/




































































































































































































































































































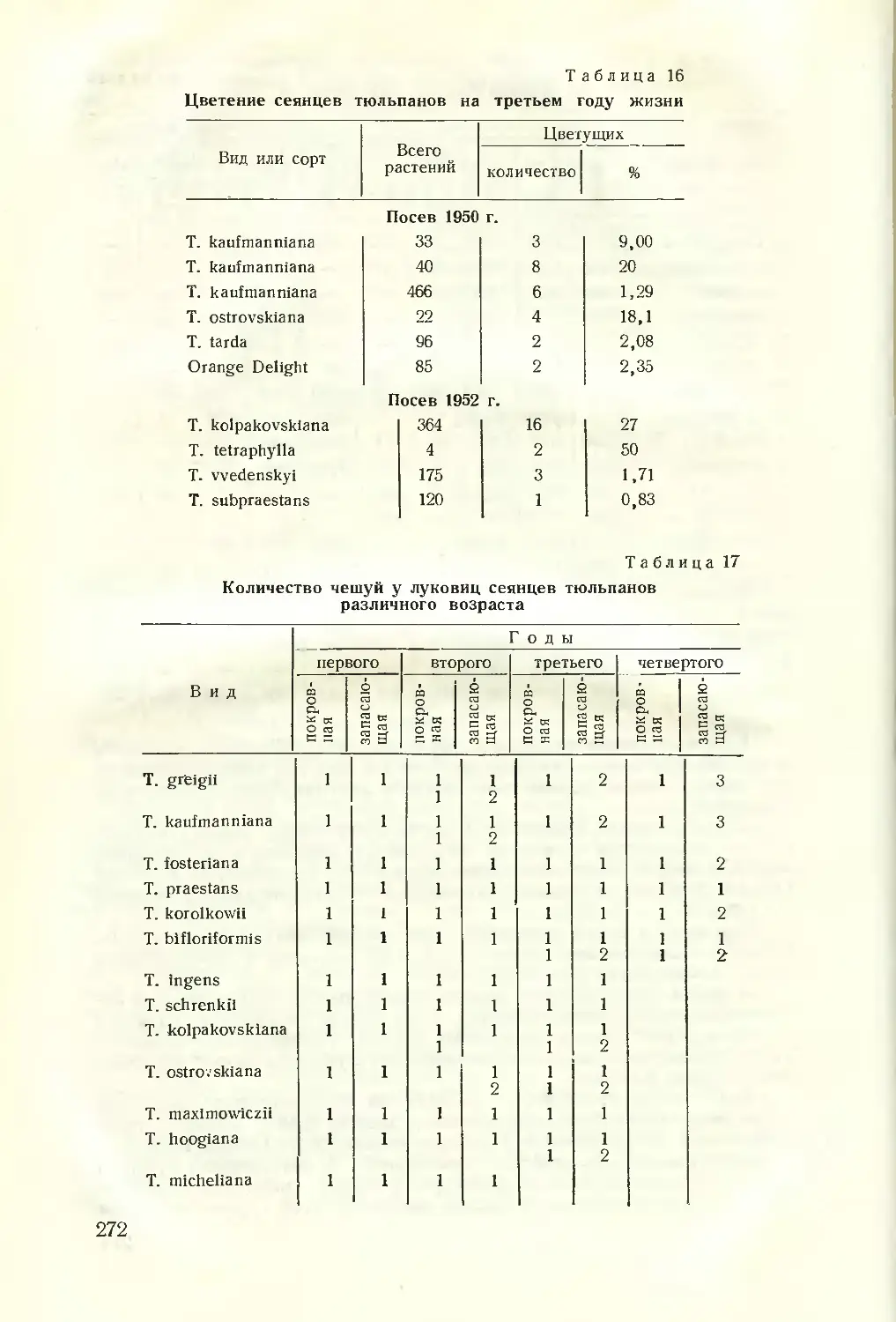








































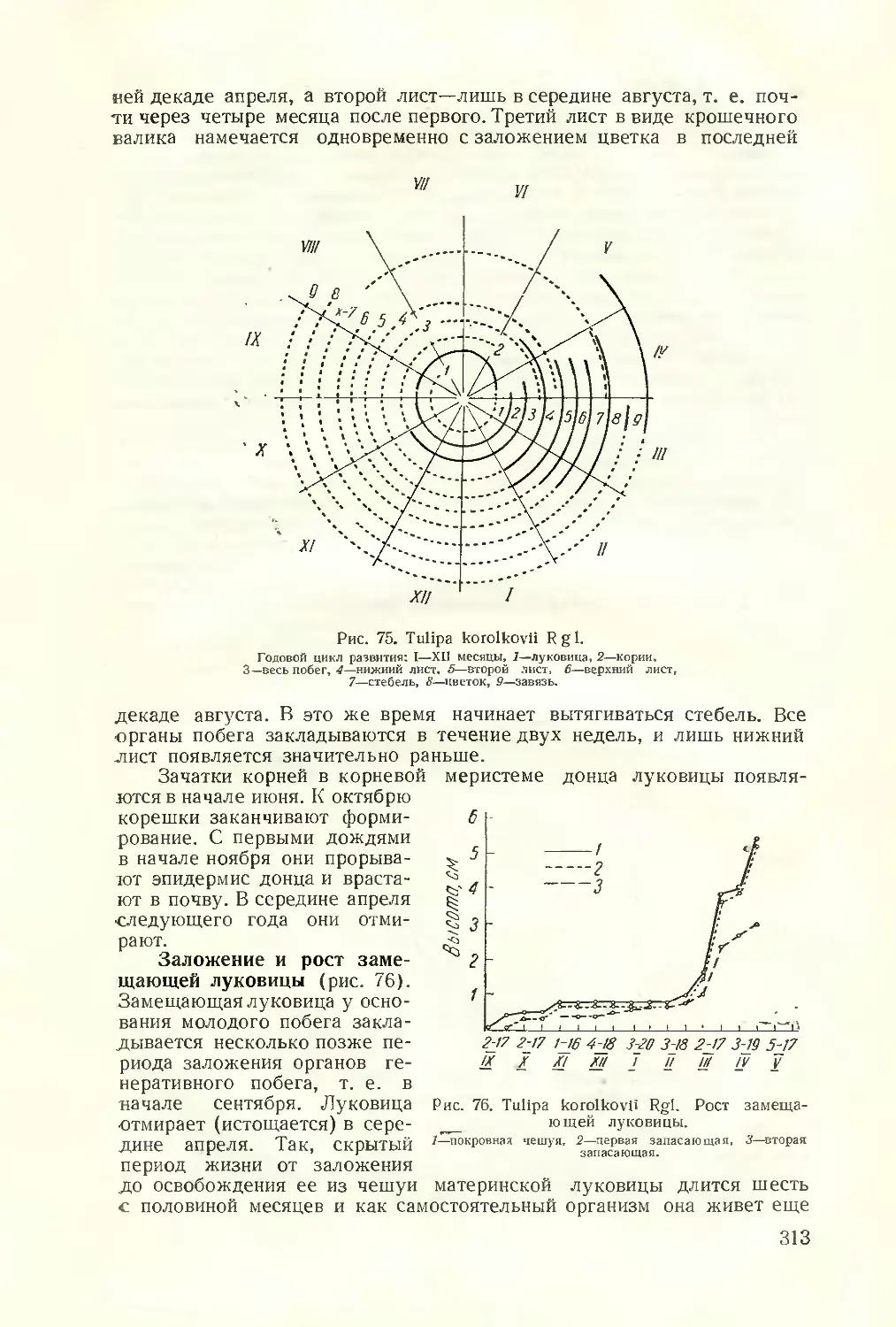





















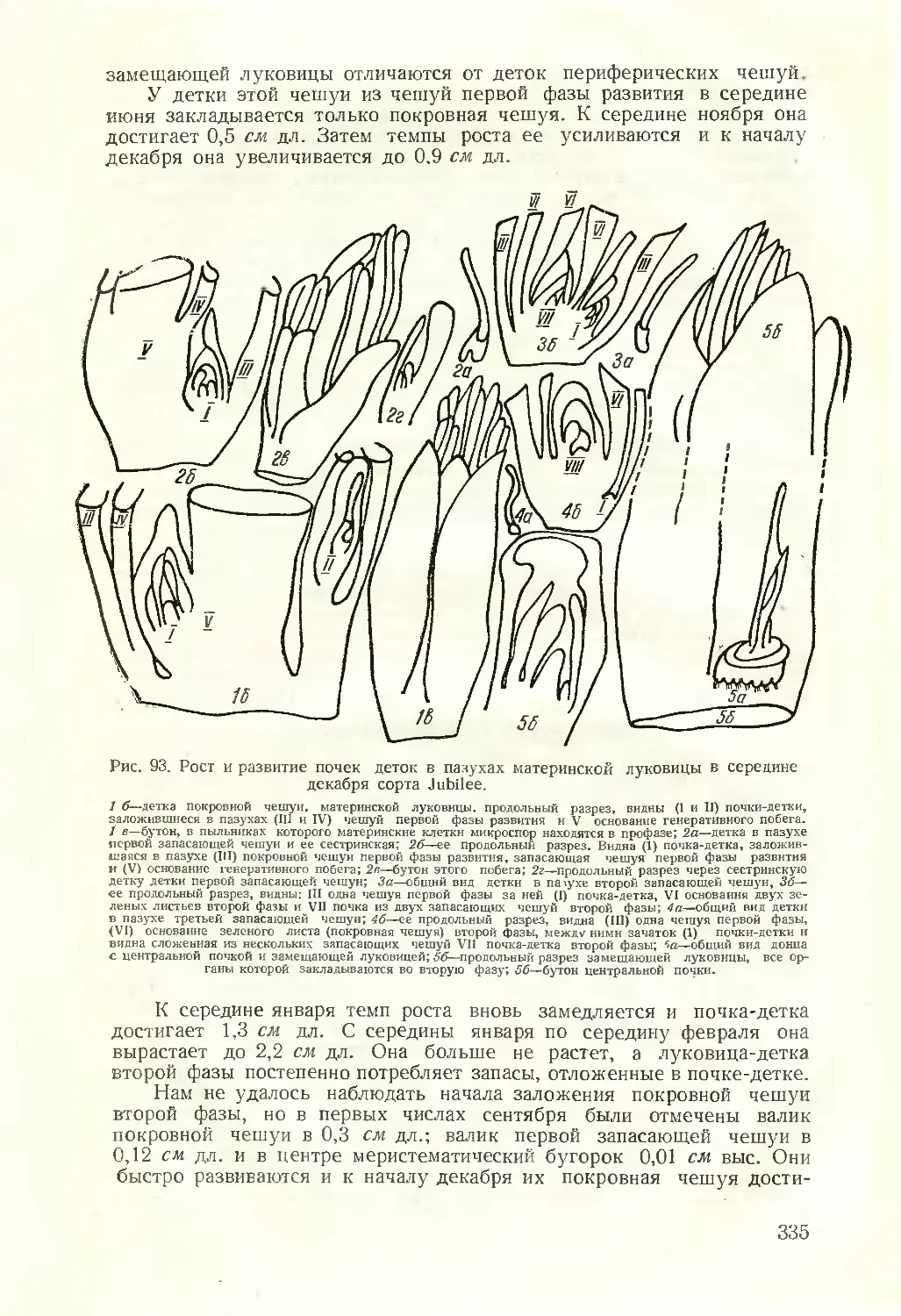




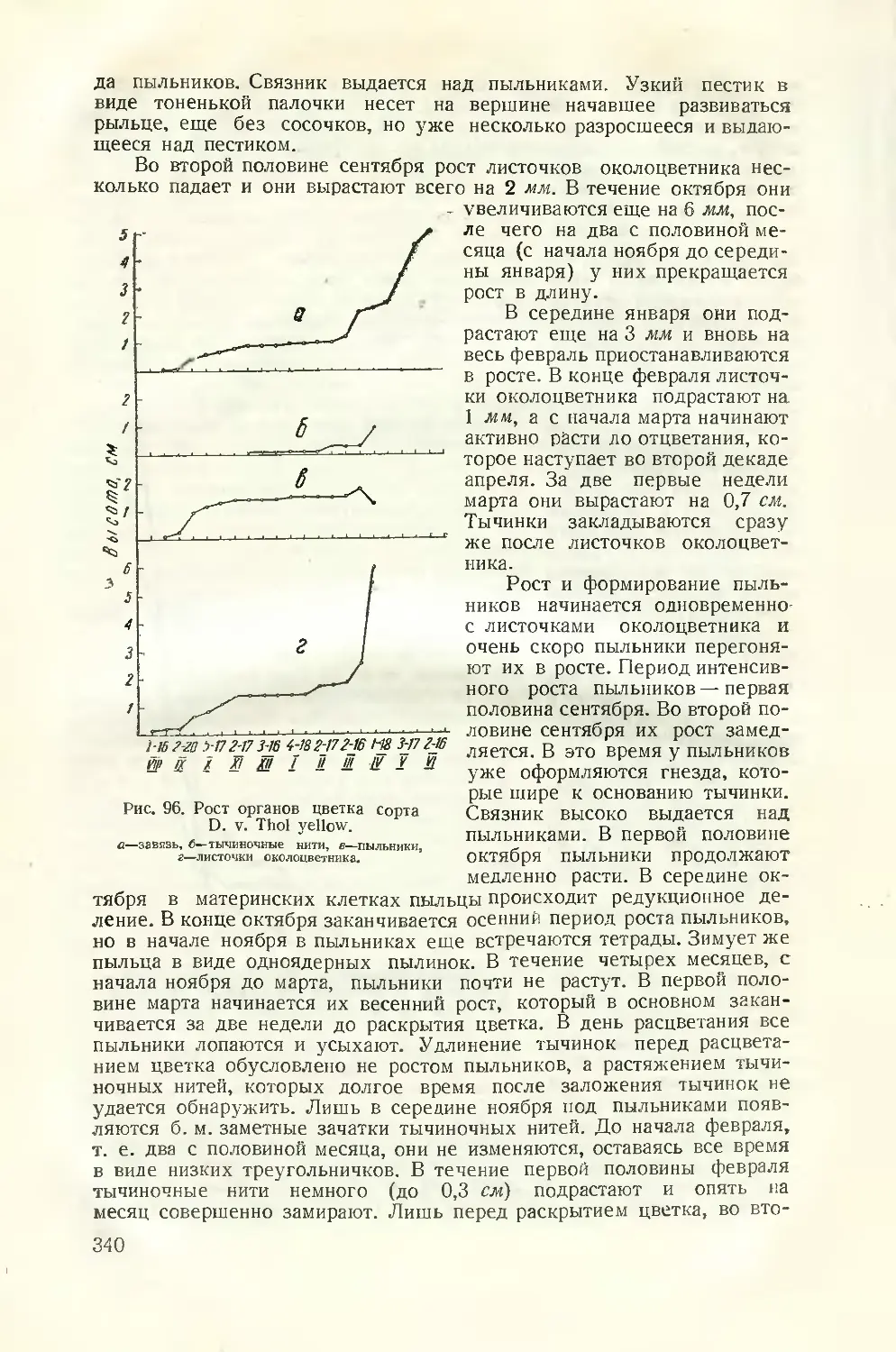







































































Текст
^ЗБЕКИСТОН ССР ФАНЛАР АКАДЕМ И Я С И
БОТАНИКА БОБИ
3. TJ. Ьочаниева
АОААААР
МОРФОЛОГИЯ, цитология
ВА БИОЛОГИЯСИ
УЗБЕКИСТОН ССР ФАНЛАР АКАДЕМИЯСИ НАШРИЁТИ
ТОШКЕНТ —1962
АКАДЕМИЯ НАУК УЗБЕКСКОЙ ССР
БОТАНИЧЕСКИЙ САД
3. TI.
Ш1Ш1
МОРФОЛОГИЯ, цитология
И БИОЛОГИЯ
ИЗДАТЕЛЬСТВО АКАДЕМИИ НАУК УЗБЕКСКОЙ ССР
ТАШКЕНТ —1962
В работе описан 61 вид в основном дикорастущих средне-
азиатских тюльпанов. В ней освещаются также география
распространения тюльпанов и вопросы их морфологии,
биологии и кариосистематики.
Книга рассчитана на ботаников и практиков-цветоводов.
Ответственный редактор—член-корр. АН УзССР
проф. И. А. РАЙКОВА
ШШШИЛ
ВВЕДЕНИЕ
Ботаническом саду Академии наук УзССР (ранее находившемся в
ведении Среднеазиатского государственного университета) с нача-
ла его существования выращивались тюльпаны местных дикорасту-
щих и культурных сортов, многие из которых высоко ценятся на миро-
вом рынке—им отводится видное место в каталогах голландских торго-
вых фирм, занимающихся, разведением и продажей луковичных растений.
Однако в нашей стране в цветниках на улицах, в парках и скве-
рах Москвы и Ленинграда только в последние годы стали появляться
тюльпаны, причем в основном иностранных сортов. Недавно тюльпаны
начали высаживаться и в цветниках Ташкента.
Многие местные дикорастущие тюльпаны по яркости, оригиналь-
ности формы, красоте и величине цветка не только не уступают за-
граничным сортам культурных тюльпанов, но часто их превосходят
(например, тюльпаны фостера, грейга, микели, кауфмана. Введенского,
альберта).
Изумительны тюльпаны грейга и микели, напоминающие по форме
вихрь пламени. Очень оригинальны тюльпаны королькова, блестящие
и розовые с прелестными небольшими цветками. Грациозны лилие-
цветные — Т. островского и близкие к нему виды.
Окраска диких видов тюльпанов исключительно разнообразна.
Широко распространен тюльпан Шренка, имеющий в своих популя-
циях все тончайшие нюансы окраски редчайших культурных сортов,
а все красные тюльпаны на Тянь-Шане обладают полихромизмом
(островского, альберта, кауфмана и др.).
Лист тюльпанов по форме, фактуре и окраске у диких видов,
безусловно, богаче и разнообразнее, чем у культурных. Дикие виды
имеют неоценимое преимущество перед культурными— они не пестреют,
т. е. они иммунны к вирусу пестрения, вызывающему инфекционное
заболевание, которому подвержены все культурные тюльпаны.
Средняя Азия с ее чрезвычайно разнообразными климатическими
и физико-географическими условиями резко отличается от других
районов СССР. Величайшие горные системы и пустыни, окружающие
ее в значительной степени определяют своеобразие этих условий.
Большая часть Средней Азии занята пустынями, с востока и юга ко-
торых находятся мощные горные системы: на юго-западе—Копет-Даг,
в центре—Памиро-Алай до границы с Афганистаном и Кашгаром, на
востоке—Тянь-Шань. Эти горные системы являются не только барье-
рами, препятствующими расселению растений, но и мостами, связы-
5
вающими флору Средней Азии с флорой сопредельных с ней стран.
Поэтому решение вопроса о филогении тех или иных групп растений,,
и в частности тюльпанов, обитающих в Средней Азии и распространен-
ных в сопредельных географических областях, связано с большими
трудностями.
В 1935 г. во „Флоре СССР", т. IV, А. И. Введенский подробно
описал тюльпаны, произрастающие на территории СССР, и дал их
классификацию. Несмотря на то, что за время, истекшее с 1935 г.,
тюльпаны* СССР изучены значительно полнее и глубже, классификация
групп рода Tulipa, предложенная А. И. Введенским, в основном может
быть сохранена.
Значительно раньше, чем тюльпаны узнали в Европе, их выращи-
вали на Востоке. В 1258 г. персидский поэт Мушрафаддин Саади в-
поэме „Гулистан" писал:
„В саду ручей с прохладной тек водой,
И пенье птиц звучало голосистых.
Румяные плоды на всех ветвях,
Тюльпанов пестрых много, роз душистых”.
Следовательно, уже в XIII в. в цветниках персидских садов было-
много „тюльпанов пестрых". Это указание поэта можно понимать
двояко: или это были пестрые тюльпаны, или—садовые тюльпаны не-
скольких видов с различной окраской.
Как известно, пестреют только сорта культурных тюльпанов, по-
этому, если последнее предположение правильно, можно сделать вы-
вод, что уже в то время на Востоке было много культурных сортов-
тюльпанов.
В XV в. в поэме „Фархад и Ширин" Алишер Навои пишет: „Вино
в зеленых пиалах смогу сравнить с тюльпаном алым на лугу", а в
поэме „Лейли и Меджнун": „Когда тюльпан узнал скорбей клеймо,,
он выжег на груди своей клеймо".
В молодости Навои жил в Самарканде и мог видеть алые с тем-
ной серединой тюльпаны Самаркандских гор. Это был Т. fosteriana
или Т. micheliana с его острыми в виде пересекающих околоцветник
лучей черными пятнами. Отсюда и сравнение глубокого горя с чер-
ным клеймом в „груди" тюльпана.
Упоминает поэт о тюльпанах и в других произведениях:
„Любила черноокая газель
Степной простор, степную пестроту,
Тюльпаны в обжигающем цвету”.
И далее, „И хлопок алый—разбросал вокруг,—смотри: тюльпаны
испещрили луг" („Семь планет").
Можно найти у поэта и указания на то, что всадах того времени
выращивались культурные сорта тюльпанов. Так, в поэме „Лейли и
Меджнун" есть такие строки:
„Камнями всех времен и всех судеб
Теперь засыпан сад, ручей ослеп...
Кайе и Лейли в толпе вошли в Ирем,
Понравились его тюльпаны всем,
Но их сердца похожи на тюльпан:
В ожогах от любовных ран”.
Алишер Навои посвящает тюльпану и такие восторженные строки:
„А кипарис изящества, тюльпан
Очарованья, нежности фазан,
Луна в айване красоты, бальзам
Струящая страдающим сердцам...”
(„Фархад и Ширин”)
6
Таким образом, в XV в. тюльпаны были широко известны на тер-
ритории современного Узбекистана, их собирали, выращивали и кра-
сота их вдохновляла поэтов.
В книге „Gardens and Gardening" (1951) есть статья Rollo Meyer
„Тюльпаны", которая начинается словами: „В русской литературе в
1123 году тюльпаны упоминаются под их русским названием „Лола".
Следует отметить, что это слово имеет тюркское, а не русское про-
исхождение, но его присутствие указывает на то, что и в России
тюльпаны были известны раньше, чем в Западной Европе, в которой
о тюльпанах узнали значительно позже 1554 г.
В Турции же в это время были известны уже довольно много-
численные сорта тюльпанов. Первые луковицы их были получены в
Вене. Позже они достигли Антверпена, в 1571 г.— Голландии и еще
позже Англии. В Россию культурные садовые тюльпаны из Европы
завезли голландцы в конце XVI и начале XVII вв. Официальное упо-
минание о садовых тюльпанах в России относится к XVIII в.
По переписи 1701 г. в Нижнем красном саду значилось 1000 тюль-
панов цветных, 3000 средних и мелких (Забелин, 1872; цит. по Мигу-
шовой, 1951).
Благодаря своей красоте и оригинальности тюльпаны быстро и
широко распространились в Европе. Громкую известность получила
вспыхнувшая в 1634 г. „золотая лихорадка", во время которой луко-
вицы тюльпанов в течение трех лет служили предметом жесточайшей
спекуляции.
По истории культуры тюльпанов в Европе начиная с 1554 г. су-
ществует обширная литература.
В нескольких работах на русском языке, появившихся за послед-
ние годы, также кратко освещается история культивирования тюль-
панов (Силина, 1953; Мигушова, 1954; Данилевская, 1955).
В результате экспорта луковиц дикорастущих тюльпанов из Сред-
ней Азии в 1929—1934 гг., организованного Ташкентским ботаническим
садом, голландские садоводы в конце прошлого десятилетия выбро-
сили на мировой рынок значительное количество совершенно новых
сортов, полученных вследствие отбора из диких видов среднеазиатских
тюльпанов и гибридизации их с культурными сортами. Особенную
славу получили в последние годы гибриды Т. fosteriana с группой
тюльпанов Дарвина.
Ботанический сад АН УзССР много .лет изучает среднеазиатские
виды тюльпанов. А. И. Введенский в 1935 г. выполнил монографиче-
скую обработку результатов этих исследований. В. К. Василевская
(1936) занималась анатомическими исследованиями, мы—цитологиче-
скими.
Для более полной морфологической характеристики растений в
настоящей работе использованы за небольшим исключением материалы
исследований только по тем видам, которые мы сами собирали в есте-
ственных условиях или выращивали из луковиц и семян, привезенных
с достоверных местообитаний. В гербариях, как известно, обычно
имеются растения или в цвету без коробочек, или с коробочками, но
без цветов. Поэтому к описаниям цветов тюльпанов добавлены опи-
сания их коробочек. Это сделано для того, чтобы раскрыть не только
видовые особенности тюльпанов во время цветения, но и морфологи-
ческие отличия их в период плодоношения. Кроме того, во время
цветения нецелесообразно заготавливать луковицы для введения их в
культуру, так как они в этот период слишком ослаблены. А позже,
не зная коробочек и их различий, легко перепутать виды.
7
В работе дано также цитологическое описание исследованных видов.
В литературе часто приводятся описания растений, выращенных в
ботанических садах Европы, без точного указания их родины, во
многих случаях садовой репродукции и гибридного происхождения;
при этом, как правило, нет гарантии соответствия исследуемого мате-
риала названию вида, присвоенному ему в цитологических или других
работах (например, тюльпаны борщева, королькова-конколер, Макси-
мовича).
Далее мы приводим данные исследований семян тюльпанов и про-
цесса их прорастания, роста и развития сеянцев.
Эти данные способствуют пониманию исторического развития и
формирования различных видов и сортов растений.
За последние годы вышло несколько работ, освещающих вопросы,
связанные с процессами годового развития взрослых луковиц куль-
турных тюльпанов на севере СССР (Силина, 1953; Мигушова, 1954;
Данилевская, 1955, 1956) и в условиях средней полосы России (Тимо-
феева, 1956). Однако ни один из перечисленных авторов не коснулся
годового цикла развития диких видов тюльпанов. При этом следует
отметить, что все они, изучая годовой цикл развития взрослого расте-
ния культурного тюльпана, почему-то под развитием растения пони-
мают только развитие луковицы.
В конце настоящей работы описаны некоторые изменения диких
видов тюльпанов под влиянием внешних условий в культуре.
В результате многолетней работы по изучению диких видов тюль-
панов Средней Азии мы получили много новых сортов путем клено-
вой селекции, межвидовой гибридизации и гибридизации дикорастущих
видов с культурными сортами.
Глава I
КРАТКИЙ ОБЗОР ИССЛЕДОВАНИЙ И ИЗУЧЕННОСТЬ
СРЕДНЕАЗИАТСКИХ ТЮЛЬПАНОВ
1873 г. Э. Регель в статье „Enumerate Specierum hucusgue cog-
nitarum generis Tulipae" („Перечисление до сих пор известных видов
рода Tulipa“) впервые описал этот род, дал ключ и объединил
в группы известные в то время 26 видов тюльпанов.
В качестве основного критерия, определяющего разделение рода
на виды, Регель использовал наличие или отсутствие опушения на
внутренних листочках околоцветника и основании тычинок. Растения
первой группы с опушенными основаниями тычинок и листочков
околоцветника он разделил на подгруппы по форме листьев, а внутри
полученных подгрупп—по опушению луковичных чешуи. Растения
второй группы с голыми основаниями листочков околоцветника и
тычинок Регель разбил на подгруппы по наличию или отсутствию
темного пятна у основания листочков околоцветника.
Таким образом, внутри первой группы он выделяет подгруппы:
1) по форме листочков околоцветника, 2) опушенности цветоножки,
3) форме листьев; внутри второй: 1) по форме листочков околоцвет-
ника, 2) опушенности цветоножки и положению цветка, 3) окаймлен-
ности листьев, 4) опушению луковичных чешуй.
По этой системе известные в то время Регелю тюльпаны распо-
лагаются в следующем порядке:
А* а. 1. Т. silvestris L.
Jb. 2. Т. biflora Pall.
с. 3. Т. celsiana Red.
А** 4. Т. saxatilis Sieber.
В* афа 5. Т. oculus solis S t. Amans.
6. T. didieri Jordan
7. T. borszczowii R g 1.
8. T. julia C. Koch.
T. montana L i n d 1.
9. T. boissieri R g 1.
10. T. greigii R g 1.
b. 11. T. euanthiae Orph.
c. 12. T. aleppensis В о i s s.
13. T. boeotica В о i s s. et H e 1 d r.
B** a-f-aao 14. T. gesneriana L.
o.o. 15. T. schrenkii R g 1.
вЬ. 16. T. lehmanniana Meckl.
c.c. 17. T. sogdiana Bunge
18. T. armena В о i s s.
19. T. clusiana Red.
20. T. montana L i n d 1.
4-+ 21. T. sibthorpiana S m..
+ + + 22. T. suaveolens Roth.
23. T. strangulata R e b о u 1.
24. T. altaica Pall.
b. + 25. T. turcica Roth.
+ + 26. T. undulatifolia В о i s s.
Регель применил все современные критерии в классификации тюль-
панов и осуществил довольно правильное распределение тюльпанов
на группы. Однако его работа осталась неизвестной ботаникам Запада.
Недостатки его классификации объясняются в основном тем, что
материал, доступный Регелю, был очень ограничен и не давал пред-
ставления об амплитуде изменчивости вида.
9
Через год после опубликования работы Регеля из печати вышла1
работа Бакера (J. Baker, 1874), получившая широкую известность.
Весь род тюльпанов Бакер делит на 2 подрода: Eutulipe с невыражен-
ным столбиком и Orithyia с хорошо развитым столбиком.
Eutulipa Бакер разделил на 5 секций: Eriobulbii, Gesnerianae, Sca-
bricapae, Saxatiles и Silvestris.
В Leiostemones у Буасье (Boissier, 1884) вошли 3 первых секции
Бакера, а в Eriostemones—2 последних.
Секция Orithyia осталась без изменений.
Такого же, что и Буасье, деления тюльпанов на секции, придер-
живается В. И. Талиев (1930), который в группе Silvestris секции
Eriostemones дополнительно выделяет две ветви—bifloraeformes и sil-
vestriformes и устанавливает промежуточные типы видов Т. praebiflo-
ra, Т. tricolor polychroma, Т. prae-Biebersteiniana, Т. praetricolor.
Талиев считает Т. turkestanica близким к прототипу видом.
На основании кариологических данных Ньютона и Тишлера Та-
лиев пишет, что число хромозом п, равное 12, встречается у „арха-
ичных групп лилейных". Это, по-видимому, указывает на то, что род.
Lilium со свитой сопутствующих родов Fritillaria и Tulipa очень бли-
зок к эволюционным первоисточникам лилейных. „Типичная для тюль-
панов одноцветковость, без сомнения, возникла из более или менее,
многоцветкового соцветия отдаленных предков, путем редукции".
Крупные цветы, наблюдаемые у некоторых тюльпанов, есть не
более „как обедневшие родичи" по отношению к цветам лилий. И
наконец, „моделировка данного рода явно вырабатывалась под влия-
нием приспособления к определенным экологическим условиям—пер-
воначально, вероятно, к высокогорному существованию, а затем, к
условиям жизни—ксерофитного весенника".
Таким образом, если бы мы стали искать примитивный вид тюль-
пана, то он, по Талиеву, должен быть многоцветным, крупноцветко-
вым и обитать в высокогорных условиях.
Талиев утверждает, что по количеству обитающих в Средней Азии
видов она занимает первое место в мире и является центром проис-
хождения тюльпанов. Для практического подтверждения его мысли,
мы думаем, в Средней Азии должны быть поставлены широкие рабо-
ты по использованию дикорастущих фондов тюльпанов и их восста-
новлению.
А. И. Введенский, изучавший род Tulipa, пишет: „Принятая в
настоящее время система Baker’a — Boissier’a весьма искусственна; пе-
реработать же ее трудно из-за скудости наших познаний о тюльпан-
ной флоре Передней Азии" (Флора СССР, т. IV).
Изменения, которые Введенский внес в систему классификации1
тюльпанов, состоят в выделении двух монотипных секций. Это — сек-
ция Spiranthera Vved. с одним видом Tulipa kaufmanniana Rgl. и
секция Lophophyllon с одним видом Tulipa regelii Krasn. Кроме то-
го, он перенес Tulipa sogdiana Bge. из секции Leiostemones в секцию
Eriostemones и обратил особое внимание на распределение тюльпанов,
внутри секций.
А. И. Введенский во „Флоре СССР" расположил виды тюльпанов
Средней Азии в порядке родства, видимо, считая наиболее древними
тюльпаны, входящие в секцию Tulipanum, и поставил их первыми во
главе всего рода. Внутри секций виды расположены по наибольшему
родству—Т. tubergeniana Hoog., Т. kuschkensis В. Fedtsch, Т. ho-
ogiana, В. Fedtsch., Т. julia С. Koch. В 1946 г. А. И. Введен-
ский описал еще один вид из этой секции—Т. subquinquifolia Vved.
10
В секции Leoistemones виды распределены в следующем порядке:'
Т. florenskyi Vo г on., Т. lanata Rgl., Т. ingens Hoog., T. fosteriana
Irv., T. carinata Wved. Это первая группа секции, включающая виды,
у которых есть определенная связь с тюльпанами секции ТиИрагшпъ
по опушению луковичных чешуй, но менее обильному, тонкому и из-
витому.
Далее следует группа из двух видов—Т. praestans Hoog. и Т.
subpraestans V v е d. А. И. Введенский сближает эти виды потому,
что они оригинальны из-за отсутствия пятна при основании листочков
околоцветника. Однако у некоторых форм есть очень небольшая рас-
плывчатая легкая оранжеватость при основании листочков околоцвет-
ника, которая, как нам кажется, связывает их с последующей группой
тюльпанов. По форме цветка они ближе к предыдущей, объединяемой
около Т. fosteriana Irv. Следующая группа тюльпанов весьма ориги-
нальна по форме цветка и близка к Т. greigii Rgl. Большинство ви-
дов, входящих в эту группу, имеет на листьях антоциановые пятна
или полосы и состоит из видов Т. alBerti RgL, Т. greigii Rgl.,
mogoltavica М. Pop. et Vved., T. micheliana Hoog.
По сложению цветка эта группа близка к видам из секции Tuli-
panum.
В следующую группу Введенский объединил 2 кавказских вида—
Т. eichleri Rgl. и Т. schmidtii Fom.
Интересной группой, состоящей из самобытных тюльпанов, близ-
ких не только по внешней морфологии, но, как показали наши дан-
ные, и по цитологическим признакам, являются 3 вида: Т. maximo-
wiczii Rgl,, Т. linifolia Rgl. и T. wilsoniana Hoog.
Следующую группу образуют тюльпаны со скорлуповидными по-
кровными чешуями луковиц —Т. lehmanniana Merci., Т. behmiana
Rgl., и Т. borszczowii Rgl., которые по сложению цветка близки к
группе тюльпанов, объединившихся вокруг Т. greigii, так как имеют
сильно продолженные луковичные чешуи. Этот признак у видов, близ-
ких к Т. greigii, может отсутствовать, а для видов группы Т. leh-
manniana он обязателен.
Группа маленьких тюльпанов Т. korolkovii Rgl., Т. nitida Hoog.
и Т. rosea Vved. совершенно естественна и состоит из близко род-
ственных видов.
Группа видов из Т. schrenkii Rgl., Т. ostrovskiana Rgl., Т. kolpa-
kovskiana Rgl. и T. zenaidae Vved. вполне естественна. Ho T. schren-
kii, по нашему мнению, в какой-то степени по сложению цветка тяготе-
ет к группе видов, объединяющихся вокруг Т. greigii, а Т. zenaidae—
вокруг Т. korolkovii. Т. brachystemon Rgl. по общему габитусу об-
наруживает родство с группой видов, близких к Т. greigii.
Однако начиная с Т. brachystemon взаимоотношения между ви-
дами тюльпанов обрисованы неясно, и все эти виды не объединены
между собой ни в какие естественные группы. У А. И. Введенского
они расположены в следующем порядке: Т. platystemon Vved., Т.
altaica Pall., Т. dubia Vved., T. iliensis Rgl., T. fergauica Vved.,
T. anisophylla Vved., T. korshinskyi Vved., T. tianschanica Rgl., T.
tetraphyIla Rgl., T. hissarica M. Pop. et Vved., T. krauseana Rgl.
Из видов этой последней группы Т. dubia Vved. на основании
тщательно проведенных наблюдений должна быть перенесена в сек-
цию Spiranthera, а Т. iliensis, Т. ferganica и Т. tetraphylla—к тюль-
панам, объединенным вокруг Т. ostrovskiana и Т. kolpakovskiana.
При этом Т. tetraphylla тяготеет к Т. korolkovii, в то же время
он очень самобытен. Т. hissarica—совершенно оригинальный тюльпан
11
и не имеет близких родичей среди перечисленных групп видов, вхо-
дящих в секцию Leiostemones.
Остальные виды этой секции мы не видели живыми и не можем
судить о том, насколько верно определено их место в системе рода.
Секция Spiranthera у А. И. Введенского содержит один вид Т.
Laufmanniana R g 1. Она выделена по способу открывания пыльников.
По нашим наблюдениям, в эту секцию необходимо перенести и Т. du-
bia, так как у него пыльники не скручиваются в спираль и открыва-
ются не сразу, как у других видов, а растрескиваются постепенно
вдоль всей длины в течение 2—3 дней, как у Т. Jcaufmanniana.
В секцию Eriostomones А. И. Введенский помещает виды: Т. vi-
olacea В о i s s. et Buhse, T. caucasica Lipsky, T. Silvestris L., T.
bieb'ersteiniana Schult., T. patens Aga rd h., T. sogdiana Bge. Пер-
вые пять видов образуют ветвь, называемую Silvestriformis (Талиев,
1930). Т. sogdiana стоит несколько особняком благодаря оригинальной
форме цветка и отсутствию у большей части особей опушения на ос-
нованиях тычиночных нитей.
Вторая группа видов этой секции, называемая в литературе Bif-
loriformis (Талиев, 1930; Holl 1940), не выделена во „Флоре СССР“,
но объединяет виды секций, которые Введенский расположил в такой
последовательности: Т. buhseana Boiss., Т. turkestanica Rgl., Т. tur-
comanica В. Fedtsch., T. bifloriformis Vved., T. biflora Pall., T. po-
lychroma Stapf., T. koktebelica Junge, T. callieri Ha lacs у et Lev.,
T. orithyioides Vved.,T. dasystemonoides Vved., T. dasystemon Rgl.,
T. tarda Stapf.
Пять первых видов образуют естественную группу. Объединение
кавказских и крымских видов в настоящее время, по-видимому, есте-
ственно, но четыре последних вида этой секции явно родственны меж-
ду собой и связывают секцию Eriostemones с секцией Orithyia.
В секцию Orithyia, по А. И. Введенскому, входят три вида—Т.
heteropetala Ldb., Т. uniflora (L.) Bess, и T. heterophylla (Rgl.) Ba-
ker.
Позже M. Г. Попов, базируясь в основном на особой форме
семян и типе сложения цветка, выделил Tulipa heterophylla в особый
род — Eduardaregelia М. Pop.
В. К. Василевская (1936) по анатомическому строению лукович-
ных чешуй делит тюльпаны на девять групп, но при этом делает ого-
ворку: „Естественно думать, что сходство видов внутри этих групп
обусловлено генетическим родством видов. Однако делать филогене-
тические построения внутри этих групп и между группами на основа-
нии только анатомических признаков было бы разумеется методоло-
гически неправильным11. В. К. Василевская по признакам строения
луковичных чешуй делит среднеазиатские тюльпаны на следующие
группы.
1--Т. heterophylla Baker, и Т. tianschanica Rgl.
• II—Т. kuschkensis В. Fedtsch., Т. tubergeniana Hoog. и Т. ho-
ogiana В. Fedtsch.
Ill—Т. lanata RgL, T. ingens Hoog. и T. sp. n.
IV—T. alberti Rgl., T. carinata Vved., T. micheliana Hoog., T.
kaufmanniana Rgl., T. dubia Vved., T. greigii Rgl., T. mogoltavica
M. Pop. et Vved. и T. schrenkii Rgl. Особое положение занимает
в этой группе Т. fosteriana Irv.
V—Т. korshinskyi Vved., Т. ferganica Vved., T. hissarica M.
Pop. et Vved., T. anisophylla Vved. и T. platystejnon Vved. Ук-
12
лоняющимися видами этой группы являются Т. praestans Н о о g. и
Т. subpraestans Vved.
VI—T.maximowiczii Rgl. и T. linifolia Rgl.
VII—T. turkestanica Rgl., T. turkomanica B. Fedtsch., T. dasys-
te„mon Rgl., T. dasystemonoides Vved., T. biflora Pall., T. biflorifor-
mis Vved., T. tarda Stapf., T. Buhseana Boiss., T. orithyioides
Vved., T. paradasystemon Vved., T. binutans Vved. и T. orthopoda
Vved. Особое положение занимает T. sogdiana Bge.
VIII—T. kolpakovskiana Rgl.. T. zenaidae Vved., T. iliensis Rgl.,
T. ostrovsjkiana Rgl., T. tetraphylla Rgl., T. nitida Hoog., T. .korol-
kovii Rgl., T. rosea Vved., T. brachystemon Rgl., T. dykesjana Vved.
и T. regelii К r a s n.
IX—T. lehmann;ana Merckl., T. behmiana Rgl., и T. borszczowii
Rgl. По морфологическому строению T. wilsoniana Hoog. резко от-
личается от видов IX группы.
Д. Холл (A. D. Hall, 1940) в книге, посвященной систематике ро-
да Tulipa, приводит следующее деление известных ему ррдов Tulipa.
Во главе рода он ставит тюльпаны секции Eriostemones,
ERIOSTEMONES
Section 1 Australes
T. australis (в широком смысле). Дополнительные вид^г: Т. austra-
lis (в узком смысле), Т. celsiana, Т. grisebachiana, Т. patens, Т. pri-
mulina, Т. urumiensis, Т. Silvestris.
Т. hageri (в широком смысле). Дополнительные виды: Т. hageri
(в узком смысле), Т. orphanidea, Т. whittallii.
Section II Saxatjles
Т. humilis (в широком смысле). Дополнительные виды: II. Т. hu-
milis (в узком смысле), Т. pulchella, Т. violacea, Т. ancheriana, Т. 1о-
wnei.
Т. saxatiljs (в широком смысле). Дополнительные виды: Т. cretica,
Т. bajceri, Т. saxatilis (в узком смысле).
Section Ill Eiflores
Т. biflora, Т. turkestanica, Т. polychroma, Т. tarda.
Мы считаем сделанное Холлом разделение тюльпанов секции Eri-
ostemones на три подсекции—Australes, Saxatiles, Biflores—правильным.
Можно, конечно, спорить с Талиевым о том, какой вид поставить
исходным в первой группе — Т. australis или Т. silyestris, а может
быть, и Т. biebersteiniana. Но дело в том, что разбивка на подсекции
проведена верно и виды, входящие в эти подгруппы, б. м. родственны
внутри групп и глубоко различны между группами..
Каждую из групп можно расширить за счет введения в нее до-
полнительных видов, неизвестных Холлу. Например, в группе Ill Bif-
lores во „Флоре СССР“ перечислены такие виды, как Т. buhseana,.
Т. turkomanica, Т. bifloriformis и др.
В конце каждой главы Холл разбирает какую-либо группу видов •
тюльпанов; часть видов он помещает в раздел „сомнительные виды“,
к которому из секции Eriostemines относит Т,- androsovii, Т. bessara-
bica, Т. bifloriformis, Т. buhseana, Т. callieri, Т. dasystemonoides, Т. kok-
tebelica, Т. orithyoides, Т. sogdiana и Т. turkomanica, собрав здесь и
четкие виды, и сомнительные, и синонимы.
13..
Секцию Leiostemones Холл делит на пять подсекций и выделяет
'еще один раздел с одиночными видами, не входящими ни в одну из
его пяти подсекций. Первая подсекция называется Clusianae. Во главе
первого раздела этой подсекции он ставит Т. aitchisonii (в широком
•смысле), а затем перечисляет дополнительные виды: Т. aitchisonii,
Т. aitchisonii cashmeriana, Т. stellata, Т. stellata chrysantha, Т. chitra-
lensis, T. clusiana.
Второй раздел этой подсекции возглавляет Т. montana (в широ-
ком смысле). Дополнительными видами раздела являются Т. linifolia,
Т. maximowiczii, Т. batalini и Т. montana.
Вторую подсекцию Холл называет Gesnerianae и также делит на
две части. Первую часть подсекции возглавляет Т. gesneriana (в ши-
роком смысле) с дополнительными видами Т. gesneriana ( в узком
смысле), Т. suaveolens, Т. armena, Т. galatica, Т. hungarica, Т. rhodo-
реа, Т. beotica.
Во втором разделе второй подсекции он перечисляет 28 видов
неотюльпанов.
Третью подсекцию секции Leiostemones Холл называет группой
Oculus-solis и разбивает ее также на два подотдела. Первый возглав-
ляет Т. oculus-solis (в широком смысле). В него входят: собственно
Т. oculus-solis, Т. systola, Т. Julia, Т. sharonensis, Т. stapfii, Т. cypria,
Т. undulatifoba. Т. praecox, Т. alepensis, Т. veneris. Второй подотдел
этой группы автор называет „Другие виды“. Сюда входят Т. kusch-
kensis, Т. lanata, Т. hoogiana, Т. sovietica, Т. amabilis.
В данной подсекции объединены виды, относящиеся, по данным
других авторов, как к секции Leiostemones, так и Tulipanum.
Подсекцию IV Холл называет Eichleres. Она включает всего один
раздел и состоит из Т. eichleri. Т. ingens, Т. tubergeniana, Т. fosteri-
ana, Т. micheliana, Т. greigii, Т. kaufmanniana, Т. praestans.
В эту подсекцию Холл включил виды трех секций флоры СССР.
Leiostemones, Tulipanum и Spiranthera; Т. eichleri он поставил рядом с
Т. ingens. Между Т. ingens и Т. fosteriana помещена Т. tubergeniana,
не имеющая с ними близких родственных связей. Рядом с Т. kauf-
manniana по неизвестным соображениям находится Т. praestans.
Подсекцию V А. Д. Холл назвал kolpakovskianae, поместив под
этим заголовком 8 среднеазиатских видов: Т. kolpakovskjana, Т. ilien-
sis, Т. altaica, Т. kesselringi, Т. corynestemon, Т. ostrovskiana, Т. ko-
rokovii, Т. lehmanniana.
В этой подсекции виды также расположены не по родству и ал-
фавиту. Т. kesselringi А. И. Введенским давно поставлен в синонимы
Т. tetraphylla, а Т. corynestemon Холла несомненно является синони-
мом этого же вида, т. е. Т. tetraphylla.
Одиночные виды Т. schmidtii Fomin и Т. sprengeri Ba.ker
А. Д. Холл объединяет в специальный раздел. Кроме того, он выде-
ляет тюльпаны, относящиеся к секции Orithyia,— это Т. uniflora,
Т. tianschanica, Т. neteropetala и Т. oxypetala.
Два вида тюльпанов из Китая и Японии — Т. edulis и Т. grami-
nifolia — Холл выделяет в особый род Amana —A. edulis A. gramini-
folia.
В конце работы в алфавитном порядке приведен список 69 видов,
которые автор не видел в живом состоянии. В работе Холла виды
описаны по растениям, выращенным в культуре, часто из семян мно-
гократной садовой репродукции. Поэтому, несмотря на тщательность
и добросовестность автора, описания не всегда отвечают действитель-
ности. Разбивка видов на группы не соответствует родству и не дает
14
представления ни об их филогенетических связях, ни о географиче-
ском местонахождении, так как часто описываемые виды куплены в
садоводствах. Так, по А. Д. Холлу, новый вид Т. sovietica несомнен-
но является Т. hoogiana, а то, что в его книге названо Т. hoogiana —
одна из форм Т. tubergeniana. Т. amabilis не имеет никаких прав
на самостоятельное название. А. И. Введенский давно поставил Т. ama-
bilis в синонимы к Т. hoogiana.
Очень интересна в филогенетическом отношении работа И. Д. Ро-
манова (1944). Он отмечает, что в группе Tulipeae род Tulipa наибо-
лее богат типами зародышевого мешка. До настоящего времени сре-
ди всех цветковых растений не найдено ни одного рода столь же
богатого в этом отношении. В роде Tulipa И. Д. Романовым обнару-
жено пять типов зародышевых мешков: Fritillaria, Druza, Adoxa, Tu-
lipa tetraphylla и Eriostemones. Наиболее широко распространен тип
зародышевого мешка Fritillaria. Он найден у 18 видов тюльпанов: у
Т. heterophylla (Тянь-Шань) из секции Orithyia; у Т. tubergeniana
(Памиро-Алай) и Т. hoogiana (Копет-Даг) — из секции Tulipanum; в
секции Leiostemones — у Т. ingens, Т. praestans, Т. maximowiczii —
все виды из Памиро-Алая; Т. alberti, T.greigii (Тянь-Шань), Т. miche-
liana (Памиро-Алай), Т. borszczowii (Сыр-Дарья), Т. lehmanniana
(Кызылкумы), Т. zenaidae (Киргизский Алатау), Т. ferganica (Фер-
ганский хр.), Т. hissarica (Памиро-Алай) — среднеазиатские виды и
Т. gesneriana с Т. ргаесох (Средиземье).
Drusa-тип зародышевого мешка имеется у четырех видов рода
Tulipa из секции Leiostemones: Т. rosea (Памиро-Алай), Т. korolkovii
(Сырдарьинская обл.), Т. wilsoniana (Горный Туркменистан) и Т. chry-
santha (Иран).
Adoxa- и Т. tetraphylla-типы зародышевого мешка И. Д. Романов
обнаружил у видов из секции Leiostemones: Т. ostrovskiana,
Т. kolpakovskiana и Т. tetraphylla, произрастающих в северном и
центральном Тянь-Шане.
Eriostemones-тип зародышевого мешка найден у видов секции
Eriostemones.
По типу зародышевого мешка виды секции Eriostemones очень
однообразны и резко отчленяются от видов других секций рода Tu-
lipa. Даже у Т. sogdjana, часто не имеющей типичного признака сек-
ции (опушенных тычиночных нитей), но габитуально сходной с ее
видами, И. Д. Романов выявил зародышевый мешок Eriostemones-ти-
па, который был открыт Гиньяром (1900) у Т. Silvestris и Т. celsiana.
И. Д. Романов нашел его у Т. bifloriformis, Т. tarda, Т. sogdiana и
Т. dasystemon. А. Д. Холл в 1939 г. привел список видов, исследо-
ванных Ньютоном. Это Т. Silvestris, Т. australis, Т. clusiana, Т. tur-
kestanica, Т. dasystemon, Т. hageri, Т. pulchella и Т. humilis.
Всего из секции Eriostemones исследовано 10 видов тюльпанов.
Из них три — Т. hageri, Т. pulchella и Т. humilis — относятся к Erio-
stemones-saxatilis, а остальные — к Eriostemones-silvestris.
Таким образом, по типу зародышевого мешка виды тюльпанов
секции Eriostemones отличаются от видов других секций. Незначи-
тельное сходство наблюдается между ними и Т. tetraphylla. Но по
кариотипу Т. tetraphylla более родственен тюльпанам из секции Erio-
stemones, чем Leiostemones (рис. 11, 17).
Среди видов других секций рода наиболее распространен Fritil-
laria-тип зародышевого мешка.
Следовательно, тюльпаны секции Eriostemones по типу зародыше-
вого мешка стоят на более высокой ступени развития, чем тюльпаны
15
других секции рода, тем более, что Fritillaria-тип зтротышевого меш-
ка, как пишет И. Д. Романов, имеет наиболее широкое распростра-
нение у близких к тюльпанам родов лилейных: Lilium, Fritillaria и др.
Не являясь примитивным типом для цветковых вообще, в данном
случае Fritillaria-тип должен считаться исходным для этой группы
родов.
Схему филогенетических отношений между родами Tulipaeae в
1937 г. дал Буксбаум.. По этой схеме род Tulipa поздно ответвляется
от общего ствола подсекции Tulipaeae, т. е. он являегся одним из
наиболее молодых родов, возникших в процессе эволюции цветковых
растений. И. Д. Романов (1944) приводит в своей работе эту схему.
Прототипом всего подсемейства Буксбаум считает род Iphigenia. Пер-
вым от него ответвляется род Gageac гаплоидным (п) числом хро-
мозом, равным 12. От ветви Platyospermum этого рода при ее возник-
новении отходит предок рода Lloydia родоначальник и современных
Lloydia, и Giraldiella (у Lloydia serotina п~12).
От прототипа рода Lloydia вначале ответвляется общий предок
для Fritillaria и Lilium (п = 12 хромозом), а несколько выше — общий
предок Erythronium-Tulipa (п = 12 хромозом).
С позиций филогенетической схемы Буксбаума рассмотрим взгля-
ды В. И. Талиева на эволюцию в семействе Liliaceae. Как известно,
представители родов Gagea и Lloydia никогда не имеют крупных цве-
тов. Род Tulipa ни в одной из известных нам филогенетических схем
ие выводится непосредственно из рода Lilium, поэтому утверждение
В. И. Талиева (1930) о том, что Tulipa относятся к Lilium, как обед-
невшие родичи, — необоснованно. Развитие этих родов происходило
совершенно независимо параллельными рядами (Lloydia — Tulipa —
Fritillaria — Lilium), и, несомненно, оно шло по линии прогрессивной
эволюции, а не регрессивной, как утверждал В. И. Талиев.
Укрупнение цветков получает большее развитие в роде Lilium.
Однако многие современные тюльпаны, имея независимое филогене-
тическое развитие, также обладают очень крупными цветами, и дан-
ный признак появляется в наиболее молодых видах рода, находящих-
ся в настоящее время в процессе становления и видообразования.
Например, группа молодых видов, распространенная в Памире-Алае,
близка к Т. fosteriana (Т. victoris, Т. affinis, Т. fosteriana и др.). Это
тем более правдоподобно, так как у тюльпанов секции Orithyia, со-
хранивших древннй признак семейства (столбик) и поэтому продолжаю-
щих в схеме Буксбаума линию предка, цветы относительно мелкие.
Такие же цветы у тюльпанов секции Eriostemones, которую Буксбаум
считает настолько примитивной, что выводит из нее секцию Leioste-
mones.
По нашему мнению, две первые секции рода не могут считаться
примитивными, так как у первой (Orithyia) прнмитгзен только длин-
ный столбик у завязи.
Ареал обеих секций на восточной границе распространения рода
фактически выключен из общего ареала рода, а их биология и карио-
логия не изучены. Все это не позволяет считать тюльпаны из Orit-
hyia и Eriostemones примитивом. Так, ветвь секции, направленная в
Среднюю Азию — Bifloraeformis, очень богата видами и содержит боль-
шое количество полиплоидов, что подчеркивает вторичный характер
как местообитаний, так и самих видов. Полиплоиды в этой секции
мозаично распределены по всему ареалу.
Со времени выхода в свет IV тома „Флоры СССР** (1935) были
установлены и описаны разными авторами 19 новых видов тюльпанов.
16
Из секции Tulipanum: Т. subquinquifolia Vved.
Из секции Leiostemones: Т. sosnovskyi Achv. etMars., T. vve-
denskyi Z. Botsch., T. mucronatha Fо m., T. kaghysmanica Fom.,
T. karabachensis A. Gross., T. butkovii Z. Botsch., T. affini?
Z. botsch.
Из секции Spiranthera: T. anadroma Z. Botsch., T. tschimganica
Z. Botsch.
Из секции Eriostemones: T. orthopoda Vved., T. binutans Vved.,
T. bpskyi A. Gross., T. ophiophylla Klok. et Zoz., T. scythica Klok.
etZoz., T. neustruevae P о b., T. alaica Vved., T. paradasystemon
Vved. и T. talievi Vved.
В СССР известно 83 вида тюльпанов. Из них 18 (отмечены звез-
дочкой) не вошли в IV том „Флоры СССР". Ниже приводим список
тюльпанов.
Tulipanum
Т. tubergeniana Hoog.
Т. kuschkensis В. Fedtsch.
Т. hoogiana В. Fedtsch.
Т. Julia С. Koch.
Т. subquinquifolia Vved.
Leiostemones
T. sosnovskyi Achv. et M i r s.
T. florenskyi W о г о n
T. kaghysmanica Fom.
T. lanata Rgl.
T. ingens Hoog.
T. fosteriana Irv.
T. carinata Vved.
T. affinis Z. Botsch (sp. n.)
T. praestans Hoog
T. butkovii Z. Botsch (sp. n.)
T. subpraestans Vved.
T. alberti Rgl.
T. greigii Rgl.
T. mogoltavica M. Pop. et
Vved.
T. brachystemon Rgl.
T. vvedenskyi Z. Botsch.
(sp. n.)
T. micheliana Hoog.
T. eichleri Rgl.
T. karabachensis A. Grossh.
T. mucronatha Fom.
T. schmidtii Fom.
T. maximowiczii Rgl.
T. linifolia -Rgl.
T. wilsoniana Hoog.
T. lehmanniana Merkl.
T. behmiana Rgl.
T. borszczowii Rgl.
T. korolkovii Rgl.
T. nitida Hoog.
T. rosea Vved.
T. schrenkii Rgl-
T. ostrovskiana Rgl.
T. kolpakoyskiana Rgl.
T. zenaidae Vved.
T. platystemon Vved.
T. altaica Pall.
T. iliensis Rgl.
T. ferganica Vved.
T. anisophylla Vved.
T. korshinskyi Vved.
T. tianschanica Rgl.
T. tetraphylla Rgl.
T. hissarica M. Pop. et Vved_
T. krauseana Rgl.
Spiranthera
T. kaufmanniana Rgl.
T. dubia Vved.
T. anadroma Z. В о t s c h. (sp. n)
T. tschimganica Z. Botsch
(sp. n.)
Lophophyllon
T. regelii К r a s n.
Eriostemones
T. violacea Boiss. et Buhse
T. caucasica Lipsky
T. lipskyi A. Grossh.
T. Silvestris L.
T. biebersteniana Schult.
T. ophiophylla Klok. et Zoz.
T. scythica Klok et Zoz.
T. patens Agarbh.
2—2229
17
T. sogdiana Bge.
T. buhseana В о i s s.
T. binutans Vved.
T. talievii Vved.
T. alaica Vved.
T. turkestanica Rgl.
T. bifloriformis Vved.
T. orthopoda Vved.
T. turkomanica B. Fedtsch.
'T. biflora Pall.
T. polychroma Stapf.
T. koktebelica Junge
T. callieri Halacsy et Lev.
T. orithyoides Vved.
T. dasystemonoides Vved.
T. dasystemon Rgl.
T, neustruevae Pob.
T. tarda Stapf.
Orithyia
T. heteropetala L d b.
T. uniflora (L.) Bess
Eduardaregelia heterophylla
M. Pop.
(T. heterophylla (R g 1) В a к er)
Из 83 видов, обитающих в СССР, в Средней Азии имеют распро “
’Странение 63 вида, т. е. 75,9% от общего количества видов. Из них
4 вида относятся к секции Tulipanum; 36— Leiestemones; 4—Spirant-
hera; 1—Lophophyllon; 16—Eriostemones и 2—Orithyia
На Кавказе встречается 14 видов тюльпанов, т. е. 16, 87%. В
секции Tulipanum имеется один вид; в секции Leiostemones —7, в
Eriostemones —5. На Кавказе род Tulipa представлен только тремя
секциями.
В Европейской части СССР наблюдаются представители двух сек-
ций рода: Leiostemones с одним видом и Eriostemones с 8 видами.
Всего 9 видов, или 10,84%.
Большое количество видов, относящихся к секции Eriostemones и
обитающих в Европейской части СССР, обусловлено тем, что Клоков
и др. один вид Т. biebersteiniana разбили на три мелких вида.
В Западной Сибири обитают представители трех секций: 2 вида
из секции Leiostemones; 3— из Eriostemones и 2— из Orithyia. Всего
7 видов, т. е. 8,43%.
В Восточной Сибири обнаружен всего один (1,21%) вид тюльпа-
на из секции Orithyia.
Таким образом, из всех географических областей, на которые
условно разделена территория СССР, Средняя Азия наиболее богата
как секциями рода Tulipa, так и количеством обитающих в ней видов
(табл. 1).
Приводим список видов тюльпанов, обитающих в СССР, но вы-
ходящих в сопредельные страны.
Секция Tulipanum Секция Eriostemones
Т. Julia С. Koch.
Секция Leostemones
Т. florenskyi W о г о п.
Т. micheliana Hoog.
Т. eichleri Rgl.
Т. schrenkii Rgl.
T. lehmanniana Merkl.
T. altaica Pall.
T. kolpakovskiana Rgl.
T. iliensis Rgl.
T. tetraphylla Rgl.
T. tianschanica Rgl.
T. biebersteiniana Schult.
T. turkomanica B. Fedtsch.
T. patens Agardh.
T. biflora Pall.
T. buhseana В о i s s.
T. silvestris L.
T. polychroma Stapf.
T. violacea Boiss et Buhse
T. dasystemon Rgl.
Секция Orithyia
T. uniflora L. Bess.
T. heterophylla (Rgl.) Baker
18
Таблица 1
Распределение видов тюльпанов по географическим областям СССР
и сопредельных стран, принятое во „Флоре СССР“
Географическая область Количество видов по секциям рода
Tulipanum 1 Leiosterno- nes Spiran thera Lophophyl- lon Eriostemo- nes Orithyia Всего %
Средняя Азия 4 36 4 1 16 2 63 75,90
Кавказ 1 8 — — 5 — 14 16,87
Европейская часть — 1 — — 8 — 9 10,84
Западная Сибирь — 2 — — 3 2 7 8,43
Восточная Сибирь — — — — — 1 1 1,21
Дальний Восток — — — — — — — —
Арктика — — — — — — — —
Джунгаро-Кашгар- ская — 5 — — 3 1 9 10,84
Иранская 1 4 — — 4 — 9 10,84
Балкано-Мало- азийская — — — — 1 — 1 1,21
Армяно-Курди- станская — 1 — — — — 1 1,21
Монгольская — — — — — 1 1 1,21
Западноевропей- ская — — — — 1 — 1 1,21
Всего за пределы СССР из обитающих у нас 83 видов выходят
22 вида, т. е. 26,5%. Но внутри сопредельных стран, конечно, есть
многочисленные виды тюльпанов, не встречающиеся в СССР.
Из 63 видов тюльпанов Средней Азии 48 (76,2%) эндемичны; рас-
пределение их по поясам или зонам Средней Азии дано в табл. 2.
В пустынном поясе, или поясе горной пустыни, на Кавказе оби-
тают два вида, которые не являются эндемами.
Таблица 2
Распределение эндемичных видов тюльпанов по поясам гор и зонам
Пояс гор или зона Количество эндемов по секциям Процент эндемов от количества
Tulipanum Leiostemo- nes Spiran thera Lophophyl- lon Eriostemo- nes Orithyia всего
эндемов и CO GJ О S '-•* Й CT u видов пояса эндемов области
Средняя Азия
Пояс
верхний — 3 1 — 3 — 7 10 70,00 20,98
средний 4 17 3 — 7 • — 31 34 91,18 64,58
нижний 4 11 1 — 3 — 18 25 76,00 52,08
Полупусты- ня и пустыня 4 8 — 1 3 — 15 25 60,00 31,25
19
Кавказ
Пояс верхний средний НИЖНИЙ — 3 1 — — 2 — 2 3 1 9 9 3 100.00 40 На Кавказе 14 видов, из них эндемов
33,33 33,33 60 20
Пустыня или горная пу- стыня — — — — — — — 2 — —
Пояс верхний Евр опейо <ая ча< сть СС СР
средний — — — — — — — — — —
нижний — — — — 2 — 2 2 100,00 50
Пустыня или горная пу- стыня — — — — — — — — —
Степь и лу- га — — — — 9 — 9 7 28,58 50
Глава II
ОБЩАЯ МОРФОЛОГИЯ ТЮЛЬПАНОВ
зрослое растение тюльпана состоит из луковицы, корней, гене-
ративного побега, стебля, листьев и цветка.
Луковица (по Серебрякову, 1952) — специализированный орган
вегетативного возобновления и размножения, в то же время запаса-
ющий орган растения. Стебель луковицы имеет дисковидную форму
и называется донцем. Запасные питательные вещества откладываются
в специализированных мясистых низовых листьях — запасающих чешу-
ях — от 1 до 6 у взрослых растений. Сверху луковица покрыта так-
же специализированной покрывной чешуей, по происхождению, дли-
тельности жизни и морфологии резко отличающейся от запасающих
чешуй. Отмершие чешуи прошлых лет, одевающие лежащую часто
на большой глубине луковицу тюльпана, предохраняют ее от внеш-
них неблагоприятных воздействий. Из верхушечной почки зрелой лу-
ковицы развивается цветоносный побег. В пазухе верхнего мясистого
листа — запасающей чешуи — растет почка замещающей луковицы,
которая на следующий год сменит материнскую. В пазухе каждой из
запасающих чешуй закладывается минимум по одной почке, из которых
могут развиваться дочерние луковицы-детки, служащие для вегета-
тивного размножения. У многих диких видов в естественных услови-
ях они обычно подавлены, но в условиях культуры часто дают кле-
новое размножение некоторых форм диких видов. Такие почки-детки
уносятся от материнских луковиц на большое расстояние столонами.
Заложение столонов происходит в'период летнего покоя и непосред-
ственно не связано с глубиной залегания луковицы.
Форма, размер и окраска луковиц весьма разнообразны и харак-
терны для определенных видов или групп. Например, луковицы сек-
ции Eriostemones и Leiostemones всегда отличны по размеру взрос-
лой луковицы, количеству чешуй в ней и покровной чешуе. Но
луковицы Т. Kaufmanniana из секции Spiranthera (в массе, а не в край-
них формах) нельзя отличить от луковиц Т. greigii из секции Leio-
stemones и т. д.
Столон — особый вырост на проростке тюльпана от места, где
находится донце сеянца. Он округлый, длинный, гладкий в узкой
части, шероховатый в нижней расширенной, в несколько раз толще
корешка, снаружи голый, внутри полый. На дне полости столона ле-
жит сместившаяся из луковицы почка, которая позже разрастается
в луковицу. Такие же выросты-столоны образуются у луковиц и в
последующие годы жизни, часто вплоть до первого цветения, и поз-
21
же — у почек-деток луковиц многих видов. Они имеют разную дли-
ну. Так, у Т. fosteriana они достигают почти 50 см. Столоны обеспе-
чивают первичное и последующее углубление луковицы, вегетативное
размножение, удаление дочерней луковицы от материнского растения
и проведение запасов питательных веществ от материнского растения
к дочерней луковице. В центре столона через всю длину проходит
воздухоносная полость от донца материнской луковицы до почечки,
заполненная волосковидными, вверх направленными жесткими выро-
стами однослойного эпидермиса. Снаружи на большей части поверх-
ности столон также покрыт однослойным, но гладким эпидермисом.
Лишь на утолщенной нижней части столона (луковице) он двухслой-
ный. После окончания роста луковицы столона в ширину эпидермис
в двухслойной части образует папиллы — клетки в виде сосочков с
утолщенными оболочками. Папиллы на луковице столона обнаружили
еще Ирмиш (Т. Irmisch, 1850) и Д. Е. Янишевский (1934). Они слу-
жат для укрепления луковицы в почве во время сокращения столона
при высыхании и защиты ее от раздавливания почвой.
В поперечном сечении стенки столона углубления имеют разную
толщину. В более толстой части столона, обращенной к донцу луко-
вицы, проходит несколько расположенных по кругу, с повернутой
внутрь круга ксилемой, сосудистых пучков. В противоположной, бо-
лее тонкой части стенки столона сосудистые пучки расположены в
один ряд. Они коллатеральны, как в листе, и обращены ксилемой
внутрь столона. Заслуживают внимания процессы длительного срас-
тания влагалища листа с осевой частью столона углубления, его мор-
фогенез, геотропически положительный рост, уносящий верхушечную
почку проростка на несколько сантиметров вниз от места заложения.
Первые этапы роста столона, во время которых одна половина донца,
обращенная в сторону выпуклой половины луковицы, начинает раз-
растаться горизонтально, а у прикрепленной к ней половины влагали-
ща листа усиленно растет основание, показывают, что в образовании
столона принимает участие не междоузлие, а узел (Бочанцева, 1956).
К видовым различиям или, наоборот, к сближению тюльпанов
может быть отнесено некоторое количество морфологических черт
столонов углубления. Так, очень тонкие, нитевидные, легко разру-
шающиеся при высыхании столоны углубления и их шаровидные луко-
вицы неполовозрелых, да и половозрелых, особей Т. praestans удиви-
тельно сходны со столонами и луковицами неполовозрелых особей
Т. fosteriana.
Различие заключается в том, что у Т. praestans верхушечная и па-
зушная (или все пазушные) почки уносятся на тонких нитевидных
столонах от места заложения, оставляя там лишь отмершие чешуи,
а у Т. fosteriana на месте старой луковицы часто остаются детки. У
замещающей луковицы у всех деток некоторых половозрелых особей
Т. praestans всегда сохраняется ювенильная способность уходить из-
старых чешуй на столоне углубления несколько вбок и вниз, что у
других представителей этого рода мы наблюдали очень редко. Эта
способность отмечена у Т. kaufmanniana и у одного сорта из группы
тюльпанов Дарвина — King Georg V., но их столоны, на которых раз-
виваются замещающие луковицы, морфологически отличны от столо-
нов Т. praestans.
У Т. fosteriana замещающая луковица стадийно зрелого растения
не углубляется, а остается на месте и в чешуях старой луковицы.
Столоны углубления в таком гнезде возникают из верхушечных по-
чек-деток, которые обычно в сильной степени омолаживаются и ча-
22
«сто после этого много лет подряд не цветут, а размножаются вегетативно.
Столоны и луковицы таких деток у Т. fosteriana и Т. praestans мор-
фологически сходны. Столоны и луковицы Т. praestans резко отличны
от подобных образований у Т. subpraestans, хотя эти виды наиболее
близки по строению цветка и другим морфологическим признакам.
У Т. subpraestans столоны значительно толще, более прочны, идут
отвесно вниз, а не в стороны, как у Т. praestans. Т. subpraestans часто
образует не углубившееся кучное гнездо луковиц. Стадийно зрелые
луковицы Т. subpaestans в большинстве случаев не углубляются и.
по форме более похожи на редьку, чем на шар.
Форма столона, длинного, толстого, мягкого, несколько суженно-
го к концам и расширенного к середине, постепенно переходящего
в основном в узкую и длинную луковицу, сближает Т. ingens с груп-
пой тюльпанов из секции Tulipanum — Т. tubergeniana, Т. kuschkensis.
Корень. Главный корень существует лишь у проростка тюль-
пана первого года жизни и к концу первого вегетационного сезона
отмирает.. Он не ветвится. Никаких других корней у тюльпана пер-
вого года жизни нет. Обычно корневая система тюльпана любого
возраста не имеет корневых волосков. Эпиблема корня их не обра-
зует. Но, как исключение, у Т. ingens, Т. maximowiczii, Т. wilsoniana
и Т. dubia в первый год жизни на главном корне нам удалось уви-
деть развивающиеся корневые волоски (Бочанцева, 1956). В после-
дующие годы на придаточных корнях у всех видов этого рода кор-
невые волоски отсутствуют.
Вся корневая система тюльпана в дальнейшем состоит из ежегод-
но сменяющихся придаточных корней, одновременно закладывающихся
в период летнего покоя в подковообразной нижней части донца на
стороне будущей замещающей луковицы. Следует особо отметить,
что корневая меристема в донце луковицы тюльпана вся расходуется
одновременно на заложение корней и к регенерации не способна.
Поэтому никакие прищипки или обрывания корней и пересадки после
укоренения луковиц невозможны. Если у растения тюльпана оборвать
корни, оно не погибнет благодаря запасам влаги и питательных ве-
ществ в луковице, но ему будет нанесен непоправимый вред на не-
сколько лет.
В условиях Ташкента корни у луковиц тюльпанов отрастают и
функционируют с осени. Сформированный корень очень эфемерен и
имеет первичное строение с эпиблемой (кожицей), первичной корой
и центральным цилиндром.
Первичная кора имеет слабо выраженную экзодерму, рыхлую,,
состоящую из тонкостенных крупных клеток, паренхимную ткань и
однослойную эндодерму.
Мелкоклетный по сравнению с первичной корой центральный ци-
линдр по периферии несет однослойный перицикл. В центре корня
находится сосудистый пучок с радиальным расположением ксилемы
и флоэмы. Корни растут своей вершиной и с оборванной точкой ро-
ста прекращают рост. Они, как и главный корень, не способны реге-
нерировать и ветвиться.
Генеративный побег развивается из верхушечной почки
луковицы. Развитие взрослого индивидуума продолжается из пазуш-
ной почки самой внутренней запасающей чешуи, т. е. симподия. Ос-
новной структурный элемент тюльпана — монокарпический побег, ко-
торый в течение многих лет до первого цветения растения развива-
ется моноподиально. Весь этот побег состоит из чередующихся
длинных и укороченных отрезков стебля.
23»
Стебель тюльпана имеет три главных формы: донце, стебель как
компонент столона и стебель генеративного побега с цветоножкой.
Однако в первые годы жизни тюльпана из семени у стебля из-
вестны только две формы. Первой из них является специфичная для
луковиц укороченная вертикально и расширенная горизонтально мо-
дификация стебля—донце, второй — часть стебля, принимающая уча-
стие в заложении, росте и развитии столона. Третья форма — стебель
генеративного побега, появляется после завершения тюльпаном ста-
дийного развития и вступления в плодоношение. Третья форма состо-
ит из отрезка стебля от нижнего листа до донца луковицы; среднего
отрезка, несущего ассимиляционные зеленые листья и верхней части
.цветоножки или плодоножки, являющейся непосредственным продол-
жением главной оси. Цветок тюльпана венчает растение. У тюльпа-
нов секции Eriostemones стебель выше верхнего листа ветвится и не-
сколько цветоножек полузонтиковидного соцветия бывают также
увенчаны конечными цветками одинакового размера с одноцветковы-
ми экземплярами.
У растений некоторых видов в природе и большего количества,
выращенных в культуре в условиях Ташкента из крупных луковиц
(Бочанцева, 1952), можно наблюдать ветвление стебля генеративного
побега ветвями первого, второго и иногда третьего порядка. На ге-
неративном побеге ветви могут вырастать из пазух всех надземных
листьев.
В луковице первым из всех органов генеративного побега закла-
дывается стебель в виде меристематического бугорка. Он длительное
время, разрастаясь горизонтально, не растет в длину. На первых эта-
пах рост стебля генеративного побега имеет много общего с ростом
донца, т. е., являясь органообразовательным элементом, закладывая все
новые и новые части побега, он почти не растет в длину. Так. сре-
динные листья на генеративном побеге тюльпана закладываются снизу
вверх на укороченной оси, а затем их венчает цветок. И только пос-
ле окончания заложения всех органов, вплоть до заложения плодоли-
стиков в цветке, начинают расти междоузлия стебля. У многих ви-
дов тюльпанов стебель генеративного побега с марта по сентябрь, не-
смотря на изменившиеся функции (заложения листьев, побега и цвет-
ка), растет, как донце луковицы; у него функционируют лишь узлы
при почти полном отсутствии роста междоузлий. В данном случае
мы устанавливаем обратную связь, т. е. заложившиеся на укорочен-
ном стебле органы, листья побега и цветок воздействуют на стебель
и вызывают у него рост междоузлий. Удаление зеленого бутона ран-
ней весной вызывает остановку роста междоузлий и цветоножки (Бо-
чанцева, 1952). Самым длинным междоузлием стебля является междо-
узлие между верхней чешуей луковицы и нижним стеблевым листом.
Это междоузлие выносит верхушечную почку тюльпана на поверх-
ность почвы. Средние междоузлия стебля в зависимости от вида и
' сорта могут быть короткими или длинными, но всегда большими,
чем междоузлия донца. Верхнее междуозлие генеративного побега
между верхним листом и цветком такой же длины, что и нижнее.
У стебля генеративного побега тюльпана первичное строение.
Его составляют эпидермис, первичная кора и центральный цилиндр.
Эпидермис стебля однослойный, покрыт кутикулой, часто с вос-
ковым налетом и простыми волосками у некоторых видов. Непосред-
ственно под эпидермисом первичная кора имеет колленхимное кольцо
из одного ряда клеток. Паренхимные клетки первичной коры, распо-
ложенные довольно рыхло, одеты в целлюлозные оболочки и содер-
24
жат хлорофилловые зерна. Периферические клетки центрального ци-
линдра в части стебля, расположенной над поверхностью почвы,
сильно склерефицированы и образуют довольно мощное кольцо ме-
ханической ткани. Это кольцо отделяет центральный цилиндр от пер-
вичной коры. Клетки основной паренхимы различны по размерам,
они крупнее в центре около сосудистых пучков. В центральном ци-
линдре разбросаны коллатеральные сосудистые пучки. На периферии
они окружены механической тканью. По направлению к центру их
форма постепенно меняется, и иногда там можно встретить биколла-
теральные сосудистые пучки.
Донце (основание луковицы) — укороченный, разросшийся в
ширину, дисковидный стебель.
Этот укороченный стебель обладает резко выраженной полярно-
стью. Сверху на нем развиваются покровная чешуя, затем запасаю-
щие чешуи, в пазухах чешуй — почки и, наконец, в центре—генера-
тивный побег. В нижней половине донца в виде полукольца, распо-
ложенного со стороны замещающей луковицы, лежит мелкоклетная
корнеродная ткань — корневая меристема, из которой возникает при-
даточная корневая система луковицы.
В маленьком по объему донце сходится большое количество
сосудисто-волокнистых пучков при переходе их из листьев, стебля
генеративного побега в чешуи луковицы, столоны и корни.
Э. Ф. Мигушова (1954) установила, что коллатеральные пучки
цветоносного надземного стебля или листьев неполовозрелых особей
при переходе в донце материнской луковицы перестраиваются в кон-
центрические пучки амфивазального типа с внутренней флоэмой (они
окружены крупноклетной паренхимой). Однако сосудистые пучки при
переходе в зону формирования корней вновь перестраиваются. При
этом концентрические сосудистые пучки преобразуются в сосудистые
пучки корневого типа.
Заложение корней в нижней зоне донца начинается у многих
видов в условиях Ташкента после окончания периода вегетации ма-
теринского растения. Уже в середине июня в донце луковиц видно
начало формирования придаточных корней. К концу периода летнего
покоя сформировавшиеся корни достигают внешнего эпидермиса дон-
ца, слегка его приподнимают и донце набухает. При избытке влаж-
ности луковица в это время способна к укоренению.
В Ташкенте на луковицах тюльпанов можно делать прививки
только в период от начала летнего покоя до середины июня, так как
позже вся меристема донца расходуется на корнеобразование и оно
не способно ни к каким регенерациям, рубцеваниям и срастаниям.
Лист. У тюльпанов имеется пять типов листьев: единственный
семядольный лист; влагалищный зеленый лист неполовозрелого расте-
ния; верховые, сидячие, зеленые листья генеративного побега; мета-
морфизированный низовой лист, или покровная чешуя луковицы; ме-
таморфизированный низовой лист, или запасающая чешуя луковицы.
В первый год жизни тюльпаны обладают всего одним семядоль-
ным листом. В последующие годы ежегодно весной вплоть до зацве-
тания над поверхностью почвы у растений появляется по одному
„черешковому“ влагалищному листу, и только при вступлении расте-
ний в пору плодоношения отрастают 2—5 зеленых, сидящих на стеб-
лях листьев. Как исключение, их бывает меньше (один лист у Т. ге-
gelii) или больше (12 листьев у Т. schmidtii).
Семядольный лист тюльпана состоит из влагалища, связника се-
мядоли и гаустории.
25
При прорастании семени верхняя бесплодная часть семядоли
остается в семени. Она бесцветная, плоская и играет роль гаустории,
никогда не зеленеет и после опустошения семени вскоре отсыхает.
Среднюю зеленую обычно цилиндрическую часть семядольного
листа И. Т. Васильченко назвал связником семядоли. Она сужена к
вершине и основанию, в поперечном сечении почти округлая, ее со-
судистые пучки распределены по кругу. Перед внедрением в почву
семядоля становится тоньше. Уменьшение объема происходит за счет
исчезновения воздухоносных полостей в паренхимной ткани. Под зем-
лей семядоля сохраняет свою округлую форму, бесхлорофильна и
может быть названа черешком.
У многих видов на одной стороне семядоли эпидермис уплощен
(Бочанцева, 1956). В нижней части связника эта уплощенная часть
образует желобок, являющийся остатком редуцированной верхней
плоскости связника (пластинки семядоли). Подходя к поверхности
почвы, желобок начинает углубляться в ткань семядоли и образует
замкнутое влагалище. Отверстие влагалища часто имеет маленький
язычкообразный вырост, прикрывающий вход в него. В верхней ча-
сти замкнутого влагалища находятся большей частью жесткие и на-
правленные вверх волоски, как и язычки, прикрывающие вход. Един-
ственная почечка проростка глубоко скрыта в основании влагалища
семядольного листа, которое сильно разрастается, по окончании веге-
тации высыхает и выполняет роль покровной чешуи луковицы пер-
вого года жизни.
Расположение листьев у ювенильных особей тюльпанов розеточ-
но-двухрядное. Главная жилка первого зеленого листа расположена
против семядоли, так как первым после семядоли листом является
запасающая чешуя. Это метаморфизированный лист с недоразвитой
листовой пластинкой и с функциями запасающего, а не ассимиляцион-
ного органа. Он закладывается в луковице первого года жизни почти
одновременно с семядолью, но отмирает лишь ко времени полного
развития первого зеленого влагалищного листа второго сезона веге-
тации. Со второго года вегетации вплоть до цветения у сеянца тюль-
пана ежегодно отрастает всего один зеленый лист с листовой пластин-
кой, у поверхности почвы сужающейся в черешок и одевающей своим
влагалищем замещающую луковицу.
Листовая пластинка зеленого листа ювенильного растения очень
похожа на сидячие листья взрослых растений. Со второго года жизни
она постепенно приобретает форму, близкую к форме нижнего листа
вида или сорта, к которому относятся сеянцы: на поверхности листа
Т. regelii появляются 1—2 выпуклых гребня; у Т. fosteriana—хрящеватый
красный или желтый край; у Т. greigii на верхней стороне листовой
пластинки—антоциановые пятна; у Т. micheliana—полосы; у Т. maxi-
mowiczii—узкие длинные листья сочетаются с красным окаймлением;
у Т. vvedenskyi на листьях возникает опушение. Таким образом, уже
на втором году жизни сеянца по морфологии листа у многих видов
можно установить видовую принадлежность. С каждым последующим
годом эти морфологические признаки проявляются все ярче.
Во влагалище зеленого листа неполовозрелой луковицы тюльпана,
преобразованном в черешок, как и у семядоли, проходит сквозной,
обычно опушенный канал. Опушение особенно развито в расширенной
нижней части листового влагалища, служащего покровной чешуей
замещающей луковицы.
Систематики тюльпанов придают большое значение строению по-
кровной чешуи и ее опушению.
26
Так, у многих видов сосудистые пучки и клетки стенок листового
влагалища одревесневают, а скопившиеся за многие годы и не разру-
шившиеся остатки покровных чешуй продолжаются до поверхности
почвы. Такие покровные чешуи наблюдаются у Т. borszczowii, Т. leh-
manniana, Т. behmiana и у некоторых форм Т. greigii. У других
видов, наоборот, покровные чешуи очень нежные и короткие (Т. schren-
kii) или имеют жесткие покровные, но не продолженные чешуи (Т. ost-
rovsjciana, Т. kolpakovskiana). У Т. korolkovii, как у и Т. greigii,
встречаются особи как с короткими, так и с продолженными покров-
ными чешуями. Опушение покровных чешуй—очень важный система-
тический признак. Так, тюльпаны, у которых покровные чешуи опу-
шены густыми, длинными, мягкими, сильно извитыми волосками, вы-
делены из секции Leiostemones и отнесены А. И. Введенским в секцию
Tulipanum, Т. tubergeniana, Т. kuschkensis, Т. hoogiana, Т. julia и
Т. subquinquifolia. У Т. ingens на покровной чешуе находятся длин-
ные жесткие волоски, собранные в косицы, у Т. wilsoniana опушение
покровных чешуй тонкими шелковистыми длинными волосками сосре-
доточено на вершине луковицы.
В большинстве случаев при наступлении у тюльпана половой
зрелости лист, одевающий своим влагалищем луковицу, перестает
развивать листовую пластинку; разрастается одно влагалище, т. е.
покровная чешуя, но его консистенция, опушение и цвет остаются
такими же, как и у ювенильных особей.
Цвет покровной чешуи — прекрасный видовой признак. Так, у
Т. bifloriformis свежие покровные чешуи окрашены в ярко-розовый
цвет; у Т. ostrovskiana и Т. kolpakovskiana они почти черные.
При наступлении половой зрелости функцию ассимиляции испол-
няют зеленые сидячие листья генеративного побега. По форме эти
листья очень напоминают листовые пластинки влагалищных листьев
неполовозрелых особей. Особенно сходен с ними нижний лист генера-
тивного побега. Чем выше по стеблю расположены листья, тем они
мельче. Следует отметить, что у них быстрее убывает ширина, чем
длина.
Обычно на листе тюльпана не видно жилок, но у Т. praestans,
Т. carinata, Т. anadroma выделяется толстая центральная жилка, а у
Т. kaufmanniana на светло-зеленой мякоти листа проступает целая
сеть из анастомозов темно-зеленых жилок. Листья на побеге тюльпана
засыхают раньше, чем созревают коробочки.
Запасающие чешуи луковицы являются метаморфизированными
органами, которые в противоположность покровной чешуе никогда не
несут зеленой листовой пластинки. Они строго приспособлены к отло-
жению в толстых паренхимных тканях запасов питательных веществ
и накоплению влаги. У большинства видов тюльпанов в луковице
первых лет жизни имеется лишь одна запасающая чешуя. Появление
второй запасной чешуи у большинства видов—предвестник зацветания
весной следующего года. На внутренней стороне запасающей чешуи
в противоположность покровной никогда не развивается опушение, и
по высыхании они всегда становятся тонкими, бумагообразными. У
зрелых луковиц наблюдается до шести запасающих чешуй.
Цветок у тюльпана циклический, т. е. все части его располо-
жены кругами в виде чередующихся мутовок. Это пятикруговой трех-
членный цветок, два круга которого приходится на околоцветник,
два—на тычинки и один—на плодолистики. Количество тычинок равно
числу частей околоцветника. Тычинки чередуются с внешним и вну-
тренним кругами околоцветника и могут быть разной длины.
27
Околоцветник тюльпана венчиковидный, ярко окрашенный, раз-
дельнолепестной, образует двойную чашу. Цветок тюльпана правильный
(актиноморфный), большей частью одиночный и крупный, сидит на
прямом упругом цветоносе и, как правило, направлен вверх. Лишь
у немногих видов и сортов тюльпанов на некоторое время поникают
бутон перед цветением и отцветший цветок. Перед окрашиванием
цветоножка бутона и плодоножка опыленной завязи быстро выпрям-
ляются. Подобное явление наблюдается у некоторых тюльпанов из
секции Eriostemones, у Т. tetraphylla из секции Leiostemones, а также
у гибридов с сортами культурных тюльпанов Due van Thol yellow и
Preserpina.
У некоторых видов (Т. ostrovskiana, Т. Silvestris) тонкая цвето-
ножка слегка сгибается под тяжестью крупного цветка. То же наблю-
дается у культурных тюльпанов из группы лилиецветных. Это свой-
ство придает цветам указанных видов и сортов большое изящество.
Форма цветка у диких видов тюльпанов очень разнообразна. Она
является видовым и даже групповым признаком. Так, у Т. greigii
внутренние листочки околоцветника образуют несколько расширенный
на вершине цилиндр, а внешние—на половине длины отогнуты на-
ружу. Цветок Т. micheliana имеет такую же форму. В цветке Т. alberti
листочки внутреннего круга околоцветника ложковидно загнуты внутрь.
Звездообразный цветок Т. maximowiezii очень оригинален и похож по
форме на цветок Т. linifolia. Тюльпаны секции Tulipanum, несмотря
на близость с тюльпанами, группирующимися вокруг Т. fosteriana, по
форме цветка сходны с тюльпанами, близкими к Т. greigii, и т. д.
Околоцветник. В устройстве, форме иокраске первого круга
околоцветника тюльпана есть некоторое отличие от второго круга.
Например, листочки внешнего круга околоцветника у Т. greigii,
Т. micheliana округло ромбические, а внутренние — обратно треуголь-
ные с б. м. внезапно оттянутыми, опушенными кончиками. Внешние
листочки околоцветника, особенно в основании, значительно толще
внутренних.
По окраске, ее интенсивности и распределению внешние листочки
околоцветника значительно отличаются от внутренних. Снаружи по
спинке они ярко окрашены или покрыты сизым налетом, иногда опу-
шены, часто с пятном у основания листочка. Края на вершине цвет-
ка одинаково окрашены. У внутренних листочков окраску другого
цвета имеют лишь жилка на спинке листочка и пятно при его осно-
вании.
Внутренняя сторона листочков околоцветника на вершине часто
одинаково окрашена у обоих кругов, но пятно на их основании всегда
разной формы, окраски и окаймления. Для тюльпанов секции Erioste-
mones характерно опушение оснований листочков околоцветника.
Таким образом, у цветка тюльпана этой секции необходимо опи-
сывать его форму и отдельно форму, окраску, опушение обоих кругов
листочков околоцветника как снаружи, так и внутри цветка.
Тычинки. В систематике тюльпанов форме тычинок придается
большое значение. Так, первичное деление тюльпанов на две глав-
нейшие секции рода проведено на основании опушения тычиночных
нитей. Виды секции Eriostemones обладают опушенными тычиночными
нитями, а у Leiostemones нити голые. В последующем из рода Tulipa
на основании других признаков были выделены более мелкие секции.
Но позже (1935 г.) на основании способа открывания пыльников
А. И. Введенский выделил Т. kaufmanniana из секции Leiostemones в
особую монотипную секцию Spiranthera.
28
У Т. kaufmanniana „пыльники постепенно и медленно от верхушки
к основанию открывающиеся, скручивающиеся* (Введенский, 1935). У
новых видов тюльпанов из Тянь-Шаня, описанных нами, пыльники
открываются постепенно, но без специфического скручивания (Т. anad-
roma, Т. tschimganica, Т. dubia А. И. Введенского).
У типичных представителей Т. kaufmanniana постепенно открываю-
щиеся пыльники не всегда скручиваются спирально.
Таким образом, сохраняя секционное деление рода, предложенное
А. И. Введенским, необходимо перенести в секцию Spiranthera из
секции Leiostemones Т. dubia и добавить в неедва наших новых вида:
Т. anadroma и Т. tschimganica.
В 1882 г. Регель описал из Джунгарского Алатау тюльпан, при-
своив ему название Т. brachistemon Rgl. — короткотычиночный.
В 1935 г. А. И. Введенский описал из Памиро-Алая новый вид
тюльпана с видовым названием Т. platystemon Vved., т. е. широко-
тычиночный (по форме тычиночных нитей). И наконец, в 1940 г. Холл
у полученных от нас тюльпанов обратил внимание на форму тычиноч-
ных нитей у Т. tetraphylla Rgl. и дал ему новое видовое название
Т. corynostemon Hall.
Тычиночные нити у видов тюльпанов весьма различной формы.
Они имеют вид низких толстых треугольников у Т. micheliana, Т. 1а-
nata; тонких колышков—у Т. borszczowii; булавовидно вздуты на вер-
шине у'Т. tetraphylla; расширены в середине и сужены к концам у
Т. platystemon к Т. zenaidae; очень массивны у Т. greigii и тонки у
Т. maximowiczii.
У тюльпанов секции Eriostemones на вершине нижней четверти
тычиночной нити часто лежит волосистое кольцо и от основания до
кольца нить расширяется, а выше к пыльнику постепенно сужается.
Если волосистое кольцо менее ярко выражено, то и расширение нити
б. м. сглажено.
Окраска тычиночных нитей тюльпанов весьма разнообразна и ха-
рактерна для вида. У Т. korolkovii, как правило, нижняя половина
нити черная, а верхняя красная, у Т. zenaidae нижняя половина нити
(формы с черным пятном при основании листочков околоцветника)
черная, а верхняя оранжевая; у светлых форм Т. ^kaufmanniana никог-
да не бывает темных тычиночных нитей, а у Т. dubia оранжевые цветки
с оранжевыми тычиночными нитями часто имеют коричневую верхнюю
заостренную часть.
Четырехгнездные пыльники тюльпанов сходны между собой по
форме, но весьма различны по размерам к окраске. У некоторых видов
над пыльником выдается связник. Он иногда окрашен в другой цвет,
чем вся тычинка. Например, у Т. buhseana при желтой тычинке вер-
шина пыльника и острие связника окрашены в черный цвет. У одних
видов пыльники более массивны, у других —мельче и тоньше. Особен-
но яркое отличие в форме пыльников проявляется по способу их
открывания. Пыльники видов секции Spiranthera открываются посте-
пенно. Стенки их гнезд поперечно ссыхаются, связник часто спирально
скручивается, иногда пыльники свернуты спирально вдоль, в верхней
части—поперек оси. Пыльники видов секции Spiranthera очень узкие
по отношению к длине—это главное отличие их, особенно заметное
у сухих растений в гербарии.
Завязь. Верхняя завязь тюльпана состоит из трех сросшихся
плодолистиков. В завязи тюльпана—три гнезда. В каждом из гнезд
на плацентах, образовавшихся из загнутых внутрь краев плодолисти-
ков, расположены два ряда семяпочек.
29
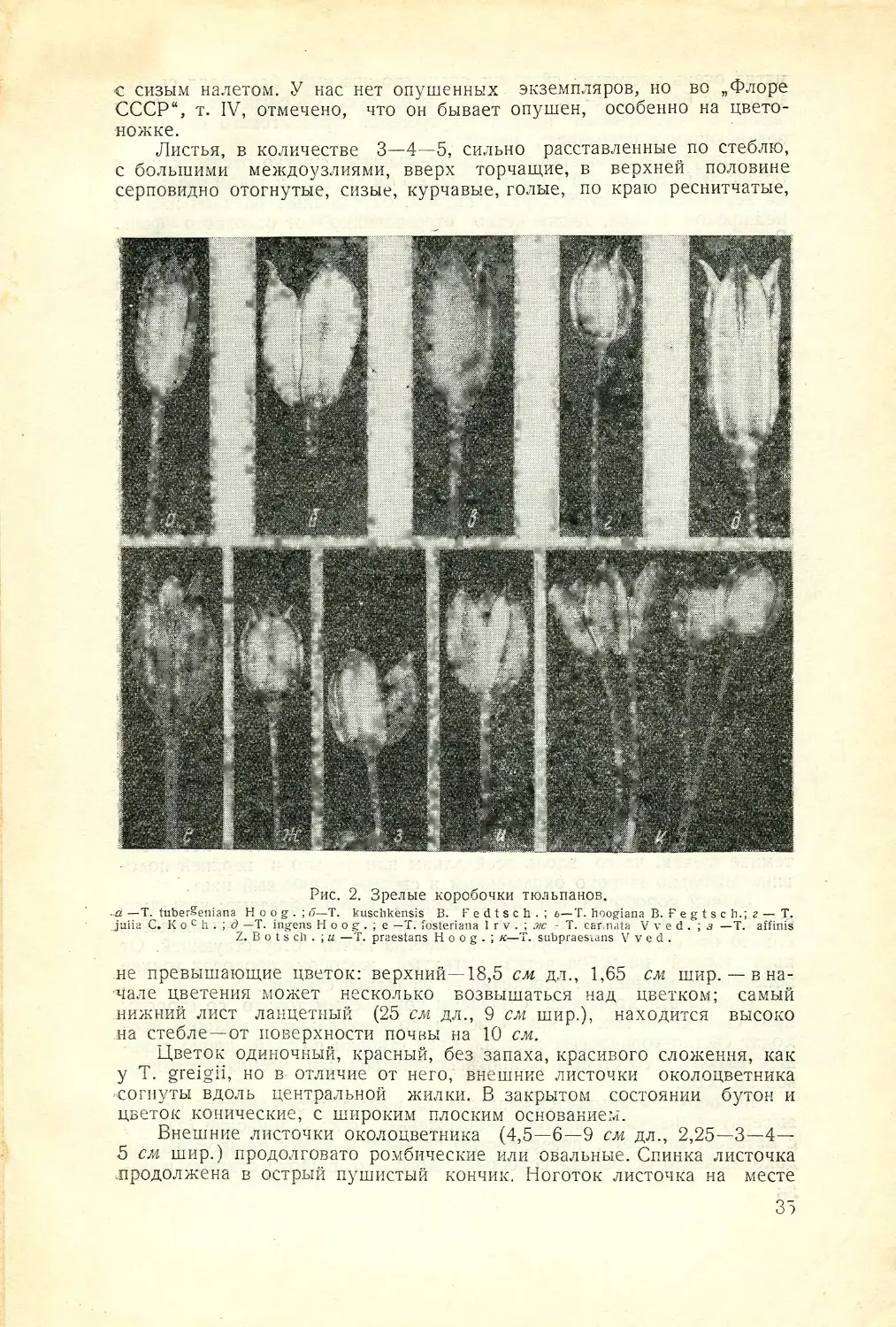
Рис. 1. Коробочка тюльпана.
а—створки коробочки; б—рыльце;
в—столбик; г—верхняя бесплодная
часть створки; д—щечки створки;
е—ребро коробочки; ж—рубец на
месте срастания плодолистиков;
з—карпофор; и—рубец от опавщих
членов околоцветника; к—плодо-
ножка.
При описании тюльпанов обращает на себя внимание размер за-
вязи относительно длины тычинок и околоцветника, форма завязи,
наличие или отсутствие столбика. Так, секция Orithyia выделена из
рода Tulipa из-за длинного столбика у входящих в нее видов.
Для завязей, имеющих маленький столбик или почти сидячее рыльце,
характерны форма перехода створки в столбики, форма рыльца и ок-
раска завязи. Она сизо-бордовая у Т. ingens, зеленая у Т. fosteriana,
сине-сизая при желтом столбике и рыльце у Т. zenaidae.
Очень часто окраска сосочков рыльца иная, чем окраска бесплод-
ных частей плодолистиков (столбика) под самым рыльцем. Например,
у Т. korolkovii рыльца желтые, а верхняя область столбика красная.
Ребра завязи часто окрашены яркими антоциановыми полосами.
Пл од тюльпана—трехгнездная коробочка, которая формируется
из верхней синкарпной завязи, состоящей из трех плодолистиков (рис. 1).
В каждом гнезде находится по два ряда
многочисленных, горизонтально расположен-
ных плоских семян, прикрепленных к завер-
нутым внутрь краям плодолистиков. Около-
плодник хорошо развит. Он характеризуется
одревеснелыми клетками, окружающими по-
лость гнезда. Кроме того, в нем присутст-
вуют механические элементы, способствую-
щие раскрыванию гнезда. Коробочки откры-
ваются тремя продольными трещинами вдоль
средней жилки каждого плодолистика. На
месте разрыва образуются характерные бах-
ромки. Не отваливающееся рыльце у многих
видов сильно разрастается. Оно сидит на
оттянутой, разной формы и величины верхней
бесплодной части створки—„столбике".
У основания коробочки бесшовным сра-
станием плодолистиков у большинства видов
образуется характерно разросшаяся бесплод-
ная часть карпофор. Коробочка отчленяется
от плодоножки часто разросшимся рубцом
от опавших членов околоцветника.
Характерные видовые признаки коробоч-
ки тюльпана—длина, диаметр и форма коро-
бочки; размер, форма и окраска карпофора;
наличие „столбика" или суженной бесплодной
верхней части коробочки; размеры, форма
и окраска столбика; форма, окраска и положе-
ние рыльца. Особое богатство признаков наб-
людается при изучении створок коробочки.
Обычно очень характерна форма створки.
Ярким признаком створки является рубец на
месте срастания плодолистиков, его углублен-
ность, характер поверхности, жилкование и
окраска. От рубца, образующего желобок
вдоль створки, приподнимаются щечки створки. Они обычно выпук-
лые, но бывают и плоские, грубые или бумагообразные, прозрачные
или матовые, морщинистые или гладкие, часто, особенно в верхней
части, поперечно полосатые. Около ребра коробочки вдоль щечек
возникает б. м. углубленный желобок, в котором коричневые полос-
ки, переходящие со створок, часто сплетаются в сложное кружево.
30
Самый край створок на ребре ярко окрашен или, наоборот, одного
цвета со всей створкой.
Раскрытие коробочки тремя продольными трещинами затрагивает
и внутренние перегородки коробочки, которые, расходясь по тяжам
пыльцепроводниковой ткани, приобретают под рыльцем на разрыве
характерную для вида форму. Глубина растрескивания внутренних
перегородок коробочки также отражает видовые особенности тюль-
панов.
Разные виды отличаются окраской, толщиной и поверхностью
створки вдоль продольного разрыва.
Степень отклонения частей столбика и рыльца, ширина и глубина
продольных щелей различны для отдельных видов тюльпанов.
Отличительные признаки—рубец от опавших членов околоцветни-
ка; окраска, толщина и относительная длина плодоножки.
Глава III
МОРФОЛОГИЧЕСКОЕ ОПИСАНИЕ
ДИКИХ ТЮЛЬПАНОВ СССР
В
настоящей работе мы придерживаемся в основном системы
секционного деления и расположения видов внутри секций рода
Tulipa, предложенной В. И. Введенским во „Флоре СССР".
Секция Tulipaniim R е b.
В IV томе „Флоры СССР" А. И. Введенский дает характеристику
данной секции: „Оболочки луковиц с внутренней стороны по всей
поверхности обильно паутинисто-мохнатые. Цветы крупные, ярко ок-
рашенные. Нити тычинок голые. Завязь с сидячим рыльцем".
Из этой секции в СССР растет 5 видов: Tulipa tubergeniana Hoog.,
Т. kuschkensis В. Fedtsch., Т. hoogiana В.^ Fedtsch., Т. julia
С. Koch, и Т. subquinquifolia Vved.
В Средней Азии находится 5 видов, на Кавказе—1 (Т. julia).
Tulipa tudergeniana Hoog.—Тюльпан тубергена (рис. 1 и 2, а)
обитает „на глинистых склонах в районах выходов пестроцветных
пород—Средняя Азия: Памиро-Алай (юго-зап. часть). Эндем" („Флора
СССР", т. IV).
Мы исследовали образцы растений из Узбекистана (Денауский
район, окрестности кишлака Сина, пестроцветы у Дербента на склоне
оврага), Таджикистана (окрестности кишлака Туткаул на лессе и
красной глине). На белых почвах он рос под кустами полыни. Луко-
вицы, обильно покрытые мохнатыми чешуями, лежали на глубине
10-15 см.
Луковицы со снятыми старыми чешуями яйцевидные, 3—5 см
в диаметре, а в чешуя.х шарообразные с бурыми или коричневатыми
(очень старые чешуи белесовато-серые) бумагообразными, с внутрен-
ней поверхности очень обильно паутинисто-мохнатыми оболочками.
Волоски опушения очень длинные, тонкие, шелковистые, сильно из-
вилистые.
Стебель вместе с цветоножкой от 5 до 30 см выс. от поверхности
почвы, опушен короткими торчащими волосками. Цветоножка вверху
красновато-розовая.
Листьев 3 (4), серповидно отогнуты, покрыты сизым налетом, б.
м. сближенные, создают впечатление розетки, лишь верхний несколько
приподнимается на стебле.
Формы этого вида из окрестностей Дербента с голыми листьями,
по краю красно или хрящевато окаймленные, реснитчатые; б. м. пу-
32
Рис. I. Т. tubergeniana Hoog. (2/3 натуральной величины).
шистые, курчавые, превышающие цветы; самый нижний—ланцетный
(18X2,8 см)\ верхний—линейный (13,7X1 см). Чем выше по стеблю
растут листья, тем глубже они сложены в желобок.
Цветок красный, один, без запаха, по форме напоминает цветок
Т. greigii, но внешние листочки околоцветника торчат вверх и отги-
баются к спинке вдоль отгиба.
Внешние листочки околоцветника (2,5—10 см дл., 1,7—7 см шир.)
острые, ромбические, с длинным, острым, опушенным кончиком, по
краю слабо волнистые, снаружи красно-оранжеватые с более белесо-
ватым отсветом, чем внутри, с сизым налетом на ноготке, с лиловым
овальным пятном на месте внутреннего просвечивающего пятна, к
основанию зеленеющие (иногда внешнее пятно зеленое), с внутренней
поверхности красно-оранжевые, с большим овальным (5,6X3 см) сплошь
черным, на вершине слегка заостренным пятном, которое окаймлено
бледно-желтой, четко отграниченной каймой; бывают формы, у кото-
рых почти незаметна кайма.
Внутренние листочки околоцветника обратно яйцевидные или
обратно продолговатые, 9—2,2 см дл., 6,3—1,5 см шир., вне-
запно оттянутые в острый, длинный пушистый кончик. Снаружи крас-
но-оранжевые, к ноготку оранжеватость несколько усиливается; на
ноготке с остро ромбическим обращенным острым углом вверх, дохо-
дящим до одной трети высоты листочка зеленым или фиолетово-бор-
довым пятном. С внутренней поверхности эти листочки красно-оран-
жевые, более блестящие, чем снаружи; имеют на ноготке черное или
черно-бордовое пятно овальной формы, к вершине несколько заост-
ренное, в нижней трети во всю ширину листочка, занимающее 3/5
длины листочка, т. е. всю толстую часть; окаймленное кругом светло-
желтой каймой, которая уже, чем на внешних листочках.
Тычинки в 2,5—3 раза короче околоцветника и немного длиннее
завязи.
Нити узкоконусовидные, тонкие, несколько сплющенные, голые,
черные, на обоих концах светлее, в 3 раза короче пыльников.
Пыльники широкие, темно-лиловые; пыльца грязно-желтая или
коричневато-лиловая.
Завязь зеленая, со светло-фиолетовым оттенком, в виде ровной
палочки.
Рыльце светло-зеленое, сидячее, шире завязи, несет желтые сосоч-
ки. Коробочка соломенно-желтая с коречневатыми полосами и штри-
хами до 4,6 см дл. и до 2,5 см шир., продолговатая, тупая на обоих
концах; у мелких растений почти шаровидная, широкий карпофор
очень короткий, 0,7 см выс., у крупных коробочек острым коротким
зубцом вдающийся в желобок створки и совсем отсутствующий у
мелких; короткий столбик, особенно ярко выраженный у мелких
коробочек (у коробочки 2,2 см дл. столбик 0,7 см дл., а у коробочки
7,7 см—всего 1 см).
Серо-коричневатое, сильно выгнутое наружу рыльце легко обла-
мывается.
Коробочка открывается узкой щелью до 0,2 см шир. с почти
параллельными краями, достигающей карпофора и образующей раст-
руб шириной в 2 см отклонением верхних суженных частей створок.
У мелких коробочек щель короткая, широкая, клиновидная.
Внутренние перегородки растрескиваются примерно на 1/3 длины,
т. е. ниже суженной верхней части створки. Внутри под рыльцем
поверхность разрыва перегородки матово-белая; от рыльца до конца
утолщения Lстворок она расширяется, затем, закругляясь, сужается
3—2229
33
и у выгиба перегородки становится соломенно-желтой и одинаковой
ширины на всем протяжении до конца разрыва.
Створки коробочки продолговатые с параллельными краями, ко-
ротко заостренным верхним концом и тупым основанием. У малень-
ких коробочек створки яйцевидной или продолговато яйцевидной
формы.
Вдоль каждой створки в центре проходит желобок—рубец, начи-
нающийся под рыльцем и кончающийся у зубца карпофора, по всей
длине широко окрашенный коричневым пигментом темнее к вершине
и светлее к основанию. Вдоль рубца у многих экземпляров в центре
проходит выпуклая гребневидная жилка, на верхней суженной части
створки часто раздваивающаяся.
Щечки створки (выпуклая часть створки) более выпуклые у мел-
ких коробочек и плоско выпуклые у крупных. Их плотность увели-
чивается с размером коробочки: у мелких коробочек они почти бу-
магообразные, полупрозрачные, у крупных—более грубые, слабо
прозрачные; от основания до вершины поперечно морщинистые. Мор-
щинки суживаются в верхней части створок. В углублениях морщи-
нок проходят тонкие ярко-коричневые жилки, связывающие коричне-
вый желобок в центре створки с желобком у ее ребра.
У мелких коробочек морщинки и полосы не так ярко выражены.
Часть их заменяется лишь неровностью выпуклой щечки створки.
Ребра коробочки у места разрыва швов окрашены в коричневый
цвет и отделены от щечек створок довольно глубоким желобком,
в котором соединяются коричневые поперечные жилки щечек. Край
разрыва ребра коробочки сильно выгнут наружу; имеет по месту
разрыва ширину до 0,5 см с поверхностью светло-палевого цвета.
Верхняя суженная часть створки более темной окраски, чем ниже-
лежащая. Под рыльцем, в начале расширенной части створки, цент-
ральный желобок ямкообразно вдавлен.
Узкий рубец от опавших членов околоцветника белесовато-ко-
ричневый, окаймлен с обеих сторон более ярко-коричневыми полоска-
ми (у мелких коробочек он просто белесовато-коричневый), сужается
к плодоножке.
Плодоножка соломенно-желтая с белесоватым оттенком, а иногда
и с коричневым отсветом, до 25 см дл.
Семя светло-коричневое, яркое, плоское, широкояйцевидное
(0,9X0,8 слг), со всех сторон окаймлено неширокой, ровной, несколько
более светлой каймой; в верхней части на скошенной стороне эндо-
сперма с темно-зеленым полумесяцем.
Зародыш прямой, лежит в центре семени, очень хорошо виден
с обеих сторон.
В условиях Ташкента зацветает в начале второй декады апреля.
Tulipa kuschkensis В. F е d ts с й,— Тюльпан кушкинский (рис II
и 2, б) встречается „на глинистых, реже на каменистых склонах,—Ср.
Азия; Горн. Туркм. (Кушкинский). Эндем“ („Флора СССР“, т. IV).
Мы изучали растения из Кушкинского района Туркмении, окрест-
ностей села Моргуновского (глинистые склоны) и Ашхабадского бо-
танического сада.
Луковица яйцевидная с бурыми, бумагообразными чешуями. По-
кровная чешуя с внутренней стороны по всей поверхности покрыта
очень густыми, тонкими, сильно извитыми, шелковистыми и длинными
волосками.
Самое высокое растение из видов этой секции. Стебель вместе
с цветоножкой 15—45 см выс. от поверхности почвы, голый, зеленый
34
Рис. II. Т. kuschkensis В. Fe d t sc h. (V2 натуральной величины).
с сизым налетом. У нас нет опушенных экземпляров, но во „Флоре
СССР", т. IV, отмечено, что он бывает опушен, особенно на цвето-
ножке.
Листья, в количестве 3—4—5, сильно расставленные по стеблю,
с большими междоузлиями, вверх торчащие, в верхней половине
серповидно отогнутые, сизые, курчавые, голые, по краю реснитчатые,
Рис. 2. Зрелые коробочки тюльпанов.
-а —Т. tubergeniana Hoog.; б—Т. kuschkensis В. Fedtsch.; 6—-Т. hoogiana В. F е g t s с h.; г — Т.
juiia С. К о с h . ; д —Т. ingens Н о о g . ; е — Т. fosteriana Irv. ; ж - Т. carmata Vved. ; з —Т. aff inis’
Z. В о t s ch . ; и —Т. praestans Hoog.; я—Т. subpraesrans V v е d .
.не превышающие цветок: верхний—18,5 см дл., 1,65 см шир. — в на-
чале цветения может несколько возвышаться над цветком; самый
нижний лист ланцетный (25 см дл., 9 см шир.), находится высоко
на стебле—от поверхности почвы на 10 см.
Цветок одиночный, красный, без запаха, красивого сложения, как
у Т. greigii, но в отличие от него, внешние листочки околоцветника
-согнуты вдоль центральной жилки. В закрытом состоянии бутон и
цветок конические, с широким плоским основанием.
Внешние листочки околоцветника (4,5—6—9 см дл., 2,25—3—4—
5 см шир.) продолговато ромбические или овальные. Спинка листочка
продолжена в острый пушистый кончик. Ноготок листочка на месте
35
пятна очень мясистый, снаружи оранжево-красный благодаря сизому
налету с белесовато-малиновым оттенком, тусклый, с как бы просвечи-
вающим свекольным цветом внутренним продолговато овальным,
зеленеющим к основанию, пятном.
Совнутри они блестяще-оранжево-красные, с четко отграниченным
большим (у листочков в 6 см дл. до 2,5 сл/дл. 1,2 см шир.) овальным,
на вершине выемчатым черным пятном, окруженным бледной желтой
неширокой каймой, также четко отграниченной от основного фона.
Внутренние листочки околоцветника обратно треугольные или обратно
дельтовидные, со спинкой на вершине (как и у внешних листочков),
продолженной во внезапно возвышающееся опушенное острие; на
ноготке они также массивные и изогнутые внутрь; вдоль листочка на
месте центральной жилки проходит глубокий желобок. Снаружи они
блестяще оранжево-красные, более краснеющие к периферии, на но-
готке несут острое, неправильно ромбическое (1,8X1,2 см), желто-
зеленое, покрытое лилово-фиолетовыми штрихами пятно; совнутри
блестящие, оранжево-красные, с большим (2,2X1,2 см) продолговатым,
усеченным, к выемчатой вершине несколько суженным, черным, жел-
то-окаймленным пятном. У растений, привезенных в виде луковиц из
Кушки, внутренние листочки околоцветника несколько шире внешних,
в то время как во „Флоре СССР" указано, что у этого вида внешние
листочки шире внутренних. Тычиночные нити короткие, массивные,
треугольные (0,5 см дл., 0,25 см шир.) в основании, к вершине пос-
тепенно суженные, черные.
Пыльники черные или черно-фиолетовые, в открытом сухом со-
стоянии 1,2 см дл., 0,37 см шир., т. е. более чем в два раза длиннее
тычиночных нитей.
Пыльца фиолетово-коричневая. Завязь на второй день цветения
выше тычинок, в виде ровного по всей длине бледно-зеленого колышка
с очень широким, сильно выдающимся над завязью такого же цвета
рыльцем.
Коробочка соломенно-желтая с легким коричнево-розоватым узо-
ром (до 4,8 см аа. и 2,2 см шир.), продолговатая, почти ровная по
всей длине, с тупым основанием и вершиной.
Карпофор очень короткий; у ребер всего 0,1 см, а у централь-
ного желобка створки, в который вдается углом, до 0,4 см выс.
Створки коробочки овальной формы с незначительными выступами
на основании в виде карпофора и столбика с рыльцем на вершине.
Вдоль створки проходит углубленный желобок, всегда несколько
темнее щечек, часто вдоль всей длины или только в верхней поло-
вине довольно широко окрашенный в светло-коричневый цвет.
Щечки коробочки выпуклые, тонкие, бумагообразные; чуть-чуть
смятые, варьируют по степени морщинистости, часто начинающейся
близко от основания коробочки, иногда почти отсутствующей. От
края створки на ребра через выгнутый желобок проходят тонкие
поперечные морщинки, переходящие на щечки створки.
Иногда эти морщинки окрашены коричневым только со стороны
края створки—в них от края створки до центрального желобка про-
ходят светло-коричневые жилки. Край створки по всей длине окра-
шен размытой со сторон желобка коричневой или узенькой малино-
вой полосой.
Коробочка открывается клиновидной щелью, вдоль которой сильно
вывернуты наружу желтовато-оранжевые, очень широкие (0,6 см)
с матовой, как фетр, поверхностью разрывы в краях створок. Щель,
доходящая до основания коробочки, довольно широко открыта и об-
36
разует широкий раструб благодаря глубокому растрескиванию ’'внут-
ренних перегородок по пыльцепроводниковой ткани. Эта ткань начина-
ется у рыльца широкой полосой во всю его ширину, на расстоянии
0,7 см от вершины рыльца, быстро сужается и подходит к выгибу
в перегородке, которая растрескивается ниже выгиба еще на 2 см.
Верхняя часть створки под рыльцем несколько оттянута, более темно
окрашена, чем вся коробочка; центральный желобок на ней уплоща-
ется, на его месте появляется что-то вроде жилки или рубчика, ко-
торый под самым рыльцем (0,4 см) раздваивается и подходит к краям
рыльца. Под рыльцем створка резко сужена и образует перетяжку
или небольшой широкий столбик 0,3 см выс. вместе с рыльцем. Рыль-
це шире столбика, к краям лопастей темнее, но вообще составляет
-единое целое со створкой.
Рубец от опавших членов околоцветника коричневый, узкий
(0,1 см), но шире, чем очень длинная, тонкая, темнее коробочки
плодоножка.
Семя светло-коричневое, округло треугольное, плоское (0,8х
ХО,55 см), окружено более светлой, в 0,1 см шир. каймой. Кайма
покрыта мелкими морщинками. Зародыш более светлый, чем семя,
хорошо виден с обеих его сторон. В условиях Ташкента цветение
этого вида начинается в начале второй декады апреля.
Tulipa hoogiana В. Fedtsch. (рис. Ill и 2, в) обитает— „Ср. Азия:
Горн. Туркм. (Центр, и Зап. Копет-Даг). Эндем. На глинистых, реже
щебнистых склонах от предгорий до степного пояса гор" („Флора
СССР", т. IV). Мы исследовали материалы из Туркмении с хр. Копет-
Даг (окрестности поселка Прохладного), из Арваза (восточный склон)
и в 1950 г. из Ашхабадского ботанического сада. Этот вид мы не-
однократно выращивали из семян.
Луковицы яйцевидные, до 5 см в диаметре, покрытые тонкими
бумагообразными, темно-коричневыми, с внутренней стороны обильно
опушенными, густыми, длинными, тонкими, шелковистыми, сильно
извитыми волосками.
Высота растений до 40 см от уровня почвы. Стебель вместе с
цветоножкой голый, часто очень сильно окрашен антоцианом (особен-
но у темных форм) до темно-бордово-коричневого цвета, покрытый
сильным сизым налетом.
У многих растений нижний лист отходит от стебля на значитель-
ной высоте (до 10 см) от уровня почвы, отчего растения имеют го-
ленастый вид.
Листья, в количестве 3—5, сильно серповидно отогнутые, по стеб-
лю расставленнные, очень сизые, голые, хрящевато окаймленные, по
краю длиннореснитчатые. Таких длинных ресничек на листьях нет ни
у одного из известных нам видов тюльпанов, но встречаются формы
•с почти голым краем. Есть среди них формы с листьями почти пря-
мыми по краю, округло желобчатыми, серповидными или с довольно
-сильно выраженной курчавостью. Часто листья превышают цветок.
Нижний лист ланцетный, при основании, как правило, стеблеобъ-
емлющий, иногда в самом низу трубковидно сросшийся, до высоты
3 см (при длине 26,5 см и ширине 10 см), верхний—узколанцетный
-(20,5 см дл., 2,5 см шир.).
Цветок одиночный, темно-малиново-красный, или светлый розова-
то-оранжевый, или просто оранжевый, очень красивой в виде двой-
ной чаши формы, в котором внешние листочки околоцветника на от-
гибе несколько отклоняются наружу, а внутренние торчат прямо вверх
37
и га вершине у mhoihx форм загнуты несколько внутрь. В начале
цветения они имеют довольно сильный своеобразный аромат.
Внешние листочки околоцветника, постепенно заостренные в пу-
шистый кончик, неправильно ромбические или широко ланцетные,
б. м. внезапно в нижней четверти оттянутые в продолговатый суженный
ноготок; по краю отгиба зубчатые, часто слегка волнистые (9,5 см дл.,
5,3 см шир.), по спинке с сильным сизым налетом, что делает их ма-
линовыми, у светлых форм с серебристым оттенком, на сторонах от-
гиба малиновые, часто становятся белесовато-оранжевыми, у самого
Рис. 3. Зрелые коробочки тюльпанов.
а —Т. Kolpakovskiana Rgl . ; б —T. brachystemon Rgl.; в —Т. ferganica Vved. ; г —Т. kauf-
manniana Rgl.; —Т. tschimganica Z. В о t s с h .; е —Т. dubia V v е d . ; ж — Т. butkovii
Z. В о t s с h з —Т. anadroma Z. В о t sc h . ; н —T. bibersteiniana Sc h uj t. ; к -T. _• buhseana
В о i s s . ; л—T. talievi Vved.
края краснеют, или краснота проступает под сизым налетом по всему
листочку; внутреннее пятно часто просвечивает; к месту прикрепле-
ния иногда зеленеет, а чаще приобретает вдоль центра массивного-
ноготка темно-бордовый оттенок; совнутри блестяще малиново-крас-
ные с часто проступающей оранжеватостью. У оранжево-розовых и
чисто оранжевых форм окраска внутри цветка более темная, видимо,
в связи с отсутствием там сизого налета. У оранжево-розовых форм
по всему листочку проходят красноватые штрихи. Совнутри на ного-
тке лежит довольно длинное (3,2X1,2 см) продолговатое пятно, час-
38
Рис. III. Т. hoogiana В. Fedtsch. (3/4 натуральной величины).
то тупое или острое, с очень меняющимся цветом—от черного до*
темно-бордового и серовато-зеленого (у светлых форм). Оно бывает
сплошным или окаймлено узкой светло-желтой полосой, различной (но
небольшой) ширины и интенсивности, часто лишь в виде светлых штри-
хов, окружающих темное пятно.
Внутренние листочки (9,2 см дл., 5,3 см шир.) обратно-продолго-
вато-яйцевидные или просто яйцевидные, б. м. внезапно оттянутые в
большой острый, на вершине опушенный кончик; часто несколько
ложкообразно наружу выпуклые; снаружи и внутри одинакового цве-
та, блестящие, темно-малиново-бордовые, малиново-красные, оранже-
вато-малиновые, оранжево-розовые или просто оранжевые; снаружи
на ноготке имеют бордовый оттенок, приподнимающийся вдоль цен-
тральной жилки, а светлые формы при самом основании несколько
желтеют. Совнутри на ноготке находится продолговатое, овальное или
остро треугольное, при основании расширенное или по всей длине
ровное, окаймленное пятно. Его окраска такая же, как и на внешних
листочках околоцветника. Пятно 3,2 см дл. и 0,7 см шир. (у формы,
взятой нами за эталон при проверке описания вида).
Тычинки выше завязи, в два, три и больше раза короче листоч-
ков околоцветника.
Тычиночные нити более длинные у внутреннего круга (1 — 1,2 см
дл.), к основанию расширенные (до 0,4 см), к пыльнику довольно
быстро суженные, в верхней половине или трети черные или бордо-
вые, у светлых форм — серовато-зеленые, в нижней половине желтые
или красноватые с постепенным переходом от темного к светлому;
под пыльником светлые.
Пыльники 1,8 см. дл., 0,4 см шир., т. е. длиннее нитей, бордово-
розово-коричневатые или желтые; часто спирально свернутые.
Пыльца коричневато-розовая, коричневато-серая или желтая. За-
вязь темно-зеленая, по ребрам к центру плодолистиков антоцианово-
бордовая, по всей длине равномерно утолщенная, или под рыльцем
чуть-чуть сужена (2,2 см дл., 0,6 см толщ.).
Рыльце чуть шире суженной части завязи, с боков бордовое, яр-
кое, сосочки желтые.
Коробочка розовато-соломенно-серая с ярким коричнево-лиловым
узором (6,7 см дл., 3,5 см шир.), треугольно-округло-цилиндрическая,,
тупая при основании, на вершине несколько более острая. Карпофор
короткий (0,6 см), широкий (при переходе в створку 1,5 см., у рубца
0,8 см), округло вершинный, без особых зубцов.
Верхняя часть створок коробочки тупо заостренная, с сидячим,
небольшим, прочно скрепленным со створками, поперечно отогнутым
коричневым рыльцем.
Коробочка растрескивается по ребрам до самого основания узко
клиновидными щелями с почти параллельными краями. На вершине,
где створки сужаются, щель несколько расширяется, образуя не очень
широкий раструб в 6 см дл., шириной на коробочке 0,2 см и у рыль-
ца 1,5 см.
Внутренние перегородки не расходятся глубоко. Растрескиваются
вглубину до конца изгиба. Совнутри под рыльцем поверхность раз-
рыва перегородки матово-белая с гребнем в центре, обратно кониче-
ская у туповерхих коробочек и вытянуто ромбическая у коробочек
с несколько продолговато заостренной вершиной.
Створки коробочки овально продолговатые или на вершине тупо
заостренные. Центральный продольный рубец створки интенсивно ок-
рашен в коричневый цвет с лиловым оттенком, немного углублен в
39>
3/5 нижней части створки, а в 2/5 ее верхней сужающейся части почти
плоский, широкий, с неровной выпуклой, в виде продолговатой мозо-
ли, поверхностью, от которой в желобке до основания рубца прохо-
дит гребневидная жилка.
Щечки створки плоско выпуклые: более выпуклые в нижней по-
ловине коробочки, грубые, толстые, матовые, морщинистые по всей
поверхности или только в верхней половине створки. В желобках мор-
щин проходят поперечные, чуть-чуть к краям створок направленные
вверх, коричнево-лиловые поперечные полосы. В глубоких продоль-
ных желобках перед выгнутым краем створок эти коричневые жилки
образуют между собой массу анастомозов.
Поверхность разрыва ребра коробочки розовато-соломенная, ши-
рокая (0,4 см) вдоль створки и суженная в верхней ее части под рыль-
цем, имеет фактуру в виде фетра. Самый край ребра створки совнут-
ри узко, а снаружи широко окрашен в темно-коричнево-бордовый
цвет, от которого на поверхность разрыва проходит легкая лиловая
или коричневая побежалость. Край ребра створки сильно выгнут на-
ружу.
Рубец от листочков околоцветника узкий (0,1 см), светло-корич-
невый, слегка выпуклый, по нему часто проходят продольные, ром-
бической формы трещинки.
Плодоножка соломенно-белая с розоватым оттенком, иногда ко-
ричневатая, не длинная (до 17 см) и довольно толстая.
Семя широкояйцевидное, светло-коричневое, окаймлено прямой
каймой, ровной по ширине на всем протяжении. В скошенном углу
семени лежит полумесяц зеленой ткани. Зародыш коротенький, ма-
ленький.
Tulipa julia С. Koch. (рис. 2, г) произрастает—„Кавказ: Южн.
Закавк. Общ. распростр.: Иранск." На сухих (каменистых) склонах"
(„Флора СССР", т. IV).
Исследованные растения получены в виде луковиц из Ереван-
ского ботанического сада от тов. Ахвердова, собранных в Армении
(Китабский район, окрестности Еревана) в среднегорном поясе на
южных открытых склонах фригана, на пестроцветных мергелистых
песчаниках в 1953 г.
Кроме того, они были выращены из семян, полученных из того
же Ботанического сада.
Луковица 1,6—3,5 см в диам., с бурыми бумагообразными (у не-
углубляющихся растений) и с жесткими (на луковицах столонов уг-
лубления) совнутри, особенно на вершине, паутинисто-мохнатыми по-
кровными чешуями.
Стебель в первый день цветения у растений, выросших в наших
условиях, невысокий: 11,5—15 cut от уровня почвы, вместе с цвето-
ножкой зеленый, с несколько коричневатым оттенком, голый.
Листьев четыре; обычно отогнуты, темно-болотно-зеленые с легким
сизым налетом, голые, курчавые, лишь слабо желобчатые, по краю
светло окаймленные, почти без ресничек, не превышающие цветок.
Нижний лист широколанцетный (13,5X3,5 см), отходит от стебля
на уровне почвы. Верхний лист отклоненный, узколанцетный, линейный,
слабо штопором завитый, вверх направленный (11 см дл., 1 см шир.).
Цветок один, в день расцветания со слабым приятным ароматом
.малиново-красный, без блеска; открывается широкой чашей.
Внешние листочки околоцветника узко-округло-неправильно-ром-
бические (5 см дл. и 3 см шир.), на вершине внезапно заканчиваю-
щиеся маленьким пушистым кончиком, снаружи малиново-бордовые,
40
к основанию с оранжеватым оттенком из-за легких желтых штрихов,
входящих на них с желто-зеленого ноготка, который занимает пло-
щадь до высоты 1,5 см. На нем в виде расплывчатого зеленоватого
пятна просвечивает внутреннее пятно.
Совнутри они красновато-малиновые, но не такие тусклые, как
снаружи, у некоторых форм бордово-оранжевые с более оранжевой
полоской вдоль центральной жилки; на ноготке расположено широко
желтоокаймленное, овальное, острое или тупое, серовато-черное пят-
но (1,1 X0,7 см). Желтая кайма пятна имеет 0,4 см, ширины в центре,
к краям шире; вдоль края листочка и пятна кайма спускается до са-
мого основания листочка.
Внутренние листочки околоцветника обратно-кругло-дельтовидные,
или обратно-вытянуто-треугольные (4,4 на 2,8 см), на вершине с вы-
емкой, в которой наблюдается острый маленький кончик, красно-бор-
довые; с обеих сторон несколько более блестящие, чем внешние;
снаружи на ноготке трехвершинное, до высоты 1,4 см, зеленоватое
пятно. От пятна желтоватые штрихи заходят выше, особенно по кра-
ям листочка. Совнутри на ноготке, на ярко-желтом фоне лежит об-
ратно-округло-треугольное (1,1X0,8 см) пятно с острым зубцом на
вершине, черное с несколько зеленоватым оттенком. Желтая кайма,
получающаяся из желтого фона ноготка, довольно широкая в центре,
до 0,4 см шир., к краям становится еще шире и заходит штрихами
на основной фон листочка, приподнимается по его краям и окаймляет
пятно с боков.
Тычинки равны завязи, имеют длинные (1,3X0,27 см), к вершине
постепенно сужающиеся, черные, при основании желтоватые нити.
Пыльники продолговатые, короче нитей (1X0,2 см), черные. Пыль-
ца коричнево-лиловая.
Завязь светло-зеленая (2,1 см дл., 0,4 см шир.), под рыльцем
суженная, с коротким столбиком.
Рыльце и область завязи под рыльцем — светло-желтые. Рыльце
шире завязи.
Коробочка беловато-соломенная (4,8 см дл., 2,3 см шир.), про-
долговатой, а в некоторых случаях почти овальной формы, при ос-
новании с длинным, до 1,4 см, карпофором. Он узкий и острым кли-
ном вдается в створки коробочки. Ребра коробочки, наоборот, в виде
невысоких, часто на вершине окрашенных, гребней спускаются на не-
го и сходят на нет на половине его высоты.
Створки овальные, на вершине б. м. внезапно сужены и оттянуты
в столбик, часто более темный, чем вся коробочка, длиною до 1 см
и короче; на нем заметны высохшие, серовато-коричневые лопасти
маленького низенького рыльца, разделенные пополам проходящим
вдоль них довольно глубоким желобком. В глубине желобка прохо-
дит окрашенная в коричневый цвет тоненькая жилка. Она несколько
расширяется у вершины щечек и в нижней части столбика, вновь су-
жаясь к его вершине,
Щечки створки выпуклые, тонкие, бумагообразные, прозрачные,
покрытые легкими поперечными морщинками. В верхней половине
коробочки по морщинкам, пересекающим поперек щечки, проходят
тонкие коричневые жилки. От края створки щечки отделяются глубо-
ким желобком, образованным выгнутым наружу по месту разрыва
ребром коробочки. По краю вдоль разрыва ребро коробочки сверху
донизу окрашено ярко-коричневой полоской. Сам же разрыв матовый,
светло-кремовый.
41
Открывается коробочка узкой клиновидной щелью, образующей
раструб отклоненными наружу частями столбика.
Внутренние перегородки растрескиваются по пыльцепроводнико-
вой ткани до выгиба в них, т. е. на всю длину столбика и немного
ниже. В верхней части внутренние перегородки по месту разрыва
имеют форму ромба, вытянутого в сторону столбика и короткого к
выгибу перегородки.
Рубец от опавших членов околоцветника очень узкий, до 0,05 см
шир., ярко-коричневый, выпуклый. Плодоножка тонкая, белее
коробочки.
Семена округло треугольные (до 0,7 см дл., 0,65 см шир.), свет-
ло-коричневые, слегка согнуты поперек, с узкой, мятой, немного-
загнутой вниз каймой. Зародыш хорошо виден с обеих сторон семени.
В условиях Ташкента этот вид цветет в середине апреля.
Секция Leiostemones В о i s s.
„Оболочки луковиц с внутренней стороны почти голые, или у
верхушки или по всей поверхности волосистые, или обильно волосис-
то шерстистые, или только у верхушки обильно паутинисто-мохнатые,
нити тыч. голые; ЗВ с сидячими рлц или с б. м. длинным столб., но
всегда более коротким, чем сама зв.“ („Флора СССР", т. IV).
По „Флоре СССР", т. IV, в СССР из этой секции находится 38
видов и, кроме того, нам известно еще 3 вида, не вошедших во Фло-
ру, т. е. 41 вид Tulipa, относимых к Leiostemones.
Tulipa florenskvi^J or on — Тюльпан Флоренского обитает — „Кав-
каз: Южн. Закавк. Общ. распр.: Иранск/ „На сухих каменистых скло-
нах" („Флора СССР“, т. IV).
Описан по гербарным экземплярам, хранящимся в БИНе СССР.
Луковица лежит на глубине до 13 см, часто с несколько про-
долженными не более чем на длину луковицы чешуями, бурыми бы-
вают только старые полуистлевшие чешуи, обычно же они коричне-
вые, тонкокожистые.
Листья, в количестве 3—4, желобчатые, сильно серповидно изог-
нутые (верхний очень узкий, торчащий), волнистые, сизые, по краю
хрящевато окаймлены.
Цветок, нам кажется, по форме несколько напоминает цветок
Т. wilsoniana.
Наружные листочки околоцветника обратно-яйцевидно-ромбиче-
ские или продолговато ромбические, часто постепенно заостренные
в короткий пушистый кончик. Пятно при основании листочков около-
цветника черное. О его форме судить трудно — оно продолговатое,
1 см дл., у листочков, 3,8 см.
Внутренние листочки обратно-продолговато-яйцевидные или об-
ратно-неправильно-узко-ромбические, иногда также с коротким пуши-
стым кончиком, пятно неясное.
Нити тычинок тонкие, к основанию чуть расширенные, темные,
при самом основании и на вершине желтые.
Пыльники желтые или коричневые, сухие, в 1,5 раза короче
нитей.
Завязь, постепенно суженная к рыльцу, достигает 2/3 высоты
тычинок. Рыльце светлое, сильно выдается над верхней суженной
частью завязи.
Из двух мест ареала есть светло-желтые формы. У них более
темные основания листочков околоцветника, тычиночные нити и пыль-
42
ники. Нити очень тонкие. Пыльники широкие и короткие, в два с
лишним раза короче нитей.
Отличается от Т. julia покровными оболочками с довольно гус-
тым опушением, состоящим из прямых жестких волосков, и формой
листьев, которые у Т. florenskyi сильно серповидно изогнуты, за ис-
ключением самого верхнего очень узкого листа (у Т. lulia - - шире)..
Рис. 4. Зрелые коробочки тюльпанов.
а — т. greigii R g 1. б —T. micheliana Hoog.; в —T. eichleri R g 1. ; г —T. vve-
denskyi Z. В о t s с b . ; д —T. maximowiczii R g 1 . ; e —T. wilsoniana Hoog. ; ж —
T. borszczowii R g 1. ; з —T. korolkovii R g 1 . ; и —T. nitida H о о g .; к —T. schrenkii
Rgl- ;
Листья, обычно в количестве 4, желобчатые и часто по краю более-
волнисты, чем у Т. julia. Они хрящевато окаймлены.
Видимо, Т. florenskyi отличается от Т. julia и формой пятна на лис-
точках околоцветника.
Tulipa lanata Rgl.—тюльпан шерстистый (рис. IV) встречается „на
глинистых, реже каменистых склонах в нижнем поясе гор. Ср. Азия:
Пам.-Алай (бассейн рр. Сурхан и Кафирниган). Эндем" („Флора СССР11,
т. IV).
Исследованные растения получены из Ходжи-Ипака Денауского
района Узбекистана, сб. А. И. Введенского, и из Душанбинского бо-
танического сада от Т. И. Рябовой. Из Шахринаузского района,
ущелье Солямич, на высоте 1600 м, восточный склон.
43-
Луковицы яйцевидные, до 4 см в диам., с нежными тонкокожис-
тыми покровными чешуями, коричневого цвета, совнутри обильно
покрытыми тонкими, длинными, шерстистыми волосками.
Стебель 13—30 см выс. от уровня почвы, вместе с цветоножкой
опушен очень густо, без антоцианового налета, бледно-зеленый, от
обильного опушения матовый.
Листья, в количестве 4, отогнутые, в начале цветения б. м. сбли-
женные, затем расставленные, курчавые, сизоватые, б. м. пушистые,
особенно верхние, часто опушены с обеих сторон, по краю часто крас-
но окаймленные, реснитчатые, в начале цветения они выше цветка,
-а затем не превышают его.
Нижний лист широколанцетный, 16,5 см дл,, 5,5 см шир. Верх-
ний — узколанцетный, 18,2 см дл. и 2,2 см шир.
Цветок одиночный, хорошего сложения, несколько напоминает
Т. greigii, снаружи сизовато-розоватый, совнутри ярко-красный, без
запаха.
Внешние листочки околоцветника (6,5 см дл., 3,2 см шир.) ром-
бические или продолговато ромбические, постепенно заостренные в
пушистый кончик; опушение с кончика переходит на внутреннюю и
внешнюю поверхность отгиба и в виде ресничек проходит по краю,
примерно до половины его высоты с наружной стороны; совнутри
опускается примерно на 1/4 высоты вниз по бокам листочка; снару-
жи серебристо-белесовато-розовые, к краям отгиба краснеющие или
оранжевеющие по спинке, особенно на вершине и ноготке; зеленова-
тые в средней части спинки, со светло-коричневатым оттенком, кото-
рый проступает и в других местах спинки; на ноготке бордовой или
черноватой тенью просвечивает внутреннее овальное пятно; совнутри
ярко-ало-красные; на ноготке лежит большое овальное или ромбиче-
ское черное, бледно, четко желтоокаймленное пятно (1,8X0,8 см),
которое на вершине может быть острым, тупым или выемчатым; в ниж-
ней части с боков ноготка основной фон листочка обесцвечен до
бледно-желтых, такого же цвета, как и кайма, штрихами входящих
в основной яркий фон листочка полосок.
Внутренние листочки околоцветника (6,7 см дл., 3,1 см шир.)
обратно-продолговато-яйцевидные или широко ланцетные, на вершине
б. м. внезапно заостренные в большой и широкий, сильно опушенный
зубец (опушение в виде ресничек переходит на края отгиба); снаружи
такие же, каки внешние, розовато-оранжево-серебристые, к краям крас-
неющие, по спинке узко с обеих сторон центральной жилки желто-
вато-зеленеющие; на ноготке с овальным, островершинным, зелено-
вато-желтоватым, сизым пятнышком (1,3 см выс. и 0,8 см шир.);
совнутри ало-красные, в пятне в нижней половине появляется и уси-
ливается малиновый, а к острому кончику на вершине заметно оран-
жевеющий оттенок.
На ноготке лежит черное, очень узко желтоокаймленное оваль-
ное пятнышко, сильно заостренное на вершине (1,3 см дл., 0,8 см шир.).
Тычинки черные, 1,8 см дл., т. е. в 2,5—3 раза короче около-
цветника; выше завязи.
Нити короткие (0,4 см дл., 0,25 см шир.), при основании в
виде черных треугольничков, под пыльниками белые. Пыльники бор-
дово-черные, 1,4 см дл„ 0,3 см шир. Пыльца бордово-лиловая.
Завязь (1,7 см дл., 0,35 см шир.) кремовая, под рыльцем сужена.
Рыльце также кремовое, сосочки бордовые.
Коробочка (4 см выс., 1,6 см в диаметре) продолговатой формы,
44
Рис. IV. Т. lanata Rgl. (3/4 натуральной величины).
при основании упиралась в короткий (0,5 см), широкий на вершине*
под створками (0,7 см шир.) карпофор.
Створка соломенно-желтая, на вершине заканчивается широким
(0,3 см), длинным (0,7 ем) столбиком, в центре которого по направ-
лению к продольному желобку створки проходит широкий окрашен-
ный в коричневый цвет тяж рубцовой ткани. Вообще же столбик
довольно темный, а рубец на нем еще темнее. На створке рубец уг-
лубляется и входит в продольный желобок створки, по всей длине
окрашенный в коричневый цвет. От желобка отходят поперечные
морщинки, пересекающие щечки створки.
Щечки створки выпуклые, по всей длине поперечно морщини-
стые, отделены от ребра створки глубоким желобком.
Ребро коробочки окрашено в темно-коричневый цвет. Поверх-
ность разрыва коробочки по ребру матовая, белая, с отходящими в
полость коробочки многочисленными белесоватыми бахромками.
Коробочка от рыльца до изгиба перегородки овальной формы,
растрескивается по пыльцепроводниковой ткани. Лопасти рыльца от-
клоняются наружу, образуя широкий раструб, в который высыпаются
семена.
Плодоножка светлее коробочки. Рубец от опавших членов око-
лоцветника широкий, светлый.
Семя 0,8X0,8 см, широко-округло-треугольное, с маленьким
прямым, достигающим только половины длины эндосперма, просвечи-
вающим с обеих сторон семени зародышем и узкой (0,1 см), несколь-
ко смятой, направленной вниз каймой.
В условиях Ташкента этот вид зацветает в середине третьей де-
кады апреля.
Tulipa ingens Hoog, — Т. Великий (рис. 2, ^встречается „на гли-
нистых склонах, до 2500 м, Ср. Азия: Пам.-Алай (Средний Зеравшан,
басе. р. Кашкадарья и к югу от Кугитанга). Эндем" (Флора СССР,,
т. IV).
Растения этого вида мы вырастили из семян растений Ботаниче-
ского сада, луковицы которых были доставлены из различных место-
обитаний естественного ареала этого вида.
Луковица продолговато яйцевидная, в культуре до 6 см в диа-
метре, с черно-бурыми или бурыми тонкокожистыми непрочными
покровными чешуями с оригинальным опушением из редких, довольно
длинных, прижатых, собирающихся концами в косицы волосков.
Растения во время цветения 25 см выс. Цветоножка зеленая, на-
верху с бордовым оттенком, опушенная. Степень бордовой окраски
и опушенность стебля сильно изменчивы у различных экземпляров с
одного и того же местообитания или из одного номера посева.
Количество листьев варьирует от 3—4 до 6.
Листья слабо приподнимаются по стеблю, изредка почти розеточно
расположены; узколанцетные, иногда по краю слабо крупно волни-
стые, желобчатые, сизые, иногда очень сильно опушенные.
Цветок открытый, формы остролучевой звезды, со слабоуглуб-
ленным центром. По форме цветок Т. ingens напоминает лилиецвет-
ный. Как правило, цветки Т. ingens тримерны, иногда димерны и тет-
рамерны, без запаха.
Внешние листочки околоцветника от 15 до 7,6 см дл. и от 6 до
2,8 см шир. У цветов от вполне сформированных луковиц листочки
околоцветника редко бывают короче 13 см дл. и 5 см шир.; широко-
ланцетные или продолговато-округло-ромбические, на вершине за-
остренные, оканчивающиеся относительно длинным, заостренным,
45
сильно опушенным кончиком; в верхней половине отгиба по краю
часто мелкозубчатые; снаружи по спинке сизо-малиновые, но чаще
палевые, разной интенсивности, с зеленоватым оттенком, который
усиливается к ноготку, а к краям листочков переходит в ало-красный
цвет; иногда сизый налет б. м. далеко заходит на края отгиба, одна-
ко самый край листочков всегда чисто ало-красного цвета.
Совнутри окраска этих листочков чисто ало-красная, в центре
листочка более густая и приобретающая несколько малиновый отте-
нок, в виде нечетко отграниченной продольной заостряющейся кверху
полосы, в нижней части которой и над пятном малиновая окраска
несколько сгущается. С боков до половины высоты пятна красная
окраска обычно исчезает и проступает основной цвет мезофилла, и
тогда нижний край листочка околоцветника белесовато-палевый.
Совнутри на внешних листочках околоцветника в нижней поло-
вине лежит черное пятно, редко окаймленное кругом узкой, до 1 мм
шир., желтой полоской, от 4,3 до 1,8 см дл. и 1 см шир., вытя-
нуто ромбической формы, у которой боковые углы сглажены, а вер-
шина, наоборот, заострена или остро треугольная, овальная и обратно
яйцевидная.
Внутренние листочки околоцветника несколько короче внешних,
от 14 до 7 см. в среднем до 12 см дл., шириной несколько большей,
чем у внешних,— от 6,9 до 4,4 см. в среднем 5,7 см; узко-обратно-
яйцевидные или округло ромбические, иногда широкояйцевидные —
все с внезапно возникающим на верхушке острым опушенным кончи-
ком, и краем, на отгибе покрытом очень мелкими зубчиками; снару-
жи обычно ало-красного цвета с сизым налетом, придающим листоч-
кам малиновый оттенок. Встречаются формы, у которых внутренние
листочки околоцветника снаружи окрашены в оранжевато-красный
цвет, причем на спинке более оранжевые, а к краям отгиба красные
с лиловым отсветом; на ноготке с внешней стороны часто с лилово-
коричневым ромбическим пятном, окаймленным желтой полоской,
которая в центре уже, а к краям шире. Эта полоса делает пятно
веерообразным; у большей части это пятно сплошь блекло-желто-
зеленое.
Совнутри внутренние листочки околопветника ало-красного цвета
с лиловыми оттенками различной интенсивности; лиловые тени сгу-
щаются в центре листочков околоцветника над пятном. На ноготке
внутренних листочков околоцветника расположено обычно чисто чер-
ное пятно. Очень редко встречаются формы с пятном в верхней час-
ти узко окаймленным желтоватой полоской. Пятно обычно узкором-
бической формы .с верхним углом более острым и как бы вытянутым,
редко более округлым, иногда трехвершинным, часто по краю чуть-
чуть зазубренным, но всегда пятно четко отграничено от основной
окраски листочка.
Тычинки от 4,5 до 2,5 см дл., в момент открытия цветка зна-
чительно выше завязи (в полтора раза). Затем пыльники быстро усы-
хают и съеживаются и завязь их перерастает. Иногда связник слегка
возвышается над пыльниками (0,3 см). Тычиночные нити кверху рез-
ко суженные, черные. Пыльники черные, коричневато-черные или
темно-бордовые.
Пыльца темно-лиловая, почти фиолетовая или темно-коричневая,
иногда коричневато-лиловая.
Завязь длинная (от 5,3 до 2,2 см дл.), узкая (0,4—0,35 см шир.),
зеленая с лиловым или бордовым оттенком, который усиливается к
рыльцу. Столбик часто зеленый.
46
Рыльце шире завязи, с боков часто красно-лиловое, иногда жел-
тое, сосочки рыльца палевые или желтые с красным.
Коробочка соломенно-желтая, очень узкая, вытянуто продолго-
ватая, заостренная на обоих концах (до 11,5 см дл. при 2,3 см шир.).
Карпофор 1,2 см выс., 0,8 см шир. над рубцом и 1,9 см шир. у
перехода в створки коробочки, коротким зубцом примыкает к жилке,
проходящей в центральном желобке створок. Грани коробочки, по
которым коробочка растрескивается, заходят на карпофор короткими
гребнями. На половине высоты карпофора эти гребни уплощаются и
исчезают. Есть коробочки, в нижней части которых семена недораз-
виваются, и тогда эта часть очень узкая и по ширине равна карпо-
фору. Столбика, как такового, у коробочек этого вида нет. Верхняя
бесплодная часть створки сильно оттянута, конически узка, но не
столбикообразна и постепенно переходит в среднюю часть створки.
Рыльце небольшое, светло-коричневое, легко отламывающееся.
Коробочка открывается узкими (до 0,1 или 0,2 см) длинными, дохо-
дящими до карпофора щелями. Верхние концы створок сильно выгну-
ты наружу, и поэтому на вершине коробочки образуется раструб до
3,5 см шир., в который высыпаются семена.
Внутренние перегородки коробочки растрескиваются значительно
глубже изгиба на перегородке, который у этого вида не такой крутой,
как у других видов. Разошедшиеся части внутренней перегородки от
рыльца до ее изгиба шире, чем лежащие ниже. Вверху перегородка
имеет ширину рыльца, ниже — это длинная б. м. плоская, постепенно
сужающаяся к изгибу створки полоска, в центре которой проходит не-
высокий острый гребень. Края перегородки до сужения ниже отхож-
дения ребер створки тоже слегка приподняты. В этой части перего-
родки Т. ingens и Т. fosteriana очень похожи.
Створки коробочки соломенно-желтые, узколанцетной или про-
долговатой, сильно вытянутой с параллельными краями, заостренной
на обоих концах формой. Вдоль створки во всю длину проходит уз-
кий желобок, в центре которого в некоторых случаях от основания
до начала сужения створки вверху проходит ясно выраженный гре-
бень, в других случаях такого сплошного гребня нет, но внизу у
карпофора он приподнимается, а выше расположен в виде выпуклой
жилки. В верхней трети коробочки эта жилка раздваивается на две и
уже в виде двух параллельных жилок подходит к основанию рыльца.
Весь желобок, а особенно жилки, окрашены в светло-коричневый
цвет; окраска усиливается после раздвоения жилок.
Щечки соломенно-желтые, плотные, тонкие, просвечивающие,
довольно выпуклые, слабо поперечно смятые и лишь в верхней части
с немногими поперечными морщинами; по всей поверхности покрыты
поперечными, светло-коричневыми тонкими полосками (к основанию
настолько бледными, что они едва заметны), которые в желобке под
стогнутым краем створки все соединяются анастомозами и образуют
сложный узор из многогранных ячеек. Край ребра коробочки окра-
шен в коричневый цвет. На вершине створки узор из анастомозов,
частые поперечные полосы, коричневая окраска ребра и желобка
сливаются и получается более темно окрашенная часть створки.
Край створки по месту разрыва на ребре коробочки 0,35 см шир.,
по отношению к ее длине очень узкий, светло-палевый, консистенции
фетра, несколько выгнут наружу, но не в такой степени, как у Т. fos-
teriana.
Рубец от опазших членов околоцветника до 0,2 см шир., корич-
47
невый. Плодоножка под ним довольно толстая, но быстро утончается;
соломенно-желтая с коричневатым оттенком, до 15 см дл.
Семя (0,8 х 0,3 слг) светло-коричневое, немного покоробленое,
т. е. не лежит плоско, а какая-нибудь часть у него приподнимается,
округло треугольное; окаймленное довольно широкой, чуть-чуть мя-
той каймой, несколько расширяющейся на его вершине, особенно с
удлиненной стороны. Прямой или в виде зяпятой зародыш хорошо
виден с обеих сторон семени, но более ясно с его верхней стороны.
В условиях Ташкента этот вид цветет в конце первой декады
апреля.
Детки у растений этого вида встречаются очень редко и только
у сильных особей.
По нашим наблюдениям, Т. ingens очень четко отграничен от
других видов, очень слабо варьирует по качественным признакам и
изменяется лишь количественно, т. е. в размерах тех или иных ор-
ганов или признаков.
Мы сравнивали растения, выросшие из семян, с вывезенными в
виде луковиц из естественных местообитаний и убедились в том, что
этот вид сохраняет в культуре видовую чистоту и не скрещивается
без принудительного опыления ни с культурными, ни с дикими вида-
ми тюльпанов.
Сеянцы в первое и второе цветение очень похожи на материн-
ские растения и отличаются в первые годы цветения лишь несколько
меньшими размерами.
Коробочки Т. ingens и Т. fosteriana очень сходны. У Т. ingens
более узкая коробочка с постепенным плавным переходом от плоду-
щей части створок к карпофору. У Т. fosteriana створки б. м. внезап-
но расширяются над ним. У Т. ingens створки плавно переходят в
бесплодную часть под рыльцем без резкого столбикоподобного или
закругленного тупого завершения сторон, как у Т. fosteriana. У Т. in-
gens 2/5 верхней части жилки, проходящей в центральной части же-
лобка, раздвоены, а у Т. fosteriana — нет. Плодоножка у Т. ingens
соломенно-желтая, с коричневатым оттенком, а у Т. fosteriana она
просто соломенно-желтая.
