
клетки.
В проточных водах, на песчинках и камнях. В СССР не обнаружена.
(Англия).
4. Protoderma frequens (Butsch.) Printz (=Ulvella frequens
Butsch.) — Протодерма частая (рис. 126, 1). — Слоевище в виде оваль-
ной пластинки 0.5—1(3) мкм в диам., состоящей из ветвящихся в плоско-
сти нитей, погруженных в слизистую обвертку. Клетки в центральной
части слоевища более или менее изодиаметрические, с закругленными
углами, 4—5 мкм шир., в распростертых по субстрату обособленных
229
веточках 4—5 мкм шир., 8—12 мкм дл. Хлоропласт с 1 часто едва
заметным пиреноидом. Размножение не прослежено.
В проточных водах. В СССР не обнаружена. (Англия).
5. Protoderma cohaerens (Wittr.) Printz (=Pleurococcus vulgaris f.
cohaerens Wittr.; Protoderma broivnii Fritsch) — Протодерма связанная
(рис. 126, 2, 3). — Слоевище одно- или многослойное, с беспорядочно
оформленными краями. Клетки изодиаметрические, многоугольно-
округлые, 5—12 мкм шир. Оболочки слизистые, толстые, бесцветные,
слоистые, средние слои слабо исчерченные. Хлоропласт с отчетливо
видным пиреноидом и крахмалом.
На скалах, фирновых полях, в окрашенном снегу, на слоевищах
Prasiola crispa. Арктические районы СССР.
Род ULVELLA Crouan - УЛЬВЕЛЛА
Слоевище микроскопическое, светло-зеленое, дисковидное или лин-
зовидное, распростертое по субстрату, вначале однослойное, позднее
в середине многослойное, состоящее из центральной группы беспорядочно
расположенных клеток, погруженных в слизь, и радиально расходя-
щихся от них коротких ползучих, беспорядочно ветвящихся нитей.
Клеи, и центральной части слоевища округлые, плотно прилегающие
друг к другу. Краевые клетки вытянутые, на конце закругленные
или вильчато рассеченные. Ядро 1 (иногда несколько?). Хлоропласт 1,
пластинчатый, цельный или сетчато продырявленный, с пиреноидом
или без него. Бесполое размножение двужгутиковыми зооспорами,
образующимися по 4, 8, 16 в зооспорангиях. Зооспорангии возникают
Рис. 127.
1, 2 — Ulvella lens: 1 — молодое слоевище, 2 — середина слоевища при большом
увеличении; 3—5 — Pseudulvella americana: 3 — фрагмент слоевища сверху,
4 — поперечный срез слоевища, 5 — зооспоры. (7, 2 — по Губеру, 3 — 5 —
по Сноу).
230
из клеток центральной части слоевища, зооспоры освобождаются через
отверстие в верхушечной части клетки. Половое размножение не про-
слежено.
В морских, соленых и солоноватых водах, для пресных вод ука-
зывается 1 вид.
1. Ulvella lens Cronan — Ульвелла линзовая (рис. 127, 1, 2). —
Слоевище 1 — 5 мм в диам. Клетки в центральной части 5—15 мкм шир.,
по краю пластинки 3—4 (5) мкм шир., (10) 15—30 мкм дл.
На раковинах моллюсков, камнях, водорослях. УССР (Березанский
лиман), Черноморское побережье Кавказа.
Род PSEUDULVELLA Wille - ПСЕВДОУ. 11>ВЕ. 1.1Л
Слоевище микроскопическое, дисковидное, в центре многослойное,
по краям однослойное. Клетки центральной части слоевища расположены
вертикальными рядами, периферической, однослойной части — более или
менее радиальными рядами. Ядро 1. Хлоропласт 1, пристенный, пластин-
чатый, иногда сетчато продырявленный, с пиреноидом. Бесполое раз-
множение четырехжгутиковыми зооспорами, образующимися по 4, 8,
16 в зооспорангиях. Зооспорангии возникают из поверхностных клеток
центральной части слоевища. Зооспоры выходят наружу в слизистом
пузыре. Половое размножение не прослежено.
Преимущественно в морях, для пресных вод указывается 1 вид.
1. Pseudulvella americana (Snow) Wille (= Ulvella americana
Snow) — Пеевдоульвелла американская (рис. 127, 3—5). —Слоевище
0.25—3 мм в диам. Центральные клетки 6.7 — 14 мкм шир., 6.7—11.7 мкм
дл., часто вытянутые в вертикальном направлении, краевые 3.4—13 мкм
шир., 10—18.4 мкм дл. В старых слоевищах на краевых клетках иногда
образуются слизистые волоски. Зооспоры яйцевидные или эллипсоидные,
8—13 мкм шир., 10—15 мкм дл.
В пресных водоемах.
Var. americana (рис. 127, 3—5). —Слоевище 1—3 мм в диам.
Центральные клетки 10—14 мкм шир., краевые 5—13 мкм шир.,
10—17 мкм дл.
На крупноклеточных водорослях. В СССР не обнаружена. (Сев. Аме-
рика, Азия).
Var. indica Philipose. — Слоевище 0.25—0.5 мм в диам. Центральные
клетки 6.7—8.4 мкм шир., 6.7—11.7 мкм дл., краевые 3.4—6.7 мкм шир.
На листьях водных цветковых растений. В СССР не обнаружена.
(Индия).
Род PRINGSHEIMIELLA Hohnel - ПРИНГСХЕЙМИЕЛЛА
(=Pringsheimia Reinke)
Слоевище микроскопическое, нитчато-пластинчатое. Нити ветвя-
щиеся, ползучие, полностью распростертые по субстрату, радиально
расходящиеся от центральной группы крупных клеток, тесно смыкаю-
щиеся в однослойную псевдопаренхимную дисковидную пластинку.
Клетки центральной части слоевища выше краевых, эллипсоидные или
булавовидные, вертикально вытянутые, беспорядочно расположенные,
краевые клетки горизонтально вытянутые в радиальном направлении,
231
Рис. 128.
Pringsheimiella scutata: 1 — взрослое слоевище с пустыми зооспорангиями
в центре, 2 — образование зооспорангиев, 3 — молодое слоевище. (По Рейнке).
с утолщенной оболочкой на внешнем конце, ветвящиеся в плоскости,
вначале как выросты, которые затем отделяются от материнской клетки
поперечной перегородкой и разрастаются. Клетки молодых слоевищ
иногда с длинными бесцветными волосками. Хлоропласт крупный,
пластинчатый, обычно с пиреноидом. Бесполое размножение четырех-
жгутиковыми зооспорами, развивающимися помногу в зооспорангиях,
в которые превращаются центральные булавовидные клетки. Половое
размножение четырехжгутиковыми гаметами, образующимися в тех же
центральных булавовидных клетках, но на других слоевищах. Гаметофит
отличается от спорофита более рыхлым расположением клеток. Оболочки
клеток у спорофита и гаметофита одинаковой толщины.
1. Pringsheimiella scutata (Reinke) Schmidt (=Pringsheimia scutata
Reinke) — Прингсхеймиелла щитовидная (рис. 128). — Пластинчатое
слоевище у спорофита до 1—2 мм в диам., у гаметофита несколько
мельче. Клетки центральной части слоевища более или менее изодиаме-
трические, но почти вдвое выше краевых, краевые клетки 4—10 (И) мкм
шир., 12—26 мкм дл. Спорангии и гаметангии эллипсоидные или
почти грушевидные, 15—22 мкм шир., 28— 38 мкм выс. Зооспоры
15 мкм шир. Гаметы 4 мкм шир.
В соленых, солоноватых и морских водах, на водных цветковых
растениях, водорослях, камнях. Побережье Балтийского, Черного, Кас-
пийского и Японского морей.
Хорошо отличается от остальных представителей подсемейства
крупными, эллипсоидными или булавовидными клетками центральной
части пластинчатого слоевища, отчетливо возвышающимися над крае-
выми, а после выхода из них зооспор или гамет характерным бесцвет-
ным центром, окруженным зеленым кольцом краевых клеток.
232
Род JAOA Fan - ЖАОА
(=Coelodiscus Jao)
Слоевище макроскопическое, пластинчатое, бляшковидное, слегка
выпуклое или почти полушаровидное, прикрепляющееся к субстрату
многочисленными простыми или разветвленными ризоидами, состоящее
из двух или трех слоев клеток, в середине пустотелое; его внутренний
слой образован крупными бесцветными клетками, наружный — мелкими
зелеными клетками. Клетки внутреннего слоя почти шаровидные,
наружного — полушаровидные до бутылкообразных. Ядро 1. Хлоропласт
пластинчатый, с цесколькими пиреноидами. Бесполое размножение
четырехжгутиковыми эллипсоидными или более или менее грушевид-
ными зооспорами со стигмой, возникающими по 4, 8, 16 в зооспо-
рангиях. Зооспорангии образуются из несколько увеличенных клеток
внешнего слоя слоевища. Половое размножение неизвестно.
Систематическое положение рода неясно. Вполне возможно, что его
следует выделить в отдельное семейство.
1. Jaoa prasina (Jao) Fan (==Coelodiscus prasinus Jao) — Жаоа
луково-зеленая (рис. 129). — Слоевище оливково-зеленое или темно-
оливковое, 0.5—6 см в диам., 1—2 (30) мм выс., одиночное или в скоп-
лениях по 2—3. Клетки внутреннего слоя 30—82 мкм шир., наружного —
(9) 10—20 мкм шир., 20—32 мкм дл. Зооспорангии 15—20 мкм дл.
В быстро текущих водах.
Редкий вид, хорошо отличается от остальных представителей
подсемейства крупными размерами выпуклого многослойного пусто-
телого слоевища.
Рис. 129.
Jaoa prasina: 1 — общий вид слоевища сверху, 2 — то же сбоку, 3 — фрагмент
верхней части слоевища (поперечный срез), 4 — то же при большем увеличении,
5 — фрагмент нижней части слоевища с ризоидами, б — зооспора, 7—9 —
стадии прорастания зооспор, 10 — молодое слоевище. (По Жао).
233
Var. prasina (рис. 129). — Слоевище 0.5—1 см в диам., 1—2 мм выс.,
обычно двуслойное. Клетки внутреннего слоя 30—70 мкм шир.,
наружного — 10—20 мкм шир., 20—32 мкм дл.
В проточных водах на скалах. В СССР не обнаружена. (Китай).
Var. bullata Jao. — Слоевище 6 см в диам., 3 см выс., обычно
трехслойное. Клетки внутреннего слоя многоугольные, 37 — 82 мкм шир.,
среднего — 14—30 мкм шир., наружного — яйцевидно-конические, 9—
15 (20) мкм шир.
В проточных водах на камнях. В СССР не обнаружена. (Китай).
Подсемейство LEPTOSIROIDEAE Воигг.
Нити прямостоячие или ползучие, ветвящиеся, реже простые,
оканчивающиеся закругленными или слегка заостренными коническими
клетками, реже клетки одиночные или объединенные в неправильные
или пакетообразные скопления. Волоски, волосковидные образования и
ризоиды отсутствуют.
Водоросли преимущественно эпифитные или эпилитные, часто
встречаются на почве или в почве и на раковинах моллюсков.
I. Хлоропласт без пиреноида или с мелким, едва заметным пиреноидом.
1. Нити не выражены, клетки одиночные или в скоплениях.
А. Клетки разобщенные или в бесформенных скоплениях, не свя-
занные друг с другом .... род Diplosphaera (с. 236).
Б. Клетки объединены обычно в пакетообразные группы, иногда
могут возникать и короткие ползучие нити.
а. Клетки делятся только в двух взаимно перпендикулярных
направлениях, пакетообразные группы клеток однослойные
...........................род Apatococcus (с. 236).
б. Клетки делятся в трех взаимно перпендикулярных направ-
лениях, пакетообразные группы клеток многослойные,
а. Пакетообразные группы клеток не всегда ясно выра-
жены, образуются обычно при старении, на ранних
стадиях развития возникают короткие малоклеточные
нитевидные группы . . . род Coccobotrys (с. 238).
р. Пакетообразные группы клеток хорошо выражены, часто
шестигранные, иногда могут возникать короткие мало-
клеточные простые или разветвленные нити..........
......................род Desmococcus (с. 238).
2. Нити хорошо выражены.
А. Нити только ползучие, разобщенные.
а. Нити малоклеточные, простые или слабо разветвленные,
а. Нити окружены слизью, клетки преимущественно ци-
линдрические, обычно легко отделяющиеся друг от друга,
зооспоры одиночные; в толще воды, эпифит..............
............................род Gloeoplax (с. 239).
р. Нити без слизи/ клетки преимущественно шаровидные
или эллипсоидные, не отделяющиеся друг от друга,
зооспоры многочисленные; в нейстоне, в свободном
состоянии.................род Streptochlora (с. 240).
б. Нити многоклеточные, обильно разветвленные.
а. Нити беспорядочно распростертые, ветвящиеся лишь
в плоскости субстрата; кроме нитей часто возникают
комковатые скопления клеток . . . род Jaagiella (с. 240).
234
р. Нити радиально распростертые, ветвящиеся не только
в плоскости субстрата, но и в вертикальном направ-
лении; комковатые скопления клеток не образуются
..................................род Diaphragma (с. 240).
Б. Нити ползучие и прямостоячие, объединенные в дерновинки.
а. Клетки прямостоячих нитей более или менее цилиндри-
ческие; аэрофиты, на- влажных субстратах.
а. Клетки цилиндрические, прямые или слегка изогнутые,
зооспоры не наблюдались; на влажных стенах . . . .
...........................род Cedercreutziella (с. 241).
р. Клетки эллипсоидно-цилиндрические, прямые, зооспоры
двужгутиковые; на влажных вулканических скалах
..................................род Zoddaea (с. 242).
б. Клетки прямостоячих нитей шаровидные, эллипсоидные
или бочонковидные; гидрофиты, а также в почвах и на
почвах............................род Leptosira (с. 243).
В. Нити только прямостоячие, обильно разветвленные, образую-
щие кустики..........................род Lochmium (с. 246).
II. Хлоропласт с хорошо выраженным пиреноидом.
1. Нити только ползучие, разобщенные.
А. Нити малоклеточные, простые или слабо разветвленные.
а. Нити заключены в слизь; короткие боковые веточки раз-
виваются только в плоскости субстрата.
а. Нити заключены в широкое слизистое влагалище,
клетки бочонковидные, прямые; эпифиты.................
............................род Hazenia (с. 246).
р. Нити располагаются в поверхностном слое общей слизи,
клетки цилиндрические, обычно искривленные; в планк-
тоне ......................род Helicodictyon (с. 248).
б. Нити без слизи; короткие боковые веточки развиваются
во всех направлениях.
а. Клетки нитей правильно цилиндрические, прямые или
слегка изогнутые, конечные клетки широко закруглен-
ные; хлоропласт не продырявленный.
* Нити легко распадаются на малоклеточные фраг-
менты или на отдельные клетки, становящиеся
эллипсоидными или шаровидными; в стоячих водо-
емах ..................род Rhexinema (с. 248).
* * Нити легко распадаются на пакетообразные скопле-
ния клеток;в почве ...............................
. род Pseudopleurococcus (частично) (с. 248).
р. Клетки нитей неправильно цилиндрические, прямые,
заметно вздутые у нижнего конца, конечные клетки
конусовидно суженные; хлоропласт сетчато продыряв-
ленный .................род Spongoplastidium (с. 250).
Б. Нити многоклеточные, обильно и беспорядочно разветвленные.
а. Клетки делятся надвое в одном поперечном направлении,
образуя только нити; в слизи колониальных синезеленых
и зеленых водорослей.
а. Клетки цилиндрические, вытянутые, зооспорангии вер-
хушечные ................род Chloroclonium (с. 251).
р. Клетки шаровидные, бочонковидные, зооспорангии рас-
сеянные ...................род Leptosiropsis (с. 253).
б. Клетки делятся в двух и трех взаимно перпендикулярных
направлениях, образуя короткие, неправильно разветвлен-
235
ные нити или гроздевидные скопления; в почве и на
почве.......................род Pleurastrum (с. 253).
2. Нити только ползучие, тесно сомкнутые в псевдопаренхимную
пластинку, иногда образуются простые гроздевидные скопления
клеток.
А. По краям пластинки отходят короткие малоклеточные веточки.
а. Веточки простые, прямостоячие, к вершинам сужи-
вающиеся; в пресных и морских водах, эпифитно ....
.......................род Pseudendoclonium (с. 254).
б. Веточки скудно разветвленные, ползучие, к вершинам не
суживающиеся; на коре деревьев и на влажных пнях
род Pseudopleurococcus (частично) (с. 248).
Б. По краям пластинки веточки отсутствуют, гроздевидные
скопления клеток не образуются . . . род Epibolium (с. 256).
3. Нити ползучие и прямостоячие, объединенные в слоевище в виде
дерновинок, подушечек или корочек.
А. Слоевище инкрустировано известью; на разных субстратах.
а. Нити состоят из клеток двух типов, более или менее
правильно чередующихся: коротких, содержащих хлоро-
филл, и длинных, без хлорофилла; слоевище в виде
плотных подушечек .... род Chlorotylium (с. 258).
б. Нити состоят из более или менее однородных клеток;
слоевище в виде плотных дерновинок, подушечек или
корочек.........................род Gongrosira (с. 260).
Б. Слоевище не инкрустировано известью; на поверхности или
внутри субстратов, главным образом известковых.
а. Прямостоячие нити разветвлены беспорядочно, спорангии
с корневидными отростками, нити подошвы обычно погру-
жены в субстрат...................род Gomontia (с. 269).
б. Прямостоячие нити разветвлены преимущественно одно-
сторонне, часто гребневидно, спорангии без корневидных
выростов, нити подошвы не погружены в субстрат ....
............................род Ctenocladus (с. 273).
. Род DIPLOSPHAERA Bial. - ДИПЛОСФЕРА
Нити не выражены. Клетки шаровидные, разобщенные, при делении
образующие разнообразные скопления. Хлоропласт 1, без пиреноида.
Зооспоры не наблюдались, иногда образуются продолговатые аплано-
споры.
1. Diplosphaera chodatii Bial. — Диплосфера Шода (рис. 130,1,2). —
Клетки 2—6 мкм в диам.
На известковых скалах. В СССР не обнаружена. (Зап. Европа).
Близка к Desmococcus vulgaris, отличается разобщенностью клеток,
а также отсутствием пиреноида.
Род APATOCOCCUS Brand emend. Geitl. - АПАТОКОККУС
Нити не выражены. Клетки шаровидные или от взаимного дав-
ления многоугольные, способные к делению только в одной плоскости
в двух взаимно перпендикулярных направлениях, в результате чего
возникают однослойные, обычно многоклеточные группы без слизистой
236
Рис. 130.
1, 2 — Diplosphaera chodatli, клетки разного возраста; 3—7 — Apatococcus
vulgaris: 3 — клетки разного возраста, 4 — спорангий, 5 — зооспоры, 6 авто-
споры, 7 — прорастание автоспор; 8—11 — Coccobotrys verrucariae: 8 — фраг-
менты слоевища, 9 — апланоспоры, 10 — образование зооспор, 11 — зооспора;
12—14 — Desmococcus vulgaris: 12 — клетки и пакеты клеток, собранные с коры,
13 — нити, выросшие на агаре, 14 — спорангий и апланоспоры. (1, 2, 8—14 —
по Вишеру, 3—7 — по Гейтлеру).
обвертки. Хлоропласт 1, без пиреноида, чашевидный, крупный, по
краям несколько изрезанный или поделенный на пластины, в старых
клетках слегка волнистый и несколько отстающий от оболочек, что
создает ложное впечатление центрального звездчатого хлоропласта. Ядро
крупное. Вегетативное размножение фрагментацией таллома. Бесполое
размножение двужгутиковыми зооспорами. Зооспоры удлиненные, не-
сколько сплющенные с боков, без стигмы, возникающие по 8—32 (64)
в клетке; они освобождаются через узкое отверстие в верхней части
зооспорангия. Часто образуются автоспоры.
Аэрофитные водоросли.
1. Apatococcus vulgaris Brand emend. Geitl. [=Pleurococcus lobatus
Chod.; Pleurastrum lobatum Printz; Apatococcus lobatus (Chod.) Boye-
Pet.] — Апатококкус обыкновенный (рис. 130, 3—7). — Клетки 8—
15 (20) мкм в диам., одиночные, по 2—4 или в однослойных много-
клеточных группах. Хлоропласт желтовато-зеленый. Оболочка клеток
237
местами едва заметно утолщенная или отчетливо бородавчатая. Зоо-
спорангии 9—20 мкм в диам. Зооспоры 2—2.8 мкм шир., 6 — 7 мкм дл.
В тенистых влажных местах в виде зеленых налетов на стенах,
скалах, пнях деревьев. В СССР не обнаружена. (Зап. Европа).
Близок к Desmococcus vulgaris, отличается более крупными клет-
ками с желтовато-зеленым хлоропластом без пиреноида, способностью
клеток делиться лишь в одной плоскости, никогда не образуя нитей,
и продуцированием зооспор и автоспор.
Род COCCOBOTRYS Chod. - КОККОБОТРИС
Нити обычно не выражены. Клетки шаровидные, способные к де-
лению в трех взаимно перпендикулярных направлениях, в результате
чего возникают объемные, обычно многоклеточные группы. Хлоропласт 1,
пристенный, без пиреноида. Бесполое размножение двужгутиковыми
зооспорами и апланоспорами, образующимися в обычных вегетативных
клетках.
1. Coccobotrys verrucariae Chod. emend. Vischer — Коккоботрис вер-
рукарневый (рис. 130, 8—11). — Клетки 5—12 мкм в диам. В молодом
состоянии иногда образуются короткие нити, переходящие при старении
в пакетообразные группы изодиаметрических клеток.
В лишайниках из рода Verrncaria. В СССР не обнаружена. (Зап.
Европа).
Род DESMOCOCCUS Brand emend. Vischer —
ДЕСМОКОККУС
(=Pleurococcus auct.; Protococcus auct.)
Нити обычно не выражены. Клетки шаровидные или угловатые,
с неослизняющимися оболочками, способные к делению в трех взаимно
перпендикулярных направлениях, в результате чего возникают правиль-
ные пакетообразные шестигранные группы, реже неправильные скоп-
ления, при некоторых условиях иногда образуются короткие, простые
или разветвленные нити. Хлоропласт пристенный, чашевидный или
поясковидный, зеленый, с 1 мелким пиреноидом без крахмальной обо-
лочки. Ядро мелкое, в живых клетках едва заметное. Размножение
исключительно вегетативное.
Аэрофитные водоросли.
1. Desmococcus vulgaris (Nag.) Brand [—Pleurococcus vulgaris
Nag.; P. naegelii Chod.; Protococcus viridis Ag.; Desmococcus viridis
(Ag.) Brand] — Десмококкус обыкновенный (рис. 130, 12—14). —
Клетки 4—8 (11) мкм в диам., обычно собранные в правильные пакето-
образные или нитевидные группы, изредка возникают короткие одно-
рядные нити. Зооспоры не наблюдались. Встречены спорангии, похожие
на клетки трохисций (fihlorococcales), в которых образуются автоспоры,
прорастающие затем в нити.
В тенистых, влажных местах, на скалах, коре деревьев, кирпичных
кладках, оштукатуренных наружных стенах строений и т. п. Повсеместно.
В естественных условиях чаще всего встречаются в виде пакето-
образных групп, в условиях культуры (преимущественно на агаре)
нередко образует нити.
238
Род GLOEOPLAX Schmidle - ГЛЕОПЛАКС
Нити ползучие, короткие, скудно разветвленные, окруженные
слизью, во взрослом состоянии легко распадающиеся на отдельные
клетки, сдвигающиеся в слизи горизонтально и частично вертикально.
Клетки тонкостенные, на ранних стадиях развития цилиндрические,
позднее округлые. Хлоропласт пристенный, в виде 1 или нескольких
мелких пластинок, без пиреноида. Продукт ассимиляции — крахмал.
Вегетативное размножение делением клеток. Бесполое размножение зоо-
спорами, образующимися по 1 в вертикально расположенных клетках.
Зооспоры освобождаются через отверстие в оболочке клеток на их
спинной стороне (по отношению к субстрату); при их прорастании
вначале образуется короткая простая нить, которая позже разветвля-
ется. Акинеты образуются в результате утолщения оболочек клеток.
Преимущественно в стоячих водоемах, в обрастаниях разных суб-
стратов.
1. Gloeoplax weberi Schmidle — Глеоплакс Вебера (рис. 131,1—3). —
Клетки на ранних стадиях развития около 5 мкм шир., по длине
в 2—4 раза больше, по мере старения более или менее округлые,
8—16 мкм в диам.
В торфяных болотах, большей частью на листьях сфагнума.
Var. weberi (рис. 131, 1—3). — Молодые клетки правильно ци-
линдрические, прямые. 1
Калининградская обл., ЭССР, ЛатвССР.
Var. suecicum Borge. — Молодые клетки неправильной формы,
вытянутые вверх, или квадратные и многоугольные, изогнутые или
раздвоенные.
В СССР не обнаружена. (Швеция).
Рис. 131.
1—3 — Gloeoplax weberi-. 1 — старое слоевище, 2, 3 — молодые слоевища; 4—
6 — Streptochlora moniliformis, нити разного возраста. (1—3 — по Шмидле,
4—6 — по Бойе-Петерсену и Гансену).
239
Род STREPTOCHLORA Boye-Pet. et Hansen —
СТРЕПТОХЛОРА
Нити свободноживущие, не связанные с субстратом, короткие,
малоклеточные, обычно однорядные, простые. Клетки на ранних
стадиях развития цилиндрические, позднее шаровидные или эллипсоид-
ные, способные делиться не только в поперечном, но и в продольном
направлении, в результате чего иногда могут возникать многорядныр
нити. Хлоропласт пристенный, без пиреноида. Продукт ассимиляции —
крахмал. Зооспоры двужгутиковые, шаровидные, со стигмой; они обра-
зуются в большом количестве (до 200) в любой шаровидной клетке
и освобождаются через боковое отверстие.
1. Streptochlora moniliformis Boye-Pet. et Hansen — Стрептохлора
четковндная (рис. 131, 4—6). — Нити большей частью 3—7-клеточные.
Молодые клетки цилиндрические, около 7 мкм шир., до 16 мкм дл.,
взрослые шаровидные, до 16 мкм в диам. Зооспоры 1—2 мкм в диам.
В нейстоне пресных водоемов. В СССР не обнаружена. (Дания).
По мнению Буррелли, может быть стадией развития Leptosira,
Gloeoplax, Rhexinema.
Род JAAGIELLA Vischer - ЯАГИЕЛЛА
Нити ползучие, ветвящиеся, в молодом состоянии при росте на агаре
короткие, однорядные, при старении дву—многорядные, переходящие
в комковатые образования, напоминающие паренхимную ткань. Клетки
более или менее изодиаметрические, врастающие в субстрат длиннее
поверхностных. Хлоропласт пристенный, без пиреноида, с мелкими
зернами крахмала. В клетках внешней части старых слоевищ образу-
ются апланоспоры и акинеты. Зооспоры не наблюдались.
1. Jaagiella alpicola Vischer — Яагиелла альпийская (рис. 132). —
Клетки 5—10 мкм шир., акинеты до 15 мкм шир.10
На влажной кислой почве, на высоте 2440 м над ур. м., в куртинках
Carex sempervirens. В СССР не обнаружена. (Зап. Европа: Швейцарские
Альпы).
Род DIAPHRAGMA Geitl. — ДИАФРАГМА
Нити ползучие, радиально расходящиеся, обильно разветвленные
в горизонтальном и вертикальном направлениях, состоящие из разоб-
щенных клеток, отделенные друг от друга длинными участками пустой
оболочки. Клетки цилиндрические с закругленными концами или эллип-’
соидные, с тонкими пектиновыми неослизняющимися оболочками. Хлоро-
пласт пристенный, пластинчатый, без пиреноида. Бесполое размножение
двужгутиковыми зооспорами. Зооспоры эллипсоидные, с 2 сократитель-
ными вакуолями и стигмой; они образуются по 4, 8 в зооспорангиях,
незначительно отличающихся от вегетативных клеток.
10 Эти размеры установлены нами по масштабной линейке на автор-
ских рисунках, в оригинальном описании они отсутствуют.
240
Рис. 132.
Jaagiella alpicola: 1 — общий вид слоевища, выросшего на агаре, 2 — автоспоры,
3 — прорастание автоспор, 4 — акинеты. (По Вишеру).
1. Diaphragms radiosum Geitl. — Диафрагма радиальная (рис. 133,
1—3). — Клетки (3.5) 4—6 мкм шир., в старых частях нитей до 8 мкм
дл., на вершинах нитей до 48 мкм дл.
На почве, среди протонемы мха. В СССР не обнаружена. (Австрия).
Род CEDERCREUTZIELLA Vischer -
ЦЕДЕРКРЕЙТЦИЕЛЛА
Нити частично ползучие, частично прямостоячие, ветвящиеся.
Клетки в ползучих нитях и в нижней части прямостоячих нитей
короткобочонковидные, эллипсоидные или почти шаровидные, к концам
ветвей постепенно становящиеся цилиндрическими, прямыми или слабо
искривленными. Хлоропласт пристенный, без пиреноида, в старых
клетках с зернышками крахмала. Размножение акинетами, прорастаю-
щими без периода покоя, или апланоспорами, возникающими по 2, 4
в слегка вздутых клетках. Зооспоры не наблюдались.
Род мало изучен, описан в основном по культуральным материалам.
Разветвленные нити образуются в культурах только на плотных средах.
16 Зеленые водоросли. Улотриксовые
241
Рис. 133.
1—3 — Dlaphragma radiosum'. 1 — общий вид слоевища, 2 — образование зоо-
спор, 3 — клетки в состоянии покоя; 4— 8 — С edercreutzlella savoniensis:
4 — молодое слоевище, 5 — образование апланоспор, 6 — акинеты, 7 — про-
растающие акинеты. Я прорастающие апланоспоры. (1 — 3 — по Гейтлеру,
4—8 — по Вишеру).
1. Cedercreutziella savoniensis Vischer — Цедеркрейтциелла савоний-
ская (рис. 133, 4—8). — Клетки 3—5 мкм шир., до 18 мкм дл., акинеты
до 18 мкм в диам.11
На влажных стенах. В СССР не обнаружена. (Финляндия).
Род ZODDAEA Borzi - ЗОДДЕЯ
Нити ползучие и прямостоячие, обильно односторонне ветвя-
щиеся. Клетки в ползучих нитях короткие, шаровидные или эллипсо-
идные, в прямостоячих — эллипсоидно-цилиндрические, тонкостенные.
Хлоропласт пристенный, пластинчатый, без пиреноида. Зооспорангии,
образующиеся на старых нитях, почти не отличаются от вегетативных
клеток. Зооспоры по 1—4, двужгутиковые, со стигмой.
1. Zoddaea viridis Borzi — Зоддея зеленая. — Слоевище 200—300 мкм
выс. Клетки в ползучих нитях 5—8 мкм шир., в молодых прямо-
стоячих — 3—5 мкм шир. *
" См.- предыдущее подстрочное примечание.
242
На влажных вулканических скалах. В СССР не обнаружена.
(Италия).
Вид изучен недостаточно, оригинального рисунка нет.
Род LEPTOSIRA Borzi - ЛЕПТОЗИРА
Нити частично ползучие, частично прямостоячие, разветвленные,
ветви отходят преимущественно от верхнего конца материнской клетки.
Клетки шаровидные, эллипсоидные, более или менее бочонкообразные,,
короткие или вытянутые, с тонкими твердыми бесцветными оболочками.
Конечные клетки иногда слегка заостренные. Хлоропласт пристенный,
пластинчатый или лентовидный, обычно без пиреноида, изредка с пире-
ноидом, иногда с зернами крахмала. Бесполое размножение зооспорами
и апланоспорами, половое — изогаметами. В спорангий или гаметангий
может превратиться любая клетка нити, слегка увеличивающаяся
в размерах. Зооспоры и гаметы развиваются в большом количестве
и выходят наружу через отверстие в оболочке, вначале окруженные
слизистым пузырем, в скором времени расплывающимся. Зооспоры
и гаметы двужгутиковые, яйцевидные или несколько заостренные
на обоих концах, со стигмой в задней части клетки. Зооспоры, про-
плавав некоторое время, прикрепляются к субстрату и прорастают
в клетки, подобные Characium, содержимое которых делится потом на
2—8 апланоспор, освобождающихся в результате полного ослизнения
материнских оболочек и прорастающих в новые нити. Гаметы слива-
ются задними, заостренными концами, образуя веретеновидную зиготу,
переходящую в состояние покоя; дальнейшее их развитие не прослежено.
Известно образование акинет.
В стоячих водоемах, преимущественно в озерах и болотах, а также
на почвах и в почвах.
I. Ветвление всестороннее, ветви прямые, оттопыренные, короткие;
клетки более 5 мкм шир.
1. Клетки цилиндрические или вздуто-цилиндрические (у ползучих
нитей большей частью шаровидные); ветви к вершине слегка
суживаются.
А. Конечные клетки заостренные; в болотах...................
.........................................L. mediciana 1.
Б. Конечные клетки обычно закругленные; гонидии лишайников
..............................................L. thrombii 2.
2. Клетки шаровидные, эллипсоидные или вздуто-бочонковидные.
А. Конечные клетки ветвей слегка вздутые, округло-конусовидные
или неправильно яйцевидные; в водоемах . . . L. obovata 3.
Б. Конечные клетки ветвей широко закругленные; в почвах,
а. Клетки ползучих нитей 10—16.7 мкм шир.....................
.......................................L. terricola 4.
б. Клетки ползучих нитей 15—40 мкм в диам................
.........................................L. terrestris 5.
II. Ветвление обычно одностороннее, ветви ровные на всем протяжении,
слегка согнутые, большей частью плотно прижатые к главной нити;
клетки 3—5 мкм шир...................................L. willeana 6.
1. Leptosira mediciana Borzi — Лептозира Медичи (рис. 134,
1—7)- — Образует маленькие ярко-зеленые дерновинки. Прямостоячие
нити обильно разветвленные, ветви короткие, прямые, оттопыренные,
243
16*
Рис. 134.
1—7 — Leptoslra mediciana: 1 — часть слоевища с зооспорангиями, частично
пустыми, 2 — прорастающая зооспора, 3,4 — образование апланоспор, 5 —
прорастание апланоспоры, 6 — гамета, 7 — копуляция гамет; 8,9 — L. thrombii:
8 — часть слоевища с зооспорангиями, 9 — зооспоры; 10 — L. obovata, часть
слоевища с зооспорангиями; 11 — L. terricola, часть слоевища с зооспорангиями,
частично пустыми; 12 — L. terrestris, часть слоевища с зооспорангиями, час-
тично пустыми; 13 — L. willeana, фрагмент слоевища. (1—7 — по Борзи, 8, 9 —
по Чермак-Вонс, 10 — по Вишеру, 11 — по Фричу, 12 — по Бристоль, 13 —
по Гансгиргу).
кверху постепенно суживающиеся. Клетки короткие, вздуто-цилиндри-
ческие, 10—20 мкм шир. Конечные клетки обычно заостренные. Хлоро-
пласт пластинчатый, с отверстием в середине и мелкими зернами
крахмала. В зооспорангии и гаметангии превращаются прежде всего
нижние клетки таллома. Зооспоры и гаметы 0.5—2 мкм шир., 2.5 мкм дл.,
по 20—60 и больше в каждом спорангии. Зиготы шаровидные или
короткоэллипсоидные, 10 мкм в диам., с толстой оболочкой и зер-
нистым содержимым, с каплями масла, вначале зеленые, позже крас-
неющие.
В болотах и на заболоченных участках, на водных цветковых расте-
ниях в виде зеленых точечек, хорошо заметных в лупу. Ленинградская,
Рязанская, Харьковская обл.
2. Leptosira thrombii Tschermak-Woess — Лептозира тромбневая
(рис. 134, 8,9). — Нити короткие, разветвленные, ползучие по субстрату
и приподнимающиеся над ним, легко переходящие в пальмеллевидное
состояние. Клетки ползучих нитей большей частью шаровидные,
12—18 (26) мкм в диам., прямостоячих — цилиндрические, 10—20 мкм
шир., в пальмеллевидном состоянии обычно эллипсоидные или шаровид-
ные. Конечные клетки обычно закругленные. Хлоропласт пластинчатый,
244
с изрезанными краями и 1 пиреноидом. Зооспоры грушевидные,
1.5—2.5 мкм шир., 7—9.5 мкм дл., с желто-зеленым хлоропластом,
I сократительной вакуолью и стигмой, по 32, 64 в шаровидных или
эллипсоидных зооспорангиях. Апланоспоры шаровидные, по 2—8 в мате-
ринской клетке.
В лишайниках, впервые выделен в культуру из Thrombium epigeum.
В СССР не обнаружена. (Зап. Европа).
Близок к Leptosira obovata.
3. Leptosira obovata Vischer — Лептозира обратнояйцевидная
(рис. 134, 10). — Образует слоевища в виде полушаровидных поду-
шечек около 1 мм в диам., состоящих из беспорядочно разветвленных
нитей. Клетки шаровидные, удлиненно-эллипсоидные или вздуто-
бочонковидные, 12—15 мкм шир. Конечные клетки ветвей вначале
округло-конусовидные, позднее неправильно яйцевидные, слегка взду-
тые, 20—30 мкм в диам. Хлоропласт темно-зеленый, изрезанный по
краям, без пиреноида. Зооспоры грушевидные, 2—8 мкм шир., 8—10 мкм
дл., со стигмой, расположенной почти в середине клетки. Апланоспоры
известны.
В стоячих и замедленно текущих водах, преимущественно в боло-
тах, на растительном субстрате. В СССР не обнаружена. (Швейцария).
4. Leptosira terricola (Bristol) Printz (=Gong rostra terricola Bris-
tol) — Лептозира наземная (рис. 134, 11). — Образует подушковидные
слоевища, состоящие из ползучих, обильно ветвящихся нитей, смыкаю-
щихся в псевдопаренхимную подошву, и прямостоячих простых и
коротких нитей. Клетки ползучих нитей почти шаровидные или не-
правильной формы, вздутые, 10—16,7 мкм шир., прямостоячих — эллип-
соидно-цилиндрические, 6 —14 мкм шир., 10—18 мкм дл., нередко расши-
ренные к одному из концов и часто изогнутые. Конечные клетки
широко закругленные. Хлоропласт пластинчатый или лентовидный,
с изрезанными округлолопастными краями, с 1—3 отчетливыми пиренои-
дами или без них; изрезанность хлоропласта часто бывает настолько
сильной, что он кажется состоящим из отдельных, неправильно округ-
лых дисков. Зооспорангии шаровидно-яйцевидные или неправильной
формы, 14—18 мкм шир., как в ползучих, так и в прямостоячих
нитях. Апланоспоры шаровидные, 8.4 мкм в диам., по 3—4 в клетке.
В обрабатываемых почвах; обнаружена в Водных культурах. Ленин-
градская, Кировская обл., БССР, УССР, КазССР, Зап. Сибирь.
5. Leptosira terrestris (Fritsch et John) Printz t==Rhextnema terres-
tris Fritsch, et John) — Лептозира почвенная (рис. 134, 12). — Нити
короткие, разветвленные, легко переходящие в бесформенные группы
клеток. Клетки шаровидные, 15—40 мкм в диам. Оболочки клеток
по мере старения слегка утолщающиеся. Конечные клетки широко за-
кругленные. Хлоропласт слегка изрезанный по краям, с 1 или несколь-
кими пиреноидами. Зооспоры яйцевидные, со стигмой; они образуются
в большом количестве в почти неизмененных вегетативных клетках.
Апланоспоры яйцевидные.
В почвах. В СССР не обнаружена. (Англия).
6. Leptosira willeana Hansg. — Лептозира Вилле (рис. 134, 13). —
Образует небольшие светло-зеленые дерновинки, инкрустированные
известью. Нити обильно разветвленные, преимущественно односторонне.
