/




















































































Текст
ЗООЛОГИЧЕСКИЙ ИНСТИТУТ АКАДЕМИИ НАУК СССР
НОВАЯ СЕРИЯ № 3
ФАУНА СССР
ГУБКИ
Т. II, выл. 2
ПРЕСНОВОДНЫЕ ГУБКИ
Составил
П. Д. РЕЗВОЙ
ИЗДАТЕЛЬСТВО АКАДЕМИИ НАУК СССР
МОСКВА • 1936 - ЛЕНИНГРАД
INSTITUT ZOOLOGIQUE DE L’ACADEmIE DES SCIENCES DE L'URSS
NOUVELLE SERIE n« 3
FAUNE DE L’URSS
Sous la direction de S. A. SERNOV, Membre de I’Academie Redige par A. A. STACKELBERG
PORIFERA
Vol. II, n° 2
Fam. SPONGILL1DAE et LUBOM1RSKIIDAE
par P. D. REZVOJ
EDITION DE I’ACADEMIE DES SCIENCES DE 1’URSS MOSCOU • 1936 • LENINGRAD
ЗООЛОГИЧЕСКИЙ ИНСТИТУТ АКАДЕМИИ НАУК СССР
НОВАЯ СЕРИЯ № 3
ФАУНА СССР
Главный редактор акад. С. А. ЗЕРНОВ Редактор А. А. ШТАКЕЛЬБЕРГ
ГУБ К И
Т. II, вып. 2
ПРЕСНОВОДНЫЕ ГУБКИ
(Сем. SPONGILLIDAE и LUBOMIRSKIIDAE)
Составил
П. Д. РЕЗВОЙ
ИЗДАТЕЛЬСТВО АКАДЕМИИ НАУК СССР
МОСКВА • 1936 • ЛЕНИНГРАД
Напечатано по распоряжению Академии Наук СССР Декабрь 1935 г.
Непременный секретарь академик Н. П. Горбунов
Редактор издания А. А. Штакельберг
Технический редактор и ученый корректор Н. Г. Редько
Сдано в набор 8 октября 1935 г. — Подписано к печати 27 декабря 1935 г.
VIII -+- 126 стр.
Формат бум. 72 X НО см. — 10 печ. л. — 50 371 зн. в печ. л. — Тираж 2170
Ленгорлит № 36758. — АНИ № 668. — Заказ № 2760
Типография Академии Наук СССР. Ленинград, В. О., 9 линия, 12
ПРЕДИСЛОВИЕ
Пресноводные губки составляют- небогатую видами группу водных животных. По своему образу жизни они принадлежат к сидячему зообентосу, который в пресных водах представлен очень бедно в сравнении с морем и состоит преимущественно из микроскопических форм. Губки вместе со мшанками являются наиболее существенным и заметным элементом пресноводного зообентоса.
Пресноводные губки у нас сравнительно мало останавливали внимание зоологов и гидробиологов. Поэтому в настоящем издании мы сочли желательным предпослать систематическому описанию губок более пространный очерк их строения и жизнедеятельности. Этот очерк составляет первую часть работы и в него входят данные по анатомии, развитию, физиологии губок и их биоценотическим отношениям; здесь же рассматривается и их практическое значение. В дополнение к этой части даются указания для собирания и исследования губок.
В систематической части описанию семейств, родов и видов предпосылаются определительные таблицы, в которые введены кроме губок, известных в пределах Союза, также и в5е европейские формы и некоторые азиатские, нахождение которых в пределах Союза является возможным. Знаком * отмечены констатированные для Союза формы.
Как дополнение к описанию современных губок и их географическому распространению, приводятся данные по ископаемым пресноводным губкам и в табличной форме даются указания по определению губок из современных и субфоссильных озерных отложений по изолированным спикулам.
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ ВИДОВ
ОТРЯД CORNACUSPONGIDA
Сем. I. Spongillidae
Род 1. Spongilla Lamark Стр.
1. Sp. lacustris L................................................................. 64
1 a. Sp. lacustris var. jordanensis Vejd. '............*............... . 66
l b. Sp. lacustris var. lacustroides Koshow................................. 66
1 c. Sp. lacustris var. paupercula Bow. . •....................................66
2. Sp. arctica Ann. . . •...................................•..................... 66
3. Sp. biseriata Welt.......................•............ • ...............•• • 67
4. Sp. fragilis Leidy.............................................................. 69
4a. Sp. fragilis var. rectituba Koshow......................................... 70
4 b. Sp. fragilis var. brevipora Grim......................................... 70
5. Sp. carteri Cart.........................’....................................... 70
5 a, Sp. carteri var. rotundacuta Rezvoj..................................... 72
5 b. Sp. carteri var. mollis Ann............................................. 72
5 c. Sp. carteri subsp. balatonensis Arndt. •................................ 72
6. Sp. stanleyi Ann.........................•.............................. ..... 73
Род 2. Ephydatia Lamouroux
1. Eph. fluviatilis L.............................................................. 75
la. Eph. fluviatilis var. teberdana Rezvoj .................................... 76
lb. Eph. fluviatilis var. hastifera Rezvoj ............................. ... 76
2. Eph. meyeni Cart.............. . ............................ •.................. 77
3. ЕрЫ miilleri Lieb............................................................. 77
3a. Eph. miilleri var. acuminata Potts ........ ................... 79
3 b. Eph. miilleri var. sibirica Koshow........................................ 79
3c. Eph. miilleri var. obtusisclera Koshow..................•.................. 80
Род 3. Heteromeyenia Potts.
1. H. baileyi Bwk.................................................................. 82
la.-H. baileyi var. stepanowi Dyb............................................. 82
lb. H. baileyi var. petri Lauterb...........................;.................. 84
1 с. H. baileyi var. repens Potts ............................................. 84
VIII
Стр.
1 d. Н. baileyi var. arndti Sqhr. ..................................... 84
le. H. baileyi var. pah tina Lauterb.................................. 85
1 f. H. baileyi var. bohemica Petr. . . . • ........................... 85
Род 4. Trochospongilla Vejd.
1. Tr. horrida Welt......................................................... 85
Сем. IL Lubomirskiidae
Род 1. Lubomirskia Dyb.
L. baicalensis Dyb.......................................................... 91
1 a. L. baicalensis m. littoralis Rezvoj............................... 92
2. L. fusifera Souk............ ..................... 93
3. L. abietina Sw. . ................................................... . 94
Род 2. Baikalospongia Ann.
1. B. bacillifera Dyb. ... ............................................... 96
2. B. intermedia Dyb............................................... ..... 98
2a. B. intermedia m. profundalis Rezvoj............................... 98
3. B. dzhegatajensis Rezvoj ............................................. 99
Род 3. Sivartschewskia Mak.
1. Sw. papyracea Dyb..................................................... 101
ОБЩАЯ ЧАСТЬ
„Жизнь этих существ не сложна и состоит главным образом в процеживании воды со взвешенными в ней пищевыми веществами".
Н. А. Холодковский.
ВНЕШНИЙ ВИД И МЕСТООБИТАНИЕ ПРЕСНОВОДНЫХ ГУБОК
Пресноводные губки имеют вид неправильных наростов или корок, покрывающих различные подводные предметы. Поверхность редко бывает более или менее гладкой; обычно она покрыта неровностями и буграми. Молодые губки представляют тонкие корочки в 2—3 мм толщиной и 10—20 мм в поперечнике. С возрастом толщина корки увеличивается и достигает нескольких сантиметров. Большинство видов встречается только в виде таких корок, но у некоторых от коркового основания поднимаются ветвистые выросты, достигающие иногда значительной длины. Таков, например, внешний вид Spongilla lacustris-, близкая ей .S'. arctica имеет более тонкие и короткие выросты.
Размеры отдельных колоний зависят от их возраста. Старые экземпляры образуют желваки до нескольких килограммов весом. Растущие по соседству колонии могут срастаться друг с другом. Таким путем в благоприятных условиях губки образуют иногда сплошные покровы; говорить тогда о величине отдельных колоний уже не приходится.
Внешность губок очень характерна, но словесное описание мало даст тому, кто не видел губок в натуре. Внимательно рассматривая поверхность губки, мы видим мелкие бугорки и тонкую полупрозрачную пленку, обволакивающую всю наружную поверхность; сквозь эту дермальную мембрану темными пятнами просвечивают многочисленные полости. На многих местах эта пленка может быть легко отделена при помощи игол-Ткань губки легко разрывается, и на разломе мы видим шероховатую, ноздреватую слизистую массу, всю пронизанную неправильными полостями и каналами. Консистенция ткани различна у различных видов и вариирует также в пределах одного и того же вида. Наиболее прочная ткань имеется у кустистых форм, например у Spongilla lacustris. В корках 5. fragilis ткань слабая и ломкая. S. carieri иногда отличается очень слабой
Фауна СССР, Губки, т. П, в. 2 1
2
ФАУНА СССР
и дряблой слизистой тканью (var. mollis). При растирании кусочка губки между пальцами всегда чувствуется некоторая шероховатость, зависящая от присутствия скелета. Этим ткань губки отличается от плюшек нитчаток или от скоплений диатомовых водорослей. На разломе или разрезе губки уже простым глазом можно заметить рассеянные в ткани мелкие шарообразные зерна около полумиллиметра в поперечнике. Это геммулы, покоящиеся почки, предназначенные для переживания неблагоприятных сезонов. У некоторых видов они собраны в основании губки, где на поверхности субстрата образуют сплошной слой. Как мы увидим в дальнейшем, присутствие геммул очень важно при систематическом определении родов и видов.
Губки, свойственные Байкалу (сем. Lubomirskiidae), сильно отличаются по внешности от повсеместно распространенных пресноводных губок или бадяг1 (сем. Spongillidae). Среди байкальских губок также преобладают корковые формы, и только один вид, Labomirskia baicalensis, образует высокорастущие, достигающие метра высоты, кустистые формы, состоящие из толстых выростов. Ткань этой губки плотная и упругая и разрывается с некоторым усилием. У других байкальских видов ткань более ломкая, но всегда более плотная, чем у бадяг. Очень существенным их отличием является также отсутствие геммул (стр. 41).
Для всех вообще губок как пресноводных, так и морских, очень характерен своеобразный резкий и неприятный запах. Чем он вызывается, пока еще не выяснено.
Почти всем пресноводным губкам, выросшим на свету, свойственна ярко зеленая окраска. Она зависит от симбиотических одноклеточных водорослей, „зоохлорелл", как их обычно называют, заключенных в протоплазме губочных клеток. У губок, выросших в тени, этой окраски не бывает. Часто на одной и той же колонии только верхняя часть оказывается зеленой. Кроме зеленого, губки могут быть грязнобелого, желтосерого, бурого или красноватого цвета. На их цвет имеет большое влияние присутствие илистых частиц, заносимых во внутренние полости. Губки, выросшие в очень мутной воде, могут быть почти черными. Цвет геммул бывает желтый, бурый или черный, реже зеленоватый.
Байкальским губкам также свойственна зеленая окраска. Lubomirskia baicalensis имеет яркий травянозеленый цвет. Различные виды, растущие в прибрежной зоне, отличаются различными оттенками зеленого. Губки, добытые с более значительных глубин, лишены зеленой окраски. До
1 Народное название пресноводных губок — бадяга или бодяга (а также водяга, виша, надожник) (Даль, 1880). К байкальским губкам, насколько мне известно, местное население не применяет этого названия (их называют „морская губа"). Слово бадяга мы будем употреблять для обозначения сем. Spongillidae, противополагая им губок сем. Lubomirskiidae. Слово бадяга привилось и в Германии (das Badiaga-Pulver, die „Badiaga Tinctur"). Это название в 1729 г. было введено в науку Буксбаумом (Buxbaum, 1729), и род Badiaga фигурирует и в современных сводках в списке синонимов.
ПРЕСНОВОДНЫЕ ГУБКИ—ОБЩАЯ ЧАСТЬ 3
последних лет не было нахождений байкальских губок глубже 200 м. В 1932 г. впервые добыты губки со значительных глубин: Baikalospongia bacillifera с 533 м, В. intermedia' (выделенная как morpha profundalis, стр. 98) с 889 м. Эти две губки, по словам сборщиков и по записям в журнале, отличались своеобразным ярким сине-зеленым цветом.
Встречаются губки в чрезвычайно разнообразных водоемах, от крупнейших озер и рек и до прудов и ручьев, следовательно, и в стоячих, и в текучих водах. Но далеко не в каждом водоеме мы обязательно найдем губки, хотя у них имеются широкие возможности расселения благодаря геммулам. Очевидно, в каждом отдельном случае их отсутствие вызывается какими нибудь особыми неблагоприятными моментами. Так, в горных речках, где по дну перекатывается галька, губки отсутствуют. Их развитию также мешает сильное заиление в мелких озерах или прудах а также обилие слизистых наростов сине-зеленых водорослей (напр. сем. Nostocaceae). Очень часто нам не ясны причины, мешающие развитию губок в том или ином водоеме. В городе Бухаре мне удалось их найти только в одном из многочисленных городских прудов („хаузов"), хотя трудно себе представить водоемы, более сходные во всех отношениях, и при этом одновременно наполняющиеся одной и той же водою.
Пресноводные губки могут уживаться не только в пресных водоемах. Они известны и в опресненных морских заливах (Финский залив), в лагунах и реликтовых озерах, а также и в континентальных солоноватых водах (оз. Иссык-куль).
Губки поселяются на очень разнообразном субстрате. Кажется, нет в воде такого твердого предмета, на котором не могли бы разрастаться губки. Прибрежные камни и скалы являются излюбленными местами, причем некоторые виды определенно предпочитают нижнюю поверхность камней. Очень часто они пышно развиваются на камнях и деревянных сооружениях, дамбах, плотинах, шлюзах, сваях мостов; поселяются таюйе на днищах барж и на плотах. Очень удобно собирать губки, поднимая затонувшие коряги и хворост.
Мы находим губки и на живых растениях, на свисающих в воду ветвях и корнях прибрежных деревьев и кустов, а также на подводной травянистой растительности. Довольно часто можно видеть губки и на раковинах моллюсков, а также на домиках ручейников.
Гораздо реже губки поселяются прямо на илистом или илисто-песчаном грунте; в таких случаях скорее всего можно думать, что губки были сорваны со своего субстрата и продолжали расти, лежа на мягком грунте. Случаи роста на грунте без прикрепления к твердому субстрату известны и среди байкальских губок. Но здесь дело, повидимому, не в случайных отрывах, а в особом способе бесполого размножения, отделения от материнского тела наружных почек, падающих на илистое дно (стр. 26).
1*
4
ФАУНА СССР
Будет ли губка расти на том или ином субстрате — зависит от того,, где прикрепится свободно плавающая личинка, или куда будет занесена геммула. Некоторые автора допускают у личинок избирательную способность. Геммулы, конечно, переносятся в то или иное место совершенно пассивно.
В крупных колониях бадяг мы в основании всегда находим старую отмершую, почерневшую ткань, покрытую молодой живой тканью. Для спонгинс?лога присутствие этой старой ткани бывает очень существенно, так как в ней всегда можно найти геммулы столь важные в систематическом отношении; в молодой же ткани их может и не оказаться.
СКЕЛЕТ И ПАРЕНХИМА
Основное студенистое вещество и клеточные элементы. Устройство ирригационной системы. Поры, приводящая и отводящая система каналов, жгутиковые камеры, оскулярные отверстия.
Рассматривая губку простым глазом, разрезая или разламывая ее, мы не находим в ней каких нибудь обособленных частей, каких нибудь заметных простым глазом органов; мы видим только шероховатое на ощупь слизистое вещество, пронизанное пустотами и каналами. Только мелкие шарики геммул останавливают наше внимание.
При изучении губки под микроскопом, при слабых увеличениях на срезах, сделанных от руки, мы отличим в ней два элемента: скелет и паренхиму. Скелет состоит из пучков, образованных кремневыми спикулами, склеенными большими или меньшими количествами спонгина— прозрачного бесструктурного органического вещества. От основания к поверхности идут главные пучки. Они соединяются друг с другом поперечными пучками, образующими между ними перемычки. Скелет пронизывает мягкое, слизистое вещество, паренхиму, и служит ей опорой. ^Кроме спикул, образующих скелет, макро с клер, у большинства видов имеются более мелкие и иначе устроенные спикулы, микро склеры, рассеянные в паренхиме или приуроченные к оболочкам геммул.
При малых увеличениях микроскопа и на срезах, сделанных от руки, мы не сможем рассмотреть структуру паренхимы. Для этого необходима более сложная гистологическая методика, приготовление срезов на микротоме, их окраска и изучение при сильных увеличениях. Рассматривая соответствующим образом приготовленный препарат, мы отличим в паренхиме основное студенистое вещество, мезоглею, и клеточные элементы, выстилающие многочисленные полости и рассеянные внутри студенистого вещества. Но можно видеть клеточные элементы и без сложной обработки. Для этого надо маленький кусочек живой или фиксированной губки расщипать иглами в капле воды на предметном стекле. При этом будут изолированы в большом количестве отдельные клетки.
ПРЕСНдвОДНЫЕ ГУБКИ— ОБЩАЯ ЧАСТЬ 5
Все тело губки, как мы уже указали, пронизано мночисленными полостями. Рассматривая поверхность разреза губки, мы получаем впечатление полного беспорядка в их расположении. Мы сможем разо
браться во взаимных связях этих полостей, и каналов только на полной серии не слишком тонких срезов. Реконструируя по срезам эти полости, мы сможем проследить их ход и соединения в теле губки, что представляет далеко не легкую задачу. Вся ткань губки сводится к сравнительно тонким перегородкам между пустотами, пронизывающими губку. Все эти полости входят в со
став важнейшей и, можно сказать, единственной системы органов губки — ирригационной системы (фиг. 1). Эта система естественно делится на две части — приводящую и отводящую.
Приводящая система начинается многочисленными порами на поверхности губки, пронизывающими дермальную мем-б рану, одевающую все тело. У бадяг эта мембрана натянута между концами скелетных пучков и почти на всем своем протяжении отделяется от ниже лежащей ткани субдермальными полостями. Только на небольших участках субдер-.мальные полости отсутствуют.
Фиг. 1. Схема строения ирригационной системы бадяги. дм — дермальная мембрана; п — поры; ск — скелетный пучок; сдп — субдермальная полость; вопк — входные отверстия приводящих каналов; пк — приводящие каналы; жк — жгутиковые камеры; ок—отводящие каналы; у — устье или оскулярное отверстие; от — оскулярная трубка. По ван-Тригту (1919).
У байкальских губок субдермальные
полости не развиты, и дермальная мембрана не отделяется от ниже лежащей ткани. Поры здесь непосредственно ведут в приводящие каналы. У бадяг приводящие каналы начинаются отверстиями на дне субдермаль-ных полостей.
Приводящие каналы представляют совокупность очень неправильных полостей и протоков, разветвляющихся и пронизывающих все тело губки. Тонкие стенки всюду отделяют приводящую систему каналов от сходной с ней отводящей системы, и между этими системами нигде нет непосредственной связи. Их сообщение достигается только при посредстве особых образований, жгутиковых камер, расположенных в толще перегородок, разделяющих обе системы (фиг. 2). Жгутиковые камеры представляют важнейший орган губок. Это очень мелкие округлые полости 25—50 р. в поперечнике,1 выстланные жгутиковыми воротничковыми клетками
1 (Л — микрон, 0.001 миллиметра
6
ФАУНА СССР
Фиг. 2. Относительное расположение приводящих каналов (пк), жгутиковых камер (жк) и отводящих каналов. По ван-Тригту (1919).
(фиг. 3). Эти полости при помощи нескольких мелких отверстий, прозопилей, соединяются с приводящей системой и при помощи одного' более крупного отверстия, апопиля, — с отводящей системой. Движение жгутов в жгутиковых камерах представляет тот двигатель, который создает непрерывный ток воды через все тело губки. О механизме этого тока мы подробно будем говорить далее.
Отводящая система, как мы сказали, сходна по своему устройству с приводящей. Отводящие каналы, постепенно сливаясь друг с другом, образуют протоки все более и более крупного сечения. Наиболее крупные каналы, часто называемые клоакальными полостями или протоками, подходят к поверхности губки, идут под. дермальной мембраной и впадают в секулярные отверстия, или устья, которыми отводящая система сообщается с наружной средой (фиг. 1).
У бадяг клоакальные протоки, просвечивающие^ сквозь дермальную мембрану, идут непосредственно под нею на значительном расстоянии. Подходя с разных сторон к оскулярным отверстиям, они образуют довольно крупные, в несколько сантиметров в поперечнике, неправильные звездообразные фигуры (табл. И, фиг. 6). Клоакаль
ные полости, проходящие под дермальной мембраной, мы можем также считать субдермальными полостями. Тогда часть субдермальных полостей будет относиться к приводящей, другая часть — к отводящей системе. Между теми и другими, конечно, нет прямого сообщения.
Края оскулярного отверстия у фиксированной бадяги не приподнимаются или только слабо приподнимаются над поверхностью губки. Но если мы живую бадягу посадим в сосуд со свежей вбдой, то через некоторое время мы увидим, что от оскулярного отверстия вытягивается оскулярная трубка до 10 мм длины, состоящая из тонкой просвечивающей пленки, подобной дермальной мембране (фиг. 4; см; также табл. II,. фиг. 4).
Совершенно другое устройство отводящей системы мы находим у байкальских губок. Оскулярные отверстия у них представляют очень неглубокие круглые ямки в 4 — 5 мм в поперечнике, затянутые тонкой пленочкой с одним или несколькими отверстиями на дне ямки. Под этой пленочкой, просвечивая сквозь нее, сходятся из глубины ткани тонкие отводящие каналы, образуя маленькую звездообразную фигуру, не выходящую за пределы оскулярной ямки (см. табл. XV, фиг. 62). Можно думать, что пленочка, затягивающая дно оскулярной ямки, не может вытягиваться в оскулярную трубку, но никто не наблюдал байкальских губок в аквариумах. На сухих экземплярах и большею частью и на.
ПРЕСНОВОДНЫЕ ГУ БКИ—ОБЩАЯ ЧАСТЬ
7
спиртовом материале, не особенно тщательно сохраненном, оскулярные пленочки оказываются сорванными, тогда отводящие, каналы прямо открываются на дне оскулярных ямок; в таком виде обычно и описываются оскулярные отверстия байкальских губок.
Фиг. 3 Жгутиковая камера ба-дяги: ch — хоаноциты; ар—апо-пиль; рг—прозопиль. По Vos-maer и Pekelharing (1898).
Фиг. 4. Оскулярные трубки Ephydatia mulleri (пруды Старого Петергофа), слабо увеличены.
КЛЕТОЧНЫЕ ЭЛЕМЕНТЫ.
Плоские клетки дермальной мембраны и полостей. Пороциты. Воротничковые клетки. Основное студенистое вещество, мезоглея. Амебоциты и их производные.
Вся‘наружная поверхность губки покрыта плоскими клетками неправильных очертаний, тесно лежащими одна около другой и образующими подобие плоского эпителия. Эти клетки носят название пинакоцитов. Ядро располагается в средней части, где клетка несколько утолщена. Пинакоциты могут сокращаться и растягиваться, а также, по некоторым указаниям, вытягивать псевдоподии в подстилающее их студенистое вещество. Они выстилают не только наружную поверхность дермальной мембраны, но и все полости и каналы внутри тела губки. Дермальная мембрана и стенки оскулярных трубок выстланы плоскими клетками и с наружной и с внутренней стороны; между этими слоями находится прослойка основного студенистого вещества со включенными в него клетками. Изолированные пинакоциты при расщипывании или протирании губки сквозь ткань принимают округлую форму, по величине м'ельче других клеток и показывают слабые амебоидные движения.
Среди пинакоцитов дермальной мембраны разбросаны клетки, несущие поры, пороциты (фиг. 5). Пора представляет округлое отверстие в клетке, от которой остается только узкий серповидный ободок, окружающий пору, более широкий около ядра и совсем узкий на противопо
8
ФАУНА СССР
ложной стороне. Кроме присутствия пор, пороциты отличаются и другими особенностями от обычных пинакоцитов: их ядро имеет меньшую величину и сильнее красится, а протоплазма более зерниста. Иногда пора образована не одним, а двумя пороцитами. Тогда две узких серповидных клетки охватывают пору и сходятся своими тонкими концами. Сжимаясь, пороциты уменьшают просвет, но, повидимому, не могут вполне замыкать поровое отверстие (Annandale, 1907 b).
Фиг. 5. Дермальная мембрана Ephydatia rrvilleri (пруды Старого Петергофа), пц— пороциты с порами; пк — пузыревидные клетки; ядр — ядра пииакоцитов.
Для всех вообще губок наиболее характерными клетками являются хоаноциты, жгутиковые воротничковые клетки, выстилающие полость жгутиковых камер (фиг. 3). Хоаноцит представляет удлиненную клетку, более широкую у основания и несколько суживающуюся к вершине, несущую воротничек и жгут. Тело клетки может менять свою форму и выпускать псевдоподии. Ядро находится в ее нижней части. Воротничек состоит из очень тонкой плазматической мембраны. У бадяг он имеет вид высокого цилиндра. От вершины клетки внутри воротничка отходит длинный подвижной жгут, беспрестанно движущийся в полости камеры.
Мы уже говорили выше об устройстве жгутиковых камер. Хоаноциты выстилают их сплошным слоем, оставляя свободными только несколько входных отверстий, прозопилей, и отсутствуя также в районе выходного отверстия, апопиля. Ниже нам придется вернуться к хоаноцитам, к их работе и механизму водного тока (стр. 16).
Очень объемистую часть паренхимы составляет основное студенистое вещество, мезоглея, в которой заключены многочисленные клетки различной величины и устройства (фиг. 6). Большинство исследователей считают мезоглею бесструктурным веществом, выделяемым погруженными в него клетками. По отношению ко всем клеткам губки мы можем сказать, что это студенистое вещество является той средой,
ПРЕСНОВОДНЫЕ ГУБКИ— ОБЩАЯ ЧАСТЬ
9
(Lieberkiihn, 1870), мезоглея
же
.ПК
Фиг. 6. Клеточные элементы паренхимы бадяги. пк — приводящие каналы, ок — отводящие каналы, жк — жгутиковые камеры, к — различные амебоциты, я — яйцевые клетки; Р — ацинета Podophrya fixa, живущая в полостях губки. По Вельт-неру (1896).
в которой они живут. Клетки наружной поверхности и полостей, пинако-цнты и хоаноциты, заключены в мезоглею своими основаниями и по многим указаниям могут уходить вглубь студенистого вещества. Для клеток губки мезоглея играет ту же роль, что и плазма крови для кровяных клеток. Но существует воззрение, отрицающее существование бесструктурной мезоглеи, если не у всех губок вообще, то по крайней мере у Spongillidae. По наблюдениям Либеркюна при нагревании, а также под влиянием давления, может нацело распадаться на клетки? с ядрами. В том же смысле высказывается Вельтнер (Weltner, 1900), а в новейшее время ван-Тригт (v. Trigt 1919). В аргументации этих трех авторов все же много неясного. С другой стороны, по их же наблюдениям, на хорошо фиксированных препаратах вполне ясно видно однородное студенистое вещество и погруженные в него клетки.
Среди клеток, находящихся в мезоглее, в зависимости от их вида и функции, различают целый ряд категорий. Большую роль при этом играет характер зернистых включений в их протоплазме.- Мы не можем здесь останавливаться на характеристике всех этих категорий. Скажем только, что многие из них, вероятно, окажутся различными состояниями одних
фичности различных клеток, выполняющих те или иные функции, также нельзя считать вполне выясненным.
Вельтнер (Weltner, 1907) доказывает, что все эти многочисленные категории клеток являются производными одной основной формы — амебоцита или (у других авторов) археоцита, содержащего пузыревидное ядро с ядрышком и зерна различной величины в протоплазме. Амебоциты являются недифференцированными клетками, обладающими, подобно яйцевой клетке, потенциальной возможностью производить все остальные элементы. Такие клетки, так наз. интерстициальные (1-клетки), известны у многих низших животных. Средняя величина амебоцитов около 20 у. Этим клеткам свойственны оживленные амебоидные движения, что легко наблюдать при их изолировании (см. ниже, стр. 14—15).
Из клеток, генетически связанных с амебоцитами, упомянем про колленциты — крупные звездчатые или веретенообразные клетки, которым приписывается выделение студенистого вещества. О с и лик о-
и тех же клеток. Вопрос о специ-
10
ФАУНА СССР
бластах и спонгобластах, строящих скелет, мы подробнее будем говорить в дальнейшем. Своеобразно измененными амебоцитами являются пузыревидные клетки, характерные для некоторых видов (Ephydatia miilleri, Е. meyeni) (фиг. 7). Эти клетки, 20—35 [л в поперечнике, заключают вакуоль с жидкостью, занимающую почти весь их объем. Протоплазма с ядром образует как бы тонкую оболочку этой вакуоли. Содер-
Фиг. 7. Пузыревидные клеткиТЁрЛу-datia (Ст. Петергоф).
жимое вакуоли окрашивается иодом в синий цвет, что указывает на присутствие какого-то углевода.
От археоцитов берут начало и половые клетки. Из их же скоплений возникают и геммулы и сориты, но о половом и бесполом размножении мы будем говорить дальше, где подробнее рассмотрим и устройство и развитие половых клеток.
Таким образом, археоциты дают начало другим клеточным элементам тела и несут функции амебоидных клеток. С другой стороны, они исполняют роль полового зачатка, передавая от поколения к поколению наследственные особенности вида.
СКЕЛЕТ
Основные элементы скелета, спикулы и спонгин. Скелетные пучки. Устройство макро- и микросклер. Образование спикул и спонгина.
Паренхима губки, состоящая из студенистого вещества и клеток, вся пронизанная полостями и каналами, не могла бы держаться, не спадаясь, без поддержки скелета. В скелете кремнероговых губок мы различаем два элемента: кремневые иглы или спикулы и склеивающий их спонгин.
На своей подошве губка выделяет базальную мембрану из спонгина, выстилающую субстрат, на котором она сидит. От этой мембраны отходят г л а в ные скелетные пучки, состоящие из склеенных спонгином спикул. Пучки пронизывают все тело губки, ветвятся и доходят до ее поверхности. У кустистых форм пучки идут по длине отростков и, дугообразно изгибаясь, подходят к их поверхности приблизительно под прямым углом. У бадяг пучки проходят через субдермальные полости и поддерживают натянутую на их концах дермальную мембрану. На поверхности дермальной мембраны уже простым глазом видны мелкие конические возвышения, к о нули, соответствующие концам скелетных пучков.
Главные пучки бывают различной толщины, с различным числом спикул на поперечном сечении. Они соединяются более тонкими попереч
ПРЕСНОВОДНЫЕ ГУБКИ— ОБЩАЯ ЧАСТЬ
11
ными пучками, толщина которых может быть очень различна. Иногда поперечный пучок состоит из немногих, иногда только из одной спикулы. Совокупность главных и поперечных пучков образует сеть или пространственную решетку в теле губки, то более, то менее правильную.
Спйкулы построены из аморфной водной, кремнекислоты, из опала,, с которым связано известное количество органического вещества, содержащего азот (тай наз. спикулин). В середине спикулы проходит осевая нить из органического вещества с небольшой примесью кремнезема. Вокруг этой нити концентрическими слоями отлагается кремнезем. Между его слоями имеются прослойки с большим содержанием органического вещества. При прокаливании вся толща спикулы буреет, что указывает на содержание органического вещества не только в прослойках, но и в плотных слоях кремнезема. Осевая нить иногда хорошо заметна, но часто' может быть рассмотрена только с трудом. При разрушении осевой нити внутри поврежденных спикул образуется осевой канал.
Все кремнероговые губки имеют одноосные спикулы, носящие общее название рабд. У пресноводных губок чаще всего встречаются спикулы в виде прямой или несколько изогнутой иглы, заостренной с обоих концов; такие спикулы называются о к с а м и или амфиоксами. Все бадяги нашей фауны имеют макросклеры в виде оке, за исключением одного вариетета Spongilia carteri (var. rotundacuid), у которого спикулы на концах закруглены (табл. VII, фиг. 28). Такие спикулы с закругленными концами называются стронгилями; эта форма спикул свойственна некоторым тропическим представителям сем. Spongillidae. У сем. Lubo-mirskiidae мы встречаем как оксы, так и стронгили (табл. X).
Обыкновенно спикула в средней части толще и к концам постепенно утончается. В таких случаях мы говорим о ее веретенообразности. Но спикула может быть и цилиндрической, с закругленными или заостренными концами.
Длина острия у оке вариирует довольно сильно даже в пределах вида. Обычно более толстые спикулы имеют более короткое острие,, у тонких острие длиннее.
Поверхность спикул может быть гладкой или покрытой шипами.. Шипы распределяются по всей спикуле, или же концы спикулы остаются свободными от шипов (Ephydatia miilleri). Иногда они концентрируются у концов, оставляя ' среднюю часть спикулы совершенно гладкой или с редкими шипами (многие байкальские губки). По величине шипы могут быть очень различны; мелкие шипы могут быть легко не замечены при небольших увеличениях микроскопа. Из бадяг очень сильными острыми шипами отличается Trochospongilla horrida (табл. VII, фиг. 29)..
У молодых спикул, еще не достигших полного роста, наблюдается часто одна особенность; преимущественно в молодых колониях на середине иглы замечается шарообразное вздутие, которому соответствует утолщение осевой нити. Такие иглы носят название цент ротилотных.
12
ФАУНА СССР
Это свойственно как бадягам, так и байкальским губкам. При дальнейшем утолщении иглы это вздутие маскируется отлагающимися слоями кремнезема, и у выросших спикул оно редко бывает заметно.
Паренхимные и геммульные микросклеры отличаются от макросклер не только величиной, но и по устройству. Они также представляют рабды с острыми или затупленными концами. Микросклеры редко бывают гладкими. В нашей фауне только один вид, Spongilla carteri, имет гладкие микросклеры. Большею частью они покрыты шипами, то мелкими и очень многочисленными, то более крупными и редкими. У одного вида Spongilla biseriata ( — proliferens), шипы микросклер имеют интересную особенность: на своей вершине шип рассечен на несколько крючочков, образующих розетку (табл. V фиг. 18). Очень часто шипы имеют разную величину в различных участках спикулы. Так, у упомянутой S. biseriata и у Heteromeyenia baileyi (= Carterius stepanowi) у паренхимных микросклер шипы в средней части склеры заметно длиннее. Но чаще бывает и обратное отношение: шипы у концов длиннее и расположены гуще. Это часто наблюдается у геммульных микросклер Spongilla lacustris, но сильнее всего это выражено из губок нашей фауны у упомянутой Spongilla- biseriata, геммульные спикулы которой, в отличие от паренхимных, в средней части несут короткие расщепленные шипы, а на своих концах спикулы снабжены простыми шипами в виде длинных, острых, изогнутых крючков.
В этом сказывается тенденция, которая нашла свое полное выражение в своеобразных геммульных микросклерах, амфидисках, свойственных нескольким родам бадяг. Амфидиск представляет стержень с двумя перпендикулярными к нему дисками на концах. Чаще всего диски имеют вид звезды с расходящимися в виде лучей шипами. Таковы амфидиски у р. Ephydatia (табл. УШ, фиг. 31). Шипы могут отходить от самого стержня, или же они сливаются своими основаниями,, образуя пластинку с зубцами по краям. Что амфидиски следует производить от покрытой шипами рабды, видно по некоторым их уклоняющимся и уродливым формам. Так, у некоторых вариететов Ephydatia miilleri наблюдается > прорастание стержня за диски в виде острых шипов. У некоторых Ephydatia (напр. Е. fluviatilis) почти постоянно имеются шипы на средней части стержня. В уродливо сформированных амфидисках шипы часто лежат не в одной плоскости, а смещаются по длине стержня; наконец, иногда среди нормальных или уродливых амфидисков встречаются на гем-мулах и настоящие рабды, покрытые по всей длине шипами. У рода Trochospongilla происхождение диска из шипов маскируется тем, что шипы совершенно слились, и диски представляют правильные цельнокрайние кружки (табл. VII, фиг. 29с).
У форм, не свойственных нашей фауне, наблюдается ряд интересных дальнейших изменений. Один из цельнокрайних дисков становится меньше другого (некоторые виды Trochospongilla). Он может быть также сведен
ПРЕСНОВОДНЫЕ ГУБКИ—ОБЩАЯ ЧАСТЬ И
к небольшой шишечке на одном конце стержня (род Tubella). Редукция может пойти и дальше: исчезает и стержень, и амфидиск сводится к одному диску с остатком стержня в виде маленького шипика в центре (род Parmula).
Из пресноводных губок микросклеры (паренхимные и геммульные) свойственны бадягам. У байкальских губок они отсутствуют. За микросклеры у них иногда принимаются более короткие и тонкие иглы, рассеянные в паренхиме. Эти спикулы представляют молодые, не достигшие полного роста макросклеры, еще не вошедшие в состав скелетных пучков. Такие спикулы можно видеть у многих губок, очень часто они встречаются и у бадяг.
Среди нормальных спикул попадаются и различные уродливости. Так среди оке встречаются спикулы с закруглениями на обоих или на одном конце; часты неправильно искривленные или сросшиеся спикулы, а также политилотные, снабженные четкообразными вздутиями. К уродливостям же должны быть отнесены и сферы, гладкие или шиповатые шарики из кремнекислоты; со сферами сходны и пикнастры, звездообразные образования, характерные для ископаемой губки Spongilla gigantea.
Склеивающий спикулы спонгин присутствует у различных форм в различном количестве. Лучше всего он заметен в местах соединения главных и поперечных пучков, где образует в углах между спикулами прозрачные скопления, склеивающие их концы (табл. IV, фиг. 13). Спонгин представляет прозрачное, бесцветное или желтоватое органическое вещество, содержащее азот и по химической природе близкое к шелку. В состав этого вещества входит и иод, присутствие которого было недавно доказано и в пресноводных губках. В морских роговых губках иод присутствует в больших количествах и известен уже давно. У различных видов бадяг количество спонгина подвержено значительным колебаниям. Чем больше спонгина, тем прочнее скелет. У кустарниковых форм Spongilla lacustris его относительно много; Spongilla fragilis и Ephydatia fluviatilis бедны спонгином, и потому эти формы являются ломкими. Исключительно большое количество спонгина свойственно 5. arctica', у этой интересной губки все пучки, и главные, и поперечные, даже отдельные спикулы целиком погружены в спонгин, образующий широкие бахромчатые мембраны, придающие ткани заметную эластичность (табл. V, фиг. 16, 17). У некоторых форм, например, у Spongilla carteri var. mollis, спонгин почти отсутствует, и спикулы пучков не плотно прилегают друг к другу. Пучки теряют свою определенность, и скелет имеет тенденцию превратиться в массу рассеянных в паренхиме спикул. У байкальских губок спонгин присутствует в больших количествах. У рода Lubomirskia он образует основу скелетных пучков (табл. XIV, фиг. 58).
В устройстве скелета имеются существенные отличия между семействами Spongillidae и Lubomirskiidae. Отдельные роды байкальских губок
14
ФАУНА СССР
в свою очередь различаются друг от друга главным образом по устройству скелета, поэтому нам придется подробнее остановиться на его устройстве в систематической части.
Нам необходимо еще коснуться образования скелетных элементов. Кремневые спикулы строятся силикобластами, особыми подвижными амебоцитами. Спикула возникает внутри материнской клетки. Прежде всего образуется осевая нить, около которой отлагаются слои кремнезема. Образование микросклер заканчивается внутри материнской клетки, но в образовании макросклер принимают участие и дополнительные силикобласты. Макросклера достигает почти полной длины в теле материнской клетки, после чего освобождается и оказывается свободно лежащей в основном студенистом веществе. К ней подходят дополнительно силикобласты, облекают со всех сторон и отлагают на ее поверхности слои кремнезема, пока спикула не дойдет до нормальной толщины (Р. Schulze 1923). Готовая спикула затем перемещается в мезоглее амебоидными клетками, которые ставят ее на нужное место в скелетном пучке. Приклеить ее к пучку составляет теперь задачу спонгобластов, амебоцитов, выделяющих спонгин.
До отложения спонгина и после его окончания спонгобласты не отличимы от обычных амебоцитов; можно сказать, что вопрос о специфичности этих клеток, так же как и других амебоцитов со специальной функцией, является еще открытым. Для отложения спонгина к спикуле подходят несколько спонгобластов и обволакивают ее сетью отростков. Плазма клеток светлеет, и тонкие отростки и псевдоподии целиком превращаются в спонгин. После его отложения тело клетки округляется и протоплазма опять становится зернистой (Р. Schulze, 1922).
ЖИЗНЕННЫЕ ОТПРАВЛЕНИЯ ГУБОК
Движение. Амебоидное движение клеточных элементов. Реакция на раздражение. О нервной функции. Механизм водного тока. Питание. Захват пищи и переваривание. Дефекация и выделение. Дыхание. .
Из всех животных губки являются бесспорно самыми неподвижными и неактивными существами. Но в противоположность такой неподвижности целого организма, все клеточные элементы губки обладают подвижностью. Наиболее энергичные движения мы наблюдаем у воротничковых клеток, неутомимо работающих своими жгутами.. Кроме того, они выпускают и втягивают псевдоподии, т. е. обладают амебоидным движением. Можно сказать, что это движение свойственно всем клеточным элементам губки, но в различной степени, и наиболее активными в этом отношении являются аМебоциты.
Амебоидное движение всего удобнее наблюдать на эмульсии, состоящей из изолированных клеток. Кусочек губки прожимается через мельничный газ, и несколько капель такой выжимки разбалтываются
ПРЕСНОВОДНЫЕ ГУБКИ—ОБЩАЯ ЧАСТЬ
15
в небольшом количестве воды. При наблюдении такой эмульсии под микроскопом мы видим массу живых клеток. Следя за какой нибудь одной из них, мы очень скоро заметим изменения формы и перемещение. При сильных увеличениях видно выпускание широких, прозрачных псевдоподий и постепенное переливание зернистой протоплазмы (фиг. 8).
Гальцов (Galtzoff, 1923) измерял скорость движения археоцитов у морских губок, главным образом у Microciona. Эта скорость варииро-вала от 0.6 у- до 3.5 у- в минуту; как максимальная им отмечена Скорость в 20 м- в минуту. У бадяг мы имеем скорости того же порядка.
Фиг. 8. Движущийся амебоцит Ephydatia miilleri (пруды Старого Петергофа): последовательные изменения вида клетки в течение 3 минут; поперечник клетки около 18 у,.
Важнейшие-жизненные функции губок связаны с амебоидным движением ее клеточных элементов. Мы уже говорили о роли этих движений при построении скелета. Как увидим ниже, они играют не меньшую роль при питании и размножении губок.
Заметить какие-нибудь общие активные движения всего тела у бадяг мы нс можем. При прекращении водного тока, тело губки несколько уменьшается в объеме благодаря спаданию клоакальных протоков и субдермальных полостей. То же происходит при вынимании губки из воды. Но н этом уменьшении объема нельзя видеть активного сокращения ТвЛа.1
У очень молодых бадяг, только что превратившихся из свободно плавающих личинок, наблюдалось медленное переползание по субстрату, пыаВЙННОО амебоидными движениями клеток подошвы. Единственным Движением у иярослых губок, заметным для простого глаза, является сокращение И нытнгинаиие оскулярных трубок. Бадяга, помещенная в аквариум со Свежей НОДОЙ, через некоторое время возобновляет свою
J Паркер (Parker, 1910) гонориг о некотором активном сокращении всего тела у морской Stylotella hoUophlla при пыпимапии ее ив воды. Заметить что-либо подобное у бадяг не удается.
16
ФАУНА СССР
жизнедеятельность. Присмотревшись к ней, мы заметим вытянувшиеся оскулярные трубки. Прикосновение к ним, а также резкие сотрясения, вызывают их медленное втягивание, длящееся несколько минут. Затем трубки снова расправляются, на что требуется еще большее время. При содержании губок в аквариуме оскулярные трубки являются хорошим показателем их самочувствия. Втянувшиеся трубки всегда указывают на какие-нибудь неблагоприятные обстоятельства, чаще всего на недостаточную смену воды.
Втягивание оскулярных трубок является не только единственным видимым простым глазом движением губки,но и единственной заметной реакцией, которою она отвечает на внешние воздействия. При сравнении губок с другими сидячими животными, мшанками, полихэтами, асцидиями, мы видим, насколько быстрее и энергичнее реагируют все эти последние на внешние раздражения.
Изучение реакций у губок, произведенное Паркером (Parker, 1910) над Stylotella heliophila, приводит его к определенным заключениям о их нервной функции. В теле губки, по его наблюдениям, почти отсутствует передача раздражения от одного пункта к другому. А та передача, которую ему удавалось наблюдать, так слаба и происходит так медленно, что в ней нельзя видеть результат действия нервных элементов. Таким образом, он подтверждает с физиологической точки зрения выводы, к которым пришли морфологи об отсутствии у губок нервных элементов.
