/
























































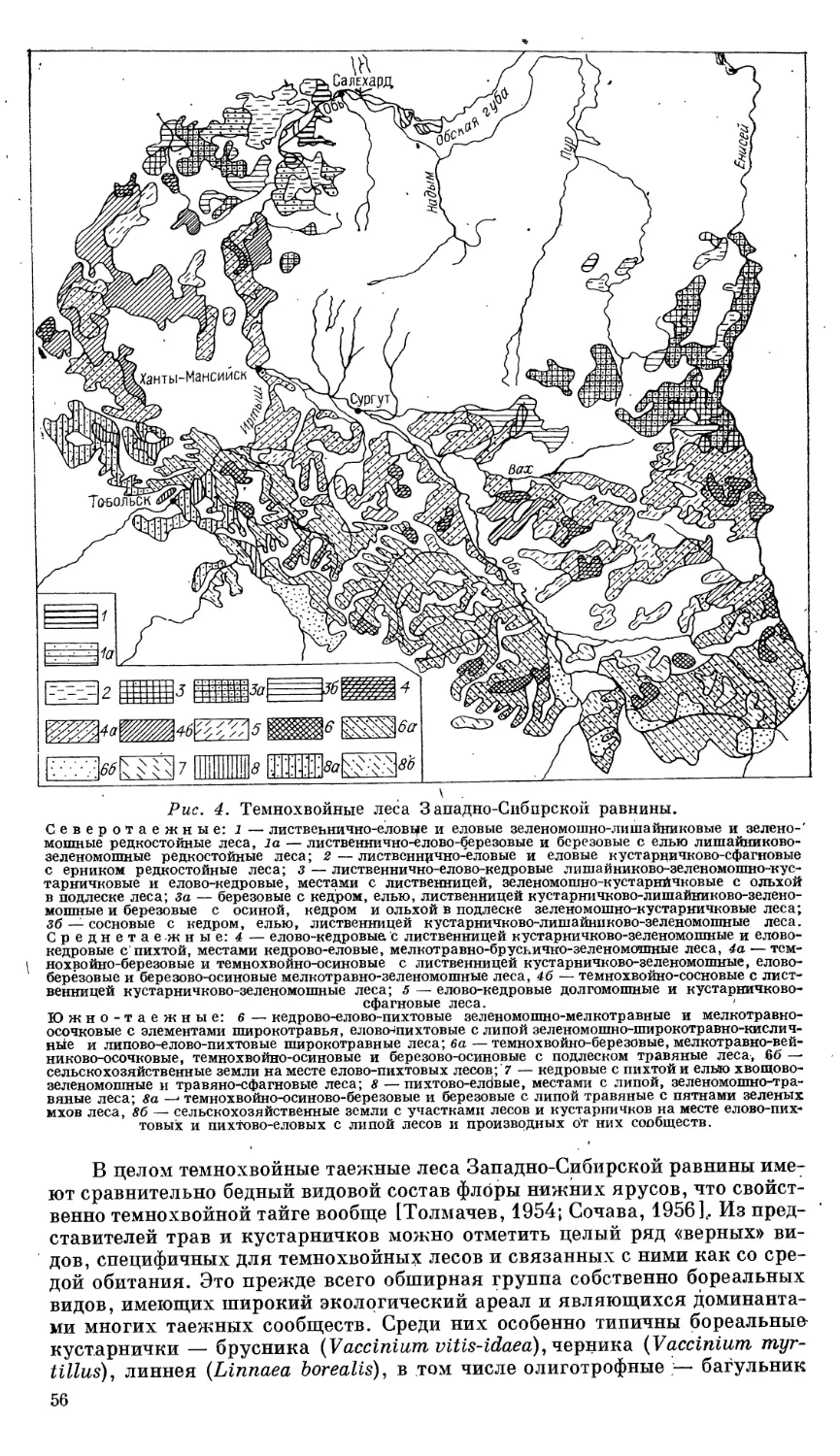

















































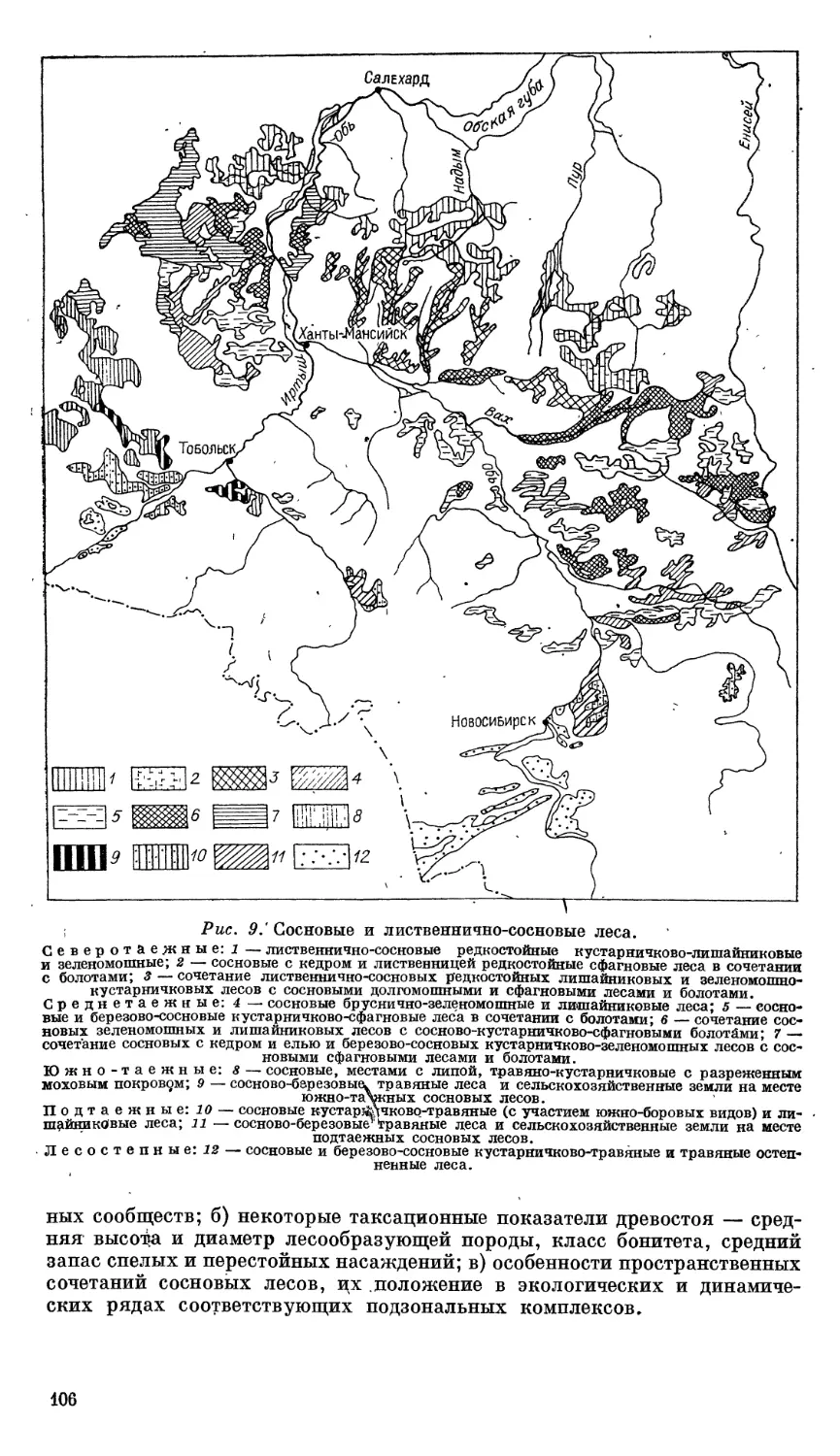

















































































































































Автор: Ильина И.С. Лапшина Е.И. Лавренко Н.Н..
Теги: картография (текстовые документы) геоботаника растительный покров
Год: 1985




Текст
РАСТИТЕЛЬНЫЙ
ПОКРОВ
Западно-Сибирской
равнины
АКАДЕМИЯ НАУКСССР
СИБИРСКОЕ ОТДЕЛЕНИЕ
ИНСТИТУТ ГЕОГРАФИИ
РАСТИТЕЛЬНЫЙ
ПОКРОВ
ЗАПАДНО-СИБИРСКОЙ
РАВНИНЫ
Ответственные редакторы
чл.-кор. АН СССР В. В. Воробьев,
канд. геогр. наук А. В. Бегов
НОВОСИБИРСК
ИЗДАТЕЛЬСТВО «НАУКА»
СИБИРСКОЕ ОТДЕЛЕНИЕ
1985
УДК 528.9 : 581.9
Растительный покров Западно-Сибирской равнины/Ильина И. С.у
Лапшина Е. И., Л а в р е н к о Н. Н. и др. — Новосибирск: Наука,
1985.
В монографии обобщены результаты многолетних исследований авторов и
новейшие литературные и картографические материалы по растительности
крупного региона — Западно-Сибирской равнины. Основная часть книги
представляет собой типологический обзор главнейших "подразделений растительного
покрова равнины ■*— тундр, темнохвойных, светлохвойных и мелколиственных
лесов, степей, а также растительности болот и долин рек. Показаны особенности
зональной и провинциальной дифференциации растительности. В то же время
монография служит пояснительным текстом к карте «Растительность Западно-
Сибирской равнины» (М., ГУГК, 1976). Рассмотрены теоретические и
методические вопросы построения карты, систематизация материалов проведена в
соответствии с ее легендой.
Книга представляет интерес для географов, картографов, геоботаников,
почвоведов, специалистов по охране и рациональному использованию
растительности.
И. С. Ильина, Е. Я. Лапшина, Н. И, Лавренко, Л. И. Мельцер,
Е А. Романова, Б'. А. Богоявленский, В. Д. Махно
Рецензенты С. А. Грибова, Г. В. Бачурин, Л. В. Попов
2091050000—790
(6) Издательство «Наука», 1985 г.
о
Посвящается светлой памяти
академика
Виктора Борисовича
СОЧАВЫ *
ПРЕДИСЛОВИЕ
Завершился большой этап в изучении растительного покрова
Западно-Сибирской равнины. Он связан с работами по созданию обзорной
геоботанической карты этого обширного региона, проводившимися более 10
лет под руководством академика В. Б. Сочавы. Были объединены усилия
большого коллектива геоботаников и картографов разных учреждений
страны. Основная часть исследований проведена сотрудниками Института
географии СО АН СССР (Института географии Сибири и Дальнего
Востока СО АН СССР), а также геоботаниками Центрального сибирского
ботанического сада СО АН СССР, Ленинградского гидрологического
института, Института географии АН СССР и Иркутского государственного
университета им. А. А. Жданова.
Многолетние экспедиционные исследования авторов, составление
серии крупно- и среднемасштабных геоботанических карт на отдельные
районы, анализ новейших литературных, картографических и фондовых
материалов, научно-теоретические разработки классификационных схем
и легенд, создание авторского макета и* издание обзорной карты
«Растительность Западно-Сибирской раввины» М 1 : 1 500 0Q0, наконец,
подготовка настоящей монографии — вот основные этапы многолетней работы.
Предлагаемая вниманию читателей монография' в основной своей
части представляет типологический обзор главнейших подразделений
растительного покрова Западно-Сибирской равнины, выполненный на основе
литературных источников и новых фактических материалов, собранных
авторами в результате экспедиционных исследований. В то же время
монография служит пояснительным текстом к карте «Растительность Западно-
Сибирской равнины» (М., ГУГК, 1976). Это обстоятельство определило
общую структуру работы и порядок расположения материала внутри
отдельных глав.
В первых главах рассматриваются общие черты природных условий
и географических закономерностей в пространственной дифференциации
растительности Западно-Сибирской равнины, ее основные зональные и
провинциальные рубежи. Здесь раскрываются принципы и методы
составления обзорной геоботанической карты; дается анализ главных источников,
использованных при составлении карты, подробно рассматриваются
методы полевых и камеральных работ, раскрываются научные принципы
построения классификации и легенды карты; приведен полный текст
легенды.
Основная часть книги посвящена Характеристике главнейших
подразделений растительного покрова равнины — тундрам, темнохЕОЙным
лесам и редколесьям, сосновым лесам, лиственничным лесам и
редколесьям, мелколиственным лесам, степям, растительности болот и долин рек.
Дан анализ растительных сообществ этих подразделений в
типологическом плане — по фитоценотическим, флористическим, экологическим,
динамическим и географическим признакам.
Материал в главах систематизирован согласно структуре легенды
карты. Основные подразделения в тексте книги соответствуют выделенным
на карте подзональвым категориям — арктическими субарктическим
тундрам, северо-, средне- и южно-таежным бореальвым типам, лесной, болот-
3
ной и пойменной растительности. Самые мелкие подзаголовки текста
раскрывают содержание непосредственно картируемых единиц, в скобках
указаны номера соответствующих выделов на карте.
Латинские названия растений, приведенные в тексте, даны в
соответствии с новой номенклатурой [Черепанов, 19811. В ряде случаев они
не совпадают с номенклатурой растений в легенде карты, поэтому в
заключительном списке латинских названий растений приведена необходимая
синонимика.
Подготовка рукописи к печати и ее научное редактирование были
начаты под руководством и при непосредственном участии академика
Виктора Борисовича Сочавы. Ему — бессменному научному руководителю,
автору основных идей, вдохновителю и организатору работы — мы, его
ученики и коллеги, с благодарностью посвящаем эту книгу.
Считаем своим долгом выразить глубокую признательность за помощь
в редактировании этой монографии старшему научному сотруднику
лаборатории географии и картографии растительности Ботанического
института АН СССР С. А. Грибовой, а также зав. лабораторией 3. В. Карамы-
шевой и научным сотрудникам Т. И. Исаченко, В. В. Липатовой, Т. К. Юр-
ковской, сделавшим ценные замечания по отдельным главам*
о
ВВЕДЕНИЕ
Западно-Сибирская равнина — крупный и уникальный по своим
природным условиям регион нашей страны, арена интенсивной хозяйственной
деятельности сотен тысяч людей. Природный потенциал его огромен:
запасы нефти и газа, богатые лесные ресурсы, крупные залежи торфа,
ценные кормовые угодья заливных лугов в поймах Оби и Иртыша. В
настоящее время Западная Сибирь — это район первоочередного хозяйственного
освоения, связанного прежде всего с разработкой нефтяных и газовых
месторождений, строительством крупных промышленных комплексов,
осуществлением мероприятий по переброске стрка сибирских рек в южные
районы страны. Освоению природных богатств края отводится
значительное место в решениях XXVI съезда КПСС, в постановлениях партии и
правительства о реализации Продовольственной программы нашей страны.
Своеобразие природы Западно-Сибирской равнины определяется
особенностями ее геолого-геоморфологического строения, палеогеографией,
а также современными гидролого-климатическими процессами.
В структурно-тектоническом отношении Западно-Сибирская равнина
представляет собой эпигерцинскую плиту со складчатым доюрским фун-
'* даментом. Современный рельеф ее характеризуется преобладанием
обширных аккумулятивных равнин, сложенных рыхлыми четвертичными
отложениями (песками, суглинками, глинами) с относительными высотами
около 100 м. Эти равнины занимают все междуречные пространства
центральных районов Западной Сибири, и только в южной ее части
располагаются невысокие (до 300 м) струйтурно-денудационные плато — Васюганское,
Чулымо-Енисейское, Приобское и др. [Николаев, 1970]. В районе
Среднего Приобья низменные зандровые равнины с запада на восток
пересечены цепью ледниковых возвышенностей — Люлцн-Вор, Сибирские
увалы. Эрозионные формы рельефа представлены долинными комплексами
крупных рек — Оби, Иртыша, Назыма, Пура, Таза и их многочисленных
притоков.
Климат Западно-Сибирской равнины характеризуется значительной
континентальностью. Он имеет существенные различия в
гидротермических показателях северных и южных районов, что связано с широтными'
изменениями прихода солнечной радиации, количества осадков,
испарения и т. д. В табл. 1 приведены некоторые данные, свидетельствующие
об этих различиях [по Мезенцеву, Карнацевичу, 1969].
Значительная протяженность Западно-Сибирской равнины с севера на
юг и широтные изменения в показателях тепла и влаги способствуют
четкому проявлению на ее территории природной зональности. Равнинность
современного рельефа усиливает это проявление. В. С. Мезенцев
и И. В. Карнацевич выделяют в пределах Западно-Сибирской равнины
восемь гидроклиматических зон, характеризующихся разными показателями
увлажнения и теплообеспеченности, что обусловливает развитие в
пределах этих зон определенных типов растительности и почв г [Мезенцев,
Карнацевич, 1969].
Наиболее выражены черты зональности в растительном покрове, на
что впервые обратил внимание Б. Н. Городков [1916а], предложив схему
дробного подзонального деления лесной и тундровой растительности
Западной Сибири. Материалы современных исследований геоботаников и
лесоводов в значительной степени уточнили и дифференцировали это деление
5
Таблица 1
Климатический показатель
Теплоресурсы климата,
ккал/(см2»год)
Осадки за вегетацией
ный период, мм
Общее увлажнение за
год, мм
Максимально
возможное испарение за
год, мм
Тундра
(I зона)
27
220
450
Тайга
(II зона)
34,5
260—325
630—770
520-630
Степь
(VIII зона)
47,4
140—170
290—320
770—820
[Смолоногов и др., 1970; Сочава д др., 1953; Лапшина, 1975; Горожанкина,;
Константинов, 1978; Ильина, 1981; и др.1.
Детально разработаны и обоснованы большим фактическим
материалом схемы зонального деления почв [Уфимцева, 19[74; Караваева, 19731,;
ландшафтов и растительности пойм [Роднянская, 1963,; 1969; Ильина,;
1976], а также болотных ландшафтов [Болота,.., 1976].
В целом, согласно современному физико-географическому делению^
территория Западно-Сибирской равнины подразделяется на пять
природных зон — тундры, лесотундры, лесов, лесостепи, степи и соответствующих .
им подзон — арктических и типичных тундр, северо-, средне- и
южно-таежных лесов, осиново-березовых лесов, северной и южной лесостепи, раз-
нотравно-типчаково-ковыльных и сухих типчаково-ковыльных стейей
[Западная Сибирь, 1963]. Таким образом, зональность — одна из главных
особенностей природы Западно-Сибирской равнины, следствие сильного
влияния гидротермического фактора в условиях равнинности территории.
Другая ее специфическая черта — исключительная- по с!воим
размерам заболоченность территории. Западно-Сибирская равнина — это
уникальный регион по широте охвата процессами болотообразования и торфо-
накопления. Средняя заболоченность ее около 50%, а в отдельных
районах (Сургутское Полесье, Васюганье) она достигает 70—75% [Болота...,,
19761.
Широкому развитию болотообразовательных процессов способствует
сочетание многих геоморфологических и гидротермических факторов *
главные из которых — равнинцость территории и ее тектонический режим
с устойчивыми тенденциями к опусканиям, особенно северных и
центральных районов [Николаев, 1970], слабая дрениробанность территории%
избыточное увлажнение, пониженная теплообеспечённость.
Болота Западной Сибири образовались еще в эпоху раннего
голоцена — более 10—12 тыс. лет назад [Нейштадт, 1977]. В их современном
развитии отмечается тенденция прогрессивного заболачивания, особенно
проявляющаяся на большей части таежных территорий [Нейштадт, 1977;
Караваева, 1973, 1982]. Как уже указывалось, болотные ландшафты несут
определенные ,черты зональности, наиболее отчетливо выраженные в их
крупных морфогенетических типах. Большинство авторов на территории
Западно-Сибирской равнины выделяют шесть болотных зон —
полигональных болот, плоско- и крупнобугристых, выпуклых олиготрофных
(сфагновых), плоских евтрофных и мезотрофных (осоково-гипновых и
лесных), вогнутых евтрофных (тростниковых) и засоленных (травяных) [Кац,;
1971; Болота..., 1976; Романова, 1977].
Трудами многих болотоведов, геоморфологов, почвоведов установлены
важные закономерности формирования болот в разных
геоморфологических условиях Западно-Сибирской равнины, выявлены механизмы
заболачивания на породах разного состава [Караваева, 1977], установлены
современные тенденции болотообразовательного процесса в разных зонах
и районах [Львов, 1976; Нейштадт, 1977; Караваева, 1982].
Третья, наиболее характерная черта природы равнины — широкое
6 '-':■'
развитие поемности и аллювиальности в долинах ее крупных рек. Б. Н.
Городков писал: «В наше время сильная заболоченность на севере Западно-
Сибирской низменности особенно обращает внимание исследователей
наряду с обширностью поемных явлений в речных долинах» [1946, с. 82].
Две крупнейшие реки Советского Союза — Обь и ее приток Иртыш —
пересекают Западно-Сибирскую равнину в меридиональном и широтном
направлениях. В их долинах особенно широко развиты поймы,
образованные сильно меандрирующим русловым потоком главного русла и его
многочисленных проток и временных водотоков. Большая ширина долин и пойм
Оби и Иртыша связана с геологической деятельностью их прарек (древних
долин) и особенностями современных русловых процессов [Петров, 19791.
На отдельных участках среднего течения Оби (у Березова,
Нижневартовска) ширина поймы достигает 40—50 км.
Гидрологический режим пойм Оби и Иртыша отличается большим
своеобразием. Это связано главным образом с прохождением длительных
весенне-летних половодий, затапливающих ежегодно большие площади
пойменных земель и обусловливающих формирование в пойме огромных
массивов естественных заливных лугов — ценнейших кормовых угодий„
требующих глубокого изучения и объективной оцецки возможностей их
рационального использования.
Отмеченные выше специфические черты природы Западно-Сибирской
равнины в полной мере проявляются в растительном покрове — в его
зонально-провинциальных особенностях, географо-генетических чертах,}
структурных и динамических свойствах.
^0
ГЛАВА I
ОСНОВНЫЕ ГЕОГРАФИЧЕСКИЕ ЗАКОНОМЕРНОСТИ
РАСТИТЕЛЬНОГО ПОКРОВА ЗАПАДНО-СИБИРСКОЙ РАВНИНЫ
Изучение растительности Западно-Сибирской равнины имеет большую
историю. Оно связано с именами многих выдающихся ботаников и
лесоводов: П. Н. Крылова, Б. Н. Городкова, В. В. Ревердатто, Л. В.
Шумиловой, Л. П. Сергиевской, Г. В. Крылова, Б. П. Колесникова, а также с
работами крупных научных коллективов Томского „университета,
Центрального сибирского ботанического сада СО АН СССР, Института географии
СО АН СССР, Института биологии СО АН СССР, Института экологии
растений и животных УФ АН СССР и других учреждений страны. Подробные
обзоры истории исследования и ботанико-географической изученности
Западно-Сибирской равнины содержатся во многих публикациях [Крылов,
1961; Западная Сибирь, 1963; Растительность..., 1963; и др."|.
Остановимся на важнейших географических закономерностях
растительного покрова равнины — его зональных и провинциальных чертах.
В. Б. Сочава писал, что «Обь-Иртышская равнина представляет собой
идеальное поле для проявления зональных явлений и их неизбежного
следствия — постепенных переходов в виде подзон» [1980, с. 451.
Зональность — это биоклиматическое явление. Как уже указывалось,
на Западно-Сибирской равнине вследствие общей равнинности территории
и ее большой протяженности с севера на юг зональность в растительном
покрове выражена очень отчетливо. Сильная заболоченность территории
и интенсивно/з развитие поемности нарушают строго широтное
расположение зональных границ и их совпадение для разных биоморфологических ?
типов сообществ — лесных, болотных, пойменных. Однако это не
затушевывает проявление в их составе и структуре тех фитоценотических
свойств, которые в большей степени обусловлены широтно-климатически-
ми факторами, что позволяет относить все эти сообщества к сходным
зональным типам.
Дифференциация растительности по зональному принципу в
Западной Сибири осуществляется уже давно. В трудах многих исследователей
[Крылов, 1919; Сочава и др., 1953; Смолоногов и др., 1970; и др.]
проводится четкое деление лесов Западно-Сибирской равнины на подзональные
категории -г- северо-, средне-, южно-таежные и подтаежные. Зональные
типы болотной.растительности выделены Е. А. Романовой [Болота...,
1976], пойменной — М. Н. Барышниковым [1933, 1961], Э. Е. Роднянскон
[1969], И. С. Ильиной [1976].
Существуют некоторые разногласия в трактовке объема и содержания
зональных единиц растительного покрова как типологических, так и
региональных. Некоторые исследователи предлагают выделять в Западной
Сибири зональные и подзональные категории только плакорных сообществ,
в наибольшей степени зависящих от широтно-климатических условий
[Шумилова, 1962; Смолоногов и др., 1970]. Другие трактуют зональность более
широко, объединяя в одно зональное подразделение экологически и
динамически связанные фитоценозы разных местообитаний. Подобный подход
для территории Западной Сибири был впервые осуществлен Б. Н. Город-
ковым. Выделенные им на огромной территории тундры и тайги зональные
и подзональные категории — арктические и субарктические тундры,
северо- и среднетаежные леса — рассматривались как ботанико-географи-
ческие единства, состоящие из разных по структуре и биоморфологическо-
8 ;
му составу сообществ лесной, болотной и пойменной растительности,
объединенных в единые взаимосвязанные ряды и серии [Городков, 1935,1946],?
Такой широкий эколого-динамический подход к выделению
зональных единиц растительного покрова впоследствии был успешно применен
А. П. Тыртиковым при изучении динамики растительности севера
Западной Сибири [1969, 1974]. Но наиболее полное воплощение этот принцип
нашел в трудах В. Б. Сочавы как структурно-динамический подход к
выделению различных типологических и региональных единиц растительного
покрова [1967, 1979].
Как будет видно из последующих глав предлагаемой монографии, при
картировании и районировании растительности Западно-Сибирской
равнины выделение зональных и провинциальных категорий было
осуществлено на основе структурно-динамического анализа-с использованием эко-
лого-фитоценотических, флористических и географических критериев•
^Рассмотрим главнейшие закономерности распределения
растительности внутри основных зональных и провинциальных подразделений,
выделенных в процессе геоботанического районирования [Ильина,1* Махно *
1976], проведенного на основе составленной геоботанической карты.
На территории Западно-Сибирской равнины проходят границы трех
геоботанических зон — тундровой, таежной (бореальной) и степной
(рис. 1).
Границы тундровой зоны совпадают с очертаниями трех
крупных полуостровов на севере — Ямал, Гыданский и Тазовский.
Южный рубеж тундровой зоны ограничивает полосу северной лесотундры
[Городков, 1935], где на плакордх преобладают кустарниковые сообщества
тундрового типа, а участки бореальных редколесий распространены
небольшими массивами по долинам рек. На территории западно-сибирских
тундр выделяются две подзоны — арктических и субарктических тундр,
что соответствует новейшей схеме районирования растительного покрова
Арктики [Александрова, 1977]. Граница между этими подзонами —
важный ботанико-географический и флористический рубе>& [Сочава,
Городков, 1956; Юрцев, 1966].
Подзоне арктических тундр свойствен определенный
подтип тундровой растительности [Александрова, 1977]. Для плакорных
местообитаний здесь характерен мозаичный покров, состоящий
преимущественно из осоково-лишайниково-моховых (с Rhacomitrium lanuginosum,
Aulacomnium turgidum, Dicranum elongatum) и кустарничково-моховых
с участием арктоальпийских кустарничков (Salix polaris, S. nummularia.
Dry as octo'petala) сообществ, приуроченных к участкам с кочковатым и
пятнистым нанорельефом. Псаммофитные варианты этих тундр связаны с
легкими почвами, 'в значительной степени подверженными процессам
ветровой эрозии и дефляции. На "дефляционны^ обнажениях здесь формируются
открытые группировки из Minuartia arctica, Armeria arctica и др.,
переходящие в местах более интенсивного делювиального сноса в сообщества
кустарничково-лишайниковых и мохово-лишайниковых (Cladonia gracilis,
CI. uncialis, Thamnolia vermicularis) полигональных тундр [Мельцер, 1980].
Заболачивание тундровых сообществ Арктики происходит разными
путями. На водоразделах заболачивание начинается в лишайниково-мо-
ховых тундрах и приводит к образованию относительно устойчивых
сообществ осоково-пушицево-моховых (Drepanocladus exannulatus, Eriophorum
polystachyon, Carex concolor) полигональных заболоченных тундр.
Собственно болотные сообщества формируются, как правило, при зарастании
различных водоемов. Наиболее характерный их арктический тип — тра-
вяно-гипновыё болота с участием Carex concolor, Eriophorum polystachyonr
Dupontia fischeri, Calliergon sarmentosum.
Помимо однородных травяно-гипновых болот в Арктике
распространены полигональные болота с ярко выраженной неоднородной
структурой. Для них характерны травяно-гипновые группировки по полигонам,
в составе которых встречаются Calliergon sarmentosum, Drepanocladus
uncinates, Tomenthypnum niiensx Carex concolor? Sphagnum fimbriatum.
9
Рис. 1. Зонально провинциальное деление растительного покрова Западно
Сибирской равнины. , .
Границы:! — геоботанических зон; 2 — геоботанических провинций; 3 — геоботанических под-
зов; 4 — Западно-Сибирской равнины. Геоботанические з о'н ы: А — тундровая, В —
Сюрреальная (таежная); В — степная. Геоботанические провинции: I — Ямальская*
II — Гыданская, III — Зауральская, IV — Обь-Иртышская, V — Приенисейская, VI —
Верхнеиртышская. Геоботанические подзоны: 5 — арктические тундры; б —
субарктические тундры; 7 — редколесья; 8 — северная тайга; 9—средняя тайга; ю — южная тайга;
11—подтайга; 12 — лесостепь; 13 — разнотравно-дерновинно-злаковая степь.
Речные долины в Арктике развиты слабо, и процессы сукцессионных
смен на речном аллювии очень замедлены. Тем не менее пойменные
сукцессии хорошо прослеживаются и здесь. Начальные их стадии — з;али-
ваемые осоковые луга из Carex concolor, которые быстро сменяются разно-
травно-кустарничковыми (Arctous alpina, Vaccinium uliginosum, Dryas oc-
topetala\ Luzula confusa) и кустарничково-осоковыми (Carex ensifolia ssp.
■arctisibirica, Vaccinium minus) с участием мхов (Ptilidium ciliare. Hyloco-
40
тщт splendens var. alaskanum) и лишайников (Cladonia macroceras, Cla-
dina arbuscula) тундровыми сообществами. Заключительными субклимак-
совыми сообществами этой гидросерии являются моховые тундры.
В подзоне' субарктических тундр в плакорных
местообитаниях распространены моховые и кустарниковые тундровые
сообщества. Гипоарктические кустарники,— ерник {Betula nana) и7
несколько видов дв {Salix glauca, S. lanata, S. pjulchra) — специфичны для
этой подзоны и характеризуют субарктический подтип тундрового типа
растительности [Александрова, 1977]. В северной части подзоны
арктические и бореальные кустарники формируют разреженные низкорослые
синузии, встречаясь отдельными небольшими группами среди сплошного
покрова моховых и лишайниковых тундр. В южной тундре они образуют
уже сомкнутые заросли, сначала низкорослые и полегающие, так
называемые низкокустарниковые тундры, а затем к югу переходящие в
собственно кустарниковые тундры — ерниковые, ивняковые, ольховниковые
[Мельцер, 1977, 1980]. -.
В пределах подзоны субарктических тундр выделяются три подзональ-
ные полосы, которые отличаются друг от друга главным образом составом
плакорных сообществ и разной степенью участия кустарников в сложении
растительного покрова. Полоса северных субарктических тундр характе-.
ризуется распространением на плакорах низко- и редкокустарниковых
с Betula папа и Salix glauca моховых тундр мозаичного сложения — бугор-
коватых и пятнисто-бугорковатых. Основу мохового покрова в них
составляют синузии зеленых мхов —Dicranum elongatum, Aulacomnium turgi-
dum, Hylocomium splendens var. alaskanum, Ptilidium ciliare.
Полоса южных субарктических тундр представлена двумя подзональ-
ными полосами II порядка. Первая более северная полоса низкокустарни-
ковых тундр характеризуется распространением низкорослых (20—30 см)
стелящихся кустарников \— Salix lanata, S. glauca, Betula nana, синузии
которых более или менее равномерно распределены среди осоково-лишай-
никово-моховых пятнисто-бугорковатых тундр. Для самой южной полосы
собственно кустарниковых субарктических тундр типичны ерниковые
и ивняково-ерниковые сообщества с лишайниково-моховым покровом,-
часто чередующиеся с группами лиственничных редколесий (Larix sibiri-
са) [Мельцер, 1980].
В местах распространения песчаных почв на всей территории
субарктической подзоны развиваются кустарничково-мохово-лишайниковые
(Cladina rangiferina, CI. stellaris,\ Cetraria cucullata, Alectoria nigricans)
полигональные и пятнисто-полигональные тундры.
При заболачивании плакорные и псаммофитные типы тундровых
сообществ переходят в травяко-кустарничково-моховые (с Drepanocladus
examvulatus, Aulacomnium palustre, Ledum decumbens, Rubus chamaemorus)'
ивнячково-ерничковые тундры с бугорковато-мочажинным микрорельефом
и далее —в болота. Для субарктической подзоны характерно широтное
развитие кустарничково-осоково-моховых валиково-полигональных и кус-
^гарничково-мохово-лишайниковых трещиновато-йолигональных
комплексных болот. Последние наиболее типичны для полосы южных
субарктических тундр.
Гидросерии долин рек субарктической подзоны развиты достаточно
широко. По субарктическому типу развивается серия пойменной
растительности приустьевых участков рек Оби, Надыма, Пура, Таза. Низкие
уровни пойм этих рек сильно заболочены, они заняты или низинными мел-
коивняково-гипново-осоковыми болотами, или кочковатыми осоковыми
(Carex acuta) и арктофиловыми (Arctophila fulva) лугами.
Заключительными стадиями этой серии являются типично тундровые сообщества —
ивняковые, ерниково-ивняковые и ольховниковые.
Таежная зона, занимающая самые большие площади на
Западно-Сибирской равнине, выделяется по преобладанию сообществ бореаль-
ного типа растительности [Сочава, 1957, 1964а]. Они образуют здесь
различные сочетания, в которых чередуются лесные, болотные и луговые фи-
11
тоценозы. Современный облик растительности этого региона во многом
определяется повышенным гидроморфизмом, обусловленным интенсивно
идущими повсеместно процессами заболачивания территории. Благодаря
этому все пространственные сочетания и ряды растительности таежной
зоны Западной Сибири в качестве обязательных компонентов имеют
сообщества заболоченных лесов и болот.
По современным представлениям, широтная дифференциация
растительного покрова внутри бореальной зоны достаточно велика. На основе
анализа обзорной геоботанической карты [Растительность..., 1976] в ее
пределах выделяются пять геоботанических подзон и восемь подзональных
полос (см. рис. 1). Остановимся кратко на отличительных особенностях
каждой из них.
Подзона редколесий находится на северном пределе
распространения лесной растительности. Вследствие суровых климатических
условий эдификаторная роль деревьев в этой подзоне ослаблена, древостой
отличаются малой сомкнутостью (0,1—0,2), небольшой высотой (4—10 м),
слабым развитием крон и усыханием вершин. Травяно-кустарничковый
ярус и мохово-лишайниковый покров имеют здесь ярко выраженное моза-
ичное сложение.
Подзона редколесий включает две подзональные полосы. Северная
полоса (лесотундровая) соответствует подзоне южной лесотундры,
выделенной Б. Н. Городковым [1935] на Западно-Сибирской равнине, Зональными
типами сообществ здесь являются елово-лиственничные (с Larix sibirica)
и лиственнично-еловые (с Picea obovata) лишайниково-зелевомошные и зе-
леномошно-кустарничковые редколесья, которые повсеместно на плако-
рах и в неплакорных местообитаниях сочетаются с кустарниковыми
тундрами — ерниковыми (Betula папа), ивняковыми (Salix glauca, S. pulchra),
ольховниковыми (Duschekia fruticosa). В травяно-кустарничковом ярусе
этих редколесий наряду с преобладанием гипоарктических кустарников
и кустарничков (Betula nana, Salix pulchra, Vaccinium uliginosum, Em-
petrum nigrum) в качестве постоянной примеси присутствуют арктоалъ-
пийские виды — Ledum decumbens, Arctous alpina. В Южной (таежной)
полосе редколесий преобладают лиственничные и лиственнично-еловые ред-
колесные сообщества, на местности чередующиеся с лиственничными
редкостойными лесами. В плакорных условиях эти редколесья имеют кустар-
ничково-зеленомошный и лишайниковогзеленомошный покров, в котором
уже нет арктоальпийских видов, а наряду с гипоарктическими
кустарничками в значительном количестве встречаются представители бореальной
флоры — брусника, линнея. Самые большие площади в этой подзональ-
ной полосе занимают лишайниковые (с Cladina, Cladonia, Cetraria),
лиственничные редколесья, приуроченные к повышенным частям водоразделов
с глубокооттаивающими песчаными почвами.
Процессы заболачивания в подзоне редколесий идут повсеместно,
везде они сопровождаются сильным промерзанием грунтов;
формирующиеся ряды заболоченных лиственничных и еловых лишайниково-сфагновых,
зеленомошно-кустарничково-сфагновых редколесий при более сильном
морозном вспучивании сменяются бугристыми заболоченными тундрами
и плоскобугристыми комплексными болотами.
Подзона северной тайги на большое расстояние
протягивается с севера на юг, и в зависимости от изменений гидротермических
условий здесь наблюдается четкая дифференциация на две широтные
полосы. Для северной подзональной полосы характерно развитие
редкостойных лиственничных, лиственнично-еловых и лиственнично-сосновых
лесов, а также плоско--и крупнобугристых мохово-лишайниковых
комплексных болот и лугово-болотно-соровых растительных сообществ в поймах
крупных рек. Леса имеют сомкнутость не выше 0,4—0,5 и среднюю высоту
10—12 м. В их покрове преобладают бореальные и гипоарктические
кустарнички {Ledum palustre, Empetrum nigrum, Vaccinium vitis-idaea, V. uli-
ginosum); моховой покров имеет сплошное распространение, состоит из
зеленых мхов (Pleurozium schreberi, Hylocomium splendens, Polytrichum al-
12
pestre), среди них пятнами встречаются лишайники (Cladina rangiferlna,
CI. arbuscula).
В южной полосе с^веротаежной подзоны на плакорах распространены
лиственнично-елово-кедровые леса. По составу нижних ярусов они
сходны с лиственничными и еловыми лесами северной полосы, но отличаются
большей сомкнутостью (0,5—0,6) и несколько большей высотой (12—14 м).
Производительность этих лесов низкая (V—Va классов бонитета),
приурочены они к суглинистым почвам с близко залегающей вечной мерзлотой.
Пространственные сочетания и ряды сообществ плакорных и непла-
корных местообитаний обнаруживают существенные различия в разных
частях северотаежной подзоны. В северной ее полосе плакорные типы ело-
во-лиственничных кустарничковых лесов чередуются с сухими
лиственничными и лиственнично-сосновыми лишайниковыми лесами и замещаются
ими на больших пространствах песчаных водоразделов^ Обь-Енисейского
междуречья. В южной полосе на песчаных почвах происходит смена
пенообразователей. Для этой территории более характерно преобладание
монодоминантных сосновых древостоев, приуроченных к зандровь!м пескам
Сургутской низины. Наиболее типичен пространственный ряд смен, который
слагают сосновые зеленомошно-лишайниковые, кустарничково-лишайни-
ковые леса и сосновые лишайниковые редколесья.
Большая часть коренных лесов дренированных местообитаний в
северной тайге испытала воздействие пожаров и в настоящее время сменилась
производными древостоями. Процессы восстановительной динамики в этих
лесах идут с обязательной сменой пород. Устойчивы и широко
распространены три типа северотаежных производных сообществ: лиственнично-бе-
резовые леса в северной полосе подзоны, -темнохвойно-березовые и
сосновые с кедром и елью в южной полосе.
Заболоченность северной тайги Западной Сибири огромна.
В'Северной полосе подзоны большие площади заняты заболоченными
лиственничными, лиственнично-сосновыми и еловыми кустарничково-долгомошными
и сфагновыми лесами. При дальнейшем заболачивании на границах с
крупными болотными массивами лиственница выпадает из состава древостоя,
формируются кустарничково-лишайниково-сфагновые еловые редколесья
и редины, переходящие в комплексы бугристых болот. Для северной
полосы подзоны особенно характерно широкое развитие кустарничково-мохо-
во-лишайниковых травяно-сфагновых и осоково-гипновых плоско- и
крупнобугристых комплексных болот [Романова, 1965]. Массивы этих болот
встречаются и в южной полосе северотаежной подзоны, однако здесь
наблюдается свой ряд заболачивания, главные компонентами которого
являются сосновые с кедром лишайниково-кустарнйчково-сфагновые леса
и олиготрофные лишайниково-сфагновые болотно-озерные комплексы.
Гидросерии долинной растительности северотаежной подзоны
обнаруживают фитоценотические связи со многими сообществами
водораздельных лесов. Их заключительные стадии (леса высоких уровней поймы
нижней Оби и надпойменных террас Надыма, Пура, Таза) по составу
близки к северотаежным типам лиственничных, еловых и кедровых лесов
[Ильина, 1976].
Подзона средней тайги Западно-Сибирской равнины
характеризуется преобладанием темнохвойных и сосновых лесов и
производных сообществ на их месте. От северртаежных типов эти леса
отличаются более высокой продуктивностью, (IV класс бонитета), большей высотой
древостоя (17—20 м) и сомкнутостью (0,6—0,7), а также возрастанием роли
таежного мелкотравья и зеленых мхов в составе нижних ярусов леса. Сред-
нетаежная подзона также делится на две подзональные полосы.
Северная — представляет переход от северной тайги и отличается
распространением на плакорах коренных сообществ елово-кедровых лесов с
участием лиственницы и кустарничково-зеленомошным покровом, в котором
ведущую роль играют Vaccinium myrtillus, V. vitis-idaea, Linnaea borealis и
бореальные виды зеленых мхов Pleurozium schreberi, Hylocomium splendens.
Для южной полосы среднетаежной подзоны характерен другой зональный
13
тип: елово-кедровые с пихтой доелкотравно-бруснично-зеленомошные леса.
Они имеют более высокий класс бонитета (III—IV), достигают высоты 20—
22 м и диаметра; стволов 30—50 см. В покрове этих лесов возрастает роль
таежного мелкотравья (Maianthemum bifolium, Gymnocarpium dryopteris,
Orthilia- secunda, Trientalis europaea).
В рядах восстановительных смен среднетаежных елово-кедровых
лесов широко представлены коротко-производные сосновые, березовые и
осиновые леса; последние более характерны для южной полосы подзоны.
Коренные и производные среднетаежные леса чаще сочетаются с
сообществами заболоченных сосняков и кедровников, а также с сухими
борами на песках. В рядах заболачивания сменяют друг друга сосново-кед-
ровые, сосново-березовые и сосновые долгомошно-сфагновые и кустарнич-
ково-сфагновые леса, обычно переходящие в сасново-кустарничково-
сфагновые залесенные* болота. Обширные болотные массивы
центральных частей междуречий представлены грядово-мочажинными,
а в центре озерково-грядово-мочажинными комплексами с характерными
для гряд багульниково-касандрово-сфагновыми (Sphagnum fuscum, Cha-
maedaphne calyculata), местами с сосной и кедром (Pinus sylvestris f. litwi-
nowii, P. sibirica f. tiirfosa) сообществами и сфагновыми с пушицей и шейх-
.церией (Sphagnum balticum. Sph. majus, Eriophdrum vaginafumx Scheuch-
zeria palustris) группировками мочажин [Романова, 1965].
К среднетаежному типу относится и растительность поймы р. Оби на
отрезке Березово—Ханты-Мансийск. На низких уровнях поймы широкое
развитие поручают осоковые (Carefx aquatilis, С acuta) луга, которые в
северной тайге встречаются на более высоких экологических уровнях.
Вместе с тем на участках средних уровней широкое развитие получают канаре-
ечниковые и разнотравно-злаковые луга, а на[ высоких пойменных уровнях
и на останцах террас среди поймы — смешанные леса из кедра, сосны
и березы. \ .
В подзоне южной тайги основным зональным типом
являются кедрово-елово-пихтовые зеленомошно-мелкотравные и мелкотрав-
но-осочковые леса, которые отличаются хорошо развитым древостоем (II —
III классы бонитета), достигающим высоты 25—28 м и диаметра 50—60 см
(в возрасте 120—150 лет), обильным подростом из темнохвойных и
лиственных пород, разнообразным по составу подлеском. В травяно-кустарнкч-
ковом ярусе ведущая фитоценотическая роль принадлежит таежному
мелкотравью, среди которого типичны Oxalis acetosella, Trientalis europaea,
Stellaria bungeana. Особую группу образуют мелкотравно-осочковые леса
с доминированием в покрове Carex macroura. Из злаков в плакорных лесах
обычны Calamagrostis obtusata, Milium effusum, Melica nutans. В качестве
обязательного элемента в покрове южно-таежных лесов присутствуют
представители таежного высоко- и широкотравъя (Aconitum septentrionale,
Cacaliahastata, Filipendula ulmaria, Paris quadrifolia). Зеленые мхи не
имеют здесь сплошного распространения.
Особую широтную Полосу в западной части подзоны южной тайги
образуют темнохвойные леса с липой (Tilia cdrdata). Эти леса отличаются
высокой производительностью (II—III классы бонитета) и богатьщ травяным
покровом, в котором значительное участие принимают неморальные
элементы— Aegopodium podagraria, Pulmonaria dacica, % Actaea erythrocarpa.
Процессы возрастных и восстановительных смен в южно-таежных
пихтовых и еловых лесах идут преимущественно со сменой пород; среди
производных типов преобладают березовые и осиновые насаждения. На
песках здесь так же, как и в средней тайге, господствуют сухие брусничные
и лишайниковые сосновые боры, но специфичным для подзоны является
характерное сочетание этих боров с собственно южно-таежным типом
сосновых с липой травяно-брусничных (с Сагех упасгожа) с разрешенным
моховым покровом (Pleurozium schreberi) лесов. '
Процессы заболачивания охватывают большие территории южной
тайги Западной Сибири. Для этой подзоны в основном характерны те ?ке, что
и для средней тайги, типы олиготрофных сфагновых болот, а в рядах сооб-
14
ществ заболоченных лесов появляются типично южно-таежные березовые
с елью, кедром и сосной вейниково-хвощовые (Calamagrostis canescens,
Equisetum sylvaticum) и осоково-сфагновые (Сагех globularis] Sphagnum
warnstorfii) леса, предшествующие образованию заболоченных сосняков
и кедровников.
Серии долинной растительности в южной тайге особенно
разнообразны. Для них характерно преобладание на низких экологических уровнях
осоковых (Сагех acuta), канареечниковых и вейниковых (Calamagrostis
langsdorffii) болотистых лугов, распространение на средних уровнях
парковых ивняков и разнотравно-злаковых полидоминантных лугов,
широкое развитде на высоких уровнях поймы березово-сосновых (на песках) t
березово-пихтовых и еловых лесов с зеленомошно-травяным покровом
[Ильина, 1968, 1976].,
Подзону, мелко лиственны хлесов (подтайгу) выделяют
исследователи западно-сибирской тайги. Это своеобразная южная полоса
таежной зоны, аналога которой нет ни в европейской, ни в
восточно-сибирской тайге. Основу растительного покрЪва составляют здесь коренные
мелколиственные (березовые и осиновые) леса, которые в плакорных условиях
характеризуются хорошо развитым злаково-разнотравным покровом
(Calamagrostis arundinacea, Brachypodium pinnatum, Aegopodium podagraria)
и участием представителей таежного мелкотравья. Эти леса часто
чередуются с участками суходольных лугов.
Для подзоны характерно значительное заболачивание, начальной
Стадией которого являются березовые вейниково-высокотравные
(Calamagrostis langsdorffii, Filipendula ulmarid) леса, в обширных понижениях
переходящие в сильно увлажненные березовые осоково-вейниковые и осоковые
(с Сагех cespitosa, С. riparia) лесные сообщества. Их дальнейшее
заболачивание ведет к образованию лесных мезотрофных и евтрофных болот — бе-
: резовых с Betula pubescens, осоковых и осоково-сфагновых. Наиболее
характерны для этой подзоны осоково-гипновые (Сагех lasiocarpa, Drepano-'
cladus vernicosus) и травяные тростниково-осоковые (Сагех juncella, Phrag-
mites australis) евтрофные болота.
Долинная растительность подтаежного типа представлена сукцессион-
ными рядами сообществ, состоящими главным образом из луговых фито-
ценозов, в южной части значительно остепненных. На высоких уровнях
прирусловых участков пойм, а также на низких террасах рек развиваются
лесные сообщества подтаежного типа — ивово-тополевые, ивово-березовые,
березово-осиновые. ^
Зона степей на территории Западно-Сибирской равнины имеет
ограниченное распространение. Она представлена северными вариантами,
среди которых наибольшую выраженность имеют растительные
сообщества лесостепного типа, характеризующиеся сочетанием степных (лу-
гово-степных) и лесных фитоценозов. В пределах этой подзоны выделяются
две широтные полосы северной и южной лесостепи. Зональная
растительность полосы северной лесостепи — это злаково-разнотравные остепненные
луга, луговые степи и остепненные травяные березово-осиновые леса.
Основу травостоя составляют мезофильное разнотравье (Filipendula
vulgaris, Lathyrus pisiformis, Galium verum, Potentilla argentea) и корневищные
злаки (Poa angustifolia, P. pratensis, Calamagrostis epigeios)..
Зональными для полосы южной лесостепи являются луговые степи
с богаторазнотравно-злаковым травостоем, состоящим в основном из
представителей корневищных и дерновинных (Stipa pennata, S. capillata, Fes-
tuca'pseudovina, Helictotrichon schellianum, Phleum phleoides) злаков и мезо-
ксерофйльного разнотравья (Artemisia latifolia, Peucedanum morisonii)'.
Березовые колки этой полосы более разрежены, чем н^ севере, и занимают
меньшие площади. Большая часть территории северной и южной лесостепи
в настоящее время занята различными сельскохозяйственными землями —
пашнями, залежами, пастбищными угодьями. Й только в местах
засоленных почв распространена комплексная растительность из галофитно-луго-
15
вых и степных сообществ (Puccinelliatenuissima, Glycyrrhiza uralensis,
Artemisia nitrosa).
Процессы заболачивания не характерны для степной зоны. Они
наблюдаются только в небольших замкнутых понижениях с близкозалегающими
грунтовыми водами.
Микропоясные ряды заболачивания прослеживаются по периферии
озерных и болотных впадин; они представлены сообществами
заболоченных и болотно-солончаковых лугов (с Carex cespitosa, Alopecurus arundina-
<ceus, Bolboschoenus maritimus, Hordeum brevisiibulatum). Болота
преобладают травяные, преимущественно осоковые и тростниковые. Их площадь
расширяется в наиболее влажные годы. К лесостепному типу относится
долинная растительность многих рек юга Западно-Сибирской равнины. На
высоких уровнях поймы Иртыша и на низких террасах более мелких рек
развиваются остецненные «луга и луговые степи, преимущественно из
корневищных злаков — мятликовые, полевицевые, костровые. Колочные
леса очень незначительны по размерам и большей частью выкорчеваны или
выж$кены при сельскохозяйственном освоении.
Подзона разнотравно-дерновинно-злаковых степей находится на
самом юге равнины. Зональными здесь являются разнотравно-красноковыль-
ные степи, которые сейчас почти полностью распаханы. Значительно
большие площади занимают сообщества галофитных (Festuca valesiaca,
Artemisia pontica, A. nitrosa), псаммофитных (Stipa pennata,Helictotrichon de-
sertorum) и кальцефитных (Stipa korshinskyi, Artemisia dracun cuius)
вариантов этих степей. . •
Провинциальные различия растительного покрова Западно-Сибирской
равнины связаны с особенностями подстилающей поверхности, рельефом,
а также близостью морей и горных систем.
Геоботанические провинции выделяются главным образом внутри зон,
пересекая последние в меридиональном направлении (см. рис. 1). В состав
каждой провинции попадают отрезки разных подзон и подзональных
полос. Они занимают б их пределах неодинаковые площади и
характеризуются разным типологическим составом растительности.
На территории тундровой геоботанической зоны выделены две
провинции — Ямальская и Гыданская. Географически они четко обособлены, а
по составу растительности отличаются преобладанием в первой сообществ
моховых и лишайникевых тундр на почвах легкого механического состава.
Состав и протяженность подзональных полос внутри этих провинций
примерно одинаковы. Однако растительность южной полосы кустарниковых
тундр Гыданской провинции разнообразнее по составу: кроме
преобладающих по площади ерниковых тундр здесь в большом количестве
присутствуют ивняковые и ольховниковые .тундровые сообщества.
Провинциальное деление растительности лесной зоны
Западно-Сибирской равнины проводилось неоднократно разными авторами [Крылов,
1961; Шумилова, 1962; Смолоногов и др., 1970]. Выделенные нами три
провинции по объему и содержанию более всего соответствуют трем
геоботаническим секторам, намеченным В. Б. Сочавой, Т. И. Исаченко,
А. Н. Лукичевой [1953] при мелкомасштабном картографировании
растительности равнины. ~
Зауральская провинция характеризуется преобладанием в подзональ-
ном спектре сообществ средней тайги, среди которых основные площади
занимают сосновые леса. Они встречаются здесь не только на легких
почвах, как это отмечалось ранее [Сочава и др., 1953], но и на суглинках пла-
корных местообитаний [Лапшина, 1975]. Другая важная особенность
Зауральской провинции — значительное распространение еловых лесов,
особенно в подзонах северной и южной тайги. Усиление роли ели (Picea
obovata) на западе региона, очевидно, обусловлено ценогенетическими
связями Западной Сибири с Уралом и частично с европейской тайгой, а также
некоторыми провинциальными особенностями климата. Флористической
особенностью этой провинции является большое участие в составе южно.
таежных темно хвойных лесов широколиственной породы липы сердцевид.
16
ной (Tilia cofdata) и целого комплекса сопутствующих ей травянистых
видов из представителей неморальной флоры.
Центральная Обь-Иртышская провинция характеризуется четко
выраженным спектром всех подзон и подзональных полос, среди ^которых
наибольшую площадь занимает северная тайга/Отличительная
особенность растительного покрова этой провинции — широкое
распространение болот. Последние занимают огромные площади в центре крупных
водоразделов, а в приречные их частях (в местах более дренированных) они
чередуется с лесными массивами. В целом растительные сообщества
заболоченных лесов и болот повсеместно образуют характерные
пространственные сочетания (эколого-динамические ряды смен) со всеми типами лесов
в пределах Обь-Иртышской геоботанической провинции.
В Приенисейской провинции хорошо представлен весь спектр таежных
подзон. Заболоченность здесь умеренная, а в составе растительности
главное место занимают сообщества коренных темнохвойных лесов и
производных, от них, преимущественно березовых и осиновых древостоев. Состав
и подзональная структура растительности этого региона имеют многие
черты сходства со среднесибирской тайгой правобережья Енисея [Сочава,
1976].
На территории Западно-Сибирской равнины собственно степная
растительность имеет очень ограниченное распространение. Ее
типологический состав и пространственная структура сходны с таковыми
североказахстанских степей, и, очевидно, провинциальное деление- этой т&ррито-
рии целесообразно проводить в соответствии с районированием
Казахстана [Карамышева, Рачковская, 1973].
Все отмеченные выше зональные и провинциальные особенности
структуры растительности Западно-Сибирской равнины имеют сложную фило-
ценогенетическую основу и подтверждаются многими
палеогеографическими данными.
История формирования растительного покрова этого региона ведет
начало с третичного времени, когда образовалась значительная часть ее
современной флоры. Во все последующие этапы четвертичного периода шла
миграция отдельных видов растений и растительных сообществ,
происходило смещение различных ботанико-географических комплексов и групп
[Крылов, 1961; Западная Сибирь, 1963; Волкова, 1965; Нейштадт, 1976].
Из всех вопросов, связанных с палеогеографическим аспектом
исследования, для понимания современного состояния растительности
существенно важно определить наиболее устойчивое генетическое ядро видов
коренных фитоценозов и проследить формирование основных
пространственных закономерностей в распределении растительности — зональных и
провинциальных.
Большинство исследователей отмечают, что современные 'коренные
сообщества темнохвойных лесов средней и южной тайги
трансформировались из биогеоценозов позднего плейстоцена и голоцена, в которых
устойчивые позиции занимали три древесные породы — кедр, ель и пихта
[Волкова, 1965; Нейштадт, 1976; Савина, 1980]. Так, в средней тайге фаза
развития растительности с господством кедра и березы отмечается более
5 тыс. лет назад [Нейштадт, 1976]. В южной тайге в один из этапов
четвертичного периода, предшествующих современному, особенно велика былач
роль пихты [Савина, 1980]. Многие исследователи отмечают также
усиление роли ели в современную геологическую эпоху; особенно в южно- и сред-
нетаежных лесах.
В целом палеогеографические данные позволяют утверждать, что
широко представленные в растительном покрове северной, средней и
южной тайги сообщества с доминированием кедра, пихты и ели — это
наиболее древнее и устойчивое ядро современных -таежных биогеоценозов.
. Зональные черты в составе и распределении растительности Западной
Сибири были выражены уже в палеогене. Как отмечает Г. В. Крылов
[1961], к концу миоцена существовали четыре зоны: хвойных лесов с
преобладанием ели, кедраг пихты, лиственницы и с участием кипарисов, гра-
2 Заказ № 297 . 17
ba, бука; хвойно-лиственных лесов с сосной, елью, пихтой и лиственными
породами — березой, буком, дубом, липой, грабом; лиственных лесов с
березой и широколиственными породами; лесостепей с «островами» степей.
В ледниковый период четвертичного времени шло формирование
тундровой зоны и ее наступление на тайгу. В эту эпоху в тайге кроме тем-
нохвойных пород широко распространились лиственничные редкостойные
леса и сосново-сфагновые «рямы». Южнее тайги располагалась зона
плейстоценовых лесостепей с сосновыми и сосново-березовыми «островными»
лесами. В послеледниковый период (голоден) при потеплении климата
тундра отступила и широко распространилась зона тайги. В современных
подзонах лесостепи и подтайги за последние 5 тыс. лат произошла смена
хвойных пород на лиственные [Крылов, 1961; Савина, 1980].
В итоге можно считать, что происходившие в разные периоды
четвертичного времени смещения на север и юг основных широтных групп
сообществ — тундровых, лесных, степных — не затушевывали существовавшей
* уже тогда четкой зональной закономерности в их пространственном
распределении.
Из регионально-генетических черт современного растительного
покрова особо следует отметить распространение темно хвойных лесов с липой.
Липа в Прииртышье была отмечена еще в конце плиоцера и в начале ран-
нечетвертичного времени [Волкбва, 1965]. В послеледниковый
климатический оптимум по долинам рек она проникала до широты Салехарда
[Петрова, 1963]. Современное распространение темнохвойных лесов с липой
приурочено к западной прииртышской части южно-таежной подзоны. Ее
проникновению на восток препятствуют сильная заболоченность и
засоленность почв, а на север — суровость климата северной и средней тайги.
- Чрезвычайно важным для понимания современного облика
растительного покрова Западно-Сибирской равнины являются изучение истории
формирования болот и выяснение тендейций их развития. Как указывалось
выше, начало формирования большей части болот следует относить к
раннему голоцену. В позднем голоцене наблюдалось уже прогрессирующее
расширение площади олиготрофных болотных комплексов, о чем
свидетельствуют данные стратиграфических разрезов [Нейштадт, 1977]. Таким
образом,' «палеогеографические корни» имеет и основная динамическая
тенденция в развитии современной растительности — отступление темно-
хвойных пород и наступление болот. Эта тенденция отмечена около 9 тыс.
лет назад и, по мнению многих исследователей [Нейштадт, 1976; Савина,
1980; и др.], будет продолжаться в предстоящем тысячелетии.
ГЛАВА II
КАРТА «РАСТИТЕЛЬНОСТЬ ЗАПАДНО-СИБИРСКОЙ РАВНИНЫ»
М:1500 000
МЕТОДИКА РАБОТ. ОСНОВНЫЕ ИСТОЧНИКИ
±{арта «Растительность Западно-Сибирской равнины» создавалась на
основе анализа и обобщения имеющихся литературных, картографических
и фондовых материалов, а также материалов многолетних экспедиционных
исследований авторов, работавших в составе западносибирских экспедиций
и стационаров Института географии СО АН СССР, Центрального
сибирского ботанического сада СО АН СССР, Государственного
гидрологического института и других учреждений страны.
Работы по составлению и изданию карты включали следующие основные
этапы и разделы: 1) сбор и анализ литературных, картографических, а
также фондовых материалов; 2) полевые экспедиционные исследования
и составление на их основе серии карт растительности крупного и среднего
масштаба на отдельные районы Западно-Сибирской равнины; 3)
разработка легенды и составление авторского макета карты «Растительность За-
18
падно-Сибирской равнины» М 1 : 1 500 000; 4) подготовка карты к изданию
и ее издание. ; . '
Анализ, проведенный на первом этапе исследований накопленных
данных, показал, что обширная и разнообразная в природном отношении
территория Западно-Сибирской равнины характеризуется различной
степенью геоботанической изученности и обеспеченности картографическими
материалами. Имеющиеся на начало работ карты растительности
характеризовали только отдельные части этого крупного региона и в
большинстве, случаев ^ыли мелкомасштабными.
Из общесоюзных мелкомасштабных карт, отражающих наиболее
полно основные зональные и провинциальные черты растительного покрова
равнины, следует отметить «Геоботаническую карту СССР (Ml: 4000000),
изданную в 1954 г. под редакцией Е. М. Лавренко и В. В. Сочавы, а также
мелкомасштабную геоботаническук^карту лесной зоны Западно-Сибирской
низменности — приложение к статье В. Б. Сочавы, Т. И. Исаченко,
А. Н. Лукйчевои, посвященной географическим закономерностям
растительного покрова этой территории [Сочава и др., 1953]. * •
На отдельные районы Западно-Сибирскод равнины, кроме
общесоюзных карт, имелись также средне- и мелкомасштабные региональные карты,
содержащие ценный фактический материал о растительности тундр, лесов
и степей этого региона. Из таких работ заслуживает особого внимания се-ч
рия карт по югу Западной Сибири сотрудников Центрального сибирского
ботанического сада СО АН СССР: Е. В. Вандакуровой — карты
растительности Кулунды и Новосибирской области, Е. И* Лапшиной — карта
растительности южной части Сибири, М. С. Кузьминой — геоботаническая
карта Барабинской низменности.
Растительность тундр и редкостойных л^сов севера Западно-Сибирской
равнины отражена на специализированных среднемасштабных картах
растительности Ямало-Ненецкого автономного округа, составленных
землеустроительными экспедициями. Ценные геоботанические данные по
тундрам Ямала содержались в рукописных материалах крупномасштабного
картографирования Б. А. Николаева.
Для таежных территорий, занимающих самые большие площади на
Западно-Сибирской равнине, имелись некоторые мелкомасштабные
лесные карты, но основным источником получения данных по распределению
и составу лесов явились фондовые материалы лесоустройства.
К моменту составления геоботанической карты Западно-Сибирской
равнины работами наземного лесоустройства была охвачена большая часть
лесных территорий Западной Сибири. Разнообразные по масштабу и
содержанию планово-картографические материалы наземного
лесоустройства — лесоустроительные планшеты, планы лесонасаждений,
схематические карты лесхозов, таксационные описания, несмотря на их
хозяйственную специфику, в настоящее время самые массовые и информативные,
позволяющие получать сведения о лесах сразу для больших территорий
[Амромин, Богоявленский, 1966; Богоявленский, Попов, 1968]. Из всех
лесоустроительных материалов в качестве главного источника при
создании обзорной карты растительности равнины использовались среднемас-
штабные планы лесонасаждений и таксационные описания к ним. Оценка
их пригодности и методы использования при полевых и камеральных
работах уже обсуждались в печати [Богоявленский, Ильина, 1970; Ильина,
1971 ] и частично освещены в последующих разделах этой главы.
Чтобы выяснить закономерности пространственного распределения,
состав и структуру растительности многих неисследованных районов, а
также для того, чтобы правильно интерпретировать имеющийся
разнокачественный по содержанию и масштабам материал, необходимо было
осуществить большой объем полевых экспедиционных исследований во многих
районах Западной Сибири. Такие исследования начались в 1964 г., когда
в Институте географии СО АН СССР была организована комплексная Обь-
Иртышская экспедиция, одной из задач которой было геоботалическое
обследование районов нового хозяйственного освоения.
2* ' . 19
В 1964—1965 гг. экспедиционные работы проводились сотрудниками
института М. Н. Баланенко, А. В. Гаращенко, И, С. Ильиной, Л. В.
Поповым в районах Нижнего Прииртышья и Среднего Приобья (бассейны рек
Демьянки, Бол. Салыма, Туртаса, Алымки); в полевой сезон 1964 г. были
начаты геоботанические исследования в бассейне Сев. Сосьвы сотрудником
Центрального сибирского ботанического сада СО АН СССР Е. И.
Лапшиной, а на Тобол-Туртасском водоразделе—сотрудником Института
географии АН СССР Е. Л. Любимовой.
В 1966 г. в Институте географии СО АН СССР под руководством
Б. А. Богоявленского был создан Октябрьский картографический отряд
Обь-Иртышской экспедиции. В его задачи входили полевое обследование
и сбор нового фактического материала для- составления геоботанической,
гипсометрической и других научно-справочных карт на территорию
Западно-Сибирской равнины. Работами этого отряда была охвачена
значительная часть Среднего Приобья (бассейны рек Назым, Атлым, Лямина, Тром-
Югана, Назинка, Югана, пойма Оби на отрезке Октябрьское — Ханты-
Мансийск—Александровское), а также районы севера — междуречья
Казыма и Надыма, Пура и Таза (рис. 2).
В составе отряда с 1966 по 1972 г. работали Б. А. Богоявленский
(картограф), И. С. Ильина и Л. И. Мельцер (геоботаники), В. Д., Махно
(картограф), Л. В. Попов (лесовод), Г. В. Пономарев (охотовед), Ю. С. Толчель-
ников (почвовед), А. П. Шведов (зоогеограф), а также
студенты-практиканты. В 1969—1971 гг. Л. И. Мельцер в составе отряда проводила
геоботанические исследования на территории западно-сибирских тундр. В этот же
период в бассейнах рек Вах, Лямин, Тым работала группа лесоводов под
руководством проф. Л. В. Попова. В районах Сосьвинского Приобья
продолжала геоботанические исследования Е. И. Лапшина. Ею же в
дальнейшем были обследованы бассейны рек Вах, Таз, Куль-Еган [Лапшина,
1978]. На юге равнины полевые исследования в подтайге и лесостепи
Омской области в 1970—1971 гг. вели Н. Н. Лавренко, а в степных районах
Курганской области — Е. И. Лапшина и В. Д. Махно.
Большой объем полевых работ был выполнен в поймах Оби, Иртыша
и их притоков. Экспедиционные исследования, начатые в этих ^районах
Октябрьским картографическим отрядом, в дальнейшем были продолжены
на базе Нищнеиртышского стационара Института географии СО АН СССР.
В них приняли участие геоботаники-картографы И. С. Ильина, В. Д.
Махно, Л. И. Мельцер, Л. П. Соколова, В. И. Инешина, а также геоморфологи
и гидрологи И. Б. Петров, К. И. Вакулищ И. Ф. Гелета. В целом
сотрудниками института за весь период экспедиционных работ были обследованы
участки Нижней и Средней Оби в районах пос. Ниж. Тугияны, Березово,
Октябрьское, Самарово, Тундрино, Соснино, в устьях рек Бол. Салым,
Юган, Назинка, в окрестностях городов Сургут и Нижневартовск
[Ильина, 1968, 1976; Петров, 1979]. Напоимте Иртыша крупномасштабное
геоботаническое картографирование и стационарное изучение растительности
проводились на пойменных массивах — Цимбельском, Моильском, Ирым-
ском, Карбинском [Ильина и др., 1977, 1980; Дружинина, 1979]. В 1978—/
1983 гг. И. С. Ильина в составе экспедиций НИИ географии ЛГУ провела
геоботанические исследования в дельте р. Оби [Ильина, 1982] и в омской
пойме Иртыша [Ильина, Скарлыгина-Уфимцева и др., 1983].
Остановимся несколько подробнее на некоторых видах
экспедиционных работ, сыгравших важную роль в создании карты растительности
Западно-Сибирской равнины.
При изучении труднодоступных малонаселенных районов,'каким
является север равнины, большое значение имели аэровизуальные
обследования. Они позволяли не только рекогносцировать территорию и
корректировать данные наземных исследований, но и давали возможность
получать значительную новую геоботаническую информацию.
В период работ Октябрьского картографического отряда были
организованы аэровизуальные маршруты с коротким десантированием для
наземных работ на обширной территории северной тайги и тундры Западно-Си-
20
Рис. 2. Схема маршрутов Октябрьского картографического отряда Института
географии СО АН СССР и Центрального сибирского ботанического сада СО АН СССР.
Маршруты: 1 — основные автомобильные и пешие; г — водные (на катерах, лодках) в сочетании
с пешими; 3 — аэровизуальные; 4 — ключевые участки.
бирской равнины (см. рис. 2). Общая протяженность этих маршрутов
7480 км. Маршруты проводились, как правило, комплексной группой
специалистов и включали различные виды работ [Богоявленский и др., 1979].
Геоботанические наблюдения, как и все прочие, велись по определенным
контрольным точкам. В результате многократной и довольно точной
привязки сведения о растительности, полученные на разных отрезках
маршрута, наносились на топографическую основу, увязывались с
географической ситуацией и таким образом приближались по детальности к
наземной геоботанической съемке. Впоследствии они дополнялись материалами
геоботанических описаний,; сделанных на точках приземления.
Аэровизуальные съемочные маршруты позволили уточнить многие геоботанические
21
рубежи, и прежде всего зональные и подзональные. С их помощью,
например, прослежены границы широтных полос лесотундровь(х и таежных
редколесий, отмечены различия в составе и производительности темнохвой-
ных лесов и болот в переходной полосе северной и средней тайги
Сургутского полесья и др. ' ,'
Другим важным видом полевых работ были маршрутные
исследования — пешие, на лодках и катерах, частично (в южных районах) на
автомашинах. В качестве географической основы при съемочных маршрутах
использовались крупно- и среднемасштабные топографические карты
и аэроснимки — черно-белые и спектрозональные [Богоявленский, 1969].
Особые трудности вызывало картографирование растительности пойм
крупных рек — Оби и Иртыша. Чрезвычайное разнообразие растительных
сообществ в пбйме и их постоянную пространственную изменчивость
трудно отразить даже в самом крупном масштабе. Тем более эта задача
усложняется при создании средне- и мелкомасштабных карт, где требуется
принять определенную степень обобщения — генерализации в показе
основных свойств и состава растительности. При картографировании пойм Оби
и Иртыша были широко использованы методы маршрутной
геоботанической съемкипо профилям высотных уровней поемности, что позволяло
выявлять пространственные смены растительности в зависимости от разной
степени заливаемости поймы водами весеннего половодья. Большое
внимание при этом уделялось среднемасштабной маршрутной съемке, которая
проводилась с борта катера или лодки путем распознавания и
дешифрирования контуров топографических, лоцманских карт и аэрофотоснимков
[Ильина, 1968]. Таким способом была составлена среднемасштабная
карта растительности среднего течения Оба (на отрезке Ханты-Мансийск —
Нижневартовск), а также отдельные участки пойм Нижней Оби и Нижнего
Иртыша. *
Особое место в полевых исследованиях отводилось крупнрмасштаб- >
ной площадной съемке ключевых (эталонных) участков. Выбор их и
количество определялись в предполевой период, особенно тщательно
подготавливалась картографическая основа — аэроснимки (контактные
отпечатки, фотопланы) и крупномасштабные топографические карты.
Ключевой участок выбирался в ландшафте, наиболее типичном для конкретного
широтного пояса, т. е. там, где ярче всего проявлялись зональные
особенности растительного покрова или его специфические региональные черты.*
Содержание'полевых работ на ключевых участках определялось задачами
картографирования растительности: выявлением^ее связи с рельефом,
изучением основных форм пространственной структуры, установлением
динамических категорий коренных, серийных, антропогенно-производных
сообществ. Методически работа на каждом из ключевых участков строилась
по трем основным этапам [Богоявленский, Ильина, 1970]: 1)
рекогносцировочное обследование территории с маршрутным дешифрированием
аэроснимков; 2) работы на опорном экологическом профиле с подробным
описанием состава растительности, сбором гербария, закладкой почвенных
и геоморфологических разрезов и т. д.; 3) составление геоботанической
карты методом площадной геоботанической съемки с использованием
аэроснимков.
На некоторых ключевых участках проводились опытные работы по
использованию материалов лесной таксации при геоботаническом
картографировании. Они проводились, совместно с таксаторами
Западносибирского лесоустроительного предприятия Леспроект, осуществлялись
методом прокладки контрольных маршрутов по просекам и повторного
дешифрирования таксационных выделов с подробными геоботаническими и
почвенными описаниями по «отработанным» таксаторами контактным
отпечаткам и лесоустроительным планшетам. Всего с разной степенью дефль,ности
было обследовано около 50 ключевых участков в отдельных частях
Западно-Сибирской равнины площадью от 5 до 20 км2 каждый.
В результате проведенных полевых исследований создавались с^)ии
крупно- и среднемасштабных карт, основанных на материалах натурных
22 "'•■- --
съемок. Крупномасштабные геоботанические карты и экологически^
профили были составлены на районы Среднего Приобья (бассейны рек Бол.
Атлым, Бол. Салым, Лямин, Назым, Назинка) Б. А. Богоявленским
и И, С. Ильиной; Омской области — Н. Н. Лавренко Шавренко, 1982];
на отдельные участки поймы Нижнего Иртыша — И. С.1 Ильиной,
В. Д. Махно, Л. И. Мельцер, Л. П. Соколовой [Ильина и др., 1977а, 1980].
Листы и блоки среднемасштабных геоботанических карт охватили
наиболее типичные районы всех природных зон и подзон Западно-Сибирской
равнины: Ямало-Гыданские тундры (Л. И. Мельцер), северную и среднюю
тайгу Обь-Иртышья (И. С. Ильина, Б. А. Богоявленский, Л. В. Попов)
и Сорьвинского Приобья (Е. И. Лапщина); южную тайгу Нижнего
Прииртышья (Е. Л. Любимова); подтаежные леса и лесостепь Омской области
(Н. Н. Лавренко), поймы Нижней и Средней Оби (И. С. Ильина).
Построение указанных средне- и крупномасштабных карт — очень
важный этап в создании обзорной карты растительности Западной Сибири.
Эти карты составлялись по новым фактическим материалам или
непосредственно по данным полевых съемок (крупномасштабные карты), или путем
геоботанической интерпретации и генерализации планов лесонасаждений
с привлечением материалов экспедиционных исследований (среднемасштаб-
ные карты). В процессе создания серии карт среднего и крупного масштаба
на различные в основном малоисследованные районы Западно-Сибирской
равнины выяснялись основные черты состава, структуры и динамики
растительного покрова, выявлялись главные закономерности его
пространственного распределения. Все это было очень важнои послужило основой
для разработки классификации растительности и построения легенды всей
геоботанической карты Западно-Сибирской равнины.
Третий этап в создании карты растительности Западно-Сибирской
равнины был определяющим. На этом этапе составлялись сводная легенда
и авторский макет карты на основе анализа, обобщения и интерпретации
литературных, картографических и экспедиционных данных.
Принципам построения легенды посвящен следующий раздел этой
главы. Здесь мы отметим только, что в основу легенды был полоясен
систематический список картируемых единиц растительного покрова, который
составлялся и уточнялся на всех этапах сбора и обобщения исходного
фактического материала. Он включал различные по объему и содержанию
подразделения растительного покрова, пространственная выраженность
которых позволяла отразить их в принятом масштабе карты.
Систематический список картируемых единиц являлся тем предварительным рабочим
вариантом легенды, с помощью которого осуществлялась геоботаническая
интерпретация исходных данных и нанесение их на географическую основу.
• Авторский макет карты составлялся в два этапа у- вначале на рабочей
основе промежуточного масштаба (1 : 300 000 — 1 : 1000 000), а затем
после уточнения легенды — на окончательной, принятой к изданию
основе. Авторские листы карты промежуточного масштаба составлялись по
принципам и методам, разработанным в современной геоботанической
картографии [Грибова, Исаченко, 1972]. Следует особо подчеркнуть большое
значение при их создании некоторых специальных отраслевых карт
приводы, сопоставление с которыми дало возможность значительно уточнить
и конкретизировать геоботаническое содержание. В частности, при
составлении авторских листов карты растительности Западно-Сибирской
равнины очень широко использовалась ландшафтно-типологическая карта
болот М 1 : 2 500 000 [Романова, 1977], в основе которой лежат материалы
многолетних экспедиционных исследований сотрудников
Государственного гидрологического института [Болота..., 1976] в этом регионе и их
данные по дешифрированию аэрофотоснимков. Из других отраслевых карт
наибольшее значение имели геоморфологическая, почвенная и карта
четвертичных отложений. С их помощью уточнялась приуроченность
геоботанических подразделений к различным типам генетических поверхностей
(поймам, террасам, водоразделам), а также к разным по составу
поверхностным отложениям и почвам.
23
После обобщения всех исходных картографических материалов по
единой легенде на карте промежуточного масштаба составлялся авторский
оригинал карты масштаба 1 : 1 500 000. Этот этап работы включал
различные виды генерализации, прежде всего геоботанической генерализации
подверглась легенда карты. В соответствии с новой нагрузкой и более
обобщенной географической основой увеличился объем многих
картируемых единиц, в основном подразделений гетерогенного растительного
покрова — типов болотных массивой и рядов сообществ долинной
растительности; некоторые малые по площади геоботанические выделы были
перенесен^ из основной легенды в число внемасштабных знаков.
Производилась и геометрическая генерализация специальной
нагрузки карты, в процессе которой уточнялись размеры и рисовка контуров,
проводилась увязка специального содержания с основными элементами
географической основы, в первую очередь с рельефом и гидрографией.
При создании авторских макетов геоботанической карты большое
значение имела также правильная разработка красочной шкалы. Система
красочно-штриховых обозначений разрабатывалась авторами на всех
этапах картосоставления и воплощалась при создании карт разного масштаба•
В соответствии с требованиями современного геоботанического
картографирования красочная шкала геоботанических карт должна отражать
фитоцецотический состав растительности, ее пространственную структуру,
динамические свойства и связь с факторами среды [Грибова, Исаченко,
1972]. Система красочно-штриховых обозначений, принятая на' карте
растительности Западно-Сибирской равнины, построена по следующим
основным принципам [Махно, 1973]:
1) качественный цветной тон отражает формационный состав
главнейших подразделений растительного покрова Западно-Сибирской
равнины;
2) оттенки цветного тона указывают на принадлежность
растительности к определенным зональным и подзональным категориям;
3) различного типа линейные штриховки по цветному тону обозначают
разные формы пространственной структуры растительности — сочетания г
ряды пойменных сообществ. На неоднородность территориальной
структуры указывают внемасштабные штриховые значки, размещенные по
цветному фону (болотная растительность, кол очные леса в лесостепи и др*);
4) линейная двухцветная заливка (типа матрац) применена для
показа антропогенной динамики (цвет узкой полосы соответствует коренной
растительности, на месте которой возникло производное сообщество, цвет
широкой — современным антропогенным модификациям);
5) сочетание цветного тона слабой интенсивности с наклонной
прерывистой штриховкой этого же тона символизирует нарушенность
растительности в результате сельскохозяйственного освоения территории.
Принципы построения системы красочно-штриховых обозначений,
разработанные в процессе составления карты, были в полной мере
реализованы при ее издании, что позволило сделать карту достаточно наглядной,
хорошо читаемой и отразить на ней комплекс фитоценотических,
флористических, физиономических, структурных и динамических свойств
растительного покрова Западно-Сибирской равнины.
Карта «Растительность Западно-Сибирской равнины» М 1 : 1 500 000
[М., ГУТК, 1976] была издана на четырех листах форматом 84x96 см.
Текстовая легенда размещена справа от основной карты на втором и
четвертом листах; она включает 144 красочно-штриховых обозначения и 26
внемасштабных знаков. Кроме основного содержания на карте помещены
пять дополнительных карт-врезок М 1 : 12 500 000.
ПРИНЦИПЫ СОСТАВЛЕНИЯ ЛЕГЕНДЫ
Составление обзорной карты «Растительность Западно-Сибирской
равнины» М 1 : 1 500 000 преследовало цель отразить на современном уровне .
геоботанического картографирования главнейшие особенности раститель-
24
ного покрова этого обширного региона'показать специфику его структуры
и динамики, фитоценотического и флористического состава. В полной мере
эта задача могла быть решена только при условии глубокого
типологического анализа растительности, что должно было найти свое отражение в
логической основе построения карты — в принципах составления ее
легенды, в определенной трактовке картируемых единиц, в их соподчинен-
ности, содержании и объеме.
При создании гёоботанических карт типологический анализ
растительности проводится на основе классификаций, разработанных для
картографирования. В основу построения карты растительности Западно-Сибирской
равнины была положена регионально-типологическая классификация
В. Б. Сочавы [1958], основные принципы х&торой неоднократно
применялись при геоботаническом картографировании крупных регионов страны
[Сочава, 1968, 1979; Белов, 1973] и земного шара в целом [Сочава, 1964а].
Согласно этой классификации выделены наиболе'е крупные типологические
подразделения растительного покрова Западно-Сибирской равнины —
типы растительности, фратрии растительных формаций и региональные
комплексы. Тип растительности, в понимании В. Б. Сочавы, объединяет
сообщества разных биоморф на основе общности их эколого-географиче-
ских, динамических и других связей* «Типу растительности присущ
определенный круг эколого^географических связей и своя
структурно-динамическая система в виде экологических и неогенетических (сукцессионных)
рядов» [Сочава, 1964, с. 12].
Растительность Западнсг-Сибирской равнины принадлежит к трем
типам — тундровому, бореальному и степному. Каждый из них представлен
набором сообществ разного биоморфологического состава — лесных
(тундровых или степных), болотных, луговых, связанных между собой
пространственно, экологически и сукцессионно, что находит свое отражение как
в соподчиненности картируемых единиц легенды, так и на самой карте —
в пространственном распределении различных подразделений
растительного покрова.
Широкая эколого-динамическая трактовка типа растительности легла
в основу содержания всех последующих типологических подразделений
растительного покрова Западно-Сибирской равнины — фратрии
растительных формаций, региональных комплексов, подзональных категорий.
При выделении фратрий формаций использован географо-генетический
принцип [Сочава, 1957, 1979].
Так, в пределах тундрового типа растительности выделены две
фратрии — арктическая и амфиатлантйческая.
Арктическая фратрия формаций в Западной Сибири объединяет
тундровые сообщества, в которых широко представлены арктические и аркто-
альпийские виды. Их генетическая однородность и обособленность в зна*-
чительной степени связаны с историей формирования арктической флоры
[Толмачев, Юрцев, 1970], а также с суровыми климатическими услрвиядои
современной эпохи, что ограждает эти территории от флористического
влияния более^ожных типичных тундр [Букс, 1973].
Амфиатлантйческая фратрия объединяет западно-сибирские
тундровые формации, приуроченные к атлантическому сектору Арктики и
обособившиеся задолго до оледенения [Буданцев, 1970]. Это многочисленные
сообщества мохово-кустарничковых и кустарниковых тундр с большим
участием бореальных видов.
К урало-сибирской фратрии формаций отнесены все. Сообщества бо-
реального типа растительности. Генетическое ядро ее образуют широко
представленные на Западно-Сибирской равнине сообщества коренных темно-
хвойных лесов из пихты, кедра, ели. Особенности структуры и динамики,;
а также своеобразие эколого-географических связей этих сообществ,
выражающиеся в формировании ими характерных пространственных
сочетаний, динамических и экологических рядов растительности, позволили
отнести эти геоботанические подразделения к особому региональному
комплексу обь-иртышских формаций. '
25
Можно отметить две главные особенности обь-иртышского
регионального комплекса формаций, отличающие его от других региональных групп
этой фратрии, например среднесибирской, южно-сибирской и др. [Белов,
1973; Сочава, 1976]:
1) формирование в пределах обь-иртышской тайги специфических
типов пространственной структуры растительности — рядов
заболачивания лесов и несопряженных сочетаний (мезокомбинаций) лесных сообществ
дренированных водоразделов с комплексами болотной растительности
[Лапшина, 1973];
2) интенсивное развитие процессов поемности и аллювиальности в
поймах крупных рек, результатом чего является широкое распространение
естественных долгопоемных *крупнозлаковых и осоковых лугов, сук-
цессионно связанных с сообществами высокие террас и водоразделов
[Городков, 1946; Роднянская, 1963; и др.].
Следующая фратрия формаций — Заволжско-Казахстанская —
выделена в пределах степного типа растительности. Ее формируют
сообщества, в которых большая фитоценотическая роль принадлежит растениям,
генетически связанным с бореальной флорой. Изучение и
картографирование ареалов этих растений дали основание обособить во флорогенетическом
и фитогеографическом отношении степь и лесостепь Западной Сибири и
Северного Казахстана и отнести их к бореальной флористической» области
[Василюхина, 1969]. Границы последней на территории степей Западно-
Сибирской равнины маркируют южные пределы распространения
сообществ заволжско-казахстанской фратрии формаций. ,
В пределах указанной фратрии выделены два региональных
комплекса — обь-иртышский и североказахстанский. Для обь-иртышских
формаций характерны богаторазнотравно-злаковые степи и остепненные л^уга
в сочетании с березовыми остепненными лесами и комплексами галофитной
растительности на солонцах и солончаках. Североказахстанские формации
представлены разнотравно-дерновинно-злаковыми степями, занимают
сравнительно небольшие территории в этом регионе (Прииртышская м Ку-
лундинская равнины).
Следуя далее эколого-динамическому принципу, в пределах
региональных комплексов все картируемые выделы растительности сгруппированы
в подзональные типы — северо-, средне-, южно-таежные и подтаежные бо-
реальные формации, арктические "и субарктические тундровые и т. д. (I —
X). Эти подзональные категории явились основными узловыми
подразделениями легенды, они отразили важнейшую черту растительного покрова
Западно-Сибирской равнины — зональность.
Как видно из содержания легенды, все подзолальные категории также
включают различные по биоморфологическому составу сообщества пла-
♦корных и неплакорных местообитаний, например лесные, болотные,
луговые (IV—VII), которые объединены на основе общности их географических
и эколого-динамическй^с связей. Так, среднетаежный тип обь-иртышских
бореальных ^формаций (V) объединяет сообщества коренных елово-кедро-
вых и производных сосновых, березовых и осиновых лесов, а также
сообщества олиготрофных сосново-кустарничково-сфагновых болот и лесо-кус-
тарниково-луговые серии сообществ пойм рек (72—87).
Возможность объединения в пределах одного подзонального типа
различных по структуре и составу растительных сообществ исходит из
широкой эколого-динамической трактовки высших единиц
регионально-типологической классификации, о чем было сказано выше. Она хорошо
согласуется также с концепцией «эпитаксона» [Сочава, 1978], согласно которой в
единое динамическое целое связываются, например, устойчивые коренные хо-
общества лесов и менее устойчивые и неустойчивые (серийные) сообщества
болот и пойм рек [Городков, 1946].
Наиболее дробные подразделения легенды объединяют
непосредственно картируемые единицы разного таксономического ранга и объема
(обозначены порядковыми номерами и буквенными индексами). При выделении
этих единиц были использованы различные классификационные подходы,
26.
что определялось главным образом структурными особенностями
растительного покрова и его динамическими свойствами. Растительность одно-
* родной (гомогенной) горизонтальной структуры — леса плакорных
местообитаний, их экологические варианты на песках и в долинах рек, степные
сообщества и другие классифицировались с помощью общепринятых фито-
ценотических единиц — формаций, классов и групп ассоциаций. При ото,-
бражении на карте они генерализовались в зависимости от их
пространственной выраженности в большинстве случаев до объема обобщенных групп
или классов ассоциаций (44, 50, 88, 89 и*др.).
Полидоминантные сообщества тундровой растительности
картировались в еще более обобщенном виде— кустарничково-моховые и моходо-
лишайниковые тундры (2, 5), ерниковые, ивняковые тундры (22, 26).
Большая часть территории Западно-Сибирской равнины имеет
растительный покров гетерогенной (неоднородной) пространственной
структуры. Для его отражения на карте использована система хорологических
единиц — типы сочетаний, эколого-динамические ряды пойменной
растительности, типы болотных массивов. Сочетания гомогенных лесных
сообществ с рядами заболачивания лесов или типами болотных массивов (55,
58, 79, 81, 82) — наиболее распространенная форма пространственной
структуры растительного покрова, характерная для территорий, где
интенсивно идут процессы заболачивания [Лапшина, 1973]. Подобные типы
сочетаний ранга мезокомбинаций [Исаченко, 1969] отражены во многих
подразделениях легенды, характеризующих растительность тундры и
тайги (3, 13, 55, 58, 79, 81, 82 и др..
Более сложными мёзокомбинадиями показана структура
растительного покрова болот, занимающих огромные пространства
Западно-Сибирской равнины. Основной картируемой единицей болот в масштабе карты
служит тип болотного массива в понимании Т.. К. Юрковской [1968],
иногда с периферийным (поясным) рядом заболоченного леса. Фитоценотиче-
ский состав картируемых единиц различных типов болотных массивов
раскрывается в соответствующих номерах легенды (83, 84, 107).
На составленной геоботанической карте Западно-Сибирской равнины
бодее дифференцированно, чем на ранее изданных картах этой территории,
показан своеобразный растительный покров обширных пойм крупных
. рек—..Оби й* Иртыша. Это также потребовало дополнительных
классификационных построений. Применительно к целям мелко- и среднемасштаб-
ного картографирования была разработана классификация растительности
островных пойм Оби и Иртыша [Ильина, 1968, 1972]..В основу ее был
положен эколого-динамический критерий.^!? качестве исходных объектов
классификации принимались дробные динамические ряды, состоящие из
сходных по формационному составу и экологии растительных сообществ,
формирующихся в одинаковых условиях поемного режима. Показ на
геоботанической карте рядов сообществ, а не отдельных фитоценозов
(обобщенных их типов) имеет большой научный и практический смысл. С одной
стороны, это увеличивает информативность карты, так как позволяет от-.
разить в пределах каждого картируемого* выдела большой диапазон
растительности, свойственной экологическим условиям данного местообитания,
при этом расположение и соподчиненность сообществ в рядах указывают
на динамические свйзи и,тенденции возможных смен растительности во
времени, с другой — картирование фитоценотических рядов наиболее
объективно отражает реально существующую пространственную структуру
пойменного растительного покрова, что чрезвычайно важно для оценки
условий его хозяйственного использования.
Установлено, что наиболее крупные различия в составе
растительности рядов пойменных сообществ связанй с их приуроченностью к
разным экологическим уровням поймы [Барышников, 1933; Роднянская,
1958, 1968; Ильина, 1968, 1976]. Эти уровни характеризуйся
определенной продолжительностью затопления, разными сроками наступления и
спада половодья, разной интенсивностью накопления аллювия и амплитудой
высот пойменного рельефа [Шенников, 1941].
27
Растительные сообщества, формирующиеся на территориях,
принадлежащих одному экологическому уровню, сгруппированы в обобщенные
ряды растительности, имеющие сходный экологический состав сообществ
и одинаковую степень их динамической устойчивости. Например, для
низких экологических уровней поймы среднего течения Оби характерны
ряды сообществ болотистых лугов — преимущественно осоковых и кана-
реечниковых, отличающихся наименьшей устойчивостью своего состава
и структуры (108а), для средних уровней типичны разнотравно-злаковые
луга и заросли кустарниковых иЪ (108г). Наиболее устойчивые
растительные сообщества формируются на высоких уровнях поймы — это
смешанные хвойно-мелколиственные леса с мохово-травяным покровом, в
котором значительное участие принимают элементы пойменного разнотравья
(108е, ж). ' .
Самые крупные подразделения классификации и легенды карты —
серии пойменной растительности. Это Типы обобщенных долных
динамических рядов, отражающих все стадии развития растительного покрова
данного участка поймы на всех экологических уровнях, начиная от открыткх
группировок соровой растительности, приуроченных к самым низким и
надолго затопляемым участкам, и кончая субклимаксовыми сообществами
долинных лесов высоких и редко заливаемых пойменных островов.
, На обзорной геоботанической карте Западно-Сибирской равнины
непосредственное отображение получили обобщенные динамические ряды,
характерные для каждого экологического уровня поймы (69а, б, в...; 108а,
б, в, г...). В легенде они объединены в серии пойменной растительности
различных участков пойм Оби и Иртыша (69, 108, 122). В составе
растительности серий, в особенностях спектров высотных поясов поемности нашли
отражение зональные черты пойменного растительного покрова. Это дало
возможность сгруппировать серии по зональному признаку и поместить
их в соответствующие (подзональные) разделы легенды.
Растительный покров долин более мелких рек, в основном притоков
Оби и Иртыша, при построении легенды также классифицирован по эко-
лого-динамическому принципу. На карте выделены серии лугово-кустар-
никово-лесных и кустарниково-лесных сообществ (109, НО, 123, 136...).
В отличие от пойм Оби и Иртыша серии долин более мелких рек включают
не только пойменные луговые сообщества, но главным образом леса и
кустарниковые заросли низких террас рек.
Более подробно принципы выделения и картирования некоторых
типов хорологических единиц обсуждались ранее в статьях Е. И. Лапшиной
[1973] и И. С. Ильиной [1968, 1976].
На карте «Растительность Западно-Сибирской равнины» нашли
отражение также динамические свойства растительного покрова. С этой целью
в процессе типологического анализа была произведена классификация
растительности jno динамическому принципу с выделением категорий
коренных, квазикоренных и серийных сообществ естественного
растительного покрова, а также коротко- и длительнопроизводных сообществ
антропогенной растительности. Указанные категории сообществ отразили
разную степень устойчивости и сформированное™ растительности.
Принадлежность картируемых единиц к той или иной динамической категории
определила порядок расположения и соподчиненность различных
геоботанических подразделений в пределах каждого раздела легенды. Так, со^т-
венно коренные [Сочава, 1963] плакорные сообщества в легенде
располагаются в самом начале каждого ее раздела (1, 10, 44, 72, 88).
Антропогенные производные категории (45, 46, 73, 74, 89, 90) подчинены коренном
(72, 88), размещены непосредственно за ними, но смещены немного вправо
в общем списке номеров легенды; далее следуют квазикоренные (4, 5,, 79*
80) и серийные (85а, б, в...). Система красочно-штриховых обозначений
также разработана с учетом динамических свойств растительности
[Махно, 1973]. '
В целом легенда карты по количеству картируемых выделов
(красочно-штриховых обозначений) в 3—4 раза превышает число подразделений
28
растительности, показанных на ранее опубликованных обзорных
геоботанических картах этой территории. Кроме основных номеров, в легенде
помещены дополнительные обозначения — внемасштабные знаки — для не-
картируемых в данном масштабе растительных сообществ или отдельных
видов растений, редких или интересных в ботанико-географическом
отношении.
В заключение остановимся на некоторых принципах построения карт-
врезок. ' -.'.-,
На карте «Растительность Западно-Сибирской равнины» помещены
пять карт-врезок М 1 : 12 500 000, которые.дают дополнительную
информацию о пространственной структуре и ресурсном потенциале
растительного покрова Западно-Сибирской равнины.
Карта-врезка «Геоботаническое районирование» (авторы И. С.
Ильина и В. Д. Махно) отражает зональную и провинциальную
дифференциацию растительности. Зональные категории — зоны, подзоны, подзональ-
ные полосы, а также провинциальные — провинции и округа — выделены
на ней по принципам структурно-динамического анализа [Сочава, 1972].
На карте нашли отражение региональные спектры эколого-динамических
рядов растительности [Ильина, 1975]. С помощью круговых
диаграмм внутри гёоботанических округов показаны соотношения площадей
плакорных, псаммофитных, болотных и долинных рядов сообществ, а
также различных динамических категорий — коренных, квазикоренных,
серийных, антропогенно-производных (последние показаны формулой
антропогенного состояния растительности).
Карта-врезка «Зональные типы болот» (автор Е. А. Романова)
составлена на основе анализа географического распространения болотных
.сообществ по карте «Растительность Западно-Сибирской равнины». На ней
выделены зональные типы болот, определяемые признаками морфологической
структуры болотных ландшафтов, их растительным покровом, способом
питания, рельефом. Чтобы подчеркнуть связь выделенных зональных
типов с климатом, на врезке нанесены границы гидрологических зон
избыточного, переменного и недостаточного увлажнения.
Карта-врезка «Фитоклимат» (автор С. И. Хомченко) составлена по
данным наблюдений за сезонными явлениями природы сети добровольных
корреспондентов-фенологов Западно-Сибирской фенологической комиссии
за период с 1959 по 1973 год. Эта карта содержит важные фенологические
.данные, позволяющие оценить продолжительность вегетационного- периода
и некоторые сроки проведения сельскохозяйственных работ.
Карта-врезка «Качественная структура земель поймы р. Оби» (автор
А. И. Асеева) составлена по данным качественной
экономико-географической оценки пойменных земель, в основу которой положена карта
«Растительность Западно-Сибирской равнины». Ступени шкалы
сельскохозяйственной ценности земель выделены по поясам поемности (экологическим
уровням), к которым приурочены соответствующие подразделения
растительности, показанные на геоботанической карте. На врезке с помощью
диаграмм отражены разные типы использования луговых ' земель
поймы Оби, требующие мелиорации, а также степень затопляемости и
процентное соотношение затопляемых площадей поймы.
Карта-врезка «Ресурсы дикорастущих пищевых; растений» (автор
Л. Н. Ильина) относится к типу оценочных карт. В ее основу был положен
анализ данных о составе, географическом распределении и средней
многолетней биологической урожайности некоторых видов пищевых растений —
ягод, грибов, орехов и др. Карта построена методом изолиний с
использованием данных статистической обработки материалов [Ильина, 1973].
В качестве географической основы для ее составленная была использована
общая геоботаническая карта Западно-Сибирской равнины. На врезке
нашли отражение некоторые общие закономерности в распределении
продуктивных сочетаний полезных растений, а также связь ресурсных
показателей растительности с ее* типологическим составом.
29
ЛЕГЕНДА ОБЗОРНОЙ КАРТЫ «РАСТИТЕ ЛЕНОСТЬ*
ЗАП4ДНО-СИБИРСКОЙ РАВНИНЫ»
Тундровая растительность
Арктическая фратрия формаций
I. Арктические западно-сибирские формации
Лишайниково-моховые, травян о-м о х о в ы е и кустарнич-
ково-моховые тундры
1. Осоково-лишайниково-моховые" (Rhacqmitrium lanuginosum, Cetraria richardsonii,
Carex ensifolia ssp. arctisibirica) кочковатые тундры в сочетании с кустарничково-
мохово-лишайниковыми (Cladonia rangiferina^, Rhacomitrium lanuginosum)1
полигональными тундрами. >
2. Кустарничково-моховые (Dicranum elongatum, Sphenolobus minutus, Salix polarisy
S. nummularia) кочковатые тундры.
3. Лишайниково-моховые и кустарничково-моховые {Rhacomitrium lanuginosum,
Dicranum elbngatum) кочковатые тундры в сочетании с пушицево-моховыми
заболоченными тундрами и осоково-гипновыми полигональными болотами.
4. Осоково-пушицево — моховые (Drepanocladus exannulatus, Eriophorum angustifo-
lium^, Carex starts) заболоченные тундры в сочетании с осоково-гипновыми
полигональными болотами.
Мохов о-лишайниковые тундры
5. Мохово-лишайниковые (Cladonia rangiferina, С, gracilis, Thamnolia vermicularis)
полигональные тундры в сочетании с открытыми группировками из Minuartia
arctica, Armeria arctica, Thamnolia vermicularis) на дефляционных обнажениях.
6. Кустарничково-мохово-лишайниковые (Cladonia rangiferina, Dactilina arctica,
Rhacomitrium lanuginosum, Cassiope tetragona) полигональные тундры в сочетании с
осоково-гипновыми болотами.
Траряные и травян о-м оховые болота
7 * Осоково-гипновые, осоково-пушицевые, злаково-осоковые (Carex stans,
Eriophorum angustifolium, Calliergon sarmentosum) болота в сочетании с участками тундр
и злаково-осоково-моховых валиково-полигональных болот (Dupontia fische-
ri, Carex stans, Drepanocladus uncinatus)2.
-Приморские луг а —«т а м п ы» и ивняково-луговые
сообщества д о л и н р е к ~
8. Приморские засоленные злаково-осоковые (Carex stans, С, subspathacea, Dupontia
fischeri, Arctophila f.ulva, Eriophorum medium) заболоченные луга («тампы»).
9. Серия разнотравно-осоковых и разнотравно-злаковых (Trisetum sibiricum,* Carex
stans, Caltha arttica) лугов и мелкоивняковых (Salix lanata) сообществ в
сочетании с участками травяно-моховых тундр и осоково-гипновых болот.
Амфиатлантическая фратрия формаций v
И. Субарктические за падно-сибирск и е формации
1
Северные субарктические
Кустарничково-моховые и лишайниково-моховые с
ивой и ерником (Salix glauca, S. lanata, Betula nana) тундры
10. Кустарничково-лишайниково-моховые (Hylocomium splendens var^ alaskanum,
Cladonia elongataA, Arctous alpina) с ивой и ерником бугорковатые тундры,
местами в сочетании с кустарничково-мохово-лишайниковыми (Cladonia rangiferina,
Arctous alpina) тундрами.
11. Кустарничковые, преимущественно дриадовые (Dryas punctata), и моховые
(Dicranum spadiceum, Ptilidium ciliare, Hylocomium splendens var. alaskanum) с ивой
и ерником бугорковатые или пятнисто-бугорковатые тундры.
12. Кустарничково-моховые (Hylocomium sp'lendens var. alaskanum, Ptilidium ciliare,
Dry as punctata) С ивой и ерником бугорковатые тундры.
13. Кустарничково-моховые и лишайниково-моховые с ивой и ерником бугорковатые
тундры в сочетании с осоково-гипновыми и осоково-сфагновыми валиково-
полигональными болотами.
Мохово-травяные тундры ч
14.* Кустарничково-мохово-травяные (Dryas punctata, Carex stans, Eriophorum angu-
stifolium, Rubus chamaemorus, Drepanocladus exannulatus, Aulacomnium turgid
dum) с ивой и ерником кочкарные заболоченные тундры в сочетании с осоково-
гипновыми болотами. '■ . ■
1 Светлым треугольником (А) обозначены виды, названия которых в тексте и в
заключительном списке латинских названий растений даны по новой номенклатуре
[Черепанов, 1981; и др.].
2 Одной звездочкой (*) обозначены сообщества, встречающиеся также в более
южных подзонах.
30 '
Кустарнпчково-мохово-лишайниковы'е тундры
15.* Кустарничково-мохово-лишайниковые (Cladonia sylvatica, С. elongata, С. ran-
giferina, S phaerophorus globosus, Cetraria cucullata, Dicranum elongatum, Arctous
alpina) полигональные тундры.
Лугово-ивняково-моховая серия сообществ долин рек
16. Серия сообществ разнотравно-злаковых лугов (Роа alpina, Ranunculus borealis^,
Alopecurus borealis), ивняков кустарниковых (Salix lanata, S. phylicifolia) и
моховых тундр в сочетании с осоково-гипновыми валиково-полигональными
болотами.
Южные субарктические .
Ерниково-ивняковые (Salix glauca, S. lanata, Betula nana)
низкокустарниковые тундры
17. Ерниково-ивняковые осоково-лишайниково-моховые (Aulacomnium turgidum,
Hylocomium splendens var. alaskanum, Dicranum elongatum, Cladonia uncialis,
C. elongata, Carex ensifolia) бугорковатые и пятнисто-бугорковатые тундры.
18. Ерниково-ивняковые, местами с ольховником (AInaster fruticosus^), кустарничково-
моховые (Dicranum elongatum, Aulacomnium turgidum, Polytrichum alpestre,
Dry as punctata, Vaccinium vitis-idaea) бугорковатые и пятнисто-бугорковатые
тундры.
19. Ерниково-ивняковые кустарничково-лишайниковые, преимущественно алекто-
риевые (Alectoria ochroleuca, A. nigricans, Ledum decumbens) заболоченные тундры
в сочетании со сфагново-лишайниковыми олиготрофными болотами.
20. Ерниково-ивняковые, лишайниково-моховые и кустарннчково-моховые
бугорковатые и пятнисто-бугорковатые тундры в сочетании с осоково-мохово-лишайнико-
выми трещиновато-полигональными болотами. ^
21. Ерниково-ивняковые травяно-кустарничково-моховые (Drepanocladus exannulatus,
Aulacomnium palustre, Ledum decumbens, Rubus chamaemorus •, EriOphorum angu-
stifolium^) заболоченные тундры в сочетании с кустарничково-мохово-лишайни-
ковыми трещиновато-полигональными болотами.
Ерниковые '(Betula nana) туидры^
22. Ерниковые и ивняково-ерниковые кустарничково-лишайниково-моховые
(Hylocomium splendens var. alaskanum, Aulacomnium turgidum, Cladonia sylvatica,
Vaccinium vitis-idaea, Ledum decumbens) бугорковатые тундры.
23. Ерниковые и ивняково-ерниковые с ольховником кустарничково-моховые
(Dicranum elongatum^ Hylocomium splendens var. alaskanum, Vaccinium vitis-idaea,
Empetrum nigrum) бугорковатые тундры.
24. Ерниковые и ивняково-ерниковые, местами с ольховником, мохово-лишайнико-
вые, преимущественно алекториевые (Alectoria ochroleuca, A. nigricans, Cetraria
cucullata, Aulacomnium turgidum, Ledum decumbens) бугорковатые тундры в
сочетании с мохово-лишайниковыми трещиновато-полигональными. болотами.
25. Ерниковые и ивняково-ерниковые с ольховником кустарничково-лишайняково-
моховые (Polytrichum alpestre, Pleurozium schrkberi, Cladonia alpestris^, Ledum
decumeens) тундры в сочетании с лиственничными (Larix sibirica) лишайниково-
кустарничковыми (Empetrum nigrum) редколесьями и плоскобугристымп тра-
вяно-мохово-лишайниковыми болотами.
Ивняковые (Salix gf 1 а и с a, S. р и 1 с b г а) тундры
26. Ивняковые травяно-моховые (Polytrichum alpestre, Aulacomnium palustre, Erio-
phorum angustifolium, Nardosmia frigida) тундры в сочетании с ивняково-ернико-
выми лишайниково-моховыми тундрами и осоково-мохово-лишайниковыми
плоскобугристыми болотами.
Ерниково-ольховниковые (A Inaster fruticosus,
Betula nan а) тундры
27. Ерниково-ольховниковые кустарничково-лишайниково-моховые (DicraAum un-
cialis, Polytrichum alpestre, Empetrum nigrum, Cladonia alpestris, C. sylvatica)
бугорковатые тундры, местами в сочетании с редкокустарниковыми
лишайниковыми тундрами и лиственничными редколесьями.
Лишайниковые редкокустарниковые тундры
28. Редкокустарниковые (Betula -nana, Salix pulchra) кустарничково-зеленомошно-
лишайниковые (Cladonia sylvatica, С* alpestris, Cetraria cucullata, Peltigera aph-
ihosa, Dicranum elongatum, Ledum decumbens) полигональные.тундры в сочетании
с открытыми группировками из Arctous alpina, Dryas punctata, Cetraria cucullata
на дефляционных обнажениях.
Лугово-кустарниковые сообщества долин рек
29. Серия гипново-осоковых* болот, болотистых осоковых лугов, ивняковых, ернико-
вых и ольховниковых тундр (устъя Оби, Надыма, Пура, Таза):
а) ряд низинных мелко ивняков о-гипново-6соковых (Carex aquatilis, Callder-
gon stramineum, Salix lanata) болот, осоково-вейниковых (Calumagrostis
purpurea3, Carex acuta) лугов и зарослей кустарниковой ивы (Salix
phylicifolia) участков низкого экологического уровня4;
3 Здесь и далее вместо Calamagroslis purpurea следует читать С, purpurea ssp.
langsdorffii (Link) Tzvel.
4 Строчными буквами обозначены последовательные стадии одной' пойменной
серии.
, ^ 31
б) ряд осоковых кочковатых (Carex caespitosa) и болотпстых (Carex aquatilis,
С. acuta) лугов и кустарниковых ивняков (Salix phylicifolia,S. dasyclados)
участков низкого экологического уровня;
в) ряд осоковых и вейниковых (Carex acuta, Сalamagrostis purpurea) лугов,
ивняковых, ивняково-ерниковых и ерниково-ольховниковых тундр участков
высокого экологического уровня.
30. Серия хвощовых (Equisetum arvense) — на песках и разнотравно-злаковых (Роа
alpigena, Alopecurus borealis, Ranunculus borealis) — на суглинках лугов,
кустарниковых ивняков (Salix dasyclados. S. phylicijolia) и ивняково-ерниковых и
ольховниковых тундр.
Субарктические болотные формации
Кустарн и чково-осоково- моховые валпково-полиго-
нальные комплексные болота
31.** Осоково-гипновые и осоково-сфагновые (Carex stans, С. rotundata, Drepanocladus
uncinatus, Calliergon sarmenthosum, Sphagnum lenense) на .полигонах и кустарнич-
ково-мохово-лпшайниковые (Rubus chamaemorus, Vaccinium vitis-idaea, Aula-
comnium turgidum, Cladonia rangiferina, Cetraria cucullata) на валиках 5.
