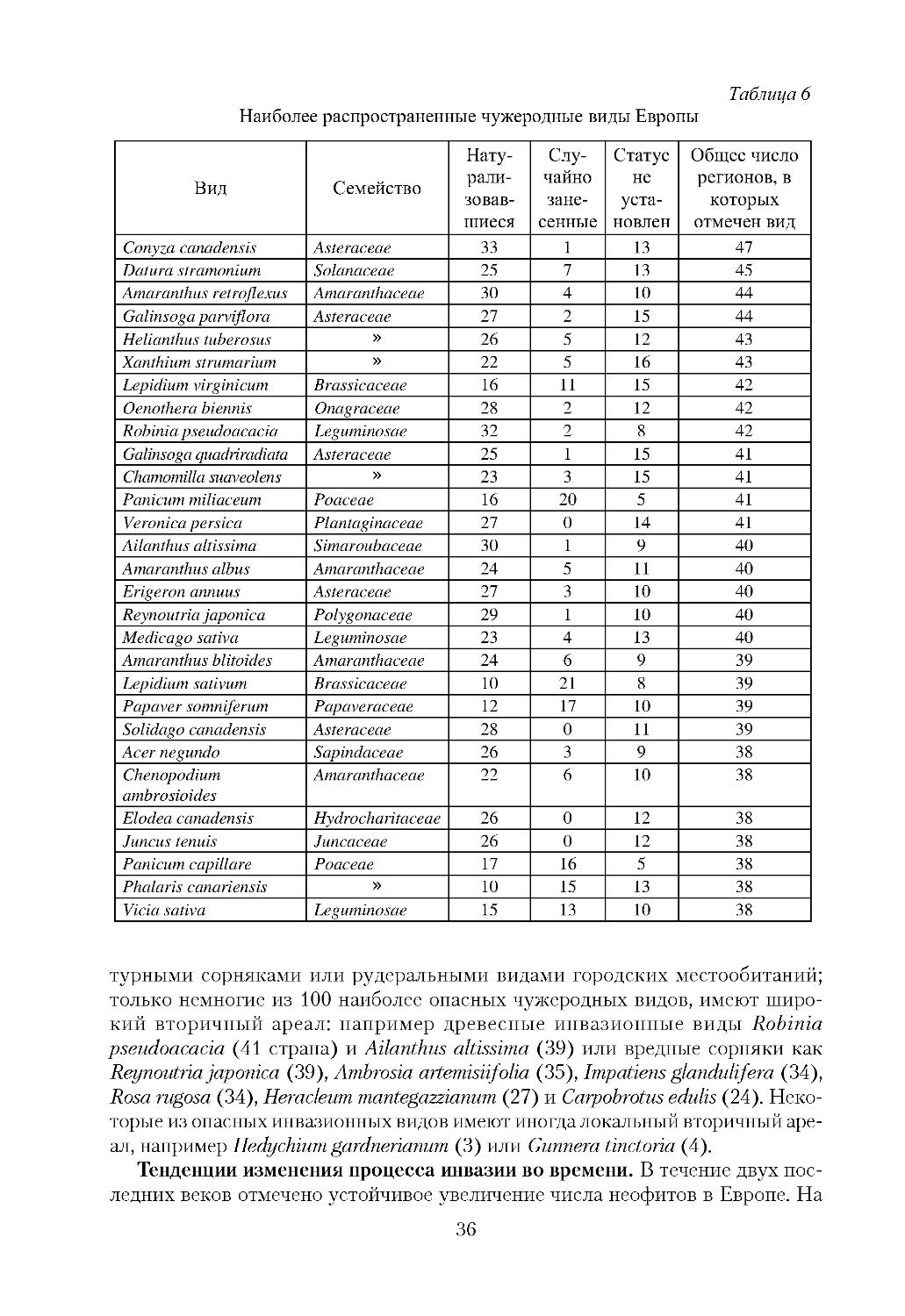
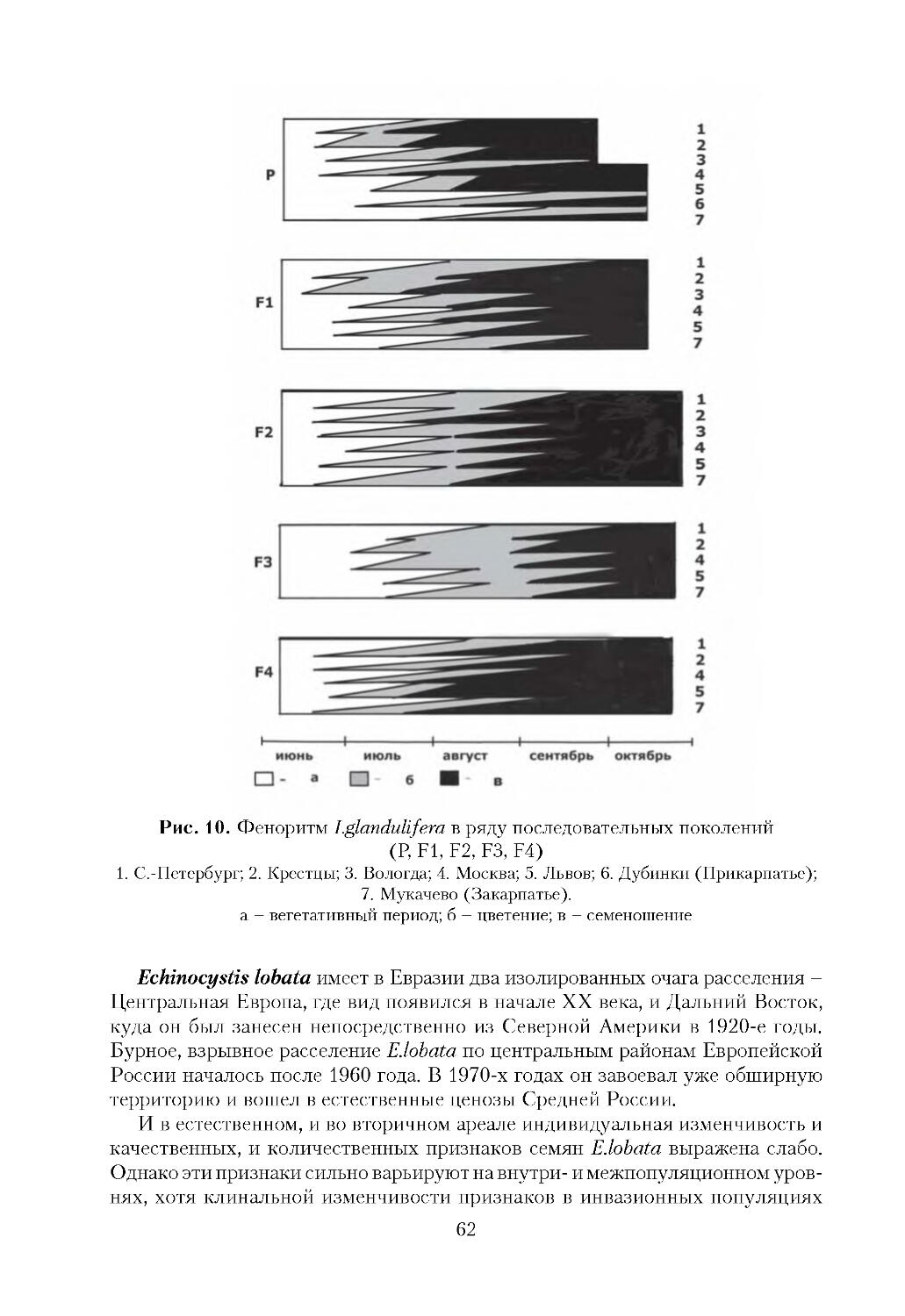
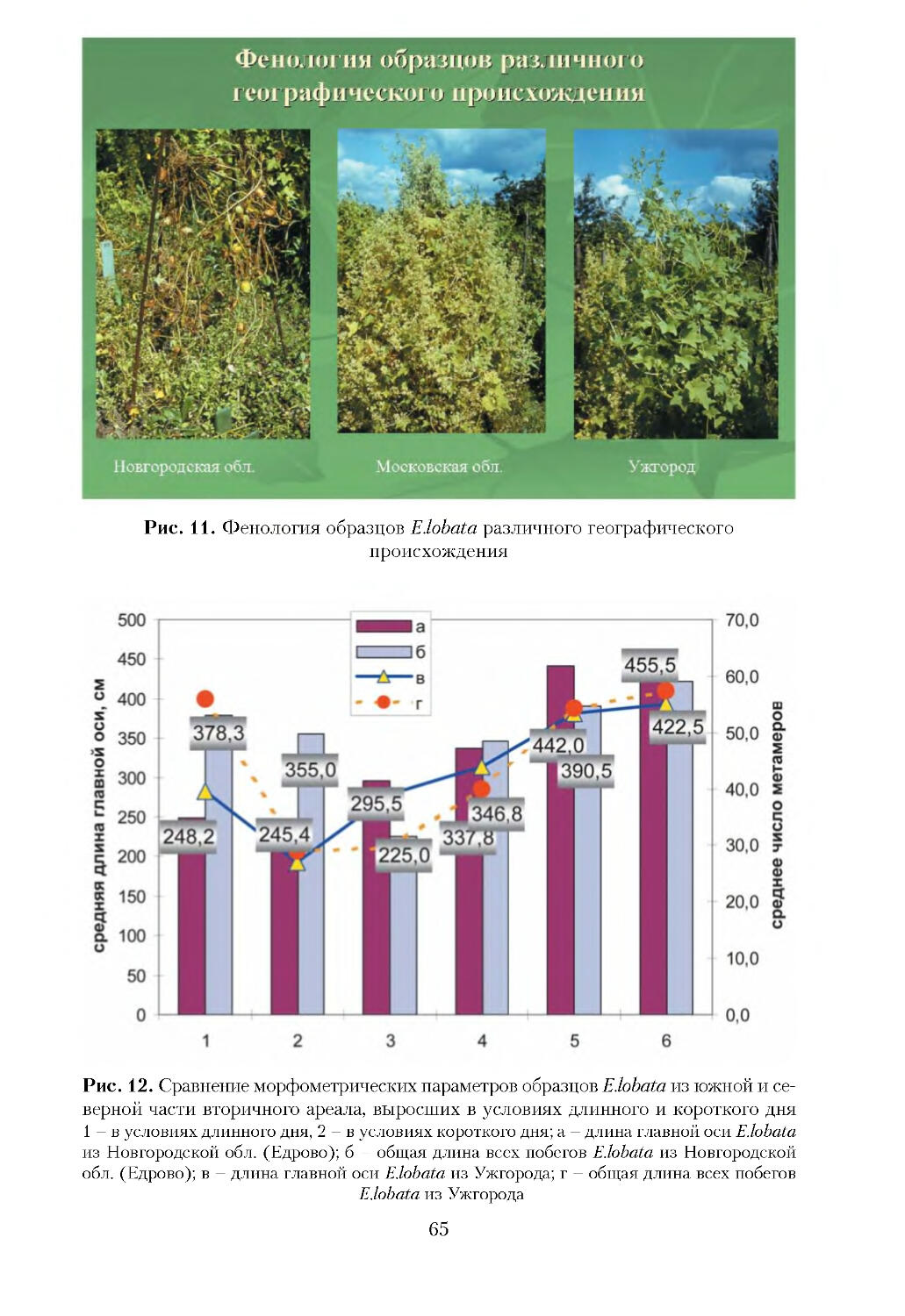
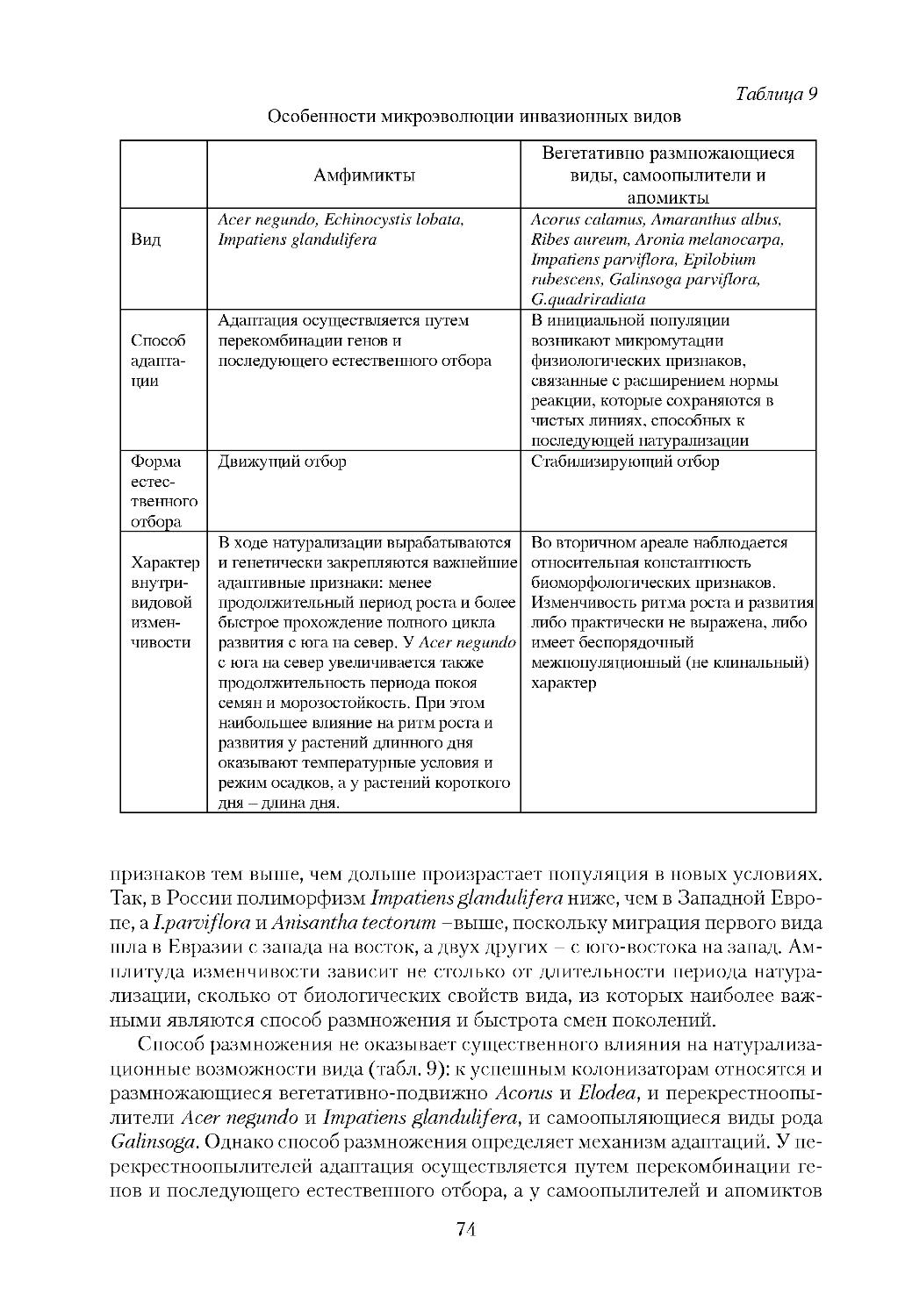
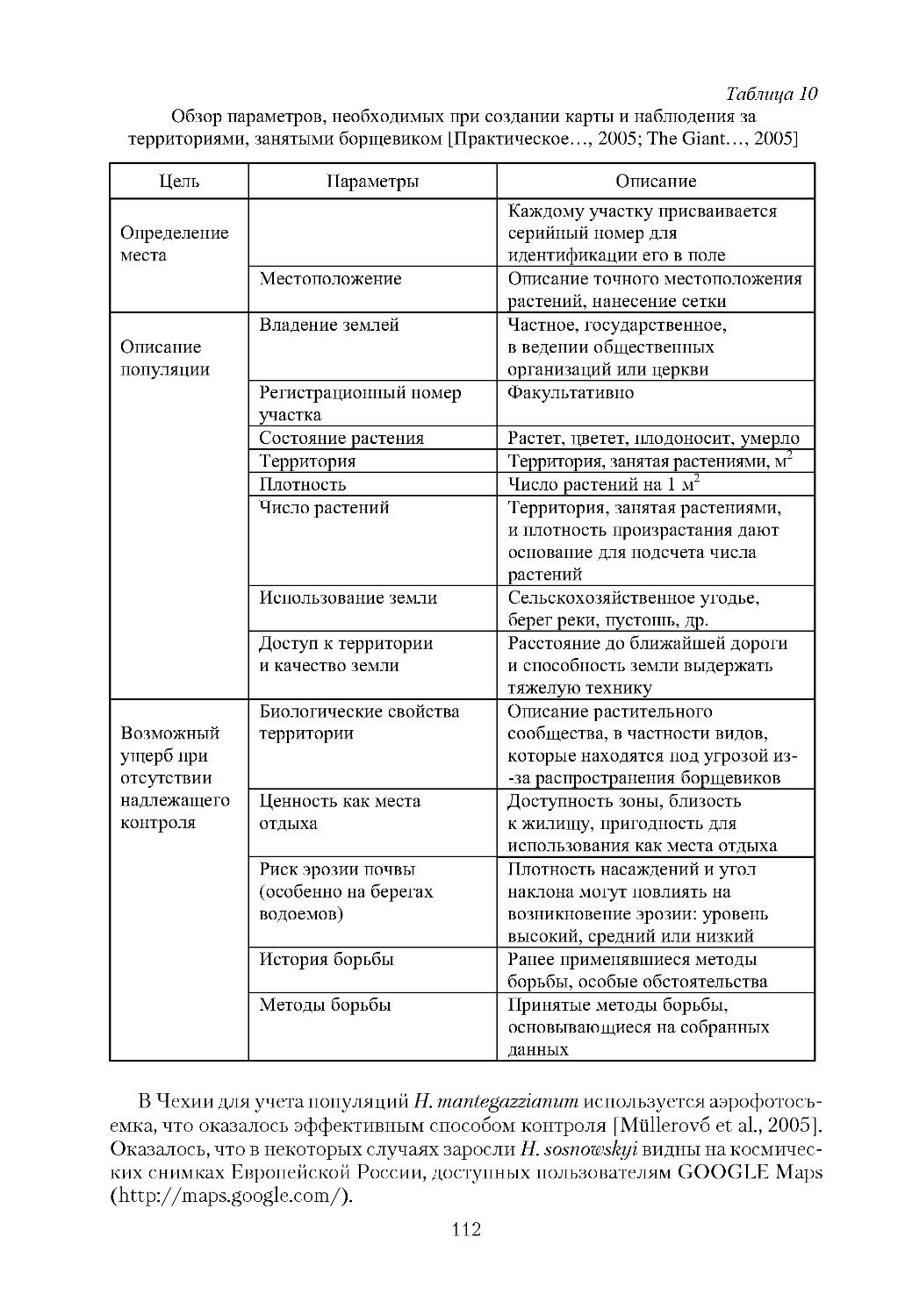
/



































































































































































































































































































































































































































































































Автор: Виноградова Ю.К. Майоров С.Р. Хорун Л.В.
Теги: джайнизм историческая геология флора
ISBN: 978-8-89119-487-9
Год: 2009




Текст
Серия «ЧУЖЕРОДНЫЕ ВИДЫ РОССИИ»
РОССИЙСКАЯ АКАДЕМИЯ НАУК
Главный ботанический сад им. Н.В. Цицина
Секция инвазий чужеродных видов Комиссии РАН по сохранению
биологического разнообразия
Программа фундаментальных исследований Президиума РАН
«Биоразнообразие и динамика генофондов»
Ю.К. Виноградова
С.Р. Майоров
Л.В. Хорун
ЧЕРНАЯ КНИГА
ФЛОРЫ СРЕДНЕЙ РОССИИ
(ЧУЖЕРОДНЫЕ ВИДЫ РАСТЕНИЙ
В ЭКОСИСТЕМАХ СРЕДНЕЙ РОССИИ)
Москва
ГЕОС
2009
2
УДК 234556
ББК 26.323
В16
Ю.К. Виноградова, С.Р. Майоров, Л.В. Хорун
Черная книга флоры Средней России (Чужеродные виды растений
в экосистемах Средней России). -- М.: ГЕОС. 2009. -- 494 с.
ISBN 978 8 89119 487 9
В книге впервые обобщены данные по биологическим особенностям 52 наи
более злостных и широко распространенных инвазионных видов флоры Сред
ней России и динамике их расселения во вторичном ареале. Даны оценка эко
номическому ущербу и рекомендации по возможному использованию инвази
онных видов. Предлагаются методы контроля чужеродных видов с целью
уменьшения их обилия, сужения ареала, а также предотвращения дальнейшего
продвижения. Представлен «black list» 100 чужеродных видов растений, кото
рые требуют первоочередного изучения и мониторинга в целях предотвраще
ния биологических инвазий.
Инвентаризация видов и составленные на основе гербарных данных карты
их ареалов призваны обеспечить квалифицированную справочную систему по
инвазионным чужеродным видам Средней России, доступной для менеджеров
по охране природы, законодателей, исследователей, студентов и всех заинтере
сованных лиц. Эта книга -- начало серьезного обсуждения необходимости уче
та биологии и экологии заносных видов для выработки эффективных мер борь
бы с фитоинвазиями.
Ответственный редактор: Ю.Ю. Дгебуадзе,
член корреспондент Российской академии наук
Научный редактор: А.С. Демидов,
директор Главного ботанического сада им. Н.В. Цицина РАН,
доктор биологических наук
Исследования проведены при поддержке научной Программы
Президиума РАН «Биоразнообразие и динамика генофондов» и гранта
РФФИ 08 04 00239а
Издание финансируется издательским проектом РФФИ 09 04 07049д
Фотографии С.Р. Майорова, Н.М. Решетниковой, Ю.К. Виноградовой,
А.Г. Куклиной, О.Н. Деминой.
© ГБС РАН, 2009
© Авторский коллектив, 2009
© ГЕОС, макет, 2009
3
ПРЕДИСЛОВИЕ
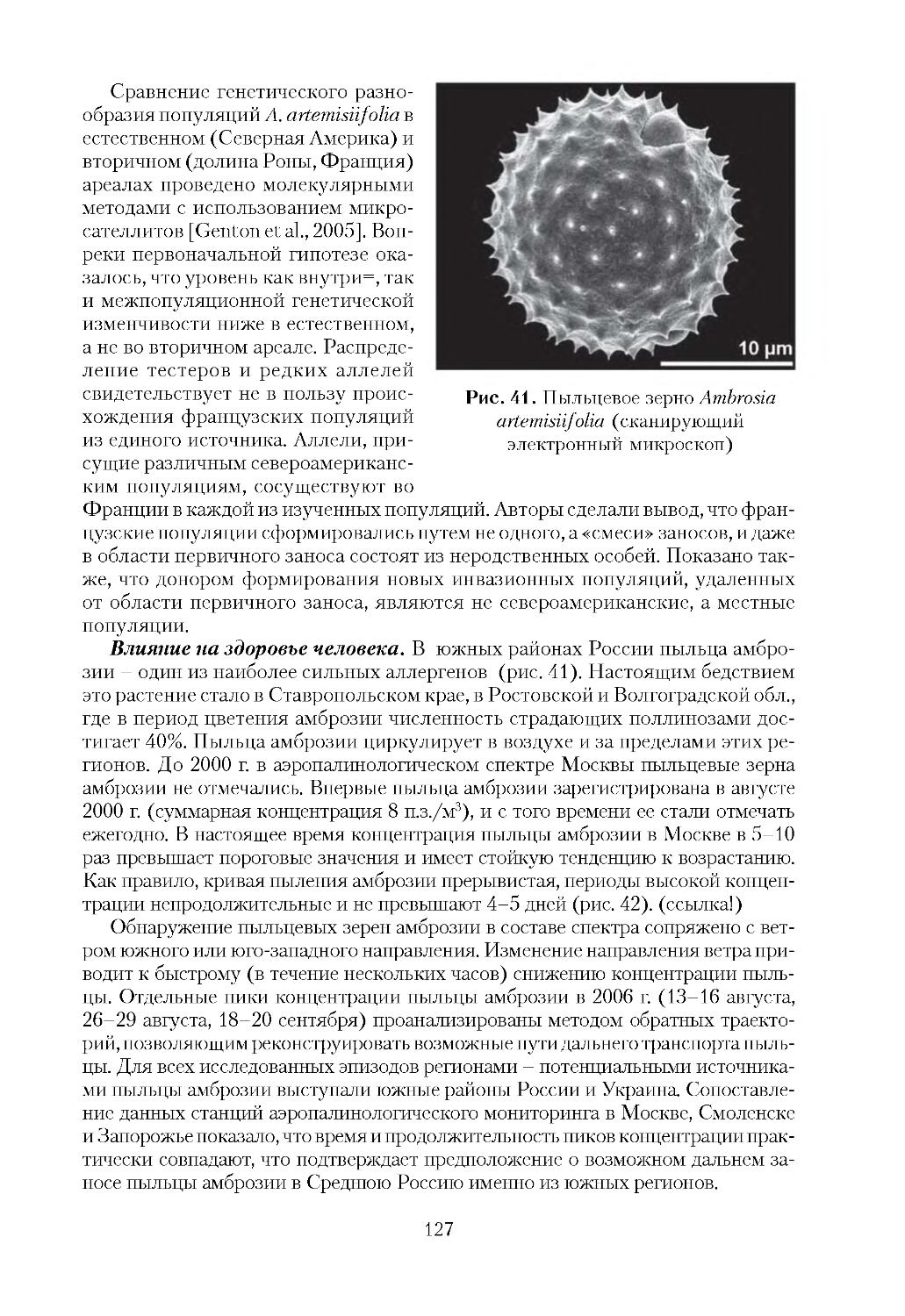
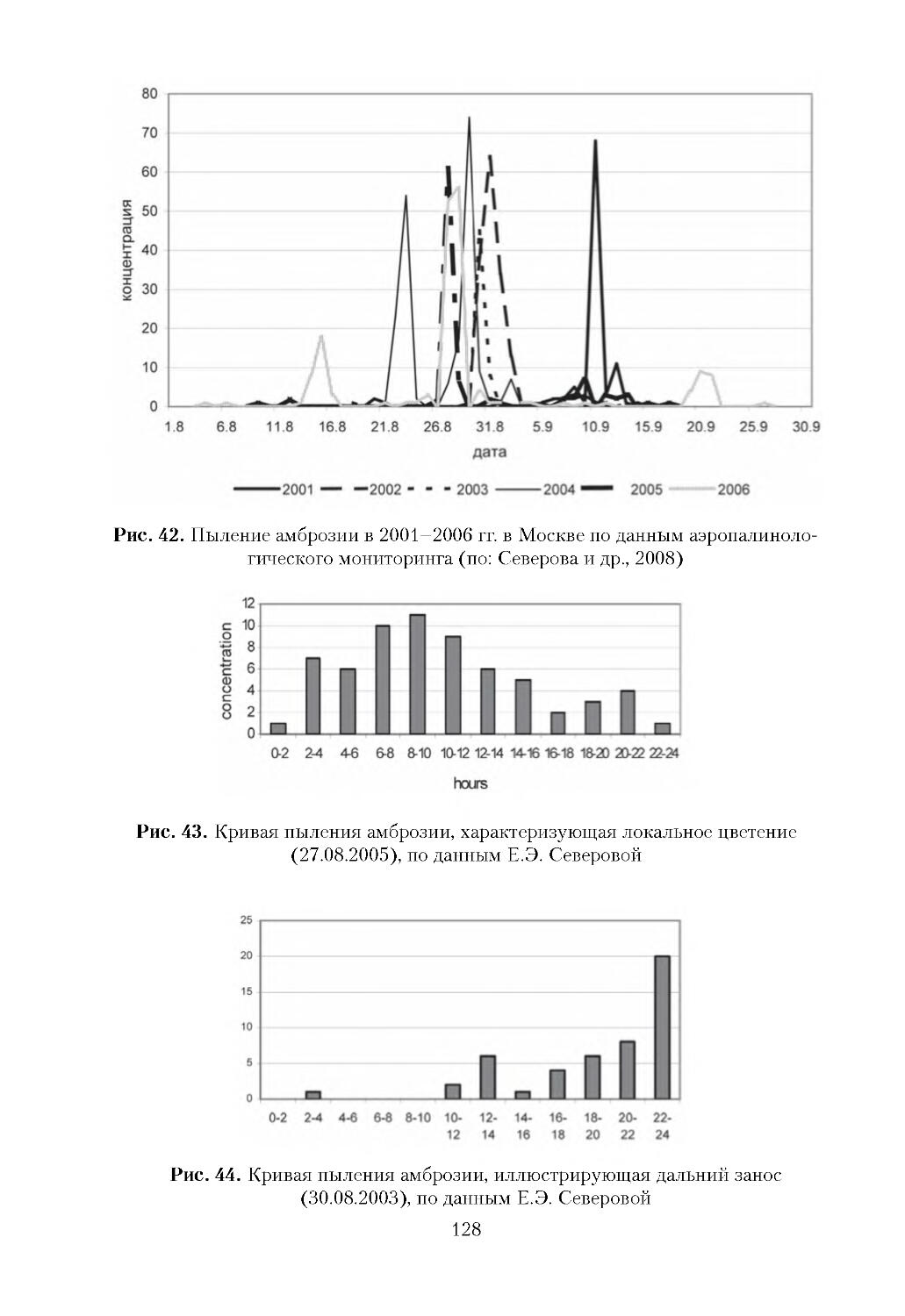
ОТВЕТСТВЕННОГО РЕДАКТОРА
Начиная с середины ХХ века во многих регионах Земного шара все чаще
стали наблюдаться существенные преобразования, связанные с проникновени
ем не свойственных для них чужеродных видов живых организмов из других
мест. Освоившись в новых для себя естественных и искусственных (антропо
генных) экосистемах, эти виды в ряде случаев поставили под угрозу безопас
ность устойчивого развития многих стран мира. Нельзя сказать, что переселе
ния организмов не происходили раньше, но интенсивность и масштабы этого
процесса в последние десятилетия столь велики, а последствия столь огромны,
что его нельзя не отнести к одной из главных экологических проблем человече
ства.