Мы уже говорили об устройстве ирригационной системы губок (стр. 5). Теперь мы рассмотрим механизм водного тока, проходящего через все тело губки, и те жизненные функции^ которые осуществляются при его помощи. Его изучением в недавние годы занимался ван-Тригт (van Trigt 1918, 1919, 1920), нашедший возможность, благодаря своеобразной методике, наблюдать работу хоаноцитов в жгутиковых камерах на совершенно не поврежденных, нормально функционирующих объектах.1 Материалом для его работ служили как раз пресноводные губки.
По его наблюдениям жгут воротничковой клетки совершает винтообразные движения, вызывающие ток воды от основания жгута к его концу. Нам уже приходилось упоминать (стр. 8), что хоаноциты отсутствуют в той части камеры, которая окружает выходное отверстие, апопиль. На боковых стенках камеры хоаноциты располагаются таким образом, что их вершины со жгутами направлены не к центру камеры, а к некоторой точке перед выходным отверстием. Маленькие токи, создаваемые отдельными жгутами, создают в сумме общий ток, проходящий через всю камеру. Вода засасывается через прозопили, проходит между телами хоаноцитов, омывает наружную сторону воротничков и направляется к выходу из камеры (фиг. 9). Таким образом каждая из бесчисленного множества камер выполняет роль насоса. Их соединенные усилия
J О методике ван-Тригта см. ниже, стр. 52.
ПРЕСНОВОДНЫЕ ГУБКИ-ОБЩАЯ ЧАСТЬ
17
ар
Фиг. 9. Схема водяных токов в жгутиковой камере. Рг — прозопиль, ар—апо-пиль; — зона уменьшенного давления; -+- зона повышенного давления. По ван-Тригту (1919).
заставляют входить воду в поры, проходить всю сложную систему каналов и выбрасываться через оскулярные отверстия.
Засасывание воды через поры очень легко наблюдать на губках в аквариумах. Если выпустить пипеткой около функционирующей губки облачко тонкой карминовой взвеси, то видно, как это облачко как бы притягивается губкой, прилипает к поверхности и исчезает. Ток воды, выходящий из оскулярных трубок легко заметить, по движению различных частиц, взвешенных в воде аквариума. Более демонстративная картина получается, если впрыснуть шприцем карминовую взвесь в тело губки. Очень скоро из ближайших оскулярных трубочек начинают бить красные струи на расстояние нескольких сантиметров. Если губка в аквариуме погружена не слишком глубоко, то присутствие оскулярных токов скажется на гладкой поверхности воды маленькими водяными бугорками в тех местах, где ток достигает поверхности.
Вместе с водой засасываются и взвешенные в ней частицы, достаточно мелкие, чтобы пройти через поры. Более крупные частицы застре
вают и скопляются на поверхности губки, образуя слой рыхлого осадка. Это скопление материала, вероятно, используется многими животными, ползающими по поверхности губки (Parker, 1910).
Частицы, прошедшие через поры, проходят вместе с водой к жгутиковым камерам. Отверстия, ведущие в эти камеры, прозопили, имеют всего 3—4 у в поперечнике и значительно меньше поровых отверстий. Далеко не все частицы, прошедшие через поры, могут попасть в камеры. Внутри камеры ток воды проходит между хоаноцитами и по наружной поверхности воротничков. При этом происходит отфильтровывание взвешенного материала. Частицы прилипают к поверхности хоаноцитов. Это происходит главным образом у основания воротничка,' но частицы могут прилипать и к воротничку. Затем движением плазмы частица передвигается по поверхности и втягивается внутрь клетки. При прибавлении кармина к воде аквариума, красные частицы очень скоро оказываются внутри хоаноцитов. Затем захваченный материал выбрасывается в мезоглею, где вновь захватывается амебоцитами.. Прибавляя к воде карминовый порошок, можно окрасить в яркокрасный цвет всю губку; ее клетки при этом оказываются переполненными кармином. Окраска эта держится несколько дней, мало по малу бледнея: клетки губки и прежде всего хоаноциты постепенно освобождаются от захваченного непитательного материала.
Фауна СССР, Губки, т. II, в. 2 2
18
ФАУНА СССР
Ван Тригт наблюдал захват частиц хоаноцитами, передачу их в мезоглею и вторичный их захват блуждающими клетками. Ему удалось также проследить судьбу и более крупных частиц, не проходящих через прозопили. Такие частицы захватываются стенками приводящих путей и оказываются втянутыми в мезоглею, где на них нападают амебоциты (у. Trigt 1919). Если частица слишком крупна, чтобы целиком поместиться в одном амебоците, то ее облегают со всех сторон несколько клеток (Cotte, 1904).
Захват губками самых различных как питательных, так и непитательных материалов (кармин, индиго, уголь, китайская тушь, крахмал, молоко), показывает, что губки лишены избирательной способности и отфильтровывают все, что взвешено в воде. От захваченных непитательных частиц губка, как мы видим, постепенно освобождается. Питательные вещества перевариваются и усваиваются.
Переваривание происходит внутри клеток, главным образом в амебоцитах, в вакуолях, которыми окружаются питательные частицы. Нелли Пурбэ (Nelly Pourbaix, 1931) наблюдала захват пищи (бактерии, зерна крахмала) и переваривание на изолированных клетках одной морской губки (Pellina semitubulosa). Ее данные, вероято, приложимы и к другим губкам. Псевдоподий охватывает частицу и втягивает ее внутрь. Еще до ее внедрения в псевдоподии образуется вакуоль, в которую и попадает втянутая частица. Сперва содержимое вакуоли кислое, затем реакция меняется, и переваривание происходит при щелочной реакции. Частица начинает растворяться, а на поверхности вакуоли появляются зерна липоидного вещества (продукт ассимиляции), все более и более многочисленные, пока частица совершенно не растворится. Таким образом, переваривание и усвоение твердых питательных веществ клетками губки может быть прослежено непосредственно под микроскопом.
На основании всех имеющихся наблюдений можно сказать, что пищей для губок служат взвешенные вещества, сестон,1 если величина частиц позволяет им проходить через поры. Питательное значение может иметь только органический сестон, слагающийся, с одной стороны, из планктиче-ских организмов, с другой, — из триптона, остатков отмерших животных и растений. Оба эти компонента, несомненно, имеют значение для питания губок.
Нелли Пурбэ (1. с.) наблюдала захват и переваривание бактерий. Ряд авторов указывают на микро- и наннопланктические водоросли, как на источник питания для губок.
Обычно принимается, что отношения между пресноводными губками и населяющими их зелеными водорослями являются- симбиотическими,
1 Сестоном называется все, что взвешено в воде и отфильтровывается через самые -гонкие номера мельничного газа, или скопляется на дие пробирок при центрифугировании-Триптон, часть сестона, состоящая из остатков отмерших животных и растительных организмов.
ПРЕСНОВОДНЫЕ ГУБКИ—ОБЩАЯ ЧАСТЬ
19
что оба организма извлекают известную выгоду из такого сожительства. По данным ван Тригта (1919) водоросли эти скорее являются пищей и перевариваются губками. Большое количество панцырей диатомей, встреч чающееся внутри ткани, вероятно, указывает на переваривание этих водорослей губками. Очень часто можно видеть панцири диатомей на поверхности геммул, иногда уложенные правильными стопками. Возможно, что эти кремневые створки после переваривания водорослей сносятся к геммулам и укладываются на их поверхности амебоцитами так же, как они это делают с амфидисками (см. ниже стр. 22). У нас нет специальных данных об усвоении губками триптона. Но судя по тем его количествам, которые имеются в воде и по его питательности, можно уверенно говорить о его значении в питании губок.
Мы должны также упомянуть о воззрениях Пюттера (1909—1914) на питание водных животных, в частности губок. Отсчитает, что количество взвешенной твердой пищи недостаточно для питания водных животных, и главным источником пищи являются растворенные органические вещества. Воззрения эти подверглись основательной критике и не были приняты большинством исследователей. Несомненно, что Пюттер недооценивает значение именно триптона, который играет существенную роль в питании водных организмов. Но, конечно, не исключается возможность усвоения и растворенных органических веществ.
У ван Тригта (1919) мы находим наблюдения и над дефекацией. В амебоидных клетках в стенках отводящих путей он наблюдал иногда крупные вакуоли, содержавшие скопления зерен. Эти вакуоли постепенно выпячивались в просвет отводящего канала и затем прорывались и изливали в канал свое содержимое. Зерна в вакуолях состояли из непереваренных остатков, а также, вероятно, из экскретов. Им описываются также довольно крупные скопления фекалий в стенках отводящих путей около жгутиковых камер, постепенный принос новых частиц к этим скоплениям и периодическое выбрасывание всего этого материала в выводящий канал. Таким путем губка быстро освобождается от избытка захваченных частиц.
Одновременно с питанием при помощи того же водного тока осуществляется и дыхание губок. Газовый обмен даже в глубоко лежащих частях губки не может представлять затруднений, так как вода всюду проникает по системе каналов и полостей, а к кислороду, растворенному в воде, еще присоединяется кислород, выделяемый на свету симбиотическими водорослями.
ПОЛОВОЕ РАЗМНОЖЕНИЕ
Образование яиц и сперматозоидов. Устройство личинок. Прикрепление личинок и метаморфоз.
Уже в конце весны или в начале лета бадяги приступают к половому размножению. Исключение составляет только один вид Spongilla biseriata. У него половое размножение или совершенно отсутствует, или наблю-2*
20
ФАУНА СССР
дается очень редко и заменяется наружным почкованием (Annandale, 1911). У байкальских губок половое размножение еще не известно, но у них описано появление свободноплавающих личинок из соритов, образований, имеющих общие черты с геммулами (Сварчевский, 1923, 1925).
Все Spongillidae являются разнополыми; одни особи продуцируют только яйца, другие только сперматозоиды. Число тех и других приблизительно одинаково, и по внешности они ничем друг от друга не отличаются. Половые клетки развиваются из амебоидных клеток, археоцитов, и образование половых продуктов не приурочено к каким нибудь определенным участкам тела. Повидимому, любой археоцит может превратиться в яйцевую клетку. Наличие недифференцированных, интерстициальных клеток стирает грань между клетками тела и половыми, между сомой и зародышевой плазмой (Исаев, 1924). Развивающееся яйцо отличается только большей величиной от археоцита, из которого оно возникло (фиг. 5). Как и остальные клетки губок, яйцо на всех стадиях своего развития лишено оболочки, и сохраняет амебоидные свойства. Яйцо растет за счет питающих амебоидных клеток, которые оно захватывает псевдоподиями. Плазма питающей клетки сливается с плазмой яйца, а ядро постепенно резорбируется. Зрелая яйцевая клетка достигает 40—50, даже 100 у. в поперечнике (Vosmaer, 1886). По мере созревания яйца к нему подходит из окружающей ткани амебоидные клетки и образуют кругом него род фолликулярного эпителия.
Мужские половые элементы, так же как и яйца, образуются из археоцитов. Археоцит растет и превращается в сперматиду, из которой путем ряда делений получаются мужские половые клетки, сперматозоид ы. К молодой сперматиде пр дходит покровная клеткаиобво-лакивает ее со всех сторон вместе с несколькими питающими клетками, образуя плазматическую капсулу, сперматоцисту. Внутри этой капсулы и происходит сперматогенез, в результате которого из одной сперматиды получается большое количество сперматозоидов (Gorich, 1904). Зрелые сперматозоиды имеют типичный вид и состоят из овальной головки и длинного жгута (фиг. 10). Они движутся внутри сперматоцисты, затем после ее разрыва освобождаются и выходят в воду, проникают через поры вместе с водным током внутрь женских особей и оплодотворяют яйца. Дробление яйца и образование личинки происходит внутри материнского тела.
У всех губок дробление полное. У кремнероговых после двух первых равных меридиональных делений происходит первое неравное экваториальное деление, при котором четыре микромера отделяются от четырех более крупных макромеров. В дальнейшем дробление приобретает беспорядочный характер и приводит к образованию стерробластулы, плотного скопления клеток, с многочисленными мелкими микромерами на анималь-ном полюсе. Гаструляция осуществляется путем эпиболии: микромеры или эктодермальные клетки от анимального полюса постепенно распро
ПРЕСНОВОДНЫЕ ГУБКИ-ОБЩАЯ ЧАСТЬ
21
страняются по всей поверхности зародыша и располагаются на ней одним сплошным слоем, окружая массу более крупных эктодермальных клеток. У наружных клеток образуются жгутики, а во внутренней энтодермальной массе начинается образование спикул и дифференциация полостей. На
этой стадии развития зародыш покидает материнское тело и превращается в свободно плавающую личинку (табл. VIII, фиг. 33).
Тело личинки у бадяг яйцеобразное, около 0.5 мм длины. На по
верхности имеется сплошной слой ’цилиндрических жгутиковых клеток. Под их покровом располагается внутренняя масса клеток, богатых питательным материалом. В более широкой части личинки, которая при плавании направляется кверху, располагается объемистая полость, выстланная уплощенными клетками. Другая половина, напра-вленная при плавании книзу,. занята компактной клеточной у массой, в которой видно до десятка прямых и тонких спикул. , 1 Здесь же можно заметить начало формирования каналов к и полостей будущей губки. Личинка в течение очень недол- / того времени оживленно плавает, вращаясь около своей про- ' дольной оси. Прикрепление личинки происходит обыкно- фиг. ю. венно в течение первых двенадцати часов после выхода из Сперматозоид Ephydatia flu-материнского тела, но иногда оно может запаздывать и до viatilis. По двух суток. Осевшая личинка уплощается и превращается Gorich (1904). в маленькое беловатое пятнышко, в котором очень скоро уже
можно узнать маленькую губочку. Об окончании развития можно судить по отслоению дермальной мембраны и образованию довольно длинной
оскулярной трубки.
При развитии губки от яйца до свободноплавающей личинки мы имеем полную аналогию с развитием других животных. Но метаморфоз личинки, начинающийся после ее прикрепления, представляет характерный для всех губок процесс, отличающий их от всех остальных многоклеточных животных. Эктодермальные жгутиковые клетки личинки после ее прикрепления начинают мигрировать внутрь тела, где они образуют клеточную
выстилку формирующихся жгутиковых камер и превращаются в хоаноциты. На поверхности тела оказываются эктодермальные клетки, дающие наружный покров губки и превращающиеся в пинакоциты. Таким образом, эктодерма и энтодерма личинки при образовании сформированной губки меняются местами. Происходит процесс „извращения зародышевых листков", открытый Делажем (Delage, 1892), отличающий губки от всех остальных
многоклеточных животных.
БЕСПОЛОЕ РАЗМНОЖЕНИЕ
Геммулы, их образование и устройство. Вылупление губочек из геммул. О геммулах у морских и байкальских губок.
У очень многих губок мы встречаем рядом с половым также и бесполое размножение. В частности у пресноводных губок оба вида размно
22
ФА УНА СС СР
жения входят в нормальный годичный цикл жизни. Половое размножение служит для образования новых колоний и расселения вида в пределах того же водоема; результат бесполого размножения, геммулы, представляет покоящуюся стадию, предназначенную для сохранения вида в течение неблагоприятных сезонов (холодных или засушливых) и восстановления старых колоний при возвращении благоприятных условий, но геммулы выполняют и другую очень важную функцию: при их помощи вид может расселяться по другим водоемам, куда геммулы могут заноситься ветром, водяными птицами или другими какими-нибудь путями.
Кроме образования геммул, мы встречаем у пресноводных губок и другие способы размножения. Мы уже упоминали о наружном почковании и образовании соритов. Все эти способы рассматриваются как бесполое размножение и противопоставляются размножению половому.
При половом размножении вообще новая особь развивается из половой клетки, из яйца, оплодотворенного или неоплодотворенного; при этом развитие всегда начинается от одной единственной клетки. При бесполом размножении начало новой особи дают соматические клетки, клетки тела. Обычно дочерняя особь создается из целого комплекса материнских клеток, но, конечно, возможны случаи, когда в этом участвует только одна соматическая клетка. Как мы уже упоминали выше, разграничение у губок соматических и половых клеток вполне условно. При развитии губки громадное число клеток остается недифференцированным и неспециализированным. Это археоциты, о которых мы уже говорили. Они дают начало половым клеткам, они же участвуют во всех видах бесполого размножения, что делает очень не резким у губок различие между половым и бесполым размножением.
Образование геммулы начинается со скопления археоцитов, богатых запасами питательных веществ. Затем к сформировавшемуся скоплению начинают собираться амебоциты, уже лишенные питательных запасов. Эти клетки располагаются одним слоем и образуют кругом скопления эпителиальную оболочку, прерывающуюся только на месте будущего порового отверстия. Клетки этой оболочки выделяют хитиновую капсулу геммулы. После образования капсулы происходит принос и укладка геммульных микросклер. Эти микросклеры развиваются в склеробластах в различных местах ткани. Готовые микросклеры захватываются особыми амебоцитами, приносятся к геммуле и втискиваются между клетками эпителиальной оболочки. Микросклеры укладываются во многих случаях с уда~ вительной правильностью. Когда геммульные спикулы уложены, клетки оболочки поднимаются, заполняя все промежутки между микросклерами зернистой или ячеистой воздухоносной тканью. Часто ими выделяется и внешняя кутикула, обволакивающая воздухоносный слой с заключенными в нем микросклерами. Во время постройки геммульных оболочек будущее поровое отверстие остается свободным. Через него проникают внутрь геммулы трофоциты, особые питающие клетки,
ПРЕСНОВОДНЫЕ Г У Б К И-—О Б Щ А Я ЧАСТЬ 23
сносящие сюда питательный материал. Когда этот процесс заканчивается, поровое отверстие затягивается пленочкой. Содержимое готовой геммулы состоит из крупных полигональных клеток, переполненных зернами запасных питательных веществ и зоохлореллами. Эта клеточная масса окружена тонкой оболочкой, плотно прилегающей к геммульной капсуле (Wierzejski, 1886, Zykoff, 1893, Evans, 1901).
Устройство и расположение геммул и характер их микросклер имеют большое значение в систематике бадяг, а потому мы должны рассмотреть некоторые детали их строения.
Форма геммульной капсулы то правильно шаровая, то несколько приплюснутая в вертикальном направлении. В ней обычно имеется одно поровое отверстие округлой формы на вершине. У некоторых видов могут иногда присутствовать и дополнительные отверстия. Край отверстия образует небольшую оторочку (Spongilla lacustris и др.), или приподнимается в виде широкоразвернутой воронки (Ephydatia, Trochospon-gilla). Во многих случаях поровое отверстие находится на вершине поровой трубки, прямой или несколько изогнутой (Spongilla fragilis, Spongilla carteri и др.). У рода Heteromeyenia на поровой трубке, несколько отступя от вершины, имеется воротник, часто снабженный по краям придатками, в виде длинных тонких выростов (Н. baileyivar. stepa-nowi — Carterius stepanowi).
Мы уже описывали геммульные микросклеры (стр. 12), которые могут иметь вид р а б д или амфидисков. Рабды у слабо вооруженных геммул ложатся тангенциально на поверхности капсулы, часто в очень небольшом количестве (Spongilla lacustris и др.), но у тех же форм геммулы могут быть снабжены и большим количеством спикул. Тогда эти микросклеры располагаются почти радиально, образуя сплошной слой, окружающий капсулу.
Амфидиски чаще всего располагаются сплошным слоем с удивительной правильностью. Один из дисков примыкает к капсуле, а стержень ориентируется радиально. Чаще всего амфидиски располагаются одним слоем. Но у некоторых видов может быть и два, и три слоя амфидисков (Ephydatia miilleri). При деградации геммульных оболочек амфидиски могут и не покрывать всей поверхности капсулы. Это обыкновенно связано и с неправильным их. расположением, и с уродливостями, которые среди них наблюдаются (стр. 12).
У одного и того же вида вариирует не только количество микросклер, но и связанная с ним толщина воздухоносного слоя. Там, где микросклеры представлены разбросанными редкими рабдами, воздухоносный слой почти или совершенно отсутствует. В других случаях он может достигать значительной толщины. Структура его у разных видов различна. В зернистом слое мы имеем пенистую ткань с мелкими пузыревидными включениями. В ячеистом слое, достигающем большой толщины, ткань состоит из правильно расположенных во много слоев полигональных ячей.
У-1
ФАУНА СССР
В разрезе такая ткань напоминает пчелиные соты (табл. IV, фиг. 14). Вещество этой ткани близко к хитину и имеет некоторую примесь кремнезема.
Геммулы могут быть рассеяны в паренхиме по одиночке (подрод Euspongilla, род Ephydatia). Но часто мы находим их в основании губки, где они уложены сплошным слоем, как булыжники в мостовой (м о с т о -видный слой). При этом воздухоносные слои отдельных геммул могут сливаться с соседними слоями, образуя один сплошной слой воздухоносной ткани с заключенными в него рабдами {Spongilla fragilis, S. starileyi). У таких форм часто можно наблюдать геммулы и в различных местах паренхимы, но они разбросаны не одиночно, а образуют более или менее крупные скопления, заключенные в общую воздухоносную ткань (S. fragilis).
Геммулы способны переносить и замораживание, и высыхание. Высохшие геммулы сохраняют жизнеспособность в течение нескольких лет. Как мы уже упоминали, это открывает бадягам широкие возможности расселения.
С наступлением весны внутри геммулы начинается развитие. Питательные запасы потребляются, начинают появляться спикулы и закладываются жгутиковые камеры. На этой стадии содержимое геммулы покидает свою оболочку. Поровое отверстие открывается и через него выползают клетки по одиночке и группами. Из освободившейся таким образом массы клеток через несколько дней формируется губочка. Если геммулы оставались на месте среди скелетных пучков материнского тела, то молодые губочки, сливаясь, образуют сплошной слой на поверхности старой отмершей ткани. У крупных желваков бадяг живым бывает только поверхностный слой. Внутри лежит старая отмершая ткань, переполненная пустыми геммулами.
Если материнское тело после образования геммул будет разрушено, геммулы рассеиваются, и каждая может дать начало новой колонии.
Геммулы известны не у всех представителей сем. Spongillidae. Среди тропического подсемейства Potamolepidinae есть формы, у которых геммулы никогда не наблюдались.
Образование геммул не является отличительной особенностью одного семейства Spongillidae. Они известны также у целого ряда морских форм из отрядов четырехлучевых и кремнероговых губок. Геммулы у морских губок изучены довольно слабо и не играют, невидимому, в их жизни значительной роли. Характерной особенностью геммул у бадяг является наличие в огромном большинстве случаев геммульных микросклер особого устройства: у морских -губок, если геммулы иногда и вооружены, то не специально приуроченными к ним спикулами.
Геммулы были описаны у одной из байкальских губок (Baikalo-spongia bacillifera-. Annandale, 1913), но Сварчевский (1923) выяснил, что здесь была допущена ошибка. Геммулы у байкальских губок до сих пор не известны; все говорит за то, что они у них и не образуются.
ПРЕСНОВОДНЫЕ ГУБКИ— ОБЩАЯ ЧАСТЬ
25
ДРУГИЕ ВИДЫ БЕСПОЛОГО РАЗМНОЖЕНИЯ
Сориты байкальских губок. Наружное почкование бадяг и байкальских губок. Редукции или редукционные почки.
В недавние годы у байкальскрх губок описан другой чрезвычайно интересный способ размножения, образование соритов и развитие из них
свободноплавающих личинок (Сварчев-ский 1923, 1925). Сориты известны у представителей различных отрядов губок—у Triaxonida, Tetraxonida и Согпа-cuspongida. Как и геммулы, они представляют шарообразные или эллипсоидальные скопления амебоидных клеток (археоцитов), но отличаются от геммул отсутствием плотных оболочек. Очень существенно, что сориты не представляют, как геммулы, покоящейся стадии, сохраняющей жизнеспособность при неблагоприятных условиях. Есть и другие существенные отличия. Содержимое геммулы состоит из многочисленных полигональных клеток, и границы между клетками хорошо заметны; содержимое соритов • представляет слившуюся син-тициальную массу. В геммуле молодая особь развивается из комплекса клеток, в сорите же только из одной, осталь
Фиг. 11. Сорит Baikalospongia bacilli-fera в капсуле из плоских клеток; в средней части видно единственное ядро; сорит переполнен зоохлореллами; по периферии видны остатки дегенерирующих ядер питающих клеток. По Сварчевскому (1923).
ные же клетки являются для нее пита-
тельным материалом. Из геммул выходит беспорядочная клеточная масса, очень быстро формирующая губку. Из соритов, поскольку известно их развитие, получаются свободноплавающие личинки, сходные с образовавшимися половым путем.
Сориты Baikalospongia bacillifera, описанные Сварчевским, представляют округлые тела 165—320 [л в поперечнике (фиг. И). На поверхности с трудом может быть обнаружена плотно прилегающая тончайшая кутикулярная оболочка. Внутри находится синцитиальная масса слившихся археоцитов, переполненная зоохлореллами и зернами питательного материала. Ядра археоцитов находятся в состоянии дегенерации. В средней части сорита находится участок протоплазмы, свободный от включений, с одним довольно крупным ядром. Образование такого сорита в общих чертах сходно с образованием геммул. После образования сорита Сварчевский предполагает для него некоторую стадию покоя, после которой начинается деление единственного ядра и формирование личинки. Очень интересно
ФАУНА СССР
указание на постепенное исчезновение симбиотических водорослей; и готовой личинке их остается очень мало. Это, конечно, указывает, что водоросли при развитии личинки потребляются как пищевой материал. Размеры образовавшейся из сорита личинки колеблются от 250 до 280 у-; форма ее тела эллипсоидальная, реже яйцевидная, причем тогда нижняя часть личинки шире верхней (у бадяг отношение обратное). В верхней части тела (более узкой у яйцевидных личинок) находится полость, не такая объемистая, как у бадяг, и занимающая ]/<t или Уз объема тела. Форма этой полости довольно неправильная. В нижней части тела рыхлым пучком лежат тонкие, слегка шиповатые спикулы. Как и у бадяг, все тело покрыто жгутиковыми клетками. Сориты часто можно наблюдать внутри ткани и у других байкальских губок {Lubomirskia baicalensis, L. fusifera, Baikalospongia intermedia).
Итак, в сорите молодая особь получается из одной единственной клетки, что сближает размножение соритами с половым партеногенетическим размножением. Мы уже говорили выше, что у губок трудно провести резкую грань между половым и бесполым размножением.
Надо отметить еще один способ бесполого размножения, встречающийся и у многих морских губок. У бадяг он описан только у Spongilla biseriata. Выше мы отмечали, что этот вид или совсем не размножается-половым путем, или крайне редко. Половое размножение заменяется здесь наружным почкованием (Annandale, 1911). На концах главных скелетных пучков, поддерживающих дермальную мембрану, непосредственно под нею, образуется скопление клеток, в состав которого входят различные элементы: паренхимные клетки, жгутиковые камеры, микросклеры; имеются и следы каналов. Такое скопление прорывает мембрану, освобождается и в дальнейшем образует новую губку.
Очень сходные явления известны и у других видов бадяг, но не как нормальный способ размножения. Почки появляются в результате неблагоприятных условий при содержании губок в аквариуме. Такие почки уже очень давно описал Лоран (Laurent, 1844) у Spongilla lacustris; их видел и Annandale (1911) у поврежденных экземпляров 5. carteri. Мне пришлось наблюдать отделение маленьких зеленых шариков около полумиллиметра в поперечнике у Ephydatia miilleri, содержавшейся в аквариуме. Через день-два шарики пристали к субстрату, уплостились и образовали молодые губочки.
Наружное почкование свойственно, повидимому, и байкальским губкам. Сварчевский (1923) находил на песчанистом илу неприкрепленные к субстрату экземпляры Lubomirskia baicalensis самой различной величины. Наиболее мелкие, 4—5 мм в поперечнике, имели округлую или несколько удлиненную форму; наиболее крупные, вытянутые в длину, экземпляры достигали нескольких сантиметров. Место прикрепления к твердому субстрату отсутствовало. Только у более крупных обозначалась по бледной окраске и по выделению спонгиновой пленки нижняя
ПРЕСНОВОДНЫЕ ГУБКИ—ОБЩАЯ ЧАСТЬ
27
подошвенная сторона. В Зоологическом институте Академии Наук имеются многочисленные экземпляры Lubomirsckia baicalensis, представляющие небольшие округлые комки или более длинные неразветвленные выросты (самый крупный имеет около 20 см длины) с закругленными концами; место прикрепления к субстрату во всех этих экземплярах отсутствует. В этих же коллекциях встречаются и маленькие вальковатые экземпляры Baikalospongia intermedia, также без поверхности прикрепления;
Сварчевский, повидимому, совершенно правильно объясняет появление таких форм роста наружным почкованием. Почки падают на дно и на каменистом грунте, вероятно, скоро прикрепляются. При попадании почки на мягкий грунт прикрепления не происходит. Губка развивается, свободно лежа на илу.
Из различных форм бесполого размножения, описанных выше, только "геммулы являются покоящейся стадией. К. Мюллер (К. Muller, 1911) описал другие образования у бадяг, по своим функциям заменяющие геммулы. Он назвал их редукциями. Если губка перед наступлением холодного времени не образовала геммул, то при наступлении неблагоприятных условий живая паренхима частью отмирает, частью сокращается и собирается в округлые клеточные комочки, которые при возвращении теплого времени восстанавливают колонию. Особенно часто это наблюдается у Ephydatia fluviatilis, у которой геммуляция может совершенно выпадать из годичного цикла.
РЕГЕНЕРАЦИОННАЯ СПОСОБНОСТЬ ГУБОК
Опыты Уильсона и Мюллера над регенерацией диссоциированных клеток. Вопрос об индивидуальности и образовании колоний у губок.
Явления бесполого размножения у губок показывают широкую способность небольших клеточных комплексов воссоздавать целый организм. Эта способность в полной мере проявляется при опытах регенерации. Мы коротко коснемся некоторых интересных наблюдений из этой области.
Губки с чрезвычайной легкостью заживляют всякие поранения их поверхности. При вырезывании кусочков ткани, рана быстро заглаживается и затягивается мембраной. То же происходит и с ходами, которые прогрызают в ткани бадяг личинки ручейников.
Отдельные кусочки губок продолжают свое существование, регенерируя дермальную мембрану и оскулярное отверстие, если оно отсутствовало на отрезанном куске. Этой способностью бадяг пользовался ван-Тригт (1919) для получения очень тонких экземпляров. Он давал регенерировать на покровном стекле маленьким кусочкам губок. Такие препараты позволяли ему изучать при сильнейшем увеличении работу и поведение
УК
ФАУНА СССР
клеток живой неповрежденной губки. Регенерационная способность губок достигает, можно сказать, крайнего предела в классических опытах Уильсона (Wilson, 1911). В его опытах над морскими губками, повторенных Мюллером (К. Muller, 1911) над бадягами, кусочки губки прожимались через мелкоячеистую ткань. Фильтрат представлял эмульсию из изолированных клеток. Несколько капель такой эмульсии размешивались в маленьком сосудике с водой. Взвешенные клетки оседали на дно в виде тонкого осадка. Изолированные клетки на дне сосудика z показывали оживленное амебоидное движение. При встрече двух клеток они слипались и продолжали двигаться,
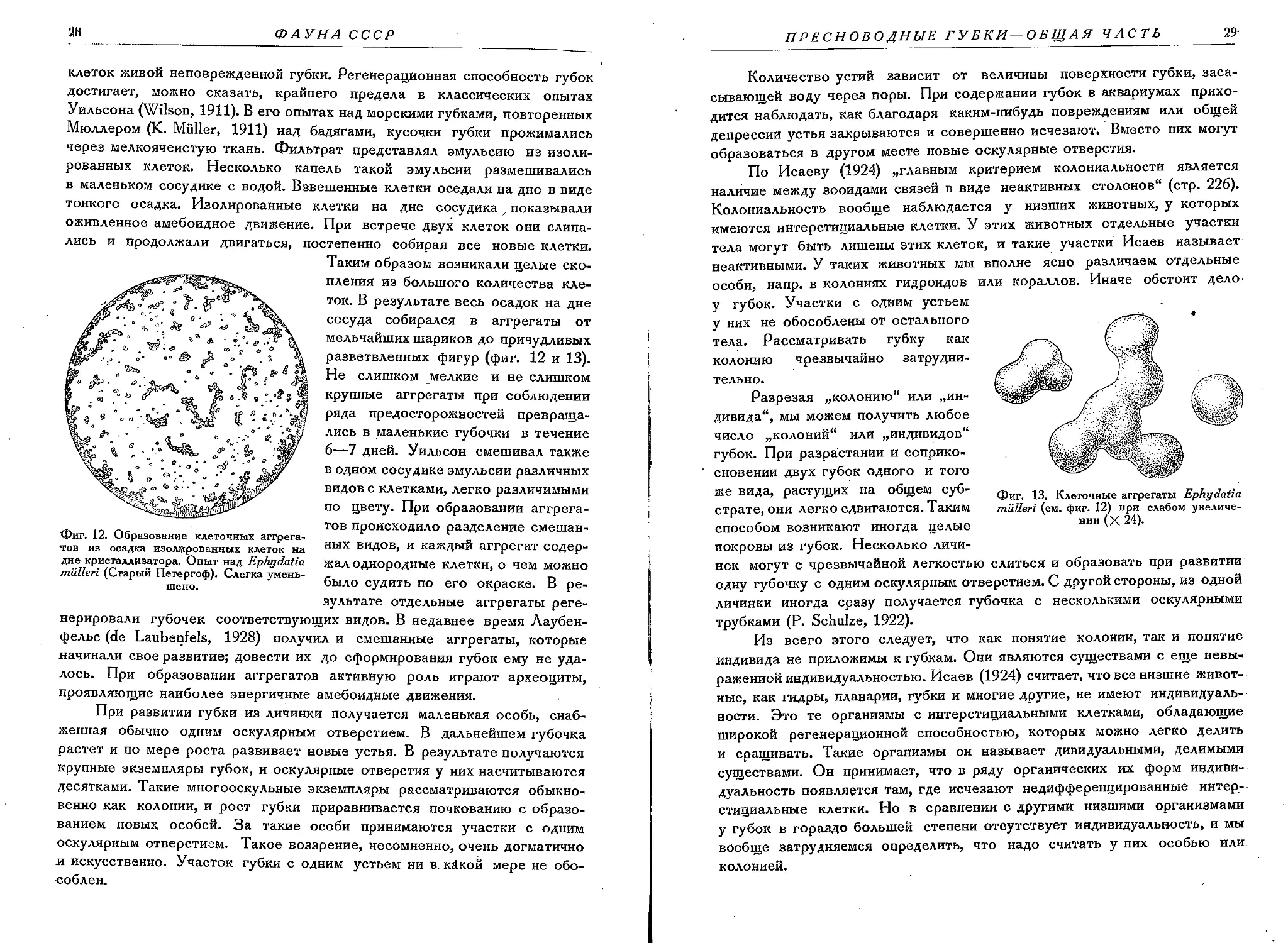
Фиг. 12. Образование клеточных аггрега-тов из осадка изолированных клеток на дне кристаллизатора. Опыт над Ephydatia miilleri (Старый Петергоф). Слегка уменьшено.
постепенно собирая все новые клетки. Таким образом возникали целые скопления из большого количества клеток. В результате весь осадок на дне сосуда собирался в аггрегаты от мельчайших шариков до причудливых разветвленных фигур (фиг. 12 и 13). Не слишком мелкие и не слишком крупные аггрегаты при соблюдении ряда предосторожностей превращались в маленькие губочки в течение 6—7 дней. Уильсон смешивал также в одном сосудике эмульсии различных видов с клетками, легко различимыми по цвету. При образовании аггрега-тов происходило разделение смешанных видов, и каждый аггрегат содержал однородные клетки, о чем можно было судить по его окраске. В результате отдельные аггрегаты реге-
нерировали губочек соответствующих видов. В недавнее время Лаубен-фельс (de Laubenfels, 1928) получил и смешанные аггрегаты, которые начинали свое развитие; довести их до сформирования губок ему не удалось. При образовании аггрегатов активную роль играют археоциты, проявляющие наиболее энергичные амебоидные движения.
При развитии губки из личинки получается маленькая особь, снабженная обычно одним оскулярным отверстием. В дальнейшем губочка растет и по мере роста развивает новые устья. В результате получаются крупные экземпляры губок, и оскулярные отверстия у них насчитываются десятками. Такие многооскульные экземпляры рассматриваются обыкновенно как колонии, и рост губки приравнивается почкованию с образованием новых особей. За такие особи принимаются участки с одним оскулярным отверстием. Такое воззрение, несомненно, очень догматично и искусственно. Участок губки с одним устьем ни в к^кой мере не обособлен.
ПРЕСНОВОДНЫЕ ГУБКИ—ОБЩАЯ ЧАСТЬ
29'
Количество устий зависит от величины поверхности губки, засасывающей воду через поры. При содержании губок в аквариумах приходится наблюдать, как благодаря каким-нибудь повреждениям или общей депрессии устья закрываются и совершенно исчезают. Вместо них могут
образоваться в другом месте новые оскулярные отверстия.
По Исаеву (1924) „главным критерием колониальности является наличие между зооидами связей в виде неактивных столонов" (стр. 226). Колониальность вообще наблюдается у низших животных, у которых имеются интерстициальные клетки. У этих животных отдельные участки тела могут быть лишены этих клеток, и такие участки Исаев называет неактивными. У таких животных мы вполне ясно различаем отдельные особи, напр. в колониях гидроидов или кораллов. Иначе обстоит дело
у губок. Участки с одним устьем у них не обособлены от остального тела. Рассматривать губку как колонию чрезвычайно затруднительно.
Разрезая „колонию" или „индивида", мы можем получить любое число „колоний" или „индивидов" губок. При разрастании и соприкосновении двух губок одного и того же вида, растущих на общем субстрате, они легко сдвигаются. Таким способом возникают иногда целые покровы из губок. Несколько личи
Фиг. 13. Клеточные аггрегаты Ephydatia miilleri (см. фиг. 12) при слабом увеличении (X 24).
нок могут с чрезвычайной легкостью слиться и образовать при развитии одну губочку с одним оскулярным отверстием. С другой стороны, из одной личинки иногда сразу получается губочка с несколькими оскулярными трубками (Р. Schulze, 1922).
Из всего этого следует, что как понятие колонии, так и понятие индивида не приложимы к губкам. Они являются существами с еще невыраженной индивидуальностью. Исаев (1924) считает, что все низшие животные, как гидры, планарии, губки и многие другие, не имеют индивидуальности. Это те организмы с интерстициальными клетками, обладающие широкой регенерационной способностью, которых можно легко делить и сращивать. Такие организмы он называет дивидуальными, делимыми существами. Он принимает, что в ряду органических их форм индиви
дуальность появляется там, где исчезают недифференцированные интерстициальные клетки. Но в сравнении с другими низшими организмами у губок в гораздо большей степени отсутствует индивидуальность, и мы вообще затрудняемся определить, что надо считать у них особью или колонией.
30
ФАУНА СССР
ЦИКЛ ЖИЗНИ И УСЛОВИЯ ОБИТАНИЯ ГУБОК
Годичный цикл жизни. Влияние озерных условий на геммуляцию. Факторы среды и их влияние: течение, глубина, свет и прозрачность, соленость, активная реакция среды, содержание кислорода. Губки в загрязненных водах. О нахождениях губок в пещерах.
В умеренных странах бадяги к зиме после образования многочисленных геммул отмирают. Зимою скелет отмершей губки обыкновенно сохраняется и геммулы остаются на своих местах в его петлях или в слое, выстилающем субстрат. С наступлением теплого времени вылупляются молодые губки из геммул и поднимаются по материнскому скелету на поверхность колонии. Здесь скопляется вся их масса, происходит их слияние и образование на поверхности старой колонии молодого функционирующего слоя. Таким образом, прошлогодняя колония воссоздается и увеличивается в объеме,. Только таким путем и образуются экземпляры бадяг, весящие иногда свыше килограмма. При разрезании такого экземпляра мы найдем внутри него старую ткань с пустыми геммулами и на поверхности сравнительно Тонкий слой молодой ткани. В старой ткани, как мы уже упоминали, можно более или менее ясно различить годичные слои нарастания. При образовании молодого слоя частично используется и старый скелет, и пространства между его петлями заполняются новой тканью.
У форм, образующих слой геммул на субстрате, иногда можно наблюдать многослойное их расположение (например, у Spongilla stanleyi, Rezvoi, 1930). Такую многослойность можно объяснить ежегодным отложением нового слоя геммул на старые отмершие слои.
Восстановившиеся колонии в конце весны и летом приступают к половому размножению. Свободноплавающие личинки в Западной Европе наблюдаются во второй половине лета, в июле, августе и до самой осени (Arndt, 1930, Schroder, 1930). Сроки эти, несомненно, сильно вариируют от условий климата и характера водоема. Там, где имеются богатые заросли губок, личинки появляются в огромных количествах и составляют заметный компонент планктона (р. Шпрее в Берлине, Lieberkuhn, 1856).
Как мы уже отмечали, личинка плавает очень непродолжительное время, от нескольких часов до двух суток. Так как они продолжают вылупляться в течение значительного периода, то и получающихся из них губочек осень застает на различных стадиях роста. Обычно к осени молодые губки представляют тонкие корочки в 2—3 см в поперечнике. Не образуя половых продуктов, они дают геммулы и отмирают с наступлением холодов, чтобы восстановиться из геммул следующей весной. Таким образом постепенно создаются объемистые колонии.
Как сроки полового размножения, так и время образования геммул зависит от климата и характера водоема. Геммуляция в умеренных широтах происходит в конце лета и в начале осени, когда в водоемах идет уже
ПРЕСНОВОДНЫЕ ГУБКИ—ОБЩАЯ ЧАСТЬ
31
•охлаждение. В окрестностях Ленинграда приходилось наблюдать образование геммул (Ephydatia miilleri) уже в конце июля. Для Москвы Зыков (1892) указывает середину августа (Ephydatia fluviatilis и Е. miilleri). Для Западной Европы приводятся более поздние сроки (Arndt, 1930).
Есть наблюдения, что в некоторых водоемах губки не образуют геммул, например Ephydatia fluviatilis в Тегельском озере в Германии •(Weltner, 1893). В этих случаях, поскольку дело касается умеренного пояса, зима переживается в состоянии редукционных почек (стр. 27).
В тропиках и субтропиках губки живут в совершенно иных условиях. Неблагоприятным временем является не холодное, а наиболее жаркое и засушливое время. В Индии Spongilla carteri процветает в течение холодного времени года с ноября по март и в конце этого периода образует геммулы. В марте, с наступлением жаркого времени, большинство губок погибает. Колонии оживают с началом периода дождей. Половое размножение происходит дважды в год при смене сезонов, в марте и в ноябре (Annandale, 1911). У нас нет данных о цикле жизни этой губки в пределах Союза. В Бухаре в городском пруде (хауз) экземпляры этой губки, а также Ephydatia fluviatilis, уже в начале мая содержали сформированные геммулы.
Другой индийский вид, Spongilla biseriata, найденный также и в Туркестане, в Индии живет активно в течение всего года. Жизнь отдельной колонии недолговечна и продолжается несколько недель, во время которых идет усиленное размножение наружными почками (стр. 26). К концу своего существования колония дает геммулы и погибает. У этой формы в любом сезоне можно наблюдать и живых губок, и находить геммулы (Annandale, 1911).
В жарких странах с сухим климатом губкам приходится проводить в состоянии геммул большую часть года. Так, у Parmula bravmi в Боливии стадия покоя длится 8—10 месяцев, у Dosilia- plumosa в реке Колорадо 9 —10 месяцев (Potts 1885; см. также Arndt, 1928, 1930; Schroder, 1932).
О жизни байкальских губок известно очень мало. Все же можно с уверенностью сказать, что они зимой не отмирают; даже в крупных ветвистых экземплярах Lubomirskia baicalensis мы не находим старой отмершей ткани. На разрезах ветвистых или корковых форм у различных видов всегда заметны слои нарастания (стр. 30), указывающие на неравномерность роста. Зимою происходит вероятно некоторая остановка или замедление в росте, но несомненно не все слои нарастания соответствуют годичным слоям. ’
О влиянии отдельных факторов среды на губки известно очень мало* В водоемах различного типа вся сумма господствующих там условий накладывает свой отпечаток на губки, но далеко не всегда мы можем связать с особенностями среды те или иные морфические изменения.
32 ФАУНА СССР
Так известно, что в некоторых крупных, а также высокогорных: озерах геммулы у бадяг недоразвивают своих оболочек и геммульных микросклер.1
Сила течения значительно влияет на внешний вид губок. Spongilla lacustris, развивающая в стоячих водах высокие кустистые формы, на течении дает корковые формы или развивает более короткие -ветви, направленные по течению. Lubomirskia baicalensis на сильном течений Ангары дает только корковые формы.
Повидимому, среди пресноводных губок нет видов, специально свойственных или только стоячим, или только текучим водам. Annandale (1911) отмечает, что Spongilla carteri в Индии встречалась ему только в стоячих водоемах. В пределах Союза, а также в Голландской Индии, она известна и из текучих вод.
Губки на желобах плотин, где они особенно хорошо развиваются, подвергаются сильному течению, которое само по себе не является помехой для их процветания. Но, конечно, перекатывание гальки в горных речках или прибой в больших озерах не дают развиваться губкам в таких местах.