Кустарн и чково-мохово-лпшайни к овые трещиновато-
полигональные комплексные болота
32. Кустарничково-зеленомошно-лпшайниковые (Ledum decumbens, Dicranum angu-
stum, Cladonia rangiferina) на полигонах и валиках и осоково-сфагновые (Carex
rotundata, Sphagnum duseniiA, S. balticum) в трещинах и мочажинах.
Б о реальная растительность
Урало-Сибирская фратрия формаций
Обь-Иртышские формации
£11. Редколесья
Лиственнично-еловые и еловые (Р i с е ^ obovata, Larix
sibirica) редколесья
33. Лиственнично-еловые зеленомошно-кустарничковые с субарктическими
элементами ( Ledum decumbens, Arctous alpina, Aulacomnium turgidum) и лишайниковые
(Cladonia mitis, С rangiferina) редколесья в сочетании с ерниковыми и
ивняковыми тундрами.
34. Лиственнично-еловые и еловые, местами с березой, лишайниково-сфагновые
(Sphagnum acutifolium, Cladonia sylvatica) редколесья в сочетании с кустарнич-
ково-мохово-лишайниковыми плоскобугристьши болотами.-
Лиственничные- и елово-лнственничные (Larix
sibirica, Picea obovata) редколесья и производные
сообщества на ихместе
35. Елово-лиственничные ерниково-лишайниково-зелевюмошные (Pleurozium schre-
beri, Cladonia sylvatica) с субарктическими элементами (Aulacomnium turgidum,
Cladonia coccifera) редколесья в сочетании с кустарниковыми, преимущественно
ерниковыми, тундрами.
36. Лиственничные, местами елово-лиственничные, кустарничково-мохово-лпшайнп-
ковые (Cladonia alpestris, Pleurozium schreberi, Vaccinium vitis-idaea) с
субарктическими элементами (Alectoria nigricans, Dry as punctata) редколесья в сочетании
с кустарничковыми и лишайниковыми тундрами..
37. Лиственничные и елово-лиственничные кустарнпчково-зеленомошные и лишайни-
ково-зеленомошные (Ledum palustre, Empetrum nigrum, Pleurozium schreberi,
Cladonia rangiferina) редколесья в сочетании с лиственничными зеленомошнымп
и лишайниковыми лесами.
38. Лиственничные и елово-лиственничные лишайниковые (Cladonia alpestris, С
rangiferina, Cetraria nivalis) редколесья в сочетании с ерниково-сфагново-лишайнико-
выми плоскобугристыми болотами.
39. Лиственнично-березовые и елово-березово-лиственничные кустарничково-зелено-
мошно-лишайниковые редколесья и редкостойные леса.
Л у г о в о - к у старни к о в о -редк о лесные сообщества долин
рек к
40. Лугово-ивняково-редколеснолиственничная (Larix sibirica, Salix glauca, Carex
aquatilis, Calamagrostis purpurea) серия сообществ.
' IV. Северотаежные
Лиственнично-еловые и еловые (Picea obovata, Larix
sibirica) редкосто й н~ы елеса и производные
сообщества на их месте
5 Двумя звездочками (**) обозначены сообщества, встречающиеся также в более
северных подзонах.
32
41. Лиственнично-еловые и еловые зеленомошно-лишайниковые и зеленомошные
(Pleurozium schreberi, Cladonia alpestris) редкостойные леса в сочетании с ерни-
ково-сфагново-лшпайниковымн плоскобугристыми болотами.
42. Лиственнично-елово-березовые иг березовые с елью лишайниково-зеленомошные
редкостойные леса в* сочетании с ерников о-сфагново-лишайниковыми
плоскобугристыми болотами.
43. Лиственнично-еловые и еловые кустарничково-сфагновые (Sphagnum warnstorfii,
Ledum palustre) с ерником редкостойные леса в сочетании с кустарничково-мо-
хово-лпшайниковыми плоскобугристыми и крупнобугристымп болотами.
Лиственнично-елово-кедровые (Pinus .sibirica, Picea
о b о v a t a, Larix sibirica) леса и производные
сообщества на их месте
44. Лиственнично-елово-кедровые лишайниково-зеленомошно-кустарничковые (Ledum
palustre, Vaccinium uliginosum, Empetrum nigrum, Pleurozium schreberi, Cladonia
alpestris) леса.
45.° Березовые с кедром, елью, лиственницей кустарничково-лишайниково-зеленомош-
ные леса 6.
46.° Сосновые с кедром, елью, лиственницей кустарничково-лишайниково-зеленомош-
ные леса.
47. Елово-кедровые,, местами с лиственницей, зеленомошно-кустарнпчковые
(Vaccinium myrtillus, V. vitis-idaea, Ledum palustre, Pleurozium schreberi) с
ольховником (Alnaster fruticosus) в подлеске леса.
48.° Березовые с осиной, кедром и ольховником в подлеске зеленомошно-кустарнич-
ковые леса.
49.° Сосновые с кедром зеленомошно-кустарнпчковые леса.
Лиственничные, елово-лиственничные п сосново-лист-
венничные (Larix sibirica, Picea obovata, Pinus
sylyestris) редкостойные леса п производные
сообщества на их месте
50. Лиственничные pi елово-лиственничные, местами с кедром, лншайниково-мохово-
кустзрничковые (Ledum palustre, Hylocomium splendens, Cladonia syluatica)
редкостойные леса.
51. Лиственничные и елово-лиственничные с примесью кедра и сосны кустарничково-
зеленомоншо-лишайниковые (Cladonia alpestris, С. rangiferina, Pleurozium
schreberi, Ledum palustre) редкостойные леса, местами в сочетании с кустарничково-
мохово-лишайниковыми плоско- и крупнобугристымп болотами.
52.° Лиственнично-березовые и елово-березово-лиственничные лишайниковые и
зеленомошные редкостойные' леса.
53. Сосново-лиственничные с кедром, местами лиственнично-сосновые зеленомошно-
кустарничковые с лишайниками (Vaccinium vitis-idaeji, V. uliginosum,
Pleurozium schreberi) редкостойные леса.
54.° Лиственнично-березовые с сосной и кедром зеленомошно-кустарничковые леса.
55. Лиственничные и елово-лиственничные с прпмесью кедра и сосны кустарннчково-
лишанниково-долгомошные и кустарничково-сфагновые (Sphagnum magellani-
сит, S. angustifolium) редкостойные леса в сочетании с кустарничково-мохово-
лишайниковыми плоско- и крупнобугристыми болотами.
Лиственнично-сосновые и сосновые (Pinus sylvestris,
Larix sibir*ica) леса и редколесья и
производные сообщества на пх месте
56. Лиственнично-сосновые лишайниково-зеленомошно-кустарничковые (Ledum
palustre, Vaccinium uliginosum, Pleurozium schreberi, Cladonia syluatica) леса в
сочетании с сосновыми лишайниковыми лесами.
57. Лиственнично-сосновые кустарничково-лишайниковые (Vaccinium vitis-iddea,
Cladonia alpestris) редкостойные леса в сочетаний с кустарничково-мохово-лишай-
никовыми крупнобугристыми болотами.
58. Сосновые с .лиственницей лишайниковые (Cladonia alpestris, С. syluatica, С.
rangiferina) леса, местами в сочетании с кустарничково-лпшайниково-сфагновыми с
сосной и кедром выпуклыми олиготрофными болотахми и сосновыми
лишайниковыми редколесьями.
5$. Сосновые лишайниковые (Cladonia alpestris, С. rangiferina, Cetraria cucullata)
редколесья в сочетании с сосновыми, с кедром и лиственницей кустарничково-
зеленомошно-лишайниковыми лесами.
60.° Сосново-березовые и березово-сосновые зеленомошно-лишайниковые леса.
61. Сосновые с кедром и лиственницей лишайниково-кустарничково-сфагновые
(Sphagnum acutifolium, S. fuscum) леса в сочетании с сосново-кустарничково-
сфагновыми олиготрофными болотами.
Кустарничково-мохово-лишайниковые п травяно-сфаг-
новые плоскобугристые комплексные болота
62.** Брниково-сфагново-лишайниковые (Cetraria cucullata, Sphagnum apiculatum,
. Betula nana, Ledum palustre) на буграх, осоково-сфагновые и пушицево-сфагно-
вые (Sphagnum balticum, Carex rotundata, Eriophorum russeolum) в мочажинах —
«ерсеях».
6 Светлым кружком (о) обозначены производные леса, возникшие на месте
коренных в результате пожаров и рубок.
3 Заказ № 297 33
63.** Багульниково-ерниково-лишайниковые (Cladonia rangiferina, Cetraria cucullata,
Sphagnum tenense, Le&um palustre) на буграх и осоково-сфагновые (Sphagnum
balticum, Carex rotundata, C. rariflora) в мочажинах —«ерсеях»— с обилием озер
и озерков.
Кустарничково-зеленомошно-л и тайников ы е, осоково-гпп-
новые и осоково-пушицево-сфагновые
крупнобугристые комплексные болота
64. Зеленомошно-лишайниковые (Cladonia deformis, Dicranum elongatum, Ledum pa-
lustre, Rubus chamaemorus) на буграх и осоково-пушицево-сфагновые (Sphagnum
balticum, Eriophorum angustifolium, Carex rotundata, C, rariflora) в мочажинах —
«ерсеях».
65. Кустарничково-зеленомошно-лшпайниковые иногда облесенные (Cetraria cucullata,
Dicranum congestum, Ledum palustre) на буграх, осоково-гипновые и осоково-
сфагновые (Sphagnum balticum, 'S. lindbergii, Drepanocladus exannulatus, Carex
rariflora, C. aquatilis) в мочажинах в сочетании с озерами.
66. Кустарничково-мохово-лишайнпковые (Cladonia rangiferina, Dicranum elongatumr
Ledum palustre) на буграх, осоково-пушицево-сфагновые и осоково-гипновые
(Sphagnum balticum, S. lindbergii) в мочажинах крупнобугристые болота в
сочетании с плоскобугристымп и грядово-мочажинными болотами.
Кустарничково-сфагновые и тр а'в яно-моховые грядо-
в о - мо ч а ж и н н ы е аапа-болота
67. Ерниково-морошково-сфагновые (Sphagnum magellanicum, Rubus chamaemorus,
Ledum palustre) на грядах/ осоково-сфагновые и осоково-гипновые (Sphagnum
mafus, Drepanocladus exannulatus, Carex rotundata, C. rariflora) в мочажинах;
западно-сибирский тип.
Лишайниково-сфагновые плосковыпуклые олиготроф-
ные болотно-озерные комплексы
68. Кустарничково-лишайниково-сфагновые (Sphagnum fuscum, Cladonia rangiferina,
Chamaedaphne calyculata) с сосной (Pinus sylvestris f. litwinowii) и кедром
(Pinus sibirica) с обилием озер и озерков, с периферийным рядом олиготрофных
комплексов и ассоциаций; сургутско-полесский тип.
Лугово-болотно-соровая растительность поймы
нижнего течения р. Оби (Салехард — Березово)
69. Серия осоково-мелкоивняковых низинных болот, сообществ соровой
растительности, болотистых лугов, ивняков кустарниковых и парковых, березовых, бере-
зово-еловых и лиственничных лесов:
а) ряд осоково-мелкоивняковых (Carex aquatilis, Salix lanata) болот и
осоковых кочковатых (Carex caespitosa, С. aquatilis) лугов, сочетающихся с ситня-
гово-полевицевыми соровыми лугами участков низкого экологического
уровня;
б) ряд группировок соровой растительности из Eleocharis acicularis, Arcio-
phila fulva, Beckmannia eruciformis и соровых лугов — арктофиловых, бек-
манниевых и полевицевых (Agrostis stolonifera) участков низкого
экологического уровня;
л в) ряд осоковых (Carex aquatilis), вейниковых (Сalamagrostispurpurea) лугов
и кустарниковых ивняков (Salix viminalis) участков среднего
экологического уровня, сочетающихся с соровыми лугами по понижениям;
г) ряд разнотравно-злаковых (Сalamagrostis purpurea, Poa palustris, Veronica
longifolia) лугов и сообществ ивняков кустарниковых и парковых (Salix
alba, S. viminalis) участков среднего экологического уровня,
сочетающихся с осоковыми лугами межгривных понижений;
д) ряд ивняков кустарниковых и парковых, ивовых и ивово-березовых
(Betula pendula, Salix alba) лесов участков высокого экологического
уровня роимы, сочетающихся с березовыми, березово-еловыми и
лиственничными разнотравно-зеленомошными лесами останцов надпойменных террас.
Кустарниково-лесная растительность долин
притоке в О б и
70. Серия ивняково-березово-еловых (Salix viminalis, Betula pendula, Picea pbovata)
с кедром и лиственницей сообществ. *
71. Серия ивняково-ольховниково-березово-лиственничных (Salix viminalis, Alnaster
fruticosus, Betula pendula, Larix sibirica) сообществ:
а) ивняково-березовый (Salix viminalis, S. alba, Alnaster fruticosus, Betula
pendula) ряд.
V. Среднетаежные
'Елово-кедровые (Pinus sibirica, Picea obovatajfleca и
производные сообщества на их месте
72. Елово-кедровые с лиственницей кустарничково-зеленомошные (Pleurozium schre-
beri, Hylocomium splendens, Vaccinium myrtillus) леса, местами в сочетании с кус-
тарничково-сфагновыми олиготрофнымп болотами.
73.° Темнохвойно-березовые с лиственницей кустарничково-зеленомошные леса.
74.° Темнохвойно-сосновые с лиственницей кустарничково-зеленомошные леса.
75. Елово-кедровые с пихтой, местами кедрово-еловые, мелкотравно-?бруснично-зе-
леномошные (Pleurozium schreberi, Hylocomium splendens, Vaccinium vitis-idaea,
Maianthemum bi folium) леса.
34
76.° Елово-березовые с пихтой и кедром мелкотравно-зеленомошные леса.
77.° Березово-осиновые и осиновые мелкотравно-зеленомошные леса.
78. Березово-сосновые и сосновые кустарничково-зелейомошные (Pleurozium schreberi,'
Hylocomium splendens, Vaccinium vitis-idaea, V. myrtillus) устойчиво производные
леса преимущественно в сочетании с сосновыми багульниково-сфагновыми
лесами и кустарничково-сфагновыми олиготрофными болотами.
79. Елово-кедровые долгомошные и кустарничково-сфагновые (Sphagnum magellanU
cum, Polytrichum commune, Chamaedaphne calyculata, Ledum palustre) леса в co-
. четании с кустарничково-сфагновыми олиготрофными болотами.
Сосновые (Pin us sylvestris) леса и. производные
сообщества на их месте
80.* Сосновые багульниково-бруснично-зеленомошные (Pleurozium schreberi) и
лишайниковые (Cladonia sylvatica, С. alpestris) леса. .
81.* Сосновые бруснично-зеленомошные и бруснично-лишайниковые (Cladonia
alpestris, С. sylvatica, С. rangiferina, Pleurozium schreberi) леса в сочетании с сосново-
кустарничково-сфагновыми олиготрофными болотами.
82.* Сосновые и березово-сосновые долгомошно-сфагновые и кустарничково-сфагновые
(Sphagnum fuscum, S. magellanicum, Polytrichum commune) леса в сочетании с
кустарничково-сфагновыми олиготрофными болотами. v
Сфагновые (Sphagnum fuscum) сосново-кустарнпчко-
вые рлиготрофные.выпуклые . б.олота
83.* Багульниково-кассандрово-сфагновые (Sphagnum fuscum, Chamaedaphne calycula-
ta) с сосной и кедром (Pinus sylvestris f. litwinowii, P. sibirica f. turfosa) на
грядах, с озерками и сфагновыми (Sphagnum balticum, S. dusenii) мочажинами, с
периферийным рядом сосново-сфагновых мезо-евтрофных лесных ассоциаций;
нарымский тип.
84.* Сосново-кустарничково-сфагновые (Pinus sylvestris, f. uliginosa и f. litwinowii,
Sphagnum fuscum, Chamaedaphne calyculata) олиготрофные болота.
Лесо-кустарниково-сорово-луговая растительность
поймы нижнего течения'р. Оби (Березово — Ханты-
Мансийск)
85. Серия сорово-луговых и ивяяково-болотисто-луговых сообществ, ивовых,
березовых, березово-кедровых и березово-сосновых лесов:
а) ряд соровых ситнягово-полевицевых, разнотравно-полевицевых (Agros-
tis stoloni.fera, Eleocharis acicularis), осоковых (Carex aquatilis) лугов,
местами сочетающихся с кочковатыми (Calamagrostis neglecta, Carex
caespitosa) лугами участков низкого экологического уровня;
б) ряд осоковых (Carex aquatilis, С. acuta) осоково-канареечниковых
(Typkoides arundinacea^, Carex acuta) и вейниково-канареечниковых
(Calamagrostis purpurea) лугов участков средне-низкого экологического
уровня, сочетающихся с осоковыми кочковатыми лугами понижений;
в) #ряд разнотравно-злаковых (Typhoides arundinacea, Calamagrostis
purpurea, Poa pratensis, Achillea ptarmica) лугов и кустарниковых ивняков
(Salix alba, S. viminalis, S. pentandra) участков средне-высокого
экологического уровня, сочетающихся с осоковыми лугами понижений и
ивняками парковыми высоких уровней7;
г) вряд ивняков паркового типа, ивово-березовых, осиново-березовых, кед-
рово-березовых (Betula pendula, Populus tremula, Salix alba) лесов
участков высокого экологического уровня поймы и березово-сосновых и
березово-кедровых мелк*отравно-зеленомошных с элементами пойменного
разнотравья (Veronica longifolia, Thalictrum simplex) лесов останцов
надпойменных террас.
Кустарниково-лесная растительность долин и
притоков Оби
86. Серия рльховниково-ивняково-березовых (Alnaster fruticosus, Salix viminalis,
Betula pendula) сообществ.
87. Серия ивово-березовых, березово-сосновых (Betula pendula, Pinus sylvestris, Salix
alba) травяно-зеленомошных и лишайниковых лесов долинных грив и сосново-
березовых осоково-сфагновых (Sphagnum girgensohnii) лесов межгривных
понижений,
VI. Южно-таеж н(ы е
Е л о в о-п ихтовые, пихтов о-е ловые и пихтово-ело-
во-кедровые (Abies sibirica, Piceaobovata, Pinus
sibirica) леса и производные сообщества на их месте
88. Кедрово-елово-пихтовые зеленомошно-мелкотравные и мелкотравно-осочковые(0;т-
lis acetosella, Maianthemum bifolium, Carex macroura, Rhytidiadelphus triquetrus)
с элементами широкотравья (Aconitum excelsum^, Cacalia hastata) леса.
89.° Темнохвойно-березовые мелкотравно-вейниково-осочковые (Carex macroura) леса.
90.° Темнохвойно-осиновые, березово-осиновые с подлеском (Caragana arborescens)
травяные леса. *
7 Залитым кружком обозначены пастбищные и сенокосные варианты сообществ.
3* ,35
91.° Березовые и осиновые молодняки с темнохвойным подростом, кипрейно-вейнп-
ковые и кустарниковые сообщества ,на месте поврежденных шелкопрядом
лесов.
92.° Сельскохозяйственные земли в сочетании с участками лесов и кустарников на
месте елово-пихтовых лесов и производных от них сообществ.
93. Елово-пихтовые с липой (Tilia cordata) зеленомошно-широкотравно-кисличные
и осочковые с неморальнымп элементами {Stellaria holostea, Actaea erythrocarpa)
леса, местами в сочетании с влажными широкотравными (Aegopodium podagra-
ria, Filipendula ulmaria) лесами.
94.° Темнохвойно-березовые и березовые с липой травяные и осочковые леса в
сочетании с влажными широкотравными лесами.
95.° Березово-осиновые с липой неморально-травяные леса.
96. Липово-елово-пихтовые широкотравные с участием неморальных видов (Aego-
podiumpodagraria, Athyrium filix-femina, Pulmonaria mollissima, Actaea ery~
throcarpa) леса.
97.° Липово-пихтово-березовые и осиновые широкотравные леса.
98. Пихтово-еловые, местами с липой, зеленомошно-травяные (Calamagrostis obtusata,
Oxalis acetosella, Milium effusum, Ptilium crista-castrensis) леса в сочетании
с влажными широкотравными (Aegop&dium podagraria, Filipendula ulmaria)
лесами.
99.° Темнохвойно-осиново-березовые и березовые с липой травяные с пятнами
зеленых мхов (Rhytidiadelphus triquetrus, Climacium dendroides ) леса.
100.° Сельскохозяйственные земли с участками ле£ов и кустарников на месте елово-
пихтовых и пихтово-еловых с липой лесов и производных от нпх сообществ.
101. Кедровые с пихтой и елью хвощово-зеленомошные (Equisetum sylvaticum, Hylo-
comium splendens) и травяно-сфагновые (Sphagnum squamosum, S. wulfianum,
Car ex globularis, Equisetum sylvaticum) леса.
102.° Березовые с пихтой, елью и кедром хвощово-вейниковые {Calamagrostis
purpurea, Equisetum sylvaticum) леса.
Сосновые (Pinus sylvestris), местам'и с липой (Tilia
cordata), леса и производные сообщества на их месте
103.* Сосновые, местами с липой, травяно-кустарничковые (Vaccinium vitis-idaea,
Carex macroura, Rubus saxatilis) с разреженным моховым покровом леса.
104.° Сосново-березовые, местами с липой, травяные леса.
105.° Сельскохозяйственные земли в сочетании с участками лесов на месте сосновых
лесов и производных от них сообществ.
Мелколиственные (Betula pubescens, Populus tr esm u 1 a)
леса
106. Березовые с елью, сосной, кедром веиниково-хвощовые (Calamagrostis canescens),
осоково-сфагновью (Carex globularis, Sphagnum angustifolium, S. wurnstorfii)
леса в сочетании с осоково-сфагновыми залесенными болотами.
С о сно в о - сфа г ню вые и сфагновые со Sphagnum fuscum
олиготрофные выпуклые болота со вторичными
озерками и мезоевтрофнымп грядово-мочажинны-
м и т о п я м и
107. Сосново-сфагновые (Sphagnum fuscum, Pinus sylvestris f. uliginosa и f. litwinowii),
кустарничково-сфагновые (Sphagnum fuscum, Chamaedaphne calyculata) с
сосной" на \грядах и осоково-сфагновые (Sphagnum balticum, Carex limosa) в
мочажинах в сочетании со вторичными озерками и осоково-гппновыми топями; юж-
но-васюганский тип.
Лесо-кустарниково-луговая растительность пойм
среднего течения Оби (Ханты-Мансийск — Пара-
бель) и нижнего течения Иртыша (ниже по с. Цинга-
л ы)
108. Серия болотистых и настоящих лугов, ивняково-мелколиственных, смешанных
и темнохвойных или сосновых лесов:
а) ряд осоковых (Carex acuta, С. aquatilis) и осоково-канареечниковых
(Typhoides arundinacea) лугов, местами переходящих в открытые
группировки соровой растительности (Equisetum limosum, Eleocharis acicu-
laris) участков низкого экологического уровня;
б) Фряд кочковатых осоковых, осоково-вейниковых и
осоково-канареечниковых (Carex acuta, С. caespitosa, Calamagrostis neglecta, Typhoides
arundinacea) лугов участков низкого экологического уровня;
в) Фряд канареечнпковых (Typhoides arundinacea), вейниковых
(Calamagrostis purpurea, С. epigeios), разнотравно-злаковых и злаково-разно-
травных (Calamagrostis epigeios, Poa pratensis, Filipendula ulmaria, Lyth-
rum salicaria) лугов участков средне-низкого экологического уровня,
сочетающихся с осоковыми лугами по понижениям;
г)#ряд разнотравно-злаковых {Calamagrostis purpurea, Alopecurus ventri-
cosus&, Lathyrus pratensis) лугов, ивняково-кустарниковых сообществ
(Salix cinerea, S. pentandra), ивняков паркового типа (Salix alba, S. vi-
minalis) участков средне-высокого экологического уровня,
сочетающихся с осоковыми и канареечниковыми лугами межгрпвных понижений;
д) #ряд ивовых (Salix alba), ивово-тополевых (Populus nigra), березовых
36
(Betula pendula), березово-осиновых (Populus tremula) разнотравных
лесов с участками разнотравно-мелкозлаковых (Роа pratensis, Agrostis
alba^, Filipendula ulmaria, Trifolium medium) лугов участков высокого
экологического уровня, сочетающихся с канареечниковыми и
осоковыми лугами понижений;
е) ряд хвойно-мелколиственных {Betula pendula, Populus tremula, Picea
obovata) травяных и кедрово-пихтово-еловых, елово-пихтовых зелено-
мошно-мелкотравных (Oxalis acetosella, Pyrola rotundifolia, Rhytidiadel-
phus triquetrus) с элементами пойменного разнотравья (Geranium sylva-
, ticum, Thalictrum simplex) лесов участков высокого экологического
уровня поймы' и останцов надпойменных террас;
ж) ряд сосново-березовых зеленомошно-травяных (Rubus saxatilis, Pleuro-
zium schreberi) и сосновых бруснично-мелкотравных (Maianthemum Ы-
folium, Vaccinium vitis-idaea) с элементами пойменного разнотравья
(Inula britannica, Lythrum salicaria) лесов участков высокого экологического
уровня поймы и останцов надпойменных террас.
Лугово-кустарничков'о-лесная растительность долин
притоков Оби и Иртышу
109. Серия лугово-ивово-тополево-темнохвойных сообществ,' преимущественно
еловых (Picea obovata) и пихтовых (Abies sibirica) травяных (Glechoma hederacea,
Geranium sylvaticum, Aconitum excelsum) лесов:
а) лугово-ивово-тополево-березовый (Calamagrostis epigeios, Salix alba,
Populus nigra, Betula pendula) ряд.
110. Серия лугово-ивняково-темнохвойных сообществ, преимущественно еловых
(Picea obovata) и пихтовых (Abies sibirica) влажнотравных (Equisetum sylvaticum,
Filipendula ulmaria, Carex globularis, Calamagrostis purpurea) лесов:
а) лугово-ивняково-осиново-березовый (Calamagrostis canescens, „ Typhoides
arundinacea, Salix pentandra, Populus tremula, Betula pendula) ряд;
б) лугово-ивняково-осиновый \Calamagrostis canescens, Salix pentandra,
Populus tremula) ряд.
VII; Подтаежные
Березовые (Betula pendula) и осиновые (Populus
tremula) леса и производные сообщества на их месте
111. Березовые, осиново-березовые, местами с липой, злаково-разнотравные
(Calamagrostis arundinacea, Aegopodium podagraria, Brachypodium pinnatum, Pul-
monaria mollissima^) с элементами таежного мелкотравья (Maianthemum bifo-
lium, Pyrola rotundifolia) леса в сочетании с осоково-злаковыми (Calamagrostis
canescens, Carex omskiana) лугами.
112.* Березовые и осиновые вейниКовые и вейниково-высокотравные (Calamagrostis
purpurea, Angelica*sylvestris, Filipendula ulmaria) леса в сочетании с лесными
суходольными лугами: ежовыми (Dactylis glomerata), мятликовыми (Роа
pratensis), овсяницевыми (Festuca pratensis).
ИЗ.0 Сельскохозяйственные земли с участками лесов и лугов на месте оспново-бе-
резовых травяных лесов.
114.* Березовые и осиново-березовые осоковые, осоково-вейниковые (Carex caespi-
tosa, С. riparia, Calamagrostis purpurea, C. canescens) леса в сочетании с
осоковыми и тростниково-осоковыми болотами.
Сосновые леса и производные сообщества на их месте
115.** Сосновые вейниковые и травяно-кустарничковые (Calamagrostis arundinacea,
С. epigeios, Rubus saxatilis, Vaccinium vitis-idaea) леса в сочетании с сосновыми
лишайниковыми лесами.
116.° Сосново-березовые разнотравно-вейниковые с орляком (Pteridium aquilinum)
леса и сельскохозяйственные земли на их месте.
Лесные-мезо- и евтрофные болота
117. Березовые, сосново-березовые осоково-сфагновые и вейниково-осоковые
(Betula pubescens, Pinus sylvestris, Carex lasiocarpa, Calamagrostis purpurea, C*
canescens, Sphagnum warnstorfii, S. angustifolium) болота.
Осоково-гипновые и травяные евтрофные'болота
118. Осоково-гипновые, ерниково-осоково-гипновые (Carex lasiocarpa, Drepanocladus
vernicosus, Betula nana) с березой и сосной (Betula pubescens, B, humilis, Pinus
sylvestris) на грядах — веретьях, осоково-гипновые (Carex diandra, C. limosa,
Drepanocladus vernicosus) в мочажинах с периферийным рядом кустарниковых
и лесных ассоциаций; южно-западносибирский тип.
119. Осоковые, осоково-гипновые, осоково-сфагновые (Carex caespitosa, С. diandra,
С. lasiocarpa, Calliergon giganteum, Sphagnum obtusum, S. warnstorfii, S. subse-
cundum) болота.'
120. Тростниково-осоковые (Phragmites communis^, Carex omskiana, C. caespitosa,
C. wiluica) в сочетании с сосново-кустарничково-сфагновыми (Pinus sylvestris,
i. litwinowii и f. uliginosa, Ledum palustre, Sphagnum fuscum) «рямами» и
периферийным рядом осоковых и осоково-вейниковых ассоциаций; барабинский
займищно-рямовый тип. '
37
Лесо-кустарниково-луговая растительность пойм, Оби
(Парабель — Кожевников о) и Иртыша (по с. Ц^нга-
л ы — Т а р а)
121. Серия разнотравно-злаковых лугов, осиновых, березовых и тополевых лесов:
а) #ряд канареечниковых и мелкозлаковых (Alopecurus pratensis, Agrostis
clavata) лугов и зарослей кустарниковых ивняков (Salix cinerea) в
сочетании с осоковыми (Carex acuta) лугами и осоковыми кочкарными (Carex
caespitosa) болотами участков низкого экологического уровня;
б) #ряд разнотравно-злаковых (Alopecurus pratensis, Agrostis clavata, Lathy-
rus pratensis) лугов, ивняков (Salix viminalis) и фарковых тополевых,
-" лесов (Populus nigra) участков среднего экологического уоовня;
в) Эряд березовых (Betula pendula), осиновых (Populus tremula), тополевых
(Populus nigra, P. alba) лесов, сочетающихся с разнотравно-злаковымп
лугами участков высокого экологического уровня.
122. Серия крупно- и мелкозлаковых лугов, низинных пойменных болот, ивово-оль-
ховых, ивово-мелколиственных, местами березово-сосновых лесов:
а) 0ряд осоково-канареечниковых и осоково-вейнркрвых (Typhoides arundi-
nacea, Carex acuta, Calamagrostis canescens) лугов участков низкого
экологического уровня;
б) #ряд разнотравно-осоковых (Carex acuta, Lythrum salicaria, Galium uligi-
nosum) лугов, сочетающихся с евтрофными осоково-тростниковыми
(Phragmites communis', Carex caespitosa) болотами участков низкого
экологического уровня;
в) Оряд разнотравно-злаковых (Calamagrostis epigeios, Agropyron repens^,
Lathyrus pratensis) лугов и сельскохозяйственных земель на их месте
участков среднего экологического уровня^ сочетающихся с разнотравно-
осоковыми лугами понижений; *
г) #ряд разнотравно-злаковых оЪтепненных (Agropyron repens, Poa pratensis,
Festuca ovina, Medicago falcata) лугов и сельскохозяйственных земель на
их месте, ивняков кустарниковых и парковых (Salix viminalis) участков
среднего экологического уровня;/
д) Фряд мелколиственных (Betula pendula, Populus tremula), ольховых'(AInus
incana) и березово-сосновых травяных (Rubussaxatilis, Filipendula ulmaria,
Maianthemum bifolium) лесов участков высокого экологического уровня.
Л у г о в о - ку старни к о в о - лесна я * растительность долин
притоков Оби и Иртыша
123. Серия лугово-ивово-тополево-березовых или осиновых (Alopecurus pratensis, Fes-
tuca pratensis, Salix alba, Populus nigra, P. tremula, Betula pendula), местами
с примесью хвойных пород (Picea obovata, Pinus sylvestris), сообществ.
VIII. Лесостепные
Березовые (Betula pendula) и осиновые (Populus
tremula) леса и сельскохозяйственные земли на их месте
124. Осиново-березовые и березово-осиновые остепненные злаково-разнотравные
(Calamagrostis epigeios, Brachypodium pinnatum,' Poa angustifolia, Medicago falcata,
Artemisia pontica) леса, местами в сочетании с ^ раздотравно-злаковыми
(Agropyron repens, Bromus inermis , Lathyrus pratensis) лугами.
125.° Сельскохозяйственные земли с участками лесов и лугов на месте березовых и
осиновых остепненных лесов.
Сосновые (Pinus sylvestris) леса и производные сооб-
. щества на их месте
126. Сосновые и березово-сосновые остепненные кустарничково-травяные и травяные
(Koeleria gracilis , Vaccinium vitis-idaea, Pulsatilla patens ■> Calamagrostis epigeios,
Carex supina, Festuca sulcata) леса.
Травяные болота и галофитные луга
127. Тростниковые и вейниково-осоковые (Phragmites communis, Carex caespitosa, С. ра-
radoxa, Calamagrostis neglecta) болота в сочетании с лисохвостными и полевице-
выми (Alopecurus ventricosus, Agrostis alba) засоленными лугами.
128. Галофитнозлаковые луга: ячменевые, лисохвостные, пырейные, бескильницевые
(Hordeum brevisubulatum, Alopecurus vehtricosus, Agropyron repens; Puccinellia
distans) в сочетании с галофитноразнотравными (Saussurea amara, Aster tripoli-
um^) лугами.
129. Галофитноразнотравные луга (Leymus paboanus, Puccinellia tenuissima) в
сочетании с открытыми группировками из галофитного разнотравья п
полукустарничков (Suaeda corniculata, Atriplex verrucifera , Salicornia herbacea, Camphorosma
monspeliacum).
Болотно-л у^г ово-лесная растительность долин рек
130. Серия разнотравно-злаковых лугов, низинных осоково-тростниковых болот и
мелколиственных лесов:
а) ряд ивняков кустарниковых (Salix viminalis), разнотравно-осоково-зла-
* ковых (Agrostis alba, Carex vesicaria, Saussurea amara) заболоченных
лугов и осоково-тростниковых (Phragmites communis, Carex caespitosa) болот;
38
б) ряд разнотравно-злаковых (Agropyron repens, Bromus inermis, Trifolium
pratense) лугов, кустарниковых ивняков, местами ивово-тополевых и
ивово-березовых травяных лесов;
в) ряд березовых и осиновых злаково-разнотравных (Agrimonia pilosa, Fili-
pendula 'ulmaria, Pulmonaria mollissima, Rubus saxatilis, Brachypodium
pinnatum) лесов.
Степная растительность
Заволжско-казахстанская фратрия формаций
Обь-Иртышские формации
IX. Луговостепные
Богато*разнотравно-злаковые луговые степи и о с т е п-
ненные луга, их галофитные варианты и
сельскохозяйственные земли на их месте
ф Разнотравно-злаковые остепненные луга и луговые степи (Calamagrostis epigeios,
Роа angustifolia, Filipendula hexapetala, Helictotrichon schellianum, Lathyrus pi-
siformis, Medicago falcata) в сочетании с участками осиново-березовых остепнен-
ных лесов 8.
131. Сельскохозяйственные земли (пашни, залежи, сбитые пастбища) на месте
разнотравно-злаковых остепненных лугов и луговых степей.
132. Разнотравно-злаковые (Роа angustifolia, Agropyron repens, Artemisia glauca, Fes-
tuca pseudovina) с галомезофильным разнотравьем (Galatella biflora, Artemisia pon-
tied) остепненные луга с полынно-бескильницевыми (Puccinellia tenuissima,
Artemisia nitrosa) группировками.
133. Волоснецовые (Leymus paboanus), разнотравно-типчаковые (Festuca pseudovina)
и бескильипцевые (Puccinellia-tenuissima) галофитные луга с бескильницево-по-
лынными (Artemisia nitrosa) и солянковыми (Kochia prostrata) группировками,
ф Злаково-разнотравные (Stipa pennata, Phleum phleoides, Libanotis intermedia■ ,
Artemisia latifolia) луговые степи в сочетании с участками осиново-березовых
остепненных лесов
134. Сельскохозяйственные земли (пашни, залежи, сбитые пастбища) на месте
злаково-разнотравных луговых степей и остепненных лугов.
135. Галофитно-разнотравные (Galatella biflora, Glycyrrhiza uralensis, Artemisia glauca)
луговые степп с полынно-типчаковыми и бескильницево-полынными (Festuca
pseudovina, Puccinellia distans, Artemisia nitrosa, A. pontica) группировками.
Лугово-степная растительность долин рек
136. Серия разнотравно-злаковых и злаковых (Festuca pratensis, Роа angustifolia, Saus-
surea amara, Galatella biflora) лугов и злаково-разнотравных (Libanotis
intermedia, Artemisia latifolia, Phleum phleoides) луговых степей.
137. Серия разнотравно-злаковых с галофитными элементами (Calamagrostis epigeios,
Bromus inermis, Leymus paboanus, Hordeum brevisubulatum) лугов и злаково-гало-
фитно-разнотравных (Phleum phleoides, Artemisia glauca, Galatella biflora)
луговых степей.
, Северо-казахстанские формации
X. Степные (настоящие степи)
Разнотравно-дерновиннозлаковые степи, их
галофитные варпанты и сельскохозяйственные земли на их
месте
О Богаторазнотравно-красноковыльные (Stipa pennata, S. rubens^, Helictotrichon
desertorum, Peucedanum morissonii) степи.
138. Сельскохозяйственные земли (пашни, залежи, сбитые пастбища) на месте богато-
разнотравно-красноковыльных степей.
О Псаммофитные разнотравно-песчаноковыльные и гемипсаммофитные разнотрав-
но-типчаково-тырсовые (Stipa pennata, S. rubens^, S, capillaia, Helictotrichon
desertorum, Artemisia marschalliana) степи.
139. Сельскохозяйственные земли на месте разнотравно-песчаноковыльных и разно-
травно-типчаково-тырсовых степей.
О Кальцефптные разнотравно-красноковыльные и разнотравно-красноковыльно-
овсецовые (Stipa rubens, S, korskinskyi, Artemisia dracunculus, Helictotrichon
desertorum) степи.
140. Сельскохозяйственные земли на месте разнотравно-красноковыльных и разно-
травно-красноковыльно-овсецовых степей.
О Разнотравно-тппчаково-красноковыльные (Stipa rubens, S. pennata, Festuca
pseudovina, Peucedanum morissonii) степи.
8 Ромбиком обозначены потенциальные сообщества, не картируемые в данном
масштабе; на их месте показаны сельскохозяйственные земли.
39
141. Сельскохозяйственные земли на месте разнотравно-типчаково-красноковыльных
степей.
О Разнотравно-типчаково-тьфсовые (Stipa capillata, S. pennata, Festuca pseudovina*
Helichrysum arenarium) степи.
142. Сельскохозяйственные земли на месте разнотравно-типчаково-тырсовых степей.
143. Галофитноразнотравно-типчаковые и ковыльногтипчаковые (Festuca pseudovina *
Stipa capillata, Glycyrrhiza uralensis, Artemisia pontica) степи с галофитностепны-
ми сообществами (Festuca valesiaca, Artemisia nitrosa, Limonium gmelimi, Kochia
prostrata).
О Разнотравно-типчаково-тырсовые и разнотравно-типчаково-ковыльные (Stipa
capillata, S. lessingiana, Festuca sulcata, Astragalus macropus) степи.
144. Сельскохозяйственные земли на месте разнотравно-типчаково-тырсовых и разно-
травно-типчаково-ковыльных степей.
Внемасштабные знаки
1. Открытые группировки из Arctous alpina, Minuartia arctica, Cetraria cucullatar
Dactilina arctica) на дефляционных обнажениях.
2. Лишайниковые (Cladonia rangiferina, Cetraria cucullata, Sphaeropfaorus globusus,
Salix nummularia) тундры.
3. Разнотравно-злаковые (Alopecurus alpinus, Agrostis borealis, Equisetum arvense ssp.
boreale, Ranunculus borealis) тундровые луговины.
4. Осоково-моховые (Carex aquatilis, Eriophorum angustifolium, Drepanocladus unci-
natus, Calliergon cordifolium) заболоченные сообщества на месте спущенных
озер — «хасыреев».
5. Участие Larix sibirica.
6. Участие Picea obovata.
7. Участие Pinus sibirica.
8. Участие Pinus sylvestris.
9. Участие Tilia cor data,
10. Участие Alnus incana,
11. Участие Calluna vulgaris.
12. Березовые (Betula pendula) и осиновые (Populus tremula) колки.
13. Заросли кустарников с единичным возобновлением березы или осины на свежих
гарях. '
14. Восстанавливающаяся растительность на вырубках (сосновые или березовые мо-
лодняки, заросли кустарников).
15. Кустарничково-пушицево (или пухоносово)-сфагновые (Sphagnum fuscum,
Eriophorum vaginatum, Trichophorum caespitosum) с редкой сосной выпуклые олиго-
трофные болота.
16. СоСново-кустарничково-сфагновые (Pinus sylvestris f. uliginosa и f. litwinowii,
Ledum palustre, Sphagnum fuscum) олиготрофные болота — «рямы» в подтайге
и лесостепи.
17. Тростниковые и вейниково-осоковые (Phragmites communis, Carex caespitosa, Ca-
lamagrostis neglecta) болота. •
18. Заросли степных кустарников (Spiraea hypericifolia, S. crenifolia, Caragana fru-
tex).
19. Участие Cerasus fruticosa в степных сообществах.
20. Остепненные разнотравно-злаковые (Роа angustifolia, Agropyron repens, A. glauca^
Festuca pseudovina) луга в комплексе с полынно-бескильницевыми (Puccinellia
tenuissima, Artemisia nitrosa) лугами.
21. Бескильницево-нитрозовополынные (Puccinellia tenuissima, Artemisia nitrosa)
степные группировки на корковых солонцах.
22» Галофитнозлаковые (Leymus paboanus, Festuca pseudovina, Puccinellia tenuissima)
луга.
23. Галофитноразнотравные (Galatella biflora, Glycyrrhiza uralensis) луговые степи
в комплексе с типчаковыми и полынными (Festuca pseudovina, Artemisia nitrosa)
группировками.
24. Галофитноразнотравно-типчаковые (Festuca pseudovina, Glycyrrhiza uralensis)
степи в комплексе е галофитнополукустарничковыми (Artemisia nitrosa, Kochia
prostrata, Limonium gmelinii) группировками.
25. Типчаковые {Festuca sulcata) и солянковые (Kochia prostrata, Camphorosma mons-
peliacum) группировки.
26. Заросли чия (Achnatherum splendens).
ГЛАВНЕЙШИЕ ПОДРАЗДЕЛЕНИЯ
РАСТИТЕЛЬНОГО ПОКРОВА
О
ГЛАВА III
ТУНДРОВАЯ РАСТИТЕЛЬНОСТЬ
Пространства Западно-Сибирской равнины, лежащие севернее
Полярного круга, относятся к тундрам. Здесь проходит граница двух
природных областей: северная оконечность материка с прилежащими
островами входит в Арктическую область, южнее этой границы расположена Обь-
Иртышская природная область [Сочава, Тимофеев, 19681, включающая
субарктические тундры.
Тундровая растительность характеризуется распространением
полидоминантных сообществ, образованных криофильными видами различных
экобиоморфологических групп (мхов, лишайников, травянистых,
кустарничков, кустарников), произрастающих в суровых климатических
условиях с коротким вегетационным периодом и низкой температурой почв
[Грибова, 1977]. Для тундровых сообществ Западной Сибири свойственна
сочетание арктических, арктоальпийских и гипоарктических видов, а
также участие в южной полосе тундровой зоны бореайьных элементов флоры.
Для кустарников и кустарничков характерны растения с гемипрострат-
ной и простратной [Полозова, 1966] формами роста. В большинстве своем
это олиготрофы, развивающиеся на бедных тундровых почвах.
Наряду с общими особенностями, определяющими тундровый тип
растительности в целом [Городков, 1935, 1946; Александрова, 1971, 1977;
Грибова, 1980; и др. J, растительный покров западно-сибирских тундр
имеет некоторые специфические черты, обусловленные положением и историей
формирования поверхности данного региона.
Территория тундр Западной Сибири расположена в области
значительных четвертичных опусканий. Поверхность ее сложена рыхлыми
четвертичными отложениями, мощность которых достигает 200—250 м. На
п-ове Ямал поверхность тундры представляет собой плоскую
аккумулятивную равнину. Берега Карского моря и Обско»й губы здесь низкие (0—2 м),.
и только в некоторых местах береговые уступы поднимаются до 20 м.
Наиболее возвышена центральная часть Ямала, где максимальная высота
достигает 98 м. Рельеф Гыданского полуострова более сложен. Северная
часть занята плоской, местами всхолмленной равниной с абсолютными
отметками 70—80 м. Центральная и юго-восточная части отличаются
наибольшей приподнятостью с преобладанием холмистых форм рельефа.
Средние абсолютные высоты юго-восточной части Гыдана составляют 150 м,.
а максимальная достигает 190 м [Рихтер, 1963].
Равнинные тундры Западной Сибири — это район сплошного
распространения многолетней мерзлоты. Ее поверхностное залегание
определяет повсеместное развитие криогенных и термокарстовых форм микро-
и мезорельефа. К повышенным водоразделам с суглинистыми торфянисто-
глеевыми почвами приурочены трещиновато-бугорковатые формы
микрорельефа. Бугорки диаметром от 0,3 до 3 м, часто с пятнами-медальонами.
На выпуклых округлых увалах с песчаными почвами развивается
полигональный микрорельеф. Полигоны тетрагональной формы со сторонами
от 10 до 20 м разделены неглубокими ложбинами шириной 0,3—0,5 м. В
условиях избыточного увлажнения с маломощным торфяным горизонтом и
неглубоким залеганием многолетней мерзлоты развиваются
полигональные болота. На юге тундровой зоны они, как правило, замещаются с лоско-
бугристыми болотами.
На формирование и распределение растительного покрова тундр
большое влияние оказывают различные климатические; факторы. Тундры
Западной Сибири располагаются в двух климатических поясах— арктическом
и субарктическом, а также в двух климатических о'бластях —
атлантического и атлантико-арктического влияния [Алисов, 1969]. Арктический
пояс характеризуется особенно резким изменением инсоляции в течение
года. Зима здесь наиболее длительная, суровая с сильными бурями и
частыми метелями.
Климат субарктического пояса западно-сибирских тундр носит
переходный характер от относительно мягкого температурного режима
европейской части к суровому восточно-сибирскому. На Ямале он умеренно
континентальный, средние температуры января —21-4-26°, июля + 3 -г- 6°.
На Гыданском полуострове климат отличается большей континенталь-
ностью. Средние температуры января —26 -f- 30°, а июля + 4ч- 11,5°.
Зимой здесь особенно сильные ветры и метели, скорость ветра достигает
30—40 м/с. Основная масса осадков, как и на Ямале, выпадает в теплый
период года в виде длительных моросящих дождей. Характерная черта
тундрового ландшафта Западной Сибири — сильная заболоченность и
обилие озер.
Значительная протяженность с севера на юг и равнинность территории
обеспечили хорошо выраженную широтную дифференциацию
растительного покрова западно-сибирских тундр.
На составленной карте [Растительность..., 1976] в качестве основных
подразделений выступают зональные и подзональные категории
растительного покрова — это тундровый тип растительности и его широтные
подразделения. Понимая тип растительности как структурно-динамическую
категорию [Сочава, 1972], в состав тундрового типа мы включаем
экологически и динамически связанные между собой коренные и серийные
сообщества плакорных и неплакорных местообитаний — собственно
тундровые, болотные, луговые. При выделении подзональных типов также
учитывается все разнообразие эколого-фитоценотических и эколого-динами-
ческих связей спонтанной растительности. Большое значение при этом
имеет выявление соотношения широтно-географических групп видов
растений, слагающих тундровые ценозы.
Ниже приводится описание основных подразделений тундровой
растительности, получивших отображение на карте. В основу положены
материалы, собранные автором во время экспедиционных исследований, а
также некоторые литературные источники.
АРКТИЧЕСКИЕ ТУНДРЫ
Арктические тундры как ботанико-географический регион высокого
ранга выделяются всеми исследователями Крайнего Севера [Поле, 1910;
Городков, 1916, 1935; Сочава, 1933, 1948; Лесков, 1947; Александрова,
1971, 1977; и др.]. Эти территории В. Б. Сочава [1948] включил в
«Арктический пояс», или «Арктическую природную область» [Сочава, Тимофеев,
1968], которая является областью распространения арктических
формаций, отнесенных к арктической фратрии [Белов и др., 1972].
Основной диагностический признак сообществ арктических тундр —
отсутствие кустарников (в первую очередь Betula nana) как в плакорных,
так и в неплакорных сообществах. В виде исключения на заболоченных
участках ближе к границе субарктики могут произрастать отдельные
угнетенные экземпляры кустарниковой ивы (Salix lanata). В
растительности плакорных местообитаний распространены кустарничковые формы
ив (Salix nummularia, S. polaris). Во флоре арктических тундр основная
роль принадлежит арктическим (Salix polaris, Alopecurus al'pinus, Cassiope
tetragona, Carexensifoliassp. arctisibirica, C. concblor, Dupontia fischeri) и арк-
тоальпийским (Dryas punctata, D. octopetala, Polygonum viviparum, Luzula
confusa) видам. Некоторые гипоарктические формы (Vaccinium minus)
проникают в Арктику, в ее южные районы, однако в незначительном коли-
42
честве. Среди болот распространены гомогенные травяно-гипновые, а
также полигональные болота. . .
Исследователи Севера [Александрова, 1977; и др.] отмечают в
качестве характерной особенности растительности арктических тундр
пространственные сочетания тундровых сообществ с участками оголенного грун^
та. Это так называемые пятнистые тундры. На Ямальском Севере,
занимающем южное положение в Арктике, пятнистыми тундрами заняты большие
пространства*: на долю пятен здесь приходится до^ 20—30% площади.
Зональным типом сообществ арктических тундр Западной Сибири можно
считать осоково-моховые и осоково-кустарничково-лишайниково-моховые
кочковатые тундры, занимающие плакорные местообитания — пологие
склоны увалов с суглинистой или супесчаной торфянисто-глеевой почвой.
Осоково-лишайниково-моховые кочковатые тундры
в сочетании с кустарничково-мохово-лишайниковыми
полигональными тундрами (1)
Сообщества лишайниково-моховых %тундр занимают плакорные
местообитания; широкое распространение они получили на Ямале, где были
описаны нами в районе фактории Тамбей и на крайнем севере полуострова
в верховьях р. Юняха.
Основу напочвенного покрова здесь образуют мхи {Rhacomitrium la-
nuginosumT, Aulacomnium. turgidum,- Polytrichum juniperinum, Dicranum
elongatum). В травяно-кустарничковом ярусе доминирует осока (Carex
ensifolia ssp. arctisibirica) с небольшим участием кустарничков (Salix пит-
miliaria, S. poldris, Arctous alpina). Кроме осоки, участие травянистых
видов в этих сообществах очень незначительно как по количеству, так и по
видовому составу. Помимо пушицы (Eriophorum polystachyon) и некоторых
злаков (Arctagrostis latifolia) отмечены Hierochloe alpina, Luzula nivalis,
KoHeria asiatica, Saxifraga foliosa. Этот номер выделен как ямальский
вариант зональных арктических тундр. По условиям масштаба данные
сообщества в большинстве случаев картируются в сочетании с
лишайниковыми полигональными тундрами.
Кустарничково-моховые кочковатые тундры (2).
Л ишайниково-моховые
' и кустарничково-моховые кочковатые тундры
# сочетании с пушицево-моховыми заболоченными тундрами
и осоково-гиггаовыми полигональными болотами (3)
В этих подразделениях легенды объединены тундровые сообщества,
характерные для плакорных местообитаний арктических тундр Гы-
данского полуострова. В их напочвенном покрове доминируют зеленые
мхи (Dicranum elongatum, Hylocomium splendens var. alaskanum) с участием
лишайников (Cladina rangiferina, Cladonia macroceras, Cetraria cucullata).
В травяно-кустарничковом ярусе гыданских тундр, в отличие от
ямальских, доминируют кустарнички (Arctous alpina, Dryas punctata), но участие
осоки (Carex ensifdlia ssp. arctisibirica) также значительно.
В данную группу включены следующие типы сообществ, описанные
нами на ключевых участках севернее р. Яруяха (северо-запад Гыданского
полуострова): осоково-бруснично-дикрановое (Dicranum elongatum, D. an-
gustum, Sphenolobus minutus, Polytrichum alpestre, Cladina rangiferina,
Cladonia macroceras, Vaccinium minus, Arctous alpina, Carex ensifolia ssp.
arctisibirica, Polygonum viviparum); дриадово-осоково-кладониево-томент-
гипново-птилидиево-гилокомиевое (Hylocomium splendens var. alaskanum,
Ptilidium ciliare, Tomenthypnum nitens, Aulacomnium turgidum, Dicranum
elongatum, Cladina rangiferina, Cladonia macroceras, Thamnolia vermicularis,
Carex ensifolia ssp. arctisibirica, Dryas punctata, Vaccinium minus, Pedicula-
ris oederi, Polygonum viviparum, Luzula nivalis, Eutrema edwardsii, Lagotis
minor); арктоусово-дикраново-гилокомиевое (Hylocomium splendens var.
43
Рис. 3. Кустарничково-моховая кочковатая тундра. Фото Л. И. Мелъцер.
alaskanum, Rhacomitrium lanuginosum, Aulacomnium turgidum, Dicranum
elongatum, Ptilidium ciliare, Cetraria cucullata, Arctous alpina, Vaccinium
minus, Pedicularis oederi); осоково-бруснично-птилидиево-ракомитриевое
{Rhacomitrium'lanuginosum, Ptilidium ciliare, Dicranum elongatum, Poly-
trichum alpestre, Cetraria cucullata, Cladina rangiferina, Thamnolia vermicu-
laris, Vaccinium minus, Carex ensifolia ssp. arctisibirica, Dry as punctata). -
В сложении плакорных сообществ арктических тундр прослеживается
неоднородность горизонтального сложения растительности, обусловленная
нанорельефом (кочки высотой 10—15 см, диаметром до 30. см) (рис. 3). На
кочках с плотным минеральным ядром преобладают Dicranum elongatum,
Sphenolobus minutus, иногда Aulacomnium turgidum, в меньшем количестве
Polytrichum alpestre, Cladonia macroceras. Среди остальных видов как вмо-
хово-лишайниковом, так и в травяно-кустарничковом ярусе определенной
приуроченности к отдельным формам нанорельефа не наблюдается.
Исключение могут составлять Vaccinium minus и в некоторой степени Dryas
punctata, которые больше приурочены к склонам кочек и их основаниям.
Описанные тундровые сообщества плакорных местообитаний на
местности часто чередуются с участками травяно-моховых заболоченных тундр
или болот. На карте по условиям масштаба они показаны характерными
сочетаниями, которые представляют собой пространственно-экологический
ряд от болот или заболоченных тундр к плакорным (коренным)
сообществам (3).
Ниже описываются некоторые экологические варианты арктических
тунд£.
Осоково-пушицево-моховые заболоченные тундры
в сочетании с осоково-гипновыми полигональными болотами (4)
В Арктике Западной Сибири на плоских возвышенных
водораздельных пространствах развиты заболоченные тундры с полигональным
микрорельефом. Полигоны (длина стороны до 10 м) плоские или слегка вогнутые,
разделены неглубокими, часто обводненными трещинами-канавками. На
полигонах хорошо развит травяной покров из Carex ensifolia ssp.
arctisibirica, Eriophorum polystachyon, с незначительной примесью Luzula con-
fusa, Saxifraga cernua. На дренированных частях полигонов произрастают
44
также кустарнички {Salix polar is, S.nummularia,Arctousalpina). В
напочвенном покрове преобладают мхи {Aulacomnium turgidum,. Dicranum angus-
tum, Sphagnum fimbriatum). Лишайники представлены целым набором
видов, но в незначительном количестве. В трещинах-канавках развиты гид-
рофитные группировки из Dupontia fischeri, Carex concolor, Drepanocladus
revolvens, С oiliergon sarmentosum.
Эти тундры представлены несколькими вариантами. В районе мыса
Дровяной (северо-восток Ямала) В. С. Михайличенко [1936] описаны
такие же плоскополигональные заболоченные тундры со слегка вогнутыми
в центре полигонами. На плоских водораздельных увалах в районе
фактории Тамбей распространены заболоченные полигональные тундры,
представляющие собой систему выпуклых полигонов, разделенных трещинами-
канавками шириной до 1,5 м [Боч и др., 1971].
В подобных местообитаниях на севере Гыданского полуострова нами
описаны заболоченные тундры.с кочковато-западинным микрорельефом.
Кочки плотные, высотой 20—30 см, диаметром от 30 до 80 см кустарничко-
во-травяно-лишайниково-зеленомошные (Dicranum elongatum, Sphenolo-
bus minutus, Aulacomnium turgidum, Polytrichum alpestre, Cladonia macro-
ceras, Dactylina arctica, Poa alpigena, Pedicularis hirsuta, Salix nummula-
ria). Межкочечные понижения влажные, осоково-моховые и пушицево-
осоково-моховые {Polytrichum jensenii, Drepanocladus f\uitans, Carex con-,
color, Eriophorum polystachyon). Иногда у основания бугорков произрастает
Salix lanata, которая имеет очень угнетенный вид с прижатыми к кочкам
оголенными стеблями.
Мохово-лишайниковые полигональные тундры
в сочетании с открытыми группировками растений
на дефляционных обнажениях (5).
Кустаршгчково-мохово-лишайниковые полигональные тундры
в сочетании с осоково-гипновыми болотами (6)
На округлых вершинах увалов с песчаными почвами развиваются
дсаммофитные варианты тундровой растительности —
мохово-лишайниковые полигональные и пятнисто-полигональные тундры. Они были описаны
на Ямале в районе фактории Тамбей и на Гыданском полуострове на
водоразделе рек Нейтаяха и Яруяха.
На полигонах преобладают кустарнички (Arcious alpina, Dryas
punctata, Salix nummularia), гораздо меньше травянистых (Arctagrostis latifolia,
Carex ensifolia ssp. arctisvbirica) и плотный мохово-лишайниковый покров
{Cladina rangiferina, CI. mitis, Cetraria islandica, C. ericetorum, Sphaeropho-
rus-globosus, Rhacomitrium lanuginosum). Ложбинки с более рыхлой
дерниной из Dicranum elongatum, Rhacomitrium lanuginosum, Polytrichum
juniperinum, Drepanocladus uncinatus, Carex ensifolia ssp. arctisibirica, Lu-
zula confusa, Arctagrostis latifolia.
Видовой состав ямальских и гыданских лишайниковых
полигональных тундр несколько отличается. На Ямале в них, обычно участвует Cas-
siope fetragona, а на Гыданском полуострове — Dryas punctata. В моховом
покрове на Ямале больше Rhacomitrium lanuginosum, а на Гыдане —
Dicranum elongatum. Несколько большее участие в сложении ямальских тундр
принимают осоки.
Лишайниковые полигональные тундры занимают относительно
небольшие площади и на карте показаны в сочетании с открытыми
группировками растительности на дефляционных обнажениях и сообществами моховых
кочковатых тундр или с участками болот.
На самых выпуклых формах рельефа, наиболее подверженного
эродирующему действию сильных, постоянно дующих ветров, появляются
участки почвы, почти Лишенные растительного покрова, — дефляционные
обнажения. Отдельные небольшие участки растительности сохранились
здесь под защитой неровностей микрорельефа. В Арктике это отдельные
экземпляры Dryas punctata, Arctous alpina, Minuartia arctica, Polytrichum
45
alpestre, Rhacomitrium lanuginosum, Cetraria cucullata, Dactylina arctica
и др. Наиболее характерны группировки растительности на дефляционных
обнажениях для туйдр Гыданского полуострова» с его более пересеченным
рельефом. На карте эти участки показаны в виде сочетаний с собственно
тундровыми сообществами или внемасштабными знаками.
Интересным образованием в тундрах являются так называемые
тундровые луговины. Они занимают достаточно крутые склоны,
ориентированные в южном направлении, с песчаными почвами. Это обычно и места но-
рения песцов, что создает дополнительное удобрение почв азотно-фосфор-
ными элементами и их дренаж, а значит, и лучшее прогревание в весенне-
летний период. На таких местообитаниях развита наиболее богатая в
условиях Арктики растительность. Разреженный кустарниковый ярус из Salix
ianata и единичных экземпляров Betula папа представлен в верхней
половине склона. Хорошо развита травянистая растительность из
преобладающей здесь осоки (Carex tripartita) и большого количества разнотравья
(Ranunculus borealis, Equisetum arvense ssp. boreale, Myosotis alpestris, Po-
lemonium coeruleum, Artemisia tilesii, Minuartia arctica). Напочвенный
покров состоит в основном из мхов (Drepanocladus uncinatus, Polytrichum
alpestre, Brachythecium austro-selebrcsum). Лишайники развиты слабо, их
пятна приурочены в основном к верхним частям склонов. Подобные
сообщества описала как нитрофильньш вариант тундровой растительности
В. Д. Александрова [1971].
На карте тундровые луговины показаны внемасштабными знаками,>
СЕВЕРНЫЕ СУБАРКТИЧЕСКИЕ ТУНДРЫ
Граница между Арктикой и Субарктикой — важный ботанико-геогра-
фический, флористический и фитоценотический рубеж [Городков, 1916,
1935; Андреев, 1932, 1954; Григорьев, 1956; Сочава, 1948, 1964; Сочава,
Городков, 1956; Александрова, 1964, 1971, 1977; Юрцев, 1966; и др].
По мнению В. Д. Александровой, «в плакорных местообитаниях иод-
зоны субарктических тундр развиты сообщества, которые можно считать
субарктическим подтипом тундрового типа растительности» [1971, с. 6].
В пределах субарктической тундры Западно-Сибирской равнины в
сложении растительных группировок плакорных местообитаний
принимают широкое участие гипоарктические кустарники — ерник (Betula nana)
и несколько видов ив (Salix glauca, S. Ianata, S. phylicifolia и др.). На
севере, в полосе северных субарктических тундр, они образуют
разреженные низкорослые синузии, при движении к югу роль кустарников в
растительном покрове субарктической тундры значите льна.. Следует отметить,
что низкорослость или полегание кустарникового яруса характерно1 дйя
возвышенных водораздельных пространств в пределах всей тундры
Западной Сибири, как результат постоянно дующих здесь ветров [Полозова,
1966]. •■ . "
В субарктической тундре Западной Сибири наряду с арктическими
элементами флоры, такими как Carex ensifolia ssp. arctisibirica, С. concolor,
Alopecurus alpinus, Dryas punctata, D. octopetala, Salix reptans и др.,
распространены бореальные виды: Ledum palustre, Comarum palustre, Trien-
talis eurdpaea, Carex chordorrhiza, Chamerion angustifolium, Epilobium
palustre, Sanguisorba officinalis.
Следует отметить, что в растительности западно-сибирских тундр
широкое участие принимает багульник, который доходит до северной
границы Субарктики.
Кустарничково-лишайниково-моховые с ивой и ерником
бугорковатые тундры, местами в сочетании
с кустарничково-мохово-лищайниковыми тундрами (.10)
На севере субарктических тундр плакорные местообитания заняты
низко- и редкокустарниковыми моховыми тундрами. Отдельные куртины
46
ивы (Salix lanata, S. glauca) и реже ерника (Betula nana) достигают высоты
10—20 см, сомкнутого покрова не образуют. Помимо зеленых мхов (Die-
ranum angustum, Aulacomnium turgidum, Rhacomitrium lanuginosum, Po-
lytrichum alpestre) в напочвенном покрове участвуют сфагны (Sphagnum le-
nense). Из лишайников наиболее часты Cladonia deformis, CI. gracilis, Cla-
dina mitis, CI. rangiferina, Cetraria cucullata, а также Alectoria ochroleuca,
Al. nigricans. ' " ■ *
В плакорных условиях севера Субарктики увеличивается по
сравнению с арктическими тундрами роль кустарничков (Vaccinium minus, Dry as
punctata), появляются морошка (Rubus chamaemorus) и багульник (Ledum
decumbens). Из травянистых помимо доминирующей здесь осоки (Сагех
■ensifolia ssp. arctisibirica) встречаются злаки (Arctagrostis latifolia) и
разнотравье (Saxifraga punctata ssp. nelsoniana, Luzula nivalis, Valeriana capitata).
Кустарничковые, преимущественно дриадовые
с ивой и ерником
бугорковатые и пятнисто-бугорковатые тундры (11)
Для плакорных местообитаний восточной части Гыданского
полуострова характерно большое участие Dry as punctata, что позволило нам выг
делить особую группу дриадовых тундр. Среди кустарничков преобладает
также Vaccinium minus, другие виды — Salix nummularia, Vaccinium
uliginosum ssp. microphyllum — встречаются в небольшом количестве. Из
травянистых наиболее широко распространена осока (Сагех ensifolia ssp.
arctisibirica) и гораздо меньше злаки (Arctous alpina, Alopecurus alpinus,
Pod arctica). Отличительная черта этих тундр, особенно в сравнении с
ямальскими,— широкое распространение разнотравья, отдельные виды
которого представлены небольшим количеством, но в общем создают аспект
цветущей тундры (Pyrola, grandiflora, Polygonum viviparum, P. bistorta,
Myosotis alpestris и др.). Мхи Aulacomnium turgidum,, Hylocomium splendens
var. alaskanum и в меньшей степени Rhacomitrium lanuginosum образуют
плотную дернину на бугорках и более рыхлую (Hylocomium splendens var.
alaskanum, Ptilidium ciliare, Aulacomnium palustre, A. turgidum) в
межбугорковых понижениях. Обилие лишайников небольшое {Cetraria
cucullata, С. hiascens, Cladonia fimbriata, CI. macroceras, Sphaerophorus globosus).
Они приурочены в основном к положительным формам микрорельефа.
Кустарничково-моховые
с ивой и ерником бугорковатые тундры (12).
Кустарничково-моховые и лишайниково-моховые
с ивой и ерником бугорковатые тундры местами
в сочетании с осоково-гипновыми и осоково-сфагновыми
валиково -полигональными болотами (13)
Эти сообщества были описаны в районе оз. Ярото (Гыданский
полуостров) и в устье р. Тадибияха. Кустарниковый ярус в них развит слабо.
Куртины ивы (Salix glauca) низкие, прижатые к земле, растут у основания
бугорков, в некоторых местах встречается ерник (Betula nana).
Бугорки высотой 0,3—0,5 м, диаметром от 0,5 до 3 м с плотным
лишайников о-моховым покровом, в котором преобладают зеленые мхи (Dic-
ranum elongatum, Sphenolobus minutus, Aulacomnium turgidum) и в
несколько меньшем количестве Rhacomitrium lanuginosum, Polytrichum juniperi-
num, Tomenthypnum nitens, Hylocomium splendens var. alaskanum, Dicranum
spadiceum. Участие лишайников на положительных формах микрорельефа
до 30% (Cetraria cucullata, С. islandica, Cladonia macroceras, CI. fimbriata,
Cladina rangiferina, Sphaerophorus globosus, Thamnolia vermicularis, Dacty-
lina arctica). В этих тундрах по сравнению с подобными сообществами на
Ямале увеличивается участие кустарничков. В основном это Dry as
punctata, Arctous alpina и несколько меньше Salix nummularia, Vaccinium minus,
V. uliginosum ssp* microphyllum. Межбугорковые понижения осоково-мо-
ховые (Carex ensifolia ssp. atctisibirica, Hylocomium splendens var. alaska-
num, Ptilidium ciliare, Tomenthypnum nitens). Разнотравье в этих тундрах
представлено несколькими видами и в небольшом количестве.
Кустарничково-мохово-травяные с ивой и ерником
кочкарные заболоченные тундры
в сочетании с осоково-гипновыми болотами (14)
Плоские, слабодренируемые водоразделы заняты мохово-травяны-
ми заболоченными тундрами. По сложению и видовому составу они
отличаются от сообществ арктических тундр, приуроченных к подобным
местообитаниям. '
Заболоченные тундры на севере подзоны были описаны на Ямале
в районе пос. Сеяха и в среднем течении р. Мордыяха, а на Гыданском
полуострове в районе оз. Ярото. В этих сообществах большое участие
принимают сфагны {Sphagnum lenense, Sph. lindbergii), образующие
постепенно разрастающиеся бугорки, на которых поселяются зеленые мхи
(Hylocomium splendens var. alaskanum, Tomenthypnum nitens, Dicranum angus-
tum, Aulacomnium turgidum), в меньшем количестве встречаются
лишайники (Cetraria cucullata, Cladonia macroceras) и цветковые (Eriophorum poly-
stachyon, E.vaginatum, Rubus chamaemorus, Luzula nivalis, Vacciniumminus),
отмечены также отдельные экземпляры угнетенной ивы (Salix lanata).
Мочажины обычно влажные или обводненные травяно-гипновые (Drepano-
cladus exannulatus, D. uncinatus, Aulacomnium palustre, Carex concolor,
С aquatilis, Eriophorum vaginatum).
Кустарничково-мохово-лишайниковые
полигональные тундры (15)
Лишайниковые полигональные тундры занимают значительные
пространства на севере Субарктики на возвышенных округлых вершинах
водоразделов с песчаными почвами. В покрове полигонов — плотный мохово-
лишайниковый (Cladina rangiferina, CI. mitis, Cetraria cucullata, Sphaero-
phorus globosus, Polytrichum alpestre, Aulacomnium turgidum) покров. Кус-
тарничковый ярус развит слабо и состоит из Dry as punctata, Arctous alpi-
na, Vaccinium minus. В трещинах-канавках рыхлая моховая (Aulacomnium
turgidum, Hylocomium splendens var. alaskanum, Rhacomitfium lanuginosum)
дернина с хорошо выраженным травяно-кустарничковым (Salix nummula-
ria, Arctous alpina, Dryas punctata, Vaccinium minus, Carex ensifolia ssp.
arctisibirica, Luzula confusa) ярусом.
Помимо описанных сообществ' для северной полосы Субарктики
характерно также распространение лишайниковых алекториевых тундр.
Значительную часть их покрова составляют Alectoria nigricans, Cornicu-
laria diver gens с примесью Alectoria ochroleuca, Cetraria cucullata. Наиболее
полно на Ямале алекториевые тундры развиты севернее оз. Нейто и р. Тиу-
тейяха [Вашкевич, 19321. На Гыданском полуострове (по сообщению
работающих здесь оленеводов) эти тундры распространены в верховьях
р. Юрибей.
ЮЖНЫЕ СУБАРКТИЧЕСКИЕ ТУНДРЫ
Ерниково-ивняковые осоково-лишайниково-моховые
бугорковатые и пятнисто-бугорковатые тундры (17)
Низкокустарниковые тундры образуют самостоятельную широтную
полосу в пределах подзоны субарктических тундр Западной Сибири [Мель-
цер, 1973]. Основное участие в сложении плакорных фитоценозов
принимают кустарники, главным образом ерник (Retula nana) и ивы (Salix
glaucax S. lanata, S. phylicifolia). Они сохраняют здесь стелющуюся форму,
48
не превышая высоты 25—30 см. Напочвенный покров имеет сложное
гетерогенное строение, что связано в основном с широким распространением
бугорковатого микрорельефа. В качестве примера проследим динамику
формирования растительности бугорков и межбугорковых понижений в
сообществах плакорньгх местообитаний, описанных на ключевом участке
у оз. Пеунто. Здесь выделяется несколько типов бугорков,
представляющих разные стадии этого процесса:
1) бугорки, основу растительности которых составляют сфагновые
мхи. Помимо сфагнов {Sphagnum girgensohnii, Sph. fuscum) в
их'напочвенном покрове участвует Polytrichum alpestre и в незначительном количестве
Hylocomium splendens var. alaskanum, Aulacomnium turgidum, а также Cla-
donia fimbriata и меньше Cladina rangiferina, CI. arbuscula. Кустарники
(Betula nana, Salix lanata) растут у основания бугорков;
2) следующая стадия — увеличение и разрастание на бугорках
зеленых мхов. В этих случаях встречаются бугорки, в покрове которых
доминируют Aulacomnium turgidum, Dicranum elongatum, Sphenolobusjninutus^
Polytrichum alpestre с небольшой примесью Hylocomium splendens var. alas-
kanum, Cetraria cucullata]
3) со временем размеры бугорков увеличиваются, основу их покрова
из мхов образуют Rhacomitrium lanuginosum, Aulacomnium turgidum,
Dicranum elongatum, Polytrichum alpestre, а из лишайников Sphaerophorus glo-
bosus, Cladonia fimbriata, Cetraria cucullata, Cladina rangiferina, CI. arbus-
cula. На бугорках поселяются кустарнички (Vaccinium minus, Dryas
punctata, D. octbpetala, Salix nummularia) и осока. Этот тип бугорков с кустар-
ничково-лишайниково-моховой ассоциацией наиболее распространен
и занимает более 40% площади в описываемых группировках;
4) бугорки с лишайниковым (Cladina rangiferina, CI. arbuscula,
Cetraria cucullata) покровом. Кустарнички (Vaccinium minus, Dryas octopetala)
и осока (Carex ensifolia ssp. arctisibirica) растут на их склонах, а у
основания — Betula nana, Salix glauca. Этот тип бугорков занимает не более
15% площади.
В межбугорковых понижениях обычно преобладает Hylocomium
splendens var. alaskanum с большим или меньшим участием Sphagnum balticum,
Sph. girgensohnii, Ptilidium ciliare, Aulacomnium turgidum. В отдельных
случаях к ним примешиваются Tomenthypnum nitens, Aulacomnium palus-
tre. В травяно-кустарничковом ярусе помимо осоки (Catex ensifolia ssp.
arctisibirica), распространенной также и на бугорках, произрастают Ledum
decumbens, Rubus chamaemorus и единично Pedicularis lapponica, Rumex
arcticus, а также кустарники (Salix lanata, S. glauca, Betula nana).
чЕрниково-ивняковые, местами с ольхой кустарничково-моховые
бугорковатые и пятнисто-бугорковатые тундры (18).,,
Ерниково-ивняковые лишайниково-моховые
и кустарничково-моховые бугорковатые
и пятнисто-бугорковатые тундры
в сочетании с осоково-мохово-лишайниковыми
трещиновато-полигональными болотами (20)
Растительные сообщества, объединенные в данную группу типов,
были описаны на п-ове Гыданский в среднем течении р. Сыдыяха, в районе
пос. Антипаюта и у оз. Ладукето.
По составу и сложению эти тундры несколько отличаются от
вышеописанных. Бугорки встречаются в основном двух типов. Одни с плотной
дерниной, осногву их растительности составляют Dicranum elongatum,
D. angustum, Sphenolobus minutus с меньшим участием других
мхов.(Aulacomnium turgidum, Polytrichum alpestre) и лишайников (Cladonia macro-
ceras, Cladina rangiferina, Cetraria cucullata, C. nivalis). На других
бугорках, более рыхлых, доминируют Aulacomnium turgidum, Hylocomium
splendens var. аЩкйпит с меньшим участием Rhacomitrium lanuginosum,
Dicranum angustum, Polytrichum alpestre. Лишайники принимают значи-
* ч
4 Заказ № 297 49
тельное (до 30%) участие в сложении описываемых группировок, среди них
преобладают Cladina rangiferina, CI. arbuscuia, Cladonia ryacroceras,
Cetraria cucullata, а другие — Cladina stellar is, Cladonia uncialis, Cetraria ery-
cetorum, Sphaerophorus globosus, Dactylina arctica, Thamnolia uermicularis,}
Alectoria nigricans, Al. ochroleuca, Nephroma arcticum — присутствуют в
меньшем количестве. Кустарнички (Vaccinium minus, Dryas punctata, Vac-
cinium uliginosum ssp. microphyllum, Ledum decumbens, Sali'x nummularia^
Empetrum hermaphroditum) более разнообразны по видовому составу в
сравнении с ямальскими тундрами. Во всех описанных плакорных сообществах
присутствует Vaccinium uliginosum ssp. microphyllum. В межбугорковых
понижениях помимо осоки (Carex ensifolia ssp. arctisibirica) встречается
пушица (Eriophorum polystachyon), реже вейник (Calamagrostis lapponica).
Неплотная моховая дернина между бугорками составлена из Hylocomium
splendens var. alaskanum, Ptilidium ciliare, иногда присутствует Drepa-
nocladus uncinatus,
В отличие от ямальских тундр в западно-гыданских отмечается
меньшее участие на плакорах сфагновых мхов, но обязательно присутствует
печеночник Ptilidium ciliare.
Ольха (Duschekia fruticosa) больше распространена в восточной части
Гыданского полуострова. < Она растет небольшими плотными куртинами
высотой 0,6—0,8 м на пологих склонах водоразделов, ближе к нижней
части склона. В восточно-гыданских тундрах увеличивается также участие
Dryas punctata по сравнению с ямальскими и западно-гыданскими, но в то
же время наблюдается значительное снижение участия дриады по
сравнению с более северными районами приенисейской части Гыданского
полуострова. ,
В плакорных местообитаниях на востоке полуострова основу
напочвенного покрова положительных форм микрорельефа составляют Aula-
comnium turgidum, Tomenthypnum nitens, Dicranum elongatum, Cetraria
cucullata, Cladina rangiferina, Cladonia macroceras и в меньшей мере tyicranum
angustum, Polytrichum alpestre, Hylocomium splendens var. alaskanum,
Cetraria hiascens,\C. islandica, Dactylina arctica. В травяно-кустарничковом
ярусе преобладают Vaccinium uliginosum var. microphyllum, Dryas
punctata, Vaccinium minus, Carex ensifolia ssp. arctisibirica., В межбугорковых
понижениях наиболее распространены Hylocomium splendens var.
alaskanum, Ptilidium ciliare, Dicranum palustre,Aulacomnium turgidum. В отдельт
ных случаях отмечены сфагны (Sphagnum lindbergif, S'ph. warnstorfii) и
лишайники (Cetraria cucullata, Peltigera dphthosa). Среди травянистых, как
и в предыдущих группировках, преобладает осока (Carex ensifolia ssp.
arctisibirica), в меньшем обилии Arctagrostis latifolia, несколько видов пушиц
(Eriophorum polystachyon, E. gracile, E. scheuchzeri) и цветковое
разнотравье (Pyrola grandiflora, Valeriana capitata, Pedicularis oederi, Nardosmia
frigida).
Вышеописанные сообщества ерниково-ивняковых моховых тундр на
местности часто чередуются с участками болот. В этих случаях по условиям
масштаба они показаны в сочетании с болотами. .....
Ерниково-ивняковые куетарничково-лишайниковые,
преимущественно алекториевые, заболоченные тундры
в сочетании со сфагново-лишайниковыми олиготрофными
болотами (19)
Для данных сообществ характерен кочкарно-бугорковатый
микрорельеф, образованный пушицей (Eriophorum vaginatum) и сфагновыми
мхами. Бугорки диаметром от-0,4 до 1,5 м, высотой 20—30 еж
плосковершинные. Помимо лишайников (Alectoria ochroleuca, A I. nigricans, Corni-
cularia diver gens, Cladina rangiferina, CI. alpestris, Cetraria cucullata),
составляющих основу напочвенного покрова, на них поселяются мхи (Dicranum
angustum, Polytrichum alpestre) и кустарнички (Vaccinium uliginosum ssp.
microphyllum, V. minus). На кочках, образованных Sphagnum lenenset
50
растут Rubus chamaemorus, Oxycoccus microcarpus, Andromeda poly folia,}
Ledum decumbens. В межбугорковых и межкочечных понижениях
распространены Sphagnum balticum, Ledum decumbens.
Подобные сообщества на Гыданском полуострове были описаны
Б. Н. Городковым [1944] и названы сфагновыми кочкарными тундрами.
По его мнению, субарктические тундры полуострова в южной своей
половине (подзона кустарниковых тундр, по Б. Н. Городкову) представляют
переходную полосу, где климаксовые тундровые сообщества Европы и
Западной Сибири на сходных местообитаниях замещаются климаксом
восточно-сибирских тундр — кочкарными тундрами [Городков, 1944].
Ерниково-ивняковые травяно-кустарничково-моховые
заболоченные тундры
в сочетании с кустарничково-мохово-лишайниковыми
трещиновато-полигональными болотами (21)
Эти сообщества приурочены к плоским слабодренируемым
водоразделам с избыточным застойным увлажнением. Кустарниковый ярус в них
более разрежен и местами угнетен. Центральные части таких водоразделов
обычно слегка вогнуты и заселены гигрофитной осоково-пушицево-гипно-
вой (Eriophorum polystachyon, lCarex concolor, Drepanocladus exannulatus,
D. aduncus) растительностью. По мере накопления органического вещества
разрастаются сфагны {Sphagnum lenense), которые образуют большие
пятна в виде плоских бугорков размером 3x5 м. На них поселяются пушица
(Eriophorum polystachyon), кустарнички (Ledum decumbens, Andromeda po-
lifolia) и морошка (Rubus chamaemorus).
Далее идет образование пушицево-сфагново-зеленомошных с ивой и
ерником сообществ на месте предыдущих. В них уже заметно проявляется
кочковатый микрорельеф. Сфагны занимают периферийные части плоских
бугорков, а в их центре произрастают зеленые мхи (Dicranum elongatum,
D. congestum, Sphenolobus minutus), образующие плотные кочки, а также
Ptilidium ciliare и в небольшом количестве Aulacomnium turgidum, Hylo-
comium splendens var. alaskanum. Небольшое участие в растительности
бугорков принимают, также лишайники (Cladina rangiferina, Cladonia macro-
ceras, Cladina arbuscula, Cetraria cucullata, Sphaerophorus globosus).
Постепенно эти сообщества замещаются ерниково-ивняковыми морошково-ба-
гульниково-пушицево-лишайниково-долгомошными. Кустарниковый ярус
(Salix lanata, S. phylicifolia, Retula nana) развивается куртинами большей
частью на склонах бугорков, достигая высоты 30—35 см. В напочвенном
покрове преобладают Polytrichum alpestre, Dicranum elongatum, Cladonia
pleurota, CI. chlorophaea, Alectoria ochroleuca, Cornicularia diver gens, Cladina
arbuscula.
Растительность плоских водоразделов Гыданского полуострова по
сложению и флористическому составу несколько отличается от подобных
сообществ, описанных для Ямала. Наиболее влажные центральные участки
заняты пушицево-гипновыми (Drepanocladus exannulatus, Calliergon sar-
mentosum, Dicranum angustum, D. elongatum, Arctagrostis latifolia, Cala-
magrostis holmii, Eriophorum polystachyon) группировками, которые дальше
сменяются злаково-пушицево-осоково-моховыми сообществами. В
напочвенном покрове их увеличивается участие зеленых мхов, образующих
кочки (Dicranum elongatum, Sphenolobus minutus, Polytrichum alpestre,
Aulacomnium turgidum). Между кочек поселяются Drepanocladus exannulatus,.
а также пушица и осока (С or ex concolor). Эти сообщества на более поздней
стадии развития сменяются низкокустарниковыми ерниково-ивняковыми
пушжцево-осоково-кустарничково-лишайниково-зеленомошными или ив-
няково-ерниковыми злаково-багульниково-лишайниково-сфагново-зелено-
мошными. Основу напочвенного покрова при этом составляют зеленые мхи
(Dicranum elongatum, Aulacomnium turgidum, Polytrichum alpestre, Conos-
tomum tetragonum, Rhacomitrium lanuginosum) и лишайники (Cladina ran-
giferina, CI. arbuscula, Cetraricr cucullata). Из кустарничков преобладает
4* 51
Ledum decumbens, меньше Vaccfnium minus, Dryas punctata. Среди
травянистых в первом случае доминируют Carex ensifolia ssp. arctisibirica, Erio-
phorum brachyantherum, во втором — Calamagrostis lapponica. Кустарники
(Betula nana, Salix glauca) низкими куртинками поселяются на бугорках
между кочек.
Брниковые и ивняково-ерниковые
кустарничково-лишайников о -моховые бугорковатые тундры (22)
Зонадьно-плакорным вариантом, характеризующим южную полосу
подзоны субарктических тундр [Мельцер, 1977], являются кустарниковые
(ерниковые, ивняковые, ольховниковые) кустарничково-лшпайниково-мо-
ховые тундры. ■ _ ' . ;
Ерниковые моховые тундры были описаны на Ямале в среднем
течении р. Хадытаяха, а также в районе нос. Яр-Сале. В сложении раститель-
♦ ного покрова наблюдается закономерность, общая для плакорных
местообитаний субарктических тундр. На бугорках распространены более мезо-
фитные кустарничковб-лишайниково-зеленомошные сообщества, а во
влажных межбугорковых понижениях — травяно-моховые. Кустарники
{Betula nana, Salix phylicifolia, S. pulchra) образуют сомкнутые синузии
высотой 0,4—0,6 м.
На одном из участков среднего течения р. Хадытаяха прослежена
следующая динамика формирования растительного покрова бугорков. На
первых этапах разрастаются сфагны (Sphagnum warnstorf и). Затем на нем
поселяются зеленые и долгомошные мхи (Aulacomnium turgidum, tDicranum
elongatum, Polytrichum alpestre), лишайники (Cladina rangiferina, CI. ar-
buscula, Cetraria cucullata), а также кустарнички (Vaccinium vitis-idaea,
V. uliginosum, Empetrum nigrum) и осока (Carex ensifolia ssp. arctisibirica).
В дальнейшем происходит разрастание зеленых мхов и лишайников
и формируются кустарничково- лишайниково-зеленомошные (Dicranum
elongatum, Aulacomnium turgidum, Polytrichum alpestre, Cladina arbuscula,
CI. rangiferina, Cetraria cucullata, Empetrum nigrum, Vaccinium vitis-idaea,
Ledum decumtens) сообщества. Межбугорковые понижения пушицево-ба-
гульниково-зеленомошно-сфагновые {Sphagnum girgensohnii, Sph. squar-
rosum, Aulacomnium turgidum, Dicranum angustum, Ledum decumbens, Erio-
phorum vaginatum, Carex ensifolia ssp. arctisibirica, Alopecurus alpinus).
Ерник и ивы произрастают, как правило, у основания бугорков.
Ерниковые и ивняково-ерниковые
с ольхой кустарничково-моховые бугорковатые тундры (23)
Ерниковые кустарничково-моховые тундры на юге Тазовского
полуострова описаны В. С. Говорухиным [1933], а на юге Гыданского
полуострова — Б. Н. Городковым [1944]. По сложению и видовому составу они.
сходны с вышеописанными, но в них меньше представлены сфагны, а
зеленые мхи (Aulacomnium turgidum, Dicranum elongatum, Tomenthypnum ni-
tens и др.) развиты более полно. Хорошо развит кустарничковый ярус
(Vaccinium vitis-idaea, V. uliginosum, Empetrum nigrum, Dryas punctata).
Среди кустарников помимо ивы и ерника произрастает ольха (Duschekia
fruticosa). • £ ,
Ерниковые и ивняково-ерниковые, местами с ольхой,
м охов о-лишайниковые, преимущественно алекториёвые,
бугорковатые тундры в сочетании с мохово -лишайниковыми
трещиновато-полигональными болотами (24)
Эти тундры выделены нами как подзональный вариант южной полосы
подзоны субарктических тундр Тазовского и Гыданского полуостровов.
Кустарники (Salix phylicifolia, S. pulchra, S. glauca, Betula nana,
Duschekia fruticosa) образуют невысокие (0,3—0,5 м),но сомкнутые синузии. В на-
52
почвенном покрове среди лишайников помимо Aulacomnium ochroteuca,
Al. nigricans, С ornicularia diver gens распространены Cladina rangiferina,
CI. arbuscula, Cetraria cucullata. Мхи (Aulacomnium turgidum, Dicranum,
elongatum, Ptilidium ciliare) произрастают, в меньшем количестве. В травя-
но-кустарничковом ярусе широко представлены кустарнички {Ledum de-
cumbens, Vactinium uliginosum, V. vitis-idaea), меньше осока (Carex ensi-
folia ssp. arctisibirica). В отдельных сообществах отмечены пушица (Erio-
phorum polystachyon) и водяника {Empetrum nigrum). Алекториевые тундры
обычно занимают небольшие пространства на дренируемых рекой
прирусловых валах и поэтому картируются в сочетании с полигональными
болотами.
Ерниковые и ивняково-ернйковые с ольхой
куетарничково-лишайниково-моховые тундры
в сочетании с лиственничными
лишайниково-кустарничковыми редколесьями
или плоскобугристыми
травяно-мохово-лишайниковыми болотами (25»)
В южной полосе подзоны субарктических тундр на плакорных
местообитаниях распространены моховые кочковатые тундры, в кустарниковом
ярусе которых помимо ёрника и ивы присутствует ольха. Эти сообщества,
как правило, занимают средние и нижние части пологих склонов увалов.
Округлые вершины увалов обычно заняты лиственничными лишайниково-
кустарничковыми редколесьями, а плоские водоразделы и депрессии —
плоскобугристыми болотами или заболоченными тундрами.
Участки лиственничных редколесий, которые сочетаются с этими
тундрами, расположены на склонах и вершинах пологих увалов с песчаными
почвами. Лиственница (Larix sibirica) высотой 8—10 м прямостоячая,
расстояние между деревьями от 3 до 20 м. Имеется подрост высотой от 0,3 до
6 м в хорошем состоянии* В травяно-кустарничковом ярусе доминируют
Empetrum nigrum, Arctous alpina, в изобилии здесь плодоносящие, а также
Equisetum palustre-.и в небольшом количестве Festuca ovina, Carex ensifolia
ssp. arctisibirica, Luzula confusa, Tanacetum bipinnatum. Лишайники
(Cetraria cucullata, Cladina arbuscula, Cladonia fimbriata, Stereocaulon pascha-
le) образуют пятна в наиболее осветленных участках.
Ивняковые травяно-моховые тундры
в сочетании с ивняково-ерниковыми
лишайников о-моховыми тундрами
и осоково-мохово-лишайниковыми плоскобугристыми болотами (26)
Ивняковые тундры выделены как ямальский вариант плакорных
сообществ подзоны субарктических тундр. В их кустарниковом ярусе
преобладают ивы (Salipc pulchra, S. phylicifolia, S. glauca) с большим или
меньшим участием ерника (Betula nana). В основе напочвенного покрова — мхи
(Aulacomnium turgidum, Dicranum angustum, Aulacomnium palustre, Poly-
trichum alpestre). Кустистые лишайники (Cladina arbuscula, CI. rangiferina,
Cetraria cucullata) растут куртинами среди густого ивняка, где они меньше
стравливаются оленями. В отличие от ерниковых тундр, в которых хорошо
представлены кустарнички, в ивняковых тундрах большее развитие
получили травы (Eriophorum polystachyon, Nardosmia frigida, Pedicularis lappo-
nica, Arctagrostis latifolia). '
В данном масштабе эти тундры картируются в сочетании с описанными
выше ивняково-ерниковыми лишайниково-моховыми тундрами или с тра-
вяно-моховыми заболоченными тундрами и болотами.
53
Ерниково-ольховниковые кустарничково-лишайниково моховые
бугорковатые тундры, местами в сочетании
с редкокустарниковыми лишайниковыми тундрами
и лиственничными редколесьями (27)
Ольховниковые тундры выделены как гыданский вариант плакорных
сообществ южной полосы подзоны субарктических тундр. Они, как
правило, приурочены к средним и нижним частям пологих склонов
дренированных водоразделов. Наиболее возвышенные участки водоразделов обычно
заняты лишайниковыми полигональными-тундрами или лиственничными
редколесьями.
Кустарниковый ярус образует как бы два подъяруса, в первом из
них -^ ольха высотой до 1 м, а во втором — ерник с примесью ив, высота
которых не превышает 0,4—0,5 м. В травяно-кустарничковом ярусе
распространены Vaccinium vitis-idaea, V. uliginosum, Empetrum nigrum, Ledum
decumberis. Напочвенный покров составлен из зеленых (Aul'acomnium tur-
gidum, Dicranum angustum, Polytrichum alpestre) мхов и кустистых
лишайников (Cladina rangiferina, CI. stellaris, Cetraria cucullata, Alectoria ochro-
leuca).
Редкокустарниковые
кустарничково-зеленомошно-лишайниковые
полигональные тундры
в сочетании с открытыми группировками растений
на дефляционных обнажениях (28)
Эти сообщества представляют собой псаммофитный вариант тундровой
растительности южной полосы подзоны субарктических тундр. Они
занимают округлые вершины увалов с песчанистой слабоподзолистой почвой.
Сомкнутость кустарникового яруса здесь сильно падает до 0,2—0,4.
На полигонах кустарничково-лишайниковая растительность, которая
лучше развита по периферии полигонов. Из кустарничков преобладает
Empetrum, nigrum, меньше Arctous alpina, Diapensia lapponica, Vaccinium
vitis-idaea, Arctous alpina, а в напочвенном покрове — Cladina arbuscula,
Cetraria cucullata, С. islandica, Thamnolia vermicular is, Rhacomitrium lanu-
ginosum. В трещинах-канавках — зеленые мхи (Hylocomium splendens var.
alaskanum, Aulacomnium turgidum) с участием Drepanocladus uncinatus.
Среди них поселяются кустарнички (Empetrum nigrum, Ledum decumbens%
Vaccinium vitis-idaea), травы (Luzula confusa, Carex ensifolia ssp. arctisibiri-
ca, Tofieldia coccinea, Campanula langsdorffiana, Hierochloe alpina) ш
низкорослые кустарники (Betula nana, Salix phylicifolia, S. pulchra, S. glauca).
ГЛАВА IV
ТЕМНОХВОЙНЫЕ ЛЕСА И РЕДКОЛЕСЬЯ
Западно-Сибирская равнина — одна из классических областей
развития темнохврйной тайги [Толмачев, 1954]. Ареал темнохвойных лесов
здесь очень обширен. Он охватывает площадь между Уралом и Енисеем,/
а на севере выходит за пределы Полярного круга. В своем
распространении темнохвойные леса как тип растительных сообществ неразрывно
связаны с природной зоной тайги, которая отличается на Западно-Сибирской
равнине умеренно холодным климатом и избыточным увлажнением
[Западная Сибирь, 1963].
Несмотря на благоприятные в целом для произрастания
темнохвойных пород условия тепло- и влагообеспеченности [Мезенцев,, Карнацевич,;
1969], на территории Западной Сибири темнохвойные лесные сообщества
не получили широкого распространения. Это объясняется рядом факторов
54
регионального порядка, среди которых главную роль играют гидролого-
геоморфологические, обусловливающие сильную заболоченность
территории и развитие на огромных пространствах центральных частей
водоразделов сообществ болотной растительности и заболоченных,
преимущественно сосновых, лесов. Распространению темнохвойных лесов на равнине
препятствуют также антропогенные воздействия, главным образом цожары
и рубки леса. Влияние пожаров на лесную растительность особенно велико.
Площади, занятые вторичными мелколиственными, сосновыми и
смешанными лесами, значительно превышают территории, на которых
сохранились коренные сообщества темнохвойной тайги [Растительность..., 1976].
В настоящее время основные массивы темнохвойных лесных сообществ
сосредоточен^ в приподнятых хорошо дренированных приречных частях
водоразделов и на высоких речных террасах. Наибольшие площади этих
лесов отмечены в центральных частях Обь-Иртышского междуречья
(Среднее Приобье, Тобольский материк, Васюганская возвышенность), а также
в приенисейской части (рис. 4).
Темнохвойные леса Западно-Сибирской равнины имеют некоторые
общие черты состава и структуры, обусловленные как внешними факторами
природной среды, так и внутренними фитоценологическими условиями,
создаваемыми самим древостоем. Лесообразователями этих лесов являются
три древесные породы — кедр (Pinus sibirica), пихта {Abies sibirica) и ель
(Picea obovata). В большинстве случаев они образуют полидоминантные
древостой с участием всех трех пород, при этом кедр наиболее типичный
здификатор в темнохвойных сообществах северной й средней тайги, пихта
выполняет функции лесообразующей породы в южно-таежной подзоне,,
а ель становится доминантом многих северо- и южно-таежных сообществ,
преимущественно в западной зауральской части равнины. Ё процессе
возрастной и восстановительной динамики, а также при незначительных
колебаниях экологических условий, доминирование в составе теми©хвойных
лесов может переходить от одной породы к другой, но при этом общие
черты состава и структуры этих лесных сообществ останутся неизмененными.
Полидоминантность — характерная чёрта почти всех лесных сообществ"
западно-сибирской тайги. Монодоминантные древостой из темнохвойных
пород в этом регионе встречаются крайне редко, занимают небольшие
площади и являются кратковременными стадиями лесообразовательного
процесса.
В составе полидоминантных темнохвойных лесов наряду с темнохвой-
ными породами обычно наблюдается примесь мелколиственных и светло-
хвойных пород. Смешанные мелколиственно-хвойные леса с участием
осины и березы типичны в основном для долинных местообитаний, где они
являются коренными сообществами; небольшая примесь березы характерна
почти для всех темнохвойных ненарушенных лесов, а для производных
(возникших после пожаров и рубок) участие березы и осины, а в отдельных
местах и сосны, возрастает до доминирующей роли на многих стадиях лесо-
восстановйтельных смен.
Специфическая особенность западно-сибирской тайги — это участие
в составе темнохвойных лесов на юге их ареала широколиственной
породы — липы сердцевидной (Tilia cordata), являющейся, по мнению
некоторых исследователей [Волкова, 1965], реликтом третичной эпохи.
Сильная эдификаторная роль древесного яруса — это одна из
характерных особенностей темнохводных лесов [Толмачев, 1954]. В Западной
Сибири-она отчетливо проявляется главным образом в средней и южной
тайге, *-где коренные темнохвойные леса достигают значительной высоты
(25—28 м) и производительности (III класс бонитета), имеют хорошо
развитые подрост, подлесок и травяной покров. В северной тайге, особенно
в полосе редкостойных лесов, эдификаторная роль темнохвойных заметно
ослабевает, в связи с чем изменяется и вся структура леса — разреженным
становится древостой, слабо развиваются подрост и подлесок, а мохово-
лишайниковый покров и некоторые виды гипо арктических кустарничков
приобретают ведущую роль в составе насаждений.
55
Северотаежные: 1 — листвеьнично-еловые и еловые зеленомошно-лишайниковые и зелено-'
мошные редкостойные леса, la — лиственнично-елово-березовые и березовые с елью лишайниково-
зеленомопщые редкостойные леса; 2 — лиственнично-еловые и еловые кустарничково-сфагновые
с ерником редкостойные леса; з — лиственнично-елово-кедровые лишайниково-зеленомошно-кус-
тарничковые и елово-кедровые, местами с лиственницей, зеленомошно-кустарнйчковые с ольхой
в подлеске леса; За — березовые с кедром, елью, лиственницей кустарничково-лишайниково-зелено-
мошные и березовые с осиной, кедром и ольхой в подлеске зеленомошно-кустарнйчковые леса;
Зб — сосновые с кедром, елью, лиственницей кустарничково-лишайниково-зеленомошные леса.
Среднетаежные: 4 — елово-кедровые с лиственницей кустарничково-зеленомошные и елово-
кедровые с пихтой, местами кедрово-еловые, мелкотравно-брускично-зеленомотиные леса, 4а — тем-
\ нохвойно-березовые и темнохвойно-осиновые с лиственницей кустарничково-зеленомошные, елово-
березовые и березово-осиновые мелкотравно-зеленомошные леса, 46 — темнохвойно-сосновые с
лиственницей кустарничково-зеленомошные леса; 5 — елово-кедровые долгомошные и кустарничково-
сфагновые леса.
Южно-таежные: 6 — кедрово-елово-пихтовые зеленомопшо-мелкотравные и мелкотравно-
осочковые с элементами широкотравья, елово-?пихтовые с липой зеленомошно-широкотравно-кислич-
Hbie и липово-елово-пихтовые широкотравные леса; 6а —темнохвойно-березовые, мелкотравно-вей-
никово-осочковые, темнохвойно-осиновые и березово-осиновые с подлеском травяные леса, &б —■
сельскохозяйственные земли на месте елово-пихтовых лесов; 7 — кедровые с пихтой и елью хвощово-
зеленомошные и травяно-сфагновые леса; 8 —пихтово-елс|вые, местами с липой, зеленомошно-тра-
вяные леса; 8а —* темнохвойно-осиново-березовые и березовые с липой травяные с пятнами зеленых
мхов леса, 86 — сельскохозяйственные земли с участками лесов и кустарничков на месте
елово-пихтовых и пихтово-еловых с липой лесов и производных от них сообществ.
В целом темнохвойные таежные леса Западно-Сибирской равнины
имеют сравнительно бедный видовой состав флоры нижних ярусов, что
свойственно темнохвойной тайге вообще [Толмачев, 1954; Сочава, 1956],. Из пред-
ставителей трав и кустарничков можно отметить целый ряд «верных»
видов, специфичных для темнохвойных лесов и связанных с ними как со
средой обитания. Это прежде всего обширная группа собственно бореальных
видов, имеющих широкий экологический ареал и являющихся
доминантами многих таежных сообществ. Среди них особенно типичны бореальные
кустарнички — брусника {Vaccinium vitis-idaea), черника (Vaccinium туг-
tillus), линнея (Ыппаеа borealis), в том числе олиготрофные ,— багульник
56
(Ledum palustre), клюква (Охуcoccus palustris), Кассандра (Chamaedaphne
calyculata), а также представители таежного мелкотравья — майник (Mai~
anthemum bifolium), седмичник (Trientalis ешчтраеа), кислица (Oxalis aceto-
- sella), папоротник (Gymnocarpium dryopteris), грушанка (Pyrola rotunda
folia) ж др.
На севере в область распространения темнохвойной тайги широко
внедряются гипоарктические виды, особенно Betula nana, Salix lanata,
Calamagrostis lapponica, а в отдельных местах и собственно арктические
и арктоальпийские элементы — Arctagrostis latifolia, Deschampsia brevi-
folia, Pedicularis lapponica, Arctous alpina.
В южных подзонах, а также в долинах рек повсеместно
распространены представители широкотравья и высокотравья — лабазник (F.ilipendula
ulmaria), борец (Aconitum septentrionale), какалия (Cacalia hastata),
папоротники (Athyrium filix-femina, Dryopteris carthusiana). Участие
неморальных видов наблюдается главным образом в западном секторе южной тайги
региона, где в составе темнохвойных лесов присутствует реликтовый
элемент флоры — липа сердцевидная (Tilia cordata). .
Из неморальных видов наиболее распространены Aegopodium podag-
raria, Daphne mezereum, Pulmonaria dacica, Viola epipsila, Milium effusum,
Melica nutans.
Таковы некоторые общие особенности состава и структуры
темнохвойных лесов Западно-Сибирской равнины. Необходимо остановиться на
некоторых фитоценотических, экологических и динамических чертах
темнохвойных лесных сообществ, формируемых каждой из основных пород —
лесообразователёй.
Кедровые леса. Темнохвойные кедровые леса Западно-Сибирской
равнины — это огромное богатство ее растительного мира. Они
распространены на площади около 8 млн. га [Леса СССР, 1969].. Полезность
кедровых лесов велика. По своей хозяйственной ценности они занимают
ведущее место среди всех видов лесосырьевых ресурсов [Крылов, 1961;
Седых, 1979]. '
Главная лесообразующая порода в темнохвойно-кедровых лесах —
кедр сибирский (Pinus sibirica). Это дерево первой величины: в равнинной
тайге Западной Сибири достигает высоты 35 м и возраста свьдпе 300 лет
[Крылов и др., 1969]. ~ v
Экологическая амплитуда условий существования сибирского кедра
достаточно широка, но экологический оптимум его произрастания и
формирование им наиболее производительных древостоев связаны с хороша
дренированными местообитаниями и развитием подзолистых суглинистых
достаточно увлажненных почв [Поварницын, 1944: Сочава, 1956; Крылов,
1961; Бобров, 1978],
Сообщества кедровых лесов достаточно хорошо произрастают на
песчаных и заболоченных почвах, о чем свидетельствуют исследования многих
авторов и наши собственные [Городков, 1916; Поварницын, 1944; Тырти-
ков, 1974; Лапшина, 1971; Аралова, 1971а; Ильина и др., 1975]. В работах
Б. Н. Городкова [1916] указывается на произрастание кедра на сфагновых
болотах, где кедр приобретает рямовую форму Pinus sibirica /. turfosaGo-
rodk..аналогично рямовой сосне Pinus sylvestris jAitwinowii. А. И. Лесков
[1940] отмечает существование полурямовой формы кедра в бассейне р. По-
луя на промерзших сухих торфяниках. А. А. Храмов и В. И. Валуцкий
[1970] описывают кустовидную форму болотного кедра, обнаруженную
ими на Иксинском болотном массиве в Новосибирской области.
Распространение кедра на песчаных субстратах отмечено многими
авторами [Поварницын, 1944; Лесков, 1940; Тыртиков, 1974; и др.]. О
приуроченности кедровых лесов к бедным песчаным почвам на северной
границе ареала кедра пишет Н. П. Мишуков [1973]. Разные типы кедровых
лишайниковых лесов на песчаных отложениях террас и междуречий северной
тайги характеризуют А. П. Тыртиков [1974] и Л. А. Аралова [1971а].
В условиях пойменного режима кедр хорошо переносит
непродолжительное затопление во время половодья, благодаря этому он широко рас-
57
пространен на высоких уровнях поймы Оби [Ильина, 1976], а также в до-
линах более мелких рек [Тыртиков, 1974; Лапшина, 1975].
Среди условий, определяющих современный ареал распространения
кедровых лесов, их состав, структуру и производительность, важное место
принадлежит процессам возрастной и восстановительной динамики.
Кедр в первые годы жизни растет очень медленно, плодоношение его
наступает уже на зрелых стадиях развития. Так, кедрачи,
сформировавшиеся после разрушения березового полога, начинают плодоносить в
возрасте 110—150 лет [Мишуков, 1973].
Процессы возрастных смен кедровых лесов имеют специфические
особенности, которые связаны с эколого-биоморфологическими свойствами
кедра как породы, обладающей большой продолжительностью жизни*
На примере лесов Зауральского Приобья и Урала эти процессы были
детально изучены Б. П. Колесниковым и Е. П. Смолоноговым [Колесников,
Смолоногов, 1960; Смолоногов, 1968; Костюченко, Смолоногов, 1976],:
а в районах Среднего Приобья — В. Н. Седых [1979].
Исследования показали, что на водоразделах в коренных лесах, не
нарушенных пожарами и рубками, первые стадии возрастного развития
примерно до 120 лет кедр проходит под пологом смешанных еловых и
пихтовых с примесью березы древостоев. Здесь он участвует в формировании
подроста и подчиненного яруса всегда в меньшем количестве, чем пихта и ель.
Только в возрасте от 160 до 200 лет кедр выходит в основной ярус и начинает
преобладать в древостое [Колесников, Смолоногов, 1960]. Аналогичные
особенности свойственны и долинным типам кедровых лесов.
Исследования В. Н. Седых [1979] в пойме р. Куль-Еган в Среднем Приобье
показали, что кедр, поселяясь и постепенно развиваясь под пологом сосновых
и березово-сосновых древостоев, только через 300—350 лет после
освобождения участка от достоянного воздействия водного потока становится
господствующей породой.
Чрезвычайно сложно проходят и восстановительные процессы в
динамике кедровых лесов. В районах Среднего Приобья, например, В. Н.
Седых [1979] выделил три периода и шесть фаз восстановления пирогенных
кедровников. Первый и второй периоды характеризуются интенсивным
заселением гарей лиственными порогами, главным образом березой и осиной.
В третьем периоде эдификаторная роль переходит к дихте, ели и кедруt
образующим через 120—140 лет от начала лесовосстановления
одноярусные лиственно-темнохвойные насаждения. Только к 160—1*80 годам*кедр
достигает максимальной величины и начинает занимать господствующее
положение в лесных сообществах.
В целом, как показывают работы многих исследователей [Колесников,
Смолоногов, 1960; Седых, 1979; Лапшина, 1975 и др.], в современном
растительном покрове западно-сибирской тайги большая часть темнохвойно-
кедровых, мелколиственно-кедровых и сосново-кедровых лесов
представляет собой не коренные насаждения, а различные стадии восстановления
пирогенных кедровников. Смешанные леса с преобладанием березы, осины
и сосны в плакорных местообитаниях также являются в большинстве своем
коротковременно производными лесными сообществами. Для них
характерно формирование под пологом подроста из темнохвойных пород —
пихты, ели, кедра. Напочвенный покров в таких лесах по составу'и структуре
сходен с покровом коренного темнохвойного леса, где наибольшее развитие
получают синузии зеленых мхов, таежного мелкотравья и бореалЬных
кустарничков.
* Таковы некоторые общие черты возрастной и восстановительной
динамики, присущие всем темнохвойным лесам с доминированием кедра
сибирского.
Пихтовые леса Западно-Сибирской равнины приурочены к южной
части таежной зоны, где они представлены как зональными и плакорными,
так и экологически замещающими типами сообществ с доминированием
Abies sibirica. В целом ареал пихты сибирской почти совпадает с ареалом
кедра и ели7/ что отмечается для всей таежной зоны нашей страны [Бобров,;
58 .
1978), и свидетельствует о тесных ценотических связях этих древесных
пород. Пихтовые леса Западной Сибири — это также полидоминантные
сообщества, в которых наряду с пихтой в сложении древостоя большую
роль в разных условиях играют кедр, ель, береза, иногда липа.
Сибирская пихта — одна из основных пород, слагающих темнохвой-
ные леса. Это дерево первой величины: высота его 30—37 м, а диаметр 50—
70 см. Пихта — одна из самых теневыносливых пород. Она имеет/ всегда
густое охвоение, слабо очищена от сучьев и успешно возобновляется под
пологом даже в самых густых насаждениях.
Предельный возраст пихты 300 лет [Крылов, 1961 ]. На Среднем Урале
и на востоке Западно-Сибирской равнины встречаются деревья в возрасте
150—200 лет [Горчаковский, 1959;Фалалеев, 1964]. Она хорошо
размножается вегетативно,-но экземпляры вегетативного происхождения обычно
не достигают большой высоты и возраста, так как очень рано поражаются
сердцевинной гнилью и отмирают.
