






































































































































































































































































































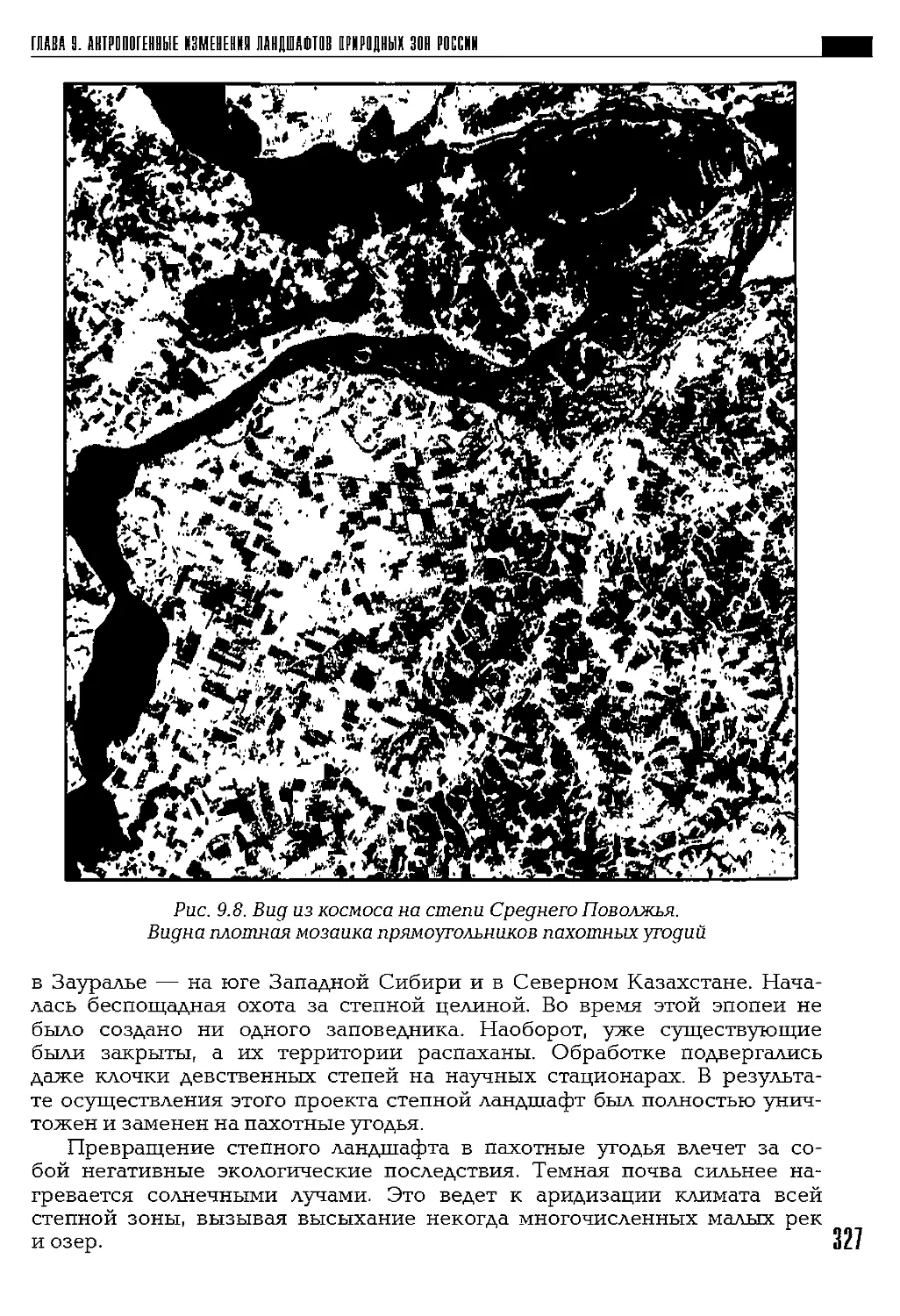




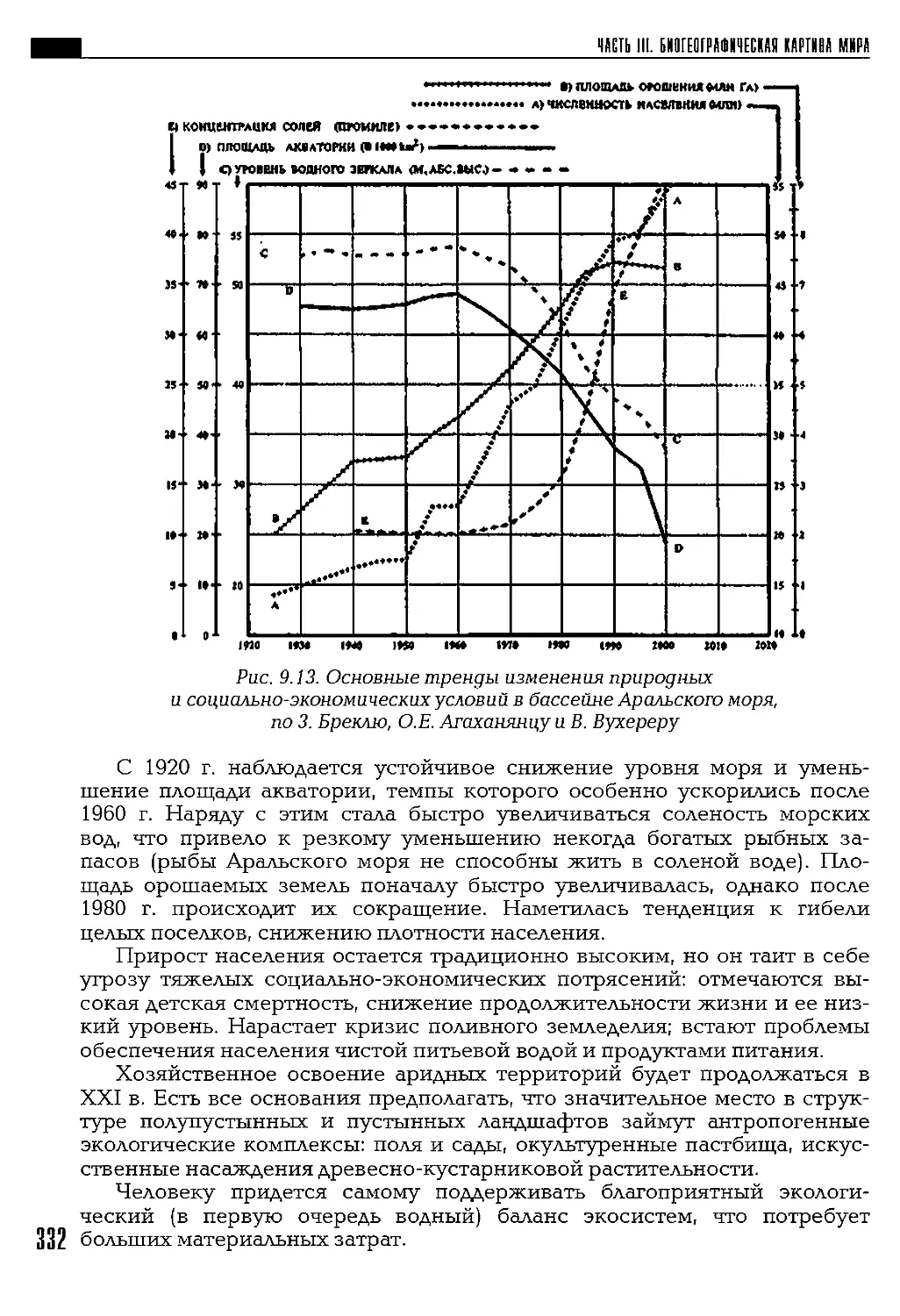



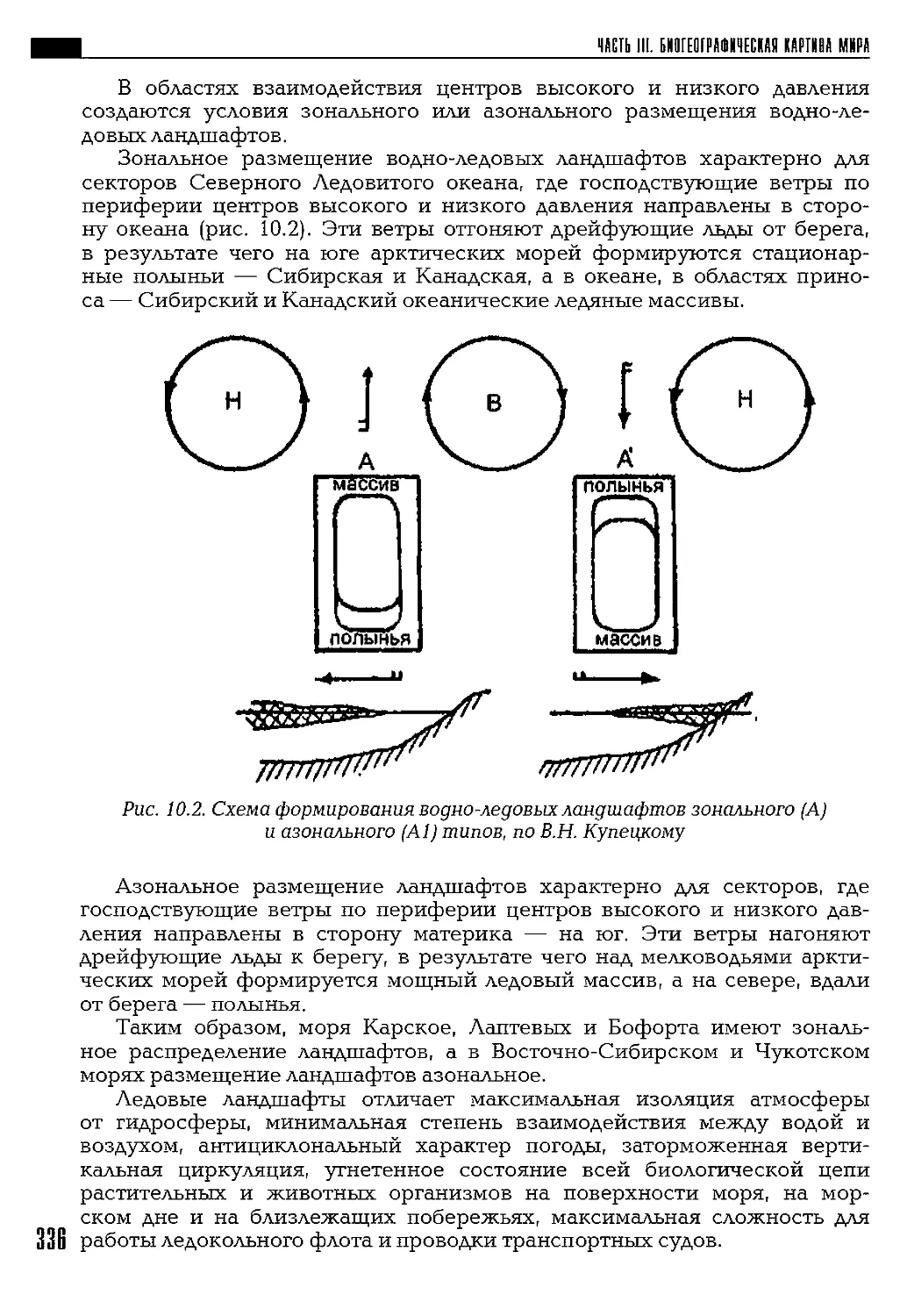










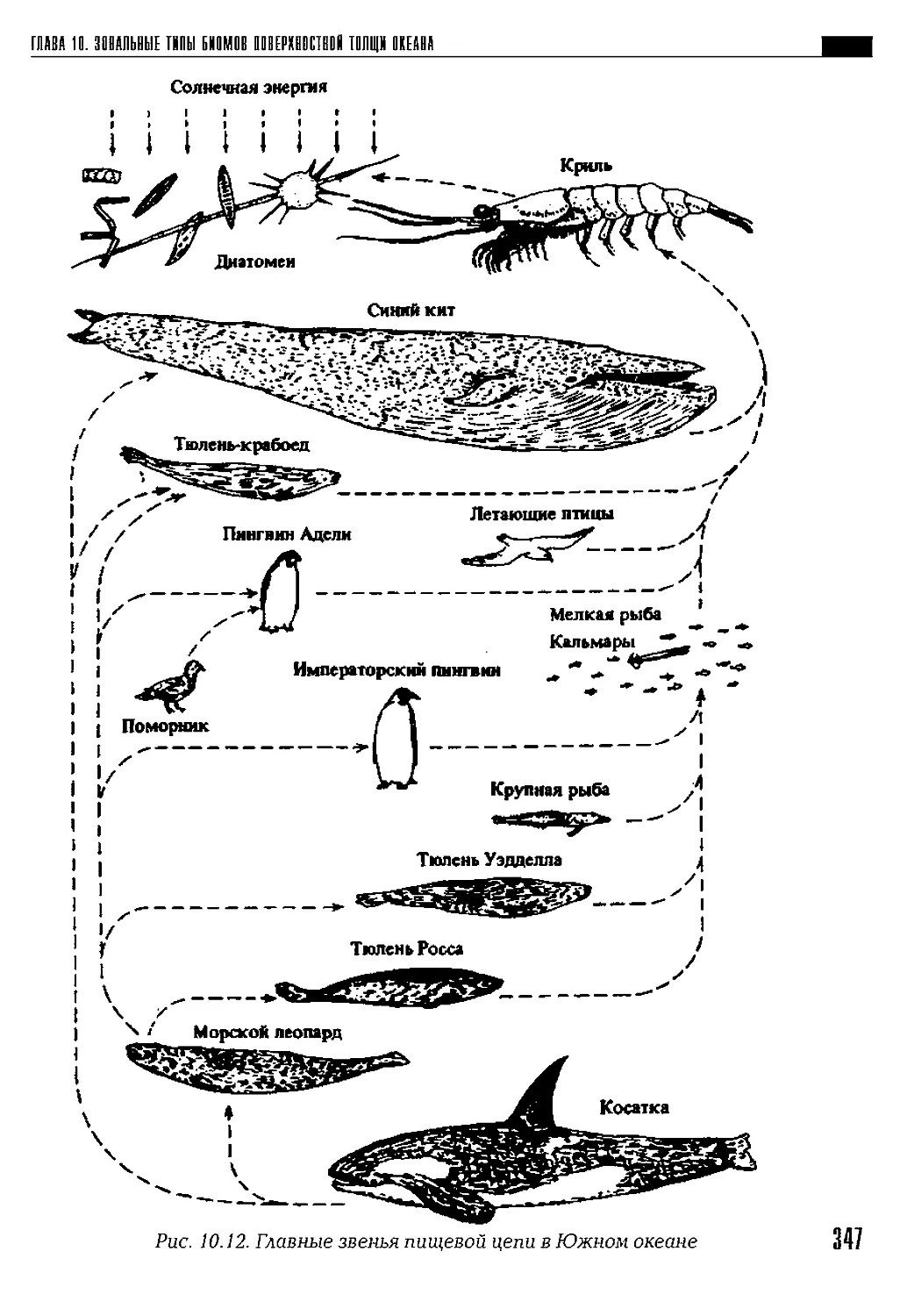




















































Автор: Петров К.М.
Теги: общая экология биоценология гидробиология биогеография экология и биогеография охрана живой природы география биология экология учебник для вузов издательство москва академический проект
ISBN: 978-5-8291-3025-1
Год: 2020




Санкт-Петербургский государственный университет
Учебник
для высшей школы
К.М. Петров
I
БИОГЕОГРАФИЯ
Рекомендовано Ученым советом
Санкт-Петербургского государственного университета
в качестве учебника для студентов,
обучающихся по географическим специальностям
Москва
Академический Проект
2020
УДК 574
ББК 28.08
П30
Рецензенты:
|Д.А. Криволуцкий |
А.К. Бродский
— кафедра биогеографии МГУ, зав. кафедрой, чл.-корр.
РАН, профессор;
— доктор биологических наук, профессор биолого-
почвенного факультета СПбГУ
Петров К.М.
ПЗО Биогеография: учебник для вузов. — М.: Академический Проект,
2020. — 400 с. — («Gaudeamus»).
ISBN 978-5-8291-3025-1
В предлагаемой книге раскрывается сущность биогеографии как науки гео-
графической. Для объяснения разнообразия жизни на Земле используется эво-
люционный подход. Обсуждаются общегеографические факторы, определяющие
подразделение арены жизни. Излагается учение об ареалах. Отмечается зави-
симость рисунка биогеоценотического покрова от экологических условий, от
структуры ландшафтов. Биогеографическая картина мира раскрывается через
описания особенностей биотических царств, зональных типов биомов суши и
океана.
Учебник предназначен для студентов вузов, обучающихся по географиче-
ским специальностям.
УДК 574
ББК 28.08
ISBN 978-5-8291-3025-1
© Петров К.М., 2006
© Академический Проект, оригинал-
макет, оформление, 2020
«Полна чудес могучая природа...»
ПРЕДИСЛОВИЕ
Своеобразие планеты Земля подчеркивается наличием на ней жиз-
ни, образующей особую оболочку— биосферу. Живое вещество (так
В.И. Вернадский назвал совокупность всех организмов) играет важную
роль в регулировании как физико-географических, так и социально-
экономических процессов. Поэтому растения и животные как элемен-
ты природных систем так или иначе рассматриваются географами.
Однако в семействе географических наук только биогеография имеет
непосредственным объектом своего изучения жизнь на Земле — рас-
тительность и животный мир как компоненты ландшафтов.
Огромную роль в функционировании экосистем играют также бак-
терии и вирусы, простейшие, цианобактерии (сине-зеленые водоросли)
и грибы. Однако в силу специфики названных групп организмов в дан-
ной книге они не рассматриваются.
Биогеография — наука о распространении на Земле растений, живот-
ных и образуемых ими сообществ в зависимости от естественно-истори-
ческих, географических, экологических и антропогенных факторов.
Объектами биогеографических исследований являются: ареалы рас-
тений и животных; биоты — исторически сложившиеся на данной тер-
ритории или акватории совокупности всех видов растений (флоры) и
животных (фауны); биомы, образованные территориальными сочетани-
ями сообществ организмов— биогеоценозов (экосистем).
Как видно из самого термина, биогеография— наука, занимающая
пограничное положение между системами биологических и географи-
ческих наук. Особенности биологического и географического подходов
в биогеографии показаны на рисунке. Основные объекты биологических
и географических исследований изображены в виде стопки дисков.
Ареалы растений
Ареалы животных
Экология
Система™ ха
Морфология и
анатомия
Физиология
Генетика
'Биота определенной территории
'(акватории)
Растительность
Животный мир
Климат
Воды
Почвы
Рельеф
(оологическое
строение
Биом определенной территории
(акватории)
Биологические и географические объекты биогеографических исследований
3
ПРЕДИЕЛОВИЕ
При биологическом подходе (а) своеобразие видов, образующих
биоту, характеризуется последовательно на генетическом, физиологи-
ческом, анатомо-морфологическом, систематическом, экологическом и
ареалогическом уровнях. При географическом подходе (б) своеобразие
биома раскрывается с учетом его растительного покрова и животного
мира, а также основных ландшафтообразующих факторов — геологи-
ческого строения, рельефа, почв, вод и климата. Секторы, вырезанные
из стопок, соответствуют территориальным (экваториальным) особен-
ностям биот (а) и биомов (б).
В данном учебнике раскрывается сущность биогеографии как
науки географической. Для объяснения разнообразия жизни на Земле
используется эволюционный подход. Обсуждаются общегеографиче-
ские факторы, определяющие подразделение арены жизни. Отмечается
зависимость рисунка биогеоценотического покрова от экологических
условий, от структуры ландшафтов. Биогеографическая картина мира
раскрывается через описания особенностей биотических царств, зо-
нальных типов биомов суши и океана. Научные основы взаимодействия
общества и природы постулируют не покорение природы, а рациональ-
ное использование и охрану ее ресурсов. Культурологический аспект
утверждает необходимость гармонии между духовным миром человека
и природой.
Императив устойчивого развития общества возможен только при со-
блюдении «принципа взаимности», как в общении между людьми, так
и в отношении человека к природе. Теория и практика биогеографиче-
ских исследований имеет целью разработку рекомендаций по охране,
воспроизводству и рациональному использованию ресурсов органиче-
ского мира. Структура книги последовательно раскрывает сущность на-
званных подходов.
Разработанный курс отвечает современным требованиям и соответ-
ствует образовательным стандартам, утвержденным Учебно-методическим
объединением университетов России. Вместе с тем ряд разделов предлага-
емого учебника выходит за рамки существующих стандартов, благодаря
чему он дополняет имеющиеся учебники, не повторяя их, и может широ-
ко использоваться в географическом и экологическом образовании.
Курс биогеографии является фундаментальным при подготовке гео-
графов всех специальностей. Не одно поколение студентов воспитано
на учебниках А.Г. Воронова и коллектива сотрудников Московского
университета (Воронов, 1963; Воронов, Дроздов, Мяло, 1985; Воронов,
1987; Второе, Дроздов, 2001 и др.). Известны монографии и учебники
по биогеографии У. Нейла (1973), П.Д. Ярошенко (1975), Ж. Леме (1976),
О.Е. Агаханянца (1992). К этому следует добавить обширную литерату-
ру по географии растений и животных.
В основу данного учебника автором положен многолетний опыт
экспедиционных исследований в разных природных зонах суши и океа-
на; работа в библиотеках научных центров России, Англии, Голландии
и Дании; чтение лекций в Российском педагогическом университете
им. А.И. Герцена и Санкт-Петербургском государственном универси-
тете. Я благодарен коллегам, прочитавшим рукопись и сделавшим ряд
ценных замечаний.
Часть I
ИСТОРИЧЕСКИЕ
И ГЕОГРАФИЧЕСКИЕ ФАКТОРЫ
БИОЛОГИЧЕСКОГО РАЗНООБРАЗИЯ
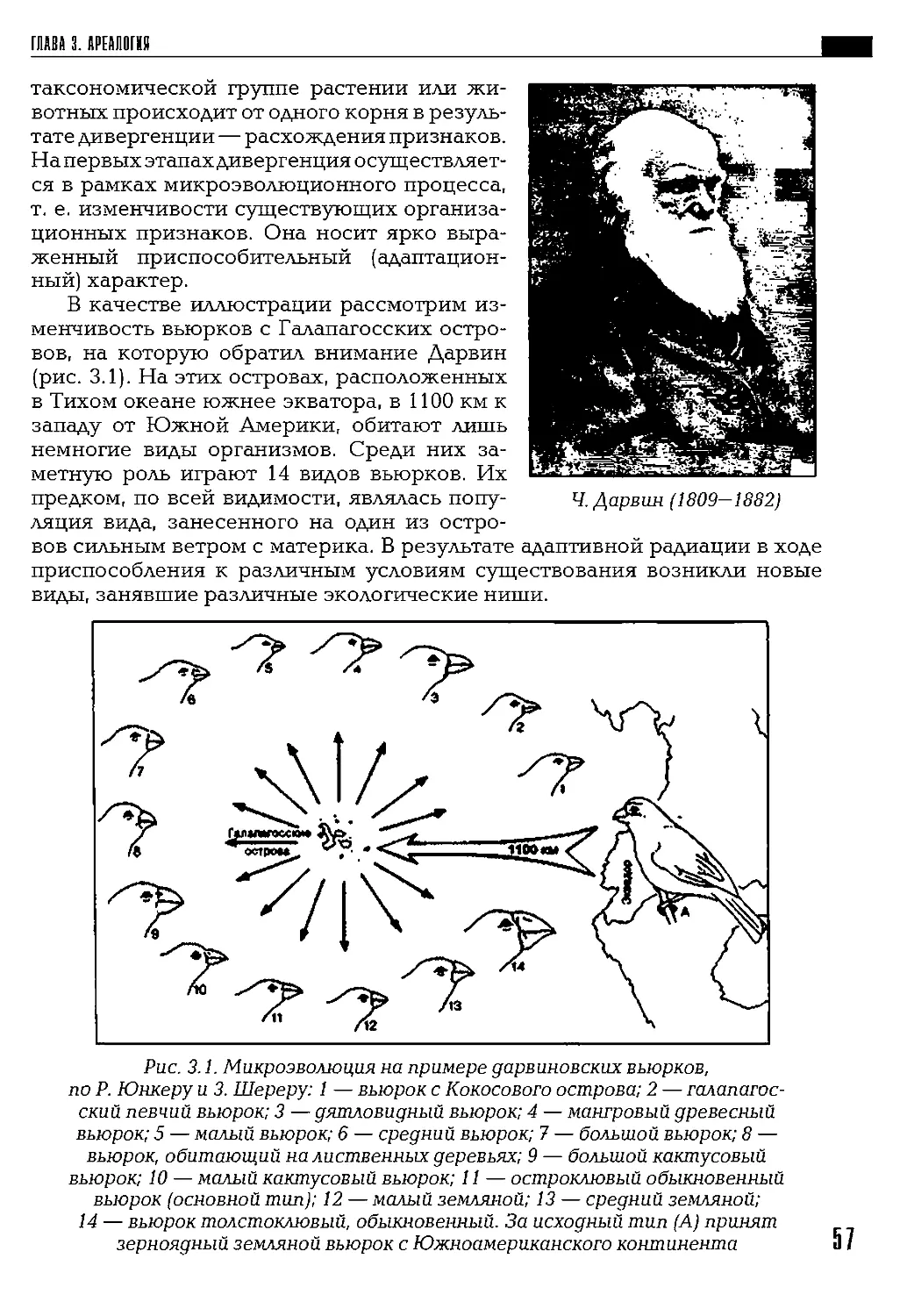
При обсуждении темы происхождения и развития жизни в центре
внимания должны находиться пять феноменов: жизнь, организм, био-
разнообразие, экосистема и биосфера.
Тайна возникновения жизни до сих пор остается неразгаданной.
В конце XVII в. голландский ученый X. Гюйгенс в книге «Космотеорос»
сделал следующее научное обобщение: «Жизнь есть космическое явле-
ние, в чем-то резко отличное от косной материи». В.И. Вернадский
назвал это обобщение принципом Гюйгенса.
Я присоединяюсь к мнению тех ученых, которые рассматривают
жизнь как космический феномен. Такое допущение не отвечает на
вопрос, как возникла жизнь, но позволяет предположить, что простей-
шие формы живых существ были занесены на Землю из космического
пространства. Таким образом, допускается, что феномен организм в его
простейшей форме появился на Земле в готовом виде.
Биоразнообразие возникло как результат микро- и макроэволюции.
Ч. Дарвин заключает свою теорию происхождения видов словами: «Есть
величие в этом воззрении, по которому жизнь с ее различными прояв-
лениями Творец первоначально вдохнул в одну или ограниченное чис-
ло форм; и между тем как наша планета продолжает вращаться согласно
неизменным законам тяготения, из такого простого начала развилось
и продолжает развиваться бесконечное число самых прекрасных и са-
мых изумительных форм» (Дарвин, 1991).
Сожительство на Земле определенного множества организмов не-
минуемо должно было привести к усложнению их отношений между
собой и с окружающей средой. Так возникли экосистемы. Экосистемы
планеты в совокупности образуют целостность высшего порядка —
биосферу, в которой живое вещество взаимодействует с элементами
литосферы, гидросферы и атмосферы.
Особенность названных феноменов состоит в том, что с начала гео-
логической истории все они существовали в неразрывной связи друг с
другом: жизнь на Земле была представлена организмами, которые эво-
люционировали в условиях конкретных экосистем и единой биосферы.
При этом каждому из названных феноменов присущи свойства устой-
чивости и саморазвития, способности меняться и приспосабливаться к
меняющимся условиям внешней среды, повышать
эффективность переработки потоков вещества,
энергии и информации, воздействовать на окружа-
ющую среду так, чтобы она приобретала все более
благоприятные качества для сохранения жизни.
Начала многих наук были заложены мысли-
телями Древней Греции. Теофраст (феофраст)
создал основы ботаники как самостоятельной
науки. В девятитомном «Исследовании о расте-
ниях» им описано около 500 видов, уделено боль-
шое внимание экологии, т. е. зависимости от
теплоты, влажности, освещенности, почв и т. п.
Как биогеографическую аксиому можно рас-
сматривать ето слова: «Своеобразие растительно-
сти создается разницей в месте».
Теофраст
(372—287дон. э.)
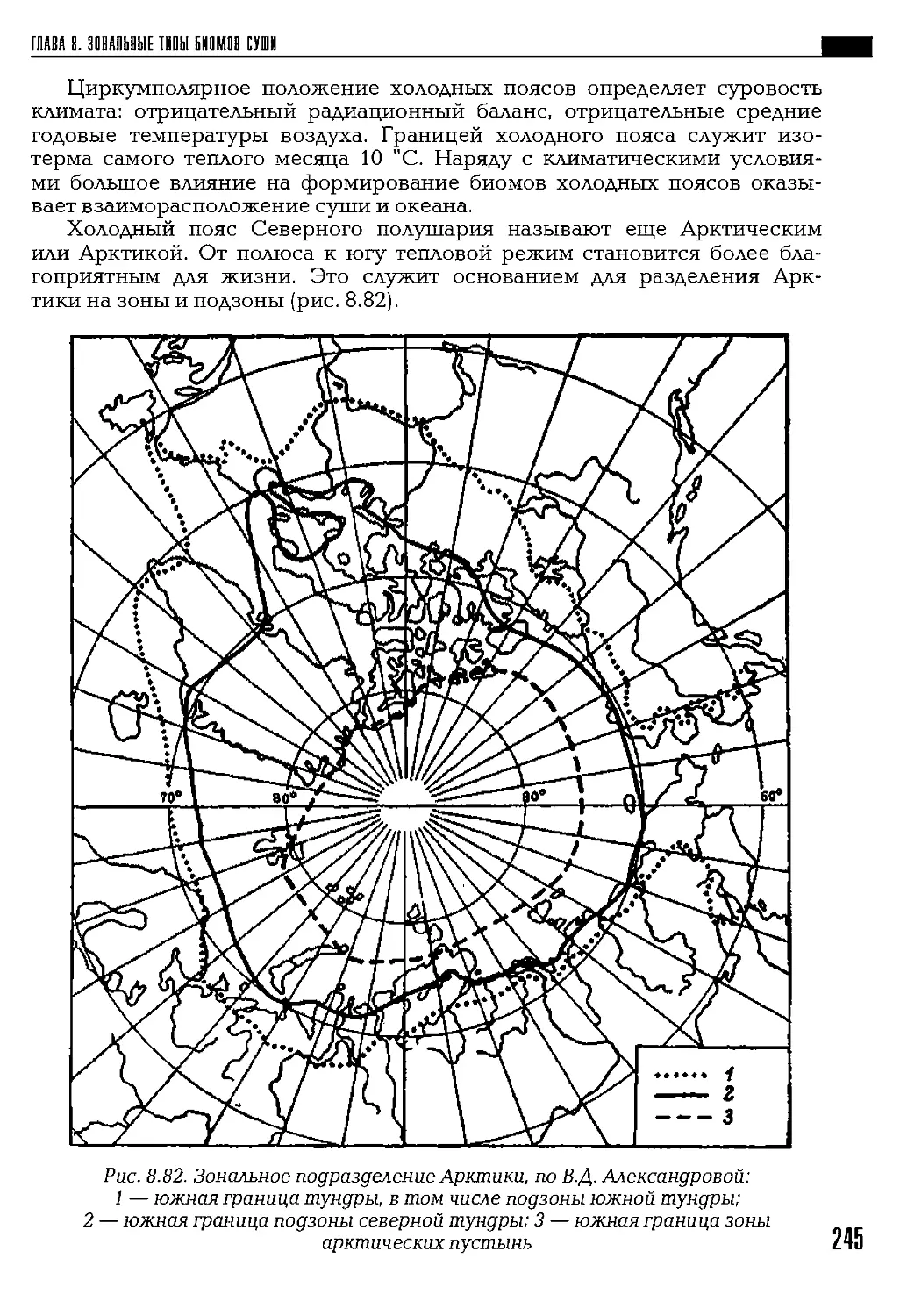
Глава 1
11ШЦ1МЫ1 П1ДХ1Д К 1БЪЯБ1Е1ИЮ
РАЗВООБРЛЗИЯ IH3IH ИА ЗЕМЛЕ
Вселенная возникла в результате взрыва, когда все спиральные туман-
ности были как бы выстрелены из небольшой части пространства, воз-
можно, из одной точки. По теории Большого взрыва вся масса и энер-
гия, а также время и пространство появились около 15 млрд лет назад.
Какую же гигантскую массу должна была иметь эта точка! Вся масса
Вселенной концентрировалась в ней: миллиарды галактик, каждая из
которых содержит по сотне миллиардов звезд. Этот сверхтяжелый за-
родыш Вселенной трудно вообразить.
Одним из парадоксальных объяснений является предположение, что
он вовсе не имел массы, а представлял собой некое средоточие инфор-
мационно-энергетического поля. Начало Вселенной мыслится как осо-
бая точка на границе сферы понятий, относящихся к физике и теосо-
фии. Любой вопрос о том, что предшествовало этому началу или какова
была его природа, уже не рассматривается физикой. Законы физики
появились вместе с рождением Вселенной.
На вопросы «Что появилось раньше — Земля или жизнь?» или «Как
произошла жизнь на Земле?» до сих пор нет однозначного ответа. Дол-
гое время считалась справедливой гипотеза А.И. Опарина о происхож-
дении первичных белков в результате химических реакций, а затем и
первых форм живых организмов на изначально безжизненной Земле.
В.И. Вернадский говорил о космической и геологической вечности
жизни, и это вовсе не метафора. Есть основания допускать, что био-
сфера Земли изначально развивалась одновременно с другими геосфе-
рами. В классическом труде «Биосфера» В.И. Вернадским еще в 1926 г.
были сформулированы эмпирические обобщения, которые в наши дни
получают новые подтверждения и завоевывают все большее признание.
Первое обобщение гласит: в течение всех геологических периодов нет
никаких следов абиогенеза (т. е. непосредственного создания живого
организма из мертвой косной материи); второе — никогда в течение
всего геологического времени не наблюдались азойные (т. е. лишенные
жизни) геологические эпохи.
В.И. Вернадский признавал справедливость принципа Реди — живое
только от живого — и допускал космическую вечность жизни. Ему при-
надлежат слова: «...жизнь зарождается не на Земле, а на Землю приносит-
ся извне, из космического пространства в готовом виде» (Вернадский, 1978).
ГЛАВА 1. ЭВОЛЮЦИОННЫЙ ПОДХОД I ОБЪЯСНЕНИЮ РЙЗНООБРЛЗИЯ ЖИЗНИ НА ЗЕМЛЕ
Согласно концепции панспермии, в космическом пространстве на-
шей Галактики присутствует огромное количество микроорганизмов.
Возможно, что это космическое живое вещество «заразило» Землю,
когда условия на ней стали благоприятными. Следы древнейшей жиз-
ни обнаруживаются на углистых метеоритах (хондритах), возраст ко-
торых 4,5 — 4,6 млрд лет (Жмур и др., 1997).
Допускается, что происхождение органического вещества в хондри-
тах связано с космической жизнедеятельностью микроорганизмов, ве-
роятно цианобактерий. Высокий уровень организации данных форм
жизни позволяет предположить, что начальная точка происхождения
жизни древнее возраста метеоритов. Следовательно, начало жизни в
Солнечной системе отодвигается дальше рубежа 4,5 —4,6 млрд лет. Воз-
раст Земли — около 4,6 млрд лет.
Таким образом, на нашей планете просто не было времени для абио-
генного происхождения жизни: с первых же этапов геологической исто-
рии на Земле существовала жизнь. Древнейшие породы водно-осадочно-
го происхождения (Гренландия, Иссуа — 3,8 млрд лет) уже содержат
остатки микроорганизмов, обладавших аппаратом фотосинтеза.
И 1.1. Космические предпосылки развития жизни____________________
Космическое живое вещество может реализовать потенциальные
возможности своего эволюционного развития только на планете, обла-
дающей благоприятными условиями. К числу космических предпосы-
лок развития жизни на Земле относятся следующие. Планета входит в
состав звездной системы. Тип звезды подобен Солнцу, излучающему
относительно равномерный поток энергии. Угасающие звезды не спо-
собны дать излучения, необходимого для развития жизни на планете.
Звезды, переживающие бурный этап своего развития, обладают слиш-
ком мощным, губительным для всего живого излучением.
Орбита вращения планеты вокруг звезды близка к круговой, поэто-
му энергетический поток от звезды оставался равномерным. Период
обращения планеты вокруг звезды (длина года) и период обращения
планеты вокруг своей оси (длина суток), по-видимому, должны быть
близки земным. Длина года определяет сезонные, а длина суток — су-
точные ритмы жизнедеятельности. Очевидно, что для жизни неблаго-
приятны ни слишком длинные, ни слишком короткие ритмы.
Масса планеты — близкая земной. Сила притяжения планет с мень-
шей массой (Луна, Марс) не способна удерживать газы атмосферы; у
планет с большей массой (Юпитер, Сатурн) образуется мощная атмо-
сфера с облачным покровом, препятствующим проникновению лучис-
той энергии от звезды к поверхности планеты. Планета должна обла-
дать магнитным полем, образующим защиту от космических частиц
высоких энергий. Температурный режим на поверхности планеты соз-
дает условия для нахождения воды в трех агрегатных состояниях: жид-
ком, газообразном и твердом (лед).
На планете Земля все перечисленные условия соблюдаются. Таким
образом, она относится к небесным телам, на которых развитие жизни 9
ЧАСТЬ I. iЕГОРИЧЕСКИЕ К ГЕОГРАФИЧЕСКИЕ ФАКТОРЫ БИОЛОГИЧЕСКОГО РАЗНООБРАЗИЯ
обусловлено благоприятными космиче-
скими предпосылками. К этому еще следу-
ет добавить важную роль, которую игра-
ла и продолжает играть тектоническая
активность Земли. Развитие жизни гало в
неразрывной связи с мощными процесса-
ми горообразования, вулканизма, выделе-
ния из недр изверженных пород, газов,
воды. Земной канал поступления в био-
сферу вещества и энергии оставался по-
стоянно открытым.
В.И. Вернадский (1931) допускал, что
жизнь на Земле изначально была разнообраз-
ной как по составу, так и по выполняемым
живым веществом геохимическим функци-
ям. Иными словами, существование и эволю-
ция организмов на Земле сразу складывались
г, . mm в условиях конкретных экосистем и биосфе-
Карл Линнеи (1707—1778) 7 „ Р
ры. Вернадский пишет: «...эволюционный
процесс, какую бы его форму мы ни взяли, всегда идет уже внутри био-
сферы, т. е. в живой природе. Логически заключить отсюда об изменени-
ях форм организмов путем эволюции вне живой природы, как часто это
делают, будет логической ошибкой, недопустимой экстраполяцией».
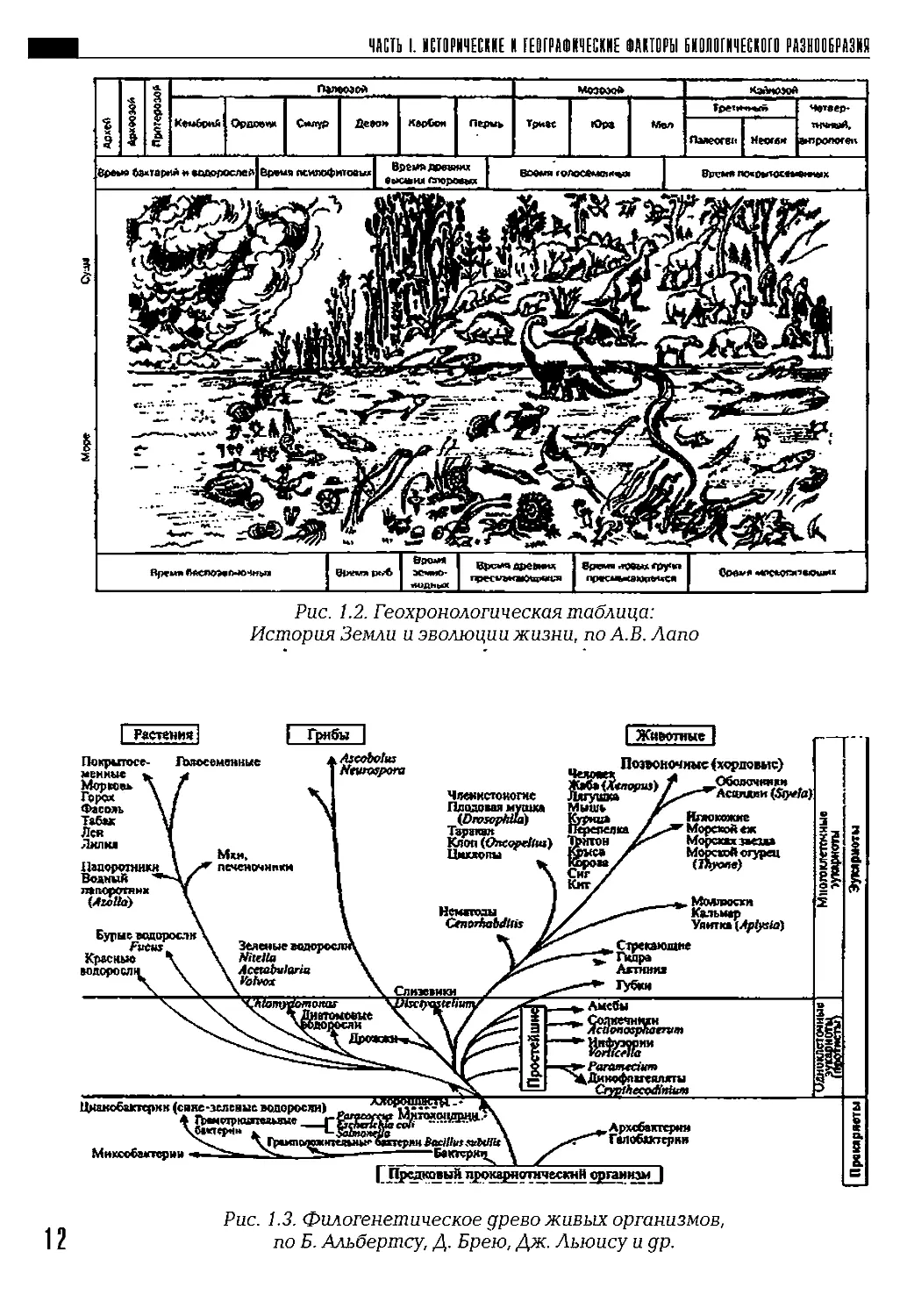
Всю историю жизни на Земле делят на два неравных отрезка: до-
кембрийский (время скрытой жизни— криптозой), его продолжитель-
ность около 3,5 — 4 млрд лет, и фанерозой (время явной жизни), начав-
шийся примерно 570 млн лет назад (рис. 1.1). Наглядное представление
о хронологии Земли и истории развития жизни дает рис. 1.2. Совре-
менная биосфера представлена огромным разнообразием видов и сооб-
ществ живых организмов. Родственные связи выражаются системой
таксономических единиц. Биоты Земли можно адекватно описать, толь-
ко если используются названия, принятые в систематике. Появлением
таксономической системы мы обязаны Карлу Линнею.
Теперь все виды обитателей суши и океана объединены в систему,
отражающую ход эволюционного процесса. Главная цель систематиче-
ского метода — присвоить каждому виду собственное единственное
наименование и показать последовательность происхождения разных
систематических групп организмов, т. е. родословное (филогенетическое)
древо (рис. 1.3).
Выделяется четыре царства живых организмов (рис. 1.4). Иерархию
таксонов одного из представителей растительного царства рассмотрим
на примере Сосны обыкновенной.
Поддарство Высшие растения
Отдел Голосеменные
Класс Хвойные
Порядок Сосновые
Семейство Сосновые
Род Сосна
1 0 Вид Сосна обыкновенная (Pinus sylvestris)
Рис. 1.1. Геохронологическая шкала
. ЭВОЛЮЦИОЯНЫЙ ПОДХОД I ОБЪЯСНЕНИЮ РАЗНООБРАЗИЯ ЖИЗНИ НА ЗЕМЛЕ
ЧАСТЬ I. iЕГОРИЧЕСКИЕ К ГЕОГРАФИЧЕСКИЕ ФАКТОРЫ БИОПОГИЧЕСИОГО РАЗНООБРАЗИЯ
$ 1 и! Палеозой М09000Й Кайнозой
Кембрмй Ораоечн Смлур Девон Kapton Пермь трмве |Ор8 Мол TpetHovw Чдтвер- ПРЙПМЙ, 8М1 ропотен
Палеоген Неоген
время бактерий и вадорослей время лсилофитовык Воемя гр/юеемсвн^о1 Время локомтосямею^х
Время Ляспоэемочньп ЗДмлля р»6 время ЭСМИО' «ЮАМЫХ Время аревмм гфеемдеаюшшця время .«рвых кручл лрес»^мдц|щ«ся время мясядопяюшнк
Рис. 1.2. Геохронологическая таблица:
История Земли и эволюции жизни, по А.В. Лапо
I Растений]
Покрытосе-
менные
Морвоеь
Горох
Фасоль
Табаг
Лее
Липка
Папоротники
Водный
77*ПОДОТННХ
(Au>Ua)
Позвоночные (хордовые)
Ободочизпш
Асами* (Styria)
Илюкожие
Морской еж
Морская змзла
Морской огурец
| Грибы ~|
[ЗКивотиые |
ГЬлоеемонные
Мши*
Слизевики
Hwwpora
Нематоды
CtnorhxMtto
Мхи»
печеночным
"ИГ
Г Предковый прокариотический организм |
Амебы
pnamtm
)ИМ
Моллюски
Кальмар
Уяитка (Aplysia)
.Архсбактджм
Галобактернв
Атгаямсмгя»
Динофпвгеялггы
Crypfacwffaiwn
Стрекающие
Гшфа
Актиния
плат
атмажие
Рис. 1.3. Филогенетическое древо живых организмов,
по Б. Алъбертсу, Д. Брею, Дж. Льюису и др.
Зеленые
Hilrila
Acetofatariti
fofvat
Бурые водоросли
Fuchj
Красные
водоросли
Цнакоб&ктсрин (сяяе-эеленые водоросли)
Миксобактсрын
Членистоногие
Плодовая мушка
(Drvsepfiua)
Тзрзют
Клоп «ТлссреЛш)
Циклопы
3
I
S
8.
3
12
ГЛАВА 1. ЭВОЛЮЦИОННЫЙ ПОДИОД I ОБЪЯСНЕНИЮ РАЗНООБРАЗИЯ ЖИЗНИ НА ЗЕМЛЕ
Рис. 1.4. Четыре царства органического мира, по «Жизни растений»
Характерными чертами естественно-исторического пути разви-
тия биосферы являются: усложнение строения организмов (аромор-
фоз); увеличение биологического разнообразия; «волны жизни» (рас-
цвет и катастрофическое вымирание видов на рубежах геологических
эпох); усложнение структуры экосистем; возрастание роли живых
организмов в организации потоков вещества и энергии в географи-
ческой оболочке.
Одна из особенностей развития жизни на нашей планете состоит в
том, что грандиозные события, знаменующие основные вехи ее исто-
рии, протекают со всевозрастающей скоростью. Если условно принять
время существования нашей планеты за 12 месяцев, то в таком кален-
даре продолжительность одного дня будет равна 12,6 млн лет, а одного
часа — 525 тыс. лет. Ускорение основных этапов эволюции в таком
масштабе времени становится особенно заметным.
1 января — догеологическая история формирования Земли.
28 марта — появление первых бактерий.
12 декабря — время расцвета динозавров.
26 декабря — исчезновение динозавров.
31 декабря, 1ч — появление общего предка обезьяны и человека.
31 декабря, 17 ч 30 мин — появление гоминоидов — австралопитеков.
31 декабря, 23 ч 54 мин — появление неандертальцев.
31 декабря, 23 ч 59 мин 46 с — начало новой эры.
31 декабря, полночь — человек шагнул на Луну.
Формирование крупнейших биогеографических подразделений
происходило в неразрывной связи с геологической историей литосфер-
ЧАСТЬ I. ИСТОРИЧЕСКИЕ К ТЕОГРАФИЧЕСХИЕ ФАКТОРЫ БИОЛОГИЧЕСКОГО РАЗНООБРАЗИЯ
ных плит. Последние вместе с впаянными в них материковыми блока-
ми испытывали глобальные горизонтальные перемещения («дрейф кон-
тинентов»), сталкивались и раскалывались. Площади и очертания мате-
риков и океанов испытывали значительные изменения. Изоляция
материков водами Мирового океана определяла неповторимые черты
эволюции биоты в их пределах; объединение континентальных блоков,
возникновение сухопутных мостов приводило к смешиванию биот, име-
ющих различные генетические корни.
И 1.2. Эволюция жизни в криптозое_________________________________
В истории образования Земли как планеты Солнечной системы око-
ло 1 млрд лет занимает догеологический этап — эпоха формирования
тонкой неустойчивой земной коры, которая легко дробилась, расплав-
лялась и возникала вновь. Изливались огромные количества магмы,
образовавшие лавовые покровы. Происходила дифференциация веще-
ства внутри планеты. Обособилось ядро, появилось магнитное поле.
Земля подверглась энергичной бомбардировке метеоритами, и ее поверх-
ность напоминала современную Луну, а точнее, Венеру, поскольку на
Земле существовала первичная атмосфера, облака которой плотной
пеленой закрывали Землю.
Когда земная кора остыла до температуры ниже кипения воды, воз-
никли первые озера и моря, потекли реки. Начались процессы разруше-
ния и переотложения продуктов разрушения изверженных пород, т. е.
стали формироваться толщи осадочных пород. Около 4 млрд лет назад
закончился догеологический этап формирования планеты и началась
геологическая история Земли. Древнейшую геологическую эпоху на-
зывают кришпозой.
Образование Мирового океана сопровождалось его минерализаци-
ей. Конденсированные воды, проходя через атмосферу, насыщенную
тогда СО2, СО, СН4, H2S, Н3ВО3, НС1, HF, сразу же становились кислы-
ми. Скатываясь по свежим поверхностям застывших первозданных
потоков лавы, они быстро минерализовались. Находившиеся в водах
крепкие кислоты оказывали сильное воздействие на изверженные по-
роды, выщелачивая из них натрий, магний, кальций, барий вместе с
катионами двухвалентного железа и марганца. На поверхности суши,
постоянно омываемой кислыми дождями, развивались энергичные про-
цессы гидролиза и гидратации разных минералов. Таким образом, хи-
мическое выветривание привело к образованию основной массы катио-
нов в водах первичного океана.
В течение криптозоя жизнь на Земле прошла длительный путь эволю-
ции, но оставалась сосредоточенной преимущественно в пределах теплых
морских мелководий. Если обратиться к истокам событий, оказавших
влияние на формирование современной биосферы, то выясняется, что
подчас они носили драматический характер. Около 2 млрд лет назад исто-
рия жизни оказалась разделенной на два почти равных отрезка. Вначале
1 4 жизнь была представлена только анаэробными одноклеточными доядер-
ГЛАВА 1. ЭВОЛЮЦИОННЫЙ ПОДХОД I ОБЪЯСНЕНИЮ РАЗНООБРАЗИЯ ЖИЗНИ НА ЗЕМЛЕ
ными организмами (прокариотами), существовавшими в условиях восста-
новительной атмосферы из углекислого газа, метана и азота. Второй этап
связан с формированием окислительной атмосферы из азота и кислорода.
Анаэробные организмы были вытеснены в ограниченные экологические
ниши. Их место занял принципиально новый ствол жизни (ядерные орга-
низмы — эукариоты), венцом эволюции которой стал человек.
В экосистеме океана четко обособились основные функциональные
звенья, состоящие из продуцентов, консументов и редуцентов. Эта
модель сохраняет постоянство в любых экосистемах Земли до наших
дней. Благодаря появлению и расцвету фотосинтезирующих организ-
мов (прежде всего цианей — сине-зеленых водорослей) в атмосфере
появился свободный кислород. Сформировался озоновый экран, защи-
тивший Землю от губительного для живых существ жесткого ультрафио-
летового излучения.
С биологической точки зрения содержание свободного кислорода в
атмосфере и гидросфере значимо потому, что позволило перейти орга-
низмам от использования энергии процессов ферментативного броже-
ния и хемосинтеза к энергетически более эффективному (в 30 — 50 раз)
окислению при дыхании. Именно это послужило причиной настояще-
го биологического взрыва.
Появились многоклеточные формы. Каким образом организмы, со-
стоящие из большого числа клеток, произошли из одноклеточных, до
сих пор непонятно. Возможно, что определенные прокариоты внедри-
лись в другие прокариоты и возникшее содружество стало функциони-
ровать как единое целое.
К концу криптозоя (в вендском
периоде) эволюция животных про-
двинулась настолько, что они уже
были представлены основными типа-
ми беспозвоночных животных, изве-
стных и в настоящее время: стрекаю-
щими (65 — 70%), червями (около 25%)
и членистоногими (около 5%). Отли-
чительная особенность фауны состоя-
ла в том, что она была образована
бесскелетными организмами. И хотя
длина некоторых из этих животных
достигала 1 м, они состояли из желе-
образного вещества подобно совре-
менным медузам. Многие из обнару-
женных групп не имеют современных
аналогов (рис. 1.5).
Возникнув в океане, организмы
коренным образом трансформирова-
ние. 1.5. Реконструкция вендской
биоты, по К.Л. и М.А. Фентонам.
ли химические свойства его вод, газовый состав атмосферы. Измени-
лось соотношение ионов: бывшая кислой морская вода приобрела ще-
лочную реакцию. Фотосинтез обогатил воду и атмосферу свободным
кислородом. Образование биогенных известняков регулировало содер-
жание углекислого газа в атмосфере. 1 5
ЧАСТЬ I. ИСТОРИЧЕСКИЕ К ТЕОГРАФИЧЕСХИЕ ФАКТОРЫ БИОЛОГИЧЕСКОГО РАЗНООБРАЗИЯ
Благодаря огромной инерционной способности океаносферы она со-
храняет константность важнейших экологических параметров — тем-
пературы, солености, состава газов — на протяжении 2 млрд лет. По-
этому гидробионты не выносят резких изменений природных условий,
в том числе вызванных антропогенной деятельностью.
Жизнь на суше, видимо, была представлена в сырых местах прокари-
отными пленками, о чем свидетельствуют палеопочвы, содержащие орга-
ническое вещество. Их возраст — около 2,4 млрд лет. Процессы разруше-
ния обнаженной поверхности горных пород на материках и выноса
терригенного осадочного материала в океан были очень интенсивными.
В основе естественного отбора всегда лежат резкие изменения фи-
зико-географических (палеогеографических) условий, определяющих
формирование новых видовых генотипов и характер распределения и
доминирования жизненных форм в биоценозах. Раскрывая тему эво-
люционного подхода к объяснению разнообразия жизни в Мировом
океане, необходимо обратить внимание на следующие важнейшие со-
бытия на протяжении фанерозоя.
И 1.3. Эволюция жизни в Данерезое_________________________________
Фанерозой делят на эры: палеозой (время древней жизни), мезозой
(время средней жизни) и кайнозой (время новой жизни). Высказыва-
ется гипотеза о зависимости развития Земли и жизни на ней от движе-
ния Солнечной системы по галактической орбите (Астафьева-Урбай-
тис, Ясаманов, 1993). Период обращения Солнечной системы вокруг ядра
Галактики образует галактический год, равный 215 — 217 млн лет. Мы
живем в эпоху зимы XXII галактического года.
Эпохи тектоно-магматической активности, экологические катастро-
фы, обусловившие вымирание больших групп организмов в разные гео-
логические эпохи, приурочены к границам галактических годов, т. е. они
были вызваны космическими факторами. Вероятно, такая связь вызвана
тем, что Солнечная система во время своего перемещения по галактиче-
ской орбите периодически проходит через газопылевые спиральные струи.
В каждом таком потоке Солнечная система пребывает несколько миллио-
нов лет, и именно в это время усиливается приток к Земле космических
лучей, а на земную поверхность выпадает 102— 103 галактических комет.
Эпохи орогенеза, активные движения литосферных плит, глубокие
изменения климата, общепланетарные трансгрессии и регрессии Ми-
рового океана, изменения состава гидросферы и атмосферы, экологи-
ческие катастрофы, вызывавшие коренную перестройку биоты, проис-
ходили в те отрезки геологического прошлого, когда наша планета
пребывала в газопылевом вихревом галактическом потоке.
Палеозой (начало — около 570 млн лет назад). Палеозой делится на
периоды. В нижнем палеозое выделяются кембрий, ордовик, силур;
в верхнем палеозое — девон, карбон, пермь.
В нижнем палеозое жизнь развивалась преимущественно в океане.
В верхнем палеозое началось активное формирование растительного
1 G покрова и животного мира суши.
ГЛАВА 1. ЭВОЛЮЦИОННЫЙ ПОДХОД К ОБЪЯСНЕНИЮ РАЗНООБРАЗИЯ ЖИЗНИ НА ЗЕМЛЕ
Кембрий. В океане встречаются представители основных типов бес-
позвоночных животных. Примечательно, что если в криптозое орга-
низмы были мягкотелые, то в начале палеозоя произошел скачок в
физиологии большинства беспозвоночных, они обрели твердые изве-
стковые или хитиновые покровы. Страницы геологической летописи
стали заполняться остатками именно этих организмов. Господствовав-
шими и самыми развитыми животными в кембрии были трилобиты,
относящиеся к членистоногим. Растения были представлены низши-
ми споровыми — водорослями: одноклеточными, входящими в состав
фитопланктона, и многоклеточными зелеными, красными, позже —
бурыми водорослями (макрофитами), образовывавшими подводные за-
росли на мелководьях (рис. 1.6).
Рис. 1.6. Жизнь на дне кембрийского моря, по И. Аугусте и 3. Буриану:
биоценоз зеленых водорослей, кремневых губок и трилобитов.
В толще воды — медузы
Все растения и животные размножались путем высева большого
количества гамет в воду, где происходило их слияние. Водная среда
являлась необходимым условием для жизни всех организмов.
Ордовик — силур. Ордовикский период является переломным эта-
пом в эволюции морских экосистем. Геологические события кардиналь-
но изменили условия жизни на Земле. Об этом свидетельствуют следы
глобального потепления в среднем и материковые оледенения в позднем
ордовике. Отмечается максимальное за всю фанерозойскую историю
Земли распространение эпиконтинентальных морей в раннем и сред- 11
ЧАСТЬ I. ИСТОРИЧЕСКИЕ К ТЕОГРАФИЧЕСХИЕ ФАКТОРЫ БИОЛОГИЧЕСКОГО РАЗНООБРАЗИЯ
нем ордовике и резкое их сокращение во время материкового оледене-
ния в позднем ордовике. Расширение морского жизненного простран-
ства в среднем ордовике можно рассматривать как благоприятный
фактор для широкого расселения бентосных сообществ. Сокращение
шельфовых морей в позднем ордовике, вероятно, было основной при-
чиной резкого снижения таксономического разнообразия, которое
контрастно проявилось в мелководных обстановках и в меньшей степе-
ни — в более глубоководных частях шельфов.
В атмосфере Земли сформировался озоновый экран, защитивший
живые организмы от жестких космических излучений и обеспечивший
экспансию бентосных и пелагических форм жизни в морских экосис-
темах, а затем выход растений и животных на сушу.
В морских экосистемах произошло крупнейшее в фанерозое быстрое
увеличение таксономического разнообразия гидробионтов. Новые эко-
логические гильдии почти мгновенно (в масштабе геологического време-
ни) осуществили глобальную колонизацию дна и водной толщи морей,
что сопровождалось формированием многочленной трофической струк-
туры экосистем. В ордовике впервые появились и, начиная со среднего
ордовика, достигли максимального расцвета многие группы фильтратор-
ных организмов с каркасным скелетом. На низшем трофическом уровне
доминирующее положение заняли остракоды. Расцвет колониальных
организмов сопровождался дроблением экологического пространства и
усложнением трофических цепей. До начала среднего ордовика в эпи-
континентальных морях доминировали строматолитовые постройки,
которые свидетельствуют о важном значении в трофических цепях
цианобактерий. Вероятно, со среднего ордовика главным компонентом
первичной продукции становится фитопланктон. Формируется зоопела-
гиаль: появляются и расцветают специализированные планктонные и
нектонные организмы.
В конце ордовика произошло резкое снижение таксономического раз-
нообразия морских биот, которое, однако, не сопровождалось полным
вымиранием каких-либо экологических гильдий в отличие от более позд-
них великих биотических кризисов (например, на границах перми и
триаса, мела и палеогена). В раннем силуре позднеордовикское снижение
таксономического разнообразия полностью компенсировалось быстрым
восстановлением новых экологических гильдий и более сбалансирован-
ными соотношениями между старыми и новыми гильдиями (рис. 1.7).
Большое распространение получили древние кораллы. По дну полза-
ли трилобиты. Одно из первых мест среди животного мира занимали
головоногие моллюски, обитавшие в прямых или слабоизогнутых укра-
шенных красивым узором раковинах. Характерными представителями
иглокожих были морские лилии, длинные стебли которых заканчива-
лись красивой чашечкой с венцом подвижных щупалец. Значительного
разнообразия достигли представители плеченогих, мшанок, брюхоно-
гих и пластинчатожаберных моллюсков. От иглокожих берет свое на-
чало мощная ветвь эволюционного древа — хордовые. Они явились пред-
ками рыб, четвероногих животных, птиц и в конечном счете человека.
В силуре получили развитие бесчелюстные рыбообразные — агнаты
(в современной фауне к ним ближе всего миноги).
ГЛАВА 1. ЭВОЛЮЦИОННЫЙ ПОДХОД I ОБЪЯСНЕНИЮ РАЗНООБРАЗИЯ ЖИЗНИ НА ЗЕМЛЕ
Рис. 1.7. Жизнь на дне силурийского моря, по И. Аугусте и 3. Буриану: среди
зарослей водорослей и морских лилий располагались ныне вымершие каравае-
образные и кубковидные кораллы. По песчаному дну ползали трилобиты,
головоногие моллюски Orthoceras (с прямой раковиной) и Cyrtoceras (со сла-
боизогнутой раковиной). В левом углу изображена небольшая группа плече-
ногих беспозвоночных, похожих на двустворчатых моллюсков
В целом весь нижний палеозой — это эпоха океанической биосфе-
ры, образованной низшими споровыми растениями и в основном бес-
позвоночными животными. Эволюция растений привела к появлению
высших споровых, обладающих приспособлениями к наземному об-
разу жизни. Начинается выход организмов из морской среды на сушу.
Первыми обитателями почв стали беспозвоночные — черви и члени-
стоногие.
Девон. С этого периода начинается верхний палеозой — начало ак-
тивного завоевания суши растениями и животными. Наземная флора
девона принадлежала к единому фитогеографическому царству суши.
В раннем и среднем девоне она была представлена однолетними водны-
ми или полуводными растениями, заселявшими мелководья морских
бассейнов, а также периодически затопляемые плоские побережья.
Развитие первых в истории Земли травянистых болот привело к обра-
зованию древнейших пластов угля.
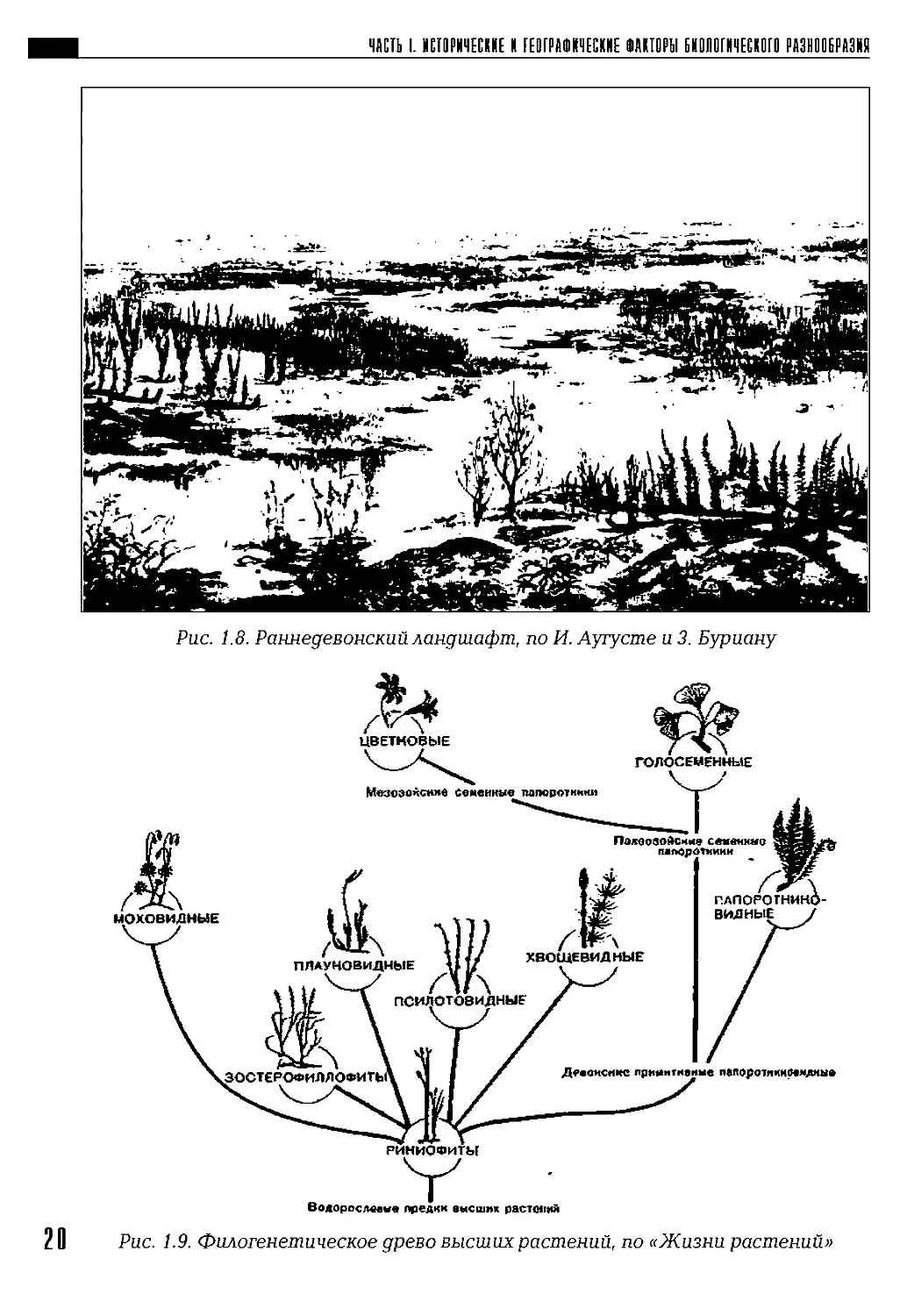
Начало формирования наземной растительности связано с высши-
ми споровыми (рис. 1.8). В конце палеозоя от них произошли голосе-
менные, а в конце мезозоя — цветковые (рис. 1.9). 19
ЧАСТЬ I. iЕГОРИЧЕСПЕ К ГЕОГРАФИЧЕСКИЕ ФАКТОРЫ БИОЛОГИЧЕСКОГО РАЗНООБРАЗИЯ
Рис. 1.8. Раннедевонский ландшафт, по И. Аугусте и 3. Буриану
2 0 Рис. 1.9. Филогенетическое древо высших растений, по «Жизни растений»
ГЛАВА 1. ЭВОЛЮЦИОННЫЙ ПОДХОД I ОБЪЯСНЕНИЮ РАЗНООБРАЗИЯ ЖИЗНИ НА ЗЕМЛЕ
Наземная растительность обитала на плоских заболоченных мор-
ских побережьях и не отличалась ни богатством жизненных форм, ни
их разнообразием. Жизнь на суше потребовала выработки целого ряда
приспособлений. Дифференцировались различные органы: корень, сте-
бель, листья, органы размножения. Обособились специализированные
ткани: основные, покровные, проводящие. Наземные растения образо-
вали органы размножения, сначала споры, позже — семена, защищен-
ные от высыхания.
Во флоре позднего девона появляются первые многолетние древес-
ные формы плауновидных, хвощевидных и папоротниковидных. С ни-
ми связано возникновение отсутствовавшего ранее лесного типа расти-
тельности. Это обстоятельство значительно ослабило эрозию суши и
поступление осадочного материала в океан. Однако развитие раститель-
ного покрова еще было ограничено приморскими равнинами. Склоны
гор продолжали интенсивно разрушаться.
Эволюция хордовых в океане дала большое разнообразие рыб. По это-
му признаку девон называют эпохой рыб. Эволюция рыб привела к появ-
лению древнейших земноводных (амфибий). Слово «амфибия» означает
«ведущий двойную жизнь». Большинство представителей этой группы
позвоночных часть своей жизни проводит в воде, а часть на суше. Свою
икру большинство амфибий откладывает в воду. Вылупляющиеся из нее
личинки являются исключительно водными существами, похожими на
рыб. Со временем они подвергаются метаморфозу, приобретают легкие и
конечности, после чего вполне способны выходить из воды и жить на суше.
Для высших споровых растений, рыб и земноводных животных по-
прежнему необходимым условием размножения являлась водная сре-
да. Это служило определенным препятствием для продвижения орга-
низмов в глубь суши.
Карбон (каменноугольный период). Облик Земли в начале карбона
определялся в Северном полушарии группой континентов, из которых
основными были Еврамерийский (Европа и Северная Америка), Ангар-
ский (Восточная Сибирь), Катазиатский (юго-восточная часть Китая и
Индокитай); в Южном — гигантским континентом Гондвана (рис. 1.10).
Рис. 1.10. Реконструкция положения кон-
тинентов в начале карбона, по Л.П. Зонен-
шайну и др.: 1 — континенты; 2 — транс-
формные разломы; 3— оси расхождения
литосферных плит. Основные континен-
ты: ЕА — Еврамерийский; А —Ангарский;
К— Катазиатский; Г— Гондвана 21
ЧАСТЬ I. iЕГОРИЧЕСПЕ И ГЕОГРАФИЧЕСКИЕ ФАКТОРЫ БИОЛОГИЧЕСКОГО РАЗНООБРАЗИЯ
Влажный и теплый климат карбона благоприятствовал формирова-
нию пышных лесов высших споровых растений: плауновидных, хво-
щевидных, папоротниковидных (рис. 1.11). Появляются семенные па-
поротники и первые голосеменные — саговники, кордаиты. Высокая
продуктивность растительности служила источником для образования
мощных пластов каменного угля.
Рис. 1.11. Каменноугольный лес, по И. Аугусте и 3. Буриану
Развиваются первые по-настоящему сухопутные животные — пре-
смыкающиеся (рептилии). В отличие от большинства амфибий эти жи-
вотные не использовали воду в процессе воспроизводства, а откладыва-
ли яйца, сохранявшие жизнеспособность в наземных условиях. Таким
образом, у рептилий появилась настоящая возможность осваивать сушу.
Широкое распространение получают наземные членистоногие —
многоножки, пауки, скорпионы, насекомые (прямокрылые — гигант-
ские стрекозы, см. рис. 1.11). Со временем насекомые станут самым раз-
нообразным классом членистоногих. Более того, они окажутся самой про-
цветающей группой во всем царстве животных. В настоящее время их
известно около 2 млн видов, что составляет 70% всех видов животных.
У семенных растений, членистоногих и пресмыкающихся живот-
ных появляется более выгодный для сухопутных условий способ раз-
множения, когда гаметы оплодотворяются, у животных в женском орга-
низме, у растений — в специальном генеративном органе.
В наземных биотах карбона впервые в истории Земли четко опреде-
лились биогеографические особенности разных континентов (рис. 1.12).
ГЛАВА 1. ЭВОЛЮЦИОННЫЙ ПОДИОД I ОБЪЯСНЕНИЮ РАЗНООБРАЗИЯ ЖИЗНИ НА ЗЕМЛЕ
Рис. 1.12. Процесс обособления основных биогеографических областей,
по СВ. Мейену
На Еврамерийском континенте (Аавразийская флористическая об-
ласть) господствовали леса лепидофитов. Низменности Гондваны покры-
вали леса глоссоптериевых. В качестве особой биогеографической об-
ласти выделялся Ангарский континент, где с конца карбона росли леса
кордаитов.
Пермь. Этот период отмечен резкими изменениями климата и релье-
фа. Активизировалась вулканическая деятельность, поднимались новые
горы (герциниды). ВЛавразии господствовал пустынный климат; в Юж-
ном полушарии в Гондване на суигу надвигались ледники. Возможно, что
причина похолодания климата крылась в том, что пышная растительность
каменноугольного периода поглотила в процессе фотосинтеза большое
количество СО2. В атмосфере с пониженным содержанием углекислого
газа и паров воды не возникало парникового эффекта, в результате чего
Земля стала отдавать тепло в космическое пространство.
Похолодание было серьезным испытанием для растений и живот-
ных. Исчезли каменноугольные леса из высших споровых растений. На
смену им пришли более приспособленные к сухому и холодному кли-
мату леса голосеменных из гинкго, тиса, пихты, кипариса, секвойи. От
холода страдали и древние ящеры. У эдафозавров появилось специаль-
ное приспособление — высокий спинной гребень (рис. 1.13). Ящеры
грелись, разворачивая гребень перпендикулярно солнечным лучам.
В конце пермского периода произошла грандиозная экологическая
катастрофа, в результате которой около 90% видов организмов вымер-
ло. В океане осталось менее 5% видов животных. Причиной кризиса,
по-видимому, была мощная вулканическая деятельность, проявившая-
ся, в частности, в образовании восточносибирских траппов. Изверже-
ния вулканов сопровождались выбросами в атмосферу диоксида серы и
оксида азота, послуживших причиной интенсивных кислотных дож-
дей. У большинства палеозойских организмов репродуктивный цикл про-
исходил в водной среде с нейтральным уровнем pH. В водоемах с кис-
лой средой гаметы и молодь организмов погибали.
ЧАСТЬ I. iЕГОРИЧЕСПЕ И ГЕОГРАФИЧЕСКИЕ ФАКТОРЫ БИОЛОГИЧЕСКОГО РАЗНООБРАЗИЯ
Рис. 1.13. Раннепермский ландшафт, по И. Аугусте и 3. Буриану
К концу палеозоя северные континенты объединились с Гондваной,
образовав единый сверхматерик Пангею, окруженный сверхокеаном
Панталассом (рис. 1.14).
Мезозой (начало около 240 млн лет назад). Делится на периоды:
триас, юра, мел. Сверхматерик Пангея, включавший в себя почти всю
сушу Земли, стал дробиться. Начался процесс формирования современ-
ных очертаний материков и океанов (рис. 1.15).
Возник океан Тетис, разделивший материки Лавразию и Гондвану.
Изоляция материков явилась предпосылкой расхождения биот. Это
находит отражение в схемах зоогеографического районирования совре-
менных материков: группа северных материков объединяется в Аркто-
гею, а группа южных — в Нотогею.
Появились Атлантический и Индийский океаны; новые обособлен-
ные материки: Североамериканский, Евроазиатский, Южноамерикан-
ский, Африканский. Австралия и Антарктида были еще едины, однако
от них откололись Новая Зеландия и Индостан.
Биогеографические особенности современных океанов объясняют-
ся их возрастом. Так, более молодая биота Атлантики противопоставля-
ется богатой и разнообразной биоте древнего Тихого океана.
В результате раскола Пангеи и образования новых океанов, соеди-
ненных проливами, циркуляция вод Мирового океана стала более сво-
бодной. Открылся доступ в высокие широты, куда поверхностные тече-
ния стали переносить тепло из тропиков. В это время вблизи полюсов
температура поверхностных вод была около 15 °C; сезонность климата —
24 слабовыраженная. В теплом океане происходило очень вялое конвекци-
ГЛАВА 1. ЭВОЛЮЦИОННЫЙ ПОДХОД I ОБЪЯСНЕНИЮ РАЗНООБРАЗИЯ ЖИЗНИ НА ЗЕМЛЕ
Рис. 1.14. Реконструкция, показывающая положение и ориентацию Пангеи
в конце палеозоя, по А.С Smith и J.C. Brinden
Рис. 1.15. Реконструкция последовательных стадий раскола Пангеи,
по Р. Дитцу и И. Холдену: показано отделениеЛавразии от Гондваны
океаном Тетис и формирование Южной Атлантики и Индийского океана.
А — 180 млн лет назад; Б — 65 млн лет назад
25
ЧАСТЬ I. iЕГОРИЧЕСПЕ И ГЕОГРАФИЧЕСКИЕ ФАКТОРЫ БИОЛОГИЧЕСКОГО РАЗНООБРАЗИЯ
онное перемешивание. Основной причиной опускания поверхностных
вод стало увеличение их солености (и плотности) в результате интен-
сивного испарения. Такой тип перемешивания слабо вентилировал глу-
бины. Недостаток кислорода в воде привел к массовой гибели глубоко-
водной фауны.
Триас. В начале триаса по сравнению с поздней пермью фитогеог-
рафическая дифференциация резко снижается. Очевидно, это явилось
следствием разрушения горных барьеров, ранее разделявших обособ-
ленные фитохоры. Глобальный климат стал более ровным и теплым
(безморозный климат). Флора раннего триаса в Лавразии оставалась еще
очень близкой палеозойской флоре. Ее отличительные признаки связа-
ны с угасанием кордаитов и возрастанием роли хвойных, папоротнико-
видных и хвощевидных. В позднем триасе флора имела уже типично
мезозойский облик (рис. 1.16).
Рис. 1.16. Мезозойский ландшафт, по И. Аугусте и 3. Буриану
Леса высших споровых сохранялись лишь на заболоченных низи-
нах. Бурный расцвет получили голосеменные: саговники с длинными
перистыми похожими на пальмовые листьями, хвойные — тисовые,
пихтовые, кипарисовые, гинкговые и др.
В триасе наступает расцвет рептилий..Филогенетическое древо пре-
смыкающихся показано на рис. 1.17. Эта схема наглядно иллюстриру-
ет значение вымерших групп для восстановления филогенеза репти-
лий в целом.
ГЛАВА 1. ЭВОЛЮЦИОННЫЙ ПОДХОД I ОБЪЯСНЕНИЮ РАЗНООБРАЗИЯ ЖИЗНИ НА ЗЕМЛЕ
Рис. 1.17. Филогенетическое древо рептилий, по А.В. Яблокову и А.Г. Юсуфову
Следует отметить появившуюся в позднем карбоне группу синап-
сидных рептилий. По мере эволюции ее представители становились все
более млекопитающеподобными. Наконец, в позднем триасе появляют-
ся самые ранние настоящие млекопитающие.
Отличительные признаки млекопитающих: теплокровность и нали-
чие у самок желез, которые выделяют молоко для вскармливания дете-
нышей. За редким исключением (однопроходные млекопитающие) они
рождают живых детенышей, а не откладывают яйца.
Родственные связи примитивных групп млекопитающих друг с другом
до сих пор остаются неясными. Однако можно предположить, что обособ-
лению основных отрядов млекопитающих способствовала изоляция ма-
териков, образовавшихся в результате раскола Пангеи. Плацентарные
возникли в Лавразии, а сумчатые и однопроходные — в Гондване.
Юра — мел. Позднеюрская и раннемеловая биоты представляют
собой единый этап развития. Они формировались в условиях влаж-
ного, безморозного, но сезонного климата. Во флоре Лавразии про-
должают господствовать представители голосеменных. Важнейшим
событием среднего мела является появление покрытосеменных (цвет-
ковых) растений. Юра и мел — время господства динозавров, кото-
рые освоили и сушу, и воду, и воздух (рис. 1.18, 1.19). С юрского
дерева совершила полет древняя птица археоптерикс (рис. 1.20). Все
создания, относимые к птицам, обладают единственным общим при-
ЧАСТЬ I. iстаРИЧЕСПЕ К ГЕОГРАФИЧЕСКИЕ ФАКТОРЫ БИОЛОГИЧЕСКОГО РАЗНООБРАЗИЯ
Рис. 1.18. Летающие и плавающие рептилии, по И. Аугусте и 3. Буриану:
летающий ящер Pteranodon. Размах его крыльев достигал 8 м.
Хищные ящеры Tylosaurus хорошо приспособлены к жизни в море —
их конечности превратились в ласты
Рис. 1.19. Гигантский травоядный ящер, по И. Аугусте и 3. Буриану:
в болотистых местах обитал ящер Diplodocus. Его длина достигала 27 м.
Большую часть жизни он проводил в воде, где питался водными растениями
ГЛАВА 1. ЭВОЛЮЦИОННЫЙ ПОДХОД I ОБЪЯСНЕНИЮ РАЗНООБРАЗИЯ ЖИЗНИ НА ЗЕМЛЕ
Рис. 1.20. Археоптерикс, по И. Аугусте и 3. Буриану:
строение первоптиц подтверждает правильности построения
филогенетического древа, выводящего птиц из пресмыкающихся.
Они имели свободные пальцы с когтями и челюсти с зубами
знаком — перьями. Перо — замечательная структура, возникшая, оче-
видно, из сильно измененной чешуи рептилий. Они служат хорошим
изолирующим покровом, помогающим сохранять тепло. Но самое важ-
ное назначение пера состоит в том, чтобы служить основой крыльев и
хвоста, необходимых для полета.
Теплокровность животных — млекопитающих и птиц — важное
приспособление, позволившее им начать заселение областей с холод-
ным климатом.
Сегодня цветковые растения, появившиеся в эпоху расцвета дино-
завров, воистину царствуют в мире. За всю их историю существовало
более четверти миллиона видов цветковых, а это гораздо больше, чем
число видов всех других групп наземных растений, вместе взятых.
Цветковые действительно превзошли все остальные группы растений
как числом видов, так и продуцируемой биомассой.
Возникновение и распространение покрытосеменных растений тес-
но связано с развитием насекомых, птиц и млекопитающих. Они пита-
ются нектаром цветков, одновременно способствуя их опылению, по-
едают листья и плоды, помогая распространению семян. Таким образом,
биота развивалась как единое целое.
С позднего мела облик растительности существенно меняется, т. к.
доминирующей группой становятся покрытосеменные, иногда сочета-
ЧАСТЬ I. ИСТОРИЧЕСКИЕ К ТЕОГРАФИЧЕСХИЕ ФАКТОРЫ БИОЛОГИЧЕСКОГО РАЗНООБРАЗИЯ
ющиеся с хвойными. Для Сибирско-Канадской фитогеографической
области позднего мела, например, характерно большое количество пла-
танолистных форм покрытосеменных, а также разнообразных папорот-
ников, хвойных, гинкговых. Эта флора существовала в условиях доста-
точно теплого и влажного климата. Поздний мел знаменует начало
формирования флоры кайнозоя.
На рубеже мезозоя — кайнозоя произошло массовое вымирание
биоты. Погибло почти две трети всех видов живых организмов, насе-
лявших Землю, полностью вымерли динозавры. Причину глобальной
катастрофы связывают с падением на Землю крупного небесного тела.
Губительной для крупных животных могла оказаться мощная ударная
волна, а для всей биоты — резкое похолодание климата, вызванное
выбросом в атмосферу большого количества пыли, которая закрыла
доступ солнечным лучам к поверхности Земли.
Кайнозой (начало 60 — 70 млн лет назад). Подразделяется на палео-
ген, неоген (раньше их объединяли в третичный период) и четвертич-
ный период. Океаны и материки приобрели современные очертания.
В наземной биоте продолжали быстро развиваться появившиеся еще
в триасе млекопитающие, в юре — птицы, а в конце мезозоя — цветко-
вые (покрытосеменные) растения. Началось вторичное освоение Миро-
вого океана: покинув сушу, к жизни в море приспособились некоторые
цветковые растения (морские травы), пресмыкающиеся (змеи, черепа-
хи), млекопитающие (киты, дельфины), птицы (пингвины) и многие
другие.
В кайнозое наземные позвоночные развивались независимо на трех
разобщенных территориях. В Лавразии господствовали плацентарные.
В Австралии — сумчатые и однопроходные. В Южной Америке, сохра-
нявшей обособленность с раннего мела, вплоть до конца кайнозоя,
неполнозубые. Именно этим объясняется своеобразие Арктогеи (Евра-
зия, Африка и Северная Америка), Нотогеи (Австралия) и Неоген (Юж-
ная Америка).
Палеоген. В Северном полушарии главные события связаны с рас-
крытием Северо-Восточной Атлантики. Нарушение устойчивой до этой
континентальной связи Евразии и Северной Америки произошло около
50 млн лет назад в результате отделения Шпицбергена от Гренландии.
Полная же изоляция Америки от Евразии установилась только в конце
палеогена. Сочленение Индостанской плиты с Азией привело к активно-
му биотическому обмену между Индией и внетропической Азией.
Климат в Северном полушарии от Индии до Арктики был тепл
географическая зональность слабо выражена. В высоких широтах Се-
верной Америки, Европы и Азии — листопадные широколиственные
леса (тургайская флора); в средних широтах — вечнозеленые тропиче-
ские леса (полтавская флора) (рис. 1.21).
В Южном полушарии произошло отделение Австралии от Антарк-
тиды. Вероятно, уже в конце эоцена прервались длительно существо-
вавшие биотические связи между Новой Зеландией и островом Н
Каледония. Это обусловило последующий высокий эндемизм биот:
островов на уровне родов и семейств, сохраняющийся и до нашего
30 времени.
ГЛАВА 1. ЭВОЛЮЦИОННЫЙ ПОДХОД К ОБЪЯСНЕНИИ) РАЗНООБРАЗИЯ ЖИЗНИ НА ЗЕМЛЕ
Рис. 1.21. Палеогеновый ландшафт, по И. Аугусте и 3. Буриану:
лесная тропическая растительность и древние хоботные животные
Неоген. Начался период активного горообразования: окончательно
выросли Альпы, сформировались Гималаи, омолодился рельеф гор гер-
цинской складчатости. Высокие горы стали преградой на пути господ-
ствующих ветров, дующих с океана. Началось иссушение внутриконти-
нентальных пространств Азии. Климат Северного полушария стал
холоднее. Площадь под вечнозелеными лесами сократилась, а под листо-
падными лесами и травянистой растительностью увеличилась. Впервые
на земле появились травянистые ландшафты саванн и степей (рис. 1.22).
Травяной биом стал единым по всей Арктогее, и каждая из ее тер-
риторий внесла свой вклад в становление фауны: из Северной Америки
пришли лошади, из Азии — полорогие (быки и антилопы), из Афри-
ки — слоны, бегемоты, носороги и жирафы. Хищные кошачьи прибы-
ли из Северной Америки, гиеновые — из Африки. 31
ЧАСТЬ I. ИСТОРИЧЕСКИЕ К ГЕОГРАФИЧЕСКИЕ ФАКТОРЫ БИОЛОГИЧЕСКОГО РАЗНООБРАЗИЯ
Рис. 1.22. Ландшафт саванн в конце неогена, по И. Аугусте и 3. Буриану:
богатая травянистая растительность давала обильный корм
стадам копытных, на которых охотились саблезубые тигры
Альпийский орогенез сопровождался разделением океана Тетис
на западную и восточную части в результате сочленения Африкан-
ской и Аравийской плит в начале неогена. Это вызвало изоляцию мор-
ских биот Индо-Пацифической и Атлантическо-Средиземноморской
областей.
В истории Средиземного моря отмечен ряд катастроф: пролив, соеди-
нявший море с Атлантическим океаном, неоднократно замыкался, и тог-
да вся морская вода испарялась. В условиях сухого и жаркого климата на
это уходило около 5000 лет. В эти периоды дно Средиземного моря пред-
ставляло собой пустыню, покрытую толстым слоем соли. Наступала об-
щая аридизация климата побережий и трансформация тропических лесс
в биомы средиземноморского типа.
Наиболее значительные вехи кайнозойской истории Северо-Восточ-
ной Азии были связаны со становлением и разрушением «Берингийск
го моста».
На Североамериканском континенте важнейшие события кайноз
происходили на Тихоокеанском побережье. Формирование прибретен-
ного вулканического пояса на рубеже мела и палеогена обусловило
последующую изоляцию внутренних районов Северной Америки и
Тихого океана и образование обширной аридной области (Мексика,
центральные районы США).
В Южной Америке сформировался своеобразный комплекс пастбищ-
ных травоядных. Он включал неполнозубых (гигантских броненосцев
и наземных ленивцев), грызунов, родственников морских свинок,
редко размером с носорога, и др. На рубеже миоцена и плиоцена Север-
ная Америка соединилась с Южной. В результате активизировался об-
3 2 мен между американскими сухопутными биотами.
ГЛАВА 1. ЭВОЛЮЦШНЫЙ ПОДИ ОД I ОБЪЯСНЕНИЮ РАЗНООБРАЗИЯ ЖИЗНИ НА ЗЕМЛЕ
Открылся пролив Дрейка, отделивший Антарктиду от Южной Амери-
ки. Вокруг Антарктиды образовалось кольцо вод Южного океана с посто-
янным дрейфовым течением, изолирующим приполярные области от про-
никновения теплых вод. Началось выхолаживание Антарктиды, приведшее
к ее полному оледенению. Отток холодных вод в глубины океанов достиг
Северного полушария, что способствовало похолоданию климата и усиле-
нию ледовитости Северного Ледовитого океана. Из-за похолодания в райо-
нах полюсов началось активное конвективное перемешивание толщи
океана. Ведущую роль играли процессы опускания холодных богатых
кислородом вод у берегов Антарктиды. Так образовались глубинные вод-
ные массы почти всех океанов. Вентиляция глубин создала благоприят-
ные условия для формирования глубоководной биоты.
Образование ледового щита Антарктиды привело к понижению
уровня Мирового океана (около 30 млн лет назад). Эта крупная регрес-
сия кайнозоя сопровождалась восстановлением и расширением многих
сухопутных связей.
В Африке, в районе Кенийского рифта, около 5 млн лет назад возник-
ла популяция древних предков человека — австралопитеков (рис. 1.23).
Рис. 1.23. Семья африканских австралопитеков
33
ЧАСТЬ I. ИСТОРИЧЕСКИЕ К ТЕОГРАФИЧЕСХИЕ ФАКТОРЫ БИОЛОГИЧЕСКОГО РАЗНООБРАЗИЯ
Современная
эпоха -1
10 000--1
Ното
sapiens
sapiens
50 000
100 000-
tiomo sapiens
(неандерталец)
Homo sapiens
(архаичный)
i
£
&
500 000
1000 000:
1 500000-
2000000-
2 500 000 4
habilis
Лет до
современной
эпохи
Рис. 1.24. Последовательные этапы формирования вида Человек разумный
Конечным звеном эволюциивчетвертичномпериоде сталЧеловекразум-
ный (рис. 1.24).
Четвертичный период (антропоген). Его начало (плейстоцен) в
Северном полушарии ознаменовалось великой ледниковой эпохой
(рис. 1.25). Вымерли гигантские млекопитающие третичного периода.
На территории Северной Америки и Евразии сформировалась своеоб-
разная мамонтовая фауна (рис. 1.26).
И все же связь четвертичной биоты с третичной, несмотря на ее
перестройку, вызванную похолоданием, сохранилась благодаря убежи-
щам, расположенным преимущественно к югу от границы ледника, в
горных долинах с более теплым климатом. Убежища, или рефугиумы,
представляли собой благоприятные для жизни островки среди враждеб-
ного моря холода. Локальные убежища могли помочь перенести клима-
тическую катастрофу ледникового периода только немногим, особенно
холодоустойчивым видам. Если говорить о третичной биоте в целом, то
главными ее убежищами были тропические и субтропические области
(рис. 1.27). В эту суровую эпоху происходила дальнейшая эволюция
гоминид, сопровождавшаяся быстрым расширением ареала (рис. 1.28).
К концу ледникового периода окончательно обособился вид — Че-
ловек разумный (Homo sapiens}. Ареал его обитания охватил всю сушу.