Толщина стенки створки в рубце у Т. ingens значительно тоньше,
чем у Т. fosteriana.
Tulipa fosteriana Iru. — тюльпан Фостера (рис. 2, е) встречается
„на мелкоземных склонах и карнизах скал. — Ср. Азия: Пам.-Алай
(Самаркандские горы). Эндем“. („Флора СССР11, т. IV).
Изученные образцы привезены из Памиро-Алая (Самаркандские
горы — западная оконечность Зеравшанского хребта). В довоенные
годы и в 1949 г. их неоднократно собирали в Агалыке и Аман-Кутане.
Луковица яйцевидная, 2—6 см по наименьшему диаметру.
Луковицы столонов углубления нередко совершенно шаровидные,
иногда репчато-плоские. Покровные чешуи черно-бурые, кожистые,
на столонах углубления часто очень крепкие. Оболочка столона уг-
лубления выше луковицы, наоборот, очень хрупкая, всегда ко вре-
мени окончания вегетации разрушающаяся. Совнутри покровная че-
шуя покрыта густыми, прямыми, жесткими, золотистыми волосками.
Опушение усиливается на вершине и при основании чешуи.
Стебель 15—50 см выс. от поверхности почвы; обычно вверху
вместе с цветоножкой окрашен антоцианом в розоватый цвет, часто
чисто зеленый; в верхней части вместе с цветоножкой опушен тор-
чащими, прямыми волосками; имеет кверху удлиняющиеся [междо-
узлия.
48
Листьев 3—5, редко 6; отклоненные, расставленные, слабо кур-
чавые, сизые или блестящие зеленые, с верхней стороны опушенные
неветвящимися торчащими прямыми волосками, по краю часто красно
или хрящевато-белоокаймленные, реснитчатые, обычно не превы-
шающие цветок.
Нижний лист, отходящий от стебля на уровне почвы, широко-про-
долговато-яйцевидный, очень варьирует по ширине, но всегда соотно-
шение длины к ширине у него меньше, чем у вышележащих листьев
(от 7 до 30 см дл., от 3 до 16 см шир.).
Второй лист часто длиннее нижнего, но значительно уже (от 7
до 32 см дл., от 1 до 12 см шир.). У вышележащих листьев резко
уменьшается длина и особенно ширина. Третий лист от 3 до 27 см
дл., от 0,4 до 10 см шир., четвертый — от 3 до 22 см дл., от 0,4 до
8 см шир.
Цветок блестящий, красный, одиночный (даже в культуре у это-
го вида трудно получить многоцветковые формы), очень крупный
(один из самых крупных и красивых цветков среди диких видов тюль-
панов), чашевидной формы, изменяющейся в широких пределах, но
свойственной лишь этому виду и типичной для него формы двойной
чаши; имеет очень слабый аромат.
Внешние листочки околоцветника (от 4,5 до 18 см дл. и от 2 до
8,5 см шир.) сильно варьируют по форме от продолговато яйцевид-
ных до узкоромбических и широко ланцетных; имеют на вершине
острый опушенный зубец; опушение от зубчика часто спускается на
верхнюю половину спинки листочка околоцветника. Край листочка в
верхней части отгиба мелкозубчатый. С внешней стороны листочки
околоцветника ярко-красные, по спинке часто с апельсиновым, сизо-
вато-малиновым или зеленоватым оттенком и окраской различной ин-
тенсивности; на ноготке снаружи зеленовато-желтое сплошное, вееро-
образное, наиболее часто одновершинное, реже выпуклое, мелкозуб-
чатое пятно.
Совнутри они ало-красного цвета с разной степенью оранжеватого
оттенка. В центре листочка к ноготку постепенно увеличивается слегка
лиловатый отсвет. Ноготок занимает черное, желтоокаймленное или
желтое, четко отграниченное от основного фона, веерообразное пятно.
Обычно оно бывает одновершинным или выпуклым, острым или ок-
руглым. Ширина желтого окаймления изменяется от легкого сияния
шириной 0,5 мм до четкой, яркой, широкой полосы.
Черное пятно чаще всего интенсивно черного цвета, но бывают
формы, у которых резкий черный цвет смягчен до коричнево-бордо-
вого разной силы и тогда темная часть пятна не сплошная, а в виде
легких штрихов на желтом фоне.
Внутренние листочки околоцветника обратно-продолговато-яйце-
видные, обычно тупые или несколько вытянутые, с коротким острым
кончиком, на самой вершине мелкозубчатым краем, снаружи и со-
внутри ярко-красного цвета, одного тона с внешними листочками
околоцветника; на ноготке снаружи пятно, окрашенное в желтый или
желто-зеленый цвет, четко отграниченное от основного фона сегмента
и повторяющее форму трехвершинного или выемчатого внутреннего
пятна; на нем может лежать острое узкотреугольное антоцианово-
лиловое, сплошное или штриховатое пятно, совнутри на ноготке чер-
ное или коричневое, обычно трехвершинное, иногда двухвершинное
с б. м. глубокой выемкой, желтоокаймленное (часто сплошь желтое,
без черного центра).
49
4—2229
Тычинки выше завязи, в открытом состоянии пыльника в 3—4
раза короче околоцветника.
Тычиночные нити тые, треугольные, расширенные к основанию
и заостренные на вершине. У цветков с темным центром тычиночные
нити черно-борцовые, лишь острый кончик желтый. У цветков с
желтым центром нити — желтые.
Пыльники черно-фиолетовые, по всей длине внезапно растрески-
вающиеся, в полтора-два раза длиннее нитей.
Пыльца коричнево-фиолетовая или желтая, очень обильная. За-
вязь зеленая, продолговато кеглевидная, под рыльцем суженная.
Рыльце желтое, почти сидячее, с сильно выступающими над сужен-
ной частью завязи лопастями.
Коробочка соломенно-желтая, часто с коричнево-лиловой или
просто светло-коричневой полосой вдоль центра створок, может быть
до 11 см дл., при 3 см шир, но чаще при значительно меньшей
длине, например 7.7 см, она имеет ту же ширину; очень изменчива
по форме, б. м. продолговатая, всегда с заостренной вершиной, часто
наподобие столбика, в который постепенно переходят ее створки. Ос-
нование у нее, наоборот, тупо закруглено, и из него выходит значи-
тельно более узкий, чем вся коробочка, карпофор.
Карпофор более светлый, чем вся коробочка; 0,9 см выс., 0,9 см
шир. у рубца и 1,6 см у перехода в створки, с заходящими в виде
коротких гребней желобками и ребрами коробочки, у желобков не-
сколько выше вдающийся в створку, чем у ребер.
Переход от карпофора коробочки в створки б. м. резкий, так как
сразу же над ним коробочка резко расширяется, а на вершине она,
наоборот, постепенно сужаясь, б. м. плавно переходит в столбик.
Рыльце широкое низкое, на вершине округлое, часто со свисаю-
щими вдоль столбика округлыми, немного выгнутыми наружу, лопа-
стями.
Коробочка открывается до самого основания ребер, чаще всего
узкой, клиновидной щелью. Иногда края щели до верха плодущей
части коробочки идут параллельно. Отклоняющиеся наружу бесплод-
ные верхние части створок создают на вершине коробочки широкий,
до 3 см раструб, в который высыпаются семена.
Внутренние перегородки коробочки растрескиваются, видимо,
только на глубину, до которой доходят специальные ткани в пере-
городках, т. е. немного ниже, чем выгиб в них. Расширение пыльце-
проводниковой ткани ниже рыльца продолговато ланцетной формы,
часто с тремя гребнями, одним в центре и двумя боковыми. Вверху
это расширение кончается у рыльца, а внизу у выгиба перегородки.
Створки коробочки широколанцетные, узколанцетные, широко
яйцевидные, несколько оттянутые, заостренные на вершине, соло-
менно-желтого цвета, часто блестящие, иногда матовые. Вдоль створ-
ки в центре, сверху донизу проходит неглубокий желобок, в центре
которого невысоким гребнем проходит жилка (есть формы, у которых
жилки не видно). В верхней трети створки желобок часто расширяет-
ся и эпидермис в нем имеет вид рубцовой ткани. Весь желобок ок-
рашен в коричневый или коричнево-лиловый цвет различной интен-
сивности, ширины и длины, часто он светло-коричневый, с едва ок-
рашенным гребнем—жилкой в центре желобка на створке; если гре-
бень и желобок темно-коричнево-лиловые, смягченные сизым налетом,
тогда желобок всегда по краю зубчатый.
Щечки створки плотные, тонкие, прозрачные, выпуклые, в нижней
половине вздутые, вверху морщинистые. Морщинистость может быть
50
только в верхней трети коробочки, или продолжается почти до ее
основания. В углублении морщинок проходят тонкие, поперечные, ко-
ричневые жилки, часто дугообразно выгнутые, с концами у ребер
коробочки чуть-чуть направленными вверх. На ребре коробочка ок-
рашена в лиловый или светло-коричневый цвет. Ребро отделяется от
щечек створки глубоким желобком, образованным выгнутой наружу
частью створки. В этом желобке соединяются анастомозами попереч-
ные жилки створок. Вывернутая наружу область разрыва ребра ко-
робочки до 0,7 см шир., соломенно-желтая с палевым оттенком у
у внешнего края, похожая на фетр.
Под рыльцем створка уплощается, сглаживаются щечки. Верхняя
часть створки до вершины плодущей части образует равнобедренный
треугольник одного цвета со всей коробочкой, со сглаженными мор-
щинками и учащенными жилками, а иногда боковые стороны тре-
угольника довольно сильно вытянуты.
Рубец от опавших листочков околоцветника светло-коричневый,
выпуклый, до 2 мм шир. и часто налегает на плодоножку округлы-
ми фистонами.
Плодоножка того же цвета, что и вся коробочка, иногда она в
верхней части немного подкрашена коричневым. Светло-коричневые
семена неправильно треугольной формы (0,9X0,8 см), чуть-чуть сог-
нуты поперек или вдоль, окаймлены широкой, направленной вниз кай-
мой. На них четко виден более светлый, в виде запятой или прямой
зародыш.
В культуре при посеве семенами этот вид долго не вступает в
плодоношение; при посадке неполовозрелых особей сильно размно-
жается вегетативно и мало цветет. Зацветает в условиях Ташкента в
начале первой декады апреля.
Интересен пушистый кончик на листочках околоцветника у тюль-
панов, в частности у Т. fosteriana. Верхний зубчик на листочках око-
лоцветника закладывается и вырастает раньше, чем крайние, боковые
части отгиба. На нем очень рано, еще в зачаточнолг состоянии, появ-
ляется опушение, которое спускается по спинке с внешней стороны
листочка околоцветника, у некоторых растений почти на треть его
длины. В то же время, на листьях тюльпанов в большинстве случаев
опушение наблюдается только на верхней стороне, но почему тогда
у листочков околоцветника оно на нижней? Листочек околоцветника—
видоизмененный лист и равен зеленому листу и чешуе луковицы, опу-
шение на внешней стороне листочка есть новообразование, видимо,
выполняющее защитную функцию в цветочной почке.
Tulipa carinata Vved. — Тюльпан килеватый (рис. V и 2, ж) встре-
чается „на каменистых склонах в среднем и верхнем поясе гор. Ср.
Азия: Пам.-Алай (Чульбаир, Ходжа-Гургур-Ата). Эндем“ („Флора
СССР", т. IV).
Образцы, которые мы исследовали, получены от В. П. Бочанцева
и А. И. Введенского из окрестностей кишлака Сины Денауского
района и выращены из семян, привезенных А. Д. Пятаевой с вер-
ховьев р. Тупаланг.
Луковицы яйцевидные, до 4 см в диаметре, с черно-коричневыми
кожистыми, с внутренней стороны волосисто шерстистыми покровными
чешуями. Опушенность покровных чешуй у разных особей очень
изменчива.
Растение 48 см выс. от уровня почвы. Стебель в надземной части
и цветоножка пушистые.
51
Листья, в количестве~3—4, слабо отклоненные, вверх торчащие,
крупно волнистые, килеватые, часто вокруг киля штопорообразно
скрученные. Нижний—отходит от стебля на 8-м см от уровня почвы;
верхние расставленные, сизовато-зеленые, пушистые, по краю хряще-
вато-желто или розовато окаймленные, реснитчатые, не превышающие
цветы. Самый нижний—широко ланцетный (21 см дл., 6,5 см шир.),
верхний—20 см дл.ХЗ см шир., т. е. листья по направлению вверх
значительно сужаются, но не укорачиваются, однако цветков не пре-
вышают, так как растения имеют довольно длинную цветоножку.
Рис. 5. Т. orthopoda Vved.
Цветок красный с розоватым оттенком, одиночный, без запаха,
острыми листочками околоцветника по форме несколько напоминает
цветок Т. ingens, но с более резко выраженной двурядностью в рас-
положении листочков околоцветника, очень изящный, хорошо держит-
ся в срезке. Листочки внутреннего круга околоцветника торчат вверх,
а внешнего б. м. отогнуты. Внешние листочки околоцветника 8,2 см
дл.Х4,2 см шир., продолговато ланцетные, или обратно продолгова-
тые, постепенно заостренные в пушистый кончик, опушение с которо-
го переходит на спинку; часто по спинке, особенно наверху и при
основании, внешние листочки околоцветника обильно опушены; на-
верху опушены и края отгиба; снаружи, во время расцветания, они
сизовато-коричневатые с легким малиново-розоватым оттенком; в даль-
нейшем они краснеют, но легкая коричневатость остается, к краям
ало-красные, на ноготке зеленеют, иногда снаружи видна просвечи-
вающаяся черная часть внутреннего пятна; совнутри они ало-красные,
очень похожие по оттенку на Т. fosteriana, при основании с неболь-
шим желтым или черным желтоокаймленным пятном. Желтое пятно
или нижняя часть его всегда веерообразная, сплошь заполняет ниж-
нюю часть листочка, на которой почти всегда расположено черное
пятно, не покрывающее, по нашим наблюдениям, всей площади ноготка.
Черное пятнышко бывает усеченно яйцеобразное, широко или про-
52
Рис. V. Т. carinata Vved. (3/4 натуральной величины).
долговато ромбическое. На макушке яйцеобразного черного пятныш-
ка есть небольшая выемка. Пятнышко занимает площадь 0,8X0,6 см.
Бледно-желтое основание листочка с краев лучами вдается в общий
красный фон. Или, наоборот, так же как и черное пятно боковые
желтые полосы четко отграничены от основного фона. Иногда жел-
тое пятно кругом окаймляет черное пятнышко. Очень маленькая пло-
щадь пятна по сравнению со всей площадью листочка околоцветника
характерна для вида.
Внутренние листочки околоцветника (до 8,8 см дл., 4,1 см шир.)
обратно-продолговато-ланцетные, узко-неправильно-ромбические, уз-
ко-вытянуто-яйцевидные, б. м. постепенно заостренные в длинный пу-
шистый кончик, снаружи розовато-белесовато-оранжевые в различной
степени интенсивности этих оттенков у отдельных особей, лишь к
краям отгиба краснеющие; на ноготке снаружи с усеченным или ром-
бическим желтым пятном, на котором часто просвечивает нижняя
часть внутреннего пятна. Совнутри они ярко-оранжево-красные, свет-
лые, довольно блестящие. Вариации окраски наблюдаются в сторону
усиления красно-малинового цвета и ослабления оранжевого или на-
оборот. На основании лежит небольшое сплошь черное ромбическое
пятно, занимающее всю ширину площади ноготка до 1,3 см выс.;
иногда это пятно бывает усеченным, т. е. обратно треугольным, иног-
да на усеченной стороне пятна в центре с острым или округлым зуб-
цом.
Формы с каймой вокруг всего пятна на внешних листочках око-
лоцветника имеют кайму у пятен и на внутренних листочках около-
цветника только в виде узкой полоски.
У этого вида желтое пятно, четко отграниченное от вышележа-
щего основного фона, всегда наверху плоское.
На внешних листочках околоцветника коричневое пятно на жел-
том фоне очень темное, на внутренних—темная часть пятна очень ос-
лаблена и проявляется лишь в виде коричневато-бордовых штрихов
на желтом фоне.
Тычинки (2,8 см) в три раза короче листочков околоцветника и
немного выше завязи. Тычиночные нити (1 см дл., 0,3 см шир.) от
основания кверху постепенно сужаются, на вершине б. м. внезапно
заострены, черные, а основание и острие под пыльником у них жел-
тые.
Пыльники (1,8 см дл., 0,28 см шир.) черные с бордовым оттен-
ком или редко желтые. Пыльца светло-коричневато-розоватая или
желтая.
Завязь (2 см дл., 0,4 см шир.) бледно-зеленая, часто кремовая с
малиновым оттенком по ребрам, или вся сплошь розовато-сизая, на
вершине под рыльцем часто суженная в длинноватый (до 0,2 см)
столбик. Рыльце с боков розовое, ярко-малиновое с лиловатым оттен-
ком или кремово-желтое и даже оранжеватое. Сосочки рыльца изме-
няются от кремовых до темно-бордовых.
Коробочка белесовато-соломенная на высокой плодоножке (5,5 см
дл., 2,2 см шир.), очень изящной коротко-округло-цилиндрической
формы, иногда широко-коротко-обратно-яйцевидной формы с длин-
ным столбиком на тупой стороне яйца. Основание коробочки вне-
запно почти на 90° сужено и переходит в округлый светлый карпо-
фор (до 0,7 смдл., 0,6 см шир.). К вершине щечек коробочка несколько
расширяется, затем выше их б. м. внезапно сужается в довольно длин-
ный столбик 1,5 см дл., 1,7 см шир. в основании и 0,4 см на вершине
лод рыльцем. Эта часть створки темнее нижележащих. Желобок на
53
ней уплощен, вместо него проходят две жилки, то идущие параллель-
но, то перекрещивающиеся на суженной части столбика; между ними
лежит узкое, более темное, чем эта часть створки, углубление. К
столбику прочно прикреплены сильно отогнутые наружу, серо-корич-
невые лопасти рыльца.
Форму створок можно было бы назвать овальной, если на учиты-
вать область карпофора и рыльца и незначительное расширение в верх-
ней части щечек.
Вдоль створки от карпофора до вершины проходит более темно
окрашенный, особенно в верхней части, желобок. Он углублен по
всей длине, особенно сильно в верхней половине створки у вершины
щечек.
Щечки створки тонкие, полупрозрачные, сильно выпуклые, осо-
бенно (почти вздутые) наверху, сверху донизу покрыты тонкими,
елочкой направленными вверх, светло-коричневыми жилками. Эти жил-
ки почти без морщинок. Лишь в верхней половине коробочки они
слабо углублены. Поперечные, косо направленные вверх жилки пере-
ходят через желобок у края створки к самому ее разрыву на ребре
коробочки, которое окрашено в розовато-коричневый цвет. Коробочка
открывается тонкой узкой клиновидной щелью, в области столбика
сильно расширяющейся, благодаря отклонению его наружу.
Внутренние перегородки коробочки имеют сильный выгиб, по ко-
торому проходит узкий, по сравнению с другими видами тюльпанов,
тяж пыльцепроводниковой ткани. Растрескиваются внутренние пере-
городки значительно ниже выгиба, примерно на две трети длины.
Рубец от опавших членов околоцветника широкий (0,2 см), яр-
ко-коричневый. Плодоножка темнее коробочки, от рубца вниз она бо-
лее тонкая—от 0,4 до 0,3 см, при длине 30 см.
Семя большое (1X0,9 см), чуть-чуть согнутое, светло-коричневое,
с широкой и волнистой каймой, с легким серпообразным потемнением
в верхнем более длинном углу эндосперма, с тонким длинным, вид-
ным с обеих сторон зародышем. Этот вид в условиях Ташкента за-
цветает в начале третьей декады апреля.
Tulipa ajfinis Z. В о t s с h. — Тюльпан родственный (рис. VI и 2, з).
Исследованные образцы мы привезли из Туркестанского хребта с бо-
кового хребта Узунгыр, против кишлака Чурбек, где их собирали в.
арчовой зоне у верхней границы каменного пояса, широким карнизом
проходящего вдоль всего хребта, среди крупных глыб камня в мел-
коземе. Эндем.
Луковица продолговато яйцевидная, 2—3—4 см в диаметре, с бу-
магообразными темно-коричневыми, часто продолженными примерно
на половину длины луковицы, чешуями. Покровная чешуя совнутри
покрыта длинными, золотистыми, прямыми, прижатыми, редкими, су-
жающимися к вершине и основанию, волосками.
По характеру опушения вид напоминает Т. ingens. Стебель вме-
сте с цветоножкой до 32 см выс. от уровня почвы, зеленый, в верх-
ней части, как и цветоножка, опушен короткими, светлыми, торчащи-
ми волосками.
Листья, в количестве трех, сизые, с верхней стороны опушенные,,
по краю белохрящеватые, реснитчатые, слабо волнистые; нижний рас-
положен на 2—3 см над уровнем почвы, верхние — б. м. расставлен-
ные. Нижний лист широко ланцетный (21X6,2 см); верхний ланцет-
ный (15X4,6 см).
Цветок одиночный, красный, очень оригинального сложения, не
похожий ни на один известный тюльпан. Дно цветка широкое, округ-
54
! I
♦♦
Рис. VI. Т. af finis Z. Botsch. (3/4 натуральной величины).
лое. У внешних листочков, на: вершине чуть-чуть отклоненных, края
отгиба вдоль спинки обращены наружу. У внутренних листочков око-
лоцветника, слегка продольно выпуклых, торчащих вверх, вершина
обращена внутрь. Цветок без запаха.
Рис. 6, Т, regelii К г a s п .
Внешние листочки околоцветника ланцетные (7,5 см дл., 3 см
шир.), острые, постепенно оканчивающиеся острым, опушенным, заг-
нутым наружу кончиком, по краю мелкозубчатые, снаружи оранже-
во-красные, по спинке с малиновым оттенком; при основании их но-
готок до высоты 1,3 см зеленый. Зеленое пятно штрихами переходит
в основной фон. От нижнего зеленого пятна с обеих сторон спинки
проходят две более или менее густые, оранжевато-зеленые штрихо-
вые полоски, сближающиеся под острым кончиком, венчающим ли-
сточки. Совнутри они блестяще-оранжевато-красные, на утолщенной
части листочка, соответствующей спинке, приобретают темно-малино-
вый оттенок, сгущающийся к пятну. На ноготке во всю ширину ле-
жит черное, выпуклое, на вершине мелкозубчатое, четко ярко-желто
кругло окаймленное пятно (черное пятно 1,3 см, желтая кайма 0,15 см).
Листочки внутреннего круга околоцветника обратно продолговато
неправильно ромбические, на вершине б. м. внезапно оттянуты в до-
55
вольно большой пушистый кончик; по краю отгиба мелкозубчатые
(7,3 см дл., 3,5 см шир.); снаружи блестяще-оранжево-красные, с
большим, во всю ширину ноготка дымчато-фиолетовым, выемчатым
или, вернее, двояко-округло-выпуклым пятном на желто-зеленоватом
фоне. Этот фон образует над пятном широкую, четко отграниченную
от основной окраски листочка, кайму. Высота дымчатого пятна 1,3 см,
ширина каймы—0,3—0,4 см.
Совнутри листочек малиново-оранжевато-красный, с большим чер-
ным, с красноватым оттенком, узко ярко-желто окаймленным пятном,
повторяющим форму внешнего пятна, но более четко, а желтая по-
лоса уже и ярче, чем снаружи. Высота черного пятна 1,6 см, во всю
ширину ноготка; желтая кайма—0,1—0,15 см шир., очень яркая.
Тычинки ниже завязи и в 3—4 раза короче листочков околоцвет-
ника. Тычиночные нити черные (лишь под пыльником их утонченные
места белые), узко-плоско-треугольные (0,7 см дл., 0,25 см шир. при
основании).
Пыльники черно-лиловые (1,3 см дл., 0,25 см шир.), линейные.
Пыльца коричнево-фиолетовая. Завязь узкая, длинная (6,5 смХ
2,5 см шир.), почти не расширена к основанию, сильно окрашена ан-
тоцианом в пурпурный цвет, под лопастями рыльца кремово-зеленая.
Рыльце кремовое, очень большое, его лопасти спускаются вдоль
завязи на 4 мм.
Коробочка в среднем 6 см дл. и 2 см шир., треугольная в попе-
речном сечении, продолговатая, на вершине оттянутая в длинный
(1,2 см), тонкий (0,4 см) столбик, на котором сидит светло-коричне-
вое, наружу слегка выгнутое, довольно крупное рыльце.
Карпофор довольно длинный (1,1 см) и узкий (0,7 см), светлее
всей коробочки и вдается острым выпуклым клином в середину створки.
Ребра коробочки растрескиваются до карпофора, на который вхо-
дят в виде коротких, высоких, в середине ее сходящих на нет, греб-
ней. Растрескиваются коробочки чаще всего узкими клиновидными
щелями, вверху коробочки образующими широкий раструб благодаря
поперечному резкому отклонению наружу верхней трети створки ко-
робочки.
Поверхность разрыва матовая, белая, с кремовым или сероватым
оттенком.
Внутренние перегородки чаще всего растрескиваются до перегиба
в них. Пыльцепроводниковая ткань растет широким тяжем от рыльца
вдоль всей области верхней суженной части створки. Там же, где
створка расширяется, образуя камеры с семенами, она начинает су-
жаться, постепенно клиновидно убывая в ширине до перегиба в створ-
ке, откуда идет вниз сплошная плацента.
Створки коробочки плотные, довольно тонкие, на щечках про-
зрачные. В центре створки вдоль всей коробочки проходит довольно
глубокий желобок. В верхней части коробочки, выше щечек, створка
уплощается и там вместо желобка видна группа темно окрашенных
параллельных жилок, исчезающих по вступлении в область собственно
столбика. Желобок в верхней трети или даже несколько ниже окра-
шен в коричневый цвет, в самом центре по жилкам темный, а с бо-
ков светло-коричневый.
Щечки коробочки соломенно-кремовые, выпуклые, гладкие, лишь
в самом верху от желобка на них заходит несколько коричневых
штрихов или поперечных полосок.
Шов створки по самому краю узко окрашен в коричневый цвет,
а все ребро однотонное кремовое, лишь чуть-чуть ярче щечек, выг-
56
нуто наружу и вдоль него у щечки узкий желобок. Верхняя часть
коробочки благодаря слиянию окраски желобка и ребер створки, со-
вершенно коричневая. Особенно ровная коричневая окраска бывает
на столбике, но есть коробочки, у которых столбик светлый, как вся
коробочка.
Рубец от опавших членов околоцветника, светлый и отделяется
от плодоножки только видом рубцовой ткани, а не цветом. Плодо-
ножка очень длинная, имеет более темную, чем коробочка, с легким
коричневатым оттенком окраску.
Коричневое округло-скошено-треугольное семя окружено каймой,
которая шире на скошенной части семени. Зародыш длинненький,
светлый, в виде запятой, ясно виден на темном фоне семени.
В условиях Ташкента этот вид зацветает в середине апреля.
Tulipa praestans Н о о g.—Тюльпан превосходящий (рис. 2, и) ра-
стет „на каменистых и глинистых склонах в среднем поясе гор.—
Ср. Азия: Пам.-Алай (ю.-в. часть). Эндем“. (Флора СССР, т. IV).
Образцы для исследования получены из Таджикистана от В. П. Бо-
чанцева в 1933 г. из окрестностей Туткаула и с горы Себистан, затем
их в течение долгих лет выращивали в Ботаническом саду.
Кроме того, в 1950 г. были получены луковицы этого вида из
Душанбинского ботанического сада. В 1956 г. мы накопали луковиц
Т. praestans Hoog. в долине р. Варзоб и в урочище Кандара в Гис-
сарском хребте. Один образец вида выращен из семян, полученных в
1950 г. из г. Люнда в Швеции.
Луковица яйцевидная, 2 см в диаметре, с толстыми, кожистыми
черно-бурыми, с внутренней стороны блестящими покровными обо-
лочками, на вершине часто покрытыми редкими, толстыми, рыжева-
тыми, прижатыми волосками. Немного волосков бывает и при осно-
вании чешуи.
Стебель от поверхности почвы 10—30—45—60 см выс.; зеленый,
часто бывает кверху очень сильно окрашен антоцианом; в верхней
части вместе с цветоножкой покрыт белесоватыми торчащими волос-
ками.
Листьев 3—4, иногда 5—6, а у многоцветных экземпляров и боль-
ше в зависимости от количества добавочных цветов. Они отклоненные,
расставленные, светло-зеленые, чуть-чуть седоватые, не курчавые, но
иногда слабо волнистые; с верхней стороны суженные, по краю часто
красно окаймленные, реснитчатые, с ясно выраженным килем, а часто
и толстой центральной жилкой, кверху постепенно уменьшающиеся, у
бесплодных экземпляров не круто штопором свернутые, вертикально
торчащие. Нижний лист продолговатый или ланцетный с острым или
тупым кончиком (22X6,1; 11X6,5; 19,5X5,3: 25ХП см) отходит от
стебля на уровне почвы.
Цветок имеет форму чаши, правда, на дне он несколько оттянут,
но не раскрывается широко звездой, как у Т. subpraestans, без запаха.
Внешние листочки околоцветника широко ланцетные или обратно
продолговато яйцевидные, на вершине постепенно или внезапно за-
остренные или почти тупые, с небольшим острым кончиком, снаружи
оранжевато-красные, по спинке с сизовато-малиновым или желтоватым
оттенком, в нижней части листочка околоцветника постепенно пере-
ходящем в желтое глубоко выемчатое пятно, которое у разных форм
различных размеров часто занимает нижнюю четверть листочка око-
лоцветника, но не менее часто поднимается от основания сегмента
всего на несколько миллиметров. Совнутри эти листочки околоцвет-
ника ровного оранжево-красноватого цвета, лишь в нижней половине
57
по краям, в местах прикрытых внутренними листочками околоцветника
проходят две обесцвеченных продольных, узких (до 1,5 мм) и длин-
ных (до 15 мм) полоски (бывают формы без этих полосок).
Внутренние листочки околоцветника чуть короче внешних
(7,2X2,5; 6,5X2,9; 4,2X2,3 см)„ ланцетные, овальные, на обоих концах
заостренные, часто обратно яйцевидные, тупые, с небольшим, слабо
опушенным кончиком на вершине; снаружи оранжево-красные, к ос-
нованию до разной высоты оранжевый оттенок усиливается и посте-
пенно переходит в чисто оранжевый или желтый цвет; совнутри
сплошь оранжево-красные, лишь к месту прикрепления несколько
светлеют, приобретая более сильный оранжевый оттенок.
Тычинки с открытыми пыльниками в два с половиной раза короче
листочков околоцветника. Нити голые, в виде заостренных на вершине
колышков, в верхней половине несколько вздутые, красные, к месту
прикрепления желтеют.
Пыльники продолговатые, желтые, фиолетовые или бордовые;
сухие, в два с половиной раза длиннее нитей. Пыльца серовато-ко-
ричневая, малиновая или красная.
Завязь фиолетово-сизого оттенка в виде ровной палочки с сидя-
чим рыльцем, лопасти которого выдаются на ней; короче тычинок.
Рыльце с боков часто оранжево-красное или красное, его сосочки
всегда красные, различной интенсивности.
Коробочка оранжево-соломенная (4,3 см дл., 2,8 см шир.), про-
долговато-округло-цилиндрическая, тупая на вершине и при основании.
Карпофор (0,6 см выс., 0,6 см шир.) вдается в створку острым
углом. На этом углу и от открытых щелей, из которых высыпаются
семена, вдоль карпофора проходят невысокие гребни.
Рыльце сидячее, маленькое, у открытых коробочек опадающее.
Коробочка открывается широкими клиновидными или полуовальными
щелями, смещенными несколько вбок благодаря скручиванию створки.
Щель широко открыта даже в самом основании. Длина щели 4 см,
ширина 1,8 в центре коробочки, 1,3 у основания верхней бесплодной
части створки и 2 см у рыльца.
Внутренняя перегородка растрескивается почти до трех четвертей
длины, имея у рыльца его ширину, затем книзу постепенно сужается.
Ширина перегородки создается плоским тяжем пыльцепроводниковой
рыхлой ткани.
Створки коробочки оранжевато-соломенные, овальные, в центре
с продольным глубоким желобком, вдоль которого они сильно выгнуты
наружу. Расплывчатая оранжевая окраска желобка проходит вдоль
всей длины створки и в его верхней части по морщинкам переходит
на створку, иногда эта окраска становится коричневатой.
Щечки створки более плоские на вершине и очень выпуклые
внизу. Створка в верхней трети настолько сильно поперечно морщи-
нистая, что сглаживается выпуклость щечки. В морщинах окраска
оранжеватая, иногда коричневая, часто под рыльцем сплошная, более
яркая, чем вся коробочка. Желобки, отделяющие створки от шва,
глубокие. Край от желобка до рубца сильно поперечно морщинистый,
более оранжеватый, чем щечки. Шов чуть-чуть коричневатый, вы-
вернутый наружу оранжеватый, фетровой консистенции, внутренний
прогиб створки широкий, ровный по всей длине до основания рубца,
сужается к рыльцу.
Совнутри створки коробочек блестящие и более яркие, чем сна-
ружи. Рубец от опавших членов околоцветника светло-коричневого
58
Рис. VII. Т. subpraestans Vved. (3/4 натуральной величины).
цвета, сплошной, в 1 мм толщины. Плодоножка тоньше рубца, со-
ломенно-оранжевая, 16 см дл.
Семена темно-коричневые (0,7X6,6 см} с каймой, особенно раз-
витой у скошенного верхнего края семени. В этом месте кайма на-
правлена вниз. Зародыш прямой и более светлый, чем семя.
В условиях Ташкента этот вид зацветает в последние дни марта.
Вид сохраняет ювенильную способность, которая выражается в
том, что у половозрелых растений замещающая луковица уходит на
столоне углубления от места, где была материнская луковица. Так
же на столонах в различные стороны уходят детки, и на месте старой;
луковицы остаются пустые чешуи.
Tulipa subpraestans Vved. — Тюльпан превосходный (рис. VII и
2, к) встречается „на каменистых и глинистых склонах в среднем;
Рис. 7. Зрелые коробочки тюльпанов.
л—Т. bifloriformis Vved.; б—Т- turkestanica Rgl.; в—Т. polychroma S t а р f .;
г—Т. dasystemon Rgl- <?—Т. tarda S t а р f.;
поясе гор. Ср. Азия: Пам.-Ал. (Себистан, Сарсаряк). Эндем“ („Флора;
СССР“, т. IV).
Луковицы для изучения получены из окрестностей Туткаула от
В. П. Бочанцева и неоднократно впоследствии выращивались из семян.
Луковицы яйцевидные, 1,5—4 см в диаметре. Покровные чешуи
кожистые, коричнево-черные, полуразложившиеся—черно-бурые с
внутренней стороны блестящие, очень редко опушенные.
Высота растения во время цветения до 40 см. Стебель вместе
с цветоножками пушистый, покрыт сизым налетом. Нижний лист часто
растет на стебле высоко (до 6 см от уровня почвы).
Листья в количестве 3—4 на главном стебле и по одному при-
цветниковидному на ветвях расставленные, серповидно отогнутые,
крупно волнистые, в условиях Ташкента часто превышающие цветы,
по краю хрящевато окаймленные, сизые, нижние—почти без ресничек;
верхние—густореснитчатые, пушистые; нижние—только с верхней
стороны, верхние—с обеих. Самый нижний лист продолговато ланцет-
ный до 43 см дл., 12 см шир.; верхний—ланцетный—22,5 см дл.,
5,3 см шир.; прицветный листочек на ветке—5,5 см дл., 0,6 см шир.
У слабых растений один, обычно же два-три цветка, раскрываю-
щихся на ветках первого порядка, сидящих в пазухах верхних листьев.
У мощных растений их 5—6, у очень сильных растений в пазухах
59'
листьев на ветвях первого порядка появляются ветви второго порядка,
увенчанные цветами.
Цветок красный, с узкими, острыми блестящими листочками око-
лоцветника в виде простой звезды, но очень приятный и изящный.
Внешние листочки околоцветника ланцетные, постепенно заострен-
ные в пушистый кончик; снаружи по всей поверхности сизо-оранжево-
красноватые, к вершине даже со светло-коричневым оттенком; лишь
у самого края листочка чистого блестящего красного цвета; на но-
готке до высоты 1 см желтые; желтое постепенно переходит в основ-
ную краску спинки листочка (есть формы, у которых листочки около-
цветника более темного цвета и тогда по спинке они как бы со
свинцовым налетом); совнутри ярко блестяще окрашенные. В их
окраске значительно меньше оранжевого, чем у Т. greigii и Т. foste-
riana и др. При основании листочка лежит желтое небольшое пятно,
обычно почти скрытое под тычиночными нитями. Оно с боков листоч-
ка, в частях, прикрытых внутренними листочками, достигает 0,7 см
выс., 0,3 см шир., затем проходит почти квадратная выемка, в которой
желтый цвет поднимается с краев до 0,3 см, в центре до 0,4 см,
образуя выпуклость, на вершине коричневеющую и незаметно пере-
ходящую в основной цвет листочка. Это пятно может почти исчезать,
особенно у мелких растений, но всегда в месте прикрепления листоч-
ка к цветоносу листочек желтеет и желтизна поднимается по его
краям на некоторую, хотя и незначительную, высоту. Полусфериче-
ской коричневатости в выемке желтого пятна также может не быть.
Внутренние листочки околоцветника (8,7 см дл. и 2,9 см шир.)
чуть короче и уже внешних, ланцетные, также постепенно оттянутые,
но в менее пушистый, чехм у внешних листочков, кончик; снаружи
блестящие, сизо-оранжево-красные, даже чуть-чуть малиновые (сизо-
ватость не от налета, а от белесоватости окраски), при основании
максимум до высоты 0,9 см желтые (желтое основание постепенно
переходит в основной цвет); совнутри очень блестящие, красные, с
чуть-чуть заметным оранжевьш оттенком, такие же как и внешние,
при самом основании на 0,15 см желтые, выемчатые в центре.
Выемку заполняет легкое, как тень коричневатое, в виде острого
угла, штриховатое пятно (у многих форм его нет).
Место прикрепления листочка околоцветника у этого вида всегда
желтое.
Тычинки (2,8 см дл.) в 2,5—3 раза короче околоцветника и выше
завязи.
Тычиночные нити (0,7 см дл. и 0,35 см шир.) от основания кверху
постепенно сужающиеся, плоские, красные или бордовые. Красные
иногда при основании желтеют, а бордовые становятся светло-крас-
ными.
Пыльники черные или лилово-черные (1,8 см дл., 0,25 см шир.),
по высыпании пыльцы спирально свернутые. Таким образом, тычиноч-
ные нити в два, два с лишним раза короче пыльников. Пыльца темно-
коричневая или серовато-коричневая.
Завязь (1,7 см дл.Х0,6 см шир.) под рыльцем чуть-чуть сужена,
сизо-зеленая.
Рыльце шире завязи, все сплошь, вплоть до верхних бесплодных
частей плодолистиков, под завязью и до сосочков на лопастях мали-
ново-красное.
Коробочка белесовато-соломенно-желтая (1,5сждл., 2,8 см ширЭ,
почти округлая, на вершине и при основании тупая; иногда продол-
.говатая (6,8X2,3 см). Карпофор белесовато-соломенный, гладкий, без
60
гребней, 0,6 см выс., в верхней части 0,8 см шир., а внизу у рубца
0,5 см, тремя короткими зубцами вдается в створку. Очень отличается
по внешнему виду от нижней части коробочки у Т. praestans. На
верху коробочки нет никакого подобия столбика или носика. Рыльце
сидячее, очень маленькое, наружу отогнутое.
Коробочка открывается клиновидной широкой щелью, чаще всего
широко открытой до карпофора. Щель 4,2 см дл., у рыльца 2 см шир.
Внутренняя перегородка растрескивается примерно наполовину
длины коробочки. Ее расширенная часть под рыльцем овальной формы,
вернее округло ромбической (1 слХ0,7 см ширины) с гребнем в
центре.
Створки коробочки широко линзовидной формы, цвета слоновой
кости. Шов на месте прикрепления внутренней перегородки широкий,
клиновидно расширяющийся кверху, с гребнем вдоль всей длины,
иногда кверху раздваивающийся, покрыт мятым эпидермисом, часто
одинакового цвета со щечками створок. Но не менее часто он окра-
шен в светло-коричневый цвет с белесовато-лиловым оттенком.
Щечки выпуклые, примерно на две верхних трети сильно морщи-
нистые, внизу гладкие, вздутые. Морщины направлены елочкой вверх.
У всех форм желобки между морщинами окрашены в сизо-коричне-
вый цвет, но разного оттенка, такого же как центральный желобок
створки. В верхней части створки морщины мельчают, иногда совсем
сминаются и под рыльцем остается мятая поверхность. У рубца раз-
рыва на ребре коробочки окраска более темно сизо-малиновая, иногда
чуть-чуть коричневая. Разрыв отделен от створки глубоким желобком
и сильно выгнут наружу. Его внутренняя сторона 0,35 см шир., фет-
ровой консистенции, палево-оранжевая или серо-соломенная.
Рубец от опавших членов околоцветника коричневатый, довольно
широкий (0,2 ему, по верхнему краю в виде фестонов видны следы
от опавших листочков и тычинок.
Плодоножка тоньше карпофора, сизовато-соломенная, до 15<дидл.
Семя темно-коричневое (1,2 X 0,9 см), остроугольное, на вершине
округлое с широкой, загнутой вниз каймой на скошенном верхнем
углу. Зародыш кривой, плохо виден. В условиях Ташкента этот вид
зацветает в начале последней декады апреля.
Tulipa butkovii Z. Botsch — Тюльпан Буткова (рис. VIII и 3, ж).
Исследованные образцы привезены в 1954 г. с песочного перевала в
Чимгане, А. Я. Бутковым — в разные годы из многих районов Запад-
ного Тянь-Шаня—урочище Большой Майдантал (мелкоземисто щебни-
стый склон—VIII-55), урочище Малый Майдантал (среди скал в мел-
коземе VIII-55), долина р. Акбулак-Карасай, щебенчатый склон,
h-1820—2000 м, 1956 г., долина р. Акбулак при слиянии с Серкели-
саем, щебенчатый склон 1956 г.
Луковицы яйцевидные (до2,5 см в диаметре) с коричневыми,
бумагообразными, несколько продолженными покровными чешуями,
совнутри по всей поверхности покрытыми короткими, прижатыми, зо-
лотистыми волосками, особенно сгущающимися к основанию и вершине.
Растения во время цветения достигают всего до 15 см выс. от
уровня почвы. Зеленовато-коричневая цветоножка опушена и покрыта
сизым стирающимся налетом. Листья в количестве 3—4, сближенные,
отогнутые, отклоненные, по краю курчавые, б. м. пушистые, реснит-
чатые, сизые, даже голубоватые, по краю светло окаймленные, верх-
ние—желобчатые, часто в начале цветения превышают цветок.
Нижний лист во время цветения своим основанием погружен в
61
•почву, широколанцетный (12,7 см дл.,4,5 см шир.); верхний лист уз-
коланцетный (10,8 см дл., 0,9 см шир.).
Цветок кирпично-красный, одиночный, открывается широко коло-
кольчато, почти звездчато и не имеет запаха.
Внешние листочки околоцветника (5,5 см дл., 2,2 см шир.) узко-
продолговатые или ланцетные, на вершине тупо заостренные или почти
округлые; иногда продолжены в острый опушенный кончик, редкое
опушение проходит вдоль всей спинки до низа; снаружи светло-бор-
довые с малиновым оттенком, особенно по спинке, к краям оранже-
веют, в самом низу желтеют и сплошь покрыты сильным сизым
налетом, часто делающим спинки почти свинцовыми; совнутри свет-
ло-бордово-оранжевые, блестящие; в нижней половине постепенно
Рис. 8. Т. dubia Vved.
темнеют и понемногу переходят в свекольно-бордовое, нечетко от-
граниченное, длинное, узкое, в середине часто с глубокой выемкой
пятно, доходящее, примерно, от 1/6 до 1/3 дл. листочка. Внутренние
листочки околоцветника продолговатые или узко яйцевидные, на вер-
шине округлые, б. м. внезапно заостренные в короткий пушистый
кончик (4,6x2,1 см и 6,2x2,3 см). Снаружи оранжево-бордовые, вдоль
центральной жилки с легким малиновым оттенком, блестящие, на но-
готке зеленовато-оранжевые; совнутри оранжевато-бордовые, в нижней
половине приобретают свекольный оттенок, который сгущается к
основанию листочка, образуя небольшое, нечетко отграниченное,
прямоугольное, коричневато-бордовое пятно; у места прикрепления на
полтора-два миллиметра они оранжевые; оранжеватость просвечивает
во всей нижней части ноготка под свекольной окраской, особенно на
его краях.
Тычинки выше завязи и в два с лишним раза короче околоцвет-
ника. Нити разных оттенков красные и бордовые, книзу расширенные,
но, вообще, тонкие, по длине часто почти равны завязи (1,1x0,11 см).
62
Рис. VIII. Т. butkovii Z. Botsch. (:i/4 натуральной величины).
Пыльники желтые или темные, сверху постепенно к основанию
«открывающиеся, в условиях Ташкента часто в течение одного дня,
но не скручивающиеся как у Т. kaufmanniana.
Пыльца желтая или бордовая.
Завязь (1,3 см дл., 0,4 см шир.), внизу расширенная, у самого
«основания б. м. внезапно суженная, к рыльцу постепенно тонко су-
жающаяся, кремовая или бледно-зеленая, по ребрам и под рыльцем
в разной степени интенсивности бордовоокрашенная.
Рыльце шире завязи, с боков часто бордовое, сосочки желтые.
Коробочка сероватая с ярким узором из коричнево-бордовых пятен
и штрихов, может быть до 7 см дл. и 2,2 см шир., продолговатая,
овальная или коническая, но почти всегда при основании шире, чем
у столбика. Карпофор варьирует по форме, длине и ширине, но он
обычно довольно тонкий, к створкам несколько расширяется, округ-
лый, отделен от створок б. м. глубоким желобком, часто острым
углом входит в центральный желобок створки.
Наверху коробочка оканчивается довольно длинным столбиком, в
который, постепенно сужаясь, переходит верхняя часть створки. Если
•створка на вершине сужается медленно, то столбик очень короткий.
На вершине столбика расположено небольшое, но значительно шире
него согнутое вдоль коричневое рыльце. Створки коробочки имеют
форму: ланцетную, овальную с выступами на вершине и основании,
широкояйцевидную, округлую, дельтовидную. От видов, обитающих
в этих районах, их отличает большая хрупкость, тонкость; лежащие
внутри семена просвечиваются сквозь щечки коробочки и делают их
«сероватыми.
В центре створки проходит довольно глубокий желобок, ярко и
широко окрашенный в коричнево-бордовый цвет; иногда окраска его
«светло-коричневая; кверху коричневая полоса желобка расширяется,
а в столбике сужается. От нее поперек щечек створки отходят яркие,
широкие или тонкие поперечные полосы, иногда, перед краем створки,
ветвящиеся, часто на краю сливающиеся со сплошной коричнево-бор-
довой полосой на ребре створки. Щечки створки прозрачные, часто
выпуклые, поперечно морщинистые. Морщинки иногда расположены
по всей поверхности, но чаще—в верхней половине створки; часто
•бывают в виде елочки направлены концами вверх. От края створки
щечки отделены довольно глубоким желобком. Самая окраина створки
за желобком окрашена в тот же цвет, что и желобок, или коричнево-
бордовый, или просто коричневый; сильно вывернута наружу. Яркая
окраска края створки, сливаясь воедино на столбике, делает его более
темным, чем вся створка.
Коробочка открывается клиновидной щелью, растрескивающейся
до основания плодолистика с большим раструбом на вершине, из-за
резкого отклонения наружу верхней бесплодной части створки вместе
со столбиком. Края створок резко выгнуты наружу. Поверхность раз-
рыва кремовая, оранжевая или палевая, 0,4, а чаще 0,35 см шир.
Рубец от опавших членов околоцветника коричневый, до 0,2 см
шир., выгнутый. Плодоножка оранжево-коричневая.
Семена крупные коричневые, округло треугольные, часто острый
угол треугольника загнут немного вбок. Их окружает пленчатая кайма,
широкая с одного скошенного бока семени, направленная вниз. Со
стороны противоположной широкому краю каймы, в эндосперме распо-
ложен узкий полумесяц темной ткани. Зародыш лежит в семени не-
сколько сбоку, смещен в сторону широкого края каймы.
63
При посадке луковицами и выращивании из семян в условиях
Ташкента этот вид начинает вегетацию позже остальных видов тюль-
панов и зацветает также позже всех среднеазиатских видов из секции
Leiostemones — во второй половине третьей декады апреля.
Tulipa vvedensky i Z Botsch. — Тюльпан Введенского (рис. IX, X
и 4, г). Исследованные растения собраны нами в Западном Тянь-Шане
в долине р. Ангрен в полутора километрах выше кишлака Тюрк; на
склонах, образованных выходами коренных пород чуть прикрытых
мелкоземистой почвой. Эндем.
Рис. 9. Т. dasystemonoides Vved.?
Луковицы яйцевидные, редко продолговато яйцевидные, до 3 см
в диаметре, с темно-коричневыми, бумагообразными, с внутренней
стороны по всей поверхности прижато рыхло, у верхушки густо во-
лосистыми оболочками.
Стебель 15—20 см выс. от уровня почвы, вместе с цветоножкой
густо опушен белыми, короткими, оттопыренными волосками, сизо-
ватый, иногда с бордовым оттенком.
Листья, в количестве 4—5, отогнутые, сближенные, сизые, кур-
чавые, иногда опушенные редкими, короткими волосками, по краю
слабо реснитчатые, постепенно кверху уменьшающиеся, обычно дости-
гающие верхушки цветка или не превышающие его; самый нижний
лист ланцетный или широколанцетный (25 см дл. и до 6 см шир.),
отходит от стебля на уровне почвы. Верхний—узколанцетный до
23,5 см дл. и 2 см шир.
Цветок одиночный, красный, иногда оранжевый и редко палевый,
очень хорошей формы, образует как бы двойную чашу, несколько
напоминающую цветок Т. greigii. Его внутренние листочки шире
отклонены, а внешние не так сильно отогнуты наружу, как у Т. grei-
gii. Основания листочков околоцветника менее массивны, чем у
Т. greigii и дно цветка менее плоское.
64
Рис. IX. Т. vvedenskyi Z. Botsch. (3/4 натуральной величины).
Внешние листочки околоцветника (до 10,5 см- дл. и 16,2-сж-шир.),
ромбические, продолговато ромбические, иногда неправильно ромбиче-
ские, на вершине округлые или заостренные, при основании б. м. по-
степенно оттянутые. На вершине все формы листочков б. м. вне-
запно оттянуты в пушистый кончик. Опушенность с кончика
часто спускается на спинку и по-
крывает ее редкими короткими
щетинками. Снаружи по спинке
листочки околоцветника белесо-
вато-оранжево-красные, края ши-
роко, ярко-ало-красные; в нижней
половине до высоты в 2,5 см
желтеют; на желтом нередко
чуть-чуть просвечивает темное
пятно; совнутри ало-красные,
часто с желтым пятном в нижней
части листочка. Желтое пятно
занимает всю площадь листочка
до 2 см выс. На вершине оно
плоское или выпуклое, штриха-
ми соединяющееся с основным
фоном; иногда в верхней части
приобретает коричневатый отте-
нок. На желтом пятне часто ле-
светло-коричне-
продолговатое, на
заостренное пятно раза
высоте превышающее
окаймление с боков.
Рис. 10. T. buhseana Boiss.
жит темно- или
вое, овально
вершине
в два по
желтое
Иногда овально продолговатое
пятно не так велико и широко, но
тогда с его боков желтая часть
листочка становится бордовой
с желтым сиянием наверху; часто
пятно в нижней части листочка
околоцветника сплошное, темно-
коричневое, почти черное, ромбической формы; часто ромбическое
темное пятно имеет как бы перетяжку в верхней половине, и несет
очень длинный зубец, округлый или заостренный на вершине.
Внутренние листочки околоцветника (11,5 см дл., 5,2 см шир.)
обратно-продолговато-яйцевидные, на вершине внезапно заостренные
в длинный пушистый кончик; по верхнему краю слегка волнистые;
снаружи оранжево-красные; в нижней части постепенно становятся
желтыми, часто до 3 см выс. Желтое основание бывает обратно тре-
угольным или ромбическим, часто на желтом фоне из б. м. густых,
темно-бордовых, сливающихся вместе штрихов образуется остроуголь-
ное или ромбическое пятно. Иногда такое пятно сплошь покрывает
желтое, которое выступает из-под него лишь в виде сияния, иногда
пятно при основании листочка не желтое, а зеленое. Совнутри внутренние
листочки околоцветника более красные и блестящие, чем снаружи;
у форм, имеющих темное пятно при основании, они ближе к пятну
приобретают малиновый отгенок. Пятно при основании листочка
бордово-черно-коричневое с преобладанием того или другого оттенка,
к краям слабее, чем внутри; иногда сплошь темное или более
светлое с желтым сиянием по краям; сияние может простираться над
5—2229
65
всем пятном; у некоторых особей пятно сплошь желтое. Желтое
пятно бывает лишь усеченным или округлым на вершине и в виде
штрихов заходит на основной фон. Темное пятно имеет форму ромба,
островершинного или округловершинного, светлеющего к бокам; или
острого треугольника, поднимающегося высоко на листочек. Часто
пятно бывает лишь в виде легкой тени.
Тычинки в три и более раза короче околоцветника и немного
выше завязи.
Нити голые, желтые или коричневые, часто в верхней части
бордово-коричневые, при основании желтые; к основанию расширен-
ные (до 0,45 см и 1,3 см дл.).' Острие под пыльником желтое.
Пыльники желтые или черно-фиолетовые; резко в нижней части
желтые, а вверху коричневые (до 2 см дл., 0,4 см шир.), бывают
равны нитям, но чаще длиннее их, одновременно открывающиеся.
Пыльца желтая, черно-фиолетовая, коричневатая и даже серо-
вато-коричневая.
Завязь (до 2,3 см дл., 0,6 см шир.) желтовато-сизоватая, бледно-
зеленая, часто по ребрам и в центре плодолистиков окрашена антоциа-
ном: обычно к рыльцу постепенно сужающаяся, но бывает и ровная
по всей длине.
Рыльце шире завязи, желтое, иногда имеет красные бока и со-
сочки, но б. ч. они желтые.
Коробочка соломенно-желтая с коричневыми узорами (9 см дл.,
2,6 см шир.), продолговатая, узкая, в поперечном сечении треуголь-
ная, на вершине и основании заостренная. Карпофор 1 см дл. у шва
и 1,5 см дл. под рубцом плаценты, 1,4 см шир.; округлый, на него
в верхней части заходит окончание рубца между створками. Вверху
коробоч ка заострена, и постепенно оттянута в довольно длинный
столбик, увенчанный крупными, отогнутыми назад светло-коричне-
выми лопастями рыльца. Рыльце светло-серовато-коричневое, крупное,
выступает над бесплодной частью коробочки.
Коробочка открывается расширяющейся кверху и доходящей
внизу до карпофора клинообразной щелью. Края шва светло-коричне-
вые, палевые, вывернуты наружу, верхняя бесплодная часть плодо-
листиков резко отклонена наружу так, что на вершине получается
широкий 3,5 см раструб. Внутренние перегородки коробочки вначале
растрескиваются на глубину бесплодной части (до 2,5 см), а затем
еще глубже на 2,5 см, по сросшимся крепко швам в перегородке
коробочки, т. е. всего на 5 см.
Створки коробочки плотные, крепкие, однако на щечках просве-
чивают наполняющие коробочку семена. Створки продолговатые,
длинно заостренные, при основании и наверху отделены от карпофора
направленным вверх остроугольным желобком. От вершины желобка
начинается продольная, идущая вдоль всей створки выемка с невысо-
ким гребешком в центре, по всей длине окрашенная широкой (0,4сж)
зубчатой, по краям светло-коричневой полосой, усиливающейся в
верхних трех четвертях коробочки. От ее зубцов отходят по жилкам
поперечные коричневые линии, их может быть больше или меньше.
Щечки створки выпуклые, поперечно-полого-морщинистые, в верхней
части морщинок больше, но, вообще, их немного, плотные, слегка
просвечивающиеся; поперечные жилки в желобке, отделяющем щечки
створок от ребра, образуют целую сеточку из остроугольных перепле-
тений. По краю ребра рубец светло-коричневый, на вывернутой ши-
рокой стороне шва створки окраска палевая, консистенции фетра. Под
рыльцем окраска сгущается и бесплодная продолговатая часть коро-
66
Рис. X. Т. vvedenskyi Z. Botsch. (2/3 натуральной величины).
бочки коричневого оттенка. При основании коробочки расположен
коричневый широкий (0,4 см) рубец от опавших членов околоцветника.
Плодоножка толстая, массивная, недлинная (15—35 см), с корич-
невым оттенком, постепенно сужается к розетке листьев. У очень
крупных коробочек она часто при высыхании растрескивается вдоль.
Семена темно-коричневые, округло треугольные, с более светлым,
в виде запятой, зародышем и узкой направленной вниз каймой.
В условиях Ташкента этот вид зацветает с конца второй декады
апреля.
Tulipa alberti Rgl. —Тюльпан Альберта'(рис. XI) встречается „на
щебнистых склонах,—Ср. Азия. Прибалх. (Таргыл, Куянкуз), Тянь-
Шань (Каратау, Чу-Илийские горы). Эндем“ („Флора СССР“, т. IV).
Исследованные растения мы впервые получили из хр. Каратау от
П. А. Гомолицкого, со ст. Чокпар в 1949 г. от В. П. Бочанцева, в
1955 г. и в 1956 г. от Алмаатинского ботанического сада из образцов,
собранных близ ст. Отар.
В 1956 г. мы пересекли хр. Каратау и среди растений Т. greigii
привезли красивейшие экземпляры Т. alberti, собранные в трещинах
среди вертикально стоящих слоев сланца, где не было никакой
другой растительности.
Луковицы яйцевидные до 4 см в диам., с кожистыми, черно-бу-
рыми, сильно продолженными покровными чешуями; совнутри в про-
долженной части чешуи очень густо, а по всей поверхности реже,
опушены прижатыми жесткими волосками.
Стебель вместе с цветоножкой до 20 см выс. над уровнем почвы,
пушистый, сизый, с легким бордовым оттенком, толстый.
Листья, в количестве 3—4, очень сизые, почти голубые, лишь
со слабым зеленоватым оттенком, опушенные, по краю хрящевато
окаймленные, реснитчатые и отогнутые, в различной степени кур-
чавые. Нижний лист отходит от стебля у уровня почвы, верхние к
нему приближены. Нижний—широколанцетный, 14,1 см дл. и 6,2 см
шир. Верхний—ланцетный, 11 см дл. и 2,3 см шир.
Цветок одиночный, часто очень красивый, особенно у растений
из Каратау. Они более приземисты, чем растения из Чокпара, но
более оригинальны по окраске и форме. Внешние листочки у них
значительно отогнуты наружу, зато внутренние сильно выпуклые.
Формы цветка и окраска этого вида очень разнообразны: от
оранжевых с красноватым оттенком у растений из Прибалхашья до
оранжевых, оранжево-розовых, золотисто-и серебристо-желтых, темно-
бордовых различной интенсивности из Каратау. Цветок без запаха.
Опишем форму этого вида, привезенного В. П. Бочанцевым из
Чокпара.
Внешние листочки околоцветника округло ромбические, б. м.
постепенно переходящие в острый опушенный кончик (8 см дл., 6 см
шир.); всегда в различной степени наружу отогнутые; снаружи по
спинке лиловато-оранжевые с сизым налетом; к краям темно-оранже-
вые с красным оттенком, к основанию листочка лиловатость сгущается
до бордового, у некоторых стеблей основание сегмента зеленеет; со-
внутри листочки околоцветника блестящие, темно-оранжевые с крас-
новатым оттенком, который усиливается к основанию и центру лис-
точка. Пятно в нижней половине листочка темное, часто черное
с лиловатым оттенком, сгущающимся к краям, округлое, к основанию
чуть удлиненное, на вершине плосковатое, с выемкой, или заострен-
ное (1,1 см выс., 1,3 см шир.).
Внутренние листочки околоцветника неправильно ромбические,
67
к основанию суженные и оттянутые, а в верхней половине расширен-
ные, на вершине округлые, в центре с выемкой, в которой находится
небольшой опушенный кончик. У некоторых форм листочки в виде
сердца, всегда б. м. выпуклые, ложковидные; с обеих сторон блес-
тящие, темно-оранжевые, только снаружи на утолщенной нижней
части они приобретают желтый цвет; на желтом листочке лежит узко-
сердцевидное пятно (2,5 см выс., 1,6 см шир,) с острыми углами на-
верху и при основании, состоящее из бордовых штрихов; совнутри
пятно более темное—черно-коричневое с бордовым оттенком, к
основанию листочка ослабевающим, такого же размера, как и снаружи,
но более округлое. Листочек у места прикрепления желтый.
Тычинки (2,9 см выс.) во время расцветания цветка выше завязи
и в 2—3 раза короче околоцветника.
Желтые тычиночные нити 1,4 см дл. и при основании 0,4 см шир.,
т. е. при основании широкие, к вершине постепенно сужающиеся,
но все же остающиеся достаточно широкими. Под пыльниками они
внезапно сужаются в острый кончик.
Пыльники по длине равны тычиночным нитям или немного длин-
нее их, темные, или изредка, у светлых форм, желтые.
Завязь светло-зеленая (2,2 см дл., 0,7 см шир.), ровная по всей
длине, под рыльцем чуть суженная.
Рыльце кремовое, шире завязи.
Коробочка 4,3 см дл., 2,5 см шир., почти яйцевидная, внизу
внезапно, почти под прямым углом, сужается к рубцу на цветоножке.
От короткого карпофора ее отделяет поперечно морщинистый желобок.
Вверху коробочка сужена в короткий, довольно широкий столбик
(9,5X0,6 см). Дольки рыльца прочно прикреплены к вершине створки
и отламываются от нее только с кусочками столбика. Довольно
крепкая коробочка растрескивается по рубцам узкими продольными
щелями, но створки не расходятся вдоль щели. Лишь верхние части
створки чуть-чуть отходят друг от друга (на 0,5 см). Внутренние пе-
регородки также растрескиваются только на глубину бесплодной
части (1,2 см) и чуть-чуть отклоняются одна от другой.
Створки коробочки широко-или узкояйцевидные, при основании
б. м. внезапно округло заканчивающиеся, наверху оттянутые в ко-
роткий столбик. В центре створки проходит желобок, в верхней поло-
вине окрашенный в сизо-коричневый цвет; коричневая окраска плотная
неровная, но от нее не отходят никакие коричневые поперечные по-
лоски.
Щечки створки грубые, но тонкие, белесовато-соломенные, вы-
пуклые, гладкие, лишь в верхней части слабо поперечно морщини-
стые. Морщинистость с них переходит на верхнюю бесплодную часть
коробочки. На ребре коробочка окрашена в серовато-коричневый
цвет, размытый к желобку, отделяющему щечки от ребра коробочки.
На ребре по размытому серо-коричневому тону, ослабевающему в
сторону щечек, виден узор из выступающих жилок. Край створок не
выгибается. Под рыльцем столбик более темного цвета, чем вся ко-
робочка.
Рубец от листочков околоцветника выпуклый, светло-коричневый,
0,15 см шир. Плодоножка довольно плотная, крепкая и короткая
(до 12 см дл.), более темная, чем коробочка, окрашена коричневым
и бордовым пигментом, вниз от рубца к листьям резко сужается.
Семена 0,6 х 0,5 см, неправильно округло треугольные, ярко
коричневые с темно-зеленым пятном в вытянутом верхнем углу с
довольно широкой направленной вниз каймой, ровной ширины по
68
Рис. XI. Т. alberti Rgl. f2/3 натуральной величины).
всему семени. В условиях Ташкента он зацветает в конце второй
декады апреля.
Tulipa brachystemon Rgl. •—Т. короткотычиночный „растет на
щебнистых склонах.—Ср. Азия. Джунг.—Тарб. (Джунгарский Алатау,
ю. ч.)“. („Флора СССР“,т. IV).
Настоящее описание сделано по образцам, хранящимся в Ботани-
ческом институте АН СССР в Ленинграде и собранных в Семиречен-
ской области (предгорья в окрестностях Копала, Заимка Маклакова
28 апреля 1908 г., Шнитников № 15); Регелем в апреле 1879 г. (почти
белый); в Семиречье Талды-Курганского уезда, верховья р. Каратала
„Солдатская щель“. Щебнистый склон на высоте 1750 м. Шипчинским
в 1928 г. в июле; Регелем в Чингильды, Лидугом в окрестностях
.Копала.
Луковица 2,5 см в диам., с черными кожистыми, совнутри у вер-
хушки опушенными прижатыми, жесткими волосками покровными
чешуями; лежит на глубине до 12 см.
Стебель 10—28 см выс. от уровня почвы. Голая цветоножка
окрашена антоцианом.
Листьев 4 (3), по стеблю расставленные (междоузлие от нижнего
до второго листа 5 см, верхний приподнят от уровня почвы на 14 см),
хрящевато окаймленные, голые или слегка реснитчатые, по краю слабо
волнистые, от стебля мало отклоненные (даже нижний лист направлен
вверх), постепенно кверху уменьшающиеся. Самый нижний лист
линейно-ланцетный (22 см дл. и 3 см шир.). Верхний—узкий злаковид-
ный (7x0,3 см).
Цветок один, желтый или кремовый, видимо, в форме простой
звезды, или неглубокой чаши.
Внешние листочки околоцветника продолговатые или ланцетные,
на вершине тупые или постепенно заостренные, голые, снаружи на
вершине по спинке с фиолетовым оттенком, совнутри однотонные
без пятен.
Внутренние листочки овально или продолговато ромбические или
обратно продолговатые, одного цвета со внешними, но без лиловой
спинки.
Тычинки в 2, почти в 3 с лишним раза короче околоцветника.
Нити голые, к основанию сужены, к пыльникам сильно расшире-
ны, под пыльниками б. м. внезапно сужены в острый кончик, об их
цвете можно сказать только, что пыльники светлее нитей, а нити
темнее всех органов цветка. Пыльники светло-коричневые или жел-
тые, почти равные нитям, при основании с глубокими выемками, в
которые входят острые концы нитей.
Завязь с заметным столбиком, светлая, формы бутылки, сильно
сужена под рыльцем, более светлая, чем пыльники.
На родине цветет в конце апреля.
Tulipa brachystemon Rgl. —Тюльпан короткотычиночный* (рис. XII
и 3, б). Исследованные растения получены из Джунгарского Алатау
от В. П. Голоскокова.
Луковица до 2,5 см в диам., с черными, кожистыми, совнутри
у верхушки опушенными прижатыми, жесткими волосками, покров-
ными чешуями.
Стебель 10—22 см выс. от уровня почвы, сизый. Цветоножка
некоторых растений слабо окрашена антоцианом, других—сильно
опушена. Есть также почти голые особи.
* Отличается от описанного во „Флоре СССР”.
69
Листьев 3, очень сизые, с верхней стороны опушенные, с белым
хрящеватым окаймлением; сильно кудрявые, округложелобчатые, сер-
повидные, особенно верхний. Нижние два сближены и чуть погруже-
ны основанием в почву; верхний—приподнимается на стебле на 4—8 см
над уровнем почвы. Самый нижний—ланцетный (22 см дл.,7 см шир.);
верхний—12X1,5 см.
Цветок желтый, одиночный. Внешние листочки околоцветника
в верхней трети по спинке чуть-чуть отклонены и верхние края их
наверху обращены внутрь цветка, а ниже по всей длине, начиная
с основания верхней трети, выгнуты наружу. Внутренние листочки
околоцветника на самом верху отгиба чуть-чуть обращены внутрь
цветка, т. е. форма цветка у этого вида приближается к форме цветка
Т. alberti Rgl.
Внешние листочки околоцветника (5,9 см дл., 3,9 см шир.) оваль-
ные, на вершине внезапно заостренные, по спинке, особенно внизу,,
массивные; снаружи по спинке, постепенно сужаясь (шир. до 1,5 см),
до самого верха—бордово-красные, при основании (до выс. 1,3 см)
и широко по краям ярко-лимонно-желтые, на кончике и верхней
трети листочка снаружи опушенные коротенькими ворсинками, со-
внутри на вершине лимонно-желтые, вдоль всей полосы, соответствую-
щей спинке, более оранжевые; при основании с овальным, на верши-
не заостренным коричневым пятном (0,2 см дл., 0,8 см шир.), состо-
ящим из легких штрихов, под которыми просвечивает основной фон
листочка. Через 0,4—0,6 см над темным пятнышком лежит бордово-
красное, обратно-сердцевидное пятно (1,2 см дл., 1,2 см шир.), четко
отграниченное от основного фона, на вершине зубчатое.
Внутренние листочки околоцветника (5,5 см дл., 4 см шир.) об-
ратно дельтовидные, на вершине с выемкой, в которой сидит маленький,
слабо опушенный зубчик; снаружи чисто лимонно-желтые; совнутри
такие же; к основанию их окраска как бы сгущается и они приобре-
тают оранжевый оттенок; в центральной части листочка, на выпуклом
месте над центральной жилкой, отступя 0,9 см от основания листочка,
лежит узкое, продолговатое, из чуть заметных коричневых штрихов^
пятно (1x0,3 см).
Тычинки 2,1 см дл., в два с половиной-три раза короче около-
цветника.
Тычиночные нити голые, длинные (1,1 см), к основанию расши-
ренные (0,2 см шир.), выше округлые, тонкие (0,1 см), на вершине
заостренные, лимонно-желтые.
Пыльники овальные (1x0,4 см), широкие, кремовые, на вершине
с острым кончиком.
Завязь 1,6 см выс. при 0,5 см шир., по всей длине почти ровная,
лишь к рыльцу постепенно очень незначительно сужается, при осно-
вании темно-зеленая, а к рыльцу почти светло-желтая. Рыльце светло-
желтое, широкое, шире завязи; плоское.
Коробочка белесовато-соломенная, покрыта сизым налетом, про-
долговато конической формы (4,5 см дл., 2 см шир.); без карпофора;
на вершине постепенно заостренная и переходит в короткий (0,4 см)
широкий столбик, на котором расположено светлое рыльце с лопас-
тями, немного выдающимися над столбиком.
Створки внизу продолговато округлые, наверху—заостренные,
треугольные, довольно плотные. В центре вдоль них проходит жело-
бок, окрашенный в бордово-коричневый, покрытый сизым налетом,
цвет. Окрашенная полоса расширяется к рыльцу и заостряется к осно-
ванию коробочки. От нее на плоские щечки створки отходят короткие
70
Рис. XII. Т. brachystemon Rgl. (2/3 натуральной величины).
оранжевые штрихи, особенно многочисленные в верхней четверти
коробочки, там удлиняющиеся и пересекающие всю щечку поперек.
Ребра коробочки, отделенные от створки желобком, по самому
краю окрашены в тускло-коричневый цвет.
Коробочка плохо растрескивается. Край створки кремовый, на
месте разрыва не выгибается наружу.
Внутренние перегородки кремовые, не растрескиваются глубоко
под рыльцем, в верхней части продолговато ромбовидно утолщены;
к выгибу створки сужаются.
Рубец от опавших членов околоцветника широкий, серовато-жел-
товатый, окаймлен вверху и внизу узкими коричневыми полосками.
Плодоножка утончается книзу от рубца, более серая, чем коробочка.
Семя (0,8X0,55 см) косотреугольной формы, светло-коричневое
с более светлой, ровной вокруг всего семени каймой.
В условиях Ташкента он зацветает в середине апреля.
Описанные растения Т. brachystemon, выращенные из луковиц,
полученных от В. П. Голоскокова, не совсем соответствуют описанию,
данному этому виду во „Флоре СССР“ и вышеприведенному нашему
по следующим признакам: по форме листочков околоцветника, по
опушенности всего растения, по форме тычиночных нитей. Кроме
того, во „Флоре СССР11 нет указания на красные и темные пятна на
листочках околоцветника. По сложению цветка и окраске листьев,
описанные здесь растения могут быть отнесены к Т. alberti, по рас-
положению пятен на листочках околоцветника и по их форме к Т. greigii.
Tulipa greigii R g 1. — Тюльпан Грейга (рис. 4, а) растет „на гли-
нистых склонах.—Ср. Азия Арало-Каспий (Кзыл-Орда), Сыр-Дарья,
Тянь-Шань. Эндем“ („Флора СССР11, т. IV).
Исследованные нами образцы взяты из массовых посадок Ботани-
ческого сада, Бостандыкского района, бассейна р. Чирчик с гор у
Каранкульсая, с хр. Каратау (старые сборы П. А. Гомолицкого и наши
1956 г.) С р. Малый Кебин из окрестностей селения Самсоновки
(наши сборы). Из Дарбазинского района около Столовой горы; Аксу
Джебаглинского заповедника из ущелья Фаинбулак; окрестностей
ст. Чокпар от Бочанцева и из Алма-Атинского Ботанического сада, из
Отарского района; с гор Актау от Сумарокова; Ангрена и т. д.
Луковица в диких условиях яйцевидная, в культуре почти репча-
тая, или округлая; часто с продолженными на значительную длину
чешуями, очень сильно варьирующими по консистенции: то грубыми
кожистыми, то нежными бумагообразными. Экземпляры, имеющие
продолженные чешуи, хорошо отличаются от луковиц Т. kaufmanniana.
Покровные чешуи у этого вида на вершине и при основании густо
опушены прижатыми, золотистыми, прямыми, грубыми волосками.
Отдельные волоски разбросаны б. м. густо по всей поверхности чешуи.
Стебли различной высоты. У некоторых растений, цветок откры-
вается, сидя в розетке листьев, или вместе с цветоножкой поднимается
до 40—45 см выс. от уровня почвы. Стебли толстые, мощные, часто
окрашены антоцианом в розовый или коричневато-бордовый цвет;
опушены густыми короткими, торчащими волосками. Листья, в коли-
честве 3—4—5, обычно б. м. сближенные и лишь самый верхний
несколько отстоит от остальных, но есть высокие формы с листьями
расставленными, обычно, отогнутыми, б. м. курчавыми, сизыми или
сизоватыми, с верхней стороны с бордовыми пятнами различной интен-
сивности и формы; у некоторых растений после цветения пятна исче-
зают; верхние листья с верхней стороны пушистые, реснитчатые.
Нижний лист прикреплен к стеблю на уровне почвы, широколан-
71
цетный, продолговатый, а у некоторых форм из Каратау почти округ-
лый, обычно остроконечный (от 10 до 32 см дл. и от 3 до 16 сл^шир.).
Самый верхний лист ланцетный, часто прицветниковидный (от 4 до
25 см дл. и от 1 до 4 см шир.).
Цветок одиночный, оранжевато-красный, но встречаются формы с
темно-красными, оранжевыми, желтыми, кремовыми или пестрыми
цветками. Этот вид имеет совершенную форму цветка, которая соз-
дается широким дном, отогнутыми наружу на половине длины
внешними листочками околоцветника и вертикально стоящими, как
стакан, широкими, с округлой вершиной, внутренними листочками
околоцветника. Очень хороши и элегантны цветы вида и в сложенном
состоянии.
Внешние листочки околоцветника округло ромбические, с острым
пушистым кончиком или яйцевидные, на вершине также заостренные
(от 3 до 14 см дл. и от 3 до 11 см шир.), по спинке, особенно в ниж-
ней половине, очень массивные, толстые, выпуклые наружу, по краям,
особенно на вершине, тонкие, волнистые, мелкозубчатые; у открытого
цветка на половине длины б. м. резко наружу отогнутые; снаружи по
спинке от острия до основания часто опушенные; у оранжевых, крас-
ных, темно-красных форм на спинке более темные с меняющейся
интенсивностью малинового оттенка; у оранжевых и от ярко-желтых
до бледно-лимонно-желтых и кремовых, по спинке окрашены в розо-
во-малиново-красный цвет различной силы и интенсивности. К основа-
нию листочки зеленеют или у них овалом просвечивает внутреннее
пятно. Из Каратау мы привезли луковицы, из которых выросли рас-
тения палевых, красноватых, ярких желтых и кремовых окрасок, у
которых спинка внешних листочков околоцветника не была окрашена
в красно-розовый цвет, присущий большинству светлоокрашенных
особей этого вида.
Совнутри внешние листочки околоцветника имеют тот же цвет,
что и края листочков околоцветника у данного растения снаружи,
только поверхность листочка более блестящая, чем снаружи и поэтому
его окраска внутри кажется более яркой; в нижней половине
лежит ромбическое или яйцевидное черное пятно; обычно островер-
шинное (2 см выс. X 1,5 см шир.). Есть формы, у которых основной
фон листочка на ноготке постепенно переходит в желтый и на этом
желтом, расплывчатом, большом пятне лежит несколько меньшее
черное различной интенсивности. Оно как бы окаймлено желтым,
которое нечетко отделяется от основного фона. Особенно велика
желтая нижняя часть листочка у растений, собранных в Чуйской
долине и в Чуилийских горах. Черное пятно там бледнеет, становится
коричневатым. У желтых, светло-оранжевых и кремовых форм черное
пятно на внешних листочках околоцветника претерпевает редукцию,
становясь коричневым, сероватым, овальным, часто не доходящим до
основания листочка, состоящим из легких коричневатых штрихов,
вплоть до полного их исчезновения, что мы и наблюдаем у растений
из северной части ареала и у некоторых представителей из Каратау.
У желтых, кремовых, светло-оранжевых форм над черным и желтым
пятном, обрисовывая верхние контуры желтого пятна, часто распола-
гается красное пятно. Это пятно очень меняется по форме, величине
и интенсивности, оно всегда бывает в нижней части вогнутым, несколько
сужается к вершине, часто на ней оно несколько неопределенно и у
разных форм достигает различной высоты. Встречаются растения, у
которых оно занимает всю среднюю часть листочка от желтого пятна
до пушистого кончика на вершине. Но из Каратау мы привезли рас-
72
тения, у которых не было черного пятна в нижней половине листоч-
ков, а красное пятно, четко отграниченное со всех сторон, имело форму
небольшого полукруга.
Внутренние листочки околоцветника обратно-продолговато-яйце-
видные или обратно-округло-дельтовидные, на вершине внезапно пере-
ходящие в острый пушистый кончик; по спинке, особенно в нижней
половине, почти такие же массивные, как и внешние (от 4 до 16 см
дл. и от 3 до 10 см шир.). Основная окраска их такая же, как и у
внешних листочков околоцветника, только более блестящая, но желтые
формы несколько более матовые, чем оранжевые и красные. При
основании снаружи они часто с желтым ромбическим или веерообраз-
ным, нечетко отграниченным пятном; у красных форм при основании
на желтом фоне или без него лежит треугольное или ромбическое
пятнышко из лиловых штрихов. У желтых форм над желтым же
большим или меньшим пятном бывает как бы сияние из красно-ма-
линовых штрихов. Это сияние состоит из штрихов различной длины,
расположенных с различной густотой, имеет форму полукруга или на
вершине образует острый угол. Центральная жилка в средней части
окрашена в красный цвет. С внутренней поверхности эти листочки
околоцветника окрашены так же, как и снаружи, только темное пятно
в нижней части листочка значительно крупнее, овальное, продолго-
ватое, почти круглое или обратно-неправильно-ромбическое. У светлых
форм оно часто очень ослаблено, состоит из коричневых штрихов,
образующих длинное, продолговатое пятнышко в самом центре нижней
половины листочка, обычно же оно очень ослаблено в нижней части,
а на вершине в виде грибной шляпки, образованной отдельными
штрихами с размытой в основании ножкой. Вообще, у желтых и пале-
во-оранжевых форм нижняя часть пятна часто обесцвечена. У них над
желтой каймой пятна лежит красный полукруг различной ширины и
интенсивности. У форм из хр. Каратау красный полукруг небольшой
я очень четкий, есть растения с оранжевыми, желтыми или кремовыми
цветками, совершенно без красных пятен внутри цветка.
Тычинки с сухими пыльниками чуть длиннее, равны или немного
короче завязи; в 3—4 раза короче околоцветника. Тычиночные нити
голые, короткие, обычно по форме широкотреугольные (0,6X0,4 см),
но есть и более узкие (0,6X0,25 см), черные или желтые, а на вершине
чернеющие.
Пыльники желтые, фиолетовые, сероватые, очень широкие
(2,5 см дл.х0,5 см шир.), открываются одновременно и в сухом виде
слабо штопорообразно свернуты.
Завязь бледно-зеленая, обычно в виде ровного по всей длине
цилиндра, 3,3х0,6 см дл.
Рыльце большое, значительно шире завязи, светло-желтое. У
некоторых форм завязь под рыльцем постепенно несколько сужается.
Коробочки соломенно-желтые с коричневатым оттенком до 11 см
дл. и 4 см шир., но бывают и меньших размеров; часто встречаются
тетрамерные, а иногда и димерные формы, продолговатые, треугольно
цилиндрические, на вершине и основании туповатые. Карпофор корот-
кий (до 1 см выс.), обычно толстый (2,2 см), отделен от верхней
плодущей части створок мятым желобком, несколько приподнимаю-
щимся в средней части створки. С боков на карпофоре в виде толстых
гребней входят продолжения боковых швов коробочки. Верхняя
бесплодная часть створки лишь незначительно сужена против всей
створки, но иногда встречаются коробочки, у которых она вытянута и
73
сужена в столбик. Такие коробочки очень сходны с коробочками
F. kaufmanniana и отличаются лишь более коротким карпофором,
Коробочка открывается широкими клиновидными щелями, сильно
расходящимися, особенно в области верхней бесплодной части. Ширина
щели образуется за счет выворачивания наружу боковых плоскостей
ребра, а щель остается, особенно в плодущей части коробочки, узкой,
лишь на вершине створки отклоняются друг от друга.
Внутренние перегородки глубоко (на 5 см) растрескиваются, часто
ниже бесплодной.верхней части коробочки. Интересна форма внутрен-
ней перегородки в верхней бесплодной части коробочки. Она бывает
равной ширине рыльца и клиновидно сужается к плодущей части, или,
начинаясь во всю ширь рыльца, на середине длины она еще больше
расширяется и, круто сужаясь, узкой лентой доходит до общей пере-
городки в средней плодущей части коробочки, внутренняя перегородка
сразу от рыльца узкая, и, не расширяясь, ровной узкой полосой
спускается к плодущей части. Глубже и шире всех растрескиваются
коробочки последнего типа.
Створки коробочки продолговатые, тупо заостренные на обоих
концах, на месте прикрепления внутренней перегородки вдоль створки
проходит мелкий широкий желобок, который часто бывает более
углублен при основании и на вершине коробочки перед бесплодной
частью. Желобок часто окрашен в коричневый цвет, иногда лишь в-
верхней части, или вдоль всей длины, но темнее вверху; иногда—лишь
вверху и внизу, а середина остается неокрашенной. Щечки створки
выпукло плоские, очень грубые, часто по всей длине мелкоморщини-
стые, иногда более сильно морщинистые лишь в верхней части; вверху
иногда поперечно коричнево полосатые; у ребра более коричневатые,
самый край ребра узко коричневый, отчленен от щечек желобком, в-
котором четко проходит разграничивающая линия. Широкие края
створок на месте бывшего ребра вывернуты наружу, палевые, конси-
стенции фетра.
Под рыльцем сходятся коричневые полосы краев створок. Корич-
невая полоса центрального желобка и вся бесплодная часть мелко
поперечно морщинисты.
Рыльце мощное, соломенно-желтое, отклоненное наружу.
Широкий рубец от опавших членов околоцветника (0,4 см) имеет
форму фистонов и окрашен в коричневый цвет.
Плодоножка мощная серо-соломенного цвета; у наиболее крупных
растений многогранная, до 16 см дл.
Семена светло-коричневые, косовато-яйцевидные (1,3X0,7 см шир.),
с широкой загнутой вниз каймой и более светлым в виде запятой
зародышем. Как коробочки, так и семена могут быть значительно
мельче вышеописанных, а иногда и крупнее.
В условиях Ташкента он зацветает в начале второй декады
апреля.
Tulipa mogoltavica М. Pop. et Vved. — Тюльпан моголтавский
(рис. XIII) распространен „на каменистых склонах — Ср. Азия: Тянь-
Шань (Моголтау). Эндем“. (Флора СССР, т. IV).
Описанные растения привезены из Моголтау, собраны с лужайки
среди камней на вершине боковых отрогов хребта. Кроме того, в
Ангрене среди типичных Т. greigii также найдены формы с призна-
ками Т. mogoltavica (окрестности кишлака Сангинак).
Луковицы яйцевидные, до 5 см в диам. (ангренские формы), из.
Моголтау они мельче, с кожистыми, черно-коричневыми чешуями.
74
Рис. XIII. Т. mogoltavica М. Pop. et Vved. (s/4 натуральной
величины).
Покровная чешуя с внутренней поверхности покрыта прижатым опу-
шением, более густым на вершине чешуи и при ее основании.
Стебель вместе с цветоножкой 19 см выс. от поверхности почвы.
Он в верхней части вместе с цветоножкой, окрашенной антоцианом,
опушен короткими волосками, переходящими с цветоножки на внешнюю
сторону наружного круга листочков околоцветника.
Листьев 4, начинаются у самой почвы, два нижних сближены, а
два верхних расставлены; нижние—отогнуты; верхний—торчащий, не
превышают цветы, покрыты сизым налетом и с верхней стороны с
многочисленными бордовыми пятнами,б. м. пушистые, по краю сильно
курчавые мелкими волнами, хрящевато окаймленные, реснитчатые.
Есть формы, у которых верхние листья опушены с обеих сторон.
Нижний лист ланцетный, 15X5,7 см, лежит на почве. Верхний —
линейный (10X1 см}.
Цветок одиночный, ярко-красный, по форме похож на цветок
Т. greigii с очень слабым ароматом.
Внешние листочки околоцветника (3—5—10 см дл. и 1,8—3—6 см
шир.) острые, неправильно ромбические, постепенно переходящие в
острый пушистый кончик, отогнутые на половине длины; с внешней
стороны листочки околоцветника опушены по всей поверхности, но
гуще на вершине и основании; снаружи оранжевато-красные, по спинке
малиновые с сизым налетом, в нижней половине просвечивает овальное,
чуть-чуть зеленоватое, черное пятно внутренней стороны листочка. С
внутренней поверхности внешние листочки околоцветника оранжево-
красные, у разных экземпляров эта окраска различной интенсивности,,
в нижней половине с овальным черным пятном, поднимающимся на
одну треть—одну четверть листочка (у листочков в 5 см дл. оно зани-
мает площадь 1,6X1 см}. Это пятно часто бывает кругом окаймлено
неширокой, от 1 до 1,5 мм шир., бледно-желтой каймой. Кайма чаще
всего исчезает на вершине пятна и тогда создается впечатление, что
ее совершенно нет; часто она остается в виде кремового расплывча-
того фона с боков при основании пятна, но есть формы из Моголтау,
у которых совершенно нет каймы.
Внутренние листочки околоцветника обратно яйцевидные или
обратно-округло-неправильно-ромбические, торчащие, между собой
сближенные (3,1—5,1 — 10,2 см дл. и 1,9; 3,1—6,2 см шир.), с неболь-
шим острым опушенным кончиком, продолжающим центральную жил-
ку; снаружи блестяще-оранжево-красные с большим зеленовато-жел-
тым, округло ромбическим, на вершине с зубчатыми краями, пятном.
На пятне в центральной части из б. м. интенсивных штрихов возникает
лилово треугольное или ромбическое пятно. С внутренней поверхности
эти листочки околоцветника такого же блестящего оранжево-красного
цвета, как и снаружи, лишь над пятном у них появляется малиновый
оттенок. На ноготке лежит большое, овальное, черное пятно, зани-
мающее до одной трети или одной четверти листочка, но всегда более
длинное, чем на внешних листочках; иногда вершина пятна бывает
плоской, а иногда с выемкой. Пятно часто окружено четко отграни-
ченной желтой полосой, более широкой, чем у внешних листочков,
до 2X2,5 мм у пятна размером 2X0,9 см (без желтого). В двух нижних
третях пятно сплошное черное и занимает всю нижнюю часть листочка.
Тычиночные нити голые, черные, лишь острый кончик под пыль-
ником красный, плоско треугольные, обращенные вершиной к пыль-
нику; 0,65X0,3 см в расширенной части. Тычинки в два с половиной-
три раза короче околоцветника, равны или короче завязи.
75
Пыльники черные или черно-фиолетовые (0,9 см дл. X 0,25 см
шир.), в один завиток штопора скрученные. Пыльца темно-коричнево-
фиолетовая.
Завязь (1,6 см дл., 0,5 см шир.) к рыльцу постепенно сужена,
колонновидная, бледно-зеленовато-желтая, под лопастями рыльца
красноватая.
Рыльце довольно большое, желто-красноватое, на завязь спуска-
ющееся, шире суженной верхней части завязи.
Коробочка (4,2 см дл., 2,2 см шир.) продолговато треугольная,
на вершине б. м. постоянно оттянута в острый носик, внизу б. м.
внезапно суженная у маленького карпофора над рубцом от опавших
членов околоцветника. Карпофор (до 0,3 см выс., 0,7 см шир.) со-
ломенно-белый, сужен к плодоножке, расширен к легкой перетяжке
у перехода в створку; на него гребнями заходят центральный жело-
бок створки и ребра, по которым она раскрывается. На вершине до-
вольно тонкий столбик, вытянутый (до 1 см дл. и 0,4 см шир.) в
бесплодную часть створки, на котором сидит небольшое светло-корич-
невое рыльце. Лопасти рыльца по средней линии сильно вытянуты
наружу. Коробочка трескается по ребрам, но не расходится и прак-
тически закрыта. Сквозь отверстие в столбике видно, что лопнула
центральная перегородка коробочки, но насколько глубоко неиз-
вестно.
Створка коробочки промежуточной формы между овальной и яй-
цевидной, так как к карпофору она несколько расширена, а к стол-
бику незначительно сужена.
В центре створки проходит довольно круто углубленный жело-
бок, ребром выходящий на карпофор и уплощающийся на бесплодной
части створки, играющей роль столбика. Он ярко окрашен, густо
расположенными елочкой ярко-коричневыми штрихами, в центре сли-
вающимися, неровной длины, частично заходящими на щечки створки.
Эти штрихи исчезают в нижней четверти желобка, там остается лишь
слабо окрашенная коричневатая жилка.
Щечки створки сильно выпуклые, плотные, довольно тонкие, проз-
рачно матовые, гладкие или лишь наверху чуть-чуть поперечно мор-
щинистые, серовато-соломенно-белые (сероватость получается из-за
просвечивающих семян), у шва ярко-коричнево окрашенные, в желоб-
ке, отделяющем щечки от ребер, с легким узором из переплетения
светло-коричневых жилок. Края створок друг от друга почти не от-
ходят. К вершине коробочки окраска ребер темнеет и сливается с
темной окраской центрального желобка, так что столбик очень тем-
ный, иногда окрашен тремя темными полосками, с просвечивающим
светлым фоном, на каждой из створок.
Рубец от опавших листочков околоцветника светло-коричневый,
2 мм шир., к плодоножке сужающийся.
Плодоножка до 10 см, короткая, светло-соломенно-белая.
Семя косо-округло-треугольное, коричневое, с широкой тонкой
каймой, слегка направленной вниз и более светлым прямым заро-
дышем.
В условиях Ташкента он зацветает в начале второй декады апреля.
Tulipa micheliana Н о о g.—Тюльпан Микели (рис. XIV и 4, б)
произрастает „на каменистых и глинистых склонах,—Ср. Азия: Горн.
Туркм., Пам.-Алай. Общее распр. Иранск.“ („Флора СССР11, т. IV).
Исследованные нами образцы получены со станции Уратюбе, Дра-
гомирово, Бахарден; из горной Туркмении (пос. Тумановского); из
Аман-Кутана (Самаркандские горы), от Ашхабадского ботанического
76
Рис. XIV. Т. micheliana Hoog. (2/3 натуральной величины).
сада, из Западного Копет-Дага с хр. Корен-Даг; из Нуратинского
хребта. Кроме того, мы выращивали растения Т. micheliana из окрест-
ностей Тегерана (Иран). Потомки большинства растений из этих
сборов выращивались из семян.
Луковицы яйцевидные от 2 до 8 см в диам., с кожистыми, черно-
бурыми, с внутренней поверхности прижато волосистыми, особенно у
верхушки и основания, часто б. м. продолженными покровными че-
шуями.
Стебель вместе с цветоножкой до 35 см выс. от уровня почвы,
пушистый.
Листья 3—5, б. м. расставленные по стеблю (нижний лист отхо-
дит от стебля на уровне почвы); сизые, по верхней поверхности с
бордовыми сплошными или прерванными продольными полосами раз-
личной интенсивности, очень яркими в начале весны и ослабевающей
окраски к концу вегетации; иногда бывают совсем без полос, б. м.
пушистые, по краю хрящевато окаймленные, реснитчатые; через один-
два дня после начала цветения, как правило, не превышающие цве-
ток; самый нижний лист широко ланцетный, до 30 см дл. и 20 см шир.
Второй — часто бывает на 1—2 см длиннее первого. Самый верхний
ланцетный, до 21 см дл., 7,5 см шир.
Цветы одиночные от ярко-красных до темно-малиновых, очень
крупные, красивой формы, близкой к цветкам Т. greigii, но совершен-
но отличающиеся от них общей гаммой малиновых оттенков и бросаю-
щимся в глаза клинообразным, черным, окаймленным светлой поло-
сой пятном с внешней стороны внутренних листочков околоцветника.
Цветы без запаха.
Внешние листочки околоцветника ромбические; продолговато ром-
бические (до 16 см %л. и 9 см шир.), на вершине острые часто с
пушистым кончиком, достигающим 1 см дл. Опушение с кончика пе-
реходит на жилку, по которой спускается до основания и переходит
на края отгиба. Есть растения, у которых опушены только верхние
части листочка, или весь листочек от вершины до основания, особен-
но густо на вершине спинки и отгиба и на нижней толстой части
ноготка. Внешние листочки околоцветника в нижней части часто по
краю реснитчатые, при основании массивные, выпуклые, на вершине
несколько наружу отклоненные с краями нежными, волнистыми, мел-
козубчатыми; от оранжево-красной до темно-малиновой окраски сна-
ружи на спинке и широко, оставляя свободным лишь узкий край, си-
зые; на нижней толстой части с продолговатым (до х/4 дл. сегмента),
нечетко отграниченным зеленым пятном, на котором как будто
просвечивает лилово-черное внутреннее пятно. С внутренней поверх-
ности той же окраски, что и снаружи, но чистой без сизого налета
и очень блестящей; в нижней части сегмента по утолщенному месту
от одной трети до половины длины листочка расположено очень
большое, черное с лиловатым оттенком, бледно-желто или бело окай-
мленное, а иногда и без окаймления, пятно разной формы: почти
прямоугольное, продолговатое, овальное или остроугольное; на вер-
шине заостренное или тупое, или усеченное, но с тремя зубчиками;
всегда четко отграниченное — кайма неширокая и достигает значи-
тельной ширины лишь в верхней части пятна. У листочка в 13 см
дл., 7,2 см шир. продолговатое пятно, на вершине усеченное, с тре-
мя небольшими зубчиками, с черной частью в 5,8X1,8 см; кайма с
боков 0,15 см; на вершине также трехзубчатая, 0,9 см шир.
Внутренние листочки околоцветника обратно дельтовидные или
обратно яйцевидные, до 15 m дл. и 9 см шир.; на вершине внезапно
77
заостренные, часто в довольно большой, до 1 см, пушистый кончик,
который является продолжением спинки листочка околоцветника, воз-
вышающейся над отгибом. Опушение с него часто переходит на спин-
ку, центральную жилку и прилегающую к нему верхнюю часть ли-
сточка. Снаружи и внутри они такого же цвета, что и внешние
листочки. Но внутри более блестящие, почти лакированные. На мас-
сивном основании листочка снаружи лежит неправильно продолговато
ромбическое, островершинное, бледно-желтое, вернее, кремово-зеле-
новатое пятно. У листочка в 13,5 см дл. и 6,7 см шир. оно доходит
до 7 см выс. В центральной части, вдоль центральной жилки оно про-
дольное, темно-фиолетовое. Эта фиолетовость может покрывать раз-
личную площадь пятна, иногда закрывая его сплошь, или частично,
оставляя лишь легкое светлое сияние вокруг. С внутренней поверх-
ности на том же месте листочка лежит длинное продолговатое пятно,
острое или усеченное, но зубчатое на вершине, окаймленное светло-
желтой полосой, часто зубчатой на вершине, ровной в верхней поло-
вине пятна и расширяющейся в нижней. Иногда у широких темных
пятен ее не бывает в нижней половине пятна, так как темное пятно
занимает всю ширину нижней части листочка. Иногда вокруг
всего пятна нет никакой светлой полосы. У вышеописанного листочка
с внутренней поверхности пятно достигает 8 см выс.; из них черная
часть занимает площадь 7x1,3 см; светлая кайма на вершине в 1 см
шир., а с боков пятно всего 0,2 см шир.
Тычинки в 2,5—4 раза короче листочков околоцветника, но не-
много выше завязи с очень подвижными пыльниками. Тычиночные
нити треугольные, короткие, в нижней части плоские, широкие
(0,3 х 0,4 см в расширенной части), голые, черные, при основании и
на самом кончике желтоватые или красноватые. Пыльники фиолето-
вые и, очень редко, желтые, большие и широкие, в открытом состоя-
нии продолговатые, у нашего растения 2,6 см дл. х0,5 см шир.
Пыльца коричневая различных оттенков, иногда лилового, вплоть
до желтого. Завязь массивная, по всей длине ровная, 3,4 см дл.,
0,7 см шир., темно-сизо-зеленая. Рыльце большое, значительно шире
завязи, более зеленое, чем завязь, сосочки рыльца желтые.
Коробочка (9 см дл., 3 см шир.) продолговато треугольная с ту-
пым основанием и вершиной. Нижняя часть коробочки внезапно за-
кругленно переходит в карпофор, имеющий 0,5 см выс. и 1,5 см шир.
Карпофор сужается к коробочке и расширяется к рубцу, и отделяется
от створок коробочки поперечной, почти прямой морщинистой пере-
тяжкой.
Верхняя бесплодная часть коробочки очень большая, 3 см дл.,
2,8 см шир. в основании и 1,5 см шир. под рыльцем.
Рыльце сидячее, очень большое (0,5 см выс., 1,3 см шир.), серо-
коричневое, плотно сросшееся со створками коробочки, на вершине
с выпуклой складкой в 0,2 см выс.; сильно выгнутое наружу, почти
под прямым углом к створке.
Коробочка открывается вдоль всей длины узкой продольной щелью,
всего 0,35 см шир.; края ребра, по которому расходятся створки ко-
робочки, выгнуты наружу. Кроме того, довольно сильно отклонены
друг от друга верхние бесплодные части створок, образующие широ-
кий раструб (2 см шир.), через который высыпаются семена. Внут-
ренние перегородки расходятся только на глубину верхней бесплод-
ной части, т. е. на 3 см. Интересен след срастания перегородок в этой
части. Ширина срастания под рыльцем была 1,4 см и постепенно су-
жалась вниз, достигая у изгиба общей перегородки 0,2 см. Поверх-
78
ность разрыва по пыльцепроводниковой ткани кремовая, матовая, с
группой волокнистых желтоватых тяжей в центре, особенно явствен-
но проступающих в месте перехода рыльца в бесплодную верхнюю
часть створки коробочки.
Створки коробочки соломенно-желтые, с параллельными краями,
очень длинные, тупые на обоих концах. Вдоль створки в центре про-
ходит пологий желобок светло-коричневый в верхних трех четвертях
•с темно-коричневыми жилками, которые в нижней части желобка
•сливаются вместе, а в верхней трети раздваиваются и между ними
располагается масса более тонких и светлых анастамозов.
Щечки створки соломенно-желтые, плоско выпуклые, гладкие с
проступающей полосатостью, но не из коричневых полосок, как у
других видов, а из чередующихся матовых и прозрачных соломенно-
желтого цвета. Желобок у ребра глубокий, округлый, заполнен сетью
из светло-коричневых жилок. Ребра створки тускло-коричневые. Шов
разрыва очень широкий, 0,7 см шир., палевый, замшевый, сильно вы-
гнут наружу. Тускло-коричневая окраска ребра створки, несколько
расширяясь, поднимается до самого рыльца и имеет одинаковый с
ним цвет, но не переходит на бесплодную, часть створки, сплошь
покрытую густой сетью ячеек из переплетающихся светло-коричневых
жилок, которые уплощаются и поперечно вытягиваются в ее нижней
половине.
Рубец от опавших членов околоцветника имеет форму коричне-
вых фестонов и выступает над расширяющимся к нему основанием
карпофора и также расширяющейся, шестигранной, у крупных экзем-
пляров, плодоножкой.
Плодоножка массивная, соломенно-белая, светлее коробочки, до
15 см дл.
Семя широко яйцевидное, коричневое (1,1 см дл., 0,9 см шир.),
с широкой плоской каймой и более светлым зародышем.
В условиях Ташкента зацветает во второй декаде апреля.
Tulipa karabachensis A. Grossh.— Тюльпан карабахский рас-
пространен в Азербайджане, Автономной области Нагорного Карабаха,
Гадрутском районе. Между пастбищами Кагныюрд и г. Зиарат. На
южных каменистых склонах Я- И. Кирпичников собрал этот вид в
цвету 29.V 1948 г. и высушил. По А. А. Гроссгейму (1949), он растет
только в Карабахе в среднем и верхнем поясах. В лесах по опушкам.
Цветет в апреле-мае. Эндем.
Луковица до 2,5 см в диам. почти шаровидная, покрыта массой
полусгнивших чешуй коричневого и светло-коричневого цвета. По-
кровные чешуи с внутренней поверхности покрыты редкими прямыми,
жесткими светлыми волосками, несколько сгущающимися к основанию
и вершине луковицы. Луковица с тремя запасающими чешуями.
Стебель от поверхности почвы до 40 см выс. Луковица лежит
под землей на глубине от 15 до 22 см. Облиствленная часть стебля
занимает 12—15 см, таким образом, цветоножка, обычно короткая,
достигает 7—9 см дл. Стебель и цветоножка голые.
Листьев 4, отклоненные, вверх направленные, не превышающие
цветок, по стеблю расставленные, голые, ланцетные. Самый нижний
(13,5 см дл. и 3,5 см шир.) в нижней части килеватый, широко лан-
цетный.
Верхний (7 см дл., 1,3 см шир.) также ланцетный, но на вершине
оттянут в длинный (1,5 см) узкий зубец. Листья этого тюльпана ко-
роткие и, относительно длины, широкие, очень оригинальны и состав-
ляют видовую особенность Т. karabachensis.
79
Цветок один. У имеющихся в гербарии АН СССР растений труд-
но судить о его форме, кроме того, там хранятся только светло-жел-
тые экземпляры этого вида, а А. А. Гроссгейм пишет, что изредка
встречаются и красные, но как те, так и другие не имеют пятна при
основании листочков околоцветника. О разнице в окраске листочков
околоцветника снаружи и с внутренней поверхности невозможно ска-
зать ничего определенного, однако, ясно, что в нижней половине они
более ярко-желтые, чем на вершине. Наружные (6,8 см дл., 3,2 см
шир.) узко округло ромбической формы, на вершине туповатые или
оттянутые в острый, окрашенный светло-лиловым, кончик.
Внутренние листочки околоцветника (6,5 см дл. и 3,2 см шир.)
узко-обратно-яйцевидной или округло-неправильно-ромбической фор-
мы, часто на вершине с выемкой, в которой находится небольшой
зубчик; по окраске они сходны с внешними, но более желтые при
основании.
Тычиночные нити длинные (до 1,5 см), довольно тонкие, к осно-
ванию слабо расширяющиеся (до 0,12 см), часто более желтые, чем
листочки околоцветника, иногда белые.
Пыльники желтые, в. сухом состоянии короткие (0,7X0,2 см). Ты-
чинки возвышаются над завязью.
Завязь длинная (2X0,3 см), узкая к рыльцу, постепенно перехо-
дящая в короткий более светлый, чем завязь, столбик.
Рыльце довольно широкое (0,4 см), светлоокрашенное, его лопасти
слегка нависают над столбиком.
Tulipa eichleri Rgl.— Тюльпан эйхлера (рис. 4, в) произрастает
„на сухих склонах и в посевах.— Кавказ: Вост. Закавк. Общ. распр.
Иранск“. („Флора СССР“, т. IV).
Исследованные растения получены от Б. В. Сердюкова из Тби-
лисского ботанического сада в виде луковиц и семян.
Луковицы яйцевидные до 4 см в диам., очень похожие на луко-
вицы культурных тюльпанов, с кожистыми черно-бурыми, б. м. про-
долговатыми, с внутренней стороны, особенно у верхушки и осно-
вания, волосистыми покровными чешу ям и.
Растения до 39 см выс. от уровня почвы. Стебель в своей над-
земной части вместе с цветоножкой очень пушистый, зеленый.
Листья в количестве 3—4, б. м. отогнутые, расставленные, сизые,
пушистые, по краю хрящевато окаймленные, реснитчатые, мелко
курчавые.
Нижний лист ланцетный, по краю волнистый (19 см дл.Х^,5 см
шир.), отходит от стебля на уровне почвы. Верхний — почти линей-
ный, по краю круто мелко курчавый (10,5 см дл.Х1,1 см шир.).
Цветок крупный, красивый, бокаловидный, иногда широко-коло-
кольчатый, в верхней трети как бы перехвачен; весь цветок цвета
краплака среднего, без запаха, дно цветка плоское.
Внешние листочки околоцветника (7 см дл.ХЗ,5 см шир.) широ-
коланцетные, наверху постепенно заостренные в широкий кончик,
снаружи светло-бордовые, покрытые сизым налетом, при основании с
бледно-зеленым, трехвершинным пятном, занимающим всю площадь
ноготка до 1,6 см выс. С внутренней поверхности они блестящие,
светло-бордовые (краплак средний), при основании с овальным, черно-
лиловым или черно-коричневым пятном (1,7 см выс. X 0,8 см шир.),
окаймленные неширокой до 0,2 см, желтой полосой. В нижней поло-
вине пятна желтая полоса покрывает весь край основания листочка.
Внутренние листочки околоцветника (6,6 см дл., 3 см шир.) об-
ратно-продолговато-ланцетные, на вершине б. м. внезапно заостренные
80
в пушистый кончик, снаружи и с внутренней поверхности блестяще-
светло-бордовые (краплак средний); но, снаружи, вдоль центральной
жилки, иногда с чуть-чуть малиновым оттенком, снаружи при осно-
вании с острым ромбическим, трехвершинным, зеленовато-желтоватым
пятном, до 1,8 см выс.; с внутренней поверхности на ноготке лежит
ромбическое черно-лиловое или черно-коричневое пятно, нижней по-
ловиной занимающее всю ширину основания листочка. Пятно окай-
млено сверху, четко отграниченной от основного фона листочка,
чуть-чуть зубчатой полосой и достигает 1,8 см выс., при кайме
0,15 см.
Тычинки в 2,5 см дл., в два, два с половиной раза короче ли-
сточков околоцветника и немного короче завязи. Тычиночные нити
черные (0,9x0,25 см шир.), к основанию расширенные, к вершине
постепенно суженные. Их основание и острый кончик под пыльниками
белые. Пыльники черные с лиловым оттенком, иногда желтые
(1,5 см дл., 0,3 см шир.), сразу открывающиеся.
Завязь (2,7 см выс. X 0,5 см шир.) ровная по всей длине, т. е.
без перетяжки под рыльцем, светло-зеленая.
Рыльце вместе с сосочками кремовое, очень широкое, больше
и значительно шире завязи.
Коробочка оранжевато-соломенного цвета (до 9,5 см дл. при
2,4 см шир.), очень напоминает по консистенции, окраске и форме
коробочки культурных тюльпанов из группы простых, ранних, а из
дикарей ближе всего подходит к коробочке Т. schrenkii.
Коробочка узкая, длинная, ровная по всей длине и заостренная
с обоих концов, сидит на довольно длинном карпофоре. Карпофор
достигает длины в остром зубце, входящем в створку, 2 см, а у
ребер коробочки он короче — 1,5 см.
У рубца от опавших членов околоцветника его ширина 0,9 см,
а у створок 1,3 см. Он гладкой окраски, светлее верхней части ко-
робочки, но темнее нижних частей щечек створок.
На вершине коробочка тупо заострена. Под самым рыльцем
створка с коротким перехватом, выше которого она вытянута в стол-
бик до 0,4 см выс., с которым срослись мощные, выгнутые наружу,
темно-коричневые остатки рыльца.
Створки коробочки почти прямоугольной, заостренной с обеих
сторон, формы. Через всю створку в центре вдоль ее длины прохо-
дит пологий желобок, иногда этот желобок совсем не углублен, а
заполнен широкой (0,3 см) полосой оранжеватой, продольно мелко-
морщинистой ткани, а чаще, вдоль всего желобка от основания
створки до вершины щечек створок, проходит невысокий гребень
или жилка.
Щечки створок выпуклые, плотные, непрозрачные, покрытые
сверху донизу массой поперечных морщинок, более крупных и реже
расположенных в нижней части коробочки, мельчающих и сгущаю-
щихся кверху; вдоль них видны оранжевато-светло-коричневые жилки.
Верхняя заостренная часть створок покрыта морщинками, направ-
ленными в виде лучей от вершины желобка у верхнего края щечек.
Под рыльцем по столбику в центре виден короткий желобок, имею-
щий вид как бы зарубцевавшегося разрыва створки.
Край створок отделен от щечек довольно широким, но пологим
желобком, в котором сходятся поперечные жилки створок, образуя
легкую поперечную морщинистость, приподнимающуюся на край. Са-
мый край у разрыва несколько темнее, чем вся створка. Разрыв осу-
ществляется по ребру коробочки очень узкой щелью, которая расши-
6-2229 81
ряется в широкий раструб на вершине коробочки, благодаря откло-
нению наружу вместе с рыльцем верхней части створок. Края створок
вместе разрыва по ребру коробочки почти не выгибаются наружу.
Они широкие (до б/б см шир.). палевые с розовато-оранжевым
оттенком, с матово фетровой поверхностью. Интересную особенность
представляет у этого вида длинный ряд волосков-бахромок, расту-
щих у внутреннего края разрыва створок и смотрящих прямо внутрь
коробочки. У этого вида волоски-бахромки очень тонкие, извитые, а
не прямые и толстые, как у других видов.
Рубец от опавших членов околоцветника несколько толще, иног-
да уже карпофора, серовато-коричневый, до 0,15 см шир.
Плодоножка до 27 см дл., у рубца белесовато-соломенная, а не-
сколько ниже с коричневатым оттенком, равной ему толщины (1 ои),
но ниже плодоножка значительно тоньше рубца (О,5 см).
Округло-косо-треугольное семя (0,8x0,65 см), коричневое, разных
оттенков, чуть-чуть согнуто поперек, имеет вокруг неширокую, до-
вольно мягкую кайму. Более светлый, чем все семя, зародыш, относи-
тельно небольшого размера, доходит до половины длины семени.
В условиях Ташкента этот вид зацветает в конце второй декады
.апреля.
Tulipa schmidtii Fom. — тюльпан шмидта растет „на склонах и в
посевах — Кавказ: Южн. Закавк., Тал. Эндем“ („Флора СССР", т. IV).
Растения описаны по гербарным экземплярам, хранящимся в гер-
барии Ботанического сада АН СССР.
Луковица яйцевидная (2,5—5 см в диам., 5,5 см выс.) с тонкими
кожистыми светло-коричневыми и бурыми при разложении чешуями.
С внутренней стороны покровные чешуи волосисто шерстистые. Опу-
шение сгущается у основания чешуи. Луковица лежит на глубине
25 см и глубже от уровня почвы.
Стебель до 40 см выс. от уровня почвы, вместе с цветоножкой
голый. Листья высоко расставлены по стеблю, выше цветка и, в
большинстве случаев, короткой до 8 см цветоножки, которая иногда
достигает 17 см.
Листья, в количестве 6—12, отклоненные, слегка отогнутые, ки-
леватые, сизые, голые, по краю реснитчатые. Самый нижний лист уз-
коланцетный (27—30 см дл., 2—4,5 см шир.), по краю крупно волни-
стый. Верхний — узкий, почти злаковидный (17—20 см рд., 0,7 см
шир.), вверх направленный.
Цветок один, красный, образующий глубокую чашу. Наружные
листочки околоцветника красные с внутренней поверхности, при ос-
новании несут продолговатое, на вершине выемчатое, черное с жел-
той каймой пятно (5—10 см дл., 1,5—2,5 см шир.), широколанцетные
или округло ромбические, постепенно заостренные в пушистый кончик.
Внутренние — обратно-продолговато-яйцевидные, на вершине не-
сколько оттянутые и, б. м. внезапно переходящие в короткий зубчик,,
несут с внутренней поверхности на ноготке широкоромбическое, на
вершине с зубчатой выемкой, желтоокаймленное, черное пятно.
Тычинки (до 2,7 см дл.) в 2—3 раза короче околоцветника.
Нити черно-бордовые (1,1 см дл.), тонкие, к основанию расши-
ренные, к вершине постепенно утончающиеся.
Пыльники в сухом состоянии (1,5 см дл. и 0,25 см шир.) почти
равны нитям, видимо, желтые, пыльца также желтая.
Завязь немного короче тычинок (2 см дл. и 0,3 см шир.), без
заметного сужения под рыльцем, в верхней части красная. Рыльце
сидячее, назначительно шире верхней части завязи.
82
Коробочка 2—2,5 см шир., 6—7 см дл.
Tulipa tnaximowiczH Rgl.— тюльпан Максимовича (рис. XV и
4, д) произрастает „на выходах пестроцветных пород (всегда?). — Ср.
Азия: Пам.-Ал. (Вахш, Кызылсу). Эндем“ („Флора СССР", т. IV).
Исследованные нами образцы собраны В. П. Бочанцевым в Тад-
жикистане в окрестностях кишлака Сангтуда. Кроме того, мы выра-
щивали их из семян, собранных с этих маточных растений и с рас-
тений, привезенных Ф. Н. Русановым с перевала Сарбо.
Луковица яйцевидная или округлая, 1—2 см в диам., с черными
•скорлуповидными жесткими, с внутренней поверхности у верхушки
обильно волосисто шерстистыми покровными чешуями.
Стебли во время цветения 15—25 см выс. от уровня почвы. Цве-
тоножка под цветком розовато-коричневая, ниже — зеленая, голая.
Листьев 5—6—8, от стебля под острым углом отходящих пример-
но на половине высоты, полого отклоняющихся, расставленных, же-
лобчатых, некурчавых, сизоватых, по краю с красным окаймлением,
реснитчатых, линейных (злаковидных), превышающих цветок. Ниж-
ний длинный (29,5 X 2,5 см шир.) прикреплен к стеблю на высоте
3—5 см от уровня почвы, верхний тоже длинный (15,5X1 см шир.).
Цветок одиночный, красный, широко открытый, почти плоский,
звездчатый, очень изящный, без запаха.
Внешние листочки околоцветника широколанцетные, с острым
•опушенным кончиком, до 6,1 см дл. и 3,2 см шир.; снаружи по спин-
ке сизо-малиновые, к краям ало-красные, при основании зеленеющие,
•с внутренней поверхности блестяще-ало-красные; имеют четко отгра-
ниченное от основного фона, синевато-черное, обратно треугольное,
на вершине несколько выемчатое пятно, занимающее нижнюю часть
листочка, до 1,5 см выс., с боков окаймленное белыми или кремовы-
ми, к вершине пятна чуть-чуть расширяющимися, полосками.
Внутренние листочки околоцветника обратно-продолговато-яйце-
видные, с довольно крупным, внезапно оттянутым кончиком (6 см
дл. и 3,5 см шир.); снаружи блестяще-ало-красные, от основания до
высоты 1,5 см желтовато-зеленые; с внутренней поверхности также
блестяще-ало-красные; с большим сине-черным пятном, 1,5см выс.,
обратно треугольным, с выемкой на вершине, с боков окаймленным
узкими, исчезающими к основанию листочка, белыми полосками.
Тычинки 1,8 см у данного экземпляра, т. е. в 3 раза короче око-
лоцветника, но есть мелкие растения, у которых они лишь в два ра-
за короче его.
Нити тычинок (0,8X0,3 см) черные, под пыльником в узкой ча-
сти белые, плоские, в нижней четверти расширенные, к основанию
и к вершине постепенно сужающиеся. Пыльники фиолетовые; есть
формы, у которых они короче нитей. Но в описываемом растении
пыльники 1,2 см дл., 0,25 см шир. Пыльца темно-фиолетовая.
Завязь в виде колышка, выше тычинок (2,3 см дл., 0,35см шир.),
кверху чуть-чуть сужающаяся, светло-зеленая, по ребрам красно-
-фиолетовая, под рыльцем красная.
Рыльце шире завязи, со свисающими над ней красно-малиновыми
лопастями.
Коробочки (от 2 до 4,5 см дл. и от 1 до 1,4 см шир.) продолго-
ватые, очень узкие, а маленькие коробочки почти шаровидные, все
с вытянутым носиком, очень изящные. У них почти нет карпофора.
Створки внезапно сходятся к рубцу от опавших членов околоцветни-
ка. Между створками и рубцом расстояние около 2 мм. Наверху ко-
робочки суженная часть получается из постепенно заостренных бес-
83
плодных частей плодолистиков, достигающих 1 см дл., у основанию
1 см шир.
Рыльце маленькое, его серовато-коричневые лопасти сидят с бо-
ку столбика и очень легко отваливаются.
Коробочка открывается продольной, довольно широкой клиновид-
ной щелью, равномерно раскрывающейся по всей длине. Вверху ко-
робочки она значительно шире, чем при основании, но не образует
раструба, так как бесплодные части створок не отгибаются так силь-
но наружу, как у других видов. Внутренние перегородки растрески-
ваются, примерно на 2/5 (1,9 см) длины створок.
Створки коробочек соломенно-желтые, очень нежные; с формой'
сильно двояковыпуклой линзы у маленьких коробочек, а у довольно-*
крупных — боковые ребра параллельны и лишь концы заострены.
Продольная выемка на месте прикрепления на внутренней поверхности!
створки плаценты едва обозначена легким желобком одного цвета-,
со всей коробочкой.
Щечки створки плоские, слабо выпуклые, их выпуклость более
выступает вверху и при основании коробочек, изредка мелко-попе-
речно-морщинистые, но часто морщинки не имеют поперечного ха-
рактера, а представляют легкую смятость эпидермиса щечек, соло-
менно-желтые в верхней трети, а часто по ребрам розоватые. Место
разрыва ребра фетровое, слабо палевое. Край створки отделен от:
щечек четко обрисовывающимся желобком, но не выгнут наружу.
Верхняя бесплодная часть створки часто окрашена с краев в розова-
тый цвет, но вообще, она несколько желтее всей створки.
Рубец от опавших членов околоцветника светло-коричневый, не
выпуклый, достигает 0,1 мм шир.
Цветоножка тонкая, белее коробочки, до 17 см дл. Большая
высота растения достигается за счет сильно вытягивающихся междо-
узлий.
Семя коричневое, округло треугольное (0,5X0,5 см) с довольно-
широкой по отношению к величине семени плоской каймой.
В условиях Ташкента этот вид зацветает в начале второй декады
апреля.
Tulipa linifolia Rgl. — тюльпан льнолистый встречается „на ка-
менистых склонах — Ср. Азия: Пам.-Ал. (Пяндж.). Эндем“ („Флора
СССР“, т. IV).
Луковица у этого вида 1 — 2 см толщ, и 1,5—2,5 см выс., яйце-
видная, лежит на глубине от 2 до 11 см, с черными или темно-ко-
ричневыми тонко кожистыми, крепкими чешуями. Покровные чешуи
с внутренней стороны у верхушки обильно покрыты волосисто шер-
стистым опушением.
Стебель в области междоузлия очень укорочен — всего до
2,5 см выс.; цветоножка во время цветения от луковицы до верши-
ны цветка имеет максимально 27 см выс. Стебель вместе с цветонож-
кой голый.
Листья, в количестве . (3) 6—8, серповидно отогнутые, сближен-
ные, курчавые, обычно не превышающие цветок. Самый нижний
лист —8 см дл. х 0,8 см шир., верхний — от 5 см дл. до 0,5 см
шир. (при семи листьях в розетке). У более мелких растений листья
совсем узкие (0,2—0,1 см).
Цветок в виде звезды, красный, одиночный.
Листочки околоцветника (2—4 см дл., 1,2—3,5 см шир.) тупые
или с коротким острым кончиком, наружные ромбически округлые-
или ромбически продолговатые, внутренние обратно продолговатые-
84 •
Рис. XV. Т. maximowiczii Rgl. (2/3 натуральной величины).
или обратно яйцевидные, или обратно-продолговато-яйцевидные, кра-
сные, при основании с синевато-черным, по бокам белоокаймленным,
на вершине плоскоусеченным, четко отграниченным пятном.
Тычинки с сухими пыльниками в два с лишним раза короче ли-
сточков околоцветника.
Нити голые, к основанию постепенно расширяющиеся.
Пыльники серовато-желтые, в полтора раза короче нитей.
Завязь красная, бутылковидная, с очень коротким столбиком и
светлым, выдающимся над ним, рыльцем.
В условиях Ташкента он цветет в середине апреля.
Tulipa wilsoniana Hoog.—тюльпан вильсона (рис. XVI) произ-
растает „на каменистых склонах в степном поясе гор. — Ср. Азия:
горн. Турки. Эндем“. („Флора СССР“, т. IV).
Исследованные растения собраны в окрестностях поселка Прох-
ладного у подножия гор и получены из Ашхабадского ботанического
сада.
Луковица 1,5—2,5 см в диам., с черно-бурыми, твердыми, кожи-
стыми, продолженными, у верхушки обильно паутинисто опущенны-
ми, длинными, мягкими извитыми волосками, покровными оболочками.
Стебель от поверхности почвы 5—15 см выс., вместе с цветонож-
кой голый. Цветоножка ^розоватая или бордовая.
Листьев 4—5, лежат на поверхности почвы, в начале цветения
их основания находятся ниже уровня почвы, сближенные так, что
образуют розетку, сильно отогнутые, желобчатые, по краю крупно
курчавые, пурпурно окаймленные, сизые. Нижний лист узколанцет-
ный (12X1,5 см). Верхний (7X0,4 см) узко-линейно-ланцетный. Раз-
мер и форма листьев постепенно меняются. Второй лист всегда длин-
нее и несколько уже первого (14X1,2 см).
Цветок по форме похож на цветок Т. korolkovii, это двойная
чаша, но дно у нее не плоское, а клиновидное; без запаха.
Внешние листочки околоцветника (5X2,5 см) продолговато ромби-
ческие или широколанцетные, наверху с маленьким острым кончиком;
•снаружи малиново-оранжевато-красные, по спинке малиновый оттенок
переходит в лиловый, на ноготке чуть зеленеют; с внутренней поверх-
ности блестяще-бордово-красные, к основанию бордовый оттенок сгу-
щается; у основания листочка находится маленькое, коричневато-чер-
ное или черно-лиловое, расширенное кверху, на вершине с выемкой,
пятнышко (0,5X0,4 см). Края листочка по бокам пятнышка ближе
к основанию бледнеют и желтеют.
Внутренние листочки околоцветника обратно-узко-яйцевидной
формы (5x2,7 см)-, снаружи блестяще-бордово-оранжевые, к основа-
нию с лиловым оттенком, к месту прикрепления сильно темнеют, по-
чти чернеют; с внутренней поверхности блестяще-оранжево-бордовые;
к основанию с лиловым оттенком, который к пятнышку сильно сгуща-
ется; при основании с черно-лиловым, во всю ширину листочка, до
высоты 0,4 см, пятном.
Тычинки в полтора раза выше завязи и в три раза короче око-
лоцветника.
Нити конические, голые (0,65x0,2 см), в нижней трети черные,
вверху — бордовые.
Пыльники (1 Х0,15 см) узкие, желтые. Пыльца желтая.
Завязь бледно-зеленая, к основанию и под рыль цем немного сужен-
ная (1,3 см дл., 0,35 см шир.), с широким сидячим желтым рыльцем.
Коробочка (2,7 см дл. и 1,7 см шир.) коротко цилиндрическая, у
^маленьких-растений она почти шаровидная, на ве ршине и у основа-
85
ния тупая. Коробочки этого вида характерны тем, что карпофор яв-
ляется естественным продолжением их створок ниже полного сраста-
ния на ребре и не отделяется от них никаким рубцом или перетяжкой.
На вершине створки б. м. закруглены и на них сидит в виде носика
маленькое, до 1 мм выс., с выгнутыми наружу лопастями, рыльце.
Коробочка растрескивается широкой клиновидной щелью вдоль по
ребрам. Размер шели 2,5 см дл., 1,4 см шир. на вершине у рыльца.
Внутренние перегородки растрескиваются глубже верхней, расширен-
ной- части перегородки. Непосредственно под рыльцем перегородка по
ширине равна с ним, но сразу же в полости под створками она рас-
ширяется в два с ли шним раза, а ниже к плодущей части постепенно
сужается, вплоть до нерастрескивающейся части перегородки.
Створки коробочки овальные, на вершине заканчивающиеся
носиком, а при основании — рубцом от опавших членов околоцветника;
тонкие, плотные, полупрозрачные, соломенно-белые, часто с нежным
розоватым оттенком. Вдоль створки в центре на месте прикрепления
перегородки коробочек проходит неглубокий, но четко обрисованный
желобок, наверху незахватывающий бесплодную часть створки, а
внизу на 2/5 не доходящий до основания коробочки, ярко окрашен-
ный в бордово-коричневый цвет. Получается расплывчатое бордово-
коричневое пятно, которое шире под рыльцем и заострено внизу. От
него по поперечным морщинкам коробочки расходится короткое ма-
линовое сияние. На макушке коробочки под рыльцем желобка нет.
Вся верхняя часть коробочки б. м. плоская и поперечно полосатая, в
центре чуть подкрашена в серовато-малиновый цвет.
Щечки створки тонкие, плотные, но просвечивающие, плоско
выпуклые, в двух верхних третях поперечно морщинистые, соломен-
но-белые, часто с розоватым оттенком. У ребра отделены от края
желобком, в котором проступает узор из жилок створки, окрашенный
в малиновый цвет. Край створки, выгнутый наружу, около 2 мм шир.,
фетровый, розовато-палевый. Под рыльцем окраска края створки
приобретает слабый коричневый оттенок.
Рубец от опавших членов околоцветника узкий, тоньше милли-
метра, сужается по направлению к белесоватой тонкой плодоножке,
8 см дл. Междоузлия стебля после отцветания сильно вытянулись и
имеют 6,8 см дл., высота растения от уровня почвы 16 см.
Семя косо-округло-треугольное (0,65 X 0,5 см), коричневое со
светло-коричневым прямым зародышем с такой же каймой; кайма
плоская.
В условиях Ташкента зацветает в конце первой, начале второй
декады апреля.
Tulipa lehmanniana Merckl. — тюльпан лемана (рис. XVII) рас-
пространен: „опесчаненные гамады и выходы пестроцветных пород—
Ср. Аз.:Кыз.-Кум., Горн. Турки., Сыр-Дар., Аму-Дар. Общее распр.:
Иран (Сев. Афганистан)" („Флора СССР", т. IV).
Описанные растения А. Я. Бутков привез в 1930 г. из окрестно-
стей Кушки.
Луковицы яйцевидные от 1,3 до 4,5 см в диам., с очень крепкими
кожистыми, на боках сильно утолщенными, почти скорлуповидными,
черно-бурыми, длинно, до поверхности почвы, продолженными чешуя-
ми. Покровные чешуи с внутренней стороны густо шерстистые. Во-
лоски опушения курчавые.
Стебель вместе с цветоножкой от 16 до 25 см выс. от уровня
почвы, голый, покрытый сизым налетом,
Листья в количестве 4, отклоненные, отогнутые, по стеблю, на
86
Рис. XVI. Т. wilsoniana Hoog. (3/4 натуральной величины).
небольшом его отрезке, расставленные, курчавые, голые, по краю
без ресничек, сизые, не достигающие цветка. Нижний лист ланцетный
(18 см дл., 2,7 см шир.), отходит от стебля на уровне почвы. Верхний
лист линейный (8 см дл. и 0,8 см шир.).
Цветки одиночные, желтые, оранжевые, красные, сложением от-
даленно напоминающие цветок Т. greigii, но уже его и значительно
мельче.
Желтая Кушкинская форма Т. lehmanniana. Внешние листочки
околоцветника округло ромбические, на вершине с заостренным, сла-
бо пушистым кончиком с наибольшей шириной 4 см, на середине
листочка при средней длине 5,6 см. Снаружи внешний листочек около-
цветника совершенно гладкий, желтый, по широкой спинке более беле-
соватый, в нижней половине чуть просвечивает внутреннее пятно; с
внутренней поверхности ярко-желтого цвета, сгущающегося к осно-
ванию, на котором лежит овальное, на вершине с острой выемкой,
черно-коричневое по краям более светлое (2 см дл., 0,9 см шир.)
пятно.
Внутренние листочки околоцветника обратно яйцевидные (6 см
дл., 4 см шир.), с обеих сторон одинакового желтого цвета; снаружи
просвечивает внутреннее темное пятно; с внутренней поверхности к
пятну основная окраска сгущается; в нижней половине лежит про-
долговатое, к вершине слабо суженное, с выемкой на вершине (1,8 см
дл., 0,7 см шир.), черное, к краю коричневеющее, пятно.
Тычинки на 1/3 выше завязи. Тычиночные нити желтые, достига-
ют по длине половины завязи. На заостренных концах нитей под
пыльником у некоторых особей наблюдается потемнение. Пыльники и
пыльца желтые.
Завязь зеленая. Рыльце светло-желтое.
Молодые бутоны склоняются, к расцветанию выпрямляются.
Коробочка белесовато-соломенная (4,8 см дл. и 2,5 см шир.), про-
долговато овальной формы, тупо заострена на вершине и основании.
Рыльце широкое (0,6 см), низкое (0,3 см), с лиловатым оттенком,
треугольное, сидит прямо на тупом конце створки.
Карпофор гладкий, широкий (0,6 см у плодоножки и 1,3 см у
коробочки), низкий (0,3 и 0,5 см у желобка^, наплывающий на плодо-
ножку.
Створки коробочки грубые, плотные, гладкие и продолговатые.
Желобок в центре створок глубокий, но пологий, в нем часто воз-
вышается центральная жилка, образует довольно глубокую ямку на
верху в сужающейся части створки под рыльцем, на всем протяжении
окрашен так же, как и створки.
Щечки створки белесые, плотные, грубые, б. м. равномерно припод-
нимаются над поверхностью створок, но внизу над карпофором и вверху
у рыльца они как бы вспухают; по всей поверхности гладкие и на них
лишь слабо просвечивают поперечные такого же цвета, как и щечки,
жилки. Край щечки отделен от ребра коробочки желобком.
Ребро коробочки чуть-чуть окрашено в лиловый цвет.
Коробочка вверху открывается короткой, широкой, клиновидной
щелью, которая книзу суживается и образуется слабым расхождением
створок друг от друга. Края створки по разрыву довольно тонкие с
матовой, слегка розоватой поверхностью и редкими, тонкими, длин-
ными бахромками.
Расхождение внутренних перегородок происходит по округло
ромбической фигуре расширения пыльцепроводниковой ткани, которая,
начинаясь вверху во всю ширину рыльца, постепенно расширяется, а
87
затем сужается к перегибу в перегородке и спускается ниже толстым
равномерным тяжом. '
Плодоножка обычно ярко окрашена антоцианом в розовый или
коричневый цвет, но всегда темнее коробочки. Рубец от опавших
членов околоцветника почти скрыт наплывом карпофора.
Семя темно-коричневое, остроугольное (0,9 X 0,55 см) с длинным
изогнутым зародышем, просвечивающим с обеих сторон, с узкой, на-
правленной вниз, каймой.
В условиях Ташкента этот вид зацветает в первой декаде апреля.
Tulipa borszczowii Rgl.—Тюльпан борщова (рис. XVIII и 4. ж)
растет,, в песчаных и песчано-глинистых пустынях,—Ср. Азия: Арал-
Касп., Кыз-Кум. (с. ч.). Эндем. Описан из Приаральских Каракумов"
(„Флора СССР", т. IV).
Описываемые образцы получены из Казахстана с песков около
р. Сыр-Дарьи у Арыси.
Луковицы яйцевидные, от 2—4 см в диам. с очень твердыми,
кожистыми, сильно, до поверхности земли, продолженными, черно-
бурыми, с внутренней стороны густо шерстистым и покровными чешуями.
Стебель до 20 см выс. над уровнем почвы вместе с цветоножкой,
сизый, голый; очень редко встречаются растения, у которых он чуть-
чуть подкрашен антоцианом; листьев 3, ланцетные, сизые, по краю
сильно волнистые, бело-хрящевато-окаймленные, гладкие или с ред-
кими ресничками (Сыр-Дарья), отогнутые, расставленные, быстро
кверху уменьшающиеся (верхний почти как прицветник), достигающие
цветок. Нижний лист прикреплен к стеблю на уровне почвы (18 см
дл., 4 см шир.). Верхний лист 7 см дл., 0,8 см шир.
Цветок одиночный, желтый, оранжевый или оранжево-красный
(с песков у Сыр-Дарьи около Арыси все растения имели цветы оран-
жево-красные, а со станции Тюря-Там — желтые и, как отклонение,
•оранжевые и оранжево-красные. По сложению приближается к цветку
Т. greigii, но внешние листочки околоцветника у него не отгибаются
поперек листочка наружу, а торчат вверх.
Описание цветка оранжево-красной формы. Внешние листочки
околоцветника (6 см дл., 4,5 см шир.) округло ромбические, на вер-
шине оттянутые в коротко пушистый кончик, по краю отгиба зубча-
тые, в нижней трети согнуты почти под прямым углом к цветоножке,
выше плоско-прямые и лишь самый кончик отогнут наружу; снаружи
оранжево-красные, в нижней половине более оранжевые, наверху
красные, по спинке с сизым налетом, при основании с просвечиваю-
щим овальным внутренним пятном, по которому (в нижней половине
несколько шире) проходят лилово-коричневые негустые штрихи.
Внутренние листочки околоцветника (7,3 см дл., 4,6 см шир.)
обратно-продолговато-дельтовидные, на вершине чуть-чуть округлые,
над спинкой с выемкой, в которой расположен маленький зубчик,
оранжево-красного цвета, который к ноготку слабеет, у пятна почти
оранжевые. До высоты 1,9 см от основания лежит во всю ширину
листочка лилово-коричневое, неправильно ромбическое пятно.
С внутренней поверхности листочек такого же цвета, как и сна-
ружи, только вдоль центральной жилки на нем появляется малиновый
оттенок; краснота спускается значительно ниже, чем снаружи и лишь с
боков над самым пятном основной фон листочка приобретает оранже-
вый цвет; пятно черное с легким лиловым оттенком, ромбическое, во
всю ширину листочка и до высоты 2 см.
Тычинки черно-фиолетовые, 1,9 см выс., т. е. в два с половиной-
три раза короче околоцветника.
38
Рис. XVII. Т. lehmanniana Merc к 1. (3/4 натуральной величины).
Нити голые, черно-фиолетовые, у основания светлеющие (1,1 см
дл.), к основанию расширены (0,2 см шир.), на вершине заостренные.
Пыльники также черно-фиолетовые, продолговатые (0,8 см дл.), до-
вольно узкие (0,3 см шир.).
Завязь ровная по всей длине, голубовато-зеленая (2,2 см дл.,
0,5 см шир.). Рыльце светло-зеленовато-желтоватое, шире завязи.
Верхняя часть плодолистиков под рыльцем ярко-зеленая.
Коробочка белесовато-соломенная (4,6 см дл. и 2,1 см шир.),
овальной формы, тупо округлая на вершине и основании, ровной
ширины по всей длине, не имеет никакого столбиковидного коротко-
го или длинного, узкого или широкого образования.
Рыльце широкое, низкое (0,6 см), такое же светлое как створки,
•сидит прямо на тупом конце створки.
Карпофора у этого вида при основании коробочки нет. Створки
подходят прямо к рубцу от опавших членов околоцветника и даже
-слегка нависают над ним, чуть-чуть наплывают на него. Рубец полу-
скрытый наплывом, узкий, имеет под створкой сероватый, а у плодо-
ножки светло-коричневый цвет.
Створки коробочки грубые, плотные, овальной формы. Желобок
в центре створки мелкий, пологий, постепенно углубляясь кверху,
образует довольно глубокую широкую ямку в сужающейся округлой
части створки под рыльцем. Обычно у тюльпанов эта часть створки
уплощена, а у этого вида, наоборот, углублена. Вдоль желобка, от-
ступя несколько миллиметров от основания, проходит низкий гребень
или жилка. У одних особей эта жилка достигает всего половины вы-
соты коробочки, у других доходит до самого рыльца. Желобок на
всем протяжении окрашен так же, как вся створка.
Щечки створки белесые, плотные, грубые, б. м. равномерно при-
поднимаются над поверхностью створки и, лишь вверху створки у
дмки, как бы вспухают; по всей поверхности мелко поперечно мор-
щинистые, морщинки сгущаются по направлению к вершине коробоч-
ки, пересекают желобок, проходящий по краю створки и отделяющий
грань ребра коробочки от щечек, доходят до самого разрыва створ-
ки.
Отсутствие коричневой или лиловой окраски на ребре коробочки
и разрывах наблюдается у всех имеющихся у нас экземпляров
этого вида.
Коробочка открывается широкой клиновидной щелью, очень
острой в основании и, постепенно, без образования особого расшире-
ния „раструба“ на верху, широко (на 1,5 см) расходящейся у рыльца.
Края створки по разрыву не очень толсты (0,3 см) для коробоч-
ки такого размера и окрашены, как створки снаружи, но с розоватым
оттенком. От них внутри коробочки растут очень тонкие светлые гру-
бые волоски, перегораживающие щель в коробочке.
Внутренние перегородки коробочки расходятся друг от друга
примерно на 3/5 длины. Расхождение перегородок происходит по
тяжу пыльцепроводниковой ткани, который начинается вверху во всю
ширину рыльца, постепенно сужаясь книзу до перегиба в перегород-
ке, опускается ниже до глубины расхождения перегородок толстым
равномерным тяжем.
Плодоножка короткая (6 см), ровной толщины как под коробоч-
кой, так и при основании, желтоватая, более темная, чем коробочка.
Семя плоское, как выглаженное, округло-косо-треугольное, тем-
но-коричневое, окаймлено кругом более светлой чем семя, неширо-
кой, кругом же чуть-чуть загнутой вниз каймой. В нем лежит длин-
89
ный, в 34 длины семени, светлей чем семя, зародыш. В условиях
Ташкента Т. borszczowii цветет во второй половине апреля.
Tulipa korolkovii Rgl. — тюльпан королькова (рис. 4, 3) произ-
растает „на песчаных и щебнистых склонах в предгорьях.—Ср. Азия:
Сыр-Дар. Эндем. Описаны из Фарижа“ („Флора СССР“, т. IV).
Исследованные образцы собраны в окрестностях ст. Дарбаза Сред-
неазиатской железной дороги.
Луковица яйцевидная 2—3 см толщ., с черными, крепкими, почти
скорлуповидными, несколько продолженными (особенно сильно у не-
плодущих экземпляров) с внутренней стороны у верхушки волосисто
шерстистыми покровными чешуями.
Стебель вместе с цветоножкой голый, зеленовато-сизый, 18—
20 см выс.
Листьев 3, голые, отогнутые, сближенные или б. м. расставлен-
ные, курчавые, серповидно изогнутые, желобчатые, зеленые с сильным
сизым налетом, кверху уменьшающиеся. Самый нижний лист стебле-
объемлющий, линейно-ланцетный или ремневидный (25 X 1,4 см-
15 х 1,6 см); верхний — узколинейный (17 X 0,6 см; 11 х 0,75 см).
Цветок обычно один (в культуре их иногда бывает больше),
красный, желтый или пестрый, других оттенков у этого вида нет.
Сложение цветка очень оригинальное: он несколько похож на цветок
Т. greigji, но у некоторых форм очень торчат вверх листочки внут-
реннего круга; без запаха.
Листочки внешнего круга околоцветника (5,1 х 3,5 см) округло
ромбической или обратно яйцевидной формы иногда с очень малень-
ким пушистым кончиком на вершине и часто сильно волнистые по
краю; обычно снаружи оранжево-красные с сизым налетом, к краям
отгиба краснеют, к основанию, наоборот, желтеют. У желтых форм
они снаружи блекло-желтые, по спинке штриховато-малиново-красные с
зелеными редкими прожилками. У обеих форм снаружи при основании
листочков чуть-чуть просвечивает внутреннее темное пятно. С внут-
ренней поверхности они оранжево-красные, с маленьким(0,6 х 0,5 см)
черным, остро ромбическим пятном, у которого нижний угол срезан;
по бокам пятна от основания до половины проходят желтые полосы.
У желтоокрашенных форм основной фон внутренней стороны внеш-
него листочка околоцветника оранжево-желтый, на нем лежит боль-
шое, длиной до трех четвертей общей длины листочка и до половины
его ширины широко ланцетное, по краю штриховатое, оранжево-
красное пятно, на котором при основании листочка расположено ма-
ленькое, черное, остро-широко-ромбическое пятно.
Листочки внутреннего круга околоцветника (5,2 X 3,4 см) обратно
треугольные, редко обратно яйцевидные, иногда на вершине выемча-
тые; в выемке помещается очень короткий, пушистый кончик; снару-
жи красные, к центральной жилке со слабым оранжевым оттенком.
На основании имеется маленькое, веерообразное, островершинное, жел-
тое, подцвеченное антоцианом, пятно. У желтых форм, снаружи по
краю листочки внутреннего круга желтые с массой розовато-красных
штрихов, по всей площади перехода от желтого к сплошному оран-
жево-красному, занимающему пять шестых всего листочка. При осно-
вании листочка лежит фиолетовое пятно, по форме повторяющее чер-
ное внутреннее пятно.
У красных форм с внутренней поверхности листочки околоцвет-
ника ярко-красные с черным, четко отграниченным, широкоромбиче-
ским (нижний угол срезан) пятном. У желтых форм с внутренней
поверхности по верхнему краю листочка они лимонно-желтые, а ниже
90
Рис. XVIII. Т. borszczowii Rgl. (3/4 натуральной величины).
сплошь красные; красное заходит острым зубцом на желтый фон,
иногда достигая вершины листочка. При основании листочка лежит
небольшое, как и у красных форм, веерообразное черное пятно.
Тычинки расположены выше завязи.
Нити плоские, широкие, толстые, в средней части несколько рас-
ширенные (0,9 см дл., 0,2 см шир.), вверху постепенно оттянутые в
острый кончик; в нижней половине желтые, в верхней — черные;
острый кончик под пыльником красный. Черная часть на тычиночных
нитях у пестрых форм занимает несколько большую площадь, чем у
красных. Пыльники желтые (0,7 см дл. х 0,2 см шир.). Пыльца жел-
тая.
Завязь в полтора раза короче тычинок, в виде колышка, кремо-
вая, подкрашена антоцианом. Рыльце сидячее, шире завязи, бледно-
желтое.
Коробочка коротко цилиндрическая, на вершине и при основании
тупая, иногда при основании имеет небольшой, внезапно оттянутый,
не более чем на 0.6 см карпофор.
Рыльце маленькое, сидячее или возвышающееся над завязью на
0,2 см. Длинные коробочки с легкой перетяжкой на середине. Коро-
бочка открывается клиновидной, не доходящей на 1/3 или 1/2 до
основания, широкой на вершине щелью. Длина щели 2,3 см, ширина
у рыльца 1 см. Внутренние перегородки растрескиваются до места
слияния суженных частей перегородок в нерастрескивающуюся ниж-
нюю часть. Перегородка под рыльцем в разрезе неправильно ромби-
ческая, более короткая к вершине и вытянутая с растреснутыми края-
ми створок, якоревидная.
Створки коробочки овальные или продолговатые, беловато-соло-
менные, часто в верхней части с желтоватым оттенком, очень тонкие,
прозрачные; сквозь них просвечивают коричневые семена, придавая
коробочкам серовато-розоватый оттенок. Желобок иа месте прикреп-
ления перегородки на створке как бы вдавлен ножом, в верхней час-
ти коробочки окрашен в коричневатый цвет, иногда раздвоен.
Щечки створки выпуклые, по всей поверхности мелко-поперечно-
морщинистые, белесовато-соломенные, некоторые в верхней части
желтоватые, часто с легким розоватым оттенком, поперечно полоса-
тые, из прозрачных и матовых полос, настолько тонкие, что сквозь
них просвечивают семена. Палево-розоватые в 1,5 мм шир. края ство-
рок вывернуты наружу. Ребра у закрытой, но сухой коробочки вы-
даются над щечками в виде коричневого невысокого гребня. Под рыль-
цем сходятся более темные полосы ребер, а кроме того, часто и
раздвоенная верхняя часть желобка окрашена двумя серо-коричневы-
ми полосками. Характерным признаком вида является ямка под рыль-
цем, возникшая на месте проходящего через всю коробочку желобка.
Карпофор, несколько оттянутый книзу; коробочка как бы нависа-
ет над узким светло-коричневым рубцом, оставшимся от опавших
членов околоцветника. Рубец Мужается в плодоножке.
Плодоножка соломенно-желтого цвета с легким коричневым от-
тенком, тонкая, невысокая (до 15 см выс.).
В условиях Ташкента он зацветает в середине первой декады
апреля.
Tulipa nitida Hoog. — тюльпан блестящий (рис. XIX и 4, ii)
встречается „на каменистых и щебнистых склонах. — Ср. Азия: Пам.-Ал.
Эндем. Описан из Бухары по культурным экземплярам без точного
указания родины" („Флора СССР", т. IV).
Исследованные нами образцы собраны в Денауском районе Узбек-
91
ской ССР, на Термезском шоссе на 578 км слева от дороги на пестро-
цветках и в низкогорьях хр. Бабатаг у Байсуна на выходах песча-
ников.
Луковица яйцевидная в 1,5—2,5 см в диам., с внутренней сторо-
ны у макушки с волосисто шерстистыми покровными чешуями.
Стебель вместе с цветоножкой до 18 см выс. от уровня почвы,
голый, желто-зеленый, покрыт сизым налетом.
Листьев 3. Они, как правило, сильно курчавые, но есть и почти
прямые, б. м. по стеблю расставленные, серповидно отогнутые, же-
лобчатые, сизые почти голубовато-зеленые, голые, по краю более
светлые.
Нижний лист (15 см дл., 2 см шир.) линейно-ланцетный, отходит
от стебля на уровне почвы. Верхний — линейный (15 см дл. х 1 см
шир.). Цветок одиночный, красный, без запаха, близкий по сложению
к цветку Т. korolkovii, но внутренние листочки околоцветника Т.
nitida более удлиненные и соотношение частей цветка нескольно иное,
чем у Т. korolkovii.
Внешние листочки околоцветника округло-продолговато-ромбиче-
ские (5,2 см дл., 3,3 см шир.), на вершине без зубца, по краю отгиба
волнистые, мелкозубчатые, снаружи красные, покрыты сизым налетом,
при основании (до 0,7 см выс.) по обеим сторонам листочка проходят
две желто-зеленых узеньких (до 1 мм. шир.) полоски, между ними
основной фон листочка темнеет; с внутренней поверхности ярко-крас-
ные, блестящие, с легким оранжевато-малиновым оттенком, при ос-
новании (до 0,5 см выс.) с черным, очень четко отграниченным от
основного фона, в центре выемчатым пятном. В этом пятне лишь
центральная часть под выемкой темно-черная, а края размытые, даже
чуть-чуть сероватые.
Внутренние листочки околоцветника узко-обратно-яйцевидные
(5,4 см дл., 2,7 см шир.), на вершине мелкозубчатые, с узкой выем-
кой, снаружи и с внутренней поверхности одинакового ярко-красного
цвета, только с внутренней поверхности более блестящие, при осно-
вании снаружи (до 0,5 см выс.) лежит глубоко-выемчатое, черно-ко-
ричневое пятно, а с внутренней поверхности черное пятно (до 0,4 см
выс.), сплошь покрывая основание листочка, очень резко прямой ли-
нией отделяется от его основного фона.
Тычинки выше завязи, 1,6 см дл., т. е. более чем в три раза
короче листочков околоцветника.
Тычиночные нити голые, на 3/4 длины внизу черные, светлеющие
к основанию, вверху ярко-красные, к основанию они чуть-чуть сужены,
часто незначительно вздуты на красном, к пыльникам заострены (0,6
см дл., 0,2 см шир.).
Бывают растения, у которых все тычиночные нити черные или
красная часть значительно больше черной.
Пыльники продолговатые (0,8 см дл., 0,2 см шир.), желтые. Пыль-
ца желтая.
Завязь (1,5 см выс., 0,35 см шир.) к столбику постепенно сужена,
зеленая, в верхней части под столбиком окрашена антоцианом. Стол-
бик до 0,15 мм дл., кремовый.
Рыльце шире столбика, равно завязи, желтое с красными сосоч-
ками.
Соломенно-белая коробочка (до 3,5 см дл., 1,5 см шир.) оваль-
ной формы, тупая, на вершине с коротким столбиком и часто несколь-
ко заостренная на основании. Карпофор белесый, непосредственно пе-
реходит в створки, округлый часто очень короткий (0,2—0,5 см выс.),
92
Рис. XIX. Т. nitida Hoog. (2/3 натуральной величины).
слегка нависает над рубцом от опавших членов околоцветника. Не-
сколько заостренная верхняя часть створки б. м. внезапно заканчива-
ется коротким (0,2 см дл. и 1,2 см шир.) столбиком, на котором рас-
положено маленькое, в ширину столбика, светло-коричневое рыльце.
Коробочка открывается до самого основания (узкой внизу и ши-
рокой на вершине) клиновидной щелью (3,2 см дл., 1,5 см шир.) на
вершине у рыльца. Расхождение створок в верхней части коробочки
настолько велико, что внутренние перегородки ниже расширенной
части, „бляшки“ над изгибом лопаются поперек, хотя и растрескива-
ются вдоль значительно ниже изгиба. Ниже столбика, сросшиеся тка-
ни центральной перегородки сильно расширяются, а к изгибу перего-
родки вновь сужаются, получается как бы „бляшка “ 0,5 см дл. и
0,35 см шир.
Створки коробочки овальные или широколанцетные, на вершине
б. м. внезапно продолженные в короткое рыльце, при коротком ос-
новании резко расширяющиеся от рубца, а при длинном карпофоре—
постепенно к середине. Вдоль центра створок проходит как бы вдав-
ленный ножом желобок, он часто окрашен в один и тот же цвет, что
и вся коробочка, а в других случаях его верхняя часть окрашена в
лилово- коричневый цвет. На бесплодной верхней части плодолистика
желобок сплющивается и вся верхняя часть плоская, ровная, чуть-
чуть желтее или коричневее, чем вся коробочка.
Щечки створки белесовато-соломенные, хотя и очень тонкие, но
плотные и твердые, плоские, очень мало выпуклые, по всей поверх-
ности поперечно морщинистые, с чрезвычайно тонкими и мелкими
морщинками без коричневых поперечных полос. На грани створки
около шва края окрашены в легкий белесовато-коричневый цвет, от
края створки эта окраска очень тонкими штрихами спускается в узень-
кий желобок, отделяющий край от щечек. Край створки в месте раз-
рыва узкий (0,15 см), по разрыву матовый, от ребра коробочки корич-
невый, глубже палевый. Область створки под рыльцем с легким ко-
ричневатым оттенком.
Рубец от околоцветника узкий (до 0,1 см), выпуклый. Плодонож-
ка от рубца книзу сильно сужена, до 13 слдл., коричневого, или со-
ломенно-белого цвета.
Семя темно-коричневое, косотреугольное (0,65 см дл. на 0,5 см'
шир.) с узкой каймой, несколько направленной вниз.
В условиях Ташкента этот вид зацветает в середине апреля.
Tulipa rosea Vved. — тюльпан розовый. „Выходы пестроцветных
пород. — Ср. Азия: Пам.-Ал. (вост, часть предгорий Туркестанского
хребта). Эндем. Описан из КИМ (Санто)“ („Флора СССР", т. IV).
Исследованные образны взяты из массовых заготовок, проведен-
ных в окрестностях кишлака КИМ Ферганской долины с возвышен-
ности Белесник в 50клг к югу от Ленинабада в 1957 г.
Луковица яйцевидная 1,5—2 см в диам., лежит на глубине 5—10
см, с крепкими кожистыми, черными,часто продолженными почти до.