Ветви слегка искривленные, в большинстве случаев тесно прилегающие
245
к главным нитям, к вершинам не суживающиеся. Вегетативные клетки
3—5 мкм шир., по длине в 1.5 (2) раза больше, у поперечных пере-
городок слегка перетянутые. Оболочки тонкие, бесцветные. Хлоропласт
широкий. Акинеты шаровидные, яйцевидные или неправильной формы,
чаще всего 6—9 мкм шир., с бледно-красным маслянисто блестящим
зернистым содержимым.
В стоячих и слабо проточных водах, большей частью в болотах,
на водных цветковых растениях. В СССР не обнаружена. (Зап. Европу).
Род LOCHMIUM Printz - ЛОХМИУМ
Нити прямостоячие, ветвящиеся, с обособленной главной нитью
и отходящими, не отличающимися от нее боковыми ветвями, обра-
зованными 1 или несколькими клетками. Клетки цилиндрические,
яйцевидные или булавовидные. Базальная клетка цилиндрическая или
булавовидная, без ризоидных выростов. Конечные клетки широко
закругленные. Оболочки тонкие. Хлоропласт пристенный, пластинчатый,
без пиреноида. Спорангии верхушечные и вставочные, не отличающиеся
от обычных вегетативных клеток. Акинеты верхушечные, яйцевидные,
крупные, с утолщенной оболочкой.
1. Lochmium piluliferum Printz — Лохмнум шариконосный (рис. 135,
1, 2). — Слоевище микроскопическое, в форме довольно плотных,
беспорядочно разветвленных кустиков. Клетки 2—3(4) мкм шир., (4)5—
6(8) мкм дл. Акинеты 6.5—8 мкм шир., 8—13 мкм дл.
В пресных или солоноватых водах, преимущественно в болотах,
на отмерших водных цветковых растениях, в виде ярко-зеленых точек,
видимых невооруженным глазом. Сибирь.
Род HAZENIA Bold - ГАЗЕНИЯ
Нити свободноживущие, заключенные в широкое слизистое влага-
лище, слабо развитые, из нескольких клеток, преимущественно одно-
рядные, слегка суженные к концам, но благодаря продольным делениям
клеток местами могут быть многорядными. Клетки шаровидные или
бочонковидные, одноядерные. Хлоропласт пристенный, пластинчатый,
с пиреноидом. Половое размножение двужгутиковыми гаметами, возни-
кающими по 2 в слегка измененных вегетативных клетках. Гаметы
шаровидные, с хлоропластом, пиреноидом и 2 сократительными ва-
куолями. Зигота шаровидная, толстостеннная; она прорастает после пе-
риода покоя, давая начало 4 клеткам.
1. Hazenia mirabilis Bold — Газения удивительная (рис. 135,
3—5). — Образует бесформенную слизистую массу 120—150 мкм шир.
Клетки 12 — 15 мкм шир. Зооспоры около 12 мкм дл.
В прудах. В СССР не обнаружена. (Сев. Америка).
Вид изучен недостаточно.
246
Рис. 135.
7, 2 — Lochmium pilulijerum.-. 1 — молодое слоевище, 2 — слоевище с акине-
тами; 3—5 — Hazenia mirabilis-, 3 — фрагмент нити в слизистой пленке, 4 —
образование зооспор, 5 — выход зооспор (гамет?). (7, 2 — по Принтцу, 3, 5 —
по Болду).
1
Рис. 13(5.
Heltcodictyon planctonicu/n: J — молодое слоевище, 2 — взрослое слоевище, 3 —
отдельная клетка. (По Уайтфорду и Буррелли).
247
Род HELICODICTYON Whitford - ГЕЛИКОДИКТИОН
Образует свободноживущие слизистые колонии эллипсоидной или
неправильной формы, содержащие нити. Нити короткие, разветвленные,
располагающиеся только в поверхностном слое слизи. Ветви малоклеточ-
ные, простые. Клетки цилиндрические, чаще искривленные, с тонкой
оболочкой. Хлоропласты по 1—3, с пиреноидами. Способ размножения
неизвестен.
1. Helicodictyon planctonicum Whitford — Гелнкоднктион планктон-
ный (рис. 136). — Клетки 8—10.5 мкм шир., 6—18 мкм дл.
В планктоне. В СССР не обнаружен. (США: Сев. Каролина).
Род RHEXINEMA Geitl. - РЕКСИНЕМА
Нити свободноживущие, короткие, не более 8—10 клеток, одно-
рядные, разветвленные, легко распадающиеся на одиночные клетки или
малоклеточные фрагменты. Клетки обычно цилиндрические, местами
с выпуклостями, у поперечных перегородок заметно перетянутые, реже
шаровидные или эллипсоидные. Хлоропласт пристенный, пластинчатый,
с пиреноидом. Оболочки клеток легко ослизняются, особенно в усло-
виях большой влажности, в результате чего возникает пальмеллевидное
скопление клеток. Бесполое размножение двужгутиковыми зооспорами.
Зооспоры грушевидные, со стигмой и 2 пульсирующими вакуолями;
они образуются по 1—4 в слегка увеличенных вегетативных клетках
и освобождаются в результате ослизнения оболочки спорангиев.
1. Rhexinema paucicellulare (Vischer) Geitl. (=Pleurastrum, pauci-
cellulare Vischer) —Рексннема малоклеточная (рис. 137, 1—5).—
Клетки 5—7 мкм шир., 7—35 мкм дл., в основании нити часто вздутые,
при старении шаровидные, легко отделяющиеся друг от друга. Зоо-
споры яйцевидные.
В стоячих водах. В СССР не обнаружена. (Зап. Европа).
Род PSEUDOPLEUROCOCCUS Snow emend. Vischer —
ПСЕВДОПЛЕВРОКОККУС
Нити ползучие, ветвящиеся, обособленные, распростертые по суб-
страту и частично приподнимающиеся над ним, разрастающиеся
в дерновинки, или тесно сомкнутые в неправильные псевдопаренхим-
ные пластинки с обособленными по краям небольшими ползучими
веточками. Клетки цилиндрические, при старении закругленные,
соединенные неплотно, благодаря чему легко распадаются на пакето-
образные группы. Хлоропласт пристенный, с пиреноидом. Размножение
акинетами, возникающими из любых клеток. Зооспоры неизвестны.
В почве и на коре деревьев.
I. Слоевище в виде разветвленных многоклеточных нитей......
............................................Р. printzii 1.
II. Слоевище в виде неправильных псевдопаренхимных образований.
1. Клетки около 6.5 мкм шир............................Р. vulgaris 2.
2. Клетки 6.5—8 мкм шир...............................Р. botryoides 3.
248
Рис. 137.
1—5 — Rhexinema paucicellulare: 1—3 — фрагмент слоевища, 4 — образова-
ние зооспор, 5 — зооспоры; 6—11 — Pseudopleurococcus printzii: 6 — молодое
разветвленное слоевище, 7—10 — фрагменты старого слоевища, 11 — акинеты;
12 — Р. vulgaris’, 13 — Р. botryold.es. (1—11 — По Вишеру, 12, 13 — по Сноу).
1. Pseudopleurococcus printzii Vischer — Псевдоплеврококкус
Принтца (рис. 137, 6—11). — Нити многоклеточные, разветвленные,
иногда переходящие в пакетообразные скопления клеток. Клетки
ветвящихся нитей цилиндрические, 5—8 мкм шир., 15—30 мкм дл., на
обоих концах нередко слегка вздутые. Конечные клетки закругленные.
Концы нитей часто распадаются на мелкие группы клеток. Хлоропласт
с изрезанными краями, занимающий лишь среднюю часть клетки. При
образовании акинет клетки делятся, округляются и окружаются толстой
оболочкой.
В почве. В СССР не обнаружен. (Швейцария).
249
Вид изучен недостаточно, описан на основании культурального
материала.
2. Pseudopleurococcus vulgaris Snow — Псевдоплеврококкус обыч-
ный (рис. 137, 12). — Нити тесно сомкнутые в псевдопаренхимные
пластинки, только по их краям обособленные, ползучие, слабо раз-
ветвленные, реже клетки одиночные. Клетки около 6.5 мкм шир. Хлоро-
пласт заполняет лишь часть клетки, иногда заметны 2 хлоропласта.
На коре деревьев. В СССР не обнаружен. (Сев. Америка).
3. Pseudopleurococcus botryoides Snow — Псевдоплеврококкус ботри-
оидный (рис. 137, 13). — Нити тесно сомкнутые в плотные много-
рядные нитевидные образования или псевдопаренхимные пластинки
с короткими обособленными ползучими нитями по краям, реже клетки
одиночные. Клетки 6.5—8 мкм шир., с тонкой оболочкой. Хлоропласт
с пиреноидом.
На коре деревьев, влажных пнях, в виде толстых желто-зеленых
налетов. Ленинградская и Калининская обл.
Род SPONGOPLASTIDIUM Vischer - СПОНГОПЛАСТИДИУМ
Нити свободноживущие, относительно короткие, разветвленные.
Ветви короткие, направленные во все стороны, к концам слегка сужи-
вающиеся. Клетки неправильно цилиндрические, прямые, заметно взду-
Рис. 138.
Spongoplastidium proliferum: 1 — общий вид слоевища, 2—4 — образование
акинет, 5 — прорастание акинет. (По Вишеру).
250
тые у нижнего конца. Конечные клетки конусовидно суженные. Хлоро-
пласт сетчато продырявленный, заполняющий большую часть просвета
клетки, в молодых клетках без пиреноида, в старых с 1 пиреноидом.
Размножение фрагментаций таллома и акинетами. Зооспоры не-
известны.
1. Spongoplastidium proliferum Vischer — Спонгопластиднум прора-
стающий (рис. 138). — Клетки 6—10 мкм шир., 10—40 мкм дл., старые
чаще всего шаровидные, 10 мкм в диам.
В теплицах. В СССР не обнаружен. (Швейцария).
Род CHLOROCLONIUM Borzi - ХЛОРОКЛОНИУМ
Нити свободноживущие в слизи других водорослей, беспорядочно
разветвленные. Ветви обычно очередные. Клетки более или менее ци-
линдрические, удлиненные. Хлоропласт пристенный, пластинчатый
с изрезанными краями, с пиреноидом. Бесполое размножение зооспо-
рами и акинетами. Зооспорангии образуются из конечных клеток
и содержат по 2—8 двужгутиковых зооспор. Зооспоры овальные или
яйцевидные, со стигмой; они освобождаются через боковое отверстие
в оболочке материнской клетки и прорастают в новые нити. Акинеты
образуются часто, одиночные или по нескольку в ряд. Вследствие
интенсивного деления акинет может возникнуть пальмеллевидная ста-
дия. В каждой клетке, образовавшейся таким путем, возникает по
1 двужгутиковой микрозооспоре. Последние, по-видимому, могут копу-
лировать.
В слизи зеленых, синезеленых и других водорослей.
I. Клетки 8—12 мкм шир............................С. elongatum 1.
II. Клетки до 5 мкм шир.
1. Клетки 4—5 мкм шир..........................С. gloeophilum 2.
2. Клетки до 4 мкм шир.........................С. parvulum 3.
1. Chloroclonium elongatum Borzi — Хлороклониум вытянутый
(рис. 139, 7—4). — Нити широко распростертые. Ветви удаленные друг
от друга, оттопыренные, длинные. Клетки 8—12 мкм шир., по длине
в 2—4 раза больше, реже более длинные. Зооспорангии образуются
из верхушечных клеток, 2—4 в ряд.
В слизи разных водорослей, большей частью зеленых и сине-
зеленых. Вост. Сибирь (Байкал).
2. Chloroclonium gloeophilum Borzi — Хлороклониум слизелюбивый
(рис. 139, 5—7). — Нити широко распростертые. Ветви рассеянные,
очень тонкие, длинные. Клетки 4—5 мкм шир., по длине от 4 до 8 раз
больше. Зооспорангии образуются из верхушечных клеток.
Преимущественно в слизи синезеленых водорослей. В СССР не обна-
ружен. (Зап. Европа).
3. Chloroclonium parvulum Borzi — Хлороклониум крошечный
(рис. 139, 8). —Нити распростертые. Ветви очень короткие. Клетки
до 4 мкм шир., по длине в 2—3 раза больше. Зооспорангии образуются
из верхушечных клеток.
В слизи колониальных водорослей (Rivularia). Вост. Сибирь
(Байкал).
251
Рис. 139.
1—4 — Chloroclonium elongatum: 1 — веточка с образующимися зооспорами,
2 — верхушка молодой веточки, 3 — акинеты, 4 — микрозооспоры, развиваю-
щиеся из акинет; 5—7 — С. gloeophilum: 5 — вегетативная нить, 6 — образова-
ние акинет, 7 — акинеты после распадения нити; 8 — С. parvulum-, 9, 10 —
Leptosiropsis torulosa: 9 — фрагмент слоевища, 10 — апланоспоры. (1—4, 8 —
по Борзи, 5—7 — по Скуе, 9, 10 — по Жао).
252
Род LEPTOSIROPSIS Jao - ЛЕПТ03ИР0ПСИС
Нити свободноживущие в слизи других водорослей, короткие, бес-
порядочно разветвленные, постепенно суживающиеся к вершине.
Ветви короткие, оттопыренные. Клетки вначале цилиндрические, позднее
бочонковидные или шаровидные, у поперечных перегородок заметно
перетянутые, с толстой слоистой оболочкой. Хлоропласт пристенный,
чашевидный, с четким пиреноидом. Бесполое размножение двужгутико-
выми зооспорами и апланоспорами, возникающими по 16, 32 в любой
клетке таллома. Зооспоры яйцевидные, без стигмы. Апланоспоры яйце-
видные, толстостенные.
1. Leptosiropsis torulosa Jao — Лептозиропеие бочонковидный
(рис. 139, 9, 10). — Клетки 11—30 мкм шир., 14—36 мкм дл. Зооспоран-
гии 27—42 мкм шир. Оболочки клеток часто достигают 10 мкм толщ.
В слизи многих водорослей (Nostoc, Gloeotrichia, Chaetophora).
В СССР не обнаружен. (Китай).
Близок к роду Leptosira, хорошо отличается наличием пиреноида
и утолщенными оболочками клеток.
Род PLEURASTRUM Chod. - ПЛЕВРАСТРУМ
Нити свободноживущие, короткие, беспорядочно разветвленные
в горизонтальной плоскости, легко переходящие в бесформенные
группы клеток. Клетки делятся обычно в двух, реже трех взаимно
перпендикулярных направлениях, часто образуя «плеврококкоидные»
колонии. Хлоропласты по 1—2 в клетке, пристенные, пластинчатые,
с пиреноидом. Бесполое размножение двужгутиковыми зооспорами и
акинетами. Зооспорангии базальные и вставочные, по форме и вели-
чине почти не отличающиеся от вегетативных клеток. Зооспоры яйце-
видные. Внешние слои оболочек клеток часто ослизняются, клетки
разъединяются и превращаются в акинеты. Они могут быть одиноч-
ными или по нескольку вместе. Покоящиеся акинеты имеют на обо-
лочках скульптурные украшения.
Аэрофитные и почвенные водоросли.
В вегетативном состоянии с этим родом весьма сходны роды Pseudo-
pleurococcus и Pseudendoclonium, однако первый отличается отсутствием
зооспор, а второй — четырехжгутиковыми шаровидными зооспорами.
I. Нити хорошо выражены, короткие, неправильно разветвленные
..................................................Р. insigne 1.
II. Нити не отчетливые, часто переходящие в более или менее грозде-
видные образования................................Р. terrestre 2.
1. Pleurastrum insigne Chod.— Плевраетрум замечательный (рис. 140,
1—3). — Нити хорошо выраженные, короткие, неправильно разветвлен-
ные. Клетки легко делятся в трех направлениях.
На почве и в почве. В СССР не обнаружен. (Зап. Европа, Америка).
Вид описан очень неполно, без указания размеров клеток.
2. Pleurastrum terrestre Fritsch et John — Плевраетрум наземный
(рис. 140, 4, 5). — Нити неотчетливые, обильно разветвленные. Ветви
довольно короткие, часто суживающиеся к концам, в некоторых слу-
чаях слегка искривленные, всегда распростертые в горизонтальном
253
Рис. 140.
1—3 —Pleurastrum insigne: 1 — фрагмент слоевища с пустыми спорангиями,
2 — зооспора, 3 — прорастающие акинеты; 4, 5 — Р. terrestre, фрагменты слое-
вища со спорангиями. (1—3 — по Шода, 4, 5 — по Фричу и Джону).
направлении, реже ветвление радиальное или вообще ясно невыражен-
ное, так как деление клеток происходит в трех взаимно перпенди-
кулярных направлениях, и короткие, тесно сближенные выросты придают
таллому характерный вид грозди, видимой на субстрате в форме темно-
зеленой крупинки («плеврококкоидный» тип роста). Часто слоевище
состоит из нескольких плотно соединенных, расположенных цепочкой
клеток, иногда представлено одиночными клетками. Клетки обычно
неправильной формы, реже шаровидные, 6.3—20.9 мкм шир., 7—
29.3 мкм дл. Хлоропласт крупнолопастный, интенсивно-зеленый, с 1 (2)
отчетливыми пиреноидами, заполняющий иногда до 4/s полости клетки,
с волнистой поверхностью. Зооспорангии базальные и вставочные, по
величине почти равные вегетативным клеткам или несколько крупнее
их. Зооспоры яйцевидные, заостренные на переднем конце, 3.3 мкм шир.,
7.3 мкм дл., с пристенным пластинчатым хлоропластом, занимающим
примерно половину клетки.
На различных почвах. Ленинградская и Кировская обл., УССР.
Весьма вероятно, что этот вид тождествен предыдущему, но в силу
крайне недостаточного описания последнего их объединение пока
невозможно.
Род PSEUDENDOCLONIUM Wille - ПСЕВДЭНДОКЛОНИУМ
Нити ползучие, короткие, беспорядочно разветвленные, тесно сомк-
нутые в однослойную псевдопаренхимную пластинку с выходящими из
нее короткими простыми прямостоячими ветвями, реже клетки одиноч-
ные или собранные в группы «плеврококкоидного» типа. Клетки
шаровидные, короткобочонковидные или цилиндрические. Хлоропласт
пристенный, пластинчатый, с 1 пиреноидом. Бесполое размножение
четырехжгутиковыми зооспорами и акинетами. Зооспорангии шаровид-
ные. Зооспоры широконйцевидные, со стигмой или без нее. Акинеты
образуются по 1 или по нескольку вместе, некоторые из них про-
растают в новые нити сразу, другие превращаются в толстостенные
покоящиеся клетки, окруженные слизью. Гаметы неизвестны.
В пресных и морских водах.
254
Рис. 141.
1—5 — Pseudendoclonium b aslliense: 1—3 — разные формы слоевища, выращен-
ные в культурах, 4 — образование зооспор, 5 — зооспоры и их прорастание;
6—9 — Р. submarinum: 6 — слоевища в клетке древесины, 7 — фрагменты слое-
вища, 8 — зооспорангии, 9 — зооспоры. (1—5 — по Вишеру, 6—9 — по Вилле).
I. В пресных водах
П. В морских водах
. Р. basiliense 1.
Р. submarinum 2.
1. Psendendoelonium basiliense Vischer — Псевдэндоклониум базель-
ский (рис. 141, 7—5). — Нити короткие, обычно примерно 10-клеточ-
ные, беспорядочно разветвленные. Ветви отходят от верхнего конца
материнской клетки. Клетки до 5 мкм шир., до 20 мкм дл., в паль-
меллевидном состоянии шаровидные, 8—10 мкм в диам., при старении
в беспорядочных скоплениях, окруженные слизью. Зооспоры по 4 в спо-
рангии, 5—7 мкм шир., 7—8 мкм дл., со стигмой, жгутики 10—
255
12 мкм дл. При прорастании зооспоры образуют 3—5-клеточную нить,
которая может перейти в пальмеллевидное скопление клеток.
Выделен из стоячей воды в окр. Базеля, описан на основании
культурального материала. В СССР не обнаружен. (Швейцария).
2. Pseudendoclonium submarinum Wille — Псевдэндоклониум подвод-
ный (рис. 141, 6—9). — Нити короткие, ползучие, образующие неболь-
шую псевдопаренхимную пластинку с отходящими вверх короткими
прямостоячими простыми, иногда слегка изогнутыми ветвями, реже
клетки одиночные. Зооспоры без стигмы. Акинеты известны.
На древесных и каменистых субстратах, вместе с другими водо-
рослями. Балтийское море, в прибрежье ЛатвССР, ЭССР.
Вид мало изучен, в оригинальном описании размеры клеток не
указаны.
Род EPIBOLIUM Printz - ЭПИБОЛИУМ
Нити ползучие, обильно и беспорядочно разветвленные, полностью
распростертые по субстрату, местами или на всем протяжении тесно
смыкающиеся боковыми сторонами в одно-, реже дву- и трехслойную
псевдопаренхимную пластинку разнообразных очертаний. Клетки обычно
цилиндрические, по длине равные ширине или до 4 раз больше, часто
неправильной формы или беспорядочно многоугольные. Хлоропласт
пристенный, кольцевидный, слегка продырявленный, с 1—2, изредка
многими пиреноидами. Зерна крахмала дисковидные, расположенные
параллельно плоскости хлоропласта. Бесполое размножение двужгутико-
выми зооспорами и акинетами. Зооспорангии верхушечные и вставочные,
образующиеся из любых клеток нитей; увеличиваясь в размерах, они
заметно возвышаются над остальными клетками. Зооспоры по 2—32
в спорангии, округло-яйцевидные, с чашевидным хлоропластом, 1 пире-
ноидом и стигмой. Акинеты с утолщенными оболочками.
В пресных и солоноватых стоячих и проточных водах, на разных
субстратах.
I. Зооспорангии только верхушечные, 10—11 мкм в диам.............
.............................................Е. dermaticola 1.
П. Зооспорангии верхушечные и вставочные, до 35 мкм в диам.
1. Зооспорангии преимущественно верхушечные, 10.8—18 мкм
в диам.; клетки нитей 5—8 мкм шир.............Е. fonticolum 2.
2. Зооспорангии преимущественно вставочные, 30—35 мкм шир.,
30—40 мкм дл.; клетки нитей 7 — 10 мкм шир...................
........................................ Е. polysporum 3.
1. Epibolium dermaticola Printz — Эпиболиум эпидермальный
(рис. 142, 1—3). — Нити широко распростертые по субстрату, беспо-
рядочно разветвленные, местами срастающиеся в псевдопаренхимные
пластинки. Клетки цилиндрические, часто неправильные, многоуголь-
ные, 5—10 мкм шир., до 40 мкм дл. Зооспорангии верхушечные,
10—11 мкм в диам. Зооспоры по 2—8, 3 мкм шир.
В пресных и солоноватых стоячих и проточных водах, на листьях
водных цветковых растений, раковинах моллюсков. УССР, Зап. Сибирь.
2. Epibolium fonticolum Starmach — Эпиболиум родниковый
(рис. 142, 4, 5). — Нити на ранних стадиях развития одиночные,
256
разрастающиеся на листьях водных мхов по поперечным перегородкам
эпидермальных клеток, позднее образующие псевдопаренхимную плас-
тинку, иногда дву- и трехслойную. Клетки по краям пластинки и на
концах ползучих ветвей цилиндрические, в центральной части слоевища
неправильной формы, укороченные, иногда вытянутые вертикально
вверх, 5—7 (8) мкм шир., до 20 мкм дл., на поверхности двуслойного
слоевища почти квадратные, 5—7 мкм шир. Хлоропласт 1, перед
делением клетки нередко их 2, с 1 или 2 пиреноидами. Оболочка
клеток тонная, иногда утолщается и частично ослизняется. Зооспорангии
верхушечные, изредка вставочные, неправильно шаровидные, 10.8—
18 мкм в диам., в зрелости окруженные толстой слоистой оболочкой,
часто ослизняющейся. Зооспоры обычно по 4, 8 в спорангии, яйцевидные
или шаровидные, 5—6 мкм шир., 7—7.7 мкм дл., с чашевидным
хлоропластом без стигмы. Акинеты в старых слоевищах, с толстой слои-
стой оболочкой.
Рис. 142.
1—3 — Epibolium dermaticola; 1 — общий вид слоевища, 2, 3 — образование
верхушечных спорангиев; 4, 5 — Е. fonticolum; 4 — молодое слоевище на
поверхности листа мха, 5 — зооспорангии и зооспоры; 6 — 8 — Е. polysporum;
6 — фрагмент слоевища, 7 — отдельная клетка, 8 — зооспоры. (7 — 3 — по
Привтцу, 4. 5 — по Стармаху, 6—8 — по Дюрингеру).
17 Зеленые водоросли. Улотриксовые
257
В горных источниках, родниках, на листьях водных мхов. В СССР
не обнаружен. (Польша).
3. Epibolium polysporum During. — Эпиболиум многоспоровый
(рис. 142, 6—8). — Нити длинные, беспорядочно разветвленные, отходят
от группы клеток, довольно плотно сросшихся между собой боковыми
поверхностями, разрастающиеся на листьях водных цветковых растений,
чаще всего по поперечным перегородкам эпидермальных клеток. Клетки
7—10 мкм шир., до 45 мкм дл. Зооспорангии большей частью вставоч-
ные, изредка верхушечные, почти шаровидные, 30—35 мкм шир.,
30—40 мкм дл. Зооспоры до 32 в спорангии, 4—5 мкм шир., 9 мкм дл.,
жгутики до 1.5 раза длиннее тела.
В стоячих и проточных пресных водах, на листьях водных цвет-
ковых растений, раковинах моллюсков. УССР (оз. Тур, р. Припять).
Род CHLOROTYLIUM Kiitz. - ХЛОРОТИЛИУМ
Нити частично ползучие, частично прямостоячие, разветвленные,
образующие маленькие плотные подушечки, инкрустированные известью.
Ползучие нити, переплетаясь, образуют подошву. Прямостоячие нити
обильно и большей частью односторонне разветвленные, плотно при-
жатые друг к другу. Клетки цилиндрические, разной длины и окраски:
длинные светло-зеленые, часто почти бесцветные, короткие ярко-зеленые;
иногда наблюдается правильное чередование 1 — 3 длинных и 3—7 корот-
ких клеток, в результате чего внутри общего слоевища возникают
более темные и более светлые слои. Конечные клетки на вершине
закругленные. Хлоропласт пристенный, широкий, пластинчатый или
лентовидный, с пиреноидом, в длинных клетках заметным не всегда.
Бесполое размножение зооспорами и акинетами. Зооспорангии конечные,
слегка вздутые по сравнению с вегетативными клетками. Зооспоры
мелкие, яйцевидные, двужгутиковые. Половое размножение не наблюда-
лось. Акинеты образуются из коротких клеток нитей, доволно толсто-
стенные, с красноватым содержимым. При ослизнении нитей в акинетах
образуются четырехжгутиковые макрозооспоры, прорастающие в новые
нити.
В стоячих и проточных пресных водах, на разных субстратах.
I. Слоевище на вертикальном срезе внутри отчетливо горизонтально
слоистое......................................С. cataractarum 1.
II. Слоевище па вертикальном срезе внутри не слоистое...........
............................................С. mammiforme 2.
1. Chlorotylium cataraetarum Kiitz. — Хлоротилиум водопадный
(рис. 143, 1—5). — Слоевище подушковидное, 0.5—0.8 мм в диам.,
ярко- или светло-зеленое, в сухом состоянии серо-зеленое или желтовато-
серое до красновато-бурого, инкрустированное известью, твердое, внутри
горизонтально слоистое. Прямостоячие нити обильно, чаще всего одно-
сторонне разветвленные. Ветви многоклеточные, расположенные более
или менее параллельно, плотно сжатые; они отходят от верхнего конца
почти каждой клетки прямостоячей нити и ветвятся повторно с той же
стороны, что и прямостоячие нити, реже беспорядочно, местами прямо-
стоячие нити ветвятся с одной стороны, а их ветви — с противо-
положной. Клетки 6—12 (18) мкм шир. Правильное чередование корот-
ких и длинных клеток выражено хорошо, обусловливая четкую горизон-
258
Рис. 143.
7 — 5 — Chlorotylium cataractarum: 1 — общий вид слоевища, 2 — вегетативная
нить, 3 — ветви с акинетами, 4 — вертикальный срез слоевища, 5 — отдельная
нить; 6 — С. mammiforme, фрагменты слоевища. (7, 4— 6 — по Кютцингу,
2, 3 — по Смиту).
17*
259
тальную слоистость внутри слоевища, видимую на его вертикальном
срезе; короткие клетки почти квадратные, по длине лишь до 1.5 раза
больше ширины, с ярко-зеленым хлоропластом, заполняющим всю их
полость, длинные, по длине в 2 — 6(10) раз больше ширины, бес-
цветные или бледноокрашенные. Зооспоры многочисленные, возникают
весной в конечных клетках прямостоячих нитей и прорастают без
копуляции. В летное время после ряда подобных генераций наступает
ослизнение прямостоячих нитей и возникает стадия, подобная Gloeocys-
tis. Внешний слой оболочек таких клеток разрывается и сбрасывается,
как у Schizochla.rn.ys. В клетках этой генерации образуются четырех-
жгутиковые зооспоры, по 4—16 в клетке. При ослизнении слоевища
в коротких клетках прямостоячих нитей могут образоваться акинеты.
Клетки при этом округляются, несколько увеличиваются в размерах,
достигая 10—16 мкм шир., окрашиваются в красный цвет и отделяются
друг от друга.
В стоячих и быстро текущих водах, па древесине и камнях. От-
дельные подушечки нередко сливаются друг с другом, образуя сплошную
бугристую корочку. Ленинградская, Московская обл., Кавказ.
2. Chlorotylium mammiforme (Balbis) Kiitz. — Хлоротилиум сосочко-
видный (рис. 143, 6). — Слоевище подушковидное, ярко-зеленое, ин-
крустированное известью. Прямостоячие нити очень обильно развет-
вленные, состоящие из длинных и коротких клеток, в оптическом
разрезе почти квадратных. Ветви короткие и длинные; они направ-
лены вертикально, чередуясь друг с другом. Клетки около 10 мкм шир.,
располагающиеся в нитях без правильного чередования коротких и
длинных, вследствие чего слоевище на срезе не слоистое.
На влажных скалах и в ручьях. Ставропольский край.
Род GONGROSIRA Kiitz. - ГОНГРОЗИРА
Слоевище в виде маленьких подушечек, дерновинок или корочек,
состоящих из подошвы и прямостоячих нитей, обычно инкрустиро-
ванное известью. Подошва образована ползучими нитями, обйльпо развет-
вленными, переплетающимися или срастающимися в псевдопаренхимную
пластинку. Прямостоячие нити короткие, обильно разветвленные, часто
плотно сжатые, с конечными клетками несуживающимися или изредка
слабо суживающимися. Ветви отходят от верхнего конца материн-
ских клеток, однако часто образование поперечных перегородок у них
запаздывает и они продолжают расти вбок, давая начало новой ветви.
Клетки обычно неправильные, очень изменчивой формы, с тонкими
целлюлозными оболочками. Хлоропласт пристенный, преимущественно
продырявленный, с 1 — 3 пиреноидами. Бесполое размножение двужгути-
ковыми зооспорами. Зооспорангии обычно расположены на концах
ветвей, реже вставочные. У одних видов они резко отличаются вели-
чиной от вегетативных клеток и в них образуется много зооспор,
у других почти не отличаются от вегетативных клеток и содержат по
2—3 зооспоры. Зооспоры двужгутиковые, яйцевидные, плоско сжатые,
со стигмой. Гаметы двужгутиковые, но в целом половое размножение
исследовано недостаточно. Акинеты и апланоспоры имеются.
В пресных и солоноватых стоячих и проточных водах, на разных
субстратах, реже в почвах.
260
I. Спорангии преимущественно верхушечные.
1. Спорангии по величине резко отличаются от вегетативных клеток.
А. Слоевище в виде корочек, большей частью инкрустированных
известью; прямостоячие нити слабо разветвленные.
а. Ползучие нити подошвы развиваются только на поверх-
ности субстрата.
а . Клетки прямостоячих нитей около 7 мкм шир., цилин-
дрические или бочонковидные . . . G. fluminensis 1.
р . Клетки прямостоячих нитей (4.4)14—50 мкм шир.,
шаровидные или цилиндрические.
* Прямостоячие нити очень короткие, почти не ветвя-
щиеся, клетки преимущественно шаровидные .
...............................G. debaryana 2.
* * Прямостоячие нити до 500 мкм дл., слабо развет-
вленные, с короткими веточками, клетки более или
менее цилиндрические . . . . G. stagnalis 3.
б. Ползучие нити подошвы развиваются не только на поверх-
ности субстрата, но и внедряются в него................
...................................... G. lacustris 4.
Б. Слоевище в виде распростертых псевдопаренхимных пластинок
с редкими одноклеточными изогнутыми веточками...............
..........................................G. prostrata 5.
В. Слоевище в виде подушечек; прямостоячие нити более или
менее многоклеточные, разветвленные.
а. Ветви направлены во все стороны; клетки прямостоячих
нитей 6 — 10 мкм шир.
а. Клетки короткоцилиндрические, правильной формы
.....................................G. scierocoecus 6.
Р. Клетки удлиненные, изогнутые, односторонне вздутые,
реже неправильной формы . . . G. trentepohliopsis 7.
б. Ветви направлены в одну сторону; клетки прямостоячих
нитей 8—20 мкм шир.
а. Клетки у поперечных перегородок не перетянутые;
(10)15—20 мкм шир......................G. pygmaea 8.
р. Клетки у поперечных перегородок отчетливо перетяну-
тые, 8—14 .мкм шир.....................G. reeurvata 9.
2. Спорангии по величине почти не отличаются от вегетативных
клеток.
А. Клетки прямостоячих нитей цилиндрические, хлоропласт
содержится во многих клетках.
а. Клетки прямостоячих нитей цилиндрические, изодиаме-
трические, или длина их до 2 раз больше ширины; слое-
вище инкрустированное известью.
а. Прямостоячие нити простые, параллельные друг другу,
плотно сжатые........................G. disciformis 10.
р. Прямостоячие нити разветвленные, радиально расходя-
щиеся, плотно сжатые.................G. burmanica И.
б. Клетки прямостоячих нитей цилиндрические или непра-
вильной формы, длина их до 6 раз больше ширины;
слоевище инкрустированное известью.
а. Клетки прямостоячих нитей цилиндрические, 6 —
11.5 мкм шир.......................G. inerustans 12.
р. Клетки прямостоячих нитей преимущественно непра-
вильной формы, 5—17 мкм шир. . . G. depauperata 13.
в. Клетки прямостоячих нитей цилиндрические, слегка или
261
значительно вздутые, длина их не более чем до 4 раз
больше ширины; слоевище не инкрустированное известью
............................G. leptotrieha 14.
Б. Клетки прямостоячих нитей с боковыми выростами, хлоропласт
содержится только в конечных клетках . . . G. schmidlei 15.
II. Спорангии преимущественно вставочные.
1. Клетки прямостоячих нитей в длину вдвое меньше или до 2.5 раза
больше ширины.................................G. tibetana 16.
2. Клетки прямостоячих нитей в длину равные ширине или до 9 раз
больше.....................................G. scourlieldii 17.
1. Gongrosira fluminensis Fritsch — Гонгрозира речная (рис. 144,
1, 2). — Слоевище в виде корочек, инкрустированное известью.
Ползучие по субстрату нити обильно разветвленные, перепле-
тенные друг с другом в псевдопаренхимную подошву, плотно срастаю-
щуюся с субстратом. Прямостоячие нити многочисленные, короткие,
плотно сжатые друг с другом, обычно простые, реже с короткими,
3—4-клеточными веточками в верхушечной части. Клетки подошвы
8—10 мкм шир., округленные или угловатые, с толстой, неравномерно
слоистой оболочкой, клетки прямостоячих нитей цилиндрические, тонко-
стенные, у поперечных перегородок слегка перетянутые, или бочонко-
видные, около 7 мкм шир., по длине в 1.5—2 раза больше. Зооспоры
неизвестны. Акинеты по 1 — 2 на прямостоячих нитях, верхушечные,
8—8.5 мкм шир.
В проточных водах, на камнях. В СССР не обнаружена. (Англия).
2. Gongrosira debaryana Babenh. — Гонгрозира де Бари (рис. 144,
3—7). — Слоевище в виде корочек, зеленое, не инкрустированное из-
вестью. Ползучие по субстрату нити образуют однослойную псевдопарен-
химную подошву. Прямостоячие нити короткие, нередко всего из не-
скольких клеток, простые или слабо разветвленные. Клетки подошвы
округлые, с толстыми оболочками, по краям слоевища сплющенные,
нередко лопастные, клетки прямостоячих нитей более или менее ци-
линдрические, (4.4)14—50 мкм шир., по длине равные ширине или до
2.5 раза больше. Зооспорангии шаровидные или булавовидные, крупные,
нередко до 150 мкм в диам. Зооспоры многочисленные. Акинеты изме-
ненной или неизмененной формы, часто заостренные. Иногда встречаются
апланоспоры; они образуются в середине материнских клеток, окру-
жаются толстой слоистой оболочкой и могут прорастать сразу после
освобождения или, округлившись, превращаются в спорангий.
В пресных и солоноватых стоячих и проточных водах, на водных
цветковых растениях, испорченной древесине, камнях, раковинах мол-
люсков, реже в почвах.
F. debaryana (=var. robustra Hansg.) (рис. 144, 3—5). — Клетки
14—50 мкм шир. Прорастающие вегетативные клетки округленные.
В реках, озерах, прудах, болотах. В разных районах СССР, нередко.
F. minor Melnik, (рис. 144, 6, 7). — Клетки 4.4—8.8 мкм шир.
Прорастающие вегетативные клетки разнообразной формы, часто заост-
ренные на одном из концов.
В почвах. ТаджССР. Обнаружена в водных почвенных культурах.
3. Gongrosira stagnalis (G. S. West) Schmidle — Гонгрозира прудовая
(рис. 144, 8). — Слоевище в виде корочек, темно-зеленое, негладкое.
Ползучие по субстрату нити обильно разветвленные, образуют плотную
псевдопаренхимную подошву. Прямостоячие нити до 500 мкм дл., скудно
262
Рис. 144.
1, 2 — Gongrosira /Zuminensis: 1 — общий вид слоевища, 2 — акинеты и про-
растающая акинета; 3—7 — G. debaryana-. 3—5 — f. debaryana (3, 4 — взрослые
слоевища, 5 — разные по возрасту зооспорангии и зооспоры), 6, 7 — f. minor
(6 — фрагмент слоевища, 7 — разные стадии прорастания вегетативных кле-
ток); 8 — G. stagnalis, фрагмент слоевища; 9 — G. prostrata, фрагмент слоевища
со спорангиями; 10 — G. sclerococcus', 11 — 13 — G. trentepohliopsis: 11 — фраг-
менты слоевища, 12, 13 — разные стадии развития спорангиев; 14, 15 — G. pyg-
таеа-. 14 — взрослое слоевище, 15 — начальные стадии развития; 16, 17 —
G. recurvata-. 16 — фрагмент верхушечной части нити, 17 — фрагмент нижней
части нити; 18, 19 — G. disciformis: 18 — вид слоевища сверху, 19 — продольный
срез слоевища. (2, 2, 18, 19 — по Фричу, 3 — по Смиту, 4, 5 — по Вилле,
6, 7 — по Мельниковой, 8 — по Весту, 9 — по Жао, 10, 14, 15 — по Кютцингу,
11 — 13 — по Шмидле, 16, 17 — по Виттроку).
263
разветвленные, с короткими веточками. Клетки ползучих нитей более
или менее цилиндрические, 16—31 мкм шир., прямостоячих — 16—
25 мкм шир., с приближением к подошве суживающиеся, по длине
в 2—6 раз больше. Зооспорангии верхушечные, шаровидные, яйце-
видные или почти грушевидные, значительно крупнее вегетативных
клеток.