В последнее время процессы, связанные с проникновением и воздействия
ми неаборигенных видов на местные виды и сообщества, принято именовать
биологическими инвазиями чужеродных видов. Под биологическими инвазия
ми понимаются все случаи проникновения живых организмов в экосистемы,
расположенные за пределами их первоначального (обычно, естественного) аре
ала. Таким образом, к биологическим инвазиям относятся вселения чужерод
ных видов, произошедшие в результате:
· естественных перемещений, связанных с флюктуациями численности и
климатическими изменениями;
· преднамеренной интродукции и реинтродукции важных в хозяйственном
и эстетическом отношении организмов;
· случайных заносов с балластными водами, с импортной сельскохозяйствен
ной продукцией, багажом, с намеренно интродуцированными видами, и т.п.
Во многих случаях чужеродные виды, вступая в контакт с популяциями ви
дов--аборигенов, существенно преобразуют структуру биоценозов, и их появ
ление имеет глобальные экологические, экономические, а иногда, и социальные
последствия.
В Российской Федерации планомерные фундаментальные и прикладные ис
следования чужеродных видов начаты сравнительно недавно. В то же время
проблема проникновения чужеродных организмов имеет исключительно важ
ное социально экономическое значение для России. Особенности нашей стра
ны (географическое положение, большие размеры, интенсивные перемещения
людей и товаров и т.п.) безусловно, способствуют инвазионному процессу. В
связи с этим число крупных экологических катаклизмов, вызванных инвазия
ми чужеродных видов, постоянно растет. Если говорить о растениях, то здесь
потрясают уже масштабы инвазий. В настоящее время только в европейской
4
части России насчитывается более тысячи чужеродных видов растений. Значи
тельно и влияние некоторых из них на растительные сообщества и экосистемы
в целом. Для нашей страны можно вспомнить целый ряд примеров нежелатель
ных и даже вредных инвазий растений: амброзия (Ambrosia artemisiifolia), бор
щевик Сосновского (Heraсleum sosnowskyi), клен ясенелистный (Acer negundo),
недотрога желёзконосная (Impatiens glandulifera), элодея канадская (Elodea
canadensis).
В последние годы в результате активности, прежде всего специалистов Рос
сийской академии наук и ряда отраслевых институтов теоретическим и при
кладным вопросам, связанным с проблемой вселения чужеродных видов на тер
риторию РФ, уделяется исключительно большое внимание. За последние 10 лет
в рамках ряда проектов (включая ФЦНТП Федерального Агентства по науке и
инновациям, программ Президиума РАН и Российского фонда фундаменталь
ных исследований) удалось выявить основные транзитные пути инвазий, со
здать базы данных по чужеродным видам, оценить их воздействие на абориген
ные экосистемы. В частности, в рамках нескольких проектов учеными Российс
кой академии наук разработана база данных по чужеродным видам РФ,
значительная часть которой касается растений. Эта база данных частично пред
ставлена на сайтах: http://www.sevin.ru/invasive/ и http://www.nobanis.org/
Factsheets.asp./
Большое значение имеет координация всех исследовательских групп, зани
мающихся проблемами инвазий чужеродных видов. Определенную роль в про
цессе такой координации в настоящее время играет секция биологических ин
вазий Комиссии Российской академии наук по сохранению биоразнообразия,
созданная в 2002 году. За годы работы члены Секции организовали две между
народные конференции и один круглый стол, на трех последних съездах Гидро
биологического общества РАН работали специальные секции по проблеме чу
жеродных видов; издано два сборника научных статей; с 2008 г. начал выходить
электронный журнал -- «Российский журнал биологических инвазий» (см. http:/
/www.sevin.ru/invasjour/); основана серия «Чужеродные виды России» (опуб
ликованы две монографии).
Следует отметить, что, несмотря на некоторые успехи, в целом по России
фундаментальные и прикладные исследования инвазионного процесса все еще
ведутся спорадически и в небольшом объеме. В частности, много еще предсто
ит сделать в исследовании уязвимости экосистем к инвазиям, оценке послед
ствий проникновения чужеродных видов и разработке мер по его контролю. Не
так хорошо обстоят дела и в информационной поддержке этих работ, что ис
ключительно важно для быстрого оповещения, оценки риска и разработки мер
борьбы с чужеродными видами, наносящими ущерб народному хозяйству. Пред
ставляется, что одним из важных шагов в исправлении сложившейся ситуации
будет публикация «Черной книги флоры Средней России».
В книге обобщены данные по биологическим особенностям 52 х наиболее
агрессивных чужеродных видов флоры Средней России и динамике их расселе
ния в новом ареале, даны оценка экономического ущерба и рекомендации по
возможному их использованию. В работе освещена современная ситуация с
инвазиями чужеродных видов растений на территории Средней России, выяв
5
лены основные их транзитные пути, разработаны прогнозы и меры по предотв
ращению инвазий и смягчению их последствий.
Настоящая монография продолжает, уже упоминавшуюся серию «Чужерод
ные виды России», издаваемую в рамках Программы Президиума РАН «Биоло
гическое разнообразие».
Есть все основания полагать, что «Черная книга» так же, как и «Красная книга
РФ» станет не только серьезным вкладом в развитие теоретических и приклад
ных вопросов, связанных с проблемой биологических инвазий чужеродных ви
дов, но и послужит основой для разработки специального законодательства по
предотвращению биологического загрязнения как на государственном, так и на
региональном уровнях и явится важнейшим аспектом обеспечения экологичес
кой безопасности страны.
Руководитель Секции биологических инвазий
Комиссии РАН по сохранению биологического разнообразия,
Член корреспондент РАН
Ю.Ю. Дгебуадзе
6
ВВЕДЕНИЕ
Экспансия чужеродных видов
Внедрение (инвазия) агрессивных чужеродных видов является в настоящее
время значительной частью глобальных природных изменений и часто ведет к
существенным потерям биологического разнообразия и экономической значи
мости экосистем, подверженных биологическим инвазиям. Иногда это внедре
ние может наносить значительный экономический ущерб и даже представлять
опасность для здоровья людей. Число крупных экологических катастроф, выз
ванных инвазиями чужеродных видов, постоянно растет. Достаточно назвать
лишь несколько примеров: амброзия, колорадский жук, борщевик Сосновско
го... Пока еще не существует универсальных способов остановить агрессивные
виды. Разработка мер по предотвращению биологических инвазий, смягчению
их последствий и мониторингу являются обязанностью всех стран, подписав
ших в 1992 году в Рио де Жанейро Конвенцию о биологическом разнообразии
[Конвенция..., 1995].
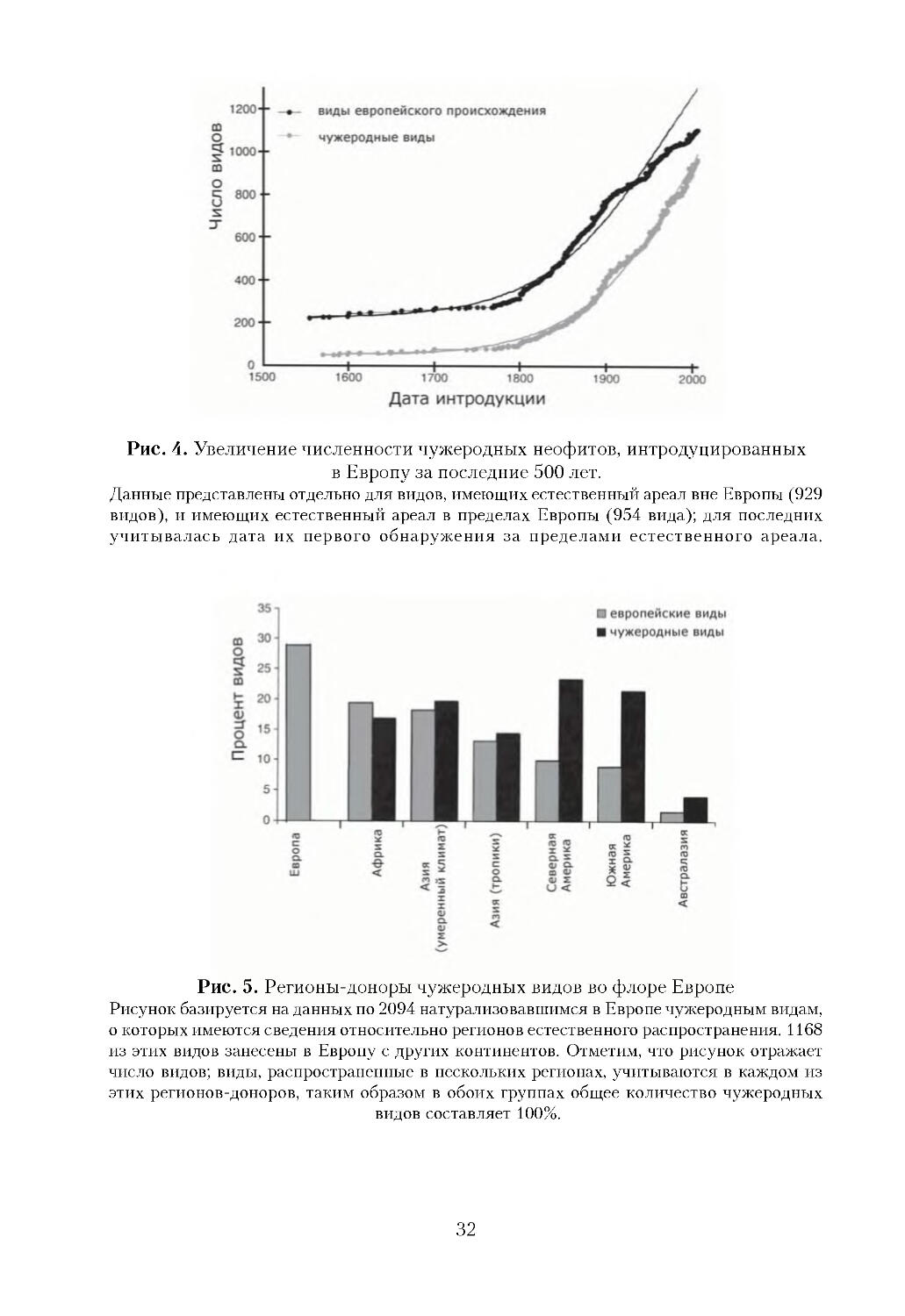
Многочисленные чужеродные виды, большинство из которых занесены все
го лишь 200 лет назад, успешно обосновались на новой родине и сформировали
широкие ареалы. Ученые предполагают, что уже в ближайшем будущем про
изойдет еще более драматическое увеличение биологических инвазий [Sala et
al., 2000; Hulme, 2007]. Глобальное потепление, повышение концентрации ат
мосферного углекислого газа и понижение содержания азота существенно из
меняют природные условия и увеличивают фрагментарность местообитаний,
что может облегчить дальнейшие инвазии [Vilа et al., 2006]. Раннее обнаруже
ние и предотвращение воздействия чужеродных видов на экосистемы является
фундаментальным требованием Стратегии ЕЭС по сохранению биоразнообра
зия в Европе до 2010 года и далее. Однако при отсутствии надежных региональ
ных исследований европейские государства не способны заняться этой пробле
мой в стратегическом плане [Miller et al., 2006; Hulme, 2007].
Экономический ущерб от биологических инвазий колоссален. Подсчитано,
что в результате инвазии заносных видов США теряют 137 млрд, Индия -- 117
млрд, а Бразилия -- 50 млрд долларов [Тишков, 2005]. Урон, наносимый биоло
гическими инвазиями только со стороны 79 наиболее злостных инвазионных
видов, в США оценивается в 97 млрд долларов [Pimentel et al., 2001]. Ниже при
ведены данные из работы Д.Пайменталя с соавт., касающиеся только ущерба
экономике США от заносных растений.
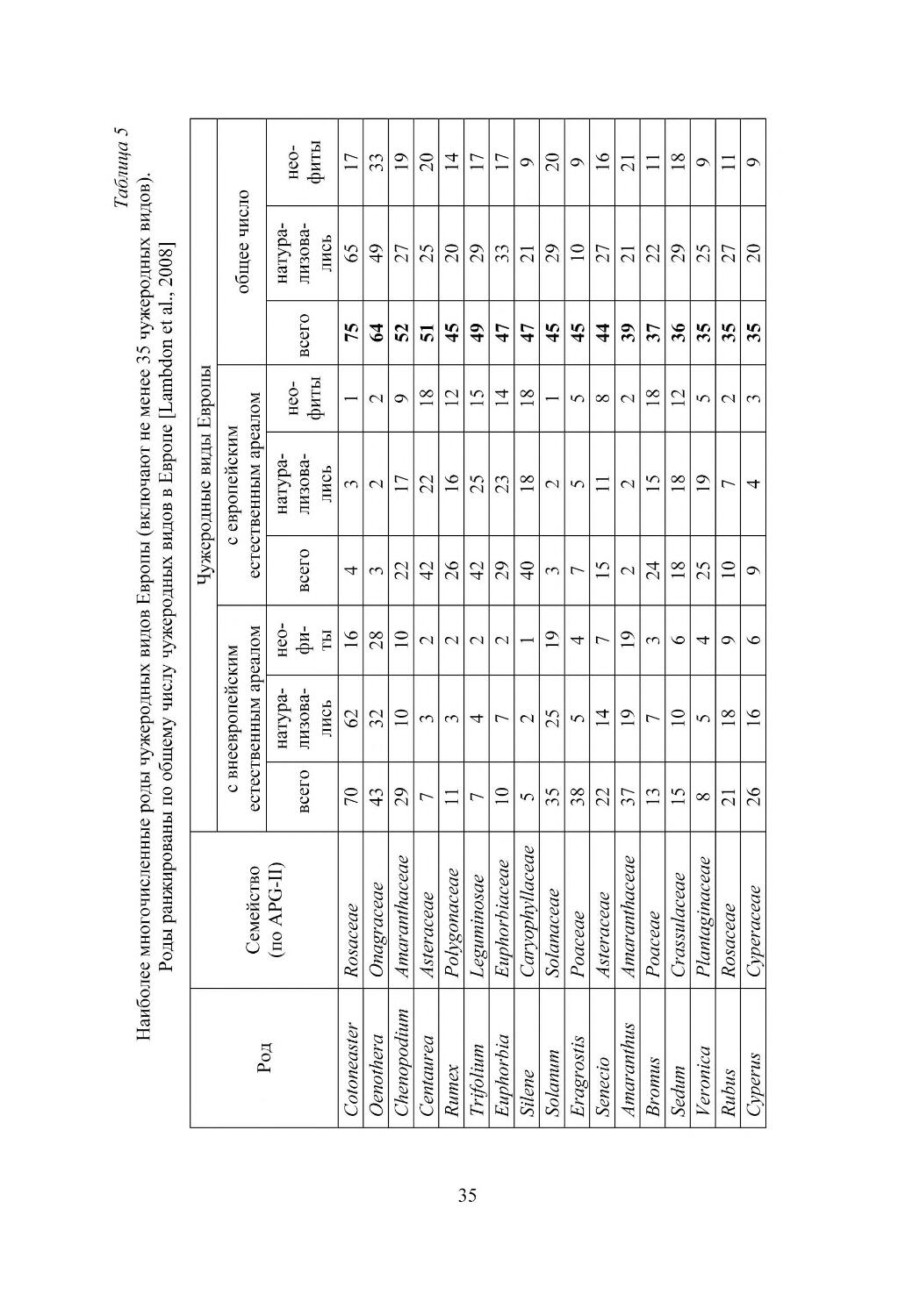
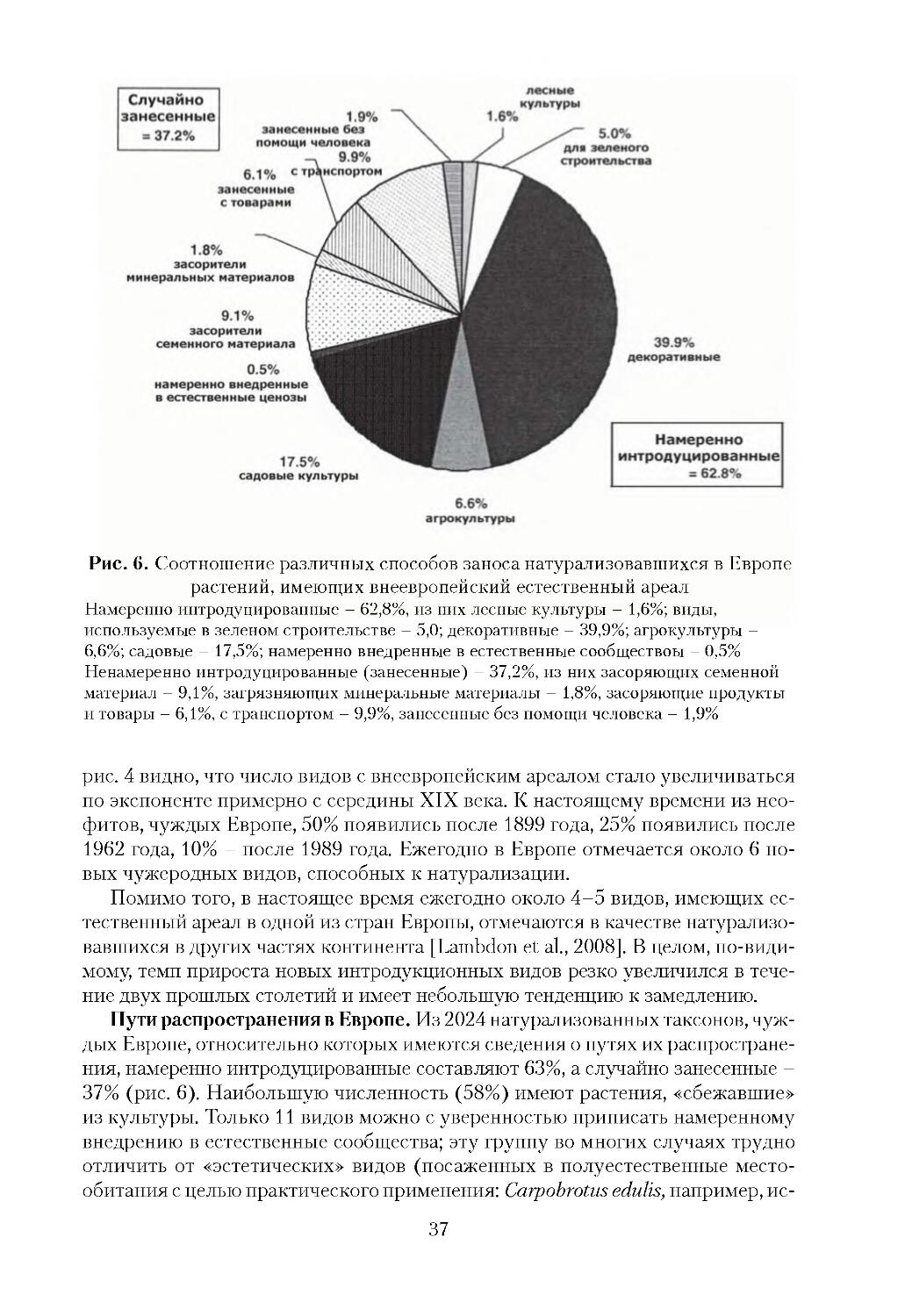
В Европе пока нет полных данных по этой проблеме, но биологические ин
вазии также приводят к экономическим потерям. Наиболее значимые свидетель
7
ства касаются ущерба, который причиняют чужеродные вредители и сорняки
сельскому хозяйству, лесоводству и водным ресурсам [Williamson, 2002]. При
мером прямого воздействия на экономику служат ущерб, наносимый поймен
ным лугам Fallopia japonica, и снижение продуктивности лесов из за поврежде
ния коры деревьев серыми белками (Sciurus carolinensis). Большое экономичес
кое значение в Европе имеют агрессивные заносные сорные виды, например,
Chenopodium ambrosioides, Paspalum paspaloides, Oxalis pescaprae (рис. 1), Conyza
canadensis. Имеются данные, что урожай сои снижается на 83%, если на 1 м2
произрастает 150 особей мелколепестника канадского Conyza canadensis. Серь
езное засорение полей мелколепестником уменьшает урожаи сахарной свеклы
в Германии на 64% и отрастание весенних почек виноградной лозы в Италии на
28%. При уборке урожая зрелые стебли C. canadensis забивают механические
комбайны, даже при низкой (1 растение/м2) плотности популяции [Weaver, 2001].
Борьба с одним из наиболее опасных заносных видов Германии -- амброзией
полыннолистной составляет 20% расходов на борьбу с сорными видами в стра
не [Sheppard et al., 2006]. Другие заносные растения служат хозяевами болез
нетворных микроорганизмов, например, Bromus catharticus является растением
хозяином желтого вируса карликовости ячменя и ржавчины пшеницы. Инвази
онные виды могут влиять на здоровье человека, вызывая фитофотодерматит при
контакте с борщевиком Heracleum sosnovskii и H.mantegazzianum, астму или сен
ную лихорадку при цветении Ambrosia artemisiifolia, а также отравление людей
ядовитыми плодами Phytolacca americana и Solanum elaeagnifolium.
Кроме того, инвазионные виды вызывают серьезные экологические послед
ствия, нанося существенный вред экосистемам [European Commission, 2004].
Экосистемы могут быть изменены вплоть до полного исчезновения природных
видов, что наблюдается при колонизации песчаных дюн видами рода Acacia. При
экспансии Carpobrotus edulis угрожающему сокращению подверглись эндемич
ные прибрежные виды (рис. 2). Однако чаще экологические изменения и био
логическая гомогенизация трудно уловимы. Например, внедрение Rhododendron
ponticum значительно сократило биологическое разнообразие приатлантических
дубрав. Тонкое, но потенциально более серьезное воздействие чужеродных ви
дов -- это возможность гибридизации с природными видами, подобно тому, что
наблюдается в Европе между заносными и природными видами рода Quercus.
Гибридизация может ввести чужие гены в естественные популяции или приве
сти к появлению сильного гибрида, способного к инвазии [Hulme et al., 2008].
Таблица 1
Урон, наносимый инвазионными видами растений [Pimentel et al., 2001]
Сорняки
Прямой ущерб,
млн долл.
Средства для контроля
инвазий, млн долл. Общие потери,
млн долл.
водоемов
10
100
110
полей
23 400
3000
26 400
пастбищ
1
5
6
газонов и садов
данных нет
1 500
1 500
ИТОГО
23 411
4605
28 016
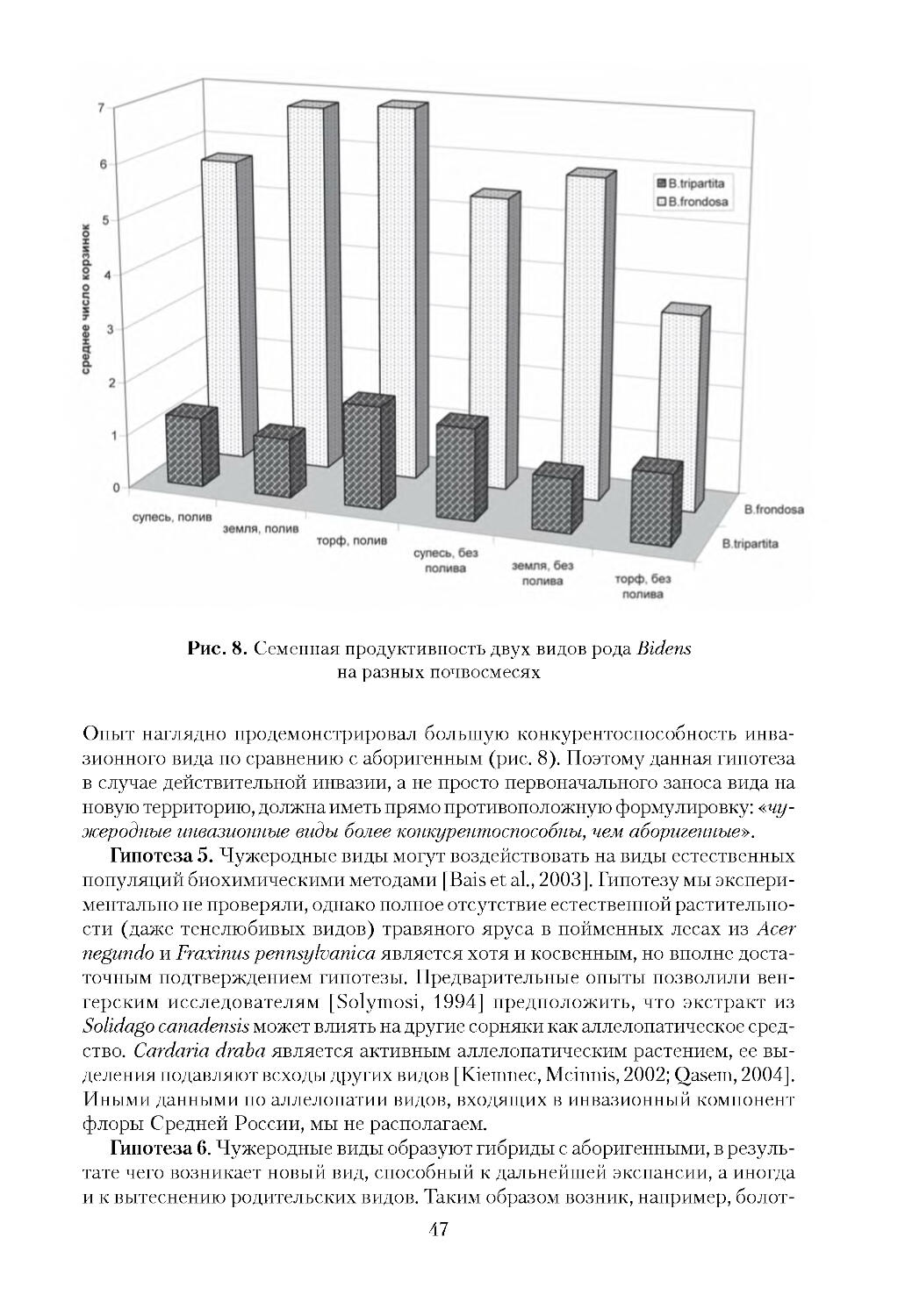
Рис. 1. Массовое цветение Oxalis pes caprae. Мальта, конец марта 2009 г.
Рис. 2. Инвазия Carpobrotus edulis в прибрежные сообщества.
Испания, Каталония, г. Салоу. 2008 г.
9
Некоторые биологические инвазии, которые в настоящее время угрожают
Европе, могли быть пресечены, если бы в отдельных странах имелся высокий
уровень понимания грозящей опасности со стороны агрессивных видов. Про
должающееся бездействие многих стран становится всё более пагубным для
сохранения биоразнообразия и экономики региона, а также здоровья населения
[Hulme, 2007]. Европейцам следует осознать опасность того, что из за их без
действия территории, находящиеся под их юрисдикцией, могут стать потенци
альным источником вторжения инвазионных видов в пределы соседних госу
дарств, а потому обязаны предпринять ряд решительных мер в попытке мини
мизировать этот риск. Для Европы такие шаги очень важны, поскольку виды,
интродуцированные на территорию одного государства, могут легко распрост
раниться на соседние, особенно те, с которыми имеются общие водные пути,
горные цепи или береговая линия. Пока же численность и воздействие вредных
инвазионных видов в Европе хронически недооценивались, особенно для ви
дов, не представляющих непосредственной опасности сельскому хозяйству или
здоровью людей.
Исторически сложилось так, что, за исключением узкого круга специалис
тов, в Европейском экономическом сообществе ранее проблеме инвазионных
видов внимания практически не уделялось. Однако в конце 1990 х годов пони
мание опасности биологических инвазий в Европе возросло в связи с явными
негативными воздействиями, сведения о которых опубликованы в региональ
ных экологических обзорах [Europe's environment..., 1995, 1998, 2003]. В 1998
году в Стратегии по сохранению биоразнообразия инвазия чужеродных видов
оценена как важная экологическая проблема [European Commission, 1998]. В
марте 2002 года Совет Европы по окружающей среде признал, что интродукция
инвазионных чужеродных видов является одной из главных зарегистрирован
ных причин потери биологического разнообразия, а также серьёзного урона эко
номике и здоровью населения [European Commission, 2002].
Совет Европы поддержал действия, включающие в качестве приоритетных
меры по предотвращению появления инвазионных видов, а также меры, при
званные уничтожать эти виды в том случае, если инвазия всё таки произошла.
В 2002 году была принята Стратегия по инвазионным видам Европы. Страте
гия по биологическому разнообразию ЕЭС [European Commission, 1998] гла
сит: «Присутствие или интродукция чужеродных видов или подвидов могут
потенциально вызвать неустойчивость и изменения в экосистемах. При этом воз
действие на природное биоразнообразие может стать необратимым ввиду гибри
дизации или конкуренции. ЕЭС должно принять меры предосторожности и пре
дотвратить пагубное влияние чужеродных видов на экосистемы, приоритетные
виды и их местообитания, а также установить, насколько это возможно, контроль
и управление для сокращения ущерба, который они причиняют».
Несмотря на эти усилия, практические программы и координация по инва
зионным видам в Европе отстают от других регионов мира. Трудности возника
ют уже при стандартизации статуса чужеродного вида. В национальных масш
табах исследователи обычно принимают во внимание региональные данные, и
классификация степени натурализации заносных видов в разных странах мо
жет различаться. Особенно это касается подвидов, гибридов, реинтродуциро
10
ванных видов, а также природных видов и популяций, естественно расширяю
щих свой ареал. Основные принципы классификации статуса вида предложе
ны не столь давно [IUCN..., 2000], однако происхождение давно интродуциро
ванных видов растений и животных часто неясно. Европейские страны разно
родны по степени подверженности биологическим инвазиям, и в ряде случаев
это ограничивает осознание ими риска, который они в состоянии причинить
соседним государствам. Наиболее агрессивные чужеродные виды в разных стра
нах Европы различны, так что согласованные акции следует планировать в суб
региональном масштабе. Наконец, чужеродный вид в одной стране может быть
аборигенным в другой стране Европы.