Расселению людей способствовало понижение уровня Мирового океа-
на в ледниковую эпоху более чем на 100 м. Оно объясняется тем, что
ГЛАВА 1. ЭВОЛЮЦИОННЫЙ ПОДХОД К ОБЪЯСНЕНИЮ РАЗНООБРАЗИЯ ЖИЗНИ НА ЗЕМЛЕ
Рис. 1.25. Карта четвертичного оледенения, по Е.В. ШонЦРУ
Рис. 1.26. Самый характерный представитель
плейстоценовой фауны — мамонт, по И. Аугусте и 3. Буриану
35
ЧАСТЬ I. ИСТОРИЧЕСКИЕ К ТЕОГРАФИЧЕСКИЕ ФАКТОРЫ БИОЛОГИЧЕСКОГО РАЗНООБРАЗИЯ
Рис. 1.27. Главнейшие убежища третичной биоты, по В. Шаферу:
1 — балкано-черноморское убежище; 2 — армяно-иранское убежище;
3—5 — среднеазиатские убежища; 6 — восточноазиатское убежище;
7—10— североамериканские убежища. Прерывистой линией обозначена
область максимального оледенения в плейстоцене
1. Австралопитеки 5 млн
2. 1,6 млн 5, 500 тыс,
3. 1 млн 6. 800 тыс,
4. 700 тыс. 10.15 000
8.33 000 12.67 000
9.12 000 13.30 000
14.30000 18.19000
16. 1 000
17.4 500
22.1600
20. 22000
21.11000
Рис. 1.28. История расселения первобытных людей по земному шару,
по ((National Geographic)'
ледники связали большое количество воды. Осушились морские мелко-
водья, и между материками появились сухопутные переходы. Остров
Тихого и Индийского океанов человек достигал на плотах и лодках
ГЛАВА 1. ЭВОЛЮЦИОННЫЙ ПОДХОД I ОБЪЯСНЕНИЮ РАЗНООБРАЗИЯ ЖИЗНИ НА ЗЕМЛЕ
Одной из причин быстрого расширения ареала людей была охота на
крупного зверя. Когда первые поселенцы попадали в новые места, оби-
тавшие там животные ничего не знали о коварных приемах охоты и не
боялись людей. Добыча доставалась легко. Но проходило время, зверей
оставалось все меньше. Люди устремлялись вперед в «край непуганых
птиц». Многие животные, ставшие легкой добычей охотников, навсег-
да исчезли с лица Земли (рис. 1.29).
Рис. 1.29. Изменение количества крупных животных (в %),
существовавших в Австралии, Северной Америке и на двух крупных
островах, обусловленное проникновением туда человека
Около 10—11 тыс. лет назад началась послеледниковая эпоха (голо-
цен). Ледник растаял, оставив на равнинах Северной Америки, Европы
и Сибири своеобразный комплекс отложений и форм рельефа. Нача-
лось потепление климата и формирование современных зональных
типов ландшафтов и вертикальной поясности в горах. Экономическое
и социальное развитие человеческого общества привело к усилению
антропогенного воздействия на биосферу.
Глава 2
ГЕОГРАФИЧЕСКИЕ ФАКТОРЫ,
ОПРЕДЕЛЯЮЩИЕ ЗАКОНОМЕРНОСТИ
ПОДРАЗДЕЛЕНИЯ АРЕНЫ ЖИЗНИ НА ЗЕМЛЕ
Ареной жизни на Земле служат ее наружные оболочки: литосфера,
гидросфера и атмосфера. Однако свойства этих оболочек крайне неод-
нородны, что определяет неоднородность пространственно-временной
структуры биосферы в целом. Глобальные физико-географические про-
цессы выступают в качестве наиболее важных экологических факто-
ров, определяющих главные закономерности распределения жизни.
В ходе эволюции в географической оболочке обособились природ-
но-территориальные комплексы планетарной и региональной раз-
мерности. Каждый такой комплекс представляет собой целостную
географическую систему (геосистему), сформировавшуюся в ходе ес-
тественно-исторического развития компонентов живой и неживой
природы, и определяет своеобразие связанных с ним растительности
и животного мира. Иными словами, наиболее крупные подразделе-
ния арены жизни на Земле контролируются физико-географическим
процессом и природно-территориальными комплексами планетарной
и региональной размерности.
К числу главных факторов, определяющих подразделение арены
жизни, относятся внутренние силы Земли, формирующие наиболее
крупные тектогенные формы рельефа, и энергия Солнца, обусловлива-
ющая действие законов географической зональности и высотной пояс-
ности. Особые условия жизни в толще воды и на дне определяют под-
разделение арены жизни в Мировом океане.
н 2.1. Подразделения арены жизни,
" обусловленные тектогенными формами рельефа
Наиболее крупные формы рельефа — материки и океанические впа-
дины, равнинно-платформенные области и горные массивы — созданы
внутренними силами Земли, проявляющимися в виде тектонических
движений и вулканической деятельности.
Материки и океаны представляют царства сухопутных и морских
обитателей. Господствующей является океаническая арена жизни,
ГЛАВА I. ГЕОГРАФИЧЕЕШЕ ФАКТОРЫ. ОПРЕДЕПЯШЩКЕ ЗАКОНОМЕРНОСТИ ПОДРАЗДЕЛЕНИЯ АРЕНЫ ЖИЗНИ НА ЗЕМЛЕ
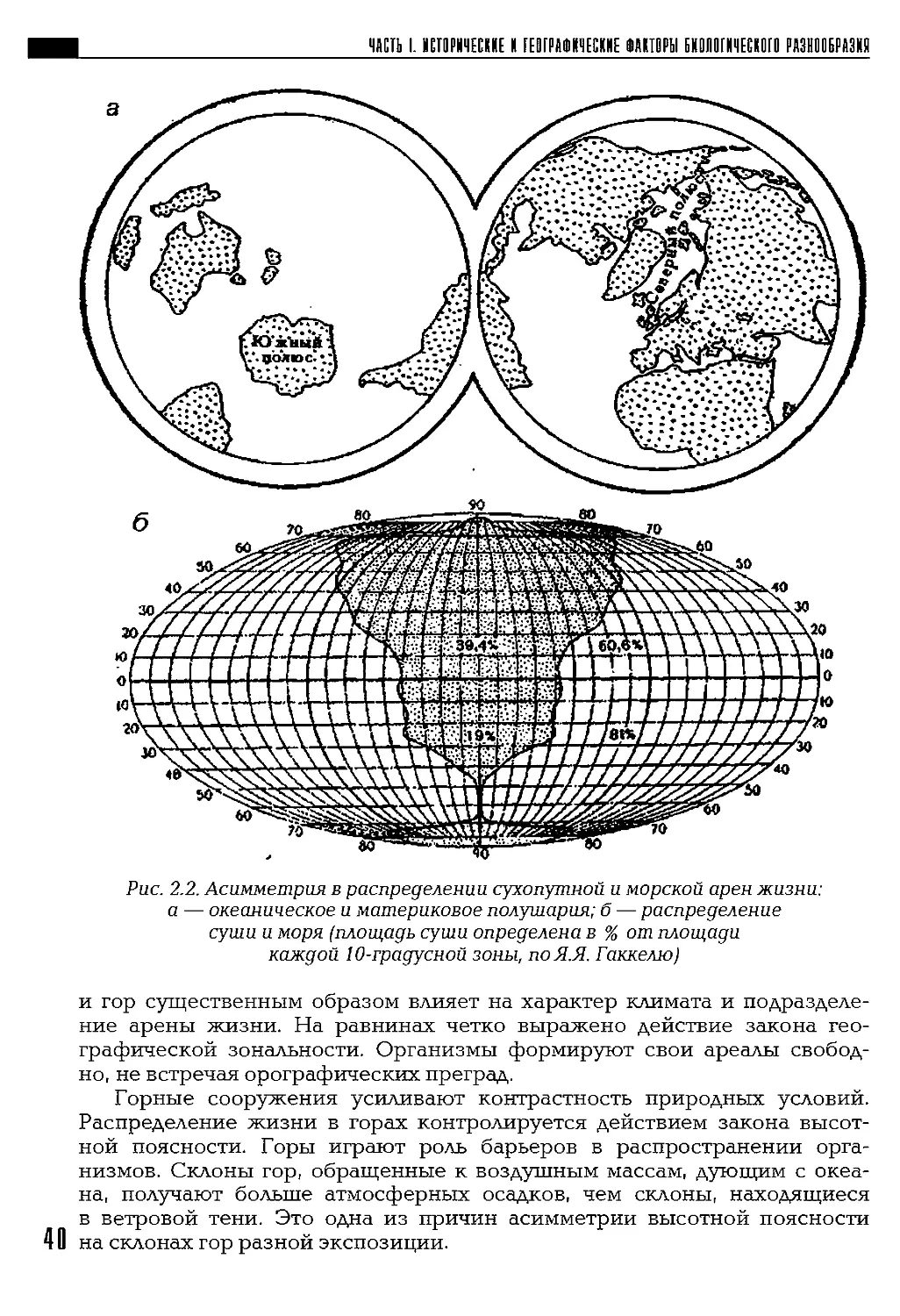
занимающая 70,8% площади всей поверхности Земли; на долю суши при-
ходится 29,2% (рис. 2.1). Океаны и моря не только самая значительная
по площади и объему часть биосферы, но и важнейшая часть географи-
ческой оболочки, регулирующая климат планеты. Распределение суши
и моря оказывает влияние н"а формирование основных типов клима-
та — морского и континентального — и связанных с климатом расти-
тельного покрова и животного мира.
Рис. 2.1. Циклограмма площадей океанов и материков, поЛ.П. Шубаеву
В Северном полушарии суши больше, ее площадь составляет 39,4%
площади полушария; в Южном полушарии площадь суши составляет
всего 19%. Соответственно сухопутная арена жизни более широко пред-
ставлена в Северном полушарии (рис. 2.2).
В Северном полушарии на обширных пространствах суши форми-
руется континентальный климат с резкими суточными и сезонными
амплитудами температур и небольшим количеством атмосферных осад-
ков. Формирование климата в Южном полушарии происходит при пре-
обладающем воздействии океана: морской климат характеризуется
пониженными амплитудами годового и суточного хода температуры,
повышенным количеством атмосферных осадков.
Важным фактором подразделения арены жизни на суше являются
крупные тектонически обусловленные формы рельефа (рис. 2.3). В пре-
делах материков на месте платформ формируются низменности, равни-
ны, плато, плоскогорья, т. е. пространства, характеризующиеся слабой
расчлененностью рельефа. Такое соответствие между тектонической
структурой и рельефом объясняется тектонической стабильностью плат-
форм в фанерозое. В областях горообразования выделяются древние
срелневысотные горы, возрожденные в процессе неотектонических дви-
жений, и молодые высокие горы. Соотношение равнинных пространств
ЧАСТЬ I. ИСТОРИЧЕСКИЕ К ТЕОГРАФИЧЕСКИЕ ФАКТОРЫ БИОЛОГИЧЕСКОГО РАЗНООБРАЗИЯ
Рис. 2.2. Асимметрия в распределении сухопутной и морской арен жизни:
а — океаническое и материковое полушария; б — распределение
суши и моря (площадь суши определена в % от площади
каждой 10-градусной зоны, по Я.Я. Гаккелю)
и гор существенным образом влияет на характер климата и подразделе-
ние арены жизни. На равнинах четко выражено действие закона гео-
графической зональности. Организмы формируют свои ареалы свобод-
но, не встречая орографических преград.
Горные сооружения усиливают контрастность природных условий.
Распределение жизни в горах контролируется действием закона высот-
ной поясности. Горы играют роль барьеров в распространении орга-
низмов. Склоны гор, обращенные к воздушным массам, дующим с океа-
на, получают больше атмосферных осадков, чем склоны, находящиеся
в ветровой тени. Это одна из причин асимметрии высотной поясности
4 0 на склонах гор разной экспозиции.
ГЛАВА I. ГЕ0ГРАФИЧЕЕК1Е ФАКТОРЫ. ОПРЕДЕПЯШЩКЕ ЗАКОНОМЕРНОСТИ ПОДРАЗДЕЛЕНИЯ АРЕНЫ ЖИЗНИ НА ЗЕМЛЕ
Рас. 2.3. Подразделения арены жизни, обусловленные тектогенными формами
рельефа, по «Миру географии»: 1,2 — равнинно-платформенные области:
1 — низкие равнины; 2— высокие равнины (плато, плоскогорья);
3, 4 — горные области: 3 — древние возрожденные горы; 4 — молодые горы;
5 — ледниковые щиты
— 2.2. Подразделения арены жизни,
" обусловленные действием закона географической зональности
Характернейшей формой подразделения арены жизни является зо-
нальность, т. е. закономерное изменение всех географических компо-
нентов и географических ландшафтов по широте (от экватора к полю-
сам). Основные причины зональности— форма Земли и положение
Земли относительно Солнца, а предпосылка— падение солнечных лу-
чей на земную поверхность под углом, постепенно уменьшающимся в
обе стороны от экватора.
С.В. Калесник (1970) подчеркивает, что по причине зонального рас-
пределения солнечной лучистой энергии на Земле зональны темпера-
тура воздуха, воды и почвы, испарение и облачность, атмосферные осад-
ки, барический рельеф и системы ветров, свойства воздушных масс,
климаты, характер гидрографической сети и гидрологические процес-
сы, особенности геохимических процессов выветривания и почвообра-
зования, типы растительности и жизненные формы растений и живот-
ных и, наконец, географические ландшафты, объединяемые в связи с
этим в систему ландшафтных зон.
Количество энергии, получаемой Землей в течение года на разных
широтах, от полюсов к экватору, существенно меняется (рис. 2.4). Из 41
ЧАСТЬ I. ИСТОРИЧЕСКИЕ К ТЕОГРАФИЧЕСКИЕ ФАКТОРЫ БИОЛОГИЧЕСКОГО РАЗНООБРАЗИЯ
Гро
Июни
июль
Догуп
Сентябрь
Ояяябр»
Ноябри
Декабрь
___________Юг
Яяоатъ
Февраль
л&кржт
Апрель
MoG
широты
Содмвчное излучение, мдеюще* за дам» не верхнюю границу ат*юеферы, вт/л1
Рис. 2.4. Трехмерная диаграмма, показывающая изменения солнечного
излучения в Северном и Южном полушариях по временам года, по «Биосфере»:
годовое изменение склонения Солнца от 23' с. ш. (тропик Рака) до 23° ю. ш.
(тропик Козерога) показано жирной кривой в широтной плоскости
диаграммы видно, что на экваторе количество солнечной энергии в те-
чение года остается примерно одинаковым; при переходе в более высо-
кие широты все отчетливее проявляются сезоны с дефицитом энергии и,
наконец, на полюсах летом устанавливается полярный день, а зимой —
полярная ночь. Особенно заметны изменения солнечной инсоляции в
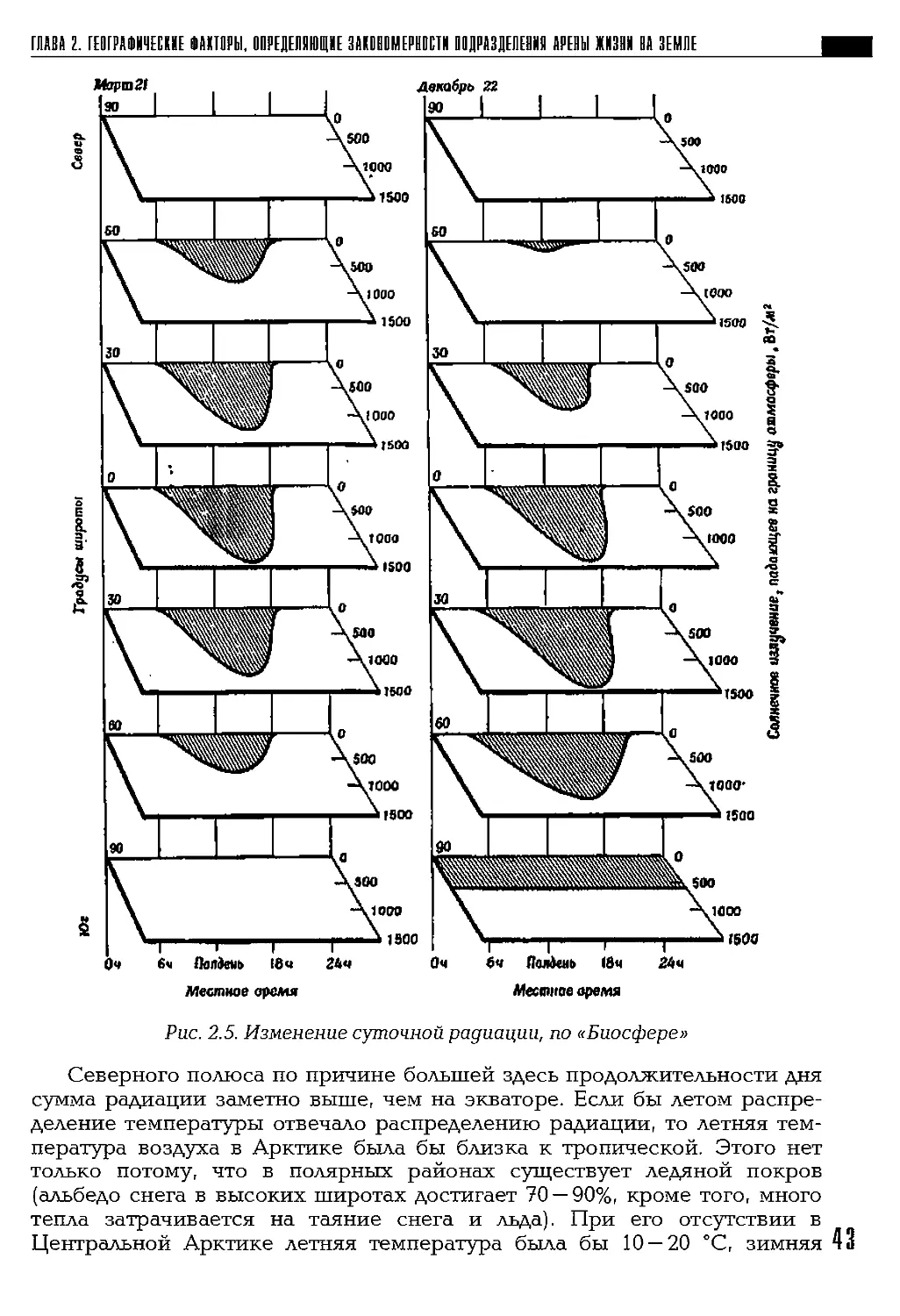
течение суток в разные сезоны и на разных широтах (рис. 2.5).
Во время равноденствия (21 марта и 23 сентября) уменьшение ин-
тенсивности солнечного излучения по направлению к полюсам симмет-
рично относительно экватора, на полюсах солнце не показывается над
горизонтом (левая диаграмма). Во время солнцестояний широтные раз-
личия между двумя полушариями достигают крайних значений. Диа-
грамма справа изображает момент летнего солнцестояния в Южном по-
лушарии — 22 декабря; в Северном полушарии — зима. Летний полюс
(внизу) освещается солнцем круглые сутки, тогда как зимний полюс
(вверху) не освещается совсем.
В распределении прихода и расхода солнечной радиации также про-
слеживается четкая зависимость от широты. В нагревании Земли Солн-
цем действуют механизм поглощения лучистой энергии, преобразова-
ния ее в тепло и излучение длинноволнового эффективного излучения
в космическое пространство. В местах, где радиационный баланс поло-
жительный, тепло накапливается; там, где он отрицательный, тепло
теряется и начинают проявляться холодные сезоны (рис. 2.6).
Связь между распределением температуры на Земле и распределе-
нием приходящей солнечной радиации совершенно отчетлива. Однако
прямая зависимость между убыванием средних величин приходящей
радиации и понижением температуры при возрастании широты суще-
42 ствует только зимой. Летом же в течение нескольких месяцев в районе
ГЛАВА I. ГЕОГРАФИЧЕСКИЕ ФАКТОРЫ. ОПРЕДЕЛЯЮЩИЕ ЗАКОНОМЕРНОСТИ ПОДРАЗДЕЛЕНИЯ АРЕНЫ ЖИЗНИ 6А ЗЕМЛЕ
Ьч 6ч flontaib 18 ч 24ч
Местное время
Оч 6ч Пшйемь 18ч 24ч
Местное время
Рис. 2.5. Изменение суточной радиации, по «Биосфере»
Северного полюса по причине большей здесь продолжительности дня
сумма радиации заметно выше, чем на экваторе. Если бы летом распре-
деление температуры отвечало распределению радиации, то летняя тем-
пература воздуха в Арктике была бы близка к тропической. Этого нет
только потому, что в полярных районах существует ледяной покров
(альбедо снега в высоких широтах достигает 70 — 90%, кроме того, много
тепла затрачивается на таяние снега и льда). При его отсутствии в
Центральной Арктике летняя температура была бы 10 — 20 °C, зимняя 43
ЧАСТЬ 1.1стаРИЧЕСКИЕ К ТЕОТРАФИЧЕСШ ФАКТОРЫ БИОЛОГИЧЕСКОГО РАЗНООБРАЗИЯ
Рис. 2.6. Годовой радиационный баланс земной поверхности
(ккал/см2 в год), по М.И. Будыке
5—10 °C, т. е. сформировался бы совсем другой климат, при котором арк-
тические острова и побережья могли одеться богатой растительностью,
если бы тому не препятствовали многосуточные и даже многомесячные
полярные ночи (невозможность фотосинтеза). То же было бы и в Антарк-
тиде, только с оттенками континентальности: лето было бы теплее, чем
в Арктике (ближе к тропическим условиям), зима— холоднее. Стало
быть, ледяной покров Арктики и Антарктики — это скорее причина, чем
следствие низких температур в высоких широтах (Будыко, 1971).
Помимо географической широты на распределение тепла на Земле
влияют характер распределения суши и моря, морские течения и господ-
ствующие ветры, рельеф, высота местности над уровнем моря. Изотермы
имеют подчас причудливые очертания, отклоняясь от параллелей (рис. 2.7).
Глобальные подразделения арены жизни, обусловленные неравно-
мерным распределением солнечной радиации и теплоты, находят выра-
жение в следующих тепловых поясах (Калесник, 1970).
Теплый, или жаркий, пояс ограничен в каждом полушарии годовой
изотермой +20 °C, проходящей вблизи 30-й северной и 30-й южной
параллелей.
Два умеренных пояса в каждом полушарии лежат между годовой
изотермой +20 °C и изотермой + 10 °C самого теплого месяца (соответ-
ственно июля или января); в Долине смерти (Калифорния) отмечена
наивысшая на земном шаре июльская температура + 56,7 °C.
Два холодных пояса, в которых средняя температура самого тепло-
го в данном полушарии месяца менее +10 °C; иногда из холодных поя-
сов выделяют две области вечного мороза со средней температурой
самого теплого месяца ниже 0°. В Северном полушарии это внутренняя
часть Гренландии и, возможно, пространство около полюса; в Южном
полушарии это все, что лежит к югу от 60-й параллели. Особенно холод-
на Антарктида; здесь зарегистрирован абсолютный минимум — 88,3 °C.
Рис. 2.7. Среднее годовое распределение температуры воздуха
науровне моря, по С.П. Хромову
В результате неравномерного нагревания Земли (океана и суши на
разных широтах) Солнцем в атмосфере формируется барический рель-
еф — центры высокого и низкого давления, которые приводят в действие
механизм движения воздушных масс. В барическом поле Земли нагляд-
но обнаруживается зональное распределение атмосферного давления,
симметричное в обоих полушариях (рис. 2.8). В соответствии с бариче-
ским рельефом складывается система постоянных ветров (рис. 2.9).
—— К&ввсргсмзад
Рис. 2.8. Схема общей циркуляции атмосферы, поЛ.П. Шубаеву:
справа меридиональный разрез. Буквами обозначены направления
господствующих ветров. Заштрихованы пояса восточного переноса.
ВиН — высокое и низкое давление
45
ЧАСТЬ I. iЕГОРИЧЕСКИЕI ГЕОГРАФИЧЕСКИЕ ФАКТОРЫ БИОЛОГИЧЕСКОГО РАЗНООБРАЗИЯ
Рис. 2.9. Глобальная система постоянных ветров по Encyclopedia of the Earth
Приэкваториальный пояс штилей — ветры сравнительно редки (т. к.
господствуют восходящие движения сильно нагретого воздуха, атмос-
ферное давление низкое).
Области затишья в субтропических поясах высокого давления (при-
чина — господство нисходящих движений воздуха).
Зоны пассатов Северного и Южного полушарий.
Зоны преобладания западных ветров в средних широтах обоих
полушарий.
Полярные ветры с восточной составляющей, дующие от полюсов в
сторону барических депрессий средних широт.
Рассмотрим механизмы, приводящие в действие систему постоянных
ветров, более подробно. Когда солнце стоит на экваторе, в зените, проис-
ходит сильный прогрев воздушных масс и их адиабатический подъем,
приводящий к падению атмосферного давления у поверхности Земли. Так
формируется приэкваториальный пояс штилей. Поднимающийся воздух
охлаждается, содержащиеся в нем водяные пары конденсируются, и вода
выпадает на землю в виде обильных дождей. Их называют зенитальными,
т. к. они обусловлены высоким стоянием Солнца над горизонтом.
Поднявшиеся на экваторе воздушные массы растекаются к северу и
югу до 30° широты, где снова опускаются. Нисходящие массы воздуха
нагреваются, становясь при этом очень сухими. Атмосферных осадков
здесь мало, атмосферное давление у земной поверхности всегда высо-
кое, погода обычно безветренная.
ГЛАВА I. ГЕОГРАФИЧЕСКИЕ ФАКТОРЫ. ОПРЕДЕЛЯЮЩИЕ ЗАКОНОМЕРНОСТИ ПОДРАЗДЕЛЕНИЯ АРЕНЫ ЖИЗНИ 6А ЗЕМЛЕ
Из субтропических зон высокого давления воздух вновь устремляет-
ся к экваториальной зоне низкого давления. Вследствие вращения Земли
ветры отклоняются в Северном полушарии вправо, а в Южном — влево,
образуя соответственно северо-восточный и юго-восточный пассаты.
Поскольку высота стояния-солнца в течение года меняется (летом в
Северном полушарии оно находится в зените над Северным тропиком,
а зимой, т. е. летом в Южном полушарии, — над Южным тропиком),
происходит смещение системы атмосферной циркуляции. В связи с пе-
ремещением тропических фронтов сдвигается экваториальная ложби-
на низкого давления: летом в Северном полушарии на север, а летом в
Южном полушарии на юг. Вместе с ней приходят зенитальные дожди
(летний период дождей); когда зимой тропические широты оказывают-
ся в зоне действия субтропических центров высокого давления, насту-
пает сухой сезон. Таким образом, формируются климатические пояса
субэкваториальных муссонов.
Под влиянием сезонных температурных контрастов между обширным
Азиатским материком и омывающими его Тихим и Индийским океанами
возникает классический режим муссонной циркуляции — зимой с суши
дуют сухие ветры, летом ветры с океана несут обильные осадки.
В умеренных широтах господствуют западные ветры, особенно
устойчивые в Южном полушарии (ревущие сороковые). В полярных об-
ластях вновь преобладает восточный перенос. Здесь, на границе между
холодным воздухом полярных широт и более теплыми воздушными
массами низких широт, непрерывно возникают мощные вихревые дви-
жения, которые в Северном полушарии формируются главным обра-
зом около Исландии и в районе Алеутских островов. Циклоны переме-
щаются с запада на восток, иногда отклоняясь к югу.
Постоянные ветры пассатов и западного переноса вызывают мощ-
ные дрейфовые течения. С глобальной системой течений связаны поч-
ти все физические, химические и биологические процессы, определя-
ющие характерные черты подразделения арены жизни в океане. В целом
распределение водных масс и связанных с ними организмов в поверх-
ностной толще Мирового океана подчинено закону географической зо-
нальности (рис. 2.10).
Солнечная радиация и температура, господствующие ветры и
атмосферные осадки (рис. 2.11) являются важнейшими факторами
климатообразования, и, как было показано, все они имеют тенденцию
к зональному распределению. Вполне естественно, что и климаты зо-
нальны. Приведем карту климатических поясов и областей, составлен-
ную Б.П.Алисовым (рис. 2.12).
Оценивая роль климатообразующих факторов в подразделении аре-
ны жизни на суше, следует подчеркнуть, что важным фактором геогра-
фической зональности является показатель увлажнения, зависящий от
соотношения тепла и влаги.
М.И. Будыко (1971) предложен радиационный индекс сухости R/Lr —
отношение среднегодового радиационного баланса земной поверхности к
количеству тепла, необходимого для испарения среднегодовой суммы осад-
ков. При радиационном индексе сухости 0,8— 1,0 тепла хватает на испаре-
ние большей части осадков, наблюдается умеренный сток, устойчивое
ЧАСТЬ I. iЕГОРИЧЕСПЕ И ГЕОГРАФИЧЕСКИЕ ФАКТОРЫ БИОЛОГИЧЕСКОГО РАЗНООБРАЗИЯ
Рис. 2.10. Распространение основных типов водных масс Мирового океана,
по В.Н. Степаному. Типы водных масс: 1 — полярный; 2 — субполярный;
3 — умеренно тропический; 4 — тропический; 5 — экваториальный;
6 — индомалайский; 7 — присредиземноморский; 8 — североатлантический
Рис. 2.11. Среднегодовое количество осадков на суше, по Г. Вальтеру
увлажнение и хорошая аэрация почвы и в целом наилучшие условия для
развития жизни, в частности лесных формаций. При радиационном
индексе сухости менее 0,8 увлажнение избыточное, тепла не хватает для
испарения осадков, происходит заболачивание. При радиационном индек-
се сухости более 1,0 увлажнение недостаточное, влага испаряется почти
полностью и избыточное тепло тратится на перегрев почвы и атмосферы.
ГЛАВА I. ГЕОГРАФИЧЕСКИЕ ФАКТОРЫ. ОПРЕДЕЛЯЮЩИЕ ЗАКОНОМЕРНОСТИ ПОДРАЗДЕЛЕНИЯ АРЕНЫ ЖИЗНИ 6А ЗЕМЛЕ
Рис. 2.12. Климатические пояса и области, по Б.П. Алисову: I — экватори-
альный пояс: 1 — континентальные, 2 — высокогорные, 3 — океанические;
11— субэкваториальный пояс: 4а — континентальные, 46 — те же, но бо-
лее засушливые, 5 — высокогорные, 6 — океанические; III — тропический
пояс: 7 — континентальные, 8 — высокогорные, 9 — западные берега кон-
тинентов, 10 — восточные наветренные берега континентов и остро-
вов, И — океанические; IV— субтропический пояс: 12 — континенталь-
ные, 13 — высокогорные, 14 — средиземноморские, 15 — муссонные облас-
ти восточных берегов материков, 16 — океанические; V— умеренный
пояс: 17а — континентальные, 176 — те же, но осадков меньше возмож-
ного испарения, 18 — высокогорные области умеренного пояса, 19 — за-
падные части материков с относительно теплой влажной зимой и уме-
ренно теплым летом, 20 — муссонные области восточных берегов мате-
риков с относительно холодной зимой и влажным дождливым летом, 21 —
океанические с относительно теплой ветреной зимой и прохладным ле-
том; VI — субарктический пояс: области сезонных перемещений аркти-
ческого фронта: 22 — континентальные, очень холодная зима и относи-
тельно теплое лето, 23 — океанические, не холодная, но очень ветреная
зима и прохладное лето; VII — арктический пояс: области арктических
воздушных масс: 24 — континентальная (внутренняя Гренландия)
с исключительно холодной зимой и морозным летом, 25 — океаническая
(Северный Ледовитый океан) с холодной зимой и сырым туманным летом;
VIII — антарктический и субантарктический пояс (объединены из-за не-
достатка данных): 26 — континентальная (внутренняя Антарктида) с
исключительно холодной зимой и морозным летом, 27 — океанические
(южные высокоширотные моря и прибрежные воды Антарктиды)
При отклонениях как в меньшую, так и в большую сторону органи-
ческий мир угнетается: величины радиационного индекса сухости ме-
нее 0,3 соответствуют зоне тундры; более 3,5 — пустыне. Между ними
располагаются остальные зоны (рис. 2.13).
ЧАСТЬ I. iСТОРИЧЕСКИЕ К ГЕОГРАФИЧЕСКИЕ ФАКТОРЫ БИОЛОГИЧЕСКОГО РАЗНООБРАЗИЯ
Рис. 2.13. Глобальный параллелизм в развитии растительных формаций,
обусловленный «периодическим законом географической зональности»,
по «Географическому энциклопедическому словарю...», 1988.
IR — радиационный индекс; относительный размер кружков отвечает
биологической продуктивности соответствующих ландшафтов
и господствующих растительных формаций
Все вышесказанное позволило А.А. Григорьеву и М.И. Будыко вы-
явить закономерность, названную ими периодическим законом зональ-
ности (Будыко, 1971). Согласно этому закону в различных широтных
поясах одним и тем же значениям радиационного индекса сухости со-
ответствуют природные зоны, сходные по ряду существенных призна-
ков. В табл. 2.1 более полно раскрываются закономерности, изображен-
ные на схеме (см. рис. 2.13).
Идеальный континент является наглядной моделью распростране-
ния зональных типов ландшафтов и биомов (рис. 2.14). Субширотное
простирание зон характерно только для холодного и умеренного поясов
Северного полушария. С увеличением радиационного баланса все оче-
виднее действие периодического закона зональности: появляются дол-
готные секторы, в которых чередование зональных типов ландшафтов
определяется изменениями радиационного индекса сухости.
Секторность — универсальная закономерность географической обо-
лочки. Она обусловлена взаимодействием океанов и материков. От соот-
ношения океанических и континентальных воздушных масс зависит сте-
пень континентальности климата, количество осадков. Это позволяет
разделить материки на физико-географические секторы — крупные регио-
нальные единицы, обычно простирающиеся в направлении, близком к
меридиональному. В каждом секторе зональность приобретает свою спе-
цифику. В приокеанических секторах зональные контрасты сглажены. Для
них характерен спектр широтных зон с доминированием лесных типов
растительности от тайги до экваториальных лесов. Континентальный
5 0 спектр зон отличается преобладанием степей, полупустынь, пустынь.
Периодический закон географической зональности суши,
по А.А. Григорьеву и М.И. Будыке
Таблица 2.1
Тепловая энерге- тическая база — радиационный баланс Условия увлажнения — радиационный индекс сухости
меньше 0 (крайне избыточ увлажнение) от Одо 1 от 1 до 2 (умеренно недостаточ- ное увлажнение) от 2 до 3 (недостаточное увлажнение) более 3 (крайне недостаточ- ное увлажнение)
избыточное увлажнение оптимальное увлажнение 0,8- 1.0
СМ о 1 о 0,2-0,4 о 1 о со о 1 о
1 2 3 4 5 6
Меньше 0 (высокие широты) I Вечный снег — — — — — — —
От Одо 50 ккал/ см2в год (южиоарктнче- ские, субарктиче- ские и средние широты) — Па Арктиче- ская пус- тыня Пб Тундра (на юге с остров- ками ред- колесий) lis Северная и средняя тайга Шг Южная тайга и смешан- ные леса Пд Лиственный лес и лесостепь III Степь IV Полупустыня умеренного пояса V Пустыня умеренного пояса
От 50 до 75 ккал/ см2 в год (субтро- пические шпроты) — — Via Районы субтропи- ческой ге- мигилей со значи- тельным количест- вом болот VI6 Дождевые субтропические леса Vila Жестколи- ственные субтропи- ческие леса и кус- тарники VII6 Субтропи- ческая степь VIII Субтропическая полупустыня IX Субтропиче- ская пустыня
Больше 75 ккал/ см2 в год (тропические широты) — — Ха Районы преобла- дания эква- ториаль- ных лес- ных болот Хб Сильно пе- реувлаж- ненный (сильно заболочен- ный) эква- ториаль- ный лес Хв Среднепе- реувлаж- ненный (среднеза- болочен- ный) эква- ториаль- ный лес Хг Экваториальный лес. переходящий в светлые тропиче- ские леса и леси- стые саванны XI Сухая саванна XII Опустыиеииая саванна XIII Пустыня тропическая
ЧАСТЬ 1.1 ЕГОРИЧЕСКИЕ К ГЕОГРАФИЧЕСКИЕ ФАКТОРЫ БИОЛОГИЧЕСКОГО РАЗНООБРАЗИЯ
южное п и к у ш a f* и е севеансс пол
Рис. 2.14. Схема идеального континента, поА.Г. Исаченке.
Секторы: I — западные приокеанические; II — восточные приокеанические;
III — слабо- и умеренно континентальные; IV — континентальные;
V — резко и крайне континентальные. Зоны: 1 — лесотундровая; 2 — при-
океанические луговые и лесо-луговые; 3 — суббореальные широколиственно-
лесные (включая переходные к субтропическим); 4 — субтропические влаж-
ные лесные; 5 — средиземноморские; 6 — субтропические лесостепные,
степные, саванновые; 7— тропические и субэкваториальные влажные лес-
ные; 8 — суббореальная полупустынная Южного полушария; 9 — бореальные
и суббореальные влажные лесные Южного полушария
ГЛАВА I. ГЕ0ГРАФИЧЕЕК1Е ФАКТОРЫ. ОПРЕДЕЛЯЮЩИЕ ЗАКОНОМЕРНОСТИ ПОДРАЗДЕЛЕНИЯ АРЕНЫ ЖИЗНИ 6А ЗЕМЛЕ
2.3. Подразделения арены жизни,
обусловленные действием закона высотной поясности
Высотная поясность, будучи функцией рельефа, обязана своим су-
ществованием понижению температуры и увеличению атмосферного
увлажнения с высотой. В самых общих чертах вертикальные клима-
тические изменения аналогичны зональным (рис. 2.15). В действитель-
ности полного тождества между широтными зонами и высотными
поясами не наблюдается. Так, высокогорный пояс парамо в Андах не
тождествен зоне тундр, точно так же как высокогорные луга и леса не
тождественны лугам и лесам соответствующих широтных зон.
► Север
Рис. 2.15. Зеркальное отображение широтных зон в высотных поясах гор,
по D. Chiras
Высотная поясность начинается с той широтной зоны, в которой
находится горная страна. Там, где высота гор позволяет, поясность за-
мыкается вечными снегами и льдами. При этом в высоких широтах
снеговая граница опускается до уровня моря; в жарких странах она
поднимается. Выше всего граница снега расположена в сухих субтро-
пиках и тропиках (в Тибете 7 км); в условиях влажного климата на эква-
торе она опускается до 4,4 км.
ЧАСТЬ I. iСТОРИЧЕСКИЕ К ГЕОГРАФИЧЕСКИЕ ФАКТОРЫ БИОЛОГИЧЕСКОГО РАЗНООБРАЗИЯ
Принято говорить о спектрах высотной поясности, которые по-раз-
ному проявляются в разных физико-географических условиях. У каж-
дой широтной зоны свой набор высотных поясов. Первым на особен-
ности высотной поясности в зависимости от географической широты
указал А. Гумбольдт (рис. 2.16).
Рис. 2.16. Спектры высотной поясности на разных широтах, по А. Гум-
больдту. Наиболее полно высотные пояса представлены на экваторе,
на широте 0—10“; в умеренных широтах 42—46° с. ш. число зон сокращается;
в холодной зоне на широте 68° с. ш. их становится еще меньше
Подобно широтной зональности высотная поясность бывает двух
основных типов: океанического и континентального. В первом случае, в
умеренных и теплых зонах, лесной пояс начинается на приморской рав-
нине; во втором — на равнине располагаются субаридные и аридные ланд-
шафты (степи, полупустыни, пустыни), а древесно-кустарниковые форма-
ции начинаются на известной высоте. На больших горных массивах, на
склонах гор разной экспозиции наблюдается асимметрия высотной пояс-
ности: спектры высотной поясности выражены по-разному (рис. 2.17).
Рис. 2.17. Схема высотных поясов растительности Закавказья,
по С.В. Колеснику. Пояса: 1 — вечного снега и льда; 2 — высокогорной пусты-
ни; 3— альпийский; 4— субальпийский; 5— высокогорных лесов с преобла-
данием восточного дуба; 6 — елово-пихтовых лесов; 7 — буковых лесов; 8 —
смешанных лиственных колхидских лесов; 9 — лесов с преобладанием груз ин-
ского дуба; 10— полупустынь, степей и аридных редколесий; 11 — пустынь
ГЛАВА I. ГЕОГРАФИЧЕСКИЕ ШТОРЫ, ОПРЕДЕЛЯЮЩИЕ ЗАКОНОМЕРНОСТИ ПОДРАЗДЕЛЕНИЯ АРЕНЫ ЖИЗНИ Ой ЗЕМЛЕ
И 2.4. Основные подразделения арены жизни в океане___________________
Толща вод Мирового океана представляет самую мощную и обшир-
ную часть биосферы — пелагиаль. Ложе океана — бенталь, арена жизни
организмов, ведущих донный образ жизни. Пелагиаль подразделяется на
верхний, освещенный солнцем фотический слой и глубоководное цар-
ство вечной ночи — афотическую толщу. В прибрежной, неритической
области условия существования отличаются от жизни в пелагиали от-
крытого океана. На морском дне выделяются глубинные пояса, связан-
ные с литоралью, батиалью и абиссалью (рис. 2.18).
ТТЕПАГИАЛь
ОКЕАНИЧЕСКАЯ
5
ШЕЛЬФ
БЕНТАЛЬ
АБИССАЛЬ
Бентос
§
Рис. 2.18. Основные подразделения арены жизни в океане
Более подробно подразделения арены жизни в океане рассмотрены
в книгах К.М. Петрова (1999, 2004).
Глава 3
АРЕАЛОГИЯ
Совокупность видов растений, встречающихся на той или иной тер-
ритории (акватории), образует флору данной территории, совокупность
видов животных — фауну. Исторически сложившаяся совокупность
живых организмов (флора и фауна) образует биоту. В задачи биогео-
графии входит районирование биоты, выявление неповторимых флори-
стических и фаунистических комплексов путем анализа особенностей
географического распространения систематических таксонов (видов,
родов, семейств и т. д.). Поэтому основополагающим разделом биогео-
графии является учение об ареалах (ареалогия).
Ареал — это часть земной поверхности или акватории, в пределах
которой встречается данный вид организма. Размеры, форма, внутрен-
няя структура, границы ареала определяются историческими, геогра-
фическими и экологическими факторами.
Методика выявления ареалов достаточно проста: на карту наносятся
места нахождения видов. Чем полнее проведены наблюдения и точнее
определена систематическая принадлежность видов, тем точнее будет
картина их географического распространения. Ареалы видов очерчива-
ют наиболее дробные единицы районирования. Чем выше таксон, тем
обширнее область его распространения. Ареалы высших таксономиче-
ских единиц, как правило, охватывают всю сушу, весь океан или всю
биосферу.
Работа по картографированию ареалов долгая и кропотливая. Требу-
ются многолетние сборы фактического материала, чтобы, соединив
периферийные точки линией, очертить границы ареала. Новые наход-
ки, уточнение систематической принадлежности организмов подчас
вносят существенные коррективы в форму ареала, заставляют по-ново-
му интерпретировать историю его формирования.
3.1. Формирование ареала
Формирование ареала связано с образованием вида и завоеванием
им пространства. Новые виды образуются путем изменчивости пред-
ковых форм, естественного отбора и сохранения наиболее приспособ-
ленных, победивших в борьбе за существование. Процесс увеличения
биоразнообразия опирается на теорию монофилитической эволюции
50 4. Дарвина. Согласно этой теории все многообразие видов в крупной
ГЛАВА 3. АРЕАЛОГНЯ
таксономической группе растении или жи-
вотных происходит от одного корня в резуль-
тате дивергенции — расхождения признаков.
На первых этапах дивергенция осуществляет-
ся в рамках микроэволюционного процесса,
т. е. изменчивости существующих организа-
ционных признаков. Она носит ярко выра-
женный приспособительный (адаптацион-
ный) характер.
В качестве иллюстрации рассмотрим из-
менчивость вьюрков с Галапагосских остро-
вов, на которую обратил внимание Дарвин
(рис. 3.1). На этих островах, расположенных
в Тихом океане южнее экватора, в 1100 км к
западу от Южной Америки, обитают лишь
немногие виды организмов. Среди них за-
метную роль играют 14 видов вьюрков. Их
предком, по всей видимости, являлась попу-
ляция вида, занесенного на один из остро-
Ч. Дарвин (1809—1882)
вов сильным ветром с материка. В результате адаптивной радиации в ходе
приспособления к различным условиям существования возникли новые
виды, занявшие различные экологические ниши.
Рис. 3.1. Микроэволюция на примере дарвиновских вьюрков,
по Р. Юнкеру и 3. Шереру: 1 — вьюрок с Кокосового острова; 2 — галапагос-
ский певчий вьюрок; 3 — дятловидный вьюрок; 4 — мангровый древесный
вьюрок; 5 — малый вьюрок; 6 — средний вьюрок; 7 — большой вьюрок; 8 —
вьюрок, обитающий на лиственных деревьях; 9 — большой кактусовый
вьюрок; 10 — малый кактусовый вьюрок; И — остроклювый обыкновенный
вьюрок (основной тип); 12 — малый земляной; 13 — средний земляной;
14 — вьюрок толстоклювый, обыкновенный. За исходный тип (А) принят
зерноядный земляной вьюрок с Южноамериканского континента
ЧАСТЬ I. iСТОРИЧЕСКИЕ К ГЕОГРАФИЧЕСКИЕ ФАКТОРЫ БИОЛОГИЧЕСКОГО РАЗНООБРАЗИЯ
Макроэволюция связана с арогенезом (ароморфозом) — крупны-
ми анатомо-морфологическими и физиологическими изменениями, в
результате которых биологический таксон поднимается на принципи-
ально новую, более прогрессивную ступень развития (рис. 3.2). Каро-
морфным изменениям относятся: появление теплокровности, четырех-
камерного сердца, возникновение перьевого покрова и приспособлений
к полету и т. п.
Рис. 3.2. Макроэволюция от простейших до человека,
по Р. Юнкеру и 3. Шереру
Примерами дивергентной макроэволюции могут быть млекопита-
ющие и птицы, произошедшие от общего корня — древних пресмыка-
ющихся.
Виды, попавшие в условия географической изоляции, дают начало вет-
вям филогенетического древа, все дальше отходящим от основного ствола
сначала на уровне микро-, а затем и макро эволюционной дивергенции.
Таким образом, формируются неповторимые черты биот, отличающиеся
не только по составу видов, но и родов, и семейств.
ГЛАВА 3. АРЕАЛОГИЯ
Т. Малыпус (1766—1834)
Каждый вид существует определенное время, его популяция растет,
увеличивается площадь распространения. Затем в силу разнообразных
причин происходит вымирание особей отдельных видов, их ареалы со-
кращаются, и, наконец, они исчезают. В одних случаях исчезновение вида
не прерывает хода эволюции, старый вид вытесняется новым; в других —
гибель вида обозначает слепую ветвь филогенетического древа.
Процесс формирования нового ареала,
как правило, монотопичен — возникнове-
ние вида происходит в одном месте, откуда
он начинает завоевывать пространство.
Новый вид происходит от одного предка
или в результате гибридизации. Эти поло-
жения являются ключевыми для понима-
ния истории формирования ареала.
В.И. Вернадский отметил огромную внут-
реннюю потенцию живого вещества к расте-
канию по земной поверхности. Он назвал это
явление давлением жизни. Оно выражается
во «всюдности» жизни, в захвате ею всякого
свободного пространства биосферы. Огром-
ная энергия давления жизни определяется
быстротой размножения. Одним из первых
количественную модель прогрессии размно-
жения разработал Томас Мальтус.
Вернадский приводит формулу геометрической прогрессии размно-
жения:
2" х D = N ,
где п — число дней с начала размножения; D — показатель прогрессии,
который для одноклеточных организмов, размножающихся делением,
соответствует числу поколений в сутки; N — число неделимых, суще-
ствующих благодаря размножению через п дней. Характерным для
каждого вида является D.
В этой формуле никаких пределов, никаких ограничений ни для л,
ни для D, ни для N не содержится. Процесс мыслится бесконечным, как
бесконечна сама прогрессия. Растекание живого вещества задержива-
ется только внешними силами. Оно замирает при низкой температуре,
при недостатке пищи, при отсутствии места для обитания, из-за конку-
ренции с другими организмами. Если нет внешних препятствий, вся-
кий вид в характерное для него время может благодаря размножению
покрыть весь земной шар, произвести потомство, равное по массе океа-
ну или земной коре (табл. 3.1).
Вернадский рассчитал теоретически возможные скорости растека-
ния жизни. Она колеблется от 33 100 см/с у бактерий, т. е. близка к
скорости звука, до долей сантиметра в секунду у крупных млекопита-
ющих— у индийского слона, например, 0,09 см/с.
Расселению организмов способствуют различные приспособления
их зачатков (диаспор): при половом размножении растений — спор, 5 9
ЧАСТЬ I. iЕГОРИЧЕСКИЕ К ГЕОГРАФИЧЕСКИЕ ФАКТОРЫ БИОЛОГИЧЕСКОГО РАЗНООБРАЗИЯ
Таблица 3.1
Скорость заселения поверхности Земли, по О.Е. Агаханянцу
Организмы Скорость заселения
Бактерии До 1,8 суток
Насекомые 203 — 366 суток
Цветковые растения (клевер) Около 11 лет
Рыбы (карп) Около 12 лет
Птицы (куры) Около 18 лет
Млекопитающие: крысы Около 8 лет
слоны Около 1000 лет
плодов, семян; при вегетативном — луковиц, клубней, корневищ, пол-
зучих побегов и т. д. У животных роль диаспор могут играть яйца, ли-
чинки, цисты.
Плоды и семена большинства водных, а равно и значительного коли-
чества наземных растений легко переносятся текучими водами, чем об-
легчается расселение растений вниз по течению рек. Многие плоды и
семена могут переноситься морскими течениями. Например, ареал коко-
совой пальмы (Cocos nucifera) охватывает берега тропических островов,
разбросанных по акваториям Тихого и Индийского океанов. Кокосовые
орехи, попадая в воду, разносятся морскими течениями. Они хорошо при-
способлены к длительным морским путешествиям. Своей плавучестью они
обязаны заполненной воздухом и непроницаемой для соленой морской
воды волокнистой наружной оболочке (рис. 3.3). Орехи не теряют способ-
ности к прорастанию после плавания в море в течение 3 — 4 месяцев. За это
время они могут быть перенесены на расстояние до 5000 км.
GO
Рис. 3.3. Кокосовая пальма, по «Жизни растений»: общий вид пальмы
на берегу океана и разрез плода, на котором видна толстая волокнистая
оболочка, обеспечивающая хорошую плавучесть ореха
ГЛАВА 3. АРЕАЛОГНЯ
Легкость спор и мелких семян множества видов обеспечивает воз-
можность переноса их на более или менее значительное расстояние
даже слабыми токами воздуха. У растений с более тяжелыми семенами
(или плодами) возможность переноса по воздуху часто возрастает бла-
годаря летательным приспособлениям — волосистым придаткам, разно-
го рода хохолкам, перепончатым лопастям, «крылышкам» и т. д., подчас
обладающим довольно сложным устройством (рис. 3.4).
Рис. 3.4. Плоды и семена, распространяемые ветром, по К. Шефферу:
I, 1а — одуванчик и его плоды; 2 — плоды чертополоха; 3 — семена сосны;
4 — плоды клена; 5 — соплодие липы; 6 — плоды вяза; 7 — плод граба;
8 — плоды березы
Перенос зачатков растений с помощью ветра может осуществлять-
ся не только по воздуху: опавшие на землю плоды и семена, при нали-
чии у них летучек, могут перегоняться ветром по поверхности почвы;
семена деревьев, опавшие зимой на затвердевшую поверхность снеж-
ного покрова, перегоняются, скользя по ней, особенно легко. Упав на
поверхность воды, снабженные летучками семена долго не тонут и под
напором ветра легко передвигаются по поверхности водоемов.
У многих растений семена или плоды переносятся животными бла-
годаря разного рода прицепкам, которыми они прикрепляются к шерс-
ти или другим покровам (рис. 3.5). То же может достигаться и при
помощи клейких выделений.
Семена растений, плоды которых поедаются животными, перено-
сятся последними в кишечнике и попадают на поверхность почвы с по-
метом. Перенос семян на значительные расстояния птицами играет В]
ЧАСТЬ I. iЕГОРИЧЕСПЕ И ГЕОГРАФИЧЕСКИЕ ФАКТОРЫ БИОЛОГИЧЕСКОГО РАЗНООБРАЗИЯ
Рис. 3.5. Плоды, распространяемые животными, по К. Шефферу:
1 — соплодие лопуха; 2 — побег подмаренника цепкого с плодами;
3 — одиночный плод череды
особо важную роль для расселения растений. Заметим, что вместе с эк-
скрементами в почву вносятся питательные вещества. А. Г. Воронов от-
мечает, что на сфагновых болотах, почти лишенных питательных ве-
ществ, семена голубики прорастают только в том случае, если они попали
сюда с экскрементами птиц. Семена многих растений обретают всхо-
жесть только после того, как пройдут через желудочный тракт живот-
ного. Бывает и так, что определенные животные делают для себя запа-
сы питательных семян, но используют их не полностью или просто
теряют часть своих «складов», которые тогда и превращаются в рассад-
ники растений соответствующего вида.
Большую роль в распространении растений играет человек. Напри-
мер, растения вводились в культуру первоначально в центрах своего
происхождения, а затем начинали культивироваться широко по земно-
му шару. Нередко в новых районах растения чувствуют себя даже луч-
ше и завоевывают большие территории, чем на родине. В качестве при-
меров можно привести плантации кофе (родина — Эфиопия), бананов
и цитрусовых (родина — Юго-Восточная Азия) в Южной Америке; кар-
тофеля, кукурузы, подсолнечника (родина — Центральная и Южная
Америка) в Европе и Азии. Многие вредные растения и сорняки разно-
G 2 сятся человеком по неосторожности.
ГЛАВА 3. АРЕАЛОГНЯ
Ярким примером формирования нового ареала является история
быстрого расселения в Европе американского по происхождению вод-
ного растения элодея (Elodea Canadensis). Она была завезена в Европу
в 1836 г. Далее ее распространение шло уже помимо воли человека
(рис. 3.6). Элодея так быстро заполняла водоемы, что это дало повод
назвать ее «водяной чумой».
Рис. 3.6. Ход расселения «водяной чумы»— элодеи (Elodea canadensis),
поА.И. Толмачеву
ЧАСТЬ I. ИСТОРИЧЕСКИЕ К ТЕОГРАФИЧЕСХИЕ ФАКТОРЫ БИОЛОГИЧЕСКОГО РАЗНООБРАЗИЯ
В способах распространения неподвижных или малоподвижных
животных много общего с растениями. Многие виды пауков летят по
воздуху благодаря выпускаемой ими паутинной нити. Ураганы и смер-
чи переносят насекомых и птиц. Сами животные разносят своих па-
разитов. Водоплавающие птицы на своих лапках переносят диаспоры
растений и беспозвоночных, земноводных и рыб, способствуя обога-
щению фауны водоемов.
Особенно велика роль морских течений в переносе диаспор живот-
ных. Многие донные беспозвоночные (моллюски, иглокожие, кишеч-
нополостные, черви, членистоногие) ведут сидячий образ жизни, при-
растая к субстрату, или передвигаются по дну в пределах ограниченной
площади. Однако их личинки проходят планктонную стадию развития
в поверхностной толще воды. Здесь они подхватываются течениями и
переносятся на многие тысячи миль.
У планктонных организмов развились замечательные приспособле-
ния для того, чтобы без лишних усилий держаться в толще воды. С помо-
щью различных выростов, щетинок они приобретают большую поверх-
ность по сравнению с массой тела (рис. 3.7). Кроме того, планктонные
организмы содержат капельки жира, которые одновременно служат
запасным веществом и облегчают массу тела в воде.
Рис. 3.7. Личиночные стадии морских беспозвоночных, проходящих
планктонную стадию развития, по Ж.П. Эрхарду и Ж. Сежену:
а —личиночные стадии морского ежа; б — личиночные стадии
многощешинкового червя; в —личиночные стадии краба
Подвижные формы животных расселяются активно. Два главных
фактора определяют их стремление к перемещению: инстинкт про-
должения рода и поиски пищи. Для мдюгих птиц районы, богатые
пищей и пригодные для выведения птенцов в летний сезон, небла-
гоприятны для жизни в зимний. Это заставляет их совершать даль-
G4 ние перелеты (рис. 3.8).
ГЛАВА 3. АРЕАЛОГНЯ
Рис. 3.8. Маршруты перелетных птиц, по Н.А. Бобринскому:
а — главнейшие маршруты ориентированы с севера, от мест гнездования,
на юг, к местам зимовок; б — распространение и перелетные пути
американской популяции длиннохвостой крачки. Черные кружочки —
места гнездования, треугольники — места зимовок, стрелки — направле-
ние и пути пролета основной массы особей на зимовку
ЧАСТЬ I. ИСТОРИЧЕСКИЕ К ТЕОГРАФИЧЕСХИЕ НАПОРЫ БИОЛОГИЧЕСКОГО РАЗНООБРАЗИЯ
Также не совпадают районы размножения и питания многих мор-
ских рыб, пресмыкающихся (рис. 3.9). Дальние миграции совершают
киты; регулярные странствования осуществляют ластоногие от райо-
нов нагула на строго определенные места размножения.
Рис. 3.9. Пути миграции трески, угрей и морских черепах, по С. Нешибе:
1 — области питания; 2 — районы нереста; 3 — район гнезд черепах;
4 — пути миграции; 5 — поверхностные океанические течения
Непреднамеренное распространение животных человеком подчас
приносит большой вред. Особенно это касается разного рода вредите-
лей и паразитов. Спутниками человека являются домовые мыши, кры-
сы, тараканы и пр. Колорадский жук, завезенный из Америки, вредит
посадкам картофеля в Европе. Европейский кролик, попавший в Авст-
ралию в 1840 г. в количестве 16 особей, вскоре размножился там настоль-
ко, что превратился в настоящее бедствие. Он стал серьезным конку-
рентом на пастбищах как местным травоядным, так и разводимым
фермерами овцам.
И 3.2. Географические и генетические элементы биоты__________________
Виды организмов, совокупность которых образует биоту, различны
в отношении их географического распространения и происхождения.
Поэтому при характеристике биоты важным аспектом исследования
является изучение закономерностей распространения видов и опреде-
G G ление мест их вероятного происхождения.
ГЛАВА 3. АРЕАЛОГНЯ
В соответствии с особенностями распространения видов выделяют-
ся географические элементы биоты. Ареалы географических элементов
в основном формируются в пределах зональных типов ландшафтов или
физико-географических регионов, обладающих единством палеогеогра-
фического развития (рис. 3.10).
Арктический (с rd')
7хт
atlId)
(ail)
all - medit
еитй
tniru
t>Or (ar) ppr <₽>
Субйорерлсный
tpeOuietfHi
mean (’*')
гкий (medit)
mMit (Ю
Понтический
fpo)
""Тиранский
Puc. 3.10. Европейские географические элементы флоры, по Г. Вальтеру:
а — схема основных географических элементов флоры Средней Европы;
б — распространение важнейших географических элементов флоры Евро-
пы. Обозначения элементов те же, что на схеме: sm — субсредиземномор-
ский элемент; submi — среднеевропейский + среднерусский элементы;
aid — арктический; Ьог (w) — бореальный западный; Ьог (о) — бореальный
восточный; atl (п) — атлантический северный; atl (s) — атлантический юж-
ный; ро— понтический; tur— туранский; medit (w)— западносредиземно-
морский; medit (о) — восточносредиземноморский. Горы не приняты во
внимание. Без обозначений — две сухие области: паннопская в Венгрии
и бассейн Эбро в северо-восточной Испании (небольшая сухая область
в юго-восточной Испании не очерчена)
ЧАСТЬ I. ИСТОРИЧЕСКИЕ К ТЕОГРАФИЧЕСХИЕ ФАКТОРЫ БИОЛОГИЧЕСКОГО РАЗНООБРАЗИЯ
Выяснение того, откуда происходят виды, как они попали в состав
изучаемой биоты, предполагает выделение генетических элементов.
Прекрасным примером такого анализа является заключение известно-
го русского ботаника А.Н. Краснова о связи тундровых кустарничков
с тропической растительностью гор Юго-Восточной Азии: «...близость
флоры горных вершин Явы к столь противоположным по свойствам
тундрам севера представляет громадный интерес, т. к. она показывает,
что флора тундры может быть непосредственным производным флоры
тропического леса» (Краснов, 1987).
Любая биота состоит из многих географических элементов. Напри-
мер, АА. Гроссгейм приводит следующее соотношение элементов фло-
ры Кавказа (в %).
Кавказские (собственно) — 20,0
Бореальные (северные) — 38,2
Средиземноморские — 13,2
Колхидские — 12,7
Переднеазиатские — 9,7
Понтические — 5,6
Туранские — 0,1
Пришлые виды — 0,5
Данный спектр элементов наглядно демонстрирует их роль в фор-
мировании флоры. Широкое участие бореальных элементов в биотах
отражает внедрение северных видов на Кавказ в ледниковую эпоху.
Свой вклад в обогащение флоры внесли соседние регионы. Однако
весьма значительна доля собственно кавказских видов, свидетельству-
ющая о том, что Кавказ представляет собой самостоятельный центр
видообразования.
И 3.3. Роль естественных барьеров________________________________
Географическое распространение видов тесно связано с их экологи-
ческой пластичностью. По способности выдерживать колебания основ-
ных экологических факторов организмы делятся на эврибионтные и
стенобионтные. Первые способны жить в условиях широкой амплиту-
ды экологических факторов, вторые выносят их изменения лишь в уз-
ком диапазоне. Соответственно эврибионтные виды способны стать
эврихорными, сформировать широкий географический ареал. Стено-
бионтные виды являются стенохорными и имеют узкие ареалы.
Заметим, что эврихорность свойственна только немногим видам-
эврибионтам. Сообщества организмов всегда более или менее стено-
хорны. Наиболее широким ареалом обладают зональные типы сооб-
ществ: дождевые тропические леса, саванны, степи, тайга и т. д. В виде
островков определенные сообщества могут заходить в соседние зоны
(явление экстразональности), но никогда вне своей зоны они не будут
доминирующими: байрачные дубовые 'леса в степной зоне, участки
широколиственных лесов в южной тайге, острова лугов в широколист-
G8 венных лесах.
ГЛАВА 3. АРЕАЛОГИЯ
На пути неограниченного увеличения численности популяций, дав-
ления жизни и стремления расширить ареал встают внешние факторы:
географические, ландшафтно-экологические и биологические.
В качестве географических факторов выступают крупные элемен-
ты строения земной поверхности, играющие роль преград на пути рас-
селения организмов. Для сухопутных растений и животных такими
преградами являются горные хребты, океаны и моря; для водных —
обширные пространства суши.
Некоторые физико-химические параметры внешней среды, играя
роль экологических факторов, в то же время выступают в качестве круп-
ных географических барьеров. На первом месте стоит распределение теп-
лоты как фактора, определяющего своеобразие биот широтных зон. На-
пример, преградой на пути распространения деревьев на север в основном
является изотерма самого теплого месяца около 10 °C. Ниже этой темпе-
ратуры деревья, как правило, расти не могут. Это одна из причин без-
лесья тундры. На втором — количество осадков и их сезонное распреде-
ление. Соотношение тепла и влаги оставляет свой отпечаток на зональных
типах биот: виды тропических лесов, живущие в условиях теплого и
влажного климата, не расселяются в жаркие и сухие пустыни. Наряду с
температурой роль географических барьеров в океане играют измене-
ния солености воды. Отклонение солености от средней океанической
(35 %о) в сторону опреснения является барьером на пути проникновения
многих морских организмов в солоноватые и пресные воды (рис. 3.11).
Рис. 3.11. Качественное обилие средиземноморской фауны и флоры и их обед-
нение с проникновением на восток, по Л.А. Зенкевичу: 1 — средиземномор-
ская биота; 2 — каспийская биота; 3 — пресноводная биота; 4 — арктические
иммигранты. Цифрами показано количество средиземноморских видов
Существуют организмы, сама природа которых ущербна с точки зре-
ния потенции к завоеванию пространства. Другие, напротив, практиче-
ски не имеют ограничений к расселению в разные жизненные среды.
Так, у иглокожих отсутствует механизм осморегуляции, поэтому их ареал
ограничен океаническими водами с полной морской соленостью. Члени-
стоногие же, обретя способность регулировать или защищать внутри-
клеточное давление, смогли завоевать океан, пресные воды и сушу. G 9
ЧАСТЬ I. ИСТОРИЧЕСКИЕ К ТЕОГРАФИЧЕСХИЕ ФАКТОРЫ БИОЛОГИЧЕСКОГО РАЗНООБРАЗИЯ
С ландшафтно-экологическими факторами связана внутренняя
структура ареала. Ни один вид не образует сплошного покрова. Внутри
ареала организмы приурочены только к определенным экологическим
условиям (местообитаниям). Характер местообитаний контролируется
структурой ландшафта. Например, географические ареалы болотных
мхов, трав и кустарничков (сфагнума, пушицы, клюквы и др.) и видов
таежного мелкотравья и кустарничков (майника, кислицы, черники и
др.) в пределах тайги совпадают. Однако первые приурочены к верхо-
вым болотам, а вторые к более сухим местообитаниям, занятым темно-
хвойными зеленомошными лесами. Таким образом, экологическая
структура ареалов названных видов образует сложное кружево, рису-
нок которого зависит от распределения морфологических единиц ланд-
шафта — верховых болот и зеленомошных лесов. Более полно пеструю
картину распределения таежных видов внутри ландшафта передает
профиль (рис. 3.12). Четко видно, что ландшафтно-экологические комп-
лексы контролируют характер местообитаний так, что определенные
виды образуют более плотные популяции там, где условия жизни для
них благоприятны. Это находит отражение прежде всего в распределе-
нии показанных на профиле растительных сообществ.
Рис. 3.12. Профиль, иллюстрирующий комбинации растительности и почв
на озерно-ледниковом моренном рельефе в Карелии, по Б.В. Виноградову:
1 — березняки злаково-разнотравные на подзолах маломощных на валунных
супесях; 2 — ельники чернично-долгомошные на подзолах среднемощных
на озерно-ледниковых суглинках; 3 — ельники-брусничники на подзолах
на моренных супесях; 4 — ельники-долгомошники на торфянисто-подзолис-
тых почвах на озерно-ледниковых суглинках; 5 — ельники-черничники на
подзолах среднемощных на моренных супесях; 6 — сосняки-брусничники на
подзолах песчаных на флювиогляциальной морене; 7 — сосняки осоково-
сфагновые на торфянистых почвах; 8 — ельники осоково-сфагновые на
торфянистых почвах болот переходного типа; 9 — сосняки сфагновые на
торфяных болотах верхового типа; 10 — березняки черничные на подзолах
среднемощных супесчаных на моренных супесях
В роли биологических барьеров выступают конкурентные отноше-
ния. На пути расселения вида встают не только географические и ланд-
шафтно-экологические преграды, но и сопротивление, которое встре-
чает новосел со стороны уже существующих видов. Чтобы завоевать
пространство, новый вид должен победить их в борьбе за существова-
ние. Расширению ареала противостоят не только отдельные организ-
ГЛАВА 3. АРЕАЛОГИЯ
мы, но и целые сообщества. Например, на границе леса и степи внедре-
нию деревьев в степное сообщество препятствует конкуренция с мощ-
ной дерниной степных трав.
И 3.4. типы ареалов
Первоначальную область распространения вида называют центром его
происхождения. Наряду с этим выделяют ту часть ареала, в которой на-
блюдается максимальная изменчивость данного вида — наибольшее ко-
личество географических форм в пределах вида, видов и подвидов в пре-
делах рода и т. п. Эту часть ареала называют центром многообразия форм
(рис. 3.13).
Рис. 3.13. Количественное распределение видов коровяка
(род Verbascum), по Ж. Леме. Цифры указывают на количество видов,
находящихся внутри зон, ограниченных линиями.
Отчетливо выделяется центр многообразия форм
Для возникновения центра многообразия форм требуется время и
подходящие условия. Как правило, такой центр тяготеет к более или
менее древней части ареала. Если он совпадает с центром происхожде-
ния вида, значит, на протяжении его истории условия существования
были благоприятными. Но вместе с тем современный центр.многообра-
зия форм того или иного вида может не совпадать с центром его про-
исхождения. Так, центр происхождения предковой формы дарвинов-
ских вьюрков лежит на материке, однако наибольшее разнообразие
видов наблюдается на Галапагосских островах.
Границы ареала могут быть статичными, расширяющимися или
сужающимися. В первом случае устойчивость границ объясняется тем,
что вид достиг естественных границ своего ареала. Границы ареала рас-
ширяются, когда молодой вид обладает потенцией к преодолению гео-
графических, ландшафтно-экологических и биологических преград. Гра-
ницы ареала сужаются, когда в силу изменения вышеназванных условий
вид теряет завоеванную территорию (акваторию). ] 1
ЧАСТЬ I. iЕГОРИЧЕСКИЕ К ГЕОГРАФИЧЕСКИЕ ФАКТОРЫ БИОПОГИЧЕСИОГО РАЗНООБРАЗИЯ
При типизации ареалов следует учитывать их размеры, непрерыв-
ность (континуальность) и прерывистость (дизъюнкции). У наземных
организмов выделяются ареалы четырех уровней размерности: конти-
нентальные, провинциальные, региональные и локальные. Ареал конти-
нентального типа охватывает большие площади одного или нескольких
континентов; провинциального — занимает большую часть физико-гео-
графической или биогеографической провинции; ареал регионального
типа ограничен площадью физико-географического или биогеографиче-
ского района; локальный тип совпадает с внутриландшафтными морфо-
логическими комплексами (рис. 3.14).
72
Рис. 3.14. Основные типы ареалов на суше, по Р. Dansereau.
Объяснения в тексте
ГЛАВА 3. АРЕАЛОГНЯ
Континуальное (сплошное) распространение обозначено на рисунке
нечетными номерами (1, 3, 5, 7), дизъюнкции ареалов — четными (2, 4, б).
Другие типы распространения обозначены буквами А, Б, В. Тип А харак-
теризуется сплошным распространением по всей площади; тип Б состоит
из одного большого и одного или нескольких небольших участков, связан-
ных между собой; тип В объединяет множество мелких участков.
Рассмотрим иллюстрации. Виды, распространенные на всех конти-
нентах, называют космополитами (что означает «распространенный по
всему миру»). Ареал, близкий к типу 1А, имеет папоротник-орляк
(рис. 3.15). Космополитами являются многие сорняки, связанные в сво-
ем распространении с деятельностью человека: мокрица (Stellaria media),
крапива (Urtica dioica), одуванчик (Taraxacum officinale). Однако, строго
говоря, контрастность экологических условий на Земле не позволяет ни
одному виду растений или животных, кроме человека, иметь истинно
космополитическое распространение.
Рис. 3.15. Ареал папоротника-орляка (Pteridium aquilinum), поА.И. Толмачеву
Возникновение разорванных ареалов объясняется событиями прош-
лых геологических эпох: исчезновением мостов-переходов на суше,
дрейфом континентов, горообразованием, изменениями климата и т. п.
Ареал вида может быть расчлененным и занимать обширные области
на разных континентах, тип 2А (рис. 3.16).
Ареал таежного кустарничка Linnea borealis в основном сплошной,
занимающий значительную часть Евразии. Наряду с этим имеются его
небольшие обособленные островки в различных местах Средней и Вос-
точной Европы, на Кавказе, на юге Западной Сибири, на Дальнем Вос-
токе (рис. 3.17). Следовательно, такой ареал можно отнести к типу 2Б.
Рис. 3.16. Ареал кизила шведского (Cornus suecica), по А.И. Толмачеву
ЧАСТЬ 1.1
со
Рис. 3.17. Ареал таежного кустарничка Linnaea borealis в пределах Евразии, по А.И. Толмачеву
Ы1
ЧАСТЬ I. ИСТОРИЧЕСКИЕ К ТЕОГРАФИЧЕСХИЕ НАПОРЫ БИОЛОГИЧЕСКОГО РАЗНООБРАЗИЙ
Рассмотрим историю формирования разорванных ареалов на следу-
ющих примерах. Дизъюнкция ареала рода Nepentes объясняется раско-
лом в мезозое материка Гондвана. В результате некогда сплошной ареал
оказался разорванным. Ныне виды Nepentes характерны для далеко
отстоящих флор Мадагаскара и Юго-Восточной Азии (рис. 3.18).
Рис. 3.18. Разорванный ареал рода Nepentes, по R. Good:
а — общий вид растения; б — черным цветом залит ареал
В верхних поясах гор Центральной и Южной Европы имеются виды,
которые отсутствуют на равнинах, но встречаются в Арктике. В данном
случае разорванность ареала объясняется похолоданием климата в плей-
стоцене. Во время ледниковой эпохи в центре Европы царил холодный
климат, и она давала приют растениям и животным, характерным для
ландшафтов тундры. В ходе потепления в голоцене эти виды остались в
арктических ландшафтах, а южнее были вытеснены в высотные пояса гор,
где они смогли выжить и стали ледниковыми реликтами. Так, заяц-беляк,
основной ареал которого охватывает тундровые и таежные ландшафты,
живет обособленно в Альпийских горах (рис. 3.19). По предложенной выше
типологии данный тип ареала можно обозначить индексом 4Б. Этими же
причинами вызван европейско-дальневосточный разрыв ареалов широко-
лиственных лесов. Иллюстрацией может служить ареал спутника широко-
лиственных лесов Hepatica nobilis (рис. 3.20); тип ареала 4А.
Рис. 3.19. Разорванный арктоальпийский ареал зайца-беляка в Европе,
по Н.А. Бобринскому
Рис. 3.20. Разорванный европейско-дальневосточный ареал Hepatica nobilis, по А.И. Толмачеву
ЧАСТЬ I. ИСТОРИЧЕСКИЕ К ТЕОГРАФИЧЕСХИЕ ФАКТОРЫ БИОЛОГИЧЕСКОГО РАЗНООБРАЗИЙ
Чтобы подчеркнуть определенные особенности ареалов континен-
тальной размерности, используют дополнительные термины. Ареалы
видов, приуроченных к арктическому поясу Северного полушария,
называются циркумполярными. Они образуют кольцо вокруг полюса
(рис. 3.21). Ареал видов, охватывающих тропический пояс, называется
пантропическим (рис. 3.22).
Рис. 3.21. Пример широко распространенного, циркумполярного,
эндемичного для Арктики вида Draba subcapitata, поА.И. Толмачеву
Особый тип образуют биполярные ареалы. Это разорванные ареа-
лы, часть которых расположена в высоких широтах Северного, а часть
в высоких широтах Южного полушария (рис. 3.23, 3.24). Возникнове-
нию биполярных ареалов способствовало-похолодание климата в лед-
никовую эпоху, когда холоднолюбивые виды севера смогли преодолеть
современный теплый пояс и проникнуть в высокие широты Южного
78 полушария.
ГЛАВА 3. АРЕАЛОГИЯ
Рис. 3.22. Пантропический ареал семейства пальмовых, по R. Good
Рис. 3.23. Биполярный ареал рода Empotrum, по R. Good
Провинциальные и региональные типы ареалов формируются внут-
ри континентов и биоклиматических зон. Например, для европейских
древесных пород давно уже установлено, что их распространение на
север и восток в наибольшей степени регулируется зимними темпера-
турами. Зависимость эта еще ярче отражается в очертаниях ареала
европейского падуба (Лех aquifolium), входящего в состав кустарнико-
вого яруса широколиственных лесов (рис. 3.25); тип ареала 5А.
Примером разорванного ареала региональной размерности (тип 6В)
может служить распространение норвежской полыни (рис. 3.26).
Ареалы локальной размерности (тип 7В) характерны для молодых
видов, освоивших специфичные местообитания, например дарвинов-
ских вьюрков. Или для реликтовых видов, сильно сузивших свой ареал
(например, ареал сейшельской пальмы ограничен долиной на одном из
островов Сейшельского архипелага). 19
ЧАСТЬ I. iЕГОРНЧЕСПЕ К ГЕОГРАФИЧЕСКИЕ ФАКТОРЫ БИОПОГИЧЕСИОГО РАЗНООБРАЗИЯ
Рис. 3.24. Биполярное распространение ламинариевых водорослей,
по Т.Ф. Щаповой
Рис. 3.25. Распространение европейского падуба (Ilex aquifolium) в Европе,
по А.И. Толмачеву: восточной границей служит январская изотерма О °C
ГЛАВА 3. АРЕАЛОГНЯ
Рис. 3.26. Разорванный ареал норвежской полыни (Artemisia norvegica),
поА.И. Толмачеву
И 3.5. Эндемики и эндемизм
Эндемики — это виды (роды, семейства, любые таксоны), свойствен-
ные только данной территории (акватории). Эндемики могут иметь
локальный ареал — встречаться только в одном конкретном местооби-
тании: в межгорной котловине или в определенном высотном поясе
горного массива и т. п. Однако территория ареала эндемика может за-
нимать и большую площадь. Например, существуют эндемики Южной
Америки, Австралии (рис. 3.27, 3.28). В открытой части озера Байкал
фауна на 60% эндемична. Полностью эндемичны в Байкале И семейств
и подсемейств, 96 родов, объединяющих почти 1000 видов.
Рис. 3.27. Характерный эндемичный род каучукового дерева (Hevea),
занимающий лесную область долины Амазонки, поА.Й. Толмачеву
81
ЧАСТЬ I. iЕГОРНЧЕСПЕ I ГЕОГРАФИЧЕСКИЕ ФАКТОРЫ БИОЛОГИЧЕСКОГО РАЗНООБРАЗИЯ
Рис. 3.28. Эндемичное панматериковое распространение рода Eucalyptus,
по R. Good
Возможны две модели формирования эндемизма. Реликтовый энде-
мизм означает сохранение вида с геологически давних времен, когда
условия для него были благоприятными, а ареал обширным. Следователь-
но, ареал реликтового эндемика находится в стадии сокращения, а сам
таксон стал эндемичным именно из-за этого уменьшения. В отличие от
реликтового молодой эндемизм связан с возникновением популяции
нового вида, поначалу занимающего ограниченный ареал. Позже, рас-
пространившись, молодой эндемик может утратить свою уникальность.
Принципиальные различия между ареалами реликтового и молодого
эндемиков иллюстрирует схема (рис. 3.29).
Рис. 3.29. Принципиальное различие между ареалами реликтового (а)
и молодого (б) эндемика, по О.Е. Агаханянцу.
Стрелки указывают состояние ареала: его сокращение (а) или расширение
(б). В первом случае вид уже эндемик (а), в другом — еще эндемик (б)
В качестве примера реликтового эндемизма рассмотрим историю
возникновения и распространения слонов Laxodonta, Mammuthus и
Elephas (рис. 3.30). Они произошли от общего предка в Центральной
Африке, откуда распространились в Европу, Азию и Северную Амери-
ку. Сегодня как реликты сохранились только два вида — африканский
82 слон (Laxodonta africand) и индийский слон (Elephas maximus).
ГЛАВА 3. АРЕАЛ ОГНЯ
Рис. 3.30. История формирования ареалов древних и современных слонов,
noJ.H. Brown, А. С. Gibson
Непременное условие существования и сохранения эндемизма —
изоляция. И чем дольше сохраняется изоляция, тем выше доля (процент
от числа таксонов) эндемизма во флоре и фауне и тем своеобразнее биота.
Неудивительно поэтому, что особенно велика доля эндемиков на остро-
вах, в высотных поясах гор. Например, на равнинах России раститель-
ных эндемиков нет совсем, т. к. виды могут мигрировать вдоль тепловых
поясов очень широко. Но в горах и на островах эндемизм высокий, и
чем древнее их изоляция, тем выше эндемизм (Global Biodiversity, 1992;
Агаханянц, 1992).
Кавказ — 25%
Горы Средней Азии — 30%
Япония — 37%
Канарские острова — 45%
Корсика — 58%
Мадагаскар — 68%
Новая Зеландия — 81%
Австралия — 80%
Гавайские острова — 82%
Остров Святой Елены — 79%
Эндемизм в высотных поясах гор объясняется экологическими при-
чинами — вид не может мигрировать ни вверх по склонам (слишком
для него холодно), ни вниз (слишком сухо или жарко) и остается в
пределах своего пояса. Чем строже экологическая изоляция, тем выше
доля эндемизма в высотных поясах. Известны виды, встречающиеся на
одной только горе: эльдарская сосна (Pinus eldarica) растет в Грузии лишь
на площади около 50 га, камчатская пихта (Abies gracilis} образует одну
рощу на восточном берегу Камчатки.
ЧАСТЬ I. ИСТОРИЧЕСКИЕ К ТЕОГРАФИЧЕСХИЕ ФАКТОРЫ БИОЛОГИЧЕСКОГО РАЗНООБРАЗИЯ
Экологическая изоляция возможна и на равнинах. Узкое субтропи-
ческое влажное побережье Южной Африки, изолированное на севере
от тропиков пустынями, имеет столь высокую долю растительного эн-
демизма (7 семейств, 210 родов), что дает основание рассматривать эту
территорию в ранге самостоятельного флористического царства (Кап-
ское царство). В юго-западной Австралии, тоже изолированной пусты-
нями, отмечена самая высокая плотность растительных эндемиков.
И 3.6. Викариат и конворгонция
Викариат в переводе с латинского означает «замещение». В биогео-
графии викаризм — это замещение одних видов другими, образовавши-
мися из одного корня, т. е. родственными, но развившимися самостоя-
тельно в разных экологических условиях. Викариат отражает процесс
дивергентной микро эволюции, когда обособление нового вида являет-
ся следствием адаптации организмов к новым условиям, отличным от
тех, в которых обитал породивший их старый вид. Классический при-
мер — ряд лиственниц в Евразии: европейская — сибирская — Гмелина
(даурская) — Каяндера — камчатская, каждая из которых занимает свой
ареал (рис. 3.31).
Рис. 3.31. Ареалы викарирующих восточносибирских
и дальневосточных видов лиственниц, по Е.Г. Боброву:
1 — Каяндера; II — Гмелина; III — Рупрехта;
IV— Ольги; V— камчатская; VI— японская
84
ГЛЙВЙ 3. ДРЕЙЛРГИЯ I
Такой же ряд образуют сосны — бореальные, средиземноморские,
гималайские, дальневосточные. Чаще наблюдаются викарные пары
видов: береза извилистая и карликовая, галка обыкновенная и альпий-
ская, тюлень каспийский и нерпа, медведь бурый и гималайский и т. д.
Но вот белый медведь и бурый не составляют викарной пары, они прои-
зошли от разного корня (Агаханянц, 1992).
Эволюция организмов протекает не только путем расхождения (ди-
вергенции), но и путем схождения (конвергенции) признаков. В по-
следнем случае в сходных условиях могут формироваться виды, очень
далекие в филогенетическом отношении, но похожие по своей орга-
низации и внешнему виду (рис. 3.32). Как пример конвергенции мож-
но привести акул из рыб, ихтиозавров из пресмыкающихся, дельфинов
из млекопитающих. В результате приспособления к сходным условиям
Рис. 3.32. Адаптивная радиация и конвергенция жизненных форм
пресмыкающихся и плацентарных млекопитающих
85
\ЧАСТЬ 1.1СТОРНЧЕСККЕI ГЕОГРАФИЧЕСКИЕ ШТОРЫ БИОПОГИЧЕБКИГО РйЗНООБРйЗИН
существования их тело, снабженное хвостовым и спинным плавника-
ми, приобрело веретенообразную хорошо обтекаемую форму и другие
черты строения, делающие их очень похожими друг на друга. Это сле-
дует учитывать при определении эндемов. Так, разнообразные формы
сумчатых животных (сумчатые волк, куница, крот, белка, медведь и др.)
отнюдь не сближают Австралийское царство с Голарктическим, для
которого характерны плацентарные волки, куницы, кроты, белки, мед-
веди, а только подчеркивают уникальность его фауны.
Часть II
ЭКОЛОГИЧЕСКИЕ
И ЛАНДШАФТНЫЕ ФАКТОРЫ
БИОЛОГИЧЕСКОГО РАЗНООБРАЗИЯ
в сфере биологических наук,
Эрнст Геккель (1834—1919)
Экология — наука, изучающая взаимоотношения организмов и об-
разуемых ими сообществ между собой и с окружающей средой.
В особую дисциплину экологию выделил в 1866 г. профессор Йен-
ского университета, активный пропагандист учения Ч. Дарвина Эрнст
Геккель для применения исключительно
главным образом зоологии.
Слово экология происходит от грече-
ских корней «эйкос» — дом и «логос» —
учение. По-русски оно обозначает «уче-
ние о доме». Понятие «эйкос» (дом) трак-
туется различно, но обычно в весьма рас-
ширенном смысле — не как собственно
дом или жилище, а как среда обитания.
Конвенция ООН (Рио-де-Жанейро,
1992) провозгласила непреходящую цен-
ность биологического разнообразия, отме-
тив его большое значение для функцио-
нирования систем, поддерживающих как
жизнь в биосфере, так и устойчивое раз-
витие общества. Биосфера и географиче-
ская оболочка представляют собой
неразрывное целое. Раздел географии,
призванный изучать закономерности
размещения любых проявлений жизни-
на Земле, — биогеография — рассматри-
вает проблему биоразнообразия как одну из наиболее актуальных.
В процессе разработки концепции биологического разнообразия сло-
жилось представление о базовых единицах (Уиттекер, 1980): альфа-раз-
нообразии — разнообразии видов; бета-разнообразии — разнообразии
сообществ; гамма-разнообразии — разнообразии видов и сообществ в
пределах ландшафтов.
Опираясь на высказывания В.Б. Сочавы (1970), можно сформулиро-
вать положение, которое необходимо учитывать при определении кри-
териев гамма-разнообразия: пространственная структура биосферы
отображает пространственную неоднородность географической обо-
лочки; система геохор адекватна соответствующей ей системе био-
хор, а точнее, фитохор. Такой подход к изучению биоразнообразия
всецело обращен к биогеографии.
Пространственная структура и параметры биоты, устанавливаемые
при изучении ландшафтов, являются хорошими косвенными индика-
торами биоразнообразия на более низких структурных уровнях. Так,
обработка спутниковых данных позволяет классифицировать раститель-
ные сообщества и по ним судить о разнообразии входящих в ландшафт
видов растений и животных.
(часть II. ШЛРГКЧЕСКИЕ И ПЙНДШАЙГНЫЕ ЙДИТРРЫ БИОПОГИЧЕЕКОГО РАЗНООБРАЗИЙ
Несмотря на всю значимость биогеографических и экологических
проблем регионального и глобального масштабов, ключ к их понима-
нию, как правило, лежит на топологическом уровне. Ему отвечают в
биогеографии биотопы и биоценозы (биогеоценозы), в физической
географии — морфологические единицы ландшафта; в социально-про-
изводственной сфере — природные угодья, конкретные объекты хозяй-
ственного использования. По Д.Н. Кашкарову (1933), проекция на мес-
тообитание есть основная характеристика экологического изучения.
Чем более дробными будут выделяемые единицы, тем ближе удастся
подойти к установлению факторов, определяющих свойства сообществ.
Глава 4
ОСНОВНЫЕ ПОНЯТИЯ И ЗАКОНЫ ЭКОЛОГИИ
Способность к размножению и расселению, которой обладают виды,
позволила бы им равномерно распространиться по всей земной поверх-
ности, если бы на их пути не существовали преграды, воздвигнутые
условиями среды и совместного обитания с другими видами. Границы
ареала вида,, а также границы территорий, на которых вид локализован
внутри ареала, определяются тем, с какими факторами среды существо-
вание вида возможно, а с какими несовместимо. Важную роль в изуче-
нии механизмов динамики биоразнообразия и закономерностей рас-
пределения организмов играют экологические исследования.
И 4.1. Организм и среда
Для жизни и процветания каждого организма требуется набор опреде-
ленных экологических факторов: абиотических, т. е. неживых, и био-
тических, т. е. живых.
Абиотические факторы. У наземных организмов к числу важнейших
прямодействующих абиотических факторов относятся климатические
(свет, тепло, влага), физико-химические свойства почвогрунтов, газы
атмосферы. Такой фактор, как рельеф, является косвенно действующим,
т. к. он перераспределяет потоки вещества и энергии в ландшафте.
Правило предварения сформулировал В.В. Алехин. По этому прави-
лу склоны северной экспозиции несут на себе растительные группи-
ровки, свойственные более северной растительной зоне, а склоны
южной экспозиции — растительные группировки, характерные для
более южной зоны. Это отклонение от зональности связано с углом
падения солнечных лучей (рис. 4.1).
Рис. 4.1. Схема правила предварения, по В.В. Алехину:
1 — северный вид, обитающий на плакоре, на юге переходит на северные
склоны и в балки; 2 — южный вид на севере встречается на южных склонах
ЧАСТЬ II. ЭКОЛВГКЧЕЕКИЕ И ЛЙНДШАЙГВЫЕ ФАКТОРЫ БИОЛОГМЧЕЕКОГО РАЗНООБРАЗИЯ
Законы ограничивающих факторов. Отношение разных видов ра-
стений и животных к условиям существования неодинаково: одним
требуется много влаги, другие живут в сухой пустыне, одни требуют
много света, другие его не выносят, одни любят тепло, другие — низ-
кие температуры и т. п.
Ход жизни организмов не нарушается, если все факторы действуют
в привычном для организма режиме. Однако картина резко меняется,
если величина одного из факторов начнет уменьшаться. Жизненность
организма будет снижаться пропорционально фактору, градиент кото-
рого стремится к минимуму.
Рис. 4.2. Модель, иллюстри-
рующая действие закона
минимума
Идея о том, что выносливость организ-
ма определяется самым слабым звеном в
цепи его экологических потребностей,
впервые была высказана Ю. Либихом и
получила название закон минимума.
Закон минимума: наибольшее влияние
на жизнь организма оказывает фактор,
напряжение которого наиболее низкое (ми-
нимальное) и приближается к пределу вы-
носливости. Действие закона минимума
наглядно показано на рис. 4.2. Высота кле-
пок бочки соответствует напряженности
экологических факторов, жидкость в боч-
ке — «жизненной силе». Как видим, «жиз-
ненная сила» вытекает через самую низ-
кую клепку, т. е. там, где напряженность
экологического фактора минимальна.
Ограничивающим процветание вида может оказаться не только
недостаток (минимум), но и избыток (максимум) фактора. Существо-
вание организма ограничено областями минимума и максимума. Меж-
ду ними располагается наиболее благоприятная (оптимальная) эко-
логическая область — экологический оптимум. Представление о
лимитирующем влиянии максимума наравне с минимумом ввел
В. Шелфорд, сформулировавший закон толерантности.
Закон толерантности: лимитирующим может быть как минималь-
ное, так и максимальное значение экологического фактора; диапазон
между минимумом и максимумом определяет область выносливости
(толерантности) организма к данному фактору.
Модель толерантности, как правило, имеет вид купола (рис. 4.3). Его
центральная часть соответствует напряженности экологического фак-
тора, при которой условия существования организма наилучшие, — это
область оптимума. Края купола отвечают или слишком низкой, или
слишком высокой напряженности экологического фактора, при кото-
рой условия существования организмов наихудшие.
Чтобы выразить степень толерантности, в экологии используют
приставку «стено» (узкий) и «эври» (широкий). Существуют стеноби-
онтные и эврибионтные виды. Первые живут в условиях узкого, вто-
рые — широкого диапазона фактора (рис. 4.4). Например, у холодолю-
бивых организмов зона температурного оптимума смещена в сторону
ГЛАВА 4. ОСНОВНЫЕ DОНЯТ1Я И ЗАКОНЫ ЭКОЛОГИИ
Рис. 4.3. Купол толерантности: существование вида определяется его вы-
носливостью по отношению к воздействию абиотических факторов.
Когда напряжение фактора слишком низкое или высокое, вид гибнет
Рис. 4.4. Типы экологических диапазонов, поЮ.И. Чернову:
А — эврибионты; Б — стенобионты; а — эврибионты с оптимумом в средней
части градации фактора; б, в — эврибионты с оптимумом в зонах низких
и высоких значений фактора; г, д, е — варианты стенобионтов
низких температур. Диапазон температур, в пределах которого могут
существовать некоторые антарктические виды рыб, составляет всего
4°С (от —2 до 2 °C). У теплолюбивых видов зона оптимума смещена в
сторону высоких температур. Коралловые рифы развиваются в морских
водах при температуре не ниже 20 °C.
Законы минимума и толерантности служат путеводной нитью для
понимания способов, позволяющих организмам переносить недостаток
или избыток определенных факторов: у организмов в процессе борьбы
за существование и естественного отбора вырабатываются приспособ-
ления, позволяющие им жить в условиях, которые поначалу были для
них неблагоприятными.
ЧАСТЬ II. ЭКРЛОГИЧЕСКИЕ И ЛДНЛШДЙГ8ЫЕ (ШШЫ БИОЛОГМЧЕБКОГО РДЭНООБРДЗИЯ
К числу важнейших экологических факторов, определяющих зональ-
ные типы биомов, относятся температурный режим и количество ат-
мосферных осадков. Диапазон значений этих факторов на Земле огро-
мен. И несмотря на существенные различия, каждому градиенту тепла
и влаги отвечает специфический набор видов (рис. 4.5).