поверхности почвы, чешуями. Покровные чешуи с внутренней сторо-
ны в верхушке луковицы и в продолженной части густо длинно-во-
лосисто-шерстистые.
Растения во время цветения достигали до 11 см выс.
Стебель голый, короткий, вместе с цветоножкой коричневато-
зеленоватый.
Листьев 3, б. м. сближенных к основанию стебля, сильно отогну-
тых, слабокурчавых, желобчатых, голых, сизых, слабо кверху умень-
шающихся, обычно превышающих цветок. Самый нижний лист линей-
93.
но-ланцетный (13 см дл., 1,8 см шир.), верхний — узколинейно-ланцет-
ный (11,5 см дл., 0,7 см шир.).
Цветок розовый, на востоке ареала бледнее, на западе — ярче,
типа Т. korolkovii с широко расставленными вверх торчащими внут-
ренними листочками околоцветника и несколько вдоль спинки нару-
жу выгнутыми наружными листочками, без запаха.
Внешние листочки околоцветника (4,4x3,2 см) ромбической фор-
мы, на отгибе мелкозубчатые и слабоволнистые, снаружи по всей
поверхности сизовато-розовато-малиновые; лишь к самому краю отги-
ба теряющие сизый налет; на ноготке зеленеющие, с просвечивающим
внутренним темным пятном; с внутренней поверхности они оранже-
вато-розоватые, причем розоватость увеличивается в нижней половине
листочка, при основании с небольшим (0,5x0,4см), не доходящим до
края, на вершине тупо заостренным, со всех сторон четко отграничен-
ным, черным пятном.
Внутренние листочки околоцветника (4,2 см дл., 2,7 см шир) про-
долговато-обратно-дельтовидной формы, на вершине мелкозубчатые,
волнистые; снаружи розовато-малиновые, к вершине с чуть-чуть оран-
жеватым оттенком, при основании просвечивает лиловым плоское, во
всю ширину листочка, до высоты 0,5 см внутреннее пятно. С внутрен-
ней поверхности они более красноватые, при основании с четко от-
граниченным тупым или трехвершинным, занимающим все основание
листочка, до высоты 0,6 см, черным пятном.
Тычиночные нити плоские, к вершине постепенно заостренные,
голые (0,75 см дл., 0,2 см толщ.), черного цвета, постепенно, но нем-
ного краснеют под пыльником и при основании.
Пыльники желтые (0,7 см дл., 0,2см шир.).
Завязь ярко или бледно-розовая (вместе с рыльцем 1,2 см выс.,
0,3 см шир.), почти не сужается под рыльцем. Рыльце и верхняя
часть завязи (столбик) окрашены в светло-желтый цвет. Лопасти рыль-
ца слегка выступают над верхней частью завязи.
В условиях Ташкента этот вид зацветает в середине апреля.
Tulipa schrenkli Rgl.— Тюльпан шренка (рис. 4, к) растет „в
степях, пустынях и полупустынях. — Европ. ч.: Заволж. (юж. ч.), При-
черн., Крым, Нижн.-Дон, Нижн.-Волж.; Кавказ: Предкавк., Даг., Зап.
(сев. ч.) и Вост. Закавк.; Зап. Сибирь: Верхн. Тоб., Ирт. (зап. ч.);
Ср. Азия: Арало-Касп. (сев. ч.). Общ. распр.: Арм.-Курд. Описан из
окрестностей Ишима“ („Флора СССР", т. IV).
В посадках у нас были растения, выращенные из луковиц, при-
везенных со станции Берчогур, собранных у горы Бохтубай среди
камней и присланных из Крыма с полуострова Камышбурун, тов. Зу-
башко. Кроме того, мы неоднократно выращивали их из семян, соб-
ранных как в естественных условиях, так и в культуре.
Луковица яйцевидная, 1,5—4 см вдиам., с черно-бурыми, некреп-
кими, кожистыми, с внутренней стороны по всей поверхности прижа-
то волосистыми покровными оболочками.
Стебель во время цветения 10—30 см выс. от уровня почвы, в
верхней части вместе с цветоножкой окрашенный антоцианом, голый
(у крымских растений часто коротко пушистый).
Листья в количестве 3—4 (5), нижние б. м. отогнутые, серповид-
ные, верхние—торчащие, расставленные, сизые, голые или опушенные,
по краю крупно волнистые, реснитчатые. Самый нижний лист отходит
от стебля на уровне почвы, ланцетный (16x6 см), а верхний — узко-
ланцетный (11X2.7 см). Бывают особи с более крупными листьями.
Цветок, обычно, одиночный, по форме очень варьирует от узко-
94
бокаловидного до широко открытой чаши почти лилейного типа,
с легким очень слабым приятным ароматом; по окраске и форме цвет-
ков самый богатый вид, есть формы белые, желтые, розовые, пестрые
и красно-бордовые.
Внешние листочки околоцветника (в среднем 6,7X3,4 см) про-
долговато ромбические, на вершине б. м. внезапно заостренные в
в пушистый кончик; окрашены в весьма разнообразные оттенки, но
снаружи их цвет бывает смягчен сизоватым налетом; снаружи по
спинке часто широко зеленовато-желтые или белесоватые. При осно-
вании с ромбическим зелено-желтым пятном; с внутренней поверх-
ности с несколько более яркой окраской, чем снаружи, могут быть
совсем без пятна, или с чуть-чуть желтеющим основанием, или при
основании с чисто желтым, заостренным на вершине, ромбическим
пятном. Это пятно не занимает больше одной четверти длины лис-
точка и бывает черным, или сероватым, иногда штриховатым, тогда
оно окаймлено желтой каймой разнообразной ширины.
Внутренние листочки околоцветника обратно-продолговато-яйце-
видные, иногда округло ромбические, или продолговато эллиптиче-
ские, в среднем 6x3 см. но бывают крупнее, а главное, шире. Они
так же, как и внешние, внезапно оттянуты, в несколько более корот-
кий чем у внешних, пушистый кончик, иногда на месте кончика на
вершине листочка имеется только выемка; снаружи более яркие, чем
внешние, часто совсем без пятна, но бледнее к ноготку, а иногда с
желтым или зеленым веерообразным пятном на ноготке, занимающим
площадь основания листочка до одной пятой его длины. С внутрен-
ней поверхности они такого же яркого цвета, как снаружи; бывают
совсем без пятна или с легким расплывчатым, а часто и с оформлен-
ным желтым пятном, которое имеет форму раскрытого веера, только
с очень выступающей вверх вершиной, увенчанной одним крупным
зубцом и многими мелкими. Это пятно чисто желтое, желтое с ко-
ричневатыми легкими штрихами,или черное, окаймленное в каждом
случае особой желтой полосой.
Тычинки в два-три раза короче околоцветника, немного выше завязи.
Тычиночные нити голые, б. м. узкотреугольные, желтые или чер-
ные (0,7 см дл.). Пыльники почти в два раза длиннее нитей (1,5 см
дл.), довольно широкие, желтые или темные.
Завязь в форме узкоцилиндрической палочки (2,2 см дл., 0,4 см
шир.), светло-зеленая, по ребрам более темная с большим, намного
шире завязи, сидячим кремовым рыльцем.
Коробочка (3,6 см дл., 1,6 см шир.) часто значительно крупнее,
с овальными створками на вершине и основании несколько оттянутая
в короткий, внезапно возникающий столбик и бесплодное основание
„карпофор", с плотными, твердыми створками, с продольным желоб-
ком вдоль створки и густой сетью поперечных морщинок; серовато-
соломенно-желтая, по ребрам створок, желобам и морщинкам розова-
то-коричневая. Рыльце часто шире столбика и тоже темное,как и тем-
ный рубец, оставшийся на месте прикрепления околоцветника. Пло-
доножка сероватая или розовато-коричневая, темнее коробочки.
В условиях Ташкента этот вид зацветает в конце первой декады
апреля.
Tulipa ostrovskiana Rgl.—тюльпан островского (рис. XX) рас-
пространен „на глинистых или глинисто-щебнистых склонах в нижнем
поясе гор. — Ср. Азия: Тянь-Шань (с. ч.) Эндем. Описан из Алма-Аты"
(„Флора ССС“, т. IV). Исследованные образцы получены из Заилий-
ского Алатау.
95
Собраны в окрестностях г. Алма-Ата и Талгара. Кроме того,
. этот вид мы неоднократно выращивали из семян разного проис-
хождения.
Луковицы яйцевидные, 1,5 3 см. в диам. с черными, крепкими,
кожистыми, с внутренней поверхности у верхушки прижато густо-во-
лосистыми покровными чешуями.
Стебель с цветоножкой 18—26 см выс. от уровня почвы.
Цветоножка гибкая, тонкая, сизо-зеленая, у некоторых форм с
^бордово-коричневым оттенком.
Листья в количестве 2—3, голые, сизые, сильно отклоненные,
сближенные, в этом их отличие от типичного Т. kolpakovskiana. Кро-
ме того, у типичных представителей этого вида они почти лежат на
земле и лишь верхний чуть приподнимается над почвой, сильно кур-
чавые, чуть-чуть серповидные изогнутые; по краю белохрящевато
окаймленные, реснитчатые. Нижний округло желобчатый, ланцетный
(15X3 гж); верхний—линейно-ланцетный (12X0,9 см), более круто свер-
нутый в желобок.
Цветок одиночный в форме звездообразной чаши, очень изящный,
оранжево-красный, оранжевый, желтый, пестрый с массой тонких пе-
реходов между этими основными оттенками, часто с запахом; в буто-
не и после цветения — поникает; очень гелиофилен.
Внешние листочки околоцветника продолговато ромбические или
широко ланцетные, с острым кончиком, со слабоволнистым краем на
вершине; снаружи по спинке сизо-малиновые, различной силы и ин-
тенсивности: иногда очень темные, или оранжево-красные, есть золо-
тисто-желтые, оранжевые или пестрые формы, в нижней трети по
краю у красных форм могут быть продольные желтые полоски; с
внутренней поверхности такого же цвета, как и края снаружи; при
основании расположено во всю его ширину до высоты 0,8—1 см жел-
тое, часто трехвершинное пятно, на котором иногда лежит черное
пятно, обратно треугольной формы, очень изменяющееся по интенсив-
ности окраски и размеру. Есть формы с пятном только на внутренних
листочках околоцветника или с черным пятном как бы окаймленным
желтым. У желтых форм часто нет никакого пятна, а у оранжевых—
оно чисто желтое. Внутренние листочки околоцветника округло-узко-
ромбические, на вершине б. м. внезапно заострены в острый кончик;
желтые, оранжевые, пестрые а чаще оранжевато-красные, снаружи на
спинке и к основанию более оранжевые; у центральной жилки по бо-
кам расположены две оранжевые, а иногда более яркие, чем основ-
ной фон, малиновых полоски. На основании во всю ширину лежит
выемчатое, оранжевое пятно, от 1 до 0,6 см выс. (в выемке); с внут-
ренней поверхности они несколько более интенсивного цвета, чем
снаружи; при основании листочка, во всю ширину лежит желтое пят-
но, которое отчленено от основного фона листочка почти прямой ли-
нией. На этом пятне, может помещаться коричневатое, разной интен-
сивности сильно уплощенное пятно (желтое пятно окаймляет коричне-
ватое в виде полосы различной ширины).
Тычинки выше завязи, в два с половиной-три раза короче около-
цветника.
Тычиночная нить (0,8X2,5 см) к основанию и вершине б. м.
сужена, но на самой вершине внезапно заострена; может быть жел-
тая, светло-коричневая, красная, бордовая, но острый кончик, на
котором прикреплен пыльник, всегда более светлый.
Пыльники продолговатые (1,2 см дл., 0,2 см шир.), от желтых
до черных. Пыльца бывает желтая, черная, коричневая, бордовая.
96
♦ ♦
♦ ♦
Рис. XX. Т. ostrovskiana Rgl. (s/4 натуральной величины).
Завязь в виде палочки (1,6 см дл., 0,4 см шир.) от сизо-зеленой
до кремово-зеленой, по ребрам более яркая, под рыльцем часто крас-
ная. Рыльце красное, сосочки на нем желтые, но область завязи под
рыльцем желтая или зеленая. Лопасти рыльца иногда коричневые.
Рыльце шире завязи.
Коробочка, Зсм дл., 1,1 см шир., узкая, продолговатая с почти парал-
лельными краями, округло прямоугольной формы; лишь слегка сужена
к карпофору, имеющему всего Злшвыс., к нему и бесплодной верхней
части коробочки—столбику — створки заканчиваются внезапно округло.
Столбик выдается над створкой в виде коричневатого зубца 0,5 см
выс. и 0,7 см шир. при основании. Характерным для этого вида яв-
ляется округло треугольная вмятина на верхнем зубце под рыльцем.
Рыльце маленькое, сидячее, легко отваливающееся.
Коробочка растрескивается вдоль всей длины до карпофо-
ра узкой щелью с почти параллельными краями (2,3 см дл., 0,45 см'
шир.), несколько расширенной в средней части коробочки. Бесплодная
верхняя часть створки, образующая носик коробочки, б. м. резко вы-
гнута наружу, поэтому в верху щели образуется широкий раструб,
0,4 см выс., 1,1 см шир., через который свободно высыпаются семена.
Внутренние перегородки коробочки растрескиваются очень неглу-
боко, всего на глубину 0,9 см. Створки коробочки соломенно-белые,
почти овальной формы, со столбиком на вершине и карпофором при
основании. В центре вдоль створки проходит неглубокий желобок,,
образующийся на месте прикрепления плаценты, под носиком он как
бы раздваивается. Раздвоенная часть до 0,5 см дл. и окрашена в тус-
клый серовато-коричневый цвет.
Шечки створки соломенно-белые, нежные, бумагообразные, про-
зрачные (сквозь них просвечивают семена), плоско выпуклые, очень-
мелко поперечно морщинистые, в верхней части под носиком, часто
очень тонко, чуть заметно, поперечно коричнево-полосатые; отгибы
на ребре приобретают легкий коричневато-розоватый оттенок; выгну-
ты, но не вывернуты наружу и отделены от створок резким неглубо-
ким желобком. Окраска краев створок сливается с коричневатой ок-
раской носика.
Коричневатый рубец, оставшийся на месте прикрепления членов
околоцветника, слегка выпуклый. Белесовато-сероватая плодоножка
от рубца книзу резко сужается, ниже она очень тонкая и изящная,
очень длинная, до 35 см дл.
Семена мелкие (0,5x0,4 см), светло-коричневые, почти без каймы.
В условиях Ташкента этот вид зацветает во второй декаде апреля.
Tulipa kolpakovskiana Rgl. — тюльпан колпаковского (рис. 3, а),
встречается „на глинистых и щебнистых склонах в предгорьях. — Ср.
Азия: Дж. Тарб. (Джунгарский Алатау, ю. ч.), Тянь-Шань (с. ч.). Об-
щее распространение: Дж. Кашгар. Описан из Алма-Аты11 („Флора
СССР11, т. IV).
Растения для исследования получены из Чуилийских гор (хр.
Сарыбастау, в окрестностях ст. Андрокай Турксиба), окрестностей ст.
Чокпар, Заилийского Алатау (долина р. Талгар, окрестности ст. Тал-
гар, Раздолье).
Луковица яйцевидная, 1,5—3,5 см в диам., с черно-коричневыми,
жесткокожистыми, с внутренней поверхности у верхушки прижато
волосистыми покровными чешуями. Волоски опушения довольно тол-
стые и прямые.
Стебель вместе с цветоножкой от поверхности почвы 10—35 см
выс., сизый, голый.
7—2229
97
Листьев 3, все вверх торчащие, ни у одного растения на земле
не лежат, отходят от стебля под углом не более 30—35°, вверху
несколько отклоненные; по стеблю на 2—3 см друг от друга расстав-
ленные, постепенно кверху уменьшающиеся, особенно по ширине, в
первый день цветения превышают цветок, сильно сизые, голые, по
краю реснитчатые, мало курчавые, но сильно желобчатые.
Нижний ланцетный (например, 18 см дл. при 2,2 см шир.), а
верхний линейный (13 см дл., 0,7 см шир.), настолько резко согнут в
желобок, что кажется еще вдвое уже. Цветок желтый, один, в буто-
не резко поникает до земли, перед цветением выпрямляется; сложен
в виде двойной чаши, со слабым медовым ароматом весенних цветов.
Внешние листочки околоцветника продолговато ромбические, на
вершине оттянуты в острый кончик (2—6 см дл., 0,7—3 см. шир,), по
спинке зеленоватые, с просвечивающим желтым фоном, сильно смяг-
ченным сизым налетом, зеленоватая спинка продолжена даже на ост-
рие на вершине листочка; по краям листочек желтый. С внутренней
поверхности в центре, соответственно внешней спинке, он более зо-
лотисто-желтый, а по краям значительно бледнее.
Внутренние листочки околоцветника обратно-продолговато-яйце-
видные или узко-неправильно-ромбические, в нижней половине удли-
ненные, на вершине оттянутые в длинный острый кончик, снаружи
желтые, более золотистые к основанию, с зеленоватой центральной
жилкой, с внутренней поверхности желтые, но более яркие к осно-
ванию.
Тычинки в 2,5—3 раза короче околоцветника и немного длиннее
завязи.
Тычиночные нити голые, ярко-желтые, в виде ровных по всей
длине палочек, на вершине б. м. округло заостренные, по длине рав-
ные пыльникам (0,8 см дл., 0,21 см шир.). Пыльники желтые, но бо-
лее бледные, чем нити, сразу по всей длине открывающиеся.
Завязь зеленая, яркая, по ребрам более темная, ровная по всей
длине в виде толстенькой палочки (1,35 см выс., 0,25—0,35 см шир.),
с почти сидячим шире завязи кремовым рыльцем. Его лопасти охваты-
вают завязь по бокам.
Коробочка (4 см дл., 1,5 см шир.) в верхней части в закрытом
состоянии продолговатая, почти цилиндрическая, вверху несколько
расширена, особенно у открытых коробочек. Следует отметить, что
у этого вида почти нет карпофора. Створки растрескиваются на 1 мм
расстояния от рубца, оставшегося от опавших членов околоцветника.
Наверху створки тупо заканчиваются и несут маленькое (0,2 см дл.,
0,2 см. шир. при основании) остроконечие, к которому были прикреп-
лены маленькие лопасти рыльца, но они при созревании коробочки
отпали.
Коробочки открываются длинной продольной узкоклиновидной
щелью, доходящей до их основания, в центральной части коробочки
несколько расширенной (0,7 см), к вершине вновь суженной (0,5 см),
наверху у рыльца несколько более широко-клиновидной. Расширение
щели в центре коробочки происходит из-за того, что края створок в
этом месте вывернуты наружу настолько, что „щечки“ створок как
бы вдавлены внутрь коробочки.
Внутренние перегородки коробочки растрескиваются всего на 1,3
см выс. от вершины. Верхние части плодолистиков отклоняются друг
от друга на 1 см, т. е. не в такой сильной степени, как у видов, име-
ющих в верху коробочки большую бесплодную часть.
Створки коробочки соломенно-желтые продолговатой формы, ту-
98
по закруглены на вершине и при основании, при котором они несколь-
ко уже, чем в верхней части. Вдоль створки на месте прикрепления
плаценты проходит неглубокий рубец, углубленный в центральной
части в связи с выгибанием краев створки наружу, в самом верху у
некоторых форм окрашен в тускло-коричневый цвет, иногда под рыль-
цем растреснут вдоль на 0,5 см.
Щечки створки слабо выпуклые, часто плоские; особенно уплоще-
ны в средней части коробочки; довольно слабые, но толще, чем у
Т. ostrovskiana, мало прозрачные, по всей длине мелкоморщинистые и
очень тонко коричнево поперечно полосатые. Край ребра, вдоль ко-
торого створки растреснулись, окрашен в светло-коричневый цвет, к
рыльцу эта окраска сгущается. Края створок, особенно в средней
части, сильно вывернуты наружу, отчего их верхняя часть несколько
отклонена, что создает оригинальную форму открытой коробочки.
Рубец от членов околоцветника коричневый, неширокий, от него
отходит плодоножка более бело-соломенная, чем створки, у которых
преобладает желтоватый тон.
Плодоножка тонкая (0,25 см и тоньше); длинная до 28 см.
Листья у растений с коробочками раздвинуты.
Светло-коричневое семя почти без каймы, 0,45 см дл. и 0,3 шир.
В условиях Ташкента Т. kolpakovskiana зацветает в конце первой
декады апреля.
Tulipa zenaidae V v е d. — тюльпан Зинаиды (рис. XXI) произра-
стает „на мелкоземных и каменистых склонах в нижнем поясе гор.—
Ср. Азия: Тянь-Шань (Киргизский Алатау). Эндем. Описан из Шекуле.
Александровский хребет". („Флора СССР", т. IV).
Мы привезли его из Тянь-Шаня, хр. Киргизского Алатау, гора
Шекуле. Кроме того, получили от Фрунзенского ботанического сада
из Киргизского Алатау; бассейн р. Джарлы-Каинды, ущелье Мокрая
Бурулю, юго-западный склон; от Р. Шпак.
Луковица яйцевидная, до 3 см в диам., с черными или сверху,
цели они очень старые, побуревшими чешуями. Жесткие покровные
чешуи с внутренней поверхности у верхушки довольно густо покрыты
прижатыми прямыми волосками.
В наших условиях во время цветения это очень низкие растения.
Цветок открывается, едва показавшись из почвы. Также в начале
цветения в почву погружены основания листьев. Стебель и цветонож-
ки голые, сизые, слегка окрашены антоцианом, затем во время цвете-
ния они вырастают до 15 см выс. от уровня почвы.
Листья в количестве трех, голые, бледно-зеленые, с более яркими
жилками, сизые, по краю реснитчатые, не курчавые, кончики листьев
часто красные, в начале цветения б. м. сближенные, выше цветка
поднимающиеся, в конце—расставленные, отклоненные, не превышаю-
щие цветок. Нижний лист продолговато ланцетный (19 см дл., 6 см
шир.). Верхний ланцетный (7 см дл., 1,5 см шир.).
Цветок одиночный, очень оригинальный (но низкая цветоножка
снижает его декоративные достоинства), склонный к образованию
изрезанности (попугайности), особенно на внутренних листочках око-
лоцветника; без запаха, образует хорошую двойную чашу, у которой
внешние сегменты в верхней трети резко отогнуты наружу, а внутрен-
ние слегка отклонены, причем эта отклоненность лишь в самом верху
более четко выражена.
Внешние листочки околоцветника широко ланцетные или узко
ромбические, заостренные (5,3 см дл., 3 см шир.), снаружи по спинке
99
широко окрашены в красновато-розовый цвет, сквозь который про-
свечивает основная желтая окраска листочков околоцветника, края
очень узко окаймлены желтым основным фоном. На основании листоч-
ка расположено такого же оттенка, как края отгиба, желтое, про-
долговато ланцетное (1,7 см дл., 0,4 см шир.) пятно. С внутренней
поверхности листочек такого же желтого цвета, как и края листочка
снаружи. На основании у него лежит маленькое, прямоугольное,
черное пятно, в центре которого возвышается зубчик. Это пятно,
вместе с зубчиком всего 0,5 см выс. и 0,5 см шир. На основании*
листочка с внутренней поверхности, особенно в части, окрашенной
в черный цвет, под лупой хорошо видны устьица.
Внутренние листочки околоцветника продолговатые или оваль-
ные с внезапно возникающим на конце острым кончиком, снаружи
и с внутренней поверхности желтые, только снаружи они чуть-чуть
с белесоватым оттенком, а с внутренней поверхности с теплым золо-
тистым, особенно сгущающимся к маленькому черному, с прямой
линией вершины, пятнышку, которое с внутренней поверхности при
основании листочка, до 0,3—0,4 см выс., занимает всю его площадь.
Тычинки 1,6 см выс. в два с половиной — три раза короче около-
цветника и в полтора раза выше завязи.
Нити голые, в форме колышка, заостренного на вершине, в ниж-
ней половине черные, вверху желтые (0,6 см дл., 0,15 см шир.);
Пыльники желтые (1 см дл., 0,15 см шир.), продолговатые.
Завязь боченочком, зеленая или фиолетовая, почти черная с силь-
ным сизым налетом (1,3 см дл. X 3,25 см шир.). Рыльце уже завязи,,
ярко-желтое.
У некоторых форм при основании листочков околоцветника нет
черного пятна, а желтое основание сливается с основной окраской*
листочков. У этих форм тычиночные нити также сплошь желтые,
Есть формы этого вида с оранжевыми или красными цветами,
но сохранившиеся у нас растения старых сборов и новые растения
желтые.
Коробочка (3,6 см дл., 1,5 см шир.) продолговатая, примерно
одной толщины по всей длине плодущей части.
В основании у рубца от опавших членов околоцветника коробочка
сужена в короткий (0,3X0,5 см) карпофор, от которого сразу б. м
внезапно расширяется плодущая часть коробочки. На вершине коро-
бочки створки заострены, их края сходятся вместе, примерно, под
прямым углом (иногда этот угол округлый), на нем сидит маленькое,
легко опадающее рыльце. Коробочка растрескивается тремя щелями,
доходящими до карпофора. У щели иногда почти параллельные края
и раструб у верхней заостренной части створок; но бывает щель от
основания широко клиновидная, внутренние перегородки растрески-
ваются на небольшую глубину, примерно на 1 см у описываемой
коробочки. Из-за растрескивания могут несколько отклониться наружу
верхние заостренные части створок.
Створки коробочки желтовато-соломенного цвета с коричневатым
оттенком, продольно прямоугольные с заостренным основанием и вер-
шиной. Вдоль створок проходит углубленный плацентарный желобок,
слегка подкрашенный часто одной или двумя продольными коричневы-
ми жилками, от него на довольно плоские, сильно морщинистые и
грубые щечки отходит около семи тонких коричневатых жилок; эти.
жилки, не доходя до желобка, отделяющего ребро створки от щечек,
образуют кружево из петель, целую сеточку у наружного края щечек*
100
Рис. XXI. Т. zenaidae Vved. (3/4 натуральной величины)
в верхней части коробочки жилки и узор сгущаются, у шва створки
темнеют, становятся коричневыми, а иногда и грязновато-розоватыми.
Края створки равномерно по всей длине сильно выгнуты наружу,
настолько сильно, что изгиб проходит по линии прикрепления цен-
тральной плаценты. Бесплодная часть коробочки под рыльцем корич-
невая. Карпофор узкий, но шире плодоножки. Рубец от членов
околоцветника коричневый, на него часто наплывает карпофор.
Плодоножка значительно светлее, чем коробочка, часто мягкая,
много тоньше карпофора. Ко времени созревания коробочка вырастает
до 32 см выс. над уровнем почвы. Листья раздвигаются на 6 см.
Нижний — остается на уровне почвы.
Темно-коричневое семя (0,5x0,4 см) имеет маленький загнутый
край.
В условиях Ташкента этот вид зацветает в начале второй поло-
вины апреля.
Tulipa ferganica Vved. — тюльпан ферганский (рис. 3, в) встре-
чается „на каменистых и мелкоземистых склонах, в среднем поясе
гор. — Ср. Азия: Памиро-Алай (Алайский хр.). Тянь-Шань (Ферганский
я Чаткальский хр.).Эндем. Описан с р. Кугарт" („Флора СССР", т. IV).
Материалы для исследования в виде луковиц и семян мы привезли
из Ферганского хребта (различные места по течению рек Кугарт и
Яссы). П. А. Гомолицкий в 1951 г. — из хребта Баубашата и Шамурат
на высоте 2700 м, Т. И. Славкина и Р. М. Мурзова — из лесничества
Аркит в районе оз. Сары-Чилек.
Луковицы яйцевидные (до 3,5 см в диам.), с черно-коричневаты-
ми, крепкими, кожистыми, с внутренней поверхности у вершины
и при основании прижато волосистыми покровными чешуями. Растение
до 25 см выс. от уровня почвы. Стебель и цветоножка пушистые,
сизые, иногда подкрашенные антоцианом, крепкие. Часто цветки
расцветают у самой почвы, но за время цветения цветоножка выра-
стает на значительную высоту.
Листьев от 3 до 5, сближенные, иногда расставленные, отогнутые,
нижние крупноволнистые, верхние б. м. курчавые, в конце цветения
не превышают цветы, сизые, пушистые (причем опушение увеличи-
вается у верхних листьев и почти отсутствует у нижних), по краю
светло-желто-хрящевато окаймленные, реснитчатые. Самый нижний
лист широко ланцетный, при основании стеблеобъемлющий (до 16 см дл.,
6,7 см шир.), верхний — узколанцетный (до 13,5 см дл., 2 см шир.).
Цветков 1—2; а иногда в культуре и больше, желтые, снаружи
сизовато-розоватые, в виде простой звездочки, несколько углубленной
в центре, почти без запаха.
Внешние листочки околоцветника (в среднем 5,3 см дл., 2,3 см шир.)
ланцетной формы, постепенно заостренные, на острие опушенные;
снаружи по спинке розовато-коричневые, с сильным сизым налетом,
на ноготке зеленые, позеленение клином вдается вверх почти до по-
ловины длины листочка; по самому краю вдоль всего листочка ярко-
желтые полосой не шире 0,4 см-, розовато-коричневое окрашивание
на спинке очень сильно варьирует в интенсивности, часто позеленение
видно и на вершине спинки у острия листочка; с внутренней поверх-
ности ярко-желтые, без всякого пятна при основании листочка.
Внутренние листочки (в среднем 5 см дл., 2,1 см шир.) обратно
продолговатые, б. м. внезапно заостренные в слабопушистый или
голый кончик.
Снаружи они такого же желтого цвета, как и с внутренней по-
верхности, лишь в нижней половине по жилке и довольно широко
101
вдоль нее зеленеющие, иногда позеленение доходит узкой полосой до
верха листочка, иногда розовеют или краснеют эти полосы и цен-
тральная жилка.
С внутренней поверхности эти листочки чисто желтые без какого-
либо пятна при основании.
Тычинки (1,6 см дл.) в два-три раза короче околоцветника, но
выше завязи.
Тычиночные нити оранжево-желтые, к основанию несколько блед-
неющие (0,7 см дл., 0,2 см шир.), при основании кверху постепенно
сужающиеся.
Пыльники оранжевые, продолговатые (0,9 см дл., 0,2 см шир.),.
вдоль всей длины одновременно открывающиеся.
Завязь короткая, толстенькая (0,9 см дл., 0,35 см шир.), желто-
зеленая, к рыльцу более темно-зеленая, под рыльцем на ребрах и
в центре плодолистиков часто с антоциановыми штрихами. Рыльце
сидячее, шире завязи, его светло-желтые лопасти чуть-чуть спуска-
ются на завязь.
Коробочка палево-желто-соломенная, тусклая (4,6с.мдл., 2с-мшир.),
продолговато яйцевидной формы. В верхней половине все створки коро-
бочки, вплоть до конца щечек, сильно поперечно морщинистые, в
морщинах — светло-коричневые. По краю створок проходит сильно
выгнутый в нижних трех четвертях створок, но пологий желобок,
как и ребро коробочки, окрашенный в светло-коричневый тусклый
цвет. Под рыльцем в плоской широкой зоне, образовавшейся из-
слившихся между собой уплощенных вверху щечек, желобка и ребер
коробочки, получается матовая светло-коричневая окраска. Створка
имеет под рыльцем до 0,7 см шир., но бывает и уже, кончается тупо
или выемкой, к ней прикреплены маленькие, темные, узкие остатки
рыльца.
Створки расходятся между собою на большую ширину, так как
в нижней части створок шель образована округло от основания выг-
нутыми створками. В средней части коробочки эта щель несколько
сужается, а наверху, наоборот, расширяется, так как створки как бы
отталкиваются друг от друга и широко расходятся. Край створки по
лопнувшему ребру коробочки сильно выгнут наружу, светло-коричне-
вый, матовый, как фетровый, вверху суживающийся.
Рубец у основания коробочки от опавших членов околоцветника
темно-тускло-коричневый, состоит из двух, параллельно идущих
выпуклых полосок.
Коробочка внезапно тупо заострена на вершине, т. е. почти под
прямым углом, ее створки сходятся к бесплодной части.
Карпофор для определения этого вида имеет важные признаки: он
белесый, невысокий, почти цилиндрический, ровный по всей длине,
почти равный по толщине плодоножке, резко отчленяется от створок:
коробочки, которые шире его и нависают над ним, вдается в створку
в области центрального желобка острым коротким зубцом.
Створки коробочки плотные, снаружи матовые, с внутренней
поверхности желтоватые, блестящие, овальные у открытых коробочек,
они, благодаря сильно загнутым наружу в нижней половине краям
створки, шире в верхней половине, чем в нижней. В центре створки
сверху вниз проходит желобок, узкий (до 0,1 см) у ее основания и
широкий (0,7 см) под рыльцем. В нижней половине створок он
углублен и сжат, в верхней — он плоский; как внизу, так и вверху
он покрыт мелкоморщинистой рубцовой тканью светло-коричневого
цвета, вдоль него в нижней половине створки проходит одна выпуклая
102
жилка, наверху она раздваивается и расходится двумя рукавами,
производя впечатление заросшего разрыва ткани, а в других случаях
мозолистого плоского нароста. От желобка на щечки створки про-
ходит масса поперечных морщинок, от которых частично свободны
лишь белесоватые, вздутые нижние части щечек. Щечки в открытой
коробочке значительно сближены между собою благодаря выгибанию
этой части створки наружу.
Длинная плодоножка темнее коробочки, покрыта заметным нево-
оруженным глазом, густым, коротким, белесоватым опушением.
Темно-коричневые, косо треугольные семена имеют довольно
широкую кайму, которая несколько смята и равномерно кругом семени
направлена вниз-
Снизу семени виден в виде короткой палочки более светлый
- зародыш.
В условиях Ташкента зацветает в начале третьей декады апреля.
Tulipa tetraphylla Rg\.—тюлъпан четырехлистный встречается
„на каменистых склонах.—Ср. Азия: Тянь-Шань (Центр. Т.-Ш.).
Общ. распр.; Дж-Кашг. Описан с р. Кочкур". („Флора СССР", т. IV).
Образцы для исследования мы привезли из Заилийского Алатау,
окрестности с. Талгар и Я. С. Весневский — с верховьев Нарына.
Луковица широко яйцевидная 1,5—3 см в диам., с коричневато-
черными, жесткими, кожистыми, с внутренней поверхности у вер-
хушки прижато волосистыми покровными чешуями. Покрывающие
чешуи — золотистые волоски, толстые и совершенно прямые.
Стебель от поверхности почвы 3—25 см выс., голый. Цветоножка
также голая, очень сизая, к цветку даже голубоватая.
Листья в количестве 3—7 (у сильных растений), но наиболее
часто, 4—5, сильно сближенные, во время цветения еще сидящие
своими основаниями ниже уровня почвы, отогнутые, серповидные,
желобчатые, часто курчавые, ремневидные, лишь на вершине посте-
пенно заостренные, сильно сизые, голые, по краю белоокаймленные,
реснитчатые, во время цветения превышающие цветок, почти равные
между собой по длине, например, самый нижний 14,7X9 см и верх-
ний 13,5X5,5 см; могут быть длиннее и шире.
Цветков 1—4; желтые, в бутоне поникают, в форме двойной
чаши, без запаха.
Внешние листочки околоцветника (2—6 см дл., 0,7—2,9 см шир.)
широколанцетные или продолговато ромбические, на половине длины
отогнутые наружу, снаружи по спинке желто-зеленые в виде длин-
ного пятна, повторяющего собой форму листочка, окаймленного бор-
дово-малиновой широкой полосой, состоящей из плотно расположен-
ных штрихов; по верхнему краю листочки желтые, с внутренней
поверхности — чисто желтые.
Внутренние листочки околоцветника узко-обратно-продолговато-
яйцевидные, на вершине несколько оттянутые в зубец (1,3—5,8 см дл.
0,8—3,5 см шир.); снаружи желтые; центральная жилка зеленая,
с обеих сторон ее расположено штриховое, большое, островершинное,
зеленое пятно, на вершине исчезающее, при основании сгущающееся
и расширяющееся. С внутренней стороны листочки чисто желтые.
Тычинки в три раза короче околоцветника, но в полтора раза выше
завязи.
Нити голые, золотисто-желтые, толстоватые, в верхней половине
расширенные, под пыльником внезапно заостренные. Пыльники желтые,
0,7 см дл., равные тычиночным нитям; иногда спирально свернутые.
Пыльца грязновато-желтая.
103
Завязь немного короче тычинок, ярко-зеленая (1,1x0,35 см),
толстенькая, округло ребристая с желтым сидячим рыльцем.
Коробочка 3,7 см дл. и 1,7 см шир. широко продолговатая,
б. м. внезапно суженная к основанию и вершине, имеет при основании
карпофор (до 0,5 см выс., 0,4 см шир.); вверху сужена в короткий
•столбик (до 0,4 см дл.), часто этого окончания не бывает и у коро-
бочки нет никакого перехвата вверху под рыльцем, т. е. створки к рыль-
цу б. м. внезапно заострены. Рыльце маленькое, легко отваливаю-
щееся. Коробочки открываются широкой (1,1 см вверху щели при
длине в 2,1 см) клиновидной щелью, с прямыми краями створок.
Ширина раскрытия зависит от того, что средние перегородки коро-
•бочки в верхней части отходят одна от другой более чем на половину
длины коробочки, часто до глубины 1,8 см.
Створки коробочки имеют очертания приближающиеся к широ-
кой линзе, но с двумя легкими перехватами у рыльца и основания.
Вдоль всей створки проходит довольно сильно углубленный рубец,
находящийся на месте прикрепления внутренней перегородки. Нет его
только на карпофоре и он уплощается вверху под рыльцем. Этот
рубец по всей длине окрашен в коричневый цвет. Коричневая полоса
по рубцу шире вверху и сходит на нет внизу.
Щечки створки, в отличие от других близких видов, сильно
выпуклые и чуть-чуть выступают над карпофором, нежные, бумаго-
образные, соломенно-белые, настолько прозрачные, что через них
просвечивают темные семена, в верхней трети поперечно морщинис-
тые. Морщинки не частые, но довольно глубокие, вдоль морщинок
проходят бледно-светло-коричневые поперечные жилки.
Края створок отделены от щечек желобком и выгнуты наружу,
имеют светло-коричневый оттенок почти вдоль всей длины, но более
интенсивно окрашены в верхней части; иногда окрашены лишь жилки,
образующие узкую зубчатую кайму вдоль выгнутого края створки.
Под рыльцем сходятся коричневые полосы желобка и краев
••створок, поэтому вся верхняя суженная зона створки окрашена в
светло-коричневый цвет.
Коричневый рубец от опавших членов околоцветника 1,5—2 мм
шир. сужается в сторону плодоножки, так как она уже бесплодной
части коробочки, имеет легкий коричневатый оттенок, т. е. она темнее
коробочки, значительно короче, чем у близких видов и в наших
условиях до 15 см. Листья, бывшие во время цветения сближенными,
довольно сильно раздвинулись.
В условиях Ташкента зацветает в конце первой декады апреля.
Tulipa korshinskyi Vved. — тюльпан коржинского обитает: „ка-
менистые склоны в среднем и верхнем поясе гор“. Ср. Азия: Пам,-
Ал. Эндем“. („Флора СССР", т. IV). Описан из Сары-Даша. Настоящее
описание сделано по растениям с классических местообитаний, храня-
щихся в гербарии Ботанического института АН СССР в Ленинграде.
Луковицы яйцевидные, до 3 см в диаметре, лежат на глубине
7—10—16 см, а иногда мельче, покрытые темно-коричневыми почти
черными чешуями. Покровные чешуи с внутренней стороны вершины
и основания опушены прижатыми, рыжеватыми, прямыми волосками.
Судя по большому количеству отмерших чешуй и округлой форме
луковицы, она лежит в рыхлом песчаном или щебенчатом грунте.
Стебель во время цветения растения не возвышается над поверх-
ностью почвы. Сближенные листья погружены основаниями в почву.
Цветок обычно расцветает у самой поверхности почвы и во время
цветения несколько приподнимается вытянувшейся цветоножкой.
104
Цветоножка вместе со стеблем голая, за время протекшее от
.зацветания до созревания коробочка вырастает до 5,5 см дл., иногда
и больше, также несколько вытягиваются междоузлия, которые почти
исчезают во время цветения,
Листья в количестве (2—3) 4 (5) сближены, сильно отклонены,
часто серповидно изогнуты, обычно голые, иногда верхние реснитча-
тые, по краю сильно курчавые, превышающие цветок. Самый нижний
лист ланцетный, во время цветения от 5 до 14 см дл. и от 1,5 до
3 см шир., а верхний от 3 до 13 см дл. и от 0,5 до 0,9 см шир.
Цветок одиночный в виде простой звезды, сидит на тонком цве-
тоносе.
Листочки околоцветника белые с внутренней поверхности, с жел-
тым пятном при основании. Внешние листочки околоцветника линейно-
ланцетные (до 3,3 см дл. при 0,6 см шир.), по спинке с фиолетовым
оттенком, внутренние(до 3 см дл. и 0,7 см шир.) — обратно-яйцевидно-
ромбические, снаружи при основании у них просвечивает желтое
внутреннее пятно, а по спинке они узко клиновидно сероватоокра-
шенные.
Тычинки на 1/4 короче околоцветника, причем пыльники на одну
треть длиннее нитей.
Нити желтые, узкие, ровные по всей длине, лишь под пыльником
они внезапно заострены. Заостренная часть нити оранжевая. Ярко-
желтый пыльник открывается одновременно. На его вершине выдается
окрашенный в желтый цвет выступ связника.
Завязь короче пыльников, со столбиком равным половине ее
длины и светлым выступающим над ним рыльцем.
Коробочки белесоватые (4—3,1 см дл., 1,6 см шир.), овальной
формы, на вершине и при основании заострены, при основании до
0,4 см выс. несколько суженный карпофор более узкий к плодоножке,
расширяющийся к створкам и вдающийся узким клином в их середину.
Ребра коробочки не заходят на карпофор.
Овальная створка коробочки на вершине б. м. постепенно сужена
и оттянута в острый зубец, часто более темный, чем сама створка
(у сухой коробочки до 0,3 см дл.). Лопасти рыльца на ней не со-
храняются.
Створки коробочки при основании оттянуты в карпофор, а на
вершине в столбик (зубец), разделены пополам проходящим вдоль
них желобком, четко проступающим от вершины до карпофора и
имеющим зубчатую форму от входящих в него со щечек створки
поперечных морщинок, часто окрашенных в ярко-коричневый, а иногда
.лишь слабо-розоватый цвет.
Щечки створки б. м. выпуклые, тонкие, бумагообразные, белесо-
вато-соломенные, в верхней части поперек всей ширины мелкомор-
щинистые, в верхней половине иногда приобретают коричневато-
розовый оттенок.
От края створки щечки отделяются пологим желобком, образо-
ванным выгнутым краем створки у места разрыва по ребру коробочки.
По краю вдоль разрыва ребро коробочки сверху донизу светло-
коричневого оттенка. Поверхность разрыва кремовая, матовая и очень
тонкая. Открывается коробочка широкой клиновидной щелью до
самого карпофора. Бахромок у разрыва почти нет.
Внутренние перегородки коробочки растрескиваются по пыль-
.цепроводниковой ткани почти до основания коробочки, в верхней
части коробочки под рыльцем до выгиба в створке места соединения
яесколько расширены, но не достигают формы ромба, как у других
105
видов. Их форма на месте разрыва может быть названа очень вытя-
нутым овалом.
Рубец от опавших членов околоцветника светло-коричневый,
почти скрыт нависшим над ним белым карпофором. Плодоножка более
темная, чем коробочка, резко сужается под рубцом.
У экземпляра гербария, хранящегося в БИНЕ, который А. И. Вве-
денский установил как тип, 2 растения двухцветковые. У левого из
них добавочный цветок выходит из пазухи второго листа и имеет на
своем стебельке маленький прицветный листик. Без этого листика
у растения всего три листа. У растения двухцветкового, на гербар-
ном листе справа —4 листа. Добавочный цветок выходит из пазухи'
третьего листа. Его стебель до высоты 1,5 см сросся со стеблем
главного цветка, а прицветного листа не видно, по-видимому, он
также сросся с верхним листом.
Интересно, что у этого экземпляра тычиночная нить имеет по
обе стороны длины параллельные узкие крылья лепесткового харак-
тера, сходящие на нет на вершине нити под пыльником. То же самое
наблюдается и у одноцветкового растения.
Tulipa hissarica М. Pop. et Vv ed. — тюльпан гиссарский обитает:
„Альпийский пояс гор. Ср. Азия: Пам.-Ал. (Гиссарский хребет). Эндем..
Описан с перевала Мура“. („Флора СССР", т. IV).
Описанные растения в 1950 г. получены из Душанбинского бота-
нического сада от Т. И. Рябовой; в 1935 г. от экспедиции Института
сухих субтропиков и в 1956 г. из Варзобского ущелья в Гиссарском
хребте (сб. 3. П. Бочанцевой).
Мы собирали их на каменистых почвах в субальпийском поясе гор.
Луковицы продолговато яйцевидные, 1 — 1,9 см толщ., с бумаго-
образными, буро-коричневыми, иногда сероватыми (при росте луко-
вицы в течение нескольких лет на одном месте) оболочками. Обо-
лочки несколько грубеют, на них сильно выступают продольные
жилки, особенно у пересаженных в культурные условия луковиц и
у столонов углубления, которые обладают свойством закладываться
как у ювенильных, так и у цветущих растений и направлены отвесно-
вниз. Покровные чешуи продолженные, с внутренней стороны при-
жато волосистые.
Стебель до 7 см выс., голый, в начале цветения скрыт в розетке,
затем, к отцветанию вытягивающийся, сизо-зеленый.
Листьев 3—4—5; сильно сближенные, образуют мутовку, отогну-
тые, слабокурчавые, коротко ремневидные, сизо-зеленые с краснова-
тым оттенком, особенно ярким с нижней стороны листьев, по краю
и на конце; бывают ярко-бордовые, гладкие или слабо реснитчатые,
до 15 мм шир., кверху постепенно уменьшающиеся и сужающиеся,
значительно превышающие цветок. Верхний лист иногда лепестко-
видный.
Цветок желтый, одиночный, имеет форму широко открытой звез-
ды с приятным весенним запахом легкого шафранного оттенка.
В культуре этот вид образует до 5 цветков на одном стебле.
Внешние листочки околоцветника ланцетные (21—30 мм дл., 7—
8 мм шир.), по краю в верхней половине часто надрезанные; снаружи
желтовато-серые или бордово-серые, в нижней трети зеленоватые,
по краю очень узкожелтые; с внутренней поверхности золотисто-
желтые.
Внутренние листочки широколанцетные, продолговатые, часто на
вершине тупые, на ноготке суженные, иногда по краю с надрезами;
снаружи золотисто-желтые, при основании покрыты зелеными штри-
106
хами, приподнимающимися вдоль центральной части зеленоватой жил-
ки вверх по листочку околоцветника; с внутренней поверхности
золотисто-желтые (22—26 мм дл. от 8 до 13 мм шир.).
Тычинки в полтора, два раза короче околоцветника, с сухими
пыльниками, до 18 мм дл. Нити желтые, голые, плоские, постепенно
от основания к вершине суженные (до 7,5 мм дл.). Под пыльником
они образуют совсем тоненький зубец. Пыльники оранжевые, линейно
продолговатые, в сухом виде до 9 мм дл. Пыльца оранжевая.
Завязь короче тычинок, зеленая (13 мм дл., 3 мм шир.), с длин-
ным (6 мм) постепенно к завязи расширяющимся столбиком, со светло-
желтым трехраздельным, сильно выемчатым, почти разрезным, чуть
шире столбика рыльцем. Цветет с середины до конца марта. В куль-
туре размножается вегетативно.
Коробочка белесовато-соломенная (3,2 см дл., 2,1 см шир.),
очень хрупкая, бумагообразная, обратно-округло-треугольная, тупая
на вершине и сужена к основанию.
Карпофор маленький (0,25 см выс., 0,2 см шир.), чуть толще
плодоножки; у мелких коробочек его нет, очень тонкий по сравнению
с нависающими над ним створками коробочки, острым зубцом вдается в
створку, белого цвета. Столбик (до 0,3 см дл., может быть и короче)
внезапно возникает на тупой вершине створки, окрашен в темно-ко-
ричневый цвет. На столбике расположено очень маленькое, коричне-
вое рыльце, лопасти которого легко отваливаются.
Створки коробочки тонкие, бумагообразные. Обратио-округло-
дельтовидные или овальные, с продольным желобком в центре, обра-
зованным на месте срастания плодолистиков. Вдоль желобка проходит
коричневая жилка, часто более темная в верхней части. От желобка
в двух верхних третях створки отходят поперечные морщинки с
ярко-коричневыми жилками в углублениях.
Щечки створки выпуклые, вверху поперечно морщинистые, внизу
продольно смятые, тонкие, бумагообразные. Края створок отделены
от щечек желобком, выгнуты наружу, окрашены в коричневый, более
темный, чем на всей коробочке, цвет.
Место разрыва очень тонкое (до 0,1 см шир.), кремовое, с ред-
кими тонкими, смотрящими внутрь коробочки, волосками. Коробочка
открывается клиновидными щелями, доходящими до основания ство-
рок. Они узкие внизу и широкие на вершине. Внутренние перегородки
растрескиваются до основания коробочки. Эти перегородки под рыль-
цем не имеют никакого расширения и тяж пыльцепроводниковой тка-
ни лишь слабо утолщен под рыльцем.
Рубец от опавших членов околоцветника темно-коричневый, бо-
лее широкий над нависающей над ним коробочкой и узкий над тон-
кой с желтоватым оттенком плодоножкой.
Семя светло-коричневое, крупное (0,7 X 0,35 см) округло тре-
угольное или в виде полумесяца, шероховатое, с узкой, неровной,
грубой каймой. Зародыш не просвечивает.
Секция Spiranthera Vved.
А. И. Введенский, устанавливая эту секцию, дает ей следующую
характеристику: „Плн. постепенно медленно от верхушки к основа-
нию открывающиеся, скручивающиеся. Нититыч. голые. Завязь с поч-
ти сидячим рлц. Оболочки луковицы с внутренней стороны более или
менее волосистые*1. („Флора СССР", т. IV).
Т. kaufmanniana Rgl. — гюльпан кауфмана (рис. 3, г) встречается
107
„на каменистых склонах в нижнем и среднем поясе гор — Ср. Азия:
Тянь-Шань (Зап. Тянь-Шань). Эндем. Описан с Чирчика“ („Флора
СССР", т. IV).
Растения для изучения собраны в районе Западного Тянь-Шаня
с гор, составляющих бассейн р. Чирчик, по долинам Пскема, У гама
и Чимгана, вдоль р. Ангрен и из Аксу Джебоглинского заповедника,
из ущелья Тавды-Булак.
Этот вид произрастает в кустарниковой зоне гор среди кустов и
крупных камней.
В ущелье Каранкульсай на склоне горы у р. Чирчик на крупной
глыбистой осыпи с мелкоземом между глыбами росли кустарники;
склоны с глубокой лессовой почвой были покрыты пыреем. Среди
кустарников в камнях мы находили большое количество коробочек
Т. kaufmanniana, а среди лужаек, заросших пыреем,—Т. greigii.
Во „Флоре СССР" А. И. Введенский пишет, что у. Т. kaufmanniana
луковица яйцевидной формы. В действительности же форма луковицы
у этого вида очень изменчива, от очень узкой, удлиненной в виде
редьки, до почти репчатой. Луковицы цветущих растений от 1,5 до
8 см в диам. (в культуре). Покровные чешуи по окраске от черно-
бурых, кожистых, до золотисто-коричневых, нежных, бумагообразных,
с внутренней стороны б. м. прижато волосистых. Золотистые волоски
•опушения сгущаются к вершине и донцу луковицы.
Стебель 10—50 см выс. от уровня почвы, в верхней части вместе
с цветоножкой б. м. пушистый, иногда голый, часто окрашен анто-
цианом в бордовый цвет различной интенсивности.
Листьев от 2 до 5, часто расставленные, сизые, с темной сеточ-
кой жилок на более светлой мякоти, обычно не курчавые, но, как
исключение, слабо волнистые, особенно у особей с узкими или круг-
лыми листьями, голые, по краю реснитчатые, постепенно кверху умень-
шающиеся, как правило, не превышающие цветок: нижние — откло-
ненные, верхние — торчащие, слегка отогнутые. Самый нижний (от 2
до 20 см шир.). ланцетный или продолговатый, или продолговато эл-
липтический, но бывают формы, особенно у особей с неразвитым ко-
ротким цветоносом, с почти круглым нижним листом.
Цветок обычно один, но у мощных растений в культуре до 5.
Цветы белые или кремовые, однако встречаются и растения с
желтыми, оранжевыми, огненными, светло-красными, кирпично-крас-
ными и почти бордовыми цветками. Все эти полихромные формы по
всем признакам, кроме окраски листочков околоцветника, являются
типичными для этого вида.
Данные наших экспериментов приводят к выводу, что предполо-
жение о происхождении полихромных форм в результате межвидовой
гибридизации несостоятельны.
Цветок по форме изменяется от совершенно плоского звездооб-
разного до лилейного или, наоборот, чашевидного, как у благород-
нейших из культурных тюльпанов. У многих форм этого вида прият-
ный весенний аромат различной силы.
Внешние листочки околоцветника (2,5—11 см дл., от 0,8 до 4,5 см
шир.). Чаще всего по форме ланцетные или широко ланцетные, иног-
да эллиптические, часто отогнутые; снаружи по спинке красные, ма-
линовые, розовые или голубые, иногда зеленоватые, очень редко бе-
лые; по краю отгиба такого же цвета, как и с внутренней поверхно-
сти цветка; при основании с желтым сильно выемчатым или овальным
пятном различной величины; с внутренней поверхности белые, кре-
мовые, желтые, красные и т. д., с большим ярко-желтым, нечетко
108
отграниченным от основного фона листочка, пятном. Над пятном окра-
ска основного фона листочка или желтеет или сгущается. Это пятно
разнообразной формы — от острого до округлого или выемчатого; под-
нимается по листочку не менее чем на две пятых его длины. На
желтом фоне пятна, отступя на одну треть высоты от вершины, часто
наблюдаются симметричные красные штрихи, которые иногда слива-
ются в v-образное или сплошное большое пятно, но часто в природе
и культуре встречаются формы без таких красных пятен.
Внутренние листочки околоцветника часто торчат вверх, продол-
говато эллиптические или обратно продолговатые, часто округло-вы-
тянуто-ромбические, на вершине тупые (10,5 до 2,5 см дл. при 5,5 до-
1 см шир.); снаружи и с внутренней поверхности одинакового цвета,
белые, кремовые, желтые, красные, иногда пестрые. Следует отметить,
что на внутренней поверхности цветка на листочках околоцветника
от пятна кверху, надвигается желтоватый оттенок, особенно у кремо-
вых и желтоватых форм; снаружи в нижней половине просвечивает
внутреннее желтое пятно; центральная жилка у некоторых светлых
форм часто окрашена в зеленый или красный цвет.
С внутренней поверхности при основании, примерно, одну треть,
длины листочка, весь ноготок, занимает выпуклое, веерообразное яр-
ко-желтое пятно, иногда с выемкой на центральной жилке. На внут-
ренних листочках околоцветника в верхней трети желтого пятна у
белых, кремовых и желтых форм наблюдаются красные пятна в виде
точек, штрихов, v-образных, округлых мелких и больших пятен.
Тычинки в полтора, два раза короче околоцветника. Нити голые,
к основанию расширяющиеся, на вершине внезапно суженные в острый
кончик, желто-оранжевые. Пыльники желтоватые, линейные, по вы-
сыпании пыльцы поперек ссыхающиеся, часто в 3—4 раза длиннее
нитей, постепенно, в течение нескольких дней, от верхушки до осно-
вания открывающиеся и в улитку скручивающиеся или сильно изги-
бающиеся.
Завязь значительно короче тычинок с целыми пыльниками и пре-
вышает их, когда пыльники скручены, зеленовато-желтая, часто по
ребрам окрашена антоцианом, с почти сидячим рыльцем, но у неко-
торых форм этого вида под рыльцем видно сужение завязи, ясно обоз-
начающее столбик. Рыльце кремовое, его лопасти выдаются над стол-
биком.
Коробочка соломенно-желтая’ (3—9 см дл., и от 1,5 до 4 см шир.),.
продолговатая, в поперечном сечении трехгранная, сильно оттянутая
при основании и на вершине.
Карпофор, особенно у крупных коробочек, до 3 см дл. и 1,5 см
толщ., толще плодоножки, но значительно уже коробочки, т. е. относи-
тельно тонкий, округлый, окрашен в более светлый цвет, чем вся
коробочка.
Столбик, которым заканчивается вверху коробочка, часто очень
длинный (до 3 см), может быть и короче, форма его зависит от того,,
насколько постепенно переходит створка коробочки в столбик. У тупо
заканчивающихся створок столбик более короткий, а у оттянутых —
очень длинный и представляет собой неотделимую часть створки. На
столбике светлоокрашенном, как и вся коробочка, находится неболь-
шое коричневое вдоль центрального желобка наружу согнутое рыль-
це, лопасти которого прочно прикреплены к столбику и лишь немного
шире его.
Створки коробочки ланцетной или широколанцетной формы. Вдоль
их центра, как и у всех видов этого рода, проходит плоский, мало-
109
углубленный желобок, сильно варьирующий у разных форм по глу-
-бине и окраске; часто в верхней половине окрашен в коричневый
цвет. Иногда в верхней половине небольшой кусочек желобка имеет
вид рубцевой ткани, от которой отходят несколько направленных вверх
в виде елочки легких морщинок. Обычно они не окрашены. Верхняя
половина створки слегка морщинистая, особенно ребра коробочки у
разрыва.
Щечки коробочки большей частью довольно плоские, иногда чуть-
чуть морщинистые, плотные, жесткие без поперечной полосатости.
Края створок отделены от щечек довольно пологим и широким же-
лобком, выгнуты наружу, по краю часто окрашены в более темный,
чем вся коробочка, цвет и поперечно морщинистые. Место разрыва
розовато-палевого цвета, фетровой консистенции, довольно широкое
(до 0,7 см).
Коробочка открывается узкими клиновидными щелями, доходя-
щими до основания створок, а вверху, благодаря отгибанию верхних
частей створок наружу, образуется довольно широкий раструб.
Внутренние перегородки растрескиваются довольно глубоко, не-
сколько ниже выгиба.
Перегородки начинаются на внутренней поверхности от рыльца в
виде длинной несколько расширяющейся книзу полосы, увенчанной
широким гребнем. В том месте, где створка под рыльцем расширяется,
тяж пыльцепроводниковой ткани сужается до выгиба в перегородке,
образуя длинную узкую линзовидную фигуру.
Рубец от опавших листочков околоцветника, часто с неровными
краями, небольшой ширины (1— 2 мм), коричневого цвета. Плодоножка
под коробочкой толстая, но уже карпофора, быстро на протяжении
4 см еще сужается, и ниже на всем протяжении одинаковой толщи-
ны. Она сильно вырастает после отцветания растения и достигает до
30 см выс. от верхнего листа.
Семена светло-коричневые, крупные (1,3X0,8 см) в виде округло
треугольных, разной длины и ширины пластинок. С широкой, особенно
в верхней части семени, каймой. Кайма несколько перекошена и за-
гнута вниз. В верхнем более длинном углу эндосперма семени лежит
темный полумесяц. Зародыш длинный, хорошо виден с обеих сторон
семени.
В условиях Ташкента он зацветает в середине февраля, но не
позже первой декады марта.
Tulipa anadroma Z. Botsch. — тюльпан вверх стремящийся
(рис. XXII и 3, з). Образцы для исследования в 1953 г. привезла
Р. М. Мурзова из Средней Азии, с Чаткальского хребта, из лесни-
чества Аркит в окрестностях оз. Сары-Чилек (под ореховыми деревь-
ями). Эндем.
Луковида яйцевидная, до 3,5 см в диам., с коричневыми бумаго-
образными чешуями. Этот вид даже в культуре вегетативно не раз-
множается.
Растение на открытом месте достигает 30 см выс. от уровня почвы.
В тени деревьев оно может быть и выше. Стебель в верхней части
вместе с цветоножкой пушистый,, зеленый, без антоциановой окраски.
Листья, в количестве 3—4, вверх стремящиеся (нижний растет на
стебле на 2—3 см от уровня почвы). Они очень слабо отклонены от
стебля; верхние — к стеблю прижатые, зеленые, с сизым налетом, сла-
бо пушистые, по краю реснитчатые, не курчавые, а очень прямые, с
ясно выраженной центральной жилкой, видной с обеих сторон листа,
по ней резко килевато согнутые.
110
Рис. ХХП. Т. anadroma Z. В о t s с h. (3/4 натуральной величины).
Цветок возвышается над прижатыми к стеблю листьями, но иногда
они превышают его. Нижний лист килеватый, ланцетный, острый
(21 см дл., 7,4 см шир.).
Цветок одиночный, желтый, изящной лилейной формы, в начале
цветения его околоцветник образует двойной бокал, а затем растре-
скивается в виде звезды, с приятным медовым запахом.
Внешние листочки околоцветника ланцетные или продолговато
ланцетные, острые (до 7,5 см дл., 2,5 см шир.), со слабо развитыми
боковыми полями отгиба и широкой спинкой; по спинке широко с
частичным захватом боковых полей отгиба — розовато-малиновые, по-
крытые сизым налетом; лишь самый край отгиба в 1,5—2 мм вдоль
всего листочка ярко-желтый, основание листочка желтое, до 1,3 сжвыс.
Желтизна поднимается вверх от него по спинке узким острым клином,
2 см выс., у основания 0,4 см шир.; с внутренней поверхности этот
листочек околоцветника ярко-лимонно-желтый, такой же, как его края
снаружи; внутри нет никакого пятна и цвет листочка к основанию не
приобретает оранжевого оттенка, а остается таким же, как и в верх-
ней части листочка. Внутренние листочки околоцветника острые, очень
узко-продолговато-неправильно-ромбические, по длине равны внеш-
ним, до 7,5 см, и такой же как у них ширины, т. е. до 2,5 см\ с
обеих сторон, как и внешние листочки внутри ярко-лимонного цвета;
не имеют никаких пятен при основании листочка с обеих сторон и
лишь снаружи в верхней половине листочка во время цветения цент-
ральная жилка и верхняя часть отгиба с обеих сторон центральной
жилки краснеют. Эта краснота проходит в виде легких узких штри-
хов вдоль желобков, сопровождающих центральную жилку или сплошь
захватывает всю верхнюю половину листочка, оставляя свободными
только его края.
Тычинки до 2,5 см дл. очень узкие. Пыльники, как у Т. kauf-
manniana, открываются постепенно сверху вниз в течение 2—3 дней,
но не скручиваются спирально, а остаются прямыми. Тычинки в два
с лишним раза короче околоцветника, но выше завязи, в закрытом
цветке сближают свои вершины над ней, а в открытом расходятся.
Нити длинные, тонкие (1 см дл., 0,15 см шир.), кажутся прозрач-
ными, светло-желтыми, от основания к вершине постепенно суженные.
Пыльники линейные (1,5 см дл., 0,15 см шир.), желтые, матовые,
но другого цвета, чем нити.
Завязь бледно чуть-чуть зеленовато-желтая, сизоватая, к верху от
основания немного сужена (2,1 см дл.,0,55 см шир.). Рыльце шире за-
вязи и чуть-чуть желтее.
Коробочка тусклая, соломенно-желтая (до4,8сждл. и 1,9 см шир.),
узкой треугольно конической формы с узким вытянутым карпофором
(0,9 см дл., 0,6 см шир.). Но есть среди образцов этого вида много
коробочек, у которых карпофор почти не развит и створки растре-
скиваются до самого рубца от опавших членов околоцветника. Все
створки коробочек у этого вида широко- и узколанце^ной формы.
Коробочка очень узкая, сильно вытянутая, в верхней части без пе-
рехвата суженая под столбиком — верхней бесплодной конической
частью, длиною в две пятых створки. Ее венчают мало заметные, слив-
шиеся со створками, маленькие лопасти рыльца, одинаковой с коро-
бочкой окраски.
Коробочка открывается до основания конической щелью, узкой
внизу и широкой (до 3 см) вверху. Это достигается тем, что верхние
части створок сильно отклоняются наружу и даже слегка, в полвитка
111
штопоровидно изгибаются. Край створки, по линии разрыва выгнутый
наружу, ярко-палевый, матовый, узкий (до 0,2 см шир.).
В центре узколанцетных створок коробочки, как и в коробочках
всех видов тюльпанов, находится желобок, который у разных видов
имеет только этому виду свойственные признаки. Так, у описываемого
вида желобок пологий; в его средней части проходит узкая жилка или
рубчик, такой же окраски, что и створка. У Т. anadroma очень
велика верхняя бесплодная часть коробочки, на каждой створке она
занимает от одной трети до двух пятых всей длины створки. На этой
бесплодной части желобок уплощается и вся она плеская, покрыта
неровным как на рубце эпидермисом. В некоторых коробочках, на-
оборот, вместо желобка появляется пологий гребень, в других, жилка
раздваивается и между ее рукавами лежит рубцовая ткань.
Щечки створки, по сравнению с общей длиной коробочек, малень-
кие, более выпуклые при основании, чем в верхней части, почти без
морщинок, лишь в верхней части при внимательном осмотре можно
отметить мелкие морщинки, окрашенные в тот же цвет, что и вся
створка. Край створки выгнут наружу и отделен от щечек пологим,
но четко очерченным, желобком. В противоположность многим видам
тюльпанов, разрыв по ребру коробочки не окрашен в коричневый
цвет, и область створки под рыльцем такого же цвета, что и коро-
бочка.
Рубец от опавших членов околоцветника несколько углублен,
одного цвета с коробочкой. Но ниже его на ткани плодоножки, от-
ступя от рубца на 1 мм., проходит очень узкая светло-коричневая ли-
ния. Такой же линией у большинства коробочек заканчивается кар-
пофор над рубцом.
Плодоножка при культуре на совершенно открытом месте до
27 см дл., под рубцом резко сужается, той же окраски, что и ко-
робочка.
Семена очень меняются по форме и размеру (от 0,8X 0,8 до
0,4x0,3 см), почти округлые, узко-косо-треугольные, светло-и темно-
коричневые, но характеризуются сильно измятой каймой вокруг семе-
ни. У большинства семян зародыш плохо виден.
В условиях Ташкента этот вид зацветает во второй декаде апреля.
От Т. kaufmanniana этот вид отличается всем габитусом готиче-
ского растения: листьями с ясно выраженной центральной жилкой, по
которой они согнуты вдоль и, вследствие этого, ясной килеватостью;
отсутствием оранжево-желтого пятна при основании листочков около-
цветника или даже просто оранжеватости в нижней части листочка;
формой семени и каймой на семени; от Т. ferganica — более ранним
цветением; формой листьев; формой цветка и листочков околоцветни-
ка; способом открывания пыльников; формой коробочек.
Tulipa dubia V v е d.—тюльпан сомнительный (рис. 3, е и 5) оби-
тает „на мелкоземных и щебнисто-мелкоземных склонах в верхнем
поясе гор.— Ср. Азия: Тянь-Шань (Зап. Т.-Ш.). Эндем. Описан с
р. Чотан" („Флора СССР", т. IV).
Описанные растения мы собирали в Западном Тянь-Шане в Чим-
гане на вершине песочного перевала на склоне восточной экспозиции
и в ущелье Большого Чимгана у снежника в цветущем состоянии;
А. Я. Бутков привез их с Ангрена. Кроме того, мы выращивали их
из семян, собранных, как в естественных условиях, так и в Саду.
Луковица 1—3 см в диам. с черными или очень темно-коричневы-
ми оболочками (старые разложившиеся оболочки бурые) бумагообразной
консистенции, с внутренней поверхности покровная чешуя прижато
112
волосистая. Этот вид имеет во время расцветания цветы, сидящие в
розетке листьев и иногда поднимающиеся над уровнем почвы на
10 см. Во время цветения цветоножка значительно вырастает до 15 см,
после цветения продолжает интенсивно расти, достигая 25 см и более
от уровня почвы, очень пушистая, часто окрашена антоцианом в ро-
зовый или коричневатый цвет, покрыта сизым налетом.
Листья, в количестве 2—4, отогнутые, даже серповидно изогну-
тые, сильно сближенные, во время раскрывания цветка два нижних
листа основаниями погружены в почву, по краю волнистые, сильно
голубовато-сизые, особенно вдоль центральной жилки, по краю густо-
реснитчатые, на острие листа с пучком ворсинок. Самый нижний лист
широколанцетный 14X6 см\ верхний — 12x0,8 см.
Цветок одиночный, желтый, снаружи с сизо-розовыми полосами,
открывающийся широко звездой, с легким приятным ароматом.
Внешние листочки околоцветника ланцетные (4X1,5 см) или узко-
неправильно-ромбические, на вершине заостренные, с боков округ-
лые, на вершине оканчивающиеся маленьким, часто пушистым кончи-
ком, от которого опушение спускается по спинке, от основания
листочка навстречу ему поднимается опушение, переходящее на ли-
сточек с цветоножки; в нижней трети внешние листочки околоцвет-
ника часто реснитчатые.
Снаружи внешние листочки околоцветника по спинке сизо-розовые,
к основанию в центральной части слегка желтеют, края лимонно-жел-
тые; с внутренней поверхности лимонно-желтые, при основании на-
ходится почти прямоугольное (1,6X0,8 см) оранжевое пятно. На вер-
шине это пятно тремя зубцами постепенно сливается с основным фо-
ном листочка, или переходит в широкую оранжевато-коричневатую
полосу, соответствующую внешней спинке, простирающуюся до крас-
ненького кончика листочка, тогда лишь края листочка остаются ли-
монно-желтыми.
Внутренние листочки околоцветника продолговатые, при основа-
нии б. м. постепенно длинно оттянутые или округло-продолговато-
ромбические, на вершине туповатые, а иногда и острые, разной интен-
сивности лимонно-желтые, постепенно в нижней половине переходящие
в оранжевый цвет; с внутренней поверхности того же оттенка, что и
снаружи, при основании до 1/3 или 1/4 выс. оранжевые с постепен-
ным переходом оранжевого цвета в основной фон листочка.
Тычинки примерно в два раза короче околоцветника. Нити мас-
сивные, к основанию расширены, на вершине в более узкой части
внезапно суженные, длиннотреугольные, голые, оранжевые.
Пыльники желтые, того же оттенка, что и листочки околоцветни-
ка, продолговатые, постепенно открывающиеся, как пыльники Т. kauf-
manniana, но не свертывающиеся спирально; на вершине с коротким
остроконечием того же цвета, что и пыльники. Внутренность пыльни-
ка более яркая, чем внешняя сторона. Пыльца такого же цвета, что и
внутренняя сторона пыльника.
Завязь в нижней части расширена, в основании внезапно сужена,
к рыльцу продолговато заострена, кремовая, покрыта сильным сизым
налетом. Рыльце желтое, сильно выступает над завязью.
Т. dubia отличается от Т. kaufmanniana более сизыми, почти го-
лубыми, узкими острыми листьями, с жилками, которые не видны в
мякоти листа, более поздним цветением, и в основном мелкими цве-
тами, с более узкими листочками околоцветника; пыльниками, откры-
вающимися так же, как у Т. kaufmanniana, но не свертывающимися
спиралью,
8—2229 113
T. dubia в Ташкенте зацветает в середине апреля, когда настоя-
щая Т. kaufmanniana и все гибриды с ней давно окончили цветение
и завязали семена.
Коробочка (6 см дл., 2,4 см шир.) продолговатая, на вершине и
при основании б. м. постепенно заостренная, белесовато-соломенная.
Карпофор коробочки несколько вздут, отделяется от створок коробочки
желобком, ниже которого имеет как бы припухлый вид и несколько
выдается над рубцом от опавших членов околоцветника; 1 см выс. и
1,5 см шир. вверху и 1 см над рубцом.
Верхняя бесплодная часть коробочки (1,7 см дл.) довольно длин-
ная, но не в виде столбика, а является б. м. постепенно заостренной
вершиной створки. Ее венчают коричневатые, несколько отогнутые
наружу лопасти рыльца. Они несколько шире суженной части створки
и легко отламываются. Створки коробочки растрескиваются по швам
и края швов несколько выворачиваются наружу, показывая внутрен-
нюю, довольно широкую розовато-желтую часть шва. Щель, в кото-
рую высыпаются семена, открыта вдоль всей плодущей части коро-
бочки, но довольно узкая, до 3 см выс. она расходится всего на 1 см,
а выше клиновидная; верхняя бесплодная часть коробочки на полови-
не длины довольно резко отклоняется наружу. Поэтому щель вверху
имеет как бы широко открытый зев в 3 см шир. Внутренние створки
коробочки отходят друг от друга в 3,5 см выс. от основания коро-
бочки.
Створки коробочки продолговатые, широколанцетные. Вдоль
створки проходит широкий углубленный рубец, в центре которого
приподнимается продолговатая складочка. Эта складочка чуть-чуть
заходит на карпофор коробочки. В верху же створки желобок несколь-
ко уплощается, а складочка исчезает, створки желтовато-соломенного
цвета. В средней части весь рубец широко окрашен в коричневый
цвет. Эта окраска не опускается на карпофор и только изредка под-
нимается на бесплодную верхнюю часть. От нее на щечки створки от-
ходят частые коричневые поперечные прожилки, сгущающиеся в верх-
ней половине.
Щечки створки довольно грубые, выпуклые, серовато-или соло-
менно-желтые; края створок грубые, по краю окрашены в коричневый
цвет, разлитый по направлению к продольному желобку, отделяющему
края от щечек. Окраска створок под рыльцем такая же, как и в верх-
ней части створок. Широкий рубец от листочков околоцветника шире
вверху и уже У тонкой соломинки. Соломина и основной фон коро-
бочки одного цвета, иногда серее.
Плодоножка выросла до 20 см выс., значительно тоньше карпо-
фора коробочки.
Семена (0,9 X 0,7 см) с широкой каймой, которая шире в верхней
части семени и круто загнута вниз.
В условиях Ташкента он зацветает во второй декаде апреля.
Tulipa tschimganica Т. Botsch.— тюльпан чимганский (рис. XXIII
и XXIV).
Исследованные образцы привезены из многих районов Западного
Тянь-Шаня — ущелье Большого Чимгана, долина Чаткала и Ак-Була-
ка; из Ташкентского заповедника.
Этот вид растет на каменистых, щебенчатых или песчаных почвах
не ниже 1400 м на ур. м. Такой предел обитания возможен в глубо-
ких долинах, где лежит скопившийся за зиму снег. Обычно он цветет
у снежников на почве, пропитанной талой водой, или на склонах,
поздно выходящих из-под снегового покрова.
114
Рис. XXIII. Т. tschimganica Z. Botsch. (3/4 натуральной
величины).
Луковицы яйцевидные, до 3,5 см в диам., с темно-коричневыми,
«бумагообразными покровными чешуями с внутренней поверхности,
•особенно у вершины и основания, покрыты редкими, длинными, пря-
мыми, прижатыми волосками.
Растения во время цветения достигают 26 см выс. от уровня
почвы. Цветоножка опушена и покрыта сизым стирающимся налетом.
Листья, в количестве 3—4, сближенные, серповидно отогнутые,
отклоненные, желобчатые, б. м. пушистые, реснитчатые, сизо-зеленые,
по краю светлоокаймленные, часто в начале цветения превышающие
цветок.
Нижний лист во время цветения основанием погружен в почву;
ъ дальнейшем отходит от стебля на уровне почвы, широколанцетный
(до 24 см дл., 7,5 см шир.). Верхний — узколанцетный (20 см дл.,
2,2 см шир.).
Цветок одиночный, красный с желтым основанием или желтый
с красными спинками внешних листочков околоцветника и красными
•обратно V-образными пятнами с внутренней поверхности, формы ши-
рокого конуса или звездочки. Есть пестрые формы.
Внешние листочки околоцветника (до 7 см дл., 2,8 см шир.) ши-
роко- или узколанцетные, на вершине постепенно заостренные. У
красных форм снаружи спинки сизо7малиновые, при основании до
1/5 выс. желтеющие, у желтых форм спинки внешних листочков око-
лоцветника красно-бордовые, при основании желтеющие, сплошь по-
крытые сизым налетом; с внутренней поверхности у красных форм
светло-бордово-оранжевые, к основанию окраска сгущается, при осно-
вании с желто-оранжевым веерообразным пятном. У желтых форм,
примерно, на одной четверти длины от основания листочка располо-
жено обратно V-образное красное пятно, варьирующее в размерах у
разных особей.
Внутренние листочки околоцветника продолговатые или узкояйце-
видные, на вершине б. м. округлые, часто с выемкой, бывают до 7 см
дл. и 3,2 см шир.; у красных форм снаружи оранжевато-бордовые,
к пятну темнеющие. При основании находится оранжевое, во всю
ширину основания, пятно. Внутренняя их окраска сходна с внешней.
У внутренних листочков околоцветника желтых форм с внутренней
поверхности более яркая окраска к основанию переходит в оранже-
вый цвет, на месте перехода желтого в оранжевый лежит V-образ-
ное красное пятнышко, несколько меньшего размера, чем на внешних
листочках околоцветника.
Тычинки выше завязи, почти в два раза короче околоцветника.
Нити желтые, с коричневатым верхним концом, внизу несколько
расширенные, вообще же тонкие, под пыльником постепенно заострен-
ные до 1,2 см дл.
Пыльники желтые или коричневатые, почти в два раза длиннее
нитей, постепенно сверху открывающиеся, но не скручивающиеся, как
у Т. kaufmanniana.
Пыльца желтая или коричневатая.
Завязь (2,3 см дл., 0,4 см шир.) желтовато-зеленая, часто по реб-
рам с коричневым оттенком, при основании сужена, под рыльцем
сильно оттянута в столбик. Рыльце шире завязи, его лопасти покрыты
желтыми сосочками, с боков иногда красные.
Коробочка соломенно-желтая (до И см дл., 2,7 см шир.). узко-
коническая, сильно вытянутая в длину, особенно велика ее верхняя
бесплодная часть — столбик. Карпофор (1,5x1,5 см) вверху довольно
115
тонкий, к створкам расширяющийся, округлый, отделен от створок;
б. м. глубо ким желобком, часто острым углом заходящий на створки.
Столбик до 3,5 см дл., в него, постепенно сужаясь, переходит
верхняя часть створки. Рыльце небольшое, согнутое вдоль, коричне-
ватое. Створки коробочки ланцетной или продолговато овальной фор-
мы. В центре створки проходит желобок, часто окрашенный в корич-
нево-бордовый цвет, который усиливается на бесплодной части.
Щечки створки выпуклые, слабо поперечно морщинистые, от края
створки отделены довольно глубоким желобком. Ребро створки иногда
окрашено в коричневый цвет. Разрыв его широкий, и края створки
как бы вывернуты наружу вдоль центрального желобка.
Коробочка открывается широкой клиновидной щелью с большим
раструбом на вершине благодаря сильному отклонению верхней
бесплодной части створки вместе со столбиком. Поверхность разрыва,
кремовая, матовая или палевая, до 0,7 см шир.
Рубец от опавших членов околоцветника коричневый, часто вы-
пуклый. Плодоножка более темная, чем коробочка, под рубцом ши-
рокая, ниже быстро утончается.
Семена крупные, коричневые, округло треугольные. Пленчатая
кайма, окружающая семя, довольно широка. Зародыш просвечивает
с одной стороны семени.
Этот вид зацветает в условиях Ташкента на 20 дней позже, чем
Т. kaufmanniana. С Т. kaufmanniana, особенно с желтоцветными и
красными формами, его сближает окраска листочков околоцветника.
Кроме того, способ открывания пыльников у него отличается от Т. ка-
ufmanniana только тем, что постепенно открывающиеся пыльники
не скручиваются спирально, как у Т. kaufmanniana, а остаются пря-
мыми, как у всех других видов тюльпанов.
В течение многих лет Т. tschimganica путали с Т. kaufmanniana»
от которого он отличается экологией, фенологией и некоторыми мор-
фологическими признаками.
Секция Lophophyllon Vved.
„Лист одиночный, на верхней поверхности с продольными курча-
выми гребнями. Оболочки луковицы с внутренней стороны волосистые.
Нити тыч. волосистые. Зв. с коротким стлб.“
Tulipa regelii К rash.— Тюльпан Регеля (рис. 6). Обитает „на
каменистых склонах — Ср. Азия: Прибалх. Эндем. Описан из Курту».
Андракай“ („Флора СССР11, т. IV).
Исследованные растения посева 1951 г. получены из семян расте-
ний, которые были выращены из луковиц, привезенных В. П. Бочан-
цевым из Казахстана со ст. Чокпар Туркестано-Сибирской железной
дороги, где они обитают на красных глинах. В 1956 г. сеянцы цвели
в первый раз.
Луковица яйцевидная, 1,5—3 см в диам., с бурыми кожистыми».,
продолженными оболочками. С внутренней стороны покровная чешуя
имеет у верхушки и основания прижатые короткие волоски. Высота
растения от уровня почвы 5—8 см. Стебель окрашен антоцианом.
Луковицы лежат на глубине 10—15 см, так как многие экземпляры
уже со второго года жизни не углубляются. Лист сизо-зеленый, по.
земле распростертый, часто одиночный (до И см дл. при 4 см шир.),
но бывают сильные растения, у которых развивается второй малень-
кий листочек. Первый лист превышает цветок, стеблеобъемлющий»,
продолговатый или широколанцетный, острый, голый, гладкий, сизый».
116
Рис. XXIV. Т. tschimganica Z. Botsch. (2/s натуральной
величины).
«с верхней стороны с продольными курчавыми, на 1—2 мм возвышаю-
щимися над плоскостью листа, гребнями, по краю листа и вершинам
гребней проходят красные, антоциановые полосы.
Цветок большей частью одиночный (у нас в культуре были экземп-
ляры с тремя цветками), белый с чуть розоватым оттенком, со слабым
весенним запахом.
Внешние листочки околоцветника ланцетные, на вершине за-
остренные (20—30 мм дл., от 5 до 8 мм шир.), снаружи по спинке
вишнево-зеленые, к краям розоватые, к ноготку коричневатые; с
внутренней поверхности фарфорово-белые с розоватым оттенком, на
ноготке несут ланцетное (16 X 4 мм) золотисто-желтое пятно.
Внутренние листочки околоцветника значительно шире внешних,
•округло-широко-дельтовидные, на вершине тупые, при основании от-
тянутые в ноготок, опушенные по краям как снаружи, так и с внут-
ренней поверхности белесоватыми ворсинками; снаружи белые, по
спинке проходит темно-зеленая жилка, которая с обеих сторон сопро-
вождается коричневато-лиловыми штрихами, т. е. листочек имеет на
белом фоне три продольных темных полоски; с внутренней поверхно-
сти фарфорово-белый, при основании с большим, широким, округло
ромбическим золотисто-желтым пятном (10x7 мм).
Тычинки с открытыми пыльниками почти в два раза короче око-
.лоцветника, но значительно выше завязи. Ширина нити почти рав-
на по всей длине, лишь на вершине внезапно сужающиеся в острый
кончик. Они опушены на 3/4 длины, при основании желтые, в верхней
трети оранжевые. Пыльники желтые, линейно продолговатые, в откры-
том виде 5—7 мм дл. Пыльца серожелтая.
Завязь (10X3 мм) зеленовато-желтая, часто по ребрам вишневая,
внизу слабо расширяющаяся, под рыльцем суженная в очень корот-
кий столбик. Рыльце желтое, чуть шире столбика.
Коробочки в 1957—1959 гг. не были получены.
Этот вид зацветает в Ташкенте в конце марта.
Секция Eriostemones В о i s s.
„Оболочки луковицы с внутренней стороны голые или волосистые
.-или паутинисто-шерстистые. Цв. у большинства видов мелкие, невзрач-
ные. Нити тыч. опушенные, у одного вида (Т. sogdiana) голые. Зв.
с почти сидячим рлц или с б. м. длинным стлб., но всегда более ко-
ротким, чем сама зв.“
Tulipa silvestris L.— тюльпан дикий встречается „в лесах; иногда
как сорное,—Общ. распр.: Западная Европа. Описан из Южной Европы.
(„Флора СССР11, т. IV).
Описываемые растения получены из Института растениеводства
Эстонской АН от В. Н. Есиновской. Они растут в Эстонии, по ее сло-
вам, на лугах.
Луковица до 2 см в диам., с черно-бурыми, кожистыми, с внут-
ренней поверхности у вершины и основания густо-прижато-волосисты-
ми оболочками.
Растение 46 см выс. от уровня почвы. Стебель вместе с цвето-
ножкой голый, в нижней части окрашен антоцианом. Цветоножка
очень изящная, тонкая, но крепкая, наверху коричневая. Листьев 3,
темно-зеленые, сизоватые, расставленные, б. м. отклоненные, желоб-
чатые, голые, по краю узко красноватоокаймленные, гладкие, иногда,
«•очень редко, реснитчатые, не превышающие цветы.
Нижний лист линейный, на вершине постепенно заостренный, от-
117
ходит от стебля на уровне почвы (24 см дл. на 2,4 см шир.). Верхний
(18,5 см дл. на 1 см шир.) линейный, также на вершине постепенно
заостренный.
Цветок в виде изящной золотистой звездочки, с острыми долями.
Внешне листочки околоцветника (5,5 см дл., 1,6 см шир.) лан-
цетные, на вершине постепенно заостренные, на ноготке реснитчатые,
снаружи по спинке широко по всей длине зеленоватые, особенно-
ярко-зеленые центральная жилка и основание листочка. Зеленое про-
дольное пятно по краям окрашивается антоцианом в розовато-корич-
невый цвет, самый край листочка по всей длине не шире 0,2 см,
чисто желтый. С внутренней поверхности они чисто желтые без вся-
кого пятна.
Внутренние листочки околоцветника такой же длины, как и
внешние (5,5 см дл., 2,6 см шир.), кажутся значительно короче внеш-
них благодаря тому, что они выпуклые, неправильно ромбические, к
вершине постепенно заостренные; с обеих сторон ярко-желтого цвета,,
лишь снаружи проступает толстая зеленая жилка, вдоль которой с
обеих сторон, особенно в нижней половине, проходят позеленевшие-
узкие полоски, иногда с краев окрашенные антоцианом; верхушка
листочка также может розоветь, особенно верх центральной жилки;
он б. м. внезапно оттянут в длинный ноготок, по краям которого гу-
сто покрыт длинными белесоватыми ресничками. Вершины листочков
околоцветника с внутренней стороны также густо и широко опушены,
длинными белесоватыми волосками, тогда как у других видов тюль-
панов кончик листочка околоцветника опушен снаружи и это опуше-
шение спускается по спинке к краям отгиба.
Тычинки (2,3 см дл.) в 2—2,5 раза короче околоцветника и
немного выше завязи. Нити (1,4 см дл, 0,3 см шир.) толстые, плоские,
ровные по всей длине, лишь при основании под густым волосистым
кольцом несколько расширенные, на вершине б. м. внезапно заострен-
ные, желтые. Пыльники оранжевые (0,9 см дл. при 0,35 см шир.),..
сразу по всей длине открывающиеся.
Пыльца оранжевая.
Завязь ярко-зеленая (1,6 см дл., 0,4 см шир.), под рыльцем б. м.
внезапно суженная в желтый столбик. Рыльце желтое, маленькое,
уже завязи, лишь едва покрывает столбик.
Коробочка у данных растений не развилась. В условиях Ташкен-
та зацветает в третьей декаде апреля.
Tulipabiebersteiniana Schult., — Тюльпан биберштейна (рис. 3, и)
произрастает „на степях, склонах, в кустарниках, на лесных поля-
нах, в долинах рек. — Европ. ч.: Верх. Волж. (ю. ч.), Сред. -Днепр.:
Волж.-Дон, Заволж. Причерн., Крым, Ниж.-Дон, Ниж.-Волж.; Кав-
каз: Предкавк., Даг., Вост. Закавк.; Зап. Сибирь: Верх.-Тоб., Ирт.(?);
Ср. Азия: Арало-Касп., Прибалх.(?). Общ. распр.: Балк.-Малоаз.(?)А
(„Флора СССР“, т. IV).
Исследованные образцы Т. А. Пономарева собрала в окрестно-
стях Берчогура и получила от Б. В. Сердюкова из Тбилисского бо-
танического сада. Кроме того, у нас были растения, случайно при-
сланные И. А. Зубашко с другими растениями с Керчинского полу-
острова (с полуострова Камышбурун).
Получены от П. А. Гомолицкого 26 апреля 1957 г. из окрестно-
стей Актюбинска.
Луковица яйцевидная, 1—2 см в диам., с кожистыми коричневы-
ми или черно-бурыми оболочками с внутренней стороны у верхушки..
118
и при основании прижато волосистыми, а часто почти голыми, с внут-
ренней поверхности блестящими, особенно у кавказских форм.
В момент зацветания растения не превышают 20 гж, но затем
они сильно растут и к концу цветения достигают 35 см. Стебель
вместе с цветоножкой голый, коричневато-бордовый.
Листья, в количестве 2—3—4, сизовато-зеленые, отклоненные или
отогнутые, разделенные длинными междоузлиями (2,5 см, 1,8 см), не
опушенные, по краю иногда красно окаймленные (у крымских форм),
гладкие, иногда реснитчатые, ремневидно-желобчатые, на вершине
заостренные, как бы сросшиеся в короткий клюв.
Нижний лист более широкий, в начале цветения еще не целиком
вынесен на поверхность почвы (13 см дл , 1,8 см шир.), верхний —
15 см дл. и 1,2 см шир.
Цветов 1—5; перед цветением всегда и во время цветения, при
пасмурной погоде, поникающие, имеют форму широко открытой золо-
тистой звезды, со слабым приятным медовым запахом.
Внешние листочки околоцветника ланцетные, острые, на ноготке
реснитчатые (2,2X0,7 см) (крымские мельче берчогурских и кавказ-
ских).
Снаружи по спинке широко коричневато-зеленые, часто с фиоле-
товым оттенком, с более яркой зеленой жилкой и желтым (до 1 мм
шир.) краем; с внутренней поверхности золотисто-желтые; при осно-
вании зеленоватые.
Внутренние листочки околоцветника широкояйцевидные, иногда
продолговато яйцевидные, на вершине б. м. постепенно заостренные,
при основании внезапно суженные и оттянутые, с внутренней поверх-
ности на суженном основании реснитчатые; снаружи — желтые с яр-
кой зеленой жилкой, около которой основной фон листочка несколь-
ко зеленеет. Жилка расширяется к основанию и несколько краснеет
на вершине. С внутренней поверхности золотисто-желтые, на нижней
суженной части с зеленоватым оттенком.
Тычинки с открытыми пыльниками в два с лишним раза короче
листочков околоцветника и немного превышают завязь. Нити желтые,
при основании светло-желтые, толстоватые, разной длины: 3 и 1 см
в одном цветке, на вершине под пыльником внезапно суженные в
острый кончик, на вершине нижней четверти расширенные и одетые
густым волосистым кольцом. Пыльники продолговатые, широкие, в
2—3 раза короче нитей (например, 0,3—0,4 см дл.х0,2 см шир.).
Завязь ярко-зеленая, чуть сужена к вершине (1,3 см дл. X 0,3 см
шир.) с почти сидячим, желтым рыльцем.
Растения из окрестностей Актюбинска отличаются более узкими
листьями и размеры всего растения у них значительно меньше. Вы-
сота растений 16 см.
Длина, см Ширина, см
Лист нижний 14,0 0,55
„ верхний 10,0 0,15
Внешний листочек околоцветника 2,7 0,60
Внутренний в „ 2,6 1,10
Тычинка 1,3 —
Нить 0,9 .—
Пыльник 0,4 0,10
Завязь 1,2 0,30
Коробочка соломенно-желтая с легким оранжевым оттенком
(3,2 см дл. и 2 см шир.) овальной формы.
119<
Почти совсем без карпофора, т. е. створка непосредственно при-
мыкает к рубцу у плодоножки. Коробочка имеет (до 0,2 см дл.) по-
добие столбика.
Рыльце коричневое, маленькое, сидячее на слегка оттянутом кон-
чике створки. У многих коробочек оно сидячее и столбиком являет-
ся короткая перетяжка на кончике створки под рыльцем.
Коробочка открывается широкой, клиновидной щелью, доходя-
щей до основания створок, без раструба на вершине (2,8 см дл.,
1,2 см шир. на вершине).
Внутренние перегородки растрескиваются на 1'3 длины, т. е. не-
сколько ниже изгиба на вершине, под рыльцем в плоскости разрыва
с ромбическим расширением (0,7X0,7 см}, ниже которого и по изги-
бу край перегородки ровной толщины.
Створки коробочки короткой широколанцетной формы, грубые,
плотные, тусклые. Желобок в центре створки неровен и извит, глуб-
же в центре створки и выпуклый на вершине и основании. Вдоль
него проходят одна или две выпуклые жилки, делающие на пути к
вершине несколько изгибов. Они одинакового цвета со створками ко-
робочки, которые светлее к основанию и темнее в области попереч-
ных морщинок.
Щечки створки нечетко выражены, их фактически нет, так как
створка несколько выпуклая над центральным желобком, без желоб-
ка перед ребром коробочки и непосредственно переходит в рубец на
краю створки. От центрального желобка к ребрам коробочки на
площади примерно двух верхних третей створки отходят поперечные
морщинки, направленные несколько вниз, а вверху — чуть-чуть вверх
Верхняя часть коробочки более оранжеватая, а морщинки светло-ко-
ричневого оттенка.
Край створки у самого ребра не выгнут, узко окрашен в свет-
ло-коричневый цвет, проступающий и с внутренней поверхности
в месте разрыва ребра, которое имеет матовую оранжеватую по-
верхность и совсем не вывернуто наружу. Несколько тупо заострен-
ная верхняя часть створки под рыльцем самая темная во всей ко-
робочке. От рыльца отходят две жилки, которые сходятся вместе в
центральном желобке створки. Между ними образуется узкое (0,1 см
шир., 0,7 см дл.) клиновидное углубление.
Рубец от опавших членов околоцветника узкий, выгнутый, туск-
ло-светло-коричневый, отделяет коробочку от тонкой длинной (26<?лг),
вверху с оранжеватый оттенком белесоватой плодоножки.
Семена темно-коричневые, округло-косо-треугольные, с очень
узкой, направленной вниз каймой. Зародыша в них не видно.
В условиях Ташкента этот вид зацветает в начале апреля.
Tulipa patens Agardh. — тюльпан поникающий растет: „На сте-
пях, каменистых склонах, солонцах. — Европ. ч.: Ниж.-Дон, Ниж.-
Волж.; Зап. Сибирь: Верх. Тоб., Ирт., Алт. (ю. -з. ч.) Эндем“. („Флора
СССР", т. IV).
Описанные растения получены в 1933 и 1954 гг. из Семипалатин-
ска, окрестности Бобровки, от Ермакова и из Ленинградского бота-
нического сада одна луковица от 3. М. Силиной.
Луковица 1—2 см в диам. с тонкокожистыми, черно-коричнева-
тыми или серовато-коричневыми, несколько продолженными, почти
на длину луковицы чешуями. Покровные чешуи луковиц с внутрен-
ней стороны у верхушки и при основании прижато волосистые, сте-
бель голый, окрашен антоцианом, 10—21—25 см выс.
Листьев 2—3 в природе, у нас — 4. Нижний лист расположен на
120
уровне почвы. Между ним и верхними листьями находятся вытянутые
междоузлия (4,2 и 15 см), т. е. листья расставленные, признак редко
наблюдаемый у среднеазиатских тюльпанов из этой секции. Он встре-
чается еще у близкого вида Т. biebersteiniana и у Т. buhseana.
Листья отогнутые, голые, по краю красно окаймленные, реснит-
чатые или гладкие, узкоремневидные, на вершине заостренные. Ниж-
ний— 24 см дл., 1,7 см шир. Верхний —22 см д,д., 0,7 см шир.
Цветок обычно одиночный, но в культуре их бывает 2—3, в бу-
тоне поникающий, во время цветения слегка выпрямляющийся, с
нежным приятным ароматом, свойственным в различных оттенках
-большинству видов тюльпанов из этой секции, произрастающих на
территории СССР. У растений, присланных из Бобровки, цветки были
белые, а из Ленинграда — нежно-сиренево-розовые.
Внешние листочки околоцветника ланцетные, заостренные (4,5 х
0,9 см), по спинке желто-зеленые, к основанию желтеют, центральная
жилка ярко-зеленая, края широко сиреневые. С внутренней поверх-
ности нежно-розово-сиреневые, при основании с большим (1,1 х0,6см)
овальным, желтым пятном.
Внутренние листочки околоцветника неправильно ромбовидные,
с сильно заостренным и оттянутым отгибом и укороченным ноготком
(3,9X1,8 см) Наиболее широкая часть листочка на вершине нижней
трети; снаружи ярко-розово-сиреневые, при основании с большим,
четко отграниченным, занимающим всю площадь основания до 1,1 см
выс. ярко-желтым, веерообразным пятном; с толстой зеленой цент-
ральной жилкой, по обеим сторонам которой на желтом пятне прохо-
дит по полосе темно-коричневых штрихов.
С внутренней поверхности листочек такого же яркого розово-
малинового цвета. При основании у него так же как и снаружи рас-
положено яркое, веерообразное, большое, до 1,3 см выс. желтое
пятно. По обеим сторонам пятна, в нижней части внутренний листо-
чек околоцветника покрыт одиночными ресничками.
Тычинки выше завязи, но в два раза короче околоцветника. Ни-
ти (1,2x0,15 см£) желтые, вверху оранжевые, плоские, при основании
в волосистой части чуть-чуть расширенные, на вершине б. м. посте-
пенно заостренные. Пыльники продолговатые, раскрытые, они в два
раза короче нитей (0,6 мм дл.).
Завязь (2 см дл., 0,3 см толщ.) ровная, палочкой, ярко-зеленая,
с почти сидячим кремовым рыльцем.
Коробочка продолговатая, соломенно-желтая, к вершине с корич-
невым оттенком (2,5 см дл., 1 см шир.), сильно сужена к основанию
и образует довольно длинную, узкую бесплодную часть при основа-
нии — карпофор, ничем не отличающийся от створок коробочки. На
вершине створки коробочки б. м. постепенно переходят в короткий
до 0,25 см столбик, который несет на своей вершине маленькое, ко-
ричневатое засохшее рыльце.
Коробочка открывается узкой клиновиднсй щелью, доходящей
до карпофора. Щель почти не образует раструба на вершине коро-
бочки. Внутренние перегородки растрескиваются до изгиба. Под рыль-
цем в плоскости изгиба они образуют продолговато ромбическое рас-
ширение. Ниже этого расширения до самого основания коробочки пе-
регородка больше не утолщается.
Створки коробочки широколанцетной формы, довольно тонкие,
блестящие или тусклые. Желобок в центре створок значительно
углублен. Он более выпуклый на карпофоре, а также и на су-
женной части, образующей столбик. Часто желобок на створке бы-
121
вает окрашен, особенно в трех верхних четвертях длины, в светло-
коричневый цвет.
Щечки коробочки плоско выпуклые, отделены очень узким же-
лобком от ребра коробочки, но ничем не отчленяются от суженного
карпофора и столбика. На них со стороны ребер коробочки из же-
лобка приподнимаются поперечные морщинки в верхней части коро-
бочки, достигающие желобка, проходящего в центре створки. На вер-
шине коробочки, особенно в суженной части створки под столбиком,
эти морщинки расположены чаще, чем в верхней части щечек и со-
вершенно исчезают в нижней. Край створки у самого ребра чуть-чуть
отогнут и окрашен узкой полоской в коричневый цвет. Поверхность
разрыва створки очень узенькая, светлая, матовая и не вывернута
наружу.
Карпофор очень изменяется по длине и ширине (достигая 0,6 см
дл., 0,2 см шир.), отличается от створки тем, что по нему в центре
проходит не желобок, а наоборот, сильновыпуклая жилка, которая,
входя в желобок створки, постепенно заостряясь и утончаясь исче-
зает.
Рубец от опавших членов околоцветника узкий, выпуклый, риф-
леный, светло-коричневый, шире нижней бесплодной части коробочки,
отделяет ее от несколько расширяющейся кверху, длинной (до 22 см),.
тонкой плодоножки, которая в верхней части темнее щечек коробочки.
Этот вид зацветает в условиях Ташкента в начале апреля.
Tulipa sogdiana В ge. ~ тюльпан согдийский растет „Впесчанныхг
реже глинистых или каменистых пустынях. — Ср. Азия: Кыз.-Кум,
Кара-Кум, Аму-Дар. Эндем“. („Флора СССР", т. IV).
Исследованные и описанные растения получены от А. Д. Пятае-
ва. Западный Копет-Даг, хребет Кюрен-Даг, окрестности родника
Доната, пески.
Луковица яйцевидная, 1 — 1,5 см в диам., с жесткими черно-корич-
невыми (более молодыми) или сероватыми (очень старыми), несколь-
ко продолженными чешуями. На покровной чешуе, с внутренней по-
верхности, особенно у верхушки в продолженной части и при осно-
вании, есть прижатые волоски.
Стебель 15 см выс. от уровня почвы, голый, сизый с двумя ли-
стьями, нижний из них прикреплен на стебле на 1—2 см от уровня
почвы; верхние —на 2—3 см над ним, т. е. листья — расставленные.
Они сизоватые, голые, по краю слегка реснитчатые или голые, линей-
но-ланцетные, отходят от стебля под острым углом и выше слабо
отклоняются. Нижний лист 15 см Rji., 1,6 см шир. Верхний — 11 см
дл. и 1 гм шир.
Цветок одиночный, фарфорово-белый с розоватыми краями, очень
красивой формы в виде чаши, несколько суженной на вершине, с до-
вольно сильным приятным ароматом.
Внешние листочки околоцветника ланцетные, всегда уже внут-
ренних, на вершине заостренные (2,6X0,8 см), снаружи по спинке
широко-коричневато-зеленоватые, в то же время, с легким розоватым
оттенком, края фарфорово-белые, у самой кромки чуть-чуть розова-
тые, при основании с овальным желтым пятном, величиной 0,7 х
0,3 см.
Внутренние листочки околоцветника весьма разнообразной фор-
мы — от узко-продолговато-дельтовидных (2,8x1,2 см) до широких
округлых (2,6X1,7 см), лишь в нижней трети б. м. внезапно узко и
длинно оттянутых, на суженной трети в нижней части можно наблюдать
реснитчатость; на вершине листочков внезапно возникает маленький,.
122
острый кончик. Как те, так и другие листочки очень выпуклые нару-
жу, что и создает оригинальную форму цветка. Снаружи и с внутрен-
ней поверхности матово-фарфорово-белые, лишь на вершине отгиба
слегка розовеющие. Снаружи вдоль всего листочка проходит зеле-
новато-желтая центральная жилка, на ноготке зеленоватость захва-
тывает прилежащие вдоль жилки участки и слегка просвечивает
внутреннее желтое пятно. С внутренней поверхности в нижней части
листочка почти до 1 см выс. во всю площадь лежит ярко-желтое
пятно, иногда оно несколько выпуклое, иногда на выпуклой вер-
шине слегка выемчатое, но всегда четко отграниченное от основного
фона листочка.
Тычиночные нити (0,6x0,15 см) ярко-желтые, к расширенному
основанию более бледные, от него вверх лишь очень слабо сужаю-
щиеся и под пыльником б. м. внезапно переходящие в тонкое острие.
У многих живых растений из этого местообитания на тычиночных
нитях мы обнаружили на расширенном основании, особенно со сто-
роны обращенной к околоцветнику, бледно-желтые, разбросано рас-
пределенные ворсинки опушения. Ничем иным эти растения от типич-
ного, т. е. описанного во “Флоре СССР" Т. sogdiana, не отличались,
видимо, существует в этом виде переход от растений с голыми тычи-
ночными нитями к растениям с нитями опушенными.
Пыльники ярко-желтые, продолговатые (0,4x0,18 см), с коротким
остроконечием. Пыльца желтая. Завязь короче (0,8 X0,3 см) тычинок
с почти сидячим рыльцем, под которым сужена в очень короткий
столбик, окрашена антоцианом в розовый цвет.
Коробочка желто-соломенная с коричневым узором, маленькая
(1,4 на 1,2 см), почти шарообразная или короткоовальная, без карпо-
фора. Створки ее круто изгибаются к рубцу от опавших членов около-
цветника, тупо заканчиваются на вершине у внезапно возникающего,
короткого (до 0,08 см) тоненького носика—столбика с сидящим на
нем коричневым маленьким рыльцем. Коробочка . растрескивается по
ребрам клиновидными щелями. Внутренняя перегородка растрескива-
ется вдоль всей длины. Она как бы не имеет тяжа пыльцепроводни-
ковой ткани.
Створки коробочки округлой формы, хрупкие, бумагообразные.
Вдоль створки проходит узенький, довольно глубокий желобок (по
всей длине он коричневого цвета), отходящие от него боковые мор-
щинки также коричневые. Край створки интенсивно коричневый.
Место разрыва узкое, кремовое, без бахромок. Рубец от опавших
членов околоцветника светлый, шире плодоножки и прижат к коро-
бочке.
Плодоножка беловато-соломенная, сразу же от коробочки резко
сужается. Семена коричневые, тусклые, разного размера (0,5 х 0,4 см),
округло треугольные, с маленькой неровной каймой. Зародыш не
виден.
Этот вид зацветает в условиях Ташкента в середине апреля.
Tulipa polychroma Stapf. —тюльпан многоцветный (рис. 7, г)
обитает: „На сухих склонах,—Кавказ: Предкавк. (вост.), Вост. Закавк.
Общ. распр.: Иранск. Описан из Ирана: Шураб“ („Флора СССР11, т. IV).
Описанные растения в количестве 150 штук получены из Ереван-
ского Ботанического сада от А. Ахвердова, который выкопал их во вре-
мя цветения в Катавском районе в окрестностях Еревана, сел. Аван,
среднегорный пояс, 700 м над ур. м. Каменистая полынная полупус-
тыня на сухих глинистых и каменистых местах, 31 марта 1953 г.
Луковица яйцевидная, 1 — 1,8 см толщ., со светло-коричневыми
123
или бурыми, тонкокожистыми, слегка продолженными, изнутри тонко
паутинисто-шерстистыми чешуями. Волоски опушения очень длинные,-
тонкие, извилистые, особенно густо собраны в верхней трети луко-
вицы, с двумя запасающими чешуями. Стебель 6—17 см выс., так. же
как и бордово-коричневая цветоножка, голый. Листьев два, б. м.
расставленные, ремневидные, желобчатые, к вершине постепенно
заостренные, серповидно отогнутые, голые, сизо-зеленые, по краю
окаймленные, узкой, красной каймой, гладкие или слегка реснитчатые.
Цветки 1—2, внутри белые с желтым дном, снаружи серовато-
,лиловые. Цветок образует изящную красивую двойную чашу с при-
ятным довольно сильным запахом.
Внешние листочки околоцветника продолговато яйцевидные,
15—35 мм дл.; 6,5—8,5 мм шир., при основании с бородкой, отогну-
тые, с ровным краем, снаружи белые, коричневато-бордовые по спин-
ке, к основанию и вершине зеленоватые; внутри—белые с золотисто-
желтым, четко отграниченным пятном при основании. Пятно на вер-
шине выпуклое, занимает всю ширину основания листочка до 8 мм выс
Внутренние листочки околоцветника широко ланцетные, на но-
готке внезапно суженные, при основании, особенно на краях, опу-
шенные густыми ресничками; на вершине заостренные, выпуклые
с ровным краем, 3—10 мм дл., 20—14 мм шир. Снаружи белые с
зелено-фиолетовой жилкой, а часто и узким зеленым (до 2 мм) пят-
ном, поднимающимся до половины высоты, на две трети высоты зе-
леного пятна во всю ширину нижней части листочка желтые, нечетко
отграниченные; с внутренней поверхности—белые, при основании с
большим, до высоты 12 мм, веерообразным, с выемкой на вершине,
четко отграниченным золотисто-желтым пятном.
Тычинки выше завязи, но с сухими пыльниками, примерно в два
раза короче околоцветника. Нити тонкие в виде ровной палочки, на
вершине внезапно заостренные, при основании с волосистым кольцом,
золотисто-желтые, до 11 мм выс. Пыльники продолговатые, до 2—7 мм
дл., желтые, с очень коротким черным остроконечием.
Завязь зеленая, 8—12 мм выс. на 1,5—2 мм шир., равномерно
утолщенная по всей длине и постепенно суженная к столбику, имеет
короткий, неясно выраженный столбик с незначительными лопастями
рыльца.
Коробочка соломенно-желтая с коричневатыми узорами, малень-
кая (1,7 х 1,3 см), овальной, коротко овальной или почти шарооб-
разной формы. У нее нет карпофора, как у коробочек других видов.
Створки коробочки при основании круто, почти под прямым углом,
изгибаются к рубцу от опавших членов околоцветника; так же тупо
она заканчивается на вершине. Ее венчает внезапно возникающий,
короткий, до 0,1 см дл., тоненький носик-столбик с сидящим на нем
неприметным рыльцем.
Коробочка растрескивается по ребрам, доходящим до основания
широкими клиновидными щелями. Внутренняя перегородка растрески-
вается более чем на половину длины. По всей длине разрыва по
краю внутренних перегородок проходит утолщенный тяж пыльцепро-
водниковой ткани. На перегородках еле-еле приподнимается изгиб.
Отсутствие изгиба во внутренней перегородке обусловливает слабое
отклонение наружу вершины створок.
Створки коробочки овальной формы, наибольшей ширины в центре,
хрупкие, бумагообразные. Вдоль всей створки проходит узенький,
довольно глубокий желобок, по всей длине окрашенный в коричневый
цвет. В верхней половине створок в нем появляется невысокий гре-
124
бель, а по боковым морщинкам от него отходят узкие, коричневые,
поперечные линии, которые на вершине коробочки сливаются в сплош-
ное коричневое пятно, иногда створка под рыльцем по желобку
раздваивается.
Щечки створки узенькие, довольно выпуклые, соломенно-желтые,
в нижней части они гладкие, а в верхней половине морщинистые,
поперечно полосатые, коричневые.
Край створки широко окрашен в коричневый цвет, слабо выгнут
наружу, желобка почти нет. Место разрыва створки широкое—у такой
маленькой коробочки оно имеет 0,15 см шир., светло-палевое.
Рубец от опавших членов околоцветника коричневый, чуть шире
плодоножки, прижат к коробочке. Плодоножка сразу ниже коробочки
резко сужается до 6 см дл., светло-коричневая. Семена разного раз-
мера, 0,5 см дл. на 0,4 см шир., но все округлые или округлотреу-
гольные, тусклые, с маленькой каймой вокруг зерна, с почти непри-
метным снаружи зародышем.
Он зацветает в Ташкенте в начале или середине марта.
Tulipa buhseana Boiss.—тюльпан бузе (рис. 3, «) растет: „На
песчаных и глинистых почвах,—Ср. Азия: Арало-Касп., Прибалх.,
Кыз.-Кум., Горн. Турки., Тянь-Шань (предгорья с севера). Общ.
распр.: Джунг.-Кашг., Иранск. Описан из Кара-Чоката". („Флора
СССР1*, т. IV).
В примечании к описанию этого вида А. И. Введенский пишет,
что у этого вида разорван ареал: растения из северной части ареала,
в общем, отличаются от растений южной части ареала.
Северную расу он хотел выделить в особый вид под названием
Т. talievii Vved. У нас есть образцы из северной и южной частей
ареала.
Описываемые растения взяты из пустынных горок Алымтау, ле-
жащих к северо-западу от Ташкента, из предгорий хребта Каратау
с западной стороны и с плоскогорья Карой в хребте Каратау, лежа-
щем между двумя цепями гор.
Луковица яйцевидная, 1—1,75 ел: толщ., с бурыми, коричневатыми,
несколько продолженными, кожистыми чешуями. С внутренней сто-
роны у верхушки ее покровная чешуя покрыта тонкошерстистыми,
белесоватыми, плоскими, закрученными штопором вокруг оси, по
слабо извитыми волосками.
Стебель 10—40 см выс., голый (у наших растений вместе с цвето-
ножкой пушистый), вверх направленный, все растение канделябро-
видное, розовато-коричневое.
Листьев 2, отходят от стебля несколько выше уровня почвы, до
5—7 см выс.; сильно расставлены по стеблю, вначале образуют
со стеблем острый угол, а затем сильно отклоняются и загибаются
вниз; значительно короче цветов, линейно ремневидные, желобчатые,
по краю красно окаймленные, голые, реснитчатые, очень узкие;
нижний несколько более широкий, 3—6—10—15 мм шир. (20 мм
редко—лишь в культуре, например: 190 мм дл. при 14 мм шир.).
Цветы в количестве (1) 2—3—6(8—10—12—16), сидят на длинных
цветоножках без прицветных листьев. Особенно характерен для этого
вида большой отрезок общего стебля, приподнимающий высоко от
листьев (3—5 см) мутовку цветоножки. Цветок сужен при основании,
затем широко открыт, но околоцветник загнут несколько внутрь, т. е.
цветок колокольчатый, но смотрит вверх. Листочки околоцветника
кремовые.
125
Внешние листочки околоцветника острые, узколанцетные, при
самом основании часто бородатые (11 — 15—20—25—30 мм дл. и от
2 до 6 мм шир.); снаружи широко по спинке розовато-коричневатые,
к краям кремовые; с внутренней поверхности кремовые, с неболь-
шим (7x3 мм) желтым овальным пятном в нижней половине.
Внутренние листочки околоцветника также острые, по форме
несколько более широколанцетные, при основании оттянутые и сужен-
ные, на ноготке реснитчато бородчатые, по длине почти равны внеш-
ним (10—14—19— 24— 29 мм)-, при ширине чуть большей, чем у
внешних (от 3 до 10 мм). Снаружи—кремовые, центральная жилка
зеленая, при основании просвечивает желтое пятно, с внутренней
поверхности также кремовые, желтое пятно основания, четко отгра-
ниченное от основного фона листочка, имеет веерообразную форму
во всю ширину листочка и занимает площадь до одной четверти его
высоты.
Тычинки с раскрытыми пыльниками почти в два раза короче
околоцветника. Нити 5—10 мм., золотисто-желтые, довольно тонкие,
на вершине внезапно суженные в острый кончик, при основании
светлеющие, расширенные под волосистым кольцом, к. месту при-
крепления вновь суженные; выше волосистого кольца часто также
волосистые.
Пыльники линейно продолговатые, 4—6 мм дл., желтые или у
верхушки темные, с коротким остроконечием. Пыльца желтая или
грязновато-желтая.
Завязь немного короче тычинок, желто-зеленая, узкая, к рыльцу
сужающаяся в короткий столбик. Рыльце бледно-зеленое чуть шире
столбика.
Коробочка белесоватая с коричневой макушкой (до 2,1 см дл. и
1,2 см шир.), продолговато цилиндрическая, реже шаровидная, на
обоих концах тупая. При основании ее створки заканчиваются непо-
средственно у рубца опавших членов околоцветника, карпофора нет;
на вершине тупая, имеет маленький (до 0,1 см выс.) внезапно возни-
кающий тоненький столбик с крошечным сухим коричневым рыльцем.
Створки коробочки прямоугольные с округлыми гранями. Вдоль них,
в центре, проходит довольно резко прорезанный желобок, на три
четверти от макушки коробочки окрашенный в светло-коричневый
цвет, в нижней четверти коробочки он такого же цвета, как и вся
створка.
Щечки коробочки очень нежные, бумагообразные, выпуклые, со-
ставляют единое целое с краем створки, так как не отделяются от
него никаким желобком. Верхняя четверть коробочки до столбика
покрыта легкими поперечными морщинками, отходящими от желобка
и окрашенными в коричневый цвет. Под столбиком эти морщинки
сливаются, образуя единое коричневое пятно. Края створок по ребрам
коробочки, разделенные узкой щелью, также окрашены коричневым.
Коробочка растрескивается, но не раскрывается. Рубец коричне-
вый, от опавших членов околоцветника плотно прижат к коробочке,
очень узкий, но шире нижележащей плодоножки, которая, выходя
из него, резко сужается.
Плодоножки желтее, даже коричневее коробочек, тонкие, длин-
ные вверх направленные с общим стеблем до 8 см дл.
Семена (0,5 X 0,4 см) светло-коричневые, треугольно округлые на
верхних углах и острые у нижнего, окаймлены узкой каймой, с очень
длинным зародышем, сдвинутым несколько вбок семени.
В условиях Ташкента зацветает в начале третьей декады марта.
126
Растения Т. buhseana отличаются от других среднеазиатских видов
этой секции стройностью, канделябровидностью, формой цветов,
общей розовато-коричневой окраской всей особи и формой нераскры-
вающихся коробочек. Мы в декабре встречали в естественных усло-
виях коробочки этого вида в закрытом состоянии.
Tulipa buhseana Во i s s.—тюльпан бузе. По всему пути вдоль
.линии железной дороги на маршруте ст. Дарбаза—Байгакум были
собраны образцы луковиц этой формы. Растения оказались однотипны-
ми, с узкими желобчатыми, сизо-зелеными, голыми листьями, с крас-
ной каймой, по краю слабо реснитчатые, очень резко отогнутые,
сильно расставленные. Нижний—15x0,7 см.
Стебель вместе с цветоножкой достигает обычно 25 см выс.,
имеет хорошо развитый общий цветонос, на котором на длинных
цветоножках сидят довольно многочисленные цветы (до 9), которые
в бутоне и при отцветании поникают.
Цветки по форме широко открытые, звездчато колокольчатые.
Внешние листочки околоцветника узколанцетные, острые (2,1х
х0,55 см), снаружи серовато-розоватые, к основанию желтеют, с
внутренней поверхности кремовые, до 4,5 мм выс. желтые. Желтое
пятно на вершине выпуклое, занимает всю ширину листочка, при
основании окаймлено густой полоской из ресничек.
Внутренние листочки околоцветника сероватые, дельтовидные
'(2,IX 1,2 см), в нижней половине б. м. внезапно сужены и оттянуты,
снаружи кремовые, с серенькой центральной жилкой, на суженной
части как бы просвечивает желтое внутреннее пятно; с внутренней
поверхности они кремовые, до 5,5 мм выс. во всю ширину с желтым,
четко отграниченным, выпукло заходящим на основной фон, пятном.
На основании этого пятна несут реснички и густую бородавку волосков.
Тычиночные нити разной длины в одном цветке (0,7 и 0,5 см),
очень расширены под волосистым кольцом, выше резко сужены, до-
вольно тонкие, на вершине постепенно заострены, внизу бледные,
вверху оранжево-желтые, выше волосистого кольца также довольно
густоопушенные.
Пыльники продолговатые, до 3 мм рл., желтые, на кончике тем-
ные. Пыльца желтая.
Завязь в два с половиной раза короче листочков околоцветника
и немного короче тычинок, в форме палочки, светло-зеленая, под
рыльцем внезапно суженная в короткий столбик, рыльце и столбик
желтые.
Коробочка соломенно-желтая (1,6 см дл., 1,1 см шир.), треугольно-
округло-продолговатая, тупо закруглена на вершине и основании,
без карпофора. На вершине с коротеньким столбиком-носиком, всего
0,05 см выс., на котором сидит такого же цвета маленькое рыльце.
Коробочка открывается узкими коническими продольными щелями,
доходящими до основания.
Верхние части створки коробочки не отгибаются наружу, как
это бывает у других видов и поэтому щель на вершине коробочки
едва достигает 0,3 см. Внутренние перегородки коробочки расходятся
лишь на глубину расширенной клиновидной проводниковой ткани.
У перегиба створки, где кончается основание этого клина, перего-
родка цельная. Клиновидная часть створки коричневого цвета, с греб-
нем посередине, широкая {0,4 см), значительно шире рыльца (0,05 см),
и короткая '(0,5 см).
Створки коробочки овальные, вернее, прямоугольно округлые,
рубец на месте срастания плодолистиков довольно узкий, но глубо-
127
кий, окрашен узкой линией в желобке в легкий коричневый цвет.
Щечки створки тонкие, но очень плотные, выпуклые, гладкие.
Небольшое количество морщинок может быть на створке под носиком,
но чаще встречаются совсем гладкие коробочки светлого желтовато-
соломенного цвета. На самой вершине на уплощенной верхней части
коробочки сплошь коричневеют. Ниже эта сплошная окраска ослаб-
ляется и часто разбивается на несколько узких коричневых полосок.
Вдоль ребра, по которому растрескивается коробочка, проходит
узкая коричневатая полоска. Желобка между краем створки и ее
щечками нет, край лишь несколько выпрямляется и натягивается,
почти не выгнут и для того, чтобы увидеть место разрыва створки,
необходимо заглянуть внутрь коробочки. Створка на ребре коробочки
в месте разрыва тонкая, всего не более 1 мм ширины в средней
части, палевого цвета. Разрыв снаружи окаймлен коричневой каймой.
Несколько выпуклый, очень узкий рубец от опавших членов
околоцветника в виде узкой коричневой муфточки прижат к коробочке.
Плодоножка в два-три раза тоньше его, прочная, такого же соломен-
ного цвета, как и коробочка.
Семена темно-коричневые, косо треугольные (о,45 см дл. на 0,25 см
шир.), с узкой толстой каймой, под которой в длинном углу семени,
на вершине лежит узкий черный полумесяц из тканей, сохраняющих
хлорофилл после созревания семени. В эндосперме через эпидермис
хорошо заметен длинный (0,3 см), для семени такого размера,
зародыш.
В условиях Ташкента эта форма вида зацветает в конце марта-
начале апреля.
Tulipa talievU V v е d.—тюльпан талиева (рис. 3, л). Растения
получены в 1949 г. из окрестностей станции Чокпар Турксиба от
В. П. Бочанцева.
Луковицы яйцевидные, с красновато-бурыми, кожистыми, несколь-
ко продолженными чешуями. Покровная чешуя с внутренней поверх-
ности сплошь покрыта длинными, прижатыми волнистыми, но не пере-
путанными волосками, сгущающимися при основании и на вершине
ее и довольно редкими в средней части.
Стебель вместе с цветоножкой 11,5 см выс. от поверхности поч-
вы, зеленый, с чуть сизовато-розоватым оттенком, опушен на цвето-
ножках редкими волосками.
Листьев 2; превышающие цветы, расставленные, отклоненные от
стебля почти под прямым углом, сизо-зеленые, голые, по краю рес-
нитчатые, линейные, желобчатые.
Нижний лист серповидно изогнут, 18 см дл, 0,9 см шир.; верхний
17 х0,7 см.
Цветы в количестве 1 — 2—3, широко-колокольчато-открытые.
Внешние листочки околоцветника очень острые, узколанцетные
(2,1x0,5 см), снаружи серые, с розоватым оттенком, к основанию
желтеющие, с внутренней поверхности кремовые с желтым овальным
пятном (0,3 х 0,2 см). В нижней половине при основании желтого пят-
на проходит узкая выпуклая полоска из коротких, густых волосков.
Внутренние листочки околоцветника тоже очень острые, ланцет-
ные (2 см дл. х 0,7 см шир.), нижняя половина широкая, при основа-
нии к месту прикрепления внезапно сужены, с обеих сторон кремо-
вые, но снаружи проходит коричневато-зеленая жилка и как бы
просвечивает в нижней четверти внутреннее желтое пятно.
С внутренней поверхности, до высоты 0,55 см от основания лис-
точка, лежит выпуклое, занимающее всю ширину ноготка, четко от-
128
граниченное пятно. При основании пятна, особенно с боков, распо-
ложены целые пучки ресничек.
Тычинки с закрытыми пыльниками почти в два раза выше завязи.
После открытия пыльники сокращаются в длину и тычинки лишь не-
значительно превышают завязь.
Нити короткие (0,5 см), в нижней трети на расширенном основа-
нии с густым волосистым кольцом; выше кольца более ярко-желтые,
конические, покрыты редкими волосками. Пыльники желтые, продол-
говатые, пыльца желтая.
Завязь бледно-зеленая, 0,8 см дл., в форме узкой палочки с
коротким столбиком и кремовым рыльцем.
Коробочка белесовато-соломенная (1,6 см дл. при 1,1 см шир.),
продолговато овальной формы, тупо заканчивающаяся на основании и.
вершине; совершенно не имеет никакого подобия карпофора, створки
коробочки отходят непосредственно от рубца у плодоножки.
В выемке на вершине коробочки сохранился маленький (0,1 см
выс.) коричневый зубчик, который являлся столбиком. На нем сохра-
нились следы маленького темного рыльца.
Створки коробочки имеют вид прямоугольника, у которого углы
закруглены. Вдоль центра створок проходит довольно глубокий жело-
бок, окрашенный в коричневатый цвет. От этого желобка, особенно
густо в верхней половине створки, отходят поперечные морщинки,
проходящие через довольно выпуклые тонкие щечки створки. На ребре
коробочки нет желобков, отделяющих щечки коробочки у разрыва
створки. Край створки у ее разрыва на ребре коробочки окрашен в
светло-коричневый цвет. Место разрыва очень тоненькое, белесовато-
соломенное, матовое. Под рыльцем створка образует как бы выемку
того же цвета, что и вся створка.
Открывается коробочка глубокой клиновидной щелью, доходящей1
до основания створок и образующей широкий раструб. Внутренние
перегородки расходятся друг от друга почти на половину длины коро-
бочки (значительно ниже выгиба на перегородках) по тяжу пыльце-
проводниковой ткани. Этот тяж возникает под рыльцем в виде доволь-
но широкой линии, которая быстро сужается и ниже начала изгиба
проходит в виде легкого утолщения по краю перегородок. Рубец от
опавших членов околоцветника светло-коричневый, узкий, почти
скрыт нависшими над ним основаниями створок коробочки. От него
отходит плодоножка, ровная по всей длине и более желтоватая, чем
коробочка. Плодоножки коробочек у некоторых наших растений до-
стигают 9 см при общем цветоносе в 3,5 см.
Семена мелкие, темно-коричневые, округло треугольные. Зародыш
в них не просвечивает.
Этот вид в условиях Ташкента зацветает в последние дни марта.
Tulipa turkestanica Rgl.—тюльпан туркестанский (рис. 7, 6}
встречается „На глинистых и каменистых склонах от предгорий до
2500 м. — Ср. Азия: Пам.-Ал., Тянь-Шань (Моголтау, Паша-ата).
Эндем“ („Флора СССР", т. IV).
Описываемые растения выращены из луковиц, привезенных в
1949 г. из лесничества Аман-Кутан, расположенного в Зеравшанском
хребте в так называемых Самаркандских горах.
Луковица широко яйцевидная, до 3,5 см толщ., с несколько про-
долженными вверх, черно-бурого цвета, жесткими кожистыми, с вну-
тренней стороны у верхушки шерстистыми чешуями. Волоски опуше-
ния довольно длинные, но грубые, толстые и слабо извитые. На че-
9—2229
129
шуях у донца также есть небольшое опушение. Луковица взрослого
растения имеет две запасающие чешуи, дает пазушные детки.
Стебель 10—30 см выс., коричневато-зеленый, в верхней части
вместе с цветоножками опушен короткими, торчащими белыми волос-
ками. Ветки отходят от стебля одновременно, как в зонтике.
Листьев 2—3, очень редко 4, сизо-зеленые с бордовым оттенком,
расставленные, отогнутые, лежащие на земле, голые по окаймленно-
му красным краю, реснитчатые, с красным кончиком, линейные или
ремневидные, слегка желобчатые, 5—25 мм шир., превышающие
цветы.
Цветы кремово-зеленоватые, в количестве 1 —12, в форме звез-
дочки, но листочки околоцветника даже в совершенно открытом цвет-
ке несколько загнуты внутрь, с сильным весенним, но не очень прият-
ным запахом, в бутоне прямые, не поникают.
Внутренние листочки околоцветника острые, ланцетные (12—25 мм
дл., 3,5—6 мм шир. при основании), с небольшим кольцом ресничек,
снаружи зелено-фиолетовые, по краешку кремовые, с внутренней по-
верхности кремовые с зеленоватым оттенком, при основании несут
овальное, на вершине заостренное желтое пятно (8—11x4 мм). Оно
поднимается по листочку на две пятых его длины.
Внутренние листочки околоцветника широколанцетные, несколько
ложковидные, на вершине заостренные, при основании по краям слег-
ка бородатые, между бородками слабо реснитчатые, чуть-чуть короче
внешних (11—24 мм дл., 4—11 мм шир.); снаружи зеленовато-кремо-
вые, с большим веерообразным во всю ширину листочка нечетко
отграниченным, желтым пятном и зеленой центральной жилкой; с
внутренней поверхности кремовые. Желтое пятно при основании ли-
сточка поднимается на две пятых его высоты и имеет ромбическую
форму (13x10 мм; 8x7 мм) во всю ширину листочка.
Тычинки в расцветшем цветке почти в два раза короче около-
цветника. Пыльники линейно продолговатые, 3—6 мм дл., желтые, на
вершине фиолетовые с коротким остроконечием.
Тычиночные нити желтые, тонкие, у бледного расширенного осно-
вания с густым волосистым кольцом, выше до самого сужения на вер-
шине редко волосистые, 77 мм дл.
Завязь желтовато-зеленая (7 мм дл.х2 мм шир.), книзу расши-
ренная, к короткому столбику (1 мм) суженная. Столбик бледно-жел-
тый, рыльце чуть шире столбика.
Соломенно-белая коробочка этого вида (до 3 см дл. и 1,8 см шир.)
продолговатая, чуть заостренная на вершине и основании. Основанием
коробочки служит очень короткий карпофор, всего 0,7 см дл., причем
половину этой длины составляет острый зубец, вдающийся в створку
коробочки. Карпофор округлый, но сбоку, со стороны створки, он
имеет форму ромба с усеченной вершиной со стороны рубца от опав-
ших членов околоцветника.
Широколинзовидная створка коротко заострена на вершине и
заканчивается коротким (до 0,1 см) носиком-столбиком, на котором
сидит крошечное, потемневшее от времени, рыльце.
Вдоль створки проходит желобок и створки открываются не отги-
банием краев у щечек, а выгибанием наружу вдоль центрального
желобка всей створки. Створки очень тонкие, бумагообразные, сверху
донизу покрыты тонкими, веерообразно расходящимися от централь-
ного желобка, морщинками. Этот желобок наиболее углублен и окра-
шен в средней части и особенно у вершины створки перед началом
ее верхней уплощенной зоны под рыльцем.
130
Щечки створки плоские, слабо очерченные, белые, блестящие,
ст края створки отделяются легкой вмятиной. Край створки по ребру
скрашен тонкой коричневой линией. Она видна и со стороны разрыва
в матовой, кремовой и лишь по краю коричневой створке.
Коробочка раскрывается глубокими щелями, широко открытыми
ст основания благодаря отгибанию всей створки, но заплетенных тон-
кой паутиной волосков, выросших на внутренней стороне створок.
Семена могут высыпаться лишь в раструб, образованный разошедши-
мися верхними частями створок.
Внутренние перегородки в месте разрыва под столбиком ромбо-
видного сечения с гребнем посередине. Они растрескиваются лишь
до основания выгиба.
Рубец от опавших членов околоцветника очень узкий, коричне-
вый, заканчивает более толстый карпофор и из него как из муфты
выходит тоненькая светлая плодоножка.
Семя коричневое, довольно крупное для видов этой секции (0,7 см
дл., 0,65 см шир.), округло-косо-треугольное, с узкой мятой каймой.
Зародыш довольно хорошо виден.
В условиях Ташкента зацветает в середине марта.
Tulipa turkomanica В. Fedt sch.—тюльпан туркменский. Место
произрастания его — „склоны гор. — Ср. Азия: Горн. Туркм. Общ.
распр.: Иранск." („Флора СССР*, т. IV).
Описанные растения получены от А. Д. Пятаевой из Туркмении,
Западный Копет-Даг, с хр. Кюрендаг.
Луковица яйцевидная, 1,5—3 см в диам., с черноватыми или бу-
рыми чешуями, с внутренней поверхности покровная чешуя густо
покрыта тонкими паутинистыми перепутанными волосками.
Стебель окрашен антоцианом, голый, 15—19—30 см выс. Листьев
2; отходят от стебля на 2 см выс. от уровня почвы; расставленные
друг от друга на 2 см, отклонены от стебля почти под прямым углом,
голые, гладкие, или чуть-чуть реснитчатые, линейные. Нижний —
17X2,2 см; верхний — 15,5X1,8 см.
Цветы, в количестве 1—2—3, сидят на длинных цветоножках,
которые отходят от двухсантиметрового общего цветоноса.
Цветок широко колокольчатый, один из лучших среди видов этой
секции в Средней Азии, нежный, с приятным запахом.
Внешние листочки околоцветника ланцетные, на вершине очень
острые (3,1x0,8 см), снаружи красновато-коричневые, по спинке к
основанию зеленеющие; с внутренней поверхности белые с продолго-
ватым (до 0,7 см выс.) зеленовато-желтым пятном, овальной, но не-
четко отграниченной формы. В центре зеленоватого пятна, вверх
почти до его вершины проходит густая полоска опушения из белень-
ких волосков.
Внутренние листочки околоцветника (3,2x1,бел:) заостренно-
яйцевидной выпуклой формы, б. м. постепенно суженные к основа-
нию; снаружи они белые с розовато-коричневой центральной жилкой
и, веерообразным, зеленеющим в нижней половине до 0,6 см выс.
пятном. По всей поверхности пятна, особенно густо в центре, разбро-
саны белые волоски. По краям листочка расположены реснички.
Волоски опушения в центре листочка часто поднимаются выше жел-
того пятна.
Тычинки и тычиночные нити разной длины (0,6х0,8). Нити в
верхней части оранжевые, в нижней — сплошь опушенно светло-жел-
тые, линейно продолговатые. На желтых пыльниках видны черные
кончики.
131
Завязь немного короче тычинок (1,5 см дл.), с коротким стол-
биком.
В условиях Ташкента этот вид цветет в начале апреля.
Tulipa bifloriformis Vved.—тюльпан ложнодвуцветковый
(рис. 7, а) растет „На глинистых и каменистых склонах, от предго-
рий до 1900 м. — Ср. Азия: Сыр-Дар., Тянь-Шань (Зап.), Пам.-Ал.
Эндем. Описан из Ташкента" („Флора СССР", т. IV).
Описанные растения собраны в Ташкенте на склонах к арыку
Анхор; близ ст. Дарбаза Ташкентской ж.-д., на глинистых холмах в
урочище Кыз-Кудук; в Западном Тянь-Шане у подножия песчаного
перевала в Чимгане.
Луковицы округлые или яйцевидные с чешуями часто продол-
женными на высоту луковицы, иногда и выше (до 2 см в диам.),
часто вегетативно размножающиеся и образующие целое гнездо моло-
дых луковиц в старых чешуях. Покровные чешуи красновато-корич-
невые или розовые, старые — бурые, жесткокожистые, с внутренней
поверхности тонко паутинисто-шерстистые.
Стебель 10—25 см выс., часто во время цветения растений окра-
шен антоцианом в бордовый цвет, который смягчен сизым опушением,,
состоящим из густых, коротких, неразветвленных белых волосков.
Листьев у взрослых растений 2, расставленных, отогнутых, часто
не волнистых, иногда окаймленных красной каймой, реснитчатых,
ремневидных или линейно-ланцетных, часто превышающих цветы.
Нижний более широкий (10—15—25 мм шир., 10-20—25 мм дл.).
Цветы в количестве от 1 до 15 на одном стебле с приятным ме-
довым весенним ароматом, в форме широко открытой звезды. Внутрен-
ние листочки околоцветника бывают несколько согнуты и направлены
вверх, а верхние отклонены под прямым углом по отношению к
стеблю.
Внешние листочки околоцветника узколанцетные, острые, при
основании бородчатые (25—30 мм дл., 6—7 мм шир.), снаружи по
спинке коричневато-зеленые, спинка окаймлена красным, к краю кре-
мовые, на ноготке желтые, иногда по спинке ярко-антоцианово окра-
шенные, с внутренней поверхности кремовые, часто с легким зелено-
ватым оттенком, при их основании лежит остро или округло треуголь-
ное пятно, обращенное вершиной вверх (7x4 мм, 4x3 мм), золотисто-
желтого цвета.
Внутренние листочки околоцветника значительно, в полтора—два
раза, шире внешних (10—13 мм при длине 24—30 мм), широко-лан-
цетной, к ноготку резко сужающейся формы, имеют форму лодочки
или ложки, при основании с бородкой; снаружи кремовые, со слабым
зеленоватым оттенком, на ноготке желтые, центральная жилка зеле-
ная; с внутренней поверхности кремовые; при основании с четко от-
граниченным ромбическим (9x7 или 4х 4 мм) золотисто-желтым пятном.
По отцветании все листочки околоцветника розовеют.
Тычинки почти в два раза короче околоцветника (10—17 см дл.),
пыльники линейно продолговатые, после открывания 5—6 мм дл.,
желтые, иногда темно-бордовые (фиолетовые). Связник продолжен в
острый кончик, возвышающийся над пыльниками. У желтых пыльни-
ков он темный. Нити желтые (7—12 мм дл.), тонкие, у расширенного
основания с густым кольцом из длинных волосков, выше обычно воло-
систые, к вершине постепенно утончающиеся.
Завязь немного короче тычинок, округло-узко-коническая, бледно-
зеленая (10—15 мм выс., 2—3 мм шир.), с коротким (в 1,5—2 мм)>
132
•бледно-желтым столбиком. Рыльце мохнатое, шире столбика, но уже
завязи.
Коробочка (4,7x1,7 см) округлая или чаще продолговатая, при
основании постепенно, а на вершине внезапно суженная в короткий
носик. Самая широкая часть коробочки лежит в основании верхней
четверти. В незрелом виде часто ребра и центральная жилка створ-
ки коробочки с антоциановыми красными продольными полосами,
от которых в верхней трети коробочки отходят сгущающиеся к вер-
шине поперечные полоски. У зрелой коробочки все эти полосы не
исчезают, а приобретают коричневатый оттенок.
Створки коробочек тонкие, нежные, вверху часто поперечно мор-
щинистые. Вскоре по созревании коробочка раскрывается по швам
почти до основания, створки часто винтообразно скручиваются и
семена высыпаются.
Семена темно-коричневые-с чуть заметным зародышем, почти без
окраины (0,6 х 0,5 см).
В условиях Ташкента этот вид зацветает в начале марта.
Tulipa orthopoda Vved.—тюльпан прямоножка (рис. 5).
А. И. Введенский в примечании к Т. bifloriformis пишет: „Р. из Кара-
Тау очень приземисты и имеют очень сильно опушенные, обычно про-
долженные оболочки луковиц: сближенные, обычно превышающие
цветы, листья; бутон и отцветшие цветы торчащие" („Флора СССР",
т. IV).
Описываемые растения взяты из Казахстана, хребет Каратау на
подъеме плоскогорья Джон из ущелья Байджонсай на крупноглыби-
стом склоне с мелкоземом, среди камней в мелкоземе, выкопаны
19 июня 1956 г. Эндем.
Луковицы продолговато округлые, с чешуями, продолженными
на 1 —1,5 см. Чешуи красновато-коричневые или розоватые, кожистые
с внутренней поверхности очень обильно покрыты длинными, тонки-
ми, сильно извитыми, паутинистыми волосками. Опушение на чешуях
настолько густое, что через него не просвечивает оболочка.
Стебель коричневато-бордовый, густо опушен не ветвистыми отто-
шыренными волосками вместе с цветоножкой до 2 см выс., а все рас-
тение возвышается над почвой во время цветения первого цветка
всего на 3,7 см.
Листьев 2, они сближены, отогнутые, почти не волнистые, жело-
бчатые, на вершине туповатые, сизо-голубовато-зеленые, по краю
окаймлены красной каймой, нижняя поверхность сильно окрашена
антоцианом, довольно короткие, во время цветения первого цветка
•еще не совсем выходят из почвы (2 см первого листа лежат еще ниже
-ее уровня); в основании чуть приподнимаются над почвой, а затем
опускаются и лежат на ней.
Цветков —1—2, а в культуре их значительно больше, со слабым
запахом. По форме это широкая чаша, у которой как внешние, так
и внутренние листочки околоцветника слегка отогнуты и направлены
•вверх.
Внешние листочки околоцветника (25 мм дл., 5,5 мм шир.) лан-
цетные, с внутренней поверхности при основании с венцом из длин-
ных, белых, нежных волосков; снаружи по спинке широко зеленые,
•спинка окаймлена розовым фоном, постепенно к краю становящимся
•белым. С внутренней поверхности листочки белые, при их основании
•(до 0,8 см выс.) расположено эллиптическое, на вершине слегка за-
остренное золотистое пятно.
133
Внутренние листочки околоцветника в полтора — два раза шире
внешних (18 мм дл., 10 мм шир.), широко дельтовидные, внизу чуть
оттянутые и на этом оттянутом месте с внутренней поверхности рас-
тут белые волоски в виде полоски сгущающихся и удлиняющихся к
краям листочка; как снаружи, так и с внутренней поверхности белые
и с обеих сторон на них лежит вытянутое в поперечном направлении
широкоромбическое (7x8 мм) пятно, желтое снаружи листочков, с
внутренней же поверхности оранжевое и несколько более угловатое.
С внешней стороны на них ярко выступает окрашенная в зеленый
цвет центральная жилка.
Тычинки с сухими пыльниками выше завязи и на одну треть ко-
роче листочков околоцветника, разной длины (14 и 12 мм), откры-
ваются неодновременно: расположенные против внутренних листочков
околоцветника, имеют более длинные тычиночные нити (7 мм) и от-
крываются раньше; у расположенных против внешних листочков-
околоцветника тычиночные нити 5 мм дл. и пыльники открываются
позже. Пыльники желтые, линейные, продолговатые, после открыва-
ния 4,5 мм дл. Связник желтый, несколько возвышается над пыльни-
ком. Нити желтые, при основании кремовые, кверху постепенно
сужающиеся в очень тонкое оранжевое острие; при основании в кре-
мовой части, где растет масса густых, белесоватых волосков, сильно
расширенные (одна пятая всей нити). Эти длинные волоски густо
особенно по краям, на две трети вверх от кольца покрывают тычи-
ночные нити.
Завязь (11 мм дл., 1,6 мм шир.) ровная, чуть сужающаяся к
столбику с почти сидячим кремовым рыльцем. Рыльце не шире стол-
бика коробочки.
Коробочка белесовато-соломенная, со светло-коричневым окаймле-
нием (2,3 см дл., 1,8 см шир.), округлой или чуть-чуть продолгова-
той формы, без карпофора.
Створки на вершине округлены и увенчаны коротким, давно вы-
сохшим коричневым столбиком до 0,15 см дл., на котором находится
крошечное, такое же желтое и узкое, как столбик, рыльце. Створки
коробочки овальной формы, вытянуто округлые на вершине и при
основании.
В отличие от Т. talievii створка коробочки этого вида без выемки
на вершине, а наоборот, чуть-чуть заострена и продолжена в корот-
кий столбик (до 0,1 см дл.). В центре створки проходят желобки,
вдоль которых створки сильно вытянуты и составляют с ними единое
целое, без каких-либо щечек и желобков у краев и т. д. У централь-
ного желобка нижняя половина неокрашена, а верхняя, вплоть до
верхней бесплодной уплощенной, часто коричневой части створки под
рыльцем, довольно сильно окрашена в ярко-коричневый цвет. Эта
окрашенная часть желобка имеет вид рубцовой ткани до 0,2 см шир.
Створки тонкие, но плотные, почти плоские, лишь чуть-чуть вы-
пуклые в верхней части; в верхней половине у желобка с рубцовой
тканью возникает много мелких и более крупных морщинок, которые
расходятся б. м. веерообразно. Жилки, проходящие вдоль морщинок,
светло-коричневые. Край створки у разрыва с коричневой полосой,
вдоль нее по створке проходит более светлая полоска. Коричневая
полоса на краю створки видна и с внутренней поверхности створки
вдоль линии разрыва. Разрыв створки, за исключением этой полосы,
матовый, белый. Край створки сильно выгнут наружу, но не вдоль
краевого желобка, а вдоль центрального желобка створки.
134
Коробочка раскрывается широкими клиновидными щелями, дохо -
дящими почти до рубца опавших членов околоцветника. От еще
более широкого отклонения створки сдерживают перепутавшиеся
концами волоски с внутренней поверхности на гранях коробочки.
Внутренние перегородки расходятся до выгиба в них. Пыльце-
проводниковая ткань и верх перегородки в месте разрыва под рыль-
цем обратно яйцевидной формы, из острого конца которой тяж пере-
городки подходит к ее выгибу. Ниже перегородка цельная. Рубец ог
опавших членов околоцветника узкий, коричневый, но шире плодо-
ножки, плотно прижат к коробочке, но ее створки не нависают над
ним как у Т. talievii. Плодоножка тонкая, желтее коробочки, до
5,5 см дл. Общая плодоножка почти не развита, ее максимальная
длина — 1 см. Темно-коричневое, узко-косо-треугольное семяедо-
вольно широкой каймой. Зародыш через эпидермис не просвечивает.
В условиях Ташкента этот вид зацветает в середине марта.
Tulipa binutans Vved. —тюльпан дваждыпоникающий А. И. Вве-
денский в примечании к Т. bifloriformis охарактеризовал следующим
образом: „Р. из Александровского хребта имеют менее опушенные
оболочки луковиц; бутон и отцветающие цветы у них поникают11
(„Флора СССР", т. IV). Впоследствии в „Ботанических материалах
АН УзССР" № 13, 1952 г. А. И. Введенский описал его как само-
стоятельный вид.
Растения выращены из семян, собранных в горах в окрестностях
станции Тюлькубас Туркестано-Сибирской железной дороги.
Луковица яйцевидная или округлая, с несколько продолженными,
лишь у вершины и немного у основания опушенными грубоватыми,
прижатыми волосками, красно-коричневыми покровными чешуями.
Замещающая луковица цветущих особей на столонах уходит в глубь
почвы.
Стебли до 12 см выс. от поверхности почвы, вместе с цветонож-
ками сильно опушены короткими, белесоватыми, оттопыренными во-
лосками, окрашены антоцианом в красно-коричневый цвет.
Бутоны и отцветшие цветы у них поникают, что, между прочим,
свойственно еще некоторым видам этой секции, например, Т. buhseana.
Листьев 2; ремневидные, желобчатые, серповидно изогнутые, на
вершине заостренные, голые, по краю и на вершине красно-окаймлен-
ные, б. м. расставленные. Цветы звездчатые, аромат не установлен.
Внешние листочки околоцветника (2,6 х 0,6 см) узколанцетные,
по спинке широко зеленые, с красновато-коричневым оттенком, по
краю очень узко кремовые, с внутренней поверхности кремовые, в
нижней половине с небольшим (0,7 х 0,3 см) овальным, желтым пят-
нышком. При основании пятна расположена полоска из белесоватых
ресничек. Незначительное количество ресничек есть на краях в ниж-
ней части пятна.
Внутренние листочки околоцветника ланцетные, на вершине за-
остренные, но несколько более широкие, чем внешние и на ноготке
б. м. внезапно оттянутые (2,5X0,65 см); снаружи кремовые, с ко-
ричневато-зеленой жилкой, до 0,6 см выс., сплошь желтые; с внутрен-
ней поверхности кремовые. Весь листочек до расширенной верхней
части, т. е. до 0,7 см выс. желтый, выпукло четко отграниченный. У
основания листочка волоски растут целыми пучками, по краям они не
только образуют реснички, но переходят на внешнюю сторону, кроме
того, они рассеяны, но более редко чем в кольце, по всему основа-
нию листочков, занятому желтым пятном.
135
Тычинки превышают завязь, но почти в полтора раза короче
листочков околоцветника, неровной длины из-за неравных тычиноч-
ных нитей, тонких в нижней трети с густым кольцом опушения. Во-
лоски опушения поднимаются вверх до сужения нити в острие под
пыльником. Нити 0,85 и 0,6 см дл. при толщине выше опушенного
кольца в 1 мм.
Пыльники желтые, линейно продолговатые, до 5 мм дл. Пыльца
желтая.
Завязь зеленая, узкая, длинная, немного короче тычинок, с почти
•сидячим кремовым, шире завязи, рыльцем.
Коробочка белесовато-соломенная, со светло-коричневыми ребра-
ми (2,8 см дл., 1,6 см шир.) овальной формы, с очень маленьким
(0,3 см) узким, в виде ножки карпофором, но у некоторых коробочек
карпофор отсутствует.
На вершине коробочки, обычно тупо законченной, нет никакого
столбика и рыльце сидит на створке в углублении или на незначи-
тельной выпуклости.
Створки коробочки овальные, вдоль их проходит сильно углуб-
ленный в центральной части, слабо окрашенный в коричневый цвет
желобок. Внизу, где он уплощен, без особой окраски. Вверху жело-
бок сильно морщинистый. Эги морщинки переходят на створки. У не-
которых коробочек по морщинкам проходят очень тоненькие корич-
невые штрихи.
Щечки створки выпуклые, слегка вздутые, очень нежные, бума-
гообразные, блестящие, отделены от края глубоким желобком. Самый
край створки (ребро коробочки) слабо подкрашен в коричневый цвет.
Коричневая окраска ребер соединяется под рыльцем. Створки коро-
бочки растрескиваются очень глубоко до рубца у основания коробоч-
ки, а наверху образуют широкий раструб. Внутренние перегородки
растрескиваются до конца выпуклой части по пыльцепроводниковой
-ткани. Верхняя часть перегородок под рыльцем ромбовидно расши-
рена, по ней проходит гребень, который опускается ниже этого рас-
ширения. Рубец от опавших членов околоцветника узкий, 0,05 см
шир., выпуклый. Плодоножка тонкая, с желтоватым оттенком.
Этот вид в условиях Ташкента зацветает в конце марта.
Tulipa biflora Р а 11.— тюльпан двуцветковый растет „на солонцева-
тых степях, солонцах, глинистых склонах. — Европ. ч. Причерн. (??),
Крым (??), Нижн. Волж.; Зап. Сибирь: Верх.-Тоб. (юж. ч.), Ирт.
(юж. ч.); Ср. Азия: Арало-Касп. Общ. распр.: Джунг.-Кашг.“ („Фло-
ра СССР", т. IV).
Луковица яйцеобразная до 2 см в диам., с коричневатыми бумаго-
образными, несколько продолженными, с внутренней поверхности
тонко паутинисто-шерстистыми покровными чешуями.
Растение 13 см выс. от уровня почвы. Стебель вместе с цвето-
ножкой голый, коричневато-зеленый, особенно цветоножка у цветка,
•сизый.
Листьев 2, расставленные, отклоненные, голые — по краю иногда
-с редкими ресничками, сизые, на концах и по краю бордовые, линей-
ные, желобчатые, превышают цветы. Нижний лист 15,5 см дл.Х0,9сти
шир., верхний—12 см дл. на 0,55 см шир.
Цветы, в количестве 1—2, широко колокольчатые. Наружные
листочки околоцветника под прямым углом от стебля отогнутые, лан-
цетные (1,9 см дл., 0,4 см шир.), с внешней стороны серовато-зеле-
новатые или серовато-фиолетовые, по краю узкоокаймленные белым;
< внутренней поверхности бело-кремовые, в нижней половине до
136
0,6 см выс. с остроугольным желтым пятном, на котором всегда рас-
положена бородка из желтых волосков.
Внутренние листочки околоцветника остро-узко-ромбические
<(1,9 см дл., 0,7 см шир.), на вершине сильно заостренные, при осно-
вании б. м. внезапно длинно оттянутые, белые снаружи и с внутрен-
ней поверхности, с зеленой центральной жилкой, желтое внутреннее
пятно просвечивает снаружи; с внутренней поверхности до 0,65 см
выс. всю площадь занимает желтое, большое, остроугольное во всю
ширину основания, с бородкой волосков пятно. По краю листочки
реснитчатые.
Тычинки желтые (0,9 см дл.), в 2 раза короче околоцветника,
почти равны завязи.
Тычиночная нить желтая, тонкая, в основном ровная, кверху
внезапно заостренная, в нижней части под густым волосистым коль-
цом расширенная; под кольцом и ниже более бледная, чем в верхней
части, такое же бледное острие нити под пыльником. Пыльники жел-
тые (0,25 см дл.), продолговатые с небольшим желтым остроконе-
чием на вершине.
Завязь зеленая (0,75 X 0,2 см), к небольшому желтому столбику
б. м. внезапно сужена. Рыльце желтое, уже завязи.
Коробочка почти шаровидная или чуть-чуть продолговатая
<1,3 X 1,3 см), без карпофора при основании, округлая на вершине,
на которой внезапно возникает короткий (0,1 см дл.), тоненький, ко-
ричневый столбик, держащий остатки коричневого, маленького, плос-
кого рыльца; открывается расширяющейся кверху клиновидной щелью,
доходящей до основания створок.
Внутренние перегородки растрескиваются до изгиба. От рыльца
до изгиба проходит узкий тяж пыльцепроводниковой ткани, по кото-
рой и происходит разрыв. Эта ткань шире к основанию столбика и
клиновидно сужается к изгибу перегородки. Створки коробочки почти
«сферической формы, блестящие и полупрозрачные. Продольный же-
лобок в центре каждой створки довольно сильно углублен, окрашен
в коричневый цвет; в нем часто видна выпуклая продольная жилка.