Экологический ареал пихтовых лесов определяется высокой
требовательностью пихты к почвенному плодородию и умеренно теплому и
влажному климату. Лучшие условия для произрастания пихты — это глубокие
(с хорошо развитым почвенным профилем) свежие суглинистые почвы [Фа-
лалеев, 1964, Поварницын, 1956]. Она не выносит заболачивания и
промерзания почв, однако в поймах хорошо переносит временное затопление
и подрост ее не гибнет даже на песчано-галечниковых сегментах молодых
пойм.
Эдификаторная роль пихты проявляется главным образом в подзоне
южной тайги, где она образует разнообразные и богатые по составу
высокопродуктивные (300—400 м3/га) лесные сообщества на дренированных
приречных частях водоразделов и высоких речных террас [Любимова, 1972;
Храмов, Валуцкий, 1977; Горожанкина, Константинов, 1978].
Разнообразие пихтовых лесов еще более возрастает в западном прииртышском
секторе южной тайги равнины, где в составе полидомйнантных елово-пихтовых
лесов появляется примесь липы [Хлонов, 1965; Ильина и др., 1975].
Фитоценотическое я экологическое разнообразие основных типов
пихтовых лесов Западно-Сибирской равнины невелико. В плакорных
условиях наибольшее распространение имеют группы ассоциаций зеленомош-
но-мелкотравных и осочковых сообществ. В них всегда обилие кислицы
(Oxalis acetosella) или осочки (Carex macroura) — показателей повышенной
трофности почв. Среди других видов преобладают представители
таежного мелкотравья, в.значительном количестве встречаются также
отдельные виды высокотравья — Aconitum septentrionale, Cacalia hastata, Athy-
rium filix-femiria, Athyrium crenatum и др. Зеленые мхи на плакорных
местообитаниях не имеют большого значения в формировании
напочвенного покрова, их обилие обычно не превышает 20—30%. Зато в разного
рода понижениях рельефа и при увеличении степени глееватостй почв
количество зеленых мхов резко возрастает (до 70—80%), главным образом
за счет распространения Hylocomium splendens и Polytrichum commune.
Как уже отмечалось, пихта не выносит сильного заболачивания, ее
крайний экологический тип в ряду застойного увлажнения —г пихтовый долго-
мошный лес [Шумилова, 1962]. Не типичны пихтовые леса и на
местообитаниях с легкими почвами. На супесях и легких суглинках в них обычно
увеличивается количество майника и линией (за счет кислицы и осочки),
а также Pleuroziuiri schreberi в составе мохового покрова.
Прииртышские варианты-пихтовых лесов с участием липы имеют
более разнообразный и богатый видовой состав травостоя. Здесь обильно
произрастает широкотравные и высокотравные виды, в большом
количестве появляются представители неморальной флоры — Aegopodium ро-
dagraria, Corydalis marchalliana, Lathyrus vernus, Paris quadrifolia. Еще
более богаты по обилию и составу всех ярусов пихтовые леса долин рек,
ложбин стока. Они достигают большой высоты (до 30 м) и бонитета (I—II кл.),;
имеют хорошо развитый подлесок и богатый высокой сомкнутости „траво-
59
стой* в котором преобладают влажно-крупнотравные виды [Крисон, Но-
люшкин, 1975]. , . ■ ■ *
Процессы возрастной и восстановительной динамики пихтовых лесов
изучены недостаточно. Известно, что пихта в молодом возрасте растет очень
медленно и достигает высоты 1 м не ранее 15—20 лет, а под густым пологом
материнских пород еще позднее — в возрасте 50—60 лет . [Фалалеев,
1964]. В дальнейшем темпы ее роста увеличиваются и к 80—100 годам она
заметно вытесняет все другие породы в благоприятных для ее
существования условиях.
Все пихтовые леса Западно-Сибирской равнины чрезвычайно
нарушены пожарами и.рубками, а на востоке южно-таежной подзоны они
поражены тутовым шелкопрядом. В связи с этим на значительных
территориях леса представлены разнообразными производными насаждениями,
в которых пихта заменена другими породами, чаще всего березой и осиной.
Еловые.леса и редколесья. В пределах Западно-Сибирской равнины
еловые леса и редколесья распространены преимущественно в западной,
зауральской, ее части. По занимаемой площади ельники (5,3 млн. га)
уступают кедровым и пихтовым лесам [Леса СССР, 1969]. Главной лесо-
образующей породой еловых лесов равнины является ель сибирская
(Picea obouata). В качестве эдификатора ель в Западной Сибири
встречается реже;, чем другие темнохвойные породы, однако в целом фитоценоти-
ческаяроль ее в строении различных типов лесных сообществ очень
велика. Основная часть ареала ели сибирской приходится на северную
тайгу и подзону редколесий [Ильина, Махно, 1976; Растительность...,
1976]. Там она образует чистые и смешанные, преимущественно с
лиственницей, древостой, отличающиеся большой разреженностью. Более
сомкнутые еловые леса на севере встречаются только в долинах рек.
В подзонах средней и южной тайгчя на большей части территории ель
в строении лесных сообществ занимает подчиненное положение. Как
сопутствующая порода она участвует в составе по ли доминантных лесов
с кедром и пихтой. В качестве примеси встречается во многих
производных мелколиственных и светлохвойных древостоях, а на некоторых
стадиях восстановления кедровых лесов приобретает доминирующее
значение [Колесников, Смолоногов, 1960]. В зауральской части средней и
главным образом южной тайги ель снова встречается в качестве
эдификатора в составе кедрово-еловых и пихтово-еловых с участием липы
лесов.
География еловых лесов Западной Сибири свидетельствует о широком
экологическом ареале их распространения. Сибирская ель
малотребовательна к температуре воздуха и почвы, она хорошо произрастает в местах
близкого залегания вечной мерзлоты, образуя там разветвленную
поверхностную корневую систему. Низкие температуры воздуха и резкие их
перепады не сказываются отрицательно на ее росте и возобновлении.
Наиболее благоприятен для произрастания еловых лесов режим проточного
увлажнения, благодаря чему по всей тайге Западно-Сибирской равнины
наблюдается ясно выраженное тяготение еловых лесов к долинам рек
и ложбинам стока. Довольно широкое распространение ели на
заболоченных почвах свидетельствует о возможности произрастания ее в местах
застойного увлажнения, но здесь она приобретает низкорослые формы,
искривленный ствол, сильную фаутность. Еловые леса одинаково хорошо
развиваются на почвах разного механического состава, наибольшей
производительности они достигают на суглинках и супесях.
В еловых лесах процессы возрастной и восстановительной динамики
протекают в зависимости от их типологического состава и условий место7
произрастания. На севере в разреженных древостоях пожары быстрее
всего уничтожают подрост и молодые ели; старые ели гибнут быстрее, чем
кедр и лиственница, так как кроны их с обилием легковоспламеняющихся
лишайников на сухих ветвях расположены почти у основания стволов и
легко охватываются огнем [Тыртиков, 1974]. На месте коренных еловых
лесов вырастают вторичные березовые древостой. Возобновление ели под
60
их пологом идет успешно даже на ранних стадиях, когда достаточное
развитие получают молодые побеги лиственниц и берез, создающие
необходимую затененность почвы. Хорошо растет подрост ели и в лесах со старым
лишайниковым покровом, где уже образовался слой лишайникового торфа
и благодаря этому установился более или менее постоянный режим
почвенной влаги [Лесков, 1940]. В долинных типах лесов восстановительные
процессы идут более интенсивно, там сомкнутость древостоя выше и
затененность больше, подрост ели развивается здесь очень обильнб, но в то же
время он сам затеняет почву, затрудняя^ ее прогревание и тем самым
способствует глубокому промерзанию почвенных горизонтов [Тыртиков, 1974].
В средней тайге некоторые коренные типы еловых лесов, встр>ечаю-
щиеся в западных районах (Сосьвинское Приобье) [Михеев и др., 1977],;
после пожаров и рубок сменяются березовыми и сосновыми древостоями.
Однако на большей части среднетаежной подзоны леса с преобладанием
ели в плакорных условиях сами являются временными стадиями в процессе
возрастной и восстановительной динамики кедровых лесов.
Для восстановительной динамики южно-таежных еловых лесов
характерно образование смешанных темнохвойно-березовых и темнохвойно-оси-
новых древостоев, постепенно переходящих в смешанно-еловые леса. В
производных южно-таежных лесах ель иногда размножается и вегетативным
путем.
Структурные и фитоценотические особенности еловых лесов различны
в разных зональных условиях. На севере эти леса имеют разреженный
древостой, слабо развитые подлесок и травяно-кустарничковый ярус, а в
напочвенном покрове наблюдается обилие мхов и лишайников.
Южно-таежные еловые леса, наоборот, отличаются высокой сомкнутостью, хорошо
развитым кустарниковым ярусом, обилием трав и почти полным отсутствием
лишайников и мхов.
Основные зональные типы темнохвойных лесов. Как уже было
отмечено, темнохвойные леса Западно-Сибирской равнины обнаруживают
очень тесную связь с зонально-климатическими условиями. Их фито-
ценотический состав, структурные особенности, а также закономерности
'географического распределения четко дифференцируются в зависимости
от приуроченности к определенным зональным полосам [Городков, 19166;
Сочава и др., 1953; Горожанкина, Константинов, 1978; Ильина, 1981].
Особенно это заметно при анализе плакорных типов лесных сообществ,;
которые в пределах разных подзон имеют резкие различия в высоте,;
сомкнутости и производительности древостоя, в составе и
горизонтальном сложении нижних ярусов леса. Специфические черты состава и
структуры, а также своеобразие в системе эколого-географических и
динамических связей присущи всем лесным сообществам сходных зональных
типов.
На карте-схеме распространения темнохвойных лесов (см. рис. 4)
показаны основные зональные типы плакорных и производных сообществ,
а также некоторые экологические варианты, главным образом
заболоченные темнохвойные леса. Все они сгруппированы по подзональным катего-
. риям — редколесья, северо-, средне- и южно-таежные леса.
В группу редколесий из темнохвойных сообществ вошли только лист-
веннично-е левые и еловые редколесья; в группу северотаежных лесов —
лиственнично-еловые и еловые редкостойные леса, характерные для
северной подзональной полосы редкостойных лесов [Идьина, Махно, 1976J,
а также лиственнично-елово-кедровые ле^а — собственно северотаежные..
В группе среднетаежных типов показаны елово-кедровые леса с участием
лиственницы (северный вариант средней тайги) и пихты (южный вариант);
в группе южно-таежных — елово-пихтовые, пихтово-еловые и пихтово-ело-
во-кедровые леса. Кроме перечисленных категорий коренных темнохвойных
лесов в указанные зональные групды вошли также производные,
преимущественно смешанные с доминированием мелколиственных и светлохвой-
ных пород, лесные сообщества, возникшие на месте коренных в результате
пожаров и рубок, а также сельскохозяйственные земли.
Далее coQTaB и структура темнохвойных лесов и производных от них
сообществ, а также особенности их географического распространения
будут характеризоваться в соответствии с содержанием геоботанической
карты — по подзональным категориям и в той последовательности, в
которой они размещены в легенде карты [Растительность..., 1976].
ЛИСТВЕННИЧНО-ЕЛОВЫЕ И ЕЛОВЫЕ РЕДКОЛЕСЬЯ
Еловые редколесья*на территории Западно-Сибирской равнины не
занимают больших площадей. Отдельными массивами они распространены
в бассейнах Оби, Полуя, Надыма. По типологическому составу и фитоце-
нотическому разнообразию еловые редколесья подразделяются на два
зональных типа — северотаежные и лесотундровые.
Лесотундровые типы отличаются наименьшей высотой (4—6 м) и
низкой сомкнутостью древостоя (0,1—0,2), участием в составе нижних ярусов
гипоарктических и арктоальпийских видов и сочетаниями на местности.
с группировками тундровой растительности (рис. 5). Распространены
лесотундровые еловые редколесья в основном в долинах рек — на склонах
высоких надпойменных террас. Северотаежные еловые редколесья имеют
близкие черты состава и структуры, но отличаются несколько большей,
высотой и производительностью древесного яруса; на местности тяготеют
к центральным возвышенным частям водоразделов, где сочетаются с
еловыми и лиственничными редкостойными лесами. В составе северотаежных
еловых редколесий наблюдается большая .примесь лиственницы и березы.
На геоботанической карте по условиям масштаба получили
отображение только некоторые типы преимущественно лесотундровых еловых
редколесий, имеющие наибольшую пространственную выражедность.
Лиственнично-еловые зеленомошно-кустарничковыё
с субарктическими элементами и лишайниковые редколеЬья
в сочетании с ерниковыми и ивняковыми тундрами (33)
Это подразделение легенды включает сочетание двух географических
типов редколесий, связанные в своем распространении с наиболее
дренированными, местообитаниями. Первый — это плакорный тип —
лесотундровые еловые редколесья с зеленомошно-кустарничковым мозаичным
покровом, в котором заметно участие представителей гипоарктической
флоры (Arctous alpina, Calamagrostis lapponica).
Для редколесий этого типа характерны очень разреженный
древостой (сомкнутость 0,1), небольшая высота деревьев (3—5 м) и диаметр не
более 10 см. У взрослых экземпляров ели в таких редколесьях часто
наблюдается сильное искривление стволов, а также характерное для этих
мест и связанное с деятельностью ветра смоление от ветвей их средних
частей, в результате чего образуются как фы двухкронные деревья
[Говорухин, 1957].
Лесотундровые типы еловых зеленомошно-кустарничковых
редколесий, описанные нами на юге Ямала недалеко от с. Панаевска, были
приурочены к суглинистйм холодным почвам, где горизонт сезонной мерзлоты
находился на глубине 30—50 см. В нижних ярусах сообщества имели
мозаичное строение. На кочках высотой не более*полуметра
концентрировались синузии гипоарктических кустарников (Betula nana, Salix pulchra)
и кустарничков \Ledum palustre, Empetrum nigrum, V actinium uliginosum\
Среди них единично встречались арктоальпийские виды — Arctous alpinaf'
A. erythrocarpa. Из травянистых растений присутствовали в небольшом
количестве Carex globularis, Equisetum sylvaticum, Calamagrostis langsdovj-
fii, а также представители гипоарктической флоры — Calamagrostis
lapponica, Pedicularis lapponica. В напочвенном покрове преобладали
зеленые мхи — Pleurozium schreberi, Dicranum elongatum, D. spadiceum^
D. groenlandicum, Aulacomnium turgidum, Polytrichum jensenii. Отдельны-
62 ^ .
Рис, 5. Лиственнично-еловое ерниковое мохово-лишайниковое редколесье. Фото
Б. А. Богоявленского.
ми пятнами среди мхов были распространены листоватые лишайники
Peltigera aphthosa\ P. canina и кустистые лишайники — Cladina arbuscula%
CL rangiferina, CI. mitis, Cladonia amaurocraea, Cetraria cucullata. В
широких межкочечных понижениях — сплошной покров из тех же видов мхов
и лишайников с единичными экземплярами кустарничков и травс
Лишайниковые типы лиственнично-еловых редколесий встречаются
на западе в бассейне р. Полуй, а также на востоке в Таз-Енисейском
междуречье. Они занимают наиболее сухие возвышенные участки с глубокоот-
таивающими песчаными почвами. Древостой их более разнообразен и
имеет сомкнутость 0,2—0,3. Ель здесь достигает высоты 10—12 м при диа-
63
метре 20—23 см, имеет сильносбежистые стволы, ветви ее покрыты эпифит-
ными лишайниками. Кроме ели обычно значительна примесь лиственницы
высотой до 15 м и березы (8—10 м). Травяно-кустарничковый ярус очень
разрежен. Отдельные виды кустарничков и трав концентрируются только
под кронами деревьев. Среди них нет арктоадьпийских видов, но зато
встречаются представители бореальной флоры— Vaccinium vitis-idaea,
Linnaea borealis. Сплошной покров из лишайников и мхов имеет
мозаичное строение. Наиболее разрастаются пятна-подушки из Cladina stellaris,
между ними — полосы Cladonia gracilis, CI. deformis, Cetraria islandica,
Cladonia amaurocraeq и группы зеленых мхов — Polytrichum jensenii, P. alpes-
tre„ Dicranum elongatum и др. В некоторых лишайниковых редколесьях
бассейна Полуя на сходных местообитаниях возрастает роль кустарничков,,
особенно Ledum palustre, и соответственно этому увеличивается количество .
зеленых мхов, которые поселяются в более затененных местах, а часто под
кустиками Betula папа и Ledum palustre [Лесков, 19401.
В восточной части ЗапаДно-Сибирской равнины еловые моховые и
лишайниковые редколесья встречаются редко. На геоботанической карте
массив этих редколесий отмечен только в левобережье р. Таз. В районе
Советских озер небольшие участки еловых багульниково-лишайниковых
редколесий описала Л. И. Мельцер. В них наряду с елью и лиственницей
отмечено большое участие березы {Betula tortuosa), а в подлеске — ерника
и ольхи (Duschekia fruticosa). Участие березы, отмеченное во многих типах
лесотундровых и таежных редколесий, по-видимому, больше всего
связано с антропогенным влиянием. В местах прошлых пожаров древостой
всегда имеет смешанный состав, а в покрове наряду с обычными видами
лишайников и зеленых мхов большую роль играют пирогенные виды (Polytrichum
juniperinurn, P. alpestre). * *
На местности все лесотундровые типы еловых редколесий сочетаются
с участками кустарниковых — ивняковых и ольховниково-ерниковых
тундр, которые занимают более возвышенные местоположения —
преимущественно северные склоны.
Лиственнично-еловые и еловые,
местами с березой лишайниково-сфагновые редколесья
в сочетании с кустарничково-мохово-лишайниковыми
плоскобугристыми болотами (34)
На севере Западно-Сибирской равнины среди массивов
плоскобугристых комплексных болот распространены отдельными участками
сообщества заболоченных лиственнично-еловых и еловых лишайниково-сфагно-
вых редколесий. Они занимают окраинные части болотных массивов и
часто чередуются с заболоченными лиственничными лесами и
редколесьями. Приурочены они к торфяно-глеевым почвам и к поверхностям с ярко
выраженным бугристым микрорельефом с торфяно-глеевыми почвами,
с чем связана сильная мозаичность напочвенного покрова этих
редколесий. Древесный ярус, как правило, очень разрежен (0,1), он состоит из
ели с примесью лиственницы, сосны и березы извилистой {Betula tortuosa).
Древостой в целом очень угнетен, его возобновление слабое. Кустарники
и кустарнички развиваются на более высоких кочках, среди них обычны
Betula nana, Ledum palustre, Enipetrum nigrum. На низких кочках к ним
примешиваются олиготрофные виды: Кассандра {Chamaedaphne calyculata),
клюква {Oxycoccusmicrocarpus),.-подбел {Andromedapolifolia). Из трав
встречаются Rubus chamaemorus, Calamagrostis neglecta, Car ex globularis.
Сплошной мохово-лишайниковый покров имеет мозаичное распределение. На
вершинах кочек преобладают мхи — Sphagnum fuscum, Sph. nemoreum,
Pleurozium schreberi, Polytrichum commune и лишайники — Cladina
stellaris, CI. rangiferina, Cladonia gracilis, Cetraria cucullata? C. islandica,
Peltigera aphthosa, Nephroma arcticum. Такие кустарничково-лишайниково-
сфагновые еловые редколесья описаны в бассейне Полуя А. И. Лесковым
" [19401. * '
64
В Ивдельском Зауралье заболоченные еловые редколесья изучала
К. Н. Игошина [1949]. Она проследила разнообразные переходы от зе-
леномошных и ягельниковых редколесий к сфагновым.
На северо-востоке Западно-Сибирской равнины в Пур-Тазовском
междуречье нами были описаны участки заболоченных еловых
редколесий с большим участием лиственницы и березы (Betula pubescens) и
ерником в подлеске. Их древостой был также сильно разрежен, угнетен и в
некоторых местах имел вид криволесья. Высота елей не превышала 10 м,,
а диаметр 15—18 см, единичные березы и лиственницы были еще ниже —
7—8.м. Возобновление очень слабое, многие экземпляры ели и
лиственницы высотой 1—3 м имели уже сухостойный угнетенный вид. Подлесок из
Betula папа был развит довольно хорошо, его покрытие в некоторых
местах доходило до 60% и высота до 1—1,5 м. В покрове, имеющем
комплексной характер, преобладали лишайниково-олиготрофнокустарничковые
группировки по кочкам и зеленомошно-сфагновые по микропонижениям.
СЕВЕРОТАЕЖНЫЕ ЛИСТВЕННИЧНО-ЕЛОВЫЕ
И ЕЛОВЫЕ РЕДКОСТОЙНЫЕ ЛЕСА
И ПРОИЗВОДНЫЕ СООБЩЕСТВА НА ИХ МЕСТЕ
Лиственнично-еловые редкостойные леса занимают значительные
площади на северо-западе и северо-востоке Западно-Сибирской равнины.
В центральной части они почти не встречаются, уступая место
лиственничным редкостойным лесам и сфагново-лишайниковым плоско- и крупно-
бургистым болотам. В Зауралье и на Казымо-Полуйском междуречье эти
леса приурочены к легким супесчаным и песчаным почвам. На востоке в
левобережье Енисея они встречаются на суглинистых Отложениях.
Наибольшие площади занимают лиственнично-еловые редкостойные леса с
покровом из зеленых мхов и лишайника. Они развиваются на
супесчаных ипесчаных подзолистых почвах, имеют очень разреженный древостой
(0,3—0,4) и местами переходят в елово-лиственничные и
лиственнично-еловые лишайниковые редколесья. От последних они отличаются большей
производительностью древостоя (V, реже IV класс бонитета), большей его,
сомкнутостью и высотой. В их'напочвенном покрове тоже преобладают
кустистые лишайники из родов Cladonia, Cladina и Cetraria, встречаются и
зеленые мхи (Pleurozium, Hylocomium). Кустарничковый ярус очень
разрежен и состоит из обычных бореальных и гипоарктических видов.
Южнее в этих лесах роль лиственницы" ослабевает, местами
встречаются почти чистые еловые древостой, а в бассейне р. Казым в еловых лерах
появляется значительная примесь сосны.
Лиственнично-еловые и еловые зеленомошно лишайниковые
и зеленомошные редкостойные леса
в сочетании с ерниково-сфагново-лишайниковыми
плоскобугристыми болотами (41)
На карте по условиям масштаба показан обобщенный тип зеленомощно-
лишайниковых и зеленомошных еловых лесов в его характерном
сочетании с заболоченными еловыми и лиственничными лесами и сфагново-ли-
шайниковыми бугристыми болотами.
Еловые лишайниково-зеленомошные леса широко распространены
в бассейне р. Казым. Они характеризуются разреженным древостоем,
состоящим из преобладающей по числу стволов ели с примесью березы и
сосны. По нашим наблюдениям, на участках таких лесов средняя высота
всех деревьев примерно одинакова — 15—16 м, а диаметр стволов у ели
20 см, у сосны 16—18 см, у березы 15—20 см. В подросте преобладает ель
высотой 1—3 м семенного происхождения, в меньшем количестве
встречается береза и единично сосна (высота 1—0,5 м). В подлеске отдельные
кусты Duschekia fruticosa высотой 2 м. Травяно-кустарничковый ярус в
• отдельных местах имеет покрытие до 50%. В нем преобладают багульник
и брусника. Группами встречаются черника, голубика; единично — во-
5 Заказ № 297 65
дяника, линнея, а из трав — осока шаровидная, хвощ лесной и морошка.
Моховой покров сплошной, состоит в основном из зеленых мхов (Pleuro-
zium schreberi, Hylocomium splendens, Polytrich um commune, P. alpestre)f
в которые равномерно вкраплены пятна кустистых (Cladina stellaris,
CI. arbuscula, CI. rangiferina) и листоватых (Peltigera aphthosa, Nephroma
arcticum) лишайников.
Сильно разреженные лишайниковые еловые леса, в которых
напочвенный покров состоит в основном из Cladina arbuscula и CI. rangiferina*
распространены отдельными массивами по наиболее повышенным местам
с песчаными почвами.
Еловые зеленомошные и лишайниковые леса восточной части
равнины (бассейны рек Пур, Таз, Енисей) имеют сходный состав и строение
всех ярусов леса. Можно отметить лишь возрастание в них роли
лиственницы, местами значительную примесь кедра и обязательное участие
ерника в подлеске.
Лиственнично-елово-березовые и березовые с елью
лишайниково-зеленомошные редкостойные леса (42)
Производные от еловых и лиственнично-еловых смешанные с
преобладанием березы леса формируются в местах, подверженных пожарам и
рубкам. Древостой этих лесов на разных стадиях восстановления имеют
различный состав. Абсолютное преобладание березы наблюдается
преимущественно на первых этапах послепожарных смен. Далее образуются
смешанные древостой со значительным участием лиственницы и ели,
впоследствии вытесняющими все другие породы. В целом встречающиеся на месте
коренных производные лиственнично-елово-березовые и березовые с елью
леса можно отнести к динамической категории короткопроизводных [Со-
чава, 19791 сообществ, относительно быстро восстанавливающихся и не
претерпевающих необратимых изменений в составе и структуре всех
ярусов леса. Современный травяно-кустарничковый ярус и мохово-лишайни-
ковый покров этих лесов имеют состав, близкий к коренному лесу.
Наиболее длительные и 'устойчивые стадии восстановления
лиственнично-еловых лишайниковых лесов характеризуются преобладанием в покрове
зеленых мхов, среди которых заметную роль играют мхи из рода Polytrichum.
Еловые зеленомошные леса восстанавливаются аналогичным путем, но в
их покрове среди зеленых мхов появляется больше гигрофильных видов
(Polytrichum commune, Sphagnum), что свидетельствует о тенденциях к
временному заболачиванию этих территорий.
Производные лиственнично-елово-березовые леса, как и коренные
типы, на местности постоянно сочетаются с плоскобугристыми болотами
и заболоченными лиственнично-еловыми и еловыми лесами.
Лиственнично-еловые и еловые кустарничково-сфагновые
с ерником редкостойные леса
в сочетании с кустарничково-мохово-лишайниковыми
плоско- и крупнобугристыми болотами (43)
Широкое распространение заболоченных еловых лесов на Западно-
Сибирской равнине в значительной степени связано с динамикой
естественных смен растительного покрова, происходящих на водоразделах и в
долинах рек под влиянием современных гидролого-геоморфологических
и криогенных процессов [Городков, 1946; Тыртиков, 1974]. Эти леса на
местности постоянно чередуются с кустарничково-мохово-лишайниковы-
ми плоско- и крупнобугристыми болотами. Ряд заболачивания при
переходе от долин рек к не дренированным водоразделам начинается с еловых
(лиственнично-еловых или кедрово-еловых) зеленомошных лесов, в
которых в связи с развитием сплошного мохового покрова и накопления
торфянистого слоя понижается температура почвы, уменьшается глубина
протаивания почвенных горизонтов и создаются условия застойного ув-
66
лажнения [Тыртиков, 1974]. В результате в покрове зеленых мхов
появляются синузии Poly.trichum commune, а затем и Sphagnum angustifolium.
Рост деревьев сильно угнетается, древостой изреживается. Во многих
местах еловые леса переходят в еловые редколесья, а затем в редины на
сфагновых болотах. Ель здесь находится на грани своего экологического
ареала, при дальнейшем заболачивании она уступает место сосне в более
южных районах [Сочава, 1956] или лиственнице в северных
[Тыртиков, 1974]. . . -■ ' •
Лиственнично-еловые и еловые кустарничково-сфагновые леса — это
наиболее распространенная и устойчивая стадия в ряду заболачивания.
Эти леса были описаны нами в бассейне р. Казым и на Пур-Тазовском
водоразделе. Они изучались Е. И. Лапшиной в левобережье Оби — на
водоразделе рек Сыни и Лесми-Югана. Для заболоченных еловых лесов
характерен разреженный древостой (сомкнутость 0,3—0,5), состоящий из
ели с примесью березы (2—3 единицы) и единично лиственницы и сосны.
Деревья угнетены, стволы их искривлены и увешаны эпифитными мхами
и лишайниками (Parmelia sp., Uznea barbata). Много сухостоя. Высота
елей 9—12 м, диаметр 12—18 см, высота берез примерно такая жа — 9—
10 м. Лиственницы несколько выше — 13—15 м. Подрост очень редкий,
угнетенный; в нем присутствует ель, единично встречается сосна, в
несколько большем количестве береза вегетативного происхождения. В
подлеске группы кустов ерника. В травяно-кустарничковом ярусе
преобладают олиготрофные кустарнички — голубика, Кассандра, багульник,
встречаются также водяника, клюква, брусника, довольно обильны осока
шаровидная и морошка.
В сплошном покрове мхов господствуют сфагны — Sphagnum angus-
tifolium, Sph. fuscum; довольно много кукушкина льна и зеленых мхов —
Pleurozium schreberi, Ptilium crista-castrensis. Лишайники встречаются
пятнами. Среди них обычны Peltigera aphthosa, Nephroma arcticum^ Cladina
rangiferina.
СЕВЕРОТАЕЖНЫЕ ЛИСТВЕННИЧНО-ЕЛОВО-КЕДРОВЫЕ ЛЕСА
И ПРОИЗВОДНЫЕ СООБЩЕСТВА НА ИХ МЕСТЕ
Эти леса встречаются преимущественно в южной полосе подзоны
северной тайги, непосредственно к югу от области распространения
редкостойных лиственничных лесов и редколесий. В этой полосе северной тайги
они отмечались Б. НГ. Городковым [1916 б, 19461, В. Б, Сочавой, Т. И.
Исаченко, А. Н. Лукичевой [1953], А. П. Тыртиковым [1974]; в районе Таз-
Енисейского междуречья различные типы северотаежных кедровых лесов
описывала Н. С. Аралова [1971], на водоразделе Надыма и Пура —
Л. Н. Тагунова [Ландшафтные индикаторы..., 1974], в левобережье
р. Оби и в бассейне р. Таз — Е. И. Лапшина [1975, 1978]. Эти леса
изучались в бассейне Казыма и на Пур-Тазовском
междуречье-Западно-Сибирской экспедицией Института географии СО АН СССР.
Лиственнично-елово-кедровые леса распространены отдельными
массивами по всей южной полосе северотаежной подзоны. В Зауральском
Приобье они встречаются на левобережье Сев. Сосьвы и в бассейнах рек
Ляпина и Сыни. В центральной части значительные массивы их отмечены
в бассейнах рек Казыма и Куновата, в верховьях Агана и Таза. Н'а
левобережье Енисея эти леса продвигаются далеко к северу, доходят до Ту-
рухана.
Коренные леса не занимают больших площадей в северной равнинной
тайге Западной Сибири. На всем протяжении своего ареала они постоянно
сочетаются с различными производными насаждениями в основном пи-
рогенного происхождения, которые находятся на различных стадиях
восстановления и образуют массивы гораздо большей площади, чем сами
коренные леса.
5* 67
Из плакорных коренных типов наибольшее распространение получили
две обобщенные группы ассоциаций северотаежных кедровых лесов.
Первая — лиственнично-елово-кедровые лишайниково-зеленомошно-кустар-
ничковые леса (44). Это основной зональный тип сообщества. *Его
отличительные фитоценотические черты — обязательное участие лиственницы в
составе древостоя, преобладание в нижних ярусах леса синузий бореаль-
ных и гипоарктических кустарничков, значительная роль лишайников в
напочвенном покрове и в целом низкая производительность древостоя
(V класс бонитета) при малой его сомкнутости и небольшой высоте
(12-15 м).
Следующая группа ассоциаций — ело$о-кедровые с лиственницей
зеленомошно-кустарничковые с ольховником в подлеске леса *(№ 47) —
несколько меньше распространена и является переходной к среднетаежно-
му типу сообщества.
Лиственнично-елово-кедровые
лишайнщсово-зеленомошно-кустарничковые леса (44)
Растительные сообщества, объединенные в эту группу, приурочены к
приречным хорошо дренированным участкам водоразделов, сложенных
суглинистыми слабоподзолистыми глееватыми оторфованными почвами.
Для таких местообитаний весьма характерно наличие слоя многолетне-
мерзлых пород, температура которых колеблется от —0,2 до ±0,1°С [Ара-
лова, 1971.)
Древостой этих лесов разрежен (сомкнутость 0,5—0,6).
Доминирующие породы кедр и ель не достигают в них большой высоты, в зрелом
возрасте (100—150 лет) они образуют полог высотой не более 12—15 м.
Лиственница в большинстве случаев превосходит по высоте эти породы,
достигая иногда 18—20 м. Как правило, в лесах наблюдается примесь
березы (Betula pubescens, В. pendula) и иногда сосны (Pinus sylvestris),
образующих более низкий полог леса. Естественное возобновление под
пологом северотаежных лиственнично-елово-кедровых лесов медленное,
подрост их угнетен, в нем из темнохвойных пород обычно преобладает ель.
Подлесок также развит плохо, единично или небольшими группами там
встречаются ольховник (Duschekia fruticpsa), можжевельник (Junipgrus
communis), рябина (Sorbus sibirica) и шиповник (Rosa acicularis).
Травяно-кустарничковый ярус образован гипоарктическими и бо-
реальными кустарничками. Их проективное покрйтие достаточно велико
(60—70%). Наиболее распространены багульник (Ledum palustre),
голубика (Vaccinium uliginosum), водяника (Empetrum nigrum), брусника (Vac-
cinium vitis-idaea), реже черника и линнея. Из трав обычны плауны, хвощ
лесной, осока шаровидная, но их участие незначительно. Мхи и пятна
лишайников образуют сплошной покров, где преобладают Pleurozium
schreberi, Polytrichum alpestre. Пятна лишайников (Cladina stellaris, Ste-
reocaulon paschale, Peltigera aphthosa) распределены довольно равномерно
и создают мозаичность напочвенного покрова.
Таковы некоторые общие особенности фитоценотического состава и
•структуры плакорных типов кедровых лесов центральной части
северотаежной подзоны;
В левобережье Оби на склонах холмистого междуречья Лесми-Югана
и Колик-Югана Е. И. Лапшиной [1971] были описаны участки коренных
лесов, в древостое которых ель местами преобладала над кедром, но в
остальном состав и структура всех ярусов этих лесных сообществ
оставались аналогичными вышеописанным типам северотаежных кедровых
лесов центральной части Обь-Енисейского водораздела. Такие колебания в
количественных соотношениях пород, не меняющие общего характера
леса, весьма характерны для западного зауральского сектора северной
тайги Западной Сибири, где роль ели в строении древостоя значительно
усилена.
68
Обследованные нами лесвййе массивы лиственнично-елово-кедровых
лесов южной части Пур-Тазовского водораздела отличались еще более
разреженным и низкорослым древостоем (11—13 м), большим участием в
его составе лиственницы, иногда отсутствием ели и значительным
развитием в напочвенном покрове синузий кустистых лишайников (Cladonia,
Cladina, Cetraria). Для таких лесов характерны почвы с близкозалегающей
вечной мерзлотой.
Описанные выше сообщества лиственнично-елово-кедровых лишай-
никово-зеленомошно-кустарничковых лесов на карте показаны
обобщенными выделами, включающими как собственно плакорные типы этих
лесов, так и многочисленные их экологические варианты, возникающие
в условиях избыточного увлажнения или, наоборот, в местах более сухих,
где происходит изменение механического состава почв. Наиболее типичны
пространственные сочетания плакорных зеленомошно-кустарничковых
лесов с различными сообществами заболачивающихся кедровников. Среди
последних, выделяются березово-кедровые и лиственнично-сосново-кедро-
вые лишайниково-зеленомошно-долгомошные и сфагновые леса. Синузий
сфагновых мхов в таких лесах появляются уже на первых стадиях
заболачивания, они первоначально концентрируются в микропонижениях и
«окнах» ~ западинах. Впоследствии в заболоченных лесах формируется
кочковатый микрорельеф и. растительный покров приобретает
мозаичное строение.
На высоких террасах рек, а ближе к северу на гривистых и холмистых
водоразделах, сложенных супесчаными и песчаными породами с глубоко-
залегающим слоем вечной мерзлоты, среди плакорных кедровых лесов
часто появляются маесивы елово-лиственнично-кедровых и березово-
лиственнично-кедровых багульниково- или бруснично-лишайниковых
лесные сообществ — псаммофитный вариант северотаежных кедровников.
Березовые с кедром, елью, лиственницей
кустарничково-лишайниково-зеленомошные леса (45)
Несмотря на то, что южная полоса северной тайги Западной Сибири
слабо заселена и мало освоена человеком, почти все леса этих территорий
в разное время подвергались пожарам [Городков, 1946; Тыртиков, 1974].
Сейчас на больших площадях они представлены короткопроизводными
березовыми и сосновыми насаждениями. Под этими лесами развивается
вечная мерзлота, хотя мощность ее обычно меньше, чем под коренными
ненарушенными лесными сообществами. По данным А. Н. Тыртикова [1974],
слой многолетнемерзлых пород в березовых лесах начинает формироваться
через 60—80 лет после пожара, когда под пологом березы появляется
подрост темнохвойных пород.
На карте показана одна из наиболее устойчивых стадий послепожар-
ных смен коренных кедровых лесов. Она представлена березовыми с
примесью хвойных пород кустарничково-зеленомошными лесами. В составе
этих лесов, описанных в бассейне Казыма, доминирование в древесном
ярусе имела береза (Betula pendula). Она вместе с лиственницей, елью и
единичной сосной образовала более сомкнутый (0,6—0,7) нижний полог
леса. В верхнем сильно разреженном пологе сохранились отдельными
группами, или «маяками», представители коренного материнского
древостоя— лиственницы и кедра. Основной восстанавливающийся древостой
имел высоту 13—15 ми средний класс бонитета'по преобладающей породе
V. Лиственницы в верхнем пологе достигали высоты 15—16 м,
имели'средний диаметр 18—20 м, а отдельные деревья в возрасте свыше 300 лет
иногда достигали диаметра 50—60 см. Повсюду в таких лесах наблюдались
следы горелого древостоя (обгоревшие старые пни, отдельные стволы
деревьев). Возобновление древесных пород происходило неравномерно. В
более затемненных местах обычно хорошо возобновлялась ель (высота под-
69
роста от 2,5 до 4 м) и в меньшем количестве кедр (высотой Q,7—1,5 м).
Сосна и береза выбирали осветленные участки, где распределялись
отдельными группами. Кустарничковый ярус в производных березовых лесах
обычно представлен группами шиповника (Rosa acicularis), можжевельника (Ju-
niperus communis), иногда карликовой березкой (Betula nana). В
напочвенном покрове доминировали синузии гипоарктических и бореальных
кустарничков, мхов и лишайников, аналогично преобладающим в покрове
коренных лесов. . ■
*Молодые стадии пирогенных кедровых лесов обычно представлены
березовыми насаждениями почти без примеси хвойных пород, В их
покрове преобладают зеленые мхи, главным образом Pleurozium schreberi, а на
осветленных местах бурно развиваются синузии политриховых мхов
(Polytrichum alpestre, P. juniperinum) и трубчатых лишайников (Stereo-
caulon paschale).
Сосновые с кедром, елью, лиственницей
кустарничково-лишайниково-зеленомошные леса (46)
Широкое развитие производных березовых лесов на месте севербта-
ежных лиственнично-елово-кедровых особенно характерно для
центральных и восточных частей Обь-Енисейского водораздела. На западе в
левобережье Оби более устойчивы и широко распространены коротко-длитель-
нопроизводные сосновые леса. В этих районах они наиболее подробно
изучены Е. И. Лапшиной [1971].
Производные сосновые северотаежные леса формируются в основном
на легких суглинках в различной степени оподзоленных. На более ранних
стадиях послепожарных смен в составе древостоя значительная примесь
березы и почти нет в подросте темнохвойных пород. В дальнейшем по мере
роста сосны береза выпадает полностью и начинают возобновляться темно-
хвойные породы и лиственница. Но часто в зрелых вторичных сосняках
происходят низовые пожары, которые уничтожают темнохвойный подрост*
и тогда вновь происходит возобновление березы и сосны. Наиболее
длительные стадии восстановления представлены сосновыми с примесью
кедра, лиственницы багульниково-зеленомошными лесами. Эти леса имеют
хорошо развитый древостой, в котором преобладает сосна, имеющая в
возрасте 100—120 лет среднюю высоту 15—17 м. Единично в верхнем пологе
встречаются ель, кедр и лиственница, они же образуют подрост разного
возраста и разной высоты — от 0,5 до 5 м. Однако большая часть экземп^
ляров подроста имеет угнетенный вид, среди него много сухостоя.
Кустарниковый ярус развит слабо, редко разбросаны отдельные кусты
шиповника и рябины. Покров из кустарников имеет неравномерное покрытие —
от 40 до 70%. В нем преобладает багульник, часто достигающий высоты
70—80 см, в меньшем количестве встречаются брусника и рассеянно
отдельными кустиками водяника. Из травянистых видов обычны только
синузии плаунов (Diphasiastrum complanatum, Lycopodium annotinum) и
осоки шаровидной (Carex globularis). Сплошной покров образуют зеленые мхи,
среди которых преобладает Pleurozium schreberi. Кроме него, отдельными
пятнами распространен Polytrichum alpestre и единично в примеси Die-
ranumpolysetum.VL3 лишайников обычны Cladina rangiferina,. CI. stellaris
и Stereocaulon paschale. ' , ■
Елово-кедровые, местами с лиственницей
зеленомошно-кустарничковые с ольховником
в подлеске леса (47)
Это вторая обобщенная группа ассоциаций полидоминантных теми о-
хвойно-кедровых лесов, которая получила отображение на
геоботанической карте. Сообщества этой группы имеют более ограниченное
распространение в северной тайге Зацадно-Сибирской равнины, чем описанные
лиственнично-елово-кедровые леса, и по ряду' фитоценотических и эколо-
70
гических свойств представляют как бы переходный вариант к лесам сред-
нетаежного типа. На карте растительности равнины [Растительность...,
1976] эти леса показаны в основном цо возвышенным массивам южных
отрогов Сибирских увалов. Древостой их образован елью и кедром,
лиственница встречается в небольшой примеси или отсутствует. Характерно
активное возобновление темнохвойных пород и хорошее развитие
подлеска, в нем следует выделить ольху кустарную {DuscheMa fruticosa), которая
встречается повсеместно, а местами образует группы зарослей. По составу
травяно-кустарничкового яруса группа ассоциаций елово-кедровых кус-
тарничковых зеленомошных лесов близка к описанному выше собственно
северотаежному т^пу лиственнично-елово-кедровых сообществ.
Несколько большее участие таежного мелкотравья (Maianthemum
bifolium, Pyrola rotundifolia, Trientalis europaea) и соответственно меньшая
фитоценотическая роль некоторых видов гипоарктических кустарничков
(Empetrum nigrum, Vaccinium uliginosum) при постоянном доминировании
Ledum palustre свидетельствуют о более южном характере этого типа
лесов. Зато почти среднетаежный облик имеет древесный ярус этих
сообществ. По описанию Е. И. Лапшиной [1978], в районах Верхнетазовской
возвышенности в составе елово-кедровых насаждений кедры достигали
высоты 16—18 м и диаметра 32—44 см, высота елей была 16—17 м при
диаметре 22—34 см, высота единично встречающихся лиственниц доходила
до 20 м. Несколько увеличились и сомкнутость древесного яруса
коренных лесов (0,6— 0,7) и их производительность (IV—V классы бонитета).
В подросте преобладали кедр и ель, которые местами формировали второй
полог древостоя высотой 7—9 м, там же встречалась и пихта,
возобновление ее шло вегетативно, путем укоренения ветвей* В кустарниковом ярусе
кроме ольхи присутствовали шиповник и* рябина.
На карте контуры этого типа северотаежных кедровых лесов также
включают, кроме собственно плакорных сообществ, многочисленные их
экологические варианты. Среди них переходными к гидроморфному ряду •
можно считать березово-кедровые насаждения с багульниково-зеленомош-
ным покровом, развивающиеся на торфянисто-подзолистых элювиально-
глеевых почвах с торфяной подстилкой мощностью 5—6 см
[Лапшина, 1978].
Различные стадии заболачивания кедровых лесов также
представлены сообществами долгомошной и сфагновой групп. В их составе постоянно
доминируют олиготрофные кустарнички, осока шаровидная, политри-
ховые (Polytrichum commune^ P. alpestre) и сфагновые {Sphagnum girgen-
sohnii, Sph. russowii, Sph. angustifolium) мхи.
Березовые с оеиной, кедром и ольховником в подлеске
зеленомошно-кустарничковые леса (48).
Сосновые с кедром зеленомошно-кустарничковые леса (49)
После пожаров елово-кедровые с ольхой леса восстанавливаются
через смену пород. Более ранней, но достаточно продолжительной
стадией, являются березовые с осиной, кедром и ольховником в подлеске
зеленомошно-кустарничковые леса. На заключительных этапах
восстановления из них формируются березово-кедровые и осиново-березово-кедровые
насаждения. Верхний полог в них часто образуют березы, осины и старые
кедры, сохранившиеся послехпожара. Во втором пологе преобладает ног
вое поколение кедра, вытесняющее мелколиственные породы. В подросте
господствуют ель и кедр, возобновление мелколиственных пород только
на осветленных местах и на участках вывалов старых деревьев. Кустар-
циковый ярус лучше развивается в производных лесах с
преобладанием березы.
На заключительных стадиях в смешанных березово-кедровых древостоях
подлесок редкий. Однако во всех типах этих лесов в подлеске обязательно
присутствует ольха кустарниковая, образующая группы различной
густоты.
71
Сосновые с кедром зеленомошно-кустарничковые леса возникают в
процессе восстановления кедровых преимущественно на почвах более
легкого механического состава — на супесях и легких суглинках. Иногда
они замещают сосново-кедровые леса и на более влажных глееватых
почвах. В. цел ом сосновые производные леса являются более устойчивой и
длительно существующей стадией и сохраняются особенно долго в местах,
подверженных часто повторяющимся пожарам.
Во всех типах производных березовых и сосновых лесов травяно-кус-
тарничковый ярус и моховой покров близки по составу к таковым в
коренных елово-кедровых сообществах. К пирогенным элементам в этих
лесах кожно отнести возрастающие по обилию синузии багульника,
водяники и политриховых мхов (Pdlytrichum alpestre, P. juniperinum).
СРЕДНЕТАЕЖНЫЕ ЕЛОВО-КЕДРОВЫЕ ЛЕСА
И ПРОИЗВОДНЫЕ СООБЩЕСТВА НА ИХ МЕСТЕ
Елово-кедровые леса наиболее типичны для подзоны средней тайги%
где занимают пологоувалистые суглинистые междуречья повышенных
равнин. Значительно реже и небольшими массивами встречаются на
супесчаных слоистых отложениях холмистых равнин, или по надпойменным
террасам и их склонам.
Наибольшие площади этих лесов и их производных сосредоточены на
Обь-Иртышском междуречье и в правобережье Оби— на Тым-Кетском и
Вах-Тымском междуречьях. Коренных елово-кедровых лесов
сравнительно немного и располагаются они разобщенными массивами среди
производных лесных сообществ, относящихся к различным стадиям послепо-
жарного восстановления. В зоне средней тайги основным фактором
нарушения естественного * хода развития растительности являются пожары*
вырубки занимают сравнительно небольшие площади и сосредоточены
преимущественно вдоль железных дорог Ивдель — Обь, Асино — Белый
Яр, Тюмень — Сургут. Относительно гомогенная растительность
местоположений плакорного типа показана на карте обобщенными выделами
коренных елово-кедровых лесов двух групп ассоциаций (72 и 75). В
северной части подзолы отчетливо проявляется преобладание лесов кустар^
ничково-зеленомошной фуппы с участием в древесном ярусе лиственницы
(72). В южной части подзоны господствуют леса мелкотравно-кустарнич-
ково-зеленомошной группы с участием в древесном ярусе пихты. В
легенде леса этой группы названы по узловой ассоциации — мелкотравно-брус-
нично-зеленомошные (75), имеющие наибольшее распространение. В
группы коренных лесов (75) объединены сообщества, наиболее близкие к
заключительной стадии восстановления,— сообщества, в которых эдифика-
торная роль принадлежит ели и кедру, а участие сопутствующих пород —
березы, осины, сосны — в составе древесного яруса минимальное.
Производные леса показаны обобщенными выделами, включающими
сообщества различных возрастных групп. Преобладают смешанные леса
(темнохвойно-березовые, темнохвойно-осиновые) со вторым ярусом .из
кедра, ели и пихты (73, 76, 77). Для восстановительного процесса елово-
кедровых лесов, формирующихся на почвах легкого механического
состава, характерно образование производных тбмнохвойно-сосновых
лесов (74, 78):
На гидроморфных местообитаниях в средней тайге развиваются
сообщества елово-кедровых лесов долгомошной й сфагновой групп,
образующих устойчивые сочетания с пространственно сопряженными
сообществами болотной растительности, формирующейся в многочисленных
микропонижениях.
Среднетаежные леса изучены недостаточно. Описания выделенных
групп елово-кедровых лесов даны в основном по опубликованным
материалам. Краткие сведения о различных сообществах этих лесов приведены
в работах М. Ф. Елизарьевой [1961], Е. П. Смолоногова и В. И.
Никулина [1963], Е. П, Прокопьева [1968], И. С. Ильиной и Ю. С. Толчельникова
72
[1968], Е. И. Лапшиной [1973], С. М. Горожанкиной, В. Д.
Константинова [1978]. Вопросам восстановительно-возрастной динамики среднетаеж-
ных кедровых лесов посвящены работы Б/ П. Колесникова, Е. П. Смоло-
ногова [I960] и В. Н. Седых [1979].
Елово-кедровые, с лиственницей
кустарничково-зеленомошные леса, местами в сочетании
с кустарничково-сфагновыми олиготрофными болотами (72)
Распространение елово-кедровых с лиственницей кустарничково-зе-
леномошных лесов отчетливо увязывается с субширотной полосой
повышенных равнин — Северного и Южного Люлимвора,,Белогорского
материка, Дубчесской холмистой равнины. .
Эти леса приурочены к пологовыпуклым водоразделам и верхним
частям пологих склонов со слабо подзолистыми элювиально-глееватыми
суглинистыми длительногсезонномерзлотными появами. [Караваева, 1973 ].
Нередко они формируются на двучленных наносах —- суглинок сменяется
супесью или опесчаненным суглинком с хрящом.
Елово-кедровые кустарничково-зеленомошные леса представляют
северный вариант среднетаежных темнохвойных лесов. Они флористически
сходны с северотаежными елово-кедровыми лесами (47), но существенно
отличаются от них более высокой продуктивностью (преимущественно
IV класс бонитета), большей высотой древесного яруса (17—20 м) и
увеличением его сомкнутости (0,6—0,8). Некоторые отличия отмечаются также
в возрастании участия видов таежного мелкотравья и зеленых мхов в
составе нижних ярусов леса.
Ключевые участки для изучения лесов этой труппы располагались в
бассейне р. Бол. Атлым на Белогорском материке и в бассейне р. Сев. Сось-
вы на возвышенностях Северного Люлимвора.
Коренных елово-кедровых кустарничково-зеленомошных лесов, не
затронутых воздействием сильных разрушительных пожаров за
последние 200—250 лет, почти не встречается. В зависимости от состояния
процесса восстановления материнского полога конкретных насаждений
соотношение пород в древостое верхнего полога сильно варьирует — от
8К2ЕедЛ до 5 — 4КЗ — 2Е2 — 1Б1 СедЛ, Ос. Лиственница, хотя и
присутствует почти во всех насаждениях, но участие ее незначительно.
Состав второго, полога более стабилен, его образует новое возрастное
поколение ели и кедра. В некоторых Насаждениях принимает единичное
участие пихта. .
Под пологом групповой подрост кедра и ели разного возраста, в более
старших возрастных группах ель заметно уступает кедру и количественно
и по состоянию. В местах вывала пихты нередко наблюдается массовое
возобновление ее за счет укоренения ветвей упавших деревьев, но
выживаемость и сохранность такого подроста низкие.
Редкий подлесок состоит из Sorbus sibirica, Salix cinerea, Rosa acicu-
laris, I)uschekia fruticosa, в некоторых насаждениях встречается Juniperus
communis.
В составе травяно-кустарничкового яруса обычно насчитывается 15—
17 видов. Доминируют бореальные кустарнички — черника, брусника,;
линнея северная. Из гипоарктических кустарников постоянно
присутствует в небольшом количестве багульник болотный.
Группами и единичными экземплярами распространены плауны
(Lycopodium anceps и L. annotinum). Из других травянистых видов
встречаются майник двулистный, седмичник европейский, марьянник луговой,,
очанка волосистая, вейник тупоколосковый, гудайера ползучая.
Проективное покрытие травянистыми растениями и кустарниками
неравномерное — от 40 до 70%. В понижениях концентрируется синузия мезогигро-
фитов из. хвоща лесного и осоки шаровидной.
Сплошной моховой покров мощностью 8—10 см сложен главным
образом Hylocomium splendens и Pleurozium schreberi с участием Dicranum
73
polysetum% Ptilium crista-castrehsis. На разложившейся древесине мелкие
скопления Ptilidium ciliare. В понижениях постоянно присутствуют Ро-
lytrichum commune, Sphagnum girgensohnii, Sph. nemoreum. Среди
моховой дернины небольшими латками распространены лишайники — Pelti-
gera aphthosa, Cladina rangiferina, CL stellaris, CI. arbuscula, Cetraria
laevigata.
С повышением увлажнения формируются гигрофитные варианты этих
лесов, приуроченных к выположенным участкам склонов с близкой
верховодкой или близким водоупорным горизонтом с хрящом — елово-кедро^-
вые леса с вороничн'о-чернично-зеленомошным покровом, или воронично-
бруснично-долгомошно-зеленомошным, или воронично-бруснично-долго-
мошно-зеленомошным покровом. Нарастание мощности моховой толщи
вызывает изменение прежде всего в обилии мезогигрофитных видов. В
число содоминирующих видов выходят, гипоарктические кустарнички —
вороника (Empetrum nigrum) и багульник (Ledum palustre), повышается
обилие Equisetum sylvaticum и Carex globularis, появляется Equisetum
scirpoides. В напочвенном покрове таких лесов преобладает Pleurozium
schreberi, увеличивается участие Polytrichum commune. Интенсивно идет
развитие сфагновых мхов — Sphagnum warnstorfii, Sph. magellanicum и»
Sph. fuscum, образующих всюду подушки разных размеров и мощности.
Обобщенный выдел елово-кедровых лесов включает мелкие контуры
заболачивающегося леса и молодых кустарничково-сфагновых сГлиготроф-
ных болот, формирующихся на вогнутых участках склонов или в логооб-
разных понижениях. На карте сочетания лесных и болотных сообществ
показаны как единый территориальный выдел.
Темнохвойно-березовые с лиственницей
кустарничково-зеленомошные леса (73)
Такие леса формируются в коротковосстановительных сменах через
промежуточную породу — березу. Наибольшие площади их отмечены нал
водоразделе рек Дубчес — Сым, небольшие массивы встречаются на
севере подзоны повсеместно.
В эту группу объединены производные насаждения разных стадий
восстановления от лиственных молодняков до смешанных насаждений с
сомкнутым вторым древесным пологом из темнохвойных пород. Молодые
березовые леса занимают относительно небольшие площади и вкраплены
# в основной фон смешанных насаждений с верхним пологом из березы с
примесью осины, иногда лиственницы. Подрост в смешанных лесах образован
группами кедра и ели, местами встречается пихта, а в более старых
насаждениях (80—100 лет) из темнохвойных пород сформирован сомкнутый
полог. В подлеске и в травяно-кустарничковом покрове, участвуют те же
виды, что и в коренных лесах. Общий видовой состав дополняется
несколькими лугово-лесньши растениями.— Solidago virgaurea, Hieracium
umbellatum, Chamerion angusiifolium, Calamagrostis langsdorffii.
, Обилие отдельных видов кустарничков в различных фитоценозах
значительно варьирует. Проективное покрытие видов неравномерное — от 30
до 70%. .
Моховой покров образован Преимущественно Pleurozium schreberi,
небольшой мощности (5—7 см), со следами пожаров, которые
прослеживаются также и в почвенном профиле. На местах свежего пала — чистые
куртины Polytrichum alpestre, в микропонижениях — Polytrichum commune
с подушками сфагновых мхов.
Темнохвойно-сосновые с лиственницей
кустарничково-зеленомошные леса (74)
Восстановление темнохвойных лесов через смешанные древостой с
сосной свойственно районам, где преобладают легкие суглинистые или
супесчаные почвы. Большие площади таких лесов сосредоточены в лево-
74
бережье Оби — на волнисто-увалистых возвышенностях Конда-Сосьвин-
ского междуречья. Широко распространены кедрово-сосновые леса с ба-«
гульниково-чернично-зеленомошным и багульниково-бруснично-зелено-
мошным покровом.
Древостой сосновые с примесью кедра, реже лиственницы и березы,
состав их< варьирует 6 — 5G3— 2К2 — 1Л 4- Б. Сомкнутость полога
неравномерная (0,5—0,6). В составе разновысотного и разновозрастного
подроста преобладают кедр и ель, сосна встречается только отдельными
экземплярами. На участках, где сомкнутость ниже 0,5, может быть
довольно большая поросль березы. В процессе восстановления многие
насаждения испытывают вторичные слабые пожары с частичным
разрушением древостоев, поэтому их структура часто носит характер «куртинно-раз-
новозрастный» [Смолоногов, 1970].
Подлесок редкий из Sorbtts sibirica, Rosa aciculqris, местами образует
группы Duschekia fruticosa. Видовюй состав травяно-кустарничкового
покрова и их обилие мало отличаются от елово-кедровых кустарничков о-
зеленомошных лесов. Заметное увеличение обилия багульника (он
становится содоминирующим видом) показательно для производных лесов, так
как его широкому разрастанию в дренированных местоположениях
способствуют низовые пожары. Несколько снижается мощность мохового
покрова (5—3 см), и вся дернина пронизана углями. Ведущую роль в составе
мохового покрова играет Pleurozium schreberi, ему сопутствуют Hyloco-
mium splendens, Dicranum polysetum.
В эти же выделы на карте включены насаждения, характерные для
ранних стадий восстановления — березово-сосновые чернично-бруснич-
но-зеленомошные леса с обильным подростом кедра и ели.
Елово-кедровые с пихтой, местами кедрово-еловые,
мелкотравно-бруснично-зеленомошные леса (75)
Леса этой группы сосредоточены в южной части подзоны, в полосе
слабо- и среднерасчлененных равнин с пологохолмисто-увалистыми
формами рельефа, где занимают относительно хорошо дренируемые1
местоположения — пологовыпуклые участки водоразделов, вершины и пологие
склоны. Почвы под ними подзолистые ^элювиально-глееватые на средних
и легких суглинках или элювиально-глееватые неоподзоленные длительно-
сезонномерзлотные на тяжелосуглинистых отложения?:.
Узловыми ассоциациями в этой группе будут елово-кедровая мелко-
травно-бруснично-зеленомошная с преимущественным распространением
на легкосуглинистых почвах и елово-кедровая мелкотравно-чернично-.
зеленомошн&я, более характерная для тяжелосуглинистых почв с
длительным сохранением почвенной верховодки. Кроме коренных ассоциаций,
обобщенный выдел этой группы включает и их экологические варианты
(елово-кедровая хвощово-бруснично-зеленомошная, линнеево-бруснично
или чернично-зеленомошная, плауново-чернично-зеленомошная),
распространенные некартируемыми выделами смежно с основными.
Небольшие массивы даких лесов наиболее распространены на Обь-
Иртышском междуречье — в бассейнах рек Салым, Ларь-Еган и Ильяк
среди преобладающих производных лесов. Реже встречаются на Вах-
Тымском и Кондо-Сосьвинском междуречьях.
Наряду с превалирующими елово-кедровыми лесами на карте в эту
же группу включены сообщества кедрово-пихтово-еловых мелкотравно-
бруснично-зеленомошных и мелкотравно-зеленомошных лесов,
встречающихся небольшими массивами на местоположениях плакорного типа в
центральной части Кондо-Сосьвинского водораздела. Детальная
характеристика таких лесов приведена в публикациях, посвященных исследованиям
на Ту греком стационаре Института географии СО АН СССР [Михеев,
Соколов, 1974; Михеев и др.,* 1977].
Среди темнохвойных среДнетаежных лесов леса этой группы
отличаются наибольшей производительностью (III и IV классы бонитета).
75
Деревья верхнего полога достигают высоты 22—24 м, а нижнего 18—20 м.
Своеобразие состава древесного яруса заключается в присутствии в нем
пихты — единично в составе верхнего полога и до 2—3 единиц в нижнем.
Соотношение между основными породами колеблется в небольших
пределах 6 — 8КЗ — 1Е1П, нередко в составе верхнего полога небольшими
куртинами или единичными деревьями находятся перестойные деревья березы
и осины, тогда состав может быть более неустойчивым (5К2— ЗЁ1 —
.— 2П1 — 20с или Б). Примесь березы и осины в плакорных лесах после-
пожарного происхождения. В подросте ель и кедр встречаются
примерно в равных соотношениях, учаотие пихты неустойчиво. В некоторых
участках леса ее подрост очень обилен, хотя сохранность и выживаемость
пихты низкая"
Подлесок развит слабо. Единично, реже группами (по 2—3
экземпляра) рассеянно встречаются рябина сибирская, шиповник иглистый, ива
козья, малина, реже жимолость.
В травяно-кустарничковом покрове преобладают бореальные
кустарнички — брусника, черника, линнея северная, образующие
самостоятельные синузии. Характерной особенностью является присутствие в его
составе группы видов, таежного мелкотравья — кислицы, майника
двулистного, седмичника евроцейского, гудиеры ползучей, ортилии однобокой —
постоянных спутников темнохвойных лесов южно-таежной подзоны. Из
других травянистых видов следует отметить бЪлыпое участие плаунов
(Lycopodium anceps, L. annotinum), папоротников, Хвощей — Gymnocarpium
dryopteris, Equisetum sylvaticum. . Единичными экземплярами
встречаются лугово-лесные виды: вейник тупоколосковый и Лангсдорфа,
марьянник луговой, золотая розга. Общее покрытие травяно-кустарничкового
яруса 60—85%.
Моховой покров сплошной из Hylocomium splendensn Pleurozium schpe-
beri с постоянным включением небольших латок Ptilium crista-castrensisf
Dicranum polysetum, Polytrichum commune. Все микропонижения заняты
сфагновыми мхами, преимущественно Sphagnum russowii, Spk. fallax.
Строению травяно-кустарничкового и мохового покрова свойственна
сильная мозаичность, вызванная неоднородностью экотопов.
Восстановительные смены ^елово-кедровых мелкотравно-бруснично-
зеленомошных лесов представляют леса со смешанным древостоем,
состоящим из темнохвойных и мелколиственных или светлохвойных пород. Наи^
более распространены елово-березовые и березово-осиновые с пихтой и
кедром леса (76> 77). Локальное распространение имеют производные
сообщества, образованные темнохвойными породами с участием сосны и
лиственницы. В таких насаждениях при часто повторяющихся пожарах
сосна может создать устойчивые производные сообщества (78).
Елово-березовые с пихтой и кедром
мелкотравно-зеленомошные леса (76)
В обобщенный выдел этой группы объединены также молодые
березовые леса (20—40 лет), встречающиеся обычно в непосредственном контакте
со смешанными лесами, но не выделяющиеся в масштабе карты.
Елово-березовые производные леса представляют одну из стадий
восстановления коренных елово-кедровых мелкотравно-бруснично-зелено-
мошных лесов (75), для которой характерны распад древостоев лиственных
пород, образующих первый ярус, и формирование сомкнутого второго
яруса из ели,^пихты и кедра с разным соотношением этих пород. Наиболее
распространены смешанные леса с преобладанием ели и пихты во втором
ярусе, что характерно для восстановительно-возрастной динамики елово-
кедровых лесов [Колесников, Смолоногов, I960]. Но пихта в возрасте
80—100 лет начинает интенсивно выпадать, и лишь немногие экземпляры
выходят в верхний ярус. Участие кедра во втором ярусе также постоянно,
но по обилию он уступает быстрорастущим пихте и ели. Доминирование
76
кедра наступает в стадии, близкой к полному восстановлению темнохвой-
ных лесов, такие участки леса вычленялись как коренные (75).
В смешанных елово-березовых лесах подрост также
полидоминантный из кедра, ели и пихты, образующих разновысотные группы. Обычно
отмечается преобладание пихты высотой 1,5—2 м (35—40 лет). Подрост
пихты часто локализован в местах разреженного полога выпавших, но
неотмерших деревьев пихты, ветви которых быстро укореняются и создают
густую заросль подроста. В западной части равнины, особенно на Кондо-
Сосьвинском водоразделе, в производных лесах этой группы принимает
участие сосна.
Напочвенный покров лесов группы существенно не отличается от
ненарушенных елово-кедровых лесов. При сохранении доминирующей роли
брусники и мелкотравья изменяются степень покрытия мхами, обилие и
встречаемость некоторых травянистых видов растений.
Березово-осиновые и осиновые
мелкотравно-зеленомошные леса (77)
Наибольшие площади производных лесов с преобладанием осины
сосредоточены на Обь-Иртышском междуречье. Они приурочены к наиболее
дренированным пологоувалистым участкам равнины, примыкающим к
речным, долинам: Почвы под ними преимущественно слабоподзолистыё
суглинистые.
Преобладают березово-осиновые и осиновые леса в фазе формирования
в них второго яруса из темнохвойных пород — кедра, ели с небольшой
примесью пихты. Береза и осина в возрасте от 80 до 120 лет образуют верхний
ярус высотой до 25 м. Наиболее старые осины имеют диаметр стволов от
50 до 76 см, березы 28—36 см. Разновысотные группы подроста образованы
елью, кедром и пихтой. Местами пихта, возобновляющаяся вегетативно*
преобладает, но из стадии подроста выходит единично. Лиственные породы
погибают от недостатка освещенности, едва достигнув высоты 0,3—0,5 м.
В редком подлеске постоянна присутствуют рябина, роза иглистая и
коричная, ива козья, реже можжевельник.
Травяно-кустарничковый покров в своей основе сходен с видовым
составом коренных темнохвойных лесов, чему способствует высокая
сомкнутость осиновых лесов (0,7—0,8). Большое участие в травяном покрове
принимают такие виды темнохвойных лесов, как Maianthemum bifolium,
Linnaea borealis, Diphasiastrum complanatum, Lycopodium clavatum, Gym-
nocarpiunt? dryopteris, Luzula pilosa. Распространение Vactinium myrtillus
и V. vitis-idaea спорадическое. Некоторые отличия определяются
большим обилием вейника тупоколоскового, хвощей лесного и лугового,
осоки шаровидной, а также сохранением таких видов, как Chamerion angus-
tifolium, Cirsium heterophyllum, свойственных ранним стадиям развития
березово-осиновых лесов.
Моховой покров не сомкнут, проективное покрытие его 40—60%.
В его сГснове те же самые виды мхов, что и в коренных лесах — Hylocomi-
ит splendens, Pleurozium schreberi, Ptilium crista-castrensis с куртинами
Polytrichum commune в понижениях.
Распределение производных и коренных лесов дает профиль в
бассейне р. Назинки (рис. 6).
Березово-сосновые и сосновые
кустарничково-зеленомошные устойчиво-производные леса,
преимущественно в сочетании
с сосновыми багульниково-сфагновыми лесами
и кустарничково-сфагновыми олиготрофными болотами (78)
В эту группу, объединены и молодые производные березово-сосновые
зеленомошно-брусничные леса с обильным подростом из кедра и ели,
характерные для ранних стадий восстановления, и длительно-производные
77
10
2\3l4\S ' 6
I 9 \1Q41\ 12 \13\141516\ 17 43
I III I I" I
19 ' 20^
900
Pub. 6. Геоботаничесдий профиль в бассейне р. Назинки (со
1 — березовый с сосной разнотравно-долгомопшый лес; 2 — вейниково-осоковое кочкарное болоте*
долгомошно-зеленомопшо-брусничный; 5 — березовый мблкотравно-долгомошно-зеленомошный;
вый хвощово-зеленомопшо-сфагновый; 8 — березовый осоково-чернично-долгомошный; 9 — кедро
никово-сфагновый; 11 — сосново-осиново-березовый княженично-долгомоптый; 12 — осиново-бе
березовый сфагново-осоковый (кочковатый); 15 — березовый вейниково-долгомошный; 16 — бере
ный; 18 — елово-березово-кедровый багульниково-долгомопшо-сфагновый; 19 — кедровый хвощово-
осиново-березово-кедровый багульниково-бруснично-зеленомопшый; 22—березово-кедровый чернич
кедрово-березовый мелкотравно-бруснично-зеленомошный; 25 — березовый с кедром, елью папо
ный; 27 — болото, облесенное сосной V6 класса бонитета, пушицево-сфагновое; 28 — сосново-кедро
и хвощом; 30 — кедровый с елью, сосной, березой чернично-долгомошный; 31 — елово-кедровый
гнумом; 33 — елово-кедровый с березой осоково-кассандрово-сфагновый; 34 — кедрово-сосновый
40 — кукушкин лен; 41 — кочкарное болото; 42 — сфагнум; 43 — зеленый мох;
сообщества сосновых кустарничково-зеленомршгаых лесов, имеющих в
составе древесного яруса и подросте темнохвойные породы. Такие сосновые
леса, формирующиеся после пожаров на местоположениях плакорного
типа с почвами суглинистого механического состава, можно рассматривать
как длительную (не менее 100-М50 лет) возрастную стадию
восстановления елово-кедровых лесов, что подтверждают и результаты анализа дан^
ных разных лет лесоустройства конкретных участков [Седых, 1979]. Их
распространение локализовано западными районами подзоны, где
занимают большие площади в бассейне верхнего течения рек Пелыма и Конды
(преимущественно в левобережье). Восточнее Иртыша встречаются
небольшими разрозненными участками в бассейне рек Югана, Куль-Егана и
Киевского-Егана.
Насаждения березово-сосновых и сосновых лесов с кустарничково-
зеленомошным покровом приурочены к вершинам и пологим склонам
низких гряд или увалистых водоразделов, сложенных двучленными
породами — легкие суглинки небольшой мощности близко подстилаются
супесью. Почвы под ними подзолисто-элювиально-глееватые, часто "маломощг
ные. Верхние горизонты и тонкая моховая подстилка обычно густо
пронизаны углями — свидетелями частых пожаров.
Длительно-производные сосновые кустарнйчково-зеленомошные леса
имеют разновозрастные древостой с высотой верхнего полога 17—19 м,
образованного сосной с единичным участием лиственницы, кедра, ели и
березы. В подросте (до 50 лет) преобладает сосна, в более старшем
возрасте на ^участках с длительным отсутствием пожаров господствуют-ели и
кедр, что и определяет дальнейшее формирование полога из темнохвойных
пород и явную тенденцию к смене сосны'как эдификатора. В лесах, часто
подвергающихся низовым пожарам, в подросте сохраняется
доминирование сосны, что приводит впоследствии к образованию нового поколения
сосны,—процесс восстановления темнохвойных лесов надолго
прерывается. J
Редкий подлесок образуют разреженно растущие можжевельник
(Juniperus communis), роза иглистая (Rosa acicularis), ива серая (Salix
caprea)]ilo склонам ольха (Duschekia frutjeosa). В составе травяно-кустар-
ничкового покрова обычные виды темнохвойных лесов. Доминируют бо-
реальные кустарнички — брусника, черника, линнея северная,
спорадически встречаются плауны, ожика бледноватая, хвощь лесной, осока
шаровидная. Общее проективное покрытие этого яруса составляет 60—70%.
Моховой покров сложен Pleuroziurri schreberi, Hylocomium splendens, Dic-
78
25025 75 м
М--' ' I ' '—I * 9
г- 1 = , f—г г-: т * *7 '«
//00 /ДО0 , 1500 м
ставлен Б. А. Богоявленским, И. С. Ильиной, Л. И. Мельцер).
с редкой березой; з — березовьгймелкотравно-зеленомошно-долгомошный; 4 — сосновый с березой
6 — елово-березово-кедровый долгомопшо-багульниково-черничный; 7 — сосново-березово-кедро-
во-елово-березовый мелкотравно-чернично-зеленомошный; ю — сосново-березово-кедровыйбагуль-
резовый княженично-зеленомошный; 13 — березовый вейниково-княженично-долгомошный; 14 —
зово-сосновый багульниково-долгомопшый; 17 — кедровоберезовый хвощово-осоково-долгомош-
зеленомошно-долгомошный; 20 — березовый с кедром бруснично-мелкотравно-зеленомопгаый; 21 —
но-зеленомошный; 23 —г- березовый с елью, кедром багульниково-чернично-зеленомошный; 24 —
ротниково-зеленомошный с кислицей; 26 — березово-кедровый хвощово-багульниково^долгомош-
вый хвощово-багульниково-сфагновый; 29 — кедровый с елью, пихтой зеленомошный с брусникой >
бруснично-зеленомошный с хвощом; 32 — елово-сосново-кедровый бруснично-зеленомошный со сфа-
чернично-багульниково-сфагновый; 35 — береза; 36 — сосна; 37 — кедр; 38 — ель; 39 — осина;
44 — осока; 45 — брусника; 46 — черника; 47 — багульник; 48 — пушица.
ranum polysetum. В понижениях развиваются Polytrichum communef>
Aulacomnium palustre, Sphagnum girgensohnii; куртинками встречаются
лишайники — Cladina arbuscula, CI. stellaris, Cetraria islandica.
Сосновые леса этой группы, расположенные вдоль железной дороги
Ивдель — Обь,, на больших площадях вырубаются. На вырубках в
возобновлении преобладают сосна и ель, а на участках, пройденных пожарами,—
густая поросль березы. Порослевые березовые леса с брусничным
покровом — категория кратковременно-производных сообществ. В результате
восстановительных процессов на их месте могут вновь сформироваться
сосновые леса или (при отсутствии пожаров) потенциальные сообщества
темнохвойных лесов.
На карте выделы этих лесов представлены в сочетании с
сообществами заболачивающегося соснового леса по уплощенным участкам
"водоразделов и кустарничково-сфагновыми олиготрофными болотами в
эрозионных ложбинах. Такие территориальные единицы характерны для
структуры растительного покрова с холмисто-депрессионным рельефом.
Елово-кедровые долгомошные
и кустарничково-сфагновые леса
в сочетании с кустарничково-сфагновыми олиготрофными
болотами (79)
Сочетание гидрофильных елово-кедровых лесов и небольших
массивов олиготрофных болот по заторфованным понижениям характерно для .
суглинисто-глинистых плоских междуречий, межгривных впадин и
ложбин древнего стока преимущественно в пределах среднетаежной подзоны.
Такие сочетания реже встречаются в северной и южной тайге.
В условиях плоского рельефа на слабодренированных поверхностях
первичными очагами заболачивания становятся многочисленные мелкие
микропонижения, где застой атмосферной влаги стимулирует начало
развития процессов поверхностного заболачивания почвенного и
растительного покрова. Механизм поверхностного заболачивания почвенного
покрова детально изучен Н. А. Караваевой [1973, 1982]. Развивается ландшафт,,
изобилующий небольшими олиготрофными болотами с прилегающими к
ним лесами, находящимися в разных стадиях заболоченности. Данный
выдел карты отражает неоднородный растительный покров такой
территории.
79
Елово-кедровые леса с доминированием Polytrichum commune
формируются на полугидроморфных минеральных почвах в начале активного
. торфонакопления (мощность торфяного горизонта 5—15 см). Они
представлены различными сообществами в зависимости от коренных тиш>в леса,;
существовавших на этих местоположениях до заболачивания. Наиболее
характерны сообщества елово-кедровых лесов с бруснично-осоково-дол-
гомошным, багульниково-чернично-долгомошным, багульниково-хвоще-
во-долгомошным покровом.
Древесный ярус таких лесов мало отличается от суходольных. В его
составе, кроме господствующего кедра в первом ярусе, обильно
представлена ель с небольшой примесью сосны, березы и единично пихты во
втором ярусе. Наиболее обычен состав 7 - 8КЗ - 2Е + Б, С едП. Высота
первого яруса 16—20 м, средний диаметр кедра 22—24 см. В подросте
также преобладает кедр, меньше ель, группами береза, единично сосна;
много сухостоя ели и пихты в'высотных группах от 1,5 до 2 м. В редком
подлеске рябина, ива (Salix cinerea), местами можжевельник.
В травяно-кустарнич^овом покрове заболоченных лесов ясно
прослеживается внедрение' болотных видов. Еще сохраняется высокое обилие
У actinium vitis-idaea тлжУactinium myirtillus, но жизненное состояние их
резко ухудшается (низкорослы, слабо плодоносят). Четко выделяются
синузии с доминированием Carex globularis, Equisetum sylvaticum ж долго-
мошно-сфагновым покровом. Всюду равномерно распространен
багульник. В*микропонижениях формируются болотные ценозы с Carex globularis
и мезотрофными сфагновыми мхами с единичным участием пушицы (ErU
ophorum uaginatum). На приствольных повышениях и валеже
группируются виды суходольных лесов — Linnaea borealis, Trientalis europaea, Ma-
ianthemum bifolium c Pleurozium schreberi, Hylocftmium splendens, Ptilium
crista-castrensis, Dicranum polysetum.
Моховой покров на выровненных участках леса состоит из мезофит-
ных видов — Polytrichum commune, Sphagnum warnstorfii, Sph. robustumf
Sph. girgerisohnii, из чистых участков олиготрофных сфагновых мхов по
понижениям — Sph. angustifolium, Sph. magellanicum и куртинок Sph.
fuscum на вершинах наиболее высоких кочек.
Елово-кедровые леса со сфагново-долгомошным покровом не
занимают больших площадей и представляют короткую стадию в процессе
интенсивного заболачивания лесов. По периферии болотных массивов они
сменяются сфагновыми елово-кедровыми лесами, испытывающими
постоянное подтопление с насыщенного влагой торфяника. Почвы под ними тор-
фянисто-глеевые с торфяной подстилкой до 35 см.
В сфагновых елово-кедровых лесах происходит существенное
изменение всех ярусов лесного ценоза. Древесный ярус разреженный,
сомкнутость 0,5—0,4, высота 13—15 м. Состав древостоя 7 - 8К1-2Е1Б + С,,
V класс бонитета. Подрост темнохвойных редкий, угнетенный: Более
жизненное возобновление составляет сосна. Подлесок чаще всего отсутствует.
Старые отмирающие кедры сплошь покрыты эпифитными лишайниками.
В травяно-кустарничковом покрове этих лесов Доминируют
багульники осока круглая. Спорадически распространены другие виды — хвощ
лесной, голубика, в меньшем обилии встречаются вороника (Empetrum
nigrum), Кассандра. На приствольных повышениях сохраняются синузии
суходольных лесов — брусника, реже черника, линнея северная с
Pleurozium schreberi% Dicranum polysetum, Ptilium crista-astrensis, Aulacomnium
palustre.
Напочвенный покров мозаичного сложения образован политриховы-
ми (Polytrichum commune, P. alpestre) и мезотрофными сфагновыми мхами
(Sphagnum robustum, Sph. fallax, Sph. squarrosum) с небольшими
участками по возвышениям олиготрофных (Sph.^ magellanicum, Sph. fuscum).
Наибольшая мощность сфагнового торфа образуется в естественных
микропони женияхг где прежде всего и начинается отпад деревьев.
80
ЮЖНО-ТАЕЖНЫЕ ЕЛОВО-ПИХТОВЫЕ, ПИХТОВО-ЕЛОВЫЕ
И ПИХТОВО-ЕЛОВО-КЕДРОВЫЕ ЛЕСА
И ПРОИЗВОДНЫЕ СООБЩЕСТВА НА ИХ МЕСТЕ
Южно-таежные темнохвойные леса распространены
преимущественно в центральной части Обь-Иртышского водораздела (рис. 7). Они
занимают широтную полосу к югу от среднего течения рек Конды и ^емьянки
в Прииртышье, а в Томском Приобье простираются южнее притоков Оби —
Васюгана и Кети. Эти леса наиболее подробно изучены на Тавда-Кумин-
ском междуречье [Южно-таежные леса...; 1972], в прииртышской части
Тобольского материка [Любимова, 1972; Ильина и др., 1975], в Восточном
Васюганье [Храмов, Валуцкий, 1977], в бассейнах рек Чулыма и Чаи
[Горожанкина, Константинов, 1978].
Южно-таежная подзона является областью экологического оптимума
темнохвойных пород, главным образом пихты и ели. Здесь леса из этих
пород достигают наибольшей производительности: II класс бонитета с
• запасом до 300 м3/га дри среднем возрасте 160—180 лет [Горожанкина,;
Константинов, 1978]. Зональные особенности этих лесов проявляются
прежде всего в составе и структуре их плакорных типов, формирующихся
на хорошо дренированных суглинистых почвах автоморфных рядов. К
таким зонально-плакорным типам в центральных районах южной тайги
Западной Сибири относятся елово-пихтовые с кедром, преимущественно
мелкотравные и осочковые леса. В зауральской части доминирование в них
чаще всего переходит к ели [Колесников, 19(72]. В северных и восточных
вариантах этих лесов, близких к среднетаежным типам, еще более
возрастает роль кедра и участие в покрове зеленых мхов. Южные прииртышские
варианты с участием липы дают повышение обилия широкотравья и
неморальных видов в составе травяного покрова. Кедровые южно-таежные
леса не образуют собственно плакорных типов и относятся большей
частью к гидроморфным рядам заболачивания. Последние представлены в
южной тайге главным образом мезо-олиготрофными сообществами кедра и
ели: хвощово-осоково-сфагновыми, осоково-сфагновыми, осоково-кустар-
ничково-сфагновыми. Гидроморфные ряды долинной растительности по-
Рис. 7. Пихтовый южно-таежный лес. Бассейн р. Четь. Фото Л. А. Игнатьевой.
6 Заказ NS 297
называют большое фитоценотическое разнообразие лесов каждой породы*
Пространственно более выражены различные типы еловых насаждений
в долинах рек и ложбинах стока. В* их покрове обилие широко- и
высокотравных мезофитов и гигромезофитов, а также представителей неморальной
флоры.
Кедрово-елово-пихтовые зеленомошно-мелкотравные,
мелкотравно-осочковые с элементами широкотравья леса (88)
Это очень обобщенный выдел легенды, включающий целую группу
растительных сообществ, в которых главными лесообразующими породами
являются пихта, ель и кедр. Участие последнего сильно варьирует от
нескольких единиц до со доминирующей роли в составе древостоя.
Лесные сообщества кедрово-елово-пихтбвой формации относятся к двум
экологически близким группам ассоциаций: зеленомошно-мелкотравной и
мелкотравно-осочковой. Географически эти леса тяготеют главным
образом к центральным и восточным частям Обь-Иртышского междуречья,;
где занимают наиболее дренированные пологосклоновьге участки
заболоченных водоразделов и высоких речных террас. Для этих местообитаний
характерны дерново-слабоподзолистые почвы в разной степени глееватые
[Уфимцева, 1974].
Сообщества зеленомошно-мелкотравной группы ассоциаций имеют
полидоминантный и разновозрастный состав древостоя (5ПЗЕ1КБ1 —
4П2ЕЗК1Б + С). Средняя высота деревьев в возрасте 150 (ели, пихты)—
220 (кедры) лет 23—25 м, что соответструет III классу бонитета, средний
диаметр стволов 30 см (у кедров он обычно больше, у ели и пихты —
меньше).
Сомкнутость, как правило, высокая — 0,7—0,8. Подрост представлен
теми же породами, что и древостой, но большая его часть приходится на
долю пихты. Подлесок выражен слабо, его сомкнутость менее 0,2, а
высота примерно 1—1,5 м. Среди кустарников преобладают жимолость (Lo-
nicera xylosteum), шиповник (Rosa cinnamomea), спирея (Spiraea media).
Хорошо развит травяно-моховой покров, неоднородный по
горизонтальному сложению. Синузии осочки, отдельных видов крупнотравья и
зеленых мхов образуют в нем микрогруппировки разного состава и величины.
В сообществах уписываемой группы ассоциаций наибольшие площади
приходятся на мелкотравные и зеленомошные группировки. В первых
доминирующая роль принадлежит Oxalis acetosella, Linnaea borealis, Stellaria
bungeana, Trientalis europaea. На более осветленных местах
присутствуют Carebc macroura и Calamagrostis obtusata, единично встречаются Tha-
lictrum minus, Aconitum septentrionale. Зеленые мха наблюдаются пятнами
и покрывают почву в общей сложности не более чем на 10—15%. Среди
них преобладает Hylocomium splendens. Зеленомошцые микрогруппировки
приурочены к более затененным местам, часто они формируются по
валежнику. Основу составляют зеленые мхи — преобладающий Hylocomium
splendens; спорадически встречаются Rhytididdelphus triquetrus и Ptilium
crista-castrensis, Pleurozium schreberi, Climacium dendrvides. Травостой
очень разрежен и состоит преимущественно из хвощей (Equisetum рта-
tense, Е. .sylvaticum), а также отдельных Представителей мелкотравья —
Oxalis acetosella, Stellaria bungeana, Orthilia secundd, Maianihemum bifo-
lium и др. В ассоциациях зеленомошно-мелкотравной группы обычны
также группировки мелкотравно-осочковые с преобладанием Carex macrourafi
приуроченные к повышенным и осветленным местам, а в увлажненных
микроцонижениях — разнотравно-папоротниковые (Dryopteris carthusia-
па, Diplazium sibiricum, Matteuccia struthiopteris, Filipendula ulmariaf)
Aegopodium podagraria) [Храмов, 1971; Храмов, Валуцкий, 1977; Горо-
жанкина, Константинов, 1978].
В целом в сообществах, принадлежащих к этой группе ассоциаций*
можно отметить несколько повышенную роль кедра и зеленых мхов, что
82
связано с более тесной пространственной их сопряженностью с
заболоченными местообитаниями и фитоценозами.
Следующая группа ассоциаций полидоминантных кедрово-елово-пих-
товых лесов представлена мелкотравно-осочковыми сообществами. Они
приурочены к повышенным частям склонов грив и увалов, чаще
встречаются на высоких террасах рек, где достигают производительности II,
а иногда I класса бонитета. Почвы под этими лесами имеют хорошо
развитый дерновой горизонт, глубокое оподзоливание и'слабое оглеение.
Древостой характеризуется абсолютным преобладанием пихты (6—8 единиц
в составе), большим участием ели (2—4) и незначительным кедра.
Сомкнутость крон 0,7—ОД Подрост и подлесок мало отличаются от
вышеописанных сообществ зеленомошно-мелкотравной группы асоциаций. JB
травяном покрове абсолютное преобладание получают синузии и
микрогруппировки осочковые (Carexmacroura), вейниково-осочковые (с Calamagrostis
obtusata), мелкотравно-осочковые, мелкотравно-папоротниковые. В них
обилие типичных представителей южно-таежной бореальной флоры: Сагех
macroura, Qxalis- acetosella, Siellaria- bungeana, Circae alpina, Dryopteris
carthusiana, Mitella nuda, Milium effusum, Cacaliahastatan^. Зеленые мхи
встречаются небольшими пятнами, главным образом по приствольным
повышениям.
Фитоценотическое разнообразие ассоциаций в пределах описанной
группы также связано со сменой экологических условий: в местах
большего проточного увлажнения (ложбинах, долинах мелких рек) в темнохвой-
ных лесах возрастает обилие влажного высокотравья и папоротников;
при тенденциях к заболачиванию увеличивается количество зеленых мхов,
главным образом Hylocomium spleridens, появляются синузии долгомошни-
ка, осоки шаровидной, вейника Лангсдорфа и иногда сфагнума [Горожан-
кина, Константднов, 1978].
Темнохвойно-березовые
мелкотравно-вейниково-осоковые леса (89)
Участки коренных елово-кедрово-пихтовых лесов на всем
протяжении южно-таежной подзоны чередуются с большими массивами темно-
хвойно-березовых, березово-осинокЬгх и осиново-березовых лесов,
являющихся лесами вторичными, производными от коренных елово-кедрово-
пихтовых, возникшими на их месте в результате восстановления расти-s
тельности после пожаров и рубок. Леса с доминированием березы
преобладают в современном растительном покрове плакорных типов
местообитаний центральной части южной тайги Обь-Иртышского водораздела.
Они представлены сообществами разных стадий лесовосстановления. На
карте показана обобщенная группа ассоциаций вторичных темнохвойно-
березовых осочковых лесйв, являющихся наиболее распространенными на
территории Обь-Иртышской тайги.
По описаниям А. А. Храмова [Храмов, Валуцкий, 1971], сделанным
в бассейне р. Чаи, древесный ярус таких лесов состоит из березы с
примесью темнохвойных пород (до 1/3 состава) и осины. Сомкнутость крон и
полнота высокие (0,7--0,8). Высота и возраст неравномерны. Основной
полог образует береза высотой 20—22 м в возрасте 70—90 лет.
Отдельными «маяками» встречаются экземпляры кедра и пихты — пород прежнего
допожарного леса. Но кроме них присутствует и молодое поколение темно-
хвойных, главным образом пихты и ели, образующее более низкий полог
'древостоя. Возобновление темнохвойных пород идет успешно, в подросте
представлены разные их возрастные поколения — от всходов до
экземпляров 60—70 лет, формирующих самостоятельный ярус. Подрост березы,
главным образом порослевого происхождения, всегда развивается в
подобных насаждениях, но иногда при хорошем развитии хвойных он бывает
сильно угнетен. Подлесок негустой и представлен обычно шиповником,,
жимолостью, караганой (Catagana arborescens). Травяной покров развит
хорошоа имеет" проективное покрытие 80—90%, по структуре он неодно-
6* 85
роден. Главную роль в нем играют синузии вейниково-оеочковые с\Carex
тасгоига и Calamagrostis obtusata. На более осветленных местах им
сопутствует большая примесь широко- и высокотравных южно-таежных видов:
Aegopodium podagraria, Aconitum septentrionale, Athyrium filix-feminafi
Dryopteris carthusiana. В затененных местах и по микропонижениям
появляются в значительном количестве синузии таежного мелкотравья и
зеленых мхов. В целом напочвенный покров темнохвойно-березовых
производных лесов очень сходен с таковым в коренных кедрово-елово-пихтовых
лесах, особенно на поздних стадиях восстановительных смен.
Темнохвойно-осиновые, березово-осиновые
с подлеском травяные леса (90)
Вторичные леса с преобладанием осины занимает меньшие площади,;
чем производные березовые леса, но и они очень характерны для лесо-
восстановительного процесса в южной тайге Западной Сибири. По
структуре древостоя они очень близки к описанным березовым лесам.
Возобновление темнохвойных пород в них происходит менее активно, так как
наряду с темнохвойным подростом здесь очень обильна поросль
мелколиственных пород. Эти леса более осветлены и более увлажнены, почвы в них
отличаются повышенной трофностью, что особенно характерно для
долинных местообитаний (высоких надпойменных террас). В подлеске
наряду с обычными для 1ожно-таежных лесов кустарниками (шиповник,
жимолость, можжевельник) в большом количестве всегда присутствует Сага-
gana arborescens, чаще встречаются черемуха, малина. В травяном покрове
обычно преобладают виды таежного широкотравья и высокотравья
(Aegopodium podagraria, Crepis sibirica, Aconitum septentrionale, Thalictrum
minus, Athyrium filix-femiria, Dryopteris carthusiana) при большом и
постоянном участии осочки и вейника (Calamagrostis obtusata).
Березовые и осиновые молодняки с темнохвойным подростом,
кипрейно-йейниковые и кустарниковые сообщества
на месте поврежденных шелкодрядом лесов (91)
На геоботанической карте особыми контурами на востоке подзоны в
бассейнах рек Кети и Чулыма показан растительный покров,
формирующийся в местах, подверженных нашествиям сибирского шелкопряда.
Этот представитель энтомофауны наносит огромный ущерб лесным
территориям, почти целиком уничтожая современную растительность. Площади,,
пораженные шелкопрядом, лишены прежнего древостоя. В настоящее
время они предствляют собой участки порослевых березовых и осиновых мо-
лодняков с темнохвойным подростом. Кустарниковый ярус от коренных
лесов на таких участках обычно не сохраняется, через несколько лет
формируется новый с преобладанием малины (Rubus idaeus), смородины
(Ribeshispidulum), йвы (Salix caprea, S. cinerea, S. pentandra), бузины (Sam-
bucus sibirica). Располагаются кустарники-куртинами вместе с валежом.
Травостой обычно очень густой и высокий. Проективное покрытие 100 %.
В нем доминируют кипрей (Chamenerium angustifolium), вейники
(Calamagrostis arundinacea, С. obtusata, С. langsdorffii), сныть (Aegopodium
podagraria). К ним иногда примешиваются Aconitum septentrionale, Delphinium
datum, Her acleumdissectum, Thalictrum minus, Geranium sylvaticum и др.
[Храмов, Валуцкий, 1978].
Елово-пихтовые с липой
зеленомошно-широкотравно-кисличные и осочковые
с неморальными элементами леса (93)
Темнохвойно-пихтовые леса — это основная зональная формация в
подзоне южной тайги Западной Сибири. На территории прииртышской
и зауральской частей равнины эти леса имеют свои специфические чертыя
84
главная из которых — значительное участие липы (Tilia cordata) в составе
большинства лесных сообществ.
Распространение и роль липы в строении лесных фитоценозов
южной тайги наиболее подробно изучены Ю. П. Хлоновым [1965].
П. Л. Горчаковский [1968] дал характеристику некоторым особенностям
северной и южной границ ареала липы, отмечая ее
произрастание в виде кустарника на северном пределе распространения и в
древовидной форме в составе второго яруса древостоя — близ южной
границы ареала. По материалам наших исследований, проведенных в
южно-таежном Прииртышье {Любимова, 1972; Ильина и др., 1975], липа
играет разную фитоценотическую роль в строении лесных сообществ.
Примерно на широте с. Горно-Слинкино проходит рубеж, севернее которого?
она встречается только в подлеске. Самое северное ее присутствие в этом
районе отмечено близ устья р. Туртас, на возвышенности Тобольского
материка [Любимова, 1972].
В южной тайге прииртышской части равницы наибольшее
распространение получили елово-пихтовые с липой-леса, относящиеся к двум
характерным группам ассоциаций — зеленомошно-широкотравно-кислич-
ной и осочковой. На карте они обозначены одним номером (93) и
показаны в районах, примыкающих к среднему и нижнему течению Иртыша.
В составе этих лесов доминирующая роль принадлежит пихте, ели,
образующим древостой средней сомкнутости (0,6—0,7), преимущественно
III класса бонитета. Липа здесь встречается главным образом в подлеске,;
но нередко в качестве небольшой примеси она присутствует и во втором
подчиненном пологе древесного яруса. Кроме липы в подлеске обычно
участвуют рябина, малина, ива. Травостой хорошо развит и имеет
типично южно-таежный облик — с обилием кислицы, осочки, различных видов
широкотравья (какалия, аконит, сныть, папоротники) и с участием
неморальных элементов. Роль мохового покрова незначительна. В составе
мхов наряду с обычными Pleurozium'schreberi, Hylocomiumsplendens
всегда встречаются южно-бо^ральные виды — Rhytidiadelphus triquetrus, Cli-
macium dendroides.
Елово-дихтовые с липой леса подробно изучались на территории Ниж-
не-Иртышского стационара Института географии СО АН СССР,
расположенного на высоком правом берегу Иртыша на водораздельной равнине
Тобольского материка в районе с. Горно-Слинкино [Южная тайга...%
1975]. Эти леса приурочены здесь к участкам плакоров небольших
водоразделов местной гидросети, где встречаются на легкосуглинистых глу-
бокооподзоленных со; вторым гумусовым горизонтом почвах [Уфимцева,:
1974]. Древостой этих лесов,, как правило, смешанный. Взрослые деревья
образуют два полога. Видовой состав господствующего полога на одном
из участков был 6П2ЕБ10 + К, подчиненного 5П4Е1Б + 0 [Крисон,
Полюшкин, 1975].
В возрасте 100 лет деревья верхнего полога имеют высоту 23—26 м,
максимальная высота пихты 32 м в возрасте 180 лет. Древостой высокопол-
нотные, сомкнутые. Запасы древесины достигают 250—300 м3/га.
Преобладает III класс бонитета. Естественное возобновление хвойных обычно
ограничено высокой полнотой материнского полога и в некоторых
насаждениях бывает очень слабым.
Подлесок образовай главным образом липой и редкими
экземплярами рябинь!. Травостой развит хорошо, его покрытие неравномерно, но
достигает 70—80%. Преобладают типичные южно-таежные виды —
кислица, осочка, цинна, звездчатка Бунге, папоротник, хвощ полевой,
кочедыжник, какалия. Наряду с ними обязательно присутствуют
представители неморальной флоры *— Aegopodium podagraria, Paris quadrijolia%
Actaea erythrocarpa.
» Моховой покров распределен пятнами по затемненным местам, он
достигает плотности 50%. Среди мхов обычны Pleurozium schreberi,
Hylocomium splendens, Rhytidiadelphus triquetrus, Climacium dendroides.
Леса с преобладанием осочки занимают несколько более выпуклые
85
поверхности с древостоем, не столь сомкнутым, как в вышеописанных
сообществах зеленомошно-кисличных лесов. Здесь верхний полог древостоя
высотой 25 м мало сомкнут, основная масса деревьев сосредоточена во
втором пологе, который имеет высоту 18 м и сомкнутость 0,5—0,6,
Возобновление древесных пород обычно слабое, в подросте больше пихты,;
чем ели, единично встречаются кедр, береза, осина. В подлеске
преобладает липа, бывает много малины, отдельными экземплярами встречаются
рябина, жимолость. Проективное покрытие травостоя от 40 до 70%.
В нем абсолютно преобладает осочка, наряду с ней встречаются кислица,,
щитовник Линнея, костяника, вейник тупоколосковый (Calamagrostis
obtusata), сныть, кочедыжник женский. Моховой покров развит слабо,;
зеленые мхи распределены группами, концентрируясь около
приствольных повышений [Ильина и др., 1975; Крисон, Полюшкин, 1975].
Темнохвойно-березовые и березовые с липой травяные
и осочковые леса в сочетании с влажными широкотравными лесами (94).
Березово-осиновые с липой неморально-травяные леса (95)
Описанные елово-пихтовые с участием липы леса в большой степени
нарушены пожарами и рубками. В результате эти коренные леса
занимают ограниченные площади и только отдельными массивами
распространены среди возникших на их месте производных мелколиственных
насаждений. Последние по составу травяно-кустарничкового яруса близки к
исходным елово-пихтовым лесам, но в них значительно шире разрастаются
осочка и вейник, в подросте и во втором ярусе всегда наблюдается примесь
темнохвойных пород. По динамическим свойствам такие леса относятся
к категории» короткопроизводных.
Эти леса на карте показаны в сочетании с длительно-производными
березовыми широкотравньгми лесами, возникшими на месте влажных
типов темнохвойных елово-пихтовых и пи*тово-еловых с липой сообществ
[Любимова, 1972]. >
Длительно-произвоДные березовые леса приурочены к подзолисто-
глеевым почвам и характеризуются густым травяным покровом, состоящим
преимущественно из представителей широкотравья — сныти, лабазника,;
папоротников.
Коротко-производные березово-осиновые с липой
неморально-травяные леса произрдстают на достаточно увлажненных хорошо гумусирован-
ных почвах. Они имеют хорошо развитый Древостой сомкнутостью 0,7—
0,8 с густым подлеском липы. В составе травяного яруса преобладают
неморальные виды, среди них наибольшее обилие дмет сныть, кроме нее
встречаются медуница (Pulmonaria dacica), зведчатка (Stellaria holosted),
воронец красноплодный (Actaea erythrocarpa).
Липово-елово-пихтовые шйрокотравные
с участием неморальных видов леса (96),
Липово-пихтово-березовые и осиновые
широкотравные леса (97)
Для южно-таежных прииртышских лесов весьма характерно широкое
распространение липы не только в подлеске, но и в древесном ярусе, где
она встречается как примесь к хвойным или мелколиственным породам
[Хлонов, 1965; Любимова, 1969; Ильина и др., 1975]. При этом часто ее
обилие возрастает настолько, что она формирует смешанные
полидоминантные сообщества: липово-елово-пихтовые, липово-пихтово-березовые
и липово-березово-осиновые. По мнению некоторых исследователей
[Любимова, 1969], такие леса следует считать реликтовыми образованиями,,
наследием эпохи более теплого климата. Им соответствуют и реликтовые
признаки почв — наличие второго гумусового горизонта? в их дерново-
подзолистых, дерново-подзолисто-глеевых и других разновидностях [Уфим-
цева, 1974].
86
На карте получили отображение липово-елово-пихтовые леса с
участием неморальных видов. Они занимают самые высокие вершины и
склоны увалов на расчлененной поверхности Тобольского материка
преимущественно в южной его части вдоль широтного отрезка Иртыша. Липа в
этих лесах образует второй ярус древостоя, достигая, высоты 14—16 м
и диаметра 16—20 см. Обильно она представлена в подлеске, где наряду
с ней типичны представители флоры темнохвойно-широколиственных
лесов — жимолость красная и калина обыкновенная. В травяном покрове
этих лесов обильны представители южно-таежных и неморальных видов:
Aegopodium podagraria, Pulmonaria dacica, Athyrium filtx-femina, Dipla-
zium sibiricum, Lathyrus vermis, Viola mirabilis, Milium effusum, Melica
nutans и др. [Любимова, 1972].
На некоторых возвышенных- участках Тобольского материка
встречаются фрагменты чистых липовых или березово-липовых лесов с шйроко-
травным, преимущественно сцьгге'вым покровом. В понижениях рельефа,
по днищам логов и небольших речек, липа встречается только в подлеске
в составе влажно-травных с Filipendula ulmaria пихтовых и еловых лесов.
Для таких лесов характерно большое участие ольхи серой (Alnus incana).
Производные липово-пихтово-берёзовые и осиновые леса (97)
представлены богатыми неморально-травяными сообществами. При увеличении
увлажнения в них возрастает количество мезофитного высокотравья и
злаков (Filipendula ulmaria, Calamagrostis ob'tusata, C. langsdorffii).
В целом большая часть пихтовых лесов с липой тяготеет к наиболее
возвышенным приречным частям древних поверхностей Тобольского
материка. Здесь эти леса образуют массивы большой площади. В отдаленных
от реки частях водоразделов, где близко подходят обширные олиготроф-
ные болота, леса быстро сменяются собственно бореальными елово-пихто-
выми без липы, но с участием кедра зеленомошно-мелкотравными лесами
и далее — рядами заболоченных л&шых сообществ. Среди последних осо-,
бенно характерны березовые вейниково-хвощовые и осоково-сфагновые
леса, которые в этом районе обычно предшествует образованию
заболоченных сосняков и сосново-сфагновых рямовых болот.
Пихтово-еловые с липой зеленомошно-травяцые леса
в сочетании с влажными широкотравными лесами (98)
Пихтово-еловые леса на карте Западно-Сибирской равнины показаны
в западной половине южной тайги на Тавда-Куминском междуречье,;
а также на высоком правом берегу Иртыша — Тобольском материке.
Наиболее подробно они были изучены в бассейне р. Тавды сотрудни-
камд Института экологии растений и животных УФ АН СССР
[Колесников, 1972; Маковский, Шадрина, 1972; Синелыцикова, 1972; Придня„
1972]. Некоторые сведения о них имеются в работе Е. П. Смолоногова и
В. И. Никулина [1963], а так^е в классификационных таблицах лесорас-
тигельных условий и типов лесЬв Свердловской области [Лесораститель-
ные условия..., 1973]. Прииртышские еловые леса частично изучались
Е. Л. Любимовой [1972] и нами [Ильина и др., 1975], краткие
упоминания об этих лесах содержатся в работах западно-сибирских ботаников
[Городков, 1916, 1924; Баранов, 1928].
По составу пихтово-еловые леса полидоминантны. Их основные лесо-
образующие породы — ель и пихта. На плакорных или условно-плакор-
ных местообитаниях соотношение между этими породами может меняться
в процессе возрастной и восстановительной динамики, а также в
зависимости от условий местообитания.
В пихтово-еловых лесах второй полог древостоя формирует в основ-,
"ном пихта, участвуя в общем его составе в количестве от 3 до 5 единиц.Иног-
да она может доминировать, это происходит в основном при возрастной
смене поколений в ненарушенных древостоях на стадии распада главной
лесообразующей породы — ели (или кедра). Из других древесных пород
в пихтово-еловых *лесах чаще всего встречается значительная примесь
4 87
березы (Betula pendula) и осины (Populus tremula), что обычно указывает
на восстановительный процесс после пожаров и рубок.
Липа в составе елово-пихтовых лесов встречается главным образом в
подлеске, иногда она выходит и во второй полог древостоя обычно на
наиболее богатых умеренно влажных дерново-подзолистых почвах со вторым
гумусовым горизонтом [Уфимцева, 1974].
При хорошем проточном увлажнении в долинах рек и в
многочисленных ложбинах стока доминирующая роль ели возрастает. Там она
образует высокобонитетные древостой. В этих лесах из сопутствующих пород
чаще встречаются береза и осина. Роль липы в подлеске незначительнах
вместо нее развивается мощный смешанный кустарниковый ярус.
В местах застойного увлажнения при увеличении глееватости почв
ель уступает место кедру, а затем сосне. .Наибольшей степени
заболачивания еловых лесов в подзоне южной тайги соответствует образование
сргры — разреженных кедрово-еловых и елово-кедровых насаждений; со
сфагново-травяцым покровом. ^ *
Состав и структура нижних ярусов леса в южно-таежных еловых
лесах сильно^ меняются в зависимости от условий местообитания. В лесах
дренированных водораздельных склонов и террас развивается довольдо
обильный травостой из представителей таежного мелко- и широкотравья.
Участие зеленых мхов значительно, хотя сплошного покрова они не
образуют. В долинных типах ельников травяной ярус особенно хорошо развит^
он состоит из мезофитных видов высодо- и широкотравья, имеющих
большое проективное покрытие. Но часто среди долинных типов встречаются
еловые леса, преимущественно молодняки, с очень разреженным
покровом и даже мертвопокровные. Такие насаждения характерны для низких
террас рек.
В легенде (98) обозначено сочетание двух наиболее характерных групп
ассоциаций южно-таежных пихтово-еловых лесов.
Основная плакорная группа представлена пихтово-еловыми лесами
с липой в подлеске или во втором пологе с зеленомошно-разнотравным
покровом. На дренированных водоразделах такие леса имеют состав
5ЕЗП2Б + Лп, Ос — ЗЕ ЗП ЗБЮс, их сомкнутость 0,8—0,7, бонитет
II—III классов. В подросте обильнее всего представлена пихта, кроме
нее в значительном количестве встречается ель, единично — кедр, береза,
липа. В подлеске средней густоты много рябины, шиповника, встречаются
также малина, черемуха, можжевельник. Травяно-кустарничковый ярус
с проективным покрытием 60—70% образован в основном
представителями таежного разнотравья — кислицей, майником, щитовником, линнеей,
костяникой, папоротником женским, плауном годовалым, зведчаткой
Бунге, снытью, медуницей, цирцеей; довольно обильны лесные злаки —
Calamagrostis obtusata, Milium effusum, часто встречается осочка Carex
macroura. Зеленые мхи расселяются пятнами по валежу, приствольным
повышениям, на кочках. Их общее покрытие 10—25%. Среди мхов
обычны Rhytidiadelphus triquetrus, Hylocomium splendens, Ptilium crista-castrensis.
Небольшие логообразные понижения на водоразделах с хорошим
проточным увлажнением заняты широкотравными пихтово-еловыми
лесами, в покрове которых преобладют лабазник (Filipendula ulmaria),
сныть (Aegopodium podagraria), герань лесная (Geranium sylvaticum),
калужница (Caltha palustris),* крупные папоротники (Athyrium filix-femina),
борец (Aconitum septentrionale), гравилат (Geum rivale), а, из злаков вей-
ник — Calamagrostis langsdorffii; зеленые мхи здесь отсутствуют.
Небольшие слабодренированные понижения с торфянисто-подзолис-
то-глеевыми почвами заселяются ельниками хвощовыми. В них снова
возрастает роль зеленых мхов (в основном Hylocomium splendens),
спорадически по микропонижениям появляются и сфагнумы (Sphagnum centra-
le, Sph. nemoreum). В травяном покрове обильны Equisetum palustre*
Е. sylvaticum, Calamagrostis langsdorffii, из разнотравья обычны кислица,
линнея, майник, грушанки, а также влаголюбивые виды — Ranunculus
repens, Nardosmia frigida, Galium boreale.
88
Темнохвойно-осиново-березовые и березовые с липой
травяные леса (99)
Во всех районах распространения пихтово-еловых водораздельных
лесов в результате многочисленных лесных пожаров на больших
площадях произошла смена коренных темнохвойных лесов на производные
березовые. В лесах Тавда-Куминского междуречья эти смены подробно
изучены 3. И. Синелыциковой [1972]. Было установлено, что в ельниках зе-
леномошно-кисличных и ельниках липняковых после пожаров чаще все-,
го Происходит смена на производные древостой мелколиственных пород,:
в основном на березовые насаждения. В процессе восстановительной
динамики в возрасте 80—120 лет в составе древостоя начинает повышаться
удельный вес ели; после 140 лет наблюдается массовое выпадение березы,
и процесс завершается превращением снова в ельник с примесью
лиственных пород. Это развитие может идти двумя путями — по типу
долговременных или кратковременных смен. Если в древостоях первых классов
возраста участие ели не достигает хотя бы одной единицы состава, то их
развитие пойдет по пути долговременной смены; если в их составе 2—3
единицы ели, -го можно предполагать кратковременный характер смены [Си-
нелыцикова, 1972]. . (
Могут быть и иные линии развития производных лесов. Однако для
Тавда-Куминского междуречья характерны именно такие пути
восстановительных смен, которые приводят к формированию на больших
территориях смешанных темнохвойно-березовых насаждений.
Их древостой имеет состав 7Б ЗЕ + П, — 9Б 1JE + Ос,1 бонитет II—
III классов. В покрове преобладает^таежное мелкотравье — майник* лин-
нея, кислица; встречается вейник Лангсдорфа и пятнами распространены
зеленые мхи — Hylocomium splendens, Climacium dendroides, Rhytididdel-
phus triquetrus.'