Рис. 4.5. Зависимость зональных типов биомов от соотношения темпера-
турного режима и количества атмосферных осадков:
Термические пояса: А — холодный; Б — умеренный; В — жаркий
Зональные типы биомов: А1 — полярные пустыни; А2 — тундра; АЗ — тайга;
Б1 — пустыни умеренного пояса; Б2 — степи; БЗ — широколиственные леса;
В! — пустыни жаркого пояса; В2— саванны; ВЗ— тропические леса
Казалось бы, характер воздействия экологических факторов проще
всего проследить у растений. Однако именно на этом примере можно
показать, насколько тонко и многообразно связаны организмы с окру-
жающей средой. Оказывается, растения как бы имеют слух, зрение,
осязание и обоняние.
Экспериментально доказано, что растения реагируют на изменение
магнитного поля, на музыкальные звуки. Растения «любят» классиче-
ские произведения, тянутся к источнику мелодичных звуков и под их
влиянием быстрее растут и лучше цветут, богаче плодоносят; ритмы
жесткого рока их угнетают. Под влиянием мелодичной музыки в два
раза увеличивается поглощение СО2 (повышается интенсивность фото-
синтеза), обогащается биохимический состав. Например, ростки пше-
ГЛАВА 4. ОСНОВНЫЕ DОНЯТ1Я И ЗАКОНЫ ЭКОЛОГИИ
ницы, выращенной под звуки классической музыки, содержали в 20 раз
больше витамина А и в 5 раз больше витаминов С и В6, чем в обычных
условиях.
Жизненные формы: в разных зонах можно выделить адаптивные
группы видов, наиболее характерные для соответствующих типов био-
мов (тундровые, таежные, неморальные, степные, пустынные и т.д.),
или группы видов, отражающие связь с конкретными местообитания-
ми: «луговые», «болотные», «прибрежные» и т. п. Все эти группы объ-
единяются понятием жизненная форма.
Жизненная форма — это прежде всего тип внешней организации,
отражающей важнейшие моменты образа жизни, отношения вида к
среде. В характеристику жизненной формы животных должны вклю-
чаться, например, особенности движения, формы тела, способа добы-
чи пищи, отношения к субстрату и т. д. (жизненные формы растений
будут рассмотрены в гл. 5).
Типы жизненных форм — объект экологической классификации,
строящейся на основе конвергентного сходства приспособительных осо-
бенностей. Такая классификация, как правило, не согласуется с система-
тикой, главный принцип которой — филогенетическая близость.
Подчас адаптивные признаки накладывают характерный отпечаток
на весь облик организма, так что по внешнему виду можно судить, в каких
именно условиях данный организм обитает. Как правило, влияет не один,
а сразу целый комплекс экологических факторов. Например, облик бело-
го медведя позволяет безошибочно судить, что он житель снежной Арк-
тики, а облик верблюда — что он обитатель жаркой пустыни; саксаул
растет в пустыне, а банан — во влажном тропическом лесу.
Приведем примеры жизненных форм рыб, свойственных двум раз-
ным биотопам мелководий Черного моря: скал и камней и подводных
песчаных равнин (рис. 4.6, 4.7).
Типы связи + — 0
+ + + ч— + 0
— —h — - 0
0 0 + 0 - 00
Биотические факторы. Все многообразие биотических отношений
может быть представлено в виде матрицы.
Обе связи положительны у организмов, взаимно нуждающихся в
совместном проживании. Например, цветковые растения и насекомые-
опылители. Связь, положительная для одного организма, отрицательна
для другого; это наиболее распространенный тип связей питания:
растительноядные животные — растения, хищник — жертва.
Нейтральная для одного организма связь может быть положитель-
ной для другого. Например, стволы и кроны деревьев служат местом
обитания для множества птиц, других растений, которые не наносят
дереву вреда, .но сами получают значительное преимущество, находя в
кроне убежище или поднимаясь ближе к свету. 9 5
ЧАСТЬ IL ЭКОЛВГИЧЕЕКИЕ И ЛАНДШАЙГВЫЕ (ВДКТОРЫ БИОЛОГИЧЕЕЮГО РАЗНООБРАЗИЯ
Рис. 4.6. Рыбы скальных грунтов, по О.Ф. Хлудовой: 1 —морская ласточка
(Chromis chromis); 2 — смарида (Smarts smarts); 3 — горбыль (Corvina umbra);
4 — каменный окунь (Serranus scriba); 5,6 — зеленушки (Crentlabrus acellatus);
7 — зеленушка рулена (Crentlabrus rulaend); 8 — бычок (Gobius niger);
9 — морские собачки (Blennius); 10 — морской налим (Gaidropsarrus
mediterraneus); 11 — морской ерш (Scorpaena)
Обе связи отрицательные у организмов, находящихся в состоянии
конкуренции; особенно сильны конкурентные отношения у особей,
относящихся к одному виду (за исключением некоторых животных, у
которых в популяции существует не только конкуренция, но и отноше-
9 9 ния взаимозависимости и взаимопомощи).
ГЛАВА 4. ОСНОВНЫЕ О О НЯТ1Я П ЗАКОНЫ 3 КОЛОГМН
Рис. 4.7. Рыбы подводных песчаных равнин, по О.Ф. Хлудовой:
1 — песчанка (Ammodvites cicerelius); 2 — морской язык (Soles lascaris);
3— ошибень (Ophidian barbatum); 4— барабуля (Mullus barbatus);
5 — морской петух (Trigla lucerna); 6 — морской дракон (Trachinus draco);
7 — морская коровка, или звездочет (Uranoscopus scaber)
Удивительная целесообразность в строении каждого организма, их
взаимная приспособленность к совместному существованию есть ре-
зультат длительной эволюции и естественного отбора в процессе борь-
бы за существование. Биоэволюция всегда носит приспособительный
(адаптивный) характер. Организмы, приспосабливаясь к абиотической
и биотической среде, меняются внешне и внутренне, становятся эко-
логически устойчивыми, приобретают четкие экологические границы.
ЧАСТЬ IL ЭКОЛПГИЧЕЕКИЕ И ЛАНДШАЙГВЫЕ (ВДКТОРЫ БИОЛОГМЧЕЕЮГО РАЗНООБРАЗИЯ
Приведем пример взаимной биологической адаптации (коадаптации).
Трематоды (плоские черви), паразитирующие в печени овцы, проходят
путь развития, меняя хозяев от муравья до овцы, где наконец приносят
потомство. Вероятность того, что овца проглотит зараженного муравья,
сама по себе очень мала, но поведение такого муравья изменяется так,
что вероятность быть проглоченным сильно возрастает.
Дело в том, что трематоды проникают в мозг муравья и вынуждают
свою жертву вести себя самым самоубийственным образом: порабощен-
ный муравей, вместо того чтобы оставаться на земле, взбирается по стеб-
лю растения и, замерев на самом кончике листа, как бы поджидает овцу.
И 4.2. Пищевые (трофические! связи________________________________
Главными биотическими отношениями, поддерживающими целост-
ность сообщества, являются трофические, т. е. питание одних организ-
мов за счет других (рис. 4.8).
Правило 10% Линдемана. В процессе питания осуществляется пере-
дача вещества и энергии с одного трофического уровня на другой
(рис. 4.9). Часть вещества и энергии при этом теряется. В среднем каж-
дое последующее звено в цепи питания содержит вещества и энергии в
десять раз меньше, чем предыдущее звено. Таким образом, трофичес-
кую структуру можно изобразить в виде экологической пирамиды,
основанием которой служат растения (первый уровень), а последую-
щие уровни — травоядные и хищные животные — образуют этажи и
вершину пирамиды (рис. 4.10).
Человечество занимает вершину экологической пирамиды, поэтому
его численность всегда будет ограничена биологической продуктивно-
стью растений и животных, которыми человек питается. Уничтожая
отдельные этажи трофической пирамиды, человек может вызывать
весьма неблагоприятные последствия (рис. 4.11).
Например, устойчивость луга может поддерживаться продуктивностью
травостоя; далее по убывающей располагаются растительноядные кузне-
чики, насекомоядные лягушки, хищные змеи; вершину пирамиды венча-
ет орел. Если человек убьет орла, это приведет к увеличению численности
змей, змеи уничтожат большую часть ля1ушек, численность кузнечиков
увеличится, а биомасса травостоя уменьшится. Потребуется немало вре-
мени, чтобы устойчивость лугового сообщества восстановилась.
и 4.3. Свойства популяций_________________________________________
Популяция — группа особей одного вида, занимающая определен-
ное пространство. Она обладает многими признаками, которые харак-
теризуют группу как целое.
Распределение особей, составляющих популяцию, в пространстве бы-
вает равномерным, случайным и кучным (рис. 4.12). Каждому виду при-
суща определенная оптимальная плотность популяции, отклонения от ко-
торой в обе стороны отрицательно сказываются на ее жизненности.
ГЛАВА 4. ОСНОВНЫЕ О ОНЯТ1Я I ЗАКОНЫ ЭКОЛОГИИ
Рис. 4.8. Пищевые отношения, связывающие членов водного
и наземного сообществ, по J.J. Moran
Рис. 4.9. Передача энергии по пищевой (трофической) цепочке, по J.J. Moran 9 9
ЧАСТЬ II. ШЛРПЧЕСКИЕII ЛДИДШДаГВЫЕ (ШТОРЫ БИОЛОГИЧЕСКОГО РДЗНООБРДЗИЯ
Рис. 4.10. Пирамида энергии, иллюстрирующая снижение количества энергии
по мере перехода от нижних уровней потребления к верхним, по J.J. Moran
Рис. 4.11. Устойчивая экосистема лугового сообщества и ее нарушение
в результате антропогенного вмешательства
100
ГЛАВА 4. ОСНОВНЫЕ О ОНЯТ1Я I ЗАКОНЫ ЭКОЛОГИИ
а — равномерное; б — случайное (беспорядочное); в — кучное
Плотность популяции определяется числом особей, приходящихся
на единицу площади (обычно так подсчитывается плотность популяций
крупных видов) или объема (так подсчитывается плотность популяций
мелких видов в объемах почвы или воды). Роль любого вида в сообще-
стве зависит от плотности его популяции.
Скорость воспроизводства популяции за счет рождаемости новых
особей определяется биологией вида. Микроорганизмы делятся пример-
но раз в минуту. Низкая плодовитость характерна для видов, которые
проявляют заботу о потомстве. Большое влияние на плодовитость жи-
вотных оказывают погодные условия, обеспеченность кормами и т. п. Бу-
дущее популяции зависит от соотношения смертности и рождаемости.
У большинства видов уровень смертности в раннем возрасте выше,
чем у взрослых особей. С наступлением биологической старости смерт-
ность резко увеличивается. Однако встречаются и такие виды, у кото-
рых смертность приблизительно одинакова во всех возрастных группах
(рис. 4.13).
Число
Рис. 4.13. Различные типы кривых выживания: 1 — высокая смертность
молоди у организмов, не проявляющих заботу о потомстве; 2 — одинаковая
смертность во всех возрастных группах; 3 — кривая выживания, характер-
ная для организмов, проявляющих заботу о потомстве, основной причиной
смертности является биологическая старость 101
ЧАСТЬ IL ЭКОЛПГИЧЕЕКИЕ И ЛАНДШАЙГВЫЕ (ВДКТОРЫ БИОЛОГМЧЕЕЮГО РАЗНООБРАЗИЯ
Факторы смертности очень разнообразны. В засушливый год гибнут
не только растения, но и многие организмы, связанные с ними трофиче-
скими связями. К мощным биотическим факторам, вызывающим повы-
шенную смертность, относятся разного рода инфекционные заболевания
как растений, так и животных; массовое развитие вредителей и паразитов
и т. п. Человек, прямо или косвенно уничтожая естественные угодья, за-
грязняя окружающую среду, способствует гибели многих организмов.
Возрастной состав популяции имеет очень большое значение для ее
существования и процветания. При благоприятных условиях в популя-
ции присутствуют все возрастные группы. В быстро растущих популя-
циях доминируют молодые особи. Популяция жизнестойка, если суще-
ствует равномерный поток особей, протекающий через все возрастные
классы данной популяции от рождения до биологической старости. Если
смертность будет превышать численность новых видов, приходящих на
смену старым, популяция деградирует; если количество молодых видов
превысит количество погибших, популяция будет расти, распростра-
няться на новые территории и вытеснять другие виды.
Сохранность того или иного вида в сообществе основана на посто-
янной борьбе жизни и смерти. Во всех организмах заложена потенция
размножения, выражающаяся геометрической прогрессией, графиче-
ским изображением которой является экспонента.
Неограниченный экспоненциальный рост популяции подобен взры-
ву, он приводит к истощению и полному разрушению ресурсов среды.
В основе существования любой популяции, подчеркивает Р. Уиттекер
(1980), лежит конфликт между свойственной организму тенденцией
увеличивать свою численность и разнообразными ограничениями, ко-
торые препятствуют такому увеличению. Если система не получает
постоянной подпитки необходимыми ресурсами извне, устойчивое со-
стояние может быть достигнуто только при условии равных значений
рождаемости и смертности особей. Рассмотрим несколько кривых, отоб-
ражающих разные типы динамики популяций (рис. 4.14).
а б
Время
Рис. 4.14. Характерные типы динамики популяций, по N.D. Levine.
Пояснения в тексте
102
ГЛАВА 4. ОСНОВНЫЕ DОНЯТ1Я И ЗАКОНЫ ЭКОЛОГИИ
Тип а — S-образная кривая — характерен для популяции, в которой
высокая рождаемость, быстро обеспечив оптимальную для данных усло-
вий численность, затем снижает ее до уровня, когда рождаемость равна
смертности, — кривая выходит на «плато», что говорит об устойчивом
существовании во времени. Тип б — куполообразная кривая — харак-
терен для популяции, быстро размножившейся, а затем так же быстро
погибшей в результате того, что все жизненные ресурсы оказались
исчерпанными. Тип в — волнообразная кривая — характерен для попу-
ляции, которая быстро восстанавливает свою численность после ее спа-
да, вызванного неблагоприятными факторами. Тип г — кривая, демон-
стрирующая, как после спада численность популяции выходит на
«плато», т. е. существование становится устойчивым.
Р. Уиттекер указывает на существование приспособлений, благода-
ря которым потери популяции сокращаются, когда ее численность и
ресурсы среды входят в конфликт. Он называет этот феномен буфер-
ностью популяции. Например, как только вид становится редким, хищ-
ничество по отношению к нему может уменьшиться. Хищники могут
забыть способ поиска подобных жертв и не считать их пищей. Многие
организмы (растения, членистоногие и др.) переносят неблагоприятный
период в форме покоящихся диаспор. Последние могут оставаться в
почве до тех пор, пока условия среды не станут благоприятными, тогда
из них вновь появляются и размножаются активные особи. Так, пус-
тынные эфемеры в виде семян переживают неблагоприятные сезоны
или, если это потребуется, даже периоды неблагоприятных лет.
«Волны жизни», или колебания численности во времени, часто но-
сят циклический характер. Быстрый подъем численности начинается
обычно под действием благоприятных погодных условий, обеспечива-
ющих пышное развитие растительности, хороший урожай плодов и
семян. Обилие корма, хорошие погодные условия способствуют быст-
рому размножению травоядных животных, например грызунов; гры-
зуны служат пищей хищников — лисиц и волков, количество которых
тоже увеличивается, но с некоторым запозданием.
В дальнейшем в процесс увеличения численности включаются регу-
лирующие факторы, имеющие характер обратной связи. Растительный
корм быстро съедается, и травоядные начинают гибнуть От голода. Тем
самым подрывается пищевая база хищников, численность которых так-
же должна естественным образом снизиться (рис. 4.15).
Существует сложная зависимость биологических циклов со средним
периодом 11,1 года от солнечной активности, связанная с образовани-
ем на Солнце пятен. На 11-летние циклы накладываются влияния мел-
ких ритмов, определяющих благоприятные и неблагоприятные дни в
течение месяца, а также ритмов с периодичностью в столетия, тысяче-
летия и даже миллионы лет. Обоснованию этих связей были посвяще-
ны труды выдающегося русского ученого А.Л. Чижевского (1897 — 1964).
Чижевский оставил и поэтическое наследие, особенно интересное
тем, что оно является продолжением научного поиска. Здесь в образ-
ной системе воплощены научные идеи о мощном влиянии космоса не
только на природные явления на нашей планете, но и на события и по-
ступки людей. Об этом, в частности, его стихотворение «Галилей»:
103
ЧДСГЬII. ШЛРГ1ЧЕСКИЕII ЛйИЛШДаГВЫЕ (ШТОРЫ БИОЛОГМЧЕЕКОГО РДЗНООБРДЗМЯ
И вновь, и вновь взошли на Солнце пятна;
И омрачились трезвые умы;
И пал престол, и были неотвратны
Голодный мор и ужасы чумы.
И вал морской вскипел от колебаний;
И норд сверкал, и двигались смерчи;
И родились на ниве состязаний
Фанатики, герои, палачи.
И жизни лик подернулся гримасой:
Метался компас — буйствовал народ;
А над землей и над людскою массой
Свершало Солнце свой законный ход.
О ты, узревший солнечные пятна
С великолепной дерзостью своей —
Не ведал ты, как будут мне понятны
И близки твои скорби, Галилей!
1940 1945 1950
Рис. 4.15. «Волны жизни» в биоценозах: а — годовой ход развития фитопланк-
тона (1) и зоопланктона (2); б — соотношение урожая семян ели (1)
и численности белки (2); в — соотношение численности зайцев (1), лисиц (2),
волков (3); соотношение количества заготовляемых лисьих шкур (1)
и больных чесоткойлис (2)
104
ГЛАВА 4. ОСНОВНЫЕ О ОНЯТ1Я N ЗАКОНЫ ЭКОЛОГИИ
И 4.4. Четыре закона общей экологии___________________________________
В формировании современной концепции экологии можно выде-
лить пять основных этапов (рис. 4.16). Сначала под экологией понима-
лись чисто биологические исследования связей каких-либо организ-
мов между собой и с окружающей средой (1). К середине 20-х годов
XX в. этот термин уже начал применяться к исследованию сообществ
организмов и включал такие понятия, как «трофическая связь» и «пи-
рамида чисел» (2). К 50-м годам XX в. были разработаны понятия «эко-
система» и «биогеоценоз», которые стали рассматриваться как основ-
ные единицы научных исследований, включающих все взаимодействия
между окружающей средой и обитающими в ней организмами (3).
Следующим шагом было признание того факта, что критической
областью исследований являются объекты, находящиеся на стыке
эко- и геосистем, что их совокупность составляет то, что мы называем
биосферой и географической оболочкой (4). И наконец, последним
событием явилось признание главенствующей роли человека в био-
сфере (5), его ответственности за ее судьбу.
Рис. 4.16. Пять основных этапов формирования современной концепции экологии.
Пояснения в тексте
Одним из результатов экологической революции стало расширение
границ экологии. Четыре основных закона современной экологии с
присущей американцам непринужденностью сформулировал известный
эколог Б. Коммонер (1974). 105
ЧДСГЬII. ШЛРПЧЕСКИЕII ЛйИЛШйаТВЫЕ (ШТОРЫ БИОЛОГМЧЕЕКОГО РЙЗНООБРДЗМЯ
1. Все связано со всем. Вред, наносимый одному компоненту экосис-
темы, может привести к большим сбоям в функционировании всей
экосистемы.
2. Все должно куда-то деваться. Ландшафты Земли, географиче-
ская оболочка в целом — в известном смысле замкнутые системы. Бы-
товые и производственные отходы, попадая в окружающую среду, не
исчезают бесследно. У природных систем остается все меньше возмож-
ностей, чтобы справляться с переработкой веществ, загрязняющих среду
обитания людей. Вокруг городов растут свалки мусора, загрязняющие
вещества разносятся далеко от мест выброса воздушными и водными
потоками.
3. Природа знает лучше. Человек, самонадеянно желая «улучшить»
природу, нарушает ход естественных процессов. Последствия разного
рода мелиорации делают среду обитания людей еще менее благоприят-
ной.
4. За все надо платить. Человек не может безвозмездно расходовать
природные ресурсы, загрязнять окружающую среду, преобразовывать
природные ландшафты в культурные и т. п. Все виды взаимодействия
человека с природой должны оцениваться экономически.
Будущее человечества зависит от того, какой станет окружающая
среда и как будут приспосабливаться к ней люди. Человек как вид мо-
жет сохраниться в том случае, если сумеет предотвратить отрицатель-
ные последствия изменения окружающей среды. Второй путь выжива-
ния — это адаптация, приспособление к неблагоприятным условиям.
Если не произойдет ни первого, ни второго, согласно биологическим
законам человечество обречено на вымирание.
Глава 5
СИНЭКОЛОГИЯ
При использовании экологического подхода к решению биогеогра-
фических задач существенно разделять общую экологию на аутэколо-
гию и синэкологию. Если первая изучает взаимоотношения между от-
дельными организмами и средой и является наукой биологической, то
вторая изучает взаимоотношение сообщества (биоценоза) со средой и
занимает пограничное положение с географией. Как утверждал
Д.Н. Кашкаров, проекция на местообитание есть основная характери-
стика экологического изучения. Такое определение задач синэкологии
сближает ее с биогеографией и ландшафтоведением. Работы синэколо-
гов, главным образом геоботаников, по сути дела явились той основой,
из которой выросли и современное ландшафтоведение, и ландшафтная
экология.
И 5.1. Биоценоз, экосистома, биогеоценоз__________________________
Проживающие совместно организмы вместе с абиотическими фак-
торами среды обитания образуют целостные природные системы.
Биоценозы. Биологический мир образован многочисленными сооб-
ществами живых существ. Еще в 1887 г. К. Мебиус на примере устрич-
ной банки в Северном море показал, как в результате взаимодействия
абиотических и биотических факторов на морском дне формируется
целостная природная система, для обозначения которой он впервые ввел
термин биоценоз. Это понятие быстро вошло в научный обиход, т. к.
отражало созревшее стремление естествоиспытателей рассматривать
совокупности организмов как целостные системы.
Всякий естественный биоценоз представляет собой исторически
сложившийся комплекс функционально связанных организмов и явля-
ется частью целостного природного образования — экосистемы. Он
занимает конкретную территорию, имеет определенный видовой состав
и внутреннюю пространственную структуру. Состав биоценоза зави-
сит от того, какие организмы встречаются в данной местности и каким
из них подходят местные условия существования.
Экосистемы. В основе концепции экосистемы лежит взаимозависи-
мость физического и биологического миров. Термин экосистема вошел
в употребление в 1935 г. Он был предложен английским ботаником 107
ЧДСГЬII. ШЛРПЧЕСКИЕII ЛйИЛШйаТВЫЕ (ШТОРЫ БИОЛОГМЧЕЕКОГО РЙЗНООБРДЗМЯ
А. Тенсли, который писал, что в экосистему входит «не только комплекс
организмов, но и весь комплекс физических факторов, образующих то,
что мы называем средой биома, — факторы местообитания в самом ши-
роком смысле. Хотя главным, интересующим нас объектом могут быть
организмы, однако, когда мы пытаемся проникнуть в самую суть вещей,
мы не можем отделить организмы от их особой среды, в сочетании с
которой они образуют некую физическую систему». Биотическую и абио-
тическую части экосистемы связывают непрерывные круговороты пита-
тельных веществ, энергию для которых дает солнечная радиация (рис. 5.1).
Экосистема
Рис. 5.1. Схема потока энергии и круговорота химических веществ
в экосистеме, по Р. Риклефсу: биологическая и физическая сферы,
представленные органическими и неорганическими соединениями,
в совокупности образуют экосистему
Теппо
Движение
В.Н. Сукачев
(1880—1967)
В современной трактовке экосистема рассматри-
ваетсякак информационно саморазвивающаяся, тер-
модинамически открытая совокупность биотиче-
ских и абиотических элементов (Реймерс, 1990).
Биогеоценозы. В.Н. Сукачев, изучая развитие
растительного покрова, пришел к выводу, что он
находится во взаимосвязи и взаимодействии с дру-
гими компонентами географической оболочки —
литосферой, атмосферой и гидросферой. Это вза-
имодействие на уровне растительных сообществ
(фитоценозов) протекает в рамках географиче-
ских комплексов. В.Н. Сукачев в своих работах
1940—1942 гг. именовал такие комплексы геоце-
нозами, однако, учитывая ведущую и активную
роль живых существ в процессах взаимодействия,
в 1945 г. он предложил использовать термин «био-
геоценоз».
Биогеоценоз — участок территории, однородный по экологическим
условиям, занятый одним биоценозом. В модели биогеоценоза Сукачев
108
ГЛАВА 5. СИНЭКОЛОГИЯ
выделяет два блока: экотоп и биоценоз (рис. 5.2). Схема взаимодействий
между организмами и средой в экосистеме или биогеоценозе показана
на рис. 5.3. Специальное внимание уделено рассмотрению функций
почвы (рис. 5.3-6).
Рис. 5.2. Структура биогеоценоза
и схема взаимодействия его компонентов, по В.Н. Сукачеву
Рис. 5.3-а. Схема взаимодействий в биогеоценозе (а),
по А.А. Молчанову и Н.В. Дылису
109
ЧАСТЬ II. ЭКОЛОГИЧЕСКИЕ И ЛАНДША(ВГ8ЫЕ (ВДКТОРЫ БИОЛОГМЧЕЕЮГО РАЗНООБРАЗИЯ
Рис. 5.3-6. Функции почвы как компонента биогеоценоза (6), по Р.А. Филенка
Несмотря на близость понятий «экосистема» и «биогеоценоз», по-
следний отличается тем, что имеет четкие пространственные размеры.
Биогеоценоз всегда приурочен к определенному небольшому по площа-
ди участку земной поверхности, однородному в экологическом отно-
шении. Экосистема — понятие безразмерное. В качестве экосистемы
можно рассматривать и грядку в теплице, и луг, и лес, и космический
корабль, и биосферу в целом.
И 5.2. Стации, местообитания, экопогическио ниши___________________
Целостные природные системы имеют сложную внутреннюю струк-
туру.
Стация. Это место, обладающее всеми условиями, необходимыми
для жизни того или иного вида. У животных это может быть часть
ареала, используемая в определенное время или для ограниченных це-
лей, — сезонные стации, стации ночевок, размножения, питания и т. д.
Существуют стации, пригодные для жизни определенных видов, но в
силу тех или иных причин ими не занятые. Это потенциальные стации,
которые еще не стали местообитаниями. Потенциальные стации пред-
ставляют хозяйственный интерес с точки зрения интродукции полез-
ных видов растений или животных. Так расширялся ареал многих ви-
дов, введенных в культуру человеком.
Потенциальные стации могут оказаться благоприятными не только
для полезных, но и дли вредных организмов, которых человек способен
занести туда по неосторожности. Стация — понятие очень широкое.
110
ГЛАВА 5. СИНЭКОЛОГИЯ
Это и наземная, водная и воздушная среды жизни, и зональные типы
биомов, и более дробные подразделения, связанные с неровностями ре-
льефа, свойствами почвогрунтов и т. п., вплоть до болотных кочек, от-
дельных валунов, организмов, являющихся стациями для других орга-
низмов, живущих с ними в симбиозе или паразитирующих на них.
Местообитание. Стация, в которой живет конкретный вид, стано-
вится его местообитанием, т. е. местом, где он живет. Например, в Ав-
стралии имеются стации, благоприятные для жизни кроликов, но до
прихода европейцев кролики там не водились. Привезенные европейца-
ми кролики быстро размножились, потеснив многие местные виды.
Потенциальные стации стали реальными местообитаниями кроликов.
Понятие «местообитание» в биогеоценологии рассматривается ши-
роко как однородная часть природного ландшафта, занятая целым со-
обществом (ландшафтно-экологическому подходу в биогеографических
исследованиях посвящена гл. б).
Экологические ниши. Внутри биоценоза в результате борьбы за суще-
ствование происходит специализация видов и расхождение их по разным
экологическим нишам. Г. Хатчинсон (Hutchinson, 1965) определяет эколо-
гическую нишу как гиперобъем в л-мерном пространстве множества эко-
логических факторов. Если в неком гиперпространстве экологических
факторов оказались виды с одинаковыми требованиями к условиям суще-
ствования, между ними начинается жесткая конкуренция, которая мо-
жет привести к трансформации этих организмов в новые виды, приспо-
собленные к жизни в разных экологических нишах (рис. 5.4).
Рис. 5.4. Схема гиперпространственной концепции экологической ниши,
по Г. Хатчинсону: Точками изображены факторы среды (температура,
источники пищи, минералы, другие организмы) в проекции на плоскость.
Неправильные фигуры включают в себя ряд факторов, которые операцио-
нально значимы для популяций определенного вида. Л — два вида занимают
неперекрывающиеся ниши; Б — ниши двух видов столь сильно перекрыва-
ются, что жестокая конкуренция за общие ресурсы может привести к вы-
теснению одного вида другим, или дивергенции ниш (показано стрелками)
Ю. Одум приводит пример, как два вида водяных клопов могут жить
совместно благодаря тому, что занимают разные трофические ниши
(рис. 5.5). Гладыш (Notonecta) — активный хищник; плавт, напротив
(Corixa), хотя внешне очень похож на гладыша, играет в сообществе
совершенно иную роль, т. к. питается в основном разлагающимися рас-
тениями.
ЧДСГЬII. ШЛРПЧЕСКИЕII ЛйИЛШйаТВЫЕ (ШТОРЫ БИОЛОГМЧЕЕКОГО РЙЗНООБРДЗМЯ
Рис. 5.5. Два водяных клопа (Hemiptera) — Notonecta (а) и Corixa (б),
которые могут жить в одном и том же месте, но занимают разные
трофические ниши из-за различий в питании, по Ю. Одуму
Каждая видовая популяция в сообществе использует конкретную
экологическую нишу, границы которой контролируются условиями
среды во времени, пространстве и в градиентах значений абиогенных
факторов. Отсюда следует, что новый вид не может образоваться, если
нет свободной ниши или если этот вид не способен ее «отобрать» у
какого-нибудь другого вида, участвующего в экосистеме. Вхождение
новых видов в устойчивую экосистему осуществляется главным обра-
зом путем открытия новых ниш, что создает тенденцию к структурно-
му усложнению, отождествляемому с морфологическим прогрессом
(Красилов, 1995).
Экологическая ниша включает не только положение вида в простран-
стве (топическую нишу), но и его функциональную роль в экосистеме.
Описание связей между растениями и животными в экологических
нишах возможно в рамках классификации В.Н. Беклемишева (1970), в
которой все многообразие связей сведено к четырем фундаментальным
типам: 1) трофические (по питанию), 2) топические (по местоположе-
нию), 3) форические (по переносу, например, пыльцы между растением
и его опылителем), 4) фабрические (по материалу, используемому жи-
вотными для обустройства гнезд, укрытий и т. п.). Эта система являет-
ся исчерпывающей и позволяет охарактеризовать любые типы взаимо-
отношений между организмами.
Узкая специализация видов позволяет им сосуществовать, макси-
мально плотно упаковываясь в пределах ограниченного пространства.
Ю. Одум (1975) приводит результаты детального исследования рас-
пределения многоножек (Diplopoda), показавшего высокую степень
специализации разных видов и соответственно узость их экологических
ниш. В качестве таковых оказались сердцевина гниющего бревна; его
наружная часть; наружная поверхность под корой; участок под брев-
ном, но на его поверхности; под бревном, но на почве; среди листьев
подстилки; под подстилкой на поверхности почвы.
Экологические ниши, объединяемые определенными типами свя-
зей, создают функциональные подсистемы (блоки) сообщества. В каче-
стве таких блоков выступают, например,, синузии и консорции расти-
тельного сообщества (см. раздел 5.3). Таким образом, экологические
ниши выступают в качестве более дробных подразделений местообита-
112 ния всего сообщества (рис. 5.6).
ГЛАВА 5. СИНЭКОЛОГИЯ
Рис. 5.6. Экологические ниши, связанные с отдельным деревом (липой),
по А.Г. Воронову: I — ниша корней: 1 — микориза, 2 — личинка хруща — потре-
бителя корней; II— ниша ствола: 3 — жук-короед; III — ниша листьев и цве-
тов: 4 — гусеница шелкопряда, питающаяся листвой дерева, 5 — жук-листоед,
6 — пчела — опылитель цветов; IV— ниша ветвей: 7 — гнездо дрозда, свитое
на ветви липы; V — ниша опушек: 8— олени — потребители веточного корма;
VI — ниша припочвенного слоя: 9 — лесная мышь — потребитель семян липы
Структуру сообщества, образованную сочетанием экологических
ниш, В.В. Жерихин (1994) уподобляет генотипу организма, в котором
записана программа, управляющая развитием системы. Пока она оста-
ется постоянной, структура сообщества и его сукцессионная последо-
вательность воспроизводятся в прежнем виде. Изменение набора про-
грамм означает отклонение от прежней нормы развития системы. Если
это изменение фиксируется и впоследствии воспроизводится, можно
говорить о переходе системы в новое состояние.
Р. Уиттекер (1980) подвергает сомнению гипотезу о том, что стабиль-
ность сообщества — результат наличия большого числа приспособленных
друг к другу видов. Представление о том, что сложность ведет к стабиль-
ности, по мнению Уиттекера, более привлекательно, чем верно. На самом
же деле увеличение количества видов и сложности взаимоотношений
скорее является причиной уязвимости сообществ при их нарушении.
Данный эффект особенно ярко проявляется при антропогенном воздей-
ствии на древние сложные по составу и структуре сообщества, например
дождевых тропических лесов или коралловых рифов. Сложные сообще-
ства оказываются более уязвимы в условиях, когда их среда резко нару-
шается. Они могут развиваться только в стабильных условиях. 113
ЧАСТЬ II. ЭКРЛОГМЧЕСКИЕIIЛДНЛШДЙГ8ЫЕ (ШШЫ БМОЛОГМЧЕБКОГО РДЭНООБРДЗИЯ
И 5.3. Растительные сообщества как каркас наземным экосистем____
Естественный растительный покров Земли является основным
средо- и ресурсообразующим фактором биосферы. Зеленые растения —
это, по словам В.И. Вернадского, живое вещество «первого порядка»,
исходное звено всех цепей питания в экосистеме. Они усваивают и
накапливают солнечную энергию в форме химических связей, поддер-
живают запас кислорода в атмосфере, поглощают углекислый газ. Рас-
тительность формирует почву с ее плодородием, выполняет водорегу-
лирующую и противоэрозионную функции, влияет на характер климата,
придает ландшафтам их неповторимый облик.
В соответствии с законом пирамиды биомасс в наземных экосисте-
мах доля растительности составляет более 90%. Растения не только слу-
жат основанием трофической пирамиды, но и благодаря своей непо-
движности они дают приют и убежище животному населению. Поэтому
с полным основанием можно говорить о том, что растительность слу-
жит каркасом всего биоценоза.
Без сохранения растительного покрова и разнообразного видового
состава растений все усилия по поддержанию на Земле условий, необхо-
димых для жизни, не могут быть эффективными. Растительные сообще-
ства выполняют свои многочисленные полезные функции в биосфере
практически «бесплатно», ибо они способны к самовосстановлению, если
нарушение функций не перешло предела, после которого самовосстанов-
ление уже невозможно.
Растительным сообществом, или фитоценозом, называют сово-
купность растений, обитающих на однородном участке земной поверх-
ности, с только им свойственными взаимоотношениями как между
собой, так и с условиями местообитания. Постулат ландшафтной гео-
ботаники гласит: один фитоценоз — один биогеоценоз — одна фация.
Однако структура фитоценозов бывает весьма сложной и динамич-
ной: фрагменты сообществ, их отдельные элементы или сочетания
иногда образуют пеструю мозаику. При использовании растительно-
сти в качестве индикатора общих биогеографических особенностей
территории необходимо рассмотреть наиболее существенные свойства
фитоценозов. Они раскрываются через описание видового состава и
ценоэлементов растительных сообществ (рис. 5.7).
Состав. Основой любого описания растительности является список
образующих данное сообщество видов. Растительные сообщества мо-
гут быть одновидовыми, бедными или богатыми по числу видов. Сте-
пень участия вида в сложении сообществ характеризуется его обилием,
численностью, биомассой, проективным покрытием, встречаемостью
и т. п. Величина названных показателей определяет фитоценотическую
мощность вида.
фитоценотипы — группы видовых популяций, характеризующие-
ся сходной ролью и фитоценотической позицией в растительном со-
обществе.
Доминанты — виды, преобладающие в разных ярусах сообщества.
Доминанты, слагающие основу сообщества и играющие главную роль
в создании фитоценотической среды, называются эдификаторами:
ГЛАВА 5. СИНЭПВЛОГИЯ
|СОСТАВ|
жизненные
формы
фитогеографииескке
к фпорогенетические
элементы
Сииморфологнческие
единицы
] ЦБНОЭЛЕМЕНТЫ |
Синэкологические
единицы
-| вертикальная ярусность]
—| горизонтальная мозаичность]
сезонные]
—фитогенная |
консорций*]
консоргеиты ( J консорты]
Рис. 5.7. Элементы структуры растительных сообществ
в лесу это деревья, на верховом болоте — сфагновые мхи и т. п. Кроме
того, выделяются второстепенные, редко встречающиеся, незначитель-
ные по биомассе и продуктивности виды.
Существуют довольно простые методы оценки обилия видов по шкале
О. Друде (табл. 5.1), а также глазомерные методы оценки проективного
покрытия. Однако наиболее перспективен количественный учет расте-
ний с последующей обработкой данных методами математической ста-
тистики.
Таблица 5.1
Шкала оценок обилия видов, по О. Друде
Обилие Баллы Примечания
Socialis (Soc) 6 Обильно
Coplosus (СорЗ) 5 Много
(Сор2) 4 Довольно много
(Copl) 3 Скорее редко, чем много
Spasus (Sp) 2 Редко, мало
SoUlanus (Sol) 1 Очень мало
Имея списки видов, можно применить к ним методы экологическо-
го анализа, т. е. установить, какими жизненными формами и экологи-
ческими типами растений образованы одни сообщества и чем они от-
личаются от других. ] ] 5
ЧАСТЬ II. ЭКОЛПГИЧЕЕКИЕ И ЛАНДШАЙГВЫЕ (ВДКТОРЫ БМОЛОГМЧЕЕЮГО РАЗНООБРАЗИЯ
Понятие о жизненной форме ввел один из основоположников эко-
логии растений Г. Варминг. Жизненные формы — это результат дли-
тельного приспособления к условиям существования. В результате виды,
сходные по внешнему облику и приспособлениям к среде, объединяют
в одну жизненную форму. Классификациям жизненных форм посвя-
щена обширная литература (Серебряков, 1964; Серебрякова, 1972).Од-
ной из наиболее известных является классификация жизненных форм
К. Раункиера (рис. 5.8).
Рис. 5.8. Жизненные формы, по Раункиеру: 1 — фанерофиты (тополь),
2—хамефиты (черника), 3—гемикриптофиты (лютик, одуванчик, злаки),
4— геофиты (ветреница, тюльпан), 5— терофиты (семя фасоли).
Выделены зимующие почки
В ее основу положена идея о том, что сходные типы приспособлений —
это прежде всего сходные способы перенесения наиболее трудных усло-
вий. В областях с сезонной периодичностью климата таковые наступают
в зимний сезон, а в аридных областях — еще и в период летней засухи.
В качестве признака, выражающего приспособленность к перенесению
неблагоприятного сезона, Раункиер использовал способ перезимовки по-
чек возобновления, т. е. их положение относительно поверхности земли.
Основные жизненные формы в системе Раункиера следующие.
Фанерофиты — деревья и кустарники, у которых почки возобнов-
ления расположены высоко над землей. Хамефиты — невысокие (20 —
30 см) кустарнички и полукустарнички с почками возобновления на
зимующих побегах, расположенных вблизи поверхности земли. На зиму
они укрываются слоем снега или опавшими листьями. Гемикриптофи-
ты — травянистые многолетники, у которых надземные органы в кон-
це вегетации отмирают, а почки возобновления находятся на уровне
почвы и защищены собственными отмершими листьями и снегом. Крип-
тофиты — растения, у которых почки возобновления скрыты: у гео-
фитов под землей, у гидрофитов на дне водоема. Терофиты — однолет-
ники, переживающие неблагоприятный период в виде семян или спор.
Экологические типы растений устанавливаются по отношению к
отдельным факторам среды (табл. 5.2). В качестве примера проанализи-
руем экологический ряд увлажнения и колебания обилия отдельных
11G видов в зависимости от условий увлажнения (рис. 5.9).
ГЛАВА 5. СИНЭКОЛОГИЯ
Таблица 5.2
Основные наземные экологические группы растений по отношению
к факторам среды, по О.Е. Агаханянцу
Экологические факторы Интенсивность воздействия
недостаточная умеренная сильная
Тепло Влага Минеральное питание Засоленность почв Песчаный фунт Обломочный фунт Скальный фунт Холодные сухие усло- вия Холодные влажные условия Солнечное освещение Ветер Микротермы Ксерофиты Олиготрофы Олигогалофиты Сциофиты (теневые) Мезотермы Мезофиты Мезотрофы Мезогалофиты Т еневыносливые Макротермы Гифофиты Эутрофы Эугалофиты Псаммофиты Хасмофиты Литофиты Криоф|ггы Психрофиты Гелиофиты Анемофиты
Рис. 5.9. Экологический ряд увлажнения на заливном лугу р. Оскол,
по Л. Г. Раменскому. Справа налево — снижение уровня поймы и возраста-
ние увлажнения и поемности: по вертикали отложены градации обилия
видов: Fs — Festuca sulcata; Kd — Koeleria delavignei; Tp — Trifolium
pratense; Tr — Trifolium repens; Tv — Taraxacum officinale; Ra — Ranunculus
acer; Pt — Poa trivialis; Rr — Ranunculus repens; Cp — Caltha palustris;
Gf— Glyceria fluitans; Cg — Carex acuta; Hp — Eleocharis palustris;
Am — Alisma gramineum; Ga — Glyceria maxima; SI — Scirpus lacustris
На рисунке отчетливо выделяются зоны оптимума увлажнения, в
которых определенные виды имеют максимальное обилие. Например,
Glyceria maxima и Scirpus lacustris — типичные гидрофиты; их экологи-
117
ЧАСТЬ II. ЭКОЛПГИЧЕЕКИЕ И ЛАНДШАЙГВЫЕ (ВДКТОРЫ БМОЛОГМЧЕЕЮГО РАЗНООБРАЗИЯ
ческий ареал ограничен левой частью ряда. Такие виды, как Роа trivialis
и Ranunculus асвг, относятся к мезофитам, они занимают среднюю
часть ряда. A Festuca sulcata — ксерофит, он замыкает ряд справа. Пре-
обладание в травостое растений определенной экологической группы
позволяет косвенно судить об условиях увлажнения данного местопо-
ложения.
Фитогеографический и флорогенетический анализ позволяет выяс-
нить, какими фитогеографическими элементами образована раститель-
ность и из каких центров происхождения пришли разные виды.
В соответствии с жизненной стратегией А.Г. Раменский (1938) под-
разделял виды растений на «львов» (виолентов), «верблюдов» (патентов)
и «шакалов» (эксплерентов). Человек иногда должен помогать «львам»
выжить в конкурентной борьбе с «шакалами».
Американские экологи (Пианка, 1981) выделяют два типа жизнен-
ных стратегий. К-стратеги — это, как правило, крупные многолетние
организмы, требующие устойчивых условий существования; их жиз-
ненная энергия расходуется главным образом на прирост биомассы, а
не на размножение. По классификации Раменского, это по преимуще-
ству «львы». R-стратеги, напротив, организмы с непродолжительным
периодом жизни, они предпочитают нестабильные местообитания и ха-
рактеризуются высокой репродуктивной способностью. По классифи-
кации Раменского, это «шакалы».
Весь ход эволюции на протяжении кайнозоя, когда благодатный
теплый и влажный климат сменялся в умеренных широтах холодным и
сухим, а в тропических широтах жарким и сухим, шел по пути замены
крупных многолетних жизненных форм (k-стратегов) на мелкие мало-
летние и однолетние (г-стратеги). «Львы» уступали место «шакалам».
Этим объясняются зачастую трудности восстановления численности
реликтовых видов, большинство из которых относится к к-стратегам.
Ценоэлементы. Известно большое разнообразие элементов струк-
туры растительных сообществ, для которых Х.Х. Трасс предложил ис-
пользовать общий термин «ценоэлементы».
Ценоэлементами называют пространственно ограниченные участ-
ки фитоценоза, отличающиеся по составу и внешнему виду. Каждый
из них занимает в сообществе свою нишу, естественную или создан-
ную человеком. Описание ценоэлементов предполагает выделение син-
морфологических и синэкологических единиц.
Синморфологические элементы сообщества — вертикальная ярус-
ность и горизонтальная мозаичность.
Вертикальная ярусность характерна для всех типов наземной
растительности: лесной (рис. 5.10), луговой, болотной и др.
Верхний ярус находится под наибольшим воздействием внешних
климатических условий. Под пологом верхнего яруса условия существо-
вания растений значительно трансформируются фитосредой. Таким
образом, верхний ярус играет эдификаторную роль в формировании
расположенных под ним ценоэлементов.
В сообществах пионеров зарастания с малой сомкнутостью ярус-
ность может быть не выражена. В моновидовых сообществах, особен-
118 но в полевых агроценозах, все растения относятся к одному ярусу.
ГЛАВА 5. СИНЭКОЛОГИЯ
о Первый
подъярус
<к
>3
3
о ~_
<о
£ Второй
ы подъярус
Рис. 5.10. Схема вертикальной ярусности лесного сообщества,
по Н.Ф. Реймерсу
Горизонтальная мозаичность, наблюдаемая в пределах почти каж-
дого фитоценоза, выражается в наличии сравнительно мелких групп ра-
стений, различающихся по составу, обилию видов, их жизненности и т. п.
Ценоэлементы горизонтальной структуры фитоценозов называют мик-
рогруппировками, или микроценозами. В растительных сообществах
выделяют экотопогенную, фитогепную и антропогенную мозаичность.
Экотопогенная мозаичность, как правило, связана с неровностями
микрорельефа или неоднородностью почвогрунтов, что особенно харак-
терно для полупустынных ландшафтов. Так, на карте ключевого участ-
ка полупустынной экосистемы в Калмыкии выделяется 12 элементар-
ных единиц (рис. 5.11).
Фитогенная мозаичность возникает как результат взаимодействия
растений в сообществе; в качестве примера можно привести распреде-
ление растений под пологом леса при его неравномерной сомкнутости
(рис. 5.12). Мозаичность может быть обусловлена особенностями био-
логии самих растений. Так, при зарастании свободного субстрата не-
равномерность в распределении видов возникает в результате случай-
ного заноса диаспор. Мозаичность может быть связана с биологией
разрастания самих видов (например, образование болотных кочек).
Антропогенная мозаичность возникает под прямым и косвенным
влиянием хозяйственной деятельности. Так, существенно меняется
структура растительного покрова под воздействием рубки леса, перевы-
паса скота; мозаичность характерна и для производной растительно-
сти, находящейся на разных стадиях развития. Приведенный выше при-
мер мозаичности в березово-лиственничном лесу можно рассматривать
и как пример антропогенной мозаичности, когда микроассоциации
березняков возникают на месте вырубленного хвойного леса.
ЧАСТЬ II. ЭКОЛВГИЧЕЕКИЕ И ЛАНДШАЙГВЫЕ (ВДКТОРЫ БИОЛОГИЧЕЕЮГО РАЗНООБРАЗИЯ
Рис. 5.11. Карта элементов мозаичной полупустынной экосистемы,
Калмыкия. Масштаб 1:3000, по Б.В. Виноградову: Элементы мозаичности:
а — ass. Stipa lessingiana + Festuca sulcata + Artemisia austriaca на лугово-
каштановых почвах; b — ass. Festuca sulcata + Artemisia austriaca на
светло-каштановых солонцеватых почвах; с — a$s. Festuca sulcata +
Tanacetum achilleifolium на светло-каштановых средпесолонцеватых поч-
вах; d — ass. Festuca sulcata + Artemisia lerchiana на степных глубоких
солонцах; е — ass. Artemisia lerchiana + Agropyron desertorum на степ-
ных средних солонцах; f— ass. Artemisiapauciflora на степных корковых
солонцах солончаковатых; д — ass. Artemisia lerchiana + A. pauciflora на
степных солонцах средних и корковых; h — ass. Elytrigia repens + Herbae
xeromesophyticae на луговых глубоких солонцах; i — ass. Agropyron
pectinatum + Elytrigia repens + Festuca sulcata + Artemisia austriaca на лу-
гово-каштановых почвах;] — ass. Agropyron pectinatum + Festuca sulcata +
Artemisia austriaca на лугово-каштановых солонцеватых почвах; k — ass.
Artemisia monogyna + Elytrigia repens на лугово-степных
средних и глубоких солонцах; 1 — ass. Camphorosma monspeliaca +
Artemisia pauciflora на лугово-степных корковых солонцах
Мозаичная структура имеет большое значение в жизни экосистемы.
Благодаря мозаичности полнее используется разнообразие условий мик-
росреды, увеличивается устойчивость фитоценоза к неблагоприятным
факторам внешней среды и конкурентным взаимоотношениям с сосед-
ними сообществами.
Синэкологические элементы сообщества — синузии и консорции.
Синузии выделяются как элементы пространственной структуры или
сезонной ритмики фитоценоза. Синузии образованы определенными
жизненными формами, или биоморфами, растений, т. е. группами ви-
дов, подчас далеких в систематическом отношении, но сходных по внеш-
120 нему виду, экологии и ритму развития. Достаточно, например, назвать
ГЛАВА 5. СИНЭПВЛОГИЯ
Рис. 5.12. Мозаичность в березово-лиственничном лесу в Верхнем Приаму-
рье (три типа микроассоциаций), по П.А. Ярошенко: а — микроассоциация
Larix dahurica + Betula platyphylla (сомкнутость 0,6) — Saussurea umbrosa +
Linnaea borealis; 6— микроассоциация Larix dahurica (сомкнутость 0,4) —
Vaccinium vitis-idaea+PyroIa incarnata; в — микроассоциация Betula
platyphylla (сомкнутость 0,3) — Calamagrostis langsdorfii + Sanguisorba
officinalis + Sanguisorba parviflora
деревья (фанерофиты), кустарнички (хамефиты), травы (криптофиты),
мхи и лишайники в таежном лесу, чтобы иметь представление об ос-
новных синузиях этого сообщества.
Сезонные синузии представлены разными биоморфами, развиваю-
щимися на одной территории в разные сроки вегетационного периода,
т. е. отграниченными друг от друга во времени. Свои сезонные аспекты
характерны для травяного яруса в лесу, на лугу, в степи и т. п. Особен-
но яркими примерами служат сезонные аспекты в степи или синузия
эфемеров в пустыне.
Разнообразное и сложное синузиальное сложение растительных
сообществ свидетельствует об их способности к наиболее полному ис-
пользованию пространственных и сезонных экологических ниш. Чем
больше в сообществе синузии, тем больше его флористическое, эколо-
гическое и биоморфное богатство. Изучение синузии помогает глубже
познать структуру сообществ и характер взаимоотношений различных
экологических групп растений. Стационарные наблюдения позволяют
выявить более жизненные (развивающиеся) синузии и менее активные
(вырождающиеся). Таким образом, разрабатывается прогноз сукцес-
сионных сдвигов растительного покрова.
Пока мы ограничивались рассмотрением структуры растительных
сообществ, абстрагируясь от связей растений с животными. Связую-
щим звеном между фито- и зооценозом является консорция.
Определяя это понятие, В.Н. Беклемишев приводит характерный
пример консорции — дерево с его микоризными грибками, с эпифит-
ными мхами и лишайниками на стволах и ветвях, с паразитическими
грибами в различных тканях, со всем множеством населяющих ее чле-
нистоногих и других животных (см. рис. 5.6).
121
ЧАСТЬ II. ЭКОЛПГИЧЕЕКИЕ И ЛАНДШАЙГВЫЕ (ВДКТОРЫ БМОЛОГМЧЕЕЮГО РАЗНООБРАЗИЯ
Консорция состоит из консоргента (ядро консорции) и консортов
(группы организмов, связанные с ядром). Консоргент играет роль эди-
фикатора, т. е. строителя главного яруса в сообществе.
Существенно также и то, что консоргент в сообществе является
автотрофом (продуцентом органического вещества). Консорты — рас-
тения и животные — связаны с ядром образом жизни и трофическими
отношениями. Типичные консорты малоподвижны, постоянно или на
определенной стадии своего развития обитают в кроне консоргента.
Разнообразие консортов, вероятно, может рассматриваться как сви-
детельство зрелости сообщества. Напротив, когда растения-эдификато-
ры лишены консортов, то это наводит на мысль о молодости биоценоза,
в котором процессы коадаптации и видообразования еще не привели к
появлению консортов, способных занять выгодные экологические ниши.
И 5.4. Сукцессии сообществ________________________________________
Последовательная смена на данной территории одних сообществ
другими называется сукцессией. Экосистемы меняются под воздействи-
ем как внутренних (ценотических), так и внешних экологических фак-
торов. Классическая схема предполагает, что в естественных условиях
смены идут к финальной стадии — устойчивому сообществу, стабиль-
ность которого поддерживается равновесием между ним и окружающей
средой. Такое сообщество называется климаксовым.
Изменения экосистем можно рассматривать как реакцию биоты на
изменения окружающей среды, в том числе на воздействие антропоген-
ных факторов. Анализ общих закономерностей динамики экосистем со-
держится в работе Ю.А. Исакова, Н.С. Казанской, А.А. Тишкова (1986).
Авторы отмечают, что экосистемы, достигшие в своем развитии терминаль-
ных стадий (климакса), часто оказываются малоустойчивыми к воздействию
антропогенных факторов. Как правило, это является отражением узкой
специализации климаксовых сообществ, следствием высокой степени сба-
лансированности свойственных им функциональных процессов.
Субклимаксовые сообщества, находящиеся на пути к терминальным
стадиям, менее специализированы и поэтому обладают большей спо-
собностью к восстановлению своей структуры. В настоящее время,
когда влияние антропогенных факторов становится почти повсемест-
ным, эта категория экосистем в силу своей адаптивности является наи-
более распространенной.
Сукцессии растительных сообществ и экосистем в целом рассмат-
риваются как реализация потенциальных возможностей биоты пере-
страивать структуру экосистемы в соответствии с изменяющимися
параметрами внешней среды. Схема типологии основных форм сукцес-
сии показана на рис. 5.13; в табл. 5.3 названы причины и характер из-
менений при различных формах динамики экосистем.
Анализ этих материалов позволяет сделать вывод, что среди много-
образных форм динамики экосистем выделяются три принципиально
различные категории: флуктуации, сукцессии и преобразования эко-
122 систем человеком.
Климатогенные
погодичные и
короткогодичные
-К
Динамика
экосистем
Направленное
преобразование
человеком
Преобразование
абиотических
компонентов экосистем
ГЛАВА 5. СШШОГИЯ
Биогенные
----Сукцессии
Перевод на положение
управляемых человеком
полуприродних
экосистем
Эндогенные
Экзогенные
Первичные
сукцессии развития
1
1
Катастрофические | Антропогенные]
1Техноэндогенные
1. * f
) Пирогенные
- {Лесоэксплуатационные
Восстановительные -* — ] Пасквальные
I Рекреационные
----- I Токсикогенные
----| Интродуционные
Создание
антропогенных
экологических
комплексов
Климатогенные .
многолетние
** изменения (естественные
или антропогенные)
.» развитие или восстановление
S изменения или развитие
^чередование противоположно
направленных изменений
* направленное преобразование
организации экосистем
человеком
Рис. 5.13. Основные формы естественной и антропогенной динамики экосистем и их взаимосвязи,
по ЮЛ. Исакову
ГО
со
I
ЧДЕТЬII. ШЛОПЧЕСКИЕ И ЛДНДШДЙГВЫЕ ШШЫ БИОЛОГИЧЕБХаГО РДЭН00БРДЗМЯ
Таблица 5.3
Некоторые основные формы погодичной,
многолетней и вековой динамики экосистем, по Ю.А. Исакову
Форма динамики Причины флуктуации и сукреций
Флуктуации биогенные климатогенные Внутренние закономерности динамики популяций организмов, корректируемые пошлинными особенностями климата Погодичные или короткоцикличные изменения гидротерми- ческих показателей
Сукцессии развития восстановления циклические со сменой режимов функционирования гологенетнческие, филоценогенетические многолетне-циклические климатогенные катастрофические Внутренние противоречия, возникающие в результате средообразующей деятельности биоты Исчезновение причин, вызвавших нарушение организации экосистем Разрешение внутренних противоречий, возникающих в процессе средообразующей деятельности биоты благодаря чередованию серийных и относительно стабильных стадий Перестройка структуры биоты в результате вековых и многовековых изменений рельефа, климата и почв, а также эволюции видов и возникновения новых форм взаимодействия между ними на консортивном уровне Многолетние изменения гидротермического режима Вековые и многовековые направленные изменения климата (глобальные, региональные) Проявления стихийных природных явлений, часто локальных
Антропогенные, в том числе способные формировать полуприродные экосистемы пирогенные лесоэксплуатационные пасквальные рекреационные техноэдоогенные токе икогенные интродукционные Пожары вызываются большей частью антропогенными причинами Вырубка коренных и формирование вторичных лесонасаждений Выпас, вызывающий изменения структуры растительного покрова Нарушения растительного покрова и почв в результате массовой рекреации Нарушения почвенного покрова и фунта, эрозия, образование отвалов и шлаков при горных разработках, создание новых форм рельефа, снижение уровня фунтовых вод Загрязнение атмосферы, водоемов и почв токсичными веществами Намеренная или случайная интродукция видов животных, растений и микроорганизмом, несвойственных данным экосистемам
Направленное и непреднамеренное преобразование абиотических компонентов экосистем Мелиорация, орошение, регулирование стока рек и другие мероприятия
Под флуктуациями фитоценозов понимаются ненаправленные их
изменения от года к году, завершающиеся возвратом фитоценоза к
исходному или, точнее, близкому к исходному состоянию. С некото-
рой долей условности к флуктуациям растительных сообществ можно
отнести изменения, вызываемые сенокошением, выпасом, лесохозяй-
ственными мероприятиями и т. п.
124
ГЛАВА 5. СИНЭКОЛОГИЯ
Сукцессионные процессы в противоположность флуктуациям более
долговременные. Развитие их происходит в определенном направлении.
Сукцессионные процессы никогда не имеют характера колебаний око-
ло какого-то среднего состояния.
Эндогенные сукцессии, определяемые внутриценотическими причи-
нами, разделяются на две главные категории. Сукцессии развития (пер-
вичные), началом которых служит освоение биотой еще незанятого суб-
страта (рис. 5.14); сукцессии восстановительные (вторичные) (рис. 5.15).
Рис. 5.14. Сукцессия развития: формирование лесного сообщества
в результате зарастания обнаженных скальных грунтов
К категории экзогенных сукцессии, вызываемых внешними факто-
рами, относятся как многолетние климатогенные, так и катастрофиче-
ские, являющиеся результатом проявления природных стихий: схода
лавин, селей в горах, извержения вулканов, наводнения и др. Особенно
большое распространение получили сейчас антропогенные сукцессии,
возникающие в результате хозяйственной деятельности. Они вызыва-
ются пожарами, вырубкой лесов, выпасом скота, рекреацией. Глубокую
трансформацию почвенно-растительного покрова вызывают строитель-
ные работы, горные выработки. Растительный покров и животный мир
меняются под воздействием загрязнения среды различными техноген-
ными веществами.
Наряду с негативным воздействием на биоту хозяйственная деятель-
ность человека может носить конструктивный характер. Природные
системы, в которых проводятся мелиоративные мероприятия, на-
правленные на повышение их продуктивности — осушение и обводнение,
ЧАСТЬ II. ШЛРПЧЕСКИЕII ЛДИЛШАаГВЫЕ ШТОРЫ БИОЛОГМЧЕЕХОГО РДЭНООБРДЗМЯ
Рис. 5.15. Восстановительная сукцессия:
последовательные стадии зарастания вырубки
удобрение, фитомелиорация, лесо-, рыбо- и охотохозяйственные рабо-
ты и др., — переводятся в категорию полуприродных. Создаются антропо-
генные экологические комплексы — сельскохозяйственные, рекреа-
ционные, водохозяйственные. Затраты на управление этими комплексами
12G человек должен брать на себя.
Глава
ЭКОЛОГИЯ И ГЕОГРАФИЯ
Растения и животные, компоненты абиотической среды, социаль-
но-производственные сферы общества объединены в территориальные
образования, которые формируют пространственную структуру био-
сферы и географической оболочки.
Методология изучения сложных объектов опирается на системный
анализ. Для обозначения множества живых существ, взаимодейству-
ющих с факторами окружающей среды, предложено понятие «экосисте-
ма». Для обозначения множества взаимосвязанных элементов живой и
неживой природы используется понятие «геосистема». Множество эле-
ментов природы, населения и производства образуют социально-про-
изводственные системы, или «социосистемы».
5.1. Геоэкологический и ландшафтно-экологический подкоды
к биогеографическим исследованиям
Биогеографические исследования должны проводиться с учетом
особенностей взаимодействия сложных территориальных образований.
Для решения этих задач на границе экологии и географии обособились
геоэкология и ландшафтная экология.
Геоэкология. Существует область исследований, где множества эле-
ментов гео-, эко- и социосистем пересекаются. Пространство, в кото-
ром они находятся во взаимодействии, составляет объект геоэкологи-
ческих исследований (Петров, 2004). Иными словами, геоэкология — это
наука о взаимодействии географических, биологических (экологиче-
ских) и социально-производственных систем (рис. 6.1). Систему гео-
графических наук можно изобразить в виде стопки дисков, где каждый
диск — частная географическая дисциплина. Осью, связующей всю
стопку, являются идеи докучаевской географии, а в современных усло-
виях — экологическая парадигма. Сегмент, вырезанный из стопки, —
это региональный объект геоэкологических и биогеографических ис-
следований. Он включает природно-территориальный комплекс (ПТК)
с присущими ему биоценозами и территориально-производственный
комплекс (ТПК) с его социально-экологическими проблемами.
Итак, геоэкологический подход к биогеографическим исследовани-
ям направлен на изучение находящихся во взаимодействии географи- 127
ЧАСТЬ II. ЭКОЛОГИЧЕСКИЕ И ЛДНДШАЙГ8ЫЕ ШШЫ БИОЛОГИЧЕЕХОГО РДЭНООБРДЗМЯ
Рис. 6.1. Объект геоэкологических исследований. Пояснения в тексте
ческих и экологических систем; особое внимание при этом обращается
на антропогенные процессы экологической дестабилизации и опусто-
шения земель, разработку рекомендаций по рациональному природо-
пользованию и охране природы.
Ландшафтная экология. Основополагающим положением ландшафт-
ной экологии является утверждение, что пространственная неоднород-
ность биосферы определяется пространственной неоднородностью гео-
графической оболочки: системе геохор (природно-территориальных
комплексов — ПТК) соответствует адекватная ей система биохор (Соча-
ва, 1970).
При средне- и крупномасштабных биогеографических исследовани-
ях синэкологические (экосистемы, биогеоценозы) и ландшафтные
объекты — ландшафты и их морфологические единицы — тесно взаи-
мосвязаны.
Учитывая неоднозначность толкования термина ландшафт, отме-
тим, что при биогеографических исследованиях предпочтительнее его
региональная трактовка.
Ландшафт — объективно существующая часть земной поверхности,
представляющая собой самостоятельный природно-территориальный
комплекс, который качественно отличается от других. Каждый ландшафт
как региональное природное образование имеет индивидуальный внеш-
ний облик и внутреннюю структуру. Он имеет конкретное положение
на земной поверхности и границы. С антропоцентрических позиций
ландшафт рассматривается как ресурсосодержащая и ресурсовоспроиз-
водящая система, как среда жизни и деятельности человека, как система,
хранящая генофонд, как природная лаборатория научных исследований,
как место отдыха и эстетического вдохновения.
Основоположник науки о ландшафтах В.В. Докучаев первым на
рубеже XIX —XX вв. обосновал необходимость изучения взаимосвязей
12В между разными природными факторами, особенно между неорганиче-
ГЛАВА G. ЭКОЛОГИЯ I ГЕОГРАФИЯ
ской и живой природой. Для обозначе-
ния целостных природных комплексов в
современной географии используется
термин «геосистема». По своей сущности
он близок понятию «экосистема». Отли-
чие состоит в том, что в первом случае
взгляд исследователя полицентричен, он
с одинаковым вниманием устремлен как
на биотические, так и на абиотические
компоненты системы. Во втором случае
взгляд исследователя биоцентричен, он
устремлен прежде всего на сообщества
живых организмов, а компоненты среды
рассматриваются им только как место-
обитание — биотоп (рис 6.2).
Проследим основные этапы становле-
ния ландшафтно-экологического метода
биогеографических исследований. Бли-
жайший ученик Докучаева Г.Н. Высоц-
В.В. Докучаев (1846-1903)
кий высказал мысль о зависимости растений и почв от одних и тех же
факторов среды — теплоты, влаги и соотношений между ними, распре-
деления их во времени, а также от типов грунта и рельефа. Он называл
эти факторы элементами местности. Подчеркивая тесную связь рас-
тительности с почвами и другими факторами среды, Г.Н. Высоцкий в
1909 г. указывал на возможность использовать растения в качестве ин-
дикатора экологических условий: «Находясь в зависимости от тех же
естественных элементов местности, первобытная дикая растительность
является, так сказать, наружным выражением той или иной комбина-
ции их». Высоцкий писал о целесообразности составления карт типов
местопроизрастаний — участков с однородными почвами и раститель-
ностью, грунтами, рельефом и климатом, для которых должны разраба-
тываться соответствующие системы природопользования.
Рис. 6.2. Схема взаимоотношения полицентрического (ландшафтного)
и биоцентрического (экосистемного) взглядов на геосистему,
по И.С. Арзамасцеву и Б.В. Преображенскому
129
ЧАСТЬ II. ЭКОЛОГИЧЕСКИЕ И ЛДНДШАЙГ8ЫЕ ШШЫ БИОЛОГИЧЕЕХОГО РДЭНООБРДЗМЯ
Д.Н. Кашкаров (1933) предложил экологические принципы дробного
подразделения арены жизни. Основной единицей он считает биотоп.
Этим термином обозначаются участки арены жизни, сходные по основ-
ному характеру жизненных условий. Д.Н. Кашкаров раскрывает также
понятие синонимичное биотопу — местообитание. Это комплекс эда-
фических (экотопических) и микроклиматических факторов.
Фундаментальное значение для разработки ландтпафтно-экологиче-
ского подхода к изучению биогеографических закономерностей на мест-
ном (топологическом) уровне имеют труды известного русского геобо-
таника Л.Г. Раменского (1938). Им заложены основы экотопологии —
учения о внешней обусловленности различных местообитаний и жиз-
ненных сред. «Долгий исторический путь развития местности, — пи-
шет Л.Г. Раменский (1971), — привел к расчленению ее на мозаику эн-
топиев, или местоположений, приуроченных к определенным элементам
рельефа и горным породам, общим гидрологическим условиям, окру-
женных другими энтониями». Энтопий в условиях данного климата
рассматривается как экотоп, характеризуемый определенным укладом
экологических режимов.
Задачу экотопологии Раменский видел в выделении однородных в
экологическом отношении участков. Поскольку распределение основ-
ных факторов существования — гидротермических показателей, почв
и растительности — в значительной степени контролируется рельефом,
Раменский разработал типологию местоположений, которую полезно
использовать в ландшафтно-экологических исследованиях (рис. б.З).
Итак, каждому сочетанию условий внешней среды соответствует своя
группировка организмов; сообщества животных и растений закономерно
повторяются всякий раз при сочетании определенных типов рельефа и
почвогрунтов с определенными же гидротермическими и другими условия-
ми жизни. Поэтому для углубленного изучения закономерностей распре-
деления организмов возникает необходимость изучать не только сами
организмы, но и ландшафты, в которых они обитают. Из трактовки тер-
минов «тип местопроизрастания» (Высоцкий, 1909), «биотоп» (Кашкаров,
1933), «энтопий» и «экотоп» (Раменский, 1938) видно, что это синонимы.
В начале 1970-х гг. В.Б. Сочава создал школу сибирских географов,
развивающих научное направление, названное им топологией геосис-
тем, или геотопологией. А.Г. Исаченко (1972) убедительно указал на
связь этого направления с классическими работами геоботаников —
Г.Н. Высоцкого, Л.Г. Раменского, В.Н. Сукачева, с одной стороны, и с
работами ландшафтоведов — с другой. К. Тролль (Troll, 1939) обозначил
науку, лежащую на рубеже географии (ландшафтоведения) и биологии
(экологии), ландшафтной экологией. Изложению основ ландшафтной
экологии посвящена монография Б.В. Виноградова (1999).
6.2. Аксиом этические о оно вы ландшафтной эко л о гии
Порядок рассмотрения теоретических основ ландшафтной экологии
устанавливается исходя из требований системного анализа: сначала
130 формулируется аксиома о целостной системе, затем даются положения
ГЛАВА 6. ЭКОЛОГИЯ I ГЕОГРАФИЯ
Пойма - — h
; >
5-/?| 13-16 ;*7-zo’
1 1 *
• 1
I | 2~—<
I СК Ась ? I
lM{ 3al
I 1 1
\Crinnf ।
---- Материк-----------
ВерхоВые местоположения
t 1a
I1
t
§1
g)
3
'б
'll!
•s!
<a>
t t
I I
I I
I ill
1 Заморезкаt
r~T"l
i*
&:
<O (
j Уклон
•около 1a
je
t
t
t
I
I
t
sg
i if ’ 6 у«
‘ iBeuieitai S 1ck/row)
' §l • t l§.‘Olg
~1
Назина
3
. <6- j 4
i
Заморозки
Грунтовая Soda
[!i калвйыю-жидким состоянии/_
Нейтряпьяна/„мерпЛыи f
ffaita S пленочной соаиаяниИ
tj
7
г
5
5
i
S
6
I
i
Обычная предельная глубина првмачиВания почбы
вддб Выщелачивание Вещестб из мчбы аросачиВающимися бодами
лл.'; Накопление ВещестВ, бмытыг ила доставленных подтоком Верхвбодки
•»т» НакоплениеВещестбиз поднятой по капиллярам Вады, испаряющейся £ поверхнос-
ти. почбы ияи В ее толще
(Движение Вод пи поверхности и б почбе. Число стрелок отражает кипиче-
стВо дВижущейся Воды
Рис. 6.3. Схема типов местоположений, по А.Г. Раменскому:
1—20 — различные типы местоположений
об ее элементах, системообразующих отношениях, структуре и иерар-
хии систем и, наконец, об их границах (Петров, 1998).
Аксиома Докучаева-Вернадского: географическая оболочка, биосфе-
ра представляют собой целостную природную систему, в которой
живое вещество взаимодействует с элементами литосферы, гидросфе-
ры, атмосферы и техносферы. Руководствуясь этой аксиомой, биосфе-
ру определяют как экосистему высшего ранга.
Эта аксиома имеет фундаментальное значение для развития тео-
рии ландшафтной экологии. Из нее вытекают важные положения об
элементах, системообразующих отношениях и структуре биосферы,
которые составляют основу познания экологических факторов окру-
жающей среды.
Положение о составе элементов. К элементам биосферы относится
множество предметов и явлений, входящих в состав биосферы, лито-
сферы, гидросферы, атмосферы и техносферы. Действительно, в любой
экосистеме — на суше или в океане — присутствуют хотя бы в незначи-
131
ЧАСТЬ II. ЭКОЛОГИЧЕСКИЕ И ЛДНДШАЙГ8ЫЕ ШШЫ БИОЛОГИЧЕЕХОГО РДЭНООБРДЗМЯ
тельных количествах и в своеобразной форме элементы любой геосфе-
ры. Например, в ландшафтах вод Мирового океана вещество литосферы
представлено растворами и взвесями минеральных веществ; в ланд-
шафтах морского дна вещество атмосферы присутствует в виде раство-
ренных в воде газов. Внешне безжизненные пустыни тропиков, поляр-
ных стран или высокогорий несут в себе множество «зародышей живого
вещества», подтверждая мысль В.И. Вернадского о «всюдности» жизни.
Все шире в биосфере распространяются элементы техногенеза — горо-
да и сельскохозяйственные угодья, горные выработки и дороги и т. д.,
оказывающие прямое или косвенное воздействие на природные экосис-
темы.
Способность природных элементов образовывать целостные систе-
мы раскрывает модель трех начал геосистемы, предложенная А.А. Крау-
Рис. 6.4. Три начала геосистемы, по А.А. Крауклису.
Стрелками обозначено взаимодействие частей геосистемы
Устойчивое (инертное) начало, к которому относятся горные поро-
ды и рельеф, выступает как «скелет» геосистемы. Оно придает ей фик-
сированное местоположение на земной поверхности и известную про-
странственную обособленность, связывая с геологическим прошлым
данного участка.
Подвижность (мобильность) в геосистему вносят, с одной стороны,
энергия Солнца, процессы, возбуждаемые силой земного притяжения,
а также скрытые в самой экосистеме источники разных видов энергии;
с другой — вещество, у которого силы молекулярного сцепления отно-
сительно слабы и которое пребывает в геосистеме в основном в виде
потоков. Это главным образом воздушные и водные массы.
Мобильная составляющая, таким образом, выполняет обменные
функции, связывает внутренние части геосистемы и объединяет по-
следнюю с ее внешним окружением. Она постоянно размывает и пере-
132 двигает пространственные границы.
ГЛАВА 6. ЭКОЛОГИЯ I ГЕОГРАЙИЯ
Совокупность живых организмов (биота) частично принадлежит
обеим рассмотренным выше составляющим, но, кроме того, еще вы-
полняет и самостоятельные функции. В деятельности биоты особую
роль играют высокая избирательность по отношению к внешним усло-
виям, способность к самовоспроизведению, размножению и росту.
Именно в активности и пластичности основная сила биоты как са-
мостоятельной части геосистемы. Взаимодействуя с остальными ком-
понентами, биота выступает как важнейший внутренний фактор само-
регуляции, восстановления, стабилизации. Оптимальное насыщение
геосистемы биотой — первостепенное условие и способ успешного
управления экосистемами и рационального использования их природ-
ного потенциала.
Системообразующая значимость элементов неравнозначна.
В.И. Вернадский ведущую роль отводил живому веществу: «На земной
поверхности нет химической силы, более постоянно действующей, а по-
тому и более могущественной по своим конечным последствиям, чем
живые организмы, взятые в целом». Оценивая роль элементов в форми-
ровании геосистем, Н.А. Солнцев ранжирует их в определенной после-
довательности. На первом месте стоит земная кора с ее структурными
формами, омоложенными новейшими тектоническими движениями,
затем следуют поверхностные воды, метеорологические условия; завер-
шают этот ряд почвенно-растительные комплексы и животный мир.
Рассматривая элементы окружающей среды как экологические фак-
торы, В.В. Мазинг предлагает следующий иерархический ряд. На пер-
вое место среди факторов, влияющих на главное звено экосистем —
растительный покров, он ставит количество теплоты (сумму солнечной
радиации) как фактор, определяющий широтную зональность. На вто-
рое место — количество осадков и их сезонное распределение; анало-
гичное значение имеет деление зон по степени гумидности (океанич-
ности) и аридности (континентальности). Третье место занимает
водный режим, зависящий от вышеназванных факторов, а также от
стока и свойств почв; одновременно с водным режимом почв отмеча-
ются их богатство и характер минерализации (засоления).
Подчеркивая значение сильных элементов, мы вместе с тем не долж-
ны забывать о роли слабых взаимодействий, без которых целостность
и устойчивость системы невозможна. Сильные воздействия не всегда
играют решающую роль в управлении системой. Одно из правил кибер-
нетики гласит, что вовремя и к месту приложенное слабое воздействие
может оказаться решающим в управлении системой.
Познание элементного состава экосистем отвечает аналитическому
пути познания. В ходе исследования из генерального множества эле-
ментов, отображающих реальное пространство биосферы, руководству-
ясь эвристическими решениями или мнениями экспертов, формируют
операционное признаковое пространство, которое представляет собой
перечень наиболее существенных элементов и их свойств, присущих
конкретным биогеоценозам. Заметим, что для разработки моделей эко-
систем главным является исследование изменчивости признаков, харак-
теризующих объекты. В этом смысле справедливо утверждение Д. Хар-
вея о том, что элементы системы суть состояние вещей, а не сами вещи.
133
ЧАСТЬ II. ЭКРЛОГИЧЕСКИЕIIЛДИЛШДИТВЫЕШТОРЫ БКОЛОГМЧЕБКОГО РДЭНООБРДЗИЯ
Сравнив элементный состав биогеоценозов, можно говорить об их
различии или сходстве в экологическом отношении. Несмотря на слож-
ность и уникальность экосистем, реальны системы, пересечение свойств
которых помогает выявлять область общих элементов. Наличие общих
свойств позволяет находить классы изоморфных экосистем, в пределах
которых действуют единые методы описания их структуры, состояния
и поведения. Инварианты структуры, присущие изоморфным систе-
мам, представляют собой наиболее ценный фонд биогеографических
исследований.
Исследование систем допускает прием сознательного отбрасывания
отдельных элементов, роль которых с точки зрения конкретной задачи
ландшафтной экологии несущественна. Этот прием отвечает принципу
ограничения разнообразия информации; его еще называют принципом
«бритвы Оккама».
Принцип «бритвы Оккама»: не следует делать посредством боль-
шего то, чего можно достичь посредством меньшего. Руководствуясь
этим принципом, процедуру описания главных свойств системы можно
существенно упростить.
Положение о системообразующих отношениях. Обязательным ат-
рибутом биосферы, как следует из аксиомы Докучаева—Вернадского,
являются связи или отношения, заданные на множестве ее элементов.
Связи между элементами составляют характерную черту экосисте-
мы любого ранга (рис. 6.5). Простейшей формой отношений являются
отношения ряда, связывающие элементы необратимой связью. Это ти-
пичный пример четких причинно-следственных отношений. Например,
сухость климата ведет к снижению биологической продуктивности.
Рис. 6.5. Формы отношений и причинно-следственных связей между
элементами экосистем, поД. Харвею: а — параллельные отношения;
б— отношения обратной связи; в — комбинирование различных форм свя-
зей; г— причинно-следственные связи типа множественности причин;
134 д — причинно-следственные связи типа множественности следствий
ГЛАВА 6. ЭКОЛОГИЯ I ГЕОГРАЙИЯ
Параллельные отношения отражают воздействие элементов х. и х. на
элемент хк (рис. 6.5, а). Например, богатство почв (х.) и благоприятный
режим увлажнения (х.) повышают биологическую продуктивность (xj.
Отношения обратной связи составляют характерную черту саморе-
гулируемых систем. Они отражают ситуацию, при которой один элемент,
влияя на другие, одновременно опосредованно воздействует сам на себя
(рис. 6.5, б]. Обратная связь может быть положительной или отрицатель-
ной. Например, высокая биопродуктивность степных экосистем способ-
ствует накоплению гумуса в почве, что повышает ее плодородие и ведет
к дальнейшему увеличению продукции растительной массы.
В основе саморегуляции экосистем лежат отрицательные обратные
связи. Положительные обратные связи, подобные рассмотренным, вы-
зывают быстрое, взрывное развитие определенных процессов. Так,
интенсивное нарастание растительной массы в степи ведет к уплотне-
нию дернины и в конечном счете к обеднению видового состава и сни-
жению продуктивности сообщества.
Высокая продуктивность степных экосистем в естественных услови-
ях поддерживается благодаря воздействию на растительный покров ко-
пытных животных. Стада травоядных постоянно обкусывают и разби-
вают дернину копытами, т. е. частично разрушают растительный покров
и открывают почву для всходов новых растений. Если поголовье станет
слишком большим, наступит ухудшение состояния пастбища, продук-
тивность растительности снизится, часть животных погибнет от голода.
Таким образом, продуктивность растительности и поголовье копытных
поддерживаются в естественных условиях на оптимальном уровне.
Комбинирование различных форм связей приближает нас к отобра-
жению структуры реальной гео- или экосистемы во всей ее сложности
(рис. 6.5, в).
Характерная черта структурообразующих связей — их причинно-
следственный характер: всякий природный процесс, хозяйственная
деятельность человека являются причиной, вызывающей изменение
связанных с ними элементов. Поэтому важным дополнением к рассмот-
ренным выше формам отношений служит анализ основных типов при-
чинно-следственных связей.
Причинно-следственные (каузальные) цепи А-^В—>С—>... задаются
последовательностью отношений ряда. Отношения ряда позволяют
выводить свойства конечного члена каузальной цепи из свойств ее
начального члена.
На этом строится практика индикации различных компонентов
окружающей среды. Например, карбонатная (известковая) горная поро-
да определяет карбонатиость отложений, на которых развиваются почвы с
высокими значениями pH, что обусловливает появление во флоре спе-
цифичных растений. Пользуясь отношениями ряда, мы можем рассмат-
ривать определенные виды растений в качестве индикатора карбонат-
ных горных пород.
Множественность причин. Этот тип отношений является компози-
цией параллельных отношений и отношений ряда (см. рис. 6.2, г). На-
пример, гидрогеологические условия (xd оказывают влияние на мине-
рализацию вод в реке (хь); сток промышленных отходов (х.) также 135
ЧАСТЬ II. ЭКОЛПГИЧЕЕКИЕ И ЛАНДШАЙГВЫЕ ЙДКТОРЫ БИОЛОГМЧЕЕЮГО РАЗНООБРАЗИЯ
оказывает свое влияние на химизм вод (xj. Объединение химических
особенностей вод, формирующихся под воздействием различных при-
чин (xb U xs), определяет общую минерализацию водоема (ху).
Множественность следствий — это отношение, представляющее
композицию ряда и параллельных отношений (рис. 6.5, д). Например,
Солнце (ха) определяет световой режим на Земле (хд); последний влияет
на радиационный баланс (xf), температурный режим (xj и интенсив-
ность фотосинтеза (хр); суммарное воздействие этих факторов обу-
словливает продуктивность растений (х;) и животных (xj.
Положение о причинно-следственном характере отношений явля-
ется фундаментальным для теории ландшафтной экологии. Связи ряда
или каузальных цепей позволяют построить наиболее простые и ясные
модели. Однако ввиду сложности структуры реальных природных сис-
тем отношения между ее элементами наиболее полно раскрываются с
помощью моделей множественных причин и множественных след-
ствий. При моделировании подобных структур невозможно проследить
и учесть абсолютно все связи, в результате характер экологического воз-
действия отдельных факторов носит неопределенный, вероятностный
характер.
Аксиома об иерархической структуре биосферы: биосфера пред-
ставляет собой систему, организованную в виде множества террито-
риальных подсистем различной размерности. В самых общих чертах
можно говорить о территориальных единицах местного (топологиче-
ского), регионального, континентального и глобального уровней. Терри-
ториальные единицы низкого уровня последовательно, как матрешки,
вкладываются в единицы более высокого ранга. Конкретные экосисте-
мы объединяются в территориальные единицы — биохоры, которые об-
разуют иерархический ряд, т. е. могут быть расположены в порядке от
низших, меньшей размерности, к высшим, более крупным (рис. 6.6, 6.7).
Иерархические уровни
Рис. 6.6. Модель иерархической структуры биосферы.
Сплошной линией разграничены биохоры континентального (I),
13G пунктиром — регионального (II), точками — топологического (III) уровней
ГЛАВА 6. ЭКОЛОГИЯ I ГЕОГРАЙИЯ
Рис. 6.7. Иерархическая структура биосферы, по Т.Т. Forman
Применительно к решению задач ландшафтно-экологических иссле-
дований, придерживаясь рекомендации Б.В. Виноградова (1999), усло-
вимся различать пять иерархических уровней биохор и отвечающих им
подразделений географической оболочки — геохор:
— микрохоры и мезохоры — биогеоценозы, входящие в состав мор-
фологических единиц ландшафта размером 10“1—10-2 км2;
— макрохоры — биогеографические (геоботанические) районы,
охватывающие территорию конкретных ландшафтов размером
10—102км2;
— мегахоры — единицы биогеографического (фитогеографическо-
го) районирования размером 103— 105 км2;
— гигахоры — главнейшие элементы биосферы и географической
оболочки; океаны и материки, зональные типы биомов размером
более 106км2.
Принцип размерности — один из главнейших в теории ландшафт-
ной экологии. Следуя ему, необходимо четко задавать территориаль-
ный уровень, на котором должны вестись исследования экосистем и
разрабатываться модели экологических связей. Переход от одного струк-
турного уровня к другому сопровождается качественным изменением
свойств системы. Практическое значение этой закономерности заклю-
чается в том, что использование модели ограничивается тем рангом
системы, для которого она разработана. 137
ЧАСТЬ II. ЭКОЛПГИЧЕЕКИЕ И ЛАНДШАЙГВЫЕ ЙДКТОРЫ БИОЛОГМЧЕЕЮГО РАЗНООБРАЗИЯ
Аксиома о границах географических, экологических и приролно-
хозяйственных систем: каждая система занимает определенную пло-
щадь и объем и отделена от соседних систем границами. Биосфера как
планетарная система обладает свойствами непрерывности (континуаль-
ности) и прерывностности (дискретности).
Границы между территориальными системами обладают барьерны-
ми свойствами, они разделяют потоки вещества и препятствуют рас-
пространению организмов. Вместе с тем латеральные потоки связывают
соседние ПТК в нарадинамические системы; континуальный характер
переходов вызывает явление экотона — краевого эффекта, обусловли-
вающего тенденцию к увеличению разнообразия и плотности организ-
мов на окраинах двух соседствующих биогеоценозов.
В каждом конкретном случае границы между территориальными
системами могут быть линейными или расплывчатыми, четко выражен-
ными или затушеванными, стабильными или подвижными, однако они
объективно существуют независимо от того, обнаружены они или нет.
Проведение границ есть начало и конец каждой ландшафтно-экологи-
ческой работы.
И 6.3. Топологический уровень биогеографических исследований______
Под топологическим уровнем подразумеваются ПТК внутриланд-
шафтной размерности, т. е. морфологические единицы ландшафта.
Остановимся на двух главных морфологических единицах — фации и
урочище — как объектах ландшафтно-экологических исследований.
Фация — наименьший элементарный природно-территориальпый
комплекс. Она занимает одно местоположение — форму микрорельефа
или одну элементарную поверхность мезорельефа; сложена одной ли-
тологической разностью покровных отношений или приурочена к од-
нородным по петрографическому составу выходам коренных пород; за-
нята одним растительным сообществом, одним почвенным контуром.
Приведенная выше схема типов местоположений Л.Г. Раменского
(рис. 6.3) может служить основанием для выделения ПТК фациальной
размерности.
Фация — первичная ячейка, в которой совершаются процессы об-
мена веществом и передачи энергии компонентами экосистемы. Глав-
ную роль здесь играют жизнедеятельность организмов, их взаимоотно-
шения между собой и со средой. С биоцентрических позиций фация
трактуется как биогеоценоз.
В качестве примера приведем карту фаций ключевого участка юж-
нотаежного сельгово-ложбинного ландшафта Северо-Западного Прила-
дожья, Карельский перешеек (рис. 6.8). Как видим, при выделении
фаций учитываются как особенности абиотических факторов, гак и
компонентов биоты — растительности.
Пространственная организация фациальной структуры территории
во многом определяется вещественно-энергетическими потоками (гео-
потоками), как вертикальными (между различными природными ком-
понентами), гак и горизонтальными, или латеральными (между сами-
ГЛАВА 6. ЭКОЛОГИЯ I ГЕОГРАЙИЯ
ми ПТК). Оба типа потоков всегда пространственно совмещены и не-
разделимы, однако в каждом случае удается выявить доминирующее зна-
чение одного из них в ландшафтной организации.
Э.Г. Коломыц (1987) подчеркивает необходимость переноса центра
тяжести ландшафтных исследований на всестороннее изучение геосис-
тем, образованных направленными латеральными геопотоками и обла-
дающих вследствие этого взаимной функциональной соподчиненно-
стью и векторной плановой структурой. М.А. Глазовская (1964) назвала
элементы такой организации ландшафтно-геохимическими звеньями;
Ф.Н. Мильков (1981) — парагенетическими и парадинамическими комп-
лексами. Парадинамические системы, в которых между подсистемами
возникают пороги перепада массы и энергии, называют также каскад-
ными. В зарубежной литературе подобное пространственное объедине-
ние фаций получило название катены.
На пестрой литогенной основе, при чередовании грунтов с различны-
ми физико-химическими свойствами, при пересеченном микрорельефе
ит. п., формируется мозаичная фациальная структура территории. Внут-
ренняя структура фаций может осложняться также образованиями, ко-
торые Б.Б. Полынов (1953) назвал предельными структурными элемента-
ми. Размеры их колеблются от нескольких сантиметров до нескольких
метров. Их особенность состоит в том, что сама природа определяет их
небольшие размеры: это сурчины или микрозападины в степи, болотные
кочки, приствольные или прикустовые бугры и т. п. Предельные струк-
турные элементы создают подчас весьма пеструю внутреннюю структу-
ру фации, растительного сообщества, биогеоценоза.
Особую категорию фациальной структуры образуют серийные фа-
ции (Сочава, 1961), которые представляют собой последовательный ряд
стадий развития фаций в условиях относительно быстрого течения гео-
графических процессов. В естественных условиях это, например, серия
фаций, сменяющих друг друга в процессе формирования поймы; в усло-
виях антропогенного воздействия — смены фаций, сопровождающие
процесс зарастания отвалов горных пород, стадии дернового процесса
на заброшенном поле и т. п.
Природные (коренные) фации испытывают сильные изменения в ре-
зультате воздействия человека. Хозяйственная деятельность особенно
сильно изменяет растительный покров и животное население фации, а
также почвы, водный режим и т. п. При сельскохозяйственном освое-
нии земель часто не учитываются мелкоконтурные фациальные разли-
чия и создаются сельскохозяйственные угодья, раскинувшиеся на боль-
ших площадях.
Однако распашка, например, не может полностью нивелировать
различия на участках, занимающих разные местоположения. Это ска-
зывается прежде всего на урожайности сельскохозяйственных культур.
Таким образом, производные варианты фаций всегда обнаружива-
ют связь с коренными типами, и эта связь должна учитываться в хо-
зяйственной деятельности.
Урочище истолковывается как участок с хорошо выраженными гра-
ницами, отличающийся от окружающей местности. Наиболее надеж-
ным является выделение урочищ по характеру литогенной основы, т. е.
мезоформам рельефа, однородным по происхождению, сложенным оди-
139
ЧАСТЬ II. ЭКОЛПГИЧЕЕКИЕ И ЛАНДШАЙГВЫЕ ЙДКТОРЫ БИОЛОГМЧЕЕЮГО РАЗНООБРАЗИЯ
140
Рис. 6.8. Карта фаций ключевого участка, поА.Г.
ГЛАВА 6. ЭКОЛОГИЯ I ГЕОГРАНИЯ
Расшифровка к рис. 6.8: I. Группа сельговых фаций: 1 — скальные фации.
Выходы гранитов; мелкозем только в трещинах и микропонижениях; увлаж-
нение резко неустойчивое; комплексный мохово-лишайниковый покров с
редкой сосной IV—V бонитета; 2 — фации крутых склонов. Уклоны свыше
10 — 15°; незначительный и неравномерный слой мелкозема; увлажнение не-
устойчивое; редкостойные сухие сосняки брусничные и вересковые; 3 — фации
выровненных вершин (уклоны до 5°) и верхних склонов (уклоны 5 — 10°).
Маломощный слой элювиально-делювиального материала; сосняки бруснич-
ные и чернично-брусничные на маломощных слабоподзолистых почвах; 4 —
фации нижних склонов южной экспозиции. Уклоны 3—10°; значительный слой
делювия; сосняки южнотаежного типа на слабоподзолистых почвах; 5 — фа-
ции нижних склонов северной экспозиции. Уклоны 5—10°; значительный слой
делювия; ельники южнотаежного типа на слабоподзолистых почвах; 6 — фации
подножий сельг. Сероольховые опушки. II. Группа фаций ложбин и низких
террас, сложенных ленточными суглинками: А. Облесенные и заболоченные
ложбины. 7 — фации облесенных ложбин. Уклоны до 2°; обильное слабопро-
точное увлажнение; сосново-березово-сероольховый лес на дерново-слабопод-
золистых (частью оглеенных) почвах; 8 — фации заболоченных ложбин. Ук-
лоны до 2°; застойное увлажнение; редкостойные сфагново-долгомошные
березняки на торфянисто-глеевых почвах. Б. Окультуренные ложбины и низ-
кие озерные террасы с сетью дренажных канав, с различными вариантами
злаково-разнотравных лугов и перелогами на дерново-скрытоподзолистых
почвах; 9 — фации приопушечные. Уклоны обычно свыше 5°; 10 — фации
склонов террас. Уклоны свыше 5°; И — фации низких террас открытых лож-
бин. Уклоны 2 — 5°; 12 — то же. Уклоны до 2°; 13 — фации узких плоских лож-
бин. Уклоны до 2°; 14 — фации замкнутых ложбин с вогнутой поверхностью.