Щечки створки поперечно-полого-морщинистые. Морщинки сгущаются
в верхней трети коробочки. У щечек нет желобка перед ребром ко-
робочки. Ребра часто бывают окрашены широкой полосой вначале в
лиловый, а затем по созревании коробочки в коричневый цвет. Край
створки, из-за отсутствия вдоль него желобков, не выгнут наружу.
Он тонкий, матовый. Под рыльцем находится самая темная часть ко-
робочки. Рубец от опавших членов околоцветника узкий, неровный,
уступом окружает основание коробочки. От него быстро сужаясь от-
ходит розоватая тонкая плодоножка.
Этот вид в условиях Ташкента зацветает в конце марта.
Tulipa dasystemonoides Vved. — тюльпан ложно-волосисто-тычи-
ночный (рис. 8). встречается „Альпийский пояс гор. — Ср. Азия: Тянь-
Шань. Эндем. Описан с Таласского Алатау: Майдантал" („Флора
СССР," т. IV).
Исследованные растения получены в 1954 г. от А. Я. Буткова с
Ангрена.
Кроме того, мы видели и выращивали растения этого вида, при-
везенные А. Я. Бутковым из окрестностей поселка Первомайского.
Луковицы яйцевидные (1 — 1,8 см в диам.) с черно-бурыми тонко-
кожистыми, изнутри густошерстистыми оболочками.
Стебель вместе с цветоножкой 5—8 см выс. от уровня почвы.
137
Цветоножка окрашена антоцианом в розовато-коричневый цвет, густо
опушена оттопыренными волосками.
Листьев 2, широколанцетные, показываются из почвы почти одно-
временно с цветами, зеленые, по краям окаймлены красным, реснит-
чатые, незначительно превышающие цветок.
Во время цветения нижний лист 4,7, 8,5 см дл., 1,3, 2,7 2,8 см
шир,; верхний лист 3,4, 5, 7,5 см дл., 0,9, 1,8, 1,5 см шир.
Цветов 1—2—3 на одном цветоносе. Общий стебель во время
зацветания растений не показывается из земли. Цветок имеет вид
узколучевой звездочки с нежным приятным ароматом, слабо напоми-
нающим шафран.
Цветы в момент раскрывания бледно-желтые, на второй день
становятся ярче.
Внешние листочки околоцветника узколанцетной формы, на вер-
шине с острым зубчиком, на ноготке реснитчатые (1,9 X 0,3 смт
12,7X0,5 см), снаружи по спинке зеленоватые, к вершине и краям
бордовые, по краю желтоватые, с внутренней поверхности желтые.
Внутренние листочки околоцветника овально ланцетные, к осно-
ванию постепенно оттянутые, реснитчатые и бородчатые (1,1x0,4 см)
и (2,7x1 см), в полтора —два раза шире внешних, снаружи желтые,
с яркой зеленой центральной жилкой, по бокам сопровождаемой
двумя узкими зелеными полосками, на вершине полоски и централь-
ная жилка розовеют, на ноготке — коричневеют; с внутренней поверх-
ности ровного желтого цвета, лишь на вершине совершенно бледные.
Желтые тычиночные нити (1X0,1 см) тонкие, нитевидные, в ниж-
ней четверти более бледные, волосистые, расширенные, так как это
свойственно нитям тюльпанов секции Eriostemones.
Пыльники узкие, линейно продолговатые, 0,6 мм ял., желтые, на
вершине с темным кончиком.
Завязь бледно-зеленая с кремовым рыльцем, на половину длины
пыльника, короче тычинок, с довольно длинным и толстым столбиком,
который зеленее завязи и отделяется от нее легкой перетяжкой.
Рыльце шире столбика.
Коробочки в условиях весны 1957 г., несмотря на искусственное
доопыление, не развились.
Tulipa neustruevae Р о b. — тюльпан Неуструевой. Во „Флоре
СССР** (т. IV) в примечании к описанию Т. dasystemon указывается,
что: „В Ферганском хребте, кроме настоящего Т. dasystemon, встре-
чается тюльпан, растущий, видимо, более низко, отличающийся более
крепкими кожистыми оболочками луковицы. Культурные экземпляры
показали, что у него ярко-желтые цветы. (Т. paradasystemon m. in
herb.)1*. Эндем.
Луковицы описанных растений были собраны Р. М. Мурзовой в
1953 г. в окрестностях оз. Сары-Чилек в Чаткальском хребте в оре-
ховом лесу. Мы выращивали их в Ботаническом саду АН УзССР.
Луковица почти шаровидная или яйцевидная (1—2 см толщ., 2 см
выс.) с черно-бурыми или бурыми бумагообразными оболочками. За
первой запасающей чешуей часто бывает детка, продолженная в мо-
мент расцветания первого цветка в узенький зелененький листочек до
9 см ял. Луковица у взрослого растения с двумя запасающими чешуями.
Стебель 10—20—30 см выс., так же как и коричневато-зеленая
цветоножка, голый.
Листьев 2, б. м. расставленные, отклоненные, но не лежащие на
земле, а приподнимающиеся под углом в 30° по отношению к стеблю,.
138
голые, по краю с красной каемкой, гладкие или слегка реснитчатые,
линейные, слабо или почти не волнистые. Нижний более широкий
(5—24 мм шир.).
Цветков 1—2—3, широко открытые звездочкой, золотисто-желтые,
с приятным медовым ароматом.
Внешние листочки околоцветника ланцетные, острые (15—30 мм
дл., 6—8 мм шир.), при основании без опушения; снаружи по спинке
коричневато-фиолетово-зеленоватые, по самому краю золотисто-жел-
тые, с внутренней поверхности яркие золотисто-желтые.
Внутренние листочки околоцветника равны по длине внешним, а
по ширине несколько шире их (1,1 —1,5 см шир.), широколанцетные,
при основании почти внезапно-длинно-оттянуты; оттянутое основание
все покрыто белесыми ресничками, особенно сгущающимися к краям;
снаружи желтые с зеленой центральной жилкой, которая от основа-
ния, примерно до одной трети высоты листочка, с боков сопровож-
дается продольными густыми зелеными штрихами, с внутренней по-
верхности ярко-золотисто-желтые.
Тычинки с раскрытыми пыльниками, почти в два раза короче
околоцветника. Пыльники продолговатые желтые до 5 мм дл., с очень
коротким желтым же остроконечием. Пыльца золотисто-желтая.
Тычиночные нити плоские, равномерно на две трети высоты по-
крыты тоненькими ресничками, в нижней трети кверху постепенно
расширяющиеся, в двух верхних третях кверху сужающиеся в очень
тоненький кончик.
Завязь (1,4X3 мм шир.) чуть короче тычинок с раскрытыми
пыльниками, постепенно к основанию расширяющаяся, зеленовато-
желтая, с довольно длинным (2 мм) бледно-желтым столбиком и не-
сколько выступающими над ним лопастями рыльца.
Коробочка соломенно-белесоватого цвета (2,7 см дл., 1,2 см шир.),
часто со створками неправильно ромбической формы с округлыми
боковыми углами.
Наиболее широкая часть коробочки лежит в основании верхней
трети, которая б. м. постепенно сужается в довольно длинный стол-
бик (до 0,5 см), но есть коробочки без столбика, а вся верхняя треть
коробочки постепенно к рыльцу сужается. Карпофор коробочки едва
достигает 1—2 мм. Створки коробочки нежные, бумагообразные. По
желобку и ребрам у нее проходят коричнево-красные, довольно тон-
кие полосы. Поперечные морщинки мелкие и более густые в верхней
части.
В условиях Ташкента этот вид зацветает в середине марта. Т. пеп-
struevae был намечен к описанию Талиевым (1930), затем А. И. Вве-
денским (1935), но описала его только Е. Г. Победимова (1949) по
выращенному из луковицы в оранжерее растению. У этого растения
в условиях оранжереи не развился цветонос. В культуре и в природе
также встречаются особи с недоразвитым цветоносом (рис. 3, а).
Вид описан по единственному растению, которое растрепали при
зарисовке; аутентичного экземпляра не сохранилось.
Tulipa dasystemon Rgl.—тюльпан волосисто-тычиночный (рис. 7, г).
Место распространения его: „Субальпийский и альпийский пояс гор.—
Ср. Азия: Пам.-Ал. Тянь-Шань. Общ. распр. Джун.-Кашг.“ („Флора
СССР", т. IV).
Описанные растения получены в июле 1955 г. от Ю. Мироновой —
Тянь-Шань, Заилийский Алатау: Алмаатинская область; выше Иссык-
ского озера; северо-восточный крутой склон. Щебень с мелкоземом,
выше осыпи, в негустом травостое.
139
В 1956 г. наблюдался большой выпад растений, зацвели они лишь
в апреле 1957 г. Кроме того, мы раньше исследовали и описали об-
разцы этого вида, собранные из тех же мест в 1935 г.
Луковица 0,9—1,5 см в диам. с черно-бурыми, а чаще со светло-
коричневатыми, бумагообразными, обычно голыми оболочками, в верх-
ней половине с отдельными белыми, прямыми прижатыми волосками.
Стебель во время цветения от поверхности почвы до 5 см выс.,
•так же как цветоножка голый. В верхней части цветоножка сильно
окрашена антоцианом в коричневато-бордовый цвет.
Листьев 2, во время цветения на половину длины еще скрыты
под землей, сизо-зеленые, красноокаймленные, гладкие. Нижний про-
долговатый, на верху б. м. внезапно оканчивающийся, 10 см дл. X
XI,1 см шир. Верхний — еще под землей, на 2 см отстоящий от ниж-
него, значительно более узкий, почти узколанцетный, более желоб-
чатый (8 см дл., 0,7 см шир.).
Цветок одиночный, довольно ярко-желтый. Во „Флоре СССР“
(т. IV) неверно указана окраска цветка этого вида. У него в действи-
тельности желтый цвет, а не бледно-желтый. На Б. Чимгане мы соб-
рали растения с белыми с серенькой спинкой и нежным приятным
ароматом цветками.
Внешние листочки околоцветника узколанцетные (1,9 см дл.,
0,4 см шир.), снаружи по спинке широко коричневато-зеленовато-
бордовые, только краешки отгиба у них желтые; с внутренней по-
верхности они чисто желтые, при основании ноготка с кольцом очень
нежных ресничек, под которыми часто лежит капля сладковатой жид-
кости.
Внутренние листочки околоцветника (1,75 см дл., 0,7 см шир.),
продолговато ромбические или широколанцетные, острые; при осно-
вании оттянуты в ноготок, который с обеих сторон по краям густо
покрыт тонкими длинными ресничками, с внутренней поверхности
кольцо ресничек смыкается; снаружи желтые, с коричневато-зеленой
центральной жилкой с обеих сторон, в нижней половине листочка
сопровождаемой продольными коричневато-зелеными штрихами, с
внутренней поверхности желтые, на ноготке чуть просвечивает цен-
тральная жилка.
Тычинки (1,6—1,1 см дл.) желтые, с закрытыми пыльниками не-
значительно короче околоцветника, с открытыми — почти в полтора
раза. Тычинки внешнего круга открываются в первый день цветения;
тычинки внутреннего круга (более длинные) — на второй день.
Тычиночные нити на протяжении всей длины почти ровной тол-
щины, даже под опушением не расширенные, лишь на вершине б. м.
внезапно суженные в острие, опушены по всей поверхности, желтые,
в нижней трети более светлые.
Пыльники (0,3—0,8 мм дл.) желтые, с коротким темным остро-
конечием, очень бросающимся в глаза у закрытых пыльников.
Завязь зеленая, с очень длинным столбиком, вся в 1 см, а стол-
бик 0,5 см, т. е. столбик равен половине ее длины. Столбик отделяет
от завязи перетяжка. Рыльце бледно-желтое, уже столбика.
Коробочка серовато, даже чуть-чуть розовато-соломенная (24 см
дл., 0,9 см шир.), обратно продолговато яйцевидная в поперечном
сечении треугольная, к основанию сильно оттянутая в узкий карпо-
фор 0,5 см дл. и 0,2 см шир. у основания; у перехода в створку
0,35 см шир. Между карпофором и створкой нет никакой перетяжки
и он является продолжением створок.
140
На вершине коробочка б. м. внезапно переходит в длинный
(0,5 см) и узкий (0,1 см) столбик. Крошечные остатки рыльца отва-
ливаются при раскрывании коробочки.
Створки коробочки серовато-розоватые, прорезаны вдоль узень-
ким желобком, который не заходит на карпофор и слабо пигменти-
рован в верхней части.
В верхней трети коробочки от желобка поперек створок отходят
5—6 тоненьких, окрашенных в коричневый цвет, морщинок. Края ство-
рок по ребрам коробочки вдоль разрыва узко, но достаточно ярко-
окрашены коричневым пигментом. Край створки не отделяется от всей
створки желобком, так же как и нижняя бесплодная часть, поэтому
нельзя говорить о щечках коробочки, так как створка представляет
собой единое целое, трудно разбиваемое на отдельные части. Она
очень нежная, бумагообразная, плоская.
Коробочка открывается узкими, кверху клиновидно расширенными
щелями, образующими на вершине нечто вроде раструба, возникшего-
из-за отклонения наружу бесплодных верхних частей створок. Внут-
ренние перегородки коробочки растрескиваются, примерно, на одну
четверть длины коробочки там, где по граням перегородки от рыльца
проходят узкие тяжи пыльцепроводниковой ткани. Часть створки, об-
разующая столбик, часто бывает окрашена так же, как и вся створка,
лишь самая верхняя часть темно-коричневая.
Рубец от опавших членов околоцветника темно-коричневый, в
0,05 см шир. Плодоножка чуть темнее коробочки, тонкая, 7,5 см pj\.
Семя темно-коричневое, округло треугольное, острое, в конце,
где лежит зародыш, с почти незаметной каймой и зародышем.
В условиях Ташкента этот вид зацветает в конце первой декады
апреля.
Tulipa tarda Stapf. — тюльпан поздний (рис, 7, д) встречается „На
скалистых и щебнистых склонах. — Ср. Азия: Тянь-Шань. Эндем. Опи-
сан из Ср. Азии: „Вероятно Туркестан“ и в примечании о нем же:
„Этот весьма интересный вид известен из немногих близко располо-
женных пунктов (Настек, Джол-Булак, Джиль-Арык)“ („Флора СССР“,
т. IV).
Исследованные образцы мы привезли из Киргизии — ущелье р.
Кастек в 10 км от селения Михайловского. Кустарниковая зона среди
камней и скал.
Луковица яйцевидная, 1,5—3 см в диам., с черно-коричневыми,
кожистыми, голыми оболочками.
Стебель вместе с цветоножкой достигает во время цветения 5—
11 см выс. от уровня почвы. Общий цветонос настолько короткий,
что кажется, будто цветы пучком выходят прямо из земли.
Цветоножки зеленые, пушистые.
Листья, в количестве 3—7, сильно сближены, б. м. отогнутые,
ярко-зеленые, голые, с бордовой каемкой по краю, часто реснитчатые,
слабо желобчатые, линейные, чуть превышающие цветы.
Нижний лист 12 х 1,5 см, верхний—-11,5X0,9 см.
Цветок белый, в форме простой широко открытой звезды, с очень
приятным нежным ароматом. На одном растении бывает до 15 цветов.
Внешние листочки околоцветника острые, широко ланцетные
(3,4 см дл., 1,3 см шир.), снаружи по спинке широко зеленые, кверху
с лиловыми штрихами, к ноготку желтеющие, по краю узкоокаймле-
ны белым, с внутренней поверхности с большим желтым, занимающим
большую часть листочка, широко ланцетным пятном (2,5 X 1,3 см).
141
Внутренние листочки околоцветника (3,5 см дл. на 1,7 см шир.)
острые, продолговато ромбические, ложковидно выгнутые, постепенно
к основанию длинно оттянутые, густо-длинно-реснитчатые, снаружи
нижняя часть листочка на две трети желтая, а верхняя — белая, пе-
реход между ними постепенный; с внутренней поверхности три чет-
верти листочка желтого цвета и лишь верхняя четверть его белая. Встре-
чаются формы с желтыми листочками околоцветника.
Тычиночные нити (0,8 см) желтые, от расширенного основания
конические. На основании находится кольцо из густых длинных блед-
но-желтых ворсинок, часть которых несколько приподнимается по
нитям.
Пыльники (0,5 X 0,15 см} желтые, окаймлены серой полоской.
Пыльца желтая или желто-зеленая.
Завязь бутылковидная, с довольно длинным столбиком, с желтым
рыльцем, которое шире столбика.
Коробочка (3,8 см дл., 1,8 см шир.) округло или продолговато
ромбическая, мелко-поперечно-морщинистая, с соломенно-белыми неж-
ными створками, при основании округлыми, на вершине оттянутыми
в столбик (до 1 см дл), который несет маленькое, не шире столбика,
рыльце. В центре створок коробочки проходит желобок, в верхней
половине окрашенный в коричневый цвет, от которого по поперечным
морщинкам расходятся тонкие коричневые штрихи, сгущающиеся к
вершине створки.
Коробочка почти без щечек. На месте щечек наблюдается вздув-
шаяся часть коробочки, под которой лежат семена. У этого вида
створки совсем плоские, но прозрачные, поперечно полосатые. Между
широким краем створки и щечкой нет привычного желобка, харак-
терного для других видов тюльпанов. Край створки отогнут, вместе
со всей половиной створки и более матовый, чем щечки. Перегиб
створки происходит по центральному желобку. Области под рыльцем
и при основании коробочки являются как бы продолжении края
створки. Место разрыва края створки на ребре коробочки сильно вы-
вернуто наружу, оно широкое (0,35 сж), матовое, бледно-соломенное.
А рубец по краю створки коричневатый. Щель между створками уз-
коклиновидная, у рыльца расширенная, благодаря сильному отклоне-
нию суженных частей коробочки (столбика) друг от друга. С внут-
ренней поверхности по столбику проходит гребень. Внутренняя пере-
городка начинается широким, крутым выступом у вершины гнезда с
семенами. Она клиновидно сужается книзу. Перегородка растрески-
вается до конца этого клиновидного сужения.
Рубец от опавшего околоцветника светло-коричневый, неровный,
выпуклый, к плодоножке суженный. Плодоножка тонкая, соломенно-
белая. У главного цветка 11 см дл.
Семена (0,6 х 0,5 см) очень темные, коричневатые, косо треуголь-
ные или широкояйцевидные, почти без каймы. Зародыш не просве-
чивает.
В условиях Ташкента этот вид зацветает во второй декаде апреля.
Глава IV
О ЦЕНТРАХ ФОРМООБРАЗОВАНИЯ ТЮЛЬПАНОВ
В СРЕДНЕЙ АЗИИ
характеризовать видообразование в роде Tulipa при помощи гео-
графического метода пытался в свое время В. И. Талиев (1930).
Он взял в основу виды этого рода, входящие в секцию Eriostemo-
Неплохо разобравшись в эволюции западной ветви секции, которая
называется Silvestriformis, В. И. Талиев не понял и не разобрался в
ветви, которая называется Bifloraeformis и направлена в Туркестан
и Арало-Каспийскую низменность. Эта ветвь в действительности со-
стоит из массы прекрасных, четко обрисованных как морфологически,
так и географически видов — Т. bifloriformis, Т. buhseana, Т. binu-
tans, Т. tarda, Т. turkestanica, Т. turkomanica, Т. sogdiana, Т. rege-
lii и др. и нескольких видов, которые еще нуждаются в географиче-
ском осознании — Т. orthopoda, Т. talievii, описанных А. И. Введен-
ским.
Кроме того, к ним примыкает целый узел новообразований на
северных склонах Памиро-Алая. В. И. Талиев не знал и не мог ви-
деть все виды в живом состоянии и нарисовать достоверную картину
эволюции секции этого рода, хотя и описывает в работе эволюцию
всего рода в целом.
В работе „Процесс видообразования в роде Tuljpa“ В. И. Талиев
обрисовал гипотетического предка тюльпанов. По его описанию при-
митивный предок в сем. Liliaceae должен быть крупным, многоцвет-
ковым, многолистным, полихромным, с крупными цветками, нектар-
никами, рыльцем, сидящим на длинном столбике, иметь большую
изменчивость в форме листа вплоть до широких сердцевидных листьев,
как у Lilium cardiocrinum.
В. И. Талиев придерживается взглядов признания в роде Tulipa
регрессивной эволюции, которая представляется ему в виде расщеп-
ления некоторого Центральноазиатского прототипа на несколько форм
и расселения их в западном и северо-восточном направлениях. „Выпа-
дение признаков при этом регулируется отбором и идет по линии
возникновения хотя и обедненных морфологически и физиологически
сочетаний признаков, но в то же время более специализированных
в смысле приспособления“ (стр. 117).
Разбирая вопрос о величине цветка, значительной у представителей
секции Tulipanum и мелкой у Eriostemones Талиев утверждает, что они
все же относятся к крупнейшим цветкам лилий как обедневшие родичи.
Это положение не вызывает особых возражений, хотя и не может
служить веским доказательством в пользу более древнего происхож-
143
дения крупного цветка у представителей рода Lilium, так как у Fri-
tillaria в широком линнеевском смысле цветы не крупнее, чем у Tulipa,
однако автор считает род Fritillaria более древним, чем Tulipa. У не-
которых видов тюльпанов из секции Leiostemones цветки более круп-
ные, чем из секции Tulipanum. Это Т. micheliana, Т. greigii, Т. foste-
riana. Особенно крупные цветы у этих растений в культуре. Дикие
виды тюльпанов очень отзывчивы на улучшения условий существова-
ния в естественных условиях и в культуре. На вспаханных участках
Т. greigji и Т. micheliana всегда крупнее, чем на целине. В условиях
культуры они часто увеличиваются в размере. Как культурные тюль-
паны, так и дикари становятся многоцветковыми (Бочанцева, 1953, 1954).
На основании опытов мы установили, что малое количество листь-
ев является действительно приспособительной реакцией тюльпанов на
условия внешней среды. Улучшением условий выращивания мы доби-
лись увеличения количества листьев у растений, привезенных в виде
луковиц из естественных условий произрастания.
Так, у Т. regelii, обычно имеющего в природе и в культуре все-
го один лист и цветок, мы получили растения с двумя листьями и
тремя цветками. У Т. kaufmanniana, Т. greigii, Т. fosteriana, Т. vveden-
skyi и многих других наблюдали заложение и развитие пазушных
почек на зеленом стебле. Из этих пазушных почек выросли побеги
с одним, двумя листьями, увенчанные пветами. При ухудшении ухо-
да в последующие годы эти же растения из пазушных почек не раз-
вивали ни добавочных цветков, ни листьев, т. е. они легко возвра-
щались в прежнее состояние.
Многоцветковость, свойственная в естественных условиях тюль-
панам секции Eriostemones, на которую указывал Талиев, по нашему
мнению, не является примитивной, так как обусловлена ветвлением
верхушечной почки, очень специализированной по типу заложения
цветов и поэтому не может быть примитивной и общей для всего
рода и почти никогда не встречается у представителей секции Leios-
temones. У видов этой секции — другой тип многоцветковости. Так,
Т. praestans часто в естественных условиях образует на ветках, вы-
ходящих из пазух зеленых листьев, до четырех верхушечных цветков
и, следовательно, в какой-то степени сохраняет древний признак рода.
Мы рассмотрели заложение цветков у имевшихся в Ботаническом
саду лилий и убедились в том, что у Lilium philippense Baker,
L. Martagon L. и L. tenuifolium Fisch, цветы на ветках выходят из
пазух листьев, и почти на каждой веточке, увенчанной цветком, один-
два листа, т. е. цветы лилий сидят на ветках, как у тюльпанов сек-
ции Leiostemones.
Развитие секций Eriostemones и Leiostemones рода Tulipa шло
разными путями. Они очень рано отделились друг от друга, но в то
же время нельзя не согласиться с В. И. Талиевым, что: „типичная
для тюльпанов одноцветковость без сомнения возникла из более или
менее многоцветкового соцветия отдаленных предков путем редукции".
Но так как усиленным питанием и поливами у тюльпанов достаточно
легко в наших условиях вызвать многоцветковость и более высокую
облиствленность, являющиеся в какой-то степени риверсиями, склон-
ность к которым сохранилась у многих видов этого рода—данные
признаки можно считать утерянными родом, видимо, в предшествую-
щую нашему времени ксеротермическую эпоху.
Не ставя перед собой такой задачи, как решение всего вопроса
о видообразовании в роде Tulipa, мы на небольшом количестве
признаков попробуем провести исследование о филогенетических свя-
144
зях распределения их по Средней Азии и изменчивости в зависимости
от местоположения вида. Для этого необходимо проследить изменчи-
вость признаков у тюльпанов в связи с их географическим распреде-
лением по Средней Азии с востока на запад.
В настоящее время не ясно, какие представители рода Tulipa мо-
гут быть найдены в Джунгарском Алатау, но по имеющимся сведе-
ниям, это тюльпаны с желтыми, а не красными цветами.
В окрестностях Бобровки у Семипалатинска растет белоцветный
Т. patens с сизыми спинками и, как свойственно всем тюльпанам этой
секции, с желтым пятном у основания листочков околоцветника.
В Джунгарском Алатау наблюдается Т. brachystemon желтая с ро-
зоватыми спинками внешних листочков околоцветника и красными
v-образными пятнами внутри; в Кетменьтау обитает Т. iliensis с жел-
тым околоцветником и тюльпан из секции Eriostemones — белый. Даль-
ше на запад в долине р. Или в песках мы встречаем Т. behmiana —
желтый и Т. talievii — белый. Южнее р. Или с востока на запад про-
стирается Заилийский Алатау, у подножия которого в невысоких гор-
ках ниже прилавков произрастает Т. kolpakovskiana с желтым и Т. ta-
lievii с белым околоцветником. Выше, в кустарниковой зоне гор,
обитает Т. ostrovskiana с красно-оранжевым, оранжевым и желтым
околоцветником. Еще выше, среди камней, растет Т. tetraphylla — ли-
монно-желтый. На вершинах боковых хребтов, часто вместе с эдель-
вейсами, встречается Т. dasystemon — бледно-желтый, а на альпийских
лужайках Т. heterophylla, или, как теперь ее называют, Eduardoregelia
heterophylla М. Pop, тоже желтый с сизыми спинками. В горах, ок-
ружающих Иссык-Куль, вплоть до верховьев р. Нарына, широко
распространен Т. tetraphylla — желтый. Там же обнаружен Т. dasys-
temon — желтый, Т. tianschanica — желтоватый и Т. heterophylla —жел-
товатый. В 1956 г. луковицы тюльпанов, выкопанные в Терскей-Ала-
тау, в зоне еловых лесов, зацвели желтыми цветами. Таким образом,
в горах, окружающих Иссык-Куль, на высоте 1607 м над ур. м. все
тюльпаны желтые или желтоватые. Первый оранжево-красный тюль-
пан на востоке ареала массового распространения этого рода отмечен
в Заилийском Алатау.
Значительно больше ярких тонов в окраске тюльпанов появляется
при дальнейшем продвижении на запад. Так, в Чу-Илийских горах и
в горках в Бетпак-Дале обитает Т. alberti с оранжево-красными цвет-
ками. Тут же, примерно несколько восточнее перевала Чок-Пар, встре-
чается Т. greigii с явным преобладанием оранжевого оттенка в осно-
вном цвете окраски листочков околоцветника и расплывчатым большим
желтым пятном на ноготках листочков околоцветника. Темное пятно
на них не черное, а коричневое или темно-коричневое, бледнеющее к
краям. Там же растет желтая Т. kolpakovskiana, кремовая, почти бе-
лая Т. talievii и Т. regelii с белыми листочками околоцветника.
На горках Алатау в 80 км от Фрунзе в сторону Алма-Аты отме-
чен Т. ostrovskiana оранжево-красный, такой же Т. greigii и один
белый тюльпанчик из секции Eriostemones. В долине р. Кемин, впа-
дающей в р. Чу, обнаружен оранжево-красный тюльпан, похожий на
Т. ostrovskiana и Т. tarda с белым околоцветником. На перевале Ка-
стек этой же горной системы мы собирали белый и желтый Т. tarda.
В долине р. Чу наблюдается красновато-оранжевый Т. greigii.
В Киргизском Алатау от Фрунзе до Джамбула (Каиндысай) встре-
чается оранжевато-красный тюльпан, близкий Т. ostrovskiana. По низ-
когорьям этой горной цепи также отмечен оранжево-красный Т. grei-
gii. На осыпях и среди камней в мелкоземе зафиксирован чаще жел-
10—2229 145
тый, чем красный тюльпан Т. zenaidae с маленьким черным четко
отграниченным пятном или без пятна при основании листочков около-
цветника. Там же есть и белые тюльпаны из секции Eriostemones.
В хребте Каратау, простирающемся с юго-востока на северо-за-
пад, мы обнаружили Т. alberti и Т. greigii с варьирующей от темно-
красновато-оранжевой, лимонно-желтой до почти кремовой окраской.
У светлых форм Т. greigii на середине внутренней стороны листоч-
ков околоцветника расположено ярко-красное пятно; у внешних ли-
сточков — красные спинки. Особенно изменяется окраска этих двух
видов на щебнях центрального плато, вытянутого параллельно двум
гребням, ограничивающим эту горную систему с востока и запада.
В Каратау растут два кремоватых тюльпана из секции Eriostemones:
Т. orthopoda — обитающий на крупноглыбистых склонах (гранитах) в
расселинах скал, заполненных мелкоземом, и Т. buhseana — на слан-
цах и щебнях в центральной и восточной части хребта.
В северной части Таласского Алатау по низкогорьям зарегистри-
рован Т. greigii — оранжево-красный, несколько более красный, чем
на р. Чу и в Актасских горах. В западной части хребта обитает кре-
мовоцветный с розовыми спинками Т. kaufmanniana, там же по низ-
когорьям распространен Т. binutans, а в высокогорьях Т. dasvstemon—
светло-желтый. На слегка солоноватых почвах вдоль Сыр-Дарьи ра-
стет Т. buhseana. В Алкакумах и других песках с обеих сторон реки
мы находили Т. borszczowii — темно-оранжевоцветный с красноватым
оттенком. До Кызыл-Орды вдоль всего восточного берега Сыр-Дарьи,
между Каратау и рекой, на юге — во всех долинах Западного Тянь-
Шаня, в нижних частях хребтов Каржантау, Угамского, Пскемского,
Чаткальского, Кураминского простирается ареал Т. greigii. Не ясно
распространение этого вида в Кураминском хребте, в частности на
склонах, обращенных к Ферганской долине.
В Моголтау обитает Т. magoltavica, сходный с Т. greigii западно-
тяньшанским, такого же ярко-красного цвета, а не красно-оранжевого,
как из восточных и северных частей ареала, с черным пятном, четко
отграниченным узкой желтой каймой. Подобный тюльпан встречается
в смеси с типичным западнотяньшанским Т. greigii на восточном бе-
регу Ангрена.
На всех хребтах Западного Тянь-Шаня растет Т. kaufmanniana,
как правило, с кремовыми, почти белыми, лишь при основании жел-
тыми, листочками околоцветника. Но часто встречаются формы этого
вида с красными, оранжевыми и желтыми цветками. Иногда попада-
ются пестрые формы, у которых по желтому фону проходят оранже-
вые или красные полосы различной интенсивности. В этих же горах,
но несколько выше по осыпям и карнизам скал обнаружен другой
вид Т. butkovii с ланцентными сизо-голубыми листьями, бордово-крас-
ными цветками, зацветающий в условиях культуры в Ташкенте на
месяц позже Т. kaufmanniana. Там же у линии снега растет Т. tschimga-
nica, относящийся к секции Spiranthera, с желтыми, оранжевыми и
красными цветами. До сих пор мы не встречали форм Т. tschimgani-
са с кремово, почти белоокрашенными цветами. Там же на высоте не
ниже 1800 м над ур. м. на рыхлых субстратах обитает желтоцветная
Т. dubia. Всюду на разных высотах встречается почти белоцветковый
Т. bifloriformis, который в некоторых местах Чаткальского хребта
дает формы, легко различающиеся в зависимости от места обитания
на различной высоте. Выше всех растет Т. dasystemon, по литератур-
ным данным, белоцветковый. Белоцветковые формы этого вида в жи-
вом состоянии мы наблюдали в 1959 г. в Чимгане. Совершенно до-
146
стоверно, что тот тюльпан, который за границей называют Т. dasyste-
mon —есть наш Т. tarda из хребтов, окружающих долину Чу. Он
действительно часто белый с большим желтым дном, но встречаются
и желтоцветные формы. Желтоватый, с нежнейшим ароматом, Т. da-
systemonoides А. Я. Бутков собрал на каменистых осыпях в верховь-
ях Ангрена. Раньше его привозили с Кызыгурта.
В бассейне Ангрена обитает ярко-красный Т. vvedenskyi, крас-
ный Т. greigii и полихромный Т. kaufmanniana. По западной окраине
Тянь-Шаня на низких холмах растет красный с черным основанием
Т. korolkovii. Там же довольно часто наблюдаются оранжево-желтые с
красным узором формы этого вида, ярко-красный Т. greigii и кремо-
вый Т. bifloriformis.
В Ферганской долине мы не встречали красных тюльпанов.
На южных склонах Чаткальского хребта обитают: Т. neustruevae—
ярко-желтый, Т. ferganica — желтый, Т. anadroma из секции Spirant-
hera— желтый. На Ферганском хребте Т. ferganica — желтый, neust-
ruevae — желтый, Т. dasystemon - желтоватый, Tulipa sp. из секции
Eriostemones — белый.
На северных склонах Алайского хребта произрастают Tulipa sp.
из Eriostemones — белый, Т. turkestanica, Т. dasystemon, Т. sp. близ-
кий к ferganica — желтый с узким листом и сильно опушенный. В
Ферганской долине, в окрестностях Намангана на низкогорьях встреча-
ется Т. sp. — желтый и Т. rosea, представляющий розовую вариацию
Т. Eorolkovii или более бледную форму его, потерявшую яркую крас-
ную окраску. Этот тюльпан известен из окрестностей кишлака Ким.
Выше Ферганской долины с верховьев Нарына (правый приток
Кекелемансая) в 1958 г. был привезен крупный цветок тюльпана
оранжево-красной окраски с желтым пятном при основании листоч-
ков околоцветника и, кроме того, с коричневыми и штриховыми пят-
нами на желтом фоне в нижней части внутренних листочков около-
цветника. Появление здесь тюльпанов с яркой окраской очень инте-
ресно. О других тюльпанах этой долины нам ничего неизвестно. На
возвышенности Белесны, как бы закрывающей вход в Ферганскую
долину, произрастает тот же Т. rosea — розовый, Т. lehmanniana —
желтый и Т. buhseana — кремово-белый.
Система Памиро-Алая резко отличается по видовому составу и
окраске листочков околоцветника населяющих его видов от преды-
дущих областей, тяготеющих к Тянь-Шаню.
На хребте Узунгыр (северная цепь Туркестанского хребта) на
высоте 200 м мы нашли красный тюльпан, похожий на тюльпан из
группы Т. fosteriana — Т. carinata, который мы называем Т. affinis. Там
же и несколько ниже рос Т. turkestanica — ярко-кремовый. По доро-
ге на Шахристан мы обнаружили Т. dasystemon — желтый. В заповед-
нике Гуралаш собрали два тюльпана: Т. dasystemon — желтый и Т. sp.
из секции Eriostemones — почти белый. Кроме того, сотрудники этого
заповедника сообщили, что в 18 км на север от опорного пункта
Кульсай есть два вида различных красных тюльпанов.
Из Мальгузарского хребта нам привозили Т. turkestanica — кре-
мовый. Возможно там находится и красный тюльпан — Т. micheliana.
В Нуратинском хребте растут: Т. micheliana —красный, Т. victoris—-
красный, вид, близкий к Т. fosteriana. Это наиболее восточное обита-
ние целой серии видов, объединяющихся вокруг Т. fosteriana. Из
всех видов, обитающих в горной системе Тянь-Шаня, к этой группе
видов, на первый взгляд, наиболее близок Т. vvedenskyi, но при вни-
147
мательном изучении он резко отличается от них, нечетко отграничен-
ными, чуть-чуть расплывчатыми пятнами на ноготках листочков около-
цветника и другими признаками.
В Нуратау встречается Т. turkestanica — кремовый. В песках у
подножья хребта и дальше в глубь пустыни растет Т. lehmanniana—
желтый и оранжево-красный, у Джизака — Т. korolkovii и Т. nitida —
красный, Т. micheliana — красный.
В западной части Зеравшанского хребта, в так называемых Са-
маркандских горах, наблюдается большое разнообразие видов тюль-
панов — красные: Т. fosteriana, Т. micheliana и Т. ingens, причем
Т. micheliana бывает ало-красного цвета, с малиновым оттенком в
красных цветах.
Т. ingens красного цвета без оранжевого оттенка, в них преоб-
ладает слегка бордовый оттенок, как в краплаке. Тут же произраста-
ет Т. turkestanica — кремовый.
В Кашкадарьинской области обитают: Т. ingens — красный, Т. ni-
tida — красный, Т. tubergeniana — красный, Т. turkestanica — кремовый.
В Гиссарском хребте — Т. lanata, Т. carinata, Т. tubergeniana и
Т. nitida — все красные, Т. turkestanica — кремовый. В центральной
части этого хребта появляются Т. praestans — красный и Т. hissarica—
желтый. Еще далее на запад мы находим Т. subpraestans — красно-ма-
линовый, Т. orithyoides — белый, в Дарвазском хребте Т. linifolia —
красный, в окрестностях кишлака Санктуда на р. Вахш Т. maximowiczii—
красный.
В Памиро-Алае обнаружено два вида желтоцветковых тюльпанов—
Т. platystemon (Сары-Бия), Т. anisophylla (Дарваз) и Т. korshinskyi —
белый.
В Памиро-Алае, возможно, находятся и другие виды, так как
в этом отношении он исследован чрезвычайно скудно. В Алайской
долине растет Т. dasystemon — желтого цвета и Т. alaica из Erioste-
mones с кремовыми цветами.
Таким образом, в Памиро-Алае мы встречаемся со следующими
видами, у которых околоцветник красного цвета, это — Т. tubergeniana,
Т. lanata, Т. fosteriana, Т. carinata, Т. affinis, Т. praestans, Т. subpraes-
tans, Т. micheliana, Т. maximowiczii, Т. linifolia, Т. victoris, Т. nitida.
Всего в настоящее время там известно 13 видов, все красные
с различным малиновым оттенком. Все формы в основном сохраняют
основной цвет вида. Только для Т. linifolia описана желтая форма,
известная в литературе под названием Т. batalini и для Т. nitida —
розовая форма — Т. rosea.
Кроме того, в Памиро-Алае обитает шесть видов тюльпанов,
листочки околоцветника которых окрашены в желтый цвет: Т. platys-
temon, Т. ferganica, Т. sp. (Северный Памиро-Алай), Т. anisophylla,
Т. hissarica, Т. dasystemon и белые: Т. korshinskyi, Т. turkestanica,
Т. alaica, Т. sp. (Северный Памиро-Алай) и Т. bifloriformis.
В Кушке обнаружен замечательный красный Т. kuschkensis с мох-
натой покровной чешуей у луковицы — признак, впервые появившийся
в Памиро-Алае. Он растет у самой границы Афганистана и, вероятно,
переходит за нее.
Продвигаясь на запад по хребту Копет-Даг, мы встречаемся с
Т. micheliana — красным с малиновым оттенком. В восточном Копет-
Даге наблюдается Т. hoogiana — малиново-красный с луковицей,
имеющей сильно опушенную покровную чешую. Эти виды растут
вдоль всего хребта. В степном поясе гор произрастает малиново-крас-
ный Т. wilsoniana и кремовато-белый Т. turkojmanica.
148
Следовательно, в области Горной Туркмении произрастает четыре
вида с красными, имеющими малиновый оттенок цветами: Т. kuschken-
sis, Т. hoogiana, Т. micheliana и Т. wilsoniana; желтый тюльпан —
Т. lehmanniana (Т. ariana) и белые — Т. buhseana и Т. turkomanica.
Особенностью самых западных среднеазиатских видов, обитающих
в Копет-Даге, является преобладание в красной окраске листочков
околоцветника малинового оттенка, а не оранжевого, как у видов из
восточных частей Средней Азии, таких как Заилийский Алатау близ
Алма-Ата, окрестности долины Чу у Фрунзе и далее до хр. Каратау
и Западного Тянь-Шаня.
Нашу мысль о преобладании малиновых оттенков у красных
тюльпанов из западной части ареала в какой-то степени подтверждают
тюльпаны, распространенные на Кавказе. Мы не наблюдали Т. schmidtii
и Т. florenskvi в живом состоянии и нам трудно судить об их окраске
по литературным и гербарным данным, но определенно Т. florenskyi
по окраске очень близок к Т. julia, а Т. schmidtii — несколько отличен.
Кроме того, описаны два кавказских вида с красными цветами,
близких к Т. florenskyi — Т. kaghismanica и Т. sosnovskyi. Т. eichleri
и Т. julia красного цвета с ярким малиновым оттенком. У Т. schrenkii
в полупустынных предгорьях на Северном Кавказе встречаются формы
бордовые, оранжевые и желтые.
На Кавказе встречаются также два тюльпана из секции Erios-
teinones с фиолетовыми листочками околоцветника: это Т. violacea
из Талыша и Т. caucasica с верховьев Кубани с окраской, неизвестной
в других ботанических областях СССР. Чудесные цветы Т. polychroma
с белыми листочками околоцветника приобретают к отцветанию розо-
вато-малиновый оттенок. На Кавказе наблюдается всего два вида с
желтыми цветами: Т. Biebersteiniana и Т. karabachensis A. G г о s s. .. '
Таким образом, к области Памиро-Алая и горной Туркмении
тяготеют тюльпаны Кавказа, хотя эти области не имеют общих видов,
но сходны по общему (малиновому) оттенку красных тюльпанов.
Несколько отличны от кавказских тюльпаны Крыма. На них ясно
видно влияние тюльпанов заволжских степей.
Т. schrenkii из степной части Крыма очень богат оттенками —
бордовые, разной интенсивности розовые, оранжевые, желто-огненные
и даже чисто белые формы, но ни в одном местообитании этот вид
не имеет таких огненно-красных цветов, как тюльпаны Средней Азии.
Остальные тюльпаны Крыма или желтые, как Т. biebersteiniana,
Т. oxypetala, или белые, как Т. callieri и Т. koktebelica.
В Европейской географической области флоры СССР (вместе
•с Крымом) обитает Т. schrenkii, сильно варьирующий в окраске. Он
бывает белый, розовый, желтый, оранжевый и почти бордовый.
Остальные виды тюльпанов желтые: Т. silvestris, Т. biebersteiniana,
Т. koktebelica, Т. callieri и два белых: Т. patens и Т. biflora.
В Сырдарьинской подгорной области произрастает три красных
тюльпана: в полупустыне — Т. greigii — красный, но он имеет и жел-
тые формы; в пустыне —Т. lehmanniana — красно-оранжевый, желтый
и оранжевый, в полупустыне — Т. korolkovii — красный и желтый и
Т. bifloriformis — кремовый, почти белый.
В Прибалхашье в полупустыне и пустыне встречается Т. alberti—
красно-оранжевый; в песках — Т. behmiana — желтые, в полупустыне
Т. altaica, Т. kolpakovskiana, Т. biebersteiniana; Т. regelii и Т. buhsea-
na — белые. В Арало-Каспийской географической области, в полупу-
стыне, распространен Т. greigii — оранжево-красный; в пустыне —
149
T. borszczowii — желтый, оранжевый и красный; в пустыне и полупу-
стыне — Т. schrenkii — белый, розовый, красный, желтый и оранжевый.
Там же обитают желтый Т. biebersteiniana и белые — Т. buhseana
и Т. biflora.
В Кызылкумской географической области в пустыне юго-запада
произрастает Т. lehmanniana — желтый, оранжевый и красно-оранже-
вый. В пустыне на северо-востоке обитает Т. borszczowii с той же
вариацией в окраске. Кроме того, там произрастают два маленьких
белых тюльпана — Т. sogdiana и Т. buhseana.
В пустынях Средней Азии с востока на запад, начинаясь у Сыр-
Дарьи и простираясь почти до Каспия, широко распространен
Т. lehmanniana, обладающий желтыми, оранжевыми, а часто и крас-
ными цветами. В Бетпак-Дале и в песках вдоль р. Или ему соответ-
ствует желтоцветный Т. behmiana; в окрестностях Кушки — желто-
цветный Т. lehmanniana (Т. ariana). Спутник Т. behmiana —Т. talievii
(восточная форма Т. buhseana). Спутником Т. lehmanniana почти на
всем значительном пространстве его ареала, кроме восточной части,
является Т. sogdiana — белый, Т. ariana — Т. turkestanica.
В Каракумской географической области обитает всего один вид
тюльпана — Т. sogdiana с белыми цветами.
В Амударьинской подгорной области в пустыне произрастает все
тот же полихромный вид Т. lehmanniana с желтыми, оранжевыми,
и красными цветами.
В Арало-Каспийской низменности, в приаральских Каракумах
и по пескам вдоль Сыр-Дарьи на юг встречается Т. borszczowii — по-
лихромный вид с желтыми, оранжевыми или оранжево-красными
цветами. В северной части этой низменности в полупустынных усло-
виях обитает Т. schrenkii также чрезвычайно полихромный с красными,
розовыми, оранжевыми, желтыми и белыми листочками околоцветника.
Этот вид, кроме того, широко распространен в Европейской части
СССР. В северной части его ареала встречаются только желтые формы
Т. schrenkii.
На юге и севере в Мугоджарах растет Т. biflora — белый. Он
распространяется далеко на юг и постепенно переходит в Т. buhseana.
Т. biflora заходит в западную часть Сибири и южную Европейскую
часть СССР. Там же, но еще более широко распространен Т. bieber-
steiniana — желтый. Т. patens белый обнаружен лишь в Европейской,
части СССР и почти не встречается в Средней Азии.
Границы двух последних видов прекрасно очерчены в работе
Талиева. Нас же интересует то, что оба эти вида являются наиболее
северно распространяющимися в пределах СССР видами и один из
них имеет желтые цветы, а другой белые.
В Западной Сибири произрастают только три желтоокрашенных
вида — Т. heterophylla, Т. altaica, Т. uniflora и два белых — Т. biflora,
и Т. patens.
В Восточной Сибири обитает один вид — Т. uniflora — желтый.
Таким образом, в Средней Азии наиболее широко распространены
виды, видимо, обитавшие на древней прибрежной пустынной суше,
а затем, после поднятия дна Тетиса, занявшие обширные пустынные
пространства. Это в первую очередь пустынный, обладающий поли-
хромными цветами Т. lehmanniana с близким к нему Т. borszczowii
и другими видами. Затем полупустынно-степной с очень обширным
ареалом, так же, как и первый, уходящий за пределы СССР — вид
с очень широкой полихромной гаммой —Т. schrenkii. К этим видам
по широте полихромной гаммы, древности местонахождений и неко-
150
торому сходству признаков примыкает также пустынный Т. alberti
и в какой-то степени предгорный и полупустынный Т. greigii. Оба
вида не уступают Т. lehmanniana и Т. schrenkii по разнообразию
окраски цветков, особенно в районе хребта Каратау.
Красные тюльпаны Тянь-Шаня отличаются оранжевым оттенком
и расплывчатыми нечетко окаймленными пятнами на основаниях ли-
сточков околоцветника. У красных тюльпанов Памиро-Алая к западу
усиливается малиновый, а не оранжевый оттенок. И если у них на
основании листочков околоцветника находится желтое пятно, то оно
четко отграничено от основного фона. Темное пятно также резко
отграничено или четко окаймлено светлой полосой. Кроме того, в.
Памиро-Алае появляются виды с сильно опушенной луковицей
Т. tubergeniana, Т. subquinquifolia. Тюльпаны Копет-Дага и Памиро-Алая
близки по окраске цветков. Среди них есть Т. hoogiana с очень
сильно опушенными покровными чешуями луковицы. Кавказские
красные тюльпаны также характеризуются малиновыми тонами и среди
них у Т. iulia сильно опушены покровные чешуи луковицы. Т. micheli-
апа и Т. greigii являются викарирующпми видами. Т. greigii характе-
рен для предгорий Тянь-Шаня, а Т. micheliana — для Памиро-Алая,
Копет-Дага и дальше Ирана. Однако цветки Т. greigii больше изме-
няются в окраске, чем Т. micheliana, который в окраске цветка варьи-
рует только от оранжево-красного до красно-малинового, а Т. greigii
из хр. Каратау — от кремовых до темно-красных. В предгорьях
Западного Тянь-Шаня встречается Т. greigii от темно-оранжевых до
темно-красных оттенков, но в другой гамме, чем в Каратау и до
оранжевых — в Чуйской долине и Чу-Илийских горах.
Т. ostrovskiana по окраске цветков, особенно в формах из Киргиз-
ского Алатау, приближается к Т. schrenkii, но в более узкой
гамме цветных оттенков. Т. korolkovii по типу является тюль-
паном Памиро-Алая. Там он широко распространен и описан как
самостоятельный вид Т. nitida. В восточной части ареала Т. korolkovii
в окрестностях ст. Дарбаза растут как красные, так и желтые формы.
А еще восточнее в Киргизском Алатау обитает желтоцветный
Т. zenaidae, отличный от типа Т. korolkovii и в то же время сходный
с ним по черным четко отграниченным пятнам при основании листоч-
ков околоцветника. Т. korolkovii — единственный вид из красных
тюльпанов, общий как для Памиро-Алая, так и Западного Тянь-Шаня.
Этот вид произрастает по низкогорьям и в основном приурочен к
осадочным породам или к продуктам разрушения коренных пород.
В Ферганской долине он розовый и выделен в самостоятельный вид
Т. rosea.
Тюльпаны восточной части ареала рода — Т. alberti, Т. greigii и
Т. korolkovii—узкой лентой заселяют пустыни и предгорья Тянь-Шаня
и в какой-то степени являются тюльпанами первичными. Тюльпаны
же собственно гор Тянь-Шаня — Т. kaufmanniana, Т. tschimganica и
Т. ostrovskiana — более молодые производные виды, заселившие новые
территории поднявшихся хребтов Тянь-Шаня, изменившиеся в новых
местообитаниях, приспособившиеся к жизни в новых условиях, но в
какой-то степени сохранившие древний тип изменчивости окраски
цветов — полихромной гаммы оттенков у разных особей вида.
В географической области Тянь-Шаня и прилегающих пустынях
мы встречаемся с шестью видами тюльпанов, обладающими большой
изменчивостью в окраске цветка — Т. alberti. Т. greigii, Т. kaufmanniana,
Т. ostrovskiana, Т. tschimganica и Т. korolkovii. Из этих видов три
первых наиболее изменчивы в окраске — вплоть до совершенно белых
151
{если не считать различно окрашенных оснований листочков около-
цветника у других видов). При этом у двух первых преобладают
красно-оранжевые формы, а у Т. kaufmanniana — кремовые. Остальные
три варьируют от светло-желтых до темно-красных.
Из тюльпанов Тянь-Шаня Т. butkovii и Т. vvedenskyi, по сравне-
нию с вышеперечисленными видами, менее изменчивы в окраске
основного фона листочков околоцветника; встречаются лишь светло-
оранжевые формы, представляющие собой ослабление обычной оран-
жево-красной расцветки околоцветника этих видов. В данном случае
мы наблюдаем изменчивость, подобную тюльпанам Памиро-Алая, ко-
торые почти не меняют окраску основного фона листочков около-
цветника.
Остальные виды секций Leiostemones и Spiranthera, обитающие
в этой области, все желтоцветные—Т. kolpakovskiana, Т. zenaidae,
Т. dubia, Т. iliensis, Т. ferganica, Т. tianschanica, Т. tetraphylla и
Т. anadroma. Из тюльпанов секции Eriostcjnones распространены
кремовые, почти белоцветные — Т. buhseana, Т. bifloriformis, Т. tarda
и Т. turkestanica (лишь в Моголтау); желтоцветные — Т. dasystemon,
Т. dasystemonoides и Т. heterophylla или, как теперь он называется,
Eduardoregelia heterophylla М. Pop.
В Памиро-Алае мы встречаемся с третьим очагом формообразо-
вания в этом роде Tulipa. Здесь, как нигде в мире, у взрослых особей
тюльпанов секции Leiostemones сохранилась способность образовывать
столоны расселения, расходящиеся не вниз, а в стороны. Из известных
нам видов этой способностью в большей степени обладают: Т. рга-
estens, Т. fosteriana, Т. carinata, Т. lanata и многие виды секции
Eriostemones. Способность к столонообразованию присуща ювенильным
растениям всех видов диких и сортам культурных тюльпанов. У раз-
личных видов ювенильная способность с возрастом исчезает в разное
время. За незначительным исключением она пропадает в фазе генера-
тивной зрелости растений (Бочанцева, 1956). Поэтому здесь целесо-
образно отметить факт образования столонов у взрослых луковиц
нескольких Памиро-Алайских видов секции Leiostemones, уносящих
луковицы в стороны от прежнего местоположения. Эта особенность
также довольно широко распространена на всей площади ареала рода
,у видов секции Eriostemones (Т. bifloriformis и др.).
В Памиро-Алае опушение луковичных чешуй тюльпанов наиболее
разнообразно. Так, у Т. ingens длинные прямые волоски образуют
редкие длинные прядки на внутренней стороне покровной чешуи.
У разных особей Т. lanata, Т. carinata опушение изменяется от силь-
ного, почти как у видов секции Tulipanum,—Т. tubergeniana, похожее
на волокно из хлопковых коробочек, до очень слабого, только у осно-
вания и у вершины чешуи.
Таким образом, по типу окраски тюльпанов и некоторым другим
признакам, характеризующим тюльпаны, ареал этого рода в СССР
можно разбить на четыре крупных округа. Первый — пустынно-степной
округ, в который входит Сырдарьинская, Прибалхашская, Аралокас-
пийская, Кызылкумская и Каракумская области; к ним тяготеют
европейские местообитания. Сибирь стоит несколько особняком. Второй
Тяньшанский округ Памиро-Алай является третьим крупным окру-
гом. К нему тяготеют Горная Туркмения и Кавказ. У тюльпанов
третьего округа, за небольшим исключением, отсутствует полихром-
ность у тюльпанов секции Leiostemones, наличие малинового оттенка
в красной окраске цветков и обильное опушение покровной чешуи
у нескольких видов.
152
В высокогорьях Средней Азии обитают виды тюльпанов только
с желтой окраской листочков околоцветника, образующие цветок
в виде звездочки и ограниченное число узких ремневидных листьев.
Высокогорные виды тюльпанов часто бывают полиплоидными, что
указывает на их относительную молодость и относительно недавнее
заселение данного местообитания видами рода Tulipa.
Крайне однообразны по окраске тюльпаны на восточной границе
ареала в Западной Сибири, где произрастают виды из секции Orithyia
и у восточной границы ареала в Семиречье. Ферганская долина также
бедна окрасками и формами тюльпанов. Один очаг современного нам
формообразования рода Tulipa лежит в пустынях Средней Азии, дру-
гой — в Западном Тянь-Шане, а третий, совершенно оригинальный, —
в Памиро-Алае.
Наши исследования видов и форм тюльпанов в природе позволяют
видеть флористическую обособленность Памиро-Алая от Тянь-Шаня.
Виды рода Tulipa, обитающие на Памиро-Алае, во всяком случае
более родственны видам Копет-Дага, Ирана и Кавказа и обособлены
от тяныпанских, которые связаны ареалами, особенностями с первым
пустынным очагом видо- и формообразования.
Предки тюльпанов, очевидно, обитали не в бедных тюльпанами
высокогорьях, тем более, что наличие там полиплоидов свидетельст-
вует о более позднем заселении этих местообитаний представителями
рода Tulipa.
Наибольшее разнообразие представителей рода Tulipa отмечено
в среднем и нижнем горном поясе. Там же строго сохраняется ха-
рактеризующее виды основное число хромозом и там, видимо, в
разных горных системах на разной высоте, но не выше 3000 м, лежат
очаги современного видообразования в роде Tulipa.
Глава V
КАРИОСИСТЕМАТИЧЕСКИЕ ИССЛЕДОВАНИЯ
В
СРЕДНЕАЗИАТСКИХ ТЮЛЬПАНОВ
Ботаническом саду АН УзССР мы провели кариосистематическое*
исследование почти всех имевшихся там в течение последних
30 лет видов и форм тюльпане в. Применение цитологического ме-
тода в систематике и филогении дало прекрасные результаты в работах
Сакамура (Sakamura, 1918); М. Г. Николаевой (1922); Кихара (Kihara,
1924); Г. Д. Карпеченко (1922); Л. Н. Делоне (1915, 1922, 1925, 1928);
Г. А. Левитского и Н. Е. Кузьминой (1927); Г. А. Левитского (1931);
Текгольм (TSckholm, 1922); Н. П. Авдулова (1931); И. Н. Свешниковой.
(1927); М. В. Сеняниновой-Корчагиной (1932) и многих других.
Особенно широкое распространение получил этот метод после
опубликования работ Г. А. Левитского (1931), посвященных отысканию-’
соответствующих фиксаторов для выявления морфологии хромозом.
Количество хромозом характеризует определенные группы растений,
например Triticum (Николаева, 1922; Т. sakamura, 1918). В других
случаях количество хромозом (24) одинаково для нескольких близких
родов семейства Liliacea, Lilium, Fritillaria, Tulipa, Erythronium (Ti-
schler, 1921; Darlington and Wylie, 1956).
В целях кариосистематики для объектов с одинаковым количе-
ством хромозом в пределах секции рода или иной таксономической
единицы необходимо использовать данные и по морфологии хромо-
зом.
Основное морфологическое отличие хромозом впервые было
описано С. Г. Навашиным (1911) и Сакамура (1915—1920) для расте-
ний и Агаром — для животных. С. Г. Навашин (1911) при изучении
Fritillaria tenella установил три типа хромозом: с медиальные сочле-
нением плеч, субмедиальным сочленением и с терминальным прикреп-
лением нитей веретена. Говоря о терминальном прикреплении нитей
веретена, С. Г. Навашин установил обязательное присутствие кинети-
ческой перетяжки у этого типа хромозом так же, как и у всех
остальных (термин „кинетическая перетяжка“ был дан позднее
Л. Н. Делоне, 1928).
Открытие „спутников" у хромозом, играющих большую роль при
составлении идиограммы, также принадлежит С. Г. Навашину (1911),
который по праву считается создателем учения о морфологии хри-
мозом.
В дальнейшем, в работах М. Черноярова (1914) и Л. Н. Делоне
(1911) были установлены членистые хромозомы, т. е. имеющие, кроме-
154
основной,—вторичные перетяжки. В 1929 г. Делоне назвал вторичную
перетяжку плеча хромозомы „акинетической".
Г. А. Левитский (1924—1931) свел в стройную систему все дан-
ные по морфологии хромозом. Он разработал также единую термино-
логию для классификации хромозом, основанную на фактическом
материале, определяющую общий план построения хромозом из двух
плеч, соединенных кинетической перетяжкой, и устанавливающую,
будет ли эта хромозома с медиальным, субмедиальным, субтерминаль-
ным или терминальным прикреплением нитей веретена.
Такое расчленение хромозом является первичным, присущим нор-
мальным хромозомам как растительного, так и животного мира (Ле-
витский, 1931; Навашин, 1936). Спутники же и акинетические переры-
вы являются признаками вторичного расчленения тела хромозомы.
При решении вопросов систематики и филогении кариосистемати-
ческий метод в систематике в связи с другими методами пользуется
вполне заслуженным признанием (Левитский и Кузьмина, 1927; Де-
лоне, 1928; Левитский, 1931; Авдулов, 1930).
Так, Делоне (1928) на основании исследований родов Muscarii,
Bellavalia, Ornithogalum и Iris пришел к заключению: „(после уста-
новления связи между кариологией и систематикой) может быть при-
дется признать, что после „элементарного вида" нет систематической
единицы более естественной, чем „род", при том именно „малый род",
как иногда выражаются, или „род кариотип", как мы предпочли бы
здесь выразиться".
Левитский же утверждает: „что морфология хромозом, хотя и
дает весьма ценные в этом отношении (филогенетическом—3. Б.}
комплексы признаков, но таксономическое их значение, вообще го-
воря, так же относительно, как это имеет место и для признаков
внешне морфологических". Это положение Левитского можно иллю-
стрировать . работами, в которых доказана изменчивость кариотипа
(Свешникова, 1927; Сенянинова-Корчагина, 1931), когда два близких
вида имеют настолько различные кариотипы, что, по Делоне, должны
быть отнесены к разным родам.
То же явление отметили мы при изучении кариотипов Т. maxi-
mowiczii и Т. linifolia. Эти виды по морфологии крайне близкие, а
по кариотипу должны быть отнесены к разным родам.
Мы наблюдали и другую картину, т. е. виды разных секций рода
Tulipa имели столь мало отличные кариотипы, что, по Делоне, они
должны быть отнесены в один „малый род". Судя по разбивке Дело-
не рода Iris, понятие „малый род" — это, видимо, секции рода.
Однако надо отметить, что работа Делоне единственная, где сде-
лана попытка подойти к понятию рода с кариологической точки зре-
ния. Вообще же, кариосистематики на основании данных только судят
о родственной близости или, наоборот, отдаленности отдельных таксо-
номических единиц в систематике и филогении, не затрагивая понятия
самих таксономических единиц.
Цитологи пытаются давать кариосистематическим признакам
определенное таксономическое значение, и выделять признаки семей-
ственные, родовые, видовые и расовые. К таким признакам Левитский
относит величину хромозом, основное количество, расчлененность, а
также и полиплоидию: „с одной стороны, каждый тип кариологических
изменений обладает известной приуроченностью к определенному так-
сономическому масштабу, с другой — эта приуроченность является
лишь преимущественной и допускает многочисленные исключения —
от высоких систематических единиц до самых низших" и, наконец, в
155-
заключение он вновь подчеркивает: „Таким образом, кариотипическая
характеристика подлежит тому же общему правилу относительности
систематических признаков, как и обычная морфологическая" (Левит-
ский, 1932, стр. 208 и 209).
Первые числа хромозом у Tulipa в результате изучения зароды-
шевого мешка и процессов оплодотворения выявили Гиньяр (Guignard,
1900), Эраст (Erast, 1901), Страсбургер (Strasburger, 1908) и Ньютон
XNewton, 1927). Эти исследователи, кроме Ньютона, не уделяли осо-
бого внимания хромозомам. Гиньяр, изучая эмбриологию четырех
видов — Т. clusiana (вероятно), Т. australis Link., Т. silvestris Linn,
и Т. gesneriana Linn., нашел, что для всех редуцированное чис-
ло хромозом равно 12. В 1925 г. Ньютон исследовал 26 видов и
5 садовых форм тюльпанов, предоставленных ему Дайксом (Dykes,
1930). К сожалению, Ньютон в работе не дал систематического опи-
сания изученных им видов и не указал на географические пункты их
происхождения, что создало большие, подчас, непреодолимые затруд-
нения при сравнении данных нашей работы с данными Ньютона. Что
касается систематической характеристики изучаемого материала, то
Ньютон, следуя Буасье, разделил весь род Tulipa на 2 секции: Leios-
temones, характеризующуюся гладкими тычиночными нитями, и Erio-
stemones, имеющую бородку волос при их основании.
Ньютон изучал хромозомы в соматическом и редукционном де-
лении у следующих видов:
Leiostemones
Т. batalini Rgl.* 24 хромозомы
Т. maximowiczii Rgl.* 24 (12) »
T. linifolia Rgl.* 24 (12) n
T. stellata Hooker 48 ff
T. chrysantha В о i s s. 48 (24) »
T. clusiana D. C. about 60 (24, 12) 1)
T. kolpakovskiana Rgl.* 24 (12)
T. greigii Rgl.* 24 fl
T. eicbleri Rgl. 24
T. kaufmanniana Rgl.* 24 (12) fl
T. praestans H о о g.* 24 (12) fl
T. sprengeri Baker 24 fl
T. armena В о i s s. 24 »
T. galatica Freyn 32 n
T. viridiflora Baker 24 (12) fl
Copper colour (hort.) 24 n
Keizerskroon (bort.) 36 fl
Massennet (hort.) 36 n
Murillo (hort.) 24 (12) fl
Due van Thol (hort.) 24 (12) fl
Eriostemones
T. dasystemon Rgl.* 24 (12) fl
T. humilis Herbert 24 (12) fl
T. pulchella Fenzl. 24 fl
T. biflora Pall.* 24
T. turkestanica Rgl.* 24 fl
T. primulina Baker 24 n
156
T. australis Link. 24 (12)
T. Silvestris Linn. 48 (24)
T. orphanidea Boiss. 24 (12)
T. whittallii Elwes. 48 (24)
T. hageri Halder 24 (12)
Виды (10), отмеченные звездочкой, растут в Средней Азии. Для
всех их Ньютон приводит соматическое число хромозом 2 п = 24.
Ньютон не ограничивается только подсчетом хромозом у тюль-
панов, он касается и морфологии хромозом, разбивая все хромозомы
тюльпанов на 3 типа: длинные с субмедиальным прикреплением,- сред-
ние с субтерминальным прикреплением;короткие с терминальным при-
креплением.
Ньютон указывает, что у Tulipa хромозомы I и II типов почти
одинаковой длины. Исключением являются Т. galatica и Т. armena, у
которых хромозомы I типа значительно длиннее, чем у II, так же, как
у Lilium и Fritillaria. У коротких хромозом III типа Tulipa и Lilium
прикрепление нитей веретена субтерминальное, а у таких же хромо-
зом Fritillaria — терминальное.
У Lilium, Fritillaria и Tulipa (за исключением 5 видов и тетра-
плоидного ряда) имеются две пары хромозом с субмедиальным при-
креплением, у 3 видов — Т. linifolia, Т. maximowiczii и Т. batalini—
все хромозомы с конечными перетяжками. Эти три вида образуют
естественную группу и отличаются от других тюльпанов цитологиче-
ски и морфологически.
У Т. kolpakovskiana нет хромозом ни с субмедиальным сочлене-
нием, ни очень маленьких, свойственных группе Т. linifolia, т. е. хро-
мозомы его промежуточные между типом Т. linifolia и нормальным
типом.
Два вида Т. armena и Т. galatica, очень близкие, отличающиеся
друг от друга только окраской цветов, имеют хромозомы I типа, бо-
лее равноплечие и, кроме того, у них разные числа хромозом: Т. ar-
mena — 24 хромозомы, а Т. galatica — 32. Добавочными являются
4 пары очень маленьких хромозом, не встречающихся у Tulipa. Нью-
тон предполагает, что они произошли путем фрагментации, а не репе-
тиций или гибридизации. Сравнение наборов хромозом Т. armena и
Т. galatica не дает для этого оснований, так как хромозомы Т. gala-
tica при сравнении с Т. armena не имеют сколько-нибудь заметных
укорочений.
Кроме того, Ньютон изучил несколько полиплоидных видов. Так,
Т. stellata и Т. chrysantha — тетраплоидные формы принимали участие
в образовании гибрида Т. clusiana. Т. whittallii — устойчивая тетра-
плоидная форма, видимо, происшедшая из форм сгруппированных под
названием Т. orphanidea.
В сводном списке по хромозомам высших растений у Тишлера
(G. Tischler, 1938) на основании работ Апкот (М. Upcott, 1935, 1936,
1937, 1957); Апкот и Лакура (М. Upcott et La Cour, 1936); Холла
(A. D. Hall, 1934, 1937); Вуда и Бамфорд (M. W. Woods and R. Bam-
ford, 1957) и Дарлингтона (Darlington, 1957) приводятся гаплоидные
числа хромозом для следующих видов тюльпанов: Tulipa altaica—12;
Т. amabilis —12; Т. armena — 12; Т. aucheriana — 12; Т. australis—12;
Т. batalini (zu linifolia) — 12; T. biflora — 12; T. boeotica — 12; T. bor-
szczowii — 12, 12+fragii; T. chrysantha (zu stellata)—12; T. cretica—12;
T. cuspidata —12; T. cypria—12; T. dasystemon (-tarda)—12; T. eich-
leri — 12; T. fosteriana — 12; T. galatica — 12 (+ frag.); T. gesnerlana—12;
157
T. greigii — 12; Т. grisebachiana — 12; Т. hageri—12; Т. hoogiana—12;
Т. humilis — 12; Т. ingens—12; Т. kaufmanniana—12; Т. kesselringii—
12; Т. kolpakovskiana — 12—24; Т. korolkovii—12—24; Т. kuschken-
sis—12; T. lanata—12—18; T. linifolia—12; T. marjolettii—12; T. mi-
cheliana — 12; T. montana — 12 (12 4- frag.); T. orphanidea — 12—18—30;
T. ostrovskiana — 12—24; T. persica — 12; T. planifolia — 12; T. poly-
chroma— 12; T. praecox (zu Oculus-solis)—12 —18; T. praestans — 12;
T. primulina —12; T. pulchella —12; T. rhodopea — 12; T. saxatilis —12—
18—18; T. schmidtii — 12; T. schrenkii—12; T. sprengeri —12; T. stap-
fii,— 12; T. stellata—12; T. suaveolens —12; T. tubergeniana — 12; T. un-
dulatifolia—12; T. urumiensis — 12; T. violacea — 12; T. wilsoniana — 12;
T. aleppensis —18; T. clusiana—18—30—12—18—24—30; T. edulis — 24;
T. Silvestris —24; T. turkestanica —24; T. whittallii —24.
Из тюльпанов, произрастающих в СССР, в этом списке приведено
27 видов. Точное происхождение многих из них неизвестно.
Только в настоящее время началось всестороннее изучение тюль-
панов, поэтому ясно, что перед исследователем ставится много проб-
лем эволюционного порядка.
МАТЕРИАЛ И МЕТОДИКА
Корешки для исследования мы получили от луковиц тюльпанов,
привозимых в течение многих лет экспедициями Ботанического сада
АН УзССР из разных районов Средней Азии.
Луковицы высаживали в питомник Сада; вырастающие из них
растения тюльпанов всесторонне изучали. Мы исследовали по 10 рас-
тений с каждого местообитания, но некоторые редкие виды, привози-
мые единичными экземплярами, изучали на меньшем количестве эк-
земпляров. У луковиц среднеазиатских тюльпанов происходит одно-
временная закладка всех корешков — это основная трудность, с которой
мы встретились при их изучении (Бочанцева, 1937).
Обрезка всех корешков вызывает в большинстве случаев гибель
луковицы, части их — ослабление луковицы, которая после этого
много лет не цветет. Чтобы изучить количество и морфологию хро-
мозом, необходимо было сохранить нормально развивающееся растение.
Мы использовали метод мазка из молодого пыльника по Левану (Le-
wan, 1931, 1933), который требует сравнительно большого количества
бутонов для предварительного отыскания стадий деления первичного
ядра пылинки. Таким образом, оба метода — корешков и мазка могут
быть использованы только при обильном материале.
Поэтому, когда материала было недостаточно, или пластинки, по-
лученные из корешков, были не удовлетворительны, а иногда для
целей сравнения, мы использовали пыльцевые трубки в момент деле-
ния генеративного ядра.
Таким образом, по некоторым тюльпанам у нас имеются хромо-
зомы, изученные в клетках корешков и на мазках из пыльника и
пыльцевых трубок.
Получение корешков от луковиц, а не из семян, вызвано тем,
что из каждого семячка тюльпанов в первый год жизни развивается
всего один корешок, быстро останавливающийся в росте. Кроме того,
семена наших тюльпанов переживают почти годовой период покоя и
прорастают только после промораживания в течение зимы. Луковицы
тюльпанов начинают трогаться в рост с октября. От одной взрослой
луковицы у крупных тюльпанов можно получить до 100 корешков, у
мелких 10—15.
158
Луковицы мы высаживали в горшочки с влажной землей, или
помещали в сосуды с водой так, чтобы вода смачивала только донце,
иначе они загнивали. Во избежание порчи луковиц и ненужной изог-
нутости корешков, с луковиц удаляли покровные чешуи.
В поисках оптимального фиксатора для корешков тюльпанов мы
использовали хромацетформол Навашина (1914) и хромформол (Левит-
ский, 1931) в нескольких вариациях.
Наилучшие результаты при фиксации хромформолом получены
при комбинации равных частей (5+5) 1-процентной хромовой кислоты
< 10-процентным формалином. Высокие дозировки формалина значи-
тельно обесцвечивают плечи хромозом у кинетического перерыва, уве-
личивая его за счет плеч. Низкие — дают картины, получающиеся при
фиксации хромацетформолом. У большинства видов Tulipa при хрома-
цетформоле кинетическая перетяжка выступает довольно явственно,
но, так как хромозомы у Tulipa большие и толстые, трудно бывает
при этом фиксаторе отыскать б. м. удачно расположенные пластинки.
В начале работ толщина срезов корешка была принята в 10 р, но
затем мы увеличили ее до 24 р, так как в тонких срезах трудно
найти во время метафазы неперерезанные хромозомы. Наличие же
неповрежденных пластинок в метафазе совершенно необходимое усло-
вие при просчете, изучении морфологии и, особенно, при измерении
хромозом. При фиксации корешков хромформолом в комбинации
5+5 плазма достаточно прозрачна даже для толстых срезов в 24 р.
Окраску микротомных препаратов производили железным гемо-
токсилином по Гейденгайну.
У видов Tulipa пыльники для мазка бывают готовы на разных
этапах развития бутона. Так, у Т. praestans следует брать очень мо-
лодые бутоны, еще не вышедшие на поверхность земли, а у Т. maxi-
mowiczii, наоборот, по достижении цветоножкой нормальной длины,
за 2 — 3 дня до раскрытия цветка.
При предварительном отыскании нужных стадий деления с уксус-
но-кислым кармином, особенно у видов с ранним созреванием пыльцы,
нарушается целостность бутона и цветок расцветает изуродованным
или часто гибнет. Однако картина настолько отчетлива (рис. 9), что
при некотором излишке растений можно несколькими бутонами по-
жертвовать, тем более, что луковица у них сохраняется. Легкость
подсчета хромозом, прекрасно выявленная морфология их (рис. 9, 10),
почти неизмененные размеры и упрощение техники по приготовлению
препаратов, по сравнению с микротомными препаратами,— все говорит
в пользу более широкого использования этого метода в цитологиче-
ской практике.
Пыльники из цветка, в котором установлено наличие соответ-
ствующей стадии, раздавливаются между двумя предметными стекла-
ми и немедленно опускаются в фиксатор следующего состава.
Первый раствор. Пикриновая кислота (насыщенный раствор)—
75 см; мочевина — 22 см; хромовый ангидрид—1,5 см.
Второй раствор. Ледяная уксусная кислота—1,5 см; формалин
(продажный) —7 см.
Первый раствор перед употреблением нагревают. Его надо взять
21 см и весь второй (т. е.—8,5 см). Соединить в момент фиксации.
Продолжительность действия от 2 до 6 час. Промывать водой, затем
приступить к окраске препарата. Капнуть на стекло 1-процентный
раствор генцианвиолет. Смыть дистиллированной водой. Капнуть на
объект J + KJ, покачать препарат; смыть дистиллированной водой. За-
тем быстро провести по спиртам возрастающей концентрации 10—20—
159
35—50—75— 95°. В 95-градусном спирте контроль окраски препарата
под микроскопом. Затем капнуть 2—3 капли смеси спирта с гвоздич-
ным маслом (АЦ< 95° + ol. Caryophillum, 1 + 1) и слить смесь обратно.
Капнуть чистое гвоздичное масло; оставить в чистом гвоздичном мас-
ле на 1/2 часа; слить гвоздичное масло обратно; капнуть 2—3 капли
смеси xylol + ol. Caryoph. в равных объемах; слить. Смыть препарат
ксилолом; капнуть смесь — 2 части xylol + 1 часть ol. cedri, слить;
капнуть смесь — 1 часть xylol + 2 части ol. cedri, слить. Поместить в
чистое кедровое масло. Закрыть покровным стеклом. При употребле-
нии кандского бальзама вместо кедрового масла происходит сжатие
пылинок и деформация их содержимого, что крайне нежелательно.
Ограниченность материала по некоторым видам Tulipa заставила
нас прибегнуть к изучению хромозом при делении генеративного ядра
в пыльцевой трубке. Этот метод по получению четкости хромозом
их морфологии и величине не уступает методу Левана (Бочанцева
1933).
При проращивании пыльцы Tulipa in vitro в висячей капле 2%.
желатина и 20% тростникового сахара через 20 мин. до 21 час. 50 мин.,
в пыльцевых трубках протекает спермиогенез.
Благодаря достаточной ширине пыльцевых трубок, хромозомы
располагаются в них вполне свободно, изменяя длину крайне редко.
В качестве фиксатора для выявления морфологии хромозом хромфор-
мол дает вполне удовлетворительные результаты. Держать в нем
вполне достаточно 2 часа, промывать водой—1 час (Бочанцева, 1937).
Затем, препараты окрашивают генцианвиолет, так же, как для окра-
шивания препарата мазка по Левану, но без предварительной провод-
ки по спиртам перед генцианвиолет. Заклейку срезов производят так-
же в кедровом масле, так как канадский бальзам и тут дает сжатие
содержимого. Приготовление препаратов по пыльцевым трубкам так-
же ускорено по сравнению с микротомными препаратами из корешка.
Уменьшенное вдвое гаплоидное число хромозом, четкость, выпрямлен-
ность и безупречное выявление морфологии хромозом — свидетель-
ствует о том, что этот метод должен найти широкое применение в
цитологической практике. Кроме того, при работе таким методом,
однажды установив срок от посева пыльцы данного вида до деления
генеративного ядра, можно смело фиксировать трубки через этот про-
межуток времени и всегда иметь на одном предметном стекле десят-
ки и сотни метафаз, из которых уже нетрудно выбрать наиболее
удачно расположенные.
Все рисунки выполнены при помощи рисовального аппарата.
Для получения более сравнимых величин хромозом у разных
видов Tulipa хромозомы были измерены по проекционному методу,
разработанному Левитским (1931).
У Tulipa иногда в одном и том же корешке встречаются нормаль-
ные и сильно укороченные хромозомы 'рис. 11, «; 12, м). Подобное
укорочение хромозом, но в менее резкой форме встречается очень
часто.
Поэтому при разбивке хромозом на разделы мы принимали во
внимание, прежде всего, соотношение их плеч и, уже внутри групп—
величину, а затем наличие акинетических перетяжек и спутников.
Хромозомы тюльпанов по соотношению плеч разбиваем на 7 раз-
делов: с соотношением плеч меньше чем 1:2; больше чем 1:2; 1:3;
1:4; 1:5; 1:6; 1:7; 1:8 и т. д.
160
2229
Рис. 11
о—Т. iRufmaniilana Rgl.; б—T. wTlsoniana Hoog.; в— T, subpraestas V v e d г — 1. kolpakovskiana; Rgl.: д—T. maximowiczii Rgl., e—T. feiganica
Vved.; on— T. tetraphylla Rgl.; з—T. greigii Rgl. и— T. linift lia R g 1 . ; к — T. wilsoiiiana il о о g . ; л — T. zcnaidae V v e d . ; м — T. maximowiczii
Rgl. н — T. praestans Hoog.; о — T. fosteriana I г у ♦
Рис. 12. Пластинки хромозом (увелич. в 1000 раз).
а - Т. bifloriformis Vved.; б — Т. bifloriformis V v е d.; в - Т. binutans Vve d.,; г — Т. binutans V v ed, д - T. Buhseana В о i s s.; с — Т. turkestanica Rgl.; ж — T buhseana
В о i s s . ; з —• T. bifloriformis Vve d ; и — T. bifloriformis Vved.; к — T. turkestanica Rgl; л — T. buliseana В о i s s . ; Л1 — T. binutans V v e d .
Секция Tulipanum R е b.
Из данной секции у нас в Союзе растет 5 видов: Т. tubergeniana
Hoog, Т. kuschkensis В. Fedtsch., Т. hoogiana В. Fedtsch. и
Т. Julia С. Koch. („Флора СССР11, т. IV). В пределах Средней Азии
обитают 3 первых вида, а Т. Julia — на Кавказе. В 1946 г. А. И. Вве-
денский описал еще один вид из этой секции Т. subquinquifolia Vved.
Число и морфология хромозом у тюльпанов этой секции впервые
были опубликованы Апкот и Лякуром (1936), для Т. kuschkensis,
Т. tubergeniana и Т. hoogiana. В 1937 г. их данные подтвердили Вудс
и Бамфорд.
Наши исследования этих видов проведены, в основном, на хро-
мозомах, полученных из корешков взрослых луковиц. Измерения и
морфологию хромозом проводили при изучении течения спермиоге-
неза в пыльцевых трубках, воспитываемых in vitro.
Соматические числа хромозом для первых трех среднеазиатских
видов одинаковы, т. е. 24 хромозомы. Если классифицировать хромо-
зомы этих видов по отношению их плеч, то все хромозомы можно
характеризовать, как резко неравноплечие, с отношением плеч около
одного к половине.
Т. tubergeniana Hoog. (рис. 13, а; 14, а). Исследованные нами
образцы были получены из Узбекистана — Денауский район, окрестно-
сти кишлака Сина; Таджикистан, окрестности кишлака Туткаул. По
вершинам холмов на лессе и красной глине.
Изучение морфологии хромозом данного вида показало, что все
хромозомы, а их в соматических клетках 24, можно разбить по со-
отношению плеч на шесть разделов.
В первую группу входит одна пара хромозом; во вторую —две
пары; в третью —одна пара; в четвертую — пять пар; в пятую — две
пары; в шестую — одна пара.
На идиограмме, составленной из хромозом при делении генератив-
ного ядра на спермин в пыльцевой трубке, хромозомы вытянуты по
сравнению с соматическими в 1,5 раза, при нормальном соотношении
плеч. Хромозомы у Т. tubergeniana по длине колеблются от 21,25 до
9,25 р.
В первом разделе наиболее короткая пара хромозом во всем на-
боре. Первая пара хромозом из второго раздела очень крупная; ее
длинное плечо равно 14,25р, на более коротком плече в нижней тре-
ти наблюдается акинетический перерыв. Вторая пара гораздо короче:
длинное плечо—9,5р. Пара хромозом третьего раздела по длинному
плечу почти равна хромозоме второй пары второго раздела, а корот-
кое — соответственно группе, уменьшено.
В первой паре четвертого раздела находятся самые крупные хро-
мозомы набора по длинному плечу, длина которого—18,75, коротко-
го—3,75 р; последующие пары постепенно убывают в длине, при этом
происходит более сильное укорачивание длинного плеча. У пятой па-
ры оно равно всего 10 р, в то время, как короткое—2,5 р. Четвер-
тая пара чуть-чуть крупнее пятой пары, а вторая и третья пары близ-
ки между собой (у второй длинное плечо равно 14 р, а у третьей —
13,25 р). Пятая группа состоит из двух пар почти равных хромозом.
У первой длинное плечо—10,62, у второй 10 р; короткие плечи этих
хромозом также мало различны—у первой—2,25 р, у второй—2 р. Ше-
стая группа представлена одной парой хромозом с длинным плечом
13,25, коротким —2 р.
163
Рис. 13. Пластинки хромозом из точки роста корня (увелич. в 1000 раз).
А—Т. tubergeniana Н о о g . ; б — Т. fosteriana 1 г v . ; в — Т. lanata R g 1 . ; г — Т. victoris Vved. ; д — Т. kusckensis В. F е d t s с h . ; е — Т. ingens Hoog.;
ж — Т, subpraestans Vved,; з — Т. praqstans Hoog.; и — Т. lioona В. Fedtsch.; к—Т. carinata Vved-
Ill Ihh
tO ft
1,1
Ю It
I
12
is
12
I
9 I
// 12
Рис. 14. Идиограмма измеренных пластинок (увелич. в 800 раз).-
ш — Т. tubergeniana Н о о g . ; б — Т. kuschkensis В. F е d t s„c ti . ; в — T. hoogiana В. Fedtsch. ;г —
Т. lanata R g 1.; д ~ T. ingens H о о g . ; e — T. carinata V v e d . ; ж - T. fosteriana 1 r v.; з — T. vic-
toris Vved.
Общая длина гаплоидного набора хромозом—175,495 р; длинных
плеч—139,995 р; коротких плеч—35,5 р; общий индекс 3,943.
Т. kuschkensis В. Fedtsh. (рис. 13, д; 14, б). Изученные нами
материалы были получены из Кушкинского района близ поселка Мор-
гуновского и окрестностей Кушки.
Разбивка на шесть разделов и распределение хромозом у Т. kusch-
kensis такое же, как у Т. tubergeniana. Однако размеры всех хро-
мозом Т. kuschkensis меньше, чем у Т. tubergeniana и они более мор-
фологически расчленены (рис. 14, б).
В первой паре хромозом первой группы в нижней трети второго
плеча также находится акинетическая перетяжка, но первое плечо ее уко-
рочено и она из второй группы перешла в первую. Длинное плечо^
равно 7,88 ц и на дистальном конце имеет акинетический перерыв,
отчленяющий головковидное образование; второе плечо —5,5 р, с аки-
нетическими перерывами в нижней трети плеча и выше его. Эти два
перерыва делят второе плечо хромозом на 3 неравных части, из кото-
рых средняя — самая маленькая.
Во второй раздел хромозом у Т. kuschkensis так же, как и у
Т. tubergeniana, входят две пары хромозом. Но, в противоположность
хромозомам этой группы у Т. tubergeniana они почти равные, длинное
плечо первой пары—8,5, второй—8,125 р. короткие одинаковые—3 р.
В третий раздел хромозом входит одна пара, почти равная по
размерам паре хромозом этого раздела у Т. tubergeniana; ее длин-
ное плечо—10 р, короткое—2,75 р. Акинетический перерыв, располо-
женный на коротком плече, делит его на две, почти равные, части,
Четвертый раздел включает пять пар хромозом; у первой из них
длинное плечо—10 р, короткое—2,125 р. Кроме того, на длинном
плече на расстоянии одной трети всей длины его от кинетического
перерыва находится акинетический перерыв. У второй пары длинное
плечо — 9, а короткое —2 р. Длинное плечо третьей пары этого раз-
дела— 8,125, а короткое—2 р, кроме того, на длинном плече эта па-
ра несет акинетическую перетяжку, расположенную чуть-чуть ближе
к проксимальному концу плеча и делящую это плечо на две, почти
равные, части. У четвертой пары длинное плечо —7,5, короткое —
1,875 р. У пятой пары длинное плечо—6,5, а короткое—1,5 р.
Пятый раздел состоит из двух почти одинаковых по размерам пар
хромозом: у первой из них длинное плечо—8, а короткое—1,5 р.
В шестом разделе одна пара хромозом; ее длинное плечо—9,375,
а короткое—1,5 р. На длинном плече у проксимального конца имеет-
ся акинетическая перетяжка, образующая шарообразный членик длин-
ного плеча у кинетического перерыва.
Общая длина гаплоидного набора хромозом—128,755 р; длинных
плеч — 101,005; коротких плеч—27,75 р; общий индекс 3,64.
Т. hoogiana В. Fedtsch. (рис. 13, и; 14,в). Исследованные
нами материалы были получены из Туркмении — окрестности поселка
Прохладного; Арваз, восточный склон.
У Т. hoogiana, как и у всех видов этой секции, количество хро-
мозом 2 п — 24. Все хромозомы по соотношению плеч могут быть
разбиты на 4 раздела (рис. 14, б).
В первый раздел входит одна пара хромозом. Ее длинное плечо —
5,5, короткое —2,75 р.
Второй раздел состоит из двух пар хромозом. У первой из них
(длинное плечо —2,5р) на коротком плече находится в нижней трети
акинетическая перетяжка. Вторая пара этой группы с длинным плё-
166
чом —6,26, коротким —2,25 у. На коротком плече имеется акинети-
ческий перерыв, делящий его на две, почти равные части.
В третий раздел входят 3 пары хромозом. У первой пары длин-
ное плечо —8,5 [л, на коротком плече (2,5 у) на расстоянии 2/5 длины
его от кинетического перерыва наблюдается акинетическая перетяжка.
У второй пары длинное плечо—6,625 и короткое—2 у; на ди-
стальном конце длинного плеча отмечен акинетический перерыв, от-
деляющий членик в форме головки.
Третья пара с длинным плечом, равным 6,25, а коротким—1,5 у
также на дистальном конце длинного плеча имеет небольшую часть,
отчлененную в виде головки акинетическим перерывом.
В четвертый раздел входит четыре пары хромозом.
Первая с длинным плечом, равным 8,5 и коротким —1,875 у с аки-
нетической перетяжкой, делящей его на две равные части на корот-
ком плече.
У второй пары длинное плечо—7,5 и короткое—1,75 у, на прок-
симальном конце длинного плеча находится акинетическая перетяжка,
отделяющая его нижнюю четверть; короткое плечо также с акинети-
ческой перетяжкой, расположенной в 2/5 всей длины плеча от его
проксимального конца.
Длинное плечо третьей пары—7 и короткое—1,625 у. Никаких
следов вторичного расчленения на этой паре хромозом мы не обна-
ружили.
У четвертой пары длинное плечо—6,5 и короткое—1,625 у без
вторичного расчленения.
Хромозом пятой группы у Т. hoogiana нет.
Шестой раздел хромозом состоит из двух пар.
Длинное плечо первой пары хромозом равно 10,5 у, короткое —
1,75 у. На дистальном конце длинного плеча лежит акинетический
перерыв, отделяющий от него продолговатый членик около 1,25 удл.
У второй пары этой группы длинное плечо—9,625, короткое —1,5 у.
Вторичного расчленения нет.
Обшая длина хромозом гаплоидного набора—110,675 у; длинных
плеч—87,75; коротких плеч—22,925 у; общий индекс—3,8.
Таким образом, виды Т. kuschkensis и Т. tubergeniana секции
Tulipanum очень близки по своему кариотипу. Большая укороченность
хромозом и морфологическая расчлененность Т. kuschkensis свидетель-
ствует о более позднем его происхождении.
Прямым и непосредственным родичем Т. kuschkensis, по-видимо-
му, является Т. tubergeniana.
Происхождение Т. hoogiana, несмотря на большое сходство его
кариотипа с кариотипами двух вышеназванных видов, неясно, однако
несомненно влияние на этот вид каких-то иных кариотипов.
Полную картину филогенеза секции Tulipanum можно восстано-
вить только после изучения тюльпанов сопредельных стран —Ирана
и Афганистана. В Средней Азии всего четыре вида этой секции и
один вид Т. julia на Кавказе.
Секция Leiostemones В о i s s.
В СССР из этой секции по „Флоре СССР“, т. IV произрастает
38 видов; 5 видов открыты после опубликования „Флоры“, т. е. в на-
стоящее время в Союзе обнаружено 42 вида Tulipa из секции Leioste-
mones. Из них мы исследовали 28 видов —все из Средней Азии.
167
Для удобства изложения при описании кариотипов видов этой
секции необходимо весь материал расположить группами, объединяя в
группу близкие виды. Таким образом, описанные в настоящей работе
виды секции Leiostemones мы разбили на восемь групп: 1) Т. lanata,
Т. ingens, Т. carinata, ,Т. fosteriana, Т. victoris, Т. affinis, Т. vvedens-
kyi; 2) Т. praestans, Т. subpraestans, Т. butkovii; 3) Т. alberti, Т. grei-
gii, Т. mogoltavica, Т. micheliana; 4) Т. maximowiczii, Т. linifolia,
Т. wilsoniana; 5) Т. lehmanniana, Т. behmiana, Т. sp. из Кушки,
Т. borszczowii; 6) Т. korolkovii, Т. nitida, Т. rosea; 7) Т. schrenkii,
Т. ostrovskiana, Т. kolpakovskiana, Т. zenaidae, Т. ferganica; 8) Т. tet-
raphylla.
Не исследованы виды — Т. eichleri, Т. schmidtii, Т. platystemon,
Т. brachystemon, Т. altaica, Т. iliensis, Т. anisophylla, Т. korshynskyi,
Т. tianschanica, Т. krauseana, Т. vvedenskyi, Т. affinis.
Из среднеазиатских видов тюльпанов, входящих в секцию Leios-
tem.ones, Ньютон в 1927 г. исследовал Т. batalini Rgl., Т. maximowi-
czii Rgl., Т. greigii Rgl., T. praestans Hoog.
А. И. Введенский во „Флоре СССР", IV Т. batalini поставил в си-
нонимы к Т. maximowiczii. Следовательно, Ньютон определил коли-
чество хромозом у пяти видов тюльпанов из этой секции, растущих
в пределах Средней Азии.
Все пять видов Tulipa, по Ньютону, в соматических клетках име-
ют 24 хромозомы. При этом, он не дает морфологического описания
растений и не указывает их родину. Так, для Т. maximowiczji Нью-
тон приводит число хромозом 2 п =24, а при нашем исследовании
образцов, полученных от экспедиции Ботанического сада ТашГУ из
из окрестностей Санктуда (Бочанцев), неоднократно установлено у
Т. maximowiczii 2 п=22.
Ньютон также указывает, что нити веретена всех хромозом это-
го вида прикрепляются терминально. Мы же как в соматических
клетках, так и в мазках по Левану наблюдали одну пару хромозом
почти с медиальным прикреплением нитей веретена. Несомненно,
Ньютон называл Т. maximowiczii совсем не тот вид, который пони-
маем мы и что был описан Регелем. Мы исследовали растения с
классических местообитаний, а откуда Ньютон и его последователи
брали материал неизвестно. Таким же недостатком, что и у Ньютона,
страдают материалы всех работ Лякура и Апкот (1935, 1936,
1937) и Вудса и Бамфорда (1937). Эти исследователи изучали не ди-
кие виды из естественных местообитаний, а потомки дикарей, вы-
ращенных в Ботанических садах Европы и Америки. Но, на основа-
нии обширного экспериментального материала мы знаем, что дикие
виды тюльпанов обладают способностью скрещиваться при благоприят-
ных условиях между собою и с культурными тюльпанами.
В работах Ньютона вызывают сомнения следующие тюльпаны:
Т. batalinii, Т. linifolia, Т. maximowiczii. В работе М. Апкот и Ля-
кура Т. ostrovskiana hort., Т. kolpakovskiana hort, Т. borszczowii hort
и др. Так, совершенно невозможно перепутать Т. hoogiana и Т. lana-
ta, однако, в работе Вудса и Бамфорда (1937) в табл. 1 написано
Т. hoogiana В. Fedtsch., а в скобках — Т. lanata.
В этих работах можно верить лишь данным по количеству хро-
мозом, так как 24 хромозомы в соматических клетках характерны не
только для рода Tulipa в целом, но и для серии близких родов, а
уклонения и полиплоды очень редки.
Таким образом, без описания растения и указания местообитания
конкретных исследуемых образцов диких видов нельзя получить до-
168
стоверных данных, которые могли бы быть использованы для каких-
либо филогенетических или иных построений. Названия, приводимые
в цитологических и эмбриологических работах обычно не внушают
доверия, так как авторы большей частью не знают изучаемых расте-
ний и, имея растения сомнительного происхождения, пишут о них, как
о достоверных видах.
Т. lanata Rgl. (рис. 13, в; 14, г). Исследованные нами образцы
выращены из луковиц сбора А. И. Введенского и В. П. Бочанцева —
Хаджиипак (Узбекистан, Денауский район). Подсчет и изучение хро-
мозом производились одновременно в корешках при соматическом де-
лении (рис. 13, в) и во время течения спермиогенеза в пыльцевых
трубках.
Все хромозомы Т. lanata по месту кинетического перерыва могут
быть разбиты на пять разделов (рис. 14, г). В первую группу входит
одна пара хромозом, ее длинное плечо равно 7.5, короткое —4 р.. На
коротком плече на расстоянии около 1 р от кинетического перерыва
лежит акинетическая перетяжка. Второй раздел состоит из двух не-
больших хромозом. У первой из них длинное плечо —5,5, короткое —
2,25 р, у второй пары этой группы длинное плечо равно длинному
плечу первой пары, а короткое — около 2 р. В третий раздел входят
три пары хромозом. Длинное плечо первой пары —8,25, короткое —
'2,66 р; длинное плечо второй пары —8; короткое —2,25 р, длинное
плечо третьей пары 5,5, короткое—1,5 р.
В четвертый раздел входят пять пар хромозом. Длинное плечо
-первой пары 9,5» короткое —2,5 р; длинное плечо второй пары 8,75,
короткое—2 р; длинное плечо третьей пары 8,5, короткое—1,75 р;
.длинное плечо четвертой пары 6, короткое—1,25 р; длинное плечо
пятой пары 5,6, короткое—1,24 р. В пятом разделе только одна пара
хромозом с длинным плечом 7,5 и коротким—1,5 р.
При изучении этого вида мы не наблюдали вторичного расчлене-
ния хромозом, кроме одной перетяжки у хромозомы первой группы.
Общая длина хромозом гаплоидного набора — 110,95 р; длинных
плеч—86,1; коротких плеч—24,85 р; общий индекс 1,875. Индекс
хромозом Ап 2,44.
Во время изучения спермиогенеза этого вида, среди нормальных
пыльцевых трубок с 12 хромозомами мы встречали пыльцевые трубки
-с 11 хромозомами. Чаще всего в таких трубках отсутствовала хромо-
зома первой группы. Только один раз мы обнаружили трубку, в ко-
торой отсутствовала другая хромозома.
Пыльцевых трубок с 13 хромозомами мы не наблюдали. Возмож-
но, пылинки с количеством хромозом п +1 не прорастают. Несбалан-
сированность хромозомального набора и вытекающие отсюда послед-
ствия, в виде гамет с разными количествами хромозом, есть показа-
тель известной молодости вида, хотя при наличии у него вегетативного
размножения он может существовать очень долгое время не изменя-
ясь и тогда возраст его будет определен ошибочно. Отсутствие среди
изученных нами 15 растений этого вида таких, у которых в сомати-
ческих тканях было бы по 22—23 хромозомы можно объяснить малым
количеством исследованных экземпляров или нежизненностью подоб-
ных форм.
Т. ingens Hoog. (рис. 13, е- 14, д'). Изученные образцы получе-
ны из Самаркандских гор (Сатуалдыбаев), окрестностей кишлака Ага-
лык, мягкие склоны (А. Я. Бутков), Аман-Кутана (3. П. Бочанцева),
Кугитанга (А. Ф. Гнездило). Т. ingens имеет 24 хромозомы (рис. 13, е),
Хромозомы изучали на корешках, полученных от взрослых луковиц.
169
Все хромозомы этого вида по отношению длины плеч могут быть
разбиты на 5 разделов (рис. 14, <?).
В первый раздел входит одна пара хромозом, длинное плечо ко-
торой 8 {л, короткое —4,5 у. На длинном плече имеется акинетическая
перетяжка, расположенная на расстоянии около 3 у от дистального
конца плеча. На коротком плече также расположена акинети-
еская перетяжка, на расстоянии около 1,25 у- от кинетической
перетяжки.
Второй раздел состоит нз двух пар хромозом. Первая из них с
длинным плечом в 7 у, коротким —2 у-; приблизительно на середине
длинного плеча может находиться акинетическая перетяжка. У второй
пары длинное плечо 5,6, короткое - 2 у-. На коротком плече, на рас-
стоянии около 3/4 от терминального конца плеча, расположена аки-
нетическая перетяжка.
В третьем разделе четыре пары хромозом. У первой пары длин-
ное плечо 7,75, короткое —2,25 у-. На длинном плече (6,25 у) второй
пары на расстоянии около 2,5 у- от терминального конца плеча поме-
щается акинетическая перетяжка; короткое плечо—2 у-. Длинное пле-
чо третьей пары 5,75, короткое—1,6 у-. Четвертая пара с длинным
плечом 6 и коротким в 1,70 у.
В четвертый раздел входят четыре пары хромозом. Первая из них
характеризуется длинным плечом в 8 и коротким в 2 у, вторая —
длинным плечом в 7,5 и коротким в 1,75 у-. Третья — длинным плечом
в 7 и коротким плечом в 1,75 у-. Кроме того, эта пара на длинном
плече имеет две акинетических перетяжки: одну у терминального
конца плеча на расстоянии около 3/4 у- от кинетической перетяжки,
другую у дистального конца плеча на расстоянии около 1,5 у. от него.
Четвертая пара с длинным плечом 7,25 и коротким 1,6 у.
В пятом разделе одна пара хромозом с длинным плечом —9,75 и
коротким — 1,75 у; на дистальном конце длинного плеча наблюдается
небольшой (около 1 у) членик, иногда при вытягивании акинетическо-
го перерыва принимаемый за спутника.
Общая длина гаплоидного набора хромозом 109,45 у-; длинных
плеч 85,35; коротких плеч 24,1 у, общий индекс—3,54.
Т. carinata Vved. (рис. 13, к; 14, е). Исследованные образцы
были получены из окрестностей кишлака Сины, сбора В. П. Бочанце-
ва и А. И. Введенского. Цветы Т. carinata в общем гораздо мельче,
чем у Т. fosteriana.
Хромозомы изучали только в корешках. Число хромозом 2 п = 24
(рис. 13, к). Все хромозомы этого вида так же, как и предыдущих,
можно по соотношению плеч разбить на 5 разделов (рис. 14, е). В
первый раздел входит одна пара хромозом; с длинным плечом 7,25 и
коротким —3,75 у. На коротком плече первой пары находятся две
акинетические перетяжки; первая на расстоянии 0,75, а вторая—1,75 у
от проксимального конца плеча.
Второй .раздел состоит из двух пар хромозом. У первой длинное
плечо—6,5 у; короткое—2,3 у- с двумя акинетическими перетяжками.
Одна из них расположена на расстоянии около 0,25 у- от прокси-
мального конца плеча, а другая около 0,625 у- от дистального конца
плеча.
У второй пары длинное плечо равно 6, а короткое — около 2,25 у.
Эта хромозома на дистальном конце длинного плеча несет спутнико-
образную головку, т. е. членик 0,4 у диаметром, который при увели-
чении акинетического перерыва приобретает вид спутника. На корот-
ком плече на расстоянии 1 у. от проксимального конца плеча лежит
170
еще одна акинетическая перетяжка, делящая это плечо на две, почти
равные, части.
В третий раздел у Т. carinata входит всего две пары хромозом.
Длинное плечо первой пары—6,25, короткое—2 р. Вторая пара с
длинным плечом —6,25 и коротким —1,75 р. Нам не удалось наблю-
дать вторичного расчленения хромозом.
Четвертый раздел состоит из шести пар хромозом, весьма различ-
ных по морфологической дифференцировке. Так, первая пара, кроме
кинетического перерыва, не имеет иных морфологических черт. Ее
длинное плечо равно 9, а короткое—2 р. Длинное плечо второй па-
ры —8,5 и короткое —2 р. На дистальном конце длинного плеча рас-
положен головковидный членик 0,75 р величины. Отделяющая его
перетяжка отличается непостоянством. Третья пара так же, как и
первая, не имеет никаких признаков вторичного расчленения. Кине-
тическая перетяжка делит ее на два плеча, из которых длинное —8,25,
короткое—1,75 р. Длинное плечо четвертой пары 8 р с акинетической
перетяжкой, расположенной почти на середине, приблизительно в
3,75 р от проксимального конца плеча, короткое—1,75 р; у пятой па-
ры длинное плечо—7,6 и короткое—1,6 р. Шестая пара с длинным
плечом в 6,25 и коротким в 1,4 р. У этой пары очень интересна фор-
ма длинного плеча, на котором находится два акинетических переры-
ва, расположенных один на расстоянии около 2,5 р, а другой —3 р ог
проксимального конца. Таким образом, длинное плечо хромозомы
имеет вид двух вытянутых члеников, соединенных между собой
овальным тельцем.
В пятый раздел входит одна пара хромозом с длинным плечом
7,75 и коротким—1,4 р. Иных признаков, кроме кинетического пере-
рыва, она не имеет.
Общая длина гаплоидного набора хромозом—111,45 р; длинных
плеч—87,6; коротких плеч—23,95 р; общий индекс 3,64.
Т. fosteriana Irv. (рис. 13, б; 14, ж). Изученные образцы соб-
раны в Агалыке и Аман-Кутане (Самаркандские горы). Зеравшанский
хребет.
Мы исследовали 6 рас этого вида, выделенных на питомнике Бо-
танического сада ТашГУ по раскраске венчика и листа. Все они в со-
матических клетках, полученных из корешков от взрослой луковицы,
имеют 24 хромозомы (рис. 13, б). Проверку числа и морфологии хро-
мозом проводили по методу Левана мазком из пыльника. Число хро-
мозом в молодых пыльниках п=12 (рис. 11, о)
По месту кинетической перетяжки хромозомы Т. fosteriana мож-
но разбить на пять разделов (рис. 14, ж).
В первый раздел входит одна пара хромозом. Длинное плечо ее
равно 7,4, короткое —5 р, на котором лежит акинетический перерыв
на расстоянии около 1,25 р от проксимального конца плеча.
Второй раздел в отличие от изученных раньше видов состоит все-
го из одной пары хромозом с длинным плечом 5,75 и коротким —2,5 р.
Третий раздел объединяет пять пар хромозом, удивительно слабо
морфологически расчлененных.
Первая пара с длинным плечом 7,6, коротким—2,25 р; вторая —
7,6 и 2,125; третья—7 и 1,9; четвертая—7 и 1,9; пятая—6,75 и 1,75 р.
Как видно из приведенных цифр, разница в их величине незна-
чительна.
В четвертый раздел входят четыре пары хромозом. У первой па-
ры длинное плечо—10,125 и короткое—2,6 р; второй—10 и 2,5;
третьей—7,6 и 1,75; четвертой 3,5 и 1,75 р.
171
В четвертом разделе у двух первых пар самые крупные хромо-
зомы в идиограмме Т. fosteriana. По размерам они равные. Две дру-
гие пары также сходны по размерам. По длине первого плеча они
приближаются к двум первым парам третьей группы.
В пятый раздел входит одна пара хромозом с длинным плечом —
10, коротким—2 р.
Общая длина гаплоидного набора хромозом—122,35 р; длинных
птеч—94,325; коротких плеч—28,025 р; общий индекс—3,36.
Т. victoris Vved. (рис. 13, г; 14, з). Новый вид, установленный
А. И. Введенским. Эндем. Морфологически очень близкий к Т. fos-
teriana. Нам неясны их различия.
Материал получен из хребта Койташ в Нуратау (сбор В. П. Бо-
чанцева). Хромозомы исследовали в корешках, взятых от взрослых
луковиц (рис. 13, г).
По соотношению плеч все хромозомы Т. victoris так же, как и
во всем цикле близких видов, объединяемых морфологически около
Т. fosteriana, можно разбить на пять разделов (рис. 14, з).
В первый раздел входит одна пара хромозом с длинным плечом
<6,26, коротким —3,57 р. На коротком плече на расстоянии 1 р от
проксимального конца плеча лежит акинетическая перетяжка.
Второй раздел состоит из одной пары хромозом. Длинное плечо
5,5 р с акинетическим перерывом, отделяющим головку около 0,5 р,
на дистальном конце и короткое —2,25, на дистальном конце которо-
го имеется головка таких же размеров.
Третий раздел объединяет 5 пар хромозом. У первой пары длин-
ное плечо 8,75, короткое—2,25 р; второй—7,75 и 2,125; третьей —
6,125 и 2; четвертой—5,5 и 1,8, пятой—5,5 и 1,75 р.
Кроме того, у второй пары на длинном плече находится акине-
тический перерыв, делящий его на 2, почти равные, части; у пятой
пары на дистальном конце длинного плеча расположен перерыв, от-
членяющий головку около 0,5 р величиной.
В четвертый раздел входят четыре пары хромозом. У первой па-
ры длинное плечо 9,5, короткое—2 р; второй—8,75 и 2; третьей —
5,75 и 1,4; четвертой—5,5 и 1,25 р.
Две первые пары хромозом самые длинные, две вторые — почти
-равные.
В пятую группу входит одна пара хромозом с длинным плечом
'б,5 и коротким—1,25 р.
Общая длина гаплоидного набора хромозом—105,2 р; длинных
плеч—81,375; коротких плеч—23,825 р; общий индекс—3,34.
Подводя итоги изучению кариотипа группы близких видов тюль-
панов, сосредоточенных в Памиро-Алае и объединяемых вокруг
Т. fosteriana, можно сделать вывод, что количество хромозом для
всей группы 2 п = 24. Отклонение в гаметах дает Т. lanata.
По месту прикрепления нитей ахроматинового веретена одна пара
у всех видов может быть названа хромозомой с субмедиальной пере-
тяжкой. У всех остальных хромозом субтерминальное прикрепление
нитей веретена. Хромозом с термальным прикреплением нитей вере-
тена у этой группы видов нет. По соотношению плеч, общими для
всей группы видов являются: 1) одна пара хромозом первого раздела
с акинетической перетяжкой на коротком плече; 2) одна пара хромо-
зом второго раздела; 3) две пары хромозом третьего раздела; 4) че-
тыре пары хромозом четвертого раздела; 5) единственная пара хро-
мозом пятого раздела.
172
Три вида — Т. lanata, Т. ingens и Т. carinata объединяются меж-
ду собой наличием второй пары хромозом во втором разделе. В то
же время у Т. lanata и Т. carinata увеличенное количество хромозом
в четвертом разделе и уменьшенное — в третьем.
Кариотип Т. ingens отличается от Т. fosteriana только лишней
хромозомой во втором разделе и недостачей одной хромозомы в
третьем.
Кариотип Т. victoris по распределению хромозом в группах вос-
производит кариотип Т. fosteriana, отличаясь большой укороченностью
и расчлененностью хромозом.
Следовательно, исходным видом этой серии, по нашему мнению,
является Т. fosteriana, очень близким к Т. fosteriana и молодым, а
может быть видом в будущем — Т. victoris, затем Т. ingens, Т. lanata
и Т. carinata. Эта группа видов тюльпанов мало дифференцирована и
специализирована, находится в процессе усиленного видообразования.
Вторая группа Leiostemones
Во вторую группу входят Т. praestans Hoog. и Т. subpraestans
Vved., Т. butkovii Z. Botsch. Луковица двух первых видов с тол-
стокожистыми черно-бурыми оболочками, с внутренней стороны бле-
стящие, почти голые.
При простом анализе видно, что эти два вида резко отличаются
друг от друга, но у них есть общие признаки, свидетельствующие об
их несомненной близости. Только у этих двух видов из наших круп-
ноцветных тюльпанов в естественных условиях, как правило, встре-
чается многоцветковость: кроме того, у обоих видов хорошо развито
вегетативное размножение.
Т. praestans Hoog (рис. 13, з; 15, а). Изученные нами образцы
получены из Таджикистана: перевал Сарбо (Ф. Н. Русанов, 1932 г.);
окрестности Туткаула (В. П. Бочанцев, 1933 г.) и с горы Себистан
(В. П. Бочанцев, 1933 г.). Долина Варзоба (3. П. Бочанцева, 1956 г.).
Хромозомы изучали на препаратах, приготовленных из корешков
(рис. 13, з) и го время спермиогенеза в пыльцевых трубках.
Исследовать хромозомы методом мазка по Левану не удалось,,
из-за рано протекающих митозов в молодой пыльце. Они происходят
в период, когда над землей еще нет никаких следов растения. В это
время цветок плотно одет листьями и извлечение пыльников вызы-
вает его гибель и сильное повреждение листьев. Таким образом, для
отыскания нужных фаз губятся сотни редких растений, в то время
как изучение хромозом во время спермиогенеза при посеве пыльцы
in vitro дает прекрасные результаты.
Для Т. praestans Ньютон (Newton, 1927) устанавливает количест-
во хромозом в соматических тканях —24. Исследованные нами 30
растений из трех местообитаний подтверждают данные Ньютона. По
соотношению плеч все хромозомы Т. praestans можно разбить на 5-
разделов (рис. 15, а; 11, н).
В первый раздел входит одна пара хромозом с длинным плечом
4,25 и коротким 2,25 р.
Второй раздел состоит из двух пар хромозом. У первой длинное
плечо—6,6, короткое—Зр; у второй—4,75 и 2,25 и..
Третий раздел объединяет две пары хромозом; первая с длинным
плечом—8,75, коротким—2,25 р; вторая—6,25 и 2 ц.
В четвертом разделе 4 пары хромозом; первая, самая крупная
хромозома набора, имеет длинное плечо 9,4 и короткое 2 р; вторая—
7,15 и 1,5 р; третья—6 и 1,4 р; четвертая—5,75 и 1,25 р.
173-
I
a
6
6
г
д
e
ж
з
8 9 Ю 11
9 10 11
Рис. 15. Идиограммы измеренных пластинок (увелич. в 800 раз).
?-Т- praestans Hoog.; б—Т. subpraestans Vved.; к—Т. alberti Rgl.; г—Т. mogoltavica М. Р о р. е t
v.v е d . ;'б—Т. greigii Rgl.; е—Т. micheliana Hoog.; ж—Т. lehmanniana М е г с k 1. ; з—Т. behmi-
ana Rgl.
В пятый раздел входят три пары хромозом; из них первая с
длинным плечом 9,25 и коротким—1,75 ц; вторая—8,25 и 1,5 р; третья—
7,5 и 1,5 р.
Нужно отметить, что хромозомы Т. praestans удивительно мор-
фологически однообразны. Никаких признаков вторичного расчленения
мы не наблюдали. Они очень толстые и укороченные, при этом длин-
ные плечи укорачиваются сильнее коротких и поэтому совершенно
меняется их внешний вид.
Попытка разбивки таких укороченных хромозом на группы дает
совершенно иные картины по соотношению плеч, чем нормальные
хромозомы.
Общая длина гаплоидного набора хромозом 108,55 р; длинных
плеч—83,9; коротких плеч—24,65 р; общий индекс—3,4.
Т. subpraestans Vved. (рис. 13, ж-, 15, б). Изученные нами об-
разцы получены из окрестностей Туткаула от В. П. Бочанцева. Хро-
мозомы исследовались в корешках, взятых от взрослых луковиц, из
пыльников, методом мазка по Левану и во время спермиогенеза. Де-
ление ядра в пыльнике на вегетативное и генеративное протекает
над землей в крупном бутоне. Число хромозом 24 (рис. 13, ж). По
соотношению плеч все хромозомы можно разбить на 6 разделов
(рис. И, в; 15, б).
В первый раздел входит одна пара маленьких хромозом с длинным
плечом в 3,25 и коротким—1,9 р.
Второй раздел состоит из двух пар хромозом. У первой длинное
плечо—6,75 и короткое—3 р. Вторая пара—самая маленькая хромозома
набора с длинным плечом 3,5 и коротким—1,25 р. На длинном плече
первой пары на расстоянии 3,6 р от кинетической перетяжки лежит
акинетический перерыв.
В третьем разделе две пары хромозом. Первая пара этой группы
с длинным плечом 8,25 и коротким—2,25 р со спутником на коротком
плече, который отличается известным постоянством и наблюдается
как в митозах, в клетках корешков, так и в митозах в молодой пыль-
це. Вторая пара хромозом с длинным плечом в 5,5 и коротким—1,75 р.
Четвертый раздел объединяет три пары хромозом: первая пара
с длинным плечом 8, коротким—1,75 р; вторая пара с длинным пле-
чом 6,75 и коротким—1,5 р, на дистальном конце длинного плеча
расположен спутник. Его нить может быть втянута в тело хромозомы
и тогда спутник сидит непосредственно на плече. У третьей пары
длинное плечо—5 и короткое—1,25 р.
В пятый раздел входят две пары хромозом. Первая с длинным
плечом—8,75, коротким—1,75 р. Вторая—8,3 и 1,5 р.
В шестом разделе две пары хромозом. У первой длинное плечо
8,25 и короткое—1,25 р, второй—6 и 1 р.
Общая длина гаплоидного набора хромозом 98,45 р; длинных
плеч—78,3; коротких плеч—20,15 р; общий индекс 3,88.
Сравнивая кариотипы Т. praestans и Т. subpraestans устанавливаем,
что первый, второй, третий хромозомальные разделы по количеству
хромозом и их пропорциям совпадают.
У обоих видов в четвертом разделе 3, а в пятом 2 одинаковых
хромозомы.
У Т. praestans по одной лишней паре хромозом в четвертом и
пятом разделе. Т. subpraestans имеет в этих разделах соответственно
меньше хромозом но компенсирует их двумя парами в шестом раз-
деле. В общем, хромозомы Т. subpraestans немного короче и более
морфологически расчленены, чем—Т. praestans.
175
При сравнении можно найти некоторые общие черты в кариотипах
Т. carinata,Т. praestans и Т. subpraestans. Так, три первых раздела
по числу входящих в них хромозом одинаковы. Правда, хромозомы
двух последних видов, особенно пара хромозом из первого раздела^
подверглись большой редукции.
Если сравнить хромозомы четвертого и пятого разделов трех
видов с другими, можно говорить вновь о большой редукции хромо-
зом Т. carinata против хромозом у близких ему видов (Т. lanata,.
Т. ingens и Т. fosteriana). О такой же редукции у Т. praestans против
Т. carinata и Т. subpraestans против Т. praestans.
Близость по кариотипу Т. carinata и Т. praestans, не беспочвенна
и макро-морфологически; близость же между собой Т. praestans и
Т. subpraestans ни кариотипически, ни макро-морфологически не вызы-
вает никаких сомнений.
Третья группа секции Leiostemones
Тюльпаны секции Leiostemones с совершенно < собым строением
цветка типа Т. greigii, антоциановыми пятнами и полосами на листьях
в Средней Азии образуют естественную группу видов, в которую'
входят 3 вида: Т. greigii, Т. mogoltavica, Т. micheliana, несколько,
особняком стоит Т. alberti, имеющая немного измененное сложение
цветка и листья без антоциановых пятен.
Из этой группы видов Ньютон исследовал лишь Т. greigii и ука-
зал для него 24 хромозомы. Изучение других близких видов, объе-
диняющихся в эту естественную группу, мы нашли в литературе
значительно позже и только относительно Т. micheliana (Тишлер, 1940).
Т. alberti Rgl. (рис. 15, в\ 16, а). Исследованные нами образцы
получены из Каратау от П. А. Гомолицкого. Хромозомы изучали в
корешках, выращенных из взрослых луковиц.
Число хромозом 2п=24 (рис. 16, а). Все хромозомы Т. alberti
можно разбить на 5 разделов (рис. 15, в).
В первый раздел входит одна пара хромозом. Длинное плечо—7,.
короткое—3,25 ц. На коротком плече на расстоянии 1 [х от кинетиче-
ской перетяжки лежит акинетическая перетяжка.
Второй раздел также состоит из одной пары хромозом, с длинным
плечом—6 и коротким—2,25 [х.
Третий раздел включает 6 пар хромозом. Первая с длинным пле-
чом 7,5 и коротким—2,5 ц; вторая—7,25 и 2 ц; третья—6,5 и 2 р;
четвертая—5,5 и 1,75 ц; пятая—5,25 и 1,75 ц; шестая—5,25 и 1,75 <х.
Четвертый раздел включает 3 пары хромозом. Первая из них
имеет длинное плечо 8 и короткое—2 [х; вторая—7 и 1,625; третья -6
и 1,5 ц.
В пятый раздел входит одна пара хромозом. Ее длинное плечо —
7,5, короткое—1,5 |х. Хромозомы Т. alberti толстые и слабо расчле-
ненные. Кроме одной акинетической перетяжки на хромозоме первой
группы, несмотря на большое количество материала, никаких перетя-
жек мы не наблюдали.
Общая длина гаплоидного набора хромозом—102,628 р-; длинных
плеч—78,75; коротких плеч—23,878 ц; общий индекс—3,346. Индекс
хромозомы Aj—2,154.
Т. mogoltamca М. Pop. et Vved. (рис. 15, г; 16. м). Изученные
нами образцы получены из Моголтау от И. И. Гранитова.
Хромозомы исследовали в корешках, полученных от взрослых
луковиц. Число хромозом 2п=24 (рис. 16, м). Все хромозомы Т. то-
goltavica по соотношению плеч можно разбить на 5 разделов (рис. 15 г).
176
12-2229
Рис. 16. Пластинки хромозом из точки роста корня (увелич. в 1000 раз).
а - Т. alberti Rgl.; 6-Т. micheliana Hoog.; в - Т. borszczowii Rgl,; г - Т. nitida Hoog.; д - Т. greigii Rgl.;e- Т. lehmanniana Merck!;ж - T. korolkovii
Rgl.; з—T. schrenkii Rgl.; и — T. behmiana; к — T. lelimaiiniana; л — Т. ostrovsKiana Rgl.; м — Т. mogoltavica М. Pop. et Vved.
В первый раздел входит одна пара хромозом. Длинное плечо~3,3
и короткое—5,5 р. На коротком плече на расстоянии 1,75 р от его
проксимального конца расположена акинетическая перетяжка.
Второй раздел состоит из одной пары хромозом. Длинное плечо—6,
короткое—2,6 р.
Третий раздел объединяет 5 пар хромозом. У первой пары
длинное плечо —11, короткое—3,25 р; второй—9 и 3 р; третьей—7,5
и 2,4 р; четвертой—6,5 и 2,25 р; пятой—6,24 и 2 р.
Кроме того, у первой пары этой группы на длинном плече на
расстоянии 4,5 р от проксимального конца плеча находится акинети-
ческая перетяжка. У второй пары также на длинном плече от прок-
симального конца на расстоянии 2 р расположена акинетическая пере-
тяжка. Третья пара имеет 2 перетяжки. Одна из них лежит на длин-
ном плече на расстоянии около 1 р от дистального конца плеча.
Другая—на коротком плече и тоже сдвинута к дистальному концу
плеча, отделяя членик в виде головки до 0,3 р величиной.
Четвертый раздел включает 4 пары хромозом: первая пара с
длинным плечом—9,75 и коротким—2 р; вторая—3 и 2 р; третья—6,5
и 1,625 р; четвертая 6 и 1,5 р.
Кроме того, вторая пара хромозом имеет на длинном плече 2
перетяжки. Одна из них расположена на расстоянии около 3 р от
проксимального конца плеча, а другая находится на расстоянии около
1 Р от его дистального конца.
В пятый раздел входит одна пара хромозом; ее длинное плечо—
7,5, короткое -1,5 р.
Общая длина гаплоидного набора хромозом—121,925 р; длинных
плеч—92,3; коротких плеч—29,625 р; общий индекс—3,456. Индекс
хромозомы Ат 1,509.
Т. greigii Rgl. (15, д\ 16, д). Исследованные нами образцы взяты
из массовых посадок Ботанического сада ТашГУ из Бостандыкского
района (сбор Сатуалдыбаева), Каратау (сбор Гомолицкого), с р. Малый
Кемин (окрестности с. Семеновки, сбор 3. П. Бочанцевой), ст. Тюль-
кубас (ущелье Файнбулак,- сбор А. Я. Буткова и В. П. Бочанцева).
Во всех случаях количество хромозом для данного вида, по
определениям Ньютона (1927) и нашим, 2п=24 (рис. 16, д).
Исследование хромозом мы проводили в корешках, полученных
от взрослых луковиц и в мазках пыльников во время формирования
двухъядерной пылинки (рис. 11, з).
Все хромозомы Т, greigii можно разбить на 5 разделов (рис. 15, д).
Анализ кариотипа проводится по хромозомам, взятым из мазка. Они
несколько короче, чем у корешков, но, как показала проверка, сох-
ранили соотношение плеч.
В первый раздел входит одна пара хромозом; длинное плечо—6,5,
короткое—3,75 р. На коротком плече в нижней трети лежит тради-
ционный акинетический перерыв.
Второй раздел имеет одну пару хромозом с длинным плечом 6,75
и коротким—3 р.
Третий раздел включает четыре пары хромозом. У первой пары
длинное плечо 7, короткое—2 р; вторая—5,75 и 1,9 р; третья—5,5
и 1,8 р; четвертая—5 и 1,5 р.
Четвертый раздел состоит из 5 пар хромозом. Первая—с длинным
плечом—7,25 и коротким—1,75 р; вторая—6,5 и 1,5 р; третья—6 и
1,5 р; четвертая—6 и 1,25 р; пятая—5,4 и 1,25 р.
В пятый раздел входит одна пара хромозом с длинным плечом—7,5
и коротким—1,5 р.
178
Исследование морфологии хромозом мы проводили у ташкентского
Т. greigii. У образцов из других местообитаний устанавливали только
число хромозом. Хромозомы Т. greigii очень толстые и вторично не
расчленяются.
Общая длина хромозом гаплоидного набора—97,8 р; длинных
плеч—75,15; коротких плеч—22,7 р; общий индекс 3,31. Индекс
хромозом А—1,733.
Т. micheliana Hoog. (рис. 15, е-, 16, б). Изученные нами образцы
получены из Каракала, со ст. Уратюбе, со ст. Драгомирове, ст. Бахар-
ден (сборы Н. Б. Никифоровой), из пос. Тумановского (сбор А. Я. Гнез-
дило).
Хромозомы исследовали в корешках, полученных от взрослых
луковиц. Число хромозом 2п=24 (рис. 16, б).
Все хромозомы Т. micheliana по соотношению плеч можно разбить
на шесть разделов (рис. 15, е).
В первый раздел входит одна пара хромозом с длинным плечом 8
и коротким—4,25 р. На коротком плече на расстоянии 1,5 р от прок-
симального конца находится кинетическая перетяжка.
Второй раздел состоит из одной пары хромозом с длинным плечом
в 5,25, коротким —2,25 р. На длинном плече на расстоянии около 2,75 р
расположен акинетический перерыв. На коротком плече на расстоянии
около 1 р от дистального конца лежит акинетическая перетяжка.
В третий раздел входят две пары хромозом. Первая из них с
длинным плечом 8 и коротким—2,6 р, на длинном плече расположены
2 перетяжки—одна на расстоянии около 2,25 р от дистального конца,
вторая почти в 3 р от первой. Таким образом, это плечо трехчленис-
тое. На коротком плече на расстоянии 1,5 р от проксимального конца
также лежит акинетическая перетяжка. У второй пары длинное пле-
чо—7,25 и короткое — 2 р; на длинном плече на расстоянии 2 р от
проксимального конца находится акинетическая перетяжка.
Четвертый раздел состоит из шести пар хромозом. Первая с
длинным плечом—9,5, коротким 2,25 р; акинетическая перетяжка рас-
положена на длинном плече на расстоянии около 2,5 р от дистального
конца плеча и на коротком плече на расстоянии около 1 р от дис-
тального конца плеча. Вторая пара хромозом с длинным плечом 8,74,
коротким—1,9 р; на длинном плече имеются две перетяжки, располо-
женные около 0,75 р от проксимального конца плеча и почти 1 р от
дистального конца плеча. На коротком плече перетяжка находится
на расстоянии около 0,74 р от проксимального конца плеча. У третьей
пары длинное плечо—7,5 и короткое—1,625 р; у четвертой—7 и 1,5 р:
в нижней половине длинного плеча с двумя перетяжками—первая на
расстоянии 0,75; вторая—2,5 р от проксимального конца плеча. Пятая
пара с длинным плечом в 6, коротким—в 1,5 р; шестая—в 5,75 и
1,4 р, признаков вторичного расчленения хромозом не наблюдается.
В пятый раздел входит одна пара хромозом с длинным плечом 6,25 р,
на котором в 1,75 р от проксимального конца плеча расположена
перетяжка, и коротким 1,925 р.
Шестой раздел составляет также одна пара хромозом с длинным
плечом—9,5 и коротким—1,5 р. На дистальном конце длинного плеча
отчленен перетяжкой членик, в 1,5 р дл.
Общая длина хромозом гаплоидного набора—123,45 р; длинных
плеч—98,75; коротких плеч—24,7 р; общий индекс 3,998. Индекс
хромозомы Ат 1,882.
Для Т. .micheliana материалы собраны в окрестностях Тегерана
{Иран). Предварительные исследования типов хромозом несколько
179
отличаются от приведенных. Отклонение наблюдается в сторону уве-
личения коротких плеч, что совершенно меняет кариотип, но материал
еще нуждается в строгой проверке. Так как изучение Т. micheliana1
из Ирана проведено в мазках, а советского—в клетках корешков, то
трудно сравнивать их кариотипы. Но, по нашему мнению, такое срав-
нение возможно, так как мы установили, что другие виды при обоих
методах исследования сохраняют соотношение плеч, изменяя только*;
абсолютную длину плеч хромозом.
Таким образом, общим для кариотипа всей группы Tulipa, объе-
диняющихся систематически в одну группу во главе с Т. greigii, яв-
ляются следующие пары хромозом: одна пара хромозом первого-
раздела; одна пара второго; две пары третьего; три пары четвертого;:
одна пара пятого.
T.mogoltavica от Т. greigji отличается большей расчлененностью
хромозом, одной лишней хромозомой в третьем разделе и отсутствием
одной пары хромозом в четвертом разделе. Следовательно, получается
так, что у Т. mogoltavica в третьем разделе пять хромозом, а в чет-
вертом—четыре; а у Т. greigii в третьем разделе четыре хромозомы,
а в четвертом—пять. Хромозомы Т. greigii вдвое толще Т. mogoltavi-
ca. У Т. micheliana в третьем разделе всего две пары хромозом, чет-
вертом—шесть пар, т. е. на одну пару больше, чем у Т. greigii и на
две—чем у Т. mogoltavica. Кроме того, у Т. micheliana одна пара
хромозом входит в шестой раздел, чего нет у других видов этой
группы тюльпанов.
У Т. alberti количество хромозом в третьем разделе увеличивается
по сравнению с Т. mogoltavica на одну пару; с Т. greigii—на две,
с Т. micheliana—на четыре пары; но в четвертом разделе у Т. albert;
меньше пар хромозом, чем уТ. mogoltavica на одну пару, у Т. grei-
gii—на две и у Т. micheliana—на три пары. Таким образом, Т. miche-
liana и Т. alberti по кариотипам наиболее отдаленные виды в пределах
группы родственных видов, что и подтверждается внешней морфологией
этих растений.
Четвертая группа видов секции Leiostemones
Четвертую естественную группу среди изученных нами видов Tu-
lipa секции Leiostemones образуют три вида: Т. maximowiczii Rgl.,.
Т. linifolia Rgl. и Т. wilsoniana Hoog. У всех трех видов луковицы
маленькие с черными кожистыми оболочками. Растения Т. maximo-
wiczii до 30 см выс., а у Т. wilsoniana мы наблюдали цветы, сидящие
прямо на земле. Стебель у этих особей можно обнаружить только'
выкопав растения. Листья у всех трех видов узкие. Листочки около-
цветника красные. Пятно черное, тупое, невысокое, во всю ширину
листочка.
Проведя сравнение признаков и наблюдая эти тюльпаны в природе,
нужно отметить большую близость Т. maximowiczii и Т. linifolia.
Т. wilsoniana занимает в группе несколько особое положение.
Из группы видов Leiostemones Ньютон исследовал: Т. maximo-
wiczii, Т. batalini и Т. linifolia. А. И. Введенский считает Т. batalini
синонимом Т. maximowiczii.
Ньютон (Newton, 1927) у всех трех видов находит один и тот же
тип хромозомы. Хромозомы с конечным прикреплением нитей веретена.
Второе плечо в виде небольшого шарика. Он отличает 'характерную
для всех трех видов единственную очень короткую пару хромозом. Мы
не могли исследовать Т. batalini, но, использовав рис. 2 из табл. 7 в
180
работе Ньютона (1927), можно сделать некоторые выводы и о нем.
Но прежде всего освещаем данные по изучению Т. maximowiczii,
Т. linifolia и Т. wilsoniana.
Т. maximowiczii Rgl. (рис. 11, м\ 17, ё) первый из трех видов
четвертой группы секции Leiostemones исследованных нами.
Изученные образцы собраны в окрестностях кишлака Санктуда
(В. П. Бочанцев, 1933).
Мы исследовали корешки, полученные от взрослых луковиц и
проверяли результаты по мазкам из молодых пыльников (рис. 11, д).
Число хромозом в клетках корешков в отличие от других видов рода
Tulipa 2п=22. Такого числа хромозом нет ни у одного из исследо-
ванных нами видов этого рода. Ньютон (Newton, 1927) определяет
2 п=24. Однако после исследования корешков 30 растений Т. maxi-
mowiczii мы получили подтверждение, что 2п=22 хромозомам (рис. 11 м\
Весной 1935 г. мы изучали хромозомы в мазках из молодых
пыльников, когда в пыльце происходило деление первичного ядра
пылинки и вновь получили тот же результат—п = 11 хромозомам. После
морфологического анализа хромозом установлено, что все хромозомы
Т. maximowiczii можно разбить по соотношению плеч на 4 раздела
4рис. 17, е).
В первый раздел входит одна пара хромозом с длинным плечом—
4,75 и коротким—4 и, хромозома почти равноплечая или, по опреде-
лению Ньютона у других видов, с медиальным прикреплением нитей
веретена.
Второй раздел состоит из одной пары хромозом с длинным пле-
чом—7,5 и коротким—2 щ В третий раздел также входит одна пара
хромозом с длинным плечом—7,75 и коротким—2 Четвертый, пя-
тый, шестой, седьмой и восьмой разделы не представлены. У восьми
пар, которые остались не разбитыми по группам, короткое плечо имеет
вид шарообразного тельца и едва достигает 0,5 Таким образом,
Т. maximoviczii содержит новый девятый раздел хромозом — головча-
тых, не встречающийся у ранее исследованных видов. В девятый раз-
дел входят 8 пар хромозом. Первая с длинным плечом — 8,75 у,, вто-
рая—8,75, третья—7,5, четвертая—5,25, пятая—5, шестая—4,25, седь-
мая—4,25, восьмая—3,5
У третьей пары хромозом на дистальных концах длинных плеч
расположен акинетический перерыв, отделяющий членики до 1 дл.
Кроме того, на коротких плечах пятой пары хромозом (наблюдалась
только в мазках) находится шаровидный спутник, сидящий на нити.
Таким образом, по морфологии хромозом, этот вид имеет три типа
хромозом (в первой, второй, третьей группах), которые Ньютон не
приводит для Т. maximoviczii, и из головчатых хромозом—одну с
акинетическим перерывом на длинном плече и одну со спутником.
Общая длина гаплоидного набора хромозом — 79,25 рь; длинных
плеч хромозом — 67,25; коротких плеч хромозом того же набора —
12; общий индекс 5,6
Т. linifolia Rgl. (рис. 11, и\ 17, ж). Исследованные нами образ-
цы получены от П. А. Баранова с перевала Кайвак (Кала-и-Хумб)
в 1934 г. Изучение хромозом проведено только на корешках, полу-
ченных от проращивания взрослых луковиц. Число хромозом 2п=
:24 (рис. 11, и).
Таким образом, данные наши и Ньютона (Newton, 1927) совпадают.
По соотношению плеч все хромозомы Т. linifolia можно разбить на
4 раздела (рис. 17, ж).
181
2 3 4
JTL
о Mi
2 3 4
13 14
4 5
5 6
5 6
15 1617 18 19 20 21
3 4 5 6 7 8
II
3
2 3 4
6 7 8 9
7 8 9 10
fO 11 12
5 5 7
4 5 6 7 8 9 10 11
4 5 6 7 8 9 1011 12
\ 8 9 Ю 11 1 2
4 5 5
Рис. 17. Идиограммы измеренных пластинок (увелич. в 800 раз).
с—Т. zenaidae Vved.: б— Г. kolpakovskiana Rgl.; e—T. ostrovskiana Rgl.; z—T. tetraphylla Rgl..
д—T. ka ufmanniana Rgl.; e—T. maximowiczii Rgl.; ж—T. linifolia Rgl.; J—T. wilsoiuana Hoog.
182
Первого раздела у Т. linifolia нет. Во второй раздел входит одна
пара хромозом, названная Ньютоном „of wery sh rt chromosomes" c
длинным плечом —2,5 и коротким — 1у; третьего раздела у Т. linifolia
также нет.
Четвертый раздел состоит из одной пары хромозом с длинным
плечом — 6,5 и коротким — 1,5 у.
В пятом разделе также одна пара хромозом с длинным плечом —
5,625 и коротким — 1 у; шестого раздела нет.
Последний раздел головчатых хромозом с шарообразными голов-
ками различной величины: первая пара с длинным плечом — 8,5 и ко-
ротким— 0,5 у; вторая — 7,4 и 0,75; третья — 6 и 0,5; четвертая — 5,75
и 0,5; пятая 5,75 и 0,4; шестая 5,75 и 0,25; седьмая 5,5 и 0,25; вось-
мая 5,4 и 0,5; девятая 5,125 и 0,5 у.
По нашим данным, у двух пар хромозом имеются короткие пле-
чи, а не головки, как у Ньютона. Вего работе (1927) на табл. 7, рис.1
изображена метафаза Т. linifolia. В ее левом нижнем углу видна хро-
мозома, короткое плечо которой никак не может быть названо шаро-
образным, так как его длина в два раза превышает ширину. Оно
может быть названо плечом. У остальных хромозом короткие плечи
приподняты, а при их незначительных размерах это незаметно.
Т. batalini по кариотипу очень похож на Т. linifolia (Ньютон, 1927,
табл. 7, рис. 2): видна одна пара маленьких хромозом; среди осталь-
ных хромозом разобраться нелегко, но можно констатировать, что ве-
личина головок весьма разнообразная, что свойственно и Т. linifolia.
Размеры хромозом у обоих видов одинаковые.
Общая длина гаплоидного набора хромозом —77,45 у; длинных
плеч —69,8; коротких плеч — 7,65 у; общий индекс—9,1.
Т. ‘wilsoniana Hoog. (рис 11, к; рис. 17, з). Исследованные нами
растения получены из Туркмении от А. И. Введенского и окрестнос-
тей поселка Прохладного у подножия гор от А. Ф. Гнездило.
Изучение хромозом проведено только в корешках, полученных от
проращивания взрослых луковиц. Число хромозом 2п=24 (рис. 11 к).
Все хромозомы у Т. wilsoniana по соотношению плеч можно разбить
на 4 раздела (рис. И, б; 17, з).
Первого раздела у Т. wilsoniana нет; во второй раздел входит одна
пара маленьких хромозом, отмеченная Ньютоном для Т. linifolia и Т.
batalini с длинным плечом — 2,25 и коротким -1 ц.
Третий раздел включает три пары хромозом. Первая с длинным
плечом—5 и коротким 1,5 у; вторая—3,5 и 1 у; третья —3 и 1 у.
Кроме того, у первой пары на коротком плече расположен аки-
нетический перерыв, делящий его на две почти равные части, у треть-
ей пары на длинном плече на расстоянии 1,25 у от проксимального
конца находится акинетический перерыв.
Четвертый раздел состоит из трех пар хромозомы. У первой длин-
ное плечо —5,6 и короткое —1,25 у; вторая —4,5 и 1; третья —4 и —2 у.
На длинном плече третьей пары имеется акинетический перерыв
на расстоянии около 1,75 у от проксимального конца плеча; свобод-
ный конец длинного плеча этой хромозомы неокруглый, как у других
хромозом, а оттянут в острие, кончающееся нитевидным образованием;
никаких спутников на ней мы не наблюдали.
Пятый, шестой, седьмой и восьмой разделы хромозом у Т. wilsonia-
na не представлены. Головчатые хромозомы многочисленны. Размеры
головок у всех хромозом б. м. одинаковы и достигают 0,75 у. Длин-
ные плечи у первой пары 6,25 у, второй 5,75, третьей 5,6, четвертой.
5,125, пятой 4,4 у.
183.
Вторая, третья и четвертая пары на длинных плечах имеют аки-
нетический перерыв на расстоянии 1 у. 1,75 и 1,5 от проксимального
конца плеча.
Общая длина гаплоидного набора хромозом — 67,47 ji; длинных
плеч — 54,975; коротких плеч — 12,5 щ общий индекс — 4,39.
Просматривая всю группу в целом, нужно отметить большую ка-
риотипическую разницу между Т. linif olia и Т. Maximowiczii. в то
время, как внешне растения морфологически слабо различаются. Оче-
видно, кариотип изменяется простой перекомбинацией хроматинового
вещества или эти виды имеют ряд существенных признаков еще не отме-
ченных систематиками. Тем более, что А. И. Введенский во „Флоре
СССР" (т. IV) в примечании к этим двум видам пишет: „Т. maximo-
wiczii и Т. linifolia очень близки друг к другу и требуют еще допол-
нительные наблюдения по живому материалу для окончательного их
разграничения". Ошибка Ньютона в числе и морфологии хромозом у
Т. maximowiczii должна быть отнесена за счет отмеченной выше
морфологической близости Т. maximowiczii и Т. linifolia и, таким об-
разом, неправильного определения вида. Проверить это невозможно,
так как Ньютон не приводит ни географических, ни систематических
данных для исследованных им видов. То же самое получается при
сравнении данных Ньютона с Т. batalini, по пластинке хромозом, при-
веденной в его работе (Newton, 1927; рис. 29). Кариотип Т. batalini
повторяет кариотип Т. linifolia.
А. И. Введенский („Флора СССР", т. IV) считает Т. batalini сино-
нимом Т. maximowiczii. Видимо, исследовали не точно определенные
растения. Т. wilsoniana, связанная с Т. linifolia одной парой малень-
ких хромозом второго раздела, одной парой хромозом четвертой груп-
пы и целым рядом головчатых хромозом, имеет более удлиненные
вторые плечи хромозом, чем у описанных выше видов, что дает
семь пар хромозом с плечиками, а не с шаровидными головками.
Кроме того, его кариотип характеризуется большим количеством,
только ему свойственных признаков. Вторичное расчленение хромозом
из всей группы близких видов наиболее ярко выражено у Т. wilsonia-
na. Морфологически Т. wilsoniana отличается от изученных нами ви-
дов этой группы.
Пятая группа видов секции
Leiostemones
Пятая группа видов секции Leiostemones в Средней Азии вклю-
чает: Т. behmiana Rg 1., Т. lehmanniana Merkl. типичный Т. lehmannia-
na из Кушки и Т. borszczowii Rgl., всего четыре вида. Мы не распо-
лагаем литературными данными о цитологическом исследовании нас-
тоящей Т. borszczowii, Т. behmiana и Т. lehmanniana.
Т. lehmanniana Merci, (рис. 15, ж\ рис. 16, е). Исследованные
образцы собраны в окрестностях ст. Малик (наш сбор 1931 г. и Т.
А. Романовича, 1933 г.).
Количество хромозом у данного вида, обычное для Tulipa, 2п—
24 (рис. 16, е}. Изучение проводили на корешках, полученных от
взрослых луковиц и во время спермиогенеза в пыльцевых трубках,
воспитанных in vitro. Все хромозомы Т. lehmanniana по соотношению
плеч можно разбить на 5 разделов (рис. 15, ж).
Первый раздел состоит из одной пары хромозом с длинным пле-
ном—7,75 и коротким—4 [х; на коротком плече расположена обычная
184
перетяжка, отделяющая членик около кинетического перерыва разме-
ром около 1/4 всей длины плеча.
Во второй раздел входят две пары хромозом. Первая с длинным
плечом 8,125 и коротким 3 [х; вторая—6,25 и 2,25 |х. У второй пары
на дистальном конце длинного плеча акинетической перетяжкой отде-
лен небольшой шарообразный членик, иногда при сильной дифферен-
цировке принимающий вид спутника.
Третий раздел составляет четыре пары хромозом: первая с длин-
ным плечом 8 и коротким—2,25 jx; вторая—6,25 и 2 [ь; третья—6 и
2 [х; четвертая — 6 и 1,75 |х.
Кроме того, на коротком плече четвертой пары находится акине-
тическая перетяжка, делящая его на две равные части.
Четвертый раздел хромозом состоит из трех пар хромозом: пер-
вая с длинным плечом 9,6 и коротким 2 [х; вторая— 7,25 и 1,75 [х;
третья —6,5 и 1,5 [X.
У третьей пары хромозом на дистальном конце длинных плеч
имеются акинетические перерывы, отделяющие шарообразные членики
такие же непостоянные, как и у второй пары хромозом из второй
группы этого вида.
Пятый раздел объединяет две пары хромозом: первая с длинным
плечом—11,75 и коротким—2,3 |х; вторая—10 и 1,75 |х.
Общая длина гаплоидного набора хромозом—120,025 |х; длинных
плеч —93,475; коротких плеч — 26,66 [х; общий индекс 3,52.
Т. lehmanniana из Туркмении (Кушка) А. И. Введенский несколь-
ко отличает от типичного Т. lehmanniana, но, видимо, из-за недоста-
точно выраженных признаков, он не выделил его в самостоятельный
вид. Мы исследовали только корешки взрослых луковиц.
Количество хромозом то же, что и у типичной Т. lehmanniana
2п—24 (рис. 16, к). У этого вида несколько иная разбивка хромозом
по пяти разделам (рис. 18, а).
В первый раздел входит одна пара хромозом с длинным плечом
5,75 и в 3,5 |х.
На длинном плече расположен акинетический перерыв, делящий
•его на две почти равные части. На коротком плече акинетический пере-
рыв отделяет членик около 1 р» в проксимальной части плеча.
Второй раздел состоит из двух пар хромозом: первая — с длин-
ным плечом — 6,5 и коротким—2,25 р; вторая —5,125 и—2 [х.
Третий раздел объединяет пять пар хромозом: у первой длинное
плечо—6,5; короткое—1,9 и; вторая — 5,25 и 1,75; третья—5,25 и
1,75; четвертая—5 и 1,6; пятая — 5 и 1,5 щ
У третьей пары этого раздела на длинном плече находится аки-
нетический перерыв, делящий его на две почти равные части. У чет-
вертой пары на длинном плече расположено два акинетических пере-
рыва—на расстоянии около 0,75 от дистального конца плеча, и 1,5 от
его проксимального конца. У этих пар на дистальном конце длинных
плеч отмечены спутники.
Четвертый раздел объединяет три пары хромозом: первая —с
длинным плечом —8,5 и коротким —2 [х; вторая — 7 и 1,5 |х; третья—
7 и 1,5 <х.
На длинном плече первой пары имеется акинетический перерыв
на расстоянии около 3,25 [х от дистального конца плеча.
В пятый раздел хромозом входит одна пара хромозом с длинным
плечом—9,25 и коротким—1,75
Общая длина гаплоидного набора хромозом — 99,125 |х; длинных
плеч—76,125; коротких плеч—23 |х; общий индекс—3,31.
185
a
б
б
г
д
е
u!V
3
I,
l!
/
23 4 5
6 7 8 9 10
6 7 8 9 10
9 10 11
9 10 И
2 3
9 5 6 7 8
8
/
/ 2
1 2
2 3
3 4 5
Рис. 18. Идиограммы измеренных пластинок (увелич. в 800 раз).
л-Т. lehmanniana Merckl.; б — Т borszczowii Rgl.; в — Т. nitida Hoog.; г—Т. korolkovii Rgl.; д—Т. rosea
Vved.; е — Т. schrenkii Rgl.; ж — Т. kolpakovskiana Rgl.; з — Т. ferganica Vved.
186
T. eehmlana Rgl. (рис. 16, «; 15, з). Исследованные образцы
собраны в окрестностях Илийска в Казахстане (сбор в 1931 г. Грани-
товой); изучали только корешки взрослых луковиц. Количество хро-
мозом 2п=24 (рис. 16, «).
Все хромозомы можно разбить по соотношению плеч на пять
разделов (рис. 15, з).
В первый раздел входит одна пара хромозом с длинным плечом—
6,25 и коротким — 3,25 у. Мы не могли установить наличия акинети-
ческой перетяжки на коротком плече.
Второй раздел состоит из двух пар хромозом: первая пара с длин-
ным плечом —6,75 и коротким —2,75 у; вторая —5,5 и 2,25 у.
На коротких плечах у первой пары хромозом на расстоянии око-
ло 1,25 у от кинетической перетяжки находится большой акинетиче-
ский перерыв. На длинном плече второй пары акинетический перерыв
отделяет продолговатый членик, расположенный на расстоянии около
1,5 у от дистального конца плеча.
Третий раздел объединяет пять пар хромозом: первая с длинным
плечом — 6,4 и коротким—1,75 у; вторая—5,5 и 1,75 у, третья —5,5
и 1,72; четвертая — 5,5 и 1,75; пятая—5,25 и 1,75 у.
Четвертая пара хромозом имеет акинетические перерывы, деля-
щие ее короткие плечи на две почти равные части и пятая пара хро-
мозом с двумя акинетическими перерывами на каждом из длинных
плеч. Перерывы пятой пары делят длинное плечо на три части, из
них средняя часть немного длиннее крайних, которые почти равны
между собой.
Четвертый раздел состоит из трех пар хромозом: у первой длин-
ное плечо —8 и короткое—2 у; у второй — 6,6 и 1,5 у- и третьей —
6 и 1,5 у
Вторая пара хромозом с акинетическими перерывами на длинных
плечах на расстоянии около 3 у. от дистального конца плеча. На дис-
тальном конце длинного плеча третьей пары акинетический перерыв
отделяет небольшой членик длиной около 0,5 у-.
В пятый раздел входит всего одна пара хромозом с длинным пле-
чом—10,25 и коротким — 2 у. На дистальном конце длинного плеча
имеется акинетическая перетяжка, отделяющая головкообразный чле-
ник длиной 0,5 у.
Общая длина гаплоидного набора хромозом—101,5 у.; длинных
плеч — 77,5; коротких плеч—24 у-; общий индекс—3,41.
Т. eorszczowii Rgl. (рис. 16, в; 18, б). Изученные нами лукови-
цы получены из Казахстана (ст. Тюрятам, сбор 1931 г. Т. С. Агане-
совой). Исследование проводили на корешках взрослых луковиц.
Количество хромозом 2п=24 (рис. 16, в). Все хромозомы Т. Borszczo-
wii Rgl. по соотношению плеч можно разбить на 5 разделов (18, б).
В первый раздел, как обычно для Tulipa секции Leiostemones,
входит одна пара хромозом с длинным плечом в 7 и коротким —4 у.
Нижняя часть короткого плеча отделена акинетическим перерывом.
Второй раздел состоит из двух пар хромозом: первая с длинным
плечом—7 и коротким—2,5 у; вторая—4,75 и 1,9 у.
Третий раздел включает пять пар хромозом: у первой длинное
плечо—6,5 и короткое—2 у; второй—6 и 2; третьей—5,5 и 1,75;
четвертой—5,5 и 1,75; пятой—5,5 и 1,5 у. У второй пары хромозом
на дистальном конце длинного плеча расположен акинетический пе-
рерыв, отделяющий продолговатый членик 1 у длины.
Четвертый раздел включает три пары хромозом: первая с длинным
плечом—8,25 и коротким—2 у; вторая — 6 и 1,5; третья — 6 и 1,25 у.
187
В пятый раздел входит одна пара хромозом с длинным плечом
8,25 и коротким 1,5 р.
Общая длина гаплоидного набора хромозом — 99,15 р; длинных
плеч—75,25; коротких плеч — 23,9 [J-; общий индекс—3,14.
Подводя итоги изучению кариотипа в V группе Tulipa секции
Leiostemones, надо отметить что общими для всей группы по числу
хромозом являются одна пара хромозом первого раздела, две пары
хромозом второго раздела, три пары четвертого, четыре пары треть-
его и одна пара пятого. По кариотипу Т. lehmanniana из Туркмении
(Кушка), более сходна с Т. Behmiana, чем с типичным Т. lehmanniana.
Кариотип Т. Borszczowii ближе к кариотипу Т. Behmiana, чем к leh-
manniana, который обладает более длинными хромозомами.
У хромозом Т. Borszczowii меньше признаков вторичного расчле-
нения, чем у других тюльпанов в этой группе.
Шестая группа видов секции Leiostemones
В VI группу видов Tulipa секции Leiostemones входят три вида:
Т. korolkovii Rgl., Т. nitida Hoog. и Г. rosea Vved. Те образцы,
которые мы изучали, были достаточно разнообразными. Так стебель
Т. nitida до 30 см дл., листья не достигали или не превышали цветок,
окраска листочков околоцветника ярко-красная, блестящая, а стебель
Т. korolkovii маленький, листья превышали цветок, окраска листочков
околоцветника красная, но более густого тона, чем у Т. nitida. А. И.
Введенский во „Флоре СССР" в примечании пишет: „Отличия Т. ni-
tida от Т. korolkovii в настоящее время не ясны и экземпляры из се-
верных частей Памиро-Алая могут с успехом относиться к тому или
другому виду11. Т. rosea отличается от этих двух видов необычайно
ярко-розовым цветом.
Т. nitida Hoog. (рис. 16, г; 18, в). Исследованные образцы со-
браны в Денауском районе. Мы изучали корешки взрослых луковиц.
Число хромозом 2п=24 (рис. 16, г). Все хромозомы Т. nitida по со-
отношению плеч можно разбить на 4 раздела.
Первый раздел состоит из одной пары хромозом. Ее длинное пле-
чо 5,5, короткое — 2,75 р. Обнаружить акинетическую перетяжку на
коротком плече не удалось.
Второй раздел объединяет 4 пары хромозом: первая пара с длин-
ным плечом —5,6 и коротким — 2 р: вторая—5,25 и 2; третья—4,75
и 2; четвертая — 4,25 и 1,9 р.