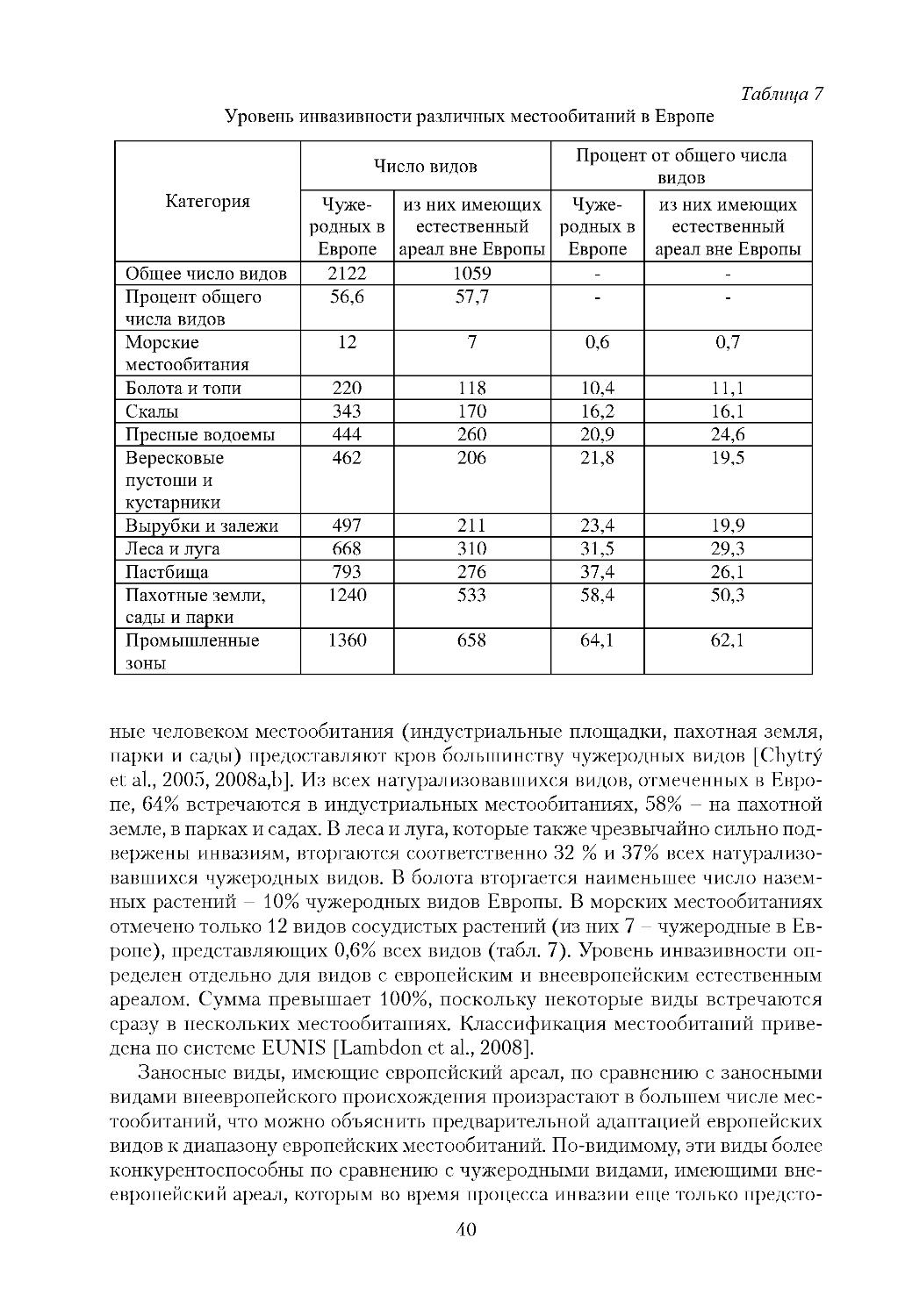
В Европе эффективному контролю за инвазией чужеродных видов препят
ствовали: 1) отсутствие мониторинга заносимых видов с достаточно частыми
интервалами; 2) отсутствие средств информирования и правильность иденти
фикации новых находок; 3) отсутствие осознания риска и оценки вероятности
превращения того или иного заносного вида в инвазионный. Информация о
распространении инвазионных чужеродных видов в Европе неполная, а то, что
доступно, рассеяно во множестве опубликованных или неопубликованных от
чётов, а также различных базах данных. Предупреждение вторжений чужерод
ных видов затруднено в связи с тем, что доступ к информации относительно их
предыдущего инвазионного потенциала (один из лучших показателей того, ста
нет ли новый вид агрессивным) в основном, малодоступен. Ключевая рекомен
дация Стратегии по инвазионным видам в Европе -- развитие региональной
инвентаризации заносных видов, зарегистрированных в природе [Council of
Europe, 2003].
Интерес к изучению чужеродного компонента флор Восточной Европы ак
тивизировался в 1970 е годы, однако долгое время исследования носили локаль
ный характер. Переход к целенаправленному изучению региональных адвен
тивных флор наметился в середине и конце 1980 х годов, продолжается он и
ныне. Более 700 адвентивных (заносных) видов выявлено в настоящее время в
составе флор стран Балтии, Беларуси, Украины и ряда регионов РФ: Ленинг
радской и Московской областей, Республики Удмуртия. 400--700 видов обна
ружено во флорах 12 регионов, 200--400 -- во флорах 30 регионов. Фрагментар
но изучены Уральская и Гурьевская области Казахстана, Архангельская область
РФ. Активные флористические исследования ведутся в Смоленской области,
ранее относившейся к малоизученным (Решетникова, 2003).
Сделано несколько крупных обзоров по вопросам антропогенной трансфор
мации флоры (Горчаковский, 1979; Малышев, 1981; Туганаев, Пузырев, 1988;
Игнатов и др., 1990; Протопопова, 1991; Березуцкий, 1999; Хмелев, Березуцкий,
2001; Миркин, Наумова, 2001; Морозова, 2003а; Папченков, 2003). Итоги изу
чения адвентивной флоры регионов Восточной Европы за последние 30 лет
подведены большой группой авторов: Н.В. Абрамов (Йошкар Ола), Л.М. Абра
мова (Уфа), О.В. Бакин (Казань), Е.А. Борисова (Иваново), М.А. Борисова, Н.А.
Тремасова (Ярославль), В.Д. Бочкин, С.Р. Майоров, А.П. Сухоруков (Москва),
З.А. Гуджинскас (Вильнюс), В.П. Дедков, И.Ю. Губарева (Калининград), О.Н.
Демина, Д.В. Вахненко (Ростов на Дону), В.А. Костина (Мурманская обл.), А.В.
Кравченко (Петрозаводск), А.П. Лактионов, В.Е. Афанасьев (Астрахань), В.А.
11
Мартыненко, Б.И. Груздев (Сыктывкар), А.А. Нотов (Тверь), С.А. Овеснов
(Пермь), Н.Н. Панасенко (Брянск), В.И. Попов (С. Петербург), В.В. Протопо
пова (Киев), А.Н. Пузырев (Ижевск), В.А. Сагалаев, Д.Е. Матвеев (Волгоград),
Е.М. Тарасова (Киров), А.С. Третьякова (Екатеринбург), Л.В. Хорун (Тула), Н.Н.
Чаадаева (Орел), Н.В. Швыдкая, Т.Г. Яненко (Краснодар) и опубликованы в
материалах международной конференции «Изучение флоры Восточной Евро
пы: достижения и перспективы» [2005].
База данных по адвентивной флоре Восточной Европы (AliS), включающая
1148 видов, создана в институте географии РАН (Морозова, 2003б). Стали дос
тупны электронные источники по адвентивной флоре Орловской области (http:/
/botanika.boom.ru, автор сайта А.В. Сотников) и Республики Татарстан. Подго
товлен электронный вариант списка адвентивного компонента флоры Тверс
кой области. Этот список доступен в интернете (Нотов, 2000). Создана основа
сайта по инвазионным видам на ботаническом сервере биологического факуль
тета МГУ (куратор сайта -- С.Р. Майоров). Тем не менее информационные базы
развиты крайне недостаточно и требуют совершенствования.
Диссертационные работы защищены по адвентивной флоре ряда регионов
Средней России: Ивановской, Калужской, Липецкой, Тверской, Тульской об
ластей и Мордовии [Малышева, 1980; Вьюкова, 1985; Борисова, 1993, 2008; Ста
родубцева, 1995; Хорун, 1999; Бармин, 2000; Березуцкий, 2000; Григорьевская,
2003; Маркелова, 2004; Крылов, 2008; Швецов, 2008]. Тверская и Ивановская
области изучены в этом отношении особенно тщательно: по результатам изуче
ния адвентивной флоры Тверской области защищены две кандидатские диссер
тации [Малышева, 1980; Маркелова, 2004], Ивановской области -- кандидатс
кая и докторская диссертации [Борисова, 2003; 2008]. В работе Е.А. Борисовой
привлечен большой материал по заносному компоненту флор Владимирской и
Костромской областей РФ.
Флора транспортных путей специально изучена в Московской и Ярославс
кой областях [Чичев, 1985; Борисова, 2002].
Адвентивная фракция флор городов явилась предметом диссертационных
исследований в Москве, Санкт Петербурге, Брянской и Ярославской областях
[Попов, 2000; Швецов, 2008]. Урбанофлоры и их динамика с характеристикой
адвентивного компонента исследованы в десятке городов [Ильминских, 1982,
1986; Григорьевская, 2000; Панасенко, 2002; Раков, 2003, и др.]. Имеются много
численные работы по адвентивной флоре заповедных территорий, парковым
интродуцированным видам, сегетальному потенциалу адвентивных видов [Гор
чаковский, Козлова, 1998; Усадебные парки..., 2003].
Данные по биологии адвентивных видов приводятся в региональных «Фло
рах...» и «Определителях...» [Александрова и др., 1996; Сосудистые растения
Татарстана, 2000; Казакова, 2004].
Адвентивная фракция флоры -- чрезвычайно динамичная система. Процесс
пополнения флористических списков новыми находками идет значительно бы
стрее, чем издаются «Флоры...» и «Определители...». Так, во «Флоре Восточной
Европы» отсутствуют Galega orientalis, Artemisia selengensis и другие виды.
Рассуждать об уровне видового богатства регионов Восточной Европы чрез
вычайно сложно, так как этот показатель определяется целым рядом объектив
12
ных и субъективных факторов. Кроме того, чем больше территория обзора, тем
меньше в составе ее флоры доли адвентивных видов. Например, М.А. Березуц
кий [1999, 2000] показал, что из 1219 видов аборигенной флоры южной части
Приволжской возвышенности на других территориях в качестве адвентивных
видов встречаются не менее 698.
Изменения показателей богатства адвентивной фракции флор зависят от сте
пени исследованности территории, ее хозяйственной освоенности и характера
взаимодействия с другими регионами, «пограничных эффектов», когда прогресси
рующие виды дают количественный всплеск на границах природных зон, и т.п.
Существует обоснованное предположение, что уровень видового богатства
региональных адвентивных флор падает с северо запада на юго восток, следо
вательно, основные направления заноса видов внутри территории Bосточной
Европы -- южное, юго восточное и юго западное, хотя существует и весьма ог
раниченная по объему группа видов, мигрирующих с севера на юг.
Отдельного внимания заслуживает проблема прогрессирующих видов. На
личие «пограничных эффектов» приводит к тому, что в одной части террито
рии ряд видов оказываются аборигенными, в другой -- адвентивными. Нередко
эти виды расселяются по антропогенно нарушенным местообитаниям, поэтому
отделить прогрессирующие виды от адвентивных не всегда представляется воз
можным [Игнатов и др., 1990].
Информации о пространственной динамике вида и его инвазионном потен
циале на настоящем этапе изучения адвентивной флоры уже недостаточно.
Необходимо перейти на более высокий этап и рассматривать процесс адвенти
зации флор в историческом аспекте путем создания ретроспективных баз дан
ных, отражающих динамику натурализации адвентивных видов за последние
200--250 лет. В историческом аспекте изучены Калужская и Московская облас
ти, Республика Мордовия, Тверская и Тульская области (Игнатов и др., 1990;
Хорун, 1999; Бармин, 2000; Маркелова, 2004; Крылов, 2008). Подобный подход
позволяет выявить многолетнюю динамику адвентивных флор, создать основу
для корректного повидового сравнения с аналогичными флорами и их истори
ческой динамикой и построения модели флорогенетических тенденций.
Разработка и реализация на национальном уровне системы мероприятий по
предотвращению неконтролируемого распространения чужеродных видов и
ликвидации его последствий отнесена к приоритетным направлениям деятель
ности по обеспечению экологической безопасности в рамках Экологической
доктрины Российской Федерации (одобрена распоряжением 1225 р Прави
тельства РФ от 31 августа 2002 г.).
Необходимость интенсификации работ по биологическим инвазиям чуже
родных видов в России определяется и рядом особенностей нашей страны, что,
в частности, затрудняет экстраполяцию результатов зарубежных исследований.
Специфика России в этом отношении состоит в следующем [Дгебуадзе, 2002]:
1. Большая территория страны при фактическом отсутствии внутреннего
контроля благоприятствует переносу организмов за пределы их естественного
ареала.
2. История России (особенно новейшая) насыщена войнами континенталь
ного и регионального характера, сопровождавшихся интенсивными перевозка
13
ми военных и гражданских грузов и перемещениями множества людей (воен
ных и беженцев) между сторонами, участвующими в конфликтах.
3. Длительное время и в больших масштабах в России (СССР) осуществля
лась политика по расселению (акклиматизации) организмов с целью повыше
ния продуктивности экосистем и получения новых продуктов.
4. Относительно высокая активность по строительству крупных сооружений
(дорог, каналов, водохранилищ, мостов, тоннелей), которые могут служить пу
тями для расселения организмов.
5. Относительно высокая активность по созданию новых местообитаний
(агроценозов, водохранилищ, мегаполисов и т.д.) для организмов, включая чу
жеродные виды.
6. Относительно высокий уровень торговых перевозок, включающих обмен
сельскохозяйственной продукцией, древесиной, жидким топливом и т.д., что
способствует обмену живыми организмами.
7. Относительно слабый контроль за переносом чужеродных организмов че
рез государственную границу.
8. Неразвитое законодательство в отношении проведения интродукций и
случайного заноса организмов из других стран.
9. Слабое развитие информационного обеспечения мониторинга инвазион
ных видов. В России создано пока 2--3 базы данных по всем группам организ
мов. Для сравнения: в США только по инвазионным растениям создано 34 базы
данных.
10. Сравнительно широкая распространенность среди населения увлечений,
связанных с домашним содержанием и разведением экзотических растений и
животных (в частности, аквариумных животных и растений), часть из которых,
оказавшись в природных условиях, превращаются в типичные инвазионные
виды.
Воздействие каждого заносного вида трудно предсказуемо ввиду множества
связанных с ним параметров. Чужеродные виды: 1) конкурируют с абориген
ными видами; 2) упрощают структуру фитоценозов; 3) выполняют роль новых
растений хозяев для различных паразитов и возбудителей заболеваний; 4) гиб
ридизируют с аборигенными видами; 5) вытесняют аборигенные виды из есте
ственных фитоценозов.
В настоящей работе обобщены данные по биологическим особенностям ин
вазионных видов флоры Средней России и динамике их расселения во вторич
ном ареале. Площадь Средней России >900 тыс. км2, то есть равна Франции,
ФРГ и странам Бенилюкса вместе взятым. В районировании, принятом во Flora
Europaea, это бульшая часть Центральной России -- Rs(C) -- и лишь правобере
жье Саратовской области входит в юго восточный район -- Rs(W) (рис. 3).
Рассматриваемая территория включает в себя области, традиционно отно
симые во флористической литературе к средней полосе Европейской России
[Маевский, 2006]. Она охватывает территорию от государственной границы
Российской Федерации на западе до Волги на востоке, Тверской, Ярославской
и Костромской областей на севере и Белгородской, Воронежской и Саратовс
кой областей на юге. Таким образом, эта территория охватывает земли следую
щих административных единиц (рис. 3):
14
Даны оценка экономическому ущербу и рекомендации по возможному ис
пользованию инвазионных видов. Цель проекта состоит в том, чтобы ознако
мить административные органы и частных лиц с базирующимися на научной
основе методами контроля наиболее злостных и широко распространившихся
чужеродных видов с целью уменьшения их обилия, сужения ареала, а также
предотвращения дальнейшего продвижения. Это начало серьезного обсужде
ния необходимости учета биологии и экологии заносных видов для выработки
эффективных мер борьбы с фитоинвазиями.
Мы надеемся, что настоящая книга на практике поможет всем, кто сталкива
ется с острой проблемой наступления агрессивных чужеродных сорняков.
Инвазионные виды Средней России авторы понимают как чужеродные
(alien) виды, которые: а) внедрились в естественные (леса, поймы рек) или по
луестественные (сбитые боровые пески, зарастающие торфяники) фитоцено
зы; б) являются кенофитами, то есть начали формировать вторичный ареал по
зднее ХVI века, и в настоящее время продолжают активное расселение; в) встре
Рис. 3. Карта территории Средней России
Белгородская область,
Владимирская область,
Воронежская область,
Ивановская область,
Калужская область,
Костромская область,
Липецкая область,
Республика Мордовия,
Московская область,
Нижегородская область,
Орловская область,
Пензенская область,
правобережье Самарской области,
правобережье Саратовской области,
Смоленская область,
Тамбовская область,
правобережье республики Татарстан,
Тверская область,
Тульская область,
Ульяновская область,
Чувашская республика,
Ярославская область
15
чаются не менее чем в 75% административных единиц, входящих в изучаемый
регион. Археофиты в число исследуемых видов не включены. Случаи, когда
инвазионный процесс наблюдается лишь на ограниченной территории, авторы
относят к локальным инвазиям; например, бурная натурализация дальневосточ
ного вида Adenocaulon adhaerescens и кавказской Veronica filiformis наблюдается
пока только в Московском регионе, а инвазионные популяции североамерикан
ской Robinia pseudoacacia отмечены лишь в южных районах Средней России.
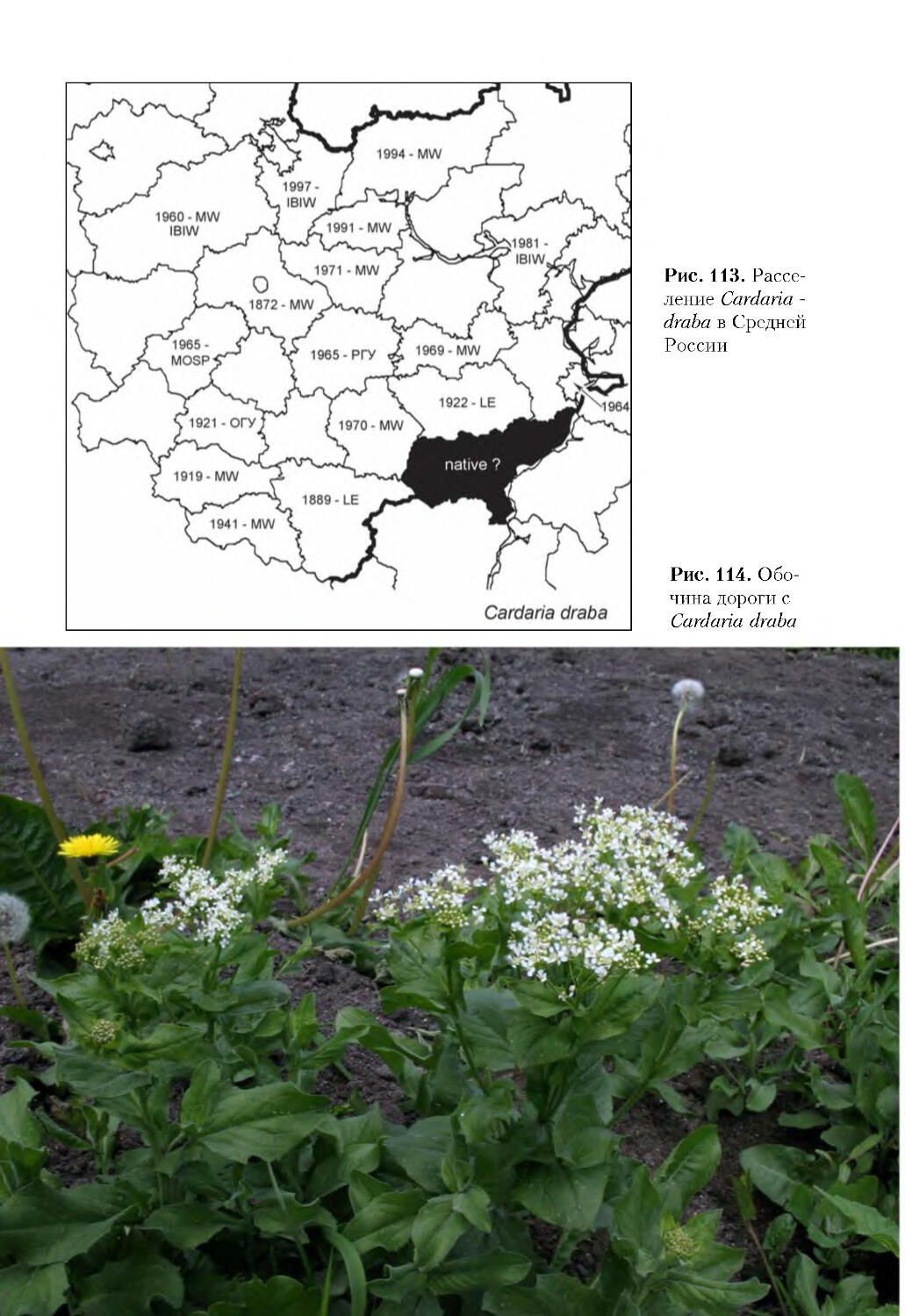
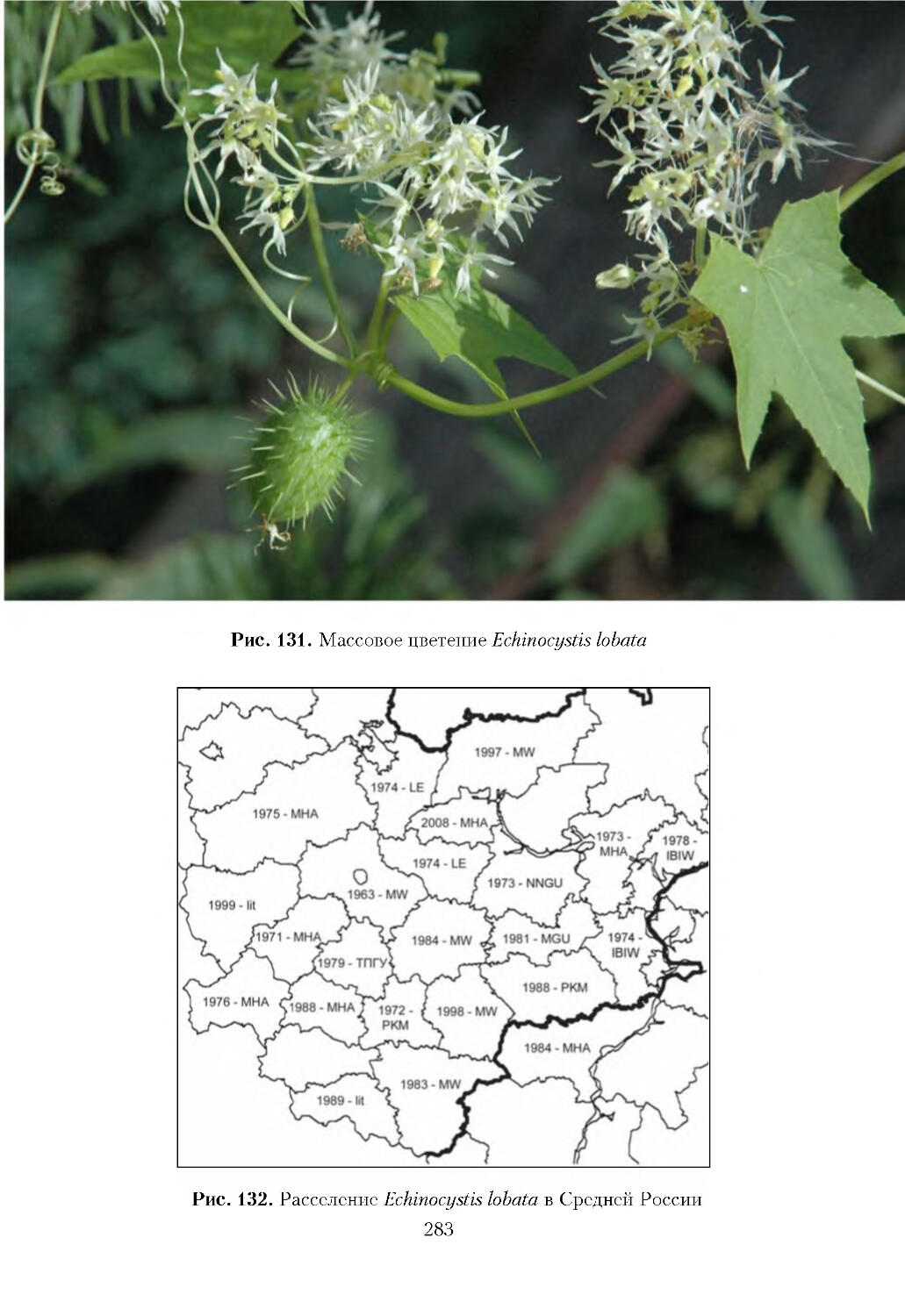
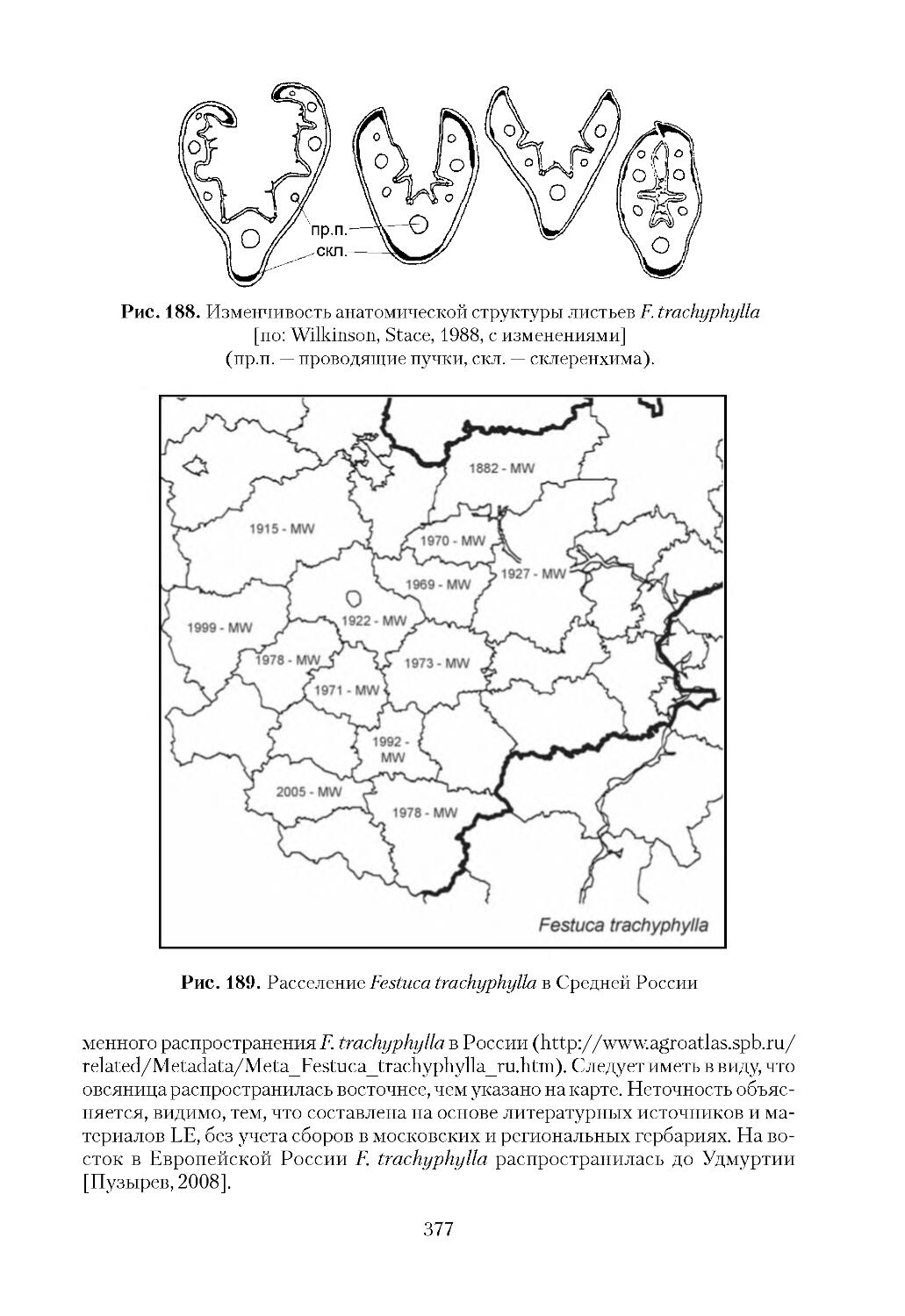
К широко распространенным инвазионным видам отнесено 52 таксона: Acer
negundo, Acorus calamus, Amaranthus albus, A.retroflexus, Anisantha tectorum, Atriplex
tatarica, Heraсleum sosnowskyi, Ambrosia artemisiifolia, Amelanchier spicata, A.
alnifolia, Aster Ч salignus, Bidens frondosa, Cardaria draba, Chamomilla suaveolens,
Crataegus monogyna, Cyclachaena xanthiifolia, Echinocystis lobata, Elaeagnus
angustifolia, Elodea canadensis, Elsholzia ciliata, Epilobium adenocaulon, E.
pseudorubescens, Conyza canadensis, Erygeron annuus, Erucastrum gallicum, Euphorbia
peplus, Festuca trachyphylla, Fraxinus pennsylvanica, Galinsoga ciliata, G.parviflora,
Helianthus tuberosus, Hippophae rhamnoides, Hordeum jubatum, Impatiens glandulifera,
I. parviflora, Juncus tenuis, Lepidium densiflorum, Lupinus polyphyllus, Oenothera
biennis, Oxalis stricta, Poa supina, Populus alba, Puccinelia distans, Reynoutria х
bohemica, R.japonica, Senecio viscosus, Solidago canadensis, S.gigantea, Sorbaria
sorbifolia, Symphytum caucasicum, Sisymbrium wolgense, Xanthium albinum.