Кедровые с пихтой и елью хвощово-зеленомошные
и травяно-сфагновые леса (101)
Кедровые южно-таежные леса на карте выступают как гидроморф-
ный вариант темнохвойной тайги. Эти леса представлены здесь двумя
характерными , группами ассоциаций — хвощово-зеленомошной и травяно-
сфагновой. Кедровые леса имеют наибольшее распространение во
внутренних более заболоченных частях водоразделов. На местности они образуют
разнообразные сложные пространственные сочетания как с лесами пла-
корного ряда — елбво-пихтовыми или пихтово-еловыми и производными
от них березняками и осинниками, так и с многочисленными
сообществами рядов заболачивания. '
Участки влажных типов кедровых лесов были описаны В. И.
Барановым [1928] в бассейне р. Урны (приток р. Демьянки) под названием бере-
зово-кедровой «коги». Широкое распространение травяно-болотных
кедровых лесов в южно-таежном Приобье отмечает И. А. Бех [1972]. На
территории Свердловской области лесоводами отмечены сфагново-травяные,
травяные и хвощово-мшистые типы кедровых лесов [Лесорастительные
условия..., 1973]. Основные массивы влажных кедровых лесов
приурочены к,окраинным частям заболоченных водоразделов, где они развиваются
на торфянисто-глеевых почвах.
Древостой этих лесов смешанный с преобладанием кедра и ели в
первом ярусе, пихты и березы — во втором. Бонитет бывает довольно
высоким — до III класса, но преобладают леса IV и V классов. Возобновление,
кдк правило, редкое — единично встречаются ель, пихта, береза и сосна.
Подлесок средней густоты, в нем присутствуют рябина, ива, черемуха,
иногда ольха серая (Alnus incana) и можжевельник (Juniperus communis).
В густом травяном покрове обильно произрастают хвощи, осокиt;
болотное разнотравье, на кочках и в понижениях встречаются сфагновые
мхи (Sphagnum squarrosum, Sph. wulfianum).
89
Березовые с пихтой, елью,
кедром хвощово-вейниковые леса (102)
Коренные кедровые зеленомошные и травяно-сфагновые леса в
местах пожаров и вырубок заменяются производными в основном березовыми
дгесами, в покрове которых заметную роль начинает приобретать вейник
Лангсдорфа (Calamagrostis langsdorffli). На карте показана одна из
наиболее распространенных стадий восстановительного ряда —
коротко-производные березовые с пихтой, елью и кедром хвощово-вейниковые леса.
Для них характерно активное участие в составе подроста древесного
яруса и темнохвойных пород. Напочвенный покров этих лесов образован
преимущественно хвощом лесным и вейником Лангсдорфа с примесью
таежного разнотравья (майник, костяника, кислица) и гидроморфных видов
зеленых мхов: Rylocomium splendens; Polytrichum commune; P. juniperinum. .
Подобные сообщества влажных кедровых лесов изучались нами вг
южно-таежном Прииртышье в бассейне р. Алымки [Ильина и др., 1975].
Один из участков такого леса был описан на плоском, слегка заболоченном
водоразделе на границе с сосново-кустарничково-сфагновым болотом.
В древесном ярусе преобладали кедры высотой 15—17 м, диаметром
стволов 12—15 см. Они имели несколько угнетенный вид, нижние ветви их
были сплошь покрыты свисающим «бородатым» лишайником (Usnea
Tbirta). Второе место пб числу стволов принадлежало березе пушистой
(Betula pubescens), имеющей* высоту 16—18 м. В подросте встречались
группы порослевых берез высотой до 4 м. Травяной покров состоял в
основном из хвоща лесного (Equisetum sylvaticum) и осоки шаровидной
(Сагёх globularis). К ним в незначительном количестве примешивались
вейник Лангсдорфа и майник двулистный, местами попадались кустики
брусники. Мхи покрывали в среднем 60% площади, среди них
преобладали Hylocomium splendens, Pleurozium schreberi, Sphagnum squamosum,
Sph. wulfianum.
ГЛАВА V
)
ЛИСТВЕННИЧНЫЕ ЛЕСА И РЕДКОЛЕСЬЯ
Лиственничные леса и редколесья на Западно-Сибирской равнине
занимают большие площади (рис. 8). На севере таежной зоны они имеют
почти сплошное распространение и образуют несколько подзональных
полос [Ильина, Махно, 1976]. В средней тайге эти леса встречаются
отдельными массивами и уже не занимают ведущее положение в сложении
древостоя. В качестве примеси к другим породам лиственница
распространяется далеко на юг, вплоть до границ с лесостепями.
Лиственничные леса и редколесья Западной Сибири обрааованы
лиственницей сибирской {Larix sibirica). Лишь на северо-востоке по
левобережью Енисея к ней местами примешивается лиственница даурская (La-
rix gmelinii), а на западе — в зауральской части — лиственница
Сукачева {Larix sukaczewii)..
Лиственница сибирская — дерево первой величины, достигающее в
оптимальных условиях высоты более 50 м и диаметра 175 см. Это самая
светолюбивая из сибирских древесных пород. Об этом свидетельствуют
высокое расположение крон, способность к быстрому очищению стволов
от, сучьев и тенденция к самоизреживанию древостоев. Почти во всех
лиственничных лесах наблюдается сильная разреженность древесного полога.
В Западной Сибири эта особенность проявляется особенно резко и
является следствием не только биологических особенностей лиственницы, но и
суровых климатических условий севера западно-сибирской тайги.
Экологический диапазон лиственничных лесов в Западной Сибири
довольно широк. Наилучшие условия для их произрастания создаются на
90
V и* ■*> и» иш^иш^ шт*
Рис. 8. Лиственничные леса и редколесья Западно-Сибирской равнины.
Редколесья: 1 — елово-лиственничные и лиственничные лишайниковые и кустарничково-зе-
леномопшые с субарктическими элементами редколесья в'сочетаний с лишайниковыми и кустар-
ничково-зеленомошными тундрами; 2 — лиственничные и елово-лиственничные лишайниковые и зе-
леномошные редколесья в сочетании с редкостойными, лиственничными лесами и плоскобугристыми
болотами; 2а— лиетвеннично-березовые кустарничкс^ю зеленомошно-лишайниковые редколесья и
редкостойные леса.
Северотаежные редкостойные леса: з — лиственничные и елово-лиственничные
кустарничковые и зеленомошно-лишайниковые леса; За — лиетвеннично-березовые местами с кедром,
сосной зеле^омошно-кустарничковые и лишайниковые леса; 4—сосново-лиственничные с кедром
зеленомошно-кустарничковые леса; 5 — лиственничные и елово-лиственничные с кедром, сосной
кустарничковые долгомошные и сфагновые леса.
суглинистых хорошо дренированных и достаточно увлажняемых почвах,
преимущественно в приречных частях водоразделов и в долинах рек. Но
такие местообитания имеют весьма ограниченное распространение.
Гораздо шире на территории севера равнины распространены лиственничные
леса с ^разреженным низкобонитетным древостоем, приуроченные к
песчаным и супесчаным почвам озерно-аллювиальных равнин и высоких
речных террас. На слабодренированных центральных частях междуречий
развиваются заболоченные лиственничники, формирующие разные по
структуре древостой (редкостойные леса — редколесья — редины) и
образующие на местности закономерные топо-экологические ряды смен
растительных сообществ по направлению от дренированных местоположений к
болотам. Под лиственничными лесами на суглинистых породах с мощно
развитым торфяным горизонтом развивается вечная мерзлота [Тыр-
тиков, 1974].
В Западной Сибири на значительных площадях лиственница образует
почти чистые древостой с небольшой примесью других пород. Чаще всего
к лиственнице примешивается ель (Picea obovata). Елово-лиственничные
леса более характерны для запада равнины, хотя небольшими массивами
они распространены повсеместно.
При продвижении к югу в составе лиственничных лесов, особенно на
песках, начинает усиливаться роль сосны, последняя уже на широте
Сибирских увалов вытесняет лиственницу и становится доминирующей лесо-
образующей породой. По-видимому, главная роль в указанной смене
принадлежит климатическому фактору и связанному с ним улучшению
теплового режима почв. Немаловажное значение в усилении роли сосны в юж-
' 91
ных районах может иметь также антропогенное влияние, главным
образом действие пожаров, в результате которых быстрее уничтожается
подрост лиственницы, чем сосны, и в целом позиция сосны, вытесняющей
лиственницу, чрезвычайно возрастает [Сочава, 1956].
Значительное участие кедра в составе лиственничных лесов для
рассматриваемого региона нехарактерно. Причина этого кроется в малой
сомкнутости лиственничных древостоев, что обусловливает повышенную
освещенность их местообитаний и в связи с этим плохое, возобновление
кедра. Тем не менее на востоке равнины на Пур-Тазовском междуречье
наблюдается постоянное участие кедра в составе лиственничных лесов и
редколесий на всем меридиональномs отрезке вдоль правого берега Пура.
Из мелколиственных пород береза— Betula pubescens, а в лесотундре
и Betula tortuosa встречаются в примеси почти во всех коренных
лиственничных древостоях, а в производных насаждениях она приобретает
доминирующее положение.
Смешанные темнохвойно-лиственнйчные и березово-лиственничные
леса в Западной Сибири распространены преимущественно в долинах рек,
где они образуют сомкнутые более производительные (III—IV классов
бонитета) древостой и представляют собой заключительные стадии
развития гйдросерий долинной растительности [Городков, 1946; Ильина, 1976].
Основная структурная особенность большинства лиственничных
лесов, как уже было отмечено,— их разреженность. Тем не менее на
территории Западной Сибири можно наблюдать все разнообразие структурных
форм лиственничного древостоя — от насаждений с высокой
сомкнутостью в долинах рек до низкорослых лиственничных редин в лесотундре и на
болотах. Наибольшее распространение и зональную приуроченность имеют
два физиономических типа лиственничных древостоев — лиственничные
редкостойные лесач свойственные северной полосе северотаежной
подзоны [Ильина, Махно, 1976], и лиственничные редколесья, образующие под-
зональные полосы на северной окраине тайги. Северная граница ареала
лиственниц на территории равнины проходит в тундровой зоне, это
северные пределы ее распространения [Крылов, 1961].
В вертикальной структуре лиственничных древостоев наблюдается
большое разнообразие. Одновозрастные и одновысотные древостой
встречаются крайне редко. Обычно лиственничные леса состоят из деревьев
различного размера и неодинакового возраста, что теснейшим образом связано
с особенностями их возрастной и восстановительной динамики.
Структура нижних ярусов лиственничных лесов меняется в
зависимости от условий местообитания. В долинных типах сообществ
развивается хороший подлесок из ольхи кустарниковой (Duschekia fruticosa), ивы
(Salix phylicifolia), шиповника (Rosa acicularis), черемухи (Padus avium),
иногда смородины (Ribes nigrum). В разреженных лиственничных лесах
водораздельных территорий подлесок развит слабо, в нем обычны
карликовая береза (Betula папа) и <5льха.
В травяно-кустарничковом ярусе коренных лиственничных» лесов
господство принадлежит бореальнымигипоарктическим кустарничкам —
багульнику, бруснике, водянике, голубике. В заболоченных
местообитаниях возрастает роль олиготрофных видов — клюквы, Кассандры. Из
трав в водораздельных лесах встречаются мезогигрофитные виды —
осока шаровидная и хвощ лесной. Роль таежного мелкотравья
очень,незначительна, а представители высокотравья встречаются только в
долинах рек. , • ■ '
Мохово-лишайниковый покров во всех типах лиственничных лесов
развит очень интенсивно. Из зеленых мхов здесь обильны Pleurozium
schreberi, Hylocomium splendens и виды Polytrichum. Из лишайников —
представители родов Cladonia, Cladina и Cetraria. Широкое
распространение в лиственничных лесах и редколесьях имеют также трубчатые
(Stereocaulon) и листоватые (Peliigera, Nephroma) лишайники.
По нашим представлениям, лиственничные редкостойные леса не
были подвержены лесным пожарам в такой степени, Как лежащие к югу
92
темнохвойные насаждения. В значительной мере это связано со слабой
заселенностью территории и ее трудно доступностью из-за большого
количества болот. Тем не менее многие районы, особенно те, которые покрыты
лиственничными лишайниковыми лесами, были подвержены пожарам,
и в настоящее время там произрастают не коренные, а производные леса.
По исследованиям многих авторов [Лесков, 1940; Сочава, 1956;
Шанин, 1965] лиственничные леса больше всего подвергаются действию
низовых пожаров, которые древостой полностью не уничтожают, но
способствуют быстрой смене нижних ярусов леса. После низовых пожаров
развивается интенсивный подрост, и, если пожары не повторяются
длительное время,* он формирует новый, полог лиственничного древостоя.
Низовые пожары стимулируют интенсивное развитие березы, которая на гарях
очень активно возобновляется вегетативным. путем и местами начинает
преобладать в подросте, а з&тем и в древостое. Гораздо медленнее в
производных лесах восстанавливается ель, подрост которой и .молодые
экземпляры после низовых пожаров уничтожаются полностью; медленно
восстанавливаются кедр и сосна.
Характерной особенностью восстановительных смен травяно-кустар-
ничкового яруса лиственничных лесов является возрастание в нем роли
багульника и водяники, а на первых стадиях — появление вейника, иван-
чая и других травянистых видов. После сплошного уничтожения
лишайникового покрова на почве сначала пбселяются зеленые мхи из рода Poly-
trichum. Восстановление лишайников в лиственничных лесах„ по данным
А. П. Тыртикова [1974], происходит через несколько десятилетий после f
пожара, если территория не подвергается вторичным антропогенным воз-4
действиям.
Лиственничные леса равнины имеют специфичные для разных
географических условий черты, что выражается в формировании характерных
групп зональных и провинциальных типов. В пределах каждой группы
намечаются свои эколого-фитоценотические ряды сообществ,
свойственные плакорным и неплакорным местообитаниям, а также характерные
флористические комплексы.
Обобщенные группы и классы ассоциаций лиственничных лесов и
редколесий отражены на составленной геоботлнической карте. По условиям
масштаба они в* большинстве случаев даны в сочетании с болотами или с
лесами других типов.
ЛИСТВЕННИЧНЫЕ И ЕЛОВО-ЛИСТВЕННИЧНЫЕ РЕДКОЛЕСЬЯ
И ПРОИЗВОДНЫЕ СООБЩЕСТВА нА ИХ МЕСТЕ
Лиственничные редколесья показаны на карте к востоку от низовий
р. Оби на всем ее протяжении до Енисея. Они образуют сплошную под-
зональную полосу, сильно расширяющуюся на Пур-Тазовском и Таз-.
Енисейском междуречьях. На правобережье Пура, по данным
аэровизуального обследования [Богоявленский и др., 1968], лиственничные редколесья
распространяются на юг почти до широты с. Тарко-Сале.
Условия существования редколесий в отличие от лесов
характеризуются более суровым климатом, в котором роль ветра, как фактора
дефляции и иссушения почв, а также влияние близко залегающей вечной
мерзлоты очень велики. Древесный полог в редколесьях сильно разрежен
(сомкнутость ОД—0,3), деревья отстоят друг от друга на расстоянии от
7 до 20 м, они имеют небольшую высоту (6—10 м), слаборазвитые кроны и
усыхающие вершины стволов. Указанные особенности структуры и
морфологических свойств редколесий приводят к резкому падению в них эди-
фикаторной роли древостоя. Вследствие этого травяно-кустарничковый
ярус и моховой покров формируются независимо от древесного полога,
испытывая наибольшее влияние общих природных условий. В этом
смысле можно говорить о полиэдификаторности редколесий [Норин, 1964].
Важнейшей особенностью, горизонтальной структуры нижних ярусов
растительности редколесий является их необычайная пестрота и неодно-
93
родность, выражающаяся в образовании различных форм гетерогенного
сложения — мозаичности, комплексности и т. п. В лиственничных
редколесьях Западной Сибири эта неоднородность проявляется повсеместно
[Норин, 1964; Тыртиков, 1970, 1974; Москаленко, 1976]. На
дренированных участках в составе фитоценозов наблюдается постоянное чередование
синузий мхов, лишайников и кустарничков в зависимости от изменений
форм нанорельефа, режима тепла и влажности почв. В районах
заболачивания структура растительного покрова еще более усложняется частой
сменой в пространстве целых фитоценозов (или их фрагментов),
приуроченных к разным по степени дренированйости формам микро- и
мезорельефа.
Деление западно-сибирских редколесий на подзональные типы —
лесостундровый и таежный — ранее было осуществлено А. И. Лесковым
[1940], детально изучившим редколесья бассейна р. Полуй. Для других
районов подобные подразделения производились Б. Н. Городковым
[1935], В. Б. Сочавой [1957], С. А. Грибовой [1977]. Однако на
геоботанической карте' эти подзональные типы редколесий получили отражение
впервые.
Для западно-сибирских лесотундровых редколесий характерна очень
низкая сомкнутость древостоя (0,1—0,2), образованного преимущественно
лиственницей, наименьшая высота деревьев (6—8 м), низкая
производительность (Va, V6 классы бонитета), слабое возобновление под пологом,,
почти полное отсутствие подлеска. Травдно-кустариичковый и мохово-
лишайниковый покров этих редколесий отличаются сильной мозаичностыо.
В их видовом составе наряду с преобладанием бореальных и бореально-
гипоарктических видов [Юрцев, 1966] в качестве постоянной примеси
присутствуют собственно гипоарктические элементы: Arctous alpina,
Ledum decumbens, Dry as punctata, Aulacomnium turgidum, Alectoria nigricans.
Северотаежные редколесья более разнообразны по составу. В каче-'
стве доминирующей породы здесь часто присутствует ель (Picea obovata)f
а на востоке — кедр (Pinus sibirica). Подлесок, состоящий в основном из
кустов карликовой березы (Betula nana), а местами из ольхи (Duschekia
fruticosa) и можжевельника (Juniperus communis), успешно развивается во
многих типах таежных редколесий. Травяно-кустарничковый и моховой
покров более гомогенны, хотя различные формы неоднородности их
структуры хорошо прослеживаются и здесь.
В географическом распределении таежных и лесотундровых
редколесий, в их приуроченности к разным элементам рельефа, в составе
образуемых ими пространственных сочетаний и топо-экологических рядов
также намечаются довольно резкие различия. Лесотундровые типы
редколесий распространены в основном на высоких террасах рек, а на
водоразделах в местах, сложенных породами легкого механического состава.
Лесотундровые редколесья на местности постоянно .чередуются с
сообществами ерниковых, ивняковых и ольховниковых тундр, а в заболоченных
частях междуречий — с участками плоскобугристых и полигональных болот.
Иное распределение имеют таежные редколесья. Они встречаются
преимущественно на водороазделах и всегда в сочетании с лесными
сообществами или сообществами бореальных болот. На вершинах крупных
грив и увалов в центральных частях Обь-Ендсейского междуречья
таежные типы редколесий образуют значительные массивы. Однако' уже на
склонах водораздельных возвышенностей они переходят в редкостойные
леса, а в понижениях — в заболоченные редины и открытые плоско- и
крупнобугристые лишайниково-сфагновые болота.
Елово-лиственничные ерниково-лишайниково-зеленомошные
с субарктическими элементами редколесья (35)
Лиственничные редколесья лесотундрового типа на территории
Западной Сибири изучены крайне слабо. Некоторые сведения о них
содержатся в работе А. И. Лескова [1940]. Более детально эти редколесья опи-
94
саны Б. Н. Нориным [1958] в бассейне р. Ныды. Отдельные участки
лесотундровых редколесий исследовались нами в окрестностях нос. Тазов-
ский, а также на юге п-ова Ямал близ дельты р. Оби.
На геоботанической карте (35) показана группа лесотундровых
лиственничных и ^лово-лиственничных редколесий с ерниково-лишайниково-
зеленомошным покровом. Сообщества этой группы встречаются среди
массивов ерниковых тундр и приурочены к суглинистым в разной степени
дренированным почвам преимущественно приречных частей водоразделов.
В древесном ярусе редколесий этого типа абсолютно преобладает
лиственница. Небольшое участие ели обычно связано с несколько повышенной
влажностью и глееватостью почв. Примесь' березы (Betula pubescens,
В. tortuosa) более значительна (до 3—4 единиц в составе) и характерна
как для коренных типов, так и для антропогенно-производных редколес-
ных березово-лиственничных древостоев.
Наиболее дренированным местообитаниям соответствует ассоциация
лиственничного лишайниково-зеленомошного редколесья, развитого на
суглинистых среднеподзолистых глееватых почвах. Древесный ярус
сомкнутостью ОД—0,2 состоит из лиственницы, имеющей высоху 6—8'м и
диаметр ствола 10—15 см, с единичной примесью березы (Betula tortuosa)
и в некоторых местах ели высотой до ,7 м, диаметром 10—12 см. Расстояние
между деревьями 10—13 м. В подлеске группы кустарников Betula nana
и единичные экземпляры Juniperus communis. Травяно-кустарничковый
ярус представлен отдельными синузиями низкорослых кустарничков,
в основном багульника, голубики, водяники, среди которых часто
присутствует гипоарктический вид Ledum decumbens. Из трав обычны Carex glo-
bularis, С. bigelowii, Eriophorum vaginatum. Хорошо развиваются
группировки зеленых мхов, занимающие 50—60% площади фитоценоза^ среди
которых преобладают Pleurozium schreberi, Polytrichum alpestre, Ptilidium
ciliare, и в небольшом количестве встречается субарктический мох Aula-
comnium turgidum. Синузии лишайников развиты в более осветленных
местах, по обилию лишайники уступают зеленым мхам. Среди них обычны
Cladinastellaris, CI. rangiferina, Cladonia uncialis, CI. cornuta, а также
трубчатые (Stereocaulon paschale) и листоватые (Peltigera aphthosa, Nephroma
arcticum) лишайники.
В целом мозаичность покрова в этом сообществе редколесий
выражена очень четко — наблюдается частое чередование синузий мхов,
лишайников и кустарничков в зависимости от разных форм нанорельефа ц
степени затенения почвы. На повышениях обычны лишайники и кусты
багульника и водяники, в микропонижениях — зеленые мхи ичкарликовая
береза. Моховой покров больше развит под низкоопущенными кронами
берез и на приствольных повышениях лиственниц и елей.
По окраинам болот в местах менее дренированных развиваются
лиственничные зеленомошные редколесья с синузиями долгомошных и
сфагновых мхов и с большим участием ерника в подлеске, местами
образующего открытые заросли. Мозаичность и комплексность покрова в таких ред-.
колесьях еще более характерны. Среди преобладающих по площади
группировок зеленых мхов (Pleurozium schreberi, Ptilidium ciliare) и
кустарничков (Vaccinium uliginosum, Empetrum nigrum. Ledum decumbens) в
небольших депрессиях концентрируются Polytrichum commune и Sphagnum gir-
gensohnii. Древостой становится еще более разреженным и угнетенным.
Средняя высота лиственниц 4—6 м. На местности сообщества подобных
редколесий обычно образуют комплексы с чистыми зарослями ерника и с
фитоценозами заболоченных долгомошно-сфагновых лиственничных редин.
В долинах рек, где ближе уровень грунтовых вод и глубже или
совсем отсутствует мерзлота, в составе лиственничных зеленомошных
редколесий усиливается роль ели и березы (Betula pubescens), разнообразнее
становятся покров и подлесок, в которых полнее представлены бореальные
виды.
- 95
Лиственничные и ел ово-лиственничные
тсустарничково-мохово-лишайниковые
с субарктическими элементами редколесья (36)
Следующая группа лесотундровых редколесий представлена
лиственничными и елово-лиственничными лишайниковыми редколесными
сообществами. На местности эти редколесья встречаются в различных
территориальных сочетаниях с группировками кустарниковых и
лишайниковых тундр. Лишайниковые редколесья из лиственницы сибирской, как
правило, приурочены к легким песчаным почвам, но нередко они
попадаются и на суглинистых грунтах, где развиваются на выдутых и обедненных
гумусом дефляционных почвах. Состав древостоя в них очень однообразен.
Лиственница абсолютно.преобладает. Ель примешивается к ней очень
редко в основном на западе [Норин, 1958]. На Пур-Тазовском водоразделе
участие ели в небольшом количестве прослеживается только вдоль рек.
Зато .обычна примесь березы — Betula tortuosa, образующей второй ярус
древостоя. Березы в сообществах таких редколесий приобретают
специфическую форму дерева-куста со множеством стволов и нередко
стелющихся ветвей. Высота берез 4—5 м с диаметром у шейки корня 7—8*см. Эта
экологическая форма берез в литературе описывается под названием «хо-
дылей» [Норин, 1958].
Сомкнутость древесного полога в лишайниковых редколесьях
наименьшая — 0,1. Лиственницы в верхнем ярусе имеют высоту 6—7 м и
диаметр стволов (на высоте груди) 14—15 см. Стволы: их часто искривлены
в сторону от господствующих ветров, имеют сухие вершины и плохо
развитые кроны, на ветках обычны «бородатые» свисающие мхи и лишайники
(Alectoria ochroleuca). В таких редколесьях почти отсутствует подлесок из
кустарников, а травяно-кустарничковый ярус развит очень слабо
(покрытие 10—20%). В нем единично встречаются багульник, водяника и
брусника, а также гипоарктические виды Arctous alpina и иногда Drtfas punctata.
Из трав обычны овсяница приземистая, золотая розга, кошачья лапка.
Синузии зелГеных мхов не имеют большого распространения. Они так же,
как и кустарнички, приурочены к более затемненным местам. Среди
зеленых' мхов обычны Pleurozium schreberi, Hylocomium splendensf Ptilium
crista-castrensis. Главную фитоценотическую роль в этом сообществе
играют лишайники, которые образуют почти сплошной покров (80—90%),}
состоящий в основном из видов Cladina и Cladonia (Cladina stellaris, CI.
rangiferina, Cladonia coccifera, CI. macroceras) и цетрарий {Cetraria cucul-
lata, C. islandica, C. nivalis), среди которых отдельными пятнами
встречаются трубчатые лишайники (Stereocaulon paschale).
Лиственничные и елово-лиственничные
кустарничково -зелецомошные
и лишайниково-зеленомошные редколесья (37)
Две группы ассоциаций, обозначенные в легенде 37 и 38, Относятся
уже к собственно бореальному типу сообществ, и на карте показаны в
характерном для них сочетании с северотаежными лесами и болотами. Эти
типы редколесий изучались нами на Пур-Тазовском междуречье
(ключевой участок в 65 км к востоку от пос. Уренгой) и в бассейне р. Надым
(водораздел Надым—Танлово).
Обобщенный выдел легенды 37 включает по существу две эдафиче-
ские группы лиственничных редколесий: первая — кустарничково-зеле-
номошная, в которой лишайники не играют доминирующей роли, связана
с суглинистыми почвами разной степени увлажнения и дренированное™;
вторая — лишайниково-зеленомошная приурочена к грунтам более
легкого механического состава (супесям и легким суглинкам), сильно
подверженным процессам дефляции.
Лиственничные кустарничково-зеленомошные редколесья
распространены на относительно дренированных пологих склонах водоразделов.
96
Они развиваются на торфянисто-глеевых почвах,, подстилаемых вечно-
мерзлыми грунтами. Древостой преимущественно чистые, лиственничные,
одноярусные. Сомкнутость крон 0,2—0,3. Высота лиственниц 8*^-10 м,
диаметр стволов 16—20 см. Расстояние между деревьями 10—18 м.
Возобновление очень слабое, встречается единичный подрост лиственниц
высотой от 0,7 до 2 м. В подлеске небольшие группы Betula nana7 а на
возвышениях отдельные экземпляры Duschekia fruticosa. Травяно-кустарничковый
ярус имеет проективное покрытие, 50—60%. В нем преобладают
багульник и^пшкша. В меньшем количестве встречаются брусника, голубика,
а из трав осока шаровидная (Carex globularis); единично попадаются
экземпляры мытника (Pedicularis lapponica), чемерицы (Veratrum lobelianum)
хвоща лесного {Equisetum sylvaticum). Мохово-лишайниковый покров
сплошной, в нем господствуют зеленые мхи Pleurozium schreberi, Hyloco-
mium splendens, Polytrichum alpestre; лишайники встречаются отдельными
небольшими группами — Cladina stellaris, CI. rangiferina, Cl.xarbuscula,
изредка попадаются пятна Peltigera aphthosa. При заболачивании
редколесий происходит изреживание древостоя (до сомкнутости 0,2), при этом
почти исчезают подрост и подлесок, в травяно-кустарничковом ярусе
увеличивается обилие олиготрофных видов —Chamaedaphne calyculata,
Oxycoccus microcarpus, Rubus chamaemorus, в покрове мхов появляются си-
нузии Polytrichum commune и Sphagnum angustifolium.
Лиственничные лишайниково-зеленомошные редколесья, как уже
указывалось, приурочены к почвам более легкого механического состава.
Они встречаются приблизительно в тех же частях водоразделов, но по
рельефу располагаются несколько выше, а потому сильнее подвергаются
процессам дефляции.
Древостой этих редколесий в районе Уренгоя преимущественно
лиственничный с небольшим участием кедра, примесь ели очень
незначительна. Высота лиственниц 8—10 м при диаметре 10—12 см. Общая
сомкнутость древесного полога 0,2—0,1. В подроете слабое возобновление
лиственницы и единично березы. Подлесок образован группами ерника
высотой до 1 м, в нем единично встречаются низкорослые кустики ив,
очевидно Salix lanata. Травяно-кустарничковый ярус разрежен — его
проективное покрытие 30—40%. Как и в предыдущих сообществах, большую
фитоценотичесуую роль играют здесь багульник, брусника и водяника.
В vменьшем количестве встречается голубика, из трав характерна осока
шаровидная. В сплошном покрове из мхов и лишайников преобладают
Pleurozium schreberi, Polytrichum alpestre, Dicranum angustum, Cladina
stellaris, CI. rangiferina, CI. arbuscula, Cetraria cucullata, Nephroma arc-
ticum, Peltigera aphthosa. Заболачивание этих редколесий происходит
аналогично вышеописанному для сообществ кустарничково-зеленомошных
лиственничников. Некоторое отличие состоит в усилении роли лишайников
{Cladina stellaris, Stereocaulon paschale) и сфагновых мхов (Sphagnum ne-
moreum, Sph. fuscum) в мозаичной структуре напочвенного покрова.
Лиственничные и елово-лиственничные
лишайниковые редколесья (38)
Лиственничные лишайниковые редколесья бореального типа были
изучены А. И. Лесковым в бассейне р. Полуй4 [1940]. В экологических
рядах, выделенных им в разных условиях местообитания, отмечены
наиболее характерные ассоциации: Cladina stellaris — Ledum palustre —
Larix sibirica; Cladina stellaris — Betula nana — Larix sibirica; Cladina
stellaris — Vaccinium uliginosum — Larix sibirica. Все они приурочены к
вершинам- материковых или долинных грив, сложенных легкими
супесчаными или суглинистыми породами. В составе древостоя, кроме
лиственницы, автор отмечает Значительное участие ели (Picea obovata), что
признает вполне закономерным для исследованного района. Отсутствие ели
он объясняет временным явлением, связанным с действием низовых
пожаров. Кроме ели в составе лиственничных редколесий бассейна Полуя
7 Заказ Ко 297 97
очень значительна примесь березы, что в большей степени связано с
антропогенным влиянием.
В подросте этих редколесий лучше всего возобновляется ель; в
кустарниковом ярусе в некоторых ассоциациях присутствует только карликовая
береза, достигающая высоты не более 0,8—1 м. Травяно-кустарничковьш
ярус принимает незначительное участие в сложении растительного
покрова (2—<10% проективного покрытия). В мохово-лишайниковом покровег
имеющем сплошное распространение, доминирующая роль принадлежит
лишайникам, прежде всего Cladina stellaris, а также CL rangiferinay
CI. arbuscula, Cetraria islandica и листоватому лишайнику Nephroma arcti-
cum. Моховая часть напочвенного покрова очень угйетена (покрытие 10—
15%).
На Пур-Тазовском водоразделе лиственничные лишайниковые
редколесья обследованы нами. Во время аэровизуальных обследований
отмечалось почти полное отсутствие в их составе ели, в то же время
значительное участие кедра, а в некоторых местах березы и сосны. Лиственничные
редколесья бассейна Пура обнаруживают четкую приуроченность к
наиболее приподнятым валообразным возвышенностям, сложенным легкими
песчаными и супесчаными породами. Лиственницы на всех описанных
участках абсолютно доминировали в древостое, создавая сомкнутость 0,1—
0,2. В возрасте 120—160 лет они имели высоту 8—12 м и диаметр 15—20 см.
Многие стволы их?были искривлены, кроны развиты плохо, на ветвях
обилие листоватых лишайников. Расстояние между деревьями в таких
редколесьях от 7,5 до 15 м. Единичные экземпляры кедра имеют угнетенный
вид, высота их 7—9 м, ветви укороченные, часто наполовину засохшие.
Возобновление древесных пород плохое, среди угнетенных с засохшими
верхушками молодых лиственниц высотой от 0,7 до 3 м встречаются
отдельные экземпляры кедрового подроста не более 1 м и иногда березы (0,5—
1 м). В подлеске можно встретить только низкорослые кусты Betula
папа, распределенные по поверхности единично или небольшими группами.
Травяно-кустарничковый ярус имел общее покрытие не более 30%.
В нем преобладали багульник, брусника, голубика, водяника, часто ветре-»
чались отдельные экземпляры и группы осоки шаровидной, а в
заболоченных местах — шгюквы, Кассандры и морошки. В Сплошном покрове-из
лишайников, образованном в основном видами кладин, кладоний {Cladina
stellaris, CI. rangiferina, Cladonia macroceras) и цетрарий (Cetraria cuculla-
ta, C. nivalis), только отдельными пятнами.главным образом в затененных
местах у стволов деревьев встречались синузии зеленых мхов — Pleurozium
schreberi, Dicranum angustum, Polytrichumalpestre. Среди мхов и кустистых
лишайников обычны пятна Stereocaulori paschale и Nephroma &rcticum.
Лиственничные лишайниковые редколесья, описанные нами на
водоразделе Надым-Танлово, имели очень сходный состав и структуру всех
ярусов. Однако в этих редколесьях наблюдается заметная примесь ели —
до 2 единиц в составе древостоя и до 5—6 единиц в подросте.
Лиственнично-березовые и елово-березово-лиственничные,
кустарничково-зеленомошно-лишайниковые редколесья
и редкостойные леса (39)
Несмотря на то, что лиственничные редколесья распространены в
труднодоступных и слабо населенных районах севера Западно-Сибирской •
равниры, многие из них были подвержены действию неоднократных
пожаров, что отразилось на составе и структуре всех ярусов этих сообществ.
На карте показана одна обобщенная группа ассоциаций, включающая
растительные сообщества, формирующиеся на разных стадиях
восстановления лиственничных моховых и лишайниковых редколесий лесотундрового
и таежного типов. В целом ряды трансформации послепожарных смен в
лиственничных редкодесьях представляются следующим образом.
При весьма типичном для данных мест воздействии низовых пожаров
смены пород в лиственничном редколесье, как правило, не происходит.
98
У лиственницы отмирают только нижние ветки и повреждаются нижние
части стволов. Гораздо сильнее страдают ели, отдельные экземпляры
которых и весь еловый подрост сгорают полностью. Повреждаются и бе-х
резы, но они сразу дают обильный подрост, который при благоприятных
условиях начинает успешно вегетировать и расселяться по окружающей
территории. После низового пожара быстро возобновляется Betula папа
и приобретают доминирующую роль кустарнички, особенно Ledum palust-
ге и Vaccinium vitis-idaea. Bf травяном покрове появляются пирогенные
виды: толокнянка (Arciostaphylos uva-ursi), хвощ (Equisetum sylvaticum),
овсяница (Festuca ouina). В моховом покрове также усиливается роль пи-
рогенных мхов из рода Polytrichum (Potytrichum alpestre.P. juniperinum).
На первых стадиях восстановления роль лишайников в покрове
невелика, после Пожаров довольно быстро расселяются отдельными небольшими
группами только трубчатые (Stereocaulori) и листоватые (Peltigera,
Nephroma) дишайники. Только на поздних более длительных и устойчивых
стадиях послепожарных смен в покрове начинают завоевывать прочные
позиции субклимаксовые синузии кустистых лишайников из родов Clado-
nia, Cladina и Cetraria.
Неоднократное действие низовых или верховых пожаров нередко
приводит к смене доминирующих древесных пород. В таких случаях из
древостоя совсем выпадает ель, и лесообразующая роль переходит к березе.
Действие пожаров может способствовать улучшению режима
влажности и гумусного состава почв, следствием чего является повышение
производительности древостоя, изменение его структуры в сторону
увеличения сомкнутости и переход лиственничных редколесий в лиственничные
редкостойные леса с хорошо развитым подлеском и травяно-кустарнич-
ковым ярусом. Однако в дальнейшем по мере восстановления коренного
растительного покрова происходит самоизреживание лиственничного
древостоя и возвращение к структуре леса, наиболее соответствующей
сложившимся экологическим условиям [Шанин, 1965].
СЕВЕРОТАЕЖНЫЕ ЛИСТВЕННИЧНЫЕ, ЕЛОВО-ЛИСТВЕННИЧЦЫЕ И
СОСНОВО-ЛИСТВЕННИЧНЫЕ РЕДКОСТОЙНЫЕ ЛЕСА И ПРОИЗВОДНЫЕ
СООБЩЕСТВА НА ИХ МЕСТЕ
Лиственничные редкостойные леса на территории Западно-Сибирской
равнины охватывают ее большую часть. Основные их массивы отмечены
на востоке равнины на междуречьях Таз—Енисей и Пур — Таз и на
западе—в левобережье Надыма^
Основная часть лиственничных лесов приурочена к легким песчаным
и супесчаным почвам, широко распространенным в северной тайге на занд-
ровых аллювиальных равнинах. Более ограниченные площади занимают
лиственничные леса на суглинках приречных частей водоразделов и
речных террас. Многие из этих лесов заболочены. Под лиственничными
лесами разной степени заболоченности, развивающимися на суглинистых
почвах, как правило, формируется вечная мерзлота [Тыртиков, 19741.
Участие ели в составе лиственничных лесов наблюдается почти
повсеместно, наиболее значительно оно на западе на междуречье Надым —
Полуй. Столь же обычна и примесь березы {Betula pubescens, В. pendula)
как в коренных, так и в производных типах лиственничных лесов.
Лиственничные и елово-лиственничные местами с кедром,
лишайниково-мохово-кустарничковые редкостойные леса (50)
Лиственничные кустарничковые лишкйниково-зеленомошные леса
распространены на дренированных частях водоразделов и высоких
речных террас преимущественно на суглинистых почвах, имеющих разной
мощности торфянистый горизонт и подстилаемых вечной мерзлотой.
7* 99
На одном из участков лиственничного багульниково-лишайниково-
зеленомошного леса, описанного нами на водоразделе Надым—Танлово,,
древостой состоял из лиственницы с небольшой примесью березы и
единичной елью (8Л2Б + Е), имел сомкнутость 0,3—0,4. Лиственницы
высотой до 12—15 м с диаметром стволов 14—16 см, ветки сплошь покрыты
эпифитными лишайниками. Во втором ярусе березы и ели высотой 9—10 м.
Возобновление лиственницы и березы очень слабое, особенно угнетена
лиственница, среди подроста ее много сухостоя; береза возобновляется
вегетативным путем, но рост ее тоже угнетен и замедлен. В подлеске
отдельные группы Betula папа высотой не более 1 м. В травяно-кустарничко-
вом ярусе преобладают бореальные кустарнички (общее покрытие 35—
50%) — багульник, брусника, черника, водяника, реже встречаются
голубика и отдельные группировки трав — осоки шаровидной, хвоща
лестного, княженики, иногда плауна (Lycopodium annotinum). Моховой
покров имеет сплошное распространение. Из зеленых мхов преобладают
Pleurozium schreberi и Hylocomium splendens, в меньшем обилии
встречаются Ptilium crista-castrensis, Ptilidium ciliare. Лишайники менее обильны,,
распространены отдельными пятнами среди мхов и кустарничков. Среди
них отмечены Cladina stellaris, CI. arbuscula, Cetraria islandica, Alectoria
nigricans, а также Stereocaulon paschale и Peltigera aphthosa.
Сообщества из группы кустарничково-зеленомошных лиственничников
формируются в условиях застойного увлажнения — в местах
заболачивающихся. Здесь увеличивается мощность торфяного горизонта до 10—
15 см, что ведет к интенсивному промерзанию почв. В покрове таких лесов
более разрежен и угнетен травяно-кустарничковый ярус, в нем
преобладают олиготрофные кустарнички — багульник, голубика, водяника, а из
трав встречаются преимущественно осока шаровидная и морошка. В
сплошном моховом покрове особенно обильны Hylocomium splendens, Polytri-
chum commune, P. alpestre и Pleurozium schreberi, отдельными пятнами по
мйкропонижениям встречаются Sphagnum nemoreum, Sph. russowii, Sph.
warnstorfii.
Лиственничные и елово-лиственничные
с примесью кедра и сосны
кустарничково-зеленомошно-лишайниковые
редкостойные леса (51)
Леса этого типа занимают большие площади на севере полосы
редкостойных северотаежных лесов. Они распространены как на водоразделах,,
так и на речных террасах и всегда приурочены к хорошо дренированным
песчаным почвам, в которых сезонно-мерзлый слой ежегодно протаивает
полностью, а вечная мерзлота не формируется совсем/
Древостой в таких лесах редкий, как и в редколесьях, слабо
затеняющий почву. Его средняя сомкнутость"0,4—0,3. Лиственницы высотой
12—14 м, диаметром 12—18 см, стволы их сильносбежистые, на ветвях
много листоватых лишайников, высота прикрепления крон 2—3 м. Иногда
в нижнем пологе произрастают лиственницы высотой 4—6 м, диаметры
стволов которых 5—6 см, а возраст около 50 лет. В лиственничных лесах
бассейна Надыма обычна небольшая примесь ели и более значительная
(часто антропогенного происхождения) — березы (Betula pubescens). Ели
(Picea obovata) высотой до 10—12 м, диаметром 10 см. Березы имеют
меньшую высоту (8—10 м) и образуют второй полог. Возобновление в таком
лесу слабое, подрост лиственницы угнетен, среди него 50% сухостоя,
однако некоторые экземпляры достигают высоты 3—5 м. Несколько лучше
возобновляется ель, хотя и в меньшем количестве, чем лиственница,
средняя высота ее 0,5—1 м. Подрост березы вегетативного происхождения,,
развивается довольно активно, достигая высоты 4—5 м. В подлеске
отмечена только Betula nana, распределяющаяся на поверхности отдельными
группами, тяготеющими к стволам деревьев. Травяно-кустарничковый
ярус разрежен (проективное покрытие 25—Й0%), он концентрируется во-
100
круг деревьев по приствольным повышениям и по валежу. В нем обильны
только кустарнички — багульник, брусника, водяника. Единично
встречается толокнянка, а из трав очень редко Carex globularis, Festuca ovina,
Antennaria dioica. Сплошной покров образуют лишайники, мощность
живого слоя их 4—5 см, а мертвого (торфянистого) — 6—9 см. Среди них
преобладают Cladina stellaris, CI, rangiferina, Cladonia uncialis, CI. macro-
ceras, в значительном количестве встречаются также Cetraria cucullatat
С. islandica, С. nivalis; из трубчатых лишайников обычен Stereocaulon
paschale, из листоватых — Nephroma arcticum. Мхи, как и кустарнички,
приурочены к более затененным местам. Среди них обычны Pleurozium
schreberi, Ptilium crista-castrensis, Polytrichum alpestre.
Лиственнично-березовые
и елово-березово-лиственничные лишайниковые
и зеленомошные редкостойные леса (52)
Лиственничные леса северной части подзоны значительно
повреждены пожарами. На их месте возникли производные леса, которые в
зависимости от эдафических условий, степени повреждения и стадии восстанави-
тельной сукцессии имеют разный состав и структуру всех ярусов. В
лиственничных редкостойных лесах, так же как и в редколесьях, в
результате низовых пожаров обычно уничтожаются сопутствующие породы — ель
и береза, лиственница только повреждается; полностью гибнут подрост,
ярус кустарничков и весь лишайниковый покров.
В лиственничных лесах на песках лишайниковый покров
возрождается через несколько десятков'лет после того, как был уничтожен полностью
и вторично сильно не повреждался [Тыртиков, 1974]. Несколько быстрее
восстанавливается покров в кустарничково-зеленомошных лесах на
суглинистых породах. Сплошной моховой покров здесь образуется уже на
первых стадиях послепожарных смен, впоследствии в основном изменяет-4
ся его видовой состав — выпадение пирогенных видов (Polytrichum juni-
perinum, P. alpestre) и возрастание обилия экологически более
устойчивых мхов — Pleurozium schreberi, Ptilium crista-castrensis и др.
Выдел 52, показанный на карте, включает обобщенные группы
ассоциаций наиболее устойчивых и широко распространенных лесов,
возникших на месте коренных типов лиственничных моховых и
лишайниковых сообществ. В качестве иллюстрации приведем одно из характерных
описаний вторичного лиственнично-елово-березового багульниково-зеле-
номошного леса. Участок этого леса расположен на пологом склоне
водораздельного увала в районе междуречья Надым — Танлово. Почва
суглинистая торфянисто-скрытоподзолистая. В горизонте Ал заметна
угольная прослойка — след гари. Древостой имеет состав 6Б4Л + Е,
сомкнутость его 0,4. Высота берез 10—11 м, диаметр 10—12 см; лиственницы
имеют высоту 14—15 м, диаметр стволов 17—20 см, ель — высоту 10 м и
диаметр 14—16. см. Лиственницы несколько угнетены, многие ветви их и
вершины засохли, березы, наоборот, в хорошем состоянии. Местами они
приобретают кустообразную форму из-за обильно разрастающегося
вегетативного подроста. В кустарниковом ярусе преобладают береза высотой 1—2 м
и ель примерно такой же высоты. В некоторых местах единично
встречаются экземпляры кедра высотой до 2 м. Подлесок образован группами
Duschekia fruticosa, концентрирующимися в осветленных местах. В травя-
но-кустарничковом ярусе преобладает багульник. Кроме него,
встречаются голубика, брусника, водяника. Изтрав заметную роль играют осока
шаровидная и хвощ .лесной. В сплошном моховом покрове преобладают
Polytrichum alpestre, P. commune, Pleurozium schreberi; из лишайников
обильно встречаются" только листоватые (Peltigera aphthosa и Nephroma
arcticum), отдельными небольшими пятнами распространены Cladina ran-
giferiria и Cetraria islandica.
101 ,
Сосново-листвённичныё с кедром,
местами лиственнично-сосновые зеленомошно-кустарничковые
с лишайниками редкостойные леса (53)
Эти леса распространены на юге полосы редкостойных лесов северной
тайги. Они не занимают больших шГощадей, встречаются в сочетаниях с
болотами и заболоченными лесами на супесях и легких суглинках в восточных
частях подзоны или с сухими лиственнично-сосновыми лишайниковыми
лесами на песках, преимущественно в западных отрогах Сибирских
увалов. В последнем случае на песчаных почвах доминирующая роль часто
переходит к сосне.
Небольшие участки этих лесов были описаны на заболоченном
водоразделе в районе истоков р. Пур. Почвы под ними легкосуглинистые сла-
боподзолисто-глеевые, подстилаемые на небольшой глубине песками.
Древостой образованы лиственницей со значительной примесью сосны и
березы (5ЛЗС2Б + К). Сосны и лиственницы имеют примерно одинаковую
высоту 13—15 м, однако по толщине стволов и производительности
лиственница превосходит сосну и имеет большое фитоценотическое значение
в строении древостоев. В таких лесах часто .наблюдается небольшая
примесь кедра (Pinus sibirica), который встречается единично также и в
подросте. Березы растут, редко, в основном только на осветленных местах,
где имеют хорошее вегетативное возобновление. В травяно-кустарничко-
вом ярусе господствуют те же бореальные кустарнички. Из трав отмечена
более или менее обильно Carex globularis, местами попадаются кустики
Ыппаеа borealis. Моховой покров развит хорошо, хотя и не имеет
сплошного распространения. Он образован зелеными мхами Pleurozium schre-
beri, Hylocomium splendens, а также Polytrichum commune и P. alpestre.
Лишайники встречаются пятнами, среди них обычны синузии Cladina
rangiferina и Peltigera aphthosa.
Лиственнично-березовые с сосной и кедром
зеленомошно-кустарничковые леса (54)
В производных лиственнично-березовых с сосной древостоях роль
лиственницы и сосны второстепенна. Леса эти более разрежены,
преобладающие в них березы не достигают большой высоты (10-*-12 м), но имеют
хорошее возобновление. В таких лесах много следов бывших верховых
пожаров в виде обгоревших стволов и пней лиственницы. Подлесок, как
правило, отсутствует, а в разреженном травяно-кустарничковом покрове
(40%) преобладают багульник, голубика и брусника. Травянистых видов
почти нет. Моховой покров чрезвычайно разрежен, местами его плотность
всего 15.-20%. Среди мхов обычны Polytrichum alpestre и Pleurozium schrе-
beri.
Лиственничные и елово-лиственничные
с примесью кедра и сосны
кустарничково-лишайниково-долгомошные
и кустарничково-сфагновые редкостойные леса (55)
Заболоченные лиственничные леса на территории Западно-Сибирской
равнины не образуют больших массивов, но распространены повсеместно
и тяготеют к окраинам плоско-и крупнобугристых болот. В
возникновении заболоченных лиственничных сфагновых лесов кроме влияния
гидротермических и геолого-геоморфологических факторов очень большая роль
принадлежит самому растительному покрову [Тыртиков, 1974]. В
динамике естественных и антропогенных смен растительности на стадии
образования лиственничных зеленомошных лесов на суглинистых почвах по
мере нарастания мхов и увеличения мощности торфяного слоя создаются
условия для появления вечной мерзлоты, которая способствует
заболачиванию растительного покрова и появлению сфагновых мхов. По данным
102
А. П. Тыртиковк [1974], лиственничники зеленомошные сменяются
сфагновыми примерно через 100 лет, т. е. за период, когда происходит
постеленное нарастание-торфянистого горизонта в почвах зеленомошных лесов
до толщины не менее 15 см. Леса лиственничные лишайниковые,
произрастающие на песчаных почвах, почти не заболачиваются, так как из-за
сухости верхних слоев почвы мхи в них не развиваются, сезонно-мерзлый
'слой почвы ежегодно протаивает полностью и вечная мерзлота не
образуется [Тыртиков, 1974]. ^
Описанный механизм заболачивания лиственничных лесов,
связанный с процессом появления вечной мерзлоты,— специфическая
особенность всех лесных сообществ северной тайги Западной Сибири, формирую*
щихся на суглинистых почвах и имеющих в своем! составе синузии^мхов.
Для западно-сибирских лиственничных лесов характерен свой ряд
заболачивания, состоящий из последовательно сменяющихся сообществ
по направлению от дренированных частей водоразделов и террас к
обширным болотным массивам. Начальной стадией заболачивания в этом ряду
можно считать описанный гидрофитный вариант лиственничных
зеленомошных лесов, в покрове которых получают развитие синузии политри-
ховых и сфагновых мхов.
На геоботанической карте получили отражение две наиболее
устойчивые и широко распространенные группы ассоциаций заболоченных
лиственничных редкостойных лесов: лиственничные кустарничково-ли-
шайниково-долгомошные и лиственничные кустарничково-сфагновые леса.
Первые были описаны нами в бассейне р. Надым. Они имели в
составе древостоя лиственницу высотой 12—14 м с диаметром стволов 10—14 см
и ветками, покрытыми эпифитными листоватыми лишайниками; ель
высотой до 10 м и небольшую примесь березы, имеющую гораздо меньшую
высоту (7—8 м), искривленный ствол диаметром 6—7 см. Древостой
разрежен (сомкнутость 0,3—0,4) и угнетен, многие деревья фаутны, некоторые
из них повалены ветром и сгнили. Подрост очень незначительный,
образован преимущественно лиственницей, высота подроста 2—3 м. В
подлеске имеются отдельные группы ерника, приуроченные к кочкам и
приствольным повышениям. Проективное пркрытие травяно-кустарничкового
яруса 30—35%. В нем преобладают кустарнички — багульник, шикша,.
брусника и черника, довольно обильна Carex globularis, единично
встречается Rubies arcticus. Мхи образуют сплошной покров, доминирует в нем
кукушкин лен (Polytrichum commune), который распространён по всем
элементам микрорельефа. Кроме него обильны Polytrichum alpestre, Pleu-
rozium schreberi, Hylocomium splendens. В микропонижениях — пятна сфаг1
нумов {Sphagnum warnstQrfii, Sph. fuscum). На кочках, где больше
сдувается снег и от этого иссушается поверхностный слой почвы, группами
поселяются синузии кустистых лишайников — Cladina stellaris, CI
arbuscula, Cladonia gracilis.
Следующая стадия заболачивания лиственничных лесов, после чего
они переходят в заболоченные редколесья, редины и открытые болота,,
отражается группой ассоциаций кустарничково-сфагновых
лиственничников. В образовании сфагнового покрове этих лесов основную роль играет
мерзлотный режим почв, создающий дополнительные условия для
избыточного застойного увлажнения и формирования торфяного слоя. Слой
вечномерзлых грунтов, по нашим наблюдениям, на водоразделе
Надым — Танлово, находился на глубине 40—50 см. Древостой лесов еще
более разрежен (сомкнутости 0,3—0,4). Лиственницы имели высоту 10—12
ми диаметры стволов 6—12 см. Кроны их были развиты слабо, вершины
ж ветки многих лиственниц засыхали, ветки покрылись листоватыми и
накипными лишайниками (Evernia thamnodes, Parmelia sulcata). Березы,;
встречающиеся в небольшом количестве на более осветленных местах „
имели искривленные в сторону от господствующих ветров стволы и
полузасохшие ветки. Высота этих берез не превышала 6—8 м. Подроста в та-
1ких лесах, как правило, нет, в подлеске единичные кусты карликовой
^березы.
103
В травяно-кустарничковом ярусе (проективное покрытие 20—30%)
преобладают олиготрофные кустарнички — Кассандра (Chamaedaphne
calyculata), подбел (Andromeda polifolia), клюква (Охуcoccus microcarpus),
а также багульник, голубика, шикша. Из трав обычны осока шаровидная
и морошка. Сфагновые мхи «образуют почти сплошной цокров (покрытие
почвы 90%), занимают все элементы микрорельефа. На кочках более
обильны Sphagnum fuscum, Sph. girgensohnii, Sph. warnstorfii, Sph. nemo-
reum, в понижениях — Sph. balticum, Sph. russowii, Sph. fuscum.
Лишайники распространены отдельными небольшими пятнами на самых высоких
кочках. Среди них встречаются Cladina rangiferina, CI. stellaris, Cladonia
gracilis, Cetraria richardsonii, C. laevigata.
ГЛАВА VI
СОСНОВЫЕ ЛЕСА И РЕДКОЛЕСЬЯ
Основные массивы сосновых лесов на Западно-Сибирской равнине
сосредоточены в таежной зоне, где площадь их дЪстигает 22,2 млн га
[Таран, 1973].
Ареал сосны обыкновенной (Pinus sylvestris) очень обширен. Уступая
в северных пределах своего произрастания лиственнице, ели и кедру, она
гораздо дальше этих пород продвигается к югу. Практически сосна
широко представлена во всех подзонах таежной зоны равнины и значительные
площади занимает.в лесостепной. Такое широкое распространение сосны
во многом объясняется широтой ее экологического ареала и
способностью в силу своих биолого-морфологических свойств расти в самых
различных местообитаниях.
Большая пластичность корневой системы сосны, быстрота ее ростаг
ветроустойчивость, неприхотливость к минеральному богатству и
влажности почв — все это обусловливает успешное ее существование, рост и
плодоношение в условиях, крайне неблагоприятных для развития других
пород. Сосна хорошо растет на бедных песчаных почвах, где она
развивает мощный, глубоко проникающий в землю, стержневой корень.
Наоборот, на торфяниках у сосны образуется поверхностная корневая
система, способная усваивать кислород и минеральное питание из верхних
горизонтов почвы. К условиям влажности и дренированности почв сосна
малотребовательна. Она одинаково хорошо растет и в очень сухих хорошо
дренируемых и аэрируемых местообитаниях и в избыточно влажных
заболоченных местах. На болотах сосна образует несколько экологических
форм. Наиболее^ распространены на равнине две ее болотные формы —
Pinus sylvestris f. uliginosa', P. sylvestris f. litwinowii.
Одним из важных факторов, определяющих распространение сосны
в Западной Сибири, является тепловой режим почв. Сосна может
переносить довольно низкие температуры воздуха и почв, но в условиях
близкого залегания вечной мерзлоты она, как правило, не произрастает. Именно
поэтому наблюдаются самые северные проникновения ее в Западной
Сибири на более теплые песчаные почвы, где слой сезонной мерзлоты
ежегодно протаивает, а вечная мерзлота не формируется [Тыртиков, 1974].
Ареал сосновых лесов находится в основном за пределами границ сплошного
распространения вечной мерзлоты.
По составу сосновые леса равнины весьма разнообразны. Чистые
сосновые насаждения формируются главным образом в экстремальных
условиях — на сухих песках и на заболоченных почвах, где с сосной не
могут конкурировать другие породы. Такие леса имеют очень широкое
распространение во всех таежных подзонах региона. Однако не менее
широко распространены и смешанные леса, в которых в качестве
сопутствующих пород вместе с сосной встречаются лиственница в северной, тайге,
кедр и ель в средней, пихта и ель — в южной. Большое количество насаж-
104
дений образует сосна и с мелколиственными породами, главным образом с
берёзой, и меньшее осиной. Среди этих лесов значительный удельный вес
принадлежит производным древостоям, возникшим как временная стадия
в процессе восстановления коренных сосновых Сообществ. Смешанные
мелколиственно-сосновые леса естественного происхождения обычно
формируются в долинах рек.
Структура и возрастной состав древостоев в сосновых лесах
неоднородны и зависят как от среды обитания, так и от степени антропогенного,
воздействия. В оптимальных условиях минерального богатства и влажное-,
ти почв при спонтанном развитии формируются разновозрастные и разно-
высотные древостой достаточно большой сомкнутости (0,7—0,8) и высокой
производительности (I —-III классы бонитета). Такие леса произрастают
в речных долинах и на водоразде/4х в южной тайге, подтайге и лесостепи.
Для сухих песчаных почв во всех подзонах характерны разреженные
сосновые леса, местами переходящие в редколесья. Также редкостойны и
заболоченные сосняки, произрастающие в районах избыточного
увлажнения во всех подзонах тайги.
Различную сомкнутость и возрастной состав имеют многочисленные
варианты коротко- и длительно-производных сосновых лесов, возникших
в результате пожаров и рубок. В целом возрастной состав сосняков в За4
падной Сибири характеризуется большим процентом молодняков и
средневозрастных насаждений, что свидетельствует об успешности возобнови-,
тельных процессов, происходящих в них [Таран, 19731. Для большинства
сосновых лесов равнины характерно их восстановление без смены пород:
Смешанные с преобладанием березы древостой обычно образуются на
первых стадиях послепожарных смен и являются кратковременными в
условиях благоприятного и естественного лесовосстановительного процесса.
Производные сосновые леса, составляющие довольно большой
процент в современном составе сосновых лесов равнины, возникли после
пожаров на месте коренных темнохвойных, главным образом кедровых и
еловых лесов, приуроченных в своем распространении к супесчаным и
песчаным почвам. Вторичные сосновые леса в большинстве случаев
представлены сробществами смешанного состава, в которых наблюдаются
значительная примесь и подрост темнохвойных пород (46, 49). Но в ряде
случаев они формируют и длительно-производные чисто сосновые насаждения,;
когда под действием периодически повторяющихся пожаров надолго
задерживается восстановление коренных темнохвойных лесов.
Типология сосновых лесов Западной Сибири разработана еще
недостаточно. В некоторых работах лесоводственного направления [Крылов^
1961; Смолоногов, Никулин, 1963; Шанин, 1965; Таран, 1973] дается
характеристика таксационных и производственных показателей различных
типов сосновых лесов. Исследований, касающихся фитоценотического,
флористического и эколого-географического анализа этой обширной
лесной формации для рассматриваемого региона, еще нет. Особенно большие
трудности вызывает обоснование зональных типов сосновых лесов. В
значительной степени это связано с тем, что на состав и структуру сообществ
сосновой формации очень сильное влияние оказывают эдафические
факторы (механический состав, заболоченность почв), которые в значительной
степени затушевывают проявление зональных фитоценотичёских
особенностей. Тем не менее в пределах Западной Сибири, где сосновые леса
распространены на огромной территории, простирающейся с севера на юг
от 65 до 51° с. ш., выявление зональных черт их состава и структуры
крайне важно.
На карте растительности Западно-Сибирской равнины получили
отображение следующие подзональные типы сосновых лесов (рис. 9): 1)
северотаежные лиственничнр-сосновые и сосновые леса и редколесья; 2) сред-
нетаежные сосновые леса; 3) южно-таежные сосновые леса с липой; 4)
подтаежные сосновые леса; 5) лесостепные сосновые леса.
Основными диагностическими признаками выделения указанных
типов явились: а) фитоценотический состав и структура всех ярусов лес-
105 .
^
;«*
^/
5(
#/
№
(Ханты-Мансийск
Щ
<?
ъ
Тобольска
V.
М(
/"
*^
Новосибирск <&Г
* EI* \
•Л 1/2
: г
; Pwc. £.'Сосновые и лиственнично-сосновые леса.
Северотае ж н ы е: 1 — лиственнично-сосновые редкостойные кустарничково-лишайниковые
и зеленомошные; 2 — сосновые с кедром и лиственницей редкостойные сфагновые леса в сочетании
с болотами; 3 — сочетание лиственнично-сосновых редкостойных лишайниковых и зеленомошно-
кустарничковых лесов с сосновыми долгомошными и сфагновыми лесами и болотами
Среднетаежные: 4 — сосновые бруснично-зеленомошные и лишайниковые леса- 5
—'сосновые и березово-сосновые кустарничково-сфагновые леса в сочетании с болотами* 6 — сочетание
сосновых зеленомопшых и лишайниковых лесов с сосново-кустарничково-сфагновьши болотами* 7
сочетание сосновых с кедром и елью и березово-сосновых кустарничково-зеленомошных лесов с
сосновыми сфагновыми лесами и болотами.
Южно-таежные: 8 — сосновые, местами с липой, травяно-кустарничковые с разреженным
моховым покровов; 9 — сосново-бзрезовыа травяные леса и сельскохозяйственные земли на месте
южно-та\нкных сосновых лесов.
Подтаежные: 10 — сосновые кустарт^чково-травяные (с участием южно-боровых видов) и
лишайниковые леса; и — сосново-березовые5 травяные леса и сельскохозяйственные земли на месте
подтаежных сосновых лесов.
Лесостепные: J2 — сосновые и березово-сосновые кустарничково-травяные и травяные остеп-
ненные леса.
ных сообществ; б) некоторые таксационные показатели древостоя —
средняя высока и диаметр лесообразующей породы, класс бонитета, средний
запас спелых и перестойных насаждений; в) особенности пространственных
сочетаний сосновых лесов, их положение в экологических и
динамических рядах соответствующих подзональных комплексов.
106
СЕВЕРОТАЕЖНЫЕ ЛИСТВЕИНИЧНО-СЬСНОВЫЕ
И СОСНОВЫЕ ЛЕСА Й РЕДКОЛЕСЬЯ
И ПРОИЗВОДНЫЕ СООБЩЕСТВА НА ИХ МЕСТЕ
Северотаежные сосновые леса занимают большие площади в средней
ш южной частях подзоны. Они приурочены к приречным частям
водоразделов, а также к высоким террасам рек, где развиваются
преимущественно на легких (песчаных и супесчаных) более теплых', лишенных слоя
мерзлоты, почвах. В составе большей части, сосновых лесов присутствует
лиственница. Ее участие возрастает по направлению к северу, особенно на
хорошо дренируемых приречных частях водоразделов.
Древостой северотаежных сосняков, как пг>авило> разрежен
(сомкнутость не выше 0,4—0,6). Средняя высота деревьев 10—14 м,
продуктивность V—IV классов бонитета. При заболачивании в условиях близкого
залегания слоя вечной мерзлоты сосновые леса сменяются
лиственничными кустарничково-сфагновыми лесами, а здтем и лишайниково-сфагновы-
ми болотами. В псаммофитных рядах, наоборот, наблюдается выпадение
лиственницы и переход чистых сосновых редкостойных лесов в сосновые
редколесья и редины на развеваемых песках.
В зональном аспекте леса этого типа имеют два варианта: северный,
представленный сосновыми лесами с большим участием лиственницы (56,
57), и южный с единичной примесью лиственницы и кедра (58, 59).
Лиственнично-сосновые
лишайниково-зеленомошно-кустарничковые леса
в сочетании с сосновыми лишайниковыми (56)
Эти леса распространены на относительно дренированных плоских
участках водоразделов и речных террас, где приурочены к супесчаным и
легкосуглинистым подзолистым почвам. Состав древостоя варьирует от
7СЗЛ + Б, Е, К до 8С2Л + Е, К. Сомкнутость 0,4—0,5, класс бонитета
V, запас спелых и перестойных насаждений 80—90 м3/га [Таран, 19731.
Высота деревьев 10—12 м, диаметр 13—20 см. Возобновление сосной
происходит удовлетворительно, но подрост довольно редкий. Кроме сосны,
в нем единично встречаются кедр, ель, берегза. Подлесок развит слабо, он
представлен небольшими груцпами ерника (Betula nana), шиповника (Ro-
за acicularis), иногда ольхи (Duschekia fruticosa) и можжевельника (Juni-
perus communis).
Кустарнички образуют фон, *их покрытие от »30 до 60%. Среди
кустарничков обильны багульник (Ledum palustre),. водяника (Empetrum
nigrum), брусника (Vaccinium vitis-idaea), голубика (Vaccinium uliginosum),
встречаются спорадически линнея (Linnaea borealis) и черника
(Vaccinium myrtillus), Из травянистых видов распространены рсока
шаровидная (Carex globularis), вейники Лангсдорфа (Calamagrostis langs-
dorffii) и лапландский (С. lapponica), марьянник (Melampyrum pratense),
майник (Maianthemum bifolium), плауны (Diphasiastrum complanatum,
Lycopodium annotinum).
В моховом покрове с общим проективным покрытием 60—70% широко
развиты Hylocomium splendens, Pleurozium schreberi, Ptilium crista-castren-
sis, в меньшем количестве встречаются Dicranum polysetum, Aulacomnium
palustre. Синузии лишайников распределены отдельными пятнами по
микроповышениям — Cladina stellaris, CI. rangiferina, CI. arbuscufa, Peltigera
aphthosa.
Лиственнично-сосновые кустарничково-лишайниковые
редкостойные леса
в сочетании с кустарничково-мохово-лишайников'ыми
крупнобугристыми болотами (57)
' Эти леса получили широкое развитие в северной тайге Западной
Сибири. Они приурочены к песчаным иллювиально-железистым подзолам
приречных частей водоразделов и высоких речных террас, где вечная мерз-
107
ю-в
с-в
13 \14\ 15 | 16
700
900
1100
25025 75ГМ
М:'. i I I I \
Рис. 10. Геоботанцческий профиль в бассейне р. Казым (состав
1 — лиственнично-сосновый кассандрово-багульниково-сфагново-зеленомошный лес; 2 — листвен
сандрово-сфагновое болото; 4—грядово-мочажинное осоково-сфагновое с сосной^ болото; 5 — Сое
нично-лишайниковый; 7 — сосновый с лиственницей багульниково-водянично-лишайниковый; 8 —
водянйчно-лишайниковый со сфагнумом лес; ю — облесенное кустарничково-зелеломошно-сфаг
лес; 12 — сосновый багульниково-бруснично-лишайниковый; 13 — облесенное кустарвичково-
новый водянично-бруснично-лишайниновый лес* 16 — сосновый багульниково-бруснично-лишайни
бруснично-зеленомошно-лишайниковый; 19 — сосновый с лиственницей» багульниковобруснично-
никово-бруснично-зеленомошный; 22 — сосновый бруснично-багульниково-зеленомошно-лишайни
березово-лиственнично-сосновый чернично-бруснично-зеленомошный; 25 — лиственнично-березово-
28 — кедр; 29 — зеленые мхи; 30 — лишайники-; 31 — сфагнум;,
лота отсутствует или залегает глубоко (рис: 10). На обследованных нами
участках в верхнем течении р. Пур и на левобережье р. Казым состав
древостоя был 8С2Л + Б, К — 9С1Л + Е, сомкнутость крон 0,3—0,5.
Леса незахламленные, хорошо проходимые. Высота деревьев 'сосны и
лиственницы в возрасте'140—150 лет 12—15 м, диаметр 20—25 см. Форма
крон узкопирамидальная. Класс бонитета V—Va, средний запас 70 м3/га.
Возобновление редкое, подрост высотой не более 1—2 м, представлен
сосной, а в некоторых местах кедром (0;5—1 м) и порослевой березой.
Подлесок развит слабо, рассеянно группами и отдельными кустами в нем
встречаются карликовая береза (Betula nana) высотой 0,5—0,7 м и шиповник
(Rosa acicularis). \
Травяно-кустарничковый ярус беден и в видовом отношении, и по
обилию, его покрытие не более 20—30%. Преобладают брусника и
багульник, единично встречаются толокнянка (Arctostaphylos uva-ursi), осока
шаровидная (Carex globularis), хвощ (Equisetum sylvaticum), овсяница (Fes-
tuca supina), майник (Maianthemum bifolium), плаун (Diphasiastrum comp-
lanatum).
Напочвенный покров из лишайников с включением отдельных сину-
зий зеленых мхов сомкнутый (90—100%). Мощность живой части
лишайниковой дернины 6—12 см. Фон создает Cladina stellaris, в примеси к ней
встречаются Cladina arbuscula, CI. rangiferina, Cetraria islandica, C.
laevigata. Из мхов здесь обычны Pleurozium schreberi, Polytrichum alpestrer
Ptilidium ciliare, Dicranum polysetum.
Сосновые с лиственницей лишайниковые леса
в сочетании с кустарничково-лищайниково-сфагновьгаи
выпуклыми болотами
и сосновыми лишайниковыми редколесьями (58)
Сосновые, преимущественно чистые или с единичной лиственницей:
лишайниковые леса наиболее характерны для Сибирских увалов и
обширной области Сургутской низины. В южной части бассейна р. Пур эти леса
описывал Дунин-Горкавич [19041, а в бассейне р. Агана — Б. Н.
Городков [19241.
108
I 26 1 27 T 28 *» 29
oo JO r, 31 $ 32 ЧзЗ
\з4 \з?
—i 1 1 1 1 1 1
1300 1500 1700 1900m
лен Б. А. Богоявленским, И. С. Ильиной, А. П. Шведовым).
нично-сосновый с березой бруснично-лишайниковый; 3 — облесенное сосново-багулышково-кас-
новый багульниково-зеленомошно-сфагновый лес; 6 — сосновый с лиственницей багульниково-брус-
облесенное багульниково-голубично-кассандрово-сфагновое болото; 9 — сосновый багульниково-
новое с пятнами лишайников болото; 11 — сосновый кассандрово-сфагновый с пятнами зеленого мха
сфагновое болото; 14—вырубка соснового леса водянично-лишайниковая; 15—лиственнично-сос-
ковый; 17 — сосновый с лиственницей бруснично-лиш^йникЬво-зеленомошный; 18— сосновый
лишайниковый; 20 — сосновый зеленомопшо-лишайниковый с брусникой; 21 — сосновый багуль-
ковый; 23 — лиственнично-березово-сосновый багульниково-зеленомопгао-лишайниковый; 24 —
сосновый багульниково-бруснично-зеленомошный с лишайниками; 26 — сосна; 27 — лиственница;
32 — брусника; 33 — багульник; 34 — осока; 35 — кассандра.
Наземные и аэровизуальные обследования лишайниковых лесов и
редколесий производились в бассейне р. Тром-Еган и на южных отрогах
Сибирских увалов в окрестностях оз. Мамонтотяй [Богоявленский и др.,
1968].
Для рельефа этих мест характерны вытянутые вдоль русла рек гряды
и дюнообразные холмы, сложенные перевеваемыми песками.
Лишайниковые сосняки приурочены к хорошо дренируемым вершинам и склонам
этих гряд и развиваются на песчаных подзолистых иллювиально-желези-
стых почвах. Древостой преимущественно чистые ЮС + А + Б, редко
в них встречаются отдельные экземпляры лиственниц или берез.
Структура одноярусная^ древесный полог разрежен (0,3—0,4). Средняя высота
деревьев 12—13 м, диаметр 18—25 см. Производительность древостоя V—
Va классы бонитета.
Естественное возобновление удовлетворительное, в подросте сосна
высотой от 1 до 4 м, но местами подрост отсутствует, видимо, как следствие
низовых пожаров. Подлеска обычно нет или он представлен единичными
кустами можжевельника и шиповника. Травяно-кустарничковый ярус
разрежен, его проективное покрытие 10—15%. Отдельные кустики
низкорослой брусники, багульника, водяники приурочены к приствольным
повышениям. Сплошной покров имеют лишайники, преобладающая среди
них Cladina stellaris образует куртины-подушки диаметром 1,5—2,5 м.
Кроме нее обильны Cladina rangiferina и CI. arbusoula. Другие виды
лишайников — Cladonia coccifera, CI. deformis и Cetraria islandica, С.
nivalis — встречаются единично или небольшими группами. Дернинки
зеленых мхов вместе с кустарничками концентрируются вокруг деревьев и
по колодинам, а также по микропонижениям. Среди них обычны Pleuro-
zium schreberi, Polytrichum alpestre. '
Сосновые лишайниковые редколесья в сочетании с сосновыми
с кедром и лиственницей
кустарничково-зеленомошно-лишайниковыми лесами (59)
Сосновые' лишайниковые редколесья показаны на геоботанической
карте Западно-Сибирской равнины впервые. Это очень специфичный для
19 \20\21\22\23\ 24- \ 25
I I I I
109
данных условий компонент растительного покрова. Его широкое
распространение на зандровых песках Сургутской низины связано с современной
деятельностью ветра. По данным А. А. Земцова [1962], проводившего
географические исследования на Аган-Пуровском междуречье, для
ландшафтов этих мест характерны песчаные гряды., которые в настоящее время
интенсивно развеваются. Ца поверхности таких гряд среди лишайниковых
боров можно встретить чистые песчаные поля, котловины выдувания,
а также засыпанные песком отдельные деревья. На поверхности бывает
много гальки и небольших валунов. Именно к таким местам наиболее
приурочены сосновые ягельниковые редколесья, которые местами занимают
обширные площади на береговых гривах вдоль рек и на развеваемых
песчаных холмах и грядах среди обширных болотных массивов.
На менее выдуваемых вершинах и оклонах сосновые ягельниковые
редколесья переходят в лишайниковые леса, а у подножий склонов и на
плоских слабодренируемых участках водоразделов и речных террас —
в заболоченные сосновые сфагновые леса и обширные лишайниково-кустар-
ничково-сфагноБые олиготрофные болота.
Состав всех ярусов сосновых лишайниковых редколесий близок та-
ковому в сосновых лишайниковых лесах (58). Но в структурном отношении
здесь наблюдаются довольно существенные отличия. Прежде всего в
редколесьях резко уменьшаются сомкнутость древесного яруса (до 0,2—0,3)у
его средняя высота (до 10—11 м) и производительность (д,о V—Va классов
бонитета). В соответствии с этим снижается эдификаторная роль
древостоя, а напочвенный покров испытывает более сильное и прямое влияние
радиационных и эдафических факторов и его горизонтальная структура
приобретает ярко выраженный гетерогенный характер.
Ценообразующую роль и наивысшее обилие сохраняют кустистые
лишайники — Cladina stellaris, CI. rangiferina, CI. arbuscula. Они занимают
наиболее выпуклые и незатененные участки f- обычно склоны и вершины
песчаных бугров, кочек и более обширных грядообразных повышений.
Плоские и неглубокие микропонижения тоже заселены лишайниками, но
более разнообразного состава с большим обилием Cladina rangiferina и с
участием Cladonia gracilis, CI. uncialis, а также Stereocaulon paschale.
Более глубокие ложбинки, располагающиеся между грядами, имеют
покров, состоящий из смеси лишайников и зеленых мхов (Hylocomium splen-
dens, Pleurozium schreberi). Здесь же можно встретить и отдельные группы
кустарничков, главным образом Ledum palustre, Vaccinium myrtillus,
V. uliginosum, а также кустики Betula nana. На плоских или слегка
выпуклых пристволовых участках, затененных кронами деревьев,
доминируют зеленые мхи, в основном Pleurozium schreberi, и хорошо
развиваются синузии брусники и черники»
Сосново-березовые и березово-сосновые
зеленомошно-лишайниковые леса (60)
Несмотря на труднодоступность и малую населенность районов
распространения сосновых северотаежных лесов, пожары там повторяются
довольно часто. Больше всего пожары бывают беглыми низовыми,
уничтожающими подрост, живой и мертвый покров, запас семян в почве. В
случае повальных верховых пожаров гибнут деревья сосны и сопутствующих
пород — лиственницы, березы, кедра.
После пожаров в места/Х, где почти полностью уничтожен
напочвенный покров и почва в связи с этим несколько сильнее минерализована,
довольно быстро появляются самосев и подрост сосны; другие породы,
особенно кедр и ель, даже при слабых пожарах почти не возобновляются.
Подрост темнохвойных пород в сосновых лесах может появиться лишь
после того, как послепожарное поколение сосны войдет в основной полог
древостоя [Шанин, 1965J. Значительно активнее в осветленных местах
развивается подрост березы, который способен чере^ несколько
десятилетий сформировать верхний полог вторичного древостоя. Однако березо-
110
йые стадии в послепожарных сменах сосновых лесов обычно очень кратко-
временны, так как при $осте под сосновым пологом береза обладает очень
слабой порослевой способностью и к возрасту спелости почти полностью
вытесняется сосной. В целом можно считать, что восстановление сосновых
древостоев после пожаров идет без смены пород.
В нижних ярусах леса при выгорании, особенно на песках, быстрее
всего разрастается брусника, в небольших микропонижениях
формируются синузии пирогенных видов зеленых мхов — Polytrichum piliferum*
P. juniperinum, P. alpestre. Позднее среди кустарничков появляются
багульник и водяника, а также некоторые типичные псаммофиты —
толокнянка и кошачья лапка. Политриховые мхи постепенно сменяются более
устойчивыми зелеными мхами — Pleurozium schreberi, Ptilium crista-
castrensis и лишайниками. Обилие последних особенно увеличивается на
песках. ,
На карте обобщенным выделом 60 показаны две стадии
послепожарных смен сосновых древостоев, которые на местности постоянно
чередуются в зависимости от давности и повторяемости пожаров.
Сосновые с кедром и лиственницей
лишайшшово-кустарничков'о-сфагновые леса
в сочетании с сосново-кустарничково-сфагновыми
олиготрофными болотами (61)
Заболоченные сосновые леса очень распространены в северной тайге
Западной Сибири. Они представлены многочисленными сообществами
разных стадий заболачивания. Как правило, все леса этого типа приурочены
к почвам с мощным торфянистым горизонтом, подстилаемым,вечномерзлы-
ми грунтами.
Начальным стадиям заболачивания соответствуют ассоциации лист-
веннично-сосновых кустарничково-зеленомошных лесов, в которых по
микропонижениям — «окнам» — активно развиваются синузии сфагнов
{Sphagnum angustifolium). При увеличении мощности торфяного слоя до
15—20 см формируются сосновые кустарничково-долгомошные с
участием сфагновых мхов и лишайников леса. Они обычно приурочены к
окраинным частям водораздельных болотных массивов. На террасах рек и при
близком залегании верховодки развиваются кустарничково-долгомошные
и осоково-кустарничково-долгомошные леса, в их древесном ярусе
наблюдается большое участие, а иногда и доминирование березы пушистой
(Betula pubescens).
В ряду заболачивания сосняки долгомошные могут индицировать и
обратный процесс, когда они возникают на месте сосновых сфагновых
лесов после пожаров, способствующих минерализации почвы и улучшению
ее гидротермического режима [Ком1ин, 1967].
При дальнейшем .заболачивании в северной части подзоны на
суглинистых грунтах формируются лиственнично-сосновые и сосново-листвен-
ничнне кустарничково-сфагновые леса. Чистые сосновые олиготрофно-
кустарничково-сфагновые леса наиболее характерны для обширной Сур-
гутскоц низины и области Сибирских увалов, где они формируются на
песчаных, супесчаных и песчано-галечных отложениях в условиях
длительного избыточного увлажнения.
На карте сосновые заболоченные леса показаны номером 61, они
описаны нами на многих участках в бассейнах рек Лямин, Тром-Еганг
Казым, Назьщ. „
Древостой этих лесов, состоящий в основном из сосны (ЮС) с редкой
примесью других пород (9С1Б + Л, К), угнетен и разрежен. Высота
сосен 10—15 м, диаметр 6—10 см. В лесу обилие сухостоя, деревья суховер-
шинны, ветви их покрыты эпифитными лишайниками (Usnea longissinea).
Возобновление слабое, единичные экземпляры низкорослой сосны иногда
с примесью кедра и березы. Подлесок образован группами кустов
карликовой березы {Betula nana), местами разрастающихся в сплошные заросли *
Покрытие травяно-кустарничкового яруса 25—40%. Травы и кустарнич-
lit
ки концентрируются на кочках, приствольных повышениях, по валежу.
Обильны олиготрофные кустарнички — багульник, Кассандра, голубика^
реже встречаются клюква (Оху coccus microcarpus), андромеда. Довольно
много трав — осока (Carex globularis), пушица (Eriophorum russeolum),
морошка: в обводненных понижениях — вахта (Menyanthes trifoliata)f
шейхцерия (Scheuchzeria palustris), росянка (Drosera intermedia), реже
осока топяная (Carex limosa), пушица (Eriophorum vaginatum).
Моховой покров сплошной (100%), состоит из сфагновых (Sphagnum
angustifolium, Sph. fuscum) и зеленых (Pleurozium schreberi, Aulacomnium
palustre, Dicranum polysetum) мхов и небольших пятен лишайников (Cla-
dina rangiferina).
СРЕДНЕТАЕЖНЫЕ СОСНОВЫЕ ЛЕСА
И ПРОИЗВОДНЫЕ СООБЩЕСТВА НА ИХ МЕСТЕ
Наибольшие площади сосновых лесов Западно-Сибирской равнины
сосредоточены в пределах среднетаежной подзоны ч преимущественно на
ее краевых частях. Преобладание этих лесов в бассейнах рек Сев. Сосьва
и Конда, Вах, Тым и Сым связано с широким распространением в этих
районах песчаных и супесчаных отложений разного генезиса. Сосновые
леса зандровых водораздельных равнин, сложенных флювиогляциальны-
ми песками, следует рассматривать как квазикоренные сообщества
спонтанной растительности [Сочава, 1963]. На аллювиальных песках и
супесях речных террас развиты сосновые леса, представляющие наиболее
устойчивую стадию долинной серии, предшествующую климаксу [Городков,
19461.
Длительно-производные сосновые леса в большинстве своем формц-
руются на более**плодородных суглинистых автоморфных почвах
(слоистых отложениях) и представляют различные ч^тадии восстановления елово-
кедровых лесов. Характеристика таких лесов дана выше (78).
Типологическое разнообразие сосновых лесов обусловлено
гидротермическим режимом пачв и в меньшей степени связано с их генезисом.
Различные группы ассоциаций в средней тайге имеют довольно устойчивые
связи с определенными подтипами почв, чего не отмечалось в условиях
северных подзон. Господствующее положение на дренированных
местообитаниях получили сосновые леса кустарничково-зеленомошной группы,
связанные в своем развитии с подзолистыми глее^атыми пбчвами: Для
лесов этой группы характерен стабильный режим влажности и /аэрации в
пределах верхних горизонтов почвенного профиля. На гидроморфных
почвах повсеместно распространены сосновые леса сфагновой группы.
Характерна совместная встречаемость сообществ сосновьгх лесов разных
экологических групп на склоновых местоположениях и понижениях, что
позволяет картировать их лишь путем территориального объединения. На
карте использованы три типа сочетаний экологически различных групп
сосновых лесов и болот '(80—82). Соотношение компонентов в этих
сочетаниях варьирует в зависимости от характера рельефа и связанного с ним
увлажнения.
Среднетаежные леса отличаются большей сомкнутостью и лучшей
производительностью древостоя. Средний класс бонитета IV, но в отличие
от северных лесов встречаются древостой II и III классов бонитета.
Возрастной состав характеризуется преобладанием перестойных
насаждений [Таран, 1973].
Древесный ярус большей частью чисто сосновый, изредка
примешиваются кедр, ель, береза, лиственница. Лиственница утратила свое
значение как содоминанта и ее участие свелось к единичным деревьям.
В травяно-кустарничковом покрове' доминирующая роль
принадлежит бореальным кустарничкам (бруснике, чернике), содоминирование бо-
реально-арктических видов ограничивается сообществами заболоченных
лесов. К югу увеличивается присутствие травянистых видов.
112
Сосновые багульниково-бруснично-зеленомошные
и лишайниковые леса (80)
* Сочетание соснового багульниково-бруснично-зеленомошного леса
и лишайникового характерно для наиболее расчлененных участков су-
песчано-песчаных равнин с крупнохолмистым и гривистым рельефом*
Вершины и верхние части склонов холмов покрыты сухими
лишайниковыми сосновыми лесами, сменяющимися в нижних частях пологих склонов и
равнинных участках сосновыми багульниково-бруснично-зеленомошными»
Такие сочетания лесов широко распространены по песчаным бугристо-
дюнным холмам и долинным зандрам кондинского левобережья, на
расчлененных возвышенных равнинах Северногоi и Южного Люлимвора,
Значительные площади они занимают на зандровых равнинах с
плоскохолмистым рельефом в восточной (приенисейской) части средней тайги —
на междуречьях и террасах Сыма, Каса, Тыма и Ваха, небольшие
массивы таких лесов встречаются и в пределах южной тайги по широким
террасам и гривам на междуречьях Кети и Чулыма.
Особенно своеобразен растительный покров зандровой равнины
низовьев Дубчеса и Сыма, где гривы тянутся на десятки километров, а
ширина их достигает 2—3 км. На всех гривах разной высоты и
горизонтальной протяженности произрастают лишайниковые сосновые леса, среди
которых в слабо выраженных депрессиях встречаются массивы зеленомош-
ных сосновых лесов. Багульниково-бруснично-зеленомошные сосновые
леса занимают также короткие пологие склоны и подножия грив,
примыкающие к болотным котловинам.
Сосновые лишайниковые леса приурочены к песчаным субстратам
с автоморфными подзолистыми иллювиально-железистыми почвами.
Древесный ярус лишайниковых лесов разреженный (сомкнутость
0,3—0,5) высотой 13—15 м. Производительность его низкая,
характеризуется V классом бонитета. По составу насаждения преимущественно
чистые, местами единично встречаются кедр или лиственница, береза.
Наряду с молодыми одновозрастными лесами ч^сто встречаются /и
разновозрастные, где возрастные поколения образуют два четко обособленных
полога. Подрост сосны редкий, часто повреждается пожарами.
Подлесок отсутствует или развит слабо из единичных кустов розы
иглистой, ивы, реже можжевельника.
Травы и кустарнички встречаются фрагментарно, видовой состав их
крайне беден (5—6 видов). Небольшие куртины образуют брусника и
толокнянка, местами Empetrum nigrum. Из травянистых видов
присутствуют вейник нкземный и тростниковидный, кипрей, майник двулистный,,
а в приенисейской части (низовья р. Дубчес) отмечается участие Koele-
ria cristata, Festuca supina, Carex macroura. На террасах р. Конды
встречается в составе травяно-кустарничкового покрова лишайниковых
сосняков вереск (Calluna vulgaris), местами образующий фон, что позволило
Е. П. Смолоногову впервые для Западной Сибири выделить как
самостоятельный тип сосняк вересковый [Смолоногов, Никулин, 1963].
В несколько более благоприятных условиях увлажнения
развиваются бруснично-лишайниковые сосновые леса, древостой которых
характеризуется IV—V классами бонитета. Древостой отличается большей
сомкнутостью (0,5—0,6), пестротой возрастной структуры. Напочвенный
покров пятнистый, с одновидовой зарослью брусники по приствольным
микроповышениям и небольшим западинкам и синузий зеленых мхов из Pleu-
rozium schreberi среди основной лишайниковой толщи.
Основу напочвенного лишайникового покрова составляют кустистые
и трубчатые формы кладоний. В зависимости от давности пожаров
видовой состав и обилие лишайников меняются. В старых ненарушенных на-
саждениях доминируют кустистые лишайники Cladina arbuscula, CI.
rangiferina, CI. stellaris с мелкими включениями Cetraria islandica. По ва-
лешу и мелким западинкам латками встречаются зеленые мхи — Pleu-
rozium schreberix Dicranum polysetumx Polytrichum juniperinutn. В лесах^ где
8 Заказ N 297 ИЗ
пожар прошел не более 15—20 лет назад, господствуют трубчатые и бо-
кальчатые лишайники — Cladonia gracilis, CI. deformis, CI. crispatai
CI. естосупсг, CI. jurcata с включениями CI. coccifera, CI. cornuta;
кустистые кладонии встречаются спорадически. В некоторых лесах чистые
агрегации образует Stereocaulon paschale.
Сосновые лишайниковые леса наиболее подвержены воздействию
пожаров, чему способствуют легко загорающийся лишайниковый покров
и общий засушливый характер местообитания. Восстановление
лишайникового покрова идет медленно (40—30 лет). На молодых гарях появляются
травянистые виды — Calamagrostis epigeios, Chamaenerion angustijolium^
но больших зарослей они не дают и быстро отмирает.
Более устойчив на молодых гарях нокров из Polytrichum iuniperimun
и различных видов трубчатых лишайников.
По пологим склонам грив и дренированным межгривным понижениям
господствуют сосновые леса с багульниково-брусни,чно-зеленомошным
покровом. Почвы под ними подзожясто-грунтово-глеевые иллювиально-
гумусово-железистые. Насаждения характеризуются довольно хорошей
жизненностью IV—V классов бонитета, сомкнутость крон 0,6—0,8. Из
других пород единично участие березы, лиственницы, кедра. Высота
сосны I полога 16—18 м при диаметре 18—22 см, II полога — 12—15 м с
диаметром 12—16 см. В подросте сосна, лиственница, редко кедр. Эти леса
реже повреждаются пожарами и под их пологом всегда много подроста и
самосева сосны. В редком подлеске чаще других видов встречаются
рябина, роза иглистая и ива козья. Травяно-кустарничковый покров густой,
довольно однородный с проективным покрытием 75—85%. Некоторая
застойность увлажнения, вызванная близостью грунтовых вод (в пределах
1,5—2 м), сказывается в доминировании олиготрофного багульника
наряду с брусникой, имеющей несколько меньшее обилие. Из других
кустарничков спорадически встречаются черника и единичными кустами
голубика. Незначительное участие травянистых видов — хвоща малого,
осоки шаровидной, вейника тростниковидного, кипрея. В сплошном моховом
покрове явно доминирует Pleurozium schreberi при постоянном небольшом
участии других зеленых мхов. Редкими пятнами по зеленым мхам
распространены лишайники — Cladina arbuscula% Peltigera aphthosa.
Сосновые бруснично-зеленомошные
и бруснично-лишайниковые леса
в сочетании с сосново-куетарничково-сфагновыми
олиготрофными болотами (81)
Закономерные сочетания таких типов сосновых лесов и болот
характеризуют растительный покров возвышенных слабо расчлененных
аккумулятивно-денудационных равнин Южного Люлимвора и Малососьвин-
ского амфитеатра; они специфичны для большей части низких эрозионно-
аккумулятивных равнин Конды-Сосьвинского и Лозьва-Пельшского
междуречий. Широко распространены они и на озерно-аллювиальных и занд-
ровых равнинах Обь-Енисейского междуречья. Наибольшие площади их
сосредоточены в бассейне рек Тыма, Сыма, Дубчеса и на междуречье Ело-
гуя и Келлога.
В этом территориальном выделе плосковершинные холмы или гривы
с пологими склонами покрыты сосновыми лесами, а ложбины или слабо-
врезанные чашеобразные понижения заняты болотами. Болота
преобладают олиготрофные, залесенные сосной с кустарничково-сфагновым
покровом. По периферии болота окаймлены узкой полосой заболоченного
соснового леса.
Наиболее дренированные местоположения — вершины и выпуклые
чдсти склонов с автоморфными подзолистыми иллювиально-железисто-
гумусовымп почвами — заняты сухими бруснично-лишайниковыми сое-.
новыми лесами, характеристика которых дана в выделе 80.
По пологим склонам распространены различные сообщества сосновых
лесов из группы кустарничково-зеленомошных. Наибольшую встречае-
114
мость и большие площади имеют сосновые бруснйчно-зеленомошные
леса. На склонах более низких грив, подверженных периодическому
переувлажнению, господствуют сосновые чернично-зеленомошные леса с
большим или меныпим участием багульника, а на вершинах — сосновые бруо-
нично-зе л еномошные.
Сосновые бруснично-зеленомошпые леса обычно связаны с
подзолистыми грунтово-глееватыми иллювиально-гумусово-жёлезистыми почвами
со слабым влиянием почвенно-грунтовых вод. В наиболее сухие периоды
уровень грунтовых вод колеблется от 3 до 2 м, поднимаясь до 1я5 м весной.
Нередко эти леса развиваются на кйнтактно-глеевых почвах, где
утяжеленные прослойки располагаются на небольшой глубине
[Караваева, 1973].
Древесный ярус лесов довольно однородный, с единичным участием
других пород — березы, лиственницы, кедра и ели. Наиболее
распространены одноярусные разновозрастные насаждения с колебаниями возраста
от 70 до 200 лет. Производительность древостоевУ, реже IV класса
бонитета. В подросте преобладает сосна, местами с участием лиственницы и ели,;
редко кедра.
Подлесок образуют единично встречающиеся Sorbus sibirica, Rosa
acicularis, Juniperus communis, Duschekia fruticosa.
Травяно-кустарничковый покров беден флористически и не имеет
«специфически боровых» видов. Общий флористический состав
насчитывает около 30 видов, а на отдельных участках встречается от 5 до 15
видов. На хорошо дренированных местоположениях доминирующее
значение имеют обычные бореально-таежные кустарнички (брусника, меньше
черника), при возрастании увлажнения и ухудшения дренажа господство
переходит к Ledum palustre, увеличивается присутствие и таких бореаль-
но-арктических кустарничков, как Empetrum nigrum и Vaccinium uligi-
nosum. Последняя обильно разрастается в осветленных лесах после пожара.
Участие трав на различных участках ассоциации незначительно.
Обычными спутниками служат Calamagrostis arundinacea, Maianthemum bifoliumt
Melampyrum pratense, Solidago uirgaured, Carex globularis, Festuca mpina,
встречающиеся единичными экземплярами.
В напочвенном покрове доминирует Pleurozium schreberi, единичное
распространение имеют другие таежные мхи — Hylocomium proliferum, Die-
ranum polysetum^ Ptilium crista-castrensis. Мощность моховой дернины 5—
12 см. Иногда редкими пятнами на валежнике и около оснований
древесных стволов растут лишайники. Только в лесах с недавними низовыми
пожарами отмечается массовое распространение трубчатых форм
лишайников -*- Cl'adonia deformis, CI. pleurota, CI. coccifera, CI. cornuta, CI. gracilis.
При восстановлении зеленые мхи вытесняют лишайники.
На склонах с хорошим транзитом влаги формируются сообщества
сосновых бруснично-зеленомошных лесов с Duschekia jruticosa в подлеске
{рис. 11). Почвы под ними хорошо увлажненные нодзолисто-грунтово-
глееватые, грунтовые воды на глубине 2—2,5 м. Насаждения IV класса4
бонитета, одноярусные с единичным участием березы, лиственницы, ели.
В подросте также участвуют темнохвойные, особенно обильна ель.
Хорошо развитый подлесок.образован ольхой кустарниковой высотой 2—2,5 м,
распространенной группами, единично встречаются рябина, ива, роза
иглистая.
Травяно-кустарничковый покров несколько богаче видами (12—15
на участке), характерно присутствие в его составе обычных спутников тем-
нохвойных лесов. Наряду с доминирующей брусникой, спорадически
распространены черника, линнея северная, плаун обоюдоострый, майник
двулистный, единично участие седмичника, марьянника лугового,
овсяницы приземистой, кипрея. Моховой покров,из Pleurozium schreberi с
участием Hylocomium splendens, Dicranum polysetum, Polytrichum commune.
На плоских поверхностях с худшим дренажем (со слоистыми
отложениями в приповерхностных толщах или при близком уровне грунтовых вод)
развиваются сосновые леса с кустарничково-зеленомошным покровом.
8* • 115
Сосновые и березово-сосновые долгомошно-сфагновые
и кустарничково-сфагновые леса
в сочетании с кустарничково-сфагновыми
олиготрофными болотами (82)
Неоднородный растительный покров плоских слабодренированных
поверхностей, испещренных мелкими/заторфованными понижениями,
показан на карте сочетанием сосновых заболоченных лесов и олиготрофных
сфагновых болот. Такое территориальное объединение различных
гидрофильных типов сообществ характерно для местоположений с торфянисто-
Рис, 11. Сосновые среднетаежные леса Белогорского материка.
^ ленского.
116
Фото Б. А. Богояв~
или торфяно-глеевъгми почвами и небольшой мощности верховыми
торфяниками в условиях длительного избыточного увлажнения.
Широкое распространение и наибольшие площади этот тип выдела
•имеет в среднетаежной подзоне, где криогенные процессы и длительное
сохранение почвенно-грунтовой верховодки стимулируют заболачивание
суходольных лесов на почвах различного механического состава. В южной
тайге такая структура растительного покрова выделена преимущественно
по периферии крупных болотных массивов, где заболачивание лесов
связано с деятельностью окружающих болот, систематически, повышающих
уровень верховодок в примыкающих к ним суходолах.
♦Березово-сосновые долгомошно-сфагновые и сосновые кустарничково-
сфагновые леса представляют конечное звено обобщенного эколого-фито-
ценотического ряда лесных сообществ разных типов суходольного
заболачивания. В картируемый выдел, названный по этим сообществам
заболоченным лесом, включены также территориально смежные им лесные
сообщества начальных стадий заболачивания, образующие в приболотной
полосе единый экологический ряд и собственно болотные сообщества, не
выраженные в масштабе карты.
При олиготрофном типе заболачивания, что свойственно
преимущественно лесам среднетаежной подзоны на супесчаных и песчаных почвах,
наиболее характерен гидрофильный ряд из лесных сообществ, следующих
по нарастанию увлажнения и увеличению мощности торфяного горизонта
в такой последовательности: сосновый багульниково-бруснично-зелено-
мошный с куртинами сфагновых мхов — сосновый багульниково-осоково-
долгомошно-сфагновый — сосновый багульниково-сфагновый. При
заболачивании сосновых лишайниковых лесов внедрение сфагновых мхов при
быстром нарастании увлажнения происходит непосредственно в
лишайниковый покров, минуя переходную зеленомошную стадию, и ряд
заболачивания образует неустойчивые сообщества из видов, контрастных по
своей экологии: сосновый кустарничково-сфагново-лишайниковый лес —
сосновый кустарничково-лишайниково-сфагновый лес — сосновый осоко-
во-кустарничково-сфагновый.
При таком типе заболачивания характерно образование сфагновой
синузии непосредственно на минеральном грунте без предшествующего
этому поселению накопления торфяного слоя, что неоднократно отмечали
разные авторы [Самбук, 1932; Горновский, 1949; Сторожева, 1960, 1968;
Глебов, 1963; Караваева, 1973]. Дальнейшее обильное развитие долгомош-
ных и сфагновых мхов приводит к их доминированию и накоплению слоя
торфа, что влечет за собой ослабление эдификаторной роли древесного
яруса и массового разрастания кустарничков — багульника, голубики,
Кассандры. На почвах с хорошо выраженным торфяным горизонтом —
торфянисто- или торфяно-глеевых — сосновые леса переходят в стадию
заболоченных.
На почвах, подстилаемых песками с прослоями супесей и суглинков,
начальные звенья ряда заболачивания образуют лесные сообщества, в
составе которых присутствуют темйохвойные породы (преимущественно кедр):
кедрово-сосновые (с единичной елью) чернично-сфагново-зеленомошные —
сосновые (С единичными кедром и елью) чернично-осоково-долгомошно-
сфагновые — сосновые осоково-кустарничково-сфагновые.
Такой тип заболачивания широко распространен в подзоне южной
тайги на Кеть-Чулымском и Кеть-Касском междуречьях [Львов, 1976:
Глебов, Горожанкина, 1979]. Для междуречий с суглинистыми
отложениями сосновые сфагновые леса представляют конечное звено
экологического ряда поверхностного заболачивания темнохвойных лесов, обычно ело-
во-кедрового бруснично-зеленомошного. Различия в начальных звеньях
этого ряда заболачивания по мере нарастания торфянистой подстилки и
развития мощного сфагнового покрова все более затушевываются, и
конечную фазу представляют лесные сообщества с господством сосны: сос-
ново-березовый (с единичным кедром) хвощово-осрково-сфагновый (Сагех
globularis, Equisetum sylvaticum, Sphagnum wulfianum, Sph. girgensohnii),
> 117
сосновый (с единичными березой, кедром) кустарничковр-кассандрово-осо-
ково-сфагновый (Sph. girgensohnii, Sph. warnstorfii), контактирующие с
периферией болотных массивов.
Собственно болотная растительность представлена кустарничково-
сфагновыми олиготрофными сообществами облесенными или открытыми.
Сосновые кустарничково-сфагновые леса, представляя одну из
крайних стадий заболачивания леса, характеризуются сочетанием признаков
лесных и болотных сообществ. Древесный ярус таких лесов разреженный
с сомкнутостью крон 0,4—0,6, разновозрастный (в пределах от 40 до
200 лет). Деревья угнетенные высотой 8—12 м в средней тайге и до 14—
15 м — в южной. В составе древесного яруса, кроме господствующей
сосны, встречается примесь кедра и березы единично или группами. Подрост
редкий из разновысотных групп: 2—3, 1—2 и и,до одного метра.
Подлесок отсутствует. Продуктивность лесов низкая: IV—V классов бонитета
в южной тайге и V—Va — в средйей. Напочвенный покров мозаичного
строения. Основную синузию составляют сфагновые мхи, образуя
специфический нанорельеф: обширные подушки, кочки или валы,
окольцовывающие приствольные повышения. Из сфагновых мхов наиболее
распространены в заболоченных лесах мезогигрофиты — Sphagnum girgensoh-
nii, Sph. warnstorfii, Sph. russowii, Sph. magellanicum, Sph. angustifolium.
На фоне основной синузии отчетливо* выделяются микроценозы прит-
стволовых повышений с кустарничками — брусникой, багульником,
Кассандрой и лесными видами мхов — Pleurozium schreberi, Dicranum poly-
setum, Aulacomnium palustre, Polyirichum alpestre.
В понижениях на минеральном грунте и небольшой мощности
сфагновой подстилке разрастаются дерновинки Carex globularis и кустики
черники, багульника, подбела. На сфагновых подушках развиты типичные
микроценозы болот с синузией стелющихся кустарничков — клюквы
болотной или мелкоплодной, багульником, с участием травянистых видов —
морошки (Rubus chamaemorus), росянок (Droserarotundifolia, D. anglica).
Общее число видов этой группы в заболоченных лесах более 30, из них
10—15 высших растений и до 15 *мхов.
ЮЖНО-ТАЕЖНЫЕ СОСНОВЫЕ ЛЕСА
И ПРОИЗВОДНЫЕ СООБЩЕСТВА НА ИХ МЕСТЕ
Основные массивы сосновых лесов сосредоточены в западной и
восточной окраине подзоны южной тайги — в районах с преобладанием
песчаных отложений. На Обь-Иртышском междуречье сосновые леса
распространены небольшими разрозненными островами по заболоченным
торфянистым отложениям и песчаным террасам.
В западной части южно-таежной подзоны, охватывающей районы
Тавдинско-Кондинской наклонной равнины и Тобол-Ишичского
междуречья, сосновые леса (103) имеют более южный облик, сближающий их с
подтаежными лесами как сибирскими, так и европейскими. Характерной
особенностью этих лесов является обогащение флористического состава
травянистыми растениями, а на достаточно плодородных почвах
присутствуют виды неморального флористического комплекса с липой в составе
древесного яруса или подлеска. Средняя продуктивность таких лесов
приближается ко II классу бонитета.
Сосновые леса этой группы давно эксплуатируются и систематически
повреждаются пожарами, поэтому значительные площади их заняты
производными березовыми и осиновыми лесами (104).
Сосновые леса, местами с липой, травяно-кустарничковые
с разреженный моховым покровом (103)
Леса этой группы наибольшее распространение имеют в бассейне
среднего течения рек Тавды и Туры (левые притоки Тобола), к востоку
встречаясь лишь небольшими массигами в бассейне р. Вагая и низовьях
118
Ишима (самое восточное их местонахождение). Занимают пологие склоны
или хорошо дренированные плоские вершины увалов с супесчаными или
суглинистыми дерново-подзолистыми почвами.
Эта обобщенная группа сосновых лесов объединяет довольно
разнообразные по составу и структуре растительные сообщества: сосновые тра-
вяно-брусничные, сосновые травяно-черничные, темнохвойно-сосновые с
липой вейниково-бруснйчные, сосновые бруснично-вересковые, сосновые
с липой.в подлеске бруснично-разнотравные.
Описания основных типов сосновых лесов среднего течения р. Тавды
и Тавда-Кумцнского междуречья даны Б. П. Колесниковым [1972], а в
прииртышск*ой части и на Тобол-Ишимсшщ междуречье — Ю. П. Хло-
новым [1965].
Наиболее высокой производительностью древостоев (II класс
бонитета) и богатством видового состава характеризуются сосновые леса с липой
сердцелистной (Tilia cordata). Они располагаются по более теплым и
умеренно увлажненным пологим склонам увалов или грив с относительно
богатыми дерново-подзолистыми почвами. Липа участвует в древесном
ярусе сосняков как сопутствующая или подлесочная nopoflaw В составе
таких сосновых древостоев всегда имеется примесь березы, осины, а на
севере подзоны — ели и пихты. Обычен состав древостоев 7С1 — 20с1 —
ЗБ ед. Лп, Е, П. Иногда липа образует второй ярус высотой 12—14м,;
но чаще она вырубается, и сохраняются лишь отдельные деревья. Подрост
того же состава, что и древостой с преобладанием сосны.
Возобновление липы порослевое. На участках с большой
сомкнутостью крон (0,6—0,8) она растет в подлеске до 20—25 лет, а затем отмирает.
Из-за часто повторяющихся низовых пожаров деревья угнетены и большей
частью не превышают высоты 2—3 м.
Подлесок неравномерной сомкнутости (0,1—0,3), разнообразный
по составу — рябина сибирская, жимолость, калина, группы розы
коричной и розы иглистой, волчье лыко, а в увлажненных
микропонижениях встречается ива пепельная, бузина.
Травяно-кустарничковый покров хорошо развит (общее проективное
покрытие 70—90%) и отличается высокой видовой насыщенностью. В его
составе более 30 видов на участке с согосподством травяных и кустарнич-
ковых видов. Доминантами в различных сообществах выступают, вейник
лесной, осока большехвостая, сныть обыкновенная, а из кустарничков
брусника, реже черника. Высокую встречаемость при небольшом обилии
имеют костяника, майник двулистный, чина весенняя, подмаренник
северный, медуница, хвощ лесной и зимующий, дудник лесной.
Моховой покров развит слабо, его слагают отдельные подушки Pie-
urozium schreberi, Dicranum polysetwn.
К этой же группе отнесены также сосновые травяно-кустарричковые
леса без участия липы, встречающиеся небольшими участками на других
экотопах, территориально объединяющихся в один контур. В древесном
ярусе таких лесов нередка примесь ели и пихты (8—9С, 2— 1Б ед.
Е, П). Подлесок редкий из единично встречающихся рябины, шиповниках
ивы козьей.
В травяно-кустарничковом покрове, кроме доминирующей брусники,, .
спорадически распространены черника, вейник лесцой, плауны,
костяника, ожика волосистая, майник, подмаренник северный, золотая розга.
Моховой покров разреженный, совершенно отсутствует в густых
зарослях кустарничков. Небольшими участками встречаются выделенные
Б. П. Колесниковым характерные для этой территории сосновые леса с
бруснично-вересковым покровом (возможно, представляют последнюю
стадию в послепожарном восстановлении сосновых разнотравно-брусничных
лесов). Почвы под такими лесами песчаные, на глубине 80—100 см
подстилаются суглинками со следами неоднократных пожаров. Древостой чисто
сосновйе III—IV классов бонитета, сомкнутость 0,5—0,7. Подлесок
отсутствует. В травяно-кустарничковом покрове согосподство Vaccinium
vitis-idaea и Calluna Vulgaris — европейский видг встречающийся за Ура-
119.
лом только в единичных местообитаниях. Участие других видов
невелико, в их числе вейник наземный, кошачья лапка, золотая розга,
багульник. На почве редкие подушки мхов из Polytrichum juniperinum, P. pili-
serum, Соснойые леса с доминированием вереска впервые выделены
Е. П. Смолоноговым [Смолоногов, Никулин, 1963] в долине р. Конды и
Б. П. Колесниковым [1972] в Тоболо-Кондинской провинции.
Сосново-березовые местами с липой травяные леса (104)
Под этим номером объединены как длительно-производные леса с
небольшими участками коренных, так и молодняки и растительность не-
возобновившихся гарей и вырубок.