Бадяги наиболее многочисленны у самого берега; далеко вглубь они обычно не идут. В более глубоких местах им часто мешает отсутствие подходящего субстрата и мягкий ил, покрывающий дно. Только в больших глубоких озерах они известны на более значительных глубинах (Иссык-куль до 30 м, Боденское озеро до 30—40 м). В оз. Бива в Японии и в Танганьике губки встречаются до исключительно больших глубин. (Бива 455 м: Kawamura and Annandale, 1916; Танганьика 640 м: Evans 1899).
В Байкале начинают встречаться с глубины в 1У2—2 м; зона от -2 до 25—30 м наиболее богато населена губками; глубже количество их убывает. Но попадаются они и на значительных глубинах. В 1932 г. добыта губка (Baikalospongia intermedia} с глубины в 889 м — глубина рекордная для пресноводных губок.
Развитие губок на больших, глубинах, а также неоднократно отмечавшиеся факты их разрастания в водопроводных трубах показывают, что свет не является необходимым условием для их существования. О том же говорит нахождение губок в пещерных водоемах.
Симбиотические водоросли могут развиваться только на свету, поэтому отсутствие света должно отражаться косвенно и на губках. Чем прозрачнее вода, тем шире будет наиболее благоприятная для губок зона.
Сильно мутные воды, содержащие большое количество минеральных частиц, помимо малой прозрачности, могут оказывать вредное влияние, забивая ирригационную систему частицами песка и ила. Тем не менее в оз. Ханке (Уссурийский край), где прозрачность всего около 0.5 м, губки встречаются, хотя особенно пышного развития и не достигают.
1 О таких морфах нам придется говорить ниже в систематической части. Некоторые авторы придавали им значение самостоятельных видов (Сварчевский, 1900, 1901).
ПРЕСНОВОДНЫЕ ГУ БКИ—ОБЩАЯ ЧАСТЬ
33
Для многих видов бадяг известны нахождения в солоноватых водах, морских и континентальных. В оз. Чилка (Chilka), обширной лагуне на восточном побережье Индии, Annandale (1915) находил бадяг вместе с чисто морскими губками. Ephydatia fluviatilis еще уживается при солености в 6% (Arndt, 1928 и 1930 а, в). В Финском заливе этот вид селится на фукусах, типичной морской водоросли (Weltner, 1893). Из реликтового озера в бухте Владимир (Японское море, Уссурийский край: Резвой, 1930) описана Е. fluviatilis var. hastifera, найденная при солености 6.89%.
Кроме опресненных участков моря, бадяги известны и из континентальных солоноватых водоемов. В Иссык-куле, например, встречается Ephydatia fluviatilis (Weltner, 1911), при солености в 5.8%, обусловленной главным образом присутствием сернокислого натра (Матвеев, Зап. Гос. Гидролог. Ин-та, № 6, 1932).
В пресных водах бадяги встречаются как в жестких и щелочных, так и в мягких и кислых водах, богатых гуминовыми веществами.
О влиянии концентрации водородных ионов имеется очень мало данных. В окрестностях Петергофа Ephydatia miilleri одинаково хорошо развивается в жестких и известковых водах при pH 8.1—8.2, так и в мягкой несколько гумифицированной воде при нейтральной или слабо кислой реакции (7.0 и несколько меньше).1 В оз. Горемыкском (Байкальский хребет, высота озера около 1000 метров), где в большом количестве встречается Spongilla lacustris, активная реакция среды немного выше 5. 15. Повидимому, к мягким, гуминовым водам специально приурочена Spongilla arctica.
Без аэрации и частой смены воды губки очень быстро погибают в аквариумах. Но их гибель может вызываться не недостатком кислорода, а отравлением воды выделениями самой губки. В водоемах губки- растут только в хороших кислородных условиях. В дневные часы еще прибавляется кислород, выделяемый зоохлореллами.
В зимнее время, в состоянии геммул, губки способны переносить не только полное отсутствие кислорода, но сильное отравление всей водной толщи сероводородом, что о<ень часто наблюдается подо льдом в таких прудах, где имеются губки.
Органическое загрязнение, и довольно значительное, если оно не сопровождается недостатком кислорода, не только не действует вредно, но скорее благоприятно для развития губок. Очень показательны в этом отношении богатые заросли губок в р. Шпрее в пределах Берлина, часто упоминаемые в литературе. Annandale (1911) отмечает, что в Индии наиболее богаты губками как раз загрязненные человеком пруды.
Так как свет не является необходимым условием существования губск, то они могут поселяться и в пещерах. Но в литературе губки очень редко указываются среди пещерного населения. В недавно появившейся
1 Данные об активной реакции по Рылову.
Фауна СССР, Губки, т. П, в. 2 3
34
ФАУНА СССР
работе Arndt (1933) приводит два нахождения губок в пещерных водоемах, а также сводит все относящиеся сюда литературные данные. Эренберг, изучавший грунты Адельсбергской пещеры (пещера Магдалины), находил в них спикулы бадяг (Ehrenberg, 1859). Но эти спикулы могли быть принесены поверхностными водами и отложены в пещере. Иозеф (Joseph, 1891) из пещеры Гурк в нижней Крайне описал бадягу, названную им Spongilla stygia. По его описанию ткань у живой губки была бесцветная и прозрачная; геммулы отсутствовали. Очень неполное описание не дает возможности составить ясное представление об этой форме. По мнению Арндта, а также и других авторов, здесь дело идет скорее всего об одном из обычных видов бадяг. В р. Эхо, протекающей через Мамонтову пещеру (Кентукки), Кофоид находил в пробах спикулы бадяг. Это дает основание полагать, что в этой пещере обитают губки (Kofoid, 1900). Арндт дает описание губок из двух карстовых пещер: из пещеры Шира-Яма в Югославии и из пещеры „ЗСЬго октября” около Сан-Канциана. В обоих случаях в губках имелись многочисленные геммулы. Обе находки Арндт относит к типичной форме Ephydatia mulleri. В первой из упомянутых пещер среди ее населения встречаются настоящие троглобионты, специально пещерные формы. Во второй пещере троглобионты отсутствуют, и ее население состоит из форм, постоянно приносящихся рекой, протекающей через пещеру.
Немногие известные случаи нахождения губок в пещерах указывают на их случайный занос. Каких-либо форм, специально приуроченных к пещерам, среди них не известно. Отсюда Арндт делает вывод, что губки среди пещерного населения должны рассматриваться как троглоксены, т. е. случайные гости пещерных биоценозов.
БИОЦЕНОТИЧЕСКИЕ ОТНОШЕНИЯ ГУБОК
Связи губок с другими организмами. Зоохлореллы и другие водоросли. Животные, питающиеся губками, комменсалисты. Поселение губок на живом субстрате. О биоценозе губок. Значение губок в жизни водоема.
Теперь мы попытаемся наметить в общих чертах те связи, которые существуют между пресноводными губками и другими водными организмами, растительными и животными. Связи эти чрезвычайно разнообразны и по своему характеру, и по степени взаимной связанности организмов.
Яркозеленая окраска свойственна большинству пресноводных губок. Она вызывается одноклеточными зелеными водорослями, „зоохлореллами”, как их обычно называют (фиг. 14). Маленькие овальные клетки этих водорослей в очень больших количествах заключены в протоплазме клеток губок, главным образом из поверхностных слоев губки. Они очень многочисленны и в скоплениях археоцитов при образовании геммул у бадяг и соритов у байкальских губок. Мы уже говорили выше в другом месте о ярко сине-зеленом цвете губок, добытых в Байкале со значитель
ПРЕСНОВОДНЫЕ ГУБКИ—ОБЩАЯ ЧАСТЬ
35
Фиг. 14. Амебоцит бадяг с зоохлореллами; сильно увеличено; изолированная клетка водоросли при большом увеличении. Видоизм. по ван Тригту (1919).
ных глубин (533—889 м). Ясно, что в этом случае окраска имеет какую-то другую природу.
Очень мало известно о цикле жизни этих растительных организмов, о способах их размножения и взаимоотношения с клетками губок, в которых они заключены. Недостаточно выяснено и их систематическое положение. Эти водоросли были выделены в чистой культуре и отнесены к роду Chlorella (Limberger, 1918), но ван-Тригт (1920) видит в них представителей рода Pleurococcus. Высказывались предположения, что в различных случаях дело идет о различных формах водорослей.
Отношение губки к водоросли обычно рассматривается как типичный симбиоз, одинаково выгодный для обоих компонентов. Губка пользуется кислородом и избытком ассимилятов, выделяемых водорослью. Водоросль же находит в клетке губки защиту, а также очевидно питательные соли и углекислоту, выделяемую губкой при дыхании. Ван-Тригт (1. с.) иначе смотрит на эти й^аимоотношения. Водоросли попадают внутрь тела губки вместе с водным tokojIh захватываются клетками, как и другие питательные частицы. Внутри амебоцитов водоросли не подвергаются немедленному перевариванию и, несомненно, способны здесь жить неко
торое время. По другим указаниям, внутри губочных клеток может* итти даже их размножение (Сварчевский, 1925). Но, по ван-Тригту, в конце концов клетки водоросли перевариваются. Именно этим он объясняет быстрое обесцвечивание губок, перенесенных в темноту, хотя вн^ губочных клеток водоросли в темноте не теряют зеленой окраски. На переваривание водорослей в клетках соритов указывает и Сварчевский (1. с.).
Известно также и нахождение в мезоглее Heteromeyenia baileyi var. palatina совершенно другой водоросли, Scenedesmus quadricauda (Lauter-born, 1902). Annandale (1911) приводит наблюдение над развитием в ткани губки (Spongilla alba) одной зеленой нитчатой водоросли. Нити водоросли пронизывают паренхиму и закупоривают каналы; при сильном разрастании водоросли губка погибает.
Животных, питающихся губками, сравнительно немного. Арндт приводит в своих работах полный их список (Arndt, 1924, 1928; см. также Р. Schulze, 1932).
Почти на каждом экземпляре бадяги можно найти на поверхности или в полостях маленькую зеленую личинку сетчатокрылого Sisyra (фиг. 15). В лупу хорошо видно, как она сосет своим двойным хоботком ткань губки, повидимому, не причиняя хозяину особого вреда.
Более заметны разрушения, производимые личинками ручейников, из которых ряд форм специально питается губками (Leptocerus senilis, 3*
36
ФАУНА СССР
fulvus, Ecnomus tenellus, Hydropsyche ornaiula, Neureclipsis bimaculatar Arndt, 1928 (фиг. 16). Ручейники протачивают в ткани канавки, иногда
Фиг. 15. Личинка Si-syra с Ephydatia miilleri. &ина личинки 6 мм (пруды Старого Петергофа).
забираясь глубоко в ткань (фиг. 17). Поврежденные части губки быстро регенерируют. Интересную картину представляют экскременты Leptocerus. Они состоят почти целиком из битых спикул. Такая пища должна оказывать самое энергичное воздействие на эпителий кишечника (фиг. 18).
В глубоких слоях губки прокладывают себе ходы личинки Chironomidae (мотыля), из группы Glyptoten-dipes. У некоторых водяных клещей (Hydracarind) личинки паразитируют внутри ткани бадяг. Иногда они буквально переполняют губку. Таков Unionicola crassipes (Wesenberg-Lund, 1918) (фиг. 19).
Есть указания, что на губок нападают и моллюски, например, Limnaea, Planorbis (Annandale, 1911). У Neritina fluviatilis наблюдались случаи, когда кишечник был переполнен спикулами (Simroth, 1902). Если иногда рыбы и нападают на губок (Annandale, 1911) и куски губок и спикулы бывали находимы в желудках рыб (Arndt, 1928), то все же вряд ли губки играют сколько-нибудь заметную роль в их питании.
На ветвистых Lubomirskia baicalensis всегда в большом количестве ползают яркозеленые рачки Acantho-gammarus parasiticus, а также некоторые другие
виды амфипод.
Помимо организмов, питающихся губками, мы найдем на их поверхности и в полостях, а также в отмершей ткани в основании колоний,
Фиг. 16. Личинка py4eaama'Leptocerus senilis, паразитирующая иа Ephydatia miillerv, к чехлику приклеены геммулы; передняя часть прикрыта растущей губкой; длина чехлика 14 мм (пруды Старого Петергофа).
довольно много животных форм. Часть из них так или иначе приурочена к губкам, некоторых можно считать комменсалистами (нахлебниками), но многие формы, в особенности из живущих в старой ткани, не имеют с губками каких-нибудь специфических связей и живут здесь, как и на других подходящих субстратах.
ПРЕСНОВОДНЫЕ ГУБКИ— ОБЩАЯ ЧАСТЬ
37
К комменсалистам можно отнести одну инфузорию из ац^нет, Podo-phria jixa, довольно постоянно встречающуюся в полостях у бадяг (фиг. 6). Такова же вероятно роль и других простейших, а также и коловраток.
Среди животных, населяющих старую ткань, иногда в большом количестве, встречаются олигохеты (виды JNais, Stylaria lacustris). Сюда же •относятся и различные Chironomidae и личинки ручейников, помимо
Фиг. 19. Нимфа гидракарины Atax (Un onicola) crassipesna Ephydatia miilleri (пруды Старого Петергофа). Увеличено.
Фиг. 17. Личинка ручейка Leptoceras senilis на Ephydatia miilleri-, чех-лик покрыт разросшейся тканью губки (пруды Старого Петергофа).
Наст, велич.
Фиг. 18. Экскременты личинки Leptocerus senilis; переполнены битыми спикулами Ephydatia miilleri, видны остатки гидракарин Atax crassipes (пруды Старого Петергофа). Длина экскремента 1.2 мм
перечисленных выше, из ракообразных — Asellus и различные Copepoda, все формы, не специально приуроченные к губкам.
Сами губки могут поселяться на живом субстрате, о чем мы уже говорили выше (стр. 3). На всякой подводной растительности, на тростниках, кувшинках, осокйх, на моховых подушках и на харах, на самых различных водяных растениях могут поселяться губки. Еще более удобный субстрат представляют обнаженные корни деревьев и свешивающиеся в воду ветви.
Губки могут селиться и на животных. Мы находим их не только на пустых, но и на живых раковинах Anodonta, Unto, Viviparus и других моллюсков. Часто можно видеть корочки губок на домиках ручейников. В последнем случае поселение на таком субстрате скорее объясняется не оседанием свободно плавающей личинки, а прикреплением ручейником геммул к своей постройке (фиг. 16).
В образе жизни губок и мшанок много общих черт. Очень часто мы их находим на одном и том же субстрате в ближайшем
38
ФАУНА СССР
соседстве. Но нередки случаи, когда трубки мшанки прорастают губоч-ную ткань.
Когда при разрастании входят в соприкосновение колонии разных видов губок, то, конечно, внутреннего их слияния произойти не может, но срастаются они настолько тесно, что только на микроскопическом препарате может быть найдена между ними граница. Очень часто один вид губки перекрывает другой и разрастается на его поверхности.
Из всего сказанного выше мы видим, насколько разнообразны те организмы, с которыми связаны губки, и насколько разнообразны и эти связи. Там, где имеются заросли губок, имеется и приуроченный к ним определенный биоценоз. Входящие в него организмы должны обладать, по крайней мере, одним общим свойством — выдерживать присутствие губок. С запахом губок, повидимому, связано выделение ими какого-то ядовитого вещества в окружающую воду. Относительно известковых губок имеются определенные наблюдения НаескеГя (1872), что мелкие организмы гибнут при приближении к губкам, после чего втягиваются водным током. Относительно бадяг Золотницкий указывает, что другие животные не уживаются с ними в аквариумах (Золотницкий, 1916).
На основании образа жизни губок мы можем составить себе представление о том месте, которое они занимают в жизни водоема. Оценивая значение губок в водоеме, приходится на первое место поставить их фильтрационную способность. Мы не знаем точно тех количеств воды,, которые за сутки пропускает весовая единица тела губки. Несомненно, что количества эти очень велики.]Из каждого оскулярного отверстия бьет струя, ясно заметная на расстоянии 15—20 см, и таких оскулярных отверстий на небольшой колонии можно насчитать свыше десятка. Большие заросли губок должны сильно влиять на очищение воды от всевозможных взвешенных частиц, в том числе измельченных гниющих органических остатков и бактерий, а также и мелких планктических организмов.
Таким образом для нас ясна роль губок в процессе самоочищения водоемов. Но проходящая вода подвергается не только механической очистке, но испытывает и химические воздействия благодаря выделению губкой продуктов распада, дыханию и деятельности зоохлорелл, продуцирующих кислород.
СИСТЕМАТИКА ГУБОК
Систематическое положение пресноводных губок. Семейства Spongillidae и Lubomirskiidae
Губки образуют отдельный тип животного царства. Среди многоклеточных животных они, несомненно, являются наиболее примитивно организованными существами. Как мы уже видели, в теле губки отдельные клетки обладают большой самостоятельностью и не образуют хорошо дифференцированных тканей и органов. Благодаря этой самостоятельности и подвижности клеточных элементов некоторые исследователи даже не признавали губок многоклеточными животными и смотрели на них, как
ПРЕСНОВОДНЫЕ ГУБКИ— ОБЩАЯ ЧАСТЬ
39
на колониальных простейших. Но дробление яйца и развитие личинки показывает, что мы имеем дело с настоящими многоклеточными животными.
Благодаря особенностям эмбрионального развития мы можем противопоставить губок остальным многоклеточным, разделив всех Metazoa (многоклеточных) на Parazoa с единственным типом и классом Porifera (губки) и Eumetazoa (все остальные типы многоклеточных: Hentschel, 1923). Принадлежность к типу Porifera определяется присутствием ирригационной системы, существенной частью которой являются жгутиковые камеры с воротничковыми клетками. Присутствие этой системы служит вполне надежным признаком для отличия губок от остальных животных.
Подразделение же типа Porifera до последнего времени подвергалось большим изменениям. В 1923 г. Хентшель (Hentschel, 1. с.) во втором издании сводки Кюкенталя предложил ряд новых перегруппировок; его система сравнительно с прежними несомненно имеет большие преимущества. По предложенной им системе тип и класс Porifera делится на пять отрядов:
1. Calcarea — известковые губки
2. Triaxonida — трехосные или шестилучевые
3. Tetraxonidae — четырехлучевые губки
4. Cornacuspongida — кремнероговые губки
5. Dendroceratida — ветвистороговые губки.
Пресноводные губки относятся к четвертому из этих отрядов, самому многочисленному по количеству форм, к Cornacuspongida (кремнероговым губкам). К этому отряду отнесено большинство прежних „одноосных" губок, Monactinellida, с которыми соединены, за некоторыми исключениями, прежние Keratoza (роговые губки).
Для отряда Cornacuspongida характерно устройство скелета, в большинстве случаев содержащего спонгин. Если спонгина мало, он только склеивает кремневые спикулы, при значительных его количествах он образует скелетные волокна, внутри которых заключены спикулы. Когда эти последние отсутствуют, скелет оказывается построенным из одного спонгина („роговые" губки). Скелетные волокна, в типичных случаях построенные из пучков спикул и спонгина, слагаются сетеобразно, реже древовидно или скелет состоит из беспорядочных скоплений спикул. Спикулы всегда одноосные (монаксонные); кроме скелетных спикул, макросклер, обычно присутствуют рассеянные в ткани микросклеры. У кремнероговых губок эти последние бывают двух типов: хелоидные, образованные изогнутым стволиком, с пластинчатыми или крючковатыми выростами, и сигмоид-н ы е, с-образно изогнутые спикулы. Микросклеры пресноводных губок, вообще имеющие вид прямых или изогнутых палочек, нужно рассматривать, как производные этого второго типа.
Строение скелета хорошо определяет отряд Cornacuspongida. Но с разделением отряда на семейства дело обстоит не так благополучно и в дальнейшем, вероятно, неоднократно будут производиться изме
40
ФАУНА СССР
нения и переделы. Надо заметить что для всех губок в ряде таксономических единиц семейство является наименее определенной единицей, для v которой обыкновенно труднее всего дать ясный диагноз. Это в особенности относится к Cornacuspongida.
Обычно всех пресноводных губок помещают в сем. Spongillidae, куда не входит ни одна морская форма. По системе Хентшеля несколько морских семейств (в том числе Celliidae и Chalinidae вместе с пресноводными Spongillidae') образуют один из подотрядов кремнероговых губок, подотряд Phthinorhabdina. Если руководиться только устройством скелета и спикул то пресноводные губки очень неясно отграничиваются от соседних семейств подотряда. Но Spongillidae в типичных случаях хорошо отличаются от соседних семейств по присутствию геммул, снабженных особо устроенными защитными микросклерами (геммульные микросклеры). Вообще геммулы известны не у одних бадяг, но у морских губок к ним не приурочены какие-либо особо устроенные спикулы. Вопрос осложняется тем, что у Spongillidae геммульные спикулы и самые геммулы могут отсутствовать. Мы уже упоминали, что и у обычных пресноводных видов при известных условиях недоразвиваются покровы геммул, а геммульные. микросклеры дают уродливые формы или отсутствуют. Известны колонии Ephydatia fluviatll-s, никогда не образующие геммул. У некоторых же родов, по всем признакам принадлежащим к настоящим Spongillidae, отсутствуют какие бы то ни было микросклеры, а иногда неизвестны и геммулы. Таковы роды Potamolepis и другие, объединяемые обычно в подсемейство Potamolepidinae, противуполагаемое подсемейству Spongillinae. Таким образом, присутствие геммул, снабженных покровными иглами, не является отличительным признаком всего сем. Spongillidae. Род Lubomirskia вместе с выделенными' из него в настоящее время р. р. Baikalospongia и Swartschewskia без каких-либо сомнений помещали и помещают и до настоящего времени в сем. Spongillidae. Байкальские губки не образуют геммул и не имеют микросклер;1 поэтому среди родов сем. Spongillidae они могли бы найти место в подсемействе Potamolepidinae.
Что байкальские губки должны быть совершенно выделены из сем. Spongillidae, об этом впервые поднял вопрос Annandale (1913, 1914). Он указал на существенное различие в устройстве канальной системы: у типичных бадяг сравнительно толстые отводящие каналы идут под дермальной мембраной коскулярному отверстию. Сходясь к нему с разных сторон и просвечивая сквозь мембрану, они образуют крупные звездчатые фигуры, для них очень характерные. У родов Potamolepis и др. (подсем. Potamolepidinae) также имеются субдермальные полости и звездчатые фигуры. У байкальских губок они совершенно отсутствуют, и канальная система имеет другое устройство. Здесь тонкие отводящие каналы подходят кпо-
1 О геммулах у Baikalospongia bacillifera, ошибочно описанных у Annandale см. стр. 24. О „паренхимных" спикулах у байкальских губок, см. стр. 13.
ПРЕСНОВОДНЫЕ ГУБКИ— ОБЩАЯ ЧАСТЬ 41
верхности из глубины ткани. Сходясь вместе, они открываются на дне неглубоких оскулярных ямок несколькими мелкими отверстиями. У бадяг от простого оскулярного отверстия вытягивается оскулярная трубка до 1 см длины. По устройству устий у байкальских губок можно думать, что таких трубок у них нет (наблюдать оскулярные трубки можно только на живом материале).
В строении скелета мы также находим большие отличия. У Lubomirskia baicalensis пучки спикул заключены в спонгин, и скелетные волокна образуют довольно правильную решетку. Главные пучки у поверхности оканчиваются коническими щетками спикул. У рода Baikalospongia спонгина меньше и строение скелета несколько менее правильное. Совершенно иначе устроен скелет у рода Swartschewskia’. в поверхностном слое имеется сеть плотных горизонтальных пучков, под которыми во внутренних частях губки спикулы располагаются беспорядочно. У байкальских губок, как и у морских Chalinidae, на разрезах видны „кольца нарастания" — полосы параллельные поверхности, с более густым расположением спикул, следы прежней поверхности, обросшей новой тканью.
У бадяг спонгина меньше, чем у байкальских губок. Большей частью он заметен только в местах соединения главных и поперечных пучков. Спонгинового волокна у них никогда не образуется. Только у одного вида бадяг, у Spongilla arctica, мы находим очень сильное развитие спонгина, но здесь он образует совершенно своеобразные перепонки, натянутые между скелетными пучками. Колец нарастания, очень характерных для байкальских губок, у бадяг не бывает. Наконец, существенным отличием можно считать образование соритов, описанных у В. bacillifera и найденных также и у L. baicalensis, заменяющих у байкальских губок геммулы. По описанию Сварчевского (1923, 1925) в сорите сохраняется одно ядро, из которого и развивается зародыш, и из сорита выходит свободноплавающая личинка. Из геммул же у бадяг выходят готовые молодые губочки, развивающиеся из целого комплекса клеток, заключавшегося в геммуле. Все эти отличия достаточно существенны, чтобы отделить байкальских губок от остальных пресноводных губок. Annandale (1914) ставит их в сем. Haploscleridae, причем род Lubomirskia им отнесен в подсем. Chali-ninae, a Baikalospongia (вместе с выделенными впоследствии р. Swart-schewskia) — в подсемейство Renierinae. По системе Хентшеля нам пришлось бы отнести р. Lubomirskia к сем. Chalinidae, a Baikalospongia и Swartschewskia — к сем. Gelliidae.
Мы не видим возможности разделять три рода байкальских губок по различным семействам или подсемействам. Мы считаем байкальских губок естественной группой, связанной родством и имеющей общее происхождение, и группа эта не может быть отнесена ни к семейству Spongillidae, ни к одному из морских семейств подотряда. Ее необходимо рассматривать как отдельное семейство Lubomirskiidae. Различия обоих пресноводных семейств, по нашему мнению, настолько существенны, что мало весьма
42
ФАУНА СССР
вероятно, чтобы семейство Lubomirskiidae произошло от семейства Spongillidae, или наоборот. Корни всей пресноводной фауны находятся в море. На байкальских губок, по нашему мнению, приходится смотреть, как на самостоятельных, независимых выходцев из моря, невидимому, более поздних, чем остальные пресноводные губки.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ ПРЕСНОВОДНЫХ ГУБОК
Если нанести на карту Союза все известные местонахождения пресноводных губок, то эти пункты расположатся очень разрозненно и между ними останутся обширнейшие, совершенно не обследованные пространства. Несмотря на такую слабую изученность территории Союза, все же и по имеющимся нахождениям мы можем составить себе общее представление о географическом распределении встречающихся у нас видов. Дальнейшее изучение фауны губок, конечно, существенно дополнит эту картину. Пресноводные губки (сем. SpongiUidae) являются космополитической группой. Наибольшего развития они достигают в тропиках и субтропиках; в умеренных широтах их фауна сильно обеднена, и только отдельные представители переходят за полярный круг.
В Палеарктике, в Европе и северной Азии, мы имеем бедную и довольно однообразную фауну бадяг. В последней сводке по Европе Arndt (1926) приводит 13 видов, а вместе с вариететами 25 европейских форм.1 Из числа этих 13 видов, нахождение в Европе Некоторых {Ephydatia robusta и Heteromeyenia tubisperma) является довольно сомнительным, для других сомнительна их самостоятельность {Spongilla helvettca) или же они с несомненностью должны быть низведены на степень вариететов других видов (5. rotundacuta). Прибавить же к списку Arndt’a можно в настоящее время только два вида (.S', arctica и З”. biseriata), что же касается форм ниже вида, то только немногим можно приписать характер подвидов; остальные же должны рассматриваться как формы или даже как уклонения, не выходящие за пределы индивидуальных вариаций. Фауну Европы можно назвать бедной при сравнении с другими странами:: Индия — 32 вида и 8 вариететов, Китай —16, Африка (чрезвычайно слабо обследованная) — 31 вид, Северная Америка — 32 вида (Arndt, 1. с ). Интересно отметить, что умеренные области Северной Америки имеют сравнительно с Европой значительно более богатую фауну.
То немногое, что известно относительно бадяг Сибири и Дальнего Востока (без Уссурийского края) указывает, что фауна этих районов сходна с европейской: ни одного вида, неизвестного в Европе, здесь до
1 Арндт приводит следующие виды: 1) Spongilla lacustris, 2) S. Helvetica, 3) б", rotund-acuta, 4) S. cart:ri, 5) & fragilis, 6) Ephydatia miilleri, 7) E. fluviatilis, 8) E. robusta, 9) Heteromeyenia ryderi, 10) H. repens, 11) Carterius stepanowi, 12) C. tubisperma, 13) Tro-chospongilla horrida.
ПРЕСНОВОДНЫЕ ГУБКИ—ОБЩАЯ ЧАСТЬ 43-
сих пор не констатировано;1 такие виды, как увидим ниже, появляются лишь в более южных частях Союза — в Туркестане и Уссурийском крае. Основное ядро фауны как в Западной Европе, так и на всей территории Союза, составляет группы широко распространенных форм: Spongilla lacustris, S. fragilis, Ephydatia miilleri, E. fluviatilis. Из этих четырех видов только Е. fluviatilis можно считать космополитом. S. lacustris, повидимому, отсутствует в Австралии и не найдена в Африке; в Индии типичная форма известна только в Гималаях, а в остальной Индии и Бирме ее заменяет хорошо выраженный подвид (subsp. reticulata Ann.). 5. fragilis не найдена в Африке; в Индии представлена хорошо выраженным подвидом (subsp. calcuttana Ann.), Ephydatia miilleri имеет голарктическое распространение, но не идет далеко к югу (вся Европа, Сибирь, Уссурийский край, Корея, Япония, Сев. Америка). В пределах Союза, в европейской части и в Сибири Е. miilleri представляет господствующую форму и. встречается так же часто, как и Spongilla lacustris. В Западной Европе она распространена повсеместно, но встречается реже, чем Е. fluviatilis и S. lacustris. Из форм, не столь широко распространенных, Troho-spongilla horrida известна из ряда европейских стран и из Северной Америки, в Азии констатирована в устье Аму-дарьи; в Китае описаны два близких к основной форме вариетета (var. sol Ann и ningpoen-sis Gee).
В недавнее время Schroder (1927) произвел ревизию рода Heteromeyenia и присоединил род Carterius к этому роду. Carterius stepanowi и ряд относившихся к нему вариететов он рассматривает, как вариететы американского вида Н. baileyi. Типичная форма встречается в Северной Америке в штате Нью-Йорк; вариететы же распространены в Европе и Азии, и, повидимому, всюду являются довольно редкими формами. Из них в пределах Союза покуда констатирована только одна форма Н. baileyi var. stepanowi на юге европейской части СССР, в окр. Харькова и в Ю. Буге в районе Николаева.2 Месяцев (1924) нашел спикулы этого вида в глубоких слоях одного из Косинских озер (окр. Москвы). Вне пределов Союза эта форма известна из Галиции и Венгрии.
Распространение Spongilla arctica еще нельзя считать достаточно выясненным. Нахождения вида в тундре Пе-мал, откуда вид описан (Полярный Урал) и иа Кольском полуострове говорят за его арктическую природу. В озерах в районе Порчнихи на Мурмане это единственный встречающийся там вид. Но имеется нахождение в Ленинградской области и еще значительно более южное нахождение в б. Рязанской губернии (озера Мещерской низменности—Глухое и Черное-Давыдовское).
1 Здесь не принимаются во внимание найденные в Байкале виды сем. SpongillidAe > описанные как новые, так как их самостоятельность более чем сомнительна (стр. 85—88).
2 Благодаря любезности М. А. Гримайловской, автор получил определенные ею препараты спикул экземпляра из Ю. Буга; данные об этом нахождении М. А. Гримайловской еще не опубликованы.
44
ФАУНА СССР
В Южно-уссурийском крае рядом с широко распространенными формами (S'. lacustris, S. fragilis, Eph. miilleri) мы находили и китайский элемент: Spongilla stanleyi в оз. Ханке и впадающих в нее реках является господствующей формой. Вид известен из Китая—из провинций Киаигсу и Чекианг. Весьма вероятно, что к китайскому элементу следует отнести и вариетет Eph. fluviatilis var. hastifera, описанный из реликтового озера в бухте Владимир. Характерный признак этой формы — копьевидные макросклеры встречается и у Eph. fluviatilis var. chui Gee, описанного из Китая, но остальные признаки не позволяют отожествить оба эти варие-тета. В фауне Средней Азии 'сказывается присутствие южноазиатского или, правильнее, тропического элемента; здесь констатирована Spongilla biseriata, найденная в дельте Аму-дарьи и в Арале в районе той же дельты. Этот вид встречается в Индии (Бенгалии), Бирме, Цейлоне, Малайском архипелаге (Ява, Целебес и Флорес), Филиппинских островах, Китае, в Ниле (окр. Каира) и в оз. Ниасса. Недавно найден и в южной Болгарии.
К южноазиатскому же элементу нашей фауны должна быть отнесена Spongilla carteri. Она широко распространена в южной Азии, а в Индии и Бирме является господствующей формой; ее ареал захватывает западную Персию, южный Китай, Малайский архипелаг; констатирована на о-ве Св. Маврикия. Африканские нахождения вида не вполне достоверны. В пределах Союза мы находим этот вид в Средней Азии (г. Бухара и Байрам-Али), в дельте Волги, в нижнем течении Дона (окр. Ростова), в Ю. Буге (район Николаева) и плавнях Днепра. Наконец, самым западным местонахождением является Балатонское озеро. Таким образом этот вид от района своего сплошного распространения в Индии через Персию, Туркестан, по устьям южнорусских рек протягивается далеко на запад. Форму из Балатонского озера Арндт рассматривает, как хорошо выраженный подвид subsp. balatonensis, основываясь, впрочем, исключительно на внешней форме балатонских экземпляров. При современных климатических условиях местонахождения вида в Балатонском озере, низовьях южнорусских рек и в Туркестане можно считать изолированными, так как между ними расстилаются обширные степные и пустынные пространства. На европейские колонии S. carteri мы вправе смотреть, как на реликты, сохранившиеся от значительно более теплой и влажной эпохи; скорее всего их можно считать третичными реликтами. Некоторые находки указывают на изменения в фауне бадяг, происшедшие в послеледниковое время. Месяцев исследовал отложения Косинских озер (окр. Москвы). Вся исследованная толща этих отложений должна быть отнесена к послеледниковому времени. В одном из озер в илу на глубине восьми метров, среди спикул различных губок им констатированы спикулы Carterius stepanowi (= Heteromeyenia baileyi vox. step anowi). Этот вид в настоящее время в районе Москвы не известен и встречается значительно южнее; нахождение его в озерных отложениях указывает на его более широкое распространение в один из более теплых периодов послеледникового времени.
ПРЕСНОВОДНЫЕ ГУ БКИ—ОБЩАЯ ЧАСТЬ 45
Совершенно другую картину географического распространения дает семейство пресноводных губок Lubomirskiidae. Все встречающиеся в Байкале виды семейства являются его эндемиками.1 Лишь в недавнее время описан вид, принадлежащий к байкальскому роду Baikalospongia из оз. Джегатай-куль в Урянхайском крае (Резвой, 1927). Высказывалось мнение, и даже в недавние годы (Макушок, 1925), что своеобразие байкальских губок может быть объяснено своеобразием экологических условий этого глубочайшего в мире озера. Нахождение Baikalospongia dzhegatajensis заставляет вопрос поставить иначе. Семейство Lubomirskiidae свойственно не одному Байкалу, и дело здесь не в экологических условиях этого „пресного моря". Только геологическая история всей, этой части Азии разъяснит нам в будущем, в каких водоемах эта группа могла сложиться, а об ее родственных отношениях мы сможем судить после детального сравнительно морфологического изучения как байкальских губок, так и бадяг, а также и некоторых морских семейств,, с которыми байкальские губки имеют несомненное сходство.
ИСКОПАЕМЫЕ ПРЕСНОВОДНЫЕ ГУБКИ
Породы, из которых известны ископаемые пресноводные губки, представляют собою видоизмененные грунты водоемов, частично или полностью потерявшие органические вещества и обогащенные минеральными • составными частями. Таковы пресноводные известняки, различные разновидности трепела или горной муки, диатомиты, полировальные и глинистые сланцы.
Микроскопическим изучением таких пород в течение ряда лет занимался Эренберг. В своей „микрогеологии" (Ehrenberg, 1854) он описывает и изображает спикулы пресноводных губок. При описании этих остатков малейшие отличия в очертаниях или какие-нибудь уродливости получают у него особые названия (Amphidiscus antediluvianus, Spongolithis furca, Lithastericus tuberculatus и т. п.). Все эти остатки, несомненно, принадлежат пресноводным губкам, но по описаниям и рисункам Эренберга нет возможности отнести его материал к каким-нибудь ныие живущим видам или составить себе ясное представление о вымерших формах. Позднейшие исследователи пересматривали материал из месторождений, изучавшихся Эренбергом, и им частично удалось дешифрировать его описания. Пресноводные губки в настоящее время констатированы во многих отложениях. Послетретичные находки отожествляются с современными видами, более же древние описываются как вымершие виды.
Всего для Европы в ископаемом состоянии (фоссильно или субфос-сильно) известно восемь видов (Arndt, 1926: Spongilla purbeckensis (-+-)-
1 Указание Дыбовского на нахождение типичной Lubomirskia baicalensis в Беринговом море не может приниматься во внимание. Более чем вероятно, что мы здесь имеем путаницу в этикетках (стр. 106—107).
>1<>
ФАУНА СССР
gigantea (•+), lachstris, fragilis, Ephydatia fossilis (•+), fluviatilis, miilleri, Heteromeyenia baileyi var. stepanowi). Самое древнее нахождение пресноводных губок было сделано в пурбекских слоях юрской системы, в пресноводном известняке южной Англии. Найдены исключительно гладкие спикулы среднего размера 370 у (и до 415 у) X 2—3 у. Эти остатки описаны как Spongilla purbeckensis Young (1887, р. 220). Из третичных слоев Чехии, относящихся к нижнему миоцену, описана Spongilla gigantea Traxler (1895, 1898), форма очень близкая к S. lacustris. Основанием для ее отделения от этого вида послужило присутствие среди геммульных спикул очень крупных игол, достигающих 200 у X 12 у и обилие пикнастров — мелких шарообразных кремневых образований, усаженных крупными шипами. Пикнастры изредка в виде уродливостей попадаются и у современных форм. К третичному же времени относится и Ephydatia fossilis Traxler (1894), найденная в глинистых сланцах и диатомитах Венгрии. Эта форма отличается очень крупными толстыми амфидисками, достигающими 41—60 у длины; диаметр диска 16—26 у, скелетные иглы частью гладкие, мелкошиповатые.
ЗНАЧЕНИЕ ПРЕСНОВОДНЫХ ГУБОК ДЛЯ ЧЕЛОВЕКА
Губки в рыбоводных прудах. Разрастание губок иа гидротехнических сооружениях и плавучих средствах. Закупорка водопроводных труб. Медицинское и косметическое применение „бадяги“. Применение губок для полировки металла.
Практическое значение губок приходится расценивать достаточно скромно. При исследовании водоемов с научно-промысловыми целями, поскольку учитывается вся фауна водоема, должны учитываться и губки. Непосредственного значения корма для рыбы они не имеют, но они должны найти себе место в общем балансе органического вещества в водоеме. В рыбоводных прудах губки иногда сильно разрастаются, и практикам приходится ставить вопрос о желательности или нежелательности такого явления. На основании всего предыдущего изложения мы можем ответить, что появление обширных зарослей губок в таких маленьких водоемах нельзя считать желательным, и необходимо бороться с их разрастанием. Такую борьбу можно вести очисткой спущенного пруда. В литературе есть указания, что спуск пруда не только уничтожает его наличное губочное население, но в некоторых случаях и без очистки после нового наполнения губки вновь не появляются.
Разрастание губок на различных гидротехнических сооружениях, шлюзах, плотинах и т. п. является, конечно, нежелательным. Появление же их в водопроводных трубах может иметь очень серьезные последствия» Отсутствие света, как мы уже говорили, не препятствует их росту. В литературе имеются довольно многочисленные указания на частичную или полную закупорку водопроводных труб, вызванную губками. При частичной закупорке уменьшается просвет и сильно увеличивается трение. При
ПРЕСНОВОДНЫЕ ГУ БКИ—ОБЩАЯ ЧАСТЬ
47
сутствие покрова из живых и отмерших губок вредно не только само по себе, но еще и тем, что способствует задержке всякого мусора, а гниение отмерших губок может дать воде отвратительный привкус и запах.
Для борьбы с заселением водопроводной сети губками единственным средством является фильтрование воды.
Губки также участвуют в обрастании судов. На деревянных баржах приходилось видеть сплошные их покровы.
Пресноводные губки находят себе, или вернее находили, некоторое применение в медицине. В русской народной (знахарской) медицине с давних пор применяется сушеная и растертая в порошок бадяга. Натирание „бадягой", сильно раздражающее кожу, применяется при ревматизме и других заболеваниях. Очень распространено и косметическое ее применение, получение „вполне естественного" румянца натиранием щек. Действие бадяги, конечно, основано на механическом раздражении кожи кремневыми спикулами. Из народной медицины бадяга перешла в прежнюю официальную медицину, и порошок бадяги еще в сравнитеььно недавние времена можно было получить в аптеках. В старой русской медицинской литературе отмечается применение бадяги после телесных наказаний (Барановский, Baranowsky, 1853). Из России употребление бадяги перешло и на Запад. Арндт (Arndt, 1930) отмечает, что кроме порошка бадяги, в недавние годы вошла в моду в гомеапатии тинктура „die Badiag-a-Tinktur", помогающая якобы от целого ряда болезней. Интересно отметить, что. бадяга получалась в Германии из России, хотя там пресноводных губок не меньше, чем у нас (Arndt, 1. с.). Благодаря этому на Западе привилось и русское народное название пресноводных губок. Война прекратила этот экспорт.
Мы можем указать некоторое применение пресноводных губок и в технике. Еще у Палласа (1'773, том III: 100) мы -находим, что местное население на Байкале употребляет губки для чистки и полировки металла.
СОБИРАНИЕ И СОХРАНЕНИЕ ГУБОК
Техника собирания и консервировки губок. Фиксация и сушка. Этикетировка.
Мы уже говорили, что бадяги являются обитателями по преимуществу береговой полосы водоемов. Здесь их приходится прежде всего искать. При сборе трудно обойтись без лодки. Самые надежные результаты получаются когда есть возможность непосредственно увидеть растущих на том или ином субстрате губок. Для этого надо пользоваться солнечной погодой и время выбирать около полудня, когда всего лучше освещается дно. При некотором навыке губок легко отличить, несмотря на их зеленую окраску, от подводной растительности. Для высматривания губок очень полезно пользоваться „морским окном" — небольшим деревянным ящиком со стеклянным дном; такой ящик плавает у борта лодки,
48
ФАУНА СССР
привязанный на веревке. Еще удобнее устроить длинный конус из листового железа, на подобие рупора, со стеклом, вставленным в его раструб.
Замеченные губки добываются руками, сачком или скребком. Если они растут на корягах или хворосте, то такой материал поднимается багром или граблями. Для добывания травянистой растительности, на которой имеются губки, удобно также пользоваться кошками, устроенными в виде маленького якорька или как-нибудь иначе; кошки своими крючьями вырывают растения. С больших камней и скал губки сдираются скребками. При помощи камнещупа Рубцова можно поднимать довольно крупные камни с растущими на них губками. На больших глубинах и в малопрозрачных водах сбор приходится вести драгами различных систем.
Все указанные выше приборы и способы пользования ими описаны в недавно изданной инструкции Гидрологического института по сбору бентоса, и мы не будем останавливаться на их детальном описании.1
Если губка добыта вместе с субстратом, то ее, по возможности, в таком виде и сохраняют. С поднятых каряг губки срезают вместе со слоем дерева. Если губку нельзя сохранить вместе с субстратом, то надо следить, чтобы на субстрате не осталось слоя геммул, без которых не может быть надежного определения собранного материала. Если добытый экземпляр слишком велик и не помещается во взятую посуду, то от него для' фиксации отрезают часть, но с таким расчетом, чтобы на взятом образце была и наружная поверхность и основа колоний со старой тканью и местом прикрепления к субстрату. Остальная часть экземпляра может быть засушена (см. ниже).
Лучшим фиксатором для систематических целей является 95 °/0 спирт-ректификат. Надо избегать спирта-сырца. Этот недоброкачественный продукт действует мацерирующим образом на животные ткани. В банку со спиртом нельзя класть слишком много губок, чтобы жидкость не потеряла необходимой крепости (должна хорошо гореть). Через день-два желательно спирт сменить и залить сбор 80—75% спиртом. Можно обойтись без смены, если губок в банке не больше Уз—% объема и спирт достаточно крепок. Нет необходимости пользоваться чистым спиртом. Вполне пригоден спирт денатурированный 1% метилового спирта („белый" денатурат). Сохранение губок в формалине совершенно недопустимо. Формалин очень быстро их мацерирует.
Для гистологических целей употребляются специальные фиксаторы, осьмиева кислота и различные смеси, содержащие сулему. На этой специальной методике мы останавливаться здесь не будем.2
Для сохранения в целости нежной дермальной мембраны надо заворачивать каждый экземпляр в тонкую материю, лучше всего в марлю.
1 Инструкция по биологическим исследованиям вод. Под ред. К. М. Дерюгина. Часть II, А. Исследование бентоса, изд. ГГИ. Лгр. 1931.
2 См. Krause Enzyklopedie d.jnikr. Technik, I, (p. 399); Mayer, 1920, Zoomicrotechnik. (p. 325); P e t e r f i, Method, d. wiss. Biol., I, 1928 (p. 1264).