Представленные в книге карты распространения инвазионных видов состав
лены на основе гербарных материалов из следующих хранилищ: LE, MW, MHA,
PKM, KLH, KOKM, IBIW, GMU, NNGU, MGU, TVBG, MOSP, ОГУ (гербарий
Орловского университета им.Н.В. Хитрово), РГУ (гербарий Рязанского универ
ситета), ИГУ (гербарий Удмурдского университета), ТГПУ (гербарий Тульско
го университета)
По сравнению со списком инвазионных видов Северо Запада России [Гель
тман, 2003], насчитывающим 22 таксона, в Средней России натурализовавших
ся видов в 2 раза больше. При этом, однако, авторы не включили в список инва
зионных видов Средней России Rudbeckia laciniata, Aronia mitchurinii и Rosa
rugosa, поскольку зарегистрированы они пока менее чем в 75% регионов.
По видимому, тенденция к увеличению числа инвазионных видов вдоль
широтного профиля действительно имеет место, поскольку на Украине зарегис
трировано 95 инвазионных видов, 66 из которых -- кенофиты, а 29 -- археофиты
[Протопопова, Мосякiн, Шевера, 2002]; в Польше -- 138 инвазионных видов
(123 кенофита и 15 археофитов) [Solarz et al., 2005].
Как и следовало ожидать, ведущее положение в инвазионном компоненте
(51%) занимают кенофиты североамериканского происхождения. Виды из Сре
диземноморья, Центральной и Восточной Азии играют меньшую роль и состав
ляют по 8--10% общего числа видов.
На Украине и в Польше североамериканские виды также преобладают
(45% и 35% соответственно), однако при этом средиземноморские и централь
ноазиатские виды играют большую роль: первые в инвазионном комплексе Ук
раины составляют 26%, вторые -- 15%.
При сравнительном анализе кенофитов Северо Запада России, Средней Рос
сии, Украины и Польши выделяется группа видов (ядро инвазионного комп
16
лекса), которые активно натурализуются во всех этих регионах. К ним относят
ся 9 видов, 7 из которых занесены в Европу из Америки: Acer negundo, Ambrosia
artemisiifolia, Conyza canadensis, Galinsoga ciliata, G. parviflora, Helianthus tuberosus,
Echinocystis lobata; 2 -- из Центральной Азии: Impatiens glandulifera и I. parviflora.
Анализируя состав инвазионных видов и их расселение, следует отметить
следующее:
1) для распространения по всей территории Средней России большей части
видов потребовалось всего несколько десятилетий;
2) начало расселения практически всех видов плохо документировано как
гербарными материалами, так и в публикациях;
3) существенная часть инвазионных видов во флоре Средней России есть
результат преднамеренной интродукции. Лишь небольшая часть видов перешли
к стремительному расселению почти сразу (Heracleum sosnowskyi). Как правило,
виды достаточно долго существовали только в культуре и лишь позднее стали
активно расселяться (например, Acer negundo или Fraxinus pennsylvanica). Среди
них немало растений, культивировавшихся в коллекциях ботанических садов:
Conyza сanadensis, Echinocystis lobata, Bidens frondosa, Impatiens parviflora,
Chamomilla suaveolens, Galinsoga parviflora и др.;
4) почти все инвазионные виды флоры Средней России проникли на эту тер
риторию из вторичных центров распространения, расселение представляет лишь
часть общего процесса их инвазии в Восточной Европе или в Старом Свете в
целом.
Авторы надеются, что инвентаризация и карты ареалов, приведенные в этой
книге, обеспечат квалифицированную справочную систему по инвазионным
чужеродным видам Средней России, доступной для менеджеров по охране при
роды, законодателей, исследователей, студентов и всех заинтересованных лиц.
Документация текущих инвазий, прогнозирование новых участков вторжения
и предотвращение инвазий жизненно важны для защиты биологического раз
нообразия в России.
Авторы выражают глубокую признательность коллегам, предоставившим
ценные сведения по распространению инвазионных видов в различных регио
нах России: В.С. Новикову, А.А. Нотову, А.В. Щербакову, Н.М. Решетниковой,
А.В. Крылову, В.Г. Папченкову, В.К. Тохтарю, А.В.Полуянову, И.Л. Мининзону.
17
Глава 1
ПОНЯТИЕ «ИНВАЗИОННЫЙ ВИД»
И ИСТОРИЯ ЕГО ФОРМИРОВАНИЯ
Приступая к рассмотрению этого вопроса, авторы решили сразу оговориться,
что термин «инвазия» и его производные -- слова для русскоязычной биологи
ческой литературы относительно новые, происходящие из западных источников.
В русском языке термин «инвазионный вид» является калькой с английского
словосочетания invasive species. История же становления терминологическо
го аппарата, включающего понятие «инвазионный вид», как в России, так и на
Западе сложна и запутанна. Авторы, не претендуя на полноту охвата бесчис
ленных источников, обращают внимание читателей на некоторые проблемы,
возникающие при определении понятия. В условиях терминологических раз
ногласий, которые имеются между западной и отечественной школами и внут
ри каждой из них, трудно надеяться, что существует исчерпывающее опреде
ление понятия «инвазионный вид», которое снимет все трудности выделения
и описания группы этих видов. Достаточно отметить, что в западной школе
изучением инвазионных видов занимается специальная дисциплина, опреде
ляемая как инвазионная экология растений, а в отечественной традиции эти
виды изучаются флористами в составе адвентивных флор регионов и отдель
но специалистами других направлений -- биохимиками, экологами и т.п. с точки
зрения биологии и экологии таких видов. Тем не менее, остаются вопросы: что
означает термин «инвазионный вид», как он определяется в отечественной и
западной школах, какова история происхождения и изменения термина?
Интересно отметить, что словосочетание «отечественная школа» для изу
чения заносных флор весьма условно. В России в начале XX века появился
интерес к этой проблеме благодаря работам немецких, польских и финских
ботаников, и именно из их работ взяты ставшие общепринятыми классифика
ционные термины [Thellung, 1905; 1918--1919; Holub, Jirasek, 1967; Schroeder,
1969]. Однако впоследствии произошло разделение исследователей на цент
рально европейских [Pyљek et al., 2004], чья школа получила свое дальней
шее развитие в России, и западных, школа которых сейчас продолжает актив
но развиваться в Европе [Williamson, 1996] и носит название инвазионная эко
логия растений. Новым является не только название: стандартизация
терминологии, которая в России в целом завершена в 1970--1980 годах, в ин
вазионной экологии происходит в настоящее время [Richardson et al., 2000].
Так как термин «инвазионный вид» относится именно к современной запад
ной школе, возникает необходимость постоянно обращаться к сравнению объе
18
ма и сути термина с категориями, применяемыми в отечественной традиции,
и таким образом попытаться уточнить место этой группы видов в принятых в
России классификационных схемах.
В самых современных российских работах термин «инвазионный вид» встре
чается чаще, чем, например, «агриофит». И именно существование такой тен
денции обусловливает актуальность теоретического осмысления термина.
Как правило, совокупность видов, определяемых как «инвазионные», яв
ляется частью обширного заносного или адвентивного элемента флоры, среди
которого они выделяются, прежде всего, агрессивностью, то есть способнос
тью быстро распространяться и внедряться в различные типы ценозов, в том
числе и ненарушенные. Таким образом, для начала нужно определить, что та
кое заносная, или адвентивная, флора.
Исторически потребность в выделении заносного, пришлого элемента фло
ры появилась с того момента, когда человек начал осознавать степень его соб
ственного влияния на флору, а последствия ускоренного им же флорогенеза ста
ли проявляться в сжатые периоды времени. Такое понимание развивалось в на
правлении выделения из всего состава флоры того ее компонента или части,
которая изменялась и адаптировалась быстрее всего. В середине XIX века в про
мышленно развитой Европе появились первые работы, касающиеся растений,
названных «синантропные», «пришлые», «чужеземцы», «колонисты» и т.д.
[De Candolle, 1855; Touchy, 1857; Smith, 1867; Watson, 1870; Martindale, 1876;
Brown, 1878]. Наглядным и проще всего фиксируемым внешним проявлени
ем процесса изменения флоры стало наличие эмиграционных и иммиграци
онных потоков видов, перемещавшихся внутри регионов и выходивших за их
пределы. Тем не менее на этапе разделения этих потоков на группы возникли
до сих пор существующие терминологические разногласия, которые касают
ся, с одной стороны, круга изучаемых объектов, а с другой -- их классифика
ции. Попытки решить эти проблемы предпринимались с начала ХХ века, од
нако до сего дня каждый исследователь в той или иной степени перестраивает
терминологическую основу в зависимости от целей и методов работы, разме
ров и изученности территории, а также субъективного понимания вопроса.
Понятие «адвентивная флора» (или «адвентивный элемент», или «компо
нент флоры») существует в системе представлений, в которую значение терми
на иерархически включено как часть флоры территориальной единицы. В та
ком понимании флора может распадаться на аборигенный (автохтонный) и ад
вентивный (аллохтонный) элементы флоры.
Автохтонный элемент флоры выражает совокупность аборигенных видов
данной территории [Юрцев, Камелин, 1991], то есть видов, возникших в той же
местности, в которой существуют в настоящее время, и составляющих наибо
лее древнее ядро ее флоры [Краткий словарь..., 1993].
В понимании западных авторов категория аборигенных видов native plants
или indigenous plants определяется как таксоны, произошедшие на данной
территории без влияния человека или распространившиеся без прямого или косвен
ного участия людей по территории, на которой они являются природными [Pyšek
et al., 2004]. Согласно мнению авторов, такое определение исключает продукты гиб
ридизации с привлечением заносных родителей из состава природной флоры.
19
Аборигенный элемент флоры формально пересекается с адвентивным с од
ной стороны, через понятие «археофит» [термин из Rikli, 1903], а с другой --
через виды, натурализация которых произошла в последнее время и была в той
или иной форме зафиксирована документально. Промежуточное положение
занимают и прогрессирующие виды в понимании Н.В. Козловской [1978], рас
ширение ареала которых могло быть связано с естественными причинами. Та
ким образом, учитывая относительность разграничения адвентивного и абори
генного компонентов, авторы понимают последний как часть флоры, заносное
происхождение видов которой не установлено для данной территории.
Определение антропофильного (синантропного, антропофитного) элемента
флоры А. Теллунгом [Thellung, 1905, 1915, 1918--1919], а вслед за ним и другими
авторами, послужило основанием для выделения более подчиненных по отно
шению к антропофильным заносных видов. Сам А. Теллунг выделял виды си
нантропных сообществ, которые делил на апофиты (синантропные растения,
происходящие из местных естественных растительных сообществ) и антропо
хоры (синантропные растения чужеземного происхождения). В общем то, та
кая точка зрения используется до сегодняшнего дня, хотя термин антропохор
во многих случаях заменяется вследствие его неоднозначного понимания, на
что указал еще Я. Ялас [Jalas, 1955].
Адвентивная часть флоры в такой системе понятий является самостоятель
но изучаемой единицей флоры. Стимулами к выделению такой категории слу
жат, с одной стороны, факт заноса на определенную территорию не обитавших
на ней ранее видов, а с другой -- степень участия человека в этом процессе и
последствия этого участия.
Адвентивный элемент флоры чаще всего определяется в отечественной ли
тературе как совокупность видов растений, не свойственных местной флоре, за
нос которых на данную территорию не связан с естественным ходом флорогене
за, а является результатом прямой или косвенной деятельности человека.
В западноевропейской литературе аналогом адвентивных флор является
совокупность видов, называемая alien plants, exotic plants, introduced plants,
non native plants, non indigenous plants и определяемая как виды растений,
которые присутствуют на данной территории благодаря прямому или кос
венному влиянию человека или распространяются без помощи человека с тер
ритории, на которой являются заносными [Pyšek et al., 2004].
Термин «адвентивный» впервые применил Де Кандолль [De Candolle, 1855],
и он получил широкое распространение изначально в Европе, а затем и в Рос
сии. Однако на Западе, в отличие от России, где его значение принимается од
нозначно, термин «адвентивный» лишь иногда используется для обозначения
разных групп видов, а в последнее время почти полностью вытеснен из научно
го обихода словосочетанием alien plants.
Разработке, оценке и уточнению классификации адвентивной фракции фло
ры посвящено значительное число работ российских и зарубежных авторов.
Практически все классификации ставят целью выделение степени и формы за
висимости расселения и распространения заносных растений от антропогенно
го влияния. Неоднозначность подходов и терминологий со всей очевиднос
тью показывает многогранность человеческого воздействия на флору и труд
20
ность отделения его от природных факторов. Тем не менее, существуют два
вполне оформленных направления классификации, основоположниками ко
торых можно считать А.Теллунга и Я.Яласа [Thellung, 1905, 1915, 1918--1919;
Jalas, 1955]. Опираясь на работы А.Теллунга, можно заключить, что первое
направление предполагает разделение преднамеренного и непреднамеренно
го способов заноса растений на новые территории, а также времени и итогов
этого заноса. Все эти характеристики первоначально укладывались в одну си
стему терминов, что и привело к двузначному определению некоторых поня
тий, таких как «неофит», «антропохор», позднее уточненных В.Крэ и Я.Яла
сом. Дальнейшее развитие более или менее обширных вариантов классифи
кационных систем привело к вычленению отдельных, наиболее важных
факторов, по которым можно оценивать процесс и итог заноса видов на новые
территории. Например, Д. Лузли [1953] выделил среди адвентивных расте
ний виды натурализовавшиеся, приспособившиеся и случайные, подойдя тем
самым к пониманию степени натурализации как отдельной характеристики
адвентивных растений.
Ф.Г. Шредер [Schroeder, 1969], обобщив такого рода данные, пришел к выво
ду о необходимости разграничения классификационных построений по трем
основным принципам: времени иммиграции, способу иммиграции и степени
натурализации. Такое разграничение послужило основой для ряда современных
классификаций и полностью оформило первое направление, оценивающее вре
менные и пространственные особенности распределения адвентивных расте
ний по вновь занятой территории.
Второе направление, основоположником которого является Я.Ялас, напро
тив, группирует растения по их способности произрастать на местообитаниях,
в различной степени видоизмененных хозяйственной деятельностью человека.
Система Я.Яласа, во первых, сыграла роль в оценке степени антропотолерант
ности различных экосистем и их компонентов, а во вторых, дала возможность
разработать экологические шкалы применительно к отдельным антропогенным
средам, например урбанизированным ландшафтам. В таком частном приложе
нии она послужила основой для классификации видов урбанофлор по их отно
шению к антропогенному воздействию [Ильминских, 1988].
Во флористических работах применимы системы первого направления, ак
центирующие внимание на особенностях современного пространственного рас
пределения заносных видов по изучаемой территории и на оценке возможных
причин такого распределения через анализ времени, путей и способов заноса
диаспор. При этом чаще всего используются построения А. Теллунга [Thellung,
1905], Я. Корнася (Kornás, 1968], Ф. Г. Шредера (Schroeder, 1969] и их модифи
кации. Мы придерживаемся системы Ф. Г. Шредера с использованием терми
нов, предложенных Н.С. Камышевым (1959], Н.А. Вьюковой (1985], А.В. Чиче
вым (1985] и выделяем по времени заноса археофиты и неофиты, по способу
заноса -- ксенофиты, ксеноэргазиофиты и эргазиофиты, по степени натурализа
ции -- эфемерофиты, колонофиты, эпекофиты и агриофиты (табл. 2). Что каса
ется времени заноса или residence status, то с ее трактовкой археофиты/неофи
ты (или кенофиты) согласны и западные исследователи. Как правило, в Евро
пе рубеж между археофитами и неофитами проводится по дате открытия
21
Америки: 1492 год. В других частях света, например в Австралии, разграниче
ние проходит между таксонами, которые распространились до или после ев
ропейской колонизации [Kloot, 1987]. На Гавайях [Webster, 1992] выделяют
виды, интродуцированные полинезийцами еще до того, как капитан Д. Кук в
1778 году открыл эти острова. Категории по способу заноса в западной клас
сификационной схеме не выделяются, их нет, по крайней мере, в работе Д.Ри
чардсона с соавторами [Richardson et al., 2000]. Англоязычная школа инвази
онной экологии растений предлагает несколько иной подход к определению
инвазионного статуса вида (invasive status), который до некоторой степени
сходен с представлением о степени натурализации [Pyšek et al., 2004]. Как и в
отечественной литературе, по степени натурализации выделяют четыре груп
пы видов, но иначе их называют: casual alien plants, naturalized plants, invasive
plants, transformers (табл. 2). Основной критерий для разграничения первых
трех групп биологический: способность вида к размножению и распростране
нию по новой территории, для четвертой (transformers) -- эколого фитоцено
тический (роль в сообществах). В оригинальном понимании А. Теллунга
(Thellung, 1918--1919] термин casual alien plants был использован для видов,
убежавших из культуры и случайно распространившихся за ее пределами.
Naturalized plants (синоним established plants) не обязательно входят в состав
природных или нарушенных сообществ. Как долго эти виды могут сохранять
ся, зависит от случайных причин, и это может быть выяснено только на прак
тике. Авторы классификационной схемы полагают, что это должен быть пери
од длительностью не <10 лет. Эти виды могут формировать самоподдержива
ющиеся популяции в течение нескольких лет, а затем угасать. Виды, которые
встречаются в местах культивирования, но после прекращения культуры пе
рестают встречаться, представляют специальную категорию, и они могут быть
классифицированы как casual или naturalized. Invasive plants -- это виды, вхо
дящие в группу naturalized plants в качестве подмножества. Их основная от
личительная особенность -- распространение на значительные территории.
Примерная оценка скорости распространения: >100 м за период длительностью
<50 лет для таксонов, которые распространяются семенами; >6 м за 3 года -- для
таксонов, распространяющихся вегетативно. Для двудомных растений, имею
щих семенную репродукцию, установление факта распространения возможно
только после заноса особей обоих полов. По мнению авторов классификации,
таксоны, распространенные прежде, но не распространенные сейчас, могут быть
названы invasive, так как локальное искоренение -- это явный пример реинва
зии. Нам трудно согласиться с этим мнением. По нашему мнению, исчезнове
ние или резкое снижение численности особей вида влечет за собой изменение
степени его натурализации в данный момент времени. В противном случае
нам придется считать инвазионными редкие или даже исчезнувшие сорняки,
которые в прошлом засоряли посевы!
Transformers -- термин исключительно экологический, поэтому сами транс
формеры ближе всего к эдификаторам. Растения относят к трансформерам по
экономическим соображениям, хотя в некоторых случаях их влияние может быть
и выгодным. Выделяют отдельные категории видов трансформеров: 1) чрез
мерные потребители ресурсов (воды -- Tamarix spp., Acacia mearnca; света --
22
Таблица 2
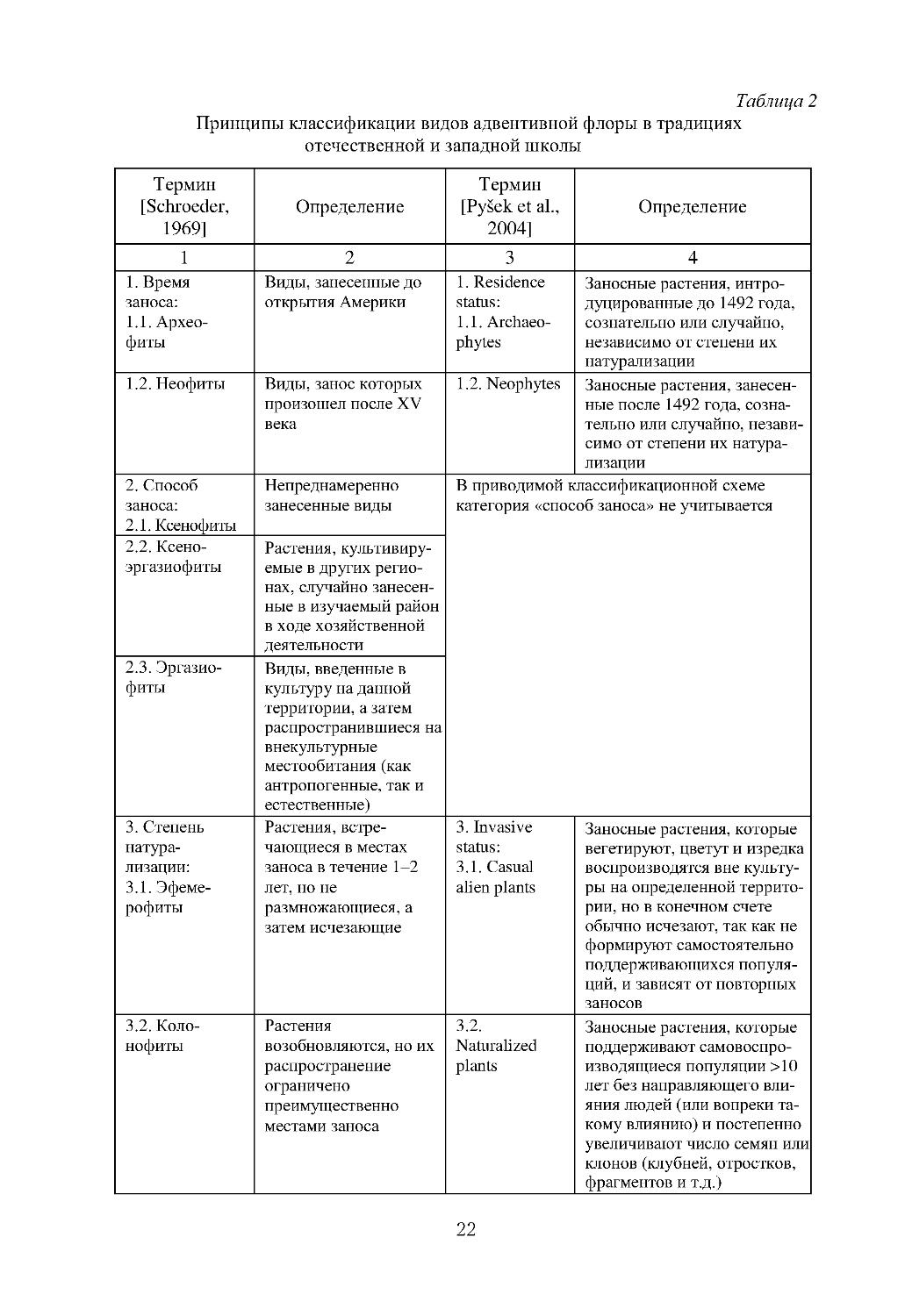
Принципы классификации видов адвентивной флоры в традициях
отечественной и западной школы
Термин
[Schroeder,
1969]
Определение
Термин
[Pyšek et al.,
2004]
Определение
1
2
3
4
1. Время
заноса:
1.1. Архео-
фиты
Виды, занесенные до
открытия Америки
1. Residence
status:
1.1. Archaeo-
phytes
Заносные растения, интро-
дуцированные до 1492 года,
сознательно или случайно,
независимо от степени их
натурализации
1.2. Неофиты Виды, занос которых
произошел после XV
века
1.2. Neophytes Заносные растения, занесен-
ные после 1492 года, созна-
тельно или случайно, незави-
симо от степени их натура-
лизации
2. Способ
заноса:
2.1. Ксенофиты
Непреднамеренно
занесенные виды
2.2. Ксено-
эргазиофиты Растения, культивиру-
емые в других регио-
нах, случайно занесен-
ные в изучаемый район
в ходе хозяйственной
деятельности
2.3. Эргазио-
фиты
Виды, введенные в
культуру на данной
территории, а затем
распространившиеся на
внекультурные
местообитания (как
антропогенные, так и
естественные)
В приводимой классификационной схеме
категория «способ заноса» не учитывается
3. Степень
натура-
лизации:
3.1. Эфеме-
рофиты
Растения, встре-
чающиеся в местах
заноса в течение 1--2
лет, но не
размножающиеся, а
затем исчезающие
3. Invasive
status:
3.1. Casual
alien plants
Заносные растения, которые
вегетируют, цветут и изредка
воспроизводятся вне культу-
ры на определенной террито-
рии, но в конечном счете
обычно исчезают, так как не
формируют самостоятельно
поддерживающихся популя-
ций, и зависят от повторных
заносов
3.2. Коло-
нофиты
Растения
возобновляются, но их
распространение
ограничено
преимущественно
местами заноса
3.2.
Naturalized
plants
Заносные растения, которые
поддерживают самовоспро-
изводящиеся популяции >10
лет без направляющего вли-
яния людей (или вопреки та-
кому влиянию) и постепенно
увеличивают число семян или
клонов (клубней, отростков,
фрагментов и т.д.)
23
Pueraria lobata, Rubus armeniacus; воды и света -- Arundo donax; света и кисло
рода -- Salvinia molesta, Eihhornia crassipes; 2) доноры ресурсов (азота -- Acacia
spp., Lupinus arboreus, Myrica faya, Salvinia molesta; 3) виды, влияющие на вы
жигание растительности (способствующие пожарам -- Anisantha tectorum,
Melaleuca quinquenervia, Melinis mimuiflora; препятствующие пожарам -- Mimosa
nigra); 4) закрепители песчаных почв (Ammophila arenaria); 5) виды, способ
ствующие эрозионным процессам (Andropogon virginicus на Гаваях, Impatiens
glandulifera в Европе); 6) закрепители прибрежных дюн (Rhizophora mangle,
Spartina spp.); 7) аккумуляторы опада и вредных веществ (Eucalyptus spp.,
Lepidium latifolium, Pinus strobus); 8) аккумуляторы солей (Mesembryanthemum
crystallinum, Tamarix spp.) [Richardson et al., 2000]. В отечественной школе по
нятию «трансформер» примерно соответствует агриофит. Но, в отличие от
transformers, агриофитом может быть только заносный вид. По нашему мне
нию, включение в группу transformers видов природной флоры не вполне удач
ное решение, так как вся классификационная схема теряет стройность, нару
шается внутренняя логика.
Основные расхождения в классификационных схемах очевидны из следую
щего лаконичного перечисления:
1) в представленной англоязычной классификации не учитывается способ
заноса видов в исследуемый регион;
2) классификация по степени натурализации (invasive status) иерархична,
по крайней мере invasive plants являются подмножеством naturalized plants, а
transformers, в свою очередь, составляют часть invasive plants. Группы колоно ,
эпеко и агриофитов строго разграничены и не являются частями друг друга;
3) группа видов, которую западные исследователи определяют как
transformers, неоднородна по составу и, согласно приведенному определению,
может включать и виды природной флоры.
Из табл. 2 видна и основная проблема, которая не позволяет корректно оп
ределить термин «инвазионный вид» по отечественной классификации, по
скольку в ней каждая группа видов, выделяемых по степени натурализации,
Таблица 2 (окончание)
1
2
3
4
3.3. Эпеко-
фиты
Заносные растения,
которые
распространяются по
одному или
нескольким типам
антропогенных мест
обитания
3.3. Invasive
plants
Инвазионные виды являются
подмножеством натурализо-
вавшихся (naturalized plants).
Они воспроизводятся, иногда
в большом количестве, на
значительном удалении от
родительских особей, и могут
потенциально распростра-
няться на значительные
территории
3.4. Агрио-
фиты
Растения,
внедрившиеся в
естественные ценозы
3.4. Trans-
formers
Подмножество инвазионных
видов (необязательно занос-
ных), которые в значитель-
ной степени преобразуют
природные экосистемы
24
не является подмножеством предыдущей. Таким образом, нельзя приравнять
invasive species к эпекофитам, как это представляется в таблице. К инвазион
ным видам можно отнести лишь часть широко распространенных эпекофитов
(голоэпекофиты) и агриофиты заносного происхождения, которые в запад
ном понимании относятся к группе transformers. Кроме того, те определения,
которые предпосланы понятию «инвазионный вид», часто оценивают эти виды
не как биологическое явление, а с экономической точки зрения. Например, на
сайте Глобальной программы по изучению инвазионных видов дается такое
определение [Гельтман, 2003]: «инвазионные заносные виды -- это чужерод
ные (non native) организмы, которые наносят или могут нанести урон окру
жающей среде, экономике или здоровью человека».
А как же быть с активно распространяющимися нейтральными или полез
ными видами? К ним относятся [по: Richardson et al., 2000] от 20 до 50% занос
ных видов. Это так называемые «мягкие инвазионные виды», чей средовой или
экономический вред весьма незначителен. На это обращают внимание многие
исследователи, призывая не вводить в определение инвазионных видов кате
горию «экономический ущерб» [Гельтман, 2003]. Для видов реально агрессив
ных, входящих в группу инвазионных, на западе предлагается применить тер
мин transformers, как подмножества заносных растений, которые изменяют ха
рактер, условия, форму и сущность экосистем на значительной территории.
Нам представляется, что для более точного вычленения группы видов, кото
рые можно рассматривать как инвазионные в нашей флоре, необходимо под
робнее рассмотреть такое понятие, как степень натурализации и его западный
аналог invasive status. Степень натурализации служит мерой «внедренности»
вида в растительный покров на данный момент или более ранние периоды, о
которых есть достоверные сведения. Понятие натурализации как итога вне
дрения вида введено А. Де Кандоллем [De Candolle, 1855], и впоследствии его
неоднократно дополняли и изменяли другие исследователи. Это понятие не
разрывно связано с флорогенезом, как способом существования флоры во вре
мени и, по видимому, является закономерным отражением ускоренных чело
веком флорогенетических преобразований. В этом смысле мы понимаем под
степенью натурализации степень адаптации растений к конкретным услови
ям на определенном временном этапе. Последовательная смена этих этапов или
отсутствие таковой в обозримом промежутке времени (например, для недича
ющих интродуцированных растений) обнаруживает способность вида к нату
рализации, то есть к нормальному возобновлению его в естественных расти
тельных сообществах вне зависимости от колебаний климата.
Рассматривая заносную флору региона в настоящее время, то есть простран
ственную ее составляющую, можно считать натурализовавшимися растения,
нормально возобновляющиеся на одном или немногих типах мест обитания,
например колонофиты, геми и голоэпекофиты, отождествляя натурализацию с
ее степенью. Это верно в том случае, если рассматривать видовой состав занос
ной флоры лишь на данный момент. Мы же имеем дело со списками видов раз
ной давности, иногда за несколько столетий. С тех пор многие виды исчезли
совсем, натурализовались или как то изменили степень натурализации. Поэто
му мы считаем правомерным подчеркнуть разделение понятий «степень нату
25
рализации» и собственно «натурализация», понимая последнюю как крайнее
проявление процесса.