Производные сосново-береэовые леса занимают те же
местоположения, что и коренные сосновые леса. Они возникли на вырубках и raj/ях,
где возобновлению сосны препятствует быстрое задернение почвы и
частые «шзовые пожары. )
* На сплошных вырубках сосновых травяно-кустарничковых лесов
в составе лесовозобновления преобладают лиственные породы, в
результате чего формируются устойчиво-производные сосново-осиново-березо-
вые и сосново-березовые леса с участием сосны от единичных деревьев до
1—2 состава древостоя. В северных районах южной тайги в составе сос-
ново-березовых производных лесов нередко отмечается примесь пихты и
ели. Эти же породы всегда присутствуют и в возобновлении. Липа в
производных лесах обычно господствует во втором ярусе, не превышая высо-.
ты 13—15 м. Лишь отдельные деревья могут выходить в первый ярус,
достигая высоты 18—20 м.
Подлесок средней густоты из липы, жимолости, рябины, шиповника,
бузины, крушины ломкой, волчьего лыка, свидины белой. Травяной
покров флористически разнообразен и насчитывает более 30 видов на 100 м2.
Наиболее обильны Aegopodium podagraria, Pteridium aquilinum, Carex
macroura, Maianthemum bifolium, Rubus saxatilis, Lathyrus vernus, Angelica
sylvestris, Equisetum sylvaticum, в незначительном количестве, но
постоянно присутствуют Melica nutans, Milium effusum, Paris quadrifolia,
Geranium sylvaticum, Thalictrum minus, Equisetum hyemale. В
микропонижениях встречаются куртинки Vaccinium vitis-idaea c Pleurozium schreberi и
Ptilium crista-castrensis.
Ю. Ц. Хлонов [1965] выделил три типа производных осиново-березо-
вых и сосново-осиновых лесов на месте травяных сосновых лесов с липой:
березняк липняковЪ-снытево-разнотравный, березняк липняково-орля-
ково-разнотравный, осинник липняково-осочково-разнотравный.
Естественный ход возобновления сосновых лесов очень длительный
и часто прерывается различными нарушениями, что создает условия для
сохранения устойчиво-производных березовых лесов с елью, подростом
сосны, осины.
ПОДТАЕЖНЫЕ И ЛЕСОСТЕПНЫЕ СОСНОВЫЕ ЛЕСА
И ПРОИЗВОДНЫЕ СООБЩЕСТВА НА ИХ МЕСТЕ
Сосновые леса, распространенные в зоне мелколиственных лесов,
где занимают местоположения с песчаными и супесчаными почвами,
объединены в группу «подтаежных».
Подтаежные сосновые леса благодаря своему положению в
переходной полосе от лесной к степной зоне существенно отличаются от типичных
остепненных и от таежных лесов, что отмечалось многими авторами [Гор-
чаковскиц, 1949а].
В древесном ярусе подтаежных лесов нередко присутствие темнохвой-
ных пород — ели, пихты и кедра, что несвойственно остепненным
сосновым лесам. В'западном секторе подзоны в составе подтаежных сосновых
лесвв широко распространена липа сердцевидная, но в* отличие от
южнотаежных лесов липа произрастает преимущественно в подлеске, в соста-
120
ве древесного полога встречается только близ северной границы [Смоло-
ногов и др., 1970]. Характерно для этих лесов и участие в подлеске
хвойного кустарника — Juniperus communis.
Переходный характер подтаежных сообществ сосновых лесов
особенно заметно проявляется в составе травяно-кустарничкового покрова —
наряду с повсеместным распространением таежных растений отмечается
широкое участие лугово-лесных, а на юге подзоны появление лугово-степ-
ных растений. Количество лугово-степных видов еще невелико (6—10),,
встречаются они и единичными экземплярами.
Флора травянистого покрова разнообразна и составляет до 40—50
видов на участке ассоциации.
Зеленые, мхи, господствующие в таежных сосняках, в подтаежных
лесах встречаются фрагментарно. Сосновые травяно-зеленомошные леса
занимают незначительные площади. Преобладают в группе подтаежных
лесов различные варианты травяных и травяно-кустарничковых лесовг
различающихся составом доминирующих видов травяно-кустарничкового
покрова и соотношением в нем растений различных экологических групп.
Таежные элементы флоры большую роль играют в сосновых лесах
восточной части подзоны. В лесах западной части их участие сокращается, и
основу растительных сообществ составляют лесо-луговые растения с
участием неморальных и лугоЬо-степных.
Подтаежные сосновые леса интенсивно используются, что
подтверждается большой распространенностью молодняков и производных сосново-
березовых лесов.
Сосновые вейниковые и травяно-кустарничковые леса
в сочетании с сосновыми лишайниковыми (115)
Основные массивы подтаежных сосновых лесов сосредоточены на
боровых террасах Пышмы, Туры, Тобола, Иртыша. Восточнее крупный
участок таких сосновых лесов находится на Обь-Томском междуречье с
примыкающими к нему высокими террасами Оби и Томи. На Обь-Иртышском
междуречье и в правобережье Оби, восточнее Томи, они имеют
ограниченное расцространение, встречаясь небольшими островками по боровым
террасам Тары, Чулыма, Кии и Яи.
Для этих территорий характерен волнисто-холмистый или
равнинный рельеф с дюнными всхолмлениями, сложенными мелкозернистыми
песками и супесями. С разнообразием экотопов в условиях расчлененного
рельефа связано и распространение различных типов сосновых лесов^
объединенных на карте в одно сочетание.
Наиболее распространены и занимают относительно большие
площади сосновые травяно-кустарничковые леса, приуроченные к вершинам и
склонам низких холмов и грив. Нижние части склонов и выположенные,-
хорошо дренированные местоположения покрыты травяными,
преимущественно вейниковыми лесами. Сосновые лишайниковые леса занимают
самые сухие местообитания — вершины дюн и холмов с относительно
глубоким размещением грунтовых вод (более 4 м) и бедными почвами.
В западном секторе подзоны на надпойменных террасах древних
долин рек Пишмы и Туры и их междуречьях в составе сосновых лесов
нередко встречается ель — единично в основном ярусе, но иногда образует
хорошо развитый второй ярус. Из лиственных пород в сосновых
насаждениях постоянна примесь березы, рерке осины и липы сердцелистной.
Липа в основных лесах этой полосы имеет подлесочную форму, едва
превышает размер кустарников и редко выходит в древесный ярус.
Наиболее распространенные сосновые травяно-кустарничковые леса
связаны в своем развитии с супесчаными и суглинистыми почвами,
подстилаемыми тяжелыми суглинками и глинами. Древесный ярус их высотой 20—
25 м имеет производительность II—III классов бонитета. Подлесок в этих
лесах редкий из единичных экземпляров шиповника иглистого и
обыкновенного, рябины, жимолости, крушины ломкой, можжевельника, чере-
121
мухи. Встречаются участки с обильно разросшимся можжевельником.
В травяно-кустарничковом покрове доминируют Vaccinium vitis-
idaea и V. myrtillus. Большим количеством лугово-лесных и боровых видов
представлены травянистые растения (45—60). Наиболее характерны из
них Chimaphila umbellata, Maianthemum bifolium, Rubus saxdtilis, Galium
boreale, Pulmonaria obscura, Geranium sylvaticum, Calamagrostis arundina-
cea, Pyrola minor, P. rotundifolia, Stellaria holostea. Моховой покров
встречается небольшими пятнами, образованными преимущественно Pleurozium
schreberi с участием других видов обычных таежных мхов — Ptilium cris-
ta-castrensis, Dicranum polysetum, Rhytidiadelphus triquetrus.
Среди травяных сосновых лесов преобладают сообщества с
доминированием вейника — Calamagrostis arundinacea. Вейниковые сосновые леса
объединяют насаждения различного генезиса как коренные, так и
длительно-производные. Общность морфологического строения их
обусловлена нивелирующим длительным воздействием хозяйственной деятельности
человека, определившим современное состояние насаждений.
Доминирование в травостое лесов Calamagrostis arundinacea связано как с частыми
низовыми пожарами, так и с постоянным выпасом скота, способствующим
задернению почвы.
Древесный ярус вейниковых сосняков характеризуется небольшой
полнотой (0,3—0,5), в его составе постоянна примесь березы (до 0,2)
и участие осины. Нередко встречаются двухъярусные древостой, в них
лиственные породы составляют второй ярус.
Подлесок редкий из одиночных кустов шиповника иглистого,, ивы
серой, крушины, можжевельника, рябины, боярышника.
Травяной покров хорошо развит (проективное покрытие 60—80%)
и отличается флористическим разнообразием. В Припышминских борах
Р. С. Зубаревой [I960, 1962] было отмечено более 100 видов (105—113)
растений различных экологических групп. Преобладают лугово-лесные
и лесные виды, небольшое участие лугово-степных (Filipendula vulgaris,
Galium verum, Pulsatilla flavescens). Наибольшее обилие имеют следующие
виды: Calamagrostis arundinacea, Rubus saxatilis, Galium boreale, Poa pra-
tensis, Trifolium medium, Maianthemum bifolium, Carex macroura,,
Vaccinium vitis-idaea, Pulmonaria obscura, Aegopodium podagraria.
Моховой покров не развит, редкие пятна мхов встречаются главным
образом в местах разложения древесных остатков.
На Обь-Томском водоразделе в.составе древесного яруса подтаежных
сосновых лесов*отмечается присутствие Pinus sibirica, Abies sibirica, реже
Picea obovata. Более обильно их участие в подросте и единично в основном
ярусе. В подлеске характерно присутствие таких видов, как Caragana
arborescens, Daphne mezereum, Spiraea media.
Широкое распространение имеют травяные сосняки с
доминированием Calamagrostis arundinacea, В г achy podium pinnatum, Carex macroura.
Древостой их преимущественно смешанные с участием березы (до 0,5),
единично лиственницы, осины. По продуктивности (I—1а классы бонитета)
эти леса превосходят насаждения других типов леса.
^ Подлесок редкий из розы иглистой, рябины, ивы.
Травяной покров хорошо развит (проективное покрытие его 80—90%)
и отличается большой видовой насыщенностью (до 50—60 видов). В его
составе кроме доминирующих растений характерно участие Maianthemum
bifolium, Antennaria dioica, Trientalis europaea, Melica nutans, Rubus
saxatilis, Equi$etum hyemale, Aegopodium podagraria, Iris ruthenica, Galium
boreale, Paris quadrifolia, а в более разреженных лесах преобладает группа
высокотравья — Delphinium elatum, Aconitum septentrionale, Heracleum
dissectum, Cirsium heterophyllum, Pleurospermum uralense.
Сосновые лишайниковые леса, встречающиеся небольшими
участками среди других типов леса подтаежных4 лесов, также отличаются
некоторым своеобразием своего строения, что было отмечено П. Л. Горчаковским
[1949а]. Древесный ярус их сосновый с неравномерным распределением
деревьев III—IV классов бонитета. Подлесок редкий из моячжевельника
122
и ракитника в лесах Притоболья и Caragana arborescens, Rosa acicularis
Spiraea media в лесах восточных районов.
Травяной покров ^разрежен (покрытие 10—30%), беден по видовому
составу. Постоянно участие Antennaria dioica, Chimaphyla umbellata, Me-
lampyrum pratense, Hieraclum umbellatum, Achyrophorus maculatus% Orthi-
lia secunda*
Лишайники покрывают почву на 40—50%, распространение их
пятнистое. Преобладает среди лишайников более ксерофильный вид Cladina
arbuscula и другие виды CI. stellaris, CI. rangijerina включениями.
Из мхов встречаются единичные куртинки Dicranum polysetum% Ple-
urozium schreberi.
Сосново-березовые разнбтравно-вейниковые
с орляком леса и сельскохозяйственные земли
на их месте (116)
Широкое распространение производных березовых лесов по всей
площади подтаежных боров связано с разными годами вырубок, не
возобновившихся сосной, лесными пожарами и выпасом скота. Наибольшие
площади среди березняков занимают сосново-березовые разнотравно-вей-
никовые леса. Они развиваются преимущественно на участках сосновых
вейников'кх и травяно-кус,тарничковых лесов, пройденных давними вы-'
борочными рубками, а позднее подвергавшихся неоднократно низовым
пожарам. Смешанные сосново-березовые древостой чаще всего двухъярусные.
Состав первого яруба 7 — 8СЗ — 2Б, второго 10Б. Среди них встречаются
и одноярусные березовые леса с очень разреженным пологом старой
сосны, под которым сформировался полог более молодой березй с примесью
осины. Средняя полнота древостоев 0,5—0,6, сомкнутость полога
неравномерная. В возрасте 80 лет средняя высота древостоя достигает 20—23 м.
Производительность II класса бонитета. Подлесок развит слабо, состоит
из единичных экземпляров ивы козьей, рябины, черемухи, шиповника
иглистого.
Травяной покров хорошо развит из большого числа видов, общих с
коренными типами сосновых лесов. Доминирует вейник тростниковидный,
равномерно распределенный по всей площади лесов с примесью
разнотравья — Rubus saxatilis, Geranium sylvaticum. Inula salicina, Maianthe-
mum bifolium, Potentilla erecta, Galium boreale и др. На склонах и
вершинах невысоких плоских холмов в составе сосново-березовых разнотравно-
вейниковых лесов большое участие принимает орляк (Pteridium aquilinum).
Его распространение приурочено к слоистым супесчаным почвам,
подстилаемым суглинкаки и глинами. Моховой покров в таких лесах не выражен,
единично встречаются на разлагающихся древесных остатках Pleurozium
schreberi, Mnium euspidatum, Dicranum scoparium.
Березовые производцые леса перспективны при использовании их
под сенокосы и пастбища.
Сосновые и березово-сосновые остепненные
\ кустарничково-травяные и травяные леса (126),
Сосновые остепненные леса распространены разобщенными
массивами в пределах лесостепи и степи. Они приурочены к песчаным отложениям
в долинах крупных рек — Оби, Иртыша, Тобола, а также к древним
лощинам стока Обь-Иртышского междуречья. В литературе эти леса
получили название островных и ленточных боров. На карте нашли отражение
более крупные из них — Притобольские, Приобские островные и
ленточные боры ' (Алеусская, Кулундинская, Касмалинская, Барнаульская
ленты). '
Для районов распространения остепненных сосновых лесов
характерен гривисто-лощинный и дюнный рельеф^создающий разнообразие
экологических условий. Занимая различное положение в рельефе, сосновые
123
остепненные леса представлены несколькими контактирующими
ассоциациями, образующими различного типа пространственные сочетания. Одно
, из самых распространенных — сочетание кустарничково-травяных и
травяных сосновых лесов. *
По составу древесного яруса чистые сосновые насаждения в группе
остепненных лесов занимают сравнительно небольшие площади и
находятся, преимущественно в южных массивах боров степной зоны. Более широко
распространены смешанные сосновые леса с участием березы (от 1 до 3
едириц состава), реже осины. Березово-яосновые леса относятся к
производным, но в условиях лесостепи могут существовать длительное время,,
чему способствуют антропогенные факторы — вырубки и пожары. Смена
(Сосны березой характерна преимущественно для травяных лесов
лесостепи. Для восстановления сосновых насаждений широко применяется метод
лесных культур. Из других древесных пород в Приобских борах
отмечалось присутствие единичных экземпляров лиственницы, ели, пихты.
В полосе лесостепи преобладают насаждения II—III классов
бонитета, реже встречаются насаждения I класса. Продуктивность боров,
расположенных в степной зоне на южном пределе распространения лесных
пород, более низкая — IV—V классов, лишь местами встречаются леса,
относящиеся к III классу бонитета.
Подлесок в остепненных сосновых лесах более или менее хорошо
развит в борах лесостепи. Из кустарников единично и группами растут
акация желтая, калина, роза коричная и иглистая, черемуха, рябина,
боярышник, ивы. В Прцтобольских борах характерно присутствие
европейских видов кустарников — дрока красильного (Genista tinctoria),
ракитника (Cytisus ruthenicus), вишни степной (Cerasus fruticosa). В степных
ленточных сосновых лесах подлесок отсутствует или единично встречаются
акация желтая, спирея городчатая и зверобоелистная, кизильник.
Травяно-кустарничковый покров остепненных сосновых лесов
отличается флористическим разнообразием, согосподством бореальных и
лесостепных видов с участием неморальных и настоящих степняков. Общий
флористический состав насчитывает более 400 видов, из которых 40%
составляют мезоксерофиты и ксерофиты, чем существенно
отличаются*остепненные сосновые леса от подтаежных. Из бореальных видов постоянно
участие Vaccihium vitis-idaea, У. myrtillus, Chimaphila umbellata, Pyrola
rotundifolia, P. chlorantha, Antennaria dioica, Melica nutans, Equisetum
hyemale, Maianthemum bifolium, Calamagrostis arundinacea, Rubus saxati-
lis и др.
Среди луговых и луговостепных видов обычны Calamagrostis epigeios,
Iris ruthenica, Veronica spicata, Lathyrus humilis, Trifolium lupinaster,
Peucedanum baicalense, Pulsatilla flavescens и др. [Горчаковский, 1949а;
Павлова, 1963]. В травяных Приобских лесах правобережья
распространен такой неморальный вид, как Aegopodium podagraria. Нередко сныть
доминирует как в коренных сосновых лесах, так и в их производных —
березовых и березово-осиновых молодняках [Игнатьева, 1966, 1973].
В южных борах увеличивается участие ксерофитов — Festuca pseudo-
vina, Koeleria cristata, Phleum phleoides, Gypsophila altissima, Scabiosa
ochroleuca, Carex supina, С duriuscula, достигая на открытых участках
76%, на что указывал еще П. Н. Крылов [1916].
Моховой"покров в остепненных лесах отсутствует или развит
фрагментарно в виде мелких куртинок из Pleurozium schreberi, Dicranum poly-
setum, реже Ptilium crista-castrensis преимущественно в лесах травяно-
кустарничковой группы сообществ^ Лишайниковый покров развит в сухих
остепненных лесах, встречающихся на вершинах холмов или дюн с
глубокими грунтовыми водами (5—8 м).
В кустарничково-травяной группе сообществ наибольшее
распространение имеют бруснично-травяные сосновые леса, с участием таежных и
боровых кустарничков и трав (Vacciniurp, vitis-idaea, V. myrtillus,
Calamagrostis arundinacea, Carex macroura, Chimaphila umbellata, Antennaria
dioica).
124
Наиболее фитоценотически разнообразна группа травяных сосновых
лесов. В этой группе выделяются сообщества сосновых лесов с
доминированием в покрове мезофитов — вейниковые (Calamagrostis arundinacea),
коротконожковые, осоковые (Carex macroura), разнотравные (Iris ruthe-
nica), снытьевые (Aegopodium podagraria) и сообщества лесов с мезоксеро-
фильной структурой травяного покрова без доминирования
определенных видов — разнотравно-злаковые (Festuca pseudovina, Calamagrostis
epigeios, Phleurn phleoides), разнотравно-лишайниковые, редкотравные с
Carex supina, С. duriuscula, Koeleria cristata, Artemisia commutata,
Veronica spicata, Cladina arbuscula.
В более мезофитных вариантах сосновых Приобских и Притобольских
лесов лесостепи характерно сочетание бруснично-разнотравных, разно-
травно-вейниковых и разнотравно-осоковых с Carex macroura сообществ;
приуроченных к выположенным вершинам и склонам различных
экспозиций, увалов и дюнных всхолмлений.
Наибольшая остепненность свойственна ленточным борам Кулундин-
ской равнины, продолжающейся до Казахстанского Прииртышья. Южные
ленты этих боров (Касмалинская, Барнаульская) расположены в степной
зоне, что и определяет своеобразие их флористического состава. Широкое
развитие имеют травянистые редкотравные и лишайниковые сосновые
леса с участием ксерофитных видов, особенно по периферии лесов с низкой
сомкнутостью древостоев, где их участие достигает до 50—7096. Гари и
поляны покрыты Stipa capillata, Spiraea hypericifolia, открытые песчаные
дюны заросли в этих лесах Leymus arenarius [Вандакурова, 1950].
Такое же строение имеют островки степных Притобольских боров
(Озернинский, Кочердынский) с разреженным травяным покровом из
единично встречающихся растений — Calamagrostis ppigeios, Agrostis
vinealis, Koeleria cristata, Carex duriuscula, Antennaria dioica, Dianthus su-
perbus и др.
Экстразональные остепненные боры — это уникальное явление
природы. Велика их водоохранная и почвозащитная роль, поэтому
восстановлению и сохранению этих лесов придается большое значение.
ГЛАВА VII
БЕРЕЗОВЫЕ И ОСИНОВЫЕ ЛЕСА
Коренные березовые леса г на юге Западно-Сибирской равнины
ограничены на северном пределе 56—57° с. ш. и 54—55° — на южном. Они
представляют собой зональное явление. Здесь, в условиях
континентального климата, березовые леса замещают полосу хвойно-широколиственных
и широколиственных лесов, характерную для европейской части СССР
[Исаченко, Лукичева, 1956].
Для ландшафтов подзоны березовых лесов — подтайги и северной
полосы лесостепи — характерна высокая степень гидроморфности и
широкое развитие элювиально-супераквальных и супераквальных
признаков — оглеения, заболачивания, осолодения и засоления почв [Полынов„
4953; Глазовская, 1964; Уфимцева, 1968]. Заболачивание и засоление почвл
происходящие во влажные климатические циклы, сменяются усыханием
и рассолением в сухие годы. Масштабы этих процессов довольно
значительны, например, в Ишимском округе в 20—40-х годах прошлого
столетия высохло 364 озера [Кириков, 1966].
JB почвенном покрове под березовыми лесами развиты зональные
дренированные почвы — дерново-подзолистые и выщелоченные черноземы^
серые и темно-серые лесные почвы. В западной, приуральскойг части боль-
1 Производные мелколиственные леса рассмотрены в главах, посвященных
коренным темнохвонньш, сосновым и лиственничным лесам.
125
шие площади занимают почвы легкого механического состава, а в лесных
сообществах велика роль сосны. Смешанные сосново-березовые леса,
чередующиеся с массивами сосняков, показательны для западного отрезка
полосы березовых лесов. В восточной приобской части ареала на
суглинках структурно-денудационных плато в составе березовых лесов обычна
примесь темнохвойных пород — кедра, ели, пихты.
Гидролого-климатические условия в полосе подтаежных
березняков характеризуются оптимальным увлажнением в средний год и
недостаточным в сухой, повторяемость которого один раз в 5 лет. Для полосы
произрастания остепненных березняков показательно оптимальное
увлажнение во влажный год один раз в 5 лет, а в средний и сухой годы
увлажнение недостаточное [Мезенцев, 19571;
Зона березовых лесов характеризуется наиболее оптимальным
сочетанием увлажнения и радиации по сравнению с тайгой и
степями/Соотношение сумм радиации и осадков, выраженное в количестве калорий,
которое нужно затратить на испарение выпадающих осадков, в полосе
березовых лесов близко к единице: в подтайге оно равняется 0,9, в
лесостепи 1,1 (в тайге составляет 0,65 и ниже, в степи — 1,5—3,0) [Григорьев^
1954].
Основная лесообразующая порода в подтайге и лесостепи Западной
Сибири — береза повислая (Betula pendula). На засоленных в разной
степени почвах, вплоть до структурных солонцов, из всех древесных
пород способна развиваться только эта. Чем глубже находится засоленный
горизонт, тем лучше развивается ее поверхностная корневая система
[Городков, 1915]. На дренированных участках береза образует
высокоствольные насаждения, на засоленных почвах древостой березы мельчает
и образует угнетенные насаждения колочного типа. Береза — одно из
самых светолюбивых деревьев, она создает осветленные изреженные
насаждения паркового характера. Ее экологическая пластичность
необычайно широка: морозоустойчива, способна переносить засуху и
длительное затопление, солевынослива и неприхотлива к почвенным условиям
' [Городков, 1915; Сукачев, 1914J. Это позволяло ей занимать
неблагоприятные для других лесообразующих пород экологические ниши не только в
современных климатических условиях, но и в течение всего антропогена
[Крашенинников, 1937; Волкова, 1977]. На периодически иссушаемых
почвах береза повислая способна создать монодоминантные фитоценозы,
в мезофитных условиях она создает смешанные с осиной и березой
пушистой сообщества разнотравного типа [Кулагин, I960* 1963; Юркевич, Гельт-
ман, 1963; Махнев, 1965].
В условиях постоянного переувлажнения и заболачивания на
окраинах болот и в понижениях береза повислая уступает доминирующее
положение березе пушистой (Betula pubescens). Эта порода более
требовательна к влаге и плохо переносит засоление почвогрунтов. Экологический
ареал березы пушистой значительно шире, чем экоареал березы повислой.
В подтайге и лесостепи роль березы пушистой в растительном покрове
достаточно велика. Наилучшего развития леса из березы пушистой
достигают в лесостепи по западинам с хорошо увлажненными осолоделыми
почвами. На переувлажненных или засоленных почвах она образует
невысокие мало продуктивные древостой колочного типа [Скворцов, Связева,
, 1980].
Исследователями рода Betula объем вида Betula pendula в Западной
Сибири оценивается по-разному. Г. В. Крылов и В. Н. Васильев выделяют
из этого вида березу Крылова (Betula krylovii) — дерево высотой 20—30 м
с гладкой серовато-белой корой даже у старых столетних и большего
возраста деревьев. Этот вид образует леса в южной части лесной области
Западно-Сибирской равнины между 57 и 58° с. ш., распространяясь от
Иртыша до восточных склонов Урала [Крылов, 1953; Васильев, 1969].
Осина на юге равнины встречается в березовых лесах в качестве
примеси, ее участие в древостоях увеличивается на богатых и хорошо
увлажненных почвах, чистые насаждения осина образует редко. Г, В. Крылов
126
отмечает два центра обилия осины — первый по линии Тара — Обь
(устье Югана), связанный с древним руслом Иртыша, второй — в
бассейне р. Чулыма. Во всей лесостепной зоне осина встречается в березовых
колках, на слабосолонцеватых суглинистых солонцах и солодях, иногда
образует чистые осиновые колки [Крылов, 1961].
Существование на юге равнины зональной полосы коренных
березовых лесов было обосновано П. Н. Крыловым и Б. Н. Городковым
[Городков, 1915; Крылов, 1916, 19191. Л. А. Гордягиным, П. Н. Крыловым и
Б. Н. Городковым довольно полно охарактеризована флора березовых
лесов. Сводный список травянистых растений, составленный по их
описаниям, насчитывает 180 видов.
Флору березовых лесов юга Западной Сибири по нашим данным
составляют около 400 видов, относящихся к 55 семействам. Наиболее
богаты видами семейства Asteraceae (60 видов), Роасеае (41 вид), Ranuncula-
ceae, Rosaceae, Fabaceae (26 видов). Десять ведущих семейств —
Asteraceae, Роасеае, Rosaceae, Ranunculaceae, Fabaceae, Apiaceae, Scrophularia-
ceae, Cyperaceae, Salicaceae, Lamiaceae содержат более 65% видов флоры.
По этому признаку, согласно А. И. Толмачеву [1974], флора березовых
лесов относится к бореальным флорам.
Самобытность флоры коренных березовых лесов проявляется при
анализе фитоценотических и флористических границ в полосе березовых
лесов юга равнины, где многие бореальные виды растений имеют
совмещающиеся южные или северные границы. Они совпадают с границами
формаций растительности. Северные границы Adonis vernalis, Anemone syl-
vestris, Elymus sibiricus, Carex riparia, Crataegus sanguinea, Galium verum,
Geranium asiaticum, Gypsophila altissima, Phleum phleoides, Pteridium aqui-
linum, Spiraea crenata расположены между 57 и 57°30' с. ш. Согласно
геоботанической карте СССР [1954] границы перечисленных видов отражают
переход от южной тайги к подтайге — полосе расдространения березовых
лесов. Южные границы Adonis vernalis, Carex riparia, Equisetum palustre,
E. pratense, E. sylvaticum, Geranium sylvaticum, Glechoma hederaceae,
Phleum phleoides, Pinus sylvestris, Pteridium aquilinum, Salix caprea, Tragopogon
orientalis располагаются между 54 и 55° с. ш. Они отражают южный
предел плакорного произрастания березовых лесов в полосе луговых степей
и остепненных лугов, т. е. южную границу лесостепи.
Основу флоры березовых лесов юга Западно-Сибирской равнины
составляют виды евразиатского (27%) и евросибирского (20%)
распространения. Видов с азиатским типом ареала всего 6%. Показательно
значительное число дизъюнктивных ареалов (9%) с разрывом ареала в
европейской части СССР, на Урале, в Заволжье, Ангаро-Саянском регионе,
на Колыме, Камчатке.
В коренных березовых *лесах представители бетулярного ценотичес-
кого и флористического элементов [Клеопов, 1941; Миняев, 1966]
должны.быть обычными и часто встречающимися видами. По
статистическим подсчетам ими являются: Vicia cracca, Rubus saxatilis, Filipendula
ulmaria, Galium boreale, Calamagrostis epigeios, Lathyrus pratensis* San-
guisorba officinalis, *Solidago uirgaurea, Poa pratensis, Cnidium dubium,
Vicia sepium, Hieracium umbellatum, Trifolium lupinaster ,*Galatella rossica,
Galium verum, Angelica sylvestris, Thalictrum simplex, Sedum telephium,
Fragaria viridis, Seseli libanotis, Pulmonaria flacica, Serratula coronata,
Achillea ptarmica, Orobus luteus, Calamagrostis arundinacea, "Artemisia pontica,
Lysimachia vulgaris, Crepis sibirica, Phlomis tuberosa, Brachypodium pinna-
turn, Filipendula vulgaris, Poa palustris, Carex disticha, Aegopodium podag-
raria, Melica nutans [Лапшина, 1963; Лавренко, Кобелева, 1973].
Флору березовых лесов составляют представители разных
фитоценотических групп — лесные, луговые, степные, болотные. Их экология
столь же разнообразна [Лавренко,' 19721. Около половины видового
состава флоры относится к мезофильным лесным (22%) и лугово-лесным (25%)
растениям. Значительно меньший вес имеют лугово-лесные ксеромезофилы
(7%) и лугово-лесные гигромезофилы (5%)/Около'40% видового состава
127
характеризуется очень пестрым экологическим спектром. Сюда входят
таежные мезофилы (5%), боровые ксеромезофилы (4%), луговые галофилы
(5%, в их числе мезо- и гигрогалофилы), лугово-болотные гигрофилы
(6%, в их числе мезо- и галогигрофилы), водные и болотные гигрофилы
(4%), лугово-степные мезоксерофилы (3%), лугово-степные
ксеромезофилы (2%), степные ксерофилы (менее 1%).
' Своеобразие березовых лесов Западно-Сибирской равнины
проявляется в экологическом составе доминантных видов травяного яруса.
На границе с южной тайгой в лесах из Betula pendula наряду с основными
синузиями подтаежных лесных злаков (Calamagrostis arundinaceae) и
разнотравья (Aegopodium podagraria, Pulmonaria dacica) представлены:
южно-таежное мезофильное разнотравье {Delphinium elatum, Aconitum
septentrionale) и таежное мезофильное мелкотравье (Maianthemum bifoli-
ит, Trientalis europaea). На юге подтайги и в лесостепи в сложении фито-
ценозов большую роль играют ксеромезофильные растения, травостой
составляют лесные мезофильные злаки (Brachypodium pinnatum), лугово-
лесные ксеромезофильные злаки (Calamagrostis epigeios, Phleum phleoidesi)r
лесное мезофильное разнотравье (Pulmonaria dacica), лугово-лесное ксе-
ромезофильное разнотравье (Filipendula vulgaris, Seseli libanotis).
Сообщества с доминированием березы пушистой (Betula pubescens)
имеют гигрофильный характер. Травяной покров состоит из
представителей синузий: подтаежных лесных гигрофильных злаков (Calamagrostis
lanceolata), лугово-болотных галогигрофильных и гигрогалофильных осок
(Carex buxbaumii, С. disticha), подтаежного мезофильного (Pulmonaria
dacica, Aegopodium podagraria) и гигромезофильного (Angelica silvestris)
разнотравья. На южном пределе распространения березовых лесов в
колках с Betula pubescens встречаются растения самой разнообразной
экологии, относящиеся к синузиям подтаежных гигрофильных лесных злаков
{Calamagrostis lanceolata), подтаежных ксеромезофильных лугово-лесных
злаков (Calamagrostis epigeios), болотных гигрофильных qcok (Carex ri-
paria), лугово-болотных гигрогалофильных осок (Carex disticha),
разнотравья: подтаежного лесного мезофильного (Pulmonaria dacica, Cnidium
dubium), подтаежного лугово-лесного мезофильного (Vicia cracca),
лугового гигрогалофильного (Tripolium pannonicum), лугово-степного ксерога-
лофильного (Gilatella biflora), лугово-болотного гигрофильного (Stachys
paludtris) и гигромезофильного (Achillea ptarmica).
Описание наиболее распространенных типов березовых лесов в
южной части тайги равнины приведено в работах Б. Н. Городкова [1915 ]t
П. Л. Горчаковского [19496], Г. В. Крылова [1953], Т. И. Исаченко^
А. Н. Лукичевой[1956], Е. И. Лапшдной [1963], Е. Л. Любимовой [1969а].
ПОДТАЕЖНЫЕ БЕРЕЗОВЫЕ И ОСИНОВЫЕ ЛЕСА
И ПРОИЗВОДНЫЕ СООБЩЕСТВА НА ИХ МЕСТЕ
Березовые и осиново-березовые, местами с липой,
злаково-разнотравные
с элементами таежного мелкотравья леса
в сочетании с осоково-злаковымн лугами (111)
Леса этой группы характерны для северной части подзоны березовых
лесов. Распространены на Зауральском плато, Вагайской и Рыбино-
Каргалинской четвертичных аллювиальных равнинах, Васюганском
плато, Обской аллювиальной террасовой равнине, Кеть7Тымском плато.
Почвы под северными березняками дерново-подзолистые. *
Древостой состоит из Betula pendula с примесью В. pubescens и Рори-
lus tremula. В ненарушенных насаждениях полотна насаждений достигает
0,7—0,9, высота деревьев 15—20 м, диаметр стволов до 50 см.
Кустарниковый ярус слагают Rosa cinnamomea, Rubus idaeusx Crataegus sanguinea9
128
Salix caprea, S. cinerea, Sorbus sibirica, Viburnum opulus, Ribes nigrum^
Spiraea media, S. salicifolia, Lonicera xylosteum.Tp&BKKoii покров высокий:
высота яруса злаков 1—1,5 м, разнотравья — 80 см, густой, степень
проективного покрытия почвы достигает 70—90%.
В ярусе травяного покрова доминируют вейники Calamagrostis
arundinacea, С. epigeios, С. langsdorffii, С. obtusata, С. canescens. В нарушенных
сообществах увеличивается видовое разнообразие злаков, в
значительном количестве появляются Brachypodium pinnatum, Elytrigia repens,
Phleum phleoides, Poa pratensis, P. trivialis, Festuca pratensis, Agrostis gi-
gantea. Основу травостоя сообществ составляют лесные виды Rubus saxa-
tilis, Carex macroura, Pulmonaria dacica, Aegopodium podagraria, Pleuros-
permum uralense, Cnidium dubium, Angelica silvestris, Crepis sibirica и
многие другие, а также лугово-лесное разнотравье — Geranium pseudosibiricum,
Galium boreale, Hieracium umbellatum, Vicia cracca, V. sepium, Lathyrus
pratensis, Gentiana pneumonanthe, Achillea millefolium, Stellaria graminea.
Характерно присутствие'в составе сообществ видов таежного
разнотравья (Delphinium elatum, Aconitum septentrionale, Rubus arcticus, Paris
quadrifolia, Trientalis europaea, Maianthemum bifolium, Pyrola rotundifolia,
P. minor, Ortilia secunda) и папоротников (Gymnocarpium dryopteris, Dryop-K
teris tcarthusiana, Athyrium filix — femina, Phegopteris connectilis, Mat-
teuccia struthiopteris, Pteridium aquilinum).
Примером состава подобных сообществ могут служить разнотравно-
вейниковая, разнотравно-злаковая, разнотравно-коротконожковая,
разнотравно-костяничная ассоциации Прииртышских березовых лесов.
В разнотравно-вейниковой ассоциации доминирует подтаежный мезо-
фильный Calamagrostis arundinacea и ксеромезофильный С. epigeios. Из
подтаежного разнотравья в большом количестве встречаются Rubus, sa-
xatilis, Aegopodium podagraria, Serratula coronata, Cacalia hastata, Poly-
gonatum humile, Castilleja pallida, Viola hirta, Pulmonaria dacica, Agri-
monia pilosa, Sedum telephium, Lathyrus pisiformis, Brachypodium pinnatum%
Chamerion angustifolium;\ немного Ciprypedium macranthon, группа
подтаежных гигромезофилов представлена Filipendula ulmaria, Angelica
silvestris. В значительном количестве в травостое присутствуют лугово-
лесные мезофилы — Geranium pseudosibiricum, Galium boreale, Hieracium
umbellatum, Viola canina, Vicia cracca, V. sepium, Lathyrus pratensis, San-
guisorba officinalis. Для ассоциации свойственно небольшое количество
таежного мезофильного (Maianthemum bifolium), гигромезофильного
(Lythrum salicaria, Parnassia palustris) и лугово-лесного ксеромезофильного
(Senecio erucifolius, Dracocephalum ruiscfiiana, Pedicularis elata, Centaurea
scabiosa) разнотравья.
В травостое разнотравно-коротконожковой ассоциации доминируют
подтаежные лесные мезофилы Brachypodium pinnatum, 'Aegopodium
podagraria, много других лесных мезофилов — Calamagrostis arundinacea,
Pleurospermum uralense, Cacalia hastata, Sedum telephium, Lathyrus pisi-
formis, Rubus saxatilis, Pulmonaria dacica, Agrimonia pilosa, обычны гигро-
мезофильные Filipendula ulmaria, Angelica silvestris. Лугово-лесные
мезофилы Geranium pseudosibiricum, Vicia cracca, V. sepium, Galium boreale и
ксеромезофилы Calamagrostis epigeios,- Filipendula vulgaris участвуют в
сложении ассоциации в довольно значительном количестве.
Травяной покров разнотравно-костяничной ассоциации представлен
видами, относящимися к различным флористическим группам. Группа
таежных мезофилов состоит из обильно встречающейся Rubus arcticus,
в небольшом количестве отмечены Paris quadrifolia и Trientalis europaea.
Обычен таежный гигромезофильный Calamagrostis canescens. Подтаежные
лесные виды представлены многочисленными мезофилами Rubus
saxatilis, Sedum telephium, Angelica silvestris и гигромезофилами Filipendula
ulmaria, Cnidium dubium. Лугово-лесные мезофилы Sanguisorba
officinalis, Trifolium lupinaster, Vicia cracca, Gilium boreale довольно обычны.
Экологический состав подобных сообществ укладывается в схему:
35—50% подтаежных мезофилов, 20—25% лугово-лесных мезофилов.
9 Заказ № 297
129
5—30% таежных мезофилов, 0—10% лугово-лесных ксеромезофилов*
В ^понижениях рельефа в полосе северной подтаиги на растительный
покров начинает оказывать влияние фактор засоления грунтовых вод,
который проявляется в их сильной минерализованное™. Березовые леса
в таких условиях перемежаются с участками первичных осоково-злако-
вых лугов на лугово-болотных и лугово-солончаковатых почвах.
Травостой лугов высокий, средняя высота достигает 70 см, степень
проективного покрытия почвы 60—75%. Доминирующее положение на этих лугах
принадлежит злакам (Calamagrostis canescens, Agrostis gigantea, Poa tri-
vialis, Alopecurus arundinaceus) и осокам (Carex omskiana, С riparia, С. ш-
ginatq,, C. tomentosa, C, cespitosa, C. disticha). Довольно пестрое разнотравье
представлено видами, мезофильной (Sanguisorba officinalis, Vicia cracca7
Filipendula ulmaria, Veronica longifolia, Inula salicina), гигромезофильной
(Achillea ptarmica, Galium uliginosum, Lythrum salicaria) и галофильной
(Plantago cornuti, Saussurea amara, Galatella rossica, Cenolofium fischeri,
Triglochin maritimum, Ostericum palustre) экологии'.
В результате неумеренного выпаса в сухих березовых лесах
развиваются разнотравно-осоковые сообщества. Они 'представляют собой коч-
коватые гигрофильные сообщества, состоящие из Carex brunnescens, S.
cespitosa, С. vaginata, С. rostrata, Calamagrostis neglecta, Agrostis gigantea,
Comarum palustre, Cnidium dubium, Vicia cracca.
К лесным почвам, бедным азотом, на месте уничтоженных березняков
приурочены разнотравно-бобово-злаковые луга. В фитоценозах богато
представлена группа бобовых трав; Lathyrus pratensis, L. pisiformis, Vicia
cracca, V. sepium, Trifolium pratense, T. lupinaster, Astragalus onobrychis,
обычны Calamagrostis arundinacea, Poa pratensis, P. trivialis и мезофиль-
ное разнотравье. При пастбищной дегрессии лугов на них увеличивается
видовое разнообразие злаков, а количество разнотравья уменьшается,
снижается высота травостоя.
Березовые и осиновые вейниковые
и вейниково-высокотравные леса
в сочетании с лесными суходольными лугами (112)
Эти леса характерны для южной части подтаиги. Они имеют
распространение на Зауральском плато, Вагайской и Рыбино-Каргалинской чет-
вертичных^аллювиальных равнинах, Иртышской аллювиальной террасной
равнине, Барабинской четвертичной аллювиальной равнине, Приобском,
Чулымо-Енисейском и Кеть-Тымском плато. Почвы под вейниковыми и
вейниково-высокотравными березняками серые лесные.
Древостой насаждений смешанный, от преобладания березы плакучей
до равного участия осины и березы плакучей, иногда осина преобладает.
Рост характеризуется в среднем II классом бонитета, высота деревьев
10—20 м, диаметр 20—30 см, полнота насаждений 0,7—0,8. В
кустарниковом ярусе встречаются Ribes nigrum, Padus avium, Rosa cinnamomea,
Cotoneaster melanocarpus, Sorbus sibirica, Viburnum opulus. Травостой
высокий (до 1 м) и густой, степень проективного покрытия почвы 50—70%.
В травяном покрове доминируют злаки: вейники — Calamagrostis
epigeios, С arundinacea, коротконожка — Brachypodium pinnatufn, ежа -г-
Dactylis glomerata, перловник — Melica nutans, прочие злаки — Poa
pratensis, Elytrigia repens, Agrostis venealis, Phleum pratensis, P. phleoides,
Festuca pratense, F. rubra и другие" представлены полнее в нарушенных
сообществах. Отличительной особенностью фитоценозов является ярко
выраженный .и богато представленный ярусбетулярного высокотравья. Его
^слагают зонтичные (Angelica silvestris, Heracleum dissectum, H. sibiricum,
Cnidium dubium, Pleurospermum uralense, Aegopodium podagraria, Carum
carvi), сложноцветные (Crepis sibirica, Centaurea scabiosa, Serratula corona-
ta, Cirsium heterophyllum, Cacalia hastata, Tanacetum uulgare}, розоцветные
(Agrimonia pilosa, Filipendula ulmaria) и представители других семейств:
Rumex crispus, Lilium martagon1 Polygonatum odoratum, Thalictrum simplex.
Сообщества богаты мезофильными видами бетулярного лесного (Pul-
monaria dacica, Viola mirabilis, Equisetum sylvaticum, Origanum vulgare,
Rubus saxatilis, Vicia megalotropis, V. sylvatica, Polemonium caeruleum)
и лугово-лесного (Hieracium umbellatum, Vicia sepium, Achillea,
millefolium, Sanguisorba officinalis) разнотравья.
Примером подобных березовых лесов может служить березняк вей-
никово-коротконожково-разнотравный с Vicia megalotropis. Насаждение
березы имеет высокоствольный характер с такими же осинами,, высота
деревьев до 15—17 м, диаметр 25—30 см. Травостой высокий, степень
проективного покрытия почвы до 95%. Бетулярное подтаежное
разнотравье представлено лесными мезофилами Thalictrum simplex, Rubus
saxatilis, Aegopodium podagraria, Angelica silvestris, Pimpinella saxifragaf
Pulmonaria dacica, произрастающими с оценкой обилия «много», обильна
Vicia megalotropis, немного Cacatia hastata, Agrimonia pilosa, из лугово-
лесных мезофильных видов отмечены Sanguisorba officinalis, Galium bo-
reale, Inula salicina, Lathyrus pratensis, Filipendula ulmaria, Solidago vir-
gaurea, в меньшем количестве экземпляров произрастают Achillea
millefolium, Centaurea scabiosa, Crepis sibirica, Hieracium umbellatum, Serrutula
coronata, Artemisia vulgaris, Viola canina, Vicia cracca, V. sepium, V. syl-
vatica, Trifolium lupinaster.
Группа ксеромезофильных подтаежных, видов представлена Libanotis
intermedia, Calium verum, Dracocephalum ruischiana. Из злаков хорошо
развиты мезофильные Calamagrostis arundinacea, Brachypodium pinnatum
и ксеромезофильный Calamagrostis epigeios, в примеси мезофил Bromopsis
inermis.
Характерны также ассоциации костянично-разнотравно-перловнико-
вого осинника.
Березовые леса склонов и понижений рельефа носят черты гидроморф-
ности. Для них показателен колочный характер и некоторое остепнение.
Они представлены мезофильно-разнотравно-злаковыми и ксеромезофйль-
но-разнотравно-злаковыми сообществами.
Примером мезофильно-разнотравно-злаковых сообществ могут
служить ассоциации с доминированием коротконожки и вейника наземного.
Их древостой состоит из Betulu pendula с примесью осины, высота деревьев
6—9 м, диаметр стволов 12—15 см, сомкнутость крон 0,8. Ярус
кустарников развит хорошо, состоит из ив, черной смородины, шиповника
обыкновенного. Травостой высокий, густой, высота первого яруса 1—1,2 м,
второго 50—60 см, степень проективного покрытия почвы 80—90%. В
травяном покрове обильны подтаежная мезофильная Brachypodium pinnatum
или ксеромезофильный Calamagrostis epigeios. Лесное разнотравье
составляют встречающиеся в большом количестве экземпляров мезофилы (Rubus
saxatilis, Pulmonaria dacica, Thalyctrum simplex, Polygonatum humile)
и гигромезофилы (Filipendula ulmaria, Lactuca sibirica). Характерны для
сообществ лугово-лесные мезофильные виды — Lathyrus pratensis,
Trifolium lupinaster, Vicia cracca, V. sepium, Geranium silvaticum, Sanguisorba
officinalis, Agrostis tenuis. Редко встречается гигрофильный Phragmites
australis.
Ксеромезофильно-разнотравно-злаковые остепненные березовые леса
представлены ассоциациями с доминированием вейника наземного или
пырея ползучего. Древостой преимущественно из Betula pendula, высота
деревьев 5—6 м, диаметр стволов 6—7 см, сомкнутость крон 0,3—0,4.
В подросте береза и осина, кустарниковый ярус из шиповника
обыкновенного. Травяной покров невысокий, первый ярус 70—80 см, второй — 40—
50 см, степень проективного покрытия почвы 70—80%. В менее
нарушенных ассоциациях доминируют злаки: мезофильный Elytrigia repens или
ксеромезофильный Calamagrostis epigeios. Представители лугово-лесного
мезофильного разнотравья (Vicia cracca, Hieracium umbellatum, Trifolium
lupinaster, Sanguisorba officinalis, Galium boreale) встречаются с оценкой
«много», также как и лесные мезофилы (Brachypodium pinnatum, Agrimonia
pilosa, Po,ygonatum humile, Thalictrum simplex, Silene nutans). Значитель-
9* . 131
ное место в ассоциации занимают лугово-лесные и луговостепные ксероме-
зофилы (Phleum phleoides, Filipendula vulgaris, Fragaria viridis, Phldmis
tuberosa, Seseli libanotis).
Экологический состав гидроморфных и галомор.фных березовых лесов
следующий: подтаежных мезофилов 40—50%, лугово-лесных мезофилов
30—40%, лугово-лесных ксеромезофилов 10—159^6, лугово-степных ксе-
ромезофилов 0—10%, лесных гигромезофилов 0—5%, гигрофилов 0—2%.
В нарушенных пастьбой колках травостой низкий — 15—20 см\
степень проективного покрытия почвы 40—50%, на выбитых участках
поселяется Festuca valesiaca. Под пологом леса остаются луговые мезофилы и-
ксеромезофилы — Elytrigia repens, Viola canina, Galium boreale, Sanqui-
sorba officinalis, Achillea millefolium, Fragaria viridis, Pimpinella saxifraga,
Phlomis * tuberosa.
В полосе южной подтайги березовые леса на сухих открытых Участках
чередуются с массивами лесных суходольных лугов. На хорошо
дренируемых территориях луга занимают 35—50% площади (Крылов, 1916; Уткин,
19351. Они богаты по видовому составу,4 их флора близка к лесным
сообществам. Доминантами и субдоминантами луговых сообществ являются
Dactylis glomerata, Poa pratensis, Festuca pratensis, F. rubra, Phleum praten-
se, Ph. phleoides, Brachypodium pinnatum, Calamagrostis epigeios, Vicia se-
pium, Trifolium pratensis, T. repens, Lathyrus pisiformis, Galium boreale,
Centaurea scabiosa, Thalictrum simplex, Th. minus, Geranium silvaticum,
Filipendula ulmaria, Sedum telephium, Argopodium podagraria, Pulmonaria
dacica, Cnidium dubium, Seseli libanotis, Phlomis tuberosa, Solidago virgau-
rea, Tanacetum vulgare, Achillea millefolium, Hieracium umbellatum и
многие другие.
Березовые и осиново-березовые осоковые,
осоково-вейниковые леса в сочетании с осоковыми
и тростниково-осоковыми болдтами (М4)
С доминированьем Betula pubescens леса широко развиты на
понижениях рельефа — на склонах грив в западинах и, котловинах, где высокое
стоярие уровня грунтовых вод вызывает заболоченность почвы. Функцию
местных центров стока выполняют неглубокие западины. При
неравномерных осадках наблюдаются значительные колебания степени их;
увлажнения.
В северной полосе подтайги гигрофильные березовые леса
развиваются на оглеенных дерново-подзолистых почвах на склонах и у подножий
грив. Древостой состоит из Betula pubescens с примесью В. pendula и Ро-
pulus tremula, высота деревьев 10—12 м. В подлеске Salixcaprea, S. cine-
rea% S. pentandra. Поверхность почвы кочковатая, кочки образованы гид-
рогалофильными и галогигрофильными осоками — двурядной, дернистой
и Буксбаума, покрыты мхами — Polytrichum alpestre, Mnium medium,
Amblistegiumr sp. Почва между кочками залита небольшим слоем воды.
Высота яруса злаков 1—1,5 м, разнотравья 70—80 м, осок 40—60 см.
Примером разнотравно-осоково-гигрофильно-злаковых * лесов могут
служить разнотравно-осоково-вейниковые с Calamagrostis canescens и
разнотравно-осоково-двукисточниковое с Phalaroides arundinacea
сообщества.
В травостое разнотравно-осоково-вейникового сообщества доминируют
^гигрофильный Calamagrostis canescens и ксеромезофильный С. epigeios.
Из других злаков в большом количестве встречаются мезофильный
Elytrigia repens и гигромезофильные Poa trivialis, Agrostis gigantea. Подтаежное
лесное разнотравье состоит из встречающихся с оценкой «много»
мезофилов (Rubus saxatilis, Aegopodium podagraria, Lathyrus pisiformis, Serratula
coronata, Chamerion ungustifolium, Crepis sibirica) и гигромезофилов
(Filipendula ulmaria, Cnidium dubium, Aconitum volubile, Angelica silvestris,
Heracleum sibiricum). Гигромезофильные Lythrum salicaria, Gxlium uli-
ginosum, Trisetum sibiricum и гигрофильный Phragmiies australis участвуют
132
в сложении ассоциаций в небольшом количестве. Довольно обычны виды
лугово-лесного мезофилъного разнотравья — Sanguisorba officinalis, Gen-
tiana pneumonanthe, Trifolium lupinaster, Lathyrus pratensis, Vicia cracca,
Galium boreale, Achillea millefolium,, Hieracium umbellatum% Geranium p$e-
udosibiricum.
В разнотравно-осоково-двукисточниковой ассоциации доминируют
гигрофильные злаки Phalaroides arundinacea и Calamagrostis canescens,
немного Phragmites australis. Из подтаежного лесного разнотравья обычны
мезофилы — Pulmonaria dacica, Aegopodium podagraria, Angelica silvestris
и гигромезофилы — Aconitum volubile, Filipendula ulmaria. Хорошо
развиты лугово-лесные мезофилы (Hieracium umbellatum, Geranium pseudosi-
biricum, Lathyrus pratensis, Vicia cracca) и гигромезофилы (Achillea ptarmi-
ca, Galium uliginosum).
Экологический спектр перечисленных сообществ укладывается в
схему: 20—30% подтаежных лесных мезофилов, 20% подтаежных лесных
гигромезофилов, 5—20% гигрофилов, 15—20% лугово-лесных мезофилов,
10—15% гигромезофилов, 10—15% галогигрофилов и гигрогалофилов.
В глубоких западинах березняки имеют характер колков, для
которых показательно наличие нескольких резко отличных микроценозов.
В центре колков в наиболее обводненных местах в воде растут Salix sibiri-
са, Alisma plantago-aquatica, Alopecurus aequalis, Equisetum fluviatile,
Gnaphalium uliginosum, Rorippa amphibia, Ranunculus repens.
На окраинах на оглеенных почвах без признаков засоления
насаждения составляет береза пушистая, ее высота достигает 12—15 м; повислая
береза и осина занимают подчиненное положение. Ярус кустарников
хорошо развит, его образуют ивы (Salix cinerea, S. starkeana, S. pyrolifolia)
калина, черная смородина. Поверхность почвы покрыта подушками
болотных мхов Нурпит sp., Aulacomnium palustre, Mnium cuspidatum.
В травостое доминируют виды гигрофильной (Calamagrostis canescens, Ag-
rostis giganteu, Phalaroides arundinacea, Glyceria lithuanica, Carex elongata,
C. orthostachys. Lysimachia vulgaris, Filipendula ulmaria, Achillea ptarnftica,
Galium palustre, Scutellaria galericulata) и мезофильной (Cnidium dubium,
Angelica silvestris) экологии. На местообитаниях, где к
заболоченности присоединяется фактор засоления, береза пушистая не образует
сомкнутых древостоев, насаждения имеют вид отдельных куртин,
составлены ивами, среди ив возвышаются отдельные экземпляры Betula pubes-
cens высотой 5—8 м. В травостое доминируют виды злаков и осок гигро-
галофильной экологии — Calctmagrostis neglecta, Alopecurus arundinaceus,
Carex disticha, C. cespitosa. В меньшем количестве встречаются
Calamagrostis phragmitoides, Poa palustris, Carex riparia. Разнотравье по видовому
составу довольно однообразно, его составляют Filipendula ulmaria,
Veronica longifolia, Cnidium dubium, Lysimachia vulgaris, Achillea ptarmica,
Inula britannica.
Куртинные ивняки и колочные березняки на обширных понижениях
чередуются с открытыми тростниково-светлуховыми займищами
{Phragmites austrialis, Scolochloa festucacea, Phalaroides arundinacea), травяными
болотами (Carex disticha, C. omskiana, C. buxbaumii\ Calamagrostis canescens,
C. neglecta), солончаковыми (Elytrigia trichophora, Calamagrostis epigeios,
Plantago cornuti, Saussurea amara, Limonium gmelinii), солонцовыми
(Festuca sulcata, Artemisia pontita, A. nitrosa) и солонцово-солончаковыми
(Puccinellia distans, P. tenuissima) лугами.
В южной полосе подтайги гигрогенные сообщества березняков
развиваются на оглеенных и оторфованных солодях в понижениях рельефа с
неустойчивым водно-солевым режийом. Они составлены березой пушистой
и ивами, представлены разнотравно-вейниковыми (с Calamagrostis canescens)
и разнотравно-осоковыми (с Carex disticha) сообществами. Древостой раз-
нотравно-вейниковых березняков состоит из Betula pubescens с примесью
В. pendula и Populus tremula, высота деревьев 10—13 м, диаметр стволов
12—14 см, сомкнутость крон 0,5—0,6. Подлесок состоит из ив (Salix caprea,
S. cinerea, S. alba, S. triandra), черной смородины и шиповника обыкно-
133
венного. Травяной покров в понижении центральной части колка
невысокий и не богат видами, преобладает гигрофильный Calamagrostis canescens
и гигромезофильные.£1/$тгас/иа vulgaris и Роа palustris. Подтаежные лес-
ные мезофилы представлены немногочисленными экземплярами Rubus
saxatilis и Pulmonaria dacica. На окраине колка травостой высокий до
1,5—2 м высоты, густой, ярус высокотравья составлен подтаёжными
лесными гигромезофилами Cnidium dubium, Heracleum sibiricum, Angelica
silvestris и гигромезофильным Phalaroides arundinacea. Лугово-лесное
мезофильное разнотравье состоит из небольшого количества Hieracium umbel-
latum, Galium boreale, Vicia cracca.
Древостой разнотравно-осоковой ассоциации представлен единичными
экземплярами березы пушистой, высота которой не превышает 4 м, между
березами разрастаются кустарные заросли ив (Salix capita, S. rosmarini-
folia, S. cinerea) высотой 2—3 м. Травяной покров хорошо развит, в нем
доминируют гигрофильные осоки: Carex disticha, С. cespitosa, Carex riparia,
С. rostrata. Значительная роль в ассоциации принадлежит лесным, лугово-
лесным и лугово-болотным гигромезофилам: Cnidium dubium, Filipendula
ulmaria, Lysimachia vulgaris, Stachys palustris, Achillea ptarmica, Galium
uliginosum. Мезофильное лугово-лесное разнотравье представлено
небольшим количеством видов — Inula britannica, Vicia cracca, Lathyrus praten-
sis. В засушливые годы состав сообщества меняется, в травостое
появляется большое количество гигрофильных и гигромезофильных злаков
(Calamagrostis canescens, Agrostis gigantea, Alopecurus arun dinaceus).
. Экологический спектр подобных сообществ состоит из лугово-болот-
ных гигрогалофилов (20—30%), лугово-болотных гигрофилов (20%),
подтаежных гигромезофилов (10—30%), лугово-лесных гигромезофилов
(2—20%), лугово-болотных гигромезофилов (10%) и лугово-лесных мезо-
филов (10%).
В лесостепи березовые и осиново-березовые осоковые и осоково-вей-
никовые леса представлены разнотравно-осоково-наземновейниковыми,
разнотравно-осоково-ланцетно-вейниковыми и разнотравно-вейниково-
осоковыми фитоценозами.
Разнотравно-осоково-наземновейниковые сообщества березовых
лесов тяготеют к более или менее глубоким западинам. В год^т с обильными
осадками дно западин заполняется водой, в травостое преобладают осоки
и гигромезофилы, в сухие — колок подсыхает, в травяном покрове
доминирует мезофильное разнотравье. Древостой состоит из Betula pendula
с примесью В. pubescens и Populus ttemula, сомкнутость крон 0,6—0,7,
высота деревьев 10—15 м, диаметр стволов 15—20 см. В подлеске —
Salix cinerea, Rosa cinnamomea. Травостой высокий, высота первого яруса
75 см, степень проективного покрытия почвы 40—50%. В травяном
покрове доминируют ксеромезофильный Calamagrostis epigeios, гигрофильная
Carex riparia и гигрогалофильная Carex disticha. Подтаежное разнотравье
представлецо небольшим числом видов: мезофильные Thalictrum simplex,
Sedum telephium, Pulmonaria dacica и гигромезофильные Cnidium dubium,
Filipendula ulmaria. Заметное участие в травостое принимают луговые
мезофилы (Crepis tectorum,Vicia cracca, Viola canina,Gentiana pneumonanthe,
Galium boreale, Inula britannica, I. salicina) и гигромезофилы (Роа trivialis,
Achillea ptarmica, Stachys palustris, Lythrum salicaria). В небольшом числе
экземпляров произрастают ксерогалофильная Galatella biflora и ксеромезо-
фильная Veronica spuria. v
Разнотравно-осоково-ланцетно-вейниковые ивово-березовые леса
занимают неглубокие западины на приозерных понижениях. Почвы под
ними — оглеенные солоди. Во влажные годы вода в колке стоит в течение
летнего периода: Древостой негустой, состоит из Betula pubescens с примесью
В. pendula, высота деревьев 6—10 м, диаметр стволов 5—10 см, в подлеске
Salix tosmarinifolia, S. cinerea. Травостой высокий, высота первого яруса
150 см, средняя высота 60 см. Степень проективного покрытия цочвы
60—70%. Видовой состав травяного покрова заметно обеднен, слагается
15—20 видами. В ассоциации доминирует гигрофильный Calamagrostis
134
lanceolata, содоминантами являются гигрофильная Carex riparia и гигро-
галофильная С. disticha. Хорошо развито подтаежное высокотравье: гигро-
мезофильные Cnidium dubium, Conioselinum vaginatum, Filipendula ulma-
ria и мезофильные Angelica silvestris, Heracleum dissectum, Thalictrum
simplex. Луговые мезофилы представлены незначительным количеством Vicia
cracca, Lathyrtts pratensis, Veronica longifolia, гигромезофилы — Poa tri-
vialis, ксеромезофилы — Calamagrostis epigeios. Отмечено небольшое
.количество экземпляров ксерогалофильной Galatella biflora.
Разнотравно-вейникодо-осоковая ассоциация развита на оторфован-
ных солодях, представляет собой кустарные заросли ив — Salix rosmari-
nifolia, S. caprea, S. cinerea, S. depressa с одиночными экземплярами
Betula pubescens высотой 3—5 м. Травостой ассоциации высокий и густой,
высота первого яруса НО—120 см, второго 70—80 см, степень
проективного покрытия почвы 75—100%. В сообществах доминируют осоки:
гигрофильная Carex riparia, галогигрофильная С. atherodes, гигрогалофильная^
С. disticha. Из злаков обилен Calamagrostis lanceolata, в небольшом
количестве произрастают гигрофильные Phragmites australis, Phalaroides arun-
dinacea, Poa palustris, Agrostis gigantea, ксеромезофильный Calamagrostis
epigeios, гигрогалофильный Alopecurus arundinaceus. Из разнотравья с
оценкой «много» встречаются гигрофильные Achillea ptarmica, Stachys
palustris, Epilobium palustre. Мезофильное разнотравье представлено
немногочисленными Sanquisorba officinalis, Lathyrus palustris, Vicia cracca,
V. septum, Veronica longifolia, Stellaria graminea, Achillea millefolium,
Inula britannica, Gentiana pneumonanthe.
Подтаежные виды — гигромезофильная Filipendula ulmaria и
мезофильные Thalictrum simplex, Sedum telephium встречаются с оценкой «ма-
ло»¥ столько же гигрогалофильной Tripolium pannonicum.
Экологический спектр сообществ очень пестрый, он слагается из луго
во-лесных мезъфилов (5—25%), гигромезофилов (5—20%), подтаежных
гигромезофилов (5—20%), луговых ксеромезофилов (5—15%), подтаежных
мезофилов (5%), галогигрофилов (0—15%),} гигрогалофилов (0—15%),
ксеромезофилов (0—5%), подтаежных гигромезофилов (0—5%).
ЛЕСОСТЕПНЫЕ БЕРЕЗОВЫЕ И ОСИНОВЫЕ ЛЕСА
И СЕЛЬСКОХОЗЯЙСТВЕННЫЕ ЗЕМЛИ НА ИХ МЕСТЕ
Осиново-березовые и березово-осиновые остепненные
злаково-разнотравные леса
местами в сочетании с разнотравно-злаковыми лугами (124)
Леса представляют собой вариант более ксерофильных лесов южной
иодтайги. Они приурочены к оподзоленным черноземным почвам,
выщелоченным черноземам и солодям. Встречаются отдельными большими
массивами в полосе остепненных лугов и луговые степей в южной частд
Зауральского плато, на Ишимской неогеновой равнине, на севере Барабин-
ской неогеновой равнины по водоразделу р. Омь-Иртыш, в южной части
Барабинской четвертичной аллювиальной равнины и на Приобском
плато полосой от оз. Чаны на восток до широты городов Новосибирск и Ка-
мень-на-Оби.
Расположение березняков сравнимо с узором кружева — леса,
чередующиеся с луговыми полянами, тянутся беспрерывно. На лугах
растут единичные экземпляры березы, осины и шиповника. Ландшафты
лесостепи носят вторичный характер, они сильно изменены антропогенной
деятельностью, березовые леса в процессе освоения территории
интенсивно вырубались. Соотношение площади лесов и безлесных участков в
прошлом были иные, леса занимали гораздо большие площади, поэтому
А. Ф. Миддендорф [1871] характеризовал нынешнюю лесоЬтепную полосу
как березовые степи, Танфильев — березовое предстепье. Сохранившиеся
массивы используются под сенокосы и пастбища.
135
Древостой в остепненных березняках представлен березой плакучей
высотой до 12—15 м, диаметр деревьев иногда достигает 50 см. В подростке
березки высотой 4—10 м, сомкнутость крон 0,7—0,8. Подлесок выражен
слабо, состоит из Rosa cinnamomea, Lonicera tatarica, Crataegus sanguinea.
Травостой высокий до 1 м, густой, степень проективного покрытия почвы
90%. В нарушенных сообществах высота деревьев достигает лишь 5—8 м
диаметром 5—10 см, травяной покров невысокий, разреженный, высота его
до 60 см, степень проективного покрытия почвы 70%.
В травяном покрове по числу видов и, обилию преобладает ксеромезо-
фильное разнотравье — Fragaria viridis, Filipendula vulgaris, Phlomis
tuberosa, Artemisia sericea, Medicago falcata, Galium veruml велика роль
злаков — Calamagrostis epigeios, Festuca ovina, Phleum phleoides, Brachy-
podium pinnatum, Poa pratensis. Из бетулярных мезофильных видов
обильна Rubus saxatilis, роль содоминантов принадлежит Sanguisorba
officinalis, Filipendula ulmaria, Vicia sepium, V. cracca, Angelica silvestris, Hera-
cleum dissectum, Pleurospermum uralense, Heiracium umbellatum, Achillea
millefolium, Pulmonaria dacica. В березовых лесах на осолоделых почвах
обычны галофилы' — Artemisia pontica, Galatella punctata.
Примером остепненных злаково-разнотравных лесов .могут служить
коротконожково-разнотравная и наземновейниково-разнотравная
ассоциации Прииртышья. Первая ассоциация встречается заметно реже, чем
вторая. Ненарушенный древостой представлен старыми экземплярами
березы плакучей высотой 12—15 м диаметром 30—50 см, в подросте березы
высотой 4—Л0 м диаметром 5—10 см. Сомкнутость крон 0,7—0,8.
Подлесок выражен ч^лабо, состоит из жимолости татарской, боярышника,
шиповника обыкновенного. Травостой высокий, высота первого яруса до
1 м, густой, степень проективного покрытия почвы 90—100%. В травяном
покрове обильны подтаежные мезофилы — Brachypodium pinnatum, Rubus
saxatilis, Polygonatum odoratum, в меньшем количестве встречаются
Pleurospermum uralense, Silene nutans, Tanacetum vulgar'e, Chamerion angusti-
folium, Lathyrus pisiformis и подтаежные гигромезофилы Filipendula ul-
maria, Cnidium dubium. Из лугово-лесных мезофилов в значительном
количестве отмечены Poa pratensis, Hieracium umbellatum', довольно обычны,
но в меньшем обилии встречаются Galium boreale, Inula britannica, Elytri-
gia repens, Plantago media, Veronica longifolia, ,Trifolium* lupinaster,
Sanguisorba qfficinalis, Solidago virgaurea; в малом числе экземпляров
произрастают Achillea millefolium, Vicia cracca, V. sepium, Lathyrus pratensis.
Ксеромезофильное разнотравье и злаки представлены встречающимися
с оценкой «много»: Filipendula vulgaris, Artemisia sericea, Phlomis tuberosa,
Medicago falcata, Seseli libanotis, Astragalus danicus, Calamagrostis epi-
geios, Festuca ovina. В сложении травостоя довольно значительное участие
принимают ксеромезогалофильная Artemisia pontica, в незначительном
количестве галоксеромезофильная Galatella rossica, гигромезофильный
Poa palustris, мезоксерофильный Stipa pennata.
Древостой разнотравно-наземновейниковой ассоциации представлен
березой плакучей высотой 5—8 м, диаметром 4—10 см, в подросте мелкая
осина, в подлеске шиповник обыкновенный. Травяной покров невысокий,
разреженный, высота первого яруса 60—70 см, степень проективного
покрытия почвы 70%. В травостое обильны ксеромезофильные
Calamagrostis epigeios, Fragaria viridis и подтаежный мезофильный 'Rubus saxatilisr
обычен подтаежный мезофильный Pleurospermum uralense, меньше
Pulmonaria dacica, Sedum telephium, Serratula coronata, Lathyrus pisiformis
и гигромезофильной Filipendula ulmaria. Обычны луговые мезофилы
Trifolium pratense, Т. lupinaster, Achillea millefolium, Veronicq, longifolia,
Sanguisorba officinalis, Inula britannica, I. salicina. Гигромезофильные
Poa palustris, Lythrum salicaria присутствуют в небольшом количестве.
В значительном числе из ксеромезофильных видов отмечены Filipendula
vulgaris, Phlomis tuberosa, Phleum phleoides, меньше встречаются Medicago
falcata, Seseli libanotis, Centaurea scabiosa, Lathyrus tuberosus, Thalictrum
minus, Tragopogon orientalis. Мезоксерофильные Peucedanum morisoniiy
136
Eryngium planum, Festuca valesiaca и ксерофильные Koeleria cristata
присутствуют в небольшом количестве.
Экологический состав ассоциаций остепненных березовых лесов
состоит из луговых и лугово-лесных ксеромезофилов (2Q—3096), подтаежных
мезофилов (20—2596), луговых мезофилов (20—25%), подтаежных тигро-
мезофилов (5—10%), лугово-степных мезоксерофилов (5—10%), ксероме-
зогалофилов (0—5%), гигромезофилов (0—5%), галоксеромезофилов
(0—5%), ксерофилов (0—5%).
В понижениях мезорельефа в неглубоких западинах с осолоделыми
почвами развиты гигроморфные разнотравно-наземновейниковые леса.
Они представляют собой ко л очные насаждения. Древостой ассоциации
составлен березЬй плакучей с примесью пушистой, высота деревьев 4—G м,
диаметр стволов 10—12 см. В подлеске немного шиповника обыкновенного
и розы иглистой. Травяной покров редкий, степень проективного покрытия
почвы 30—:40%, невысокий (высота первого яруса 50—70 см, второго —
20 см). В травостое выделяется по обилию группа подтаежных мезофилов
и гигромезофилов — Rubus saxatilis, Thalictrum simplex, Pleurospermum
uralense, Agrimonia pilosa, Pulmonaria dacica, Lathyrus pisiformis, Fili-
pendula ulmaria, Cnidium dubium. Обилен ксеромезофильный Calamagros-
tis epigeios. Заметная роль в ассоциации принадлежит гигромезофилам
Galium, uliginosum, Lythrum salicaria, Poa palustris. Мезофильное лугово-
лесное разнотравье — Vicia cracca, V. sepium, Sanguisorba officinalis,
Geranium pratense, Poa pratensis, Viola canina — встречается в небольшом
количестве.
Экологический спектр гидроморфных лесов состоит из подтаежных
мезофилов (40%), гигромезофилов (20%), йодтаежных гигромезофилов
(10%), луговых мезофилов (10%), ксеромезофилов (10%),гигрофилов(10%).
Разнотравно-злаковые остепненные луга, составляющие сочетания
с массивами лесов, сильно изменены антропогенными воздействиями —
сенокошением, палами, пастьбой, часто они имеют вторичный характер,
развиваются на месте сведенных лесов. Травостой лугов довольно высокий
(высота первого яруса 70 см, второго — 40 см), густой, степень
проективного покрытия почвы 75%. В нем доминирующее положение*занимают
злаки — Elytrigia repensr Bromopsis inermis, Calamagrostis epigeios, Poa an-
gustifolia, Phleum phleoides. Характерно присутствие небольшого
количества бетулярных мезофилов —Rubus saxatilis, Heracleum sibiricum, Crepis
sibirica. Обычны ксеромезофильные (Filipendula vulgaris, Seseli libanotis,
Phlomis tuberosa, Fragaria viridis, Veronica spuria, Campanula sibirica,
Ranunculus polyanthemos, Dianthus versicolor) и мезофильные (Galium bo-
• reale, Achillea millefolium, Solidago virgaurea) виды. Хорошо развита
группа бобовых трав, представленная ксеромезофильными Astragalus danicus
и Medicago falcata и мезофильными Trifolium lupinaster, Lathyrus praten-
sis, Vicia cracca. Степняки -*■ Peucedanum morisonii, Veronica spicata,
Plantagostepposa, Allium lineare, Stipa pennata — на таких лугах
немногочисленны.
Наиболее благоприятная в природном отношении полоса березовых
лесов юга Западной Сибири издавна является зоной хозяйственной
деятельности. Описания естественных ландшафтов и их растительности
имеются в многочдсленных работах. О Барабе первой половины XIX в.
А. Ф. Миддендорф писал: «Беспредельно раскинувшиеся местности носят
на себе характер красиво расположенных парков. Местности,
отличающиеся таким своеобразным характером, я предлагаю называть березовыми
степями. В таком только случае может быть оправдано название Барабы
степью» [1871, с. 16].
История эксплуатации лесных ресурсов юга Западной Сибири *в
XVIII и XIX в. была рассмотрена В. А. Энгельфельдом. Он отмечает, что
березовые леса вблизи населенных пунктов Омского и Петропавловского
уездов вырублены полностью или представляют собой молодняки, «в глубь
степей они полнее, а местами встречаются нетронутые насаждения с 50%
строевого леса» [Энгельфельд, 1888, с. 53].
137
С развитием промышленности и строительством Транссибирской
железной дороги росли масштабы вырубки леса для нужд промышленности,
построек и топлива, расчистки лесных площадей под пашню, луговых
пожаров-палов и скотобоя. К 20-м годам нынешнего столетия безлесные
пространства шагнули к северу на 1—1,5° [Крылов, 1919].
О Масштабах уничтожения березовых лесов в прошлом говорят
различия в описании полосы их южного распространения у-разных авторов.
Л. А. Гордягин наметил южную границу «более сплошных березовых
лесов» между 54 и 55° с. ш.— от Троицка южнее Кургана, Петропавловска,
Омска к южному берегу оз. Чаны [Гордягин, 1900]. Через 50 лет
П. Л, Горчаковский эту полосу охарактеризовал как лесостепь с березо-
вымп колками [Горчаковский, 19496]. Сведение лесов в конкретных
условиях юга подтайги вызывает потепление климата и остепнение территории
[Рутковский, 1948; Ступила, 1965].
Материалы свидетельствуют, что в течение последнего столетия
южная граница распространения березовых лесов претерпела смещение
севернее своего естественно-исторического положения.
ГЛАВА VIII
РАСТИТЕЛЬНОСТЬ БОЛОТ
Болота на Западно-Сибирской равнине занимают огромные площади
(рис. 12). Их фитоцендтический состав, структура и динамика их
растительного покрова очень сложны. В главе будут обсуждены только
основные типологические особенности структуры и динамики болотной
растительности.
Болота рассматриваются как сообщества биоморф соответствующих
типов растительности — тундрового и бореального. Основной
картируемой единицей болотнрй растительности в масштабе карты служит тип
болотного массива в понимании, близком соответствующие
подразделениям зональных типов болотных ландшафтов в ландшафтных
классификациях [Е. А. Галкина, 1946; Е. А. Романова, 1973]. При его выделении
анализировались закономерности распределения растительных сообществ на
территории болот, их фитоценотический и флористический состав,
экологические связи, особенно обусловленные такими гидрометеорологическими
факторами как степень увлажнения, вечная мерзлота, обводнение и про-
точность болотных вод и другое, так же геоморфологические условия
залегания болот, структура болотных систем и массивов, микрорельеф их*
поверхности. Были выделены различные зональные и подзональные
болотные комплексы — полигональные, плоско- и крупнобугристые, грядово-
мочажинные и грядово-озерковые, олигомезотрофные и т. д. (табл. 2),
наряду с типичной некомплексной болотной растительностью.
Таблица 2
Приуроченность болотных зон к различным зонам увлажнения
Гидрологическая
зона
Избыточного
увлажнения
Переменного
увлажнения
Недостаточного
увлажнения
Болотная зона
Полигональных, плоско- и
крупнобугристых
Выпуклых олиготрофных
(сфагновых) болот
Плоских ев- и мезотрофных
(осоково-гипновых и
лесных) болот
Вогнутых евтрофных
(тростниковых) и засоленных
(травяных) болот
Особенности
минеральных
грунтов болотной
зоны
Мерзлые
Талые
Засоленные
138
Рис. 12. Схема распределения болот на территории Западно-Сибирской равнины.
1 — болота; 2 —реки.
Для более четкого отражения зональности, картируемые единицы
болотной растительности были сгруппированы в подзональные типы на
основании обусловленных гидроклиматическими факторами
индикаторных синузий и структурно-морфологических признаков растительных
сообществ. Так, в широко распространенных Обь-Иртышских формациях
выделены севербтаежные типы — кустарничкоро-зеленомошно-лишайни-
ковые осоково-гипновые и осоково-пушицево-сфагновые крупнобугристые
комплексные болота (64—66), лишайниково-сфагновые плосковыпуклые
139
олиготрофные болотно-озерные комплексы (68), а также среднетаежные
болота — сфагновые {Sphagnum fuscum) сосново-кустарничковые
олиготрофные выпуклые болота (83, 84). Аналогичным образом болота
тундровой зоны по характеру и структуре растительности, общим климатическим
и геоморфологическим условиям, глубине забегания вечной мерзлоты и
другим признакам подразделены на арктические, северные
субарктические и южные субарктические типы.
АРКТИЧЕСКИЕ ТРАВЯНЫЕ И ТРАВЯНО-МОХОВЫЕ БОЛОТА
Общая заболоченность территории подзоны арктических тундр
составляет в среднем на п-ове Ямал 16% и на Гыданском полуострове 25 %Ч
Особенностью растительности болот этой подзоны является отсутствие
древесной растительности, комплексность травяно-мохового покрова,
незначительная мощность торфяной залежи (20—30 см), близкое залега-
нпе вечной мерзлоты, малая мощность снежного покрова.
Наиболее распространены в подзоне арктических тундр травяные
и травяно-моховые болота в сочетании с валиково-полигональными
комплексами [7]..
Осоково-гипновые, осоково-пушипевые,
злаково-осоковые болота в сочетании с участками тундр
и злаково-осоково-моховых валиково-полигональных
болот (7)
Эти болота приурочены к днищам ложбин стока, речным долинам,
приозерным понижениям.
В подзонах арктической и северной субарктической тундр широко
распространены осоково-гипновые и осоково-пушицевые обильно
увлажненные болота с господством осоки прямостоячей (Carex concolor), пушицы
узколистной (Eriophorum polystachyori) с примесью злака дюпонтии (Dupon-
tia fischeri). В моховом покрове господствуют гипновые мхи (Calliergori
sarmentosum, Drepanocladus uncinatus, D. exannulatus, D. revolvens). Болота
имеют мелкую торфяную залежь, мощность которой в подзоне
арктических тундр не превышает 30 см. Залежь сложена осоковыми и осоково-
гипновыми торфами. Н. Я. Кац [1948] называет эти болота «арктическими
минеральными осоковыми болотами».
Болота этого типа часто встречаются в сочетании со злаково-осоково-
моховыми валиково-полигональными болотами, которые достигают
наибольшего развития на молодых морских и речных террасах.
Валиково-полигональные болота имеют своеобразный микрорельеф,
который образован полигонами, валиками и трещинами. Полигоны имеют
правильную 4-х, реже 5—6-угольную форму, разделены они между собой
трещинами шириной 0,3—0,5 м, вдоль которых тянутся валики высотой
15—20 см и шириной 1—4 м. Растительность на полигонах и в трещинах
состоит из сплошного покрова гипновых мхов (Drepanocladus uncinatus,
Calliergon sarmentosum), в трещинах встречается Sphagnum girgensohnii.
В травяном ярусе преобладают дюпонтия (Dupontia fischeri), осока
прямостоячая (Carex concolor), пушица (Eriophorum medium) и др. На валиках
растительность злакозо-осоково-морошково-сфагновая с лишайниками.
Господствуют дюпонтия, пушица, ожика (Luzula wahlenbergii), осока
прямостоячая, морошка (Rubus chamaemorus), В сфагновом и мохово-лишай-
никовом покрове встречаются Sphagnum fimbriatum, Sph. balticum% Cet-
raria hiascens, Cladonia elongata, Dicranum elongatum. Торфяная залежь
на полигонах травяно-гипновая, на валиках верхний слой торфа
сфагновый, ниже сфагново-травяной. Мощность торфа на валиках 20 см,; в .центре
мочажин 30 см. Торф сильно минерализован, примесь песка составляет
50—60%. Вечная мерзлота находится на глубине около 30 см.
1 Под заболоченной территорией имеется в виду площадь, занимаемая только
болотами, не включая заболоченные тундры.
140
На морских и речных террасах встречаются сочетания
полигональных болот с низинными, заросшими кустарником, которые носят местдое
название «лапты». Огромных размеров достигают болотные «лапты» в
низовьях рек Надуйяха, Мордыяха, Юрибей, Нейтаяха и др.
Болота арктических тундр описывались многими исследователями
Севера — Б. Н. Городковым [1932, 1938, 1944], В. Н. Андреевым [1934,
В. Н. Андреев, А. Л. Панфиловский, 1938], В. С. Михайличенко [1936],
М. Н. Аврамчиком [1969], М. С. Боч [1971] и др.
СУБАРКТИЧЕСКИЕ КУСТАРНИЧКОВО-ОСОКОВО-МОХОВЫЕ
И КУСТАРНИЧКОВО-МОХОВО-ЛИШАЙНИКОВЫЕ
ПОЛИГОНАЛЬНЫЕ КОМПЛЕКСНЫЕ БОЛОТА
Северные субарктические тундры. Общая заболоченность полосы
неравномерная, и в среднем составляет на п-ове Ямал 22%, а на Гыданском
полуострове 15%.
Болота расположены в тех же условиях рельефа, что и в арктической
подзоне, но отличаются по структуре микрорельефа и растительности.
Здесь широко распространены кустарничково-осоково-моховые валиково-
полигональные. комплексные болота.
О происхождении полигональных болот большинство современных
исследователей (Гусев, 1938; Попов, 1953; Пьявченко, 1955 и др.)
высказывают следующие предположения. В результате суровых климатических
условий и маломощного снежного покрова переувлажненные участки
молодых болот аллювиальных плоских террас, спущенных озерных котло-.
вин и морских побережий покрывались сетью взаимно перпендикулярных
трещин. В трещины поступала вода, которая замерзала, образуя ледяной
клин. Ледяные клинья при расширении сжимали прилегающие к ним
окраины полигонов и придавали им вид валиков. Возникал своеобразный
микрорельеф трещиновато-полигональных и валиково-полигональных
болот, состоящий из трех элементов — трещин, валиков и полигонов
(рис. 13).
Рис. 13. Полигональное болото (Гыданскип п-ов). Фото Б. А. Богоявленского-
. 141
Валико-полигональные болота стадийно являются наиболее старыми,,
по сравнению с современными валиково-полигональными болотами
арктических тундр, рассмотренными выше. Они имеют мощную реликтовую
торфяную залежь до 3-х м и несколько иной состав растительности на
валиках и полигонах, с присутствием гипоарктических видов кустарничков
и осок (ерника, осоки кругловатой и осоки редкоцветковой). Их
происхождение и структура освещены в работах В. Н. Андреева [1934, 1954]>
Н. Я. Каца [1939,1948], Н. И. Пьявченко [1955], А. П. Тыртикова [1969],
М. С. Боч [4971].
Осоково-гипновые и осоково-сфагновые на полигонах
и кустарничково-мохово-лишайниковые
на валиках болота (31)
Расположены на плоских слабо дренированных участках
водоразделов, в долинах рек и котловинах спущенных озер. Они имеют
полигональную структуру микрорельефа, состоят из полигонов квадратной или
5-угольной формы длиной 15—20 м и валиков высотой до 40 см и шириной
3—9 м, трещин шириной 1—3 м. Отличаются от полигональных болот
подзоны арктических тундр тем, что валики более широкие, полигоны в
центре имеют термокарстовые просадки, в которых образуются мочажины.
Растительность в центральной части полигонов и и. мочажинах осоко-
во-гипновая и осоково-сфагновая, господствуют: осоки — прямостоячая
(Carex concolor), редкоцветковая (С. rariflora), кругловатая (С. rotundata);
гипновые мхи — Drepanocladus uncinatus, Calliergon sarmentosum; сфагяё-
вые мхи — Sphagnum balticum, Sph. lenense. Растительность на валиках
кустарничково-зеленомошно-сфагновая с господством кустарнпчков —
морошки, брусники (Vaccinium vitis-idaea)f подбела ^Andromeda polifolia),
ерника (Betula папа), в моховом покрове — Dicranum angustum и
Sphagnum lenense. Встречаются кустистые лишайники (Cladina rangiferina,
Cetraria cucullata). В трещинах преобладают осоки — редкоцветковая
(Carex rariflora) и кругловатая (С. rotundatd), Sphagnum balticum.
В северной и южной полосах субарктических тундр на Ямальском и
на Гыданском полуостровах встречаются некомплексные мохово-травя-
ные осоково-гипновые болота. Больших массивов они не образуют, часто
встречаются в сочетании с тундрами и тундровыми заболоченными
ивняками. Приурочены к долинам рек, озерным котловинам и пониженным
депрессиям водоразделов. Поверхность болот ровная, слабовогнутая,
иногда закочкаренная. Болота постоянно сильно обводнены. В травяном
ярусе господствуют осоки — водяная {Carex aquatilis), прямостоячая
(С. concolor), редкоцветковая (С. rariflora), тонкая (С. ten,ella),
встречаются пушица узколистная (Eriophorum polystachyon), вахта (Menyanthes
frifoliata), сабельник (Comarum palustre), единично — злаки и хвощ.
Моховой покров составляют гипновые (Calliergon, Drepanocladus) и
сфагновые (Sphagnum squamosum, Sph. balticum) мхи.
Вокруг тундровых озер и низких заболоченных приозерных террас
широкими полосами встречаются осоко,во-злаковые болота —«хаеыгреи».
В травостое господствуют вейники, осоки (прямостоячая водяная),
Торфяная залежь их маломощная 5—20 см, в связи с чем многие авторы
относят эти болота к лугам. «Хасыреи» имеют небольшую площадь, и на'кар-
те отмечены внемасштабным знаком.
Южные субарктические тундры. Общая заболоченность территории
в среднем 29 %. В широтном направлении она изменяется следующим об-
разом: на п-ове Ямал 24%, на Малом Ямальском 37%, в Гйгданской
тундре 27%.
Особенность всех болот- этой полосы — господство кустарничков
(багульник, брусника, ерник и др.).
Среди болот преобладают кустарничково-мохово-лишайниковые
трещиновато-полигональные болота (32), которые часто встречаются в
сочетании с некомплексными кустарничков о-сфагново-лишайниковыми
142
болотами. В южной части подзоны, наряду с указанными болотами,
имеются еще плоскобугристые, которые характерны для лесотундры и
северной тайги. •
Кустарничково-зеленомошно-лишайниковые
на полигонах и валиках
и ороково-сфагновые в трещинах и мочажинах болота (32)
Наибольшего развития эти комплексные болота получили в полосе
южных субарктических тундр, но встречаются также и в северной полосе.
Обычно они образуют крупные болотные массивы.
Названные болота приурочены преимущественно к плоским
водоразделам, террасам, речным долинами и котловинам спущенных озер.
Поверхность болот прорезана сетью трещин на полигоны 6- или 4-гранной
формы, размером 20x25 м. Полигоны имеют плоскую или слегка
вогнутую в центре поверхности" и резкие, почти отвесные склоны к трещинам.
Трещины шириной около 1 м и глубиной 0,8 м.
Растительность на полигонах и валиках багульниково-морошково-
мохово-лишайниковая с брусникой (Vaccinium vitis-idaea), подбелом
(Andromeda polifolia), ближе к трещинам — с ерником. Мохово-лишайни-
ковый покров сложен в основном гипновыми мхами с господством Dicra-
питп angustum, Polytrichum strictum, а также сфагновыми Sphagnum an-
gustlfolium, Sph* lenense и лишайниками Cladina rangiferina, Cetraria cu-
cullata. В тех случаях, когда центр полигона имеет термокарстовую
просадку (мочажину), развивается более влаголюбивая растительность —
пушицево-сфагновая (Eriophorum russeolum и Sph. balticum) или осоково-
сфагновая с осокой кругловатой (Carex rotundata) и Sphagnum, majus,
Sph. lindbergii, последняя характерна и для трещин. Используются эти
болота как летние оленьи пастбища.
Некомплексные кустарничково-сфагново-лишайниковые болота
расположены в южной субарктической тундре. Они также заходят далеко в
лесотундру и северную тайгу, где широко распространены на контакте
болотных зон полигональных и плоскобугристых болот. Приурочены к
плоским депрессиям на водоразделах древних террас и водоразделов в
условиях избыточного, но слабопроточного увлажнения* Образуют
отдельные массивы или входят в сочетание с другими типами болот, по
отношению к которым занимают периферийные слабо обводненные участки.
Характеристику названных болот приводят в своих работах Б. Н.
Городков [1944 J для Гыданского полуострова ж А. П. Тыртиков [1969] для
междуречий рек Надым—Пур—Таз.
Кустарничково-сфагново-лишайниковые болота приходят на смену
осоково-сфагновым и сфагновым болотам и являются в настоящее время
заключительной стадией в развитии некомплексных травяных болот при
заболачивании водоемов и суши [Городков, 1944; Тыртиков, 1969].
Микрорельеф их кочковатый от дернин пушицы и моховых подушек, кочки
занимают 60—-70% поверхности, "высота их 20—60 см, диаметр до 1 м. В
растительном покрове господствуют болотные кустарнички: багульник (Ledum
decumbens), брусника (Vaccinium vitis-idaea), морошка, ерник (Betula nana),
водяника (Empetrum nigrum), клюква (Oxy coccus palustris). Из травянистых
рдстений — пушица влагалдщная (Eriophorum uaginatum), в понижениях
между кочками пушица узколистная (Eriophorum polystachyori), осоки
кругловатая (Carex rotundata) и топяная (С. limosa) и др. В
мохово-лишайниковом покрове господствуют лишайники (Cladina alpestris, С.
rangiferina, Cetraria islandica, С. cucullata), в понижениях сфагновые мхи
(Sphagnum angustifolium, Sph. warnstorfii и др.).
В случае самоосущения болота, например при поднятии поверхности
в результате накопления торфа и пучеция грунтов при промерзании,
сфагновые мхи постепенно отмирают и их место занимают лишайники. Тогда
багульниково-сфагново-лишайниковые болота переходят в багульниково-
лишайниковые. Такой тип болот часто встречается в северной тайге, где
143
занимает довольно большие площади [Тыртиков, 1969], в частности — на
водоразделах рек Надым, Пур, Таз.
Используются эти болота как летние и осенние оленьи пастбища.
Характеристику субарктических болот приводят в своих работах
В. Л. Андреев, А. Л. Панфиловский [1938]; Б. Н. Городков [1944J;
Н. И. Пьявченко [1955]; А. П. Тыртиков [1969], М. С. Боч [1971].
СЕВЕРОТАЕЖНЫЕ КУСТАРНИЧКОВО-МОХОВЪ ЛИШАЙНИКОДЫЕ
ПЛОСКО- И КРУПНОБУГРИСТЫЕ
И ЛИШАЙНИКОВО-СФАГНОВЫЕ ОЛИГОТРОФНЫЕ БОЛОТА
Средняя заболоченность северной тайги 40%, а на междуречье Надыма
и Пура она возрастает дох70%.
Слабодренйрованные водоразделы покрыты плоско- и
крупнобугристыми комплексными и кустарничково-сфагново-лишайниковыми
некомплексными болотами, в долинах рек и приозерных котловинах развиты
травяные и осоково-сфагновые болота.
Плоскобугристые комплексные болота (рис. 14) — один из зональных
типов болот северотаежной подзоны. К северу они заходят в лесотундру,}
где занимают отдельные участки в понижениях на водоразделах, а также
встречаются в крмплексе с другими типами болот, редколесий и тундр.
К югу плоскобугристые болота распространены до Сибирских увалов,
образуя крупные массивы на водоразделах средних течений рек Надым,:
Пур, часто в сочетании с крупнобугристыми и другими болотами.
Плоскобугристые болота представляют собой чередование плоских
торфяных бугров с обводненными мочажинами —«ерсеями», с различным
соотношением их площадей. Высота бугров 0,5—1 м, диаметр от
нескольких до десятков', иногда сотен метров.
На плоских буграх господствуют кустарнички и лишайники, в
мочажинах («ерсеях») — осоки, сфагновые и гипновые мхи. По преобладанию
тех или иных кустарничков на плоских буграх выделяются два варианта
плоскобугристых болот: ерниково-сфагново-лишайниковые (62) и багуль-
никово-ерниково-лишайниковые (63).
144
Рис. 14: Крупнобугристое болото. Фото Л. И. Усовой.
Ерниково-сфагново-лишайниковые на буграх,
осоково-сфагновые и пушицево-сфагновые
в мочажинах — «ерсеях» болота (62)
Эти плоскобугристые комплексные болота характерны для северной
полосы подзоны. Они встречаются большими площадями в пониженных
котловинах на водоразделах, на надпойменных террасах рек и по берещм
озер. На буграх кустарничковый ярус густой и высокий, господствуют:
ерник {Betula папа), багульник {Ledum palustre), морошка (Rubus chamae-
morus), в меньшем количестве встречается пушица влагалищная {Erio-
phorum vaginatum). Моховой покров пятнистый состоит из сфагновых мхов —
Sphagnum lenense, Sph. apiculatum, в понижениях — Sph. vftarnstorfii,
Sph. balticum и лишайников — Cladina rangiferina, CL sylvatica, Cetraria
cucullata; довольно часто встречаются гипновые мхи —Dicranum,*Polyt-
richum. В мочажинах (ерсеях) преобладают -осоки кругловатая (Сагех
rotundata), струнокоренная {С. chordorrhiza), в отдельных случаях —
пушица рыжеватая (Eriophorum russeolum). В моховом покрове господствуют
сфагновые {Sphagnum balticum, Sph. lindbergii) и гипновые мхи. Среди
мочажин-«ерсеев» встречается довольно много озер п озерков.
Багульниково-ерниково-лшнайниковые на буграх
и осоково-сфагновые в мочажинах — «'ерсеях»,
с обилием озер и озерков болота (63)
Плоскобугристые болота этого типа широко распространены от Урала
до Енисея в подзоне северной тайги. Они занимают огромные
пространства на водоразделах рек Надым, Пур, Таз, приурочены к депрессиям
водоразделов, склонам водоразделов и притеррасным частям речных долин.
Бугры плоские высотой 0,5—2 м, поперечный размер их 5—15 м, местами
до 20 м и более., они овальной или вытянутой формы. Мочажины вытянуты
вдоль уклона поверхности болота. Соотношение площадей бугров и
мочажин различное.
Растительность на буграх кустарничково-лигпайниковая, иногда
центральная часть их покрыта корковыми лишайниками. Кустарнички
угнетены, часто имеют карликовую форму. Преобладают багульник
{Ledum palustre), морошка {Rubus chamaemorus), постоянно присутствуют
ерник {Betula nana), брусника {Vaccinium vitis-idaea). Господство
принадлежит лишайникам {Cladina rangiferina, Cladonia mitis, Cetraria cucullata,
C. nivalis) и дикрановым мхам {Dicranum elongatum), в понижениях на
буграх встречаются сфагновые мхи {Sphagnum lenense и др.). В мочажи-
v нах-«ерсеях» господствуют осоки: струнокоренная {Сагех chordorrhiza),
кругловатая (С. rotundata), редкоцветковая {С. rariflora) и сфагновые мхи
{Sphagnum balticum, Sph. majus, Sph. subsecundum и др.), встречаются
гипновые мхи {Drepanocladus revolvens). Среди мочажин-«ерсеев» много
озерков и крупных озер. Используются огромные площади
плоскобугристых болот как летние пастбища для оленей.
Подробное описание растительности плоскобугристых болот приводят
в своих работах В. С: Говорухин[1933], В. Н. Андреев [1934], В. Н.
Андреев, К. Н. Игошина, А. И. Лесков [1935], Н. Я. Кац [1939], Б. Н.
Городков [1944], Н. И. Пьявченко [1955].
Крупнобугристые комплексные болота для южной полосы
северотаежной подзоны являются зональным типом. Они наиболее характерны
для области распространения спорадической мерзлоты, где на
водораздельных болотах чередуются крупные торфяные бугры с мерзлым
минеральным ядром и мокрые мочажины —«ерсеи», на которых мерзлота
отсутствует или находится на большой глубине. Южная граница
крупнобугристых болот проходит около 64° с. ш., затем в верховьях рек Надым,
Пур она опускается до 62° с. ш.
Приурочен этот тип болот, к депрессиям водоразделов и террасам
древних озерных котловин. Бодрта образуют крупные массивы, часто в соче-
10 Заказ JVfi 297 * -. 145
тании с другими типами болот (плоскобугристыми, грядово-мочажинны-
ми). На водоразделе рек Надым—Пякупур они уступают свое место
плоскобугристым.
Крупнобугристые болота представляют комплекс торфяных бугров,
понижений мочажин-«ерсеев» и термокарстовых озер. Бугры расположены
единично или группами, размер их самый различный, от десятков до
сотен метров в поперечнике. Высота бугров 6—8 м, ближе к лесотундре
уменьшается до 2—4 м. Бугры куполообразной формы, склоны их от
пологих до обрывистых, чем они и отличаются от плоскобугристых болот.
Мочажины между буграми имеют вытянутую форму (различной ширины),
как правило, соединены между собой в единую систему, по которой талая
вода сбрасывается в. озера и речную сеть.
По форме бугров и растительности на них и в мочажинах выделяются
два варианта крупнобугристых комплексных болот (обозначены в легенде
64, 65).
Зеленомошно-лишайниковые на буграх
и осоково-пушицево-сфагновые
в мочажинах- «ерсеях» болота (64)
Такие крупнобугристые болота имеют довольно широкое
распространение, приурочены к плоским депрессиям водоразделов и террасам
древних озерных котловин. Они занимают огромные пространства
водоразделов и являются характерным элементом ландшафта.
Растительный покров бугров довольно однообразен. Вершины их
покрыты лишайниками и дикрановыми мхами (Dicranum elongatum,
D. congestum, Cladonia deformis, Ochrolechia tartarea). На склонах
господствуют кустарнички: багульник {Ledum palustre), голубика [Vaccinium
uliginosum), брусника (Vaccinium vitis-idaea), морошка (Rubus chamaemo-
rus), пушица \Eriophorum vaginatum), ерник (Betula nana). Около
основания бугров ерник и багульник образуют сплошные заросли. Иногда
встречаются единичные деревья лиственницы (Larix sibirica), березы (Betula
pubescens) и сосны (Pinussylvestris). На открытых участках склонов
сплошной покров образуют лишайники (Cladina sylvatica, CI. rangiferina, Cet-
raria cucullata и др.)- При переходе к мочажинам-«ерсеям» вокруг бугров
часто образуется (от подтаивания мерзлого ядра) кольцо открытой водной
поверхности шириной 4—5 м. Растительность в «ерсеях» осоково-сфагно-
вая, осоково-пушицево-сфагновая, господствуют осоки — кругловатая
(Carex rotundata), струнокоренная (С. chordorrhiza),и пушица (Eriophorum
polystachyon), всегда присутствуют вахта (Menyanthes trifoliata), сабельник
(Comarum palustre), хвощи. В моховом покрове преобладают сфагновые
мхи (Sphagnum balticum, Sph. lindbergii), в меньшей мере гипновые^мхи
(Drepanocladus, Calliergon).
Кустарничково-зеленомошно-лишайниковые,
иногда облесенные на буграх, осоково-гипновые
и осоково-сфагновые в мочажинах
в сочетании с озерами болота (65)
Эти крупнобугристые болота также широко распространены на
водоразделах рек Надым, Пур, Таз.
Растительность на вершинах крутых склонов и в верхних частях
бугров кустарничково-зеленомошно-лишайниковая. В кустарничковом
ярусе господствуют багульник (Ledum palustre), брусника (Vaccinium
vitis-idaea), морошка (Rubus chamaemorus), водяника (Empetrum nigrum) $
ерник (Betula nana), в мохово-лишайниковом ярусе — дикрановые мхи
(Dicranum elongatum, D. undulatum, D. congestum) и лишайники (Cladina
stellaris, CI. rangiferina, CI. sylvatica, Cladonia deformis, Cetraria cucullata).
Ниже по склону растительность сменяется кустарничково-лишайниково-
сфагновой и кустарничково-сфагновой На нижних склонах бугров пыш-
146 ч
но развиваются заросли кустарничков, ерника и багульника. У подножия
при переходе к мочажинам-«ерсеям» иногда расположено кольцо с
открытой водной поверхностью шириной 3—4 м. Бугры обычно безлесны,
встречаются отдельные угнетенные деревья лиственницы (Larix sibirica), кедра
(Pinus sibirica), березы (Betula pubescens), ели (Picea obouata), редко сосны.
К западу от Оби, вдоль восточного склона Полярного Урала, в
бассейне реки Сев. Сосьва [Говорухин, 1947] и на водоразделе р. Таз—Енисей
[Городков, 1916] выделяется особый тип крупнобугристых болот с
буграми, облесенными кедром и березой высотой 3—5 м (кедровые «рямы» на
торфу). Бугры отделены друг от друга обводненными мочажинами-«ерсе-
ями» значительных размеров, площадь их нередко превышает территорию,
занятую буграми. Мочажины-«ерсеи» заняты осоково-гипновой, осоково-
и пушицево-сфагновой растительностью, преобладают осоки (Сагех^ го-
tundata, С. limosa, С, chordorrhiza), пушицы (Eriophorum polystachyon,
Е. russeolum), всегда присутствуют вахта (Menyanthes trifoliata),
сабельник (Comarum palustre), хвощ. В моховом покрове сфагновые (Sphagnum
majus, Sph. balticum, Sph. lindbergii, Sph. teres) и гипновые (Drepanocladus,
Calliergon) мхи. Среди мочажин часто протекают малозаметные, но
чрезвычайно опасные глубокие водотоки, по краям которых пышно развиты
кустарнички ивы и ерника. Среди мочажин-«ерсеев» много крупных
термокарстовых озер глубиной 0,5—1 м. Бугры сложены сверху торфом
мощностью 0,5—0,6 м (нередко 1—1,5 м).
Крупнобугристые болота используются в качестве зимних пастбищ и
являются потенциальными сенокосными угодьями.
Крупнобугристые болота подробно описывают в своих работах
Б. Н. Городков [1928, 1932], В. Н. Андреев [1932], Н. И. Кузнецов
[1932], Н. Я. Кац [1948]. Довольно полный обзор различных гипотез
образований бугров содержится в монографиях Н. И. Пьявченко [1955*
1967], А. П. Тыртикова [1969] и др.
Кустарничково-мохово-лишайниковые на буграх
осоково-пушицево-сфагновые й осоково-гипновые
в мочажинах крупнобугристые болота в сочетании с плоскобугристыми
и грядово-мочажинными болотами (66)
Эти сложные системы болотных массивов занимают огромныь
пространства на водоразделах рек в подзоне северной тайги. Они образуют
довольно широкую полосу на контакте двух зон крупно- и плоскобугристых
болот, граница между которыми выражена не четко. Максимального
своего распространения достигают на водоразделах и в верховьях рек
Надым, Пякупур и Айваседапур, где заболоченность территории доходит
до 70% и более. Приурочены к плоским центральным частям с малыми
уклонами поверхности, где речная сеть развита слабо.
Подробное описание растительности крупно- и плоскобугристых
болот приведено выше (62—65).
Грядово-мочажинные аапа болота встречаются редко, небольшими
массивами, в подзоне северной тайги, в полосе контакта крупнобугристых
и сфагновых олиготрофных болот. Иногда они входят в комплекс с плоско-
и крупнобугристыми болотами, где занимают по сравнению с ними более
пониженные элементы рельефа.
Ерниково-морошково-сфагновые на грядах,
осоково-сфагновые и осоково-гипновые в мочажинах болота;
западно-сибирский тип (67)
Приурочены к неглубоким котловинам, расположенным на
водоразделах и склонах. Отдельные массивы этих болот имеют веерообразную
форму с вогнутой поверхностью и общим уклоном в сторону водотока,,
по которому осуществляется сток воды с болота.
Основная площадь болота занята грядово-мочажинными и грядово-
10* 147
озерковыми комплексами. Для них характерны дугообразные гряды,
чередующиеся с вытянутыми сильно обводненными мочажинами с
озерками в центре. Площадь мочажин значительно превышает площадь гряд
(до 70—80%). Растительность на грядах кустарничково-сфагновая,
преобладает ерник (Betula папа), морошка (Rubus chamaemorus), единично —
пушица, осочки, в моховом покрове сфагновые (Sphagnum magellanicum,
Sph: angustifolium и др.) и гипновые (Dicranum, Polytrichum strictum) мхи,
единично встречаются лишайники. Мочажины сильно обводнены, в
травостое осока кругловатая (Carex rotundata) и топяная (С. limosa), пушица
рыжеватая (Eriophorum russeolum), вахта (Menyanthes trifoliata),
сабельник (Comarum palustre). Моховой покров из сфагновых и гипновых .мхов
с примесью печеночнико'в.
Названные болота по своему характеру и растительности близки к
грядово-мочажинным северным болотам типа «аапа». Н. И. Пьявченко
[1967] упоминает о болотах типа «аапа» в подзоне лесотундры
Таз-Енисейского водораздела.
Олиготрофные болотно-озерные комплексы — своеобразный тип
сфагновых олиготрофных болот. Они распространены в южной полосе
северной тайги Западной Сибири, на водоразделах рек Лямин, Пим, Тромьеган,;
Аган. Общая заболоченность территории здесь более 70%, суходолы
тянутся "лишь узкими полосами вдоль рек.*
Кустарничково-лишайниково-сфагновые с сосной и кедром,
с*обилием озер и озерков,
. е периферийным рядом олиготрофных комплексов
и ассоциаций болота; сургутско-полесский тип (68)
Болотно-озерные макрокомплексы представляют собой сочетание кус-
тарничково-лишайниково-сфагновых болот с крупными озерами и
озерками. Озера занимают до 50% и более всей площади комплекса. В
северной части района, у Сибирских увалов, болота соединяются вместе и
образуют единую сложную гигантскую болотно-озерную систему,
которую Л. П. Шубаев [1956] назвал «Сургутским полесьем».
Крупные болотные системы имеют плосковыпуклую поверхность.
В Центральной части системы до 85% площади занято грядово-озерными
комплексами, а на остальной приходящейся на склоны (обращенные к
рекам), расположены грядово-мочажинно-озерковые комплексы. На
грядах господствует кустарничково-лишайниково-сфагновая растительность
с отдельными деревьями сосны и кедра (Pinus sylvestris, P. sibirica). В кус-
тарничковом ярусе преобладают хамедафна (Chamaedaphne calyculata)t
багульник (Ledum palustre), ерник (Betula nana), морошка (Rubus
chamaemorus), реже встречается пушица влагалищная (Eriophorum vaglnatum).
В мохово-лишайниковом ярусе плотный покров образует Sphagnum fuscumt
на повышениях встречаются обильно лишайники (Cladina rangiferina,
CI. sylvatica). В грядово-мочажинных комплексах гряды обычно густо
облесены сосной (Pinus sylvestris, f. litwinowii, f. wilkommii) с примесью
кедра (Pinus sibirica, f. turfosa). Высота деревьев 1—2,5 м.
Эти болота в течение лета оттаивают полностью. Лишь редко
встречаются на грядах небольшие линзы мерзлого торфа на глубине 1—1,5 м.
Продольные уклоны поверхности болота близки к уклонам рек, порядка
0,0003—0,0008. Высота берегов и поверхности болот над урезом воды в
реках всего 1—3 м.
Формирование и развитие этих болот освещены в работах Е. А.
Романовой [1965, 19671, К. Е. Иванова [1969, 1974].
СРЕДНЕТАЕЖНЫЕ^СФАГНОВЫЕ
и сосново-кустарничково-сфагновые;олиготрофные болота.
Общая заболоченность территории в подзоне^средней тайги около
50%. Наиболее заболочены водоразделы крупных рек Оби, Иртыша и
Конды (55—70%). Менее заболочена территория на западе — в верховьях
148
рек Лозьвы, Пелым, Сев. и Мал. Сосьвы и на востоке — в верховьях рек
Вах, Таз, Елогуй (5—10%).
Зональным типом болот в подзоне средней тайги являются выпуклые
олиготрофные сфагновые болота с господством Sphagnum fuscum. Болота
занимают обширные водораздельные пространства рек первого и второго
порядков, имеют атмосферное водное питание и выпуклую форму
поверхности.
Для крупных болотных систем характерно господство в центральной
части грядово-мочажинных и грядово-озерковых комплексов. На
дренированных* склонах расположены грядово-мочажинные комплексы с кус-
тарничково-сфагновыми грядами и менее обводненными мочажинами (83).
В бассейнах рек Юконды и Камы встречаются менее обводненные
некомплексные кустарничково-пушицево-сфагновые болота в сочетании с
сосново-кустарничково-сфагновыми болотами и сосновыми бруснично-
лишайниковыми и лишайниковыми борами.
По всей территории подзоны прослеживаются небольшие по площади
сосново-кустарничково-сфагновые олиготрофные болота (84) и кустарнич-
ково-пушицево- или пухоносово-сфагновое с редкой сосной. В долинах
рек и на первых надпойменных террасах отмечаются евтрофные осоковые
и осоково-гипновые болота, которые в этой подзоне не имеют большого
значения.