В стоячих, реже слабо проточных водах, на отмерших водных цвет-
ковых растениях, раковинах моллюсков, камнях. УССР, Кавказ.
4. Gongrosira lacustris Brand — Гонгрозира озерная. — Слоевище
в виде мелких темно-зеленых, почти гладких корочек, часто инкру-
стированное известью. Ползучие нити срастаются в псевдопаренхимную
подошву и часто врастают в субстрат. Прямостоячие нити скудно
разветвленные, плотно сжатые. Клетки ползучих нитей (6)11 —14 мкм
шир., погруженных в субстрат отростков не более 6 мкм шир., клетки
прямостоячих нитей частично укороченные, слегка вздутые, около 14 мкм
шир., частично удлиненно-цилиндрические, пустые или почти лишен-
ные содержимого. Конечные клетки округлые, многие из них, значи-
тельно увеличиваясь в размерах, превращаются в спорангии. Акинеты,
освобождаясь в результате ослизнения оболочек материнских клеток,
сразу прорастают. Бесполое и половое размножение не просле-
жено.
В стоячих и проточных водах, на водных цветковых растениях,
древесине, раковинах моллюсков, камнях, а также на почве глинистых
пустынь (такыров). УССР, ТаджССР.
Вид изучен недостаточно, авторский рисунок отсутствует.
5. Gongrosira prostrata Jao — Гонгрозира распростертая (рис. 144,
9). — Слоевище темно-зеленое, в виде беспорядочно распростертых пла-
стин. Ползучие нити плотно переплетаются, образуя псевдопарен-
химную подошву. Прямостоячие нити редкие, всегда одноклеточные.
Клетки разнообразные по форме и размерам, часто беспорядочно
вздутые, выпуклые на верхней стороне и уплощенные на нижней,
7 — 8 мкм шир., 13—15 мкм дл., 9—15 мкм выс. Зооспорангии одиноч-
ные, обычно верхушечные, изредка на ползучих нитях вставочные,
шаровидно-яйцевидно-почковидные, треугольные, с толстой оболочкой.
Зооспоры по 8, 16, 32 в каждом зооспорангии.
В пресных проточных водах, эпифитно на листьях валиснерии.
В СССР не обнаружена. (Китай).
6. Gongrosira sclerococcus Kiitz. (=G. viridis Kiitz.) —Гонгрозира
еклерококковая (рис. 144, 70). —Слоевище в виде подушечек, ярко-
зеленое, 0.5 — 1 мм в диам., инкрустированное известью. Подошва
псевдопаренхимная. Прямостоячие нити обильно разветвленные, с вет-
вями, направленными во все стороны. Клетки прямостоячих нитей
цилиндрические, 6.6 — 10 мкм шир., по длине равные ширине или
несколько больше, в веточках последнего порядка клетки шаровидные.
Размножение не исследовано.
В стоячих и слабо проточных водах, на раковинах моллюсков,
камнях, а также на скалах, орошаемых водой родников. ЛатвССР,
УССР, Кавказ.
7. Gongrosira trentepohliopsis Schmidle — Гонгрозира трентеполие-
видная (рис. 144, 11—13).— Слоевище в виде подушечек, 0.5—5 мм
в диам., твердое, инкрустированное известью. Ползучие по субстрату
264
нити обильно разветвленные, срастающиеся в псевдопаренхимную плас-
тинку, образующую подошву. Прямостоячие нити обильно разветвлен-
ные, плотно сжатые. У вершины нитей боковые ветви слегка сог-
нутые, у основания нитей расположенные почти параллельно им; они
отходят обычно от верхнего, изредка от нижнего конца материнской
клетки. Клетки изогнутые или вздутые с одной стороны, реже пра-
вильно цилиндрические, 6—9.9 мкм шир., по длине в 2—3 раза больше.
Спорангии верхушечные, двух форм: крупные, до 22 мкм шир., булаво-
видные с узкой шейкой и шаровидные, в которых образуются 32, 64 зоо-
споры. Зооспоры двужгутиковые, эллипсоидные или шаровидные, спе-
реди заостренные, сзади закругленные, с чашевидным хлоропластом
с пиреноидом у заднего конца; они освобождаются через округлое
отверстие на вершине спорангия. Клетка, лежащая под верхушечным
спорангием, после его опадения также превращается в такой же верху-
шечный спорангий. Кроме того, на свободных концах нитей или под
верхушечным спорангием могут образовываться небольшие изодиаметри-
ческие, преимущественно шаровидные клетки, в которых возникает по
2, 4 зооспоры, по форме и величине не отличающиеся от описанных
выше. Наконец, на верхушке свободных нитей часто образуется не-
сколько коротких веточек, состоящих из 2—3 клеток, каждая из которых
может превратиться в вышеописанный маленький спорангий.
В проточных и стоячих водах, на водных цветковых растениях,
раковинах моллюсков, камнях, в виде черных блестящих точек. УССР,
Кавказ, Ср. Азия.
8. Gongrosira pygmaea Kiitz. — Гонгрозира карликовая (рис. 144,
14, 15). — Слоевище в виде подушечек, бледно-зеленое. Ползучие нити
образуют псевдопаренхимную подошву. Прямостоячие нити до 200 мкм
дл., обильно односторонне разветвленные. Боковые ветви многоклеточ-
ные, восходящие, почти все заканчивающиеся на одном уровне. Клетки
цилиндрические, (10)15—20 мкм шир., по длине равные ширине, у осно-
вания ветвей вытянутые, в 2—3 раза длиннее. Размножение не ис-
следовано.
В слабо проточных водах, на растительном субстрате. УССР (р. Че-
репаха) .
9. Gongrosira recurvata (Wittr. et Nordst.) Printz (= Trentepohlia
recurvata Wittr. et Nordst.) — Гонгрозира вниз-отогнутая (рис. 144,
16, 17). — Слоевище почти шаровидное, до 3 мм в диам., инкрустиро-
ванное известью. Ползучие нити образуют подошву. Прямостоячие нити
односторонне разветвленные, отходящие от подошвы радиально. В ниж-
ней части нитей веточки преимущественно прямые, направленные вверх,
односторонне разветвленные, в верхушечной части ветви и концы
прямостоячих нитей отогнуты вниз. Клетки в нижней части прямо-
стоячих нитей цилиндрические, 8—14 мкм шир., по длине в 2—3 раза
больше, в верхушечной части изодиаметрические, шаровидно вздутые.
В пресных водоемах, на крупных водорослях. В СССР не обна-
ружена. (Зап. Европа).
10. Gongrosira diseiformis Fritsch — Гонгрозира дисковидная
(рис. 144, 18, 19). — Слоевище в виде подушечек около 1 мм в диам.,
инкрустированное известью. Ползучие нити обильно разветвленные,
образующие псевдопаренхимную пластинку подошвы. Прямостоячие
нити короткие, простые, плотно сжатые. Клетки ползучих нитей упло-
щенные, прямостоячих — правильно цилиндрические, 9—12 мкм шир.,
265
Рис. 145.
1—3 — Gongroslra burmanica: 7 — общий вид слоевища, 2, 3 — образование
акинет; 4—7 — G. Incrustans: 4 — часть молодого слоевища, 5 — вертикальный
разрез слоевища, 6 — ветвь прямостоячей нити, 7 — нить с акинетой; 8—10 —
G. depauperata: 8 — фрагмент слоевища, 9,10 — акинеты (?); 11—15 — G. lepto-
tricha: 11 — ползучая нить, 72 — прямостоячая нить, 13—15 — фрагменты слое-
вища. (1—3 — по Скуе, 4—7 — по Батчеру, 8—70 — по Воронихину, 11—15 —
по Рейнери).
по длине равные ширине или до 2 раз больше, со слегка утолщенными
оболочками.
В пресных водах, в обрастании урути. В СССР не обнаружена.
(Англия).
11. Gongrosira burmanica Skuja — Гонгрозира бирманская (рис. 145,
1—3). — Слоевище в виде полушаровидных подушечек до 750 мкм толщ.,
инкрустированное известью. Подошва не выражена. Прямостоячие нити
266
более или менее разветвленные, особенно в верхушечной части, от
основания расходящиеся радиально, плотно сжатые и частично срастаю-
щиеся между собой боковыми сторонами. Клетки цилиндрические или
слегка бочонковидные, 10—20 мкм шир., подлине вдвое меньше или до 1.5
раза больше, в нижних частях старых нитей частично отмирающие
и тогда пустые, или превращающиеся в акинеты. Зооспорангии верху-
шечные, эллипсоидные, чуть больше вегетативных клеток. Акинеты
эллипсоидные, 18—20 мкм шир., 22—27 мкм дл.
В обрастании камней. В СССР не обнаружена. (Зап. Европа,
Бирма).
12. Gongrosira incrustans (Reinsch) Schmidle (—Chlorotylium incru-
stans Reinsch) — Гоигрозира инкрустированная (рис. 145, 4—7). —
Слоевище в виде ярко- или темно-зеленой корочки до 1 мм толщ.,
инкрустированное известью. Подошва не выражена. Прямостоячие нити
простые или скудно односторонне разветвленные, плотно сжатые.
Ветвление гребневидное, чаще в основании нитей, реже в верхней
их части. Клетки цилиндрические, 6—11.5 мкм шир., по длине равные
ширине или до 3 раз больше. Зооспорангии верхушечные, по величине
почти не отличающиеся от вегетативных клеток. Размножение не про-
слежено.
В стоячих и проточных водах, на водных цветковых растениях,
крупноклеточных нитчатых водорослях, раковинах моллюсков. ЛатвССР,
УССР, Кавказ.
13. Gongrosira depauperata Woronich. — Гоигрозира обедненная
(рис. 145, 8—10). — Слоевище в виде ярко- или темно-зеленой корочки,
инкрустированной известью. Ползучие и прямостоячие нити не разли-
чаются между собой. Прямостоячие нити короткие, 100—260 мкм выс.,
параллельные друг другу, скудно и односторонне разветвленные,
с 3—4 короткими (из 2—5 клеток) ветвями, отходящими подряд от
клеток нити, или ветвление беспорядочное и более обильное, приводящее
к образованию сложных сплетений, иногда встречаются совершенно
простые нити или с едва заметными веточками. Клетки разнообразной
формы: от почти правильно цилиндрических до раздутых или непра-
вильно искривленных, 5—17 мкм шир., по длине равные ширине или
до 3(6) раз больше, часто разных размеров на протяжении одной
нити. Оболочки клеток обычно тонкие, реже утолщенные и слоистые,
поперечные перегородки часто утолщенные, что приводит к образованию
целлюлозных пробок. Очень часто встречаются клетки без пигмента,
иногда вытягивающиеся в гифоподобные нити. Размножение не просле-
жено. Клетки с утолщенной оболочкой, возможно, акинеты.
В горных, быстро текущих водах, в известковых налетах на камнях
и гальке. Крым (рр. Альма, Сары-Су).
Вид очень интересный, но изучен слабо, по некоторым признакам
напоминает род Gomontia.
14. Gongrosira leptotrieha Reineri — Гоигрозира тонковолоеиетая
(рис. 145, 11—15). — Слоевище в виде поросли, не инкрустированной
известью. Ползучие нити симподиально ветвящиеся, образующие рыхлую
псевдопаренхимную подошву. Прямостоячие нити короткие, слабо раз-
ветвленные. Клетки ползучих нитей неправильно эллипсоидные или
многоугольные от взаимного давления, уплощенные с нижней стороны
и сосковидно вытянутые вверх, прямостоячих — неправильно цилиндри-
ческие, 4—6(7) мкм шир., по длине не более чем до 4 раз больше.
267
Рис. 146.
1—4 — Gongrosira schmtdlei: 1 — общий вид слоевищ на субстрате (нат. вел.),
2—4 — фрагменты слоевища; 5 — G. tibetana, общий вид слоевища (при слабом
увел.); 6—9 — G. scourfieldii'. 6 — фрагмент слоевища, 7—9 — фрагменты нитей
с пустыми спорангиями. (7, 4 — по Мошковой, 2, 3 — по Воронихину, 5 —
по Скуе, б—9 — по Весту).
Конечные клетки округленные или бутылковидно вытянутые. Вегетатив-
ное размножение распадением нитей на отдельные шаровидные или
яйцевидные клетки или группы клеток. Бесполое и половое размно-
жение не прослежено.
В горных ручьях, на камнях. В СССР не обнаружена. (Швейцария).
15. Gongrosira schmidlei Р. Richter (=G. koppei Budde) — Гонгро-
зира Шмидле (рис. 146,1—4). — Слоевище в виде плоской или полушаро-
видной подушечки до 4 мм шир., 2 мм выс., инкрустированное из-
вестью. Ползучие нити образуют псевдопаренхимную подошву. Прямо-
268
стоячие нити ооильно, преимущественно односторонне разветвленные,
окруженные слизью, из которой во все стороны торчат концы ветвей.
Клетки ползучих нитей без ризоидов, обычно лишены содержимого,
прямостоячих — искривленно-цилиндрические, часто с боковыми выро-
стами в верхней части, 9—13(15) мкм шир., по длине до 5 раз
больше. Хлоропласт размещается главным образом в конечных клетках.
Оболочки клеток довольно тонкие, неслоистые. Зооспорангии конечные,
почти ничем нс отличающиеся от вегетативных клеток; в коротких
клетках образуются по 2—4 зооспоры, в длинных — 16 зооспор. Часто
перед образованием зооспор верхушечные клетки прямостоячих нитей
интенсивно делятся на ряд коротких клеток, в каждой из которых воз-
никает ио 2 зооспоры. Зооспоры около 4 мкм дл., освобождаются
через отверстие, возникающее в оболочке материнской клетки. Акинеты
не наблюдались.
В стоячих и проточных водах, на водных цветковых растениях,
древесине, раковинах моллюсков, камнях. ЛатвССР, УССР, Кавказ.
16. Gongrosira tibetana (Skuja) Printz — Гонгрозира тибетская
(рис. 146, 5). — Слоевище в виде подушечек 0.3—0.4 мм выс., не ин-
крустированное известью. Ползучие нити разветвленные, ризоидо-
образные. Прямостоячие нити обильно всесторонне разветвленные, моло-
дые веточки обычно простые, к концам слегка суживающиеся.
Клетки цилиндрические, 7—13 мкм шир., по длине вдвое меньше или
до 2.5 раза больше, слегка вздутые, с тонкими бесцветными оболочками.
Зооспорангии почти ничем не отличаются от вегетативных клеток, только
слегка шире. Зооспоры многочисленные, освобождаются через отверстие
в боковой стенке спорангия.
В горячих источниках. В СССР не обнаружена. (Центр. Азия).
17. Gongrosira scourfieidii G. S. West — Гонгрозира Скурфильда
(рис. 146, 6—9). — Слоевище в виде ярко-зеленого ложа 4—9 мм толщ.,
инкрустированное известью. Ползучие нити сетевидно переплетенные,
тесно сжатые, анастомозирующие. Прямостоячие нити скудно разветвлен-
ные, с верхушками, выдающимися за пределы известковой корочки.
Клетки ползучих нитей беспорядочно вздутые, 12—16 мкм шир., прямо-
стоячих — цилиндрические, 7.5—9.5 мкм шир., по длине равные ширине
или до 9 раз больше; короткие и длинные клетки размещаются в нитях
беспорядочно. Конечные клетки всегда тупо закругленные. Зооспорангии
вставочные, относительно короткие, 16—19 мкм шир. Зооспоры осво-
бождаются через боковое отверстие в оболочке спорангия.
В быстро текущих водах, на старой древесине. В СССР не обна-
ружена. (Англия).
Род GOMONTIA Born, et Flah. - ГОМОНТИЯ
(—Gongrosira Chod. p. p.; Foreliella Chod.)
Слоевище в виде беспорядочно разветвленных, иногда радиально
расходящихся ползучих нитей, полностью или частично погруженных
в субстрат, где они образуют рыхлые сплетения или плотные псевдопа-
ренхимные пластинки, и отходящих от них относительно коротких,
простых или беспорядочно разветвленных прямостоячих нитей (послед-
ние иногда бывают выражены неотчетливо). Клетки неправильной
формы, одно- или многоядерные. Хлоропласт преимущественно 1, при-
стенный, дисковидный или лентовидный, иногда сетчато анастомозирую-
269
щий, с 1 или несколькими пиреноидами. Нити таллома часто на зна-
чительном протяжении распадаются на отдельные клетки, продолжаю-
щие делиться и расти дальше. Бесполое размножение зооспорами и
апланоспорами. Зооспорангии вставочные, одиночные или по нескольку
в ряд; они образуются из коротких, слегка вздутых клеток. Зооспоры
по 2—4, яйцевидные, со стигмой, с 2 или 4 жгутами. Некоторые
клетки нитей превращаются в шаровидные, овальные или клиновидные
спорангии (гаметангии ?) с характерными корневидными отростками
и утолщенными оболочками, которые со временем отчленяются от
нитей. В одних из этих спорангиев образуется большое количество
грушевидных двужгутиковых зооспор разной величины (возможно,
гаметы, хотя копуляция не наблюдалась), в других, таких же по форме
и величине, — большое количество шаровидных неподвижных клеток —
апланоспор. Апланоспоры непосредственно прорастают в вегетативные
нити или повторно образуют апланоспорангии. Часто вследствие ослиз-
нения оболочек клеток наблюдается пальмеллевидное состояние, при
котором клетки также могут превращаться в спорангии, варьирующие
по форме и величине.
В морских, соленых и пресных стоячих и проточных водах, на рако-
винах моллюсков, скалах, камнях и других субстратах.
I. В морских водах, на раковинах моллюсков . . . . G. polyrrhiza 1.
II. В пресных водах, на разных субстратах.
1. Клетки в ползучих и прямостоячих нитях преобладающе много-
угольные.
А. Ветви плотно сжатые; на камнях............G. codiolifera 2.
Б. Ветви не сжатые; на раковинах двустворчатых моллюсков .
.............................................G. holdenii 3.
2. Клетки в ползучих и прямостоячих нитях цилиндрические или
бочонковидные.
А. Клетки 8—10 мкм шир......................G. aegagropilae 4.
Б. Клетки свыше 20 мкм шир.
а. Клетки цилиндрические; в древесине . . . G. lignicola 5.
б. Клетки бочонковидные; на скалах, в реках..............
..........................................G. rupicola 6.
3. Клетки нитей на поверхности субстрата неправильной формы,
вздутые и изогнутые, врастающих в субстрат — тонкие, длинные
........................................... . . . G. perforans 7.
1. Gomontia polyrrhiza (Lagerh.) Born, et Flah. (=Codiolum polyrrhi-
zum Lagerh.) — Гомонтия многокорешковая (рис. 147, 1, 2). — Ползу-
чие нити беспорядочно разветвленные, срастающиеся в псевдопаренхим-
ную пластинку. Прямостоячие нити выражены неотчетливо. Клетки
нитей 5—9 мкм шир. Апланоспорангии толстостенные, крупные, SO-
125 мкм шир., 150 — 250 мкм дл., с 1 или несколькими простыми или раз-
ветвленными ризоидными или корнеобразными выростами, располо-
женными венцом на конце клетки, обращенном к субстрату.
В морях, большей частью в загрязненных местах, на раковинах
моллюсков. Сев. Ледовитый океан, Черное (лиманы Одесской и Нико-
лаевской обл., Крымское и Кавказское побережья) и Каспийское
моря.
2. Gomontia codiolifera (Chod.) Wille — Гомонтня кодиолиевая
(рис. 147, 3, 4). — Ползучие нити короткие, внедряющиеся в извест-
ковый субстрат. Прямостоячие нити короткие, малоклеточные, плотно
270
Рис. 147.
1,2 — Gomontia polyrrhiza, разные по возрасту апланоспорангии; 3, 4 —
G. codiolifera: 3 — фрагмент нити, 4 — отдельный спорангий; 5, 6 — G. holden.it:
5 — фрагменты нитей, 6 — акинеты с апланоспорами; 7, 8 — G. aegagropilae:
7 — разные формы нитей, 8 — акинета. (7 — по Лагергейму, 2 — по Линдстедту,
3, 4 — по Шода, 5, 6 — по Коллинсу и Смиту, 7, 8 — по Актону).
сжатые. Клетки нитей многоугольные, с толстой оболочкой, у прямо-
стоячих нитей иногда переходящие в цилиндрические. Хлоропласт
с 1—2 пиреноидами. Спорангии шаровидные или овальные, 20 мкм
шир., 45 мкм дл., с корнеобразными выростами утолщенной оболочки,
обращенными к субстрату (стадия Codiolum).
В пресных быстро текущих водах, в обрастании камней. ЛатвССР,
УССР (Крымский заповедник).
3. Gomontia holdenii Collins — Гомонтия Холдена (рис. 147, 5,6). —
Ползучие нити беспорядочно разветвленные. Прямостоячие нити
выражены неотчетливо. Клетки толстостенные, очень изменчивой формы
и величины, многоугольные, эллипсоидные или цилиндрические,
12—50 мкм шир. Конечные клетки цилиндрические или суженные.
Спорангии яйцевидные, с удлиненным основанием, 30 мкм шир.,
100 мкм дл.
В пресных водах, на раковинах двустворчатых моллюсков. В СССР
не обнаружена. (Америка).
4. Gomontia aegagropilae Acton — Гомонтия эгагропильная
(рис. 147, 7, 8). — Ползучие нити радиально отходящие от центра,
с тонкими веточками, врастающие в субстрат. Прямостоячие нити
выражены неотчетливо. Клетки ползучих нитей многоядерные (от 1
до 6 ядер), цилиндрические, более или менее бочонковидные или
яйцевидные, 8—10 мкм шир., 10—16 мкм дл., клетки врастающих
в субстрат нитей цилиндрические, сильно вытянутые, 2 мкм шир.,
до НО мкм дл., обычно бесцветные. Конечные клетки от слегка-заост-
ренных до широко закругленных. Хлоропласт с 1—6 пиреноидами.
Вегетативное размножение отчленением от таллома конечных клеток,
иногда и некоторых срединных клеток, сразу прорастающих в новые
271
Рис. 148.
1—6 — Gomontla ligntcola: 1,2 — вегетативные клетки, 3 — зооспоры, 4, 5 —
стадии прорастания зооспор, 6 — прорастание акинеты; 7, 8 — G. rupicola:
7 — фрагмент нити, 8 — апланоспорангий. (1—6 — по Муру, 7, 8 — по Жао).
нити или превращающихся вначале в покоящиеся акинеты. Подвиж-
ных клеток не найдено.
В отмерших клетках кладофоры эгагропильной. В СССР не обнару-
жена. (Англия).
5. Gomontia lignicola Moore — Гомонтия древесинная (рис. 148,
1—6). — Ползучие нити обычно простые, изредка разветвленные. Пря-
мостоячие нити выражены неотчетливо. Клетки неправильные, в осно-
вании цилиндрические, 25—45 мкм шир., 100— 200 мкм дл., с 1—6 ядрами.
Хлоропласт с многочисленными пиреноидами, хорошо развитый лишь
в конечных клетках, тогда как в нижележащих клетках он редуци-
272
руется или исчезает вовсе. Зооспоры двужгутиковые, шаровидные или
эллипсоидные, 10 — 12 мкм шир., 12 15 мкм дл.; они сразу прорастают
в новые нити или вначале превращаются в покоящиеся акинеты, про-
растающие затем в ползучие разветвленные нити.
В клетках древесной паренхимы. В СССР не обнаружена. (Сев.
Америка).
6. Gomontia rupicola Jao — Гомонтия наскальная (рис. 148, 7,8).—
Ползучие нити беспорядочно разветвленные, плотно переплетенные.
Прямостоячие нити выражены неотчетливо. Клетки разнообразной
формы и величины, чаще неправильно бочонковидные, вздутые, (16)20—
60(100) мкм шир., 38—117 мкм дл., с 1—4 ядрами. Оболочка клеток
толстая, слоистая. Аплаиоспорангии верхушечные и вставочные,
неправильной формы, 40—70 мкм шир., 50—80 мкм дл., часто с 1 или
многими короткими выростами оболочки. Апланоспоры многочисленные,
шаровидные, около 10 мкм шир.
В реках, на скалах, постоянно омываемых водой. В СССР не обна-
ружена. (Китай).
7. Gomontia perforans (Chod.) Acton [—Foreliella performs Chod.;
Tellamia perforans (Chod.) Wille] — Гомонтия пронзающая (рис. 149,
1—3). — Ползучие нити на поверхности субстрата немногочисленные,
искривленные, скудно ветвящиеся, нити, врастающие в субстрат,
обильные, более или менее прямые, дихотомически разветвленные.
Прямостоячие нити выражены неотчетливо. Клетки наружных ползу-
чих нитей относительно короткие, неправильной формы, вздутые,
изогнутые, вильчато разделенные, иногда слагающиеся в псевдопа-
ренхимные образования, клетки врастающих в субстрат нитей длинные,
тонкие, 4—5 мкм шир., постепенно переходящие в ризоидные выросты,
разнообразно вздутые на концах. Хлоропласт с пиреноидом. Продукт
ассимиляции — крахмал, в ризоидных выростах еще и масло. Вегета-
тивное размножение отчленением клеток, растущих на наружной
поверхности субстрата. Бесполое размножение зооспорами и акинетами.
В зооспорангии превращаются клетки, растущие на внешней поверхности
и внутри субстрата. Акинеты с толстыми оболочками.
В пресных водах, на раковинах живых двустворчатых моллюсков.
В СССР не обнаружена. (Зап. Европа).
Род CTENOCLADUS Borzi - КТЕНОКЛАДУС
(—Lochmiopsis Woronich. et Popova)
Слоевище в виде плотных дерновинок или бородавчатых корочек
0.2—10 мм выс., состоящих из подошвы, образованной ползучими
нитями из округлых или более или менее многоугольных клеток,
и отходящих от них прямостоячих нитей. Прямостоячие нити в нижней
части простые, в верхней обильно разветвленные, преимущественно
односторонне, часто гребневидно. Ветви неодинаковой длины, одно-
сторонние или очередные; они всегда отходят от верхнего конца мате-
ринских клеток, у которых перед поперечной перегородкой обычно
образуются боковые выпячивания, продолжающиеся в ветвь. Клетки
прямостоячих нитей большей частью цилиндрические, конечные клетки
закругленные. Хлоропласт пристенный, пластинчатый, с 1 — 6 пиренои-
дами. Бесполое размножение зооспорами и акинетами. Зооспорангии
18 Зеленые водоросли. Улотриксовые
273
Рис. 149.
1—3 — Gomontta perforans: 1, 2 — срез через наружные слои раковины мол-
люска, 3 — срез через внутренние слои раковины моллюска; 4—9 — Ctenocladus
clrcinatus: 4 — фрагмент слоевища с зооспорангиями, 5 — вегетативная ветвь,
6, 7 — фрагменты слоевища со вставочными и верхушечными акинетами, 8 —
прорастание акинет, 9 — слоевище в пальмеллевидном состоянии. (1—3 —
по Шода, 4, 8 — по Борзи, 5—7,9 — по Воронихину и Поповой).
яйцевидные или бочонковидные, верхушечные или вставочные, одиноч-
ные или иногда по нескольку в коротких цепочках, мало отличаю-
щиеся от вегетативных клеток. Зооспоры двужгутиковые, двух разме-
ров — крупные и мелкие; они освобождаются через неправильно округлое
отверстие на вершине или сбоку спорангия. Акинеты шаровидные или
яйцевидные, верхушечные или вставочные, с толстой слоистой оболочкой
и зернистым содержимым, зеленые, с большим количеством крахмала;
274
'они развиваются на прямостоячих нитях, одиночно или по нескольку
в ряд. Половое размножение двужгутиковыми грушевидными гаметами.
Зигота шаровидная.
1. Ctenocladus circinatus Borzi [=С. fastigiatus Borzi; Lochmiopsis
sibirica Woronich. et Popova; L. printzii Woronich. et Popova; L. chodatii
(Reineri) Printz; Gongrosira circinata (Borzi) Schmidle; G. fastigiata
(Borzi) Schmidle] — Ктенокладус закрученный (рис. 149, 4^9). —
Слоевище до 10 мм выс. Подошва сетчато распростертая или клубко-
видно утолщенная, псевдопаренхимная. Ползучие нити беспорядочно
разветвленные. Прямостоячие нити слегка дуговидно изогнутые, плотно
сжатые, в верхушечной части преимущественно односторонне гребне-
видно разветвленные. Клетки цилиндрические, у поперечных перего-
родок слегка перетянутые, 10—15 мкм шир., почти такой же длины
или в 1.5 — 2 раза больше, к вершинам нитей слегка суживающиеся.
Оболочки клеток утолщенные (до 2—2.5 мкм), неясно слоистые. Хлоро-
пласт с пиреноидом. Макрозооспорангии чуть больше вегетативных
клеток. Макрозооспоры по (4)8—32, реже больше, двужгутиковые,
со стигмой, яйцевидные или овально-яйцевидные, 5—7 мкм шир.
Акинеты шаровидные или эллипсоидные, 12.5—20 мкм шир.; возрас-
тание солености среды вызывает массовое развитие акинет. Известно
и пальмеллевидное состояние, наступающее после образования макрозоо-
спор и ослизнения оболочек клеток. При этом во многих клетках
могут возникать микрозооспоры 2—3 мкм шир.; они образуются по 4—16
в клетке, но дальнейшее их развитие не выяснено. Из других клеток
пальмеллевидной стадии может развиться подошва нового организма,
состоящая из очень длинных, беспорядочно разветвленных нитей,
от которых вверх отходят одиночные клетки или нити, богатые хлоро-
филлом. При неблагоприятных условиях эти клетки могут снова
переходить в пальмеллевидное состояние и образовывать микрозооспоры.
Некоторые клетки пальмеллевидной стадии не ослизняются, а увеличи-
ваются до 80 мкм шир. и превращаются в акинеты с толстыми оболоч-
ками; длительное время они находятся в состоянии покоя, после чего
в них образуется 30—60 гамет. Гаметы до 3 мкм шир., похожие на
микрозооспоры; вышедшие из одного спорангия, они не копулируют
друг с другом. Развитие зигот не исследовано.
В солоноватых стоячих или медленно текущих водах, часто в содовых
озерах, в свободном Состоянии или на камнях, видных цветковых
растениях, древесине; обнаружена также в культурах, засеянных
почвой. Кировская обл., Ханты-Мансийский авт. окр., Омская обл.,
Алтайский край (Кулундинская степь), Хакасская авт. обл. (Качинская
степь), КазССР, ТуркмССР, УзССР.
Семейство TRENTEPOHLIACEAE De-Toni -
ТРЕНТЕПОЛИЕВЫЕ
Слоевище нитевидное, разрастающееся на субстрате в форме
рыхлых порошковатых налетов или войлочных дерновинок. Нити
простые или ветвящиеся, полностью распростертые по субстрату или
частично прямостоячие. У некоторых распростертых по субстрату
форм нити плотно смыкаются друг с другом, образуя однослойные
псевдопаренхимные пластинки, над поверхностью которых возвышаются
одиночные короткие членики; у некоторых видов имеются ризоиды
и волосовидные выросты. У тропического рода Cephaleuros, приспо-
18*
275
сооившегося к паразитным условиям существования, слоевище состоит
из многослойной подошвы или псевдопаренхимного, иногда эндо-
фитного ложа, от которого к субстрату отходят ризоиды, а над поверх-
ностью возвышаются спорангии и многоклеточные волосовидные
выросты. Клетки шаровидные, цилиндрические, бочонковидные,
нередко многоугольные, с целлюлозной, преимущественно толстой,
слоистой оболочкой, соединяющиеся между собой плазмодесмами.
Ядро 1, реже их несколько. Хлоропласты многочисленные, дисковидные
или лентовидные, без пиреноидов. Продукт ассимиляции — масло,
в клетках накапливается также Р-каротин, благодаря чему слоевище
обычно окрашено в желтый, красный или оранжевый цвета. Рост
нитей верхушечный или, нередко, вставочный. Вегетативное размно-
жение фрагментацией встречается редко. Бесполое размножение
четырехжгутиковыми зооспорами без стигмы, возникающими в особых
крючковидных или воронковидных зооспорангиях, а также акинетами
и апланоспорами. Половое размножение двужгутиковыми изогаметами
без стигмы, образующимися в шаровидных сидячих гаметангиях; они
нередко прорастают партеногенетически.
Аэрофитные водоросли, обитающие на коре деревьев, стеблях
и листьях кустарниковых и травянистых растений, влажных валунах
и скалах. Встречаются в северных широтах, в горах доходят до субаль-
пийского пояса. Имеются эндофитные и эпифитные формы, встре-
чающиеся преимущественно в тропических и субтропических районах,
но легко акклиматизирующиеся в оранжереях умеренных широт.
I. Слоевище в форме разветвленных ползучих, приподнимающихся
или прямостоячих обособленных нитей, разрастающихся на субстрате
в порошковатые налеты или войлочные подушковидные дерновинки,
изредка на слоевищах образуются одноклеточные волосовидные вы-
росты ..................................род Trentepohlia (с. 276).
II. Слоевище в виде псевдопаренхимных пластинок.
1. Псевдопаренхимные пластинки дисковидные, однослойные, без
ризоидов и волосовидных выростов...........................
..................................род Phycopeltis (с. 297).
2. Псевдопаренхимные пластинки разнообразных очертаний, одно-
или многослойные, с ризоидами и многоклеточными волосовид-
ными выростами.......................род Cephaleuros (с. 299).
Род TRENTEPOHLIA Marti us - ТРЕНТЕПОЛИЯ
(=Chroolepus Ag.)
Слоевище нитевидное. Нити ветвящиеся, частично ползучие,
частично приподнимающиеся или прямостоячие, образующие спутанные,
большей частью подушковидные дерновинки, реже порошковатые на-
леты. Ползучие по субстрату нити без ризоидов, обычно короткие,
нередко распадающиеся на отдельные клетки, прямостоячие нити
длинные или короткие, простые или ветвящиеся, с веточками, направ-
ленными во все стороны, отходящими как от верхнего конца мате-
ринских клеток, так и от середины. Ветвление нитей моноподиальное.
Клетки шаровидные, эллипсоидные, цилиндрические, бочонковидные,
реже многоугольные. Конечные клетки прямостоячих нитей в боль-
шинстве случаев оканчиваются своеобразными пектозными колпачками,
которые по мере роста клеток постепенно сдвигаются ца их боковую
поверхность. Оболочки клеток толстые, целлюлозные, слоистые. Хлоро-
276
{пласты многочисленные, дисковидные, без пиреноидов, нередко соеди-
ненные в короткие ленты или цепочки. Вегетативное размножение
фрагментацией слоевища выражено слабо. Бесполое размножение четы-
рехжгутиковыми зооспорами и акинетами. Зооспоры образуются
в шаровидных, эллипсоидных или яйцевидных спорангиях, возникающих
на прямых или крючковидно согнутых субспорангиальных клетках.
В спорангий преобразуется верхушечная часть трубчатого выроста
субспорангиальной клетки, отделяющаяся от остальной части клетки
2 кольцевидными утолщениями. Отверстие для выхода зооспор горло-
видное, короткое, образуется сбоку или на вершине спорангиев, которые
открываются чаще всего после их отпадения, обычно в сухую погоду.
Акинеты известны лишь для Г. umbrirur, при их образовании клетки
округляются, заполняются запасными питательными веществами
и освобождаются после ослизнения внешних слоев оболочки, прорастая
затем в новые нити. Половое размножение двужгутиковыми, удли-
ненно-эллипсоидными или яйцевидными, плоско сжатыми изогаметами,
образующимися по 16, 32, 64 в шаровидных или яйцевидных верху-
шечных, боковых или вставочных гаметангиях, возникающих на ползу-
чих или прямостоячих нитях. Освобождаются гаметы через круглое
или удлиненное отверстие на горловидном выросте гаметангия.
На стволах деревьев, полусгнивших пнях и каменистом субстрате.
I. Слоевище без одноклеточных волосовидных выростов.
1. Клетки цилиндрические, по крайней мере в прямостоячих нитях.
А. Прямостоячие нити простые или очень скудно ветящиеся.
Зооспорангии не образуются, известны только гаметангии,
а. Прямостоячие нити 5—12 мкм шир., к вершине отчетливо
^суживающиеся, прямые.
а. Гаметангии одиночные или по нескольку в ряд, пре-
имущественно на прямостоячих нитях, окутанных буро-
ватым веществом.......................Т. calamicola 1.
р. Гаметангии многочисленные, преимущественно на пол-
зучих по субстрату нитях, реже на базальных частях
прямостоячих нитей или на коротких веточках, отхо-
дящих от ползучих нитей....................Т. effusa 2.
б. Прямостоячие нити 16—20 мкм шир., к вершине не сужи-
вающиеся, обычно изогнутые.................Т. jucunda 3.
Б. Прямостоячие нити ветвящиеся (иногда простые). Известны
зооспорангии и гаметангии.
а. Гаметангии шаровидные, эллипсоидные или яйцевидные,
со слабо развитым горловидным выростом.
а. Зооспорангии воронковидные, поперечно-эллипсоидные,
с прямой субспорангиальной клеткой ..................
...................................Т. annulata 4.
р. Зооспорангии шаровидные, овальные или яйцевидные,
часто с крючковидной субспорангиальной клеткой.
* Зооспорангии одиночные, реже по 2—3, на вздутой
верхушечной клетке прямостоячей нити.
4- Оболочки клеток толстые, продольно-слоистые,
пектозные колпачки имеются; прямостоячие нити
длинные, многоклеточные.
X Прямостоячие нити не более 10 мкм шир.,
пектозные колпачки развиты слабо.............
............................Т. abietina 5.
277
XX Прямостоячие нити в большинстве свыше
10 мкм шир., пектозные колпачки развиты
хорошо.................... . . . Т. аигеа 6.
+ + Оболочки клеток топкие, не слоистые, пектоз-
ные колпачки отсутствуют: прямостоячие нити
короткие, малоклеточные . . . . Т. uncinata 7.
* * Зооспорангий по 2—8 на вздутой верхушечной
клетке прямостоячей нити.
+ Нити и ветви довольно ровные на всем про-
тяжении, клетки 13—28 мкм шир................
.............................Т. агЬогшп 8.
+ + Нити и ветви к концам суженные, бичевидно
удлиненные, клетки 17—32 мкм шир.
...........................Т. hunanensis 9.
б. Гаметангии бутылковидные, с довольно хорошо развитым
горловидным выростом...................Т. lagenifera 10.
2. Клетки шаровидные, эллипсоидные, яйцевидные или неправиль-
ные, иногда асимметричные — выпуклые на спинной и уплощен-
ные на брюшной стороне, изредка цилиндрические.
А. Оболочки клеток косослоистые, с перекрещивающимися на-
ружным и внутренним слоями; преимущественно на каменис-
том субстрате..............................Т. jolithus И.
Б. Оболочки клеток продольно- или концентрически слоистые,
с чешуйчато расходящимся наружу внешним слоем; преиму-
щественно на стволах деревьев.
а. Нити образуют дерновинки; зооспорангии имеются, часто
с крючковидной субспорангиальной клеткой, открыва-
ющиеся внизу.
а. Нити большей частью ползучие, длинные, разнообраз-
но изогнутые, слегка ветвящиеся; клетки асимметричные
или неправильно цилиндрические ......................
....................................Т. rigidula 12.
р. Нити преимущественно прямостоячие; клетки цилиндри-
ческие, эллипсоидные или неправильно шаровидные.
* Клетки эллипсоидные, неправильно шаровидные,
реже цилиндрические, с толстой слоистой оболоч-
кой, чешуйчатой с поверхности, пектозные колпачки
мелкие, гаметангии с простым выводным отвер-
стием ........................... . Т. odorata 13.
* * Клетки цилиндрические, тонкостенные, пектозные
колпачки отсутствуют, гаметангии со сложным
выводным отверстием на тупом носике..............
....................................Т. gobii 14.