Третий раздел составляет пять пар хромозом: у первой пары длин-
ное плечо — 5,75, короткое—1,5; у второй—5,75 и 1,5; третьей—5,75
и 1,5; четвертой—5,25 и 1,75; пятой—5,25 и 1,5 р. Четвертая пара
хромозом на длинном плече имеет акинетическую перетяжку, распо-
ложенную на расстоянии около 3 р от проксимального плеча.
Четвертый раздел не представлен.
В пятый раздел входят две пары хромозом: первая с длинным
плечом—9 и 1,75, вторая—7 и 1,25 р.
Общая длина гаплоидного набора хромозом —90,5 р; длинных
плеч—69,1; коротких плеч—24,4 р; общий индекс—3,22.
Т. korolkovii Rgl. (рис. 16, ж; 18 г). Исследованные образцы со-
браны в окрестностях ст. Дарбаза Среднеазиатской железной дороги.
Мы изучали хромозомы в корешках взрослых луковиц. Количе-
ство хромозом у Т. korolkovii 2п=24 (рис. 16, ж). Все хромозомы
этого вида могут быть разбиты на 4 раздела (рис. 18, г).
488
В первый раздел входят две пары хромозом: длинное плечо пер-
вой пары—7,75 и 3,75 и.; второй — 4,5 и 2,25 р.. На коротком плече
первой пары, очень похожей по величине на подобную хромозому
других видов, мы не обнаружили акинетическую перетяжку. Необыч-
но появление второй пары хромозом в первом разделе, чего не было
в кариотипах, описанных выше видов.
Второй раздел хромозом состоит из трех пар: первая с длинным
плечом —7,25 и коротким —3 р.; вторая —6,25 и 2,25; третья —5,2
и 1,9 [а.
Третий раздел объединяет пять пар хромозом: длинное плечо пер-
вой пары—8,5 и короткое—2,25 р.; второй—8,5 и 2,25; третьей—7 и
2; четвертой—6,25 и 2; пятой—6,25 и 1,9 р.
Четвертый раздел хромозом у Т. korolkovii так же, как у Т. ni-
tida, отсутствует.
Пятый раздел включает две пары хромозом: первая с длинным
плечом—12 и 2,25 р; вторая—9,75 и 1,7 р.
Общая длина гаплоидного набора хромозом—113,2 р; длинных
плеч—85.2; коротких плеч —28 р; общий индекс— 3,03.
Т. rosea Vved. Грис. 18, д). Исследованные образцы собраны в
окрестностях Намангана.
Изучение хромозом мы проводили во время спермиогенеза в пыль-
цевых трубках, воспитанных in vitro.
Число хромозом 2п=24. Все хромозомы можно разбить на четы-
ре раздела.
В первый раздел входят две пары хромозом: у первой длинное
плечо 8,25 и короткое 3,125 р; второй—5,5 и 3,125 р.
Второй раздел состоит из трех пар хромозом: длинное плечо пер-
вой—6,25 и короткое — 2,25 р; второй—5,5 и 2; третьей—4,25 и 1,5 р.
. Третий раздел объединяет пять пар хромозом: первая с длинным
плечом 8,5 и коротким 2,5 р; вторая—7,25 и 2,25; третья —6 и 1,75;
четвертая — 5,75 и 1,5; пятая —5,5 и 1,5 р.
Хромозом четвертого раздела нет, а в пятый раздел входят две
пары. У первой пары длинное плечо 9,5 и короткое 2,15 р, у второй—
9,25 и 1,75 р.
Признаков вторичного расчленения хромозом у Т. rosea не наб-
людается.
Общая длина гаплоидного набора хромозом —106,9 р; длинных
плеч—81,5; коротких плеч—25,4 р; общий индекс—3,2.
Кариотипы трех близких видов Т. korolkovii Rgl., Т. nitida.
Hoog. и Т. rosea Vved. также близки, как и их морфология. Почти
единственное их различие в том, что у Т. korolkovii и Т. rosea в пер-
вом разделе имеется одна маленькая лишняя пара хромозом. Но, ес-
ли сравнить эту пару хромозом с четвертой парой во втором разделе
Т. nitida, ясно видна их близость, так как это самые мелкие хромо-
зомы в идиограммах этих видов. Хромозомы Т. korolkovii и Т. rosea
несколько крупнее, чем хромозомы Т. nitida. Кариотип Т. rosea
полностью повторяет кариотип Т. korolkovii.
Седьмая группа видов Leiostemones
В седьмую группу видов секции Leiostemones входит пять ви-
дов: Т. schrenkii Rgl., Т. kolpakovskiana Rgl., Т. ferganica Vved.,
T. zenaidae Vved. и T. ostrovskiana Rgl., T. kolpakovskiana, T. fer-
ganica, T. zenaidae и T. ostrovskiana — близкие виды. Так, долинные
Т. kolpakovskiana отличаются от Т. ostrovskiana, взятой из кустарни-
189
ковой зоны гор, но на месте стыка местообитаний различить их поч-
ти невозможно.
А. И. Введенский во „Флоре СССР11 дает примечание к описанию
этих двух видов: „Многочисленные естественные гибриды между
Т. kolpakovskiana и Т. ostrovskiana, известные, например в окр. Алма-
Ата, перекомбинируя окраску цветов и габитуальные свойства этих
видов, совершенно стирают границу между ними и делают определе-
ние их чрезвычайно затруднительным
В пределах этой группы Ньютон изучил хромозомы только у од-
ного Т. kolpakovskiana и число хромозом определил равным 24. По-
следующие исследователи изучили садовые формы Т. ostrovskiana.
Т. schrenkii Rgl. (рис. 16, з; 18, е). Исследованные образцы при-
везены экспедицией Ботанического сада из Бер-Чогура в 1930 г.
Хромозомы изучали в корешках взрослых луковиц. Количество хро-
мозом 2п=24 (рис. 16, з).
Хромозомы Т. schrenkii по соотношению плеч можно разбить на
четыре раздела (рис. 18, е).
В первый раздел входят две пары хромозом: первая пара с длин-
ным плечом—7,5 и коротким—3.75 р.: вторая — 4,6 и 2,6 р.
Второй раздел состоит из одной пары хромозом с длинным пле-
чом - 6 [1 и коротким—25 р. Третий раздел объединяет пять пар хро-
мозом: длинное плечо первой пары 10,25 и короткое—3 р; второй
—8,5 и 2,25 р; третьей—6,75 и 1,75 р; четвертой—6 и 1,75; пятой
—5,25 и 1,6 р.
Четвертый раздел составляет четыре пары хромозом: первая с
длинным плечом —10,5 и коротким —2,6 р; вторая - 9,5 и 2,25; третья—
6 и 1,5; четвертая — 5,75 и 1,25 р. Мы не обнаружили признаков вто-
ричного расчленения хромозом.
Общая длина гаплоидного набора хромозом—113,5 р; длинных
плеч—86,6; коротких плеч—26,55 р; общий индекс—3,22.
Т. kolpakovskiana Rgl. Исследованные образцы собраны в Сары-
Бастау (Чу-Илийские горы) в окрестностях ст. Андрокай Туркестано-
Сибирской ж.-д. (сборы 1932 г. 3. П. Бочанцевой); Заилийском Ала-
тау; долина р. Талгар — окрестности станции Талгар, Раздолье (сбор
3. П. Бочанцевой); бассейн р. Малый Кемин, в верховьях р. Карта-
Булга (сбор 3. П. Бочанцевой).
Изучали корешки взрослых луковиц. Количество хромозом 2п=24.
Ньютон (Newton, 1927 г.) для Т. kolpakovskiana приводит то же ко-
личество хромозом.
Он находит, что хромозомы Т. kolpakovskiana по форме, вернее
по длине вторых плеч, занимают промежуточное место между типом
хромозом, характерным для Т. linifolia и нормальным типом рода.
Ньютон отмечает, что среди хромозом Т. kolpakovskiana отсутствуют
хромозомы с субмедиальной перетяжкой, а также пара маленьких
хромозом, характерных для Т. linifolia.
Мы при исследовании Т. kolpakovskiana по морфологии хромозом
выделяем два кариотипа.
Первый кариотип обнаружен у Т. kolpakovskiana из Чу-Илийских
гор и Заилийского Алатау (рис, 11, г). Хромозомы в этом кариотипе
по соотношению плеч можно разбить на пять разделов (рис. 17, б).
В первый раздел входит одна пара хромозом с длинным плечом—
4,5 и коротким — 2,25 р.
Второй раздел состоит из трех хромозом: первая с длинным пле-
чом—6,25 и коротким—3 р; вторая—4,5 и 1,75; третья ^4,5 и 1,6 р.
190
У первой пары на длинном плече находится акинетический перерыв
на расстоянии около 2 |х от кинетической перетяжки.
Третий раздел объединяет две пары хромозом: первая с длинным
плечом — 6,75 и коротким—2 |х, вторая — 6 и 2 |х.
В четвертый раздел входят четыре пары хромозом: у первой длин-
ное плечо —8 и короткое—2 |х; второй—5 и 1,25; третьей—4,75 и 1;
четвертой—4,5 и 1 |х.
У второй пары хромозом на проксимальном конце длинного пле-
ча расположен акинетический перерыв, отделяющий головкообразный
членик до 0,5 jx.
Пятый раздел составляет две пары хромозом: длинное плечо пер-
вой пары—8 и короткое —1,5 jx; второй—7,75 и 1,5 р-. У второй па-
ры на дистальном конце длинного плеча акинетическим перерывом
отделен продолговатый членик длиной немного больше 1 jx.
Общая длина гаплоидного набора - 91,35 р-; длинных плеч—70,5;
коротких плеч —20,85 |х; общий индекс 3,38
Второй кариотип Т. kolpakovskiana из Малого Кемина в сомати-
ческих клетках также имеет 24 хромозомы, но разбивка по группам
несколько иная.
Все хромозомы данного типа можно по соотношению плеч раз-
бить на четыре раздела (рис. 18, ж),
В первый раздел входят две пары хромозом: первая с длинным
плечом —6,5 и коротким—3,7 р-; вторая—4 и 2,6 [х. Первая пара хро-
мозом имеет два кинетических перерыва — один на расстоянии около
2,5 р, от дистального конца длинного плеча и второй —на коротком
плече около 1,5 н от его проксимального конца.
Второй раздел хромозом состоит из одной пары хромозом с длин-
ным плечом —7 и коротким - 2,75 fx.
Третий раздел объединяет четыре пары хромозом: первая—с длин-
ным плечом—7 и коротким—2 jx; вторая—5,75 и 1,9; третья—5,75 и
1,5; четвертая—5,5 и 1,5 |х.
В четвертый раздел входят пять пар хромозом: первая—с длин-
ным плечом—10,5 и коротким—2,4 р-; вторая 9,35 и 2; третья—8 и 2;
четвертая—7,9 и 1,5; пятая—6,25 и 1,25 р-.
Общая длина гаплоидного набора хромозом—107,6 jx; длинных
плеч—83,5; коротких плеч—24,1 |х; общий индекс—3,28.
Таким образом, Т. kolpakovskiana имеет два различных кариотипа
—кариотип Т. kolpakovskiana с Малого Кемина, близок к кариотипу
Т. schrenkii и Т. korolkovii, а кариотип из окрестностей Алма-Ата и
Чу-Илийских гор приближается к Т. zenaidae и Т. ostrovskiana.
Т. ferganica Vved. (рис. 11, е\ 18, з). Исследованные образцы
собраны в Ферганском хребте (различные места по течению Кугарт и
Яссы; сбор 1933 г. 3. П. Бочанпевой).
Мы изучали корешки взрослых луковиц (рис. 11, в) и, кроме того,
хромозомы во время деления генеративного ядра в пыльцевых труб-
ках, воспитанных in vitro. На основании такого комплексного иссле-
дования хромозомы Т. ferganica можно по соотношению плеч разбить
на шесть разделов (рис. 18, з).
В первый раздел входит одна пара маленьких хромозом с длин-
ным плечом —7,5 и коротким—2,25 [х.
Второй раздел состоит из двух пар хромозом: длинное плечо пер-
вой—8,25 и короткое—3,5 [х; вторая—4,5 и 1,6 [х.
Третий раздел объединяет четыре пары хромозом: длинное плечо
первой пары—8 и короткое—1,9 [х; второй—6,5 и 1,75; третьей—6 и
1,9; четвертой—5,75 и 1,75 [х.
191
Четвертый раздел составляет две пары хромозом: первая с длин-
ным плечом—9,75 и коротким—2,25 у; вторая—7,75 и 1,6 у..
В пятый раздел входят две пары хромозом с плечами 9 и 1,5;
8,5 и 1,5 у.
В шестом разделе одна пара хромозом с длинным плечом — 9 и
коротким—1,4 у. Никаких признаков вторичного расчленения хромо-
зом у этого вида не обнаружено.
Общая длина гаплоидного набора хромозом—114,1 у; длинных
плеч—90,5; коротких плеч—23,6 у, общий индекс—3,84.
Т. zenaidae Vved. (рис. 11, л-, 17, а). Исследованные образцы
привезены из Александровского хребта (гора Шекуле, сбор 1932 г.,,
3. П. Бочанцевой).
Мы изучали корешки взрослых луковиц.
Количество хромозом 2п=24 (рис. 11, л). Все хромозомы по соот-
ношению плеч можно разбить на шесть разделов (рис. 17, а).
Первый раздел включает одну пару маленьких хромозом с длин-
ным плечом—4 и коротким—2,25 у.
Второй раздел состоит из двух пар хромозом: длинное плечо пер-
вой—7 и короткое—3 у; второй—6,5 и 2,25 у.
В третий раздел входит две пары хромозом: первая —би 1,6; вто-
рая—5,6 и 1,5 у.
Четвертый раздел включает 4 пары хромозом: первая с длинным
плечом—7,75 и коротким—1,75 у; вторая—7 и 1,4; третья—6 и 1,25;
четвертая—6 и 1,25 у.
В пятый раздел входят две пары самых длинных хромозом набо-
ра: первая с длинным плечом—10,25 и коротким—2 у; вторая—9,25
и 1,6 у.
Шестой раздел включает одну пару хромозом с длинным плечом —
7,75 и коротким—1 у. Признаки вторичного расчленения на хромозо-
мах этого вида не выявлены.
Общая длина гаплоидного набора хромозом—103,95 у; длинных
плеч—83,1; коротких плеч—20,85 у; общий индекс—4,02.
Т. ostrovskiana Rgl. (рис. 16, л-, 17, в). Исследованные образцы
Т. ostrovskiana получены из Заилийского Алатау, окрестности с. Тал-
гар (ущелье Байбулак). Розарий. Глинисто-щебнистый склон (сбор
1933, 1934, 1936 гг., 3. П. Бочанцевой).
Мы изучали хромозомы в точках роста корешков и прорастаю-
щей пыльце во время спермиогенеза. Число хромозом 4п=48 (рис. 16, л)
постоянно для данного вида при всех окрасках цветка от бледно-жел-
того, оранжевого, оранжево-красного и ярко-темно-красного. Мы про-
смотрели хромозомы 16 форм (по окраске цветка) Т. ostrovskiana и
все они дали количество хромозом —48 в соме и 24 в генеративной
сфере. Все хромозомы Т. ostrovskiana по соотношению плеч можно
разбить на пять разделов (рис. 17, в).
Первая группа состоит из одной пары хромозом с длинным пле-
чом—6,5 и коротким—3,75 у с акинетическим перерывом на коротком
плече на расстоянии 1 у от проксимального конца.
Второй раздел хромозом объединяет пять пар: длинное плечо
первой—6,9 и короткое—2,5 у; второй—6,75 и—2,5; третьей—6 и 2,25;
четвертой—5 и 2; пятой—4,5 и 1,5 у.
Третий раздел включает девять пар хромозом: первая с длинным
плечом—9,4 и коротким—2,5 у; вторая—8,5 и 2,25; третья—7 и 2;
четвертая—6,25 и 2; пятая—5,5 и 1,9; шестая—5,25 и 1,75; седьмая—
5,25 и 1,5; восьмая—5,25 и 1,5; девятая—4,4 и 1,3 у.
В четвертый раздел входят семь пар хромозом: первая с длинным
192
плечом—9,5 и коротким—3,25 а; вторая—8,25 и 1,9; третья—8 и 1,75;
четвертая—6 и 1,4; пятая—6,5 и 1,25; шестая—6,5 и 1,25; седьмая—
5 и 1,25 [х.
В пятый раздел входят всего две пары хромозом: длинное плечо
первой—8,75 и короткое—1,5 [а; второй —8,25 и 1,70 [а.
Общая длина гаплоидного набора хромозом —205,9 у-; длинных
плеч хромозом—159,7; коротких плеч—46,2 и; общий индекс—3,24.
Подвергая тщательному разбору Т. schrenkii, Т. kolpakovskiana,
Т. ferganica, Т. zenaidae и Т. ostrovskiana, объединяющихся в одну
естественную группу, надо отметить, что Т. schrenkii непосредственно
связывает данную группу с кариотипом Т. korolkovii. В то же время
кариотип Т. kolpakovskiana с Малого Кемина распределением хромо-
зом на группы почти повторяет кариотип Т. schrenkii.
Первый, второй, пятый и шестой разделы хромозом Т. ferganica
и Т. zenaidae по количеству входящих в них хромозом одинаковы.
В третий раздел Т. ferganica входят четыре пары хромозом, а в чет-
вертый —две пары, третий раздел Т. zenaidae состоит из двух пар
хромозом, четвертый—из четырех.
Сравнивая кариотипы Т. ostrovskiana и близких видов, можно
утверждать, что Т. ostrovskiana не простой тетраплоид, а амфидип-
лоид. Одним из компонентов, участвовавших в создании Т. ostrovskia-
na является Т. kolpakovskiana типичный или, как мы его называем,
равнинный с кариотипом Т. kolpakovskiana из Раздолья (Талгар, Заи-
лийский Алатау). Особенно ярки и показательны укороченные хромо-
зомы Т. kolpakovskiana из второго и четвертого разделов; они, несом-
ненно, имеются в кариотипе Т. ostrovskiana. Другой компонент нам
неизвестен. Данная группа видов, по-видимому, в настоящее время
находится в стадии видообразования. Изучение большого количества
видов и внутривидовых форм с учетом экологических условий место-
обитания в настоящее время должно пролить свет на процессы видо-
образования в естественных условиях.
Принимая во внимание неполное исследование кариотипов данного
раздела, степень родства между видами данной группы можно пред-
ставить в виде следующей цепочки: Т. schrenkii — Т. kolpakovskiana
(Малый Кемин) —Т. ferganica — Т. zenaidae — Т. kolpakovskiana (Алма-
Атг) — Т. ostrovskiana.
Т. tetraphylla Rgl. (рис. И, ж-, 17 г). Исследованные образцы
получены из Заилийского Алатау, из окрестностей с. Талгар (сбор
1933—1936 гг., 3. П. Бочанцевой).
По хромозомальному набору Т. tetraphylla не менее оригинален,
чем по внешнему виду (рис. 11, ж). Количество хромозом обычное:
2 п = 24, но бывают и тетраплоидные формы 4 п = 48; их разбивка по
группам совершенно необычна для Tulipa секции Leiostemones. Все
хромозомы Т. tetraphylla по соотношению плеч можно разбить на 5
разделов (рис. 17, г).
В первый раздел входят три пары хромозом: первая с длинным
плечом —8,5 и коротким—4,5 [а; вторая—7,5 и 4,25; третья—7 и 4,1 [а.
У первой пары на середине короткого плеча расположен акинетиче-
ский перерыв. У второй пары на коротком плече также находится
акинетический перерыв на расстоянии около 1 у- от проксимального
конца плеча.
Второй раздел состоит из трех пар хромозом: первая с длинным
плечом—8,75 и коротким—3,25 р.; вторая 6,5 и 3 [а; третья—6,25 и
2,5 [а. У третьей пары на коротком плече имеется акинетический пере-
рыв, делящий его на две равные части.
13—2229
193
Третий раздел объединяет три пары хромозом: длинное плечо
первой—7,4 и короткое—2 р.; второй—6,25 и 1,75; третьей—5,75 и 1,5 р..
В четвертый раздел входят две пары хромозом: первая с длинным
плечом—9 и коротким—2,25 р.; вторая—7 и 1,75 р.
В пятый раздел входит пара хромозом с длинным плечом в 7 и
коротким в 1,4 р.
Общая длина гаплоидного набора хромозом—119,75 р.; длинных
плеч—86.9; коротких плеч—32,25 р; общий индекс—2,69.
Следовательно, кариотип Т. tetraphylla не имеет себе подобных
среди изученных нами тюльпанов этой секции.
По трем парам хромозом с отношением плеч меньше чем 1:2 этот
вид более близок к некоторым разновидностям видов из секции Erio-
stemones, чем Leiostemones. Из видов секции Leiostemones Т. tetrap-
hylla по кариотипу приближается к Т. schrenkii или Т. korolkovii;
имеет большое вторичное расчленение хромозом; часто встречаются
полиплоидные формы.
Возможно, после исследования Т. iliensis Rgl. и Т. dykesiana
удастся связать Т. tetraphylla с тюльпанами, объединяющимися вокруг
Т. kolpakovskiana и с самим Т. kolpakovskiana, так как в природе и
культуре мы наблюдали растения, образующие целую гамму перехо-
дов между этими видами и являющимися несомненными гибридами
между ними.
Секция Spiranthera Vved.
Во „Флоре СССР“ (т. IV) к секции Spiranthera Vved. отнесен один
вид. Однако по способу открывания пыльников в эту секцию необхо-
димо перенести из секции Leiostemones Т. dubia и включить два
новых вида Т. anadroma с Сары-Чилека и Т. tschimganica из Бостан-
дыкского района.
Т. kaufmanniana Rgl. (рис. 11, а; 17, д). Изученные нами образ-
цы были взяты из массовых посадок Ботанического сада ТашГУ.
Материал для них брали из Западного Тянь-Шаня по долинам рек
Пскем и Угам, из Казахстана— ст. Тюлькубас, ущелье Тавды-Булак,
второй правый приток (сбор В. П. Бочанцева и А. Я. Буткова).
Число хромозом для соматического митоза Т. kaufmanniana было
впервые установлено Ньютоном (Newton, 1927 г.) — 24 хромозомы.
При подсчете в меристеме корешка количество хромозом под-
тверждается — их 24, в молодой пыльце —12.
При изучении морфологии были использованы хромозомы из ме-
ристемы корешка и молодых пылинок (рис. 11, а).
Все хромозомы Т. kaufmanniana по соотношению плеч можно
разбить на шесть разделов (рис. 17, д').
В первом разделе одна пара хромозом с длинным плечом —8,5 и
коротким—4,375 р. На коротком плече расположена акинетическая
перетяжка, присутствующая на всех хромозомах этого типа. Отлича-
ется она лишь тем, что сдвинута ближе к проксимальному концу пле-
ча и находится от него на расстоянии одной четверти всей длины
плеча.
На длинном плече также имеется акинетическая перетяжка, отде-
ляющая продолговатый членик около 1,75 и дл.
Во второй раздел входит одна пара хромозом с длинным плечом—
7,25 и коротким—2,75 р. Вторичное расчленение выявить не удалось.
Третий раздел хромозом состоит из шести пар хромозом, слабо
разнящихся по длине плеч.
194
Первая пара с длинным плечом — 8,5 и коротким —2,75 р. На
коротком плече наблюдается акинетическая перетяжка, чуть-чуть сдви-
нутая с середины ближе к проксимальному концу плеча. Длинное
плечо второй пары —7,75, короткое—2,375 р. Никаких перетяжек,
кроме кинетической, у этой хромозомы выявить не удалось. Третья
пара с длинным плечом—7,25 и коротким—2 р; четвертая пара—7 и
4 р с акинетической перетяжкой на длинном плече на расстоянии
около 2 р от проксимального конца. Пятая пара, по размерам равная
четвертой, на дистальном конце длинного плеча имеет небольшой
шарообразный спутник, который отличается непостоянством и не на
всякой пластинке может быть обнаружен. Шестая пара, по размерам
равная двум предыдущим, если и меньше по длинному плечу, то на
десятую долю микрона с непостоянным крошечным шарообразным
спутником на коротком плече.
Четвертый раздел состоит из двух хромозом. Первая—с длинным
плечом—7,25 и коротким—1,75 р с акинетической перетяжкой у прок-
симального конца длинного плеча, отделяющей небольшой (до 1 р)
членик около кинетического перерыва. Вторая — с длинным плечом в
7 и коротким в 1,75 р; акинетических перерывов не несет.
В пятый раздел входит одна пара хромозом с длинным плечом —
7,25 и коротким—1,25 р. На длинных плечах хромозом этой пары
находятся акинетические перетяжки, немного сдвинутые с середины
ближе к проксимальному концу плечей.
Шестой раздел состоит из одной пары хромозом с длинным пле-
чом—9,5 и коротким—1,5 р; на длинных плечах этой хромозомы име-
ются акинетические перетяжки, расположенные на расстоянии около
-3 р от дистального конца плеча.
Общая длина гаплоидного набора хромозом—118,75 р; длинных
плеч—91,25; коротких плеч—26,5 р; общий индекс—3,44.
Других видов этой секции мы не изучали. Единственный ее пред-
ставитель по кариотипу из изученных нами видов других секций при-
«ближается к тюльпанам III группы секции Leiostemones.
Секция Lophophyllon Vved.
Это монотипная секция с одним видом Т. regelii Krasn. В лите-
ратуре нет указаний на цитологическое исследование данного вида.
Т. regelii Krasn. (рис. 19, ж-, 20, ж). Мы изучали семена Т.re-
gelii с гербарных экземпляров из Казахского филиала АН СССР, со-
бранные на Андракае в 1935 г. Дмитриевым. Кроме того, в 1947 г. их
привез со ст. Чокпар В. П. Бочанцев.
Число хромозом соматических клеток 2п = 24.
Все хромозомы Т. regelii по месту кинетического перерыва могут
быть разбиты на пять разделов (рис. 19, ж-, 20, ж).
В первый раздел входят две пары хромозом: первая с длинным
.плечом—7 и коротким—5,8 р. На коротком плече на расстоянии около
1,5 р от кинетического перерыва расположена
тяжка; вторая пара хромозом с длинным
2,625 р.
Второй раздел состоит также из двух
пары длинное плечо —7, короткое —2,5 р-;
Третий раздел включает четыре пары
длинное плечо—7, короткое—2,125 р-; у второй—6,5 и2ц; у третьей—
6,25 и 1,75 р; у четвертой—6 и 2 р.
Четвертый раздел объединяет три пары хромозом: у первой пары
длинное плечо—10 и короткое—2,25 р, на котором на расстоянии
акинетическая пере-
плечом —5 и коротким —
пар хромозом. У первой
у второй —5,75 и 2,25 р.
хромозом: у первой пары
195
Рис, 19. Пластинки хромозом.
в-Т. biebersteiniana S с h u i t.; б^-Т. patens A g а г d if в—Т. biflora ДаП.. JJ^asys&oid^f V ’v/d?1
ndrossovii L i t w. ; e—T. turKomanica B. F e d s c h , ; ж—T. regelii
Рис. 20. Идиограммы измеренных пластинок (увелич. в 800 раз).
л—Т. ’biebersteiniana Schult.; б—Т. patens A g а г d h . ; п—Т. biflora Pall .; г—Т. sogdiana В g е.,
д—Т. androssovii L i t w . ; e—T. turcomanica Rgl.; ж -T. regelii Krasn.
около 0,75 p. от кинетического перерыва находится акинетическая
перетяжка. Вторая пара с длинным плечом —9,25 и коротким —2,25 р-
и третья—6,625 и 1,5 р..
В пятый раздел входит одна пара хромозом с длинным плечом—
7,4 и коротким —1,125 р.
197
Таким образом, кариотип Т. regelii характеризуется наличием
двух пар хромозом в первом разделе. Большая из них имеет пере-
тяжку, характерную для этой хромозомы в роде Tulipa.
Первая хромозома четвертого раздела также с перетяжкой характер-
ной для некоторых кариотипов видов Tulipa из секции Eriostemones.
Общая длина всех хромозом гаплоидного набора—111,65; длинных
плеч—83,775; коротких плеч—27,875 и; общий индекс—3,005. Индекс
хромозомы А—1,27. Индекс Ai—1,86.
Секция Eriostemones Boiss.
Во „Флоре СССР11 (т. IV) А. И. Введенский дает следующую ха-
рактеристику секции: „Оболочки луковиц с внутренней стороны голые
или волосистые или паутинисто-шерстистые. Цветы у большинства
видов мелкие, невзрачные, нити тычинок опушенные, у одного вида
(Tulipa sogdiana) голые. Завязи с почти сидячими рыльцами или с
более или менее длинным столбиком, но всегда более коротким, чем
сама завязь11.
В книге Холла (1940) секция Eriostemones в мировом масштабе
представлена 21 видом, которые он считает достаточно наделенными
видовыми отличиями и разбивает их на три подсекции.
В СССР произрастает 18 видов тюльпанов из секции Eriostemo-
nes: Т. violaceae Boiss. et Buhse, T. caucasica Lipsky, T. Silves-
tris L., T. biebersteniana* Schult, T. patens* Agardh, T. sogdiana*
Bge, T. Buhseana Boiss., T. turkestanica* Rgl., T. turkomanica*
B. Fedtsch., T. bifloriformis* Vved., T. biflora* Pall., T. polyc-
hroma Stapf., T. koktebelica Junge, T. callieri Holacsy et Lev.,
T. orithyioides Vved., T. dasystemonoides* Vved., T. dasystemon*
Rgl., T. tarda* St a pf.
Виды, отмеченные звездочкой, исследованы нами. Кроме того, мы
изучали виды этой секции, поставленные во „Флоре СССР11 в приме-
чаниях — Т. androssovii L i t w., T. neustruevae P о b., T. binutans Vved.,
T. talievi Vved., и T. orthopoda Vved.
Таким образом, мы исследовали 16 видов тюльпанов этой секции
из различных местообитаний.
При сравнении списков видов, приведенных Холлом (1940) и
А. И. Введенским (1935), видно, что в Советском Союзе произрастают
в основном виды, относимые Холлом к III подсекции, называемой Bif-
lorae. Однако эта подсекция представлена Холлом всего тремя видами,
не вызывающими у него сомнения, а девять видов, относимых дру-
гими исследователями к этой подсекции, расцениваются им как со-
мнительные.
Из видов первой подсекции (по Холлу) в Советском Союзе встре-
чается три вида: Т. biebersteiniana, Т. patens и Т. Silvestris. Мы ис-
следовали только Т. biebersteiniana и Т. patens. Т. biebersteiniana Холл
считает синонимом Т. australis, что вызывает сомнение, так как эти
виды имеют слишком разобщенные ареалы, и этот вопрос можно
решить лишь после дальнейшего глубокого исследования. Тем более,
что виды секции Eriostemones отличаются большой пластичностью
в морфологических признаках (и в признаках, характеризующих ка-
риотип), нет резких признаков перехода от одного к другому и по-
этому при описании вида необходимо точно указывать на географи-
ческое происхождение изучаемого образца. Тюльпаны этой секции
удивительно разнообразны по кариотипу. Часто представители одного»
вида секции из разных точек ареала имеют разный кариотип. Из II
198
подсекции в Советском Союзе произрастает Т. violacea. По „Флоре
СССР" Т. violacea может быть встречена в Талыше.
Т. biebersteiniana Schult. Исследованные образцы собраны в
окрестностях ст. Берчугура (сбор 1932 г., Пономаревой). Изучали
корешки взрослых луковиц. Количество хромозом 2п = 24 (рис. 19, а).
Все хромозомы Т. biebersteiniana по соотношению плеч можно разбить
на пять разделов (рис. 20, а).
В первый раздел входит одна пара хромозом с длинным плечом—
6,75 и коротким —4,25 р. На коротком плече на расстоянии 1 р от
кинетического перерыва расположена акинетическая перетяжка. Вто-
рая такая же перетяжка делит длинное плечо этой пары на две почти
равные части, в 3 р от кинетической перетяжки.
Второй раздел состоит из двух пар хромозом, длинное плечо
первой пары—7,5 и короткое—2,75 р; вторая пара 7 и 2,5 р. У первой
пары хромозом на длинном плече на расстоянии 3 р от кинетической
перетяжки находится акинетическая перетяжка. У второй пары на ко-
ротком плече также имеется кинетическая перетяжка, расположенная
на расстоянии около 1 р от кинетического перерыва.
Третий раздел объединяет четыре пары хромозом: первая с длин-
ным плечом—8 и коротким—2,5 р; вторая—6 и 2; третья—6 и 1,74 и
четвертая—6 и 1,75 р. На длинном плече первой пары на расстоянии
2 р от дистального конца плеча лежит акинетическая перетяжка.
Четвертый раздел включает три пары хромозом: длинное плечо
первой пары—8,25 и 2 р; второй—7 и 1,75; третьей—6 и 1.25 р.
У второй пары хромозом на длинном плече на расстоянии 2,25 р
от кинетической перетяжки расположен акинетический перерыв. В
пятый раздел входят две пары хромозом: первая с длинным плечом—
7,5 и коротким—1,5 р; вторая—7,25 и 1,25 р.
Вторая пара хромозом на длинном плече имеет три акинетических
перетяжки: первая на расстоянии 0,75 р от дистального конца хромо-
зомы; вторая—1,85 р от первой и третья—0,75 р от кинетического
перерыва.
Общая длина всех хромозом гаплоидного набора —108,5 р; длин-
ных плеч—83,25; коротких плеч—25,25 р; общий индекс—3,297. Ин-
декс хромозомы Ai —1,588.
Т. patens Aga rd h. (рис. 19, б; 20, б). Исследованы корешки
взрослых луковиц из Семипалатинска окр. Бобровки от Ермакова в
1933 г. Количество хромозом у данного вида 2 п = 24. Все его хро-
мозомы по соотношению плеч можно разбить на пять разделов. В
первый раздел входит одна пара хромозом с длинным плечом—6,75 и
коротким—5 р дл. с одной акинетической перетяжкой на длинном
плече на расстоянии 3 р от дистального конца плеча и второй — на
коротком плече в 1 р от кинетического перерыва.
Во второй раздел входит три пары хромозом: первая с длинным
плечом—6,25 и коротким—2,75 р: вторая—6 и 2,25; третья — 5,5 и
2,25 р. Первая пара хромозом имеет акинетические перетяжки на
обоих плечах: на длинном плече в 0,75 рот кинетического перерыва,
на коротком в 0,5 р. Вторая—имеет акинетическую перетяжку на ко-
ротком плече на расстоянии 1 р от кинетической перетяжки.
Третий раздел объединяет пять пар хромозом: первая с длинным
плечом—8,25 и коротким—2,9 р; вторая—6,25 и 2 р; третья—6,25 и 2;
четвертая—5,75 и 1,75 и пятая—5,5 и 1,75 р. Первая и пятая пары
хромозом имеют акинетические перетяжки на длинных плечах. У пер-
вой пары эта перетяжка расположена в 3,75 р от кинетической пере-
тяжки. У пятой пары перетяжка находится ближе к проксималь-
199'
ному концу плеча, перешнуровывая хромозому на расстоянии 1,75 р
от конца плеча.
Четвертый раздел включает две пары хромозом: длинное плечо
первой пары—8,25 и короткое—2 р; вторая—5,75 и 1,25 р. У первой
пары на длинном плече на расстоянии 4 р от кинетической перетяжки
лежит акинетическая перетяжка. У второй — на расстоянии 2 р от
проксимального конца плеча также расположена акинетическая пере-
тяжка.
В пятый раздел входит всего одна пара хромозом с длинным пле-
чом—6,75 и коротким —1,25 р.
Общая длина всех хромозом гаплоидного набора у этого вида —
104,4 р; длинных плеч—77,25; коротких плеч—27,15 р; общий индекс—
2,845; индекс хромозомы Ai—1,35.
Т. biflora Pall. (рис. 19, в; 20, в). Исследованные образцы лу-
ковиц получены в 1932 г. от Ф. Н. Русанова с Черного Иртыша.
Изучали корешки взрослых луковиц. В соматических пластинках из
точки роста корня установлено 2 п = 24.
Все хромозомы данного вида по соотношению плеч можно раз-
бить на пять разделов. В первый раздел входят две пары хромозом:
первая с длинным плечом- 6,75, коротким—5 р, а вторая—5,25 и 3 р.
У первой хромозомы на коротком плече в 1 р от кинетического пере-
рыва лежит первая акинетическая перетяжка и в 1,75 р от нее по
направлению к дистальному концу плеча—вторая.
Второй раздел объединяет четыре пары хромозом. Первая пара
с длинным плечом—5,5 и коротким—2,5 р; вторая—5,5 и 2,25; третья
5,25 и 2; четвертая—4,5 и 1,75 р. У первой пары хромозом на длин-
ном плече на расстоянии 1,75 от кинетического перерыва расположена
акинетическая перетяжка.
Третий раздел состоит из трех пар хромозом. Первая пара с длин-
ным плечом—6,75 и коротким—1,75 р; вторая пара—6 и 2; третья —
€ и 1,75 р. У третьей пары на длинном плече в 2,75 р от кинетиче-
ского перерыва находится ахроматиновая перетяжка.
Четвертый раздел представлен двумя парами хромозом; первая с
длинным плечом —9,25 и коротким — 2,25 р; вторая —7 и 1,75 р.
По соотношению плеч в пятый раздел не входит ни одна пара, а
в шестом одна пара с длинным плечом —6 и коротким —1 р.
Общая длина всех хромозом гаплоидного набора—100,75 р; длин-
ных плеч—73,75; коротких плеч—27 р; общий индекс—3,73. Индекс
Ai-1,35; Ап -1,75.
Т. sogdiana Bge (рис. 19, г; 20, г). Исследованные образцы по-
лучены в 1931 г. от И. В. Андросова с отметкой „начало песков близ
Ашхабада".
Кончики корешков мы отрезали от корней взрослых луковиц при
проращивании; 2п=24.
Все хромозомы Т. sogdjana по соотношению плеч можно разбить
на шесть разделов (рис. 20, г). В первом разделе нет хромозом с
соотношением плеч больше чем 1:2. Во второй раздел входит одна
пара хромозом с длинным плечом—3 и коротким — 1,5 р.
Третий раздел состоит из двух пар: первая с длинным плечом —
7,5 и коротким —2,5 р, на котором в 1 р от проксимального конца
лежит акинетическая перетяжка. У второй пары длинное плечо —5,25
и короткое—1.75 р.
Четвертый раздел объединяет четыре пары: первая пара с длин-
ным плечом—9 и коротким—2,25 р; вторая—7 и 1,5; третья — 5,5
и 1,25 и, наконец, четвертая — 4,5 и 1 р. У первой пары на коротком
200
плече на расстоянии 1 р от проксимального конца плеча находится
акинетическая перетяжка.
Пятый раздел состоит из двух пар хромозом. длинное плечо пер-
вой—10 и короткое—1,75 р; вторая—8,75 и 1,5 у..
Шестой раздел представлен двумя парами хромозом: первая с
длинным плечом — 8,25 и 1,25 р и вторая—6,5 и 1 у..
В седьмой раздел входит одна пара хромозом с длинным плечом—
7 и коротким—1 р.
Таким образом, образцы данного вида, полученные из Ашхабада
от Н. В. Андросова, обладают хромозомами с более высоким соотно-
шением плеч, чем вышеописанные виды.
Общая длина всех хромозом гаплоидного набора у этого вида —
100,5 р; длинных плеч —82,25; коротких плеч —18,25 р; общий индекс—
4,5. Индекс А—2.
Т. androssovii L itw. (рис. 19, <?; 20, д). Образцы тюльпана мы полу-
чили под наименованием Т. androssovii Litw. от Н. В. Андросова из
Сераке, собранные на глинистых местах около р. Тедгасан. У этого
тюльпана 2п=-24 хромозом, которые можно разбить на пять разделов.
В первый раздел входит одна пара хромозом с длинным плечом —
7,5 и коротким —4,25 р, на котором расположена характерная для
хромозом этого раздела акинетическая перетяжка на расстоянии око-
ло 1 р от кинетического перерыва.
Второй раздел состоит из трех пар хромозом. Первая с плечами—
6 и 2,75 р несет акинетическую перетяжку на коротком плече в 1 р-
от проксимального конца и другую на расстоянии 1 р от кинетиче-
ского перерыва на длинном плече.
Вторая пара с длинным плечом — 5,5 и коротким —2 р, имеет две
перетяжки — на длинном плече на расстоянии 0,75 р от его прокси-
мального конца и на коротком—в 0,5 р от кинетической перетяжки.
Третья пара хромозом с длинным плечом—5 и коротким—2 р.
В третий раздел входит три пары хромозом. Первая с плечами—
-5,75 и 1,75 р, на расстоянии 2,5 р от дистального конца длинного
плеча имеется акинетическая перетяжка, а на коротком плече наблю-
дается сферический спутник на тонкой нити. Вторая—5,25 и 1,75 р,
третья пара—5 и 1,5 р с двумя акинетическими перетяжками на
длинном плече. Первая из этих перетяжек расположена на расстоянии
около 1,75 от кинетической перетяжки, вторая—около 1 р от нее по
направлению к проксимальному концу плеча.
В четвертый раздел входит две пары хромозом: первая с длинным
плечом—9, а коротким—2,25р; вторая—5 и 1,25 р. У первой пары
хромозом на длинном плече в 0,75 р от кинетического перерыва ле-
жит акинетическая перетяжка.
Пятый раздел представлен одной парой с длинным плечом —8,75
и коротким—1,5 р. На длинном плече в 3.75 р от кинетического пе-
рерыва находится акинетическая перетяжка.
Шестого, седьмого, восьмого и девятого разделов нет.
Две оставшиеся пары хромозом относятся к 10 разделу и могут
быть отнесены к головчатым. Они одинакового размера с длинным
плечом —7,5 и коротким — 0,75 р. У первой пары на длинном плече
расположены два акинетических перерыва —первый на расстоянии
3,75 р от кинетического перерыва, а второй —1,5 р от него по направ-
лению к проксимальному концу плеча.
Общая длина всех хромозом гаплоидного набора у этого вида —
100,25; длинных плеч —77,75; коротких плеч —22,5 р; общий индекс —
.3,456. Индекс А —1,764.
201
T. turkomanica В. Fedtsch. (рис. 19, е; 20, е). Исследованные об-
разцы получены из различных мест Восточного Копет-Дага (сбор А. Ф.
Гнездило). Этот вид с четкими морфологическими признаками.
Хромозомы изучали в корешках взрослых луковиц, в мазках из мо-
лодых пыльников и в пыльцевых трубках в момент спермиогенеза.
Соматическое число хромозом у этого вида равно 24 (рис. 20, е).
Все хромозомы этого вида можно разбить на пять разделов.
В первый раздел входят две пары хромозом. Первая с длинным
плечом —6,5 и коротким—5,75 у. Вторая пара—6,25 и 3,75 у. На ко-
ротком плече этой хромозомы имеется спутник.
Второй раздел представлен одной парой хромозом с длинным
плечом—5,75 и коротким—2,5 у.
Третий раздел объединяет три пары хромозом, первая с длинным
плечом—7,75 и коротким—2,1 у; вторая—7 и 2 и третья—6,25 и 2 у.
Четвертый раздел состоит из пяти пар хромозом. Первая пара с
длинным плечом—11 и коротким—2,75 у; вторая—8,5 и 2; третья —
7,75 и 1,75 у со спутником на коротком плече; четвертая — 7,75 и 1,75,
пятая—6,25 и 1,65 у.
В пятый раздел входит одна пара хромозом с длинным плечом —
11,25 и коротким—2,25 у.
Общая длина всех хромозом гаплоидного набора у этого вида —
121,25 у; длинных плеч—92; коротких плеч—29,25 у; общий индекс—
3,124. Индекс Ai—1,130, индекс Ап —1,66.
Т. dasystemonoides Vved. (рис. 19, з; 21, а) Исследованные об-
разцы получены в 1935 г. из окрестностей поселка Первомайского от
В. П. Бочанцева. Изучали корешки взрослых луковиц. Этот вид в
соматических клетках имеет 2п—24. Все хромозомы этого вида могут
быть разбиты на шесть разделов.
В первый раздел входит одна пара с длинным плечом — 7,75 и ко-
ротким—6 у, на котором в 1,75 у от кинетического перерыва распо-
ложена акинетическая перетяжка, характерная для хромозом этого*
раздела большинства видов рода Tulipa.
Второй раздел состоит из трех пар хромозом: первая с длинным
плечом — 7,75 и коротким—3,75 у; вторая—7,25 и 3,25; третья—6,75
и 2,5 у. У второй пары хромозом на расстоянии около 1 у от дисталь-
ного конца короткого плеча имеется акинетическая перетяжка, аки-
нетическая перетяжка третьей пары ближе сдвинута к проксимальному
концу короткого плеча и находится в 1 у от кинетической перетяжки.
Третий раздел объединяет пять пар хромозом: первая пара с пле-
чами — 10,675 и 3,55 у; вторая—8,25 и 2,75; третья—8,05 и 2,5; чет-
вертая—7,75 и 2,6; пятая—7,6 и 2,25 у.
Четвертый раздел представлен одной парой хромозом с длинным
плечом—12,5 и коротким—2,75 у.
В пятом разделе нет хромозом с отношением плеч больше чем 1:4.
Шестой раздел включает одну пару хромозом с длинным плечом —
9 и коротким—1,6 у.
Седьмой раздел имеет одну пару хромозом с длинным плечом—
10,62 и коротким — 1,75 у.
Общая длина всех хромозом гаплоидного набора у этого вида —
140,145 у; длинных плеч — 104,945; коротких плеч—35,2 у; общий ин-
декс—2,98. Индекс А—1,291.
Tulipa sp. (рис. 22, з; 23, а). Образцы привезены в 1933 г. В. П.
Бочанцевым и А. Я- Бутковым с гор Кызыкурт. Исследовали кореш-
ки взрослых луковиц.
202
т
а 1 i / lil: Il 2 3 4 5 1 11 lli 17 8 9
6 1 1 1 6 s lilt i 1!
f 2 3 4 5 6 7 В
e I'll Illi / llllll II 3 4 5 6 llllll 7 8 9
Рис. 21. Идиограммы измеренных пластинок (увелич. в 800}раз).
. dasystemonoides Vved.; б—Т. neustruevae Р о b . ; в—Т. dasystemon Rgl.; г—Т. dasystemon’
Pg 1 .; д— Т. dasystemon Rgl.; Т. dasystemon Rgl.; ж—Т. bifloriformis Vved.
Рис. 22. Пластинки хромозом.
° - iienstrnevae Р и b . ; б — Г. dasystemon R g 1 . ; в — Т. iLisystemou Rgl,; г — Т. dasystemon Rgl.; д — Т. dasystemon R g 1 . ; е — Т. biflori forints Rgl,; ж — Т. bitloriformts
Vved.; з—T.f sp.; и ~T. bifloiiformis Vved.
Illll
4 5 6 7 8
Рис. 23. Идиограммы из^меренных пластинок (увелич. в 800 раз).
а—Т. buhseana В о i s s . ; б—Т. bifloriformis Vved.; в—Т. bifloriformis Vved.; г—Т? bifloriformis
Vved.; <?—Т. binutans Vved.; e—T. buhseana В о i ss . ; ж—T. turkestanica Rgl.
В соматических клетках этого вида 2п=24 (рис. 22, з).
Все хромозомы по соотношению плеч можно разбить на семь раз-
делов.
В первый раздел входит одна пара хромозом с длинным плечом—
6,5 и коротким — 4,25 р.
Второй раздел состоит из двух пар хромозом: первая с длинным
плечом—7 и коротким — 2,73 р: вторая—5 и 1,75 р.
Третий раздел состоит из трех пар хромозом: первая с длинным
плечом —6,75 и коротким—1,75 р; второй—6,62 и 2,06 и третья—6,33
и 1,69 р.
Четвертый раздел представлен двумя парами хромозом, длинное
плечо первой—7,45 и короткое—1,57 р; вторая—6,53 и 1,62 р.
В пятый раздел входит одна пара хромозом с длинным плечом—
6,7 и коротким—1,5 р; на дистальном конце длинного плеча находится
акинетический перерыв, отчленяющий конец плеча длиною до 1,25 р.
Шестой раздел объединяет две пары хромозом, первая с плечами
11,44 и 1,86 р, а вторая—9,18 и 1,52 р.
Седьмой раздел с плечами 8,75 и 0,78 р.
Общая длина всех хромозом гаплоидного набора у этого образ-
ца—111,25 р; длинных плеч—88,25; коротких плеч—23 р; общий ин-
декс—3,84. Индекс Ai —1,529.
Т. neustruevae Pob. (рис. 21, б; 22, а). Мы исследовали образцы
из Джалалабадского района. Левый берег р. Кугарт, против с. Дмит-
риевского. Верхняя речная терраса. Галька с мелкоземом.
Хромозомы изучали в корешках и в пыльцевых трубках, воспи-
танных на искусственной питательной среде в период деления генера-
тивного ядра на спермин.
У данного вида 2п=24. Все его хромозомы по соотношению плеч
могут быть разбиты на шесть разделов (рис. 21, б).
В первый раздел входит две пары хромозом. Первая пара с длин-
ным плечом—13,125 и коротким—8,125 р. На длинном плече на рас-
стоянии 5,25 р от кинетического перерыва расположена акинетическая
перетяжка. На коротком плече в 1,25 р от кинетического перерыва
находится акинетическая перетяжка, свойственная хромозомам этой
группы большинства видов тюльпанов. Вторая пара хромозом с длин-
ным плечом —9,5 и коротким —4,4 р.
Второй раздел состоит из трех пар хромозом: первая с плечами —
12 и 5,4 р; вторая —9,5 и 4,25 р, на коротком плече второй пары в
1,5 р от дистального конца плеча лежит акинетический перерыв; третья
пара с длинным плечом 9,5 и коротким —3,5 р.
Третий раздел объединяет три пары хромозом: первая с длинным
плечом—12 и коротким—3,125 р; вторая—11,25 и 3,5 р. Почти по-
середине длинного плеча на расстоянии 5,75 р от кинетического пере-
рыва лежит акинетическая перетяжка; третья пара с длинным плечом —
9,5 и коротким — 2,75 р. На коротком плече на расстоянии 1,25 р от
кинетического перерыва находится акинетическая перетяжка.
Четвертый раздел представлен одной парой хромозом с длинным
плечом—19,8 и коротким—4,1 р.
В пятый раздел входит две пары хромозом с плечами —17,5 и
3,6 р; 11,25 и 2,25 р.
В шестом разделе не встречаются хромозомы с отношением плеч
больше чем 1:5.
В седьмой раздел входит последняя 12-я пара хромозом с длинным
плечом—18,28 и коротким—1,5 р.
Общая длина всех хромозом гаплоидного набора у этого вида —
206
195,375 р; длинных плеч— 148, 875; коротких плеч—46,5 р; общий
индекс — 3,206. Индекс А( —1,618. Индекс Ап—2,159.
Т. dasystemon Bge. (рис. 21, е-, 22, ж). Этот вид, собранный
нами в разных частях ареала, имеет весьма изменчивый кариотип.
Исследованные образцы мы привезли из Ферганского хребта с
перевала Кугарт (задернованный серозем), изучали корешки взрослых
луковиц.
Число хромозом у этого вида с данного местообитания в сомати-
ческих клетках бывает 24 и 48. Таким образом, у него имеется как
диплоидные, так и полиплоидные формы. У диплоидной формы все
хромозомы можно разбить на шесть разделов.
Первый раздел состоит из двух пар хромозом. Первая с длинным
плечом —7,5 и коротким —4,75 р.
На длинном плече у этой хромозомы на расстоянии 3 р от кине-
тического перерыва расположена акинетическая перетяжка. На корот-
ком плече в 0,75 р от кинетического перерыва также находится аки-
нетическая перетяжка, характерная этой хромозоме почти всего рода.
Вторая пара с плечами — 6,25 и 3,25 р.
Второй раздел состоит из двух пар: первая с длинным плечом —
7 и коротким — 3 р: вторая—5,2 и 2, на коротком плече на расстоя-
нии 0,5 р от дистального конца плеча лежит акинетическая перетяжка.
Третий раздел объединяет четыре пары хромозом: первая с длин-
ным плечом—8,75 и коротким—2,5 р; вторая—7 и 2 р. На длинном
плече у хромозом этой пары на расстоянии 3,5 р от кинетического
перерыва расположена акинетическая перетяжка. Третья пара с длин-
ным плечом—6,25 и коротким—2 р; четвертая—6,25 и 1,9 р. На ко-
ротком плече этой хромозомы лежит акинетическая перетяжка на
расстоянии 0,5 р от кинетического перерыва.
В четвертый раздел входит одна пара хромозом с длинным плечом—
8,75 и коротким —2 р.
Пятый раздел также представлен одной парой хромозом с длин-
ным плечом — 8,25 и коротким —1,5 р.
Шестой раздел составлен двумя парами хромозом: первая с длин-
ным плечом—9,25 и коротким —1,25 р. Вторая —7,5 и 1,25 р.
На длинном плече этой хромозомы на расстоянии 3,25 р от кине-
тического перерыва расположена акинетическая перетяжка.
Общая длина всех длинных плеч гаплоидного набора у диплоид-
ных экземпляров Т. dasystemon этого местообитания равна —87,95 р;
коротких плеч—28,4; всех хромозом—116,35 р. Индекс Aj —1,579.
Индекс Ап —1,923. Общий индекс—3,1.
Т. dasystemon Rgl (рис. 21, в; 22, б). Образцы собраны в районе
Алайской долины (восточная часть). Урочище Арча-Булак, типчаковые
степи. Сбор 1935 г. М. Величко. Мы отнесли эти растения к Т. dasys-
temon, но у А. И. Введенского во „Флоре СССР" при описании Т.
turkestanica есть следующее примечание: „Растения из Алайской до-
лины заслуживают дальнейшего изучения: по словам Величко, они име-
ют бледно-желтые цветы".
В клетках кончиков корня у этих образцов мы определили 24
хромозомы.
Все хромозомы этого вида можно разбить на пять разделов. В
первый раздел входят две пары хромозом. Первая с длинным плечом—
6 и коротким—4 р. На коротком плече на расстоянии 1,75 р от кине-
тического перерыва лежит акинетическая перетяжка; вторая — с длин-
ным плечом —5,25 и коротким —2,75 р.
207
Второй раздел объединяет три пары хромозом: первая с плечами—
6,75 и 2,5 р; вторая—5,25 и 2,25; третья—4,75 и 2,25 р. Третий раз-
дел состоит из четырех пар хромозом: первая—6,5 и 1,625 р; вто-
рая—6,25 и 1,75; третья—5,25 и 1,75; четвертая—5 и 1,625 р.
В четвертый раздел входит две пары хромозом: первая с длин-
ным плечом—7,75 и коротким—2,25 р; вторая—6,5 и 1,5 р.
В пятом и шестом разделах хромозом не наблюдается.
К седьмому разделу отнесена одна пара хромозом с длинным пле-
чом—9,5 и коротким — в 1 р.
Общая длина всех длинных плеч хромозом у этого вида —74 р;
коротких плеч —25,625; хромозом гаплоидного набора данного образ-
ца—99,925 р; общий индекс—2,88. Индекс Ai —1,5. Индекс Ап —
1,9 (09).
Т. dasystemon Rgl. (рис 21, г; 22, в). В Казахстане (Заилийский
Алатау, озеро Иссык) встречается два типа растений этого вида: дип-
лоиды — 2п=24; тетраплоиды — 2п=48.
Все хромозомы этого вида можно разбить по соотношению плеч
на шесть разделов. В первый раздел входит одна пара хромозом с
длинным плечом —6,25 и коротким —4,75 р.
Второй раздел состоит из четырех пар хромозом: первая пара с
плечами — 6,5 и 2,6 р; вторая —6,5 и 2,25; третья —5,75 и 2,6; четвер-
тая —5,4 и 2,4 р.
Третий раздел представлен двумя парами хромозом: первая пара
с длинным плечом—6 и коротким—2 р, вторая пара—5,4 и 1,75 р.
Четвертый раздел содержит три пары хромозом: первая с плеча-
ми—10,75 и 2,6 р; вторая—9,9 и 2,25; третья — 6,75 и 1,5 р.
В пятом и шестом разделах по одной паре хромозом с длинными
плечами—8,25 и 10 и короткими —1,6 и 1,5 р.
Общая длина всех хромозом гаплоидного набора образцов дан-
ного местообитания— 125,25 р; длинных плеч—99,625; коротких плеч—
25,625 р; общий индекс—3,887. Индекс Aj—1,316.
Т. dasystemon Rgl. (рис. 21, д; 22 г, д). Терскей Алатау. Дже-
ты-Огуз (сбор 1931 г. А. И. Введенского). Данные образцы состоят
из двух форм: тетраплоидной 2п=48 (рис. 22, г) и диплоидной 2п=24
хромозомы (рис. 22, d).
Все хромозомы диплоидной формы по соотношению плеч можно
разбить на пять разделов (рис. 21, д).
В первый раздел входит одна пара хромозом с длинным плечом—
7,5 и коротким —4 р. На коротком плече — в 1,5 р от кинетического
перерыва расположена акинетическая перетяжка.
Второй раздел представлен также одной парой хромозом с длин-
ным плечом —7 и коротким —3,25 р.
Третий раздел хромозом Т. dasystemon включает шесть пар хро-
мозом: первая пара с длинным плечом—9,25 и коротким 2,75 р; вто-
рая—8,25 и 2,5; третья—6,75 и 1,75; четвертая — 6,25 и 1,75; пятая—
6,1 и 1,75 и шестая—5,75 и 1,75 р.
Четвертый раздел объединяет три пары хромозом: первая пара с
плечами—9,25 и 2,25 р, вторая—8,5 и 2; третья—6,75 и 1,5 р.
В пятый раздел входит одна пара хромозом с длинным плечом—
6,75 и коротким—1,25 р.
Сумма всех длинных плеч у этой формы —97,05; коротких плеч-
27,8; длина всех хромозом — 124,25 р; общий индекс —3,49. Индекс
Ai-1,875.
7. bifloriformis Vved. Этот вид в различных точках ареала дает
совершенно несходные картины.
208
T. bifloriformis Vved. (рис. 22, И; 23, б). Таласский Алатау. Га-
лечниковые выступы в конусе выноса сая у сел. Новониколаевка (сбор
1930 г. А. Я. Буткова и В. П. Бочанпева). Мы исследовали хромозо-
мы в точках роста корня и в период спермиогенеза. Количество хро-
мозом в соматических клетках этого вида равно 24 (рис. 22, и) и со-
ответственно —12 в пыльцевых трубках. Все хромозомы по соотношению
плеч можно разбить на шесть разделов (рис. 23, б).
В первый раздел входит одна пара хромозом с длинным плечом—
8,7 и коротким —4,5 р.
Второй раздел объединяет три пары хромозом. У первой пары
каждый компонент с длинным плечом —13,62 и коротким —5 р; вто-
рой —8,5 и 3,25 и третьей —8,25 и 3,25 р.
Третий раздел состоит из двух пар: длинное плечо у первой па-
ры—8,25 и короткое—2,75 р, второй—8,15 и 2,32 р. •
Четвертый раздел объединяет три пары хромозом: первая с длин-
ным плечом—8,75 и коротким —1,95 р; вторая—8,62 и 1,87; третья—
8 и 1,87 р.
Пятый раздел состоит из двух пар хромозом: первая с длинным
плечом—13,8 и коротким—8,5 р; вторая—9,75 и 1,87 р.
Шестой раздел представлен одной парой хромозом с длинным
плечом—12 и коротким—1,75 р.
Общая длина всех хромозом данного вида—147,27 р; длинных
плеч—115,39; коротких плеч—31,88 р; общий индекс 3,619. Индекс
А,-1.955.
Т. bifloriformis Vved. (рис. 21, да?; 22, ё). Ферганский хребет, ле-
вый берег реки Кугарт у сел. Димитриевского. Склон над мельницей.
Кустарниковая зона (сбор 1933 г. 3. П. Бочанцевой).
Хромозомы исследованы в момент спермиогенеза в пыльцевых
трубках при выращивании их на искусственной питательной среде.
Гаплоидное число хромозом для данного вида установлено п=12
(рис. 22, е), т. е. 2 п = 24.
Все хромозомы этого вида по соотношению плеч можно разбить
на шесть разделов (рис. 21, ж).
В первый раздел входит одна пара хромозом с длинным плечом—
5,25 и коротким—3,38 р.
Второй раздел представлен также одной парой хромозом с длин-
ным плечом —4,25 и коротким—1,35 р.
Третий раздел объединяет шесть пар хромозом: первая пара с
длинными плечами—6,75 и короткими—1,94 р; вторая— 5,5 и 1,75;
третья—5,5 и 1,7; четвертая—5,35 и 1,625; пятая—5 и 1,5 и шестая—
4,3 и 1,35 р.
Четвертый раздел состоит из трех пар хромозом: первая пара с
длинным плечом—7,5 и коротким —1,8 р; вторая —6,375 и 1,625; тре-
тья —5 и 1,25 р.
В пятом и шестом разделах хромозом нет.
Седьмой раздел включает одну пару хромозом с длинными пле-
чами—5,35 и короткими—0,69 р.
Общая длина всех хромозом этого образца —75,845 р; длинных
плеч—66,085; коротких плеч—19,76 р; общий индекс—3,344. Индекс
хромозомы Ai —1,553.
Т. bifloriformis Vved. (рис. 12, а-, 23, в). В 1935 г. этот образец
взят из коллекции тюльпанов Института сухих субтропиков, собран-
ных в окрестностях Намангана. Исследование проведено на хромозо-
мах в пыльцевой трубке в процессе спермиогенеза при проращивании
пыльцы на искусственной питательной среде.
14—2229
209
Число хромозом для данного образца п = 12; следовательно, в
соме 24 (рис. 12, а).
Все хромозомы этого образца по соотношению плеч можно раз-
бить на шесть разделов (рис. 23, в).
В первый раздел входит одна пара хромозом с длинными плеча-
ми—10 и короткими—5,5 р.
На длинном плече в 1,25 р от кинетического перерыва ясно видна
акинетическая перетяжка, на таком же расстоянии от перерыва не
заметная обычно на коротком плече.
Во второй раздел входят три пары хромозом: первая пара с длин-
ными плечами—6,125 и короткими—2,4 р; вторая—6 и 2,4; третья —
5,125 и 2,35 р.
Третий раздел объединяет две пары хромозом: первая с плечами—
'8,75 и 2,75 р; вторая —7 и 2 р.
В четвертом разделе нет ни одной пары.
Пятый раздел представлен двумя парами: первая с длинным пле-
чом—6,625 и коротким—1,25 р; вторая—6,25 и 1,125 р.
В шестом разделе нет ни одной пары.
Седьмой раздел содержит три пары хромозом: первая пара с пле-
чами—11 и 1,9 р; вторая—10,55 и 1,75 р; третья—9,9 р и 1,35 р.
В восьмой раздел входит одна пара хромозом с длинными пле-
чами —7,55 и короткими —0,833 р.
Общая длина гаплоидного набора всех хромозом этого вида —
120,375 р; длинных плеч—94,875; коротких плеч—25,5 р; общий ин-
декс—3,799. Индекс хромозомы Ai—1,81.
Т. bifloriformis Vved. (рис. 12, б; 23, г). Этот образец взят из
коллекции тюльпанов, выращиваемых Институтом сухих субтропиков
на загородном участке Ботанического сада. Этикетки о происхожде-
нии образца не было. Но экспедиции Института сухих субтропиков
того года были в окрестностях Самарканда, Денау и Душанбе.
Исследовали хромозомы в пыльцевых трубках во время спермио-
тенеза при проращивании пыльцы на искусственной питательной среде.
Число хромозом для этого образца установлено п=12 или при
пересчете на диплоид 2 п = 24 (рис. 12, б).
Все хромозомы можно разбить на шесть разделов (рис. 23, г).
В первый раздел входит одна пара хромозом с длинными плечами
у каждого компонента — 5,25 и короткими—4,125 р.
Второй раздел состоит из трех пар хромозом: первая пара с длин-
ным плечом—5,75 и коротким—2,5 р; вторая—5,75 и 2,15; третья —
4,95 и 1,75 р.
Третий раздел представлен тремя парами хромозом: первая с
длинным плечом—5,75 и коротким—1,8; вторая—5,25 и 1,5 и третья—
5,125 и 1,625 р.
Четвертый раздел также включает три пары хромозом: первая с
длинными плечами—9,75 и короткими—2,25 р; вторая—7,75 и 1,575 р;
третья—5,75 и 1,1р. У второй пары хромозом на длинных плечах в
1,55 р от кинетического перерыва расположена акинетическая пере-
тяжка.
В пятый раздел входит одна пара с длинным плечом —8,25 и ко-
ротким—1,5 р.
В шестом разделе нет хромозом.
Седьмой раздел содержит одну пару с длинным плечом —6,875 и
коротким —0,95 р.
210
Общая длина всех хромозом гаплоидного набора этого образца —
$9,025 у; длинных плеч—76,2; коротких плеч—22,825 у; общий индекс—
3,347, индекс Aj —1,27.
Т. euhseana Boiss. (рис. 12, ж-, 24, а). Этот образец получен от
В. П. Бочанцева и А. Я. Буткова с этикеткой: „Равнина со слабыми
признаками засоления у Чар-Дары“ в 1933 г.
Хромозомы исследовали в кончиках корешков взрослых луковиц
и в пыльцевых трубках в период спермиогенеза при проращивании
пыльцы в искусственных условиях.
Число хромозом для данного вида установлено 2 п = 24 и п=12
(рис. 12, ж).
Все хромозомы данного вида можно по соотношению плеч раз-
бить на пять разделов (рис. 24, а).
В первый раздел входит три пары хромозом: первая пара с длин-
ными плечами—8,5 каждая, а короткие— 5 у. На коротком плече в
1,75 [х от кинетического перерыва расположена акинетическая пере-
тяжка. Вторая пара с длинными плечами—6,25 и короткими —4 у, на
которых в 1,3 у расположен акинетический перерыв. Третья пара с
длинным плечом—5,75 и коротким—3 у.
Во второй раздел входят три пары хромозом: первая пара с пле-
чами—7,5 и 3,25 у; вторая—7,25 и 3; третья—7,5 и 2,7 у.
Третий раздел включает четыре пары хромозом: первая с плеча-
ми —9,25 и 3 у; вторая —7,5 и 2,5; третья —6,75 и 2,25; четвертая —
6,75 и 1,75 у.
В четвертом разделе хромозом нет.
В пятом разделе одна пара хромозом с длинными плечами—11,25
и короткими—2,25 у. На длинных плечах эта пара хромозом имеет
акинетический перерыв, делящий плечо на две равные части.
В шестой раздел входит одна пара хромозом с длинным плечом —
3,5 и коротким —1,25 у.
Общая длина всех хромозом гаплоидного набора равна—125,7 у;
длинных плеч —92,75; коротких плеч—32.95 у; общий индекс—2,815.
Индекс Ai—1,7. Индекс Ап—1,56. Индекс Аш 1,92.
Т. bifloriformis Vved. (рис. 12, з, и; 24, б). Исследованные об-
разцы произрастают в Ботаническом саду под деревьями и на склонах
к Анхору. Первичное происхождение их установить не удалось.
Они очень похожи на образцы этого вида из Акташа и Хумсана. Мы
изучали корешки, полученные при проращивании взрослых луковиц,
хромозомы из молодых пылинок в момент деления первичного ядра
пыльника на вегетативное и генеративное ядра и хромозомы в пыль-
цевых трубках при спермиогенезе.
Этот образец имеет две формы: наиболее часто встречается поли-
плоидная форма—триплоид. Она хорошо цветет, крупными цветами и
не дает семян. У этой формы сильно развито вегетативное размноже-
ние. Растет она большими колониями — клонами. Изредка встречается
диплоидная форма. При изучении морфологии хромозом мы устано-
вили, что у триплоида Т. bifloriformis из этого местообитания мы
имеем дело с автополиплоидией.
Все хромозомы этого вида как диплоида (2 п =24), так и трип-
лоида, прекрасно разбиваются по соотношению плеч на четыре раздела.
В первый раздел входит три пары хромозом: первая с длинным
плечом—4,5 и коротким—4 у; вторая—4,25 и 2,75; третья—4 и 2,25 у.
Второй раздел состоит из четырех пар хромозом: первая с длин-
ными плечами—6,25 и короткими—2,75 у; вторая—4,75 и 2; третья —
4,5 и 1,75 и четвертая —4 и 1,5 у.
211
6 II Illi
/ 2 3
1 llllll
12 3
3 4 5 6 7
78 9 10
8 9 /0
8 9 10
IB
77
7/ 12
fl 12
В
f2
i I
/7 12
Рис. 24. Идиограммы измеренных пластинок (увелич. в 800 раз).
а—Т. buhseana В о i s s . ; б— Т, bifloriformis Vved.; е—Т. turkesianica Rgl.; г—T.fbuhseana В о i s s.j
o'—-T. binutans V v e d . ; г—T. buhseana В oiss.; Ж—T. lurkesianica Rgl.
212
Третий раздел представлен тремя парами хромозом: первая с
длинными плечами—5,25 и короткими—2 р; вторая—6,25 и 1,75; тре-
тья —5 и 1,5 р.
В четвертом разделе нет хромозом.
Пятый раздел содержит две пары: первая с длинными плечами —
6 и короткими—1,1; вторая—5,25 и 1 р.
Общая длина всех хромозом гаплоидного набора этого образца—
85,35 р; длинных плеч—61; коротких плеч—24,35 р; общий индекс —
2,5. Индекс Ai —1,13. Индекс Ац —1,58. Индекс Ат —1,7 (7).
Т. turkestanica Rgl. (рис. 12, к-, 24, в). Исследованный образец
происходит из гор Нуратау, хребет Койгаш. Лужайка, верховья сая
около кишлака Каракон (сбор 1934 г. В. П. Бочанцева). Хромозомы
мы изучали в пыльцевых трубках и корешках.
Этот образец является диплоидом, 2 п хромозом =24.
Все хромозомы по соотношению плеч можно разбить на четыре
раздела.
В первый раздет входит три пары хромозом:' первая с длинным
плечом—4,75 и коротким—3,75; вторая—5 и 2,25; третья —3,75 и 2р.
Второй раздел объединяет четыре пары хромозом: первая с длин-
ным плечом—6,75 и коротким—2,25 р; вторая—5,75 и 2,1; третья—5
и 2; четвертая—4,5 и 1,9 р.
Третий раздел составлен тремя парами хромозом: первая с пле-
чами—7 и 2 р, вторая—5,75 и 1,75; третья—5,25 и 1,5 р.
Четвертый раздел включает две пары хромозом: первая с длин-
ным плечом—6,6 и коротким—1,5 р; вторая—5,5 и 1,25 р.
Общая длина всех хромозом гаплоидного набора этой географи-
ческой формы данного вида равна—89,85 р; длинных плеч—65,6; ко-
ротких плеч—24,25 р; общий индекс—2,76. Индекс хромозомы Ai —
1,27, Индекс хромозомы Ац —2,2. Индекс хромозомы Аш—1,87.
Т. euhseana Boiss. (рис. 12, л-, 24,г). Этот образец в 1935 г.
мы привезли с разъезда 146 Ташкентской ж.-д. Равнина с явными
признаками засоления, кое-где переходящая в пухлые солончаки с
тамариксами.
Исследовали хромозомы в корешках взрослых луковиц и в пыльце-
вых трубках.
Данная географическая форма является диплоидом и имеет 2п=24.
Все хромозомы этого вида можно разбить на шесть разделов.
В первый раздел входят три пары хромозом. Первая пара с длин-
ными плечами у обоих компонентов пары—9,25 и короткими—6 р;
вторая пара—8,87 и 5,25; третья—6,75 и 4 р.
Второй раздел включает три пары хромозом: первая с длинным
плечом—11,65 и коротким—4,5 р; вторая—10 и 4,25; третья —9,25 и
3,87 р.
Третий раздел представлен одной парой с длинным плечом —12,5,
коротким —3 р.
Четвертый раздел объединяет три пары хромозом: первая с длин-
ными плечами каждого компонента—14,5 и короткими—в 3,55 р; вто-
рая—12,5 и 2,75; третья—9,75 и 2,25 р.
Пятый, шестой и седьмой разделы не имеют хромозом.
Восьмой раздел состоит из одной пары хромозом с длинным пле-
чом—12,87 и коротким—1,78 р.
В новый 13-й раздел отнесена одна пара с длинным плечом—15,12
.и коротким в 1,12 р.
Общая длина хромозом гаплоидного набора этого образца равна—
213
175,33 р; длинных плеч—133,01; коротких плеч—42,32 р; общий ин-
декс—3,145. Индекс Ai —1,54. Индекс Ап—1,69. Индекс Ащ—1,69.
Т. binutans Vved. (рис. 12, в; 23, д). Исследованные образцы
мы привезли из Киргизии, на 7-м км выше слияния р. Аламедин с
р. Чонкурчак у подножья нижней гряды скал.
Хромозомы исследованы в кончиках корешков взрослых луковиц
и в молодых пыльниках в момент деления первичного ядра на гене-
ративное и вегетативное (рис. 12, г).
Этот образец является диплоидом и количество хромозом в его
соматических тканях равно 24, а в генеративных—12.
Все ее хромозомы можно разбить на пять разделов.
В первый раздел входит одна пара хромозом с длинным плечом
каждого компонента этой пары—5,75 и коротким—4 р.
Второй раздел состоит из четырех пар хромозом: первая пара с
длинным плечом каждого компонента пары—6,75 и коротким—2,75 р,
на коротком плече имеется акинетическая перетяжка, расположенная
в 0,75 р от кинетического перерыва. Вторая пара с длинным плечом —
6,25 и коротким—2,5 р; третья—5,5 и 2,25; четвертая—5,5 и 2 р.
Третий раздел составлен тремя парами хромозом: первая пара с
плечами—5,4 и 1,75 р; вторая—5,25 и 1,75; третья пара—5,25 и 1,5 р.
В четвертый раздел входит три пары хромозом, первая с плечами—
8 и 2; вторая—8 и 1,75; третья—6 и 1,5 р, у второй и третьей пар к
дистальным концам коротких плеч прикреплены тоненькими нитями
округлые, небольшие спутники.
В пятом разделе хромозом нет.
К шестому разделу относится последняя пара хромозом с длин-
ными плечами 6 и короткими—1 р.
Общая длина всех хромозом гаплоидного набора этого образца
равна—98,4 р; длинных плеч—73,65; коротких плеч—24,75 р; общий
индекс —2,985. Индекс хромозомы Ai —1,44.
Т. binutans Vved. (рис. 12, м; 24, <?). Образец получен из Се-
верной Киргизии с хребта Киргизский Алатау из ущелья Чулак Ка-
инды (сбор 1934 г. Красовского).
Хромозомы изучены в пыльцевых трубках во время спермио-
генеза.
Установлено, что гаплоидное число хромозом этой географической,
формы п=12, соматическое 2 п=24.
Все его хромозомы можно разбить на семь разделов.
В первый раздел входят две пары хромозом: первая пара с длин-
ным плечом у каждого компонента—8,75 и коротким—4,5 р; вторая—
7,5 и 4,12 р.
Второй раздел состоит из двух пар хромозом: первая с плечами—
6,25 и 2,25 р; вторая—5,75 и 2,7 р.
Третий раздел представлен одной парой хромозом с длинным пле-
чом—7,12 и коротким—2,12 р.
В четвертый раздел отнесены четыре пары хромозом. Первая с
плечами—10,75 и 3 р; вторая—9 и 2,5; третья 8,75 и 2; четвертая —
6 и 1,5 р.
В пятом одна пара с длинным плечом—11,5 и коротким—2,25 р.
В шестом разделе одна пара с плечами—11,07 и 2 р.
В седьмом разделе одна пара с плечами 8 и 1,125 р.
Общая длина хромозом гаплоидного набора равна—130,84 р; длин-
ных плеч —100,19; коротких плеч—30,065 р; общий индекс—3,ЗОН
Индекс хромозомы Ai —1,72. Индекс хромозомы Ац —1,82.
214
T. euhseana Boiss. (T talievii Vved) (рис. 24, e; 25, а). Иссле-
дованный образец получен из окрестностей г. Джамбула с горы Тек-
турмас, среди камней (сбор 1934 г. А. Пономаревой). Хромозомы изу-
чали в кончиках корешков взрослых луковиц и из молодых пылинок
при делении первичного ядра пылинки на генеративное и вегетатив-
ное ядра.
У этого образца количество хромозом в соматических тканях рав-
но 24, в генеративных—12.
Все хромозомы можно разбить на четыре раздела.
Первый раздел объединяет две пары хромозом. Первая с длинным
плечом каждого компонента—6,25 и коротким—4,5 вторая —7 и 3,75 р.
На коротком плече второй пары имеется маленький шарообразный
спутник.
Второй раздел состоит из четырех пар: первая с плечами—8 и 3 р;
вторая 7 и 3,25; третья—6,25 и 2,25; четвертая—6 и 2,25 р.
Третий раздел содержит две пары хромозом: первая с плечами —
8,75 и 2,5 р; вторая—7 и 2,25 р.
В четвертый раздел входят четыре пары хромозом: первая с пле-
чами—9 и 2 р; вторая—8и 2; третья—7и 1,75; четвертая—5,5 и 1,25р.
Общая длина всех хромозом гаплоидного набора равна —117,5 р;
длинных плеч—85,75; коротких плеч—31,75 р; общий индекс—2,89.
Индекс Ai —1,4. Индекс Ап —1,9.
7. euhseana Boiss. (рис. 12, д\ 23, е). Этот образец из окрестно-
стей Кушки в 1935 г. привезен А. Пономаревой.
Хромозомы исследовали из точек роста корешков при проращива-
нии взрослых луковиц. Установлено, что соматическое число хромо-
зом для этого вида равно 24.
Все хромозомы можно разбить на четыре раздела. Хромозомы
первой группы отсутствуют при относительно большой величине ко-
ротких плеч.
Во второй раздел входит пять пар хромозом: первая пара с длин-
ным плечом каждого компонента пары—8,5 и коротким—3,75 р. На
коротком плече этой хромозомы на расстоянии 1 р от кинетического
перерыва находится акинетическая перетяжка. Таким образом, по мор-
фологии это типичная хромозома первого раздела, но соотношение
плеч у нее нарушено. Возможно из-за увеличения длинного плеча
или соответствующего уменьшения короткого, что мало вероятно, так
как акинетическая перетяжка делит короткое плечо на отрезки, меж-
ду которыми сохраняется правильное соотношение длины, свойствен-
ное этой хромозоме.
Вторая пара хромозом с длинным плечом— 8,25 и коротким—3,75 р;
третья —8 и 3,45; четвертая —7 и 2,75; пятая —6,25 и 2,75 р.
Третий раздел включает три пары хромозом: первая с плечами —
10 и 2,75 р; вторая—8 и 2,25; третья—7 и 2,25 р. У третьей пары на
коротких плечах в 0,75 р от кинетического перерыва расположена
акинетическая перетяжка.
Четвертый раздел состоит из хромозомы: первая с плечами—10 и
2,6 р; вторая—9,35 и 2,95; третья—7,4 и 1,75 р.
Пятый раздел представлен одной парой с длинным плечом каж-
дого компонента—6,4 и коротким —1,25 р.
Общая длина всех хромозом гаплоидного набора равна—126,865 р;
длинных плеч—95,65; коротких плеч—31,215 р; общий индекс—3,08.
Т. turkestanica Rgl. (рис. 12, е-, 13, ж). Исследованный образец
привезен с горы Кунгус-Бучак, северный склон к роднику Ташбуч
(сбор 1935 г. А. Ф. Гнездило).
215
216
Рис. 25. Пластинки хромозом.
а-Т. binutans Vved.; б- Т. turkestanica Rgl.; в - Т. turkestanica Rgl.; г - Т. tarda S t а р f.; д - Т. tarda S t а р f.; е - Т. orthopoda V v е d. ж — Т. turkestanica Rgl.
Хромозомы изучали во время спермиогенеза при посеве пыльцы
на искусственной среде.
Данный образец содержит в соматических клетках 2 п хромозом —
24 и генеративных п=12.
Все хромозомы этого образца можно разбить на шесть разделов.
В первый раздел входят две пары хромозом: первая с длинным
плечом у каждого компонента—12 и коротким—8 р; вторая—8 и 4,87 р.
На коротком плече этой хромозомы на тонкой нити расположен ша-
ровидный спутник.
Второй раздел представлен одной парой хромозом с длинным пле-
чом—9,2 и коротким—3,62 р.
Третий раздел объединяет пять пар хромозом: первая с длинным
плечом—11,12 и коротким—2,87 р; вторая—10,5 и 3; третья—9,5 и 3;
четвертая—9 и 3; пятая—9 и 2,7 р.
В четвертом разделе нет хромозом.
В пятый раздел входят две пары хромозом: первая с длинным
плечом —16,5 и коротким—3 р; вторая- 15,37 и 3 р.
Шестой раздел состоит из одной пары с длинными плечами—18 и
короткими—3 р.
В седьмом разделе также одна пара с плечами—11,75 и 1,75 р.
Общая длина всех хромозом гаплоидного набора равна—181,55 р;
длинных плеч—139,74; коротких плеч—41,81 р, общий индекс—3,33.
Индекс хромозомы Ai —1,5. Индекс хромозомы Ап —1,643.
Т. turkestanica Rgl. (рис. 24, ж-, 25, б, в). Хребет Кугитанг ок-
рестности зимовки Майдана в зарослях Акантофиллюма (сбор 1932 г.
А. Ф. Гнездило). Исследовали хромозомы в корешках взрослых луко-
виц, в молодой пыльце в момент деления первичного ядра пылинки
на генеративное и вегетативное ядра и в пыльцевой трубке при деле-
нии генеративной клетки на два спермия.
В соматических клетках данного образца установлено 2п=24 хро-
мозомам, встречаются формы с 36 хромозомами, т. е. триплоидные.
При изучении гаплоидных наборов мы нашли формы с п=12,
п=17 и п=18, чаще всего с первым и последним. Среднее число
хромозом мы наблюдали всего один раз.
Все хромозомы как диплоидного (рис. 25, <5), так и триплоидного
набора (рис. 25, в) по соотношению плеч можно разбить на 5 разде-
лов. Мы установили, что триплоидные формы этого образца являются
автополиплоидами (рис. 24, ж).
Приводим разбивку хромозом на типы по диплоидному образцу.
Первый раздел содержит две пары хромозом: первая пара с длин-
ным плечом—6 и коротким 4,75 р. На коротком плече в 1,5 р от
кинетического перерыва расположена акинетическая перетяжка, кроме
того, на дистальном конце этого плеча, на тонкой нити прикреплен
маленький шарообразный спутник. Вторая пара с длинным плечом —
6,25 и коротким—3,25 р. На длинном плече этой хромозомы в 1,75 р
от дистального конца плеча находится акинетическая перетяжка. На
коротком плече в 0,75 р от кинетического перерыва также наблюда-
ется акинетическая перетяжка.
Второй раздел объединяет пять пар хромозом: первая с длинным
плечом—7,5 и коротким—2,9 р; вторая—6,75 и 2,75; третья —7 и 2,5;
четвертая—6,5 и 2,5; пятая—6,25 и 2,25 р. У пятой пары на корот-
ком плече посередине отмечен акинетический перерыв.
В третий раздел входят две пары хромозом: первая с длинным
плечом—8,75 и коротким—2,5 р. На коротком плече посередине рас-
положен акинетический перерыв. Вторая пара с длинным плечом-
217
6,95 и коротким—2,25 р. У этой пары короткие плечи также разде-
лены акинетическим перерывом на две равные части.
В четвертом разделе хромозом нет.
Пятый раздел включает одну пару хромозом с длинными плеча-
ми—8,75 и короткими—1,75 р. На дистальных концах коротких плеч
находятся маленькие спутники, а длинное плечо разделено акинети-
ческим перерывом на две равные части. В шестой раздел входят две
пары хромозом: первая с плечами 7,25 и 1,25 ри вторая—7 и 1 р. На
коротких плечах хромозом второй пары расположен спутник.
Общая длина всех хромозом гаплоидного набора равна—123,3 р;.
длинных плеч—84,25; коротких плеч—39,05 р; общий индекс—2,16.
Индекс хромозомы Ai —1,264. Индекс хромозомы Ап—1,923.
Т. turkestanica Rgl. (рис. 25, ж-, 26, а). Мальгузарский хребет.
Вершина в восточной части хребта. Среди камней (сбор 1934 г.
П. А. Гомолицкого). Исследовали хромозомы в точках роста корней,,
полученных от взрослых луковиц, и во время спермиогенеза, проте-
кающего в пыльцевых трубках при посеве их на искусственной пита-
тельной среде.
Рис. 26. Идиограммы измеренных пластинок (увелич. в 800 раз).
а—Т. turkestanica Rgl.; б—Т. tarda S t а р f . ; в— Т. orthopoda Vved.
Для соматических тканей установлено, что 2 п хромозом =24.
Для генеративных п=12.
Все хромозомы по соотношению плеч можно разбить на пять
разделов.
В первый раздел входят две пары хромозом: первая с длинным
плечом каждого компонента пары -7,5 и коротким--4,25 р; на корот-
ком плече в 0,75 р от кинетического перерыва расположена акинети-
218
ческая перетяжка; вторая пара с длинным плечом—7,5 и коротким —
4,12 {л.
Второй раздел состоит из четырех пар хромозом: первая с плеча-
ми—8,25 и 2,87 р.; вторая —7,87 и 2,70; третья —7 и 2,5; четвертая —
6,6 и 2
Третий раздел содержит три пары хромозом: у первой длинное пле-
чо —8,65, и короткое —2,52 р.; второй —8,48 и 2,37 р. третьей —8,4 и 2,23 р..
Четвертый раздел включает две пары хромозом: первая с плеча-
ми—11,32 и 2,27 р.; вторая—8,7 и 2 и.
В пятом и шестом разделах у этого образца нет хромозом.
Седьмой раздел представлен одной парой с длинными плечами —
9,75 и короткими—1,27 р..
Общая длина всех хромозом гаплоидного набора равна —131,22 р-;
длинных плеч—100,02; коротких плеч—31,2 р.; общий индекс—3,23
Индекс хромозомы Ai —1,764. Индекс хромозомы Ац—1,82.
Т. tarda Stapf. (рис. 25, г, д; 26, б). Исследованные образцы
мы привезли в 1932 г. из ущелья р. Кастек, в 10 км от сел. Михай-
ловского. Кустарниковая зона с Primus prostrata и многочисленными
выходами коренных пород в виде скал и щебня. И р. Малый Кемин.
Гряда на юг от перевала Джол-Булак. Южный скалистый склон среди
крупного щебня, засыпанного мелкой, сильно рыхлой почвой от само-
го низа до верха гряды (сбор 1932 г. 3. П. Бочанцевой). Растения со
значительным вегетативным размножением. Хромозомы исследовали в
корешках взрослых луковиц и в молодой пыльце в момент деления
первичного ядра пылинки на вегетативное и генеративное.
Диплоидные и тетраплоидные формы растут совместно; 2п=24;
4 п=48; п=12.
Все хромозомы гаплоидного набора можно разбить на шесть раз-
делов.
Первый раздел включает две пары хромозом: первая с длинным
плечом—9,25 и коротким—6,75 р. На коротком плече в 2,25 у- от ки-
нетического перерыва находится акинетическая перетяжка, характер-
ная для хромозом этой группы; вторая 6,5 и 3,75 р..
Второй раздел объединяет пять пар хромозом: первая с плечами —
8,75 и 4,125 р.; вторая—8,5 и 3,125; третья—7,79 и 3,95; четвертая —
7,5 и 3; пятая—6,75 и 2,4 р..
Третий раздел состоит из двух пар хромозом: длинное плечо пер-
вой—12 и короткое—3,75 р.; второй—8,25 и 2,4 р..
В четвертом разделе нет хромозом.
В пятый раздел входит одна пара с плечами—12,5 и 2,5 р-. На
длинном плече у компонентов этой пары в 5,75 р. от кинетического
перерыва расположена акинетическая перетяжка.
Шестой раздел представлен одной парой с длинным плечом—9,125'
и коротким —1,5 р.. По соотношению плеч в девятый раздел входит
одна пара с длинными плечами—15,25 и короткими—1,6 р.. На длин-
ном плече в 1,5 р. от кинетического перерыва лежит акинетическая
перетяжка.
Общая длина всех хромозом гаплоидного набора равна—150,215 р.;.
длинных плеч—112,65; коротких плеч—38,05 р-; общий индекс—2,975.
Индекс Ai —1,37. Индекс Ац —1,73 (3).
Т. orthopoda Vved (рис. 25, е; 26, в). Исследованные образцы
получены из хребта Каратау. Сай Джинаган-Ата, и др. (сбор 1930 г.
П. А. Гомолицкого). В Каратау встречается также северная форма
Т. Buhseana. Хромозомы Т. orthopoda мы исследовали в точках роста
корня и в молодой пыльце во время спермиогенеза; 2 п хромозом=24
219
или 48 и п=12 или 24. Все хромозомы по соотношению плеч можно
разбить на шесть разделов.
В первый раздел входят две пары хромозом: первая с плечами—
7 и 3,875 р; вторая—6,625 и 3,35 р.
Второй раздел состоит из двух пар хромозом: длинное плечо пер-
вой—7,25 и короткое—2,44; второй —6,75 и 2,625 р.
Третий раздел объединяет четыре пары хромозом: первая с пле-
чами—10 и 2,69 р; вторая—6,75 и 1,75; третья—6,75 и 1,625; четвер-
тая—5,75 и 1,75 р.
Четвертый раздел представлен одной парой с длинными плечами —
7,75 и короткими —1,9 р.
В пятый раздел отнесено две пары хромозом: первая с длинным
плечом—9,4 и коротким—1,94 р; вторая—9,25 и 1,625 р.
В шестой раздел входит одна пара с плечами—7,94 и 1,14 р.
Общая длина всех хромозом гаплоидного набора равна—126,565 р;
длинных плеч —90,855; коротких плеч —25,71 р; общий индекс —3,5.
Индекс Ai —1,824. Индекс Ап —1,948.
Несмотря на цитологическое исследование большого количества
видов из секции Eriostemones рода Tulipa в настоящее время трудно
сделать какие-либо серьезные выводы о направлении путей эволюции
кариотипа или филогении у видов этой секции.
Мы исследовали 14 видов из 18, приведенных во Флоре СССР. Из
одного местообитания изучены Т. biebersteiniana 2п=24, Т. patens 2п=24,
Т. biflora 2n=24. Т. turkomanica 2n=24, Т. orthopoda 2п=24, 36, 48; из
двух—Т. sogdiana 2n=24, Т. tarda 2п=24, 36, 48, Т. binutans 2п=24;
из трех—Т. turkestanica 2п=24, 36; из четырех—Т. dasystemon 2п=24,
36, 48; из пяти—Т. Buhseana 2п—24, Т. bifloriformis 2п=24, 36, 48.
Каждое местообитание этих видов при сохранении основного
числа хромозом п=12 и кратных ему чисел у полиплоидов дает но-
вый кариотип, который трудно увязывается как у особей из других
местообитаний, но относящихся к этому виду, так и с другими вида-
ми, из близких мест, где обитает исследованный вид.
Встречаются виды с тремя парами хромозом и с отношением плеч
меньше чем 1:2 в первом разделе хромозом — Т. Buhseana (Чердара),
Т. bifloriformis (Ташкент), Т. turkestanica (Койташ). Эти виды из дру-
гих местообитаний в том же разделе имеют лишь по одной паре хро-
мозом—Т. Buhseana, Т. bifloriformis (Наманган).
У хромозом тюльпанов из северной части ареала секции Т. Bie-
berstejniana, Т. patens, Т. biflora наблюдается значительная вторичная
расчлененность. У кариотипа Т. sogdiana из окрестностей Ашхабада
имеется тенденция к резкому укорочению второго плеча хромозом.
У этого вида существуют также кариотипические расы. Вторая раса
этого вида из Серакса с сильным вторичным расчленением хромозом.
Т. dasystemon из Памиро-Алтая отличается от Т. dasystemon из
Тянь-Шаня более длинными вторыми плечами хромозом и большей
склонностью к образованию вторичных перетяжек. Возможно для того,
чтобы составить хотя бы относительно верную картину филогенеза
у видов, входящих в секцию Eriostemones нужно охватить большое
количество видов из различных местообитаний, чем это было сделано
в настоящей работе.
Несомненно, что эволюция кариотипа в секциях Lophophyllon и
Eriostemones шла иным путем, чем в секциях Tulipanum, Leiostemo-
nes и Spiranthera.
Пути эволюции первых двух секций с тремя последними разош-
лись на очень ранних этапах филогенеза.
Гл аеа VI
ПОЛИПЛОИДИЯ У ТЮЛЬПАНОВ
примитивности или прогрессивности видов, секций рода и родов
семейства трудно судить, не имея кариологических данных.
Основным гаплоидным числом хромозом в роде Tulipa является 12.
Это число свойственно большинству представителей семейства Liliaceae
и, видимо, — первичное основное число и древний семейственный
признак. В группе Tulipeae этого семейства оно является основным
гаплоидным числом для родов: Lilium, Fritillaria, Gagea (Darlington,
1956). Кроме того, для одного вида из рода Lloydia, именно для
Lloydia serotina, по данным Тишлера, оно установлено Saki в 1935 г.
Эти данные важны потому, что род Lloydia поставлен Буксбаумом
в ключевое положение для родов Erythronium, Tulipa s. 1., Fritillaria s. 1.
и Lilium s. 1. Для Cardiocrinum cordatum и C. glehi Тишлер (1938)
приводит, по данным Oikava (1937), гаплоидное число хромозом,
равное двенадцати. То же число открыто в 1936 г. Дарлингтоном для
Noimocharis suluensis.
Уклонение в сторону умножения основного числа хромозом —
явления вторичного порядка, т. е. всякое умножение основного числа,
полиплоидия и анеуплоидия, указывают на относительную молодость
данного рода, вида или сорта по сравнению с диплоидами. То же
можно сказать и об уменьшении основного числа хромозом. Виды
с изменением основного числа хромозом также являются производны-
ми видами.
В роде Tulipa имеются две тенденции, ведущие к изменению
основного числа хромозом. Первая из них представляет отклонения
в сторону уменьшения основного числа хромозом, а вторая —его
увеличения.
С первой из них мы встречаемся у Т. maximowiczii из Памиро-
Алая с красных песчаников в окрестностях кишлака Сангтуда. У этого
вида 2га=22 хромозомам. В редукционном делении у него наблюдает-
ся 11 отдельностей, т. е. п хромозом равно 11.
Для Т. maximowiczii в литературе приводились другие числа
хромозом. Но наши неоднократные исследования хромозом у растений,
выращенных из луковиц с естественных местообитаний, показали не-
правильность данных, впервые приведенных Ньютоном (1925), полу-
ченных от растений из ботанических садов, а за ним повторяемых
массой ученых.
И. Д. Романов (1944) исследовал зародышевые мешки этого вида
и также подтвердил наши данные о том, что 2 п хромозом равно 22.
221
Кроме того, как видно из приведенного выше описания кариотипа
этого вида, он имеет очень короткие вторые плечи хромозом. Данный
признак, т. е. уменьшение числа хромозом, у Т. maximowiczii соче-
тается с уменьшением коротких плеч хромозом.
Уменьшение основного числа хромозом мы встретили также
в пыльцевых трубках Т. lanata во время изучения спермиогенеза
(Бочанцева, 1937). Луковицы его были привезены из Памиро-Алая
(окрестности кишлака Сина). Во время деления первичного ядра на
два спермин в некоторых пыльцевых трубках в метафазах генератив-
ного ядра наблюдалось по 11 хромозом. В других пыльцевых трубках
имелись метафазы с 12 хромозомами, но мы не обнаружили 13-хромо-
зомных метафаз. Видимо, пылинки с 13 хромозомами у вида Т. lanata
отмирали. Этот образец плохо завязывал семена, но неплохо размно-
жался вегетативно. Оба примера показывают, что у представителей
рода Tulipa из Памиро-Алая отмечается тенденция к уменьшению
основного числа хромозом. Кроме того, как исключение, уменьшение
основного числа хромозом в группе Tulipeae известно в роде Fritilla-
ria. У Fritillaria ruthenica 2 п—18, а у Fritillaria pudica 2«=26. В послед-
нем случае наблюдается увеличение основного числа хромозом.
В роде Erythronium у Е. albidium, по данным Купера. «=11.
Таким образом, на фоне общей, хотя и не ярко выраженной, тен-
денции некоторых представителей группы Tulipeae к уменьшению
основного числа хромозом исключение рода Calochortus из группы
Tulipeae представляется нам неверно обоснованным. В этом роде наб-
людаются гаплоидные числа хромозом: 6, 7, 8, 9 и 10 и кратные им.
Возможно, что в противоположность взглядам Ньютона (1937)
и Беля (Beal, 1939) основным и исходным числом этого рода все же
являлось 12 хромозом, но затем их число претерпело уменьшение,
что безусловно нашло отражение так же и во внешней форме расте-
ний. Хромозомы с почти терминальным прикреплением нитей веретена
у представителей рода Calochortus более резко выражены, чем у
многих представителей этой группы семейства. У Т. maximowiczii
уменьшение основного числа хромозом привело к изменению морфо-
логии кариотипа или, наоборот, изменение морфологии кариотипа
привело к уменьшению основного числа хромозом. Кроме Т. maxi-
mowiczii еще у нескольких тюльпанов из Памиро-Алая есть хромозомы
с почти терминальным прикреплением нитей веретена (Т. linifolia,
Т. wilsoniana) и другие.
Род Calochortus можно считать наиболее далеко филогенетически
продвинувшимся, чем любой другой представитель группы Tulipeae
и поэтому получивший новые арены жизни и новые морфологические
признаки. Это тем более правдоподобно, так как такое же явление,
но не затрагивающее основное число хромозом кариотипа, имеется
на востоке ареала рода Tulipa. Так, в Китае и Японии произрастают
Т. eduljs и Т. graminifolia с линейными б. м. парно расположенными
листьями, пониклыми цветами и округлыми семенами. Первый из них—
полиплоид. Оба они настолько морфологически отличны от тюльпанов,
что А. Д. Холл нашел нужным выделить их в особый род Amana.
На востоке же, но ближе к основному ядру рода, в Тянь-Шане, произ-
растает Т. heterophylla из секции Orithyia. М. Г. Попов на основании
•б. м. ребристой округлой формы семян выделил его в особый род
Eduardaregelia, несмотря на то, что он имеет мало измененный ка-
риотип и даже не является полиплоидом. Необходимо исследовать
Т. uniflora и Т. heteropetala из этой секции на форму семян, число
и морфологию хромозом. Возможно, всю секцию Orithyia необходимо
222
будет выделить в особый род, хотя кариотипически Eduardaregelia
ничем особенным от представителей рода Tulipa не выделяется.
В роде Gagea, поставленном Буксбаумом в схеме филогенетиче-
ских отношений в группе Tulipeae наиболее близко к ключевому
предку группы, встречаются как круглые, так и плоские семена.
В этом роде форма семян позволяет выделить подроды: I) Eugagea —
семена шаровидные; II) Platyspermum — семена плоские; секция
Foliatae — семена вальковатые; секция Euchloranthae — семена плоские.
Увеличение основного числа хромозом в роде Tulipa наблюдалось
значительно реже и не было так ярко выражено. Мы встречали его
у одного тюльпана, присланного из Тегерана (Т. sp. — 4«=52). Это
тетраплоид с четырьмя экстра-хромозомами. Кроме того, Ньютон
сосчитал в соматических клетках Т. galatica 32 хромозомы, т. е. также
четыре лишних хромозомы.
У Т. lanata мы наблюдали в пыльцевых трубках гаметы с 11 и
12 хромозомами, но не встречали пыльцевых трубок с 13 хро-
мозомами. Однако в некоторые годы гаметы могут быть более
жизненными и тогда у этого вида также возникает форма с 26 хро-
мозомами. Таким образом, в роде Tulipa мы встречаемся как с уве-
личением основного числа хромозом, так и уменьшением. Это тем
более вероятно, если вспомнить, что увеличение основного числа
наблюдается и у Fritillaria pudica. Особый интерес представляют числа
хромозом у видов рода Gagea. Они известны у трех видов: Gagea
spathcea «=102, G. lutea «=36, G. minima «=12. Если принять 12 хро-
мозом за основное число в этом роде, то у первых двух видов имеется
слишком высокая полиплоидия. Это обстоятельство заставляет нас
серьезно усомниться в верности отводимого Буксбаумом (1937) ключе-
вого положения для рода Gagea, так как подобная полиплондность не
свойственна всему подсемейству и не укладывается в общее направление
•его эволюции. Тем более, что всякая полиплоидия — явление вторич-
ного порядка и тогда едва ли возможно ставить род Gagea близко
к прототипу всей группы Tulipeae.
Полиплоидные формы тюльпанов давно известны в литературе.
Де Моль (De Moll, 1925, 1927) первый открыл полиплоидию у тюльпа-
нов, найдя у садовых тюльпанов 36 хромозом. Он объяснил полип-
лоидию отсутствием редукции хромозом. Ньютон (1927) при первом
цитологическом исследовании этого рода обнаружил два тетраплоида:
Т. silvestris и Т. whittiallii, Т. silvestris считают тетраплоилом, прои-
зошедшим от Т. australis. Но Т. australis распространен в Апеннинах,
Южной Франции, Испании и Португалии; Т. silvestris — вдоль по
Средиземью, сорничая в виноградниках, в Англии и Иране в окрестно-
стях Табриза. Считают, что Т. silvestris произошел в результате авто-
тетраплоидии. Так же автотетраплоидом является и Т. whittallii. Пред-
полагают (Hall, 1940), что он происходит из очень изменчивого
греческого вида Т. orphanidea. Кроме того, Ньютоном (1927) была
установлена тетраплоидная Т. turkestanica без точного указания родины.
Т. saxatilis — триплоид и давно введен в культуру. Т. cretica —
тетраплоид с острова Крита (Hall, 1940).
Интересным видом с широким ареалом и большим размахом
полиплоидии является Т. clusiana. Ньютон (1927) у этого вида из
Средиземья нашел тетраплоидный набор хромозом, 5 «=60. А у пред-
ставителя Т. clusiana из Читраля найдена тетраплоидия — 4 «—48;
в коллекции Садовою института в Англии, у экземпляров без точного
указания родины, обнаружена диплоидная форма, 2«—24. У Т. chry-
.santha, которого отделяет от Т. stellata только желтая окраска около-
223
Цветника, найдено 48 хромозом, т. е. он — тетраплоид. Близкий к
Т. chrysantha морфологически, но мелкий, желтый тюльпан из Кашмира
оказался диплоидом.
Кроме того, за границей широко известна форма Т. korolkovii,
Т. korolkovii concolor, которая имеет тетраплоидное число хромозом
(4га = 48). Среди большого количества растений этого вида, взятых нами
из естественных условий, мы не наблюдали тетраплоидов и это дает
нам основание предполагать, что полиплоидная форма Т. korolkovii
получена в культуре, как и многие другие полиплоиды среднеазиат-
ских видов, указания на которые встречаются виностранной литературе.
Тетраплоидный шарлаховокрасный Т. kolpakovskiana, полученный
за границей из Советского Союза, несомненно, является естественным
гибридом между Т. kolpakovskiana и Т. ostrovskiana. Он был выве-
зен нашим Садом из окрестностей Талгара в Заилийском Алатау и
экспортирован за границу.
Кроме вышеперечисленных полиплоидных видов, Холл еще ука-
зывает на Т. ргаесох — триплоид. Он сообщает, что в коллекции Садо-
вого института среди видов близких к Т. ргаесох находятся еще две
полиплоидные формы из Малой Азии и Палестины.
Т. Oculus-solis, которого как пишет Холл, в продаже называют
Т. borszczowii, имеет несколько фрагментов. Видимо, их „Т. borszczowii“
действительно близок к Т. oculus-solis, так как произрастающий в
Средней Азии и исследованный нами Т. borszczowii диплоид с пра-
вильным редукционным делением без каких-либо фрагментов. Холл
считает, что полиплоиды обладают способностью столонообразования,
способствующего их вегетативному размножению, взамен полового —
подавленного или вовсе отсутствующего.
На основании большого количества работ, касающихся приурочен-
ности тетраплоидов к определенным экологическим условиям, в лите-
ратуре создалось мнение, что с изменением физиологического баланса
тетраплоиды и, вообще, полиплоиды часто имеют другие ареалы, чем
диплоиды. Полиплоиды способны заселять другие области, чем дип-
лоиды, хотя это не всегда может быть направлено в сторону небла-
гоприятных условий. Изменения, произошедшие в результате полип-
лоидии, могут позволить полиплоидам существовать в совершенно'
других условиях, чем те. в которых живут и процветают исходные
виды или формы. Это подтверждается на примерах Т. australis —
Т. silvestris; Т. wittallii — Т. orphanidea; Т. clusiana, которые в разных
местообитаниях имеют неодинаковое число хромозом. Из всего выше-
сказанного можно сделать вывод, что Средиземноморье, в узком
смысле, может считаться вторичным местообитанием для представите-
лей рода Tulipa, так как там слишком широко представлено умножение
основного набора хромозом. Явления полиплоидии у видов, произра-
стающих на берегах современного Средиземного моря, встречаются
чаще, чем где бы то ни было в других странах. Среди культурных
тюльпанов, нашедших себе вторую родину в Западной Европе, также
много полиплоидов.
Среди исследованных нами среднеазиатских тюльпанов наибольшее
число полиплоидов найдено в секции Eriostemones. Полиплоидия у
тюльпанов этой секции встречается у Т. bifloriformis из окрестностей
Ташкента и из долины р. Угама в Западном Тянь-Шане. В обоих
местообитаниях были найдены как диплоидные (2га=24), так и тетра-
плоидные (4га=48 хромозом)формы. В данном случае, как показывает
анализ кариотипа, наблюдается авто-тетраплоидия.
224
У Т. turkestanica из Кугитанга найдены диплоидные (2 /г=24 хро-
мозом) и триплоидные (2 п = 36) формы. Гаметы, продуцированные
триплоидами, обычно содержат по 18 хромозом. В некоторых гаметах
встречаются отклонения в числе хромозом: иногда с 17 или с 19 хро-
мозомами. Подсчет хромозом у этого вида производится как в клет-
ках кончиков корней, так и во время течения спермиогенеза в
пыльцевых трубках.
В хребте Каратау растет два вица из секции Eriostemones —
Т. buhseana (2 га=24) и Т. orthopoda — тетраплоид (4га=48 хромозом).
Т. tarda с перевала Кастек имеет как диплоидные, так и тетрап-
лоидные формы. У тетраплоида образуются гаметы с 14, 20 и 24 хро-
мозомами. Широко распространенный по высокогорьям Средней Азии
Т. dasystemon, обычно диплоидный, но образцы этого вида с перевала
Кугарт и с Терскей Алатау и Заилийского Алатау являются тетрап-
лоидами. В Терскей Алатау встречаются как диплоидные, тетраплоидные,
так и триплоидные формы этого вида; все они являются автополип-
лоидами. Таким образом, полиплоидные тюльпаны секции Eriosetemones.
в Средней Азии встречаются в Памиро-Алае, Тянь-Шане. За исклю-
чением Т. bifloriformis из окрестностей Ташкента среди долинных
тюльпанов этой секции нет полиплоидов.
Полиплоиды Т. dasystemon, Т. tarda б. м. высокогорные тюльпаны,
а Т. turkestanica и Т. orthopoda — тюльпаны среднего пояса гор.
Среди исследованных нами представителей других секций рода
Tulipa очень интересен Т. ostrovskiana из секции Leioste^nones.
Т. ostrovskiana имеет 48 хромозом и является амфидиплоидом. В его
идиограмме соединен один полный набор хромозом Т. kolpakovskiana
и другой неизвестного нам вида (рис. 17, в). Диплоидных форм
у Т. ostrovskiana в Заилийском Алатау не было встречено.
В том же Заилийском Алатау у Т. tetraphylla найдены как дип-
лоидные (2 га=24), так и тетраплоидные формы (4 га=48) хромозом. На
основании тщательного исследования кариотипа Т. tetraphylla установ-
лено, что 48-хромозомная форма этого вида является автотетраплоидом.
Она встречается в Заилийском Алатау не менее часто, чем диплоидная.
Таким образом, из семи видов, у которых мы обнаружили полип-
лоидию, шесть обитают в Тянь-Шане и один в Памиро-Алае. Тянь-
шанские полиплоидные виды распределяются следующим образом:
один вид в Каратау, один — в Западном Тянь-Шане и четыре в
Заилийском Алатау. Из них Т. tarda обитает на склонах хребта,
обращенных к долине р. Чу. А остальные три можно найти в окрест-
ностях г. Алма-Аты или Талгара.
Большой интерес представляет распределение этих видов по
вертикали, хотя бы при грубом разделении на зоны, — пустыня,
предгорья, кустарниковый пояс гор (розарий), еловый лес и альпий-
ские луга. В пустыне — Чуилийские горы — растет Т. greigii — 24 хро-
мозомы, Т. kolpakovskiana — 24, Т. alberti — 24, Т. regelii — 24 и
Т. buhseana-T. talievii — 24 хромозомы. На песках при впадении р. Тал-
гар в р. Или растет Т. behmiana — 24 хромозомы. Вверх по течению
р. Талгар по берегам в широкой долине между невысоких гор, покры-
тых травянистой растительностью, обитает Т. kolpakovskiana с 24 хро-
мозомами.
По этим горкам Т. kolpakovskiana слегка поднимается выше и там
встречается с Т. ostrovskiana, имеющей 48 хромозом (из них один
набор хромозом идентичный набору Т. kolpakovskiana). Эти два вида
здесь образуют массу естественных гибридов с 24—36 и 48 хро-
мозомами.
15—2229
225
Выше в кустарниковой зоне, например, в ущелье Акбулак растет
чистый Т. ostrovskiana с 48 хромозомами. При подъеме еще выше по
ущелью в зону смешанного леса до верхних границ еловой зоны
встречается Т. tetraphylla с 24 и 48 хромозомами. На сухих альпий-
ских лужайках и в верхней зоне елового леса обитает Т. dasystemon
с 24—48 хромозомами. Еше выше растет Eduardaregelia с 24 хромо-
зомами.
Таким образом, в пустыне и в нижнем поясе гор нет полиплоидов.
Все они сосредоточены в среднем и верхнем поясе Заилийского
Алатау, очерчивая своеобразную высотную и восточную границы
обитания полихромных представителей рода. Следовательно, наиболь-
шее количество полиплоидов имеется на западной и ближе к восточ-
ной границе ареала рода. Центр же ареала почти свободен от них.
Исключение представляет Т. turkestanica из Кугитанга. Но, вообще,
это исключение лишь подтверждает правило, так как тюльпаны секции
Eriostemones по кариотипу, зародышевому мешку, полиплоидии и
внешней морфологии глубоко отличны от тюльпанов секции Tulipa-
num и Leiostemones.
Глава VII
СЕМЯ ТЮЛЬПАНА И ЕГО ПРОРАСТАНИЕ
лоские семена тюльпанов лежат в трехгнездной коробочке шестью
стопками. В каждом из трех гнезд у коробочки находятся по две
стопки плоских семян, образовавшихся из обратных семяпочек
(рис. 27, б). Семена тюльпанов, разных оттенков, коричневые, округло
треугольной формы несколько напоминают перекошенный сектор круга,
Рис. 27. Семя тюльпана.
д;_обратная семяпочка тюльпана перед созреванием; (7—поперечный
разрез микропиллярной части молодых семян тюльпана у микропиле; в—
поперечный разрез завязи Т. kaufmanniana; г—извлеченный из созрева-
ющего семени Т. fosteriana зародыш; д—зародыш, извлеченный из созреваю-
щего семени Т. kaufmanniana.
заключенный между двумя неравными радиусами и отрезком окруж-
ности в одну шестую ее длины (рис. 27. а). Сосудистый пучок, вхо-
дящий из плаценты через ножку в семяпочку, проходит вдоль узкого
края, обращенного внутрь гнезда и заканчивается в халазе у нуцел-
люса. Этот пучок при созревании семени полностью облитерируется.
Однако ткани семяпочки, лежащие несколько внутрь гнезда и снару-
жи от места окончания сосудистого пучка у халазы (рис. 27, а, б)
при основании нуцеллюса, очень долго остаются живыми. В них со-
храняются хлорофилловые зерна. У многих видов и сортов в сухом се-
227
сухого
ном сечении состоит из
рассматривании
Рис. 28. Продольный разрез через
семя тюльпана Т. subpraestans.
«—халазальиый конец, б—мнкропилярный.
пени на месте этих тканей просвечивает темно-зеленое, почти черное,,
серпообразное пятно. Коричневый цвет семян зависит от цвета стенок
покровов семени. Высохшая оболочка одевает семя прозрачно корич-
невой пленкой. Она образует широкую до 1 мм шир. тонкую слегка
загнутую вниз кайму (рис. 27, б). Кайма способствует рассеиванию
семян из коробочки. Клетки эпидермиса покрова семени над эндо-
спермом вытянуты поперек семени, а в кайме — параллельно краю
семени.
Стенки клеток эпидермального слоя интегуметов, слагающие обо-
лочку, при созревании почти не видоизменяются. Оболочка семян
очень тонкая, в зрелом семени состоит из двух рядов клеток, пред-
ставленных внешним и внутренним эпидермисом интегумента (рис. 28, б).
Находящиеся в ней клетки внутренних слоев интегументов облитери-
руются и сплющиваются в тоненькую пленочку, в которой почти не-
возможно найти их следы.
Разрушение клеток, заполняющих оболочку семени по краям, об-
разующих кайму, происходит задолго до полного созревания, т. е.
когда эндосперм не приобрел роговидной консистенции и легко про-
калывается иглой.
В нормальных семенах эндосперм обычно не заполняет всей се-
менной оболочки. Молочно-белый, полупрозрачный, целлюлярный эн-
досперм тюльпанов, имеющий роговидную консистенцию, в попереч-
в очень толстостенных клеток. При
а под микроскопом видно слоистое
строение его клеточных стенок, а
полости в клетках представлены
лишь незначительными щелевидны-
ми просветами неправильной фор-
мы. Их протопласты в сухих клет-
ках образуют аморфные комочки,,
прижатые к стенкам (рис. 28).
Следовательно, запасные пита-
тельные вещества в семенах тюль-
панов откладываются в виде утол-
щенных стенок в клетках эндоспер-
ма. От хлорцинкиода эти стенки не
окрашиваются. Но когда, после дей-
ствия хлорцинкиодом, на срезы эн-
досперма наносилась капля глице-
рина, они энергично набухали и-
приобретали вид мошных слизистых
образований, во много раз превы-
шающих размеры полостей, в ко-
торых они лежат. Следовательно,
запасные питательные вещества в
эндосперме семени тюльпанов пред-
ставлены слизями.
Слоистые стенки клеток эндосперма состоят из слизей. Зародыш
и протопласт регулируют их растворение и преобразование в легко
растворимые и транспортабельные сахара. Без такого регулирования
при избытке влажности и при соответствующих температурах семя-,
может разрушиться и не прорасти. Подобные картины мы наблюдали
неоднократно при попытках прорастить семена, у которых развился:
только один не совсем полноценный эндосперм и часто отсутствовал
зародыш.
228
Запасные вещества, отложенные в стенках эндосперма, при помощи
протопластов поглощаются растущим зародышем. Когда стенки кле-
ток эндосперма становятся совершенно тонкими, зародыш своими вы-
делениями не только стимулирует растворение протопластом клеточ-
ных стенок, но разрушает собственно протопласт поглощает его.
Следовательно, зародыш для питания не может непосредственно
использовать запасы питательных веществ, отложенные в виде утол-
щенных оболочек в клетках эндосперма. Лишь после того, как запас-
ные питательные вещества растворяются под воздействием ставшего
вновь жизнедеятельным протопласта, пробуждающегося к жизни за-
родышем, они переходят в состояние, доступное для зародыша.
На поверхности сухого семени наблюдается небольшое углубле-
ние, где находится зародыш. Прямой или в виде запятой вальковатый
зародыш просвечивает сквозь эндосперм и тончайшую оболочку семе-
ни (рис. 27, а). Он особенно хорошо виден с плоской верхней стороны
семени, и у многих видов едва просвечивает с нижней.
При смачивании семян для проращивания набухший зародыш
слегка приподнимает эндосперм и оболочку семени, выступая над его
поверхностью в виде чуть приподнятого валика. В сухом семени за-
родыш лежит в его остром конце, примерно на половину не дости-
гая до вершины эндосперма (рис. 27, а).
Зародыш сухого семени состоит из двух неравных частей (рис.
27, г, д). Верхняя — в 6—12 раз длиннее и больше нижней и состоит в
основном из гаусториальной части единственной семядоли зародыша.
В нижней части семядоля образует влагалище, сообщающееся узким
каналом с наружной средой и плотно охватывающее почечку.
Почечка, зачаток стебля и корня составляют меньшую нижнюю
часть зародыша. Стеблевая часть зародыша — будущее донце всего
в несколько микрон, расположена непосредственно под почечкой, по-
степенно переходит в такой же маленький зачаток корня. Несмотря
на значительные размеры зародыша, без большого увеличения лупы и
специального окрашивания невозможно различить его составные
части.
Зародыш сухого семени довольно свободно лежит в полости эн-
досперма. Чем в лучших условиях растут растения, тем меньше в
них пустых и беззародышевых семян. К вершине и основанию коро-
бочки семена всегда мельчают и теряют характерную форму. Среди
семян тюльпанов бывает много пустых, особенно у основания и на
вершине коробочки. Пустые семена имеют вид светло-коричневой
или белой, прозрачной, тоненькой пленочки, под которой нет ни эн-
досперма, ни зародыша. Кроме того, на вершине и при основании
коробочки часто встречаются недоразвитые семена (жесткие,) и такого
же коричневого цвета, как и нормальные. В этих семенах имеется
неокончивший развитие эндосперм, но нет зародыша. Запасные веще-
ства в них не откладываются, как обычно, в виде слоистых утолщен-
ных стенок клеток эндосперма, а остаются в полостях клеток в виде
слизей и легко растворимых сахаров. При проращивании семян даже
при низких температурах такие семена в начале в результате высо-
кой концентрации сахарного раствора внутри клеток всасывают боль-
шое количество воды и вспухают, а затем быстро поражаются пле-
сенью.
Отложение в эндосперме семени запасов питательных веществ
протекает при наиболее высоких температурах активного периода
жизни взрослого растения тюльпана (май, июнь), а их растворение и
преобразование в формы, доступные для зародыша, могут происходить
229
только при наиболее низких температурах первого активного периода
в жизни (декабрь—февраль) нового растения (Бочанцева, 1951).
Прорастание семян. Семена тюльпанов прорастают только при
пониженных температурах от 0° до 10° и повышенной влажности
осенью и зимой (Бочанцева, 1951). Но даже при благоприятных усло-
виях зародыш длительное время растет и формируется внутри семени.
Этот срок тесно связан с условиями, при которых формировали нас-
ледственность исследуемые виды.
Во время прорастания семян зародыш постепенно заполняет всю
полость в эндосперме и плотно прижимается к его клеткам. Семена
тюльпанов прорастают длительное время — осенью, зимой и ранней
весной. Когда протопласты клеток эндосперма оживают, набухает их
плазма, заполняющая все увеличивающуюся полость клетки. Ядра
клеток эндосперма растут, в них хорошо вырисовываются ядрышки и
глыбки хроматина. В ядре наблюдаются явления морфологически
подобные телофазам.
В данном случае ярко обрисовывается сушествующая взаимосвязь
между начавшим развитие зародышем и протопластами клеток эндо-
сперма. Чем ближе к зародышу расположены клетки эндосперма в
прорастающем семени, тем крупнее в них протопласты. Растущий
зародыш стимулирует развитие протопласта в клетках эндосперма.
Протопласты этих клеток растворяют утолщенные клеточные стенки,
являющиеся хранилищами запасных питательных веществ.
В первое время развития зародыша внутри семени интенсивно
растет верхняя часть семядоли, представляющая гаусторию, испол-
няющая функцию всасывающего органа. Она совершенно не содержит
хлорофилловых зерен и некоторое время растет прямо внутрь семени
(это особенно заметно на крупных семенах садовых сортов тюльпа-
нов). Не дойдя приблизительно одной трети до конца эндосперма по
направлению к вершине семени, она слегка изгибается в виде буквы
„г“ и растет вбок. Лишь после завершения роста гаустории у заро-
дыша начинает расти нижняя часть семядольного листа, которая раз-
двигает и прорывает клетки эндосперма под корешком и выходит из
семени с верхушечной почкой и зачатком корня.
Зародыш долго питается запасными питательными веществами,
отложенными в эндосперме семени. За счет этих веществ развивается
семядоля, корешок, почечка и мешковидный вырост — „столон11, уно-
сящий эту почечку далеко от места заложения.
Пустая семенная оболочка впоследствии спадает с гаустории. Еше
до опадения семенной кожуры семядольный лист бывает продолжи-
тельное время зеленым, а его гаусториальная часть остается белой и
вскоре после опадения семенной оболочки отсыхает.
Мы установили, что проростки тюльпанов проходят стадию яро-
визации в течение длительного роста зародыша внутри семени и еще
некоторое время по выходе из него до позеленения и выпрямления,
приблизительно так, как при проращивании семян косточковых (Ро-
дионов, 1959).
Требования проростков тюльпанов к условиям внешней среды
после их позеленения и выпрямления меняются — им необходимы свет
и невысокая температура.
При посеве семян тюльпанов осенью они обычно всходят весной.
Однако в особенно теплую зиму семенам многих горных видов и не-
которых сортов культурных тюльпанов для прорастания не хватает
дней с пониженными температурами. Тогда, высохнув весной и летом
и намокнув следующей осенью, они добирают недостающее им для.
230
прорастания и прохождения первой ступени стадии яровизации коли-
чество дней с пониженными температурами и всходят на следующий
год весной.
При изучении прорастания семян особенно заметна роль среды в
формировании наследственной природы растения. Однако сведения по
этому вопросу для семян тюльпанов очень бедны. Только Е. И. Про-
скоряков (1926) исследовал в 1921 —1923 гг. температурные условия
прорастания семян весенних многолетников у трех видов тюльпанов:
Т. biflora, Т. schrenkii и Т. biebersteiniana.
Позже он (Проскоряков, 1952) изучил большое количество семян
представителей различных семейств. В том числе и род Tulipa.
МАТЕРИАЛЫ ИССЛЕДОВАНИЯ
В 1949 г. мы проращивали семена тюльпанов следующих видов и
сортов. Секция Tulipanum—Т. tubergeniana, Т. kuschkensis, Т. hoogia-
na; секция Leiostemones—Т. ingens, Т. fosteriana, Т. praestens, Т. sub-
praestens, Т. alberti, Т. greigii, Т. micheliana, Т. maximowiczii,
Т. wilsoniana, Т. korolkovii, Т. kolpakowskiana, Т. dubia, Т. vveden-
skyi, Т. hissarica; секция Spiranthera—Т. kaufmanniana; секция Lop-
hophyllon—T. regelii; секция Eriostemones—T. turkestanica, T. biflori-
formis. Из культурных тюльпанов, относимых к Т. gesneriana (дар-
винские тюльпаны) изучены сорта Jubilee, Elephant, Kathleen Parlow,
King Georg Y.
Из культурных тюльпанов, относимых старыми садоводами к
Т. suaveolens, исследованы сорта Due van Thol yellow maximus, van
der Erden, Thomas Moore, а из группы Cottage—Mathers Day.
Происхождение исследованных семян тюльпанов. Семена куль-
турных тюльпанов собраны с растений, выращиваемых в Ботаническом
саду АН УзССР, полученных в виде луковиц в 1932 г. из Голландии
и Главного ботанического сада АН СССР.
Т. tubergeniana. Исследовали семена с экземпляров, выращивае-
мых с 1933 г. в Ботаническом саду АН УзССР. Луковицы тюльпана
собраны в 1933 г. В. П. Бочанцевым на лессовидных склонах г. Се-
бистан в Памиро-Алае. С 1933 г. они росли без пересадки и были вы-
копаны в 1940 г. с 60 см глубины, посажены на глубину 15—20 см.
Т. kuschkensis. Проращивали семена растений, которые росли на
горке в Ботаническом саду ТашГУ с 1931 г. Луковицы тюльпана вы-
копаны в 1931 г. А. А. Мали на лессовых холмах в окрестностях
села Маргуновки Кушкинского района. В 1949 г. их выкопали и пере-
несли на удобренные гряды, в последующем выкапывали и пересажи-
вали каждые два года.
Т. hoogiana. Исследовали семена растений, росших в 1935 г. в
Саду. Луковицы тюльпана в 1935 г. собраны А. Ф. Гнездило на щеб-
нистых склонах в Западном Копет-Даге. С 1935 по 1949 гг. они рос-
ли на горке в Ботаническом саду. С 1949 г. каждые два года их вы-
капывали и пересаживали.
Т. fosteriana. Мы собрали в 1949 г. в Зеравшанском хребте в
районе Аманкутанского лесничества, начиная с высоты около 1500 м
и выше. Он растет главным образом по осыпям из крупных глыб
мрамора или мраморовидного известняка, в щелях между глыбами, в
расселинах скал, заполненных мелкоземом, и реже, на мелкоземис-
тых склонах со щебенкой. Обычно последнее местообитание приуро-
чено к вершинам хребтов. Луковицы лежат на глубине до 35 см.
Т. subpraestans. Исследовали семена экземпляров, выращиваемых
в Ботаническом саду с 1933 г. Луковицы собраны В. П. Бочанцевым
231
на глинистых склонах западных отрогов Памиро-Алая в окрестностях
кишлака Туткаула. обладают хорошей способностью размножаться
вегетативно, лежат группами все в одной лунке на глубине 20 см.
Этот вид часто дает многоцветковые стебли.
Т. greigii. Мы собрали семена в 1949 г. в Дарбазинской щели
близ ст. Дарбаза Ташкентской ж. д. Этот тюльпан обитает на глини-
стых склонах холмов и в долинках между ними. Особенно густые,
почти сплошные, заросли образуют в долинках, где почва сильно
гипсирована. Луковицы в таких местах залегают до глубины 35 см.
Т. korolkovii. Семена собраны нами в 1949 г. близ ст. Дарбаза
Ташкентской ж. д. в долине Кыз-Кудук, на карнизах холмов из пес-
чаника, покрытых неглубоким слоем песчаной почвы. По мере увели-
чения мощности почвы этот тюльпан исчезает, уступая место Т. gre-
igii. Его луковицы расположены на глубине до 12 см.
Т. kaufmanniana. Семена собраны нами в 1949 г. в Западном
Тянь-Шане, в долине р. Сукок в среднем поясе гор в расселинах
скалистых карнизов, заполненных мелкоземом. Луковицы находятся
на глубине до 25 см.
Т. turkestanica. Мы собрали семена в Зеравшанском хребте в
районе Аманкутанского лесничества при начале подъема из долин, на
северных, западных и восточных каменистых склонах. Этот вид в ос-
новном встречается ниже чем Т. fosteriana, хотя верхние границы его
местообитаний налегают на нижние границы обитания Т. fosteriana.
Луковицы лежат на глубине до 10 см.
Т. bifloriformis. Семена собраны нами в 1949 г. близ ст. Дарбаза
Ташкентской ж. д. на глинистых склонах и на пустынных горках
Алымтау, лежащих к северо-западу от Ташкента, на красных глинах,
имеющих большую примесь гипса. В обоих этих местообитаниях лу-
ковицы расположены на глубине 10—16 см.
ЭКСПЕРИМЕНТЫ ПО ПРОРАЩИВАНИЮ СЕМЯН ТЮЛЬПАНОВ
В практике садоводства семена, обладающие жесткой оболочкой
или твердым эндоспермом, для ускорения проращивания заваривают
кипятком.
В сентябре 1948 г. , мы пытались прорастить семена двух видов
диких тюльпанов—Т. bifloriformis и Т. greigii. Семена взяты в двух
повторностях—сборы 1941 г. и 1948 г.
Сухие семена 2 сентября заварили кипятком, 3 сентября, слив
воду, половину заваренных семян вновь залили кипятком, а другую
половину свежей водой комнатной температуры; 4 сентября раздели-
ли заваренные 3 сентября семена на две части и одну из них вновь
заварили. Таким образом, у нас были семена, заваренные один, два
и три раза. Все семена собрали в марлевые мешочки и 7 сентября
посеяли в ящики. Контролем служили посевы семян, смоченных во-
дой комнатной температуры. Посевы поливали каждый день, но осенью
семена не взошли. В сентябре и октябре преобладали положительные
температуры, в зимние месяцы — отрицательные до полного замерза-
ния ящиков с посевами. Только в феврале 1949 г. после оттаивания
почвы появились всходы обоих видов.
Семена Т. greigij и Т. bifloriformis сбора 1941 г. и 1948 г., зава-
ренные по одному разу, взошли на 50%. Семена обоих видов, зава-
ренные по два и три раза совершенно не взошли. Контрольные семе-
на обоих видов взошли на 100%.
2 32
Кроме описанных посевов заваренных семян в теплое время года,
проводили посев семян диких и культурных тюльпанов в парнике,
оранжерее и комнате. При тщательном уходе эти семена не взошли.
Высевали семена (Т. korolkovii, Т. greigii, Т. bifloriformis, Т. fos-
teriana), смоченные растворами биостимуляторов в термостате при 22°.
После двухмесячного содержания в термостате у семян даже не на-
бухли зародыши.
При посеве семян тюльпанов в грунт в августе или сентябре
всходы появляются только следующей весной.
Некоторые исследователи считают, что в семенах растений содер-
жатся особые вещества, тормозящие их прорастание (Исип, 1939; Яр-
кова, 1939).
И. Н. Исип указывает, что при намачивании семян в большом
количестве воды „защитные вещества11 растворяются и при сливании
воды могут быть частично удалены. Подсушивание намоченных семян,
особенно при высокой температуре, еще больше разрушает „защит-
ные вещества11. Для разрушения „защитных веществ" И. Н. Исип
предлагает семена некоторых культур намачивать и подсушивать дву-
кратно.
Учитывая указание М. Г. Николаевой (1948) на тормозящее дей-
ствие воды комнатной температуры на прорастание семян Ferula kara-
tavica — растения, обитающего в сходных с тюльпанами условиях сре-
ды, мы смачивали семена в воде при температуре 0° и 18°.
Семена Т. korolkovii, Т. greigii, Т. kaufmanniana и трех сортов
культурных тюльпанов — Jubilee, Kathleen Parlow и Due van Thol
yellow maximus — партиями no 100 штук подвергали:
1) осторожному перетиранию с промытым речным песком в фар-
форовой ступке для удаления семенной оболочки, трехкратному сма-
чиванию в воде при 0° и трехкратному высушиванию;
2) однократному намачиванию в воде при температуре 0° и одно-
кратному высушиванию;
3) двукратному намачиванию в воде при температуре 0° и дву-
кратному высушиванию;
4) трехкратному намачиванию в воде при 0° и трехкратному вы-
сушиванию;
5) трехкратному намачиванию в воде при 18° и трехкратному вы-
сушиванию.
Контрольные семена не подвергались никакому воздействию.
Каждое смачивание и высушивание семян продолжалось 24 часа.
Перед посевом, произведенным 25 января, каждую партию семян
мы разделили пополам и высеяли в чашки Коха на фильтровальную
бумагу, находящуюся на смоченном водой лигнине. Половину семян
поместили в комнате при температуре 16°—18°, остальные семена
проращивали при низких температурах от 2° до 4- 3°.
У Т. korolkovii прорастание началось на 25-й день одновременно
на всех вариантах опыта, проводимого при низких температурах
^табл. 3).
Из табл. 3 трудно сделать вывод о наличии или отсутствии в се-
менах тюльпанов защитных веществ, • так как прорастание начинается
одновременно у контроля и во всех вариантах опыта. Однако эти
данные представляют интерес с точки зрения биологии прорастания
семян растений, приспособленных к климату Средней Азии. Семена
всех видов тюльпанов требуют для прорастания длительного периода
пониженных температур при наличии значительной влажности. Ни в
каких других условиях они не прорастают. Но в окрестностях Таш-
233
кента, откуда происходят растения Т. korolkovii, с которых взятье
для этого опыта семена, часто в течение зимы и осени бывает смена
влажного периода большой сухостью, низких температур — длитель-
ным периодом высоких. Для изучения характера прорастания семян
в таких резко изменяющихся условиях и интересны данные настоя-
щего опыта. Однако воздействие влажности и высушивания было*
слишком кратковременным, но и оно дало, несомненно, интересные
результаты.
Прорастание семян, % Таблица 3
Номер варианта 21.11 23.11 25.11 27.11 1.III З.Ш 6.III 8.II1 10.111 13.111 Всего
Т. кого 1 к о v I 1
1 2 0 0 0 6 0 4 0 2 4 18
2 6 12 32 26 4 4 14 2 0 0 100
3 4 32 16 10 14 14 0 0 6 4 100
4 22 16 28 4 0 8 2 4 0 0 84
5 20 8 24 26 4 8 8 2 0 0 100
Контроль 2 20 8 8 22 0 0 0 0 4 64
Т. g г е i g i i
2 — 12 6 30 9 10 14 4 4 2 84
3 .— 12 12 12 12 4 6 18 2 8 86
4 — 14 14 16 6 4 10 0 0 2 66
5 — 4 6 20 8 14 8 2 8 18 68
Контроль — 12 18 0 14 6 6 10 2 0 72
Предварительное намачивание и высушивание семян не ускоряет
начала прорастания семян, но несколько сокращает его длительность
во 2, 4 и 5-м вариантах и немного изменяет ход. Прорастание прохо-
дит неодинаковыми темпами—сразу дает резкий скачок в 4 и 5-м ва-
риантах, постепенно нарастая во 2-м. В 3 и 2-м вариантах картина
прорастания в первые 10 дней наиболее сходная. Чем больше мы
предварительно вымачивали семена Т. korolkovii в воде низкой тем-
пературы или же увеличивали ее температурм и высушивали, тем
раньше они начинали прорастать. Но во 2 и 3-м вариантах всходы
были дружнее, чем в 4, 5 и 6-м (контроль).
Очень низкий процент проросших семян мы наблюдаем в 1-м
варианте опыта на семенах с удаленной семенной оболочкой, что про-
изошло, видимо, в результате механического повреждения семян и их
зародышей.
Семена Т. greigii (табл. 3), проращивавшиеся в таких же условиях,
что и Т. korolkovii, начали наклевываться на 27-й день одновременно
во всех вариантах опыта. Совершенно не взошли семена с удаленной
оболочкой. В данном опыте также трудно говорить о наличии или
отсутствии в семенах ускорения против сроков прорастания семян
Т. greigii, посеянных в горшках ранней весной 1949 г. Даже одно-
кратное подсушивание изменило характер прорастания семян этого
вида. В 3-м варианте максимальное прорастание семян началось 8 мар-
та, а в 5-м—13 марта, а не 27 февраля. В 4-м варианте после трех-
кратного вымачивания в воде при 0° и подсушивания семена Т. greigii,
как и Т. korolkovii, взошли очень плохо. Вообще же повторное нама-
чивание и подсушивание семян значительно меняет характер прора-
стания.
234
Семена всех видов и сортов как подвергавшиеся многократному
намачиванию и высушиванию, так и не подвергавшиеся ему, но вы-
ращиваемые при температуре в 16—18° или в термостате при 22°—25°,
дней через десять становились темно-коричневыми и, примерно, на
25-й день поражались плесневыми грибами. Особенно ярко это на-
блюдалось у семян 1-го варианта со снятой семенной оболочкой. Кроме
того, очень легко повреждались плесенью семена Due van Thol yellow
maximus; меньше—Jubillee. Появление плесени мы предупредили
опылением семян порошком древесного угля, но и при этом семена
не проросли. Увеличившиеся при намачивании зародыши приостановили
развитие.
Таким образом, никакого ускорения в прорастании семян подоб-
ного типа методом, предложенным И. Н. Исипом, получить нельзя.
В семенах тюльпанов не удалось также обнаружить действия за-
щитных веществ, которые могли бы тормозить или приостанавливать
их прорастание. Видимо, природа длительности прорастания семян
тюльпанов является физиологической, но более сложной, чем та, на
которую указывает Т. Д. Лысенко (1943), описывая семена сорных
трав: „Семена овсюга без перезимовки обычно всходят не все, а при-
мерно, только на 10—30%. Остальные же не прорастают летом, кажут-
ся мертвыми, а в действительности они живые и ожидают весны, чтобы
дать всходы. Пока семенная оболочка у семян овсюга не промерзнет—
воздух к зародышу семени мало проходит. Поэтому семена и нахо-
дятся в состоянии, так называемого, покоя“.
Повторные намачивания семян в воде с последующим высушива-
нием должны значительно разрушить семенную оболочку и обеспечить
более сильные притоки воздуха к эндосперму семени. Однако взаимо-
отношения среды и семени не так просты и объяснить ускорение или
торможение в прорастании семян одним нарушением целостности
семенной оболочки невозможно.
Данные опытов свидетельствуют о разнокачественности семян у
исследованных видов. Одни из них в природе при повторяющихся
оттепелях, засухах и морозах ускоряют прорастание, а другие, наобо-
рот, тормозят, обеспечивая большую растянутость прорастания и воз-
можность появления части всходов лишь через год. Чем больше коле-
баний во влажности, т. е. чем чаще в естественных условиях с осени
влажность почвы будет сменяться сухостью, тем более это отразится
на последующем ходе прорастания семян. Теплые осенние дожди с
последующими засухами также могут изменить характер и скорость
прорастания семян. Приспособленность семян к изменению особенно-
стей прорастания в разные годы с различным температурным режимом
способствует сохранению вида. Так, в годы со сменами дождей и
засух у некоторых семян происходит торможение роста. Они замедляют
развитие и не успевают прорасти в год рассеивания из коробочки.
В дальнейшем они высыхают и при наличии благоприятных условий
всходят через год. Эти опыты еще раз показывают единство растения
и среды.
Темпы прорастания семян, количество всходов весной определяет-
ся всем ходом погодных условий данного года.
В конце февраля 1949 г. мы посеяли необработанные семена
Т. bifloriformis, Т. greigii, Т. fosteriana и Т. kaufmanniana в горшки и
вынесли их на воздух; поливали по мере необходимости. Температура
воздуха в это время падала ночью до—1°, а днем поднималась до4-12,5°.
При таких колебаниях температуры первые семена Т. biflori-
formis проросли через 22 дня, а Т. greigii через 25 дней, т. е. почти
235
в тот же срок, что и в предыдущем опыте. С третьей декады марта
температура начала повышаться и семена Т. kaufmanniana и Т. foste-
riana, которым необходимо для прорастания более длительное воздей-
ствие низких температур, взошли только весной 1950 г. вместе с по-
сеянными осенью 1949 г.
Обрезка эндосперма. М. Т. Николаева (1948) утверждает, что у
семян Ferula karatavica при обрезке эндосперма, покрывающего кончик
корешка зародыша, ускоряется прорастание. При температуре от 0 до
10° для их прорастания требуется 45 дней. Руководствуясь этим мы
повреждали оболочку семени и обрезали часть эндосперма, прилегаю-
щую к точке роста корешка зародыша. Проращивание обрезанных
семян проводили во влажной камере на влажной фильтровальной бу-
маге при температуре 18—20°. В таких условиях поврежденные семе-
на легко загнивают. Часть семян, обрезанных без повреждения заро-
дыша, избегает заражения плесневыми грибками и дней через пять
начинает „прорастать". На самом деле, здесь происходит только вы-
талкивание зародыша набухшим эндоспермом.
Молодое растение тюльпана долго живет за счет питательных
веществ, отложенных в эндосперме семени поступающих в проросток
’через верхнюю бледную бесхлорофилловую часть семядольного листа
и являющуюся как бы гаусторией. Связь молодого растения с семе-
нем при нормальном прорастании не прекращается очень долго.
Если при обрезке оставить тонкий слой эндосперма над точкой роста
корня, то семя прорастает при комнатной температуре (18—20°С).
Но из-за твердости эндосперма и тонкого слоя над корешком заро-
дыша обрезать эндосперм без повреждения корешка так, чтобы над
корешком остался тоненький слой, очень трудно. Этот способ значи-
тельно ускоряет прорастание семян тюльпанов. Мы получили несколько
проростков Т. fosteriana в течение 35 дней. По внешнему виду эти
проростки мало отличались от нормальных.
Более широкие опыты по изучению оптимальных условий для
прорастания семян тюльпанов мы провели осенью 1949 г. Предвари-
тельными данными, указывающими на необходимость постановки опы-
тов по проращиванию семян тюльпанов при низких температурах,
явились все наши опыты осенью 1948 г. и весной 1949 г.
Е. И. Проскоряков (1926) провел наблюдения над проращиванием
семян трех видов: Т. biflora—характерное растение чернополынной
формации комплексной полупустыни юго-востока Средней Азии;
Т. schrenkii—обитатель широких степных пространств Заволжья, про-
израстает на тяжелых глинистых почвах; Т. biebersteiniana имеет более
широкое географическое распространение, встречается нередко в степях
и лесах.
Е. И. Проскоряков проращивал семена тюльпанов при температу-
рах от 0 до —6°. В его опытах прорастание продолжалось при темпе-
ратуре —0,5°. Семена Т. biflora начали прорастать на восемнадцатый
день после посева, и на стопроцентное прорастание потребовалось
31 день (с 20. III по 20. IV).
У Т. schrenkii в течение 150 дней, или 5 месяцев проросло всего
47% семян. У Т. biebersteiniana за период в 176 дней проросло 96%
семян. Из работы Е. И. Проскорякова видно, что свет тормозит про-
растание семян. При высоких температурах семена тюльпанов не всхо-
дили ни на свету, ни в темноте.
При исследовании прорастания семян тюльпанов мы отобрали
хорошо выполненные и обладающие развитым зародышем семена по
500 каждого вида и сорта на один сосуд (чашки Коха или глиняный
236
горшок) и высеяли в три срока. Первую партию семян посеяли в
песок (в чашках Коха) в начале августа.
Вторую партию семян высеяли 29 августа в маленькие горшки и
поместили в песок; гретые—1 октября в чашки Коха на фильтроваль-
ную бумагу, лежащую на лигнине и сверху укрытую фильтровальной
бумагой, затем лигнином. Мы покрывали семена песком или лигнином,
чтобы предохранить их от тормозящего, по Проскорякову, действия
света. Все посевы поместили в электрохолодильник системы ЭКД-120,
и по мере необходимости поливали.
В первые дни работы холодильника (с 4 по 29 августа) мы вклю-
чали его с 8 час. 30 мин. до 17 час. За это время температура в хо-
лодильнике падала на верхней полке (у холодильной камеры) до—5°,
на средней—до -Ь2‘. К утру она поднималась до + 18°. В помещении,
где находился холодильник, температура доходила до 4-30 , а иногда
и выше. Высокие температуры способствовали появлению на горшках
и чашках Коха с семенами грибков и сине-зеленых водорослей. С 29
августа мы удлинили время работы холодильника—с 7 час. до 22—23
час. При этом минимальная температура на верхней полке была—8
и максимальная, утром перед включением -г7 . Эти температуры более
благоприятствовали прорастанию семяи и тормозили появление снне-
зеленых водорослей иа горшках и чашках.
В опьпах, результаты которых приведены в табл. 4, мы пытались
приблизить эксперимент к естественным условиям и учесть естествен-
ные требования семян тюльпанов к условиям внешней среды. Для
этого мы давали переменные температуры от высоких до очень низких.
В естественных условиях Средней Азии особенно резкие колеба-
ния дневных п ночных температур наблюдаются поздней осенью, зи-
мой и ранней весной, когда семена тюльпанов прорастают, а растения
(луковицы) живут скрытой жизнью.
Таким образом, для прорастания семян Т. bifloriformis, Т. Jcorol-
kovii и Т. greigii (нз которых Т. bifloriformis принадлежит к другой
секции рода), обитающих в одинаковых климатических условиях "гли-
нистой полупустыни, хотя и растущих на различных почвах в окре-
стностях ст. Дарбаза требуется почти одинаковое количество холод-
ных дней. В первых двух опытах для прорастания семян Т. korolkovii
и Т. bifloriformis потребовалось по 40 дней, а в третьем (для Т. bu-
hseana и Т. korolkovii)—по 24 дня (табл. 4). Некоторую роль здесь
по-видимому, играла среда, в которой они проращивались.
Семена Т. greigii в первом опыте были недоброкачественными,
почернели и не взошли. Во втором и в третьем опытах получены
близкие цифры.
Т. turkestanica, Т. fosteriana и Т. kaufmanniana привезены из гор.
Т. kaufmanniana и Т. fosteriana взяты из сходных экологических
условий и требования их семян к пониженным температурам при про-
растании одинаковы. Т. turkestanica, произраставший в более низком
горном поясе, требует для прорастания меньшее количество холодных
дней (табл. 4).
Т. tubergeniana, обитая невысоко в горах на лессовых склонах,
требует меньше холодных дней, чем Т. fosteriana и Т. kaufmanniana
(табл. 4), обитающие в более высоком горном поясе, среди скал, в
щелях, заполненных мелкоземом.
Культурные тюльпаны прорастают быстрее при более высоких
температурах.
Самый поздний по срокам цветения в наших условиях тюльпан
Kathleen Parlow и самый ранний из культурных тюльпанов D. v. Т_-
237
Таблица 4 Проращивание семян тюльпанов в холодильнике
Вид или сорт Дата посе- ва, август Начало прора- стания Количе- ство дней от посева до начала прораста- ния Конец про- раста- ния Количе- ство дней от посева до конца прораста- ния Растяну- тость прора- стания, дни Примеча- ние
Пос ев в н а чале аг г у с т а
Т. bifloriformis 5 14.IX 40 19.IX 76 36
Т. korolkovii 4 15.IX 42 Ю.Х 76 34
Т. turkestanica 6 29. IX 54 2.XII 118 64
Т. kaufmanniana Т. fosteriana — — — — — — 18. I про- растание еще не закончи- тесь
Т. tubergeniana Jubilee 4 27.Х 84 11.XI 99 15 Проросло всего 50% семян
Elephant Kathleen Parlow 5 5 29. IX ю.х 54 67 10.XI 4.XI 97 91 43 25
D. van Thol yellow maximus 5 10.x 67 4.XI 91 25
П осев в конце августа
T. bifloriformis 28 8.Х 40 26.Х 58 18
T. korolkovii 28 8.Х 40 25.X 57 17
T. greigii T. fosteriana* 29 29 8.Х 10.1 40 104 25.Х 57 17
Jubilee* 29 10.1 104 — — —
T. kaufmanniana* 29 10.1 104 — — —
По сев в начале oj < т я б р я
T. buhseana из 1.Х 24.Х 24 10.XI 41 17
г. Алымтау
T. korolkovii Т. greigii 1.Х 1.Х 24.Х 24.Х 24 43 10.XI 41 17 до 1.III проросло около 80% семян
Т. kaufmanniana 1.Х 20.XII 80 -—• — — —
Т. subpraestens Т. fosteriana 1.Х 1.Х 1.Ш** 151 17.II 140 — —
Т. tubergeniana 1.x — нет — — — —
* До 14. XI еще не окончили прорастания и были заморожены, оттаяли под
снегом к 8.1 и к ЮЛ проросли.
** Был вынесен и засыпан снегом, после оттаивания пророс Эти данные
требуют уточнения.
238
yellow требуют для прорастания семян одинаковое количество холод-
ный дней. Этот факт подчеркивает значение внешних условий, в ко-
торых создаются сорта культурных растений. Сорта Kathleen Parlow
и D. V. T-yellow были созданы в Голландии и привезены в виде вполне
сформированных взрослых луковиц.
Семенам голландских тюльпанов для прорастания необходимо
почти то же количество холодных дней, что и семенам тюльпанов
из горных областей (табл. 4).
При проращивании семян культурных тюльпанов в более низких
температурах удлиняется срок начала прорастания и приближается к
срокам начала прорастания семян горных тюльпанов.
Требования тюльпанов к необходимому количеству холодных дней
определяются не видовой принадлежностью семян, а условиями, из
которых они происходят.
В июне 1953 года мы продолжили опыты по проращиванию семян
тюльпанов. К восьми видам исследованным в 1949 г. добавили еще
тринадцать.
Из секции Tulipanum взяли испытывавшийся в 1949 г. Т. tuber-
geniana и два новых—Т. kuschkensis и Т. hoogiana.
Из секции Leiostemones вновь проращивали Т. fosteriana. Т. grei-
gii и Т. subpraestens для сравнения прорастания семян с близкими к
ним видами и исследовали новые — Т. ingens, Т. praestans, Т. alberti,
Т. micheliana, Т. maximowiczii, Т. wilsoniana, Т. kolpakovskiana,
Т. dubia, Т. vvedenskyi и Т. hissarica.
Кроме того, вторично проращивали семена Т. kaufmanniana из
секции Spiranthera. Впервые исследовали семена Т. regelii из монотип-
ной секции Lophophyllon. Таким образом, в 1953 г. испытывали семена
18 видов тюльпанов. Мы посеяли также семена четырех сортов, от-
носящихся к трем группам садовых тюльпанов: Van der Erden, Mot-
her’s Dav, King George Y и Thomas Moore.
В 1953 г. мы раскладывали семена в чашки Петри на влажную
фильтровальную бумагу, прикрывали бумагой и в закрытых чашках
ставили в холодильник. Температура в холодильнике колебалась от—8
до 4-7°. Через каждые 8 дней выбирали и просчитывали проросшие
и наклюнувшиеся семена. Их высаживали в горшки, а непроросшие
вновь ставили в холодильник до полного прорастания всех семян. Во
время просмотра из чашек удаляли все загнившие или испортившиеся
по какой-либо причине семена. Бумагу меняли по мере необходи-
мости.
Приводим результаты исследований видов через каждые пять дней
с момента посева (табл. 5).
Хорошо характеризуя темпы прорастания внутри вида, абсолютные
цифры не дают возможности сравнивать отдельные виды между собой.
Процент проросших семян мы высчитывали для каждого вида отдель-
но, принимая за 100% не количество посеянных семян, а количество
семян данного вида, проросших в настоящем эксперименте. Дикие
виды расположены в этих таблицах соответственно срокам прораста-
ния семян. Они образуют в них интересные столбцы цифр, идущие с
левого верхнего угла таблицы к правому нижнему ее углу. При этом
виды из пустыни или низкогорья—Т. regelii, Т. alberti, Т. kuschkensis,
Т. tubergeniana—расположились в верхнем левом углу таблицы, а виды
несколько более высокогорные—Т. fosteriana, Т. kaufmanniana, Т. sub-
praestans, Т. dubia—в нижнем правом углу. Что же отличает в данном
случае первые виды от вторых? Виды из пустынь и низкогорья про-
ходят более короткий срок от посева семян до прорастания первого
239
to
Интенсивность прорастания семян диких видов тюльпанов при проращивании
их в холодильнике
Таблица 5
Растение Июль Август Сентябрь Октябрь
30 5 10 15 20 25 30 5 10 15 20 25 30 5 10 15 20 25 30 5 10 15 20 25 30
Т. regelii 3,1 — 3,1 6,2 9,3 15,6 28 15,6 9,3 — 3,1 3,1 — — 3,1 — — — — — — — — — —
Т. alberti 2,2 4,4 8,8 13,6 13,6 15,8 31,2 6,6 6,6 6,6
Т. tubergeniana — 2,2 15,0 17,0 24,0 16,5 11,6 3,3 2,5 1,5 1,3 1,4 — 0,8 0,7 — 1,3 1,3 — — — — —
Т. ingens — 2,8 — 2,8 2,8 — 2,8 8,3 16,6 — 8,3 16,6 — 14,0 — 2,8 — 2,8 — — 5,6
Т. wilsoniana — 0,5 3,3 2,3 1,9 3,75 4,7 3,75 3,76 19,7 15,5 13,1 3,3 15,5 5,6 6,1 2,5 — 0,9 — 0,5 — 0,9 — —
Т. kuschkensis — — 7,1 13,6 22,4 14,4 8,2 8,0 7,2 10,4 4,8 — — 3,2 — — — — — — — — — —
Т. micheliana — — 0,7 1,4 3,5 4,9 11,9 4,9 7,0 8,7 7,7 7,0 2,8 1,4 4,2 7,0 1,7 4,2 6,3 6,8 1,4 — — — 1,1
Т. greigii — — — — — 1,7 8,6 6,9 6,9 12 3,4 5,2 6,9 8,6 6,9 5,2 1,7 1,7 3,4 3,4 5,2 6 9 3,4 1,7
Т. hoogiana — — — 0,9 2,6 7,1 15,1 2,5 5,0 7,1 18,7 10,7 2,6 1,8 3,6 — 0,9 — — —
Т. vvedenskyi — — — — — — 0,5 1,5 7,8 9,9 4,0 4,6 7,8 9,3 3,6 1,0 4,1 3,1 1,5 — — 0,5 — —
Т. fosteriana — 1,5 1,5 2,2 4,3 1,5 3,7 2,2 1,5 1,5 2,2 2,9 0,75 0,75 2,2 4,3
Т. kaufmanni- ana 1,0 1,0 2,0 8,0 10,0 7,0 3,0 1,0 3,0 — — 1,0
Т. subpraeslans — — — — — — — — — — — — 1,2 — — 1,2 6,2 6,2 9,9 13,6 12,3 6,2 2,4 1,2
Т. maximowi- czii — — — — — — — — — — — — — — — — 2,0 — 2,0 — 6,0 — 3,0
16 -2229 241
Растение Ноябрь
5 10 15 20 25 30 5
Т. regelii — — — — — — —•
Т. alberti — —• — — — —• —
Т. tubergeniana — — — — — — —
Т. ingens — — — 5,6 — — 5,6
Т, wilsoniana — — — — — — —
Т. kuschkensis — — —• — — — —
Т. micheliana 1,4 1,4 1,4 — 1,4 —
Т. greigii — — — — — — —
Т. hoogiana — — — — — — -
Т. vvedenskyi — — 1 — — 1,0 10,2
Т. fosteriana 8,6 13,2 5,1 2,9 0,75 0,75 1,5
Т. kaufmanni- ana 2,0 6,0 11,0 3,0 11,0 — 1,0
Т. subpracstans 1,2 2,4 3,7 6,2 3,7 24,0 1,2
Т. maximowi- czii — 3,0 3,0 8,0 9,0 12,0 20,0
Продолжение таблицы b
Декабрь Январь Февраль
10 15 20 25 30 5 10 15 20 25 30 5 10 15 20
— — — — — — — — — — — — — — —
— — — — — — — — — — — — — —
— — — — — — — — — — — — — —
—
0,7 — — — — — — — — — — — — — —
— — — — — — — — — — — — — —
10,2 20 — — — — — — — — — — — — —
1,5 2,2 4,3 6,6 8,6 4,3 2,9 1,5 1,5 — — — — — —
3,0 6,0 12 9,0 1 ,о —- —
— 1 Л 1,2 1,2 2,4 1,2 1,2 1,2 1,2 1,2 1,2 1,2 1,2 2,4 2,4
14 7 6 5,0 4,0 3,0 2,0 4,0 5,0 2,0 — — — —
семени и процесс прорастания протекает в более сжатые сроки (рис.