Понятие «натурализация» и связанные с ним концепции активно дискути
руются и на Западе. Здесь принят несколько иной подход к определению того
же биологического процесса, который можно назвать концепцией барьеров. При
этом используется представление о лимитирующих факторах, задерживающих
перемещение вида на большие расстояния и расселение его на новой террито
рии. От числа преодоленных барьеров зависит invasive status. Например, если
вид преодолел межматериковый барьер, и встречен единично на другом конти
ненте, он является casual alien, то есть эфемерофитом в нашем понимании. Пре
одоление барьера размножения в новых условиях делает его naturalized, а ак
тивное расселение переводит в подмножество invasive или даже transformers.
Что касается самого термина «натурализация», имеющего богатую историю, то
сейчас европейские авторы склонны именовать это явление invasion process,
имея в виду последовательную смену видами invasion status или степеней нату
рализации. Уход от самого термина натурализация в современных европейских
источниках вызван его расхожестью и крайне неоднозначным пониманием. Эта
проблема подробно обсуждается в работе «Натурализация и инвазия чуже
земных растений: концепции и тенденции» [Richardson et al., 2000].
При сравнении подходов становится очевидным, что даже в отсутствие стан
дартизированной терминологии по степени натурализации видов подразумева
ется тот же процесс и те же четыре группы видов, названных иначе.
Несмотря на многочисленные попытки описать сущность процесса натура
лизации, его внутренний механизм до сих пор не вполне изучен, что не позво
ляет объективно подойти к прогнозам относительно поведения отдельных ви
дов в будущем. Перспективы натурализации в каждом случае оцениваются
субъективно и ориентировочно.
Г. Бейкер [Baker, 1959] определяет четыре стадии жизненного цикла, в ре
зультате прохождения которых растение может считаться натурализовавшим
ся -- вегетативный рост, цветение и образование семян, расселение зачатков на
подходящие местообитания, возобновление на том же или на новом местооби
тании. Тем не менее трудно заранее определить, сможет ли растение пройти все
эти стадии а, если сможет, то через какой промежуток времени это произойдет.
Исследователями приводятся многочисленные факты самого разнообразного
поведения растений после заноса их на определенную территорию. П.И. Бело
зеров [1957, 1960], Е.В. Дорогостайская [1972] приводят примеры видов, «...на
турализация которых происходит очень легко даже на Крайнем Севере, или, мо
жет быть, им не нужна акклиматизация, а натурализация их сводится к одному
только расселению». Ряд исследователей [Назаров, 1927; Erkamo,1946; Бочкин,
1991 и др.] указывают на влияние периодов нестабильности и войн на расселение
растений. Однако последствия такого расселения различны и, как указывают те
же авторы, могут привести к натурализации или, наоборот, исчезновению вновь
расселившихся видов после прекращения военных действий.
Ридли [Ridley, 1930], М.С. Игнатов [1989], А.К. Скворцов, Ю.К. Майтули
на (Виноградова) [1989] и ряд других авторов обращают внимание на значи
тельную разницу во времени между первым появлением растения и началом
26
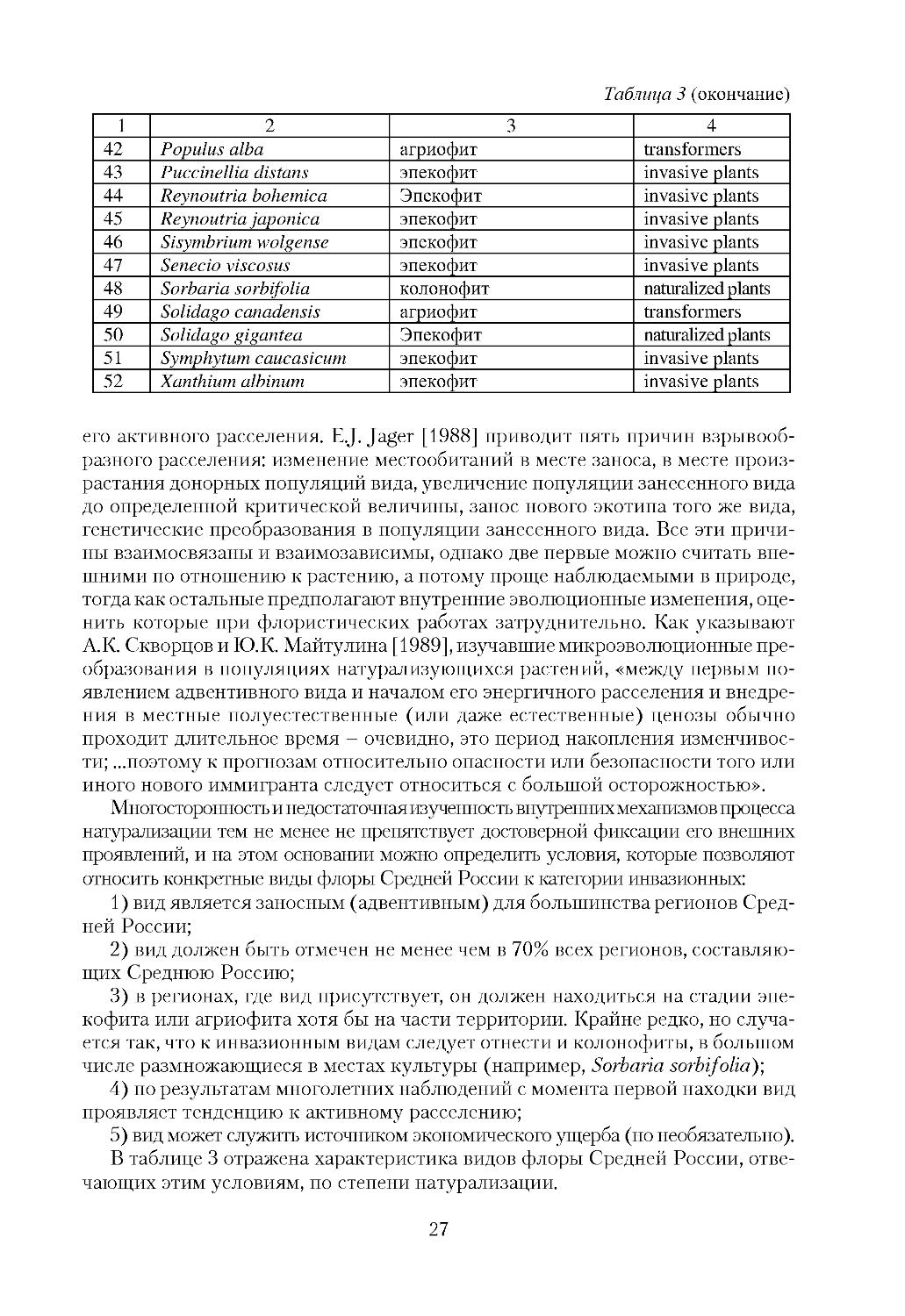
Таблица 3
Характеристика инвазионных видов флоры Средней России
по степени натурализации и инвазионному статусу (invasive status)
No
Вид
Степень натурализации Invasive status
1
2
3
4
1 Acer negundo
агриофит
transformers
2 Acorus calamus
агриофит
transformers
3 Amaranthus albus
эпекофит
invasive plants
4 Amaranthus retroflexus
эпекофит
invasive plants
5 Ambrosia artemisiifolia
эпекофит
invasive plants
6 Amelanchier spicata
агриофит
transformers
7 A.alnifolia
эпекофит
invasive plants
8 Anisantha tectorum
эпекофит
invasive plants
9 Aster x salignus
эпекофит
invasive plants
10 Atriplex tatarica
эпекофит
invasive plants
11 Bidens frondosa
агриофит
transformers
12 Cardaria draba
эпекофит
invasive plants
13 Chamomilla suaveolens
агриофит
transformers
14 Conyza сanadensis
агриофит
transformers
154 Crataegus monogyna
эпекофит
invasive plants
16 Cyclachaena xanthiifolia
эпекофит
invasive plants
17 Echinocystis lobata
агриофит
transformers
18 Elaeagnus angustifolia
колонофит
naturalized plants
19 Elodea сanadensis
агриофит
transformers
20 Epilobium adenocaulon
агриофит
transformers
21 Epilobium pseudorubescens эпекофит
invasive plants
22 Elsholtzia ciliata
эпекофит
invasive plants
23 Erigeron annuus
агриофит
invasive plants
24 Erucastrum gallicum
эпекофит
invasive plants
25 Euphorbia peplus
эпекофит
invasive plants
26 Festuca trachyphylla
колонофит
naturalized plants
27 Fraxinus pennsylvanica
колонофит
naturalized plants
28 Galinsoga ciliata
агриофит
transformers
29 Galinsoga parviflora
агриофит
transformers
30 Helianthus tuberosus
эпекофит
invasive plants
31 Heracleum sosnowskyi
агриофит
transformers
32 Hippophaë rhamnoides
эпекофит
invasive plants
33 Hordeum jubatum
эпекофит
invasive plants
34 Impatiens glandulifera
агриофит
transformers
35 Impatiens parviflora
агриофит
transformers
36 Juncus tenuis
агриофит
transformers
37 Lepidium densiflorum
агриофит
transformers
38 Lupinus polyphyllus.
агриофит
transformers
39 Oenothera biennis
агриофит
transformers
40 Oxalis stricta
эпекофит
invasive plants
41 Poa supine
агриофит
invasive plants
27
его активного расселения. E.J. Jager [1988] приводит пять причин взрывооб
разного расселения: изменение местообитаний в месте заноса, в месте произ
растания донорных популяций вида, увеличение популяции занесенного вида
до определенной критической величины, занос нового экотипа того же вида,
генетические преобразования в популяции занесенного вида. Все эти причи
ны взаимосвязаны и взаимозависимы, однако две первые можно считать вне
шними по отношению к растению, а потому проще наблюдаемыми в природе,
тогда как остальные предполагают внутренние эволюционные изменения, оце
нить которые при флористических работах затруднительно. Как указывают
А.К. Скворцов и Ю.К. Майтулина [1989], изучавшие микроэволюционные пре
образования в популяциях натурализующихся растений, «между первым по
явлением адвентивного вида и началом его энергичного расселения и внедре
ния в местные полуестественные (или даже естественные) ценозы обычно
проходит длительное время -- очевидно, это период накопления изменчивос
ти; ...поэтому к прогнозам относительно опасности или безопасности того или
иного нового иммигранта следует относиться с большой осторожностью».
Многосторонность и недостаточная изученность внутренних механизмов процесса
натурализации тем не менее не препятствует достоверной фиксации его внешних
проявлений, и на этом основании можно определить условия, которые позволяют
относить конкретные виды флоры Средней России к категории инвазионных:
1) вид является заносным (адвентивным) для большинства регионов Сред
ней России;
2) вид должен быть отмечен не менее чем в 70% всех регионов, составляю
щих Среднюю Россию;
3) в регионах, где вид присутствует, он должен находиться на стадии эпе
кофита или агриофита хотя бы на части территории. Крайне редко, но случа
ется так, что к инвазионным видам следует отнести и колонофиты, в большом
числе размножающиеся в местах культуры (например, Sorbaria sorbifolia);
4) по результатам многолетних наблюдений с момента первой находки вид
проявляет тенденцию к активному расселению;
5) вид может служить источником экономического ущерба (но необязательно).
В таблице 3 отражена характеристика видов флоры Средней России, отве
чающих этим условиям, по степени натурализации.
Таблица 3 (окончание)
1
2
3
4
42 Populus alba
агриофит
transformers
43 Puccinellia distans
эпекофит
invasive plants
44 Reynoutria bohemica
Эпекофит
invasive plants
45 Reynoutria japonica
эпекофит
invasive plants
46 Sisymbrium wolgense
эпекофит
invasive plants
47 Senecio viscosus
эпекофит
invasive plants
48 Sorbaria sorbifolia
колонофит
naturalized plants
49 Solidago canadensis
агриофит
transformers
50 Solidago gigantea
Эпекофит
naturalized plants
51 Symphytum caucasicum
эпекофит
invasive plants
52 Xanthium albinum
эпекофит
invasive plants
28
Глава 2
ЧУЖЕРОДНЫЕ И ИНВАЗИОННЫЕ ВИДЫ
В ЕВРОПЕ
База DAISIE
Консорциум ведущих исследователей биологических инвазий в Европе,
включающий 18 учреждений из 15 стран, в феврале 2005 года начал проект, по
лучивший название DAISIE (Delivering Alien Invasive Species Inventories for
Europe). Проект продолжался в течение трех лет и потребовал затрат 2,4 млн
евро. Основные цели проекта:
1) создание кадастра чужеродных видов, которые угрожают наземным, пре
сноводным и морским биоценозам в Европе;
2) структурирование кадастра как базы для контроля и предотвращения био
логических инвазий посредством учета экологических, социальных, экономи
ческих и других факторов;
3) оценка и суммирование экологических, экономических и влияющих на
здоровье человека воздействий наиболее широко распространенных и агрес
сивных видов в Европе;
4) использование данных по распространению видов и опыт отдельных го
сударств как основу для создания индикаторов, способствующих раннему об
наружению видов --агрессоров.
Для достижения этих целей DAISIE стремится сделать одну общую базу по
информации о биологических инвазиях в Европе.
Стратегия по Инвазионным видам Европы [Council of Europe, 2003] одоб
рила проведение инвентаризации агрессивных чужеродных видов в Европе и
мобилизовала экспертов для составления обзоров и инвентаризации видов,
объединения существующих баз данных, включения потенциально инвазион
ных видов, которые имеют высокую вероятность спонтанного расширения аре
ала из близлежащих стран, и выявление приоритетных видов. Информация, По
возможности, информация включает: таксономию и биологические особеннос
ти вида, дату и место интродукции, пути внедрения и распространения, диапа
зон и динамику распространения, угрозу экспансии в близлежащие страны, втор
жение в природные ценозы, размер популяций и основные тренды, зарегистри
рованные случаи инвазии и уровень опасности вида, другие данные, важные для
анализа угрозы и для системы раннего обнаружения, методы предотвращения,
уменьшения или восстановления и их эффективность, ссылки и контактные
данные. Основываясь на этих требованиях, программа DAISIE сосредоточилась
на трех основных блоках сбора и распространения информации: 1) регистра
29
ция европейских экспертов по чужеродным видам: справочник исследовате
лей и исследований; 2) база данных чужеродных видов, включающая все изве
стные чужеродные виды в Европе; 3) информационная система инвазионных
чужеродных видов в Европе: описание ключевых агрессивных чужеродных
видов, действительно или потенциально опасных, ввиду их воздействия на
окружающую среду или на экономику с картами ареалов [Hulme et al., 2008].
Эти три блока объединены на интернет портале www. europe aliens.org.
Блок 1. Регистрация европейских экспертов по чужеродным видам: спра
вочник исследователей и исследований
Текущей экспертной оценкой биологических инвазий занимается в Европе
ряд исследовательских организаций, и она финансируется, главным образом,
национальными программами. Регистрация в программе DAISIE является фун
даментальным шагом к объединению этих организаций и исследователей.
Регистр содержит информацию относительно области исследований экс
пертов (распространение, сохранение, экология, экономика, генетика, законо
дательство, управление, способы распространения, физиология, оценка сте
пени риска и таксономия). Уже через год после начала программы справочник
содержал информацию о 1500 экспертах из 90 стран почти для 3000 высших
таксонов (уровень семейства или выше), и с тех пор их число устойчиво уве
личивается. Эти данные отражают общий недостаток экспертов в восточноев
ропейских регионах, а также экспертов по чужеродным грибам, мхам и бес
позвоночным животным, особенно насекомым.
Блок 2. База данных по чужеродным видам в Европе
Современная инвентаризация всех чужеродных видов, известных в Евро
пе, является основой построения ранней системы обнаружения и предупреж
дения инвазий для менеджеров по охране окружающей среды. Этот шаг пред
ставлял главную сферу деятельности DAISIE и включал компилирование и
обзор национальных списков сотен видов грибов, растений, беспозвоночных,
рыб, амфибий, рептилий, птиц и млекопитающих. Были сопоставлены дан
ные по всем 27 государствам -- членам ЕЭС и, кроме того, Андорры, Ислан
дии, Лихтенштейна, Молдовы, Монако, Норвегии, европейской части России,
Швейцарии, Украины, а также прежних югославских государств на Балканах.
Для каждого вида была собрана информация относительно естественного
ареала, времени интродукции, среды обитания, известных случаев инвазии и
статуса популяций. Потребовались значительные усилия по сведению синони
мики. К февралю 2008 года в базу данных включены сведения по 11 000 чуже
родным видам; большинство из них относятся к сосудистым растениям и бес
позвоночным [Olenin, Didziulis, 2009].
Блок 3. Информационная система инвазионных чужеродных видов в Европе
База включает сведения о 100 наиболее вредоносных инвазионных видах,
вторгающихся в естественные и полуестественные местообитания Европы. Эти
100 видов включают: 3 гриба, 18 наземных растений, 16 наземных беспозво
ночных, 15 позвоночных животных, 16 водных и 32 прибрежноводных вида.
30
Выбор обосновывался широким спектром жизненных форм и функциональ
ных типов, большим диапазоном экосистем, подверженных вторжению, и чет
кими примерами различных воздействий на биоразнообразие, экономику и
здоровье населения. Ключевое требование для эффективного контроля инва
зионного чужеродного вида -- возможность идентифицировать его, нанести
на карту и отслеживать инвазии, чтобы оценить их степень и динамику.
Заносные сосудистые растения Европы. Сосудистые растения являются
наиболее интенсивно исследуемой таксономической группой относительно
биологии инвазий. По крайней мере 95 инвазионных видов были предметом
детального изучения в глобальном масштабе, что составляет 44% всех изучен
ных инвазионных таксонов. После Северной Америки Европа -- наиболее ин
тенсивно исследуемый континент, по крайней мере по 80 инвазионным видам
растений [Pyšek et al., 2008]. Однако, хотя и имеется значительный объем ин
формации по основным инвазионным видам в Европе [Weber, 2003], ситуация
является значительно менее удовлетворительной, поскольку в полной мере на
циональные инвентаризации чужеродных видов не проведены. До проекта
DAISIE только немногие страны имели проверенную информацию о составе их
адвентивных флор, доступную в специализированных чек листах: Австрия [Essl,
Rabitsch 2002], Чешская Республика [Pyšek et al., 2002], Германия [Klotz et al.,
2003; Kühn, Klotz, 2003], Ирландия [Reynolds, 2002] и Великобритания
[Clement, Foster, 1994; Preston et al., 2002, 2004]. Эта ситуация напрямую от
ражала недостаточную изученность биоты европейского континента. Един
ственный доступный анализ инвазионных видов в Европе [Weber, 1997] был
основан на данных Flora Europaea [Tutin et al., 1964 -- 1980] -- единственного
синтетического источника информации о флорах отдельных стран, включая
заносные виды. Этот источник, однако, является в настоящее время устарев
шим и содержит многочисленные погрешности [Pyšek, 2003]. Вообще, инфор
мация о присутствии и расселении чужеродных видов растений в большин
стве европейских стран была рассеяна во множестве опубликованных и нео
публикованных отчетов и баз данных; так же обстоит дело и на других
континентах [Meyerson, Mooney, 2007].
Территория Европы, включенная в программу DAISIE, определяется, в ос
новном, границами, установленными во Flora Europaea [Tutin et al., 1964--1980]
в отношении северных и центральных районов (восточная граница проходит
по Уральским горам к берегу Черного моря, исключая Кавказ). На юго востоке
границей включенного в изучение региона был Кипр, включена была также Тур
ция. Всего в рассматриваемую территорию входит 49 стран или регионов.
По инвазионному статусу различали натурализовавшиеся и случайно зане
сенные чужеродные виды, а по продолжительности инвазии различали архео
фиты и неофиты, следуя критериям, предложенным в работах [Richardson et al.,
2000; Pyšek et al., 2004]. В тех случаях, если для определения точной катего
рии (археофит или неофит, натурализовавшийся или случайно занесенный)
данных не хватало, таксон регистрировался как чужеродный без детального
уточнения. Подобное происходит обычно в отношении стран, имеющих недо
статочно детальную регистрацию заносных видов (например, Беларусь, Бол
гария, Молдова) и включающих в базы данных только инвазионные виды.
31
Заносные виды Европы разделены на две группы. Первая включает виды,
естественно произрастающие в одних регионах Европы, но чуждые для дру
гих ее регионов. Вторая группа включает виды с естественным ареалом, рас
положенным вне Европы [Lambdon et al., 2008].
Разнообразие чужеродных видов в Европе. База данных DAISIE содержит
описания 5789 заносных растений Европы, из которых 2843 вида имеют внеев
ропейское происхождение (для 275 видов статус происхождения не установ
лен, поскольку нет возможности определить их естественный ареал). Из этих
заносных видов соответственно 1507 и 872 относятся к категории случайно
занесенных во всех регионах, в которых они были отмечены. Таким образом,
всего в Европе насчитывается 3749 натурализовавшихся чужеродных видов
растений, из которых 1780 имеют внеевропейское происхождение. Общее чис
ло натурализовавшихся неофитов не определено, так как результат этот был
бы основан на данных только 19 стран, для которых имеются данные по инва
зионному статусу и периоду интродукции, что привело бы к недооценке чис
ла натурализовавшихся неофитов, которые в настоящее время произрастают
в Европе [Lambdon et al., 2008].
В опубликованном 11 лет назад [Weber, 1997] обзоре адвентивной флоры
Европы сообщается о 1568 натурализовавшихся в Европе видах растений, что
намного меньше, чем зарегистрировано DAISIE, и нужно иметь в виду два ас
пекта, объясняющих это различие. Обзор Е.Вебера [Weber, 1997] основан на
издании Flora Europaea, которое, в свою очередь, опиралось на данные 1960 --
1970 х [Tutin et al., 1964--1980]. Начиная с этого периода, однако, в отдельных
странах шел непрерывный приток чужеродных видов [Pyšek et al., 2003]. Ког
да публикация Flora Europaea была завершена, многие чужеродные виды, дол
жно быть, уже присутствовали в Европе (рис. 4), но не были включены в изда
ние. Другой принципиальной причиной низкого числа чужеродных видов, о
которых имеются данные во Flora Europaea, был довольно низкий уровень де
тализации, используемой для скрининга заносных видов. Повышение внима
ния к проблеме чужеродных видов и возросшая интенсивность исследований
в прошлые десятилетия [Pyšek et al., 2006] резко увеличили число видов, вклю
ченных в базы данных DAISIE, которое намного ближе к действительности.
Происхождение. Для чужеродных видов Европы основным ареалом до
нором являются другие регионы этого же континента. 29% интродуцирован
ных видов (рис. 5) имеют естественный ареал в одних странах Европы и инва
зионные популяции в других странах. Объединение их с заносными видами
азиатского происхождения (31%) иллюстрирует основной вклад (60%) Евра
зии в богатство чужеродных видов в Европе. Северная и Южная Америка вме
сте являются естественным регионом произрастания для 19% интродуциро
ванных чужеродных видов в Европе.
Раздельное рассмотрение видов внеевропейского происхождения отдель
но приводит к другой картине (рис. 5): среди заносных видов Европы 34% ин
тродуцированных видов имеют азиатское происхождение (регионы Азии с уме
ренным климатом дают больше видов, чем тропики), 23 и 22% происходят со
ответственно из Северной и Южной Америки, а 17% -- из Африки. Эти числа
согласуются с распределением происхождения в национальных флорах [Kühn,
32
Рис. 5. Регионы доноры чужеродных видов во флоре Европе
Рисунок базируется на данных по 2094 натурализовавшимся в Европе чужеродным видам,
о которых имеются сведения относительно регионов естественного распространения. 1168
из этих видов занесены в Европу с других континентов. Отметим, что рисунок отражает
число видов; виды, распространенные в нескольких регионах, учитываются в каждом из
этих регионов доноров, таким образом в обоих группах общее количество чужеродных
видов составляет 100%.
Рис. 4. Увеличение численности чужеродных неофитов, интродуцированных
в Европу за последние 500 лет.
Данные представлены отдельно для видов, имеющих естественный ареал вне Европы (929
видов), и имеющих естественный ареал в пределах Европы (954 вида); для последних
учитывалась дата их первого обнаружения за пределами естественного ареала.
33
Klotz, 2003; Pyšek et al., 2002] и подтверждают закономерности, о которых со
общает Е.Вебер [Weber, 1997] для Европы.
Таксономия. Представители заносной флоры преобладают во многих круп
ных семействах (табл. 4); самое высокое их число отмечено в Asteraceae (692
чужеродных представителя), Poaceae (597), Rosaceae (363), Fabaceae (подсемей
ство Leguminosae, 323) и Brassicaceae (247). Эти семейства имеют тенденцию к
сорничанию [Daehler, 1998; Pyšek, 1998], а в умеренных областях обладают
значительным видовым разнообразием, за исключением Rosaceae, в котором
большинство интродуцированных в Европу видов принадлежат к умеренно
бореальным деревьям и кустарникам. Единственным семейством с преобла
данием древесных чужеродных видов является Pinaceae (53 чужеродных вида).
Отмеченные в Европе чужеродные виды относятся к 213 семействам [Lambdon
et al., 2008], что почти в 2 раза больше, чем сообщил Е.Вебер [Weber, 1997].
Возможно, в некоторых случаях успех может быть связан с частотой интро
дукции, поскольку некоторые особенности семейства делают виды ценными
для использования человеком (например, Rosaceae используются как плодо
вые растения, Pinaceae -- для древесины, а Lamiaceae -- в качестве приправ и
как декоративные растения).
Отмечено 1567 родов, представленных в Европе по меньшей мере одним
чужеродным видом (табл. 5). Наиболее обычны роды с центрами разнообразия,
расположенными в Европе или Евразии, с высоким естественным разнообрази
ем (Centaurea, Silene, Euphorbia, Rumex) или экстенсивно используемые людьми
(Trifolium, Vicia, Rosa). Весьма различны закономерности для чужеродных ви
дов с внеевропейским происхождением; заносные растения Европы в большин
стве случаев принадлежат к глобально разнообразным родам, включающим, глав
ным образом, городские и агрикультурные сорняки (Amaranthus, Chenopodium и
Solanum), а также наиболее часто культивируемые виды (самый большой род
Cotoneaster, насчитывающий 70 чужеродных видов, состоит почти исключитель
но из видов, интродуцированных в декоративных целях, или другие декоратив
ные растения, такие как Sedum или Narcissus). Особый случай -- род Oenothera, в
котором гибриды после нескольких поколений изоляции начинают нормально
скрещиваться. Только в некоторых крупных успешно инвазионных родах (на
пример, Oxalis, Panicum, Helianthus) преобладают виды внеевропейского проис
хождения.
Насчитывается 128 видов, зарегистрированных более чем в половине стран
[Lambdon et al., 2008]. Самый распространенный европейский чужеродный вид --
Conyza canadensis, уроженец Северной Америки, обнаружен в 47 странах (94%).
Другие виды, отмеченные более чем в 80% изученных регионов, включают
(табл. 6): Datura stramonium, Helianthus tuberosus, Robinia pseudoacacia (все они
уроженцы Северной Америки), Amaranthus retroflexus, Lepidium virginicum
(Северная и Центральная Америка), Galinsoga quadriradiata и G. parviflora
(Центральная и Южная Америка), Xanthium strumarium (Евразия), Panicum
miliaceum, Veronica persica (Азия) и Oenothera biennis (этот вид, вероятно, воз
ник в Европе, но в большинстве стран считается чуждым).
Однако широко распространенные виды необязательно являются самыми
агрессивными. Многие таксоны, перечисленные в табл. 6, являются агрикуль
36
турными сорняками или рудеральными видами городских местообитаний;
только немногие из 100 наиболее опасных чужеродных видов, имеют широ
кий вторичный ареал: например древесные инвазионные виды Robinia
pseudoacacia (41 страна) и Ailanthus altissima (39) или вредные сорняки как
Reynoutria japonica (39), Ambrosia artemisiifolia (35), Impatiens glandulifera (34),
Rosa rugosa (34), Heracleum mantegazzianum (27) и Carpobrotus edulis (24). Неко
торые из опасных инвазионных видов имеют иногда локальный вторичный аре
ал, например Hedychium gardnerianum (3) или Gunnera tinctoria (4).
Тенденции изменения процесса инвазии во времени. В течение двух пос
ледних веков отмечено устойчивое увеличение числа неофитов в Европе. На
Таблица 6
Наиболее распространенные чужеродные виды Европы
Вид
Семейство
Нату-
рали-
зовав-
шиеся
Слу-
чайно
зане-
сенные
Статус
не
уста-
новлен
Общее число
регионов, в
которых
отмечен вид
Conyza canadensis
Asteraceae
33
1
13
47
Datura stramonium
Solanaceae
25
7
13
45
Amaranthus retroflexus Amaranthaceae
30
4
10
44
Galinsoga parviflora Asteraceae
27
2
15
44
Helianthus tuberosus
»
26
5
12
43
Xanthium strumarium
»
22
5
16
43
Lepidium virginicum Brassicaceae
16
11
15
42
Oenothera biennis
Onagraceae
28
2
12
42
Robinia pseudoacacia Leguminosae
32
2
8
42
Galinsoga quadriradiata Asteraceae
25
1
15
41
Chamomilla suaveolens
»
23
3
15
41
Panicum miliaceum
Poaceae
16
20
5
41
Veronica persica
Plantaginaceae
27
0
14
41
Ailanthus altissima
Simaroubaceae
30
1
9
40
Amaranthus albus
Amaranthaceae
24
5
11
40
Erigeron annuus
Asteraceae
27
3
10
40
Reynoutria japonica
Polygonaceae
29
1
10
40
Medicago sativa
Leguminosae
23
4
13
40
Amaranthus blitoides Amaranthaceae
24
6
9
39
Lepidium sativum
Brassicaceae
10
21
8
39
Papaver somniferum Papaveraceae
12
17
10
39
Solidago canadensis
Asteraceae
28
0
11
39
Acer negundo
Sapindaceae
26
3
9
38
Chenopodium
ambrosioides
Amaranthaceae
22
6
10
38
Elodea canadensis
Hydrocharitaceae 26
0
12
38
Juncus tenuis
Juncaceae
26
0
12
38
Panicum capillare
Poaceae
17
16
5
38
Phalaris canariensis
»
10
15
13
38
Vicia sativa
Leguminosae
15
13
10
38
37
рис. 4 видно, что число видов с внеевропейским ареалом стало увеличиваться
по экспоненте примерно с середины ХIХ века. К настоящему времени из нео
фитов, чуждых Европе, 50% появились после 1899 года, 25% появились после
1962 года, 10% -- после 1989 года. Ежегодно в Европе отмечается около 6 но
вых чужеродных видов, способных к натурализации.