б. Нити образуют порошковатые налеты; зооспорангии от-
сутствуют или имеются.
а. Зооспорангии отсутствуют.
* Клетки (7)12—27(38) мкм шир., с бесцветной тол-
стой, грубо концентрически слоистой оболочкой;
содержимое клеток красное или красно-бурое . . .
...............................Т. umbrina 15.
* * Клетки (5)8.6—15 мкм в диам., со светло-серовато-
желтой тонкой плотной, четко очерченной оболоч-
кой; содержимое клетки желтовато- или оранжево-
бурое ..............................Т. piceana 16.
278
р. Зооспорангии имеются, открывающиеся на вершине .
................................. . Т. hygrophila 17.
II. Слоевище с одноклеточными волосовидными выростами.
1. Клетки шаровидные или эллипсоидные, 5—15 мкм шир., с 1 — 3
волосовидными выростами (2)3—6.5 мкм шир., 26—120 мкм дл. .
.............................................Т. bogoriensis 18.
2. Клетки цилиндрические или бочонковидные, 7 — 15 мкм шир., с 1
волосовидным выростом немного уже клетки, по длине до 10 раз
больше.......................................Т. lagerheimii 19.
1. Trentepohlia calamicola (Zeil.) De-Toni et Levi — Трентеполия
тростниковая (рис. 150, 1—3). — Образует дерновинки, во влажном
состоянии зеленоватые, при высыхании желтоватые. Нити перепутан-
ные; прямостоячие нити вильчато ветвящиеся, ветви к вершинам сужи-
вающиеся, почти всегда направленные в одну сторону; те и другие
покрыты бурым веществом, образующим на них своеобразную сеточку.
Клетки ползучих нитей 9 мкм шир., по длине до 1.5 раза больше,
прямостоячих — 7 — 12 мкм шир., по длине до 2—4 раз больше. Гаме-
тангии одиночные или по нескольку в ряд, шаровидные или эллипсоид-
ные, 12 мкм шир., сидячие, боковые, реже верхушечные.
На листьях тростника. В СССР не обнаружена. (Англия, тропи-
ческая Азия).
2. Trentepohlia effusa (Kremp.) Hariot [=T. setljera (Farlow)
Hariot] — Трентеполия разметанная (рис. 150, 4—9). — Образует
тонкие буровато-оранжевые дерновинки. Прямостоячие нити довольно
короткие, до 300—400 мкм, отчетливо суженные к вершине, обычно
простые, иногда скудно ветвящиеся. Клетки ползучих нитей цилин-
дрические или более или менее вздутые, прямостоячих — цилиндри-
ческие, 5—10(11.4) мкм шир., 18—22 мкм дл. Гаметангии многочислен-
ные, шаровидные или яйцевидные, 11—20 мкм шир., 22—30 мкм дл.,
с простым круглым выводным отверстием;. они сидячие, возникают
чаще всего непосредственно на ползучих нитях, реже на базальных
клетках прямостоячих нитей или на верхушках их одноклеточных
боковых веточек, изредка на верхушках особых коротких малоклеточных
прямостоячих нитей, отходящих от ползучих нитей.
Во влажных местах, на каменистых и древесных субстратах. УССР
(горный Крым).
Близка к Т. аигеа, известна пока из немногих мест, но несомненно
распространена значительно шире.
3. Trentepohlia jucunda (Cesati) Hariot — Трентеполия приятная
(рис. 150, 10). — Образует тонкие светло-желтые дерновинки, мало
меняющие цвет при высыхании. Прямостоячие нити длинные, скудно
ветвящиеся, несколько изогнутые, боковые веточки вытянутые, изогну-
тые. Клетки прямостоячих нитей цилиндрические, 16—20 мкм шир.,
по длине равные ширине или до 2—3 раз больше, клетки веточек 9—
11 мкм шир., по длине в 2—4 раза больше. Гаметангии шаровидные,
28—36 мкм в диам., верхушечные или боковые.
На коре Populus nigra. В СССР не обнаружена. (Сев. Италия).
Малоизученный вид, близкий к Т. аигеа.
Ь. Trentepohlia annulata Brand (=T. maleiformis Gutw.) — Трен-
теполия кольчатая (рис. 151). — Образует бархатистые, во влажном
состоянии желто-оранжево-зеленые или буроватые дерновинки 1 — 1.5 мм
279
1—3 — Trentepohlia calamicola'. 1 — базальная часть слоевища с прямостоячими
нитями, 2, 3 — фрагменты прямостоячих нитей с сидячими гаметангиями;
4—9 — Т. effusa'. 4—6 — ползучие и прямостоячие нити, 7—9 — фрагменты
слоевища с сидячими и верхушечными гаметангиями; 10 — Т. juciinda (1 3 —
но Весту, 4—9 — по Вильдеману п Фарлоу, 10 — но Харкоту).
280
Рис. 151.
Trentepohlia annulata: 1 — общий вид слоевищ, 2—4 — начальные стадии раз-
вития зооспорангия, 5 — зрелый зооспорангий, 6 — зооспора. (7, 5, 6 — по
Скуе, 2—4 — по Бранду).
выс., при высыхании серые. Прямостоячие нити скудно разветвленные,
отходящие от псевдопаренхимной подошвы или четковидных, скудно
ветвящихся обособленных ползучих нитей. Клетки прямостоячих нитей
цилиндрические или слегка бочонковидные, 9.5—19(24) мкм шир.,
по длине в 1.5—3 раза больше, в базальных участках иногда очень
вздутые, до 38 мкм шир. Оболочки клеток бесцветные, до 2 мкм толщ.,
косослоистые,12 преимущественно с довольно гладкой поверхностью
12 Слоистость оболочки хорошо заметна при обработке нитей едким
калием.
281
и лишь местами чешуйчатые; на одной из сторон поперечных перего-
родок иногда появляются крапинки. Зооспорангии воронковидные,
поперечно-эллипсоидные, 30—58 мкм шир., 17—38 мкм дл., одиночные,
на верхушках прямостоячих нитей. Зооспоры удлиненно-цилиндри-
ческие, на концах округленные, 2.5 мкм шир., около 13 мкм дл.,
по 32, 64 в спорангии; иногда внутри спорангия зооспоры одеваются
оболочкой, превращаясь в апланоспоры. Гаметангии шаровидные или
эллипсоидные, 16—30 мкм шир., 29—52 мкм дл., с толстой, концентри-
чески слоистой оболочкой 6—7 мкм толщ., боковые или верхушечные,
на ползучих нитях.
В затененных сыроватых местах, на лежащих на земле полуразру-
шенных стволах деревьев, на боковых и торцовых поверхностях пней
и т. п., преимущественно в горных районах до высоты 950 м над ур. м.
ЛатвССР, УССР (Карпаты), Абхазская АССР (окр. оз. Рица).
5. Trentepohlia abietina (Flotow) Hansg. — Трентеполия пихтовая
(рис. 152). — Образует тонкие дерновинки, во влажном состоянии крас-
новатые, золотисто-желтоватые или желтоватые, при высыхании серые.
Нити беспорядочно ветвящиеся, веточки искривленные или согнутые.
Клетки ползучих нитей и оснований прямостоячих нитей вздутые,
на остальных участках слоевища преимущественно цилиндрические,
3.5 — 10 мкм шир., по длине равные ширине или до 4 раз больше. Обо-
лочки клеток слоистые, с расходящимися слоями, у верхушечных
клеток на конце довольно толстые; пектозные колпачки развиты слабо.
Зооспорангии шаровидные или яйцевидные, 9—20 мкм в диам. Гаме-
тангии более или менее шаровидные или яйцевидные, боковые, сидячие
или верхушечные, особенно обильные на прямостоячих нитях.
В районах с высокой влажностью воздуха, в тенистых лесах,
па коре елей и сосен, реже на стволах широколиственных деревьев.
F. abietina (рис. 152). —Клетки прямостоячих нитей 4—10 мкм
шир. Зооспорангии 12—20 мкм в диам.
Ленинградская, Калининская обл., окр. оз. Селигер, ЛатвССР,
ГрузССР, Абхазская АССР (окр. оз. Рица).
F. minor Wild. — Клетки прямостоячих нитей 3.5—5.5 мкм шир.
Зооспорангии 9—15 мкм в диам.
УССР (Карпаты).
6. Trentepohlia aurea (L.) Martius (—Bissus aurea L.; Chroolepus
aureum Kiitz.; C. velutinum Kutz.; Trentepohlia velutina Kiitz.; T. ger-
manica Gluck) —Трентеполия золотистая (рис. 153). — Образует дер-
новинки до 6 мм выс., во влажном состоянии золотисто-желтые, красно-
вато-оранжевые или буровато-красные, при высыхании желтовато- или
зеленовато-серые. Ползучие нити довольно длинные, беспорядочно
ветвящиеся, плотно прилегающие к субстрату или слегка приподни-
мающиеся. Прямостоячие нити простые или обильно разветвленные,
часто переплетенные, прямые, изогнутые или волнистые. Клетки ползучих
нитей вздутые, четковидные, реже цилиндрические, 10—15 (24) мкм
шир., с довольно толстой оболочкой, прямостоячих — длинные, цилин-
дрические, слегка вздутые, у поперечных перегородок иногда перетяну-
тые, 7.5 — 30 (32) мкм шир., по длине в 1.5—3 раза больше. Конечные
клетки обычно широкоокруглые, реже слегка суженные. Оболочки
молодых клеток довольно тонкие, гладкие, у старых клеток толстые,
слоистые, местами чешуйчатые, часто с пектозными колпачками,
достигающими иногда 10 мкм шир., 30 мкм дл. Зооспорангии на концах
прямостоячих нитей; сначала на расширенной конечной клетке нитей
282
Рис. 152.
Trentepohlia abietina: J — нити с гаметангиями, 2 — фрагменты слоевища
с лектозными колпачками на верхушечных клетках прямостоячих нитей, 3 —
начало образования верхушечного спорангия. (J — по Вильдеману, 2 — по
Блюму, 3 — по Кютцингу).
образуются 1 — 3 крючковидные субспорангиальные клетки 20—25 мкм
дл. и уже на них по эксцентрически расположенному эллипсоидному
спорангию 25—30 мкм шир., 27—40 мкм дл. Зооспоры 5—6 мкм шир.,
13 — 15 мкм дл. Гаметангии шаровидные, эллипсоидные, иногда кувши-
яовидные, 15—25 (55) мкм шир., по длине в 1.5 раза больше; они
образуются в большом количестве как на ползучих, так и на прямо-
стоячих нитях. Гаметы 4—5 мкм шир., 13—15 мкм дл.
На камнях, реже коре многих широколиственных и некоторых
хвойных деревьев.
Var. aurea (=var. genuina Hansg.) (рис. 153).—Дерновинки
маленькие, позднее сливающиеся. Прямостоячие нити более или менее
прямые или слегка согнутые, ветвящиеся. Клетки (7.5) 10 — 30 (32) мкм
шир., по длине в 1.5—3 раза больше. Конечные клетки обычно широко-
округлые. Зооспорангии 25—30 мкм шир., 27—40 мкм дл. Гаметангии
15—35 мкм в диам.
На скалах, крупных валунах, реже стволах деревьев. ЭССР,
ЛатвССР, Карельская АССР, Ленинградская, Московская обл., УССР,
Краснодарский край (Хостинская тиссо-самшитовая роща), Кавказ,
Полярн. Урал, Бурятская АССР.
283
Рис. 153.
Trentepohlia аигеа: 1, 2 — конечные части веточек, 3, 4 — пектозные колпачки.
5 — зооспорангий, 6 — субспорангиальная клетка, 7 — базальная часть слое-
вища с верхушечными и сидячими гаметангиями, 8 — разные формы гаметан-
гиев. (7, 2, 5, 6, 8 — по Мошковой, 5, 4 — по Весту, 7 — по Бранду).
Одна из распространенных водорослей, встречается как в равнинных,
так и в горных районах.
Var. tomentosa Kiitz. — Дерновинки подушковидные, 4—6 мм выс.,
с плотно переплетенными прямостоячими нитями.
Преимущественно в горных районах, на скалах и крупных валунах.
УССР (Карпаты).
Var. lanosa Kiitz. — Нити извитые, курчавые. Прямостоячие нити
слегка переплетенные, с многочисленными, несколько утончающимися
к концам сближенными ветвями.
Преимущественно на скалах и крупных валунах. В СССР не обна-
ружена. (Зап. Европа).
Var. polycarpa (Nees et Mont.) Hariot. — Прямостоячие нити ветвя-
щиеся, переплетенные, к концам не суживающиеся. Клетки цилин-
дрические, 14—26 мкм шир., по длине в 2—3 раза больше. Гаметангии
шаровидные или слегка эллипсоидные, 20—45 (55) мкм в диам., боковые,
одиночные или по нескольку в ряд, часто супротивные, или верхушеч-
ные, почти всегда группами по 2—5, реже одиночные.
Преимущественно в горных районах, на скалах, крупных валунах.
УССР (горный Крым).
Var. subsimplex (Caspary) De-Toni. — Прямостоячие нити простые
или только с 1 — 2 короткими веточками.
Преимущественно на скалах и крупных валунах. ГрузССР (окр.
Боржоми), Абхазская АССР (окр. Гагры).
Var. chinensis (Harv.) Printz. — Клетки прямостоячих нитей 16 —
32 мкм шир., 20— 48 мкм дл. Гаметангии 24 мкм в диам.
В СССР ие обнаружена. (Китай).
284
7. Trentepohlia uncinata (Gobi) Hansg. (—Chroolepus uncinatum
Gobi; Trentepohlia polymorpha Deck.) — Трентеполия крючковатая
(рис. 154). — Образует тонкие дерновинки, во влажном состоянии
красновато-бурые, оранжеватые, желтоватые, реже желто-зеленые.
Ползучие нити беспорядочно ветвящиеся. Прямостоячие нити много-
численные, простые или ветвящиеся, короткие, из небольшого числа
клеток. Клетки ползучих нитей вздутые, почти шаровидные, реже
вытянутые, 12—25 мкм шир., по длине равные ширине или до 3.5 раза
больше, прямостоячих — удлиненные, цилиндрические или неправильно
цилиндрические, у поперечных перегородок едва заметно перетянутые,
6.4 — 10 мкм шир., 11 — 21.5 мкм дл. Конечные клетки прямостоячих
нитей или их ветвей часто несколько расширены на верхнем конце
и несут субспорангиальную клетку со спорангием. Субспорангиальные
клетки крючковидные, 25—30 мкм дл., с расширенной нижней частью
и отходящим сбоку вверх узким, несколько изогнутым выростом
(«шейка»), на вершине которого и образуется по зооспорангию. Обо-
лочки клеток тонкие. Зооспорангии шаровидные или эллипсоидные,
15—24 мкм шир., 24—28 мкм дл. Отверстие для выхода зооспор возни-
кает в нижней части спорангия, с вогнутой стороны субспорангиальной
клетки. Зооспоры красноватые, эллипсоидные, 5—8 мкм шир., 12—
15 мкм дл., по 8—16 в спорангии. Гаметангии шаровидные или эллип-
соидные, 15 мкм шир., 17.2 мкм дл., с простым выводным отверстием;
они возникают как на ползучих, так и на прямостоячих нитях. Гаметы
5 — 7 мкм шир., 10—12 мкм дл. После выхода гамет гаметангии сдви-
гаются в сторону, так как лежащие рядом клетки прорастают в новые
нити.
На стволах различных деревьев, преимущественно у их основания,
в равнинных и горных районах. ЛатвССР, Ленинградская обл., окр.
Москвы, Горьковская обл., УССР (Карпаты, горный Крым), Красно-
Рис. 154.
Trentepohlia uncinata-. 1 — верхушечная часть ветви, 2,3 — ветви с зооспоран-
гиями, 4—в — образование зооспор, 7 — ветвь с гаметангием. (По Мошковой).
285
дарский край (Хостинская тиссо-самшитовая роща), Кавказ (окр. оз.
Рица, Батумский бот. сад).
8. Trentepohlia arborum (Ag.) Hariot — Трентеполия древесная
(рис. 155). — Образует дерновинки. Прямостоячие нити длинные, ветвя-
щиеся. Веточки отходят от материнских клеток под прямым углом,
к концам едва заметно суживаются. Клетки цилиндрические, у попе-
речных перегородок не перетянутые или слегка перетянутые, 13—
20(28) мкм шир., по длине до 3 раз больше. Конечные клетки веточек
очень часто вытянутые и заостренные. Оболочки клеток бесцветные
или слегка желтоватые, довольно твердые, слоистые, слои сходящиеся,
ровные или морщинистые. Зооспорангии яйцевидные или шаровидные,
18—24 мкм шир., 24—35 мкм дл., очень редко одиночные; обычно
от сильно вздутой конечной клетки прямостоячей нити отходит по 2—8
крючковидных субспорангиальных клеток с 1 спорангием каждая.
Иногда субспорангиальные клетки могут прорастать в нити, которые
на конце вновь образуют зооспорангии. Гаметангии боковые, сидячие,
одиночные или по 2—3, нередко на коротких боковых веточках.
На стволах деревьев, скалах. Широко распространена в тропических
районах, случайно завезена в европейские оранжереи, где обитает
на листьях некоторых растений и на стенах. В СССР не обнаружена.
(Зап. Европа: окр. Вены).
Без зооспорангиев этот вид трудно отличить от Т. аигеа.
9. Trentepohlia hunanensis Jao — Трентеполия хунаньская
(рис. 156). — Образует рыхлые дерновинки, состоящие из ползучих
по субстрату и прямостоячих нитей. Прямостоячие нити бичевидные,
длинные, беспорядочно ветвящиеся, к вершине постепенно суживаю-
щиеся. Веточки вытянутые, слабо разнообразно изогнутые, иногда
отогнутые назад. Клетки цилиндрические, ближе к основанию слоевища
слегка вздутые, бочонковидные, в главных нитях 17—32 мкм, в веточках
Рис. 155.
Trentepohlia arborum: 1 - фрагмент слоевища, 2—4 — верхушки прямостоячих
нитей с зооспорангиями. (7, 2, 4 — по Вильдеману, 3 — по Фишеру).
286
Рис. 156.
Trentepohlia hunanensis-. 1 — нижняя часть слоевища, 2 — верхняя часть слое-
вища, 3 — зооспорангии при большом увеличении. (По Жао).
10—15 мкм шир., по длине равные ширине или до 3 раз больше.
Конечные клетки вытянутые, суженные, по длине в 12—15 раз больше.
Оболочки клеток толстые, до 5.5 мкм, слоистые, слои расходящиеся,
что делает поверхность нитей лохматой. Зооспорангии шаровидные,
яйцевидные, сидячие, обычно по 2—7 вместе, реже одиночные, (14)25—
37 мкм в диам. Гаметангии неизвестны.
На скалах и стволах деревьев. В СССР не обнаружена. (Китай).
10. Trentepohlia lagenifera (Hild.) Wille [—Chroolepus lageniferum
Hild.; C. gracile Rabenh.; C. tenue Zell.; Trentepohlia tenuis De-Toni;
T. phyllophila W. et G. S. West; T. gracilis (Rabenh.) Iyengar] — Трен-
теполия бутыльчатая (рис. 157). — Образует тонкие дерновинки,
во влажном состоянии оливково-желто-зеленые до оранжево-желтых.
Нити довольно длинные, обильно и беспорядочно разветвленные,
ползучие по субстрату и прямостоячие. Клетки шаровидные, эллип-
соидные, яйцевидные, бочонковидные или цилиндрические, 2.5—20 мкм
287
Trentepohlia lagenifera: 1 — 6 — var. lagenifera (1 — фрагмент слоевища, 2, 3 —
бутылковидны^ гаметангии, 4, 5 — гаметы, 6 — веточка слоевища с зооспоран-
гиями), 7, 8 — var. hansgirgit (7 — молодое слоевище, 8 — зооспорангии),
9 — var. hildenbrandtli, зооспорангий, 10, 11 — var. africana, фрагменты слое-
вища с бутылковидными гаметангиями. (1—9 — по Мейеру, 10, 11 — по
Принтцу).
шир., по длине до 4(8) раз больше. Зооспорангии встречаются редко,
па концах особых, обильно разветвленных спорангиеносных нитей,
состоящих из длинных цилиндрических клеток, к вершине несколько
укорачивающихся; на конечных клетках этих ветвей образуются
1 — 2 субспорангиальные клетки с 1 спорангием каждая. Субспоран-
гиальные клетки цилиндрические, крючковидно изогнутые. Зооспо-
рангии эллипсоидные или почти шаровидные, (12)15.7—17.8 мкм шир.,
(15)17.5—20 мкм дл. Отверстие для выхода зооспор возникает в нижней
части спорангия с вогнутой стороны субснорангиальной клетки или
на вершине спорангия. Гаметангии верхушечные, реже вставочные,
по 1—2, бутылковидные, 9—24 мкм гпир., с довольно хорошо развитым
горловидным выростом, иногда в 1.5—2 раза превышающим длину
самого гаметангия. Гаметы зеленоватые, с капельками оранжевого масла,
удлиненно-эллипсоидные, слегка искривленные, 4.8—5.8 мкм шир.,
8—10 мкм дл.
Тропический вид, в умеренных широтах обычно встречается
в оранжереях и теплицах, реже в природных условиях.
Var. lagenifera (рис. 157, 1—6). — Нити длинные, беспорядочно
разветвленные. Клетки до 20 мкм шир. Зооспорангии 15.7 — 17.8 мкм
шир., 17.5—20 мкм дл. Гаметангии до 24 мкм шир., одиночные или
по 2 в ряд.
288
В оранжереях, на листьях некоторых тропических растений,
стенах, досках. В ботанических садах Риги, Ленинграда, Москвы,
Харькова, Батуми, Краснодарский край (Хостинская тиссо-самшитовая
роща).
Var. hansgirgii Meyer (рис. 157, 7, 8). —Нити короткие, беспо-
рядочно разветвленные, легко распадающиеся. Клетки почти шаровид-
ные, 7.5—12 мкм в диам., при достаточной влажности цилиндрические,
2.5—4 мкм шир. Зооспорангии 12—15 мкм шир., 15 — 17(20) мкм дл.,
с пунктированной оболочкой и верхушечным выводным отверстием.
Субспорангиальные клетки 20—25 мкм дл. Гаметангии 10 —11 мкм шир.,
до 35—40 мкм дл., обычно в большом количестве.
В оранжереях, на стволах некоторых тропических деревьев. Бот.
сад МГУ, Краснодарский край (Хостинская тиссо-самшитовая роща).
Var. hildenbrandtii Meyer (рис. 157, 9). — Нити короткие, беспо-
рядочно разветвленные, легко распадающиеся. Клетки 7—8 мкм шир.
Зооспорангии группами или одиночные. Субспорангиальные клетки
35 — 40 мкм дл. Гаметангии 16—20 мкм шир., до 42 мкм дл. Гаметы
4.8—5.8 мкм шир., 8—10 мкм дл.
В оранжереях, на стволах некоторых тропических деревьев, нередко
вместе с var. hansgirgii. Бот. сад МГУ.
Var. africana Printz (рис. 157, 10, 11). — Нити короткие, ползучие,
разнообразно ветвящиеся, нередко образуют псевдопаренхимную
подошву. Клетки преимущественно эллипсоидные или яйцевидные,
4—7 мкм шир., по длине в 1.5—2 раза больше, реже цилиндрические,
по длине в 3—4 раза больше. Гаметангии 12—18 мкм шир.
На стволах деревьев. Краснодарский край (Хостинская тиссо-
самшитовая роща), Кавказ (окр. оз. Рица).
11. Trentepohlia jolithus (L.) Wallr. — Трентеполия Жолита
(рис. 158). — Образует дерновинки, во влажном состоянии красновато-
или кроваво-бурые, при высыхании серовато-желтые или серовато-
зеленые, тонкие или до 2—3 мм толщ., войлочные или в виде плотного
порошковатого налета. Ползучие и прямостоячие нити обильно и беспо-
рядочно ветвящиеся, прочные. Веточки отходят от верхнего конца
материнских клеток и располагаются обычно параллельно основным
нитям, к вершине слегка суженные. Клетки эллипсоидные или цилин-
дрические, несколько вздутые в средней части, у поперечных пере-
городок слегка перетянутые, (9)12 — 35 мкм шир., по длине обычно до 2,
а в конечных клетках нередко до 6 раз больше. Оболочки толстые,
косослоистые; наружные и внутренние слои перекрещивающиеся,
расходящиеся наружу. Зооспорангии шаровидные, 20—48 мкм в диам.,
эллипсоидные, 17.2—23.4 мкм шир., 18—23 мкм дл., или яйцевидные,
36 — 40 мкм шир., 45 — 54 мкм дл., субспорангиальные клетки крючко-
видно согнутые. Гаметангии шаровидные, 34—42 мкм в диам., верху-
шечные, вставочные или боковые, одиночные, реже по нескольку вместе.
Преобладает в высокогорных районах, преимущественно на каме-
нистом субстрате.
Var. jolithus (рис. 158, 1—8).— Дерновинки до 2—3 мм толщ.,
войлочные. Клетки 12—35 мкм шир., по длине до 2 раз больше. Зооспо-
рангии шаровидные, 20 — 40 мкм в диам., или яйцевидные, 36—40 мкм
шир., 45 — 54 мкм дл., преимущественно на прямостоячих нитях. Гаме-
тангии шаровидные, 34 — 42 мкм в диам.
ЛатвССР, Ленинградская обл., УССР, Кавказ (Абхазия).
Var. bovina (Flotow) Rabenh. (рис. 158, 9, 10). — Дерновинки
тонкие, в виде плотного порошковатого налета. Клетки 9—26 мкм
19 Зеленые водоросли. Улотриксовые
289
Рис. 158.
Trentepohlia jolithus: 1—8 — var. jolithus {7 — фрагмент слоевища, 2—5 —
схема развития зооспорангия, 6 — 8 — зооспорангии при разных увеличениях),
9, 70 — var. bovina (9 — фрагмент слоевища, 70 — зооспорангий). (7, 6 — 10 —
по Мошковой, 2—5 — по Бранду).
шир., 20—32 мкм дл. Зооспорангии шаровидные или эллипсоидные,
17.2—23.4 мкм шир., 18—23 мкм дл., с простым выводным отверстием
на верхушке, образуются как на ползучих нитях, так и на коротких
приподнимающихся веточках.
УССР (Карпаты), Кавказ (окр. оз. Рица).
12. Trentepohlia rigidula (Mull.) Hariot — Трентеполия жестковатая
(рис. 159, 1, 2). — Образует тонкие дерновинки, во влажном состоянии
оранжево- или красно-бурые, при высыхании бледно-зеленовато-
желтые. Нити преимущестенно ползучие, длинные, жестковатые,
разнообразно изогнутые, слабо ветвящиеся, местами приподнимающиеся
над субстратом. Ветви вытянутые, отстоящие. Клетки неправильно
цилиндрические или эллипсоидные, часто более или менее асимметрич-
ные, с выпуклой дорсальной и сплющенной вентральной стороной,
16—24 мкм шир., по длине равные ширине или до 2 раз больше,
у поперечных перегородок заметно перетянутые, с довольно тонкими
плотными бесцветными оболочками. Зооспорангии шаровидные, 20—
25 мкм в диам., с крючковидной субспорангиальной клеткой, в нижней
части яйцевидной, около 20 мкм шир., в верхней вытянутой в длинный,
290
слегка изогнутый горловидный вырост 10 мкм шир. Гаметангии шаро-
видные, 14—25 мкм в диам., боковые, верхушечные или вставочные.
На стволах широколиственных и хвойных деревьев, преимущест-
венно в горных районах. УССР (Карпаты), Кавказ (окр. оз. Рица,
Батумский бот. сад).
13. Trentepohlia odorata (Wigg.) Wittr. — Трентеполия душистая
(рис. 159, 3, 4). — Образует войлокообразные дерновинки, во влажном
состоянии оранжевые, красновато-желтые или красновато-бурые.
Нити разветвленные, прямые или изогнутые, чаще прямостоячие,
реже ползучие. Клетки эллипсоидные, неправильно шаровидные,
нередко цилиндрические или бочонковидные, у поперечных перегоро-
док перетянутые, (10)15—31 мкм шир., по длине равные ширине
или до 2(4) раз больше, с толстыми слоистыми оболочками. Внешние
слои оболочки расходящиеся, вследствие чего поверхность нитей чешуй-
чатая, шероховатая. Пектозные колпачки мелкие. Зооспорангии шаро-
видные или широкоэллипсоидные, (15)30 — 40(50) мкм в диам., верху-
шечные, боковые и вставочные. Гаметангии подобны зооспорангиям.
На коре широколиственных и хвойных деревьев, реже на скалах
и обработанной древесине.
Вид известен пока из немногих мест, но несомненно распространен
значительно шире.
Рис. 159.
7, 2 — Trentepohlia rigidula: 1 — вегетативная нить, 2 — зооспорангий; 3, 4 —
Т. odorata: 3 — var. odorata, 4 — var. elongata; 5—11 — T. gobii: 5, 6 — прямо-
стоячие нити с зооспорангиями, 7—9 — опадание зооспорангия, 70, 77 —
ползучие нити с гаметангиями. (7, 2 — по Мошковой, 3, 4 — по Фишеру,
5—11 — по Мейеру).
19*
291
Var. odorata (рис. 159, 3). — Дерновинки оранжевые или красновато-
желтые. Прямостоячие нити к вершине почти не суживающиеся.
Клетки эллипсоидные или неправильно шаровидные, реже более или
менее цилиндрические, 15—30 мкм шир., по длине равные ширине
или до 2 раз больше. Гаметангии (15)30—40 мкм в диам.
На стволах деревьев, реже скалах. Ленинградская обл., УССР
(Карпаты), Камчатка.
Var. elongate (Bleisch) Fischer (рис. 159, 4). — Дерновинки красно-
вато-бурые, шелковисто блестящие. Прямостоячие нити нередко к вер-
шине суживающиеся, скрученные. Клетки бочонковидные, 22—31 мкм
шир., по длине до 2 раз больше, а также цилиндрические, тонкие,
10—20 мкм шир., по длине до 4 раз больше. Гаметангии до 45 мкм
в диам.
На стволах деревьев. В СССР не обнаружена. (Зап. Европа).
14. Trentepohlia gobii Meyer — Трентеполия Гоби (рис. 159,
5—11). — Образует дерновинки, во влажном состоянии темно-красные,
с неровной бугорчатой поверхностью. Ползучие нити короткие, четко-
видные, обильно ветвящиеся, с многочисленными короткими приподни-
мающимися веточками, растущими в косом направлении, от которых
вырастают вверх нити иного строения. Прямостоячие нити короткие
или длинные, разветвленные. Клетки ползучих нитей шаровидные,
17.5—25 мкм в диам., с толстыми слоистыми оболочками, прямосто-
ячих — более или менее цилиндрические, 12.5—18 мкм шир., тонко-
стенные, оканчивающиеся обычно зооспорангиями, одиночными или
пе 3 вместе. Зооспорангии шаровидные, около 25 мкм в диам., или
эллипсоидные, 18—25 мкм шир., 25—27.5 мкм дл., с довольно толстой
оболочкой, субспорангиальные клетки 48—65 мкм дл., крючковидно
изогнутые, их нижняя, расширенная часть сильно удлиненная, иногда
почти цилиндрическая или слегка вздутая, верхняя, узкая изогнутая
часть приблизительно в 1.5 раза короче широкой части. Зооспоры
7.5 мкм шир., 13 мкм дл. Гаметангии шаровидные, реже слегка эллип-
соидные, 25 — 30 мкм в диам., с небольшим тупым носиком, в котором
образуется сложное выводное отверстие, представляющее собой короткий
канал, проходящий в утолщении оболочки; они образуются в боль-
шом количестве на ползучих нитях главным образом из конечных
клеток, нередко бывают вставочными, поодиночке или по 2 подряд.
Гаметы 7.5—8 мкм шир., 12—15 мкм дл.
На стволах деревьев до высоты 2—3 м. РСФСР, местами.
Близка к Т. uncinata, хорошо отличается формой субспорангиальной
клетки, а также строением выводного отверстия гаметангиев.
15. Trentepohlia umbrina (Kiitz.) Born. [=Chroolepus umbrinum
Kiitz.; C. quercinum Kiitz.; Trentepohlia umbrina (Kutz.) Born. var.
quercina (Kiitz.) Rabenh.] —Трентеполия теневая (рис. 160). — Обра-
зует порошковатый налет до 2 мм толщ., во влажном состоянии красно-
бурый до кирпично-красного. Нити короткие, беспорядочно разветвлен-
ные, легко распадающиеся, ползучие и слегка приподнимающиеся.
Клетки шаровидные, эллипсоидные, яйцевидные или неправильной
формы, (7)12—27(38) мкм шир., по длине равные ширине или до 2 раз
больше. Оболочки клеток толстые, бесцветные, концентрически слоистые,
Содержимое клеток красное или красно-бурое. Зооспорангии не обра-
зуются. Гаметангии 18—38 мкм в диам., по форме не отличающиеся
от вегетативных клеток, обычно с простым выводным отверстием, реже
вытянутым в короткую шейку. Гаметы 4—8 мкм шир., 9—13 мкм дл.
292
Рис. 160.
Trentepohlia umbrina: 1,2 — нити разного строения, 3,4 — фрагменты нитей
при большем увеличении, 5 — образование гамет, 6 — выход гамет из гаметан-
гия, 7 — гаметы, 8 — копуляция гамет. (1, 2 — по Гоби, 3, 4 — по Мошковой,
5, 6 — по Карстену, 7, 8 — по Вилле).
На стволах разных деревьев, досках, реже на скалах и железо-
бетонных блоках, часто в нескольких метрах от земли.
Близка к Т. kygrophila и Т. -piceana. От первой отличается отсут-
ствием крючковидных спорангиев и ксерофитными условиями обитания,
от второй — крупными клетками с кирпично-красным содержимым
и толстыми концентрически слоистыми бесцветными оболочками. Неко-
торыми авторами рассматривается как разновидность Т. odorata.
Встречена в трех формах.
F. umbrina (рис. 160). — Клетки шаровидные, неправильно шаро-
видные или слабо эллипсоидные, 7—27 (38) мкм шир.
Повсеместно.
F. elongate Reinsch. — Клетки 24—36 мкм шир., по длине в 2 раза
больше.
УССР (Харьковская обл.).
F. unilateralis (Cribb) Moschk. (-Т. odorata var. unilateralis
Cribb). — Клетки шаровидные, эллипсоидные, яйцевидные или непра-
вильной формы, 7—17 мкм шир., подлине равные ширине или до 1.5 раза
больше, обычно заметно вытянутые в дорсивентральном направлении,
с оболочкой, особенно сильно утолщенной на вершине клеток, нередко
в виде притупленного или округленного сосковидного выроста 3.5—
7 мкм выс.
На стволах деревьев. Краснодарский край (Хостинская тиссо-
самшитовая роща).
Близка к типовой форме, отличается более мелкими клетками
с утолщенной оболочкой на вершине.
293
Рис. 161.
Trentepohlia piceana, разные формы слоевища. (По Мейеру).
16. Trentepohlia piceana Meyer — Трентеполия еловая (рис. 161). —
Образует порошковатый налет, во влажном состоянии желтовато- или
оранжево-бурый. Нити короткие, четковидные, беспорядочно разветвлен-
ные, разнообразно изогнутые, ползучие и слегка приподнимающиеся,
реже они переходят в простые скопления клеток разной величины,
тесно прилегающих друг к другу. Клетки шаровидные, (5)8.6—15 мкм
в диам. Оболочки клеток довольно толстые, до 1 мкм, светло-серо-
вато-желтые, плотные, четко очерченные, обычно неслоистые, у молодых
клеток тонкие, бесцветные. Содержимое клеток желтовато- или оран-
жево-бурое. Зооспорангии не образуются. Гаметангии 16—20 мкм
в диам., по форме не отличающиеся от вегетативных клеток, с простым
выводным отверстием.
На стволах деревьев, преимущественно хвойных. Московска'я
и Горьковская обл., УССР (Карпаты), Кавказ (Абхазия), Ср. Азия.
Формой клеток и внешним видом слоевища напоминает Т. umbrina,
однако хорошо отличается меньшими размерами клеток, плотными,
четко очерченными оболочками и желто-бурым цветом клеточного
содержимого.
17. Trentepohlia hygrophila Moschk. — Трентеполия влаголюбивая
(рис. 162). — Образует тонкий порошковатый налет, во влажном
состоянии красновато-бурый. Нити короткие, беспорядочно ветвящиеся,
ползучие и слегка приподнимающиеся. Клетки почти шаровидные,
11.4—24.4 мкм в диам. Оболочки тонкие у молодых клеток, толстые,
концентрически слоистые у старых. Содержимое клеток красное.
Зооспорангии шаровидные, 12.4—19.8 мкм в диам., реже эллипсоидные,
17—20 мкм шир., 21—22.8 мкм дл., с простым отверстием на вершине;
они образуются на крючковидных субспорангиальных клетках 22.8—
30.4 мкм дл., внизу расширенных, 12.4—15.5 мкм шир., кверху сужен-
ных, 5—7 мкм шир., и вытянутых в несколько изогнутый вырост,
эксцентрично отходящий от основания, возникающих как на ползучих,
так и на коротких приподнимающихся веточках, нередко состоящих
всего из 1—2 клеток. Гаметангии шаровидные, 19.5—35 мкм в диам.,
по форме подобные вегетативным клеткам, на вершине с простым
выводным отверстием 4.4 мкм в диам.
У основания деревьев, растущих во влажных условиях — по берегам
рек, ручьев, водоемов в равнинных и горных районах. УССР (Карпаты,
правобережная лесостепь).
294
Trentepohlia hygrophila: 1 — общий вид слоевища, 2—4 — зооспорангии разной
степени зрелости. (По Мошковой).
Очень напоминает Т. umbrina, отличается более всего наличием
крючковидных спорангиев и экологией. Отверстием для выхода зооспор
на вершине спорангия хорошо отличается от Т. aurea, Т. uncinata,
Т. arborum и др.
18. Trentepohlia bogoriensis Wild. — Трентеполия богорская
(рис. 163). — Образует тонкие рыхлые дерновинки. Нити короткие,
ветвящиеся, ползучие и приподнимающиеся. Клетки более или менее
шаровидные или эллипсоидные, (5)7.5—15 мкм шир., по длине равные
ширине, реже до 2 раз больше. Оболочки клеток довольно толстые,
слоистые, шероховатые. От спинной поверхности многих клеток отходят
1 — 3 одноклеточных удлиненно-цилиндрических волосовидных выроста
(2)3—5.5(6.5) мкм шир., 26—56(120) мкм дл., слабо или значительно
головчато расширяющихся на вершине, бесцветных или с оранжевым
содержимым. На концах волосовидных выростов нередко возникают
своеобразные вздутия, постепенно превращающиеся в вегетативные
клетки, на спинной поверхности которых вновь образуются длинные
цилиндрические одноклеточные выросты или возникают короткие нити
из нескольких клеток, также с волосовидными выростами. Зооспо-
рангии шаровидные или полушаровидные, 12—20 мкм в диам.; они
изредка возникают на коротких, до 110 мкм выс., прямостоячих нитях,
на вершине которых сначала образуются одиночные крючковидные
субспорангиальные клетки. Гаметангии шаровидные или яйцевидные,
15 мкм шир., 17.2—19.5 мкм дл., сидячие, боковые или верхушечные.
На коре деревьев с северо-северо-западной стороны, на высоте
20—50 см от поверхности земли. Краснодарский край (Хостинская
тиссо-самшитовая роща), Кавказ (окр. оз. Рица, Батумский бот. сад).