ПРЕСНОВОДНЫЕ ГУБКИ—ОБЩАЯ ЧАСТЬ
49
Это желательно делать на месте, при фиксации. При таком способе в одну банку можно класть губки, собранные на разных станциях в том же водоеме или даже в разных водоемах, так как при этом этикетка кладется внутрь марлевого пакетика, который перевязывается веревочкой. Пакетики не должны болтаться в банке, поэтому все свободное место заполняется каким-нибудь материалом — ватой, паклей или тонкой упаковочной стружкой. Сухой материал вполне пригоден ддя систематических целей, за исключением некоторых случаев, когда нужна проверка на присутствие пузыревидных клеток (см. выше, стр. 10 и ниже стр. 52). Поэтому при сушке желательно хотя маленький кусочек, около 1 кб. см, сохранить в спирту.
Для сушки губки кладут где-нибудь на сквозняке или около печки, но не на солнце. Более крупные экземпляры перед сушкой полезно вымочить несколько часов в 3— 4% формалине (продажный формалин разводится в 10—15 раз водой). За такое короткое время формалин не может оказать вредного действия на ткань.
Сухие губки очень ломки и легко растираются в порошок. Их приходится упаковывать очень тщательно. Имея дело с сухими губками, надо беречь глаза и дыхательные пути от пыли, которая легко от них получается.
Без правильно составленной этикетки материал теряет свою научную ценность. В отношении этикеток сборщики часто допускают досадные пробелы, а потому мы здесь перечислим те требования, которым этикетка должна удовлетворять:
1. Этикетка должна сама по себе достаточно характеризовать материал, независимо от каких-либо записей, журналов или объяснительных записок.
2. В этикетке указывается место сбора, время сбора и к е м собран материал.
3. Место сбора указывается так, чтобы оно легко могло быть положено на карту, хотя бы и приблизительно. Для мелких малоизвестных водоемов, кроме его названия, надо дать направление и расстояние до ближайшего населенного пункта, который легко отыскать на карте. Для крупных общеизвестных водоемов таким же образом определяется место на водоеме, где произведен сбор.
4. Необходимо указывать глубину (в особенности в более крупных озерах) и субстрат, с которого снята губка, а также цвет живой губки, так как губки в спирту и прй сушке обесцвечиваются.
5. Записывая месяц и число сбора, надо проставить и год, о котором часто забывают.
6. Фамилия сборщика непременно должна быть указана. Без нее материал является анонимным и не внушающим доверия.
7. В экспедициях, где ведется правильная нумерация взятых станций, на этикетке, кроме всего указанного, отмечаются эти номера. По ним могут быть получены дополнительные данные о месте сбора, температурах, химизме и прочих условиях местонахождения.
8. При единичных и случайных сборах полезно, если все Дополнительные данные сборщик даст в краткой записке, которую приложит к материалу.
9. Очень существенно качество бумаги для этикеток. Этикетка обязательно кладется внутрь банки и должна сохраняться в жидкости, не превращаясь в труху. Пишут мягким, черным карандашом (не химическим), сильно нажимая. При вовможности этикетка заменяется другой, написанной на пергаментной бумаге тушью (карандашом писать на пергаменте нельзя. Проверить, не расплывается ли тушь в жидкости).
Фауна СССР, Губки, т. В, в. 2 4
50
ФАУНА СССР
ПРЕПАРОВКА ГУБОК
Препаровка губок для микроскопического исследования. Препараты изолированных -спикул. Ручные срезы для изучения скелета и геммул. Окраска спонгина. Препараты пузыревидных клеток.
Определение губок ведется под микроскопом, для чего необходимо приготовление постоянных или временных микроскопических препаратов.
Для систематических целей руководящее значение имеют спикулы, для чего их надо получить в изолированном виде.
Для ориентировочного просмотра, когда не требуется приготовление постоянных препаратов, удобно пользоваться жавелевой водой на холоду.1 Очень маленький кусочек губки (2—3 мм в поперечнике), содержащий геммулы, кладется на предметное стекло в каплю жавелевой воды. В несколько минут ткань должна разрушиться. Тогда ее осторожно промывают, прибавляя капли воды и отсасывая жидкость фильтровальной бумагой. После некоторой промывки покрывают покровным стеклом. Операцию с жавелевой водой можно вести и в часовом стекле. После разрушения ткани несколько раз прибавляют и отсасывают пипеткой воду. Нагревать или кипятить спикулы в жавелевой воде нельзя: едкая щелочь при нагревании разъедает кремнезем.
Для постоянных препаратов кусочки губки с геммулами вывариваются в крепких кислотах, азотной, соляной или серной. Серная всего удобнее, так как не выделяет едких паров. Вместо серной кислоты удобно пользоваться хромовой смесью — серной кислотой, к которой прибавлена двухромокалиева соль. Для вываривания губок крепкая серная кислота (или крепкая хромовая смесь) разбавляется вдвое водой. В химическую пробирку опускают кусочек губки (около 5 мм в поперечнике) и очень осторожно (беречь глаза и платье!) кипятится в 2—3 куб. см жидкости. Пробирку полезно заткнуть рыхлым тампоном ваты. В 1—2 минуты кусочек губки распадается, разрушаются также и геммулы.
Содержимое из химической пробирки переливается в центрифужную коническую пробирку, добавляется дестиллированная вода до определенной метки, пробирка затыкается пальцем и перебалтывается, чтобы вся жидкость была однородной и на дне не осталось бы тяжелой хромовой смеси.
Центрифугируют короткое время, 1—2 минуты, после чего жидкость с осадка спикул отсасывается пипеткой. Теперь осадок надо промыть, для чего вновь наливают дестиллированную воду, перебалтывают и центрифугируют снова и повторяют это раза два или три. При очень мягкой водопроводной воде (например, в Ленинграде) можно обойтись без дестил-лированной воды. Отсосав лишнюю воду, берут пипеткой несколько
1 Жавелевая вода — крепкий раствор едкого натра, насыщенный хлором. Обращаться с осторожностью; прожигает платье и очень вредно действует на металлические предметы, лежащие поблизости.
ПРЕСНОВОДНЫЕ ГУ БКИ—-ОБЩАЯ ЧАСТЬ
51
капель вместе с осадком и выпускают их на предметное стекло. Выпущенная на стекло капля высушивается над пламенем спиртовки.
Хорошо промытые спикулы на высушенном стекле должны лежать прозрачным, еле заметным осадком. Получение мутного белого налета показывает, что промывка была недостаточной. Спикулы на препарате должны лежать не слишком густо, чтобы не затруднять просмотра.
Если губка была добыта из богатых известью вод, то при выварке в серной кислоте (и хромовой смеси) на препарате вместе со спикулами окажутся и микроскопические кристаллики гипса. Этого легко избежать, если после первого центрифугирования и удаления жидкости с осадка прибавить немного соляной кислоты, после чего продолжать промывку. Описанный способ получения препаратов может показаться сложным и мешкотным. Тем не менее он удобен и надежен, в особенности когда обрабатывается большой материал. Тогда все операции ведут последовательно: сразу отделяют кусочки губок в целый ряд пробирок, в раструбы которых вставляют написанные на бумажках этикетки. Затем идет выварка в кислоте и центрифугирование по четыре или по две, в зависимости от устройства центрифуги. Само центрифугирование 'во много раз ускоряет оседание спикул и дает гарантию, что не будут потеряны микросклеры.
На высушенный препарат опускают каплю канадского бальзама и накрывают покровным стеклом без ножек.
Пипетки и пробирки необходимо тщательно мыть, чтобы не занести на препарат посторонних спикул.
Для изучения скелета и геммул приготовляются срезы от руки. Их можно готовить по методу, принятому у ботаников: режется бритвой маленький кусочек губки, зажатый в сердцевину бузины. Большею частью удается сделать достаточно тонкие срезы и скальпелем, положив губку на кусок пробки. Получаются срезы от 0.5 — до 1 мм.
Срезы обезвоживаются в абсолютном спирту (минут 10) и просветляются в гвоздичном масле (до полного просветления геммул). Заделывают их в канадский бальзам, покрывая стеклышком с восковыми ножками.
В некоторых случаях для более детального изучения скелета, в особенности для выяснения количества спонгина, приходится делать препарат мацерированной губки. Иногда на материале удается найти такие мацерированные участки. Для мацерации кусочки губки или ручные срезы продолжительное время (неделями) вымачиваются в водопроводной воде. Рекомендуются также растворы поташа или соды, а также выдерживание в аммиаке в термостате (около 30°). Есть указания на применение трипсина (см. литерат. указания на стр. 48).
Для изучения спонгина его необходимо окрасить. Хорошие результаты дает кислый фуксин (Fuksin S) с пикриновой кислотой: в часовое стеклышко наливают около 1 куб. см насыщенного спиртового раствора пикриновой кислоты и прибавляют 5—10 капель 1°/0 водного фуксина.
4*
52
ФА УНА СССР
Окраска длится около 10 минут. Окрашенные срезы промываются в спирту,,, обезвоживаются в абсолютном спирту, просветляются и заделываются в бальзам.
Для рода Ephydatia требуется еще одна дополнительная операция, проба на пузыревидные клетки (стр. 10). Кусочек ткани, свежей или фиксированной (2—3 мм в поперечнике) расщипывается иглами в капле воды на предметном стекле и исследуется под микроскопом.
Для изучения гистологической структуры специально фиксированный материал заливается в целлоидин-парафин и режется на микротоме вместе со спикулами. В зависимости от цели исследования применяются те или иные окраски. Этой гистологической методики мы здесь касаться не можем (см. указанную выше литературу по микроскопической технике).
Для изучения строения живой функционирующей губки ван-Тригт (1919) выработал своеобразную и остроумную методику, основанную на широкой регенерационной способности губок. Он брал .цилиндрический отросток Spongilla lacustris (этот вид наиболее удобен, так как обладает быстрым ростом) и разрезал его вдоль по всей длине. Затем эти половинки, нарезались на кусочки длиною в 1 см. Такой кусочек клался плоской стороной на большое покровное стекло и помещался в аквариуме. Скоро ткань губки прирастала к стеклу, и от кусочка по поверхности стекла распространялась тонкая прозрачная мембрана. Через неделю эта мембрана увеличивалась в толщине и в ней появлялись жгутиковые камеры, и каналы. Регенерированная губка на покровном стеклышке помещалась стеклом кверху на стеклянных ножках в маленький сосуд с водой таким образом, чтобы. вода не покрывала поверхности стекла. Такой живой препарат можно изучать под микроскопом при любых увеличениях до масляной иммерзии включительно.
О СОДЕРЖАНИИ ПРЕСНОВОДНЫХ ГУБОК В АКВАРИУМАХ
Для целого ряда ценных наблюдений и легко доступных экспериментов, имеющих большое значение и для систематики, пресноводные губки держат в аквариумах.
Даже кратковременные наблюдения над губкой, принесенной с экскурсии и посаженной в сосуд с водой, могут оказаться очень полезными. При этом можно рассмотреть устройство оскулярных трубок, познакомиться с различными паразитами и сожителями губок, а также иногда получить и свободноплавающих личинок.
При продолжительном содержании губок возможны более систематические наблюдения и эксперименты, но для этого надо создать им подходящие для жизни условия. Держать губки в аквариумах не легко, благодаря их высокой требовательности. Здесь мы дадим некоторые практические указания по технике этого дела (см. Schroder, 1930, Золотницкий, 1916).
ПРЕСНОВОДНЫЕ ГУБКИ—ОБЩАЯ ЧАСТЬ ;53
Губки для содержания в аквариумах берутся вместе с субстратом. При добывании их лучше не вынимать из воды, а под водой же поместить в сосуд достаточной вместимости. При переноске надо избегать сильного нагревания, . для чего сосуд обертывается мокрой тряйкой. Их можно переносить и без воды, завернутыми вместе с водяными растениями в мокрую тряпку. Такую упаковку губки выдерживают в течение нескольких часов.
Размеры аквариума могут быть и не велики, если держать в них небольшие колонии. Но, конечно, чем больше аквариум, тем легче в нем приживаются губки. Вместимость от 3 до 15 л можно считать достаточной. Слой воды не должен быть слишком глубок, чтобы не затруднять аэрации. При непроточных аквариумах Золотницкий рекомендует наливать слой воды не выше двух вершков.
Основными вопросами являются проточность воды и аэрация. Держать губки продолжительное время можно только в проточной воде, причем предпочтительнее вода прудовая или озерная, а не водопроводная.
Для питания аквариума проточной водой устраивают достаточных размеров бак, куда наливается или накачивается вода и откуда она по трубкам распределяется по аквариумам. Можно воспользоваться и водопроводом, если вода в нем не слишком богата железом.
Даже при хорошем обмене воды трудно обойтись без аэрации (продувания воздуха). Для этого в аквариальной технике применяются специальные аппараты, состоящие из резервуара, куда ручным насосом накачивается воздух. По резиновым трубкам воздух проводится к аквариумам. На конец трубки насаживается специальный наконечник из пемзы или пористого камня, сквозь который воздух проходит в виде мельчайших пузырьков.
Но и хорошая аэрация не уменьшает необходимости постоянного возобновления воды. Губки, как нам уже приходилось говорить, не столько страдают от недостатка кислорода, сколько от отравления воды продуктами их обмена. При непроточных аквариумах Золотницкий рекомендует менять воду каждый час.
Прямой солнечный свет и перегревание вредно действуют на губки, поэтому аквариум надо ставить так, чтобы он не освещался Солнцем. Шредер считает, что 20—25° С являются наиболее благоприятной температурой, Золотницкий рекомендует более низкие температуры (17—19° С). Если условия обмена и аэрации не слишком благоприятны, то лучше, если температура будет более низкой.
При хорошем обмене воды обыкновенно нет надобности заботиться б питании губок: сама вода приносит достаточное количество пищи. Но водопроводная вода может оказаться недостаточно питательной. Чтобы улучшить условия питания, на дне аквариума устраивается грунт и засаживается не слишком густо водяной растительностью. Это способствует развитию микрофлоры и фауны. Полезно также подсадить в аквариум
54
ФАУНА СССР
различных животных, рыбок или моллюсков, которые сами обычно губкам не вредят, а их экскременты обогащают воду питательным детритом. Для питания губок рекомендуют также настои сена или салатных листьев,, в которых развивается богатая бактериальная флора. Такие настои прибавляются к воде небольшими порциями. Шредер прибавлял также прудовой плактон, выловленный тонкими номерами мельничного газа. Такая подкормка может > давать хорошие результаты, но прибавка гниющих настоев сопряжена с загрязнением аквариума, с появлением плесеней и может вредно отозваться на губках.
Ван-Тригт (v. Trigt, 1919), много экспериментировавший над губками, держал их в аквариумах 50—100 л вместимости. Аквариумы были проточные, питались постоянно протекавшей водопроводной водой и полный обмен воды происходил в течение 5—10 часов. В аквариумах не было ни грунта, ни растений, не давалось также никакой подкормки. Все это ван-Тригт считает вредным. Губки помещались на металлических сетках в 10—20 см от дна и на 15—20 см под поверхностью воды. Иногда губки воспитывались и в непроточных сосудах 3—5 л вместимости, с возобновлением воды раз в сутки.
ГЛАВНЕЙШАЯ ЛИТЕРАТУРА ПО ГУБКАМ
Гримайл1вська, М. А. 1927. До фавни Spongillidae та Bryozoa П1вд. Бог. Сбирник: Б1олог. станц., Киев: 267—276.
Даль, В. 1880. Толковый словарь живого великорусского языка. 2-е изд., I.
Д ы б о в с к и й, В. И. 1884 а. Заметка о бодягах южной России. Тр. Общ. исп. пр. Харьк. ун., 17.
---- 1884 б. Дополнительные сведения к познанию пресноводной губки Dosilia stepanowi. Тр. Общ. исп. пр. Харьк. ун., 18: 201—208.
Золотницкий, Н. Ф. 1916. Аквариум любителя, изд. 4-е.
Зыков, 1892. Дополнения к фауне окрестностей Москвы. С. R. Bull. Soc. NatMoscou., n. s., 6.
Исаев, В. M. 1924. Этюды об органических регуляциях. Тр. Ленинград. Общ. Ест., 53: 176-275.
К о ж о в, М. М. 1925. Очерк по фауне пресноводных губок Иркутской губ. и Прибайкалья. Изв. Биол. геогр. Н.-исс. инет. Ирк. ун., 2: 27—65.
---- 1931. К познанию фауны Байкала, ее распределения и условий обитания. Изв. Биол. Геогр. Н.-и. инет. Ирк. ун., 5: 3—170.
Маку ш ок, М. Е., 1925. К вопросу о происхождении спонгиофауны озера Байкала. Русск. зоол. жури., 5: 50—78.
---- 1927. К систематике байкальских губок. I, II. Русск. зоол. журн., 7: 79—124. Матвеев, В. П. 1932. О солености оз. Иссык-куль. Зап. Гос. Гидролог. Ин-та, № 6. Месяцев, И. И. 1924. Ископаемая фауна Косинских озер. Раб. Косинск. биол. ст., I: 35. Пр ен дель, А. Р. 1915. Заметка о губках Днестровских плавней. Тр. Бессарабск. Общ.
ест., 1915, 6: 1—7.
Резвой, П. Д. 1922. Заметка о пресноводных губках, собранных в районе Окской биологической станции. Раб. Окск. биол. ст., 2: 12—18.
—- 1923. К спонгиофауне реки Волги. Раб. Волжск, биол. ст., 7: 2—5.
1926. К фауне пресноводных губок нижней Волги. Раб. Волжск, биол. ст., 9: 65—69.
ПРЕСНОВОДНЫЕ ГУБКИ—ОБЩАЯ ЧАСТЬ
55
Рылов, В. М. 1929. Некоторые наблюдения над концентрацией водородных ионов в водоемах окрестностей Петергофского естественно-научного института. Русск. гидроб. журн., 8.
Сварчевский, Б. А. 1900. О губках Байкальского озера. Зап. Киевск. общ. ест., проток., 17: IX—XIII.
---- 1901. Материалы по фауне губок Байкальского озера. Зап. Киевск. общ. ест., 17: 329—359.
---- 1923. Спонгиологические очерки. I, II. Тр. Ирк. общ. ест., 1: 329—352.
---- 1925. Спонгиологические очерки. III. Изв. Биол. Геогр. Н.-и. инет. Ирк. ун., 2:1—30.
Совинский. 1890. О фауне пресноводных губок Днепра. Вест, ест.: I—II.
Сукачев, Б. 1895. Несколько новых данных о губках оз. Байкала. Тр. СПб. общ. ест., 25: 1—18.
Ч е р н а й, А; 1870. Об иглах губок, встречающихся в иле озера Лиман и Чайка Змиев-ского уезда Харьковской губ. Тр. Харьк. общ. исп. пр., 1: 1—4.
Annandale, N. 1907 a. Notes on the freshwater fauna of India. IX. Jour. Asiatic Soc. Bengal, n. s., 3: 15—26.
---- 1907 b. The nature of the pores in Spongilla. Rec. Ind. Mus., I: 269—271.
---- 1911. Freshwater Sponges, Hydroids et Polyzoa. Faun. Brit. Ind.: 1—126.
---- 1913. Notes on some sponges from Lake Baikal. Ann. Mus. Zool. Ac. Imp. Sc. St. Pet., 18:96-101.
—— 1914. Further notes on the Sponges of Lake Baikal. Rec. Ind. Mus., 10: 137—148.
---- 1915 a. Description of a freshwater sponge from the North-West of Siberia. Mem. Ac.
Imp. Sc. Petrogr., s. 8, 28: 1—3.
---- 1915 b. Fauna of the Chilka Lake. Mem. Ind. Mus., 5: 1—146.
---- 1916. Freshwater Sponges from the T’ai Hu (Great Lake) of the Kiangsu Province of China. J. N. China Br. Roy As. Soc., 47: 49—52.
---- 1918. Zoological Results of a Tour in the Far East. Sponges. Mem. As. Soc. Bengal, 6: 193—216.
Annandale, N. and Kawamura. 1916. The’sponges of Lake Biwa. Tokyo Journ. Coll. Sci, 39, Art. I: 1—27.
Arndt, W. Bau und Leben der deutschen Siisswasserschwamme. Mikrosk. fiir Nat.-freund., Jahrg. 4, H. 5: 1—14.
---- 1923. Balkanspongillen. Zool. Anz., 56: 74—81.
---- 1926. Die Spongillidenfauna Europas. Arch. Hydrobiol., 17: 337—365.
---- 1928 a. Lebensdauer, Alter und Tod der Schwamme. Sitz. Ber. Ges. Naturf. Fr. Berlin: 23—44.
---- 1928 b. Porifera, Schwamme, Spongien. In Dahl, Tierwelt Deutschlands, 4: 1—94.
---- 1930a. Schwamme. In Oppenheimer und Picussen, Tabulae Biologicae, 6, suppl. 2: 39—797.
---- 1930 b. Schwamme und Gesundheitswesen. Zeitschr. Desinfect. Gesundh., Jahrg. 23, H. 3: 1—25.
---- 1933. Ephydatia miilleri (Liebk.) als Bewohnerin von der Hercegovina und des Kar-stes. Mitt. Hohlen- u. Karstforsch.: 19—31.
Baranowsky. 1853. Die Heilkrafte der Spongia fluviatilis. Med. Ztg. Russlands. St. Pet., 10: 123.
Bower bank, J. S. 1863. A Monograph of the Spongillidae. Proc. Zool. Soc. London, Nov. 24: 440—472.
Buxbaum J. C. 1729. Nova Plantarum genera (Badiaga) etc. Comm. Ac. Petropol., 2: 343.
Carter, H. J. 1849. A descriptive Account of the Freshwater Sponges in the Island of Bombay, etc. Ann. Mag. N. H., s. 2, 4: 81.
---- 1859. On the Identity in Structure and Composition of the so called Seed-like Body of Spongilla with the Winter-egg of the Bryozoa. Ann. Mag. Nat. Hist., s. 3, 3: 331—343.
56 ФАУНА СССР
Cotte, J. 1904. Des phenomenes de la nutrition chez les Spongiaires. C. R. Ass. Franc. Av. Sc., 32: 776—780.
Delage, J. 1892. Embryogenie des Eponges. Arch. Zool. exp. et gen., s. 2: 10.
Dybowski, W. 1878. Mitteilung fiber Spongien. Zool. Anz., 1: 30—32 und 52—54.
---- 1880 a. Studien fiber die Spongien des Russischen Reiches etc. Mem. Ac. Imp. Sc. St. Petersburg, s. 7, 27: 1—69.
---- 1880 b. Einige Bemerkungen uber die Veranderlichkeit der Form und Gestalt von Lubo-mirskia baicalensis. Bull. Ac. Imp. Sc. St. Petersburg, Mel. biol., 9: 41—47.
---- 1882. Studien uber die Siisswasser-Schwamme des Russischen Reiches. Mem. Ac. Imp. Sc. Petersburg, s. 7, 30.
---- 1884. Notiz iiber die aus Siid-Russland stammenden Spongillen. Sitz. ber. Naturf. Ges. Dorpat, 4: 507—515.
---- 1886. Mitteilung iiber einen neuen Fundort des Schwammes Lubomirskia baicalensis. Sitz. ber. Naturf. Ges. Dorpat, 7: 44—45.
Ehrenberg, 1846. Ueber die geformten uncristallinischen Kieselteile von Pflanzen, beson-ders iiber die Spongilla erinaceas etc. Ber. Ac. Berlin: 96—101.
---- 1854. Mikrogeologia.
---- 1859. Ueber die mit Proteas anguinus zusammenlebenden Tierformen. Mon. Ber. Ak. Berlin: 758—777.
Evans, R. 1899. A description of two new species of Spongilla from lake Tanganyika. Quart-Journ. Mier. Sei. (n. s), 41.
Galtzof, P. S. 1901. A description of Ephydatia blembingia with an account of the formation and structure of the gemmules. Quart. Journ. Mier. Soc., 44.
---- 1923. The amoeboid movement of dissociated sponge cells. Biol. Bull. Woods Hal. 45: 153-161.
Gee Gist, N. 1926. Freshwater sponges. The China Journ. of Sc. a. Arts, 4: 180—184.
---- 1927. Chinese freshwater sponges. The Lingnaan Agricult. Rev., 4: 1—10.
Gee Gist, N. and W u C. F. 1925. Description of some freshwater sponges from China 1. 3, № 11: 609—610.
;--- 1927—28. Chinese Fresh-water Sponges. A Synopsis etc. Bull. Peking Soc. N. H., 2:1—14.
G о r i c h, W. 1904. Zur Kenntnis der Spermatogenese bei den Poriferen und Coelenteraten etc. Zeitsch. wiss. Zool., 76: 523—543.
Haeckel, E. 1872. Die Kalkschwamme. 1: 376.
Hentschel, E. 1923. Parazoa-Porifera in Kiikenthal und 'Krumbach, Handb. Zool. 1.
Joseph, G. 1881. Mitteilungen iiber einen Grottenschwamm (Spongilla stygia n. sp.). Jahr. ber. Schles. Ges. Vaterl. Cult., 59: 253.
Kirkpatrick, R. 1915. Notes on Fresh-water Sponges from the Volga-Basin. Раб. Волжск. Биол. Ст., 5, № 2: 102—106.
Kofoid, Ch. A. 1900. The plankton of the Eche River Mammoth Cave. Trans. Am. Mier. Soc. 21: 113-126.
Koshow, M. 1930. Material zur Spongillidenfauna Ostsibiriens. Zool. Anz., 90: 155—168.
L a u b e n f e 1 s, de M. W. 1928. Interspecific grafting using sponge cells. Journ. Elisha Mitchell Soc., 44: 82—86.
Laurent, J. L. M. 1844. Recherches sur 1’Hydre et 1’Eponge d’eau douce. Paris.
Lauterborn, R. 1902. Ein fiir Deutschland neuer Susswasserschwamm etc. Biol. Zentr.
BL, 22: 519.
L e i d y. 1851. On Spongilla etc. Pr. Ac. Nat. Sc. Philadelfia, 5: 278.
Lieberkuhn, N. 1856 a. Beitrage zur Entwickelungsgeschichte der Spongilla. Arch. Anat. Physiol.: 1—19.
-- 1856 b. Zusatze zur Entwickelungsgeschichte der Spongilliden. Arch. Anat. Phys.: 496—514.
“ 1870. Ueber Bewegungserscheinungen der Zellen. Schr. Ges. Bef. Ges. Naturwiss. Marburg, 9.
ПРЕСНОВОДНЫЕ ГУБКИ—ОБЩАЯ ЧАСТЬ
57
Limberger, А. 1918. Ueber die Reinkulturen der Zoochlorella aus Euspongilla lacustris etc. Sitzungsber. Ak. Wiss. Wien, Math.-Naturw. Kl.
Linne, C. (Linnaeus) 1759. Systema Naturae, 10 ed., 2: 1348.
M i к 1 u c h o-M a с 1 a y, 1870. Ueber einige Schwamme des nordlichen Stillen Oceans und des Eismeeres. Mem. Ac. Imp. Sc. St. Petersburg, s. 7, 15: 1—24.
Muller, K. 1911a. Reductionserscheinungen bei Susswasserschwammen. Arch. Entw. Meeh., 32. ---- 1911 b. Das Regerationsvermogen der Siisswasserschwamme. Arch. Entw. Meeh., 32.
---- 1911 c. Beobachtungen iiber Reduktionsvorgange bei Spongilliden, nebst Bemerkungen
zu deren ausseren Morphologie und Biologie. Zool. Anz., 37: 114.
Pallas, P. S. 1772—1773. Reise durch verschiedene Provinzen des Russischen Reiches, 3: 710.
Parker, G. H. 1910. The reactions of Sponges etc. Contr. Zool. Lab. Mus. Compar. Zool. Harvard Coll. n° 204: 765—804.
Petr, Fr. 1886. Dodatky ku taune ceskych hub sladkowodnich. Sitz. ber. Bohm. Ges. Wiss. Prag: 147—174.
Potts, E. 1880. On Fresh-Water Sponges. Proc. Ac. N. Sc. Philadelphia: 356—357.
---- 1881 (1882). Some new Genera of Fresh-water" Sponges. Proc. Ac. N. Sc. Philadelphia: 149—150.
---- 1887. Fresh-water sponges, a monograph. Proc. Ac. N. Sc. Philadelphia: 1—279.
P'ourbaix, Nelly. 1931. Contribution a I’etude de la nutrition chez les Spongiaires. Bull. Stat. Ocean. Salammlo, n° 23.
P:u tter, A. 1909. Die Ernahrung der Wassertiere. Ztsch. Allgem. Physiol., 7.
---- 1914. Der Stoffwechsel der Kieselschwamme. Ztsch. Allgem. Physiol., 16.
Rezvoj, P. 1926. Note on freshwater Sponges from Turkestan. C. R. Ac. Sc. URSS: 107— 110.
__— 1927. Note on Sponges from Dzhegataj-kul in the Urjankaj region. C. R. Ac. URSS: 296—300.
—— 1928. Zur Spongillidenfauna Russlands. Zool. Anz., 76: 221.
---- 1929 a. Siisswasserschwamme aus der russischen Arktis. Zool. Anz., 85: 283.
—1929 b. Ueber Spongilla proliferens Annandale und S. sectospina mihi. Zool. Anz., 84: 158.
---- 1930. Siisswasserschwamme aus dem Ussuri-Gebiet. Zool. Anz., 87: 175.
.Schroder, K. 1927 a. Spongilliden-Studien. II. Zool. Anz., 70: 75—82.
---- 1927 b. Spongilliden-Studien. HI. Zool. Anz., 73.
---- 1930. Haltung und Aufzucht von Susswasserschwammen. In Abderhalden. Hand-buch der biol. Arbeitsmeth., Abt. 9, T. 2/II: 1563—1576.
---- 1932. Spongilliden-Studien. V. Zool. Anz., 98: 161—170.
---- 1933. Spongilliden-Studien. VI. Zool. Anz., 104: 113—119.
Schulze, P. 1922 a. Ueber die Bildung der Spongielins bei den Susswasserschwammen. Zool. Anz., 55: 213-215.;
---- 1922 b. Spongiaria etc. Biol. d. Tiere Deutschl., 1—12.
----- 1923. Beitrage zur Kenntnis der Kieselnadelbildung besonders bei den Spongilliden. Arch. Zellforsch., 17: 105—130.
Simroth, H. 1902. Ueber den Ursprung der Wirbeltiere, der Schwamme und der gesch-lechtlichen Fortpflanzung. Verb. Deutsch. Zool. Ges., Jahrg. 12: 152.
Traxler, L. 1894. Ephydatia fossilis, eine neue Art der fossilen Spongilliden. Foldtan. Kozlony, 24: 234.
---- 1895. Beitrage zur Kenntnis der Siisswasserschwamme. Fold. Kozl., 25: 241.
---- 1898. Spongilla gigantea n. sp. Foldt. Kozl., 28: 186.
Van Trigt, H. 1918. A contribution to the physiology of the fresh-water Sponges. Proc. Wet. K. Ak. Amsterdam, 20, № 7: 1061—1075.
---- 1919. A contribution to the physiology of the freshwater Sponges. Leiden Tijdschr. Ned. Dierk. Ver., s. 2, 17: 1—220.
---- 1920. A contribution to the physiology of the freshwater Sponges. Biol. Zentralbl., 40.
58
ФАУНА СССР
Vejdovsky, F. 1877. О ceskych hubach sladkovodnich. Vesmir. Obrc., 67. 2: 212.
---- 1883 a. Die Siisswasserschwamme Bohmens. Abh. Bohm. Ges. Wissensch. Prag., S. 6, 12,. n° 5: 26—31.
---- 1883 b. Pzispevky к znamostem о houbach sladkovodnich. Sitzber. K. Bohm. Ges. Wiss. Prag, S. 6, 12: 331.
---- 1887. Diagnosis of the European Spongillidae. In Potts: A Monograph: 172.
Vosmaer, G. C. J. 1887. Porifera. In Broun, Die Klassen und Ordnungen des Tierreichs, 2.
Weber. 1890. Spongillidae des Indischen Archipels. Zool. Erg. Reise Niederl. Ost-Indien., 1: 30—47.
Weltner, W. 1893 a. Ueber die Autorenbezeichnungen von Spongilla erinaceus. Sitz. ber. Ges. Nat. Fr. Berlin: 7—13.
------ 1893 b. Spongillidenstudien. II. Arch. Naturg., 59, I: 245—282.
---- 1895. Spongillidenstudien. III. Arch. Naturg., 66: 114—144.
---- W. 1900. Siisswasserschwamme. In: R. S e m о n. Zool. Forschnngsreisen in Australien etc. 5. Denkschr. Med. Natw. Ges. Jena 8.
---- 1907. Spongillidenstudien. V. Arch. Naturg., 73.
---- 1909. Spongillidae, Siisswasserschwamme. In Brauer, Die Susswasserfauna Deutscb-lands, 19: 177—190.
--- 1911. Spongilliden des Issyk-Kul-See und de? . Baches bei Dschetu-Ogus. Tp. СПб.
О. Ест. 42: 57—97.
W esenber g-L und, C. 1918. Contribution to the knowledge of the postembryonal development of the Hydracarina. Viedensk. Medd. fra Dansk. nsturh. Foren., 70.
W i e r z e j s к i, A. 1886. Le developpement des gemmules des eponges d’eau douce d’Europe.
Arch. Slaves Biol., I, fasc. I: 26—46.
Wilson, H. V. 1911. Development of Sponges from dissociated tissue cells. Bull. Bureau Fish., 30, n° 750: 1—30.
Young, J. T. 1878. On the occurence of a Freshwater Sponge in the Purbeck Limestone. Geol. Mag., 5: 220.
Z у к о f f, W. 1893. Die Entwickelung der Gemmulae bei Ephydatia fluviatilis auct Bull. Soe.
Imp. Nat. Moscou, n. s., 6: 1—16.
СПЕЦИАЛЬНАЯ ЧАСТЬ
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА СЕМЕЙСТВ ПРЕСНОВОДНЫХ ГУБОК
1 (2). Дермальная мембрана имеет вид тонкой легко отделяющейся пленочки, сквозь которую просвечивают хорошо развитые субдермаль-ные полости. К простым оскулярным отверстиям радиально сходятся просвечивающие сквозь мембрану отводящие каналы, образующие довольно большие (до 2—3 см) звездообразные фигуры. У созревших экземпляров или в старой отмершей ткани в основании губки обычно имеются геммулы, снабженные микросклерами особого устройства .
.........................................Сем. Spongillidae.
2 (1). Субдермальные полости отсутствуют, дермальная мембрана не отделима от ниже лежащей ткани. Крупные звездчатые фигуры, состоящие из просвечивающих отводящих каналов, не образуются. Геммулы отсутствуют. Бесполое размножение при помощи соритов .....
.....................'...............Сем. Lubomirskiidae.
Сем. SPONGILLIDAE —БАДЯГИ
Субдермальные полости развиты хорошо; дермальная мембрана легко отделяется от остальной ткани. У живой губки от оскулярных отверстий поднимаются довольно длинные оскулярные трубки. Отводящие каналы, сходясь к оскулярным отверстиям, образуют в совокупности характерные крупные звездчатые фигуры. Скелет построен главными и поперечными пучками, но часто правильность его нарушается. Спонгин присутствует в различных, в общем, незначительных, количествах. Микросклеры диактины, чаще всего амфиоксы. Нет особых дермальных спикул. Микросклеры различного устройства; никогда не бывает хелоидов. Бесполое размножение при помощи геммул, из которых выходят готовые губки; геммулы снабжены плотной хитиноидной капсулой, воздухоносным слоем и особыми защитными геммульными микросклерами.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ СЕМ. SPONGILLIDAE
1(2). Геммульные микросклеры представлены рабдами, прямыми или изогнутыми, гладкими или шиповатыми..................1. Spongilla.
2(1). Геммульные микросклеры представлены амфидисками.
60
ФАУНА СССР
3 (4). Диски у амфидисков цельнокрайние в виде правильных кружков, гладкие или с тонкой радиальной исчерченностью . . 4. Trochospongilla.
4(3). Край диска рассечен более или менее глубоко на лучи, и диск имеет вид звезды или зубчатого колесика.
5(6). Длина стержня амфидисков колеблется около одной средней величины и амфидиски не распадаются по величине на две группы . . .
............................................2. Ephydatia.
6(5). Амфидиски распадаются по длине стержня на две группы—длинные и короткие .................. 3. Heteromeyenia.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ СЕМ. SPONGILLIDAE ПРИ !
ОТСУТСТВИИ ГЕММУЛ
Экземпляры, лишенные геммул, обычно не могут быть определены с полной достоверностью. Для ориентировки в безгеммульном материале мы приводим настоящую таблицу, заранее указывая на условность получаемых по ней определений. Надо, однако, заметить, что и при отсутствии геммул иногда удается находить рассеянные в паренхиме геммульные спикулы, например в паренхиме у Ephydatia. можно встретить рассеянные амфидиски. В таких случаях (за некоторыми исключениями) определение можно вести, как и в присутствии геммул.
1 (2). В паренхиме имеются пузыревидные клетки. Микросклеры шиповатые, за исключением концов, спикул, или рядом с шиповатыми имеются и гладкие спикулы; в редких случаях гладкие микросклеры преобладают или еще реже шиповатые отсутствуют...............
.......................................* Ephydatia miilleri.
2(1). Пузыревидные клетки отсутствуют.
3 (12). Микросклеры имеются.
4(11). Микроскеры шиповатые.
5 (6). У микросклер шипы расположенные в средней части спикулы, рассечены на своих вершинах на два или несколько крючков ....
.....................................* Spongilla biseriata.
6(5). Микросклеры не несут шипов, рассеченных на своих вершинах.
7 (10). Паренхимные микросклеры сплошь покрыты тесно расположенными мелкими шипиками.
8(9). Устройство скелета своеобразное: спонгин не только обволакивает как главные, так и поперечные пучки и даже отдельные спикулы, но и образует широкие прозрачные мембраны, натянутые между пучками. Скелетные элементы тонкие (макросклеры от 1 до 11 р. толщины) .
................................... . . * Spongilla arctica.
9(8). Устройство скелета обычное; спонгин иногда присутствует в значительном количестве, в особенности ближе к основанию губки, но не образует широких мембран. Скелетные элементы более толстые (толщина макросклер 7 у —17 у).............* Spongilla lacustris.
SPONGILLIDAE—Б АДЯГИ 61:
10(7). Шипы на паренхимных микросклерах сидят не так густо, с промежутками; наиболее длинные палочкообразные шипы расположены в средней части спикулы; они посажены перпендикулярно и имеют Обыкновенно несколько утолщенную вершину. Макросклеры гладкие или с редкими острыми шипиками......................................
. . . . . . . . . . . * Heteromeyenia baileyi (var. stepanowi etc.).
11(4). Микросклеры гладкие: в паренхиме среди гладких слабо веретено- образных макросклер в небольшом количестве встречаются спикулы меньшей длины (100—300 р. X 3—8 р.), отличающиеся сильной вере-тенообразностью, тонкими и острыми концами, обычно более сильно изогнутыми; эти спикулы тожественны со спикулами, покрывающими геммулы у снабженных ими экземпляров ... * Spongilla carteri.1
12(3). Паренхимные микросклеры отсутствуют.2
13(14). Микросклеры покрыты многочисленными длинными и острыми шипами.................................... * Trochospongilla horrida.
14(13). Макросклеры гладкие или гладкие и шиповатые с редкими и мелкими шипиками.
15(16). Рядом с гладкими, обычно построенными макросклерами встречаются в значительном количестве гладкие спикулы с копьевидновздутыми концами. Макросклеры крупные, достигают 440 р. длины.
...........................* Ephydatia fluviatilis var. hastifera.
16(15). Копьевидные спикулы отсутствуют.
17 (18). Микросклеры в среднем крупнее, чем у других видов; средний их размер больше 300 р. (максимальный размер до 550 р.). Рядом с гладкими микросклерами обычно присутствуют (иногда в очень малом количестве) слабо шиповатые спикулы, с малочисленными мелкими шипиками........................................* Ephydatia fluviatilis..
18 (17). Средний размер макросклер значительно ниже 300 р, обычно и максимальные размеры спикул не достигают этой величины. Шиповатые макросклеры совершенно отсутствуют.
19(20). Толщина макросклер 5—15 р.. Консистенция губки очень рыхлая и ломкая. Скелет из слабых, плохо отграниченных пучков, образующих рыхлую сеть...................................* Spongilla fragilis.
20(19). Толщина макросклер 11—18 р.. Консистенция губки сравнительно плотная. Скелет состоит из довольно беспорядочных, плохо отграниченных пучков, образующих плотную сеть . . . * Spongilla stanleyi.
1 Spongilla carteri с трудом может быть узнана при отсутствии геммул, так как гладкие микросклеры всегда могут быть приняты за молодые макросклеры. Отметим, что в пределах СССР преимущественно описывались формы этого вида с очень короткими, почти закругленными или даже явственно закругленными концами. Длина макросклер 215— 330 р. X 11—16 р..
2 Виды, лишенные паренхимных микросклер, ие могут быть сколько-нибудь надежно-определены при отсутствии геммул. Trochospongilla horrida составляет исключение.
62
ФА УНА СССР
1. Род SPONGILLA LAMARK
Геммульные микросклеры представлены прямыми или изогнутыми рабдами, палочкообразными спикулами с острыми или затупленными концами, шиповатыми или в редких случаях гладкими. Паренхимные микросклеры имеются или отсутствуют. Макросклеры, за исключением немногих тропических видов, гладкие.
Род Spongilla подразделяется на три подрода: Euspongilla, Eunapius, Sfratospongilla.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДРОДОВ РОДА SPONGILLA
1 (2). Геммульные микросклеры образуют вокруг геммульной капсулы один или несколько плотных слоев тангенциально расположенных спикул, соединенных в типичных случаях кожистыми мембранами. Кругом этих слоев беспорядочная масса микросклер, заполняющая всю толщу воздухоносного слоя и маскирующая его структуру . .
....................................3. Подрод Stratospongilla.
2(1). Геммульные микросклеры не образуют плотных слоев тангенциально расположенных спикул. Геммульные микросклеры заключены в воздухоносном слое, имеющем зернистую или ячеистую структуру. Воздухоносный слой может быть очень тонким или отсутствовать.
3(4). Воздухоносный слой зернистый, различной толщины. При толстом воздухоносном слое геммульные микросклеры располагаются в нем почти радиально; при тонком воздухоносном слое они лежат тангенциально на поверхности геммульной капсулы, большею частью в небольшом числе, не образуя сплошного плотного слоя.................
.......................................1. Подрод Euspongilla.
4(3). Воздухоносный слой ячеистый, значительной толщины, состоящий из правильно расположенных полиэдрических ячей. Геммульные микросклеры расположены внутри воздухоносного слоя беспорядочно.
......................................... 2. Подрод Eunapius.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ И ВАРИЕТЕТОВ РОДА SPONGILLA
1 (20). Микросклеры (геммульные или паренхимные) шиповатые. 2(15). Есть паренхимные микросклеры.
3 (4). Шипы в средней части микросклер (паренхимных и геммульных) на своих вершинах рассечены на 2—4 крючка, загнутых в разные стороны. У концов спикулы шипы простые, прямые или изогнутые . .
..............................................* 3. S. biseriata.
4(3). Шипы у микросклер простые, не рассеченные. Паренхимные микросклеры покрыты частыми мелкими шипиками. Геммульные микросклеры с более редкими и крупными шипами.
SPONGILLIDAE —БАДЯГИ
63
5(6). Все скелетные пучки и отдельные спикулы совершенно погружены в спонгин, образующий широкие перепонки между скелетными пучками. Все спикулы, и макро- и микро-склеры очень тонкие ....
.....'....................................... * 2. S. arctica.
6(5). Спонгин развит нормально; иногда присутствует в значительном количестве и в основании губки обволакивает пучки, но не образует широких мембран между ними.
7 (8). Геммульные спикулы постоянно отсутствуют, так же как и воздухоносный слой геммул. У геммул большею частью несколько поровых отверстий...............................................S. Helvetica.1
8(7). Как правило, имеются и геммульные спикулы, и воздухоносный слой.2 Обычно только одно поровое отверстие . . * 1. S. lacustris. 9(14). Геммульные микросклеры шиповатые; только изредка среди них встречаются гладкие.
10(11). Паренхимные микросклеры присутствуют в массе, как в дермальной мембране, так и внутри ткани. Воздухоносный слой сильно развит и снабжен большим количеством геммульных спикул..................
..................................la. S. lacustris v. jordanensis.
11 (10). Паренхимные микросклеры присутствуют в малых или средних количествах.
12(13). Геммульные микросклеры присутствуют в исключительно малых: количествах............................* 1с. S. lacustris v. paupercula.
13(12). Геммульные микросклеры в средних количествах.
14(9). Среди геммульных микросклер преобладают гладкие, большей частью с характерным перегибом недалеко от концов спикулы; вместе с гладкими встречаются и шиповатые; приблизительно х/10 всех геммул имеет по 3—6 дополнительных поровых отверстий ...................
.................•....................S. lacustris forma rbenana.
15(2). Нет паренхимных микросклер.
16(19). Геммулы собраны в основании губки, где образуют мостовидный слой, и заключены в общий толстый слой воздухоносной ткани, состоящей из крупных полиэдрических ячей; или же геммулы собраны группами в толще паренхимы и также заключены в общую воздухоносную ткань. В воздухоносной ткани многочисленные беспорядочно расположенные геммульные микросклеры, довольно тонкие с мелкими многочисленными шипами; геммулы снабжены поровыми трубками ..................................................* 4. S. fragilis.