Помимо того, в настоящее время ежегодно около 4--5 видов, имеющих ес
тественный ареал в одной из стран Европы, отмечаются в качестве натурализо
вавшихся в других частях континента [Lambdon et al., 2008]. В целом, по види
мому, темп прироста новых интродукционных видов резко увеличился в тече
ние двух прошлых столетий и имеет небольшую тенденцию к замедлению.
Пути распространения в Европе. Из 2024 натурализованных таксонов, чуж
дых Европе, относительно которых имеются сведения о путях их распростране
ния, намеренно интродуцированные составляют 63%, а случайно занесенные --
37% (рис. 6). Наибольшую численность (58%) имеют растения, «сбежавшие»
из культуры. Только 11 видов можно с уверенностью приписать намеренному
внедрению в естественные сообщества; эту группу во многих случаях трудно
отличить от «эстетических» видов (посаженных в полуестественные место
обитания с целью практического применения: Carpobrotus edulis, например, ис
Рис. 6. Соотношение различных способов заноса натурализовавшихся в Европе
растений, имеющих внеевропейский естественный ареал
Намеренно интродуцированные -- 62,8%, из них лесные культуры -- 1,6%; виды,
используемые в зеленом строительстве -- 5,0; декоративные -- 39,9%; агрокультуры --
6,6%; садовые -- 17,5%; намеренно внедренные в естественные сообществоы -- 0,5%
Ненамеренно интродуцированные (занесенные) -- 37,2%, из них засоряющих семенной
материал -- 9,1%, загрязняющих минеральные материалы -- 1,8%, засоряющие продукты
и товары -- 6,1%, с транспортом -- 9,9%, занесенные без помощи человека -- 1,9%
38
пользуется для озеленения, а Robinia pseudoacacia часто применяется для ук
репления почвы). К примерам преднамеренной интродукции относится насе
комоядное растение Sarracenia purpurea, которое специально интродуцирова
но ботаниками в болотные трясины Великобритании и Ирландии, а также
Spartina alterniflora, интродуцированная на солонцах.
Засорители семян, минеральных материалов и других предметов потребле
ния ответственны за 403 интродуцированных в Европу вида (17% всех видов),
и предполагается (рис. 6), что 235 видов прибыли как «безбилетники» (непос
редственно связаны с перевозками людей и прибывают независимо от товара
[Hulme et al., 2008a]. Однако число «безбилетников» почти наверняка недооце
нивается из за технически трудной систематической регистрации этого пути
распространения, например, разбора семян и примесей к ним [Mack, 2000].
41 заносный вид внеевропейского происхождения (2% видов, чуждых Евро
пе), является продуктом спонтанной гибридизации с вовлечением одного или
обоих чужеродных родителей (рис. 6).
Биогеографические закономерности. Оценка уровня инвазии европейс
ких регионов по числу натурализующихся заносных видов не обязательно от
ражает инвазибельность региона [Lonsdale, 1999; Richardson, Pyšek, 2006;
Chytrý et al., 2008a]. На континенте самое высокое богатство чужеродных ви
дов связано с высокоиндустриальными северо западными странами, имеющи
ми давние традиции хорошей ботанической регистрации или интенсивного
недавнего исследования. Самая высокая численность всех чужеродных видов,
независимо от их статуса, отмечена в Бельгии (1969), Великобритании (1779),
Чешской Республике (1378), во Франции (1258), в Швеции (1201) и Австрии
(1086), что объясняется детальным включением всех случаев заноса. Более
200 натурализовавшихся неофитов зарегистрировано в Великобритании (857),
Германии (450), Бельгии (447), Италии (440), на Украине (297), в Австрии (276),
Польше (259), Литве (256), Португалии (250) и Чешской республике (229).
Выделено [Lambdon et al., 2008] пять основных типов распространения:
1) северо--западный, включающий Скандинавию и Великобританию; 2) запад
но--центральный, простирающийся от Бельгии и Нидерландов до Германии и
Швейцарии; 3) балтийский, включающий только прежние советские респуб
лики; 4) центрально--восточный, включающий остальные страны Централь
ной и Восточной Европы; 5) южный, покрывающий всю Средиземноморскую
область. Некоторые чужеродные инвазионные виды даже представляют эти
биогеографические зоны: например, Rhododendron ponticum -- северо западный,
Heracleum sosnowskyi -- балтийский, Echinocystis lobata -- центрально восточ
ный, Opuntia ficus indica -- южный тип распространения.
Не исключено, что наблюдаемые соотношения возникают частично из за
региональных различий в подходе к ботанической регистрации, почти навер
няка имеется значительное влияние климата и культуры. Валовой внутрен
ний продукт, низкий уровень осадков и температура воздуха, по видимому,
играют роль в дифференцировании чужеродной фракции флоры, но их ин
терпретировать трудно, так как эти факторы сильно зависят от широтного или
меридионального градиента. Площадь отдельных стран и численность насе
ления не объясняли вариабельность, и это позволило предположить, что фак
39
торы, связанные с плотностью населения (например, урбанизация), незначи
тельно влияют на флористический состав заносных растений. По видимому,
биоклиматические условия, устанавливающие приспособленность видов к
физическим параметрам окружающей среды, наиболее важны и играют глав
ную роль в формировании фракции чужеродных видов в Европе. Культурные
факторы, например региональные торговые связи, традиции выращивания
сельскохозяйственных культур или декоративных растений, также могут ока
зывать сильное влияние на состав интродуцированных видов [Lambdon et al.,
2008]. Кюн с соавторами [Kühn et al., 2003] показали, что по крайней мере в
Германии чужеродные виды в значительной степени повторяют биогеографи
ческие закономерности, свойственные природным видам.
У всех пяти типов распространения доминирующие семейства и географи
ческие области происхождения существенно не отличались от параметров, об
щих для всей Европы, сходны были и доминирующие роды. Однако южная аг
ломерация имеет множество отличий и содержит более низкое число видов,
относящихся к родам умеренных широт, таких, как Chenopodium, и более значи
тельную представленность родов тропического или американского происхож
дения (например, Acacia, Opuntia). То, что эта категория совпадает со Среди
земноморским регионом, имеющим по сравнению с остальной частью Европы
особенно контрастный климат, подтверждает важную роль климата в формиро
вании фракций чужеродных видов. Многие из обычных заносных сорняков сель
скохозяйственных культур в Европе также имеют средиземноморское проис
хождение [Pyšek et al., 2004], а потому исключены из числа чужеродных видов.
Кроме того, страны Средиземноморья расположены на перекрестке трех конти
нентов, что делает их доступными для биотических элементов, происходящих
из множества источников, что также способствует флористической обособлен
ности южной агломерации заносных видов. По степени распространения чуже
родные виды в Европе формально можно разделить на четыре группы: 1) широ
ко распространенные -- натурализовавшиеся на большей части континента (448
видов, 76 из которых отмечены во всех биогеографических зонах); 2) обычный
на региональном уровне -- натурализовавшиеся в какой либо одной биогеогра
фической зоне (196 видов); 3) спорадические -- появляющиеся редко, но в не
скольких биогеографических зонах; 4) локальные -- натурализовавшиеся толь
ко в отдельных регионах Европы. Две последние категории включают большин
ство оставшихся видов [Lambdon et al., 2008].
Наиболее инвазибельные местообитания. В большинстве региональных
Флор описания местообитаний относительно грубы [Chytrý et al., 2008a,b].
Информация о сходстве местообитаний имеется для 30 стран/регионов. В це
лом для Европы зарегистрировано 57% натурализовавшихся растений с евро
пейским ареалом (2122 вида) и 58% -- с внеевропейским ареалом (1059 видов),
которые классифицированы по степени встречаемости в том или ином место
обитании (табл. 7). Показано, что тип местообитания является более важным
фактором, чем климат и количество семязачатков, как для уровня инвазионнос
ти вида [по Hierro et al., 2005], так и для инвазибельности фитоценозов [Chytrý
et al., 2008a]. Поэтому информация об уровне инвазий в отдельные местооби
тания крайне важна для управления чужеродными видами в Европе. Создан
40
ные человеком местообитания (индустриальные площадки, пахотная земля,
парки и сады) предоставляют кров большинству чужеродных видов [Chytrý
et al., 2005, 2008a,b]. Из всех натурализовавшихся видов, отмеченных в Евро
пе, 64% встречаются в индустриальных местообитаниях, 58% -- на пахотной
земле, в парках и садах. В леса и луга, которые также чрезвычайно сильно под
вержены инвазиям, вторгаются соответственно 32 % и 37% всех натурализо
вавшихся чужеродных видов. В болота вторгается наименьшее число назем
ных растений -- 10% чужеродных видов Европы. В морских местообитаниях
отмечено только 12 видов сосудистых растений (из них 7 -- чужеродные в Ев
ропе), представляющих 0,6% всех видов (табл. 7). Уровень инвазивности оп
ределен отдельно для видов с европейским и внеевропейским естественным
ареалом. Сумма превышает 100%, поскольку некоторые виды встречаются
сразу в нескольких местообитаниях. Классификация местообитаний приве
дена по системе EUNIS [Lambdon et al., 2008].
Заносные виды, имеющие европейский ареал, по сравнению с заносными
видами внеевропейского происхождения произрастают в большем числе мес
тообитаний, что можно объяснить предварительной адаптацией европейских
видов к диапазону европейских местообитаний. По видимому, эти виды более
конкурентоспособны по сравнению с чужеродными видами, имеющими вне
европейский ареал, которым во время процесса инвазии еще только предсто
Таблица 7
Уровень инвазивности различных местообитаний в Европе
Число видов
Процент от общего числа
видов
Категория
Чуже-
родных в
Европе
из них имеющих
естественный
ареал вне Европы
Чуже-
родных в
Европе
из них имеющих
естественный
ареал вне Европы
Общее число видов 2122
1059
-
-
Процент общего
числа видов
56,6
57,7
-
-
Морские
местообитания
12
7
0,6
0,7
Болота и топи
220
118
10,4
11,1
Скалы
343
170
16,2
16,1
Пресные водоемы
444
260
20,9
24,6
Вересковые
пустоши и
кустарники
462
206
21,8
19,5
Вырубки и залежи
497
211
23,4
19,9
Леса и луга
668
310
31,5
29,3
Пастбища
793
276
37,4
26,1
Пахотные земли,
сады и парки
1240
533
58,4
50,3
Промышленные
зоны
1360
658
64,1
62,1
41
ит адаптироваться к характеру европейских местообитаний [Lambdon et al.,
2008]. Другой причиной может быть более продолжительный период произ
растания заносных видов, имеющих естественный ареал в Европе, дающий им
возможность колонизации более широкого диапазона местообитаний [Pyšek
et al., 2003].
Влияние на экологию и экономику. Исчерпывающие данные о воздействии
на экологию и экономику существуют в базе данных DAISIE только для тех
стран (Латвия, Литва и Великобритания), где, как считается, ~20% натурализо
вавшихся видов оказывают подобное влияние. Наилучшие данные имеются в
Швейцарии, где из 97 натурализовавшихся чужеродных видов 50 оказывают
влияние на экологию и экономику. Однако можно получить некоторое пред
ставление о различных воздействиях, причиной которых являются чужеродные
виды, из обзора списка 100 самых вредных чужеродных видов. Из 18 включен
ных в этот список растений 17 сокращают местообитания природных видов, а 8
приводят к разрушению естественных сообществ. Carpobrotus edulis [Vilа et al.,
2006] и Heracleum mantegazzianum [Ecology..., 2007] являются примерами чуже
родных видов, вызывающих серьезное снижение биоразнообразия сообществ,
в которые они вторгаются. Однако наблюдается это не всегда, отмечено, напри
мер, что в некоторых случаях инвазия Impatiens glandulifera приводит не к поте
ре разнообразия всех сообществ, в которые она вторгается, а лишь к изменению
состава рудеральных видов и видов, требующих повышенного содержания азо
та [Hejda, Pyšek, 2006; Hulme, Bremner, 2006].
Заносные виды растений воздействуют на экологию и экономику на мно
жественных уровнях и прямо, и косвенно. Из выделенных 22 типов воздей
ствия [Binimelis et al., 2007], в группе растений, включенных в 100 наиболее
опасных чужеродных видов, на каждый вид приходится в среднем более чем
по четыре типа воздействия, что ставит эту группу на 2 е место по разнообра
зию оказываемых воздействий после наземных млекопитающих. Относитель
но воздействия на экономику широко известны Oxalis pes caprae, Opuntia
maxima, Paspalum paspalodes и Rhododendron ponticum, которые негативно вли
яют на урожайность сельскохозяйственных культур и лесозаготовки. Robinia
pseudoacacia, Ailanthus altissima и Cortaderia selloana являются типичным при
мером чужеродных видов в Европе, наносящих серьезный ущерб инфраструк
туре и общественным местам. Уникальная генетическая природа аборигенных
видов или даже эндемиков с высоким уровнем редкости может быть потеряна
ввиду интрогрессии с широко распространенными чужеродными видами [Vilа
et al., 2000]. В чешской флоре [Pyšek et al., 2002] гибриды составляют 13% об
щего числа чужеродных видов, и гибридизация более часта среди археофитов
(19%), чем среди неофитов (12%). Вероятно, самый известный пример недав
ней эволюции инвазионного вида относится к североамериканскому виду
Spartina alterniflora, который при скрещивании с европейским S.maritima в
Англии и во Франции образовал стерильный S. Ч townsendii. Этот гибрид от
мечен в Европе только в двух местах и не проявлял способности к натурализа
ции до появления аллотетраплоидной формы S. x anglica, возникшей в 1890 х.
Эта форма, способная расти в более широком диапазоне условий, стала агрес
сивным инвазионным видом в Европе и других регионах [Williamson, 1996]. В
42
Рис. 7. Ailanthus altissima на территории раскопок античного г. Танаис,
Ростовская обл.
43
Германии при гибридизации заносной Rorippa austriaca с аборигенной R.
sylvestris образовался комплекс гибридных форм с уровнями плоидности 3x --
5x, размножающихся половым путем и распространяющихся в регионах, где
родительские виды не встречаются [Bleeker, 2003].
Имеются сообщения, что заносные виды сокращают опыление природных
видов, как, например, в случае Impatiens glandulifera [Chittka, Schürkens, 2001].
На Балеарских островах аборигенный вид Lotus cytisoides реже посещается опы
лителями в сообществах, в которые вторгся южноафриканский Carpobrotus spp.,
чем в сообществах, в которых этот вид не отмечен [Traveset, Moragues, 2004].
Инвазионные растения могут также изменить структуру сообщества на более
высоких трофических уровнях. Показано [Pétillon et al., 2005], что Elymus
athericus влияет на популяционную динамику пауков на засоленных маршах
во Франции. К инвазионным видам, которые изменяют функционирование
экосистемы (трансформерам в понимании Richardson et al., 2000], относится,
например, Robinia pseudoacacia, которая нарушает цикл азота в почве. Другие
виды влияют на режим пожаров; примером служат Cortaderia selloana и
Ampelodesmos mauritanica, которые увеличивают риск возникновения пожа
ров в средиземноморских кустарниковых сообществах, поскольку являются
хорошим топливом и разлагаются более медленными темпами, что приводит
к формированию толстых слоев опада в тех зарослях, куда вторглись эти виды
[Vilа et al., 2001; Grigulis et al., 2005]. Наконец, некоторые чужеродные расте
ния, по видимому, затрагивают культурные аспекты цивилизации, изменяя
восприятие человеком естественных пейзажей (например, Opuntia maxima,
Rosa rugosa, Oxalis pes caprae), уменьшая площадь рекреационных природных
участков (например, Heracleum mantegazzianum) или произрастая в изобилии
на древних руинах (например, Ailanthus altissima; рис. 7).
Ближайшее будущее -- тенденции и возможность управления инвазиями.
Базы данных DAISIE представляют собой первую общеевропейскую подроб
ную сводку заносных видов растений на этом континенте. Объем иммиграции
новых чужеродных видов растений в Европе замедляется довольно незначи
тельно (рис. 4). Как и в других частях света, в Европе в ближайшем будущем
угроза растительных инвазий вряд ли уменьшится.
44
Глава 3
ГИПОТЕЗЫ УСПЕШНОСТИ
ИНВАЗИОННЫХ ВИДОВ
Дискуссия о факторах, способствующих натурализации, продолжается и
поныне. Изучение этой проблемы связано с адаптивными стратегиями вида,
каждая из которых характеризуется соответствующим набором морфологичес
ких и биологических признаков. Детально этот вопрос освещен в работе Б.Н.
Головкина [1988], поэтому нам остается добавить лишь несколько гипотез, не
вошедших в его монографию.
Интересна гипотеза о влиянии ареала растения на его способность к коло
низации. Установлено [Smith, 1981], что виды, относящиеся к родам, распрос
траненным в обоих полушариях, имеют большую способность к колонизации,
чем виды, относящиеся к родам с более ограниченным распространением. Ги
потеза о преимуществе видов, сочетающих вегетативный и генеративный спо
собы размножения [Considine,1986], не вполне приемлема, поскольку подоб
ных видов не так уж много. Гипотеза о конкурентном превосходстве видов,
обладающих перекрестным опылением [Russell, Levin, 1988], как нашими, так
и другими исследованиями [Pernes, 1987; Виноградова, 1992] не подтвержда
ется. Широко распространено также мнение, что адвентивные виды должны
иметь на семенах или плодах специальные приспособления для распростра
нения животными, ветром или водой. Однако в Окленде (Новая Зеландия) из
615 натурализовавшихся видов 368 никаких явных приспособлений не имеют
[Esler, 1988].
Чтобы экспериментально исследовать эту проблему необходимо сравнить
биоморфологические признаки: или 1) адвентивного вида и активно вытесняе
мого им аборигенного вида из этого же рода, или 2) нескольких близкородствен
ных видов, из которых только один два стали инвазионными.
Примеров применения первой методологии очень мало. Практически пол
ное вытеснение адвентивным видом аборигенного произошло лишь в двух слу
чаях: в Центральной Европе Impatiens parviflora выдворила I.noli tangere, а в
Австралии Cakile maritima вытеснила C.edentula. Евразиатский Myriophyllum
spicatum, появившийся в Северной Америке около ста лет назад, в настоящее
время в значительной степени вытеснил аборигенный M.exalbescens, однако изу
чение темпов роста и продуктивности обоих видов в различных экологических
условиях не выявило заметных преимуществ заносного вида [Aiken, Picard, 1980].
При сравнении устойчивости в условиях полупустыни штата Юта (США)
аборигенного Agropyron spicatum и интродуцированного из Евразии в начале ХХ
века A.desertorum выявлены признаки, показывающие преимущества последне
45
го [Pyke, 1990]. У A.desertorum формируется в 70 раз больше семян (причем не
все они после созревания сразу опадают, что позволяет избежать уничтоже
ния вредителями и грызунами); всхожесть семян в 2 раза выше, выживаемость
проростков также выше, чем у A. spicatum.
Совершенно редкий случай вытеснения одним заносным видом другого,
ранее занесенного вида наблюдается на Британских островах [Simpson, 1990],
где интродуцированная из Америки в 1966 году Elodea nuttallii активно заме
щает E.canadensis. Опыты по выращиванию обоих видов при разных экологи
ческих условиях показали, что E. nuttallii растет вдвое энергичнее и образует
более длинные боковые побеги, что приводит к угнетению E.canadensis в при
родных водоемах.
Применение второй методологии исследования можно найти в ряде работ.
Опыты с пятью видами Xanthium в Австралии [Martin, Carnahan, 1983], два из
которых распространились там очень широко, а три имеют ограниченный аре
ал, показали, что последние обладают более сильным врожденным и индуциро
ванным покоем семян.
Сравнение репродуктивного потенциала аборигенной на востоке США
Celastrus scandens и быстро расселяющейся интродуцированной C.orbiculatus
показало [Dreyer et al.,1987], что последняя имеет более высокие жизненность
пыльцы и всхожесть семян.
Два южноамериканских перечных дерева Schinus molle и S.terebinthifolius
были интродуцированы в Калифорнию более ста лет назад. Действительно
натурализовался там только S. molle. Изучали прорастание семян обоих видов
с целью определения механизма, препятствующего натурализации [Nilsen,
Muller, 1980]. Различия обнаружены по степени холодостойкости, по отноше
нию к воздействию фенольных кислот и во времени, протекающем между на
буханием и прорастанием. Именно медленное прорастание семян
S.terebinthifolius тормозит его возобновление, так как проростки не успевают
развить корни во время коротких периодов обильного увлажнения почвы.
При детальном изучении натурализовавшихся растений Новой Зелан
дии были сформулированы [Esler, 1988] параметры «идеального сорняка», ко
торые включают: экологическую многосторонность, быстрое созревание, пло
довитость, эффективность диссеминации, вегетативное размножение и конку
рентоспособность.
Таким образом, однозначного объяснения причин того, что чужеродные виды
более обильны в регионах вторичного ареала, чем в регионах естественного
ареала, до сих пор нет. В работе, посвященной этой проблеме [Hierro et al., 2005]
приведено восемь основных гипотез для объяснения успеха чужеродных рас
тений, внедряющихся в естественные сообщества. Первые четыре гипотезы
можно проверить путем сравнения аборигенных и инвазионных популяций,
последующие четыре можно оценить при изучении чужеродных видов только
во вторичном ареале. Мы протестируем эти гипотезы, используя данные, полу
ченные нами при изучении инвазионных видов Средней России.
Гипотеза 1. Чужеродные виды не имеют естественных врагов, которые
контролируют рост их популяций [Элтон, 1960]. Подтверждение гипотезы мы
получили при исследовании Echinocystis lobata. В естественном ареале его попу
46
ляции сильно поражают вредители и болезни, что обусловливает мозаичность
распространения этого вида, тогда как в Средней России подобного явления
не наблюдается [Виноградова, 2007]. Не отмечена и поражаемость вредителя
ми у большинства других инвазионных видов. Инвазионный вид флоры Мос
квы Adenocaulon adhaerescens Maxim. повреждается вредителями слабо и лишь
в конце вегетационного периода, тогда как в его естественном ареале на Даль
нем Востоке из за сильного поражения этого вида болезнями и вредителями с
трудом можно собрать растения для гербария. Из видов, входящих в инвази
онный компонент флоры Средней России, сильно поражается только Fraxinus
pennsylvanica (и только в Москве), его вредителем является ясеневая изум
рудная узкотелая златка, личинки которой вызывают усыхание деревьев. Од
нако подобное исключение только подтверждает правило, поскольку этот вид
златки также инвазионный.
Гипотеза 2. У чужеродных видов быстро эволюируют генетические при
знаки, связанные с давлением естественного отбора в новых условиях среды.
Гипотеза подтверждена нами при изучении Acorus calamus, Amaranthus albus,
Ribes aureum, Aronia mitschurinii, Acer negundo, Impatiens glandulifera, I.parviflora,
Epilobium adenocaulon, E.rubescens, Echinocystis lobata, Bidens frondosa, Conyza
canadensis, Chamomilla suaveolens, Galinsoga parviflora, G.quadriradiata [Виногра
дова, 1992]. Выявлена общая для этих видов черта -- все они на родине произра
стают вне зоны экологического оптимума и потому обладают широкой нор
мой реакции. Подробнее гипотеза рассмотрена в главе 4.
Гипотеза 3. Чужеродные виды приспособлены к типу и интенсивности на
рушений природной флоры, которые являются новыми для аборигенных ви
дов [Baker, 1974]. Думается, гипотеза представляет собой «более ученый» ва
риант предыдущей гипотезы -- чужеродные виды действительно лучше при
способлены к высокому уровню «беспорядочности», поскольку имеют более
широкую норму реакции генотипа.
Гипотеза 4. Чужеродные виды используют ресурсы, неиспользуемые або
ригенными видами [Элтон, 1960]. На начальном этапе внедрения на новые
территории чужеродные виды действительно поселяются в открытых место
обитаниях. Однако инвазионные виды именно вытесняют аборигенные виды,
то есть конкурируют с ними за имеющиеся ресурсы. Эксперименты по срав
нительной оценке конкурентоспособности близких видов, один из которых
инвазионный, другой -- аборигенный, проведены нами на видах рода Bidens.
Было подготовлено шесть образцов, представляющих собой смесь из 50 семян
североамериканской B.frondosa и 50 семян аборигенной B.tripartita, которые
высеяли в пикировочные ящики, вкопанные в грядки. В двух ящиках была су
песь, в двух -- садовая земля, в двух -- торф. Три ящика (с супесью, землей и
торфом) через день два усиленно поливали, в остальных трех растения росли
без искусственного полива [Виноградова, Возна, 2008]. По всхожести семян
B.frondosa получила преимущество лишь в четырех вариантах. В самых небла
гоприятных условиях (в ящике с супесью без полива и в ящике с торфом при
поливе) взошло на 30% больше семян B.tripartita. Однако, несмотря на это, во
всех шести вариантах опыта B.frondosa имела преимущество по приросту в вы
соту и по биомассе (в 2--3 раза) и по семенной продуктивности (в 3--5 раз).
47
Опыт наглядно продемонстрировал большую конкурентоспособность инва
зионного вида по сравнению с аборигенным (рис. 8). Поэтому данная гипотеза
в случае действительной инвазии, а не просто первоначального заноса вида на
новую территорию, должна иметь прямо противоположную формулировку: «чу
жеродные инвазионные виды более конкурентоспособны, чем аборигенные».
Гипотеза 5. Чужеродные виды могут воздействовать на виды естественных
популяций биохимическими методами [Bais et al., 2003]. Гипотезу мы экспери
ментально не проверяли, однако полное отсутствие естественной растительно
сти (даже тенелюбивых видов) травяного яруса в пойменных лесах из Acer
negundo и Fraxinus pennsylvanica является хотя и косвенным, но вполне доста
точным подтверждением гипотезы. Предварительные опыты позволили вен
герским исследователям [Solymosi, 1994] предположить, что экстракт из
Solidago canadensis может влиять на другие сорняки как аллелопатическое сред
ство. Cardaria draba является активным аллелопатическим растением, ее вы
деления подавляют всходы других видов [Kiemnec, Mcinnis, 2002; Qasem, 2004].
Иными данными по аллелопатии видов, входящих в инвазионный компонент
флоры Средней России, мы не располагаем.
Гипотеза 6. Чужеродные виды образуют гибриды с аборигенными, в резуль
тате чего возникает новый вид, способный к дальнейшей экспансии, а иногда
и к вытеснению родительских видов. Таким образом возник, например, болот
Рис. 8. Семенная продуктивность двух видов рода Bidens
на разных почвосмесях
48
ный злак Spartina x anglica -- естественный гибрид афроевропейского S.maritima
с американским S.alternifolia, завезенным на южное побережье Англии и зак
репившимся там в начале XIX века. Сейчас S.anglica стал почти космополи
том, захватив колоссальные пространства на побережье Англии, Франции, Се
верной и Южной Америки, Австралии и Новой Зеландии [Guйnйgou,
Levasseur, 1993]. Из видов, входящих в инвазионный компонент флоры Сред
ней России, гибридогенным является Reynoutria x bohemica -- гибрид между
R.japonica и R.sachalinensis. И, действительно, гибрид в экспериментальной
посадке дает больший прирост по биомассе, чем родительский вид R.japonica.
В Москве высокую активность проявляет Symphytum x uplandicum, гибрид
между местным S. officinale и кавказским S. asperum.
Amelanchier spicata, по видимому, также является естественным гибридом
между североамериканской A. сanadensis, интродуцированной в 1590 году из
провинции Квебек (Канада) для королевского ботанического сада в Париже, и
введенной в культуру в 1596 году A. ovalis, являющейся единственным видом
ирги с европейским ареалом [Куклина, 2006а].
Гипотеза 7. Многовидовые сообщества более стойки к вторжению чуже
родных видов, чем маловидовые [Элтон, 1960]. Теоретически увеличение ви
дового богатства значительно снижает возможности инвазии, видимо потому,
что более разнообразные сообщества полнее и эффективнее используют име
ющееся пространство, ограничивая ресурсы для местных видов. Снижение био
разнообразия, таким образом, создает благоприятные условия для инвазий в
эту систему, потенциально ускоряя потерю разнообразия и гомогенизацию биот
на земном шаре. Эта гипотеза, разработанная на основе теоретического моде
лирования, впоследствии была подтверждена экспериментально в лаборатор
ных условиях в микрокамерах с контролируемыми гомогенными условиями
обитания. Однако правомерность этой гипотезы для естественных сообществ
остается дискуссионной, поскольку экспериментальных данных, полученных
именно в природных неконтролируемых условиях, до сих пор мало. В связи
со спорностью вопроса нами был проведен эксперимент по искусственному
внедрению Bidens frondosa в естественные сообщества с варьирующим чис
лом слагающих их видов и с различными доминантами [Виноградова, 2003].
Вопреки теории оказалось, что на участках без явных доминантов и с
минимальным числом видов в фитоценозе всхожесть семян B.frondosa была са
мой низкой и составляла 10%. Вовсе не проросли семена на участке без доми
нантов с максимальным числом видов в фитоценозе. Зато 100% ная всхожесть
наблюдалась на двух участках: на первом монодоминантом была Zizania aquatica
и росло еще шесть видов, а на втором -- монодоминантом выступала Carex rostrata
и росло еще пять видов. В полидоминантных сообществах независимо от числа
слагающих их видов всхожесть составляла 10--40%. Следовательно, помимо
биоразнообразия фитоценозов, не последнюю роль играет и их структурирован
ность. Сообщество противостоит натиску чужеродных видов, если состоит из
2--3 выраженных доминирующих и 6--10 сопутствующих видов.
Гипотеза 8. Изменчивость уровня инвазибельности различных естественных
сообществ зависит от числа чужеродных видов, вторгающихся в сообщество
[Williamson,1996]. Думается, эта гипотеза вполне правомерна. Во всяком слу
49
чае, число адвентивных видов, отмеченных по берегам рек, обочинам дорог и
на пустырях, значительно превышает их число в лесных или степных фитоце
нозах. Соответственно имеется прямая корреляция и между числом адвентив
ных и инвазионных видов. Разработка методики экспериментальной провер
ки этой гипотезы станет для нас очередной задачей.
Очевидно, что успешность расселения заносных видов невозможно объяс
нить лишь одной причиной. Процесс натурализации задействует целый комп
лекс факторов, и, вероятно, для разных видов определяющие факторы в этом
комплексе будут различными.