19. Trentepohlia lagerheimii Wild. — Трентеполия Лагерхейма
(рис. 164). — Образует тонкие дерновинки. Нити ползучие, реже слегка
приподнимающиеся, скудно ветвящиеся. Клетки более или менее
цилиндрические или бочонковидные, 7—15 мкм шир., по длине равные
ширине или до 2 раз больше. От некоторых клеток ползучих и припод-
нимающихся нитей отходят одноклеточные, волосовидные или шланго-
видные выросты, по ширине лишь незначительно уступающие веге-
тативным клеткам, на верхнем конце иногда головчато вздутые, по длине
до 10 раз больше ширины. Зооспорангии шаровидные, сидячие. Субспо-
рангиальные клетки крючковидные,' внизу расширенные, кверху
295
Рис. 163.
Trentepohlia bogoriensis: 1—3 — фрагменты слоевища, 4 — прямостоячая нить
с зооспорангием, 5—7 — ползучие нити с гаметангиями. {1, 4, 6, 7 -— по Виль-
деману, 2, 3, 5 — по Мошковой).
Рис. 164.
Trentepohlia lagerheimii: 1,2 — фрагменты слоевища с зооспорангиями, 3, 4 —
фрагменты слоевища с гаметангиями. (По Вильдеману).
296
суженные и вытянутые в слегка изогнутую шейку. Гаметангии шаро-
видные, боковые и сидячие.
Преимущественно на коре деревьев, реже на скалах. Кавказ (окр.
оз. Рица).
Род PHYCOPELTIS Mill. - ФИКОПЕЛЬТИС
Слоевище пластинчатое, образованное нитями. Нити ползучие,
рыхло или плотно смыкающиеся друг с другом в однослойные, пра-
вильно дисковидные или вееровидные псевдопаренхимные пластинки
разнообразной формы или сетевидные сплетения. Волосовидные
выросты и ризоиды отсутствуют. Клетки неправильные, часто много-
угольные, интенсивно окрашенные. Хлоропласты дисковидные, эллип-
соидные, без пиреноидов. Бесполое размножение зооспорами. Зооспо-
рангии шаровидные или яйцевидные, сидящие на довольно искривлен-
ных субспорангиальных клетках, возвышающихся над поверхностью
слоевища. Зооспоры яйцевидные, двужгутиковые, без стигмы; они обра-
зуются в большом количестве и освобождаются из спорангиев после
их опадения^ Гаметангии по форме не отличаются от вегетативных
клеток. Гаметы яйцевидные, двужгутиковые, без стигмы.
Большинство видов рода обитает в тропических, реже субтропи-
ческих районах, в умеренных широтах встречается обычно в теплицах
или оранжереях.
I. Слоевище из беспорядочно разветвленных нитей, образующих
рыхлые или плотные псевдопаренхимные пластинки разнообраз-
ной формы......................................Р. irregularis 1.
II. Слоевище из радиально расходящихся разветвленных нитей, обра-
зующих четко очерченные псевдопаренхимные пластинки.
1. Слоевище правильно дисковидное; нити плотно смыкаются
друг с другом, клетки 3—4 мкм шир..............Р. epiphyton 2.
2. Слоевище неправильной формы; нити неплотно смыкаются друг
с другом, часто образуя сетевидные сплетения, клетки 5—
7 (10) мкм шир.................................Р. flabelligera 3.
1. Phycopeltis irregularis (Schmidle) Wille {—Hansgirgia irregularis
Schmidle) — Фикопельтис неправильный (рис. 165). — Слоевище в виде
цельных или продырявленных псевдопаренхимных пластинок разно-
образных очертаний. Нити беспорядочно разветвленные, рыхло или
плотно смыкающиеся друг с другом. Клетки неправильной формы,
местами вздутые, часто многоугольные, (4)6—10 мкм шир., по длине
равные ширине или до 1.5 раза больше, с довольно плотной, иногда
слегка буроватой оболочкой. У плотных дисковидных слоевищ краевые
клетки отличаются от многоугольных центральных клеток выемчатым,
дольчато изрезанным внешним краем. Зооспорангии редкие, на короткой
субспорангиальной клетке. Гаметангии шаровидные или неправильные,
13—18 мкм в диам., верхушечные, вставочные либо боковые, одиночные
или по нескольку рядом.
На коре деревьев. Краснодарский край (Хостинская тиссо-самши-
товая роща), Кавказ (окр. оз. Рица, Батумский бот. сад).
2. Phycopeltis epiphyton Mill. — Фикопельтис эпифитный
(рис. 166). — Слоевище дисковидное, до 130 мкм в диам., зеленое,
желто-бурое или золотисто-желтое, под влиянием неблагоприятных
297
Рис. 165.
Ph-ycopeltis Irregularis, разные формы слоевища. (По Шмидле).
Рис. 166.
Phycopeltis eplphyton: 1 — общий вид слоевища, 2—4 — схема образования
краевых клеток слоевища, 5 — образование зооспорангия, 6 — зрелый зоо-
спорангий, 7 — гаметангий, 8 — гамета. (7—4, 7, 8 — по Милларду, 5, 6 —
по Воронихину).
условий становится нитевидным, с короткими боковыми ответвлениями.
Клетки в периферических участках большей частью прямоугольные,
часто дольчато поделенные, около 4 мкм шир. Зооспорангии изучены
недостаточно, шаровидные. Гаметангии до 14 мкм шир. Гаметы яйце-
видные, 5 мкм дл., по 20—30 в гаметангии, освобождаются через круглое
отверстие, образующееся на верхней поверхности клетки. Копуляция
не наблюдалась.
На листьях плюща, малины, самшита, на иглах лиственницы
в виде пунктирных пятнышек. Кавказ (ГрузССР).
ппо
Рис. 167.
1, 2 — Phycopeltis flabelligera; 1 — общий вид слоевища, 2 — фрагмент слое-
вища со вставочным спорангием; 3—5 — Cephaleuros endophyticus: 3 — по-
перечный срез однослойного слоевища, 4 — трехслойное слоевище, 5 — одно-
слойное слоевище со спорангием. (7 — по Принтцу, 2 — по Вильдеману, 3 — 5 —
по Фричу).
3. Phycopeltis flabelligera (De-Toni) Hansg. — Фикопельтис вееро-
носный (рис. 167, 1, 2). —Слоевище пластинчатое, неправильной
формы, более или менее изрезанное по краям, в центральной части
плотное, к периферии с радиально расходящимися, рыхло располо-
женными нитями, часто образующими сетевидные сплетения. Клетки
5—7(10) мкм шир., в периферической части слоевища более вздутые,
чем в центре. Гаметангии вставочные, шаровидные или эллипсоидные,
более вздутые, чем вегетативные клетки.
В теплицах, на листьях многих тропических растений. В СССР
не обнаружен. (Зап. Европа).
Род CEPHALEUROS Kunze - ЦЕФАЛЕУРОС
(=Mycoidea Cunning.; Phylloplax Schmidle;
Weneda Racib.; Chrooderma Fritsch)
Слоевище пластинчатое, образованное нитями, эпифитное или
эндофитное. Нити ветвящиеся, ползучие, плотно смыкающиеся в одно-
и многослойные псевдопаренхимные пластинки или беспорядочные
скопления. У эндофитных форм от клеток верхней части слоевища
отходят прямостоячие, обычно простые нити, пронизывающие ткани
растения-хозяина и оканчивающиеся на его поверхности волосовидными
выростами или спорангиями, а от клеток нижней части слоевища —
обильно разветвленные ризоидообразные нити, глубоко проникающие
в ткани пораженного растения. Хлоропласты пристенные, мелкие,
пластинчатые, или сетчатые, без пиреноидов, часто полностью зама-
скированные красящими веществами. Бесполое размножение зооспо-
рами. Зооспорангии крючковидные, на длинных ножках, обычно груп-
пами по нескольку. Зооспоры двужгутиковые. Гаметангии шаровидные;
они образуются из вегетативных клеток, расположенных глубоко
в слоевище. Гаметы двужгутиковые, многочисленные.
299
Преимущественно в тропических и субтропических областях, явля-
ется возбудителем заболеваний культурных цветковых растений.
В умеренной зоне обнаружен 1 вид.
1. Cephaleuros endophyticus (Fritsch) Printz (=Chrooderma endo-
phytica Fritsch) —Цефалеурос эндофитный (рис. 167, 3—5). — Слое-
вище эдифитное или эндофитное, одно- и многослойное, 0.5—2 мм толщ.,
в виде бледных желто-коричневых корочек, нередко сливающихся
в крупные диски. Клетки изодиаметрические, 6.5—15 мкм шир., на попе-
речных перегородках часто с утолщениями в виде крапинок. Зооспо-
рангии на малоклеточных коротких нитях, приподнимающихся над
поверхностью псевдопаренхимного диска. Гаметангии яйцевидно-
шаровидные; они образуются из обычных округлившихся клеток
подошвы диска.
На листьях малины. В СССР не обнаружен. (Англия).
Семейство MICROTHAMNIACEAE West -
МИКРОТАМНИЕВЫЕ
Слоевище нитевидное. Нити ветвящиеся, прямостоячие, без слизи,
образующие плотные или рыхлые кустики, прикрепляющиеся к суб-
страту базальной клеткой. Волосовидные образования на концах
ветвей отсутствуют. Клетки цилиндрические, с тонкой оболочкой,
продолжающие расти вбок перед верхними поперечными перегородками,
что и дает начало боковым ветвям. Ядро 1. Хлоропласт 1, пристенный,
пластинчатый, без пиреноида. Продукт ассимиляции — маслоподобное
вещество. Бесполое размножение двужгутиковыми зооспорами, возни-
кающими во всех клетках слоевища, становящихся зооспорангиями,
за исключением базальной, и акинетами.
По своеобразному заложению поперечных перегородок занимает
изолированное положение в системе. В пресных водах, выделен также
из почв.
Род MICROTHAMNION Nag. - МИКРОТАМНИОН
Нити прямостоячие, ветвящиеся, с беспорядочно отходящими боко-
выми ветвями, образованными 1 или несколькими клетками, на ранних
стадиях развития прикрепленные к субстрату, позднее нередко отры-
вающиеся и свободноплавающие. Главный ствол и боковые ветви друг
от друга неотличимы. Клетки, за исключением базальной, более или
менее цилиндрические, оболочки тонкие. Хлоропласт бледно- или ярко-
зеленый, в форме полуцилиндра, прилегающий к одной из продольных
сторон клетки. Зооспоры удлиненно-грушевидные, со стигмой, по 4, 8
в зооспорангии. Акинеты образуются редко, главным образом при
подсыхании нитей.
В проточных и стоячих водах, в болотах, преимущественно на
растительных субстратах, реже в толще воды; найден также в почвах.
I. Конечные клетки ветвей прямые.
1. Кустики плотные, ветви короткие, отстоящие; длина клеток
превосходит ширину до 4 раз..........М. kuetzingianum 1.
2. Кустики рыхлые, ветви длинные, более или менее прижатые;
300
Рис. 168.
Microthamnion kuetzingianum: 1 — общий вид слоевища, 2— 4 — образование
зооспор, 5— прорастание зооспор. (По Рейсу).
длина клеток превосходит ширину в (2) 5 раз и более..........
......................................М. strictissimum 2.
II. Конечные клетки ветвей изогнутые...............М. curvatum 3.
1. Microthamnion kuetzingianum Nag. — Микротамнион Кютципга
(рис. 168). — Кустики плотные, чаще всего 100—200 мкм дл., реже
больше. Нити обильно разветвленные. Ветви одно- или многоклеточные,
более или менее отстоящие, нередко слегка искривленные. Клетки
цилиндрические, иногда кверху булавовидно расширенные, 3—5 мкм
шир., по длине обычно в 2—4 раза больше. Конечные клетки прямые,
на верхушке широко закругленные.
В стоячих и проточных водах, на водных цветковых растениях,
древесных субстратах, раковинах моллюсков, встречается одиночными
кустиками или образует дерновинки. Выделен также из почв с помощью
водных культур. Во многих районах СССР, нередко.
2. Microthamnion strictissimum Rabenh. — Микротамнион сжатый
(рис. 169, 7—3). — Кустики рыхлые, до 0.5—0.75 мм дл. Нити умеренно
разветвленные. Ветви прямые, из 1 или нескольких клеток, более или
менее прилегающие, вертикально направленные, к концам слегка
суживающиеся или не суживающиеся. Клетки удлиненно-цилиндриче-
ские, 2.5—4 мкм шир., по длине в (2)5—6(12) раз больше. Конечные
клетки прямые, на верхушке закругленные.
В стоячих и проточных водах, на водных цветковых растениях,
мхах, раковинах моллюсков, одиночными кустиками или довольно
плотными дерновинками. Выделен также из почв с помощью водных
культур.
301
Рис. 169.
1—3 — Microthamnion strictissimum: 1, 2 — var. strictissimum, 3 — var. macro-
cystis, 4, 5 — M. curvatum. (1, 2 — по Рейсу, 3 — по Газену, 4, 5 — по Весту).
Var. strictissimum (рис. 169, 1, 2). — Клетки 3—4 мкм шир.,
по длине обычно в 5—6;раз больше. Конечные клетки ветвей к концам
не суживаются.
В реках, ручьях, канавах, болотах, нередко. ЭССР, ЛатвССР, РСФСР
(Европ. ч., Зап. и Центр. Сибирь), БССР, УССР, ТуркмССР.
Var. macrocystis Schmidle (рис. 169, 3). — Клетки 2.5—3 мкм
шир., по длине в 6—8(12) раз больше. Конечные клетки ветвей к концам
суживаются.
В сфагновых болотах, одиночными кустиками на листьях Sphagnum.
УССР, Сев. Урал.
3. Microthamnion curvatum W. et G. S. West — Микротамнион
изогнутый (рис. 169, 4, 5). — Кустики рыхлые, до 0.1 мм дл. Нити
умеренно разветвленные.,Ветви из 1 или 2—3 клеток, прямые или слабо
изогнутые. Клетки 1.6—2.2 мкм шир., по длине в 9—10 раз больше.
Конечные клетки сильно изогнутые, на верхушке закругленные.
В стоячих и проточных водах, чаще в болотах, на растительных
субстратах, обычно одиночными кустиками. УССР, РСФСР (Центр.
Сибирь).
302
Семейство APHANOCHAETACEAE (Printz) Bourr. —
АФАНОХЕТОВЫЕ
Слоевище нитчатое. Нити простые или ветвящиеся, обособленные,
полностью распростертые по субстрату или частично приподнимающиеся,
реже прямостоячие. Клетки шаровидные или цилиндрические, часто
неправильной формы, в большинстве с волосками. Волоски преимуще-
ственно одноклеточные, большей частью луковицеобразно расширенные
у основания, обычно без влагалища, отделяющиеся поперечной перего-
родкой от материнской клетки еще на ранней стадии развития, реже
не отделяющиеся. Ядро 1. Хлоропласт 1, пластинчатый, дисковидный,
реже кольцевидный, цельный или сетчато продырявленный, с 1 или
несколькими пиреноидами. Вегетативное размножение случайным раз-
рывом нитей. Бесполое размножение четырехжгутиковыми зооспорами,
образующимися по 1—4 в почти не измененных вегетативных клетках,
реже апланоспорами, возникающими по 1 в клетке, и акинетами.
Половое размножение гетерогамное или оогамное.
В пресных проточных и стоячих водах, на различных субстратах,
а также в слизи преимущественно красных, реже некоторых зеленых
водорослей.
Семейство искусственное, охватывающее в морфологическом отно-
шении разные роды. Общим признаком, объединяющим их, является
образование волосков.
I. Волоски длинные, одно-, реже многоклеточные, у основания обычно
луковицеобразно вздутые, реже ровные.
1. Нити ползучие, многоклеточные, длинные или короткие, простые
или ветвящиеся.
А. Нити на поверхности субстрата.
а. Нити обособленные, не инкрустированные известью.
а. Конечные клетки нитей без волосков................
..............род Aphanoehaete (частично) (с. 304).
р. Конечные клетки нитей с волосками.................
............................род Gonatoblaste (с. 308).
б. Нити слагаются в подушковидное слоевище, слегка или
значительно инкрустированное известью.
а. Клетки нитей неправильно цилиндрические, часто
вздутые, почти шаровидные; волоски отделяются от
клеток поперечными перегородками....................
............................род Chaetomnion (с. 309).
(3. Клетки нитей только удлиненно-цилиндрические; во-
лоски не отделяются от клеток поперечными перего-
родками ..........................род Fridea (с. 310).
Б. Нити внутри субстрата.
а. В слизи зеленых и красных водорослей.
а. Нити только стелющиеся...........................
...........род Aphanoehaete (частично) (с. 304).
р. Нити стелющиеся с прямостоячими ветвями .....
.............................род Chaetonema (с. 310).
б. В пустых клетках и в оболочках зеленых нитчатых водо-
рослей, преимущественно кладофоры....................
..............................род Ectochaete (с. 313).
В. Нити на поверхности или в толще воды..................
...............род Aphanoehaete (частично) (с. 304).
303
2. Нити прямостоячие, малоклеточные, обычно из 3—6 клеток,
простые или скудно ветвящиеся . . . род Thamniochaete (с. 315).
II. Волоски короткие, одноклеточные, у основания не вздутые, только
слегка расширенные..........................род Micropoa (с. 316).
Род APHANOCHAETE А. Вт. - АФАНОХЕТЕ
Нити простые или беспорядочно ветвящиеся, слегка суживающиеся
к концам, ползучие, полностью распростертые по субстрату, реже
свободноплавающие. Клетки в средней части нитей большей частью
шаровидные, ближе к периферии бочонковидные, или цилиндрические.
Конечные клетки конусовидные, закругленные. От спинной, выпуклой
поверхности многих клеток отходят длинные бесцветные одноклеточные
волоски, чаще всего луковицеобразно расширенные у основания, очень
ломкие, особенно в нижней части. Хлоропласт 1, пристенный, дисковид-
ный, с 1—4 пиреноидами. Бесполое размножение четырехжгутиковыми
зооспорами и апланоспорами. Зооспорангии несколько крупнее вегета-
тивных клеток и расположены обычно в средней части слоевища.
Зооспоры изменчивы по форме и величине, чаще всего грушевидные,
с 2 пульсирующими вакуолями, стигмой и хлоропластом с 1 пиреноидом,
нередко с каплей масла или зернами крахмала; они выходят через
круглое отверстие, образующееся в зооспорангии, окруженными вначале
тонким слизистым пузырем, прикрепляются к субстрату и прорастают,
но иногда, потеряв жгутики, переходят к амебоидному движению. Апла-
носпоры встречаются реже, по 1 в клетке. Половой процесс гетерогамный,
водоросли гомоталличны. Оогонии расположены главным образом в цент-
ральной части слоевища, крупные, шаровидные, лишенные волосков.
Женские гаметы (оосфера, или макрогаметы) четырехжгутиковые, круп-
ные, темно-зеленые, с большой каплей масла внутри, по 1 в оогонии.
Антеридии образуются из конечных клеток нитей в результате их
повторных делений; они представляют собой маленькие бесцветные
клеточки. Мужские гаметы (микрогаметы) четырехжгутиковые, мелкие,
грушевидные, по 1—2 в антеридии. Гаметы выходят из материнских
клеток окруженными слизистым пузырем, быстро растворяющимся,
женская гамета некоторое время свободно плавает, затем останавли-
вается, втягивает жгутики и оплодотворяется подвижной мужской
гаметой. Зигота выделяет толстую оболочку, наполняется красно-
ватым маслом и переходит в состояние покоя. Прорастание зигот не про-
слежено.
Преимущественно в пресных стоячих или проточных водах, на
водных цвс >вых растениях, крупноклеточных водорослях, древесном
субстрате, раковинах моллюсков, камнях, реже внутри субстрата или
свободноживущие.
I. Волоски у основания луковицеобразно расширенные.
1. Волоски одноклеточные, нити распростертые по субстрату.
А. Клетки шаровидные или эллипсоидные, их длина может только
слегка превышать ширину.
а. Волоски по 1(2) почти на каждой клетке . . A. repens 1.
б. Волоски по 2—6 на каждой клетке.
а. Слоевище заключено в слизь, волоски у основания
окружены коротким влагалищем с бахромчатым верхним
краем..............................A. pilosissima 2.
р. Слоевище без слизи, волоски без влагалищ.........
.................................A. polychaete 3.
304
Б. Клетки цилиндрические, их длина до 2—3 раз превышает
ширину...................................A. vermiculoides 4.
2. Волоски многоклеточные, нити свободноплавающие...............
.............................................A. globiferum 5.
II. Волоски на всем протяжении ровные................A. pascheri 6.
1. Aphanochaete repens A. Br. [= Herposteiron confervicolum Nag.;
H. repens (A. Br.) Wittr.; H. braunii Nag.; H. bertholdii Hub.] — Афано-
хете ползучая (рис. 170, 1—5). — Нити простые или ветвящиеся, обособ-
ленные, распростертые по субстрату. Клетки эллипсоидные или почти
шаровидные, 4.2.—12.5 мкм шир., 7.8—18 мкм дл., несущие 1(2)
одноклеточных волоска до 200 мкм дл., луковицеобразно расширенных
у основания. Оболочка клеток несколько утолщенная. Зооспоры и
апланоспоры известны. Женские гаметы в неподвижном состоянии
шаровидные, 14.5 — 20 мкм в диам.
В стоячих и проточных водах, на водных цветковых растениях,
древесном субстрате, раковинах моллюсков, часто на нитчатых водо-
рослях, лишенных слизи. Повсеместно.
2. Aphanochaete pilosissima Schmidle — Афанохете волосистая
(рис. 170, 6). — Нити простые или разветвленные, полностью распро-
стертые по субстрату, окруженные толстой бесцветной гомогенной
массой слизи, из которой торчат лишь верхние части волосков. Клетки
шаровидные или эллипсоидные, в средней части нитей 4 мкм шир.,
6 мкм дл. Конечные клетки 2 мкм шир., 5 мкм дл. От каждой клетки
отходит 2—6 длинных одноклеточных волосков, у основания луковице-
образно расширенных и окруженных коротким отчетливым влагалищем
с бахромчатым верхним краем, отстающим от волосков. Оогонии
шаровидные, 12—19 мкм в диам. Антеридии нередко несколько крупнее
вегетативных клеток и возникают вблизи оогониев. Внешняя оболочка
зигот с едва заметной грануляцией.
На нитях Mougeotia, Zygnema, Oedogonium. ЭССР, ЛатвССР.
3. Aphanochaete polychaete (Hansg.) Fritsch (==Herposteiron po-
lychaete Hansg.) — Афанохете многощетинковая (рис. 170, 7—9). —
Нити прямые или слегка изогнутые, простые или скудно разветвленные,
полностью распростертые по субстрату. Клетки шаровидные или
эллипсоидные, 9—12 мкм шир., по длине в 1.5 раза больше. От каждой
клетки отходит 2—6 длинных одноклеточных волосков, почти прямых
или слегка искривленных, у основания луковицеобразно расширенных,
реже без расширения, к вершине постепенно суживающихся. Размноже-
ние не исследовано.
В стоячих и слабо проточных водах, на нитчатых водорослях,
водных цветковых растениях, древесном субстрате. В ряде районов
СССР, часто вместе с A. repens.
Вид малоизученный. Близок к A. repens.
к. Aphanochaete vermiculoides Wolle [=Herposteiron vermiculoides
(Wolle) Collins; H. hyalothecae Hansg.; Aphanochaete hyalothecae
(Hansg.) Schmidle inkl. var. mucicola Schmidle] — Афанохете червеобраз-
ная (рис. 170, 10—11). — Нити длинные или короткие, иногда всего
из нескольких клеток, обычно обильно разветвленные, с короткими
веточками, прилегающие к клеткам водоросли-хозяина полностью или
отдельными участками, часто оплетающие их. Клетки цилиндрические,
иногда неправильных очертаний, у поперечных перегородок не перетя-
20 Зеленые водоросли. Улотриксовые
305
Рис. 170.
1—5 — Aphanochaete repens'. 1 — общий вид слоевища, частично в профиль,
2 — слоевище с оогониями и аятеридиями, 3 — оосфера, 4 — микрогаметы,
<5 — оплодотворение; 6 — A. pilosissima; 7—9 — A. polychaete; 10, 11 — A. ver-
miculoides. (1—5 — по Губеру, 6, 10, 11 — по Шмидле, 7—9 — по Весту).
306
нутые или слегка перетянутые, 4—6 мкм шир., 5—12 мкм дл., обычно
без волосков, иногда на отдельных клетках с одноклеточными волосками
до 50 мкм дл., луковицеобразно расширенными у основания.
В стоячих и проточных водах, преимущественно в слизистых
обвертках десмидиевых и зигнемовых водорослей. УССР (Южн. Буг).
5. Aphanochaete globiferum (Hansg.) Printz (=Herposteiron globife-
rurn Hansg.) — Афанохете шароносная (рис. 171, 7, 2). — Нити беспо-
рядочно ветвящиеся, ползучие. Клетки цилиндрические до шаровидных,
3—4(6) мкм шир., по длине равные ширине или до 3 раз больше,
в старых нитях сплошь шаровидные, 6—12 (15) мкм в диам., с толстыми
оболочками. От многих клеток отходят 1—2 длинных многоклеточных
волоска, у основания слегка луковицеобразно расширенных. Зооспоры
шаровидные, по 2, 4 в спорангии. Половой процесс не исследован.
В пресных водоемах, нередко плавает по поверхности воды. Калинин-
ская обл. (оз. Селигер).
Вид изучен еще очень недостаточно, положение его в роде Apha-
nochaete сомнительно.
Рис. 171.
1, 2 — Aphanochaete globiferum’, 3—6 — A. pascheri: 3 — общий вид слоевища,
4 — выход амебоидной зооспоры, 5 — зооспора, 6 — прорастание зооспоры;
7—9 — Gonatoblaste rostrata; 7 — взрослое слоевище, 8 — зооспора, 9 — про-
растающая зооспора. (7, 2 — по Гансгиргу, 3—6 — по Геерингу, 7—9 — по
Губеру).
20*
307
6. Aphanoehaete pascheri Heering. — Афанохете Пашера (рис. 171,
3—6). — Нити ветвящиеся, ползучие, полностью распростертые по суб-
страту. Клетки шаровидно-цилиндрические, 8—16 мкм шир., 9—12 мкм
дл. Конечные клетки удлиненно-конусовидные. От многих клеток
отходит по 1 длинному волоску без луковицеобразного расширения
у основания, длина волоска нередко в 10—16 раз больше его ширины.
Зооспоры четырехжгутиковые, грушевидные, со стигмой и пульсирующей
вакуолью в переднем конце и с блюдцеобразным хлоропластом с 1 —
2 пиреноидами. После короткого периода движения зооспоры оседают
на субстрат, теряют грушевидную и приобретают амебоидную форму,
затем, одеваясь оболочкой, переходят в состояние покоя, после чего
прорастают. Вместо грушевидных зооспор в клетках нитей могут
развиваться амебоидные споры. Половой процесс не исследован.
В стоячих и слабо проточных водах, на водных цветковых растениях,
нитчатых водорослях, раковинах моллюсков. УССР, РСФСР (Сибирь),
нечасто.
Малоизученный вид. Гееринг с сомнением относит его к роду
Aphanoehaete.
Род GONATOBLASTE Huber - ГОНАТОБЛАСТЕ
Нити короткие, скудно ветвящиеся, полностью распростертые по
субстрату, окруженные слизистым влагалищем. От спинной поверхности
клеток отходит по 1—3 длинных одноклеточных волоска, слегка расши-
ренных у основания и выходящих верхушечной частью из слизи.
У конечных клеток нитей волоски отходят сбоку и вначале расположены
по направлению роста нити, позднее, слегка изогнувшись, поднимаются
вверх и постепенно смещаются на спинную поверхность клетки,
поскольку образование волосков не приостанавливает ее дальнейшего
роста и деления. Хлоропласт пластинчатый, с 1 или несколькими
пиреноидами. Бесполое размножение четырехжгутиковыми зооспорами.
Зооспоры грушевидные, без стигмы, с 2 пульсирующими вакуолями и
1—3 пиреноидами; они возникают по 1—2 в клетке, выскальзывают
наружу, окруженные вначале слизистым пузырем, в скором времени
расплывающимся, прикрепляются передним концом к слизистой
обвертке нитчатых водорослей, теряют жгутики и начинают прорастать,
наискось внедряясь в слизь; соприкоснувшись с твердой оболочкой
водоросли-хозяина, проросток слегка отгибается и образует бесцветный
бугорок, который постепенно удлиняется, отделяется поперечной
перегородкой и превращается в волосок.
Малоизученный род. По внешнему виду и характеру бесполого
размножения очень сходен с родом Aphanoehaete. Отсутствие данных
о половом размножении делает его систематическое положение весьма
неясным. Достоверен только 1 вид.
1. Gonatoblaste rostrata Huber — Гонатобласте носатый (рис. 171,
7—9). — С признаками рода. Размеры клеток в исходном описании
не указаны.
На нитях Zygnema, Oedogonium. Новгородская обл. (Валдайское
озеро).
308
Род CHAETOMNION Skuja - ХЕТОМНИОН
Нити обильно ветвящиеся, частично ползучие по субстрату, частично
приподнимающиеся, слагающиеся в мелкие подушкообразные слоевища,
иногда слабо инкрустированные известью. Приподнимающиеся ветви
беспорядочно ветвятся и в большинстве заканчиваются одноклеточным
длинным бесцветным волоском, обособленным от клетки. Клетки более
или менее цилиндрические, часто слегка вздутые. Хлоропласт кольцевид-
ный, с 1—2 пиреноидами. Бесполое размножение акинетами и аплано-
спорами. Акинеты грушевидные или эллипсоидные; они образуются
преимущественно из верхушечных клеток веточек. Апланоспоры шаро-
видные или эллипсоидные.
В пресных водоемах, на водных цветковых растениях и крупно-
клеточных водорослях.
I. Волоски отходят от верхней части конечных клеток приподнимаю-
щихся ветвей . ...................................С. pyriferum 1.
11. Волоски отходят от спинной поверхности конечных клеток ползучих
нитей..............................................С. nakaoi 2.
1. Chaetomnion pyriferum Skuja — Хетомнион грушеносный
(рис. 172). — Слоевище шаровидное или подушкообразное, около 1 мм
в диам., из беспорядочно переплетающихся нитей. Клетки нитей
большей частью неправильно цилиндрические, 8—17 мкм шир., по длине
равные ширине или до 2.5 раза больше. Волоски около 4 мкм шир.
у основания, до 600 мкм дл. Акинеты 20—30 мкм шир., 35—48 мкм дл.
На листьях Potamogeton. В СССР не обнаружен. (Китай).
2. Chaetomnion nakaoi Hirano — Хетомнион Накао (рис. 173).—
Слоевище подушковидное, из тесно сближенных, лучисто расходящихся
нитей. Клетки нитей в центральной части слоевища угловатые, 7.3 мкм
шир., 10—11 мкм дл., в средней части неправильно цилиндрические,
вздутые. Конечные клетки конусовидные. Апланоспоры шаровидно-
эллипсоидные, 17—24 мкм в диам.
На клетках Hydrodictyon. В СССР не обнаружен. (Непал).
309
Рис. 173.
Chaetomnion nakaoi. (По Хирано).
Род FRIDEA Schmidle - ФРИДЕЯ
Нити ветвящиеся, ползучие и прямостоячие, слагающиеся в полу-
шаровидные слоевища, инкрустированные известью; сливаясь, они
образуют мелкобугорчатую известковую корочку до 0.5 мм толщ, или
известковый налет. Ползучие нити образуют подошву. Прямостоячие
нити короткие, плотно сжатые, повторно ветвящиеся. Боковые ветви
отходят от верхних концов материнских клеток. Конечные клетки
прямостоячих нитей часто очень длинные, в верхушечной части несколько
расширенные. Многие клетки вытягиваются в длинный бесцветный
одноклеточный волосок с расширенным основанием. Ядро 1. Оболочки
слоистые. Хлоропласт пристенный, с 1—4 пиреноидами и многочислен-
ными зернами крахмала; он присутствует обычно только в 2—3 конечных
клетках прямостоячих нитей, размещаясь преимущественно у верхнего
их конца, и только в самых верхушечных клетках полностью выстилает
продольные стенки. Бесполое размножение зооспорами, образующимися
в большом количестве в пузыревидно вздутых спорангиях, -возникающих
на коротких веточках.
1. Fridea torrenticola Schmidle — Фридея быстроводная (рис. 174). —
Прямостоячие нити (без волосков) до 200—300 мкм дл. Клетки нитей
подошвы 9.5 мкм шир., клетки прямостоячих нитей 6—16 мкм шир.,
по длине до 2.5 раза больше. Некоторые верхушечные клетки оканчи-
ваются 1—2 волосками, второй волосок обычно отходит сбоку, несколько
ниже верхушки. Волоски 2—3 мкм шир., до 700 мкм дл.; они лишены
содержимого, легко обламываются.
В быстро текущих пресных водах, на камнях. Крым.
Род CHAETONEMA Nowak. - ХЕТОНЕМА
Нити ветвящиеся, ползучие, обособленные, с приподнимающимися
простыми или повторно разветвленными ветвями. Клетки цилиндриче-
ские, несущие волоски и в этом месте слегка вздутые. Волоски однокле-
точные, бесцветные, у основания слегка расширенные, по 1—4, высту-
пающие из слизи растения-хозяина как ветви, направленные в одну сто-
310
Рис. 174.
Fridea torrenticola: 1 — фрагмент прямостоячей нити, 2 — нить со споран-
гиями, 3 — верхушечные клетки в старых нитях. (По Смиту).
рону; они могут возникать повторно, вследствие чего на старых клетках
находится много базальных частей обломанных волосков. Вегетативное
размножение своеобразным почкованием увеличенных в размерах
вегетативных клеток в период перехода нитей в пальмеллевидное
состояние. Почки, вначале небольшие, полушаровидные, постепенно
увеличиваясь в размерах, округляются, отделяются от материнской
клетки перегородкой и освобождаются вследствие ослизнения оболочек;
они могут вновь начать почковаться или непосредственно прорастают
в нить. Некоторые из обособленных клеток образуют многочисленные
апланоспоры. Бесполое размножение четырехжгутиковыми зооспорами
и апланоспорами. Зооспоры со стигмой, по 2 в клетке. Прикрепившись
к слизи водоросли-хозяина, зооспоры образуют выросты, проникающие
сквозь слизь к нитям, обвивают их и растут далее вдоль этих нитей.
Половой процесс оогамный, особи гетероталличны. Оогонии обра-
зуются из конечных клеток боковых ветвей; содержимое такой клетки
311
преобразуется в крупную яйцеклетку, которая выскальзывает наружу
через образующуюся в оболочке клетки верхушечную пору. Антеридии
бочонковидные, с желто-зеленым мелкозернистым содержимым; они раз-
виваются из клеток коротких боковых ветвей или из отдельных клеток
главных нитей, по нескольку в коротких рядах, в каждом из них обра-
зуется по 8 гамет. Мужские гаметы без стигмы, двужгутиковые, эллип-
соидные, с 2 пульсирующими вакуолями в переднем конце и редуциро-
ванным желто-зеленым хлоропластом в заднем конце. Оплодотворение
происходит вне оогония.
В проточных, реже стоячих водах, в слизи зеленых и красных водо-
рослей.
I. Длина вегетативных клеток в 2—4 раза больше ширины, волоски
длинные, оогонии грушевидные......................С. irregulare 1.
II. Длина вегетативных клеток равна ширине или до 3 раз больше,во-
лоски короткие, оогонии почти шаровидные............С. ornatum 2.
1. Chaetonema irregulare Nowak. — Хетонема неправильная
(рис. 175). — Клетки нитей 9—18 мкм шир., по длине до 4 раз больше.
Волоски одиночные, длинные, у основания 4—6 мкм шир., часто согнутые.
Оболочки клеток бесцветные или, иногда, ржаво-красные. Зооспоры
5 мкм шир., 8—10 мкм дл. Оогонии 20—25 мкм шир., 40—45 мкм дл.,
яйцеклетка 20—25 мкм шир., 35 мкм дл.
Преимущественно в проточных водах.
Var. irregulare (рис. 175). — Вегетативные клетки одинаковой
ширины, 9—15 мкм шир., по длине в 2—4 раза больше. Волоски длин-
ные, у основания 4—5 мкм шир., часто согнутые.
В стоячих и проточных водах, в слизи Tetraspora, Schizochlamys,
Chaetophora, Batrachospermum и других водорослей. ЛатвССР, РСФСР
(Московская обл., Вост. Сибирь: оз. Байкал), УССР, местами.
Var. subvalidum Hansg. — Вегетативные клетки разные по ширине,
чаще 15—18 мкм шир., 18—48 мкм дл. Волоски очень длинные, в 10—
Рис. 175.
Chaetonema Irregulare: 1 — вегетативная нить на слоевище Batrachospermum,
2 — нить с оогониями, 3, 4 — освобождение яйцеклетки, 5 — нить с антери-
диями, 6 — мужские гаметы. (7 — по Губеру, 2—6 — по Мейеру).
312
Рис. 176.
Chaetonema ornatum, фрагменты слоевища. (По Трансо).
20 и более раз длиннее несущей их клетки, у основания около 6 мкм шир.,
в средней части 3 мкм шир., у вершины 1 мкм шир.
В слизи Chaetophora elegans. В СССР не обнаружена. (Зап. Европа).
2. Chaetonema ornatum Trans. — Хетонема украшенная (рис. 176). —
Клетки нитей 7 — 10 мкм шир., 15—30 мкм дл., нередко односторонне
вздутые. Волоски по 1—4, довольно короткие. Оогонии почти шаровид-
ные, 18—22 мкм в диам., с нежной мелкоямчатой оболочкой, с терми-
нальным волоском или без него.
В слизи водорослей Тetraspora, Draparnaldia. В СССР не обнаружена.
(Сев. Америка).
Род ЕСТОСНАЕТЕ (Huber) Wille - ЭКТОХЕТЕ
Нити беспорядочно разветвленные, ползучие, обособленные или
образующие паренхимные пластинки внутри пустых клеток других
водорослей. Клетки цилиндрические, вытянутые или почти шаровидные.
Волоски тонкие, длинные, одноклеточные, не отделяющиеся поперечными
перегородками от материнских клеток, у основания слегка вздутые
и перетянутые, без влагалищ. Хлоропласт пристенный, пластинчатый,
цельный или продырявленный, со многими пиреноидами. Бесполое
размножение двужгутиковыми зооспорами. Зооспоры без стигмы; они
образуются в большом количестве в обычных более крупных клетках.
Половое размножение изогаметами, возникающими в неизмененных
клетках — гаметангиях, как и зооспоры.
Виды преимущественно морские, реже встречающиеся и в пресных
водах.
I. Клетки 8 — 20 (30) мкм шир., с длиной, в несколько раз превосходя-
щей ширину........................................Е. endophytum 1.
II. Клетки 5—15 мкм шир., с длиной, равной ширине или несколько
большей ее........................................Е. leptochaete 2.
313
Рис. 177.
1—5 — Ectochaete endophyturn: 1 — слоевище в пустой клетке Cladophora,
2—5 — стадии формирования слоевища на поверхности нитчатой водоросли;
6,7 — Е. leptochaete, фрагменты слоевища. (По Губеру).
1. Ectochaete endophytum (Mob.) Wille (=Bulbocoleon endophytum
Mob.; Endoderma endophytum Huber; E. jadinianum Huber) — Эктохете
эндофитный (рис. 177, 1—5). — Клетки 8 — 20 (30) мкм шир., по длине
в несколько раз больше. Волоски многочисленные, заполненные цито-
плазматическим содержимым.
В солоноватых, соленых и морских водах, на нитчатых водорослях,
преимущественно в пустых клетках базальных частей кладофоры.
УССР (Сиваш), Азовское море.
При усиленном росте таллома у этого вида образуются подушечки,
под давлением которых оболочки клеток растения-хозяина разрываются
и водоросль, оказавшись снаружи, продолжает расти дальше. В резуль-
тате ее интенсивного разрастания на поверхности нитей кладофоры
возникают крупные плотные налеты, которые постепенно переходят
на камни и скалы.