Багульниково-кассандрово-сфагновые с сосной и кедром
на грядах, с озерками и сфагновыми мочажинами,
с периферийным рядом сосново-сфагновых мезо-евтрофных
лесных ассоциаций болота; нарымский тип (83)
Эти болота достигают большого развития в подзоне средней тайги,
а также далеко заходят в подзону южной тайги. Они образует большие
сложные болотные системы на водоразделах крупных рек и их притоков,
особенно на Обь-Иртышском водоразделе. Здесь болота единым массивом
занимают все водоразделы рек первого и второго порядков. Характерная
особенность их — выпуклая форма прверхности с более или менее ровной
слегка волнистой центральной частью и пологими длинными склонами,
с широкой довольно ровной окраиной, занимающей основание склонов и
нижнюю часть болот. Центральная часть возвышается над окраиной до
7—10 м, в большинстве случаев на 4—5 м. По геоморфологическим
условиям залегания и по характеру стока воды с них болота такого типа
относятся к водораздельным плакорного залегания.
Болота Нарымского типа в сущности представляют собой огромную
сложную по своей структуре систему болотных массивов, которая
образовалась в результате слияния по мере накопления торфа отдельных,
изолированных очагов заболачивания. Эти процессы наиболее интенсивно
проходили в ложбинах стока древнеледниковых вод и депрессиях озерных
котловин [Бронзов, 1930]. Об этом свидетельствуют неровности рельефа
дна и напластования торфяных отложений болот. В процессе торфо-
накопления на болотных системах выработался; сложный рельеф
поверхности, состоящий из ряда местных выпуклостей, в понижениях между
которыми сформировались внутриболотные вторичные озерки, озера,;
ручьи и топи [Бронзов, 1930; Романова, 1967; Романова Е. А.,
Усова Л. И., 1969].
Растительные сообщества этих болот имеют определенную
закономерность в'расположении и строгую приуроченность к местным элементам
рельефа болот. На более плоских центральных частях болот, вследствие
малых уклонов и переобводнения их поверхности, образовались
вторичные озерки, которые в сочетании с вытянутыми кустарничково-сфагновы-
ми грядами создают грядово-озерковые комплексы. На склонах, имеющих
относительно большие уклоны и лучшие условия стока поверхностно-
149
фильтрационных вод, расположены менее обводненные грядово-мочажин-
ные комплексы с сфагново-шейхцериевыми мочажинами., На участках,,
где обеспечен хороший естественный дренаж, на крутых склонах, вокруг
озер расположены сосново-кустарничковые и сфагново-кустарничково-
сосновые ассоциации. Менее дренированные плоские участки,
преимущественно примыкающие к окраинам болот, заняты моховыми редко
облесенными сообществами (кустарничково-пушицево-сфагновыми, сфагново-
пушицевыми редко, облесенными сосной, а*в некоторых восточных и
западных районах кедром).
Наиболее распространены грядово-мочажинные и грядово-озерковые
комплексы, которые занимают 50—70% площади болот. Соотношение
площадей гряд, мочажин и озерков различное, оно зависит от уклонов
поверхности. Чем они меньше, тем болыйая площадь занята мочажинами и
озерками [Романова, 1961, 19671. Растительность на грядах кустарничково-
сфагновая с редким облесением сосной, в некоторых восточных и западных
частях подзоны с примесью кедра. Из кустарничков преобладают
багульник {Ledum palustre), хамедафне (Chamaedaphne calyculata), подбел
(Andromeda polifolia), в моховом покрове — Sphagnum fuscum. В мочажинах
растительность зависит от их обводненности. В менее обводненных
мочажинах преобладает шейхцерия (Scheuchzeria palustris), в более
обводненных осока топяная (Carex limosa), очеретник (Rhynchospora alba).
Центральные части мочажин, как правило, с открытой водной поверхностью или
обнаженным торфом. В моховом покрове преобладают сфагновые мхи
(Sphagnum balticum, Sph. majus),
В сложных' болотных системах не менее распространены сосново-
кустарничково-сфагновые сообщества, которые занимают от 20 до 45%
всей площади болота. Древесный ярус сомкнутостью 0,4—0,6 состоит из
сосны (Pinus sylvestris. f. litwinowii и f. uliginosa) высотой 4—7 м (до 10 м).
В кустарничковом ярусе господствует багульник (Ledum palustre),
Кассандра (Chamaedaphne calyculata), голубика (Vaccinium uliginosum), на
более осветленных участках встречаются брусника (Vaccinium, vitis-idaea),-
морошка (Rubus chamaemorus), черника (Vaccinium myrtillus). Моховой
покров хорошо развит, в нем преобладают сфагновые мхи — Sphagnum
fuscum, Sph. magellanicum, в' западинах — Sph. angustifolium.
В ложбинах стока и в понижениях между выпуклостями на болотных
системах расположены олиготрофные и мезотрофно-евтрофные. топи,
которые широкими полосами тянутся от центра к периферии болот и часто
являются истоками многих рек.
' Огромная территория средней тайги, занятая сильно обводненными
комплексными болотами, в народном хозяйстве не используется. Эти
болота служат потенциальным торфяным фондом, который может быть
использован в различных отраслях промышленности, сельского хозяйства и
для топлива.
Некомплексные кустарничково-пушицево-сфагновые в сочетании с
сосново-кустарничково-сфагновыми болота занимают сравнительно
небольшую площадь в бассейне р. Конды, между ее левыми притоками Юкон-
да и Кама.4 Кустарничково-пушицево-сфагновые болота редко облесены
сосной. Кустарничково-травяной покров-состоит из пушицы ^Eriophorum
vaginatum) и угнетенных кустарничков багульника (Ledum palustre) %
Кассандры (Chamaedaphne calyculata); «подбела (Andromeda polifolia),
клюквы (Охуcoccus palustris). В моховом ярусе господствуют сфагновый
мох Sphagnum fuscum, в понижениях Sph. angusti folium.
Более дренированные участки некомплексных болот заняты сосново-
кустарничковой растительностью. Древесный ярус состоит из сосны
(Pinus sylvestris f. uliginosa и f. litwinowii), высота деревьев 4—7 м,
сомкнутость крон 0,3—0,5. В пышно развитом кустарничковом ярусе
преобладают багульник (Ledum palustre), хамедафне (Chamaedaphne calyculata),
голубика (Vaccinium uliginosum). Моховой покров сплошь состоит из
сфагновых мхов.
150
Сосново-кустарничково-сфагновые
. олиготрофные болота (84)
Болота широко распространены в западной и восточной частях
средней тайги — в Зауралье, в приенисейской части и на водоразделах рек
Кеть—Чулым. Они часто.представлены небольшими по площади
массивами, или занимают большие участки на крупных болотных системах с гря-
дово-мочажинными комплексами, где расположены на наиболее
дренированных склонах, и полосами вокруг озер. В литературе сибирские
ученые такие болота называют «рямами» [Жаркова, 1963; Шумилова, 1969;
и дрЛ- Мы считаем «рямами» только те сосново-кустарничковые болота,
которые встречаются в подтаежной и лесостепной зонах среди тростнико-
во-осоковых болот «займищ» (120).
Растительность этих болот довольно однообразна. Древесный ярус
сострит из сосны (Pinus sylvestris, f. litwinowii и f. uliginosa). Высота
деревьев от 4 до 10 м. Хорошо развит кустарничковый ярус из багульника
„(Ledumspalustre), хамедафне (Chamaedaphne calyculata), в межкочечных
понижениях встречаются мелкие вересковые кустарнички и пушица
(Eriophorum vaginatum). В моховом покрове господствует Sphagnum fuscum.
Кустарничково-пушицево или пухоносово-сфагновые болота с редкой
сосной и периферийным рядом мезотрофных и евтрофных ассоциаций
образуют небольшие по площади отдельные массивы в сточных котловинах на
водоразделах рек, а также полосами по-периферии крупных олиготрофных
болот. Они встречаются.более часто в северо-западной и северо-восточной
частях подзоны, на водоразделах рек Вах—Тым и в верховьях рек Мал.
Сосьвы и Конды. Эти болотные массивы имеют слабо выпуклую и плоскую
поверхность. В растительности господствует пушица (Eriophorum
vaginatum) или пухонос (Baeothryon ceespitosum), на повышениях угнетенные
кустарнички: подбел (Andromeda polifolia), Кассандра- (Chamaedaphne
calyculata), багульник (Ledum palustre), клюква (Oxicoccus palustris).
В моховом покрове преобладают сфагновые мхи (Sphagnum angustifolium,
Sph. magellanicum, реже Sph. fuscum). Торфяная залежь-мелкозалежная,
различная как по глубине, так-и по составу.
Сведения по болотам различных районов подзоны средней тайги
приводят в своих работах А. Я. Бронзов [1930, 1936], Н. Я. Кац [1948, 1971],
Е. А. Галкина [1956], С. Н. Тюремнов [1957]; С. Н. Тюремнов, О. Л. Лисе,
Г. Г. Куликова [1971], П. Е. Логинов [1958], Ю. А. Львов [1959, 1966],
Е. А. Романова [1965, 1967, 1976], Е. А. Романова, Л. И. Усова [1969],
Ф. 3. Глебов [1969], Г. Г. Куликова [1971], О. Л. Лисе, Н. А. Березина
[19771, А. А. Храмов, В. И. Валуцкий [1977].
ЮЖНО-ТАЕЖНЫЕ СОСНОВО-СФАГНОВЫЕ ОЛИГОТРОФНЫЕ
- И ОСОКОВО-ГИПНОВЫЕ МЕЗОТРОФЦО-ЕВТРОФНЫЕ БОЛОТА
Общая заболоченность подзоны южной тайги 50%. Болота
представляют собой сложные выпуклые системы, расположенные на водоразделах
рек Тавда — Конда, Иртыш — Демьянка и на южных склонах огромного
Васюганского болотного массива. Особенность болот этой подзоны —
выпуклая поверхность, большая облесенность сосной, наличие сильно
обводненных мезотрофно-евтрофных (грядово-мочажинных) топей,
которые занимают до 30% и более площади болотных система
Сосново-сфагновые, кустарничково-сфагновые
с сосной на грядах и осоково-сфагновые в мочажинах,
в сочетании с вторичными озерками
и осоково-гипновыми топями болота;
южно-васюганский тип (107)
Эти болота занимают более половины площади водоразделов рек
Кондо—Тавда и Демьянки-— Иртыш, а также полностью южные склоны
Васюганского болота. Они представляют собой сложные болотнь!е систе-
151
мы, которые состоят из ряда крупных выпуклых олиготрофных болотных
массивов и огромных мезотрофно-евтрофных топей, и образуют так
называемые «болотные макрокомплексы» [Романова, Усова, 1977] г.
Выпуклые олиготрофные болотные массивы в наиболее
дренированных местах заняты гомогенной сосново-сфагновой и сосново-кустарнич-
ково-сфагновой растительностью; менее дренированные — комплексами
грядово-мочажинных и грядово-озерковых болот.
Участки с сосново-сфагновой и сосново-кустарничково-сфагновой
растительностью на склонах и полосах вокруг озер имеют древесный ярус
сомкнутостью 0,3—0,5 высотой 3—5 м. Хорошо развит кустарничковый
ярус из багульника (Ledum palustre), Кассандры (Chamaedaphne calyculata),
клюквы (Оху coccus palustris), пушицы (Eriophorum vaginatum). Моховой
покров сплошной, состоит из сфагновых мхов (Sphagnum fuscum, Sph.
magellanicum) с незначительной примесью лишайников.
Основную площадь (до 60%) выпуклых болотных массивов занимают
олиготрофные грядово-мочажинные и грядово-озерковые комплексы.
В грядово-мочажинных комплексах соотношение площадей гряд и
мочажин различное, как правило, преобладают мочажины. Гряды шириной
2—5 м возвышаются над мочажинами на 20—30 см. Растительность на
грядах кустарничково-сфагновая с сосной (Pinus sylvestris f. litwinowii)%
реже с кедром (Pinus sibirica f. turfosa). Высота деревьев 1—2,5 м. В кус-
тарничковом ярусе преобладают подбел (Andromeda polifolia), багульник
(Ledum palustre), хамедафне (Chamaedaphne calyculata), морошка (Rubus
chamaemorus), в понижениях — пушица (Eriophorum vaginatum).
Моховой покров плотный, из сфагновых мхов (Sphagnum fuscum, Sph.
magellanicum, Sph. angustifolium) с примесью лишайников. Мочажины
вытянутые, длиной от десятков до сотен метров, сильно обводненные, с озерками
в центре.
По составу травостоя и степени обводненности выделяются три
варианта мочажин: шейхцериево-сфагновые, шейхцериево-осоково-сфагновые
и осоково-очеретниково-сфагновые. В моховом покрове повсюду
преобладают Sphagnum balticum, Sph. majus. Однако с увеличением обводнения
мочажин первой исчезает шейхцерия (Scheuchzeria palustris), затем —
сфагновые мхи, дольше всего сохраняется очеретник (Rhynchospora alba);
на оголенном торфяном субстрате остаются только единичные экземпляры
печеночников. В грядово-озерковых комплексах, расположенных на
площадях с малыми уклонами и на контакте с мезотрофно-евтрофными
топями, растительность на грядах аналогична таковой на грядах в грядово-
мочажинных комплексах.
Мезотрофно-евтрофные (грядово-мочажинные) топи занимают
значительную площадь болот (более 30%), располагаются между выпуклыми
олиготрофными массивами. В этом комплексе обширные осоково-гипно-
вые мочажины занимают до 80% площади, а узкие и редкие кустарничко-
во-сфагново-гипновые гряды занимают всего 20% площади. Гряды
(«веретья») достигают в длину несколько километров, имеют ширину 2—5 м с
превышением над мочажинами 20—40 см. На грядах редко встречаются
отдельные угнетенные деревья березы (Betula pubescens) и сосны (*Pinus
sylvestris f. litwinowii, f. wilkommii), высотой 1—2 м. Травяно-кустарнич-
ковый покров угнетенный, состоит из багульника (Ledum palustre), xa-
медафны (Chamaedaphne calyculata), ерника (Betula nana), на более низких
грядах обильно растет вахта (Menyanthes trifoliata), сабельник (Сотагит
palustre). Моховой покров сплошной из сфагновых мхов (Sphagnum
magellanicum, Sph. angustifolium, Sph. warnstorfii, Pleurozium schreberi, Campto-
thecium nitens). Мочажины топкие, трудно проходимые, в центральной
части залиты водой, с пятнами оголенного торфа. Растительность в
мочажинах имеет определенную приуроченность: .ближе к грядам полосой
осоково-сфагновая, далее к центру — осоково-гипновая с обилием вахты
1 А. Я. Бронзов [193G] называл .пх «южные болотные комплексы», Н. Я. Кац
[1948, 1971] — «гетеротрофные макрокомплексы».
152
(Menyanthes trifoliata), в центральной части — гипновая (Scorpidium
scorpioides) с открытыми озерками и пятнами торфа. Травостой разрежен,
представлен осоками: топяной (Carex limosa), двутычинковой (С. diandra)x
\ волоеистоплодной (С. la&ocarpa) и вахтой (Menyanthes trifoliata). Моховой
покров рыхлый из сфагновых мхов (Sphagnum obtusum, Sph. subsecundum),
ближе к центру из гипновых Calliergon stramineum, Drepanocladus vernico-
sus, Scorpidium scorpioides.
В настоящее время описанные болота в народном хозяйстве не ис
пользуются. В перспективе — это торфяной фонд и лугово-пастбищные
угодья.
Болота на водоразделах рек Туртас — Демьянка описаны М. К.
Барышниковым [1929]*.Е. А. Романовой, Л. И. Усовой [1977], южные
склоны Васюганского болота описаны А. Я. Бронзовым [1930, 1936 J, Г. Г. Яс-
нопольской [1965], Ю. А. Львовым [1966, 1959], А. А. Храмовым,
В. Н. Валуцким [1977], Q.. Л. Лисе, А. А. Березиной [1977].
ПОДТАЕЖНЫЕ ЛЕСНЫЕ
И ОСОКОВО-ГИПНОВЫЕ МЕЗОТРОФНО-ЕВТРОФНЫЕ БОЛОТА
Заболоченность территории всей подзоны в среднем около 20%
[Романова, 1976]. Заболоченность неравномерная, возрастает с юго-запада
на северо-восток. Относительная площадь болот северной и центральной
частей Барабинской низменности составляет 31,6% общей площади
территории [Кузьмина, 196Ц. На северо-востоке подзоны заболоченность
достигает 60% площади. Наиболее заболочены водоразделы крупных рек
Ишим—Иртыш, на востоке верховья рек Тара, Тартас, Омь, Каргат, Бакса.
Зональным типом болот в подтайге являются водораздельные осоково-
гипновые и травяные евтрофные болота с грядово-мочажинным
комплексом (118) и тростниково-осоковые («займища») в .сочетании с сосново-
кустарниково-сфагновыми «рямами» (120). Болота расположены на
водораздельных пространствах рек и имеют плоскую и слабовогнутую
поверхность.
Зональная особенность этих болот — наличие среди них участков,,
занятых выпуклыми олиготрофными рямами. Последние встречаются
только на западно-сибирских болотах, их нет на болотах европейской
территории СССР.
Крупные болотные массивы, занимающие обширные водораздельные
пространства, имеют богатое водно-минеральное питание.
В подтайге хорошо выражена широтная смена растительности болот,
которая прослеживается с севера на юг в следующем порядке. В северной
части подзоны проходит узкая полоса лесных мезо- и евтрофных болот с
господством березы и примесью сосны, кедра, реже ели. Это березовые,
сосново-березовые осоково-сфагновые и вейниково-осоковые болота ч(117).
К югу от них расположейа полоса осоково-гипновых и травяных
евтрофных болот с грядбво-мочажинными комплексами (118), ближе к южной
границе подтайги проходит полоса тростниково-осоковых займищ в
сочетании с рямами (120).
По всей территории подтайги встречаются отдельными небольшими
массивами осоковые, осоково-гипновые и осоково-сфагновые болота (119).
Лесные мезо- и евтрофные болота подтайги изучены слабо, некоторые .
сведения приведены в работах А. Д. Панадиади [1953] и С. Н. Тюремнова
[19571. Лесные болота, как правило, встречаются по окраинам евтрофных
и олиготрофных болот, расположенных на древних и надпойменных
террасах рек, в условиях смешанного (грунтового и поверхностного) водно-
минерального питания.
Березовые, сосново-березовые осоково-сфагновые
и вейниково-осоковые болота (117)
По составу древесной, травяно-моховой растительности и степени
богатства водно-минерального питания лесные болота можно расположить в
следующий ряд: березовые вейниково-осоковые ->■ березово-осоко-
153
вые -> березово-осоково-гипновые -»- сосново-березово-осоково-сфагновые.
Березовые вейниково-осоковые болота отдельных массивов почти не
образуют, приурочены к плоским сильно минерализованным понижениям
и представляют собой одну из начальных стадий заболачивания лесов.
В древесном ярусе доминирует береза (Betula pubescens) в травяном
ярусе — вейник Лангсдорфа (Calamagrostis langsdorffii) и осоки волосисто-
плодная (Carex lasiocarpa).
Березово-осоковые болота встречаются по окраинам евтрофных и
мезотрофных болот, расположенных на боровых террасах приобья, в
Барабинской низменности они сплошным или прерывистым кольцом
окружают рямы. В древесном ярусе доминирует Betula pubescens высотой 4—-
8 м, встречается сосна (Pinus sylvestris). В кустарниковом ярусе имеются
Betula humilis и Salix rosmarinifolia. В травяном ярусе господствуют осоки:
Carex appropinquata, С. cespitosa, С. juncella, С. lasiocarpa, С. vesicaria,
С. diandra. Из болотного разнотравья встречаются вахта (Menyanthes
trifoliata), сабельник (Comarum palustre), хвощ и др. В моховом ярусе на
кочках распространены гипновые мхи — Aulacomnium palustre, Sphagnum
warnstorfii, между кочками — Drepanocladus ve/nicosus и виды Calliergon.
При дальнейшем развитии мохового покрова и наличии его сплошной
сомкнутости эти болота переходят в стадию березово-осоково-гипновую.
В березово-осоково-гипновых болотах древостой становится сильно
разреженным и угнетенным. В травостое выпадают дернистые осоки,
господствующая роль принадлежит гипновым мхам. Эти болота
встречаются обычно в условиях напорного грунтового питания и отличаются
значительной обводненностью. Они занимают окраины притеррасных
евтрофных и мезотрофных болот приобья и окраинные участки гипновых
болот, расположенных на водоразделах рек Тары—TapTacj [Тюрем-
нов, 19571.
Сосново1березово-осоково-сфагновые болота распространены на
вторых террасах рек Кеть, Тым. Они являются последующей стадией
развития описанных болот в связи с уменьшением грунтового питания и проточ-
ности. В разреженном древесном ярусе, кроме березы {Betula pubescens)f
в значительном количестве присутствует сосна (Pinus sylvestris) высотой
4—6 м, иногда кедр (P. sibirica). Довольно хорошо развит подлесок из
березы приземистой (Betula humilis) с примесью ив (Salix rosmarinifolia,
S. lapponum). Микрорельеф кочковато-бугристый. На кочках хорошо
развиты сфагновые мхи Sphagnum warnstorfii, Sph. centrale, между
кочками — Sph. angustifolium, Sph. obtusum, Sph. subsecundum. Гипновые
мхи уступают свою роль сфагновым. Травяной ярус представлен'теми же
видами, что и в предыдущих болотах, но сильно угнетен и разрежен,
преобладает хвощ топяной (Equisetum heleocharis).
Торфяная залежь описанных болот преимущественно древесно-осо-
ковая, глубина на водоразделах до 2 м, в долинах ред 4—5 м.
Осоково-гипновые и травяные евтрофные болота в Западной Сибири
достигают особенно широкого развития на водораздельных пространствах
и древних надпойменных террасах крупных рек Туры, ,Тавды, Тобола,
Ишима, Иртыша. В северной части Барабинской низменности они
занимают депрессии водораздельных пространств и надпойменных террас,
образуя отдельные массивы и сложные болотные системы с грунтовым водно-
минеральным питанием.
В растительности этих болот доминируют осоки и гипновые мхи,
однако в различных вариантах болот изменяется их количество и видовой
состав. Древостой почти отсутствует или встречаются отдельные деревья.
По составу и структуре растительности выделены три типа болот:
комплексные осоково-гипновые, ерниково-осоково-гипновые с березой и
сосной на грядах (веретьях) и осоково-гипновые в мочажинах с
периферийным рядом кустарниковых и лесных ассоциаций (118); осоковые, осоково-
гипновые, осоково-сфагновые болота (119); тростниково-осоковые в
сочетании с сосново-кустарниково-сфагновыми «рямами» и периферийным
рядом осоковых и осоково-вейниковых ассоциаций (120). **
154
Осоково-гипновые, ерниково-осоково-гшшовые
с березой и сосной на грядах-веретьях, осоково-гипновые
в мочажинах с периферийным рядом кустарниковых
и лесных ассоциаций;
южно- и западно-сибирский тип болот (118)
В западно-сибирской подтайге евтрофные осоково-гипновые болота
с грядово-мочажинными комплексами являются зональным и
господствующим типом-болот. Они были описаньь Н. И. Пьявченко [1963] в районе
р. Чулым, А. М. Жарковой [1963] на водоразделах рек Ишим — Иртыш
(болота Гледенское и Артевское), Е. А. Романовой [1964] на
надпойменной террасе левобережья р. Туры (болото Тарманское) и в верховьях
р. Баксы.
Болота образуют сложные системы, огромной площади в несколько
сот и даже тысяч квадратных километров с ровной и слабовогнутой
поверхностью. Характерная их особенность — обилие на них крупных озер,
заполненных сапропелем. В северных районах Б арабы среди евтрофных
болот встречаются островки олиготрофных выпуклых сосново-кустарнич-
ково-сфагновых «рямов». В размещении растительности на болотных
массивах большой площади наблюдается определенная закономерность.
Ближе к суходолам и минеральным островам расположены фитоцено-
зы с господством кочкарных осок Carex cespitosa, С omskiana и др. По мере
удаления otv суходолов кочкарные осоки сменяются корневищными осо-
ками — Carex lasiocarpa, С. vesicaria, С. diandra, C/chordorrhiza. Моховой
покров развит слабо, состоит из гипновых мхов. На более дренированных
окраинах болот расположены травяно-лесные и лесные ассоциации.
Ближе к центру болот расположены осоково-гипновые фитоценозы,
в которых доминируют корневищные осоки с участием болотного
разнотравья (вахты, сабельника). Моховой покров плотный из гипновых мхов
(Drepanocladus aduncus, D. sendtneri, D. verrucosus и виды Calliergon).
В центральной более обводненной части болотных систем с
уплощенной или вогнутой поверхностью, там, где уклоны поверхности
незначительные, развиваются болотные комплексы с расчлененным микрорельефом
на гряды и мочажины. Эти комплексы занимают до 50—70% всей
площади болотной системы. По структуре микрорельефа и физиономичности
растительности среди них выделяются грядово-мочажинные, мозаичные,
пятнистые комплексы с различным составом, растительности на грядах и
в мочажинах.
В грядово-мочажинных комплексах гряды облесены сосной и березой
(Betula pubescens), в северо-восточной части Б арабы'встречается
лиственница и кедр. Деревья высотой 2—4 м и более, они тянутся на грядах4
цепочкой вдоль морозобойных трещин-канавок. Эти трещины весной служат
мельчайшей внутриболотной сетью поверхностного стока, тем самым
улучшают дренаж болота и оказывают благоприятное влияние на развитие
древесной растительности на грядах. В травяно-кустарничковом ярусе
доминируют осоки Carex lasiocarpa, С diandra и карликовая береза (Betula
папа). В моховом ярусе хорошо развиты гипновые мхи, господствует
Tomenthypnum nitens с участием Drepanocladus vernicosus, D. sendtneri,
около стволов деревьев Pleurozium schreberi пятнами встречается Sphagnum
warnstorfii. Мочажины более обводнены, древесная растительность
отсутствует, иногда встречаются мелкие экземпляры ив (Salix lapponum,
S. rosmarinifolia). В травостое доминируют в основном те же осоки, что и
на грядах, с большой примесью вахты (Menyanthes trifoliata), сабельника
(Comarum palustre), 'с небольшой примесью шейхцерии, хвоща топяного
(Equisetum fluviatile), подмаренника (Galium palustre) и др. В моховом
ярусе больше Drepanocladus vernicosus, D. sendtneri, виды Calliergon и др.
Мозаичный микрокомплекс является следующей генетической стадией'
развития грядово-мочажинного комплекса. В отличие от предыдущего
комплекса облесенные сосной и березой гряды соединены здесь между
собой и оконтуривают необлесенные осоково-гипновые мочажины, что в
155
плане напоминает мозавду- Растительность на грядах мало отличается от
таковой грядово-мочажинного комплекса — доминируют осоки Сагех
lasiocarpa, С. diandra, С. buxbaumii, карликовая береза, пятнами
встречается сабельник. Мочажины, заключенные между грядами, более
обводнены, вода часто стоит на поверхности мохового покрова. Древесная
растительность отсутствует полностью. В травяном покрове господствует
осока Сагех buxbaumii, с примесью С. lasiocarpa и С. vesicaria и болотного
разнотравья из вахты, пушицы узколистной (Eriophorum polystachyori).
В моховом ярусе Сомкнутостью 60—80%, кроме Drepanocladus vernicosus,
D. aduncus, в более обводненных участках появляется Scorpidium
scorpioides.
Пятнистый комплекс занимает центральную, почти бессточную часть
болотного массива. Характерным для него является более сильная
обводненность и обилие озер, по сравнению с предыдущими комплексами.
Своеобразное строение пятнистого комплекса в плане представляет собой
чередование относительно повышенных полос — гряд шириной 5—10 м,
которые оконтуривают переобводненные участки — зыбуны различной
конфигурации. В связи с увеличением обводненности и уменьшением про-
точности воды, древесная растительность на грядах отсутствует,
доминирует карликовая береза (Betula nana), из осок встречаются Сагех chordor-
rhiza, С. lasiocarpa, С. buxbaumii, С limosa, а также вахта, хвощ. Моховой
покров разрежен, состоит в основном из гипновых мхов Drepanocladus
vernicosus, D. sendtneri видов Calliergon. Понижения (зыбуны) составляют
80—90% площади комплекса, обводненность их настолько велика, что
моховой покров, состоящий из Scorpidium scorpioides находится во
взвешенном состоянии. В травяном покрове редкие осоки {Сагех chordorrhiza,
С. limosa, С. теуегапа) чередуются с пятнами вахты.
Торфяная залежь в комплексах глубиной от 2 до 4 м сложена
осоковым и осоково-гипновым торфами. Местами в пятнистом комплексе, где в
моховом покрове господствует Scorpidium scorpioides, верхний горизонт
торфа мощностью 1—2,5 м сложен скорпидиум-торфом.
В осоково-гипновых болотах с грядово-мочажинными комплексами^
расположенных на водоразделе рек Ишим — Иртыш, залегает мощный
слой (6—9 м) сапропеля, над которым слой торфа составляет всего 0,5—
1,5 м [Жаркова, 1963].
Водораздельные осоково-гипновые болота занимают огромные
площади и являются потенциальным фондом для народного хозяйства. Они
могут быть использованы для торфодобычи, а также под сенокосные луга,
пастбища и другие сельскохозяйственные угодья.
Осоковые, осоково-гипновые,
осоково-сфагновые болота (119)
Наиболее распространены в поймах рек. Они не занимают больших
площадей п обычно встречаются в сочетании с олиготрофными и мезотроф-
ными болотами как на периферии, так и в центре их. Иногда они
представлены небольшими массивами среди заболоченных лесов и лугов.
В связи с малой площадью большинство указанных болот не отражены на
карте, но они вошли в контуры заболоченных лесов, лугов или
верховых болот.
Осоковые болота, как правило, безлесны, иногда попадаются редкие
кусты ив (Salix cinerea, S. rosmarinifolia). Основной фон составляют осоки
корневищные или кочкарные с примесью болотного разйотравья, редкими
пятнами встречаются гипновые или сфагновые мхи.
В зависимости от господства в травостое того или* иного вида осоки
С. Н. Тюремнов [1957] выделяет среди осоковых болот три варианта:
lasiocarpa-ъд^идлт, cespifosa-вариант, оттгз&штга-вариант. Описание их
приводится по указанному первоисточнику.
Первый из них £бшосаг/?а-вариант осоковых болот чаще всего
встречается на периферийных участках водораздельных олиготрофных болот с
156
грунтовым славо минерализованным водным питанием. Травостой
образован корневищными осоками, доминирует Carex lasiocarpa с примесью
С. vesicaria с большим количеством разнотравья, в котором основную
массу составляют вахта, сабельник, хвощ, вербейник (Lysimachia vulgaris).
В моховом покрове в незначительном количестве присутствуют евтрофные
гипновые и сфагновые мхи.
Второй cesp^osa-вариант осоковых болот занимает довольно большие
площади на окраинах болот в зоне переменного увлажнения и граничат,
с одной стороны, с тростниковыми «займищами», а с другой — с
солончаковыми лугами. В Барабе такие болота концентрической полосой
окаймляют рямы. Формируются всегда в условиях переменного увлажнения,
обильно обводняются талыми поверхностно-сточными и полыми речными
водами в весенний период и в значительной мере пересыхают во вторую
половину лета. Микрорельеф сильно кочковатый, кочки высотой 25—
30 см образует С. cespitosa. На кочках и между кочками произрастают
С. buxbaumii, вейник (Calamagrostis neglecta), калужница (Caltha palust-
ris), подмаренник (Galium palustre)1 сабельник (Comarum palustre). По
данным многих исследователей, по мере усыхания болота кочки с Сагех
cespitosa быстро минерализуются и способствуют капиллярному поднятию
к поверхности засмоленных почвенно-грунтовых вод.
Третий оттгз/аатга-вариант осоковых болот встречается в широких
долинах рек и прилегающих к ним полосах водораздельных плато, в
условиях поверхностно-сточных и грунтовых вод с повышенным
содержанием минеральных солей. В травостое господствует Carex omskiana и
С. riparia. Болота характерны дЛя западной части Барабинской
низменности, где они расположены на придолинных полосах водоразделов,,
прилегающих к долинам рек. В растительном покрове доминирует осока
омская (С. omskiana), в меньшей степени осока береговая (С. riparia) с
небольшой примесью болотного разнотравья.
Моховой покров во всех вариантах осоковых болот развит слабо или
отсутствует.
Торфяная залежь неглубокая 1—2 м, осоковая, осоково-гипновая.
Дальнейшее развитие гипнового мохового покрова обеспечивает
переход евтрофных болот из осоковой стадии в стадию осоково-гипновую, где
в травяном покрове доминируют осоки (Carex lasiocarpa, С. diandra,
С. vesicaria) и гипновые мхи (Drepanocladus aduncus, D. sendtneri, виды
Calliergon и др.). В последующем евтрофная осоково-гипновая стадия
при обеднении водно-минерального питания переходит в мезотрофную
осоково-сфагновую, из осок доминируют Carex diandra, С. chordorrhiza,
в болотном разнотравье появляется шейхцерия (Scheuchzeria palustris).
Моховой покров образован сфагновыми- мхами Sphagnum warnstorfii,
Sph. centrale, Sph. papillosum, в понижениях Sph; obtusum, Sph. subsecun-
dum, гипновые мхи играют незначительную роль.
Троетниково-осоковые ( «займища»)
в сочетании с сосново-кустарничково-ефагновыми «рямами»
и периферийным рядом осоковых
и осоково-вейниковых ассоциаций;
барабинский займищно-рямовый тип болот (120)
Болота в основном сосредоточены на водоразделах верхних течений
Оби и Иртыша, где они простираются на десятки километров и являются
основным элементом ландшафта центральной части Барабинской
низменности. Заболоченность здесь достигает 25—30% общей площади. Болота
центральной части Барабы приурочены к депрессиям водоразделов,
современным или древним ложбинам стока и имеют характерную вытянутую
форму с СВ и ЮЗ. Напротив, болота, расположенные к западу на
водоразделах рек Иртыш—Ишим—Тобол, Занимают незначительные
площади и имеют более округлую форму.
157
Основные заболоченные пространства весьма значительной площади
занимают тростниковые и тростниково-осоковые болота — «займища».
Растительный покров составляют тростник (Phragmites australis) и осоки
(Carex cespitosa, С. appropinquata, С. riparia), в большом количестве
встречаются светлуха (Scolochloa festucacea), вейник (Calamagrostis neglecta).
Древесный покров на займищах отсутствует, он начинает появляться
только у самых границ рямов, встречающихся островкдми среди займищ.
По окраинам займищ и на подсушенных участках произрастает светлуха
{Scolochloa festucacea). Ближе к периферии болота к тростнику и светлухе
примешивается вейник {Calamagrostis neglecta), такие участки примыкают
к суходолу, образуя высокопродуктивные сенокосы.
По данным М. С. Кузьминой [1949] приведем две схемы экологичен
ских рядов,растительности займищ.
1..Схема эволюции растительности при естественном усыхании
займища
Ассоциация тростниково-светлуховая
Лисохвостово-со-
лончаковая
Солончаково-
разнотравная
Шслковицево-разнотравная
2. Схема эволюции растительности при искусственном осушении
займища
Ассоциация
тростниковая
Тростниково-вейнико-
во-светлуховая
\
Тростниково-светлу-
хово-вейниковая
1
Светлуховая
Светлуховая
вейниковая
I Л
Вейниковая с примесью
лисохвоста
Вейниковая с примесью
солончакового разнотравья
Необходимо фтметить, что как на осушенном, так и неосушенном болотных
массивах происходит смена экологических типов болотных сообществ —
от гидрофитных до галофитных, причем на неосушенном массиве этот
процесс протекает более интенсивно.
Как уже отмечалось, отличительной особенностью западно-сибирских
тростниково-осоковых займищ является наличие рямов. Нами этот ^тип
болот назван «бдрабинский займищно-рямовый тип».
, Рямы — это типичные олиготрофные выпуклые сосново-кустарнич-
ково-сфагновые болота, которые разбросаны среди обширных пространств
евтрофных тростниково-осоковых займищ и выделяются среди них густо
облесенной поверхностью. Древесный ярус на рямах состоит из сосны
{Pinus sylvestris f. litwinowii, реже /. uliginosa). Высота сосны 1,5—3 м,
на склонах 5—8 м. В кустарничковом ярусе доминируют багульник {Ledum
palustre), хамедафне {Chamaedaphne calyculata), брусника {Veccinium vitis-
idaea). Моховой ярус состоит из Sphagnum fuscum.
Ближе к склонам ряма в значительном количестве появляется морош-
158
ка (Rubus chamaemorus) и подбел {Andromeda polifolia), в моховом ярусе
преобладает Sphagnum fuscum.
У подножья хклонов ряма расположен сосново-березово-пушицевый
пояс, где в древесном ярусе доминирует сосна (Pinus sylvestris) и береза
(Betula pubescens), в травяном ярусе преобладает пушица (Eriophorum
vaginatum), в моховом ярусе развиты Sphagnum magellanicum и Sph.
recurvum. Далее при переходе к займищам идет полоса березово-осоково-
сфагновой растительности с Carex lasiocarpa и редким ярусом тростника,
которая сменяется полосами сначала березового, а затем безлесного
осокового кочкарника с Carex cespitosa, С. appropinquata.
Тростниково-осоковые болота — займища являются потенциальными
кормовыми угодьями, однако использование их в большинстве случаев
возможно лишь после осушения.
^ Подробное описание болот подтаежной зоны приведено в работах
Б. Н. Городкова ([1915], М. И. Нейштадта [1936], А. А. Генкеля и
П. Н. Красовского [1937], Н. Я. Каца [1948], М. С. Кузьминой [1949,
1957,1961], К. Е. Иванова, Л. В. Котовой [1964], Е. А. Романовой [1978].
ЛЕСОСТЕПНЫЕ ЕВТРОФНЫЕ ТРАВЯНЫЕ БОЛОТА
И ГАЛОФИТНЫЕ ЛУГА
Заболоченность лесостепной зоны Западной Сибири незначительная,,
Волнистый рельеф, наличие многочисленных пресных озер и понижений с
богатым грунтовым водным питанием способствуют развитию
высокотравных (тростниковых, камышовых, рогозовых) осоковых, реже осоково-
гипновых болот. Эти болота и являются зональным типом для лесостепной
подзоны. '
Высокотравные евтрофные болота приурочены к периферии пресных
озер или днищам озерных котловин, а также к центральным частям
ложбин, соединяющих озера в период весенних разливов. Для травяных болот
характерно полное отсутствие древесной растительности. Наиболее
распространенными являются тростниковые и вейниково-осоковые болота в
сочетании с лисохвостными (Alopecurus arundinaceus) и полевицевыми
(Agrostis gigantea) засоленными лугами (127).
Тростниковые и вейниково-осоковые болота
в сочетании с лисохвостными
и полевицевыми засоленными лугами (127)
Они занимают небольшие участки в замкнутых озерных котловинах
или располагаются полосами вокруг озер в старицах и долинах рек»
По площади ^болота настолько малы, что на карте помечены в основном
внемасштабным знаком, или вошли в контуры галофитных лугов.
В травяном ярусе болот господствует тростник (Phragmites australis)
высотой 1—2 м, ближе к суходолу во втором ярусе появляются кочкар-
ные осоки — Carex cespitosa, С. appropinquata. В тех случаях, когда
происходит самоосушение болота и увеличивается засоленность, появляется
вейник вытянутый (Calamagrostis neglecta), который постепенно вытесняет
тростник и болото переходит в вейниково-осоковое. Таким образом в
зависимости от увеличения засоленности грунта и удаления от уреза воды
динамика смены болотной растительности происходит в следующем
порядке: тростниковые —>- тростниково-осоковые ->■ вейниково-осоковые
болота ->■ галофитные луга.
В южной части лесостепи развитие этих болот заканчивается стадией
осоковых кочкарников из Carex omskiana, С. cespitosa, С. appropinquata.
Более подробное описание болот лесостепной зоны приводится в
работах М. Д. Спиридонова [1928], М. И. Нейштадта [1936], А. А. Генкеля
и П. И. Красовского [1937], М. С. Кузьминой [1949, 1957], А. А. Смирен-
ского [1949].
159
Галофитно-злаковые луга — ячменевые, лисохвостные,
пырейные, бескильницевые
в сочетании с галофитно-разнотравными лугами (128)
Галофитные луга широко распространены в лесостепи Барабинской,
Кулундинской равнины и на междуречьях Иртыша и Тобола — Ишим-
ской степи. Они формируются по пониженным элементам рельефа в
условиях постоянного или периодического действия сильно минерализованных
грунтовых вод преимущественно карбонатного типа засоления.
Наибольшие площади галофитные луга занимают в межгривныХ
понижениях северной лесостепи, в южной — неширокими поясами
обрамляют болотные и озерные котловины или сырые березовые колки. Почвы под
ними — луговые солончаки, лугово-болотные солончаковые, торфянисто-
болотные солончаковые, сформировавшиеся на озерно-аллювиальных
отложениях. В связи с особенностями гидрологического режима,
засоленностью почв, грунтовых вод, развиваются и различные сообщества гало-
фитных лугов, образующие в совокупности микропоясные экологические
ряды. На торфянисто-болотных и лугово-болотных солончаковых почвах,
периодически промываемых опресненными талыми водами, преобладают
менее солеустойчивые лисохвостовые луга с Alopecurus arundinaceus и
постоянным участием Bolboschoenus maritimus, Agrostis stolonifera, Triglochin
maritimum, T. palustre, Eleocharis palustris. При увеличении засоленности
на лугово-солончаковых почвах с близкоЬтоящими минерализованными
грунтовыми водами в травостое лугов господство переходит к Hordeum
brevisubulatum, а затем к наиболее солестойкому злаку Puccinellia distans.
В составе ячменевых и бескильницевых лугов до 40% галофильных видов:
Juncus gerardii, Plantago cornuti, P. salsa, Glaux maritima, Saussurea amaraf
Cirsium esculentum, Carex diluta, Trypolium vulgare, Artemisia rupestrisf
A. laciniata, Taraxacum bessarabicum. Во влажные годы, когда обильными
осадками вредные соли вымываются из верхнего корнеофгтаемого слоя
почвы, в травостое этих лугов увеличивается обилие луговых и лугово-
болотных видов Alopecurus arundinaceus, Poa palustris, Phragmites austra-
lis, Elytrigia repens, Potentilla anserina, Lathyrus palustris. Видовой состав
галофитно-злаковых лугов беден (не более 15—20 видов на 100 м2), высота
травяного покрова 60—70 см, проективное покрытие составляет 80—
100%. Во влажные годы луга становятся ценными сенокосами с
урожайностью 15—20 ц/га [Вагина, 1962].
Среди галофитно-злаковых лугов небольшими участками на более
засоленных солончаковых почвах и солончаках распространены гало-
разнотравные фитоценозы, объединенные со злаковыми лугами в один
территориальный выдел. В их травяном покрове преобладает галофильное
разнотравье — Saussurea amara, Plantago cornuti, P. salsa, Claux maritima,,
Triglochin maritima, Artemisia nitrosa, Atriplex littoralis, Salicornia europaea,
Suaeda corniculata. Площади таких фитоценозов неизменно расширяются
за счет контактирующих с ними галофитно-злаковых лугов, травостой
которых деградирует при неумеренном выпасе скота. Уплотнение почвы при.
выпасе усиливает засоление, что ведет к постепенному выпадению Злаков
из травостоя лугов и внедрению сорнолуговых галофильных видов —
Atriplex littoralis, Halimione pedunculata, Taraxacum bessarabicum, Lepidi-
um latifilium. На выбитых участках с засоленными почвами поселяются
сочные солянки — Salicornia europaea, Suaeda prostrata, S. corniculata.
Растительность галофитных лугов освещена в работах Т. А. Вагиной
[1962, 19741,'К. А. Куркина [1952, 1976], М. С. Кузьминой [1953].
Галофитно-разнотравные луга
в сочетании с открытыми группировками
из галофитного разнотравья и полукустарничков (129)
В многочисленных бессточных понижениях, по днищам высохших
или усыхающих озер, по периферии болот на сильно засоленных
солончаковых почвах развит комплексный растительный покров из галофитно-
160
разнотравных лугов в сочетании с однолетними солянками и галофильны-
ми полукустарничками. Такое сочетание сообществ галофитов характерно
для районов южной лесостепи, где наиболее широко распространены
комплексы луговых солонцов и. солончаков хлоридно-сульфатного и: со-
дово-сульфатно-хлоридного типа засоления. Большие площади галофиль-
ная растительность занимает в сухой долине Камышловского лога и на
обширной низменной равнине Сумы-Чебаклинского понижения с
множеством усыхающих мелких озер. Южнее в степных районах Кулундинской
равнины галофильная растительность распространена небольшими
очагами на террасах солевых озер Кучукско-Кулундинской котловины и др.
Комплексный растительный покров солонцово-солончаковых почв
с близким уровнем грунтовых вод высокой минерализации (20—30 г/л)
формируется разнообразными вариантами волоснецовых, полынных,,
лебедовых и солеросовых фитоценозов. К корковым солонцам и луговым
солончакам приурочены во л оснецово-лебеДовые, во лоснецово-по лынные
и бескильницево-полынные сообщества с преобладающими растениями
Leymus paboanus, Puccinellia tenuissima, Halimione verrucifera, H. peduncu-
lata, Artemisia nitrosa, Saussurea salsa, Limonium gmelinii. Травяной покров
разрежен, покрытие растениями почвы неравномерное от 15 до 40%. Коли-4
чество видов на участке не превышает 8—12..
На участках с максимальной концентрацией солей солончаки покрыты
выцветами солей и лишены растительности, во влажные годы на них
развиваются сообщества сочных солянок — Salicornia europaea, Suaeda
corniculata, S. linifolia, S> prostrata/Ofaiston monandruml Растительность
открытых солончаков озерных террас и днищ высохших соленых озер
состоит из галофитных кустарничков — Halocnemum strobilaceum, Nitra-
ria sibirica, реже Kalidium foliatum, между которыми встречаются
однолетние солянки Salicornia europaea, Suaeda corniculata, S. liriifolia, Petrosimo-
nia litwinowii.
ГЛАВА IX
СТЕПНАЯ РАСТИТЕЛЬНОСТЬ
Степная растительность на юге Западной Сибири распространена
широкой полосой от предгорий Урала до Оби, занимая плоские равнины
(Ишимскую, Барабинскую, Кулундинскую), а также холмисто-увалистые
плато — Зауральское и Приобское.
Водораздельные пространства рек Тобола и Иртыша — Ишимская
степь — представляют собой плоскую равнину с абсолютными высотами
100—130 м и с общим наклоном к северу. Поверхность рассечена
широкими и неглубокими долинами рек Тобола, Ишима, Иртыша, почти не
имеющими притоков в пределах степного региона. Гидрографическая сеть
имеет слабодренирующее воздействие, большие площади водораздельных
пространств лишены поверхностного стока. Характерно наличие крупных
и мелких ббссточных впадин с многочисленными озерами разных размеров.
В пределах Обь-Иртышского междуречья расположена Барабинская
степь — плоская равнина с абсолютными высотами 105—145 м и гривно-
ложбинньш рельефом. Межгривные ложбины часто заняты озерами и
болотами. Крупные озера Барабы — Чаны, Убинское, Сартлан.
Южнее Барабы на Обь-Иртышском водоразделе находится Кулундин-
ская степь. В центральной своей части она представляет собой
слабовогнутую чашу с абсолютными отметками 120—140 м. Кулунда широко
открыта на север в сторону Барабы и ограничена с востока и юго-востока
пологими склонами увалов Приобского плато. На ее поверхности
расположены бессточные озера, часто значительных размеров. В строении
рельефа Кулунды большая роль принадлежит системе древних ложбин стока „
11 Заказ К* 297 161
расчленяющих поверхность на серию параллельных сравнительно узких и
платообразных увалов, имеющих наклон в сторону Иртыша.
Приобское плато расположено на юго-востоке Западно-Сибирской
равнины между предгорьями Алтая и долиной Оби. Его поверхность
расчленена на ряд плоских гряд-увалов (от 15 до 50 км ширины),
вытянутых с северо-востока на юго-запад и отделенных друг от друга широкими
и глубокими ложбинами. Дно ложбин длоское,лоно занято долинами рек
или озерами и болотами [Николаев, 1970].
Степная растительность связана с выщелоченными, обыкновенными
и южными черноземами, а также каштановыми почвами различного
механического состава. Существенное влияние на развитие почвенного и
растительного покрова степей оказывает засоленность грунтов и грунтовых воду
уровень которых р послеледниковое время был, очевидно, выше
современного. Развитию процессов засоления содействовали равнинность рельефа
и отсутствие поверхностного стока. На водораздельных понижениях
равнин широко распространены комплексы и сочетания обыкновенных г
солонцеватых черноземов, луговых солонцеватых и солончаковых почв,
солонцов и солончаков. В низких недренируемых западинах лутово-чер-
ноземные почвы сочетаются с солонцами, солончаками и солодями. При-
оз'ерные понижения и древние ложбины стока занимают солончаки разной
степени засоленности. В северных районах распространения степной
растительности преобладает содовое засоление, в южных — хлоридами и
сульфатами с участием соды [Егоров, Базилевич, 1976].
Степная растительность юга равнины представлена заволжско-казах-
станской фратрией формаций в восточной части ее ареала. Формации этой
фратрии распространены в пределах геоботанической провинции того.же
названия, согласно ботанико-географическому делению Евразиатской
степной области Е. М. Лавренко [1970, 1980], давшему и исчерпывающую
характеристику особенностей ее степной растительности. .
Сообщества степного типа растительности состоят «преимущественно
из многолетних микротермных и ксерофилвеых (морозо- и
засухоустойчивых) травяных растений, большей частью дерновинных злаков»
[Лавренко, 1956, с. 595]. Корневищные злаки в настоящих степях принимают
небольшое участие, но характерны для луговых степей и остепненных
лугов. Большинство степных сообществ имеет полидоминантный состав с
тремя или большим числом доминирующих видов.
Доминантными и эдификаторными видами формаций, объединенных в
Заволжско-Казахстанскую фратрию, являются дерновинные злаки —
Stipa pennata, S. zalesskii, S. capillata, S. lessingiana, S. korshinskyi, Fes-
tuca pseudovina, F. valesiaca, реже Koeleria cristata, Helictotrichon deser-
torum. Кроме дерновинных злаков в качестве постоянных доминантов
выступают представители лугово-степного и степного разнотравья —
Peucedanum morisonii, Seseli libanotis, S. ledebourii, Filipendula vulgaris,
Pulsatilla flavescens, Galatella biflora, Artemisia glauca, A. latifolia, A. seri-
cea и полукустарнички — Artemisia austriaca, A. frigida, A. nitrosa, Thymus
marschallianus. Нередко в степньъх сообществах доминируют мезоксеро-
фильные корневищные осоки — Carex supina, С. duriuscula. Другие
многочисленные виды многолетних растений разных жизненных форм
встречаются в меньшем обилии, но характерны для различных плакорных
сообществ степей и их эдафических вариантов — Artemisia pontica, А. таг-
schalliana, Potentilla bifurca, Phlomis tuberosa, Galium verum, Veronica incanar
Gypsophila paniculata, Goniolimon speciosum. В составе степных сообществ
также присутствуют степные кустарники, встречающиеся единично или
образующие заросли,— Spiraea crenata, S. hypericifolia, Caragana frutex,
а в бассейне Тобола европейские виды — Cerasus fruticosa и Cytisus
ruthenicus. Коротковегетирующие многолетники (эфемероиды) и
однолетники (эфемеры) для степей Сибири не типичны и представлены лишь
единичными видами — Androsace septentrionalis, A. turczaninovii^ Gagea
granulosa, G. minima, Alyssum desertorum, Erophila verna.
Формации степной растительности, составляющие Заволжско-Ка-
16 2
захстанскую фратрию, распространены в различных геоморфологических
и климатических условиях, что оказывает существенное влияние на
флористический состав формаций и их структуру. На территории равнинн
выявились два региональных комплекса формации, отражающих
экологические связи растительности соответствующих регионов — обь-иртыш-
ские и северо-казахстанские. '
Обь-иртышский комплекс объединяет формации луговых степей, остеп-
ненных лугов и эволюционно связанных с ними — галофильных лугов ^
Отличительная особенность фитоценотического состава комплекса
группы — преобладание Галофитных вариантов зональных типов луговых
степей и остепненных лугов, комплексирующихся с галофитно-разнотрав-
ными сообществами солонцов и солончаков. Наиболее широко формации
комплекса распространены в северной подзональной полосе степей, Обь-
Иртышского и Тобол-Иртышского междуречий, где засоленные почвы
занимают значительные площади. На карте «Растительность Западно-
Сибирской равнины» обь-иртышские формации представлены
разнотравно-злаковыми остепненными лугами и злаково-богаторазнотравными
луговыми степями плакоров, а также различными галофильными
вариантами их сообществ — разнотравно-полынно-ковыльными, галоразнотравно-
злаковыми, полынно-типчаковыми солонцеватыми степями, комплекси-
рующимися с сообществами галофильных лугов.
Северо-казахстанский комплекс объединяет формации разнотрав-
но-дерновинно-злаковых (разнотравно-типчаково-ковыльных) степей*
включающих как плакорные сообщества, так и разнообразные эдафиче-
ские варианты, а также комплексную растительность из галофитных
разностей дерновинно-злаковых степей с галофитно-степными и гадофитными
сообществами на солонцеватых черноземах или каштановых почвах и
солонцах. Формации, объединенные в комплекс, распространены в двух
подзональных полосах: разнотравно-типчаково-ковыльных и типчаково-
ковыльных степей. Они связаны, главным образом, с территорией Ку-
лундинской и Прииртышской равнин и южной частью Приобского плато*
На карте «Растительность Западно-Сибирской равнины» разнотравно-
дерновинно-злаковые степи показаны дифференцированно. Выделяются
следующие картируемые подразделения: богаторазнотравно-красноко-
выльные степи плакоров (138), псаммофитные (139) и кальцефитные (140)
и их варианты, характерные для более северной полосы подзоны разно-
травно-типчаково-ксвыльных степей; разнотравно-типчаково-тырсовые
степи (142) и их галофитные варианты (143), широко распространенные
в южной части подзоны; ксерофитно-разнотравно-типчаково-тырсовые и
полынно-типчаково-ковыльные степи (144), встречающиеся локально в
районе восточной границы типчаково-ковыльных степей.
БОГАТОРАЗНОТРАВНО-ЗЛАКОВЫЕ ЛУГОВЫЕ СТЕПИ
И ОСТЕПНЕННЫЕ ЛУГА
Луговые степи и остепненные луга, распространенные в северной
части степной зоны и «сочетающиеся с сообществами остепненных осиново-
березовых лесов, образуют полосу лесостепи. Эта широтная полоса
охватывает Ишимскую и Барабинскую равнины, центральную часть
Приобского и Зауральского плато. Растительный покров характеризуется
неоднородностью сложения, преобладанием галофитных вариантов луговых
и степных сообществ, образующих разнообразные комплексы с галофит-
ными лугами на луговых солонцах и солончаках.% *•*
Зональные сообщества луговых степей и остепненных лугов [131 %
135] занимали до распашки плакорные местоположения —
выровненные вершины грив и повышенные участки равнины с выщелоченными и
обыкновенными черноземами. Ныне эти территории все распаханы.
Описания их даны в работах А. Я. Гордягина [1900, 1901], П. Н. Крылова
. [1916, 1919], В. И. Баранова [1923, 1927], И. М. Крашенинникова,
Н. А. Ивановой [1934], В. В. Ревердатто [1931]. Большая часть террито-
11* - 163*
рии была распахана еще в начале XX в., интенсивной распашке они
подвергались в 30-е годы. В настоящее время остатки таких степей можно
встретить лишь по обочидам пашен, на склонах увалов, а также на
засоленных равнинах, где степные сообщества встречается в комплексе с
солонцеватыми лугово-степными и луговыми сообществами.
Сравнивая плакорные луговые степи я остепненные луга Западной
Сибири с восточно-европейскими, Е. М. Лавренко [1956] отмечал их
большую мезофильность, флористическую обедненность, отсутствие в их
составе многих европейских. степных видов, участие восточных видов,
основной ареал которых связан с Северным Казахстаном, Южной и Вое- .
точной Сибирью,— Peucedanum morisonii, Iris ruthenica, Geranium bifo-
lium, Artemisia glauca, A. lacinata и др., бедность эфемероидами,
представленными лишь единичными видами гусиного лука — Gagea granulosa,
G* minima.
Галофитные варианты луговых степей и остепненных лугов
отличаются от плакорных меньшим фитоценотическим разнообразием,
обеднением видового состава, большей ксерофильностью, постоянным участием
в их составе видов галофильного разнотравья — Artemisia pontica, A.
glauca, Galatella biflora, Limonium gmelinii, Glycyrrhiza uralensis и др.
Солонцеватые степи и остепненные луга генетически связаны с
луговыми сообществами солончаков, что отчетливо выявляется в условиях,Ба-
рабинской низменности, где процессы формирования и развития
современного почвенного и растительного покрова идут более интенсивно.
Здесь можно наблюдать множество переходов от солончаков к типично
степным почвам. Прогрессирующая ксерофитизация луговых
солончаковых сообществ, обусловленная постепенным понижением уровня
грунтовых вод и изменением водно-солевого режима почв, ведет к доминированию
на их месте солонцеватогстепных сообществ, образующих единый
генетический ряд от галофитных лугов к солонцеватым степям [Вагина, 1982].
Площади солонцеватых степей и остепненных лугов невелики и на
карте 'они показаны территориальными выделами (132, 135) —
комплексами степных, луговых и галофитных сообществ.
На почвах солонцово-солончакового комплекса, широко
распространенного в межгривных низинах по территории озерных и болотных
впадин, развита комплексная растительность, образованная сообществами
солонцеватых степей и остепненных лугов, галофитных лугов и
однолетних солянок (133). Распределение сообществ в комплексном растительном
покрове на Причановской пониженной равнине показано на рис. 15.
Ниже приводится краткая характеристика растительности
выделенных на карте территориальных сочетаний сообществ лугово-степных
формаций (131—135). Распаханные плакорные степи показаны на карте как
сельскохозяйственные земли — пашни, залежи с небольшими участками
вклинивающихся между ними естественных лугово-степных сообществ,
используемых как.сенокосы и пастбища. Описания даны для коренной
растительности, определяющей ^экологический потенциал этих земель.
Разнотравно-злаковые остепненные луга и луговые степи
в сочетании с участками
осиново-березовых остепненных лесов (131)
Сочетание растительных сообществ остепненных лугов, луговых
степей и остепненных осиново-березовых лесов характеризует
растительность плакоров полосы северной лесостепи. При картографировании сб-
общества этих формаций объединены в единый территориальный выдел.
В северной лесостепи (дернисто-луговой подзоне [по П. Н. Крылову,
1919]), являющейся переходной от лесной зоны к настоящим степям, ос-
тепнение выражено лишь на повышенных формах рельефа. Зональными
для этой полосы считаются остепненные разнотравно-злаковые луга,,
в прошлом занимавшие гривы и высокие водоразделы с выщелоченными
черноземами и лугово-черноземными почвами, богаторазнотравно-злако-
164
3* ffiB^
9 ^Ш\Ю\11-
11 \Z~~
12
15
Рис. 15. Лугово-степной микрокомплекс межгривного понижения на Причановской
депрессии [по Вагиной, 1974].
Ассоциации: 1 — ковыльная степь со Stipa capillata на черноземовидной почве; 2 — разно-
травно-пырейно-типчаковая степь с Festuca pseudovina, Elytrigia repens на лугово-степном средне-
столбчатом солонце; 3—полынно-тонконогово-типчаковая степь с Festuca pseudovina, Koeleria cris-
tata, Artemisia pontica на лугово-степном высоком солонце; 4 — злаково-разнотравный остепненный-
луг с Calamagrostis epigeios, Festuca pseudovina, Peucedanum morissonii на лугово-степном глубоком
солонце; 5 — вейниково-типчаково-пырейный луг с Elytrigia repens, Festuca pseudovina,
Calamagrostis epigeios, на черноземно-луговой солонцевато-солончаковой почве; 6 — разнотравно-вейниковый
луг с Calamagrostis epigeios, Galatella biflora, Medicago falcata, Saussurea amara, Cirsium esculentum на
черноземно-луговой солонцевато-солончаковой почве; 7 — разнотравцо-ячменевый луг с Hordeum
brevisubulatum, Elytrigia repens, Artemisia rupestris, Artemisia laciniata на луговой осолоделой
солонцевато-солончаковой почве; 8 — бескильницевый с Puccinellia tenuissima луг на высоком луговом
солойце; 9 — бескильницевый с Puccinellia distans, Saussurea salsa, Tripolium vulgare луг на луговом
корковатом солонце; 10 — светлухово-лисохвостный луг с Alopecurus ventricosus, Scolochloa festuca-
cea, Carex disticha, Phragmites communis на солончаковато-солонцеватых почвах; 11 — осоковое с Са-
rex gracilis, С. disticha и С. ruparia болото на торфянисто-болотных осолоделых солонцевато-солон-
чаковатых почвах; 12 — светлуховое с Scolochloa festucacea болото на торфянисто-болотной
сильноосолоделой почве; 13 — открытые фитоценозы на луговых солончаках с Sueda maritimat Saussurea
salsa, Puccinellia distans.
вые луговые степи и осиново-березовые остепненные леса. Луговые степи
приурочены к обыкновенным черноземам, в прошлом они занимали
небольшие участки наиболее повышенных поверхностей водоразделов или
их южных склонов. В настоящее время сохранились лишь на почвах с
ясно выраженным засолением, как галофитовый вариант плакорных
луговых степей.
Леса также занимали гривы и другие повышенные элементы рельефа
с выщелоченными черноземами илихерыми лесными почвами, отличались
высокой продуктивностью и отсутствием болотных видов. В наши дни
облесенность сократилась до 15—25%, осиново-березовые леса (колки)
сохранились лишь в западинах с солодями и осолоделыми почвами или по
склонам и днищам логов и балок.
Целинные остепненные луга й начале XX в. еще существовали и были
описаны в работах П. Н. Крылова [1916, 1919], В. И. Баранова J1923,
1927], В. Ф. Семенова [1924], В. В. Ревердатто [1931]. Для таких остеп-
ненных лугов характерно преобладание в травостое ксеромезофильных
луговых корневищных злаков — Calamagrostis epigeios, Poa angustifoliaf
Bromopsis inermis, а из\ рыхлодерновинных — Phleum phleoides, Helictotri-
chon schellianum. Участие степных плотнодерновинных злаков — Stipa
165,
pennata, Festuca pseudovina — незначительно. Из бобовых обильно
представлены Lathyrus pratensis, L. pisiformis, Trifolium lupinaster, Vicia cracca.
Среди разнотравья наиболее распространены мезофильные и ксеромезо-
фильные растения — Filipendula vulgaris, F. stepposa, Seseli libanotis,
Inula salicina, Achillea millefolium, Ranunculus polyanthemos, Pulsatilla
flavescens, Artemisia latifolia, A. sericea, Galium verum, Fragaria viridis,
Anemone sylvestris, Adonis vernalis.
Травостой остепненных лугов густой и высокий (средняя высота
70 см), общее проективное покрытие в ассоциациях 90—100%, количество
Видов на 100 м2 70—80 экз.
Небольшие площади разнотравно-злаковых лугов, сохранившиеся
до настоящего времени, занимают межколочные поляны или окаймляют
колки среди пашен и используются под сенокосы. Нередко встречаются
остепненные луга вторичного происхождения на месте уничтоженных
березовых лесов или образовавшиеся в результате обсыхания травяных
болот. Вторичные остепненные луга на участках, вышедших из-под леса,
характеризуются преобладанием мезофильных растений и меньшим
участием ксерофильных. В их составе преобладают из злаков Calamagrostis
epigeios, Poa angustifolia, P. pratensis, Bromopsis inermis, Elytrigia repens,
среди разнотравья наряду с обычными видами остепненных лугов
сохранились лугово-лесные Serratula coronata, Solidago virgaurea, Hieracium
umbellatum, Galium boreale.
Остепненные луга, и'спользуемые под интенсивный выпас, имеют
низкий, разреженный травяной покров, представлены обычно разнотравно-
мятликовыми лугами с доминированием наиболее устойчивого к выпасу
Poa angustifolia и розеточным разнотравьем с участием различных сорных
одно- и двулетников.
Сельскохозяйственные земли на месте остепненных лугов заняты
под посевы зерновых культур, преимущественно пшеницы, и зернофураж-
но-овсяных смесей, ячменя и др.
Разнотравно-злаковые с галомезофильным разнотравьем
остепненные луга
в комплексе с полынно-бескильницевыми группировками (132)
Этот выдел на карте характеризует комплексный растительный
покров, образованный сообществами галофитных вариантов остепненных
лугов, развитых на лугово-черноземных солоццеватых почвах, средне-
столбчатых лугово-степных солонцах и полынно-бескильницевых
галофитных лугов на корковых солонцах. Галофитные остепненные
(солонцеватые) луга широко распространены в северной полосе лесостепи, где
занимают нижние части склонов грив, низких увалов и слабоприподнятые
участки межгривных низин и древних ложбин. В южных райо.нах
лесостепи они нередко приурочены к плоским понижениям-блюдцам на месте
обсохших заболоченных лугов и травяных болот в процессе
прогрессирующей ксерофитизации лесостепи [Вагина, 1962].
Остепненные солонцеватые луга по сравнению с плакорными
отличаются большим участием лугово-степных и степных видов, преобладанием
в травяном покрове ксеромезофитов, значительным распространением ксе-
рогалофитов (до 10%) и незначительным участием (менее 1 %) лесных
мезофитов [Вагина, 1962]. Общее число видов на участке ассоциации этих
лугов (100 м2) 46—60. Проективное покрытие травяного покрова 65—85%.
Обычно доминирующими видами выступают Calamagrostis epigeios, Poa
angustifolia, Festuca pseudovina. Из других злаков постоянное участие
имеют Elytrigia repens, Phleum phleoides, Koeleria cristata, Agrostis vinealis.
Среди группы разнотравья наибольшим постоянством и обилием
отличаются Filipendula vulgaris, Peucedanum morisonii, Pulsatilla flavescens,
Artemisia glauca, A. latifolia, Seseli libanotis, Plantago stepposa, Fragaria
viridis, Galium verum, Achyrophorus maculatus, Achillea millefolium.
Характерно для этих лугов постоянное участие видов галофильной экологии
166
Galatella biflora, Artemisia pontica, A. laciniata, A. nitrosa, Plantago
maxima. Особенно увеличивается их обилие в сухие годы, когда мезофиль-
ные злаки выпадают. Из других видов, имеющих небольшое обилие, но
встречающихся повсеместно следует отметить ксеромезофильные растения
Carex praecox, Astragalus danicus, Vicia tenuifolia, Medicago falcata, Onobry-
chis sibirica.
В обобщенную разнотравно-злаковую группу ассоциаций
объединено большое количество луговых сообществ, из которых наиболее
широко распространены разнотравно-пырейно-мятликовые, вейниково-типча-
ково-пырейные, разнотравно-пырейно-типчаковые солонцеватые луга.
Другим постоянным компонентом комплекса являются фрагменты
ассоциаций галофитных лугов полынно-бесшщьницевые фитоценозы.
В их составе преобладают ксерогалофильные виды Puccinellia tenuissima
и Artemisia nitrosa, постоянно участие других видов растений,
встречающихся единично — Festuca pseudooina, Elytrigia repens, Plantago cornuti,
P. salsa, Limonium gmelinii, Artemisia laciniata, A. rupestris. Травяной
покров редкий, проективное покрытие колеблется от 40 до 60%.
Встречаются пятна почвы, лишенной растительности или $ единичными
однолетними солянками — Salicornia europaea, Sueda corniculata, S. linifolia.
Территории, занятые галофитными вариантами остепненных лугов,,
издавна используются под выпас сельскохозяйственных животных. *JIpn
интенсивном постоянном выпасе в составе травяного покрова
увеличивается обилие пастбищеустойчивых растений Festuca pseudoiina, Poa angu-
stifolia, Plantago media, Taraxacum officinale, Trifolium repens,
формирующих пастбищные модификации остепненных лугов — разнотравно-осоч-
ково-мятликовые (Carex praecox), типчаково-вейниково-осочковые, разно-
травно-вейниково-типчаковые. В засушливые годы неумеренный выпас
увеличивает засоление почв, что ведет к расширению площадей под по-
лынно-бескильницевыми галофитными лугами.
Волоснецовые, разнотравно-типчаковые
и бескильницевые галофитные луга
с бескильницево-полынными
и солянковыми группировками (133)
Наиболее сложный комплексный растительный покров представляет
территориальный выдел, объединяющий различные сообщества
остепненных галофитных лугов или их фрагментов, и открытые группировки
эвгал офитов. Такой комплексный растительный покров широко
распространен в подзоне лесостепи по древним лощинам стока, межгривным
понижениям, террасам крупных озерных впадин на почвах солонцово-солонча-
кового комплекса.
Состав компонентов комплексного растительного покрова и его
состояние определяется водно-солевым режимом местообитаний и находится
в прямой зависимости от глубины стояния грунтовых вод. Наибольшие
площади в этом сложном комплексе занимают волоснецовые и
бескильницевые луга на луговых солончаках, корковых солонцах и
солонцах-солончаках, где уровень грунтовых вод в годы с обильными осадками может
подниматься до 20—30 см от поверхности, а в годы сильных засух —
опускаться на глубину до 200 см. Наилучшее развитие волоснецовые луга
имеют на луговых солончаках сульфатного типа засоления. В их составе
преобладают Leymus paboanus, Puccinellia giganteaf Galatella bifloraf/
Plantago cornuti, в меньшем обилии, но постоянно присутствуют Limonium
gmelinii, Saussurea amara, Artemisia nitrosa. Под воздействием постоянного
выпаса волоснец усиленно разрастается, образуя иногда почти чистые
заросли [Куркин, 1957].
На солончаках с повышенным содержанием хлоридов доминирующим
растением становится Artemisia nitrosa, а волоснец изреживается. Для
таких местоположений характерны волоснецово-полынные и бескильнице-
во-полынные галофитные луга. Травяной покров их разрежен (проектив-
167
ное покрытие составляет не более 40%), видовой состав беден. Кроме
господствующих Artemisia nitrosa, Puccinellia tenuissima, единично
встречаются Limonium %meliniix Plantago salsa, P. cornuti, Halimione verrucifera.
На корковых содовых солонцах-солончаках из многолетних галофитов
произрастают только бескильницы — Puccinellia dolicholepis или
Puccinellia tenuissima. В годы сильных засух бескильница целиком выгорает и
почва оказывается лишенной растительности. На луговых* солончаках в
засушливые годы в связи с усилением засоления верхних горизонтов
почвы создаются благоприятные условия для поселения и развития
однолетних галофитов — Salicornia europaea, Suaeda corniculata, S. prostrataf
Atriplex litoralis, Halimione pedunculata, Petrosimonia litwinowii,
образующих сообщества неустойчивого состава.
Участки типичных солончаков с группировками из тех же видов
однолетних солянок и куртинами полукустарничков Halimione verrucifera^
Camphorosma monspeliaca, а на юге лесостепи с участием кустарника
Halocnemum strobilaceum, занимают небольшие площади, но также
являются постоянными компонентами этого комплекса.
Разнотравно-типчаковые остепненные солонцеватые луга, входящие
в состав комплексного растительного покрова, связаны в своем
распространении со средне- и высокостолбчатыми солонцами. В травяном
покрове разнотравно-типчаковых лугов наряду с доминирующим степным дер-
новинным злаком Festuca pseudovina большое участие принимают лугово-
степные и луговые злаки Calamagrostis epigeios, Poa angustijolia, Agrostis
vinealis, Phleum phleoides. Почти всегда присутствует корневищная осока
Carex praecox. Среди разнотравья преобладают галомезо- и ксерогалофиль-
ные растения — Artemisia glauca, A. pontica, A. latifolia, A. nitrosa, Gala-
tella biflora. * ,
Злаково-разнотравные луговые степи
в сочетании с участками
осиново-березовых остепненных лесов (134)
Луговые степи были i широко распространены в южной части
Зауральского плато — на водоразделе рек Миасс — Юргамыш — Курта-
мыш, на Ишимской неогеновой равнине — Тобол-Ишимском и Ишим-
Иртышском междуречьях, в Барабинской равнине и на Приобском
структурно-денудационном плато. Почвы под луговыми степями —
обыкновенные, выщелоченные и солонцеватые черноземы, легкосуглинистые и
карбонатные. Они занимали плакорные местоположения в полосе южной
лесостепи (раздотравно-луговой подзоны по П. Н. Крылову) и южные склоны
в северной лесостепи. По описаниям П. Н. Крылова [1916] и Е. В. Ванда-
куровой [1950] луговые злаково-разнотравные степи отличались высоким
травостоем (50—60 см), большой видовой насыщенностью (60—70 видов
на участке ассоциации), полидоминантностью состава — в них
доминировало несколько видов злаков и разнотравья. Обычными доминантами из
злаков были эври- и мезоксерофильные плотнодерновинные растения —
Stipa pennata, Koeleria cristata, Festuca pseudovina, ксеромезофильные
рыхлодерйовинные Phleum phleoides, Helictotrichon schellianum и
корневищные Calamagrostis epigeios, Poa angustijolia. Для сообществ луговых
степей характерно преобладание разнотравья, представленного большим
количеством видов преимущественно ксеромезофильной экологии —
Adonis vernalis, Galium verum, Filipendula vulgaris, Seseli libanotis, Phlomis
tuberosa, Pulsatilla flavescens, Artemisia latifolia, A. sericea, A. scoparia%
A. dracunculus, Peucedanum morisonii, Achillea nobilis, Plantago stepposa*
Участие ксерофильных видов небольшое — единично встречались Poten-
tilla humifusa, Seseli ledebourii, Salvia stepposa, Artemisia austriaca.
Вблизи лесных массивов, на межколочных полянах с выщелоченными
черноземами степные сообщества имеют наиболее мезофильный характер
(рис. 16). Из злаков доминируют Stipa pennata, Festuca pseudovina„
Phleum phleoides, немного корневищных Bromopsis inermis, Poa angusti-
168
Рис. 16. Разнотравно-злаковая луговая стень. Фото Н. Н. Лавренко.
folia у Elytrigia repens и ксерофильного нлотнодерновинного Koeleria
cristata. Разнотравье составлено из большого количества хорошо
развивающихся лугово-степных, луговых и отчасти лесных растений мезофиль-
ной и ксеромезофильной экологии — Solidago virgaurea, Gentiana pneumo-
nanthe, Hieracium umbellatum, Stellaria graminea, Achillea millefolium,
Inula salicina, Silene nutans, Dianthus versicolor, Seseli libanotis, Galium
verum, Fragaria viridis.
• Группа бобовых также представлена ксеромезофилами —- Astragalus
danicus, Lathyrus tuberosus, Onobrychis sibirica. Из степных видов
многочисленны Plantago stepposa, Peucedanum morisonii, P. alsaticum.
Количество видов на 100 м2 достигает 50—60. Проективвгое покрытие составляет
90-95%.
На всхолмленных равнинах с суглинистыми обыкновенными
черноземами разнотравно-злаковые луговые степи более ксерофильны.
Доминирующее положение занимают наряду с Stipa pennata, Siipa capillata,
полыни — Artemisia latifolia и A. sericea и другие виды разнотравья
Peucedanum morisonii, Filipendula vulgaris. Богаче представлена группа
степных видов, чем в сообществах луговых степей на выщелоченных
черноземах. Наиболее часто встречаются такие мезоксерофилы, как
Astragalus onobrychis, Eryngium planum, Onosma simplicissima, Thymus marschal-
lianus, Carex supina, Artemisia dracunculus.
Небольшие участки целинных луговых степей сохранились лишь на
склонах балок, террас, по неудобьям среди пахотных земель. Луговые
степи, используемые под постоянный выпас вблизи населенных пунктов,
претерпели значительные изменения, такие участки обычно находятся на
«типчаковой» стадии дигрессии — здесь сформировались вторичные мелко-
дерновинные типчаково-полынные или осочково-тонконогово-типчаковые
степи. Травяной покров их низкий, сильно выбит скотом, в его составе
преобладают устойчивые к вытаптыванию травы — типчак, полыни
(Artemisia frigiada, A. austriaca), Plantago stepposa, Potentilla bijurca.
Флористический состав их беден, не более 16—20 видов на 100 м2.
169
Березовые леса, также интенсивно используемые под выпас,
приобрели вид разреженного мелколесья с низкорослыми (5—7 м) угнетенными
деревьями и остепненным травяным покровом со следами явной дигрессии.
Галофитно-разнотравные луговые степи
с полынно-типчаковыми
и бескильницево-полынными группировками (135)
Галофитно-разнотравные и галофитно-разнотравно-злаковые луговые
(солонцеватые) степи развиты на сильно солонцеватых черноземах,
средне- и глубокостолбчатых солонцах по склонам и шлейфам грив, межгрив-
ным приподнятым участкам. Для этих степей характерно многообразие
экологических и жизненных,форм. Среди доминирующих видов дерновин-
ные, рыхлодерновинные и корневищные злаки — Festuca pseudovina^
Koeleria cristata, Poa stepposa, Elytrigia repens, Calamagrostis epigeios.
Другие злаки — Stipa pennata, Poa angustifolia — встречаются единично.
Значительное обилие имеют корневищные осоки — Сагех ргаесох, С. su-
pina. Среди многочисленного разнотравья наряду с обычными видами
луговых степей постоянно присутствуют растения галофильной экологии
(лугово-степные и лугово-солончаковые) — Galatella biflora, Glycyrrhisa
uralensis, Limonium gmelini, Plantago salsa, Artemisia pontica, A. nitrosa,
A, laciniata, A. rupestris. Флористический состав солонцеватых степей
насчитывает 30—35 видов на участке ассоциации, проективное покрытие
составляет 55—70%. Наиболее распространены на глубокостолбчатых
солонцах злаково-разнотравные солонцеватые степи (Glycyrrhisa uralensis,
Peucedanum morisonii, Artemisia austriaca, A. pontica), на среднестолбчатых
солонцах — типчаково-разнотравные (Galatella biflora, Artemisia austriaca,
A. nitrosa, A, glauca). Полынно-типчаковые солонцеватые степи
формируются на высокостолбчатых солонцах. Они встречаются небольшими
участками как постоянные компоненты сложного комплексного
растительного покрова. Для них характерны бедность видового состава и
разреженность травяного покрова (проективное покрытие составляет не более
30—40%). В их составе преобладают ксерофильные злаки и ксерогало-
фильные виды разнотравья (Festuca pseudovina, Koeleria cristata, Artemisia
nitrosa, A. rupestris, Galatella biflora, Plantago salsa, Limonium gmelini)
[Вагина, 1982].
Сообщества луговых солонцеватых степей комплексируются с гало-
фильно-луговой растительностью на корковых солонцах, образуя единый
территориальный выдел. С корковыми солонцами связаны фитоценозы
бескильницевых и бескильницево-полынных ксерогалофитных лугов,
отличающихся также бедностью видового состава и изреженностью
травяного покрова (проективное покрытие составляет 30—40%). В составе
травяного покрова преобладают Puccinellia tenuissima, Artemisia nitrosa. Обилие
других растений незначительно. Из постоянно присутствующих видов
следует назвать Limonium gmelinii, Plantago salsa, Festuca pseudovina,
Artemisia rupestris, Saussurea salsa, Salicomiaherbacea, Sueda corniculata, Halimio-
ne verrucifera.
Площадное соотношение компонентов комплексного растительного
покрбва этого выдела варьирует, но площади солонцеватых степей занима-
• ют не менее 70% комплекса.
Комплексная растительность солонцеватых степей сохранилась
благодаря приуроченности к почвам, не пригодным для распашки, и повсеместно
используется для выпаса скота. Урожайность этих степей неустойчива,
а в засушливые годы низкая и це превышает 2—3 ц/га сухой травяной
массы.
РАЗНОТРАВНО-ДЕРНОВИННО-ЗЛАКОВЫЕ СТЕПИ
Формации разнотравно-дерновинно-злаковых степей формирует
растительность подзоны разнотравно-типчаково-ковыльных степей,
простирающихся по южной окраине Западно-Сибирской равнины. Широкое рас-
170
пространение эти степи имели на Прииртышской и в особенности на
Кулундинской равнинах, где занимали обширные площади. Эти степи
связаны с южными черноземами различного механического состава.
Описания зональных типов степей даны в работах П. Н. Крылова
[1916, 1919], выделившего настоящие степи в подзону, названную им
«ковыльно-кипцовой», В. И. Баранова [1923, 1927], В. Ф. Семенова [1924],
Е. В. Вандакуровой [1950].
Основные признаки плакорных степных сообществ этой подзоны —
доминирование дерновинных ксерофильных и мезоксерофильных злаков
(Stipa zalesskii, S. capillata, *S. pennata, Festuca pseudovina, F. ualesiaca),
полидоминантность, постоянное участие богатой видами группы
разнотравья. Виды разнотравья особенно характерны для сообществ разнотрав-
но-дерновинно-злаковых степей — Peucedanum morisonii, Seseli ledebouri,
Artemisia latifolia, ACdracunculus, A. glauca, а для некоторых эдафических
вариантов этих степей — обильное развитие полукустарничковых
полыней — Artemisia marschalliana, A. austriaca, A. frigida. Наибольшим
числом видов представлена группа разнотравья в сообществах богаторазно-
травно-ковыльных степей, преобладающих в северных районах подзоны
{138). В южных районах подзоны распространены сообщества,
отличающиеся большей 'ксерофитностью и уменьшением роли разнотравья —
разнотравно-типчаково-тырсовые и разнотравно-типчаково-ковыль-
ные (144).
Значительное разнообразие экологических условий в пределах
подзоны обусловливает широкое распространение экологических вариантов
разнотравно-дерновинно-злаковых степей, связанных с условиями
местообитания, отличными от, плакорных. Так, на маломощных карбонатных
черноземах в южной части Ишим-Иртышского междуречья были развиты
кальцефитные разнотравно-ковыльно-овсецовые степи с господством
Stipa korshinskyi, S. zalesskii, Helictotrichon desertorum и довольно
разнообразным ксерофильным разнотравьем. Эти степи распаханы и показаны
на карте сельскохозяйственными землями (140). Псаммофитные и геми-
псаммофитные степи (139), приуроченные к супесчаным и
легкосуглинистым черноземам, показаны в основном по приречным увалам |Иртыша и
Тобола, где они занимали большие площади. Широкое распространение
таких степей в Кулундинской равнине не нашло отркжения на карте из-за
малых площадей, не выраженных в данном масштабе.
Большое разнообразие растительных сообществ встречается на южных
разностях черноземов и каштановых почв .различной степени солонцева-
тости и на солонцах. Наиболее распространены галофитные варианты
разнотравно-типчаково-ковыльных степей — ковыльно-типчаковые,,
гал офитно-разнотравно-типчаковые, полынно-типчаковые сообщества,
комплексирующиеся с галофитно-степныки сообществами солонцов —
солончаков. Такая комплексная растительность занимает большие
площади по склонам к многочисленным озерным котловинам, по древним
ложбинам стока преимущественно в центральной части Кулундинской
равнины (на карте показана одним территориальным выделом 143).
В пределах разнотравно-типчаково-ковылышх степей расположены
наиболее крупные массивы посевных площадей — озимой и яровой
пшеницы, зернобобовых и других культур. Широко возделываются картофель,
сахарная свекла, кормовые корнеплоды и огородные культуры.
Распаханные степи показаны на карте сельскохозяйственными землями,
включающими и небольшие участки степей, используемых под выпас.
Богаторазнотравно-ковыльные степи (138)
Занимали на юге Западно-Сибирской равнины широкую широтную
полосу в южной лесостепи. Они были распространены на равнинах
Зауральского плато, на озерно-аллювиальных четвертичных и неогеновых
равнинах — в южной части Ишимской, Барабинской и северной части
Кулундинской степи, представляющий восточный • отрезок ареала этих
171
степей. На равнинах с гривно-западинным. и лощинным рельефом степи
приурочены к плакорным местоположениям с обыкновенными сред-
негумусными черноземами. Степи занимают промежуточно© положение
между луговыми и настоящими степями, являясь самыми мезофильными
среди настоящих степей. Для них показательно отсутствие перибда
летнего покоя в течение вегетационного сезона, большое количество синузий,
быстрая смена многочисленных красочных аспектов [Исаченко, Рачков-
ская, 4961].
Злаки и разнотравье в рассматриваемых степях представлены видами
различной экологии. Доминирующими злаками являются корневищные
эвримезофилы (Bromopsis inermis, Elytrigia repens), корневищные ксеро-
мезофилы (Calamagfostis epigeios, Poa angustifolia), рыхлодерновинный
ксеромезофильный вид Phleum phleoides, плотнодерновинные мезоксеро-
фильные Stipa zalesskii, S. pennata n эвриксерофильные Koeleria cristatat
Festuca pseudovina.
Разнотравье состоит из большого количества мезофитов — Sanguisor-
Ъа officinalis, Lathyrus pratensis, Vicia cracca* Plantago media, Inula britanni-
ca, I. salicina, Achillea millefolium, Achyrophorus maculatus, ксеромезофи-
лов — Filipendula stepposa, Seseli libanotis, Phlomis tuberosa, Galium verumr
Fragaria viridis, Adonis vernalis и мезоксерофилов — Thymus marschallianus^
Gypsophila altissima, Hieracium echioides, Peucedanum morisonii.
Доминирующим среди разнотравья является Peucedanum morisonii*
который в богаторазнотравно-красноковыльных степях Ишим-Иртышского
междуречья, Барабы и Кулунды играет наряду со Stipa zalesskii рол]ь содо-
минанта. Брлыпое участие в составе травостоя принимают также осоки —
Сагех ргаесох и С. supina. В мезофильных вариантах сообществ этих
степей, произрастающих по окраине опушечной полосы колков, роль содо-
минантов играют Achillea millefolium, Seseli libanotis, Galium verum*
Plantago media.
В сообществах степей хорошо развита группа полыней,
представленная значительным числом видов разной экологии: ксеромезофильных —
Artemisia laiifolia, A. pontica, A. sericea, мезоксерофильных — Artemisia
macrantha, A. dracunculus, A. glauca. Более ксерофильный характер носят
богаторазнотравно-овсецово-красноковыльные сообщества с
доминированием Helictotrichon desertorum. В них многочисленны степные виды —
Сагех supina, Peucedanum morisonii, Potentilla humifusa, Salvia stepposa f
Aster alpinus, Onosma simplicissima, Seseli ledebourii, Thymus marschallia-
nus, Jurinea multiflora. Из ксеромезофилов в большом числе произрастают
Seseli libanotis, Medicago falcata, Galium verum.
На пониженных равнинных участках с солонцеватыми черноземами
развиты разнотравно-полынно-красноковыльные степи. Их основу
составляют Stipa zalesskii, S. capillata, Festuca pseudovina с небольшой примесью;
Calamagrqstis epigeios, Phleum phleoides, Bromopsis inermis, Poa stepposa.
Большим числом видов представлены полыни — Artemisia latifolia*
A. glauca, A. pontica, A. nitrosa, A. sericea, A. austriaca. Хорошо развиты
представители галофильного разнотравья — Glycyrrhisa uralensis, Galatella
biflora, G. hauptii. Богаторазнотравно-ковылыше степи полностью
распаханы, на карте на их месте показаны сельскохозяйственные земли.
Псаммофитные разнотравно-песчаноковыльные
и гемипсаммофитные
разнотравно-типчаково-тырсовые степи (139)
Эти степи довольно широко распространены в полосе настоящих раз-
нотравно-типчаково-ковыльных степей. Приурочены они к
легкосуглинистым и супесчаным разностям черноземов и песчаным почвам,
распространенным на современных и древних речных террасах Тобола и Иртышаt
а также на аллювиальных равнинах Обь-Иртышского междуречья.
Особенности псаммофильных степей связаны с условиями гидрологического
режима почв легкого механического состава — хорошей водойроницае-
172
мостью при малых потерях влаги через испарение. Поэтому растительный
покров легких почв имеет мезофильный характер по сравнению с плакор-
ными участками, В то же время однообразие физических и
гидрологических свойств легких почв в пределах зоны вызывает сходство видового
состава и структуры псаммофильных степных сообществ. В тех случаях,,
когда песчаные почвы подстилаются засоленными глинами или близко к
поверхности залегают сильно минерализованные грунтовые воды,
песчаные почвы носят признаки засоления, которые отражаются на составе
степных группировок [Калинина, 1961].
К супесчаным и легкосуглинистым разностям черноземов приурочены
сообщества, относящиеся к формациям ковыля песчаного (Stipa pennata)
и тырсы (Stipa capillata). Отличительной особенностью псаммофил ьных
сообществ является участие в них группы псаммофильных и псаммопетро-
фильнЦх видов — Onosma simplicissima, Artemisia frigida, A. marschalliana,
Helichrysum arenarium, Gypsophila paniculata, Jurinea cyanoides, Potentilla
acaulis, Cleistogenes squarrosai Spiraea hypericifolia. Среди злаков в
псаммофильных степях доминантную и эдификаторную роль играют те же
растения, как и в плакорных сообществах,— Stipa pennata, S. capillata, Festuca
valesiaca, Koeleria cristata.
В условиях , лучшего увлажнения супесчаных почв вследствие
верховодки или близкого к поверхности залегания грунтовых вод
усиливается роль рыхлодерновинных и корневищных злаков в сложении псаммо-
фитных степей. Нередко соэдификаторами в них наряду с ковылем
песчаным являются тимофеевка степная и вейник наземный. Такие сообщества
имеют более мезофильный облик и отличаются большей разнотравностью.
В разнотравно-тырсово-песчаноковыльных сообществах
доминирующее значение принадлежит Stipa pennata, S. capillata, Festuca valesiaca,
Koeleria cristata, наряду с ними в большом количестве развивается Phleum
phleoides, примешиваются Роа angustifolia, Bromopsis inermis. В группе
разнотравья наряду с типичными псаммофилами (Gypsophila paniculata,
Artemisia marschalliana) многочисленны виды, характерные для
плакорных местообитаний, Peucedanum morisonii, Potentilla humifusa, Achillea
nobilis, Artemisia latifolia.
На легкосуглинистых почвах возрастает значение ковыля Залесского,
и псаммофитно-разнотравно-песчаноковыльные сообщества замещаются
разйотравно-ковыльными с ковылем Залесского. Основа их состоит из
Stipa zalesskii, S. pennata, Festuca valesiaca, в небольшом количестве
примешиваются Phleum phleoides, Bromopsis inermis. В, травостое много геми-
(полу)псаммофилов и псаммофилов — Silene parviflora, Gypsophila
paniculata, Pulsatilla flavescens, Thymus marschalianus, Jurinea cyanoides. Хорошо
развиты полыни — Artemisia austriaca, A. latifolia, A. glauca, A. dracuncu-
lus, A. sericea, A. marschalliana .
Гемипсаммофитно-разнотравно-типчаково-тырсовые степи на легких
разностях черноземов встречаются в различных подзональных полосах
вследствие широкой, экологической амплитуды эвриксерофила Stipa
capillata. В разнотравно-типчаково-тырсовых сообществах эдификаторами
и доминантами среди злаков остаются те же растения, как и в плакорных
ассоциациях,— Stipa capillata, S. zalesskii, Festuca valesiaca, Koeleria
cristata. В примеси к ним встречаются эвримезофилы и ксеромезофилы —
Elytrigia repens, Bromopsis inermis, Calamagrostis epigeios, Poa angustifolia
и плотнодерновинный мезоксерофильный Helictotrichon desertorum. Содо-
минантами служат Artemisia sericeaf Seseli ledebourii, Carex supina.
Многочисленна группа гемипсаммофильных и псаммофильных видов —
Artemisia marschalliana, A. frigida, Centaurea sibirica, Pulsatilla flavescens,,
Jurinea cyanoides, Onosma simplicissima, Silene parviflora, Alyssum obova-
tum, Kochia prostrata, Potentilla acaulis.
173
Кальцефитные разнотравно-ковыльные
и разнотравно-ковыльно-овсецовые степи (140)
Кальцефитные варианты настоящих разнотравно-красноковыльных
степей встречаются на Ишим-Иртышском водоразделе на склонах увалов
и высоких террасах озер, н& плоских равнинных пространствах между
ними. Почвы под ними — карбонатные черноземы тяжелого механического
состава.
Индикаторное значение в сообществах кальцефитных степей
принадлежит ковылю Коржинского (Stipa korshinskyi) и овсецу пустынному
(Helictotrichon desertorum), являющимися содоминантами и соэдификато-
рами. Овсец пустынный — Helictotrichon desertorum — имеет широкий
ареал, но степные сообщества с его преобладанием встречаются редко.
В большом количестве он произрастает на карбонатных, супесчаных и
щебнистых почвах. В Заволжье, Западной Сибири и Северном Казахстане
Helictotrichon desertorum в значительном количестве встречается в степных
сообществах с Stipa penhata, Stipa zalesskii, S. lessingiana, S. korshinskyi
на карбонатных почвах [Исаченко, Рачковская, 1961].
Степи с господством ковыля Коржинского связаны только с
карбонатными черноземами. Ареал Stipa korshinskyi невелик. Этот типичный за-
волжско-казахстанский вид довольно широко распространен на юге
Западной Сибири и в Северном Казахстане в полосе настоящих разнотравно-
ковыльных и сухих типчаково-ковыльных степей.
В полосе разнотравно-ковыльных степей на юге Западной Сибири
кальцефитные степи представлены сообществами разнотравно-овсецово-
ковыльных (с ковылем Залесского и Коржинского) и разнотравно-ко-
выльно-овсецовых (с ковылями Залесского и тырсой) степей.
В разнотравно-овсецово-ковыльных степях с ковылями Залесского
и Коржинского основу травостоя составляют ксерофильные и мезоксеро-
фильные злаки — Stipa zalesskii, S. korshinskyi, Festuca valesiaca, Helicto- "
trichon desertorum, к ним примешиваются Koeleria cristata, Stipa lessingiana,
Phleum phleoides и корневищные мезофильные Elytrigia repens, Bromopsis
inermis. Группа разнотрарья состоит в основном из мезоксерофилов и
ксерофилов, типичных для разнотравно-ковыльных степей. В нем наиболее
обычны Veronica incana, Potentilla humifusa, Artemisia dracunculus, Seseli
ledebourii, Peucedanum morisonii, Pulsatilla flavescens, Galium verumy
Phlomis tuberosa, Artemisia latifolla, Eryngium planum, Achillea nobilisr
Salvia stepposa. Характерно участие в травостое растений более южных
степей Dianthus leptopetalus, Phlomis agraria, Linosyris vilosa. Бобовые
представлены Astragalus austriacus, A. onobrychis, Hedysarum gmelinii7
Medicago falcata, встречающихся в небольшом обилии.
В разнотравно-ковыльно-овсецовых сообществах из злаков
доминируют Helictotrichon desertorum, Stipa zalesskii, S. capillata, Festuca valesiaca*
в небольшой примеси имеются Koeleria cristata, Phleum phleoides, Poa
stepposa. Среди разнотравья преобладают мёзоксерофилы Peucedanum
morisonii, Seseli ledebourii, Veronica spicata, Galatella hauptii, Artemisia
latifolia, A. glauca. В небольшом обилии постоянно присутствуют
корневищные осоки — Carex supina, С. ргаесох. Степные эвксерофилы
представлены Goniolimon speciosum, Dianthus leptopetalus, Potentilla humifusa.
Из гемиэфемероидов встречаются Astragalus testiculatus, Nepeta ucrainicay
Serratula nitida. Степень проективного покрытия почвы в этих степных
сообществах 70—80%.
Разнотравно-типчаково-ковыльные степи (141)
Эти степи по сравнению с богаторазнотравно-ковыльными являются
более ксерофильными.-Их распространение приурочено к южной окраине
Прииртышской равнины и к центральной ча^ти Кулундинской степи.
В отличие от богаторазнотравно-ковыльных сообществ в разнотравно-
типчаково-ковыльных степях значительно большее развитие имеют плот-
174
нодеряовинные злаки. К ним относится основной эдификатор Stlpa zales-
skii, Festuca valesiaca, Koeleria cristata и встречающиеся в примеси Helecto-
trichon desertorum и % Stipa capillata. Рыхлодерновинные и корневищные
злаки Phleum phleoides, Bromopsis inermis имеют незначительное участие,
В группе разнотравья доминирует мезоксерофильный Peucedanum mori-
sonii или ксеромезофильные Seseli libanotis и Filipendula vulgaris. Богата в
видовом отношении группа полыней — Artemisia austriaca, A. glauca,
A. pontica, A. sericea, A. latifolia, A. marschalliana.
В некоторых сообществах разнотравно-красноковыльных степей
наряду со Stipa zalesskii роль соэдификатора осуществляет S. capillata.
Для таких степей характерно развитие полыней — Artemisia latifolia,
A. dracunculus, A. glauca, A. austriada. В мезофильных вариантах разно-
травно-тырсово-красноковыльных степей доминируют Phleum phleoides,
Filipendula vulgaris, Peucedanum morisonii, P. ruthenicum, Astragalus
onobrychis, в ксерофильных — преобладание остается за ковылями и
типчаком. В травостое вместе со Stipa capillata появляется значительное
число типично степных ксерофильных видов Dianthus leptopetalus, Iris scariosa,
Sisymbrium polymorphum, Jurinea multiflora, Crinitaria villosa, Calatella
hauptii и мезоксерофильных Peucedanum alsaticum, Adonis wolgensis,
Potentilla nudicaulis, Astragalus testiculatus, A. buchtormensis.
Разнотравно-типчаково-тырсовые степи (142)
Степи со Stipa capillata и S. pennata на юге Западной Сибири имеют
распространение на плакорах Кулундинской равнины и на Иртыш-Омском
междуречье. Почвы под ними южные и обыкновенные черноземы от
тяжелого до легкого механического состава. Эдификаторная роль в этих степях
принадлежит плотнодерновинным ксерофилам Stipa capillata и Festuca
valesiaca, в качестве постоянных доминантов выступают Koeleria cristata .
и мезоксерофильный Stipa pennata. Характерно присутствие
разнообразных полыней — Artemisia glauca, A. dracunculus, A. scoparia, A. pontica,
A. latifolia, A. sericea, A. marschalliana, A. austriaca, степных
кустарников — Spiraea hypericifolia, S. crenata.
На легкосуглинистых и супесчаных разностях южных черноземов
среди злаков в большом количестве появляется Helictotrichon desertorum.
Из корневищных злаков в таких степях присутствуют * Bromopsis inermis
и Calamagrostis epigeios, в некоторых, ценозах довольно хорошо
развиваются рыхлодерновинные Phleum phleoides и Cleistogenes squarrosa —
показатель легких почв, В разнотравье доминирующая роль принадлежит
Artemisia austriaca, меньше^ развитие имейт Artemisia glauca и A.
marschalliana, единично встречаются Potentilla bifurca, Seseli ledebourii, Veronica
spicata, Onosma simplicissima, Thymus marschallianus, Hieradium virosum%
H. echioides, Helichrysum arenarium, Orostachys spinosa. Из бобовых
постоянно участие Medicago falcata, Astragalus onobrychis [Вандакурова, 1950].
Среди сообществ тырсовых степей наибольшее распространение
имели разнотравно-тонконогово-тырсовые и полынно-типчаково-тырсовые.
В разнотравно-тонконогово-тырсовых степях основу составляют плот-
нодерновинные ксерофилы Stipa capillata, Koeleria cristata, Festuca
valesiaca и луговые мезофильные и ксеромезофильные корневищные злаки
Bromopsis inermis, Festuca pratensis, Poa angustifolia. Травостой составлен
в основном степными мезоксерофильными видами, произрастающими в
небольшом количестве, Carex supina, Peucedanum alsaticum, Salvia stepposaf
Astragalus testiculatus, Plantago stepposa, Calatella hauptii, Erysimum
hieracifolium, Seseli ledebourii. Род полыней представлен в них большим
количеством видов — Artemisia austriaca, A. dracunculus, A. glauca, A.
latifolia, A. pontica, A. rupestris,A. sericea, встречающихся единично.
Характерно присутствие в составе сообществ галофильных видов — Plantago
cornuti, Kochia prostrata, Calatella biflora и ксеромезофильного степного
кустарника Spiraea crenata. Степень проективного покрытия почвы
травостоем в сообществах 40—55 %, количество видов на 100 м2 достигает 50—60,
175
- Полынно-типчаково-тырсовые степные сообщества более галофиль-
ны. Их основа составлена доминирующими Festuca valesiaca и Stipa capilla-
ta с участием Calamagrostis epigeios. В разнотравье большое развитие
получают полыни разной экологии — Artemisia latifolia, A. pontica, A. glauca,
A. sericea, A. austriaca, A. frigida, A. nitrosa. Единично встречаются Fili-
pendula vulgaris, Inula salicina, Phlomis tuberosa, Seseli libanotis, Peuceda-
num morisonii, Phlomis tuberosa, Seseli libanotis, Peucedanum morisonii.
Количество видов на 100 м2 составляет 30—40.
В юго-восточной части Кулунды разнотравна-типчаково-тырсовые
степи носят черты ксерофилизации: эдификаторами являются Stipa capil-
lata, S. zalesskii, Festuca valesiaca, соэдификаторное значение принадлежит
Artemisia austriaca. Среди разнотравья появляются ксерофильные Ferula
caspica, Dianthus ramosissimus, Allium clathratum, Phlomis agraria, Paeonia
hybrida, Iris scariosa, Nepeta ucrainica, Syrenia siliculosa, Astragalus stenoce-
ros, Ephedra distachya. Ксеромезофильные виды, как Seseli libanotis,
Filipendula vulgaris, Pulsatilla flavescens, приурочены к понижениям
микрорельефа, Описание таких степей дано Е. В. Вандакуровой [1950].
Небольшие участки целинных тырсовых степей можно встретить
узкими полосами среди'пахотных земель. Вблизи населенных пунктов эти
степи в результате постоянного интенсивного выпаса превратились в тип-
чаковые и типчаково-полынные с Artemisia austriaca и A. frigida, Festuca
pseudovina и постоянно присутствующей Carex supina. При еще большей
пастбищной нагрузке угнетаются и эти растения, заменяясь сорными
однолетниками Polygonum aviculare, Atriplex tatarica, а на легких почвах
Ceratocarpus arenarius.
Галофитно- разнотравно -типч&ковые
и коЬыльно-типчаковые степи
с галофитно-степными сообществами (143)
В полосе распространения зональных разнотравно-типчаково-ко-
выльных и типчаково-ковыльных степей на Кулундинской равнине
целинная степная растительность сохранилась лишь в депрессионных формах
мезорельефа в пределах комплексного почвенного покрова с
солонцеватыми разностями темно-каштановых почв или южных черноземов и солонцов.
На солонцеватых почвах и глубоких солонцах Е. В. Вандакурова
[1950] выделила галофитную разность зональных степей — разнотравно-
типчаковые степи с Glycyrrhiza uralensis. В их составе преобладают Festuca
valesidca, Stipa capillata, Koeleriacristata, в небольшом количестве
присутствуют Phleum phleoides, Calamagrostis epigeios, Leymus secalinus. Богато
' видами разнотравье с господством полыней — Artemisia frigida, A. glauca,,
A. campestris, A. nitrosa, A. austriaca,' A. dracunculus и постоянным
участием галоксерофилов -— Calatella hauptii, G. macrosciadia, Polygonum
pabulum, Limqnium gmelinii, L. coralloides и др. Пятнистость травостоя создает
солодка Glycyrrhiza uralensis, образующая небольшие куртины.
На средне- и особенно высокостолбчатых солонцах, которые
постоянно входят в комплекс с солонцеватыми темно-каштановыми почвами%
развиваются типчаково-полынные сообщества с участием
пустынно-степных полукустарничков и южных ксерогалофильных растений — Kochia
prostrata, Artemisia schrenkiana, A. gracilescens,A. nitrosa, Goniolimon speci-
osum, Limonium gmelini,r L. coralloides.
Разнотравно-типчаково-тырсовые
и разнотравно-типчаково-ковылковые степи (144)
Сочетание ксерофитно-разнотравно-типчаково-тырсовых и ксерофит-
но-разнотравно-типчаково-ковылковых степей характерно для юго-
вападной части Кулундинской равнины в каштановыми почвами преиму-
176
щёственно супесчаного и легкосуглинистого механического состава. С
легкими почвами связано широкое распространение сообществ тырсовых
степей, описанных в Кулундинских степях Е. В. Вандакуровой [1950J. *
Развитию тырсовых степей в прошлом способствовал также и постоянный
рыпас. В условиях интенсивного выпаса сохранились наиболее стойкие
растения — Stipa capillata и Festuca valesiaca.
Разнотравно-типчаково-тырсовые степи в южной полосе их
распространения отличаются большей ксерофитностью, уменьшением роли
разнотравья, меньшей видовой насыщенностью фитоценозов (20—35 видов
на 100 м2), участием эфемеров, эфемероидов и гемиэфемероидов. Травяной
покров низкий и разреженный, проективное подрытие 30—50%. Основу
травостоя этих степей составляют плотнодерновинные злаки — Stipa
capillata и Festuca valesiaca, в примеси встречаются Stipa lessingiana,
S. sareptana, Koeleria cristata и рыхлодерновинная Cleistogenes squarrosa.
Среди разнотравья и полукустарничков преобладают полыни — Artemisia
frigida, A. austriaca, A. glauca, A. marschalliana, другие виды встречаются
единично — Dianthus ramosissimus, D. leptopetalus, Hieracium echioides,
Potentilla acaulis, Ephedra distachya. Несколькими видами представлены
коротковегетирующие многолетники (геми эфемероиды) — Astragalus
testiculatus, A. buchtormensis, Serratula nitida, Ferula caspica, весной
появляются эфемероиды — Gagea pusieea, Valeriana tuberosa.
В годы с влажным вегетационным периодом отмечалось развитие
малолетников. Среди однолетников представлены как эфемеры Ally sum
desertorum, Erophiea verna, Androsace septentrionalis, так и летне-осенние
однолетники Ceratocarpus arenarius, Bassia sedoides. Из двулетников
присутствуют Trinia ramosissima, Erysimum marschallianum, Syrenia siliculosa.
Характерно постоянное произрастание кустарников, образующих
небольшие куртины в микропонижениях, Spiraea hypericifolia, S. crenata, реже
Caragana frutex.
На фоне разнотравно-типчаково-тырсовых степей отдельными
островами были распространены ксерофитно-разнотравно-типчаково-ковылко-
вые степи с доминированием Stipa lessingiana. Они занимали небольшие
плоптди на плоских увалах между боровыми лощинами (Коростелевская
и Узкая степь) с каштановыми суглинистыми почвами. Эти районы —
восточный предел распространения ковылковых степей в Западной Сибири,,
являющихся зональным типом в подзоне сухих типчаково-ковыльных
степей. Эдификаторная роль в ковылковых степях принадлежит Stipa
lessingiana, S. capillata, Festuca valesiaca; Stipa zalesskii принимает лишь
небольшое участие. Среди немногих видов разнотравья характерно
присутствие таких эвксерофилов, как Phlomis agraria, Dianthus leptopetalus,
D. ramosissimus, а из полукустарничков — Artemisia austriaca, Kochia
prostrataf Ephedra distachya. Бобовые представлены гемиэфемероидами —
Astragalus testiculatus, A. buchtormensis. Травяной покров ковылковых
степей,, разреженный, проективное покрытие не превышает 25—30%,
количество видов на 100 м2 не более 25—35 [Вандакурова, 1950].'
ГЛАВА X
РАСТИТЕЛЬНОСТЬ РЕЧНЫХ ДОЛИН
Растительность долин рек Западно-Сибирской равнины отличается
чрезвычайным разнообразием. Многочисленные ряды пойменных
ассоциаций слагают различные по составу и структуре сообщества древесной*,
кустарниковой и луговой растительности. Основная черта пойменных
местообитаний — их динамичность, т. е. быстрые смены во времени и
пространстве. Отсюда главное свойство растительности пойм —
неустойчивость и слабая сформированность ее состава и структуры. Большую
часть пойменных сообществ мы относим к динамической категорий
серийных [Сочава, 1963] наименее устойчивых, формирующихся в условиях
12 Заказ № 297 177
постоянно изменяющегося, большей частью избыточного увлажнения;
в условиях меньшего увлажнения, например на высоких уровнях поймы?
формируются более устойчивые сообщества — субклимаксовые
[Городков, 1946], преимущественно лесные, приближающиеся по своему составу
к водораздельным типам соответствующей зональной растительности
(климаксовым типам).
Основными объектами изучения и картографирования пойменной рас*
тительности равнины являлись поймы двух крупнейших рек — Оби и
Иртыша, а также долинная растительность их притоков — Тобола, Иши-
ма, Салыма, Васюгана и др.
Поймы Оби и Иртыша по сравнению с поймами других рек
Советского Союза имеют свою специфику. Их своеобразие заключается прежде все-
то в огромных размерах поймы, в исключительном по продолжительности
и величине весенне-летнем паводке, в формировании чрезвычайно
сложного по структуре и динамическим * свойствам растительного
покрова.
Чтобы понять закономерности "территориального распределения
пойменной растительности, необходимо выявить те факторы среды, которые
определяют его пространственную выраженность, основные фитоценотиче-
ские, экологические и динамические черты. Как показал опыт изучения и
картирования пойменной растительности, наибольшее влияние на ее-
состав, структуру и пространственное распределение оказывают режим
половодья рек, относительная высота, возраст и генезис элементов
пойменного рельефа [Барышников, 1933; Роднянская, 1968; Ильина и др.,
1977; и др.]. Рассмотрим несколько подробнее вл'ияние этих природных
компонентов на растительность Обь-Иртышской поймы.
Пойма в геоморфологическом смысле — это часть дна речной долины,
затапливаемой в половодье. Все особенности ее строения хорошо
прослеживаются на разных типах пойменных массивов х. Массивы пойм Оби и
Иртыша в основном представлены системой валов и межваловых
понижений, характер и расположение которых меняются на разных участках
поймы. Для Нижней и Средней Оби, а также для низовий Иртыша
характерно формирование островной поймы [Еленевский, 1936]. Пойменные
массивы островной поймы отделены друг'от друга, кроме основного русла,
множеством проток и вторичных водотоков и образуют целую сеть
островов, осередков, останцов обтекания и т." п. Внутри пойменных массивов
преобладают обширные плоские и слабовогйутые соровые понижения,
чередующиеся с прирусловыми гривами вдоль небольших проток. Для
среднего и верхнего течения Иртыша и для Верхней Оби характерен
другой морфологический тип поймы — сегментно-гривистый [Еленевский,
1936; Петров, 1979]. Основную часть пойменных массивов здесь занимают
сочетания грив и межгривных понижений разной высоты, ширины и
глубины вреза. Боле§ или менее плоские поверхности гораздо меньшей
площади встречаются только в притеррасных частях или в старых сегментах
центральной поймы (рис. 17).