50
Глава 4
МИКРОЭВОЛЮЦИЯ ИНВАЗИОННЫХ ВИДОВ
Успешному прогнозированию результатов инвазионного процесса препят
ствуют две проблемы: 1) невозможность предугадать вероятных «захватчи
ков»; 2) степень нарушенности естественной растительности не играет такой
определяющей роли, которая отводилась ей ранее. Если бы можно было точно
предсказать, что конкретный вид или вид, обладающий определенным набо
ром биологических признаков, будет самостоятельно завоевывать новые тер
ритории, задача состояла бы в усилении карантинной службы. Однако про
блема осложняется тем, что заносные виды могут долгое время произрастать
на ограниченной территории, и лишь затем происходит взрывное расширение
их ареала. К сожалению, моделей, прогнозирующих вторжение заносных ви
дов, не так много, и затрагивают они, в лучшем случае, только узкий диапазон
организмов. Между тем ботанические сады и садоводческие фирмы постоян
но вводят в культуру как новые дикорастущие виды, так и виды, успешно куль
тивируемые в других регионах, тем самым приводя в действие пусковой меха
низм микроэволюции. Начальная популяция интродуцированного вида неус
танно расширяется (благодаря усилиям по его размножению), но имеется
возможность постоянного возникновения и новых центров (например, за счет
введения в культуру других сортов того же вида на смежной территории).
Интродукция растений -- процесс, как правило, многократно повторяющий
ся; чаще всего он дублируется в разных точках континентов или стран.
Что же происходит после введения вида в культуру? Большинство интро
дуцированных растений произрастают в ботанических садах лишь благодаря
действиям человека. Однако некоторая часть видов доходит до стадии коло
низации, то есть способности размножаться самосевом, а половина из этой
части достигает стадии натурализации, то есть «сбегает из культуры» в есте
ственные ценозы. Таким образом, ботанические сады являются как бы резер
вуарами, подпитывающими естественные ценозы новыми видами, а конкрет
ные флоры постоянно обогащаются «сбежавшими экспонатами».
При изучении истории интродукции широко натурализовавшихся в Рос
сии в настоящее время растений установлено, что все из исследованных одно
летних видов являются беженцами из ботанических садов [Виноградова, 1992].
Так, мелколепестник канадский Conyza canadensis в 1646 году был завезен в
ботанический сад Нюрнберга как редкостное заморское растение. В XVIII веке
он был обнаружен в Южной Германии уже в качестве адвентивного вида. Эхи
ноцистис шиповатый Echinocystis lobata сбежал из культуры в ботаническом
саду в Румынии в 1904 году. Из ботанического сада Потсдама в 1896 году сбе
51
жала североамериканская Bidens frondosa. Недотрогу мелкоцветковую
Impatiens parviflora в конце XIX века выращивали в ботанических садах Льво
ва, Санкт Петербурга, Тарту, Риги и Пярну, откуда она и сбежала через 50 лет.
Chamomilla suaveolens сбегала дважды: из ботанического сада Гельсингфорса
(Финляндия) в 1849 году и из Краковского ботанического сада в 1886 году.
Южноамериканскую галинзогу мелкоцветковую Galinsoga parviflora культи
вировали в 1785 году в Парижском ботаническом саду, откуда и ведут свое
происхождение большинство европейских популяций этого вида. Перечень
беглецов можно пополнить и такими широко распространенными видами, как
Amaranthus retroflexus, Veronica persica, Cyclachaena xanthiifolia, которые через
ботанические сады разбрелись по всей Европе и стали злостными сорняками.
По счастью, не все заносные виды способны к натурализации. Подсчитано,
что только 10% заносных видов успешно доходят до стадии колонизации и
лишь 5% -- натурализации [Considine, 1986]. К тому же иммиграционный пе
риод (лаг фаза) длится обычно довольно долго. На примере ряда однолетних
растений нами установлено, что между первым появлением вида на новой тер
ритории и его внедрением в естественные ценозы может пройти до 100 смен
Таблица 8
Cравнительная оценка факторов микроэволюции в естественных фитоценозах
и в ботанических садах
Естественные фитоценозы
Ботанические сады
Мутационный процесс
Преимущество выживания
доминантной аллели А1
В новых условиях может получить
преимущество рецессивная аллель а2
Рецессивная аллель а2 встречается
редко, в основном в гетерозиготном
состоянии
Искусственное сохранение и
размножение рецессивной аллели а2
Поток генов
Скорость потока генов невелика
Скорость потока генов возрастает.
Увеличивается вероятность появления
рецессивной аллели а2 за счет
многократной интродукции
Отбор
Только естественный отбор
Естественный и искусственный отбор
Идет стабилизирующий отбор,
дающий преимущество генотипам, не
отклоняющимся от нормы
В новых условиях среды возрастает
роль ведущего отбора
Возникшие макромутанты обычно
нежизнеспособны
Макромутанты размножаются
искусственно
Дрейф генов
Минимален, так как в многочисленной
популяции есть возможность
восполнить потерю утраченного
генотипа
Низкая численность особей в
интродукционной популяции. Кража
или повреждение ведут к утрате
генотипа
52
поколений. Epilobium adenocaulon, имеющий естественный ареал в Северной
Америке, собран впервые в Британии в 1891 году и произрастал там в течение
полувека, не расширяя ареала. В Финляндии этот вид стал бурно расширять
ареал через 20 лет после первого появления. В Европейской России появился
не позднее 1909 года, но стал обычным растением только через 50 лет. Бурное
расселение Bidens frondosa началось лишь после Второй мировой войны -- че
рез 50 лет. Гималайский вид Impatiens glandulifera культивировался в России с
конца XIX в., а бурная его натурализация началась лишь через 60--70 лет. Сред
неазиатская I.parviflora появилась в Западной Европе в 1830 х годах, дичать
cтала через 10--15 лет, а стремительное расширение ее ареала и натурализация
начались лишь через 120 лет. Galinsoga parviflora интродуцирована в Англию
в 1796 году, найдена там вне ботанического сада через 13 лет, а массовое ее
распространение началось только через 125 лет! Североамериканский
Amaranthus albus впервые был отмечен в Италии в 1723 году, в Средней Евро
пе появился в конце XIX века, натурализовался только к 1960 м годам, то есть
через 250 лет! За этот период в первоначальной генетически обедненной ин
вазионной популяции происходит накопление генетической изменчивости.
Какими же путями происходит изменение генофонда интродуцированных
растений в ботанических садах? Здесь имеют место все четыре основных фак
тора эволюции, два из которых (мутационный процесс и поток генов) создают
изменчивость, а два (отбор и дрейф генов) регулируют эту изменчивость (табл.
8). Мутационный процесс в ботанических садах поддерживается благодаря ис
кусственному сохранению растений с рецессивной аллелью а2, нередко пред
ставляющих интерес с декоративной точки зрения. В новых условиях среды
эти растения могут даже получить преимущество выживания. Напротив, в ес
тественных устойчивых популяциях преимущество выживания имеет доми
нантная аллель А1, тогда как рецессивная а2 встречается крайне редко, в ос
новном в гетерозиготном состоянии.
Скорость потока генов в ботанических садах намного выше, чем в естествен
ных популяциях, поскольку многократная интродукция декоративных видов
из разных источников (из разных природных популяций или разных ботани
ческих садов) увеличивает вероятность многократного поступления рецессив
ной аллели а2 в интродукционные популяции. Процесс этот усугубляется еще
и тем, что в последнее время в России в связи со значительным сокращением
экспедиционных поездок ботанические сады стали друг для друга основными
поставщиками исходного интродукционного материала. В природных усло
виях возникшие макромутанты (в основном рецессивные) обычно нежизнес
пособны и, как правило, погибают. В ботанических же садах, применяя искус
ственное вегетативное размножение растений, макромутанты можно сохранять
сколь угодно долго. Этим способом в конечном счете созданы практически
все сорта широко распространенных вегетативно размножающихся декоратив
ных растений: розы, тюльпаны, сирень и т.д.
Значительные сдвиги частот аллелей А1 и А2 могут происходить в интро
дукционных популяциях случайным образом. Действие этого чисто случай
ного компонента в изменении генных частот, известного под названием дрей
фа генов, в естественных популяциях практически сведено к нулю, поскольку
53
в многочисленной популяции всегда есть возможность восполнить потерю ут
раченного генотипа. Однако в ботанических садах из за низкой численности
культивируемых особей любое случайное повреждение или кража десятка ра
стений может привести к полной утрате какого либо гена в интродукционной
популяции. Этот фактор сильно влияет на генофонд декоративных растений,
поскольку именно они чаще всего становятся объектами подобного варварс
кого отношения со стороны посетителей ботанических садов.
Таким образом, в интродукционных популяциях, по сравнению с природ
ными, все факторы эволюционного процесса (кроме естественного отбора) дей
ствуют интенсивнее и, помимо того, прибавляются еще два мощнейших фак
тора -- искусственный отбор и искусственная гибридизация. Факторы, созда
ющие изменчивость, дают начало процессу микроэволюции, а факторы,
регулирующие изменчивость, продолжают этот процесс, что приводит к уста
новлению новых частот вариантов. Генофонд интродукционных популяций
столь сильно преобразуется, что можно говорить о наличии процесса видооб
разования в ботанических садах. К сожалению, работ по сравнению генофон
да естественных и интродукционных (переживших несколько смен поколе
ний) популяций одного и того же вида слишком мало. Однако многочислен
ные примеры значительного расширения ареала «сбежавших из культуры»
видов являются неоспоримым свидетельством микроэволюции растений во
вторичном ареале.
Из всего вышеизложенного вытекают три проблемы. Во первых, встает воп
рос: можно ли считать, что интродукционные (особенно натурализовавшие
ся) и естественные популяции относятся к одному и тому же виду? Эта про
блема особенно остро касается агамоспермных видов. Если последовательно
применять критерии выделения видов, можно столкнуться со множеством
трудностей. Для современной систематики вид есть понятие морфолого эко
лого географическое, а по всем этим признакам дикорастущие и культивируе
мые особи сильно различаются. Так, широко распространенная в России чер
ноплодная арония, отличающаяся от родоначального вида Aronia melanocarpa
по числу хромосом, способу опыления, количеству и размерам цветков и пло
дов, форме листовой пластинки, зимостойкости, была описана в качестве но
вого вида Aronia mitschurinii [Скворцов, Майтулина, 1982]. Во вторых, уста
новление новых частот аллелей в интродукционных популяциях является важ
нейшим препятствием для успешности процесса репатриации видов в
естественные условия произрастания. Требуется такое же (если не большее)
число смен поколений для возврата генофонда в исходное состояние, незави
симо от того, изменились или нет условия среды в «родоначальном» место
обитании репатриируемого вида. И в третьих, «растением захватчиком» по
тенциально может стать любой интродуцированный вид. Следовательно, ана
лиз генетических ресурсов ботанических садов на предмет выявления видов,
которые размножаются самосевом и в ближайшем будущем могут превратиться
в сорняки, должен стать важной задачей на ближайшее десятилетие.
Внутривидовая изменчивость заносных видов выявлена у многих расте
ний, имеющих обширный вторичный ареал: у Zizania aquatica отмечена из
менчивость количественных признаков в северном Саскачеване, куда она за
54
несена была в 1935 году. [Peden, 1982]; Echium vulgare, занесенный из Старого
Света в Северную Америку в начале ХIХ века, обладает настолько высокой
амплитудой изменчивости, что некоторые авторы описывают его разновиднос
ти [Gleason, 1963]; мультивариантный анализ по 64 морфологическим призна
кам, проведенный в популяциях Bromus japonicus в штате Вашингтон, показал,
что экземпляры из мезофитных местообитаний имеют более значительную меж
популяционную изменчивость, чем из ксерофитных [Heidel et al., 1982].
Широкую вариабельность проявляют виды, интродуцированные в Авст
ралию в ХIХ веке -- Trifolium glomeratum [Woodward, Morley, 1974], три вида
рода Avena [Whalley, Burfitt, 1972] и др.
Изучение внутривидовой изменчивости интродуцированного в Северной
Америке Avena barbata на уровне генома [Allard, 1975] позволило выделить
у этого вида 35 вариаций генома, три из которых распространены довольно
широко.
Изучение внутривидовой изменчивости Trifolium hirtum показало, что раз
новидность Wilton, завезенная в Калифорнию, претерпела значительные эво
люционные преобразования, причем уже в самых ранних по времени пастбищ
ных посевах. Межпопуляционные различия отмечены и по морфологическим,
и по количественным, и по ферментативным признакам, однако большинство
генетических изменений проявлялось в виде изменения частот аллелей, а не в
виде появления новых аллелей. У калифорнийских популяций отсутствуют
две аминокислоты, имеющиеся в родоначальных турецких растениях. Разно
видности были более гомозиготными, чем естественные популяции, но они
также проявили большое генетическое разнообразие. Внутрипопуляционная
изменчивость отмечена для количественных признаков и времени цветения.
Авторы сделали вывод, что расширение ареала вида на новой территории обес
печивают быстрые микроэволюционные изменения, но не путем изоляции и
дрейфа, а, более вероятно, силами отбора [Martins, Jain, 1980].
При изучении генетической структуры 19 дикорастущих и культивируе
мых популяций Pennisetum typhoides было показано [Pilate Andre et al., 1987],
что отдельные локусы различаются по уровню полиморфизма аллелей. Уро
вень изменчивости дикорастущих популяций везде выше, чем культивируе
мых, причем наблюдается уменьшение градиента изменчивости популяций с
востока на запад африканского континента.
При изучении изменчивости аллозимов у трех видов Aeschynomene показа
но [Carilli, Fairbrothers, 1988], что заносный сорняк A. indica имеет большую,
по сравнению с аборигенным видом A.virginica, вариабельность.
Повышение генетической изменчивости в ходе колонизации растениями
новых местообитаний отмечено и у других интродуцированных видов: Ricinus
communis [Harland, 1947] и Avena fatua [Jain, Marshall, 1967].
Эволюция генетической пластичности наглядно изучена на примере
Sarracenia purpurea. Первая популяция вида возникла от посаженного на сфаг
новом островке в штате Огайо одного экземпляра. С помощью метода элект
рофореза аллозимов была изучена изменчивость популяции -- «основателя»
и 10 других популяций в США. В популяции на сфагновом островке изменчи
вость была самой низкой -- найден лишь один полиморфный локус (в некото
55
рих популяциях половина локусов была полиморфной), а гетерозиготность
на особь на 50% ниже, чем средняя по виду [Schwaegerle, Schaal, 1979].
По данным электрофореза ферментов, уровни генетической изменчивости
у Helianthus bolanderi ssp. exilis и сорных ssp. bolanderi и H.annuus существенно
не различаются [Rieseberg et al., 1988]. Различия наблюдаются в распределе
нии изменчивости: у H. bolanderi ssp. exilis -- в пределах популяции, а у сорных
таксонов -- между популяциями.
При изучении изменчивости 10 ферментных систем методом электрофо
реза у двух интродуцированных в Америку видов клевера установлено, что у
обоих видов изменчивы 8 локусов из 15 (у Trifolium hybridum в среднем 1,73
аллели на локус, а у T.pratense -- 1,93 аллели на локус). Межпопуляционная
изменчивость у обоих видов составляет в среднем 10%, а внутрипопуляцион
ная изменчивость довольно высокая [Hickey, Vincent, Guttman, 1991].
Определение среднего значения признаков, меры их пластичности, харак
тера фенотипической пластичности для видов в пяти родах и подсчет расхож
дений между видами по каждому из этих измерений показало, что фенотипи
ческая пластичность признаков может эволюировать независимо от самих при
знаков и что дивергенция видов по пластичности признаков вполне реальна.
Растения в одной популяции могут различаться по фенотипической пластич
ности; возможен отбор по этому признаку [Schlichting, Levin, 1986].
Интереснейшая работа проведена с Lupinus succulentus -- американским од
нолетником, который в Калифорнии стал поселяться на рудеральных местах.
Сравнивали генные частоты в естественных и рудеральных популяциях этого
вида и изучали полиморфизм в четырех локусах. Три локуса (ВВ, РР и DD)
отвечают за окраску цветков, а локус SS определяет рисунок семенной кожу
ры. Большинство популяций L. succulentus оказались мономорфными по ло
кусам ВВ, РР и DD. Лишь в одном из каньонов найдены популяции, полимор
фные по локусу D/d, здесь наблюдалась клинальная изменчивость -- от осно
вания к вершине каньона уменьшалось количество голубых цветов. Локус S/
s оказался полиморфным в подавляющем большинстве популяций, но чаще
это наблюдалось в популяциях, растущих в нарушенных местообитаниях. По
лиморфизм в локусах D/d, B/b и P/p найден только в нарушенных местообита
ниях. Таким образом, небольшие популяции, начинающие колонизировать но
вые местообитания, не только не являются генетически однородными, но, на
оборот, имеют большую генетическую изменчивость, чем естественные
популяции.
Данные о лабильности генетического аппарата видов колонизаторов име
ются в ряде других работ [Kornas, 1982; Oka, 1983]. Однако различия между
таксонами по пластичности отдельных признаков изучены недостаточно.
Экспериментальное изучение внутривидовой изменчивости инвазионных
растений впервые начал Дохальд, который отметил у адвентивного в Австралии
Trifolium subterraneum существование нескольких рас, различающихся по срокам
созревания при выращивании их в однородных условиях. Позднее изучение внут
ривидовой изменчивости этого же вида позволило выделить в Австралии более
130 его рас, различающихся не только по морфологическим признакам, но и по
устойчивости к болезням [Morley, Katznelson, 1965; Kellock et al., 1978].
56
Наиболее детально изучена вариабельность Chondrilla juncea -- облигатно
го апомикта, появившегося в Австралии в 1917 году. Исследованиями McVean
[1966] установлено, что в Австралийском регионе этот вид морфологически
единообразен. Но уже через 6 лет обнаружены три формы Ch. juncea, различа
ющиеся по морфологии соцветия, характеру плодов и форме розеточных лис
тьев [Hull, Groves, 1973]. По меньшей мере, в двух генерациях растений, выра
щенных в однородных условиях, эти морфологические различия сохранялись.
Формы различались также по продолжительности префлорального периода и
способности к вегетативному размножению. Среди растений формы А отме
чена клинальная изменчивость префлорального периода, который увеличи
вается с запада на восток.
Все три формы (А, В и С) произрастают вместе на площади ~3,3 млн
га. Вокруг этого участка имеется концентрический ареал, в котором встреча
ются только формы А и С. На внешней стороне этого второго ареала найдены
растения только формы А, которые широко распространены и встречаются в
Юго Восточной Австралии в районах с разным климатом и типом почв. Ареа
лы всех трех форм продолжают увеличиваться.
Применение электрофоретического метода позволило установить, что 8 эн
зимов из исследованных 14 дают для каждой из трех форм различные изозим
ные пятна [Burdon et al.,1980]. Дальнейшие исследования касались влияния
трех популяций клеща Aceria chondrillae на три формы Ch. juncea [Cullen, Moore,
1983]. Выяснилось, что первая популяция клеща поражает только форму А, а
две другие -- только форму В.
У двух близких видов Hordeum leporinum и H.glaucum, естественно произ
растающих в Европе, Западной Азии и Северной Америке, во вторичном аре
але в Австралии (где комплекс этих видов появился в 1840--1850 х годах), от
мечена межпопуляционная изменчивость времени вступления в фазу цвете
ния [Cocks et al., 1976]. Признак этот находится в коррелятивной зависимости
от длины вегетационного периода в пункте сбора семян, причем у H.leporinum
коррелятивная связь была более тесной, чем у H.glaucum.
Выявлен диапазон внутривидовой изменчивости Bromus mollis -- адвентив
ного вида североамериканской флоры [Allard, 1965]: количество побегов и вы
сота растений изменяются незначительно, а период развития от всходов до
колошения тем меньше, чем южнее произрастает популяция.
Имеются также экспериментальные данные об отличиях между популяци
ями различного географического происхождения адвентивного в Америке
Rumex crispus по требованиям к яровизации и фотопериодизму [Hume, Cavers,
1983].
Таким образом, в более или менее заметных масштабах изучение внутри
видовой изменчивости инвазионных видов ведется лишь последние 30 лет. За
это время были получены данные об изменчивости не более чем сотни видов
колонизаторов, причем экспериментальными методами изучено В 50 из них.
Наши исследования, проведенные в Главном ботаническом саду им. Н.В.
Цицина РАН [Виноградова, 1992], охватили 16 заносных видов, принадлежа
щих к различным жизненным формам, с различной биологией размножения,
в разное время и разным путем занесенные в нашу страну: Anisantha tectorum,
57
Acorus calamus, Amaranthus albus, Ribes aureum, Aronia mitschurinii, Acer negundo,
Impatiens glandulifera, I.parviflora, Epilobium adenocaulon, E.pseudorubescens,
Echinocystis lobata, Bidens frondosa, Conyza canadensis, Chamomilla suaveolens,
Galinsoga parviflora, G.quadriradiata. Общим для них было только то, что они
претерпевают процесс активной натурализации и расширения ареала.
Тем не менее следует понимать, по сравнению с громадным числом занос
ных видов, встречающихся в России, это капля в море. Требуются вовлечение
в исследования гораздо более широкого набора объектов и изучение их по сход
ной методике. В ходе исследования должны быть зафиксированы достовер
ные микроэволюционные сдвиги, произошедшие в натурализующихся видах
за более или менее точно определимый срок. Под микроэволюционным сдви
гом мы понимаем такое изменение генофонда популяции, которое приводит к
достаточно ясному изменению адаптивных свойств этой популяции. Появле
ние отдельных мутаций или каких либо индивидуальных отклонений, адап
тивный смысл которых неясен, в счет не идет. В связи с этим считаем целесо
образным привести, хотя бы схематично, методологию исследования внутри
видовой изменчивости и микроэволюции заносных видов:
· изучение истории интродукции и составление карты естественного и вторич
ного ареалов вида по литературным источникам и гербарным образцам, что позво
лит определить продолжительность периода эволюционной дифференциации;
· выявление диапазона модификационной изменчивости морфологических
признаков в природных условиях путем обследования естественных и инва
зионных популяций изучаемых видов, причем особое внимание уделяя при
знакам, имеющим диагностическое значение;
· изучение биологических признаков семян различного географического
происхождения посредством проращивания семян в лабораторных при раз
личных температурных режимах и сроках стратификации, а в условиях пи
томника -- при различных сроках посева: подзимнем, весеннем и летнем;
· выращивание растений в однородных почвенно климатических условиях
из семян различного географического происхождения (создание интродукци
онных популяций). Сбор семенного материала проводится вдоль широтного
профиля вторичного ареала изучаемых видов -- от крайних северных до край
них южных точек произрастания вида на территории России. Для более пол
ного изучения полиморфизма растений можно использовать также семена, при
сланные по делектусам. Для определения эндогенной изменчивости семена
собирают с каждого экземпляра отдельно. Внутрипопуляционную изменчи
вость оценивают в интродукционных популяциях, сформированных из се
мян, собранных в 4--6 микропопуляциях с 10--15 растений в каждой, в двух
повторностях. Выращивание образцов всех сравниваемых популяций в од
нородных условиях питомника позволяет оценить действительно генотипы,
а не вариации фенотипов;
· изучение в интродукционных популяциях темпов и продолжительности
роста, фенологии и соответствующих фенофазам морфологических измене
ний, способа опыления; для некоторых видов необходимо изучение вопроса
об озимости/яровости, а также фотопериодической реакции сравниваемых
образцов;
58
· сравнение диапазона и характера изменчивости изучаемых видов в пер
вичном и вторичном ареалах;
· выявление различий изучаемых видов в естественном и вторичном ареа
лах молекулярно генетическими методами;
· в очень редких случаях сравнение конкурентоспособности близкород
ственных аборигенного и заносного видов.
Целью подобных исследований должны стать выявление основных законо
мерностей микроэволюции, происходящих при инвазии растений, и определение
факторов, влияющих на процессы микроэволюции, а также диагностика призна
ков, способствующих широкой натурализации видов колонизаторов, и установ
ление зависимости процессов микроэволюции от способа размножения, жизнен
ной формы, отношения к длине дня и других биологических признаков.
Результаты изучения амфимиктных видов
Acer negundo появился в России почти двести лет назад, но на большей
части ареала он произрастает лишь с конца XIX -- начала ХХ века, а первые
упоминания о внедрении клена в аборигенную флору и размножения его са
мосевом относятся к 1920 м годам. Учитывая, что клен вступает в стадию пло
доношения на 6--7 м году жизни, можно утверждать, что эволюция приспосо
бительных признаков затронула лишь 9--11 поколений вида.
В естественном ареале у A.negundo наблюдается клинальная изменчивость
количественных признаков плодов: от севера к югу происходит постепенное
уменьшение длины, ширины и массы крылатки. Длина крылаток северного
происхождения превышает длину крылаток южного происхождения в 1,5 раза,
а масса крылаток -- в 2 раза [Williams, Winstead, 1972; Виноградова, 2006]. Во
вторичном ареале изменчивость плодов изучена в инвазионных популяциях в
Ашхабаде, Астрахани, Москве и Вологде. Внутрипопуляционная изменчивость
оказалась довольно высокой: длина крылатки варьировала у ашхабадских об
разцов от 3,7 до 4,6 см, у московских -- от 3,3 до 4,5 см, а у астраханских и
вологодских -- от 2,9 до 4,6 см (коэффициент вариации 20--25%). Наибольшая
амплитуда изменчивости (коэффициент вариации 30--36%) отмечена для сред
ней массы крылаток, которая варьировала от 20 до 88 мг. Однако по среднему
значению длины, ширины и массы крылаток исследованные инвазионные по
пуляции практически не различались. Таким образом, во вторичном ареале
внутривидовая изменчивость количественных признаков плодов A.negundo не
носит четко выраженного географического характера и является в основном
внутрипопуляционной (рис. 9).
В лабораторных условиях семена различного географического происхож
дения проявили сходство по требованиям к определённой температуре прора
стания и сроку стратификации, однако семена A.negundo из южных частей вто
ричного ареала обладали меньшим периодом покоя, чем семена из более се
верных районов.
Интродукционная популяция была заложена из семян, собранных в Ашха
баде, Астрахани, Москве и Вологде. Семена московского происхождения име
ли наиболее высокую всхожесть -- 69%; всхожесть астраханских семян -- 40%.
Семена, собранные в крайних точках ареала -- Вологде и Ашхабаде, имели очень
59
низкую всхожесть (вологодские -- 23%, ашхабадские -- 26%), а также низкую
энергию прорастания, которая вычислялась за первые 15 дней.
Установлено наличие клинальной изменчивости ритма развития однолет
них сеянцев. Чем севернее находится пункт сбора семян, тем раньше распус
кается соответствующая пара листьев у выросших из них растений; различие
между образцами вологодского и ашхабадского происхождения составляет 2
недели. Сеянцы московской репродукции в основном закончили свой рост в
первой декаде сентября, вологодской и астраханской репродукций -- на неде
лю позже, а ашхабадской репродукции -- на месяц позднее. В связи с продол
жительным периодом роста даже к концу ноября хорошо сформированной вер
хушечной почки не было у 44% сеянцев московской репродукции, у 71% сеян
Рис. 9. Внутривидовая изменчивость крылаток
A.negundo во вторичном ареале
60
цев вологодской репродукции, у 77% сеянцев астраханской и у 92% сеянцев
ашхабадской семенных репродукций.
Однолетние сеянцы в большой степени пострадали от мороза в суровую
зиму 1978/79 г. Полностью вымерзли 6% сеянцев ашхабадской репродукции,
остальные 94% сильно обмерзли (в среднем на 5--6 междоузлий, то есть на 24 см).
Вымерзли также 2% сеянцев вологодской и 1% сеянцев астраханской репро
дукций, соответственно 12 и 3% из них существенно обмерзли. Среди сеянцев
московского происхождения полностью вымерзших экземпляров не было, 23%
сеянцев совершенно не пострадали от морозов, а у остальных подмерзли 1--2
междоузлия (6--7 см). К началу второго вегетационного периода средняя вы
сота растений московского происхождения в 3 раза превышала высоту расте
ний ашхабадского происхождения и в 1,5 раза -- высоту растений вологодско
го и астраханского происхождения.
За три периода вегетации наибольшим приростом по высоте, диаметру и
биомассе отличались деревца местной (московской) семенной репродукции.
У них отмечены также наименьшая продолжительность периода роста и наи
большая морозостойкость. К концу 3 го года жизни деревья московского про
исхождения были в 1,5 раза выше деревьев ашхабадского происхождения; пос
ледние к тому же из за ежегодного обмерзания побегов имели кустообразную
форму.
Таким образом, за 9--11 смен поколений у A.negundo уже выработалась ге
нетически закреплённая клинальная географическая изменчивость некоторых
биологических признаков: продолжительности периода покоя семян, морозо
стойкости деревьев, прироста по диаметру стволика и по биомассе растений, а
также ритма их развития.
Impatiens glandulifera в качестве декоративного растения выращивали в
садах Европы с середины XIX века, в Северной Америке -- с начала ХХ столе
тия. Первые случаи дичания отмечены через 20 смен поколений, внедрение
в естественные ценозы началось через 60, а бурная экспансия -- через 100
смен поколений.
Интродукционная популяция была заложена при осеннем посеве семян,
собранных в городах Санкт Петербург, г.Крестцы Новгородской области, Во
логда, Москва, Львов, Дубинки (Прикарпатье), Мукачево (Закарпатье).
Установлено наличие клинальной изменчивости ритма развития недотро
ги: чем севернее находится пункт сбора семян, тем быстрее развиваются рас
тения. В интродукционной популяции растения из семян закарпатского про
исхождения вступают в генеративный период развития на 5 недель позднее,
чем вологодские, причем 2/3 экземпляров не успевают даже вступить в ста
дию плодоношения, погибнув от первых заморозков еще в цвету. Все растения
северных образцов полностью проходят жизненный цикл и заканчивают его в
третьей декаде сентября еще до заморозков.
Отмечена клинальная изменчивость прироста по высоте и по биомассе ра
стений: от севера к югу у сравниваемых образцов наблюдается достоверное
увеличение средней высоты главного побега, среднего числа метамеров на глав
ном побеге, биомассы надземной части, а также продолжительности периода
роста растений. Северные образцы раньше закончили рост, и к концу вегета
61
ционного периода высота их была невелика: особи крестцовской репродук
ции в среднем достигли высоты 63 см и имели 10 метамеров, петербургской --
72 см (12 метамеров), московской -- 90 см (10 метамеров) и вологодской -- 107
см (12 метамеров). Южные образцы росли дольше, и к концу вегетации львов
ские экземпляры имели среднюю высоту 122 см (13 метамеров), а мукачевские --
111 см (13 метамеров).
Таким образом, за 150 смен поколений у I.glandulifera во вторичном ареале
выработалась генетически закреплённая географическая изменчивость ритма
роста и развития растений.