' 17 (18). Поровая трубка короткая и прямая, длиной от 38 до 60 [л, диаметром 57—76 [л, оканчивающаяся нешироким раструбом......................
.................................... * 4 а. S. fragilis v. rectituba.
1 S. helvefica известна только из Женевского озера. Самостоятельность этого вида вызывает сомнения; очень возможно, что он представляет морфу б’, lacustris.
2 В крупных озерах часто встречаются экземпляры, у которых все или часть геммул лишены геммульных микросклер и воздухоносного слоя.
64
ФАУНА СССР
18 (17). Поровая трубка прямая и еще более короткая,- длиной 22 и, дия-метром 49 у-.............................* 4Ь. S. fragilis v. brevipora.
19(16). Геммульные спикулы крупные и очень толстые (до 13 у. толщины), снабженные редкими грубыми шипами, располагаются вокруг каждой геммулы тангенциальными слоями, образуя плотную многослойную мозаику; кроме того все промежутки между геммулами заполнены сплошной массой геммульных спикул, совершенно маскирующих воздухоносный слой; длинные изогнутые поровые трубки.....................
• • °.......................................... * 6. S. stanieyi.
20(1). Микросклеры гладкие, однотипные; геммульные не отличаются от паренхимных. Они в небольшом количестве рассеяны в ткани и образуют скопления у геммул, располагаясь как на поверхности, так и внутри хорошо развитого толстого ячеистого воздухоносного слоя. Короткая прямая боченкообразная поровая трубка • . * 5. S. carteri.
21 (22). Микросклеры с длинными, средними или короткими остриями . .
............................................* 5. S. carteri typica.
22(21). Макросклеры почти исключительно с закругленными концами; в остальном сходна с типичной формой.................................
..................................* 5 а. S. carteri v. rotundacuta^
1. Подрод Euspongilla Vejdovsky ♦
Воздухоносный слой зернистый и может достигать зничительной толщины; при развитом воздухоносном слое геммульные спикулы располагаются внутри него почти радиально, на его же поверхности — тангенциально. При мало развитом слое микросклеры лежат тангенциально на поверхности геммульной капсулы.
* 1. Spongilla lacustris L. (Табл. I, фиг. 1, 2, т. П, ф. 4, т. IV, ф. 12—14).
Linnaeus, 1759, 10 ed., 2 :1348; Vejdovsky, 1887: 172; Potts, 1887 :186, pl. 5; Weltner, 1895: 118; Weltner in Brauer, 1909 :181, f. 305—308; Arndt, 1926 : 342.
Колонии ветвистые или в виде неправильных наростов или корок. Скелет из главных и поперечных пучков, образующих неправильную решетку; спонгин в значительном количестве. Макросклеры гладкие с длинными и тонкими остриями; реже — острия средней длины. Паренхимные микросклеры в виде тонких веретенообразных мелкошиповатых слабо изогнутых иголочек, обычно с длинными острями. Геммульные спикулы более толстые, часто сильно изогнутые с более редкими и крупными шипами, несут короткие острия или затуплены.
Вид вариирует очень сильно в отношении всех своих признаков. Абе-рантные формы неоднократно описывались, как новые виды; установлено много вариантов, из которых некоторые не выходят в сущности из пределов видовой изменчивости. Мы можем сказать, что природа большинства
SPONGILLIDAE—БАДЯГИ
65
этих отклонений от нормы еще не известна. Мы не знаем, какие из этих отклонений являются наследственно закрепленными и какие носят морфи-ческий характер, находясь в зависимости от условий местообитания. Относительно описанных местных рас нельзя с уверенностью сказать, что они имеют подвидовой характер и не могут быть встречены в совершенно иных районах и совместно с типичной формой.
Кустистые формы представляют совокупность длинных ветвистых отростков, достигающих до 10 мм в поперечнике и поднимающихся от корковой основной части. Описаны экземпляры до 1 м высоты. Такие формы развиваются только в спокойной воде; на течении губки дают корковые формы или образуют неправильные объемистые наросты; впрочем, корковые формы встречаются и в стоячих водах. Цвет губки зависит от условий освещения: губка, выросшая на полном свету, имеет темнозеленый цвет. У теневых экземпляров цвет вариирует от грязно-белого до коричневого, красноватого и бурого. Спикулы вариируют и по величине, и по форме. Длина макросклер чаще всего колеблется в пределах от 225 до 300 [J. длины, при 7—17 [х толщины. Длина часто переходит за 300 у-: в литературе указываются экземпляры с иглами до 414 р. максимальной длины; толщина макросклер может достигать 25 [л. Вариации формы касаются степени изгиба и длины острия. Преобладают то почти прямые, то более сильно изогнутые; острия чаще длинные, и спикулы веретенообразные; реже острия средней длины или короткие, тогда обычно и вере-тенообразность выражена слабо. Паренхимные микросклеры веретенообразные, постепенно утончаются к концам и заканчиваются остриями, реже самые концы бывают притуплены; на всем протяжении они густо покрыты мелкими шипиками. Средние размеры 70—80 у. X 3—7 р., пределы колебаний приблизительно 45—117 у.. Количество этих спикул может сильно вариировать. Геммульные микросклеры сильнее паренхимных вариируют по внешности и величине. В типичных случаях они толще паренхимных, цилиндрические, с тупыми концами, довольно сильно и неправильно изогнутые; шипы более редкие и крупные; обычные размеры 50 —80 X 4—8 у; бывают и совершенно прямыми; иногда они несут короткие острия; количество шипов может сильно уменьшаться; встречаются и совершенно 1 ладкие спикулы. Обычно капсула геммулы покрыта нетолстым зернистым воздухоносным слоем с небольшим количеством тангенциально лежащих геммульных спикул. Реже воздухоносный слой развит сильно, достигает значительной толщины; иногда он снабжен наружной кутикулой; количество геммульных микросклер может быть очень велико; располагаются они в слое или совершенно неправильно или более или менее радиально; снаружи на поверхности воздухоносного слоя спикулы лежат тангенциально. Очень часто воздухоносный слой отсутствует совершенно. Могут отсутствовать и геммульные микроскеры. В типичных случаях порус один, реже геммула имеет дополнительные порусы. Он имеет вид простого круглого отверстия или снабжен низкой развернутой воро~
Фауна СССР, Губки, т. II, в. 2. б
66
ФАУНА СССР
ночкой с плоской оторочкой. Поперечник геммульной капсулы от 330 до 800 (д.
Распространение: вся Европа (от Кольского полуострова до Португалии и Болгарии); вся Сибирь до Камчатки включительно; Закавказье, Туркестан, Уссурийский край, Япония, Китай (Киангсу, Юнань, Манчжурия), Корея, Гималаи и северо-западная Индия (в остальной Индии и Бирме subsp. reticulata Ann.), Сиам. Вид не известен из Африки, Индо-Малайского архипелага, Австралии и Новой Зеландии.
Сильная изменчивость всех элементов послужила основой для описания ряда вариететов, из которых мы упомянем главнейшие.
la. S. lacustris var. jordanensis (Vejdovsky)
Vejdovsky, 1877; Vesmir, 67, 2 :212 (<S. jordanensis).
Паренхимные микросклеры в громадных количествах. Геммулы с толстым воздухоносным слоем и большим количеством беспорядочно расположенных микросклер.
Распространение: Зап. Европа (Чехословакия, Г ермания, Франция, Ирландия). В пределах Союза не отмечался, но несомненно встречается.
* lb. S. lacustris morpha lacustrioides Koshow
Кожов, 1925, 45 (S. lacustroides)', Koshow, 1930 :158.
Близка к предыдущему вариетету. Паренхимные микросклеры в очень большом количестве. Толстый воздухоносный слой с радиально расположенными внутри него геммульными спикулами; хорошо выраженная внешняя кутикула, покрывающая воздухоносный слой, и на ней большое скопление беспорядочно расположенных геммульных микросклер.
Распространение: Прибайкалье.
1с. S. lacustris var. paupercula (Bowerbank)
Bowerbank, 1863:470, pl. 38 (5. paupercula)', Potts, 1887 :189.
Вариетет характеризуется отсутствием воздухоносного слоя и очень малым количеством геммульных микросклер, которые обычно слабо шиповаты. У паренхимных микросклер шипы также не так густо покрывают спикулу.
Распространение: С. Америка (Бостон), Франция (горные озера Оверни).
* 2. Spongilla arctica Annandale. (Табл. V, фиг. 15—17, т. VI, ф. 19).
Annandale, 1915 :1—3, f. 1—4; Rezvoj, 1928 :219, f. 1—3; Rezvoj, 1929: 283, f. 1—6.
От корки, покрывающей субстрат, поднимаются тонкие и короткие отростки, иногда ветвистые. Макросклеры короткие и тонкие, гладкие амфидиски, сильно вариирующие по длине и в особенности по толщине. Паренхимные микросклеры сходны с таковыми S. lacustris, но тоньше и короче этих последних. Геммульные микросклеры прямые или слабо изогнутые с малочисленными крупными шипами. Скелет характерного для вида
SPONGILLID АЕ — Б АДЯГИ
(Я
устройства: как главные, так и поперечные пучки и отдельные спикулы заключены в широкие спонгиновые мембраны, имеющие часто округлые отверстия. Геммулы рассеяны в ткани, снабжены очень небольшим количеством геммульных микросклер; поверх тонкого зернистого слоя имеется спонгиновая оболочка, соединяющаяся со спонгином скелетных пучков, соприкасающихся С геммулой.
Типовой экземпляр представляет обломки тонкой корки на листьях водяного растения. Большинство экземпляров с Мурманского берега представляют корки от 0.5 до 1 см толщиной очень темного зелено-бурого цвета; от корки поднимаются светлые изумрудно-зеленые отростки 2— 4 мм толщины и 5—6 см длины;, отростки иногда разветвляются. Некоторые экземпляры с Мурмана и из Рязанской губернии представляют тонкие налеты на дерновинках подводного мха. Все известные экземпляры итого вида отличаются малыми размерами спикул. Все типы спикул в среднем короче, а главное тоньше соответствующих спикул S. lacustris, но их размеры сильно колеблются, и отдельные спикулы по длине даже превышают средние размеры таковых «S', lacustris. Макросклеры 101— 303 р. X 1—И паренхимные микросклеры 30—128 р. X 1.5—4 р, гем-мульные — 24—161 р. X 1.5—5 р.. Строение скелета совершенно своеобразно. Главные пучки тонкие, из плотно расположенных спикул; ветвятся и соединяются друг с другом поперечными пучками, часто из 1—2 рядов спикул. Много отдельных макросклер. Все эти скелетные элементы заключены в спонгиновые мембраны, натянутые между пучками. В мембранах этих часто видны крупные округлые или овальные отверстия. Мембраны пронизывают ткань во всех направлениях. На их поверхности приклеено большое количество паренхимных микросклер. Геммулы рассеяны в ткани. Их поперечник колеблется от 300 до 748 р.. Поровое отверстие одно, реже их два; порус окружен низкой оторочкой. Геммульных микросклер очень мало, они плоско лежат на поверхности капсулы. Зернистый воздухоносный слой довольно тонкий, сверху он перекрыт гиалиновым спонгиновым слоем, соединяющимся со спонгином подходящих к геммуле пучков. Эти пучки образуют около геммулы род корзинки.
Распространение: вид описан с Полярного Урала (тундра Пе-мал, долины р Пидераты). Найден на Кольском полуострове (Мурманский берег, озера в районе Порчнихи; Терский берег — оз. Масловское, бассейн р. Чапомы), в Сяберских озерах юго-зап. части Ленинградской области и в озерах Мещерской низменности (б. Рязанской губ.).
* 3. Spongilla biseriata Weltner. (Табл. V, фиг. 18, т. VI, ф. 24).
Weltner, 1895: 117 et 138; Annandale, 1907 а: 15, f. 1. (*У. proliferens); Annandale, 1911: 72, f. 9 (S’. pr.); Rezvoj, 1926: 107 (S’, sectospina); Rezvoj, 1929, b: 159, f. 1, 2 (S. proliferens var. sectospina)-, Schroder, 1933: 113, f. 1—4.
Образует небольшие корочки на листьях водяйых растений; цвет зеленый; оскулярные отверстия снабжены коническими оскулярными труб-5*
68
ФАУНА СССР
ками. Иногда наблюдаются короткие выросты, но чаще они отсутствуют. Скелет развит слабо, спонгин имеется в малом количестве. Макросклеры гладкие, веретенообразные с длинными и тонкими остриями; размеры 315—350 р. X 10—12 [л. Паренхимные микросклеры в большом количестве как в дермальной мембране, так и внутри ткани, веретенообразные, с притупленными концами; размеры 59—79 X 2—3 сильно изогнутые,
острые шипы сидят преимущественно у концов спикулы; в средней части спикулы шипы имеют характерное для вида строение: они рассечены на своих вершинах на 2, 3, 4, реже больше, крючечка, образующих маленькую розетку. Геммульные микросклеры в общем сходны с паренхимными, но они цилиндрические; наиболее крупные шипы, сильно изогнутые острые крючья, собраны у самых концов спикулы; в средней части располагаются такие же рассеченные шипы, как и у паренхимных спикул; размеры бб— 77 [л X 3 |л. Геммулы расположены в основном слое губки, сильно варии-руют по величине и снабжены толстым воздухоносным слоем с большим количеством микросклер и толстой изогнутой поровой трубкой. Размножение происходит при помощи наружного почкования и геммул; половое размножение или совершенно, или почти отсутствует.
Распространение: Азия—Ьенгалия, Бирма, Цейлон, Ява, Целебес, Флорес, Филиппины, Китай, Арал и Аму-дарья; Африка — Нил у Каира, оз. Ниасса, Европа — южн. Болгария.
Как вариетет настоящего вида описана из дельты Аму-дарьи S. рго-liferens {—biseriata v. sectospina Rezvoj). Этот вариетет отличается от типичной формы только строением паренхимных микросклер. Веретенообраз-ность у них выражена сильнее, чем у типичной формы; концы микросклер острые, и около них концентрируются мелкие острые простые шипики; средняя часть спикулы занята характерными рассеченными шипами, причем самые длинные шипы сидят в середине спикулы, и к концам их длина уменьшается. Вариетет описан по очень небольшому материалу. В настоящее время из Аральского моря, из района дельты Аму-дарьи, имеется большой материал по S. biseriata, относящийся к типичной форме. Это сильно уменьшает значение описанного вариетета и не позволяет придавать ему характер подвида. Вследствие неполноты, а иногда и ошибочности первых описаний и изображений этого вида, его синонимика значительно усложнилась. Вельтнер (Weltner, 1895) кратко описал вид из окрестностей Каира, не приложив рисунков. В 1907 г. вид был описан из Калькутты, как новый (S. proliferens Annand. 1907), но приложенные рисунки микросклер не имеют ничего общего с действительностью и попали в печать, очевидно, по какому-то недоразумению, что выяснилось при изучении котипа S. proliferens из Калькутты (Rezvoj, 1929). Вследствие ошибочности рисунков Эннендала материал из Аму-дарьи был описан как новый вид, S. sectospina (Rezvoj, 1926), причем впервые была подмечена наиболее характерная особенность вида — его рассеченные шипы на микросклерах. В результате изучения вышеуказанного котипа, 5. sectospina
SPONGILLIDAE — БАДЯГ И
69
была низведена на степень вариетета *У. proliferens (Rezvoj, 1929). В самое последнее время Шредер (Schroder, 1933) доказал идентичность S', proliferens и S, biseriata. К этому же последнему виду, по мнению этого автора, вероятно, относятся также S', semispongilia Ann. и S. micron Ann.
2. Подрод Eunapius Gray.
Толстый воздухоносный слой из хорошо развитой, правильно построенной крупноячеистой ткани. Геммулы лежат сплошным слоем на субстрате (мостовидный слой) или собраны группами в ткани губки. Геммульные микросклеры лежат беспорядочно как внутри слоя, так и на его поверхности.
* 4. Spongilla fragilis Leidy. (Табл. VI, фиг. 21, т. VII, ф. 26).
Leidy, 1851: 279; Dybowski, 1878: 53 (Д. sibirica)-, Dybowski, 1882: 10, f. 12 (Д. sibi-rica); Vejdovsky in Potts, 1887: 176; Potts 1887: 197, pl. 5 et 8; Weltner, 1895: 177, Weltner in Brauer, 1909: 183; Кожов, 1925: 58 (Д. rectiiuba); Arndt, 1926: 342; Гримайл1вська, 1927: 273, 6—8 (Д. fragilis var. brevipora); Arndt in Dahl, 1928: 60.
Форма корковая, co слабо развитым скелетом. Макросклеры гладкие. Паренхимные микросклеры отсутствуют. Геммульные спикулы прямые или слабо изогнутые, с притупленными концами, реже с короткими остриями. Геммулы снабжены поровыми трубками и заключены в толстый воздухоносный слой с многочисленными геммульными спикулами. Они образуют в основании губки сплошной мостовидный слой на субстрате или же собраны в ткани группами, заключенными в воздухоносную ткань.
Губки образуют тонкие корки или более массивные наросты, иногда с короткими, неразветвленными выростами. Цвет беловатый, грязно-серый, буроватый, реже зеленый. Ткань мягкая и ломкая. Спонгин развит слабо. Скелет из рыхлых главных и поперечных пучков. Микросклеры гладкие с длинными остриями, короче и тоньше, чем у S. lacusfris, 140— 260 у X 5—15 р. Паренхимные микросклеры отсутствуют. Геммулы небольшие, часто уплощенной формы, снабжены довольно длинной, слабо изогнутой поровой трубкой, торчащей из воздухоносного слоя. Геммулы лежат сплошным „мостовидным" слоем на субстрате; кроме того, они часто встречаются и в ткани, образуя группы по две, по три и значительно более. Воздухоносный слой покрывает слой геммул на субстрате, а также образует толстую оболочку около геммульных групп, разбросанных в ткани. Он состоит из крупных полигональных ячей и заключает многочисленные геммульные спикулы. Эти последние представляют прямые или слабо изогнутые иголочки, притупленные или снабженные короткими остриями, покрытые мелкими острыми шипами. Размеры геммульных спикул 60—140 р X 4—12 р.
Распространение: Европа(европ. ч.СССР, Финляндия,Польша, Венгрия, Румыния, Болгария, Германия, Чехословакия, Бельгия, Боденское озеро, Англия, Ирландия, Франция), азиатск. ч. СССтКЗакавказье, При
70
ФАУНА СССР
байкалье, Уссурийский край, Камчатка), Япония, Китай (Киангсу и Чекианг). С. и Ю. Америка, Австралия. Из Индии (Калькутта) известна var. calcut-tana Ann.; var. decipiens описана из Индо-Малайского архипелага (Целебес); var. rectituba из Прибайкалья (р. Култучная); var. brevipora из Буга и р. Синюхи (район г. Николаева).
Из Голландской Индии была описана S. fragilis subsp. decipiens Weber. Эта форма отличается от S. fragilis короткой и прямой трубкой (Weber, 1890, Annandale, 1911). Форма очень близка свариететами 51. fragilis, описанными из Прибайкалья и Ю. Буга.
* 4а. S. fragilis var. rectituba Koshow.
Поровая трубка прямая, 38—60 р. длины и 57—76 р. в диаметре.. „Трубка эта развернута на конце, почему иногда делается похожей на воронку" (Кожов, стр. 60). Макросклеры 172—190 р, X 5—7 р. Геммульные микросклеры 60—165 р X 8—12 р. Геммулы 247—437 р в диаметре.
Распространение: Прибайкалье.
* 4b. S. fragilis var. brevipora Grimailowskaja.
Поровая трубка еще более короткая, 22 р вышины, при 49 р диаметра. Верхний край несколько развернут. У М. А. Гримайл1вськой на верхнем крае поровой трубки описывается . „облямивка", имеющая вид „ком1рчика“, т. е. неширокая оторочка с зубчатым краем. Просмотр препарата этой формы по типу заставляет меня считать, что такой зубчатой закраинки не имеется. На поровой трубке имеются следы полигональных ячей воздухоносного слоя. При просмотре трубки сбоку, на ней получается зубчатая линия; при просмотре сверху эта зубчатая линия видна как замкнутая зубчатая фигура, что и дало вероятна основание описать зубчатую оторочку, „комтрчик". Микросклеры 205—240 р X 11—14 р.. Геммульные спикулы 80—120 р X 4—6 р, геммулы 320—450 р.
Распространение: Ю. Буг.
* 5. Spongilla carteri Carter. (Табл. VI, фиг. 23, т. VII, ф. 28).
? Carter, 1849:81 (.S', fragilis)-, Carter, 1859: 334 pl. 8 (5. carteri)-, Bowerbank, 1863: 469; Annandale, 1911: 87; Arndt, 1923:74 (S. c. var. melli et subsp. balatonensis)- Резвой, 1923: 65 (.S', rotandacata)-, Rezvoj, 1926: 107 (5. c. var. rotandacata)-, Гримайл1вська 1927; Rezvoj, 1928: 219.
Корковые или массивные формы. Макросклеры гладкие; микросклеры гладкие с длинными остриями; образуют покровные иглы геммул; встречаются в небольшом каличестве и в паренхиме. Геммулы снабжены толстым ячеистым воздухоносным слоем и короткой прямой боченкообразной трубкой.
Типичная форма по Annandale (Индия) характеризуется следующими Признаками: встречается в виде массивных наростов, с гладкой поверх-MOQTIHO, или с неправильными морщинами, принимающими иногда вид пе
SPONGILLIDAE—БАДЯГИ
71
тушьих гребней. Цвет сероватый, иногда с зеленым налетом на поверхности. Геммулы по одиночке рассеяны в ткани. Экземпляры S. carteri из Байрам-Али (Ср. Азия) представляют массивные наросты с многочисленными геммулами. В Старой Бухаре' находились исключительно корковые формы с рассеянными геммулами. Цвет губок зеленый, такой же, как и у Eph. fluviatilis, с которой S. carteri росла рядом в том же пруде. Все экземпляры из дельты Волги, а также найденные Гримайловской в Ю. Буге, представляют тонкие корки с геммулами, собранней в „мостовидном" слое в основании губки. Размер макросклер колеблется мало (Волга 215— 305 р- X 12—13 р-; Байрам-Али и Бухара до 250 р. X 9—16 у.; Дон 280— 330 р. X 11—14 [л, Ю. Буг 290—300 у X 12—16 [л), но длина острия довольно значительно. У бухарских экземпляров преобладают длинные острия, но встречаются и очень короткие. Экземпляры из Байрам-Али имеют более короткие острия; часто спикулы почти закругленные. То же надо сказать и про донской и волжские экземпляры, за исключением одного, выделенного в особый вариетет (см. ниже).
Микросклеры отличаются от макросклер своей величиной, сильно выраженной веретенообразностью и всегда длинными и тонкими остриями. Для них характерна более сильная изогнутость и некоторый перегиб недалеко от концов спикулы. Их размеры колеблются мало: 145—196 у. X 4 р. (Волга), 155—190 р. X 5—8 р. (Дон), 95—190 р. X 3—4 р. (Бухара и Байрам-Али), 126—288 р. X 4—5 р (Ю. Буг). Обгцие пределы колебаний от 95— 155 р до 190—288 р, при толщине в 3—8 р. Эти спикулы играют роль геммульных микросклер, встречаясь как внутри воздухоносного слоя, так и на его поверхности. Те же спикулы рассеяны в небольшом количестве в паренхиме. Эти спикулы Annandale не считает настоящими микросклерами, а недостигшими полного роста скелетными спикулами. Действительно, в каждом экземпляре можно подобрать немногочисленные переходные формы между микро- и макросклерами, но все же эти обе группы хорошо отличаются друг от друга, и считать спикулы, лежащие в воздухоносном слое геммул, за молодые скелетные спикулы, нет основания.
Поперечник геммульной капсулы 300—445 р, воздухоносный слой около 50—90 р, длина поровой трубки около 75 р.
Распространение: Индия (с Цейлоном), Бирма, вост. Персия (Сеистан), Китай (Фукиен, Квантунг), Сиам, Малайский архипелаг (Ява, Мадура), о-в Св. Маврикия, Африка (Виктория, Нианза, Танганьика, Сахара); Туркестан (г. Бухара, Байрам-Али), дельта Волги, низовья Дона (Ростов н/Д), Днепра и Ю. Буга; Балатонское озеро (Венгрия). Географическое распространение этого вида представляет значительный интерес. Вид господствует в Индостане и Бирме. В Туркестане является, повидимому, обычной формой. Далее на северо-запад он идет по устьям южно-русских рек — Волги, Дона и Ю. Буга. Самый западный пункт его распространения— Балатонское рзеро в Венгрии; венгерская форма выделена
ТЛ ФАУНА СССР
как subsp. balatonensis, но отличается от типичной формы только по внешнему виду.
* 5а. Spongilla carter! var. rotundacuta Rezvoj отличается от основной формы закругленными цилиндрическими макросклерами несколько меньшего размера, 220—265 (л X 13—17 [>. Эта форма была описана из дельты Волги, как отдельный вид, когда основная форма еще не была известна ни из Астрахани, ни из Туркестана. После нахождения основной формы в Астрахани и Туркестане, у которой ясно выражена тенденция укорачивать острия или почти их закруглять, приходится расценивать нашу форму не выше чем вариетет S. carteri.
* 5b. S. carteri var. mollis Annandale. Под этим названием Annandale отличает разновидность со слабо развитым скелетом. Спикулы относительно очень малочисленны; скелетные пучки тонкие, ткань мягкая, дряблая и очень ломкая. Эта форма обычна для Калькутты. В г. Бухаре в одном и том же „хаузе" (пруде) рядом с основной формой встречались экземпляры, вполне отвечающие описанию этого вариетета.
5 с. S. carteri subsp. balatonensis Arndt. Arndt отличает форму «S’, carteri из Балатонского озера (Венгрия), как подвид S. carteri balatonensis. Различия этого подвида от основной формы касаются только внешней формы тела. Балатонские экземпляры представляют корки, от которых поднимаются многочисленные вальковатые или несколько уплощенные отростки, достигающие 2.5 см длины и 1 см в поперечнике, обычно несущие на вершинах оскулярные отверстия. Строение спикул и геммул как у типичной формы. По слабому развитию скелета балатонские экземпляры сходны с var. mollis. От индийской var. lobosa Annandale, также снабженной короткими выростами, балатонская форма отличается лишь тем, что у var. lobosa оскулярные отверстия открываются не на вершинах отростков, а на базальной части.
Экземпляры из Кантона (Китай) также снабжены маленькими короткими отростками (4 мм длины и 2—3 мм в поперечнике); широкие оскулярные отверстия открываются на вершинах возвышений. Эта китайская форма описана, как var. melli Arndt.
3. Подрод Stratospongilla Annandale
Геммульные спикулы лежат на геммульной капсуле тангенциально, образуя один или несколько слоев, в которых спикулы плотно уложены на подобие мозаики. В типичных случаях каждый слой этих спикул связан плотной хитиноидной мембраной, окутывающей геммулу. Этих мембран по числу слоев спикул может быть один или несколько. Кроме слоев с мозаично уложенными спикулами, геммулы окружаются беспорядочным скоплением микросклер.
SPONGILLIDAE—БАДЯГИ
73
* 6. Spong'lia Stanley! Annandale. (Табл. VII, фиг. 25, т. VIII, ф. 30).
Annandale, 1916: 50; Gee et Wu, 1925: 609; Gee 1927, 3; Gee et Wu, 1927: 9, f. 13; Rezvoj, 1930: 177, f. 1-4.
Образует гладкие корки; макросклеры гладкие, толстые, с остриями короткими или средней длины. Паренхимные микросклеры отсутствуют. Геммулы собраны мостовидным слоем в основании губки. Каждая геммула снабжена длинной изогнутой поровой трубкой и окружена несколькими плотными слоями геммульных микросклер, расположенных тангенциально; поверх этих слоев имеется масса беспорядочно расположенных микросклер, окутывающих весь слой геммул. Микросклеры толстые, с малочисленными грубыми шипами.
Губки образуют гладкие корки без каких-либо отростков или возвышений и обыкновенно покрывают мертвые или живые раковины Cri-staria plicata (сем. Unionidae-, вид распространен по бассейну Амура и в Китае). Один экземпляр найден на Unio murchisonianum-, многочисленные колонии были найдены на погруженных в воду ветвях ивы. Толщина корок вариирует от немногих мм до 1 см. Консистенция ткани плотная. Цвет живых губок на раковинах Cristaria светлый, серовато-зеленый. Экземпляры на ветвях ивы темнобурые, почти черные. Скелет составлен из коротких довольно рыхлых пучков, образующих неправильную полигональную сеть; лишь у поверхности дифференцируются главные и поперечные пучки. Макросклеры гладкие, слабо изогнутые, или почти прямые; острия обычно средней длины, но встречаются и длинные; у некоторых экземпляров преобладают короткие острия. Размеры 196— 286 р. X 11—18 [>.. Геммульные микросклеры представляют очень толстые иглы с короткими остриями или затупленные с малочисленными грубыми, широкими у основания и острыми шипами; встречаются почти гладкие спикулы. Размеры 64—121 [л X 8—13 [л. Кроме этих микросклер встречаются и более тонкие, обыкновенно затупленные склеры, с многочисленными мелкими шипами, 70—72 [7. X 4—7 р.. Обе группы вполне соединяются промежуточными формами. Геммулы лежат плотным мостовидным слоем на субстрате; иногда этих слоев несколько, что, вероятно, объясняется отложением новых слоев геммул на старые, уже отмершие, слои. Геммулы окружены сплошной массой микросклер, и очень трудно видеть, что находится между ними. На срезах иногда видно какое-то желтоватое вещество, точно склеивающее все эти спикулы. Из просмотренных сотен геммул лишь на 2—3, снабженных меньшим количеством микросклер, удалось увидеть довольно хорошо образованный воздухоносный слой. Нахождение воздухоносного слоя у данного вида не покажется таким неожиданным, если принять во внимание, что типом подрода Stratospongilla является S. bombayensis Ann., и что этим же автором описан вариетет этого вида, var. pneumatica, у которого геммулы снабжен» -толстым воздухоносным слоем и крупными неправильными ячеями. Попе* речник геммульной капсулы 300—400 [/., поровая трубка около 100 !*-•
74
ФАУНА СССР
Распространение: Уссурийский край, южная часть оз. Ханка,, рр. Лефу и Мо при их впадении в оз. Ханка; Китай, оз. Т’аи-ху, остров Си-донг-динг, провинция Киангсу, долина Янтсе, Сучоу.
2. Род EPHYDATIA LAMOUROUX.
Геммульные спикулы представлены амфидисками, радиально располагающимися на поверхности геммулы. У амфидисков оба диска одинаковой величины, и края их рассечены более или менее глубоко на зубцы. По величине и устройству амфидиски не распадаются на две группы. Паренхимные микросклеры отсутствуют.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ И ВАРИАНТОВ РОДА EPHYDATIA
1 (8). Пузыревидные клетки имеются.
2 (7). Стержень амфидиска короче или равен диаметру диска. Диск глубоко рассечен на небольшое число лучей (4—13). Макросклеры шиповатые и гладкие, чаще с преобладанием шиповатых, реже гладких;, в редких случаях присутствуют только одни или только другие
..........................................* 3. Eph. miilleri.
3 (4). Стержень амфидисков около 12 (л X 8 [J-; диаметр диска около 36 [J-. Макросклеры шиповатые и гладкие; у шиповатых спикул концы остаются гладкими; мелкие, но хорошо заметные острые шипы густо покрывают всю среднюю часть иглы . * За. Eph. miilleri forma typica.
4 (3). Имеются значительные отклонения от типичной формы.
5 (6). Макросклеры в значительно преобладающем числе или исключительно шиповатые. В большом количестве встречаются амфидиски со стержнем, продолженным за диски и оканчивающимся с каждой стороны острым шипом; много уродливо построенных амфидисков.. На поверхности геммулы амфидиски не образуют сплошного слоя и расположены беспорядочно ... * Зв. Eph. miilleri v. acuminata..
6 (5). Макросклеры двух типов: нормальные с острыми концами, шиповатые в средней части и гладкие у концов, и спикулы с закругленными концами, толстые и короткие, гладкие или слабо шиповатые. . . .
. ...........................* Зс. Epb. miilleri v. obtusosclera.
7 (2). Стержень амфидиска длиннее диаметра диска. Диск глубоко рассечен: на тонкие и острые лучи. Макросклеры исключительно гладкие.
............................................2. Eph. meyeni.
8 (1). Пузыревидные клетки отсутствуют.
9 (10). Амфидиски с толстым стержнем, более длинным, чем диаметр амфидиска. Диски с грубыми, толстыми лучами, часто неправильно построенными. На стержне обычно сидят шипы такой же длины как лучи на диске............................................Eph. robusta..
10 (9). Стержень амфидиска длиннее радиуса диска, тонкий, часто сужен в средней части; иногда несет 1—2 шипа, более коротких чем лучи
SPONGILLIDAE — БАДЯГ И
75
диска. Диск обычно неглубоко рассечен на многочисленные лучи; обычно несколько более глубоких вырезок делят диск на секторы с зубчатыми краями. Преобладают или исключительно имеются гладкие макросклеры; шиповатые несут мелкие малочисленные шипики
.................................................* 1. Eph. fluviatilis.
11 (12). Макросклеры исключительно гладкие; рядом с нормально построенными, в преобладающем количестве имеются копьевидные спикулы, снабженные вздутиями перед концевыми остриями ....
....................................* lb. Eph. fluviatilis v. hastifera.
12 (11). Среди амфидисков преобладают уродливые формы; у построенных более или менее нормально длина стержня может быть меньше диаметра диска; встречаются амфидиски с шипами на продолжении стержня за диски. На геммулах амфидиски не образуют сплошного слоя и располагаются беспорядочно ........................................
...................................*la. Eph. fluviatilis v. teberdana.
* 1. Ephydatia fluviatilis (L.). (Табл. VIII, фиг. 31, 32, т. IX, ф. 37).
Linnaeus, 1759: 1348 (Spongilla); Dybowsky, 1882: 13 f. 3, 9 (Meyenia n° 1); Vej-dovsky in Potts, 1887: 178; Potts, 1887: 219; Weltner, 1895: 122; Weltner in Brauer, 1909: 185, f. 314—317; Weltner, 1911: 59, f. 1—39; Kirkpatrick, 1915: 102, f. 2 (E. miilleri var. behningi); Резвой, 1922: 16 (E. miilleri var. 1); Re svoj, 1928: 221, f. 8—10 (E. fl. var. teberdana); Rezvoj, 1930: 185, f. 7 (E. fl. var. hastifera).
Преобладают гладкие макросклеры, к которым обычно присоединяются слабо шиповатые спикулы. Амфидиски с длинным стержнем, более длинным, чем диаметр диска. Диски в виде пластинок, рассеченных по краям на большое число зубцов (20 и более). Пузыревидные клетки отсутствуют.
Формы корковые или массивные, с поверхностью ровной или снабженной буграми и невысокими выростами. У экземпляров, выросших на свету, цвет яркозеленый; при росте в тени окраска грязнобелая, светло-бурая, иногда розоватая. Скелет развит сравнительно слабо, и ткань губки мягкая, ломкая. Спонгин в небольшом количестве. Вместе с гладкими микросклерами встречаются и шиповатые, с немногочисленными мелкими шипиками, иногда шиповатые спикулы совершенно отсутствуют. Размеры макросклер колеблются в пределах 175—550 X 9—25 [л; чаще всего размеры лежат в пределах 250—350 [/. X 10—15 Паренх. мнЫе микросклеры отсутствуют. У амфидисков длина стержня превосходит диаметр диска. Стержень часто имеет сужение в средней части и может нести один или несколько зубцов. Диски имеют вид пластинок, рассеченных по краям на зубцы, число которых около 20 и более; часто пластинка диска делится более глубокими зарубками на несколько секторов с зубчатыми краями. Длина стержня 19—35 [л, диаметр диска 10—22 [л. В типичных случаях радиально расположенные амфидиски покрывают геммулу одним сплошным слоем. Шарообразные геммулы по одиночке встречаются в ткани или сосредоточены ближе к основанию губки. Поперечник геммул
76
ФАУНА СССР
колеблется в пределах 260—700 [л. Поровое отверстие несет небольшую плоскую оторочку.
Распространение: космополит. Вся Европа, Дальний Восток, Сибирь, Туркестан (дельта Аму-дарьи, г. Бухара), оз. Иссык-куль, Закавказье, Сирия (var. syriaca Topsent), западные Гималаи (subsp. hima-layensis Annandale), Китай (Джили, Шантунг, Киангсу, Чекианг), Япония, С. и Ю. Америка, Ю. Африка (var. capensis Kirkpatrick), Австралия; есть указания на нахождения на Малайском архипелаге (Weber, 1890), оспариваемое Annandale (1911).
Eph. fluviatilis вариирует в своих признаках значительно слабее, чем Eph. miilleri. Мы укажем для нее два вариетета. ‘
* la. Eph. fluviatilis var. teberdana Rezvoj.
Отличается от типичной формы строением амфидисков и их расположением на геммуле. Среди амфидисков значительно преобладают уродливые формы: то диски развиваются неравномерно, то спикула принимает вид шиповатой палочки или совершенно неправильного тела; часто стержень амфидиска продолжается за диски двумя острыми шипами, как у Е. miilleri v. acuminata. Правильно построенные амфидиски имеют стержень обычно равный по длине поперечнику диска, иногда даже несколько более короткий; длина стержня 14—18 р, диска 14—19 Диски представляют пластинки с неглубоко вырезанными зубцами, числом 9—12, иногда и больше; иногда число зубцов всего 5—8 и зубцы глубоко-вырезанные, как у Е. miilleri. На геммуле диски разбросаны, не образуют сплошного слоя и лежат на капсуле во всевозможных положениях, что также напоминает Eph. miilleri v. acuminata. Остальные признаки, как у типичной Eph. fluvialilis: пузыревидные клетки отсутствуют, среди макросклер преобладают гладкие; шиповатые покрыты немногочисленными мелкими шипиками. Размер гладких макросклер 250— 285 р. X10 [л, шиповатых 195—245 р. X 5—7 ;л.
Как указано выше, некоторые признаки (строение амфидисков) у этого вариетета сходны с Eph. miilleri, но остальные признаки и прежде всего отсутствие пузыревидных клеток заставляют с уверенностью отнести вариетет к Eph. fluviatilis. Var. teberdana по отношению к основной форме занимает такое же место, как var. acuminata по отношению Eph. miilleri.
Распространение: Теберда, оз. Кара-кель, б. Кубанская область, Сев. Кавказ.
* lb. Ephydatia fluviatilis var. hastifera Rezvoj.
Вариетет отличается от типичной формы строением макросклер. Рядом с нормально заостренными макросклерами в преобладающем количестве встречаются копьевидные спикулы, имеющие хорошо заметные вздутия около концов; все микросклеры гладкие. Размеры нормально
S PON GIL LI DAE — БАДЯГИ
77
построенных спикул — 285—440 [л X 10—17 копьевидных 220— 310 X 10—15 [л. В остальном вариетет сходен с основной формой. Пузыревидные клетки отсутствуют. Длина стержня амфидиска 20—26 |л, поперечник диска 18—22 [>. Геммулы с одним сплошным слоем правильно расположенных амфидисков, поперечник геммул 350—460 [J.
Копьевидные макросклеры характерны также для Eph. fluviatilis var. chui Gist Gee, описанного из Китая, но этот последний вариетет отличается от v. hastifera и строением, и размерами макросклер, и особенно устройством своих амфидисков.
Распространение: Уссурийский край,реликтовое озеро в бухте Владимир, с солоноватой водой (содержание хлора 3.8 °/0).
2. Ephydatia meyeni (Carter).
Carter, 1849: 33 (Spongilla}- Annandale, 1911: 108.
Макросклеры исключительно гладкие. Амфидиски со стержнем, более длинным, чем диаметр диска. Число лучей (зубцов) на диске около 16. Пузыревидные клетки встречаются в большом количестве.
Губки образуют корки, не достигающие значительной толщины, часто с невысокими толстыми выростами, на вершинах которых видны отводящие каналы, сходящиеся к малозаметным оскулярным отверстиям. Ткань губки' довольно твердая, но ломкая. Цвет беловатый, иногда зеленоватый. Многочисленные пузыревидные клетки в паренхиме. Скелет плотный и неправильный, количество спонгина значительное. Скелетные спикулы все без исключения гладкие, довольно тонкие, слабо изогнутые, с длинными остриями. Амфидиски со стержнем более длинным, чем диаметр диска. Стержень часто несет 1—3 толстых прямых шипа. Диск рассечен то более, то менее глубоко на большое число (около 16) лучей. Геммулы с толстым воздухоносным слоем и с одним до трех рядов амфидисков.
Е. meyeni не встречается в пределах Союза. Мы помещаем здесь его описание, так как этот тропический вид как бы соединяет признаки двух наших обычнейших видов, Е. fluviatilis и Е. miilleri. По форме макро-и микросклер он ничем существенным не отличается от Е. fluviatilis', присутствие пузыревидных клеток, а также наблюдающееся иногда расположение амфидисков, более, чем в один ряд у него обще с Е. miilleri.
Распространение: Индия, Суматра, Китай (Киангсу).
* 3. Ephydatia miilleri (Lieberkiihn^. (Табл. VIII, фиг. 33, т. IX, ф. 36).
Lieberkuhn, 1856: 510 (Spongilla}-, Dybowski, 1882: 15. 18 (Meyenia n° 2, n° 3 var. a, var. ff)-, Vejdovsky in Potts, 1887: 177; Potts, 1887: 222, pl. 9, f. 2 (Meyenia fluviatilis var. acuminata}-, Weltner, 1895: 125; Weltner in Brauer, 1909: 186; Роавой, 1922: 17 (E. m. var. II); Arndt, 1926: 343; Кожов, 1925: 29 и 36 (E. sibirica, E. sol’da)-, Koshow, 1930:162 (E. m. var. sibirica)- Кожов, 1925: 38 (E. obtusosclera); Koshow, 1830, 164 (E. m. obtusosclera).
Макросклеры шиповатые и гладкие. Амфидиски короткие, стержень короче диаметра диска, с небольшим числом (4—13) рассеченных до стержня лучей (шипов). Пузыревидные клетки в большом количестве.
ФАУНА СССР
Корки или массивные наросты с гладкой или неровной поверхностью, покрытой буграми или короткими выростами. Цвет зеленый, иногда беловатый, желтый, буроватый или красноватый. Консистенция ткани более плотная, и спонгин развит сильнее, чем у Eph. fluviatilis. Шиповатые макросклеры покрыты мелкими острыми шипиками, за исключением концов, которые остаются гладкими. Между шиповатыми и гладкими спикулами можно найти переходы: спикулы с меньшим количеством шипов и почти гладкие — с единичными мелкими шипиками, разбросанными по длине спикулы. Обычно шиповатые макросклеры встречаются вместе с гладкими, но преобладают в числе. Иногда гладкие спикулы совершенно отсутствуют, но описываются также формы, имеющие только гладкие макросклеры. Указание на такие формы мы находим в диагнозе Вейдов-ского (1887: 178). Совинский описывает для Днепра под Киевом экземпляры с преобладанием гладких спикул над шиповатыми рядом с типичными, где шиповатые преобладают (Совинский, 1890: 7). По Пренделю (1915: 5) экземпляры Е. millleri из Днепровских плавней имели гладкие спикулы в преобладающем количестве и лишь в ничтожном количестве шиповатые спикулы. Мною была описана как вариетет Е. millleri форма с исключительно гладкими макросклерами [Резвой, 1922: 17 — „Е. millleri (Lieb.) var. И“]. В перечисленных выше случаях строение амфидисков типично для Е. millleri, но указания на присутствие пузыревидных клеток, к сожалению, отсутствуют. Несмотря на этот пробел, все же можно считать, повидимому, установленным, что существуют формы Е. millleri с преобладанием или исключительным присутствием гладких макросклер. Размеры макросклер колеблются очень широко. Общие пределы колебаний, указываемые в литературе: 132—411 [7. X 6—20 р. Чаще всего раз" меры лежат между 200 и 300 [л при 13—17 р толщины. Для вида характерны короткие амфидиски со стержнем более коротким, чем диаметр диска. Сам диск в типичных случаях состоит из небольшого числа лучей, глубоко рассеченных до стержня. Размеры амфидисков колеблются очень значительно: стержень 6—30 длины, диаметр диска б—40 |л, число лучей 4—13 (иногда и более). Иногда лучи несут шероховатость или большее или меньшее число зубчиков или шипиков. Крупные шарообразные геммулы рассеяны в ткани или в большем количестве скопляются ближе к основанию губки. Капсула одета воздухоносным слоем с одним, двумя или тремя слоями радиально расположенных амфидисков. Поровое отверстие с небольшой воронкообразной оторочкой. Поперечник геммул 300—670 р. Пузыревидные клетки встречаются в большом количестве.
Распространение: Голарктика: Европа (европ. часть СССР, Финляндия, Эстония, Польша, Венгрия, Румыния, Болгария, Югославия, Чехословакия, Австрия, Германия, Швейцария, Бельгия, Голландия, Англия, Ирландия, Франция), вся Сибирь до Камчатки включительно, Уссурийский край, Корея, Манчжурия, Япония, С. Америка.