Уклоны до 2°. III. Группа фаций зарастающих озер: 15 — переходные фации при-
брежных гигрофитов (вдоль меженного уреза воды); 16 — фации хвощевые.
Глубины до 0,5 м; растительный покров сомкнутый; 17 — фации хвощевоосо-
ковые. Глубины 0,5—1,2 м; растительный покров сомкнутый; 18 — фации кув-
шинковокубышковые. Глубины от 1,2 до 2 — 3 м; покров несомкнутый
наковыми по составу породами. Урочища могут выделяться на равнине,
где мезоформы выпуклостей и вогнутостей рельефа отсутствуют. В этом
случае разные урочища будут формироваться на отличающихся физико-
химическими свойствами породах (рис. 6.9).
Литогенная основа является важным экологическим фактором.
Физико-химические свойства пород влияют на характер почвообразо-
вательного процесса, режим увлажнения, аэрацию, богатство почв и т. п.
От рельефа зависят распределение стока, экспозиция склонов, энергия
процессов денудации, микроклимат.
Урочище — более устойчивое к антропогенному воздействию обра-
зование. Даже уничтожение почвенно-растительного покрова и замена
его на техногенный покров может не изменить существа литогенной
основы того или иного урочища, например речной террасы, на которой
построен жилой массив города. Вместе с тем человек, вооруженный
современной техникой, может преобразовывать или уничтожать целые
урочища, создавать новые рукотворные искусственные террасы, карье-
ры горных выработок, терриконы.
141
ЧАСТЬ II. ЭКОЛПГИЧЕЕКИЕ И ЛАНДШАЙГВЫЕ ЙДКТОРЫ БИОЛОГМЧЕЕЮГО РАЗНООБРАЗИЯ
Рис. 6.9. Типы урочищ, по АЛ. Макуниной. Смена урочищ в пространстве,
предопределенная: I — разной мощностью рыхлых отложений;
И — тем же с дополнительным влиянием грунтовых вод; Ш — разным лито-
логическим составом рыхлых отложений; TV — разным литологическим со-
ставом коренных пород; V— положением на элементах рельефа разной кру-
тизны и экспозиции и выходами разных пород; 1—5 — разные типы урочищ
Морфологические ПТК не всегда четко выделяются в ландшафте.
С постепенными переходами между морфологическими единицами
ландшафта связано свойство континуальности (непрерывности) геогра-
фической оболочки. В этом случае между соседними ПТК выделяется
переходная полоса — экотон.
Фации, как уже отмечалось, могут образовывать мозаику,, не подчи-
няющуюся какой-либо пространственной закономерности. Раститель-
ный покров в этом случае будет характеризоваться комплексностью,
сочетанием разнородных по составу и строению фрагментов раститель-
ных сообществ. Таковы, например, мелкобугристые пески в пустыне.
В этом случае бывает трудно выделить урочище как систему определен-
ным образом организованных фаций.
Размеры фаций и урочищ могут сильно варьировать. На однородном
субстрате формируются значительные по площади фации (фации тростни-
ковых плавней по плоским отмелым берегам Северного Каспия). В то же
время четко выраженный бэровский бугор, возвышающийся над ильме-
нями, даже небольших размеров, относится к рангу урочища. Иногда
природно-территориальные комплексы образуют постепенный ряд от
небольших до значительных. Переход от солончакового понижения в
Прикаспийской полупустыне или прирусловой гривы речной поймы фа-
циальной размерности к размерности солончака или гривы, представляю-
щих собой полноценные урочища, зачастую совершается постепенно.
ГЛАВА 6. ЭКОЛОГИЯ I ГЕОГРАЙИЯ
В практической работе следует обращать внимание прежде всего на
выделение однородных по природным и экологическим условиям участ-
ков земной поверхности (иногда повторяющееся сочетание разнород-
ных элементов может рассматриваться как однородность). Определе-
нием же ранга ПТК — фация это или урочище можно пренебречь.
Природно-хозяйственные системы — морфологические ПТК — луга,
леса, болота и т. п., рассматриваемые человеком как разного рода природ-
ные угодья. На их месте создаются сельскохозяйственные угодья: пашни,
сады, лесопосадки, сенокосы и пастбища, а также разного рода техноген-
ные комплексы: города, промышленные предприятия, транспортные ма-
гисчрали и т. д. Хозяйственное использование природного угодья может
меняться, но принадлежность его к определенному природному типу ос-
тается неизменной, за исключением тех случаев, когда техногенное воз-
действие коренным образом изменяет его природу. Таковы, например,
открытые горные выработки, терриконы, крупные водохранилища и т. д.
При изучении природно-хозяйственных систем традиционные био-
географические объекты — естественные биомы — дополняются тре-
мя типами антропогенно-преобразованных экосистем.
Первый тип преобразования состоит в повышении биологической
продуктивности экосистем без изменения их типа. Примером может
служить внесение удобрений на естественных пойменных лугах, руб-
ки ухода в лесу, исключающие, однако, превращение одного типа эко-
систем в другой.
Второй тип антропогенных преобразований преследует цель заме-
ны одного типа экосистем другим, что бывает обусловлено конкретны-
ми хозяйственными задачами. В качестве примера можно привести
вырубку леса с последующей заменой его лугом или пашней, превраще-
ние лугов в пахотные земли и т. п.
Третий тип преобразований приводит к полной или частичной де-
струкции естественных экосистем, что имеет место при отводе земель под
города и села, заводы и фабрики, горнорудные разработки, транспортные
артерии и т. п. Каждому виду человеческой деятельности соответствуют
различные пространственные структуры. Это положение привело
Г.И. Швебса (1987) к разработке концепции природно-хозяйственных
систем (ПХС), согласно которой в зависимости от природных условий, вида
хозяйственных объектов, их плотности, интенсивности обмена веществ и
других факторов формируются вторичные по отношению к исходным ланд-
шафтам природно-хозяйственные системы разного ранга. В современных
условиях, когда биосфера сильно трансформирована человеком, биогео-
граф чаще имеет дело не с естественными экосистемами и ландшафтами,
а с природно-хозяйственными системами. В объекте исследования биогео-
графа неизбежно сливаются воедино природная и социальная среды.
На топологическом уровне Г.И. Швебс обосновывает выделение трех
категорий природно-хозяйственных (ПХ) территориальных единиц:
контуров, массивов, местностей.
Природно-хозяйственный контур — элементарная единица, приуро-
ченная преимущественно к одному элементу рельефа, однородная по
технологии использования техногенного покрова и хозяйственного
функционирования. Примеры: селитебный контур (квартал городской
ЧАСТЬ II. ЭКРЛОГИЧЕСКИЕIIЛДИЛШДИТВЫЕШТОРЫ БКОЛОГМЧЕБКОГО РДЭНООБРДЗИЯ
застройки); рекреационный контур (городской сквер); промышленный
контур (заводская территория); сельскохозяйственный контур (пахот-
ные, садовые, сенокосные угодья — рис. 6.10).
Рис. 6.10. Контуры сельскохозяйственных угодий в окрестностях поселка,
по И. Шмитхюзену
Природно-хозяйственный массив — группа ПХ контуров, приурочен-
ная к смежным элементам рельефа, образующая единую систему с
преобладанием одного типа техногенного покрова и хозяйственного
функционирования. Примеры: селитебный массив (жилой район); сель-
скохозяйственный массив (группа пахотных угодий).
Природно-хозяйственная местность — сочетание природно-хозяй-
ственных массивов, объединяемых общими историко-ландшафтными
предпосылками развития, со своим типом архитектурно-планировоч-
ной организации, функционирования, однотипным подходом к опти-
мизации природопользования. Примеры: селитебно-промышленная
местность (планировочный район); лесохозяйственная местность (со-
четание лесхозов и лесничеств); лечебно-рекреационная местность (са-
наторно-курортная зона города).
Хозяйственная деятельность вызывает появление новых ПХ систем.
Однако природные границы при этом не исчезают. Они рассекают хозяй-
ственные контуры на участки с разными свойствами. При переходе через
естественную границу меняется весь комплекс природных условий.
Часть III
БИОГЕОГРАФИЧЕСКАЯ
КАРТИНА МИРА
Внешние оболочки Земли — лито-
сфера, гидросфера и атмосфера обо-
значают наиболее существенные чер-
ты лика нашей планеты. Но там, по
словам А. Гумбольдта, где миросозер-
цание стремится подняться до более
возвышенной точки зрения, эта кар-
тина была бы лишена своей наиболее
очаровательной прелести, если бы она
не представила нам и сферу органи-
ческой жизни в разных степенях ее
типичного развития.
Жизнь на Земле отличается огром-
ным разнообразием как систематиче-
ского состава флоры и фауны (биоты),
так и сообществ живых организмов
(биомов).
Расхождение признаков (диверген-
ция) в ходе микро- и макроэволюци-
онных процессов определило своеоб-
разие биот биогеографических регионов разного ранга. В ходе
естественно-исторического развития под влиянием физико-геогра-
фических факторов складывались биомы природных зон на суше и
в Мировом океане. Внутри ареалов распределение видов и сообществ
контролировалось ландшафтно-экологическими условиями.
Общее представление о количестве видов и их распределении по
основным систематическим группам дает таблица.
Обратим внимание на особенности в соотношениях видов растений
и животных в современном органическом мире. Численность видов
животных примерно в четыре раза больше числа видов растений; насе-
комые составляют 75% от числа видов животных. На суше растет 92%
всех видов растений и живет 93% всех видов животных. На суше биомас-
са растений составляет 2400 х 109т (99,2%); животных 20 х 109т (0,8%). Это
соотношение отражает фундаментальный экологический закон (закон
пирамиды биомасс) и показывает ведущую роль растительности в орга-
низации и распределении как биоценозов, так и биомов любого ранга.
Гумбольдт пишет: «Хотя характер различных местностей земного шара
определяется одновременно всеми внешними явлениями, хотя очерта-
ния гор, физиономия флоры и фауны, синева неба, строение облаков и
прозрачность воздуха влияют на общее впечатление, тем не менее нельзя
отрицать, что главное, определяющее это впечатление обусловливается
растительным покровом». На формирование биот большое влияние ока-
зали четвертичные оледенения Северного полушария. Похолодание кли-
мата в меньшей степени затронуло тропические широты. Поэтому здесь
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ КАРГИ0А МИРА
Количество видов основных групп живых организмов в биосфере,
по В. Groombridge
Организмы Растения Животные Другие царства и подцарства
Растения:
Покрытосеменные 346 000
Голосеменные 600
Высшие споровые:
Плауновидные 1000
Хвощевидные 25
Папор отниковидны е 10 000
Моховидные 27 000
Водоросли 40 000
Животные:
Позвоночные 40 000
Моллюски 70 000
Ракообразные Паукообразные 40 000
и клещи 75 000
Насекомые 950 000
Круглые черви Грибы 15 000 100 500
Бактерии 4 000
Одноклеточные 40 000
Вирусы 5 000
Всего по группам Всего в биосфере: 1 764 125 424 625 1 190 000 149 500
располагаются наиболее древние и богатые по составу биоты материков.
Иллюстрацией сказанному может служить карта флористического раз-
нообразия суши (рисунок). Отчетливо видно резкое уменьшение числа
видов растений по мере продвижения в высокие широты.
Б. Грумбридж (Global Biodiversity, 1992) приводит следующие данные
о количестве видов высших растений на континентах.
Южная Америка.....85 000
Северная Америка. 17 000
Африка.................45 000
Тропическая Азия..50 000
Австралия..............15 000
Европа................ 12 500
148
Показатели флористического разнообразия материалов, по Е.В. Вульфу
Глава 7
БИОТИЧЕСКИЕ ЦАРСТВА СУШИ
Биогеограф разделяет биоты, опираясь на различия в систематиче-
ском составе растений и животных. Главный упор делается на выявление
регионального своеобразия флор и фаун. Например, чтобы иметь представ-
ление о биоте Австралии, необходимо прежде всего составить список видов
и более высоких таксонов организмов, обитающих на этом континенте.
Наличие в этом списке эндемичных семейств, родов и видов служит осно-
ванием для выделения Австралии в особое биогеографическое царство.
Согласно эволюционной модели различия состава биот становятся
тем больше, чем дольше сохраняется изоляция. Теория дрейфа конти-
нентов позволяет объяснить особенности самых крупных биогеогра-
фических подразделений суши (см. гл. 1). По этой теории современные
континенты представляют собой осколки некогда единой Пангеи, су-
ществовавшей в конце палеозоя. Южные континенты прошли стадию
разделения раньше, чем северные; с этим связаны их значительные био-
географические различия. Биоты северных континентов, между кото-
рыми еще недавно существовали мосты суши, обладают многими чер-
тами сходства.
Названные особенности исторического развития явились предпо-
сылкой первого подразделения материковых фаун на царства: Нотогею
(сумчатые и однопроходные), Неогею (неполнозубые) и Арктогею (пла-
центарные) .
В последующем границы территорий со сходной историей разви-
тия, с самым высоким рангом эндемизма (на уровне семейств и родов),
высокой долей эндемиков более низкого таксономического ранга, осо-
бым набором географических элементов флоры и фауны стали уточ-
няться и выделяться в качестве высших таксономических единиц био-
географического районирования суши — царств. Ботаники выделяют
флористические, а зоологи фаунистические царства1.
Однако выделение даже самых крупных биогеографических подраз-
делений встречает трудности. Во-первых, многие систематические груп-
пы имеют очень широкое распространение и, следовательно, малопри-
годны для районирования.
’ Первоначально зоологи высшие единицы районирования материков называли фау-
нистическими царствами, однако позже более употребительным стал термин «фаунисти-
ческие области». Для единообразия я сохраняю для всех высших единиц биотического
районирования термин «царство».
ГЛАВА 1. БИОТИЧЕСХЙЕ ЦАРСТВА СУША
Во-вторых, различные систематиче-
ские группы организмов (например, цвет-
ковые и высшие споровые растения,
млекопитающие, птицы, рептилии, чле-
нистоногие) дают подчас несовпадаю-
щие картины своего распространения.
Какую именно группу взять за основ-
ную — не ясно и крайне спорно.
Флористическое и фаунистическое
районирование суши имеет давнюю ис-
торию. Своеобразие основных флори-
стических царств было обозначено в
конце XIX — начале XX в. А. Энглером
и Л. Дильсом. Последняя схема флори-
стического районирования принадле-
жит А.Л. Тахтаджяну (1978; рис. 7.1, А).
Фаунистическое районирование впер-
вые было выполнено в XIX в. П. Склете-
ром и А. Уоллесом. В России схема фау-
нистического районирования уточнена Н.И. Вавилов (1887—1943)
В.Г. Гептнером (1936; рис. 7.1, Б).
Вопрос о том, можно ли выделять единые биогеографические цар-
ства или следует рассматривать их отдельно по флористическому и фау-
нистическому принципу, до последнего времени дискутируется. Од-
нако если пренебречь частностями, то мы увидим, что обобщенные
схемы флористического и фаунистического районирования во многом
совпадают (рис. 7.1, В). Следует, однако, подчеркнуть, что выделенные
границы не бесспорны.
Своеобразие биот определяется набором характерных для каждой
из них полезных видов растений и животных, в том числе и тех, кото-
рые человек ввел в культуру. Это положение легло в основу учения
Н.И. Вавилова о центрах происхождения культурных растений (рис. 7.2).
Перейдем к краткой характеристике биогеографических царств суши.
Описания составлены на основании работ А.Г. Воронова, В.Г. Гептнера,
Е.М. Лавренко, И.И. Пузанова, А.Л. Тахтаджяна, А.И. Толмачева и других.
И 7.1.Голарктичоскоо царство__________________________________________
Голарктическое царство занимает большую часть суши к северу от
тропика Рака: Европу, внетропическую Азию (Палеарктику) и Север-
ную Америку (Неарктику). В палеозое вся суша в Северном полушарии
образовывала единый материк Лавразию. Отчленение Северной Аме-
рики от Европы в результате образования Атлантического океана про-
изошло в мезозое.
Благодаря общей геологической предыстории биоты Голарктиче-
ского царства характеризуются большим сходством, причем оно быстро
возрастает при продвижении с юга на север, доходя почти до полного
тождества в зоне тундры, опоясывающей северные побережья Европы, 151
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ КАРГИ0А МИРА
Рис. 7.1. Царства суши, выделяемые по флористическому, фаунистическому
и общим биотическим признакам. А. Флористические царства,
по А.Л. Тахтаджяну: 1 — Голарктическое; 11 — Палеотропическое; III — Кап-
ское; IV — Неотропическое; V— Австралийское; VI — Голантарктическое.
Б. Фаунистические царства, по В,Г. Гептнеру: I — Голарктическое:
1, а — Палеарктическая область; 1,6 — Неарктическая область; II — Эфиоп-
ское; III — Индо-Малайское (Восточное, или Ориентальное); W— Неотропиче-
ское; V — Австралийское; VI— Мипарктическое: В. Биогеографические царства:
I — Голарктическое: 1,а — Палеарктическая область, 1,6 — Неарктическая об-
ласть; II — Эфиопское; III — Индо-Малайское (Восточное, или Ориентальное);
IV — Неотропическое; V — Австралийское; VI — Голантарктическое
ГЛАВА 1. БИОТИЧЕСХЙЕ ЦАРСТВА СУША
Рис. 7.2. Центры происхождения культурных растений, по Н.И. Вавилову:
I — Юго-Западная Азия; II — Юго-Восточная Азия;
III — Средиземноморское побережье; IV — Абиссиния и Эритрея;
V— горные районы Южной Америки и Мексики
Азии и Америки. Это сходство отчасти объясняется и тем, что зональ-
ные типы ландшафтов (тундры, тайги, широколиственных лесов, сте-
пи) в одинаковой последовательности повторяются в высоких и сред-
них широтах Восточного и Западного полушарий.
В составе голарктической флоры более 30 почти эндемичных се-
мейств. К их числу относятся: магнолиевые, лавровые, лютиковые, бар-
барисовые, березовые (береза, ольха, граб, орешник и др.), буковые (бук,
дуб), кленовые (клен), сосновые (сосна, ель, пихта, лиственница и др.).
Преимущественно к флоре Голарктического царства относятся гвоздич-
ные, розоцветные, зонтичные, гречишные, первоцветные и др.
Голарктическое царство — родина многих культурных растений,
ставших основой для формирования древнейших земледельческих ци-
вилизаций. Средиземноморский центр дал вишню, сливу, маслину, лавр,
капусту, морковь, тмин, свеклу, петрушку, спаржу, лук, чеснок, лен, мак;
Юго-Западноазиатский центр (Средне- и Переднеазиатский) — вино-
град, грушу, персик, алычу, фисташку, миндаль, абрикос, яблоню, ин-
жир, гранат, дыню, пшеницы однозернянку и двузернянку, ячмень,
рожь, грецкий орех, бобовые (горох, чечевицу, маш, нут), коноплю,
морковь, лук, чеснок; Восточноазиатский (Китайский) центр — просо,
гречиху, сою, хурму, чай, шелковицу, местные сорта абрикоса, перси-
ка, сливы, вишни, айвы, капусты, моркови.
Большое влияние на формирование биоты Голарктики оказали со-
бытия третичного и четвертичного периодов. В палеогене суша была
занята двумя лесными флорами (а по сути дела — биомами) — тургай-
ской и полтавской. Первая состояла главным образом из листопадных 153
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ К АРТИ В А МИРА
и хвойных пород, свойственных теплоумеренному климату; к ним при-
мешивались некоторые субтропические растения (например, Sequoia,
Taxodium). Эта флора занимала обширные пространства на севере, да-
леко проникая в современную безлесную Арктику.
Полтавская флора состояла преимущественно из вечнозеленых де-
ревьев и кустарников, характерных для тропического и субтропическо-
го климата (фикусы, лавровые, миртовые, пальмы и др.). Ее северная
граница проходила приблизительно по линии, соединяющей устья рек
Вислы и Янцзы.
По берегам древнего океана Тетис — в Передней, Средней и Цент-
ральной Азии, но побережью современного Средиземного моря, а так-
же во внутренних пустынных областях Северной Америки сложилась
ксерофильная флора Древнего Средиземья. Она образовывала расти-
тельность степей, пустынь, сухих редколесий и жестколистных лесов с
засухоустойчивой флорой из эфедровых, тамариксовых, ворсянковых,
маревых, губоцветных и других растений.
Начиная с неогена, тургайская флора, в связи с охлаждением клима-
та, стала вытеснять полтавскую. Последовавшие затем покровные чет-
вертичные оледенения привели к глубокому преобразованию раститель-
ного покрова. Расчленению древней третичной флоры способствовало
также возникновение альпийской системы складчатости на Европей-
ско-Азиатском материке и в Северной Америке.
Современная биота Голарктики отличается относительной бедностью
вследствие как суровости современного климата, так и опустошения,
вызванного четвертичными оледенениями. Вместе с тем оледенения
повлекли за собой возникновение столь характерных для Голарктики
холодолюбивых тундровых и таежных форм животных и растений.
Ледники покрывали большую часть территории Евразии и Север-
ной Америки. В Евразии свободными ото льда оставались равнины и
предгорья Южной Европы и Кавказа, Южный Урал, предгорья Алтая и
Саян. В этих «убежищах жизни» (рефугиумах) сохранялась лесная
флора, и из них началось новое расселение растительности на освобо-
дившуюся ото льда голоценовую сушу. Изучение ареалов основных
древесных лесных пород показывает, что широколиственные леса с
буком, грабом, ясенем, дубом и др. распространялись на восток из
Центральной Европы и последовательно захватывали Восточно-Европей-
скую равнину на восток вплоть до Волги и на север до Онеги. Далее всех
на север и восток продвинулись липа, остролистный клен, вяз, в мень-
шей степени орешник, дуб, граб, менее всего — бук. Вместе с широко-
лиственными породами проникли с запада еще ель и сосна.
Главным ледниковым «убежищем жизни» сибирской тайги были
предгорья Алтая и Саян, а также Южный Урал. Из этих мест таежные
породы (вначале сибирские ель и лиственница, а также сосна, затем
сибирская пихта и позже всех сибирский кедр) распространились к
северу и западу — за Урал, до бассейна рек Северной Двины и Онеги.
Вместе с проникшими сюда с запада европейской елью и сосной они
образовали широкий пояс темнохвойной тайги, простирающийся на
север до зоны тундры. К востоку от Енисея (в Забайкалье и всюду в
154 Якутии) распространились светлохвойные леса из даурской лиственни-
ГЛАВА 1. БМОТИЧЕСХЙЕ ЦАРСТВА СУШИ
цы и сосны. Эти породы во время ледниковых эпох сохранялись в Вос-
точной Сибири и на Дальнем Востоке с третичного периода, т. к. по-
кровное оледенение здесь отсутствовало.
В течение ледникового периода территория современных равнин-
ных степей располагалась по окраинам ледниковых щитов, в так назы-
ваемой перигляциальной тундро-степной гиперзоне, и подвергалась всем
губительным влияниям этого соседства. Окончательно степной тип ра-
стительности смог сложиться только в послеледниковое время.
Современная тундровая зона в основном сложилась в позднечетвер-
тичное время; в особенности это относится к западной части Европей-
ско-Азиатского материка. На севере Центральной и Восточной Сибири
тундровая флора сохранилась до наших дней, по-видимому, с раннечет-
вертичного времени.
Основными эндемичными группами позвоночных животных в Гол-
арктическом царстве являются: млекопитающие — бобры, кроты, туш-
канчики; птицы — тетеревиные, гагары, чистиковые; рыбы — осетро-
вые, лососевые, щуковые, колюшковые и др.
Голарктическое царство делится на Палеарктическую и Неарктиче-
скую области, характеризующиеся чертами как сходства, так и разли-
чия. Общности биот, в частности фаун Палеарктики и Неарктики, спо-
собствовала связь Северо-Восточной Азии с Северной Америкой через
Берингию, прекратившаяся лишь в послеледниковое время. По мосту
суши между Аляской и Восточной Сибирью происходила широкая
миграция видов (рис. 7.3).
Рис. 7.3. Обмен фаун Азии и Северной Америки, по Дж. Симпсону.
Положение животных соответствует направлению движения данных видов 155
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ КАРТИ В А МИРА
Америка получила с Европейско-Азиатского материка горного бара-
на, мамонта (вскоре вымершего), лося, оленя вапити, бурого медведя и
др. В свою очередь Америка дала Европейско-Азиатскому материку
северного оленя, овцебыка и др.
Кроме того, существовала постоянная связь Палеарктики с биотами
тропической Африки и Индии. Связь Неарктики с Южной Америкой
осуществляется через Панамскую сушу. Это обстоятельство способство-
вало вселению в Северную Америку многих южноамериканских видов.
Остановимся на характерных особенностях фаун Палеарктики и
Неарктики.
Палеарктическая область (Палеарктика) включает всю Европу,
внетропическуто часть Азии (с прилежащими островами) и Северную
Африку. Среди млекопитающих много эндемичных родов и видов, на-
пример выхухоль (из насекомоядных), барсуки (из хищных), хомяки
(из грызунов), верблюд, як, косуля, сайга и серна (из копытных). Среди
птиц также имеются эндемики: семейство усатых синиц, роды соловь-
ев, тетеревов, глухарей, горных индеек, турухтанов, серпоклювов.
Фауна Палеарктики на ее громадной территории обладает существен-
ными различиями (рис. 7.4, 7.5). В Арктической подобласти она настоль-
ко близка к фауне тундры Неарктической области, что иногда говорят
о единой циркумполярной зоне, населенной такими характерными
формами, как песец, пеструшки, белая и тундряная куропатки.
Южная половина Палеарктики имеет разнообразную теплолюбивую
фауну, пополненную выходцами из Африки и Индии.
Неарктическая область (Неарктика) занимает весь материк Север-
ной Америки с прилежащими островами: Алеутскими, Гренландией,
Ньюфаундлендом и Багамскими. Фауна Неарктики отличается от фау-
ны Палеарктики наличием ряда эндемичных групп животных (рис. 7.6).
Среди млекопитающих животных эндемичны четыре семейства: ви-
лорогие антилопы, бобровые белки, мешотчатые крысы (или гоферы),
мешотчатые мыши. Характерны снежная коза, олени рода Odocoileus,
пекари, скунс, пума, медведь-барибал, бизон, древесный дикобраз. Носо-
роги, верблюды и дикие лошади, населявшие Неарктику в четвертичный
период, в современной фауне не сохранились. Полностью отсутствуют
обезьяны, ежи, виверры и даже настоящие мыши (Mus).
Из птиц по преимуществу эндемичными родами представлены ку-
риные, например луговой тетерев, дикий индюк. Большим разнообра-
зием отличаются дятлы, а также корольки, свиристели, крапивники.
Характерны стервятники. Встречается много семейств птиц южноаме-
риканского происхождения, проникающих в Неарктику, в частности
12 видов колибри.
И 7.2. Палеотропические царства - Эфиопскее и Индо Малайское_____
Во флоре и фауне большей части Африки, тропической Азии (Индо-
стан, Индокитай и Малайский архипелаг) много общего. Определенные
черты сходства биот объясняются общностью естественно-историче-
ского развития: принадлежностью в прошлом к единому Гондванскому
15G материку, а также положением в теплых тропических широтах.
ГЛАВА 1. БМОТИЧЕСХИЕ ЦДРЕТВА СУШИ
Рис. 7.4. Животные Палеарктической области (западная часть)
Голарктического царства, по БСЭ: 1 — лисица; 2 — лесная куница;
3 — белобрюхий глухарь; 4 — филин; 5 — волк; 6 — бурый медведь; 7 — выд-
ра; 8 — бобр; 9 — барсук; 10 — зубр; 11 — выхухоль; 12 — косуля; 13 — заяц-
русак; 14 — серна; 15 — благородный олень; 16 — сайгак; 17 — байбак; 18 —
хомяк; 19 — бесхвостый макак; 20 — дикообраз; 21 — кабан
157
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ КАРТИ В А МИРА
Рис. 7.5. Животные Палеарктической области (восточная часть)
Голарктического царства, по БСЭ: 1 — рысь; 2 — росомаха; 3 — белка;
4 — снежный баран; 5 — колонок; 6 — бурундук; 7 — соболь; 8 — черный
медведь; 9— темнобрюхий улар; 10— антилопа-оронго; 11 — енотовидная
собака; 12 — сибирский козерог; 13 — горал; 14 — кеклик; 15 — длиннохвостый
сурок; 16 — як; 17 — пятнистый олень; 18 — суслик; 19—кулан;
158 20 — джейран; 21 — варан; 22 — уссурийский тигр
ГЛАВА 1. БМ0ТИЧЕЕ1ИЕ ЦДРЕТВА СУШИ
Рис. 7.6. Животные Неарктической области Голарктического царства,
по БСЭ: 1 — толсторогий баран; 2 — снежная коза; 3 — овцебык; 4 — бобр;
5 — древесный дикобраз; 6 — пума; 7 — енот; 8 — индейка; 9 — ондатра;
10 — гофер; 11 — медведь orjyec барибал; 12 — скунс; 13 — койот; 14 — оцелот;
15 — вилорог; 16 — луговая собачка; 17 — луговой тетерев;
18 — виргинский олень; 19 — бизон 159
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ КАРТИ В А МИРА
П. Склетер и А. Уоллес, составившие первую карту зоогеографиче-
ского районирования суши в конце XIX в., выделяли единое царство Па-
леогея, объединявшее названные пространства суши. На современной
карте флористического районирования А.Л. Тахтаджяна (1978) пример-
но в тех же границах показано единое Палеотропическое царство.
Несмотря на определенное сходство, биоты крупных, ныне удален-
ных друг от друга регионов обладают своеобразными чертами, что дало
основание А.Г. Воронову (1987) выделить два самостоятельных биогео-
графических царства: Эфиопское и Индо-Малайское. Прежде чем оста-
новиться на характеристике особенностей этих царств, отметим черты
их флористического сходства.
Флора Палеотропического царства очень богата. Ее характеризует
около 40 эндемичных семейств. Только в Юго-Восточной Азии встреча-
ется около 45 тыс. видов. Помимо преобладающих во флоре Палеотро-
пического царства пантропических и палеотропических элементов в
пределы этой области заходят с севера (преимущественно в горах) го-
ларктические элементы, а с юга — капские (в Африке), голантарктиче-
ские и австралийские (на островах Тихого океана). Количество голарк-
тических элементов во флоре Палеотропического царства, видимо,
увеличилось в конце третичного периода и в плейстоцене в связи с аль-
пийским горообразованием и четвертичным оледенением.
К семействам, свойственным преимущественно флоре Палеотро-
пического царства, относятся пять наиболее известных: двукрыло-
плодные (Dipterocarpaceae), панданусовые (Pandanaceae), непентесовые
(Nepenthaceae), банановые (Musaceae), флагелляриевые (Flagellariaceae).
Кроме того, здесь широко представлены виды рода Ficus и семейства,
характерные для всего тропического пояса (пальмовые, молочайные,
мареновые, ароидные, аноновые, мимозовые, цезальпиниевые, мирто-
вые, бигнониевые, акантовые и многие другие).
Дифференциация флоры внутри Палеотропического царства во
многом определяется климатом и формированием зональных типов
биомов: дождевых тропических лесов, листопадных тропических лесов,
саванн и пустынь.
Палеотропическое царство — родина большого числа культурных
растений.
В Эфиопском центре — сорго, просо, кофейное дерево, твердая
пшеница, клещевина, алоэ; в Индийском центре — рис, цитрусовые,
сахарный тростник, хлопчатник, манго, огурцы, баклажаны; в Индоне-
зийском центре — хлебное дерево, таро, ямс, кокосовая пальма, бана-
ны, дуриан; пряные растения: черный перец, кардамон, корица, гвоз-
дичное дерево, мускатный орех.
Основанием для разделения царств служит прежде всего своеобра-
зие их животного мира.
Эфиопское царство. Располагается в тропических широтах и занима-
ет большую часть Африки (к югу от Сахары), юго-восток Аравийского
полуострова и близлежащие острова в Индийском океане. Рельеф Афри-
канского континента характеризуется обширными нагорьями и плоско-
горьями. В распределении осадков наблюдается закономерный переход
от обильно и постоянно увлажненной экваториальной зоны дождевых
тропических лесов к сезонно-влажным субэкваториальным зонам лис-
ГЛАВА 1. БМОТИЧЕСХЙЕ ЦАРСТВА СУШИ
топадных лесов и саванн и постоянно сухим зонам тропических пустынь.
Характерные представители фауны Эфиопского царства изображе-
ны на рис. 7.7. Особенно богато представлены млекопитающие: встре-
чаются 38 семейств, в том числе 13 эндемичных (жирафы, бегемоты,
трубкозубы, земляные волки, долгоноги, тенреки, златокроты, шило-
хвостые белки и др.). Представители шести семейств характерны и для
Индо-Малайского царства: человекообразные обезьяны, лемуры, слоны,
носороги, оленьки, панголины. Из копытных многочисленны и разно-
образны антилопы, зебры.
Птицы представлены 89 семействами, в том числе 15 эндемичными:
бананоеды, лесные удоды, мышанки, китоглавые цапли, секретари и др.
Представители 12 семейств свойственны и Индо-Малайскому царству:
птицы-носороги, питты, медоуказчики и др. Куриные представлены
только цесарками и африканским павлином. Попугаев мало.
Капское флористическое царство. Занимает территорию внетропи-
ческой Южной Африки. Несмотря на небольшие размеры (около 80 км
шириной и до 800 км длиной — вдоль берега океана), капская флора
настолько своеобразна, что выделяется в самостоятельное царство.
Эндемичными являются 7 семейств, 280 родов, 7000 видов.
Корни капской флоры связаны с растительностью Гондваны. Когда
Африка обособилась, условия существования на ее южной оконечно-
сти оказались изолированными от основной территории континента
безводными пустынями.
Растительность Капского царства напоминает безлесные ландшаф-
ты вересковых пустошей. На каменистых плоскогорьях господствуют
вечнозеленые кустарники, представленные протейными, вересковыми,
теребинтовыми, истодовыми, кисличными, рутовыми. Капская фло-
ра — неисчерпаемый источник декоративных растений, в том числе ама-
риллисов, кливий, гладиолусов и др.
Индо-Малайское (Восточное, или Ориентальное) царство. Занима-
ет тропическую часть Азиатского материка с примыкающими к нему
островами. Северная граница области пересекает пустыню Тар в Запад-
ной Индии, затем проходит по южному склону Гималаев, пересекает
горные хребты на севере полуострова Индокитай и выходит к Тихому
океану примерно по водоразделу рек Хуанхэ и Янцзы.
Из всех тропических царств Индо-Малайское ближе всего стоит к
Эфиопскому, с которым обмен видами происходил вплоть до конца
третичного периода. Столь характерные для современной Африки
жирафы и антилопы являются выходцами из миоценовой и плиоцено-
вой Индии. Вместе с тем из Африки перешли в Индию слоны. Менее
тесны связи с фауной Австралийского царства. Лишь некоторые остро-
ва, в миоцене находившиеся в кратковременной связи с австралийской
сушей, получили оттуда сумчатых (кускусов), большеногих кур и дру-
гих птиц и дали Австралии водяных мышей (Hydromyinae).
Включая в себя страны весьма древней культуры, Индо-Малайское
царство дало ряд ценных домашних животных; например, из млекопита-
ющих — буйвола, горбатого быка зебу, балийский скот, отчасти свинью
и собаку; из птиц — домашнюю курицу и павлина; из насекомых — ин-
дийскую пчелу. Индо-Малайское царство характеризуется весьма разно-
образными природными условиями и неоднородной фауной (рис. 7.8).
1G1
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ КАРТИ В А МИРА
Рис. 7.7. Животные Эфиопского царства, по БСЭ: 1 — гвереца; 2— мышон-
ка; 3 — нектарница; 4 — прыгунчик; 5 — иглохвостая летяга; 6 — окапи;
7 — буйвол; 8 — горилла; 9 — шимпанзе; 10 — бананоед; 11 — долгоног;
12 — антилопа-орикс; 13 — леснойудод; 14 — даман; 15 — носорог; 16 — го-
лого; 17 — страус; 18 — павлин; 19 — оленек; 20 — зебра; 21 — ящер; 22 —
слон; 23 — фенек; 24 — лев; 25 — жирафа; 26 — златокрот; 27 — трубко-
зуб; 28 — антилопа-гну; 29 — земляной волк; 30 — бегемот; 31 — мандрил;
32 — пятнистая гиена; 33 — бородавочник; 34 — шпорцевая лягушка;
35 — птица-носорог; 36 — китоглав; 37 — виверра; 38 — выдровая земле-
ройка; 39 — крокодил; 40 — ламантин; 41 — протоптерус; 42— секретарь;
1G2 43 — цесарка
[ЛАВА 1. БМОТИЧЕСХИЕ ЦДРЕТВА СУШИ
Рис. 7.8. Животные Индо-Малайского царства, по БСЭ: 1 — панда;
2 — мангуст; 3 — малайский медведь; 4 — павлины; 5 — тигр;
6 — птица-носорог; 7— орангутан; 8 — черная пантера; 9 — королевская
белка; 10 — летучая лисица; 11 — макак; 12 — сиаманг; 13 — бантенг;
14 — нильгау; 15 — аксис; 16 — хохлатый павиан; 17 — двуцветный тапир;
18 — бабирусса; 19 — индийский слон; 20 — кобра; 21 — однорогий носорог
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ К АРТИ В А МИРА
Типичны представители тропической, весьма богатой и красочной
фауны. Из копытных встречаются индийский буйвол, гаур, разнообраз-
ные олени, серповидные антилопы, два вида оленьков, тапиры, три вида
носорогов, индийский слон. Из хищных характерны тигр и пантера.
Среди грызунов особенно многочисленны белки, летающие белки и
дикобразы. Весьма обычны громадные рукокрылые (калонги, или лету-
чие собаки). Как и везде в тропиках, разнообразны приматы: долгопяты
(Tarsiidae), лемуры; семейство низших узконосых обезьян представлено
тонкотелыми обезьянами IColobinae) и макаками; человекообразные —
гиббонами и орангутаном. Из птиц характерны: павлины, фазаны, на-
стоящие куры, яркие зимородки, щурки и похожие на колибри нектар-
ки; сравнительно малочисленны попугаи.
И 7.3. Неотропическее царство______________________________________
Неотропическое царство занимает Южную Америку, большую часть
Центральной Америки и прилегающие острова. Четко выраженные
индивидуальные черты неотропиков обусловлены длительной изоляци-
ей континента. Однако на севере границы царства выражены не резко,
т. к. установившаяся в третичном периоде сухопутная связь с Север-
ной Америкой обусловила смешение биот.
Богатству и разнообразию флоры и фауны Южной Америки способ-
ствует широкий спектр природных зон и высотных поясов. На севере и
юге располагаются зоны субтропиков. На западе в Андах, поднявшихся
в конце мелового периода, в горах и на обширных плоскогорьях сформи-
ровались ярко выраженные высокогорные флора и фауна.
Бассейн Амазонки, плоскогорья Бразилии и Гвианы располагаются
в зоне дождевых тропических лесов. Для Мексики и прибрежных райо-
нов Перу и Чили характерны пустыни. Равнины Аргентины заняты
необозримой саванной и степью (пампой).
Материк Южной Америки, являющийся основной территорией раз-
вития неотропической флоры, входил в состав Гондваны, которая к концу
мезозоя распалась. Палеоботанические данные показывают относитель-
ное постоянство развития неотропической флоры в течение всего третич-
ного периода. Соединение Южной Америки с Северной произошло толь-
ко в плиоцене. Это определило, с одной стороны, наличие в Неотропическом
царстве многих пантропических элементов, а с другой — возможность
проникновения по горной цепи Анд некоторых голарктических элемен-
тов, роль которых сравнительно невелика. Наличие антарктических эле-
ментов, особенно во флоре Анд, объясняется тем, что на юге Южной
Америки издавна господствовали элементы антарктической флоры.
Во флоре Неотропиков представлено несколько генетических эле-
ментов: 1) пантропические, характерные для всей тропической зоны;
2) неотропические, свойственные только тропикам и субтропикам Юж-
ной Америки; 3) антарктические, встречающиеся преимущественно в
южных Андах и сближающие флору Неотропиков с флорой Антаркти-
ческого царства; 4) голарктические, также представленные преимуще-
1G4 ственно в Андах.
ГЛАВА 1. БМОТИЧЕСХЙЕ ЦАРСТВА СУШИ
Для Неотропического царства эндемичны семейства Нутепо-
phyllopsidaceae, Marcgraviaceae, Caryocaraceae, Pellicieraceae, Quiina-
ceae, Peridiscaceae, Bixaceae, Cochlospermaceae, Tovariaceae, Lissocar-
paceae, Brunelliaceae, Dulongiaceae, Columelliaceae, Julianiaceae, Picro-
dendraceae, Goupiaceae, Desfontainiaceae, Plocospermataceae, Dialype-
talanthaceae, Nolanaceae, Calyceraceae, Heliconiaceae, Cannaceae,
Thurniaceae, Cyclanthaceae.
Поражает исключительное богатство флоры — около 45 тыс. видов. Из
них почти четверть эндемична для Бразилии. Ни одно тропическое цар-
ство не сравнится с Неотропическим по численности видов и необыкно-
венной красоте орхидей. В Мексиканской области огромное разнообра-
зие кактусовых (около 1,5 тыс. видов), агав, древовидных лилейных — юкк.
Неотропическое царство — родина культурных растений, большин-
ство из которых разводится не только в жарком, но и в умеренных
поясах (рис. 7.9).
Рис. 7.9. Основные пищевые растения, введенные в культуру в Централь-
ной и Южной Америке: 1 — томаты; 2 — кукуруза; 3 — земляная груша
(топинамбур); 4 — какао; 5 — подсолнечник (дикорастущая форма);
6 — картофель; 7 — арахис; 8 — батат
Из Южной Мексики и Центральной Америки произошли: кукуру-
за, некоторые виды фасоли (Phaseolus vulgaris, Ph. multiflorus и др.),
тыквы {Cucurbita pepo, С. moschata и др.), батат [Ipomaea batatas), мек-
сиканский перец (Capsicuum аппиит), некоторые виды хлопчатника
(Gossypium hirsutum), мелкоплодный томат [Lycopersicum crasifonne),
какао (Theobroma), а также другие тропические культурные растения:
анона (Апопа), дынное дерево (Carica papaya), авокадо [Persea americana),
гуайява [Psidium guajava) и др. 1G5
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ КАРТИ В А МИРА
Горные страны Южной Америки — родина многих клубненосных ви-
дов картофеля [Solarium andigenum, S. ajanhuiri, S. juzepczukii и мн. др.),
клубненосных видов кислицы (Oxalis tuberosa и др.), арахиса (Arachis),
крахмалистой кукурузы, томата (Lycopersicum esculentum pyriforme и др.),
а также тыквы (Cucurbita maxima], южноамериканских видов перца, куль-
тивируемых только на родине, некоторых видов хлопчатника (Cossypium
barbadense и др.), хинного дерева (Cinchona), табака и многих др. Обык-
новенный картофель (Solanum tuberosum) происходит из Чили.
Тропические леса равнин — родина фейхоа (Feijoa sellowiana), кау-
чукового дерева — гевеи (Hevea brasiliensis), маниока (Manihot), анана-
са (Ananas comosus), парагвайского чая (Ilexparaguariensis) и др.
По климатическим условиям Южная Америка сходна с Африкой,
между тем их фаунистические различия весьма существенны. В пер-
вую очередь в глаза бросается отсутствие больших стад копытных. Уди-
вительно, что в современной фауне Южной Америки вообще нет особо
крупных четвероногих, подобных слонам, бегемотам, буйволам и др. в
Африке. Хотя в Южной Америке в третичный период жили гигантские
формы ленивцев и броненосцев.
В целом фауна Неотропического царства весьма своеобразна и чрез-
вычайно богата (рис. 7.10). Основными эндемичными группами позво-
ночных животных в Неотропическом царстве являются: из сумчатых —
опоссумы, из отряда неполнозубых — броненосцы, ленивцы, муравье-
ды; характерны кровососущие рукокрылые — вампиры. Из отряда гры-
зунов эндемичны цепкохвостые дикобразы, морские свинки, агути,
вискаши, шиншиллы.
Из отряда хищных встречается несколько видов псовых, один вид
медведей, а из кошачьих — ягуар, оцелот, пума и некоторые другие.
Копытные малочисленны: из непарнокопытных — три вида тапиров, а
из парнокопытных — пекари и мелкие олени (Mazama, Odocoileus,
Pudua); эндемичный род лам.
В реках Южной Америки обитают крупные водные млекопитающие —
ламантины и речные дельфины. Все обезьяны Неотропиков принадле-
жат к подотряду широконосых, включающему как мелких белкообраз-
ных игрунок, так и цепкохвостых обезьян (например, сапажу и ревуны).
Фауна птиц насчитывает 26 эндемичных семейств (виды лишь 9 се-
мейств успели проникнуть в Неарктическую область). Имеются трехпа-
лые страусы — нанду, тинаму (похожие на куропаток), весьма много-
численны и разнообразны древолазы, колибри, попугаи (выделяются
крупные ара и зеленые амазонские попугаи). Из хищных характерны
кондоры и гарпии; интересен единственный представитель отряда —
гоацин, птенец которого лазает по ветвям с помощью когтей на перед-
них конечностях. Из дятловых характерны туканы.
Богатство и разнообразие фауны Неотропического царства объяс-
няется как физико-географическими, так и историческими причина-
ми. Ландшафты Южной и Центральной Америки чрезвычайно пестры.
Большая их часть занята дождевыми тропическими лесами, условия
жизни в которых особенно благоприятны для жизни как растений, так
и животных. Кроме того, здесь имеются аридные редколесья и степи,
166 пустыни и огромные горные массивы и плоскогорья.
ГЛАВА 1. БМ0ТИЧЕЕ1МЕ ЦДРЕТВА СУШИ
Рис. 7.10. Животные Неотропического царства, поБСЭ: 1 —капуцин;
2 — игрунка; 3 — опоссум; 4 — древесный дикобраз; 5 — ленивец;
6 — игуана; 7 — ара; 8 — колибри; 9 — носуха; 10 — большой броненосец;
И — муравьед; 12 — тукан; 13 — нутрия; 14 — водосвинка; 15 — ягуар;
16 — тапир; 17 — кондор; 18 — пекари; 19 — вискаша; 20 — тинаму;
21 — гуанако; 22 — шиншилла; 23 — нанду 1В7
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ КАРТИ В А МИРА
И все же фауна Неотропического царства представляет собой еди-
ный комплекс, связанный общей историей. Единство фауны, несмотря
на различия в ландшафтах, объясняется тем, что животное население
Неотропиков в течение очень долгого геологического времени развива-
лось изолированно.
В геологическом прошлом в формировании фауны Неотропиков
участвовали фауны других континентов, с которыми Южная Америка
некогда образовывала единое целое. Древнее всего связи с Австралий-
ским и Эфиопским царствами. Весьма возможно, что сумчатые попали
в Австралию из Южной Америки через Антарктиду, где в меловой пе-
риод был субтропический климат. На древнюю связь с Эфиопской
областью и Мадагаскаром указывает родство антильских щелезубов с
тенреками, а также наличие в пресных водах обеих областей сирено-
вых (ламантинов) и близкородственных двоякодышащих — Lepidosiren
и Protopterus. Хищные (ягуар, оцелот, медведь) и копытные (олени, ламы
и др.) проникли в Неотропики из Неарктики через Панамский переше-
ек, образовавшийся лишь в плиоцене. В свою очередь фауна Неотропи-
ков обогатила биоту Неарктики.
И14. Австралийское царство
Биота Австралийского царства отличается глубокой древностью и
замечательным своеобразием своего состава. Между Австралийским
материком и Индо-Малайским царством на севере располагается боль-
шая группа островов Малайского архипелага. А. Уоллес первым обозна-
чил границу, отделяющую Индо-Малайское царство от Австралийско-
го. Ее называют линией Уоллеса. Перекрытие биот происходит между
островами Ява и Новая Гвинея. Оно объясняется сближением конти-
нентальных шельфов Юго-Восточной Азии и Австралии (рис. 7.11) и
существовавшим здесь в недавнем прошлом мостом суши.
Рис. 7.11. Граница между Индо-Малайским и Австралийским царствами,
по С. Barry Сох: Жирная линия обозначает границу Уоллеса, которая прохо-
дит по краю шельфа Юго-Восточной Азии; пунктирная линия обозначает
168 границу австралийского шельфа
ГЛАВА 1. БМОТИЧЕСХЙЕ ЦАРСТВА СУШИ
Значительная часть Австралии представляет собой сухое плато со
скудной растительностью (скрэб); многие места совершенно пустын-
ны. Обильно увлажнены только северные и юго-восточные берега, здесь,
как и в Тасмании, развиваются роскошные леса. Новая Гвинея и сосед-
ние острова гористы и покрыты тропическим лесом.
Австралийская флора богата и своеобразна. Здесь растет 12 049 ви-
дов сосудистых растений, из которых 75% эндемики. Почти весь род
эвкалиптов, содержащий 600 видов, филлодийные акации (280 видов),
казуарины (около 25 видов), банксии (46 видов), так называемые травя-
ные деревья (Xanthorrhoea — И видов) и многие другие не встречаются
за пределами Австралийского флористического царства.
В конце мезозоя — начале кайнозоя, во время бурного распростра-
нения на земле цветковых растений, Австралия, Тасмания, Новая Гви-
нея составляли изолированный материк. Древние роды, характерные
для тропического и умеренного климата IFlindersia, Brachychiton,
Acacia, Grevillea, Melaleuca, Eucalyptus, Casuarina, Banksia, Tristania и
др.), составили основное ядро эндемической флоры Австралии. Пред-
ставители этих родов распространены по всей Австралии, но в наибо-
лее чистом виде эндемическая флора представлена в юго-западной и
центральной частях континента.
Вместе с тем в формировании флоры Австралии принимали участие
представители других биогеографических царств (рис. 7.12). В третич-
ном периоде происходило вторжение в северо-восточные провинции
Австралии множества представителей флоры Индо-Малайского царства
(Podocarpus amarus, Elaeagnus latifolia, Aleurites moluccana и др.). Так
сложился один из основных элементов флоры Австралии — малезий-
ский. Многие его представители составляют главную массу влажных
тропических лесов Австралии.
При всей самобытности австралийская флора в ряде случаев обнару-
живает замечательную общность с антарктической флорой, представи-
тели которой распространены в Новой Зеландии, южной оконечности
Южной Америки и Южной Африки (Libocedrus, Nothofagus, Drimys,
Araucaria, Fitzroya, Casuarina, древовидные папоротники, многие протей-
ные и др.). Представители антарктической флоры встречаются в юго-
восточной горной Австралии и на острове Тасмания, что объясняется
существованием в мезозое единого материка, включавшего Антарктиду.
В Австралии почти нет растений с сочными плодами. Австралийская
флора лишена дикорастущихрастений, обогативших культурную фло-
ру ценными пищевыми культурами. По-видимому, это послужило при-
чиной того, что австралийские аборигены не смогли освоить земледе-
лие и перейти к оседлому образу жизни. До прихода европейцев они
оставались бродячими охотниками-собирателями. Вместе с тем Авст-
ралия — родина большого числа засухоустойчивых деревьев и кустар-
ников, распространяемых отсюда в культурах по всему свету (эвкалип-
ты, акации, казуарины и др.).
Фаунистические особенности Австралийского царства складывались
также под влиянием, с одной стороны, изоляции, а с другой — древних
связей с соседними материками. В мезозое существовала связь Австра-
лии с прилегающими частями Азии и с Южной Америкой. В это время 169
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ КАРГИ0А МИРА
ПРОШЛЫЕ И СОВРЕМЕННЫЕ СВЯЗИ ФЛОРЫ АВСТРА*
ЛИИ С ФЛОРАМИ ДРУГИХ ОБЛАСТЕЙ И ПУТИ МИГ»
РАЦИЙ ФЛОР
СВЯЗИ С ДРЕВНЕЙ АНТАРКТИЧЕСКОЙ
ОБЛАСТЬЮ (ПлМООкЕАисний элеыемп
____Дроаиио «моэодскио, pa*» яарушанми» cam
~ е Каясао* флоое* Юж*- Афоинм
МмОВоАсМи» САМ** АйСТМЛИа
с Ат доннмеквй флорой
СВЯЗИ С ПАЛЕОТРОПИЧЕСКОЙ
флористической областью
Ф»»а«*^^> моловиа фж»е«мч»смм* сам» м пу*м м«гоаамк
ТРЕТИЧНЫЕЧГЛ. ОБР. МДЛЕЭМЙСНМЕ
связи австралийской ФЛОРЫ
Т&пнмирс (до ммоцом*) сами е маеЫ 3о»амдм(й
(до лтаца**) cam < Тасмииа*
напрасен* (мгмцм* иаламааЫсиро аанамта
Флоов» Ак'раямм
напрытка диграф аашлааЫсмаго >ммх<»
флоеы
Главии* цамгрч экминаоа аЧХаидаммчаехмя
ыам а цамрах (п© E.B Вуяъфу и H.K.Ky»wu»wy)
Число видов а ивстмих флова» Аосгоаянм
(по ЕВ. Эуавфу)
Рис. 7.12. Пути формирования флоры Австралии, по НА. Комарницкому
сюда переселились низшие представители млекопитающих — клоачные
(из Азии) и сумчатые (из Южной Америки.). Дальнейшая изоляция Ав-
стралии способствовала тому, что населявшие ее животные и растения
продолжали развиваться собственным, независимым от остального
170 органического мира путем (рис. 7.13).
[ЛАВА 1. БМОТИЧЕСХМЕ ЦДРЕТВА СУШИ
Рис. 7.13. Животные Австралийского царства, по БСЭ: 1 — утконос;
2 — ехидна; 3 — сумчатый кускус; 4 — сумчатый барсук; 5 — сумчатый крот;
6 — проехидна; 7 — лирохвост; 8 — райская птица гоби; 9 — большеног, или
сорная курица; 10 — эму; И — динго; 12 — сумчатая белка; 13 — сумчатый
прыгун; 14 — большой кенгуру; 15 — древесный кенгуру; 16 — попугай корелла;
17 — медосос; 18 — чешуеног; 19 — вомбат; 20 — сумчатый волк; 21 — коала
(сумчатый медведь); 22 — черный какаду; 23 — какаду-инка; 24 — безногая
райская птица; 25 — молох; 26 — плащеносная ящерица; 27 — беседковая пти-
ца (шалашник); 28 — казуар; 29 — гигантский зимородок; 30 — сумчатая ку-
ница; 31 — сумчатый мурашеед; 32 — свинья папуасская; 33 — черный лебедь 171
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ КАРТИ В А МИРА
Бросается в глаза почти полное отсутствие высших млекопитающих,
или плацентарных. Распространенная в Австралии собака динго проник-
ла сюда в древности с человеком и впоследствии одичала. Свиньи Новой
Гвинеи и Тимора также представляют собой одичалых потомков домаш-
ней свиньи, некогда завезенной сюда человеком. Зато необычайно
развита фауна низкоорганизованных млекопитающих. Особого расцве-
та достигли сумчатые — более 130 видов.
Эндемичные формы сумчатых весьма многообразны и соответству-
ют различным биологическим типам высших млекопитающих (кон-
вергенция жизненных форм): аналоги хищных — сумчатый волк
{Thylacinus) и сумчатая куница (Dasyurus); насекомоядных — сумчатый
крот {Nothoryctes); грызунов (типа байбака) — вомбат {Phascolomys), типа
белок и летяг — сумчатая белка {Petaurus). Лазящие сумчатые — коала,
или сумчатый медведь {Phascolarctus). Аналоги копытных — многочис-
ленные кенгуру.
Для Австралийского царства характерны единственные ныне живу-
щие однопроходные (клоачные) млекопитающие — утконос {Omithor-
hynchus) и два рода ехидн {Echidna).
Из 286 родов птиц, гнездящихся в Австралийском царстве, 95 ро-
дов эндемичны. В их числе отряды казуаров {Casuarius) и эму
(Dromaeus). Далее ряд семейств, например беседковые {Amblyomis)
и райские птицы с роскошным причудливым оперением; лирохво-
сты (Menuridae); 30 видов какаду {Cacatuinae); нитеязычные попу-
гаи; большеногие куры {Megapodiidae), которые не высиживают яиц,
а закапывают их в кучи гниющих растительных остатков; черный
лебедь {Chenopais) и др.
И 7.5. Голантарктическое царство____________________________________
А.Л. Тахтаджян (1978) выделяет Голантарктическое флористическое
царство, относя к нему южную оконечность Южной Америки с Огнен-
ной Землей и Патагонией, острова Южного океана и Антарктиды, Но-
вую Зеландию.
Сходство флор удаленных друг от друга участков суши объясняется
тем, что в меловом периоде Южная Америка, Австралия и Антарктида
образовывали единый материк, характеризовавшийся теплым климатом
и пышной растительностью.
Современная флора Голантарктического царства включает 10 энде-
мичных семейств, 1600 видов сосудистых растений, из которых 1200,
т. е. 75%, эндемичны. Например, виды южного бука {Nothofagus), фиц-
рои {Fitzroya), араукарии {Araucaria), дримиса {Drimys winteri), древовид-
ных папоротников {Blechnum), трава туесок {Роа flabellatd) и др. Харак-
терной особенностью флоры является биполярность — наличие ряда
родов и видов растений, обитающих в умеренных и высоких широтах
Северного полушария, но отсутствующих в тропических поясах Земли.
Таковы, например, виды сфагнума, красная водяника {Empetrum rubrum),
а также луговик извилистый, тимофеевка альпийская, примула, яскол-
ка, генциана и др.
172
ГЛДВД1. БИОТИЧЕЕКИЕ ЦДРЕТЯД СУШИ|
Наиболее северные теплые районы по склонам Анд и на западе Ог-
ненной Земли заняты лесной растительностью. Тундры охватывают ост-
рова Южного океана. Полярные пустыни занимают собственно Ан-
тарктический материк. Южная полярная граница распространения
цветковых растений — антарктического луговика и колобата [Colobanthus
crassitolius, Aira antarctica) — проходит приблизительно по 64" ю. ш. (Земля
Грейама). Далее к югу встречаются только мхи (5 видов, в их числе эн-
демичный род Sarconeurori), лишайники (около 90 видов) и водоросли.
Голантарктическое царство животных занимает побережье Антарк-
тиды и прилежащие острова. Фауна отличается бедностью видового
состава. Здесь очень мало наземных животных, нет летающих насеко-
мых, сухопутных млекопитающих, пресноводных рыб. Жизнь позво-
ночных животных полностью связана с океаном. Поэтому большин-
ство зоологов не выделяет Антарктического царства на суше, а относит
эту фауну к океанической.
К числу немногих связанных с сушей птиц принадлежат три энде-
мичных вида: из куликов — белая ржанка, или футляронос (Chionis alba),
поедающая яйца пингвинов, своеобразный конек (Anthus antarcticus) и
небольшая утка (Anas georgica), обитающая на острове Южная Георгия.
Самыми типичными птицами Голантарктического царства следует
считать пингвинов, среди которых наиболее крупный императорский
пингвин (длина тела до 115 см, масса до 42 кг), немного уступающий ему
в величине королевский пингвин и мелкий пингвин Адели. Пингвины
остаются в Антарктиде круглый год, устраивая свои гнездовые колонии
на побережье. Летом очень многочисленны буревестники (антарктиче-
ский, капский, снежный, серебристо-серый и др.), нередко встречают-
ся альбатросы, качурки, чайки, крачки и поморники.
Класс млекопитающих представлен ластоногими и китами. У бере-
гов Антарктического материка, около островов и в поясе дрейфующих
льдов, обитает 5 видов настоящих тюленей (Phocidae), из которых тес-
нее всего связан с сушей морской слон (Mirounga leonina). В зоне не-
подвижных прибрежных льдов обитает тюлень Уэдделла (Leptonychotes
weddelli) и частично тюлень Росса {Ommatophoca rossi); в поясе плаву-
чих льдов встречается тюлень-крабоед (Lobodon carcinophagus) и мор-
ской леопард (Hydrurga leptonyx). Довольно многочисленные в Южном
полушарии ушастые тюлени были представлены в Антарктическом цар-
стве морским котиком (Arctocephalus austral is), полностью истреблен-
ным к началу XX в.
Обитателями вод Антарктики в летнее время, когда создаются наи-
более благоприятные условия для откорма, являются киты: из усатых
китов — синий кит, финвал, сейвал, горбач, малый полосатик и в еди-
ничных экземплярах гладкий кит; из зубатых китов — кашалот, бутыл-
конос и касатка.
Глава
ЗОНАЛЬНЫЕ ТИПЫ БИОМОВ СУШИ
Биом представляет собой сочетание конкретных экосистем. Зональ-
ные типы биомов формируются при определенном соотношении тепла
и влаги, включают растительные сообщества и животное население,
неразрывно связанные друг с другом. Как крупные экологические еди-
ницы биосферы они, в свою очередь, могут быть подразделены на более
мелкие типы — региональные типы биомов.
Зональные типы биомов — совокупности естественных экосистем
со сходной структурой и обликом сообществ, сформировавшиеся в
процессе исторического развития и отражающие гидротермический
потенциал территории. Их границами являются рубежи, за пределами
которых экосистемы данного зонального типа не могут существовать
из-за несоответствия условиям среды.
Биогеографическая картина мира, основанная на выделении биоти-
ческих царств суши, отражает уникальность флоры и фауны, являю-
щуюся следствием дивергентного пути эволюции (см. гл. 7). Выделение
зональных типов биомов подчеркивает черты сходства органического
мира, формирующегося в условиях одинаковой обеспеченности теплом
и влагой. Об общей картине распределения зональных типов биомов
суши дают представление карты растительности и почв (рис. 8.1, 8.2).
Наглядной моделью распространения зональных типов ландшафтов
служит гипотетический материк (см. рис. 2.14). Сопоставление с кар-
той растительных и почвенных зон Земли показывает, что эта модель
достаточно полно отображает географию зональных типов биомов в це-
лом (табл. 8.1). Зоны в таблице перечислены в том же порядке, в каком
дано их описание в тексте.
Перейдем к характеристике зональных типов биомов суши. В каче-
стве основы взяты представленные в табл. 8.1 системы зональных типов
ландшафтов и растительных зон Земли. При описании биомов исполь-
зовались работы В.В. Алехина, Л.В. Кудряшова, В.С. Говорухина (1961),
Н.А. Бобринского (1951), Г. Вальтера (1968), А.Г. Воронова, Н.Н. Дроздо-
ва, Е.Г. Мяло (1985), П.П. Второва, Н.Н. Дроздова (2001), А.Г. Исаченко
174 и А.А. Шляпникова (1989), П.Д. Ярошенко (1975) и др.
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ 5Н0М0В СУШИ
Рис. 8.1. Зональные тины биомов суши. А — схема растительных зон Земли,
по Г. Вальтеру: Тропические и субтропические зоны: 1 — вечнозеленые тро-
пические дождевые леса низменностей и горных склонов (леса пояса тума-
нов); 2 — тропические полувечнозеленые и листопадные леса; 2, а — сухие
леса, естественные саванны и заросли колючих кустарников, частично зла-
ковники зоны тропиков и субтропиков; 3 — тропические и субтропические
полупустыни и пустыни, простирающиеся в сторону полюсов только до 30°
(35°) с. ш. и ю. ш. (в других местах — растительность, отмеченная в пунк-
те 7, а). Умеренная и арктическая зоны: 4 — переходная область жестколи-
стной растительности (с зимними дождями), включая аридные области с
зимними дождями; 5 — вечнозеленые лиственные леса умеренно теплой
зоны или зон с постоянно влажным прохладным климатом; 6 — летнезеле-
ные лиственные леса умеренной зоны с зимним периодом покоя (немораль-
ная зона); 7— злаковники умеренной зоны (степи, прерии, пампа); 7, а— по-
лупустыни и пустыни с холодной зимой (простираются в сторону полюсов
от 30—35° с. ш. и ю. ш.), в Тибете — холодная пустыня; 8 — бореальная зона
хвойных лесов Северного полушария; 9 — арктическая зона тундры Северно-
го полушария; 10 — горы с различными поясами растительности, включая
альпийский и нивальный пояса. Б — наземный покров (land cover), по данным
космического мониторинга в рамках программы Seasonal Land Covers Regions
175
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ К АРТИ В А МИРА
17G
Рис. 8.2. Зональные типы почв, по Earl D. Shaw
Таблица 8.1
Сопоставление основных зональных типов ландшафтов
на идеальном континенте, растительных и почвенных зон Земли
Схема основных зональных типов лан- дшафтов на идеальном континенте, по АГ. Исаченко (см. рис. 2.14) Схема основных растительных зон Земли (см. рис. 5.1) и зональных типов почв (см. рис. 5.2)
А Жаркий пояс
Экваториальный лес Вечнозеленые тропические дождевые леса; ферраллит- ные почвы
Тропические и субэкваториальные сезонно влажные лесные Тропические листопадные леса; ферраллитные почвы сезонно влажных тропических лесов
Тропические и субэкваториальные саванны Сухие леса, естественные саванны и заросли колючих кустарников; коричнево-красные альферритные и черные тропические почвы
Пустыни Тропические пустыни; почвы примитивные песчаные, щебнистые, пустынные сероземы
Переходная субтропическая зона
Субтропические влажные лесные Вечнозеленые субтропические леса; красноземы и желтоземы
Средиземноморские сухие Жестколистная растительность; коричневые почвы
Б Холодный пояс
Арктические и антарктические пустыни
Лесотундровая и тундра Арктическая зона тундры Северного полушария; тундровые глеевые и болотные почвы
Умеренный пояс
Тайга Бореальная зона хвойных лесов Северного полушария; подзолы, подзолистые и болотные почвы
Широколиственно лесные Летнезеленые лиственные леса умеренной зоны с зимним периодом покоя (неморальная зона); бурые и серые лесные почвы
Лесостепь и степи Злаковники умеренной зоны (степи, прерии, пампа); черноземы и каштановые почвы
Суббореальная полупустыня и пустыня Полупустыни и пустыни с холодной зимой; бурые полупустынные и серые пустынные почвы
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ ВИН МОН СУШИ
Рис. 8.3. Девственный тропический лес в Бразилии.
Иллюстрация к книге «Жизнь растений» А. Кернера
177
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ КАРТИ В А МИРА
И 8.1. Бирмы жаркого пояса_____________________________________
Жаркий пояс Земли простирается к северу и югу от экватора в
тропические широты; в каждом полушарии его ограничивает годовая
изотерма 20 °C. Важнейшим фактором зональной дифференциации в
жарком поясе является увлажненность (см. гл. 2). В соответствии с за-
кономерностями атмосферной циркуляции и режимом выпадения ат-
мосферных осадков формируется зональный ряд биомов: дождевых
тропических лесов, тропических листопадных лесов, саванн и пустынь.
Переходное положение между биомами жаркого и умеренного поясов
занимают биомы субтропических зон.
8.1.1. Дождевые типические песо
Биомы дождевых тропических лесов — одни из самых древних и
богатых на Земле (рис. 8.3). Их ареал охватывает несколько царств:
Неотропическое, Эфиопское, Индо-Малайское, Австралийское. Коли-
чество видов растений и животных здесь больше, чем во всех других
биомах, вместе взятых.
По климатическим особенностям различают леса экваториального
и субэкваториального поясов, муссонных дождей, горных склонов. Наи-
более полно свойства вечнозеленых дождевых тропических лесов (ги-
лей) выражены в бассейне Амазонки, по берегам Гвинейского залива и
в бассейне Конго, на Малаккском полуострове, островах Малайского
архипелага, на северо-востоке Австралии (рис. 8.4). Главный фактор,
определяющий формирование вечнозеленых дождевых тропических
лесов, — сохраняющийся на протяжении миллионов лет благоприят-
ный теплый и влажный климат. Обилие тепла и влаги обеспечивает
круглогодичную вегетацию растений. Сезонная ритмика нечеткая.
Средние месячные температуры воздуха от 25 до 35 °C, их годовая
амплитуда всего 2 —4 °C (суточные амплитуды больше годовых). Атмо-
сферных осадков выпадает от 1500 до 4000 мм, а на наветренных склонах
гор до 10 000 мм и более. Для существования гилей, однако, более важно
не общее количество осадков, а распределение их в году. В субэкватори-
альном поясе дождевой климат имеет летний пик осадков, когда солнце
стоит в зените (субэкваториальный муссон). Эти зенитальные дожди
вызывают резкий подъем уровня воды в реках и затопление низменно-
стей. С переходом в тропические широты продолжительность зимнего
сухого сезона увеличивается. Климатической границей дождевых лесов
является продолжительность периода засухи более двух месяцев.
Длительность дня и ночи вблизи экватора в течение года приблизи-
тельно одинакова. Это постоянство имеет большое значение для суточ-
ного ритма жизни растений и животных. Красочное описание суток
во влажном тропическом лесу дает А. Уоллес: «Рано утром, в первые два
часа после восхода солнца, небо было неизменно безоблачным, термо-
метр стоял на 22 — 23 °C, тяжелая роса или следы ночного дождя, отя-
гощавшие листья, быстро испарялись под палящими лучами солнца, под-
178 нимавшегося как раз на востоке и быстро приближавшегося к зениту.
Рис. 8.4. Ареал дождевых тропических лесов, по П. Ричардсу
со
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ БИОМОВ СУШИ
I
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ КАРТИ В А МИРА
Вся природа дышала свежестью, молодая зелень и почки цветов быстро
распускались... Жара усиливалась с каждым часом и около 2 ч дня
достигла 33 — 34 °C, к каковому времени умолкали голоса всех птиц и
зверей. Листья, такие свежие и сочные утром, блекли и свешивались
вниз, лепестки цветов осыпались. В июне и июле около полудня обык-
новенно выпадает сильный дождь, приносящий желанную прохладу.
Туча приближается всегда одинаковым, весьма любопытным маневром.
Сначала прохладный морской бриз, появляющийся около 10 ч утра
и усиливающийся по мере того, как поднимается солнце, начинает осла-
бевать и, наконец, прекращается совершенно. Тогда жара и электриче-
ское напряжение воздуха становятся просто невыносимыми. Тяжелое
расслабление овладевает всеми, даже обитатели леса выдают это своими
движениями. Но вот на востоке показывается белый туман и начинает
собираться в кучевые облака, нижний край которых становится все тем-
нее и темнее. Весь восточный горизонт внезапно чернеет; эта мгла рас-
пространяется все больше и больше, заслоняя под конец и солнце. Затем
мощный порыв ветра с туманом проносится по лесу, раскачивая верши-
ны деревьев, сверкает ослепительная молния, раздается удар грома и
разражается стремительный ливень. Такие грозы быстро проходят и
оставляют за собой неподвижные исчерна-синие тучи, остающиеся на
небе до самой ночи. Вместе с тем вся природа освежена; лишь лепестки
цветов и листья кучами валяются под деревьями. К вечеру природа про-
буждается к новой жизни, которой полон каждый куст, каждое дерево.
На другой день солнце снова появляется на безоблачном небе, возобнов-
ляя, таким образом, круговорот тропического дня, в течение которого
перед нашими глазами как будто проходят весна, лето и осень...»
Почвы влажных тропических лесов формируются на мощной бесплод-
ной ферраллитной (А1203, Fe203) коре выветривания. Основной запас
питательных веществ сосредоточен в живых растениях. Полагают, что
растения получили этот запас миллионы лет назад в процессе биохими-
ческого разложения материнских горных пород богатого минералогиче-
ского состава. В конечном счете в коре выветривания сосредоточились
наиболее устойчивые, но бесплодные соединения алюминия и железа.
Продуктивность лесов огромна. Запас древесины свыше 5000 ц/га
(в Бразилии — до 17 000 ц/ra), а годовой прирост 350 — 500 ц/га и более.
Парадокс влажных тропических лесов заключается в том, что пышная
растительность развивается на маломощных почвах. Это становится
возможным благодаря очень быстрому круговороту биогенных солей.
Отмершая масса ветвей и листьев быстро, в течение нескольких не-
дель, разлагается и минерализуется. В лесах мало опавшей листвы и
нередко видны пятна оголенной земли. В разложении опада большое
участие принимают термиты; главная же роль, по-видимому, принад-
лежит грибам.
Корневая система деревьев не проникает на большую глубину, вся
она сосредоточена в первых десятках сантиметров маломощного слоя
почвы, где образует фильтр, поглощающий растворимые питательные
вещества. Важную роль в питании растений играет микориза. Высокие
деревья с поверхностной корневой системой малоустойчивы. Досковид-
ные корни — специальное приспособление, подобно контрфорсам у
готических соборов они поддерживают высокие стволы (рис. 8.5).
ГЛАВА 8. ЗОВАЛЬНЫЕ ТИПЫ 5И0М0В СУШ
Рис. 8.5. Сосковидные корни, придающие большую устойчивость высоким
стволам деревьев с поверхностной корневой системой, по А. Кернеру 181
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ КАРТИ В А МИРА
Особенности почв влажных тропических лесов следует учитывать
при их хозяйственном освоении. Издавна здесь применялась подсечно-
огневая система земледелия: лес вырубался, сжигался, участок обраба-
тывался для выращивания тех или иных сельскохозяйственных куль-
тур (рис. 8.6).
Рис. 8.6. Выжигание тропического леса
под сельскохозяйственное угодье и его последствия.
182 Внизу — эрозия склонов после уничтожения лесной растительности
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ БИО МОН СУШИ
Но запасы питательных веществ в почве ограничены, и они быстро
вымываются дождями. Через пару лет возделанный участок приходит-
ся забрасывать. Губительна сплошная вырубка лесов, особенно на скло-
нах, которая ведет к смыву почв и интенсивной эрозии. Около
100 000 км2 тропических лесов Подвергаются деградации ежегодно в ре-
зультате антропогенного воздействия. Обнажение бесплодной феррал-
литной коры выветривания на больших площадях — угроза того, что
влажные тропические леса могут утратить способность к самовосста-
новлению и на их месте возникнут вторичные растительные сообще-
ства с более бедным видовым составом и низкой продуктивностью.
Богатство биома влажных тропических лесов находит выражение
в огромном разнообразии видов, жизненных форм и в сложной струк-
туре лесного сообщества. 70% видового разнообразия растений состав-
ляют деревья (цветковые). Участие трав незначительно, обычно они
растут на опушках леса. Преобладают травы, своими размерами не
уступающие деревьям, бананы достигают высоты б м. Их подземные
органы живут неопределенно долго.
Число видов деревьев на 1 га может превышать 100; переходя от
одного дерева к другому, исследователь каждый раз обнаруживает но-
вый вид. Лесные сообщества не отделены друг от друга четкими грани-
цами, а связаны постепенными переходами. Такое явление называется
горизонтальной континуальностью растительного покрова.
Также сложен вопрос о вертикальном сложении леса: между 30 —
40-метровыми гигантами, образующими верхний полог, и деревьями в
3 —5 м существуют виды деревьев, образующие кулисообразные полога,
перекрывающие друг друга (рис. 8.7). Это дает основание говорить о
вертикальной континуальности, которая усиливается благодаря массе
внеярусных растений — лиан и эпифитов.
Рис. 8.7. Вертикальная структура дождевого тропического леса,
по Г. Вальтеру: разные виды деревьев высотой от 4—5 до 30—35 м
образуют переходы, затрудняющие выделение четких элементов
вертикальной структуры лесного сообщества
183
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ К АРТИ В А МИРА
Рис. 8.8. Орхидеи-эпифиты, по А. Кернеру
Число видов эпифитов и лиан в два-три раза превосходит число видов
деревьев (рис. 8.8). Особую жизненную форму представляют деревья-
душители, большинство из которых относится к роду Ficus. Благодаря
птицам их семена попадают на ветви крон деревьев верхнего яруса. На
первой стадии развития росток ведет жизнь эпифита. Однако сразу
начинается быстрый рост корней: один корень устремляется в землю
и снабжает растение водой и питательными солями; ответвления воз-
душных корней охватывают ствол хозяина и душат его (рис. 8.9).
Влажный тропический лес создает под своим пологом своеобразный
микроклимат. Кроны деревьев смыкаются, в лесу царит зеленый сум-
рак, редкий луч солнца пробивается сквозь густую листву. Здесь влаж-
но и душно, как в оранжерее. Условия обитания накладывают отпеча-
ток на строение растений. Стволы гладкие и стройные, кора тонкая и
светлая. Корка, как правило, не образуется. У многих деревьев доско-
видные и ходульные корни. Почки защищены очень слабо, почечные
чешуи отсутствуют. Форма и размер листьев в нижних ярусах леса мало
варьируются: обычно они большие и цельнокрайние. В более высоких
ярусах леса листья вырабатывают приспособления для защиты от ярко-
го солнца и потери влаги — уменьшаются в размерах, они становятся
кожистыми, блестящими, с толстой кутикулой. Представление о «веч-
нозелености» тропических лесов создается потому, что смена листьев
в течение года происходит постепенно: одни листья разворачиваются
из почек, другие опадают. Наряду с этим у некоторых деревьев может
происходить одновременное массовое сбрасывание листвы, хотя рядом
184 стоящие деревья того же вида могут оставаться зелеными.
ГЛАВА 8. ЗОНАЛЬНЫЕ ИЛЫ БИОМОВ СУШИ
Рис. 8.9. Деревья-душители: а — схема развития фикус а-душителя,
по «Жизни растений»: 1 — факус развивается как эпифит в кроне
дерева-хозяина, 2 — воздушные корни достигают почвы, 3, 4 — ответвле-
ния воздушных корней оплетают и сдавливают ствол дерева-хозяина;
б — общий вид, по A. Scliimper
В нижнем ярусе влажность воздуха постоянно высокая. Защиты от
испарения растениям здесь не требуется. Напротив, у них есть ряд
приспособлений, которые облегчают транспирацию: тонкие листья с
большой поверхностью и большим количеством устьиц; листья, в кото-
рых отдача влаги обеспечивается капельными устьицами на концах
листовых жилок, из которых выдавливаются капли воды.
Многие тропические деревья цветут и плодоносят круглый год. На
одном дереве имеются цветочные почки и цветки, зеленые и спелые
плоды. Иногда деревья зацветают дружно раз в 10 лет. Учитывая, что
деревья одного вида в тропическом лесу растут на большом удалении
друг от друга, приходится только догадываться, благодаря какому сиг-
налу они зацветают одновременно, обеспечивая тем самым возможность
перекрестного опыления. Одной из особенностей плодоношения у не-
которых видов растений является каулифлория, т. е. образование цвет-
ков, а затем и плодов непосредственно на стволах или толстых ветвях.
Наиболее известный пример — шоколадное дерево (рис. 8.10).
Животный мир влажных тропических лесов характеризуется огром-
ным богатством и разнообразием: более 10 000 000 видов, в основном
насекомых и других беспозвоночных (рис. 8.11). Благоприятные уело- 185
ЧАСТЬ III. БИОГЕОГРАШИЧЕСШ КАРТАВА МАРА
Рис. 8.10. Плоды шоколадного дерева (какао) на стволе и ветвях дерева.
Рядом продольный разрез плода
Рис. 8.11. Мир насекомых обладает приспособлениями, делающими их
трудноразличимыми среди зелени листьев и цветов тропического леса
вия обитания — тепло, влажно, обилие зеленого корма, масса убежищ —
способствуют формированию сообществ со сложными биотическими
отношениями. Круглый год климатические условия благоприятны для
размножения и развития. Основная жизнь сосредоточена в кронах
деревьев; у земли лес кажется безжизненным.
В Неотропическом царстве дождевые тропические леса занимают
огромные пространства на Амазонской низменности, где их описал еще
18G А. Гумбольдт, введший в науку термин «гилея» (рис. 8.12).
ГЛАВА 8. ЗОНАЛЬНЫЕ Т1ПЫ БИОМОВ СУШИ
Рис. 8.12. Амазонская гилея, по А. Кернеру.
На переднем плане монстера с воздушными корнями
187
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ КАРТИ В А МИРА
А.Л. Тахтаджян (1978) выделяет особую Амазонскую флористиче-
скую область, которая включает 500 эндемичных родов и не менее
3000 видов. Здесь центр распространения пальм (до 200 видов), из кото-
рых 60 эндемичны. Из деревьев нужно назвать цейбу (Ceiba) — дерево,
возвышающееся над всем лесным пологом; бертоллецию (Bertolletia
excelsa), плоды которой называют бразильскими или американскими
орехами; гевею (Hevea brasiliensis), дающую каучук; деревья с ценной
красной древесиной — махагонию и цезальпинию. К нижнему ярусу
относится шоколадное дерево (Theobroma cacao); дынное дерево (Carica,
около 40 видов), сладкие плоды которого содержат фермент папаин,
благоприятствующий пищеварению. Деревья всех ярусов переплетают
лианы. Среди эпифитов выделяются своим разнообразием орхидеи,
папоротники, виды из семейства Bromeliaceae. В заводях Амазонки
растет знаменитая гигантская кувшинка виктория-регия (рис. 8.13).
Рис. 8.13. Виктория-регия на Амазонке, по А. Кернеру
Кроны густого тропического леса являются средой обитания мно-
жества зверей и птиц с ярким оперением (рис. 8.14). Почти все они
проводят большую часть жизни на деревьях, не опускаясь на землю.
Приспособление, характерное для животных неотропиков, — цепко-
хвостость. Цепкие хвосты обезьян (рис. 8.15), опоссумов, древесных
муравьедов и дикобразов, енотов-носух помогают им ловко передвигать-
188 ся с ветки на ветку.
ГЛАВА 8. ЗОНАЛЬНЫЕ Т1ПЫ БИОМОВ СУШИ
Рис. 8.14. Характерные представители фауны дождевого тропического
леса. Бразильская область Неотропического царства (рисунки В.А. Ватаги
на, по Н.А. Бобринскому): 1 —ленивец; 2 — средний муравьед; 3 — цепко-
хвостый дикобраз; 4 — попугай-ара; 5 — тукан; 6 — игрунка; 7 — опоссум
8— большой муравьед; 9— ревун; 10— броненосец-гигант; 11— тапир;
12 — водосв инка 189
ЧАСТЬ III. БИ0ГЕ11ГРЙ1ШЕСШ КАРТИ В А МИРА
Рис. 8.14 (окончание): 13 — пекари; 14 — ягуар; 15 — носуха; 16 — щелезуб;
17 — нутрия; 18 — колибри; 19 — паукообразная обезьяна; 20 — удав боа;
21 — бразильский попугай; 22 — гоацин; 23 — игуана;
190 24 — кракс, или шлемоносная курица
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ 8I0M08 Ml
Рис. 8.15. Виды широконосых обезьян с длинными цепкими хвостами,
по К. Шефферу
Типичными представителями фауны Неотропического царства в
гилее являются неполнозубые — ленивцы. Их когти срослись в серпо-
видные крючья, с помощью которых животные висят на ветвях без
всякого напряжения мускулов (рис. 8.16).
Впрочем, представление о том, что в тропическом лесу на каждом
шагу встречаются интересные птицы и звери, не соответствует действи-
тельности. Девственный лес кажется мертвенно тихим: в густой чаще
трудно передвигаться и в то же время легко спрятаться. Лишь невиди-
мые стаи обезьян и птиц производят подчас невероятный шум. Изредка
встречаются равнинный тапир и похожие на кабанов пекари. Крупные
грызуны замещают здесь копытных; наибольших размеров достигает
капибара [Hydrochoerus capybara), до полутора метров в длину при массе
до 60 кг. Хищники лредставлены кошачьими — ягуаром и оцелотом.
Важную роль в жизни влажных тропических лесов играют терми-
ты — главные пожиратели отмершей растительной массы, благодаря
чему она быстро разлагается и минерализуется. Термиты строят свои
термитники, начиная с глубоких слоев почвы и кончая ветвями круп-
ных деревьев. Основательницей сообщества является «царица» — круп-
ная, до 12 см в длину, самка, непрерывно продуцирующая яйца (рис. 8.17). 191
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ КАРТИ В А МИРА
Рис. 8.16. Трехпалый ленивец
Рис. 8.17. Царица в термитнике
Количество термитов на 1 м2— от 500 до 10 тыс. Переваривание клет-
чатки в кишечном тракте термитов осуществляется с помощью одно-
клеточных жгутиковых, без симбиоза с которыми термиты существо-
вать не могут. Поэтому появившуюся на свет личинку рабочий термит
подносит к анальному отверстию царицы, чтобы новорожденная обо-
гатила флору своего кишечника.
Тропический лес от почвы до крон самых высоких деревьев запол-
няет множество видов муравьев. У ряда растений выработались особые
симбиотические отношения — мирмекофилия (любовь к муравьям): в
стволах и ветвях деревьев возникают специальные пустоты, в которых
живут хищные муравьи, уничтожающие насекомых-вредителей. Му-
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ БИО МОН СУШИ
равьи-листорезы ведут иной образ жизни. Они искусно вырезают ку-
сочки листьев, переносят их в свои подземные гнезда, пережевывают и
на этой разлагающейся массе выращивают определенные виды грибов,
которыми выкармливают личинок. Когда самка муравья-листореза от-
правляется в брачный полет, она берет в рот кусочки гифов гриба, что-
бы создать плантацию на новом месте.
Дождевые тропические леса, более подробно рассмотренные на
примере Амазонской гилей, сохраняют свои главные черты и в других
областях своего распространения. Индивидуальный отпечаток накла-
дывает их принадлежность к разным биогеографическим царствам.
В Эфиопском царстве дождевые тропические леса с мощным древес-
ным ярусом, обилием лиан и эпифитов занимают большую часть бас-
сейна Конго, а также побережье Гвинейского залива. В составе флоры
13 000 видов; более 250 родов растений эндемичных или преимуществен-
но свойственных этим лесам. Широко представлены виды из тутовых,
бобовых, пальмовых, стеркулиевых, панданусовых и других семейств. В д-
ревесном ярусе господствуют роды Ficus, Treculia, Chlorophora, Coula,
Albizzia, Copaifera, Khaya, Ceiba, Cola, Omphalocarpum.
Обитатели тропических лесов (см. рис. 8.8) — гориллы, шимпанзе и
различные мелкие обезьяны; из грызунов — длиннохвостая летяга; из
копытных — малый оленек и окапи; из хищников — леопард. Из птиц
следует упомянуть серого попугая, длинноклювого токо, птицу-носоро-
га, цесарку и африканского павлина.
Характерным приспособительным навыком животных, позволяю-
щим им постоянно обитать в кронах деревьев, является планирование.
Способностью к планирующему полету с дерева на дерево обладают не
только птицы, но и белки, змеи, ящерицы и лягушки.
В Индо-Малайском царстве дождевые тропические леса занимают
преимущественно полуостров Малакку и острова Суматра, Ява, Борнео,
Целебес, Новая Гвинея и соседние более мелкие острова.
Флора подобласти одна из наиболее богатых на Земли (около 45 тыс.
видов растений); эндемичных родов около 500; видовой эндемизм чрез-
вычайно велик. Так, например, в Новой Гвинее около 12% родов и 85%
видов являются эндемичными для острова; из 2546 видов орхидей 2534
встречаются только здесь.
В состав древесного яруса входят Dipterocarpus, Sborea, Caesalpinia,
Canahum, Epicharis, фикусы и другие; многочисленны пальмы из родов
Corypha, Areca, Cyrtostachys. Огромное количество эпифитов и лиан
(в том числе пальма Calamus). В горных гилеях встречаются хвойные —
Podocarpus, Libocedrus. Индо-Малайское царство — родина многих пря-
ных растений: гвоздичного дерева {Eugenia aromaticus), мускатного
ореха {Myristica fragrans), коричного дерева {Cinnamomum zeylanicum).
Виды рода Rafflesia растут на Больших Зондских островах. Это па-
разиты, питающиеся за счет деревьев тропического леса. Растение про-
растает из-под коры растения-хозяина в виде бутона, развертывающе-
гося в пятилепестковый мясистый красный с пятнами цветок. Rafflesia
arnoldii на острове Ява достигает более 1 м в диаметре, это величайший
в мире цветок (рис. 8.18). Он издает сильный трупный запах, привлека-
ющий мясных мух-опылителей.
193
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ К АРТИ В А МИРА
Рис. 8.18. Цветок раффлезии —растения, паразитирующего
на деревьях тропического леса
Растения дождевых тропических лесов испытывают недостаток ми-
нерального питания. Некоторые из них компенсируют дефицит био-
генных солей благодаря ловле и перевариванию мелких животных. К их
числу относится род Nepentes, часто встречающийся в тропических
лесах на островах Малайского архипелага. Это большей частью кустар-
никовые или полукустарниковые лианы, поднимающиеся по стволам
деревьев на десятки метров. Наряду с обычными листьями у них раз-
виваются видоизмененные листья-кувшины с крышечкой, представля-
ющие собой своеобразные ловушки длиной от 2,5 до 30 см (рис. 8.19).
Яркие кувшины, висящие между ветвями деревьев, привлекают к себе
не только насекомых, но даже мелких птиц и зверьков. Кувшин непен-
теса выполняет функции желудка у животного: количество жидкости в
нем может достигать двух литров, причем само растение выделяет фер-
менты, играющие роль желудочного сока.
Высоко в кронах деревьев в тропическом лесу Юго-Восточной Азии
произрастают виды Myrmecodia. Стебель этих растений образует круп-
ный клубень, пронизанный многочисленными галереями и ходами, в
которых живут муравьи (рис. 8.20). Как уже говорилось, это явление
называется мирмекофилией: симбиоз с муравьями помогает растению
194 защититься от насекомых-вредителей.
ГЛАВА 8. ЗОНАЛЬНЫЕ ШЫ БИНИОН СУШИ
Рис. 8.19. Непентес (Nepentes hybridd), по «Жизни растений»
Рис. 8.20. Мармекофилия — симбиоз растения с муравьями, по К. Шаффе-
ру. Мармекодия колючая (Myrmecodia echinata); на продольном разрезе клуб-
ня видны полости, в которых обитают муравьи
195
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ К АРТИ В А МИРА
Весьма разнообразны виды фикусов, среди которых выделяется
баньян — дерево Будды, или священная смоковница [Ficus bengalensis).
По мере роста его развесистой кроны от ветвей выходят прямые воз-
душные корни, которые, достигнув земли, укореняются. Со временем
они приобретают вид настоящих стволов, от них также начинают от-
ходить облиственные ветви. Так постепенно из одного дерева выраста-
ет целая роща (рис. 8.21).
Рис. 8.21. Баньян (Ficus bengalensis). Ходульные корни несут огромную крону
Индусы увидели в непрерывно растущем дереве олицетворение веч-
но творческой силы природы. Согласно легенде, именно в тени баньяна
Будда познал ничтожество земной жизни и вечность мироздания. Для
разрастания кроны требуются особенно благоприятные условия, поэто-
му священные смоковницы растут обычно вблизи храмов, где за ними
ухаживают монахи.
Животный мир дождевых тропических лесов Индо-Малайского цар-
ства наиболее типичен для царства в целом (см. рис. 7.8). Млекопитаю-
щие представлены эндемичным отрядом шерстокрылов. Из человекооб-
разных обезьян обычны гиббоны; на островах Борнео и Суматра
встречаются орангутаны. Из копытных только здесь обитают чапрач-
ный тапир, три вида носорогов, дикий бык бантенг. Из хищных харак-
терны тигры, леопарды. Из птиц замечателен великолепный фазан ар-
гус. Из пресмыкающихся гигантский варан с острова Комодо.
В Австралийском царстве влажные тропические леса имеются на
северо-восточном побережье, флористически эти гилей близки к индо-
малайским. Здесь растут фикусы, пальмы, тропические хвойные
(Agathis), но есть и араукарии и подокарпусы, что сближает эти гилей
с амазонскими. Много древовидных папоротников (рис. 8.22). Обиль-
19G ны эпифиты и лианы.
ГЛАВА 8. ЗОНАЛЬНЫЕ Т1ПЫ БИОМОВ СУШИ
Рис. 8.22. Заросли древовидных папоротников
на северо-восточном побережье Австралии
Для животного мира австралийских гилей обычны лазающие сум-
чатые (см. рис. 8.11): древесный кенгуру, сумчатые куницы, сумчатые
мыши, сумчатые барсуки. Богато представлены плотоядные крыланы,
гроздьями висящие днем на высоких деревьях. Скрываясь в густых
лесах, живут страусоподобные казуары. Интересны большеногие, или
сорные, куры, замечательные тем, что они не насиживают яйца, а сно-
сят их в кучи растительного сора, где они развиваются под влиянием
теплоты от гниения. Украшением леса являются райские и беседковые
птицы, попугаи.