Высота элементов пойменного рельефа — важный показатель,
влияющий на распределение пойменной растительности. В зависимости от
высоты рельефа разные участки поймы попадают в различные условия поем-
ности и аллювиальностп и неодинаково формируют свой растительный
покров. Превышение относительных высот только на 0,5 м существенно
меняет состав и свойства пойменной растительности. По относительной
высоте над меженным урезом воды на участках поймы Оби и Иртыша
выделяются ярусы (уровни) рельефа — высокий, средний, низкий [Кузьмин,
Русаков, 1978; Петров, 1979].
Для понимания пространственных различий в составе растительности
при изучении рельефа необходимо учитывать возраст и положение
«отдельных его элементов по отношению к русловым потокам. Последнее в
1 Под пойменным массивом понимается участок поймы, заключенный в дугу
излучины русла и формирующийся по мере роста ее кривизны [Петров, 1979].
178
Рис. 17. Растительность низкой и средней поймы р. Оби. Фото В. А. Богоявленского.
значительной степени определяет интенсивность осадконакопления —
состав и мощность аллювия, являющегося здесь основной почвообразую-
щей породой.
Существует деление поймы по поперечному профилю на
экологические зоны — прирусловую, центральную, притеррасную [Вильяме,
1946], которое хорошо отражает ее морфологическое строение и широко
используется всеми геоботаниками, изучающими поймы рек. Это деление
оказывает сильное дифференцирующее влияние на растительность пойм
Оби и Иртыша, главным образом в их средних и верхних течениях, где
реки теряют многорукавность и их растительный покров находится под
воздействием основного руслового потока.
Возраст рельефа и история его формирования накладывают отпечаток
на режим поемности и, следовательно, на состав растительности.
Достаточно сказать, что на одинаковых по высоте элементах рельефа формируются
разные по составу и динамическим свойствам растительные сообщества.
Например, на высоких гривах в молодых частях пойменных массивов
Нижнего Иртыша произрастают березово-ивовые молодые и
средневозрастные злаково-разнотравные леса, а на таких же по высоте гривах
старых частей — зрелые и перестойные березово-сосновые с шиповником
разнотравные леса. В зависимости от возраста, степени сформированности
и устойчивости на разных массивах Обь-Иртышской поймы
выделяются молодые, Средневозрастные и старые (зрелые) сегменты поймы
[Петров, 1979].
Режим поемности — основной гидроклиматический фактор,
определяющий состав и свойства пойменной растительности и ее связь с
особенностями рельефа и осадконакопления. Частота и длительность заливания
водами весеннего половодья различны на разных отрезках
Обь-Иртышской поймы [Петров, Бачурин, 1976], а внутри пойменных массивов
они имеют определенную амплитуду колебаний в соответствии с высотой
пойменного рельефа и другими факторами геоморфологического строения.
Характеристика режима поемности разных участков поймы — задача
чрезвычайно сложная и трудоемкая. В настоящее время еще нет достаточ-
12* 179
но подробных, хорошо сопоставимых данных для всей Обь-Иртышской
поймы. Наиболее пригодные для решения поставленных задач
гидрологические сведения о режиме поемности содержатся в работах А. А.
Максимова и Е. П. Мерзляковой [1972], И. Б. Петрова и Г. В. Бачурина [1976]
к И. Б. Петрова [1979].
Материалы этих работ свидетельствуют, что длительность заливания
и сроки начала — конца половодья сильно отличаются на разных отрезках
пойм Оби и Иртыша (от устья вверх цо течению). Кроме того, эти
показатели имеют резкие различия и внутри пойменных массивов в зависимости
от приуроченности к высотным ярусам рельефа. В конечном итоге
особенности режима половодья целиком определяют условия вегетации
пойменной растительности —* продолжительность прохождения ею отдельных
фаз развития. Это существенно влияет на ее продуктивность и возможные
сроки сельскохозяйственного использования (сенокошение, выпас).
Приведенные сведения о гидролого-геоморфологических условиях
Обь-Иртышской поймы указывают на тесную связь, существующую между
рельефом и режимом поемности в пойменных ландшафтах. Если
рассматривать эти факторы с точки зрения условий формирования растительности,
то целесообразно лх объединить и привести к широко применяемым в
ботанической географии понятиям «экологический уровень», или «высотный
пояс поемности» [Шенников, 1941]. Экологические уровни выделяются
в зависимости от продолжительности затопления и соотношения высот*
основных элементов рельефа по соответствующим экологическими
диапазонам растительности — группам (рядам) пойменных сообществ,
свойственных данным условиям поемно-аллювиального режима. В поймах
Оби и Иртыша исследоватеди выделяют 3—4 экологических уровня
[Барышников, 1933; Роднянская, 1963, 1968; Ильина, 1976].
Дифференциация пойменного растительного покрова по
экологическим уровням, связанным в таежной зоне главным образом с режимом
поемности и рельефом,— это наиболее важная особенность его
пространственной структуры, которая должна найти отражение на картах. Другая
не менее важная особенность, которую также необходимо выявить и
показать на карте — это зональность пойменной растительности, ее связь
с общими биоклиматическими различиями. Весь растительный покров
Западно-Сибирской равнины четко Дифференцируется по зональным и
подзональным типам, что нашло отражение на карте [Растительность...,
1976]. Пойма — интразональное явление, однако в особенностях фито-
ценотического состава, экологических и динамических свойствах ее
растительного покрова достаточно четко проявляются зональные черты.
Наиболее характерными показателями зональных типов пойменной
растительности являются сообщества заключительных стадий пойменных серий.
Они приурочены к высокий* редко заливаемым поверхностям старых,
преимущественно притеррасных частей пойм. Это полностью
сформировавшиеся относительно устойчивые (субклимаксовые) сообщества, которые по
фитоценотическому составу близки к зональным типам водораздельной
растительности, но отличаются от последних присутствием более
влаголюбивых луговых элементов. Определенные черты зональности
прослеживаются в спектрах высотно-экологических уровней разных отрезков
поймы; в их пределах изменяются состав сообществ эколого-динамических
рядов каждого уровня, соотношения площадей луговых, кустарниковых
и лесных фитоценозов, экологические свойства пойменных сообществ
северной и средней тайги — они занимают более высокие экологические
уровни,чем в поймах южной тайги и т. п.
Описание растительного покрова пойм рек Западно-Сцбирской
равнины построено в соответствии со структурой легенды карты — по
наиболее крупным подзональным типам серий пойменной растительности, в
пределах которых характеризуются картируемые эколого-динамические
ряды сообществ, приуроченные к поверхностям разного экологического
уровня.
180
ТУНДРОВЫЕ АРКТИЧЕСКИЕ И СУБАРКТИЧЕСКИЕ
БОЛОТНО-ЛУГОВО-КУСТАРНИКОВЫЕ СЕРИИ
РАСТИТЕЛЬНЫХ СООБЩЕСТВ
Для долинных серий рек крайнего севера Западно-Сибирской
равнины характерно преобладание болотисто-луговой и болотной
растительности на низких эколргических уровнях и развитие тундровых моховых
и кустарниковых сообществ на высоких поверхностях поймы и
прилегающих участках надпойменных террас.
Особенность пойменных серий тундровой растительности — тесная
связь их растительных сообществ с процессами промерзания и протаива-
ния грунтов. Последовательные смены растительности, происходящие в
поймах северных рек, сопровождаются постепенным уменьшением
глубины протаивания и понижением температуры почвы. Так, по данным
А. П. Тыртикова [1974], глубина протаивания под ерниково-лишайниково-
моховыми сообществами высоких уровней поймы в 2,5—4 раза меньше,,
чем под зарослями арктофилы на низкой пойме. В результате на
заключительных стадиях сукцессии нередко нижние слои почвы переходят в вечно-
мерзлое состояние.
Динамика растительного покрова в долинах рек тундровой зоны
изучалась Б. Н. Городковым [1932], ,А. П. Тыртиковым [1970, 1974],
Л. И. Мельцер [1977].
Арктическая серия разнотравно-осоковых
и разнотравно-злаковых лугов
и мелкоивняковых сообществ в сочетании с участками
травяно-моховых тундр и осоково-гипновых болот (9)
Начальными стадиями большинства пойменных серий в подзоне
арктических тундр Западной Сибири являются заливные осоковые луга,
которые быстро сменяются разнотравно-злаковыми и разнотравно-кустарнич-
ковыми сообществами и участками травяно-моховых тундр. В пойме
р. Яруяха, например, осокой (Carex concolor) занята узкая полоса
прирусловой части, затем идут чередующиеся между собой участки
разнотравно-осоковых (Carex concolor, Caltha arctica, Ranunculus borealis, Pedicularis
sudetica) и разнотравно-злаковых (с Trisetum sibiricum ssp. litorale) лугов
с редкими кустиками угнетенной ивки, местами на повышенных участках
они переходят в мелкоивняковые сообщества (Salix lanata). В
притеррасных частях поймы на месте зарастающих стариц и пойменных озер
развиваются осоково-гипновые болота. Помимо осоки (Carex concolor, С. ade-
lostoma) в травяном покрове присутствует Caltha arctica. Основу мохового
покрова составляют Calliergon sarmentosum, Drepanocladus exannulatus,
Aulacomnium palustre. По периферии на кочках сфагнума поселяются
зеленые мхи и печеночники (Dicranum angustum, Ptilidium ciliare), а также
некоторые травянистые виды и редкие экземпляры угнетенной ивы (Salix
lanata). Эти болота обычно небольших размеров, но в общей сложности в
долинах арктических рек они занимают значительные площади.
Наиболее возвышенны^ и хорошо дренированные участки поймы
р. Яруяха заняты разнотравно-кустарничковыми (Potentilla uniflora%
Pedicularis lapponica, Luzula confusa, Arctous alpina, Dryas punctata) и
кустарничково-осоковыми (Carex ensifolia ssp. arctisibirica, Vaccinium
minus, Dryas punctata, Arctous alpina) сообществами с хорошо развитым
моховым покровом (Dicranum angustum, Ptilidium ciliare).
Заключительными стадиями пойменной серии являются Относительно
устойчивые субклимаксовые сообщества травяно-моховых тундр (Carex
concolor).
181
Приморские засоленные злаково-осоковые
заболоченные луга ( «тампы» ) (8)
На низких побережьях Карского моря, особенно в устьях крупных
рек Арктики и Северной Субарктики, развиты засоленные приморские
луга — «тампы», затопляемые во время приливов слабо солонцеватой водой.
Как и в речных долинах, здесь развивается серийная растительность и
сукпессионные\смены идут по типу гидросерий пойм рек. Однако по
составу эта растительность-отличается от долинной. Приморские тампы на
побережье пролива Малыгина изучались В. С. Михайличенко [1936], в
заливе Шаронов Шар — М. Н. Аврамчиком [1969]. Наиболее полно сук-
цессионные серии низинных побережий Гыдана описаны Б. Г. Городко-
вым [1932J. Начальная стадия их зарастания — заросли Arctophila fulva,
Eriophorum medium, с участками Drepanocladus uncinatus. Дальше от воды
они сменяются болотистыми пушицево-злаково-о'соковыми лугами из
Carex concolor, С. subspathacea, Dupontia fisheri, Arctophila fulva,
Eriophorum medium, E. polystachyon. На более дренированных и менее засоленвГых
местообитаниях появляются Alopecurus alpinus, Calamagrostis neglecta,
разреженно — Salix reptans, пятнами встречаются мхи, главным образом
Drepanocladus uncinatus. Заключительной стадией этой серии, так же как
и в долинах рек, являются сообщэства. кустарничково-моховых тундр,
которые на всем протяжении сочетаются с участками полигональных осо-
ково-гипновых бойот.
Субарктическая серия сообществ разнотравно-злаковых лугов,
ивняков кустарниковых и моховых тундр
в сочетании с осоково-гшгаовыми
валиково-полигональными болотами (16)
Эта серия долинной растительности отнесена к субарктическому
типу. Она характерна для рек полосы северных (типичных) моховых 'тундр
и описана нами в долинах рек Сеяха (п-ов Ямал) и Сыдыяха (Гыдадский
полуостров). Начальные ее стадии представлены или пионерными
группировками из Equisetum arvense, Veratrum lobelianum, Hedysarum arcticum
на песчаном аллювии, или хвощово-пушицево-злаковыми {Calamagrostis
neglecta, Poa alpigena, Eriophorum polystachyon, Equisetum arvense)
сообществами на зарастающих илистых наносах. Более устойчивы и широко
распространены разнотравно-злаковые луга из Alopecurus alpinus, Poa
alpina, Calamagrostis holmii, Ranunculus borealis, Pedicularis sudetica, Та-
nacetum bipinnatum. На следующей стадии появляются
низкокустарниковые ивняки из Salix lanata, S. reptans, S. phylicifolia; в их покрове кроме
разнотравья и злаков обычны пятна зеленых мхов (Aulacomnium turgidum,
Pleurozium schreberi) и сфагнов {Sphagnum warnstorfii).
На более .высоком пойменном уровне развиваются ивнячково-ернич-
ковые злаково-осоково-кустарничково-моховые {Aulacomnium turgidum,
Rhacomitrium lanuginosum, Empetrum hermaphroditum, Dryas punctata%
Carex acuta, Alopecurus alpinus) кочковатые тундры, которые являются
заключительными сообществами пойменной серии.
Большие пространства в поймах рек занимают осоково-гипновые
болота, развивающиеся в результате зарастания многочисленных пойменных
озер и стариц. Основу их напочвенного покрова образуют Drepanocladus
revolvens, D. uncinatus, Calliergon stramineum, Aulacomnium palustre, в
травяном ярусе преобладают Carex concolor, C. chordorrhiza, Eriophorum polys-
tachyon. ' * ,
Субарктическая серия хвощовых на песках
и разнотравно-злаковых на суглинках лугов,
кустарниковых ивняков и ивняково-ерниковых
ольховниковых тундр (30)
«
В поймах рек полосы южных (кустарниковых) субарктических тундр
распространена серия сообществ, начальные стадии которой по составу
очень близки к вышеописанным для серий долин рек северной тундровой
182 ' -
полосы. В отличие от последних здесь менее выражена луговая стадия и
уже на низких уровнях появляются группировки ивняков-
кустарниковых (Salix lanata, «S. reptans, S. phylicifolia), которые занимают
значительные площади в центральных частях пойм. Наиболее распространены
ивняки травяные, в покрове которых преобладают, вейник (Calamagrostis
langsdorffii), арктагростис (Arctagrostis arundinacea), хвощ (Equisetum pra-
tense), подмаренник (Galium palustre), пушица (Eriophorum polystachyon),
незабудка (Myosotis palustris), мятлик (Poa pratensis), синюха (Polemonium
acutiflorum), горец (Polygonum viviparum), пиретрум (Tanacetum bipinna-
tum), лютик (Ranunculus repens), княженика (Rubus arcticus), чемерица
(Veratrum lobelianum), вероника (Veronica longifolia), фиалка (Viola epip-
sila). В ивняках накапливается достаточно снега, чтобы предохранить
почву от глубокого промерзания. Сезонно-мерзлый слой ее обычно ежегодно
протаивает полностью, и вечная мерзлота под ивняками деградирует [Тыр-
тиков, 1974J. В таких ивняках развивается подрост ольхи, и на следующей
стадии ивняки сменяются ольшаниками.
На периодически заливаемых участках поймы формируются
ольшаники травяные, сходнее по составу с вышеописанными ивняками, но
имеющие более разреженный травяной покров. В местах, где почти
прекращается отложение наилка в половодье, на высоких экологических уровнях
развиваются ольшаники, а затем ерники зеленомошные, в которых
постепенно формируется слой мерзлоты.
По описаниям А. П. Тыртикова [1974], в ерниках зеленомошных
карликовая береза высотой до 60 см образует редкий ярус (сомкнутость
полога 0,2—0,5), в котором в значительном количестве присутствуют ивы
(Salix glauca, S. pulchra). В травяно-кустарничковом ярусе преобладают
водяника, морошка,- голубика; обильны багульник, брусника; единично
встречаются арктагростис, вейник. Сплошной покров образуют зеленые
мхи и сфагнумы — Hylocomium splendens var. alaskanum, Aulacomnium
turgidum, Polytrichum alpestre, Sphagnum angusti}olium,Dicranum spadiceum.
Заключительными и наиболее устойчивыми стадиями этой пойменной
серии являются подобные вышеописанным ивняково-ерниковые с
ольхой (Betula nana, Dryas punctata, Salix lanata) кустарничково-лишайнико-
во-зеленомошные (Hylocomium splendens var. alaskanum, Drepanocladus
uncinatus, Dicranum elongatum, D. angustum, Cladina rangiferina)
сообщества, близкие по составу к тундровым водораздельным типам полосы
южных субарктических тундр. Они были описаны нами в долине р. Анти-
Паюта (Гыдан).
Дальнейшее развитие и увеличение обилия мхов ведет, по мнению
А. П. Тыртикова -[1974], к смене кустарниковых тундровых сообществ в
долинах рек кустарничково-дикрановыми тундрами. Этот процесс
сопровождается формированием вечной мерзлоты, понижением температуры
почв, уменьшением глубины протаивания.
Серия гипново-осоковых болот, болотистых осоковых лугов,
ивняковых, ерниковых и ольховниковых тундр (29)
Эта наибольшая по площади серия пойменной растительности
тундрового субарктического типа представлена в дельте р. Оби, а также в устке-
вых участках крупных северных рек ~ Надым, Пур, Таз.
Дельта Оби — это огромный архипелаг различных по величине
островов, площадь которых колеблется от 0,6 до 163 км2 (о. Хаманельский).
Началом дельты считается о. Бол. Яры, делящий Обь на два больших
рукава — надымскую Обь и хаманельскую Обь. Ширина дельты у мыса
Ямсале достигает почти 60 км.
На территории дельты Оби выделяются участки трех пойменных
уровней — низкого, среднего и высокого (рис. 18).
Низкая пойма наиболее широко представлена в пределах дельты.
Относительная высота ее над меженью не превышает 1,5—1,8 м. Сложена
она обычно чередующимися между собой слоями суглинков и супесей*
183
ю-с
10-
20-21 I 22 25
I И
1\2Ъ 4 \5\6\1\8\9\10\11\12\ 13
I I
| I I I I I I I I
nil L\.J J i
i
Рис. 18. Геоботанпческий профиль в дельте р. Оби, район
Растительные сообщества: 1 — пушицево-юсоковое с кустами ивы и ерника; 2 — за
ково-сфагновое; 5 — багульниково-сфагновое; 6 — багульниково-лишайниковое; 7 — багульниково-
ково-морошково-сфагново-зеленомошное; 10 — комплекс низкокустарничково-сфагново—зеленомош
лишайниково-зеленомошных сообществ по буграм и багульниково-зеленомошно-сфагновых по за
травно-осоковым покровом; 14,— отдельные экземпляры пушицы и осоки в воде (протока); 15 — ба
ково-сфагновое; 18 — багульниково-зеленомошно-лишайниковое; ,19 — багульниково-морошково-
ково-сфагновое; 22 — багульниково-пушипево-лишайниковое с ерником; 23 — осоково-пушицево-
вых и осоковых группировок по трещинам; 25 — багульниково-пушицево-сфагнбвое; 26 — осоково-
но-лишайниковое; 29 — лишайниково-багульниково-морошково-сфагновое; 30 — осоково-пушице
цевое со сфагнумом; 33 — багульниково-морошково-сфагновое с лишайниками; 34 — бугристо-за
ерниково-багульниково-сфагновое; 36 — ерниково-ивняковое; 37 — ерниково-багульниково-сфаг
Доминирующие виды: а — ольха; б — ива; в — ерник; г — пушица; д — осока; е — ба
новые мхи; н — лишайники
На поверхности низкой поймы развит кочкарный микрорельеф. Кочки
имеют высоту 0,5—0,7 м, диаметр 2—5 м. Между ними микрозападины
шириной не более 1 м. В обширных понижениях дельтовых островов,
часто образованных руслами «бездействующих» проток, формируются
болота. В центральных частях островных массивов дельты наблюдаются также
соровые понияюния, они имеют овально-округлую форму и глубину до
2—3 м. В половодье соры заливаются водой, а при низких уровнях
осушаются. С рукавами дельты соры обычно соединяются неглубокими
(до 3 м) узкими протоками. Вдоль берегов современных проток и рукавов
имеются прирусловые валы, по режиму поемности относящиеся к
среднему пойменному уровню, высота их над поверхностью низкой поймы 0,6—
1 м. Между валами понижения, к которым приурочены небольшие озера
или болота. Высокая пойма (3—4 м) развита в основном в западной
(вершинной) части дельты, но нигде не образует больших массивов; сильно
изрезана системой протоков и ложбин.
Вся дельта Оби находится в области распространения вечной
мерзлоты. Мощность многолетнемерзлых пород от 10 до 300 м. Минимальные
мощности наблюдаются в низкой пойме (от 0 до 50 м). Выше слоя многолетне-
мерзлых пород находится слой сезонно-талых грунтов. Максимальное про-
таивание характерно для песчаных пород (до 2 м), которыми сложены
прирусловые гривы в пойме [Трофимов, 19751.
Низовья Оби характеризуются сильно растянутым весенне-летним
половодьем. В период половодья подъем уровня в дельте достигает 2 м.
Наиболее многоводные годы повторяются 1 раз в 18—20 лет, тогда
заливается высокая пойма. Средние уровни обычно затапливаются в годы со
средними паводками, низкие — ежегодно и на длительный срок (с конца
мая до начала сентября). Указанные различия в рельефе, длительности в
сроках .половодья обусловливают четкую дифференциацию растительного
покрова по экологическим уровням. . -
Для участков низкого экологического уровня, расположенных в
центре дельтовых островов, характерны ряды пойменных сообществ, главными
компонентами которых являются низинные болота и соровые болотистые
луга (29а).
Болота здесь образуются на месте бессточных проток при их
обмелении и постепенном зарастании. Они, как правило, сильно обводнены и за-
кочкарены. Кочки образованы осокой водной (Carex aquatilis) и дернистой
(Carex cespitosa), можкочечные понижения заняты травяно-гипновыми
184
О 50М
i i
\ a W Ч в
} г I д / е
/®Ж $ 3 $ U
с. Панаевска (составлен И. С. Ильиной, А. В. Васильковым).
росли ерника с одиночными кустами ивы; г — осоково-пушицевое; 4 — ерниково-ольховое морош*
бруснично-лишайниковое; 8 — низкокустарничково-зеленомошно-лишайниковое; 9 — кустарнич-
ных сообществ по полигонам и багульниково-сфагновых по трещинам; 11 — комплекс багулышково-
падинам; 12 — заросли ерника с покровом из багульника и морошки; 13 — заросли ивняка с разно-
гульниково-морошково-сфагновое; 16 —нушицево-оеоково-сфагновое; 17 — багульниково-морош-
сфагновое; 20—пушицево-багульниково-зеленомопшо-лишайниковое; 21 —морошково-багульни-
сфагновое; 24 — комплекс багульниково-морошково-лишайниковых сообществ по буграм, сфагно-
пушицевое; 27 — багульниково-морошково-сфагновое; 28 — багульниково-морошково-зеленомош-
вое со сфагнумом; 31 — багульдаково-морошково-зеленомршно-лишайниковое; 32 — осоково-пуши-
падинный комплекс кустарничково-морошково-сфагновых и осоково-пушицевых сообществ; 35 —
новое.
гульник; ж — морошка; з — брусника; и — вейник; к — разнотравье; л — зеленые мхи; м — сфаг-
0 — глубина залегания сезонной мерзлоты.
(Carex chordorrhiza, Eriophorum polystachyon, Calliergon stramineum)
группировками; торфяной горизонт под ними мощностью 30—50 см. Из ив
абсолютно преобладает ива шерстистая (Salix lanata) высотой 1,5 м.
Так как низкая пойма заливается водами половодья почти на весь
вегетационный период, то все встречающиеся здесь виды растений
приспособлены к длительному затоплению, они способны не только вегетировать^
но даже цвести в воде. Для самых обводненных участков дельцы
характерны группировки арктофилы и хвоща. Во время половодья генеративные
побеги арктофилы возвышаются над поверхностью воды на 30—40 см.
Arctophila fulva иногда образует довольно сомкнутые сообщества с
проективным покрытием 65—70%. Монодоминантные группировки Equisetum
fluviatile также очень типичны, но обычно они приурочены к менее
глубоким местам и песчаным наносам. Наибольшие площади на участках
низкого уровня, главным образом с соровыми понижениями, заняты
кочковатыми осоковыми лугами, где доминируют Carex aquatilis, С. acuta (296).
Проективное покрытие этих лугов 75—100%. Кроме осоки обычно
встречаются представители гигрофитного разнотравья — Comarum palustre,
Alisma plantago-aquatica, Naumburgia thjfrsiflora. В сезоны особенно
большой воды на таких местообитаниях сначала поселяются соровые бекман-^
ниевые (Beckmannia eruciformis) и арктофилловые группировки, которые
затем сменяются осоковыми лугами. По обводненным западинам с близким
залеганием сезонно-мерзлых пород обычно встречаются пушицево-осоко-
вые луга с господством Carex aquatilis и Eriophorum polystachyon. Берега
зарастающих озер, как правило, имеют микропоясный растительный
покров, в котором последовательно сменяют друг друга группировки
гигрофитного е разнотравья (A lisma plantago-aquatica, Naumburgia - thyrsiflora,
Menyanthes trifoliata), хвоща болотного (Equisetum fluviatile) и кочковатых
осок (Carex aquatilis, С. cespitosa).
В низкой пойме, кроме травянистых сообществ, представлены
кустарниковые группировки, образованные Salix lapponum, S. lanata, S. phyli-
cifolia. Они распределены одиночно или небольшими группами зарослей
по мезоповышениям вдоль небольших проток.
На поверхностях, принадлежащих к среднему и высокому
экологическим уровням преимущественно молодых участков Обской дельты,
определенный ряд смен образуют сообщества осоковых и вейниковых лугов,;
ивняковых, ивняково-ерниковых и ерниково-ольховдиковых тундр (29в).
По рельефу эти местообитания обычно представляют скопления невысоких
185
24 2526\2829303132х 35 I 34 \ JS 36 37 >\
I ' 27 ' ! I ' '
. грив с плоскими вершинами или более высоких грив, расположенных вдоль
русла. Такие местообитания не каждый год заливаются водами
половодья, а если и заливаются, то на небольшой срок (8-^—10 дней).
На нижних частях склонов грив обычно распространены осоковые
и осоково-вейниковйе луга среднего уровня с Carex acuta и
Calamagrostis langsdorffii. Выше они сменяются разнотравно-вейниковыми лугами,
среди которых часто встречаются отдельные экземпляры и небольшие
группы низкорослых кустов ивы (Salix phylicifolia) и ольхи (Duschekia fruti-
cosa). Травостой хорошо развит, в нем доминирует Calamagrostis
langsdorffii (40—60%), содоминантами являются представители разнотравья —
^Achillea ptarmica, Veronica longifolia, Comarum palustre. В значительном
количестве присутствуют Роа sublanata, Calamagrostis neglecta, из
бобовых — Vicia sepium,\ Lathyrus palustris. Единично встречаются Galium
uliginosum, Geranium sylvaticum, Lactica sibirica, var. integrifoliax Stella-
ria graminea, Festuca rubra, Carex aquatilis.
На высоких гривах прирусловой части наиболее молодых сегментов
поймы распространены ивняковые сообщества с разнотравно-вейниковым
докровом. В древесно-кустарниковом ярусе обычны Salix viminalis и
S. dasydados, иногда достигающие высоты 6—7 м, но в основном
образующие группы кустарниковых зарослей, в которых кроме ив отдельными
экземплярами встречаемся ольха (Duschekia fruticosa).
Во внутренних частях дельтовых островов, вдалеке от действующих5
проток, находятся небольшие массивы древней высокой поймы. По
рельефу это едва заметные сглаженные вершины грив, заливаются они очень
редко — только в годы катастрофических паводков (таким был паводок
1978 г.). Растительный покров этих участков по составу близок к
зональным тундровым фитоценозам. Наиболее типичны сообщества ивняково-
ерниковых и ерниково-ольховниковых тундр. Первые обычно имеют
более сомкнутый кустарниковый ярус (до 60—70% проективное покрытие)г
Betula nana достигает здесь высоты 0,8—1 м. В сообществах ерниково-
ольховниковых тундр Betula nana распределена по площади единичными
экземплярами и более равномерно, высота ее бывает до 2—2,5 м.
Травяной ярус разрежен (проективное покрытие до 30%). В значительном
количестве здесь представлены Ledum palustre, Vaccinium uliginosum, Cha-
maedaphne calyculata. Единично встречаются Vaccinium vitis-idaea, Arctag-
rostis latifolia, Calamagrostis langsdorffii, Rubuschamaemorus, Diphasiastrum
complanatum, Empetrum nigrum, Oxycoccus microcarpus. Моховой покров
неравномерен, но местами его плотность достигает 60%. Преобладают в
нем^зеленые мхи Dicranum spadiceum (30%), Polytrichum jensenii (20%)r
Aulacomnium turgidum (10%). Единично встречаются Aulacomnium pa-
lustre и Ceratodon purpureum. Из сфагновых мхов отмечен только
Sphagnum girgensohnil (10%).
СЕВЕРОТАЕЖНЫЕ ЛУГОВО-БОЛОТНО-СОРОВЫЕ
И КУСТАРНИКОВО-ЛЕСНЫЕ СЕРИИ РАСТИТЕЛЬНЫХ СООБЩЕСТВ
Серия осоково-мелкоивняковых низинных болот,
сообществ соровой растительности,
болотистых и торфянистых лугов,
ивняков кустарниковых и парковых, березовых,
березово-еловых и лиственничных лесов (69)
• Эта серия эколого-динамических рядов пойменной растительности
характерна для поймы нижнего течения р. Оби на отрезке от Салехарда
до пос. Березово. Река на этом участке распадается на множество
протоков и рукавов, по своей ширине и мощности не уступающих иногда
основному руслу (Мал. Обь). Пойма в связи с этим приобретает
проточно-островной характер [Еленевский, 1936]. Ее ширина от 10 до' 45 км. Наиболее
важной особенностью пойменных островов этого участка поймы является
186
наличие в их центре «соров». Площадь соров на этом отрезке поймы очень
велика (до 40% от общей площади).
По данным А. С. Герасимовой, С. Б. Ершовой и др. [19671, пойма
Нижней Оби (ниже нос. Ванзетур до Салехарда) представляет типичную
аккумулятивную террасу с мощным чехлом (26—28 м) аллювиальных
отложений, среди которых преобладают суглинистые. Ширина поймы от 10
до 45 км (максимум 65).
Проточно-островной характер поймы почти полностью стирает
обычные для большинства рек различия в горизонтальном ее делении на
прирусловую, центральную и притеррасную части [Вильяме, 1946 J. Для
такой поймы более характерна вертикальная ее дифференциация по
экологическим уровням, или высотным поясам, в зависимости от рельефа и
режима поемности [Шенников, 1941]. Участки каждого уровня имеют
специфический набор растительных сообществ, сходных по экологическим й
динамическим свойствам и составляющих определенные эколого-динами-
ческие ряды. В пойме Нижней Оби на участке Салехард — Березово
выделено три экологических уровня [Барышников, 1961; Ильина, 1976].
Низкий уровень поймы охватывает обширные соровые понижения,
площадь которых иногда достигает нескольких сотен квадратных
километров и увеличивается вниз по течению Оби. Относительная высота поверх^
ности соров над уровнем средней многолетней межени составляет 2—3,5 м.
Соры заливаются на самое продолжительнее время — 80—100 дней
[Барышников, 1961; Петров, 1979].
Участки среднего уровня поймы лежат? на высоте 3,5—5 м над
меженью [Петров, 19791. Они меньше заливаются, чем поверхности соров
(70—75 дней). Участки этого уровня представляют собой пологие
растянутые склоны к соровым понижениям или небольшие плоские гривы,
окаймляющие мелкие протоки, или нижние части склонов высоких
прирусловых грив.
Пойма высокого уррвня занимает прирусловые валы по берегам
больших проток, вершийы и верхние части склонов высоких грив. Средняя
высота ее над меженным уровнем превышает 5 м. Почти ежегодно она
заливается на 25—40 дней [Барышников, 1961; Петров, 1979].
Состав растительности этого отрезка обской поймы обнаруживает
очень четкую связь с высотными уровнями поемности.
Для участков низкого экологического уровня самых северных районов
поймы Ниж. Оби характерен ряд пойменных сообществ с участием
мелкоивняковых болот (69а). Мелкоивняковые болота Обской поймы описаны
М. К. Барышниковым [19611 и отражены в материалах
Землеустроительной экспедиции МСХ РСФСР, работавшей в 1960—1963 гг. на территории
, Ямало-Ненецкого автономного округа под руководством Е. Н. Широков-
ской. Они занимают центральные части пойменных островов, поверхность
их плоская или слегка вогнутая. Болота сильно кочковаты и обводнены,
имеют торфяной слой мощностью 30—50 см. Из видов ив абсолютно
преобладает Salix lanata высотой 0,5—1,5 м. Кочки образованы осокой водной
и дернистой (Carex aquatilis, С. cespitosa). Между кочками в небольшом
количестве встречается Petasites frigidus, сабельник (Comarum palustre).
На более возвышенных, но плохо дренируемых плоских участках
низкого уровня мелкоивняковые болота переходят в заболоченные сильно
кочковатые осоковые луга, образованные теми же кочкарными осока-
ми — Carex cespitosa, С. aquatilis. По составу растительности они
довольно сходны с болотами, отличаются от них отсутствием кустарниковых ив
и большим проективным покрытием осок.
Примерно к югу от Полярного круга в пойме Нижней Оби площадь
болот сильно сокращается. Здесь они все чаще сочетаются с участками со-
ровых лугов, которые на севере попадаются среди болот только
отдельными массивами.
Южнее соры начинают преобладать по площади, их растительный
покров образует самостоятельные ряды смен сообществ, картируемые в
данном масштабе (696). В северных районах Йижнеобской поймы, ближе к
187
дельте Оби, встречаются огромные соры, почти лишенные растительности,;
с отдельными пятнами разреженных группировок сорового .разнотравья и
злаков из арктофилы (Arctophila fulva), ситнягов игольчатого (Eleocharis
acicularis) и болотного (Е. palustris), бекмании (Beckmannia eruciformis)
и с обширными мелководными озерами, иногда зарастающими сусаком
зонтичным (Butomus umbellatus). На большей части характеризуемого
отрезка Обской поймы преобладают соры, зарастающие и в разной степени
покрывающиеся соровой луговой растительностью. После спада воды
на самых низких участках в центральных частях таких соров сначала
поселяются разреженные группировки ситняга, хвоща, бекмании,
полевицы. По мере обсыхания на них формируются вполне сомкнутые
группировки соровых лугов — арктофиловых, бекманиевых,полевицевых.
Наибольшие площади занимают полевицевые луга из полевицы побегообра-
зующей (Agrostis stolonifera). Они обычно имеют сомкнутый травяной
покров с проективным покрытием ВО—90%, высота генеративных
побегов не превышает 15—20 см. В составе травостоя на долю полевицы
приходится 70—80%. Кроме нее в небольшом количестве присутствуют бек-
мания {Beckmannia eruciformis), хвощ иловатый (Equisetum fliiviatile)f]
чистец болотный (Stachys palustris), лютик едкий (Ranunculus acris), же-
рушник (Rorippa sylvestris), подмаренник болотный (Galium palustre),
чихотная трава (Achillea ptarmica), девясил британский (Inula britannica),
частуха (Alisma plantago-aquatica), иез&букка. болотная (Myosotispalustris).
Соровые понижения на всех массивах островной поймы окружены
обширными слабонаклонными поверхностями, которые заливаются на
менее длительный срок и по режиму поемности относятся к среднему
экологическому уровню. Для них характерен ряд ассоциаций осоковых и вей-
никовых лугов и ивняков кустарниковых, сочетающихся с соровыми
лугами по понижениям (69в).
Центральной ассоциацией этого ряда является закочкаренный осоч-
ник из осоки водной (Carex aquatilis), приуроченной к обширным
слабонаклонным поверхностям/ обращенным к сорам. Кочки высотой 25—30 см
диаметром 20 см занимают почти 1/3 площади. Травостой высокий — до
100 см, проективное покрытие 90%. В составе абсолютно преобладает
осока водная (Carex aquatilis), единично между кочками встречаются
подмаренник болотный (Galium palustre), сердечник луговой (Cardamine praten-
sis), полевица стелющаяся (Agrostis stolonifera).
Почти такие же местообитания занимают кочковатые вейниковые
луга, отличающиеся, однако, более разнообразным составом травостоя,
среди которого преобладают вейник Лангсдорфа (Calamagrostis langsdorf-
fii), осока водная (Carex aquatilis) и мятлик, луговой (Роа pratensis). Мел-
кокочкарные водноосоковые и вейниковые луга занимают самые большие
площади на участках среднего уровня.
Бескочкарные мятликово-вейниковые, а в более южнь!х районах ка-
нареечниково-вейниковые луга также распространены довольно
широко, приурочены к широким плоским гривам вдоль небольших проток,
где чередуются с группами ивняйов кустарниковых из Salix viminalis и
S. cinerea.
Луга среднего уровня образуют сплошные массивы большой
площади только на крупных пойменных островах. На более мелких, а также
в местах гривистого рельефа вдоль небольших проток они чередуются с
соровыми лугами понижений. Поэтому на карте ряд лугов среднего
уровня показан в характерном сочетании с участками лугов низкого уровня,
последние занимают до 30% площади.
Для более расчлененных гривистых участков островной поймы
среднего уровня, расположенных вдоль основного русла и больших протоков
Оби, характерен ряд сообществ разнотравно-злаковых лугов и ивняков
кустарниковых и парковых, сочетающихся с осоковыми лугами межгрив-
ных понижений (69г). .
Начальным звеном этого ряда являются луговые фитоценозы. Среди
них разнотравно-злаковые \ полидоминантные луга М. К. Барышников
188
[1961] считает переходными от лугов'среднего уровня. В их составе
значительное участие принимают вейник Лангсдорфа {Calamagrostis langsdorf-
/it), мятлик (Роа palustris, P. pratensis), лисохвост (Alopecurus pratensis),
бекмания (Beckmanniaeruciformis), полевица гигантская (Agrostis gigantea),
канареечник (Phalaroides arundinacea).
Вейниковые из Calamagrostis langsdorffii, особенно мятликовые
(Роа pratensis), луга занимают довольно большие площади на верхних
частях склонов прирусловых грив и принадлежат уже к лугам высокого
уровня. Вейниковые луга чаще чередуются с сообществами кустарниковых ив.
Последние образуют группы довольно густых зарослей из Salix viminalis,
S. cinerea высотой до 2—3 м.
Наиболее возвышенные участки, преимущественно плоские вершины
грив, покрыты вейниковыми ивняками паркового типа, а в более южных
районах — ивняками канареечниковыми.
Участки высокого экологического уровня занимают самую малую
площадь на данном отрезке Нижней Оби. Для высоких прирусловых грив,
расположенных ближе к воде характерны ряды сообществ ивняков
кустарниковых, парковых и ивовых травяных лесов (69д). В глубине пойменных
массивов развиваются ивово-березовые, березовые, местами смешанные
травяные леса.
По данным М. К. Барышникова [19611, а* также по нашим
наблюдениям, лиственные, а тем более хвойные леса очень редко встречаются на
собственно пойменных участках этого отрезка Нижней Оби. Наиболее
значительные площади подобные сообщества занимают только на
останцах надпойменных террас, расположенных среди островов поймы и
хорошо картирующихся в данном масштабе. Эти лесные массивы останцов
также включаются в описываемую серию на том основании, что они в
своем составе четко отражают влияние окружающих пойменных экотопов„
и в то же время, имея все свойства устойчивых лесных сообществ
водораздельного типа, представляют как бы потенциальную заключительную
стадию рассматриваемой пойменной сукцессии (серии).
В смешанных березово-лиственничных и березово-еловых лесах
высоких уровней поймы и останцов надпойменных террас наряду с таежными
видами мхов (Pleurozium schreberi, Polytrichum commune) и
кустарничков — брусники (Vaccinium vitis-idaea), багульника (Ledum palustre)
встречаются представители пойменного разнотравья — мята (Mentha аг-
vensis), молокан (Lactuca sibirica), подмаренник (Galium boreale), вероника
(Veronica longifolia).
' Лугово-ивняково-редколеснолиственничная
серия сообществ (40)
Указанная серия долинной растительности характерна для отдельных
участков нижних течений северных таежных рек Западной Сибири —
Енисей, Надым, Пур, Таз, а также рек тундровой зоны — Хадытаяха,;
Мессояха и др. В составе рядов растительных сообществ здесь много
общего с дельтой и низовьями Оби. Луговые стадии низких уровней также
представлены зарослями арктофилы, хвоща (Equisetum fluviatile) и
осоки водной. На более высоком экологическом уровне развиваются
кустарниковые разнотравно-злаковые с редкими деревьями лиственницы (Larix
sibirica) сообщества. В них хорошо развит кустарниковый ярус (Duschekia
fruticosa, Salix hastata, S. phylicifolia, Juniperus sibirica, Betula nana)
густой, высокий разнотравно-злаковый покров из Alopecurus pratensisf)
Роа sublanata, Festuca ovina, Veratrum lobelianum, Equisetum pratense и др.
На высоких участках старых притеррасных частей поймы, а в основном на
склонах и в прибровочных частях первой надпойменной террасы
распространены лиственничные редколесья. Лиственница (Larix sibirica)
прямоствольная высотой 15—17 м в хорошем состоянии. В кустарниковом ярусе
преобладают ерник и ивы (Betula nana^ Salix hastata, S. phylicifolia) f
меньше ольхи (Duschekia fruticosa) и можжевельника (Juniperus sibirica).
189
Основу травяно-кустарничкового яруса составляют Vactinium uliginosum.
Rubus arcticus, в меньшем количестве встречаются Festuca ovina, Equise-
trum pratense и некоторые виды разнотравья. В напочвенном покрове
господствуют зеленые мхи: Hylocomium splendens, Drepanocladus uncinatus,
Pleurozium schreberi, Polytrichum commune.
Для лиственничных редколесий долин низовий Таза и Пура характер- „
на значительная примесь березы, ели и хорошее развитие кустарникового
яруса, в котором встречается много ольхи (Duschekia fruticosa).
Пойменные озера и старицы, которые типичны для притеррасных час-
. тей долин многих рек, имеют свои «микросерии» — микропоясные ряды
сообществ. Первая стадия их зарастания представлена осоково-гипновы-
ми группировками. Среди погруженной в воду растительности
преобладают Carex aquatilis, Equisetum fluviatile. Дальше от воды идет пояс
осоковых кочковатых лугов, а следом за ним, в некоторых местах и прямо от
воды, широкая полоса низинных мелковдняковых (Salixhastata, S. lanata),
травяно-моховых (Carex rariflora, Eriophorum polystachyon, Aulacomnium
turgidum, Hylocomium splendens var. alaskanum, Sphagnum subsecundum)
болот.
Серия ивняково-березово-еловых с кедром
и лиственницей сообществ (70).
Серия ивняково-ольховшшово-березово-листвеыничных
сообществ (71, 71а)
Для нижних и средних течений рек Надым, Пур, Таз, а также для
северных притоков Оби — р. Полуй и других характерны серии
растительных сообществ, заключительными стадиями которых являются
лиственничные и еловые леса северотаежного типа (рис. 19). Начальные
стадии этих серий здесь представлены арктофиловыми и осоковьГми лугами,
но эти группировки занимают очень незначительные площади,
.протягиваясь узкими полосами вдоль русел. Гораздо'большую пространственную
выраженность имеют стадии ивняков кустарниковых из Salix phylicifolia,
£. lanata, S. viminalis. По составу они очень разнообразны. На хорошо
увлажняемых прирусловых гривах развиваются ивняки травяные— вейни-
ковые и разнотравно-злаковые. В местах слабопроточных они переходят
в заболоченные варианты — вейниково-осоковые, осоково-сфагновые, ер-
никово-сфагновые. Последние характеризуются наличием слоя вечной
мерзлоты на глубине 40—70 см и связанным с этим ухудшением условий
произрастания и постепенным отмиранием ив [Тыртиков, 1974]. На дре-
ННВВН^ИННЯННННННнНШ
Рис. ./^Растительность долины р. Таз (нижнее течение). Фото Б. А. Богоявленского.
190
нированных местообитаниях долинная сукцессия после стадии ивняков
травяных ведет к образованию лиственных древостоев — ивово-березовых
и березовых лесов. В них преобладает береза Betula pendula, а из ив
типичны древовидные Salix viminalis и S. alba, В подлеске много
кустарников: Rosa qcieularis, Duschekia fruticosa, Sorbus sibirica, Salix phylicifolia.
Среди трав господство принадлежит зл&кам — Calamagrostis langsdorffii
(С. epigeios н& легких почвах), Роа pratensis, P. palustris; встречаются
также различные представители мезофитиого разнотравья — Ranunculus
repens, Veronica longifolia, Galium boreale, Viola epipsila и др.
Мелколиственные травяные леса занимают значительные площади в
долинах среднего течения Пура и Таза, -поэтому они нашли свое отражение
на составленной карте как отдельные стадии пойменной серии (71а).
Заключительными в сукцессиях долинных гидросерий северных рек
являются относительно устойчивые (субклимаксовые) 'сообщества
смешанных, темно- и светлохвойных лесов. На этих стадиях сукцессионные
смены протекают по-разному, £ зависимостп от характера субстрата. На
глинистых отложениях формируются березово-елрвые и еловые травяно-
зеленомошные леса (70). \
Для почв более легкого механического состава характерны березово-
^иственничные и лиственничные леса (71). Для всех лесов очень
характерно преобладание в подлеске ольхи. В покрове господствуют таежные
виды — Vaccinium vitis-idaea, Cymnocarpium dryopteris, Trientalis europaea,
Viola epipsila. Влияние пойменного ,режима проявляется в присутствии
вейника (Calamagrostis langsdorffii) и некоторых представителей
пойменного мезофитного разнотравья: Geranium sylvaticum, Anemonidium dicho-
tomum, Aconitum septentrionale. Хорошо развит в таких лесах и моховой
покров, хотя он, как правило, не имеет сплошного распространения.
Типичны таежные виды мхов^— Polytrichum commune, Hylocomium splen-
dens, Climacium dendroides.
СРЕДНЕТАЕЖНЫЕ ЛЕСО-КУСТАРНИКОВО-СОРОВО-ЛУГОВЫЕ
СЕРИИ РАСТИТЕЛЬНЫХ СООБЩЕСТВ
Серия сорово-луговых
и ивняково-болотистолуговых сообществ,
ивовых, березовых, березово-кедровых
и березово-сосновых лесов (85)
К этой серии нами отнесены пойменные ряды сообществ,
распространенные на отрезке Оби от г. Ханты-Мансийска до пос. Березово.
Для данного участка поймы Оби Э. Е. Роднянская [1958, 1968]
выделяет четыре экологических уровня:
1) ежегодно заливаемый исключительно долгопоемный пояс, или
низкий экологический уровень. Он охватывает участки, занятые соровыми
понижениями, а также слабонаклонные поверхности, обращенные к со-
рам. Продолжительность их затопления 2,5—3 мес;
2) ежегодно заливаемый долгопоемный пояс, или средненизкий
экологический уровень. Участки средненизкого' экологического у^овяя,
затопляются на 1—2, реже 2,5 мес; '
3) периодически заливаемый слабопоемный пояс, или средневысокий
экологический уровень. Он занимает средние гривы, высотой 6—7 м,
заливаемые 1 раз в 2—4 года на срок 1,5—2 мес;
4) редко заливаемый краткопоемный пояс, или высокий
экологический уровень. Участки этого уровня заливаются 1 раз в 7—8 лет на срок
до 1—1,5 недель. - • - ,
Характеризуемый участок Обской поймы можно отнести к типу
проточно-островных пойм, хотя в отличие от более северных районов здесь
ширина многих проток и рукавов, отчленяющих пойменные острова,
значительно меньше.»
191
Для участков самого низкого экологического уровня характерен ряд
сообществ соровой растительности и кочковатых осоковых и вейниковых
лугов (85а). На рассматриваемом участке Обской поймы соры
распространены очень широко, но в отличие от соров более северных районов поймы
они не имеют больших размеров. Поэтому на карте сообщества соровых
понижений показаны главным образом в сочетании с болотистыми
осоковыми лугами. Большинство соров после спада воды высыхает довольно
быстро и заселяется типичными сорово-разнотравными луговыми
сообществами.
Один из типичных соров был описан нами на пойменном острове,
расположенном в 12км от с. Полноват вверх по протоке Токсика. Сор
занимал обширное слабовогнутое понижение с небольшим зарастающим
озером, в центре которого еще сохранились группы зарослей сусака
зонтичного, частухи и манника водного. На такыровидных понижениях,
недавно освободившихся от воды, поселились открытые фитоценозы из ситняка
игольчатого и хвоща поникающего. Но большая часть сора заросла
сплошным ковром полевицы ползучей (Agrostis stolonifera), создающей
характерный красно-бурый аспект. Средняя высота травяного покрова 20—-30 см,
покрытие 60—70%. В составе абсолютна преобладает полевица ползучая
(Agrostis stolonifera), в значительном количестве встречается ситняг
болотный (Eleocharis palustris) и пятнами — звездчатка (Stellaria gramijiea).
Остальные виды имр*от небольшое обилие — незабудка болотная (Муо-
sotis palustris), птармика (Achillea ptarmica), хвощ (Equisetum fluviatile)f
лютик ползучий (Ranunculus repens), мята (Mentha arvensis), чистец
болотный (Stachys palustris), поручейник (Sium latifoliuni), подмаренник
болотный (Galium palustre) 1. Подобные соры наблюдались в окрестностях
пос. Нижние Тугияны, сел Березово, Октябрьское.
На описываемом отрезке Обской поймы вокруг соров
распространены сообщества водно- и остроосоковых лугов. Эти луга большей частью
закочкарены, особенно когда в них преобладает осока водная (Carex aqua-
tilis). Кроме нее на таких лугах обычны птармика, поручейник,
подмаренник болотный, хвощ болотный, девясил (Inula britannica).
В эко лого-динамический ряд низких уровней включены также осоко-
во-вейниковые крупнокочковатые луга. Они часто встречаются
небольшими участками на плоских обводненных местах вокруг соров. Высота ко-
"чек достигает 50 см, кочки образованы вейником незамечаемым (Calamag-
rostis^neglecta), осокой дернистой (Carex cespitosa) и мятликом болотным
(Роа palustris).
Второе место по величине занимаемой территории на этом участке
Обской поймы принадлежит динамическому ряду сообществ средненизкого
экологического уровня (856). Здесь в составе растительности абсолютно
преобладают осоковые и осоково-канареечниковые луга. Типичен
следующий эко л ого-динамический ряд их растительных сообществ:
водно-остроосоковая ->- остроосоковая —>• остроосоково-вейниковая —>• остроосоково-
канареечниковая ->- вейниково-канареечниковая.
Водно-остроосоковые луга занимают наиболее пониженные и выполо-
женные участки этого экологического уровня. В районе с. Октябрьское
на таких лугах высота травостоя была 70—80 см, проективное покрытие
до 100%. В составе преобладали осока острая (Carex acuta), осока водная
(Carex aquatilis) и канареечник (Phalaroides drundinacea); единично
встречались звездчатка (Stellaria graminea), незабудка (Myosotis palustris)^
хвощ полевой (Equisetum amense), птармика (Achillea ptarmica), чина
болотная (Lathyrus palustris), подмаренник болотный (Galium palustre)ft
чистец болотный (Stachys palustris), лютик ползучий (Ranunculus repens).
Сообщества вейниково-осоковых (с Calamagr.ostis neglecta) канарееч-
никово-осоковых и осоково-канареечниковых лугов занимают склоновые
1 Здесь и далее при описании конкретных участков гербарные сборы были
сделаны И. С. Ильиной и Л. И. Мельцер; видовой состкв растений определен сотрудниками
«Гербария им. М. Г. Попова» Института физиологии и биохимии растений СО АН СССР.
192
местообитания, причем канареечниково-осЪковые луга характерны для
их нижних частей. На одном из типичных участков осоково-канареечнико-
вого луга на пологом склоне низкой гривы в окрестностях нос. Ниж. Ту-
гияны в составе травостоя преобладали канареечник (Phalaroides агип-
dinacea), осока острая (Carex acuta), вейник Лангсдорфа (Calamagrostis
langsdorffii); повсеместно, но в небольшом количестве были
распространены звездчатка (Stellaria graminea), бодяк (Cirsium heterophyllum) t
лютик (Ranunculus acris), калужница (Caltha palustris), незабудка.
(Myosotis palustris)-.
Вейниково-канареечниковые луга приурочены к наиболее
возвышенным местам — вершинам мелких грив и верхним частям склонов. Их
проективное покрытие 80—90%, высота травостоя до 100 см. Доминируют
канареечник и вейник Лангсдорфа, в значительном количестве встречается
вейник ланцетовидный (Calamagrostis canescens), присутствуют также
чина луговая (Lathyrus pratensisf, дербенник (Lythrum salicaria), бодяк (Cir-
siumheterophyllum), пт&фмжка. (Achillea ptarmicd), лютик ползучий
(Ranunculus repens), незабудка (Myosotis palustris), подмаренник болотный
(Galium palustre).
Луга средненизкого экологического уровня — преимущественно грив- »
ные, хотя приурочены они к гривам низким с пологими растянутыми
склонами. На местности гривы сочетаются с неглубокими понижениями,
занятыми кочковатыми водноосоковыми и вейниковыми из Calamagrostis neg-
lecta лугами. Именно в этом сочетании и показаны луга средненизкого
уровня на геоботанической карте.
Основными растительными сообществами средневысокого
экологического уровня являются настоящие и болотистые луга и заросли
кустарников (85в) [Шенников, 1941].}
Разнотравно-злаковые луга на местообитаниях этого уровня
занимают пологие склоны и вершины грив высотой 5—6 м. Травостой всегда
густой (80—100% проективное покрытие). Высота генеративных побегов
до 170см, вегетативных — 100—110 см. В составе преобладают
канареечник (Phalaroides arundinacea) и вейник (Calamagrostis langsdorffii),
остальные виды встречаются в небольшом количестве — мятлик луговой (Роа
pratensis), осока острая (Carex acuta), лютик ползучий (Ranunculus repens)%
василистник (Thalictrum simplex), девясил (Inulat britannica), чистец
болотный (Stachys palustris), подмаренник болотный (Galium palustre),
молокан (Lactuca sibirica), звездчатка (Stellaria graminea), чина луговая
(Lathyrus pratensis), незабудка (Myosotis palustris) г.
Чисто верниковые луга из Calamagrostis langsdorffii встречаются по
вершинам и верхним частям склонов грив, расположенных вдоль
небольших проток, вблизи от русел. Именно эти луга,в большинстве случаев,,
чередуется с зарослями кустарниковых ив. Кусты ивы — Salix viminalis,
S. pentandra, S. cinerea — образуют группы сообществ сомкнутостью
0,7—0,8, высотой от 1,5 до 4 м.
Топо-экологические ряды лугов и кустарников средневысокого
уровня на местности включают также участки более низких уровней,
занимающие склоны грив и межгривные понижения. На долю последних
приходится около 20% всей площади контура.
Высокие уровни поймы этого Отрезка Нижней Оби заселяются
древесной растительностью — мелколиственными и смешанными лесами (85 г).
В целом такие леса не занимают большой площади, но являются очень
характерным элементом пойменного ландшафта.
Динамический ряд начинается ивняками паркового типа,
приуроченными к выположенным склонам и вершинам высоких прирусловых грив.
Древостой их очень разрежен — сомкнутость не более 0,1—0,2. Деревья
часто многоствольные, напоминающие огромные разросшиеся кусты,
высота их 8—12 м, диаметр .у одноствольных до 40 см, у кустообразных 10—
1 Это описание сделано на участке поймы в 30 км вверх по течению Оби от с. Бол.
Атлым.
13 Заказ № 297 193
JiiijIfUywigTuKy in; uHli
jliiili
10 , 11 \ 12 \ 13
Рис. 20. Геоботанический профиль в пойме р. Оби, район пос. Верх
Растительные сообщества: 1 — ивовый лес с березой и осиной канареечниково-вей
ковый; 4 — полевицево-осоковый кочковатый луг; 5 — осоково-разнотравно-вейниковый луг;
9 — открытая группировка с ситнягом, хвощом, частухой; 10 — ситняговая группировка; 11 —
нотравно-полевицевый луг; 15 — открытые группировки из ситняга, хвоща, полевицы, звезд
Доминирующие виды: а — осока; б — канареечник; в — вейник; г — полевица стелю
никовый; и — ива
15 см. В ивняках парковых обычно преобладает один вид ив — ива белая
(Salix alba), в примеси могут быть кустарники из Salix cinerea, S. beb-
biana, S. lapponum. Травостой хорошо развит, представлен или вейниковы-
ми сообществами с Calamagrostis langsdorffii или разнотравно-злаковыми,
где кроме вейника присутствуют в значительном количестве мятлик
луговой (Роа pratensis), полевица гигантская (Agrostis gigantea), канареечник
(Phalaroides arundinacea), а из разнотравья представители бобовых —
клевер (Trifolium medium), чина луговая (Lathyrus pratensis), мышиный
горошек (Vicia cracca) и др. Обычно луговые массивы ивняков парковых
наиболее интенсивно используются под выпас, так как имеют хороший
травостой ж меньше заливаются, чем другие участки поймы.
Следующая стадия этого эколого-динамического ряда представлена
ивово-березовыми пойменными лесами. Такие леса занимают наиболее
возвышенные участки прирусловых грив вдоль больших проток и
основного русла Оби. На одном из участков леса, расположенном на верпщне
прирусловой гривы близ пос. Верх. Рязаны, в составе древостоя (7БЗИ)
сомкнутостью 0,5—0,6 преобладали березы (Betula pendula) высотой 17—
18 м, диаметром 20—25 см и ивы {Salix alba) высотой 14—15 м и
диаметром 15—18 см. В подросте встречались порослевая береза и единично
поросль ивы. Подлесок густой, состоял из шиповника (Rosa cihnamomea),
черной смородины (Ribes nigrum), черемухи (Radus avium), свидины (Swi-
da alba). В составе травяного яруса с проективным покрытием 70—80%
преобладали рейник (Calamagrostis langsdorffii), мятлик луговой (Роа
pratensis), анемона вильчатая (Anemonidium dichotomum), льнянка
Urinaria vulgaris), звездчатка (Stellaria graminea), в меньшем количестве
встречались вероника (Veronica longifolia), хвощ полевой (Equisetum
arvense), мышиный горошек (Vicia cracca), молокан (Lactuca sibirica),
бодяк (Cirsium heterophyllum), подмаренник болотный (Galium pali^tre),
чистец (Stachyk pulusiris).
Ивово-березовые леса высоких уровней поймы местами переходят в
ивово-березово-осиновые (Populus tremula) с более богатым травостоем,
в котором преобладают обычно не злаки, а разнотравье (василистник,
аконит, айемона вильчатая, герань полевая, вероника), но в целом осиновые
лера на этом отрезке Обской поймы встречаются довольно редко и в
основном на юге, ближе к Ханты-Мансийску. Смешанные хвойно-мелколиствен-
ные лесные сообщества появляются только на самых высоких опесчанен-
ных гривах: редко небольшими участками встречаются пойменные леса с
преобладанием хвойных — сосны и кедра.
Мелколиственно-хвойные и чисто хвойные с пойменными элементами
леса значительные площади занимают только на останцах надпойменных
террас. Они характеризуются более высокой производительностью
древостоя и хорошим развитием нижних ярусов леса. Так, на одном из останцов
цадпойменной террасы, расположенном среди участков поймы около
с. Березово, в составе древостоя были отмечены 4Б20сЗК1С. Высота
берез 20—25 м, диаметр 25—30 см; высота осин 22—28 м, диаметр 35—
40 см; высота кедров 18—20 м, диаметр 18—20 см; высота сосен 16—18 м,
194
±kl±SllS±
14 15 /tf /7
Тугияны. (Составлен И. С. Ильиной, Л. И. Мельцер).
виковый; 2 — рдестово-частуховые группировки; 3 — березовый лес с осиной разнотравно-вейни-
6 — осоковый луг; 7 — звездчатково-полевицевый луг; 8 — полевицо-ситняговая группировка;
ситнягово-полевицевый луг; 12 — осоково-канареечниковый луг; 13 — осоковый луг; 14 — раз-
чатки; 16 — канареечниково-вейниковый луг; 17 — ивняк парковый канареечниково-вейниковый.
шаяся; д — ситняг игольчатый; е — соровое разнотравье; ж — влажнотравье; з —ивняк кустар-
древовидная; к — береза. ' »
диаметр 20—25 см. В подросте поросль берез, осин и единично кедр, а в
подлеске рябина и шиповник. Хорошо развит напочвенный покров, проек
тивное покрытие которого 80%. В его составе в значительном количестве
распространены брусника, хвощ лесной, кислица, костяника, папоротник
Линнея,-майник, плаун булавовидный, какалия, бор развесистый,, ане-
нона вильчатая.
Моховой покров встречается только пятнами, его покрытие 10—30%.
Из мхов отмечены Ptilium crista-castrensis, Rhytidiadelpkus triquetrus.
Описанные- ряды растительных сообществ среднетаежной пойменной
серии можно проиллюстрировать эколого-геоботаническим профилем,;
заложенным на одном из пойменных массивов в районе пос. Верх.
Тугияны (рис. 20).
Серия ивово-березовых,
березово-сосновых травяно-зеленомошных
и лишайниковых лесов долинных грив
и сосново-березовых осоково-сфагновых лесов
межгривных понижений (87).
^ерия ольховниково-ивняково-березовых сообществ (86)
Названные серии долинной растительности характерны для
большинства притоков Оби, протекающих по Сургутской низине (реки Лямин,
Пим, Аган и др.)- Эти реки проложили свои русла среди зандровых
песчаных полей, чередующихся с обширными болотными массивами, что
наложило существенный отпечаток на формирование их растительного
покрова. Наиболее устойчивые конечные стадии долинных серий
представлены березово-сосновыми и сосновыми травяно-зеленомошными (Maianthe-
тит bifolium, Equisetum sylvaticum, Rubus arcticus, Rubus saxatilis, Hylo-
comium splendens, Pleurozium schreberi, Cladina rangiferina, CI. arbuscula)
и лишайниковыми лесами, которые приурочены к старым выположенным
гривам притеррасных частей пойм и первым надпойменным террасам.
В составе лесов сосны и березы в зрелом возрасте (около 100 лет)
достигают высоты 20—25 м и значительного запаса древесины (до ЗЙО м3/га).
В травяно-кустарничковом ярусе преобладают среднетаежные виды
мелкотравья и зеленые мхи. Чисто лишайниковые сосняки характерны
только для наиболее высоких грив, сложенных целиком песчаными
породами. Их древостой разрежен, сосны высотой 15—18 м и диаметром 20—
25 см прямоствольные с высоким прикреплением крон, возобновление
слабое. Зеленые мхи и кустарнички, преимущественно брусника и линцея^
сконцентрированы на приствольных повышениях. Основное пространство
занято лишайниками из родов Cladina и Cladonia. В целом эти типы
сосняков ничем не отличаются от аналогичных водораздельных сообществ.
Процессы заболачивания на участках высоких уровней пойм и
низких террасах идут очень интенсивно, они приурочены ко всем
отрицательным формам рельефа — микропонижениям, ложбинам между рривами,
западинам. Сначала в покрове заболачивающихся лесов исчезают предста-
13* 195
вители таежного разнотравья, а среди мхов появляются кукушкин лен,;
осоки. Далее заболачивание идет по мезоолиготрофному типу с
образованием осоковых и осоково-сфагновых (Carex globularis, С. lasiocarpat
Sphagnum angustifolium, Sph. girgensohnii) лесов, преимущественно с
березовым из Betula pubescens древостоем.
Долинные серии рек восточной части Обь-Иртышского водораздела
(реки Вах, Тым, Васюган и др.) на заключительных стадиях своего
развития имеют несколько иной состав лесных сообществ, что связано с
характером почвообразующих пород, которые имеют здесь суглинистый
механический состав. К таким почвам приурочены смешанные мелколиственно-
темнохвойные, преимущественно березово-кедровые и еловые леса, в
покрове которых преобладают зеленые мхи (Hylocomium splendens, Rhytidia-
delphus squarrosus), таежное мелкотравье (Trientalis europaea, Rubus arcti-
cus, Maianthemum bifolium) и вейник Calamagrostis langsdorfii. Наиболее
широко представлены стадии этих серий, сформированные берёзовыми
(Betula pendula), зеленомошно-травяными (Rubus arcticus, Hylocomium
splendens, Veronica longifolia, Solidago virgaurea) лесами, которые в
основном и получили картографическое отображение вместе со стадиями более
молодых ивняковых и ольховниковых (Salix uiminalis, Duschekia fruticosa)
сообществ (86).
В целом обе анализируемые серии долинной растительности имеют
очень сходный состав начальных стадий пойменной сукцессии. Это в
основном порослевые ивняки на прирусловых отмелях, участки сильна
сомкнутых молодых ивовых лесов «бамбукового типа» или разреженные
ивово-березовые местами с осиной древостой.
ЮЖНО-ТАЕЖНЫЕ ЛЕСО-КУСТАРНИКОВО-ЛУГОВЫЕ СЕРИИ
РАСТИТЕЛЬНЫХ СООБЩЕСТВ
Серия болотистых и настоящих лугов,
ивняково -мелколиственных, смешанных,
темнохвойных и сосновых лесов (108)
Эта серия дшдов растительных сообществ охватывает наибольший по
протяженности отрезок Обь-Иртышской поймы: Ханты-Мансийск—Пара-
бель;, Ханты-Минсийск—Цингалы. Пойма здесь в целом еще сохраняет
проточно-островной характер,, но морфологическое ее строение, главным
образом высота и форма поверхности, заметно меняется на разных участках
[Петров, Бачурин, 1976]. В местах значительно расчлененного рельефа
(на сургутском и александровском отрезках) среди массивов островной
проточной озерно-соровой поймы появляются участки сегментно-гривис-
того типа с характерным веерообразным расположением проток, ложбин
и грив. .
В бе различия в морфологическом строении поймы проявляются на
фоне общих; ее гидролого-геоморфологических особенностей,
обусловленных в первую очередь режимом поемности и высотой элементов
пойменного рельефа. Отсюда основная, наиболее крупная дифференциация
пойменного растительного покрова здесь, как и на других отрезках поймы,,
может быть прослежена по экологическим уровням или высотным поясам
поемности.
Большинство авторов на рассматриваемом отрезке Обь-Иртышской
поймы «выделяют четыре экологических уровня [Роднянская, Самойлова^
1971; Ильина, 1968, 1976; Хромых, 1973]: .
1) участки низкого экологического уровня имеют высоту
относительно меженного уровня до 5 м и заливаются на срок от 1,5 до 3 мес;
2) участки средненизкого экологического уровня ежегодно
заливаются на длительный срок от 1 до 2 мес, они располагаются в интервале
относительных высот 5—7 м;
196
3) участки средневысокого экологического уровня заливаются
периодически один раз в 2—4 года сроком от 0,5 до 1,5 мес, находятся на
высоте 7—8 м;
4) участки высокого экологического уровня относятся к короткопоем-'
ному поясу и заливаются редко — один раз в 8—10 лет на период от 7 до
15 дней. Это наиболее высокие поверхности, принадлежащие пойме.
Особенности растительности каждого экологического уровня
отражены в содержании номеров легенды, соответствующих картируемым
подразделениям.
Ряд осоковых, осоково-канареечниковых лугов и открытых
группировок соровой- растительности характерен для участков самого низкого
экологического уровня (108а). Эти выделы включают в себя обширные
Острова, главным образом центральной части поймы, в которых основные
площади (80—90%) заняты плоскими или слабоволнистыми равнинами
низкого экологического уровня. В середине таких островов обычны соровые
понижения. Но на этом участке Обь-Иртышской поймы соры еще меньших
размеров, чем на предыдущем отрезке Нижнеобской поймы
(Ханты-Мансийск — Березово). Кроме того, их количество постоянно уменьшается по
мере продвижения вверх по течению Оби.
Растительный покров этих участков поймы представлен одним
типичным рядом серийных ассоциаций остро- и водноосо^овых и канареечнико-
вых лугов и открытых соровых группировок. По составу растительности
среднеобские соры несколько отличаются от нижнёобских. В них не
произрастают бекмания и арктофила, гораздо меньше встречается полевицы
ползучей, в сложении травостоев основная роль принадлежит соровому
разнотравью (Eleocharis acicularis, Equisetum palustre, Polygonum scabrum,
Juncus compressus), значительно распространена семенная поросль ив,
хорошо переносящих седиментацию (Salix dasyclados, S. cinerea).
Основные площади в ряду сообществ низкого уровня занимают
болотистые луга, среди которых абсолютно преобладают остроосоковые (Сагех
acuta). Они распространены па плоских пониженных участках, иногда
на днищах старых соров., В таких сообществах еще значительна примесь
сорового разнотравья — Eleocharis acicularis, Juncus compressus, CaWa
palustris, Ranunculus lingua. Чисто осоковые луга на периферийных
участках пойменных островов в местах, несколько приподнятых над центром
или слегка волнистых (от наличия низких наносных гряд), сменяются ка-
нареечниково-осоковыми и осоково-канареечниковыми лугами. В них
обилие осоки и канареечника часто варьирует в зависимости от условий
обводнения [Дыдина, 1961; Хромых, 1973]. '
Для поймы Средней Оби на ее восточном отрезке, расположенном
выше цо течению в пределах Томской области, характерен несколько
другой ряд сообществ низкого экологического уровня (1086).
По данным М. Ф. Елизарьевой [1951], луга низкого уровня поймы
здесь представлены осоковыми кочковатыми вариантами, разнообразие
видового состава в них не превышает 10—12 видов. Обычно среди осоки
острой много вейника, чихотной травы, лютика ползучего, вероники,
подмаренника, василистника, мышиного горошка. В притеррасной пойме
часто на низких уровнях развиваются чистые заросли хвоща' (Equisetum
fluviatile). Осоковые луга низкого уровня переходят в осоковые
кочковатые болота, в которых возрастает обилие Сагех aquatilis, Calamagrostis
neglecta. Местами в наиболее обводненных местах появляются заросли
Phalaroides arundinacea с влаголюбивым разнотравьем — Caltha palustris*
Mentha arvensis, Lythrum salicaria. На низких уровнях центральной поймы
(в низовьях р. Пайдугина Парабельского района) Н. Ф. Вылцан [1964aJ
описаны полевицево-болотномятликово-кайареечниковая, канареечни-
ковая, вейниковая и вейниково-осоковая ассоциации. В широко
распространенных в этом районе вейниковых и канареечниковых лугах она
отмечает, кроме Calamagrostis langsdorffii, Phalaroides arundinacea, обилие
Poa palustris, Equisetum fluviatile, Juncus filiformis, Caltha palustris.
На широтном отрезке Средней Оби широко представлены также
197
участки средненизкого экологического уровня (108в). Они наиболее
типичны для периферийных частей центрально-пойменных островов*
Низкие гривы и межгривные пространства здесь заняты канареечниковыми
лугами. Канареечник тростниковидный (Phalaroides arundinacea) —
основной эдификатор этих лугов, образует в большинстве случаев почти чистые с
небольшой примесью других видов травостои высокой сомкнутости. Осо-
ково-канареечниковые' и осоковые сообщества характерны для межгрив-
ных понижений и нижних частей склонов грив. Выше по течению Оби,,
на томском отрезке Обской поймы, такие сообщества распространены и на
низких гривах центральной поймы [Хромых, 1973].
Относительно высокие местоположения этого экологического уровня
образуют сообщества настоящих лугов. По площади они уступают
болотистым, даже в местах своего преобладания (в краевых частях пойменных
островов) всегда чередуются с болотистыми лугами межгривных
понижений; последние занимают около 30% площади (среди сообществ настоящих
лугов). Сукцессионный ряд сообществ настоящих лугов по мере
уменьшения увлажнения образуют канареечниково-вейниковые и вейниковые с
вейником Лангсдорфа (Calamagrostis langsdorffii) и вейником наземным
(С. epigeios) злаково-разнотравные с ястребинкой зонтичной (Hieracium
umbellatum), хвощом полевым (Equisetum arvense) и бобовыми (Vicia crac*
са, Trifolium medium, Lathyrus pratensis) луга; разнотравно-злаковые с
мятликом полевым (Роа pratensis), вейником (Calamagrostis epigeios) и
лисохвостом (Alopecurus pratensis) луга. На участках, занятых разнотравно-
злаковыми и злаковыми лугами, появляются отдельные группы
кустарниковых ив, главным образом пяти- (Salix pentandra) и трехтычинковой
(S. triandra)*
Вот одно из описаний разнотравно-злакового луга на участке,
расположенном в устье Салымской протоки. Плоская поверхность со слабым
наклоном к центру поименного острова. Хорошо выражены два яруса
травостоя высотой 100—120 см (канареечник) и 50 см (разнотравье).
Проективное покрытие 60—80%. Из злаков преобладают вейник
(Calamagrostis langsdorffii) и канареечник (Phalaroides arundinacea). В значительном
количестве встречаются представители разнотравья: подмаренник (Galium
Ъогеа1е)л анемона вильчатая (Anemonidium dichotomum), вербейник (Lysi-
machia vulgaris), хвощ полевой (Equisetum arvense), таволга вязолистная
(Filipendula ulmaria), девясил (Inula britannica\, дербенник (Lythrum sa-
licaria), птармика (Achillea ptarmica), вероника (Veronica longifolia).
Участки поймы, относящиеся к среднёвысокому экологическому
уровню, представляют собой или группы мелких островов, или значительные
по площади пойменные массивы, где наблюдается большое скопление грив,
чередующихся с понижениями! Для таких местообитаний характерен ряд
растительных сообществ (108г), начальной стадией которого являются
разн9травно-злаковые луга с одиночными кустами ивы. По составу и
структуре они мало отличаются от аналогичных лугов средненизкого уровня.
Выше по склонам эти луга сменяются зарослями ивняков
кустарниковых с разнотравно-вейниковым покровом. Среди ив преобладают Salix
viminalis, S. ЪеЪЫапа, S. pentandra. Под пологом этих ив в густом
травяном покрове преобладают вейники Лангсдорфа (Calamagrostis langsdorffii)
и наземный (С. epigeios), а из разнотравья — подмарейники (Galium bo-
reale, G. palustre), лютики (Ranunculus repens, R. acris), вероника
(Veronica longifolia), герань (Geranium sibiricum), дербенник (Lythrum salicaria)x
таволга вязолистная (Filipendula ulmaria). Сообщества ивняков
кустарниковых не образуют сплошных массивов и на всем протяжении
чередуются с участками разнотравно-злаковых лугов.
Следующее звено этого ряда составляют сообщества ивняков
паркового типа. Древовидные ивы, белая и корзиночная (Salix alba, S.
viminalis), пяти- (Salix pentandra) и трехтычинковая (S .triandra) высотой 10—
12 м, с диаметром стволов до 1,5 м, образуют разреженные насаждения
на плоских вершинах прирусловых грив. Наиболее распространены
ивняки парковые разнотравно-злаковые с преобладанием в покрове вейника
198
Calamdgrostis canescens, мятлика лугового (Роа pratensis), лисохвоста
(Alopecurus arundinaceus) и разнотравные, часто василистниковые (Tha-
lictrum simplex). В годы сильных разливов в покрове парковых ивняков
начинают преобладать осоки и канареечцик.
Для участков высокого экологического уровня характерен ряд
сообществ лесной растительности (108д), р котором значительная рольчпринад-
лежит ивовым и тополевым древостоям, сосредоточенным главным образом
в более приподнятой прирусловой части пойменных массивов. На участке
Средней Оби в пределах Томской области Е. П. Прокопьев [1974] выделяет
следующий ряд ассоциаций ивовых и тополевых лесов, отражающий
постепенное возрастание влажности почв: осокорник кустарникобо-разно-
травный — осокорник и ивняк разнотравный — осокорник и ивня:к разно-
травно-канареечниково-вейниковый — осокорник разнотравно-вейниково-
осоковый — ивняк осоковый — ивняк наумбургиевый — ивняк чистецо-
вый — ивняк хвощовый. Наиболее высокие прирусловые гривы с
аллювиальными песчаными почвами заняты злаково-разнотравными ивовыми
лесами с хорошо развитым подлеском из Swida alba, Ribes nigrum, Rosa
acicularis, R. cinnamomea, Lonicera xylosteum, Sambucus sibirica. В
травяном покрове из злаков преобладают Phalaroides arundinacea и Calamagros-
tis langsdorffii, а из разнотравья — Lysimachia vulgaris\ Thalictrum flavum%
Filipendula ulmaria, Anamonidium dichotomum, Cerastium holosteoides,
Veronica longifolia.
Тополевые леса также приурочены к высоким гривам прирусловой
поймы. Наибольшие по площади массивы образуют осокорники из Populus
nigra, среди которых преобладают разнотравные и кустарниково-разно-
травные [Алехина, 1971]. Древостой обычно смешанные с примесью ивы
и местами белого тополя (Populus alba) I—II классов бонитета. К 30—35-
летнему возрасту насаждения достигают высоты 20 м при среднем
диаметре 22—30 см [Прокопьев, 1974].
Мелколиственные леса, расположенные по рельефу выше ивняков
и топольников, представлены березняками и осинниками (часто
смешанными между собой) кустарниково-вейниково-разнотравными.
Встречаются они небольшими массивами, обычно вытянутыми по вершинам грив»
Сомкнутость древостоев различна — от 0,2 до 0,6. Высота деревьев 20—
22 м при среднем диаметре стволов 18—25 см. Подлесок хорошо развит и
представлен разными видами кустарников: ивами (Salix viminalis, S. beb-
biana), жимолостью алтайской (Lonicera altaica), шиповником (Rosa
cinnamomea), смородиной черной (Ribes nigrum) и ольхой (Duschekia fruticosa).
Травяной покров средней густоты состоит из вейников Calamagrostis
langsdorffii, С. epigeios, С. arundinacea со значительной примесью
костяники (Rubus saxatilis), хвоща лесного (Equisetum sylvaticum), полевицы (Ag-
rostis tenuis), мятлика лугового (Роа pratensis). Менее обильны лютик
ползучий (Ranunculus repens), вероника длиннолистная (Veronica longifolia)f/
вербейник (Lysimachia vulgaris), подмаренник северный (Galium boreale)t
а также злаки — Melica nutans, Milium effusum.
Эколого-динамические ряды растительности, включающие только
лесные сообщества, относятся к поверхностям самых высоких
экологических уровней. Из них к собственно пойменным можно отнести смешанные
хвойно-мелколиственные леса (108е), главным образом сосново-березовые
и елово-пихтово-березовые. По составу и структуре верхних ярусов они
сходны с мелколиственными лесами, однако в травяном покрове их
появляются типично таежные виды — майник, костяника, папоротник Линнея.
Следующие за ними сообщества хвойных лесов четко различаются в
зависимости от характера подстилающих пород. На песчаных массивах,,
в основном примыкающих к правому берегу Оби, встречаются сосновые
зеленомошно-М(Влкотравные и брусничные леса, иногда с пятнами
лишайников (108ж). На останцах центральной части поймы и ближе к
левобережью распространены темнохвойные — елово-пихтовые и кедрово-пнхто-
во-еловые зеленомошно-мелкотравные леса. По составу они близки к
водораздельным, но их подлесок несколько богаче — рябина, шиповник%
199
ю-в -~с-з
50 м
i I
Рис. 21. Геоботанический профиль в пойме р. Оби, район
Растительные сообщества:! — ивовый лес с березой с подлеском ивы, черной^сморо
березовый лес с подлеском ивы, черемухи, калины, жимолости; 4 — заросли кустарниковой ивы;
нареечниково-осоковый луг; 9— вейниково-канареечниковый луг; ю—разнотравно-вейниковыи
13 — канареечниково-осоковый луг; 14 — осоковый закочкаренный луг; 15 — соровое понижение
осоково-канареечниковый луг; 18 — разнотравно-осоковый луг; 19 — молодой
Доминирующие виды: а — осока; б — канареечник; в — вейник; г —^полевица стелю
i видная;
ивы« жимолость, черемуха* Травостой состоит из типично лесных видов:
майника (Maianthemum bifolium), седмичника (Trientalis europaea)f rpy-
шанок (Pyrola rotundifolia^ Orthilia secunda), но в напочвенном покрове
еще нет сплошного яруса мхов, единично встречаются представители пой-
менного разнотравья — Cacalia hastata, Solidago virgaurea.
В заключение краткого описания растительности южно-таежного
отрезка Обской поймы приводим геобот'анический профиль через пойму
Оби в районе пос. Тундрино (рис. 21).
Серия лугово-ивово-тополево-темнохвойных сообществ,
преимущественно еловых и пихтовых травяных лесов (109).
Серия лугово-ивняково-темнохвойных сообществ,
преимущественно еловых и пихтовых
влажнотравных лесов (110)
Долинная растительность наиболее крупных притоков Оби и Иртыша
(реки Демьянка, Салым, Юган, Чулым, Васюган, Кеть и Парабель)
отражена на карте двумя южно-таежными сериями растительных сообществ
(109, 110), а в ряде случаев их отдельными стадиями (109а, 110а, 1106).
В долцнах нижних течений этих рек формируются ряды и серии
сообществ, в которых наиболее полно представлены луговые и ивняково-кус-
тарниковые стадии низких и средних пойменных уровней. В средних
течениях большую пространственную выраженность имекУг поверхности,
высоких уровней поймы, а также участки первой надпойменной ^террасы,
к которым приурочены лесные сообщества. Заключительными стадиями
этих долинных серий являются темнохвойные пихтовые и еловые леса
южно-таежного типа.
Один из наиболее полных динамических'рядов долинной
растительности показан в нижнем течении р. Демьянки. В этом ряду получила
отражение серия одного из южно-таежных типов (109), представленная на
низких уровнях болотистыми, преимущественно осоковыми'и канареечни-
ковыми, лугами и ивняками кустарниковыми (Salix viminalis, S. rosmari-
nifolia); на средних уровнях — ивовыми (Salix alba, S. viminalis) и
тополевыми (Populus nigra) молодыми лесами и рощами и на участках высокого
уровня — березовыми, смешанными и пихтовыми лесами с хорошо
развитым подлеском (свидина, черемуха, шиповник, калина, жимолость) и
высокотравным покровом из щжно-таежных видов (Aconitum septentrionale/
Cacalia hastata, Calamagrostis obtusata) с примесью пойменного
разнотравья (Glechoma hederacea, Geranium sylvaticum, Filipendula ulmaria). Ряд
ассоциаций, показанный в долине среднего течения р. Демьянки (109а),
уже не образует полной серии. Он заканчивается на высоких уровнях со-
200
L •?
f Г,
Г г ' Y д V e
V* $, фв
пос. Тундрино. (Составлен И. С. Ильиной).
дины, свидины; 2 — разреженные группировки из хвоща, мяты, девясила; 3 — молодой осиново-
6 — разнотравно-вейниковый луг; 6 — осоково-канареечниковый луг; 7 — осоковый луг; 8 —
калуг с кустами ивы; 11 — водно-разнотравно-осоковый луг; 12 — вейниково-канареечниковый луг;
с открытыми группировками из ситняга, хвоща, лютика; 16 — осоковый закочкаренный луг; 17 —
березовый лес с кустами ивы и шиповника вейниково-разнотравный.
щаяся; д— влажнотравье; е — соровое разнотравье; ж — ивняк кустарниковый; з—ива древо-
и — береза.
обществами березовых травяных лесов, приуроченных к почвам более
легкого механического состава, в покрове которых много вейника (Calamag-
rostis epigeios). Луговые стадии этого ряда, как правило, не являются
длительными и не занимают больших площадей. Прирусловьш участки поймы
заняты порослевыми ивняками, быстро сменяющимися ивовыми и
тополевыми ленточными лесами, переходящими в притеррасных частях в
молодые березовые и осиновые древостой.
Долинная серия растительных сообществ, свойственная притокам
Оби (110), развивается в несколько иных условиях. Большая часть рек
здесь имеет йониженные заболоченные долины. В центральной пойме и на
низких притеррасных понижениях распространены заболоченные
березовые леса (согра) и болота [Елизарьева, 1951; Прокольев, 1974], а хорошо
дренированные участки высоких уровней заняты смешанными и темно-
хвойными еловыми и пихтовыми разнотравными лесами. Луговые стадии
низких уровней этой серии сходны почти во всех рядах по составу
входящих в них сообществ, но, как и в предыдущей серии, они имеют широкое
распространение только в нижних течениях рек и занимают очень
ограниченную площадь (узкую полосу или отдельные участки) в их. средних и
верхних частях.
В нижнем течении р. Кеть, по данным Н. Ф. Вылцан [19646],
господствуют осоковые, канареечниковые и вейниковые луга, занимающие
наиболее низкие участки прирусловой и центральной поймы. На средних
уровнях их сменяют луга с преобладанием лисохвоста и бобового разнотравья,,
а на высоких — овсяницевые, полевицевые и разнотравно-злаковые
луговые сообщества. В верхних течениях рек Тым и Васюган луговая
растительность отсутствует. На низком уровне там господствуют заросли
ивняков, а прирусловые гривы заселяются всходами березы (Betula pendula)
и сосны сразу же после того, как они выйдут из полосы ежегодного
затопления полыми водамц [Прокощ>ев, 1974].
В средних и верхних течениях большинству описываемых рек
основная площадь поймы и первой надпойменной террасы занята
мелколиственными, смешанными и темнохвойными лесами, образующими различные
по составу нижних ярусов ассоциации в зависимости от услов'ий поемности
и аллювиальности. Наиболее молодые прирусловые и Центральные части
поймы на заливаемых гривах с мощным чехлом аллювиальных отложений
заселены мелколиственными лесами. Среди них наиболее распространены
кустарниково-разнотравные березовые и осиновые насаждения. Это леса
высокой производительности I—1а классов бонитета, сомкнутостью 0,6—
0,8; средняя высота берез и осин 20—25 м при диаметре ствола 20—50 см.
В подросте обычна примесь темнохвойных пород—пихты, ели, кедра. До-
вольдо густой подлесок, состоящий из Padus avium, Sambucus sibirica, Ru-
201
bus idaeus, Sorbus sibirica, Lonicera sibirica, Swida alba. Густой травяной
покров неоднородного сложения. В ложбинах преобладают Veratrum lobe-
Напит, Calamagrostis langsdorffii, Equisetum syluaticum, Galium septentrio-
nale, Athyrium filix-femina; на высоких участках — Aconitum septentrio-
nale, Cacalia hastata, Paris quadrifolia, наблюдается примесь таежного
мелкотравья и зеленых мхов.
Менее заливаемые высокие прирусловые грцвы заняты смешанными
и темнохвойными (из пихты, ели, кедра) .травяными лесами. Сомкнутость
древостоя здесь 0,5—0,6, производительность II—Ш классов бонитета.
Подлесок хорошо развит и состоит из шиповника, рябины, черемухи,
бузины, жимолости, свидины. Травяной покров с проективным покрытием
60—90%, в нем преобладают Calamagrostis obtusata, Solidago virgaureax
Cacalia hastata, Aconitum septentrionale, Senecio nemorensis. В местах
наибольшей сомкнутости древостоя начинают господствовать срнузии
таежного мелкотравья из Oxalis acetosella, Pyrola rotundifolta, Linnaea borealis,
пятнами появляются зеленые мхи.
Выположенные гривы высоких уровней и плоские участки во
внутренних частях поймы и на первой надпойменной террасе заняты темно-
хвойными кустарниково-мелкотравно-зеленомошными лесами, часто
заболачивающимися [Прокопьев, 1974].
В поймах Салыма и Югана на высоких уровнях и на низких первых
надпойменных террасах также распространены темнохвойные и елово-пих-
товые зеленомошно-мелкотравные леса. Для высокой поймы среднего
течения Салыма характерно широкое распространение кедровых сильно
сомкнутых мертвопокровных лесов, приуроченных к вершинам грив с
почвами легкого механического состава [Ильина, Толчельников, 1968].
Описанная серия растительных сообществ (НО) показана на карте в разных
своих стадиях, соответственно пространственной выраженности тех или
иных сообществ. Полная серия (110) видна в долинах Васюгана, Кети и
Чулыма. Ряд сообществ, заканчивающийся березовыми лесами (110а),
отражен в долинах Салыма, Югана и Конды; ряд сообществ с преобладанием
осиновых лесов закартирован в среднем течении Чулыма и Кети (1106).
ПОДТАЕЖНЫЕ ЛЕСО-КУСТАРНИКОВО-ЛУГОВЫЕ
СЕРИИ* СООБЩЕСТВ
Серия разнотравно-злаковых лугов, осиновых,
березовых и тополевых лесов (121)
Эта серия рядов растительных сообществ в основном представлена на
участке Среднеобской поймы Парабель — Кожевникове. Большая часть
пойменных массивов, формирующихся на этом отрезке, может быть
отнесена к сегментно-гривистому типу, для которого характерен
волнообразно-расчлененный рельеф с серией грив, межгривных понижений и
стариц. Здесь яснее, чем на участках Нижней Оби проявляется деление поймы
по поперечному профилю на прирусловую, центральную и притеррасную.
При этом наибольшую площадь занимают участки центральной поймы,
отличающиеся относительно сглаженным рельефом и смешанным составом
поверхностных отложений. В неширокой полосе прирусловой поймы
наблюдается более сильное расчленение — чередование высоких наносных
грив, имеющих большой мощности наилок, и глубоких, узких межгривных
ложбин. Притеррасная часть характеризуется обычно ровной и слабо-
вогнутой*поверхностью с обилием озер и.заболоченных понижений.
Большинство авторов на этом отрезке поймы выделяют три
экологических уровня Шлизарьева, 1951; Вылцан, 1964а; Бокк, 1968]. По данным
Э. Н. Бокк [1968], на участке Верхней Оби от с. Кожевникова до устья
р. Томь низкий уровень поймы с глубиной залегания грунтовых вод в
межень до 1—1,5 м характеризуется периодом затопления более 2 мес,
средний — 1—2 мес при глубине грунтовых вод 1,5—2,5 м, высокий — зали-
202
вается только в годы с максимальным половодьем, и уровень грунтовых
вод здесь опускается ниже 2,5 м/Средние многолетние данные,
приведенные для Колпашевского района в работе А. А. Максимова и Е. П. Мерзля-
•ковой [1972], соответственно характеризуют период затопления разных
высотных уровней поймы*, низин (63-,8 дня), грив (17,3 дня) и всей поймы»
включая наиболее высокие поверхности (2 дня).
Участки низкого экологического уровня на этом отрезке поймы
занимают, как правило, отрицательные формы рельефа. В прирусловой части
к ним принадлежат межгривные понижения и песчаные отмели по берегам
русел и проток. В центральной пойме — наиболее глубоковрезанные
ложбины, а также нижние части склонов невысоких грив, в притеррасной
пойме к участкам низкого уровня относятся плоские заболоченные понижения
и берега пойменных озер и стариц.
В эколого-динамическом ряду растительных сообществ низкого
уровня, получивших отображение на карте (121а), наиболее пониженные и
сильноувлажненные местообитания занимают остроосоковые из Carex
acuta луга, чередующиеся по берегам речек и озер с зарослями Equisetum flu-
viatile, Scolochloa festucacea. На нижних частях склонов грив и в неглубоких
ложбинах Эти луга сменяются осоково-канареечниковыми и канареечни-
ковыми. В Колпашевском районе Н. Ф. Вылцан [1964а, 1968. 1969] на
низких уровнях отмечает распространение канареечниковых, канареечнико-*
во-осоковых, лабазниково-канареечниково-вейниковых. На более южном
кривошеинском участке поймы, по данным Е. Ф. Пеньковской [Лапшина
и др., 1971], канареечниковые луга занимают средние части склонов грив
всех экспозиций. В качестве содоминанта в этих лугах часто выступает
Calamagrostis langsdprffii, в значительном количестве встречаются Сагех
acuta, С. vulpina, Ranunculus repens, Thalictrum baikalense, Lysimachia
vulgaris, Sanguisorba officinalis, Stachys palustris, Veronica longifolia. Мелко-
злэковые луга из лисохвоста (Alopecurus pratensis), полевицы (Agrostis
clavata) и мятлика (Роа pratensis) с большой примесью канареечника и
осоки острой характерны только для вершин наиболее низких грив на
участках центральной поймы и не занимают больших площадей. Гораздо шире
на гривах и в неглубоких понижениях центральной поймы
распространены группы зарослей кустарниковых ивняков из лугово-травяной группы
[Бокк, 1968]. Основной полог в них сложен ивами Salix cinerea и S. ros-
marinifolia с примесью S. dasydados. Из других кустарников к ним
примешиваются Rosa cinnamomea, Ribes nigrum, Viburnum opulus, Lonicera tata-
rica. Травяной покров с общим проективным покрытием от 10 до 60%
составляют в основном Rubus caesius, Filipendula ulmaria, Phalaroides arun-
dinacea, Carex acuta, Cirsium setosum.
Луга среднего уровня занимают большие площади на массивах
центральной поймы. Там они приурочены к вершинам и склонам грив, к
широким выположенным межгривным понижениям. Эколого-динамический ряд
разнотравно-злаковых лугов (1216) образуют ассоциации, состав которых
несколько меняется в северных и южных районах. В Колпашевской пойме
доминантном больщинства луговых сообществ является лисохвост
(Alopecurus pratensis). Образуемые им ассоциации можно расположить по
степени увлажнения в такой последовательности: осоково-канареечниково-
лисохвостная, бобово-осоково-лисохвостная, осоково-мятликово-лйсо-
хвостная, бобово-разнотравно-полевицево-лйсохвостная [Вылцан, 1^64а].
В составе этих лугов кроме лисохвоста доминируют Agrostis clavata, Poa
pratensis, Carex cespitosa, Lathyrus pratensis, Vicia cracca. Из разнотравья
обычны Hieracium umbellatum, Equisetum arvense, Sanguisorba officinalis,
Galium boreale, Ranunculus polyanthemos и некоторые другие мезофиты.
Южнее, • в районе Кривошеино—Кожевникову, ведущее положение
среди луговых сообществ занимает овсяницевая формация (Festuca
pratensis), которая образует полидоминантные ассоциации с участием лисохвоста
(Alopecurus pratensis), пырея (Elytrigia repens), мятлика (Роа pratensis) и
других злаков. Из разнотравья в них обычны представители бобовых (Ьи-
thyrus pratensis) (Vicia cracca, V. sepium) и другие мезофитные виды.
203
Завершающие этот динамический ряд ивовые и тополевые леса
приурочены, как правило, к участкам прирусловой, реже центральной
поймы. Они не занимают больших площадей, и поэтому на карте показаны
только в сочетании с вышеописанными рядайи луговых сообществ.
Эколого-динамический ряд пойменной растительности участков
высокого экологического уровня (121в) включает лесные и луговые
сообщества. Залес&нность и закустаренность пбймы на рассматриваемом отрезке в
Колпашевском районе ниже, чем в Кривошеинском, но и в последнем она
не превышает 30—40% [Лапшина и др., 1971]. На карте получили
отражение в основном наиболее приподнятые участки центральной поймы, а
также останцы надпойменных террас, где под влиянием эрозионной
деятельности водного потока на подмываемых берегах формируются ленты и
полосы ивовых (Salix alba и S. viminalis), тополевых (Populus nigra, P. alba)
и молодых березовых (Betula pendula) древостоев. В этих лесах хорошо
развивается кустарниковый ярус из Salix rosmarinifolia, Swida alba% Ribes
nigrum, R. atropurpureum, Padus avium. Наиболее высокие гривы
центральной поймы покрыты тоже молодыми осиновыми из Populus tremula
и березовыми из Betula pendula лесами, местами чередующимися с
зарослями Rosa cinnamomea или на опушках с группами кустов Viburnum opulus
и Spiraea salicifolia. Более старые выположенные гривы, а также останцы
надпойменных террас, встречающиеся среди поймы, заселены березовыми
(Beш1а pendula), осиновыми, а иногда сосново-березовыми лесами. В таких
лесах обычно хорошо развив подлесок из Rosa acicularis,^ Rhamnus catha-
rtica, Padus avium, Viburnum opulus. Травяной покров состоит в основном
из представителей мезофитного высокотравья (Thalictrum simplex, T.
minus, Lythrum salicaria, Filipendula ulmaria, Cnidium dubium) и злаков
(Agrostis gigantea, Poa pratensis, Calamagrostis canescens). Хвойные йеса
на этом отрезке поймы встречаются исключительно редко. У с. Батурино
Э. Н. Бокк [1972] отмечает на высоких прирусловых гривах отдельные
участки еловых и кедровых древостоев. Участки лесов обычно чередуются
с луговыми сообществами на более выположенных гривах высокого
уровня. В целом луга здесь занимают довольно значительные площади. По
описанию Н. Ф. Вылцан [1964а] на Колпашевском участке поймы наиболее
распространены ассоциации мятликово-овсяницевых, мятликово-полеви-
цевых, лисохвостно-овсяницево-мятликовых лугов.
Серия крупно- и мелкозлаковых лугов,
низинных пойменных болот, нвово-ольховых, n
двово-мелколиственных, местами березово-сосновых лесов (122)
Выделенная обобщенная серия сообществ характеризует
растительность поймы Иртыша на большом отрезке его среднего и нижнего течения
от пос. Цингалы до пос. Брлыперечье. На рассматриваемом отрезке ИртыЩ
формирует двухстороннюю сегментно-гривистую пойму шириной от 5 до
10 км. Эта пойма состоит из множества разновозрастных и разновысотных
поименных массивов, представляющих собой системы валов и понижений
[Петров, Бачурин, 1976]. По высоте над меженным урезом Иртыша на
разных его отрезках выделяется несколько ярусов рельефа. На одном из
пойменных массивов в нижнем течении (ИрымсКий пойменный массив), по
данным И. В. Петрова, нижний ярус рельефа располагается в интервале
высот 4,5 м от уреза межени, средний — от 4,5 до 8,5 и высокий — выше
8,5 м над урезом средней многолетней межени [Ильина и др., 1977].
Режим поёмности резко меняется на участках с разнрй высотой пойменного
рельефа. Для Тобольского.района поймы Иртыша средняя
продолжительность заливания грив высокого уровня, которые покрываются водой только
в многоводные годы, 39 дней, среднего уровня (также в
многоводные годы) — 90 дней и низкого уровня — 118 дней [Петров, 1979].
Участки с преобладанием поверхностей низкого экологического
уровня (122а) выделены в основном в северной части рассматриваемого отрезка
Нижнеиртышской поймы. Этот район находится в области Ханты-Мая-
204
сийской впадины, которая испытывает прогибание [Чернышева, 1970L
Отсюда широкое распространение на многих пойменных массивах
обширных выровненных поверхностей, по высоте соответствующих уровню
невысоким половодьям [Петров, Бачурин, 1976]. Местами сюда проникают
с севера участки проточно-островной соровой поймы, но в целом по типу
строения эта пойма также относится fc сегментно-гривистому типу.
Растительность молодых прирусловых частей поймы не нашла отображения на
карте, на ней показаны преимущественно средневозрастные и старые
центральные части пойменных массивов, занятые выположенными низкими гри-
вами> участками плоских или слабовогйутых равнин, а также обширными
заиливающимися зарастающими староречьями. Последние характеризуют
самые обводненные местообитания. Образующиеся здесь фитоценозы по
составу и структуре часто близки к соровым. Местами они сформированы
отдельными экземплярами хвоща топяного (Equisetum fluviatile), сусака
зонтичного (Butomus umbellatus), осоки острой (Carex acuta) и других
видов, хорошо выносящих длительное затопление. Их сменяют чисто
осоковые луга с высоким проективным покрытием (100%), состоящие из осоки
острой (Carex acuta) с примесью других видов осок: дернистой (С. cespitosa)f
пузырчатой (С. lesicaria), ситниковой (С. juncella). Высота генеративных
побегов осок 110—120 см. Представители разнотравья встречаются
единично: хвощ полевой {Equisetum arvense), лютик (Ranunculus linqua),
незабудка (Myosotis palustris), дербенник (Lythrum salicaria), подмаренник
северный (Galium boreale).
Наиболее устойчивые сообщества этого ряда образуют осоково-кана-
реечниковые и осоково-вейниковые луга, приуроченные к плоским гривам
и выровненным участкам центральных частей пойменных массивов. По
составу эти луга очень близки к аналогичным канареечниковым и вейнико-
вым лугам среднего уровня, описанным для более северных среДнеобских
участков поймы, но в этом районе они приурочены к местообитаниям более
низкого экологического уровня; отличаются несколько меньшим видовым
разнообразием (рис. 22).
Для южных районов рассматриваемого отрезка Иртыша наиболее
характерен ряд растительных сообществ разнотравно-осоковых лугов,
сочетающихся с евтрофными осоково-тростниковыми болотами (1226).
Большие площади поверхностей низкого уровня здесь свойственны
слабовогнутым равнинам притеррасных частей поймы, а также широким выположен-
ным долгопоемным межгривным ложбицам центральной поймы.
Характерно, что значительная часть этих низких поверхностей заболочена.
Например, в Тевризском районе Омской области на долю заболоченных земель
приходится около 22% общей площади [Прокопьев, 1978].
Наиболее дренированным местообитаниям низкого экологического
уровня свойственны разнотравно-осоковые луга с господством осоки
острой, нередко образующей кочки, и со значительным участием
гидрофильных видов разнотравья: Myosotis palustris, Galium uliginosum, Lythrum
salicaria, Ranunculus linqua. В более обводненных местах с высоким стоянием
грунтовых вод развиваются осоково-тростниковые (Phragmites australis,
Carex cespitosa, C. acuta) болота, иногда с большим участием манника (Gly-
ceria maxima) и тростянки (Scolochloa festucacea). В притеррасной части
преобладают чисто осоковые кочкарные болота из Carex cespitosa, С. juncellaf
нередко закустаренные. Встречаются также залесенные березово-осоковые
и сосново-сфагновые болотные фитоценозыч [Прокопьев, 19781. На южном
лесостепном отрезке Верхнеиртышской поймы обширные притеррасные
понижения Ьбычно заняты солончаковатыми лисохвостными (Alopecurus
arundinaceus) лугами и галофитно-разнотравно-бескильницевыми (PuccU
nellia distans, Plantago maritima) группировками по солончакам [Ильина,,
Скарлыгина-Уфимцева и др., 1983]. [
Показанные на карте ряды растительных сообществ участков среднего
экологического уровня (122в, г) характерны в основном для центральной
поймы Среднего Иртыша и колыванского отрезка Верхнеобской поймы.
Здесь широко распространены разнообразные по видовому составу .
205
ю-з ►с-в
4
О
J. I l i I I I I U
1 2 J 4 5 6 7 8 9
Рис. 22. Геоботанический профиль в пойме р. Иртыш, Цимбельский поймен
Растительные сообщества: 1 — разреженный ивовый лес разнотравно-канареечнико
заболоченный луг; 5 — ивняк кустарниковый высокотравный; 6 — осоковый луг; 7 — ивняково-
ивняково-кустарниково-бобово-разнотравно-злаковый луг; 11 — осоковый кочковатый заболочен
ренное болото; 14 — ивняково-кустарниково-бобово-разнотравно-злаковый луг; 15 — осоковый луг;
няк с кустами ивы злаково-
Доминирующие виды: а — осока; б — канареечник; в — вейник; г — мятлик, пырей;
ива корзиночная; к — ивняки кустарничковые
разнотравно-злаковые луга, приуроченные к пологим склонам и плоским
вершинам грив, а также к широким днищам неглубоко врезанных ложбин.
В пойме Нижнего Иртыша состав этих лугов более мезофитен — здесь
доминируют канареечник, мятлики (Роа pratensis, P. palustris), вейник (Са-
lamagrostis epigeios), пырей (Elytrigia repens) и бобовое разнотравье (Lathy-
rus pratensis, Trifolium medium). На Среднем Иртыше, ближе к Болыпе-
речью, наблюдается заметная ксерофитизация и галофитизация лугового
травостоя. На лесостепном отрезке Омской поймы, где в настоящее время
нарушен естественный режим поемности и обширные пространства
центральных и притеррасных частей в течение многих лет не заливаются
паводковыми водами, процессы засоления и иссушения идут особенно
интенсивно. Здесь широко распространены ксерофитно-разиотравно-злаковые^
преимущественно тиичаковые с Festuca valesiaca степные сообщества в
комплексе с группировками полыни (Artemisia nitrosa, A. austriaca, A.schren-
kiana) no солонцам [Ильина, Скарлыгина-Уфимцева и др., 1983].
Большая часть лугов среднего уровня используется в качестве
сенокосов и пастбищ, в связи с этим на карте получили отображение в основном
вторичные фитоценозы этих лугов.
На высоких экологических уровнях пойменных массивов Иртыша
распространены преимущественно лесные сообщества. Среди них
абсолютно преобладают мелколиственные березовые и осиновые леса; небольшими
участками встречаются также березово-сосновые древостой (122д).
Все они приурочены к наиболее старым сегментам поймы и
развиваются на дерново-подзолистых суглинистых или супесчаных почвах. Наиболее
типичны березовые и осиново-березорые с густым подлеском шиповника
злаково-разнотравные леса. Древостой их высокой сомкнутости (0,7—0,8)
и высокой производительности (II—III классы бонитета). Хорошо
выражены подрост и подлесок. Травостой с проективным покрытием 60—
70% имеет в составе злаки: Роа pratensis, Agrostis alba, Festuca ovina, Phle-
um pratense и мезофитное разнотравье Rubus saxatilis, Lythrum salicaria,
Anemondium dichotomum. Geranium sylvaticum.
Смешанные березово-сосновые леса завершают сукцессионные ряды
смен растительности на участках пойм, сложенных легкими песчаными
отложениями. По составу и структуре эти леса являются наиболее
устойчивыми [Ильина и др., 1977]*
Серия лугово-ивово-тополево-березовых или осиновых,
местами с примесью хвойных пород, сообществ» (123)
Долинная растительность малых рек подтаежного типа показана
одной серией лугово-кустарниково-лесных сообществ. В ее составе набор
растительных группировок разного эколого-фитоценотического состава,
приуроченных к различным поясам поемности. Значительные площади в
долинах Томи, Верхней Оби, а также иртышских притоков — Тобола, Тавды
и Ишима занимают луга. Они распространены преимущественно на
низких и средних экологических уровнях и приурочены к прирусловым и цен-
206
О 50м
Уд v e fж
Уз Ъи Фн Фл
10 11 12 13 П 15-16 17 18
ный массив. (Составлен И. С. Ильиной, И. Б. Петровым, Л. П. Соколовой).
вый; 2 — осоковснканареечниковый луг; 3—вейниково-осоковый луг; 4 — осоковый кочковатый
кустаряиково-разнотравно-злаковый луг; 8 — разнотравйо-злаковый луг; 9 — осоковый луг; 1 о —
ный луг; 12—ивняково-кустарниково-бобово-разнотравно-злаковый луг; 13— осоковое закочка-
16 — бобово-разнотравно-злаковый луг; 17 — гигрофитно-разнотравно-осоковый луг; 18 — берез-
костянично-разнотравный.
д — бобовое разнотравье; е — кустарнички; ж — соровое разнотравье; з — высокотравье; и —
(смешанные); л — ива древовидная; ж — береза.
тральным частям поймы. Ведущее место среди луговых сообществ
занимают овсяницевые (Festuca pratensis), лисохвостные (Alopecurus pratensis)%
ежовые (Dactylis glomerata) и полидоминантные
разнотравно-мелкозлаковые луга центральной поймы [Елизарьев&, 1951; Вылцан, 1969], на всем
протяжении чередующиеся с участками зарослей кустарниковых ив и то-
полево-березовых рощ. В межгривных понижениях господствуют осоковые
(Carex acuta, С. cespitosa) и крупнозлаковые (Phalaroides arundinacea, Poa
palustris, Calamagrostislangsdorffii)jiyT8L. В прирусловых участках поймы,
главным образом на прибрежных участках отмелей, узкими полосами
распространены хвощовые (Equisetum arvense), костровые (Bromopsis inermis)%
полевицевые (Agrostis gigantea) луга.
В описываемой серии долинной растительности очень большая роль
принадлежит сообществам ивняков, образующих как кустарниковые
заросли, так и лесные насаждения. И те и другие свойственны наиболее
молодым прибрежным и центральным частям поймы. В прирусловой пойме
у самых берегов рек образуются заросли молодых порослевых ивняков,
главным образом Salix viminalis. Несколько дальше от русла на
пониженных, сильно затопляемых (до 2 м в половодье) участках формируются
высокоствольные (до 20 м) ивовые древостой из Salix alba высокой
сомкнутости (0,7—0,8), но с разреженным травяным покровом
преимущественно из крупнотравья, без подлеска и подроста. Далее на средних элементах
рельефа с уровнем затопления около 1 м господствуют ежевичные ветля-
ники, осокоре-ветляники {Salix alba,- Populus tremula) и топольники (Ро-
•pulus nigra, P. alba) с хорошо развитым подлеском из смородины красной
и черной, крущины ломкой, калины обыкновенной, шиповника и др.
[Бокк, 1972].