SPONGILLIDAE —БАДЯГИ
79
Вид вариирует чрезвычайно сильно во всех своих признаках. Многие аберантные формы описаны как вариететы, часто по очень несущественным отклонениям от типичной формы. Благодаря близости Е. millleri и Е. fluviatilis их' часто смешивали. Такие вариететы, как Eph. millleri var. A, var. В и var. astrodiscus Vejdovsky 1883 не заслуживают выделения и относятся нами к типичной форме, как это делает и сам Вейдовский в монографии Поттса (Potts, 1887). Вариететы Е. millleri var. behningi Kirkpatrick и E. m. var. I. Rezvoj ошибочно отнесены к E. millleri и ничем существенным не отличаются от типичной Е. fluviatilis.
Ниже мы даем описание трех вариететов, известных в пределах Союза. Два из них, var. acuminata и var. sibirica, имеют несомненно морфический характер. У первого из них наблюдается недоразвитие геммульных оболочек, у второго, наоборот, геммулы снабжены тройным слоем амфидисков, — явление, известное и для Зап. Европы. Недоразвитие геммульных оболочек, невидимому, связано с условиями больших, а также высокогорных озер. Переразвитие оболочек характерно для Прибайкалья, что, вероятно, связано с суровостью его климата. Третий вариетет, var. obtusosclera, представляет своеобразную аберантную форму.
* За. Ephydatia mul'eri (Lieb.) var. acuminata (Potts)
Макросклеры исключительне' шиповатые, размеры 210—235 у. X 10— 17 [л. Пузыревидные клетки около 22 у в поперечнике. Амфидиски часто уродливой формы; среди более правильно построенных много амфидисков со стержнем, продолженным за диск в виде острого шипа. Длина стержня (от диска до диска) 9—11 у., диаметр диска 12—18 у, число лучей 5—14; общая длина амфидисков с продолженным за диски стержнем 20—24 у-. Воздухоносный слой на геммуле развит сл^бо. Амфидиски немногочисленны, не образуют сплошного слоя, разбросаны по капсуле геммулы и расположены на ней самым различным образом — и радиально, и косо, и тангенциально. Устройство амфидисков и их расположение сходно с тем, что мы имеем у Е. fluviatilis var. teberdana, а также у таких форм, как описанные Сварчевским из Байкала Е. olchonensis, Е. goriaevi и др. (см. ниже стр. 86—88).
Распространение: Северо-американские Соед. Штаты, Франция (?), б. Рязанская губ. (оз. Сокорево), оз. Имандра.
Зв. Ephydatia millleri (Lieb.) var. sibirica Koshow
Под этим названием M. М. Кожов описывает форму, распространенную в Прибайкалье и характеризующуюся очень сильным развитием всех покровных элементов у геммул.
Макросклеры типичного строения с большим преобладанием шиповатых над гладкими. Размеры 250—280 у. X 15—20 у.. Амфидиски 14—15 у, длины, диск 18—20 у, число лучей 4—8. Толстый воздухоносный слой с тремя рядами амфидисков и внешней кутикулой. Поперечник геммул 300—670 у.
80
ФАУНА СССР
Как видно из этого описания, в сибирской форме нет каких-либо особенностей, не известных и для других мест. Многослойное расположение амфидисков неоднократно описывалось, но встречается оно не так часто. Интересно отметить, что столь сильное развитие геммульных покровов мы встречаем в Прибайкальи в водоемах, очень близко расположенных у Байкала, в самом же Байкале описанные для него формы Ephydatia характеризуются противуположными чертами—редукцией и дегенерацией геммульных покровов.1
Распространение: старицы и озера в ближайших окрестностях южн. Байкала, окрестности Иркутска, Верхняя Лена.
* Зс. Ephydatia mulleri (Lieb.) var. ohtusosclera Koshow.
Отличается плотной консистенцией и сильным развитием спонгина. Рядом с типично построенными шиповатыми спикулами в большом количестве встречаются спикулы с закругленными концами, короткие, толстые,, совершенно гладкие или с редкими мелкими шипиками. Размер нормальных спикул 230—-240 w. X 16 и, закругленных —180 р. X 18 у. Амфидиски и геммулы нормального строения, два слоя амфидисков.
Распространение: оз. Чертово, окр. Иркутска.
3. Род HETEROMEYENIA POTTS
Амфидиски распадаются на две группы или только по величине, или по величине и строению. Края дисков зубчатые или снабжены более длинными, изогнутыми к средине амфидиска лучами. Обычно имеется поровая трубка, большею частью снабженная на своей вершине Воротником, без придатков или с придатками.
До настоящего времени в пределах Союза известен только один вид (или вариетет одного вида), относящийся к этому интересному роду.2
В определительные таблицы введены и некоторые другие представители рода из числа встречающихся в Зап. Европе. Краткие же описания даются только для тех из этих форм, которые объединяются Шредером (Schroder, 1927) как вариететы Н. baileyi и к которым принадлежит и встречающийся в Союзе представитель рода, Н. baileyi var. stepanowi.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ И ВАРИЕТЕТОВ РОДА HETEROMEYENIA.
1(10). У больших амфидисков диски рассечены на длинные лучи с загнутыми под углом концами, направленными к средине иглы.
2(7). Поровая трубка отсутствует; поровое отверстие не несет каких-либо придатков, возвышающихся над уровнем воздухоносного слоя.
1 Такова Е. goriavei Sw., по всей вероятности морфичееки измененная Е. miilleri, а также и другие описанные Сварческим виды (см. стр. 86—88).
2 Отметим здесь, что описанный Сварчевским (1923) Carterias primitivus никакого отношения к р. Carteritis или Heteromeyenia не имеет и представляет недостаточно полно описанную форму Ephydatia (см. стр, 87).
SPONGILLIDAE—Б АД ЯГИ
81
У больших амфидисков небольшое число лучей с острыми, перегнутыми под углом концами................................Н. ryderi.
3 (4). Все спикулы отличаются значительной толщиной; макросклеры густо шиповатые, толщиной до 25 ]л. Консистенция губки жесткая ....
...................................Н. ryderi forma pictovensis.
4 (3). Спикулы не отличаются особенной толщиной. Консистенция губки мягкая.
5 (6). Толщина макросклер около 10—13 р. при длине в 200—330 р. Разница между большими и малыми амфидисками значительная (40—55 р-и 30—35 р) .*.................................Н. ryderi forma typica.
6 (5). Все спикулы отличаются своею тонкостью; макросклеры не толще 5 [л при длине в 160—260 р. Между двумя размерами амфидисков сравнительно мало разницы.....................И. ryderi forma baleni.
7 (2). Имеется короткая или длинная поровая трубка.
8 (9). Поровая трубка тонкая и длинная, цилиндрическая; по длине может достигать поперечника геммулы; на своей вершине несет несколько тонких нитевидных придатков различной длины. Загнутые концы лучей больших амфидисков закругленные; сами лучи несколько изгибаются по направлению к средине иглы И. tubisperma.
9 (8). Поровая трубка короткая^ на вершине несет воротник в виде диска или широкой воронки. Загнутые концы лучей больших амфидисков закругленные, сами лучи почти прямые и сильно наклонены к средине иглы.......................................1с. Н. baileyi var. repens.
10 (1). Концы лучей у больших амфидисков не загнуты.
11 (16). Между большими и малыми амфидисками значительная разница в величине; лучи дисков представляют небольшие, неровные зубцы; более глубокими вырезками диск может быть рассечен на лопасти с зубчатыми краями.
12 (15). Поровая трубка снабжена округлым или четырехугольным воротником, от краев которого отходят различной длины придатки.
13 (14). Воротник округлый; его край несет до 10 придатков неправильной формы.........................................lb. Н. baileyi v. petri.
14 (13). Воротник четырехугольный; от каждого угла отходит по 4— 7 придатков; придатки разной длины, тонкие, оканчиваются остро, иногда развилены..........................*la. Н. baileyi v» stepanowL
15 (12). Воротник отсутствует; поровая трубка несет на вершине 4—6 или более лопастных или червеобразных придатков, неправильно изогнутых, иногда разветвленных, на концах закругленных..................
..................................... 1с. И. baileyi v. palatina.
16 (11). Разница в величине больших и малых амфидисков выражена слабо; все же большие амфидиски выступают из воздухоносного слоя. Короткая поровая трубка снабжена воротником в виде широкого, Фауна СССР, Губки, т. П, в. 2 6
82
ФАУНА СССР
неровного диска или неглубокой воронки. Диски у амфидисков довольно правильно рассечены на многочисленные лопасти, несущие по краям мелкие ровные шипики .... If. Н. baileyi v. bobemica.
1. Heteromeyenia baileyi (Bwk.) forma typica
Bowerbank, 1863: 451 (Spongilla)', Schroder, 1927:108
Макросклеры с длинными остриями, гладкие или с мелкими шипи-ками, 200—315 у X 8—10 (л. Паренхимные микросклеры с заостренными концами, покрыты многочисленными шипами, более длинными в средней части иглы и уменьшающимися к ее концам 70—80 /АХ 2—4 у. Крупные амфидиски имеют диски в виде 8—12 лучей или отростков, сильно наклоненных к средине иглы и перегнутых недалеко от концов; длина стержня 95—130 Мелкие амфидиски снабжены дисками с неправильными зубчатыми краями, неглубоко рассеченными на треугольные зубцы, длина стержня 75—80 /и,. Стержни амфидисков обоих размеров снабжены шипами. Между обоими размерами амфидисков многочисленные промежуточные формы. Округлые геммулы до 650 у в поперечнике снабжены толстым мелкоячеистым воздухоносным слоем, покрытым наружной мембраной. Имеется поровая трубка.1
Распространение: Сев. Америка, шт. Нью-Йорк.
* la. Heteromeyenia baileyi (Bwk.) Var. stepanowi (Dybowski)
Dybowski, 1884: 507 (Dosilia ? stepanowi)', Дыбовский 1884: 289, pl. 18, 201; Petr.: 1886: 147 (Carterius stepanowi)-, Weltner 1895: 129 (C. s.); Weltner in Brauer, 1909:187, •f. 324, 325, 328 (C. s.); Arndt, 1926: 334 (C. s.); Schroder, 1927 a: 75 (H. repens f. stepanowi); Schroder, 1927: 101 (H. baileyi var. stepanowi).
Макросклеры гладкие или с редкими мелкими шипиками. Паренхимные микросклеры веретенообразные с длинными, перпендикулярно сидящими шипами в средней части иглы; к ее концам длина шипов сильно убывает. Геммулы с толстым воздухоносным слоем и амфидисками двух размеров, около 40 у. и около 25 у. длины, с шиповатым стержнем и дисками в виде пластинок с зубчиками по краям, рассеченных на несколько секторов более глубокими вырезами. Поровая трубка несет у своей вершины пленчатый воротник четырехугольных очертаний, от углов которого отходят длинные тонкие, иногда развиленные придатки.
Губка образует тонкие корки на подводных растениях и на древесных остатках, попавших в воду — коре, листьях и хворосте. Цвет голубовато-зеленый. Макросклеры тонкие с длинными остриями, несут редкие очень мелкие шипики, разбросанные по длине спикулы, встречаются и совершенно гладкие; размеры 104—200 у. X 4—6.5 у.. Паренхимные микросклеры представляют веретенообразные прямые или слабо
1 Характеристика вида дана по Шреберу (1. с.), исследовавшему единственно сократившиеся микроскопические препараты типа. Устройство поровой трубки по этим препаратам не могло быть прослежено.
SPONGILLIDAE — БАДЯГИ
83
изогнутые иглы, снабженные характерными шипами. На концах спикула усажена мелкими, острыми шипиками, изогнутыми к средине иглы. Ближе к средине острые шипы становятся крупнее, они слабее изогнуты или сидят перпендикулярно. Наиболее длинные шипы расположены в средней части спикулы; они посажены перпендикулярно к игле, на конце затуплены или закруглены и в свою очередь покрыты мелкими шипиками. Иногда вершина шипа несколько вздута или несет палочковидные образования в виде буквы Т; размеры 40—50 [л X 2.5—10 |л. Амфидиски распадаются по размерам на две группы, длинные и короткие. Как те, так и другие несут на стержне редкие, перпендикулярно посаженные крупные шипы, и имеют диски в виде пластинок с мелкими зубчиками по краям, глубоко рассеченные на несколько секторов; диаметр дисков 8—12 [z, стержень длинных амфидисков 40 р X 2 р, коротких 24— 28 р X 2—4 р. Шарообразные геммулы, 300—500 р. в диаметре, окружены довольно толстым 26—30 р толщины воздухоносным слоем, покрытым тонкой наружной кутикулой. В этом слое радиально расположены амфидиски; короткие целиком погружены в слой и не выходят за внешнюю мембрану, более длинные прободают ее и выходят своими дистальными концами за ее пределы. Короткие амфидиски более многочисленны, чем длинные. Промежутки между амфидисками заполнены пузыревидными ячеями в 2— 4 р- в поперечнике, не имеющими полигональных очертаний. Геммулы снабжены цилиндрической поровой трубкой 80 р. длины. Несколько отступя от вершины поровая трубка несет тонкий пластинчатый придаток, воротник. Воротник имеет вид четырехугольника, от углов которого отходят придатки в числе 2—5 и более от каждого угла. Придатки эти представляют тонкие, различной длины нитевидные выросты, на концах острые или тупые, иногда развиленные. Геммулы собраны группами в основании губки на субстрате.
Эта форма описана Дыбовским в 1884 г. и провизорно отнесена им к роду Dosilia. В 1881 Potts (1881) установил род Carterella для трех американских видов, имеющих мелкие придатки на поровой трубке. В 1886 г. Petr показал, что под именем Carterella уже ранее описана одна ископаемая губка, дал роду новое название Carterius и отнес к нему форму, описанную Дыбовским. До последнего времени род Carterius принимался для всех форм, снабженных придатками на поровой трубке. В 1927 г. Schroder (1927в: 101) пересмотрел взаимоотношения родов Carterius Petr., Astromeyenia Annandale и Heteromeyenia Potts. Основываясь на устройстве спикул он’слил первые два рода с р. Heteromeyenia. Этот последний род, на основании присутствия и устройства паренхимных микросклер, он разделил на три подрода (Qxyheteromeyenia, Astroheteromeyenia, Anhetero-meyenia). Schroder произвел также ревизию видов. Н. repens и Carterius stepanowi с причислявшимися к ним вариетстам он отнес к Н. baileyit сохранив за ними лишь значение вариететов этого последнего вида. Таким образом, Н. baileyi, известная лишь из окрестностей Нью-Йорка, прини-6*
84
ФАУНА СССР
мается теперь за широко распространенный вид с многочисленными вариететами (подвидами). Выше, в зоогеографическом очерке, мы касались соотношения этих вариететов и их таксономического значения.
Распространение: европ. часть СССР: б. Харьковская губ.,, оз. Великое и озерко в долине Донца у дер. Кочеток; Ю. Буг в районе г. Николаева; в озерных отложениях Косинских озер (окрести. Москвы) найдены спикулы в толще ила в слоях от 4 до 17.5 м; Польша (вост. Галиция), Венгрия.
1в. Heteromeyenia baileyi (Bwk.) var. petri (Lauterborn).
Lauterborn, 1902: 528 (Carterius stepanowi f. petri)', Arndt, 1925: 344 (C. s. f. p.); Schroder, 1927a: 75 (H. repens f. petri)-, id., 1927 b: 101 (H. baileyi var. petri)-, Arndt in Dahl, 1928: 74 f. 87, 88 (H. b. f. p.).
Корковые формы до 1 см толщины и 15 см в поперечнике. Цвет зеленоватый или желтоватый. Макросклеры гладкие или с мелкими шипиками, 170—310 у X 6—10 у.. Паренхимные микросклеры сходны с Н. Ъ. var. stepanowi, 65—95 у X 5—8 у. Диски на амфидисках рассечены более глубокими вырезками на несколько лопастей с мелкошиповатыми краями. Длинные амфидиски 60—80 у. X 4—6 у, диски 18—19 у; короткие —22— 50 у, диски 18 у. Геммулы 400—500 у. в поперечнике. Вариетет отличается от /7. b. v. stepanowi главным образом устройством поровой трубки. На ее вершине имеется воротник, от краев которого отходят до 8 неправильных придатков.
Распространение: Чехословакия, Германия, Китай (Чжили),. Япония (оз. Бива).
* 1с. Heteromeyenia baileyi (Bwk.) var. repens (Potts)
Potts, 1880: 357 (Spongilla repens); 1887: 237, pl. 11 (H. r.); Schroder, 1927a: 75 (H. r.); id., 1927 b: 107; Arndt in Dahl, 1928: 72, f. 85, 86.
, Макросклеры тонкие с редкими шипиками, 200—280 у. X 5—10 у. Паренхимные микросклеры 54—76 у. X 2.5 у, строение как у Н. b. stepanowi. Диски длинных амфидисков снабжены длинными отростками-лучами, наклоненными к средине иглы с сильно перегнутыми закругленными концами. Стержень длинных амфидисков 54—80 у X 3—5 у, диски с отростками до 23 у. Короткие амфидиски 38—44 у X 4 у, диски 17—18 у, рассечены по краю на треугольные зубцы с затупленными вершинами. Поровая трубка короткая, иногда не превышает толщины воздухоносного слоя; воротник в виде небольшого диска или воронки.
Распространение: Америка — Канада и Соед. Штаты;Европа — Галиция, Германия.
Id. Heteromeyenia baileyi (Bwk.) var. arndti Schroder.
Schroder, 1927 a: 79 (H. repens v. a.); Schroder, 1927 b: 107; Arndt, 1928: 75, fig. 89.
Рассматривается Шредером, как промежуточная форма между Н. b. var. repens и petri. У длинных амфидисков лучи на дисках не перегнуты
SPONGILLIDAE—БАДЯГИ
85
и заканчиваются остро. У коротких амфидисков лучи на дисках треугольные. Поровая трубка короткая, снабжена небольшим воротником угловатых очертаний, иногда с короткими неправильными зубцами, без длинных придатков, иногда трубка не выражена, и поровое отверстие расположено на небольшом возвышении с дискообразным воротником. Макросклеры — 220—330 [J. Х8—12 паренхимные микросклеры—54—90 jx X 2.5—
4 (л, амфидиски — длинные и короткие вместе:—35—62 fx X 2.5—4 [х, диски 16—21
Распространение: Германия, округ Ротенбург, пруды в долине Эльбы.
le. Heteromeyenia baileyi (Bwk.) var. palatina (Lauterborn)
Lauterborn, 1902: 528 (Carterius stepanowi f. palatina}-, Schroder, 1927 a: 81 (H. repens f. palatina}-, Schroder, 1927 b: 107.
Поровая трубка довольно длинная, коническая, 80—100 [х длины; воротник отсутствует; непосредственно от трубки отступя от ее вершины, отходят 4—6 червеообразных, неправильно изогнутых отростка. Макросклеры слабо шиповатые, 150—320 [х X 6—10 [х; паренхимные микросклеры 50—80 jx X 8—10 амфидиски (оба размера) 35—80 у, X 4—6 р.
Распространение: Германия (Рейнпфальц).
If. Heteromeyenia baileyi (Bwk.) var. bohemica (Petr)
Petr, 1886:92 (Ephydatia bohemica)- Weltner in Brauer, 1909: 189 (Carterius stepanowi var. bohemica)-, Schroder, 1927 a: 81 (H.'repens f. bohemica); Schroder, 1927 b: 107 (H. b. var.).
Макросклеры гладкие. Слабое различие по длине между обеими группами амфидисков. Диски рассечены на лопасти с мелкими шипиками по краям. Устройство поровой трубки в общем сходно с Н. Ъ. var. arndti.
Распространение: Чехия, Польша.
4. Род TROCHOSPONGILLA VEJDOVSKY
Диски у амфидисков цельнокрайние в виде тонких кружков с гладким краем. Оба диска на амфидиске одинаковой величины или верхний несколько меньше нижнего, прилегающего к капсуле геммулы. Иногда диски имеют тонкую радиальную исчерченность; в Европе и в пределах Союза только один вид.
* 1. Trochospongilla horrida Weltner
Ehrenberg, 1846: 96 (Spongilla erinaceus); Vejdovsky in Potts, 1887: 177 (Trocho-spongilla erinaceus); Weltner, 1893 a: 7; Arndt, 1926, 344; Rezvoj, 1926: 109 (T. polysclera); Rezvoj, 1928: 226, f. 6, 7.
Макросклеры с многочисленными крупными и острыми шипами; острия длинные. С этими спикулами связаны постепенными переходами более короткие макросклеры с более короткими остриями и более мелкими, не такими многочисленными шипами. Эти спикулы образуют «густые скопления в основании губки над слоем геммул. Геммулы
Н6
ФАУНА СССР
собраны в мостовидный слой, плотно прирастающий к субстрату. Каждая геммула окружена толстым воздухоносным слоем, в основании которого на капсуле располагается слой амфидисков. Амфидиски с дисками одинаковой величины, цельнокрайными, иногда радиально исчерченными. Паренхимные микросклеры отсутствуют.
Образует корки или более массивные наросты; поверхность губки ровная, шероховатая. Цвет беловатый, желтоватый или буроватый. Макросклеры густо покрыты сильными острыми шипами. Величина шипов вариирует у одного и того же экземпляра, но шипы, можно сказать, всегда крупнее чем у спикул Ephydatia miilleri. Макросклеры могут быть разделены на две группы, связанные переходами: более длинные, 215—246 p. X 2—9 сильно шиповатые, с длинными остриями, и более короткие, 99 (и менее)—188 р. X 4—7 р., с меньшим количеством более мелких шипов, с короткими остриями. У амфидисков оба диска одинакового диаметра; стержень 7—10 р X 2.5—4 р., диски 9—12 р.. Иногда на дисках заметна тонкая радиальная исчерченность. Амфидиски образуют на геммуле один сплошной слой, облекающий геммульную капсулу. Этот слой амфидисков лежит в основании толстого, построенного из рядов полигональных ячей воздухоносного слоя, окружающего каждую геммулу. Капсула геммулы с несколько приплюснутой верхней поверхностью, на которой располагается поровое отверстие, приподнимающееся над капсулой в виде широкой воронки. Геммулы собраны в основании губки и плотно прирастают к субстрату, образуя сплошной мостовидный слой. Над слоем геммул, а также пронизывая их воздухоносные оболочки, скопляются макросклеры, и именно более короткие, с короткими остриями (см. выше).
Распространение: Европа (европ. СССР, Латвия, Румыния, Германия, Франция), Арал (дельта Аму-дарьи), Китай (var. sol Annandale 1918 и var. ningpoensis Gee 1926), С. Америка.
СОМНИТЕЛЬНЫЕ ИЛИ НЕДОСТАТОЧНО ПОЛНО ОПИСАННЫЕ ВИДЫ
Сварчевским впервые были констатированы в Байкале представители сем. Spongillidae, описанные им как новые виды. К сожалению, его описания содержат досадные пробелы, а приложенные рисунки сделаны без рисовального аппарата и не дают удовлетворительного представления об этих формах. Нахождение в Байкале настоящих бадяг рядом со специфическими для Байкала видами сем. Lnbomirskiidae представляет большой интерес, но принадлежность их к новым самостоятельным видам, как увидим ниже, вызывает большие сомнения.
* 1. Ephydatia oichonensis Swarczewsky
Сварчевский, 1900: 3; Сварчевский, 1901 : 14, тб. 4.
Спикулы двух родов: гладкие и сплошь покрытые'мелкими шипиками, очень тонкие, с длинными и тонкими остриями; размеры 174—205 р X са
SPONGILLIDAE — БАДЯГ И
ST
4 |л. Амфидиски очень мелкие—6—И р, диски 11—14 [Л, число лучей от 4 до 8. Среди, амфидисков преобладают уродливые формы. На поверхности геммулы амфидиски имеются в очень небольшом количестве и ориентированы различным образом. Диаметр геммул 418—560 р..
Распространение: Байкал, Малое море у мыса Кабанья голова на о-ве Ольхоне (гл. 3—5 сажен).
* 2. Ephydatia goriaevi Swarczewsky
Сварчевский, 1901: 16, тб. 5.
Макросклеры шиповатые, покрытые мелкими шипами, за исключением концов, остающихся гладкими. Острия короткие. Большею частью в средней части спикулы имеется одно или два вздутия. Размеры 187—249 [л X 27 [л. Среди амфидисков преобладают совершенно уродливые формы. Расположены они на геммуле, как и у Eph. olchonensis, неправильно и не образуют сплошного слоя. Длина стержня амфидисков 21—23 р. Геммулы 338—444 [л.
Распространение: Байкал, Чевыркуйский залив, у берега.
* 3. Spongilla microgemmata Swarczewsky
Сварчевский, 1901: 18, тб. 5.
Спикулы шиповатые и гладкие. Первые несут шипы по всей длине, но ближе к концам шипы длиннее и расположены гуще: размеры 275— 311 р. X до 27 [л. Гладкие спикулы более тонкие, 294 p. X 14 р. Очень мелкие геммулы, 178—187 р в поперечнике, образуют скопления около основания губки и в ткани. Геммульные спикулы отсутствуют.
Описанная как Spongilla, эта аберантная форма несомненно принадлежит к р. Ephydatia.
Нахождение: Байкал, Малое море, 20 саженей.
* 4. Carterius1 primitivus Swarczewsky
Сварчевский, 1923: 2, тб. 1.
Макросклеры шиповатые, острия длинные. Количество шипов небольшое, иногда спикулы почти гладкие. Размеры 200—250 [л X 4—10 [л. Амфидиски с очень неправильно развитыми дисками, длина стержня 9—11 р, диск до 15 р. (таким образом амфидиски не могут быть разделены на две группы по величине). Геммулы от 200 до 500 [л в поперечнике. Амфидиски не образуют сплошного слоя и располагаются на поверхности геммулы во всевозможных положениях. При описании этой формы Сварчевский главное внимание обращает на устройство порового отверстия геммул „Отверстие геммулы округлой формы, и края его слегка приподняты над общей поверхностью геммулы. Вокруг отверстия, на некотором рас
1 Сварчевский пишет всюду „Carierias"
88
ФАУНА СССР
стоянии от него, находится невысокая тонкая оторочка, приподнимающаяся с поверхности геммулы". Сварчевский сравнивает эту оторочку с придатками, имеющимися на поровой трубке у р. Carterius {Heteromeyenia). Без просмотра типового материала трудно сказать, что представляет эта оторочка у описанной Сварчевским формы. Из данного им рисунка ясно, что она ничего общего не имеет с теми „воротниками" и придатками, которые имеются у рода Heteromeyenia. Скорее всего это часть наружной оболочки воздухоносного слоя, спускающаяся к поровому отверстию в виде воронки и прирастающая к капсуле геммулы. Форма, несомненно, принадлежит к р. Ephydatia (Е. miilleri?)
Нахождение: Байкал, Ольхонские ворота при входе в Базарную бухту (у берега).
У всех четырех форм, описанных Сварчевским, имеются общие черты. Характерно уродливое и неполное развитие и даже отсутствие геммульных микросклер. Как это и указывает Сварчевский (1923), такое явление наблюдается у форм, живущих в крупных озерах. Можно прибавить, что дело здесь не только в величине водоема, так как эти же явления известны для форм и из небольших озер (так, Eph. fluviatilis var. teberdana описана мною из небольшого горного озера на Сев. Кавказе; вариетет этот характеризуется теми же особенностями). Редукция защитного покрова у геммул должна рассматриваться, как морфологическое изменение, вызванное еще неясными для нас особенностями в характере водоемов. Эти явления редукции защитных покровов геммул не дают основания для выделения особых видов, между тем у Е. olchonensis, Е. goriaevi и У. microgemmata мы не находим каких-либо других характерных признаков. К сожалению, Сварчевский не упоминает о пузыревидных клетках. Их присутствие или отсутствие могло бы пролить свет на эти формы. Относительно .S’. microgemmata надо заметить следующее: она описана по единственному экземпляру, у которого может быть случайно совершенно отсутствовали геммульные микросклеры. По устройству шиповатых макросклер, ясно, что это не Spongilla, а по всей вероятности тоже Ephydatia, так как шиповатые макросклеры известны только у очень немногих тропических видов рода Spongilla. Малая величина геммул также указывает на общую их дегенерацию, вызванную внешними условиями. Carterius primitivus Сварчевскбго, несомненно, не имеет ничего общего с родом Carterius, и, как мы указали уже выше, оторочка у порового отверстия не имеет сходства с воротником р. Heteromeyenia. Относительно этой формы можно с уверенностью сказать, что она, так же как и предыдущие, относится к роду Ephydatia, к одному из двух обычных видов, морфически видоизмененных. Аналогичное мнение о трех первых формах высказано Annandale (1913, 1914), а относительно Carterius primitivus имеется указание у Schroder, что эта форма не может быть отнесена к роду Heteromeyenia ( — Carterius) и требует дополнительного описания (Schroder, 1927:jlll) .
LUBOMIRSKIIDAE—БАЙКАЛЬСКИЕ ГУБКИ 89
2. Сем. LUBOMIRSKHDAE— Байкальские губки
Субдермальные полости отсутствуют. Дермальная мембрана не отделяется от ниже лежащей ткани. Отводящие каналы идут к устьям из глубины ткани, не проходя под самой поверхностью и не образуя кругом устья крупных звездообразных фигур. Оскулярные трубки отсутствуют.1 Устройство скелета различное у различных родов семейства. Спонгин всегда присутствует в значительных количествах. Макросклеры амфиоксы или стронгили, шиповатые или реже гладкие. Микросклеры отсутствуют. Геммулы отсутствуют. Бесполое размножение при помощи соритов.
Несмотря на различия в строении скелета у трех составляющих семейство родов, надо признать их близко родственными и имеющими общее происхождение. Наиболее примитивным является скелет рода Baikalospongia. Он состоит из довольно беспорядочных главных и поперечных пучков. В поверхностном слое всегда имеется большое скопление спикул, и при просматривании тангенциальных срезов с поверхности мы видим концы вертикальных пучков, соединяющихся поперечными пучками; их совокупность образует более или менее правильную полигональную сеть, в каждой петле которой располагается пора. От строения р. Baikalospongia можно перейти к строению скелета р. Swartschewskia. У него тенденция образовывать скопления спикул в поверхностном слое находит свое полное отражение. Здесь в поверхностной корке имеется довольно правильная сеть толстых пучков из тангенциально лежащих спикул. С другой стороны, от скелета р. Baikalospongia можно перейти и к скелету р. Lubomir-skia. Здесь пучки, главные и поперечные, образуют более правильную пространственную решотку. Главные пучки на своих концах несут щетки или кисти расходящихся своими концами спикул. У Baikalospongia мы также находим эти концевые щетки, но не столь ясно дифференцированные. Характер спикул дает также основание считать все семейство естественной группой. У Baikalospongia имеются стронгилы и оксы. У В. intermedia обычно имеются оксы, но встречаются экземпляры со стронгилями. У В. bacillifera обычны стронгилы, но изредка попадаются спикулы с короткими остриями. Количество шипов у всех видов во всех родах сильно вариирует. У одних и тех же экземпляров рядом с шиповатыми спикулами могут встречаться почти гладкие или даже совершенно гладкие. Иногда почти гладкие и гладкие спикулы преобладают над шиповатыми. До известной степени связующим звеном для отдельных родов является В. dshegatajensis, единственный вид, встречающийся вне Байкала. Характер скелета заставляет его отнести к Baikalospongia’, по характеру спикул он имеет сходство с В. intermedia, по некоторым же их особенностям —
1 Против существования оскулярных трубок говорит устройство устий у всех байкальских губок, которые мы рассмотрим при описании отдельных видов. Наблюдений над живыми байкальскими губками не имеется.
90
ФАУНА СССР
с Lubomirskia fusifera’, образование цилиндрических отростков сближает его с L. baicalensis’, по форме оскулярных отверстий он приближается к р. Swartschewskia.
Разнообразие в устройстве скелета не позволяет поместить байкальские губки ни в одно из известных морских семейств. О существенных анатомических различиях с пресноводными сем. Spongillidae мы уже говорили выше (см. стр. 40—42).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ СЕМ. LUBOMIRSKIIDAE
1 (2). Скелет не дифференцирован на главные и поперечные пучки; поверхностный слой резко обособлен в виде корочки с толстой полигональной сетью пучков тангенциально расположенных спикул; скелет внутренней ткани из изолированных макросклер. Спикулы короткие и толстые, стронгили шиповатые или гладкие..............
.......................................... 3. Swartschewskia.
2 (1). Скелет дифференцирован на главные и поперечные пучки. Резко обособленная наружная корочка с тангенциальными спикулами отсутствует.
3 (4). Спонгин присутствует в большом количестве и равномерно распределен по длине главных пучков, образуя сплошное волокно, обволакивающее спикулы. Спикулы в пучке лежат с известной правильностью, параллельно длине пучка или несколько отклоняясь своими дистальными концами. Главные пучки оканчиваются на поверхности губки хорошо выраженными коническими щетками спикул..............
..............................................1. Lubomirskia.
4 (3). Количество спонгина может быть значительным, но распр>еделяется он по главным пучкам неравномерно, образуя неправильные скопления и не образуя сплошного волокна вокруг главных пучков. Спикулы в главных пучках располагаются без большой правильности; часто видны группы тесно склеенных параллельных друг другу спикул, лежащих по отношению к оси пучка в различных направлениях^ Правильно сформированных конических щеток на вершинах главных пучков не образуется........................ 2. Baikalospongia.
1. Род LUBOMIRSKIA DYBOWSKI
Скелет состоит из хорошо дифференцированных главных и поперечных пучков. Спикулы в главных пучках лежат параллельно длине пучка или несколько отклоняются в наружные стороны своим дистальными концами. Главные пучки оканчиваются на поверхности коническими щетками расходящихся своими концами спикул. Спонгин сильно развит и образует сплошные волокна, заключающие главные и поперечные пучки спикул. Спикулы исключительно амфиоксы. Ткань губки упругая и не ломкая, разрывается с некоторым усилием.
LU BOMIRSKIID АЕ—Б АИКАЛЬСКИЕ ГУБКИ
91
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ И ВАРИЕТЕТОВ РОДА LUBOMIRSKIA
1 (2). Спикулы в главных пучках располагаются под некоторым углом к оси пучка, несколько отклоняясь наружу своими дистальными концами. Скелет образует почти исключительно одни главные пучки; поперечные пучки развиты слабо и состоят из одиночных спикул . .
...............................................* 3. L abietina.
2 (1). Спикулы в главных пучках располагаются параллельно оси пучка. 3 (4). Спикулы представлены амфиоксами с сильно выраженной веретено-
образностью; много центротилотных спикул. Шипы мелкие. Значительно преобладают спикулы с шипами, собранными только у концов, и с совершенно гладкой всей средней частью спикулы; встречаются спикулы и совершенно гладкие. Спикулы крупные, 225 [J- X 20 р. в среднем......................................* 2. L. fusifera»
4 (3). Спикулы с шипами по всей длине. Веретенообразность выражена
слабо. Центротилотность, если и встречается, то только у небольшой части спикул.
5 (6). Высокорастущие ветвистые формы. Спикулы сравнительно тонкие и короткие, 193 u. X 13 и в среднем . . . * 1. L. baicalensis typica.
6 (5). Корковые прибрежные формы. Спикулы короткие и очень толстые, покрыты на всем протяжении толстыми шипами. Размеры в среднем 190 [л X 18 р.....................* la. L. baicalensis morpha iittoralis.
* 1. Lubomirskia baicalensis (Pallas). (Табл. II, фиг. 5, т. Ill, ф. 7, 9 —10, т. X, ф. 38, 40, т. XII, ф. 47, т. XIII, ф. 54, т. XIV, ф. 58, т. XV, ф. 62).
Pallas, 1772—1773: 710 (Spongia): Miklucho-Maklay, 1871: 8 (Veluspa polymorpha var. baicalensis)' Dybowsky, 1880a: 12; Dybowsky, ibid.: 19 (L. b. var.); Сварчевский, 1901: 19 (Veluspa ); Annandale, 1913: 98 (Veluspa); Annandale, 1914: 142.
Высокорастущие ветвистые формы. Устья рассеяны по всей поверхности губки и представляют округлые неглубокие ямки 3—4 мм в поперечнике, затянутые на- дне тонкой мембраной, с одним или несколькими отверстиями; сквозь мембрану просвечивают отверстия тонких отводящих каналов. Спикулы амфиоксы с короткими или средней длины остриями, цилиндрические или слабо веретенообразные, сравнительно тонкие, в сред-, нем 13 р. толщины и 193 р- длины. Шипы расположены по всей длине спикулы. Центротилотность, как правило, отсутствует.
Чаще всего губка состоит из более или менее цилиндрических отростков, поднимающихся от коркового основания. Отростки могут дихотомически ветвиться и срастаться с соседними отростками при тесном их соприкосновении. По длине отростков в большинстве случаев имеются пережимы, разделяющие отростки на четкообразно расположенные вздутия; вершины отростков закругленные. Высота достигает 30—60 см; Дыбовский указывает экземпляры до 1 м высотой. Толщина отростков .вариирует
92 ,
ФАУНА СССР
значительно, от 1—2 см до 3—4 см. Некоторые экземпляры отличаются уплощенной формой отростков, имеющих вид треугольных лопастей, расширяющихся к вершине; ширина у вершины может достигать 5 см, при толщине в 1 см. Более редкую форму роста представляют гребенчатые экземпляры: от стелющегося по субстрату цилиндрического стержня отходит кверху ряд цилиндрических отростков. Губка своим корковым основанием прикрепляется к подводным предметам — к скале, крупной неподвижной гальке, или к затонувшему дереву. Толщина базальной корки колеблется около 1 см. Как на особую форму роста надо указать на экземпляры, лишенные базальной корки и места прикрепления. Такие экземпляры в виде длинного отростка с обоими закругленными концами растут свободно лежа на мягком грунте (о причинах появления таких форм см. стр. 26 — 27). Устья встречаются как на базальной корке, так и на отростках и расположены довольно часто, в 2—3 см друг от друга, иногда намечается их рядовое расположение по длине отростков. Ветвистая типичная форма L. baicalensis характеризуется сильным развитием спонгина и вместе с тем тонкими спикулами. Характер спикул, довольно постоянный у отдельного экземпляра, значительно меняется от экземпляра к экземпляру. Изменения касаются главным образом величины и количества шипов, всегда располагающихся по всей длине спикулы. Преобладают короткие острия, реже они средней длины. Спикулы цилиндрические, реже слабо веретенообразные. Средний размер спикул 193 p. X 13 р, пределы колебаний 145—233 р X 9—18 р. Кроме спикул, склеенных в скелетные пучки, на срезах видны более тонкие и короткие спикулы, свободно расположенные в ткани. Они представляют гладкие тонкие амфиоксы с длинными остриями. Между ними и спикулами пучков можно найти все переходы. Это молодые, не достигшие полного роста макросклеры, еще не вклеенные в пучки; их ни в коем случае нельзя рассматривать как паренхимные микросклеры. На этих молодых спикулах у L. baicalensis, в противоположность другим представителям байкальских губок, только очень редко наблюдается в средней части вздутие (центротилотность). Цвет живых губок ярко зеленый.
Распространение: оз. Байкал. Ветвистая форма L. baicalensis начинает попадаться с глубины 3—4 м. Ее заросли достигают максимального развития на глубине 5—8 м (Кожов, 1931). Нижний предел ее распространения точно не установлен, но имеются ветвистые экземпляры с 50 м глубины. По берегам Байкала она распространена всюду, где имеется подходящий субстрат.
* la. Lubomirskia baicalensis (Pallas) morpha littoralis nova. (Табл. X, фиг. 40).
Dybowski, 1880: 46; Кожов, 1931: 25 („Прибрежная корковидная морфа“).
Форма губки корковая, толщина колеблется около 1 см. Отличается от типичной формы не только отсутствием выростов, но и характером
LUBOMJRSKIIDAE — БАЙКАЛЬСКИЕ ГУБКИ
93
спикул. Длина спикул колеблется в тех же пределах, что и у ветвистой формы, но толщина значительно больше, в среднем 18 [л. Веретенообраз-ность выражена слабо, центротилотность так же редка; как у ветвистой формы. Крупные грубые шипы распределяются по всей длине спикулы... Цвет живых губок яркозеленый.
Распространение: оз. Байкал. Встречается начиная' с глубины в 1.5 м. и до 3—6 м.
Указания на эту форму мы находим уже у Дыбовского (1. с.). s Кожов (I. с.) выделяет „прибрежную корковидную морфу", приспособленную к прибрежной зоне. Верхние 1.5 м недоступны и для этой формы, так как сила прибоя здесь слишком велика. Но оба цитированные автора не указывают на отличие в толщине спикул. Кожов отмечает, что обе формы, прибрежная корковая и ветвистая, по характеру спикул друг от друга не отличимы. Большая толщина спикул у m. littoralis, вероятней должна быть поставлена в связь, так же как и ее корковая форма, с условиями прибойной зоны. Средний размер спикул 190 [л X 18 [л, пределы колебаний: 174—229 X 11—-24 [л. Вниз по Ангаре по Дыбовскому (1. с.) встречается L. baicalensis, но исключительно в виде корок. О характере спикул ангарских экземпляров у нас нет данных.
* 2. Lubomirskia fusifera Soukatschoff. (Табл. X, фиг. 42).
Dybowski, 1830а: 20—22 (L. baicalensis var. var. 3, у, 8);. Сукачев, 1895: 7 (L. fusifera). Сукачев, ibid. (Z,. baicalensis var. e)
Корковая форма от 1 до 2 см толщины. Отличается от L. baicalensis, кроме отсутствия отростков, только по величине и характеру спикул: средние размеры спикул 225 (J. X 20 и.; форма их веретенообразная, очень часто наблюдается центротилотность; малочисленные шипы собраны, у концов спикул и большая часть их поверхности гладкая.
Ткань губки довольно рыхлая, мягкая и эластичная. Спикулы L. fusifera длиннее, чем у L. baicalensis, толще, чем у типичной формы и даже превосходят толщиной морфу littoralis. Веретенообразность хорошо выражена. Острия средней длины или короткие. Поверхность спикулы,, как правило, гладкая, и малочисленные шипы собраны у концов. Изредка встречаются спикулы с малочисленными шипиками и в средней части спикулы. Много спикул почти гладких или совершенно гладких. Очень большой процент центротилотных спикул с неясным вздутием в средней части и резко обозначенным вздутием осевого канала. Молодые макросклеры, рассеянные в ткани, имеют тот же характер, как у L. baicalensis,. но большинство из них имеют резко обозначенное вздутие в средней части. Средняя величина для всех исследованных экземпляров 225 р-Х20 [л, колебание длины от 161 до 282 |л, толщины — от 12 до 26 [л.
Распространение: оз. Байкал. Сукачев указывает для исследованных им экземпляров глубины от 4 до 44 м. В материалах Зоологического института Академии Наук имеются экземпляры с глубин от 6 до»
94
ФАУНА СССР
50 м. Таким образом, L. fusifera начинает встречаться у верхней границы распространения губок в Байкале и идет до значительных глубин, все время сохраняя характер корковой формы. Нижний предел ее батиметрического распространения нельзя считать точно установленным.
L. fusifera чрезвычайно близка к L. baicalensis, и самостоятельность этого вида далеко нельзя считать бесспорной. Несомненно, что вариететы L. baicalensis Дыбовского тожественны с описанным Сукачевым видом, хотя он их в своей работе и не отожествляет. Отделение L. fusifera базируется только на строении спикул, вообще сильно вариирующйх у байкальских губок. Только при просмотре значительного материала можно убедиться, что параллельно с L. baicalensis littoralis и typica, приуроченными к определенным глубинам и постепенно сменяющими друг друга, имеется еще какая то особая форма, довольно стойко сохраняющая свои особенности независимо от глубины и имеющая значительное батиметрическое распространение. Вопрос несомненно требует переисследо-вания на основе большого, специально собранного материала.
* 3. Lubomirskia abietina (Swarczewsky). (Табл. X, фиг. 39, т. XII, ф. 48, т. XIII, ф. 56, т. XIV, ф. 60).
Сварчевский, 1901 : 10 (Veluspa); Annandale, 1914: 193.
Корковая форма, достигающая 0.5—1 см толщины. Наружная поверхность имеет характерный вид и покрыта сетью мелких бороздок. Скелет состоит почти исключительно из главных пучков характерного строения. Спикулы в них несколько отклоняются своими дистальными концами от оси пучка, что придает пучкам сходство с молодыми побегами ели. Главные пучки местами соединяются горизонтально лежащими одиночными спикулами, представляющими поперечные пучки.
Развитие спонгина такое же, как и у других корковых форм Lubomirskia. Спикулы оксы с короткими остриями, с хорошо выраженной вере-тенообразностью. Средние размеры 186 (л X 17 у.. Шипы собраны у концов, и остальная поверхность спикулы гладкая, реже встречаются спикулы с шипами в средней части. Главные пучки поднимаются от основания губки к ее наружной поверхности и дихотомически ветвятся. К наружной поверхности пучки постепенно расширяются и оканчиваются щеткой торчащих спикул. Поперечные пучки развиты слабо, состоят часто из одной или двух-трех спикул, соединяющих главные пучки. Количество спонгина значительное, он образует волокна, из которых торчат в стороны дистальные концы спикул. Средние размеры спикул исследованных экземпляров 186 р. X 17 р., пределы колебания 145—240 р. X 13—21 р.. По своему характеру они очень сходны со спикулами L. fusifera, но меньших размеров. Веретенообразность хорошо выражена. Острия короткие или средние. Шипы малочисленны и собраны у концов; реже имеются более редкие шипы и в средней части спикулы; встречаются почти гладкие и совершенно гладкие спикулы.