На влажном восточном побережье Австралии господствуют эвкалип-
товые леса (рис. 8.23). Они довольно сильно отличаются от дождевых
тропических лесов других царств. Несмотря на то что деревья достига-
ют внушительных размеров, листовые пластинки у них располагаются
вертикально. Поэтому лес светлый, с хорошо развитым травяным по-
кровом. В кронах эвкалиптов обитают питающиеся их листвой сумча-
тые медведи коала (рис. 8.24). 197
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ КАРТИ В А МИРА
Рис. 8.23. а — влажный эвкалиптовый лес; б — основание ствола эвкалипта
8.1.2. Тропические листападные леса и саванны
На материках, лежащих в тропических широтах к северу и югу от
экватора, где устанавливается режим субэкваториальной муссонной
циркуляции, осадки в течение года выпадают неравномерно. Вследствие
198 более сильного нагревания летом суши соответствующего полушария в
ГЛАВА 8. ЗОНАЛЬНЫЕ ШЫ 8I0M08 Ml
Рис. 8.24. Сумчатый медведь коала, поА.Э. Брему
субэкваториальный пояс смещаются от экватора низкое атмосферное
давление и влажный воздух; лето — сезон дождей. Зимой в субэкватори-
альный пояс из тропических широт смещаются высокое атмосферное
давление и сухой воздух; зима — сезон засухи. У экватора бездождье
может продолжаться не более одного месяца; чем ближе к субтропиче-
ским поясам высокого атмосферного давления, тем отчетливее начина-
ют проявляться сезоны бездождья и дождей, тем меньше выпадает осад-
ков и тем дольше сухой сезон.
Периодичность выпадения атмосферных осадков четко выражена так-
же в областях с муссонным климатом. Вся Юго-Восточная Азия находит-
ся во власти муссонных ветров, дующих зимой с Азиатского материка
(сухой сезон), а летом с Тихого и Индийского океанов (сезон дождей).
Неравномерность выпадения атмосферных осадков, уменьшение их
количества при сохранении высокого радиационного баланса является
главной причиной перехода биома дождевых тропических лесов (гилей)
в биомы листопадных лесов и саванн. Животный мир тропических ле-
сов уступает место животным саванн. В конечном счете направленная
аридизация климата приводит к формированию биома тропических
пустынь, завершающего ряд биомов жаркого пояса (рис. 8.25).
Таким образом, причиной формирования листопадных тропических
лесов является сезонность климата, в которой выражены два времени
года — первое очень сухое и жаркое, второе — более прохладное, но
дождливое. Переход от типичного дождевого леса к лесу, где выражены
сезонные явления, осуществляется весьма постепенно. В условиях, где 199
ЧАСТЬ III. БАОГЕОЕРАФАЧЕСААЯ КАРТА НА МАРА
8-10 10-11
и
50
40
X
№
to
о
Число сухих
месяцев
1-3
4-6 6-8
600—500 300-200
12
менее 100
Вечнпеленый ВечнпелмнЛ Нмдвечшшткый Сухы лиане- Суше лйаммв- Щмнрмшявая
OaitMeS Ate сезонный лес тонный нее iwnuS лее ные мроаш шуме/пыня Йцеошня
। । нуста^нимв
Вшомтравные саванны Ниттравные саванны
Рис. 8.25. Зависимость развития растительных формаций в тропическом
поясе от продолжительности периода засухи, по Г. Вальтеру.
Стрелки внизу указывают пути деградации лесов в саванны после пожаров
засушливый период длится 1 — 2,5 месяца, уже намечается известная
периодичность в развитии растений. Она проявляется в одновремен-
ном сбрасывании деревьями листвы в сухой сезон и появлении новых
листьев с началом сезона дождей. При этом листопадные породы обра-
зуют верхний ярус, в то время как в нижних ярусах сохраняются веч-
нозеленые виды. К числу приспособлений, защищающих деревья от
засухи, относятся почечные чешуи и толстая кора на стволах. У вечно-
зеленых растений листья становятся мелкими и жесткими.
Другим приспособлением является суккулентность. Суккуленты
(стеблевые и листовые) обладают специальными запасающими воду
тканями, которые наполняются после дождей и затем длительное вре-
мя удовлетворяют потребность растений в воде. Транспирация у сукку-
лентов крайне слабая; растут они медленно. Стеблевыми суккулентами
являются кактусы, древовидные молочаи; листовыми суккулентами —
агавы, алоэ и др.
При уменьшении количества атмосферных осадков и увеличении
продолжительности сухого сезона число листопадных пород увеличи-
вается. Поскольку листву сбрасывают деревья верхних ярусов, вглубь
леса начинает проникать все больше солнечного света, благодаря чему
деревья нижнего яруса, кустарники и травы получают бурное разви-
тие. Увеличение продолжительности сухого сезона до б — 8 месяцев и
уменьшение годового количества осадков до 600 мм и менее приводит
к замещению тропических лесов биомами саванн.
Саванновые формации разных биогеографических царств имеют фло-
ристические различия, но объединены общими чертами строения: на-
личием травяного покрова (преимущественно из ксерофильных злаков)
и несомкнутого верхнего яруса из деревьев и кустарников, стоящих по-
одиночке или группами. Важную роль в переработке растительной мас-
сы играют термиты; термитники составляют характерный элемент ланд-
200 шафтов листопадных тропических лесов и саванн (рис. 8.26).
ГЛАВА 8. ЗОНАЛЬНЫЕ Т1ПЫ БИОМОВ СУШИ
Рис. 8.26. Редкостойный листопадный лес в облиственном состоянии
в сезон дождей. На переднем плане — четырехметровый термитник
В сокращении площади тропических лесов и увеличении площади
саванн важную роль играет деятельность человека. После массирован-
ной вырубки девственные тропические леса уже не восстанавливают-
ся. Кроме того, в период засухи часто случаются пожары. Происходит
деградация лесов, ведущая к формированию редколесья и саванн на их
месте. Можно предположить, что еще первобытный человек, вызывая
пожары, сознательно или по неосторожности, способствовал формиро-
ванию обширной зоны африканских саванн.
Процесс трансформации саванн связан также с неумеренным выпа-
сом скота, который ведет к разрастанию колючих кустарников. Колюч-
ки у растений появились в результате естественного отбора как защита
от пасущихся диких животных. Колючие деревья и кустарники осо-
бенно характерны для африканской саванны, где издавна паслись мно-
гочисленные стада диких копытных. В Австралии только кенгуру могут
быть отнесены к пастбищным животным, поэтому здесь почти совер-
шенно отсутствуют колючие растения. Для борьбы с кустарниками,
которыми зарастают пастбища, широко применяется огневая чистка,
т. е. те же самые палы. К отрицательным последствиям выжигания
растительности относятся усиление процессов эрозии на склонах и
дефляции легких песчаных почв. 201
ЧАСТЬ III. БАОГЕОЕРАФАЧЕСААЯ КАРТА НА МАРА
Следует отметить, что многовековое, регулярное воздействие огня
привело к появлению растений-пирофитов, приспособленных и даже
нуждающихся в воздействии высоких температур. К числу таких адапта-
ций относятся запас спящих почек, способность к корневому возобновле-
нию, устойчивость к нагреву стволов и семян. Высокая всхожесть семян
на открытых участках позволяет растениям успешно переживать пожары.
Особый интерес представляют более специфические адаптации: стимуля-
ция цветения обгоранием и прорастания семян водными вытяжками из
обугленной древесины. Может быть, самый яркий пример зависимости
от пожаров — некоторые австралийские и южноафриканские протейные
с одревесневающими толстостенными плодами. Такие плоды по нескольку
лет сохраняют жизнеспособные семена и хорошо защищают их, но для
раскрывания плода и рассеивания семян необходим сильный нагрев.
Если следствием антропогенного воздействия на тропические леса
является превращение их в саванны, то следствием воздействия челове-
ка на саванны является их опустынивание. «Сахельская трагедия» слу-
жит тому примером. Сахель — переходная полоса между саваннами и
пустыней Сахара, населенная народами, занимающимися в основном
скотоводством и ведущими кочевой образ жизни. Начиная с 1960-х гг.
эта область испытывает сильнейшие засухи. Растительный покров де-
градирует, чему немало способствуют неумеренный выпас скота и
использование древесно-кустарниковой растительности на топливо. Ли-
шенная растительности почва отражает больше солнечных лучей, что
меняет термодинамику атмосферы так, что условия для формирования
атмосферных осадков становятся еще менее благоприятными. Саванна
превращается в пустыню.
В Эфиопском царстве биомы листопадных лесов и саванн занимают
обширные пространства в средней части Африки к югу от Сахары, а
также всю восточную часть Южной Африки, окаймляя экваториальную
область распространения вечнозеленых дождевых тропических лесов.
Саванны образованы сомкнутым злаковым покровом высотой до 3 —
4 м, на фоне которого равномерно распределены группы деревьев и
кустарников. Переход от леса к открытым пространствам постепенный.
Кажется, что ты стоишь на лесной поляне с одиночными деревьями,
окруженной со всех сторон густым лесом. Однако достичь леса никог-
да не удается, ибо поляна перемещается вместе с идущим (рис. 8.27).
Вся жизнь в саванне подчиняется сезонному ритму выпадения атмо-
сферных осадков. В период зимней засухи местность выглядит пустынно
и печально. Небо всегда безоблачно, но воздух полон пыли. В полдень под
иссушающими лучами солнца травы трещат, как будто охваченные огнем.
Очень жарко. Температура в тени поднимается до 35 — 44 °C. Насыщенная
пылью атмосфера пылает желтым цветом; желтым кажется и море трав, а
деревья выглядят серыми, безлистными. В сухой сезон большая часть
населения голодает. Но самая острая проблема — вода. Вереницы женщин
несут воду от редких колодцев, преодолевая десятки километров пути до
своего селения. К концу сухого сезона травяной покров съеден дикими и
домашними животными и имеет высоту всего 20 — 30 см. Эту траву мест-
ные жители выжигают, расчищая землю для пашни. Сгорая, трава превра-
202 щается в пепел, используемый как удобрение.
ГЛАВА 8. ЗОНАЛЬНЫЕ Т1ПЫ БИОМОВ СУШИ
Рис. 8.27. Африканская саванна
С приближением лета облик ландшафта меняется. Небо проясняет-
ся, дымка исчезает, на ярко-синем небосводе появляются кучевые обла-
ка. Летом солнце поднимается в зенит. Белые кучевые облака уплот-
няются и превращаются в темные угрожающие облачные скопления.
Их вал надвигается подобно черной стене. Издалека, раскатываясь над
боязливо примолкнувшей местностью, доносятся первые удары грома.
С молнией и громом на жаждущую влаги землю обрушивается силь-
нейший дождь. В короткое время растрескавшаяся высохшая почва ока-
зывается напоенной и размягченной. Все расцветает. Быстро растут
травы. На кустах и деревьях появляется молодая листва. Торопятся на
водопой звери (рис. 8.28).
Травянистый покров саванн образован главным образом злаками из
родов Pennisetum, Andropogon, Imperata, Aristida и др. На их фоне раз-
бросаны деревья с опадающей на бездождное время листвой: баобабы
(Adansonia, сем. Bombacaceae) (рис. 8.29), многочисленные виды ака-
ций, часто с характерной зонтиковидной кроной (рис. 8.30), пальмы,
древовидные суккулентные молочаи, виды Aloe.
Мощная корневая система злаков способствует наиболее полному
улавливанию атмосферных осадков и обеспечивает успешную конку-
ренцию с корневой системой деревьев. Ежегодное отмирание большой
массы травянистых растений, чередование сухих и влажных сезонов
обусловливают формирование специфичных богатых перегноем черных
тропических почв.
203
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ К АРТИ В А МИРА
Рис. 8.28. Тысячные стада антилоп гну совершают миграции, покидая саванну
в период засухи и возвращаясь в период дождей
Рис. 8.29. Баобаб (Adansonia digitata) и масличная пальма (Elaeis guinensis)
в сухой сезон в саванне нижнего Конго, по Г. Вальтеру
204
ГЛАВА 8. ЗОНАЛЬНЫЕ ШЫ БИОМОВ СУШИ
Рис. 8.30. Зонтиковидные акации, по Г. Вальтеру.
Внизу— вертикальная структура растительного покрова
Обычное дерево саванн Западной Африки — масличная пальма
(Eiaeis guinensis). Унее развивается два типа соцветий — мужские и
женские, из которых вторые характеризуются мясистой осью и мно-
гочисленными цветками. Мякоть околоплодника содержит масло (от 22
до 70%), используемое в пищу и как сырье для промышленности. Семя
также содержит масло, отличающееся более высокими вкусовыми и
питательными качествами. Из сочной оси соцветий путем надреза по-
лучают сладкий сок, идущий на приготовление пальмового вина.
Животный мир африканских саванн отличается прежде всего оби-
лием разнообразных копытных. Чтобы избежать конкуренции, в ходе
естественного отбора образовались виды, занявшие различные ниши
потребления растительной пищи — от высоко расположенных ветвей
деревьев и кустарников до мелкотравья (рис. 8.31). Обычными обитате-
лями саванны являются антилопы (канна, орикс, газели, гну), зебры,
буйволы, слоны и носороги, жирафы (рис. 8.32, 8.33, 8.34); в водоемах —
бегемоты и крокодилы. К жизни в открытых ландшафтах саванн при-
способился нелетающий, но быстро бегающий африканский страус.
205
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ К АРТИ В А МИРА
20G
Рис. 8.31. Распределение копытных зверей по ярусам питания
в африканской саванне, поА.Г. Воронову: 1 —жираф;
2— антилопа геренук; 3— антилопа дик-дик; 4— носорог; 5— слон;
6 — зебра; 7 — гну; 8 — газель: 9 — антилопа бубал
Рис. 8.32. Африканский
слон
Многообразие экологических ниш по-
зволяет поддерживать высокую биологиче-
скую продуктивность животных в саванне.
Исходя из этого, экологи дают рекоменда-
цию не разводить крупный рогатый скот, а
заниматься умеренной охотой, тем более
что коренные виды животных менее вос-
приимчивы ко многим тропическим пара-
зитам и болезням, которые сильно поража-
ют домашних животных.
Обилие травоядных в саванне определяет
разнообразие хищных животных — это львы,
леопарды, гепарды, гиены и др. Они регули-
руют численность травоядных, а также выполняют санитарные функции,
поедая падаль, уничтожая прежде всего слабых и больных животных.
Рис. 8.33. Жирафы
Наиболее могучими представителями отряда хищных являются львы
(рис. 8.35). Они не любят лесных местностей, предпочитая открытые
саванны. Нередко львы прячутся у водопоев. Когда наступает ночь,
ГЛАВА 8. ЗВНАЛЬНЫЕ ШЫ БИОМОВ СУШ
Рис. 8.34. Белый носорог
антилопы, зебры, буйволы спешат к воде. Все время нюхая воздух, при-
слушиваясь и зорко вглядываясь в темноту, ведет стадо передовая анти-
лопа. Если из-за сильного желания утолить жажду антилопы пренебре-
гут осторожностью, львы настигнут свою добычу.
Рис. 8.35. Берберийский лев, поА.Э. Брему
207
ЧАСТЬ III. БАОГЕОТРАФАЧЕСААЯ КАРГИ0А МАРА
В обыденной жизни гиены, как правило, вызывают ассоциацию чего-
то грязного и мерзкого. Эти ночные животные отличаются большой жад-
ностью и прожорливостью, имеют пронзительный голос и отвратитель-
ный запах. Только после заката солнца они начинают рыскать; тогда
повсюду слышится их вой. Всего охотнее гиены идут на запах падали;
пожирая ее, они выполняют функции санитаров саванн (рис. 8.36).
Рис. 8.36. Полосатые гиены, поА.Э. Брему
В Индо-Малайском царстве большая часть Индостанского полуост-
рова покрыта листопадными тропическими лесами и саваннами. Рас-
пространены насаждения тикового дерева [Tectona gran dis) и сала
(Shorea robusta). Во внутренних частях Индокитая преобладают тропи-
ческие листопадные леса и отчасти саванны. Среди листопадных лесов
доминируют леса с Tectona grandis, Dipterocarpus tuberculatus, Diospyros
birmanica, видами Terminalia и др.
Саванны Индостана и Идокитая на большей части территории вто-
ричны по происхождению. Выжигание растительности, вырубки силь-
но тормозят рост и возобновление деревьев, травы же при этом хорошо
сохраняются благодаря подземным корневищам и, быстро отрастая в
сезон дождей, прочно удерживают за собой территорию (рис. 8.37).
В Неотропическом царстве ксерофитные тропические редколесья
(каатинга) занимают восточную часть Бразильского нагорья (рис. 8.38).
В начале сухого периода большинство деревьев сбрасывает листья;
однако цветут деревья именно в сухой период. Деревья Chorisia
crispiflora, Cavanillesia arborea и др. (сем. Bombacaceae, к этому же семей-
ству относятся баобабы) имеют вздутый бочкообразный ствол. В рас-
тительном покрове аридных областей Неотропического царства господ-
ствуют суккуленты, ксерофитные кустарники и полукустарники.
Большую роль играют кактусы: многочисленные виды Opuntia, Cereus,
Mamillaria, Echinocactus и др. (рис. 8.39); крупные агавы и лилейные
208 (Yucca, Dasylirion).
ГЛАВА 8. ЗОНАЛЬНЫЕ ШЫ БИОМОВ СУШИ
Рис. 8.37. Саванна Раджастана
Рис. 8.38. Ландшафт каатинги: а — бутылочные деревья, по A. Schimper; б — цве-
тущее бутылочное дерево Cavanillesia arborea, по Г Вальтеру
В Центральной Америке, в южной части Бразилии и Уругвае огромные
пространства заняты саваннами, называемыми здесь кампосами (рис. 8.40).
Так же как в саваннах Африки, основу травяного покрова в них составля-
ют многолетние злаки, правда, они не достигают столь больших размеров
(обычно не более 1 м высотой) и редко образуют сомкнутый травостой. На-
ряду со злаками здесь много сложноцветных, бобовых, вербеновых, ама-
рантовых и др. На фоне трав разбросаны 209
ЧАСТЬ III. БИОГЕОГРАШИЧЕСШ КАРТАВА МАРА
Рис. 8.39. Заросли кактусов весной
невысокие листопадные и вечнозеленые деревья и кустарники. По до-
линам рек — галерейные леса с большим участием пальм. Особенно
типичны леса из восковой пальмы (Copernica cerifera) (рис. 8.41). Листья
пальмы покрыты толстым восковым налетом, который собирают как
ценное сырье.
Рис. 8.40. Саванна Центральной Америки
Характерными обитателями кампосов являются многочисленные
грызуны — капибара, пака, агути, а также питающиеся термитами
большой муравьед и броненосцы (рис. 8.42, 8.43); из птиц — американ-
ский страус нанду. Важную роль в опылении цветков играют колибри
210 (рис. 8.44).
ГЛАВА 8. ЗВНАЛЬНЫЕ ШЫ БИОМОВ СУШИ
Рис. 8.41. Роща восковой пальмы (Copernica cerifera)
Рис. 8.42. Большой муравьед, поА.Э. Брэму
211
ЧАСТЬ III. БИОГЕОГРАШИЧЕСШ КАРТАВА МАРА
Рис. 8.43. Шестипоясный броненосец, поА.Э. Брему
Рис. 8.44. Колибри, поА.Э. Брему
В бассейне Ориноко, в пределах Венесуэлы и отчасти Колумбии,
распространены саванны, называемые льянос. Мощные разливы рек во
время периода дождей приводят к затоплению огромных пространств,
а после ухода воды остаются труднопроходимые болота с зарослями
осоковых и водных растений. Для льяносов характерна маврикиева
пальма (Mauritia flexuosa), которая, так. же как масличная пальма для
жителей Африки, является «деревом жизни». Сердцевина ее ствола
наполнена крахмалистым веществом, плоды съедобны, ось соцветий
212 содержит сладкий сок, идущий на приготовление вина.
ГЛАВА 8. ЗОНАЛЬНЫЕ ИВЫ БИОМОВ СУШИ
В Австралийском царстве в областях, где резко выражен сухой се-
зон, распространены редколесья и саванны, но в отличие от других
тропических царств листопадные деревья в них отсутствуют. Здесь
распространены вечнозеленые эвкалипты, а у многочисленных видов
цсаций вместо листьев образуются филлодии — расширенные черешки
цистьев, принимающие облик листовой пластинки (рис. 8.45).
а б
Рис. 8.45. Виды филлодийных акаций Австралии, по К. Шефферу:
а— акация окаймленная (Acacia marginata); б— акация обманчивая
(Acacia decipiens); в— акация крылатая (Acacia alata)
в
Характерный элемент ландшафта — термитники (рис. 8.46). Стада
копытных, обычные для африканской саванны, в Австралии замеща-
ются крупными кенгуру (рис. 8.47).
Рис. 8.46. Эвкалиптовое редколесье с многочисленными термитниками
213
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ KAPIK8A МИРА
Рис. 8.47. Гигантский рыжий кенгуру
По мере продвижения в глубь континента, где климат становится
все суше, эвкалиптовые и акациевые редколесья замещаются жестколист-
ными вечнозелеными кустарниками (скребами). Различают несколько
типов скребов: малли-скреб с господством кустарниковых эвкалиптов,
мульга-скреб с господством акаций (рис. 8.48).
214
Рис. 8.48. Мульга-скреб в период дождей
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ БИО МОН СУШИ
8.1.3. Пустыни
Ведущим фактором, определяющим формирование биомов пустынь,
является жаркий и сухой (аридный) климат; пустынные биомы называ-
ют также аридными. Они характеризуются разреженным растительным
покровом и своеобразным животным миром. И растения, и животные
приспособлены к жизни в условиях жары и дефицита воды. Биологиче-
ские процессы почвообразования замедлены. Почвы примитивные, обыч-
но их разделяют по механическому составу на пустынные щебнистые,
галечные, песчаные, глинистые; на засоленных грунтах — солончаковые.
В более благоприятных условиях формируются пустынные сероземы.
Области с аридным климатом располагаются симметрично по обе
стороны тропического пояса и находятся под воздействием субтропи-
ческих центров высокого атмосферного давления. Они тяготеют к за-
падным окраинам континентов, где зарождаются пассаты (см. гл. 2).
В Азии и Северной Америке пустыни заходят в континентальные сек-
торы умеренного пояса, мало доступные для воздушных масс, несущих
влагу с океана (рис. 8.49).
Рис. 8.49. Аридные области земного шара, по Г. Вальтеру:
а — крайне засушливые; б — засушливые; в — полузасушливые
(саванны, полупустыни, степи). Пустыни: 1 — Сонора; 2 — Намиб;
3 — чилийско-перуанская прибрежная; 4 — Карру; 5 — засушливые области
Центральной Австралии; 6 — Сахара с египетско-аравийской пустыней
Парадокс образования пустынь в приокеанических секторах, где
близость океана, казалось бы, должна смягчать сухость климата, объяс-
няется главным образом тем, что пассаты, дующие с континента, в
«корнях» своих — сухие ветры. Кроме того, они отгоняют от берега
теплую воду, на место которой поднимается с глубины холодная. Так
образуются холодные вдольбереговые течения. Формирующиеся над
океаном воздушные массы, приходя на горячую сушу, умеряют жару,
но не дают дождя. 215
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ К АРТИ В А МИРА
В условиях благоприятного режима тепла и влаги облик биомов
определяется характером растительного покрова. В пустынях же, где
растительный покров отсутствует или очень разрежен, внешний вид
ландшафта связан прежде всего с характером рельефа и составом по-
верхностных отложений. Выделяют пустыни аккумулятивные и дену-
дационные. Аккумулятивные пустыни представляют собой равнины,
сложенные толщей рыхлых отложений. Под воздействием ветра песча-
ные отложения, не защищенные растительностью, перевеиваются,
образуя эоловые формы: барханы, бугры, гряды и т. п. (рис. 8.50). На
отложениях тяжелого механического состава формируются глинистые
пустыни; на засоленных грунтах и залегающих близко к поверхности
сильно минерализованных грунтовых водах — солончаковые.
21G
Рис. 8.50. Барханы в пустыне Намиб. Внизу— снимок из космоса
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ ВИН МОН СУШИ
Второй тип пустынь — денудационные. Это возвышенные терри-
тории, на которых обнажаются коренные породы или развит плащ
щебнисто-галечниковых отложений. Соответственно денудационные
пустыни могут быть каменистыми, щебнистыми и галечниковыми.
В облике этих пустынь в условиях аридного климата, как это ни пара-
доксально, широко развиты эрозионные формы рельефа, образован-
ные временными водотоками (рис. 8.51). Отдавая должное работе во-
дотоков, возникающих в пустыне после ливневых дождей, подчеркнем,
что многие ныне аридные области в четвертичном периоде пережили
влажные (плювиальные) эпохи, когда осадков выпадало больше и дея-
тельность поверхностных вод, как эрозионная, так и аккумулятивная,
была весьма значительной.
Рис. 8.51. Сильно расчлененная временными водотоками
поверхность низкогорий. Западная Туркмения
Само название «пустынные биомы» отражает скудость органиче-
ской жизни, ее низкую биологическую продуктивность. Однако следует
заметить, что биоты пустынь начали формироваться еще в неогене,
25 — 30 млн лет назад, когда установились глобальные закономерности
океанической и атмосферной циркуляции, а вслед за формированием
горных цепей, преградивших путь воздушным массам, дующим с океана,
последовала аридизация климата внутриконтинентальных областей. 217
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ КАРТИ В А МИРА
Ввиду своей древности пустынные биомы характеризуются большим
биологическим разнообразием. Они порождены биотами тропических
лесов. За миллионы лет постепенной аридизации климата появились
новые формы, прекрасно приспособленные к жизни в условиях дефи-
цита влаги и высоких температур. Заметим, что многие адаптации пу-
стынных растений и животных оказались полезными и для жизни в
условиях похолодания, наступившего в конце неогена.
Пустынные растения представлены как травами (эфемерами, эфе-
мероидами, многолетниками), так и переходными формами от полудре-
весных (полукустарников) к древесным — кустарникам и деревьям.
Формы приспособления растений пустынь к ограничивающим факто-
рам представлены в табл. 8.2.
Таблица 8.2
Формы приспособления растений пустынь к ограничивающим
факторам, по В.К. Василевской и М.П. Петрову
Формы приспо- собления Ограничивающие факторы
Пер иоди ’ г н ость увлажнения и перегрева Постоянный недостаток влаги и высокие температуры воздуха и почвы Подвижность субстрата Засоленность субстрата (солончаки)
Жизненные формы и экологические типы Однолетние и мно- голетние травы. Мезофиты, мезоксе- рофиты Преимущественно кустарники и полу- кустарники. Ксерофиты: склерофиты, суккуленты Преимущественно деревья и кустар- ники, иногда мно- голетние и одно- летние травы. Ксерофиты — псам- мофиты Кустарники, полукустарники, многолетние и одно- летние правы. Ксерофиты — галофиты
Биологические (ритм развития) Эфемеры и эфеме- роиды. Вегетация в благоприятный период увлажнени51 при сравнительно низких температурах. В знойный, сухой период— г г р одолж ител ьн ы й, иногда многолетний, покой подземых органов. Озимость Продолжительная вегетация как в благоприятное,так и жаркое, сухое время года. Летний листопад и веткопад. Смена весенних листьев на летние,очень мелкие Продолжительная вегетация как в благоприятное, так и в жаркое, сухое время года. Образование придаточных корней на засыпае- мых ветках и ство- лах и отпрысков на выдутых корнях. Ускоренный рост засыпанных веток Продолжительная вегетация как в благоприятное, так и в жаркое, сухое время года. Хорошая влагообеспечен- носгь
физиологи- ческие Низкое осмотическое давление. Преимущественно высокая интенсив- ность транспирации Жаро- и засухо- устойчивость. Наличие в клетках водо- удержи вающи х соединений, эфирных и жирных масел, дубильных веществ, алкалоидов. У склерофитов — высокая или сниженная интен- сивность транспирации, повышенное осмотическое давление в летний период. У суккулентов — снижен- ная интенсивностьтранс- пиранни, низкое осмоти- ческое давление. Редукция листьев — микрофиллия, афиллия То же, что и у склерофитов Солеусгойчивосгь и солевыносли- восгь. Жароустойчи- вость. Высокое осмоти- ческое давление. Пониженная транспирация
218
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ БИО МОН СУШИ
Таблица 8.2 (окончание)
Формы приспособле- ния Ограничивающие факторы
Периодичность увлажнения и перегрева Постоянный недостаток влаги и высокие температуры воздуха и почвы Подвижность субстрата Засоленность субстрата (солончаки)
Формативные Листья сравнительно крупные; период роста продолжитель- ный. Строение дор- зи вентральное, сла- бое развитие механи- ческой н проводящей ткани, крупные меж- клетники. Отно- сительно слабо раз- витые корневые системы Метенный и короткий период роста листа. Толстая кутикула, воско- вой налет, лакообразные покрытия, опушение, сгруженные устьица, изопалисадносты плотная сомкнутость клеток. Сильное развитие ме- ханической н водопрово- да щей ткани у склерофи- тов; редукция этих тканей и сильное развитие водоза- пасающей ткани в стеблях или листьях суккулентов. Длинные горизонтальные и глубокие вертикальные корни. Наличие на корнях утолщений, запасающих волу и питательные веще- ства, корневые чехлы Корневые сис- темы тс же. что у склерофитов, но преимущественно гори зонтал ьн ые. Корневые чехлы. Высокая парус- ность семян Стеблевые и лис- товые галофнльные суккуленты. Мик- рофиллия и афил- лия. Из оп ал и садн ость, плотная сомкнутость тканей,развитие во доза паса юшей ткани в листе или стебле. Солевыде- ляющие клетки или ткани
Адаптации животных к недостатку влаги, высоким температурам
также выработались в процессе длительного естественного отбора.
Они проявляются в способности ограничиваться небольшим количе-
ством питьевой воды и заменять ее влагой, находящейся в растениях,
в крови и тканях поедаемых животных; использовать метаболиче-
скую воду; в зимней и летней спячке; в уходе в убежища для сокра-
щения расхода воды на физиологические процессы.
Наиболее приспособленным к
крайним тепловым воздействиям
и условиям недостатка воды в пу-
стыне можно считать верблюда
(рис. 8.52). Механизмы регулиро-
вания водного и температурного
режимов этого самого крупного
животного пустыни очень своеоб-
разны (Shmidt-Nilsen, 1964). В отли-
чие от мелких животных, которые
могут укрыться в жаркое время в
тени или в норах, верблюд всегда
находится под прямым воздействи-
ем солнца.
Верблюды относительно мало
потеют и выделяют с мочой и
калом ограниченное количество
влаги. Важное значение имеет
изоляционный слой шерсти, бла-
Рис. 8.52. Верблюды в пустыне Сахара 219
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ КАРТИ В А МИРА
годаря которому расход воды на охлаждающее испарение сокращается.
Сравнение количества воды, испаряемой стриженым и нестриженым
верблюдом, показало, что первый испаряет на 50% больше воды, чем
второй.
Объем плазмы крови верблюда даже при остром недостатке воды ме-
няется мало, что обеспечивает нормальное кровообращение. Это объяс-
няется тем, что потеря жидкости происходит за счет той ее част, кото-
рая содержится в клетках и тканях животного. Верблюд выдерживает
очень сильное обезвоживание организма, теряя в жаркие дни без водопоя
до 20,5% массы тела и более. Дефицит воды при обильном водопое быстро
восстанавливается. Так, верблюд после длительной жажды может выпить
за 10 мин до 100 л воды. Эта вода быстро всасывается из желудка и уходит
на восстановление запасов воды в тканях. Кроме того, у верблюда имеется
еще одно замечательное свойство, позволяющее ему приспосабливаться в
условиях пустыни как к недостатку воды, так и к дефициту пищевого белка.
И известно, что основной объем воды выделяется почками с продуктами
обмена, главную часть которых составляет мочевина. У верблюда же боль-
шая часть мочевины всасывается в кровь. Далее она поступает в предже-
лудки. Микрофлора последних синтезирует из мочевины полноценный
белок, который утилизируется организмом. Таким образом, одновремен-
но резко снижается количество воды, затрачиваемой на выведение моче-
вины, и восполняется дефицит белка. Подобным свойством в какой-то мере,
вероятно, обладают и другие жвачные животные пустынь.
В пустыне туманов Намиб почти не выпадает дождей. Жуки-черно-
телки в предрассветные часы поднимают вверх заднюю часть тела и
ожидают, пока капелька росы не скатится к ним в рот (рис. 8.53).
Рис. 8.53. В ожидании капли конденсационной влаги. Пустыня Намиб
Не менее важны адаптации животных к жизни в условиях высоких
температур. Особенно сильно раскаляется в дневные часы поверхность
почвы. Поэтому большинство животных пустыни живет в норах и ведет
ночной образ жизни. На рис. 8.54 показано, что температуры понижа-
ются как при переходе в приземные слои воздуха, так и при опускании
в норы. Кроме того, система нор строится таким образом, что в них
возникает естественный ток воздуха и .они хорошо проветриваются.
Большие уши пустынных животных, покрытые частой сеткой
кровеносных сосудов, служат своеобразными охлаждающими радиато-
220 рами (рис. 8.55).
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ ВИН МОН СУШИ
Рис. 8.54. Распределение температур в Каракумах в мае. Цифры —темпера-
тура в предрассветные часы; цифры в кружках — дневная температура, °C
Рис. 8.55. Пустынный заяц. Сахара
Одной из форм адаптации к условиям открытых пространств и раз-
реженной растительности в пустынях является способность животных
к быстрому перемещению. Быстрый бег и полет помогают как в поис-
ках пищи и воды, так и в спасении от врагов.
В Северной Африке расположена одна из величайших пустынь
мира — Сахара. Она представлена песчаными, щебнистыми и галечни-
ковыми равнинами, среди которых возвышаются островные горы. По-
лоса пустынь простирается еще дальше на восток по направлению к
Красному морю, захватывает Аравию и доходит до пустыни Тар на
северо-западе Индии.
Климат Сахары экстрааридный, обусловленный ее положением в
тропических широтах в зоне зарождения сухих пассатных ветров. Это
одна из наиболее жарких пустынь Северного полушария. Средние го- 221
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ КАРТИ В А МИРА
довые температуры воздуха более 20 °C, а июльские доходят до 37 °C;
январские — более 10 "С. Годовые суммы осадков на большей части
территории составляют менее 70 мм. Наиболее сухой является цент-
ральная Сахара, здесь осадки не выпадают по нескольку лет подряд.
Испаряемость же очень высокая — 4500 — 5000 мм в год.
Приведем описание начала дня в Сахаре: «В ранние утренние часы
с поражающей ясностью видны все подробности окружающего мира.
Однако по мере того, как усиливается прогревание, воздух на горизон-
те начинает мерцать и становится мутным. Спавший до этого ветер со
свежими силами спешит в далекий путь. Он вздымает пыль и песок и
вскоре окутывает горы и равнины пеленой мглы. Ветер пустыни не
приносит прохлады. Наоборот, он безмерно усиливает жару, делая ее
нестерпимой. Облака песка и пыли заслоняют солнце, превращая его
диск в мутно-красное пятно. В полуденные часы пустыня превращается
в огромный спящий мир без тени. Мы как будто в печи или внутри
вогнутого зеркала. Колышущаяся, горячая, раскаленная страна...»
Органический мир Сахары крайне скуден. Редкие куртины кустар-
ников, полукустарников и трав приурочены к временно обводняемым
сухим руслам и долинообразным понижениям; останцевые горы, по-
крытые черным «пустынным загаром», лишены видимых признаков
жизни (рис. 8.56).
Рис. 8.56. Песчаная пустыня с редкими куртинами Zygophyllum album,
ZiUa spinosa, Fagonia glandulosa, Stipagrostis ciliata в долинообразном
понижении между останцевыми горами. Западная Сахара
В Южной Африке расположены три относительно небольшие по
площади пустыни: Намиб, Калахари и Карру. Пустыня Намиб занимает
222 полосу суши вдоль побережья Атлантического океана протяженностью
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ БИО МОН СУШИ
около 1500 км и шириной от 50 до 150 км. Холодное Бенгальское тече-
ние создает температурный контраст с раскаленной сушей, поэтому
дожди здесь выпадают крайне редко (количество осадков составляет
менее 10 мм в год; в течение ряда лет дождей не бывает совсем), но
отмечается большое число дней с туманами. Холодное течение несколь-
ко умеряет жару: средняя температура самого теплого месяца 19 — 24 °C.
В северной части Намиба скопления песков образуют огромный мас-
сив до 450 км в длину (рис. 8.50). Из-за сильной сухости пески лишены
растительности; отдельные ее экземпляры встречаются лишь по глубо-
ким межгрядовым понижениям.
Наиболее известным растением подгорной равнины в пустыне
Намиб является вельвичия удивительная (Welwitschia mirabilis) —
единственный вид семейства, порядка и подкласса Welwltschiidae
отдела голосеменных растений (Жизнь растений, 1978). Увельвичии
похожий на пень толстый ствол, от которого отходят два ремневидных
листа. Последние разорваны на множество узких ремешков, волнооб-
разно стелющихся по земле (рис. 8.57). Возраст растения может дости-
гать 100 лет.
Рис. 8.57. Вельвичия удивительная (Welwitschia mirabilis)
Другое необычное растение обитает в пустыне Карру. Это Lithops —
род листовых суккулентов, внешне напоминающих камни. Основная
часть растения находится в почве, над поверхностью выступают только
так называемые «оконные листья», между которыми появляется краси-
вый цветок (рис. 8.58).
В Северной Америке пустыни охватывают В1гутреннюю часть Мек-
сиканского нагорья и на западе выходят к берегам Тихого океана. Наи-
более крупной является пустыня Сонора (точнее, Сонорская группа
пустынь). Средняя годовая температура воздуха здесь около 20 °C; наи-
высший абсолютный максимум 56,7 °C отмечен в Долине Смерти.
Средняя температура самого холодного зимнего месяца — положитель- 223
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ К АРТИ В А МИРА
Рис. 8.58. Суккулент Lithops: Видны «оконные листья» и цветок.
Справа — положение растения в почве
ная, хотя возможны кратковременные заморозки. Количество атмосфер-
ных осадков колеблется от 50 мм в Долине Смерти до 400 мм у восточ-
ной границы гор. Таким образом, зональные типы биомов меняются от
пустынных до полупустынных.
Отличительной особенностью Сонорской группы пустынь является
сложный характер рельефа — чередование сильно расчлененных эрозией
равнинных плато, невысоких горных хребтов и широких межгорных
котловин. В растительном покрове проявляются флорогенетические связи
как с Голарктическим царством — северная часть пустыни занята полуку-
старниковыми полынными пустынями с участием терескена (Eurotia
lanatd) и лебеды (Atriplex canescens, A. confertifolid), так и с Неотропиче-
ским царством — на юге господствуют кактусы. В том числе эндемичный
для Соноры гигантский канделябровидный кактус [Carnegiea gigantea)
высотой до 10— 12 м, живущий более двухсот лет (рис. 8.59).
Рис. 8.59. Общий характер растительного покрова пустыни Сонора,
образованного кактусами Carnegiea gigantea, Ferocactus wislizenii и др.,
224 по Г. Вальтеру
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ БИО МОН СУШИ
Все кактусы обладают поверхностной, далеко распространяющей-
ся в радиальном направлении корневой системой, расположенной на
глубине всего нескольких сантиметров (рис. 8.60). С ее помощью бы-
стро улавливаются атмосферные осадки, пока верхние слои почвы еще
увлажнены. В период засухи растения вынуждены обходиться только
собственными запасами влаги. Крупные экземпляры Camegiea запа-
сают до 2000 — 3000 л воды и могут существовать без ее пополнения
больше года.
Рис. 8.60. Корневая система Ferocactus wislizenii, по Г. Вальтеру.
Вверху— корни простираются в горизонтальном направлении на глубине
всего 2 см; внизу — план корневой системы того же экземпляра
В Южной Америке пустыни вытянуты вдоль гористого тихоокеанско-
го побережья Чили и Перу. Подобно тому как холодное Бенгальское тече-
ние создает особые климатические условия у берегов Юго-Западной
Африки (пустыня Намиб), Перуанское течение обусловливает особенно-
сти климата полосы пустынь тихоокеанского побережья Южной Амери-
ки: без дождей, но с приходящими с океана туманами. В прибрежной
полосе среднее годовое количество осадков не превышает 5—10 мм. Мес-
тами осадки не выпадают по 20 лет. Океан умеряет летнюю жару до 20 —
22 °C; средняя температура самого холодного зимнего месяца 13— 16 °C.
Растительность очень скудная. Ее распределение в предгорьях тесно
связано с воздействием туманов, а выше в горах — с увеличивающимся
количеством атмосферных осадков (рис. 8.61).
На прибрежной равнине встречаются преимущественно сине-зеле-
ные водоросли (Nostoc commune) и лишайники [Cladonia). По мере уси-
ления туманов на склонах гор появляются многочисленные эфемеры,
полукустарники и злаки. Из всех пустынь тихоокеанского побережья
Южной Америки наиболее известна пустыня Атакама. Она тянется
узкой полосой почти на 1000 км. Это одно из самых сухих мест на зем-
ном шаре. Старожилы не помнят, чтобы здесь когда-либо выпадали 225
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ К АРТИ В А МИРА
Рис. 8.61. Ландшафтный профиль через приморскую пустыню до высокого-
рий Кордильер на широте г. Лимы, по Г. Вальтеру: 1 — море и устья рек;
2 — пустыни; 3 — поля гуано на островах; 4 — полоса густого тумана;
5 — растительность на холмах прибрежной полосы; 6 — пустыни с покро-
вом из Tillandsia; 7 — прибрежный лес; 8 — субнивальный пояс растений-
подушек; 9 — злаковники; 10 — злаковая степь с редкими кустарниками;
И — скалистые склоны, покрытые видами бромелиевых; 12— редкостой-
ный вечнозеленый лес; 13 — кустарниковая степь; 14 — склоны долин, по-
крытые летнезеленым травостоем; 15 — заросли Carica uJatropha;
16— заросли колоннообразных кактусов; 17—ледники и фирновые поля
дожди. По данным метеорологических станций, среднее годовое коли-
чество атмосферных осадков составляет 3 — 5 мм. Пустыня занимает
обширное нагорье, постепенно поднимающееся от побережья к подно-
жью Анд. Растительность практически отсутствует (рис. 8.62).
22G
Рис. 8.62. Пустыня Атакама
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ БИО МОН СУШИ
Омывающие пустынные берега воды одни из самых продуктивных
в Мировом океане, поэтому на побережье гнездятся многочисленные
птицы (пеликаны, бакланы и др.), кормящиеся морской рыбой. В усло-
виях сухого климата накопились многометровые толщи птичьего поме-
та — гуано. Это прекрасное богатое фосфатами сельскохозяйственное
удобрение.
В Австралии пустыни занимают около 45% ее площади. Они распо-
лагаются на поверхности Западно-Австралийского плато на высоте 200 —
600 м. Обнажения красноцветной коры выветривания придают ланд-
шафту характерный цвет, благодаря которому эта область получила
название «красного» или «мертвого сердца» Австралии. Над плоской
поверхностью плато возвышаются островные горы красно-бурой окрас-
ки (рис. 8.63).
Рис. 8.63. Островные горы в песчаной пустыне
Располагаясь в тропических и субтропических широтах, пустыни
Австралии характеризуются жарким климатом: средняя месячная тем-
пература самого теплого летнего месяца (января) 28 — 30 °C, самого
холодного (июля) 12—14 °C. Основным источником влаги являются
восточные ветры, дующие с Тихого океана, однако большая ее часть
задерживается Большим водораздельным хребтом. Наименьшее годо-
вое количество осадков (около 120 мм) выпадает во впадине оз. Эйр.
Испаряемость достигает 3000 мм.
Растительный покров представлен ксерофитными травяно-кустар-
никовыми сообществами. В пустынях Австралии известно 2600 видов
высших растений. Из эндемичных пустынных растений следует отме-
тить ряд кустарников (Накеа, Eucalyptus, Casuarina, Acacia и др.), обра-
зующих низкорослые заросли — скрэбы (рис. 8.64). Весной пустыня
покрывается ковром цветущих эфемеров (рис. 8.65).
Но наряду с эндемами здесь встречаются рода и виды, характерные
для азиатских пустынь: Nitraria, Atriplex, Kochia, Salsola, и др. В живот-
ном мире много общего с фауной австралийских саванн. 227
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ КАРТИ В А МИРА
Рис. 8.64. Скрэб
Рис. 8.65. Весна в австралийской пустыне
8.1.4. Бнвмы невехедных субтремческих зен
Субтропические зоны располагаются в Северном и Южном полу-
шариях преимущественно между 30 — 40° и являются переходными от
жаркого пояса к теплым зонам умеренных поясов. С учетом количе-
ства и режима выпадения атмосферных осадков различают биомы влаж-
228 ных субтропиков и сухих субтропиков средиземноморского типа.
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ БИО МОН СУШИ
Биомы влажных субтропиков. Средняя температура самого холод-
ного месяца 4—10 °C, годовое количество осадков 1600 — 4000 мм, засухи
не выражены. Влажные субтропики охватывают восточную окраину
Азии, острова Японии, субтропические широты на восточном побере-
жье Северной Америки; а в Южном полушарии — ряд районов Южной
Америки, южной половины Австралии и Новую Зеландию.
Субтропики Восточной Азии, к югу от р. Янцзы, характеризуются
влажным муссонным климатом. Лето совпадает с влажным сезоном; его
продолжительность 7 — 8 месяцев. Годовое количество осадков около
1500 мм. Зима теплая, но сухая.
Флора отличается богатством видового состава, высокой степенью
эндемизма на уровне семейств и родов. Среди голосеменных известны
мезозойские и третичные реликты: гинкго (Ginkgo biloba), головчатотис
(Cephafotaxus], катайя (Cathayа), кетелеерга (Keteteeria\, торрета боль-
шая (Torreya grandis), лжетис (Pseudotaxus chienii), фокиения (Fokienia
hodginsii), лжелиственница миловидная (Pseudolarix amabilis], криптоме-
рия японская (Cryptomeria japonica), куннингамия (Cunninghamia), мета-
секвойя (Metasequoia gliptostroboides), лжетсута (Pseudotsuga), кипарисо-
вик тупой (Chamaecyparis obtusa), саговник (Cycas revoluta), подокарпус
(Podocarpus macrophylla) и др. Древние реликты есть и среди вечнозеле-
ных древесных пород, например три вида тунга (Aleuritesfordii, A, montana,
A. cordata), камфарный лавр (Cinnamomum camphora) и др.
Зональный тип растительности — полидоминантные вечнозеленые
леса сложного состава. Характерны представители семейств лавровых,
лаковых, чайных (чайный куст Thea chinensis) и др. Многими видами
представлены дубы (в том числе вечнозеленые), рододендроны, которые
с некоторыми другими вечнозелеными кустарниками встречаются в
подлеске. Много лиан, в том числе лимонник (Schizandra sphaenantha),
смилакс (Smilax glabra], плющ (Hedera nepalensis] и др.
Коренных лесов на материке сохранилось мало. Равнинные участки
освоены под сельскохозяйственные угодья. На склонах холмов и гор
распространены вторичные леса и заросли кустарников.
В фауне субтропических биомов Восточной Азии много эндемиков
и реликтов: бамбуковый медведь (рис. 8.66) и др.
Рис. 8.66. Бамбуковый медведь
229
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ КАРТИ В А МИРА
В субтропических лесах Китая встречаются три вида южных оленей,
белки, летяги, мангуст, леопард, ящер, макаки-резусы, тропические виды
птиц. Очень богата фауна беспозвоночных. В почве много термитов,
поедающих отмершую древесину, листья, хвою. У многих насекомых
наблюдается зимний (с декабря по февраль) перерыв в активности.
Биологический круговорот во влажных субтропических лесах проте-
кает очень активно. Органическое вещество интенсивно разлагается и
минерализуется на протяжении всего годового цикла, и в почве накапли-
вается не более 1,5 —2,0% гумуса. Желтоземы и красноземы, присущие
данной зоне, характеризуются низким содержанием азота и фосфора.
В Японии влажные субтропические леса состоят нередко из смеси
лиственных — бука, дуба, клена, ясеня, листопадной магнолии и хвой-
ных пород — тсуги, пихты, кипарисовика (Chamaecyparis) и др. Очень
своеобразна зонтичная пихта, или зонтичная сосна, относимая к осо-
бому роду (Sciadopitys) и являющаяся японским эндемиком, хотя в
миоценовую эпоху она была распространена в Западной Европе. Тре-
тичные реликты сохранились в Японии, т. к. там (в особенности на
южных островах) не было резкого плейстоценового похолодания. Ме-
стами по вырубкам встречаются заросли бамбука. В подлеске представ-
лены вечнозеленые виды.
Богатству растительности явно уступает разнообразие животного
мира. И все же японская фауна включает по крайней мере 140 видов
млекопитающих, 450 видов птиц, большое разнообразие рептилий, зем-
новодных и рыб. Из приматов — единственный вид, японский макак,
обитает на острове Хонсю. Хищники представлены черными медведя-
ми, лисами, барсуками. Из пушных зверей следует назвать куницу,
норку, выдру, ласку. Многочисленны грызуны: зайцы, белки, крысы и
мыши, хотя обычная домовая мышь отсутствует. Из двух видов оленей
чаще встречается маленький японский олень. Среди птиц обычны ку-
кушка, дятел, фазан; особенно почитаемы у японцев два вида соловья.
В Северной Америке на атлантическом побережье располагаются
влажные субтропики, на тихоокеанском наблюдается переход от влаж-
ных к сухим субтропикам средиземноморского типа.
На востоке биомы влажных субтропиков занимают часть Приатлан-
тической и Примексиканской низменностей. Летом 25 — 28 °C. Зимние
температуры меняются от 2 — 4 °C на севере до 12—14 °C на юге. Годо-
вое количество осадков 1200—1500 мм. В отличие от Восточной Азии
дожди здесь выпадают довольно равномерно в течение года. Большие
запасы тепла и обилие влаги определяют высокую активность геохими-
ческих и микробиологических процессов. Почвы представлены красно-
земами и желтоземами.
Лиственные леса образованы многими видами дуба, клена, липы,
белой акации (Robinia pseudoacacia} и др. Третичные реликты — тюль-
панное дерево (Liriodendron tulipifera] и великолепная вечнозеленая
магнолия (Magnolia grandiflora} с крупными душистыми белыми цвет-
ками, широко культивируемая в субтропиках на черноморском побере-
жье. Много деревьев второй величины, вечнозеленых и листопадных
кустарников, лиан. В составе лесов отмечается также широкое присут-
230 ствие местных видов сосен.
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ БИО МОН СУШИ
К югу от Аппалачских гор леса постепенно обогащаются тропиче-
скими элементами: появляются пальмы, бамбуки, саговники, вечнозе-
леные лианы, эпифитные бромелиевые, кактусовые, ароидные и др.
Весьма специфичны пойменные леса с болотным кипарисом
(Taxodium distichum}. Палеоботанические находки свидетельствуют, что
в третичном периоде подобные леса были обычны в Северной Америке,
Европе и Азии. Современные деревья достигают 45 м в высоту, облада-
ют толстым (до 12 м в обхвате) конусовидным стволом и своеобразной
корневой системой, помогающей удерживать дерево в зыбкой болот-
ной почве. Расходящиеся горизонтально корни образуют оригинальные
выросты — пневматофоры, возвышающиеся на 1—2 м над уровнем
почвы (рис. 8.67).
Рис. 8.67. Группа болотных кипарисов, ветви которых увешаны
«испанским мхом» (Tillandsia, сем. бромелиевых).
Из почвы выступают почти на метр пневматофоры
Животный мир восточного субтропического побережья Северной
Америки принадлежит обширной Сонорской фаунистической подобла-
сти Голарктического царства. Ее характерные представители показаны
на рис. 8.68.
На узкой полосе тихоокеанского побережья южнее 42° с. ш. зима
теплая: средняя температура января 8—10 °C. Однако лето прохладное,
что обусловлено влиянием холодного Калифорнийского течения. Сред-
няя температура самого теплого месяца (сентября) 14—18 "С. Годовая
сумма осадков 500 — 600 мм. Но в северных районах на горных склонах
она увеличивается до 1000 мм и более. Основная часть осадков (60%
годовой нормы) выпадает зимой — с декабря по февраль, тогда как в
июле — августе дождей практически нет.
В субтропических биомах запада наблюдается переход от влажных на
севере к сухим средиземноморского типа на юге. Отличительной чертой
биомов первого типа являются леса из секвойи (рис. 8.69). На тихо-
океанском побережье продольные цепи Кордильер создали убежище для 231
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ КАРТИ В А МИРА
Рис. 8.68. Характерные представители фауны Сонорской области
Голарктического царства, по А.Г. Воронову: 1 — гризли; 2 — пекари;
3 — индюковый гриф; 4 — индюк; 5 — луговая курочка; 6 — скрытножабер-
ник; 7 — аллигатор; 8 — грифовая черепаха; 9 — койот; 10 — луговая со-
бачка; 11 — каролинская утка; 12 — вилохвостый лунь; 13 — рисовка; 14 —
232 вилорог; 15 — виргинский олень; 16 — белоголовый орлан; 17 — бизон
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ ВИН МОН СУШИ
Рис. 8.69. Лес из секвойи гигантской.
Справа — туннель, выпиленный в стволе секвойи
многих реликтовых родов и видов растений. Особую известность полу-
чили секвойя гигантская [Sequoiadendron giganteum), называемая так-
же мамонтовым деревом, и секвойя вечнозеленая [Sequoia sempervirens).
Это величественные и монументальные деревья, достигающие высоты
80—100 м, со стволом до 10 м в диаметре. Вопрос об их предельном
возрасте до сих пор остается нерешенным: называют и 3, и 4 тыс. лет.
Биомы южного тина имеют средиземноморский облик. Раститель-
ный покров образуют жестколистные вечнозеленые дубовые леса (10 ви-
дов дуба: Quercus chrysolepis, Q. vislizeni, Q. leriocarpus и др.) и заросли
кустарников чапарраль. Известно свыше 100 видов кустарников. Наи-
более характерны аденостома, или чамиз (Adenoetoma fasciculatum], —
ксерофит с игловидными листьями, толокнянка [Aictostaphylos, 18 ви-
дов), жестер шафранный [Rhamnus сгосеа), близкий к жестеру цеанотус
[Ceanothus cuneatus, С. oliganthus, С. spinosus, С. divaricatus и др.), лав-
ровишня падуболистная [Laurocerasus = Prunus ilicifolia), кустарнико-
вые формы дубов. На песчаных и каменистых почвах — сосновые рощи.
Труднопроходимые заросли чапарраля замещают дубовые леса пос-
ле пожаров или вырубки. По мере аридизации климата чапарраль усту-
пает место пустынно-степным сообществам из шалфея, полыни, юкки,
кактусов. Естественные и производные субтропические биомы посте-
пенно уступают место плантациям цитрусовых и виноградникам.
В Южной Америке влажные субтропические биомы распространены
в приатлантической части. Годовое количество осадков 1600 — 2500 мм.
Зимние температуры около 12— 13 °C. Растительность представлена влаж-
ными высокоствольными многоярусными вечнозелеными лесами с гус-
тым кустарниковым ярусом. В верхнем ярусе (40 — 55 м) господствуют
вечнозеленые эукрифия (Eucryphia cordifolia), антарктический бук 233
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ К АРТИ В А МИРА
(Nothofagus dombeyi}, из хвойных — фицройя (Fitzroya patagonica] и др.;
во втором ярусе — дримис (Drimys winteri) и др.; в третьем — протейные
и древовидные папоротники (Blechnum magellanicum]. Деревья обвиты
лианами и покрыты эпифитами.
В горах встречаются чисто араукариевые (Araucaria brasiliana,
A. angustifolia) леса с наземным злаковым покровом. Редко стоящие
деревья араукарии достигают высоты 25 — 40 м при диаметре ствола у
основания до 9 м и имеют характерную плоскую зонтиковидную
крону (рис. 8.70).
Рис. 8.70. Лес из араукарии
В сообществе с араукарией растут листопадная имбуйя (Phoebe porosa)
и вечнозеленая йерба-мате, или парагвайский чай (Ilex paraguaiensis), из
листьев которого приготовляют напиток. Имбуйя местами составляет
до 20% древостоя, а иногда она, как и йерба-мате, образует чистые мас-
сивы с густым подлеском. Ареал распространения йерба-мате несколь-
ко шире, чем араукариевых лесов. Кроме того, парагвайский чай широ-
ко культивируется на плантациях.
К числу достопримечательностей относятся древний род дримис
из магнолиевых, агатис и подокарпус из хвойных, пальма из рода арека
(Агеса), древовидные лилейные (Cordyline), напоминающие американ-
ские юкки, а также лианы, эпифиты (последние главным образом из
папоротников).
Биомы влажных субтропиков на севере Новой Зеландии представ-
лены богатыми лесами, имеющими облик субтропической дождевой
гилей. В лесах много древовидных папоротников (рис. 8.71).
Флористически Новая Зеландия ближе к субантарктической Аме-
рике (к Голантарктическому царству), чем к Австралии. Здесь нет эвка-
липтов, акаций, ксанторрей, казуарин; мало протейных. Но широко
234 распространены леса из вечнозеленых южных буков (Nothofagus).
ГЛАВА 8. ЗОНАЛЬНЫЕ ШЫ БИНИОН СУШИ
Рис. 8.71. Лес древовидных папоротников, Новая Зеландия
Специфика животного населения Новой Зеландии носит резко вы-
раженный островной характер. Отсутствует ряд групп, например, почти
нет млекопитающих, зато много нелетающих птиц: киви, пастушок узка,
совиный попутай. Из рептилий совсем нет змей, крокодилов и сухопут-
ных черепах. Еще в исторически обозримое время здесь обитало 7 родов
и около 20 видов гигантских страусов моа, истребленных после прибы-
тия в Новую Зеландию первых полинезийцев (рис. 8.72, 8.73).
Рис. 8.72. Охота полинезийцев на гигантскую птицу моа
235
ЧАСТЬ III. БИОГЕОГРАШИЧЕСШ КАРТАВА МАРА
Рис. 8.73. Характерные представители новозеландской фауны.
Рис. В.А. Ватагина, по Н.А. Бобринскому: 1 — киви; 2 — совиный попугай; 1
3 — попугай нестар; 4 — гуйя (слева — самец, справа — самка); 5 — нелетающая
23G ултанская курица; 6 — нелетающий пастушок узка; 7 — моа; 8 — гаттерия
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ БИО МОН СУШИ
Биомы сухих субтропиков. Эти биомы характеризуются жарким и
сухим летом, прохладной и дождливой зимой. Такой климат получил
название средиземноморского, а биомы — общее наименование «био-
мы средиземноморского типа» (БСТ). Они располагаются в Северном и
Южном полушариях между субтропическим поясом высокого атмо-
сферного давления и областями зарождения циклонов в умеренном поя-
се — это Средиземноморье, Калифорния, Среднее Чили, Южная Аф-
рика, юг Австралии. Летом господствующим является воздействие
центров высокого атмосферного давления, поэтому стоит жаркая и
сухая погода; зимой в субтропики спускаются из умеренного пояса
центры низкого давления, что сопровождается усилением циклониче-
ской деятельности, дождливой погодой.
В БСТ преобладают формации жестколистных (склерофильных)
вечно- или зимнезеленых кустарников. Из многочисленных местных
названий БСТ наиболее известны маквис, гаррига, фригана и шибляк
в Средиземноморье, чапарраль в Америке, финбош в Южной Африке и
малли-скрэб в Австралии.
Происхождение вечнозеленых жестколистных кустарниковых
формаций БСТ связывают с подлеском субгумидных лавролистных
лесов. По мере усиления аридности климата древесный ярус этих
лесов редуцировался, а кустарниковый подвергался изменениям в на-
правлении увеличения числа склерофильных видов и их усиливаю-
щейся адаптации к сезонной сухости. В сухих лесах обычны низо-
вые пожары, и адаптации к огню также могли начать формироваться
уже под пологом леса.
Свидетельства о существовании типичных БСТ появляются не ра-
нее позднего миоцена-плиоцена. Таким образом, БСТ представляются
довольно молодыми биомами. В плейстоцене и голоцене ареал БСТ неод-
нократно то сокращался, то расширялся. Климатогенные пульсации в
голоцене усугубились действием антропогенных факторов.
В качестве примера БСТ остановимся на биомах Средиземноморья.
В начале третичного периода на равнинах господствовали тропиче-
ские и субтропические леса, после поднятия гор сменявшиеся пояса-
ми широколиственных (платан, грецкий орех, каштан, тополь, дуб, бук
и др.) и хвойных (сосна, ель пихта) лесов, отчасти местных, отчасти
проникших с востока по гималайскому пути миграций. Межледнико-
вые эпохи в плейстоцене вызвали формирование засушливых центров
на Пиренейском полуострове, в Северной Африке и Малой Азии. Они
обусловили ксерофитизацию и сильное обеднение когда-то богатой
средиземноморской флоры. Тем не менее именно флора третичного
Средиземья явилась ядром, из которого развилась в основном на мес-
те своего происхождения, т. е. автохтонно, современная средиземно-
морская флора.
Формирование современных БСТ происходило в условиях субтро-
пического климата с жарким сухим летом и влажной зимой. Сред-
ние температуры зимы 7—14 °C, хотя абсолютные минимумы могут
достигать —15 °C. Годовая сумма осадков 400 — 700 мм. Дефицит вла-
ги (превышение испарения над выпадающими осадками) достигает
1000— 1500 мм.
237
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ КАРТИ В А МИРА
Берега Средиземного моря гористые. В нижней части склонов гос-
подствует вечнозеленая жестколистная (склерофильная) древесная и
кустарниковая растительность на коричневых почвах. Характерный
вид — маслина (Olea europea); ее ареал почти точно совпадает с ареалом
БСТ (рис. 8.74, 8.75). у
238
Рис. 8.74. Роща маслин
Рис. 8.75. Ареал маслины (Olea europea), по В.В. Алехину
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ БИО МОН СУШИ
Господствующими формациями являются маквис, гаррига, фригана
и шибляк.
Маквис — заросли с преобладанием небольших вечнозеленых дере-
вьев и кустарников с мелкими жесткими листьями. Наиболее обычны
здесь земляничное дерево (Arbutus uneda), мирт (Myrtus communis), ла-
данники (Cystus), фисташка (Pistacia lentiscus), дикая маслина (Olea
europea var. oleaster), вечнозеленый каменный дуб (Ouercus ilex) (рис. 8.76).
Лианы — сассапариль (Smilax asperd), ломонос (Clematis flammuld), плющ
(Hedera helix) — делают заросли труднопроходимыми. На юге Испании
распространены заросли карликовой пальмы (Chamaerops humilis) — един-
ственного вида пальм, имеющего свой естественный ареал в Европе.
Рис. 8.76. Маквис (Ouercus ilex, Pistacia lentiscus, Euphorbia dendroides,
Rhamnus alaternus, Spartium juncus) на средиземноморском побережье
Гаррига развивается на сухих каменистых почвах по южным скло-
нам гор. Деревья отсутствуют, господствуют вечнозеленые кустарники
с примесью дуба (Ouercus coccifera), называемого на юге Франции «га-
рулия» (отсюда гаррига).
Фригана формируется в условиях сильной и продолжительной лет-
ней засухи, а также в местах усиленного выпаса как крайний вариант
дигрессии растительного покрова.
Растения растут разреженно, всюду видна скелетная почва. Господ-
ствуют колючие, пахучие, часто сильно опушенные полукустарники и
многолетники с одеревеневшими основаниями; весной обильно цветут
эфемеры и эфемероиды. В Испании на гипсовых плато в районе Сара-
госы фригана представлена ассоциациями с господством солнцецвета
(Helianthemum), розмарина (Rosmarinus officinalis), тимьяна (Thymus
vulgaris), полыни белой (Artemisia herba-alba) с участием степных зла-
ков: ковыля (Stipa parviflora), типчака (Festuca valesiaca) и др. (рис. 8.77).
Подобный ландшафт, казалось бы, не оставляет места для занятия
земледелием. Однако на аэрофотоснимке этого района виден своеоб-
разный членистый рисунок сухих русел, отображающий рукотворные
терраски, удерживающие мелкозем и весеннюю влагу. Они использу-
ются для выращивания ячменя (рис. 8.78). 239
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ КАРТАВА МАРА
Рис. 8.77. фригана на гипсовом плато в Испании
Рис. 8.78. Террасовое земледелие в Испании:
а — фрагмент аэрофотоснимка. Масштаб 1:5000. Виден членистый
рисунок террасированных сухих русел; б — земледельческие террасы
в сухом русле, прорезающем поверхность гипсового плато
240
ГЛАВА 8. ЗОНАЛЬНЫЕ ШЫ БИО МОН СУШИ
Рис. 8.79. Шиблякна Южном берегу Крыма
Шибляк представляет северный вариант средиземноморской расти-
тельности на черноморских склонах гор Крыма и Кавказа (рис. 8.79).
Он образован зарослями листопадных форм деревьев и кустарни-
ков: дуба пушистого (Ouercus pubescens), грабинника (Carpinus orientalis],
держидерева (Paliurus spina-christi) и др. в сочетании со степными зла-
ками и разнотравьем.
Животный мир Средиземноморья (рис. 8.80) сильно изменился под
влиянием многовековой деятельности человека. В настоящее время ведет-
ся большая работа по его охране; единственный вид европейских обезьян
живет в естественных условиях в заповеднике на Гибралтарской скале.
В БСТ растительная масса слабо используется консументами как в
пастбищных, так и в детритных пищевых цепях (Жерихин, 1995). Эта
особенность хорошо объясняется климатическим режимом. Для фито-
фагов прямое климатическое ограничение связано с пиком прироста
фитомассы в наиболее прохладный и дождливый период, когда неблаго-
приятная погода не позволяет растительноядным беспозвоночным не
только достичь высокой численности, но и поддерживать постоянно
высокую активность. Растительный опад, напротив, накапливается пре-
имущественно в сухой и жаркий сезон, когда дефицит влаги лимитирует
активность сапрофагов. Большинство почвенных беспозвоночных про-
водит лето в состоянии покоя или в фазе низкой численности. Таким
образом, период наибольшей массы корма и период благоприятной по-
годы для потребителей не совпадают. В результате биологическое разру-
шение подстилки происходит медленно, за 3 — 5 лет. Существенным
фактором, ограничивающим деятельность консументов, является жест-
колистность. Жесткие листья малопитательны, защищены механически
и обычно слабо поедаются как фито-, так и сапрофагами.
241
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ КАРТИ В А МИРА
Рис. 8.80. Характерные представители фауны Средиземноморской области
Голарктического царства, поА.Г. Воронову: 1 — муфлон; 2 — протей;
3 — тосканская землеройка; 4 — белоголовый сип; 5 — канарейка; 6 — лань;
7 — гюрза; 8 — желтопуз; 9 — геккон; 10 — шакал; 11 — греческая черепа-
ха; 12 — смирнский белый зимородок; 13 — пятнистая кукушка; 14 — ди-
кобраз; 15— франколин; 16— дикий кролик; 17—леопардовый полоз;
242 18 — горная курочка; 19 — генетта; 20 — бесхвостый макак
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ БИО МОН СУШИ
Из-за низкого уровня потребления первичной продукции в БСТ обра-
зуется большой запас сухой подстилки, на 70 — 80% состоящей из лис-
тового опада. Эта подстилка, препятствуя возобновлению, должна была
бы делать мертвопокровные сообщества БСТ неустойчивыми во време-
ни. Этого, однако, не происходит, поскольку БСТ повсюду характеризу-
ются высокой частотой пожаров, которые в в данном случае являются в
значительной мере природным, а не антропогенным феноменом. При
снижении частоты пожаров они делаются более сильными и разруши-
тельными из-за увеличения запаса горючего материала. Значение пожа-
ров для поддержания БСТ подтверждается тем, что их искусственное ис-
ключение в течение 60—100 лет приводит к почти полной остановке
возобновления доминантой. Историческая типичность пожаров в БСТ,
так же как в саваннах, подчеркивается широким распространением адап-
тации к воздействию высоких температур у их обитателей.
Таким образом, в БСТ пожары являются постоянно действующим
фактором и играют важнейшую роль в функционировании экосистем,
обеспечивая возврат в оборот биогенов, связанных в мортмассе. По
существу, огонь выполняет в этих экосистемах функцию своеобраз-
ного небиологического редуцента, действующего эпизодически, но
чрезвычайно быстро и придающего обороту системы своеобразную
многолетнюю ритмику. Без пожаров устойчивое существование БСТ
немыслимо. Возврат биогенов в круговорот веществ оплачивается це-
ной высоких потерь, главным образом азота, в результате вымывания
атмосферными осадками. Важнейший биологический компенсатор-
ный механизм — широко распространенная в БСТ азотфиксация, к
которой способны не только такие классические азотфиксаторы, как
бобовые, но и многие другие растения.
И 8.2. Биомы колодных и умеренных поясов_________________________
От полюсов к субтропическим широтам простираются холодные и
умеренные пояса. Если в жарком поясе ведущим фактором формирова-
ния зональных типов биомов является возрастающая сухость климата,
то в холодных и умеренных поясах главную роль играет первичный
фактор географической зональности — температурный режим. Различия
в климате и особенности палеогеографии накладывают свой отпечаток
на своеобразие ландшафтов. Наряду с биполярностью органического
мира отмечаются существенные черты отличия зональных типов био-
мов холодных и умеренных поясов Северного и Южного полушарий.
В Северном полушарии приполюсное пространство занято океаном,
полярные пустыни представлены прерывисто в основном на островах.
Биомы тундры и тайги имеют субширотное простирание, протягиваясь
в виде концентрических полос через континенты Северной Америки и
Евразии, принадлежащие к одному Голарктическому биогеографиче-
скому царству. Но по мере перехода в низкие широты, где климат ста-
новится более теплым, все отчетливее проявляется роль увлажнения, и
в распределении зональных типов биомов находит выражение закон
географической секторности (рис. 8.81).
243
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ К АРТИ В А МИРА
450
льоЗ*и
^_^-410
Лестцнац
бОО
+2
60
rjH^—-150°
850'^7
Знеи^-гэО
190-
-э ккал/см^гоЗдо
^С^-65-—
+11
♦5^
Лесостепь
Степь /)«/стьия
ПоЭзояы и подзолис-
тые долотные почвы
Почвы Северные
тундры подзолы
Серые лесные
почвы Черноземы
и мощные и каштоно-
черноземы вые почвы Сероземы
Рис. 8.81. Схема изменения климата, растительности и почв на профиле
Восточной Европы в направлении с северо-запада на юго-восток к Прикас-
пийской низменности, поА.П. Шенникову и Г. Вальтеру. Зачернен гумусо-
вый горизонт; мощные черноземы соответствуют лесостепи.
Заштрихован иллювиальный горизонт
В Южном полушарии приполюсное пространство занято Антаркти-
ческим континентом, лежащим в зоне полярных пустынь. Субантарк-
тические и умеренные широты заняты в основном океанами. Назем-
ные биомы формируются на островах и окраине Южной Америки.
Типичные тундры, тайга и широколиственные леса, характерные для
Голарктического царства Северного полушария, здесь отсутствуют. Зо-
нальные типы биомов Голантарктического царства обладают своими спе-
цифическими свойствами.
8.2.1. Тундра
Астрономической границей холодных поясов, где формируются
биомы полярных пустынь и тундр, являются полярные круги — север-
ный и южный, располагающиеся на 66° 33' с. ш. и ю. ш. У полюсов сол-
нце встает и садится лишь один раз в год. Полярный день и полярная
ночь, начинающиеся и заканчивающиеся в дни равноденствий, длятся
по шесть месяцев только на самих полюсах. У полярных кругов с нача-
лом весеннего равноденствия продолжительность дня увеличивается
ежедневно на 8 мин. Только одни сутки, в день летнего солнцестояния,
солнце не заходит за горизонт; после осеннего равноденствия продол-
244 жительность дня быстро уменьшается.
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ БИО МОП СУШИ
Циркумполярное положение холодных поясов определяет суровость
климата: отрицательный радиационный баланс, отрицательные средние
годовые температуры воздуха. Границей холодного пояса служит изо-
терма самого теплого месяца 10 "С. Наряду с климатическими условия-
ми большое влияние на формирование биомов холодных поясов оказы-
вает взаиморасположение суши и океана.
Холодный пояс Северного полушария называют еще Арктическим
или Арктикой. От полюса к югу тепловой режим становится более бла-
гоприятным для жизни. Это служит основанием для разделения Арк-
тики на зоны и подзоны (рис. 8.82).
Рис. 8.82. Зональное подразделение Арктики, по В.Д. Александровой:
1 — южная граница тундры, в том числе подзоны южной тундры;
2 — южная граница подзоны северной тундры; 3 — южная граница зоны
арктических пустынь 245
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ К АРТИ В А МИРА
Тундры — главный зональный тип биомов холодного пояса Голарк-
тического царства. Если весь холодный пояс называют Арктикой, то
тундры с расположенной южнее лесотундрой относят к Субарктике.
Тундра в переводе с финского языка означает «плоское безлесное
пространство». Формирование тундровых биомов определяется суровы-
ми климатическими условиями. Зима длинная, до 8 месяцев. Средняя
годовая температура ниже нуля. Однако не зимние холода (полюс хо-
лода Северного полушария располагается, как известно, в Восточной
Сибири в таежной зоне), а холодное и короткое лето определяют осо-
бенности среды обитания в тундре. Заморозки возможны в любой лет-
ний день, но бывают и жаркие дни, когда температура воздуха подни-
мается до 25 — 30 °C. Атмосферных осадков (в основном в виде снега)
выпадает немного — 300 — 400 мм. Снеговой покров маломощный, но он
играет важную защитную роль, предохраняя находящиеся под ним
растения, а также животных от сильных морозов, ветра и поземки.
В связи с низкими температурами испаряемость низкая, что приводит,
несмотря на небольшое количество осадков, к избыточному увлажне-
нию: заболачиванию и образованию многочисленных озер.
От расположенной южнее зоны тайги тундру отделяет граница леса.
Одним из условий формирования таежной растительности является сред-
няя месячная температура самого теплого месяца выше 10 °C (рис. 8.83).
Обращает внимание асимметрия границ тундры у западных и вос-
точных окраин Евразии. В Атлантическом секторе, где сказывается
отепляющее влияние Гольфстрима, она поднимается выше Полярного
круга, в Тихоокеанском секторе под влиянием холодных течений опус-
кается к 60° с. ш.
В тундрах Голарктического царства еще свежа память о недавнем
оледенении: почти повсеместно распространены многолетнемерзлые
породы («вечная мерзлота»). Их мощность изменяется от 300 — 400 м в
арктических тундрах до первых метров в южных тундрах. Глубина
сезонного оттаивания колеблется от 30 — 40 см в северной полосе до 0,5 —
2 м в южной. Постоянно мерзлый слой обусловливает охлаждение поч-
вы и замедление биохимических процессов, препятствует фильтрации
поверхностных вод и способствует заболачиванию; он резко ограничи-
вает грунтовое питание рек и их эрозионную деятельность. С мерзло-
той связаны специфические геоморфологические процессы и формы
мезо- и микрорельефа. Наиболее крупные формы — гермокарстовые —
возникают в результате оседания грунта при вытаивании подземных
льдов. Морозное растрескивание грунтов и образование ледяных клинь-
ев в трещинах ведут к формированию полигонального микрорельефа
(рис. 8.84). С мерзлотным пучением грунтов связан мелкобугристый
рельеф. Криогенная сортировка твердого материала имеет следствием
пятна-медальоны, структурные грунты.
Стабильность многолетнемерзлых грунтов, колебания верхней гра-
ницы и даже само существование мерзлоты зависят от многих обстоя-
тельств, и в первую очередь от растительного покрова, имеющего значе-
ние важнейшего стабилизирующего фактора. Растительная дернина
затрудняет теплообмен между мерзлым слоем и воздухом (поддерживает
24G тепловое равновесие), скрепляет почву, препятствует сортировке грунта.
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИЛЫ 510МОН СУШИ
Рис. 8.83. Полярная граница древесной растительности, ограничивающая
субарктическую зону на юге, по Б. А. Тихомирову. Границу образуют:
1 — Betula tortuosa; 2 — В. ermani; 3 — Picea obovata; 4 — P. mariana;
5 — Larix sibirica; 6 — L. dahurica
Puc. 8.84. Полигональная тундра. Аэрофотоснимок
247
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ КАРТИ В А МИРА
Недостаток тепла и избыток влаги, мерзлота, низкая емкость биоло-
гического круговорота, небольшое количество растительного опада, его
замедленная гумификация и минерализация — все эти факторы опре-
деляют главные черты геохимических процессов и почвообразования.
Химическое выветривание протекает слабо, высвобождающиеся осно-
вания вымываются из почвы, так что она обеднена кальцием, натрием,
калием (но обогащена железом и алюминием). Широкое распростране-
ние в Субарктике, особенно в южной ее части, имеют тундровые глее-
вые почвы с торфянисто-перегнойным горизонтом, под которым нахо-
дится сравнительно однородная глеевая минеральная вязкая толща.
В связи с низкой продуктивностью растительности торфообразова-
ние слабое; мощность торфа редко превышает 1 м. Почвенный по-
кров, как и растительный, характеризуется мозаичностью.
Арктика — это нивальные ландшафты, мир снега и льда. Снежный
покров играет важную роль в жизни растений и животных. Продолжи-
тельность залегания снежного покрова до 220 — 280 дней — главный
отрицательный фактор в жизни большинства животных и растений.
Толщина снежного покрова зависит от рельефа (с возвышений снег
сдувается, в понижениях накапливается), а также от континентально-
сти климата — к западу от Таймыра она больше, к востоку меньше. В то
же время снег играет и роль положительного экологического фактора.
В качестве хорошего теплоизолятора он защищает почву, растения и
животных от зимних морозов. Кроме того, слой снега предохраняет
растения от снежной корразии: ветви, выступающие над снегом, сре-
заются или сильно повреждаются зимней поземкой, поэтому высота
растений полностью зависит от мощности снегового покрова.
Тундровые биоты Палеарктики и Неарктики близки между собой.
Многие виды растений (куропаточья трава (Dryas octopetala), брусни-
ка, водяника, альпийская толокнянка, альпийский мятлик и др., как и
животных: северный олень, белый медведь, песец, белая сова и др.),
имеют циркумполярный ареал. Общность тундровых биот можно
объяснить общностью истории развития северных районов Евразии
и Северной Америки, претерпевших оледенение. Биоты тундр харак-
теризуются молодостью и бедностью видового состава: теплолюбивые
формы третичного периода погибли или были коренным образом пре-
образованы во время четвертичного оледенения. Вместе с тем для
Арктики характерен значительный процент эндемиков на родовом и
видовом уровнях (рис. 8.85).
В тундре отсутствует древесная растительность. По выражению
М.В. Ломоносова, «тундрами называют места мохом зарослые». В ра-
стительном покрове кроме мхов и лишайников распространены низ-
корослые психрофитные и криофитные формы, приспособленные к
существованию в условиях низких температур и высокой влажности
почв, к использованию тепла приземного слоя и защитных функций
снежного покрова.
Растения тундр характеризуются низкой интенсивностью фотосин-
теза, медленным накоплением органической массы, малой энергией
роста. Годичные кольца растений нередко состоят из одного ряда кле-
248 ток. Побеги полярной ивы удлиняются за год на 1—5 мм. Годичный
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИЛЫ ВИН МОН СУШИ
Рис. 8.85. Характерные представители фауны тундры. Рис. В.А. Ватагина,
по Н.А. Бобринскому: 1 — тундровая куропатка; 2 — белая куропатка;
3 — мохноногий канюк; 4 — лапландский подорожник; 5 — пуночка;
6 — белая сова; 7 — белолобая казарка; 8 — черная казарка; 9 — черноша-
почный сурок; 10 — краснозобая казарка; 11 — копытный лемминг;
12 — обский лемминг; 13— снежный баран; 14 — северный олень; 15 — песец 249
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ КАРТИ В А МИРА
прирост ягеля составляет около 4 мм. Однолетники в Арктике — явле-
ние редкое; они не успевают завершить жизненный цикл в одно лето.
Наиболее распространенная жизненная форма — карликовые кустар-
нички и многолетние травы (рис. 8.86).
Рис. 8.86. Растения тундры, по Т.Н. Гордеевой и О.С. Стрелковой:
А — крупка волосистая (Draba pilosa); Б — ива полярная (Salix polaris);
В — карликовая береза (Betula папа); Г — мак подушковидный (Papaver
pulvinatum); Д — дриада точечная, или куропаточья трава (Dryas punctata)
Стебли у них короткие, листья малых размеров и у многих собраны
в прикорневую розетку. Обычны здесь и подушковидные формы, возни-
кающие при отмерзании верхушечных почек. В таком случае трогаются
в рост нижние пазушные почки, что приводит к обильному ветвлению
и образованию коротких стелющихся побегов. Густо расположенные мел-
кие ветви с листьями на верхушках побегов придают кусту обтекаемую
форму, которая хорошо противостоит ураганным ветрам. Под пологом
такой подушки создается более мягкий микроклимат, а нижние отмер-
шие листья, постепенно разрушаясь, служат дополнительным источни-
250 ком азота для скудной им почвы.
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ БИО МОП СУШИ
У животных тундры также выработался целый ряд приспособлений,
облегчающих существование в суровых условиях. Например, у песца бе-
лая покровительственная окраска (рис. 8.87.). Он быстро нагуливает толс-
тый слой жира. Зимний мех его пушист и тепел, песец — ценный пушной
промысловый зверь. Уши короткие. Подошвы густо покрыты мехом,
который и греет лапы, и препятствует их скольжению на плотном снегу.
Рис. 8.87. Песец
Рассматривая экологические черты фауны полярных стран, нужно
подчеркнуть, что жизнь многих животных связана с морем. Это ки-
тообразные и ластоногие, а также охотящиеся главным образом за тю-
ленями и рыбой белые медведи. За рыбой также охотятся птицы: чи-
стиковые, чайки и другие обитатели больших птичьих базаров на
приморских скалах.
По мере продвижения на юг тепловой режим холодного пояса ста-
новится более благоприятным. В зоне тундр различают три подзоны:
северную, или арктическую, тундру; субарктическую (типичную) мо-
хово-лишайниковую тундру и южную кустарниковую тундру. Чередо-
вание центров высокого и низкого давления в Арктике определяют осо-
бенности атмосферной циркуляции; последняя служит причиной
разделения Арктики на секторы. Отображением географической сек-
торности является разделение растительности тундр России на провин-
ции: Кольский полуостров, восток Русской равнины, Западно-Сибир-
ская низменность, Средняя Сибирь (от Енисея до Лены), Восточная
Сибирь (от Лены до Колымы) и Дальний Восток.
Остановимся на краткой характеристике подзональных особенно-
стей тундровых биомов Палеарктической области.
Подзона арктических тундр. Расположена по берегам Ледовитого
океана. Под влиянием крайне сурового климата растительный покров
не сплошной: растительность занимает до 60% площади. Остальное
пространство суши занято россыпями щебня, голыми пятнами выпу-
ченного мерзлотой суглинистого субстрата, каменными многоугольни-
ками, пятнистыми тундрами и т. п. На участках, занятых растительно-
стью, преобладают дриадовые тундры с Dryas punctata, камнеломкой
(Saxifraga) и др. В травянистом покрове много пушиц {Erioph.orum
angustifolium, Escheuchzeri), злаков {Роа arctica, Dupontia fischeri,
Arctagrostis и пр.), полярных маков (Papaver radicatum]. 251
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ КАРТИ В А МИРА
Среди мхов преобладают дикрановые и политриховые, а также зеле-
ные мхи и печеночники. Много лишайников (роды Cetraria, Alectoria,
Comiculaha, Dactylina, Cladonia и др.); на камнях и скалах— накипные
лишайники.
Обилие микронеровностей рельефа — бугров и впадин, пятен-
медальенов, структурных грунтов — создает характерную мозаичную
структуру тундровых биомов (рис. 8.88).
Рис. 8.88. Пятнистая тундра, по Н. Walter, S.-W. Breckle.
Горизонтальная проекция и вертикальный разрез площадки 1x2 м:
1 — обнаженная почва; 2 — трещины; 3 — моховой покров в понижениях;
4 — лишайники; 5 — мятлик (Роа alpina); 6 — мак (Papaversp.);
7 — ожика (Luzula nivalis); 8—11 — камнеломки: Saxifraga platysepala,
S. caespitosa, S. nivalis, S. polaris; 12 — лисохвост (Alopecurus alpinus)
Подзона типичной мохово-лишайниковой тундры. На плакорах
преобладают мохово-лишайниковые тундры комплексного характера.
Моховая дернина состоит из зеленых мхов: Aulacomnium turgidum,
Dicranum elongatum, Hylocomium alascanum, Rhacomitrium canescens и
др. Обычны также политриховые мхи и печеночники (Polytrichum
hyperboreum, Ptilidium ciliare). В моховом пркрове почти всегда имеют-
ся лишайники (главным образом кустистые кладонии и цетрарии, а
также листоватые — нефрома и др.). В верхнем ярусе мохово-лишайни-
252 ковых сообществ представлены: осока гиперборейская, мятлик аркти-
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ БИО МОП СУШИ
ческий, горец живородящий, дриада — куропаточья трава, мелкие ивы,
иногда голубика, брусника и др. На песчаных грунтах распространены
лишайниковые тундры. В южных окраинах подзоны они напоминают
лишайниковый покров сосновых боров и лиственничных редколесий.
В западном секторе Арктики (до Енисея) преобладают лишайнико-
вые тундры с кладониями (олений мох) и исландской цетрарией. В вос-
точном секторе — в восточной части Таймыра и в полярной Якутии —
распространены алекториевые (Alectoria) и цетрариевые (Cetraria) тунд-
ры. Характерные для них кустистые лишайники растут в условиях
бесснежной арктической зимы и легко выносят жесткость погодных
условий, губительных для ягелевых формаций.
Ягельники требуют для перезимовки глубокого снежного покрова,
почему и распространены главным образом в западных тундрах, нахо-
дящихся под влиянием влажных ветров с Атлантического океана. Рас-
пределение лишайниковых тундр имеет большое значение для олене-
водства. Отсутствие в арктической Якутии ягельных формаций не
благоприятствует широкому распространению оленеводческих хо-
зяйств. Напротив, к западу от Таймыра сосредоточено наибольшее
поголовье оленей.
Обширные плоские понижения заняты низинными плоско-, мелко-
бугристыми и полигональными болотами. В понижениях преобладают
гипновые мхи; на грядах и невысоких буграх — политриховые и зеле-
ные мхи. Сфагновые мхи распространены мало.
Подзона южных кустарниковых тундр. Характерна для западного
сектора Палеарктики. В растительном покрове преобладают трехъярус-
ные сообщества из кустарников, кустарничков и трав и мохово-лишай-
никового напочвенного покрова. Кустарники находятся под снежным
покровом, и холодные зимние ветры не уничтожают их молодые побеги
и почки. Именно вымерзание и иссушение чаще всего губят кустарники
в восточном секторе Арктики и в более северной тундровой подзоне.
Из кустарников особенно характерна карликовая береза (Betula папа),
которую к востоку от Енисея замещает близкий вид железистой березы
(В. exilis). Много карликовых ив: круглолистная, полярная, травянистая,
ледниковая. Более крупные ивы растут по речным долинам и болотным
окраинам: Salix lanata, S. lapponum, S. glauca и многие другие. По доли-
нам рек местами встречается кустистая ольха (Alnus barbata). На между-
речных пространствах распространены водяника (Empetrum nigrum) и
ягодники: альпийская толокнянка, голубика, брусника; различные верес-
ковые кустарнички (багульник, подбели др.), много морошки.
Зимой тундра пустынна. Подавляющее число зверей и птиц откоче-
вывает на юг. Среди немногих видов, остающихся в тундре на зиму,
следует выделить леммингов (рис. 8.89).
Лемминги образуют большие колонии, прокапывая многочисленные
норы в почве. В зимний период они не только сохраняют активность,
но даже приносят потомство. В отдельные годы после благоприятных
летних сезонов, когда накапливается большой запас растительной пищи,
происходят вспышки размножения леммингов. Летом, выев большую
часть растений, миллионные полчища зверьков начинают мигрировать
в поисках новых мест обитания. Они двигаются в одном направлении, 253
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ К АРТИ В А МИРА
Рис. 8.89. Лемминг
невзирая на водные преграды. Многие тонут. Когда лемминги подходят
к берегу моря, они, видимо, воспринимают его как берег очередной реки,
которую стараются переплыть. Всех их ожидает гибель.
Лемминги составляют основу питания хищных птиц и животных
тундры. Установлена прямая зависимость между численностью попу-
ляции леммингов и песцов.
На короткий летний сезон тундра меняет свой облик. Под лучами
незаходящего солнца быстро сходит снег, в многочисленных водоемах
появляются в неисчислимом количестве личинки двукрылых насеко-
мых. Возвращаются перелетные птицы — кулики и водоплавающие: гуси,
утки, лебеди и др. Обильный корм ягодников и личинок насекомых,
долгий полярный день благоприятны для выведения и откорма птенцов
(рис. 8.90). Прикочевывают олени, спасаясь от кровососущих насе-
комых, которых на открытых пространствах тундры отгоняет ветер
(рис. 8.91). За оленями приходят волки.
В Неарктической области тундровые ландшафты занимают всю
северную окраину Северной Америки и значительную часть прилега-
ющих островов. Положение их южной границы у западной и восточ-
ной окраин материка так же, как в Палеарктике, асимметрично. В Ти-
хоокеанском секторе она проходит вблизи 60° с. ш., а в Атлантическом
секторе на полуострове Лабрадор под влиянием холодного течения
опускается почти до 50° с. ш. Как уже говорилось, у биомов тундр
Голарктического царства много общего. Почвенно-растительному по-
крову присущи типичные субарктические черты и соответствующая
дифференциация тундры на подзоны.
Из крупных животных, свойственных тундрам Неарктики, назовем
мускусного овцебыка (рис. 8.92). В прошлом он обитал и в Европе, ив
Азии, но в результате неумеренной охоты его ареал сильно сократился.
В настоящее время предпринимаются попытки восстановить популяцию
этого животного.
Несмотря на суровость природных условий, тундра была издавна
254 освоена и заселена человеком. На протяжении тысячелетий коренные
ГЛАВА 8. ЗОНАЛЬНЫЕ ШЫ БИНИОН СУШИ
Рис. 8.90. Белые гуси (а) и черные казарки (б)
Рис. 8.91. Северные олени
255
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ К АРТИ В А МИРА
Рис. 8.92. Овцебык, поА.Э. Брему
народы Севера выработали такие навыки хозяйствования, которые
поддерживали устойчивость хрупких тундровых экосистем. Индустри-
альное преобразование природы Крайнего Севера оборачивается серь-
езными отрицательными экологическими последствиями.
Голантарктическое царство. На покрытом льдом Антарктическом
континенте растительность отсутствует. В свободных ото льда и снега
береговых оазисах растут мхи и лишайники. Из цветковых растений
обнаружены два вида: Colobanthus crassifolius (сем. гвоздичных) и злак
Deschampsia antarctica. Заметим, что род Deschampsia имеет биполяр-
ный ареал, он широко распространен в Голарктическом царстве.
В омывающие со всех сторон Антарктиду моря вкраплено несколь-
ко субантарктических островов, образующих прерывистый пояс суши,
лишь немного выступающий к северу за 50° ю. ш. Острова безлесны,
хотя по своему положению они соответствуют в Северном полушарии
Британским островам и Камчатке. Климат Субантарктики, с одной
стороны, находится, под охлаждающим влиянием ледяного континен-
та, а с другой — смягчается водами океана. При постоянно ветреной
и туманной погоде в течение лета температура не поднимается выше
10 °C. Зимы мягкие, малоснежные. Средняя температура самого хо-
лодного месяца 2 — 3 °C.
Субантарктические биомы представлены не тундрами, а лугами.
Широко распространены туссоковые (кочкарные) злаковники (виды
Festuca и Роа), а также подушковидные формы растений: Azorella,
Donatio, Gaismardia, Astelia (рис. 8. 93).
Низины заняты обширными, в основном осоковыми болотами со
сфагновыми мхами, росянкой. На острове Кергелен растет Pringlea
antiscorbutica (сем. крестоцветных), «кергеленская капуста», крупные
листья которой моряки раньше употребляли как противоцинготное
25G средство (рис. 8.94).
ГЛАВА 8. ЗОНАЛЬНЫЕ Т1ПЫ БИОМОВ СУШИ
Рис. 8.93. Растительность острова Кергелен, по Г. Вальтеру.
На переднем плане — подушки Azorella selago;
на склоне слева — сомкнутый покров Асаепа adscendes и Azorella
Рис. 8.94. Кергеленская капуста
На Фолклендских островах обильны туссоковые злаковники; во
внутренних областях господствуют вересковые пустоши с зарослями
вересковых, водяники, азореллы и др. Много болот. Всего на Фолкленд-
ских островах 135 видов цветковых растений, из них 20% эндемики.
Много биполярных видов. 257
ЧАСТЬ III. БИОГЕОГР АШИЧЕСШ КАРТИ В А МИРА
8.2.2. Taira
В Северном полушарии от северной границы леса на юг простира-
ется умеренный пояс. Из-за огромной протяженности он подразделя-
ется на умеренно холодную (бореальную) и умеренно теплую (немо-
ральную) зоны. Таежные биомы определяют облик бореальной зоны
Голарктического царства. Подобные зональные типы биомов не ха-
рактерны для умеренного пояса Южного полушария.