2. Ectochaete leptochaete (Huber) Wille (—Endoderma leptochaete
Huber) — Эктохете тонкощетинистая (рис. 177, 6, 7). — Клетки 5—
15 мкм шир., 10—30 мкм дл., почти изодиаметрические в плотных
псевдопаренхимных участках таллома и более вытянутые в свободно
распростертых нитях. Волоски длинные, многочисленные. Спорангии
с клювовидно утолщенной оболочкой на верхушке.
В соленых и морских водах, на нитях кладофоры между наружными
и внутренними оболочками ее клеток. Каспийское море.
314
Род THAMNIOCHAETE Gay - ТАМНИОХЕТЕ
Нити прямостоячие, короткие, простые или ветвящиеся, прикреплен-
ные к субстрату неправильно цилиндрической, прямой или слегка
изогнутой, несколько суженной книзу базальной клеткой. От верхушеч-
ной клетки и от некоторых клеток, расположенных ниже, отходит по
1 — 3 длинных бесцветных одноклеточных волоска, слегка расширенных
у основания. Хлоропласт пластинчатый, с 1 пиреноидом. Бесполое
размножение зооспорами и акинетами. Зооспоры образуются по
1 в клетке. Акинеты с толстыми буроватыми оболочками; при их
образовании нити распадаются.
В пресных водоемах, на нитчатых водорослях.
I. Нити разветвленные, волоски отходят от верхушечной части конечных
клеток ......................................... Т. huberi 1.
Рис. 178.
1—4 — Thamniochaete huberi: 1, 2 — общий вид нитей, <?, 4 — акинеты; о, 6 —
Т. aculeata, общий вид нитей; 7—9 — Micropoa leptochaete: 7 — фрагмент
слоевища, 8 — образование зооспор, 9 —• зооспора. {1 — 4 — по Ге, 5, 6 —
по Весту, 7—9 — по Мёвусу).
315
II. Нити простые, волоски отходят от боковой поверхности конечных
клеток.......................................Т. aculeata 2.
1. Thamniochaete huberi Gay — Тамниохете Губера (рис. 178,
1—4). — Нити (без волоска) около 38 мкм дл., из 4—6 клеток. Клетки
шаровидные или эллипсоидные, 7—8 мкм шир. Волоски длинные.
На нитях Oscillatoria, Tribonema, нередко на зеленых водорослях.
В СССР не обнаружена. (Зап. Европа).
2. Thamniochaete aculeata W. et G. S. West — Тамниохете шипова-
тая (рис. 178, 5, 6). — Нити из 3—4 клеток. Клетки более или менее
цилиндрические, 5.5—13 мкм шир., 25—31 мкм дл. Верхушечная клетка
10 — 13 мкм шир. Волоски короткие.
На нитях Gloeotrichia natans, В СССР не обнаружена. (Англия).
Род MICROPOA L. Moev. - МИКРОКОД
Нити беспорядочно ветвящиеся, ползучие. Клетки вначале более
или менее цилиндрические, нередко несколько вздутые и искривленные,
позднее становятся шаровидными. Волоски слегка расширенные у осно-
вания, без влагалища. Хлоропласт с 1 пиреноидом. Размножение
двужгутиковыми зооспорами, прорастающими без периода покоя.
1. Micropoa leptochaete L. Moev. — Микропоа мелковолосистая
(рис. 178, 7—9). — С признаками рода.
В обрастаниях на Lemna minor. В СССР не обнаружена. (Зап. Ев-
ропа) .
В условиях культуры образует подушковидные налеты около
100 мкм в диам. Вид изучен недостаточно и описан кратко и схематично.
Семейство COLEOCHAETACEAE -
КОЛЕОХЕТОВЫЕ
Слоевище нитчатое или пластинчатое. Нити ползучие, ветвящиеся,
полностью распростертые по субстрату, обособленные или тесно соеди-
ненные в однослойную псевдопаренхимную пластинку округлых или
неправильных очертаний или образующие подушковидные слоевища
с подошвой и отходящими вверх от ее клеток простыми или разветвлен-
ными короткими ветвями. Клетки различной формы, преимущественно
цилиндрические. От спинной, свободной поверхности многих клеток
отходит по 1 цитоплазматической длинной бесцветной, легко обламы-
вающейся щетинке, длительное время связанной с содержимым материн-
ской клетки. Ризоидные выросты отсутствуют. Ядро 1. Хлоропласт
с 1—2 пиреноидами. Бесполое размножение двужгутиковыми зооспо-
рами. Половой процесс оогамный.
В пресных водах.
Род COLEOCHAETE ВгёЬ. - КОЛЕОХЕТЕ
Нити ползучие, ветвящиеся, обычно радиально расходящиеся от
центральной группы клеток и тесно смыкающиеся боковыми сторонами
в однослойную плоскую, более или менее правильно дисковидную
пластинку, реже слоевище подушковидное или представлено обособлен-
316
ними, беспорядочно ветвящимися нитями. Кроме эпифитных слоевищ
встречаются эндофитные, обитающие внутри оболочек клеток некоторых
харовых водорослей, преимущественно лишенных коры. Клетки изо-
диаметрические или несколько удлиненные, многие со щетинками.
Щетинки образованы внутренним слоем оболочки, у основания окружены
своеобразным воротничком — трубчатым внешним слоем прорванной
оболочки. Хлоропласт пристенный, пластинчатый, с 1—2 крупными
отчетливыми пиреноидами. Вегетативное деление свойственно лишь
краевым клеткам дисковидных слоевищ и верхушечным клеткам
прямостоячих нитей подушковидных слоевищ. Бесполое размножение
двужгутиковыми зооспорами и апланоспорами. Зооспоры грушевидной
формы, без стигмы; они образуются по 1 в любой клетке слоевища,
реже в конечных клетках прямостоячих нитей и освобождаются через
небольшое круглое отверстие, возникающее в стенках материнских
клеток. Прорастание зооспор различно у разных видов. Апланоспоры
с толстыми оболочками; они возникают изредка, по 1 в клетке. Половой
процесс оогамный. Двужгутиковый мелкий подвижный сперматозоид
оплодотворяет неподвижную крупную яйцеклетку, находящуюся в оого-
нии. Оогонии или бутылковидные с длинной шейкой (трихогиной),
заполненной бесцветной плазмой, или шаровидные и яйцевидные без
шейки; в подушковидных слоевищах они возникают обычно в результате
преобразования отдельных конечных клеток прямостоячих нитей, в пла-
стинчатых слоевищах — из некоторых краевых клеток радиально ползу-
чих нитей и могут размещаться 1 или несколькими концентрическими
кругами. Женские гаметы по 1 в оогонии, шаровидные. Антеридии
бутылковидные, бесцветные; в подушкообразных слоевищах они обра-
зуются на укороченных веточках, в пластинчатых — из любых вегета-
тивных клеток в результате их повторных делений. Мужские гаметы
по 1 в антеридии, бесцветные, шаровидные или эллипсоидные, дву-
жгутиковые. Вследствие разбухания и ослизнения верхушечной части
шейки оогония возникает отверстие, через которое проникает мужская
гамета. После оплодотворения у основания шейки оогония образуется
поперечная перегородка. Зигота некоторое время увеличивается в раз-
мерах, и от клетки, несущей оогоний, и от соседних с ней клеток
начинают отрастать веточки, плотно прилегающие к поверхности оогония,
в итоге чего вследствие их обильного ветвления оогоний оказывается
окруженным своеобразным плотным псевдопаренхимным коровым слоем.
В пластинчатых слоевищах, где оогонии полушаровидно выдаются над
поверхностью, коровым слоем оплетается преимущественно их верхняя,
выступающая часть. Между корой и поверхностью ооспоры возникает
толстая буроватая оболочка, выделяемая одновременно с обеих сторон.
После длительного периода покоя (зимний период) в ооспоре происходит
редукционное деление ядра, после чего образуется поперечная пере-
городка, затем возникают продольные перегородки и в каждой полусфере
образуется по 8—16 клиновидных клеток. С окончанием деления
оболочка ооспоры разрывается в области поперечной перегородки и
из каждой клиновидной клетки выходит двужгутиковая зооспора,
называемая карпозооспорой. Прорастая, она дает начало новому слое-
вищу.
В стоячих и проточных пресных водах, на водных цветковых
растениях, крупных нитчатых водорослях, древесине, раковинах моллюс-
ков, камнях, а также в оболочках клеток харовых водорослей.
Водоросли микроскопические, однако нередко слоевища, распола-
гаясь тесно одно возле другого, образуют пятна налета, достигающие
нескольких миллиметров в поперечнике.
317
I. Слоевище плоское, пластинчатое.
1. Нити разветвленные, обособленные, радиально отходящие от цент-
ральной группы клеток и не срастающиеся боковыми сторонами
в плотную пластинку; преимущественно на водных цветковых
растениях.
А. Нити расположены рыхло, клетки разнообразной формы,
(6)10—25 мкм шир., краевые клетки большей частью глубоко
вильчато рассеченные или закругленные . . . .С. soluta 1.
Б. Нити расположены более или менее плотно, клетки преимуще-
ственно цилиндрические, 6—10 мкм шир., краевые клетки
цельные, широко закругленные .... С. pseudosoluta 2.
2. Нити простые или разветвленные, обособленные или участками
образующие псевдопаренхимные пластинки; на оболочках и в обо-
лочках харовых водорослей.....................С, nitellarum 3.
3. Нити плотно срастающиеся боковыми сторонами в псевдопарен-
химную пластинку; на разных субстратах.
А. Пластинка неопределенных очертаний, клетки 7.4—15.2(25)
мкм шир., растение однодомное..............С. irregularis 4.
Б. Пластинка правильно дисковидная; растение двудомное,
а. Клетки 8—12(20) мкм шир.................С. orbicularis 5.
б. Клетки 14—48 мкм шир....................С. sculata 6.
II. Слоевище выпуклое, подушковидное, с вверх приподнимающимися
от клеток нитей подошвы или прямостоячими нитями.
1. Нити прямостоячие, расходящиеся радиально; слоевище полу ша-
ровидное, однодомное..........................С. pulvinata 7.
2. Нити прямостоячие, расходящиеся беспорядочно; слоевище полу-
шаровидное, двудомное...........................С. disjuncta 8.
3. Нити распростертые или приподнимающиеся; слоевище слегка
выпуклое.
А. Клетки цилиндрические, 7—14 мкм шир. . . .С. sampsonii 9.
Б. Клетки удлиненные, неправильной формы, (7.6) 11.4—25 мкм
шир........................................С. divergens 10.
1. Coleochaete soluta Pringsh. — Колеохете разделенная (рис. 179,
7, 2). — Слоевище пластинчатое, округлое в очертаниях, однодомное.
Нити ползучие, радиально расходящиеся от 2 —4 центральных клеток,
разобщенные, ветвящиеся, иногда заключенные в хорошо развитую
слизь. Клетки цилиндрические, вздутые в средней части, реже шаровид-
ные, бочонковидные или неправильных очертаний, (6)10—25 мкм шир.,
по длине равные ширине или до 3.5 раза больше. Краевые клетки
большей частью глубоко вильчато рассеченные. Зооспоры при прораста-
нии делятся вертикальной перегородкой на 2 клетки, от которых
позднее отходят прямостоячие нити. Оогонии развиваются из некоторых
конечных клеток нитей, вначале бутылк'овидные, с отчетливым горло-
видным выростом, позднее, сплетаясь корой, становятся шаровидными,
30 — 220 мкм в диам. Ооспоры 140—150 мкм в диам. Антеридии обра-
зуются вблизи оогониев из вегетативных клеток, маленькие, бесцветные.
В стоячих и проточных водах, на различных субстратах.
Var. soluta (рис. 179, 7). — Клетки цилиндрические или непра-
вильные, 10—25 мкм шир., по длине в 2—3.5 раза больше. Краевые
клетки вильчато рассеченные. Оогонии 200—220 мкм в диам., ооспоры
140—150 мкм в диам.
В реках, прудах, озерах, болотах, на водных цветковых растениях,
реже раковинах моллюсков. В различных районах СССР, нередко.
318
1
3
Рис. 179.
1,2 — Coleochaete soluta' 1 — var. soluta, 2 — var. brevicellularis; 3 — C. pseudo-
soluta, фрагмент слоевища с оогониями и антеридиями. (7 — по Прингсгейму,
2 — по Шмидле, 3 — по Готье-Ли евр).
Var. minor Hansg. — Клетки цилиндрические или неправильные,
6 — 9(12) мкм шир., по длине равные ширине или до 3 раз больше.
Краевые клетки вильчато рассеченные. Оогонии 30—36(60) мкм в диам.
В стоячих и слабо проточных водах, на раковинах моллюсков.
УССР, Кавказ.
Var. brevicellularis Schmidle (рис. 179, 2). — Клетки бочонковидные
или шаровидные, 18—24 мкм шир., по длине равные ширине или
в 1.25 раза больше. Краевые клетки закругленные. Ветви косо приподни-
мающиеся. Оогонии 80—110 мкм в диам., плотно оплетенные корой.
В водоемах с замедленным течением, на водных цветковых расте-
ниях. УССР (пойменные водоемы Горыни).
2. Coleochaete pseudosoluta Gauth.-Liev. — Колеохете ложноразде-
ленная (рис. 179, 3). —Слоевище пластинчатое, более или менее
округлое в очертаниях, двудомное. Нити ползучие, ветвящиеся в плоско-
сти, радиально расходящиеся от центральной группы клеток, плотно
расположенные, но не срастающиеся боковыми сторонами в псевдо-
паренхимный диск. Клетки цилиндрические, 6—10 мкм шир., 10—
25 мкм дл. Краевые клетки широко закругленные. Оогонии 30—50 мкм
шир., 50—60 мкм дл., покрытые корой лишь с поверхности. Антеридии
образуются по 4 в вегетативных клетках.
В пресных водоемах, на водных цветковых растениях. В СССР
не обнаружена. (Сев. Африка).
319
Рис. 180.
1,2 — Coleochaete nitellarum: 1 — общий вид слоевища, 2 — оогоний; 3—5 —
С. irregularis-. 3 — фрагмент слоевища, 4 — антеридий, 5 — оогоний; 6 — С. orbi-
cularis, общий вид слоевища с оогониями. (7, 2, 4, 5 — по Прескотту, 3 —
по Принтцу, 6 — по Прингсгейму).
3. Coleochaete nitellarum Jost — Колеохете нителловая (рис. 180,
1, 2). — Слоевище нитчатое, однодомное. Нити ползучие, простые или
ветвящиеся, обособленные или участками тесно смыкающиеся друг
с другом и образующие псевдопаренхимные пластинки. Клетки разно-
образные по форме и величине. Влагалища щетинок чаще всего отвер-
девшие. Зооспоры, прикрепившись к поверхности клеток харовых
водорослей, образуют вырост, внедряющийся в оболочку клеток;
разрастающиеся нити расслаивают стенки клеток, создавая в них поло-
сти. Оогонии шаровидные или яйцевидные, с поверхности полностью
или частично покрытые корой. Антеридии в виде мелких угловатых
клеток, развивающиеся в большом количестве по углам вегетативных
клеток.
В пресных водоемах, на оболочках и в оболочках клеток харовых
водорослей, лишенных коры, преимущественно из рода Nitella. ЛатвССР,
РСФСР (Карельская АССР, Ленинградская и Московская обл.),
АрмССР.
Вид описан недостаточно, без указания размеров, но легко узнается
благодаря своеобразному местообитанию.
4. Coleochaete irregularis Pringsh. — Колеохете неправильная
(рис. 180, 3—5). — Слоевище пластинчатое, неопределенных очертаний,
однодомное. Нити ползучие, более или менее радиально расходящиеся,
ветвящиеся в плоскости, тесно смыкающиеся боковыми сторонами
в однослойную пластинку. Клетки 7.4—15.2(25) мкм шир., по длине
равные ширине или до 2 раз больше. Щетинки отходят сбоку, нередко
по нескольку. Оогонии без горловидного выроста, шаровидные или
яйцевидные, 60—100 мкм шир., 60—120 мкм дл., без корового слоя или
частично оплетенные корой лишь сверху.
В стоячих и слабо проточных водах, на различных субстратах.
Var. irregularis (рис. 180, 3—5). — Клетки 11.4—15.2(25) мкм шир.,
по длине равные ширине или до 2 раз больше.
В прудах, озерах, болотах, реже реках, на водных цветковых
растениях, раковинах моллюсков. В разных районах СССР, нередко.
Var. minor Skvortz. — Клетки 7.4—11 мкм шир., 14.8 мкм дл.
На водных цветковых растениях. В СССР не обнаружена.
(Китай).
5. Coleochaete orbicularis Pringsh. — Колеохете округлая (рис. 180,
6). — Слоевище пластинчатое, дисковидное, двудомное. Нити ползучие,
радиально расходящиеся, ветвящиеся в плоскости, тесно смыкающиеся
боковыми сторонами в однослойную округлую пластинку. Клетки
8—12(20) мкм шир., по длине до 2—2.5 раза больше. Оогонии яйцевид-
ные, без горловидного выроста, 50—66 мкм шир., 60 —86 мкм дл.,
с нижней стороны голые, сверху частично одетые коровым слоем, реже
без коры.
В стоячих и проточных водах, на растительных субстратах, ракови-
нах моллюсков. Повсюду, нередко.
6. Coleochaete scutata Breb. — Колеохете щитовидная (рис. 181). —
Слоевище обычно правильно дисковидное, двудомное. Нити ползучие,
радиально расходящиеся, ветвящиеся в плоскости, тесно смыкающиеся
Рис. 181.
Coleochaete scutata f. scutata: 1 — общий вид слоевища, 2 — фрагмент слоевища
со зрелыми оогониями, 3 — зооспора, 4, 5 — стадии развития слоевища, 6 —
молодое слоевище. (1 — по Кютцингу, 2—в — по Прингсгейму).
1/а21 Зеленые водоросли. Улотриксовые
321
320
боковыми сторонами в округлую однослойную пластинку, лишь в самом
центре более толстую. Клетки прямоугольные, реже многоугольные,
у женского растения 14—24(48) мкм шир., в центральной части слое-
вища преимущественно 18—19 мкм шир., по длине равные ширине
или почти до 3 раз больше, у мужского растения 15—20 мкм шир.,
15—25 мкм дл. Зооспоры, прикрепившись к субстрату, делятся горизон-
тальной перегородкой на 2 клетки, из которых верхняя теряет способ-
ность к дальнейшему делению и на ее спинной поверхности образуется
щетинка, а нижняя в результате двух перекрестных делений образует
диск, состоящий вначале из 4 клеток, разрастающихся в дальнейшем
в однослойное пластинчатое слоевище. Оогонии яйцевидные, без горло-
видного выроста, 60—140 мкм шир., покрытые корой лишь сверху.
Ооспоры до 120 мкм шир., до 160 мкм дл. Антеридии образуются
по 4 из одной вегетативной клетки, 6—10 мкм шир., 6—10 мкм дл.,
с 1 зеленой гаметой каждый.
В стоячих и проточных водах, на различных субстратах.
F. scutata (рис. 181). — Вегетативные клетки 24—48 мкм шир.
Оогонии до 140 мкм шир.
В озерах, прудах, реках, на водных цветковых растениях, древесном
субстрате, раковинах моллюсков. Повсеместно.
F. minor Mob. — Вегетативные клетки 14—22 мкм шир. Оогонии
60—70(100) мкм шир.
В пресных водоемах, на раковинах моллюсков, часто вместе с типом.
В СССР не обнаружена. (Зап. Европа).
7. Coleochaete pulvinata А. Br. — Колеохете подушковидная
(рис. 182). — Слоевище полушаровидное, до 2 мм выс., или почти
дисковидное, однодомное. Нити прямостоячие, ветвящиеся, радиально
расходящиеся от клеток подошвы, образованной ползучими по субстрату
нитями. Клетки цилиндрические, реже многоугольные, 10—43 мкм шир.,
по длине равные ширине или до 2 (4) раз больше. Щетинки встречаются
часто. Зооспоры образуются лишь в верхушечных клетках прямостоячих
нитей; при прорастании они делятся вертикальной перегородкой на 2
клетки, от которых позднее отходят прямостоячие нити. Оогонии
на ранней стадии развития бутылковидные, с отчетливым горловидным
выростом, позднее, оплетаясь корой, становятся шаровидными, до
150 мкм в диам. Ооспоры до 110 мкм в диам. Антеридии бутылковидные,
7—17 мкм шир., по длине до 2.5 раза больше, расположенные на веге-
тативных клетках вблизи оогониев, преимущественно по 2—3.
В стоячих водах — озерах, болотах, реже проточных водах, на
водных цветковых растениях, камнях, раковинах моллюсков.
Var. pulvinata (рис. 182, 1—4). — Слоевище полушаровидное,
с хорошо развитыми прямостоячими нитями. Клетки 20—43 мкм шир.,
по длине до 2 раз больше. Оогонии до 150 мкм в диам. Ооспоры 80—110
мкм в диам. Антеридии 17 мкм шир., по длине до 2.5 раза больше.
В озерах, реках, болотах. ЛатвССР. Ладожское и Валдайское озера,
Московская обл., УССР, оз. Байкал. УзССР.
Var. minor Pringsh. (рис. 182, 5). — Слоевище почти дисковидное,
со слабо развитыми прямостоячими нитями. Клетки 10—17 мкм шир.,
по длине большей частью равные ширине. Оогонии 58—65 мкм в диам.
Ооспоры 28—41 мкм в диам.
В озерах. ЛатвССР, оз. Имандра и Нотозеро, Московская обл.,
часто вместе с типовой формой.
Var. baileyi (Mob.) Printz (=C. baileyi Mob.) (рис. 182, 6). — Слое-
вище полушаровидное, с довольно хорошо развитыми прямостоячими
322
Рис. 182.
Coleochaete pulvinata'. 1—4 — var. pulvinata (7 — фрагмент слоевища, 2 —
зрелый оогоннй, покрытый корой, 3 — стадии прорастания ооспоры, 4 — карпо-
споры), 5 — var. minor, 6 — var. baileyi. (1—5 — по Прингсгейму, 6 — по
Мёбиусу).
нитями. Клетки 14—20 мкм шир., по длине в 1.5—2(4) раза больше.
Оогонии со слабо развитой корой. Антеридии 7 мкм шир., 12 мкм дл.
На водных цветковых растениях. В СССР не обнаружена. (Зап.
Европа).
8. Coleochaete disjuncta Tiff. — Колеохете разобщенная (рис. 183,
1, 2). — Слоевище подушковидное, двудомное. Нити прямостоячие,
ветвящиеся, беспорядочно расходящиеся от клеток ползучих по субстра-
'/221*
323
Рис. 183.
1,2 — Coleochaete disjuncta-. 1 — фрагмент слоевища с оогонием, 2 — фрагмент
слоевища с антеридиями; 3 — С. sampsonii; 4 — С. divergens. (1, 2 — по Тиф-
фани, 3 — по Трансо, 4 — по Прингсгейму).
ту нитей подошвы. Клетки 16—26 мкм шир., 12—40 мкм дл. Оогонии
шаровидные, 45—65 мкм в диам., покрытые корой. Ооспоры 26—40 мкм
в диам. Антеридии грушевидные, 10—13 мкм шир., 22—26 мкм дл.
В прудах, на водных цветковых растениях. В СССР не обнаружена.
(Пуэрто-Рико).
9. Coleochaete sampsonii Trans. — Колеохете Сампсона (рис. 183,
3). — Слоевище более или менее подушковидное, однодомное, неопреде-
ленных очертаний. Нити ползучие по субстрату, беспорядочно ветвя-
щиеся, с отходящими вверх ветвями. Клетки цилиндрические, 7—14
мкм шир., 14—36 мкм дл., нередко слегка вздутые в средней части или
беспорядочно изогнутые. Оогонии без горловидного выроста, шаровид-
ные, 35—48 мкм в диам., полностью покрытые корой. Ооспоры 29—
36 мкм в диам. Антеридии шаровидно-грушевидные.
В различных пресных водах, в обрастаниях красных водорослей,
преимущественно Batrachospermum. В СССР не обнаружена. (Сев. Аме-
рика) .
10. Coleochaete divergens Pringsh. — Колеохете расходящаяся
(рис. 183, 4). — Слоевище подушковидное, неправильной формы, одно-
домное. Нити ползучие, ветвящиеся, не сливающиеся в псевдопарен-
химную пластинку, распростертые по. субстрату и приподнимающиеся,
разрастающиеся дерновинками. Клетки большей частью удлиненные,
неправильной формы, (7.6) 11.4—25 мкм шир., по длине равные ширине
или до 2.5—3 раз больше. Зооспоры непосредственно прорастают
324
в обособленные, беспорядочно ветвящиеся нити. Оогонии бурые, шаро-
видные или эллипсоидные, 38—135 мкм шир., полностью оплетенные
корой. Ооспоры 23—95 мкм шир. Антеридии маленькие, грушевидные,
по 3—4 и больше на вегетативных клетках вблизи оогониев.
В стоячих и проточных водах, на водных цветковых растениях,
древесине, раковинах моллюсков.
Var. divergens (рис. 183, 4). — Клетки около 25 мкм шир., по длине
до 3 раз больше. Оогонии до 135 мкм шир. Ооспоры до 95 мкм шир.
В реках, озерах, пойменных водоемах, прудах. ЛатвССР, РСФСР
(Карельская АССР, Новгородская, Московская, Владимирская, Горь-
ковская обл.), УССР, нередко.
Var. minor Hansg. — Клетки (7.6)11.4—22 мкм шир., по длине
равные ширине или до 2.5 раза больше. Оогонии 38—60(80) мкм шир.
Ооспоры 23—54(70) мкм шир.
Преимущественно в реках и пойменных водоемах. УССР
(рр. Южн. Буг, Самара).
Семейство CHAETOSPHAERIDIACEAE Bourr. -
ХЕТОСФЕРИДИЕВЫЕ
Слоевище обычно многоклеточное, дисковидное, чаще всего в виде
шаровидных шли полушаровидных подушечек, прикрепленных к суб-
страту, реже одноклеточное. Клетки шаровидные, эллипсоидные, непра-
вильных очертаний, реже более или менее цилиндрические, с гладкой
тонкой или толстой оболочкой, с 1 или несколькими щетинками,
обычно рыхло соединенные в короткие нити или беспорядочные
скопления, погруженные в слизистую массу. Щетинки в основании
обычно окружены цилиндрическим или конусовидным, одинарным или
двойным гомогенным или слоистым влагалищем, реже оно отсутствует.
Ядро 1. Хлоропласт пристенный, чашевидный или пластинчатый,
иногда их бывает по 2, с 1 или несколькими пиреноидами или без них.
Размножение преимущественно вегетативное. У некоторых родов
(Chaetosphaeridium, Conochaete) известны двужгутиковые яйцевидные
зооспоры, возникающие по 2—8 в несколько видоизмененных клетках
и выходящие наружу в слизистом, быстро расплывЩрщемся пузыре.
Акинеты и апланоспоры неизвестны.
В. пресных водах, на растительных субстратах.
I. Щетинки в основании окружены слизистым влагалищем.
1. Влагалище у щетинок одинарное (однослойное).
А. Слоевище многоклеточное, бесформенное, клетки шаровидные
или полушаровидные.
а. Влагалище у щетинок цилиндрическое, клетки соединены
длинными или короткими слизистыми тяжами .....
род • Chaetosphaeridiu'm (с. 326).
б. Влагалище у щетинок конусовидное, клетки без слизистых
тяжей............................род Conochaete (с. 329).
. Б. Слоевище многоклеточное, нитчатое, клетки более или менее
цилиндрические ......................род Chaetotheke (с. 331).
2. Влагалище у щетинок двойное (двуслойное)...................
......................................... род Dicoleon (с. 331).
П. Щетинки без слизистого влагалища.
1. Клетки с 2—4 щетинками . . . род Oligochaetophora (с. 333).
2. Клетки с 8—12 щетинками . . . род Polychaetpphora (с. 333).
325
Род CHAETOSPHAERIDIUM Kleb. - ХЕТОСФЕРИДИУМ
Слоевище в разросшемся состоянии многоклеточное, иногда окру-
женное слизью. Клетки шаровидные или полушаровидные, соединенные
друг с другом длинными иди короткими слизистыми цилиндриче-
скими, лишенными содержимого тяжами, благодаря чему слоевище
принимает вид ползучих или слегка приподнимающихся нитей; иногда
тяжи очень короткие и клетки располагаются близко друг возле друга.
От спинной стороны почти каждой клетки отходит по 1 длинной
щетинке с расширенным основанием, окруженным коротким цилиндри-
ческим слизистым влагалищем. При вегетативном размножении клетки
делятся горизонтально, нижняя дочерняя клетка поворачивается на бок
и продолжает расти в сторону в виде рукавообразного цилиндрического
тяжа, протопласт при этом перемещается к концу тяжа; в дальнейшем
кеглевидно расширенный конец тяжа отделяется поперечной перегород-
кой, а тяж остается, связывая клетки. Иногда наблюдается верти-
кальное деление клеток без образования цилиндрических тяжей.
Хлоропласты в числе 1—2, каждый с 1 пиреноидом. Бесполое размноже-
ние двужгутиковыми зооспорами, возникающими по 4 в зооспорангии.
Половое размножение неизвестно.
В пресных стоячих и проточных водах.
I. Клетки шаровидные или полушаровидные, с 1 хлоропластом.
1. Клетки заключены в общую, хорошо выраженную слизистую
обвертку и соединяются друг с другом очень короткими цилиндри-
ческими тяжами, благодаря чему размещаются плотно...........
...............-.........................С. giobosum 1.
2. Клетки не заключены в общую слизистую обвертку и соеди-
няются друг с другом длинными цилиндрическими тяжами,
благодаря чему размещаются рыхло ... С. pringsheimii 2.
II. Клетки яйцевидные, с 2 хлоропластами............С. ovale 3.
III. Клетки цилиндрические, неправильные, с 1 хлоропластом ....
...........................................С. huberi 4.
1. Chaetosphaeridium giobosum (Nordst.) Kleb. (=Aphanochaete
globosa Nordst.) — Хетосферидиум шаровидный (рис. 184, 1—5). —
Слоевище с общей слизистой обверткой, до 0.4 мм в диам. Клетки
плотно размещенные, шаровидные, иногда сплющенные в дорсивент-
ральном направлении и тогда эллипсоидные, (1)12—20 мкм в диам.,
при делении полушаровидные, окруженные хорошо развитой слизистой
обверткой. Щетинки длинные, 2—3 мкм шир., 16—17 мкм дл., окружен-
ные у основания короткими влагалищами. Оболочки клеток тонкие
или толстые. Соединительные тяжи 4—9 мкм толщ., очень короткие,
на некоторых участках слоевища трудно различимые. Хлоропласт 1.
В стоячих или слабо проточных водах, преимущественно в сфаг-
новых болотах, на растительном субстрате.
F. giobosum (рис. 184, 1, 2).— Клетки шаровидные, 12—18 мкм
в диам., с тонкими оболочками.
ЭССР, ЛатвССР, УССР, Центр. Сибирь.
F. incrassatum Kleb. (рис. 184, 3,v 4). — Клетки шаровидные,
13—20 мкм в диам., с толстыми, до 4—6 мкм,оболочками.
В СССР не обнаружена. (Зап. Европа).
F. depressum G. S. West (рис. 184, 5). —Клетки эллипсоидные,
сплюснутые в дорсивентральном направлении.
В СССР не обнаружена. (Зап. Европа).
326
Рис. 184.
1—5 — Chaetosphaeridium giobosum: 1, 2 — f. giobosum (1 — вид клеток с по-
верхности слоевища, 2 — вид клеток сбоку), 3, 4 — 1. incrassatum (3 — вид
слоевища сверху, 4 — слоевище в разрезе), 5 -» f. depressum; 6—8 — С. pring-
sheimii: 6, 7 — f. pringsheimii (6 — общий вид слоевища, 7 — образование
и выход зооспоры), 8 — f. confertum. (1—4, 6—8 — по Клебану, 5 — по Весту).
F. microscopicum Playf. — Клетки шаровидные, около 1 мкм в диам.
На нитях Oedogonium, Melosira. В СССР не обнаружена. (Австра-
лия).
2. Chaetosphaeridium pringsheimii Kleb. — Хетосферидиум Прингс-
гейма (рис. 184, 6—8). — Слоевище без общей слизистой обвертки.
Клетки рыхло размещенные, шаровидные или полушаровидные, 9 —
12 мкм в диам., соединенные друг с другом хорошо развитыми ползучими
или крючковидно приподнимающимися цилиндрическими тяжами.
Щетинки длинные, до 300 мкм. Влагалища вокруг основания щетинок
около 2 мкм шир., 11 —18 мкм дл. Хлоропласт 1.
В стоячих и слабо проточных водах, нередко в болотах, на раститель-
ном субстрате.
F. pringsheimii (рис. 184, 6, 7). — Клетки шаровидные или полу-
шаровидные, до 12 мкм в диам., соединительные тяжи длинные.
В озерах, прудах, болотах, малых реках. ЭССР, Кольский п-ов,
Ленинградская и Московская обл., УССР, Центр. Сибирь.
F. confer turn Kleb. (рис. 184, 8). — Клетки шаровидные, 9 мкм
в диам., соединительные тяжи сравнительно короткие.
В болотах, прудах, малых реках, часто вместе с типовой формой.
ЛатвССР, Новгородская обл., УССР.
3. Chaetosphaeridium ovale Smith — Хетосферидиум овальный
(рис. 185, 1). — Слоевище без ясно выраженной слизи и тяжей. Клетки
327
Рис. 185.
1 — Chaetosphaeridium ovale; 2, 3 — C. huberl (1 — по Смиту, 2, 3 — по Мё-
биусу) .
яйцевидные, 13—15 мкм шир., 20—22 мкм дл. Щетинки до 125 мкм
дл., у основания окруженные слизистым влагалищем 3 мкм шир.,
15 мкм дл. Хлоропластов 2.
В пресных водоемах, па зеленых нитчатых водорослях. В СССР не
обнаружен. (Сев. Америка).
4. Chaetosphaeridium huberi Mob. — Хетосферидиум Губера
(рис. 185, 2, 3). — Слоевище нитчатое, без общей слизи. Нити беспо-
рядочно ветвящиеся, ползучие, частично приподнимающиеся. Клетки
обычно цилиндрические или многоугольные, слегка вздутые, иногда
несколько изогнутые или односторонне выпуклые, 8—10 мкм шир.,
по длине равные ширине или до 2 раз больше. Щетинки тонкие,
нежные; они отходят от верхушечных клеток веточек и в основании
окружены влагалищем. Хлоропласт 1, с 1 пиреноидом. Размножение
неизвестно.
В пресных водоемах, в слизи Chaetophora tuberculosa. В СССР
не обнаружен. (Австралия).
Вид изучен недостаточно, весьма близок к роду Dicoleon.
328
Род CONOCHAETE Kleb. - KOHOXETE
Слоевище с общей слизистой обверткой или без нее, шаровидное
или полушаровидное. Клетки разобщенные, преимущественно шаровид-
ные. От спинной стороны каждой клетки отходит по нескольку щетинок.
Щетинки очень длинные, окруженные у основания конусовидным
гомогенным или слоистым влагалищем. Хлоропласты по 1—2, с 1—2
пиреноидами в каждом. Продукт ассимиляции — крахмал, встречается
также масло. Вегетативное размножение делением клеток в двух
направлениях; после деления дочерние клетки вначале окружены разор-
ванными оболочками материнских клеток, которые вскоре исчезают
в общей массе слизи. Бесполое размножение двужгутиковыми зооспо-
рами, образующимися по 4, 8 в увеличенных по размерам клетках;
они освобождаются через отверстие, возникающее в оболочке на спинной
стороне клетки.
В пресных водах, преимущественно на растительном субстрате.
I. Влагалища у щетинок образованы плотной слизью, окружающей
клетки.
1. Клетки 8 — 12 мкм в диам.
А. Число щетинок 5 и больше, их влагалища на вершине закруг-
ленные ....................................С. klebahnii 1.
Б. Число щетинок не больше 5, их влагалища на вершине
заостренные..............................С. lichenicola 2.
2. Клетки 22—26 мкм в диам......................С. comosa 3.
II. Влагалища у щетинок образованы внешним слоем клеточной оболочки
...............................................С............................................... polytricha 4.
1. Conochaete klebahnii Schmidle — Конохете Клебана (рис. 186,
1—3). —Слоевище в виде небольших, довольно плоских колоний 20—
40 мкм в диам., окруженных тонким общим слоем слизи, хорошо
заметной лишь при окрашивании. Клетки шаровидные, 8 — 12 мкм
в диам., с тонкой оболочкой и длинными щетинками в числе 5 и более.
Влагалище у щетинок 2—4 мкм шир., 5 — 7 мкм дл., конусовидное,
на вершине обычно широко округленное, образованное слизью, окру-
жающей клетки. Над центральной клеткой колонии влагалище колпа-
ковидно утолщенное, без щетинки.
В торфяных болотах, на листьях Sphagnum. Калининградская обл.,
ЛатвССР.
2. Conochaete lichenicola Suessen. — Конохете лишайниковая
(рис. 186, 4—7). — Слоевище в виде более или менее шаровидных или
полушаровидных колоний. Клетки более или менее шаровидные,
8.4—10.5 мкм в диам., окруженные плотной слизью, образующей
влагалища щетинок. Щетинки немногочисленные, нежные, около 1 мкм
толщ., 6—15 мкм дл. Влагалище у щетинок конусовидное, на вершине
заостренное, иногда слегка изогнутое.
На слоевищах Peltigera aphtosa. В СССР не обнаружена. (Зап.
Европа).
Малоизученный вид, возможно являющийся разновидностью
Conochaete klebahnii.
3. Conochaete comosa Kleb. — Конохете хохлатая (рис. 186,
8). — Слоевище в виде более или менее полушаровидных или диско-
видных слизистых колоний 0.2—0.5 мм в диам. Клетки шаровидные,
22—26 мкм в диам., с оболочками 2—8 мкм толщ., с 2—3(5) отходя-
22 Зеленые водоросли. Улотриксовые
329
Рис. 186.
1—3 — Conochaete klebahnii: 1 — вид сбоку, 2, 3 — вид сверху; 4—1 — С. liche-
nicola: 4 — вид сбоку, 5—1 — отдельные клетки; 8 — С. comosa, фрагмент
слоевища; 9, 10 — С. polytricha: 9 — молодое слоевище, 10 — группа клеток,
оставшаяся после распадения слоевища. (1—3 — по Шмидле, 4—1 — по Сес-
сенгуту, 8 — по Нордстедту, 9, 10 — по Клебану).
щими вверх щетинками. Щетинки длинные, у основания 1.5—2 мкм
толщ., окруженные слизистыми влагалищами. Влагалище у щетинок
удлиненно-конусовидное, в нижней части 6.5—8 мкм шир., в верхней
2.5 мкм шир.
В пресных водах, на растительных субстратах. В СССР не обна-
ружена. (Англия, Новая Зеландия).
4. Conochaete polytricha (Nordst.) Kleb. (=Aphanochaete polythricha
Nordst.) — Конохете многоволосковая (рис. 186, 9, 10). — Слоевище
в виде шаровидных или почти шаровидных подушечек 0.2—0.5 мм
в диам. Клетки шаровидно-клиновидные, 10—16 мкм в диам., с несколь-
кими щетинками. Щетинки длинные, отходящие во все стороны,
в основании 1.5 мкм толщ. Влагалище у щетинок конусовидное, 6—7 мкм
шир. в нижней части, 2 мкм шир. в верхней, 9 — 13 мкм дл., образованное
внешним слоем клеточной оболочки, утолщающимся и расщепляющимся
на большое количество неясных слоев-пластинок, благодаря чему оно
кажется многослойным.
В пресных водах, на растительных субстратах. ЛатвССР.