29, 30, 31). Исключением является Т. ingens. У видов с сильно задер-
жанным началом прорастания (табл. 5 и рис. 29, 30, 31)—Т. fosteriana,
Т. kaufmanniana, Т. kolpakovskiana, Т. dubia, Т. praestans и Т. hissari-
ca—семена всходят вяло и в значительно удлиненные сроки.
По данным табл. 6 мы сравнили виды тюльпанов, ареалы которых
в какой-то части накладываются друг на друга, а экология не слиш-
ком различается. Семена таких видов имеют сходный тип прорастания,
Таблица 6
Прорастание семян тюльпанов
Вид Дата Количе- ство дней от посева до про- растания Дата конца прора- стания Количе- ство дней от посева до конца прора- стания Длитель- ность прора- стания
посе- ва, июнь начала прора- стания
Секция Tulipanum Т. tubergeniana 8 7.VII 30 7.IX 95 65
т. kuschkensis 8 7.VII 30 10. IX 94 64
Т. hoogiana 8 5. VIII 58 9.Х 123 65
Секция Leiostemones Т. ingens 8 3.VII 26 З.ХП 178 152
Т. fosteriana 30 19.IX 81 13.11 229 147
Т. praestans 8 28.Х 142 13.11 250 108
Т. subpraestans 8. 29.VIII 82 13.11 250 168
Т. alberti 8 7.VII 30 28. IX 112 82
Т. greigii (Каранкульсай) 30 5.VIII 36 23.1 207 161
Т. micheliana 8 7.VII 30 9.XII 183 153
Т. wilsoniana 30 21. VII 22 6.XII 159 137
Т. maximowiczii 30 25. IX 87 13.11 228 141
Т. korolkovii — — 42 —• 76 34
Т. kolpakovskiana 8. VI 19.IX 103 13.11 250 147
Т. vvedenskyi 8. 5.VIII 58 12.1 218 160
Т. dubia 30 28.IX 90 13.11 228 138
Т. hissarica 30 13.11 229 — — —
Т. Секция Spiranthera kaufmanniana 30 14 IX 76 13.и 228 152
Т. Секция Lophophyllon regelii 8 29.VI 21 9.IX 93 72
Т. Секция Eriostemones turkestanica 54 118 64
Т. bifloriformis — — 40 — 76 36
хотя систематически могут быть очень далеки друг от друга— Т. al-
berti из секции Leiostemones и Т. regelii из монотипной секции Lop-
hophyllon. Луковицы обоих видов получены из окрестностей станции
242
Чокпар Туркестано-Сибирской ж. д. Т. alberti обитает на осыпях, а
Т. regelii на каменистых мелкоземистых склонах. Они были одновре-
менно высажены на соседние гряды на старой территории Ботаниче-
ского сада АН УзССР. Мы собрали с них семена и проращивали. У
Т. regelii период от посева до прорастания семян длится 21 день, у
Т. alberti—30. Растянутость прорастания от первого наклюнувшегося
семечка до последнего у первого—72 дня, а у второго—82.
Семена Т. regelii раньше всех прорастают и заканчивают рост.
Т. regelii по морфологическим и биологическим признакам ближе к
видам секции Eriostemones, чем Tulipanum или Leiostemones.
По характеру прорастания семян близки также Т. greigii Т. korol-
Lovii из одной секции, но морфологически очень далекие виды. В
окрестностях станции Дарбаза Ташкентской ж. д. Т. greigii растет на
глубоких, часто сильно гипсированных, сероземах, а Т. korolkovii чаще
всего—на прикрытых песком карнизах из песчаника или на песчаных
склонах холмов. Семенам Т. greigii необходимо 40 холодных дней
ют посева до прорастания первого семени, Т. korolkovii —42—24 дня.
У Т. greigii проходит 57 дней от посева до конца прорастания, а у
Т. korolkovii—41—76 дней.
Всего от начавшего прорастать первого семени до последнего для
-семян тюльпанов из окрестностей станции Дарбаза Т. greigii нужно
17 дней, а Т. korolkovii от 17 до 34 дней. На весь срок от посева до
конца прорастания семян у Т. greigii необходимо 57 дней, а семенам
Т. korolkovii—17, в другом случае 76. У этих видов секции минималь-
ная растянутость прорастания семян. Однако следует отметить, что мы ис-
следовали в этом отношении относительно небольшое количество видов.
Семенам Т. greigii со склонов Каранкульсая в долине Чирчика от
посева до начала прорастания требуется 36 дней, а до прорастания
всех семян уже не 57, а 207 дней, т. е. прорастание семян длится не
17, а 161 день.
Луковицы всех растений, с которых мы собрали семена для
проращивания, были выкопаны и выращены на грядах в Ботаническом
саду в Ташкенте.
Характер прорастания семян Т. greigii из окрестностей ст. Дарбаза
и Каранкульсая (рис. 30) различен. Насколько первая дает резкий
скачок и "быстрый спад, настолько вторая имеет вялый и постепенный
ход. Здесь ярко проявляется влияние внешних условий на создание
довольно устойчивых требований растений к этим условиям. Один
произрастает в глинистой полупустыне с осадками 360 мм в год; дру-
гой в невысоких горах (до 800—1200 м над ур. м.) с осадками до
600 мм. В этих условиях наблюдаются различные годовые и суточные
температуры.
В результате длительного развития поколений в неодинаковых
условиях наблюдается глубокое различие при проращивании семян
растений, росших после переноса луковиц несколько лет в одинаковых
условиях. Внешняя среда в процессе ближайшего исторического раз-
вития многих поколений оказала большое влияние на наследственность
организмов, вследствие чего получились различные типы прорастания
семян в пределах одного вида, сложившиеся под влиянием внешних
условий и закрепившиеся наследственно. Однако кривая прораста-
ния семян у Т. greigii из Каранкульсая по характеру сходна с кривой
Т. micheliana (рис. 30) из Аманкутана (Самаркандские горы). Оба
вида произрастают в одинаковых условиях. Следовательно, близкое
систематическое родство при сходстве экологических условий обусло-
вило сходство кривых прорастания семян. Другой пример близкого
243
систематического родства и относительной однотипности в прорастании,
семян можно наблюдать у видов Т. tubergeniana, Т. kuschkensis, Т. ho-
ogiana секции Tulipanum, встречающихся в различных районах Сред-
ней Азии с довольно сходной экологией (табл. 5, 6, 7 и рис. 29).
Сходство кривых прораста-
ния семян и относительная
близость по всем трем сро-
кам характеризует не
только сходство их эколо-
гии, но и их филогенети-
ческую близость. Причем,
Т. kuschkensis ближе Т. tu-
bergeniana, чем Т. hoogiana.
Но кривые прорастания
семян у этих видов очень
близки (табл. 7).
Т. ingens по развитию
проростка, ювенильному
возрасту, опушению чешуй,
является связующим звеном
между видами секции Tuli-
panum и Leiostemones, близ-
кими кТ. fosteriana.
Семена Т. ingens начи-
Рис. 29. Ход прорастания семян у тюльпанов
из секции Tulipanum.
а—Т. hoogiana, 6—Т. kuschkensis, в—Т. tubergeniana.
нают проростать вскоре после посева, даже быстрее, чем семена видов
из секции Tulipanum, а прорастание проходит более вяло, почти так же^
Таблица 7
Продолжительность прорастания семян тюльпанов в днях
Вид Начало Вид конец Вид Растяну-] тость I
Т. regelii 21 Т. regelii 93 Т. kuschkensis 64
Т. wilsoniana 22 Т. kuschkensis 94 Т. tubergeniana] 65
Т. ingens 26 Т. tubergeniana 95 Т. hoogiana 65
Т. kuschkensis 30 Т. alberti 112 Т. regelii 72
Т. tubergeniana 30 Т. hoogiana 123 Т. alberti 82
Т. alberti 30 Т. wilsoniana 159 Т. praestans 108
Т. micheliana 30 Т. ingens 178 Т. wilsoniana 137
Т. greigii 36 Т. micheliana 183 Т. dubia 138-
Т. hoogiana 58 Т. greigii 207 Т. maximowiczii 141
Т. vvedenskyi 58 Т. vvedenskyi 218 Т. fosteriana 147
Т. kaufmanniana 76 Т. kaufmanniana| 228 Т. kolpakovskiana 147
Т. fosteriana 81 Т. fosteriana 228 Т. ingens 152
Т. subpraestans 82 Т. maximowiczii 228 Т. kaufmanniana 152
Т. maximowiczii 87 Т. dubia 228 Т. micheliana 153-
Т. dubia 90 Т. kolpakovskiana 250 Т. vvedenskyi 161
Т. kolpakovskiana 103 Т. praestans 250 Т. greigii 161
Т. praestans 142 | Т. subpraestans 250 Т. subpraestans 168,-
Т. hissarica 228
244
'хак у семян видов секции Leiostemones, близких к Т. fosteriana
*(рис. 30).
В 1952 г. мы исследовали всхожесть семян Т. kolpakovskiana из
'Талгара, росшего несколько десятков лет в Ботаническом саду Таш-
кента. Т. kolpakovskiana в естественных условиях растет в предгорь-
ях, но тип прорастания семян его более свойствен горным видам.
Однако температура ниже, а количество осадков в предгорьях Заилий-
ского Алатау больше, чем в предгорьях Западного Тянь-Шаня в бас-
сейне р. Чирчик.
д
О 10 20 30 10 20 30 10 20 30 10 20 30 10 20 30 Ю 20 30
е
в 10 20 30 10 20 30 10 20 30 Ю 20 30 Ю 20 30 10 20 W 10 20 30 Ю 20
Л О ГО hi
Рис. 30. Ход прорастания семян у тюльпанов из секции Leiostemones в холодиль-
нике.
с—Т. alberti, <5—T. greigii, е—T. micheliana, г—Т. subpraestans, д— Т. ingens, е—Т. fosteriana.
Таким образом, экология Т. kolpakovskiana из Заилийского Алатау
соответствует экологии более высоких районов Западного Тянь-Шаня,
не говоря уже о предгорьях Копет-Дага или Памиро-Алая характер
прорастания семян у этого вида вполне соответствует его экологии/
При исследовании было выделено два вида Т. wilsoniana и Т. maxi-
mowiczii. Семена Т. wilsoniana начинают прорастать первыми. Через
22 дня после посева в холодильнике наклевываются первые всходы
(табл. 7). Прорастание же растянуто на 137 дней. У Т. maximowiczii
лервые всходы появляются через 87 дней, весь процесс прорастания—
245
141 день, почти 5 месяцев (табл. 6 и 7). Процесс прорастания семян
Т. maximowiczii и Т. wilsoniana имеет незначительное сходство (рис. 31).
Однако кривая Т. wilsoniana в такой же степени сходна с кривой
Т. korolkovii, хотя последняя значительно компактнее. Как Т. vilso-
niana, так и Т. maximowiczii не имеют близких родичей на территории
Средней Азии среди представителей рода Tulipa за исключением Т. li-
о ' io 'го ' зо ‘ io ' 20 30 ip' го' зо 10 20 30 1 io 'го зо io ' го зо' io ' 20' зо
5
в ю го зо 'io ‘го *~зо io ' го' зо' io 20 зГ '10^20
зо
10 20 30 10 20 30
О 10 20 30 10 20 30 10
. о io ' го^зо ' io ‘го 'за'io 20 ' зо 'io '20^30^10^?о
Рис. 31. Ход прорастания семян у тюльпанов из секции Leiostemones и Spiranthera
в холодильнике.
а—Т. kaufmanniana, б—Т. wilscniana, в—vvedensky i, Т. korolkovii, d—Т. maximowiczi.
S
nifolia, семена которого мы не проращивали. Их ближайшие родичи на-
ходятся в Иране и Афганистане. Особенности прорастания семян
Т. wilsoniana и Т. maximowiczii объясняются наследственностью, чуж-
дой большинству исследованных нами видов.
На примере Т. korolkovii мы убедились, насколько велико влия-
ние экологических условий на ход прорастания семян.
В табл. 6 виды тюльпанов размещены в соответствии с их рас-
положением во „Флоре СССР“. Для наших целей подобное распо-
ложение, возможно, в связи с малым количеством исследованных видов,
не дает целостной картины ни по вопросам филогенетического, ни
экологического родства. В табл. 7 виды расположены в соответствии
со сроками начала, конца и энергии прорастания. Каждый из трех
столбцов этой таблицы в той или иной степени подтверждает выше-
описанные взаимоотношения между видами.
В 1953 г. кроме перечисленных выше семян диких видов, мы
впервые проращивали семена пяти сортов культурных тюльпанов —
Van der Erden, Mothers Day, King George V, Thomas Moore и Elephant.
Исследованные сорта относились к четырем группам садовых тюльпанов
Van der Erden — к группе Менделя; Thomas Moore — к простым ран-
ним; Mother's Day — к группе коттеджей; a King George Y и Eleph-
ant—к Дарвиновским. Начало прорастания семян в холодильнике при
низких температурах у этих видов и сортов колеблется от 42 до 58 дней.
Разница в сроках начала прорастания колеблется в пределах 15 дней.
246
Быстрее прорастают семена сорта Van der Erden, медленнее — Thomas
Moore, который заканчивает прорастание вперед начавшего прорастать
сорта Van der Erden, т. е. имеет большую энергию прорастания. По-
следовательно кончают прорастание Mother's Day, King George V и
Thomas Moore. В основном прорастание семян у всех перечисленных
сортов тюльпанов укладывается в сжатые и близкие между собой
сроки. И лишь у сортов более ранних тюльпанов — Van der Erden и
Thomas Moore позже прорастания основной массы семян, наклевы-
вается небольшая часть семян, задержавшая прорастание у первого—
на два, а у второго на три с прловиной месяца. Эти семена несколь-
ко сближают сорта ранних тюльпанов с дикими видами и свидетель-
ствуют о двойственной природе данных сортов. По-видимому, среди
предков этих гибридов были дикие виды.
Процесс прорастания семян у сортов Van der Erden и Thomas
Moore протекает почти одинаково (рис. 32). Это, вероятно, связано с
тем, что оба сорта относятся к группам ранних тюльпанов, которые
происходят от Т. suaveolens. Прорастание семян у Mother's Day и
King George V более сходно между собой (рис. 32), чем Van der
О 10 20 30 Ю 20 30 Ю 20 ,30 10 20 30 Ю 20 30 10 20 30
о. да го- за да го зо да го зо ю го зо ю го зо 1& го зо
о да го зо ю го зо да го зо да го зо да го зо ю го зо
о да го зо ю го л ю го 36 го го зо ia го зо ю го зо ю го зо да га зо
Лоты
Рис. 32. Ход прорастания семян у культурных тюльпанов.
а—Van der Erden, б— Mothers Day, e—King George V, z—Thomos Moore.
Erden и Thomas Moore. Между Mother's Day и King George V суще-
ствует некоторое филогенетическое родство, так как происхождение
обоих связано с Т. gesneriana.
Проращивание недозрелых семян. В 1953 г. мы проращивали
недозрелые семена тюльпанов до перехода их роговидного эндоспер-
ма в твердое состояние из следующих районов.
На участке Ботанического сада АН УзССР в Ташкенте среди
растений Т. greigii из Каранкульсая выбрали зеленую коробочку с
зелеными створками, в которой лежали начавшие желтеть семена
размером h = 9,19 лиг и d = 7,69 мм. Микропилярный конец семени
и путь сосудистого пучка до халазы были еще зелеными. На верши-
не гребня горы у перевала в Ташкентский заповедник у кишлака
Сангинак Паркентского района собрали зеленые коробочки Т. greigii
Семена были крупные, в среднем h = 8,98 мм, d = 6,97 мм, но совсем
мягкие и свободно прокалывались острым концом пинцета. С участка
247
Ботанического сала АН УзССР в Ташкенте из зеленой коробочки
взяли семена сорта Kathleen Parlow. Там же собрали семена из зеле-
ных коробочек сорта Elephant. Контроль — сухие семена Elephant,
там же, но сбора 1951 г. Семена сорта Elephant из той же коробочки,
что и зеленые, но только подсушенные в половине створок на столе
посеяли 3 августа 1953 г.
Семена Т. greigii из Сангинака в холодильнике долго лежали
зелеными, затем начинали желтеть. На их поверхности начали выде-
ляться какие-то белые кристаллы, что не наблюдалось ни на каких
семенах тюльпанов, посеянных сухими и зрелыми. К 10 августа семе-
на потемнели. В некоторых стал просматриваться крупный зародыш,
принимавший постепенно характерную изогнутую форму. Но позже
на семенах появились какие-то темные пятна (заболевание?) и к
28 августа все семена сгнили.
Семена Т. greigii, начавшие перед посевом желтеть, но с зеленой
халазой и микропилярным концом, поставленные в холодильник 13 ию-
ня, начали прорастать 5 августа, т. е. через 53 дня после посева, на
17 дней позже чем сухие и зрелые семена, происходящие из тех же
мест. По-видимому, это время понадобилось для дозревания семян.
Следовательно, семена тюльпанов могут дозревать и при низких тем-
пературах. Однако до начала прорастания половина семян сгнила, а
до конца прорастания (25 января) также выбросили значительную
часть испортившихся семян.
Семена Kathleen Parlow ко времени посева были зелеными. Раз-
мер их h =3,5; d = 7 мм. Только к 10 августа у одного зерна про-
свечивал зародыш. Затем все зерна были поражены черной пятнистой
болезнью и к 29 августа все сгнили.
Семена сорта Elephant высеяли из зеленой коробочки, взятой с
растения со времени цветения которого, так же, как и у сорта Kath-
leen Parlow, прошло не менее 54 дней. Размеры семян h = 12,95,
d = 8,65 мм. В них за все время проращивания мы не наблюдали
ни одного увеличенного зародыша. К концу августа семена все
сгнили.
Посев семян сорта Elephant 3 августа 1954 г. из половинки той
же коробочки, что и в предыдущем опыте. С 13 июня до времени
посева (3 августа) она лежала на столе в Лаборатории и высохла.
Эти семена 14 сентября дали первые всходы. Таким образом, в дан-
ном опыте прошло всего 30 дней от посева до появления первых
всходов. Отмечаем, что посев недозрелых, но подсушенных семян
ярче выявляет их разнокачественность. В данном случае проявляется
новое свойство растений — у них появляется большая разнокачествен-
ность. Одним из них для прорастания нужно всего 30 холодных дней,
а другим не менее 160, т. е. семена растений, попавших в конце веге-
тации в засуху, получают свойство растянуть прорастание на ряд лет.
Следовательно, условия созревания семян безусловно влияют на
течение последующего прорастания. Это свойство семян тюльпанов, а
возможно, и многих других растений, является биологически полез-
ным свойством.
ПРОРАСТАНИЕ СЕМЯН ТЮЛЬПАНОВ, ПОДВЕРГНУТЫХ ВОЗДЕЙСТВИЮ
УЛЬТРАЗВУКОМ
Способность семян в естественных условиях прорастать только
при низких температурах и необычайная растянутость прорастания
является хорошим приспособлением к специфике климата с осадками
248
в наиболее холодное время года. Это давний признак, ставший исто-
рически сложившейся необходимостью в развитии тюльпанов. Однако
при выращивании тюльпанов в культуре из семян, которым в наших
условиях не хватает количества холодных дней и поэтому они всходят
через две, а иногда и три зимы, необходимо нарушить их консерватизм
и добиться более быстрого прорастания.
В 1952 г. руководитель одной из лабораторий Института им. Мен-
делеева в Москве Н. Н. Долгополов предложил нам в целях ускоре-
ния прорастаниия подвергнуть воздействию ультразвуком семена двух
видов тюльпанов Т. fosteriana и Т. kaufmanniana.
Помещенные в химические стаканы семена заливали водой ком-
натной температуры и обрабатывали ультразвуком в течение 1 мин.
30 сек.; 1 мин. и 30 сек. при частоте колебаний в 30000 к. г. у. При
этом для Т. fosteriana были взяты два первых варианта, а для Т. kauf-
manniana — все три. Контролем служили не озвученные семена сбора
того же года и тех же образцов. В 1952 г., получив пакеты с озву-
ченными семенами, мы не могли поставить их на проращивание в
холодильник, а проращивание в чашках Петри на влажной фильтро-
вальной бумаге в условиях Ташкента летом не дало положительного
результата. В 1953 г. 30 июля мы поставили в холодильник чашки с
озвученными семенами и контроль к ним.
Чем дольше подвергались семена обоих видов воздействию ультра-
звука, тем быстрее они начали прорастать (табл. 8). Так, у Т. kauf-
.manniana при экспозиции в 1 минуту 30 секунд они начали прорастать
Таблица 8
Длительность прорастания семян тюльпанов, подвергавшихся действию
ультразвука при частоте колебаний 30000 в 1 сек.
Вид Длитель- ность экспозиции Количество дней от Растя- нутость прорас- тания Пророс- шие семена, %
мин. сек. посева до начала прорас- тания начала до конца прорас- тания
т. fosteriana 1 30 76 158 82 81,40
т. fosteriana 1 0 90 129 40 89,00
т. fosteriana (конт- роль) 0 0 81 144 63 87,00
т. kaufmanniana 1 30 41 168 127 36,70
т. kaufmanniana 1 0 54 164 НО 68,75
т. kaufmanniana 0 30 87 141 54 90,40
т. kaufmanniana (кон- троль) 0 0 87 141 54 98,80
через 41 день, одну минуту—через 54 дня, а в 30 секунд так же,
как и в контроле, через 87 дней. В данном опыте экспозиция в 30 се-
кунд не дала никакого эффекта.
При ускорении начала прорастания с увеличением продолжитель-
ности экспозиции растянулся период прорастания семян, т. е. здесь
так же, как и в опытах с семенами из срезанных за две недели до
созревания и затем подсушенных коробочек — проявилась большая
разнокачественность семян.
С увеличением продолжительности экспозиции количество пророс-
ших семян падало (табл. 8).
249
Почти то же самое мы видим у семян Т. fosteriana. Разница
только в том, что при воздействии ультразвуком в течении 30 сек. у
этого вида несколько затягивается (на 9 дней) начало прорастания
семян, сокращается растянутость прорастания (на 13 дней против конт-
роля) и с 87 до 89 — повышается процент проросших семян.
На основании наших данных и опыта Е. Л. Рубан и И. А. Кома-
рова по озвучиванию семян жимолости и бирючины можно сделать-
вывод, что ультразвук можно использовать как фактор, в какой-то сте-
пени ускоряющий прорастание весьма разнообразных семян. Практи-
ческое применение ультразвука для этой цели должно быть специально-
разработано для каждого вида или группы биологически близких,
видов. Кроме того, необходимо проследить развитие растений, выра-
щиваемых из озвученных семян, так как возможны нарушения обще-
го развития, а также проявление всевозможных уродств, особенно
при применении очень большой частоты колебаний или повышенной
длительности экспозиции.
А. В. Попцов (1954) высказывает предположение, что в семенах
во время стратификации, происходит процесс прорастания, а не после
уборочного дозревания.
Все семена, нуждающиеся в стратификации, относятся к типу
семян, обладающих низким температурным оптимумом прорастания..
Необходимо выяснить, находится ли зародыш в состоянии покоя или
затрудненность прорастания обусловливается другими причинами.
Существует мнение, что большое влияние на прорастание семян ока-
зывают семенные покровы, эндосперм и перисперм.
Многие исследователи получили прорастание зародышей, выде-
ленных из нестратифицированных семян и на основании этого они
пришли к выводу, что зародыши семян не находятся в покое и в пос-
леуборочном дозревании не нуждаются. Другие указывают, что про-
растают далеко не все зародыши, выделенные из семени. При нару-
шении покровов семени и эндосперма в области корешка зародыша
Николаева (1950) наблюдала ускорение прорастания; но неизвестно
как протекало бы дальнейшее развитие растений. Иногда зародыш не
способен к быстрому прорастанию и тогда прорастание проходит
очень медленно, растения плохо растут и поздно вступают в плодо-
ношение (Радионов, 1950).
Наблюдения Николаевой показали, что затрудненность продвиже-
ния воды в зародышах, видимо, связана с недостаточной проницае-
мостью их тканей для воды.
Покровы семени, эндосперм, перисперм, плотно окружающие заро-
дыш, тормозят его прорастание. А. В. Попцов (1954) делает вполне
обоснованный вывод, что это торможение не является чисто механи-
ческим, но представляет препятствие физиологическое, выражающееся
в затруднении газообмена и дыхания. Зародыш с недостаточно прони-
цаемыми тканями, также может осложнить дыхание.
Выделенные из нестратифицированных семян зародыши, как пра-
вило, прорастают и при более высоких температурах, и низкие тем-
пературные оптимумы стратификации не являются лучшими при про-
ращивании выделенных из семян зародышей.
А. В. Попцов считает, что основные моменты, позволяющие до
известной степени понять, почему какие-либо семена могут прорастать
только при определенной и при том низкой температуре, следующие:
1) необходимый для дыхания кислород поступает в семя только в
растворенном в воде виде; 2) растворимость кислорода в воде с пони-
жением температуры увеличивается; 3) интенсивность дыхания (а*
250
следовательно, потребность зародыша в кислороде) с понижением
температуры значительно уменьшается (термический коэффициент
дыхания равен 2—3); 4) хотя проницаемость тканей также уменьша-
ется, но в значительно меньшей степени, так как эта проницаемость
связана с процессами диффузии и осмоса, имеющими термический
коэффициент 1,1 —1,3.
Таким образом, потребность зародыша в кислороде, по мере сни-
жения температуры уменьшается в значительно большей степени, чем
проницаемость тканей семени, окружающих зародыш. В результате
этого определенное сочетание условий может обеспечить доступ к
зародышу достаточного количества кислорода для начала его жизне-
деятельности.
В классификации жизненных форм Е. И. Проскоряков (1952)
принял систему Раункиера. Признавая, что эта система является весь-
ма искусственной и соглашаясь с ее критикой, М. В. Культиасов (1950)
пишет, что в настоящее время нет никакой другой системы жизнен-
ных форм, которую можно было бы использовать в работе. А умело
примененная система Раункиера, дает хорошие результаты. Пользуясь
этой системой он помещает тюльпаны в „геоксерофитные эфемероиды*
и дает им следующую характеристику: „Геоксерофитные эфемероиды—
хорошо очерченная в биологическом отношении жизненная форма
Средиземья — независимо от систематического положения и морфоло-
гической выраженности, обладают семенами, имеющими оптимум про-
растания при низкой температуре, что является хорошим приспособ-
лением к специфике климата, так как позволяет растению лучше ис-
пользовать небольшое количество выпадающих здесь осадков44.
Таким образом, одни семена в природе при повторяющихся от-
тепелях, засухах и морозах ускоряют прорастание, а другие, наоборот,
тормозят, обеспечивая большую растянутость прорастания и возмож-
ность появления части всходов лишь через год.
Приспособленность семян к изменению особенностей прорастания
в разные года с различным температурным режимом способствует со-
хранению вида.
Темпы прорастания семян, количество всходов весной определяет-
ся всем ходом погодных условий данного года.
Из опытов по проращиванию недозрелых семян мы выяснили, что
зеленые семена тюльпанов, снятые за две недели до созревания,
имеющие мягкий эндосперм — не прорастают. Семена, начавшие жел-
теть прорастают на 50%.
Снятые с растений и подсушенные зеленые семена прорастают,
несколько ускоряя начало прорастания, но сильно его растягивая.
Семена, захваченные в естественных условиях засухой в конце веге-
тации, получают свойство растянуть прорастание на много лет — в них
ясно выявляется разнокачественность.
Виды из пустыни или низкогорья (Т. regelii, Т. alberti, Т. kusch-
kensis и Т. tubergeniana) имеют более короткий срок от посева до
начала прорастания семян, да и само прорастание проходит у них
более интенсивно и в короткий срок.
У видов с задержанным началом прорастания (Т. fosteriana. Т. kauf-
manniana, Т. kolpakovskiana, Т. dubia, Т. praestens и Т. hissarica) —
оно протекает более вяло и в значительно удлиненные сроки.
Виды, с налегающими ареалами и близкой экологией имеют сход-
ный тип прорастания (Т. regelii, Т. alberti; Т. korolkovii, Т. greigii).
251
Семена с особей одного вида, взятые из различных мест ареала,
хотя и не очень отдаленных, но с разной экологией, резко от-
личаются между собой по характеру прорастания и эти различия за-
креплены у них наследственно (Т. greigii из Капланбека и Коранкуль-
сая по долине р. Чирчика выше ст. Барража). Филогенетическая
близость также находит отражение в характере прорастания семян.
Из данных настоящего исследования по прорастанию семян вытекает,
что исследованные виды секции Tulipanum имеют сходный тип про-
растания семян, а так же, что Т. kuschkensis из этой секции значи-
тельно ближе стоит к Т. tubergeniana, чем к Т. hoogiana; что Т. in-
gens занимает промежуточное положение между видами, относящи-
мися к секции Tulipanum и видами из секции Leiostemones, группи-
рующихся вокруг Т. fosteriana.
Глава VIII
РОСТ И РАЗВИТИЕ СЕЯНЦЕВ
роростки. Растения просмотренных нами 17 видов и многих сортов?
тюльпанов в первый год жизни имеют всего один главный корень.
Только однажды мы встретили проросток первого года жизни.
Т. bifloriformis с разветвленным корнем. В конце периода вегетации
первого года жизни растения главный корень отмирает.
В 1953 г., проращивая семена
тюльпанов во влажной камере в
холодильнике, через три недели
после наклевывания семян мы
обнаружили у проростков Т. wi-
lsoniana Т. inqens, Т. maximo-
wiczii и Т. dubia корневые волос-
ки длиной до 3 мм (рис. 33). У
сеянцев других видов этого рода
корневых волосков мы не наблю-
дали. На корнях тюльпанов более
старшего возраста также нет кор-
невых волосков. Вместо отмер-
шего главного корня в донце
луковицы в конце периода жаро-
покоя первого года жизни зак-
ладываются придаточные корни.
На второй год жизни у различ-
ных видов и сортов отмечено
неодинаковое количество совер-
шенно не ветвящихся придаточ-
ных корней.
В условиях Ташкента всходы
тюльпанов появляются весной
при наступлении в середине дня
первых положительных темпера-
тур. Этот срок колеблется в пре-
делах месяца—с конца первой
декады февраля до первой дека-
ды марта. При ранних всходах,
когда по ночам значительно па-
Рис. 33. Корневые волоски на корешках
у проростков тюльпанов.
а—Т. ingens, б—Т. wilsoniana, в—T. maximowiczii,
г—Т. dubia.
дает температура, они долго не
поверхности почвы, а затем стоят
зеленеют и вначале лежат на'
ярко окрашенные антоцианом.
253-
Но когда бы тюльпаны не взошли, при наступлении в апреле высоких
температур, обычно в конце второй половины месяца, они оканчивают
вегетацию. Из выращиваемых на участках Сада сеянцев диких видов,
дальше всех вегетируют сеянцы Т. subraestans.
Растение любого вида или сорта тюльпана в первый год жизни имеет
один семядольный лист. При прорастании семян тюльпанов верхняя бес-
цветная часть семядольного листа долгое время остается в семенной ко-
журе, играя роль гаустории. Эта часть семядоли сплющена, на вершине
тупо закруглена, проходящие в ней сосудистые пучки расположены
не по кругу, как в ассимиляционной части семядоли, а плоско.
Средняя зеленая часть семядольного листа, названная И. Т. Ва-
сильченко (1945) „связником семядоли11, почти цилиндрически округ-
лая в поперечном сечении, к середине более расширенная, постепенно
к вершине и к поверхности почвы суженная—служит ассимиляцион-
ной листовой пластинкой семядоли.
Ассимиляционная часть семядоли, например у Т. greigii (рис. 34)
или у Т. Kaufmanniana, покрыта эпидермисом, прямоугольные клетки
Рис. 34. Эпидермис со средней
части связника семядоли ~
greigii.
т.
которого вытянуты в продольном направле-
нии. Наружние стенки эпидермиса сильно
утолщены — покрыты кутикулой. Устьица
на эпидермисе мелкие с бобовидными за-
мыкающими клетками. Их щели также
расположены в продольном направлении.
На поперечном срезе (рис. 35, 36)под эпи-
дермисом находится крупноклетная рыхлая
не дифференцированная паренхимная ткань,
в которой очень развиты воздухоносные
полости (рис. 35,я). Сосудистые пучки, глу-
боко погруженные в паренхимную ткань,
расположены по кругу (рис. 35,6). У Т. ka-
ufmanniana чаще всего бывает пять или
семь пучков (рис. 36)—три крупных и два
или четыре мелких. У Т. greigii — пять;
у Т. korolkovii — два. Распределение со-
судистых пучков в семядоле
kaufmanniana
самых крупных
расположены в
имеет одну
Т.
ось симметрии. У
(рис. 36, а) три
сосудистых пучка
одной половине семядоли, два мел-
’Ч
Рис. 35. Поперечный срез через среднюю
часть связника семядоли Т. greigii.
а—воздухоносные полости, б—сосудистые пучки.
254
Рис. 36. Поперечный срез через нижнюю
часть связника семяаоли Т. kaufmanniana
а сосудистые пучки, б—краевые сосудистые пучки,
6—верхняя плоскость связника семядоли.
ких—в противоположной половине. Между ними часто лежат два бо-
лее мелких пучка (рис. 36, б). Сосудистые пучки б. м. равномерно
распределены по кругу. Округлая форма семядоли, насколько можно
судить по ее анатомическому строению, образовалась в результате
редукции верхней плоскости семядольного листа.
Об этом свидетельствует расположение ксилемы и флоэмы в со-
судистых пучках семядоли (рис. 36, а), которые повернуты флоэмой
к периферии, а ксилемой внутрь семядоли. Ксилемная часть пучка
представлена всего несколькими кольчатыми сосудами. Флоэма более
развита. Между флоэмой и ксилемой в пучках лежат мелкие, округлые,
тонкостенные клетки. Сосудистые пучки окружены двумя—тремя ря-
дами более мелких, чем остальная часть мезофилла, паренхимных
клеток. Никакой склеренхимной ткани или отдельных склеренхимных
клеток в семядоле нет (рис. 36)
У Т. kaufmanniana со стороны двух самых мелких, рядом лежа-
щих, сосудистых пучков 8—10 рядов клеток эпидермиса образует
слабо углубленный желобок. Этот желобок, видимо, и есть остаток
верхней плоскости листовой пластинки или связника семядоли
(рис. 36, д).
Перед внедрением в почву семядоля становится значительно
тоньше, так как в паренхимной ткани исчезают воздухоносные по-
лости. Под землей семядоля сохраняет округлую форму, безхлоро-
фильна и может быть названа черешком, но вернее, это влагалище.
Первичное углубление верхушечной почки сеянца тюльпана пер-
вого года жизни происходит обычным среди лилейных способом.
Проростки углубляются при помощи геотропически положительного
роста черешковой (влагалищной) части семядольного листа. Она вра-
стает в почву и погружает почку проростка часто на глубину несколь-
ких сантиметров. Углубление проростка тюльпана в первый год жизни
у большинства исследованных видов превышает углубление при по-
мощи столона, который закладывается у тюльпанов всех видов и
сортов.
Недалеко от места прикрепления семядоли к донцу и с противо-
положной стороны от корешка в черешке семядоли имеется сквозное
отверстие влагалища, которое доходит до верхушечной почки стебля.
Отверстие входа во влагалище у Т. kaufmanniana помещается со сто-
роны двух мелких, рядом расположенных сосудистых пучков в па-
ренхимной ткани между ними, т. е. со стороны редуцированной верх-
ней плоскости семядоли.
У Т. fosteriana и Т. kaufmanniana над входом во влагалище
листа иногда мы наблюдали возвышающийся незначительный вырост
в виде маленького язычка, который состоял из четырех—шести рядов
клеток и напоминал эпибласт злаков.
У Т. greigii, как у большинства других видов тюльпанов, это
отверстие образуется следующим образом: в нижней части черешка
семядоли со стороны ее редуцированной верхней плоскости появляется
желобок, который постепенно углубляется, его края сближаются и,
замыкаясь он входит в черешок, образуя полость влагалища.
Столоны. Со стороны входа во влагалище семядоли от места на
проростке, где должно быть донце, у сеянцев всех видов тюльпанов
отходит особый округлый, внутри полый, в несколько раз толще ко-
решка, но без извитости вырост. Развитие его заметно невооружен-
ным глазом уже в то время, когда у сеянца начинают расти, но еще
не функционируют как органы, семядоля и корешок.
255
Этот вырост не получил в литературе общепризнанного специаль-
ного названия, но неоднократно описывался многими авторами. Ирмиш
(1850), а еще раньше Сент Пьер и Фабр называли его — шпорой
(„schporen"); А. Д. Холл (1929)—„dropper", если вырост углублялся
отвесно вниз, и „stolones"—при горизонтальном росте. Д. Е. Янишев-
ский (1934)—„выводковым выростом", „ризоморфным выростом", „ри-
зоморфным корневищем"; Н. Г. Серебряков (1952) —„луковичными
столонами", „своеобразными столоновидными органами" и „столонами
углубления". Во „Флоре СССР" (т. IV) он назван „подземным побе-
гом".
Рис. 37. Заложение и рост столона
на проростке тюльпана.
а—во время выхода проростка из семени,
в, г—последовательные фазы роста сто-
лона, д—столон в период окончания
роста в длину.
Мы, следуя за знатоками морфологии растений, с полным правом
можем продолжать называть этот вырост столоном.
Морфологически и по происхождению он отличается от столонов
у других высших растений, но по выполняемым функциям—вегетатив-
ное размножение, удаление дочерней луковицы, отдаление ее от мате-
ринского растения и проведение питательных веществ от материнско-
го растения к дочерним луковицам — сходен с ними.
Столон углубления на проростке тюльпана закладывается через
несколько дней после выхода сеянца из покровов семени.
Вначале, в результате одностороннего роста тканей, т. е. первого
узла проростка и разрастания одной стороны влагалища семядоли,
верхушечная почка растения начинает смещаться в сторону отверстия
во влагалище листа. Эта фаза хорошо заметна на проростках тюль-
панов, во время прорастания семени еще до позеленения семядолей,
и на первых этапах роста внешне выражается легкой припухлостью
с одной стороны проростка при основании семядоли (рис. 37, а). За-
тем благодаря интеркалярному росту в стебле и листовом влагалище
образуется все удлиняющийся мешко-
видный вырост (рис. 37, б, в, г), изме-
няющий отрицательную геотропическую
реакцию на положительную и растущий
подобно корню (рис. 37, а). При этом
стебель растет вершиной, а семядоля —
основанием.
Верхушечная почка, смещенная в
столон, в это время представлена зачат-
ком запасающей чешуи (рис. 38) формы
цилиндрического выроста, более мощ-
ного со стороны противоположной семя-
доли (рис. 38, а) и несколько меньше с
прилежащей. Зачаток является влагалищ-
ной частью первого листа с недоразви-
той листовой пластинкой. Этот лист пре-
вращается в первую запасающую чешую
луковицы. Его центральная жилка, обоз-
начающаяся несколько позже со сто-
роны противоположной семядоли (рис.
38, в) показывает, что проросток сохра-
няет тип двурядного листорасположения
это время нет никакого маристематичес-
лилейных. В центре почки в
кого бугорка. Размеры почки остаются почти неизменными во всё
время роста столона в длину.
В первый год жизни сеянцев всех видов тюльпанов столон растет
как побег углубления, играя важную роль в перемещении молодых
луковиц (табл. 9). Так как легкое, крупное и плоское семя тюльпана
256
не имеет никаких приспосоолении для зарывания в почву, семена
тюльпанов обычно прорастают на поверхности почвы или заносятся
ветром в щели. Однако первичное углубление верхушечной почки
Рис. 38. Верхушечная почка проростка
Т. Kaufmanniana, смещенная в столон.
а—спинка зачатка запасающей чешуи,
— брюшная часть зачатка запасающей чешуи,
в—система сосудистых пучков донца.
Рис. 39. Проводящая сосудистая система
проростка Т. kaufmanniana при ее переходе
из листа и корня в столон углубления.
а—крупные проводящие пучки семядоли, б— сосуди-
стые пучки корня, в—сосудистые пучки, проходящие
в столон из донца, г—краевые мелкие пучки семя-
доли, входящие непосредственно в столон.
проростка происходит благодаря положительному геотропическому
росту „ черешка “ семядоли и только в дальнейшем—столона.
Столон углубления тюльпана, как это хорошо показал Янишев-
ский (1934) для взрослых особей Т. schrenkii и Т. biebersteiniana, а
в дальнейшем для многих видов различного возраста Бочанцева (1956),
образован из срастания тканей двух органов — стебля и влагалища
семядоли или листа. Количество сосудистых пучков, входящих из
стебля в столон проростка, колеблется от двух до семи; У Т. korol-
kovii и Т. bifloriformis— два —три; Т. greigii и Т. fosteriana — пять, у
Т. kaufmanniana — пять — семь пучков.
У Т. kaufmanniana три или пять проводящих пучков из семядоли
(три крупных и два мелких, рис. 39, а) сливаются в донце с прово-
дящими пучками корня (рис. 39, б) и проходят в столон (рис. 39, в).
Два же тонких краевых пучка непосредственно из влагалища входят
в столон без слияния с другими пучками (рис. 39, г).
Таким образом, в образовании столона принимает участие не меж-
доузлие, как у двудольных растений, а узел, так как часть листовых
сосудистых пучков входит в стебель на месте донца, а часть, проходя
через весь столон, присоединяется к ним значительно позже только'
у основания почки.
Таким образом, у ювенильных и омоложенных растений тюльпана
наблюдается оригинальное поперечное разрастание узла, достигающее
от 10,5 до 30 и 50 см дл. (у деток Т. fosteriana) при полном отсут-
ствии роста междоузлий.
Над отверстием при входе во влагалище семядоли иногда удается
увидеть небольшой язычковый вырост, подобный эпибласту злаков
(рис. 40, а) очень похожий на рудимент второй семядоли. Но если
согласиться с этим, то расположение спинки первого листа сеянца
17—2229 257
Таблица 9
Средние размеры сеянцев тюльпанов первого года жизни, см
Вид Длина гау- стории Пластинка семядоли Углубление почки че- решком се- мядоли Диаметр черешка Углубление почки сто- лоном Диаметр столона Общее уг- лубление луковицы в Ivoj Hi4ad9u
длина диа- метр
Т. kaufmanniana 0,7 8,5 0,30 4,3 0,10 2,5 0,10 6,8
Т. greigii 0,7 8,0 0,18 3,0 0,10 5,0 0,12 8.0
7. alberti 0,5 4,7 0,10 4,0 0,05 4,0 0,07 8,0
7. micheliana 0,7 6,0 0,16 6,0 0,12 1,5 0,18 7,5
Т. subpraestans 0,7 10,5 0,15 4,0 0,11 4,5 0,14 9,5
Т. praestans 0,6 6,5 0,13 3,6 0,05 2,6 0,05 6,2
7. ostrovskiana 0,4 7,0 0,15 4,0 0,08 3,7 0,16 7,7
7. kolpakovskiana 0,35 6,0 0,12 5,0 0,08 3,5 0,10 8,5
7. schrenkii 0,6 5,0 0,10 4,5 0,03 1,5 0,10 6,0
7. korolkovii 0,5 6,5 0,12 5,0 0,06 3,0 0,16 8,0
7. bifloriformis 0,3 4,5 0.08 5,0 0,05 3.0 0,07 8,0
7. maximowiczii °,4 4,5 0,075 2,5 0,05 1,5 0,07 4,0
7. hoogiana 0,7 6,0 0,15 3,5 0,10 1,5 0,10 5,0
7. ingens 0,6 6,5 0,15 4,0 0,08 4,0 0,80 8,0
7. fosteriana 0,5 7,0 0,30 3,0 0,10 3,0 0,10 6.0
(40, б) со стороны этого „рудимента* нарушает представление о дву-
рядном типе листорасположения у лилейных (табл. 9).
Язычковый вырост состоит всего из 6 рядов клеток, если считать
вместе с эпидермисом внешней и внутренней стороны. В выросте и
стенке столона под ним нет никаких проводящих пучков.
В центре столона, через его длину, с начала отверстия во вла-
галище проходит воздухоносная полость. На дне полости находится
верхушечная почка сеянца, сместившаяся туда благодаря поперечному
разрастанию узла проростка и основания влагалища семядольного
листа. Почка воздухоносной полостью связана с внешним воздухом
(рис. 40, в). Эта полость у многих видов почти от донца до почечки
заполнена длинными неветвящимися волосками. Концы этих волосков
направлены вверх (рис. 41, а).
Растущий столон с наружной стороны покрыт гладким эпидерми-
сом, однако, если вынуть из влажной земли еще не начавший утол-
щаться столон сеянца, то вдоль всей длины он как футляром будет
покрыт приставшими комочками почвы. Свободным от них будет
только растущий кончик столона.
По окончании роста столона в длину на его конце образуется
утолщение, указывающее, что наступил следующий период в росте
столона — начала расти верхушечная почка сеянца (табл. 10).
Рост почки в этот период происходит за счет укрупнения уже
заложенных органов. На месте утолщения столона характер эпидер-
миса меняется, некоторые его клетки становятся бутылковидными и
значительно выступают над соседними, производя впечатление корот-
ких корневых волосков, но покрытых толстыми оболочками (рис. 41,6).
258
Под ними лежит еще один слой клеток с утолщенными наружными
и поперечными стенками. .
Многие исследователи тюльпанов, находившие на концах столо-
нов углубления удлиненные в виде сосочков клетки, предполагали
что это корневые волоски. На самом
же деле эти удлиненные крепкие
клетки появляются на конце сто-
лона, когда он заканчивает рост в
длину и начинает утолщаться. Эти
клетки служат для укрепления лу-
ковицы в почве во время сокраще-
ния столона при высыхании и защи-
ты ее от раздавливания почвой.
Следовательно, столон прохо-
дит три этапа роста: первый — сме-
щение почки сеянцев в сторону,
второй — рост столона в длину, а
третий — утолщение столона, т. е.
образование луковицы из смещен-
ной в столон верхушечной почки
(табл. 11).
С первого дня возникновения
столон углубления сеянца тюльпа-
на является хранилищем запасных
питательных веществ. Крахмал в
клетках столона появляется до по-
зеленения семядоли, т. е. из запа-
сов, отложенных в семени.
Стенки столона с момента его
заложения являются покровом поч-
ки, которая в дальнейшем превра-
тится в луковицу. До окончания ро-
ста в длину почка столона несколь-
ко увеличивается в продольном
направлении, но остается очень уз-
кой. В этот период почка не со-
держит крахмала. Нет крахмала и
Рис. 40. Столон углубления Т. fosteriana
с язычковым выростом.
о—язычковый вырост у влагалища семядоли, в—
спинка первого (запасающего) листа сеянца, б—
возд у хо ноская полость.
Рис. 41. Т. greigii.
а эпидермис, покрывающий полость столона с волосками, на правленными
вверх, б—клетки внешнего эпидермиса с утолщенными стенк а ми на конце
столона во время его утолщения.
259
Таблица 10
Рост столона углубления у тюльпанов первого года жизни
в полевых условиях
Вид Появление столона Окончание роста в длину и начало утол- щения Конец веге- тации Количество дней роста Всего дней вегетации растения
в длину в тол- щину
Т. bifloriformis 28.11 10.III 15.IV 10 36 46
Т. fosteriana 28.11 23. III 18.IV 23 26 49
Т. kaufmanniana 28 II 20.1 II 20.IV 20 31 51
Т. greigii 28.11 26.111 19.IV 26 24 54
в группе клеток, составляющих основание почки — будущее донце,
во внешнем и внутреннем эпидермисе столона; в клетках, окружаю-
щих сосудистую систему, и в продолговатых клетках, из которых
впоследствии развиваются сосуды (в проваскулярных пучках). Осталь-
ные клетки столона, особенно на уровне почечки, густо заполнены
крахмалом. Он лежит долго в одних и тех же клетках столона, а
вместе с его углублением переходит все в новые и новые клетки.
Луковица первого года жизни. Во время утолщения столона у
сеянца тюльпана первого года жизни новые органы не закладываются,
а интенсивно растет первый и единственный низовый лист (сочная
луковичная чешуя), заложившийся в молодой луковице первого года.
Этот лист — запасающий орган луковицы первого года жизни. В се-
редине или конце апреля у сеянпев диких видов в наших условиях
отмирает корешок, засыхает семядоля. Из покровной чешуи луковицы,
образовавшейся из нижней части столона, крахмал перемещается в
запасающую чешую, а покровная чешуя также засыхает. Наступает
период летнего покоя.
Несмотря на то, что растение тюльпана в первый период жизни
имеет только один зеленый орган — семядолю, незначительного, но
различного у разных видов размера (табл. 9) и один единственный
корешок, — оно успевает накопить в луковице необходимые запасы
питательных веществ и воды (табл. 11). Этих запасов достаточно для
того, чтобы однолетняя луковица пережила период жаропокоя или
сухопокоя, заложила органы ассимиляции будущего года и начала
расти в конце периода жаропокоя. Луковицы первого года жизни
тюльпанов не более 180 мг у видов с крупными луковицами и даже
10 мг —у видов с мелкими луковицами (табл. 12).
В середине мая у покоящейся луковицы Т. greigii первого года
жизни под коричневой покровной чешуей наблюдается всего одна
запасающая чешуя, густо заполненная крахмалом. Никаких зачатков
других органов в такой луковице еще нет. В чешуе различаются
два, примерно равных по толщине, слоя: периферический и внутрен-
ний (рис. 42, а). Периферическая половина чешуи густо заполнена
крахмалом. Во внутренней — составляющие ее клетки лежат более
рыхло и крахмала значительно меньше.
В центре запасающей чешуи проходит узкий канал, выстланный
одним слоем тонкостенного, не содержащего крахмала эпидермиса.
При заложении первого зеленого листа эпидермис канала не проры-
вается. Клетки эпидермиса, видимо, сохраняют не только способность
260
Таблица 11
Соотношение роста верхушечной почки и почки Детки столона углубления
Вид Второй год Третий год Четвертый год Пятый год
нали- чие 1 углубле- ние почки глубина зале- гания лукови- цы нали- чие углубле- ние почки глубина зале- гания лукови- цы нали- чие углубле- ние почки глубина зале- гания лукови- цы нали- чие углубле- ние почки глубина зале-! гания лукови- цы
столона ! i детки 1 верху- шечной детки столона детки верху-^ ' шечной детки 1 столона 1 детки верху-^ шечной детки столона 1 детки верху- 1 шечной , детки
Т. fosteriana 4- 4- 3,0 0 9,0 + 4- 15,0 12,0 23,5 4- 4- 9,0 8 25,0 4- + 16 8 21
т. carinata 4- 4- 3,5 1,5 10,5 4- 4- 3,7 3,3 13,0 + 4- 3,5 4 12,5 4- + 29 20 33
т. kaufmanniana — 4” 5,0 0 10,5 4- + 6,5 0 14,0 + + 8,0 0 16,5 — — — — —
т. greigii + + 4,0 0 14,0 4- 4- 7,5 0 16,0 + + 10,0 0 28,0 — — — — —
т. subpraestans 4- 4- 12,7 4 17,0 + 4- 7,7 5,4 13,0 4- 4- 13,0 10 26,0 — — — — —
т. praestans 4- 4- 5,5 0 16,0 + 4- 11,5 0 18,0 4- + 7,0 5 15,5 — — — — —-
т. korolkovii 4- — 3, 0 0 12,0 4- 4- 4,5 1,8 14,5 0 0 0 0 16,5 — — — — —•
т. kolpakovskiana 4- 4- 9,0 0 13,0 + 4- 16,0 2,0 20,0 0 0 0 0 20,0 — — — — —
т. hoogiana 4- 4- 3,0 0 11,0 4- 4- 11,0 0 17,0 3,0 15,0
т. bifloriformis 4” + 2,0 0 9,0 4- 4- 3,4 1,0 11,0 4- + 3 — — —
т. ingens + — 4,0 0 12,0 4- + 11,4 0 20,0 — — — — — —- —
т. ostrovskiana 4- 4- 6,0 0 10,5 4- + 11,0 0 16,5 — —
т. schrenkii + — 4,0 0 15,0 + + 14,5 0 17,5 — — —- —- — — — — —
т. maximowiczii 4- — 3,3 0 15,0 4- 4- 11,0 4- 17,0 — — — — — — — — — —
т. micheliana 4- 4- 10,0 0 20,0 4- +
го
Примечание. После второго и четвертого вегетационных сезонов луковицы
были пересажены на глубину 7—10 см в зависимости от их величины
Таблица 12
Луковицы сеянцев тюльпанов
Вид и сорт Первый год жизни Второй год
вес, мг углубле- ние, % детки, % вес, мг углубление, % детки, %
крупная средняя к СЗ ч CJ крупная средняя мелкая
Т. tubergeniana 130 73 30 100 0 782 612 550 100 5
Т. kuschkensis 125 90 32 100 0 717 580 440 100 6
Т. hoogiana 140 75 35 100 0 985 651 450 100 ЗО
Т. ingens 100 60 25 100 0 625 512 420 99 0
Т. fosteriana 120 77 30 100 0 550 412 250 100 50
Т. praestans 155 100 50 100 0 795 595 230 100 100
Т. subpraestans 160 122 60 100 0 1000 325 80 100 90
Т. alberti 80 66 25 100 0 1220 726 390 94 0
Т. greigii 145 80 24 100 0 1050 220 31 100 16
Т. micheliana 140 100 40 100 0 725 526 360 100 2
Т. eichleri — — — 100 0 460 332 265 73 0
Т. maximowiczii 30 20 10 100 0 200 125 80 100
Т. wilsoniana 45 33 15 100 0 370 293 200 10 8
Т. korolkovii 60 43 20 100 0 370 280 160 31 0
Т. schrenkii 80 62 25 100 0 — — — — 0
Т. ostrovskiana 130 72 20 100 0 770 466 290 98 13
Т. kolpakovskiana 135 90 25 100 0 450 329 106 92 5
Т. dubia 120 94 27 100 0 970 636 380 100 0
Т. vvedenskyi 150 100 40 100 0 1350 984 635 100 500—
100
Т. kaufmanniana 180 104 37 100 0 1900 1160 65 100 20
Т. regelii 54 28 16 100 0 474 134 43 40 32
Т. bifloriformis 27 18 10 100 0 77 43 18
D. V. Thol yellow 80 58 20 100 0 1400 97 35 60 10
Jubilee 77 50 28 100 0 372 167 72 20 27
Elephant 80 55 25 100 0 327 133 15 20 5
Kathleen Parlow 138 104 57 100 0 372 162 59 30 14
к растягиванию, но и к росту оболочек. Судя по внешнему виду,
клетки внутреннего эпидермиса могут даже делиться (рис. 42, б), так
как в них нет никаких признаков старения.
Клетки внутреннего эпидермиса запасающей чешуи луковицы
первого года жизни в мае не вакуолизированы, в их ядрах и плазме
нет признаков отложения каких-либо запасных веществ, разрушения
или отмирания. По строению протопласта они сходны с клубком ме-
ристематических клеток в основании луковицы (рис. 42, б). В толще
клубка меристематических клеток при основании луковицы во второй
половине жаропокоя, примерно с конца июля, начинают закладывать-
ся первые придаточные корешки. У основания щели в запасающей
чешуе полукольцом поднимается зачаток первого зеленого листа а
второго низового — запасающего.
262
Лист. Расположение листьев у тюльпанов розеточно-двурядное.
Главная жилка первого зеленого листа находится со стороны семядо-
ли, так как между ними расположен единственный лист первого года
жизни тюльпана с недоразвитой листовой пластинкой и с функциями
запасающего, а не ассимиляционного органа. Второй запасающий лист
появляется через год, имеет главную жилку со стороны первого запа-
сающего листа, но отделен от него на длину столона углубления.
Таким образом, влагалищные листья тюльпана до закладки более
мощной луковицы из нескольких чешуй в течение всех предшествую-
щих цветению лет находятся с одной стороны луковицы.
Первый зеленый лист долго растет внутри луковицы и поздно
Рис. 42. Продольный разрез луковицы в сере чине мая.
а—общий вид луковицы, б—донце этой луковицы при большом
увеличении (видны живые клетки внутреннего эпидермиса).
осенью выходит из нее, но только весной он показывается над поч-
вой. Этот лист — единственный зеленый орган второго года жизни
тюльпана и состоит из листовой пластинки зеленого цвета, белого
подземного черешка и влагалища.
Размеры листовой пластинки первого ассимиляционного листа
тюльпана весьма разнообразны (табл. 13). Но соотношение длины ли-
ста к ширине для листьев неплодоносящих особей изучаемого вида
наблюдается и у сеянцев второго года жизни. Этот лист является пер-
вым настоящим листом тюльпана и, несмотря на небольшие размеры,
консистенция, форма и окраска характерны для листьев изучаемого
вида. Так, на первом настоящем листе Т. micheliana появляются ан-
тоциановые полосы; у Т. greigii — пятна; Т. regelii над верхней пло-
скостью листа выступают ребра; край листьев f. fosteriana имеет хря-
щеватую окраину, часто окаймленную красным; края листочков
Т. korolkovii приобретают характерную извитость; у Т. kaufmanniana
на белесоватой мякоти просвечивают не только более ярко-зеленые
продольные жилки, но и сеточка из их боковых ответвлений и т. д.
Листовая пластинка обильно снабжена устьицами и является ор-
ганом дыхания и транспирации. Она воспринимает воздействие света,
необходимое для прохождения возрастной ступени световой стадии;
в ней протекает фотосинтез.
Черешок листа — единственный транзиторный орган. Он выносит
листовую пластинку на поверхность почвы. Длина черешка колеблет-
ся в зависимости от глубины залегания луковицы: при глубокой по-
садке— удлиняется, при мелкой — укорачивается.
263-
Таблица 13
Размеры листьев у нецветущих сеянцев тюльпанов, см
Вид Второй год
длина ширина
макси- мальная i 1 средняя мини- мальная макси- мальная средняя мини- мальная ।
Т. hoogiana 14,0 9,0 6,0 0,65 0,44 0, Ь
Т. ingens 16,0 12,7 10,0 1,0 0,8 0,4
Т. fosteriana 12,0 9,5 6,0 2,0 1,0 0,5
Т. carinata 9,0 8,0 7,0 0,6 0 5 0,4
Т. praestans 13,2 7,0 6,0 0,5 1,0 0,6
Т. subpraestans 16 2 12,7 11,6 1.2 0,7 0,4
Т. greigii 10,0 8,2 6,0 1,4 1,1 1,0
Т. micheliana 10,5 8,0 7,0 4,1 0,9 0,7
Т maximowi- czii 16,0 14,2 13,0 0,3 0,2 0,12
Т. korolkovii 9,0 7,0 5,5 0,5 0,3 0,15
Т. schrenkii 9,2 6,0 4,5 0,8 0,6 0,3
Т. ostrovskiana 15,0 13,0 9,0 0,7 0,43 0,35
Т. kolpakovski- ana 7,5 4,5 3,0 1,75 0,7 0,15
Т. kaufmanni- ana 17,1 9,0 6,0 2,2 1,8 1,3
Т. biflorifor- mis 9,0 8,0 6,0 0,4 0,3 0,15
18,0
13,0
17,0
Третий
длина
я я
। я я
я я я
(J я J=t
я я и
я я о.
3 3 о
15,0 11,0
15,0 12,2
15,4 12,3
11,0 9,4
14,0 11,6
21,0 19,4
15,0 10,0
16,0
12,0
я
я
я
3
я
я
я
я
я
8,7
8,8
6,0
9,0
6,0
18,0
9,2
14,0
9,0
год
и 1ирина
я я я
1 я я я
Я я я д я
о я я я
я я (D Я t-
я я О. я я
U 3 3
2,2 1,5 0,5
3,9 2,6 0,6
6,5 4,0 1,4
1,0 0,7 0,6
1.9 1,3 0,9
2,5 1,8 0,5
4,3 2,8 1,7
0,5 0,4 0,3
0,9 0,4 0,3
1,9 1,2 0,6
частично
я
о
я
я
3
я
я
я
я
я
19
25
Пятый год
год
длина ширина длина ширина
к
к
о
а
я
я
3
я
я
я
я
я
я
о
я
я
я
я
я
я
я
3
я
я
я
1=1
о
л
я
я
я
3
я
я
я
Я
я
я
3
я
о
я
я
3
к
Я
я
я
я
я
3
Л
к
л
я
я
я
я
я
я
я
я
о
я
Я
3
я
я
я
я
я
я
3
я
Я
я
я
О
мини-
мальная
15,0
22,3
16 12,5
25 |17,0
Цветет
25 14,8
13,5
19,0
12,0
7,0
6,2
6,0
3,8
4,8
3,8
3,5
2,8
3,2
2,7
1,5
1,4
1,2
19,0 16,0
19,5'15,0
10
8
16,0
5,3
10,0
4,9
5,0
1,3
частично
12,0 8,0
4,6
3,1
11ветст
В1 сь
Цветет частично
21
15,2
12,0
1,6
1,2
0,7
Цветет
17,0 14,0 11,9 2,0 1,4 0,9 Цветет
19,0 13,0 10,0 2,5 1,0 0,5 Цветет
Цветет частично
16,4 10,9 5,8 7,8 4,0 2,1 18 16,4
13,0 10,0 7,0 0,8 0,5 0,3 17 15,0
<4,0
13,0
8,5
6,3
4,0
Плодоносит весь
1,2
1,0
0,7
Плодоносит весь
столона уг-
года жизни.
ЧеТ“ а—кольцо сосудистых пучков, вошедших в столон
из стебля, о—листовые сосудистые пучки, в—‘цент-
ральная полость столона.
хождения, повернутые
Рис. 44. Смещение верхушечной
почки однолетней луковицы Т.
kaufmanniana в сторону выпук-
лой половины луковицы"
в октябре'
/7—смещенная верхушечная почка, б—
разрыхление ткани чешуи перед сме-
щающейся почкой, в—заложение поч-
ки-детки, г—зачатки корней.
Листовое влагалище ассимиляционного листа неполовозрелых
особей играет роль покрова, одевающего почку в ранние этапы ее
развития и позже. Совместно со
стеблевой частью растения в период
как роста, так и покоя оно являет-
ся покровной чешуей результирую-
щей луковицы. При углублении
верхушечной почки влагалищная
часть первого зеленого листа, сра-
стаясь со стеблем, образует столон
углублениятак же, как у семядоли.
Двойная природа столона уг-
лубления более отчетливо видна на
столонах, формирующих луковицу
второго и последующих лет жиз-
ни, чем на подобных же образо-
ваниях сеянцев первого года.
Столоны неполовозрелых осо-
бей. На поперечном сечении столо- рис. 43. поперечный разрез
на углубления, например у Т. grei- дубления Т. greigii второго
gii более старшего возраста,
ко выражена картина распределе-
ния сосудистых пучков (рис. 43).
Ясно видны лежащие кольцом сосудистые пучки стеблевого проис-
ксилемой внутрь, а флоэмой наружу. По обе
стороны кольца, образованного сосудисты-
ми пучками, проходят сосудистые пучки,
вошедшие в столон непосредственно из
листа.
Подобную картину распределения про-
водящей системы мы наблюдали у всех
исследованных нами видов тюльпанов, под-
твердив этим общность особенностей для
всех видов. Следовательно, в образовании
столона углубления принимает участие не
междоузлие, а узел (рис. 43, 44).
Из 22 видов тюльпанов, изученных нами
(табл. 18), у Т. ingens, Т. alberti, Т. ost-
rovskiana, Т. kolpakovskiana, Т. dubia,
Т. regelii и Т. bifloriformis часть растений
не закладывает столонов углубления в кон-
це первого года жизни.
По степени столонообразования в юве-
нильном возрасте все исследованные виды
тюльпанов можно разбить примерно на че-
тыре группы. К первой группе относятся
виды, у которых все луковицы дают сто-
лоны углубления — Т. tubergeniana, Т. ku-
schkensis, Т. hoogiana Т fosteriana, Т. рга-
estans, Т. subpraestans, Т. greigii, Т. miche-
liana, Т. maximowiczii, Т. dubia, Т. vveden-
skyi и Т. kaufmanniana (табл. 15, 16, 17,
18, 19). Вторую группу составляют виды, у которых не более 10%
растений не закладывают столонов углубления — Т. ingens, Т. alberti,
Т. ostrovskiana и Т. kolpakovskiana. Третья группа представлена ви-
265
дами Т. eichleri и Т. regelii, у которых количество особей с неуглу-
бившимися луковицами второго года жизни составляет 25—40%,
К четвертой группе относятся виды Т. wilsoniana, Т. korolkovii, у
которых углубившиеся луковицы второго года жизни составляют
менее 40%.
Половозрелые экземпляры первой группы имеют крупные луко-
вицы. Исключение составляет Т. maximoviczii Rgl., по некоторым
признакам стоящий несколько особняком в роде Tulipa. Ко второй
группе относятся тюльпаны с мелкими луковицами и некоторые сорта
культурных тюльпанов.
Третья и четвертая группы состоят также из видов с мелкими
луковицами и некоторых сортов садовых тюльпанов (табл.'12, 14.)
Столон углубления у тюльпанов в условиях Ташкента в конце
периода жаропокоя как первого, так и последующих лет жизни (ча-
ще всего до стадийной зрелости растения) развивается из основания
зачатка влагалища первого ассимиляционного — единственного зеле-
ного листа второго года, позже—из ежегодно сменяющихся листьев
последующих лет жизни и вставочного роста подпочечного узла
(донца).
Прекращение углубления луковицы есть один из признаков наи-
более раннего вступления в плодоношение.
Столон углубления у растений всех возрастов, кроме сеянцев
первого года жизни, в условиях Ташкента начинает расти с конца
сентября или начала октября; у выкопанных луковиц —еще до по-
садки их в грунт.
В это время зачаток столона снаружи луковицы не заметен и
первоначально состоит только из смещенной верхушечной почки
тюльпана, растущей в направлении выпуклой стороны луковицы
(рис. 44). Стенка влагалища зачаточного листа, одевающего почку,
при этом разрастается в сторону смещения верхушечной почки. У
основания верхушечной почки образуется небольшая выпуклость
(рис. 44, а).
В прилегающей к выпуклости утолщенной половине луковичногг
чешуи, в месте соприкосновения с выпуклостью, клетки разрыхляют-
ся, крахмал в них растворяется и исчезает (рис. 44, б).
Клетки в смещенной части узла, усиленно делясь и разрастаясь,,
сдвигают верхушечную почку в сторону разрыхления клеток в лу-
ковичной чешуе.
Донце луковицы является укороченным побегом, один узел ко-
торого у запасающей чешуи и настоящего листа как у неполовозре-
лых особей, так и у деток половозрелых луковиц сохранил способ-
ность к усиленному боковому одностороннему росту, достигающему
у некоторых видов 10 см и более (табл. 14). Разница между столо-
ном углубления и побегом половозрелой особи заключается в том,
что у столона углубления в поперечном направлении разрастается
узел верхушечной почки неполовозрелых особей, обладающий поло-
жительным геотропическим ростом, а у половозрелых особей узел
не растет, а вытягиваются междоузлия, которые растут отрицательно
геотропично, образуя побег.
Положительный геотропический рост вышеуказанного узла непо-
ловозрелых особей продолжается у большинства видов и сортов
тюльпанов до наступления стадийной зрелости. У культурных сортов
и некоторых видов тюльпанов иногда после первых двух-трех лет
жизни этот узел теряет способность к вставочному росту вплоть до
наступления половой зрелости. Среди диких тюльпанов такое явление
266
Таблица 14
Длина столонов углубления и общая глубина залегания луковиц верхушечных почек у тюльпанов
Вид и сорт Первый год Второй год Третий год Четвертый год
Средняя длина столона Общая глубина длина столона Общая глубина длина столона Общая глубина длина столона Общая глубина
наи- большая средняя меньшая наи- | большая средняя меньшая наи- большая средняя меньшая
Т. tudergeniana 8,0 22,0 7,0 4,0 2 29 29 9,2 4 40
Т. kuschkensis 4,0 16,0 3,5 2,7 2 20 8 4,6 2 20 — —
Т. hoogiana 1,6 5,0 7,0 4,2 2 21 7 6,1 3 18 —- — — —
Т. ingens 4,0 8,0 7,0 4,0 2 20 — — — — —. — ——
Т. fosteriana 3,0 6,0 5,0 3,0 2 16 — — — — —— —
Т. psaestans 3,6 6,2 5,0 3,0 1 15 8 5,4 3 15 — — — —
Т. subpraestans 4,0 8,5 8,0 5,4 3 20 — — — — 9 5 4 24
Т. alberti 4,0 8,0 10,0 8,0 5 10 — — — — — — —
Т. greigii 3,0 8,0 7,0 4,3 2 20 7 5,7 5 17 15 6,7 4 37
Т. micheliana 1,7 7,5 6,5 2,8 2 27 — — — — — —
Т. eichleri 3,0 13 4,0 2,0 0 17 — — — — — — .
Т. maximowiczii 1,5 4,0 2,5 15,0 0 17 — — — — —. — —
Т. wilsoniana 1,5 8,0 5,0 0,0 0 13 — — — — — — — .
Т. korolkovii 3,0 8,0 4,5 4,0 3 11 7 0 0 10 5 2 0 17
Т. schrenkii 1,5 6,0 — — — — И 7 4 16 — __ —
Т. ostrovskiana 3,7 7,7 8,0 5,4 4 21 4 3,6 3 22 — — —
Т. kolpakovskiana 3,5 8,5 5,0 3,5 3 20 — —— — — — — — —
Т. dubia 2,5 4,0 7,0 5,5 4 10,6 — — — — — — — ——
Т. vvedenskyi 5,0 13,0 7,0 4,3 2 20 — — — — — — —. —
Т. kaufmanniana 2,5 6,8 7,0 4,6 3 15,1 5 4,1 3 23 12 6,4 4 34
Т. regelii 9,5 — 18,5 5,5 1,9 0,75 — — — .— — — — —.
D. V. Thol yellow 4,0 8,0 5,0 4,0 3 13 — — — — 7 5 3 19
Jubilee 2,0 6,0 6,0 4,2 3 10 —— 9 6,7 18 6 4,3 3 19
Kathleen Parlow — — 10,0 6,0 3 16 9 8 5,9 23 8 7 5 17
Рис. 45. Разрыв тканей
запасающей чешуи про-
растающим столоном Т.
kaufmanniana (в фев-
рале).
/г—столон углубления прорвал
чешуи луковицы, б—-столон
углубления и его присоедине-
ние к донцу (чешуи сняты).
наблюдается у видов с мелколежащими луковицами (табл. 14). Чем
раньше растение вступает в пору цветения, тем раньше из верхушеч-
ной почки молодой луковицы появляются неуглубляющиеся особи
(табл. 12, 14).
Сдвинутая вбок верхушечная почка, одетая сросшимися тканями
стеблями листа, имеет вид очень толстого корешка. Она разрывает
ткани запасающей и покровной чешуи. На за-
пасающей чешуе (если их несколько, то на всех)
образуется продольная щель, в которую и вы-
ходит столон углубления (рис. 45). Разрыв тка-
ней запасающих чешуй подготовлен разрыхле-
нием клеток в направлении роста столона уг-
лубления.
К видовым отличиям относятся и некоторые
морфологические черты столонов углубления.
Так, очень тонкие, нитевидные, легко разруша-
ющиеся по высыхании столоны углубления вер-
хушечных почек и шаровидные луковицы непо-
ловозрелых особей Т. praestans (рис. 46. А) уди-
вительно сходны с идентичными столонами и
луковицами Т. fosteriana (рис. 46, Б). Различие
заключается в том, что у Т. praestans верхушеч-
ная почка или все пазушные уносятся на тон-
ких нитевидных столонах от места заложения,
оставляя лишь отмершие чешуи. А у Т. foste-
riana на месте старой луковицы довольно часто
остаются детки (рис. 46, Б). У некоторых по-
ловозрелых особей Т. praestans всегда у заме-
щающей луковицы и всех деток сохраняется
ювенильная способность уходить из старых
чешуй на столоне углубления несколько в бок
и вниз (рис. 46, А), что у других представите-
лей этого рода мы наблюдали очень редко.
Кроме Т. praestans, этой особенностью обладают
Т. kaufmanniana (рис. 45) и один сорт из груп-
пы тюльпанов Дарвина — King George V. Однако
столоны их, на которых развиваются замещаю-
щие луковицы, морфологически отличны от сто-
лонов Т. praestans.
У Т. fosteriana замещающая луковица ста-
дийно зрелого растения не углубляется, остается на месте и в чешуях
старой луковицы. Столоны углубления в таком гнезде возникают из
верхушечных почек деток этой же особи, которые обычно в значи-
тельной степени омолаживаются и часто после обособления от мате-
ринского растения много лет не цветут, а размножаются вегетативно.
Столоны таких деток, а также сеянцев Т. fosteriana и Т. praestans
морфологически сходны. Различие между ними заключается в том,
что сеянцы и детки Т. fosteriana (рис. 46, Б) способны оставлять на
месте и в чешуях старой луковицы не углубившиеся детки, а у Т.
praestans со старого места столонами уносятся все почки.
Столоны и луковицы Т. fosteriana и Т. praestans резко отличаются
от подобных образований у Т. subpraestans (рис. 46, В), по цветку и
другим морфологическим признакам наиболее близкого к Т. praestans.
У Т. subpraestans столоны значительно толще, более прочные,
расположены почти отвесно вниз, а не в стороны, как у Т. praestans.
268
T. subpraestans часто образует не углубившееся кучное гнездо луко-
виц: стадийно зрелые луковицы его в большинстве случаев не углуб-
ляются и по форме более похожи на редьку, чем на шар.
Форма столона длинного, толстого, мягкого, несколько суженного
к концам и расширенного к середине, постепенно переходящего в уз-
Рис. 46. Столон углубления с луковицей: А—Т. praestans
а—луковниа прошлого года, б—замещающая луковица, в—луковица-детка; £—Т. fosteriana
(двулетка)—а—замещающая луковица, б—луковица детка; В—Т. subpraestans (двулетка)—
а—замещающая луковица углубилась, детка осталась в чешуях старой луковицы, б—-серия
коллатеральных не углубившихся деток, в—все луковицы углубились, г—углубились заме-
щающая и две детки, а одна детка осталась на месте старой; Г— Т. ingens Hoog. (двулет-
ни)—а—расширенная мягкая трубка столона луковицы; Д—Т. kuschkensis В. Fedtsch. (трех-
летка)—а—расширенная мягкая трубка столона, б—почка детки, в—замещающая луковица.
269
кую и длинную луковицу, сближает Т. ingens (рис. 46, Г) с группой
тюльпанов из секции Tulipanum—Т. tubergeniana, Т. kuschkensis (рис.
46, Д), а не с Т. fosteriana.
1 В луковице тюльпана любого возраста (от первого года) всегда
имеется не меньше двух почек — верхушечной и пазушной (рис. 47).
В пазухе запасающей чешуи у всех видов тюльпанов разных возрас-
тов всегда закладывается почка (рис. 47, 48), но часто она не разви-
вается дальше меристематического
бугорка, к которому подходит мощ-
ный сосудистый пучок. У многих
видов тюльпанов формирование па-
зушных почек связано с питанием
луковицы в предшествующий год.
Но у некоторых видов Т. korolko-
Рис. 47. Заложение почки-детки в пазухе ^ис- 48. Заложение почки-детки в пазухе
запасающей чешуи у однолетней луковицы запасающей чешуи у четырехлетней луко-
Т. greigii Rgl. вицы Т. fosteriana I г v.
а — верхушечная почка, начавшая смещаться в сто- а -поперечный разрез через луковицу в начале сме-
лой, б— почка-детка. шения центральной почки в столон, 6—продольный
разрез в то же время, в—почка-детка.
vii, Т. michelliana, Т. dubia, Т. maximowiczii, и формы Т. schrenkii
(табл. 12, 15), несмотря на благоприятные условия для роста луковицы,
обычно пазушные почки не развиваются в почки-детки.
У неполовозрелых особей тюльпанов при наличии заложившейся
детки в большинстве случаев верхушечная почка уходит на столоне
углубления, а детка остается на месте и в чешуях старой луковицы.
У этих особей покровная оболочка детки-луковицы состоит только из
тканей влагалища листа, но не из срастания его со стеблем, как у
•столонов углубления. Покровная оболочка — чешуя не углубившихся
деток, блестящая, более тонкая и яркая, чем у тех, которые, как и
верхушечная почка, углубились. У некоторых особей одного и того
же вида иногда встречаются как углубившиеся, так и не углубившие-
ся детки. Например, у Т. subpraestans мы наблюдали коллатерально
заложившиеся детки (рис. 46, В). У некоторых экземпляров все
детки остались на месте старой луковицы (рис. 46 В, б), у других,
так же как и верхушечная почка, углубились (рис. 46 В, в), у тре-
тьих — некоторые детки углубились, а другие остались на месте старой
луковицы (рис. 46 В, г), Вегетативное размножение дикого тюль-
пана превышает не только наше представление о размножении боль-
шинства диких видов, но и садовых сортов тюльпанов. При таком
интенсивном размножении возможно было ожидать, что этот вид позд-
но вступит в плодоношение, но в действительности 25,3% растений
этого вида из посевов 1949 г. зацвело в 1953 г., т. е. после 3 лет
вегетации.
-270
Вес луковиц сеянцев тюльпанов, мг
Таблица 15
Вид и сорт Третий год Четвертый год
крупная средняя из 100 мелкая j углубле- ние, % детки, % крупная средняя из 100 мелкая углубле- ние, % X
"Т. tubergeniana 3100 1000 412 100 26 — — — — —
Т. kuschkensis 2150 1372 900 100 47 —• — — — —
Т. hoogiana 4080 2275 1100 100 47 — — — — —
Т. fosteriana 1050 674 400 100 — 4500 3606 800 55 —
Т. ingens — — — — — — — — — —
Т. praestans 2000 1050 370 100 100 — — — — —
Т. subpraestans 870 730 50 100 100 10440 2826 50 100 90
Т. subpraestans Т. greigii i 650 [другая 348 [ форм; 32 0 100 17 15900 26500 10450 16500 4737 9400 4 100 20
Т. korolkovii 1100 400 100 17 5 1700 720 280 10 0
Т. schrenkii 6400 2765 800 100 100 — — — — —
Т. ostrovskiana Т. ostrovskiana 5500 i 2301 другая 500 [ форм; 100 а) 100 8260 6036 4290 100 —
Т. kaufmanniana 1760 1311 600 100 24 11220 5500 3073 100 —
Т. bifloriformis 205 63 19 100 76 5020 2464 860 — —
Jubilee 1630 800 250 — — — — — — —
Elephant 1920 910 500 25 25 — — — — —
Kathleen Parlow 1250 820 500 — — — — — — —
T. ta da 2250 539 320 100 100 5600 400 100 37 26
Скороплодность тюльпанов. Д. Холл (1929) пишет, что тюльпаны
зацветают при посеве семенами на 5, 6, 7-й год. Но, вполне возможно,
что виды с малым количеством чешуй в луковице могут зацвести на
3 или 4-й год.
Среди растений, зацветших у нас на третьем году жизни (табл.16),
«есть представители видов с крупными луковицами и относительно
большим количеством запасающих чешуй — Т. Kaufmanniana и Orange
Delight, а также со средними луковицами — Т. ostrovskiana и мелкими
Т. tarda.
Все виды и сорта тюльпанов при первом цветении имеют не
более 2—3 запасающих чешуй (табл. 15) и лишь в последующие
годы их количество увеличивается.
У видов, относящихся к секции Leiostemones, развитие двух за-
пасающих чешуй свидетельствует о том, что в следующем году эта
луковица будет цвести (табл. 16, 17, 18). Исключение составляет
Т. fosteriana, у которого часто не зацветают луковицы даже с четырьмя
запасающими чешуями.
В 1953 г. из посевов осени 1949 г. на четвертом году жизни за-
цвело значительно больше видов, чем в посевах 1948 г. Кроме Т. kauf-
manniana, Т. ostrovskiana и Т. tarda и одного сорта Orange Delight, в
этом году зацвели Т. greigii, Т. korolkovii, Т. subpraestans, Т. biflo-
riformis, сеянцы нескольких культурных сортов (табл. 18) и гибриды
между садовыми сортами и дикими видами. К этой группе относятся
271
Таблица 16
Цветение сеянцев тюльпанов на третьем году жизни
Вид или сорт Всего растений Цветущих
количество %
Посев 1950 г.
Т. kaufmanniana 33 3 9,00
Т. kaufmanniana 40 8 20
Т. kaufmanniana 466 6 1,29
Т. ostrovskiana 22 4 18,1
Т. tarda 96 2 2,08
Orange Delight 85 2 2,35
Посев 1952 г.
Т. kolpakovskiana 364 16 27
Т. tetraphylla 4 2 50
Т. vvedenskyi 175 3 1,71
Т. subpraestans 120 1 0,83
Таблица 17
Количество чешуй у луковиц сеянцев тюльпанов
различного возраста
Вид Год ы
первого второго третьего четвертого
покров- ная запасаю- щая покров- 1 ная запасаю- веТп покров- ная запасаю- щая ПОКрОВ’ . пая запасаю- щая
Т. grfeigii 1 1 1 1 1 2 1 3
1 2
Т. kaufmanniana 1 1 1 1 1 2 1 3
1 2
Т. fosteriana 1 1 1 1 1 1 1 2
Т. praestans 1 1 1 1 1 1 1 1
Т. korolkowii 1 1 1 1 1 1 1 2
Т. bifloriformis 1 1 1 1 1 1 1 1
1 2 1 2
Т. ingens 1 1 1 1 1 1
Т. schrenkii 1 1 1 1 1 1
Т. kolpakovskiana 1 1 1 1 1 1
1 1 2
Т. ostrovskiana 1 1 1 1 1 1
2 1 2
Т. maximowiczii 1 1 1 1 1 1
Т. hoogiana 1 1 1 1 1 1
1 2
Т. micheliana 1 1 1 1
272
виды, различные как по размерам листьев, луковиц (табл. 17, 18, 21),
так и по степеням вегетативного размножения. Такие виды как Т.
korolkovii, Т. greigii, Т. bifloriformis, Т. subpraestans зацвели одновре-
Таблица 18
Первое цветение сеянцев тюльпанов на четвертом году жизни
Вид или сорт Всего растений Из них цветущих Процент цветущих растений
Посев 1949 г.
Т. korolkovii 351 1 0.28
Т. sudpraestans 207 7 3,38
Т. subpraestans 75 19 25,3
Т. kaufmanniana 238 34 14,2
Т. greigii 77 57 48
Т. greigii 106 30 28,3
Т. binutans 39 1 2,6
Т. sp. Cykok 210 10 4,8
Т. sp. Eriostemones 72 7 9,7
Т. bifloriformis 202 9 1
Т. tarda IS 3 18,75
Т. tarda 39 12 30,8
Preserpina 122 2 1,64
D. v. Thol yellow 702 2 0,28
Keizerskronn 51 1 1,06
Kathleen Parlow 1005 5 0,49
Elephant 248 8 3,2
Jubilee 900 10 1,1
Посев 1951 г.
T. tubergeniana 580 8 1,38
T. kuschkensis 940 3 0.32
T. hoogiana 679 28 4,13
T. julia 67 1 1,49
T. ingens 800 40 5,00
T. praestans 400 35 8,75
T. alberti 131 11 8,48
T. micheliana 482 54 11,20
T maximowiczii 300 10 3,34
T. wilsoniana 640 4 0,62
менно, несмотря на существенные различия в степени их развития 1
иную морфологию луковиц.
При посеве тюльпанов семенами не все особи вида зацветают од
новременно — наблюдаются скороплодные и тугоплодные растения.
18—2229
27;
Из трех исследованных нами видов тюльпанов посева 1948 г.
большое количество растений не зацвело даже на 5-м году жизни, в
то время как некоторые экземпляры цвели уже по второму разу
(табл. 19).
Таблица 19
Первое и второе цветение сеянцев тюльпанов (посев 1948 гЛ
Вид 1952 г. 1951 г.
всего расте- ний из них цветущих всего растений из них цветущих в абсо- лютных цифрах цветение замеща- ющей луковицы без деток
абсолют- ные цифры %
1953 г. 1952 г.
Т. kaufmanniana 273 68 24.9 211* 128 60,7 —
Т. kaufmanniana 667 127 19,1 760 599 78,8 89,8
Т. kaufmanniana 40 21 52,5 46 41 89,1 102,5
Т. kaufmanniana 60 20 33,3 68 58 85,3 113,3
Т. greigii 15 5 30 15 12 80,0 80,0
Т. greigii 16 1 6,2 18 15 83,3 93,7
Т, bifloriformis 20 3 15 21 15 71,3 75,0
Т. fosteriana 100 0 0 200 0 0 0
* Часть луковицы была в 1953 г. уничтожена слепышом.
Но, если процент цветения в 1953 г. высчитать не по отношению
к числу растений этого года, так как в это число вошли и детки-лу-
ковички, а от количества луковиц 1952 г., тогда он значительно по-
высится и у Т. kaufmanniana превысит 100 %, а у Т. greigii приблизит-
ся к 100 % (табл. 19).
Растянутость периода от посева до первого зацветания сеянцев в
условиях Ташкента характерна не только для диких видов, но и для
садовых сортов. Поэтому при посеве тюльпанов семенами без соот-
ветствующего отбора в течение многих лет трудно получить декора-
тивный эффект от сеянцев. .
Биологически это можно объяснить тем, что растение приспосо-
билось к жизни в жестких условиях существования, где не каждый
год внешняя среда благоприятствует их росту и развитию. Историче-
ски сложившееся растягивание всхожести семян на несколько лет и
неодновременное вступление растений в фазу цветения и плодоноше-
ния — приспособительная реакция рода, обеспечивающая его процве-
тание в естественных условиях.
Как виды, так и все садовые сорта тюльпанов в ювенильном воз-
расте до первого цветения и во взрослом состоянии закладывают в
пазухах запасающих чешуй почки, которые могут развиться в дет-
ки-луковицы. Эти детки, как органы вегетативного размножения, пре-
терпевают некоторое омоложение, по сравнению с верхушечной поч-
кой ювенильного растения и замещающей луковицей взрослого. Омо-
ложение у разных видов и сортов тюльпанов происходит с неодина-
ковой степенью глубины.
Детки луковиц Т. fosteriana после обособления от материнского
растения несколько лет размножаются вегетативно, давая столоны уг-
лубления как из верхушечной, так и пазушных почек и не переходят
274
к цветению, хотя часто состоят из нескольких больших запасающих
чешуй.
Детки луковицы цветущих растений Т. kaufmanniana или Т. grei-
gii с диаметром по наибольшему сечению не менее 1,5 см, в услови-
ях Ташкента зацветают на следующий год после обособления от ма-
теринского растения.
При работе с сеянцами диких видов тюльпанов можно отобрать
«формы со значительно развитой способностью к вегетативному разм-
ножению. Путем воспитания и последующих отборов вполне возмож-
но из диких видов создать сорта подобные садовым тюльпанам.
Детки-почки — органы вегетативного размножения. У многих
сортов садовых тюльпанов вегатативное размножение заменило поло-
вое. Широко известный среди садоводов сорт садового тюльпана из
группы одинаких ранних — Keizerskroon существует около 200 лет. Он
почти не завязывает семян и не имеет никаких признаков вырожде-
ния. При хорошем уходе его цветы достигают крупных размеров. Он
размножается вегетативно при помощи почек-деток. В данном случае
сорт сохранился без вырождения потому, что при заложении луко-
виц-деток происходит частичное их омоложение. А. А. Авакян (1948)
отмечал, что у лука снятие стадийности происходит при заложении
любых органов размножения — полового или бесполового. Но у тюль-
панов при вегетативном размножении происходит лишь частичное
омоложение луковиц, специфичное для разных видов. Детки Т. kauf-
manniana и Т. greigii омолаживаются слабо, а Т. fosteriana — сильно.
У тюльпанов с возрастом растения наблюдается смена форм во-
зобновления от моноподиального к симподиальному.
У любого вида тюльпана в течение первых лет онтогенеза до на-
ступления стадийной зрелости луковица нарастает и сменяется моно-
подиально с помощью верхушечной точки роста. При достижении
растением тюльпана половой зрелости цветоносные побеги всегда
образуются из верхушечной почки луковицы, а в случае многоцвет-
ковости третьего типа (Бочанцева, 1953, 1954) —из верхушечных по-
чек-деток. Индивидуум продолжает жи:ь за счет развития пазушной
почки самой внутренней чешуи, т. е. симподиально.
Развитие тюльпана из семени в зависимости от внешних ус-
ловий. Первое цветение тюльпана при посеве его семенами происхо-
дит через несколько лет. Каждый год растение начинает естественный
рост осенью при пониженных температурах и влажности и продолжает
его под землей всю зиму. При первых теплых днях в конце зимы
или начале весны над поверхностью почвы появляется почка, которая
в зависимости от зрелости тюльпана развертывается в один лист или
розетку листьев. По диаметру показавшейся из земли почки можно
судить о том, зацветет ли тюльпан весной. Цветущие растения имеют
розетку листьев, а не цветущие—один лист. Растения из семян в пер-
вый год цветения бывают мельче, чем в последующие годы. Видовые
признаки у сеянцев диких видов, собранных при свободном цветении,
проявляются с большой четкостью. Они точно повторяют материнские
растения, за исключением сеянцев Т. alberti, Т. tuBergeniana и Т. vve-
denskyi. Наследственный консерватизм диких видов очень велик. Се-
янцы же культурных сортов из семян от свободного опыления весьма
изменчивы. Их потомство совершенно не сходно с материнским расте-
нием.
Мы установили, что во время вегетации для нормального развития
растений должна существовать определенная взаимосвязь между дли-
275
ной светового дня и температурой, при которой развиваются растения-
Чем ниже температура, тем длиннее должен быть день.
В 1952 и 1953 гг. были поставлены опыты с укорочением дня для
нескольких видов различного возраста тюльпанов (табл. 20, 21). Уко-
рочение дня влияет на вес луковиц. Так, средний вес двулетних лу-
Таблица 20
Средний вес 100 двулетних луковиц тюльпанов
при различной длине дня, мг
Вид Длина дня
нормальная 8-часовая 4-часовая
Т. korolkovii 280 280 —
Т. vvedenskyi 984 736 —
Т. hoogiana 651 487 —
Т. fosteriana 412 217 180
Т. kaufmanniana 1160 633 —
ковиц Т. vvedenskyi, Т. hoogiana, Т. fosteriana и Т. kaufmanniana
при 8-часовом световом дне в течение всего периода вегетации мень-
ше среднего веса луковиц при нормальной для Ташкента длине
дня. Нет уменьшения в весе только у Т. korolkovii. Почти в два раза,
уменьшился вес у Т. kaufmanniana.
Таблица 21
Средний вес 100 луковиц (лг) и процент цветения
четырехлетних растений при различной длине дня
Вид Нормальный день 8-часовой день
средний вес луковиц процент цветения средний вес лу- ковиц процент цветения
Т. korolkovii 1700 0,3 561 0
Т. fosteriana 2605 0 1253 0
Т. kaufmanniana 9740 17,1 7000 18
Т. greigii 12650 48 7950 25
Вес четырехлетних луковиц растений, выращиваемых при 8-ча-
совом световом дне, также значительно меньше веса луковиц расте-
ний, развивающихся при нормальной длине дня.
Из четырех видов три зацвели на четвертом году жизни. У рас-
тений Т. korolkovii, воспитывавшихся при нормальной длине дня, цве-
тение было очень незначительное, всего 0,3%. Среди растений этого
вида, выращиваемых при 8-часовом световом дне, цветущих экзем-
пляров не было (табл. 21).
У Т greigii при нормальной длине дня на 4-м году жизни впер-
вые зацвело 48% растений (рис. 49), а при 8-часовом дне в течение
двух сезонов зацвело 25%. Таким образом, при 8-часовом дне умень-
276
шается процент цветущих особей почти в два раза. Формирование
цветка тюльпана начинается в конце периода покоя в год, предшест-
вующий цветению. Таким образом, если при 8-часовом световом дне
1953 г. не было прямого влияния на заложение цветов, то 8-часовой
световой день 1952 г. должен был подействовать на цветение в 1953 г.
Рис. 49. Первое цветение Т. greigii при посеве семенами на четвертом
году жизни.
Однако у Т. kaufmanniana укорочение дня не повлияло на количе-
ство зацветших растений, т. е. мы наблюдаем некоторое увеличение
зацветших растений, бывших на укороченном световом дне.
Опыты по изучению влияния на растения тюльпанов укороченно-
го светового дня дали совершенно неожиданные результаты — укороче-
ние дня у тюльпанов резко снижает их холодостойкость, т. е. сопро-
тивляемость утренним заморозкам и холодным ночам.
В начале первой декады апреля ночью на нашем участке мороз
доходил до —6°. На подопытном участке сеянцы всех возрастов и
взрослые растения, росшие при нормальной длине дня, не пострадали
от мороза. У растений укороченного дня, крытых картонными короб-
ками, померзло от 30 до 80% листьев. Особенно пострадали сеянцы
4-часового светового дня.
Из двух- и четырехлетних сеянцев от мороза пострадали Т. grei-
gii, Т. kaufmanniana, Т. korolkovii, Т. fosteriana. Не замерзли толь-
ко сеянцы Т. vvedenskyi. Снижение устойчивости растений к понижен-
ным температурам произошло потому, что продукты фотосинтеза листа
ко времени наибольшего суточного понижения температуры в предут-
ренние часы, видимо, успели переместиться из листьев в луковицу.
Концентрация клеточного сока и плазмы листьев к этому времени
возможно была значительно слабее, чем у листьев при нормальной
длине дня, что и способствовало их более легкому замерзанию. Наи-
более сильно концентрация клеточного сока уменьшается у растений
с наименьшим запасом ассимилятов, т. е. в первую очередь у растений
четырехчасового светового дня, а затем восьмичасового дня. У этих
277
растений промежуток между концом ассимиляции и наступлением
ночного понижения температуры был более длительным, чем у расте-
ний при нормальной длине дня. Это и привело к их замерзанию.
Результаты наших опытов не противоречат данным опытов Мош-
кова (1937), в которых укорочение дня приводило к концу периода
вегетации к большему вызреванию древесины подопытных растений
и вследствие этого повышало их зимостойкость. Тем не менее, полу-
ченные нами данные чрезвычайно интересны для решения вопроса
о холодостойкости тюльпанов и расселении тюльпанов в широтном'
направлении.
Тюльпаны в культуре прекрасно растут и размножаются как в
Ташкенте, так и в Ленинграде. В то же время в Ленинграде во время
роста и развития они значительно чаще, чем в Ташкенте, подвергают-
ся утренним заморозкам, но там они не вымерзают, так как в Ленин-
граде тюльпаны вегетируют при более длинном световом дне.
Наши опыты с изменением длины дня показали, что здесь имеется
прямая связь между длиной дня и температурными условиями — при
длинном световом дне растение не вымерзает, а при коротком свето-
вом дне и низких температурах растения погибают.
Глава IX
РОСТ ЮВЕНИЛЬНЫХ ЛИСТЬЕВ И ГЕНЕРАТИВНЫХ ПОБЕГОВ
У Т. KAUFMANNIANA И Т. GREIGII ПОСЛЕ ПОЯВЛЕНИЯ ИХ
НАД УРОВНЕМ ПОЧВЫ
аадземные органы неплодоносящего тюльпана представлены всегда
одним листом. Появление розетки означает перелом, происшед-
ший в их развитии, — переход их во взрослое состояние.
Надземная часть стадийно зрелого растения тюльпана является
генеративным побегом и после появления ее над поверхностью почвы
быстро отмирает, т. е. она эфемерна. У тюльпана вырабатываются при-
способления к исключительно короткому периоду вегетации. Так, за-
ложение, формирование и первоначальный рост происходят во время
длительного развития почки внутри луковицы в осений и под землей
в зимний периоды.
В появившейся над поверхностью почвы почке сформированы все
органы, в дальнейшем они лишь увеличиваются в размерах и стареют,
т. е. переходят из ювенильных структур во взрослые. Наибольшие
изменения в это время происходят в генеративных органах — окраска
листочков околоцветника; раскрывание цветка; созревание пыльников,
пыльцы и зародышевых мешков; оплодотворение; рост и развитие за-
родыша; заложение, формирование, развитие и созревание семян.
В 1954 г. мы провели наблюдения над надземными органами двух
видов тюльпанов — Т. greigii и Т. kaufmanniana — с момента появле-
ния над поверхностью почвы маленькой почки или листочка, еще ин-
тенсивно окрашенных антоцианом.
Для исследований было выделено по 10 растений каждого образ-
ца 2, 3, 4-го года жизни и 4-го года начавших плодоношение. Все
растения в наступивший сезон вегетации развили над поверхностью
почвы по одному ассимиляционному листу, а растения, начавшие 4-й
год жизни, — по розетке листьев и цветку.
Данные изучения темпов роста почти окончивших формообразо-
вание органов позволили установить интересные закономерности в
развитии растений и их взаимосвязь с внешней средой.
Измерение длины и ширины листа у неполовозрелых особей и
всех листьев розетки бутона, цветка, завязи и цветоплодоножки у
взрослых проводилось с интервалами в 5 дней для получения досто-
верных и сравнимых данных одновременно для всех образцов.
Первые измерения мы провели 20 февраля 1954 г. и установили
стеклянные этикетки около каждого из подопытных растений. К вече-
ру пошел дождь и позже снег. До середины марта установилась хо-
лодная погода. В начале этого периода температура доходила до
— 23°. Значительное потепление наступило 15 марта, снег сошел и мы
,27»
возобновили наблюдения за подопытными растениями. Температура
постепенно повышалась, а с 25 апреля наступила жара и тюльпаны
начали гореть. Эти резкие климатические колебания нашли яркое от-
ражение в росте и развитии исследуемых в настоящем эксперименте
растений. Так, понижение температуры в течение 25 дней затормози-
ло рост растений (рис. 50, 51).
Рис. 50. Рост листьев неполовозрелых особей Т. kaufmanniana.
а—в ширину, б—в длину; 7—чвулетка, 2—трехлетка, 3—четырехлетка.
За 25 дней листья неполовозрелых растений Т. kaufmanniana 4-го
года жизни всего в два раза увеличили длину и немного больше ши-
рину. Растения 2-го и 3-го года жизни выросли еще меньше (рис. 50).
У растений Т. greigii такими колебаниями температуры в наиболь-
. шей степени затронуты неполовозрелые растения 4-го года. У сеянцев
Т. kaufmanniana меньше всего затронуты растения этого возраста (рис.
50, 51). Длительное понижение температуры у обоих видов значитель-
но задержало рост в длину и почти не отразилось на росте в ширину
(растения Т. greigii 2-го и 3-го года). Чем старше растения обоих ви-
дов, тем более интенсивно растут их листья. И чем они крупнее, тем
менее страдают от жары. У слабых растений всякого возраста вначале
отмирают надземные части. Оставшиеся растения продолжают рост,
а так как они были крупнее, то и средний прирост у них больше. С
25 апреля рост их резко увеличивается. В действительности же, в
связи с засыханием некоторых особей он должен снизиться.
280
Любое молодое растение тюльпана непрерывно растет до начала
пожелтения вершины листа. С наступлением жары начинает желтеть
и засыхает более старая верхняя часть листа, а нижняя часть остает-
ся свежей и интенсивно зеленой. Рост листьев неполовозрелых осо-
бей обоих видов в ширину прекращается на пять дней раньше, чем
в длину (рис. 50, 51).
На 4-м году жизни у растений, впервые вступающих в плодоно-
шение, мы изучали рост листьев розетки, бутона и цветка, цветопло-
доножки и завязи. У всех этих органов измеряли длину и ширину
через каждые 5 дней в течение всего периода вегетации. Характер
роста листьев розетки и неполовозрелых особей сходен у обоих ви-
дов (рис. 50, а; 51, а, б-, 52 и 55).
Рис. 51. Рост листьев неполовозрелых особей Т. greigii.
а—в ширину, б—в длину, 1—двулетка, 2—трехлетка, 3—четырехлетка.
У Т. kaufmanniana в 1954 г. листья в длину росли равномерно до
10 апреля, лишь верхний лист остановился в росте на 5 дней раньше
нижних (рис. 52).
Все листья розетки во время бутонизации с цветением и в тече-
ние 10 дней после цветения растут в основном равномерно. Верхний
лист в начале периода распускания цветов резко снижает прирост.
Но во вторую половину этого периода темпы роста листа восстанав-
ливаются. Однако он прекращает рост в длину раньше нижележащих
листьев. В первую пятидневку после массового цветения падает темп
роста среднего листа. В следующую пятидневку он вновь возрастает,
281
затем вновь падает и совершенно прекращается. На росте в длину
нижнего листа фенофазы развития растения почти не отражаются.
Все листья розетки в ширину (рис. 53) растут равномерно в пе-
риод бутонизации и цветения, после цветения развитие нижнего лис-
Рис. 52. Рост листьев розетки впервые зацветшего
растения Т. kaufmanniana в длину.
7—нижннй лист, 2—средний лист, 3—верхний лист (уменьшено в 2 раза).
та резко снижается. У среднего и верхнего листьев оно продолжается
еще пятнадцать дней.
У Т. greigii листья розетки в длину у особей, впервые на 4-м:
году жизни вступающих в плодоношение, в 1954 г. росли^в среднем
до 25 апреля.
Рис. 53. Рост листьев розетки впервые зацветшего
растения Т. kaufmanniana в ширину.
1—нижний лист, 2—средний лист, 3—верхний лист.
Самый нижний и второй лист растут дольше, чем верхние. Все-
листья розетки в первую пятидневку с 15 марта после стаивания сне-
га и наступления теплой погоды росли очень интенсивно как в длину,
так и в ширину (рис. 54). Во вторую пятидневку их рост по обоим
измерениям несколько упал, оставаясь наиболее интенсивным у пары
нижних листьев. В третью пятидневку после стаивания снега листья
вновь начинают усиленно расти до зацветания. С наступлением перио-
да цветения, особенно в первую пятидневку, листья растут еще ин-
тенсивнее. В дальнейшем пара нижних листьев в длину продолжает
неуклонно увеличиваться, а в ширину темп роста незначительно па-
дает.
У третьего и четвертого (рис. 55) листьев во вторую пятидневку
после наступления периода цветения резко сокращается прирост в.
282
длину. Четвертый лист на одну пятидневку растет дольше третьего.
Это объясняется тем, что у Т. greigii четвертый лист обычно нахо-
дится высоко на стебле и, видимо, перехватывает часть питательных
веществ, поступающих по стеблю в интенсивно растущую в этот пе-
риод. плодоножку.
Рис. 54. Рост листьев розетки впервые зацветшего расте-
ния Т. greigii в ширину.
1—нижний лист, 2—второй лист, о—третий лист и 4—верхний лист.
Во время цветения третий и четвертый листья в ширину растут
равномерно, но очень незначительно (рис. 54).
Однако все листья розетки как в длину, так и в ширину рано
прекращают рост— до окончания периода цветения (рис. 54, 56).
Бутон тюльпана, появляясь на поверхности почвы, бывает уже
значительных размеров. До расцветания он интенсивно растет в дли-
Рис. 55. Рост листьев розетки впервые зацветшего растения
Т. greigii в длину.
7—-нижний лист, 2— второй лист, 2—третий лист, 4—верхний лист.
ну и лишь незначительно в ширину. У Т. kaufmanniana на поверхно-
сти почвы бутон в большинстве случаев быстро окрашивается и рас-
крывается. В этот период резко возрастает его рост в ширину и не-
сколько уменьшается в длину (рис. 57).
В 1954 г. в связи с неблагоприятной зимой многие тюльпаны не дали
семян, а у Т. kaufmanniana завязи были значительно мельче, чем в
обычные годы. Рост завязей наиболее интенсивно протекал во время
283
•бутонизации и периода цветения и резко упал сразу после цветения.
Цветоплодоножка интенсивно росла во время цветения и после него
(рис. 57). Хотя темп роста ее в последний период несколько снизился,
но за 25 дней цветоплодоножка выросла лишь на 0,5 см меньше, чем
за 19 дней цветения и бутонизации. Ее рост продолжался до 25 ап-
реля и после цветения сохранял удивительно ровный ритм.
Рис. 56. Рост бутона и цветка
Т. greigii.
1—в длину, 2—в ширину.
Рис. 57. Рост бутона и
цветка Т. kaufmanniana.
1—высота, 2—диаметр.
Цветы Т. kaufmanniana часто расцветают как только бутон пока-
жется в розетке. Особенно часто это наблюдается у раннецветущих
•форм — из земли обычно показывается цветок с окрашенными лис-
точками околоцветника. Изредка встречаются формы, у которых цве-
ток в первый и второй день открытия листочков околоцветника и рас-
трескивания пыльников лишь на половину длины выходит из земли.
После цветения плодоножка таких растений бурно растет и выносит
завязь на 15—20 см над уровнем почвы.
Среди особей Т. kaufmanniana также часто встречаются формы
с еще зеленым бутоном, высоко поднятым над почвой.
Между формой листа, темпом роста и длиной цветоплодоножки
Т. kaufmanniana существует некоторая зависимость. При широких округ-
лых листьях цветы обычно расположены на коротких цветоножках;
узкие длинные листья сопутствуют бутонам на длинных цветоножках
и цветам, раскрывающимся высоко над почвой.
Растениям Т. kaufmanniana требуется меньший срок от появления
бутона над почвой и его зацветанием, чем Т. greigii (рис. 56, 57, 58,
•59). Бутон Т. kaufmanniana (рис. 57) в длину растет интенсивнее, чем
цветок, и энергичнее, чем бутон Т. greigii (рис. 56).
Бутон Т. greigii при появлении над почвой длительное время
очень слабо растет, а затем — интенсивно, как цветок.
Резкий скачок в росте в ширину обусловлен у обоих видов рас-
крытием бутонов, но бутон и цветок у этих видов развиваются по-
разному (рис. 56, 57).
284
У Т. kaufmanniana (рис. 58) завязь сильно растет во время буто-
низации, слабее — в период цветения и быстро прекращает рост после
цветения; у Т. greigii (рис. 59)—во время цветения и после него до 5
мая включительно. Таким образом, у Т. kaufmanniana долго растет
плодоножка, а завязь быстро пре-
кращает рост, у Т. greigii, на-
оборот (рис. 59), — плодоножка
кончает развитие почти одновре-
менно с отцветанием, но на пол-
месяца дольше, чем у Т. kauf-
manniana, растет завязь.
В заключение следут отме-
тить, что у тюльпанов с увели-
чением возраста растений наблю-
дается смена форм возобновления
от моноподиальных к симподи-
альным. У любого вида или сорта
тюльпана в течение первых лет
онтогенеза до наступления ста-
дийной зрелости луковица нара-
стает при ПОМОЩИ верхушечной рире 58. Рост плодоножки и завязи Т. ка-
точки роста, т. е. моноподиально. ufmanniana.
В ЭТОЙ точке роста В течение 7—длнна плодоножки, 2—длина завязи, 3—ширина
многих лет происходят ступен-
чатые изменения, приводящие р астение к половой зрелости.' f При
достижении растением зрелости цветоносные побеги всегда заклады-
Рис. 59. Рост плодоножки и завязи Т. greigii.
1—длина плодоножки, 2— длина завязи, 5—ширина завязи.
ваются из верхушечной почки луковицы, а в случае в многоцветково-
сти — из верхушечных почек-деток-луковичек.
Жизнь взрослого растения продолжается обычно за счет развития
285
пазушной почки самой внутренней запасающей чешуи, т. е, симподи-
ально. Основньш структурным элементом тюльпана, таким образом,
является монокарпический пибег, который до первого зацветания рас-
тения развивается моноподиально в течение многих лет. Побег состо-
ит из чередующихся длинных и укороченных отрезков стебля.
В первый год жизни тюльпана особенно сильно растет ось перво-
го узла, которая, срастаясь с брюшной стороной влагалища семядоли,
растет геотропически положительно и уносит верхушечную почку на
несколько сантиметров вниз от места ее заложения на проростке.
Длительное срастание влагалища семядоли с осевой частью сто-
лона углубления заставляет признать, что данное образование являет-
ся не вытянутым междоузлием, а односторонне вытянутой, разросшей-
ся в поперечном направлении узловой частью побега. Анатомическая
структура стеблевой части данного оригинального образования более
сближает его с междоузлием, чем с узлом. А боковые листовые со-
судистые пучки, например у сеянца Т. kaufmanniana, свободно про-
ходящие вдоль всего столона до почки, не сливаются со стеблевыми
у более взрослых растений и убеждают нас в том, что в данном слу-
чае мы имеем поперечное разрастание узла, а не междоузлие.
Поперечное разрастание узла в сторону выпуклой половины дон-
ца и его геотропически положительный рост являются ювенильной
особенностью тюльпанов, которой обладают сеянцы, не завершившие
стадии развития до образования плодущих побегов, и органы вегета-
тивного размножения—почки-детки.
Первая розетка листьев тюльпана состоит из семядоли и единст-
венного низового запасающего листа — запасающей чешуи.
В конце периода покоя следующий узел вновь растет" односторон-
не в ширину, а по выходе из луковицы получает геотропически по-
ложительный рост. Он, как и в первый год жизни растения, плотно
срастается с влагалищем первого зеленого листа.
Вторая розетка листьев состоит из первого настоящего зеленого
листа и второго низового листа (запасающей чешуи). На поверхности
почвы ежегодно из розетки листьев тюльпана вырастает один зеленый
ассимиляционный лист.
Заложение в луковице сеянца почки, состоящей из одного асси-
миляционного листа и двух или больше запасающих, показывает про-
исшедший перелом в развитии сеянца и его зацветание на следующий
за этим год.
Приближение к завершению развития побега у большинства ви-
дов характеризуется прекращение.м образования столонов углубления
(за исключением Т. kaufmanniana, Т. praestans и сорта King George
V) и заложением розетки из одного зеленого листа и не менее двух
запасающих листьев (чешуй).
Луковица такого растения в наступивший период жаропокоя за-
канчивает развитие формирование.м из верхушечной почки генератив-
ного побега, который характеризуется геотропически отрицательньш
ростом, длинными междоузлиями, безвлагалищными, сидячими сере-
динными листьями и конечным цветком.
С этого момента кончается моноподиальный рост полицикличе-
ского побега, и жизнь индивидуума продолжает почка в пазухе само-
го внутреннего запасающего листа луковицы.
Глава X
ГОДОВОЙ ЦИКЛ РАЗВИТИЯ ВЗРОСЛОЙ ЛУКОВИЦЫ
ТЮЛЬПАНА
rjg зучение годового цикла развития растения тюльпана из взрослой
Е луковицы представляет интерес, особенно в связи с кратковре-
менным активным периодом жизни зеленой части растения.
В работах 3. М. Силиной (1953) и Э. Ф. Мигушовой (1954) уде-
ляется значительное внимание годовому циклу развития луковиц
у культурных тюльпанов (только иностранные сорта) в условиях
Ленинграда и Ленинградской области.
3. М. Силина в течение 1949, 1950 и 1951 гг. проводила наблю-
дения над ростом и развитием пяти сортов тюльпанов—Бин I, Turner,
William Copland, Розовый Шарик и Prince of Austria. Три первых, по
Силиной, принадлежат к группе тюльпанов Дарвина, Розовый Шарик-
к группе Cottage, a Prince of Austria—к простым ранним тюльпанам.
Однако сорт Бин I по всем признакам принадлежит к тюльпанам из
группы Cottage, а сорт, называемый 3. М. Силиной „Розовый Шарик11,
на самом деле является одним из наиболее старых сортов Дарвинов-
ских тюльпанов и называется Margaret (Gretchen).
Наиболее полно в работе 3. М. Силиной описаны луковицы сорта
William Copland, типичного для группы тюльпанов Дарвина, и изредка
сравниваются с луковицами сорта Бин I. О развитии луковиц прочих
сортов только упоминается в начале раздела. Таким образом, прихо-
дится верить автору, что в годовом цикле развития луковиц из раз-
ных групп садовых тюльпанов в условиях Ленинграда нет никакого
различия.
По ритму развития надземных органов, по данным 3. М. Силиной,
сорта Turner и Prince of Austria несколько различаются. Так, растения
Turner зацветают 25—30 мая, Prince of Austria на несколько дней
раньше—23—26 мая. В условиях Ташкента (1954 г. и последующие
годы) различие в сроках начала зацветания у этих сортов значительно
более резкое. Так, сорт Turner зацвел 21 апреля, а первый цветок
у сорта Prince of Austria раскрывался 12 апреля —на 9 дней раньше,
чем у Turner. Сорт Prince of Austria в условиях Ташкента является
одним из наиболее поздних из ранних сортов.
Э. Ф. Мигушова (1954) отмечает, что заложение дочерних луко-
виц у сортов ранних тюльпанов в условиях Ленинграда наблюдается
позже, чем у сортов поздноцветущих. Так, у сорта Zwanenburg (груп-
па Дарвина) замещающая луковица закладывается в мае, а у сорта
Marchal Niel (группа Махровые ранние) —через месяц. Э. Ф. Мигу-
шова изучила морфогенез у 14 сортов тюльпанов, но не распределила
287
их по группам. Судя по работе, она исследовала сорт Kansas из груп-
пы Triumph, Zwanenburg из группы Дарвина и Marchal Niel из группы
ранних Махровых тюльпанов. Остальные сорта неизвестны. Выводы
о развитии луковицы тюльпанов в годовом цикле у обоих авторов
весьма не сходны. Для наиболее полного понимания этих процессов,
видимо, необходимо изучить их сравнительно с развитием диких видов.
В работах 3. М. Силиной и Э. Ф. Мигушовой основное внимание
уделено развитию органов вегетативного размножения—замещающей
луковицы и почек-деток.
Развитие годового цикла у взрослых луковиц диких видов тюль-
панов еще никем не изучено.
МАТЕРИАЛ И МЕТОДИКА
В течение 1950—1955 гг. мы проводили наблюдения над ростом
и развитием половозрелых луковиц пяти диких видов и двух куль-
турных сортов тюльпанов.
В 1950 г. по 150 луковиц высшего разбора Т. fosteriana и сорта
Jubilee из группы тюльпанов Дарвина мы высадили на отдельном
участке и для исследования через каждые две недели в течение года
выкапывали не менее двух луковиц каждого вида. Однако у Т. foste-
riana оказалось много неполовозрелых особей. Поэтому и в последую-
щие годы мы продолжали изучение этого вида.
При выкопке производилось описание и измерение всего растения
от надземных органов до корешков. Обычно с луковицы тут же
последовательно снимали, измеряли и зарисовывали чешуи. Извле-
ченные пазушные почки раскладывали в порядке заложения на влаж-
ную фильтровальную бумагу, затем резали и изучали. Растущие мери-
стематические ткани луковицы выделяли от запасающих при помощи
раствора йода в йодистом калии (J+KJ). В центральной почке после-
довательно измеряли все органы, а развитие генеративных органов
изучали ускоренными методами при помощи ацетокармина. Пути сле-
дования сосудистых пучков проявляли флероглюцином с соляной
кислотой. Однако этим методом распределение сосудистой системы
могло быть изучено только в области донца луковицы и в тех орга-
нах, где прошло одревеснение. Из-за отсутствия отложений крахмала
в окружающей сосудистые пучки паренхиме, в чешуях и почках лукови-
цы в молодом состоянии сосудистую систему выделяли раствором йода в
йодистом калии, а в старых луковицах, наоборот, ее выделяли фут-
ляры из клеток, заполненных крахмалом, как правило, одевающих
сосудистые пучки. Сахара изучали с помощью фелинговой жидкости.
Целлюлозные оболочки исследовали хлорцинкйодом, жиры—Суданом III.
В последующие годы мы исследовали и другие виды тюльпанов
и несколько изменили принцип отбора растений для анализа.
У культурных тюльпанов (сорта Jubille, D. v. Т. yellow) все лу-
ковицы одного сорта являются вегетативно размноженными потомками
одной единственной луковицы-родоначальницы или нескольких, но
очень немногих родичей, т. е. клоном, поэтому, исследуя несколько
луковиц одного сорта и размера, выращиваемых в одинаковых усло-
виях, можно получить сравнимые данные. У диких же видов при
слабом вегетативном размножении в естественных условиях каждая
луковица вырастает из семени и представляет организм с особой
наследственностью, оригинальным путем развития, сложившимся под
влиянием разнообразных внешних условий, и разным возрастом. В
первые годы мы не могли обеспечить все этапы исследования диких
288
видов вегетативно размноженными луковицами, потомками единствен-
ной родоначальной луковицы.
Поэтому годовой цикл у Т. kaufmanniana мы изучали на луковицах,
выросших из семян одной коробочки посева 1948 г. Эти луковицы
впервые зацвели в 1952 г. Из них в 1954 г. отобрали для исследования
150 луковиц первого сорта и высадили на особые гряды.
У Т. korolkovii взяли луковицы цветущего размера из посева
1950 г., впервые зацветшие в 1954 г.
У Т. subpraestans мы исследовали луковицы первого разбора,,
полученные в результате вегетативного размножения. Каждый месяц
(2-го и 16-го) луковицы исследовали. Если в назначенный срок наблю-
дение не было проведено, то луковицы и растения измеряли и фик-
сировали в спирте для того, чтобы в более свободное время провести
дальнейшее изучение.
В условиях Средней Азии каждый год вегетация тюльпанов начи-
нается в разное время, но всегда значительно раньше, чем в более
северных районах Советского Союза.
В Ташкенте ранней весной, в феврале, а иногда в конце января,
отрастают розетки листьев тюльпанов. Ранние тюльпаны начинают
цвести в первой декаде февраля или марта, иногда в начале апреля.
Первыми зацветают тюльпаны секции Eriostemones, затем Spiranthera
и Lopholhyllon, а виды секций Leiostemones и Tulipanum—почти
одновременно. Сроки окончания вегетации и рассеивания зрелых
семян у видов всех секций тюльпанов более сближены.
В развитии тюльпана можно отметить три узловых момента: от-
растание корней, появление почки над поверхностью почвы и уход
в период летнего покоя. Как в первом, так и во втором этапе мы встре-
чаемся с тем, что органы как вегетативного, так и генеративного
размножения растений уже заложились и в значительной степени
выросли. Период летнего покоя для растений тюльпанов наступает
в конце вегетации, когда начинается подсыхание кончиков листьев.
В этот период сплющенная часть стебля, проходящая около луковицы,
становится вялой и мягкой, но стебель, листья и коробочка еще зеле-
ные. Чешуи материнских луковиц уже вполне опустошены, а на за-
мещающей луковице и детках покровная чешуя утончена, прозрачна
и начинает подсыхать. Гнездо еще не распадается. Внутри луковиц
происходит развитие почек. В условиях Ташкента этот период на-
ступает очень рано, вскоре после цветения—для Т. fosteriana—в на-
чале мая. Таким образом, с момента начала увядания надземных
зеленых органов листьев, стеблей и подземных корней, у тюльпана
начинается летний период покоя, но в различное время у растений
неодинакового возраста.
Окончание периода вегетации связано с принадлежностью тюль-
пана к определенной систематической единице, возрастом особи и с
ее состоянием. В условиях Ташкента тюльпаны секции Eriostemones
кончают вегетацию раньше видов других секций. Все дикие виды
среднеазиатского происхождения, за исключением Т. subpraestans,
созревают раньше всех культурных тюльпанов. Особи, не вступившие
в пору плодоношения, оканчивают вегетацию раньше половозрелых
представителей группы. Раньше заканчивают вегетацию растения
с удаленной завязью.
Циклы развития отдельных органов в течение года так перепле-
таются, что разорвать, разъединить их без ущерба для целостности
представления о жизни растения невозможно. В недрах старой луко-
вицы, находящейся в полной силе, зарождаются новые луковицы,
19—2229 289
сменяющие на будущий год старую. Обычно еще до засыхания зеле-
ного побега и окончания роста органов, в период, когда разверты-
вается ассимиляционная поверхность листьев, у основания побега в за-
мещающей луковице в виде крошечного валика уже закладывается
первый лист побега будущего года. С этого момента мы и начнем
списание годового цикла развития тюльпанов.
Мы долго искали пути графического изображения годового жиз-
ненного цикла роста и развития тюльпанов. Нужно было изобразить
их органы во взаимосвязи как в последовательности, так и в одновре-
менности роста, т. е. скрытого роста органов внутри луковицы и откры-
того роста надземного побега. Ход годового развития тюльпанов мы
отобразили графически с помощью круговой диаграммы (рис. 60, 64,
70, 75, 79, 83, 94), на которой оказалось возможным изобразить во
времени и взаимосвязи рост всех главнейших органов растения. Не-
которая отвлеченность и схематичность круговых диаграмм искупается
тем, что они дают яркое представление об организме в целом. Диа-
грамма дополняется графиками в плоской прямолинейной проекции,
которые дают представление о росте отдельных органов.
Круг мы разбиваем на 12 секторов, соответствующих 12 месяцам
года, и обозначаем каждый сектор римской цифрой. Спиральные круги
соответствуют количеству органов, которые представлены на диаграмме.
Функционирующие органы растения изображены сплошной линией,
а находящиеся в зачаточном состоянии и еще не выполняющие своих
функций—пунктиром.
Начальный отрезок от центра круга спирали, обозначенный циф-
рой 1, изображает процесс с момента заложения луковицы до ее обо-
собления в самостоятельный организм и отмирания. Далее (цифра 2)
показан рост корней (сравнить круговую диаграмму со схематичным
изображением растения на рис. 59); затем (цифра 3)—рост побега со
стеблем, цветком и листьями; 4—нижнего листа по направлению к
периферии круга; 5—второго листа; 6—третьего листа. У некоторых
видов добавляются четвертый и пятый листья, их заложение и рост
откладываются на соответственных местах по направлению к периферии
круга, но обычно на 7-м месте — стебель, 8 — цветок, завершаю-
щего цикл развития, и 9 — завязь. Эмбриональное состояние растения
на графике отмечено точками, а активное—сплошной линией.
Tulipa kaufmanniana Rgl.
Замещающая луковица (рис. 60) закладывается в середине июня.
До второй декады мая следующего года она растет в недрах мате-
ринской луковицы. После отсыхания чешуй материнской луковицы
замещающая луковица обособляется и до второй половины мая треть-
его года существует как самостоятельный организм. Таким образом,
луковица тюльпана развивается и растет в недрах материнской лу-
ковицы в течение И месяцев, и 12 месяцев живет самостоятельно.
Всего с момента заложения до отмирания луковица существует 23
месяца.
В третьей декаде июня, почти одновременно с заложением почки
замещающей луковицы, в донце материнской луковицы начинается
заложение корней (рис. 60, 2), которые медленно развиваются и к
началу октября подходят к эпидермису донца. При пониженных темпе-
ратурах и увеличивающейся влажности почвы корни очень быстро
растут, прорывают эпидермис и немедленно начинают функционировать,
подавая в луковицу воду и питательные вещества из почвы.
290
Корни луковицы, как и сама луковица, отмирают во второй дека-
де мая.
Меристема донца, в которой закладываются зачатки корней, не-
долговечна. Заложение корней вначале происходит в центральной
части подковы, образуемой донцем, затем корни распространяются по
направлению к периферии подковы донца. Чем мощнее луковица, тем
больше рядов корней закладывается в донце.
При прорастании луковицы сохраняется тот же порядок в отра-
Рис. 60. Т. kaufmanniana. Годовой цикл развития.
I—XII месяцы, 1—луковица, 2—корни, 3—весь побег, 4—нижний лист.
5—средний лист, 6—верхний лист, 7—стебель, 8—цветок.
станин корней, что и при закладке: вначале прорывают эпидермис
донца корни центральной части дуги, а затем через одну-две недели
показываются и периферические. Если осенью во время прорастания
луковицы обрезать некоторые корни, то часть донца, с которой они
были срезаны, останется совершенно голой, так как меристема, зало-
жившая их, вся расходуется одновременно и в дальнейшем не спо-
собна регенерировать. У тюльпанов очень ограничено время деятель-
ности меристем и совершенно отсутствуют явления регенерации. По-
этому агротехнические или биологические приемы—подрезка или при-
щипка корней—невозможны. А пересадка вегетирующих растений с
повреждением хотя бы части корневой системы наносит им непопра-
вимый ущерб на много лет. Период заложения и роста корней в
донце у Т. kaufmanniana продолжается три с половиной месяца. Период
активной жизни, когда они функционируют как органы, длится семь
месяцев и около десяти—двадцати дней. При засушливой осени корни
позже отрастают и увеличивается период скрытого роста, а при влаж-
ной осени—уменьшается. Корни с начала заложения в меристеме
донца до отмирания живут около 11 месяцев. Один месяц в годовом
цикле растения не имеют ни старых, ни зачатков новых корней.
А в течение четырех с половиной—пяти месяцев после отмирания кор-
ней старой луковицы и отсутствия функционирующих новых растение
291
не имеет никакой корневой связи с почвой. Это и есть период покоя,,
т. е. период наиболее возможного при сохранении жизни разрыва
связи растения со средой.
Верхушечная почка растения (рис. 60, 3) закладывается в тот
период, когда замещающая луковица находится в чешуях материнской
луковицы, у которой на побеге едва развертывается первый лист.
Поздней весной это происходит даже до появления побега материнской
луковицы над поверхностью почвы. В нашем опыте 1954 г. первые
бугорки побега следующего года заложились в замещающих лукови-
цах в начале марта или в последние дни февраля. Заложение, внутри-
луковичный и подземный рост побега продолжались одиннадцать меся-
цев и несколько дней. Активный надземный рост до полного усыхания
длился почти четыре месяца. Следовательно, побег от заложения до
полного усыхания существует 15 месяцев.
Нижний лист (рис. 60, 4) в виде едва заметного валика на мери-
стематическом бугорке побега впервые становится заметным через
две недели после заложения, обычно в середине марта. Время от за-
ложения до начала развертывания листа мы считаем скрытым периодом
его роста. У этого вида он длится десять месяцев. После появления
над поверхностью почвы и началом развертывания в середине февра-
ля до усыхания в конце мая, т. е. три месяца и несколько дней,
нижний лист живет активной жизнью. Всего с момента заложения до
засыхания нижний лист существует немного больше четырнадцати
месяцев. Второй лист (рис. 60, 5), средний, у большинства форм этого
вида закладывается значительно позже нижнего. Появление валика
этого листа мы наблюдали в третьей декаде мая, во время отмирания
его на побеге материнского растения. В первых числах марта следую-
щего года второй лист обычно показывался над поверхностью почвы
и вскоре развертывался. Он отмирал на несколько дней раньше ниж-
него листа.
Жизнь второго листа короче первого. Скрытый период его роста
от момента заложения до развертывания длится почти девять месяцев
и десять дней. Активный период—с момента развертывания до засы-
хания—примерно два месяца двадцать дней, т. е. средний лист на
побеге от момента заложения до засыхания живет двенадцать ме-
сяцев.
Верхний лист закладывается значительно позже двух первых.
Впервые валик этого листа можно наблюдать в середине июля, на
четыре месяца позже, чем валик первого листа, и почти на два месяца
позже среднего. Но над поверхностью почвы он появляется почти
одновременно со вторьим листом и вегетирует почти такое же коли-
чество дней (рис. 60, 6). Подземный скрытый рост его с момента
заложения до выхода над поверхностью почвы продолжается семь
с половиной месяцев. Активный период жизни у этого листа длится
немного менее двух с половиной месяцев, т. е. с момента заложения
верхнего листа до засыхания проходит 10 месяцев—на два месяца
меньше среднего и на четыре—нижнего листа. Однако активный
период его жизни немногим короче, чем у нижних листьев. В данном
случае развитие органа ускоряется в связи с положением на побеге.
Чем выше на побеге закладывается лист, тем короче период его скры-
того роста и развития и тем быстрее он созревает для выполнения
своих функций.
Стебель (рис. 60, 7) начинает расти значительно позже листьев.
В начале марта закладывается весь побег и стебель, который долго
не растет в длину, но функционирует как орган. На стебле в это
292
время закладываются листья. Первой заметно растет часть стебля от
луковицы до нижнего листа. Начало ее роста, скрытого внутри луко-
вицы, наблюдается в первых числах сентября. Следующей весной в
первой декаде марта стебель выносит цветочную почку на поверхность
почвы и продолжает вытягиваться до конца мая, т. е. зачаток стебля
функционирует как орган почти с момента закладки. Но расти он
начинает в последней декаде сентября после закладки всех листьев
и венчающего побег цветка. Таким образом, развитие стебля слага-
ется из трех периодов: органообразовательный—с начала марта по
двадцатые числа сентября (примерно, 195 дней), скрытого подземного
роста—с середины сентября по десятые числа марта (около 180 дней)
и выхода бутона на дневную поверхность почвы в десятых числах
марта до созревания коробочки и засыхания в конце мая (около 75
дней). Весь период жизни стебля длится 450 дней.
Соотношение роста листьев и стебля в длину с момента их зало-
жения до отмирания четко выявляет: 1) интенсивный рост листьев
у этого вида наступает задолго до укоренения растения; 2) в зимние
месяцы—с начала декабря до первых чисел февраля—рост как стебля,
так и листьев падает или очень замедляется; 3) при выходе из зим-
него покоя раньше наступает интенсивный рост стебля, а затем и
листьев.
Более детальные наблюдения за листьями и стеблями проведены
после того, как они появляются над поверхностью почвы. Цветок
закладывается после листьев (рис. 60, 5). В середине августа стано-
вятся заметными три валика, соответствующие трем листочкам внеш-
него круга околоцветника. Формирование цветка заканчивается к
началу сентября. В конце октября в пыльниках в материнских клетках
пыльцы происходит редукционное деление. Пыльца зимует в виде
одноядерных пыльцевых зерен. У исследованных в данном опыте
растений цветок показался над поверхностью почвы в середине марта
и к первому апреля отцвел. Завязь созрела в конце мая. Таким об-
разом, генеративные органы развиваются с момента заложения первых
листочков околоцветника до появления их над поверхностью почвы
в течение 7 месяцев. Цветение продолжается 15 дней, после чего
завязь созревает еще два месяца.
Надземные органы Т. kaufmanniana после опадения листочков
околоцветника не прекращают роста. Цветоножка, став плодоножкой,
значительно вырастает. Развивающаяся завязь стимулирует рост плодо-
ножки и листьев. Завязь во время цветения и после него сильно рас-
тет в длину и соответственно увеличивается в ширину.
Так, у растений, зацветших в середине марта, завязь при распу-
скании цветка была 2 см pji. и 0,4 см шир., а в момент взятия пробы
1 апреля, через неделю после опадения листочков околоцветника,—3,5 см
дл. и 0,7 см шир. Это было приземистое сильное растение с листьями
-следующих размеров: нижний—11,3 см pji., 8,4 см шир.; второй-
11,5 см дл., 5,9 см шир. и третий — 11 см дл., 4,8 см шир. Нижняя
часть стебля слагается из двух отрезков: первый—от донца луковицы
до уровня почвы (6 см) и второй—от уровня почвы до места при-
крепления нижнего листа (2,3 см). А. Д. Холл (1940) выделяет виды,
у которых нижняя часть стебля б. м. высоко приподнимает листья
над поверхностью почвы. Из среднеазиатских видов растений такое
листорасположение имеет Т. hoogiana. У Холла в каждом диагнозе
-тюльпана отмечено, на какой высоте от уровня почвы отходит первый
лист растения.
293
У Т. kaufmanniana мы исследовали заложение почек в луковицах
с момента появления верхушечной почки над землей до усыхания
листьев (табл. 22).
Следовательно, надземные органы тюльпана этого вида функцио-
нируют недолго.
Красно-зеленая верхушечная почка тюльпана 16 февраля поднялась,
над землей на 3 см. С 1 марта развернулась вершина нижнего листа
Таблица 22
Рост генеративного побега Т. kaufmanniana
Время на- блюдения Лист Стебель (низ)
нижний средний верхний дл. шир.
дл. шир. дл. шир. дл. шир.
16.11 5,7 2 5 1,3 4,35 0,9 6 0,5
1.Ш 8,5 4 8,5 2,5 7,5 2 6,5 0,5
16.III 10,4 5,8 10,5 3,4 9,8 1,8 8,5 0,7
Цветение
1.IV 11,3 8,4 11,5 5,9 И 4,8 8,9 0,7
18.IV 12,5 9 12 ' 6 12 4,5 10,5 0,7
Начали > <елтеть -
3.V 11,5 7 11,2 4 11 3 8 0,8
16.V Посохли и сломан ы 11 0,7
2. Засохло все растение, но сез лена еще i лажные.
Время на- блюдения Междоузлие Плодоножка Завязь Рыльце
нижнее верхнее дл. шир. дл. шир. дл. шир.
дл. шир. дл. шир.
16.11 0,5 0,4 0,4 0,35 1 0,27 0,08 0,08 0,12 0,2
1.1II 0,6 0,48 0,5 0,45 1,3 0,4 1,4 о,з 0,4 0,4
16.Ш 1,2 0,55 0,9 1,45 3 0,5 1,9 0,4 0,2 0,6
Цветение
1.IV 1,3 0,65 1,5 0,55 8,9 0,5 3,5 0,7 0,5 0,7
18.1V 2,1 0,6 2,5 0,50 12,6 0,5 5,3 1,5 0,2 0,55-
Начали желтеть
3.V 2,3 0,6 2,5 0,58 11,3 0,4 6,5 2,6 0,8 0,7
16.V 2 0,4 1,9 0,4 10 0,4 6,3 2,4 — —
2
и стал виден свернутый в трубку второй лист. К 16 марта у растения
наблюдалась розетка из трех листьев и начавший распускаться бутон,,
к 1 апреля растение отцвело.
Растение с неудаленной завязью 18 апреля еще зеленое, 3 мая
верхняя треть листьев пожелтела и усохла, коробочка в это время
зеленая. Листья засохли 16 мая, коробочка—бледно-зеленая. Все над-
земные органы 2 июня засохли, но семена еще влажные.
294
Таким образом, надземная часть растения Т.kaufmanniana со вре-
мени появления верхушечной почки над поверхностью почвы в 1955 г.
жила 107 дней.
В первые две недели после выхода почки из земли (табл. 22)
нижняя часть стебля растет очень слабо, междоузлия — несколько
активнее и цветоножка почти совсем не растет. В последующие две
недели вплоть до зацветания растения резко увеличивается рост ниж-
ней части стебля, равномерно—междоузлий, интенсивно—цветоножки..
Во время цветения рост нижней части стебля несколько снижается.
Цветоножка в это время увеличивает размеры интенсивнее других
зеленых органов растения; по-прежнему медленно растут междоузлия-
По выходе из почвы междоузлия растут равномерно, на них не влияет,
как на другие части оси, зацветание растения и только в первые две
недели после отцветания наблюдается относительно резкое увеличение
их роста, после чего до конца вегетации рост междоузлий прекра-
щается.
В течение двух недель после отцветания растений значительно»
растет цветоножка и чуть заметно нижняя часть стебля, затем все
части оси резко останавливаются в росте.
В первые две недели после выхода почки из почвы наиболее
интенсивно растет в длину нижний лист, несколько сбавляя темпы в
следующие две предшествующие цветению недели. Рост нижнего
листа медленно увеличивается во время цветения и в течение двух
недель после его окончания, а через месяц — совершенно прекра-
щается. Пожелтение кончиков листьев свидетельствует о начале
прекращения их роста. Вполне возможно, что основание листа про-
должает расти, когда верхняя часть листа уже увядает. Завязь интен-
сивно растет в первые две недели после выхода бутона на поверхность
почвы и почти прекращается за две недели перед цветением. После
раскрытия цветка до отцветания рост завязи вновь увеличивается,
особенно в длину. В последующие недели завязь непрерывно увели-
чивается. После цветения завязь продолжает интенсивно расти теми
же темпами, как и во время цветения, и лишь несколько меньшими,
чем в первые две недели после отцветания. В это время все другие
органы — подземная часть стебля, междоузлия, цветоножка, ставшая
плодоножкой, и листья—совершенно прекращают рост. Таким образом,
завязь Т. kaufmanniana растет на две недели дольше всех других
надземных органов тюльпана.
Заложение и рост замещающей луковицы и луковиц-деток.
Весна и осень 1954 г. для изучения вегетативного размножения диких
видов в условиях Ташкента были неудачными. Однако начальные этапы
заложения деток в крупных луковицах были обеспечены предшест-
вующим вегетационным сезоном и поэтому о начальных этапах роста
и развития деток можно составить верное представление (рис. 61). У
Т. kaufmanniana, как правило, отсутствуют детки покровной чешуи.
Детка первой запасающей чешуи закладывается в первых числах мая
во время обособления замещающей луковицы из чешуи материнской
луковицы. После заложения полтора месяца детка первой запасающей,
чешуи не увеличивается в размере. Позже, с начала июня до сере-
дины июля, она постепенно увеличивается, достигая к концу периода
0,5 мм дл. Во вторую половину июля и начале августа рост ее пре-
кращается. Во второй половине августа она вновь начинает увеличи-
ваться и растет весь сентябрь. В октябре ее рост резко возрастает до
середины ноября. До середины декабря темпы роста луковицы несколь-
ко снижаются, с середины декабря для этой детки наступает зимний
295
период покоя. Однако рост детки луковицы первой запасающей чешуи
до вступления в период зимнего покоя был недостаточным, достигнув
всего 1 см, она больше не растет. Почти одновременно с вышеописан-
ной деткой в пазухе второй запасающей чешуи закладывается детка,
которая с момента заложения до середины сентября растет очень слабо.
С середины сентября в течение двух месяцев она усиленно растет. В
середине ноября, достигнув размера детки первой запасающей чешуи,
она совершенно прекращает развитие.
Рис. 61. Заложение и рост замещающей луковицы и почек-деток
Т. kaufmanniana.
1—детка первой запасающей чешуи, 2—детка второй запасающей чешуи, <?-—дет-
ка третьей запасающей чешуи, 4—детка четвертой запасающей чешуи, 5—замещаю-
щая луковица.
Детка третьей запасающей чешуи закладывается только в середи-
не июля. У нее не наблюдается такого длительного периода микро-
скопического роста — полуанабиоза, как у деток первых двух чешуй,
так как в июле состояние запасных питательных веществ в чешуях
совершенно иное, чем в момент заложения деток первых двух чешуй.
Непосредственно после заложения детка третьей запасающей чешуи
начинает активно расти. С середины сентября до середины ноября
темпы роста этой детки возрастают, достигая максимума в период с
середины ноября по середину декабря, после чего прирост у детки
этой чешуи резко падает, однако, в период покоя она не вступает и
равномерно увеличивается в размерах до конца вегетации. У некото-
рых сильных луковиц в середине ноября наблюдается закладка детки
четвертой запасаюшей чешуи. До середины января она быстро, но
равномерно растет, после чего дает резкий скачок в росте в длину.
Дальнейшее развитие мы не могли проследить, так как эта детка
наблюдалась у ограниченного количества растений.
Рост замещающей луковицы (рис. 62). Замещающая лу-
ковица закладывается в середине июня у основания генеративного
побега (раньше, чем детки в пазухах третьей и четвертой запасающих
чешуй) и до первых чисел августа почти не растет. Затем до 1 сен-
296
тября она медленно растет. В течение двух последующих недель ее
прирост падает. С двадцатых чисел сентября запасающая луковица
интенсивно растет вплоть до зимнего покоя. С первых чисел ноября
до начала февраля она находится в покое. В середине этого периода
наблюдается вспышка роста, видимо, обусловленная таянием почвы и
возобновившейся деятельностью корней. С середины января или начала
I6J46 1-Ю 2-20549 146 2-16447 146 1-Ю 1-18 3462-16 1-16 146 2-26543 146246447146 !46 148 3-Ю 246
п т т 1х х х/ м _/ х х[ хи -1 1L HL S £ ™
Рис. 62. Рост замещающей луковицы Т. Kaufmanniana.
1—в длину, 2—в ширину.
февраля растение вступает в весенний рост. Значительный рост отме-
чен с 1 по 16 февраля, в дальнейшем до 16 марта рост замещающей
луковицы уменьшается, но не прекращается, усиливается с середины
марта. Замещающая луковица Т. kaufmanniana непрерывно растет до
первых чисел мая, после чего наблюдается как бы уменьшение ее
размера, обусловленное усыханием покровной чешуи.
В данном случае мы проследили рост замещающей луковицы в
целом, не затрагивая органообразования, протекающего внутри нее во
время этого роста.
Бугорок почки замещающей луковицы закладывается в середине
июня. На этом бугорке лишь в середине августа явственно обозначает-
ся валик покровной чешуи. При разрезе этой почки в начале сентября
внутри валика зачатка покровной чешуи уже заметен второй валик —
зачаток первой запасающей чешуи. За ним немедленно поднимаются
валики второй и третьей запасающих чешуй. Несколько позже закла-
дывается валик четвертой запасающей чешуи, которой у многих поло-
возрелых особей этого вида может и не быть. Таким образом, зало-
жение чешуй в замещающей луковице Т. kaufmanniana происходит
через два-три месяца после заложения меристематического бугорка
почки, однако, к моменту наступления активного периода осеннего
роста в ней уже закладываются все луковичные чешуи. К зиме заме-
щающая луковица имеет в зачаточном состоянии все органы.
Заложение и рост органов цветка (рис. 63). Заложение органов
цветка у Т. kaufmanniana проходит снизу вверх. Первыми в начале
297
августа закладываются в виде полулуний три валика внешних листоч-
ков околоцветника. Вскоре также в виде валиков намечаются зачатки
листочков внутреннего круга околоцветника. Тычинки закладываются
в такой же последовательности. Бугорки тычинок внешнего круга
бывают уже достаточно обрисованы, а внутреннего круга—едва наме-
чаются. В первых числах сентября про-
Нр 2-20 5-W /16216 4/7 Мб /16 /18
Ш /X XX/ ХП 7 и /// iy
Рис. 63. Заложение и рост орга-
нов цветка у Т. kaufmanniana.
а—завязь, б—пыльники, в—бутон и цве-
ток; 1—длина, 2—ширина.
исходит заложение плодолистиков, кото-
рые поднимаются в виде трех отдельных
валиков. Эти валики на вершине свобод-
ны и несколько позже сливаются ниже-
лежащими частями.
Рост всех органов цветка протекает
почти однотипно (рис. 63), т. е. все они
сразу после заложения интенсивно ра-
стут до начала октября. С начала до
середины октября рост частей цветка за-
медляется. Затем, видимо, в связи с на-
чавшимся ростом корней и поступле-
нием через них в луковицу из почвы
воды и растворенных в ней солей, рост
вновь возобновляется, причем наиболее
интенсивно у тычинок. Но к первому
ноября развитие органов цветка падает.
Завязь растет не так быстро как тычин-
ки, но продолжается до начала декабря.
У листочков околоцветника рост в пер-
вых числах октября не прекращается, но
значительно падает и в таком темпе про-
должается до началя декабря. С этого
времени, а у пыльников на две недели
раньше, в органах цветка на зимний пе-
риод рост прекращается.
Первым из зимнего покоя выходит
пестик, рост которого начинается с пер-
вых чисел января и в медленном темпе продолжается до начала мар-
та, когда наблюдается резкий скачок в темпе роста пестика.
На полтора месяца позже завязи в середине февраля из периода
покоя выходят листочки околоцветника и очень интенсивно растут до
конца цветения.
Последними в начале марта начинают расти тычинки. Рост пыль-
ников почти заканчивается осенью, а весной перед цветением лишь
вытягиваются тычиночные нити.
Tulipa fosteriana J г *v. (рис. 64)
Заложение и рост замещающей луковицы. В середине или конце
июля у основания генеративного побега, который зацветет на буду-
щий год, закладывается замещающая луковица (рис. 64). Десять
месяцев, до середины мая следующего года, она растет в недрах
старой луковицы. Освободившись от чешуи материнской луковицы,
замещающая луковица еще год живет как самостоятельный организм.
Вся жизнь луковицы Т. fosteriana от заложения до отмирания
длится 22 месяца.
Меристематический бугорок почки замещающей луковицы у осно-
вания генеративного побега становится заметным лишь в третьей
298
декаде июля (рис. 65)’. До начала сентября он слабо растет и на нем
незаметно заложение чешуй. В начале сентября на бугорке появля-
ются валики зачатков покровной чешуи первой и второй запасающих.
Причем, когда первые две обозначились уже достаточно отчетливо,
третья едва-едва намечается. Через две недели, в середине сентября,
IX
X
VI
Рис. 64. Т. fosteriana. Годовой цикл развития.
I—XII месяцы: 1—луковица, 2—корни, 3—весь побег, 4—нижний
лист, 5—второй лист, 6—третий лист, 7—верхний лист, 8—стебель, 9—цветок,
10—завязь.
набухают зачатки еще двух чешуй. Таким образом, в почке замещаю-
щей луковицы в течение, примерно, двадцати дней закладываются
Рис. 65. Рост замещающей луковицы Т. fosteriana.
7—покровная чешуя, 2—первая запасающая чешуя, 3—вторая запа-
сающая чешуя, 4—третья запасающая чешуя.
зачатки всех чешуй. Чешуи этой почки до осеннего укоренения
материнской луковицы почти не растут. Исключение представляет
299
покровная чешуя, которая в течение двух недель после заложения
сильно увеличивается в размере, а затем с середины сентября до
середины ноября (время выпадения осадков и укоренения материнской
луковицы) больше не растет.
С начала до середины ноября немного вырастают первая и третья
запасающие чешуи. С середины ноября постепенно увеличивается
покровная чешуя. Ее ровный без резких скачков и спадов рост
продолжается без перерыва до середины января, резко увеличиваясь
во второй половине января и уменьшаясь в начале февраля. С сере-
дины февраля до середины апреля покровная чешуя растет очень
интенсивно. Ее рост в длину заканчивается на две недели раньше, чем
рост запасающих чешуй, которые со времени заложения до середины
февраля увеличиваются в размере очень незначительно. Так, первая
запасающая чешуя со времени заложения до середины февраля дости-
гает всего 0,9 см выс., а третья—0,5 см. С середины февраля до
начала мая все запасающие чешуи растут ровно, быстро и дольше,
чем покровная чешуя. Таким образом, для этого вида характерно,
что замещающая луковица в основном растет весной до периода лет-
него покоя растения.
Корни (рис. 64, 2). В первых числах июля в зоне корневой ме-
ристемы донца луковицы ясно видно начало формирования корней,
заканчивающих формирование к октябрю. При наличии влажности в
почве корни могут прорасти, или задержать прорастание до конца ноября
и позже. Отмирают корни в мае, а у бесплодных особей—часто в конце
апреля.
Побег (рис. 64, 3). В середине марта, когда рост замещающей
луковицы еще не закончен и луковица тесно связана с материнским
растением, в центре ее закладывается генеративный побег. В это время
будущий побег представлен весьма незначительным меристематическим
бугорком. Вскоре на нем закладывается валик нижнего листа. А к
началу апреля совершенно четко вырисовывается валик второго листа.
На этом закладка листьев временно прекращается и возобновляется
после наступления у материнского растения периода летнего покоя.
В середине июня закладывается третий лист, в начале июля— четвер-
тый. На этом заложение листьев обычно заканчивается. Взрослые
растения имеют 3 листа, особи с большим количеством листьев редки
среди представителей этого вида. Между заложением первого и вто-
рого листьев проходит меньше времени, чем между заложением вто-
рого и третьего. Заложение третьего и четвертого листьев также б. м.
сближено, т. е. листья как бы парами закладываются на побеге.
В начале июля начинается закладка цветка. В октябре побег
выходит из луковицы. С середины февраля до начала марта он появ-
ляется над поверхностью почвы. А в середине мая заканчивает жизнь.
Коробочки засыхают значительно позже, чем листья на стебле.
Рост листьев (рис. 64, 4, 5, 6; 65). До обособления в самостоя-
тельный организм и завершения роста в замещающей луковице за-
кладываются нижний и второй листья побега, которые два с половиной
месяца почти не увеличиваются. Но в начале июня после окончания
формирования роста и обособления замещающей луковицы оба листа
внезапно дают небольшой прирост. К середине июня их рост прекра-
щается приблизительно на два с половиной месяца.
Третий лист после заложения в начале июня растет толь-
ко в начале августа. В начале сентября он по длине догоняет нижние
листья и все три листа до середины августа растут энергично. Но
так как в Ташкенте в это время нет осадков, то посаженные лукови-
300
цы не могут укорениться и рост их листьев вновь падает. Третий
лист первым на месяц (до середины октября) прекращает рост. Два
нижних листа менее интенсивно продолжают рост. С начала до сере-
дины октября останавливается в росте второй лист, а нижний продол-
Рис. 66. Рост листьев и стебля Т. fosteriana.
1—нижний лист, 2—средний лист, 3—верхний лист, 4—стебель.
жает увеличиваться. Со второй половины октября до начала ноября
все три листа вновь активно растут. В связи с зимним покоем рост
двух верхних листьев прекращается до начала января; нижний лист
301
в это время медленно, но беспрерывно растет. У двух верхних листь-
ев может быть незначительный прирост во второй половине этого
периода.
С начала января все три листа опять начинают энергично расти.
С середины января до начала февраля рост листьев несколько сни-
жается, что связано с усиленным ростом стебля. Во второй половине
февраля и начале марта насту-
Рис. 67. Заложение и рост органов цветка
Т. fosteriana.
а—завязь, б—тычинки, в—бутон, а затем цветок.
пает активный период в росте
листьев, а рост стебля значи-
тельно снижается. В начале
апреля наступает весенний ак-
тивный период в росте стебля,
а рост листьев вновь падает.
Затем он прекращается, а
стебель продолжает активно
расти. Следовательно, чем бы-
стрее растут листья, тем мед-
леннее увеличивается стебель
и, наоборот (рис. 66).
Цветок (рис. 64, Р; 67). В
начале июня одновременно с
заложением корневых мери-
стем и верхнего листа на вер-
шине стебля начинается зак-
ладка цветка. Закладываются
валики листочков околоцвет-
ника и почти одновременно
бугорки тычинок, но до нача-
ла сентября они очень мало
увеличиваются. В первых чис-
лах сентября у растения насту-
пает активный период роста
всего побега и органов цвет-
ка листочков околоцветника и
пыльников тычинок (рис. 67).
Закладываются и растут пло-
долистики.
Этот первый осенний рост
тычинок и плодолистиков весь-
ма непродолжителен. Он за-
тихает на две недели в се-
редине сентября, а в начале
октября начинается с прежней
интенсивностью у пыльников
и в значительно меньшей сте-
пени у плодолистиков. Во вто-
рой половине октября соотно-
шение их роста меняется—ин-
тенсивнее растут плодолистики и несколько замедляется рост пыльни-
ков. Листочки околоцветника продолжают активно расти, несколько
снижая темпы роста к началу ноября.
С начала ноября до первых чисел января рост органов цветка
совершенно прекращается на период зимнего покоя. В начале января,
видимо, в связи с оттаиванием почвы и активизацией деятельности
корневой системы, органы цветка дают незначительный прирост, а во
302
вторую половину января рост их вновь приостанавливается. В начале
февраля органы цветка окончательно выходят из периода зимнего
покоя. С этого времени, постепенно нарастая, их рост не прекращается
до конца цветения во второй половине апреля.
Тычинки, впавши в период зимнего покоя, во второй половине
ноября не выходят из него и не растут до начала февраля. Весь
февраль они медленно, но непрерывно увеличиваются в размере,
давая резкий скачок в росте в первой половине марта. До зацветания
в конце марта или начале апреля они немного вырастают за счет
вытягивания тычиночных нитей. После раскрытия цветка их пыльники
лопаются и сморщиваются.