Понятие «тайга», широко используемое в ботанической и географи-
ческой литературе, привнесено в науку из языка народов, проживаю-
щих в Сибири. Тайга — это густой хвойный лес (рис. 8.95). По заклю-
чению большого знатока таежной растительности А. И. Толмачева, во
внетропических широтах Северного полушария (в Голарктическом цар-
стве) тайга является самым распространенным типом растительности.
И ни один тип растительности на Земле не занимает столь обширного
цельного пространства, как она. Тайга является наиболее ярким выра-
зителем зонального распределения растительности в современную гео-
логическую эпоху.
Рис. 8.95. Западносибирская тайга.
Фотография с борта дирижабля «Граф Цеппелин», сделанная во время
кругосветного перелета в августе 1929 г., по Н. Walter, S.-W. Breckle
Таежная (бореальная) зона располагается на севере умеренного
пояса. Здесь наиболее четко выражены все четыре сезона года: лето,
осень, зима и весна. Средняя температура самых холодных месяцев
от —10 до —40 °C; самых теплых 13—19 °C. Зимы суровые, особенно в
Восточной Сибири, где располагается полюс холода Северного полуша-
258 рия —71 °C. Лето теплое, местами жаркое.
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ БИО МОН СУШИ
Господствующий тип атмосферной циркуляции — западный пере-
нос, сопровождающийся прохождением циклонов, несущих атмосфер-
ные осадки. Поэтому в Евразии (Палеарктике) четко выражена сектор-
ность — с удалением от Атлантического океана континентальность
климата усиливается. Соответственно уменьшается количество осадков
от 600 — 700 мм на западе до 400 мм в Восточной Сибири. Большая их
часть выпадает в виде снега. Испаряется 50 — 70% осадков. Избыток вла-
ги стекает в полноводные реки, пополняет грунтовые воды, способству-
ет образованию болот.
Зима в таежной зоне более морозная и более снежная, чем в зоне
тундр. Снежный покров играет защитную роль. Под мощным слоем сне-
га почва сохраняет тепло и часто не промерзает. Благодаря этому травы
и кустарнички, а также многие животные хорошо переносят лютые
морозы.
В Палеарктической области типичным проявлением таежного типа
растительности является темнохвойная тайга — тенистый вечнозеленый
лес, представляющий собой яркий пример олигодоминантной, бедной по
видовому составу растительности (вспомним богатство биоты дождевых
тропических лесов). В классической области своего развития — Запад-
ной и Средней Сибири — она образована различными сочетаниями ели,
пихты, сибирского кедра (Pinus sibirica) и лиственницы. Лесообразовате-
лем темнохвойной тайги на территории Восточно-Европейской равни-
ны служит в основном ель. На песчаных отложениях господствуют
сосновые леса. В производных насаждениях, возникающих на месте
уничтоженных коренных лесов, значительное участие принимают
мелколиственные породы: береза, осина, ольха.
Типичной темнохвойной тайге свойственна не только олигодоминант-
ность древостоя, но и бедность состава таежной флоры, обычно не более
двух десятков видов сопутствующих растений. С ограниченностью ви-
дового состава гармонирует общая скудность растительного покрова,
развивающегося под пологом леса. Если древостой тайги поражает нас
интенсивностью своего развития, значительностью растительной мас-
сы, сосредоточенной в верхнем ярусе, то растения, живущие под поло-
гом темнохвойного леса, образуют в совокупности лишь ничтожную
часть биомассы. Отмечается зависимость растений от тайги как среды
обитания. Существование многих видов почти неотделимо от темнохвой-
ного леса. Обитая под его пологом, они практически не встречаются в
других типах растительности. С другой стороны, растения нетаежных
фитоценозов проникают под полог тайги также весьма ограниченно.
Говоря о главнейших чертах таежной флоры, следует обратить осо-
бое внимание на значительное участие вечнозеленых растений в ее
составе — это не только хвойные деревья, но и вечнозеленые кустар-
нички и мхи. Таким образом, можно сказать, что самые большие про-
странства суши, занятые «вечнозелеными лесами», располагаются не в
тропиках, а во внетропическом поясе Северного полушария — это тем-
нохвойная тайга.
Животный мир тайги от берегов Балтийского моря до Тихого океа-
на однороден. Характерные представители палеарктической таежной
фауны показаны на рис. 8.96. 259
ЧАСТЬ III. БИОГЕОГР АШИЧЕСШ КАРТИ В А МИРА
Рис. 8.96. Характерные представители таежной фауны Палеарктической
области Голарктического царства. Рис. В.А. Ватагина, по Н.А. Бобринскому:
1 — трехпалый дятел; 2 — желна; 3.— глухарь; 4 — кедровка;
5 — свиристель; 6 — бородатая неясыть; 7 — белокрылый клест;
8 — мохноногий сыч; 9 — соболь; 10 — колонок; 11 — белка; 12 —лось;
2ВД 13 —летяга; 14 — бурундук; 15 — кабарга; 16 — росомаха
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ БИО МОН СУШИ
Отметим, что один из важнейших кормовых ресурсов в тайге —
семена хвойных деревьев. Ими питаются многие птицы, грызуны и даже
хищные звери. Однако урожайность ели, пихты, кедровой сосны силь-
но колеблется по годам. С этим связаны значительные колебания чис-
ленности птиц и зверей, как питающихся семенами, так и хищных.
Для тайги зональными процессами почвообразования являются под-
золистый и болотный. В условиях избытка влаги и кислой среды про-
исходит глубокий распад минеральной части почвы. В подзолистом
горизонте на песчаных почвах сохраняется наиболее устойчивое со-
единение — кремнезем SiO2. Соединения железа, марганца и гумус
выносятся в нижележащие горизонты вмывания. В производных мел-
колиственных лесах с развитым травяным покровом формируются дер-
ново-подзолистые почвы. Подзолистый процесс почвообразования от-
тесняется дерновым также на карбонатных материнских породах.
В условиях обильного увлажнения идет болотный процесс почвообра-
зования, характеризующийся накоплением слоя торфа.
Таежный тип растительности сформировался в послеледниковое вре-
мя, в голоцене, когда похолодание сменилось потеплением и климат
умеренных широт приблизился к современному. Темнохвойная тайга
умеренных широт — молодой биом, возраст которого не превышает
10 тыс. лет. О молодости тайги говорит и ее простая биогеоценотическая
структура, и бедность видового состава. Вместе с тем генетические кор-
ни таежного биома весьма древние. Наиболее вероятна гипотеза проис-
хождения тайги на равнинах Палеарктической области, высказанная
А.И. Толмачевым. Он полагает, что темнохвойная тайга как особый биом
сложилась в горах на юге Восточной Сибири еще в третичный период.
Горная тайга является, таким образом, биомом значительно более древ-
ним, чем хвойные леса равнин. В свою очередь, горная тайга имеет еще
более древние связи с растительностью высотных поясов Гималаев и гор
Юго-Восточной Азии, расположенных в тропических широтах.
Огромная протяженность таежных ландшафтов с севера на юг и с
запада на восток определяет зональные и провинциальные изменения
растительных сообществ. С севера на юг тайга делится на подзоны лесо-
тундры, северной, средней, южной тайги и подтайги (смешанных ле-
сов). С запада на восток выделяются провинции: Западно-Европейская
(Скандинавская), Восточно-Европейская, Западно-Сибирская, Средне-
Сибирская (с господством светлохвойной лиственнично-сосновой тай-
ги) и Притихоокеанская.
Остановимся более подробно на характеристике господствующих в
темнохвойной тайге Палеарктической области типов растительности:
лесного и болотного.
Леса. В.Н. Сукачев выделяет шесть главных типов мест произраста-
ния, которые определяют формирование основных типов лесных сооб-
ществ: 1) водоразделы (плакоры) с суглинистыми или супесчаными
среднебогатыми почвами, хорошо дренированные, режим увлажнения
устойчивый; 2) такие же почвы, но хуже дренированные, заболачива-
ющиеся, режим увлажнения переходный к обильно-застойному; 3) по-
нижения в рельефе, почвы еще хуже дренированные, более заболочен-
ные, режим увлажнения обильно-застойный. Эти три типа условий 2G1
ЧАСТЬ III. БАОГЕОЕРАФАЧЕСААЯ AAPIASA МАРА
обитания составляют один эколого-топологический ряд, отражающий
усиление процесса заболачивания. Далее: 4) дно узких долин рек и
ручьев (лога) с обильным увлажнением проточными водами; 5) выпук-
лые формы рельефа, сложенные песчаными отложениями, почвы бед-
ные, режим увлажнения резко-переменный; 6) плакоры, имеющие не
свойственные таежной зоне богатые почвы, чаще всего связанные с
близким залеганием известковых пород или с карбонатной мореной,
режим увлажнения устойчивый.
В.Н. Сукачев составил классическую схему деления еловых лесов,
связав их с названными условиями местопроизрастания (рис. 8.97).
_ В*рхо«ов
(Еу- сфагновое болото
Рис. 8.97. Экологический крест В.Н. Сукачева. Пояснения в тексте
Схема имеет вид креста, где центральное место представляют усло-
вия произрастания первого типа, занятые ельниками-зеленомошника-
ми (кисличниками, черничниками и брусничниками). Направления
осей указывают на последовательные смены сообществ, вызванные из-
менениями условий обитания. Вверх (к А) условия приближаются к ха-
рактерным для местопроизрастаний пятого типа, где еловые леса сме-
няются борами-беломошниками; налево (к Б) — к условиям второго и
третьего типа, где ельники сменяются верховыми сфагновыми болота-
ми; вниз (к Г) — к условиям местопроизрастаний четвертого типа, за-
нятым травяными ельниками; вправо (кВ) — к условиям шестого типа,
где формируются сложные ельники. Схему Сукачева можно использо-
вать и при типизации сосновых лесов, экологический ареал которых во
многом перекрывается с ельниками (рис. 8.98).
Сосняки вместе с мелколиственными породами (березой и осиной)
часто выступают в качестве производных лесов на месте ельников,
уничтоженных пожарами и вырубками.
Елъники-зеленомошники и сосняки-зеленомошники занимают пер-
вый тип местопроизрастаний. В верхнем ярусе господствует ель, в про-
изводных лесах сосна и мелколиственные породы (рис. 8.99).
Травяной покров обычно не сплошной (в производных лесах роль
трав увеличивается). Моховой покров хорошо развит. Господствуют так
называемые блестящие зеленые мхи (Hylocomium splendens, Pleurozium
sehreberi, Rhytidiadelphus triquetrus, Ptilium crista-castrensis, Dicranum
2G2 undulatum, D. scoparium).
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ БИО МОН СУШИ
Рис. 8.98. Основные типы условия произрастания еловых
и сосновых лесов и положение сообществ на профиле.
А. Еловые леса: 1 — ельник-брусничник; 2 — ельник-кисличник;
3— ельник-черничник; 4— ельник травяной; 5— ельник-долгомошник;
6 — сфагновое болото с низкорослой сосной.
Б. Сосновые леса (боры): 1 — бор-беломошник; 2 — бор-брусничник;
3 — бор-черничник; 4 — бор-долгомошник;
5 — сфагновое болото с низкорослой сосной
Ель в коренных лесах данной группы является подлинным создате-
лем (эдификатором) лесного фитоценоза. Она сильно воздействует как
на процессы почвообразования, так и на микроклимат под своим поло-
гом, и тем самым влияет на видовой состав и экологию остальных чле-
нов образуемых ею сообществ.
Широта экологического ареала ельников-зеленомошников позволя-
ет расчленить их на ряд групп.
Ельники-кисличники занимают обычно лучшие, хорошо дрениро-
ванные почвы. Они характеризуются значительным участием в тра-
вяно-кустарничковом ярусе таежного мелкотравья: кислицы {Oxalis
acetosella), майника (Maianthemum bifolium), седмичника (Trientalis
еигораеа), грушанки (Pyrola rotundifolia) и др. Из зеленых мхов часто
преобладают Rhytidiadelphus triquetrus, но обильны также Pleurozium
schreberi, реже Hylocomium splendens.
263
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ К АРТИ В А МИРА
Рис. 8.99. Производный елово-сосновый лес зеленомошной группы.
Виден обильный подрост ели. Почва покрыта свежевыпавшим снегом
Ельники-черничники и сосняки-черничники занимают более влаж-
ные, чем кисличники, с худшей аэрацией почвы. Они характеризуются
преобладанием в травяно-кустарничковом ярусе черники (Vaccinium
myrtillus). Из мхов преобладают Hylocomium splendens, реже Pleurozium
schreberi, Ptilium chsta-castrensis.
Ельники-брусничники и сосняки-брусничники приурочены к более
сухим и бедным песчаным почвам. Даже в коренных ельниках наблюда-
ется большая примесь сосны. Таежное мелкотравье (кислица, майник и др.)
мало распространено или вовсе отсутствует. В моховом покрове преобла-
дает Pleurozium schreberi. Продолжением экологического ряда ельников-
брусничников является пятый тип местообитаний — выпуклые формы
рельефа, сложенные песком, характеризующиеся резкопеременным ре-
жимом увлажнения. В этих условиях коренные зеленомошные еловые леса
уступают место сосновым борам-беломошникам с наземным ярусом из
лишайников, по преимуществу видов Cladonia и Cetraria.
Ельники-долгомошники и сосняки-долгомошники приурочены к
местопроизрастаниям второго типа. Они характеризуются господством
в древесном ярусе ели или сосны, к которым примешивается в большей
или меньшей степени береза. Травяной покров беден как в отношении
видов, так и по числу особей. Зато чрезвычайно характерен мощный
сплошной ковер мха «кукушкин лен» (Polytrihum commune). В таком лесу
эдификаторная роль древесного яруса постепенно переходит ко мхам.
Мощный покров кукушкина льна неблагоприятно влияет на возобнов-
ление ели и на большинство представителей таежного мелкотравья.
В формировании почв начинает проявляться болотный процесс — фор-
мируется торфянистый горизонт.
Ельники сфагновые и сосняки сфагновые приурочены к третьему
типу местопроизрастаний. Общими чертами лесов этой группы явля-
2В4 ются: угнетенный рост ели, примесь березы и сосны, значительное уча-
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ БИО МОН СУШИ
стие в моховом покрове сфагнума. В травяно-кустарничковом ярусе
преобладают болотные формы: голубика (Vaccinium uliginosum), багуль-
ник (Ledum palustre), морошка (Rubus chamaemorus), пушица (Eriophorum
vaginatum).
Если в предыдущей группе роль созидателя сообщества оспаривается
у древесного яруса кукушкиным льном, то здесь в роли эдификатора все
заметнее выступает сфагнум. В почвенном покрове хорошо выражен
значительный по мощности торфянистый горизонт. По мере увеличе-
ния мощности слоя торфа ель начинает вытесняться сосной. Продолже-
нием экологического ряда ельников сфагновых является заболоченный
сосновый лес и далее верховое сфагновое болото.
Влажно-травяные ельники приурочены к четвертому типу местопро-
израстаний. Древостой негустой из ели с примесью березы. Подлесок
развит довольно хорошо. Травяной покров мощный, густой, высокий, в
видовом отношении очень богатый, образованный мезогигрофитами и
типичными гигрофитами: дудником лесным (Angelica sylvestris], грави-
латом речным (Сеит rivale), вахтой (Menyantes trifoliatd), сабельником
(Comarum palustre), мятликом болотным (Роа palustris), калужницей (Calta
palustris), белокрыльником (Calla palustris) и др. Моховой покров, как
правило, не развит, но по кочкам обычно находятся лесные мхи, а также
кукушкин лен и редко сфагнум.
Сложные ельники приурочены к шестому типу местопроизраста-
ний. Они характеризуются участием в составе сообщества широколи-
ственных пород. По этому признаку сложные ельники являются свя-
зующим звеном между темнохвойными лесами таежной зоны и
сообществами смешанных и широколиственных лесов. Весьма харак-
терен густой и разнообразный подлесок. Заметные изменения проис-
ходят в травяно-кустарничковом ярусе. В нем таежные элементы-ку-
старнички, таежное мелкотравье уступают место видам, характерным
для широколиственных лесов (дубравные или неморальные элементы
флоры). Травяной покров получает значительное развитие, моховой
ковер слабо развит или вовсе отсутствует.
Характерными представителями неморальной флоры в травяном
ярусе сложных ельников являются следующие виды: воронец (Actaea
spicata), копытень (Asarum еигораеит), сочевичник (Orobus vernus), ме-
дуница (Pulmonaria obscura), звездчатка (Stellaria holostea), сныть
(Aegopodium podagrarid), перловник (Melica nutans).
Болота. Это особые природные комплексы, характеризующиеся
своеобразными условиями существования: обильным увлажнением и
накоплением полуразложившихся растительных остатков (торфа). В за-
висимости от гидрологического режима, определяющего характер пи-
тания, болота таежной зоны можно разделить на низинные, переход-
ные и верховые.
Низинные болота, связанные в своем развитии с грунтовыми во-
дами, обогащенными элементами минерального питания, являются
эвтрофными (богатого питания). Это моховые (гипновые и сфагновые),
осоковые и лесные или кустарниковые болота. Мощность торфяной
толщи низинных болот обычно небольшая — менее 1 м. В связи с по-
стоянным торфонакоплением поверхностный слой болота повышает- 265
ЧАСТЬ III. БАОГЕОТРАФАЧЕСААЯ КАРГИ0А МАРА
ся, и роль грунтовых вод, питающих живой растительный покров, по-
степенно уменьшается: уже не все корни достигают минерального грун-
та. В снабжении растений водой все большую роль начинают играть
атмосферные осадки. Низинные болота вступают в стадию переходных
мезотрофных (средних по богатству минерального питания) болот. Это
преимущественно сфагновые болота с мощностью торфа более 1 м.
При значительном повышении поверхности болота, приводящем к
полному отрыву корневых систем растений от грунтовых вод, живой
растительный покров начинает питаться преимущественно атмосфер-
ными осадками, бедными минеральными солями, и болото переходит в
стадию верхового сфагнового олиготрофного (бедного питания) боло-
та. Мощность торфа сфагновых болот в среднем 3 — 4 м.
Рассмотрим процесс формирования верхового болота в подзоне се-
верной тайги. Обратим внимание на то, что вместе с торфяником воз-
никает толща многолетне-мерзлых грунтов (рис. 8.100).
5ГЦЧ
Рис. 8.100. Схема заболачивания низины и роста торфяника в подзоне се-
верной тайги, по В.В. Крючкову: 1 — направление стока воды; 2 — граница
многолетнемерзлых (вечномерзлых) грунтов; 3 — погребенные в торфяни-
ках стволы деревьев; 4 — торфяные кочки; 5 — торфяники; 6 — бугры пу-
чения с новообразованиями мерзлоты. Пояснения в тексте
266
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ БИО МОН СУШИ
Нарастание мохово-торфянистого слоя ведет к ухудшению гидро-
термического и аэрационного режимов почвы, в которой не могут
существовать корни деревьев (7). На месте низины поднимается
выпуклый торфяник, с которого сметается снег, что способствует интен-
сивному промерзанию торфяника и формированию под ним веч-
ной мерзлоты мощностью в несколько десятков метров (II). Между
торфяником и лесом образуется понижение, в которое стекает вода
не только со стороны леса (расположенного в верхней части равни-
ны), но и с торфяника. Это ведет к формированию переувлажнен-
ной полосы на границе между торфяником и лесом — мохово-осо-
кового болота (БГЦ-5), которое, в свою очередь, подтапливает и
заболачивает полосу, примыкающую к лесу, и способствует фор-
мированию на ней кочковато-бутристого болота (БГЦ-46). Растущий
выпуклый торфяник вызывает заболачивание примыкающего к нему
леса. Конечной стадией заболачивания является выпуклый кустар-
никово-лишайниковый сухой мерзлый торфяник (III). Сток идет
только в одном направлении — с торфяника в сторону леса. Рост тор-
фяника вверх прекращается, он расползается вширь, вследствие чего
резко увеличивается скорость заболачивания и гибели лесов, а так-
же формирования многолетне-мерзлых грунтов.
Сфагновые мхи являются растениями-эдификаторами, полностью
контролирующими весь комплекс экологических условий: гидрологи-
ческий и газовый режимы, температуру корнеобитаемого горизонта,
образование форм мезо- и микрорельефа, процессы почвообразования,
точнее, формирования торфяной залежи.
Сфагнум впитывает атмосферные осадки, как губка. Среди клеток,
составляющих его листья и стебель, имеются крупные мертвые клетки
с отверстиями в оболочке, которые легко наполняются водой. В сухой
сезон года головки сфагнума могут высыхать до воздушно-сухого состоя-
ния. Однако, если вырвать пучок мха из болотной кочки, мы увидим,
что в своей нижней части он напитан водой. Сжав сфагнум в руках,
можно выдавить из него воду.
Погрузим руку в толщу сфагнума. Рука свободно уходит в мох. Раз-
двинув куртину, видим, что стебельки мха внизу теряют свою инди-
видуальность, все сливается в сплошную коричневую массу полураз-
ложившихся частей болотных растений — это торф. Почвы в обычном
понимании здесь нет. Все растения укореняются или в еще живом
сфагнуме, или в торфе.
Ощущается быстрое понижение температуры с глубиной. Если тем-
пература воздуха 25 °C, то на глубине 20 см она может уменьшиться
вдвое, а на глубине 50 см опуститься до 5 —6 °C. Резкое снижение тем-
пературы с глубиной объясняется плохой теплопроводностью как жи-
вого сфагнума, так и торфа.
Торфяные сфагновые болота характеризуются высокой степенью
кислотности, значительно более высокой, нежели почвы хвойного леса.
Низкая температура корнеобитаемого горизонта вместе с высокой кис-
лотностью, плохой аэрацией и малым содержанием питательных мине-
ральных солей создают весьма специфические условия существования
болотных растений.
2В7
ЧАСТЬ III. БАОГЕОЕРАФАЧЕСААЯ КАРГИ0А МАРА
Прежде всего они страдают от физиологической сухости. Несмот-
ря на то что корнеобитаемый горизонт насыщен влагой, она с трудом
всасывается корнями. Недостаток минерального, прежде всего азот-
ного, питания, физиологическая сухость являются причиной ксеро-
морфизма болотных кустарничков: мелколиственности, жестколист-
ности, образования плотной кутикулы, чешуек, волосков, глубокого
расположения устьиц.
Растения холодных и влажных местообитаний образуют особую
экологическую группу — психрофиты. Они характерны для раститель-
ного покрова тундровых биомов. Таким образом, облик верховых бо-
лот таежной зоны как бы переносит нас в расположенный в арктиче-
ских широтах тундровый ландшафт. Как для верхового болота, так и
для тундры, кроме ковра из мхов и лишайников, характерны мелкие
кустарнички: подбел (Andromeda polifolia), вереск обыкновенный
(Calluna vulgaris), багульник болотный (Ledum palustre), два вида клюк-
вы (Oxycoccus), голубика (Vaccinium uligi-nosum), болотный мирт
(Chamaedaphne calyculata) и др. Это все вечнозеленые растения,
относящиеся к одному семейству вересковых (Ericaceae).
Любопытны флорогенетические связи болотных кустарничков. Их
родина — горы тропических широт Юго-Восточной Азии. На это обра-
тил внимание русский географ А.Н. Краснов, исследовавший в нача-
ле века растительность высотных поясов островов Зондского архи-
пелага. Вот как описывает он свои впечатления о растительности
субальпийского пояса одного из вулканов на острове Ява: «...любопыт-
но, что дошедшие до вершин формы слагаются в ассоциации, большей
частью составленные теми же родами или семействами, которые идут
далеко на север и образуют зону тундры. Род Agapetes заменяет нашу
голубику. Это Ericaceae — по облику его можно назвать тропическим
деревом голубикой... Его плоды и цветы так напоминают чернику, что
вы сразу признаете в нем Vaccinium, а черные губы ваших спутников-
малайцев напомнят вам ваши детские прогулки по болотам и лесам
родины» (1987, с. 315).
А.Н. Краснов делает вывод, что «флора сфагновой тундры, защищен-
ная от мороза снежным покровом... есть прямой продукт вырождения
тропической растительности в полярную под влиянием понижения тем-
пературы при сохранении прочих условий, т. е. влаги и, возможно,
меньшего колебания температуры».
Тундровый облик растительности верховых болот хранит следы
послеледниковья. Сфагновые болота начали формироваться по перифе-
рии отступавшего ледника около 10 тыс. лет назад. Специфические
условия существования растительности на верховых болотах позволи-
ли им сохранить свой тундровый облик в таежной зоне до наших дней.
Из древесных пород на верховых болотах растет преимущественно
сосна (Pinus sylvestris), образующая здесь ряд экологических форм, рез-
ко отличающихся от высокоствольных деревьев, растущих в лесу на
минеральном грунте. Болотная сосна никогда не достигает больших
размеров: мутовки ветвей близко расположены одна к другой, что ука-
зывает на малый годичный прирост. Возраст сосенки высотой около
268 метра может достигать 50 — 70 лет. На торфяниках с быстрым прирос-
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ БИО МОН СУШИ
том сфагнума (3 — 4 см в год) встречается своеобразная кустарниковая
форма сосны, у которой на поверхности торфяника видны только мо-
лодые побеги, покрытые хвоей.
На поверхности мохового покрова растет росянка (Drosera rotundi-
folia) — небольшое травянистое растение с розеткой прикорневых ли-
стьев. Это удивительное растение заинтересовало Ч. Дарвина. Поста-
вив ряд экспериментов, он установил, что ввиду недостаточного
минерального питания росянке необходима белковая пища. Поверх-
ность листа росянки покрыта волосками. На каждом волоске имеется
капля жидкости, которая на солнце блестит, как роса. Под действием
сока, выделяемого волосками, росянка переваривает попавших на лист
мелких насекомых. Питаясь таким образом, росянка получает с бел-
ками недостающий ей азот. Содержащиеся в росянке элементы мине-
рального питания постепенно переходят в торф, способствуя повыше-
нию его плодородия.
Растительности крупных массивов верховых болот присуща ярко
выраженная комплексность, обусловленная хорошо развитым болотным
микрорельефом (рис. 8.101). В образовании микрорельефа (атакже зна-
чительной части мезорельефа) болота большую роль играет сама расти-
тельность.
Рис. 8.101. Грядово-мочажинный комплекс, по СМ. Горожанкиной
и В.Д. Константинову. Растения: 1 — Rubus chamaemorus;
2 — Eriophorum vaginatum; 3 — Ledum palustre; 4 — Drosera rotundifolia;
5 — Betula папа; 6 — Chamaedaphne calyculate; 7 — Andromeda pollfolia;
8 — Sphagnum fuscum; 9 — Cladonia alpestris; 10 — Cladonia sylvatica;
11— Pleurozium schreberi; 12— Dicranum polysetum; 13— Sphagnum
balticum. Виды торфа: 14 — фускум; 15 — мочажинный
Верховое болото имеет выпуклую форму (вид перевернутой тарел-
ки), поэтому более обводненными являются его центральная и перифе-
рийная части. Сильно обводненная окраинная часть болота занята осо-
ковыми комплексами с шейхцерией (Scheuchzeria palustris). Вместе с 2В9
ЧАСТЬ III. БАОГЕОЕРАФАЧЕСААЯ КАРТАВА МАРА
шейхцерией произрастают осока топяная {Carex Hmosa), сабельник
[Comarum palustre}, вахта {Menyanthes trifoliata). Встречаются кустарни-
ки: береза карликовая {Betula папа), ива {Salix myrtilloides) и др. Из
сфагновых мхов обычны Sphagnum fallax, Sph. riparium, образующие
рыхлый моховой покров. Выше, в нижней половине склона с наиболее
активным стоком, развит окраинный сосново-кустарничково-сфагно-
вый комплекс. Густой кустарничковый ярус образуют карликовая бе-
реза, багульник, голубика, вереск.
Еще выше, в верхней половине склона, развивается грядово-мочажин-
ный комплекс. Вытянутые поперек склона гряды чередуются с узкими
обводненными мочажинами. Они резко отличаются друг от друга по струк-
туре и видовому составу. На грядах, сложенных сфагновыми мхами, об-
разуется плотный покров Sphagnum fuscum, Sph. magellanicum, растут бо-
лотные кустарнички, сильно угнетенные формы сосны {Pinus sylvestris f.
wilkommii). В мочажинах обычны сфагновые мхи, отличающиеся рыхлы-
ми дернинами {Sphagnum cuspidatum, Sph. balticum, Sph. fallax).
При переходе от склона к более выровненной верхушке сфагнового
торфяника возрастает обводненность; грядово-мочажинный комплекс
сменяется грядово-озерковым, в котором отдельные мочажины запол-
нены водой и принимают вид узких и длинных озер.
На плоской вершине верхового болота сильно увеличивается обвод-
ненность. Плоские гряды едва возвышаются над широкими, очень
топкими мочажинами, заросшими сфагнумами. На повышениях —
Sphagnum fuscum, печеночники, местами пятна лишайников, редкие
кустарнички, пушица, осоки, а также единичные сосенки высотой
30 — 50 см {Pinus sylvestris f. pumila), у которых все стволики погружены
в торф и возвышаются только концы ветвей. Местами в центре болота
расположены озера, к которым часто невозможно подойти из-за боль-
шой топкости и обводненности берегов.
В Неарктической области хвойные леса имеют свои отличительные
особенности. Они занимают в Северной Америке очень большие тер-
ритории: большую часть полуострова Лабрадор, значительные части
Аляски и широкую полосу, связывающую эти два полуострова.
Вдоль берегов хвойные леса продвигаются далеко на юг: на западе
вдоль тихоокеанского побережья, особенно по горным цепям до самой
Центральной Америки (Скалистые горы, Каскадные, Сьерра-Невада
и др.); на востоке по приатлантической прибрежной равнине до Мекси-
канского залива. Здесь они образуют особые формации, входящие в
состав субтропических биомов.
По сравнению с европейскими леса Неарктики отличаются боль-
шим разнообразием хвойных пород: многочисленны виды сосны, ели,
пихты, лиственницы, можжевельника и пр. Кроме того, встречаются
Tsuga, Pseudotsuga, Thuja. Основной причиной этого богатства явля-
ются благоприятные условия миграций во время ледникового перио-
да: все эти породы свободно отступали в убежища на юг, а затем снова
возвращались после таяния ледника. Как и в Европе, в более холод-
ных частях Северной Америки хвойные широко распространены на
равнинных территориях, в более же южных частях они переходят в
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ БИО МОН СУШИ
Наиболее далеко на север заходит белая ель (Picea alba), а также
лиственница (Larix americanu); на севере Канады преобладающее дерево
хвойных лесов — сосна (Pinus banksiana), к которой присоединяются
некоторые ели, бальзамическая пихта (Abies balsamea), лиственница и
мелколиственные — осина, береза. На Аляске растут леса ситхинской
ели (Picea sitchensis), тсуги (Tsuga heterophylla, Т. mertensiana), аляскин-
ского «кипариса» (Cbamaecyparis nutkatensis). В области Великих озер и
реки Св. Лаврентия леса состоят главным образом из веймутовой со-
сны (Pinus strobus), к которой присоединяются туя (Thuja occidentalis),
тсуга (Tsuga occidentalis).
Вдоль восточного побережья хвойные леса далеко спускаются к югу,
и, например, в штатах Вирджиния и Каролина леса состоят главным
образом из сосны (Pinus rigida, Р. taedd). На западном побережье хвой-
ные леса представлены особыми видами: так, для Британской Колумбии
наиболее характерны туя, дугласова пихта, сосна (Pinus murreyand) и др.
Животный мир Неарктики имеет много общего с таежным биомом
Палеарктической области. Отличия прослеживаются на видовом, реже
на более высоком таксономическом уровне (рис. 8.102). Своеобразие
таежной фауне Неарктики придают древесный дикобраз, ондатра,
скунс; панамериканские формы — колибри, пума.
8.2.3. Иимквпиствевные леса Севернеге нелушация
Широколиственные леса располагаются в приокеанических секто-
рах умеренно-теплого пояса Европы, Восточной Азии и Северной Аме-
рики. Основу биомов образуют летнезеленые (листопадные) широко-
лиственные породы деревьев. Такие леса называют еще неморальными,
а саму зону широколиственных лесов неморальной зоной. В Северном
полушарии она полностью принадлежит Голарктическому царству.
Неморальная зона отделена от тайги переходной зоной смешанных
хвойно-широколиственных лесов. Подобные зональные типы биомов
не характерны для умеренного пояса Южного полушария.
Как уже говорилось, по мере перехода в низкие широты, где климат
становится более теплым, все отчетливее проявляется роль увлажнения,
и закон географической секторности в распределении зональных ти-
пов биомов находит здесь свое более яркое, чем в холодном поясе,
выражение. Так почти в одном интервале широт, но на разном удале-
нии от океана формируются биомы широколиственных лесов, степей
и пустынь.
Летнезеленые леса одеты листвой лишь в благоприятное летнее вре-
мя года, осенью листья опадают, и зимой деревья стоят без листьев.
Почки на ветвях деревьев покрыты почечными чешуями, а стволы толс-
той коркой — это защита не только от мороза, но и от зимней засухи,
когда вода замерзает и становится малодоступной для метаболизма.
Широколиственные леса отличаются от тайги богатством видового
состава и сложной структурой. Наиболее богаты леса Северной Амери-
ки и Восточной Азии, поскольку здесь прослеживается их тесная связь
с арктотретичной флорой плиоцена. 271
ЧАСТЬ III. БАОГЕОТРАФАЧЕСААЯ AAPIABA МАРА
Рис. 8.102. Представители таежной фауны, по БСЭ: 1 — щур; 2 — клест-
еловик; 3 — кукша; 4 — трехпалый дятел; 5 — желна; 6 — обыкновенный
гоголь; 7 — обыкновенный свиристель; 8 — обыкновенная чечетка; 9 — кед-
ровка; 10 — снегирь; И — каменный глухарь; 12 — дикуша; 13 — рябчик; 14 —
воробьиный сыч; 15 — длиннохвостая неясьипь; 16 — кабарга; 17— росомаха;
18 — обыкновенная бурозубка; 19 — соболь; 20 — лесной лемминг; 21 — север-
ная пищуха; 22 — темная полевка; 23 — красносерая полевка;
24 — красная полевка; 25 — летяга; 26 — бурундук; 27 — лось; 28 — рысь;
29 — бурый медведь; 30 — северный кожанок; 31 — колонок; 32 — живородя-
щая ящерица; 33 — обыкновенная гадюка; 34 — таежный клещ;
272 35 — еловый дровосек; 36 — сибирский шелкопряд и его гусеница (а)
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ БИО МОН СУШИ
Палеарктика. В Евразии биомы широколиственных лесов тяготеют
к океаническим секторам Атлантического и Тихого океанов. Широко-
лиственные леса занимают большую часть Западной Европы; в Восточ-
ной Европе, где заметно усиливается континентальность климата, леса
выклиниваются. Здесь их северная граница проходит приблизительно
по линии Калининград —Москва —Уфа, а южная — по линии Киши-
нев—Киев—Тула—Уфа. Таким образом, область распространения этих
лесов на Европейском континенте напоминает по форме треугольник,
вершиной обращенный к Уралу. В западной части ареала широколист-
венные леса располагаются к северу от биомов средиземноморского
типа; в восточной — к северу от степей.
Соотношение тепла и влаги в неморальной зоне близко к оптимально-
му. И хотя здесь, так же как в тайге, выражены все четыре времени года
и зимой происходит спад биологической активности, продолжительность
вегетации удлиняется до 6 — 7 месяцев против 4 — 5 в бореальной зоне.
В Западной Европе биом широколиственных лесов относительно
однороден и образует сплошную зону, ширина которой местами дости-
гает 1500 км (от Южной Скандинавии до северной части Пиренейского
п-ова). Температура самого теплого месяца 15—18 °C на севере и 18 —
22 °C на юге. Зимы мягкие. Средняя температура января от —5 °C до
положительной 4 — 7 °C. Длительность безморозного периода 150 —
200 дней. Среднее годовое количество атмосферных осадков порядка
600 — 800 мм. Испаряемость примерно равна этой величине.
Европейские широколиственные леса намного беднее по составу и
структуре, чем биомы данного типа, распространенные в Восточной
Азии и Северной Америке. Эта бедность есть следствие четвертичных
оледенений. Сохранению в этот период богатой третичной лесной рас-
тительности на юге в убежищах препятствовали альпийские ледники и
Средиземное море. И все же состав и ярусная структура европейских
широколиственных лесов намного богаче и сложнее, чем у таежных
биомов. В их сложении, как правило, принимают участие три яруса —
древесный, в котором различается до трех подъярусов, кустарниковый
(до двух подъярусов) и травяной (с двумя-тремя подъярусами). Обиль-
ная опавшая листва образует мощный слой подстилки. Моховой ярус
выражен плохо.
Умеренно-теплый и влажный климат, активная деятельность почвен-
ных организмов, грибов и животных способствуют быстрому разложе-
нию опавших листьев и накоплению гумуса. Господствующий тип
почв — бурые лесные.
Лесообразующими породами служат четыре вида дуба (основные —
черешчатый Ouercus robur и скальный Q. petraed), бук лесной (Fagus
sylvatica), а также ясень обыкновенный [Fraxinus excelsior}, граб обыкно-
венный (Carpinus betulus), несколько видов липы (Tilia], клена (Acer), вяза
(Ulmus). В Северной Америке и Восточной Азии почти все перечисленные
рода деревьев представлены десятками, а дуб и клен даже сотнями видов.
Несмотря на такой сравнительно небольшой набор эдификаторов,
европейские широколиственные леса представлены довольно многочис-
ленными вариантами — зональными, провинциальными, высотно-по-
ясными, эдафическими. 273
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ К АРТИ В А МИРА
В материковой части приатлантического сектора в его северной под-
зоне распространены буковые леса (рис. 8.103). По составу и структуре
это наиболее бедные леса. Часто в них развит только один древесный
ярус. Почва, покрытая толстым слоем опавших листьев, почти лишена
яруса трав.
Рис. 8.103. Ареал буковых лесов, по В.В. Алехину: 1 — границы ареала;
2 — области наибольшего распространения; 3 — области меньшего
распространения; 4 — искусственные посадки; справа — буковый лес
В Южной и Центральной Европе буковые леса перемещаются в
предгорья и на склоны гор. Здесь же произрастают каштан (Castanea
sativa), клен ложноплатановый (Acer pseudoplatanus), липа крупно-
листная (Tilia platyphyllos). В приатлантических лесах встречаются веч-
нозеленые виды — падуб остролистный (Hex aquifolium), морозник
(Helleborus), плющ (Hedera helix).
Для равнин центральноевропейского сектора типичны мезофильные
леса из дуба черешчатого (Quercus robur), которому сопутствуют липа
сердцелистная (Tilia cordata), клен платановидный (Acerplatanoides), граб,
ясень. В подлеске присутствуют лещина (Corylus avellana), виды боярыш-
ника (Crataegus), бересклет бородавчатый (Ей опуши s verrucosa), волчье
лыко (Daphne mezereum) и другие кустарники. Для травяного покрова
характерны подснежники, а также виды, которые успевают закончить
вегетацию до облиствения деревьев, например хохлатка полая (Corydalis
cava), гусиный лук (Садеа lutea), ветреница дубравная (Anemone
nemorosa), или отцвести — сочевичник весенний (Lathyrus vernus), меду-
ница лекарственная (Pulmonaria officinalis), пролесник многолетний
(Mercurialis perennis) и др. Из злаков типичен перловник (Melica).
Травянистые растения широколиственных лесов, имеющие широ-
кие нежные листовые пластинки, относят к экологической группе дуб-
равного широкотравья. Некоторые из них проникают в таежные био-
мы, где их называют неморальными элементами. В частности, они
2/4 составляют характерную черту сложных (неморальных) ельников.
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ БИО МОН СУШИ
В юго-восточной части Центральной Европы (внутри горной дуги
Карпат) происходит некоторое усиление сухости и континентальности
климата при повышенной теплообеспеченности. В широколиственных
лесах начинают доминировать дубы австрийский, или цер {Ouercus
cerrts), и венгерский (О. frainetto), намечая переход, с одной стороны, к
субсредиземноморским ландшафтам, а с другой — к лесостепным.
В восточноевропейских широколиственных лесах состав пород ме-
няется. Здесь нет бука, дуба скального, клена явора (Acer pseudoplatanus].
Граб распространен только до левобережья Днепра. Ясень не переходит
за Волгу. Основной эдификатор — дуб черешчатый (Quercus robur),
дерево, способное переносить летнюю засуху и зимние морозы. Воз-
раст отдельных деревьев может достигать 2000 лет. Их по праву называ-
ют лесными патриархами (рис. 8.104). На западе в качестве соэдифика-
тора выступает граб, к востоку усиливается роль липы.
Рис. 8.104. Ареал дуба черешчашого, по В.В. Алехину. Точками обозначена
граница ареала ясеня; справа — опушка дубового леса, по П.Е. Васильковскому
Животный мир широколиственных лесов намного богаче, чем в
таежных биомах. Типичные представители показаны на рис. 8.105. Есть
виды, общие с фауной тайги. Из млекопитающих это лесная куница,
лесная кошка, рысь, черный хорек, барсук, белка, заяц-русак, заяц-
беляк, еж. В горных лесах встречается медведь.
Небольшая мощность снежного покрова или даже полное его отсут-
ствие, обилие растительного корма благоприятствуют распространению
копытных — зубра, благородного оленя, косули, кабана. Из птиц мож-
но отметить дятла, синицу, дрозда, зяблика и др. Почва широколиствен-
ного леса отличается высокой насыщенностью фауной беспозвоноч-
ных — насекомыми, дождевыми червями.
Важную часть пищевого рациона зверей и птиц составляют зеле-
ная растительная масса, сочные плоды, семена, желуди и лесные орехи
(рис. 8.106). 275
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ КАРТИ В А МИРА
Рис. 8.105. Характерные представители, фауны европейского широко-
лиственного леса. Рис. В.А. Ватагина, по Н.А: Бобринскому: 1 —лесная куница;
2 — черный хоры 3 — садовая соня; 4 — полчок; 5 — иволга; 6 — квакша;
7 — орешниковая соня; 8 — зеленый дятел; 9 — дубонос; 10 — европейская
27G неясыть; И — белка; 12 — зубр; 13 — европейская косуля
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ БИОМОВ СУШИ
Рис. 8.106. Пищевая сеть в экосистеме широколиственного леса,
по Н.А. Бобринскому
В Средние века европейская зона широколиственных лесов начала
активно осваиваться под пахотные угодья. В настоящее время — это
сильно преобразованные человеком территории, густозаселенные, с
мощной индустриальной и городской инфраструктурой.
Дальневосточные биомы широколиственных лесов расположены на
юге Приамурья, Сихотэ-Алиня, Приморья, Восточно-Маньчжурских
гор, на Корейском полуострове, на островах Хоккайдо и Хонсю. Их отли-
чия от европейских аналогов определяются положением на восточ-
ной окраине обширного континента, муссонным режимом атмосфер-
ной циркуляции, а также длительным развитием, не прерывавшимся
материковыми оледенениями в плейстоцене. Северная граница немо-
ральной зоны здесь значительно сдвинута на юг, до 36 — 37° с. ш. (в Европе
северная граница широколиственных лесов благодаря теплому течению
Гольфстрим достигает 57° с. ш.). В восточном секторе теплобореаль-
ной зоны в соответствии с муссонным типом климата зима сухая и
холодная, лето теплое и дождливое. На материке средняя температура 277
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ КАРТИ В А МИРА
самого холодного месяца — 16 °C; самого теплого около 20 °C. За год вы-
падает примерно столько же осадков, сколько и в европейских широко-
лиственных лесах (500 — 700 мм), но по сезонам они распределяются
крайне неравномерно. Большую часть осадков приносит летний мус-
сон; всю осень и зиму, с октября по апрель, — сухой сезон. В январе-
феврале выпадает всего 5—10 мм осадков, снежный покров — маломощ-
ный. На островах климат значительно мягче, контраст сезонов сглажен.
Температура самого холодного месяца около —2 °C. Годовая сумма осад-
ков 1100 — 1800 мм, что более чем в два раза превышает количество осад-
ков на материке. Зимы снежные.
Леса многоярусные, увитые лианами (рис. 8.107). Широколиственные
породы деревьев представлены видами дуба, клена, ясеня, липы, ильма,
орехом маньчжурским (Juglans mandschuricd) и др., а также рядом ви-
дов, отсутствующих в Европе: софора (Sophora japonica), гледичия
(Gleditschia sinensis), айлант (Ailanthus glandulosa), магнолия (Magnolia
kobus), камелия [Cornelia japonica), бархат амурский (Phellodendron
amurense) и др. Многие из названных растений культивируются в дру-
гих странах как декоративные.
Рис. 8.107. Уссурийский лес
Очень богат подлесок, состоящий из жимолости (Lonicera), сирени
(Syringa), рододендрона (Rhododendron), аралии (Aralia elata) и др. Боль-
шинство этих родов представлено многочисленными видами, отсутству-
ющими в Европе. Помимо обильного подлеска хорошо представлены
лианы: амурский виноград (Vitis amurensis), актинидия (Actinidia
kolomikta), лимонник (Schisandra chinensis). На юге Дальнего Востока в
лесах растет легендарный женьшень (Panax ginseng) (рис. 8.108).
Животный мир наиболее богат и разнообразен в материковом сек-
торе, где сочетаются северные и южные виды и присутствуют многие
дочетвертичные реликты и эндемики. В переходной полосе смешанных
хвойно-широколиственных лесов наряду с представителями обычной
278 сибирской фауны (бурундук, барсук, колонок, горностай, ласка, бурый
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ БИО МОН СУШИ
Рис. 8.108. Женьшень
ориентацию. Это происходит
медведь, волк, лисица, косуля, кабан
и др.) обитают такие южные или энде-
мичные виды, как изюбр, амурский
тигр, гималайский медведь, харза, лес-
ной маньчжурский кот, много птиц
(втом числе утка-мандаринка), из пре-
смыкающихся — полозы Шренка и
узорчатый, дальневосточный щито-
мордник, из земноводных — древесная
лягушка. К югу постепенно исчезают
такие сибирские экземпляры, как со-
боль, лось, росомаха, северный олень,
и появляются южные виды млеко-
питающих (пятнистый олень, леопард,
амурский горал, гигантская бурозуб-
ка). Характерные представители фау-
ны Амуро-Уссурийского края показа-
ны на рис. 8.109.
Неарктика. В Евразии зональные
типы биомов имеют субширотное про-
стирание. В Северной Америке грани-
цы зон холодного пояса и тайги также
имеют направление, близкое к субши-
ротному, однако к югу картина меня-
ется (рис. 8.110). От Атлантического по-
бережья на запад к Скалистым горам
зоны приобретают субмеридиональную
оттого, что благодаря теплому течению Гольфстрим северные границы
зон смещаются в более высокие широты. Атмосферные осадки прино-
сятся с Атлантического океана и из Мексиканского залива. В глубь
континента их попадает все меньше. Поэтому с востока на запад про-
исходит уменьшение увлажненности и смена зон от широколиствен-
ных лесов к степям (прериям) и пустыням умеренного пояса.
Вдоль атлантического побережья развиты буковые леса из американ-
ского бука (Fagus americana), а также сахарного клена (Acer saccharum).
В составе древесного яруса встречается до 40 видов клена (рис. 8.111). Эти
леса замечательны своей окраской — все оттенки красного и желтого
осенью, которую в России называют золотой, в Канаде и США индейской.
Американские буковые леса менее тенисты, здесь хорошо развит
травянистый ярус и лианы: «дикий виноград» (Parthenocissus quinquefolia)
и несколько видов настоящего дикого винограда (Vitis). Дубовые леса,
так же как в Европе, занимают более континентальные районы даль-
ше от побережья Атлантического океана. Но в отличие от европей-
ских они обладают очень богатым видовым составом. Кроме несколь-
ких видов дубов многочисленны виды кленов, орехов (Juglans), гикори
(Сагуа), тюльпанное дерево (Lihodendron tulipifera). фауна широколи-
ственных лесов Северной Америки относится к обширной Сонорской
области Голарктического царства, включающей субтропические биомы
Неарктики. 279
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ КАРТИ В А МИРА
Рис. 8.109. Характерные представители фауны Амуро-Уссурийского края.
Рис. В.А. Ватагина, по Н.А. Бобринскому: 1 — харза; 2 — маньчжурский
заяц; 3 — тигр; 4 — дальневосточный лесной кот; 5 — крот-могера;
6 — красный волк; 7 — енотовидная собака; 8 — черный медведь;
9 — восточный широкорот; 10— пятнистый олень; 11 — наземный три-
280 тон; 12 — утка-мандаринка; 13 — черепаха-трионикс; 14 — горал
Рис. 8.110. Карта растительности восточной части Северной Америки,
по Г. Вальтеру: 1 — районы, покрытые льдом; 2 — северная тундра;
3 — южная тундра; 4 — лесотундра; 5 — бореальные хвойные леса; 6 — се-
верные лиственные леса с примесью отдельных видов хвойных пород; 7 —
буково-сахарнокленовые лиственные леса; 8 — смешанные дубовые леса;
9 — дубовые леса с большой примесью сосны (частично доминирующей);
10 —пойменные и заболоченные леса, частично субтропического характе-
ра; И — дубово-гикориевыелеса; 12 — северные прерии с осиновыми колка-
ми; 13 — высокозлаковые прерии; 14 — смешанные и низкозлаковые прерии
(Великие равнины) 281
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ КАРТИ В А МИРА
Рис. 8.111. Широколиственный лес (дубы, клены, гикори) в долине Миссури
8.2Л. Биомы умереш теплы» эм Шжюга во дуиарн
Умеренно-теплые зоны Южного полушария располагаются пример-
но между 38° и 50° ю. ш. Наземные биомы формируются здесь на раз-
деленных океанами окраинах Южной Америки и Австралии и островах
Тасмания, Новая Зеландия и др. А.Л. Тахтаджян (1978) относит их к
Голантарктическому царству.
Внетропическая часть Южной Америки известна под названием
Патагония. Ботаники выделяют ее как Чилийско-Патагонскую область
Голантарктического царства, а зоологи как Патагонско-Чилийскую
область Неотропического царства. Благодаря влиянию океана и горно-
му барьеру Анд климат здесь очень влажный и ровный. Температура
самого холодного месяца 4 — 8 °C, но лето — прохладное 13— 16 °C. Годо-
вое количество осадков 2000 — 3000 мм.
Почвообразование характеризуется интенсивным промывным ре-
жимом. На кислых осадочных породах формируются бурые лесные
почвы. На значительных площадях склонов и подножий гор, покрытых
мощным слоем вулканического пепла, образуются своеобразные вулка-
нические почвы — «трумао», с большим содержанием гумуса. Местами
в северных районах на древней коре выветривания пепловых отложе-
282 ний почвы близки к красноземам.
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ 510МОН СУШИ
В растительном покрове наблюдается постепенный переход от влаж-
ных субтропических лесов к умеренно-теплым листопадным. Раститель-
ность представлена так называемыми магелланскими субантарктиче-
скими лесами из вечнозеленых нотофагусов (Nothofagus betuloides),
дримиса, либоцедрусов, подокарпусов и др. Здесь мало лиан и эпифи-
тов. В наземном покрове господствуют мхи, покрывающие также ство-
лы и ветви деревьев (рис. 8.112).
Рис. 8.112. Лес из южного бука
К югу и в верхней части склонов вечнозеленые буки замещаются
листопадными видами (Nothofagus pumilio, N. antarctica), лавролистным
тиком (Aextoxicum punctatum) и хвойными из либоцедрусов, подокарпусов
и саксеготеи (Saxegothea conspicua). В нижнем ярусе густые заросли вы-
сотой до 10 м образуют бамбуки (Chusquea quila, Ch. colihue), древовид-
ные папоротники (Blechnum magellanicum), вечнозеленые кустарники
мирта, барбариса и др. В наземном покрове — красная водяника, яскол-
ка, папоротники и обильные мхи. В этих летнезеленых лесах много тор-
фяных болот.
Лесная растительность сильно нарушена. Особенно интенсивно сво-
дится лес на восточных, защищенных от ветра склонах хребтов и на
равнинах. Земли после вырубки и выжигания подвергаются сильной
эрозии, на склонах образуются оползни. Расчищенные от леса участки
используются главным образом под пастбища и посевы кормовых трав
для крупного рогатого скота.
Животный мир Патагонии сохранился преимущественно в горах.
Он неотделим от Неотропического царства. Характерные представите-
ли фауны Патагонско-Чилийской области показаны на рис. 8.113.
283
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ КАРША МИРА
Рис. 8.113. Характерные представители фауны Патагонско-Чилийской обла-
сти Неотропического царства. Рис. В.А. Ватагина, по НА. Бобринскому:
1 — кондор; 2 — гуанако; 3 — олень пуду; 4 — шестипоясный броненосец;
5 — вискаша; 6 — шиншилла; 7 — тинаму; 8 — хохлатая паламедея;
284 9 — пума, 10 — олень пампасов; 11 — мара; 12 — нанду
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ БИО МОН СУШИ
Юг Австралии, остров Тасмания и Новая Зеландия располагаются
южнее 38° ю. ш. в пределах умеренно-теплой зоны. Климат влажный,
океанический. Средняя температура самого холодного месяца 5 — 8 °C;
лето прохладное — 14—18 °C. Годовое количество осадков превышает
1000 мм; распределяются они-по сезонам равномерно. Благодаря теплой
зиме вегетационный период продолжается практически круглый год.
В растительном покрове много вечнозеленых видов, что придает
биомам зональных типов субтропический облик. Их характерные чер-
ты наиболее ярко выражены в Новой Зеландии. Отличительной чертой
растительности являются леса из южного бука (Nothofagus), аналогич-
ные Чилийско-Патагонским.
Это дает основание ботаникам относить Новозеландскую область к
Голантарктическому царству. Зоогеографы выделяют Новозеландскую
область в составе Австралийского царства.
8,2.5. Степи
В умеренно-теплых зонах по мере усиления засушливости климата
лесные биомы сменяются травянистыми безлесными сообществами.
В травостое господствуют многолетние зимостойкие, способные перено-
сить длительную засуху злаки и ксерофитное разнотравье. Этот тип рас-
тительности в Северном полушарии в Евразии называют степью, в Север-
ной Америке — прерией, в Южном полушарии в Аргентине — пампой,
в Новой Зеландии — туссоком. Учитывая господствующую роль злаков
вбиомах этого типа, употребляют и общее название — злаковники. Обра-
щает внимание, что такие роды злаков, как ковыль (Stipa), овсяница
(Festuca), мятлик (Роа), житняк (Agropyrum) и др., доминирующие в рас-
тительном покрове злаковников, имеют биполярное распространение.
Северное полушарие. От венгерских пушт через всю центральную
часть Евразии до Алтая и далее через Забайкалье в Монголию и Северо-
Восточный Китай простирается Палеарктическая степная область
Голарктического царства. Граница степных биомов с более северными
лесными не резкая. Переходом служит лесостепь, представляющая со-
бой мозаичный макрокомплекс, в котором массивы леса и простран-
ства луговых степей резко отграничены друг от друга. Этим лесостепь
отличается от саванны, где отдельно стоящие деревья естественно вкрап-
лены в травянистый покров.
Биомы степей геологически сравнительно молоды. В пределах со-
временного ареала они стали формироваться в основном уже в после-
ледниковое время. В течение ледникового периода в так называемой
перигляциальной зоне по окраине ледникового щита располагалась об-
ширная тундростепь.
Степная флора, однако, существовала и ранее. В доледниковое вре-
мя степные формации уже отмечались в пределах древнего Средиземья
(в Передней и Средней Азии и в Иране), а также в горах Карпат, Балкан-
ского полуострова, на Кавказе и в Средней Азии. Все эти центры дали
основную массу ксерофитной флоры современных степей Западной
Евразии (рис. 8.114).
285
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ КАРТИ В А МИРА
28G
условные
I Арктическая флора
Эоарнтичесная флора вне области лоиров-
него оледенения
Древняя эоаритнческая суша, соединявшая
Таймыр и Якутию с Канадским архипелагом
Пути обогащения Арктики альпийскими
элементами и пути расселения альпийской
(горной) флоры
вторжение лесных флор в тундру е «осле*
ледниковое время
Расселение лесной и полярной флоры из
Древней Берингии
Флора гольцов и горной лесотундры
Северные и южные пределы современной
лесной древесной флоры и растительности
Направления обеднения -полярных флор
эоарктическими элементами
Поэдиетретичные связи Западной Арктики с
флорами Гренландии и Канадского
архипелага
БОЗНАЧЕния
II флора Еераэиатсной степной подобласти
Первичные (внеледникоеые) и вторичные
—1 центры происхождения степной флоры
(. Древний центр плейстоценовой горной
У лесостепи (по И.М. Крашенинникову)
Пути расселения степной флоры
Южная граница степной подобласти (по
* Е. М. Лавреико)
II! Флора Древнего Средиземья
xs. Предполагаемые центры происхождения
чрА пустынной флоры
t.... Третичные н дотреткчные пути расселения
флор Сев- Вост. Африки, Ирана и Монголии
.... Пути расселения флоры песчаных пустынь
Средней Аэж
ш Пути вторжения пустынной галофитной
" флоры в область степей
Е
О
2
Рис. 8.114. Схема путей миграций и центров распространения флор сте-
пей, пустынь и тундр Евразии, по В.В. Алехину и В.С. Говорухину
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ БИО МОН СУШИ
Степную растительность образуют виды, сформировавшиеся в
основном в теплых и сухих условиях средиземноморского климата. С на-
ступлением похолоданий в плейстоцене эта ксеротермофитная расти-
тельность трансформировалась в криоксерофитную перигляциальную.
Северные более влаголюбивые виды (широкотравье, корневищные лу-
говые мезофитные злаки и осоки северных степей) проникли в степи
из лесной зоны.
Огромная протяженность евразийских степных ландшафтов с севера
на юг и с запада на восток определяет зональные и провинциальные
изменения растительных сообществ. С запада на восток выделяют про-
винции: Европейско-Казахстанскую, Забайкальскую, Центрально-Ази-
атскую. С севера на юг степи делятся на подзоны. Так, в Европейско-Ка-
захстанской провинции выделяют подзоны северных луговых, типичных
и южных сухих степей (рис. 8.115).
Рис. 8.115. Деление степной зоны на подзоны, по А.А. Чибилеву:
1 — лесостепь; 2— северная луговая степь; 3 — типичная степь;
4 — южная степь
По мере увеличения засушливости климата меняется облик расти-
тельности (рис. 8.116).
Усиление континентальности климата в степной зоне связано с дей-
ствием Сибирского антициклона. Мощный барический центр устойчи-
вого высокого давления формируется осенью и зимой над просторами
Центральной Азии вследствие активного выхолаживания территории и
формирования масс плотного и тяжелого воздуха. Напомним, что в
умеренном поясе над Атлантическим океаном располагается барический
центр низкого давления. Воздушные массы устремляются из Азии в
западном направлении. Отрог изобар высокого давления, тянущийся
на запад от Сибирского антициклона, был впервые описан в начале XX в.
климатологом А.И. Воейковым и получил название «оси Воейкова». Эта
ось служит ветроразделом: к северу от нее дуют теплые, влажные, несу-
щие осадки западные ветры (западный перенос); к югу преобладают сухие 287
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ КАРТИ В А МИРА
Б — типичная ковыльная степь; В — южная типчаково-ковыльная степь
и холодные северо-восточные и восточные ветры. По мере продви-
жения на запад воздействие Азиатского антициклона ослабевает.
Степная зона Восточной Европы доступна теплым и влажным воз-
душным массам, приходящим с Атлантики. Средние температуры янва-
ря в Молдавии —4 °C (абсолютный минимум до —25 °C); в Туве, нахо-
дящейся во власти зимнего антициклона, до —35 °C (абсолютный
минимум до — 50 °C).
Весной Сибирский антициклон ослабевает, а затем исчезает. На его
месте вследствие прогревания земной поверхности и нижних слоев атмо-
сферы устанавливается область низкого давления. Значения температу-
ры июля мало различаются по мере движения по долготе:,в Придунай-
ской низменности 23 °C, на Южном Урале 22 °C, в Забайкалье 20 °C.
Важным фактором формирования степных биомов является коли-
чество и ритмика выпадения атмосферных осадков. В европейской лесо-
степи выпадает до 500 мм, к югу и востоку количество осадков снижает-
ся — в Туве 215 мм. Испаряемость в Восточно-Европейском секторе
около 1000 мм.
Сезонный ход выпадения осадков определяется в Европе западным
переносом, в Центральной Азии — муссонами. В европейских степях
осадки выпадают в основном весной и осенью; летом царит засуха,
процессы жизнедеятельности замирают. В степных областях Централь-
ной Азии наблюдается летний пик увлажнения на фоне засушливых и
холодных весны и осени (рис. 8.117).
Здесь под действием летнего муссона дожди выпадают в течение 1 —
1,5 месяца (в июле — августе). Поскольку в этот же период наблюдается
288 и максимум тепла, интенсивность жизнедеятельности высокая.
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ ВИН МОН СУШИ
Центральная Азия
Весна
Лето
Осена
Рис. 8.117. Сезонная ритмика выпадения атмосферных осадков в степных
биомах Восточной Европы и Центральной Азии, по В.Г. Мордковичу
Настоящим феноменом степных биомов являются черноземы —
самые богатые почвы на планете. Классическая область их распростра-
нения — причерноморские и приазовские степи. Карта типов почв степ-
ной области Восточной Европы и прилегающих лесных районов дает
наглядное представление о пространственной структуре степных био-
мов в целом (рис. 8.118).
Высокая продуктивность травянистой растительности, засушливое
лето и известьсодержащие почвообразующие и материнские породы
(лесс, мел, известняк) — вот три определяющих условия формирования
типичного чернозема.
В структуре растительных сообществ мощно развиты как надзем-
ная, так и подземная части (рис. 8.119). Степные травы ежегодно про-
дуцируют десятки тонн зеленой и корневой массы на гектар. Эта масса
потребляется растительноядными животными; значительная ее часть
отмирает и разлагается редуцентами. Накопление гумуса в чернозе-
мах является следствием неполного цикла деятельности микроорганиз-
мов, которые из-за летней засухи и зимних морозов успевают осущест-
вить лишь первые.стадии трансформации органического вещества.
Это приводит к резкому доминированию в черноземах процессов гу-
мификации органических остатков над процессами их минерализации.
Известьсодержащие материнские породы способствуют связыванию
гуминовых кислот в труднорастворимые гуматы кальция. Таким обра-
зом, осуществляется главный процесс формирования черноземов — гу-
мусонакопление.
289
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ КАРТИ В А МИРА
I г 3 4 5 8 7 8 9 ГО II 12 13
Рис. 8.118. Карта типов почв степной области Восточной Европы
и прилегающих лесных районов, по Г. Вальтеру: 1 — подзолистые почвы;
2 — луга и болота; 3 — карбонатное засоление; 4 — серые лесные почвы;
черноземы: 5 — северные; 6 — мощные; 7 — обыкновенные; 8 — южные;
9 — каштановые и бурые пустынно-степные почвы; 10 — песчаные дюны;
И — каменистые почвы; 12— поды (бессточные степные блюдца);
13 — засоленные почвы (солончаки)
По мере продвижения на юг условия для образования черноземов в
связи с повышением температуры и усилением засушливости климата
становятся все менее благоприятными: уменьшается мощность гумусо-
во-аккумулятивного горизонта и иллювиального горизонта (горизонта
вмывания). Последний формируется под действием нисходящего потока
веществ. Поднимается граница непромачиваемого почвенного горизон-
та, в котором наличие карбонатов устанавливается с помощью соляной
кислоты, — от капли кислоты начинается бурное выделение углекислого
газа, почва «вскипает».
Подзональные различия степных биомов находят отражение в поч-
венном покрове: от луговых степей к опустыненным последовательно
сменяют друг друга мощные, обыкновенные и южные черноземы, тем-
290 но- и светло-каштановые почвы (рис. 8.120).
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ ВИН МОН СУШИ
Рис. 8.119. Вертикальная структура степного сообщества, по М.С. Шалы-
ту: 1 — Euphorbia segueriana; 2 — Festuca sulcata; 3 — Salvia nutans;
4 — Stipa lessingiana
Богатство почв предопределило судьбу степных биомов: почти по-
всеместно они превращены в пахотные угодья. Некоторое представле-
ние о первозданной степи дают картины художников, произведения
поэтов и писателей XIX в. (рис. 8.121) и заповедники. 291
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ КАРТИ В А МИРА
Темно- Светло-
Типичный Обыкновенный Южный нашта- кашта—
Рис. 8.120. Широтно-зональная смена почв, по ВТ. Мордковичу:
1 — гумусово-аккумулятивные горизонты; 2 — иллювиальные горизонты;
3 — материнская порода; 4 — потолок вскипания карбонатов
под действием соляной кислоты
Рис. 8.121. Ковыль в степи (Южная Россия).
Иллюстрация к книге «Жизнь растений» А. Кернера
292
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ БИО МОН СУШИ
Выше уже были названы подзоны восточноевропейских степей.
При движении с севера на юг отмечаются следующие изменения в рас-
тительном покрове: 1) густой травостой луговых степей сменяется раз-
реженным на юге; 2) красочность степей уменьшается; 3) на севере
преобладают многолетники, к югу значительно усиливается роль одно-
летников; 4) широколистные злаки к югу уступают место злакам уз-
колистным — ковылям, типчаку и др.; 5) видовая насыщенность (чис-
ло видов на определенную площадь) убывает от 50 — 80 видов на севере
до 10—12 на юге. Ксерофитизация степей к югу все более усиливает-
ся, и степи постепенно переходят в полупустыни.
В самых общих чертах можно говорить о северных и южных типах
степных биомов. В северных степях доминируют красочное разнотравье
(Salvia pratensis, Filipendula hexapetala, Anemone sylvestris и др.) и широ-
колистные злаки (костер — Bromus riparius, дикие овсы — Helictotrichon
pubescens, И. schellianum, полевица— Agrostis). Из ковылей— ковыль
обыкновенный (Stipa pennata) и узколистный (S. stenophylla).
Южные степи — ковыльно-типчаковые с южным разнотравьем и
узколистными злаками. Из ковылей — ковыль Лессинга (Stipa lessingiana),
ковыль-тырса (S. capillata); кроме того, типчак (Festuca valesiaca), келе-
рия (Koeleria cristata); полыни (Artemisia).
«Курской растительной аномалией» назвал в начале XX в. профес-
сор В.В. Алехин степи под Курском. Но эта «аномалийность» связана не
с тем, что нетипична растительность, а с тем, что здесь сохранились в
условиях сплошной распашки одни из немногих нетронутых целинных
участков луговой степи. На этих землях организован Центрально-Чер-
ноземный государственный заповедник им. В.В. Алехина.
Степная растительность заповедника характеризуется богатством
видового состава — до 220 видов трав — и большим числом сезонных
аспектов: с начала весны до осени их насчитывают до одиннадцати.
Общая картина чередования весенне-летних аспектов в луговой степи
изображена на рис. 8.122.
Луговые степи необыкновенно красочны. В течение вегетационно-
го периода они постоянно изменяют свой облик. Ранней весной степь
бурая, но уже в апреле, как по волшебству, раскрывает нежные лиловые
бутоны сон-трава, за ней спешит адонис весенний. Его крупные сол-
нечно-желтые цветки появляются еще до развития листьев. Во время
цветения адониса степь постепенно одевается в зеленый наряд. За-
цветают незабудки, ирис безлистный, ветреница лесная. Начало лета
знаменует цветение лугового шалфея.
Это период наибольшего буйства красок. Но вот зацветает перистый
ковыль. Все остальные цветущие виды разнотравья скрываются под его
пушистыми перьями. И степь вновь преображается: теперь она похожа
на седое бескрайнее море. Проходят дни, постепенно степь одевается
ярко-желтыми соцветиями подмаренника. Живокость клиновидная и
чемерица черная — последние цветущие растения уходящего лета.
Животный мир евразийских степей, так же как растительность,
обладает своими подзональными и провинциальными особенностями.
Например, различия в фауне Европейско-Казахстанских и Забайкаль-
ских степей можно увидеть, сравнив рис. 8.123и8.124. 293
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ К АРТИ В А МИРА
Рис. 8.122. Весенне-летние аспекты в луговой степи. Вертикальная
проекция, квадраты дециметровые, по Г. Вальтеру и В.В. Алехину
294
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ БИО МОН СУШИ
Расшифровка к рис. 8.122: а — начало апреля: бурый аспект с лиловыми
пятнами Pulsatilla patens; желтые — мужские колоски Carex humiJis. Первые
цветки Adonis; б — вторая половина апреля: желтый аспект от Adonis
vernalis. Кроме того, цветет нежно-голубой Hyacinthus leucophaeus. Степь
еще не зазеленела; в — конец-мая: голубой аспект от Myosotis sylvatica,
отдельные цветущие экземпляры Anemone Silvestris и желтые пятна Senecio
campestris. Зацветает ковыль; г — июнь. Слева направо: Trifolium repens, Salvia
pratensis, Festuca sulcata, Viola arenaria, Trifolium montanum, Stipa pennata,
Carex caryophyllea, Ranunculus polyanthemus, Leucanthemum vulgare,
Hypochoeris maculata, Anthoxanthum odoratum, Arenaria graminifolia, Filipendula
hexapetala, Carex montana, Agrostis vinealis, Viola canina, Scorzonera purpurea,
Euphorbia gracilis, Potentilla opaca, Festuca rubra, Astragalus danicus, Phlomis
tuberosa, Koeleria delavignei, Galium boreale, Echium rubrum, Carex montana.
Ограничимся краткой характеристикой животного мира Европей-
ско-Казахстанской степной провинции. Ее границы хорошо очерчива-
ют ареалы байбака и большого тушканчика (рис. 8.125).
Фауна провинции представляет собой пестрый комплекс, переход-
ный от биомов широколиственного леса к азиатским пустыням. Тем не
менее он имеет эндемичное ядро, в состав которого входят несколько
видов сусликов, байбак, слепыш, большой тушканчик, обыкновенный
хомяк, степная мышовка. К этой же группе относится антилопа сайгак.
Из наиболее характерных птиц следует назвать стрепета, дрофу, малого
журавля, степного луня, степного орла; из пресмыкающихся — степ-
ную гадюку, разноцветную ящурку.
Для естественных ландшафтов открытых степных равнин характерны
крупные стадные копытные и грызуны. Из-за отсутствия естественных
укрытий степные копытные обладают быстрыми ногами и хорошим зре-
нием, а грызуны обзаводятся норами. Суровая зима заставляет многих
зверей и птиц откочевывать в более южные районы.
Следует подчеркнуть, что названные группы животных не только
потребляют растительную массу, но и активно воздействуют на почвен-
но-растительный покров, разбивая дернину и роя многочисленные
норы. Высокая биопродуктивность растительности составляет основу
цепей питания в степных экосистемах (рис. 8.126).
Наблюдения в заповедниках показали, что умеренные нарушения
являются необходимыми для устойчивого развития степных сообществ:
благодаря им появляются открытые участки почвы, доступные для се-
менного возобновления. Если же биопродуктивность растений не регу-
лируется, накапливается мощный слой ветоши, дернины злаков уплот-
няются, так что семенное возобновление становится невозможным,
многие виды не выдерживают конкуренции, и биоразнообразие сооб-
ществ резко падает. Следует сказать, что в естественных условиях чис-
ленность животных и биопродуктивность растительности саморегули-
руются и степные биомы развиваются устойчиво.
Степные биомы Европейско-Казахстанских степей испытали особен-
но сильное воздействие человека. В Восточной Европе практически все
степи распаханы, рассечены густой сетью дорог, заняты городами и про-
мышленными предприятиями. Вместе с растительным покровом исчезли
многие виды растений и животных, у других резко сократились ареалы.
295
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ К АРТИ В А МИРА
Рис. 8.123. Характерные представители фауны Европейско-Казахстанских
степей: 1 — степная пищуха; 2 — заяц-русак; 3 — крапчатый суслик;
4 — степная пеструшка; 5 — слепыш; 6 — сайгак; 7 — полоз желтобрюхий;
8 — дрофа; 9 — черный жаворонок; 10 — степной жаворонок;
2 9 G Н — обыкновенный хомяк; 12— стрепет
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ ВИН МОН СУШИ
Рис. 8.124. Характерные представители фауны
Забайкальских степей: 1 — монгольская песчанка; 2 — заяц-толай
3 — даурский суслик; 4 — монгольский жаворонок; 5 — манул;
6 — монгольская ящурка; 7 — даурский цокор; 8 — дзерен;
9 — тушканчик-прыгун; 10 — корсак
297
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ К АРТИ В А МИРА
Рис. 8.125. Ареалы животных Европейско-Казахстанских степей,
по Н.А. Бобринскому
Рис. 8.126. Цепи питания в степных экосистемах, поН.А. Бобринскому
Неарктическая область прерий Голарктического царства в Север-
ной Америке располагается вдали от атлантического океанического
сектора, занятого лесными биомами. Данные об изменениях климата,
растительности и почв прерий с востока на запад к подножию Скали-
298 стых гор отображены на профиле (рис. 8.127).
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ 510МОН СУШИ
3 *
пн
ИЮ-
300-
200-
100-
О
м
Средняя годовая
температура 8
Абсолютный
минимум 37__
уменыыенив
Великие равнины (низкозляко- Смешанная
бая прерия) прерия
У&миченцд
Абсолютный
минимум 34'
Высокозлаковая
прерия
В
700
гббобая Г-600
температура 11* .эду
Йл^вст
400
о
Мертвая,
постоянна сухая почва
м
1
бурые пустынно- Темные черноземы
степные почвы бурые почвы IT)
<-----Уменьшение содержания гумуса Нарастание,^тенсшню
Рис. 8.127. Схематический профиль через район прерий
с востока на запад к подножию Скалистых гор, по Г. Вальтеру
Доминантами растительного покрова являются дерновинные злаки,
на востоке — высокие, а на сухом западе — низкие. Общая картина
ареалов прерий изображена на карте (рис. 8.128).
Рис. 8.128. Ареалы прерий, по Т.Н. Гордеевой и О.С. Стрелковой:
Прерии: 1 — настоящая высокотравная восточная и южная; 2 — низкотрав-
ная и разнотравная; 3 — северные из овсяницы (Festuca); 4 — северо-запад-
ная; 5 — пустыни и полупустыни; 6 — территории с преобладанием засолен-
ных почв, растительный покров несомкнутый 299
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ КАРТИ В А МИРА
В высокозлаковой, или настоящей, прерии господствуют дерновин-
ные злаки, образующие травостой высотой 0,5—1 м. К числу доми-
нантов относятся: ковыль {Stipa spartea), овсяница {Festuca scabrella),
житняк {Адгоругит smithii), бородач, или трама (Bouteloua curtipendula),
тонконог {Koeleria cristata); обильно разнотравье. Эти прерии почти
сплошь распаханы.
Низкозлаковая прерия покрывает пространства Великих равнин.
Здесь абсолютное господство переходит к плотнодерновинным злакам:
бизоновой траве {Buchloe dactyloides), траве трама {Bouteloua gracilis)
(рис. 8.129). Эти прерии используются под пастбища или орошаемое
земледелие.
Рис. 8.129. Доминанты низкозлаковых прерий: а — бизонова трава
(Buchloe dactyloides); б— трава срама (Bouteloua gracilis)
Прерии относятся к обширной Сонорской области Неарктики, вклю-
чающей широколиственные леса и субтропические биомы. Символом
животного мира мог бы служить степной бизон, поголовье которого до
колонизации Северной Америки европейцами, вероятно, достигало 50-
60 млн (рис. 8.130).
Индейцы не могли нанести ему существенного урона, т. к. охоти-
лись с копьями или луком и стрелами. С 1872 по 1874 г. белыми было
убито более 5 млн бизонов, а к концу 1889 г. их осталось всего около
тысячи. Для степного бизона роковой оказалась постройка транскон-
тинентальной железной дороги. Отстрел животных происходил прямо
с проходящих поездов (рис. 8.131).
Южное полушарие. Основные площади злаковников располагают-
300 ся в Южной Америке и Новой Зеландии.
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ 510МОН СУШИ
Рис. 8.130. Стадо степных бизонов
Рис. 8.131. Охота на бизонов с поезда трансконтинентальной
железной дороги в конце XIX в.
Пампа — южноамериканский вариант субтропических степей, за-
нимает равнины Параны — Ла-Платы (рис. 8.132). Рельеф плоский, рав-
нинный. Климат субтропический. Зима теплая, температура самого
холодного месяца 8—12 °C; летние температуры 22 — 25 °C. Годовое
количество осадков возрастает от 500 — 600 мм на западе до 1000 мм у
атлантического побережья. Испаряется влаги несколько больше, чем
выпадает. Два-три раза в течение каждых 10 лет случаются очень за-
сушливые годы. Основная почвообразующая порода в пампе — лёсс. На
лёссе формируются черноземовидные почвы. 301
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ КАРТИ В А МИРА
Рис. 8.132. Ареал пампы в Южной Америке, по Г. Вальтеру
Безлесие пампы — явление во многом вторичное, возникшее еще до
прихода европейцев в результате выжигания растительности индейца-
ми. К причинам безлесия относятся также периодические засухи, сла-
бая дренированность и засоление грунтов.
В настоящее время это наиболее густонаселенные сельскохозяй-
ственные районы Южной Америки. Первичная растительность пампы
практически уже отсутствует. Однако в результате ее реконструкции
установлено, что доминантами являлись злаки, многие рода которых
характерны для лугов, степей и прерий Северного полушария: ковыль
(Stipa), мятлик (Роа), аристида (Aristida), перловник (Melica), костер
(Bromus) и др. (рис. 8.133); кроме того, для пампы характерны свои
рода — Paspalum, Ischaemum, Cortaderia, Gynerium и др. Разнотравье пред-
ставлено люпином (Lupinus), горошком (Vicia), вербеной (Verbena), пор-
тулаком (Portulaca), видами паслена.
Фауна пампы относится к Патагонско-Чилийской области Неотро-
пического царства. В прошлом здесь обитали стада пампасского оленя и
гуанако. В настоящее время многочисленны лишь грызуны — вискаша;
302 встречаются броненосец, страус нанду.
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ ВИН МОН СУШИ
Рис. 8.133. Пампа с ковылем (Stipa brachychaeta), по Г. Вальтеру
В Новой Зеландии злаковники известны под названием туесок. Они
распространены на Южном острове в дождевой тени Южных Альп от
морского побережья до высоты 1500 м. Климат умеренно-теплый. Годо-
вое количество осадков 600 — 500 мм. Почвы буро-серые.
Естественный растительный покров представлен злаковыми кочкар-
никами. Доминирующие рода — овсяница (Festuca, F. novae-zelandiae),
мятлик (Роа, Р. caespitosa], житняк (Адгоругит, A. scabrum) — характер-
ны для степей Голарктического царства, т. е. имеют биполярный ареал.
Это так называемый низкий туесок (рис. 8.134). Всего в группу форма-
ций низких злаковников входит около 40 видов злаков и 35 видов сложно-
цветных. На влажных торфянистых почвах произрастает высокий туесок,
образованный дантонией (Danthonia raoulii). Флористически эта форма-
ция беднее, т. к. высокие и плотно расположенные дернины дантонии
подавляют другие виды.
Рис. 8.134. Туссоковый злаковникна о. Южный, Новая Зеландия,
Вальтеру: доминирует овсяница (Festuca novae-zelandtcae)
303
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ К АРТИ В А МИРА
Со времен первых переселенцев злаковники Южного острова ис-
пользовались как пастбища, поэтому девственных, нетронутых европей-
цами и домашними животными злаковых биомов здесь не сохранилось.
8.2.6. Внетрепипсие пустыни
В континентальном секторе Евразии к юго-востоку от зоны степей
располагаются пустыни. Они занимают обширные пространства уме-
ренного пояса Средней и Центральной Азии (рис. 8.135). На юге они
смыкаются с пустынями жаркого пояса.
Рис. 8.135. Области пустынь умеренного пояса, по М.П. Петрову.
Среднеазиатские пустыни: I — Ирано-Туранская; И — Джунгаро-Казах-
станская; центральноазиатские пустыни: HI — собственно центрально-
азиатские; IV— Тибетские высокогорные
Переход от пустынь жаркого пояса к внетропическим пустыням
происходит постепенно. Он связан прежде всего с понижением зим-
них температур. Устойчивый морозный период определяет своеобра-
зие биомов. Границей, разделяющей тропические и внетропические пу-
стыни, могут служить пределы возделывания финиковой пальмы,
которая не переносит длительных заморозков.
Общие сведения о рельефе пустынь, формировании пустынных
биомов, адаптациях растений и животных к жизни в условиях высоких
температур и дефицита влаги рассмотрены при описании пустынь
жаркого пояса (см. раздел 8.1.3). Остановимся на некоторых особенно-
стях биомов пустынь Средней Азии и Казахстана.
Южные степи и северные пустыни объединены полосой динами-
ческого контакта, выделяемой в качестве полупустыни. По мере про-
движения на юг дерновинные злаки постепенно уступают место пус-
тынным полукустарникам: видам полыни (род Artemisia, подроды
Seriphidium и Dracunculus), прутняку [Kochia prostrata), разнообразным
солянкам и т. п. Следует подчеркнуть, что наряду с полынью семейство
маревые, или лебедовые {Chenopodiaceae), куда входят все виды, объ-
единяемые термином «солянки», является ландшафтообразующим. Аре-
ал саксаула (род Haloxylon, сем. Chenopodiaceae) довольно точно очер-
304 чивает границы пустынь Средней Азии и Казахстана (рис. 8.136).
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ БИО МОН СУШИ
Рис. 8.136. Ареал рода саксаул, по «Жизни растений»
Пустынные ландшафты Туранской низменности простираются на
10—12° по широте. На этом протяжении зональные изменения терми-
ческих условий и режима увлажнения приводят к формированию двух
подзон — северных и южных пустынь. Климатическими условиями
образования внетропических пустынных биомов, общими для пустынь
жаркого пояса, являются: большая солнечная радиация, исключитель-
ная засушливость на протяжении всего вегетационного периода, высо-
кие летние температуры (25 — 30 °C), малое количество атмосферных
осадков (в среднем 100—150 мм).
Испаряемость в 10 — 25 раз превышает сумму выпадающих осадков.
Зимние температуры отрицательные (—10, — 15 °C), лишь на юге Туран-
ской низменности они несколько выше 0 °C; абсолютные минимумы
на севере Казахстана достигают —50 °C, в Ташкенте — 30 °C.
В жизни пустынь в условиях дефицита атмосферных осадков боль-
шое значение имеют подземные воды, в частности грунтовая верховод-
ка. В пустынях, сформированных на месте древних аллювиальных рав-
нин, они образуют сплошное зеркало. Грунтовые воды засолены, часто
залегают неглубоко (3 —4 м) и доступны для корней растений; местами
располагаются линзы пресной верховодки.
Засушливый климат обусловил особый тип почвообразования: для
южных пустынь характерны различного тина сероземы, для северных —
серо-бурые почвы. На огромных территориях песчаных пустынь поч-
вообразовательный процесс находится в начальной стадии. Значитель-
ное участие в почвообразовании принимают водорастворимые соли —
карбонаты, сульфаты и поваренная соль. Они переходят в почву из
сильно минерализованных материнских пород или поднимаются по ка-
пиллярам из грунтовых вод. В то же время в пустынных почвах очень
мало гумуса (менее 1%). Это связано с тем, что растительность бедна и
в почву поступает небольшое количество органических остатков, кото-
рые к тому же быстро минерализуются. Пестрота почвогрунтов, раз-
ная степень засоления, неодинаковые условия увлажнения — причины
комплексности растительного покрова. 305
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ КАРТИ В А МИРА
В основе классификации пустынь Средней Азии и Казахстана лежит
характер поверхностных отложений и почвогрунтов. Основными типа-
ми пустынь являются песчаные, глинистые, гипсовые, лёссовые; своеоб-
разные природные комплексы представлены такырами, солончаками и
поймами рек; особый тип ландшафтов связан с горами (рис. 8.137).
Рис. 8.137. Пустыни Средней Азии и Казахстана, по «Пустыням»
В тесной связи с почвогрунтами находится растительность. Выделя-
ются следующие типы пустынной растительности (вне речных пойм):
псаммофитный древесно-кустарниковый на песках (Haloxylon persicum,
Н. aphyllum, Calligonum caput-medusae, С. setosum, Salsola richteri,
Ammodendron conollyi, Ephedra strobilacea и др.); гипсофильный мелко-
кустарничковый на щебнистых и каменистых третичных плато (Salsola
arbuscula, S. rigida, S. gemmascens, S. laricifolia, Anabasis salsa, Hammada
leptoclada}; эфемерово-эфемероидный на лёссах; полукустарниковый
полынный на суглинках (Artemisia semiarida, A. kemrudica, Kochia
prostrata, Eurotia ceratoides с участием многолетних злаков, а также
эфемеров и эфемероидов); водорослево-лишайниковый на такырах со
своеобразными группировками пустынных лишайников и сине-зеленых
водорослей (Петров, 1973).
В качестве иллюстрации зависимости растительности от почвогрун-
тов приведен аэрофотоснимок песчаной пустыни Каракумы (рис. 8.138).
На нем отчетливо видны гряды песков, заросшие псаммофитами, и
ЗОБ солончаковая депрессия с сообществами галофитов.
ГЛАВА 8. ЗОНАЛЬНЫЕ ШЫ ВИН МОН СУШИ
Рис. 8.138. Грядовый рельеф песков с сообществами псаммофитов;
серое овальное пятно (а) — солончаковая депрессия с сообществами галофи-
тов. Аэрофотоснимок
Фауна пустынь Средней Азии и Казахстана относится к Арало-Кас-
пийской области Палеарктики (рис. 8.139). Она имеет свои специфи-
ческие особенности и вместе с тем носит переходный характер между
сахара-средиземноморской и центральноазиатской фаунами.
Адаптации растений и животных к жизни в условиях пустыни вы-
рабатывались на протяжении более 100 млн лет, с мелового и третично-
го периодов. Они шли по пути приспособления к засухе, жаре, большой
амплитуде температур, бедным и засоленным грунтам, к сыпучим пес-
кам и т. п. Главное направление адаптации у растений привело к обра-
зованию многочисленной группы ксерофитов (см. табл. 8.2).
Основными приспособительными признаками ксерофитов являют-
ся особенности формирования их листа, ведущие к уменьшению раз-
меров листовой пластинки — микрофилии и афилии (Василевская, 1954).
Продукция надземной части пустынных растений ничтожна по срав-
нению с продукцией подземной: на корневую систему приходится до
95% всей фитомассы. Корни очень глубоко проникают в землю, дости-
гая капиллярной каймы грунтовых вод на глубине до 20 — 25 м.
Весьма своеобразны приспособления псаммофитов — растений,
живущих на рыхлых, иногда даже подвижных песках. Их корни протя-
гиваются по склонам барханных цепей на 25 — 30 м и уходят в глубину
на 2 — 2,5 м, достигая капиллярного подпора грунтовых вод и обеспечи-
вая растения достаточным количеством влаги. Во время передвижения
песков ветер выдувает из-под корней грунт, они частично обнажаются,
но не гибнут, т. к. хорошо защищены от палящих лучей солнца. Засы-
панные песком кусты не погибают. Они дают массу придаточных кор-
ней, способствующих закреплению песка (рис. 8.140). 307
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ КАРТИ В А МИРА
Рис. 8.139. Характерные представители фауны Арало-Каспийских пус-
тынь. Рис. В.А. Ватагина, по Н.А. Бобринскому: 1 — тонкопалый суслик;
2 — саксаульная сойка; 3 — гребнепалый тушканчик; 4 — ушастая круглого-
ловка; 5 — цинковый геккон; 6 — варан; 7 — каракал; 8 — белобрюхий рябок;
9 — барханный кот; 10 — сырдарьинский фазан; И — степная черепаха;
12 — полуденная песчанка; 13 — ушастый еж; 14 — джейран;
308 15 — пегая землеройка; 16 — кулан
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ БИО МОН СУШИ
Рис. 8.140. Ярусная корневая система Ephedra strobilacea, сформировавшаяся
в результате засыпания растения песком, по Н.Т. Нечаевой
Плоды и семена многих псаммофитов имеют приспособления для
переноса ветром (рис. 8.141). У песчаной акации они пропеллерообраз-
ной формы, семена кандыма и джузгуна снабжены упругими щетинка-
ми, у саксаула — перепонками, у селина — летучками и т. п. Благодаря
этому парусность плодов и семян увеличивается, и они разносятся по
пустыне на большие расстояния.
Растения солончаков — галофиты безлистны или мелколистны, с
толстыми членистыми веточками и листьями, обычно сочными и мя-
систыми. Клеточный сок их насыщен водорастворимыми солями —
сульфатами и хлоридами. Иногда содержание солей доходит до 45% от
абсолютно сухого веса листьев. Обилие водорастворимых солей повы-
шает осмотическое давление клеточного сока, что увеличивает сосущую
силу корней и уменьшает транспирацию.
Одной из характернейших черт приспособления растений к суро-
вым условиям пустынь является их способность уменьшать продолжи-
тельность вегетационного периода или впадать в состояние анабиоза.
Это связано с периодичностью в годовом ходе выпадения атмосферных
осадков, их малым количеством, летней жарой и зимними холодами.
В результате этого в пустынях сформировались две оригинальные жиз-
ненные формы растений — эфемеры и эфемероиды.
Эфемеры — травянистые однолетние растения с коротким вегета-
ционным периодом; они быстро, за 1—2 весенних месяца, проходят все
стадии развития и, образовав семена, погибают. Неблагоприятные пе- 309
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ КАРТИ В А МИРА
Рис. 8.141. Приспособления плодов и семян псаммофитов для переноса
ветром, по М.П. Петрову: 1 — солончаковый саксаул; 2 — джузгун;
3 — кандым; 4 — черкез Рихтера; 5 — песчаная акация; 6 — селин;
7 — джузгун безлистный; 8 — кумарчик широколистный
риоды летней жары и зимних холодов они переносят в виде семян.
Эфемероиды — многолетние травянистые растения с коротким перио-
дом вегетации. Они проходят все стадии развития и образуют семена
также весной в течение 1,5 — 2 месяцев. С наступлением летней засухи
наземная часть растений высыхает, подземные же органы (корневища,
луковицы, клубни и т.п.), расположенные на глубине 20 — 25 см, пере-
ходят в стадию покоя. !
Ранней весной пустыня покрывается густым зеленым покровом ярко
цветущих трав (рис. 8.142). Благодаря тому что развитие эфемеров и
эфемероидов происходит в благоприятных гидротермических услови-
ях, они лишены ксероморфных признаков. Многие из них — выходцы
из северных лугов.
В формировании растительного покрова песчаных пустынь умерен-
ного пояса принимает участие много разнообразных жизненных форм
(Нечаева, Василевская, Антонова, 1973), благодаря чему сообщества
характеризуются достаточно высокой видовой насыщенностью, слож-
ной структурой и высокой продуктивностью. Фитомасса белосаксауль-
ников — около 270 ц/га, с ежегодным приростом 75 ц/га; фитомасса
черносаксаульников — 540 ц/га.
В Каракумах в черно- и белосаксауловой формациях четко обособ-
ляются два или даже три надземных яруса; соответственно этому ярус-
310 ное распределение имеют и корневые системы (рис. 8.143, 8.144).
ГЛАВА 8. ЗОНАЛЬНЫЕ ШЫ ВИН МОН СУШИ
Рис. 8.142. Сообщество эфемеров и эфемероидов, по Е.П. Коровину:
1 — Carex pachystylis; 2 — Роа bulbosa var. vivipara; 3 — Ranunculus
pinnalifidus; 4 — Scaligeria alloides; 5 — Haplophyllum perforatum;
6 — Phlomis thapsoides; 7 — fxiolirion tatahcum; 8 — Gentiana olivieri;
9 — Eremostachys labiosa
Puc. 8.143. Заросли черного саксаула (Haloxylon aphyllum) с богатой травянис-
той растительностью на слабозасоленной влажной почве, по М.П. Петрову 311
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ КАРТИ В А МИРА
Рис. 8.144. Распределение корневых систем в зарослях черного саксаула
(Haloxylon aphyllum), по М.П. Петрову: 1 — Haloxylon aphyllum;
2— Calligonum turkestanicum; 3— Ephedra strobilacea; 4— Carex physodes;
5— эфемеры и эфемероиды
Сезонный ритм жизненных процессов в пустыне определяется свое-
образным сочетанием теплового и водного режимов, при котором са-
мый теплый период оказывается и самым сухим, а относительно влаж-
ная зима недостаточно теплой. Поэтому наибольшая активность биоты
приходится на переходные сезоны, особенно на весну.
Зимой погода неустойчивая. Вторжение теплого воздуха с Иранского
нагорья приводит к резким повышениям температуры на юге, при холод-
ных вторжениях возможно ее понижение до —30° (на севере до —50°).
В южных пустынях устойчивый снежный покров бывает редко. На се-
вере устойчивый покров наблюдается с конца ноября до марта. Мощ-
ность его невелика — 5—15 см.
В южной подзоне в марте — апреле господствует зеленый аспект эфе-
меров и эфемероидов, создаются максимальные запасы зеленой массы.