330
Род CHAETOTHEKE During. - ХЕТОТЕКЕ
Слоевище нитчатое. Нити ползучие, беспорядочно ветвящиеся,
окруженные нежной слизистой обверткой. Клетки более или менее
цилиндрические, со слоистыми оболочками, у поперечных перегородок
слегка перетянутые. От спинной стороны некоторых клеток отходит
по 1 щетинке, окруженной в основании слизистым влагалищем. Хло-
ропласт 1, пластинчатый, с 1 пиреноидом. Бесполое размножение
двужгутиковыми зооспорами. Зооспоры шаровидные, довольно метабо-
личные, по 1—2 в конечных клетках нитей, без стигмы, с пластинчатым
хлоропластом с 1 пиреноидом и приблизительно с 6 пульсирующими
вакуолями.
В пресных водах.
1. Chaetotheke reptans During. — Хетотеке ползучая (рис. 187). —
Клетки около 7.5 мкм шир., 7.5—19.5 мкм дл. Щетинки до 700 мкм дл.,
в нижней части 2—2.5 мкм шир., в верхней 0.5—1 мкм шир. Зооспоры
около 12 мкм дл.
Найдена в аквариумах на листьях водных цветковых растений.
В СССР не обнаружена. (Австрия).
Род DICOLEON Kleb. - ДИКОЛЕОН
Слоевище в разросшемся состоянии многоклеточное, нитчатое,
погруженное в полушаровидную или шаровидную слизистую массу.
Нити беспорядочно ветвящиеся, ползучие. От ползучих нитей отходят
вверх одноклеточные веточки. Клетки более или менее шаровидные.
От спинной стороны почти каждой клетки берет начало длинная
щетинка, окруженная двойным влагалищем, внутренним длинным и
наружным очень коротким. Размножение не изучено.
Рис. 187.
Chaetotheke reptans-. 1 — общий вид, 2 — участок нити, окруженной толстой
слизистой обверткой, 3 — образование зооспор, 4 — зооспоры. (По Дюрингеру).
22*
331
Рис. 188.
1, 2 — Dicoleon nordstedtii: 1 — фрагмент слоевища, 2 — отдельная клетка;
3 — Oligoch.aetoph.ora simplex, вид клеток сбоку; 4 — Polychaetophora lamellosa,
вид клеток сверху. (1, 2 — по Клебану, 3, 4 — по Весту).
1. Dicoleon nordstedtii Kleb. — Диколеон Нордстедта (рис. 188,
1, 2). — Слоевище слизистое, округлое, 250 — 500 мкм в диам. Клетки
плотно собранные, 18—29 мкм в диам., с двуслойной оболочкой до
5 мкм толщ. Щетинки длинные, на концах загнутые, одетые двойным
влагалищем. Внутреннее влагалище у щетинок в 3—4 раза длиннее
клетки, 60 —80 мкм дл., у основания 5 мкм шир., вверху 3 мкм шир.,
образованное внутренним слоем ее оболочки, наружное влагалище корот-
кое, 5—7 мкм, образованное выпячиванием наружного слоя оболочки.
В торфяных лужах, болотах, на слоевищах Nostoc pachydermum,
Tolypella и других водорослей, а также на побегах рогоза. В СССР
не обнаружена. (Норвегия).
332
Род OLIGOCHAETOPHORA G. S. West -
ОЛИГОХЕТОФОРА
Слоевище в виде 2—3 рыхло соединенных клеток или одноклеточное.
Клетки почти шаровидные или яйцевидные, с тонкой гомогенной
оболочкой. От спинной стороны каждой клетки отходит по 2—4 щетинки.
Щетинки простые, длинные, очень тонкие, слегка изогнутые, без вла-
галищных расширений клеточной оболочки вокруг их основания.
Хлоропласт 1, чашевидный, преимущественно в нижней части клетки,
без пиреноида, с 2—3 маленькими зернышками крахмала. Размножение
не прослежено.
1. Oligochaetophora simplex G. S. West (=Polychaetophora simplex
G. S. West) — Олигохетофора простая (рис. 188, 3). — Клетки 15—
20 мкм в диам. Щетинки у основания 0.6 мкм шир., 50—120(210) мкм дл.
В пресных водах, на цветковых растениях. В СССР не обнаружена.
(Ирландия).
Род POLYCHAETOPHORA W. et G. S. West -
ПОЛИХЕТОФОРА
Слоевище в виде 2—6 рыхло соединенных в нити клеток или
одноклеточное. Клетки почти шаровидные, эллипсоидные или яйцевид-
ные, с очень толстой, явственно слоистой оболочкой. С боков и от
спинной стороны каждой клетки отходит по 8—12 щетинок. Щетинки
простые, длинные, тонкие, слегка искривленные, без влагалищных
расширений клеточной оболочки вокруг их основания. Хлоропласт 1,
дисковидный, без пиреноида. Вегетативное размножение делением
клеток в двух направлениях. Зооспоры неизвестны.
1. Polychaetophora lamellosa W. et G. S. West — Полихетофора
пластинчатая (рис. 188, 4). —Клетки 19—35 мкм в диам. Оболочка
2.8 — 10.8 мкм толщ. Щетинки 86—183 мкм дл.
В пресных водах, на нитях Tolypothrix pygmaea. В СССР не обна-
ружена. (Зап. Европа, Сев. Америка).
Подпорядок PRASIOLINEAE —
ПРАЗИОЛОВЫЕ
Слоевище однорядно и многорядно нитевидное или однослойное,
лентовидное и пластинчатое, с ризоидами при основании или, реже,
на всем протяжении. Клетки округленно-угловатые, с 1 эксцентрично
расположенным ядром, в пластинчатых слоевищах сближенные друг
с другом группами, вследствие чего все слоевище кажется разбитым
на отдельные участки, состоящие из 4 клеток или из числа, кратного 4.
Хлоропласт 1, центральный, звездчатый, с 1 пиреноидом. Запасные
питательные вещества точно не установленной природы (возможно,
крахмал). Вегетативное размножение фрагментацией слоевища, при этом
нити распадаются или на сегменты из 1—4 клеток, сходные с нитями
из рода Stichococcus, или иногда на массу шаровидных клеток, легко
отделяющихся друг от друга, пластинчатые слоевища чаще распадаются
на узкие отпочковывающиеся отростки. Бесполое размножение акине-
тами. Акинеты образуются непосредственно из клеток слоевища,
333
или их образованию предшествует специальное деление клеток, в резуль-
тате чего возникают двурядные участки, клетки которых могут непосред-
ственно превращаться в акинеты или каждая клетка вновь делится
на 4 дочерние, преобразующиеся затем в акинеты; прорастают акинеты
или непосредственно в новое растение, или, иногда, превращаются
в апланоспорангии, содержащие по нескольку апланоспор. Половой
процесс гетерогамный, но он известен лишь для нескольких видов.
Гаметы двужгутиковые.
На влажной земле, влажных стенах, скалах, камнях, чаще всего
в местах, богатых аммиачными соединениями, а также в пресных,
быстро текущих холодных водах, реже на морских берегах, на влаж-
ных камнях возле воды. Встречается, во всех частях света, в том числе
в арктических и антарктических районах.
Семейство PRASIOLACEAE (Rabenh.) Borzi —
ПРАЗИОЛОВЫЕ
Характеристику см. в описании подпорядка.
I. Слоевище во взрослом состоянии нитевидное.............
.............................род Schizogonium (с. 334).
II. Слоевище во взрослом состоянии лентовидное или пластинчатое
разнообразных очертаний; нити встречаются преимущественно на
ранних стадиях развития................род Prasiola (с. 335).
Род SCHIZOGONIUM Kiitz. - СХИЗОГОНИУМ
Слоевище нитевидное. Нити простые, неограниченной длины,
окруженные бесцветным плотным гиалиновым влагалищем. Клетки
цилиндрические или короткоцилиндрические, с довольно толстыми
оболочками. Иногда встречается ложное ветвление, возникающее
вследствие отмирания 1 или 2 клеток нити и прорастания прилегающих
частей сквозь влагалище. Изредка можно наблюдать продольное деление
клеток, в результате которого слоевище приобретает лентовидную
форму в 2 или несколько клеток шириной. Ризоидные выросты
отходят беспорядочно.
Рис. 189.
Schizogonium. murale, нити разного строения. (По Кютцингу).
334
Редкий малоизученный род, многими авторами рассматривается
как стадия роста представителей рода Prasiola.
1. Schizogonium innrale Kiitz. — Схизогониум стенной (рис. 189). —
Нити 9—18 мкм шир., одно-, иногда двурядные, слегка жестковатые,
прямые или разнообразно изогнутые.
На сырой земле, скалах, сырых стенах, пнях образует ярко- или
темно-зеленые войлокообразные дерновинки. В разных районах СССР,
редко.
Род PRASIOLA Ag. - ПРАЗИОЛА
Слоевище прикрепляющееся к субстрату, лентовидное или пластин-
чатое, однослойное. Нити встречаются преимущественно на ранних
стадиях развития, в дальнейшем клетки их начинают делиться не
только в поперечном, но и в продольном направлении, в результате
чего образуются 2 параллельных ряда клеток, затем 3 и т. д.; постепенно
расширяясь, слоевище приобретает вид узкой ленты, переходящей
в дальнейшем в широкую пластину. Клетки в лентовидных и пластин-
чатых слоевищах,округленно-угловатые, на поперечном срезе палочко-
видно вытянутые; они располагаются довольно правильными квадрат-
ными участками, отграниченными друг от друга утолщенными частями
оболочек. Оболочка клеток двуслойная, внутренний слой целлюлозный,
наружный кутикулярный. Ризоиды короткие, одно- или многоклеточные,
большей частью простые, отходящие от любой клетки, чаще всего
в основании слоевища.
На влажной почве, старых досках, стенах, скалах, увлажняемых
брызгами, особенно в местах, богатых органическими загрязнениями,
многие виды обитают в воде, преимущественно в пресных водах,
но встречаются также на морских берегах возле воды.
I. Слоевище прикрепляется к субстрату без обособленного стебелька
или базального диска.
1. Слоевище прикрепляется непосредственно своей базальной ните-
видной частью, состоящей из одного ряда клеток.................
.............................................Р. calophylla 1.
2. Слоевище прикрепляется ризоидами.
А. Ризоиды развиваются вдоль боковых краев слоевища или
отсутствуют.
а. Слоевище широкое, пластинчатое, цельное или разорван-
ное на лопасти, курчавое; аэрофит . . . Р. crispa 2.
б. Слоевище узкое, лентовидное, волнистое или спирально
скрученное; гидрофит......................Р. velutina 3.
Б. Ризоиды развиваются только в основании слоевища.
а. Высота клеток на поперечном срезе равна ширине или до
2 раз больше.
а. Клетки с поверхности слоевища более или менее квадрат-
ные; слоевище до 20 см дл.................Р. japonica 4.
р. Клетки с поверхности слоевища округлые или непра-
вильно угловатые; слоевище до 2—3 см дл.................
.....................................Р. nevadensis 5.
б. Высота клеток на поперечном срезе в 3—4 раза больше
их ширины.
а. Клетки многоугольные, расположенные беспорядочно
.............................................Р. sinica 6.
335
р. Клетки округлые, треугольные, почковидные, располо-
женные обычно группами по 2—4 . Р. formosana 7.
в. Высота клеток на поперечном срезе в 5—6 раз больше их
ширины.
а. Клетки с поверхности слоевища почковидные, округлые;
разделение слоевища на группы клеток едва заметно .
......................................Р. subareolata 8.
р. Клетки с поверхности слоевища округло-многоугольные;
разделение слоевища на группы клеток выражено отчет-
ливо ......................................Р. junnanica 9.
II. Слоевище прикрепляется к субстрату обособленным стебельком.
1. Стебелек короткий, быстро исчезающий, после него развивается
широкая стопа..................................Р. furfuracea 10.
2. Стебелек длинный, не исчезающий, оканчивается подошвой . . .
.................................. . . . Р. stipitata И.
III. Слоевище прикрепляется к субстрату особыми образованиями.
1. Слоевище с широкой стопой с плотно расположенными ризоидами
.............................................Р. fluviatilis 12.
2.Слоевище с подошвой, образованной вытянутыми булавовидными
клетками.......................................Р. mexicana 13.
1. Prasiola calophylla (Carm.) Menegh. — Празиола красиволистная
(рис. 190, 7). —Слоевище лентовидное, 1 мм шир., свыше 2 см дл.,
нередко короче и уже, с ровными или волнистыми краями, к вершине
незначительно расширяющееся, закругленное или оборванное, в основа-
нии однорядное, непосредственно прикрепляющееся к субстрату,
выше многорядное. Клетки обычно располагаются отчетливыми продоль-
ными рядами.
На камнях и скалах во влажных затененных местах.
F. calophylla (рис. 190, 1). — Края слоевища ровные. Клетки
с поверхности слоевища в его основании удлиненные, 3—5 мкм шир.,
9—11 мкм дл., в верхней части слоевища изодиаметрические, 5—6 мкм
шир., на поперечном срезе 8—10 мкм выс. Продольные ряды клеток
выражены отчетливо.
Во влажных местах, на камнях, часто по берегам морей. В СССР
не обнаружена. (Зап. Европа, Сев. Америка).
F. antarctica Wille. — Края слоевища беспорядочно изогнутые
или волнистые. Клетки мелкие. Продольные ряды клеток выражены
нечетко.
Во влажных местах. В СССР не обнаружена. (Зап. Европа,
Антарктика).
2. Prasiola crispa (Lightf.) Menegh. (—Hormidium, parietinum
Kiitz.; Schizogonium crispum Gay) — Празиола курчавая (рис. 190,
2—6). — Слоевище на начальных стадиях развития нитевидное или
лентовидное, затем пленчатое, пузыревидно-чепцеобразное, позднее
разорванное на лопасти, складчато-курчавое, по краям иногда с корот-
кими ризоидами или выростами, прикрепляющееся без обособленного
стебелька. Клетки с поверхности слоевища квадратные или вытяну-
то-прямоугольные, на углах закругленные, 3 — 13 мкм шир., у нитевидных
и лентовидных форм 6—20 мкм шир., располагающиеся чаще всего
правильными, реже неправильными участками.
В тенистых, влажных, часто загрязненных местах, на почве,
коре деревьев, скалах, стенах и т. п. Во многих районах СССР.
Вид очень изменчивый, образующий много экологических, ростовых
336
Рис. 190.
1 — Prasiola calophylla-, 2—6 — P. crispa: 2, 3 — общий вид слоевищ, 4,5 —
фрагменты слоевища, 6 — акинеты. (7 — По Йенсену, 2—4 — по Мошковой,
5, 6 — по Весту).
и возрастных форм, незаметно переходящих друг в друга и поэтому
имеющих небольшую таксономическую ценность.
3. Prasiola velutina (Lyngb.) Wille — Празиола бархатистая
(рис. 191, 1—3). — Слоевище лентовидное, иногда пленчатое, 30—
70(110) мкм шир., до 2 см дл., изогнутое, волнистое, часто слегка
спирально скрученное, на ранней стадии развития однорядное, нитчатое,
прикрепляющееся ризоидами. Клетки 4—12 мкм шир., в широких
частях слоевища располагающиеся отчетливыми группами по 4, 8 или
в числе, кратном 4.
В быстро текущих холодных горных источниках. Земля Фран-
ца-Иосифа.
4. Prasiola japonica Yatabe — Празиола японская (рис. 191,
4—6). — Слоевище пластинчатое, овальное, удлиненно-линейное, лан-
цетовидно вытянутое или беспорядочно дольчатое, с волнистыми или
курчавыми краями, 0.5—4 см шир., 1—20 см дл., без стебелька, прикреп-
ляющееся удлиненными бесцветными ризоидами, образующимися при
основании слоевища. Клетки с поверхности слоевища более или менее
квадратные, расположенные группами по 4, на поперечном срезе
вертикально вытянутые, 7.5—10 мкм шир., 17.5—22.5 мкм дл., сближен-
ные по 2. Наблюдалась копуляция гетерогамет с 2 длинными жгутиками.
Зиготы шаровидные.
337
Рис. 191.
1—3 — Prasiola velutina.'. 1 — общий вид слоевищ, 2 — вид клеток с поверхности
слоевища, 3 — однорядная нить; 4—6 — Р. japonica: 4 — общий вид слоевища,
5 — вид клеток с поверхности слоевища, 6 — поперечный срез. (7—3 — по
Вилле, 4—6 — по Ятабе).
В быстро текущих холодных горных ручьях на скалах. В СССР
не обнаружена. (Япония).
5. Prasiola nevadensis Setch. et Gardn. — Празиола невадская
(рис. 192, 1—3). — Слоевище пластинчатое, удлиненное до ланцето-
видного, с волнистыми или курчавыми краями, 0.5—1 см шир., 2—
3 см дл., 14—25 мкм толщ., без стебелька, с широким основанием,
прикрепляющееся к субстрату вначале маленьким диском, позднее
ризоидами. Клетки с поверхности слоевища округлые или неправильно
угловатые, 2.7—4.1 мкм шир., 4.1—8 мкм дл., расположенные группами
по 2, 3, 4, на поперечном срезе от округлых до слегка вертикально
вытянутых, сближенные по 2.
В быстро текущих водах. В СССР не обнаружена. (Сев. Америка).
338
6. Prasiola sinica Jao — Празиола китайская (рис. 192, 4, 5). —
Слоевище пластинчатое, яйцевидное, ланцетовидное, почковидное, по
краям часто глубоко изрезанное, 3 см шир., до 9 см дл., 30—33 мкм толщ.,
с очень коротким стебельком, образованным бесцветными спутанными
ризоидами, с помощью которых прикрепляется к субстрату. Клетки
с поверхности слоевища многоугольные, полукруглые или неправильных
очертаний, 5—8(12) мкм шир., 9—13(15.5) мкм дл., расположенные
беспорядочно, иногда сближенные по 2 (4), на поперечном срезе цилинд-
рические, 24—26 мкм выс.
В горных ручьях и канавах. В СССР не обнаружена. (Китай).
7. Prasiola formosana Okada — Празиола формозская (рис. 192,
6—8). — Слоевище пластинчатое, удлиненно-ланцетовидное, клино-
видное или лентовидное, по краям волнистое или дольчатое, 0.2—2.6 см
шир., 2—10.7 см дл., 25—30 мкм толщ., на вершине закругленное или
оборванное, в основании клиновидно суженное, без стебелька, прикре-
пляющееся к субстрату ризоидами, образующимися при основании
слоевища. Клетки с поверхности слоевища округлые, треугольные или
почковидные, 2—5 мкм шир., 3—7 мкм дл., расположенные обычно
Рис. 192.
1—3 — Prasiola nevadensis-. 1 — общий вид слоевищ, 2 — вид клеток с поверх-
ности слоевища, 3 — поперечный срез; 4, 5 — Р. sinica: 4 — вид клеток с поверх-
ности слоевища, 5 — поперечный срез; 6—8 — Р. formosana: 6 — общий вид
слоевищ, 7 — вид клеток с поверхности слоевища, 8 — поперечный срез. (7—3 —
по Сетчеллу и Гарднеру, 4, 5 — по Жао, 6—8 — по Окада).
339
Рис. 193.
1,2 — Prasiola subareolata: 1 — общий вид слоевища, 2 — поперечный срез;
3, 4 — Р. yunnanica: 3 — вид клеток с поверхности слоевища, 4 — поперечный
срез. (7, 2 — по Скуе, 3, 4 — по Жао).
группами по 2—4, на поперечном срезе вертикально вытянутые,
2—6 мкм шир., 16—21 мкм выс.
В быстро текущих горных ручьях, на камнях. В СССР не обнару-
жена. (Япония).
8. Prasiola subareolata Skuja — Празиола субареолированная
(рис. 193, 1,2). — Слоевище пластинчатое, на ранних стадиях развития
продолговато-яйцевидное, позднее лентовидное, узколанцетное или
листовидное, по краям волнисто-курчавое, 0.5 см шир., до 6 см дл.,
45—54 мкм толщ., на вершине заостренное, прикрепляющееся к субстрату
ризоидами, отходящими от нижних клеток слоевища. Клетки с поверх-
ности слоевища округлые, почковидные, 6—8 мкм в диам., обычно
располагающиеся группами по 2 или 4, слабо обособленными друг
от друга, на поперечном срезе вертикально вытянутые, 28—38 мкм выс.
В ручьях, на камнях. В СССР не обнаружена. (Юго-восток Китая).
9. Prasiola yunnanica Jao — Празиола юиьнаньская (рис. 193,
4) . — Слоевище узколанцетовидное, кверху и книзу клиновидно
заостренное, по краям волнистое или гофрированное, 4 см шир., до 20 см
дл., 51—87 мкм толщ., прикрепляющееся к субстрату бесцветными
340
ризоидами, образующимися при основании слоевища. Клетки с поверхно-
сти слоевища округло-многоугольные, 5—7 мкм шир., 7—12 мкм дл.,
расположенные по 2—4 правильными плотными четырехугольными
участками, на поперечном срезе цилиндрические, вертикально удлинен-
ные, по высоте около 6 раз больше ширины. Гаметы по 64, 128 в клетке.
В горных ручьях, на скалах. В СССР не обнаружена. (Китай).
10. Prasiola furfuracea (Mert.) Menegh. (=P. leprosa Kiitz.) —
Празиола шелушащаяся (рис. 194, 1—3).— Слоевище пластинчатое,
лопатообразное, ложкообразное, ковшеобразное или, иногда, пузыревид-
ное, с волнистыми или изогнутыми краями, около 2 мм дл., на ранних
стадиях развития прикрепляющееся к субстрату коротким стебельком,
позже переходящим в широкую стопу, сложенную из удлиненных
клеток нижней части слоевища. Клетки неправильно многогранные,
4—6 мкм шир., 14—16 мкм дл., расположенные в нижней и нередко
в верхней частях слоевища беспорядочно или неправильными участками.
На морских берегах и во влажных затененных местах, где образует
на камнях и древесных субстратах плотные дерновинки. В СССР
не обнаружена. (Зап. Европа).
И. Prasiola stipitata Suhr — Празиола длинноногая (рис. 194,
4—6). — Слоевище узкое, ланцетовидное или ширококлиновидное,
часто овальное или почковидное, до 12 мм дл., прикрепляющееся
к субстрату длинной стебельковой частью с подошвой. Клетки с поверх-
ности слоевища довольно правильно четырехугольные, почти всегда
расположенные отчетливо выраженными участками.
На морских берегах, на камнях возле воды. В СССР не обнаружена.
(Зап. Европа).
Рис. 194.
1—3 — Prasiola furfuracea: 1,2 — общий вид слоевищ, 3 — вид клеток с поверх-
ности слоевища; 4—6 — Р. stipitata: 4, 5 — общий вид слоевищ, 6 — вид клеток
с поверхности слоевища. (7 — 3 — по Мошковой, 4—6 — по Блидингу).
341
Рис. 195.
1—3 — Prasiola jluviatilis: 1 — общий вид слоевища, 2 — вид клеток с поверх-
ности слоевища в средней части, 3 — то же в нижней части; 4—8 — Р. mezicana:
4—6 — общий вид слоевищ, 7 — вид клеток с поверхности слоевища, 8 —
поперечный срез. (1—3 — по Стармаху, 4—8 — по Сетчеллу и Гарднеру).
12. Prasiola fluviatilis (Sommerf.) Aresch. (~P. sauteri Menegh.) —
Празиола речная (рис. 195, 7—3). — Слоевище лентовидно-пластинчатое,
в нижней части суженное и переходящее в стопу, кверху постепенно
расширяющееся, с ровными или слегка волнистыми краями, 0.35—6.75 см
шир., до И см дл., 13—19 мкм толщ., обычно по 3—10 пластинок
вместе, расходящихся от места прикрепления в форме розетки, прикреп-
ляющееся к субстрату плотно расположенными ризоидами; верхушка
узкого слоевища чаще всего округлая, у более широкого слоевища
тупая или волнисто зазубренная. Клетки с поверхности слоевища
квадратные, 4—6 мкм шир., на поперечном срезе 8—13 мкм выс.,
расположенные в верхней части слоевища неправильными участками,
в нижней — продольными рядами.
В холодных горных источниках, ручьях, в арктических районах.
Кавказ, Полярн. Урал, Ср. Азия.
342
13. Prasiola mexicana J. Ag. — Празиола мексиканская (рис. 195,
4—8). — Слоевище пластинчатое, листоватое, по краям сильно изрезан-
ное, до 10 см дл., 25—30 мкм толщ., прикрепляющееся к субстрату
подошвой, образованной булавовидными клетками, вытянутыми в виде
ризоидов. Клетки с поверхности слоевища более или менее прямоуголь-
ные, угловатые, 6—16 мкм шир., расположенные довольно тесно
правильными участками.
В быстро текущих ручьях,. на камнях. В СССР не обнаружена.
(Центр, и Сев. Америка).
Редкий и несколько сомнительный вид, описанный недостаточно.
Таксономические преобразования —
Nomen novum
Chlorhormidium korsehikovii Hollerb. nom. nov. — Hormidium muco-
sum Korsch. Arch. Protistenk. 1941, 95, 1 : 40, fig. 13a, 14, non H. muco-
sum Boye-Pet. Det Kgl. Danske Videnskab. Selsk. Skrift., 7. Raekke,
Math.-Nat. Afd. 1915, 12, 17 : 340, tab. Ш, fig. 38-40.
343
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ 1
А
abietina (Flotow) Hansg. (Trente-
pohlia) 282
— f. abietina 282
— f. minor Wild. 282
aculeata W. et G. S. West (Tam-
niochaete) 316
acuta (Ag.) Kiitz. (Draparnaldia)
201
acuta Pasch. (Elakatothrix) 104
aegagropilae Acton (Gomontia) 271
aequalis Kiitz. (Ulothrix) 38
— f. aequalis 39
— i. crassior Borge 41
aestivale Hazen (Myxonema) 152
aestivale (Hazen) Collins (Stigeo-
clonium) 152
africanum Borge (Uronema) 46
ahlneriana Blid. (Enteromorpha)
130
alpicola Vischer (Jaagiella) 240
alpina (Koi) Hind. (Koliella) 98
alpina Koi (Raphidonema) 98
alpinis Smith et Klyver (Draparnal-
diopsis) 207
americana (Snow) Wille (Pseudul-
vella) 231
— var. americana 231
— var. indica Philipose 231
americana Snow (Ulvella) 231
amoenum (Kiitz.) Hazen (Myxo-
nema) 171
amoenum Kiitz. (Stigeoclonium)
171
— var. amoenum 174
— f. biforme Collins 171
— var. insigne (Nag.) Islam 175
amorphum Collins (Monostroma)
117
Ankistrodesmus Corda 90
annulata Brand (Trentepohlia) 279
Apatococcus Brand emend. Geitl.
236
Aphanochaetaceae (Printz) Bourr.
303
Aphanoehaete A. Br. 304
appressa Moschk. (Draparnaldia)
201
arborum (Ag.) Hariot (Trentepoh-
lia) 286
arenaria Meyer (Draparnaldia) 193
arenaria (Meyer) Meyer et Ska-
bitsch. (Draparnaldiella) 193
arnoldii Meyer (Draparnaldia) 194
arnoldii (Meyer) Meyer et Ska-
bitsch. (Draparnaldiella) 194
askenasii Schmidle (Stigeoclonium)
189
atomus Skuja (Stichococcus) 82
attenuata Hazen (Chaetophora) 184
attenuatum Hazen (Myxonema) 155
attenuatum (Hazen) Collins (Sti-
geoclonium) 155
aurea L. (Bissus) 282
aurea (L.) Martius (Trentepohlia)
282
— var. aurea 283
— var. chinensis (Harv.) Printz 284
— var. genuina Hansg. 283
— var. lanosa Kiitz. 284
— var. polycarpa (Nees et Mont.)
Hariot 284
— var. subsimplex (Caspary) De-
Toni 284
— var. tomentosa Kiitz. 284
aureum Kiitz. (Chroolepus) 282
autumnale Collins (Stigeoclonium)
152
1 Курсивом обозначены синонимы.
344
в
bacillaris Nag. s. str. (Stichococcus)
84
baicalensis Meyer (Draparnaldia)
194
baicalensis (Meyer) Meyer et Ska-
bitsch. (Draparnaldiella) 194
baileyi Mob. (Coleochaete) 322
balticum (Aresch.) Wittr. (Mono-
stroma) 125
basicladiae Jao (Endoderma) 223
basicladiae (Jao) Starmach (Ento-
cladia) 223
basiliense Vischer (Pseudoclonium)
255
basiramosa Fritsch (Enteromorpha)
137
beesleyi (Fritsch) Printz (Proto-
derma) 228
beesleyi Fritsch (Ulvella) 228
bernina (Koi) Hind. (Koliella) 93
berninum Koi (Raphidonema) 93
bertholdii Huber (Herposteiron) 305
biasolettianum Kiitz. (Draparnal-
dia) 154
biasolettianum (Kiitz.) Kiitz. (Sti-
geoclonium) 154
Binuclearia Wittr. 77
biplex (Nyg.) Hind. (Elakatothrix)
104
Blidingia Kylin 113
bogoriensis Wild. (Trentepohlia)
295
botryoides Snow (Pseudopleurococ-
cus) 250
braunii Nag. (Herposteiron) 305
brevirostre Scherff. (Raphidonema)
87
— var. brevirostre 87
— var. canadense Koi 87
brownii Fritsch (Protoderma) 230
bullosum (Roth) Wittr. (Mono-
stroma) 117
burmanica Skuja (Gongrosira) 266
C
Caespitella Vischer 212
calamicola (Zell.) De-Toni et Levi
(Trentepohlia) 279
caldaria Kiitz. (Gloeotila) 63
caldaria Collins (Ulothrix) 63
calophylla (Carm.) Menegh. (Pra-
siola) 336
— f. antarctica Wille 336
— f. calophylla 336
capitata Tiff. (Cloniophora) 189
carolinianum Islam (Stigeoclo-
nium) 165
cataractarum Kiitz. (Chlorotylium)
258
Catena Chod. 89
cateniformis Kiitz. (Ulothrix) 38
Cedercreutziella Vischer 241
Cephaleuros Kunze 299
chadefaudii (J. Feldm.) Blid. (Bli-
dingia) 115
chadefaudii J. Feldm. (Entero-
morpha) 115
Chaetomnion Skuja 309
Chaetonema Nowak. 310
Chaetophora Schrank 179
Chaetophoraceae (Harv.) De-Toni et
Levi 143
Chaetophorineae 141
Chaetophoroideae Bourr. 144
Chaetosphaeridiaceae Bourr. 325
Chaetosphaeridium Kleb. 326
Chaetotheke Diiring. 331
champlainensis Cook (Draparnal-
dia) 200
chlorelloides Grintz. et Peterfi
(Stichococcus) 84
Chlorhormidium Fott 49
Chloroclonium Borzi 251
Chlorophyta 24
Chlorospira Korsch. 65
Chlorotylium Kiitz. 258
Chodatia Koi 86
chodatii Bial. (Diplosphaera) 236
chodatii Bial. (Diplosphaera) 82
chodatii (Koi) Hind. (Koliella) 93
chodatii (Reineri) Printz (Loch-
mtopsis) 275
Chodatii Koi (Raphidonema) 93
chodatii (Bial.) Heering (Stichococ-
cus) 82
Chrooderma Fritsch 299
chroolepiforme Szym. (Endoclo-
nium) 214
chroolepiforme (Szym.) Heering
(Stigeoclonium) 214
Chroolepus Ag. 276
circinata (Borzi) Schmidle (Gon-
grosira) 275
circinnatus Borzi (Ctenocladus)
275
cladophorae Hornb. (Endoderma)
222
23 Зеленые водоросли. Улотриксовые
345
cladophorae (Hornb.) Starmach
(Entocladia) 222
clathrata Roth (Conferva) 135
clathrata (Roth) Grev. (Entero-
morpha) 135
clavata Horn. (Chaetophora) 184
Cloniophora Tiff. 187
closterioides (Kuff.) Hind. (Ko-
liella) 98
closterioides Kuff. (Raphidonema)
98
Coccobotrys Chod. 238
codiolifera (Chod.) Wille (Gomon-
tia) 270
Coelodiscus Jao 233
eohaerens (Wittr.) Printz (Proto-
derma) 230
Coleochaetaceae (Nag.) Pringsh.
316
Coleochaete Rreb. 316
comosa Kleb. (Conochaete) 329
compressa (L.) Grev. (Entero-
morpha)
— var. maeotica Pr.-Lavr. 129
— p. prolifera (Ag.) Grev. 134
— var. racemosa Ahln. 130
compressa L. (Ulva)
— p. prolifera Ag. 134
conferta W. West (Cylindrocapsa)
108
confervicolum Lagerh. (Uronema)
44
— var. africanum (Borge) Printz 46
— var. confervicolum 44
— var. uncinatum Popova 44
confervicolum Nag. (Herposteiron)
305
confervoides Bory (Draparnaldia)
201
confervoideum Lyngb. (Diplonema)
122
conjunctivum Schmidle (Radiofi-
lum) 72
Conochaete Kleb. 329
contorta Chod. (Gloeotila) 59
corcontica Hind. (Koliella) 98
cornu-dame (Roth) Ag. (Chaeto-
phora) 183
comurdame Roth (Rivularia) 183
crassa Hind. (Koliella) 96
crassa Korsch. (Chlorospira) 65
crassa Kjellm. (Ulva) 139
crassimembranacea Meyer (Myxo-
nemopsis) 187
crassiseta (W. et G. S. West) Printz
(Pseudochaete) 213
crassisetum W. et G. S. West (Her-
posteiron) 213
crassum Chod. (Hormidium) 53
crenulata Kiitz. (Ulothrix) 63
crenulatocollis Prescott (Geminella)
71
crenulatum (Kiitz.) Heering (Hor-
midiopsis) 63
crenulatum Kiitz. sec. Brand (Hor-
midium) 63
crinita (Roth) J. Ag. (Entero-
morpha) 137
crispa (Lightf.) Menegh. (Prasiola)
336
crispum Gay (Schizogonium) 336
cryophilum R. Chod. et F. Chod.
(Raphidonema) 91
Ctenocladus Borzi 273
curta Skuja (Gloeotila) 61
curvatum W. et G. S. West (Micro-
thamnion) 302
cyclostoma Waern (Entocladia) 221
cylindrica Hind. (Fottea) 101
cylindrica Wichmann (Ulothrichop-
sis) 79
Cylindrocapsa Reinsch 107
Cylindrocapsaceae Wille 107
D
daphnieola Fott (Protoderma) 228
debaryana Rabenh. (Gongrosira)
262
— f. debaryana 262
— f. minor Melnik 262
— var. robusta Hansg. 262
depauperata Woronich. (Gongro-
sira) 267
dermaticola Printz (Epilobium) 256
Desmococcus Brand emend. Vischer
238
Diaphragma Geitl. 240
Dicoleon Kleb. 331
Diplosphaera Bial. 236
diplosphaera (Bial.) Chod. (Sticho-
coccus) 82
disciformis Fritsch (Gongrosira) 265
disjuncta Tiff. (Coleochaete) 323
dissectum (Gay) Fareoqui (Chlor-
hormidium) 56
dissectum (Gay) Chod. (Hormi-
dium) 56
dissectus Gay (Stichococcus) 56
346
distans Kiitz. (Draparnaldia) 205
divergens Pringsh. (Coleochaete)
324
— var. divergens 325
— var. minor Hansg. 325
Draparnaldia Bory 196
Draparnaldiella Meyer et Skabitsch.
190
Draparnaldiopsis Smith et Klyver
207
dubius Chod. (Stichococcus) 84
E
Ectochaete (Huber) Wille 313
effusa (Kremp.) Hariot (Trentepoh-
lia) 279
Elakatothrix Wille 102
elegans Roth (Rivularia) 180
elegans (Roth) Ag. (Chaetophora)
180
elegans Woronich. (Thamniolum)
216
ellipsoideum Prescott (Hormidiop-
sis) 64
elliptica Volk. (Enteromorpha) 129
elongata Hass. (Draparnaldia) 155
elongatum Borzi (Chloroclonium)
251
elongatum (Hass.) Kiitz. (Stigeoclo-
nium) 155
elongatum Hodg. (Uronema) 46
endiviifolia Roth (Rivularia) 183
endiviifolia (Roth) Ag. (Chaeto-
phora) 183
Endoclonium Szym. 214
endophytica Fritsch (Chrooderma)
300
endophyticus (Fritsch) Printz (Ce-
phaleuros) 300
endophytum Mob. (Bulbocoleon)
314
endophytum (Mob.) Wille (Ecto-
chaete) 314
endophytum Huber (Endoderma)
314
Enteromorpha Link. 123
Entocladia Reinke 218
Epibolium Printz 256
Epicladia Reinke 226
epiphyton Mill. (Phycopeltis) 297
epiplancticus Skuja (Stichococcus)
82
eriensis Tiff. (Binuclearia) 77
exiguus Gern. (Stichococcus) 86
F
falclandicum Kiitz. (Stigeoclonium)
155
— var. longearticulatum Hazen 149
farctum Berth. (Stigeoclonium)
178
— var. farctum 179
— var. rivulare Butcher 179
— var. simplex Fritsch 179
fasciculare Kiitz. (Stigeoclonium)
167
fastigiata (Borzi) Schmidle (Gon-
grosira) 275
fastigiatus Borzi (Ctenocladus) 275
fenestrata P. et R. (Ulva) 139
fischeri Borge (Monostroma) 117
— var. fischeri 117
— var. minor Borge 117
flabelligera (De-Toni) Hansg. (Phy-
copeltis) 299
flacca (Dillw.) Thur. (Ulothrix) 43
flaccida (Kiitz.) Lagerh. (Hormis-
cia) 50
— var. montana Hansg. 52
— var. nitens (Menegh.) Hansg. 52
flaccida Kiitz. (Ulothrix) 50
flaccidum (Kiitz.) Fott (Chlorhor-
midium) 50
— var. crassum Chod. 53
— var. flaccidum 50
— — f. aquaticum Heering 52
— — f. flaccidum 52
— — f. montanum Heering 52
— — f. tumidum Heering 52
— — f. typica Heering 52 •
— var. lubricum Chod. 52
— var. nitens Menegh. emend.
Klebs 52
— — f. nitens 53
— — f. tenuis Boye-Pet. 53
flaccidum, A. Br. s. str. (Hormi-
dium) 50
flaccidum (Kiitz.) A. Br. s. str.
(Hormidium) 50
flaccidum (Kiitz.) Silva, Mattox et
Blackwell (Klebsormidium) 50
flaccidus (Kiitz.) Chod. (Hormococ-
cus) 50
flaccidus Hazen (Stichococcus) 52
flaccidus (Kiitz.) Gay (Stichococ-
cus) 50
flagellifera Kiitz. (Chaetophora) 184
flagelliferum Kiitz. (Stigeoclonium)
175
23*
347
flavescens G. S. West (Radiofilum)
75
flexuosa Wulf. (Conferva) 132
flexuosa (Wulf.) J. Ag. (Entero-
morpha) 132
— subsp. flexuosa Blid. 132
— subsp. pilifera (Kiitz.) Blid. 131
flexuosa Wulf. (Ulva) 132
fluitans (Gay) Starmach (Chlorhor-
midium) 53
fluitans (Gay) Heering (Hormi-
dium) 53
fluitans Gay (Stichococcus) 53
fluminensis Fritsch (Gongrosira)
262
flustrae Reinke (Epicladia) 226
fluviatilis (Sommerf.) Aresch. (Pra-
siola) 342
fonticolum Starmach (Epibolium)
256
Foreliella Chod. 269
formosa Meyer (Ireksokonia) 187
formosana Okada (Prasiola) 339
Fottea Hind. 101
fragilis Korsch. (Gimenellopsis) 77
fragilis Gay (Stichococcus) 86
frequens Butsch. (Ulvella) 229
frequens (Butsch.) Printz (Proto-
derma) 229
Fridea Schmidle 310
furfuracea (Mert.) Menegh. (Pra-
siola) 341
fuscum (Post, et Rupr.) Wittr.
(Monostroma) 121
G
gelatinosa Wille (Elakatothrix)
105
— f. biplex Nyg. 104.