В центральной пойме на средних уровнях преобладают заросли
ивняков кустарниковых, главным образом Salix cinerea, S. dasyclados с
примесью S. pentandra и S. rosmarinifolia. Они постоянно чередуются с
большими [участками лугов. На высоких-уровнях прирусловой поймы
поселяются ветловые и осокоревые леса, чередующиеся с березняками и
осинниками. Средняя высота древостоя в ивовых лесах 20—22 м и сомкнутость
0,5—0,6. В подросте много березы, попадаются единичные экземпляры
елей и кедров. Эти леса имеют густой подлесок из акации желтой (Caragana
arborescens), ивы (Salix dasyclados), черемухи (Padus auium), рябины (Sor-
bus sibirica). В хорошо развитом травяном покрове обильно встречаются
дягиль сибирский (Angelica decurrens), ежевика (Rubus caesius), хвощ
луговой (Equisetum pratense), какалия (Cacalia hastata), папоротник (Matteuccia
struthiopteris), будра (Glechoma hederacea). Такие ветляники уже через 50—
60 лет 'сменяются елово-березовым древостоем [Бокк, 1972]. Березовые
и осиновые леса в долинах этих рек занимают довольно большие площади
как на высоких прирусловых гривах, так и во внутренних притеррасных
частях пойм и на первых надпойменных террасах. На прирусловых
гривах они имеют наиболее высокую производительность (I класс бонитета) ^
высокую сомкнутость, хорошо развитый подлесок и густой,
преимущественно высокотравный [и крупнозлаковый, покров. Во внутренних частях
207
пойм на притеррасных равнинах эти леса менее производительны, более
разрежены и часто заболочены. Местами они переходят в березово-сосно-
вые осоковые болота или березовое заболоченное редколесье — согру
[Хромых, 1973].
ЛЕСОСТЕПНЫЕ КУСТАРНИКОВО-ЛУГОВО-СТЕПНЫЕ СЕРИИ
РАСТИТЕЛЬНЫХ СООБЩЕСТВ
Серия разнотравно пз лаковых лугов,
низинных осоково-тростниковых болот
и мелколиственных лесов (130)
Выделенная серия сообществ характерна в основном для долин
крупных притоков Иртыша — Тобола, Дшпма, Вагая в их среднем и верхнем
течении. Некоторые ряды сообществ, принадлежащие этой серии,
показаны такя^е в.пойме Верхней Оби\— выше Новосибирского водохранилища.
Долинные системы грив и межгривных понижений на этих участках
поймы четко связаны с главным, более спрямленным [и почти [не меандри-
рующим руслом речного потока. Поэтому они развиваются в направлении,,
параллельном главному руслу реки, и обнаруживают ясно выраженное
деление по поперечному профилю на горизонтальные зоны: более высокую
и расчлененную гривисто-ложбинную прирусловую часть- поймы, полого-
гривистую центральную и равнинно-западинное притеррасье. В
последнем часто сохраняются отдельные старые массивы высокой пологогриви-
стой поймы, почти незаливаемой. • '
В составе и зональных особенностях долинной растительности здесь
тоже есть свои специфические черты. Указанные реки пересекают
водораздельные пространства, покрытые лесостепью. Влияние зональных
факторов сказывается в формировании степных фитоценозов на высоких
плоских и удаленных от реки гривах [Пеньковская, 1963а], а широко
распространенные на высоких уровнях мелколиственные сообщества
лесостепного типа — березовые колочные леса являются субклимаксовыми
заключительными стадиями сукцессионных рядов, что и позволило нам отнести
рассматриваемую серию к лесостепному типу долинной растительности.
Первый эколого-динамическпй ряд (130а) характеризует
растительность тех участковЪоймы,на которых преобладают пониженные заозерен-
ные и заболоченные поверхности, т. е. наиболее хорошо выражены
притеррасные части. Однако начинается этот ряд молодыми сообществами
прирусловых грив, представленными в основном зарослями ивняков
кустарниковых (Salix viminalis), далее переходящих в ивовые (Salix alba) и
тополевые (Populus nigra) леса. В цритеррасной части поймы, кроме
отмеченной выше заболоченности, очень характерна также галофитность
растительности. Встречающиеся здесь солончаковые луга имеют в качестве
доминирующих видов Agrostis gigantea, A. alba и Alopecurus arundinaceus;
в составе их'травостоя присутствуют представители галофитного
разнотравья — Saussurea amara, Juncus gerardiif Scorzonera paruifloray Plantago
cornuti, Triglochin maritima. Низкие берега временных водотоков и
зарастающих стариц часто покрыты тростниковыми (Phragmites australis)
и тростниково-осоковыми (Carex cespitosa) евтрофными незасоленными
болотными сообществами.
Ряд растительных сообществ, в котором широко представлены
сообщества разнотравно-злаковых лугов, в значительной мере остепненных
и засоленных (1306)^ характерен для тех участков поймы, где хорошо
выражены поверхности среднего и высокого экологических уровней. Это
преимущественно центральные части пойменных массивов.
Одной из наиболее распространенных является ассоциация разнотрав-
но-пырейная с господством Elytrigia repens, значительным участием Вго~
mopsis inermiSy Poa iangustifolia и представителей мезофитного
разнотравья — Trifolium pratense,. Ranunculus repens, Cnidium dubium, Filipen-
dula ulmaria% Cirsium ^setosum.
208
На многих гривах центральной поймы, особенно вблизи глубоковре-
занных ложбин, появляются заросли ивняков кустарниковых, главным
образом Salix alba. Если эти гривы более старые и пологие, то они
заселяются лесными сообществами, в основном березняками. Более
производительными и богатыми по составу являются леса высоких грив
прирусловой зоны. В описываемый ряд сообществ включены в основном
ленточные ивово-березовые и ивово-тополевые леса, которые широкими полосами
окаймляют русла главного потока реки и характеризуются хорошо
развитым древостоем высокой сомкнутости 0,7—0,8, разнообразным
подлеском (смородина, шиповник, калина, жимолость, черемуха, ивы1 и богато-
разнотравным покровом, в котором встречаются Thalictrum minus, Th.
simplex, Lythrum salicaria, Geranium sylvaticum, Fragaria viridis, Rubus saxati-
lis, Achillea millefolium и другие виды.
На старых, удаленных от основного русла реки возвышенных
массивах центральной и притеррасной поймы, а также на первой надпойменной
террасе встречаются довольно большие массивы лесов. Среди них наиболее
распространены березовые (Betula pendula) леса (130в). Они чаще всего
разрежены и остепнены. Сомкнутость этих лесов не превышает (1,4—0,5,
высота деревьев до 18—20 м и диаметр стволов 25—30 см. В подлеске
обычно встречаются шиповник (Rosa cinnamomea) и кусты ив (Salix cine-
rea, S. caprea). Покров довольно разрежен (50—60), в нем кроме злаков
Роа angustifolia, Festuca ovina, Bromopsis inermis распространены лугово-
степные, в том числе галофитные виды FiUpendula vulgaris, Galatella Ъ\-
flora, Agrimonia pilosa, Brachypodium pinnatum, Eryngium planum. Более
мезофитный вариант этих лесов обычно имеет вейниково-разнотравный
покров с Calamagrostis epigeios, Pulmonarid dacica, Rubus saxatilis,
FiUpendula ulmaria, Hieracium umbellatum.
Серия разнотравно-злаковых и злаковых лугов
и злаково-разнотравных луговых степей (136).
Серия разнотравно-злаковых с галофитными элементами лугов
и злаково-галофитноразнотравных луговых степей (137)
Эти серии долинной растительности характеризуют растительный
покров поймы верхнего течения Обгг, а также растительность верховий
некоторых притоков Иртыша — Тобола, Тавды и др.
Детальные геоботанические исследования поймы Верхней Оби в этом
районе проводились Е. Ф. Пеньковской [1963а, б J, а еще раньше В. И.
Барановым и В. А. Шелудяковой [1929], Г. Я. Бронзовой [1929]. По
морфологическому строению пойма верхнего течения Оби также относится к сег-
ментно-гривистому типу.
Для наиболее выположенных и занимающих большую часть долины
участков центральной поймы выделена серия сообществ
разнотравно-злаковых лугов и злаково-разнотравных луговых степей (136). Самые большие
площади в центральной пойме занимают разнотравно-злаковые с мятликом
узколистным (Роа angustifolia), овсяницей (Festuca pratensis) и полевицей
(Agrostis tenuis) луга. Они приурочены к склонам и вершинам невысоких
выположенных грив, заливаемых в половодье почти ежегодно, но на
непродолжительное время.
Наиболее зысокие гривы центральной поймы, редко заливаемые во
время весенних разливов, обычно бывают заняты более ксерофитными
лугово-степными сообществами — костровыми (Bromopsis inermis) или
осоково-кострово-типчаковыми с Festuca pseudovina, Bromopsis inermis,
Carex duriuscula. В них значительна примесь чисто степных ксерофитов —
Koeleria cristata, Artemisia frigida [Пеньковская, 19631.
Следующая серия растительных сообществ (137) выделена для тех
участков поймы, где, -с одной стороны, хорошо выражена расчлененная
высокогривистая прирусловая часть, но с другой — хорошо
представлены центральные и притеррасные, более пониженные и засоленные, ее
массивы. Закартированный обобщенный эколого-динамический ряд состоит
14 Заказ № 297 ' 209
из сообществ костровых и вейниковых (Calamagrostis epigeios) лугов,
типичных для прирусловой поймы и, как правило, сочетающихся там с
ленточными зарослями ивняков кустарниковых по берегам проток. Но
большую часть территории занимают здесь галофитно-разнотравно-злаковые
с Hordeum brevisubulatum, Leymus paboanus, Limonium gmelinii, Phleum
phleoides луга, злаково-галофитно-разнотравные с Galatella biflora, Arte-
misia glauca луговые степи, приуроченные к выположенным низким гривам
притеррасной поймы и к шлейфам гривообразных склонов останцов
надпойменных террас.
*
В приводимом в конце книги «Списке латинских названий растений»
дается перечень основных видов растений, зарегистрированных на
территории Западно-Сибирской равнины, с распределением их по группам
формаций. Цифрами в таблице показываются подзональные подразделения,
в которых данный вид имеет распространение: 1 — арктических тундр,
2 — субарктических тундр, 3 — редколесий, 4 — северотаежная, 5 —
среднетаежная, 6 — южно-таежная, 7 — подтаежная, 8 — лугово-степ-
ная (северная), 9 — лугово-степная (южная), 10 — разнотравно-дерновин-
но-злаковых настоящих степей.
Латинские названия видов сосудистых растений даны по «Флоре СССР»
с некоторыми изменениями, согласно последней сводке по синонимике
видов в книге С. К. Черепанова [1981J. Старые названия растений,
упомянутые в легенде карты «Растительность Западно-Сибирской равнины»,
приводятся в списке в качестве синонимов *.
-о
ЛИТЕРАТУРА
Аврамчик М. Н. К подзональной характеристике растительното покрова тун|и>ы,
лесотундры и тайги Западно-Сибирской низменности.— Ботан. журн., "1969,
т. 54, № 3, с. 410-420.
Александрова В. Д. Арктические тундры СССР. Автореф. докт. дис. Л., 1964. 43 с.
Александрова В. Д. Принципы зонального деления растительности Арктики.— Ботан.
журн., 1971, т. 56, № 1, с. 3—21.
Александрова В. Д. Геоботаническое районирование Арктики и Антарктики. Л.:
Наука, 1977. 188 с.
Алехина А. Ф. Топольникп поймы р. Оби. Автореф. канд. дис. Свердловск, 1971. 23 с.
Алисов Б. П. Климат СССР. М.: Высш. шк., 1969. 104 с.
Амромин П. Д., Богоявленский Б. А. Об использовании материалов лесоустройства
при съемке. М 1 : 10 000 и 1 : 25 000.— Геодезия и картография, 1966, № 7,
с. 51-53.
Андреев В. Н. Типы тундр запада. Большой атлас Земли.— Тр. Ботан. музея АН СССР,
1932, вып. 25, с. 121—268.
Андреев В. Н. Кормовая база Ямальского оленеводства.— Сов. оленеводство, 1934,
вып. 1, с. 99—164.
Андреев В. Н. Растительный покров Восточно-европейской тундры и мероприятия по
его использованию. Автореф. докт. дис. Л., 1954. 39 с.
Андреев В. Н., Игошина К. И., Лесков А.И.Оленьи пастбища и растительный покров
Полярного Приуралья.— Сов. оленеводство, 1935, вып. 5, с. 171—406.
Андреев В. Н., Панфиловский А. Л. Обследование тундровых пастбищ с помощью
самолета. Вып. 1. 1938. 172 с.
Аралова Н. С. Растительность — индикатор состава и свойств грунтов северной тайги
Западной Сибири. Автореф. канд. дне. М., 1971. 23 с.
Атлас ареалов и ресурсов лекарственных растений СССР. М., ГУГК, 1976. 340 с.
Баранов В. И. Очерк растительности Калачинского уезда Омской губ.— Тр. Сиб.
с.-х. акад., 1923, вып. 2, с. 77—137.
Баранов В. И. Растительность черноземной полосы Западной Сибири.— Зап. Зап.-
Сиб. отд-ния РГО, 1927, т. 39, №3, с. 1—162.
Баранов В. И. Почвы и растительность Демъян-Р1ртышского водораздела в бассейнах
рек Урны и Утугаса.— В кн.: Материалы по изучению Тарского Васюганья.
(Районное переселенческое управление Сибири. Исследование колонизационных
фондов). Новосибирск, 1928, с. 1—70.
Баранов В. И., Шёлудякова В. А/Материалы к познанию лугов и болот долины
р. Оби.— В кн.: Труды Сибирского ин-та сельского хозяйства и лесоводства.
' Т. X, вып. 1-6. Омск, 1928,' с. 463-510.
Барышников М. К. Осоково-гигшовые болота Западного Васюганья (Нарымский
край).— Бюл. Ин-та луговой и болотной культуры им. В. Р. Вильямса, 1929,
№ 2, 38 с.
Барышников М. К. Луга Оби и Иртыша Тобольского Севера. М.: Изд-во Гос. землобъ-
единения, 1933. 95 с.
Барышников М. К. Луга низовьев р. Оби, их характеристика и перспективы
использования.— Тр. НИИ с.-х. Крайнего Севера, 1961, т. 10, с. 115—158.
Белов А. В. Карта растительности юга Восточной Сибири.— В кн.: Геоботаническое
картографирование. Л.: Наука, 1973, с. 16—30.
Белов А. В., Букс И. И., Ильина И. С. Вопросы составления карты растительности
Азиатской России (структура легенды, методы и принципы ее составления).—
Докл. Ин-та геогр. Сибири и Дальнего Востока СО АН СССР, 1972, вып. 36,
с. 30—40.
Бобров Е. Г. Лесообразующие хвойные СССР. Л.: Наука, 1978. 188 с.
Богоявленский Б. А. Картографирование таежной растительности по спектрозональ-
ным аэроснимкам. Материалы к симпозиуму на IV совещании географов Сибири
и ДВ. Иркутск: изд. ИГСиДВ СО АН СССР, 1969.
Богоявленский Б. А., Ильина И. С. О методике составления обзорной гсоботанической
карты Западно-Сибирской равнины.— В кн.: Проблемы, тематического
картографирования. Иркутск: изд. ИГСиДВ СО АН СССР, 1970, с. 236—244. %
Богоявленский Б. А., Попов Л. В. Об использовании материалов лесоустройства при
создании геоботанических карт.— В кн.: Вопросы тематического
картографирования. Материалы к III научно-технической конференции по
картографированию. Иркутск, 1968.
14*
211
Богоявленский Б. А., Дибцев Е. Н., Ильина И. С. и др. Западная Сибирь1 с самолета
(аэровизуальное обследование для обеспечения мелкомасштабного
картографирования Западной Сибири).— Там же, с. 146—148.
Богоявленский Б. А., Дибцев Е. Н., Михайлов Ю. П. и др. Комплексное
аэровизуальное обследование Западной Сибирп.— В кн.: История развития речных долин
и проблемы мелиорации земель. Западная Сибирь и Средняя Азия. Новосибирск:
Наука, 1979, с. 169—192.
Бокк Э. Н. Ивняки поймы Верхней Оби. Автореф. канд. дис. Новосибирск, 1968. 28 с.
Бокк Э. Н. Общие черты динамики прирусловых ивовых лесов в пойме Оби.— Тр.
Биол. пн-та СО АН СССР, 1972, вып. 15, с. 124—132.
Болота Западной Сибири, их строение и гидрологический режим. Л.: Гидрометеоиздат,
1976. 446 с. '
Боч М. С, Герасименко Т. В., Толчельников Ю. С. Болота Ямала.— Ботан. журн.,
, 1971, т. 56, № 10, с. 1421—1435.
Бронзов А. Я. Верховые болота Нарымского края (бассейн р. Васюган). М., 1930. 99 с.
Бронзов А. Я. Гинновые болота на южной окраине Западно-Сибирской равнинной
тайги.— Почвоведение, 1936, № 2, с. 224—245.
Бронзова Г. Я. По обским лугам (предварительное сообщение).— Изв. Гос. лугового
ин-та, 1929, № 4—6, с. 72—92.
Букс И. И. Принципы составления легенды мелкомасштабной обзорной карты
растительности Азиатской России (на примере тундровой растительности).— В кн.:
Почвы и растительность мерзлотных районов СССР. Магадан: изд. ДВНЦ
АН СССР, 1973, с. 110—114.
Буданцев Л. Ю. Некоторые проблемы изучения флоры Арктики эпохи раннего кайно-
фита.*— В кн.: Северный Ледовитый океан и его побережье в кайнозое. Л.:
Гидрометеоиздат, 1970, с. 76—86.
Вагина Т. А. Луга Барабы. Новосибирск: изд. РИО СО АН СССР, 1962. 197 с.
Вагина Т. А. Растительность.— В кн.: Структура, функционирование и эволюция
системы биогеоценозов Барабы. Т. 1. Новосибирск: Наука, 1974, с. 189—204.
Вагина Т. А. Генезис и структура солонцеватых степей Барабы.— В кн.: Стенная
растительность Сибири и некоторые черты ее экологии. Новосибирск: Наука, 198^,
с. 108-111.
Вандакурова Е. В. Растительность Кулундинской степи. Новосибирск: изд. Зап.-Сиб.
фил. АН СССР, 1950. 128 с.
Васильев В. II. Байкало-Саянская область как один из главнейших центров
формирования рода Beiula L.— Тр. Ин-та леса АН СССР, 1958, т. 37, с. 120—141.
Васильев fe. Н. Березы Урала,— Тр. Ин-та экологии раст. и живот. УФ АН СССР,
1969, вып. 69, с. 59—140. .
Василюхина М. А/Южные пределы распространения элементов бореальной флоры на
юге Западной Сибири и в Северном Казахстане.— В кн.: Ареалы растений
флоры СССР. Вып. 2. Л.: Изд-во Ленингр. ун-та, 1969, с. 195—228.
Василюхина М. А., Лавренко И. Н. О возможности применения фитохорологических
данных в геоботаническом картографировании.— Вестн. Ленингр, ун-та, 1977,
, № 9, с. 29-33. s
Вашкевич Ф. В. Изучение пастбищ оленей на Тобольском Севере,— В кн.: Сборник по
оленеводству, тундровой ветеринарии и зоотехники. М.: Власть Советов, 1932,
с. 63-97.
Вильяме В. Р. Почвоведение. М.: Сельхозгпз, 1946. 456 с.
Вдовин В. В. Становление западно-сибирской эпигерцинской плиты и развитие ее
поверхности в мезозойскую и в начале кайнозойской эры как предыстория
формирования ее современного рельефа.— В кн.: Западно-Сибирская равнина. М.:
Наука, 1970, с. 10—44.
Волкова В. С. История развития растительности и основные этапы палеогеографии
низовья Иртыша в четвертичное время.— В кн.: Основные проблемы изучения
четвертичного периода. М.: Наука, 1965, с. 342—354.
Волкова В. С. Стратиграфия и история развития растительности Западной Сибирп в
позднем кайнозое. М.г Наука, 1977. 238 с.
Вылцан Н. Ф. Материалы к изучению пойменных лугов р. Оби,— Учен. зап. Томск,
ун-та, 1964а, № 49, с. 158—167.
Вылцан Н. Ф. Пойменные луга р. Кети (Томская обл.).— Изв. Томск, отд-ния Всесо-
юз: ботан. о-ва, 19646, т. 5, с. 49—59.
Вылцан Н. Ф. Пойменные луга долины р. Оби северных районов Томской обл.— В кн.:
Природа и экономика Александровского нефтеносного района. Томск: Изд-во
Томск, ун-та, 1968, с. 212—225.
Вылцан Н. Ф. Луга Томской обл. Автореф. канд. дис. Томск, 1969. 21 с.
Галкина Е. А. Болотные ландшафты и принципы их классификации.— В кн.: Сборник
научных работ Ботанического ин-та АН СССР, выполненных в Ленинграде за
три года Великой Отечественной войны (1Ш—1943). Л., 1946, с. 139—156.
Галкина Е. А. Сфагновые болота.— В кн.: Растительный покров СССР. Пояснительный
текст к геоботанической карте СССР. Т. II. М.— Л.: Изд-во АН СССР, 1956,
с/553-572.
Генкель А. А., Красовский П. Н. Материалы по изучению озер, займищ, болот и
торфяников Западно-Сибирской лесостепи.— Учен. зап. Пермск. ун-та, 1937, т. 3,
вып. 1, с. 3 — 75.
212
Геоботаническая карта СССР/Под ред. Е. М. Лаврснко,В. Б. Сочавы. М.: ГУГК,« 1954.
8 с.
Герасимова А. С, Ершова С. Б. Значение нсотектонлки при инженерно-геологическом
районировании Западной Спбпрп.— В кн.: Тектонические движения и новейшие
структуры земной коры. М.: Недра, 1907, с. -128—136.
Глазовская М. А. Геохимические основы типологии и .методики исследований
природных ландшафтов. М.: Изд-во Моск. ун-та, 1064. 230 с.
Глебов Ф. 3. Болота и заболоченные леса восточной части бассейна р. Дубчес.— В кн.:
Заболоченные леса и болота Сибири. М.: Изд-во АН СССР, 1963, с. 33—64.
Глебов Ф. 3. Болота и заболоченные леса лесной зоны Енисейского левобережья. 3VL:
Наука, 1969. 131 с.
Глебов Ф. 3., Горожанкина С. М. Влияние ландшафтной структуры на особенности бо-
лотообразовання.— В кн.: Исследование таежных ландшафтов дистанционными
методами. Новосибирск: Наука, 1979, с. 109—134.
Говорухин В. С. Очерк растительности лесных пастбищ северного оленя в тундрах
Обско-Тазовского п-ова.— Землеведение, 1933, т. 34, выи. 1, с. 68—92.
Говорухин В. С. Бугристые болота Северной Азии и потепление Арктики (Западная
Сибирь, бассейн р. Сев. Сосьвы).— Учен. зап. Моск. обл. нн-та, 1947, т. 9,
вып. 4, с. 106—124.
Говорухин В. С. Редколесья на полярном пределе лесов Малою Ямала и в Приобской
лесотундре.— Землеведение, 1957, т. 44, № 4, с. 214—228.
Гордягин А. Я. Материалы для познания почв и растительности Западной Сибири.—
Тр. о-ва естествоиспыт. при Казан, ун-те, 1900, т. 34, вып. 3. 222 с.
Гордягин А. Я. Материалы для познания почв и растительности Западной Сибири.—
Тр. о-ва естествоиспыт* при Казан, ун-те, 1901, т. 35, вып. 2. 528 с.
Горновский К. В. Растительность Кондо-Сосьвпнского заповедника.— Науч.-метод,
зап., 1949, вып. 12, с. 55—57.
Городков Б. Н. Подзона лиственных лесов в пределах Ишпмского уезда Тобольской
губ.— Тр. лочв.-ботан. экспедиций Переселен, vnp. по псслед. колониз. районов
Азиатской России в 1912 г., Пг.,1915, ч. 2, 3* с. 102—133.
Городков Б. Н. Опыт деления Западно-Сибирской низменности на ботанико-географи-
ческие области.— Ежегод. Тобольск, губ. музея, 1916а, т. 27, с. 1—56.
Городков Б. Н. Наблюдения над жизнью, кедра в Западной Сибири.— Тр. Ботан.
музея, 19166, вып. 16, с. 153—172.
Городков Б. И. Западно-Сибирская экспедиция Российской Академии Наук и
Русского Географического Общества.— Природа, 1924, Д° 7—12, с. 3—32.
Городков Б. II. Круннобугристые торфяники и их географическое распространение.—
Природа, 1928, № 6, с. 599—601.
Городков Б. Ы. Почвы Гыданской тундры.— Тр. Полярной комис. АН СССР, 1932,
вып. 7, с. 1—78.
Городков Б. Н. Растительность тундровой зоны СССР.— М.— Л.: Изд-во АН СССР,
1935. 142 с.
Городков Б. Н. Растительность Арктики и горных тундр СССР.— В кн.:
Растительность СССР. Т. ,4. М.— Л.: Изд-во АН СССР, 1938, с. 297—354.
Городков Б. Н. Тундры Обь-Енисейского водораздела,— Сов. ботан., 1944, №'3—5,
с. 20-31.
Городков Б. Н. Движение растительности на севере лесной зоны Западно-Сибирской
низменности.— В кн.: Проблемы физической географии. Вып. 12. М.—Л ,
1946, с. 81-106.
Горожанкина С. М., Константинов В. Д. География тайги Западной Сибири.
Новосибирск: Наука, 1978. 190 с.
Горчаковский П. Л. Сосновые леса Приобья как зональное ботанико-географическое
явление.— Ботан. журн., 1949а, т. 34, № 5, с. 524—538.
Горчаковский П. Л. Таежные и лесостепные березняки Приобья.— В кн.: Сборник
труДов но лесному хозяйству. Вып. 1. Свердловск, 19496, с. 62—100.
Горчаковский П. Л. Темнохвойная тайга Среднего Урала и прилегающей части
Северного Урала.— В кн.: Материалы по классификации растительности Урала.
(Тез. докл.). Свердловск, 1959, с. 18—22.
Горчаковский П. Л. Растения европейских широколиственных лесов на восточном пре-
» деле их ареала. Вып. 59. Свердловск: изд. УФ АН СССР, 1968. 207 с.
Грибова С. А. Шпротная дифференциация растительного покрова тундр Европейской
равнины.— В кн.: Проблемы экологии, геоботаники, ботанической географии
и флористики. Л.: Наука, 1977, с^ 37—46.
Грибова С. А., Исаченко Т. И. Картирование растительности в съемочных масштабах.—
В кн.: Полевая геоботаника. Вып. 4. Л.: Изд-во АН СССР, 1972, с. 137—331.
Григорьев А. А. Географическая зональность и некоторые ее закономерности.— Изв.
АН СССР. Сер. геогр., 1954, № 5, с. 17—39.
Григорьев А. А. Субарктяка. Опыт характеристики осповных типов географической
среды. М.: Географгиз, 1956. 223 с.
Гусев Л. И. Тетрагональные грунты в арктической тундре.— Изв. Гос. геогр. о-ва,
1938, т. 70, вып. 3, с. 377—386.
Дружинина Н. П. Фптомасса растительных сообществ пойменного масеива Нижнего
Иртыша (на примере Моильского массива).— В кн.: Географические аспекты
проблемы перераспределения стока в Западной Сибири. Иркутск, 1979, с. 76—88.
213
Дунин-Горкавич А. А. Тобольский Север. Общий обзор страны, ее естественных
богатств и промышленной деятельности населения. Т. 1. Снб., 1904, с. 1—75.
Дыдина Р. А. Обь-Иртышские луга в пределах Ханты-Мансийского округа.— Тр.
НИИ с.-х. Крайнего Севера, 1961, т. 10, .с. 159—250.
Егоров В. В., Базилевич Н. И. Карта типов химизма засоления почв СССР. М
1 :2 500 000. М.: ГУГК, 1976.
Еленевский Р. А. Вопросы изучения и освоения иойм. М.: Изд-во ВАСХНИЛ, 1936.
100 с.
Елизарьева М. Ф. Луговая растительность Томской области как кормовая база
животноводства.— Тр. Томск, ун-та, 1951, т. 114, с. 135—149.
Елизарьева М. Ф. Растительность зандрового ландшафта вне ледниковой зоны на
восточной окраине Западно-Сибирской низменности.— Учен. зап. Красноярск,
пед. ин-та, 1961, т. 20, вып. 1, с. 27—45.
Жаркова А. М. Генетическая схема болотных форхмаций Западно-Сибирской
низменности по ландшафтным зонам.— Изв. Омск, отд-ния ВГО, 1963, вып. 5(12),
с. 61—68.
Земцов А. А. Развевание песков на севере Западно-Сибирской низменности.—В кн.:
- Вопросы географии Сибири. Сб. 4. Томск: Изд-во Томск, ун-та, 1962, с. 81—92.
Западная Сибирь. М.: Изд-во АН СССР, 1963. 488 с.
Зубарева Р. С. Лесная растительность Припышмпнских боров Зауралья.— В кн.:
Природные условия и леса лесостепного Зауралья. Вып. 19. Свердловск: изд.
УФ АН СССР, I960, с. 97—124.
Зубарева Р. С. Пастбищное использование лесной растительности Припышминского
борового массива Зауралья.— В кн.: Материалы по изучению флоры и
растительности Урала. Свердловск: изд. УФ АН СССР, 1962, с. 57—80.
Иванов К. Е. Эрозионные явления на болотах и их роль в формировании озерно-
болотных ландшафтов Западной Сибири.— Тр. Гос. гидрол. ин-та, 1969,
вып. 157, с. 78—97.
Иванов К. Е. Болотно-озерные системы и их устойчивость при преобразовании
избыточно-увлажненных территорий.—Учен. зап. Ленингр. ун-та, 1974, № 376, вып.
23, с. 5—81.
Иванов К. Е., Котова Л. В. Вопросы динамики развития и гидроморфологические
характеристики рямов Барабииской низменности.— Тр. Гос. гидрол. ин-та, 1964,
вып. 112, о. 35—53.
Игнатьева Л. А. Продуктивность травостоя лесных ценозов Приобья.— Ботан. журн.,
1966, т. 51, № 2, с. 196—209.
Игнатьева Л. А. Определение обилия видов травяного покрова приобских боров.—
В-кн.: Растительность Приобья и ее хозяйственное использование. Новосибирск:
Наука, 1973, с. 176- J90.
Игошина К. Н. Редколесье и мерзлые болота Ивдельского Зауралья.— Ботан. журн.,
1949, т. 34, № 5, с. 493-506.
Ильина И. С. Картографирование растительности поймы р. Оби на отрезке
Ханты-Мансийск — Нижневартовское.— Докл. Ин-та геогр. Сибири и Дальнего Востока
СО АН СССР, 1968, До 17, с. 12—19.
Ильина И. С. Использование материалов лесоустройства при геоботаническим
картографировании (на примере картирования растительности Западной Сибири).—
В кн.: Геоботаническое картографирование. Л.: Наука, 1971, с. 37—48.
Ильина И. С. Классификация растительности островных иойм Оби и Иртыша для
целей картографирования.— В кн.: Растительность речных иойм, методы ее
изучения и вопросы рационального использования. (Тез. докл.). Уфа, 1972, с. 47—
50.
Ильина И/С. Некоторые вопросы объективизации геоботанического районирования.—
Докл. Ин-та геогр. Сибири и Дальнего Востока СО АН СССР, 1975, вып. 48,
с. 49-55.
Ильина И. С. Обзорное картографирование растительности поймы р. Оби.— Сиб.
геогр. сб., 1976, вып. 12, с. 161—,182.
Ильина И. С. Вопросы географии растительности темнохвойной тайги
Западно-Сибирской равнины.— География и природ, ресурсы, 1981, № 4, с. 120—127.
Ильина И. С. Современное состояние растительного покрова дельты р. Оби.— В кн.:
Проблемы охраны природы и рационального использования природных
ресурсов в Северных регионах. (Тез. докл.).. Архангельск, 1982, с. 55—56. -
Ильина И. С, Махно В. Д. Геоботаническое районирование. Врезка на карте
«Растительность Западно-Сибирской равнины». М.: ГУГК, 1976.
Ильина И. С, Толчельников Ю. С. Бассейн р. Бол. Салыма (почвенно-гсоботанический
очерк).— Докл. Ин-та геогр. Сибири и Дальнего Востока СО АН СССР, 1968,.
вып. 18. с. 11—21.
Ильина И. С, Крисон X. П., Полюшкин Ю. В. Растительность.— В кн.: Южная тайга
Прииртышья. Новосибирск: Наука, 1975, с. 81—93.
Ильина И. С, Махно В. Д., Мельцер Л. И. Геоботаническое картографирование
пойменных земель Нижнего Иртыша для сельскохозяйственного использования.—
География и природ, ресурсы, 1980, № 2, с. 83—97.
Ильина И. С, Петров И. Б., Соколова Л. П. Закономерности пространственного
распределения растительности погшы Нижнего Иртыша в связи с гпдролого-геоморфо-
214
логическими условиями.— В кн.: Региональные биогео1рафпческпе
исследования в Сибири. Иркутск: изд. ИГСиДВ СО ЛН СССР, 1977, с. 19-40.
Ильина И. С, Лапшина Е. И., Махно В. Д., Романова Е. А. Структура растительности
Западно-Сибирской равнины.— В кн.: Пятое Всесоюзное совещание по
классификации растительности. (Тез. докл.). Новосибирск, 1977а, с. 24—27.
Ильина И. С, Лапшина Е. И., Махно В. Д., Романова Е. А. Принципы составления
обзорной «Карты растительности Западно-Сибирской равнины».— В кн.: Гео-
ботанпческое картографирование. Л.: Наука, 19776, с. 41—58.
Ильина И. С, Скарлыгина-Уфимцева М. Д., Кузьмин А. И., Русаков В. Н. Проблема
Омской поймы Иртыша (ботанико-географпческий аспект).— Вести. Ленингр.
ун-та. Сер. геол. те геогр., 1983, № 12, с. 60—69.
Ильина Л. II. Некоторые аспекты оценки урожайности, ягодников тайги Западной
Сибири.— Матер. ВННИОЗ, Киров, 1973, с. 202-204.
Исаченко Т. И. Сложение растительного покрова и картирование.— В кн.: Геобота-
нпческое картографирование. Л.: Наука, 1969, с. 20—23.
Исаченко Т. И., Лавренко Е. М. Ботанико-географпческое районирование.— В кн.:
Растительность европейской части СССР. Л.: Наука, 1980, с. 10—20.
Исаченко Т. И., Лукичева А. Н. Перезовые и осиновые леса.— В кн.: Растительный
покров СССР. Т. II. М.— Л.: Мзд-во АН СССР, 1956, с. 319-345.
Исаченко Т. И., Рачковская Е. И. Основные зональные типы степей Северного
Казахстана.— Тр. Ботан. ин-та АН СССР. Сер. 3, Геоботаника, 1961, вып. 13,
с. 133-397.
Калинина А. В*. Песчаные степи Северного Казахстана.— Там же, с. 398—443.
Караваева Н. А. Почвы тайги Западной Сибири. М.: Наука, 1973. 167 с.
Караваева Н. А. Типы и механизмы современного заболачивания почв в лесной зоне.—
В кн.: Научные предпосылки освоения болот Западной Сибири. М.: Наука,
1977, с. 124—137.
Караваева Н. А. Заболачивание и эволюция почв. М.: Наука, 1982. 296 с.
Карамышева 3. В., Рачковская Е. И. Ботаническая география степной части
Центрального Казахстана. Л.: Наука, 1973. 278 с.
Кац Н. Я. Болота низовьев реки Оби.— В кн.*: Президенту АН СССР акад. В. Л.
Комарову к 70-летпк>£О дня рождения и 45-летию научной деятельности. Л., 1939,
с. 372—405.
Кац II. Я. Типы болот СССР и Западной Европы и их географическое распространение.
М.: Географгпз, 1948. 315 с.
Кац Н. Я. Болота земного шара. М.: Наука, 1971. 290 с.
Кац II. Я., Кац СВ. Стратиграфия торфяников Приобского Севера.— Тр. Комис. по
изучению четвертичного перпода, 1948, т. 7, вып. 1, с. 15—54.
Кириков С. В. Промысловые животные, природная среда и человек. М.: Наука, 1966.
348 с.
Клеопов Ю. Д. Основные черты развития флоры широколиственных лесов европейской
части СССР.— В кн.: Материалы по исторпн флоры п растительности СССР.
Вып. 1. М.-Л.: Изд-во АН GCCP, 1941, с. 215—249.
Колесников Б. П. Тины южно-таежных лесов среднего течения р. Тавды и Тавды-Ку-
мннского междуречья.— В кн.: Южно-таежные леса Западно-Сибирской
равнины. Свердловск: изд. УНЦ АН СССР, 1972, с. 66—98.
Колесников Б. П., Смолоногов Е. П. Некоторые закономерности возрастной и
восстановительной динамики кедровых лесов Зауральского Приобья.— Тр. по лесн.
. хоз.-ву, 1960, вып. 6, с. 21—31.
Комин Г. Е. Влияние пожаров на возрастную структуру н рост северотаежных
заболоченных сосняков Зауралья.— Тр. Ин-та экологии раст. и животн. УНЦ АН
СССР, 1967, вып. 53, с. 207—222.
Крашенинников И. М. Анализ реликтовой флоры Южного Урала в связи с историей
растительности и палеогеографией плейстоцена.— Сов. ботан., 1937, № 4,
с. 16-45.
Крашенинников И. М., Иванова II. А. К истории развития растительных ландшафтов
Западной Сибири.— Землеведение, 1934, т. 36, вып. 1, с. 1—38.
Крисон X. П., Полюшкин Ю. В. Структура растительного покрова.— В кн.: Южная
тайга Прииртышья. Новосибирск: Наука. 1975, с' 165—185.
Крылов П. Н. Степи западной части Томской губ.— Тр. почв.-ботан. экспедиций
Переселен, упр. по исслед. колониз. районов Азиатской России. Ч. II. Пг., 1916,
139 с.
Крылов П. Н. Очерк растительности Сибири.— Стат. экон. бюл., 1919, № 17, с. 1—24.
Крылов Г. В. Березовые леса Томской области и их типы. Новосибирск, 1953. 123 с.
Крылов Г. В. Леса Западной Сибири. М.: Изд-во АН СССР, 1961. 255 с.
Кузнецов Н. М. Лайды в низовьях р. Енисея, их строение, образование и место в
классификационной схеме болотно-лесных образований.— Тр. Полярной комис.
АН СССР, 1932, '№ 12, с. 5—40.
Кузьмин А. И., Русаков В. Н. Влияние геоморфологии на водный режим пойменных
земель.— Науч. тр. Омск. с.-х. ин-та, 1978, т. 178, с. 33—37.
Кузьмина М. С. Эволюция растительности болот Барабинской низменности в связп с
пхл осушением и естественным усыханием.— В кн.: Труды юбилейной сессии,
посвященной 100-летию со дня рождения В. В. Докучаева. М.— Л., 1949,
С. 589-596.
215
Кузьмина М. С. Растительность Барабы.— Тр. Почв, пн-та АН СССР, 1953, т. 36,
с. 106-171.
Кузьмина М. С. Принципы типизации болот Барабы.— Изв. Новосиб. отд-ния ВГО
СССР, 1957, вып. 1, с. 23-32.
Кузьмина М. С. Болота.— В кн.: Растительные богатства Новосибирской области.
Новосибирск: пзд. РИО СО АН СССР, 1961, с. 65—72.
Кулагин 10. 3. Особенности водного режима листьев различных форм березы.—
' Изв. РИО СО АН СССР, 1960, № 3, с. 108—115.
Кулагин 10. 3. Экология березы бородавчатой и березы пушистой в связи с
особенностями их водного режима.— Тр. Ин-та бнол. УФ АН СССР, 1963, вып. 35,
с. 7-45.
Куликова Г. Г. К динамике растительного покрова водораздельных болот
левобережного Васюгана в голоцене.— Вести. Моск. ун-та. Сер. бнол. и почв., 1971,
№ 1, с. 115—116.
Куркин К. А. Луга Барабы и их улучшение. Щ.: Изд-во АН СССР, 1957. 145 с.
Куркин К. А. Системные исследования динамики лугов. М.: Наука, 1976. 284.с.
Лавренко Е. М. Степи и сельскохозяйственные земли на месте степей.— Ц кн.:
Растительный покров СССР. Т. 2. М.— Л.: Изд-во АН СССР, 1956, с. 595—730.
Лавренко Е. М. Провинциальное разделение Причерноморско-Казахстанской
подобласти степной области Евразии.— Ботан. журн., 1970, т. 55, с. 609—625.
Лавренко Е. М. Степи..— В кн.: Растительность европейской части СССР. Л.: Наука,
1980, с. 203-242, 254—262.
Лавренко Н. И. Фитонндикационные связи подтаежных геосистем Прииртышья и их
картографирование.— В кн.: Геоботаническое'картографирование. Л.: Наука,
1982, с. 3—15. •
Лавренко Н. Н., Кобелева И. В. Применение метода корреляционных плеяду
использованием ЭВМ для выявления сопряженности видов в растительных
сообществах.— Докл. Ин-та гсогр. Сибири и Дальнего Востока СО АН СССР, 1973,
№ 41, с. 33-45.
Ландшафтные индикаторы инженерно-геокриологических условий севера Западной
Сибири и их дешпфровочные признаки. М.: Недра, 1974. 133 с.
Лапшина Е. И. Карта растительности Сибири в масштабе 1 : 2 500 000». Автореф. канд.
дис. Новосибирск, 1952. 16 с.
Лапшина Е. И. Березовые леса лесостепи юго-востока Западной Сибири.— В кн.:
Растительность стенной п лесостепной зон Западной. Сибири. Новосибирск:
пзд. РИО СО АН СССР, 1963, с. 103—131.
Лапшина Е. И. Северная тайга левобережья р. Оби.— Изв. Новосиб. отд-ния ВГО
СССР, 1971, вып. 5, с. 156—171.
Лапшина Е. И. Геоботапическбс картографирование таежной зоны Западной
Сибири.— В кн.: Геоботаническое картографирование. Л.: Наука, 1973, с. 49—60.
Лапшина Е. И. География растительного покрова Сосьвпнского Прнобья.— В кн.:
Сосьвпнское Прпобье. Иркутск: изд. ИГСпДВ СО АН СССР, 1975, с. 278—352.
Лапшина Е. И. Растительность бассейна р. Таз в верхнем течении.— В кн.: Геобота-
нпческне исследования в Западной и Средней Сибири. Новосибирск: Наука,
1978, с. 47—70. ,
Лапшина Е. И., Пеньковская Е. Ф., Храмов А. А. Характерные черты растительного
покрова долины р. Оби от Барнаула до с. Кривошехшо. Новосибирск: изд.
ЦСБС СО АН СССР, 1971. 35 с.
Леса СССР. Т. 4. М.: Наука, 1969, с. 157—248.
Лесков А. И. Фитоценотическпй очерк редколесий бассейна р. Полуй.— Тр. Ботан.
ин-та АН СССР, 1940, сер. 3, вып. 4, с. 253—276.
Лесков А. И. Арктическая тундровая область, Б. Европейско-Снбирская
кустарниковая (лесотундровая) область.— В кн.: Геоботаническое районирование СССР.
М.— Л.: Изд-во АН СССР, 1947^ с. 14—22.
Лесорастительные условия, и типы лесов Свердловской области. Свердловск: изд.
УНЦ АН СССР, 1973. 175 с.
Лисе О. И., Березина М. А.. Характеристика торфяной залежи4 п растительного
покрова.— В кн.: Природные условия центральной части Западно-Сибирской
равнины. М.: Изд-во Моск. ун-та, 1977, с. 120—138.
Львов М. А. К характеристике Иксинского водораздельного болота.— Изв. Томск,
отд-ния ВГО, 1959, т. 4, с. 59—62.
Львов Ю. А. Растительность Васюганского района.— В' кн.: Природа и экономика
Привасюганья. Томск: Изд-во Томск, ун-та, 1966, с. 190—201.
Львов Ю. А. Характеры механизма заболачивания территории Томской области.—
В кн.: Теория и практика лесного болотоведения и гндролесомелиорации.
Красноярск, 1976, с. 36—44.
Любимова Е. Л. Некоторые вопросы палеогеографии центральных районов Западной
Сибири.— Материалы Моск. фил. геогр. о-ва СССР, 19696, вып. 3
(биогеография), с. 19—20.
Любимова Е. Л. Растительность Обь-Иртышского междуречья.— В кн.: Прпродпые
условия освоения междуречья Обь—Иртыш. М.: изд. Ин-та гидрол. All СССР,
1972, с. 302—321.
Маковский В. И., Шадрина Н. И. Типология заболоченных и болотных лесов Тавды-
216
Куминского междуречья.— Тр. Ин-та экологии раст. и животн. УНЦ АН СССР,
1972, вып. 83, с. 131—163.
Максимов А. М., Мерзлякова Е. П. Характеристика половодий в пойме р. Оби.— В кн.:
Биологические ресурсы поймы Оби. Новосибирск, 1972, с. 362—391.
Марков К. К., Гричук В. П., Чеботарева Н. С. Взаимоотношения леса и степи в
историческом освещении.— В кн.: Вопросы географии. М.: изд. Ин-та гидрол.
АН СССР, 1950, с. 85—119.
Материалы межвузовской конференции по геоботаническому районированию. М.:
Изд-во Моск. ун-та, 1967. 288 с.
Махнев А. К. О взаимоотношении березы бородавчатой и пушистой.— Изв. вузов.
Лесн. журн., 1965, № 3, 29—33.
Махно В. Д. Способы отображения растительности на обзорной геоботанической карте
Западной Сибири.— В кн.: Исследование по проблемам геодезии и картографии.
Иркутск, 1973, с. 114—123.
Мезенцев В. С. Зоны увлажнения и теплообеспеченности Западно-Сибирской
равнины.— Тр. Томск, ун-та. Сер. геог., 1957, т. 146, с. 113—121.
Мезенцев В. С., Карнацевич И. В. Увлажненность Западно-Сибирской равнины. Л.:
Гидрометеоиздат, 1969. 168 с.
Мельцер Л. И. Основные закономерности распределения растительного покрова
западно-сибирских тундр.— В кн.: Природа и природные ресурсы Тюменской обл.
Тюмень, 1973, с. 159—162.
Мельцер Л. И. Вопросы классификации и картографирования растительности
западносибирских тундр.— В кн.: Региональные биогеографические исследования в
Сибири. Иркутск: изд. ИГСиДВ СО АН СССР, 1977, с. 40^-59.
Мельцер Л. И. Отображение гетерогенной структуры растительности западно-сибир-
скихтундр при среднемасштабном картографировании.— В кн.: Геоботанлческое
картографирование. Л.: Наука, 1980, с. 11—24.
Мельцер Л. И. Ботанико-географический анализ тундр Западной Сибири. Автореф.
канд. дне. Свердловск, 1982. 18 с.
Миддендорф А. Ф. Бараба. Спб., 1871. 123 с.
Миняев Н. А. История развития флоры северо-запада европейской части РСФСР с
конца плейстоцена. Л.: Изд-во Ленингр. ун-та, 1966. 37 с.
Михайличенко B.C. Естз рослинност1 швшчносх1дно1 окраши п-ова Ямалу.— Журн л.
1нст. ботан. Укр. АН СССР, 1936, № 7 (15), с. 71—103.
Михайлов Ю. П., Ильина Л. Н., Червяков В. А. Картографирование природных
ресурсов (методы и результаты работ в Сибири)"— В кн.: 'Тематическое
картографирование в целях охраны природы. М.— Братск, 1976, с. 69—75.
Михеев В. С, Соколов Н. А. Изучение элементарных геотопологических структур:
описание, анализ, локализация.— В кн.: Методы комплексных исследований
геосистем. Иркутск, 1974, с. 37—51.
Михеев В. С, Полюшкин Ю. В., Сазонов А. Г. Объекты исследования и исходные
характеристики.— В кн.: Природные режимы среднейчтайги Западной Сибири.
Новосибирск: Наука, 1977, с. 30—51.
Мишуков П. Н. Кедр сибирский на северном пределе распространения в бассейнах
• Пура и Таза.— В кн.: Природа тайги Западной Сибири. Новосибирск:
Наука, 1973, с. 22—37.
Москаленко Н. Г. Горизонтальная структура некоторых болотных и тундровых фито-
ценозов севера Западной Сибири.— Бюл. МОИП. Отд-ние биол., 1976, т. 81 (3),
с. 129—143.
Мусаев И. Ф. Северные пределы распространения характерных компонентов Туран-
ской пустынной флоры.— В кн.: Ареалы растений флоры СССР. Л.: Изд-во
Ленингр. ун-та. 1965, с. 93—139.
Нейштадт М. И. Торфяные болота Барабпнской лесостепи.— Тр. Центр, торф,
опытной стадции, 1936, т. 1, с. 75—87.
Нейштадт М.И. О смене природных условий'в средней тайге Западной Сибири в
голоцене.— В кн.: Палинология в СССР. М.: Наука, 1976, с. 156—162.
Нейштадт М. И. Возникновение и скорость развития процессов заболачивания.—
В кн.: Научные предпосылки освоения болот Западной Сибири* М.: Наука,
1977, с. 39-47.
Николаев В. А. Геоморфологическое районирование Западно-Сибирской равнины.—
В кн.: Западно-Сибирская равнина. М.: Наука, 1970, с. 226—255.
Норин Б. Н. К познанию семенного и вегетативного возобновления древесных пород в
лесотундре.— В кн.: Растительность Крайнего Севера СССР и ее освоение.
Вып. 3. Л.: Изд-во АН СССР, 1958, с. 154—244.
Норин Б. Н. Проблема лесотундры и задачи ее комплексного стационарного
изучения.— В кн.: Проблемы Севера. Вып. 8. М.— Л.: 1964, с. 58—66.
Носова Л. М. Флорогенетический анализ северной степи европейской части СССР.
М.: Наука, 1973. 187 с.
Определитель лишайников CCGP/Отв. ред. И. И. Абрамов. Т. 1—5. Л.: Наука. Т. 1,
1971. 411 с; Т. 2, 1974, 283 с; Т. 3, 1975. 275 с; Т. 4, 1976. 343 с; Т. 5, 1978.
304 с.
Павлова Г. Г. Сосновые леса в лесостепной и степной зонах Приобья.— В кн.:
Растительность степной и лесостепной зон Западной Сибири. Новосибирск: изд.
РИО СО АН СССР, 1963, с. 131—162.
15 Закаа № 297
217
Панадиади А. Д. Барабинская низменность. М.: Госгеографиздат, 1953. 232 с.
Пеньковская Е. Ф. Растительность поймы Оби.— В кн.: Растительность степной и
лесостепной зон Западной Сибири. Новосибирск: изд. РИО СО АН СССР, 1963а.
с. 211—263.
Пеньковская Е. Ф. Разногодичная изменчивость луговой растительности в Колыван-
ской пойме Оби.— Там же, 19636, с. 264—277.
Петров И. Б. Обь-Иртышская пойма (типизация и качественная оценка земель).
Новосибирск: Наука, 1979. 136 с.
Петров И. Б., Бачурин Г. В. Гидролого-геоморфологические условпя освоения пойм
Средней Оби и Нижнего Иртыша.— Сиб. геогр. сб., 1976, вып.- 12, с. 75—112.
Петрова Е. П. Встречаемость пыльцы широколиственных пород в спорово-пыльцевых
голоценовых торфяниках севера Западной Сибири на широте г. Салехарда.—
Бюл. Моск. о-ва испыт. природы. Отд. геол., 1963, № 3, с. 147.
Поварницын В. А. Кедровые леса СССР. Красноярск, 1944. 221 с.
Покровская И. М. О нижней границе четвертичной системы по палеоботаническим
данным.— В кн*: Материалы по четвертичной геологии и геоморфологии СССР.
Вып. 4. М.: Госгеолтехиздат, 1965, с. 45—53.
Поле Р. Программа для ботанико-географических исследований <гундры.— В кн.:
Программы для ботанико-географических исследований. Вып. 2. Спб., 1910,
с. 29—65.
Полозова Т. Г. К биологии п экологии карликовой березки в восточно-европейской
лесотундре.— В кн.: Приспособление растений Арктики к условиям
среды. М.— Л.: Наука, 1966, с. 166—225.
Полынов Б. Б. Учение о ландшафтах.— В кн.: Вопросы географии. Сб. 33. 1953,
с. 30-44.
Попов А. И. Вечная мерзлота в Западной Сибири. М.: Изд-во Моск. ун-та,^1953.
229 с.
Придня М. В. Естественное возобновление на концентрированных вырубках ельников
зеленомошниковых Тавды-Куминского междуречья.— В кн.: Южно-таежные
леса Западно-Сибирской равнины. Вып. 83. Свердловск: изд. УНЦ АН СССР,
1972, с. 194—217.
Прокопьев Е. П. Типы леса левобережной части Александровского района.— В кн.:
Природа и экономика Александровского нефтегазоносного района (Томская
область). Томск: Изд-во Томск, ун-та, 1968, с. 194—211.
Прокопьев Е. П. Пойменные леса рек северной части Томской области.— Тр. НИИ
биол. и биофиз. при Томск, ун-те, 1974, т. 3, с. 104—116.
Прокопьев Е. П. Топографическое распределение растительности поймы Среднего
Иртыша.— Там же, 1976, т. 7, с. 33—51
Прокопьев Е. П. Очерк растительности поймы Иртыша в пределах Омской области
в связи с геоморфологией поймы.— В кн.: Природное районирование Омского
Прииртышья. Омск, 1977, с. 83—92.
Прокопьев Е. П. Проблемы охраны, улучшения и рационального использования
растительности поймы Иртыша.— В кн.: Проблемы охраны природы Сибири. Томск:
Изд-во Томск, ун-та, 1978, с. 29—46.
Пьявченко Н. И. Бугристые торфяники. М.: Изд-во АН СССР, 1955. 280 с.
Пьявченко Н.И. Особенности растительного покрова и строения некоторых болот
Красноярского края.—Учен. зап. Тартуского гос. ун-та, 1963, вып. 145, с. 232—240.
Пьявченко Н. И., Федотов С. С. Природа лесотундры Таз-Енисейского междуречья.—
В кн.: Растительность лесотундры и ее освоение. Л.: Наука, 1967, с. 157—163.
Растительность степной и лесостепной зон Западной Сибири. Новосибирск: изд. РИО
QO АН СССР, 1963. 440 с.
Растительность Западно-Сибирской равнины. Карта М 1 : 1 500 000/Ред. И. С. Ильгс-
на. Авт.: Ильина И. С, Лапшина Е. И., Махно В. Д., Романова Е. А. М.: ГУГК,
1976. 4 л.
Ревердатто В. Вт Растительность Сибири'.— В тга.: Естественно-исторические условия
сельскохозяйственного производства Сибири. Ч. 3. Новосибирск, 1931. 176 с.
Ревердатто В. В. Основные моменты развития послетретичной флоры Средней
Сибири.— Сов. ботан., 1940, № 2, с. 48—64.
Рихтер Г. Д. Рельеф и геологическое строение.— В кн.: Западная Сибирь. М.: Изд-во
АН СССР, 1963, с. 22—69. •
Роднянская Э. Е. Растительность поймы р. Оби в пределах Березовского района.—
Науч. докл. высш. школ! Геол-геогр. науки, 1958, № 4, с. 90—98.
Роднянская Э>. Е. Типология пойменных ландшафтов на примере р. Оби в пределах
таежной зоны. Автореф. канд. дис. Л., 1963. 13 с.
Роднянская Э. Е. Особенности растительности поймы р. Оби в таежной зоне.— Вестн.
Ленингр. ун-та, 1968, № 24. Сер. геол. и геогр., вып. 4, с. 127—134.
Роднянская Э. Е. Характеристика зональных особенностей растительности поймы
р. Оби в таежной зоне.—Вестн. Ленингр. ун-та, 1969, N° 12. Сер. геол. и геогр.,
с. 115—125.
Роднянская Э. Е.,* Самойлова Г. С. Роль гидрологического режима ландшафтов
р. Оби.— В кн.: Природные условия Западной Сибири. М.: Изд-во Моск. ун-та,
1971, вып. 1, с. 136—144.
Романова Е. А. Геоботанические основы гидрологического изучения верховых болот.
Л.: Гидрометеоиздат, 1961. 214 с.
218
Романова Е. А. Типы болотных микроландшафтов как показатели видов торфа в
верхних слоях залежи низинных болот.-— Тр. Гос. гидрол. ин-та, 1964, вып. 112,
с. 54-81.
Романова Е. А. Краткая ландшафтно-морфологическая характеристика болот
Западно-Сибирской низменности.— Тр. Гос. гидрол. ин-та, 1965, вып. 126, с. 96—112.
Романова Е. А. Л андшафтно-морфо логическая характеристика болот в бассейне р. Кон-
ды (Западно-Сибирская низменность).— Тр. Гос. гидрол. ин-та^ 1967, вып. 145,
с. 27—51.
Романова Е. А. К структуре обзорно-типологической карты болот Западно-Сибирской
равнины.— Докл. Ин-та геогр. Сибири и Дальнего Востока СО АН СССР, 1973,
вып. 40, с. 25—31.
Романова Е. А. Зональные типы болот. Врезка М 1 : 1 250 000 на карте «Растительность
Западно-Сибирской равнины». М.: ГУГК, 1976.
Романова Е. А. Типологическая карта болот Западно-Сибирской равнины. М.: ГУГК,
1977.
Романова Е. А. Динамика растительного покрова и торфяной залежи некоторых
низинных болот лесостепной зоны Западной Сибири.— В кн.: Генезис и динамика
болот. Вып. 2. М.: Изд-во Моск. ун-та, 1978, с. 34—39.
Романова Е. А., Усова Л. И. Геоботаническая и краткая гидрологическая
характеристика болотных ландшафтов водораздела рек В ах и Ватинский Еган Западной
Сибири.— Тр. Гос. гидрол. ин-та,^ 1969, вып. 157, с. 98—122.
Романова Е. А., Усова Л. И. О структуре болотного микрокомплекса междуречья Де-
мьянки и Бол. Туртаса.— Тр. Гос. гидрол. ин-та, 1977, вып. 236, с. 3—12.
Ронгинская А. В. Степи юго-востока Западно-Сибирской низменности.— В кн.:
Растительность степной и лесостепной зон Западной Сибири. Новосибирск: изд.
РИО СО АН СССР, 1963, с. 77—103.
Рутковский В. И. Климатическая и гидрологическая роль леса.— Тр. Второго все-
союз. геогр. съезда, 1948, с. 27—31.
Савина Л. Н. Тенденция развития лесной растительности Сибири в голоцене (по
данным спорйво-пыльцевого анализа).— В кн.: Динамика лесных биогеоценозов
Сибири. Новосибирск: Наука, 1980, с. 28—53.
Савич-Любицкая Л. И., Смирнова 3. Н. Определитель листостебельных мхов СССР.
Л.: Наука, 1970. 824 с.
Самбук Ф. В. Печорские леса.— Тр. Ботан. музея АН СССР, 1932, вып. 24, с. 63—250.
Седых В. Н. Формирование кедровых лесов Приобья. Новосибирск: Наука, 1979. 110 с.
Семенов В. Ф. Краткий очерк растительности Омской губернии.— Изв. Зап.-Сиб.
отд-ния РГО, 1924, т. 4, № 1, с. 13—46.
Синелыцикова 3. И. Развитие елово-березовых древостоев на междуречье Тавда-Кон-
да.— В кн.: Южно-таежные леса Западно-Сибирской равнины. Вып. 83.
Свердловск: изд. УНЦ АН СССР, 1972, с. 99—130.
Скворцов А. К., Связева О. A. Betula pubescens Ehrh.— В кн.: Ареалы деревьев и
кустарников СССР. Т. 1. Л.: Наука, 1980, с. 97—99.
Смиренский А. А. Торфяные районы Казахской ССР.— В кн.: Труды юбилейной
сессии, посвященной 100-летию со дня рождения В. В.' Докучаева. М.— Л.:
Изд-во АН СССР, 1949, с. 597—602.
Смолоногов Е. П. Особенности динамики и строения темнохвойных кедровых лесов
северной тайги Урала и прилегающей части Западно-Сибирской равнины.—
В кн.: * Материалы по динамике растительного покрова. Владимир, 1968,
с. 39—40.
Смолоногов Е. П., Никулин В. И. Природные экономические условия эксплуатации
лесов в южной части Уральского Приобья. Свердловск: изд. УФ АН СССР,
1963. 119 с.
Смолоногов Е. П., Вегерин А. М., Колесников Б. П. Лесорастительное районирование
Тюменской области.— В кн.: Ботанические исследования на Урале. Вып. 5.
Свердловск, 1970, с. 34—57.
Сочава В. Б. Тундры бассейна р. Анабар.— Изв. Гос. геогр. о-ва, 1933, т. XV, вып. 4,
' с. 340—365.
Сочава В. Б. Географические связи растительного покрова на территории СССР.—
Учен. зап. Ленингр. гос. пед. ин-та, 1948, т. 73, с. 3—52.
Сочава В. Б. Основные положения геоботанического районирования.— Ботан. журн.,
1952, т. 37, с. 349—361.
Сочава В. Б. Принципы и задачи геоботанической картографии.— В кн.: Вопросы
ботаники. Т. 1. М.— Л., 1954, с. 259—272.
Сочава В. Б. Темнохвойные леса.— В кн.: Растительный покров СССР. Т. 1. М.— Л.:
Изд-во АН СССР, 1956, с. 139—216.
Сочава В. Б. Тайга на северо-востоке Среднесибирского плоскогорья.— Ботан. журн.,
1957, т. 42, № 9, с. 1408—1415.'
Сочава В. Б. Пути построения единой системы растительного покрова.— В кн.:
Делегатский съезд Всесоюзного ботанического общества. (Тез. докл.). Вып. 4.
Секция флоры и растительности. Л.: изд. Всес. ботан. о-ва, 1958, с. 40—50.
Сочава В. Б. Перспективы геоботанического картографирования.— В кн.:
Геоботаническое картографирование. М.— Л.: Изд-во АН СССР, 1963, с. 3—10.
Сочава В. Б. Растительность. В атласе: Физико-географический атлас Мира. М.:
• ГУГК, 1964а, с. 66—67.
15*
219
Сочава В. Б. Классификация и картографирование высших подразделений
растительности Земли.— В-кн.: Современные проблемы географии. М*.: Изд-во АН СССР,
19646, с. 167-173.
Сочава В. Б. Макет новой карты растительности мира.— В кн.: Геоботанпческое
картографирование. М.— Л.: Изд-во АН СССР, 1964в, с. 3—16.
Сочава В. Б. Структурно-динамическое ландшафтоведение и географические проблемы
будущего.— Докл. Ин-та геогр. Сибири и Дальнего Востока СО АН СССР,
1967, вып. 16, с. 18—31.
Сочава В. Б. Ближайшие задачи геоботанического картографирования в мелком
масштабе и перспективы создания обзорной карты растительности СССР.— В кн.:
*' Геоботаническое картографирование. Л.: Наука, 1968, с. 72—77.
Сочава В. Б. Классификация растительности как иерархии динамических систем.—
В кн.: Геоботаническое картографирование. М.— Л.: Наука, 1972, с. 3—18.
Сочава В. Б. Логические основы и пути повышения информативности карт
растительного покрова.— В кн.: Геоботаническое картографирование. Л.: Наука, 1976,
с. 12—18.
Сочава В. Б. Введение в учение о геосистемах. Новосибирск: Наука, 1978. 319 с.
Сочава В. Б. Растительный покров на тематических картах. Новосибирск: Наука,
1979. 189 с.
Сочава В. Б. Географические аспекты Сибирской тайги. Новосибирск: Наука, 1980.
256 с.
Сочава В. Б., Городков Б. Н. Арктические йустыни и тундры.— В кн.: Растительный
покров СССР. Т. 1. М.— Л.: Изд-во АН СССР, 1956, с. 61—138.
Сочава В. Б., Исаченко Т. И., Лукичева А. Н. Общие черты географического
распространения лесной растительности Западно-Сибирской низменности.— Изв.
ВГО, 1953, № 2, с. 126—139.
Сочава В. Б., Тимофеев Д. А. Физико-географические области Северной Азии.— Докл.
Ин-та геогр. Сибири и Дальнего Востока СО АН СССР, 1968, вып. 19, с. 3—19.
Спиридонов М. Д. Материалы к изучению-озер и болот в южной части Западной
Сибири.— Изв. Зап.-Сиб. музея, 1928, № 1, с. 5—25.
Сторожева М. М. Материалы и характеристика болот восточного склона Северного
Урала и Зауралья.— Тр. Ин-та биологии УФ АН СССР, 1960, вып. 20, 55 с.
Сторожева М. М. О положении южной границы северотаежной подзоны*на восточном
склоне Уральского хребта.— Зап. Свердлов, бтд-ния всесоюз. ботан. о-ва,
1968, вып. 4, с. 65—73.
Ступина Н. М. Западно-Сибирская лесостепь. Автореф. канд. дис. М., 1965. 16 с.
Сукачев В. Н. О Betula pubescens Ehrh. и близких к ней видах в Сибири.— Изв. Акад.
наук. Сер. 3, 6, 8, 1914, с. 219—239.
Таран И. В. Сосновые леса Западной Сибири. Новосибирск: Наука, 1973. 291 с.
Толмачев А. И. К ^истории возникновения и развития темнохвойной тайги. М.— Л.:
Изд-во АН СССР, 1954. 156 с.
Толмачев А. И. Введение в географию растений. Л.: Изд-во Ленингр. ун-та, 1974. 244 с.
Толмачев А. И., Юрцев Б. А. История арктической флоры и ее связь с историей
Северного Ледовитого океана.— В кн.: Северный Ледовитый океан и его побережья
в кайнозое. Л.: Гидрометеоиздат, 1970, с. 87—101.
' Троицкий В. А. Гидрологическое районирование СССР. М.— Л.: Изд-во АН СССР,
1948. 112 с.
Трофимов В. Т. Полуостров Ямал (инженерно-геологический очерк). М.: Изд-во Моск.
ун-та, 1975. 278 с.
Тыртиков А. П. Влияние растительного покрова на промерзание и протаивание
грунтов. М.: Изд-во Моск. ун-та, 1969. 192 с.
Тыртиков А. П. Смена редколесий тундрами в Западной Сибири.— Вестн. Моск. ун-та.
Сер. биол.-почв., 1970, № 4, с. 46—52.
"Тыртиков А. П. Динамика растительного покрова и развитие вечной мерзлоты в
Западной Сибири. М.: Изд-во Моск. ун-та, 1974. 197 с.
Тюремнов С. Н. Растительный покров.— В кн.: Торфяные месторождения Западной
Сибири. М., 1957, с. 30—44.
Тюремнов С.Н., Лисе О. Л., Куликова Т. Г. Торфяные отложения левобережного При-
обья северной части Томской области.— В кн.: Природные условия Западной
Сибири. Вып. 1. М.: Изд-во Моск. ун-та, 1971, с. 65—75.
Уткин Л. А. Ботанико-географический очерк Причулымского края.— Ботан. журн.,
1935, т. 20, № 6, с. 646—669.
Уфимцева К. А. Современные и реликтовые свойства почв Западно-Сибирской
низменности.— Почвоведение, 1968, № 5, с. 24—33.
Уфимцева К. А. Почвы южной части таежной зоны Западно-Сибирской равнины. М.:
Колос, 1974. 203 с.
Фалалеев Э. М. Пихтовые леса Сибири и их комплексное использование. М.: Лесн.
пром-сть, 1964. 166 с.
Хлонов Ю. И. Липы и липняки Западной Сибири. Новосибирск: изд. РИО СО АН
СССР, 1965. 155 с.
Храмов А. А. Южно-таежный Западно-Сибирский стационар.— В кн.: Геоботаническпе
исследования в Западной.и Средней Сибири. Новосибирск: Наука, 1971, с. 191—
203.
220
Храмов А. А., Валуцкий В. И. Необычная форма Pinus sibirica на верховом болоте.—
Ботан. журн., 1970, т. 55, № 2, с. 280—284.
Храмов А. А., Валуцкий В. И. Лесные и болотные фитоденозы восточного Васюганья.
Новосибирск: Наука, 1977. 219 с. '
Хромых В. С. Влияние поемности на формирование ландшафтов поймы Средней Оби.—
В кн.: Географический сборник. Томск: Изд-во Томск, ун-та, 1973, с. 49—59.
Чернышева 3. С. Отражение новейшей морфоструктуры Западной Сибири в
продольных профилях рек.— Тр. Зап.-Сиб. НИГНИ, 1970, вып. 37, с. 162—174.
Черепанов С. К. Сосудистые растения СССР. Л.: Наука, 1981. 509 с.
Шанин С. С. Строение сосновых и лиственничных древостоев Сибири. М.: Лесн.
пром-сть, 1965. 106 Сг ,
Шенников Н. П. Луговедение. Л.: Изд-во Ленингр. ун-та, 1941. 512 с.
Шостакович В. П. Климатический очерк Сибирского края. Естественно-исторические
условия сельскохозяйственного производства Сибири. Ч. 1. Новосибирск, 1931.
62 с.
Шубаев Л. П. Сургутское Полесье Западно-Сибирской низменности.— Изв. ВГО
СССР, 1956, т. 88, вып. 2, с. 167—169,
Шумилова Л. В. Ботаническая география Сибири. Томск: Изд-во Томск, ун-та, 1962.
439 с.
Шумилова Л. В. Болотные регионы Западной Сибири в пределах Тюменской области.—
Докл. Ин-та геогр. Сибири и Дальнего Востока СО АН СССР, Иркутск, 1969,
№ 23, с. 14—20.
Энгельфельд В. А. О лесах Западной Сибири. 1-е приложение к «Лесному журналу»
за 1888 г.* Спб., 1888. 71 с.
Южная тайга Прииртышья. Новосибирск: Наука, 1975. 248 с.
Южно-таежные леса Западно-Сибирской равнины (бассейны рек Тавды и Конды).
Вып. 83. Свердловск: изд. УНЦ СО АН СССР, 1972. 255 с.
Юрковская Т. К. О некоторых принципах построения легенды карты растительности
болот.— В кн.: Геоботаническое картографирование. М.— Л.:»Изд-во АН СССР,
1968, с. 44—5Д.
Юрцев Б. Н. Гипоарктический ботанико-географический пояс и происхождение его
флоры. Комаровские чтения, вып. XIX. М.— Л.: Наука, 1966. '93 с.
Яснопольская Г. Г. К характеристике растительности и торфяной залежи Васюганско-
го болота.— Учен. зап. Томск, ун-та. Сер. биол. и почвовед., 1965, № 51,
с. 49—63.
вхокоа
ипзхэ
со*
91ЧННЭНПЭХЭО
8WHXH(|K)irBJ
энннэрщоц £
I*
со"
i^oo «о
<Ms# CO
иякнаи 2
suaosdqnd '3
ermoeaddg
энаоеэй
-Э9-0ЯОНИЭО
t^ r-
5
m
©
о
ft I
&
К
eo
О
eo
cd
И
И
s
ев
ЭНЯ
-оххип-оаовгэ
ЭТЧЗ
юайэи-oaoira;
эиаовгэ-он
-ьиннэаьэик
[
со
эняонэоэ
00 00
СО *>*
00
I
ЭНН
-ьиннэахэвд
в
с
п<Шн&х
5
I
S
{*
© НН
• ев
© Ф 'в Ф •
ft J3"fce3rt
-§
^ I
«о
со ^
•с а,
I
S §*
СО
R 5?
О Й eo<tiH^
и О -g ^^ 2
Я«*ч" J2
ft Р-Н
<ь
3
«8 &
&5
со §*
"US
© о
л со
ГЙ
о
Сб
ft*5 •=*
К в
~^£
Q Q
5ft
;^5
5* ^! *- —"
C51
CD
.2 £ 3
л со o*
ft Pr!
О TW
I
•5
05
* I
ft 32 *"■* *»\
222
1>
stf t>- l>- t- !>■ !>•
■* -I I I II
CO vf vf vf vf Vf
0000 О
00 00
I I
00
I
I
vf
*l I
Ю Vf Vf
со со
I I
vf vf
,лсо ^ со со
•l".! I
LO Vf Vf Vf Vf
CO
I
vf
Ю
I
CO
r-co t^oo
*: i i i i
CO Ю Vf 1Л) Vf
I
vf
vf
CO**
.i
^o
tense Eh
poides M
— pra
— scir
a
mticum
i
о
cea
rt
T$
copo
5
.) Holub — Ly
Wallr.
%tum (L
anceps
tinum
compl
L.—
. о
. Й- й
Й 5
dium a
iastrum
planati
Lycopo
Diphas
com
о
• cd
CD
о
Pina
ф ф Ф
ф ф Ф
g
^co<«
ill
§ Й s
151
Ph
ё
•a
и
со со со во
v«a «r^> »ю «го
Jk, ?^ Ik. )ч.
a i
*?>!
Co JO
й-о
S CO
►Й
Ф
cea
ев
Hi
>»
H
\
л>-
sch
Я
о
со
•
ngustifolia L.
lia L.
mnii Lepech.
zagifiii Kryl. et
52 «£s ¥
£i i r
ф
aniac
9°
cd
Ph
CO
Й
3
•~ g
й -5.
со J
фй .
Si
-is в
аб л. 1
л ж е ни е т
I
о
С
1П
**
со
S3
3
о
OS
оо
t>
со
ю
**
5-7
**
-л
00 t-
1
00
1
ем
6-9
cd
i
41*
О
1
1
ю
00
CD
1
00
Vf
CD CD
00 00
о о о о
О ОО ^
CD oi C5 CD
00
00 00 00 0000 00
о
^ CD
G* 00"
о о
.,00 00 00 |
CD ^ ч^
'
00 00
оооо оо
оооо t>- ^ ^
CD
00 °°^
CQCSJ
CSI
CD
ocf
CD
CD
00
of
CD
^CD
oo"
1 1 J*
Ю
00
00 00
^oo
t^4 l>^
oo oo
■8
I я
^1
>
Ф
3
о
CO
а ь
сб
О
Рн
J < 5
3
со
si
a-a
в' a.
S
"5
05
-4 Ф
5^ ^ ф
«о Й
Ф %
Л-
1^ »•
5 13 §IHI
В
со
Ph
to
s4
^r>
3
.
со
1
'Wilensky
— zalesskii
Milium effusum L.
4^»
?4
Сб
1^4
eoides (L.)
L.
ф
.Рн
.9 «°
н S
ее 'Г*
« ?
Й £
О .
^ -^
■яг !
aequalls S
Schmith -
iceiw Poir.
Phleum phi
— pratense
Alopecurus
— alpinus
— arundina
J
— pratensis
сб
Ф
ffl
Й
£Л
^^
8Й
arundinac
(R. Br.)
rate Trin.
Arctagrostis
— lati folia
Agrostis cla
«j
з
«
«
►o
в
4
Roth. —
J
P
II
л
ч-а
£
CO
&
*ю
Й '
Й»
ibth. —A
Schreb.
— tenuis S
— vinealis
224
05
00~
со
С5
оо"
о
О*
О
о
а?
об
CD
со"
С75 **н t>- !>•
| | СМ | СМ |
Ю СО СМ т-1
оо~
1 1
СО vji
со ^
со
со
со"
1^ гн ОО^О^
J*»N I III
со со со vf* *•#
со со
I I
со см
00 00
со
t^4
00
00 00 СО
СО
1^*
со со
со сосооо
*? COCO
I *tf ^ .
oo oo
I I
oo
I
00
I
CO
00
CO
CO
^^
л
3
о
О «3
3
s
>
-я
..о
Я-1 ?Ч
43
3
,еф.« ^^5 8-^5
в3 Э-.^яТ|ь«
3
ё и g1
О О О)
& 8 9 g '. ф
'allele -
а
1
3.
g 3'
5^s
?Рч
i.S"8W§'
's.s г 5«
.S
.9.« d^
.?г
s gj^tf
о
о
Р5
3 * я iiii
-* -J33 S 8*2
<3 I I I I. I | I I 3 I I £ I I *§
■3 1
s^3£
225
«
се 1
н
о
Я
И
Ф
14
о
fcl
о
•А
е
»о
<*
со
(М
„00 „ т*
осГоо оо
о
ста
ОЭ OS OS OS OS
I .. ..
00 COCO 0000
ОЬ О* Ot> OS OS OS
^i.00 00 CO 0000
OS
00
ooo
<«r-l rrl rH OS
I .I I
0Q OSOO t^
*N OS CO OOOO^f* 00 «чч
-I .* .II* I I
-чгч С— С— t-CO^ !>. 00
о
-ЧТИ *tf
I
OS
I
OS
со
CO
1>г
00
СО 00 00 00 00
„00 1>- 00 ^ ^ ^ СО
V
СО
со |
со
со*
3
-•а 8
£ ф
аз
" S .СО
ей
Si.
3"ёдд-< "3-е '
8 8-»
3:SJ
Ф
, се
Л CD
sz;
?
се
82« 5
bo
'So J;
.a
s^ij-s
'За*© кЬй
3 6*^1
. ** 4 S
со £j
GQ
0^"§ 8
-I I V) f-^>
9 J* fc. J*
-•S*
rVJ ч SJ £.
« 3 a e-o
бэ ' ' ' " ■ • P S* »i & as »5 *° « ^
ft.-1 I I II ttiBQuq^-ftJ I ^ I I ft,i4
Зим
226
t>. CO ЧР ФС5С50000505 „000000О5
00 -*
СО 00 N 00 t^ ^-л] t^ О t^ ^ О" ОС
I .I I 17 - -~ II I I
SttNMlOsJ< I ^С —ГСМ СМ СС СО
t^-00 Г-
I I I
о
cT
OS 05 СЛ CO
CD
00**
Oi OS Ot>
.1 I
oo t-r^
0000 00Ю
II II
vF-tf CM CM
ooi> oo
I I I-
CM CM sj*
CO
ю*4
CM t>-
- I
чч CM
CO
00 t"»
I I
CO 00
00 00 t^
. - I
t^ t^ ю
00 00
00
I
CO
I
CO
I
G3
I
00 !>»
I I
CO
CM
CM ~
.8
ft
& 1
IIIй*
I l£
3
I
CO
ft
1 >
.• о
5 &%
< <ъ
«о.. • a
Й"
«H •
ffie
О * S-?
d|
Id
CD
,§3 2
it
0g
d3
lMli
,—J Д • О- ф
-а ведь
^ •
I I eg | |£ }5
л «J S» S) 3
!<§<3 I ! I I I I
S3
1-й
г1б
•s
a
о
со
e *S
ill
fe § e
й S .2
1 < ф
•в ■хз -в 'в 43
CD
53 J
§■, .2
. О Ф ,д
СО Гг. ^^ *^
•§ 2 в « S4
о -S оЗ а ^>
»***» j) S л .о
? »- а ^ '
^ 4} V *<^» t
I I
227
ю
сб
Н
00'М
00 Г*- *vF G5 СЭ 00 v}< <*н (М 05 00 00 СЭ 00 00 С5 С" С5 Oi t— !>•
- . I «f I I I I I I ..III II . .... I
Ь- Ю CN1 »■* t- CNJ CS1 00 ■riOO ^СОЮСО CD vj< 00 ОС °0 00 CD LO
oo
05 05
о
о
tc
о
ft
00
оо~
о
1
оо
05 СЭ
1 -
^•00
9, 1
CSI
«гн <л
t*
00 05
*- 1 1
t^co
OS t>-
1 1
г*» со
05 OS
оо" оо"
00
OOs^COsf !>.
00
00
0*00
00"t^
00
I
CD
LO00
.1
4t<
со РЧ'ЗЗ ю
•9 е- g .9
; «is
Ъо
S
I 5?
и»
1Нл
нЗ*Э
а»
Я
.о
9
1
и
■8
•Я
Is
Р.9
i^jjS
3
S «о
$.3 S>
9& лЗз^«5й-||з
л g с* %э *> fi
^ g g g I
£2 a
ft, ft.
ft, ft, ft,
5i g ц £ fi -^ *? «
SS'-S -г»1 P P *5
ft,.©
kkkk^^^eo-w
I I I II II I II I II I I I II I I I I I I
И
CO *•
о ES
2 g> ^
fc ^ 3 «o S* '"*
45 ^ .s <*> F AS «o
e a r» *■> *- I3 3
J^ ^ 3 ot2 JJ ^?
s I I f I I I
228
CM
CO
2 °
о о
см сооь of со"4
со"
00 CO 00*
CO** ^
CO CD
GO 00 CO CO
00
со*
• CD
О Рч
•"U CO
a _
feo
К ^> О "—"'О
о eg <j -5*- s»
2? ro
5 • «e
S jj1j
sill
Ь. CO CO &
cs
S 3 5 •- is.
3 I I I I I t
3
о
•2-й i
С Й К
« .4 * £
3
^
I
229 ,
а
ш
ф
«
о
«I
о
р. I
G
со~
00 00СЛ t^Cfc
.«-II -I
t*» CD CO CD CO
о* о> ел t>- чр см
I I » | |еч | „
ю со чреч еччн
t-oo со
оо
00 00 Г» !>- t-v?
t- | | 00 | | ЧНСМ vF I I
ю ю s#csi in см
00 00 00
1>Г|>Г|>Г
00
oo
00 00 00
о
CD
CO
1
*tf
CO
Ю |
vF
00 00
CD
t>-
CD"
00
00
1
CO
i>-00
1 1
CD
2
со
CO
О f*
coo
.tf-S
ocr;
S •
c3^
33
PQ <s
§ fi i
•SO «oJ?e "»
SI
§
-S J £ ss s -~
^§.2
OHM g
CO . О
© .03
о • CO
:й g
sfep-cl-ggilJI
- S
45 g3 §^ gi^-8 Sis?
1 33 g « § a» g | | 42 % §
to
№
§ g «
'a fc «
ft.~;2
a
со
v Л
QJ
В
1-3 « to J
-ч-ал
^Sil-St*?
& й 2.« 2 «
55
3* ч> «■*-. о >->
230
о
00 OO
05
ocT
00 CM C5 00 00
CO -чн СО CO rM
t^ 00 CO OS 00 Oi 05 OS CM OS 00 CD тг- GO G^ CO
II. I I I *» I - -I I I I I I I
vfI>.LO 00 CO CO I> 00 ^ SF CO Np l*-sj«st«CM
я
I
00
I
CO
00
00 00
.1
t-co
00
Oi00
I .
00
00
oo
ю«с> со
|>CO 00CO Ю
.III
CO VPIO vt< stf
CO CO CO**
Ф
a
Ф
Ф
CO
£ J.
о
CO
^ Ф
I Ф
ft
?!
PS
Ф-
••go
a. £
о
t е., -„.
-si
05
<s fc й Ь
- ^ Й ^J
^ '-So
• I hxs J—)
8.3 3
Pd^.S
8 Si
I I I I I II 6 1
is.
3
Ф
О
• о
Ctf Co
о
О
? S е
^Ss
3-Я'Й.
S
ft!
2£ 5 g*
'SJ
[sSSe^
Mil
Q«.*l .
I I I I I I3£ I II
231
оо~
а
и
о
* i
о
« I
о
л I
о
с?
Oi 00O5
оо* оГсхГ
o^qSSSo 32222o2Ss 2°22 ^
оо оо сл со оо оГ аъоъсъаъоъаъоо оо аГ оо аъоъаъ °о
С5 СЭ
i i
(М СО
00
о
в
Q
^
«
со
.3
о .
Ел
^-^ *->
• О
Я bd
Рч :^
«.о •
sal?
Mill
я s §
ЯРч^ S
Рч § g-
«^ S g
Ь О со О
С^ со *к> ?s»
s
§1
Q Ьс^ 1—1
a ш g «
Si ■ S5 I I I I
■S
! s^J
232
00
„
t^
о
"чгН
OS
cs
«»
00
OS OS OS OS
V
о
"-о*
OS
о
os^
os
о
OS"*"4
OS
^ч os^ О
OS
oo
OS
OS ^ OS
00
oooot>.
I I ,
OS
I
00
I
00
OS
I
CO 00
00
00
00
00
00 OS
00
OS
od
os
00 ~00
00
00
2 .
о
N
PS^M
42*^
42
eo
?н rv,
co,^
ил
о о
•Se
CO 5
« fan
i
s
о
.2
#«o
l«S I
14^
fo
7нЛ и
$o
►2 дЗ
«O i-S^
•2 b/ R»
tuo О «
£* fe
Ui ^j *J
IP
<D
со
PQ
! e~ ■
'eg
3
a.
Cj
M
о M
a.
£**o
ije
"o
i-H Ф
со .3
I ^£ I I I I I 1*34
-*l£
^
42 3 M
£- 1 >
• ° *^4
CO R^ С
•§■13 1
5 II I I I
5 Ki.C
3 3 5?
Ь. со Л
О
CQ
!_; tf
: as»
§j2
55.2 'I
^ I 6ч
-s
.2
^> в 2 ^
233
I
I
00
н
м
ф
ч
о
*=С I
о
е-1
оо
С5 С5
О
сГ
05 -:s
С5
оо~
оо
оооо
о
35
I
00
00
1
1
со
1>
1
1
00
1
1
оо
1
1
СО
00
t>*
00
t^4
) 00 t>-00 00 00 00 СЪ С5 С5
.111.1 II.
<t>-rHCDNl> Ю ЮЮ00
00
!
О
^O
I
I
00
00 .^00
t^
>£
00
00 00
„t*-
iCt-Г
00 00 00
r^t^t-T
oooo
tCtC
00
о
Рч
-a
о о
Й
pQ
со о
й
Q
cu r-i со 12
§ « ^
■♦■* Т^ 'is •
£ Й Й Й1-1
* ^ S 5 й
СО r-i Си Си *-
Й Й
I I ^ ^ '
3
3
N3
О
со
cq
о
г-н* • в
>>.-н
fts
1
о
есп
О
"5
PU
в
i/ios
й
So!
б-g.
$3
"2 S3
j .12
9Sf°
Й
,rl*
, Й
» .—и **
^ to 8 ^ ^ • -S
cu^^goa
| g. 8 8 S
S5^ йО |
coO
r3 ал
со rf* t2 "*
-^ « ^ S Й «о •
cu в Й О в 'J? Й
"r* cu bo Й J^ °* ^
is й й о "Je р сх,
- - й s^ а, с
й
Q. >
СО
5 I I
I £ I ^ I I
-й
<0
234
00 ОС
I I
оо
оо о
(Л
оо"
° 2
CS О 00 00 О СО ОО
. I «г ■<• I I II I
00 ^3 - "NsD vjDvt* ^O
оо оа
I .
<М 00
О4
оо"
00
00 00 С5 00 00
I I
"3
л- 8'
E3
1
ui g о .ь» «к» a*
о о 2 g
eg со "* 3s *, eo ■ Ь
• ~ • >, Ф
"*■* ^Q ф d "^ 9?
© 4 Ф -; ^^ Л С£
!аз
• Ф
за
® § и
(-3 8Д
8
я I
8
ф.
о
§..5
Й
^ a ^
о
Ь- *■> с»
s S
8 ss *й £•■•? 5> 'зд се ^ м "* 2? « S *й 'й в S "§
§>§г§
I 15
.- ^ ~ ^ сб .* '
jtt S§ о -5 СЛ ^
« г3
*-» •« ^?
g
«з
235
I
со
00
оо~
'«
ф
о
«I
о
а
о
оо
аГоГ
оооо
t>CO
оо
i
t*»Ob 00 00 !>• 00 СО О* t^OO
II . .1 II I I I
NflCO t- l>»vO СОч}«СО ЮЮ
00
00
I
CO
00
00
00
00
0000 00 00
!>• 00 ^t>«00l>- ^t-
со со \om>
LI . .'
SP Np tf CO
I
со
!>• Sf CO 00
» «»I I I .
Ю CO *tf t-
о
04
•?ij
^1
5 K^
со
« о ^ a
sfSgg
- й
-
a
ев
-PL,
I I I §6
S°5
S I
ф
о
a
a
3
a
«
3
•a
•Г—Э
о
N
О
О
!3 Рч
a J §
•S a a
a
. о
5fc:s.
ь. a^r
^S 8
a <й
s^ 8
gS-SPS .
SJS3 8
Ф
a
236
!>• CD CD
| . 00
CM 00 00
1
9,10
CD
~ 00
00
CD00 00
1 1 1
CO Ю CM
00
f- 00
00 00 00
1>Г|>Г|>Г
CO
CO
oooo
1 1
СОЮ
CM
О о °
"^ CD4-* ^CDCDS
CD CD* CD*
CD CD CD CD CD
_ 00 ^ ^ ^
00 00000000
CD
00
о
00 CD CD 00 -^ С
..I.I 1
N00 N N 00 1Л
00 000000 I>>00
i>~ 1>^ьГ|>Г oo t-~
♦%
00 t-000000 00
CD
CD
00
> CD
00 00
CO
CO
oo oo
-*H
CO
CM
^CM
9, 10
CD
00
CD
1
со
00
00
oo
CO
CO
9, 10
CD
00*
CD
CD
00*
00 00 CD
1 1 1
юсосо
со ^i>-
CO*
t>
oo oooooooo
oooo ^ „ ^ fc
CO CO CO
COCO
1 .«
vf ю
oooooooo
..11^-
CM
CD
CD
00
00
l>
00
о о
CD CD
CD CD
ж „ CD CD
oooo
CD
CD 00 00
oooooo
l>
237
та 6 л
ЗИН
о
Прод
Н
-* 1
со
<NI
*■"' 1
°
Од
00
с-
со
ю
**
О
о
CD CD
CDCD 0000 CD
оо~оо~
t
/
oo°?cDCp°?
.1.11»
г*» со оо со со
!>.
0000 0000 °0
csTiC |>Г|>Г|>Г
со
СО
00 00
1
t> СО
cd
CD
1
СМ
9-10
9, 10
9, 10
cdooo
о
1
00
с^
00 !>•
i>^oo~
оооо
10
9, 10
9, 10
cdcdcd
е
а?
CD
1
00
СО
00
о
оо"
00
00
о
о"
cdoo
cd _ л
00 CD
о
8-10
9, 10
5-8
00
00
00
о
<м
о
CD CD CD
CD
„CD00 00 00
оо"
CD 00
-1
оо со
00
1
00
оооооо
со
1
оооо
coco
оо оо V
^00 00 00 00
i з
"3 .|J
•S 8 S i §
S9
gd
о x
.л
■» 3 о 3
i£. i i
J 4 «J
'Saga
Hal
i 13.1
? ев
31'
9
Iftfl
< Ъ
8^
' «ft
! * 3
§ ~ S g.*c
a *r
I ! I I I I I I I I
5»
id.
о
CO
■§3ta
PQ
Q
«o
<*>!
i ids i
J CD
238
35
00
I
CM
сг сэ
о
of
q 00O5 О
^00 00 00
оо" оГоо* ос*
оо'оо"4 оо"
О О С5
оо"*оо~ сю
00*
о
с?
I .
с^оо
I -I
С5 О500
I I I
I
00 C5 00
I I I
о ю со
00
I
00
00 00 00^° OOOOCiOO
000000 00
00 00 00
00
со
to
0000 00 00
1> i>« t>- CO
00 00 CO
00 ^ ^00 I
tCt^r со
00
CO
I
00
CO
. £
3-
Ф
Ф
'О
.s *
Ф
ее
ф
о
се
нз
се
О
ф
се
ф
о
се
S-t
се
Й
ел
"Й **
.№СЛ
tm
IS*3
. « g<
• ■'•3 « «
•3 l 1.1 I I
a
й g
«
ce
SW4
CtJ
fl^SH^s*
I
CO Co
й ?rS
.5J й
3 I
ее "тт
I
О
ф- оо
ф «
О со
ее о
'се §
Й8
1
а
р
S3
*§
ila hibri
О
о.
irica L
- sib
1
ф
■3 1
о .©
-С tS3
Eup
edeb.
ak) С
J *o
& a
»*■*
, 8
Co • •«
r&to di
osa L.
Idsteini
t.
s I 1
bq 1 1
ф
cea
CO
Empetr
rup
ф
Hag
ditum
g
aph
S
^
гит /ге
гши L
•к» Go
^
1
Ф
cea
ее
Balsamin
tangere L.
•Л
tens no
«3
>■<
Ф
ce
mna<
ce
PS
Д
Я
со
a
da aln
rangi
C«N
239
05
осГ
о
I
05
и
о
ч
о
«I
о
Он |
00
стГ
об* оо*
00 „. 00
00
о
ЧГ-»
I -
оооо
00
I
со
ОЪ Cfr 05 *<Н
г^юоо ем
00
I
со
о
оооо
I .
СО**
00
оооо
I .
СО t-
00 OS
t-^ оо~
оооо
оо t^
со*
о*
оо"
чО*
ф
сб
ф
ф
сб
ф
II
^^1
Ф
Ф
SS
Ф
о
сб
■a
2
EC «A
J5 . , ,
"a
•к»
It
'? /si
IS'73
со «J
а
«
«
£
а.
ф
сб
ф
о
сб
ф
.сб
ф
а
>»
л
н
ф
сб
ф
о
сб
agn
©
Л J
и te
ф
сб
ф
о
сб
Р>а
1-1
"55 J ьД «^
• Л9
si
fop.
.&SS
I I I I и
■a*
I*
I I
8-
«
^
8^
4
240
i
00~
00
OS OS
oo"* t^
oo
of of
о о о
О ^"* О ^"* "^
С5 Ot) C5
00* 00** 00~00 ОО**
оо
О* С»
00* 00*
оо"*
ОЭ 00 t*- С5 СТ> 00 О О500 O^tt-O^OO
J I I II I 1 I . I 1 -I
I I -I I
ю юоо«ою
I
00
00
00
t^*
00
00000000 00 0000
00
00
oooo
i>*i>r
CD со
**ю*
I
oooooo
1>Г|>Г1>Г
I
00 Oi
tr-Too"*
oo*
d
2 *
d 4~"
о
а о
a»
со
M
ф
.d
43
ев
d
о.
5
<
d -.a
«а а^
О ч-н и—ч
ииа
•■*■• е+Ч
1-3 .в
й <е <s *-»
е fc Й «
fi^"£ g
*5
e £
a.*
g§
t3 «O
tUO Ik
g
a »
; Й о s .^ .
со •
££ в
bq^
eo -^
_u к. О ^* ?3
8 g §>§ S^
H
^^^
a
ф
d
о
M
л
о
со
ф
fa
•
£•§
''со. а
-Spd eo •
«43
1 2? © .^-5
a —- о»
: g T ^ о, .2 "s/
3 &JD43 t^
^ Ф в ^o «R 8 '
■J* *» • CS
со jP «o
^ со ^ § ss»^ b.
g^^C^co g
to к | I to I
241
as
H
CM |
СО N
I I
CM vP
CD
4
о
о
ft I
И
00
I
CO
00
i
CO
00
g-j CO COCO CO CO
..II.
CO
I
CO
I
«©со со
00 «ON 00 r^ 00 N N OCOCO CO OOO 00COCO
I . .►--1 .1 . I I.I . II III
Ю Ю CO ~lOCOvF CO Nj* ЧРЮ^ Ю stf •<!< VF sf< v}<
со со со со
о
с
•8
Й
о
cd
й
й -^
^ Й
cd |
я ж-
о
ft
о
43
Со
Й
!>>
о
► 3-Й
g too-5 S
^ I I I
>A4
co^:
23-
, Й C3 V
4l д
О Й со
■sag
£2£
43
Й
о
cd
ьЙ "£
X
©
43
43
о
«J
3
^1 1
о
■ l§
й S ft
о • со _•
Q "^S
W « ^43 , ^ _
^—v "i Й со »i—( <^ -i
w-3 s s^rio-—
, о
Й
43. 43
s ^ s Л -s ^J>
4 *9 -ST'g/e S a
f
•Й
ft
й
ф
cd
a»
о
cd
i
«O S « У ^ ^
43
в |
О О 43
й й-~
й
S§
от S з з "S
s * *
cS 1
I
1
*3
242
CD CD
I I
i
CM
о
CD**
CD CD
oo~oo~
CD
oo~
CD
00*
CD
00
о
oo"
00
о
cd~
^n I (
CD CD 00
о
cT
CD
oo~
CD 00 ^н CD
.1 It- I
00 Vf* St» ^
о
I
00
CD CD
I I
t^ CO
00 CD
I -
VP 00
00
t>**
I>»00
oo"i>^
CO
I
41*
00
I
CO
a-
2
PS
j e
-Л со
© 5
&^
£!■
eegCS
«£«*£**■
S ^s «S be t,
s • * -3 g -s~§
я.
о
i-h сб
2 я
о
PL.
2 J
О
.9
3 ■;
243
vo
се
н
о
В
К
00
05
об*
О5 05
оо*оо*
оГ
ОЭ СТ>
о
«I
о
Рч |
G
С5
00**
05
^
00
ОЭ
°о ^
00
05 О 05 ОЭ
Од ^^ »» .» »»°
об* об* об*об*
Од Од ОЪ СЪ
об* оо
оэ
об*об «Г
Од
об*
оооо t^ оэ
III I
сою vj< ю
00
I
00 OS t^ 00
I II I
Од t^
I I
Od
I
Ю
.1
cocsi
00
I
Ю
00
I
Ю
00
t^*
Od
I
Ю
00
CO*
0©
00 00
f- ^00
00 t^OOOd 00 00
oo oo ^ „ ^ ^t'-oo
t>Too t^*oo* t>T i>T
00
I
CD
00
о
о
s
г»
s
I
. 4
* й £*
I
.2
Q
a>
<л .S3 _
*> I—I
CQ
о
со
,ю
со П2 t* **
§ £ §
3 e §
S2 e e
£3 ■+* ►»*
§.§*•
1 IЛ
••I л J
•S .2 «
•£ S j
в « 2
^ о s «j
3 § *§
CO ^
**^ 5
52 « "3 ^
•8 I S §
g|||l
cot^jQ II
. й
PQ
•2
v. ^ .
.2 I
Sg
IS S
с*ь
о ■; s*
^^ S
О « 25
со ^^
J eg ^ <o
* § * I 5
S S |
з а £
S3 * e
II-
S
3
t3-
а
244
cdcd
i i
CD
00~
CD
00
I
CD
00
cd
i
ю
CDCD
1 -
l>-00
00
cd
1
Ю
00
00 00
«ЧН
cd
00
CD
00*
со
1
4*
1
cd
CD
„00
00
CD
CD
00**
CDCD
00 00
I>
©0
тН
00
Ю
Ю
•*<
CO*
<M
00 OCT
1 1
сою
t>
00
<N(
о о oo
^CD ^CD *чН'44
CD CD CD CD
CD CD CD CD _.
00 00 00 0000
CDt^OOCD 00 CDCDO
1 1 II . .11
00
CO
00 CD
00 l>» t^ 00 „00 „
CD
00 0000 00
00
1
CD
CD
oo"*
о
1
00
00
<D
g ЧГ
J 53
'•8:
§.
CO J
на а
g dtl
SI el
'•S Js o, 8<
I l^
ft
о
a
PQ
i ф
§co
ьр
I S«5 l l I
о "й^
« s ь.
в Q SJ
.2 «
§ S«*E
ft* ^ «o
I I I I I I
со
1
&
I
3 И
О
3
s
245
OS
об4
О4
00*
к
ф
ч
о
о
л |
С
о
стГ
(хГсхГ оо"
о о
о
05 05 тн 05
ММ
1>- Ю 00 СО
t^ ОЗ 00
- I I
CD Ю СО
О
"Ч-Н
О?
СП
00*
г^оо
I .
05
оо~
05
I
о
сГ
оо~ оо"4
СО
I
00 CD
t^co*
СО
I
00
t^*
00
l>*
00 00
00
00
СО СО СО СО СО СО
со*
00 СО
СО | | СО
СО -SP
л
о
<Т>
»—«
л
о
сд
§3
8
&
^
^
"Q
J
к
i*5
ф
СО
СО
2
*-8
S S S
г-Н
Рч
§
<»<»
§
сб
*-i
а,
~1
*
1
£
во
О
О
СО
а
о 3 v. Sfi о
'«* о Ончо to л а
3 I I Г II !
Q
I I I
Ъ
246
OOiw'
oo'oo"00
со"
Oi C5 35 ОЭ СЪ
О ОО
"^ ^ч "^ ^ С
C5 G5 05
о
oooo
Ci CD C5 G5
C5 C5 05
00 00 00
05
oo~
о
oo~ «Г
C5 OS 05
oo~oo~oo~
oo
о
стГ
I -
CM 00
I
CM
^ да
OS СП) OS
I I I
00 00 00 G5 05
I I II I
to vt< vf Ю Ю
00
GOOO 00
00
00
00 00 00
00
00 00 t^
t^
со
t^
t^
oooooo
i> t>-1^
00 00
со"
Ф
^5
ев
a
a
.Q
CD
Ф
|
jj
o.-
Ъ0 i» CO $
• i i 43
О
>
о
о
-А Н Д
ей
Н
§ - ч.
ад «о
о $з Ji
Ьо Gl—I
со,—I
1-3 о CD -2
•JS *> 2 в
^^з
о ь-
>5
2
9Ъ-
^р
Р *S
6 1
>>0§
3 J §1
СО
£<Л
■Д- .cq
о
5 §в
О в
,g-SU S
1Й .
со С fc
oq
1 ^
со -IS
в S» в
Й в в
В § |
'ад^
<Ъ >^. s~> s^
** в а а,
о g -^ -^>
о '
^ g
1 &J
s I §.«
О
со
3 «
з- S .•» •§ *s?
6^^ 1^
247
оо~
оо оэ
Я
я
cd
я
о
К
О
о> ^
о—^
ОО ООО С5 О
О "гЦ ^ О ^ ^ чеН О "гН <
-^ „ Л^1 ^ ^ «.-*■< о* •."
ОЪ Oi Oi Oi OS TiOi
СЭ
oo~
О*
oo~
05
06*
OS CT>
СЭС5
00 оэ ст> 00 о
oTod" 00" a* 00**
OS
00"
<J5 OOO
°0 оГ аГсгГ
05 00
1 1
00 CD ОЭ 4f OS
I I I I I
Ю <M CO CSI Ю
о <5^
■чН 05 -чН
I - I
00 00 00
О Oi t>* 00 G5 05
I MM.
ю юсо^ t^oo
00
00
00 00
0000
1>Г|>Г
00
CD
I
•8
о
о
о
•J .J
Ю
12
JO S
s «
-s
к
a. -S ее
fcf) <M
.§ 1 g ^ a
s» X a в "й
b4 о
""PQ
« .2 §
I I
I k^^ I Q
"'* 11
.1
. • И
a t> Рн :2
' § в «з *й
43 «£. Ьс Ьс
-S .со
S-- .-§ х
Mill
CD
<D
^ CO «
§•££
""3 ^ '2?
•a
•1—»
'o
. . и
со g •
s « •
ed о ЙЛ
« ^ .S J .2
<j 4j 3 •< «
CO «O =0 Ч«» й>
«
^
cd
U-
M "-^ -: jo Д
q со •—' .^ as
a? .^^
3 <Ц в HJ г
248
CO
СО**
со
со"
о о
СО ^ ~"
О 00 о
00 С5 ОС
о о
со ^ *~*
СО СО
СО СС СО 00 -о СО СП)
-1.1 .1 1
00 vf 00 Ю 1-« f«- SD
оо . со
00
СО
.со
о
СО
СО "^ """ ^ "*~ "*"' СО
СО с: С", г:
со со со r ^
зо"сс" х' -с"
со" _/гГ
Г- ОС 00 СО СО СО С5 С: СТ.
!..!!! .11
Ю Г"- £>. Ю Г>- О У С^О
СО
I
h-
00 °°
о
to
00
00
00 00 СО ОС
со
00 00 00 00
оо"
со
-
~" ст.
с^
1-
со
ОС*" CO*"
"Л
i
00
1>^
сс
со
с: О
со со ^
со" со"
со со
ос" ос"
С- Ci ОЭ
! : J
'- -г- О
00
ОС ОС 00
СО
со оо оо
CQ
1
е
о
«
и
«ф
:§*
"8 Z*
ф
i«fi
~3£
g
Si-Э
^ Се So
sills
! 15 IS
• ffi
8
I
ф
3
^8 5
л
ф
О 4*
со -яр
3 *
•^ 3 В*
3
[Its
liilll
IG I I M
Afl.il
Ulit
if|||
isJi.§
If litis
' « x с
о
ОГЛАВЛЕНИЕ
Предисловие 3
Введение (И. С. Ильина) . , s . s . . . s й . * .. . 5
Глава I. Основные, географические закономерности растительного покрова
Западно-Сибирской равнины (И. С. Ильина) 8
Глава II. Карта «Растительность Западно-Сибирской равнины» . М 1 : 1 500 000 18
Методика работ. Основные источники (Б. А. Богоявленский, И. С. Ильина,
В, Д. Махно) . —
Принципы составления легенды (Я. С. Ильина) 24
Легенда обзорной карты «Растительность Западно-Сибирской равнины» -
{И. С. Ильина, Е. И. Лапшина, Л. И. Мельцер,' Е. А. Романова) .... 30
ГЛАВНЕЙШИЕ ПОДРАЗДЕЛЕНИЯ РАСТИТЕЛЬНОГО ПОКРОВА
Глава III. Тундровая растительность (Л. И. Мельцер) . 41
Арктические тундры (Л. И. Мельцер) 42
Северные субарктические тундры (.'7. И. Мельцер) 46
Южные субарктические тундры (Л. И. Мельцер)^ 48
Глава IV. Темнохвойные леса и редколесья (И. С. Ильина) 54
Лиственничио-еловые и еловые редколесья {И. С. Ильина) ....... 62
Северотаежные лиственнично-еловые и еловые редкостойные леса и
производные сообщества на их месте (Я. С, Ильина) 65
Северотаежные лиственнично-елово-кедровые леса и производные
сообщества на их месте (И. С, Ильина) 67
Среднетаежные елово-кедровые леса и производные сообщества на их месте
(Е. И. Лапшина) ." . 72
Южно-таежные елово-пихтовые, пнхтово-еловые и пихтово-елово-кедровые
леса и производные сообщества на их месте (И. С. Ильина) . \ 81
Глава V. Лиственничные леса и редколесья (Я. С. Ильина) .......'.. 90
Лиственничные и елово-лпственничные редколесья и производные
сообщества на их месте (И. С. Ильина) 93
Северотаежные лиственничные, елово-лиственничные и сосново-листвен-
ничные редкостойные леса и производные сообщества на их месте
(Я. С. Ильина) 99
Глава VI. Сосновые леса и редколесья (И. С. Ильина) 104
Северотаежяые лиственнпчно-сосновые и сосновые леса и редколесья и
производные сообщества на их месте (И. С. Ильина) • • • ^7
Среднетаежные сосновые леса и производные сообщества- на их месте
(Е. И. Лапшина) .' .:. 112
Южно-таежные сосновые леса и производные сообщества на их месте
(Е. И. Лапшина) ♦ • • • • 118
Подтаежные и лесостепные сосновые леса н производные сообщества на их
месте (Е. И. Лапшина) -1 • ; *^
Глава VII. Березовые и осиновые леса (Я. Я. Лавренко) 125
Подтаежные березовые и осиновые леса и производные сообщества на их
месте (Я. Я. Лавренко) . . . . .... . ."• • ••• "8
Лесостепные березовые и осиновые леса п сельскохозяйственные земли
на их месте (Я. Я. Лавренко) . .*'. ... . . ■ . • • ■• • • '•'. *35
Глава VIH. Растительность болот (Е. А. Романова) ............ 138
Арктические травяные и травяно-моховые болота (Е. А. Романова) . . . . с 140
Субарктические кустарничково-осоково-моховые и кустарничково-мохово-
лишайниковые полигональные комплексные болота (Е. А. Романова) ... 141
Северотаежные кустарничково-мохово-дишайниковые плоско- и крупно-
бугристые и лишайниково-сфагновые олиготрофные болота (Е. А.
Романова) ... 144
Среднетаежные сфагновые и сосново-кустарничково-сфагновые
олиготрофные болота (Е. А. Романова) 148
Южно-таежные сосново-сфагновые олиготрофные и осоково-гипновые мезо-
трофно-евтрофные болота (Е, А. Романова) 151
Подтаежные лесные и осоково-гипновые мезотрофно-евтрофные болота
(Е. А, Романова) 153
Лесостепные евтрофные травяные болота и галофитные луга (Е. А.
Романова, Е. И. Лапшина) 159
Глава IX. Степная растительность (Е. И, Лапшина, Я. Н. Лавренко) 161
Богаторазнотравно-злаковые луговые степи и остепненные луга (Е. Я.
Лапшина, Я. Я. Лавренко) 163
Разнотравно-дерновинно-злаковые степи (Я. Я. Лавренко, Е. И. Лапшина) 170
Глава X. Растительность речных долин (Я. С. Ильина) «... 177
Тундровые арктические и субарктические болотно-лугово-кустарниковые
серии растительных сообществ (И, С: Ильина, Л. И. Мелъцер) .... 181
Северотаежные лугово-болотно-соровые и кустарниково-лесные серии
растительных сообществ (Я. С. Ильина) 186
Среднетаежные лесо-кустарниково-сорово-луговые серии растительных
сообществ (Я. С, Ильина) 191
Южно-таежные лесо-кустарниково-луговые серии растительных сообществ
(Я. С, Ильина) 196
Подтаежные лесо-кустарниково-луговые серии сообществ (Я. С. Ильина) 202
Лесостепные кустарниково-лугово-степные серии растительных сообществ
(Я. С. Ильина) 208
Литература (Я. Я. Лавренко) 211
Список латинских названий растений (Я. Я. Лавренко, Е. И. Лапшина) 222
Правда Степановна Ильина, Евгения Ивановна Лапшина,
Наталья Николаевна Лавренко, Лия Ионовна Мелъцер,
Ефросинья Андреевна Романова, Борис Александрович Богоявленский,
Валентина Дмитриевна Махно
РАСТИТЕЛЬНЫЙ
ПОКРОВ
ЗАПАДНО-СИБИРСКОЙ РАВНИНЫ
Утверждено к печати
Институтом географии СО АН СССР
Редактор издательства Л. И. Шпаковская
Художественный редактор Е. Ф. Гришин
Художник Я. А. Пискун
Технический редактор А. В. Сургаиова
Корректоры В. В. Борисова, С. В. Блинова
ИБ № 23689
Сдано в набор 10.07.84. Подписано к печати 28.01.85. МН-02013.
Формат 70XI08Vie. Бумага типографская № 3. Обыкновенная
гарнитура. Высокая печать. Усл. печ. л. 21,7. Усл. кр.-отт. 22,1. Уч.-изд. л. 25.
Тираж 1000 экз. Заказ № 297. Цена 4 р. 10 к.
Ордена Трудового Красного Знамени издательство «Наука», Сибирское
отделение. 630099, Новосибирск, 99, Советская, 18.
4-я типография издательства «Наука». 630077, Новосибирск, 77g
Станиславского, 25.