Плодолистики, как и околоцветник с тычинками, во второй поло-
вине ноября вступают в период зимнего покоя, но он более короткий,
чем у периферических органов. Плодолистики вместе с листочками
околоцветника выходят из состояния покоя в середине января и не
прекращают микроскопически малого роста до начала марта. В первой
половине марта перед выходом на поверхность почвы их рост затор-
маживается. Во второй половине марта рост плодолистиков и листоч-
ков околоцветника резко увеличивается и вновь падает в конце марта
перед распусканием цветка. После этого усиленно начинает расти
завязь. Ее рост продолжается дольше всех органов цветка и длится
до второй половины мая.
Заложение и рост почек-деток (рис. 6§). Среди изученных нами
луковиц Т. fosteriana очень редко наблюдалось заложение почки-детки
в пазухе покровной чешуи, поэтому мы не могли изучить развитие
этой почки-детки в годовом цикле.
Рис. 68. Заложение и рост почек-деток Т. fosteriana.
1—детка первой запасающей чешуи, 2—детка второй запасающей чешуи,
о—детка третьей запасающей чешуи.
При вступлении растения Т. fosteriana в период покоя после со-
зревания семян и по обособлении замещающей луковицы в начале
июня, когда нет заложения никаких других органов, в пазухе первой
запасающей чешуи на центральной жилке возникает бугорок меристе-
матической ткани. Он появляется на клиновидной меристеме, лежащей
между чешуями и сильно разрастающейся к этому времени (рис. 69).
После заложения бугорок начинает медленно расти, достигая за 1
месяц нескольких миллиметров. Через полмесяца после заложения
бугорка в пазухе первой запасающей чешуи в пазузе второй запа-
сающей чешуи также закладывается меристематический бугорок поч-
ки-детки и растет в начальные этапы значительно медленнее, чем
первый. После заложения этих двух почек проходит полтора месяца
до заложения почки в пазухе следующей чешуи.
303
Между заложением почек второй и третьей запасающих чешуй у
растений этого вида закладывается масса различных органов — корни,
верхние листья и органы цветка,—после чего в первых числах августа
закладывается почка в пазухе третьей запасающей чешуи. Она закла-
дывается позже зачатка замещающей луковицы, являющейся пазушной
почкой четвертой запасающей чешуи (рис. 68).
В августе меристемы почек-деток дают незначительный прирост,
но быстро останавливаются в росте. Такое же незначительное увели-
чение их роста наблюдается
во время укоренения луко-
виц во второй половине ок-
тября до середины ноября.
Почка-детка третьей за-
пасающей чешуи, заложив-
шаяся последней, первой из
всех пазушных почек-деток
начинает развитие в середи-
не декабря и за полмесяца
увеличивается в два раза.
Однако ее рост замедляется
равномерно, продолжаясь до
начала мая, но не дает того
прироста, как у почек дру-
гих чешуй.
Почка-детка второй че-
шуи начинает активный рост
со второй половины января
и в быстром темпе продол-
жает его до второй поло-
вины апреля. Позже всех
вступает в период активного
роста почка в пазухе пер-
вой запасающей чешуи. Она
выходит из состояния зим-
него покоя в начале февра-
ля и со второй половины
февраля активно растет в
длину до начала апреля,
т. е. до зацветания растения.
Таким образом, из зим-
него покоя почки-детки вы-
ходят постепенно от центра
к периферии, а рост прекра-
щается раньше у внешних
деток и позже у внутрен-
ние. 69. Заложение почки-детки в пазухе первой
запасающей чешуи луковицы Т fosteriana.
I—ткани запасающих чешуй, заполненные крахмалом, 2— кли-
новидная меристема, д—зачаток почки-детки.
НИХ.
Заложение и развитие
чешуй в детках проходит
в следующем порядке. В
начале июля на меристематическом бугорке детки первой запасаю-
щей чешуи намечается валик покровной чешуи. В конце ок-
тября — первой запасающей; в начале ноября явно обозначается
зачаток второй запасающей; в начале декабря—третьей, а в конце —
четвертой. У детки второй запасающей чешуи также в начале июля
было отмечено заложение валика покровной чешуи. Длительное время
304
не было заложения других частей луковицы, только в начале ноября
отмечены валики двух чешуй: крупный—первой запасающей чешуи и
едва наметившийся валик—второй запасающей чешуи. В начале дека-
бря появляется валик третьей чешуи, а в начале января—четвертой
чешуи детки из пазухи второй запасающей чешуи.
У детки третьей запасающей чешуи, заложившейся позже всех, к
первому сентября на меристематическом бугорке появляется валик
зачатка покровной чешуи; первого ноября — первой запасающей. В
начале декабря отмечены зачаточные валики второй и третьей запа-
сающих чешуй. На этом заложение чешуй у этой детки заканчи-
вается.
Коллатеральные почки-детки у этого вида наблюдаются очень
редко, а сериальные почки-детки могут закладываться у каждой поч-
ки за любой чешуей.
Обычно сериальная почка-детка закладывается у основания, вер-
нее на сосудистом пучке почки-детки после заложения меристемати-
ческого валика покровной чешуи, но до заложения зачатка первой
запасающей чешуи. Такие почки-детки мы наблюдали также у заме-
щающей луковицы этого вида.
В течение года только в мае и феврале не происходит заложения
никаких органов. Распределение заложения различных частей расте-
ния по месяцам представлено в табл. 23 . Весной во время вегетации
Время заложения органов Т. fosteriana
Таблица 23
Условные обозначения. I л—первый лист, II л—второй лист, III л—тре-
тий лист, IV л—четвертый лист, пп меристематический бугорок, л—листочки около-
цветника, т—тычинки, птс—пестик, I—покровная чешуя, II —первая запасающая чешуя,
III—вторая запасающая чешуя, IV—третья запасающая чешуя.
растения закладывается побег с двумя листьями. В период летнего
покоя происходит интенсивное органообразование; закладываются вы-
шележащие листья, весь цветок, корни, почки-детки, у которых обра-
20—2229 305
дуется валик покровной чешуи, замещающая луковица со всеми че-
шуями. Во время осеннего роста и зимнего покоя закладываются все
запасающие чешуи почек-деток. Причем, в зимние месяцы (ноябрь,
декабрь и январь), когда совершенно приостанавливается рост частей
побега, происходит активное органообразование в почках-детках. По-
ражает взаимосвязь и взаимообусловленность всех процессов роста и
развития в этом растении.
Отрицательной особенностью вегетативного размножения Т. fos-
teriana является способность почек-деток к сильному омоложению. У
половозрелой луковицы этого вида почки-детки обычно не уносятся
на столонах углубления, а остаются в чешуях материнской луковицы.
После обособления они также закладывают одну—две детки, которые
образуют столоны углубления, отходящие в стороны и несколько вниз,
а иногда и вверх, и уносят почки-детки на расстояние от 1 до 40 см
от материнской луковицы. Замещающая луковица остается на месте.
Но растение, образовавшееся из пазушной почки, сильно омолажива-
ется и обычно не цветет, а образует всего один лист. На рост и
построение таких длинных столонов у него уходит много сил, и ре-
зультирующие луковицы мельчают. На следующий год наблюдается
та же картина, и так каждый год. Очень редко удается среди массы
особей отобрать такие, которые почти не образуют столонов углубле-
ния, а дают кучное гнездо деток, как культурные тюльпаны. Размно-
жая такие растения, из этого вида можно создать сорта. Видимо, от-
бором среди растений, полученных в виде луковиц из СССР, в Гол-
ландии был создан из этого вида сорт „Red Empirior“, „Madam Lefebr“
и другие. Простым отбором красивоцветущего и хорошо вегетативно
размножающегося, но слабо образующего столоны растения мы выде-
лили сорт „Материнская слава“.
Tulipa subpraestans (рис. 70,71).
Заложение и рост замещающей луковицы. В начале июля у ос-
нования нового побега в центре только что обособившейся молодой
луковицы закладывается бугорок замещающей луковицы, который
полтора месяца (с начала июля до середины августа) не растет. С се-
редины августа до середины сентября он незначительно увеличивается,
после чего плавно растет до середины ноября. Период роста связан с
началом деятельности корней материнской луковицы. С середины ноя-
бря до начала февраля продолжается период зимнего покоя и моло-
дая замещающая луковица не растет. Но с начала до середины фев-
раля эта луковица начинает интенсивно расти. С середины февраля
до середины марта ее рост вновь приостанавливается. С середины
марта луковица вновь быстро растет. В начале апреля рост ее заме-
дляется, а в начале мая—продолжается с новой силой. В середине мая
развитие луковицы заканчивается и в связи с усыханием покровной
чешуи размеры ее уменьшаются. В этот период прерывается связь
луковицы с окончившей существование материнской луковицей и на-
чинается жизнь самостоятельного организма.
Порядок заложения чешуй влуковице. В начале июля
закладывается бугорок замещающей луковицы. Валик зачатка покров-
ной чешуи появляется на нем в середине августа. Валик первой запа-
сающей чешуи обозначается через месяц, т. е. в середине сентября.
Дальше закладка чешуй проходит более быстрыми темпами — через
две недели вслед за первой появляется валик второй запасающей че-
шуи и почти одновременно третьей.
306
Таким образом, закладка всех чешуй замещающей луковицы про-
текает до начала ее роста перед зимним покоем. Ко времени укоре-
нения материнской луковицы в ее замещающей—закладываются все
чешуи. Органообразовательный период новой замещающей луковицы
проходит в период летнего покоя материнской луковицы.
Рис. 70. Tulipa subpraestans Vved.
Годовой цикл развития: I—XII месяцы, 1—луковица, 2—корни, 5—весь побег,
4—нижний лист, —второй лист, 6—третий лист, 7—четвертый лист, 8—пятый тист,
9—шестой лист, 10—стебель, 77—цветок, 12—завязь.
Скрытый период жизни замещающей луковицы, т. е. период, когда
сна тесно связана с материнской луковицей, продолжается с середины
июля до середины или конца мая следующего года (305—320 дней).
С момента обособления из чешуй материнской луковицы замещающая
луковица живет еще год. Полтора-два месяца со времени обособле-
ния ее протекает до начала за-
ложения корней в меристеме
донца, четыре с половиной, а
иногда и пять месяцев до их
укоренения в середине октяб-
ря. В среднем, со времени
заложения до отмирания, за-
мещающая луковица живет 20
-с половиной месяцев.
Корни. Заложение корней
в меристеме донца луковицы
начинается в середине июля
одновременно с замещающей
луковицей. В скрытом состоя-
2-J8 г~18 3~ls г21§ 2d? 4-22 5-J8 2J7
т ш № х я/ т Г я « /г v у/
Рис. 71. Рост и развитие замещающей
луковицы Т. subpraestants Vved.
нии внутри донца корни находятся, примерно, до середины, а иногда
и до конца октября, т. е. до первых осенних осадков. По выходе из
307
донца они начинают всасывающую деятельность, отмирают в середине
или конце мая, начале июня следующего за заложением года. Весь
период со времени заложения до отмирания корней длится 10 меся-
цев, из них более трех месяцев приходится на период скрытого роста
и развития внутри донца (рис. 70).
Побег. В третьей декаде апреля, во время цветения растения, в
центре замещающей луковицы закладывается бугорок побега буду-
щего года (рис. 70). Два с половиной месяца этот бугорок не увели-
чивается в размере и на нем не закладываются органы. В конце июня
или в начале июля на бугорке появляется валик первого листа, через-
два-три дня с противоположной стороны обозначается валик второго
листового бугорка и только в середине августа появляется валик
третьего листа, а вскоре за ним и четвертого. В середине августа
закладывается верхушечный цветок, а в пазухах двух верхних листь-
ев обозначаются бугорки двух или более ветвей, на которых в конце
августа почти одновременно закладывается по листу.
В начале сентября на вершинах боковых ветвей закладывается
по цветку. Несмотря на то, что эти цветки появляются позже верху-
шечного, они лишь незначительно отстают в развитии и часто зацве-
тают почти одновременно. В первой половине марта красная вершина
побега показывается над поверхностью почвы. Вскоре развертываются
листья побега. А в конце мая или начале июня побег высыхает, соз-
ревают семена и начинается самостоятельная жизнь нового организма,
развивающегося из замещающей^луковицы у основания этого побега.
Таким образом, и у растений Т. subpraestans все главные органо-
образовательные процессы в целом закончились до его выхода из
периода летнего покоя внутри луковицы, т. е. во время наибольшего
отрыва от внешней среды, когда у растения нет функционирующих
листьев и всасывающих корней.
Рост листьев и стебля (рис. 72). На обозначившемся в се-
редине апреля бугорке побега в начале июля закладывается в виде
продолговатого валика нижний лист и почти одновременно второй. До
середины августа они плохо растут. В это время закладывается тре-
тий и чуть позже—четвертый. Первый и второй листья со второй по-
ловины августа начинают понемногу удлиняться. Во второй половине
сентября нижний лист очень вырастает, значительный прирост дают
также второй, третий и четвертый листья. В это же время заклады-
ваются пятый, шестой и т, д. листья на боковых побегах в пазухах
уже заложившихся листьев главной оси. Чем выше на оси располо-
жен лист, тем быстрее после заложения он начинает расти и тем,
сравнительно, меньший прирост он дает. С первых чисел октября рост
листьев несколько приостанавливается, а в середине октября вновь,
усиливается. Это свидетельствует о том, что растения, подготовлен-
ные к укоренению, в условиях жаркого и сухого климата Ташкента
не могут укореняться.
Часть запасов материнской луковицы используется начавшими рост
чешуями замещающей луковицы. Наиболее незначительный прирост у
нижнего и четвертого листьев. Кроме того, в этот период заклады-
ваются пазушные веточки с зачаточными листьями, что, видимо, от-
нимает много сил у растения и сказывается на последующем росте
листьев. Во второй половине октября вновь начинается усиленный
рост всех листьев, затухающий у четвертого и пятого листьев в пер-
вой половине ноября. У второго, третьего и четвертого листьев он
приостанавливается во второй половине ноября. Нижний лист все это
время непрерывно растет до начала января, после чего совершенно
308
прекращает рост до начала февраля. С начала февраля нижний лист
растет до конца жизни, особенно сильно в апреле.
Вышележащие листья по-разному реагируют на внешние условия.
Нижний из них в середине ноября в период прекращения роста лис-
тьев дает лишь легкое снижение темпа роста; у второго листа период
покоя длится всего две. недели; у третьего — месяц; у четвертого —
Рис. 72. Рост стебля и листьев Т. subpraestans Vved.
1—нижний лист, 2—второй лист, 3—третий лист, 4—четвертый аист,
5—пятый лист, 6—стебель.
почти полтора месяца; у пятого—наступает раньше, а к началу дека-
бря уже снижается, но в течение последующих двух месяцев он ра-
стет очень вяло до начала наиболее сильного роста нижележащих
листьев.
После выхода из ноябрьского покоя второй лист до начала января
растет активней, чем нижний; они одновременно впадают в период
зимнего покоя и дальше развиваются одинаково. Третий лист, лежа-
щий внутри почки, на две недели позже выходит из ноябрьского покоя
309
и в начале января не прекращает роста» а только тормозит его до
начала марта.
Четвертый лист, лишь в начале января выходящий из периода
осеннего покоя, в середине января вновь приостанавливается в росте
до середины февраля, после чего все листья розетки усиленно растут,
особенно в апреле. Исключением является пятый лист, который с
первых чисел марта до начала апреля почти не растет. Это происхо-
дит, видимо, потому, что вышележащий цветок и более сильные
нижние листья захватывают большую долю передвигающихся из лу-
ковицы по стеблю запасов воды и питательных вешеств. С первых
чисел апреля и он начинает активно расти, но не достигает размера
нижележащих листьев.
Стебель у генеративного побега, заложившись в середине апреля,
очень медленно растет до середины сентября, т. е. времени, когда
заканчивается закладка всех органов этого побега. С середины сен-
тября по середину ноября он интенсивно растет, а затем до середи-
ны января впадает в период зимнего покоя с полным прекращением
видимого роста. В течение второй половины января стебель начинает
медленно расти, а в первой половине февраля дает резкий скачок в
росте. Во второй половине февраля, видимо из-за похолодания, его
рост, как и нижнего листа, падает, но не прекращается. Однако у
всех листьев в середине марта начинается наиболее активный период
роста, а стебель приступает к нему лишь в начале апреля, опережая
в этом отношении лишь пятый лист.
В 1954 г., когда проводились эти наблюдения, условия для роста
диких видов тюльпанов были неблагоприятными. Но приведенное опи-
сание взаимозависимости и взаимообусловленности роста отдельных
органов генеративного побега в связи с большим объемом изученного
материала в основном соответствует действительной картине.
Цветок (рис. 73). У Т. subpraestans стебель, как правило, ветвит-
ся и кроме цветка, расположенного на главной оси, на ветвях, выхо-
дящих из пазух листьев, наблюдаются добавочные верхушечные
цветы. Бугорок цветка на главном стебле закладывается в середине
августа. Через две недели на нем ясно обозначаются валики листоч-
ков внешнего круга околоцветника, вслед за ними листочки внутрен-
него круга. В начале сентября околоцветник начинает разрастаться и
намечаются бугорочки тычинок, в заложении которых существует та
же последовательность, что и листочков околоцветника: вначале в
виде овалов обозначаются бугорки внешнего круга, а за ними внут-
реннего. Почти одновременно с тычинками внутреннего круга закла-
дывается пестик в виде трех продолговатых, обособленных друг от-
друга валиков.
В начале сентября листочки околоцветника через две недели после'
заложения интенсивно развиваются до середины ноября. Так же ра-
стут пыльники тычинок, в отличие от листочков околоцветника, давая
лишь некоторое снижение темпов роста в середине октября и пре-
кращение роста к середине ноября. Медленнее всех растет пестик.
Достигнув к началу ноября весьма незначительных размеров, он на
две недели раньше других органов цветка впадает в зимний покой.
У листочков околоцветника период покоя продолжается три с
половиной месяца, во второй половине которых отмечается незначи-
тельный рост листочков околоцветника. У пыльников период покоя
длится четыре месяца -— со второй половины ноября по вторую поло-
вину марта. За это время только в начале февраля наблюдается не-
значительный прирост длины. Зимний покой пестика продолжается с
310
Z*' / й/ 27/ 7 il ш (V
Рис. 73. Заложение и рост органов
у цветков с одного растения.
1—верхушечный цветок, 2—второй цветок,
3—третий цветок; а—завязь, б—тычиночные нити,
в—пыльники, г—листочки околоцветника.
начала ноября по конец первой половины марта, т. е. четыре с поло-
виной месяца.
В начале декабря обозначаются тычиночные нити, представленные
маленькими треугольничками. Только во второй половине февраля
в связи с наступившим потеплени-
ем они дают незначительный при-
рост.
Выход из зимнего покоя начи-
нается одновременно во всех частях
цветка во второй половине марта.
За две недели окончательно вы-
растают в длину пыльники. В пер-
вой половине апреля они уже не
прибавляют в длину и удлинение
тычинок в это время происходит за
счет интенсивного роста тычиноч-
ных нитей. На две недели раньше
пыльников начинают расти листоч-
ки околоцветника, наиболее интен-
сивный рост которых наблюдается
во второй половине марта и начале
апреля, когда уже не растут пыль-
ники,
Пестик одновременно с пыль-
никами выходит из периода покоя
и начинает расти. С середины мар-
та по середину апреля он растет
непрерывно, но не очень активно.
И лишь после раскрытия цветка
до начала мая пестик вырастает до
размеров взрослой завязи.
Таким образом, раньше всех
органов цветка прекращают рост
пыльники. Тычиночные нити и ли-
сточки околоцветника прекращают
его одновременно. Позже всех за-
канчивает рост пестик, после опыле-
ния развивающийся в завязь. Боко-
вые цветки растут и развиваются,
как главный (рис. 73).
Почки-детки (рис. 74). Расте-
ние Т. subpraestans обладает спо-
собностью, наряду с заложением
почек-деток в пазухах запасающих
чешуй, подобно культурным тюльпанам, закладывать их в пазухе
покровной чешуи.
Заложение деток обеспечивается предшествующей вегетацией и
поэтому, несмотря на неблагоприятные условия 1954 г., мы могли
исследовать их.
Во второй половине апреля в пазухе покровной чешуи обнаружи-
вается меристематический бугорок несколько меньшего размера. Бу-
горок наблюдается и в пазухе первой запасающей чешуи. В начале
мая закладывается меристематический бугорок в пазухе второй запа-
сающей чешуи. Затем в заложении почек наступает длительный пере-
рыв и лишь в середине сентября через четыре с половиной месяца
311
после заложения последнего бугорка в пазухе второй запасающей
чешуи набухает бугорок в пазухе третьей запасающей чешуи.
Детка покровной чешуи начинает расти сразу после заложения
и через две недели (в середине мая) она достигает 0,2 см дл., мед-
леннозрастет и к середине сентября составляет всего 3 мм дл. С се-
174-22 548 247 248 248 2/8 348 348 246 448 24? /4? 221 548 247
Ш [У V W VU № к X Af Ж 1 И Щ /V V V/
Рис. 74. Рост деток одной лу кори цы Т. subpraestans Vved.
1—детка покровной чешуи, 2—детка первой запасающей чешуи, 3—дет-
ка второй запасающей чешуи.
редины сентября по середину октября она активно растет и достигает
0,6 см. Затем темп роста снижается и возобновляется в середине
января. К середине марта детка покровной чешуи равна 1 см выс. С
середины марта она активно растет, достигая к июню 3,2 см дл.
Заложившаяся одновременно с деткой покровной чешуи, детка
первой запасающей чешуи до середины августа почти не растет в
длину. В течение последующих двух месяцев она растет, но медленно,
составляя к середине октября 0,35 см выс., к середине января —
0,5 см, к середине апреля—0,7 см. Но с середины апреля она резко
увеличивается в длину, часто достигая к июню 3 см выс.
Рост заложившейся в начале мая детки второй запасающей чешуи
становится заметным только в середине сентября: к началу октября
юна вырастает до 0,2 см и вновь останавливается в росте. До начала
февраля она прибавляет всего 0,1 см дл. Весь февраль растет активно
и достигает к первому марта 0,7 см дл. Дальнейший рост детки зави-
сит от весенних условий—если весна достаточно влажная и теплая,
детка развивается очень активно и достигает размера замещающей
луковицы. Так же развиваются детки третьей и четвертой запасающих
чешуй. Они закладываются в середине сентября, примерно, через пять
месяцев после заложения первых деток луковицы. У них не наблю-
дается длительного летнего анабиоза. После заложения они немедлен-
но начинают расти и в первый же месяц достигают 0,5—0,7 см выс.
Так же как и детки, заложившиеся раньше, они почти не растут зи-
мой. Весной активный период роста наступает у них несколько рань-
ше, чем у нижележащих деток. С первого марта они трогаются в
рост и при благоприятных условиях догоняют по размеру замещающую
луковицу.
Tulipa korolkovii (рис. 75)
В середине апреля после отцветания или во время цветения в
замещающей луковице растения этого вида, точнее, в ее верхушеч-
ной точке роста, происходит закладка нового побега или побега бу-
дущего года. А в середине или конце мая следующего года на этом
шобеге можно собирать зрелые коробочки. Таким образом, побег жи-
вет 13 месяцев. Характерным в развитии этого побега является раз-
рыв в сроках заложения органов. Так, нижний лист закладывается
вскоре после появления меристематического бугорка побега в послед-
512
лей декаде апреля, а второй лист—лишь в середине августа, т| е. поч-
ти через четыре месяца после первого. Третий лист в виде крошечного
валика намечается одновременно с заложением цветка в последней
V//
VI
XII
Рис. 75. Tulipa korolkovii Rgl.
Годовой цикл развития: I—XII месяцы, Г—луковица, 2—корни,
3— весь побег, 4—нижний лист, 5—второй лист, 6—верхний лист,
7—стебель, 8—цветок, 9—завязь.
декаде августа. В это же время начинает вытягиваться стебель. Все
органы побега закладываются в течение двух недель, и лишь нижний
лист появляется значительно раньше.
Зачатки корней в корневой
ютсяв начале июня. К октябрю
корешки заканчивают форми-
рование. С первыми дождями
в начале ноября они прорыва-
ют эпидермис донца и враста-
ют в почву. В середине апреля
•следующего года они отми-
рают.
Заложение и рост заме-
щающей луковицы (рис. 76).
Замещающая луковица у осно-
вания молодого побега закла-
дывается несколько позже пе-
риода заложения органов ге-
неративного побега, т. е. в
начале сентября. Луковица
•отмирает (истощается) в сере-
дине апреля. Так, скрытый
период жизни от заложения
до освобождения ее из чешуи
меристеме донца луковицы появля-
Рис. 76. Tulipa korolkovii Rgl. Рост замеща-
ющей луковицы.
1—покровная чешуя, 2—первая запасающая, 3—вторая
запасающая.
материнской луковицы длится шесть
с половиной месяцев и как самостоятельный организм она живет еще
313
почти год. Следовательно, луковица Т. korolkovii живет 1 год 6,5
месяца.
Замещающая луковица поздно закладывается и в начале растет
медленно. К началу сентября она достигает всего 0,05 см выс., через
две недели—0,18 см. В этот период закладывается первая запасающая
чешуя. Рост покровной чешуи замедляется и первая запасающая че-
шуя почти догоняет ее к началу октября, составляя 0,17 см дл. С се-
редины октября до начала ноября покровная и запасающая чешуя
незначительно вырастают. В этот период закладывается вторая запаса-
ющая чешуя и все они до начала февраля едва прибавляют по 0,05
см в месяц. С середины февраля они растут быстрее, особенно увели-
чиваясь во второй половине марта. В это время замещающая луко-
вица достигает 1,9 см выс. В период цветения рост чешуй падает, а
с начала апреля опять усиливается. Покровная чешуя в конце пери-
ода роста с вытянутым на вершине луковицы носиком имеет длину
2,6 см-, первая запасающая—1,5 см и вторая запасающая—1,2 см. На
этом развитие заканчивается.
Следовательно, заложение каждой новой чешуи несколько задер-
живает рост предшествующей. После того как новая чешуя почти
догоняет предшествующую, они растут параллельно друг другу. Вто-
рая запасающая чешуя, замедляя рост в длину во время цветения,
уже не восстанавливает его и после цветения. А покровная и первая
запасающая чешуи и после цветения растут с неменьшей силой.
Рост побега. В период, когда молодая замещающая луковица
останавливается в росте, в ее верхушечной точке роста закладываются
побег и первый лист побега будущего года (рис. 77). Кроме первого листа
Рис. 77, Tulipa korolkovii. Рост листьев и стебля.
1—нижний лист, 2—средний лист, 3—верхний лист, 4—стебель.
и побега в последующие четыре месяца нет заложения других орга-
нов. С середины августа первый лист начинает медленно расти. В
начале сентября закладываются второй и третий листья и£несколько
вытягивается нижнее междоузлие стебля.}
314
Все листья и стебель активно растут до середины октября. В это
время начинают работу корни. Из-за отсутствия осадков рост листьев
во вторую половину октября значительно падает, а в первую поло-
вину ноября прекращается. Стебель продолжает расти. Во второй
половине ноября выпадают осадки, отрастают корни и рост всех ор-
ганов поэтому восстанавливается, но только на две недели. С начала
декабря до начала февраля рост всех листьев приостанавливается на
два месяца. Стебель прекращает рост во второй половине декабря,
а с начала до середины января вырастает на 2 см. За две недели до
выхода из покоя листьев стебель вновь приостанавливается в росте
на полтора месяца: с середины января до начала марта. С начала
февраля листья усиленно растут и при благоприятных температурных
условиях появляются над поверхностью почвы. К середине марта
листья заканчивают рост, около полутора месяцев остаются зелеными,
но больше не растут. За две недели до прекращения роста листьев
вновь наступает период роста стебля. С начала до середины марта он
растет медленно. Но со времени раскрытия цветка (с середины
марта до середины апреля) он вырастает на 4 см. Раннее прекращение
роста листьев является особенностью этого вида.
Цветок (рис. 74, 8, 9). В начале сентября на верхушке побега, ле-
жащего внутри луковицы, закладывается цветочная почка—бутон. За-
ложение всех органов цветка происходит почти одновременно. Так,
в середине августа при анализе мы не
заложение цветка, а в начале сентября
уже были зачатки всех его органов
(рис. 78). Первые фазы роста листочков
околоцветника, тычинок и пестика толь-
ко внешне б. м. одинаковы.
С начала сентября по первые числа
октября листочки околоцветника выра-
стают на 8 мм, после чего их рост рез-
ко снижается и за две недели они увели-
чиваются только на 1 мм. С середины
октября до середины ноября рост ли-
сточков околоцветника прекращается.
После укоренения растения до конца
ноября они вырастают на 2—3 мм и
рост их прекращается до начала февра-
ля, т. е. на два месяца. С начала фев-
раля до середины марта они вырастают
на 3 мм. Следовательно, за весь период
роста с начала сентября по середину
марта они увеличились в размере на
1,3 см, а за две последних недели мар-
та—на 1,5 см.
Тычинки закладываются одновре-
менно с листочками околоцветника и
наблюдали даже намека на
Рис. 78. Tulipa korolkovii Rgl.
Рост органов цветка.
а—завязь, б—нить, в—чыльник.г—листочки
околоцветника.
непрерывно растут до середины ноября. В это время в пыльни-
ках происходит редукционное деление. Пыльца зимует в виде одно-
ядерных пылинок. Затем с середины ноября до начала февраля ты-
чинки не растут. В начале февраля наблюдается слабый рост и за
месяц они вытягиваются на 2 мм. За вторую половину марта тычин-
ки увеличиваются еще на 0,5 см, т. е. половину всего роста пыль-
ника, лопаются во время цветения и прекращают жизнь.
315
Завязь, заложившаяся одновременно с тычинками в начале сен-
тября в виде трех раздельных, а затем слившихся бугорков, медлен-
но всего на 4 мм растет до начала ноября. На 3,5 месяца до сере-
дины февраля ее рост прекращается. С середины февраля она вновь
начинает расти — в первые две недели медленно, с начала марта более
активно; с середины марта до начала апреля ее рост почти прекра-
щается. С начала до середины апреля (период открытия цветка до
начала подсыхания листьев) завязь быстро растет, а с середины ап-
реля больше не увеличивается.
Tulipa bifloriformis (рис. 79)
В середине апреля в молодой замещающей луковице Т. biflori-
formis, только что получившей самостоятельность в период, когда
Рис. 79. Tulipa bifloriformis.
Годовой цикл развития; I—XII месяцы, 1—луковица, 2—корни,
«3—весь побег, 4—нижний лист, 5—верхний лист, 6—стебель,
7—цветок, 8—завязь.
листья материнского растения наполовину высохли, но не кончили
вегетацию, закладывается меристематический бугорок нового замеща-
ющего побега.
В середине мая на нем уже обозначается валик нижнего листа.
В начале июня в корневой меристеме этой луковицы закладываются
зачатки корней.
В середине июня у основания центральной почки четко обрисо-
вывается еше несколько плосковатый меристематический бугорок—за-
чаток будущей замещающей луковицы. Зачатки корней, заложившие-
ся в середине мая, к этому времени уже связаны общей сосудистой
системой донца.
В начале июня на плоском зачатке побега закладывается второй
и последний лист. В начале августа зачаток побега вытягивается в
длину. А при анализе в середине августа на нем отмечается форми-
316
рование двух бутонов—верхушечного—крупного и бокового—более
мелкого. К этому времени у более крупного бутона закладываются
валики листочков околоцветника и бугорки тычинок.
С начала августа заметно начинает вытягиваться нижняя часть
стебля, находящаяся между последней луковичной чешуей и нижним
листом побега.
Завязь в верхушечном цветке закладывается в начале сентября.
Позже у основания молодого побега формируется луковица. В нача-
ле октября, при достаточной влажности и невысоких температурах
почвы, укореняются луковицы и после этого происходит формирова-
ние новой замещающей луковицы, заложившейся у основания молодо-
го побега в пазухе последней чешуи. В ней сразу можно отметить
наложившиеся покровную и первую запасающую чешуи; а в центре,
за первой запасающей чешуей, довольно крупный бугорок—будущая
вторая запасающая чешуя. В это же время закладывается детка в
пазухе первой запасающей чешуи материнской луковицы.
В середине октября сбоку у замещающей луковицы появляется
коллатеральная почка-детка, состоящая из одного центрального бу-
горка и одевающей его мясистой чешуи.
На этом органозаложение у данного вида в годовом цикле разви-
тия заканчивается. У почки-детки первой запасающей чешуи лукович-
ные чешуи закладываются
в начале декабря, а у
коллатеральной почки-
детки замещающей луко-
вицы—в конце октября.
У этого вида имеется
ограниченное количество
листьев и чешуй в луко-
вице. На стебле генера-
тивного побега всегда два
листа, а чешуй в полово-
зрелой луковице — три:
одна покровная и две за-
пасающие, т. е. в луковице
больше видоизмененных
листьев, чем настоящих
на побеге. Таким образом,
нарушается двурядное
расположение органов,
свойственное лилейным.
Рост листьев и стеб-
ля (рис. 79, 80). Нижний
лист побега у всех видов
тюльпанов отличается по
строению, морфологии и
функции от вышележа-
щих листьев. Нижний
лист всегда вначале вы-
полняет защитную функ-
Рис. 80. Рост листьев и стебля T. bifloriformis Vved.
в длину.
1—нижний лист, 2—верхний лист, 3—стебель.
цию, покрывая более
верхушечные листья и генеративные органы побега, защищая их в
почве от трения и воздействия неблагоприятных факторов ранней
весной до развертывания почки над почвой, т. е. по функции он в
какой-то степени сходен с покровной чешуей луковицы. Но нижний
317
лист выполняет защитную функцию только до выполнения им основ-
ной его функции ассимиляционного органа, а покровная чешуя вна-
чале служит органом запасающим, а после отмирания—органом за-
щиты нежных запасающих чешуй от неблагоприятных внешних воз-
действий. Еще больше различаются эти органы по росту и развитию
(рис. 79).
Меристематический бугорок оси в замещающей луковице первым
закладывается в середине апреля (рис. 80). Через месяц на нем по-
является валик зачатка первого листа, через три месяца—валик вто-
рого листа. За это время первый лист успевает лишь охватить почку,
но почти не вырастает в длину. Через две недели после заложения вто-
рого листа первый вырастает до 0,5 см дл., второй—до 0,4 см. Не-
смотря на период летнего покоя растения (наибольшего отрыва его
от внешней среды), нижний лист в новой почке не прекращает рос-
та. В августе и сентябре он плавно растет, достигая к началу октяб-
ря 1,35 см дл. Второй лист с первых чисел августа прекращает рост
и до начала октября почти не изменяется в длине.
Стебель со времени заложения в середине апреля до середины
сентября вырастает до 0,2 см выс. Во второй половине сентября—еще
на 2 мм и опять прекращает развитие на полмесяца.
С начала октября до середины февраля первый лист растет более
быстрыми темпами, чем два предыдущих месяца, достигает около 4,5
см выс. Период активного роста у этого листа длится семь месяцев.
Следовательно, каждый месяц 1-й лист в среднем вырастает на
0,64 см.
В период максимального роста этот лист увеличивается на
11,8 см, т. е. в 37 раз больше, чем в период активного роста.
Рост второго листа проходит в другом темпе. На его росте силь-
но сказывается как влияние внешних условий, так и состояние дру-
гих органов побега.
Энергичный осенний рост второго листа начинается в начале ок-
тября, а в середине октября падает, так как начинается рост стебля.
В первой декаде ноября равновесие восстанавливается и оба листа и
стебель активно растут. В середине ноября из-за неблагоприятных
внешних условий (видимо, затормаживается работа корневой системы)
вновь приостанавливается рост второго листа и стебля. Стебель на-
чинает активно расти в начале декабря, а лист—в середине декабря.
В начале января рост листа сильно замедляется и до начала февраля
он увеличивается всего на 0,4 см. Стебель же с середины января до
начала февраля дает прирост в 3 см, а в середине февраля резко
на полмесяца до выхода почки на дневную поверхность затормажи-
вает рост. С середины февраля до начала марта листья этого вида
достигают максимальных размеров и больше до конца вегетации (в
середине апреля или начале мая) не растут.
Стебель растет на две недели дольше листьев.
Цветок (рис. 79, 7, 8 и 81). Цветок закладывается в конце июля
или начале августа, так как при анализе почки в начале августа мы
наблюдаем в ней зачатки листочков околоцветника, тычинок и завя-
зи (рис. 81). До середины сентября они не увеличиваются в размере,
после чего листочки околоцветника и тычинки начинают расти и фор-
мироваться. Листочки околоцветника растут до середины ноября, а
тычинки—до начала. Тычинки в это время вполне сформированы и в
пыльниках лежат материнские клетки пыльцы. Как те, так и другие
больше до начала февраля не растут. Пестик после одновременного
с околоцветником и тычинками заложения начинает расти позже них
318
Рис. 81. Рост органов цветка Т. bi-
floriformis.
и растет всего две недели, а с середины октября не увеличивается
до начала февраля.
В начале февраля мы наблюдали резкий перелом в росте и раз-
витии всех частей цветка. В пыльниках в это время происходило ре-
дукционное деление, причем в метафазах мейозиса были определены
хромозомы—по 24 пары в диакинезе и по 24 отдельности в метафазе.
Следовательно, форма Т. bifloriformis из Ташкента являлась тетра-
плоидом.
Все питательные вещества мобилизуются на снабжение генератив-
ных органов, которые начинают усиленно расти, а рост таких органов,
как стебель, на это время прекращается
(рис. 80). С начала февраля после дли-
тельного перерыва начинают расти око-
лоцветник, тычинки и завязь. Но во
второй половине февраля, во время бур-
ного роста листьев, стебля и собственно
генеративных органов—тычинок и за-
вязи, рост околоцветника на две недели
задерживается, с начала марта (вре-
мя раскрытия цветка) околоцветник
вновь усиленно растет.
Тычинки, у которых лопнули пыль-
ники, сморщиваются, усыхают и первы-
ми прекращают рост, через две недели
заканчивают рост листочки околоцвет-
ника, но значительно дольше всех орга-
нов растения растет завязь—до начала
мая. Созревает завязь в середине мая.
Ташкентская формаТ. bifloriformis пре-
красно завязывает коробочки и дает мно-
го нормально ВСХОЖИХ семян. У него/-завязь, 2-тычинки, 3-бутон, 4-цветок,
неплохо представлено и вегетативное
размножение. Несмотря на то, что этот вид имеет одну пазушную
почку-детку в пазухе первой запасающей чешуи, у основания
замещающей луковицы закладывается ее коллатеральная почка-дет-
ка, которая вырастает в крупную луковицу. В старых ташкентских
садах по берегам арыков и под деревьями встречаются большие
куртины из вегетативно размножавшихся особей вида Т. biflori-
formis. Из нескольких сотен растений, произрастающих в таких кур-
тинах, обычно зацветает до десятка особей, а остальные растут и ве-
гетативно размножаются.
Заложение и рост замещающей луковицы(рис. 79, 1 и 82). Заме-
щающая луковица закладывается в середине июня, а в середине сен-
тября она все еще состоит из зачатка покровной чешуи и меристе-
матического бугорка в центре и едва достигает 2 мм выс. Только в
начале октября в ее центре дифференцируется валик зачатка первой
запасающей чешуи. В начале ноября начинается рост покровной че-
шуи, которая за две недели увеличивается до 0,6 см выс. Первая
запасающая чешуя в течение месяца медленно охватывает луковицу
с флангов, а к середине ноября она несколько превышает 0,35 см. В
середине ноября появляется зачаток второй и последней запасающей
чешуи у зрелой луковицы Т. bifloriformis.
Покровная чешуя в это время, видимо, из-за пониженных темпе-
ратур в почве, замедляет рост и к 1 февраля вырастает до 1 см д.л.
В первую половину февраля она прекращает рост. В это время сте-
319
бель и листья, начавшие активный рост, также его притормаживают.
Вызвано это заморозками, наступившими после длительной весенней
теплой погоды. Лишь органы, скрытые глубоко внутри растения, час-
ти цветка или внутренние чешуи замещающей луковицы, т. е. не
Рис. 82. Рост замещающей луковицы и детки Tulipa
Bifloriformis Vved.
1— покровная чешуя, 2—первая запасающая чешуя, 3—вторая
запасающая чешуя; а—замещающая луковица, б—сравнение
роста 1) замещающей луковицы, 2) ее детки, 3) коллатеральной
почки.
подвергающиеся б. м. не-
посредственному воздей-
ствию внешних усло-
вий,—не приостанавлива-
ют начавшийся несколько
ранее рост.
Первая запасающая
чешуя начинает вытяги-
ваться со второй полови-
ны января и к середине
февраля достигает 0,6
см дл.
Вторая запасающая
чешуя только через месяц
после заложения начина-
ет расти и ко второй по-
ловине декабря состав-
ляет 0,2 см выс. и опять
прекращает рост. Это выз-
вано, видимо, тем, что
запасные питательные ве-
щества перехватываются
начавшей расти коллате-
ральной почкой-деткой.
С начала февраля покровная чешуя перестает расти, а вторая за-
пасающая чешуя начинает вытягиваться. У всех трех чешуй луковицы
активный период роста в длину приходится на вторую половину фев-
раля. В начале марта наступает цветение и чешуи растут медленно.
Вторая запасающая чешуя в середине марта первой прекращает
рост в длину. Она значительно короче (на 2 см} как покровной, так
и первой запасающей чешуи. Рост покровной и первой запасающей
чешуи постепенно затухает до конца марта, после чего месяц или
полтора происходит накопление новых запасных веществ и пере-
мещение запасов из покровной чешуи в запасающие. После отмирания
листьев и корней луковица вступает в период покоя. Покровная
чешуя замещающей луковицы у этого вида закладывается позже, чем
у других исследованных нами видов тюльпанов, но на три с полови-
ной месяца раньше, чем другие чешуи или детки. После заложения
в луковице длительное время не закладываются никакие органы.
Заложение всех прочих органов сосредоточено в осенние месяцы, в
которые меристема донца проявляет наибольшую активность. Так,
в начале октября закладывается первая запасающая чешуя в заме-
щающей луковице. Одновременно с ней в пазухе первой запасающей
чешуи материнской луковицы закладывается детка (рис. 82, б).
В середине октября рядом с замещающей луковицей закладывается
коллатеральная почка-детка, к середине ноября уже заложившая
все три чешуи.
В середине ноября закладывается вторая запасающая чешуя
в замещающей луковице, а в конце ноября — начале декабря форми-
руется детка первой запасающей чешуи материнской луковицы, в ко-
торой закладываются первая и вторая запасающие чешуи.
320
На этом образование органов вегетативного размножения закан-
чивается. В дальнейшем они только увеличиваются в размере и в них
происходят процессы внутренней организации. Органы вегетативного
размножения в некоторой степени повторяют рост замещающей луко-
вицы, но часто не достигают ее размера, и на две—три недели раньше
останавливаются в росте. У почки-детки первой запасающей чешуи
материнской луковицы активный период роста наступает на две неде-
ли позже, чем у замещающей и ее коллатеральной луковиц.
Луковица Т. bifloriformis десять с половиной месяцев растет в
недрах старой луковицы, а затем, освободившись от ее чешуй, еще
год живет как самостоятельный организм. Покровная чешуя луковицы
закладывается в период летнего покоя в середине июня, а засыхает
в конце вегетации — начале мая следующего года, т. е. она живет
всего десять с половиной месяцев.
Первая запасающая чешуя этой луковицы закладывается в начале
октября. До конца апреля — начала мая следующего года она нахо-
дится в чешуях материнской луковицы, а год живет самостоятельно,
т. е. всего 18 месяцев. В первые месяцы она растет и не является
запасающим органом. В последние—она истощается и также не имеет
запасов.
Вторая запасающая чешуя закладывается на полмесяца позже
первой и живет 17,5 месяца. До начала вегетации в январе и феврале
первого года жизни она уже является органом запасающим, куда
перемещаются избытки крахмала и других питательных веществ из
чешуй материнской луковицы. В последние месяцы (февраль, март
и апрель) она истощается и не имеет запасов.
Сорт Дарвиновского тюльпана Jubilee (рис. 83)
У этого сорта обычно в начале марта, а иногда и в конце февраля
над поверхностью почвы показываются головки отрастающих побегов.
К середине марта (в начале развертывания розетки листьев) в центре
сидящей у их основания еще не завершившей рост молодой заме-
щающей луковицы начинается закладка меристематического бугорка
побега будущего года (рис. 83). На бугорке в это время наблюдается
валик зачатка нижнего листа. Меристематический валик второго листа
появляется в первых числах мая. Третий лист закладывается в начале
июня, а четвертый — в середине июня. Стебель, как осевой орган,
закладывается раньше листьев, но нижнее междоузлие у него вытя-
гивается в начале сентября, а закладка цветка начинается в середине
июля. Но одновременно с закладкой в середине июня на молодом
побеге третьего листа, у его основания появляется зачаточный буго-
рок из меристематической ткани. В дальнейшем из этого бугорка
развивается новая замещающая луковица. Она почти год существует
как часть материнского организма.
Рост замещающей луковицы (рис. 84). Обычно в середине июня
на меристематическом бугорке у основания молодого побега закла-
дывается в виде полулуния зачаток покровной чешуи замещающей
луковицы. В центре его возвышается бугорок из меристематической
ткани, на котором в начале августа четко обозначается валик первой
запасающей чешуи, а в начале сентября — второй запасающей. В на-
чале октября один за другим закладываются валики третьей и четвер-
той запасающих чешуй.
Таким образом, весь процесс заложения отдельных органов
замещающей луковицы у сорта Jubilee протекает в период летнего
21—2229
321
покоя и длится примерно три с половиной месяца (середина июня по
.первые числа октября).
4 - Покровная чешуя замещающей луковицы в течение первых двух
месяцев, прошедших со времени заложения, почти не растет. В сере-
дине августа она начинает очень слабо расти. С первых чисел
•сентября темпы ее роста значительно усиливаются, а с середины
4У
Рис. 83. Сорт Jubilee.
Годовой цикл развития: 1— луковица, 2—корни, 3—побег, 4—нижний лист,
5—второй лист, 6—третий лист, 7—четвертый лист, 8—стебель, 9—цветок,
10—завязь.
сентября до первых чисел октября — резко возрастают. К началу
октября зачаток покровной чешуи замещающей луковицы достигает
0,9 см дл. Затем в течение месяца покровная чешуя почти не растет,
что обусловлено, по-видимому (как и у других видов), отсутствием
влаги в почве и пассивностью корневой системы. К началу ноября
в связи с осенними осадками происходит укоренение луковиц и на-
ступает активный рост покровной чешуи, вырастающей до начала
декабря до 4,5 см дл., т. е. почти до нормальной длины луковицы.
В первых числах декабря рост покровной чешуи луковицы, возможно,
в связи с резким понижением температур воздуха и почвы, несколько
приостанавливается. С середины декабря рост покровной чешуи уси-
ливается и не прекращается до конца вегетации, так как у заме-
щающих луковиц этого сорта покровная чешуя, обычно, продолжена
в листовую пластинку. Таким образом, покровной чешуей у замещаю-
щей луковицы является расширенное влагалище зеленого листа и
характер роста ее сходен с ростом листа, а не чешуи (рис. 84, 85).
322
Первая запасающая чешуя у этой луковицы закладывается вскоре
после заложения покровной, и первое время (4,5 месяца) характер
их роста одинаков. Но с первых чисел ноября в период активного
роста верхней листовой части покровной чешуи наблюдается различие
в их развитии.
Первая, вторая, третья и четвертая запасающие чешуи почти не
растут до начала февраля. Третья и четвертая чешуи через месяц
после заложения в течение первой половины ноября несколько под-
растают до размера двух первых запасающих чешуй.
Рис. 84. Рост почки замещающей луковицы сорта Jubilee.
I—покровная чешуя, 2—первая запасающая, 3—вторая запасающая, 4—третья запасающая,
а—четвертая запасающая чешуя.
Из периода относительного зимнего покоя первая запасающая
чешуя выходит в начале февраля, активно растет и за месяц дает
прирост 1,3 см. Одновременно с ней начинает рост вторая запасающая
чешуя, но медленнее, чем первая, и прибавляет в росте 0,6 см.
В середине февраля выходят из покоя третья и четвертая запасающие
чешуи. До начала марта они растут медленно, а с середины марта
так активно, что несколько подавляют рост первой запасающей чешуи.
С середины марта до начала июня все запасающие чешуи замещающей
луковицы растут непрерывно, быстро и равномерно. В конце первой
декады июня, когда усыхает покровная чешуя, все чешуи несколько
уменьшаются и в таком виде остаются до времени начала осеннего
отрастания корней. На втором году жизни в середине или конце июня,
т. е. в конце периода вегетации, замещающая луковица обособляется
от чешуи и донца материнской луковицы. Таким образом, луковица
сорта Юбилей один год две недели живет в чешуях за счет запасов
материнского растения; 12 месяцев—как самостоятельный организм,
т. е. 2 года с двумя — тремя неделями, а затем уступает место
323
зреющей в ее недрах новой замещающей луковице. В период летнего
покоя в середине июля второго года жизни луковицы в корневой
меристеме донца закладываются корни. Осенью в октябре или ноябре
в зависимости от количества осадков и влажности почвы корни про-
рывают эпидермис донца и выходят наружу. У посаженных в почву-
Рис. 85. Рост нижнего листа и стебля сортатЛиЬПее.
1—лист, 2—стебель.
луковиц они входят в почву и начинают функционировать. Корневая
система культурных тюльпанов так же, как и диких видов, начинает
функционировать с осени. Она активно действует всю зиму, за исклю-
чением периодов вынужденного покоя, обусловленных низкими тем-
пературами почвы, и отмирает в начале следующего года во время
окончания вегетации вместе с отмиранием листьев и стеблей. Коро-
бочка с семенами долгое время после отмирания корней, увядания
листьев и засыхания стеблей не созревает.
Рост побега (рис. 83). Первый лист закладывается на побеге в
в середине марта, растет очень медленно. Ко времени появления за-
324
чатка второго листа зачаток первого листа сильно вырастает и в нем
появляются проваскулярные пучки, ближе к центру крупные, а пери-
ферические значительно мельче. С середины мая до середины июня
первый лист почти не растет, прибавляя за месяц всего 1 мм в длину.
С середины июня до начала августа у этого зачатка прирост не наб-
людается. В августе он начинает энергично расти до начала сентября,
достигая к этому времени 1 см дл. С начала до середины сентября
его рост прекращается, а во вторую половину сентября — увеличи-
вается в три раза. С начала октября до начала ноября рост зачатка
останавливается из-за отсутствия влаги и температуры. Рост органов
тормозится внешними условиями. На листьях, лежащих в центре
почки и выросших в меньшей степени, это не проявляется так резко,
как на нижнем листе, служащем покровом всей почки.
Мы наблюдали, что у тюльпанов быстро отрастают корни (до
6 см дл.), если ранней осенью при высоких температурах воздуха
и почвы их полить, но затем почва перегревается настолько, что они
'больше не растут. У них сильно утолщалась вся зона роста корня.
В дальнейшем эти корни в течение всего последующего периода веге-
тации растения при самых благоприятных условиях уже не росли.
Правда, растения не погибали, но были малорослыми и хилыми.
Результирующие луковицы таких растений были значительно мельче,
чем у нормальных, и они развивались в меньшем числе.
При понижении температур и выпадении осадков в начале ноября
резко увеличивается рост листа, до середины ноября он вырастает
•еще на 2 см и показывается из луковицы. При дальнейшем понижении
•температуры во второй половине ноября и первой половине декабря
темп роста нижнего листа постепенно падает, но не прекращается.
Лист в течение месяца увеличивается еще на 0,7 см. Во второй по-
ловине декабря и первой половине января темп роста этого листа
вновь возрастает до 1,5 см в месяц, а с середины января до конца
второй декады апреля (более трех с половиной месяцев) рост нижнего
листа протекает бурными темпами без спадов и торможений, достигая
к цветению максимальной длины.
Рост второго, третьего и четвертого листьев происходит почти
как и у нижнего листа, отличаясь большей плавностью, более поздним
заложением и меньшими размерами (рис. 85).
Стебель закладывается в виде меристематического бугорка на
вершине замещающей луковицы раньше заложения первого (нижнего)
листа (рис. 85). Однако зачаточный стебель бывает настолько мал,
что обычно не отмечается, или наблюдается как зона мелких меристе-
матических клеток, расположенных у основания самой внутренней
чешуи. Трудно назвать срок, когда это вначале довольно плоское
образование можно назвать зачатком стебля. Началом заложения
стебля мы считаем время, когда на нем начинает выступать валик
зачатка первого листа. С этого момента стебель существует как ось
(в зачаточном состоянии), на которой закладываются листья.
Во время заложения всех листьев и цветка стебель не увеличи-
вается в длину. Лишь через полтора месяца после закладки цветка,
когда все его части видны невооруженным глазом, т. е. к середине
августа, начинает расти стебель. До середины сентября осенний рост
стебля заканчивается и со второй декады октября по начало декабря
он не изменяется в длину. С начала декабря до середины марта
стебель выходит из состояния покоя и начинает расти по 0,5 см в
месяц. До середины марта стебель вырастает еще на 3 см. С середи-
ны марта до начала третьей декады апреля стебель вырастает до
325
Рис. 86. Рост органов цветка сорта Jubilee.
а—завязь, д’—тычинка, —нить, в—листочки около-
цветника.
64 см выс., т. е. прирост в день в среднем почти 1,7 см. В три дня*
он вытягивается на весь прирост за предшествующий год. В следую-
щее за цветением время перед созреванием семян весь стебель у
этого сорта вырастает только на 5 см. Рост на последнем этапе про-
текает за счет вытягивания цветоплодоножки.
Таким образом, первые этапы роста стебля происходят за счет
вытягивания самого нижнего междоузлия, лежащего между верхней
чешуей луковицы и нижним ли-
стом побега. Средние междоуз-
лия во время подземного роста
стебля увеличиваются слабо. Ак-
тивно, но значительно слабее,
чем нижнее междоузлие, в это
время растет цветоножка. После
выхода розетки листьев на по-
верхность почвы нижнее междо-
узлие почти перестает расти.
В это время активно растут сред-
ние междоузлия и особенно цве-
тоножка (рис. 85).
Цветок (рис. 86). Цветочная
почка закладывается в середине-
июля на вершине побега, на ко-
тором уже заложились все ли-
стья, не позже, чем через месяц
после обособления замещающей
луковицы из чешуй материнской
луковицы. Листочки околоцвет-
ника после заложения начинают
расти и непрерывно, хотя и мед-
ленно, растут до середины но-
ября, достигая к этому времени
1 см дл. В середине ноября в.
связи с укоренением луковиц
темп роста листочков околоцвет-
ника несколько возрастает. В се-
редине декабря он вновь падает
до прежней интенсивности, а в
середине февраля начинает постепенно увеличиваться до начала ап-
реля. За последние две недели листочки околоцветника вырастают
на значительно большую величину, чем за все предыдущее время с
момента заложения.
Бугорки тычинок закладываются в середине июля почти одновре-
менно с зачатками листочков околоцветника и сразу же начинают
интенсивно расти, достигая за две недели 0,5 см. До середины ноября
они растут более медленными темпами, вырастая за это время еще
на 0,5 см. С середины ноября до середины декабря пыльники, как
и листочки околоцветника, растут несколько более активно, затем
равномерно до раскрывания цветка. После того, как пыльники
лопаются, величина тычинок резко падает. С середины марта перед
цветением начинается усиленный рост тычиночных нитей и создается
впечатление сильного увеличения роста тычинок. Однако этот рост
продолжается всего один месяц. За это время тычиночные нити,
тюльпана сорта Jubilee достигают 1,3 см дл.
326
Завязь, закладываясь одновременно с околоцветником и тычин-
ками в середине июля, до середины марта следующего года растет
беспрерывно и б. м. равномерно. Затем она резко увеличивается в
размере, достигая в течение месяца 3,7 см, а за все предшествующее
время (8 месяцев) она выросла всего на 1,2 см. С середины апреля
почти на две недели полностью прекращается ее рост. С начала мая
завязь вновь растет более интенсивными темпами и за последние две
недели после цветения вырастет еще на 2,5 см. На этом ее рост
заканчивается.
Рост и развитие пазушных почек луковицы (рис. 87). Для
сравнения с дикими видами тюльпанов, на которые жесткие условия
существования в континентальном климате Средней Азии наложили
Рис. 87. Рост и развитие почки-детки покровной чешуи
сорта Jubilee.
I—первая фаза роста, II—вторая фаза роста;
1—первая чешуя, 2—вторая чешуя, 3—побег, 4—чешуя луковицы
второй фазы роста.
известный отпечаток, интересно изучить рост и развитие пазушных
почек у культурных сортов. Забота человека о них, условия суще-
ствования с момента их зарождения в течение веков значительно'
327
смягчены; кроме того, человек вел среди них сознательный отбор на
вегетативно размножающиеся формы.
За последние годы у нас в СССР вышло в свет много работ, по-
священных вегетативному размножению культурных сортов тюлпанов.
Первое место среди них занимает работа 3. М. Силиной (1952).
Хотя она и посвящена годовому циклу развития тюльпана, но расте-
ния тюльпана, как такового, в этой работе нет. В ней описано только
развитие луковицы и ее вегетативное потомство. О развитии побега
у тюльпанов упоминается только на одной странице и общими сло-
вами для всех сортов. То же самое можно сказать и о работах
Э. Ф. Мигушовой (1954), О. Н. Данилевской (1955) и Л. С. Тимо-
феевой (1956). Жизнь тюльпана они подменили жизнью его части, так
как луковица, при всей ее значимости, лишь в период летнего покоя
включает в себе все растение, а осенью, зимой и весной она является
только его частью. Исследования вышеназванных авторов проведены
в северной части СССР, и их данные о развитии луковиц культурных
тюльпанов, их размножении для сравнения с дикими видами мы
можем использовать с большой натяжкой. Поэтому мы исследовали
культурные сорта параллельно с дикими видами, выращиваемыми в
одинаковых условиях.
Почки-детки, развивающиеся в пазухах луковичных чешуй,
являются почками первого порядка главного стебля, укороченного
у луковицы и называемого донцем. Эти почки могут развиться в
боковые побеги, или накопить в чешуях (листьях) запасы питательных
веществ, обособиться от луковицы во время ее усыхания и прорасти
в самостоятельный организм, т. е. они являются органами вегетатив-
ного размножения.
Почка-детка покровной чешуи. Первой у этого сорта заклады-
вается детка покровной чешуи (рис. 87): у замещающей луковицы
очень рано—в начале февраля, когда луковица еще крепко связана
с материнским организмом и составляет с ним единое целое. В это
время у детки покровной чешуи закладывается покровная чешуя,
которая интенсивно растет и к началу мая, когда происходит закладка
первой запасающей чешуи, достигает 0,5 см дл. На этом часто ее рост
и заканчивается. Но у большинства изучавшихся особей мы наблю-
дали еще одну фазу роста у покровной чешуи, протекающую во
второй половине июля. За указанное время покровная чешуя этой
детки вырастает еще на 1 см и в дальнейшем уже не растет. В пер-
вых числах мая, во время цветения материнской луковицы, в детке
покровной чешуи закладывается первая запасающая чешуя. Она
беспрерывно и активно растет, достигая к середине июля размера
покровной чешуи. На этом первая фаза развития детки покровной
чешуи заканчивается и эти чешуи больше не растут.
В середине июня из верхушечной точки роста этой почки-детки
закладываются сразу два органа—побег и покровная чешуя ее заме-
щающей луковицы. Долгое время их развитие проходит параллельно.
Так, со времени заложения до начала августа они почти не увели-
чиваются. За полтора месяца до середины сентября они вырастают
до 6—8 мм. С середины сентября до середины ноября они растут
очень интенсивно, но в одинаковом темпе, достигая к этому времени
2 см выс. С середины ноября побег растет более интенсивно, а по-
кровная чешуя новой замещающей луковицы детки покровной чешуи
более медленно, но также непрерывно. На их росте не сказывается
период зимнего покоя, так как они получают питание от материнской
луковицы и поглощают запасы, отложенные в двух первых чешуях,
328
которые совершенно истощаются и уже к началу зимы представлены
тоненькими пленочками без запасов крахмала и других веществ, оде-
вающими основание побега и его замещающей луковицы.
В июле на побеге центральной почки этой детки уже видны все
три листа, а в начале сентября—все органы цветка.
В пазухах чешуй детки покровной чешуи первой фазы в период,
когда она находится еще в полной зависимости от материнского
растения, также закладываются детки. В начале июля у чешуй первой
•фазы развития мы наблюдали детку покровной чешуи второй фазы
развития и заложение бугорка детки первой запасающей чешуи.
Кроме этих деток, у детки, заложившейся в пазухе покровной
чешуи первой фазы, часто закладывается сестринская детка. Она обыч-
но расположена со стороны материнской луковицы на том же сосуди-
стом пучке, что и детка покровной чешуи, которая скрывает сестрин-
скую детку.
Обычно замещающая луковица детки покровной чешуи, нахо-
дящаяся у основания генеративного побега второй фазы развития
этой луковицы, не имеет большого количества чешуй. У ее основания
всегда можно обнаружить в виде сухих пленок остатки чешуй первой
•фазы. Она одета покровной чешуей и имеет под ней две, максимум
три запасакмцие чешуи.
За покровной чешуей детки покровной чешуи первой фазы в
10 случаях из 24 мы обнаружили заложившуюся и развивающуюся
детку покровной чешуи.
Не менее часто мы наблюдали у нее и детку первой запасающей
чешуи. Сестринскую детку за все время исследования мы встретили
трижды. Кроме того, у каждого цветоносного побега, а их мы наб-
людали 10 раз из 24 анализов, имелась своя замещающая луковица.
За покровной чешуей у крупных луковиц сорта Юбилей почти
у половины растений развивается цветоносный побег, заложившийся
во вторую фазу роста, т. е. обыкновенный пазушный побег, оканчи-
вающийся цветком. Вегетативное размножение в данном случае
осуществляется за счет почек-деток, заложившихся в пазухах чешуй
первой фазы развития этой детки, которые грубо могут отождеств-
ляться с почечными чешуями других растений и сестринской детки.
Такиги образом, в пазухе покровной чешуи можно обнаружить
четыре луковицы: три луковицы и один побег, или у более слабых
материнских луковиц всего одну, на крупную луковицу.
г; i-гв зчд 2-17 i-jg г-я 7 не зчз г-ю з-к зя i-wt-js зчд
Ш. 12 2 и п т к х м к! 7 и щ tv г у/
Рис. 88. Рост и развитие детки в пазухе первой запа-
сающей чешуи сорта Jubilee.
1—первая фаза роста, чешуя; 2—вторая фаза роста, покровная
чешуя,
Почка-детка первой запасающей чешуи (рис. 88). Покровную
чешую детки первой запасающей чешуи в виде меристематического
лимонно-желтого валика мы наблюдали в первых числах мая в период,
когда у детки покровной чешуи уже заложилась первая запасающая
329
чешуя первой фазы развития. В первые дни роста до середины мая
она выросла до 8 мм дл. За следующие два месяца (до середины
июля) она прибавила всего 2 мм.
В это время наметилось заложение следующего органа. Однако
это не была первая запасающая чешуя первой фазы развития, как
у детки покровной чешуи. Как показало дальнейшее развитие, первая
фаза развития детки первой запасающей чешуи ограничилась всего
одной покровной чешуей, видимо, в связи с ее более поздним зало-
жением. А заложившаяся за ней в середине июля чешуя вновь была
покровной чешуей, но уже относилась ко второй фазе развития. Одно-
временно с покровной чешуей у детки первой запасающей чешуи во
второй фазе наметился валик первой запасающей чешуи. В середине
сентября были отмечены валики еще двух чешуй.
В девяти случаях из 24, исследованных в течение года, первая
фаза состояла из двух чешуй—покровной и запасающей, в остальных
15 случаях первая фаза состояла из заложения одной чешуи, которая
росла как покровная, вернее, запасающая без зеленой листовой пла-
стинки. В ней откладывалась масса крахмала. Росла она до середины
февраля и достигала 2,5 см дл., но в декабре ее переросла запасающая
чешуя этой же детки второй фазы развития, которая к середине
декабря достигала максимальной длины (4 см), а в дальнейшем и
ширины.
В середине ноября за покровной чешуей первой фазы развития
детки первой запасающей чешуи мы обнаружили детку, состоящук>
из двух чешуй. Примерно через месяц она состояла уже из четы-
рех чешуй.
В середине декабря у детки первой запасающей чешуи на сосу-
дистом пучке со стороны второй чешуи мы наблюдали сестринскую
(сериальную) детку из двух чешуек и меристематического бугорка.
В середине января у экземпляра, имевшего в первой фазе развития
у детки первой запасающей чешуи, кроме покровной чешуи, еще-
и одну запасающую, была обнаружена, кроме детки покровной чешуи,
детка за первой запасающей чешуей. Она к этому времени имела
зачаток покровной чешуи и двух запасающих.
В одном случае из 24 установлено, что у луковицы, разрезанной
в середине января в первой фазе развития, было две чешуи, а во-
второй фазе уже развился цветонос. При нем наблюдалась детка по-
кровной чешуи, а детка первой запасающей чешуи была его заме-
щающей.
Из 24 исследованных луковиц у трех деток первой запасающей
чешуи не было второй фазы развития. Одна из этих деток засохла
в зачаточном состоянии.
Таким образом, за первой запасающей чешуей материнской луко-
вицы может развиться всего одна детка. Обычно за этой чешуей
у луковиц сорта Юбилей бывает одна детка, развивавшаяся из вер-
хушечной точки роста второй фазы развития; вторая—в пазухе по-
кровной чешуи первой фазы развития; третья—в пазухе первой запа-
сающей чешуи первой фазы развития и сестринская (сериальная)
основной детки, расположенная на том же сосудистом пучке, что
и основная детка или даже комплекс деток, но не в их чешуях и на
несколько миллиметров ниже от их места заложения.
Следовательно, за первой запасающей чешуей у луковиц сорта
Юбилей наблюдается иногда 'комплекс из четырех деток разных сро-
ков заложения и различного происхождения.
330
Почка-детка в пазухе второй запасающей чешуи (рис. 89).
В пазухе второй запасающей чешуи у замещающей луковицы в начале
мая закладывается детка. Первым виден лишь валик зачатка ее по-
кровной чешуи, но тут же закладывается и валик первой запасающей
чешуи. Зачаток покровной чешуи за две недели после заложения
вырастает до 7 мм дл. Первая запасающая чешуя в это время не
растет. Покровная чешуя после двух недель довольно активного
роста на два с половиной месяца прекращает рост. В первой половине
августа покровная чешуя вырастает на 2 мм, за три последующие
Ю -
8-
21 F28 4-Ю 349 247 1-Ю 247 7_ 148 348 249 348 3-2! 1-28 449 349
Щ IV 2 VI Hi £// IX X Z XII _/ // III IV V V/
Рис. 89. Рост и развитие детки в пазухе второй
запасающей чешуи сорта Jubilee.
Первая фаза развития: 1—покровная чешуя, 2—запасающая чешуя;
вторая фаза развития; 3—покровная чешуя, на вершине развившая
листовую пластинку.
месяца—еще на 3 мм. На этом ее рост заканчивается. В течение пер-
вых 6,5 месяцев после заложения покровная чешуя достигает всего.
12 мм дл. Первая, вернее, единственная запасающая чешуя первой
фазы развития начинает расти в длину с начала августа. Через три
месяца после заложения она растет очень медленно плавно и к
середине января достигает максимальной величины—9 жгдл. И позже
обе эти чешуи очень слабо растут. В течение июня из центральной
меристемы этой детки закладывается желто-зеленый меристематиче-
ский валик луковицы второй фазы развития.
В начале июля появляется один валик, а в начале ноября мы
наблюдали еще два валика зачатков чешуй у детки-луковицы второй
фазы.
Таким образом, зачаток детки луковицы за второй запасающей
чешуей в первой фазе состоит из двух чешуй, а во второй фазе—из
пяти: одной покровной и четырех запасающих. Покровная чешуя
второй фазы закладывается очень рано и до второй половины ноября
растет очень медленно. К этому времени она имеет всего 3 мм дл.
Со второй половины ноября в ее росте наступает резкий перелом: за
две недели она увеличивается на 0,5 см. С начала до середины декабря
она быстро растет и прибавляет еще 3,5 см. В дальнейшем она не-
сколько снижает темпы роста и к середине марта достигает 11,6 см дл.
331
На вершине покровной чешуи закладывается и развивается пластинка
зеленого листа.
Появившаяся в начале июля дужка зачатка первой запасающей
чешуи второй фазы развития вначале так же долго не растет, как и
ее покровная чешуя. С первой половины ноября она начинает мед-
ленно расти в длину. К началу декабря дужка достигает 4 мм дл.
До середины января она не увеличивается, а к середине февраля
достигает уже 1,2 см дл. В дальнейшем она растет беспрерывно.
Валики двух следующих чешуй, впервые отмеченные в начале ноября,
вступают в период роста одновременно с двумя первыми и растут в
дальнейшем, не снижая темпа.
Из 24 пар образцов, исследованных в течение года, лишь у одной
детки второй запасающей чешуи в верхушечной точке роста мы обна-
ружили заложение побега. В остальных случаях это были луковицы.
У луковиц второй фазы развития с тех пор, как наступил активный
период роста, т. е. с декабря, мы почти у всех обнаружили, что
покровная чешуя продолжена в зеленый лист. Раньше это трудно
сделать, так как на ранних фазах роста почти невозможно различить
листовую пластинку от влагалища листа.
У второй запасающей чешуи во второй половине ноября мы обна-
ружили одну сестринскую (сериальную) детку. В это же время в па-
зухе покровной чешуи первой фазы развития была отмечена детка из
двух чешуй: покровной и первой запасающей. В середине декабря
наблюдалось уже три чешуи. В середине января у экземпляра, имев-
шего в первой фазе развития две чешуи, кроме детки покровной
чешуи, в пазухе ее первой и единственной запасающей обнаружен
меристематический бугорок детки первой запасающей чешуи. В даль-
нейшем в начале февраля эта детка в пазухе первой запасающей
чешуи первой фазы развития у детки второй запасающей чешуи за-
ложила бугорки покровной и первой запасающей чешуй.
Таким образом, у луковиц этого сорта за второй запасающей
чешуей может лежать также четыре детки, как и за первой и за пок-
ровной чешуями. Однако обычно их бывает меньше. Так, очень редко
развивается сестринская (сериальная) детка.
Почка-детка в пазухе третьей запасающей чешуи (рис. 90). В
пазухе третьей запасающей чешуи замещающей луковицы в начале
мая закладывается валик покровной чешуи почки-детки. За первые
две недели после заложения он вырастает на 2 мм, после чего три
месяца не растет. С конца августа начинает постепенно расти. За
первую половину сентября он вырастает до 4 мм-, за вторую половину
сентября — еще на 6 мм. В дальнейшем его рост протекает очень
медленно, и лишь ко второй половине января он достигает 1,5 см дл.
и больше не растет. На этом часто останавливается развитие первой
фазы, так как первая запасающая чешуя первой фазы развития в поч-
ках-луковицах, заложившихся за третьей запасающей чешуей заме-
щающей луковицы, закладывается редко. Мы наблюдали ее заложение
в шести случаях из 24. Она закладывается в начале июня и в развитии
повторяет этапы роста покровной чешуи; служит хранилищем запасных
питательных веществ, которые потребляются в период сильного роста
луковицей второй фазы и ее пазушной деткой. Заложение валика покров-
ной чешуи луковицы второй фазы развития мы наблюдали в середине
июля. В двух случаях в пазухе третьей запасающей чешуи в течение
года не было заложено чешуй первой фазы развития и рост почки-
детки начался позже, но сразу со второй фазы.
332
Покровная чешуя второй фазы развития после заложения с сере-
дины июля до середины сентября не растет. Затем начинает посте-
пенно прибывать в росте, достигая к концу первой декады октября
4 мм дл. С этого момента она растет непрерывно, достигая к середине
января 4,7 см дл. Затем покровная чешуя приобретает очень высокие
темпы роста, достигая во вторую половину февраля 19 см дл. Все
Рис. 90. Рост и развитие детки в пазухе третьей
запасающей чешуи сорта Jubilee.
/—первая фаза, заложилась одна чешуя, 2—вторая фаза, заложился
и растет побег.
особи этого сорта у детки третьей запасающей чешуи имеют покров-
ную чешую второй фазы в виде влагалища зеленого листа, в связи
с чем все запасающие чешуи луковицы сильно отстают в росте от
покровной чешуи. Так, в середине января покровная чешуя имеет
4,7 см дл.; первая запасающая — 0,47 см дл.; вторая —0,37; третья —
0,26; четвертая — 0,15 см дл. Затем они очень быстро растут, но ни-
когда не догоняют в росте покровную чешую, которая имеет пластин-
ку зеленого листа.
У почки-детки, заложившейся в пазухе третьей запасающей
чешуи, как и у всех исследованных нами почек более периферических
чешуй может заложиться: сестринская (сериальная) почка-детка; почка-
детка покровной чешуи за покровной чешуей первой фазы развития
и почка-детка за первой запасающей чешуей почки первой фазы раз-
вития, если эта чешуя закладывается. Кроме того, в середине января
за покровной чешуей луковицы второй фазы развития мы увидели
333
меристематический бугорок—зачаток ее пазушной детки 0,02x0,02 см.
Таким образом, мы можем наблюдать четыре почки-детки и за третьей
запасающей чешуей замещающей луковицы. К этим почкам-деткам
добавляется крупная почка-детка за покровной чешуей главной почки-
детки этой чешуи.
Почка-детка в пазухе четвертой запасающей чешуи (рис, 91,
92, 93). Рост и развитие детки в пазухе четвертой запасающей чешуи
Рис. 91. Рост и развитие детки в пазухе четвер-
той запасающей чешуи сорта Jubilee.
1—первая фаза, заложилась одна чешуя, 2—вторая фаза,
заложилась покровная чешуя.
Рис. 92. Рост и развитие почек-деток в пазухах материнской луковицы в начале июля
сорта Jubilee.
7—детка покровной чешуи, 2—детки первой запасающей чешуи, 3—детка второй запасающей чешуи,
4-детка третьей запасающей чешуи, 5—верхушечная ночка. Все детки заложили чешуи только первой
фазы развития.
334
замещающей луковицы отличаются от деток периферических чешуй,
У детки этой чешуи из чешуй первой фазы развития в середине
июня закладывается только покровная чешуя. К середине ноября она
достигает 0,5 см дл. Затем темпы роста ее усиливаются и к началу
декабря она увеличивается до 0,9 см дл.
Рис. 93. Рост и развитие почек деток в пазухах материнской луковицы в середине
декабря сорта Jubilee.
1 6—детка покровной чешуи, материнской луковицы, продольный разрез, видны (1 и II) почки-детки,
заложившиеся в пазухах (III и IV) чешуй первой фазы развития и V основание геиеративиого побега.
/ в—бутон, в пыльниках которого материнские клетки микроспор находятся в профазе; 2а—детка в пазухе
первой запасающей чешуи и ее сестринская; 26—ее продольный разрез. Видна (1) почка-детка, за ложив-
шаяся в пазухе (III) покровной чешуи первой фазы развития, запасающая чешуя первой фазы развития
и (V) основание генеративного побега; 2/?—бутон этого побега; 2г—продольный разрез через сестринскую
детку детки первой запасающей чешуи; За—общий вид детки в па чухе второй запасающей чешуи, 36—
ее продольный разрез, видны: III одна чешуя первой фазы за ней (I) почка-детка, VI основания двух зе-
леных листьев второй фазы и VII почка из двух запасающих чешуй второй фазы; 4 а—общий вид детки
в пазухе третьей запасающей чешуи; 46—ее продольный разрез, видна (III) одна чешуя первой фазы,
(VI) основание зеленого листа (покровная чешуя) второй фазы, между ними зачаток (1) почки-детки и
видна сложенная из нескольких запасающих чешуй VII почка-детка второй фазы; общий вид донца
с центральной почкой и замещающей луковицей; 56—продольный разрез замещающей луковицы, все ор-
ганы которой закладываются во вторую фазу; 56—бутон центральной почки.
К середине января темп роста вновь замедляется и почка-детка
достигает 1,3 см дл. С середины января по середину февраля она
вырастает до 2,2 см дл. Она больше не растет, а луковица-детка
второй фазы постепенно потребляет запасы, отложенные в почке-детке.
Нам не удалось наблюдать начала заложения покровной чешуи
второй фазы, но в первых числах сентября были отмечены валик
покровной чешуи в 0,3 см дл.; валик первой запасающей чешуи в
0,12 см дл. и в центре меристематический бугорок 0,01 см выс. Они
быстро развиваются и к началу декабря их покровная чешуя дости-
335
гает 3,5 см дл. За покровной чешуей в это время расположены за-
чатки еще четырех чешуй. В детке-луковице, заложившейся за
четвертой запасающей чешуей запасающей луковицы, может быть
одна покровная чешуя и пять запасающих. Максимального размера
покровная чешуя детки достигает в конце апреля. В конце апреля за
чешуями луковицы второй фазы развития детки, заложившейся в
пазухе четвертой чешуи замещающей луковицы, мы обнаружили
меристематические бугорки будущих деток следующих размеров: за
покровной чешуей — 0,25x0,05 см-, за первой запасающей чешуей —
0,2x0,07 см и за второй запасающей чешуей — 0,1x0,085 см. Детка,
которая закладывается в пазухе покровной чешуи первой фазы у
детки четвертой запасающей чешуи, мы в этом исследовании не
встретили, но в предыдущее изучение этого сорта мы ее неоднократ-
но наблюдали, как и сестринскую детку.
У материнской луковицы, имеющей пятую запасающую чешую,,
детка закладывается после первой фазы развития деток и наступления
второй. Мы не наблюдали у детки за этой чешуей заложения чешуй
первой фазы развития, а следовательно, и их деток.
Таким образом, в результате вегетативного размножения одной
сильной луковицы сорта Юбилей с шестью запасающими чешуями
в течение одного года можно иметь гнездо луковиц, из 21 почки-
детки различного возраста, размера и происхождения. Но их бывает
меньше за счет развития цветоносных побегов из многих почек.
Образование побегов вместо почек-деток является риверсией
к отдаленным предкам тюльпанов, когда не была так специализиро-
вана почка-детка, когда вместо почки-детки из пазухи чешуи, а тогда
листа, развивался настоящий побег. Таким образом, развитие побегов
вместо почек-луковиц помогает нам понять вопрос о происхождении
органов растения тюльпана. Даже из такой почки, как почка заме-
щающей луковицы, может развиться побег. Подобное явление неод-
нократно встречается у описываемого нами сорта (Бочанцева, 1954).
Сорт Due van Thol yellow (рис. 94)
Цветение сортов группы Ван Толей протекает одновременно с
дикими видами, поэтому годовой цикл роста и развития диких видов
интересно сравнивать именно с этим сортом.
Сорт D. V. Т. yellow обычно зацветает в условиях Ташкента в
конце марта и не позже пятого апреля. В это время в его замещаю-
щей луковице еще не наблюдается заложение никаких новых органов.
Лишь в конце второй декады апреля во время отцветания растения,
а часто после опадения листочков околоцветника в центре молодой
замещающей луковицы намечается валик зачатка первого листа буду-
щего года. А у некоторых луковиц примерно в середине апреля ясно»
вырисовывается не только зачаток побега — меристематический буго-
рок, но и валик нижнего листа. Валик листа после заложения быстро
приобретает характерную форму — возвышается с одной стороны и
сходит на нет с другой. Через полтора месяца после заложения,
в первых числах июня, с противоположной от зачатка первого листа
стороне почки на центральном меристематическом бугорке, уже на
3/4 охваченном основанием зачатка первого листа, закладывается
полулуние зачатка второго листа. В середине июля происходит зало-
жение третьего листа, а в конце — зачатков всех органов цветка.
Зачаток луковицы появляется одновременно с валиком второго
листа в первых числах июня. Корни в донце обособившейся заме-
щающей луковицы закладываются в середине июля.
336
Рост листьев и стебля (рис. 94, 3, 7). Стебель начинает функ-
ционировать в виде незначительного плоского меристематического
бугорка одновременно с заложением первого листа, который после
заложения в середине апреля до середины августа почти не растет;
так же не растут до середины августа в течение 2,5 месяцев после
Рис. 94. Сорт Due van Thol yellow.
Годовой цикл развития: /—луковица, 2—корни, 5—весь побег,
4—нижний лист, 5—второй лист, б—верхний лист, 7—стебель,
8—цветок, 9—завязь.
заложения второй лист и 1 месяц — третий лист. Таким образом, не-
зависимо от срока заложения органа на зачатке побега его рост
начинается в середине августа, т. е. начало роста листьев в длину
не зависит от их возраста, а связано с определенным состоянием
внутри луковицы, обусловленным определенным календарным сроком.
Луковица, в которой происходят указанные процессы, обычно на-
ходится или в сухой почве, или на полке хранилища. Все три
листа сразу начинают интенсивно расти и равномерно растут до
середины сентября. В середине сентября на вершинах двух нижних
листьев появляются волоски опушения. В это время у нижнего
листа наблюдаются некоторые неровности в темпе роста: в середине
сентября его рост несколько снижается, но уже в начале октября
выравнивается. Такая же картина падения интенсивности роста наблю-
дается у нижнего листа в середине октября, но к началу ноября —
вновь восстанавливается
Два других листа все это время растут ровно и непрерывно.
С середины ноября наступает зимний период покоя и все три листа
прекращают рост в длину. Нижний и второй листья не растут с сере-
дины ноября по середину декабря. Третий лист вступает в период
покоя одновременно с первыми двумя листьями, но выходит из покоя
в середине января.
22—2229
337
Два нижних листа в середине декабря выходят из покоя и начи-
нают активно расти, за две недели увеличиваясь на 5—6,6 мм, но к нача-
лу января, в связи с падением температуры почвы, их рост вновь
падает, и к середине января они вырастают всего на 4 мм. У нижнего
листа с середины января вновь начинается активный рост, и во второй
половине февраля он достигает максимума. Средний лист с середины
января до начала февраля полностью прекращает рост, а с начала фев-
раля начинает активно расти, также достигая максимума во второй
половине февраля. С начала марта, видимо, в связи с активным ростом
цветка и его частей на весь месяц прекращается рост обоих нижних
листьев. С начала апреля они вновь активно растут. К концу цвете-
ния и по отцветании, т. е. с середины апреля до начала мая, рост ли-
стьев вновь усиливается, а позже, из-за возрастающей сухости почвы
и высокой температуры воздуха, он прекращается.
Несколько иная картина роста, чем у двух нижних листьев,
наблюдается у третьего верхнего листа побега. Можно предположить,
что изменение в характере его роста по сравнению с двумя нижними
.листьями обусловлены его относительной близостью к верхушечной
почке побега, т. е. к цветку. С начала до середины ноября его рост
ничем не отличается от роста нижних листьев. Затем наступает по-
кой, более глубокий и длительный, чем у двух нижних листьев, так
как лишь к середине января, когда средний лист вновь прекращает рост,
верхний лист начинает расти. В начале февраля верхний лист опять
почти прекращает рост. С середины февраля, в период наиболее актив-
ного роста нижних листьев, он также довольно активно растет. В начале
марта, когда оба нижних листа останавливаются в росте, верхний
хотя несколько и задерживает его, но не прекращает. Во второй по-
ловине марта у верхнего листа наблюдается активный рост, и за 2 не-
дели он вырастает на 3 см, в то время как нижние два совсем не
растут. Возможно, это обусловлено тем, что во второй половине марта
растет верхушечный цветок, который мобилизует все запасы пита-
тельных веществ растения, и верхний лист, расположенный на пути
этих веществ, перехватывает часть запасов, поступающих в цветок;
или тем, что он использует для собственного роста продукты ассими-
ляции, так как два нижних листа вполне обеспечивают цветок. В по-
следующие две недели рост верхнего листа падает более чем в три
раза и почти равен росту двух нижних листьев. С середины апреля
он, как и у нижних листьев, усиливается, а в первых числах мая
прекращается. Таким образом, рост листьев связан как с влиянием
внешних условий, так и с внутренним состоянием растения.
Стебель, заложившийся в середине апреля, начинает вытяги-
ваться, расти в длину одновременно с листьями лишь в начале августа,
т. е. через три с половиной месяца после заложения. Его рост вначале
очень медленный. Так, за первый месяц он вырастает всего на.2 мм.
Начинает рост самое нижнее междоузлие между верхней чешуей
луковицы и нижним листом. С середины сентября стебель интенсивнее
вытягивается в длину, но медленнее, чем листья. Особенно замедляет-
ся его рост в первой половине октября перед укоренением луковиц,
затем в первой половине ноября он восстанавливается. Во второй по-
ловине ноября и в первой половине декабря стебель почти не растет.
Во второй половине января рост его восстанавливается. Весь январь
стебель растет медленно — в первой половине его прирост равен Змм,
а во второй — всего 2 мм. Но в феврале вновь наступает активный пе-
риод роста. Если за 5,5 месяцев роста он вытягивается всего на
2,7 см, то за февраль — на 5 см. В первые две недели марта, когда
338
начинают активно расти члены цветка, рост стебля несколько падает.
За это время он увеличивается на 1,1 см, но в середине марта актив-
ность роста стебля восстанавливается и не прекращается до середины
мая, т. е. стебель растет непрерывно как во время появления бутона
над поверхностью почвы,
так и во время расцвета-
ния цветка и роста завязи.
Заложение и рост
цветка (рис. 94, 8, 9\
96). В конце июля — на-
чале августа верхушечная
почка побега разбивается
на несколько бугорков и
начинается заложение
цветка. Первыми закла-
дываются в виде продол-
говатых полулуний три
валика листочков наруж-
ного круга околоцветни-
ка. Вслед за ними и од-
новременно с тычинками
закладываются широкие и
более короткие, чем внеш-
ние, валики внутреннего
круга листочков около-
цветника. При заложении
бугорков тычинок с мень-
шей вероятностью можно
установить, какие из них
принадлежат внешнему и
внутреннему кругам (впо-
следствии это можно уста-
новить безошибочно).
Завязь закладыва-
ется в виде трех оваль-
ных бугорков, которые
вначале свободны, но по
мере роста сливаются ос-
нованиями.
Листочки околоцвет-
ника после заложения в
последних числах июля
до начала сентября вы-
растают всего на 2 мм дл.
С начала сентября в те-
Ю Н8 347 246 146 1-Ю 2-20 547 247 346 448 2-17 246 146 347
III IV V VI VII VIII IX X X! хп 7 п Ш IV V
чение двух месяцев они
активно растут и в пер-
вую половину сентября
Рис. 95. Рост листьев и стебля сорта Due van
Thol yelow.
—нижний лист, 2—средний лист, 3—верхний лист, 4—стебель,
увеличиваются на 6 мм.
В этот период на листочках околоцветника уже наблюдается жил-
кование. На их вершинах появляются волоски опушения. Внутренние
листочки околоцветника с выемкой на вершине короче внешних. Про-
тив внутренних коротких листочков располагаются более высокие
тычинки, а против внешних—короткие. У тычинок хорошо видны гнез-
339
116 226 6-172-17 3-16 4-18247 2-16 148 347 246
да пыльников. Связник выдается над пыльниками. Узкий пестик в
виде тоненькой палочки несет на вершине начавшее развиваться
рыльце, еще без сосочков, но уже несколько разросшееся и выдаю-
щееся над пестиком.
Во второй половине сентября рост листочков околоцветника нес-
колько падает и они вырастают всего на 2 мм. В течение октября они
увеличиваются еще на 6 мм. пос-
ле чего на два с половиной ме-
сяца (с начала ноября до середи-
ны января) у них прекращается
рост в длину.
В середине января они под-
растают еще на 3 мм и вновь на
весь февраль приостанавливаются
в росте. В конце февраля листоч-
ки околоцветника подрастают на
1 мм, а с начала марта начинают
активно расти до отцветания, ко-
торое наступает во второй декаде
апреля. За две первые недели
марта они вырастают на 0,7 см.
Тычинки закладываются сразу
же после листочков околоцвет-
ника.
Рост и формирование пыль-
ников начинается одновременно
с листочками околоцветника и
очень скоро пыльники перегоня-
ют их в росте. Период интенсив-
ного роста пыльников—первая
половина сентября. Во второй по-
ловине сентября их рост замед-
ляется. В это время у пыльников
уже оформляются гнезда, кото-
рые шире к основанию тычинки.
Связник высоко выдается над
пыльниками. В первой половине
октября пыльники продолжают
медленно расти. В середине ок-
тября в материнских клетках пыльцы происходит редукционное де-
ление. В конце октября заканчивается осенний период роста пыльников,
но в начале ноября в пыльниках еще встречаются тетрады. Зимует же
пыльца в виде одноядерных пылинок. В течение четырех месяцев, с
начала ноября до марта, пыльники почти не растут. В первой поло-
вине марта начинается их весенний рост, который в основном закан-
чивается за две недели до раскрытия цветка. В день расцветания все
пыльники лопаются и усыхают. Удлинение тычинок перед расцвета-
нием цветка обусловлено не ростом пыльников, а растяжением тычи-
ночных нитей, которых долгое время после заложения тычинок не
удается обнаружить. Лишь в середине ноября под пыльниками появ-
ляются б. м. заметные зачатки тычиночных нитей. До начала февраля,,
т. е. два с половиной месяца, они не изменяются, оставаясь все время
в виде низких треугольничков. В течение первой половины февраля
тычиночные нити немного (до 0,3 см) подрастают и опять на
месяц совершенно замирают. Лишь перед раскрытием цветка, во вто-
Рис. 96. Рост органов цветка сорта
D. v. Thol yellow.
а—завязь, б—тычиночные нити, в—пыльники,
г—листочки околоцветника.
340
рой половине марта, они вырастают до нормальной длины. После цве-
тения тычинки отваливаются вместе с листочками околоцветника.
Завязь тремя отдельными меристематическими бугорками закла-
дывается почти одновременно с тычинками в середине августа. В про-
цессе роста в течение первых двух недель бугорки сливаются и по-
ручается треугольная завязь. В начале сентября завязь усиленно
растет, достигая 0,5 см. Затем в течение двух месяцев она растет
очень медленно и увеличивается всего на 0,4 см. С середины декабря
до середины февраля завязь не растет. С середины февраля до начала
марта она постепенно начинает вытягиваться и за две недели вырас-
тает еще на 2 мм. Усиленный весенний рост завязи приходится на
первую половину марта. Во время цветения и по отцветании она рас-
тет очень быстро. Ко времени пожелтения и созревания ее рост пос-
тепенно замедляется. Завязь живет дольше всех членов цветка. Наи-
более интенсивно растет завязь в период, когда остальные органы
цветка заканчивают рост и отмирают.
Заложение и рост замещающей луковицы (рис. 94, 7; 97, г).
.Замещающая луковица у этого сорта закладывается поздно. Впервые
Рис. 97. Рост замещающей луковицы и почек-деток
сорта Due van Thol yellow.
a—почка-детка третьей запасающей чешуи, д'—почка-летка второй
запасающей чешуи, t—почка-летка первой запасающей чешуи,
г—замещающая луковица.
меристематический бугорок почки замещающей луковицы мы наблю-
дали в середине июля. Полтора месяца с момента заложения до на-
чала сентября бугорок не увеличивался в размере. За первую поло-
вину сентября он подрос на 0,3 см, а во второй половине сентября
перестал расти. В октябре он вновь стал прибавлять по 0,1 мм в не-
делю. С начала ноября до первых чисел февраля замещающая луко-
вица почти не растет. За ноябрь, декабрь и январь луковица вырастает
на 0,2 см и к началу весеннего роста она имеет всего 0,8 см дл. За
февраль она увеличивается в длину на 0,5 см. В первой половине
341
марта при развернутой розетке листьев и действующей корневой сис-
теме замещающая луковица за две недели вырастает в длину на 1,7 см.
Во второй половине марта —еще на 1,2 см. В апреле, видимо, в связи
с цветением и ростом завязи рост луковицы замедляется. За месяц
(апрель) б. м. равномерного роста она увеличивается всего на 0,4 см.
С начала мая в связи с усыханием покровной чешуи луковицы не-
сколько уменьшаются.
Рост и развитие пазушных почек луковицы (рис. 97, а, б, в).
Весна и осень 1954 г. были неблагоприятными для роста и развития
пазушных почек-деток. Но детки у этого сорта обычно закладываются
во время летнего покоя, т. е. в период, когда существует наиболь-
ший разрыв между растением и средой.
Детки заложились и хотя большею частью не достигли нормаль-
ных размеров, но основные показатели их роста и развития нами были
изучены и описаны.
У сорта Due van Thol yellow детка в пазухе покровной чешуи
не закладывается.
Детка в . пазухе первой запасающей чешуи закладывается в на-
чале мая и до середины сентября почти не растет. В середине сен-
тября до начала октября она вырастает до 0,4 см выс. и опять пре-
кращает рост на 3 месяца —с начала октября до начала января. В
первую половину января детка подрастает на 0,3 см, а с середины
января до первых чисел апреля не растет. С начала апреля в зависи-
мости от состояния листовой и корневой системы растения эта детка
начинает б. м. усиленно расти. Ее рост у растения с развивающейся
завязью продолжался до середины апреля. Затем покровная чешуя
детки становится прозрачной и крахмал из нее транспортируется в
запасающие чешуи, после чего покровная чешуя утончается, ссыхает-
ся и коричневеет.
Детка в пазухе второй запасающей чешуи в замещающей луко-
вице закладывается так же, как и детка первой запасающей чешуи,
в начале мая, когда замещающая луковица еще не получила само-
стоятельности и не отделилась от чешуй и донца материнской луко-
вицы. Со времени заложения до начала сентября, т. е. четыре меся-
ца, детка не растет. В начале сентября за две недели она увеличи-
вается всего на 0,2 мм, а с середины сентября до начала ноября ее
рост вновь прекращается. С первых чисел ноября она растет до начала
декабря. Но в год исследования в связи с малым количеством осадков
материнские луковицы осенью не укоренились и за осенний период вто-
рая запасающая чешуя выросла всего на 0,2 мм. С начала декабря до
середины марта детка не растет. С середины марта на две недели
раньше, чем у детки первой запасающей чешуи, у детки второй за-
пасающей чешуи наступает активный период роста. За две недели
апреля детка второй запасающей чешуи вырастает на 1,6 см, в три
раза больше, чем за весь период роста с момента заложения до се-
редины марта. Больше она не растет в длину и во время окончания
вегетации обособляется от высохших чешуй материнской луковицы.
Детка третьей запасающей чешуи, расположенная с той же сто-
роны луковицы, что и детка первой запасающей чешуи, закладывает-
ся в начале июня, т. е. на месяц позже деток первой и второй запа-
сающих чешуй. Луковица детки третьей запасающей чешуи начинает
расти через две недели после заложения (без большого периода по-
коя), т. е. с середины июня и увеличивается за две недели на 0,2 см.
С начала июня до начала октября у нее наступает период покоя, за-
тем эта детка вновь растет до начала ноября, увеличиваясь всего на
342
0,2 см и опять прекращает рост на два с половиной месяца зимнего
покоя — с начала ноября до середины января. В январе она еще не-
много (на 0,1 см) вытягивается, а в первую половину февраля опять
замирает. Со второй половины февраля у детки начинается активный
весенний рост и во второй половине марта она вырастает на 1,6 см,
достигая общей длины в 2,6 см. В дальнейшем она растет более рав-
номерно.
Рассматривая заложение и рост почек-деток у сорта D. v. Т. yel-
low, следует отметить, что чем выше по донцу закладывается почка,
тем позже происходит ее заложение. Самой последней закладывается
почка замещающей луковицы. Но чем выше закладывается меристе-
матический бугорок почки-детки, тем быстрее она вступает в первую
фазу роста и тем раньше наступает у нее осенняя и весенняя фазы
роста. Это особенно ярко проявляется в наиболее сильно выраженной
весенней фазе роста.
У детки первой запасающей чешуи она наступает в первых чис-
лах апреля; у детки второй запасающей чешуи—в середине марта;
у детки третьей запасающей чешуи —в середине февраля, а у почки
замещающей луковицы—в начале февраля. Таким образом, централь-
ные или верхушечные почки находятся в наиболее благоприятных
условиях, чем почки, расположенные по периферии донца.
Тюльпаны по экологическому типу являются геоксерсфитными эфе-
мероидами, т. е. многолетними весенними растениями, быстро разви-
вающимися, цветущими и плодоносящими ранней весной, с повыше-
нием сухости, температуры воздуха и почвы уходящими в период
летнего сухо- или жаропокоя. Причем периодом покоя у живого ор-
ганизма нельзя считать полную приостановку жизненных отправлений,
так как это уже будет не покой, а смерть. Период покоя—истори-
чески сложившаяся необходимость в жизни живого организма, без
которой не может быть его дальнейшей жизни и нормального разви-
тия. Период покоя обусловлен наиболее возможным при сохранении
жизни разрывом между организмом и средой. Так, у тюльпана во
время летнего периода покоя отмирают ассимиляционные зеленые
листья и корни. Растение не ассимилирует, не всасывает с водой раст-
воренные в ней минеральные вещества; очень слабо дышит, так как
на внешнем эпидермисе луковичных чешуй тюльпана нет устьиц. Ра-
стение тюльпана в этот период все заключено в луковице. Оно как бы
оторвалось от внешней среды и поддается только температурным
факторам. В это время в растении протекают глубочайшие перестрой-
ки; происходит заложение и развитие всех органов побега будущего
года, а в корневой меристеме закладываются и растут корни.
В Средней Азии особенно распространены многолетние растения
с двумя периодами покоя—обычным у растений умеренного климата
зимним или хладопокоем и летним сухо-или жаропокоем, обусловлен-
ным высокими температурами воздуха и почти полным отсутствием
осадков в летние месяцы. К растениям, имеющим в годовом ритме
развития два периода покоя в Средней Азии, относятся многолетние
растения—эфемероиды. Это представители родов: Ferula, Rheum, Rin-
dera, Tulipa, Iris, Eremurus, Crocus и многие другие.
У них при первых осенних осадках развиваются питающие корни.
Почки надземных органов или почки возобновления усиленно растут
под покровом почвы и отмерших листьев. Осенью они обычно не
343
выкидывают розетку зеленых листьев. Рост почек возобновления бы-
вает скрытым и по сложившейся теперь терминологии называется эм-
бриональным.
Для роста и развития почки возобновления в осенне-зимний пе-
риод развития требуют пониженных температур, а при отсутствии их
заложившиеся в период жаро- или сухопокоя почки, особенно цветоч-
ные, могут отмереть. Это мы проверили экспериментально при выра-
щивании растений в оранжерейных условиях (Бочанцева, 1952). То же
можно сказать и о периоде летнего сухо- или жаропокоя.
Если растениям тюльпанов не давать пережить период сухопокоя,
то вызвать вегетацию у этих растений бывает невозможно, так как у
тюльпанов во время жаропокоя происходит перераспределение и пре-
образование запасных питательных веществ, отложенных в лукович-
ных чешуях, заложение корней, заложение и развитие почек. Эти
процессы протекают обязательно в период летнего покоя растений.
Таким образом, явления покоя, обусловленные специфическими
"особенностями климата, стали исторически сложившейся необходимо-
стью в жизни растений.
Но следует различать покой, без которого растение не может
продолжать развития, и вынужденный покой, вызываемый неблаго-
приятными условиями временного порядка,
Во время необходимого периода жаропокоя в отложенных в че-
щуях луковицц запасных питательных веществах происходит преоб-
разование одних веществ в другие, благодаря которому пазушные
почки приобретают способность проходить дальнейшие более высокие
ступени развития (сорт Jubilee).
В наступившее затем осенне-зимнее похолодание, сопровождае-
мое осадками, эти почки проходят недостающую им ступень развития.
Влияние осеннего света (Лысенко, 1942) в данном случае исключается,
так как эти почки глубоко скрыты внутри луковицы, а сама лукови-
ца лежит под толстым, не менее 10 см. слоем почвы. Весной при по-
мощи листьев, выросших из верхушечной почки, растения проходят
необходимую ступень световой стадии. Данный тип развития, видимо,
свойствен всем луковичным растениям с симподиальным типом раз-
вития луковицы. Оригинальным в этом типе развития растений явля-
ется то, что боковая почка первого порядка, не имея зеленых листьев,
заканчивает развитие при помощи листьев верхушечного, увенчанного
цветком, побега и, вступая в период покоя, закладывает в верхушеч-
ной почке цветоносный побег. В это время запасы питательных ве-
ществ, отложенные в специальных органах—-чешуях, вновь преобра-
зуются, переходя из одного состояния в другое, и пазушные почки
второго порядка, омолаживаясь, подготовляются к прохождению не-
достающей ступени развития.
Годовой цикл развития тюльпана, вырастающего из взрослой лу-
ковицы, интересен в связи с кратковременным активным периодом
жизни зеленой части растения.
В советской литературе годовым циклом развития взрослого расте-
ния у садовых сортов тюльпана занимались 3. М. Силина (1953),
Э. Ф. Мигушова (1954), О. Н. Данилевская (1955), Л. С. Тимофеева
(1956), Е, Н. Зайцева (1959). В основе их работ лежит жизнь луко-
вицы, а не растения в целом.
Мы исследовали годовой цикл развития у видов Т. kaufmanniana,
Т. fosteriana, Т. subpraestans, Т. korolkovii и Т. bifloriformis.
Для того чтобы выяснить, какие изменения произошли в расте-
ниях в результате их культивирования в течение четырех веков по
344
сравнению с видами окультивированными недавно, исследованы два
сорта из двух резко различных групп садовых тюльпанов: из группы
простых ранних тюльпанов—сорт Due van Thol yellow и из группы
тюльпанов Дарвина—сорт Jubilee.
Циклы развития отдельных органов тюльпана необходимо ото-
бразить во взаимосвязи—в последовательности и одновременности
роста, т. е. скрытого роста органов внутри луковицы и под землей и
•открытого роста надземного побега.
Заложение замещающей луковицы у диких видов тюльпанов про-
исходит в период жаропокоя, но у различных видов в разное время.
В начале июня—у Т. bifloriformis, в середине июня—у Т. kauijman-
niana, в начале июля—у Т. fosteriana, в середине июля—у Т. subpraestans,
в середине августа—у Т. korolkovii. У сорта Jubilee замещающая лу-
ковица закладывается в начале июня, а у сорта D. v.Thol yellow — в
середине июля. Таким образом, заложение замещающей луковицы у
диких видов и сортов культурных тюльпанов приурочено к середине
периода жаропокоя и занимает время для рода в целом с начала июня
по середину августа—всего 2,5 месяца (табл. 24).
Луковица дикого вида тюльпана живет от 20 (Т. korolkovii) до
23 (Т. kaufmanniana) месяцев. Активный период жизни, т. е. период
со времени обособления луковицы из чешуй материнской луковицы
до ее отмирания, равен 12 месяцам. Разница в продолжительности
жизни луковицы у видов зависит от продолжительности эмбрионально-
го, а не активного периода их развития (табл. 24).
Мы установили, что у сортов садовых тюльпанов жизнь луковицы
продолжается от 22,5 месяца у ранних сортов и до 24,4—у поздних.
Меристематические бугорки почек-деток закладываются в пазухах
чешуй у всех видов.
Однако заложение и рост почек-деток в пазухах чешуй диких
видов протекает более вяло, чем у сортов культурных тюльпанов, у
Т. korolkovii их развитие обычно не продвигается дальше меристема-
тических бугорков. У многих видов почка-детка не развивается в па-
зухе покровной чешуи. У Т. bifloriformis она совершенно не встре-
Та б л и ц а 24
Продолжительность жизни органов тюльпана, месяцы
Луковица Корни Побег Лист
НИЖНИЙ
Вид или сорт О oS И $ О 6 E К И О О 6 ’S К и о О о 1ВНЫЙ иод О
Ф ф g я* ф ф
CO ctf c и co «3 с и со со С и со со С со
Т. Kaufmanni- ana 11 12 23 3,5 7,6 11,0 11,0 4,0 15,0 10,0 3,8 13,8
Т. fosteriana 10 12 22 3,6 7,0 10 6 11,0 3,8 14,8 10,8 3,4 12,2
Т. subpraestans 10,5 12 22,5 3,5 6,5 10,2 10,5 2,7 13,2 8,4 2,4 10,8
Т. korolkovii 8 12 20 5,0 5,5 10,5 10,0 3,5 13,5 9,8 2,7 12,5
Т. biflorifor- mis 10,5 12 22,5 4,5 6,8 н.з 10,0 2,2 12,2 9,1 2,0 Н,1
Jubilee 12,4 12,2 24,6 2,6 8,0 10,6 11,5 3,4 14,9 11,5 4,0 15,5
D. v. Thol yel- low 10,5 12 22,5 4,5 6,5 11,0 10,0 2,0 12,0 9,6 2,4 9,6
345
Продолжение таблицы 24
Вид или сорт
Завязь
Т. kaufmanni-
ana
Т. fosteriana
Т. subpraestans
Т. korolkovii
Т. biflorifor-
mis
Jubilee
D. v. Thol yel-
low
м 2,6 12,0 7,5 2,5 10,0 12,5 2,5 15,0 7,5 2,5 10,0
8,4 2,5 10,9 8,1 2,4 10,5 П,6 2,3 13,9 9,0 1,0 10,0
7,2 1,8 9,0 6,0 1,7 7,7 11,0 2,5 13,5 8,0 0.7 8,7
5,7 2,3 8,0 5,6 2,2 7,8 9,9 2,1 12,0 6 8 0,4 7,2
-— __ —- П,1 8,1 10,6 10,4 1,7 12,1 7,1 0,5 7,6
9,1 2,9 12,0 9,3 2,7 12,0 11,5 3,5 15,0 8,5 1,3 9,8
9,0 2 0 |и,о 9,3 1,8 9,1 9,5 2,5 12,0 8,5 0,7 9,2
6,5 2,5 9,0-
7,0 2,0 9,0
7,4 1,6 9,0
6,6 1,8 8,4
6,1 1,8 7,9
9,3 2,3 11,6
8,0 1,5 9,5-
чается; У Т. kaufmanniana и Т. fosteriana—наблюдается довольно часто;
у Т. subpraestans—почти у каждой особи. У тюльпанов из группы простых
ранних, например у D. v. Thol yellow, она редко развивается, но-
всегда закладывается и развивается у тюльпанов из группы Дарвина.
В пазухах запасающих чешуй почти у всех диких видов и сор-
тов культурных тюльпанов обязательно закладываются почки-детки.
В развитии почек-деток как у культурных сортов, так и у диких
видов, имеются некоторые закономерности. Так, заложение почек-
деток начинается от периферических чешуй к внутренним.
Процесс роста почки-детки слагается из четырех периодов: 1) за-
ложения и первоначального роста меристематического бугорка,
2) валика покровной чешуи, 3) длительного органообразовательного-
периода со слабым ростом, 4) быстрого весеннего роста без заложе-
ния каких-либо новых органов. Чем позже закладывается детка, тем
быстрее протекают у нее три первых периода и тем меньше срок
между заложением чешуй, особенно между заложением покровной и
первой запасающей чешуей.
При вступлении в период зимнего покоя в почке-детке диких ви-
дов обычно заложены все чешуи (почки-детки культурных тюльпанов
развиваются несколько в ином плане).
В зимний покой первыми вступают периферические детки, а вес-
ной раньше пробуждаются центральные.
У сортов культурных тюльпанов, особенно из группы Дарвина,
мы наблюдаем две фазы в развитии почек-деток.
В первой фазе развития заложение органов происходит в то вре-
мя, когда замещающая луковица, в пазухах чешуй которой происхо-
дит заложение почек, еще не обособилась от материнской луковицы,
живет всецело за счет накопленных в ней запасов, так как побег ма-
теринской луковицы в это время едва показывается из земли и лишь
начинает ассимилировать. С наступлением периода жаропокоя рост
почек-деток первой фазы развития прекращается и в пазухах их че-
шуй закладываются почки-детки второй фазы развития. Покровная
чешуя деток первой фазы никогда не заканчивает морфологического
развития и после использования отложенных в ней запасов почками-
346
детками второй фазы имеет вид пленки, характерной для истощенной
запасающей чешуи. Чем позже закладывается почка-детка первой фа-
зы развития, т. е. та почка, которая расположена ближе к центру лу-
ковицы, тем она имеет меньшее количество чешуй и меньше почек-
деток второй фазы может заложиться в пазухе ее чешуй и тем быст-
рее они закладываются, но тем большее число чешуй они имеют.
В пазухах чешуй детки покровной чешуи первой фазы развития
во второй фазе закладываются почка-детка покровной чешуи, почка-
детка первой запасающей чешуи и, часто, цветоносный побег из вер-
хушечной почки-детки первой фазы развития. На месте этого побега
иногда закладывается замещающая луковица. У детки покровной че-
шуи может быть и сестринская (сериальная) детка.
В результате двухфазного вегетативного размножения одной за-
мещающей луковицы сорта Юбилей с шестью запасающими чешуями
в течение одного года можно иметь гнездо луковиц, состоящее из 21
почки-детки различного возраста, размера и происхождения. Обычно
их бывает меньше. Уменьшение происходит за счет развития цвето-
носных побегов из части почек. Образование побегов вместо почек-
деток является риверсией к тем отдаленным предкам тюльпанов, когда
не была так специализирована почка-детка и вместо почки-детки из
пазухи чешуи, а тогда листа, развивался настоящий побег. Таким об-
разом, развитие побегов вместо почек-деток дает объяснение о про-
исхождении органов растения тюльпана.
Корни в донце замещающей луковицы закладываются в период
жаропокоя после обособления луковицы из чешуи и от донца мате-
ринской луковицы. Из исследованных видов раньше всех в середине
мая зачатки корней наблюдались у Т. bifloriformis, в начале июня —
у Т. korolkovii, а в середине июня — у Т. kaufmanniana и сорта
D. v. Thol yellow, в начале июля—у Т. fosteriana и в середине июля —
у Т. subpraestans и сорта Jubilee.
В заложении корней намечается совершенно определенная зако-
номерность — чем раньше дикий вид или сорт тюльпана вступает в
период покоя, тем раньше обособляется его замещающая луковица и
в ее донце происходит заложение корней. Заложение корней в донце
замешающей луковицы диких видов или культурных сортов тюльпа-
нов приурочено к периоду жаропокоя и у рода в целом занимает
период с середины мая до середины июля, т. е. растянуто на 2 месяца.
Корни с момента заложения в донце луковицы до отмирания жи-
вут как у диких видов, так и у сортов садовых тюльпанов, от 10,2 до
11,3 месяца (табл. 24). Их жизнь также слагается из двух периодов:
эмбрионального — с момента заложения до выхода их из донца в поч-
ву и активного периода —с момента выхода их из донца до отмирания.
У некоторых видов эмбриональный период развития корней очень ко-
роткий (Т. subpraestans—1,7 месяца, сорт Jubilee—2,6 месяца); у других
видов он длиннее (Т. korolkovii—8 месяцев, сорт D. v. Thol yellow—
4,5 месяца). Виды и сорта, имеющие более длинный период эмбрио-
нального роста корней, имеют более короткий период их активной
жизни (табл. 24).
Период в жизни донца от обособления луковицы до заложения
корней в его корневой меристеме может считаться периодом, когда
на донце луковицы возможно производить хирургические операции —
прививки, пересадки и прочее.
Это обусловлено тем, что только в данный период донце заме-
щающей луковицы освобождается от закрывавших его тканей мате-
ринского растения и меристематическая ткань донца в какой-то сте-
347
пени еще способна к новообразованиям. Позже она вся расходуется
на корнеобразование и уже не способна ни к каким регенерациям.
Следует отметить полярность, глубокую специализированность и крат-
ковременность действия меристематических тканей тюльпанов, т. е. их
быстрое старение. Закладка генеративного побега в замещающей лу-
ковице у каждого вида и сорта приурочена к строго определенному
времени.
В замещающей луковице Т. kaufmanniana в конце февраля или
начале марта закладывается меристематический бугорок генеративного
побега будущего года, у Т. fosteriana и сорта Jubilee—в середине
марта, у Т. korolkovii—в начале апреля, у Т. biiloriformis и Т. sub-
praestans — в середине апреля, у сорта D. v. Thol yellow — в сере-
дине мая.
Для рода в целом период заложения побега занимает время от
начала до конца вегетации побега материнской луковицы, т. е. у всех
видов рода его закладка происходит в период, когда замещающая
луковица еще прочно соединена с материнским растением.
Меристематический бугорок—зачаток осевого органа-стебля дол-
гое время после заложения не вытягивается в длину, несколько раз-
растается в ширину и вступает в органообразовательный период. У
сорта Jubilee валик первого листа закладывается на меристематичес-
ком бугорке немедленно после его появления, а у других исследо-
ванных видов и сортов—в различное время, от нескольких дней
(Т. fosteriana, Т. korolkovii, сорт D. v. Т. yellow—10 дней, Т. kaufman-
niana—15 дней) до месяца (Т. bifloriformis—30 дней) и даже больше
двух месяцев (Т. subpraestans—75 дней).
Во время следующей фазы — органообразования, которое длится
от 70 дней у Т. subpraestans, 80—у сорта D. v. Т. yellow, ПО —у
Т. bifloriformis, 120 —у сорта Jubilee, 150—у Т. Korolkovii и Т. kauf-
manniana и 170— у Т. fosteriana,— стебель побега развивается так же,
как донце луковицы. Вначале на нем закладываются от периферии к
центру один за другим валики листьев, а затем части цветка, кото-
рого нет у донца, и в этом их различие. В период органообразования
стебель не растет в высоту. Лишь после заложения всех частей цвет-
ка стебель вступает в следующий период развития (рис. 66, 72, 77,
80, 85, 95). В это время (осень и зима) органы уже не закладываются
и происходит довольно плавный рост как стебля, так и заложившихся
на нем органов. Но в декабре и январе бывает приостановка роста
стебля. В начале февраля наступает бурный период роста стебля,
который продолжается и после отцветания растений вплоть до засы-
хания листьев.
Взаимосвязь и взаимообусловленность в развитии органов тюль-
пана особенно ярко выявляются в развитии стебля и цветка. Мы уста-
новили, что до заложения цветка стебель растет как донце. После
оформления цветка он начинает вытягиваться. Удаление бутона пре-
кращает рост стебля (Бочанцева, 1952). Стимулятором роста стебля
-тюльпана в длину является цветок или его части. При вступлении
стебля в период роста осенью первой начинает расти самая нижняя
его часть—от луковицы до первого листа. Но лишь следующей весной,
в феврале или марте, в зависимости от вида растения и погоды, ниж-
нее междоузлие выносит розетку листьев на поверхность почвы. Из
частей стебля дольше всех растет верхняя часть — цветоплодоножка.
У разных видов в росте стебля есть характерные особенности
(рис. 66, 72, 77, 80, 85, 95). В росте листьев так же, как и стебля
существует три периода: 1) с момента заложения валика листа до
348
начала роста, 2) плавного роста с осени до конца зимы, 3) бурного^
роста с февраля до пожелтения кончиков листьев.
Заложение листьев у тюльпанов очередное, но (табл. 25) часто
между заложением на генеративном побеге первого и второго листь-
ев проходит меньше времени, чем между заложением второго и
третьего. Заложение же третьего и четвертого листьев (если послед*
ний существует) также б. м. сближено, т. е. листья как бы пара-
та б л и ц а 25
Количество дней от заложения одного органа тюльпана до другого
Растение От стебля до 1-го листа От 1-го до 2-го листа От 3-го до 4-го листа Ог 3-го до 4-го листа От 3-го или 4-i о листа до цветка От цветка до завязи Or побега до луко- вицы О г 1-го ли- ста до лу- ковицы От побега до корней От корней до луко- вицы От побега । до завязи
Т. kaufmanni- ana 15 60 60 — . 30 0 105 90 105 0 165
Т. fosteriana 10 20 75 15 0 60 135 135 105 30 165
Т. subpraestans 75 5 40 5 5 15 90 15 85 0 130
Т. korolkovii 10 130 10 — 0 10 135 125 60 75 150
Т. bifloriformis 30 45 — — 35 30 50 20 30 15 110
Jubilee 0 45 30 45 , 0 0 75 75 115 -45 120
D. v. Thol yel- low 10 30 35 ; 0 15 75 65 45 15 9S
ми закладываются на побеге. В последующем листья на побеге не-
сколько раздвигаются разросшимися междоузлиями и тогда труднее
заметить их парное расположение.
Чем активнее растут листья в период плавного роста, тем слабее
увеличивается рост стебля и, наоборот, когда начинается сильный
рост стебля, приостанавливается рост листьев. Однако в период бур-
ного роста у растения хватает сил для обеспечения бурного роста
как листьев, так и стебля. Чем позже, т. е. чем выше по стеблю за-
кладывается лист, тем короче первый период его жизни (период эм-
брионального развития без заметного роста) и тем быстрее он созревает
для выполнения своих функций и короче его жизнь. Укорочение
периода жизни верхних листьев возникает в основном благодаря уко-
рочению первых фаз развития, т. е. ускорению развития. Заложение
частей цветков тюльпанов обычно протекает с июля по сентябрь:
т. е. оно приходится на вторую половину периода жаропокоя. Сроки
и темпы заложения частей цветка у разных видов следующие.
Первыми из исследованных тюльпанов в начале июля закладывают
цветок Т. fosteriana и сорт Jubilee. В начале августа закладываются
меристематические бугорки листочков околоцветника у двух видов —
Т. bifloriformis, Т. kaufmanniana и сорта D. v. Т. yellow. В середине ав-
густа появляются валики листочков околоцветника на зачатках побега
у Т. subpraestans и, наконец, в начале сентября - у Т. korolkovii.
В заложении и развитии цветочных почек тюльпанов есть общие
черты для всех видов, но есть и видовые различия.
У некоторых видов—Т. bifloriformis, Т. fosteriana, Т. subpraestans
и сорта D. V. Т. yellow—части цветка непосредственно после заложе-
ния в течение некоторого времени не растут. У Т. fosteriana — в те-
чение двух месяцев (с первых чисел июня по первое сентября), у
349
T. biflorifor,mis — полтора месяца (с начала августа по середину сен-
тября), у Т. subpraestans — всего две недели (с середины августа по
начало сентября) и у сорта D. v. Т. yellow — один месяц (с первых
чисел августа до начала сентября).
У этих видов в росте частей цветка можно различить следующие
фазы: заложения органов с микроскопически малым ростом, осеннего
роста, зимнего покоя и бурного весеннего роста.
Зимнюю остановку в росте частей цветка имеют Т. subpraestans
(4,5 месяца), Т. bifloriformis (3,5 месяца) и Т. fosteriana (3 месяца).
У сорта D. V. Т. yellow остановка в росте пыльников длится 4 месяца,
пестика — 3, а у листочков околоцветника в середине зимы покой
прерывается наступившей оттепелью и потом продолжается дальше.
У других видов — Т. kaufmanniana, Т. korolkovii и сорт Jubilee-
части цветка после заложения немедленно начинают расти. Подобное
явление нельзя объяснить поздними сроками их заложения, так как
цветок сорта Jubilee закладывается в середине июня, у Т. kaufman-
niana — в первых числах августа и у Т. korolkovii — в первых числах
сентября.
У двух последних видов в развитии можно различить три фазы:
постепенного осенне-зимнего роста, зимнего покоя и бурного роста.
У Т. kaufmanniana 3,5 месяца в течение зимы, а у Т. .korolkovii 4 ме-
сяца полностью приостанавливается рост частей цветка.
Цветки сорта Jubilee имеют всего две фазы в развитии — посте-
пенного осенне-зимнего роста и бурного весеннего роста. Остановки
в росте частей цветка зимой в ташкентских условиях у них не наблю-
дается. Специфическими для каждого вида являются сроки редукци-
онного деления и характер роста листочков околоцветника. Например,
у Т. fosteriana раньше всего вырастает и стареет их острый опушен-
ный кончик, а у Т. korolkovii долго и сильно растет центральная
часть листочка—„спинка“ и лишь позже вырастает отгиб. На ногот-
ке листочка околоцветника развиваются устьица.
Тычиночные нити закладываются довольно рано, вырастают у
всех видов лишь перед самым раскрытием цветка.
Глава XI
ИЗМЕНЕНИЕ ТЮЛЬПАНОВ ПОД ВЛИЯНИЕМ
а ВНЕШНИХ УСЛОВИЙ
правление развитием растений позволяет экспериментатору соз-
давать новые формы растений как из сеянцев гибридов, так и
из чистых видов. От гибридных сеянцев легче добиться по-
ложительных результатов, чем от сеянцев и растений чистых видов.
Для получения положительного результата нужно познать биоло-
гию растения и соответственно представлять условия, воздействуя кото-
рыми можно изменить их наследственность. В дальнейшем для нормаль-
ного развития этих растений необходимо наличие новых условий среды.
Консерватизм растений, особенно диких видов, очень велик. Усло-
вия Средней Азии с резко континентальным климатом, с холодной
зимой и жарким летом, значительными суточными и годовыми колеба-
ниями температур, с обилием солнечного света и малым количеством
осадков создали среди многолетних травянистых растений особый тип,
названный эфемероидным. Эти многолетние растения с осени после
первых дождей дают массу молодых корешков, интенсивно развиваю-
щихся под землей в течение зимы. Весной растения очень быстро
развивают буйную зелень, быстро зацветают и к началу лета закан-
чивают вегетацию, вступая в период летнего сухо- или жаропокоя.
К таким растениям относятся тюльпаны. Луковица тюльпана с
осени после наступления пониженных температур и первых незначи-
тельных осадков дает из донца массу молодых корешков. Ее цент-
ральная почка при благоприятных условиях уже в октябре показы-
вается над вершиной покровной чешуи. У стадийно зрелого тюльпана
в этот период в верхушечной почке имеются все листья. Нижний
лист стебля покрывает почку и морфологически значительно отличает-
ся от верхних листьев. На цветоножке длиной в несколько милли-
метров обычно располагается один цветок со всеми органами, еще не
окончившими развитие, но уже определимыми. В это время листочки
внешнего круга околоцветника своим жилкованием и опушением верх-
ней части резко отличаются от листочков околоцветника внутреннего
круга. Тычинки лишь на три четверти длины прикрыты листочками
околоцветника. Тычиночные нити не развиты, их зачатки в виде треу-
гольных подставочек видны под хорошо развитыми пыльниками с
ясно очерченными гнездами и ярко выделенным связником, у которо-
го на вершине пыльника находится вырост, характерный для вида и
не обращающий на себя внимание в сформированных цветках. У видов,
обладающих несколькими цветками на одном стебле, в этот период
351
все уже налицо и в материнских клетках пыльцы крупных цветков у
некоторых видов протекает редукционное деление.
Многоцветковость в семействе лилейных. Многоцветковость.
более свойственна семейству лилейных, чем одноцветковость и, не-
сомненно, является более древним признаком, тогда как одноцветко-
вость появилась в результате приспособления растений к жестким
условиям среды.
В подсемействе Lilioideae большинство видов рода Lilium и Fri-
tillaria на стебле имеет много цветков.
Многоцветковые роды подсемейства Lilioideae: Lloydia, Scilla,
Ornithogalum, Bellevalia, Hyacinthus, Hyacintella, Leopoldia, Muscari-
mia и Muscari („Флора СССР", 1935 г.).
Многоцветковость диких видов тюльпанов в естественных усло-
виях. Среди диких видов тюльпанов Средней Азии в естественных
условиях многоцветковость в основном свойственна тюльпанам секции,.
Eriostemones. В литературе, при ссылках на многоцветковость в есте-
ственных условиях, всегда цитируют найденный М. Г. Поповым близ-
ст. Дарбаза Ташкентской ж. д. экземпляр Т. bifloriformis Vved. с
И цветками на одном стебле. Большинство видов этой секции, расту-
щее в Средней Азии, обладает способностью давать в естественных
условиях два, три цветка на одном основном стебле.
А. И. Введенский (1935) во „Флоре СССР" приводит следующие
данные по многоцветковости среднеазиатских видов секции Erioste-
mones1:
-1 (5)
-1 (2)
- 1-2-3 (6)
- 1—7
1 (2-3)
-(1) 2-3 (11)
— 1-2 (6)
- 1
Т.
Т.
Т.
Т. biebersteiniana
Т. sogdiana
~ buhseana
turkestanica
turco manica
T. bifloriformis
T. biflora
T. orithyioides
T. dasystemonoides — 1 (2)
T. dasystemon — 1
T. tarda - 1-8
Из И видов, перечисленных в данном списке, 10 видов (кроме
Т. orithyioides) мы выращивали из луковиц, собранных в естественных
местообитаниях. Виды этой секции в культуре не только сохраняют
способность давать многоцветковые соцветия, но при улучшении
условий культуры эта способность значительно усиливается.
У растений секции Orithyia, Tulipanum, Spiranthera и Lophophyl-
lon, собранных в естественных условиях, многоцветковости не наблю-
дается. Секция Leiostemones, богатейшая секция рода не только по
количеству входящих в нее видов, но и по красочности и декоратив-
ной ценности их, содержит значительное количество многоцветковых
видов. Виды этой секции по морфологическим признакам легко рас-
падаются на естественные группы. Некоторые из них с многоцветко-
выми видами. А. И. Введенский (1935) указывает, что Т. praestans
имеет 1 —2 цветка. Этот вид давно пользуется заслуженным внимани-
ем среди садоводов за яркие, безупречно чистого тона, цветки. Т. prae-
stans составляет естественную группу с Т. subpraestans, который,
1 В скобках указано количество цветков, редко встречающееся у растений этого
вида.
352
как и предыдущий вид, растет в Памиро-Алае, но не обладает много-
цветковостью.
Многоцветковость часто встречается в группе относительно близ-
ких видов секции Leiostemones, у растений которой в основном жел-
тые, не очень красивые, средней величины цветки: уТ. platystemon—
2 цветка; Т. ferganica — 1 (2); Т. anisophylla — 1 (2); Т. korshinskyi —
1 (2) и Т. tetraphylla — 1 — 2 (4).
Из этой группы тюльпанов мы выращивали два вида — Т. fergani-
ca и Т. tetraphylla. Некоторые экземпляры имели многоцветковые
стебли.
Многоцветковость у садовых тюльпанов. А. И. Тютюнников (1932),
много лет занимавшийся культурой и выведением новых сортов тюль-
панов, обратил внимание на их многоцветковость. В 1923 г. выведен-
ный им сорт тюльпана „Русский богатырь" дал из одной луковицы
стебель, несущий два цветка. В 1924 г. у одной особи этого сорта наблю-
дался стебель с четырьмя цветками. В дальнейшем старые сильные
луковицы давали многоцветковые стебли. Одиночные цветы появля-
лись на слабых или впервые зацветших растениях. Тютюнников пи-
шет, что при выращивании 112 садовых форм тюльпана в питомнике
наблюдалась многоцветковость у Махрового красного (название утеря-
но) в 1923 и 1924 гг.— по одной особи с двумя цветками; Прозерпины
(одинакий) в 1926 и 1931 гг.—то же; Махрового белого (название
утеряно) в 1926 г.—несколько особей с двумя-тремя цветками;
Гоббемы (одинакий) в 1928 г.— одна особь с двумя цветками и
одна —с четырьмя; Колер Кардинала (одинакий) в 1929 г. — одна
особь с тремя цветками; Бридера тюльпан Фелицитата в 1930 г. —
одна особь с тремя цветками; Лихтфайера (махровый) в 1930 г.—одна
особь с тремя цветками; Бульденеж (махровый) в 1931 г.— одна
особь с двумя цветками; Дарвин тюльпан Нора Вар в 1931 г.—одна
особь с двумя цветками.
Таким образом, у А. И. Тютюнникова с 1923 по 1931 г. из 112
культурных сортов тюльпанов, в том числе и сорт „Русский бога-
тырь66, многоцветковость дали десять сортов—пять из группы махро-
вых тюльпанов; три из одинаких тюльпанов; один — из тюльпанов
Дарвина и один из тюльпанов группы Бридер.
У А. И. Тютюнникова было 30 сортов тюльпанов Дарвина, полу-
ченных от Ван дер Шота, и лишь одна особь из сорта Нора Вар дала
два цветка на одном стебле. Сеянцы же тюльпанов из этой группьц
выращиваемые А. И. Тютюнниковым в питомнике, ежегодно давали
особи с двумя — четырьмя цветками. У сеянцев „Русского богатыря"
при первом цветении после посева также наблюдались многоцветко-
вые особи.
Холл (1929) считает, что среди культурных сортов есть в прода-
же один сорт „ М. Scipio Motet64, имеющий многоцветковость. Под
многоцветковостыо у тюльпанов он понимает следующее: два или
более стеблей могут возникнуть из большой детки, которая не может
обособиться. В данном случае Холл (1929) говорит о многоцветко-
вости несколько иного типа, чем многоцветковость, встречающаяся в
естественных условиях у диких видов тюльпанов, полученная экспери-
ментально А. И. Тютюнниковым у культурных сортов. Говоря о много-
цветковости у диких видов тюльпанов, Холл (1940) и многие другие
указывают на пример с так называемым Т. turkestanica, приводимый
под таким названием в работе В. И. Талиева (1930) и под названием
Т. bifloriformis Vved. во „Флоре СССР66.
23—2229
353
Методика получения многоцветковости. В условиях культуры и
определенного воспитания на участках Ботанического сада АН УзССР
многие дикие среднеазиатские тюльпаны можно вырастить многоцветко-
выми. Для этого обычно осенью (в сентябре или октябре) половозрелые
особи тюльпанов, привезенные из экспедиций, высаживаются в гряды,
на которых обычная почва (культурнополивной ташкентский серозем)
омешана с песком и листовым перегноем из расчета по 15—16 кг то-
го и другого на 4,5 ж2 и удобрена аммиачной селитрой из расчета
30 г/ж2 при посадке и 30 г под первый осенний дождь.
Весной по мере оттаивания почвы на гряды под рыхление мы
вносили еще 50 г аммиачной селитры на ту же площадь и в момент
цветения один раз подкармливали.
По мере отцветания на растениях острой бритвой или ножом
удаляли завязи, а цветоножки оставляли. Тогда все пластические
вещества, накопленные разросшейся ассимиляционной поверхностью
листьев и стеблями для воспитания массы семян, и продукты даль-
нейшей ассимиляции вынуждены направиться в луковицу. Удаление
€утона не дает нужного эффекта, так как в этом случае междоузлия
перестают расти и получается розеточная форма, у которой листья
затеняют друг друга.
В результате раннего удаления завязей луковица увеличивается
в размерах. Эгу луковицу по вступлении в летний покой выкапывают
и хранят до посадки в ящиках, а затем вновь сажают на гряды, при-
готовленные вышеуказанным способом.
Пересаженным экспериментальным луковицам дают четыре под-
кормки аммиачной селитрой. По мере отцветания у растений вновь
удаляют завязи. При отсутствии дождей осенью или в период весен-
ней вегетации гряды с тюльпанами по мере необходимости поливают.
Особенно важным мы считаем осенний полив, так как луковицы с
осени должны хорошо укорениться с тем, чтобы к весне их почки
прошли необходимую для цветения на следующий год ступень раз-
вития.
Одни виды и сорта тюльпанов легко отзываются на улучшенные
условия и уже на второй год дают многоцветковость, а другие—
лишь на третий год, и то в очень небольшом количестве. Некоторые
сорта или целые группы сортов или видов вообще не бывают много-
цветковыми (Бочанцева, 1953, 1954).
Как культурные сорта, так и дикие виды тюльпанов—все подда-
ются влиянию условий культуры, но по-разному. Это влияние прежде
всего сказывается на увеличении размеров растений.
Размеры растений Т. greigii R g 1 в культуре. Дикие виды не-
обычайно чутко реагируют на улучшение условий культуры. Их цве-
ты в культуре часто достигают значительной величины.
Сопоставляя описания растений Т. greigii из естественных усло-
вий и культуры в различных местах Советского Союза, мы убедились,
что условия выращивания сильно влияют на размеры растений этого
вида
Во „Флоре СССР“ (1938) приводятся данные для диких особей
вида: высота растений 20—40 см, количество листьев—3—4; нижний
лист —3,5—14 см шир.; листочки околоцветника 3—10 см дл.
При культуре этого вида в Ленинграде (Сааков, 1950) он имеет
4 листа при высоте растений 25 — 40 см. Внутренний листочек около-
цветника 5,6—8 см дл., с 2,5 до 3,2 см шир. Наружный листочек
околоцветника 5—7,5 см дл., от 2,3 до 3 см шир.
354
В 1952 г. мы измерили растения Т. greigii непосредственно в поле
«а холмах близ ст. Дарбаза Ташкентской ж. д. (табл. 26).
Весна была влажной, благоприятной для роста растений. Высота
растений в среднем—18,1 см. Самое высокое растение достигало
24,5 см, низкое—13 см. На растениях было по 4, изредка 3 листа.
Нижний лист—в среднем 9,6 см дл. и 4,1 см шир. Самый длинный
Таблица 26
Размеры растений Т. greigii в естественных условиях
(Дарбазинская щель, Келесский район)
Высота растений Количество листьев Длина и ширина листьев, м* Листочки около- цветника, см Количество членов цветка
1 2 3 4 внешние внутрен- ние
18 4 10X4 10X2,8 8,5X1,5 7,2X0,9 5,4x3,6 5,2x3,8 6x6x3
14,5 4 13,6x5,6 13x3,3 12x1.7 10.5x9,2 5,4x4,5 5,4x4,8 6x6x3
.20 4 9x4,6 9,5x3,2 7X1,4 7X0,8 5,6x3,8 6,3X3,9 6x6x3
18,5 4 7,5X4,5 5,5x3,1 6,5X1,7 6,3x0,9 4,3X3,6 4,6x3,4 6x6X3
20,5 3 7X3,6 6,3x1,6 5,5хС,9 — 5,5X4,3 3,6x3,3 6x6x3
14 3 7x3 6,7X1,7 6,5x0,7 — 4,7X3,2 4,6x3,5 6x6x3
13 4 7X3,9 6x2,3 4,5x1 4x0,6 3X2,1 3,2x2 6x6x3
19 4 10X4,2 9,6x2,5 7,7x1,1 5,7x0,7 5,9X3,8 6,2x3,3 6x6x3
•22 4 10,5x3,8 10x2,6 10x1,6 7x0,8 5,2X4,1 5,5x3,6 6x6x3
21,5 4 12x5,2 12x3,4 9x1,3 6,7x0,8 4,7x3,2 4,5x3,4 6x6x3
24,5 4 13x4,7 12x3,9 9,5x1,5 7,5x0,9 5,5X3,9 5,9x4,3 6X6x3
13,5 4 9X3,6 8x1,9 6,5x1 5,2x0,5 4,5x2,8 4,5x3,1 6x6X3
19 4 11,5x3,7 9x2,2 8x1 6,7x0,7 4,5x3 4,8x3,2 6x6x3
16,5 4 8X3,6 7x2,6 4,3x0,9 4,5X0,7 3,7x2,7 4,3x3,6 6x6X3
17 4 9,5x3,3 9X2,4 8x1 6,3x0.7 4,5x3,2 4,5x3.4 6x6x3
18,1 9,6x4,1 8,9X2,6 5,6x1.2 6,5x0,8 4,5x3,4 4,9X3,6
Примечание. В данной таблице и во всех последующих в итоге приводятся сред-
ние цифры.
из нижних листьев у исследованных растений был 13,6 см, а широ-
кий—7 см. Листочки наружного круга околоцветника в среднем 4,5 см
дл. и 3,4 см шир. при наибольшей длине 5,9 см и ширине 4,5 см.
Мелкий цветок имел листочки наружного круга околоцветника 3 см
дл. и самые узкие из них были 2,1 см шир. Листочки внутреннего
круга околоцветника в среднем—4,9 см дл. и 3,6 см шир. при наи-
большей длине 6,2 см и ширине 4,8 см. Короткий листочек внутрен-
него круга околоцветника 3,2 см дл., а узкий—2 см шир.
Весной 1952 г. (2 апреля) мы измерили растения Т. greigii, кото-
рые были перенесены в виде луковиц с холмов близ станции Дарбаза
и первый год росли и цвели в культуре. Эти растения измерили вес-
ной 1952 г. Высота их—16,4 см. из них самое высокое—19 см, а низ-
кое—12 см. На растениях было по 4 листа. Нижний лист в среднем—
11,8 он дл. и 5,4 см шир. Самый длинный из нижних листьев—13 смдл.,
а короткий—всего 10 см. По ширине они изменялись от 7 до 3 см.
Листочки наружного круга околоцветника в среднем—5,2 см дл.
355
и 4,6 см шир. Максимальная длина и ширина наружных листочков
околоцветника не превышала 7 см. Самые мелкие—4 см дл. и такой
же ширины.
Листочки внутреннего круга околоцветника в среднем—6,6 см дл.
и 4,6 см шир. Их максимальная длина—9 см, а ширина—6 см. Мини-
мальные размеры—5 см дл. и 4 см шир. Листочки внутреннего круга
околоцветника были несколько больше, чем у растений из естествен-
ных условий (табл. 26 и 27).
В эксперименте при усиленном кормлении азотными удобрениями
и направлении потока ассимилянтов не на развитие семян, а в луко-
вицу резко изменяется величина растения. В наших опытах растения
несколько более приземисты (табл. 28), что обусловлено выращива-
нием растений на открытом участке, не защищенном от избытка
солнечного света и подверженного сильным восточным ветрам.
Высота растений в среднем—25,2 см. Наибольшая высота—45 см,
наименьшая—10 см. На стебле было 4 листа, а иногда и 5. Средние
размеры нижнего листа—19 см дл., 10,5 см шир. при максимальной
длине 32 см и ширине 16см. Самый маленький нижний лист—13 см
дл., а узкий—6 см шир.
Самый маленький нижний лист эксперимента по длине равен
самому крупному листу в естественных условиях и в первый год
после перенесения в культуру (табл. 26 и 27).
Длина листочков наружного круга околоцветника в среднем—
10,67 см, а ширина—8,5 см. Крупный—14 см дл., И см шир., самый
маленький—8 см дл., а узкий—6 см. Длина листочков внутреннего
круга околоцветника в среднем равна 10,9 см, а ширина—8 см. Самый
крупный из них—16 см дл., а широкий—10 см шир. Самый маленький
всего 8 см дл., а узкий 6 см шир. Следовательно, самый мелкий цве-
ток в эксперименте значительно больше крупного цветка как из ес-
тественных условий, так и из растений в первый год культуры у нас
и в Ленинграде (табл. 26, 27, 28).
Таблица 27
Размеры растений Т. greigii в первый год цветения после перенесения
из естественных условий в культуру
Высота ра- стений, см Количество листьев Длина и ширина листьев, см Листочки около- цветника, см Количество членов цветка
1 2 3 4 внешние внутрен- ние
19 4 12x5 8X5 8x3 5x1 6x6 7X6 бхбхз
14 4 11x7 юхб 8X5 4X2 5x5 7X5 6x6x3
19 4 13X6 11X4 7X2 7X1 7x4 9X4 6x6x3
12 4 13x3 11x2 8X1 5X1 4x4 5x4 6x6x3
18 4 10x5 9x4 7x2 5X1 4x4 5X4 6x6x3
11,8x5,4 9,8x4,2 7,4x2,6 5,2x1,2 5,2X4,6 6,6X4,6
Размеры Tulipa micheliana Hoog. Во „Флоре СССР" (1935) для
Т. micheliana приводятся размеры: высота растений 15—40 см; листья
в количестве 4—5; нижний лист—2—6 см шир.; листочки околоцвет-
ника имеют 2,5—10 см дл.
В работе С, Г. Саакова (1950) приводятся следующие данные по
размерам растений этого вида в условиях культуры в Ленинграде:
356
Размеры растений Tulipa greigii в эксперименте (в Ташкенте)
Таблица 2Й
Высота ра- стений, см Количество листьев Листья, длина и ширина, см Листочки околоцветника, см Количество членов цветка
1 2 3 4 5 внешние внутренние
28 5 18x11 20л9 21x6 13x4 13X4 12x7 14X9 6x6x3
19 4 21x13 17x9 18X3 13X3 — 10X6 11X7 6X6X3
24 4 16x6 15X6 12x4 11X4 —* 9X9 10x7 6x6x3
18 5 18X10 17X9 17x3 13x3 12x3 11X11 10X9 6x6x3
12 4 14X6 14X5 13x4 11X4 •*- 9X7 9X9 6x6x3
24 4 18x11 16x7 15x3 11X3 — 9X9 10x7 8X7x3
26 4 20x8 17x5 17X3 15X3 — 13X10 12X7 6X6X3
31 4 18X11 22X8 18x5 18X3 —* 11X9 13X8 6x6x3
31 4 19X13 19X9 16X4 11x3 — 8X8 8x7 6X6X3
24 4 19x8 14X6 16X4 14x3 — Пхб 9x6 6x6x3
10 4 14x10 15x7 14X4 9x3 — 10x9 9x9 6X6x3
10 4 13X9 12x9 10X4 5X2 — юхю 10x9 6x6x3
45 4 32x16 зохп 26ХЮ 25X4 — 14 9 16X10 6X6x3
38 4 20X12 22X12 20хЮ 16x4 — юхю 10x9 бхбхз
38 4 26X12,7 26ХИ 23X7 22x4 — 13X8 13x8,4 6x6x3
25,2 19,07x10,45 18,06x8,2 17,06X4,93 13,8x3,33 12,5x3,5 10,67x8,53 10,93x9,09 6x6x3
высота растений 25 — 35 см; количество листьев—4; внутренние лис-
точки околоцветника от 5,3 до 8 см дл. и от 2,2 до 2,9 см шир.; на-
ружные листочки околоцветника от 5 до 7 см дл. и от 2,3 до 3 см шир.
Сравнивая данные „Флоры СССР" (1935) сданными С. Г. Саакова
(1950) и с приведенными нами в табл. 29, мы видим, что растения
Т. micheliana в условиях культуры в Ташкенте несколько приземи-
стые, но с более широкими листьями. В нашем опыте самый узкий
из нижних листьев превышает самый широкий из приведенных во
„Флоре СССР" (1935) в 1,5 раза, а самый широкий —почти в 5 раз.
По размерам цветка растения из наших опытов (табл. 29) даже
по средним данным значительно превышают как ленинградские, так
и приводимые во „Флоре СССР" (1935). В среднем листочки внешнего'
круга околоцветника—12,9 см дл. и 7,5 см шир. и внутренние—12,75 см
дл. на 7,3 см шир. Средняя величина листочков околоцветника в на-
ших опытах почти равна их длине в ленинградских. Количество листь-
ев основного стебля и соотношение частей цветка почти не измени-
лось. Из 20 растений лишь у двух увеличено число листочков около-
цветника, что нельзя отнести за счет влияния улучшенных условий
культуры. Мы наблюдали у тюльпанов, выращенных из семян, при
первом цветении отклонение в числе листочков околоцветника и со-
хранение этого изменения в течение ряда лет.
Размеры Tulipa ingens Hoog. Количество листьев, приводимое
для Т. ingens во „Флоре СССР" (1935), в Ленинграде, по данным
С. Г. Саакова (1950) и в наших опытах (табл. 30) одно и то же.
Растения в наших опытах более приземисты. Однако неизвестно, что
считали эти авторы высотой растений,—весь стебель от луковицы или
его надземную часть, т. е. приблизительно на 1 см ниже места при-
крепления нижнего листа.
Непонятна фраза в работе С. Г. Саакова (1950) на стр. 281: „Цве-
тет обильно в открытом грунте". Как может растение, имеющее на
стебле всего один цветок, цвести обильно или слабо? При перенесе-
нии из Средней Азии в Ленинград в связи с изменением внешних
условий, требующихся для прохождения необходимой ступени ста-
дийного развития, оно может не цвести, или зацветут в связи с полу-
чением из семян неоднородного материала только половозрелые особи,,
а молодь не будет цвести в течение нескольких лет. Тогда можно го-
ворить о проценте цветения к определенному количеству особей с
определенным размером луковиц или их возрастом.
Во „Флоре СССР" самый нижний лист Т. ingens 2—5—(9) см
шир., по С. Г. Саакову, 29—32 см дл., 6 см шир. В наших опытах
(табл. 31) нижний лист Т. ingens имеет в среднем 21,3 см дл., 10,8
шир. Максимальная длина 28 см и ширина 13 см. Самый короткий
из нижних листьев—16 см дл., а узкий—6 см шир. Во „Флоре СССР"
А. И. Введенский приводит лишь ширину листьев, а длину считает
признаком не характерным. Однако мы убедились, что для многих
видов тюльпанов соотношение длины и ширины листьев является ха-
рактерным признаком.
Нижние листья Т. ingens в условиях наших опытов несколько
короче, чем в условиях Ленинграда, но они значительно шире, чем
в Ленинграде и в природе.
Длина листочков околоцветника этого вида во „Флоре СССР" рав-
на 4—11 см. В Ленинграде (Сааков, 1950) внутренние листочки около-
цветника от 6,2 до 8,4 см дл. и от 3,1 до 4,3 см шир., наружные-
листочки околоцветника от 6 до 8 см дл. и от 3 до 4 см шир.
В наших опытах (табл. 30) средние размеры листочков околоцвет-
358
359’
Размеры одноцветковых растений Tulipa micheliana в эксперименте (в Ташкенте)
Таблица 29
Высота растений, см Количество листьев Длина и ширина листьев, см Листочки околоцветника, см Количество членов цветка
1 2 3 4 1 5 внешних внутренних
24 4 24X14 25X10 22x9 22X5 — 15x8 14x8 6x6x3
19 5 24X14 24X11 20x7 19x4 10X4 11X9 11X9 6x6X3
18 4 26X12 26X10 20x4 18x4 — 11X8 12X8 6x6x3
29 5 30/14 27X13 25X3 23x6 20x4 14x7 14X6 бхбхз
26 4 24X17 22x12 23X9 2СХ6 — 14X8 14x8 6x6x4
28 5 27X13 23ХЮ 20X8 23X6 18X4 14X8 13x6 6x6x3
16 4 23X9 17X7 16X5 15x4 — 10X5 11 >8 6x6x3
17 5 23X11 23X9 21X7 16X5 15X4 10X7 9X6 6x6x3
22 5 22x16 21X13 21X9 18X5 19X3 11X7 12X7 7x6x4
12 4 19X11 19X8 18x5 15X4 — 9x6 9x6 6x6x3
22 3 27 + 14 27X12 20X5 — — 13X8 14X7 6х 6x3
28 5 24X18 28X15 26X10 22X8 23 - 4 16X9 15X8 6x6x3
27 6 25X15 24X10 22X6 25x6 21x5 14x8 13x9 8/.6ХЗ
23 4 21X16 25x12 21x9 18x7 23x3 14x8 14X8 5X6X3
27 5 27x20 26X15 24X8 16'Х9 21X8 14x9 15x8 6X6x3
26 4 29; 18 25x8 21x7 24X4 — 13x7 14x7 6 6x3
2-1 4 27X12 26+10 24x7 23X5 — 13x8 13X6 6X6X3
24 4 24X16 24X12 22x8 18x4 — 13x8 13X6 6x6 \3
26 4 24X20 24X14 22X10 18X4 — 13X7 12X7 6X6X3
27 5 30x20 31X14 26X7 21x5 — 16X8 13x8 6x6X3
23,25 1 25X15 24,35X11,25 I 21,7X7,4 | 19,69X5,31 | 18,2x4,5 I 12,9X7,5 12,7x7,3 |
ника. Т. ingens равны: для наружного круга—по длине 12,7 см при
ширине 5,6 см и для внутреннего круга—12,1 см при ширине 5,2 см.
Максимальные же размеры внешнего круга —15 см дл. и 7 см шир.;
для внутреннего круга максимальная длина несколько меньше—всего
14 см, а максимальная ширина остается такой же, как и у внешних.
Таблица 30
Размеры растений Tulipa ingens в эксперименте (в Ташкенте)
3S сз ~ •lecTBO ев Длина и ширина листьев, см Листочки около- цветника, см О и cj g м «
о X g “
Л сз й £ 12 3 4 внешние внУтРен- л ° ° О £Д
m л ние sr cf
15 3 20x6 19X4 15X4 - 11x6 12X7 6x6x3
25 3 21x10 20X8 22x6 — 11X6 12x5,5 6x6X3
-30 4 22x 13 30x11 27X9 13x1 14X5 13x6 7x6x3
20 3 20x10 22 x 8 22X6 — 13x5 13x5 6x6x3
17 4 19X8 21X6 19x6 17X3 11X5 9x6 7x6x3
18 3 28x10 24x6 24X6 13x5 13X5 бхбхз
20 3 24x13 23x11 17x7 — 13X6 12x7 8X7X4
18 3 24X13 22x9 20X 7 - 14x7 12X6 6X7x4
15 4 22X10 22x9 18x5 20x4 12x6 11x6 бхбхз
-35 3 22x11 22X9 22x8 — 15x6 14X6,5 бхбхз
-27 5 22x11 24x8 27X4 24x3—24X4 13x6 11X5 6x6x3
18 3 16X11 19X9 19x7 — 13x6 12X6 6X5X3
.15 4 16x10 18X11 16X10 20x5 12x7 12x6 бхбХЗ
24 4 22x11 24x8 24X4 24x3 13x5 13x5,5 6x6x3
20 4 22ХЮ 27x6 25x3 25X2 13x5 13x5 бхбХЗ
21,1 1 1 21,3x10,8 |22,5х8,2'21,1x6,1| 20,4x3 |12,7X5,б|12,1x5,2) 1
Из приведенного сравнения видно, насколько увеличились отдель-
ные части растений, особенно ширина листьев и цветы, по сравнению
< растениями в естественных условиях и в Ленинграде.
Особенных нарушений в отношении членов цветка у этого вида
не наблюдается. Из 15 измеренных растений у двух было по 7 лис-
точков околоцветника при нормальном количестве остальных членов
цветка, у одного растения—8 листочков околоцветника, 7 тычинок и
4 плодолистика; у другого растения при нормальном количестве лис-
точков околоцветника и плодолистиков—всего пять тычинок.
Размеры Tulipa fosteriana. Т. fosteriana, по данным „Флоры СССР“
<1935), имеет в природе растения 15—35 см выс. По С. Г. Саакову
(1950), высота растения при выращивании его из луковиц в открытом
грунте в Ленинграде —20—40 см. В наших экспериментах высота рас-
тения от 25—44 см и в среднем—32,6 см (табл. 31). Ширина нижнего
листа, по данным „Флоры СССР,,, колеблется от 3 до 15 см. В Ле-
нинграде длина нижнего листа—12—14, ширина—6 см. В наших опы-
тах в среднем длина листьев 22,3 см, а ширина —13,65 см, максималь-
ная длина—30, ширина—16 см.
Длина листочков околоцветника в природе 4,5—12,5 см. В Ленин-
граде члены внутреннего круга листочков околоцветника имеют 8,5 см
360
Размеры растений Tulipa fosteriana в Эксперименте (в Ташкенте)
Таблица 31
Высота ра- стений, см Количество листьев Длина и ширина листьев, см Листочки около- цветника, см Количество членов цветка Размеры листьев у деток
1 2 3 4 5 внешних внутренних 1 2
23 4 18X12 19X11 18x7 16x6 — 12X7 11X7 бхбхз — —
25 3 18X11 18X9 15X5,5 — —> 11X5 11X5 5x5x3 — —
27 4 19X16 18X11 17X7 14X7 — 14X6 12x6 8X6X4 ж» —
43 4 24X16 24X12 20x8 15x7 — 14X6,5 14x7 6x6x3 8X2 —
40 5 24X14 23X10 21X10 19x7 19X3 12X5 ЮХ7 бхбхз — —
30 3 22X15 22X12 19ХЮ — — 12X7 12X6 6x6x3 — —
34 3 24X16 27X12 23X9 — — 15X6 14X6 6X6X4 20X2 —
38 4 22X18 26X9 22X6 20x6 14X6 14X6 6X6X3 15X3 —-
28 4 18x12 19X11 19X7 14X5 — 14X5 14x5 бхбхз 5X1 —
30 4 22X10 24X10 21X8 юхб — 12X6 15x5 бхбхз 20x6 18X2
34 3 25X10 20x8 15x5 — — 12X6 11X6 6x6x3 16X4 —
33 3 24X16 24X11 21X7 мя — 13X6 12x5 бхбхз 15X3 —
31 4 24X16 25X12 23X7 22x6 — 13X7 12x6 6x6x3 17X8 16X2
35 3 21X16 22X10 18/8 — — 15X7 15X7 6x6x4 — —
44 3 30X15 31X11 27X9 — — 15X7 15x7 бхбхз 25X3 22X2
28 3 22x14 21X10 22X7 — — 14X7 12x6,5 бхбхз — —
33 3 29X13 27x9 21X5 — — 12X6 12,5X6 6x6x3 20X2 —
27 3 21X13 20Х Ю 14X5 — — 13X6 12X7 бхбхз 11X2 —
30 3 21X11 20X8 15X6 — — 13X6 12x7 бхбхз 14X2 6x1
33 5 18x14 19x10 19X8 16X4 16X4 12X6 10X5 бхбхз 14X3 —
32,6 | 1 22,3x13,65 22,7X10,3 | 18,5X7,2 16,2X6 | 17,5X3,5 13,1X6,2 | 12,5x6,1 15,4X3,2 15,5X1,75
дл., 4 см шир.; у наружного круга —8 см дл., 3,8 см шир. В наших
опытах средняя длина листочков околоцветника наружного круга—
13,1 см. а ширина —6,2 см при максимальной длине—15 см и ширине—
7 см. Средняя длина внутреннего круга листочков околоцветника —
12,5 см. ширина —6,1 см, максимальная длина их—15 см и шири-
на—7 см.