LUBOMIRSKIIDAE-БАЙКАЛЬСКИЕ ГУБКИ 95
Распространение: оз. Байкал; границы вертикального распространения нельзя указать с точностью, так же как и распространение-по Байкалу. Губка описана по нахождению у о-ва Ольхона на глубине 60 м. Известны нахождения на глубинах 20—40 м.
12. Род BAIKALOSPONGIA ANNANDALE
Корковые или массивные формы, за исключением одного вида, дающего анастомозирующие отростки. Расположение спикул в главных и поперечных пучках довольно беспорядочное; они лежат большею частью в различных направлениях по отношению оси пучка, собранные в тесные группы параллельных друг другу спикул. Спонгин присутствует в значительном количестве, но не образует сплошного волокна, обволакивающего спикулы. Спикулы представлены оксами и стронгилями. Ткань не упругая и ломкая, и смотря по видам то жесткая, то мягкая и довольно дряблая.
Корковые или массивные формы; только один вид дает короткие пальцеобразные выросты, обычно анастомозирующие (В. dzhegatajensis). Консистенция ткани различна у разных видов, но не достигает эластичности и прочности таковой Lubomirskia. Строение оскулярных отверстий различное. Скелет, сравнительно с родом Lubomirskia, отличается меньшей правильностью, и главные и поперечные пучки слабее дифференцированы. Когда главные пучки достигают значительной толщины, они представляют довольно беспорядочное скопление спикул. В таком скоплении намечаются отдельные, плотные пакеты тесно соединенных спикул; эти пакеты располагаются в пучке в различных направлениях. Степень развития скелета очень различна у различных видов и даже в пределах одного и того же вида. У некоторых- экземпляров В. intermedia скелет очень слабый; иногда он напоминает скелет р. Reniera. Главные пучки оканчиваются на поверхности то лучше, то хуже выраженными щетками спикул, но не такими, как у р. Lubomirskia. Эти щетки вместе со связывающими их поперечными пучками обыкновенно образуют в поверхностном слое значительное скопление спикул, имеющее при рассматривании с поверхности вид менее или более правильной полигональной сети. Благодаря скоплению спикул в поверхностном слое на разрезах хорошо видны „кольца нарастания", следы прежних наружных слоев с более частым расположением спикул. Спонгин, сравнительно с р. Lubomirskia развит слабее и не образует сплошного волокна, целиком охватывающего спикулы пучка. Значительные массы спонгина образуют неравномерные скопления, склеивающие главным образом концы спикул. Спикулы с заостренными или закругленными концами, с шипами по всей длине спикулы или собранными только у концов; встречаются и совершенно гладкие. Спикулы в общем крупнее, чем у рода Lubomirskia.
%
ФАУНА СССР
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ РОДА BAIKALOSPONGIA
1(4). Главные скелетные пучки сравнительно тонкие с немногими спикулами в поперечном сечении; по толщине они мало отличаются от поперечных пучков. Спонгина меньше, чем у остальных видов семейства и он заметен главным образом в местах соединения главных и поперечных пучков. Спикулы амфиоксы, реже стронгили с шипами по всей длине. Шипы чаще мелкие. Средний размер спикул 226 jU X 15 .................... . . . * 2. В. intermedia»
2(3). Спикулы обычно амфиоксы, реже стронгили. Средний размер спикул 226 [J. X 15 [J., пределы колебаний 172—266 p. X 9—22 [л.............
.......................................*2. В. intermedia typica.
3(2). Спикулы стронгили с мелкими шипиками по всей длине. Средний размер спикул 405 [7. X 22 [л, пределы колебаний 330—470 у X 20— 25 [л......................... *2а. В. intermedia morpha profundaiis»
4(1). Главные пучки толстые с многочисленными спикулами в поперечном сечении; они значительно толще поперечных пучков. Спонгин в значительных количествах.
5 (6). Спикулы — крупные толстые стронгили с шипами только на конп&х; шиповатые по всей длине спикулы встречаются только изредка. Средние размеры спикул 262 [7. X 27 у...............* 1. В. bacillifera»
6 (5). Спикулы — амфиоксы с мелкими шипами по всей длине, встречаются и гладкие спикулы, у некоторых экземпляров даже в преобладающем количестве. Средние размеры спикул 245 [а X 19 р.................
........................ •................3. В. dzhegatajensis.
* 1. Baikalospongia bacillifera (Dybowski). (Табл. Ill, фиг. 8, т. XI, ф. 43, т. XII, ф. 49, т. XIII, ф. 55, т. XIV, ф. 61, т. XV, ф. 63).
Dybowski, 1880: 22 (Lubomirskia b. et L. b. var a, ft, 7); Сукачев, 1895: 6 (Lubomirskia tsherskii); Annandale, 1913: 100 (Veluspa); Annandale, 1914: 144; Сварчевский, 1923: 12 (Veluspa); Макушок, 1927 : 79 и 124 (Baicalolepis Jungiformis, bacillifera и tscherskii).
Корковые или массивные формы. Скелет из толстых беспорядочных главных и более тонких, не всегда хорошо дифференцированных, поперечных пучков. Спонгин в значительных количествах беспорядочными скоплениями склеивает спикулы, представленные грубыми, толстыми строителями с шипами на закругленных концах; среди таких спикул иногда встречаются спикулы с шипами в средней части. Средний размер спикул 262 у X 27 у.
Ткань жесткая и ломкая. Толщина корок достигает нескольких сантиметров, массивные формы имеют вид шаров или эллипсоидов в 5— 6 см в поперечнике. Часто встречается грибовидная форма роста, описанная Макушком как отдельный вид под названием Baicalolepis fungiformis
LUBOMIRSKIIDAE —БАЙКАЛЬСКИЕ ГУБКИ
97
(Макушок, 1927). Подошва В. bacillifera покрыта толстой спонгиновой мембраной, на которой хорошо заметны кольца нарастания. Образование грибовидных форм можно объяснить себе тем, что в некоторых случаях корка перестает разрастаться в ширину по субстрату и рост идет только в вышину. Такие экземпляры несколько расширяются к верхнему концу и получают большое сходство с грибом-дождевиком (Макушок, 1. с.). Устья располагаются не только у верхнего края боковой поверхности, как это описывает Макушок, но и по всей верхней поверхности. Возможно, что грибовидные формы развиваются тогда, когда губка стеснена растущими по соседству другими губками, или ее разрастанию по субстрату мешает осевший ил. Боковая поверхность грибовидных экземпляров одета такой же спонгиновой пленкой, как и подошва. Между этой формой роста и обычной корковой можно найти все переходы. Никаких отличий во внутреннем устройстве подметить не удается. Устройство оскулярных отверстий В. bacillifera сходно с таковыми L. baicalensis. Здесь также на дне плоского углубления в 3—4 мм в поперечнике, затянутого пленочкой, имеется одно или несколько отверстий. Сквозь пленку просвечивают радиально сходящиеся тонкие отводящие каналы. Отметим, что на фиксированном материале пленочка обычно не сохраняется, и устье имеет вид маленькой звездчатой фигуры в 3—4 мм, образованной открытыми отводящими каналами. Ткань плотная, довольно твердая, нд ломкая и совершенно не упругая. Скелет состоит из толстых главных пучков, представляющих большое скопление довольно неправильно расположенных спикул. Ясно выраженные щетки или кисти на концах пучков здесь отсутствуют.
Главные пучки соединяются друг с другом довольно неправильными поперечными пучками. Спонгин образует значительные скопления, склеивающие спикулы, главным образом их концы, но сплошное спонгиновое волокно, окутывающее весь пучок, отсутствует. Спикулы представляют крупные стронгили с шипами, собранными на закругленных концах.
Остальная поверхность спикулы обычно совершенно гладкая; среди обычных спикул шиповатые в средней части встречаются довольно редко и не у всех экземпляров. У некоторых экземпляров среди стронгилей попадаются спикулы, которые можно считать амфиоксами с очень короткими остриями. Есть спикулы с очень малым количеством шипов на концах, но совершенно гладкие не были констатированы. Средний размер спикул 262 (л X 27 [л, пределы колебаний 170—365 X 16—40 |л. Цвет живых губок зеленый, но не такой яркий, как у L. baicalensis.
Распространение: озеро Байкал. Встречается начиная с глубины немногих метров. Недавно добыты с глубины 533 м два массивных экземпляра, росших на железистых конкрециях. Эти экземпляры имели своеобразный яркий сине-зеленый цвет. Распространена всюду по берегам Байкала, где имеется подходящий грунт.
Фауна СССР, Губам, т. П, в. 2 1
98 ФАУНА СССР
* 2. Baikalospongia intermedia (Dybowski). (Табл. XI, фиг. 46, т. XII, ф. 50, 51, т. XIII, ф. 57).
Dybowski, 1880а: 28 {Lubomirskia i. et L. i. var. а); Сукачев, 1895:4 (L. i. var. /8); Annandale, 1913 :100 {Veluspa); Annandale, 1914 :144.
Тонкие корковые формы. Скелет довольно беспорядочный и слабый, состоит из главных и поперечных пучков, то лучше то хуже обозначенных. Спонгин развит относительно слабо. Спикулы чаще всего тонкие оксы с остриями средними или короткими, по всей длине покрытые мелкими шипами. Реже встречаются экземпляры со стронгилями с редкими шипами в средней части и с концентрацией их на закругленных концах. Средний размер спикул 226 р X 15 р. Ткань мягкая и ломкая.
Корки имеют около 4—5 мм толщины. Сравнительно с другими байкальскими губками ткань более мягкая и ломкая. Сухая ткань легко растирается между пальцами. Устья в виде небольших плоских углублений с несколькими мелкими отверстиями на дне. Скелет развит значительно слабее, чем у В. bacillifera, л степень его развития сильно колеблется у различных экземпляров. Главные пучки и короткие поперечные, состоящие из одной или нескольких спикул, образуют не слишком правильную решетку. У экземпляров со слабым развитием скелета главные пучки тонки, состоят в поперечном сечении всего из 2—3 спикул и заканчиваются на поверхности несколькими торчащими спикулами. Эти пучки соединяются- поперечными, состоящими из одиночных спикул; в таких случаях скелет получает сходство со скелетом р. Reniera. При более развитом скелете главные пучки толще и оканчиваются на поверхности беспорядочными щетками, образующими в поверхностном слое значительное скопление спикул. Спонгин развит слабее, чем у других байкальских губок. Его скопления склеивают спикулы главных пучков; он хорошо заметен в местах соединения главных пучков с поперечными (см. табл. XIII, фиг. 57). Спикулы сильно вариируют по толщине и строению. Часто они довольно тонкие с остриями средними или дтже длинными. Средние размеры 226 X 15 р-, пределы колебаний 172 —266 р, X 9 —22 р. Шипы всегда рассеяны по всей длине, но часто они гуще расположены у концов. В некоторых случаях, наоборот, самые концы спикул свободны от шипов. У некоторых экземпляров преобладают более толстые стронгили.
Распространение: оз. Байкал. Встречается от верхнего предела распространения губок в Байкале (1.5—2 м) до глубины нескольких десятков метров. Нижняя граница не может быть указана с достоверностью.
*2а. Baikalospongia intermedia (Dyb.) morpha profcndalis nova. (Табл. XI, фиг. 44).
Отличается от основной формы размерами спикул, средняя величина которых 405 р. X 22 р (у основной формы 226 р. X 15 р). Пределы колебаний размеров 330—470 р X 20—25 р.. Спикулы представляют слабо изогнутые или прямые стронгили по всей длине покрытые мелкими шипи-
LU BOMIRSKIIDAE—БАЙКАЛЬСКИЕ ГУБКИ 99
ками (табл. XI, фиг. 44); на закругленных концах густое скопление мелких точечных шипиков.
Единственный экземпляр представляет несколько мелких обрывков мягкой тонкой корочки зеленоватого цвета (спирт). Сохранность плохая. На плотной базальной спонгиновой мембране имеется слой ткани 3—4 мм толщиной. Наружная поверхность повреждена. Вследствие плохой сохранности не удалось получить хорошего препарата скелета. Насколько позволяет судить препарат, скелет слабый, состоит из неправильной сети спикул, склеенных своими концами.
Распространение: оз. Байкал. Губка добыта летом 1932 г. у Б. Котов с глубины 889 м. По записи в журнале и по словам сборщиков, М. Я. Базикаловой и Д. Н. Талиева, губка была ярко голубовато-зеленого цвета; цвет сильно отличав гея от травянисто-зеленого цвета, обычно свойственного байкальским губкам с малых глубин. Такой же цвет был и у Baikalospongia bacillifera, добытой теми же сборщиками с глубины 533 м. Глубина в 889 м является рекордной для пресноводных губок. Находка несомненно является редкой, так как на такой глубине развиты илистые грунты и трудно найти подходящий твердый субстрат. Эту интересную глубинную форму мы здесь описываем как новую морфу В. intermedia, основываясь на величине спикул. По ничтожности имеющегося материала такое выделение надо считать провизорным. Только более обильный ' материал позволит установить правильные взаимоотношения этой формы. В. intermedia вариирует сильнее других байкальских губок. Обычно мы встречаем у нее спикулы в виде амфиоксов со средними или длинными остриями. Но встречаются также экземпляры со стронгилями. Может быть в дальнейшем, когда в распоряжении исследователя будет более богатый и систематически собранный материал, придется форму со стронгилями выделить в самостоятельный вид, к которому будет отнесена и наша морфа profundalis.
3. Baikalospongia dzhegatajensis Rezvoj. (Табл. IV, фиг. 11, т. XI, ф. 45, т. ХШ, ф. 53, т. XIV, ф. 59, т. XV, ф. 64).
Rezvoj, 1927: 296, f. а—d (В. d. et В. d. var. lissosclera)
Тело массивное, клубневидное или состоит из цилиндрических анастомозирующих, образующих клубок отростков. Устья малочисленные в виде небольших отверстий на дне маленьких ямок. Субдермальные полости отсутствуют. Главные пучки со значительным числом спикул в Поперечном сечении, расположенных под различными углами к оси пучка. Спонгин в значительном количестве, но не образует сплошного волокна, охватывающего спикулы. Спикулы амфиоксы с короткими или средней длины остриями, с мелкими шипиками по всей длине или у концов; встречаются и совершенно гладкие. Средний размер 245 |л X 19 [>.
Материал по этому виду состоит из 7 полных экземпляров и некоторого числа обломков. Размеры от 2 до 4 см в поперечнике; диаметр 7*
100
ФАУН А СССР
отростков у ветвистых экземпляров от 3 до 10 мм. Цвет (в спирту) белый, желтоватый или сероватый. Поверхность ровная, слегка шероховатая на ощупь. Консистенция плотная, ткань довольно эластичная. У всех экземпляров очень небольшое число устий, открывающихся небольшими отверстиями на дне ямок в 2—3 мм в поперечнике. Отводящие каналы подходят к устью из глубины ткани в количестве 1—2. Кольца нарастания также хорошо заметны, как и у других представителей семейства. Скелет представляет довольно беспорядочную решетку. В главных пучках хорошо заметно расположение спикул плотными группами под различными углами к оси пучка. Спонгина больше, чем у В. bacillifera, но все же он не образует сплошного волокна, как у Lubomirskia. Спикулы варии— руют довольно сильно у одного и того же экземпляра и от экземпляра к экземпляру. Типичными можно считать амфиоксы с короткими или средней длины остриями, равномерно покрытыми по всей длине мелкими шипами. Иногда наблюдается более частое расположение шипов у концов. Очень часто уменьшается размер шипов и от шиповатости может оставаться только тонкая шероховатость. Встречаются и совершенно гладкие спикулы. Среди нормальных встречаются также спикулы своеобразных очертаний, толстые и короткие, цилиндрические, с резко обозначенными конусообразными концами. Центротилотность встречается почти так же часто, как и у L. fusifera. Таковы спикулы у экземпляров, снабженных отростками. У двух клубневидных экземпляров без отростков спикулы в среднем крупнее и среди них в значительно преобладающем количестве встречаются гладкие или почти гладкие спикулы. Эти же два экземпляра отличаются несколько более темным, сероватым цветом. При описании вида эти два экземпляра были провизорно, выделены как var. lissosclera. Средний размер спикул для всех экземпляров 245 [л X 19 [л, пределы колебаний 170—320 [л X 13—25 [л.
•Распространение. Материал по этому виду добыт А. Я; Тугариновым в 1915 г. на берегу Джегатай-куля (Танну-тувинская республика, Урянхайский край). Губки собраны на скоплениях хары; выброшенных прибоем; они еще не успели высохнуть и все хорошей сохранности. Все экземпляры росли на харах, следовательно, губка живет на незначительных глубинах. Эта находка представляет единственный пока случай нахождения байкальских губок вне самого Байкала и Н. Ангары. По устройству скелета и характеру распределения спонгина этот вид должен быть отнесен к р. Baikalospongia, но стоит ближе к р. Lubomirskia, чем остальные виды Baikalospongia. В устройстве спикул больше всего сходства с В. intermedia. Часто встречающаяся центротилотность сближает В. dzhegatajensis с L. fusifera, наличие же цилиндрических отростков с L. baicalensis. По устройству устий вид может быть сопоставлен с р. Swar-tschewskia. Таким образом, В. dzhegatajensis занимает как бы промежуточное положение среди различных представителей семейства байкальских губок.
LU BO MI RS К11D АЕ—Б АИ К АЛЬС КИЕ ГУБКИ
101
III. Род SWARTSCHEWSKIA MAKUSCHOK.
Скелет не дифференцирован на главные и поперечные пучки. В дермальном слое обособляется полигональная сеть пучков, состоящих из толстых тангенциально расположенных спикул. Внутри губки рассеяны малочисленные не связанные в пучки макросклеры. Спонгин имеется в большом количестве в поверхностной корке. Спикулы исключительно стронгили, шиповатые или гладкие. Консистенция наружной корочки жесткая и ломкая, внутренняя ткань мягкая и рыхлая. Один вид. •
* 1. Swartschewskia papyracea (Dybowski). (Табл. X, фиг. 41, т. XII, ф. 52, т. XV, ф. 65).
Dybowski, 1880а: 13 (Lubomirskia)-, Сварчевский, 1901: 10 (Lubomirskia р. var. а); ibid.: 13 (Lubomirskia irregularis)-, Annandale 1914: 144 (Baikalospongia p. et irregularis)-, Макушок, 1927: 79 (Swartschewskia p. et irregularis).
Характеризуется приведенным выше описанием рода. Зеленая окраска всегда отсутствует. Форма тела корковая или до некоторой степени массивная. Малочисленные устья рассеяны по поверхности или расположены на вершинах низких, уплощенных конусообразных выростов. Губка чаще всего имеет вид небольшого невысоко поднимающегося сосочка с одним оскулярным отверстием на вершине. Устья расположены в неглубоких ямках и большею частью окружены то более низким, то более высоким валиком. Отверстие устья круглое. Поверхность тела совершенно гладкая. Для невооруженного глаза она равномерно покрыта мелкими точками, представляющими просвечивающие петли полигональной скелетной сети с порами посредине каждой петли. На поверхности дифференцирован наружный слой в виде плотной гладкой корочки, под которой лежит рыхлая и мягкая внутренняя ткань. Скелет корки в виде многоспикульной полигональной сети, богатый спонгином; скелет внутренней ткани из изолированных спикул. Спикулы короткие и толстые, дугообразно изогнутые в виде колбасок стронгили. Средний размер спикул 133 р. X 21 р, пределы колебаний 112—150 р. X 13—29 р.. Поверхность спикул покрыта мелкими шипиками, гуще расположенными на концевых закруглениях. Иногда наблюдается расположение шипиков группами. Реже встречаются экземпляры с совершенно гладкими или почти гладкими спикулами. Такая форма была описана по одному экземпляру Сварчевским (1923) как „Lubomirskia papyracea var. d“. К форме с гладкими макросклерами относится и Lubomirskia irregularis Swarczewsky (1. с.), также описанная по одному экземпляру, отличающаяся от типичной формы только тем, что скелетная сеть в поверхностной корке не имеет обычной правильности.
Распространение: оз. Байкал. Вид попадается в общем реже остальных байкальских губок. По Кожову (1931) он довольно многочисле-нен в районе Б. Котов. Встречается от 2 до 80 м глубины.
Приложение
ГУБКИ В ОТЛОЖЕНИЯХ СОВРЕМЕННЫХ ВОДОЕМОВ
Определение губок по изолированным спикулам в современных и субфоссильных озерных отложениях. ,
В работах, посвященных изучению грунтов современных озер, указываются обыкновенно и спикулы пресноводных губок. Чернай (1870) исследовал грунты озера Лимана и Чайки (б. Харьковская губ.) и нашел в них спикулы Spongilla lacustris. В работе Месяцева (1924) даны результаты исследования грунтов трех Косинских озер (б. Московский уезд)— Черного, Белого и Святого. Спикулы губок найдены во всех трех озерах. Часто на спикулах были заметны следы разъедания с поверхности и со стороны осевого канала. Самым богатым губками оказалось Черное озеро, где во всех пройденных буром горизонтах ила от 4 до 17.5 м были найдены спикулы, особенно в слоях на 8 и 9 метрах. Констатированы Spongilla lacustris, S. fragilis, Ephydatia millleri, Heteromeyenia baileyi var. stepanowi. Интересно, что в настоящее время губки в Черном и Белом озерах не живут, a Heteromeyenia — в живом состоянии вообще не была встречена в окрестностях Москвы и известна только по значительно более южным нахождениям. й
Спикулы губок сохраняются не только в современных пресноводных грунтах, но также и в сапропелях, подстилающих торфяники, отчасти и в самом торфе, а также в известковых туфах, диатомитах и других отложениях, иногда в значительных количествах. Сохранность может быть достаточно хорошей, позволяющей рассмотреть все детали строения спикулы. В более древних отложениях органическое вещество частично или полностью исчезает, и отложение обогащается минеральными составными частями, в том числе кремневыми спикулами губок и створками диатсмовых. Целые слои оказываются сложенными из такого кремнеземистого материала (пресноводные трепелы, инфузорные земли, диатомиты).
Определение губок и в современных, и в более древних отложениях представляет значительный интерес, но только в редких случаях оно может быть более или менее надежным. В помощь тем, кто обрабатывает подобные отложения, мы попробуем сопоставить морфологические особенности спикул с теми видами, у которых они встречаются.1 Дать настоя
1 Мы будем иметь в виду только формы, вошедшие в настоящий определитель. В более древних отложениях вполне возможно встретить формы вымершие, а из ныне живущих — виды, в настоящее время не встречающиеся в нашей фауне. Учесть эти пос--ледние мы здесь не можем.
ПРИЛОЖЕНИЕ
103
щую определительную таблицу не представляется возможным. Мы будем исходить из наиболее трудного случая, когда имеются изолированные макро-.или микросклеры. При большом обилии спикул и совместном нахождении макро- и микросклер, можно делать более определенные заключения.
I. Макросклеры
1. Гладкие амфиоксы. Размеры не ниже 100 р. Гладкие амфиоксы встречаются у большинства видов. Исключается Trochospongilla horrida И‘ с большой долей вероятия Ephydatia millleri. Возможны все виды р. Spongilla, а также Ephydat'.a fluviatilis. При обилии и однородности спикул некоторые указания дают размеры: средняя длина больше 300 р указывает на Ephydatia fluviatilis", средняя длина меньше или до 300 р — возможны все остальные виды с гладкими амфисксами.
2. Шиповатые амфиоксы:
а) шипы длинные и острые, густо покрывают спикулу; от осевого канала спикулы часто отходят под прямым углом боковые канальцы и идут к более крупным шипам. Trochospongilla horrida;
б) шипы мелкие и острые, густо по срывают спикулу, за исключением концов, которые остаются гладкими Ephydatia miilleri;
в) шиповатость выражена слабо, малочисленные мелкие шипики ,раэ-бросаны по длине спикулы Ephydatia fluviaiilis (возможно и Е. тйНёЬЛ.
II. Микрбсклеры
Размеры в большинстве случаев <С 100 р.
1. Шиповатые р а б д Ы, заостренные или притуплен* ны е у концов
а) веретенообразность хорошо выражена, спикула постепенно утончается к обоим концам, изгиб слабый. Концы острые, на всей поверхности тесно прижатые друг к другу, густо сидят мелкие шипики. Размеры обычно 70—80 р при 3—7 р толщины; пределы колебаний 30—128 р: паренхимные спикулы Spongilla lacustris или S. arctica.
б) Веретенообразность не выражена, спикулы цилиндрические с различным изгибом, от прямых до сильно и неправильно изогнутых, концы с короткими остриями или притупленные. Шипы немногочисленные, сравнительно крупные и острые, иногда гуще расположены ближе к концам. Обычно в пределах 50—80 р, в редких случаях длиннее (до 160 р): геммульные спикулы Spongilla lacustris или S. arctica.
в) Шипы в средней части спикулы рассечены на своих вершинах на 2—4 крючка; ближе к концам шипы простые, острые. Концы спикулы тупые и несут наиболее длинные, изогнутые острые шипы, или же концы острые, и наиболее длинные шипы сидят в средней части спикулы: геммульные и паренхимные спикулы Spongilla biseriata.
104 ФАУНА СССР
г) Слабо изогнутые цилиндрические шиповатые рабды, 60—140 у. X X 4—12 [л. Концы притупленные или коротко заостренные. Шипы мелкие, многочисленные; расположение шипов не настолько частое, чтобы между ними не оставалось промежутков. Геммульные спикулы Sp. fragilis.
д) Грубые, толстые микросклеры 64—120 р X 8—15 р. Острия короткие, или концы притуплены. Спикула усажена малочисленными толстыми острыми шипами: геммульные спикулы З'. sianleyi.
е) Тонкие, прямые микросклеры, в средней части иглы сидят длинные затупленные или вздутые на вершинах шипы; к концам спикулы шипы укорачиваются.- Концы спикулы заострены. Размеры: 40—50 р X 2.5—10 р: паренхимные спикулы Heteromeyenia baileyi.
III. Амфидиски
1. Диск в виде звездообразной фигуры, глубоко рассеченной на лучи; число лучей небольшое, 6—13 или несколько больше. Стержень короткий, короче диаметра диска. Размеры: стержень 6—30 у, диск 6—40 р.... Ephydatia miilleri.
2. Число лучей на диске более значительное, от 15 до 35; диск имеет вид пластинки с зубчатым краем; большею частью пластинка делится более глубокими надрезами на несколько „лепестков" и диск получает сходство с цветком гвоздики; в более редких случаях лучи более глубоко рассечены и диск имеет сходство с диском Е. miilleri, но число лучей всегда более значительное. Стержень длинный, длиннее диаметра диска; очень часто он несколько сужен в средней части и иногда несет 1—3 шипа по своей длине. Размеры: стержень 19—35 у, диск 10—22 у...........
......................... . • . . . ,.........Ephydatia fluviatilis.
3. Стержень диска относительно очень длинный в 2.5 до 3.5 раз длиннее диаметра диска. Он цилиндрический и большею частью имеет немногочисленные короткие шипы по своей длине. Диски в виде пластинок с зубчатым краем, зубцы короткие, треугольные, часто более глубокие вырезки придают диску неправильную форму. Эти амфидиски на одной и той же геммуле бывают двух размеров, более длинные, около 40 р и более короткие, 24—28 у. У некоторых вариететов (Н. baileyi var. repens) длинные амфидиски отличаются от коротких и своим устройством: диск представляет звездообразную’ фигуру с немногочисленными длинными лучами, загнутыми к середине амфидиска, с сильно перегнутыми в том же направлении закругленными концами . . . Heteromeyenia b ailei. (несколько вариететов, в том числе известный в Союзе v. stepanovi).
Диски в виде цельнокрайних тонких пластинок, имеющих вид правильных кружков. Иногда з'аметна на таком кружке тонкая лучистость. Стержень короткий: 7—10 р, диск 9—12 у-. Оба диска одной и той же величины....................................Trochospongilla horrida.
ZUSAMMENFASSUNG
Der vorliegende Band der Fauna der U. d. S. S. R. ist den Siisswas-serschwammen, die innerhalb der Union leben, gewidmet. Ungeachtet dessen, dass unsere Spongiofauna noch recht wenig erforscht ist, fehlen bei uns nur wenige Formen, die in Westeuropa bekannt sind. Andererseits konnen wir unter den echten Spongilliden nur eine einzige Art verzsichnen, die nur innerhalb der Grenzen der Union vorkommt. Ein ganz eigenartiges Element unserer Fauna stellen Formen aus dem Baikalsee dar, und im vorliegenden Auszuge werden wir deren systematische Stellung betrachten und Beschrei-bungen einzelner Arten gcben. Eine Anzahl Arten aus dem genannten See, die gegenwartig auf drei verschiedene Gattungen bezogen werden, unter-scheiden sich schon ihrem ausseren Habitus nach schroff von den gewohnlichen Susswass.erschwammen oder Badiaga’s, wie sie russisch genannt werden. Die Baikalschwamme haben schon langst die Aufmerksamkeit der Spongiologen auf sich gelenkt, und ihnen ist eine Anzahl Arbeiten gewidmet. Trotzdem darf es gesagt werden, daSs sie bis auf den heutigen Tag noch bei weitem nicht genugend erforscht sind und dass inbezug auf ihre systematische Stellung sowie ihr Verhaltn's zu anderen Schwammen noch gar Manches unauf-geklart verbleibt. Die Baikalschwamme wurden gewohnlich und werden auch bisjetzt zu der einzigen Siisswasserfamilie Spongillidae gerechnet. So wird in einer Reihe von Zusammenfassungen die Gattung Lubomirskia in dieser Familie neben den anderen Siisswassergattungen gestellt (Vosmaer 1887, Weltner 1895, Hentschel 1923, Arndt 1926). Von den erwahnten Autoren scheidet Weltner die Gattung Lubomirsk'.a (zusammen mit Arten, die nachher in besondere Gattungen ausgeschieden wurden) in die Unterfamilie £zz6omz‘r~ skiinae aus, neben der Unterfamilie Spongillinae und Meyeninae. In den letzten Jahren hat sich fur eine innigste Verwandtschaft der Baikalschwamme zu den ubrigen Siisswasserschwammen Makuschok (1925) ausgesprochen. Neben den eigenartigen Formen sind aus dem Baikalsee ©auch typische Spongilliden bekannt, die vorwiegend in dessen Buchten (,,Ssoren“) vorkommen, aber auch in der offenen Kiistenzone des Baikalsees selbst vorgefunden wurden. Diese Schwamme wurden von Swarczewsky als selbstandige Arten der Gattung Spongilla, Ephydatia und Carterlus beschrieben. Weiter unten werden wir noch zur Frage uber diese seind Arten zuruckkommen. Hier sei nur bemerkt, dass die Beschreibungen von Swarczewsky recht unvollstandig und ringenugend sind. Aus denselben erhelit nur, dass samtliche seine neue Arten
106
FAUNA URSS
(Sp. microgemmata, Eph. olchonensis, Eph. goriaevi, Carterius primitivus} zur Gattung Ephydatia und zwar, aller Wahrscheinlichkeit nach, zu der einen oder zu den beiden gewohnlichsten Arten, Eph. miilleri oder Eph. fluviatilis, angehoren. Bei alien diesen Formen wird eine unregelmassige Ausbildung oder eine Degenaration der Gemmulahiillen sowie der Belagsnadeln beo-bachtet, was in grossen Seen oft vorkommt.
Diese morphisch abgeanderten Vertreter der Fam. Spongillidae geben nach Makuschok den Schlussel zum Verstandnis der Genese der Baikal-Spongiofauna. Nach seiner Meinung sollen im Baikalsee gewisse eigenartige physikalisch-chemische Bedingunen existieren, welche in kurzer Zeit auf die fortwahrend in den Baikalsee geratenden Vertreter der weitverbreiteten Spongillidenarten einwirken. Schon in der zweiten oder dritten Generation erscheinen nach seiner Meinung abweichende Formen mit degradierenden Gemmulahiillen und Belagsnadeln. Und eine langandauernde Einwirkung der Baikalbedingungen fiihrt zu einer Verwandlung der Einkommlinge in typische Baikalschwamme. „Meiner Meinung nach, haben wir es bei der Erforschung der Spongien-Fauna des Baikalsees meist mit feststehenden Formen, sondern mit einer Masse von einzelnen Momenten der Formbildung zu tun“. Die ganze Baikal-Spongiofauna wird von ihm in 5 Gruppen eingeteilt, welche diese einzelnen Etappen der Umwandlung darstellen, von typischen Spongilliden (1-te Gruppe) bis Lubomirskia baicalensis (5-te Gruppe).
Wir mochten beziiglich dieser Anschauungen folgendes bemerken: 1) Eine Degradation der Gemmulahiillen, derer Belagsnadeln, sowie, zuweilen auch ein Verschwinden der Gemmulae ist bei Schwammen aus zahlreichen Seen bekannt. 2) Es sind uns im Baikalsee keinerlei besonders eigenartige physikalisch-chemische Bedingungen bekannt, nach welcher dieser See sich schroff von anderen grossen Qewassern unterscheiden sollte (es sei daran erinnert dass die Schwamme vorwiegend auf geringen Tiefen des Baikalsees, angefangen mit 1—1.5 m, vorkommen). 3) In den Auseinandersetzungen von Makuschok sind, wie wir weiter unten sehen werden, die wesentlichen anato-mischen Besonderheiten, welche die Baikalschwamme von den gewohnlichen Vertretern der Fam. Spongillidae unterscheiden, nicht beriicksichtigt worden. 4) Den Anschauungen von Makuschok wiederspricht dis Vorfinden von einem Vertreter der Gattung Baikalospongia ausserhalb des Baikalsees, in einem kleinen und offenbar seichten See des Urjanchai-Gebietes.
Neben den Tendenzen einer Annaherung der Baikalschwamme an die Fam. Spongillidae find an wir in der Literatur auch Versuche vor, einen Zusammenhang der Baikalschwamme mit der Meeresfauna nachzuweisen. Den ersten Versuch in dieser Richtung hat Miklucho-Maklai (1870) gemacht, welcher aus dem nordlichen Teil des Stillen Ozeans die ,,Art“ Veluspa polymorpha mit 10 ,,Varietaten“ beschrieben hat. Als die elfte Varietat seiner Art betrachtete er den einzigen damals aus dem Baikalsee bekannten Schwamm—Spongia baicalensis. Die von Dybowsky (1880) durchgefiihrte Revision des Materials von Miklucho-Maklai hat gezeigt, dass diese Art mit
susswasserschwammen
107
Varietaten ein Konglomerat von Schwammen darstellt, die den verschieden-sten Familien angehoren. Dybowsky stellte die neue Gattung Lubomirskia auf und bfachte dieselbe in die Nahe der Siisswassergattung Spongilla. Spater (1886) aber veroffentlichte er eine Mitteilung, die bis auf den heutigcn Tag zu grossen Misverstandnissen fiihrt. W. Dybowsky erhielt von seinem Bruder B. Dybowsky spongiologisches Material aus dem Behringsmeer, von der Insei Medny. In diesem Material fand er einen Schwamm vor, der sich in nichts von der typischen buschartigen Lubomirskia baicalensis unterschied. Seit dieser Mitteilung von Dybowski wird in einer Reihe von Zusammenfas-sungen angenommen, dass die Baikal-Art sowohl im Baikalsee als auch im. Behringsmeere vorkommt. Es sei aber daran erinnert, dass B. Dybowski, ehe er zum Behringsmeere gelangte, eine langero Zeit am Baikalsee arbeitete, von wo er seinem Bruder die Schwamme iibersandte. Das Vorfinden von L. baicalensis auf der Insel Medny muss nur als ein Misverstandnis erklart werden, das infolge der Verwechselung der Etiquetten entstanden ist. Daher muss die erwahnte Angabe vollig vernachlassigt werden. In diesem Sinne spricht sich mit Recht auch Makuschok (1. c.) aus.
1901, teilweise unter dem Einfluss der Notiz von Dybowski uber das Vorfinden von L. baicaensis im Behringsmeer, wird von Swarczewsky die Gattung Veluspa wiederhergestellt, zu welcher eine Anzahl Baikalarten bezogen wird. Bei Makuschok (1. c.) finden wir eine berechtigte und griind-liche Kritik der nomenklaturischen Reform voh Swarczewsky vor.
Einen wichtigen Schritt in der Erkenntnis der Baikalschwamme stellt die Arbeit von Annandale (1914) dar. Er weist auf einen wesentlichen Unterschied der Baikalschwamme von der Fam. Spongillidae hin, welcher in dem. Fehlen von subdermalen R’aume besteht, sowie auf Verschiedenheiten in der Einrichtung des Kanalsystems, der Oscularoffnungen und des Skelets. Er schlagt zum ersten Mai vor auf Grund bestimmter anatomischer Daten die Baikalschwamme von dar Fam. Spongillidae abzugliedern und findet fur die-selben Platz in der marinen Familie Haplosc’eridae. Die Gattung Lubomirskia mit den Arten L. ba:calensis und L. abietina stellt er in die Unterfamilie Chalninae ( = Fam. Chalntdae nach dem System von Hentschel 1923), und die von ihm abgesonderte Gattung Baikalospongia — in die Unterfam. Renie-rinae ( = Fam. Gelliidae nach dem System von Hentschel).
Obwohl wir eine voile Abgliederung aller drei Gattungen der Baikalschwamme von der Fam. Spongillidae fiir ganz berechtigt erachten, halten wir es doch fiir unmoglich dieselben in irgendeine andere Familie der Ordnung unterzubringen. Wie wir im weiteren zu zeigen versuchen, stellen samtliche spezifischen Baikalschwamme eine aus naheverwandten Formen bestehende Gruppe dar, welche in drei Gattungen zerfallt. Diese drei Gattungen konnen nur als Representanten einer und derselben Familie betrachtet werden. Zugleich ist aber der Bau des Skelets bei verschiedenen Gattungen so verschiedenartig, dass es keine einzige Familie existiert, die dieselben vollig in sich aufnehmen konnte. Wir sehen, dass auch Annandale genotigt
108
FAUNA URSS
war die beiden damals bekannten Baikalgattungen in zwei verschiedene Unterfamilien (jetzt Familien) unterzubringen. Der einzig mogliche Angang ist die Absonderung der Baikalgattungen in eine besondere Familie Lubomirskiidae, welche ihre Stellung in der Untsrordnung Phthinorhabdina sowohl neben den marinen Familien, als auch neben der Susswasserfamilie Spongillidae nimmt. Es sei bemerkt, dass eine derartige Absonderung der Baikal-schwamme noch nicht uber die Frage von ihrer Genese entscheidet.
Betrachten wir nun etwas genauer die Frage fiber die Unterschiede der Baikalschwamme von der Familie Spongillidae.
Bei den Spongilliden ist der Korper fast auf seiner ganzen Ausdehnung von einer diinnen von Poren durchsetzten Dermalmembran iiberzogen. Unter dieser Dermalmembran befinden sich weite Subdermalraume welche dem zuffihrenden System angehoren. Die zuffihrenden Kanale nehmen ihren Anfang am Grunde dieser Raume. Die Dermalmembran wird durch die Enden der Hauptskeletbiindel, welche mit wenigen Spicula enden, unterstiitzt. Im abfuhrenden System finden wir Haupt-(Kloakal-)kanale von, welche aus der Tiefe des Gewebes herkommen und gerade unter der Oberflache des Schwammes zu der Oskularoffnung hinziehen. Sie werden von dem Aus-senmilieu durch eine ebensolche dfinne Dermalmembran, wie auch im zuffihrenden System abgegrenzt, nur ist diese Membran nicht von Poren durchsetzt. Diese abfuhrenden Kanale, die sich unter der Dermalmembran befinden, konnen wir ebenfalls als Subdermalraumen, die aber zu dem abfuhrenden System gehoren, betrachten. Von verschiedenen Seiten gegen die Oskularoffnung zusammentretend, bilden diese, durch die Membran durchschimmernde Kanale, die fur die Familie Spongillidae charakteristischen grossen (2—3 cm.) unregelmassigen sternformigen Figuren. Beim lebenden Schwamm ist die Oskularoffnung mit einer bis 1 cm langen Oskularrohre versehen.
Ganz andere Verhaltnisse finden w‘r bei der Familie Lubomirskiidae var. Subdermale Raume sind hier nicht vorhanden. Die Poren fiihren unmittelbar in die zuffihrenden Kanale, die sich in die Tiefe des Gewebes begeben. Die Dermalmembran ist nicht abgesondert und von dem darunterliegenden Gewebe abtrennbar. Die diinnen abfuhrenden Kanale treten aus dem Inneren des Gewebes zu den Oskularoffnungen ohne ihren Weg unter der Dermalmembran zu nehmen. Die Oscula stellen kleine (3—4 mm.) flache Griibchen dar, auf deren Boden, der von einem diinnen Hautchen iiberzogen ist, sich eine oder zwei Offnungen befinden. Durch das Hautchen sind radial gelagerte Kanale sichtbar, welche hierher aus dem Inneren des Gewebes treten. Das Hautchen bleibt an einem nicht besonders gut fixierten Material nur seiten erhalten. Auf Taf. XV, Fig. 64 ist ein Osculum von B. dzhegatajensis dargestellt; hier ist das erwahnte Hautchen offenbar nicht erhalten geblieben. Aller Wahrscheinlichkeit nach sind die Oskularrohren bei der Fam. Lubomirskiidae nicht entwickelt; jedenfalls lasst es sich schwer vorstellen, dass solche vorhanden sein konnten, falls man fixiertes Material in Betracht zieht. Es sei dabei
SOSS WASSERSCH WAMMEN
109
bemerkt, dass noch niemand lebende Baikalschwamme in Aquarien beo-bachtet hat.
Zweifellose Unterschiede zwischen den beiden Familien liegen im Ban des Skelets vor. Jedoch ist das Skelet auch innerhalb der Fam. Lubomirskiidae selbst recht verschie denartig. Bei der Gattung Lubomirskia haben wir ein ziemlich regelmassiges Gitterwerk aus Haupt- und Querbiindeln. Die Hauptbiindein enden an der Schwammoberflache mit gut ausgesproche-nen kegelformigen Spiculabiirsten. Das Spongin ist reichlich entwickelt und umgibt die Spicula der Hauptbiindel, sowie in bedeutendem Grade auch die jenigen der Querbiindel. Bei der Gattung Baikalospongia ist das Skelet weniger regelmassig. Bei B. dzhegatajensis und B. bacillifera-. finden wir noch die kegelformigen Spiculabiirsten vor, jedoch sind dieselben weniger gut als bei Lubomirskia ausgesprochen. Am unregelm assigsten ist das Skelet bei B. intermedia entwickelt, wo sich eine gewisse Annaherung an die Verhaltnisse die der Fam. Spongillidae zeigt. Bei B. bacillifera und B. dzhegatajensis ist viel Spongin vorhanden, jedoch bildet es hier nicht, wie bei Lubomirskia, eine kontinuierliche Faser aus. B. intermedia ist bedeutend armer an Spongin (Taf. XIII, Fig. 57). Am eigenartigsten erscheint das Skelet bei der Gattung Swartschewskia. Der Schwammkorper ist hier scharf dif-ferenziert in die zerbrechliche harte Kruste und das weiche, lose innere Gewebe. In der Kruste finden wir vor ein polygonales Netz von kurzen dicken Biindeln mit einer grossen Quantitat von Spongin. Im inneren Gewebe sind relativ wenige Spicula entwickelt und sie sind daselbst ohne merkliche Ordnung zerstreut. Fiir samtliche Baikalschwamme sind „Zuwachsringe" charakterisch, die auf Schnitten durch den Schwamm beobachtet werden konnen. Die genannten Ringe stellen Schichten mit einer dichteren Lage-' rung der Spicula dar — Spuren der friiheren Aussencchishten, die beim Wachstum des Schwammes von neuem Gewebe iiberwachsen wurden. Derartige ,,Zuwachsringe“ werden bei den Spongilliden niemals vorge-funden.
In der Fam. Spongillidae sind Belagsnadeln vorhanden.sowie meistens auch im Gewebe zerstreuten parenchymale Microskleren. Bei den Lubomirskiidae fehlen jegliche Microskleren. Fiir parenhymale Microskleren werden bie den Baikalschwammen zuweilen junge, noch nicht in Biindel eingeschlossene Skeletspicula gehalten.
Sehr bedeutende Unterschiede zeigt die Art und Weise der ungeschlechtli-chen Vermehrung in beiden Familien. Bei den Spongilliden entwickeln sich Gemmulae, die von einer komplizierten harten, mit eigenartigen Microskleren bewaffneten Hiille umgeben sind. Aus dem in der Gemmula eingeschlossenen Zellkomplex entwickelt sich der junge Schwamm, der durch die Porenoffnung nach aussen gelangt.