Но ко второй декаде мая (в сухие годы — в апреле) эти растения полно-
стью засыхают. В северной подзоне вегетация начинается во второй
половине марта — начале апреля. Зацветают ранние эфемеры, затем
эфемероиды, появляется листва на кустарниках и полукустарниках.
Основной период вегетации — апрель —май. В июне растения выглядят
угнетенными; в первой декаде июня заканчивается плодоношение боль-
шинства эфемеров и эфемероидов, у полукустарничков перестают рас-
ти побеги и опадает часть листьев.
Лето — время длительной засухи. На юге с июня по сентябрь расти-
тельный покров выгорает. На севере в июле —августе большинство ра-
стений пребывает в состоянии покоя, но в сентябре начинается осен-
няя вегетация. Наблюдается цветение и максимальный прирост у
полукустарничков, в октябре они уже плодоносят и к концу месяца пре-
кращают вегетировать. Во влажные годы в это время может возобно-
виться вегетация эфемеров и эфемероидов. Если затем последует теп-
лая зима, в южной подзоне эфемероиды могут вегетировать непрерывно
(как озимые) почти до 200 дней, тогда как в сухие годы лишь около
312 60 дней (как яровые).
ГЛАВА 8. ЗОНАЛЬНЫЕ ТИПЫ БИО МОН СУШИ
* * *
Завершая обзор зональных типов биомов суши, подчеркнем огром-
ную потенцию биоты к завоеванию жизненного пространства. На про-
тяжении геологической истории эволюция биоты шла по пути дивер-
генции и экологической адаптации. По мере усиливающегося давления
жизни, в результате борьбы за существование и естественного отбора
возникали все новые формы, способные захватывать территории, где
напряженность экологических факторов находится как в минимальных,
так и в максимальных пределах. В настоящее время практически все по-
верхностные оболочки Земли представляют собой арену распростране-
ния жизни.
Процессы адаптации к новым условиям жизни протекают непрерыв-
но. На строении одних организмов лежит печать древнейших геологи-
ческих эпох, другие сформировались на протяжении последнего чет-
вертичного периода. При этом наряду с расхождением признаков
наблюдается конвергентный путь эволюции, когда далекие в система-
тическом отношении организмы благодаря сходству условий существо-
вания приобретают сходный облик. Своеобразие природных зон опре-
деляет особенности жизни человека, характерные приемы ведения
хозяйства и мероприятия по сохранению и приумножению биологиче-
ских ресурсов.
Антропогенное воздействие на биосферу достигло в наши дни гло-
бальных масштабов. Зональные типы биомов в естественном виде почти
не сохранились. Проблема изменения биосферы в результате антро-
погенного воздействия требует специального рассмотрения.
313
Глава
АНТРОПОГЕННЫЕ ИЗМЕНЕНИЯ
ЛАНДШАФТОВ ПРИРОДНЫХ ЗОН РОССИИ
На разнообразии ландшафтов России лежит печать географиче-
ской зональности. Основоположник отечественной географии В.В. До-
кучаев отмечал, что «человек зонален во всех проявлениях своей
жизни: обычаях, религии (особенно в нехристианских религиях), в
красоте, в одежде, во всей житейской обстановке; зональны — домаш-
ний скот, так называемая культурная растительность, постройки,
пища, питье». Антропогенное воздействие ведет к экологической деста-
билизации ландшафтов, проявляющейся по-разному в разных природ-
ных зонах.
9.1. Зональные особенности распределения экологического потенциала
ландшафтов
Одним из фундаментальных понятий эколого-географической тео-
рии взаимодействия человека и природы, по утверждению А.Г. Иса-
ченко (2001, 2003), является экологический потенциал ландшафта
(ЭПЛ) — совокупность условий, необходимых для жизни и воспроиз-
водства населяющих данную территорию организмов. В этом понятии
отражена одна из важнейших социальных функций ландшафта — спо-
собность обеспечивать человечество необходимыми средствами к су-
ществованию.
В любой зоне экологический потенциал ландшафта не беспределен,
он способен прокормить ограниченное количество животных. Возни-
кает понятие емкость ландшафта — количество домашних или диких
животных (иногда тех и других суммарно), способных жить и нормаль-
но размножаться на единице площади данного ландшафта в течение
неопределенно долгого времени.
Экологическая емкость ландшафта — это также численность насе-
ления, которую ландшафт способен поддерживать своими естественны-
ми экологическими ресурсами без ущерба для собственного функцио-
нирования и
биологической
моотношениях
314 пряженность.
экологического потенциала. Когда использование
продукции ландшафта превышает его емкость, во взаи-
между человеком и природой возникает растущая на-
та 8. АНТРОПОГЕННЫЕ ИЗМЕНЕНИЙ ЛДНДШДЙТОВ ПРИРОДНЫХ ЭОН РОССИИ I
Всесторонняя характеристика ЭПЛ требует учета сотен показате-
лей, однако его сравнительная оценка может быть основана на двух
определяющих факторах — тепле и влаге, от которых в первую очередь
зависит биологическая продуктивность. Картографическое изображе-
ние ЭПЛ России показано на карте (рис. 9.1).
Рис. 9.1. Экологический потенциал ландшафтов России, по А.Г. Исаченко:
1 — наиболее высокий; 2 — относительно высокий; 3 — средний; 4 — низ-
кий; 5— очень низкий; 6— экстремально низкий; 7— горные территории
На распределении ЭПЛ ясно прослеживается влияние закона гео-
графической зональности (Исаченко, 2001). В пределах европейской тер-
ритории России хорошо выражена зона экологического оптимума, в ко-
торую входят широколиственно-лесные и лесостепные ландшафты.
К северу ЭПЛ снижается по мере сокращения теплообеспеченности, к
югу — с уменьшением водообеспеченности при одновременном увели-
чении запасов теплоты. Крайние ступени зонального ряда — арктиче-
ские ландшафты на севере и пустынные на юго-востоке. В Предкавказье
и на Северном Кавказе, где наблюдаются признаки перехода к субтро-
пикам и увеличение как тепла, так и влагообеспеченности, формирует-
ся вторая область экологического оптимума. В Западно-Сибирском и
Восточно-Сибирском секторах в связи с усилением континентально-
сти ЭПЛ снижается. С переходом к Дальневосточному сектору ЭПЛ
вновь увеличивается.
Н.П. Базилевич (1976) отмечает следующие общие закономерности
в функционировании зональных типов экосистем на территории Рос-
сии: наибольшие запасы живого и мертвого органического вещества
присущи экосистемам, развивающимся в условиях радиационного ба-
ланса около 35 — 40 ккал/год при индексе сухости, близком к единице.
Это биомы лесостепи и луговой степи, наиболее удобные для пахотного
сельскохозяйственного освоения. При таком же радиационном балан- 315
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ КАРТИ В А МИРА
се, но индексе сухости больше 1 формируются биомы степей, в кото-
рых мертвое органическое вещество (до 95%) накапливается в виде гу-
муса. Плодородие почв возрастает, однако сухость климата делает эту
зону рискованной для земледелия.
Только в таежных биомах отношение между живым и мертвым орга-
ническим веществом складывается в пользу живого органического ве-
щества — древесной растительности. Однако почвы бедны гумусом и
малоплодородны. На базе этих биомов развивается лесное хозяйство.
Наименьшие запасы живого органического вещества характерны для
биомов тундры и пустыни. Традиционным видом хозяйственной дея-
тельности местного населения является пастбищное животноводство —
оленеводство в тундре и овцеводство в пустыне.
Интенсивность биотического круговорота проявляется в скорости
разложения органических веществ в ландшафте, о чем свидетельствует
отношение массы подстилки к массе зеленой части опада. Чем интен-
сивнее протекают в ландшафте процессы разрушения органических ве-
ществ, в том числе антропогенного происхождения, тем больше этот
коэффициент. В тундре на разложение 10 ц опада требуется около 100 лет,
в лесотундре и северной тайге количество опада увеличивается более чем
в два раза, а скорость его разложения — более чем в четыре раза и
составляет около 20 лет, снижаясь в южной тайге до 10 лет, что на
порядок выше, чем в тундре. В степных ландшафтах наземный опад
разлагается за 1 — 3 года, а в сухих степях и пустынях менее чем за год
(Кочуров, 1988).
Под воздействием человека природные ландшафты преобразуются в
антропогенные. Ф.Н. Мильков (1973) предложил выделять особую их
категорию — естественно-антропогенные ландшафты. Их особенность
состоит в том, что в результате уничтожения коренных растительных
сообществ активизируются такие природные процессы, которые в нена-
рушенных ландшафтах были выражены слабо или не проявлялись вовсе.
Отмечаются зональные закономерности в распределении естественно-
антропогенных ландшафтов. Так, максимальное развитие термокарста
наблюдается в тундре и лесотундре; вторичных лесов, суходольных и
пойменных лугов, пустошей — в зоне тайги и смешанных лесов; вторич-
ных болот — от лесотундры до смешанных лесов; оврагов — от лесо-
степи до полупустыни; вторичных солончаков и развеваемых песков —
в полупустыне и пустыне (рис. 9.2).
С антропогенной деятельностью связана деградация почв, сниже-
ние их биологической продуктивности. В широком смысле говорят об
опустошении земель, или детериорации. Антропогенному опустошению
подвержены почвенный и растительный покровы, животный мир. Оно
возникает как под воздействием деструктивных физических сил, так и
в результате химического загрязнения воздуха, воды и почвы. Антро-
погенной детериорации могут быть подвержены земли любой природ-
ной зоны. Угроза опустошения возрастает по мере усиления засушли-
вости климата. Особенно резко эти процессы проявляются в сухих,
субгумидных областях, в аридной и субаридной зонах. Здесь они извест-
31G ны под названием опустынивание.
ГЛАВА 9. АНТРОПОГЕННЫЕ ИЗМЕНЕНИЯ ЛАНРАФТОВ ПРИРОДНЫХ ЗОН РОССИИ
Тип естественно- антропогенных ландшафтов Географические зоны
Тундре Лесо- тундра Тайга Смеш. леса Лесо- степи Стели Полу- пустыни Пустыни
Термокарст
Л
вторичные леса
Пустоши
Луга суходольные
поименные
вторичные болота — ш ж
Овраги
вторичные солончаш
Развеваемые лески
Рис. 9.2. Зональное распределение естественно-антропогенных
ландшафтов, по Ф.Н. Милькову
Обсуждая вопросы трансформации зональных типов биомов в про-
цессе их хозяйственного освоения, следует иметь в виду глобальные
изменения ландшафтов в XX —XXI вв., связанные с потеплением кли-
мата. По мнению А.А. Величко (1992), мы живем в межледниковую
эпоху: естественный тренд — понижение температуры.
Потепление полностью обусловлено хозяйственной деятельностью
человека, сопровождающейся все увеличивающимся выбросом парни-
ковых газов в атмосферу. И потепление, и похолодание климата по-раз-
ному проявляются в ландшафтах природных зон. Нельзя сказать одно-
значно, что лучше для человека — сохранить естественный тренд
похолодания и бороться с выбросами парниковых газов или, напротив,
сохранять действие факторов, ведущих к повышению температуры. По
мнению специалистов, в начале XXI в. под воздействием антропоген-
ных факторов среднеглобальная температура может повыситься при-
близительно на 1 °C.
Основные типы экологической дестабилизации ландшафтов суши
рассматриваются на примере территории России. Они обусловлены
зональными типами ландшафтов и особенностями природопользования
в разных природных зонах. При характеристике функционирования и
изменения зональных типов экосистем использованы данные Н.И. Ба-
зилевич и др. (1976) и Ю.А. Исакова и др. (1986). Особенности вероят-
ных изменений ландшафтов в связи с потеплением климата даются по
работе А.А. Величко (1992).
317
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ КАРТИ В А МИРА
И 9.2. Зона тундры___________________________________________________
В XIX в. тундра была малообитаема. Местное население занималось
кочевым оленеводством и пушным промыслом, а также рыболовством
и охотой на морского зверя. В начале XX в. возникли немногочислен-
ные и малонаселенные поселки, расположенные в долинах рек. С сере-
дины XX в. быстрыми темпами стала развиваться геологоразведка, до-
быча нефти и газа, минерального сырья, строительство дорог, нефте- и
газопроводов, городов и поселков.
Наличие в тундре многолетней мерзлоты — важнейший фактор, сдер-
живающий техногенное освоение территории. Растительная дернина, в
том числе моховой покров и торфяной слой, являются хорошим есте-
ственным теплоизолятором. Разрушение этого слоя способствует таянию
подземных льдов, образованию просадок и провалов. Поэтому необходи-
мо исключительно бережное отношение к почвенно-растительному по-
крову. Достаточно гусеничному транспорту проехать по тундре и разру-
шить мох, чтобы вскоре следы от гусениц превратились в канаву, а затем
и в глубокую промоину (рис. 9.3). Расчищенная грунтовая площадка через
несколько лет может стать провальным озером.
Рис. 9.3. След от гусеничного транспорта, превращающийся в овраг
Попадающие в грунт нефтепродукты разлагаются очень плохо. Низ-
кие температуры и незначительное содержание кислорода в почве спо-
собствуют длительному сохранению нефтяного загрязнения. Самоочи-
щения в зоне многолетней мерзлоты практически не происходит.
Растительность является чутким индикатором общего состояния тунд-
ровых экосистем. Восстановление экологического потенциала тундро-
вых ландшафтов полностью определяется процессами демутации рас-
тительного покрова.
Антропогенная трансформация флоры. Арктические виды, облада-
ющие специфическими адаптациями к суровым условиям среды, не
могут произрастать южнее. Когда в результате антропогенного воздей-
ствия естественные местообитания уничтожаются, они могут рассе-
ляться только в северном направлении; на место арктических видов при-
318 ходят бореальные: идет процесс «бореализации» флоры Севера. Однако
ГЛАВА 9. АНТРОПОГЕННЫЕ ИЗМЕНЕНИЯ ЛАНРАФТОВ ПРИРОДНЫХ ЗОН РОССИИ
он наблюдается только в подзоне южных тундр. Вопрос о том, насколько
возможно внедрение бореальных видов в типичные (а тем более в аркти-
ческие) тундры, остается открытым.
В результате хозяйственной деятельности происходит также синан-
тропизация локальных флор Севера. В первую очередь этот процесс
связан с увеличением во флорах числа рудеральных синантропных
видов, что происходит за счет как заноса рудеральных видов с юга, так
и переселения части аборигенных видов на антропогенные пустоши.
Ядро аборигенной части синантропных флор образуют виды неплакор-
ных местообитаний, приуроченные в естественных условиях к эроди-
рованным, слабозадернованным участкам — береговым обнажениям,
речным и озерным отложениям, осыиным склонам и т. п. Такие виды
называются апофитами. В антропогенно-нарушенных местообитани-
ях может расти от 40 до 60% видов-апофитов, причем подчас обилие и
жизненность растений здесь выше, чем в естественных экотопах.
Несмотря на существенные компенсационные возможности природ-
ной флоры, она несет значительные потери, невозместимые увеличе-
нием разнообразия антропофитов. Систематическая и географическая
структура флоры претерпевает при этом существенные изменения.
Синантропизация флоры ведет к ослаблению ее зональных черт, по-
скольку зарастание свободных субстратов происходит с участием не-
многих широко распространенных рудеральных видов.
Список основных ценозообразователей растительных сообществ
антропогенных местообитаний включает около 120 видов. Их разнооб-
разие возрастает с севера на юг: в полярных пустынях выявлено 8 ви-
дов, в арктических тундрах 28, в северных гипоарктических 46, в юж-
ных гипоарктических — 91 вид. Характерно большое количество общих
видов для всех пунктов, где есть антропогенные нарушения. Иными
словами, в антропогенных сообществах главная роль принадлежит ви-
дам с широким географическим ареалом либо географически замеща-
ющим видам.
Антропогенная трансформация растительности. Под влиянием че-
ловека доминанты типичных тундровых сообществ снижают обилие и
исчезают. В то же время происходит увеличение обилия и видового
разнообразия злаков, этот процесс называют «озлаковением» или «олу-
говением» тундры.
Видовой состав сообществ антропогенных местообитаний зна-
чительно отличается от состава естественных сообществ. Иногда
антропогенные сообщества могут быть даже одно-, маловидовыми,
что крайне редко встречается в естественных экотопах. Простран-
ственная (вертикальная и горизонтальная) структура растительно-
го покрова также меняется. Уменьшается сомкнутость, общее про-
ективное покрытие растительности на сильно нарушенных участках
может быть менее 10%, тогда как в естественных сообществах тундр
сомкнутость варьирует от 50 до 90—100%. Ярусы плохо выражены,
некоторые ярусы вообще отсутствуют (неполный набор ярусов).
В результате происходит «унификация» растительного покрова: фи-
зиономически антропогенные сообщества в различных районах
Крайнего Севера незначительно отличаются друг от друга. 319
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ КАРТИ В А МИРА
Специальные исследования и анализ литературных источников по-
казывают, что на обширных пространствах тундр ведущую роль в фор-
мировании их облика играл выпас северных оленей — сначала диких,
а в последние столетия — и домашних (рис. 9.4).
Рис. 9.4. Оленеводство — традиционное занятие местного населения
Оленеводство развито практически на всем протяжении тундровой
зоны, лесотундры и северной тайги на площади более 3 млн км2. Дикие и
домашние олени используют, особенно в снежный период года, одни и те
же пастбища. Они одинаково адаптированы к пастбищным условиям Се-
вера, но выпас домашних оленей имеет ряд особенностей, определяемых
системой пастбищной «стратегии» и традициями местного оленеводства.
Домашние олени круглый год содержатся в стадах по 500 — 2000 го-
лов и более. Угодья, освоенные домашним оленеводством, периодиче-
ски испытывают нагрузки, превышающие их биологическую емкость.
В большей части районов развития оленеводства пастбищные ресурсы
используются на 80 — 90%. В структуре пастбищ основная роль принад-
лежит продукции злаков, осок, разнотравья и кустарников, тогда как
доля продукции мхов, лишайников и кустарничков относительно не-
велика. Потери лишайников и зеленых частей кустарничков могут
составлять в местах концентрации оленей 60—100% от общего запаса.
Ежегодный прирост надземной фитомассы варьируется на пастбищах
тундровой зоны от 0,3 —0,5 до 2 — 3 т/га в год. В местах расположения
стоянок растительность представлена луговинами, и ее естественный
облик восстанавливается крайне медленно.
Лишайниковые пастбища на значительных территориях уже выби-
ты. Обращает внимание общий процесс делихенизации — деградации
лишайниковой тундры. Помимо неумеренного выпаса исчезновение
лишайников вызывается пожарами, а также воздействием вездеходно-
го транспорта.
Пастбищной дигрессией в настоящее время охвачены огромные
территории тундровой зоны и лесотундры. Прямое и косвенное унич-
тожение растительности на пастбищах, разбивание мохового и торфя-
320 нистого покровов приводят к трансформации гидротермического режи-
ГЛАВА 9. АНТРОПОГЕННЫЕ ИЗМЕНЕНИЯ ЛАНРАФТОВ ПРИРОДНЫХ ЗОН РОССИИ
ма почв и замещению климаксных кустарниково- и кустарничково-
моховых, лишайниковых и других тундр злаково-моховыми, травяно-
моховыми и луговинами.
Наблюдения на полигонах, заложенных в 1930 г. в печорских тунд-
рах, показали, что за прошедшее время в результате перевыпаса возрос-
ли разнообразие и проективное покрытие разнотравья и особенно зла-
ков, увеличилась мощность мохового покрова, но проективное покрытие
лишайников, кустарников и кустарничков снизилось наполовину,
островки леса исчезли, наметилась тенденция к смещению нижней
границы активного слоя.
Назовем основные стадии пастбищной дигрессии тундровых эко-
систем:
1) выпадение из растительного покрова видов, хорошо поедаемых
животными, но обладающих узкой толерантностью, и преоблада-
ние в растительном покрове видов, менее поедаемых или характе-
ризующихся более широкой экологической амплитудой;
2) снижение продукции коренных экосистем, а на поздних стадиях и
их видового разнообразия. Процесс пастбищной дигрессии вызы-
вает также изменение абиотических компонентов экосистем: по-
вышение уровня залегания мерзлоты, развитие солифлюкции и
образование пятен голого грунта. Эти процессы, часто имеющие
необратимый характер, усиливают невысокую устойчивость тундро-
вых экосистем к выпасу скота.
9.3. Зона тайги
Характерными ландшафтами самой обширной по площади таежной
зоны являются хвойные леса и верховые болота.
Таежные леса. Территория средней и особенно северной тайги ста-
ла интенсивно осваиваться в России сравнительно недавно. Основу
хозяйственной деятельности населения долгое время составляли лес-
ные промыслы: заготовка древесины, сбор дикорастущих, охота. Пре-
обладали коренные, не измененные человеком ландшафты.
В первой половине XIX в. население было немногочисленным и кон-
центрировалось преимущественно в долинах рек. Именно здесь стало
развиваться животноводство, а затем и земледелие, поскольку почвы пойм
более плодородны. На месте вырубленных лесов появились пастбища и
пашни. Принципиальные изменения в структуре таежной зоны произо-
шли во второй половине XX в. Объем заготовок древесины необычайно
возрос. Расширилось техническое вооружение лесозаготовителей, появи-
лась сеть автомобильных и железных дорог, благодаря чему значитель-
ная часть территории тайги стала сравнительно легкодоступной. Круп-
ные районы освоения лесных ресурсов охватили практически всю
европейскую часть зоны тайги, значительную часть Урала, большие пло-
щади в Западной, Средней и Восточной Сибири, а также Дальнего Вос-
тока. В связи с расширением геологоразведочных работ, добычей полез-
ных ископаемых даже в труднодоступных местах таежные ландшафты
стали страдать от хозяйственной деятельности человека (рис. 9.5). 321
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ КАРТИ В А МИРА
Рис. 9.5. Покорение тайги. Западная Сибирь.
Сверху вниз: сооружение газопровода; лесозаготовки
Значительный урон тайге наносят пожары, выжигающие большие
площади леса, ценные охотничьи, ягодные и грибные угодья. Вблизи
больших городов леса все больше замещаются лугами и пашнями.
Леса играют важную почвозащитную и водоохранную роль: там, где
они уничтожаются, увеличивается поверхностный сток и как следствие
усиливается смыв почвы, возникают овраги, уменьшается пополнение
подземных вод, реки мелеют.
Следует иметь в виду, что таежная зона играет огромную роль в
насыщении атмосферы кислородом и поглощении углекислого газа.
Один гектар леса выделяет за один год около 1 тыс. м3 кислорода, что
удовлетворяет годовую потребность в нем человека. Подсчитано, что
кислорода, выделяемого лесами тюменского Севера за год, достаточно
для обеспечения потребностей в нем населения всей Тюменской обла-
322 сти, Урала, Казахстана в течение этого времени.
[ЛАВА 9. АНТРОПОГЕННЫЕ ИЗМЕНЕНИЙ ЛАНДШАФТОВ ПРИРОДНЫХ ЗОН РОССИИ
Для примера скажем, что леса США уже не могут обеспечить кис-
лородом население этой страны. В промышленно развитые страны, где
площадь лесов сильно сокращена, кислород приносится воздушными
потоками из сохранившихся лесных ландшафтов, в том числе с терри-
тории таежной зоны России.
Верховые (сфагновые) болота таежной зоны. Этот уникальный при-
родный комплекс (рис. 9.6) долгое время рассматривался с хозяйствен-
ной точки зрения как источник торфа на топливо или (после осуше-
ния) как сельскохозяйственные угодья. На протяжении последних
десятилетий на болота шло массированное наступление. Однако оказа-
лось, что экономическая польза от осушения болот очень небольшая, а
отрицательные последствия огромны.
Рис. 9.6. Грядово-озерковые болота западносибирской тайги. Аэрофотоснимок
Присущая болотам естественная растительность через несколько лет
после осушения погибает, и болота превращаются в бесплодные пусто-
ши. Слой торфа быстро срабатывается. За год его толщина уменьшает-
ся в среднем на 1 — 2 см; максимальная скорость достигает 12 см в год.
Двух-трехметровый торфяной слой срабатывается до минерального
грунта через 10 — 20 лет после осушения. В Белорусском Полесье, быв-
шем крае болот, теперь случаются пыльные бури, в воздух поднимают-
ся черные тучи пересушенного торфа.
Верховые болота — настоящие кладовые чистой воды. Пройдя через
сфагновый ковер, вода становится стерильной, поскольку эти мхи —
хорошие антисептики. Кроме того, экосистема болот является геохи-
мическим фильтром, задерживающим различные вещества, в том числе
тяжелые металлы (свинец, ртуть, кадмий и т. п.).
Особенно велика фильтрующая роль болот вблизи поселений и в
зонах отдыха. Вода болот — это огромный экологический и экономи-
ческий потенциал, ценность которого со временем будет возрастать.
Осушение болот ведет к нарушению режима питания рек: мелеют и
исчезают вытекающие из болот ручьи и малые реки — истоки крупных 323
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ К АРТИ В А МИРА
рек. Снижается уровень грунтовых вод на прилегающих к осушенным
болотам территориях. Усыхают леса, сокращается разнообразие болот-
ных растений, животных, количество перелетных птиц. Прекращаются
традиционные промыслы — сбор ягод, лекарственных трав, охота.
В XXI в. тенденции роста лесозаготовок, отчуждения ландшафтов
таежной зоны на строительство городов и промышленных объектов бу-
дут сохраняться. Осушение и загрязнение болот приведет к обмелению
рек, ухудшению качества природных вод. Чтобы избежать неблагопри-
ятных экологических последствий, человек должен взять на себя затра-
ты на восстановление ресурсного потенциала таежных ландшафтов.
И 9.3. зона смешанных и широколиственных лесов______________________
Эта зона обладает плодородными почвами и мягким климатом. Не
случайно многие дворянские усадьбы, в том числе родовые имения ве-
ликих русских писателей, поэтов и композиторов, располагались именно
в этой зоне. Она была заселена раньше и с большей плотностью, чем
другие зоны России. Уже в начале прошлого века пашни и пастбища за-
нимали здесь большую площадь, чем лесные угодья (рис. 9.7).
Рис. 9.7. Сельскохозяйственные угодья на месте ландшафтов смешанных
и широколиственных лесов
В XX в., особенно во второй его половине, разрушение естествен-
ных ландшафтов нарастало. Значительное развитие получили техноген-
ные промышленно-городские комплексы. Реликты естественных эко-
систем сохранились лишь в немногих заповедниках.
Отдаленная перспектива этой зоны рисуется в следующем виде:
господствующее место займут антропогенные комплексы, на которые
будут переложены как ресурсовоспроизводящие функции, так и средо-
324 регулирующие. Поскольку в противоположность естественным экоси-
ГЛАВА 9. АНТРОПОГЕННЫЕ ИЗМЕНЕНИЯ ЛАНДШАФТОВ ПРИРОДНЫХ ЗОН РОССИИ
стемам, где эти функции выполнялись «бесплатно», антропогенные
комплексы полностью управляются людьми, заботу о поддержании
благоприятного экологического режима в антропогенных ландшафтах
человеку придется взять на себя. Долю экологических затрат в эконо-
мике нужно будет резко увеличить.
И 9.5. Лесостепная и степная зены________________________________
Заселение лесостепной и степной зон произошло очень давно, поч-
ти одновременно с освоением зоны широколиственных лесов. Вся
послеледниковая история степей — это в то же время и история отно-
шений между человеком и природой. Начало земледелия было положе-
но в широколиственном лесу, потому что в степях первобытный чело-
век с его примитивными орудиями не мог бороться против травянистой
растительности, заглушающей посевы. Оседлые племена использовали
древесину в качестве строительного материала, топлива, для обжига
керамики и плавки металла. Основу сельского хозяйства составляло
подсечно-огневое земледелие. В конце голоцена произошло значитель-
ное сокращение площади лесов, их граница отступила на север.
Ко времени образования Киевской Руси девственные лесостепные
ландшафты уже отсутствовали. В то же время леса служили местами
укрытия от набегов кочевников и играли огромную оборонительную
роль. Поэтому в пограничных районах лесные массивы всячески обере-
гались. Кочевники, наоборот, выжигали леса, чтобы подорвать оборону
русских и расширить участки с травянистой растительностью для ско-
та. В результате противоположного отношения к растительности рус-
ских племен и кочевников леса и луга (степи) в лесостепи неоднократно
сменяли друг друга.
С середины XVI в. после освобождения от монголо-татарского ига
начались укрепление и рост Московского государства. Интенсивность
хозяйственной деятельности и вырубка лесов резко возросли.
Конец XVIII в. стал поворотным этапом в освоении южных окраин
России. В результате присоединения Крыма страна получила свободный
выход к Черному морю и начала осваивать богатства вновь обретенного
края. Быстрая колонизация южнорусских степей сопровождалась пересе-
лением в новые районы крестьян и городского населения из северных
губерний, постройкой городов и селений, распашкой бескрайних степных
просторов. Естественных местообитаний становилось все меньше. Исче-
зали многие характерные представители фауны степных ландшафтов.
Изучению процессов восстановления широколиственных лесов или
степной растительности в местах, где они были уничтожены или силь-
но трансформированы в результате хозяйственной деятельности чело-
века, посвящена обширная литература. Однако вопрос о том, насколько
полно может возродиться биоразнообразие биома коренного леса, лу-
говых или настоящих степей, остается открытым.
О возможности восстановления лесов можно судить исходя из того,
какое место занимали леса в естественных ландшафтах юга Русской
равнины. Оригинальную картину естественных ландшафтов и динами- 325
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ КАРТИ В А МИРА
ки растительного покрова под влиянием естественных и антропоген-
ных факторов рисует Ф.Н. Мильков (1995). Оригинальность подхода со-
стоит в том, что среди естественных ландшафтов южнорусских степей
выделяются саванны. Ф.Н. Мильков считает обоснованным рассматри-
вать саванны как мегатип ландшафта, состоящий из трех типов: тро-
пических, субтропических и северных (неморальных) саванн. В пользу
этого, по мнению Ф.Н. Милькова, говорят:
1) общность фитоценотической структуры: и там и здесь покров ксе-
рофильных злаков с одиночными деревьями, реже — группами де-
ревьев;
2) общность в сезонной ритмике ландшафтов: и там и здесь летний
сезон — пора активной вегетации растительности; зимний — время
покоя, когда деревья теряют листву, вызванное в тропиках отсутстви-
ем осадков, в умеренном поясе — низкими температурами воздуха;
3) высокое плодородие почв;
4) большая, часто ведущая роль в формировании тропических и север-
ных саванн антропогенного фактора.
Разнотравно-луговые саванны являются одной из стадий сукцес-
сионного ряда, ведущего к восстановлению лесной растительности.
В 1968—1969 гг. под руководством Ф.Н. Милькова было осуществлено
ландшафтное картирование Каменной степи, в том числе ее степных
залежей. Почти через 20 лет, в 1986— 1987 гг., по той же методике было
проведено повторное картирование. Практически на всех некосимых
залежах сформировались участки настоящих лесов, местами непрохо-
димых чащ, высотой до 15—18 м, почти или полностью лишенных тра-
вяного покрова. В древостое принимают участие клен остролистный
{Acerplatanoides), клен американский {Acernegunda), вяз гладкий {Ulmus
laevis), ясень обыкновенный {Fraxinus excelsior), груша обыкновенная
{Pyrus communis), яблоня лесная {Malus silvestris), черемуха {Padus
racemosa), рябина {Sorbus aucuparia), жимолость татарская {Lonicera
tatarica), крушина обыкновенная (Rhamnus catbartica), боярышник
однопестичный {Crataegus monogina), боярышник коричный {Crataegus
oxycantha), шиповник {Rosa canina).
Коренным типом растительности плакоров типичной и южной сте-
пи были плодовые саванны: одиночные деревья груш и яблони в сопро-
вождении колючих кустарников — терна, боярышника, шиповника.
Деревья и кустарники как наиболее уязвимая структурная часть есте-
ственной растительности исчезали при сенокошении и неумеренной
пастьбе скота, не говоря уже о распашке земель. Консервативной сину-
зией оказывалась степь. Однако на защищенных от антропогенного
воздействия участках степи и даже залежах плодовая саванна начинает
возрождаться.
Черноземы — самые плодородные почвы. Из 300 млн га чернозем-
ных почв мира 100 млн га находится в России. Они дают 80% всей сель-
скохозяйственной продукции. В настоящее время степная зона стала
полностью земледельческой. В некоторых районах пашни занимают 70%
территории, а местами и более (рис. 9.8).
Роковым для степных ландшафтов было принятое в 1954 г. решение
32В советского правительства, направленное на освоение целинных земель
[ЛАВА 9. АНТРОПОГЕННЫЕ ИЗМЕНЕНИЙ ЛАНДШАФТОВ ПРИРОДНЫХ ЗОН РОПСИН
Рис. 9.8. Вид из космоса на степи Среднего Поволжья.
Видна плотная мозаика прямоугольников пахотных угодий
в Зауралье — на юге Западной Сибири и в Северном Казахстане. Нача-
лась беспощадная охота за степной целиной. Во время этой эпопеи не
было создано ни одного заповедника. Наоборот, уже существующие
были закрыты, а их территории распаханы. Обработке подвергались
даже клочки девственных степей на научных стационарах. В результа-
те осуществления этого проекта степной ландшафт был полностью унич-
тожен и заменен на пахотные угодья.
Превращение степного ландшафта в пахотные угодья влечет за со-
бой негативные экологические последствия. Темная почва сильнее на-
гревается солнечными лучами. Это ведет к аридизации климата всей
степной зоны, вызывая высыхание некогда многочисленных малых рек
и озер. 327
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ КАРТИ В А МИРА
К неблагоприятным явлениям в степной зоне относятся засухи, су-
ховеи и пыльные бури. Массированная распашка земель, потеря гумуса
вызывают истощение почв. Плодородный гумусный слой может быть
унесен с пашни одной сильной бурей (рис. 9.9). Поистине планетарный
характер имели пыльные бури середины 60-х годов XX в., пронесшиеся
над степями СССР.
Незащищенная почва
Защищенная почва
Разбрызгивание
Пахотный слой
Подпочва
Вегер
Поверх ностный
сток
Рис. 9.9. Роль степной растительности в защите почвенного слоя:
тонкие частицы гумуса, глины и пыли легко уносятся потоками воды
или ветром, оставляющими только грубый материал.
Растительный покров защищает почву от всех видов эрозии
Отсутствие степного покрова подорвало, а местами полностью унич-
тожило запасы целинных охотничье-промысловых видов фауны. Пре-
данием стала увлекательная охота на степную дичь: дрофу, стрепета,
серую куропатку, перепела, кречетку. Некогда сплошной ареал сурка
распался на ряд изолированных популяций. Сайгак оттеснен в полупус-
тынную зону.
К неблагоприятным социально-экономическим последствиям освое-
ния целины следует отнести бедственное положение зерновых хо-
зяйств, созданных на востоке страны в сухостепной зоне. Урожайность
зерновых составляет здесь 3,5 —8,5 ц с гектара и неравномерна по го-
дам, в то время как экономически оправдана урожайность — 7,5 ц с
328 гектара. Большинство зерновых хозяйств в этой зоне убыточны. С на-
та 8. АНТРОПОГЕННЫЕ ИЗМЕНЕНИЯ ЛАНДШДЙТаВ ПРИРОДНЫХ ЭОН РОССИИ I
чалом проведения экономических реформ положение хозяйств стало
еще более тяжелым, что вызвало волну миграции населения из целин-
ных районов.
Немаловажный моральный аспект освоения целины — это отсут-
ствие у переселенцев естественно-исторических традиций степного при-
родопользования. Уничтожение степного ландшафта, истребление фло-
ры и фауны поставили целинные регионы на грань экологической
катастрофы. Так расправляться с природой может только чужеземец.
За несколько десятилетий освоения степная природа Заволжья, Урала
и Казахстана деградировала сильнее, чем средняя полоса России за
столетия хозяйственного освоения.
И 9.6. Полупустынная и пустынные зоны
Прикаспийская низменность — единственная область в европей-
ской части России, где степная зона сменяется зоной полупустыни; по-
следняя широкой полосой продолжается к востоку, образуя северное
обрамление азиатских пустынь. Именно в сухих (аридных) зонах наи-
более ярко проявляются признаки опустынивания.
Аридные ландшафты всегда оставались малозаселенными. Большая
часть населения группировалась в оазисах, долинах рек и в предгорьях.
Традиционной для полупустынь и пустынь формой хозяйства явилось
пастбищное скотоводство.
В настоящее время в структуре аридных ландшафтов произошли
заметные изменения. Осуществлены крупные программы по орошению
засушливых и пустынных земель. Пастбища стали использоваться с
большей нагрузкой. Разрушительное воздействие на пустынные ланд-
шафты оказывают геологоразведка, добыча газа и нефти, минерально-
го сырья, строительство дорог, нефте- и газопроводов, новых городов и
поселков (рис. 9.10).
Рис. 9.10. На строительстве газопровода
329
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ КАРТИ В А МИРА
В ландшафтах полупустыни и пустыни процессы опустынивания
происходят особенно быстро. Остановимся на некоторых примерах.
Уничтожение растительного покрова. Пустынные пастбища, харак-
теризующиеся низкой продуктивностью, особенно сильно страдают от
длительного выпаса скота. При перегрузке пастбищ происходит обедне-
ние растительности и как следствие — опустынивание ландшафта.
Ветровая эрозия. Ветер развевает песчаные толщи, лишенные рас-
тительного покрова. Рыхлый материал переносится и отлагается на
огромных пространствах. Ветровая эрозия может иметь площадное
равномерное и очаговое проявления.
В обоих случаях ее результатом являются разнообразные формы
аккумулятивного рельефа (барханы). Последние характеризуются под-
вижностью, тенденцией захвата новых территорий (рис. 9.11).
Засоление почв. Наблюдается в районах интенсивного орошаемого
земледелия. В результате длительного использования одних и тех же
земель с избыточным поливом поднимается уровень грунтовых вод. При
прохождении воды через грунты содержание солей в ней увеличивает-
ся до 3—10 г/л, и после испарения влаги соль остается на поверхности.
На орошаемых землях в условиях жаркого климата пустыни с одного
гектара испаряется до 10 000 м3 воды, при этом в верхнем слое почвы
может накопиться до 20 т солей.
Рис. 9.11. Барханы наступают на поселок
330
[ЛАВА 9. АНТРОПОГЕННЫЕ ИЗМЕНЕНИЙ ЛАНДШАФТОВ ПРИРОДНЫХ ЗОН РОЕСИН
В бывшем СССР проблема Аральского моря явилась одной из пер-
вых, всколыхнувших общественное мнение. Вслед пропагандистской
шумихе о могуществе советского человека, способного создавать ру-
котворные моря и поворачивать течение рек, пришлось признать, что
неумелое хозяйствование привело к одной из крупнейших экологиче-
ских катастроф.
Питающие Аральское море реки Амударья и Сырдарья берут начало
в горных ледниках. Далее, от подножия гор на север, реки несут свои
воды через пустынные ландшафты Средней Азии. С давних пор долины
Амударьи и Сырдарьи были цветущими оазисами. Воды хватало и на
орошение полей, и на поддержание баланса вод в Аральском море. Но
десятилетия социалистического строительства сопровождались бурным
развитием орошаемого земледелия. Длина оросительных каналов до-
стигла тысяч километров. Сток рек в Аральское море практически пре-
кратился. Некогда цветущие дельты стали подвергаться процессам опу-
стынивания. На дневную поверхность вышли тысячи квадратных
километров бывшего морского дна. Сравнение карты Аральского моря,
помещенной в Большой советской энциклопедии в 1970 г., с космиче-
ским снимком, сделанным в 1996 г., показывает, как сильно сократи-
лась площадь моря и увеличилась площадь солончаковых пустынь по
его берегам (рис. 9.12).Основные тренды изменения природных и
социально-экономических условий в бассейне Аральского моря показаны
на рис. 9.13.
Рис. 9.12. Усыхание Аральского моря:
а — карта Аральского моря, по БСЭ, 1970 г;
б — Аральского моря из космоса, 8 октября 1996 г.
331
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ КАРТИ В А МИРА
Рис. 9.13. Основные тренды изменения природных
и социально-экономических условий в бассейне Аральского моря,
по 3. Бреклю, О.Е. Агаханянцу и В. Вухереру
С 1920 г. наблюдается устойчивое снижение уровня моря и умень-
шение площади акватории, темпы которого особенно ускорились после
1960 г. Наряду с этим стала быстро увеличиваться соленость морских
вод, что привело к резкому уменьшению некогда богатых рыбных за-
пасов (рыбы Аральского моря не способны жить в соленой воде). Пло-
щадь орошаемых земель поначалу быстро увеличивалась, однако после
1980 г. происходит их сокращение. Наметилась тенденция к гибели
целых поселков, снижению плотности населения.
Прирост населения остается традиционно высоким, но он таит в себе
угрозу тяжелых социально-экономических потрясений: отмечаются вы-
сокая детская смертность, снижение продолжительности жизни и ее низ-
кий уровень. Нарастает кризис поливного земледелия; встают проблемы
обеспечения населения чистой питьевой водой и продуктами питания.
Хозяйственное освоение аридных территорий будет продолжаться в
XXI в. Есть все основания предполагать, что значительное место в струк-
туре полупустынных и пустынных ландшафтов займут антропогенные
экологические комплексы: поля и сады, окультуренные пастбища, искус-
ственные насаждения древесно-кустарниковой растительности.
Человеку придется самому поддерживать благоприятный экологи-
ческий (в первую очередь водный) баланс экосистем, что потребует
332 больших материальных затрат.
Глава 10
ЗОНАЛЬЛЫЕ ТИПЫ БИОМОВ
НООЕРХНВЕТНПЙ ТОЛЩИ ОКЕАНА
Зональные типы биомов — характерные сочетания биогидроцено-
зов в поверхностной толще океана — формируются при определенном
комплексе — климатических, гидрологических, биологических и других
характеристик. Их границами являются рубежи, за пределами которых
экосистемы данного зонального типа не могут существовать из-за не-
соответствия условиям среды.
В системе единиц географической зональности поверхностной тол-
щи Мирового океана так же, как на суше, различаются пояса и зоны
(см. гл. 7). В каждом полушарии выделяются холодный и умеренный
пояса, между ними располагается жаркий пояс. Различный температур-
ный режим названных поясов обусловливает своеобразие флоры и
фауны в их пределах.
Жаркий пояс. В экваториальных широтах положительный в тече-
ние всего года радиационный баланс дополняется большим притоком
тепла конденсации. В тропических широтах становится заметным се-
зонное колебание радиационного баланса.
Умеренные пояса. Их границей с жарким поясом служит годовая
изолиния радиационного баланса 100 ккал/см2 в год. Резко выражен
сезонный ход радиационного баланса. Зимой радиационное выхолажи-
вание тормозится отдачей тепла океанами, адвекцией тепла и освобож-
дением тепла при выпадении осадков. Среди названных источников
нагревания атмосферы главенствующее положение занимают океаны:
приток явного тепла от водной поверхности составляет 50% радиацион-
ного баланса.
Холодные пояса. Их границей с умеренными поясами служит годо-
вая изолиния радиационного баланса 20 ккал/см2 в год. Над полярными
областями годовой радиационный баланс отрицательный. Баланс тепла
поддерживается благодаря горизонтальному переносу тепла воздушны-
ми массами. Перенос тепла океаническими течениями сильно ослаблен
ледяным покровом.
Каждый пояс характеризуется атмосферной циркуляцией с выра-
женной широтной составляющей скорости ветра: в жарком поясе пре-
обладает восточная составляющая ветра — пассаты, в умеренном по-
ясе — западный перенос, в холодном вновь появляется восточная
составляющая ветра — полярный восточный перенос.
333
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ К АРТИ В А МИРА
Перейдем к краткой характеристике географических зон и свой-
ственных им биомов на поверхности Мирового океана. За основу взята
система единиц, выделенная по термическим условиям (см. гл. 7). Опи-
сания составлены на основании работ Д.В. Богданова (1961, 1963, 1978,
1985), В.Г. Богорова (1960), С.В. Калесника (1970), Атласов океанов (1974,
1977, 1980)и др.
И 1D.1. Биомы полярных по довитых морей___________________________
Ведущим фактором формирования биомов полярных ледовитых мо-
рей служит холод, обусловленный малым количеством солнечной радиа-
ции (баланс менее 10 ккал/год/см2), падением солнечных лучей под косым
углом, большой отражающей способностью льда и снега, длительной по-
лярной ночью. В летнюю половину года солнце не заходит за горизонт, но
из-за малого угла падения и большой отражающей способности подстила-
ющей поверхности около 90% солнечной радиации отражается, не преоб-
разуясь в тепло. В зимнюю половину года солнечная радиация вообще не
поступает в полярные зоны. Океан и суша теряют тепло, излучая его в
пространство. Таким образом, в целом за год радиационный баланс отри-
цательный; приход тепла значительно меньше его расхода.
Северная зона полярных ледовитых морей. Зона, которая называ-
ется еще арктической, занимает среднюю часть Северного Ледовитого
океана, так называемый Центральный полярный бассейн, а также боль-
шую часть эпиконтинентальных морей Евразии (Карского, Лаптевых,
Восточно-Сибирского, Чукотского) и Северной Америки. В Южном
полушарии соответствующие широты заняты антарктической зоной.
Рассмотрим физико-географические особенности арктической
зоны. Это бассейн, окруженный материками, своего рода тупик, окон-
чание циркуляционных потоков Мирового океана. Он имеет более или
менее широкую связь с Атлантическим океаном через Гренландское и
Норвежское моря. Однако и здесь имеются мелководные пороги, за-
трудняющие циркуляцию. Связь Северного Ледовитого океана с Тихим
осуществляется через узкий и мелководный Берингов пролив.
Зима в Арктике суровая, особенно в восточном секторе, между Тай-
мыром и морем Бофорта. Температура воздуха от —25 до —30 °C. Часты
морозы в — 40, — 50 °C. Температура летом обычно около 0 °C. Однако глу-
бокого выхолаживания Северного Ледовитого океана не происходит, по-
тому что в него проникают из Атлантики самые дальние струи теплого
Гольфстрима. С этим связан один из парадоксов Центрального полярно-
го бассейна: его поверхностные воды холодны, скованы льдом, а на глу-
бине 200 — 800 м лежит теплая (0 — 2,5 °C) и соленая (35%) толща атлан-
тических вод.
В целом в температурной и соленостной вертикальной структуре
Северного Ледовитого океана выделяется поверхностный слой холод-
ной (до —1,7 °C) и распресненной (до 30%, в Азиатском секторе до 15%)
воды, промежуточный слой более теплой и соленой атлантической воды
и, наконец, более холодный придонный слой. Наличие распресненного
334 поверхностного слоя обусловливает устойчивую стратификацию (в дан-
ГЛАВА 10. ШАЛЬНЫЕ ТИПЫ БИОМОВ ПОВЕРХНОСТНОЙ ТОЛЩИ ОКЕАНА
ном случае — несмешиваемость) вод даже зимой, когда образование
льда несколько повышает соленость. Плохое конвективное перемеши-
вание затрудняет подъем из глубин богатых питательными солями вод.
Низкое плодородие наряду с суровыми климатическими условиями яв-
ляется причиной низкой продуктивности вод арктической зоны, круп-
ные рыбы отсутствуют— «безрыбное море».
С наступлением зимних холодов происходит интенсивное ледообразо-
вание. Каждый год появляются новые льды толщиной около 3 м. Летний
прогрев воды невелик. Среднегодовая температура воды ниже О °C. В ре-
зультате ежегодно льда образуется больше, чем тает. В течение всего года
океан покрыт тяжелыми многолетними льдами: паковыми, торосистыми.
Избыток льда выносится течениями за пределы арктической зоны. Льды
движутся с востока на запад и выносятся в Северную Атлантику, главным
образом Восточно-Гренландским течением.
Казалось бы, все физико-географические процессы в Северном Ледо-
витом океане должны строго подчиняться формуле зональности, из кото-
рой следует, что на севере ледовитость должна быть больше, а на юге
меньше. Однако оказывается, что наряду с зональным размещением ланд-
шафтов, гармонично вписывающимся в эту схему, выделяются секторы с
азональным размещением водно-ледовых ландшафтов, что создает пара-
доксальные контрасты, объяснение которым нашел В.Н. Купецкий (1961).
В Северном Ледовитом океане расположены четыре центра действия
атмосферы (рис. 10.1). Два центра высокого давления— один между
полуостровами Ямал и Чукотка, второй между Гренландией и Аляской.
По периферии этих центров ветры дуют по часовой стрелке. Два центра
низкого давления — между Гренландией и полуостровом Ямал и между
Чукоткой и Аляской. Здесь ветры дуют против часовой стрелки.
Рис. 10.1. Четыре центра действия атмосферы в Северном Ледовитом океане:
два — высокого (В) и два — низкого (Н) давления, по В.Н. Купецкому 335
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ КАРТИ В А МИРА
В областях взаимодействия центров высокого и низкого давления
создаются условия зонального или азонального размещения водно-ле-
довых ландшафтов.
Зональное размещение водно-ледовых ландшафтов характерно для
секторов Северного Ледовитого океана, где господствующие ветры по
периферии центров высокого и низкого давления направлены в сторо-
ну океана (рис. 10.2). Эти ветры отгоняют дрейфующие льды от берега,
в результате чего на юге арктических морей формируются стационар-
ные полыньи — Сибирская и Канадская, а в океане, в областях прино-
са — Сибирский и Канадский океанические ледяные массивы.
Рис. 10.2. Схема формирования водно-ледовых ландшафтов зонального (А)
и азонального (А1) типов, по В.Н. Купецкому
Азональное размещение ландшафтов характерно для секторов, где
господствующие ветры по периферии центров высокого и низкого дав-
ления направлены в сторону материка — на юг. Эти ветры нагоняют
дрейфующие льды к берегу, в результате чего над мелководьями аркти-
ческих морей формируется мощный ледовый массив, а на севере, вдали
от берега — полынья.
Таким образом, моря Карское, Лаптевых и Бофорта имеют зональ-
ное распределение ландшафтов, а в Восточно-Сибирском и Чукотском
морях размещение ландшафтов азональное.
Ледовые ландшафты отличает максимальная изоляция атмосферы
от гидросферы, минимальная степень взаимодействия между водой и
воздухом, антициклональный характер погоды, заторможенная верти-
кальная циркуляция, угнетенное состояние всей биологической цепи
растительных и животных организмов на поверхности моря, на мор-
ском дне и на близлежащих побережьях, максимальная сложность для
33В работы ледокольного флота и проводки транспортных судов.
[ЛАВА 10. ШАЛЬНЫЕ ТИПЫ БИОМОВ ППВЕРХННСТНОЙ ТОЛЩМ ОКЕАНА
Водным ландшафтам среди льдов — полыньям — свойственна мини-
мальная изоляция атмосферы от гидросферы, максимальная степень
взаимодействия между водой и воздухом, циклональный характер по-
годы, наиболее интенсивная вертикальная циркуляция вод вплоть до са-
мых больших глубин, наилучшая их аэрация, расцвет всех звеньев био-
логической цепи от микроорганизмов до самых крупных на планете
позвоночных — китов, максимальная продуктивность органической
жизни на поверхности моря, на морском дне и на близлежащих побе-
режьях, наиболее благоприятные условия для ледокольного флота.
Флора и фауна Северного Ледовитого океана, несмотря на весьма
обширные его размеры, как по видовому составу, так и по биомассе,
бедны. Особенно заметно обеднение жизни при продвижении к вос-
току от Атлантического океана, к берегам Азии и Канады. Мелково-
дья лишены литоральной фауны и флоры. Из-за длительной полярной
ночи и ледового покрова процесс фотосинтеза возможен в течение
крайне непродолжительного периода и только для диатомовых водо-
рослей. Своеобразным биологическим приспособлением является
обрастание диатомовыми водорослями нижней поверхности многолет-
них льдов. В целом в арктической зоне процессы первичного продуци-
рования выражены в минимальной для Мирового океана степени —
«биологический минимум».
В арктической зоне расположены остров Гренландия и несколько
архипелагов. Большая их часть покрыта ледниками, а свободные ото льда
участки представляют собой арктические пустыни. Обитающие по бере-
гам животные— птицы, ластоногие, полярный медведь (рис. 10.3) —
питаются преимущественно морскими организмами. Основные взаимо-
связи компонентов природы в арктической зоне показаны на рис. 10.4.
Рис. 10.3. Полярные (белые) медведи
Бедность жизни вод арктической зоны объясняется не только суро-
востью природных условий, но и особенностями ее палеогеографиче-
ского развития и географического положения. В третичный период арк-
тические широты Северного полушария характеризовались теплым
климатом, океан был свободен ото льда. Резкое похолодание и оледене- 337
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ КАРТИ В А МИРА
Рис. 10.4. Основные взаимосвязи компонентов природы в арктической зоне
ние, наступившие в начале четвертичного периода, стали экологической
катастрофой для теплолюбивых биоценозов пелагиали, шельфа и бере-
гов арктических морей. К этому нужно добавить, что в четвертичном
периоде Северный Ледовитый океан подвергался опреснениям. Значи-
тельная часть древней фауны вымерла. Формирование биоты арктиче-
ской зоны началось практически заново около 2,5 млн и закончилось
500 тыс. лет назад, в плейстоцене. Тогда возникли эндемичные аркти-
ческие виды. Центральная часть Северного Ледовитого океана не игра-
ла никакой роли в обогащении пелагических и донных биоценозов мел-
ководной арктической зоны.
Южная зона полярных ледовитых морей. Зона, которую еще назы-
вают антарктической, охватывает воды Южного океана, омывающего
Антарктический материк. Асимметрия Северного и Южного земного
пространства (на севере океан обрамлен материками, на юге материк
окружен океаном) накладывает свой отпечаток на особенности приро-
ды зон в Северном и Южном полушариях.
Антарктический континент является «главным холодильником»
планеты и сильно охлаждает все Южное полушарие. Поэтому термиче-
ский экватор расположен севернее географического, а все зоны на по-
верхности океанов в Южном полушарии оказываются сдвинутыми в
338 более низкие широты по сравнению с зонами Северного полушария.
ГЛАВА 10. ШАЛЬНЫЕ ТИПЫ БИОМОВ ПОВЕРХНОСТНОЙ ТОЛЩИ ОКЕАНА
Акватория Южного океана, лежащая в антарктической зоне, покры-
та морскими льдами. Большие пространства материковой отмели зани-
мают шельфовые ледники, простирающиеся иногда до континенталь-
ного склона. У края шельфовых ледников зарождаются обширные
столообразные айсберги. Объем воды, заключенной в самых больших
айсбергах, так велик, что мог бы обеспечить годовой сток Волги.
В результате осенне-зимнего охлаждения воды и ледообразования
формируются большие массы переохлажденной воды с соленостью,
близкой к нормальной. Эти воды погружаются и движутся в северном
направлении вдоль дна, заполняя глубокие котловины Атлантического,
Индийского и Тихого океанов. На движение антарктической придон-
ной воды большое влияние оказывает рельеф (рис. 10.5.). По мере дви-
жения в северном направлении вода скапливается у преград, образуе-
мых подводными хребтами, находит расселины и устремляется по ним
бурными подводными потоками.
Рис. 10.5. Влияние рельефа Срединно-Атлантического хребта на движение
антарктической придонной воды, по С. Нешибе
339
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ КАРТИ В А МИРА
Роль Северного Ледовитого океана в питании глубин остальных океа-
нов холодными водами невелика, т. к. он имеет очень узкий выход в
Тихий океан и отделен от Атлантического океана мелководным порогом.
Антарктические воды несут в глубины океанов кислород, что делает
возможным существование животных на дне самых глубоководных
впадин. Вместе с тем они несут холод — температура придонных вод во
всех океанах, даже в тропических широтах, колеблется около 1 °C.
Низкие температуры над Антарктидой способствуют формированию
здесь области высокого атмосферного давления. С ледяного купола сто-
ковые ветры дуют на север. Под влиянием силы Кориолиса они откло-
няются на запад. Их воздействие распостраняется на восточное антар-
ктическое прибрежное течение. Динамика вод в антарктической зоне
осложняется рядом циклонических вихрей, из которых наиболее выра-
жен циклонический круговорот моря Уэддела. Конвективное переме-
шивание вод, циклонический апвеллинг способствуют подъему глубин-
ных вод и обогащению зоны фотосинтеза биогенными солями, что
определяет высокую продуктивность антарктической зоны.
Несмотря на низкие температуры воды, жизнь в океане богата и
разнообразна. Это объясняется также палеогеографическими условия-
ми и особенностями географического положения антарктической зоны.
Начало образования Южного океана — открытого водного простран-
ства, окружающего Антарктиду, относится к раннему олигоцену (36 млн
лет назад), когда в процессе распада Гондваны стал обособляться Антар-
ктический континент. Последней, 14 млн лет назад, от него отделилась
Южная Америка, в результате чего открылся пролив Дрейка.
Кольцевая система течений, сформировавшаяся в Южном океане,
изолировала южное полярное пространство от проникновения тепла с
севера, что способствовало его выхолаживанию и оледенению Антарк-
тиды. Таким образом, формирование пелагических и донных сообществ,
приспособившихся к суровым условиям Антарктической зоны, проис-
ходило на протяжении более десятка миллионов лет.
Южный океан широко открыт в Атлантический, Индийский и Ти-
хий океаны. Это способствует проникновению в его воды представите-
лей фауны и флоры из богатых жизнью тропических широт. В резуль-
тате длительной адаптации к суровым условиям антарктической зоны в
Южном океане сформировались относительно разнообразные в видовом
отношении и богатые по биомассе пелагические и донные сообщества.
Антарктическая зона на побережье ледяного материка бедна жиз-
нью, однако благодаря своеобразию она относится к особой Антаркти-
ческой биогеографической области. Животный мир Антарктиды сосре-
доточен в редких оазисах, участках суши, летом свободных ото льда и
снега, и на кромке прибрежных льдов. Именно здесь живут единствен-
ные млекопитающие — тюлени и гнездятся пингвины; самый распро-
страненный — пингвин Адели; самый крупный — императорский пинг-
вин (рис. 10.6).
Поразительным в образе жизни императорских пингвинов является
то, что период их размножения приходится на суровую антарктиче-
скую зиму. В полной темноте, в ледяную пургу стоят эти птицы на снегу,
340 согревая яйца, из которых лишь после двух месяцев насиживания вы-
[ЛАВА 10. ШАЛЬНЫЕ ТИПЫ БИОМОВ ППВЕРХННСТНОЙ ТОЛЩМ ОКЕАНА
Рис. 10.6. Императорские пингвины
лупляются птенцы. Самки в это время вынуждены голодать по мень-
шей мере месяц, самцы — три, а часто и дольше. Зимой императорский
пингвин — единственная птица Антарктиды. Летом здесь гнездятся
поморники, чайки, крачки, качурки, буревестники. Все позвоночные
Антарктиды добывают пищу из моря, которое, в противоположность
суше, характеризуется богатством и высокой продуктивностью.
и 10.2. БИОМЫ субполярным ЗОН_______________________________________
Северная субполярная (субарктическая) зона включает Северную
Атлантику, южную часть Баренцева моря и северную окраину Тихого
океана — Берингово море. В южном океане субполярная, или субантарк-
тическая, зона лежит между 50 — 70° ю. ш. Характерным признаком суб-
полярной зоны является сезонный характер ледового покрова: зимой
море сковано льдом, летом частично или полностью освобождается ото
льда. Температура воды на поверхности летом колеблется от 0 до 10 °C.
Зимние погодные условия в субполярных зонах сходны с теми, что
наблюдаются в полярных зонах. Температура воздуха падает от —20
до —40 °C. При замерзании морской воды большая часть содержащих-
ся в ней солей вытесняется: морской лед пресный, с включениями ка-
пель высокосоленого рассола. В результате изъятия из океана пресной
воды, связываемой в виде льда, соленость поверхностных вод повыша-
ется. Итак, зимой поверхностные воды в субарктической зоне охлаж-
даются до —1, —2 °C, их соленость повышается до 35%о, увеличивается
плотность. Это вызывает погружение поверхностных вод. Так развива-
ется зимнее конвективное их перемешивание. Оно способствует пере-
носу кислорода с поверхности в глубины океана и выносу биогенных
солей из глубины к поверхности.
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ КАРТИ В А МИРА
Своеобразная циркуляция вод, характерная для субарктической
зоны, особенно ярко выражена на севере Атлантики. Теплые течения
системы Гольфстрим, прижимающиеся к берегам Скандинавии, взаи-
модействуют здесь с холодными течениями, идущими из Северного
Ледовитого океана и прижимающимися к берегам Гренландии и Кана-
ды (Восточно-Гренландское, Восточно-Исландское, Лабрадорское). В ре-
зультате в океане образуется циклонический круговорот (против ча-
совой стрелки), порождающий токи воды из глубины к поверхности.
Аналогичная картина наблюдается на севере Тихого океана, в Беринго-
вом море. Циклонический апвеллинг вместе с конвективным переме-
шиванием вод еще больше способствует обогащению поверхностных
вод в субарктической зоне биогенными солями.
Южная граница субарктической зоны в Атлантическом секторе
резко перекошена. На западе она доходит до Нью-Браунсунка, лежаще-
го почти на широте северной границы субтропиков в восточной части
Атлантики; на востоке граница между субполярной и умеренной зона-
ми, напротив, сильно сдвинута к северу. Причина этого — течения:
холодные воды на востоке и теплые на западе.
В Северном Ледовитом океане и в Северной Атлантике природные
условия субарктической зоны существенно различаются в западном и
восточном секторах. В западный сектор Субарктики (Норвежское и
Баренцево моря) входят мощные струи теплых течений, порождаемых
системой Гольфстрим. Они значительно смягчают термические усло-
вия, препятствуют замерзанию моря. Здесь создаются особенно бла-
гоприятные условия для формирования богатых и разнообразных пе-
лагических и донных биоценозов на мелководьях. По своему составу
они ближе к биоценозам расположенной к югу бореальной зоны. Та-
ким образом, в западном секторе Субарктики благодаря теплым тече-
ниям ярко выражено явление экстразональности — присутствие в
ландшафтах черт, свойственных более южной зоне.
Из Тихого океана в Северный Ледовитый поступление теплых вод
течения Куросио затруднено из-за узости Берингова пролива. Поэтому
в восточном секторе (моря Лаптевых, Восточно-Сибирское, Чукотское,
шельф Северной Америки) субарктическая зона резко сужается, места-
ми полностью вытесняясь арктической зоной ледовитых морей.
В ноябре и декабре в субполярных зонах — полярная ночь; с мая по
июль — полярный день. Поверхность океана весной и летом получает
относительно много солнечного тепла. Постепенно исчезают морские
льды, температура тонкого поверхностного слоя воды поднимается до
5 °C, на юге до 10 °C. В результате прогрева и интенсивного таяния льдов
летом в субполярных зонах образуется относительно теплая, опреснен-
ная водная масса, обогащенная питательными веществами, резко от-
граниченная от более плотных глубинных вод слоем скачка.
Фауна пелагиали и шельфа морей субполярной зоны характеризу-
ется большим разнообразием (рис. 10.7).
Расшифровка к рис. 10.7: Млекопитающие: 1 — гренландский тюлень;
2 — нарвал. Рыбы: 3 — нельма; 4 — корюшка; 5 — семга; 6 — треска; 7 —
342 пикша' 8 — европейская навага; 9 — сайка; 10— морской окунь; 11 — поляр-
[ЛАВА 10. ШАЛЬНЫЕ ТИПЫ БИОМОВ ППВЕРХННСТНОЙ ТОЛЩМ ОКЕАНА
Рис. 10.7. Характерные животные морей субполярной зоны, по БСЭ
ный ликод; 12 — полярная камбала; 13 — липарис; 14 — четырехрогий бычок;
15 — морская лисичка. Губки: 16 — Tethyu llncurlum; 17 — Polymastia
mammillaris. Стрекающие: 18 — актиния Metridium dlanthus; 19 — гидроид
Sertularia plumosa; 20 — медуза Cyanea arctlca; 21 — медуза Aurelia aurlta;
22 — гребневик Mertensla ovum. Мшанки: 23 — Flustra foliacea; 24 —
Reterpora cellulose. Кольчатые черви: 25 — стрелка Sagitta elegans; 26 —
нереис Nereis vlrens; 27 — полихета Amphltrita cirrata; 28 — полихета
Harmothoe imbricata. Моллюски: 29 — хитон Tonicella rubra; 30 — морское
блюдечко Acmaea testudinalis; 31 — клион Clio borealis; 32 — трубач Buccinum
undatum; 33 — россия Rossia glaucopis; 34 — морской гребешок Pecten
islandicus; 35 — сердцевидка Serripes groenlandicus; 36 — иольдия Joldla
hyperborea. Ракообразные: 37 — веслоногий рачок Calanus finmarchicus; 38 —
краб Hyas araneus; 39 — бокоплав Gammarus locusta; 40 — морской таракан; ЭДЗ
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ К АРТИ В А МИРА
41 — морской желудь Balanus balanoides; 42 — креветка Pandalus borealis.
Иглокожие: 43 — морской еж Strongyllocentrotus droebuchiensis; 44 — мор-
ской еж Brlsaster fragilis; 45 — морская звезда Ctenodiscus crispatus; 46 —
морская звезда Crossaster papposus; 47 — морская звезда Astertus rubens; 48 —
офиура Ophiucantha bidentata; 49 — морская лилия Heliometra glacialis; 50 —
голотурия Cucumaria frondosa. Оболочник: 51 — асцидия Pyura aurantium.
По мере таяния льда и увеличения освещенности.усиливается «цве-
тение» воды в результате размножения диатомовых водорослей. Вслед
за ним наступает массовое размножение зоопланктона (рис. 10.8). Оби-
лие корма привлекает летом в Субарктику многих рыб — сельдь, треску,
мойву; у дна держатся камбала, палтус, морской окунь. Скопления сель-
ди бывают столь велики, что плотные косяки растягиваются в длину на
многие десятки миль.
Рис. 10.8. Годовой цикл развития жизни в субарктической зоне, по С. Нешибе
Обилие зоопланктона, рыб, богатая донная фауна служат надежной
пищевой базой для миллионов птиц. На труднодоступных прибрежных
скалах шумные птичьи базары образуют чайки-моевки, чистики, гагар-
ки, кайры, глупыши (рис. 10.9). В море кормятся тюлени, несколько
видов китов, на мелководье — моржи.
Рассмотрим более подробно пищевую цепь моржей. С. Нешиба (1991)
анализирует ее на примере экосистемы Берингова моря. Шельф послед-
него, со средними глубинами 75 м, имеет ширину около 500 м. В весен-
ние месяцы, когда льды отступают к северу, наблюдатели насчитывают
вдоль края пакового льда до 40 тыс. моржей. Каждый взрослый морж
съедает в день более 100 кг двустворок. Животное использует свои два
344 длинных клыка, чтобы, вспахивая ими дно, выкапывать моллюсков.
ГЛАВА 10. ШАЛЬНЫЕ ТИПЫ БИОМОВ ВВВЕРШСШЙ ТОЛЩМ ОКЕАНА
Рис. 10.9. Пищевая сеть и пирамида животных субарктической зоны
Каким же образом поддерживается огромная популяция моллюсков,
питающая моржей? Первым звеном пищевой цепи (продуцентами) слу-
жат высокопродуктивные заросли морской травы на мелководье. От-
мерший растительный материал разлагается и превращается в детрит,
служащий пищей моллюскам-фильтраторам, которыми и питаются
моржи (рис. 10.10).
Рис. 10.10. Пищевая цепь моржей, по С. Нешибе
345
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКДЯ КАРША МИРА
Основные взаимосвязи компонентов природы в субарктической зоне
показаны на рис. 10.11.
________________I
В «сеяно-легняй период вспышка
продуктивности фитопланктона, бла-
годаря уснлевию солнечной радиации
н высокому плодородию морских иод
Вслед за вспышкой
фитопланктона про-
исходит вспышка
зоопланктона
Обилие корма
привлекает массу
рыб, в том числе
промысловых
Рис. 10.11. Основные взаимосвязи компонентов природы в субполярной зоне
На суше, вдоль северных берегов Евразии и Северной Америки, из-
за холодного лета формируются тундровые ландшафты — безлесные
пространства с господством мохово-лишайниковой и кустарничковой
растительности. Наземные биоценозы менее продуктивны, чем океа-
нические, поэтому значительная часть животных находит пищу в море.
Субполярная, или субантарктическая, зона в Южном полушарии
охватывает воды Южного океана, располагающиеся кольцом к северу
от антарктической зоны. Это южная часть области западных ветров.
Основной характерной чертой Южного океана является гигантское
западное Антарктическое циркумполярное течение, опоясывающее
материк Антарктиды.
В отличие от Северного полушария течения меридионального направ-
ления в Южном океане практически отсутствуют. Здесь не наблюдается
вхождения мощных теплых струй, подобных Гольфстриму в Атлантиче-
ском и Куросио в Тихом океане, что, как уже отмечалось, способствует
еще большему выхолаживанию приполярных пространств Южного по-
лушария.
С севера субантарктическая зона ограничена линией антарктиче-
ской конвергенции, лежащей в среднем на 53 — 54° ю. ш. Вдоль этой ли-
нии холодные воды субарктической зоны сходятся с теплыми водами
умеренного пояса Южного полушария. На сравнительно небольшом
расстоянии по меридиану резко изменяются температура и соленость
воды, а также температура воздуха: к югу заметно холоднее, а жизнь
обильнее.
Весной, с усилением солнечной радиации, начинается массовое раз-
витие диатомовых водорослей, потребляющих биогенные соли. Фито-
планктоном кормится зоопланктон. Исключительно важное значение в
пищевых цепях Южного океана имеют довольно крупные рачки эвфа-
узииды — криль длиной 2 — 4 см. Им питаются рыбы (нототения, путас-
34G су), киты, ластоногие, птицы (рис. 10.12).
ГЛАВА 10. ЗАВАЛЬНЫЕ ТИПЫ БИОМОВ ПЛВЕРХНВСТНВЙ ТОЛЩВ ОКЕАНА
Солнечная энергия
Рис. 10.12. Главные звенья пищевой цепи в Южном океане
347
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКДЯ КАРША МИРА
И ю-з. Биомы умеренных зон________________________________________
Умеренные зоны располагаются между холодными и жаркими поя-
сами Мирового океана. В Северном полушарии их принято называть
бореальными, в Южном — нотальными.
Умеренные зоны — это акватории больших сезонных изменений тем-
пературы воды в верхних слоях, области интенсивной циклонической
деятельности, сильных штормов, преобладающих западных ветров («реву-
щие сороковые» в Южном полушарии). Все, вместе взятое, — температур-
ная конвекция, ветровое перемешивание, циклонический апвеллинг —
способствует подъему к поверхности богатых биогенами глубинных вод.
Умеренные зоны — самые продуктивные в Мировом океане. Они
дают основной улов рыбы, беспозвоночных, водорослей.
Климат, гидрологические условия умеренных широт весьма контраст-
ны у северных и южных границ. Это дает основание подразделять
бореальную суперзону на холодно- и теплобореальные зоны. Первая
располагается между зимними изотермами воздуха 0 — 5 °C, воды 5 —
10 °C, летними изотермами воздуха 10—15 °C, воды 10—15 °C; вторая —
между зимними температурами воздуха 5 — 10 °C, воды 10—15 °C, лет-
ними температурами воздуха 15 — 20 "С, воды 15 — 20 °C. Этот же прин-
цип положен в подразделение нотальной суперзоны в Южном полуша-
рии: холодно- и теплонотальную.
Зимой благодаря понижению температуры стратификация поверх-
ностных вод нарушается: происходит температурная конвекция — опус-
кание охлажденных поверхностных вод. Океан, за исключением неко-
торых внутренних морей, не покрывается льдом. Осенью и зимой
перемешиванию вод способствует сильное ветровое волнение. Подъем
глубинных вод обогащает зону фотосинтеза биогенными солями.
Летом, особенно во второй половине, вода на поверхности хорошо
прогревается и отделяется от более глубоких холодных вод резким сло-
ем скачка (термоклином). В это время подъем глубинных вод к поверх-
ности затруднен. Поэтому вслед за весенней вспышкой продуктивно-
сти планктона, после того как все питательные вещества использованы,
продуктивность вод падает. Однако осенью с началом штормов и пони-
жением температуры происходит обогащение поверхностных вод био-
генами, поднимающимися из глубин. В это время наблюдается вторая
(более слабая) вспышка продуктивности планктона. Весенний макси-
мум образуется за счет диатомей, а осенний — за счет перидиней.
Климатическим и гидрологическим сезонам умеренных зон соот-
ветствуют четко выраженные биологические сезоны; с сезонной дина-
микой продуктивности планктона связаны сезонные миграции рыб,
китов и др. Сезонный ход температур обусловливает циклы размноже-
ния и роста организмов нектона, фито- и зообентоса на мелководьях.
Бореальная зона Северного полушария. Характерной особенностью
умеренной зоны в Северной Атлантике и в северной части Тихого океа-
на являются системы течений, образующих кольца циклонических
круговоротов. С последними, как известно, связаны крупные апвеллин-
348 ги, обогащение поверхностных вод биогенными солями, повышенная
[ЛАВА 10. ШАЛЬНЫЕ ТИПЫ БИОМОВ ПЛВЕРХНОСТНЛЙ ТОЛЩИ ПИТАНА
биологическая продуктивность. В Северной Атлантике циклонический
круговорот (против часовой стрелки) образован у берегов Европы иду-
щими с юго-запада струями теплого Северо-Атлантического течения, у
берегов Северной Америки идущими с северо-востока струями холод-
ного Восточно-Гренландского- и Лабрадорского течений. В северной
части Тихого океана он образован теплым Аляскинским течением на
востоке и холодными Камчатским и Курильским на западе.
Теплые струи течений создают в умеренных широтах мощные тепло-
вые аномалии, центры низкого атмосферного давления — Исландский
минимум в Атлантике и Алеутский в Тихом океане. Вступая во взаимо-
действие с холодными арктическими воздушными массами, они посто-
янно генерируют циклоны. Под воздействием вращения Земли циклоны
движутся на восток. Умеренные широты характеризуются преобладани-
ем западных ветров — это область так называемого западного переноса.
Благодаря теплым течениям на востоке и холодным на западе струк-
тура бореальной зоны в Атлантическом и Тихом океанах резко асим-
метрична. Например, вдоль границ Америки арктическое влияние про-
никает на юг до 45° с. ш., а вдоль берегов Европы оттесняется на север
почти до 73° с. ш. Аналогичная картина наблюдается в умеренных ши-
ротах Тихого океана.
Бореальная зона характеризуется весьма богатыми биоценозами на
каменистых грунтах морских мелководий. Это заросли крупных бурых
водорослей — ламинарии (морской капусты), гигантского макроцисти-
са, разнообразных красных водорослей, сообщества беспозвоночных —
моллюсков, ракообразных, иглокожих, губок, мшанок и др.
Назовем характерных представителей промыслового комплекса хо-
лоднобореальной зоны: сельдь, морские окуни, камбала, треска, палтус;
кроме того, в Атлантическом океане — пикша, сайра, зубатка, бельдю-
га, мойва, семга и др., в Тихом океане — минтай, навага, лососевые и
др. Характерными представителями промыслового комплекса теплобо-
реальной зоны являются скумбрия, ставрида, сардина; кроме того, в Ат-
лантическом океане — макрелещука, кефаль, шпрота и др.; в Тихом океа-
не — анчоус, акулы, сайра и др.
Основные взаимосвязи компонентов природы в умеренных зонах
Северного полушария показаны на рис. 10.13.
Нотальная зона в Южном полушарии охватывает кольцо вод Южного
океана, опоясывающих Антарктиду. Преобладающие западные ветры со-
здают общее течение поверхностных вод на восток — Западный дрейф,
или Антарктическое циркумполярное течение. В нотальной зоне нахо-
дится лишь та часть течения, что лежит севернее антарктической кон-
вергенции.
Назовем характерных представителей промысловых комплексов
холодно- и теплонотальных зон. К первому относятся мерлуза, шпрота,
макрорус, ко второму — анчоус, скумбрия, ставрида, мерлуза, морские
караси, луфарь и др.
На суше в умеренном поясе формируется широкий спектр зональ-
ных типов ландшафтов, наиболее полно представленных в Северном
полушарии. На широтах, отвечающих холоднобореальной зоне, раски-
нулись ландшафты хвойных таежных лесов. 349
ел
Сезогаые изме-
нения темпера-
туры
В теплый сезон — стратифика-
ция вод: отделение теплых по-
верхностных вод от холодных
глубин слоем термоклина
Затрудненное вертикальное
перемешивание вод
^Низкое плодородие вод
В холодный сезон - темпера-
турная конвекция вод
Интенсивна»
циклоническая
деятельность
атмосферы
Зяпедный перенос
Температурная конвекция, ветровое
перемешивание, циклонический апвел-
линг вызывают подъем к поверхности
глубинных вод, богатых биогенными
солями
Циклонический
(против часовой
стрелки) круго-
ворот течений в
океане
Теплые течения у восточных
окраин океана (у западных
окраин материков)
Холодные течения у запад-
*ных окраин океана (у вос-
точных окраин материв»)
Асимметрия хода изотерм
воды у западных и восточ-
ных окраин океанов; асим-
метрия границ природных
зон у западных и восточных
окраин материков
Высокое плодородие вод
Высокая биологическая про-
дуктивность морских вод, в
ток числе промысловыми ор-
ганизмами *
Рис. 10.13. Основные взаимодействия компонентов природы в умеренных зонах Северного полушария
[ЛАВА 10. ШАЛЬНЫЕ ТИПЫ БИОМОВ ПЛВЕРХНОСТНЛЙ ТОЛЩИ ПИТАНА
В теплобореальной зоне проявляется секторность ландшафтов, связан-
ная с аридизацией климата по мере удаления в глубь материков. Так, на
атлантическом побережье Европы представлены ландшафты широколист-
венных лесов, на северных берегах Черного моря — ландшафты лесостепи
и степи, на берегах Каспия — ландшафты полупустыни и пустыни.
И 10.4. Биомы субтропнчвскмх ЗОН__________________________________
Биомы этих зон занимают промежуточное положение между уме-
ренными поясами и жарким. Океанские субтропики располагаются
вдоль широты 30° в Северном и Южном полушарии. Температуры зи-
мой: воздуха 10— 15 "С, воды 15 — 20 "С; летом: воздуха и воды 20 — 25 °C.
Характерной чертой субтропических зон является приуроченность
к ним областей высокого атмосферного давления. Здесь преобладают
нисходящие потоки воздуха, поступающего с антипассатами. В Север-
ном полушарии в Атлантическом океане субтропическая зона испыты-
вает воздействие Азорского максимума атмосферного давления, а в
Тихом океане — Гавайского. Отсюда воздух растекается в более высо-
кие и низкие широты. Это область зарождения пассатов. Течения сла-
бые, переменных направлений. Аналогичная картина наблюдается в со-
ответствующих широтах Южного полушария.
Летом в субтропических зонах обычно ясное небо, ничтожное ко-
личество осадков, сухой воздух. За год в океане испаряется слой воды
толщиной более двух метров. Поэтому здесь максимальная для океана
соленость: в Атлантике 37,5%о, в Средиземном море до 39%о, в Тихом
океане 35%о.
В результате сильного нагрева поверхностных вод создается устой-
чивая стратификация, когда поверхностные воды отделяются от хо-
лодных глубинных вод термоклином. В осенне-зимний период темпе-
ратура воды несколько понижается и благодаря высокой солености
возникает плотностная конвекция.
Однако погружение поверхностных вод не компенсируется подъ-
емом глубинных; дефицит восполняется поступлением вод с севера и юга.
Воды субтропиков бедны питательными биогенными солями, что огра-
ничивает продуктивность фитопланктона, а в конечном счете опреде-
ляет небольшие запасы рыбы в море.
Характерные представители промыслового комплекса субтропиче-
ской зоны являются переходными от теплобореального к тропическо-
му и экваториальному биомам. Они будут названы ниже.
Промежуточное положение субтропических зон проявляется, в част-
ности, в том, что в зимнее время области высокого атмосферного давле-
ния смещаются к экватору и субтропические зоны захватываются атмо-
сферной циркуляцией, характерной для умеренных зон, — циклонической
деятельностью, западным переносом. В холодный сезон усиливается ве-
тер, волнение, увеличивается количество атмосферных осадков.
На суше зональные типы ландшафтов представлены влажными и
сухими субтропиками. Более южные области, по западным окраинам
континентов, отвечают окраинам великого пояса тропических пустынь.
351
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКДЯ КАРША МИРА
И 10.5. Бирмы тропическим ЗОН_____________________________________
Тропические зоны обоих полушарий — это прежде всего области
пассатных ветров и течений в океане. Устойчивая система ветров под-
держивается движением воздушных масс от областей высокого давле-
ния, располагающихся в субтропиках, к области низкого давления, вы-
тянутой вдоль экватора. Под воздействием силы Кориолиса ветры,
дующие в сторону экватора, отклоняются в Северном полушарии впра-
во: пассаты имеют северо-восточное и восточное направление; а в Юж-
ном полушарии — влево: пассаты дуют с юго-востока, востока.
Устойчивая система ветров создает пассатные течения, движущиеся
на запад. В «корнях» пассатов, у западных окраин материков, происхо-
дит сгон поверхностных теплых вод в открытый океан и подъем холод-
ных, богатых биогенами глубинных вод. Поэтому в тропиках, у запад-
ных окраин материков, формируются холодные течения, продуктивность
вод высокая.
На суше, в тропических широтах в «корнях» пассатов располагают-
ся наиболее сухие и жаркие пустыни. Это связано с тем, что зарожде-
ние пассатов происходит из опускающихся в субтропических зонах
воздушных масс антипассата. Опускаясь, воздух сжимается и нагре-
вается, его относительная влажность понижается. Кроме того, омыва-
ющие материк течения холодные, испарение ограничено, океан дает в
атмосферу мало водяных паров.
У противоположных берегов океана пассаты осуществляют нагон теп-
лых поверхностных вод. Проходя над океаном, воздушные массы насыща-
ются влагой, поэтому западные побережья океанов характеризуются влаж-
ным климатом. Однако биологическая продуктивность вод низкая.
Постоянный нагон теплой воды у западных берегов океанов, напри-
мер у Центральной Америки в Атлантическом океане или у Юго-Вос-
точной Азии в Тихом океане, является причиной образования мощных
компенсационных течений, направленных в сторону полюса. В Атлан-
тическом океане — это Гольфстрим, в Тихом — Куросио.
Под воздействием силы Кориолиса течения отклоняются на восток.
Основные струи, достигнув восточных берегов океана, устремляются
на север, но другие отклоняются на юг, где постепенно усиливаются
благодаря действию пассатов.
Таким образом формируются гигантские круговороты, направлен-
ные по часовой стрелке в Северном полушарии и против — в Южном.
Например, в Северном полушарии в Атлантическом океане такой кру-
говорот образован в «корнях» пассатных ветров у берегов Африки хо-
лодным Канарским течением, далее теплыми течениями: Северным
пассатным, Гольфстримом, Северо-Атлантическим; в Тихом океане —
Калифорнийским, Северным пассатным, Куросио. Аналогичные кру-
говороты, захватывающие воды субтропических и тропических широт,
формируются в Южном полушарии.
Тип циркуляции в таких круговоротах антициклонический: вода с пе-
риферии собирается к центру и затем погружается. Нисходящие токи
уносят с поверхности органические остатки и вместе с ними питательные
352 вещества. В результате воды тропических широт — одни из самых бедных
[ЛАВА 10. ШАЛЬНЫЕ ТИПЫ БИОМОВ ПЛВЕРХНОСТНЛЙ ТОЛЩИ ПИТАНА
жизнью в Мировом океане. Это океанические пустыни. Они характери-
зуются высокой прозрачностью, до 40 — 60 м, и глубоким синим цветом.
Температуры тропической зоны зимой: воздуха 15 — 20 °C, воды 20 —
25 °C. Теплые поверхностные воды отделяются от холодных глубинных
вод резко выраженным слоем-температурного скачка. Благодаря высо-
кой испаряемости, соленость воды в целом повышенная — от 33 до
37,5%о. Максимальная соленость отмечается в полузамкнутых морях и
заливах: Красном, Персидском. Ярко выражена секторность, связанная
с различной увлажняемостью.
Как уже говорилось, пассат в своих истоках, т. е. в восточной части
океанов, — сухой ветер. В связи с этим у западных окраин континентов
в тропических широтах располагаются величайшие пустыни мира, су-
хие саванны. По мере движения над океаном пассат насыщается вла-
гой, и климат восточных окраин материков становится более влажным.
Здесь располагаются постоянно- и сезонновлажные леса.
В связи с сезонной миграцией центров атмосферного давления ле-
том в тропическую зону входит экваториальный воздух, отличающий-
ся большой влажностью и облачностью, идут частые дожди. Зимой в
тропические широты смещается центр высокого атмосферного давле-
ния. Устанавливается сухая жаркая погода. Таким образом, в тропи-
ках два сезона: сухая зима и дождливое лето.
Сезонный ход метеорологических явлений особенно ярко выражен
в Индийском океане, в азиатском секторе Тихого океана, находящихся
под воздействием муссонной циркуляции. В Южной Азии летом уста-
навливается очень высокая температура и необычайно низкое давле-
ние. Над Индийским и Тихим океанами, в Южном полушарии в это
время зима, температура низкая и давление высокое. Поэтому летом
возникает устойчивый поток воздуха с океана на сушу — летний мус-
сон. Он приносит влажный воздух, большую облачность, обильные
дожди.
Зимой в результате выхолаживания суши над Азией формируется
область высокого давления. Над теплыми водами океана давление ниже.
В результате возникает зимний муссон — поток воздуха с суши в оке-
ан. Он приносит сухой воздух, солнечную погоду.
Смена ветра оказывает влияние на направление муссонных течений,
особенно заметное в северной части Индийского океана: Сомалийское
течение вдоль берегов Африки летом направлено на северо-восток (в это
время оно сгонное, сопровождается апвеллингом, высокой продуктив-
ностью и рыбностью), зимой — на юг (в это время оно нагонное, про-
дуктивность и рыбность падает).
Устойчивый характер основных метеорологических процессов в
тропических широтах нарушается лишь во время ураганов (тайфунов).
Это чрезвычайно опасные разрушительные циклонические вихри, воз-
никающие в конце лета у западных теплых окраин океанов.
Промысловый комплекс тропических широт характерен для всего
теплого пояса — это сардинелла, анчоус, рифовые окуни, акулы, скаты,
тунцы, марлины, корифэны, летучие рыбы. Типичные представители
фауны теплых морей приведены на рис. 10.14. Основные взаимосвязи
компонентов природы показаны на рис. 10.15. 353
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ КАРША МИРА
8
Рис. 10.14. Фауна теплых морей, поА.Г. Воронову: 1 —мадрепора; 2 — пиросо-
ма; 3 — красный коралл; 4 — сифонофора; 5 — фунгия; 6 — парусник;
7 — мечехвост; 8 — ципрея; 9 — сростночелюстная рыба балистес;
354 10 — жемчужница; 11 — морская змея; 12— акула гетеродонтус
C-FI
C-FI
Рис. 10.15. Основные взаимосвязи компонентов природы в тропических зонах
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ КАРТИ В А МИРА
И 10.6. Биомы экваториальной зоны____________________________________
Основные взаимосвязи компонентов природы в экваториальной зоне
показаны на рис. 10.16.
Рас. 10.16. Основные взаимосвязи природы в экваториальной зоне
35G
Вдоль термического экватора располагается ложбина низкого
атмосферного давления — это зона затишья между пассатными ветрами
Северного и Южного полушарий. Из-за сильного нагрева штилевой
поверхности океана возникают восходящие токи воздуха, питающие
антипассаты. Они увлекают влажный воздух, содержащаяся в нем вла-
га конденсируется, образуются мощные облака, выпадают обильные
дожди.
Сильная инсоляция и прогрев воды препятствуют распостранению
растений и животных на рубеже суши и моря. Почти на нет сходят
литоральные заросли водорослей, слабо развита литоральная фауна.
Характерной особенностью экваториальной зоны является мощ-
ное Межпассатное течение, часто называемое Экваториальным про-
тивотечением. Оно является компенсационным по отношению к пас-
сатным течениям, формирующимся к северу и югу от экватора, и
имеет противоположное направление. В этой зоне на громадном про-
странстве происходит подъем глубинных вод. Это хорошо видно по
более низкой температуре воды, обогащению ее биогенами, богатству
жизни.
Подъем глубинных вод обогащает зону фотосинтеза питательными
солями — фосфатами и нитратами. В результате увеличивается количе-
ство фитопланктона, зоопланктона, рыб. Обилие жизни в экваториаль-
ной зоне влияет на прозрачность и цвет воды: прозрачность меньше на
10 м, чем в пассатной зоне, цвет воды становится зеленоватым.
Промысловый комплекс экваториальной зоны по составу аналоги-
чен названному выше при характеристике тропических зон, однако по
количеству рыбы намного его превосходит. Экваториальная зона —
один из важнейших районов мирового рыболовства.
Экваториальная зона на суше характеризуется влажным климатом.
Секторность, обусловленная разной увлажненностью, выражена сла-
бо. Благодаря тому что годовые амплитуды температур меньше суточ-
ных, мало заметна сезонная ритмика ландшафтов. Господствуют два
ГЛАВА 10. ЗОВДЛЬНЫЕ ТИПЫ БИОМОВ ППВЕРХНВСТНОЙ ТОЛЩИ ОКЕАНА
типа ландшафтов: постоянно влажные вечнозеленые леса (гилей) и влаж-
ные леса с коротким засушливым периодом. Последние тяготеют к во-
сточной периферии материков, где экваториальный тип климата ослож-
няется муссонной циркуляцией.
И 10.7. Мангровые Солота и коралловые рифы________________________
На мелководьях жаркого пояса (тропической и экваториальной зон)
формируются два типа биомов, заслуживающих специального рассмот-
рения. Это мангровые болота и коралловые рифы.
Мангры, или мангровы, образованы наземными растениями, кото-
рые выдерживают соленость открытого моря. Зеленой стеной встают
они из морских вод вблизи побережий. Во время прилива над водой
возвышаются лишь кроны деревьев, а при отливе обнажается густая сеть
корней, в том числе своеобразные дыхательные корни (рис. 10.17).
Рис. 10.17. Мангры: густая сеть корней-подпорок;
внизу— дыхательные корни (пневматофоры)
357
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ КАРТИ В А МИРА
Пышные заросли образованы немногими видами, относящимися к
трем основным семействам: Rhizophoraceae, Combretacecte и Verbenaceae.
Мангровы развиваются в местах, защищенных от сокрушительных уда-
ров волн коралловыми рифами или прибрежными островами. В зависи-
мости от высоты и продолжительности затопления во время прилива в
структуре растительности формируется поясность (рис. 10.18).
Рис. 10.18. Поясность прибрежной мангровы, по Г. Вальтеру.
Африканское побережье Индийского океана
Рис. 10.19. Плоды ризофоры
с проростками на ветке дерева,
358 по «Жизни растений»
Наиболее характерному представите-
лю мангр — ризофоре — свойственно
живорождение. Семена прорастают еще
на дереве: на ветвях образуется готовая
рассада с сигаровидными корнями, до-
стигающими в длину десятков сантимет-
ров (рис. 10.19).
Во время отлива такой проросток
вонзается в илистый грунт, быстро за-
крепляется и дает начало новому расте-
нию. Унесенные течением проростки
могут совершать длительные (до года)
путешествия, сохраняя при этом жизне-
способность. Этим объясняется широ-
кий ареал мангр на тропических побе-
режьях.
Коралловые рифы являются одной
из наиболее ярких достопримечатель-
ностей морей жаркого пояса. Их ареал
ограничен за пределами вод, температу-
ра которых не опускается ниже 20 °C
(рис. 10.20).
Рифообразующие, или, иначе, гер-
матипные кораллы, — это колониаль-
ные кишечнополостные животные
ГЛАВА 10. ШАЛЬНЫЕ ТИПЫ БИОМОВ ПОВЕРИОСПОЙ ТОЛЩИ ОКЕАНА
(тип Coelenterata), относящиеся к отряду мадрепоровых (Madreporaria).
Риф строится благодаря биологическому связыванию карбоната каль-
ция; скелет мадрепоровых кораллов — чистая известь (Наумов, Пропп,
Рыбаков, 1985). В связи с этим следует подчеркнуть роль герматипных
кораллов в утилизации излишков углекислого газа из атмосферы и
морской воды. В огромных количествах он связывается в известковых
скелетах рифостроящих организмов.
Живой коралловый риф — это сочетание биологической и геологи-
ческой структур, образовавшихся в результате жизнедеятельности
многих организмов. Он имеет вид отмели, которая поднимается с глу-
бины к поверхности моря.
Этот сложный биологический комплекс должен быть достаточно
прочным, чтобы противостоять воздействию океанических волн; весь-
ма крупным, чтобы обеспечивать существование целостного биома и
влиять на экологическое своебразие окружающих акваторий.