Несмотря на значительное увеличение размеров всех частей рас-
тения, в том числе и цветка (табл. 31), следует указать на сохране-
ние соотношения членов цветка. Небольшие уклонения в ту или дру-
гую сторону показывают лишь консерватизм данного явления, но не
его изменение.
Из всех среднеазиатских тюльпанов Т. fosteriana имеет самые
крупные цветы. Из сравнения размеров наиболее крупных тюльпанов
Т. micheliana (табл. 29), Т. ingens и Т. fosteriana (табл. 31) видно, что
цветы первых двух видов не намного, но все же в среднем мельче,
чем у Т. fosteriana.
В работе Б. П. Сердюкова (1952) приводятся данные по увеличе-
нию размеров растений Т. eichleri в течение многих лет под влиянием
перенесения их из естественных условий в культуру. Но Сердюков
дает им слабые дозы азотных удобрений, применяемые при полевых
работах, а не для садовых культур. Поэтому в его опытах медленно
увеличиваются размеры растений. Наши работы (Бочанцева, 1953, 1954)
и С. М. Брагилевской (1952) с цветочными однолетниками показыва-
ют, что в цветоводстве повышенными дозами азотных удобрений до-
стигается колоссальный эффект.
В результате выращивания растений на высоком агрофоне с до-
статочной влажностью в период укоренения и вегетации, высокими
температурами очень длительного периода летнего жаро- или сухопокоя
и удаления молодых завязей у многих диких и культурных тюльпанов
появляется многоцветковость.
В. И. Талиев (1930) в работе, посвященной роду Tulipa, назвал
тюльпаны обедненными родичами лилий, указывая, однако, что в
секции Eriostemones рода Tulipa многоцветковость преобладает над
одноцветковостью. Группе близких видов секции Leiostemones, расту-
щих в основном на Памиро-Алае и Тянь-Шане,—Т. platystemon, Т. fer-
ganica, Т. anisophylla, Т. korshynskyi, Т. tetraphylla—присуща много-
цветковость, вернее двухцветковость.
Способность давать многоцветковость в естественных условиях
отмечена и у такого вида, как Т. praestans.
ТИПЫ МНОГОЦВЕТКОВОСТИ
Многоцветковость у тюльпанов в естественных условиях и в ре-
зультате воздействия условий культуры может быть трех типов. У
видов секции Eriostemones на одном растении наблюдается несколько
верхушечных цветков. Они возникают на стебле несколько выше
верхнего листа, сидят на длинных цветоножках и имеют лишь один
прицветный лист (рис. 98). Эти цветки расположены в виде развилки,
кисти одно-, двух- или трехъярусного зонтика. У разных экземпля-
ров одного и того же вида этой секции можно встретить всевозмож-
ные типы соцветий. Такой тип многоцветковости возникает благодаря
ветвлению верхушечной почки стебля. Он свойственен только видам
секции Eriostemones и поэтому назван „типом Eriostemones41 или пер-
вым типом многоцветковости, так как первым отмечен в литературе
о тюльпанах.
362
У тюльпанов секции Leiostemones наблюдается иной тип много-
цветковосги—„тип Leiostemones", или второй тип многоцветковости.
Добавочные цветки у растений, относящихся к этому типу, возникают
на облиствленных побегах из почек, сидящих в пазухах зеленых
листьев (рис. 99). На пазушных побегах в основном бывает лишь
у некоторых культурных сортов тюльпанов
почти
глав-
сорта
100).
. б
Рис. 98. Tulipa bifloriformis Vved.
а—четырнадцатицветковое растение; верхушечная
почка ветвится и дает зонтик цветков с одним при-
цветным листом; из пазухи второго листа развивается
3 цветка также без прицветных листьев; их цветонож-
ки в нижней части срастаются со стеблем; б—четы-
рехцветковое растение этого же вида; цветоножка
пазушного цветка свободна.
одна из другой,
коллатеральные
один прицветный лист, но
пазушные побеги несут
столько же листьев, что и
ный стебель. Например, у
„Русский богатырь" (рис.
Второй тип многоцветковости
мы иногда отмечали и у видов
секции Eriostemones (рис. 98, б).
У второго типа многоцветковости
тюльпанов, относящихся к обеим
вышеназванным секциям рода, в
пазухе одного листа может раз-
виться не одна, а две или три поч-
ки (рис. 98, б). Они располага-
ются рядом или
Сериальные и
почки.
У растений, относящихся к
секции Leiostemones, каждый по-
бег, возникающий из такой почки,
несет один или несколько ли-
стьев и заканчивается цветком
(рис. 100, 101). Пазушные по-
беги растений из секции Eriostemones не имеют листьев. Дальнейшее
развитие второго типа многоцветковости секции Leiostemones приводит
к получению из пазух листьев на побегах первого порядка почек и раз-
витию из них побегов второго порядка с листом и цветком (рис. 101).
Третий тип многоцветковости возникает в результате ускорения
развития замещающей луковицы и деток. В год заложения в естест-
венных условиях замещающая луковица дикого вида и культурного
сорта тюльпана является спящей почкой. Обычно она дает корни
и листья только следующей весной.
В условиях же культуры при избыточном кормлении материн-
ской луковицы замещающая луковица трогается в рост в год форми-
рования (вторая фаза развития) и часто в этот же год дает облиствлен-
ный побег, увенчанный цветком.
Многоцветковость первого типа—„Eriostemones"—не встречается
у видов и сортов, относимых к секции Leiostemones. Иногда у тюль-
панов секции Leiostemones мы наблюдаем картины ветвлений на вер-
шине стебля, напоминающие собой тип ветвления у тюльпанов секции
Eriostemones (сорта Kathleen Parlow, Proserpina, La Tulipa noire и
Elephant).
В данном случае побег развивается из пазушной почки самого
верхнего листа основного стебля тюльпана. Главный цветок этого
стебля как бы лишается прицветного листа, который всегда сохраняется
у цветка боковой ветви в том случае, если ветвь не дает ветвь сле-
дующего порядка.
Отличить основной цветок стебля от цветка, который развивается
на ветке первого порядка, удается легко, даже в случае сращения
основного стебля с веткой. Для этого следует проследить по стеблю
363
ют места срастания цветков до узла, от которого боковая ветвь начи-
нает расти. Ветвь первого порядка будет ближайшей к центральной
жилке листа данного узла. Она обычно несет прицветный лист.
Основная ось побега удалена от листа и у ее цветка нет такого
листа.
Рис. 99. Tulipa praestans Hoog.
а—двухцветковое растение; ветвь первого порядка с цветком и листом вы-
росла из пазухи второго листа; трехцветковое растение; из пазух второго и
третьего листьев выросло по ветви первого порядка; в—четырехцветковое
растение; из паяух третьего, четвертого и пятого листьев выросло по вет-
ви первого порядка; пятый лист главного стебля, несущий в пазухе ветвь
первого порядка, является прицветным листом для главного цветка.
Первый тип многоцветковости. В 1951 г. мы установили, что
как у диких видов, так и у культурных сортов тюльпанов три типа
многоцветковости (Бочанцева, 1951, 1954). Однако без соответствую-
щего воспитания растений они проявляются редко и обычно проходят
незамеченными.
Для видов, относящихся к секции Eriostemones, в литературе для
полевых условий отмечено по 2—3 цветка. Под Ташкентом был най-
ден Т. bifloriformis с 11 цветками, вошедший во все сводки о тюль-
панах. Найденное растение имело тип многоцветковости, свойственный
лишь видам этой секции и обусловленный ветвлением верхушечной
почки роста стадийно зрелого побега.
У видов секции Eriostemones как пучок цветков, оканчивающий
главный стебель, так и добавочные цветы, возникшие на побегах
первого порядка из пазухи второго листа (рис. 98), не имеют при-
цветных листьев. Отсутствие прицветных листьев и развитие цветов в
виде зонтика на вершине стебля — существенное отличие первого ти-
па многоцветковости от второго. Это различие свидетельствует о край-
ней редукции некоторых органов у представителей секции Eriostemo-
nes, об утере ими не только листьев на главном стебле (половозре-
лые экземпляры видов этой секции имеют в основном по два листа),
но и прицветных листьев как на цветоносах, венчающих побег цветов,
так и на ветвях первого порядка, а не о примитивности видов этой
секции, как утверждает Талиев (1930). Если бы венчающие стебель
цветки сохранили прицветные листья, то получилось бы соцветие ти-
па соцветия Fritillaria с ветвистым стеблем, сериальные ветви перво-
го порядка у которого были бы также облиствлены. Уменьшение
364
размеров растений, количества листьев и поверхности листовой плас-
тинки — результат приспособления растений к жизни в условиях резко
континентального климата.
Многоцветковость нового типа
ких видов.
Секция и вид
Eriostemones
Т. bifloriformis
Т. turkestanica
Lophophyllon
Т. regelii
Leiostemones
Т. ferganica
была получена у следующих ди-
Количество
в экспери- менте 1951 г. 1952 г.
100 75 75
40 37 37
75 2 2
20 0 1
У Т. regelii, относящегося к секции Lophophyllon также получе-
на многоцветковость первого типа. Т. regelii раньше относили к сек-
ции Eriostemones, но А. И. Введенский на основании необычной фор-
мы листьев выделил его в особую секцию. С секцией Eriostemones
Т. regelii сближает наблюдавшийся нами
новый признак -- добавочный цветок,
возникший путем ветвления верхушеч-
ной почки и не имеющий никакого при-
цветного листа. Здесь развилась только
цветоножка и цветок, а не ветка. Сле-
довательно, у Т. regelii — Eriostemones
тип многоцветковости.
Особого внимания заслуживает факт
появления в 1952 г. трех цветков у од-
ного из двух растений Т. ferganica
Vved., ставших многоцветковыми. При
этом все ветки были без добавочных
листьев (другой многоцветковый экземп-
ляр этого вида был обычного для ви-
дов секции Leiostemones второго типа
многоцветковости). Появление растения
с ветвлением, но без листьев на ветках
у вида, относящегося к секции Leioste-
mones, как бы сближает этим совершен-
но обособленные секции. Тем более,
Рис. 100. „Русский богатырь". Че-
тырехцветковое растение, в па-
зухах четвертого и пятого листьев
заложились ветви первого поряд-
ка, несущие по одному цветку, но
по три листа; в пазухе пятого
листа выросло два побега рядом;
боковые побеги срослись с глав-
ным стеблем до первого, лежаще-
го над местом их заложения, листа.
что по внешнему виду Т. ferganica и
близкие к нему виды несколько отли-
чаются от других видов своей секции.
Второй тип многоцветковости у
диких видов тюльпанов. Второй тип
многоцветковости, названный нами „Lei-
ostemones типом“, встречается преиму-
щественно у растений, относящихся к
секции Leiostemones.
От предыдущего типа многоцвет-
ковости он отличается тем, что добавочные почки развиваются на
ветках, выходящих из пазух верховых зеленых листьев. В пазухах
этих листьев на всех побегах тюльпанов часто можно обнаружить за-
чатки почек. Они хорошо видны в середине зимы на сильном моло-
дом побеге, недавно показавшемся из луковицы. Почки большей ча-
365
стью представлены в виде довольно крупных недифференцированных
меристематических бугорков. Такие почки, из которых вырастут по-
беги или луковицы, в это время бывают сильно развиты.
Добавочные цветы у многоцветковых растений сидят на ветках,
несущих небольшое количество листьев — один, два или три листа
(рис. 99). Эти листочки обычно небольшого размера и приблизитель-
но равны верхнему листу данного экземпляра.
В пазухах стеблевых листьев тюльпанов может прорасти несколь-
ко почек, так как тюльпаны склонны к коллатеральному почкообра-
Рис. 101. Kathleen Parlow.
а—трехцветковое растение. Ветвление первого и второго узлов; б—трехцветковое
растение. Ветвление второго узла. В этом узле вырос побег с листком и цветком.
В пазухе листа на побеге первого порядка вырос побег второго порядка, также, с
листом и цветком; в—пятицветковое растение- Ветвление второго и третьего узлов.
§ обоих развилось по две коллатеральных почки, лежащих рядом. Каждая из них
дала по стеблю с листом и цветком.
зованию. У сортов культурных тюльпанов и диких видов в культуре
мы часто наблюдаем по нескольку облиствленных веток, увенчанных
цветами, выходящих из пазухи одного листа. Подобное явление часто
встречается у Т. subpraestans, Т. praestans, сортов Elephant, Kathleen
Parlow, Mothers Day, „Русский богатырь", Murillio и многих других.
В пазухах листьев, растущих на ветвях первого порядка, также
развивается одна или несколько почек и тогда вырастают ветви вто-
рого порядка с листьями и цветами. Ветви третьего порядка особенно
часто встречаются у Т. subpraestans. Эти ветви обычно несут лишь
по одному цветку и листу. Чем ветвление более высокого порядка,
тем мельче листья на ветках. Из культурных сортов только сорт Kat-
hleen Parlow имеет растения с ветвями второго порядка. У Т. sub-
praestans наблюдается настолько сильное ветвление, что при неболь-
шой высоте растений их кусты во время цветения производят впечат-
ление красных шаров, лежащих на земле.
В 1951 г. второй тип многоцветковости был получен у предста-
вителей 5 видов секции Leiostemones. В 1952 г. он появился у 14
среднеазиатских видов (табл. 32) —11 видов секции Leiostemones, два
вида секции Tulipanum и один вид секции Spiranthera.
У одного экземпляра Т. greigii (рис. 102) пробудились почки
второго и третьего узлов зеленого стебля. Почка во втором узле не
до развивалась. Она дала два небольших листочка и недоразвитый,
неокрашенный цветок, у которого, однако, можно было различить
366
Таблица 32
Дикие виды с многоцветковостью второго типа
Секция и вид Количество луковиц в эксперимен- те 1951 г. 1952 г.
Leiostemones
Т. fosteriana 500 1 3
Т. ingens 30 0 1
Т. praestans 15 11 11
Т. subpraestans 28 22 39
Т. greigii 400 1 11
Т. micheliana 130 0 1
Т. korolkovii 300 0 2
Т, vvedenskyi 450 0 1
Т. zenaidae 4 0 2
Т. ferganica 20 0 1
Т. hissarica 50 2 2
Spiranthera
Т. kaufmanniana
Tulipanum 1000 0 27
Т. kuschkensis 50 0 3
Т. hoogiana 75 0 3
пестика. В
пазухе
одним
третьего
зеленым
главно-
все основные элементы, за исключением
листа этого растения развился нормальный побег с
листом и цветком, который по размерам и окраске подобен
му цветку этого экземпляра. У
Т. fosteriana (рис. ЮЗ) мощный по7
бег первого порядка развился из
пазухи первого листа. Он нес
один лист и был увенчан круп-
ным цветком, по величине и ок-
раске сходным с главным цветком
этого растения. Из 63 растений
Т. subpraestans 22 экземпляра
под влиянием воспитания стали
многоцветковыми (рис. 104). Так
же, как у сортов культурных
тюльпанов Proserpine и La Tulipe
noire, у этого вида ни у одного
экземпляра не проснулась ни одна
почка в пазухе первого узла. У
12 экземпляров Т. subpraestans
проснулись почки в пазухе вто-
рого листа, у 20 экземпляров—в
пазухе третьего листа; У 8 ЭК- рис Ю2. Т. greigii Rgl. Двухцветковый
земпляров—в пазухе четвертого экземпляр.
листа и у 5 экземпляров — в
пазухе пятого листа. Т. praestans имеет, как отмечали многие
исследователи (Тютюнников, 1938; Холл, 1939; Бочанцева, 1956),
367
склонность к ветвлению стебля и многоцветковости. Перенесе-
ния из естественных условий в культуру оказалось достаточно, чтобы
из 61 экземпляра, росшего в питомнике, 14 дали многоцветковые
стебли. Кроме того, следует отметить, что несколько самых крупных
растений было сорвано случайными людьми и вследствие этого не уч-
тено. У Т. praestans (рис. 99), так же как и у Т. subpraestans, не
просыпаются пазушные почки первого узла. У первого вида наиболь-
шее количество пробудившихся почек появляется на втором узле —
у 13 из 14 многоцветковых растений. У Т. subpraestans пробудивших-
ся почек больше на третьем узле. В третьем узле просыпаются почки
у 9 растений Т. praestans.
При этом почки просыпались у растений, развивших почки вто-
рого узла, за исключением одного растения. В четвертом и пятом уз-
Рис. 103. Tulipa fosteriana
Jrv. Двухцветковый экзем-
пляр.
Рис. 104. Tulipa subpraestans Vved. Се-
мицветковое растение. Ветвление в третьем,
четвертом и пятом узлах стебля. В треть-
ем и четвертом узлах развилось по две
сериальных почки. В пятом—одна почка.
На одном из побегов первого порядка,
выросших из пазухи третьего листа, есть
побег второго порядка, возникший из пазу
хи его единственного листа.
лах проснулись почки у нескольких экземпляров, но растения были
сорваны. Сохранился только экземпляр, представленный на рис. 99.
Мы не приводим измерения всех многоцветковых растений, полу-
ченных у тюльпанов. Только у Т. greigii в 1952 г. мы измерили все
многоцветковые растения (табл. 33). У других видов тюльпанов из-
мерялись лишь отдельные многоцветковые особи (табл. 34).
Из сравнения данных по измерениям многоцветковых растений
Т. greigii и других видов с размерами раскормленных не многоцвет-
ковых растений (табл.28, 29, 30, 31) видно, что у многоцветковых рас-
тений размеры цветов и других частей не уменьшаются. По сравне-
нию же с растениями из естественных условий, описанными во „Флоре
СССР" (1935), измеренными нами в поле, а также выращенными
С. Г. Сааковым в Ленинграде, многоцветковые растения являются зна-
чительно более крупными.
На примере Т. greigii трудно говорить о преимущественном пре-
обладании развития почек в пазухе какого-либо одного узла зеленого
стебля. Из данных табл. 33 видно, что процент заложения и развития
почек в пазухе самого нижнего и лежащего над ним узла совершенно
368
24 —22 29 3 69
Размеры многоцветковых растений Tulipa greigii в эксперименте Таблица 33
Количество Длина и ширина листьев, СМ
Высота растений, см цветов на растении листьев основного стебля 1 2 3 4 5 6 7 8
31,5 4 6 1 27,2X16 26,3x13,5 1 22,7X10,3 22,5x6,1 21,7X4,2 17,5x3,5 — —
16 2 6 18X11 19X8 13X5 17X3 13X3 12X2,5 1 —
31 3 4 25X15 22X10 21X7 20X5 —• — — —
34 2 5 31X21 26X14 25X7 23X5 4X2 — — —
34 2 6 27X23 28X14 24хЮ 25x8 24X5 24x3 — —
23 3 6 24X10 24x9 22X7 19X2 8X1 4X1 — 1—
35 2 6 24X15 25X12 19x10 19X7 18x4 8X2 —
29 2 8 | 28X16 29X13 27X7 25X4 21X6 22x3 25x3 20X2
25 2 6 19X13 17X12 15X7 14X5 12X3 12x5 — —
36 2 6 32X10 28X19 22x6 24x5 24x3 19X2 — —
33 2 4 22X13 22X10 17X5 19X3 — — — —
29,8 5,7 ^25,3X14,8 24,2ХИ,3 20,7X7,5 20,7X4,8 16,2x3,5
о Высота растений, см Основной цветок Ветви и их части Добавочный цветок
количество членов цветка длина и ширина ли- сточков околоцвет- ника, см количество веток узел зало- жения длина вет- ки, см длина и ширина листьев, см количество членов цветка Длина и ширина ли- сточков околоцвет- ника, см
внешние внутренние 1 2 3 4 внешних внутренних
31,5 5X5X3 12X8 12x7,9 3 3 18 10,4X1,8 — — — 5x5x3 9X5,9 8,5X4,9
4 23 11X1,9 — — — 5x6x3 8,8x5,5 9X5,5
5 17,5 8,2 • 1,4 — — 5x5x3 6,4x3 5 6,3X4,2
16 5X5X3 6X4 7X5 1 3 20 10 '2 — — — 5X5X3 5x3,5 5,5> 4
31 6X6X4 10X7 11X8 2 1 16 5X1 — — — 6X6X3 бхб 5X4
2 10 5X2 — — — 6х6> 2 8x6 7X6
34 6x6x3 6X6 7X6 1 1 24 4X2 - — —- 6x6x3 5/5 5x5,5
34 6x6x3 11x8 12x8 1 2 30 14 6 10X3 8X3 — 6x6x3 9X6 8,5X5
23 бхбхз 12X7 13X8 2 4 18 4X1 3X1,5 3X1 2X1 6x6x3 9, .6,5 8,6X5,1
5 12 3; 1 1,5x0,5 — — бхбхз 7x4,5 7X5
35 бхбхз 10X5 12x6 1 2 29 8X3 — —- — 6X6X3 9X6 8X5
29 6x6x3 10x6 12X6 1 1 29 14X3 10X2 — — 6X6X2 10X4 9X5
25 6x6x3 9x9 11X7 1 3 20 13X2 6X3 —« — 4X4X4 10X6 9X6
36 6X6X2 8x8 8X7 1 2 24 24x5 19X2 .— — 6X6X4 8X7 7X7
33 бхбхз 9X9 9x8 1 1 24 2X1 2у 1 2X1 — 5x5x2 8X4 8x4
29,8 9,1X7 10,4X6,8 21 9X2,3 7,3X1,9 4,3x1,7 —— 7,9X5,3 7,4x3
одинаков, что в пазухе третьего, четвертого и пятого узлов они мо-
гут также заложиться. В наиболее верхних узлах почки закладывают-
ся при наличии их в нижележащем узле.
Как у раскормленных одноцветковых растений (табл. 28, 29, 30,
31), так и у многоцветковых (табл. 33, 34) нет никаких признаков
махровости.
Мы пришли к заключению, что ветвление стебля и многоцветко-
вость— это развитие признака, которым обладали растения в недале-
ком прошлом. Но в связи с тем, что они вынуждены были занять
новые, более ксерофитные арены жизни или благодаря некоторой
ксерофитизации уже занятых ими пространств они почти утратили
свойство ветвления надземного побега в естественных условиях.
При обеспеченности таких растений влагой и необходимыми запа-
сами питания у них начинает проявляться бывшая до того скрытой
способность к ветвлению. Таким образом, ветвистость тюльпанов есть
древний признак в данном роде, как это показал и К. Н. Тараканов
(1952) для некоторых гетерокарпных видов солончаковых растений.
Он обрисовал, как разная приспособленность морфологически различ-
ных семян к условиям прорастания сложилась в результате историче-
ского развития свойств растений под воздействием внешних условий
и, приобретая новое свойство, не утратили, а удержали в себе старое.
Махровость же предполагает совершенно иную перестройку ор-
ганизма— изменение направления развития одних органов и превра-
щение их в другие. Она образуется в результате глубочайшего пот-
рясения организма, в результате нарушения корреляций в развитии
органов цветка и в действительности может быть расценена как та-
Таблица 34
Размеры многоцветковых растений семи видов тюльпанов в эксперименте
Вид Высота растений, см Количество Длина и ширина листьев, см
| цветов на ра- стении листьев основно- го стебля 1 2 3 4 5
Т. micheliana 32 2 4 26x19 25x19 22x19 20x18 —
Т. hoogiana 10 2 4 12x11 20x5 12x3 12x4 —
Т. hoogiana 15 2 4 25x8 19x3 19x3 19X3 —
Т. kuschkensis 32 2 5 25X10 24x10 24x8 22x8 13X4
Т. ingens 30 2 4 22X12 24x8 22x8 5X1 —
Т. fosteriana 33 2 5 22x20 18X10 22x8 14x7 14x6
Т. fosteriana 34 2 4 22x18 21X12 16x8 10X5 —
Т. kaufmanniana 25 3 3 20x8 17X6 16,5x4 —. —
Т. kaufmanniana Т. kaufmanniana 40 5 4 16x17,5 16x6 15x5,5 12x4 —
Т. korolkovii 37 3 3 20x18 17x6,5 17X8 — —
16 2 4 20x3 20x3 21X2 20X1 —
371
Продолжение табл. 34
Вид Основной цветок Ветви и их части [ Добавочный цветок
количество ' членов цветка длина и ширина листочков око- лоцветника, см а о и _> т S 3 й узел заложения длина ветки, см ширина и дли- на листа количе- ство цветка длина и ширина листочков около- цветника, см
внешних внут- рен- них внешних внутрен- них
Т. micheliana 6x6x3 13x5 13x6 1 3 13 5x2 4x6x3 5x4 5X3
Т. hogianna 6x6x3 6X3 6x3 1 2 8 11X4 6x6x3 6x4 6X3
Т. hoogiana бхбхз 7X3 7x3 1 2 12 16x4 6x6x3 7X4 7X3
Т. kuschken- sis бхбхз юхз 9x4 1 3 16 15x4 6x6x3 10x4 9X4
Т. ingens 6x6x3 14x5 13x6 1 2 18 5X1 6x6x3 7X4 6,5X3,5
Т. fosteriana 6x6x3 13x8 11X5 I 3 25 14x6 6x6x3 10x8 10x8
Т. fosteriana бхбхз 11X6 11X6 1 1 16 10x5 6x6x2 5X5 5X6
Т. kaufinan- niana бхбхз 8,5x2,7 8x2,6 2 3 16 7,5x1,4 6x6x3 6,5X2,3 5,2x2
3 16 1,5x0,5 6x6x3 3x0,9 3X1
Т. kaufman- niana 6x5x3 12X3 11X4 4 1 12 8x2 5x5x3 9x3 8X3-
Т. kaufman- niana 2 20 7x2 6x6x3 7X3 5X3
9 25 13x3 5x4x3 7x2 7X3
3 10 8X1 6x6x3 6x2,5 6X3
Т. korolkovii 5x5x3 8X3 6X3 2 1 19 17X2 5x5x3 юхз 6X3
2 15 5X1 5x5x3 3X2,5 2X2
6x6x3 5X6 4,5x6 1 1 12 21X1 6x6x3 4X5 3X5
ратологическое явление. У тюльпанов махровость появляется обычно
у особей гибридного происхождения.
Второй тип многоцветковости у культурных тюльпанов. У куль-
турных тюльпанов многоцветковость второго типа в 1951 г. была по-
лучена у 9 сортов. В 1952 г. — у 22 сортов (табл. 35). При этом все
сорта, бывшие в 1951 г. многоцветковыми, в 1952 г. также дали мно-
гоцветковые экземпляры. Представителей сортов со вторым типом
многоцветковости мы получили почти во всех группах культурных
тюльпанов. Однако в одних группах их было больше, в других—
меньше.
Во втором типе многоцветковости у разных сортов пробуждаются
почки, относящиеся к разным листьям, т. е. к разным узлам зеленого
стебля тюльпана. У сорта Kathleen Parlow (рис. 101) наблюдается
преобладание прорастания почек в пазухе первого зеленого листа.
Они проросли у четырнадцати экземпляров этого сорта, что составля-
ет почти 50% от числа всех пробудившихся у него пазушных почек:
во втором узле — у 8, в третьем — у 7 растений.
Преобладание прорастания почек во втором узле при полном его
отсутствии в первом мы наблюдали у сорта Proserpine.
У сорта La tulipa noire (рис. 105) так же, как и у Kathleen Par-
low в пазухе зеленого листа не пробуждается ни одной почки. В па-
зухе второго листа они проросли у трех растений, в пазухе третьего
372
Таблица 35
Культурные сорта тюльпанов с многоцветковостью второго типа
Группа и сорт тюльпана 1951 г. 1952 г.
количество луковиц многоцвет- ковых количество луковиц много- цветко- вых
Single early Tulips
Proserpine 98 20 не считали 8
Keiserskroon 200 0 не считали 2
Double early Tulips
Murillo 30 10 не считали 15
Mr. v. d. Hoef 7 1 13 1
Double late Tulips
Русский богатырь 4 1 8 4
Breeder Tulips
Medee 8 2 9 3
Красный с желтым основанием 55
(из Ленинграда) 5 0 10 1
Золотистый 60 (из Адлера) 1 0 3 1
Orahge Delight 7 0 16 1
Goud Lake 4 0 8 1
La Danee 3 0 10 1
Single late Cottage Tulips
.Mothers Day 2 0 2 1
Marchal Victor 4 0 6 2
Arethusa 7 1 11 0
Golden Harvest 10 0 24 4
Lily-flowered Tulips
Solfatare 4 0 6 1
Darin Tulips
Philip Snowden 1 0 17 1
Kathleen Parlow 175 26 не считали 55
La Tulipe noire 270 28 не считали 34
Elephant 532 29 не считали . 90
Princess Elisabeth 1 0 1 1
Фиолетовый (из Ленинграда) 2 0 6 1
листа —у 15, в пазухе четвертого листа — у трех. Таким образом, у
этого сорта в основном прорастают почки третьего узла.
У сорта Elephant в пазухах первого листа пробуждаются почки
у 7 растений, второго — у 3 растений, третьего — у 15 растений и
четвертого — у 5 растений. У этого сорта могут прорастать почки
373
всех узлов главного стебля, но большее количество их все-таки &
пазухах третьего узла.
Таким образом, у бывших в эксперименте сортов культурных
тюльпанов с большим количеством луковиц наблюдается прорастание
почек у одного сорта в первом узле, у другого—во втором и у двух —
в третьем. Сорта, представленные малым количеством луковиц в экс-
Рис. 105. La Tullpe noire.
с—двухцветковое растение. Типичная форма ветвления этого сорта с очень высо-
ким срастанием ветви со стеблем, ветка’ заложи тасъ в третьем узле; б—треховет-
ковос растение, ветви развились в третьем < четвертом узлах, срастание стеблей
высокое; в—махровая форма, возникшая в результате полнен о срастания ветви с
основным побегом. Цветок имеет 12 листочков околоцветника, 12 тычинок и завязь
с шестилопастным рытьцем.
перименте, так же, как Medea и Arethusa, дали прорастание почек &
третьем узле. У „Русского богатыря*, имеющего богато облиствленный
стебель, прорастают — только верхние почки в 4-м и 5-м узлах.
Следовательно, увеличением увлажнения питающего субстрата в
критически важные моменты жизни растения, усилением кормления
и лимитированием плодоношения при высоких температурах во время
заложения почек можно вызвать у этих растений изменение в ритме
развития роста, морфологические изменения, направленные как в сто-
рону появления новых листьев, так и на увеличение их поверхности
и увеличение количества цветов.
Листья и п азу ш ные почки тюл ьпанов. Большинство тюль-
панов на главном стебле половозрелой особи имеет от двух до шес-
ти листьев. У известного вида Т. regelii Krasn., отнесенного А. И.
Введенским в особую секцию рода Lophophyllon, у половозрелых эк-
земпляров обычно наблюдается всего одни лист. Известны также сор-
та культурных тюльпанов, главный стебель которых несет более
шести листьев, например, сорт „Русский богатырь*. Но это исключе-
ние. Обычно же на главном стебле находится от двух до шести уз-
лов, на которых могут закладываться почки пазушных побегов. Таким
образом, по-видимому, в зависимости от сорта или вида можно полу-
чить тюльпаны с одной, двумя, тремя, четырьмя, пятью или шестью
ветвями первого порядка. На самом деле пробуждение и развитие па-
зушных почек в побеги первого порядка у каждого вида и сорта
происходит по-разному. У некоторых просыпаются почки первого узла.
У других в этих же условиях — почки второго узла и т. д. Но инте-
ресно то, что у сортов Kathleen Parlow, La Tulipe noire, Proserpine,
„Русский богатырь" и видов Т. subpraestans и Т. bifloriformis в пазу-
374
хах отдельных листьев просыпается не по одной, а по две или даже
по три почки. Это значительно увеличивает количество цветков на
одном растении и дает возможность получить букеты от одной луко-
вицы (рис 101, 105). Почки, из которых развиваются подобные доба-
вочные побеги, мы называем коллатеральными. Они сидят в пазухе
листа одна рядом с другой, но никогда не располагаются одна над
другой (рис. 101). Подобное ветвление в результате развития несколь-
ких почек в пазухе одного листа отмечено также и для тюльпанов
секции Eriostemones. Например, у Т. .bifloriformis из Сукока на тре-
тий год культуры (рис. 98).
У всех описанных сортов и видов тюльпанов у ветви первого по-
рядка, как правило, наблюдается один лист и заканчивается цветком.
Но у нескольких экземпляров сортов Elephant и Kathleen Parlow вет-
ви первого порядка, возникающие в пазухе первого листа стебля,
имеют по два листа; у сорта Elephant одна ветвь первого порядка в
первом узле имела три листа; у сорта „Русский богатырь" ветви пер-
вого порядка в пазухе четвертого и пятого листьев — по три листа. В
пазухе пятого листа у этого сорта две коллатеральные почки, обе
несут по одному цветку и три листа (рис. 100). У сорта „Русский
богатырь" отмечается тенденция к увеличению облиствленности стеб-
ля и к умножению количества членов цветка. У Т. greigii недоразви-
тая ветвь в пазухе второго листа также состоит из двух узлов и двух
крошечных листочков (рис. 102). Многоцветковые растения Т. greigii мы
встречали в горах Каратау в 1959 г.
С увеличеним количества листьев на ветвях первого порядка уве-
личивается зеленая, т. е. ассимилирующая, поверхность растения.
Кроме того, на ветвях первого порядка в пазухах листьев могут воз-
никнуть ветви второго порядка, что мы и наблюдали в 1951 г. у сор-
та Kathleen Parlow (рис. 101) и у Т. subpraestans (рис. 104),
В результате выращивания на высоком агрофоне, поливов в кри-
тические моменты жизни растения, удаления завязей немедленно по-
сле отцветания и высоких температур в период летнего покоя были
созданы для растений такие условия, при которых обычно неветви-
стые сорта и виды тюльпанов дали ветви первого и второго порядков.
В данном случае мы отмечаем факт полной перемены привычных
морфологических черт растения под влиянием измененных внешних
условий.
Срастание ветвей. У тюльпанов часто наблюдается срастание
ветвей первого порядка с основным стеблем. Это явление встречается
среди сортов садовых тюльпанов и дикарей, ставших под влиянием
условий культуры многоцветковыми. Крайнюю степень срастания стеб-
ля с веткой первого порядка мы наблюдаем у сорта La Tulipe noire
(рис. 105, а, б, в). Срастание происходит настолько полно (рис. 105, е),
что стебель округляется, а цветки осевой ветви и ветви первого по-
рядка сливаются. Образуется цветок с 12 листочками околоцветника,
12 тычинками и завязью из 6 плодолистиков. В этом случае мы отме-
чаем кажущуюся махровость в результате полного срастания двух
цветков. Срастание стеблей у этого сорта чаще всего происходит
только до цветков, а цветки на очень коротеньких цветоножках си-
дят скученно на вершинах растений (рис. 105). Однако не всегда,
даже у этого сорта, стебли срастаются полностью. Главный стебель
сорта Proserpine обладает такой же ярко выраженной склонностью к
срастанию с ветвями первого порядка. У других сортов хотя и на-
блюдается срастание стеблей, но в значительно меньшей степени. При-
растание ветви первого порядка до второго узла над узлом, где за-
375
вкладывается ветка, отмечается у сорта Kathleen Parlow. Ветка прира-
стает к главному стеблю в основном только до лежащего над ней
узла, а дальше растет свободно (рис. 101). Такую же форму роста
имеет и ветвь второго порядка, заложившаяся у одного из растений
этого же сорта (рис. 101, б). Подобную форму роста часто имеют
ветви сортов Elephant, Русский богатырь (рис. 100), Т. subpraestans
(рис. 104).
У сорта Elephant ветви первого порядка, заложившиеся в пазу-
хе первого листа, не срастаются выше места выхода из влагалища
листа и растут свободно. Такую же форму роста ветви мы наблюда-
•ем иногда у сортов Kathleen Parlow (рис. 101.),Medea, Arethusa и ви-
дов Т. greigii и Т. fosteriana. Эта форма роста дает пышные кусты
< крупными свободными ветвями и цветами, не сближенными в плот-
ные головки, как у Tulipe noire. Срастание ветвей с главным стеб-
лем очень напоминает фасциацию, так как большей частью при по-
добном срастании, кроме вышеописанного случая с La Tulipe noire
(рис. 105.), оно не бывает полным и стебель уплощается. В действи-
тельности это не полное срастание, а как бы прирастание ветви к
-стеблю, которое появляется у растений, не бывших в действительно-
сти многоцветковыми, а ставших ими в результате направленного вос-
питания. Как известно, бутон у тюльпана закладывается с осени и
до ранней весны остается внутри луковицы. Самая внутренняя чешуя,
внутри которой закладывается побег с листьями и цветами, очень ма-
ла и играет роль своеобразной повязки, прижимающей один побег
•со всеми его элементами к другому. В результате длительного пре-
бывания в таком связанном состоянии побеги срастаются. Следова-
тельно, чем крупнее у луковицы тюльпана ее внутренняя чешуя, тем
«свободнее у него растут ветви; чем быстрее побеги после своего
заложения выходят из луковицы, тем меньше степень их сраста-
ния.
Воздушные луковицы. Количество видов и сортов тюльпа-
нов, которые при хорошем уходе и обеспеченности питанием вновь
обретают утерянные родом в целом свойства давать ветвящиеся стеб-
ли, может быть значительно увеличено, если добавить виды и сорта,
у которых в пазухе чаще всего нижнего стеблевого зеленого листа
развились почки, разросшиеся в луковицу.
В 1951 г. у нас было всего две таких луковицы — сорта Kathleen
Parlow и Jubilee. Сорт Kathleen Parlow в 1951 г. дал значительное
количество особей, у которых заложились и выросли в ветви почки
в пазухах верховых листьев (табл. 37); у сорта Jubilee ни в 1951, ни
в 1952 гг. при большом количестве растений не было ни одной ветви
в пазухах верховых листьев. Почки низовых листьев (чешуй) у это-
го сорта обладают способностью к ускоренному прорастанию. Ветвле-
ние надземного стебля в сорте Jubilee осуществляется лишь при по-
средстве воздушных луковиц, вырастающих в пазухе нижнего листа
надземного побега (табл. 36).
В 1952 г. мы наблюдали образование луковицы в пазухе верхо-
вого зеленого листа у растений тюльпанов, которые дали в экспери-
ментах ветвление надземного побега, и у растений, еще не проявив-
ших этой особенности (табл. 36).
Из 14 видов и сортов исследованных нами растений у 9 не было
ветвления надземного побега (табл. 36). В основном воздушная луко-
вица закладывалась в пазухе первого верхового листа. Лишь в двух
случаях мы наблюдали заложение воздушных луковиц в пазухах вы-
шележащих листьев.
376
У сорта Marchal Victor одно растение имело воздушные луковицы
в пазухах первого и второго листьев. У Т. kaufmanniana в пазухе
первого листа развилась довольно крупная луковица (рис. 106). В
пазухе второго листа у этого же растения луковица проросла побе-
гом с толстым зеленым листочком и недоразвитым цветком. В пазухе
третьего листа развился зеленый пенек — ни луковица, ни побег.
Таблица 36
Образование луковиц в пазухах зеленых листьев
Сорт или вид Количество ра- стений с доба- вочными луко- вицами Многоцвет- ковость II типа Добавочное число на одном растении Узел зало- жения лу- ковицы
Keizerskroon 4 + 1 1
Marchal Niel 1 — 1 1
Orange Nusau 1 — 1 1
Amidonate 1 — 1 1
1 — 1 1
Marchal Victor 4 + 1;2 1;2
Miss Holland 1 — 1 1
Venus 1 — 1 1
Afterglow 2 — 2 1
Elephant 2 — 1 1
Jubilee 5 — 1 1
T. kaufmanniana 1 + 3 1;2;3
T. fosteriana 1 + 1 1
Kathleen Parlow 1 1 1
У экземпляра Т. fosteriana в 1951 г. из пазухи первого листа вы-
росла ветвь с листом и крупным цветком (Бочанцева, 1958). В 1952 г.
у этого растения, выращенного из замещающей луковицы, на месте
побега в пазухе первого листа наблюдалась воздушная луковица. Та-
ким образом, у одного и того же растения в один год в пазухе пер-
вого верхового зеленого листа может образоваться побег, а в дру-
гой — воздушная луковица.
Следует отметить, что воздушные луковицы у тюльпанов закла-
дываются на стадийно зрелом побеге, который вырастает из почки,
закончившей стадийное развитие. Достигнув вершины стадийного раз-
вития, почка изменяет форму роста.
Междоузлия этого побега б. м. вытягиваются, а рост побега за-
канчивается с появлением на нем цветка. Самым длинным в таком
побеге является междоузлие между чешуями луковицы и первым зе-
леным листом. Это междоузлие выносит прошедшую стадийное раз-
витие почку на поверхность почвы. Средние междоузлия могут быть
в зависимости от вида или сорта короткими или длинными, но никог-
да не бывают такими короткими, как в донце.
Верхнее междоузлие, или цветоножка, между самым верхним ли-
стом и цветком почти такое же, а в иных случаях и более длинно,
чем нижнее. Следовательно, у половозрелого растения изменяется
377
форма роста, стебля; форма, размеры и консистенция листьев. У ра-
стений, не достигших половой зрелости, имеются один влагалищный
и низовые запасающие листья, а у половозрелого растения на над-
земном побеге появляются зеленые верховые листья. В пазухе вер-
хового зеленого листа на стебле, который в связи с пройденной ста-
дийностью изменяет форму роста и становится конечным против
бесконечного роста побега, не закончившего стадийного развития,
появляется луковица. Луковица моложе, чем побег минимум на одну
ступень стадийного развития, и зацветает при посадке ее в грунт на
будущий год. Следовательно, при зало-
Рис. 106. Растение Tulipa kauf-
manniana с луковицами в пазухах
верховых листьев стадийнозрелого
побега.
жении этой луковицы происходит омо-
ложение — снятие стадийности.
Снятие стадийности сходно с тако-
вым же у луков, описанных А. А. Ава-
кяном (1948), при образовании воздуш-
ных луковиц, но оно несколько отлично
от имеющегося у луков, так как здесь
нет полного снятия и омоложения.
На примере с Т. kaufmanniana (рис.
106) можно сделать вывод, что чем ни-
же по стеблю залегает почка, тем пол-
нее снимается стадийное развитие. Са-
мая нижняя почка-луковица, сидящая
в пазухе первого верхового ассимиляци-
онного листа, зацветает только на сле-
дующий год. А у почки-луковицы из
второго междоузлия лишь первый мясис-
тый лист приобретает форму и функ-
цию запасающего органа, а следующие
листья остаются зелеными, на вершине
находится, хоть и недоразвитый, цветок.
В пазухе третьего листа развивается лишь стеблевая часть ветки.
Многоцветковость второго типа обусловлена ветвлением стебля
тюльпана. Луковицы, получаемые в пазухах зеленых листьев, также
являются ветвями, но несколько видоизмененными. Ветвление второго
типа получено нами в 1952 г. у 30 сортов и 14 видов тюльпанов.
На примере изучения второго типа многоцветковости мы можем
сказать, подтверждая выводы Н. А. Шахова (1952), что развитие ра-
стений, а в наших опытах тюльпанов, исторически слагается из по-
следовательных типов развития. Особенно ярко в данном явлении
подтверждается его положение о том, что „переход от одного типа
развития к другому не всегда обозначает утрату более раннего и
характерного при иных экологических условиях типа развития*4. Бо-
лее древним признаком у тюльпанов является кустовая форма надзем-
ного побега. Но в связи с ксерофитизацией условий у них преобла-
дают одноосные неветвящиеся одноцветковые надземные побеги.
Изменение условий в опыте вызывает проявление более раннего
типа развития. Вместо привычных одноосных одноцветковых неветвя-
щихся растений тюльпанов мы получили ветвящиеся кустовые мно-
гоцветковые формы с ветвями до третьего порядка.
Третий тип многоцветковости, или тип многоцветковости куль-
турного тюльпана. Выделенный нами третий тип многоцветковости ча-
ще всего встречается у культурных тюльпанов, поэтому он назван ти-
пом многоцветковости культурного тюльпана.
378
От двух вышеописанных типов с развитием почек в пазухах зе-
леных листьев он отличается тем, что здесь наблюдается ускоренное
прорастание почек-деток, закладывающихся в пазухах лишенных хло-
рофилла листьев — луковичных чешуй.
Как и два предыдущих типа, этот тип многоцветковости вызы-
вается усиленным раскормом тюльпанов и обусловлен ускорением
развития результирующих луковиц на один год против обычного.
В наших экспериментах раскорм состоит из удобрения почвы по-
вышенными дозами аммиачной селитры и направления потока асси-
милянтов из листьев и стебля не в завязывающуюся коробочку, а в
луковицу. Изменение направления потока ассимилянтов достигается
срезанием завязи немедленно после опадения членов цветка. К это-
му времени ассимиляционная поверхность растения тюльпана дости-
гает максимальной мощности. При удалении завязи растение вынуж-
дено направить поток ассимилянтов в замещающую луковицу и детки.
Количество деток в год удаления завязей не увеличивается, так как
все детки в луковице закладываются в период жаропокоя предше-
ствующего года и первое время растут за счет накопленных в преды-
дущие сезоны питательных веществ. В результате проведенной опера-
ции в сезон удаления завязей увеличивается лишь размер луковиц и
выполненность чешуй крахмалом.
Обычно после окончания вегетации наступает жаропокой, во вре-
мя которого происходит закладка и первоначальный рост почек. Бла-
годаря большим отложениям пищи в запасающих чешуях луковицы
рост закладывающихся в пазухах чешуй почек происходит более ин-
тенсивно — в один год проходит две фазы развития почек-деток.
В чистом виде третий тип многоцветковости мы получили у сор-
та Jubilee, относящегося к группе тюльпанов Дарвина.
При хорошей подготовке этого сорта в предшествующий год,
особенно после удаления завязей и подкормки аммиачной селитрой
после посадки, весной, в период осеннего роста корней, из земли, где
была посажена одна луковица, показывается группа ростков. По фор-
ме, соотношению длины и ширины этих ростков можно наблюдать,
что из одной луковицы вырастает по нескольку цветоносных побегов.
Обычно из земли вырастает до 6 побегов. У трех-четырех из них
можно предположить наличие бутона. Однако цветение всех этих по-
бегов не всегда осуществляется. Четыре цветка из одной луковицы,
каждый на отдельном стебле, встречаются у данного сорта не чаще
одного раза на 100 луковиц. Чаще наблюдается три цветка и как массо-
вое явление — два. Одноцветковые экземпляры растений этого сорта
на наших экспериментальных грядах представляют такое же исключе-
ние, как и четырехцветковые. Весной при развертывании листьев по-
бегов очень часто удается установить, что цветет основной побег,
побег замещающей луковицы и побег детки покровной чешуи. Если
пронумеровать все чешуи луковицы этого сорта, начиная с покровной
по направлению в глубь луковицы, то у половозрелой луковицы будет
шесть таких чешуй. В пазухе каждой чешуи всегда закладывается
почка. Некоторые из почек при наличии соответствующих условий
вырастают до значительных размеров, иногда равных замещающей
луковице.
Обычно почка-детка служит органом вегетативного размножения
и до полного обособления от материнской луковицы не прорастает ни
корнями, ни листьями. То же можно сказать и о замещающей луко-
вице. Но при наличии соответствующих условий некоторые детки
второй фазы могут прорасти в год формирования и листьями и кор-
379
ними. Эти условия определяются выращиванием луковицы преды-
дущего года и ее питанием в год изучения. Если же эта детка про-
ходит в год заложения и формирования вторую фазу развития, тогда
получаются многоцветковые растения типа культурного тюльпана.
Задержка в развитии деток по сравнению с главной почкой обуслов-
ливается недостатком притока питательных веществ и температурны-
мы условиями периода жаропокоя. Первое очень легко обосновать,
если проследить расход запасных питательных веществ в луковичных
чешуях.
В период жаропокоя происходит глубокая перестройка внутри
луковицы. Под влиянием высокой температуры почвы в луковице
происходит уменьшение количества воды, а центральная почка начи-
нает медленно развиваться. Ее развитие происходит за счет накоп-
ленных в запасающих чешуях питательных веществ. Вначале расхо-
дуются запасы, накопленные в самой внутренней чешуе, непосредст-
венно одевающей главную почку и почку замещающей луковицы,
затем следующей за ней и так далее к периферии луковицы. В каждой
чешуе вначале беднеет крахмалом субэпидермальный слой, прилегаю-
щий к внутреннему эпидермису, потом — следующий за ним и так
далее. При продвижении этого процесса в глубь чешуи трансформа-
ция крахмала уже не захватывает целые слои, а происходит преры-
вистыми участками и чем дальше вглубь, тем эти участки становятся
меньше. В результате получаются участки пустых клеток в форме
усеченных пирамид, обращенных основанием внутрь луковицы. Этими
участками является паренхимная ткань, лежащая в чешуе между со-
судистыми пучками. Сосудистые пучки остаются долгое время со
всех сторон окруженными обкладкой из клеток, густо заполненных
крахмалом.
При растворении крахмала внутренней чешуи в объеме около
одной трети ее толщины начинается растворение крахмала в следую-
щей за внутренней периферической чешуе. Очень медленный процесс
трансформации крахмала и опустошения луковичных чешуй не заме-
тен снаружи луковицы до тех пор, пока в начале сентября не начнет
набухать донце и не лопнет покровная чешуя, а верхушечная и за-
мещающая почки, растущие внутри луковицы, заполняют появившую-
ся в результате опустошения чешуй полость. Из всех почек лукови-
цы эти почки пробуждаются первыми и на них расходуется основная
масса запасных питательных веществ, после чего начинается рост
почки второй чешуи, затем — третьей. При поперечном разрезе луко-
вицы можно наблюдать, что чем ближе к периферии, тем мельче за-
ложившиеся детки. Если луковица мощная, то питательных веществ
хватает на начальные этапы развития не только центральной почки и
ее замещающей, которая является пазушной почкой самой внутрен-
ней чешуи, но и для почек более периферических чешуй. При недо-
статке запасных питательных веществ у культурных сортов они пе-
рехватываются ушедшими вперед в развитии перед остальными поч-
ками—верхушечной почкой и ее замещающей, и тогда развитие всех
прочих почек подавляется. У некоторых диких видов, несмотря на
избыток запасов в луковице, развития почек-деток не наступает.
При избытке запасных питательных веществ и длительном периоде
жаропокоя замещающая почка догоняет центральную и также закла-
дывает несколько листьев и цветок.
На более позднем этапе развитие ближайших к центру деток от-
стает от периферических, так как во внутренних чешуях, в связи с
их небольшим объемом, меньше запасных питательных веществ, чем
380
в периферических, и начавшийся к этому времени рост центрального*
комплекса почек успевает значительно опорожнить внутренние чешуи
прежде чем их собственные почки придут в силу.
Не то происходит с детками периферических чешуй. Пробудив-
шись несколько позже они, однако, находятся у более мощных запа-
сов питательных веществ, кроме того вскоре после их пробуждения
у луковицы отрастают корешки и она начинает получать питание извне.
Поэтому периферические детки, вначале слабо развитые перегоняют
деток внутренних чешуй. Особенно далеко в росте и развитии про-
двигается детка покровной чешуи. Эта детка после отслаивания по-
кровной чешуи, очень нежной у большинства культурных сортов^
непосредственно соприкасается с почвой и дает корни. У сорта Jubilee
периферические детки развивают цветоносные побеги при условии
посадки мощных луковиц в богатую почву. При некоторой бедности
почвы иногда вместо цветоносных стеблей развиваются лишь облист-
вленные стебли, а цветочная почка не развивается (рис. 107, г). По-
кровная чешуя деток, за-
ложившихся у внутрен-
них чешуй, вырастает в
лист. Эти почки-детки
оказываются по сравне-
нию с другими почками
в наименее обеспеченном
положении. Но в связи с
относительно большими
запасами пищи и мощной
корневой системой мате-
ринской луковицы они все
же более продвинуты в
развитии, чем детки в
обычных условиях куль-
туры.
Сериальность зало-
жения почек и их разви-
тие, описанные выше у
второго типа многоцвет-
ковости, встречается и в
Рис. 107. Сорт Jubilee, многоцветковый экземпляру
а—направо основной побег, налево проросшая побегом заме-
щающая луковица; б,в—детки 2 и 3-й чешуй имеют один лист и
сериальную почку-детку; г—детка третьей чешуи имеет несколь-
ко листьев и сестринскую детку, цветок не развился; д—детка
покровной чешуи имеет стебель с листьями и цветком.
этом типе многоцветно-
вости. Однако вторая почка в пазухе запасающей чешуи в данном
случае никогда не прорастает в год заложения и формирования ни
побегом, ни листом, она вырастает только в почку-детку среднего
или малого размера (рис. 107). Это происходит в связи с тем, что
вторая почка закладывается значительно позже, чем главная и даже
ее сестринская и, в связи с этим, отстает от них на одну ступень
стадийного развития.
Массовое цветение деток покровной чешуи мы наблюдали у мно-
гих культурных сортов, воспитывавшихся в эти годы на наших участ-
ках, как у тюльпанов с многоцветковостью второго типа, так и у не-
которых из группы „одинаких ранних1* —Due van Thol yelow maximus,
D. v. Thol white, D. v. Thol red, Proserpine и Keizerskroon; из „мах-
ровых ранних11—Murillo и Mr. van Tubergen; из „Тюльпанов Дарвина11 —
Jubilee, Elephant, Kathleen Parlow и La Tulipe noire; из диких видов—
часто у Т. subpraestans, у одного растения Т. greigii и у одного есте-
ственного гибрида Т. ingens х Т. fosteriana.
381
Примененный нами метод выращивания растений дает способ не
только по управлению их цветением, но и вегетативным размно-
жением. У нескольких диких видов мы уже получили особи с боль-
шим количеством деток, что дает возможность размножать их как
сорта новых тюльпанов.
Многоцветковость тюльпанов культурного типа еще раз подтвер-
дила наш вывод о том, что все почки в половозрелой луковице—ста-
дийно одновозрастные. Но почки-детки, являясь спящими и не про-
растая ни корнями, ни листьями, не способны к прохождению стадий-
ных изменений и в связи с этим отстают от центральной почки на
одну ступень стадийного развития. При избытке питательных веществ
в луковице почки-детки, просыпаясь почти одновременно с централь-
ной почкой, догоняют ее в развитии и могут зацвести одновременно.
Внешние условия и развитие почек в луковице. У мощных луко-
виц закладка почек происходит раньше вступления их в период жа-
ропокоя. Это начинается еще до окончания растением последней
ступени стадии яровизации, не говоря уже о световой стадии.
Мы проследили процесс заложения и развития почек у мощных
луковиц Т. fosteriana, Т. kaufmanniana, Т. korolkovii, Т. subpraestans,
Т. bifloriformis и сортов D. v. Т. yellow и Jubilee в условиях Таш-
кента. Насколько вяло протекает этот процесс у Т. fosteriana и вообще
у большинства дикарей (исключение составляют немногие виды), на-
столько он интенсивен у сорта Jubilee и несколько подавлен у D. v.
Т. yellow.
В условиях Ташкента жаропокой для массы тюльпанов наступает
в середине мая. Но сорт Jubilee относится к позднецветущим и в
конце мая еще вегетирует. Разрезая крупные луковицы этого сорта
при наличии еще зеленых листьев, мы видим в пазухах низовых ли-
стьев или луковичных чешуй заложившиеся почки.
Верхушечная почка в это время состояла из меристематического
бугорка и двух полукруглых валиков. Почка замещающей луковицы
еще не наметилась. В пазухах всех чешуй луковицы заложились
детки размером —за второй покровной чешуей 0,4x0,31; за первой
запасающей 0,4x0,2; за второй запасающей 0,2x0,15 (за третьей
запасающей в данном случае лежала верхушечная почка и ее запа-
сающая). Обычно запасающих чешуй у этого сорта бывает больше.
Все детки луковицы в мае представляют лишь меристематические
бугорки. В начале июля в детке покровной чешуи обычно видны две
чешуи, зачаток листа и в центре меристематический бугорок. У детки
первой запасающей чешуи намечается одна чешуя и в центре буго-
рок. Детка второй запасающей чешуи представлена лишь одним мери-
стематическим бугорком. В верхушечной почке в это время уже зак-
ладывается три листовых валика и меристематический бугорок. В двух
периферических валиках намечается проваскулярная система. Почка
замещающей луковицы состоит лишь из одного недифференцирован-
ного еще весьма плоского меристематического бугорка.
В течение июля в мощной луковице прорастают все заложив-
шиеся почки. Наступает второй период в жаропокое крупных луко-
виц у культурных сортов тюльпанов.
У детки покровной чешуи луковицы, состоящей из двух белых
низовых запасающих чешуй, верхушечная почка начинает интенсивно
расти, чаще всего преобразуясь в побег. В пазухе ее двух белых
низовых чешуй закладывается по почке. Одна из них вырастает из
верхушечной почки этой детки и будет замещающей луковицей по-
382
бега, а другая более периферическая, чаще всего дальше не разви-
вается.
В почках запасающих чешуй, как правило, к этому времени бы-
вает по одной, максимум две (у более периферических из них) белых
запасающих чешуи первой фазы развития.
Во второй фазе развития у некоторых из этих почек из верху-
шечной точки роста начинает формироваться побег, но значительно
чаще закладывается и растет влагалищный лист. Расширенное осно-
вание вышеуказанного влагалищного листа будет покровной чешуей
для закладывающейся из верхушечной меристемы почки-детки луко-
вицы будущего года.
В пазухах чешуй почек-деток первой фазы развития закладыва-
ются новые почки-детки второй фазы развития.
Почка замещающей луковицы закладывается и развивается самой
последней, значительно позже верхушечной, при вступлении луковицы
во второй период жаропокоя. У мощных луковиц ее развитие обычно
начинается со влагалищного листа, свойственного луковицам второй
фазы, минуя первые запасающие листья,—чешуи первой фазы разви-
тия, свойственные всем нижележащим почкам. Если этот влагалищ-
ный лист замещающей луковицы закладывается раньше наступления
второго периода жаропокоя, то под ним может заложиться еще один
влагалищный лист.
Двупокровные луковицы. Замещающие луковицы с двумя зало-
жившимися и развившимися из оснований влагалищных листьев пок-
ровными чешуями мы наблюдали, кроме сорта Jubilee, еще у многих
сортов (табл. 37, рис. 108).
Таблица 37
Двупокровные луковицы у культурных сортов тюльпанов
Сорт Количество мощных луковиц
всего в том числе с двумя покровными оболочками
абс. ! %
Proserpine И 2 18,2
Medea 25 2 8
Красный с желтым 18 2 И
Золотистый 3 2 66,7
Orange Delight 23 2 8,7
Бин 1 27 1 3,7
Good W. Leak 8 1 12,5
Orange King 20 5 25
Golden Harwest 25 2 8
Venus 11 1 9,2
Jubilee 50 25 50
Elephant 50 20 40
Kathleen Parlow 50 10 20
Двупокровные луковицы бывают самыми мощными луковицами
в сорте. У этих луковиц как под первой, так и под второй покровной
оболочкой часто закладываются коллатеральные детки. Они бывают
под одной из покровных оболочек, или под обеими.
383
луковицы развивается
Рис. 108. Двупокровная луковица тюль-
пана сорта Jubilee.
а—первая покровная чешуя, б'—пазушная почка,
в—вторая покровная чешуя, г—коллатеральные
почки в пазухе второй покровной чешуи, д—детки
в пазухах запасающих чешуй, е—почка замещаю-
щей луковицы, ж—верхушечная почка с бутоном
и зачатками ассимиляционных листьев.
В пазухах других низовых чешуй у бывших под нашим наблю-
дением сортов и видов тюльпанов коллатеральные детки закладыва-
ются очень редко.
Обе покровные чешуи мощных луковиц могут быть низовыми или
влагалищными листьями, которые встречаются значительно чаще, чем
низовые. Иногда, а у сорта Jubilee достаточно часто, вместо заме-
етоносный побег (рис. 109). Зало-
жение его начинается несколько
позже побега из верхушечной почкщ
но почти одновременно с побегом
почки покровной чешуи. Побег
развивающийся вместо замещающей
луковицы в начале августа не-
сколько менее развит, чем побег
из верхушечной почки и почти
равен побегу покровной чешуи.
Иногда у основания этого побега
на том же сосудистом пучке чешуи,
на котором сидит этот побег, но
несколько ниже его, закладыва-
ется в ноябре—декабре сериальная
почка. Заложение подобных почек
мы наблюдаем довольно часто в
мощных луковицах у деток дру-
гих чешуй и не только у сортов
культурных тюльпанов, но несколь-
ко реже и позже и у диких видов
(рис. ПО).
К наступлению периода пони-
женных температур верхушечные
точки роста пазушных почек тюль-
панов вполне сформированы для
прохождения возрастно необходи-
мой ступени стадии яровизации. Пазушные почки второй фазы раз-
вития, нижние сериальные почки и боковые коллатеральные, не ус-
певают заложить, а в некоторых случаях едва начинают формировать
отдельные части к наступлению осенне-зимнего комплекса понижен-
ных температур и отстают от почек-деток первой фазы на одну ступень
стадийного развития.
В мощной луковице тюльпана находятся почки трех степеней
развития: 1) закончившие стадийное развитие и готовые зацвести на
будущий вегетационный сезон; 2) закончившие формирование зачат-
ков чешуй луковиц с точками роста, готовыми к прохождению ста-
дийных изменений; 3) детки второй фазы, формирующиеся и не зало-
жившие зачатков чешуй, еще не подготовленные к восприятию влия-
ния внешних условий.
Первые зацветают одновременно с верхушечной почкой луковицы;
вторые—в первый год после обособления от материнской луковицы;
третьи—на третий год от момента закладки. В третью группу попа-
дают луковицы, развивающиеся из почки второй фазы, которые явля-
ются пазушными почками пазушных почек первой фазы материнской
луковицы, следовательно, ее внучатые потомки, сериальные и колла-
теральные, т. е. сестринские к пазушным почкам и являющиеся пря-
мыми потомками—детками первой генерации материнской луковицы.
384
или
Рис. 109. Растение
тюльпана сорта Ju-
bilee с ускоренным
развитием замеща-
ющей луковицы.
а—основной побег, б—
побег замещающей лу-
ковицы (имеет влага-
лищный лист с расши-
ренным основанием).
треть ей группе (если можно для вегетативно размножаемых расте-
ний говорить слово генерация) будут детки двух генераций.
Ускоренный рост почек в мощных луковицах в период жарочо-
коя, их соответствующее морфологическое состояние ко времени отра-
стания корней луковицы обеспечивает прохождение точками роста
заложившихся почек необходимой ступени стадии яровизации. В дан-
ном случае с необыкновенной ясностью выступает связь питания
(обеспеченность закладывающихся почек запасными питательными
веществами) и прохождения точками роста соответствующих ступе-
ней стадийного развития.
В слабых луковицах заложение почек несколько задерживается.
Их эмбриональное развитие в период летнего покоя
происходит очень медленно и ко времени осеннего
отрастания корней они недостаточно сформированы
и не могут воспринять влияние комплекса осенних
условий.
Тюльпаны, которые прорастают с осени корнями
и дают под землей усиленный рост верхушечной поч-
ки, могут быть названы озимыми растениями. Одна-
ко они не образуют с осени розетки зеленых листьев
над поверхностью почвы, так что при прохождении
их точками роста стадии яровизации исключается
влияние осеннего света.
Луковица тюльпана расположена достаточно глу-
боко под землей и ее растущие почки защищены от
влияния света не только живыми запасающими че-
шуями текущего года и мертвыми оболочками преды-
дущих лет, но и толстым слоем почвы.
Набухающие семена тюльпанов также проходят
стадию яровизации в то время, когда растущий в
них зародыш еще не окрашен в зеленый цвет
(3. П. Бочанцева, 1951). Таким образом, в комп-
лексе условий требуемых озимым растениям тюль-
панов для прохождения стадии яровизации исключа-
ется влияние осеннего света, выдвинутое академиком
Т. Д. Лысенко (1952) как необходимое условие для
прохождения стадии яровизации у озимых растений.
Озимость тюльпанов несколько отличается от
пшеницы или ржи. К озимым растениям типа пше-
ницы из многолетников относятся многие растения
средиземноморского типа—Anemone coronaria, Cycla-
men ponticum или Sternbergia lutea. Тюльпаны же—
Ferula, Leontice и другие многолетние растения-
эфемероиды Средней Азии отрастают с осени под
землей, но их побеги не выходят до весны на поверх-
ность почвы и не подвергаются воздействию осен-
них солнечных лучей.
В условиях Ташкента цветение тюльпанов на
открытом воздухе обычно начинается в конце первой декады марта
и заканчивается в первой декаде мая, продолжаясь в среднем не бо-
лее двух месяцев.
У взрослого растения любого вида или сорта тюльпанов во время
цветения наблюдается несколько стеблевых зеленых листьев. Самый
крупный и широкий из этих листьев—нижний лист. К вершине стебля
листья уменьшаются, становятся значительно уже и короче (табл. 26,
25-222Э
385
27, 28, 29, 30, 31). Эти листья, окрашенные в зеленый цвет, у разных
видов и сортов сильно различаются по форме, окраске, размерам и
распределению их по стеблю. Они могут располагаться мутовкой при
основании цветоножки (Т. tetraphylla); подниматься по стеблю почти
до цветка (Т. maximowiczii); могут быть почти округлыми (некото-
рые формы Т. kaufmanniana); широколанцетными (Т. praestans); рем-
невидными (Т. subpraestans) или злаковидными (Т. maximowiczii). По-
верхность листьев бывает гладкой или покрытой выступающими реб-
рами (Т. regelii); антоциановыми пятнами (Т. greigii); полосами (Т. mi-
cheliana); сизой (Т. alberti) или покрытой опушением (Т. vvedenskyi).
Рис. 110. Сериальная почка
детки первой чешуи Tulipa
fosteriana в марте.
а—основная почка, б—ее сериаль-
ная.
Различаясь морфологически, все зеленые ли-
стья имеют одну функцию—это основные ор-
ганы ассимиляции растений. Несмотря на то,
чго зелеными у растений являются стебель,
молодой бутон и завязь, основой основ для
ассимиляции и восприятия фотопериодическо-
го влияния света являются зеленые листья.
У взрослого растения они обычно располо-
жены на побеге, увенчанном цветком и за-
кончившем стадийное развитие. Эти листья,
кроме функции ассимиляции, воспринимают
и влияние света для прохождения световой
стадии. Для каких же почек эти листья вос-
принимают влияние света? Или они только
ассимилируют и не способны к восприятию
фотопериодических воздействий. Мы думаем,
что зеленые листья взрослого растения тюль-
панов прежде всего работают для заверше-
ния развития завязи и воспринимают влияние
света для почки замещающей луковицы, ко-
торая заложилась в предыдущий период жа-
ропокоя в пазухе верхней запасающей чешуи
несколько позднее верхушечной и является
ее сестринской почкой.
В предыдущий, осенне-зимний период эта почка прошла ступень
стадии яровизации и теперь при помощи листьев, выросших из вер-
хушечной почки, проходит необходимую для вступления в генератив-
ную фазу ступень световой стадии. В наступивший затем период жа-
ропокая замещающая луковица, выросшая из этой почки, в верхуше-
чной почке заложит цветок. Обычно верхушечная почка замещающей
луковицы заканчивает стадийное развитие при полном отсутствии
собственных листьев еще до обособления замещающей луковицы от
материнского растения.
То же произойдет и с почкой покровной чешуи материнской
луковицы, которая закладывается одновременно с меристемой верху-
шечной почки в конце вегетации, перед периодом жаропокая в пре-
дыдущем году. Эта почка зацветет одновременно с верхушечной по-
чкой материнской луковицы.
Также развиваются и другие пазушные почки, если они заклады-
ваются в луковице, которая достаточно обеспечена запасными пита-
тельными веществами. В этом случае их развитие ускоряется и они
подходят к осени достаточно сформированными для восприятия осен-
не-зимних условий. При ускорении развития как замещающей луко-
вицы, так и любой из пазушных почек мы наблюдаем ускорение раз-
вития, названное третьим типом многоцветковости или многоцветко-
386
востыо культурного типа (Бочанцева, 1954). Этот тип многоцветко-
вости обычен для культурных тюльпанов, но встречается в наших
опытах и у диких видов.
Когда ускоряется развитие почки самой внутренней чешуи, из-
которой должна развиться замещающая луковица, в этом случае функ-
цию замещающей луковицы исполняет детка ближайшей запасающей
чешуи. Часто побег, выросший на месте замещающей луковицы, имеет
свою замещающую луковицу, развивающуюся из сериальной почки
основания побега. В этом случае отмечается развитие двух замещаю-
щих луковиц. У таких луковиц цветущая детка покровной чешуи
часто также закладывает замещающую луковицу, развивающуюся из-
коллатеральной почки или же из детки ее покровной чешуи. Таким
образом, мы можем получить в одном гнезде несколько замещающих
луковиц—от одной до двух-трех, а иногда и больше, но все они бу-
дут разного происхождения, разновозрастны и не равны между собой
стадийно.
У луковиц растений, получивших описанное выше воспитание,,
наблюдается раннее заложение почек, их интенсивный рост до вступ-
ления в жаропокой и вторичный рост — в период жаропокоя. Этим
точкам роста обеспечено прохождение необходимой для цветения
ступени стадии яровизации, Они зацветают одновременно с верхушеч-
ной почкой.
Детка покровной чешуи. При изучении третьего типа многоц-
ветковости особенный интерес представляет почка покровной чешуи.
Эта почка у раскормленных луковиц закладывается одной из первых.
У диких видов она обычно даже не закладывается. Только иногда у
очень мощных луковиц этих видов мы наблюдаем ее заложение и
рост.
У культурных тюльпанов после применения нашего способа куль-
туры в течение предшествующих лет детка покровной чешуи не
только закладывается, но очень часто и зацветает одновременно с
верхушечной почкой луковицы.
В некоторых группах тюльпанов ускорением развития детки по-
кровной чешуи обладают многие сорта (табл. 38).
В группе лилиецветных у обоих сортов зацвели детки покровной
чешуи. Внутри сортов процент зацветших деток от общего количества
зацветших луковиц не очень высок.
Большой процент ускоренного зацветения детки покровной чешуи
отмечается у махровых ранних тюльпанов. В группе махровых ран-
них 73% сортов дало ускоренное развитие этой детки. У некоторых
сортов детки покровной чешуи зацвели у 62% цветущих луковиц.
Особый интерес в этом отношении представляет группа одинаких
ранних тюльпанов. Детки тюльпанов этой группы небольшого размера,
однако в последующий год после обособления от материнской луко-
вицы они зацветают почти на 100%. Однако всего у одной четверти
имевшихся в нашем распоряжении сортов было получено ускорение
цветения детки покровной чешуи.
Внутри сортов группы самый высокий процент ускорения развития
едва достигал 25%. У сортов, относящихся к группе одинаких ранних,
существует недоразвитие деток периферических чешуй. Луковицы их
в наших условиях не так крупны, за небольшим исключением сортов,
которые мы культивируем более полутора десятка лет—Keizerskroon
и Proserpine. Но у этих сортов мы не провели учет вегетативного
размножения и цветения, так как в 1951 г. их не пересаживали.
387'
Некоторая слабость развития тюльпанов из группы одинаких ран-
них, возможно, обусловлена неудачными грядами, на которых они
росли в 1952 г. Эти гряды были приподняты, а между ними проходили
канавы для полива методом инфильтрации. Приподнятые гряды быстро
иссушались. На стороне гряды, обращенной на юг, растения зацветали
Таблица 38
Многоцветковость третьего типа у культурных сортов тюльпанов
Группа сортов Количество сортов Сорта с цветущей деткой пок- ровной че- шуи, % Цветущие детки в сортах, %
всего с цветущей деткой по- кровной чешуи с цветущей замещающей луковицей
Юдинакие ранние 24 7 0 25 25
.Махровые ранние 11 8 0 73 62
Махровые поздние 2 1 0 50 25
Гоночные (Менделя) 21 7 0 33 50
Бридер 18 13 1 72 40
Расписные 3 1 1 33 13
Попугайные 4 3 0 75 20
Лилиецветные 2 2 0 100 22
Триумф 29 5 0 18 30
Котедж 24 15 5 63 60
.Дарвина 44 23 4 52 33
Всего | 182 1 85 | 1 11 47 34
на 3—5 дней раньше, чем на противоположной. Гряды несколько
перегревались и растения быстро заканчивали вегетацию. На грядах
же несколько углубленных поливные воды очень уплотняли почву.
В 1951 и 1952 гг. на углубленных грядах тюльпаны дали лучшее цве-
тение, чем на приподнятых.
Самый малый процент сортов с ускорением развития детки дали
-тюльпаны группы Котедж—всего 17,6%.
Среди сортов, относящихся к тюльпанам Дарвина,—63% имеют
склонность к многоцветковости третьего типа. В 1952 г. почти у трех
четвертей сортов из групп махровых ранних, Бридер, попугайных
детка покровной чешуи ускорила развитие. Однако внутри сортов,
относящихся к этим группам, процент детки с ускорением развития
очень меняется. Максимальный процент ускорения дают махровые
ранние—61,5% и минимальный—попугайные—всего 20%.
Зацветание детки покровной чешуи у диких видов. У диких
видов тюльпанов на опытных грядах также было получено зацветание
детки покровной чешуи одновременно с верхушечной почкой. Однако
в связи со слабым развитием почек в пазухах низовых «листьев (че-
шуй), у диких видов это явление наблюдалось в значительно меньшей
степени, чем у культурных сортов (табл. 39).
Цифры для Т. kaufmanniana значительно занижены в связи с не-
удачной популяцией этого вида, попавшей на экспериментальную гряду
‘(табл. 39). Среди других посадок Т. kaufmanniana третий тип много-
цветковости в 1952 г. проявился значительно ярче.
388
Вегетативное размножение видов Т. micheliana и Т. korolkovii
практически равно нулю. Эти виды не дают цветущих деток. Мы
раскормили луковицы Т. micheliana до 130 г, но деток они не зало-
жили. У этих видов наблюдалось подавленное развитие почек в пазу-
хах луковичных чешуй.
Т. micheliana очень изменяется морфологически при укорочении
дня. Мы надеемся преодолеть его консерватизм и получить формы с
вегетативным размножением.
Таблица 39
Многоцветковость третьего типа у диких видов
тюльпанов
Вид Количество луковиц В том числе давших цветение
детки покровной чешуи заме- щающей луковицы
количе- ство %
Т. kaufmanniana 46 1 2 1
Т. greigii 33 5 15 0
Т. ingens 18 1 5 0
Т. fosteriana 74 3 4 0
т. subpraestans 28 9 32 0
т. praestans 15 4 27 0
т. turkestanica 22 11 50 0
т. micheliana 20 0 0 0
т. hoogiana 75 2 3 0
У. korolkovii 300 0 0 0
Г. ferganica 20 1 5 0
Некоторые формы Т. fosteriana неплохо размножаются вегетатив-
но, но его детки после обособления от материнской луковицы не у
всех особей одинаково быстро зацветают. Видимо, в связи с такой
направленностью в их развитии, наблюдается сравнительно небольшой
процент зацветания деток покровной чешуи.
Наивысший процент цветения третьего типа дали Т. praestans и
Т. subpraestans. Это наиболее пластичные виды, их изменчивость при
улучшении условий выращивания равна изменчивости сортовых тюль-
панов.
Развитие почек-деток культурных тюльпанов. Почти у всех видов
и сортов тюльпанов при хорошем уходе можно усилить луковицы для
получения из одной луковицы по несколько цветов, за счет ускорен-
ного цветения деток и ветвления главного стебля.
Зацветание замещающей луковицы в 1952 г. было получено из
тюльпанов Дарвина: у большого количества особей сорта Юбилей,
у нескольких растений сорта Слон; по одному экземпляру у сортов
Turner и Clara Butt у пяти сортов из группы Котедж: Бин I, Miss
Н Hand, Marchal Victor, Mother’s Day и Golden Harwest.
У одного экземпляра сорта Kroschel из группы Бизарр зацвела
389
замещающая луковица и все три детки. Все они имели свои замещаю-
щие луковицы. То же наблюдалось у одного экземпляра сорта Turner.
У сорта Lucifer из группы Бридер зацвела не только детка
покровной чешуи, но и детка первой запасающей. Зацветание несколь-
ких деток одновременно с верхушечной почкой луковицы чаще всего
и в большом количестве отмечалось у сорта Юбилей.
Следовательно, все детки луковицы в потенциале способны к за-
цветанию одновременно с ее верхушечной почкой, т. е. все почки-
детки луковицы, заложившиеся в один сезон с верхушечной почкой,
стадийно одновозрастны с ней. Это подтверждается цветением детки
покровной чешуи, т. е. нижней детки на луковице во время цветения
верхушечной—самой верхней почки луковицы. У сильных луковиц к
большом количестве случаев (табл. 38) зацветает замещающая луко-
вица до ее обособления от материнского растения. Прочие детки не-
однократно цветут одновременно с цветением верхушечной почки.
Отставание в развитии почек обусловлено необеспеченностью ма-
теринской луковицы запасными питательными веществами, задерживаю-
щими их закладку. Дальнейшее их развитие находится также в пря-
мой зависимости от количества запасных питательных веществ в лу-
ковице.
Отсутствие надлежащего питания в первые моменты роста почек
у диких видов чаще всего ведет к их отмиранию. Это становится по-
ложительным фактором в борьбе за сохранение жизни индивидуума
в условиях тощих почв и слабой влажности. У многих тюльпанов-
пустынь и полупустынь даже неполовозрелые особи почти не размно-
жаются вегетативно.
При решении вопросов вегетативного размножения необходимо
было выяснить, что же наблюдается при размножении тюльпанов дет-
ками от цветущей луковицы: отставание—незаконченность стадийного*
развития деток-луковиц против развития верхушечной почки растения,
или снятие стадийности—стадийное омоложение деток?
В предыдущих работах мы (1953) придерживались взгляда, что
детки отстают стадийно от верхушечной точки роста луковицы на одну
ступень стадийного развития. На основании последующего изучения
развития растений разных возрастов, процессов заложения деток как.
в пазухах низовых листьев в луковице, так и в пазухах срединных
листьев стадийно зрелого плодоносящего побега, мы пришли к выво-
ду, что при заложении деток-луковиц в пазухах низовых листьев или
срединных зеленых листьев на стебле, как и при заложении воздуш-
ных луковиц на стебле лука, описанном А. А. Авакяном (1948), про-
исходит снятие стадийности. Но при заложении воздушных луковиц,
на вершине цветочной стрелки лука происходит полное снятие ста-
дийности и развитие растений начинается заново как из семени. При
заложении деток в пазухах чешуй луковицы происходит лишь частич-
ное омоложение на одну-две ступени стадийного развития. Это утверж-
дение обосновано заложением меристемы почек-деток одновременно'
с меристемой верхушечной почки после прохождения как стадии
яровизации, так и световой стадии в период жаропокоя; зацветанием
у сильных луковиц всех деток, заложившихся в пазухах низовых
листьев (чешуй) одновременно с верхушечной почкой, как у сортов
Jubilee, Kroschel и Turner; зацветанием детки покровной чешуи, т. е.
самой нижней детки, сидящей при основании донца (стебля) и как бы
наиболее стадийно молодой одновременно с верхушечной (т. е. с наи-
более стадийно старой) почкой луковицы в очень большом количестве
случаев (табл. 38, 39). Вышеизложенные факты свидетельствуют о
390
стадийном равенстве между верхушечной почкой и пазушными почками
луковицы тюльпана. Стадийное отставание пазушных почек, даже в
сильных луковицах, обусловлено их частичным омоложением за счет
некоторой ограниченности поступления запасных питательных веществ
в момент их заложения, так как превращение крахмала в сахар в
чешуях луковицы лишь начинается в период жаропокоя и слабо на-
растает к периоду отрастания корней.
У культурных тюльпанов, особенно у сильных луковиц, развитие
пазушных почек происходит следующим образом. Они закладываются
в пазухах почек-деток первого порядка, расположенных между чешуя-
ми материнской луковицы. Эти почки-детки второго порядка всегда
закладываются как луковицы с низовыми чешуями. Вместе с верху-
шечной точкой роста почки-детки первого порядка проходят период
пониженных температур, составляя единое целое с материнской луко-
вицей и находясь под покровом ее чешуй. При достаточном развитии
почки-детки второго порядка с помощью листьев материнской луко-
вицы проходят световую стадию. Почка-детка первого порядка, в
пазухах чешуй которой находятся почки-детки второго порядка в конце
периода вегетации обособляется от материнской луковицы. В период
жаропокоя в почке-детке первого порядка начинается еще одна фаза-
заложение органов. После прохождения последней ступени стадийного
развития почти перед началом заложения генеративных органов в
этой почке закладывается верхушечная почка и, несколько позже, за-
мещающая луковица.
Вторую фазу заложения органов можно наблюдать у всех взрос-
лых луковиц тюльпанов как культурных сортов, так и диких видов.
Во время закладки верхушечной почки обычно у сильных луковиц
прорастают ранее заложившиеся почки-детки, наступает вторая фаза
развития, а у слабых луковиц и многих диких видов почки-детки
только начинают закладываться.
Прорастание почек-деток, бывших почками-детками второго по-
рядка, а после обособления почки-детки первого порядка и превраще-
ния ее в самостоятельную луковицу, ставших почками-детками
первого порядка, выражается в том, что в пазухах их чешуй закла-
дывается новая серия почек-деток, а их верхушечная почка образует
луковицу или цветоносный побег.
Новая луковица второй фазы, возникшая из верхушечной почки,
часто морфологически отличается от той почки луковицы, из которой
она развилась —покровная чешуя новой луковицы второй фазы явля-
ется расширенным влагалищем ассимиляционного листа с развитой в
той или иной степени листовой пластинкой (табл. 40, 41). Количество
запасающих чешуй этой почки луковицы больше, чем той, из которой
она развилась. Чем мощнее гнездо деток, тем большее количество их
вместо обычной низовой покровной чешуи имеет влагалищный лист.
У одних видов эта особенность проявляется значительно слабее, у
других—сильнее.
Л *
-5-
1. В работе по направленному воспитанию растений было обнару-
жено сильное влияние условий культуры на растения тюльпанов. Дикие
виды значительно увеличили размеры всех частей растений.
2. Под влиянием внешних условий; подкормки азотными удобре-
ниями, поливов и удаления завязей как у диких, так и у культурных
тюльпанов, увеличилось количество цветов от высадки одной луковицы.
Появились многоцветковые формы тюльпанов трех типов.
391
Таблица 40
Развитие влагалищного листа у почек-деток диких видов
тюльпанов
Вид Количество луковиц В том числе давших в гнезде деток с влага- лищным листом Растения, давшие в материнском гнезде вла- галищные листья у де- ток, %
одну две три и более
Т. kaufmanniana 46 5 2 0 15,21
Т. greigii 33 5 4 0 27,63
Т. ingens 18 1 3 0 22,22
Т. fosteriana 74 12 12 5 39,18
Т. subpraestans 28 15 10 3 100
Т. praestans 15 10 2 0 80
Т. turkestanica 22 22 0 0 100
Т. micheliana 20 1 0 0 5
Т. hoogiana 75 10 1 0 8,8
Т. korolkovii 300 1 0 0 0,33
Т. ferganica 20 1 0 0 5
3. Первый тип многоцветковости—Erjostemones тип обусловлен
ветвлением верхушечной точки роста стебля.
4. Второй тип многоцветковости—Leiostemones тип появляется
благодаря пробуждению и развитию обычно недоразвитых, спящих
пазушных почек надземной части тюльпана. Получены тюльпаны с
Таблица 41
Развитие влагалищного листа у деток культурных сортов
тюльпанов до их обособления
Группа сортов Количество сортов В том числе давших в гнезде деток с влага- лищным листом
одну две три и более
Одинакие ранние 24 12 11 5
Махровые ранние 13 3 3 —
Махровые поздние 2 — — —
Гоночные (мен- деля) 21 5 5 2
Бридер 18 11 11 4
Бизарр 3 — —. —
Попугайные 4 — — —
Лилиецветные 2 1 1 1
Триумф 29 6 6 6
Котедж 24 12 12 7
Дарвина 44 24 17 21
392
облиствленными и увенчанными цветами ветвями первого, второго и
третьего порядков.
5. Третий тип многоцветковости—тип культурного тюльпана пред-
ставляет ускорение развития пазушных деток в луковице и зацветание
их одновременно с верхушечной почкой луковицы.
6. В цветущей луковице тюльпана имеется две фазы заложения
деток. Первая начинается до вступления растения в покой, а вторая—
в период покоя. Первая фаза протекает до окончания растением стадии
яровизации и последней ступени световой стадии и тогда все закла-
дывающиеся почки являются вегетативными; вторая фаза —во второй
половине жаропокоя, когда ранее заложившиеся почки прорастают.
На базе их верхушечных меристем закладываются новые луковицы
или цветоносные побеги, а в пазухах их чешуй новые детки. В это
же время происходит развитие верхушечной почки луковицы в цве-
тоносный побег, а немного позже закладывается почка замещающей
луковицы, иногда в сильных луковицах переходящая в побег. Почки
луковицы, заложившиеся во вторую фазу, на базе верхушечных ме-
ристем пазушных почек морфологически иные, чем почки первой фазы.
Их покровной чешуей служит основание влагалищного листа, впослед-
ствии развивающего зеленую пластинку над поверхностью почвы. Ко-
личество чешуй таких почек всегда больше, чем у почек из верху-
шечных меристем, из которых они развились.
7. В слабых луковицах культурных тюльпанов и у большинства
диких видов начало заложения почек задерживается до вступления
растения в период покоя и прорастание почек с заложением влагалищ-
ных листьев выпадает.
8. Изменение условий культуры тюльпанов при избыточном удо-
брении азотом, полив их и направление запасов питательных веществ,
не в молодую завязь, а в луковицу, в наших опытах при высоких
температурах жаро- или сухопокоя вызвало у тюльпанов проявление
раннего типа развития—ветвление стебля и ускорение зацветания деток.
При появлении у тюльпанов ветвления стебля и многоцветковости
мы наблюдаем развитие признака, которым обладали растения в неда-
леком историческом прошлом, но в связи с тем, что они вынуждены
были занять новые более ксерофитные арены жизни или же благодаря
некоторой ксерофитизации уже занятых ими пространств, они почти
утратили свойство ветвления надземного побега в естественных место-
обитаниях. Таким образом, ветвление стеблей и кустовая форма—древ-
ний признак в данном роде. Одностебельность же и одноцветковость—
это результат исторического развития свойств растений под воздей-
ствием внешних условий. Однако, приобретая новый тип развития,
тюльпаны не полностью утрачивают старый.
ЛИТЕРАТУРА
Авакян А. А. Цветение двухлетних культур в первом году жизни, „Агробиоло-
гия 1950, № 2.
Авакян А. А. Некоторые вопросы индивидуального развития растений, „Агроби-
ология", 1948, № 2.
Авакян А. А. Стадийные процессы и так называемые гормоны цветения, „Агро-
биология", 1948, № 2.
Авдулов Н. П. Кариосистематическое исследование сем. Gramineae, Труды по
Прикладной ботанике, генетике и селекции, приложение, Л., 1931.
Авдулов Н. П. О сверхкомплектных хромозомах у кукурузы, Труды по приклад-
ной ботанике, генетике и селекции, сер. 2, Л., 1033, № 2.
Авдулов Н. П. и Титова Н. Н. Сверхкомплектные хромозомы у Pospalum sto-
loniferum Bosco, Труды по прикладной ботанике, генетике и селекции,
сер. 2, Л., 1933, № 2.
А вдулов Н. П. Дополнительные кариологические данные к систематике злаков,
Труды по прикладной ботанике, генетике и селекции, сер. 2, Л., 1933, №2.
Алкиперова С. Г. Новый способ вегетативного размножения нарциссов, „Совет-
ские субтропики", 1940. №11—12.
Алферов В. А. К развитию цветочных луковиц в советских субтропиках, 1934, №1—2
Алферов В. А. Луковичные цветочные растения, М., Сельхозгиз, 1956.
Артюшенко 3. Т. К вопросу о морфологической сущности нижней завязи у не-
которых представителей Caprifoliaceae, „Ботанический журнал", 1948.
Арутюнова Л. Г. Морфология хромозом некоторых видов клевера, ДАН СССР,
т. 27, 1940, № 8.
Ахвердов А. А. Биология некоторых декоративных геофитов флоры Армении,
Автореферат диссертации на соискание ученой степени кандидата биоло-
гических наук, Ботанический институт АрмССР, Ереван, 1954.
Ахвердов А. А. Биология некоторых декоративных геофитов флоры Армении,
Бюллетень Ботанического сада АН Армянской ССР", 1956, № 15.
Ахвердов А. А. и Ми рзоев Н . В. Новый вид тюльпана из Ю. Армении Т.
Sosnovskyi, Труды Ботанического института АН СССР, т. 7. Ереван, 1950.
Базилевская И. А. Декоративное цветоводство СССР, „Сад и огород", 1957,
№ 11
БеклемешевЕ. А. О культуре луковиц на Кавказе, „Вестник Император-
ского Российского Общества Садоводства*, СПб., 1898, № 1.
Беляева В. Промышленное разведение цветочных луковиц в Англии и Голландии
„Советские субтропики", 1935, № 12.
Б е р г Л. С. Закономерность в образовании органических форм. Труды по приклад-
ной ботнике, генетике и селекции. Л., 1924, 14.
Блументаль И . X. Геоботанический очерк западной оконечности Заилийского
Алатау и Чуилийских гор, Л., изд. ЛГУ, 1937.
Бородин И. П. Курс анатомии растений, третье издание, Л„ 1904.
Бочанц’ева 3. П. К биологии цветения Гуаюлы Parthenium argentatiim Gray, Тру-
ды САГУ, сер. 8, вып. 15, Ташкент, 1933.
Бочанцева 3. П. К методике цитологического исследования растений, „Социали-
стическая наука и техника", 1934, № 3—4.
Бочанцева 3. П. Спермиогенез у тюльпанов и его изучение для целей кариоси-
стематики, Бюллетень САГУ, ^ып. 22, Ташкент, 1937.
394
Бочанцева 3. П. Преодоление нескрещиваемости при межродовой гибридизации
в семействе маковых (Papaveraceae), Бюллетень АН УзССР, 1945, № 5.
Бочанцева 3. П. Миксоплоидия у некоторых растений. Бюллетень АН УзССР,
Ташкент, 1946, № 12.
Бочанцева 3. П. К вопросу о прорастании семян тюльпанов, Труды Ботаничес-
кого сада АН УзССР, т. 2, Ташкент, 1951.
Бочанцева 3. П. К вопросу о стадийном развитии многолетних растений, Труды
Инстинута ботаники АН УзССР, т. 1, Ташкент, 1952.
Бочанцева 3. П. Многоцветковость тюльпанов, полученная экспериментально,
Труды Ботанического сада АН УзССР т. 3, Ташкент, 1953.
Бочанцева 3. П . Выступления участников совещания представителей ботаничес-
ких садов в 1952 г. в Москве, Бюллетень Главного Ботанического сада,
М., 1953, № 15.
Бочанцева 3. П. Многоцветковость тюльпанов, Труды Ботанического сада АН
УзССР. вып. 4, Ташкент, 1954.
Бочанцева 3. П. Новый вид тюльпана из Западного Тянь-Шаня, Ботанические
материалы Гербария Института ботаники АН УзССР, вып. 14, Ташкент
Бочанцева 3. П. Тюльпаны и их культура в Ташкенте, Реферат работ АН
УзССР, внедряемых в народное хозяйство, Ташкент, Изд-во АН УзССР,
1954.
Бочанцева 3. П. Прорастание семян тюльпанов, подвергнутых воздействию уль-
тразвука ДАН УзССР, 1955, № 4.
Бочанцева 3. П. Онтогенез тюльпанов, Труды Ботанического сада АН УзССР
вып. 5, Ташкент, 1956.
Бочанцева 3. П. Отдаленная гибридизация у тюльпанов, Тезисы докладов и со-
общений на юбилейной научной сессии, посвященной 40-летию Великой
Октябрьской социалистической революции, Ташкент, АН УзССР, 1957.
Бочанцев 3. П. Отдаленная гибридизация у тюльпанов, Тезисы докладов сове-
щания по отдаленной гибридизации растений и животных, М., 1958.
Бочанцева 3. П. Лола, .Фан ва Турмуш", 1958, № 5.
Бочанцев В. П. и Б утков А. Я. Дикорастущие декоративные растения Узбе-
кистана, Ташкент, Узгиз, 1937.
Брагилевская С. М. Применение удобрений в цветоводстве, Бюллетень „Обмен
опытом по озеленению городов УзССРКиев 1949, № 1.
Бреславец Л., Медведева Г. и Магитт М. Цитологические исследования
лубяных растений, II. кендырь, рами, кенаф, канатник, сап, бамия, Труды
Института нового лубяного сырья, 2, М., 1933.
Бреславец Л. Цитологические исследования лубяных растений I Cannabis saliva
(конопля), Труды Института нового лубяного сырья, 2, М., 1933.
Б v г а е в В. А. Климат Средней Азии и Казахстана, Изд-во АН УзССР, Ташкент,
1946.
Бы л да А. 3. О годичном цикле развития у плодовых растений, „Природа", 1952,
№ 2.
В а к а р Б. А. Цитологическое исследование гибридов Triticum peisicum Vav. с дру-
гими видами пшеницы, Труды съезда по генетике,селекции, семеновод-
ству и племенному животноводству, т. 2, 1930.
В а кар Б. А. Пшенично-пырейные гибриды (Гилогенетическое исследование), Труды
по прикладной ботанике, генетике и селекции, сер. 2, Л., 1935, № 8.
В а к а р Б. А. Цитология пшенично-пырейных гибридов, Омск, Омское областное
издательство, 1935.
Вальнев Е. М. Тюльпаны Средней Азии, „Социалистическая наука и техника*,
Омск, 1937, № 9.
Василевская В. К. Значение анатомических признаков в чешуе луковиц средне-
азиатских тюльпанов для их систематики, Труды САГУ, сер. 8, Ташкент.
1936.
Василевская В. К. О значении анатомических коэффициентов, как признаке за-
сухоустойчивости растений, т. 23, Ашхабад, 1938, № 4.
Васи льев А. В. Перспективы развития промышленного садоводства на юге и в
субтропиках СССР, „Субтропики", 1930, № 7, 12.
Введенский И. А. Тюльпан— Tulipa, Флора СССР, т. 4, 1935.
Введенский И. A. Tulipa binutans Vved., Ботанические материалы АН УзССР,
1952, № 13.
Введенский А. И., Д р о б о в В. П., К о р о в и н Е. П., К у л ь т и а с о в М. В., По-
пов М. Г., Райкова И. А. Определитель растений окрестностей
Ташкента, вып. I, Ташкент, изд. САГУ, 1923.
Верушкин С. М. Пшенично-пырейные гибриды, М., Сельхозгиз, 1933.
В е ру ш к ин С. М. Основные закономерности в отдаленной гибридизации, Пособие
по селекции, вып. I., М. 1936.
395
Вильсон Э. Клетка и ее роль в развитии и наследственности, т. 2, М.—Л., Изд-во*
АН СССР, 1940. Влияние климата и условий на развитие цветочных лу-
ковичных растений, „Опыт зарубежных субтропиков", 1938, № 7.
Воробьев А. И. Кратные хромозомные числа у родственных форм, Труды по при-
кладной ботанике, 1енетике и селекции, сер. I., Л., 1934, № 6.
Вульф Е. В. Историческая география растений, М.—Л., 1944.
Г а б е Д. Р. Наследование пола у Mercurialus annua в связи с цитоплазматической
теорией наследования пола, ДАН, т. 23, 1939. № 5.
Герасимова Е. Н. О размерах спутников хромозом, Известия АН СССР, Отдел
биологических наук, М., 1940, № 1.
Герасимова-Навашина Е. Н. Развитие зародышевого мешка, двойное опло-
дотворение и вопрос о происхождении покрытосемянных, Ботанический"
журнал, т. 39. 1954, № 5.
Глотов В. Диморфизм кариогнпа у Chrysanthemum coronarium L., ДАН., т. 24, 1939?.
№ 2.
Глотов В. Действие колхицина из Colchicum urnbrosum Stev. на камфарный бази-
лик, ДАН, т. 24, 1939, № 5.
Глотов В. В. Амфидиплоидная плодовитая форма Mentha piperita L., полученная
после обработки колхицином, ДАН, т. 28, 1940, № 5.
Голландские цветочные луковицы, Иллюстрированное описание. 1947.
Голубинский И. Н. и Шлосе М. С. К вопросу о портеногенезе у Humulus Lu
pulus L., ДАН, т. 23, 1932, № 5.
Гроссгейм A. A. Tulipa L.—Флора Кавказа, т. II. изд. 2, Баку, 1940.
Гущин Г. Н. Итоги отбора диких луковичных растений на Денауском опытном по-
ле ВНИПСС и их хозяйственно-декоративные качества, Бюллетень по
культурам сухих субтропиков, Душанбе, 1940, № 4 (56).
Даева О. В. Ритм развития некоторых среднеазиатских растений в условиях Моск-
вы, Труды Главного Ботанического сада, т. 2, М., 1951.
Даева О. В. Среднеазиатские виды лука и опыты их культуры в Москве, Бюлле-
тень Главного Ботанического сада, вып. 31, Изд-во АН СССР, 1958.
Д а е в а О. В . Биоморфологические типы лука Средней Азии, Бюллетень Главного
Ботанического сада АН СССР, вып. 33, Изд-во АН СССР, 1959.
Данилевская О.Н. Размножение тюльпанов на Севере, Автореферат диссерта-
ции на соискание ученой степени кандидата биологических наук, Л., 1955.
Данилевская О. Н. Культура тюльпанов, Л., Лениздат, 1956.
Дарвин Чарльз. Сочинения, тт. 6—7, М., 1950.
Делоне Л. Н. Сравнительно-кариологическое исследование нескольких видов Mus-
cat! Mill, записки Киевского общества естествознания, Киев, 1915.
Делоне Л. Н. Тетраплоидная особь Muscat! latifolium. Протокол заседания Киев-
ского общества естествоиспытателей, Киев, 1915.
Делоне Л. Н. Сравнительно-кариологическое исследование видов Muscarl Mill, u
Bellevalia Lopeyr, Вестник Тифлисского ботанического сада, сер. 2, вып. 1,
Тбилиси, 1922.
Делоне Л. Н. Хромозомы видов Ornithogallum L., Труды Государственного Тими-
рязевского исследовательского института, Отдел экспериментальной эво-
люции. сер. 1, отд. 2, вып. 3, 1925.
Делоне Л. Н. Хромозомная теория наследственности у некоторых лилейных, „Вест-
ник Тифлисского ботанического сада", сер. 2, вып. 2, 1926.
Делоне Л. Н. Превращения клеточного ядра в наследственной изменчивости, Тру-
ды с/г ботаники т. I, вып. 1, 1926.
Делоне Л. Н. Применение кариологического анализа к решению вопросов спе-
циальной систематики, Сборник им. С. Г. Навшина, М., 1928.
До роган ев ск ая Е. А. О связи географического распространения растений с их
обменом веществ, М., Изд-во АН СССР, 1951.
Есиновская В. И. Тюльпаны в Эстонской ССР, „Известия АН Эст. ССР“ т. 7,
1958, № 4.
Ефремова Л. Д. Эмбриологическое исследование декоративных тюльпанов и их
значение в садоводстве, Автореферат диссертации на соискание ученой
степени кандидата биологических наук, М., 1951.
Жадовский А. Е. Гормоны клеточных делений (раневые гормоны и нейрогормо-
ны) и их роль в развитии нормальных и придаточных зародышей у рас-
тений, „Успехи экспериментальной биологии", 1924, № 3—4.
Жадовский А. Е. Типы развития пыльцы и их филогенетическое значение, Труды
по прикладной ботанике, генетике и селекции, 14,5, Л, 1924—1925.
Забелин Н. А. Пестролепестность тюльпанов в зависимости от сорта и, среды
разведения, Труды государственного Никитского Ботанического сада,
т. 24, вып. 3, 1948.
396
Зайцева Е. Н. Дикорастущие виды тюльпанов и их садовые формы в коллекции
Главного ботанического сада, Бюллетень Главного ботанического сада»
вып. 26, М., 1956.
Зайцева Е. Н. Коллекция садовых форм тюльпанов в Главном Ботаническом са-
ду, Бюллетень Главного ботанического сада, вып. 27, Изд-во АН СССР.
1957.
-Зайцева Е. Н. Тюльпаны, М., 1958.
Затворницкий Г. Ф. Степной тюльпан, Бюллетень Главного ботанического сада
АН СССР, вып. 18, 1954.
Золотарев И. И. Флора садоводства, Иллюстрированный указатель растений.
М, 1896.
ИльинМ. М. К происхождению флоры пустынь Средней Азии, „Советская Бота-
ника", 1937, № 6.
Исип И. Н. Влияние защитных веществ в растительном организме, „Советская
Ботаника", 1939, № 3.
Казарян В. О. Стадийность развития и старения однолетних растений, Ереван,
Изд-ro АН АрмССР, 1952.
К а п и но с Г. Е. К тератологии цветка тюльпана (Tulipa Gesneriana L.), ДАН
АзербССР, т. 3, 1947, № 6.
К а п и н о с Г. Е. Из наблюдений по фенологии тюльпана на апшероне, Бюллетень
Главного ботанического сада, вып. 4, М., 1949.
Капинос Г. Е. К тератологии цветка тюльпана, сообщение II, „Известия АзербССР*,
1951, № 11.
Капинос Г. Е. Выращивание гиацинтов и тюльпанов из семян, ДАН АзербССР,
т. 8, 1952, № 11.
.Карпеченко Г. Д. Числа хромозом и генетические взаимоотношения у культур-
ных Gruciferae, Труды по прикладной ботанике и селекции, т. 13, Л.»
1922—1923.
Карпеченко Г. Д. Кариологический очерк рода Trifolium, Труды по прикладной
ботанике и селекции, т. 14, Л., 1924—1925.
Карпеченко Г. Д. Полиплоидные гибриды Raphanus saiivus L. и Brassica olera-
ceae L., Труды по прикладной ботанике, генетике и семеноводству, вып.
3, т, 17, Л., 1927.
Карпеченко Г. Д. К синтезу константного гибрида из трех видов, Труды съезда
по генетике, селекции, семеноводству и племенных животных, т. 2, Л.,
1930.
.Карпеченко Г. Д. Теория отдаленной гибридизации, Теоретические основы се-
лекции, т. I, 1935.
Каспарян А. С. Обзор работ по полиплоидии и амфидиплоидии за последние го-
ды, Труды по прикладной генетике и селекции, сер. 2, Л., 1934, № 6.
Кахидзе Н. Т. Хромомерное строение хромозом пшениц в митозе, ДАН, т. 26,
1940, № 5.
Киселев Г. Е. Опыт цветоводов-москвичей, М„ Изд-во Московский рабочий, 1950.
Кичунов Н. И. Лучшие луковичные растения для грунта, Садовая библиотека,
СПб, 1905.
Кичунов Н. И. Красиво цветущие грунтовые луковичные растения, М., 1909.
Кичунов Н. И. Многолетники, М.—Л., 1935.
К лимочкина Л. В. Морфология хромозом Hellianthus annus L., ДАН, т. 27,
1940, № 6.
Кожевников А. В. Весна и осень в жизни растений, М., Изд-во Московского
общества испытателей природы, 1950.
Кожин А. Е, Пути селекции цитрусовых в новых районах их культуры, „Селекция
и семеноводство", 1950, № 2.
Комаров В. Л. Ученье о виде растений, М.— Л., Изд-во АН СССР, 1940.
Комаров В. Л. Избранные сочинения, М.— Л., 1945—1953.
Коновалов И. Н. О причинах возникновения явлений пролификации цветов с
точки зрения стадийного развития растений, ДАН СССР, 66, 1949.
Коновалов И. Н. и Артюшенко 3. Т. Морфология плодов типа коробочка и
ягода, В сб. „Морфология и анатомия растений", сер. 7, вып. 2, Л., 1951.
Коровин Е. П. Род Scaligeria D. С. (Umbeliferae) и его филогения, Труды САГУ,
сер. 8, вып. 2, Ташкент, 1928.
Коровин Е. П. Растительность Средней Азии, Ташкент, 1934.
Коровин Е. П. Основные пути естественной акклиматизации теплолюбивой флоры,
Цитрусовые и субтропические культуры Узбекистана, Ташкент, Изд-во
АН УзССР, 1949.
Коровин Е. П. и Шувалов С. А. О биогенной комплексности почвенно-расти-
тельного покрова в оридной зоне, Бюллетень Московского общества ис-
пытателей природы, Отдел биологии, т. 53 (I), М., 1948.
397
Красинский Н. И. Методы ускоренной выгонки цветочных растений, М., Сель-
хозгиз, 1937.
Ереван П. Еще к вопросу о культуре гиацинтов и тюльпанов в СССР, «Сад и
огород 1928, № 10.
К р е н к е Н. И. Хирургия растений, М., Изд-во Новая деревня, 1928.
К р е н к е Н. И. Теория циклического старения и омоложения растений, М., Сель-
хозгиз, 1940.
К р е н к е Н. И. Регенерация растений, М.-Л., Изд-во АН СССР, 1950.
Крокер В. Рост растений, М., Изд-во Ил, 1950.
Кудряшов С. Растительность района гор Хабдун-Тау и Карача-Тау, Труды САГУГ
сер. 8 в, вып. 13, Ташкент, 1930.
Кузьмина Н. Е. О хромозомах Beta vulgaris L., Труды по прикладной ботанике,,
генетике и селекции, т. 17, Л., 1927, № 3.
Кузьмина Н. Е. Цитология культурного мака (Papaver Somniferum L.) в связи с
его происхождением и эволюцией, Труды по прикладной ботанике, гене-
тике и селекции, сер. 2., Д., 1935, № 8.
КультиасовМ. В. Проблема становления жизненных форм у растений. Сб. Проб-
лемы ботаники, М.-Л., Изд-во АН СССР, 1950.
К у п е р м а н Ф. М. Морфофизиологический метод исследования на службу селекции
растений, „Селекция и семеноводство", 1951, № 9.
Куперман Ф. М. Биологические основы культуры пшеницы, М., изд. МГУ, 1953»
Куперман Ф. М., Дворянкин С. А., Ростовцева 3. П., Р ж а н о в а Е.
Этапы формирования органов плодоношения злаков, 1, М., Изд-во МГУ
1955.
Лавренко Е. М. Памяти Валерия Ивановича Талиева, Ботанический журнал, т. 42^
9, М.-Л., 1957.
Лапин П. И. Опыт управления цветением декоративных растений, Диссертация на
соискание ученой степени кандидата биологических наук, Л., 1936—1938.
Лапин П. И. К вопросу о расширении периода цветения луковичных растений,
„Зеленое строительство", Л., 1939, № 5—6.
Лапин П. И. Весенний показ цветов, „Природа", 1956, № 8.
Лебедев Д. В. Эволюционные взгляды Валерия Ивановича Талиева, Ботанический
журнал, т. 42, М.-Л., 1957, № 9.
Левитин А. Н. Дикорастущие луковичные растения Средней Азии и их исполь-
зование в культуре, Бюллетень Главного ботанического сада АН СССР,,
вып. 17, 1954.
Левитин А. Н, Гущин Г. Н. Краткое описание сортов массового и группового
отбора из дикорастущих луковичных растений Средней Азии, Бюллетень
по культурам сухих субтропиков, Душанбе, 1953, № 11—12.
ЛевитскийГ А. Материальные основы наследственности, Киев, 1924.
Левитский Г. А. О цитологическом методе в систематике, Труды Всесоюзного
съезда по генетике, селекции, семеноводству и племенному животновод-
ству, т. 2., Л., 1929.
Левитский Г. А. Морфология хромозом, Труды по прикладной ботанике и се-
лекции, т. 27, вып. I, Л., 1931.
Левитский Г. А. Опыт цитологического анализа фиксационного действия хром-
ацет-формола. Труды по прикладной ботанике, генетике, селекции, т. 27,.
вып. 1, 1931.
Левитский Г. А. Морфология хромозом и понятие кариотипа в систематике на
.основе исследования подсем. Helleboreae, Труды по прикладной генетике
и селекции, т. 27, вып. I, Л., 1931.
Левитский Г. А. Цитологический метод в селекции, „Социалистическое растение-
водство", 1933, № 5/6.
Левитский Г. А. Новые фиксационные смеси, проявляющие морфологию хромо-
зом, ДАН СССР. т. 4, 1934, № 3.
Левитский Г. А. Фиксационные изменения тела хромозом, ДАН СССР, т. 4Г
1934, № 4.
Левитский Г. А. Цитологический метод в селекции, Теоретические основы в се-
лекции, т. I. Л., 1935.
Левитский Г. А. Очерк генетической цитологии, Пособие по селекции, вып. 1,„
М., 1936.
Левитский Г. А. Цитологические основы эволюции, „Природа", 1939, № 5.
Левитский Г. А. и Араратян А. Преобразование хромозом под влиянием
рентгеновский лучей, Труды по прикладной ботанике, генетике и селек-
ции, Л., 1931.
Левитский Г. и Бенецкая Г. О кариотипе Solan urn tuberosum L., Труды по
прикладной ботанике, генетике и селекции, т. 17, вып. 3, 1927.
Левитский Г. А. и Бенецкая Г. К. Цитологическое исследование константно-
промежуточных ржано-пшеничных гибридов, Список докладов и тезисов-
398
Всесоюзного съезда по генетике, селекции и т. д., Л., 1929.
Левитский Г. А. и Б е нецка я Г. К. Цитология пшенично-ржаных амфидиплои-
дов, Труды по прикладной ботанике, генетике и селекции, т. 27, вып. I,
Л., 193L
Левитский Г. А. и Кузьмина Н. Е. Кариологический метод в систематике и
филогенетике рода Festuca, Труды по прикладной ботанике, генетике и
селекции, т. 17, вып. 3, 1927.
Левитский Г. А. и Сизова М. А. О закономерностях в преобразованиях хро-
мозом, вызываемых х-лучами, ДАН СССР, т. 4, 1934, № 1-2.
Левитский Г. А., Шепелева Е. М., Титова Н. Н. Новые кариотипические
расы, выведенные из потомства рентгенезированных растений, „Социа-
листическое растениеводство", Л., 1937. № 11.
Липшиц С. Ю. Заметка о Tulipa Krauseana Rgl. Известия Ботанического сада
АН СССР, т. 30, вып. 5-6, Л., 1932.
Литовченко А. Г. Значение трикотеледонии (трехдольности) в растениеводстве,
ДАН, т. 27, 1940, № 8.
Лодкина М. М. Особенности развития тычинок пшеницы и лилии в связи с общей
Физиологией цветка, Труды Ботанического института АН СССР, сер. 7,
вып. 4, М.-Л., 1957.
Лопотт М. С. и О з о л и н П. К. Практические указания по грунтовому цвето-
водству в Средней Азии, Душанбе, 1941.
Лучник 3. И. Декоративные растения* Горного Алтая, М.» Сельхозгиз, 1951.
Лысенко Т. Д. Работы в дни Великой Отечественной войны, М., Огиз-Сельхоз-
гиз, 1943.
Л ы с е н к о Т. Д. Агробиология, М„ 1948.
Львова И. Н. Некоторые вопросы цитологии оплодотворения у злаков, „Селекция
и семеноводство", 1950, № 9.
Л ю б и ч Ф. П. Разнокачественность плодов и семян у растений и ее значение в
жизни вида, „Агробиология", 1951, № 5.
Магешвари П. Эмбриология покрытосемянных. М., 1954.
Маевский П. Ф. Флора Средней полосы Европейской части СССР, 8-е изд., М.-Л.,
Сельхозгиз, 1954.
Макарова Н. А. Некоторые экологические особенности в анатомическом строе-
нии и физиологии тюльпанов юго-востока, „Советская ботаника", Л„
1934, № 3.
М а м п о р и я Ф. Д. Особенности воспроизведения, роста, развития и формообразо-
ния цитрусовых и некоторых других померанцевых, Тбилиси, 1951.
Марков А. Г. Отечественные сорта декоративных растений, М., 1955.
МашинскийЛ. О. Промышленное освоение культуры дикорастущих тюльпанов в
садоводствах Ленинграда, „Зеленое строительство", 1939, № 11 — 12.
Международный кодекс номенклатуры для культурных растений, М.-Л., Изд. АН
СССР, 1957.
Мигушова Э. Особенности садового тюльпана, Автореферат диссертации на соис-
кание ученой степени кандидата биологических наук, Л., 1954.
Мигушова Э. Особенности биологии садового тюльпана, Автореферат диссерта-
ции на соискание ученой степени кандидата биологических наук, Л.,
1954.
Минина Е. Г. Смещение пола у растений воздействием факторов внешней среды,
М., Изд-во АН СССР, 1952.
М и ч у р и н И. В. Выведение новых улучшенных сортов плодовых и ягодных расте-
ний, М., Сельхозгиз, 1933.
Мичурин И. В. Сочинения, т. I, М.-Л., Огиз-Сельхозгиз, 1941.
Мищенко П. Дикие виды Tulipa (тюльпан) Scilla Кавказа, Крыма и Средней Азии
как материал для культуры. Труды Бюро по прикладной ботанике, СПб.,
т. 5, 1912, № 2.
Модилевский Я. С. Об аномальных случаях развития зародышевого мешка у
высших растений, Записки Киевского общества естествоиспытателей,
вып. 20, Киев. 1910.
Модилевский Я. С. Жиночий гаметофит у Angiosperma „Украинский ботаниче-
ский журнал". 1929, № 5.
Молчанов Л. А. Климатические условия наиболее теплых районов Средней Азии,.
Сб. „Субтропические культуры Средней Азии", Ташкент, Госиздат
УзССР, 1935.
Мурзаев Э. М. Средняя Азия, М., Географгиз, 1957.
Мыт аре в И. М. К вопросу о культуре гиацинтов и тюльпанов в СССР, „Сад и
огород", 1929, № 1.
Навашин М. С. Сб изменении числа и формологических признаков хромозом
у межвидовых гибридов, Труды по прикладной генетике и селекции,,
т. 27, вып. 3, Л., 1927.
399
Н а в аш и н М. С. Морфология клеточного ядра у видов Crepis в связи с вопросом
видообразования, Сборник им. С. Г. Навашина, М., 1928.
Навашин М. С. Новые данные по вопросу о самопроизвольных мутациях, „Биоло-
гический журнал", т. 2, вып. 2/3, М., 1933.
Навашин М. С. Методика цитологического исследования для селекционных целей,
М., Сельхозгиз, 1936.
Навашин М. С. и Герасимова Е. Н. Получение полиплоидов из листьев обра-
ботанных колхицином, ДАН, т. 24, 1939.
Навашин С. Г. О некоторых явлениях при слиянии ядер в зародышевом мешке
после оплодотворения и замечания о типическом делении ядер у Fritil-
laria tenella, Протокол зеседания Киевского общества естествоиспытате-
лей, Киев, 1910
Навашин С. Г. Об индивидуальных и видовых отличиях хромозом, Протокол
заседания Киевского общества естествоиспытателей, 1911.
Навашин С. Г. О диморфизме ядер в соматических клетках у Galtonia candicans.
Известия Импер. акад, наук, 1912.
Навашин С. Г- Новые наблюдения над превращением клеточного ядра у Galtonia
candicans. Протокол Киевского общества естествоиспытателей,
Киев, 1912.
Навашин С. Г. Гетеро и идиохромозомы у растений в связи с вопросом об опре-
делении пола у животных, Протокол Киевского общества естествоиспы-
тателей, Киев, 1913.
Н а в а ш и н С. Г. Заметка о числе хромозом в клеточном ядре у Najas Major, Про-
токол Киевского общества естествоиспытателей, Киев, 1914.
Навашин С. Г. О некоторых признаках внутренней организации хромозом, Сбор-
ник статей, посвященных К. А. Тимирязеву, М, 1914—1916.
Навашин С. Г. Резюме возражений на доклад Л. Н. Делоне, „Русское ботаниче-
ское общество", т. 6, 1921.
Навашин С. Г. Природа и порядок возбуждения ядерного и клеточного деления
в меристеме корешка и митогенные лучи проф. А. Г. Гурвича, „Биоло-
гический журнал*, т. 5, вып. 2. М., 1936.
Навашин С. Г. О диминуции хроматина Ascaris megalocephala, .Биологический
журнал", т. 5, вып. 2, 1936.
'Навашин С. Г. Избранные труды, т. I, М.—Л., Изд-во АН СССР, 1951.
Навои Алишер. Пять поэм, М., 1948.
Н е п о р о ж н ы й Г. Д Гладиолус, М., Сельхозгиз, 1950.
Никитине А. Пески Западного Прибалхашья, Труды Почвенного института
АН СССР, т. 11, М., 1936,
Николаева А. Г. Применение цитологическо!о метода при решении некоторых
вопросов генетики, Труды 111 Всероссийского съезда по селекции и семе-
новодству в Саратове, Саратов, 1920.
Николаева А Г. Цитоло!ические исследования рода Triticum, Труды по приклад-
ной ботанике и селекции, т. 13, Л., 1922—1923.
Николаева А. Г. Материалы по цитологии ржано-пшеничных гибридов. Селек-
ционная станция Сельскохозяйственной Академии, М., 1924.
Николаева М. Г. К биологии прорастания некоторых видов Ferula, Труды БИН,
сер. 4, Л., 1948, № 6.
Николаева М. Г. Зависимость прорастания семян некоторых зонтичных от
состояния покровов зародыша (на примере видов Ferula), Труды Бо-
танического института В. Л. Комарова АН СССР, сер. 4, вып. 6, Л
1948.
Николаева М. Г. Значение стратификации семян для дальнейшего развития сеян-
цев европейского бересклета, ДАН, т. 71, 5, 1950.
Новиков Г Н. и Николаева М. Г. О прорастании семян некоторых видов
Ferula, „Советская ботаника", 1940. № 5—6.
Опыты по культуре декоративных луковичных растений в Англии, „Опыт зарубеж-
ных субтропиков", М., 1937, Ks 4.
Павильон цветоводство и озеленение, Путеводитель, М., Госкультпросветиздат, 1954.
Павильон цветоводство и озеленение, ВСХВ, Путеводитель, М , 1955.
Павлов Н. В. Литературные источники по флоре и растительности Казахстана,
Казахский филиал, Труды, 13, 19, М. — Л., Изд-во АН СССР, 1940.
Павлов Н. В. Растительное сырье Казахстана, М.—Л., 1947.
Павлов Н. В. Растительные ресурсы Южного Казахстана, М.—Л., 1947.
Павлов Н. В. Рассказы о диких цветах Казахстана, Алма-Ата, Изд-во АН КазССР,
1959.
Петров Д. Ф. Генетическое и эволюционное значение различных типов зародыше-
вых мешков, Журнал Общей Биологии, 1942, № 1—2.
Победимова Е. Г. Новый вид тюльпана из Средней Азии Tulipa Neustruevae
Pob. sp. novae, Ботанические материалы гербария, т. Xl, М. — Л.у 1949.
400
Поддубная-Арнольди В. А. Эмбриологический метод в систематике покрыто-
семянных растений, „Известия ассоциации научно-исследовательских
институтов*, т. 3, М., 1930, № 1а.
Поддубная -А рнольди В. А. Эмбриогенез при отдаленной гибридизации в роде
Taraxacum, ДАН, т. 24, 1939, № 4.
Поддубная-Арнольди В. А. К морфологии хромозом твердых пшениц, ДАН,
т. 48, 1945, № 4.
Поддубная-Арнольди В. А. Успехи советской эмбриологии растений, Бюлле-
тень Московского обшества испытателей природы, Отдел биологии,
т. 52 (5), М., 1947.
Полиплоидия, Сборник статей под редакцией П. А. Баранова и Б. Л. Астаурова,
М„ 1956.
Попов В. И. Тюльпаны в США, „Советские субтропики", М., 1937, № 8.
И о п о в М. Г. Флора пестроцветных толщ Краснопесчанниковых низкогорий,
Ташкент, 1923.
П о п о в М. Г. 10 лет работы в Средней Азии, Известия Института почвоведения и
геоботаники САГУ, вып. 1, Ташкент, 1925.
Попов М. Г. Основные черты истории развития флоры Средней Азии, Бюллетень
САГУ, Ташкент, 1927, № 15.
Попов М. Г. Опыт монографии Eremostachis Bge., Мемуары Московскою общества
испытателей природы, т. 19, 1940
Попов М. Г. Растительный покров Казахстана, вып. 18, Филиал Каз АН СССР,
Алма-Ата, 1940.
Попов М. Г. О применении ботанико-географического метода в систематике расте-
ний, Сб, „Проблемы ботаники", М.—Л., Изд-во АН СССР, 1950.
П о п о в М. Г. О взаимоотношениях и истории родов Papa ver и Roemeria, „Ботани-
ческий журнал", т. 42, М. — Л., 1957, № 9.
Грозина М. Н. Сравнительное кариологическое исследование у подсолнечника, 1,
Соматическое деление, „Русское ботаническое общество", М., 1923.
Прозина М. Н. Опыт построения филогенетического развития рода Eremurus,
„Биологический журнал", М., 1937, № 3.
Прозоровский А. Б. О биологических типах растений пустыни, „Биологический
журнал СССР", т. 21, 1936, № 5.
Проскоряков Е. И. Температурные условия прорастания семян весенних многолет-
ников, „Известия Главного ботанического сада СССР", т. 25, 1926.
Проскоряков Е. И. Приспособления прорастающих семян растений среднеазиат-
ской флоры, Автореферат на соискание ученой степени доктора биоло-
гических наук, УзГУ, Самарканд, 1952.
Проскоряков Е. И., Джалилов А. С. К биологии одного из среднеазиатских
тюльпанов, Труды Ботанического сада АН УзССР, вып. 5, Ташкент, 1956
Пыл ков Н. А. Комнатная культура лучших луковичных растений (практическое
руководство), Садовая библиотека, изд. Сойкина, М., 1915.
'Г а б от нов Г. А. Работы в области изучения жизненного цикла многолетних травя-
нистых растений в естественных ценозах, Сб. „Вопросы ботаники",
М. — Л., Изд-во АН СССР, 1954, № 2.
Тегель Э. Л. Туркестанская флора по материалам собранным О. А. Федченко,
Карелиным и Кириловым, Каильбарсом, Корольковым, Краузе, Кушаке-
вичем, Потаниным, Семеновым, Северцевым, Шарнгорстом, Шренком
и др., СПб., 1876 (6).
Регель Э. Л. Вестник Российского общества садоводства, 1873, 1877, 1878, 1879,
1884, СПб.
Тегель Э. Л. Журнал „Gartenflora" с 1877 по 1885.
Р е г е л ь Э. Л. Весенние красивоиветущие многолетние и луковичные растения, их
содержание и воспитание в садах, СПб., 1888.
Родионов А. И. Изучение стадии яровизации у персика и песчаной вишни, Изве-
стия АН СССР, серия биологическая, 1950, № 2.
Розанов М. А. О генотипическом различии рас, Rubus caesius L., ДАН, 27,
1940, № 6.
Розанов М. А. Генетические основы современной систематики растений, Успехи
современной биологии, 2,4 5, М., 1933.
Романов И. Д. Изменчивость и направление эволюции зародышевого мешка в роде
Gagea, Автореферат диссертации на соискание ученой степени кандидата
биоло! ических наук, Ташкент, 1940.
Романов И. Д Новая форма зародышевого мешка Adoxa у Т. tetraphylla и
Т. Ostrovskiana, ДАН, т. 19, 1938, № 1—2.
-Романов И. Д. К морфогенезу хлопчатника, Вопросы цитологии, эмбриологии и
анатомии хлопчатника, Ташкент, СоюзНИХИ, 1936.
Романов И. Д. Две новые формы зародышевого мешка в роде Tulipa, ДАН, т. 22,
1939, № 3.
.26—2229
401
Романов И. Д. Эволюция зародышевого мешка цветковых растений, Автореферат
диссертации на соискание ученой степени доктора биологических наук,.
Ташкент, 1944.
Романов И. Д. К анатомии реликтовых растений, Анатомия листа Tulipa Regelii,
Бюллетень Московскою общества испытателей природы, Отдел биологии,,
т. 57 (5), М„ 1952.
Романов И. Д. Зародышевый .мешок в роде Tulipa, ДАН, т. 115, 1957, № 5.
Руб а ше в ск ий А. А. Философское значение теоретического наследства И. В. Ми-
чурина, М., 1949.
Рубцов Л. И. Продление жизни срезанных цветов, „Зеленое строительство", Л.г
1939, № 7, 8.
Рябова Т. И. Вегетативное размножение эремурусов, „Известия отделения естест-
венных наук АН ТаджССР", вып. 14, 1956.
Рыбин В. А. Гибриды терна и алычи и проблема происхождения культурной сливы,.
Труды по прикладной ботанике и селекции, сер. 2, Л., 1936, № 10.
Рыбин В. A/Цитологический метод в плодоводстве иогородничестве, Достижения
и перспективы в области прикладной ботаники, генетики и селекции, 1929,.
Саади Мушрфаддин Гулистан, М., 1951.
Сааков С. Г. Дикорасту.щие тюльпаны Средней Азии и опыты их практического
использования, „За зеленый Ленинград", Л., 1935, № 6.
Сааков С. Г. Опыт использования в цветоводстве диких тюльпанов Средней Азии,
Труды Ботанического института АН СССР, сер. 6 М. — Л., 1950, № 1.
Сааков С. R Цветоводство в СССР и перспективы его развития, Труды Ботаниче-
ского института АН СССР, сер. 6, вып. 4, М. — Л., 1955.
Светозарова В. В. и Эленгори А. Е. Процесс оплодотворения у растений^,
„Селекция и семеноводство", 1950, № 9.
Свешникова И. Н. Кариологический очерк рода Vicia, Труды по прикладной
ботанике, генетике и селекции, т. 17, вып, 3., Л., 1927.
Свешникова И. Н. Vicia Sativa L. u Vicia cracca L. (к вопросу о значении ка-
риотипа в систематике), „Известия сельскохозяйственной академии
им. Тимирязева", кн. 4, 1929.
Свешикова И. Н. и Белехова Ю„ Проблема транслокации у гибридов, Труды
по прикладной ботанике, генетике и селекции, сер. 2, Л., 1936, № 9.
Свешникова И. Н., Белехова Ю. Изменение хромозом в результате межви-
довой гибридизации у Вики и их филогенетическое значение, М., Изд-во
ВАСХНИЛ, 1936.
Свешникова И. Н. Новый метод изучения эволюция вида, Бюллетень Москов-
ского общества испытателей, Отдел биологии, т. 52 (3), М.. 1947.
Свешникова И. Н. Изучение мутационной изменчивости в связи с проблемой
эволюции у растений, Бюллетень Московского общества испытателей,
Отдел биологии, т. 53 (1), М., 1948
Северцов А. Н. Морфологические закономерности эволюции, М., 1939.
Сеня н инов а-К орчагина М. В. Кариосистематическое исследование рода
Aegilops, Труды по прикладной ботанике, генетике и селекции, т. 28,
Л, 1931.
Сеняни нов а-К орчагина М. В. Кариологическое исследование вопроса о
происхождении Vicia Faba, Труды по прикладной ботанике, генетике и
селекции, т. 28, Л., 1931.
Сердюков Б. В. К вопросу акклиматизации и введения в культуру цветочных
растений дикой флоры Грузии, Вестник Тбилисского ботанического сада,
вып. 59, 1950.
Сердюков Б. В. Введение в культуру некоторых дикорастущих цветочных рас-
тений Грузии, Вестник Тбилисского ботанического сада, вып. 60, 1952.
Серебряков И Г. Морфология вегетативных органов высших растений, М.. 1952-
Сизова М. А. Структурные изменения хромозом в пределах видаTriticum durum,
ДАН, т. 25, 1939, № 1.
Силина 3. М. Тюльпаны в Ленинграде (биология и селекция), Автореферат дис-
сертации на соискание ученой степени кандидата биологических наукг
Л., 1952
Силина 3. М. Опыты по гибридизации тюльпанов, Труды Ботанического институ-
та АН СССР, сер. 6, вып. 2, Л, 1952.
Силина 3. М. Культура тюльпансв в Ленинградской области. Труды Ботаниче-
ского института АН СССР. сер. 6, вып. 3. Л , 1953.
Силина 3. М. Описание сортов тюльпана, Труды Ботанического института АН
СССР, сер. 6, вып. 4, 1955.
Смирнова В. А. О передвижении ядер при мозаичных заболеваниях некоторых
однодольных, Бюллетень Московского общества испытателей природы.
Отдел биологии, т. 59 (5), 1954.
О культуре тюльпанов, „Советские субтропики", 1935, N? 12.
Соколов В. С. Алкалоидоносные растения СССР, М. —Л., Изд-во АН СССР, 1952
Соколовская А. Н. и Стрелкова О. О. Кариологическое исследование
402
высокогорной флоры Главного Кавказского хребта и проблема геогра-
фического распределения полиплодов, ДАН, т. 29, 1940, № 5-6.
Сорокина О. Н. О хромозомах видов Aegilops, Труды по прикладной ботанике,
генетике и селекции, т. 19, вып. 2, 1928.
Сорокина О. Н. Гибридизация эгилопс с пшеницей, Труды по прикладной бота-
нике, генетике и селекции, сер. 2, 1936, № 6.
Талиев В. И. Процесс видообразования в роде Tulipa секция Eriostemones,Труды
по прикладной ботанике, генетике и селекции, т. 24, вып. 2, Л., 1930.
Тараканов К. Н. Опыт историко-экологического изучения гетерокарпии у не-
которых видов солончаковых растений, Сб. „Растение и среда’1, Л., Изд-
во АН СССР, 1952.
Тахтаджан А. Л. Морфологическая эволюция покрытосемянных растений, изд.
Московского общества испытателей природы, 1948.
Тахтаджан А. Л. Вопросы эволюционной морфологии растений, Л., 1954.
Терновский М. Ф. Межвидовая гибридизация Nicotiana получение полиплоидных
и гаплоидных растений, Труды по прикладной ботанике, генетике и се-
лекции, сер. 2, вып. 9, 1936.
Терпило А. И. Анатомический атлас лекарственных растений, Харьков, 1948.
Тимофеева Л. С. Динамика роста и развития садового тюльпана как основа
рациональной агротехники, Автореферат диссертации на соискание уче-
ной степени кандидата сельскохозяйственных наук, М., 1956.
Транковский Д. А. Метод цитологического исследования пыльцевых трубок и
его перспективы, Труды съезда генетики, селекции, семеноводства и пле-
менного животноводства, т. 2, 1930.
Трофимов Т. Т. Вегетативное размножение хохлатки, Галера плотной (Coridalis
Halleri Willd.), Бюллетень Московского общества испытателей природы,
Отделение биологии, т. 57 (3), 1952.
Трофимов Т. Т. О подснежном росте и развитии ранневесенник растений, „Природа",
1954, № 1.
Турбин Н. В. Генетика с основами селекции, М., 1950.
Турбин Н. В. О биологической роли чужеродного доопыления, Вопросы биологи-
ческого оплодотворения, Л, изд. ЛГУ, 1954.
Турбин Н. В. Вопросы биологии оплодотворения, Л., изд. ЛГУ, 1954.
Тут а юк В. X. Образование микро- и мегаспор в одном споролистке у покрыто-
семянных, „Ботанический журнал", т. 34, 1949, № 3.
Тута юк В. X. Анатомо-морфологический анализ махровости покрытосемянных рас-
тений, Труды Ботанического института АН СССР, сер. 7, М.-Л., 1952,№ 3.
Тютюнников А. И. О тюльпане дазистемон (Т. dasystemon Rgl.) и тюльпане
тарда (Т. tarda Stapf.), „Советская ботаника", т. 13, 1945, № 5.
Тютюнников А. И. Многоцветковый дернистый тюльпан, „Известия ботаниче-
ского сада АН СССР", т. 30, вып. 5—6, 1932.
Устинова Е. И. Ритм развития конуса нарастания в луковица?; пролески (Scilla
sibirica) в осенне-зимнее время, ДАН, т. 64, 1949, № 6.
Фаворский М. В. Новый метод дифференцировки препаратов, окрашенных же-
лезным гематоксилином, ДАН, т. 26, 1940, № 5.
Фаворский М. В. Роль гвоздичного масла в процессе дифференцировки препаратов
окрашенных генцианой по Ньютону и возможности его замены, ДАН, т. 26,
1940, № 5.
Федоров А. А. Случай аномалии в строении цветка тюльпана, „Советская бота-
ника", 1947, № 5.
Федоров А. А. О некоторых аномалиях у тюльпанов и о знчении этих аномалий
для незнания морфологической структуры Tulipa, Ботанический журнал,
т. 41, 1956, N2 5.
Федоров А. А., Кирпичников М. Э. и Артюшенко 3. Т. Атлас по
описательной морфологии высших растений: „лист", М. -Л., Изд-во АН
СССР, 1956.
Федоров А. И. О дифференциации конуса нарастания, „Агробиология", 1953, № 1.
Федченко Б. А. Растительность Туркестана, Петроград, 1915.
Федченко Б. А. Красивоцветущие растения Средней Азии, Труды по прикладной
ботанике, генетике и селекции, т. 23, вып. 5, Л., 1929—1930.
Федченко Б. А. Красиво цветущие растения, Растительные богатства СССР,
кн. 3, Лесоматериалы и декоративные растения, 1932.
Филимонова 3. Н. Период летнего покоя у диких луков. Сборник работ аспи-
рантов АН УзССР, Отделение биологическик наук, вып. 2, Ташкент.
1958.
Филимонова 3. Н. Морфология и биология сеянцев первого года развития у
видов рода Allium L., „Узбекский биологический журнал", 1958, № 6.
Филимонова 3. Н. Морфология луковицы некоторых видов рода Allium L.,
„Узбекский биологический журнал", 1959, № 4.
403
Флора Киргизской ССР, т, 3, Фрунзе. 1951.
Флора Туркмении т. 1, вып. 2, Л., 1932.
Флора Узбекистана, т. 1, Ташкент, 1941.
Ха рузин Д. П. Культура и выгонка нарциссов и тюльпанов, „Сад и огород*,
1948, № ‘3.
Шахов А. А. О типах развития растений в процессе исторического приспособле-
ния к засушливым условиям, Сб. „Растение и среда", Л., Изд-во АН СССР,
1952.
Шмальгаузен И. Флора Средней и Южной России, Крыма и Северного Кавказа,
т. 2, М., 1897.
Штамм В. А. К биологии гусиного лука, Бюллетень Главного ботанического сада
АН СССР, вып. 19, М., 1954.
Чернова Н. М. Изменчивость тюльпана Шпенка в Крыму, Бюллетень Главного
ботанического сада, вып. 5, М., 1950.
Цингер Н. В. О причинах мешенного прорастания семян пионов, Труды Главного
ботанического сада, 2-е изд. АН СССР, М., 1951.
Цицин Н. В. Отдаленная гибридизация растений, М., Сельхозгиз, 1954.
Щербаков Н. И. Фитоклиматические аналоги лесных зон Узбекистана и соот-
ветствующих по широте частей Таджикистана и т. д., ТИЗ УзССР, Таш-
кент, 1932.
Эленгорн Я. Е. О различии хромомеров у аллеломорфных спутничных хромозом
Allium сера, ДАН, т. 27, 1940, № 4.
Эленгорн Я. Е. Критический пересмотр митоза в связи с учением о спиральной,
структуре хромозом Ботанический журнал СССР, т. 25, 1940, Ns 3.
Эмме Е. К. Результат цитологического исследования некоторых видов Aegilops
Русское общество, т. 8, 1923.
Эмме Е. К. Материалы к цитологи ячменей, „Русское ботаническое общество*,
т. 9, 1924.
Эмме Е. К Материалы к цитологии рода Secale L., Труды по прикладной ботанике
генетике и селекции, т. 17, вып. 3, Л.. 1927.
Эмме К. А. Триплоидные гибриды стойких к фитофторе видов Solatium Antipo-
viczii Вис. sp. coll., „Биологический журнал*, т. 1, вып. 5, 1936.
Юрцев В. Н. Особенности новообразований и стадийного развития придаточных
почек и побегов у многолетних травянистых и древесных растений. Тру-
ды по прикладной ботанике, генетике и селекции, т. 32, вып. 1, Л., 1955.
Яковлев М. С. Однодольность в свете данных эмбриологии, „Советская ботани-
ка*,, т. 14, 1946, К° 6.
Яковлев М. С. Структура эндосперма и зародыша злаков как систематический
признак, Труды Ботанического института Аг! СССР, сер. 7, вып. 1,
М.-Л., 1950.
Яковлева С. В. Кариологические исследования некоторых видов Salvia, Труды
по прикладной ботанике, генетике и селекции, сер. 1,5, Л., 1933.
Я нише век ий Д. Е. Из жизни тюльпанов на нижней Волге, „Советская ботани-
ка*, 1934, № 3.
Яркова А. С. Гипотеза защитных веществ в растительном организме, „Советская
ботаника*, 1939, № 3.
Abraham A. Chromosome structure and the mechanicus of mitosis and me iosis,
Annals of Botany, vol. 3,11, 1939.
Anderson Edgar. Intregressive hibridisation, London, 1949.
Bailey L. N. The standard cyclopedia of horticulture, London, 1927.
Baker J. D. Revision of the genera and species of Tulipeae, Journ. Linn. Soc. Bot.,
London, 14, 1874.
Barth T., Weinhausen К., Parey B. Die kulture der blumenzwiebeln und knol-
len, ihre vermehrung, Anzucht und Behandlung, 1954.
Beal J. Cytologikal studies in relation to the classification of the genus Calachortus,
Bot. gas. 100, 1939.
Belling J. The iron-aceto-carmine method of fixing and staining chromosomes, Biol.
Bull., 50, 1926.
Bergner A. D. Geographikal, distribution of ,chromosome types in Datura MeteP
American Journ. of Botany, vol. 30, 3 1944.
Boissier E. Flora Orientalis, V, Geneva, Basle and Lyons, 1882.
Brittingham W. Cytological studies on some genera of the Iridaceae Americ., Journ
of Botany, vol 21,1, 1934.
Bruun H. D. Cytological studies in Primula, Uppsala, 1932.
Burrell P. C. Root tip metod for difficult material, Stain Tech. 14, 1939.
Buxbaum F. Die Entwicklungs linien der Liliodeas., 11, Die Wurmbacoideae, Bot.
Archiv. 38, 1936.
Buxbaum F. Die Entwicklungs linien der Lilioideas. 11, Die Sistematische stellung
der Gattung Gagea. Bot. Archiw. 38, 1937.
404
Crane M. В., Lawrence W. J. C. The genetics of Garden Plants, London, 1952.
Dark S. O. S. Crotnosome studies in Scilleae. 11, Journ. Genet., 29, 1934.
Darlington C. D. The bechavior of polyploids, Nature, 1927.
Darlington C. D. Chromosomes and plant breding, Macmillan, 1932.
Darlington C. D. Recent Advances in Cytology, Churchile, 2, 1937.
Darlington C. D. The Handling of chromosomes. London, 1942.
Darlington С. B. and J a n a к i A m in a 1 K. Chromosome atlas of cultivated plants
London, 1945.
Darlington C. D. and Wylie A. P. Chromosome atlas of flowering plants,.
London, 1956.
D у c e s W. R. Notes on tulip species, London, 1930.
Everett T. H. The american Gardeners Book of Bulbs, New York, 1954.
Fedtschenko B. A. Species of wild flowers of the USSR, Moscow, Lektechsyrio
corporation, 1935.
Fernandes A. Estudos nos cromosomas das Liliaces e Amarilidaceas, Bol. de Soc.
Broteriana 7. 1930.
Foerster Karl. Der steingarten der sieben Yahreszeiten, Leipzig, 1954.
Frylink Adrian. The 1 ulip, Part 1, Its early history, The Garden Journal of the-
New York Bot. Garden, vol. 4, 1, 1954.
Frylink A drian. The Tulip, Part II, The development of the modern Tulip, The
Garden Journal of the New York Bot. Gard., vol. 4,2, 1954.
G a i s e r L. O. A list of chromosome numbers in angiosperms, Genetica, 8, 1926.
Gai s er L. O. Chromosome numbers in angiosperms II, Bibliographia genetica, 6
1930.
G a i s e r L. O. Chromosome numbers in angiosperms III, Genetica, 10, 1930.
G a i s e r L. O. Chromosome numbers in angiosperms IV, Bibliographia Genetica, 10,
1933.
Goodsped T. H., UberF. M. and Avery P. Chromosomes structurein Lilium
longiflorum Univ. Calif. Pub. Bot., 28, 3, 1935.
G n e у C. N. Hardy bulbs, vol. Ill, London, 1938.
Griffiths D. Tulips, U. S. Deport, of Agric. Wash. D. C. Cirk., 372, 1936.
Guiliermond A. Mangenot G., Plantefol L. Traite De Cytologie vegetale,
Paris, 1933.
G i 11 i A. Neue Liliaceen und Iridaceen an Afghanistan. Feddes Repert. spec. Now
regni weget. 2, 1954—1956.
Hagerup O. Morphological and Cytological studies of Bicornes, Dansk Botanick
Arkiv, 6 bind, i928.
Hagerup O. Uber Polyploidie in beziehung zu Klima, Okologie und Phylogenie
Chromosom Zahlenaus timbuctu. Hereditas, 16, 1932.
Hagerup O. Studies on polyploid ecotipes in Vaccinium fuliginosum L. Hereditas
(Lund) 18, 1933.
Hall A. D. The book of the Tulip, London, 1929.
Hall A. D. Polyploidy in tulips, Gard. Chron. Ill, 1930.
Hall A. D. Polyploidy in Tulipa, The Linnean Soc. Journ, Bot. vol. 50, 335, 1937.
Hall A. D. The genus Tulipa, London, 1940.
Hargrave J., Tompson and Wood. Size as a factor in the chemical composition
and morphological structure of tulips bulbs, 18 (4), 1941.
Heilborn O. Cromosome numbers and taxonomy, Proc, internal. Congress of plant
sei Itaca, 1929.
Heitz E. Der Nachweis der Chromosomen, Zeitsch. f. Bot. 18, 1926.
H el Iyer A. G. Tulips with a difference, Amateur Gardening, 73, № 3740, 4, 1956,
Hsu Slang. Structure of somatic chromosomes in Lilium tigrinum, La Cellula X1L
1932.
Logansen D. A. Plant Embriology Waltham, Mass., USA, 1950.
Ingwersen Will. Using Bulbs in the Rock Garden, Gardens et Gardening, London
et New York, 1951.
Irmisch Thilo . Zur Morphologie der monokotylischen knollen und zweibelgewa-
chse, Berlin, 1850.
Ishikawa M. A list of the numbers of chromosomes, Bot. Mag. 30, Tokio 1916.
Kimura Yojiro. Sisteme et phylogenie des monocotyledones, Rapp, et commons
Huiteime congr. internal, bot., Paris, sec. 2, 1954.
Kirchner O., Loew E., SchrOter C. Lebensgeschichte der Bliitenpflanzen Mitte-
leuropas, Stutgart, 1914.
Kermit Olson. Want early spring color, plan your bulb planting now, Minnesota,
Horticulturist, 81, № 8, 1953.
Let о u r L. Improvements in everyday technique in plant cytology, Journ. Roy. Mic-
rosc. Soc. 51, 1931.
Lei er Emile. Les Tulips de Г Europe, 1884.
Levan A. Cytological studies in Allium, A preliminary note. Hereditas, 15, 1931.
405
Levan A. Cytological studies in Allium II, Hereditas, 16, 1933.
Matsura H. On karyo-ecotypes of Fritillaria camschatkensis (L) Ker. Gawler. Journ.
of the Faculty of Science, ser. Y, vol. Ill, № 5, 1935.
Meyer Canon H. Rollo. The tulip. Gardens and Gardening, London, 1951.
Mol W. E. de-. Het celkundig erfellyk on der zoek in dienst van de veredeling der
hyacinten, narcissen en tulipen. Genetica. 7, 1925.
Mol W. E. de-. Duplication and quadriplication of the generative nuclei of Tulips.
Genetica, 9, 1927.
Mol W. E. de-. Zusammenfassung der cytologlschen und genetischen Ergebnisse des
Versuche sur Dublizierung und Quadriplizierung von Sexualkernen bei
Hyacinten und Tulipen, Lehre, 48, 1928.
Mol W. E. de-. The originating of diploid and tetrapioid pollen grains in Due van
Thol-Tulips (Tulipa suaveolens) dependandent on themetod of culture app-
lied, Genetica, XI, 1928.
Mol W. E. de Producing at will of fertile diploid and tetrapioid gametes in Due
van Thol, Scarlet (Tulipa suaveolens Roth.). Tetschrift, Haus Schinz, 1928.
Al ol W. E. de-. Die Entstehung weise anormaler Pollenkorner bei Hyazinthen, Tuli-
pen und Narzissen, Cytologia (Tokyo), 5, 1933.
Aloor Nicolas. Historic tulips. Amateur Gardening, 73, № 3743, 4, 1956.
M о r i n a g a T., F u к u s h i m a E„ К a no T., M a r ny a m a Y. and Yamasaki.
Chromosome numbers in cultivated plants, Bot. Mag. 43, 1929, Tokyo.
Miintzing, Arne. Hybrid incompatability and the origin polyploidy, Hereditas, 18,
1933.
Nakajama G. The chromosome numbers in the cultivated and wild angiospermes,
Bot. Mag., Tokyo, 45, 1931.
Newton W. C. F. Chromosome studies in Tulipa and some related genera, Journ.
Linn. Soc. 47, 1927.
Newton W. С. T. and Darlington C. D. Meiosis in a triploid tulip, Nature, 120,
№ 3009, 1927.
Newton W. С. T. and Darlington C. D. Meiosis in polyploids, Part I Triploid.
and pentapioid and tulips Part II. Aneuploid hyacinth es. Journ. Gene-
N e w t о n W. C. F. and Darlington C. D. Meiosis in polyploid Tulips, Journ
Gen. 19. 1929.
Nisbet A. Bulb propagation in the Hebrides, Commerc. Grower, 3, 1956.
O’ Mara J. Division of the generative nucleus in the polen tube of Lilium, Bot. Gas.
94, 1933.
Pamela Ai a n d e F. A list of chromosome numbers of species of British flowering
plants, New Phytol., 38, 1935.
R e g e 1 E. Descriptiones plantarum novarum in regionibus Turkestanicis ad viris Fed-
chenko, Korolkow, Kuschakewicz, et Krause collectis cum adho tationibus ad
plantas vivas in Horto botanico Petropolitano cultas, Fasc. I, Act. Hort. Petr,
vol. II, 2, 1874.
Regel Enumerate specierum hucusque cognitarum generis Tulipae, Act. Hort. Petr.
2, 1873.
Regel E. Descriptions plantarum novarum et minus cognitarum vol. VIII, Fasc. IX, 3,
1884.
Regel E. Description plantarum nonularum. Horti botanici in statu vivo examinatarum,
vol. X, 1, 1887.
Regel E. Descriptiones et emendationes plantarum in Horto botanico Petropolitano
cultarum. Acta Hort. Petrop., vol. X, 2, 1889.
Sakamura T. Kurze Mitteilung fiber die Chromosomenzahlen und die Verwandtsc-
haftsverhaltnisse der Triticum — Arten, Bot. Magaz., Tokio, vol. 32, 1918.
Schlechtendal D. T. L. Uber die Monstrositat der gemeinen Garten tulipen (Tuli-
pa Gesneriana L.)— Linnea Easter Band. 1826.
Selden M. AL The ups and downs of tulips, bulbs, Nation horticult. Mag. 21, 1, 1942.
Square Vincent. A classified list of Tulips names. The Royal Horti cultural Socie-
ty, London, s. w. 1. Krelagehnis, Haarlem. Holland. The Generaldutsch
bulbgrowers Society. 1952.
Scharp Lester W. Fundamentals of Cytology. N.—Y. London. 1943.
Simons ArthurJohn. Bulb growing, London, 1955.
Stephen G. Cutting, Forcing bulbs in Homme, The Garden Gournal of the New
York Botanikal Garden, vol. 4, 6. 1954.
Stewart Robert N. The morphology of somatic chromosomes in Lilium, American
Journal of Botany, vol. 34, № 1, 1947.
Stout A. B. Tulip droppers. Journ. of the International Garden Club, vol. Ill, 3, 1919,
New York.
Sugiura T. A list of chromosome numbers in Angiospermous plants, Botan. Magaz.,
vol. 45, 1931.
406
Sugiura T. A list chromosome numbers in Angiospermous plants III, Botan. Magaz.,
vol. 51, № 606, 1937.
Syngle Patrick M. Dictianary of Gardening, Oxford, 1956.
T а у 1 о r W. R. Chromosome morphology in Fritillaria, Alstroemeria, Silphyum, and
other genera. Amer. Journ. Bot. Xiii. 1926.
Tischler G. Allgemeine Pflanzen Karyologi, Berlin, 1921.
Tischler G. Pflanzenche Chromosomen zahlen IV. Tabule Biological, vol. XVI, 8,
1938.
Thibodean Jean. Adventures with, the species tulips, Horticulture 34, № 9, 1956.
Trankowshy D. Zytologische Beobachtungen uber die Entwicklung der Pollenscha-
cheli higer Angiospermen, Arch. fur. wiss. bot. Abt. E. 12, 1930.
Troll W. Praktische Einfuhrung in de Pflanzenmorphologie, Yena, 1954.
Upcott M. Chromosome of the Tulip in mitosis, Nature, vol. 135, 1935.
Upcott M. The mechanics of mitosis in the pollen-tube of Tulipa, Proceding of the
Royal Society of London, ser., В № 822, vol. 121, 1936.
Upcott M. The genetic structure of Tulipa, II Structural Hvbridity, Journ. of genet.,
vol. 34, 2, 1937.
Upcott M. and La Cour L. The genetic structure of Tulipa, I A chromosome sur-
vey., Journal of Genetics., vol. 33, № 2, 1936.
Upcott M. and Philip James. I'he genetic structure of Tulipa, IV balance section
and fertility, Journ. of Genet, vol. 38, N 1—2, 1939.
Vvedensky A. Tulipae et Junones novae, B3, 1935.
Wansch er J. H. The basic chromosome numbers of the higher plants, The New
Phytologist, 33, 1934.
W a r m к e H. E. A permanent root tip smear metod, Stain Tech., 10, 1935.
Wiedmer T. Wild tulipen Zanber. Schweliz, Carten 24, № 9, 1954.
Woods M. W. The nucleolus in Tulipa, Amer. Journ. Bot. 24, 1937.
"Woods M. and Bamford R. Chromosome Morphology and number in Tulipa, Amer.
Journ. of Botan., vol. 24, № 4, 1937.
Wright Murray. The garden of ten million bulbs, The Garden Journ. New York
Botan. Garden, vol. 7, № 2, 1957.
ОГЛАВЛЕНИЕ
Введение........................................ _...................... 5
Глава I. Краткий обзор исследований и изученность среднеазиатских тюльпанов 9
Глава 11. Общая морфология тюльпанов..................................... 21
Глава III. Морфологическое описание диких тюльпанов СССР................. 32
Глава IV. О центрах формообразования тюльпанов в Средней Азии .... 143
Глава V. Кариосистематические исследования среднеазиатских тюльпанов . . 154
Глава VI. Полиплоидия у тюльпанов....................................... 221
Глава VII. Семя тюльпана и его прорастание........................... . 227
Глава VIII. Рост и развитие сеянцев . •................................. 253
Глава IX. Рост ювенильных листьев и генеративных побегов у Т. kaufman-
niana и Т. greigii после появления их над уровнем почвы................. 279
Глава X. Годовой цикл развития взрослой луковицы тюльпана............... 28Z
Глав а XI. Изменение тюльпанов под влиянием внешних условий............. 351
Литература........................................................... 394-
Зинаида Петровна
Бочанцева
Тюльпаны
Редактор
М. Рахманова
Оформление художника
В- Гий
Цветные рисунки выполнены
Г. А. Семеновым и М. К. Луканиной
Художественный редактор
А. Т. Шепельков
Технический редактор
3. П. Горъкоеая
Корректор
Ф. М. Денисова
* *
¥
Р04282. Сдано в набор 21/ХЦ-61 г.
Подписано к печати 22/1П-62 г. Бумага
70xl08I/ie=12-75 бум. л. -34,94 печ. л.
Уч.-изл. л. 38,3 (24 вкл.). Изд. № 2807.
Тираж 2000. Заказ № 2229.
Цена 3 р. 4b к. Переплет 20 к.
* , #
*
Типография Объединенного издательства
„Цизил ^збекистон", „Правда Востока"
и „Узбекистони Сурх". Ташкент —1962.