Bei den Lubomirskiiden bilden sich Sorite aus, — fester Hiillen entbehrende Ansammlungen von Zellen, welche miteinander zusammen-fliessen und ein Syncytium bilden. Die Entwicklung der Soriten wird
110 FAUNA URSS
durch die Teilung eines einzigen Kernes eingeleitet. Aus dem Soriten entsteht eine freischwimmende Larve (Swartschewsky 1923, 1925).1
Die angefiihrten Unterschiede geniigen unserer Meinung nach vollkom-men zur Trennung der beiden Familien. Es wurde schon bemerkt, dass eine derartige Trennung noch keineswegs uber die Frage der Genese der Baikalschwamme entscheidet. Wir verfijgen im keinerlei Anhaltspunkte fur eine Annaherung der Lubomirskiiden zu irgend einer marinen Familie, von der wir die Baikalschwamme herleiten konnten. Bemerkt sef, dass die Familien, aus denen sich die Unterordnung Phth'norhabdina zusammensetzt, wohin sowohl die Lubomirsklidae als auch die Spongillidae und auch eine Anzahl von marinen Familien gehorten, im allgemeinen ziemlich nahe einander stehen. Wir sind aber vollig berechtigt eine folgende Schlussfolgerung negativen Charakters zu ziehen. Die Unterschiede zwischen den beiden Siisswasserfamilien sind so bedeutend, dass die Moglichkeit einer Ableitung der Lubomirskiiden von den Spongilliden ausgeschlossen bleibt. Wie es fiir die gesammte Siisswas-serfauna angenommen wird, so darf es auch von den Siisswasserschwammen gesagt werden, dass deren Vorfahren einst aus dem Meere hergekommen sind. Analysiert man den Bau der Spongilliden so wie den der Lubomirskiiden, so muss angenommen werden, dass sie von verschiedenen marinen Vorfahren entstanden sind und wohl auch zu verschiedener Zeit, wobei aller Wahrscheinlichkeit nach die Lubomirskiiden von einer spateren Herkunft sind. Wir haben kein Material um zu entscheiden, wo, in welchem G ewasser das stattfinden konnte. In dieser Frage gehort das Wort den Geologen, welche nicht nur die Geschichte deg Baikalsees selbst, sondern auch diejenige eines betrachtlichen Teiles des asiatischen Kontinents, der den Baikalsee umgibt, aufklaren miissen. Nach dem Funde von B. dzheg itajensis ist es klar, dass die Frage uber die Fam. Lubomirskiidae nicht nur mit dem Baikalsee allein verkniipft werden darf.
Fasst man das Gesagte zusammen, so konnen fiir die beiden Siisswas-serfamilien folgende Diagnosen gegeben werden:
A. Fam. Spongi.lidae Gray. Subdermalraume gut entwickelt; Dermal-membran von dem ubrigen Gewebe leicht abtrennbar. Beim lebendigen Schwamm erheben sich von den Oscularoffnungen ziemlich lange Oscular-rohren. Abfiihrende Kanale, gegen die Oscularoffnungen zusammentretend, bilden in ihrer Gesamtheit grosze sternformige Figuren. Das Skelet besteht aus Haupt-und Querbiindeln, seine Regelmassigkeit wird jedoch oft beeintrachtigt. Das Spongin ist in verschiedenen, im allgemeinen in unbedeu-tenden Quantitaten vorhanden. Macroskleren durch Diactinen dargestellt,
1 Von Annandale (1913, 1914) wurden Gemmulae bei einem Exemplar von B. bacilllfera beschrieben. Swarczewsky (1923) bezweifelt die Richtigkeit dieser Beachtung AnnandaL’s. Wie es aus der Beschreibung, sowie den Abbildungen Annandale's erhellt, sind die von ihm beschriebenen Gemmulae unter der Basalmembran des Schwammes gelegen und stellen nichts anderes dar, als Eier verschiedener Tiere, die vom Schwammgewebe iiberdeckt sind. Somit griindet sich die Beschreibung von Annandale auf einer irrtiimlichen Beobachtung-
sOsswasserschwAmmen 111
meist durch Amphioxen. Microskleren in Gestalt von Rhabden oder deren Abanderungen; Cheloide niemals vorhanden. Ungeschlechtliche Vermehrung durch Gemmulae, aus denen fertige Schwamme heraustreten. Die Gemmulae sind mit einer festen chitinoiden Kapsel, einer Luftkammerschicht sowie besonderen schiitzenden Belagsnadeln versehen.
B. Fam. Lubomirskiidae. Subdermalraume fehlen. Dermalmembran vom darunterliegenden Gewebe nicht abtrennbar. Die abfuhrenden Kanale treten an die Oscula aus der Tiefe des Gewebes ohne ihren Gang unmittelbar unter der Oberflache zu nehmen und ohne Bildung von grossen sternformigen Figuren um das Osculum. Oscularrohren nicht vorhanden. Skeletbau bei verschiedenen Gattungen der Familie verschiedenartig. Spongin in verschiedenen, jedoch meist in betrachtlichen Quantitaten vorhanden. Macroskleren sind Amphioxen oder Amphistrongylen bedornte oder seltener glatte. Microskleren fehlen. Gemmulae fehlen. Ungeschlechtliche Vermehrung mittelst Soriten, aus denen freischwimmende Larven erscheinen.
Die Familie Lubomirskiidae besteht aus drei Gattungen — Lubomirskia, Baikalospongia und Swartschewskia. Trotz ihrer Verschiedenheiten betrach-ten wir sie als eine natiirliche Gruppe von einer gemeinsamen Herkunft. Die Frage iiber ihre gegenseitige Verwandtschaft betrachten wir weiter unten, aachdem wir uns mit den einzelnen Vertretem naher bekannt gemacht haben.
Gattung LUBOBIRSKIA DYBOWSKI.
Das Skelet besteht aus gut differenzierten Haupt-und Querbiindeln. Die Spicula liegen in der Hauptbiindeln parallel der Biindellange oder auch etwa aussenwarts mit ihren Distalenden divergierend. Die Hauptbiindel enden an der Oberflache mit kegelformigen Bursten von Spicula, deren Enden von einander divergieren. Das Spongin ist stark entwickelt und bilden kontinuier-liche Fasern, awelche sowohl die Haupt-als auch die Querbiindel in sich einsshliessen. Die Spicula stellen ausschliesslich Amphioxen dar. Das Schwamm-gewebe ist elastisch und nicht zerbrechlich, kann nur mit einiger Kraftan-wendung zerrissen werden.
Lubnmirskia baicalensis (Pallas)1 forma typica. (Taf. II, Fig. 5, Taf. Ш, Fig, 7, 9, 10, Taf. X, Fig. 38, Taf. XII, Fig. 47, Taf. ХШ, Fig. 54, Taf. XIV, Fig. 58, Taf. XV, Fig. 62).
Hochwachsende verzweigte Formen. Die Oscula sind auf der ganzen Schwammoberflache zerstreut und stellen rundliche wenig tiefe Griibchen von
1 Die Synonymik und die hauptsachliche Litcratur Я. im russischen Text.
112 FAUNA URSS
3—4 mm. im Durchmesser dar, welche an ihrem Boden von einer diinnent mit einer oder mehreren Offnungen versehenen Membran iiberzogen sind; durch die Membran schimmern die Offnungen der diinnen abfiihrenden Ka-nale durch. Die Spicula sind bedornte Amphioxen, mit kurzen oder mittellan-gen Spitzen; sie sind von zylindrischer oder schwach spindelformiger Gestalt, relativ diinn und kurz, durchschnittlich 13 [ь dick und 193 lang. Die Dor-nen sind auf der ganzen Spiculalange verteilt. Die zentrotyloten Skleren feh-len in der Regel.
Samtliche hochwachsende Formen beziehen wir auf die forma typica, obwohl unter ihnen sich mehrere Wachstumsformen gut unterscheiden lassen. Am gewohnlichsten trifft man Exemplare mit zylindrischen, sich verzweigen-den Auswiichsen, welche von einer krustenartigen Basis ausgehen. Die Dicke der Auswuchse schwankt von 1—2 bis 3—4 cm im Durchmesser. An den Auswiichsen bemerkt man perlschnurartige Einschniirungen. Die Hohe erreich, >sq 60 cm, in der Literatur werden Exemplare bis 1 m hoch erwahnt. E’ne geringere Hohe erreichen facherformige Exemplare, welche aus abgeflachten, gegen das Ende breiter werdenden dreieckigen Auswiichsen bestehen, die ihren Anfang von einem kurzen Stiel nehmen. In ihrer Gesamtheit bilden diese Auswuchse eine Art von kegelformigen, in Segmente zerteilten Becher (Taf. Ill, Fig. 7). Seltner kommen kammformige Exemplare vor (Taf. Ill, Fig. 10). Von dem an das Substrat befestigten Basalteil entspringt ein horizontal liegender Auswuchs, von dem sich diinne vertikale Zweige erheben. Als eine besondere Wuchsform konnen Exemplare betrachtet werden, welche auf einem weichen Grunde ohne Anheftung ans feste Substrat wachsen. Derartige Exemplare stellen im Jugendzustande runde Korper von der Grosse einer Erbse bis zu der einer Nuss dar. Mit dem weiteren Wachstum verlangern sie sich und werden zu unverzweigten Auswiichsen mit abgerundeten beiden Enden (Taf. II, Fig. 5)-Swarczewsky (1923) erklart eine solche Wuchsform wohl mit Recht durch aussere Knospung. Wenn eine aussere Knospe nach dem Abtrennen vom Mut-terorganismus auf ein festes Substrat gelangt, wachst sie dort an und ergibt gewohnliche Wachstumsformen. Gelangt sie dagegen auf einen weichen schlammigen Boden, so entwickelt sie sich auf dem Schlamm freiliegend. Alle soeben erwahnten Wuchsformen lassen keinen Unterschied in dem Bau ihrer Spicula erkennen. Lebendige Schwamme haben eine grelle gras-griine Farbung.
Die verzweigte Form beginnt vorzukommen in einer Tiefe von 3—4 m, und auf 5—8 m erreichen die Bestande der Schwamme ihre maximale Entwicklung. Verzweigte Exemplare sind aus Tiefen von 3 bis 50 m bekannt.
Die Form ist entlang der ganzen Kiistenzone des Baikalsees verbreitet.
Lubomirskia baicalensis (Pallas) morpha llttoralis nova. (Taf. X, Fig. 40).
Die Gestalt des Schwammes ist krustenformig, von etwa 1 cm Dicke. Sie unterscheidet sich von der typischen Form nicht nur durch das Fehlen
SUSSWASSERSCHWAMMEN
11$
von Auswiichsen, sondern auch durch den Charakter ihrer Spicula. (Taf. X, Fig. 40). Lange der Spicula wie bei der typischen Form oder etwas grosser, Breite jedoch bedeutend grosser; durchschnittlich 190 (л lang und 18 (J. breit. Die Dimen-sionen schwanken zwischen 174—229 |л resp. 11—24 jj.. Die Spindelformig-keit nur schwach ausgesprochen, centrotylote Spicula ebenso selten, wie bei. der typischen Form. Grosse und grobe Dornen verteilen sich langs der ganzen Spiculalange.
Kommt in der Kiistenzone des Baikalsees vor, angefangen von etwa 1.5 m und bis 3—6 m Tiefe.
Lubomirskia fusifera Soukatschoff. (Taf. V, Fig. 42).
Krustenform von 1—2 cm Dicke. Unterscheidet sich von L. baicalensis,. ausser dem Fehlen von Auswachsen nur durch die Grosse und die Beschaf-fenheit der Spicula (Taf. X, Fig. 42). Mittlere Dimensionen der Spicula 225 X 20 [л. Ganzen der Grossenschwankungen 161—282 [л X 12—26 a. Gestalt der Spicula spindelformig, sehr oft werden centrotyloten Spicula beobachtet. Eine geringe Anzahl Dornen ist bei den Enden der Spicula kon-zentriert so dass der grossere Teil defen Oberflache glatt bleibt. Ofi^werden fast ganz glatte oder vollig glatte Spicula beobachtet. Das Schwammgewebe ist ziemlich lose, weich und elastisch.
Diese Art steht ausserst nahe zu L. baicalensis. Nur bei Durchsicht eines umfangreicheren Materials kann bemerkt werden, dass neben der kru-stenformigen und der verzweigten Form von L. baicalensis, die mit zunehmen-der Tiefe allmahlich einander ersetzen, noch eine Form existiert, die sich durch ihren Spiculabau unterscheidet und ihre Krustenform auch auf relativ bedeutenden Tiefen standig beibehalt. Es sind Exemplare aus 4 bis 50 m Tiefe bekannt (Baikalsee).
Lubomirskia abietina (Swarczewsky). (Taf. X, Fig. 39, Taf. XII, Fig. 48, Taf. XIII, Fig. 56, Taf. XIV, 60).
Eine Krustenform von 0.5—1 cm Dicke. Die aussere Oberflache hat ein charakteristisches Aussehen und ist mit einem Netz von kleinen Rinnen bedeckt. Das Skelet setzt sich fast ausschliesslich aus Hauptbiindeln von einen charakterischen Geprage zusammen. Die Spicula darin sind mit ihren Distalenden von der Biindelachse etwas abgelenkt, was den Biindeln eine Anlichkeit mit jungen Tannentrieben gibt. Die Querbiindel sind schwach entwickelt und werden meist durch vereinzelte Spicula dargestellt. Die Spicula sind amphioxen mit kurzen Spitzen und gut ausgesprochener Spindelfor-migkeit. Mittlere Dimensionen 186 |) X 17 <л, Schwankungsamplitude 145— 240 [л X 13—21 [7.. Die Dornen befinden sich nur bei den Enden der Spicula, wahrend die iibrige Obeflache glatt ist; seltener werden Spicula mit Dornen in ihrem mittleren Teil angetroffen.
Die bekannt gewordenen Fundstellen in Tiefen von 20—40 m (Baikalsee).
Фауна СССР, Губки, т. II, в. 2
8
114
FAUNA URSS
Gattung BAIKALOSPONGIA ANNANDALE
Krustenformige oder massive Formen mit Ausnahme von einer Art, die anastomosierende Auswiichse ergiebt. Die Lagerung der Spicula in den Haupt-sowie an den Querbiindeln ist eine ziemlich regellose; oft liegen sie in ver-schiedenen Richtungen im Verhaltnis zur Biindelachse, indem dichte Gruppen einander parallel liegenden Spicula bilden. Das Spongin ist in bedeutender Quantitat vorhanden, ohne jedoch eine kontinuierliche, die Spicula iiberzi ehende Faser zu bilden. Die Spicula sind durch Amphioxen und Amphis trongylen reprasentiert. Gewebe nicht elastisch und zerbrechlich, je nach der Art bald derb, bald weich.
Baikalospongia bacillifera (Dybowski). (Taf. Ill, Fig. 8, Taf. XI, Fig. 43, Taf. XII, Fig. 49, Taf. XIII, Fig. 55, Taf. XIV, Fig. 61, Taf. XV, Fig. 63).
Krustenformige oder massive Formen. Skelet aus dicken regellosen Haupt- und dunneren, immer gut differenzierten Querbiindeln. Das Spongin in bedeutenden Mengen und in unregelmassigen Anhaufungen klebt die Spicula zusammen, welche durch grobe dicke Strongylen mit Dornen an ihren abgerundeten Enden reprasentiert sind, unter derartigen Spicula werden zuweilen Spicula mit Dornen in ihrem mittleren Teil angetroffen. Mittlere Dimensionen der Spicula 262 д X 27 ft, Grossenschwankungen 170—365 Д x 16—40 fi. Gewebe derb und zerbrechlich.
Kommt im Baikalsee angefangen mit wenigen Metern und bis 533 m tief vor. Aus der letzterwahnten Tiefe wurden vor kurzem zwei massive Exem-plare erbeutet, die auf Eisenkonkretien wuchsen. Sie batten im lebenden Zu-stande (nach Angabe der Sammler) eine eigenartige grell blaugriine Farbe. Schwamme aus geringen Tiefen sind grasgriin.
Baikalospongia intermedia (Dybowski). (Taf. XI, Fig. 46, Taf. XII, Fig. 50, 51, Taf. XIII, Fig. 57).
Dunne Krustenform. Skelet ziemlich regellos und schwach,aus bald besser, bald schlechter differenzierten Haupt-und Querbiindeln bestehend. Spongin, relativ schwach entwickelt. Spicula meistenteils Amphioxen mit mittelangen oder kurzen Spitzen, langs ihrer ganzen Ausdehnung mit kurzen Dornen bedeckt; selfener werden Exemplare angetroffen mit Strongylen mit sparlichen Dornen in ihrem mittleren Teil und konzentrierten Dornen auf ihren abgerundeten Enden. Mittlere Dimensionen der Spicula 226 fi X 15 fi. Dimensionenschwan-kungen 172—266 fi X 9—22 fi. Gewebe weich, nicht elastisch und zerbrechlich.
Farbung der lebenden Schwamme grim mit einer grau-oliven Tonung.
Kommt im Baikalsee vor angefangen mit der oberen Verbreitungs-grenze der Schwamme (1.5—2 m) bis zu einer Tiefe von mehreren Dutzend Meter.
SOS'S WASSERSCH WAMMEN
115
Baikalospongia intermedia (Dyb.) morpha profundalis nova. (Taf. XI, Fig. 44).
Unterscheidet sich von der Hauptform durch die Dimensionen der Spicula, die durchschnittlich 405 ц X 22 Ц betragen und zwischen 330—470 X 20—25 [л schwanken. Die Spicula stellen schwach gebogene oder gerade Strongylen dar, die langs ihrer ganzen Ausdehnung mit kleinen Dornchen bedeckt sind; an den abgerundendeten Ende je eine dichte Anhaufung von punktartigen Dornchen (Taf. XI, Fig. 44).
Es wurde nur ein schlecht erhaltenes Exemplar in der Form von Frag-menten einer diinnen Kruste aus einer Tiefe von 889 m erbeutet. Die Farbung des lebenden Schwammes war (nach Angaben der Sammler) grell blaugriin.
Vielleicht ware es berechtigt, die erwahnte Morphe samt den Schwam-men aus geringen Tiefen, die anstatt von Amphioxen im Besitz von Strongylen sind und die von uns nicht von der typischen Form B. intermedia abge-sondert werden, als selbstandige Art abzusondern.
Baikalospongia dzhegatajensis Rezvoj. (Taf. IV, Fig. 11, Taf. XI, Fig. 45, Taf. XIII, Fig. 53, Taf. XIV, Fig. 59, Taf. XV, Fig. 64).
Korper massiv oder aus zylindrischen, meist ahastomosierenden, einen fiunregelmassigen Knoten bildenden Auswiichsen bestehend. Skelet gut in ziemlich dicke Haupt- und Querbundel differenziert. Die Hauptbundsl enden bald mit deutlicher, bald mit schlechter ausgesprochenen kegelformigen Bur-sten von Spicula. Recht deutlich sind die Zuwachsringe sichtbar. Spongin in bedeutender Quantitat, bildet jedoch keine kontinuierliche Fasern. Spicula Amphioxen mit kurzen oder mittellangen Spitzen und mit kleinen Dornchen jangs ihrer ganzen Lange oder bei den Enden; es kommen vor und bei man-chen Exemplaren dominieren sogar fast ganz oder vollig glatte Spicula (var. lissosclera). Mittlere Spiculadimensionen 245 (л X 19 (л. Variationsgrenzen 170—320 [л X 13—25 ,u. Schwammgewebe fest, ziemlich elastiach und nicht sprode.
Das Material wurde von Dr. A. J. Tugarinov am Ufer des Sees Dzhegataj-kul im Urjanchai-Gebiet (Tannu-Tuwinsche Republik, Becken des oberen Jenissej) gesammelt, — im ganzen 7 voile Exemplare und mehrere Fragmente. Samtliche Exemplare wuchsen auf Characeen, mit denen zusammen sie durch die Brandung ans Ufer herausgeschleudert waren. Der See Dzhegataj-kul hat etwa 15 km im Durchmesser und besitzt niedrige flache sandige Ufer und augen-scheinlich eine geringe Tiefe.
Gattung SWARTSCHEWSKIA MAKUSCHOK,
Skelet nicht in Haupt- und Querbundel differenziert. In der Dermalschicht, welche eine harte Kruste darstellt, ist ein polygonales Netz aus kurzen BUndoln abgesondert, die aus dicken, tangential gelagerten Spicula bestehen. Im inneren Gewebe sind wenig zahlreiche nicht zu Biindelh verbundene Spi-8*
116
FAUNA URSS
cala zerstreut. In der Oberflachenkruste sind die Spicula durch eine bedeu-tende Menge von Spongin zusammengeklebt, wahrend im inneren Gewebe das Spongin nicht zu bemerken ist. Die Spicula stellen relativ kurze und sehr dicke bogenartig gekrummte Strongylen dar mit kleinen Dornchen auf ihrer ganzen Ausdehnung und einer Anhaufung derselben an den Enden, oder kon-nen auch glatt sein.
Oscularoffnungen in geringer Anzahl, in der Form von kleinen Griibchen, die bald von einen hoheren, bald von einem niedrigen Wall umgeben sind. Aussere Kruste derb und sprode, inneres Gewebe weich und lose.
Eine Art mit den Merkmalen der Gattung.
Swartschewskia papyracea (Dybowski). (Taf. X, Fig. 41, Taf. XII, Fig. 52, Taf XV, Fig. 65).
Eine griine Farbung wird niemals beobachtet, mittlere GrossenVerhalt-nisse der Spicula 133 [л X 21 ji, Grossenschwankungen von 112—150 F 13—29 II, Kommt im Baikalsee in Tiefen von 2 bis 80 m vor.
Trotz der Verschiedenartigkeit der ausseren Gestalt und den Unterschieden im Bau des Skelets bei den drei Gattungen der Baikalschwamme, betrachten wir sie als eine aus naheverwandten Formen sich zussammensetzende natiir-liche Gruppe, die sich in eine einzige Familie zusammenfassen lasst.
Ein am regelmassigsten gebautes Skelet finden wir in der Gattung Lubomirskia vor. Hier haben wir gut differenzierte Haupt- und QuerbiindeL In der Oberflachenschicht sind gut ausgesprochene kegelformige Bursten von Spicula, welche die Hauptbiindel abschliessen, entwickelt. In derselben Gattung ist das Spongin in der maximalen Quantitat vorhanden und bildet eine ununterbrochene Skeletfaser.
Bei der Gattung Baikalospongia ist das Skelet weniger regelmassig obwohl bei B. dzhegaiajensis und teilweise bei B. bacillifera wir dennoch kegelformige, wenn auch nicht so regelmassig gebaute Endbiirsten von Spicula vorfinden. Bei samtlichen Baikalschwammen wird eine starkere Anhaufung von Spicula in der Oberflachenschicht beobachtet. Bei Lubomirskia geschieht das infolge der Anwesenheit von kegelformigen Bursten die mit ihren Randern fast einander beriihren; bei Baikalospongia (B. bacillifera) vereinigen sich in der Oberflachenschicht die Enden der Hauptbiindel durch machtigere Querbundel, was bei der Ansicht von der Oberflache ein polygonales Netz von kurzen Biindeln ergiebt (Taf. XII, Fig. 55). Dank der Anhaufung von Spicula in der Dermalschicht ergeben sich an Schnitten durch den Schwamm-korper „Zuwachsringe" — d. h. Spuren der friiheren ausseren Oberflachen., Die Struktur der Oberflachenschicht von Baikalospongia kann an die Skelet-struktur der Gattung Swartschewskia gekniipft werden, falls man sich vor-stellt, dass die Dermalschicht sich nach der Anzahl der Spicula nach schrof-
sOsswasserschwAmmen
117
fer von dem inneren Gewebe unterscheidet und die oberflachlichen Querbiin-del eine grossere Machtigkeit und Festigkeit erlangen.
Fiir die Frage von der gegenseitigen Verwandtschaft der Baikalschwamme gibt sehr wichtige Hinweise die einzige ausserhalb des Baikalsees vorkommende Art, — B. dzhegatajensis. Von den Schwammen die im Baikalsee vorkommen, besitzt nur die typische Form von L. baicalensis Auswiichse von verschiedener Gestalt. Exemplare mit Auswiichsen liegen auch in dem aus zur Verfiigung stehenden Material von B. dzhegatajensis vor. Nach der Skeletstruktur zeigt B. dzhegatajensis Ziige, fiir die Gattung Baikalospongia typisch sind, namlich die Lagerung der Spicula in den Hauptbiindeln in dich-ten Paketen unter verschiedenen Winkeln zu der Biindelachse. Daneben fin-den wir bei dieser Art ziemlich gut ausgesprochene Endbiirsten von Spricula und eine grosse Quantitat von Spongin, wodurch die Konsistenz des Schwam-mes sich der Konsistenz von L. baicalensis nahert. In dem Bau der Oscularoffnungen hat ,B. dzhegaiajensis eine Ahnlichkeit mit 5. paryracea. Nach dem Spiculabau dagegen nahert sich die urjanchaische Art der L. fusifera sowie L. abietina.
In dem vorliegenden Auszuge aus unserer Arbeit uber die SiissWasser-schwamme der Sovietunion werden wir nicht genauer die Vertreter der Fam. Spongillidae betrachten. Wir bleiben nur etwas bei ihrer geographischen Verteilung in den Grenzen der Union stehen.
Es kann gesagt werden, dass auf dem gesamten Territorium der Union die Gruppe der weit verbreiteten Formen verherrscht und zwar: Spongilla lacustris, Ephydatia miilleri, E. fluviatilis und S. frag lis. Die zweitgenannte Art, E. miilleri, erweist sich als eine dominierende Form vorwiegend in den mehr nordlich gelegenen Bezirken. S. fragilis wird seltener als die ubrigen Arten angetroffen. Heteromeyenia baileyi var. stepanowi (Carterius stepanowi) hat eine sporadische Verbreitung im Siiden des europaischen Teils der U. d. S.S.R. im friiheren Gouv. Charkow (See Welikoje und im Tale des Donetz-Flusses) und im Siidlichen Bug. In subfossilem Zustande wurden Spicula dieser Art in tiefen Schlammschichten in den Kossino-Seen bei Moskau ent-deckt. Trochospongilla horrida ist ebenfalls nach Einzelfunden aus dem europaischen Teil der Union bekannt. Von ihren asiatischen Fundstellen muss der Aralsee und zwar im Bezirk des Amu-Darja-Deltas erwahnt werden. Spongilla arctica stellt die einzige Art vor, die nur aus dem Bereich der Union bekannt ist. Diese Art steht zweifelsohne S. lacustris sehr nahe, unterscheidet sich jedoch scharf durch den sonderlichen Bau ihres Skelets, welches diinne, breite Sponginmembranen darstellt, die Skeletbiindel verbinden (Taf. V, Fig. 15, 16, 17). Die Art wurde aus dem Polar-Ural (Pe-mal-Tundra, Tai des Pideruta-Flusses) beschrieben und ist auch von der Kola-Halbinsel (Seen der Murman- und der Terskij-Kiiste) bekannt; als deren siidlichere Fundstellen seien genannt die Sjaberschen Seen des Leningrader Ge-bietes sowie die Seen der Meschtscherskaja-Niederung (der friihere Gouv. Rjasan). Die Art ist an oligotrophe und dystrophe Gewasser
118 FAUNA URSS
mit sehr salzarmien Wasser gebunderi. Wie bereits erwahnt, in das Bereich unserer Union das siidasiatische oder tropische Element ein. Im Aralsee, im Bezirk des Amu-Darja-Deltas, herrscht .S', biseriata (S. proliferens} vor, welche uniangst auch in Bulgarien vorgefunden wurde. Zu demselben Element gehort auch .S', carteri, die in Turkestan (Buchara, Bairam-Ali), in dem Wolga-Delta, im Unterlauf der Fliisse Don, Dnjestr und Siidl. Bug nachgewiesen ist. In dem Ussuri-Gebiet, im Chanka-See und den sich darin ergissenden Fliis-sen herrscht die chinesische S'. stanleyi vor, und aus der Bucht St. Wladimir (an der Kuste des Japanischen Meeres) ist eine interessante Varietat von E. fluviatilis, var. hastifera beschrieben, welche manche Besonderheiten mit der chinesichen Varietat derselben Art, var. chui, teilt.
Es ist schon friiher die Rede fiber die im Baikalsee vorgefundenen Ver-treter der Spongillidae gewesen. Die Beschreibung dieser Formen, welche von Swarczewsky gegeben wurde, ist ausserst ungeniigend. Aus derselben geht hervor, dass im Baikalsee sowohl S', fragilis, als auch eine oder beide der gewohnlichen Ephydatia-Axien. (E. miilleri und E. fluviatilis) vorkom-men. Alle diese j Formen tregen Spuren einer Degeneration der Gemmula-hiilen sowie der Belagsnadeln.
ОБЪЯСНЕНИЕ ТАБЛИЦ
Т а б л. I.
1. Spongilla lacustris. Ветвистый экземпляр. Наст. вел. По Шредеру.
2. Spongilla lacustris. Экземпляр с тонкими выростами. Уменьшено. По Шредеру.
3. Ephydatia miilleri. Наст, величина. По Шредеру.
Табл. ,11.
^. Spongilla lacustris. Снимок с живой губки в аквариуме. Видны оскулярные трубки и сходящиеся к устьям отводящие каналы. Наст, величина. По К. Мюллеру.
5. Lubomirskia baicalensis. Мелкие неприкрепленные экземпляры с илистого грунта. Уменьшено.
6. Trochospongilla horrida. Сквозь дермальную мембрану просвечивают звездчатые фигуры, сходящиеся к устьям отводящих каналов. Наст, величина. По Шредеру.
Табл. III.
7. Lubomirskia baicalensis. Веерообразная форма роста. Уменьшено.
8. Baikalospongia bacillifera. Массивная форма роста.
9. Lubomirskia baicalensis. Типичная кустистая форма роста. Уменьшено.
10. Lubomirskia baicalensis. Гребенчатая форма роста. Сильно уменьшено.
Табл. IV.
11. Baikalospongia dzhegatajensis. а, Ь, с — экземпляры с выростами; d — экземпляр без выростов (var. lissosclera). Наст, величина.
12. Spongilla lacustris. а — макросклеры; b — паренхимные микросклеры; с — геммульные микросклеры (р. Лефу, Уссур. край).
13. Spongilla lacustris. Скелет; окрашен (Карелия).
14. Spongilla lacustris. а) геммула с тонким зернистым воздухоносным слоем и малым количеством микросклер; тоже: воздухоносный слой толстый, микросклер мало (озеро, сев. Прибайкалье).
Табл. V.
15. Spongilla arctica: а — макросклеры (типовой экземпляр из Пе-мала); Ь — тоже (оз. Мещерской низм.); с — паренхимные микросклеры (тип); геммульные микросклеры (тип).
16. Spongilla arctica. Скелет со спонгиновыми мембранами (тип).
17. Spongilla arctica. Скелет со спонгиновыми мембранами (Мещерск. озера).
18. Spongilla biseriata. а — макросклеры: Ь — паренхимные микросклеры; с — геммульные микросклеры (Арал, район дельты Амударьи); d—паренхимная спикула var. sectospina (дельта Аму-дарьи).
120
ФАУНА СССР
91
Табл. VI.
19. Spongilla arctica. Геммула (тнпичн. экземпляр).
20. Ephydatia miilleri. Геммула с двумя слоями амфидисков (Сев. Прибайкалье).
21. Spongilla fragilis. Геммулы (Уссур. край).
22. Trochospongilla horrida. Геммулы (Арал, дельта Аму-дарьи).
23. Spongilla carteri. Геммула в разрезе (в середине изображен участок ячеистой ткани с поверхности). (Волга).
24. Spongilla biseriata. Геммула (дельта Аму-дарьи).
Табл. VII.
25. Spongilla Stanleyi. Геммула (Уссур. кр.).
26. Spongilla fragilis. а — макросклеры; Ъ — геммульные микросклеры (Уссурийский край).
27. Heteromeyenia baileyi var. stepanowi. Геммула. По Дыбовскому.
28 Spongilla carteri. а — макросклеры (Бухара): Ь — тоже (Днепр, плавни): с — тоже (Дон у Ростова); d — тоже (Волга); е — тоже (Байрам-Али); f — тоже (дельта Волги): var. rotundacuta; g — микросклеры (Бухара); h — тоже (Волга); i — тоже (Дон), j — тоже (Байрам-Али).
29. Trochospongilla horrida. a, b — микросклеры, с — амфидиски (Арал, район дельты Аму-дарьи).
Табл. VIII.
30. Spongilla stanleyi. а — макросклеры, геммульные микросклеры (Ханка).
31. Ephydatia fluviatilis. а — макросклеры (Бухара), Ь — тоже (б. Харьковск. губ.); с — амфидиски (б. Харьковск. губ.).
32. Ephydatia fluviatilis var. teberdana. a — макросклеры, b — амфидиски, с — геммула (Теберда, Кавказ).
33. Ephydatia miilleri. .Личинка (Петергоф). По живому материалу.
Табл. IX .
34. Heteromeyenia baileyi. Поровые отверстия геммул различных вариететов: а — v. bohe-mica; b — v. repens-, d, g — v. palatina-, f, e — v. petri; c — амфидиски v. repens. По Шредеру.
35. Heteromeyenia baileyi var. stepanowi. a — макросклеры, b — паренхимные микросклеры: (a, b — Днепровск. плавни); с — амфидиски (с — по Дыбовскому).
36. Ephydatia miilleri. а — макросклеры; b —• амфидирки; с — микросклеры молодой губочки (возр. 1 неделя); d — спикулы личинки (Петергоф).
37. Ephydatia fluviatilis var. hastifera. a — макросклеры, b — концы макросклер при большем увеличении, с — амфидиски (Уссур. край).
Табл. X.
38. Lubomirskia baicalensis. Скелетные иглы кустистой формы: а — с глуб. 50 м.; b — с 28 м, с — с 10 м.
39. Lubomirskia abietina. Скелетные спикулы.
40. Lubomirskia baicalensis morpha littoralis. Скелетные спикулы.
41. Swartschewskia papyracea. Скелетные спикулы.
42. Lubomirskia fusifera. Скелетные спикулы (тип Сукачева).
Табл. XI.
43. Baikalospongia bacilliferd. Скелетные спикулы (тип Lubomirskia tsherskii Сукачева).
44. Baikalospongia intermedia morpha profundalis. Скелетные спикулы.
ОБЪЯСНЕНИЕ ТАБЛИЦ
121
45. Baikalospongia dzhegatajensis. Скелетные спикулы, а — vwrjfjlissosclera. (Джегатай-куль, Урянх. край). *
46. Baikalospongia intermedia. Скелетные спикулы.
Табл. XII.
47. Lubomirskia baicalensis typica. Скелет.
48. Lubomirskia abietina. Скелет.
49. Baikalospongia bacillifera. Скелет.
50. Baikalospongia intermedia. Экземпляр co слабо развитым скелетом.
51. Baikalospongia intermedia. Экземпляр с сильно развитым скелетом.
52. Swartschewskia papyracea. Скелет наружной корки с поверхности.
Табл. XIII.
53. Baikalospongia dzhegatajensis. Скелет.
54. Lubomirskia baicalensis. Расположение скелетных пучков.
55. Baikalospongia bacillifera. Скелет наружного слоя с поверхности: видны выходы главных пучков, соединенных поперечными , пучками; в петлях скелетной сети видны поры.
56. Lubomirskia abietina. Расположение скелетных пучков.
57. Baikalospongia intermedia. Строение скелета (см. объясн. фиг. 58).
Табл. XIV.
58. Lubomirskia baicalensis. Строение скелета. Спонгин окрашен; на рисунке спонгин, покрывающий спикулы, обозначен точечной тушовкой; сплошные массы спонгина зачерчены.
59. Baikalospongia dzhegatajensis. Строение скелета (см. объясн. фиг. 58).:
€0. Lubomirskia abietina. Строение скелета (см. объясн. фиг. 58).
61. Baikalospongia bacillifera. Строение скелета (см. объясн. фиг. 58).
Табл. XV.
62. Lubomirskia baicalensis. Оскулярное отверстие.
63. Baikalospongia bacillifera. Оскулярное отверстие.
64. Baikalospongia dzhegatajensis. Оскулярное отверстие.
65. Swartschewskia papyracea. Оскулярное отверстие.
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ!
Стр.
Acanthogammarus parasiticus......... 36
Amphidiscus antediluvianus . • . . . 45
Anheteromeyenia..................... 83
Anodonta . ............................. 37
Asellus................................ 37
Astroheteromeyenia ........ 83
Astromeyenia........................ 83
Badiaga........................ . . . 2
Baicalolepis bacillifera............ 96
„ fungiformis .... 96
„ tscherskii.................... 96
Baikalospongia .......... 90, 95
„ bacillifera ...... 96
„ dzhegatajensis .... 96, 99
v. lisso-
sclera ...... . . 99
„ intermedia....................96, 98
„ „ m. profun-
dalis....................96, 98
„ irregularis.............. 101
„ papyracea................ 101
Calcarea................................. 39
Carterella .............................. 83
Carterias primitives..................... 87
Carterius................................ 80
„ primitives......................... 87
Carterius stepanowi...................... 82
„ „ v. bohemica ... 85
„ „ v. palatina .... 85
„ „ v. petri ..... 84
„ tubispermum........................ 42
Chalinidae............................ 40, 41
Chalininae............................... 41
Chironomidae . •..................• . 36
Cblorella ............................. 35
Copepoda................................. 37
Cornacuspongida.......................... 39
Стр.
Cristaria plicata......................... 73
Dendroceratida........................... 39
Dosilia............................. . 83
Dosilia (?) stepanowi.................... 82
Ecnomus tenellus.......................... 36
Ephydatia ..............................60, 74
bohemica ....... 85
fluviatilis ..............61, 75
fluviatilis v. chui......... 77
„ v. hastifera . 61, 75, 76
„ v. teberdana . . 75, 76
fossilis (-»-).............. 46
goriaevi.................... 87
meyeni . . . ...... 74, 77
mUlleri ......... 6(f 74
„ v. acuminata ... 74, 79
„ „ v. astrodiscus ... 79
„ „ v. behningi .... 75
„ „ v. obtusosclera 74, 77, 79, 80
„ obtusosclera ....... 77
„ olchonensis.................. 87
„ robusta...................... 74
„ sibirica.................. 77, 79
„ solida..................... ... 77
Eumetazoa ...... ........... 39
Eunapius ......... . 62, 69
Euspongilla............................. 62
Gelliidae............................40, 41
Glyptotendipes.......................... 36
Haploscleridae.......................... 41
Heteromeyenia................... . . 60, 80
„ baileyi...................... 82
„ „ v. arndti ... 84
„ „ v. bohemica . 82
„ „ v. palaiina . . 81
„ baileyi v. petri . . . 81,84
„ „ v. repens . . . 81,84
1 Наявания, введенные в определительные таблицы, напечатаны курсивом.
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
123
Стр.
Heteromeyenia „ v. stepanowi 61, 81, 82
» repens 84
a if v. arndti . . . 84
f. bohemica . . 85
и f. palatina . . 85
a if f.' petri .... 84
ft и v. stepanowi . 82
Heteromey enia ry deri . 81
" » f. baleni .... 81
>> » f. pictovensis . . 81
» tubisperma....................... 81
Hydracarina................................ gg
Hydropsyche ornatula....................... 36
Keratosa................................... 39
Leptocerus senilis . . •................. 35 36
» fulvus............................ *36
Limnaea.................................... 3g
Lithastericus tuberculatus................. 45
Lubomirskia................................ qq
» abietina...................... 91, 94
» bacillifera................ 96
» baicalensis................ 91
” >> m. littoralis . 91, 92
>, fusifera.......................91 93
>, intermedia................. 98
» irregularis............... 101
» papyracea.............
» tscherskii............
Lubomirskiidae...................
Metazoa.......................
Meyenia.......................
Meyenia fluviatilis v. acuminata ....
Microciona ...................
Monactinellidae...............
Nais..................
Neritina fluviatilis...............
Neureclipsis bimaculata..............
Oxyheteromeyenia...................
Parazoa.......................
Parmula............................
>> browni ......................
Pellina semitubulosa.................
Phthinorhabdina....................
Planorbis..........................
Pleurococcus .................
Podophria fixa.......................
Porifera...........................
Potamolepidinae......................
Potamolepis........................
Renierinae...........................
Scenedesmus quadricauda
101
96
59,89
39
75
77
15
39
37
36
36
83
39
13
31
18
40
36
35
9,37
39
40
40
41
35
Стр.
Sisyra.................................... 35
Spongia baicalensis........................ 91
Spongilla........................... . 59, 62
„ arctica . . . . . . . 60, 63, 66
„ biseriata ..;... 60, 62, 67
,, bombayensis..................... 73
» v. pneumatica 73
„ carteri.......................61, 64, 70
„ carteri subsp. balatonensis . 70, 72
„ „ v. melli . . 70
v. rotunda-
cuta ........ 64, 70, 72
ii »
V if
»
j, »
» » » a
»
» if
n if
erinaceus ........ л 85
fluviatilis........................ 75
fragilis....................61, 63
„ v. brevipora . 64, 69, 70
„ subsp. calcuttana . 70
fragilis subsp. decipiens . . 70
„ v. rectituba..............63, 70
fragilis........................ 70
gigantea (ч-)................... 46
Helvetica ........ 63
jordanensis..................... 66
lacustris............... 60, 63, 64
„ v. jordanensis . 63,66
„ m. lacustroides . 66
„ v. paupercula . . 63, 66
„ subsp. reticulata 66
„ f. rhenana ... 63
lacustroides.................... 66
meyeni.......................... 77
microgemmata.................... 87
micron.......................... 69
miilleri........................ 77
paupercula . ................... 66
proliferens.................67, 68
„ v. sectospina . 67
purbekensis (ч-)............... 45
rectituba...................... 69
repens....................... 84
rotundacuta.................. 70
sectospina..................67, 68
semispongilla................... 69
sibirica........................ 69
stanleyi ....... 61, 64/73
„ stygia................................ 34
Spongillidae................. . ... . 59
Spongillinae................................ 40
Spongolithis furca (ч-)..................... 45
Stratospongilla ........... 62, 72
Stylaria lacustris.......................... 37
124
ФАУ.НА СССР
Стр.
Стр.
Stylotella heliophila .................... 16
Swarischewskia.........................90,101
„ irregularis .... 101
„ papyracea ............... 101
Tetraxonida............................... 39
Triaxonida................................ 39
Trochospongilla....................... 60, 85
„ erenaceus................. 85
„ erinaceus................. 85
„ horrida ....... 61, 85
„ polysclera ... . . 85
Tubella ..... ................ 13
Unio . ...........................'. . 37
„ murchisonianum......................... 73
Unionicola crassipes ...................36, 37
Unionidae................................. 73
Veluspa abietina . .................... 94
„ bacillifera •.................... 96
„ baicalensis .................... 91
„ intermedia........................ 98
„ polymorpha v. baicalensis ... 91
Viviparus................................. 37
СОДЕРЖАНИЕ
Стр.
Предисловие............................................................... V
Систематический список видов........................................... VII
Общая часть
Внешний вид и местообитание пресноводных губок........................... 1
Скелет и паренхима........................................................ 4
Клеточные элементы........................................................ 7
Скелет.................................................................. 10
Жизненные отправления губок.............................................. 14
Половое размножение.........................._.......................... 19
Бесполое размножение (геммулы)........................................... 21
Другие виды бесполого размножения ...................................... 25
Регенерационная способность губок........................................ 27
Цикл жизни и условия обитания губок . . ................................ 30
Биоценотические отношения губок........................................ 34
Систематика губок........................................................ 38
Географическое распространение пресноводных губок........................ 42
Ископаемые пресноводные губки............................................ 45
Значение пресноводных губок для человека................................ 45
Собирание и сохранение губок...................................... .... 47
Препаровка губок......................................................... 50
Содержание пресноводных губок в аквариумах.............................. 52
Главнейшая литература по губкам.......................................... 55
Специальная часть
Определительная таблица семейств......................................... 59
Сем. Spongillidae — Бадяги .............................................. 59
Сем. Lubomirskiidae — Байкальские губки.................................. 89
Приложение: Губки в отложениях современных водоемов..................... 102
Резюме................................................................ 105
Объяснения таблиц....................................................... 119
Алфавитный указатель................................................... 122
Таблица
иг. 3
к р
и
к Г
Фиг. 2
Таблица 11
Фиг. 4
Фиг. 5
Фиг. 6
I
Фиг. 7
Фиг. 8
Фиг. 9
Фиг. 10
Таблица IV
Фиг. 11
Фиг. 12
Фиг. 14а
Таблица V
о
о0' о
Фиг. 15
9 ’ д1»
Фиг. 16 /
л*.
Фиг. 17
Фиг. 18
Таблица VI
Т а б л й Ц a VII
Таблица VIII
Таблица IX
Фиг. 35
Фиг. 36
Таблица
Фиг. 38
РИГ.
39
Фиг. 41
Фиг. 40
Фиг. 42
Таблица XI
Фиг. 45
300Ц
Фиг. 46
Таблица XII
Фиг. 47 Фиг. 48
300^
Фиг. 50
Фиг. 49
Фиг. 52
Фиг. 51
Таблица XIII
I
Фиг. 53
Фиг, 54
Dnr. 57
Таблица
XIV
г
Фиг
60
иг.
IQQJL
иг. 61
Таблица XV
Фиг. 62
Фиг. 64
Фиг. 63
ФиМВ