К настоящим коралловым рифам относятся сообщества, в которых
главную роль на морском дне по общей биомассе и продуктивности
играют рифостроящие кораллы. Наряду с кораллами важную роль в
образовании рифа играют известковые красные водоросли, так назы-
ваемые литотамнии. Таким образом, коралловый риф в действительно-
сти представляет собой кораллово-водорослевое образование.
Ч. Дарвин впервые выделил три типа рифов: барьерные, окаймляю-
щие и атоллы. Он же выдвинул гипотезу происхождения атоллов, кото-
рая в наши дни блестяще подтвердилась. Атоллы — рифы, имеющие вид
кольца или подковы с центральной лагуной внутри. Подобно жемчуж-
ным ожерельям, они светятся в темно-синих водах океана (рис. 10.21).
Стадии образования атолла показаны на рис. 10.22.
Согласно Ч. Дарвину, развитие атолла начинается с берегового рифа,
окаймляющего остров. Последний обычно представляет собой вулка-
нический конус, поднимающийся из глубин океана. В результате жиз- 359
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ КАРТИ В А МИРА
Рис. 10.21. Атолл
Рис. 10.22. Образование атолла, по Ч. Дарвину
недеятельности герматипных организмов на береговом склоне риф на-
чинает разрастаться в сторону моря. Однако после того, как активная
вулканическая деятельность прекращается, остров под действием силы
тяжести начинает погружаться в земные недра. Кораллы же по мере
опускания дна успевают надстраивать риф до уровня поверхности моря.
На этой стадии коралловые постройки окружают остров сплошным или
прерывистым кольцом, т. е. барьерным рифом, между которым и ост-
ровом образуется лагуна. Дальнейшее опускание острова приводит к его
полному исчезновению, и тогда на поверхности остается лишь рифо-
вое кольцо — атолл с внутренней лагуной.
Коралловые рифы — настоящие оазисы жизни среди пустынных
тропических вод (рис. 10.23). Наряду с дождевыми тропическими леса-
ми они относятся к наиболее продуктивным экосистемам Земли. Лю-
бопытен парадокс богатства жизни в водах тропических морей, бедных
питательными солями. Он объясняется симбиозом коралловых поли-
пов с водорослями.
Расшифровка к рис. 10.23: Мадрепоровыо кораллы: 1 — поритес; 7 — моз-
говик; 9, 14, 21 — ветвистые кораллы Асторота; 17 — лобофеллия; 39 — фун-
гия; 2, 8 — горгоновые кораллы. Водоросли: 22 — галимеда. Черви: 24 — труб-
3G0 чатый червь серпула. Моллюски: 28 — тридакна; 32 — осьминог Octopus
ГЛАВА 10. ШАЛЬНЫЕ ТИПЫ БИОМОВ ПОВЕРХНОСТНОЙ ТОЛЩИ ОКЕАНА
Рис. 10.23. Характерные обитатели коралловых рифов
(Индо-Западно-Тихоокеанская область), по БСЭ
pictus; 34 — мурекс; 36 — каури; 37 — рог тритона; 38 — элизия;
40 — конус. Ракообразные: 19 — креветка стенопус; 30 — рак-отшельник
Dardanus; 31 — краб Atergates. Иглокожие: 23 — голотурия Cucumaria tricolor;
18, 25, 35 — морские звезды линкия (18); ореастер (25) и терновый венец
Acanthaster (35); 33 — офиура Opluotrichoides; 20, 29 — морские ежи гетеро-
центротус (20) и диадема (29). Рыбы: 3, И, 16 — разные виды рыб-бабочек
рода Chaetodon; 4 — рогатый занкл; 5 — вымпельный щетинозуб Heniochus;
6 — рыба-единорог Osbeckia; 10 — пигоплит; 12 — ринекант; 13 — рыба-хи-
рург Acanthurus; 15 — Никобарский голакант; 26 — амфиприон; 27 — хромис. 3G1
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ КАРТИ В А МИРА
Жизнь грандиозных рифов полностью зависит от одноклеточных
пирофитовых водорослей [Symbiodirtium microadriaticum), селящихся в
тканях кораллов-рифостроителей. Продукты фотосинтеза водорослей
напрямую используются полипами. Ночью коралловый полип, как жи-
вотное, переходит на питание планктоном. Тогда продукты пищеваре-
ния служат источником биогенных солей для водорослей.
Физиологическая связь между герматипными кораллами и симбио-
тическими водорослями настолько велика, что ни те ни другие в при-
родных условиях не могут жить самостоятельно. Эти кораллы так же
немыслимы без водорослей, как немыслим без водорослей гриб в теле
лишайника.
Поэтому симбиотическое сочетание «герматипные кораллы-водорос-
ли» следует рассматривать как целостный организм, служащий един-
ственным примером животных-продуцентов. Недаром К. Линней вклю-
чал кораллы в отряд Lithophyta, что значит «камни-растения».
Известный эколог Ю. Одум пишет, что человек многому может на-
учиться на примере коралловых рифов, в частности как повторно ис-
пользовать те или иные продукты и как преуспеть в мире со скудными
ресурсами.
Глава 11
УЯЗВИМЫЕ ЗВЕНЬЯ ЭКОЛВГИЧЕСКОИ
СИСТЕМЫ МИРОВОМ ОКЕАНА
В начале 1960-х гг., когда стало ясно, что «неисчерпаемые» запасы
сырья на суше стали быстро таять, взоры и мысли обратились к Миро-
вому океану. Имея ввиду прежде всего его огромные размеры, загово-
рили об «океане возможностей» — колоссальных запасах биологиче-
ских, минеральных, энергетических и других ресурсов, гигантской
емкости как резервуара для сброса отходов производства. Однако вско-
ре стало ясно, что действие мощного антропогенного пресса сделало
океан весьма уязвимым. И уже сейчас видно, что не с «океаном воз-
можностей», а скорее с «океаном проблем» будут иметь дело наши
потомки в XXI в., причем проблем в основном экологических.
Человечество наносит два удара по природе: во-первых, истощает
ресурсы, во-вторых, загрязняет ее. Оба эти удара поражают не только
сушу, но и океан. Мировой океан является перспективным источником
ресурсов. Значительный энергетический потенциал заключен в океа-
нических волнах, приливах, термальных градиентах, морских течени-
ях. Дно океана дает около четверти суммарной мировой добычи нефти.
Предполагается, что в начале XXI в. морская добыча нефти повысится
еще на 50%. По оценкам специалистов, морские ресурсы магнетита,
ильменита, рутила, олова, золота, платины и других металлов сопоста-
вимы с их запасами на суше. В железо-марганцевых конкрециях, усти-
лающих дно океана, содержится около 30 элементов таблицы Менделе-
ева. Океан обеспечивает 90% мировой добычи брома, 60% магния, 30%
поваренной соли, большую часть алмазов.
Океан служит одним из важнейших источников пищевых ресурсов
для человечества, поставляя 25% белков животного происхождения.
Велико его транспортное значение — более 75% мирового грузооборота
приходится на его долю.
Возрастающая эксплуатация ресурсов Мирового океана уже сама по
себе оказывает все более сильное воздействие на его экосистему. Одна-
ко имеются и мощные внешние источники загрязнения — это атмо-
сферные потоки и материковый сток. В результате на сегодня можно
констатировать наличие загрязняющих веществ не только в зонах,
прилегающих к материкам, и в районах интенсивного судоходства,
но и в открытых частях океанов, включая высокие широты Арктики и
Антарктики.
3G3
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ К АРТИ В А МИРА
В Мировой океан ежегодно сбрасывается более 30 тыс. различ-
ных химических соединений, общая масса которых составляет
1,2 млрд т. Загрязнение его вод отдельными веществами приобрело гло-
бальный характер. Самыми опасными загрязнителями, поражающи-
ми экосистему океана из-за их отравляющего, мутагенного либо кан-
церогенного действия на морские организмы, являются углеводороды
(пестициды, нефть и нефтепродукты и др.), токсичные металлы
(ртуть, свинец, кадмий, мышьяк и др.) и радиоактивные вещества.
Все более опасным для прибрежных районов является микробиоло-
гическое загрязнение. Патогенные организмы, в частности кишеч-
ная микрофлора, начинают проникать в открытые акватории неко-
торых морей. Загрязняющие вещества не только непосредственно
воздействуют на экосистему океана, но и изменяют физико-химиче-
ские свойства вод, определяющие газообмен, потоки солнечной радиа-
ции и теплоты через его поверхность.
Сгущения и пленки жизни. Жизнь распределена в океане очень
неравномерно. В.И. Вернадский выделял две формы концентрации
живого вещества: жизненные пленки на границе газообразной и жидкой
сред — планктонная, на границе жидкой и твердой сред — донная —
и сгущения жизни. Остальная часть океаносферы представляет собой
зону разрежения живого вещества.
Между распределением биомасс планктонной и донной пленок
жизни в открытом океане существует тесная корреляционная зависи-
мость: акваториям с высокой продуктивностью планктона соответству-
ют участки с повышенной биомассой бентали («принцип соответствия»
Л.А. Зенкевича).
Сгущения жизни В.И. Вернадский разделял на три типа: прибреж-
ные, саргассовые, рифовые. Позже были открыты еще два: апвеллинго-
вые и гидротермальные рифтовые (рис. 11.1).
В обеспечении экологического равновесия биосферы эти пленки и
сгущения жизни в океане играют ключевую роль. Живое вещество
океана вносит свой вклад не только в продуцирование биомассы, но и
в поддержание газового состава атмосферы, регулирование химического
состава океанических вод, процессов образования осадочных горных
пород, формирование берегов и подводного рельефа.
Мировой океан как единая система. Научный подход к решению
проблем океана заключается в изучении закономерностей функциони-
рования его как единой природной системы, определении простран-
ственной и функциональной неоднородности, выяснении характера и
интенсивности антропогенного воздействия на разные блоки океани-
ческой природно-хозяйственной системы. Чтобы понять целое, необ-
ходимо рассмотреть взаимодействие его частей.
Океан неоднороден по физико-географическим условиям. Элемен-
ты его структуры, отличающиеся по характеру природных процессов,
разделяют так называемые активные граничные поверхности, являю-
щиеся в то же время областями бурно протекающих биологических
процессов: 1 — контакт поверхностной толщи океана с атмосферой; 2 —
контакт океана с сушей (морские мелководья, шельф); 3 — контакт
364 океана с дном (рис. 11.2).
ГЛАВА 11. УЯЗВИМЫЕ ЗВЕНЬЯ ЭКОЛОГИЧЕСКОЙ СИСТЕМЫ МИРОВОГО ОКЕАНА
/ E3w E]/v
Рис. 11.1. Пленки и сгущения жизни в Мировом, океане, по А.В. Лапо:
I — планктонная пленка жизни; II — донная пленка жизни; III — сгущения
жизни: 1 — прибрежное; 2 — саргассовое; 3 — рифовое; 4 — апвеллинговое;
5 — гидротермальное рифтовое; IV— апвеллинг;
А — область разрежения живого вещества
Рис. 11.2. Модель физико-географической и природно-хозяйственной
структуры океана: 1 — контакт поверхностной толщи океана — гидросферы
(Г) с атмосферой (А), зона рыболовства и мореплавания; 2 — контакт оке-
ана (Г) с дном —литосферой (Л), в перспективе добыча металлоносных
осадков; 3 — контакт океана с сушей (Г, Л, А), прибрежная зона освоения
биологических, минеральных, рекреационных и других ресурсов,
интенсивного строительства
365
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ К АРТИ В А МИРА
Деятельность человека уже охватывает практически всю акваторию
океана, но локализуется в тонких контактных зонах: поверхностная
зона служит для рыболовства и мореплавания; прибрежная зона — для
получения биологических, минеральных, энергетических, рекреацион-
ных ресурсов, интенсивного жилого и промышленного строительства;
дно океана — в перспективе зона добычи полезных ископаемых.
Океаны представляют собой огромные впадины в рельефе Земли.
В эти впадины устремляется практически весь сток поверхностных вод
суши, промывая самые отдаленные участки земной поверхности. За
исключением небольшого числа замкнутых бессточных котловин, зем-
ная поверхность промывается атмосферными осадками. Ручейки впа-
дают в реки, реки несут свои воды в моря и океаны, и таким образом
в загрязнении океана участвуют почти все города и поселки. Поскольку
скорость течения в реках довольно высока (в среднем вода из верховьев
рек достигает океана всего за 12 дней), отбросы, попавшие в реку у ее
истоков за тысячи километров от берега, очень скоро стекут в океан.
Площади речных бассейнов огромны (рис. 11.3). Реки вбирают в себя и
несут в океан не только сточные воды городов, но также ядохимикаты
и удобрения, которые в больших количествах вносятся на поля.
Рис. 11.3. Водосборные бассейны материков, по С. Нешибе
|\* Амктпмжогискмм
Га, (мимам смямАЛяМКТЫ*)
Устье реки — это точка в береговой зоне. Однако оказываемое сто-
ком рек загрязнение выходит далеко за пределы прибрежных вод. Бла-
годаря мощным циркуляционным процессам океан представляет собой
единое целое. Воды океанов и морей связаны течениями. Даже внут-
ренние моря (например, Черное) обмениваются своими водами с от-
366 крытым океаном. Течения способны многократно увеличивать площадь
ГЛАВА 11. УЯЗВИМЫЕ ЗВЕНЬЯ ЭКОЛОГИЧЕСКОЙ СИСТЕМЫ МИРОВОГО ОКЕАНА
загрязнения и переносить ядохимикаты, смытые с полей, за тысячи
километров, например к берегам Антарктиды. Здесь они, пройдя по
пищевой цепи, обнаруживаются даже в теле пингвинов.
Существуют глубоководные котловины и желоба, которые открыто
или тайно используют для захоронения вредных, в том числе радиоак-
тивных, отходов производства. Несмотря на кажущуюся стагнацию вод
в котловинах, доказано, что существует медленное перемешивание всей
толщи вод Мирового океана. Это должно служить основанием для меж-
дународного соглашения, запрещающего захоронение ядовитых и ра-
диоактивных отходов в глубинах океана.
Единство океана поддерживается целостностью его экологической
системы. Океан заполнен жизнью от поверхности до максимальных глу-
бин. Однако весь процесс биологического круговорота начинается в
верхнем, освещаемом Солнцем слое воды. Главным производителем пер-
вичной биологической продукции в океане является фитопланктон —
микроскопические водоросли (продуценты), плавающие в толще воды.
Дальнейшие звенья круговорота состоят из мелких животных, питаю-
щихся фитопланктоном, различных хищников, поедающих свои жерт-
вы, детритофагов и микроорганизмов — редуцентов. В цепочке пище-
вых связей биомасса каждого последующего звена приблизительно в
10 раз меньше, чем предыдущего.
Одна из особенностей техногенеза — активное рассеяние металлов.
Происходит интенсивная «металлизация» общего фона геохимической
среды. Более 30 химических элементов опасны для морских организмов
и человека, причем большую часть составляют переходные и тяжелые
металлы. Многие из них биофильны, т. е. морские организмы накапли-
вают их в своих тканях, в концентрациях, в десятки, сотни тысяч раз
превышающих содержание металлов в морской воде.
Опасность заключается в том, что содержание тяжелых металлов
увеличивается с продвижением по трофической цепочке. В 1 кг круп-
ной хищной рыбы содержится металлов примерно столько же, сколько
их сконцентрировано в 10 000 кг фитопланктона. По сравнению с пира-
мидой биомасс пирамида содержания тяжелых металлов в экосистеме
оказывается перевернутой. Об этом следует знать, поскольку человек
питается преимущественно крупными хищными морскими рыбами.
Металлы, рассеиваемые человеком в процессе техногенеза, возвраща-
ются к нему в концентрированном виде вместе с морской пищей.
Живое вещество в океане выполняет важную функцию насоса, от-
качивающего СО2 из атмосферы. Как известно, сжигание углеводород-
ного топлива обусловливает выброс в атмосферу огромного количества
СО2. Повышение его концентрации в атмосфере вызывает парниковый
эффект, глобальное потепление климата. В результате начинают таять
ледяные шапки на полюсах, повышается уровень океана. Трансгрессия
океана уже сейчас вызывает озабоченность государств Микронезии, на-
селение которых живет на атоллах, возвышающихся над поверхностью
океана на считанные метры. Океан захватывает из атмосферы и захо-
ранивает С02 в осадочных породах типа известняков и доломитов, а
также в толще морской воды благодаря образованию биомассы живого
вещества и растворенной органики.
367
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ КАРТИ В А МИРА
Уязвимость этого звена в том, что темпы антропогенного накапли-
вания С02 в атмосфере выше, чем скорость откачивания его морскими
организмами. К тому же многие экосистемы, особенно интенсивно свя-
зывающие СО2, расположены на мелководьях или в поверхностной
толще океана. Они страдают в первую очередь от антропогенного воз-
действия и снижают свою продуктивность, поэтому злободневен вопрос
об охране коралловых рифов — одной из наиболее продуктивных эко-
систем.
Загрязнение океана разрушает тонкие биохимические связи между
его обитателями. Разные группы организмов, особенно бактерии и бес-
позвоночные животные, выделяют растворенные органические веще-
ства, играющие роль пищи и биостимуляторов для членов сообщества.
Физиологические системы многих морских организмов являются от-
крытыми, а все сообщество объединяется биохимическими связями по
принципу дополнительности: гидробионты выделяют вещества, необ-
ходимые для других организмов, и сами включают в свой обмен выде-
ления соседей. Морская экосистема представляет собой физиологиче-
скую целостность. Гибель определенных групп организмов в такой
системе можно уподобить удалению железы внутренней секреции, про-
изводящей гормоны у высшего животного, а введение в воду ядовитых
веществ — инъекции яда.
Есть еще одна важная сторона экологического метаболизма — это
роль прижизненных выделений в общении гидробионтов. Подобно спе-
цифическим ароматическим веществам, выделяемым животными на
суше, эти вещества помогают морским обитателям общаться между
собой: самцам и самкам находить друг друга, особям определенных
видов собираться в стаи, хищникам находить жертву, а жертве избе-
гать хищника. Вторжение чужеродных веществ в океан способно вы-
звать хаос в поведении гидробионтов, разрушить всю систему биоцено-
тических отношений.
Итак, в структуре океана выделяются контактные зоны, для кото-
рых одновременно характерны максимальная напряженность физико-
географических и биологических процессов и деятельность человека,
обусловливающая истощение ресурсов и загрязнение. Граничные по-
верхности при этом не являются непреодолимыми барьерами. Напро-
тив, здесь вещество и энергия интенсивно переходят из одной природ-
ной системы в другую. Циклы массоэнергообмена не только связывают
географическую оболочку планеты (атмосферу, гидросферу, часть ли-
тосферы и биосферу), материки и океаны в единое целое, но и поддер-
живают устойчивость океана как единой природной системы.
Рассмотрим своеобразие природных процессов в контактных зонах
океана, выделяя антропогенные нарушения, которые имеют негатив-
ные последствия. Характерной особенностью функционирования глав-
нейших экосистем океана, формирующихся на разделах геосфер, явля-
ется единство противоположностей. С одной стороны, каждая из этих
систем относительно автономна. С другой стороны, каждая система от-
крыта и обладает широкими связями с контактными системами. По-
следнее определяет ее вклад в целостность структуры океана и всей гео-
368 графической оболочки.
ГЛАВА 11. УЯЗВИМЫЕ ЗВЕНЬЯ ЭКОЛОГИЧЕСКОЙ СИСТЕМЫ МИРОВОГО ОКЕАНА
Контакт океана с сушей. В прибрежных мелководьях, занимающих
13% площади океанов, продуцируется 40% органического вещества; здесь
вылавливается более 90% рыбы. Вместе с тем это зона наиболее активно-
го антропогенного воздействия. Около 70% населения Земли живет в
пределах 500 км от морских побережий; около 50% городов с населением
более 1 млн человек расположено около устьев рек и заливов океанов
(рис. 11.4). Поэтому огромная масса веществ, в том числе промышлен-
ных, сельскохозяйственных и бытовых отходов, поступает на мелково-
дья со стоком с суши, замыкающим большой круговорот воды.
Каждая точка соответствует 10 000 человек. Затушеваны территории,
где плотность превышает 75 чел./км
Твердый сток при впадении рек в море осаждается лавинно, так что
до 90% веществ, выносимых с суши, остается в пределах мелководий и
в океан не поступает. Таким образом, морские мелководья являются
глобальной геохимической ловушкой, задерживающей большую часть
веществ, сносимых с континентов. Морские мелководья — область сгу-
щения жизни, и на их долю приходится основная масса рыбы и море-
продуктов, употребляемых человеком в пищу. Относительная замкну-
тость биохимической системы морских мелководий делает ее особенно
уязвимой как в отношении нарушения экологии гидробионтов, так и в
отношении биоаккумуляции веществ, опасных для человека.
В пределах морских мелководий выделяется еще более узкая контакт-
ная полоска — береговая зона. Важную роль в формировании берегов
играет вдольбереговой поток наносов: там, где он не насыщен, образу-
ется дефицит наносов — берега разрушаются морем. Казалось бы, на
современном этапе тектонического развития планеты, когда «раствор 369
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ КАРТИ В А МИРА
гипсометрических ножниц» достиг максимальной величины, можно
было бы ожидать поступления с суши в береговую зону огромного
количества обломочного материала, насыщения потока наносов и ши-
рокого развития пляжей. Сама природа создала гигантский цех
строительных материалов, который, как надеялись, с лихвой может
обеспечить нужды быстро развивающихся портово-промышленных
комплексов и песком, и гравием, и галькой. Однако на изъятие чело-
веком рыхлых наносов из береговой зоны море ответило катастро-
фически быстрым разрушением берегов. Возникла необходимость
проведения дорогостоящих мероприятий по защите инфраструкту-
ры портово-промышленных комплексов от волн.
Оказывается, для стока рек характерен «точечный» вынос осадоч-
ного материала. Устья рек — это, как правило, ловушки осадков. Из этих
ловушек наносы не попадают на соседние участки береговой зоны.
Современный этап развития береговой зоны океана характеризует-
ся дефицитом рыхлого материала, в связи с чем преобладают процессы
волнового разрушения берегов. Поэтому так уязвим баланс наносов в
береговой зоне. Если человек выбирает рыхлый материал, это неминуе-
мо ведет к разрушительным последствиям: исчезают пляжи, а вместе с
ними дороги и строения.
Особую проблему создает переудобрение морских мелководий,
вызываемое избытком растворенных в воде питательных солей. Послед-
ние попадают в море в результате смыва с полей минеральных удобре-
ний. Парадокс состоит в том, что, в то время как биопродуктивность
большинства морских экосистем сдерживается недостатком биогенных
солей, человек навязывает традиционно малоплодородным морским
экосистемам избыточное минеральное питание. Экосистемы отвечают
на это сначала бурной вспышкой биопродуктивности специфических
групп гидробионтов, а затем их гибелью. Уязвимым звеном становится
быстрое потребление кислорода. Без растворенного в воде кислорода
гибнут практически все животные морских мелководий. Разложение
трупов ведет к сероводородному заражению, которое завершает превра-
щение прибрежных экосистем в черную зловонную мертвую зону.
Контакт океана с атмосферой. Эта граничная поверхность может
рассматриваться широко, тогда к ней относится вся фотическая зона,
или узко, тогда это поверхностная пленка и первые сантиметры воды.
Ю.П. Зайцев (1974) выделил особую зону жизни в океане (поверхность
раздела океан — атмосфера), назвав ее гипонейсталь. Эта зона исключи-
тельно богата так называемым неживым органическим веществом. Его
роль для всех обитателей океана весьма велика — это готовая пища, ис-
точник биостимуляторов или ингибиторов. Гипонейсталь хорошо осве-
щается солнцем, в том числе биологически активными инфракрасными
и ультрафиолетовыми лучами; она насыщена кислородом. Особое сти-
мулирующее влияние на рост и развитие живых организмов оказывает
взбиваемая ветром на поверхности моря пена. В верхних 5 см морской
воды наблюдается скопление своеобразных форм жизни; свыше 90%
беспозвоночных и рыб, обитающих в пределах мелководий и в открытой
части океана, используют поверхностную пленку как своеобразный ин-
370 кубатор для откладывания икры и выращивания молоди (рис. 11.5).
ГЛАВА 11. УЯЗВИМЫЕ ЗВЕНЬЯ ЭКОЛОГИЧЕСКОЙ СИСТЕМЫ МИРОВОГО ОКЕАНА
Рис. 11.5. Обитатели поверхности раздела океана и атмосферы,
по Ю.П. Зайцеву: а — физалия, или португальский кораблик, и сопутствующие
ей рыбки номеус; б — парусник; в — медуза-порпита; г — малек кефали;
д — икринка кефали; е — личинка краба
Главные загрязнители поверхности океана — нефть и нефтепродукты.
Экологической катастрофой грозят аварии супертанкеров, перевозящих по
несколько сотен тысяч тонн нефти. Но основная масса нефтепродуктов
сносится в океан с суши. Около 60% загрязнений дает автотранспорт. Это
мытье в ручьях автомашин, слив бензина и масла. Капля бензина создает
на поверхности воды крут пленки диаметром 30 см. Мальку рыбы, едва он
появился из икринки, чтобы заполнить плавательный пузырь, необходи-
мо сделать глоток воздуха. Малек поднимается к поверхности, но там неф-
тяная пленка. Малек делает глоток и погибает.
Нефтяные пленки, образуемые на поверхности моря разного рода
нефтепродуктами, — настоящее бедствие для обитателей гипонейстали.
Эти пленки препятствуют газообмену между океаном и атмосферой, что
ведет к дефициту кислорода в воде, экранируют солнечные лучи, практи-
чески лишая организмы биологически активных инфракрасных и ультра-
фиолетовых лучей. Многие углеводородные соединения являются силь-
нодействующими ядами, к которым особенно чувствительны икра и
молодь организмов, составляющих основу жизни в гипонейстали. 371
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ КАРТИ В А МИРА
В.И. Вернадский (I960), говоря о поверхностной пленке жизни, имел
в виду более мощный, освещаемый Солнцем (фотический) слой океана
глубиной около 100 м. С ним связана жизнедеятельность фитопланкто-
на — первого звена трофической цепочки в океане.
А.П. Лисицын коренным образом пересмотрел значение живого
вещества для осадочного процесса. Если прежде основная роль отводи-
лась обломкам горных пород, выносимым в океан материковым стоком,
то теперь установлено, что морские мелководья захватывают практи-
чески весь твердый сток с суши. Ведущую роль в питании дна откры-
того океана осадочным материалом А.П. Лисицын отводит его верхне-
му слою, где благодаря жизнедеятельности организмов, главным
образом диатомовых водорослей, ежегодно возникает огромное коли-
чество взвеси, во много раз превосходящее массу осадочного материа-
ла, поступающую с континентов. Вместе с осадками из зоны жизне-
деятельности фитопланктона уносятся на дно загрязняющие вещества.
Это очищает поверхность океана, но загрязняет его дно.
Следует выделить особую роль биофильтра, которую выполняет зоо-
планктон. Для удовлетворения суточного рациона организмы зоопланк-
тона отфильтровывают около 100 км3 воды в сутки. Вся зона планктона
в океане отфильтровывается всего за 20 суток. Образуемые фильтрато-
рами пищевые комочки — пеллеты — оседают на дно. Понятно, что на-
рушение экологического равновесия в фотической зоне, даже частичная
гибель зоопланктона, может привести к сбоям в механизме самоочище-
ния вод океана, а это еще больше усугубит кризисные явления.
В последние годы доказано участие атмосферных переносов в за-
грязнении Мирового океана. Так, вещества континентального проис-
хождения попадают в океан в результате переноса ветром пыли. По экс-
периментальным данным, среднегодовой поток пыли на поверхность
Тихого океана в его экваториальной части, например, составляет 15 —
30 мкг/м2, что соизмеримо со среднегодовой скоростью осаждения
взвешенных частиц в морской воде (50 мкг/м2). Это подтверждает поло-
жение о том, что атмосферные потоки играют большую роль в процес-
сах морской седиментации и формировании донных отложений в от-
крытой части Мирового океана.
Из атмосферы на поверхность океана оседают вещества, которые
необходимы для развития бактерионейстона — «живого фильтра».
Общий поток важнейшего биогенного элемента фосфора в океан до-
стигает 1,4 — 103 т/год.
Свинец, ртуть и другие тяжелые металлы, а также используемые в
сельском хозяйстве ядохимикаты, низкомолекулярные нефтяные угле-
водороды и иные органические вещества в газообразной фазе или во
взвешенном состоянии переносятся атмосферными потоками на десят-
ки тысяч километров.
Контакт океана с дном. Срединно-океанические хребты, а точнее,
рифтовые разломы, образующиеся в результате расхождения литосфер-
ных плит, занимают ключевое положение в геохимии океана. Здесь на
глубине около 4000 м при давлении 400 атм происходит постоянный кон-
такт расплавленной магмы с холодными придонными водами. Средин-
372 но-океанические хребты — это мощные химические реакторы, где во
ГЛАВА 11. УЯЗВИМЫЕ ЗВЕНЬЯ ЭКОЛОГИЧЕСКОЙ СИСТЕМЫ МИРОВОГО ОКЕАНА
взаимодействие вступают атомы всех известных находящихся в земной
коре элементов. О масштабах геохимических процессов можно судить
по тому, что протяженность этого «реактора» около 800 тыс. км, а весь
объем океанической воды проходит через него в течение 3 млн лет.
Глубокие трещины — рифты можно сравнить с ранами в тонкой зем-
ной коре, через которые сочатся вязкие потоки лавы. Застывая в холод-
ной воде, они превращаются в причудливые нагромождения. Холодная
придонная вода (ее температура даже в тропических широтах едва пре-
вышает 0 °C), проходя сквозь пористые вулканические породы в недра,
нагревается, растворяет содержащиеся в горных породах металлы, обо-
гащается вулканическими газами. Горячая вода под большим давлением
устремляется вверх и выбрасывается в виде подводных гейзеров (гидро-
терм), температура которых достигает 300 — 400 °C. Благодаря высокому
содержанию сульфидов металлов вода гейзеров окрашена в черный цвет.
Она похожа на черный дым. Поэтому глубоководные рифтовые гейзеры
окрестили «черными курильщиками».
Гидротермы напоминают промышленный ландшафт с лесом труб ме-
таллургических и химических заводов, из которых валит ядовитый дым.
Такая картина на суше стала символом экологической катастрофы. Од-
нако в глубинах океана именно гидротермы в условиях полного мрака
и холода служат животворными источниками, создающими оазисы
жизни (рис. 11.6).
Рис. 11.6. Оазисы жизни на дне океана, по СВ. Галкину: а— край скалы
в зоне гидротермальной активности (стрелками показано истечение
флюидов). Видны сообщества вестиментифер (погонофор), двустворчатых
моллюсков и др. Грунт покрыт слоем бактериальных обрастаний;
б—гидротермальные постройки (их высота достигает 100 м),
склоны которых обросли разнообразными животными.
Стрелками показано просачивание флюида
Вблизи горячих источников кишат бесчисленные черви и червеоб-
разные существа — вестиментиферы; лежат «морские огурцы» (куку-
марии), ползают крабы, кружат рои креветок. Скалы облеплены круп-
ными, величиной с большую тарелку двустворчатыми моллюсками.
Ключ к разгадке тайны богатой органической жизни находится в
высоких концентрациях в водах гидротерм сероводорода и метана.
Миллиарды хемосинтезирующих бактерий питаются этими ядовиты-
ми соединениями. Высокая биологическая продуктивность обусловле- 373
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ К АРТИ В А МИРА
на прежде всего симбиозом животных с бактериями. Такой способ пи-
тания дает ряд преимуществ: бактерии, синтезирующие органические
соединения, вводят питательные вещества непосредственно в клетки.
Когда горячий источник иссякает (он существует несколько десятков
лет), ледяные, погруженные в вечный мрак глубины вновь становятся
безжизненными.
С «черными курильщиками» связано формирование обширного
класса рудных гидротермальных месторождений. Поднимаясь из недр,
гидротермы насыщаются не только метаном, сероводородом, но и сер-
нистыми соединениями металлов, главнейшими из которых являются
железо, цинк, медь, молибден, серебро, свинец, кобальт, марганец, ва-
надий, мышьяк и др. По сути дела, «дым» — это настоящая жидкая руда.
При контакте гидротермальных растворов с холодной придонной
водой происходят интенсивные геохимические реакции, в результате
которых многие минералы выпадают в осадок. В итоге гидротермаль-
ный источник обрастает осадочными породами, образующими гигант-
ские конусы высотой с 20-этажный дом (до 50 — 70 м). Отложения гид-
ротерм представляют собой весьма богатые полиметаллические руды.
Итак, дно океана — это мощный геохимический фильтр, связываю-
щий тяжелые металлы, поступающие из недр. Инженеры пытаются
решить вопрос о промышленной разработке металлоносных осадков.
Возникает опасение, что такого рода деятельность нарушит работу гео-
химического фильтра, и тогда концентрация тяжелых металлов в водах
океана быстро возрастет, что представляет опасность как для гидроби-
онтов, так и для человека.
Проблемы охраны океана — прежде всего задача международного
сотрудничества. Об этом писал известный ученый и путешественник Тур
Хейердал: «Я призываю отказаться от близоруких личных и нацио-
нальных мерок, призываю осознать огромную ответственность перед
нынешним и грядущим поколениями. Морские течения не считаются с
политическими границами. Государства могут делить между собой
сушу, но океан — не знающий неподвижности океан, без которого
невозможна жизнь, — всегда будет общим и неделимым достоянием
человечества».
Вместо предисловия
УЧЕНИЕ В БИОСШЕРЕ ОХРЯ» ПРИРОДЫ
В бескрайних далях Вселенной голубая
планета Земля остается пока единствен-
ной известной науке планетой, на кото-
рой имеются условия, благоприятные для
существования жизни и формирования
биосферы.
Особая роль в развитии учения о био-
сфере принадлежит великому русскому
ученому В.И. Вернадскому. «Своеобраз-
ным, единственным в своем роде, отлич-
ным и неповторяемым в других небесных
телах представляется нам лик Земли — ее
изображение в космосе, вырисовывающе-
еся извне, со стороны, из дали бесконеч-
ных небесных пространств» — так начи-
нает Вернадский свою книгу. Вся она
пронизана идеей взаимодействия не толь-
ко земных, но и космических тел и явле-
ний. И главную роль среди них играют живые
В.И. Вернадский
(1863-1945)
организмы, «живое ве-
щество» планеты.
Небольшая книга В.И. Вернадского «Биосфера» вышла в свет в 1926 г.
Он написал ее в возрасте 63 лет. Понимание идей Вернадского пришло
только в 60-е годы XX в. Оно крепло по мере осознания человечеством
угрозы экологического кризиса. Решение глобальных экологических
проблем невозможно без понимания законов, управляющих живыми
организмами в биосфере.
Вернадский раскрывает ведущую роль живых организмов в транс-
формации солнечной энергии и преобразовании веществ, слагающих
наружные оболочки Земли: «По существу, биосфера может быть рас-
сматриваема как область земной коры, занятая трансформаторами,
переводящими космические излучения в действенную земную энер-
гию, — пишет Вернадский. — ...Лучи Солнца обусловливают главные
черты механизма биосферы... Солнцем в корне переработан и изменен
лик Земли, пронизана и охвачена вся биосфера».
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ КАРТИ В А МИРА
Живое вещество, по словам Вернадского, выполняет космическую
функцию, связывая Землю с космосом: «Вещество биосферы благодаря
им (солнечным лучам. — К.П.) проникнуто энергией; оно становится
активным, собирает и распределяет в биосфере полученную в форме
излучений энергию, превращает ее в конце концов в энергию в земной
среде свободную, способную производить работу... Лик Земли ими
меняется, ими в значительной мере лепится. Он не есть только отраже-
ние нашей планеты, проявление его вещества и ее энергии — он одно-
временно является и созданием внешних сил космоса». Развитием уче-
ния о биосфере стала концепция ноосферы, в которой заключена мысль j
о том, что человечество благодаря своему разуму становится мощным
фактором, преобразующим лик Земли.
Что ожидает людей — благоденствие, опирающееся на достижения
научно-технического прогресса, или глобальная экологическая катаст-
рофа в результате замены естественных процессов техническими?
Чтобы переход из биосферы в ноосферу не оказался несбыточной уто-
пией, коллективный разум людей должен быть направлен на охрану
природы и создание общественных мер экологической безопасности.
Биосфера — главная арена жизни живых существ, а также жизни и
хозяйственной деятельности человека. Биосфера включает нижнюю
часть воздушной оболочки (атмосферы), всю водную оболочку (гидро-
сферу) и верхнюю часть твердой оболочки (литосферы) (рис. 1).
37G
Рис. 1. Распределение живых организмов в биосфере
ВМЕСТО ПРЕДИСЛОВИЙ. УЧЕНИЕ 0 01 ОСЛЕРЕ И ОХРАНА ПРИРОДЫ
Вернадский четко обозначает границы биосферы — верхний и ниж-
ний пределы распространения жизни. Верхний — обусловливается
приходящей из космоса лучистой энергией, губительной для живых
существ. Речь идет о жестком ультрафиолетовом излучении; оно задер-
живается озоновым экраном, нижняя граница которого проходит на
высоте около 15 км — это верхняя граница биосферы.
Нижний предел жизни связан с повышением температуры в зем-
ных недрах. На глубине 3 — 3,5 км температура достигает 100 "С. Наи-
большую мощность биосфера имеет в океане: от поверхности до мак-
симальных глубин в нем обитают живые существа. *
Для биосферы характерно не только присутствие живого вещества.
Она обладает также следующими тремя особенностями: во-первых, на
нее падает мощный поток солнечной энергии; во-вторых, в ней в зна-
чительном количестве содержится жидкая вода; в-третьих, в биосфере
проходят поверхности раздела между веществами, находящимися в трех
фазах — твердой, жидкой и газообразной. Все это служит предпосыл-
кой для активного обмена веществом и энергией, в котором большую
роль играют организмы.
И 1. Функции живого вещества и механизмы устойчивости биосферы_____
Все виды устойчивости, наблюдаемые в живых организмах и био-
сфере в целом, не являются статическими, а достигаются за счет непре-
рывных процессов, активно препятствующих любой тенденции к нару-
шению этого постоянства. Устойчивость всего живого есть непрерывная
борьба за существование. Ключ к загадке, которую представляет для
человека красота и гармония органического мира, заключается в одном
слове — это слово «смерть». Смерть, рано или поздно пресекающая все
уродливое, все бесполезное, все несогласное с окружающими условия-
ми. И если эта вечная борьба, это бесконечное истребление невольно
вселяют в душу ужас, то мы не должны забывать, что
...у гробового входа
Младая будет жизнь играть;
И равнодушная природа
Красою вечною сиять.
А. С. Пушкин
Ключевое положение в понимании механизмов устойчивости био-
сферы занимает изучение функций живого вещества.
Функции живого вещества. Чтобы более доходчиво раскрыть сущ-
ность процессов, протекающих в биосфере и поддерживающих ее устой-
чивость, рассмотрим основные функции живого вещества: энергетиче-
скую, разрушительную, накопительную и средообразующую (Лапо, 1987).
Энергетическая функция выполняется прежде всего растениями,
которые в процессе фотосинтеза аккумулируют солнечную энергию в
виде разнообразных органических соединений. По словам Вернадско-
го, зеленые хлорофилльные организмы, зеленые растения являются
377
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ КАРТИ В А МИРА
главным механизмом биосферы, который улавливает солнечный луч и
создает фотосинтезом химические тела — своеобразные солнечные
консервы, энергия которых в дальнейшем становится источником
действенной химической энергии биосферы, а в значительной мере —
всей земной коры.
Внутри экосистемы эта энергия в виде пищи распределяется между
животными. Частично энергия рассеивается, а частично накапливает-
ся в отмершем органическом веществе и переходит в ископаемое состоя-
ние. Так образовались залежи торфа, каменного угля, нефти и других
горючих полезных ископаемых, служащие в настоящее время энерге-
тической базой для жизни и работы людей. Растения — главный источ-
ник пищи для людей и животных.
Разрушительная (деструктивная) функция состоит в разложении,
минерализации мертвого органического вещества, химическом разло-
жении горных пород, вовлечении образовавшихся минералов в биоти-
ческий круговорот.
Мертвое органическое вещество разлагается до простых неорганиче-
ских соединений (углекислого газа, воды, сероводорода, метана, аммиака
и т. д.), которые вновь используются в начальном звене круговорота. Этим
занимается специальная группа организмов — редуценты (деструкторы).
Особо следует сказать о химическом разложении горных пород. Бла-
годаря живому веществу биотический круговорот пополняется минера-
лами, высвобождаемыми из литосферы. Например, плесневый грибок в
лабораторных условиях за неделю высвобождал из вулканической гор-
ной породы 3% содержащегося в ней кремния, 11% алюминия, 59% маг-
ния, 64% железа. Пионеры жизни на скалах — бактерии, сине-зеленые
водоросли, грибы и лишайники — оказывают на горные породы силь-
нейшее химическое воздействие растворами целого комплекса кислот —
угольной, азотной, серной и разнообразных органических. Разлагая с их
помощью те или иные минералы, организмы избирательно извлекают и
включают в биотический круговорот важнейшие питательные элемен-
ты — кальций, калий, натрий, фосфор, кремний, микроэлементы.
Общая масса зольных элементов, вовлекаемая ежегодно в биотиче-
ский круговорот только на суше, составляет около 8 млрд т. Это в не-
сколько раз превышает массу продуктов извержения всех вулканов мира
на протяжении года. Благодаря жизнедеятельности организмов-деструк-
торов создается уникальное свойство почв — их плодородие.
Накопительная (концентрационная) функция заключается в из-
бирательном накоплении организмами в процессе их жизнедеятельно-
сти атомов веществ, рассеянных в природе. Способность концентри-
ровать элементы из разбавленных растворов — это характерная
особенность живого вещества. Наиболее активными концентратора-
ми многих элементов являются микроорганизмы. Например, в продуктах
жизнедеятельности некоторых из них по сравнению с природной средой
содержание марганца увеличено в 1 200 000 раз, железа — в 65 000,
ванадия — в 420 000, серебра — в 240 000 раз.
Морские организмы активно концентрируют рассеянные минера-
лы для построения своих скелетов или покровов. Существуют, напри-
мер, кальциевые организмы (известковые водоросли, моллюски, корал-
378
ВМЕСТО ПРЕДИСЛОВИЙ. УЧЕНИЕ 0 01 ОСЛЕРЕ И ОХРАНА ПРИРОДЫ
лы, мшанки, иглокожие и т. п.) и кремниевые (диатомовые водоросли,
кремниевые губки, радиолярии). Особое внимание следует обратить
на способность морских организмов накапливать микроэлементы,
тяжелые металлы, в том числе ядовитые (ртуть, свинец, мышьяк), ра-
диоактивные элементы. Их концентрация в теле беспозвоночных и рыб
может в сотни тысяч раз превосходить содержание в морской воде
(рис. 2). Вследствие этого морские организмы полезны как источник
микроэлементов, но вместе с тем употребление их в пишу может гро-
зить отравлением тяжелыми металлами или быть опасным в связи с
повышенной радиоактивностью.
Рис. 2. Пирамида чисел в экосистеме океана, показывающая процесс
биогенной концентрации загрязняющих веществ, по CM. Woodwell:
а — пирамида биомасс; б — перевернутая пирамида концентрации
загрязняющих веществ (ДДТ) в звеньях пищевой цепи
Средообразующая функция состоит в трансформации физико-хи-
мических условий среды (литосферы, гидросферы, атмосферы) в усло-
вия, благоприятные для существования организмов.
Можно сказать, что средообразующая функция есть совместный
результат всех рассмотренных выше функций живого вещества: энер-
гетическая функция обеспечивает энергией все звенья биотического
круговорота (в ходе фотосинтеза растения выполняют газовую функ-
цию: поглощают углекислый газ и выделяют кислород); деструктив-
ная и концентрационная способствуют извлечению из природной
среды и накоплению рассеянных, но жизненно важных для организ-
мов элементов.
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ КАРТИ В А МИРА
Средообразующие функции живого вещества создали и поддержива-
ют баланс вещества и энергии в биосфере, обеспечивая стабильность
условий существования организмов, в том числе человека. Вместе с тем
живое вещество способно восстанавливать условия обитания, нарушен-
ные в результате природных катастроф или антропогенного воздействия.
Эту способность живого вещества к восстановлению благоприятных
условий существования выражает принцип Ле Шателье, заимствован-
ный из области термодинамических равновесий. Он заключается в том,
что изменение любых переменных в системе в ответ на внешние возму-
щения происходит в направлении компенсации производимых возмуще-
ний. В теории управления аналогичное явление носит название отрица-
тельных обратных связей. Благодаря этим связям система возвращается
в первоначальное состояние, если производимые возмущения не превы-
шают пороговых значений. Например, на повышение содержания угле-
кислого газа в атмосфере биосфера отвечает усилением фотосинтеза,
который:снижает концентрацию СО2. Таким образом, устойчивость био-
сферы оказывается явлением не статическим, а динамическим.
В результате средообразующей функции в географической оболоч-
ке произошли следующие важнейшие события: был преобразован газо-
вый состав первичной атмосферы; изменился химический состав вод
первичного океана; образовалась толща осадочных пород в литосфере;
на поверхности суши возник плодородный почвенный покров (также
плодородны воды океана, рек и озер).
Чистота морских вод — результат фильтрации, осуществляемой
разнообразными организмами, но особенно зоопланктоном. Большин-
ство из этих организмов добывает пищу, отцеживая из воды мелкие
частицы. Работа их настолько интенсивна, что весь океан очищается от
взвеси за 4 года. Озеро Байкал исключительной чистотой своих вод во
многом обязано веслоногому рачку эпишуре, который за год трижды
процеживает его воду.
Вернадский объясняет парадокс: почему, несмотря на то что общая
масса живого вещества — пленка жизни, покрывающая Землю, — нич-
тожно мала, результаты жизнедеятельности организмов сказываются
на составе и литосферы, и гидросферы, и атмосферы?
Если живое вещество распределить на поверхности Земли ровным
слоем, его толщина составит всего 2 см. При такой незначительной массе
организмы осуществляют свою планетарную роль благодаря весьма
быстрому размножению, т. е. весьма энергичному круговороту веществ,
связанному с этим размножением.
Масса живого вещества, соответствующая данному моменту вре-
мени, с трудом сопоставляется с тем грандиозным ее количеством,
которое производило свою работу в течение сотен миллионов лет
существования организмов. Если рассчитать всю массу живого веще-
ства, воспроизведенного за это время биосферой, она окажется равной
2,4 х Ю20т. Это в 12 раз превышает массу земной коры.
На земной поверхности нет химической силы, более постоянно дей-
ствующей, а потому и более могущественной по своим конечным резуль-
татам, чем живые организмы, взятые в целом. Глины, известняки, доло-
380 миты, бурые железняки, бокситы — это все породы органогенного
ВМЕСТО ПРЕДИСЛОВИЙ. УЧЕНИЕ 0 01 ОСЛЕРЕ И ОХРАНА ПРИРОДЫ
происхождения. Наконец, свойства природных вод, соленость Мирово-
го океана и газовый состав атмосферы определяются жизнедеятельно-
стью населяющих планету существ.
Рассмотрим влияние средообразующей функции организмов на
содержание кислорода и углекислого газа в атмосфере. Напомним, что
повышение концентрации СО2 в атмосфере вызывает «парниковый
эффект» и способствует потеплению климата. Свободный кислород
выделяется при фотосинтезе. Впервые на Земле массовое развитие
фотосинтезирующих организмов — сине-зеленых водорослей — имело
место 2,5 млрд лет назад. Благодаря этому в атмосфере появился кисло-
род, что дало импульс быстрому развитию животных. Однако интен-
сивный фотосинтез сопровождался усиленным потреблением С02 и
уменьшением его содержания в атмосфере. Это привело к ослаблению
«парникового эффекта», резкому похолоданию и первому в истории
планеты (гуронскому) оледенению. В наши дни накопление в атмосфе-
ре углекислого газа от сжигания углеводородного топлива рассматрива-
ется как тревожная тенденция, ведущая к потеплению климата, таянию
ледников и грозящая повышением уровня Мирового океана более чем
на 100 м. В связи с этим следует отметить функцию захвата и захоро-
нения избыточной углекислоты морскими организмами путем перево-
да ее в соединения углекислого кальция, а также путем образования
биомассы живого вещества.
Биотические круговороты. Устойчивое функционирование экосис-
тем и выполнение ими разнообразных функций возможно только при
условии соблюдения закона постоянства вещества и энергии, который
реализуется в биотических круговоротах. Глобальные циклы миграции
химических элементов в биосфере связывают наружные оболочки на-
шей планеты (атмосферу, гидросферу и литосферу) в единое целое,
обеспечивая, с одной стороны, ее устойчивость, а с другой — непре-
рывную эволюцию ее состава.
К. Бэр установил закон бережливости. Вернадский очень образно
формулирует этот закон.
Закон бережливости: атомы, вошедшие в какую-нибудь форму жи-
вого вещества, захваченные единичным жизненным вихрем, с трудом
возвращаются, а может быть, и не возвращаются назад, в косную
материю биосферы.
Благодаря закону бережливости можно говорить об атомах, оста-
ющихся в пределах живой материи в течение геологических периодов,
все время находящихся в движении и миграции, но не выходящих назад
в косную материю. Иными словами, основу функционирования живо-
го вещества составляет биотический круговорот веществ.
Биотический круговорот обеспечивается взаимодействием трех основ-
ных групп организмов: 1) продуцентов — зеленых растений, осуще-
ствляющих фотосинтез, и бактерий, способных к хемосинтезу; они
создают первичное органическое вещество; 2) консументов, потребля-
ющих органическое вещество; это растительноядные и хищные живот-
ные; 3) редуцентов, разлагающих мертвое органическое вещество до ми-
нерального; это в основном бактерии, грибы и простейшие животные
(рис. 3).
381
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ КАРТИ В А МИРА
Растителаноядные
(консументы {порядка)
образные бещ^
|Л?ллгг««ая
’ НгО^СОз '
Зеленые растения
(продуценты)
Хищники
Л________1_______
|gca<?owwg породы]
Рис. 3. Модель биотического круговорота веществ
Хищники
(консументы II порядка)
органические
остатки
В восходящей части биотического круговорота, основанного на вы-
полнении энергетической функции зелеными растениями, происходит
накопление солнечной энергии в виде органических веществ, синтези-
руемых растениями из неорганических соединений — углекислого газа,
воды, азота, зольных элементов питания. Нисходящая часть биотиче-
ского круговорота связана с потерями органического вещества. Важней-
ший процесс — дыхание растений, при котором наполовину ассимили-
рованное при фотосинтезе органическое вещество окисляется до С02
и возвращается в атмосферу. Второй существенный процесс расходова-
ния органического вещества и накопленной в нем энергии — это по-
требление растений животными. Запасаемая с пищей энергия также в
значительной мере расходуется на дыхание, жизнедеятельность, раз-
множение, выделяется с экскрементами.
Таким образом, биотический круговорот представляет собой непре-
рывный процесс создания и деструкции (разрушения) органического
вещества. Он реализуется при участии представителей всех трех групп
организмов: без продуцентов невозможна жизнь, поскольку лишь они
производят основу жизни — первичное органическое вещество; консу-
менты разных порядков, потребляя первичную и вторичную продук-
цию и переводя органическое вещество из одной формы в другую, спо-
собствуют возрастанию многообразия форм жизни на Земле; наконец,
редуценты, разлагая органическое вещество до минерального, возвра-
щают его к началу круговорота. В биотическом круговороте помимо
образующих органическое вещество элементов (кислород, углерод, во-
дород) принимает участие большое число биологически важных элемен-
тов (азот, кальций, натрий, калий, кремний, фосфор, сера) и минераль-
ных веществ (бром, йод, цинк, серебро, молибден, медь, магний, свинец,
кобальт, никель). Список элементов, поглощающихся живым веществом,
можно значительно расширить, причем в него входят даже ядовитые
элементы (ртуть, мышьяк) и радиоактивные. Глобальные циклы мигра-
382
ВМЕСТО ПРЕДИСЛОВИЙ. УЧЕНИЕ 0 01 ОСЛЕРЕ И ОХРАНА ПРИРОДЫ
ции химических элементов не только связывают три наружные обо-
лочки нашей планеты в единое целое, но и обусловливают непрерыв-
ную эволюцию ее состава (рис. 4).
Рис. 4. Круговорот углерода, кислорода, водорода, азота и серы в биосфере,
по «Биосфере»
Отметив циклический характер массоэнергообмена, ответим на во-
прос о скорости круговорота различных веществ в биосфере. Все живое
вещество биосферы обновляется в среднем за 8 лет. В океане циркуля-
ция идет во много раз быстрее: вся масса живого вещества обновляется
за 33 дня, а масса фитопланктона — каждый день. В атмосфере смена
кислорода происходит за 106 лет, углекислого газа за 6,3 года. Процесс
полной смены вод в гидросфере осуществляется за 2800 лет, а время,
необходимое для фотосинтетического разложения всей массы воды,
исчисляется 5 — 6 млн лет.
И 2. Экологические проблемы человечества__________________________
Человечество постепенно приходит к осознанию того, что индустриа-
лизация, если ее осуществлять без оглядки на биосферные процессы,
порождает явления, разрушительный потенциал которых сопоставим с
последствиями применения ядерного оружия. Экологический фронт
проходит по самому переднему краю борьбы за выживание человече-
ства наравне с регулированием региональных конфликтов, преодолени-
ем экономической отсталости и т. п.
В 1992 г. в Рио-де-Жанейро на уровне глав государств и правительств
проходила Конференция ООН по окружающей среде и развитию, ко-
торая констатировала невозможность движения развивающихся бед-
ных стран по пути, которым пришли к своему благополучию развитые
богатые страны. Признано, что эта модель ведет к катастрофе: Земля не 383
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ К АРТИ В А МИРА
выдержит столь мощного потребления ее ресурсов и загрязнения окру-
жающей среды.
В связи с этим провозглашена необходимость перехода мирового
сообщества на рельсы устойчивого развития (sustainable development).
Заметим, что широко вошедшее в обиход понятие «устойчивое разви-
тие» — весьма неточный перевод англоязычного термина. Sustain —
поддерживать, подкреплять; так что более точно sustainable development
переводится как поддерживающее или сбалансированное развитие.
Однако, учитывая, что в русскоязычной литературе уже укоренилось
понятие «устойчивое развитие», будем продолжать пользоваться этим
термином.
Международной комиссией по окружающей среде и развитию в
1987 г. дано следующее определение.
Устойчивое развитие — это такое развитие, которое удовлетворяет
потребности настоящего времени, но не ставит под угрозу способность
будущих поколений удовлетворять свои собственные потребности.
Особенности экологической ситуации, в которую постепенно втяги-
вается человечество, начали привлекать внимание широкой общественно-
сти в конце 60-х годов XX в. В популяризации этих проблем большую роль
сыграл так называемый Римский клуб — группа бизнесменов, юристов,
ученых, объединенных общим интересом к перспективам развития че-
ловеческого сообщества, проблемам, порождаемым неконтролируемым
ростом населения, разрушением биосферы и непрерывным уменьшени-
ем природных ресурсов. Начало исследованиям было положено работой
Дж. Форрестера «Мировая динамика». Затем последовала целая серия
публикаций, среди которых наибольшую известность получили работы
«Пределы роста» и «За пределами роста», выполненные под руководством
Д. Медоуза (Meadows etc., 1972, 1992). Они превратили глобальную эколо-
гию в одну из самых популярных научных проблем.
Компьютерный анализ мировых трендов (рис. 5) показал, что при
сохранении современных тенденций развития общества какое-то вре-
мя еще будут продолжаться рост численности населения, промышлен-
ности и производства продуктов питания, а также будут возрастать
загрязнение окружающей среды и сокращаться природные ресурсы
(рис. 5, а). Затем численность населения начнет быстро уменьшаться.
Это типичная ситуация развития популяции, когда источники благо-
приятных для жизни условий ограничены.
На рис. б изображена гипотетическая картина роста народонаселе-
ния (силуэт человека) на фоне сокращающихся ресурсов биосферы
(силуэт дерева) и производства сельскохозяйственных продуктов (ко-
лос злака). Уже в наши дни биосфера и сельское хозяйство не в состоя-
нии обеспечить продуктами питания все население: каждый четвертый
житель планеты голодает. Понятно, что ситуация, при которой народо-
население в несколько раз будет превосходить ресурсы биосферы, не-
возможна.
Если человечество не остановит быстрый рост рождаемости, приро-
да сделает это решительно и жестко. Вслед за демографическим взры-
вом неизбежно наступит коллапс: народонаселение резко сократится
384 вследствие того, что ресурсы будут исчерпаны, а среда загрязнена. Затем
ВМЕСТО ПРЕДИСЛОВИЙ. УЧЕНИЕ 0 01 ОСЛЕРЕ И ОХРАНА ПРИРОДЫ
Рис. 5. Компьютерный анализ мировых трендов, по D.H. Meadows:
а — модель развития при сохранении современных тенденций развития
общества: по инерции население и загрязнение окружающей среды будут
еще некоторое время увеличиваться после пика индустриализации, однако
в конце концов наступит драматически быстрое сокращение численности
населения; б— модель развития, предполагающая нулевой прирост населе-
ния: в этом случае возможен переход на рельсы устойчивого развития:
1 — ресурсы; 2 — производство продуктов питания на душу населения;
3 — численность населения; 4 — производство промышленной продукции
на душу населения; 5 — загрязнение
Рис. 6. Гипотетическая картина роста народонаселения с 1980 по 2020 г.
на фоне сокращающихся ресурсов биосферы
и производства сельскохозяйственных продуктов
начнется постепенный процесс восстановления возобновимых природ-
ных ресурсов и самоочищения окружающей среды; численность насе-
ления стабилизируется на более низком уровне. Ученые говорят о «зо-
лотых четырех миллиардах» — количестве жителей, при котором
возможно управляемое взаимодействие общества и природы.
Переход на рельсы устойчивого развития невозможен в условиях
дальнейшего увеличения численности людей и роста потребления. Вы-
ход трендов на плато произойдет только при уменьшении численности
и нулевом приросте населения, применении ресурсо- и энергосберегаю-
щих чистых технологий (рис. 5, б). Однако сокращение невозобнови-
мых природных ресурсов будет продолжаться, что грозит грядущим
обострением ситуации. 385
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ КАРТИ В А МИРА
И 3. Ноосфера иди техносфера?______________________________________
Ноосфера (от греч. noos — разум) — сфера разума, буквально «мыс-
лящая оболочка». Термин введен в науку в конце 20-х годов XX в. Одна-
ко до сих пор представления о ноосфере остаются крайне противоречи-
выми. Ноосферное учение признается, с одной стороны, как величайшее
научное достижение, более того, как основной закон социальной эколо-
гии, с другой — как светлая, но зыбкая мечта об управляемой челове-
ческим разумом окружающей среде.
В концепции ноосферы сложно переплелись материалистические и
религиозно-философские взгляды на роль и предназначение человече-
ства, человеческой мысли в окружающем мире. Для христианского
мировоззрения, веками формировавшегося под знаком безусловного
права человека на владение всеми богатствами природы, дарованного
свыше, она естественна и закономерна.
Немаловажную роль в зарождении ноосферных идей сыграли рус-
ские философы-космисты, особенно Н.Ф. Федоров и С.Н. Булгаков.
Первый из них в своем труде «Философия общего дела», опубликован-
ном в 1906 г., заявляет, что главная цель общего дела человечества состо-
ит в управлении слепыми, хаотичными силами природы: «...нет в приро-
де целесообразности... ее должен внести сам человек, и в этом заключается
высшая целесообразность». Средством для наведения порядка в природе
должна стать хозяйственная деятельность.
Эту мысль развивает в 1912 г. С.Н. Булгаков в работе «Философия
хозяйства». Он пишет, что хозяйственный труд есть уже как бы новая
сила природы, новый мирообразующий, космогонический фактор,
принципиально отличный от всех остальных сил природы. Человек
создает как бы новый мир, новые блага, новые знания, новые чувства,
новую красоту — он творит культуру. Рядом с миром естественным
создается мир искусственный, творение человека, и этот мир новых сил
и новых ценностей увеличивается от поколения к поколению.
Главным творцом ноосферной концепции стал В.И. Вернадский.
Двадцатилетним студентом Санкт-Петербургского университета, рабо-
тая над рефератом, он записывает мысль, ставшую впоследствии одной
из ведущих в его научном поиске: «Человек настоящего времени пред-
ставляет из себя геологическую силу; сила эта все возрастает и предела
ее возрастания не видно».
Появление Человека разумного стало итогом направленного разви-
тия живой природы. Современник Дарвина североамериканский уче-
ный Д.Д. Дана пришел к выводу, что эволюция живого вещества идет в
направлении обособления и формирования головного отдела централь-
ной нервной системы. Это явление он назвал цефализацией (от греч.
kefale — голова). Дана указал, что в ходе геологического времени на-
блюдается скачкообразное усовершенствование центральной нервной
системы (мозга), начиная от ракообразных и моллюсков и кончая че-
ловеком (принцип Дана).
Ключевые положения концепции В.И. Вернадского следующие: а) че-
ловечество — великая геологическая сила; б) эта сила есть разум и воля
38G человека как существа социально организованного; в) лик планеты из-
ВМЕСТО ПРЕДИСЛОВИЙ. УЧЕНИЕ 0 01 ОСЛЕРЕ И ОХРАНА ПРИРОДЫ
менен человеком настолько глубоко, что оказались затронутыми ее био-
геохимические круговороты; г) человечество эволюционирует в сторо-
ну обособления от остальной биосферы.
Закономерный и неотвратимый характер перехода биосферы в стадию
ноосферы лежит в основе социального оптимизма Вернадского. Он счи-
тал науку той великой силой, которой удастся сделать то, что не удалось
философии, религии, политике, — объединить человечество. «Мы пережи-
ваем не кризис, волнующий слабые души, — писал Вернадский, — а вели-
чайший перелом научной мысли человечества, совершающийся лишь раз
в тысячелетия, переживаем научные достижения, равных которым не
видели долгие поколения наших предков... Стоя на переломе, охватывая
взором раскрывающееся будущее, мы должны быть счастливы, что нам
суждено это пережить, в создании такого будущего участвовать».
Вместе с тем Вернадский видел и предсказывал отрицательные по-
следствия антропогенного воздействия на природу. В геологической
истории биосферы перед человеком открывается огромное будущее,
если он поймет это и не будет употреблять свой разум и свой труд на
самоистребление. Лик планеты, пишет Вернадский, резко меняется
человеком сознательно и главным образом бессознательно: меняются
физически и химически почва, воздушная оболочка Земли, природные
воды суши, прибрежные моря и части океана.
Эволюция человеческого общества предстает как совокупная эволю-
ция умственных способностей человека, науки и культуры, освоения
все более эффективных источников энергии, орудий и технологий тру-
да; биосфера замещается техносферой (рис. 7).
С момента выделения предков человека из животного состояния на
протяжении почти 2 млн лет развивалось первобытное общество. Для
него были характерны: охота и собирательство, поддержание огня,
появление членораздельной речи. Численность населения была неболь-
шой. Человеческое общество еще являлось частью вмещающего ланд-
шафта. Разрушение биосферы носило узколокальный характер.
Около 6 — 8 тыс. лет назад люди перешли к производящему хозяй-
ству — земледелию и животноводству. Появляются высокоразвитые
цивилизации древней Месопотамии, Египта, Средиземноморья, Индии,
Китая и Центральной Америки. Возрастает численность населения.
Растут крупные города. Разрушение биосферы увеличивается от локаль-
ных до крупных региональных размеров.
Промышленный период, сопровождавшийся преобразованием био-
сферы в техносферу, начался примерно 200 лет назад. Развитию науки
и культуры способствовало изобретение книгопечатания. Появились
электронные средства связи — радио и телевидение. Сбор и обработка
информации стали осуществляться с помощью портативных, но мощ-
ных по своим возможностям компьютеров. Произошел демографиче-
ский взрыв. Экологический кризис достиг глобальных масштабов.
Итак, ноосфера или техносфера? Это предмет дискуссии, в которой
активное участие принимал крупнейший специалист в области этноге-
неза Л.Н. Гумилев. Он был противником идеи ноосферы (Гумилев, 1990).
Это несогласие заключено в его вопросе: так ли уж разумна «сфера
разума»? Водь ее развитие ведет к замене живых процессов. По словам
387
ЧАСТЬ III. БИОГЕОГРАФИЧЕСКАЯ КАРТИ В А МИРА
§
з
3
S
^Деструкция биосферы '^-.Ьиоинжёмеряя
L; Компьютеризация
2
пом
£.Хямнзацм
Ауд’ ;. Наука .
Пар-техяогенез
•**«*•**•
t-ЬЮигопеяат'эяиф-.?
20Э net
е
6-Втыс. лет
£
«г
г
7 о
2 е?
Ш О
з °
н
2
° Л
ъ «
<в £
8 S
t- Прямохождение
V Письмо
Культурная
эволюция-
обучение
L Пахота
Металлурги*»
L Рабы -скот
Мотыжное
земледелие
Адаптация Нопто-
формирование рас
LЗагонная охота
Огонь, 1,4 млн
лег
Гекетическая эволюция -
рост мозга
S
о
I
।
।
I
Рис. 7. Периодизация истории обществ — перехода от генетической
эволюции (биогенеза) к культурной (ноогенезу), поВ.А. Зубакову
Гумилева, человечество вырывает из природы частицы вещества и ввер-
гает их в оковы форм. Камни превращаются в пирамиды или Парфе-
нон, шерсть — в пиджаки, металл — в сабли и танки. А эти предметы
лишены саморазвития. В итоге, кроме развалин прежних цивилизаций
и мусорных свалок вокруг современных городов, ноосфера не дала че-
388 ловеку ничего.
ВМЕСТО ПРЕДИСЛОВИЙ. УЧЕНИЕ О БИОСФЕРЕ И ПХРАНД ПРИРОДЫ
Если Вернадский подчеркивал роль человече-
ства как единого целого, то Гумилев, наоборот,
обращает внимание на его пространственно-вре-
менную неоднородность, разделение человече-
ства на этносы — естественно сложившиеся
общности людей, характеризующиеся одинако-
выми нормами поведения, противопоставляю-
щие себя всем другим коллективам, исходя из
положительного или отрицательного ощущения
других этносов.
Население нашей планеты, разделенной на
государства, трудно назвать единым и разум-
ным целым. Поэтому вполне уместно сравне-
ние человечества с кораблем, терпящим круше-
ние: в спасательных шлюпках находятся жители
богатых стран, они затеряны среди бушующих
волн и окружены многочисленными тонущи-
ми народами — жителями бедных стран. Эти-
Л.Н. Гумилев
(1912—1992)
ка «спасательной шлюпки» столь же очевидна, сколь беспощадна: что-
бы кого-то спасти, его нужно взять на борт шлюпки, вместимость
которой ограничена. Спасти чью-то жизнь — значит пожертвовать
своей, бросившись в волны.
— 4. Причина кризиса - противоречив во взаимоотношениям
и общества и природы
Рассмотрим упрощенную модель взаимодействия общества и при-
роды (рис. 8).
Рис. 8. Обобщенная модель взаимодействия общества и природы
389
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ КАРТИ В А МИРА
Население (А), промышленное и сельскохозяйственное производство
(Б) имеют положительные связи. Население — это трудовые ресурсы,
необходимые для роста промышленности и сельского хозяйства; про-
мышленность обеспечивает человека материальными благами и содей-
ствует интенсификации сельского хозяйства; сельское хозяйство дает
продукты питания и сырье для промышленности.
Положительные связи блока А и Б являются причиной быстрого безу-
держного роста как населения, так и промышленности и сельского хозяй-
ства. Современная эпоха развития человеческого общества характеризу-
ется тенденцией к экспоненциальному (взрывному) росту численности
населения, потребления энергии, ресурсов, продуктов питания и т. п.
Отрицательные обратные связи, усиливающиеся по мере воздей-
ствия человека на окружающую среду, служат причиной ограничения
роста населения и хозяйства. Природные ресурсы положительно влия-
ют на развитие общества, но интенсивное потребление отрицательно
сказывается на их запасах и воспроизводстве. Воздействие общества на
природу вызывает загрязнение окружающей среды, отрицательно влия-
ет как на условия жизни людей, так и на качество природных ресур-
сов — чистоту почвы, воды, воздуха.
Одной из причин возникшего напряжения во взаимодействии обще-
ства и природы является укоренившееся в сознании людей потребитель-
ское отношение к природным ресурсам. Не одно тысячелетие человек
вырубал лес, охотился на диких зверей, использовал созданное природой
плодородие почв, ловил рыбу, расходовал чистую пресную воду. Сама
природа восстанавливала нарушенное в экосистемах равновесие, и это
породило у человека представление о безграничности природных ресур-
сов, о возможности брать их у природы, ничего ей не возвращая.
Экологические проблемы человечества имеют своими истоками
различия в процессах, совершающихся в природных и социально-эко-
номических системах. Для естественного ландшафта характерны про-
цессы самоуправления, ведущие к устойчивости (гомеостазу) экосисте-
мы — сохранению внутренних и внешних связей, обеспечивающих ее
устойчивость. Гомеостаз природных систем поддерживается, с одной
стороны, в результате постоянных контактов с внешней средой, из
которой система черпает энергию и вещество для своего функциониро-
вания. Это первый блок управления, отвечающий за устойчивость сис-
темы в условиях разнообразных внешних воздействий.
С другой стороны, гомеостаз системы поддерживается благодаря
внутренним процессам функционирования, осуществляющимся в виде
круговоротов вещества при примерно постоянных энергетических за-
тратах, преимущественно за счет рассеянных источников энергии. Это
второй блок управления, отвечающий за внутреннюю целостность сис-
темы. Оба блока управления — внешний и внутренний — находятся
«в руках» экосистем. Ведущая роль в гомеостазе природных систем
принадлежит функциям живого вещества, без которого системы быст-
ро деградируют; в них усиливается роль механических форм движения
вещества, структура их упрощается.
Если применить к прогнозу поведения природных систем второй
закон термодинамики, то можно сказать, что в них работают естествен-
390
ВМЕСТО ПРЕДИСЛОВИЙ. УЧЕНИЕ 0 01 ОСЛЕРЕ И ОХРАНА ПРИРОДЫ
ные механизмы чрезвычайно экономной передачи энергии с одного
уровня потребления на другой. Количество энергии, выделяемой сис-
темой в окружающую среду, незначительно. Природные системы бла-
годаря механизмам саморегулирования постоянно стремятся перейти
из состояния менее организованного в более организованное, из состоя-
ния менее устойчивого в более устойчивое. Энтропия (мера неупорядо-
ченности) в них естественным образом убывает.
В техногенных системах — городах, промышленных предприятиях —
массоэнергообмен резко отличается от процессов, протекающих в при-
роде. Ресурсные циклы производства и потребления открыты. Поток
вещества и энергии через производственную систему почти не имеет
обратной положительной связи. На входе — это все возрастающее по-
требление ресурсов и энергии, ведущее к деградации природных систем;
на выходе — огромное количество отходов и отбросов, поступающих в
окружающую среду, что является главной причиной ее загрязнения.
Энтропия техногенных систем имеет тенденцию к росту: возника-
ют аварии, нарушаются связи управления, усиливается хаос. Для под-
держания этих систем в рабочем состоянии, как показывает истори-
ческий опыт, требуются все увеличивающиеся энергетические затраты,
причем энергия нужна преимущественно в высококонцентрированном
виде. Рост производства, увеличение населения городов (урбанизация)
ведут к обострению экологических проблем. Сравнение основных про-
цессов, протекающих в биосфере и в обществе, представлено в табл. 1.
Таблица 1
Основные особенности процессов, протекающих в биосфере и обществе
Биосфера Общество
Регулирование чис; Борьба за существование. Жесткие, порой безжалостные механизмы, предотвраща- ющие вспышки численности отдельных видов Потреблен Эволюция биосферы от простей- ших форм до современного разно- образия жизни совершалась при относительно постоянном потоке рассеянной солнечной энергии М а с с о В основе функционирования эко-сис- тем лежат квазизамкнутые круговороты вещества. При этом ресурсный потенциал ландшафта не истощается, а окружающая среда не загрязняется Поддержание Природные системы обладают способно- стью самоорганизовываться, самовоостанавливаться, саморазвиваться. Энтропия уменьшается ценности популяций Гуманные принципы развития циви-ли- зованного общества. Жизнь челове-ка — высшая ценность. Быстрый рост народона- селения ие энергии Развитие цивилизации требует быстро- го роста потребления энергии. Главные энергетические ресурсы — нефть и газ — быстро истощаются. Надвигается угроза энергетического кризиса обмен В процессе производства человек исполь- зует природные ресурсы, постепенно подрывая их запасы. Отходы производства и жизнедеятельности людей загрязняют окружающую среду устойчивости Техногенные системы не обладают способ- ностью саморазвиваться. Предоставленные самим себе, они разрушаются. Энтропия увеличивается
391
ЧАСТЬ III. БИОГЕОГРАШИЧЕСКАЯ КАРТИ В А МИРА
Коренное отличие природных систем от техногенных заключается в
их способности к самоорганизации и саморазвитию; энтропия в природ-
ных системах естественным образом уменьшается. Техногенные систе-
мы могут сохранять устойчивость только при условии постоянных
материальных и энергетических затрат со стороны общества; предо-
ставленные сами себе, они разрушаются, энтропия растет (рис. 9).
Рис. 9. Применение второго закона термодинамики к оценке состояния
техногенных и природных систем, по Р.К. Баландину: 1 — возрастание
энтропии в техногенных системах (часы); 2 — ритмичное изменение
энтропии, характерное для обычных часов: когда часы заводят, энтропия
уменьшается; в целом — повышение энтропии в результате трения
деталей, износа механизма; 3 — уменьшение энтропии в природных
системах (античасы); 4 — ритмичное изменение энтропии с общим
ее уменьшением, идет как бы обновление старых часов
Установка на активное силовое преобразование природы признается
в настоящее время ошибочной. Вместе с тем человек не может существо-
вать на Земле, не нарушая ее природы. Важным критерием рациональ-
ного природопользования является сохранение принципа географической
392 и экологической эквивалентности (Алпатьев, 1978).
ВМЕСТО ПРЕДИСЛОВИЙ. УЧЕНИЕ 0 01 ОСЛЕРЕ И ОХРАНА ПРИРОДЫ
Для пояснения существа этого принципа можно привести следую-
щий пример.
Вырубка леса сопровождается отчуждением из лесной экосистемы
углерода, кислорода, азота, фосфора, калия, воды при одновременном
изменении баланса углекислого газа и кислорода в атмосфере. Проис-
ходит увеличение скорости разрушения земной поверхности и сопря-
женной с ней интенсивности выноса из ландшафта веществ. Корен-
ным образом перестраиваются фауна и микробиологические процессы.
При рациональном природопользовании новая экосистема на месте вы-
рубленного леса, например луговое сообщество, должна быть эквива-
лентна исходной прежде всего по массе продуцируемого органического
вещества и энергии.
Следует признать, что создаваемые человеком динамические равно-
весия в природно-хозяйственных системах, видимо, никогда не будут
полностью эквивалентны средообразующим функциям естественных
биоценозов. Тем не менее эти управляемые равновесия можно лишь
тогда назвать нормальными, когда они будут адекватны скорости при-
способления человека к изменяющейся окружающей среде. При несо-
ответствии темпов изменения среды и приспособления человечества к
этим изменениям последнее обречено на вымирание. Возможны три
пути обеспечения экологической безопасности на Земле:
1) сохранение расширенного воспроизводства естественных экосис-
тем, прежде всего растительных сообществ, средообразующая функ-
ция которых является гарантией нормальных динамических равно-
весий в среде обитания человека;
2) управление природно-хозяйственными системами, средообразую-
щие функции которых эквивалентны функциям естественных эко-
систем, место которых они заняли;
3) внедрение экологически чистых технологий, устраняющих послед-
ствия хозяйственной деятельности, неблагоприятные для жизни че-
ловека.
393
Литература
Агаханянц О.Е. Биогеография. Минск, 1992.
Александрова В.Д. Геоботаническое районирование Арктики и Антарктиды. А., 1977.
Алехин В.В., Кудряшов Л.В., Говорухин В. С. География растений. М., 1961.
А4патьевА.М.Опринципиальныхосновахохраныприроды//Вопросыохраныприро-
ды ирациональногоиспользования природных ресурсов / Под ред. В. С. Жекулина.
Л., 1978.
Астафьева-Урбайтис К.А., Ясаманов НА. Биотические катастрофы на галактиче-
ской орбите Земли // ДАН. 1993. Т. 332. № 6.
Аугуста И., Буриан 3. По путям развития жизни. Прага, 1961.
БазилевичН.И.,ГребенщиковА.Н.,ТишковА.А. Закономерности структурыэкосистем.
М., 1976.
Баландин Р.К. Время, земля, мозг. Минск, 1979.
Беклемишев В.Н. Биоценотические основы сравнительной паразитологии. М., 1970.
Биосфера: Пер. с англ. / Под ред. чл.-кор. АН СССР М.С. Гилярова. М., 1972.
Богданов Д.В. Карта природных зон океана // Океанология. 1961. Т. 1. Вып. 5.
Богданов Д.В. География голубого континента. М., 1963.
Богданов Д.В. География Мирового океана. М., 1978.
Богданов Д.В. Региональная физическая география Мирового океана. М., 1985.
БогоровВ.Г. Биогеоценозы пелагиали океана // Программа и методика изучения био-
геоценозов водной среды. М., 1970.
БогоровВ.Г. Проблемы зональности Мирового океана //Советская география. Итоги
и задачи. М., 1960.
Будыко М.И. Климат и жизнь. Л., 1971.
Булгаков СИ. Философия хозяйства. 1912.
Вальтер Г. Растительность земного шара. Т. I — III. М., 1968 — 1975.
Василевская В.К. Формирование листа засухоустойчивых растений. Ашхабад, 1954.
Величко А А. Возможное будущее земных ландшафтов // Природа. 1992. № 12.
Вернадский В.И. Живое вещество. М., 1978.
ВернарскийВ.И.ОбусловияхпоявленияжизнинаЗемле//Изв.АНСССР.Сер.7.1931.
№5?
Вернадский В.И. Биосфера// Избр. соч. Т. V. М., 1960.
Виноградов Б.В. Основы ландшафтной экологии. М., 1999.
Воронов А.Г. Биогеография. М„ 1963.
Воронов А.Г. Биогеография с основами экологии. М., 1987.
Воронов А.Г, Дроздов Н.Н. Биогеография мира. М., 1985.
Воронов А.Г, Дроздов Н.Н., МялоЕ.Г. Биогеография мира. М., 1985.
Воронов А.Г, Дроздов Н.Н., Мяло Е.Г. Биогеография с основами экологии. М., 1999.
Второе П.П., Дроздов Н.Н. Биогеография. М., 2001.
Высоцкий ГН. О фитотопологических картах, способах их составления и их практи-
ческом значении // Почвоведение. 1909. Т. II. № 2.
Гана Ф. Африка // Всемирная география / Под общ. ред. В. Сиверса. СПб., 1905.
Гептнер В.Г. Общая зоогеография. М.; Л., 1936.
Глазовская МА. Г еохимические основы типологии и методики исследований природ-
ных ландшафтов. М., 1964.
394
ЛИТЕРАТУРА
Гордеева Т.Н., Стрелкова О.С. Практический курс географии растений. М., 1968.
Горшков В.Г. Энергетические потоки биосферы и их потребление человеком //
Изв. ВГО. 1980. №5.
Гумбольдт А. География растений. М.; Л., 1936.
ГумилевЛ.Н. Этногенез и биосфера Земли. Л., 1990.
Дан илов-Дан плянВ.И., ГоршковВ.Г.,АрскийЮ.М.,Лосев/С С. Окружающаясредамеж-
ду прошлым и будущим: Мир и Россия // Эко-Информ. 1994. №5 — 6.
Дарвин Ч. Происхождение видовпутем естественного отбора, илиСохранениеблаго-
приятных рас в борьбе за жизнь: Пер. с 6-го изд. (Лондон, 1872). СПб., 1991.
Дарлингтон ф. Зоогеография. М., 1966.
Докучаев В.В. Избр. соч. Т. 3. М., 1949.
ЖерихинВ.В. Эволюционная биогеоценология:проблемавыборамоделей// Экосис-
темные перестройки и эволюция биосферы / Отв. ред. А.Ю. Ровенов, М.А. Семи-
хатов. М., 1994.
Жизнь растений. Т. 1-6. М., 1978—1982.
Жмур СИ., Розанов А.Ю., Горленко В.М. Следы древнейшей жизни в космических
телах Солнечной системы // Природа. 1997. № 8.
Зайцев Ю.П. Жизнь морской поверхности. Киев, 1974.
Зедлаг У. Животный мир Земли. М., 1975.
Зоненшайн Л.П., Кузьмин М.И., Моралев В.М. Глобальная тектоника, магматизм и
металлогения.М, 1976.
Ипатов В.С., Кирикова Л.А. К вопросу о континууме и дискретности растительного
покрова // Вотан, журн. 1985. № 7.
Исаков Ю.А., КазанскаяН.С., ТишковА.А. Зональные закономерности динамики эко-
систем. М„ 1986.
Исаченко А.Г. Экологическая география России. СПб., 2001.
Исаченко А.Г. Введение в экологическую географию. СПб., 2003.
Исаченко А.Г, Шляпников А.А. Природа мира. Ландшафты. М., 1989.
Исаченко ГА. Ландшафтно-динамическое картографирование — настоящее и буду-
щее//Изв. РГО. 1994. Т. 126. Вып. 3.
Колесник СВ. Общие географические закономерности Земли. М., 1970.
КашкаровД.Н. Среда и сообщество (основы синэкологии). М., 1933.
Коломыц Э.Г Ландшафтные исследования в переходных зонах. М., 1987.
Колчинский Э.И. Эволюция биосферы. Л., 1990.
Коммонер Б. Замыкающийся круг. Л., 1974.
АрасиловВ.А.Общаямодельэволюцииэкосистем//Эволюцияэкосистем.Тезисы.М.,
1995.
Краснов Н.А. Подтропиками Азии. М., 1987.
Крючков В.В. Чуткая Субарктика. М., 1976.
Купецкий В.Н. О морских ландшафтах в Арктике // Изв. ВГО. 1961. Т. 93. Вып. 4.
ЛапоА.В. Следы былых биосфер. М., 1987.
Лархер В. Экология растений. М., 1978.
ЛемеЖ. Основы биогеографии. М., 1976.
МазингВ.В.Теоретическиеиметодическиепроблемыизученияструктурыраститель-
ности. Тарту, 1969.
Мейен СВ. Основы палеоботаники. М, 1987.
Мильков Ф.Н. Физическая география. Современное состояние, закономерности,
проблемы. Воронеж, 1981.
Мильков Ф.Н. Человек и ландшафты. Очерки антропогенного ландшафтоведения.
М., 1973.
Наумов Д.В., Пропп М.В., Рыбаков С.Н. Мир кораллов. Л., 1985.
Нейл У. География жизни. М., 1973.
Неш иба С. Океанология. Современные представления о жидкой оболочке Земли. М.,
1991.
395
Л1ТЕРЛТУРА
Одум Ю. Основы экологии. М., 1975.
Петров К.М. Общая экология. Взаимодействие общества и природы. СПб., 1998.
Петров К.М. Биогеография океана. Биологическая структура океана глазами геогра-
фа. СПб., 1999.
Петров К.М. Экология человека и культура. СПб., 2000.
Петров К.М. Биогеография с основами охраны биосферы. СПб., 2001.
Петров К.М. Биономия океана. СПб., 2004.
Петров К.М. Геоэкология. СПб., 2004.
Петров М.П. Пустыни земного шара. Л., 1973.
ПианкаЭ. Эволюционная экология/Под ред. М.С. Гилярова. М., 1981.
Полынов Б.Б. Учение о ландшафтах // Вопр. географии. 1953. Сб. 33.
Пригожий И., Стенгере И. Время, хаос, квант. М., 1994.
Пригожий И., Стенгере И. Порядок из хаоса: новый диалог человека с природой.
М„ 1986.
РаменскпйЛ.Г.Введениевкомплексноепочвенно-геоботаническоеисследованиезе-
мель. М., 1938.
РаменскийЛ.Г.Проблемыиметодыизучениярастительногопокрова//Избр.работы.
Л., 1971.
Реймерс Н.Ф. Природопользование. М„ 1990.
Ричарде П. Тропический дождевой лес. М., 1961.
Серебряков И. Г. Жизненныеформырастенийиих изучение//Полевая геоботаника.
Т. III. М., 1964.
Серебрякова Т.П. Учение о жизненных формах на современном этапе // Итоги науки
и техники. Ботаника. М., 1972.
Сочава В.Б. Вопросы классификации растительности, типологии физико-географи-
ческих фаций и биогеоценозов // Труды ин-та биологии Уральск, фил. АН СССР.
1961. Т. 27.
Сочава В.Б. География и экология. Л., 1970.
Сочава В.Б. Классификация растительности как иерархия динамических систем //
Геоботаническое картографирование. Л., 1972.
Сочава В.Б. Переменные состояния и инварианты растительного покрова // Совре-
менные проблемы биогеографии. Л., 1980.
Сочава В.Б. Растительный покров на тематических картах. Новосибирск, 1979.
СтруминскицВ.В. Каки зачем возниклажизньнаЗемлеи других план етахКосмоса//
Вести. РАН. 1995. Т. 65. № 1.
Сукачев В.Н. Биогеоценология и фитоценология // ДАН СССР. 1945. Т. 47. № 6.
Тахтаджян А.А. Флористические области Земли. М., 1978.
Толмачев А.И. Основы учения об ареалах (Введение в хорологию растений). Л., 1962.
Уиттекер Р. Сообщества и экосистемы. М., 1980.
Федоров Н.Ф. Философия общего дела // Русский космизм. М., 1993.
Фентон К.Л., Фентон М.А. Каменная книга. Летопись доисторической жизни. М.,
1997.
Хакен Г. Синергетика: Пер. с англ. / Под ред. Ю.Л. Климонтовича. М, 1994.
ХарвейД. Научное объяснение в географии. М., 1974.
Хромов СП. Метеорология и климатология. Л., 1968.
Чернов Ю.И. Природная зональность и животный мир суши. М., 1975.
Швебс Г.И. Концепция природно-хозяйственных территориальных систем и вопро-
сы рационального природопользования // География и природные ресурсы. 1987.
№4.
Шмитхюзен И. Общая география растительности. М„ 1966.
ШубаевЛ.П. Общее землеведение. М., 1977.
Экотоны в биосфере / Под ред. В.С. Залетаева. М„ 1997.
ЭрхардЖ.П., СеженЖ. Планктон. Л., 1984.
39В Юнкер Р., Шерер 3. История происхождения и развития жизни. Кайрос. М., 1992.
ЛИТЕРАТУРА
Юрцез БЛ. Основные направления современной науки о растительном покрове //
Ботан. журн. 1988. Т. 73. № 10.
Ярошенко П.Д. Общая биогеография. М., 1975.
Brown J., Gibson А.С. Biogeography. The С. V. Mosby Company, 1983.
Clements F.E. Nature and Structure of the Climax // Ecology: 1936. Vol. 24. Ne’l.
Сох C.B., Hecleyl.N., Moore P.D. Biogeography // An Ecological and Evolutionary
Approach. Blackwell Sci. Pub., 1973.
Global Biodiversity. Status of the Earth's Living Resources. A Report compiled by World
Conservation Monitoring Centre / Editor: Briari Grdbmbndge. Chapman & Hall, 1992.
Meadows D.H., Meadows D.L. RahdersJ. Beyond the Limits. London, 1992.
Good. R. The Geography of the Flowering Plants. Logmans, 1953.
Hutchinson G.E. The Niche: an Abstractly Inhabited Hypervolume // The Ecological
T Theatreand/EvolutionaryPlay.:NewHaveri, 1965.
Moran J.J., Morgan M.D., Wiersma J.H. Introduction to Environmental Science. San
Francisco, 1980.
Polunin N. Introduction to Plant Geography and some Related Sciences. Logmans, 1965.
ShawEarlB. Fundamentals of Geography. John Wiley and SonsV1965. ""’«r * —
Tansley A: The Use and Abuse of Vegetational Concepts and Terms // Ecology. 1935.
Vol. 16. №4.
TivyJ. Biogeography. A Study of Plants in the Ecosphere. Oliver and Boyd, 1971.
Troll C. Luftbildplan und okolologishe Bodertforschung. 1939. Zeister. Ges. Erkunde zu
Berlin. № 7/8.
Walter H., Breckle S.-W. Ecological Systems of the Geobiosphere. Vol. 3. Temperate and
Polar Zonobiomes of Northern Eurasia. Springer-Verlag, 1986.
Содержание
Предисловие...................................................3
Часть I. Исторические и географические факторы
биологического разнообразия
Глава 1
ЭВОЛЮЦИОННЫЙ ПОДХОД К ОБЪЯСНЕНИЮ
РАЗНООБРАЗИЯ ЖИЗНИ НА ЗЕМЛЕ..........................8
1.1. Космические предпосылки развития жизни.....................9
1.2. Эволюция жизни в криптозое................................14
1.3. Эволюция жизни в фанерозое................................16
Глава 2
ГЕОГРАФИЧЕСКИЕ ФАКТОРЫ, ОПРЕДЕЛЯЮЩИЕ ЗАКОНОМЕРНОСТИ
ПОДРАЗДЕЛЕНИЯ АРЕНЫ ЖИЗНИ НА ЗЕМЛЕ......................38
2.1. Подразделения арены жизни, обусловленные тектогенными формами
рельефа...........................................................38
2.2. Подразделения арены жизни, обусловленные действием закона
географической зональности........................................41
2.3. Подразделения арены жизни, обусловленные действием закона
высотной поясности................................................53
2.4. Основные подразделения арены жизни в океане..................55
Глава 3
АРЕАЛОГИЯ.............................................ПО
3.1. Формирование ареала........................................56
3.2. Географические и генетические элементы биоты...............66
3.3. Роль естественных барьеров.................................68
3.4. Типы ареалов...............................................71
3.5. Эндемики и эндемизм........................................81
3.6. Викариат и конвергенция....................................84
Часть II. Экологические и ландшафтные факторы
биологического разнообразия
Глава 4
ОСНОВНЫЕ ПОНЯТИЯ И ЗАКОНЫ ЭКОЛОГИИ.....................81
4.1. Организм и среда............................................91
4.2. Пищевые (трофические) связи.................................98
4.3. Свойства популяций..........................................98
4.4. Четыре закона общей экологии...............................105
Глава 5
СИНЭКОЛОГИЯ............................................107
5.1. Биоценоз, экосистема, биогеоценоз............................107
5.2. Стации, местообитания, экологические ниши....................110
5.3. Растительные сообщества как каркас наземных экосистем .......114
5.4. Сукцессии сообществ..........................................122
СОДЕРЖАНИЕ
Глава
ЭКОЛОГИЯ И ГЕОГРАФИЯ..................................127
6.1. Геоэкологический и ландшафтно-экологический подходы
к биогеографическим исследованиям...............................127
6.2. Аксиоматические основы ландшафтной экологии................130
6.3. Топологический уровень биогеографических исследований......138
Часть III. Биогеографическая картина мира
Глава 7
БИОТИЧЕСКИЕ ЦАРСТВА СУШИ..............................150
7.1. Голарктическое царство......................................151
7.2. Палеотропические царства — Эфиопское и Индо-Малайское.......156
7.3. Неотропическое царство......................................164
7.4. Австралийское царство.......................................168
7.5. Голантарктическое царство...................................172
Глава 8
ЗОНАЛЬНЫЕ ТИПЫ БИОМОВ СУШИ..........................174
8.1. Биомы жаркого пояса......................................178
8.2. Биомы холодных и умеренных поясов.......................243
Глава
АНТРОПОГЕННЫЕ ИЗМЕНЕНИЯ ЛАНДШАФТОВ ПРИРОДНЫХ ЗОН РОССИИ.314
9.1. Зональные особенности распределения экологического
потенциала ландшафтов..........................................314
9.2. Зона тундры................................................318
9.3. Зона тайги.................................................321
9.4. Зона смешанных и широколиственных лесов....................324
9.5. Лесостепная и степная зоны.................................325
9.6. Полупустынная и пустынные зоны.............................329
Глава 10
ЗОНАЛЬНЫЕ ТИПЫ БИОМОВ ПОВЕРХНОСТНОЙ ТОЛЩИ ОКЕАНА......333
10.1. Биомы полярных ледовитых морей............................334
10.2. Биомы субполярных зон.....................................341
10.3. Биомы умеренных зон.......................................348
10.4. Биомы субтропических зон..................................351
10.5. Биомы тропических зон.....................................352
10.6. Биомы экваториальной зоны.................................356
10.7. Мангровые болота и коралловые рифы.........................357
Глава 11
УЯЗВИМЫЕ ЗВЕНЬЯ ЭКОЛОГИЧЕСКОЙ СИСТЕМЫ
МИРОВОГО ОКЕАНА.......................................363
Вместо заключения................................................375
Литература.......................................................394