Geminella Turp. 65
geminella Wolle (Cylindrocapsa)
109
— var. geminella 110
— var. minor Hansg. 110
Geminellopsis Korsch. 76
genevensis Reverd. (Ankistrodes-
mus) 104
genevensis (Reverd.) Hind. (Elaka-
tothrix) 104
germanica Gliick (Trentepohlia)
282
gigas Vischer (Uronema) 46
globiferum (Hansg.) Printz (Apha-
nochaete) 307
globiferum Hansg. (Herposteiron)
307
globosa Nordst. (Aphanochaete) 326
giobosum (Nordst.) Kleb. (Chaeto-
sphaeridium) 326
— f. depressum G. S. West 326
— f. giobosum 326
— f. incrassatum Kleb. 326
— f. microscopicum Playf. 327
gloeocystiformis Korsch. (Elakato-
thrix) 103
gloeophilum Borzi (Chloroclonium)
251
Gloeoplax Schmidle 239
Gloeotila Kiitz. 58
Gloeotilopsis Iyengar et Philipose
80
glomerata (Vauch.) Ag. (Draparnal-
dia) 204
— var. acuta Ag. 201
— var. distans (Kiitz.) Hansg. 205
— var. genutna Kirchn. 205
— var. glomerata 205
— var. groenlandica (Bachm.)
Printz 206
— var. remota Rabenh. 205
glomeratum Hazen (Myxonema) 167
glomeratum (Hazen) Collins (Sti-
geoclouium) 167
gobii Meyer (Trentepohlia) 292
Gomontia Born, et Flah. 269
Gonatoblaste Huber 308
Gongrosira Kiitz. 260
Gongrosira Chod. 269
goroschankinii Meyer (Draparnal-
dia) 193
goroschankinii (Meyer) Meyer et
Skabitsch. (Draparnaldiella) 193
gracile Rabenh. (Chroolepus) 287
gracile Skwortz. (Stigeoclonium)
160
gracilis Hansg. (Entocladia) 219
gracilis (Hansg.) De-Toni (Endo-
derma) 219
gracilis W. et G. S. West (Pseudo-
chaete) 213
gracilis (Rabenh.) Iyengar (Trente-
pohlia) 287
gracillima G. S. West (Entero-
morpha) 136
— f. breviramosa G. S. West 137
— f. gracillima 136
grevillei Thurp. (Enteromorpha)
120
grevillei (Thurp.) Wittr. (Mono-
stroma) 120
groenlandica Bachm. (Draparnal-
dia) 206
348
н
Hazenia Bold 246
Helicodictyon Whitford 248
Helvetica (Koi) Hind. (Koliella) 91
helveticum Koi (Raphidonema) 91
helveticum Vischer (Stigeoclonium)
171
Heterothrichopsis Iyengar et Kan-
thamma 81
holdenii Collins (Gomontia) 271
Hormidiella Iyengar et Kanthamma
48
Hormidiopsis Heering 63
Hormidium Kiitz. emend. Klebs 49
Hormospora Nag. 65
huberi Mob. (Chaetosphaeridium)
328
huberi Heering (Stigeoclonium) 178
huberi Gay (Thamniochaete) 316
hunanensis Jao (Trentepohlia)
286
hyalothecae (Hansg.) Schmidle
(Aphanochaete) 305
— var. mucicola Schmidle 305
hyalothecae Hansg. sec. Schmidle
(Herposteiron) 305
hygrophila Moschk. (Trentepohlia)
294
I
idiospora G. S. West (Ulothrix) 34
immane Schmidle (Endoderma) 223
immane (Schmidle) Starmach (En-
' tocladia) 223
implexa Rabenh. (Hormiscia) 43
implexa (Kiitz.) Kiitz. (Ulothrix) 43
implexum Kiitz. (Hormidium) 43
incrassata (Hudson) Hazen
(Chaetophora) 183
— f. clavata (Horn.) Rabenh. 184
— f. crassa (Ag.) Kiitz. 184
— f. crystallophora Kiitz. 184
— f. elongata Kiitz. 184
— f. genuina 184
— f. incrassata 184
— f. incrustans Rabenh. 184
— f. linearis Kiitz. 184
— f. polyclados Kiitz. 184
incrassata Hudson (Ulva) 183
incrustans Reinsch (Chlorotylium)
267
incrustans (Reinsch) Schmidle
(Gongrosira) 267
indica Bharadwaja (Draparnaldiop-
sis) 207
insigne Chod. (Pleurastrum) 253
insigne' Nag. (Stigeoclonium) 175
intermedia Obuch. (Draparnaldiop-
sis) 210
intermedia Blid. (Enteromorpha)
132
intermedium Bourr. (Uronema) 46
interrupta (Turp.) Lagerh. (Gemi-
nella) 67
intestinalis (L.) Link. (Entero-
morpha) 125
— f. attenuata Ahln. 125
— var. capillaris Kiitz. 131
— f. elliptica Volk. 129
— f. intestinalis 125
— f. ramosa Vinogr. 128
— f. saprobia Vinogr. 127
intestinalis L. (Ulva) 125
involuta Reinsch (Cylindrocapsa)
109
Ireksokonia Meyer 186
irregulare Kiitz. (Stigeoclonium)
160
irregulare (Wille) Brunnth. (Radio-
filum) 75
irregulare Nowak. (Chaetonema)
312
— var. irregulare 312
— var. subvalidum Hansg. 312
irregularis Korsch. (Chlorospira)
65
irregularis Pringsh. (Coleochaete)
321
— var. irregularis 321
— var. minor Skvortz. 321
irregularis Schmidle (Hansgirgia)
297
irregularis (Schmidle) Wille (Phy-
copeltis) 297
irregularis Wille (Hormospora) 75
Iwanoffia Pasch. 210
J
Jaagiella Vischer 240
jadinianum Huber (Endoderma) 314
Jaoa Fan 233
japonica Yatabe (Prasiola) 337
jolithus (L.) Wallr. (Trentepohlia)
289
— var. bovina (Flotow) Rabenh.
289
— var. jolithus 289
349
jucunda (Cesati) Hariot (Trente-
pohlia) 279
judayi Prescott (Draparnaldia) 206
jugoslavica Blid. (Enteromorpha)
133
К
klebahnii Schmidle (Conochaete)
329
klebsii (tenue) Heering (Stigeoclo-
nium) 160
Klebsormidium Silva, Mattox et
Blackwell 49
Koliella Hind. 90
koppei Budde (Gongrosira) 268
korschikovii Hollerb. (Chlorhormi-
dium) 58, 343
kuetzingianum Nag. (Microtham-
nion) 301
L
lactuca L. (Ulva) 139
— d. rigida Le Jobs 139
lacustris Korsch. (Elakatothrix) 104
lacustris Brand (Gongrosira) 264
lacustris Chod. (Stichococcus) 84
lagenifera (Hild.) Wille (Trente-
pohlia) 287
— var. africana Printh 289
— var. hansgirgii Meyer 289
— var. hildenbrandtii Meyer 289
— var. lagenifera 288
lageniferum Hild. (Chroolepus) 287
lagerheimii Wild. (Trentepohlia)
295
lamellosa W. et G. S. West (Poly-
chaetophora) 333
lauterbornii (Schmidle) Pr.-Lavr.
(Binuclearia) 77
lauterbornii Schmidle (Plancto-
nema)~ 77
lebelii Islam (Stigeoclonium) 163
leibleinii Kiitz. (Schizomeris) 110
lens Crouan (Ulvella) 231
leprosa Kiitz. (Prasiola) 341
leptochaete Huber (Endoderma) 314
leptochaete (Huber) Wille (Ecto-
chaete) 314
leptochaete L. Moev. (Micropoa)
316
Leptosira Borzi 243
Leptosiroideae Bourr. 234
Leptosiropsis Jao 253
leptotricha Beineri (Gongrosira)
267
lichenicola Suessen. (Conochaete)
329
lignicola Moore (Gomontia) 272
limnetica Lemm. (Ulothrix) 36
— var. limnetica 36
— var. minor Lemm. 36
linza (L.) J. Ag. (Enteromorpha)
— f. c'rispa J. Ag. 130
— f. lanceolata J. Ag. 130
linza L. (Ulva) 130
lobatum Printz (Pleurastrum) 237
lobatus (Chod.) Boye-Pet, (Apato-
coccus) 237
Lochmiopsis Woronich. et Popova
273
Lochmium Printz 246
longearticulatum (Hansg.) Heering
(Stigeoclonium) 149
longipilum Kiitz. (Stigeoclonium)
155
— var. lacustre Chod. 152
— var. longipilum 157
— var. minus Hansg. 157
longiseta (Vischer) Hind. (Koliella)
95
longiseta Vischer (Raphidonema) 95
lubrica Dillw. (Conferva) 171
lubrica Meyer (Draparnaldia) 196
lubrica (Meyer) Meyer et Skabitsch.
(Draparnaldiella) 196
lubricum (Dillw.) Fries (Myxo-
nema) 171
— var. varians Hazen. 171
lubricum Chod. (Hormidium) 52
lubricum (Dillw.) Kiitz. (Stigeoclo-
nium) 171
— var. lubricum 171
— var. varians (Hazen) Collins 171
M
macrocladia (Nordst.) Bourr. (Clo-
niophora) 190
macrocladium (Nordst.) Schmidle
(Stigeoclonium) 190
maeotica Pr.-Lavr. (Enteromorpha)
129
malleiformis Gutw. (Trentepohlia)
279
mammiforme (Balbis) Kiitz. (Chlo-
rotylium) 260
marginata (J. Ag.) Dang. (Blidin-
gia) 115
350
— f. marginata 116
— subsp. subsalsa (Kjellm.) Blid.
116
— f. ramulosa Vinogr. 116
marginata J. Ag. (Enteromorpha)
115
mediciana Borzi (Leptosira) 243
membranaceum W. et G. S. West
(Monostroma) 118
membranaefaciens Chod. (Sticho-
coccus) 84
mesomorphum Skuja (Radiofilum)
75
mexicana J. Ag. (Prasiola) 343
micrococca Kiitz. (Enteromorpha)
micrococca Ahln. (Enteromorpha)
— f. subsalsa Kjellm. 114
— var. subsalsa Kjellm. 116
micrococca (Kiitz.) Gobi (Ulva) 115
Micropoa L. Moev. 316
Microthamniaceae West 300
Microthamnion Nag. 300
minima (Nag.) Kylin (Blidingia)
113
— f. minima 114
— var. ramifera Blid. 114
— f. subsalsa (Kjellm.) Vinogr. 114
— var. subsalsa (Kjellm.) Scagel
114
minima Nag. (Enteromorpha) 113
— f. glacialis Kjellm. 114
minor (Nag.) Heering (Geminella)
68
minor Nag. (Hormospora) 68
minor Nag. s. str. (Stichococcus) 84
minutissimus Skuja (Stichococcus)
82
minutus Grintz. et Peterfi (Sticho-
coccus) 84
mirabilis Bold (Hazenia) 246
mirabilis Lagerh. (Stichococcus) 86
moebiusianum Collins (Endoclo-
nium) 175
moebiusianum (Collins) Printz (Sti-
geoclonium) 175
moniliformis Boye-Pet. et Hansen
(Streptochlora) 240
moniliformis Kiitz. (Ulothrix) 38
— var. braunii (Kiitz.) Rabenh. 38
— var. moniliformis 38
monospora J W. G. Lund (Gemi-
nella) 68
Monostroma Thur. 116
Monostromataceae Kunieda 112
mucosa Kiitz. (Gloeotila) 62
mucosa Thur. (Ulothrix) 36
mucosum Boye-Pet. (Hormidium)
56
mucosum (Boye-Pet.) Starmach
(Chlorhormidium) 56
mucosum Korsch. (Hormidium) 58
murale Kiitz. (Schizogonium) 335
mutabilis Roth (Conferva) 200
mutabilis (Roth) Cedercr. (Drapar-
naldia) 200
mutabilis (Nag.) Wille (Geminella)
68
mutabilis Nag. (Hormospora) 68
— var. minor Hansg. 68
Mycoidea Cunning. 299
Myxonemopsis Meyer 187
N
naegelii Chod. (Pleurococcus) 238
najadeanum Skvortz. (Stigeoclo-
nium) 178
nakaoi Hirano (Chaetomnion) 309
папа (Sommerf.) Sjostedt (Entero-
morpha) 114
— var. minima (Nag.) Sjostedt 114
— var. subsalsa (Kjellm.) Sjostedt
114
папа Dillw. (Conferva) 158
nanum (Dillw.) Hazen (Myxonema)
158
nanum (Dillw.) Kiitz. (Stigeoclo-
nium) 158
— f. subsimplex Collins 158
nelsonii Prescott (Stigeoclonium)
169
nevadensis Setch. et Gardn. (Pra-
siola) 338
nitellarum Jost (Coleochaete) 320
nitens (Kiitz.) Rabenh. (Hormi-
dium) 50
nitens Manegh. (Ulothrix) 52
nitens (Menegh.) emend. Klebs
(Hormidium) 52
nivale Lagerh. (Raphidonema) 89
— var. nivale 89
— var. taylori Koi 89
nivale (Lagerh.) Chod. sensu Printz
(Raphidonema) 89
nivale Chod. (Raphidium) 91
nivalis (Chod.) Brunnth. (Ankistro-
desmus) 91
nivalis (Chod.) Hind. (Koliella) 91
nordstedtii Kleb. (Dicoleon) 332
351
norvegica Hind. (Koliella) 96
nudiusculum Kiitz. (Stigeoclonium)
152
О
obovata Vischer (Leptosira) 245
obscura (Kiitz.) Gayral (Ulvaria)
121
odorata (Wigg.) Wittr. (Trentepoh-
lia) 291
— var. elongata (Bleisch) Fischer
292
— var. odorata 292
— var. unilateralis Gribb 293
Oligochaetophora G. S. West 333
opposite (Lyngb.) Ag. (Draparnal-
dia) 203
orbicularis Pringsh. (Coleochaete)
321
ordinata (W. et G. S. West) Hee-
ring (Geminella) 66
ordinata W. et G. S. West (Hormo-
spora) 66
ornatum Trans. (Chaetonema) 313
oscillarina Kiitz. (Ulothrix) 36
ovale Smith (Chaetosphaeridium)
327
oxyspermum (Kiitz.) Doty (Mono-
stroma) 118
P
pallescens Chod. (Stichococcus) 86
pallida Kiitz. (Gloeotila) 63
pamirica (Wille) Printz (Ulothrix)
36
paradoxa (Chod. et Topali) Printz
(Geminella) 75
paradoxum Chod. et Topali (Inter-
filum) 75
paradoxum (Chod. et Topali) Printz
(Radiofilum) 75
parietinum Kiitz. (Hormidium) 336
parvula (Arch.) Hind. (Elaka-
tothrix) 105
parvula Iyengar et Kanthamma
(Hormidiella) 49
parvula Arch. (Spirotaenia) 105
parvulum Borzi (Chloroclonium)
251
pascheri Heering (Aphanochaete)
. 308.
pascheri Vischer (Caespitella) 212
pascheri (tenue) Heering (Stigeo-
clonium) 160
paucicellulare Vischer (Pleurast-
rum) 248
paucicellulare (Vischer) Geitl.
(Rhexinema) 248
pelagica (Nyg.) Skuja (Gloeotila)
61
pelagica Nyg. (Ulothrix) 61
percursa (Ag.) Kjellm. (Dipion etna)
122
percursa Ag. (Enteromorpha) 122
percursa (Ag.) Bory (Percursaria)
. 122
Percursaria Bory 122
perforans Huber (Endoderma) 222
perforans (Huber) Levr. (Entocla-
dia) 222
perforans Chod. (Foreliella) 273
perforans (Chod.) Acton (Gomon-
tia) 273
perforans (Chod.) Wille (Tellamia)
273
phlllophila W. et G. S. West (Tren-
tepohlia) 287
Phycopeltis Mill. 297
Phylloplax Schmidle 299
piceana Meyer (Trentepohlia) 294
pilifera Kiitz. (Enteromorpha) 131
pilosa Meyer (Draparnaldia) 196
pilosa (Meyer) Meyer et Skabitsch.
(Draparnaldiella) 196
pilosissima Schmidle (Aphanochae-
te) 305
piluliferum Printz (Lochmium)
246
pisiformis (Roth) Ag. (Chaeto-
phora) 182
pisiformis Roth (Rivularia) 182
pithophorae G. S. West (Endo-
derma) 221
pithophorae (G. S. West) Starmach
(Entocladia) 221
planctonica Bolochonz. (Gloeotila)
68
planctonica (Bolochonz.) Tiwary et
Pandey (Geminella) 68
— var. indica Tiwary et Pandey
69
— var. planctonica 68
planctonica Iyengar et Philipose
(Gloeotilopsis) 80
planctonica Hind. (Koliella) 93
planctonicum Whitford (Helicodic-
tyon) 248
platyzonata Hazen (Draparnaldia)
198
352
Pleurastrum Chod. 253
Pleurococcus auct. 238
plumosa (Kutz.) Bourr. (Clonio-
phora) 189
plumosa (Vauch.) Ag. (Draparnal-
dia) 200
— var. opposite, Lyngb. 203
— var. plumosa 200
— var. pulchella Kiitz. 200
plumosum Kiitz. (Stigeoclonium)
189 ,
plumosum Vauch. (Batrachosper-
mum) 200
polychaete (Hansg.) Fritsch (Apha-
nochaete) 305
polychaete Hansg. (Herposteiron)
305
Polychaetophora W. et G. S. West
333
polymorpha G. S. West (Endo-
derma) 222
polymorpha (G. S. West) Starmach
(Entocladia) 222
polymorpha Deck. (Trentepohlia)
285
polymorphum Franke (Endoclo-
nium) 214
polymorphum (Franke) Heering
(Stigeoclonium) 214
polyrrhiza (Lagerh.) Born, et Flah.
(Gomontia) 270
polyrrhizum Lagerh. (Codiolum)
270
polysporum During. (Epibolium)
258
polythricha Nordst. (Aphanochaete)
330
polytricha (Nordst.) Kleb. (Cono-
chaete) 330
pontica Roehl. (Epicladia) 227
prasina (Jao) Fan (Jaoa) 233
— var. bullata Jao 234
— - var. prasina 234
prasinus Jao (Coelodiscus) 233
Prasiola Ag. 335
Prasiolaceae (Rabenh.) Borzi 334
Prasiolineae 333
Pringsheimia Reinke 231
Pringsheimiella Hohnel 231
pringsheimii Kleb. (Chaetosphaeri-
dium) 327
— f. confertum Kleb. 327
— f. pringsheimii 327
printzii Woronich. et Popova (Loch-
miopsis) 275
printzii Vischer (Pseudopleurococ-
cus) 249
procera Ahln. (Enteromorpha) 130
— var. denudata Ahln. 130
prolifera (O. F. Miill.) J. Ag. (En-
teromorpha) 134
— f. prolifera 134
— subsp. prolifera Blid. 134
— f. simplex Vinogr. 135
prolifera O. F. Miill. (Ulva) 134
proliferum Vischer (Spongoplasti-
dium) 251
prostrata Jao (Gongrosira) 264
prostratum Fritsch (Stigeoclonium)
177 - ..
protensum (Dillw.) Kutz. (Stigeo-
clonium) 163
— var. subuligerum (Kiitz.) Ra-
benh. 160
Protococcus auct. 238
Protoderma Kiitz. 227
protogenita Kutz. (Gloeotila) 61
Pseudendoclonium Wille 254
Pseudochaete W. et G. S. West 212
pseudoflacca Wille (Ulothrix) 43
pseudogelatinosa Korsch. (Elakato-
thrix) 106
Pseudopleurococcus Snow emend.
Vischer 248
pseudosoluta Gauth.-Liev. (Coleo-
chaete) 319
pseudostichococcus (Heering) Fott
(Chlorhormidium) 56
pseudostichococcus Heering (Hor-
midium) 56
Pseudulvella Wille 231
pulchella Kiitz. (Draparnaldia) 200
pulchra Skuja (Gloeotila) 61
pulvinata A. Br. (Coleochaete) 322
— var. baileyi (Mob.) Printz 322
— var. minor Pringsh. 322
— var. pulvinata 322
pumila .Meyer (Draparnaldia) 191
pumila (Meyer) Meyer et Skabitsch.
(Draparnaldiella) 191
punctiformis Kutz. (Chaetophora)
182
pusilia Lyngb. (Conferva) 169
pusilium (Lyngb.) Kiitz. (Stigeo-
clonium) 169
pygmaea Kiitz. (Gongrosira) 265
pygmaeum Hansg. (Endoderma)
215
pygmaeum Hansg. (Stigeoclonium)
215
353
pyrenoidifera (Korsch.) Hind. (Ko-
liella) 100
pyrenoidifera Korsch. (Raphido-
nema) 100
pyriferum Skuja (Chaetomnion) 309
Q
quaternaria Playf. (Ulothrix) 35
— f. angustissima Playf. 36
— f. quaternaria 35
quaternarium Desm. (Monostroma)
118
quercinus Kiitz. (Chroolepus) 292
R
radiata J. Ag. (Enteromorpha) 116
Radiophilum Schmidle 72
radiosum Geitl. (Diaphragma) 241
ramulosa Borge (Enteromorpha)
116
ramulosa (Engl. Bot.) Hooker
(Enteromorpha) 137
Raphidium Kiitz. 90
Raphidonema Lagerh. emend. Hind.
87
Raphidonema Lagerh. 90
ravenelii Wolle (Draparnaldia) 203
recurvata (Wittr. et Nordst.) Printz
(Gongrosira) 265
recurvata Wittr. et Nordst. (Tren-
tepohlia) 265
reineckei Schmidle (Endoderma)
225
reineckei (Schmidle) Starmach
(Entocladia) 225
repens A. Br. (Aphanochaete) 305
repens (A. Br.) Wittr. (Herpo-
steiron) 305
reptans During. (Chaetotheke) 331
Rhexinema Geitl. 248
rigida Ag. (Ulva) 139
rigidula (Miill.) Hariot (Trente-
pohlia) 290
rivulare (Kiitz.) Starmach (Chlor-
hormidium) 55
rivulare Kiitz. (Hormidium) 55
rivularis Kiitz. (Ulothrix) 55
rivularis De-Toni (Hormiscia) 55
rivularis (Kutz.) Hazen (Stichococ-
cus) 55
rostrata Huber (Gonatoblaste) 308
rupicola Jao (Gomontia) 273
354
S
sabaudum Koi (Raphidonema) 87
— var. minus Koi 89
— var. sabaudum 89
saccharophila Coker et Shanor (Sap-
rochaete) 216
salina Kutz. (Enteromorpha) 134
salishensis Prescott (Draparnaldiop-
sis) 210
sampsonii Trans. (Coleochaete)'324
Saprochaete Coker et Shanor 216
sauteri Menegh. (Prasiola) 342
savoniensis Vischer (Cedercreut-
ziella) 242
Schizogonium Kiitz. 334
Schizomeridaceae Smith. 110
Schizomeris Kiitz. 110
schmidlei P. Richter (Gongrosira)
268
sclerococcus Kiitz. (Gongrosira) 264
scopulina (Hazen) Heering. (Gloeo-
tila) 60
scopulinus Hazen (Stichococcus) 60
scourfieldii G. S. West (Gongrosira)
269
scutata Breb. (Coleochaete) 321
— f. minor Mob. 322
— f. scutata 322
scutata Reinke (Pringsheimia) 232
scutata (Reinke) Schmidt (Prings-
heimiella) 232
segare Islam (Stigeoclonium) 163
sempervirens (Chod.) Hind. (Ko-
liella) 97
sempervirens Chod. (Raphidonema)
97
setifera (Farlow) Hariot (Trente-
pohlia) 279
setigerum Kiitz. (Stigeoclonium)
151
sibirica Woronich. et Popova (Loch-
miopsis) 275
sigmoidea Hind. (Koliella) 97
simplex Meyer (Draparnaldia) 192
simplex (Meyer) Meyer et Ska-
bitsch. (Draparnaldiella) 192
simplex Jao (Draparnaldiopsls) 210
simplex G. S. West (Oligochaeto-
phora) 333
simplex G. S. West (Polychaeto-
phora) 333
sinica Jao (Prasiola) 339
smithii Bourr. (Geminella) 68
soluta Pringsh. (Coleochaete) 318
— var. brevicellularis Schmidle 319
— var. minor Hansg. 319
~ var. soluta 318
sparsifasciculata Prescott (Drapar-
naldia) 202
spicata (Schmidle) Islam (Clohio-
phora) 189
spicatum Schmidle (Stigeoclonium)
189
spiculiforme Vischer (Raphido-
nema) 94
spiculiformis (Vischer) Hind. (Ko-
liella) 94
spinosa Kiitz. (Draparnaldia) 201
spiralis Chod. (Gloeotila) 59
spirochroma Reverd. (Ankistrodes-
mus) 105
spirochroma (Reverd.) Hind. (Ela-
katothrix) 105
spirochroma Reverd. (Raphidium)
105
spiroides (G. S. West) Printz
(Gloeotila) 59
spiroides G. S. West (Ulothrix) 59
spirotaenia G. S. West (Ankistro-
desmus) 96
spirotaenia (G. S. West) Hind. (Ko-
liella) 96
spirotaenia (G. S. West) Korsch.
(Raphidonema) 96
spirulinoides Hind. (Koliella) 98
spongophila Web.* van Bos. (Clado-
phora) 225
spongophila (Web. van Bos.) Printz
(Endoderma) 225
spongophila (Web. van Bos.) Star-
mach (Entocladia) 225
spongophila (Web. van Bos.) Wille
(Gongrosira) 225
Spongoplastidium Vischer 250
stagnalis (G. S. West) Schmidle
(Gongrosira) 262
stagnalis Hind. (Koliella) 93
stagnatile Hazen (Myxonema) 163
stagnatile (Hazen) Collins (Stigeo-
clonium) 163
stellare Kutz. (Stigeoclonium) 160
Stichococcus Nag. 81
Stigeoclonium Kutz. 145
stipitata Suhr (Prasiola) 341
Streptochlora Boye-Pet. et Hansen
240
strictissimum Rabenh. (Microtham-
nion) 301
— var. macrocystis Schmidle 302
— var. strictissimum 302
subacuta Korsch. (Elakatothrix)
103
subareolata Skuja (Prasiola) 340
subconstricta (G. S. West) Printz
(Gloeotila) 62
subconstricta G. S. West (Ulothrix)
62
subflacca Wille (Ulothrix) 43
‘submarina Kutz. (Ulothrix) 43
submarinum Wille (Pseudendoclo-
nium) 256
subsecunda Kiitz. (Conferva) 149
subsecundum (Kutz.) Hazen (My-
xonema) 149
subsecundum (Kiitz.) Kiitz. (Sti-
geoclonium) 149
— var. subsecundum 149
— var. tenuis Nordst. 149
subspinosum Kiitz. (Stigeoclonium)
159
subtile (Kiitz.) Starmach (Chlor-
hormidium) 55
subtile Heering (Hormidium) 55
subtilis (Kiitz.) De-Toni (Hormis-
cia) 55
— var. tenerrima De-Toni 32
— var. variabilis (Kiitz.) Krrchn.
34
subtilis Kleriker (Stichococcus) 55
subtilis Kiitz. (Ulothrix) 55
— f. subtilissima Rabenh. 32
— var. tenerrima Kirchn. 32
— var. variabilis Kirchn. 34
subtilissima (Lagerh.) Printz (Ge-
minella) 66
subtilissima Lagerh. (Hormospora)
66
subtilissima Rabenh. (Ulothrix) 32
subuligerum Kiitz. (Stigeoclonium)
160
subuligerum (Kiitz.) Hazen (Myxo-
nema) 160
T
tatrae Koi (Ankistrodesmus) 98
tatrae (Koi) Hind. (Koliella) 98
— var. bratislavensis Hind. 98
— var. fogarasensis (Koi) Hind. 98
— var. tatrae 98
tatrae Koi (Raphidonema) 98
— var. fogarasensis Koi 98
tatrany Wittr. (Binuclearia) 79
tectorum (Kiitz.) Beger (Binuclea-
ria) 79
355
tectorum Kiitz. (Gloeotila) 79
tenerrima Kiitz. (Conferva) 32
tenerrima Kiitz. (Ulothrix) 32
— f. dentatospora Hollerb. 34
— f. tenerrima 34
tenue Zell. (Chroolepus) 287
tenue (Ag.) Rabenh. (Myxonema)
160
tenue (Ag.) Kiitz. (Stigeoclonium)
160
— var. genuinum Kirchn. 160
— var. gracile Kiitz. 160
— var. irregulare (Kiitz.) Rabenh.
160
— var. lubricum Rabenh. 171
— var. tenue 160
— var. iniforme (Ag.) Kiitz. 160
tenuis Kogan (Binuclearia) 77
tenuis Ag. (Draparnaldia) 160
tenuis Kylin (Entocladia) 223
tenuis De-Toni (Hormiscia) 38
tenuis De-Toni (Trentepohlia) 287
tenuis Kiitz. (Ulothrix) 32, 38
tenuissima Kiitz. (Ulothrix) 38
terrestre Iwanoff (Stigeoclonium)
212
terrestre Fritsch et John (Pleurast-
rum) 253
terrestre Mitra (Uronema) 46
terrestris (Iwanoff) Pascher (Iwa-
noffia) 212
terrestris (Fritsch et John) Printz
(Leptosira) 245
terrestris Fritsch et John (Rhexi-
nema) 245
terricola Boye-Pet. (Geminella) 71
terricola Bristol (Gongrosira) 245
terricola (Bristol) Printz (Lepto-
sira) 245
testarum Kylin (Entocladia) 223
tetrallantoidea Koi (Chodatia) 87
Thamniochaete Gay 315
Thamniolum Woronich 216
thermale (A. Br.) Hazen (Myxo-
nema) 149
thermale A. Br. (Stigeoclonium) 149
thrombii Tschermak-Woess (Lepto-
sira) 244
tibetana (Skuja) Printz (Gongro-
sira) 269
torrenticola Schmidle (Fridea) 310
torta Mert. (Conferva) 125
torta (Mert.) Reinb. (Entero-
morpha) 125
torulosa Jao (Leptosiropsis) 253
356
transilvanica (Koi) Hind. (Ko-
liella) 96
transilvanica Koi (Raphidonema) 96
trentepohlia Martins 276
Trentepohliaceae De-Toni 275
trentepohlioides Schmidle (Gongro-
sira) 264
tribonematoideum (Skuja) Star-
mach (Chlorhormidium) 56
tuberculosa (Roth) Ag. (Chaeto-
phora) 180
tuberculosa Roth (Rivularia) 180
tubulosa Kiitz. (Enteromorpha) 132
turfosa Skuja (Gloeotila) 61
U
Ulothrichopsis Wichman 79
Ulothrichopsis Iyengar et Kant-
hamma 81
Ulothrix Kutz. 31
Ulotrichaceae Kiitz. 27
Ulotrichales 25
Ulotrichineae 26
Ulotrichophyceae 24
Ulva L. emend. Thur. 138
Ulvaceae Lamour. 122
Ulvella Cronan 230
Ulvelloideae Bourr. 218
Ulvineae 112
umbrina (Kiitz.) Born. (Trentepoh-
lia) 292
— f. elongata Reinsch 293
— var. quercina (Kiitz.) Rabenh.
292
— f. umbrina 293
— f. unilateralis (Cribb) Moschk.
293
umbrinum Kiitz. (Chroolepus) 292
incinata (Gobi) Hansg. (Trentepoh-
lia) 285
uncinatum Gobi (Chroolepus) 285
uniforme (Ag.) Rabenh. (Stigeoclo-
nium) 160
uniformis Ag. (Draparnaldia) 160
Uronema Lagerh. 43
V
4
variabile (Nag.) Elenk. emend. (Sti-
geoclonium) 175
variabilis W. et G. S. West (Sticho-
coccus) 86
variabilis Kutz. (Ulothrix) 34
velutina (Lyngb.) Wille (Prasiola)
337
velutina Kiitz. (Trentepohlia) 282
velutinum Kiitz. (Chroolepus) 282
vermiculoides Wolle (Aphanochae-
te) 305
vermiculoides (Wolle) Collins (Her-
posteiron) 305
verrucariae Chod. emend. Vischer
(Coccobotrys) 238
villosa Meyer (Draparnaldia) 195
villosa (Meyer) Meyer et Skabitsch.
(Draparnaldiella) 195
viretii Chod. (Raphidonema) 93
viretii (Chod.) Brunnth. (Ankistro-
desmus) 93
viretii (Chod.) Hind. (Koliella)
93
viretii R. Chod. et F. Chod. (Raphi-
donema) 93
viride Kiitz (Protoderma) 228
viridis Chod. (Catena) 89
viridis Kiitz. (Gongrosira) 264
viridis (Reinke) Lagerh. (Endo-
derma) 220
viridis Reinke (Entocladia) 220
viridis lyngar et Kanthamma (Hete-
rothrichopsis) 81
viridis Ag. (Protococdus) 238
viridis (Ag.) Brand (Desmococcus)
238
viridis Borzi (Zoddaea) 242
vulgaris Brand emend. Geitl. (Apa-
tococcus) 237
vulgaris (Nag.) Brand (Desmococ-
cus) 238
vulgaris Nag. (Pleurococcus) 238
— f. cohaerens Wittr. 230
vulgaris Snow (Pseudopleurococ-
cus) 250
W
weberi Schmidle (Gloeoplax) 239
— var. suecicum Borge 239
— var. weberi 239
Weneda Racib. 299
westii (tenue) Heering (Stigeoclo-
nium) 160
willeana Hansg. (Leptosira) 244
willei Tiff. (Cloniophora) 189
wittrockii Born. (Monostroma) 119
yunnanica Jao (Prasiola) 340
Z
zaisanica Skwortz. (Binuclearia) 79
Zoddaea Borzi 242
zonata Web. et Mohr (Conferva) 41
zonata (Web. et Mohr) Aresch.
(Hormiscia) 41
zonata (Web. et Mohr) Kiitz.
(Ulothrix) 41
— var. attenuata (Kiitz.) Rabeh. 41
— var. inaequalis (Kiitz.) Rabenh.
41
— var. pectinalis (Kiitz.) Rabenh.
41
— var. rigidula (Kiitz.) Hansg. 41
— var. zonata 41
357
СОДЕРЖАНИЕ
От редакции ................................................
I. Общая часть..............................................
1. Общая характеристика улотриксовых водорослей.............
Основные признаки улотриксовых водорослей (7). Проис-
хождение и родственные связи (8). Распространение (9).
Практическое значение (10). Методы сбора и изучения (10).
2. Морфология улотриксовых водорослей.................. . .
Строение клетки (12). Нитчатые формы (13). Пластинчатые
формы (14). Разнонитчатые, или гетеротрихальные,
формы (15). Размножение (17). Смена форм развития (19).
л *
3. Система улотриксовых водорослей..........................
4. Основные систематические сводки..........................
12
20
22
л.
II. Специальная часть............................................. 24
Отдел Chlorophyta ................................................. 24
Класс Ulotrichophyceae......................................... 24
Порядок Ulotrichales ...................................... 25
Подпорядок Ulotrichineae ........ ... 26
Семейство Ulotrichaceae.............................. 27
Роды: Ulothrix (31). — Uronema (43). — Hor-
midiella (48). — Chlorhormidium (49). —
Gloeotila (58) i — Hormidiopsis (63). — Chlo-
rospira (65). — Geminella (65). — Radiofi-
lum (72). —' Geminellopsis (76). — Binuclea-
ria (77). — Ulothrichopsis (79). — Gloeotilop-
sis (80). — Heterothrichopsis (81). — Sti-
chococcus (81). — Chodatia (86). — Raphido-
nema (87). — Catena (89). — Koliella (90). —
Fottea (101). — Elakatothrix (102).
Семейство Cylindrocapsaceae ......... 107
Род Cylindrocapsa (107).
Семейство Schizomeridaceae . ... . .... 110
Род Schizomeris (110).
Подпорядок Ulvineae.....................................112
Семейство Monostromataceae.................... . 112
Роды: Blidingia (113). — Monostroma (116).
358
Семейство Ulvaceae ............
Роды: Percursaria (122). — Enteromorpha
(123). - Ulva (138).
ПодпЪрядок Chaetophorineae.........................
Семейство Chaetophoraceae . ...................
Подсемейство Chaetophoroldeae . ... . . .
Роды: Stigeoclonium (145). — Chaetophora
(179). — Ireksokonia (186). — Myxonemopsis
(187). — Cloniophora (187). — Draparnal-
diella (190). — Draparnaldia (196). — Drapar-
naldiopsis (207). — Iwanoffia (210). — Caespi-
tella (212). — Pseudochaete (212). — Endoclo-
nium (214). — Thamniolum (216). — Sapro-
chaete (216).
Подсемейство Ulvelloideae ...................
Роды: Entocladia (218). — Epicladia (226). —
Protoderma (227). — Ulvella (230). — Pseudul-
vella (231). — Pringsheimiella (231). — Jaoa
(233).
Подсемейство Leptosiroideae ........
Роды: Diplosphaera (236). — Apatococcus
(236). — Coccobotrys (238). — Desmococcus
(238). — Gloeoplax (239). — Streptochlora
(240). — Jaagiella (240). — Diaphragma
(240). — Cedercreutziella (241). — Zoddeae
(242). — Leptosira (243). — Lochmium
(246). — Hazenia (246). — Helicodictyon
(248). — Rhexinema (248). — Pseudopleuro-
coccus (248). — Spongoplastidium (250). —
Chloroclonium (251). — Leptosiropsis (253). —
Pleurastrum (253). — Pseudendoclonium
(254). — Epibolium (256). — Chlorotylium
(258). — Gongrosira (260). — Gomontia
(269). — Ctenocladus (273).
Семейство Trentepohliaceae.....................
Роды: Trentepohlia (276). — Phycopeltis
(297). — Cephaleuros (299).
Семейство Microthamniaceae .........
Род Microthamnion (300).
Семейство Aphanochaetaceae . . . . . . . .
Роды: Aphanochaete (304). — Gonatoblaste
(308). — Chaetomnion (309). — Fridea
(310). — Chaetonema (310). — Ectochaete
(313). — Thamniochaete (315). — Micropoa
(316).
Семейство Coleochaetaceae......................
Род Coleochaete (316).
Семейство Chaetosphaeridiaceae ........
Роды: Chaetosphaeridium (326). — Conochaete
(329). — Chaetotheke (331). — Dicoleon
122
141
143
144
218
234
275
300
303
316
325
359
(331). — Oligochaetophora (333). — Poly
chaetophora (333).
Подпорядок Praslolineae...........................
Семейство Prasiolaceae........................
Роды: Schizogonium (334). — Prasiola (335)
Таксономические преобразования — Nomen novum...............
Алфавитный указатель латинских названий....................
Нонна Алексеевна Мошкова
Максимилиан Максимилианович Голлербах
Определитель
пресноводных водорослей СССР. Вып. 10 (1)
УЛОТРИКСОВЫЕ
Утверждено к печати
Ботаническим институтом им. В. Л. Комарова
Академии наук СССР
Редактор издательства Г. Т. Никифорова
Технический редактор Н. Ф. Соколова
Корректоры М. В. Ед ем скал, Н. Г. К а ц е н к о
и Э. Г. Рабинович
ч
ИБ № 21516
Сдано в набор 29.07.85. Подписано к печати 13.01.86.
М-18508. Формат 84Х1081/з2- Бумага для глубокой печати.
Гарнитура обыкновенная. Фотонабор. Печать офсетная.
Усл. печ. л. 18.90. Усл. кр.-отт. 18.90. Уч.-изд. л. 23.89.
Тираж 1000. Тип. зак. 270. Цена 1 р. 70 к.
Ордена Трудового Красного Знамени
издательство «Наука». Ленинградское отделение
199164, Ленинград, В-164, Менделеевская линия, 1
Ордена Трудового Красного Знамени
Первая типография издательства «Наука»
199034, Ленинград, В-34, 9 линия, 12
360
333
334
343
344