Экспериментально установлено, что в результате процесса адаптации и сво
бодного переопыления растений в интродукционной популяции, в ряду пос
ледующих поколений микроэволюция адаптивных признаков идет в сторону
нивелирования различий между географическими образцами и сближением
их с московскими экземплярами. Одновременное вступление в одну и ту же
фенофазу и одинаковый темп роста наблюдались уже у растений 5 го поколе
ния (рис. 10).
Изучено влияние длины дня на рост и фенологию образцов различного гео
графического происхождения. Для этого сравнивали темпы роста и фенорит
мы растений, которые на стадии проростков в течение трех недель росли при
искусственном коротком дне (9 часов) с контрольными растениями, произра
ставшими в течение того же периода при естественной длине дня.
Особи вологодской, крестцовской и львовской репродукций, росшие при
коротком дне, вступили в соответствующие фенофазы на 3 недели позднее,
чем контрольные растения. У московского образца опытные растения вступи
ли в генеративный период развития на неделю позднее контрольных. У даль
невосточного образца опытные растения зацвели практически одновременно
с контрольными.
У московского и владивостокского образцов влияние длины дня на высоту
растений и на число метамеров достоверно не сказалось. У образцов крест
цовского, вологодского и львовского происхождения, напротив, отмечался
более медленный рост опытных растений.
Следовательно, в eвропейской части вторичного ареала недотрога желез
коносная является длиннодневным растением, а в дальневосточной части --
растением, нейтральным по отношению к длине дня. В качестве гипотезы мож
но выдвинуть предположение, что длительное произрастание I.glandulifera в
Европе до начала его стремительной натурализации связано с тем, что это ко
роткодневное в естественном ареале растение не было способно эффективно
обсеменяться в новых условиях. Однако, как только в результате рекомбина
ции генов появились экземпляры, способные полностью завершить жизнен
ный цикл в условиях длинного дня, началось продвижение именно этой длин
нодневной формы на север и стремительное расширение ареала недотроги.
Наша гипотеза противоречит выводам английских ученых [Willis, Hulme, 2002]
о ведущей роли температуры в комплексе факторов, лимитирующих расши
рение вторичного ареала недотроги. Нам она, однако, кажется предпочтитель
нее, поскольку в естественном ареале I.glandulifera достаточно холодостойка --
ведь она растет в горах до 3500 м над уровнем моря!
62
Echinocystis lobata имеет в Евразии два изолированных очага расселения --
Центральная Европа, где вид появился в начале ХХ века, и Дальний Восток,
куда он был занесен непосредственно из Северной Америки в 1920 е годы.
Бурное, взрывное расселение E.lobata по центральным районам Европейской
России началось после 1960 года. В 1970 х годах он завоевал уже обширную
территорию и вошел в естественные ценозы Средней России.
И в естественном, и во вторичном ареале индивидуальная изменчивость и
качественных, и количественных признаков семян E.lobata выражена слабо.
Однако эти признаки сильно варьируют на внутри и межпопуляционном уров
нях, хотя клинальной изменчивости признаков в инвазионных популяциях
Рис. 10. Феноритм I.glandulifera в ряду последовательных поколений
(P, F1, F2, F3, F4)
1. С. Петербург; 2. Крестцы; 3. Вологда; 4. Москва; 5. Львов; 6. Дубинки (Прикарпатье);
7. Мукачево (Закарпатье).
а -- вегетативный период; б -- цветение; в -- семеношение
63
не прослеживается. Выявлено 17 дискретных вариаций семян, встречающих
ся и в естественном, и во вторичном ареале [Виноградова, 2007]. Степень реа
лизации фенофонда в 90% изученных популяций составляла 100%; лишь в
двух популяциях на северном пределе вторичного ареала отсутствовало 2--3 фена.
Таким образом, в инвазионных популяциях перекрестноопыляющихся видов, рас
ширяющих вторичный ареал (к которым, несомненно, относится E.lobata), сохра
няется высокий уровень генетического разнообразия, и именно это обеспечивает
успешную адаптацию видов к новым условиям произрастания.
Интродукционные популяции закладывали неоднократно из семян, собран
ных от Закарпатья до Владивостока и от Вологды до Таганрога; всего испыта
ны 43 локальные популяции.
а) Изменчивость периода покоя семян. При весенних сроках посева не от
мечено заметных отличий в темпах прорастания как нестратифицированных,
так и стратифицированных семян различного географического происхожде
ния: семена начинали прорастать на 5--6 е сутки после посева и полностью
прорастали на 10 й день. Однако при осеннем посеве проявляется тенденция к
более быстрому прорастанию семян, собранных в северной части ареала. На
пример, в 1984 году к 18 апреля взошло 26% семян северного происхождения
(из Ленинградской, Новгородской и Вологодской областей), 15% семян мос
ковского происхождения и только 8% семян закарпатского происхождения.
б) Изменчивость фенологического ритма развития и темпов роста. В 1978
году растения московского и закарпатского происхождения вступили в гене
ративный период развития на 1,5 мес. раньше, чем владивостокского, причем
последние к концу вегетационного сезона не успели образовать полноценные
всхожие семена. Интенсивность и продолжительность роста убывали в следу
ющей последовательности: владивостокские образцы (длина главной оси в
конце вегетации 2,5--3 м), московские и закарпатские (1--2 м). Разница в сро
ках вступления в одну и ту же фенофазу между московскими и закарпатски
ми экземплярами составляла всего 1 неделю, а по длине главной оси образцы
достоверно не различались. Это дало нам основание утверждать [Виноградо
ва, 1992], что внутри восточноевропейской части ареала адаптивная изменчи
вость биологических признаков эхиноцистиса четко не выражена, а имеются
лишь два биотипа, отличающихся по темпам роста и фенологическому ритму
развития -- восточноевропейский и дальневосточный.
Суровая зима 1978/79 наложила серьезный отпечаток на распространение
эхиноцистиса. Многие из наблюдавшихся нами популяций вымерзли совсем,
а оставшиеся насчитывали всего по 3--5 особей. Именно после этой зимы бла
годаря естественному отбору наиболее морозостойких экземпляров началось
стремительное продвижение E.lobata в северные регионы.
Бурное расширение ареала сопровождалось значительным темпом адаптив
ной эволюции: эхиноцистис выработал и генетически закрепил важнейшие
адаптивные признаки: более короткий период роста и быстрое прохождение
полного цикла развития (на 2--3 месяца скорее) по направлению с севера на
юг. В интродукционной популяции у самых северных новгородских образцов
семена прорастают на 2--3 недели раньше, период роста заканчивается в конце
июля, главная ось практически не ветвится, общая длина всех побегов не пре
64
вышает 1--2 м, цветение наступает в середине июня, плоды завязываются в
пазухе 5--13 го листа. У закарпатских образцов, напротив, период роста про
должается до первых заморозков, растения сильно ветвятся, образуя побеги
II, III и даже IV порядка, общая длина всех побегов достигает 9--13 м, цве
тение наступает в конце августа, плоды завязываются в пазухе 60--80 го
листа, семена практически не вызревают (рис. 11). Внутрипопуляционная
изменчивость изученных адаптивных признаков значительно ниже меж
популяционной.
Сумма эффективных температур к моменту начала той или иной фенофа
зы во все годы наблюдений различалась столь сильно, что мы констатируем
отсутствие корреляции между этими величинами.
в) Изменчивость фенологии и темпов роста в ряду последовательных по
колений. Сравнение фенологического ритма родительских и дочерних особей
проводили путем одновременного высева Р и F1 семян. При свободном опы
лении в интродукционной популяции уже в 1 м поколении происходит сбли
жение феноритмов дочерних особей как южных, так и северных образцов с
образцами местного, московского происхождения: дочерние особи вологодс
кого происхождения зацветают на неделю позднее родительских, а дочерние
особи закарпатских образцов -- на 2 недели раньше родительских. Однако по
количественным признакам, характеризующим нарастание биомассы, роди
тельские и дочерние экземпляры не различаются. Следовательно, рост расте
ний -- показатель менее пластичный, чем феноритм. При изменении условий
существования растения в течение по меньшей мере 1 года сохраняют темпы
прироста вегетативной массы, тогда как феноритм их изменяется в сторону
сближения с феноритмом образцов местного происхождения.
г) Влияние длины дня. Поскольку мы не выявили зависимости между фено
ритмом растений и суммами эффективных температур, был поставлен опыт по
выращиванию образцов различного географического происхождения при разных
световых режимах. В середине июня, когда на растениях полностью раскрылся 1
й настоящий лист, опытные экземпляры накрывали плотной черной бумагой, со
здавая короткий 8 часовой световой день. Контрольные экземпляры росли при
полном световом дне, который в июне на широте Москвы составляет 17,3 часа.
Многолетние наблюдения показали, что растения одного и того же места
сбора зацветали в одно и то же время, несмотря на различия суммы эффектив
ных температур к дате начала цветения.
У растений северных репродукций длина главной оси и число ее метаме
ров выше в условиях короткого дня, у растений же южных репродукций, наобо
рот, длина главной оси и число ее метамеров выше в условиях длинного дня
(рис. 12), у московского образца достоверных различий между опытными и
контрольными экземплярами не зафиксировано. Ритм развития растений в
условиях короткого дня с севера на юг убыстряется практически в 2 раза.
У растений северных репродукций феноритм контрольных и опытных эк
земпляров не различается; растения московской репродукции при коротком
дне вступают в генеративный период развития на 2 недели раньше, а карпатс
кой репродукции -- на 1--1,5 месяца раньше, что дает последним возможность
на широте Москвы завязать плоды, а иногда и дать зрелые семена.
65
Рис. 11. Фенология образцов E.lobata различного географического
происхождения
Рис. 12. Сравнение морфометрических параметров образцов E.lobata из южной и се
верной части вторичного ареала, выросших в условиях длинного и короткого дня
1 -- в условиях длинного дня, 2 -- в условиях короткого дня; а -- длина главной оси E.lobata
из Новгородской обл. (Едрово); б -- общая длина всех побегов E.lobata из Новгородской
обл. (Едрово); в -- длина главной оси E.lobata из Ужгорода; г -- общая длина всех побегов
E.lobata из Ужгорода
66
Рис. 13. Изменчивость количественных признаков образцов
Chamomilla suaveolens в интродукционной популяции
Рис. 14. Феноритм Bidens frondosa в интродукционной популяции при
различной длине дня
1 -- образцы московского происхождения, естественная длина дня; 2 -- короткий день;
3 -- образцы из Приморья, естественная длина дня; 4 -- короткий день: а -- вегетация, б --
бутонизация, в -- цветение, г -- плодоношение
67
Таким образом, сформированный во вторичном ареале и генетически зак
репленный феноритм E.lobata в значительной степени обусловлен продолжи
тельностью длины дня в местах его произрастания.
Результаты изучения самоопылителей, апомиктов
и вегетативно размножающихся видов
Chamomilla suaveolens появилась в Европе в середине XIX века и начала
внедряться в нарушенные ценозы как «беженец» из ботанических садов. В юж
ной Финляндии и в Петербурге этот процесс занял 20 лет, в Москве -- 10 лет, в
Томске -- 4 года. Бурная натурализация вида наблюдалась в начале ХХ века.
Следовательно, между первым появлением Ch. suaveolens в Европе и натура
лизацией вида прошло не менее 20 смен поколений. В настоящее время в Рос
сии произрастает по меньшей мере 100 ое поколение этого вида.
Изменчивость морфометрических признаков Ch. suaveolens изучали в инт
родукционной популяции, сформированной из семян, собранных в Сыктыв
каре, Твери, Москве, Орле и в Казахстане в окрестностях Алма Аты [Виног
радова, 2001].
У образцов сыктывкарской и орловской репродукций цветоносные побеги
начали появляться в середине июня, у растений алма атинского происхожде
ния -- в конце июля, а у образца из Твери -- только в середине августа. Экземп
ляры сыктывкарского происхождения зацвели в начале июля, орловского -- на
2 недели позднее, алма атинского -- на месяц позднее и тверского -- на 2 месяца
позднее. У образца из Твери 25% растений так и не образовали ни одного цвето
носного побега и погибли от осенних заморозков на стадии розетки.
Период роста сыктывкарских растений закончился к началу августа, об
разцов из Орла и Медео -- к началу сентября, а тверской образец продолжал мед
ленно расти до начала октября. К концу вегетационного периода тверской обра
зец имел самые короткие побеги, наименьшее число листьев и корзинок и досто
верно отличался от всех остальных образцов (рис. 13). Отмечено и достоверное
различие по длине побега между сыктывкарскими и орловскими образцами.
Таким образом, у ромашки душистой в ее вторичном ареале выявлено наличие
нескольких биотипов, различающихся по фенологическому ритму развития. При этом
изменчивость феноритмов носит неупорядоченный, а не клинальный характер.
Bidens frondosa в естественном ареале является высокополиморфным ви
дом. Различные разновидности B.frondosa заносились в Европу с 1762 года,
однако натурализовались только две из них: B.frondosa var. frondosa с тройча
тосложными листьями, короткими листочками обертки и коричневыми боро
давчатыми семянками и B.frondosa var. anomala, которая отличается от типич
ной разновидности вверх направленными зубцами на ости семянки. Началом
формирования вторичного ареала B.frondosa var. frondosa в Европе следует
считать 1896 год, а B.frondosa var. anomala -- 1954 й.
На территории бывшего СССР существует три изолированных очага рас
пространения B.frondosa: Приморье, куда она была занесена из Америки не
позднее 1950 года, запад европейской части, где этот вид стал расселяться с
1955 года, и Черноморское побережье Кавказа, куда череда занесена в конце
1970 х годов. Все растения представляют собой типичную разновидность.
68
Единственная находка B.frondosa var. anomala сделана в 2003 году в Москве.
За последние 15--20 лет B.frondosa начала активно расширять вторичный аре
ал и внедряться в естественные фитоценозы по берегам водоемов, вытесняя
аборигенную B.tripartita [Виноградова, 2009].
Интродукционная популяция заложена из семян приморского (окрестнос
ти оз. Ханка), ростовского (Ростов на Дону) и московского происхождения
[Виноградова, 2003]. Экземпляры, выросшие из дальневосточных семян, на
чали бутонизировать в середине сентября и погибли от осенних заморозков,
так и не начав цвести. Экземпляры московского происхождения образовали
бутоны в середине июля, зацвели в начале августа и дали плоды в конце сен
тября (рис. 14).
Отмечен более быстрый рост растений московского происхождения, мак
симальная интенсивность которого приходится на июль. Максимальная ин
тенсивность роста дальневосточных образцов приходится на август. Поэтому
в начале августа московские экземпляры были значительно выше дальневос
точных и имели больше метамеров, а в начале октября оба образца по высоте
уже не различались, а по числу метамеров дальневосточные растения даже до
стоверно перегнали московские.
Внутрипопуляционная изменчивость ритма роста и фенологии B.frondosa
оказалась очень низкой. Коэффициент вариации даже самого изменчивого
признака -- высоты растений -- составлял всего 8--12%.
Влияние длины дня на рост и фенологию череды в интродукционной попу
ляции изучали путем сравнения темпов роста и феноритмов опытных расте
ний, произраставших в стадии проростков в течение 3 недель при искусствен
ном коротком дне (9 часов) с контрольными растениями, произраставшими
при естественной длине дня [Майтулина, 1990]. Опытные растения и москов
ского, и дальневосточного происхождения начали бутонизацию в конце июня
и полностью прошли весь жизненный цикл до середины октября. Таким обра
зом, при коротком дне московские образцы вступают в генеративный период
развития на месяц раньше контрольных, а дальневосточные -- на 2,5 месяца
раньше контрольных, что дает им возможность сформировать на широте Мос
квы зрелые семена. По темпу роста опытные растения различного географи
ческого происхождения друг от друга не отличались. Следовательно, в даль
невосточной части ареала B.frondosa является короткодневным растением и
на широте Москвы обнаруживает более медленный рост и неспособность всту
пить в стадию цветения. Наоборот, в европейской части ареала произрастает
длиннодневная форма череды, отличающаяся более быстрым ростом и спо
собностью полностью проходить жизненный цикл при естественной длине дня
на широте Москвы.
Между образцами, выросшими из семян, собранных внутри изолирован
ных очагов вторичного ареала, не отмечено различий ни по темпам роста, ни
по фенологическому ритму развития.
Impatiens parviflora появилась в Европе в 1830 е годы, начала дичать че
рез 10--15 лет, стремительное расширение ее ареала и натурализация начались
через 120 лет. В Европейской России и Балтии культивировалась с 1870 х го
дов, дичать стала через 50 лет, превратилась в массовый сорняк через 100 лет.
69
Интродукционная популяция закладывалась и при осеннем, и при весен
нем посеве семян [Майтулина, 1988; Виноградова, 1992]. Семена желтоцвет
ковой формы собраны в Санкт Петербурге (4 образца), Вологде (1 образец),
Москве (2 образца, один из них имел почти белые лепестки), Алма Ате (1 об
разец), Таджикистане (3 образца с Памира) и Туркмении (1 образец с хребта
Кугитанг); семена лиловоцветковой формы собраны в Туркмении (1 образец с
Кугитанга) и Таджикистане (6 образцов).
При осеннем посеве единичные всходы появились во второй декаде, а мас
совые всходы -- в третьей декаде апреля. Практически одновременно (в 20 х
числах мая) растения вступили в фазу бутонизации, в первой декаде июня -- в
фазу цветения. В середине июня все растения образовали плоды, а в начале
июля стали разбрасывать семена. Исключение составили желтоцветковые об
разцы из Рушана и Кугитанга, у которых долгое время часть особей не вступа
ла в стадию цветения, а также лиловоцветковые образцы из Комсомолабада и
Калаи Хумба, плоды которых стали быстро растрескиваться и рассеивать се
мена. Однако эти различия не превышали 2 х недель. Следует отметить, что у
особей, находящихся в стадии плодоношения и семеношения, новые бутоны и
цветки продолжали образовываться вплоть до осенних заморозков.
Уже в середине июля после семеношения часть особей стала засыхать, но
только лиловоцветковый образец из Кугитанга полностью выпал в конце сен
тября. У остальных образцов часть особей отмирали, а дркгие продолжали раз
виваться до середины октября.
Амплитуда изменчивости высоты растений была невысокой. Имели место
достоверные межпопуляционные различия между образцами по этому пара
метру, но они не носили клинального характера.
Число метамеров главного побега, число боковых побегов, цветков и плодов
оказались весьма изменчивыми как внутри образца, так и между образцами.
При весеннем посеве нестратифицированными семенами к середине мая
проросли лишь единичные экземпляры из Кугитанга и Таджикистана, кото
рые начали бутонизировать в середине июня. Следовательно, в естественном
ареале имеются отдельные особи, не нуждающиеся в стратификации семян.
Таким образом, у недотроги мелкоцветковой наблюдается сильная внутри
популяционная изменчивость по числу метамеров побега, числу цветков и пло
дов и по степени ветвления, сильная межпопуляционная изменчивость (не но
сящая, однако, закономерного характера) по высоте, числу метамеров главно
го побега, числу цветков и плодов и по степени ветвления. Существенных
различий по темпам роста и фенологии как внутри образцов, так и между об
разцами, не отмечено.
Amaranthus albus в 1860 х годах был занесен в порты Черного моря и на
чал распространяться по южным районам Российской империи. Однако в бо
танических садах Москвы и Санкт Петербурга его культивировали еще в 1812
году [Виноградова, 2002].
При осеннем посеве семян, собранных в Московской области, Узбекиста
не, Абхазии и Киргизии, всходы у всех образцов появились в последней декаде
мая. Растения дружно прошли цикл развития и дали зрелые семена: образцы из
Ферганы, Сухуми и Подмосковья -- в конце июля, а образец из Бишкека --
70
на неделю позднее. Никаких других различий между образцами не наблюда
лось -- все представляли собой типичную форму A.albus f.albus. Более того,
количественные признаки (высота, число побегов II и III порядков, размеры
листовой пластинки) также варьировали незначительно.
При весеннем посеве нестратифицированных семян, собранных в Москов
ской и Ростовской областях, Узбекистане, Абхазии, Грузии и Киргизии, всхо
ды появились через 12 дней после посева, цветение наступило через 40 дней
после прорастания и продолжалось до осенних заморозков, семена образова
лись через месяц и стали осыпаться через 1,5 месяца после начала цветения.
Все эти «весеннепроросшие» растения образовали побеги II, а некоторые -- и
III порядка; ни стелющихся, ни подушкообразных особей отмечено не было.
Никаких различий ни в морфологии, ни в сроках прохождения фенофаз нами
не выявлено -- все особи относились к типичной A.albus f.albus. Наибольшей
высотой отличался подмосковный образец (рис. 15), он же содержал наиболь
шее число особей (45%), имеющих побеги III порядка. Самыми низкими и сла
боветвящимися оказались оба образца из Ростовской области (средняя высо
та 23 см).
В июне -- начале июля начали прорастать оставшиеся в почве семена всех
образцов. Выросшие из них «летнепроросшие» неотеничные особи скорее всту
пили в генеративный период развития, чем весенние, но росли очень медлен
но, в конце вегетации не превышали 10 см в высоту, совершенно не ветвились
и представляли собой A.albus f.minimus. К осени образцы из Средней Азии на
Рис. 15. Средняя высота двух форм Amaranthus albus в интродукционной
популяции
71
22--26% состояли из A.albus f.minimus; образец подмосковного происхождения --
на 33%, грузинского -- на 44--62%, а в образцах ростовского происхождения
эти карликовые экземпляры превалировали и составляли от 46 до 75%. Наи
меньшая высота экземпляров A.albus f.minimus отмечена в подмосковном об
разце (средняя высота 3 см) и образце из Рустави (4 см). У остальных образ
цов высота A.albus f.minimus составляла 8--10 см.
Таким образом, описываемые в литературе формы A.albus являются отра
жением модификационной изменчивости вида. Генетически закреплены лишь
две формы: типичная A.albus f.albus, семена которой прорастают весной, и нео
теничная A.albus f.minimus, семена которой прорастают в середине лета. Об
щая генетическая изменчивость А.albus во вторичном ареале очень низка, что
объясняется обедненным генофондом первоначальных малочисленных инва
зионных популяций в сочетании с самоопыляемостью вида. При этом измен
чивость феноритмов не выражена, а изменчивость ритма роста носит неупо
рядоченный характер.
Epilobium adenocaulon занесен в северо западные районы СССР в конце XIX
века, однако встречался крайне редко вплоть до 1960 х годов [Скворцов, 1976].
При осеннем посеве семян, собранных в Архангельске, Москве, Сыктывка
ре и Вологде, не отмечено достоверных различий между образцами ни по тем
пам роста, ни по фенологическому ритму развития. Растения зацвели одно
временно в конце июля, через 4 недели образовали семена и оставили зимую
щие розетки, которые на следующий год так же дружно вступили в
генеративный период развития и погибли.
При осеннем посеве из большего числа пунктов сбора семян -- от Ростова
на Дону до Вологды -- различий в феноритме также не наблюдалось. Однако
замечены различия по высоте побегов: чем севернее находилось место сбора
семян, тем выше были растения. Исключение составили экземпляры тульс
кой репродукции, имеющие такую же среднюю высоту, что и растения из Рос
това на Дону. Высота ростовских растений варьировала от 6 до 53 см (в сред
нем 30 см), липецких -- от 7 до 59 см (в среднем 37 см), тульских -- от 10 до 66
см (в среднем 30 см), московских -- от 28 до 72 см (в среднем 46 см), вологод
ских-- от22до62см(в среднем49см).
Таким образом, во вторичном ареале в Европе E.adenocaulon имеет очень
низкую генетическую вариабельность. Различий по феноритму не наблюда
ется, а изменчивость ритма роста выражена недостаточно отчетливо.
Galinsoga parviflora интродуцирована в Англию в 1796 году, найдена там
вне ботанического сада через 13 лет; массовое ее распространение началось
только через 125 лет! В Дерптском и Санкт Петербурском ботанических са
дах G.parviflora стала расширять свой ареал лишь через 135 лет! Поскольку
этот вид является однолетником, можно считать, что период его натурализа
ции на территории бывшего СССР насчитывает не менее 50 смен поколений.
Интродукционные популяции этого вида закладывали неоднократно пу
тем как осеннего, так и весеннего посева из семян, собранных в 1981 году в
Санкт Петербурге и Москве; в 1983 году в Санкт Петербурге и Таджикистане
(Хорог, Рушан); в 1985 году в Москве и Узбекистане (Коканд, Фергана); в 1987
году на Украине (Львов, Славяногорск), в Армении (Эчмиадзин), Грузии
72
(Тбилиси) и в Киргизии (Бишкек). При любых вариантах образцы практи
чески не различались по темпу роста и фенологии. Выращивание образцов
при различной длине дня также не оказало влияния на феноритмы: и опыт
ные, и контрольные экземпляры изучаемых образцов начали цвести в конце
июля, а плодоносить -- в конце сентября [Виноградова, 2002].
Следовательно, популяции G.parviflora на всем протяжении вторичного аре
ала в нашей стране являются гомогенными по важнейшим адаптивным при
знакам: ритму роста и фенологическому ритму развития. Однако наблюдает
ся значительная межпопуляционная изменчивость по степени опушенности
верхней части стебля, цветоложа и обертки.
Galinsoga quadriradiata культивировалась в ботаническом саду Санкт Пе
тербурга с 1842 года. В одичавшем виде стала встречаться в Европейской Рос
сии с 1920 х годов. Массовое распространение на Украине и в Белоруссии на
чалось в послевоенные годы, а в центральных и северо западных районах Рос
сии и в Средней Азии -- в 1970 е годы. Поэтому можно считать, что в настоящее
время в Восточной Европе и Средней Азии произрастает по меньшей мере 40
е поколение вида.
Интродукционные популяции закладывали неоднократно путем как осен
него, так и весеннего посева из семян, собранных в 1982 году в Вологде, Мос
кве и Таганроге; в 1983 году в Санкт Петербурге и Душанбе; в 1985 году в
Москве, Узбекистане и Киргизии; в 1987 году на Западной Украине и в Грузии
[Виноградова, 2003].
При весеннем посеве стратифицированными семенами разрыв между на
чалом цветения северных и южных образцов составляет всего 5 дней. При осен
нем посеве наиболее заметные различия в феноритме отмечены в 1985 году: обра
зец из Киргизии начал цвести на 2 недели позднее остальных образцов. Других
вариаций по ритму роста и фенологическому ритму развития не отмечено.
Acorus calamus интродуцирован в Европу в ХVI веке. Период произраста
ния инвазионных популяций аира в России насчитывает, по видимому, более
300 лет.
Интродукционная популяция сформирована путем посадки фрагментов
корневищ, собранных во Пскове, в Московской области, Армении, Грузии, Эс
тонии, Таганроге и Хабаровске [Виноградова, 2004].
В середине апреля начинают развертываться 3--4 весенних листа, которые
постепенно отмирают к середине мая и окончательно засыхают в середине
июня, достигнув средней длиы от 6--7 (Таганрог и Псков) до 11 см (закавказс
кие и подмосковные образцы). Коэффициент вариации длины весенних лис
тьев очень высок (не ниже 83%)!
Одновременно с отмиранием весенних листьев начинают отрастать летние
листья, развертывание которых завершается в конце июля. У подмосковных и
таганрогских образцов образование летних листьев начинается в первой дека
де мая, у остальных на 1--1,5 недели позднее. За вегетационный период на од
ном растении образуется 6--7 летних листьев, длина которых составляет в сред
нем от 50 (Псков) до 114 см (Хабаровск). Различия по длине летних листьев
практически между всеми образцами достоверны; коэффициент вариации это
го признака -- от 6% (Тарту) до 60% (Псков).
73
Летние листья у растений закавказского происхождения начали засыхать
на 2 недели раньше, чем у остальных, более северных, образцов. Полное отми
рание летних листьев отмечено в начале сентября. В середине августа начина
ют расти листья (по 4--5 на особь), которые уходят под снег и в следующем
вегетационном сезоне развиваются как весенние листья.
У подмосковных и таганрогского образцов соцветия появились в первой
декаде июня, остальные образцы зацвели на 1--1,5 недели позднее. На каждом
растении образовалось только по 1 початку, средняя продолжительность цве
тения составляла 3 недели. Длина покрывала всегда превышает длину листь
ев и составляет в среднем от 73 (Грузия) до 133 см (Подмосковье). Длина соц
ветия у всех образцов 6--8 см, диаметр 1,1 см. Зрелых семян после отцветания
не обнаружено.
Таким образом, во вторичном ареале аира на территории европейской час
ти России и Закавказья выявлена генетически закрепленная межпопуляци
онная изменчивость (не носящая клинального характера) по темпам роста как
весенних, так и летних листьев, однако изменчивость фенологического ритма
развития практически не выражена.
Заключение
Изученные виды являются наиболее яркими представителями видов ко
лонизаторов. Они имеют обширный вторичный ареал (в большинстве случа
ев значительно превышающий площадь естественного ареала), в котором на
блюдается более широкая амплитуда изменчивости природно климатических
факторов. Все эти виды на родине произрастают вне зоны экологического оп
тимума, а вследствие этого обладают довольно широкой нормой реакции. Ни
один из видов, доминирующих в естественных ценозах, не стал инвазионным!!!
Общей чертой натурализующихся видов является относительная генети
ческая бедность инвазионных популяций при широкой норме реакции. Отно
сительное генетическое обеднение обусловлено происхождением инициаль
ной популяции от одного или нескольких семян или кусочков побегов. Поэто
му между появлением вида на новой территории и его внедрением в
естественные ценозы проходит длительное время -- период накопления мик
ромутаций и рекомбинаций, продолжающийся у некоторых видов до 100 смен
поколений.
Высокая фенотипическая пластичность позволяет генетически однородным
популяциям приспосабливаться к различным местообитаниям. Вследствие
формирующего эффекта, вызванного различием местообитаний, идет генети
ческое дифференцирование популяций и наступает период бурного, иногда
взрывного, расселения, поэтому к прогнозам относительно опасности или бе
зопасности того или иного нового иммигранта относиться следует с превели
кой осторожностью.
При расширении ареала период между первым появлением вида на новой
территории и началом его натурализации стремительно сокращается и может
продолжаться всего 3 года.
В популяциях натурализовавшегося вида формируются ряды изменчивос
ти неадаптивных морфологических признаков. Амплитуда изменчивости этих
74
признаков тем выше, чем дольше произрастает популяция в новых условиях.
Так, в России полиморфизм Impatiens glandulifera ниже, чем в Западной Евро
пе, а I.parviflora и Anisantha tectorum --выше, поскольку миграция первого вида
шла в Евразии с запада на восток, а двух других -- с юго востока на запад. Ам
плитуда изменчивости зависит не столько от длительности периода натура
лизации, сколько от биологических свойств вида, из которых наиболее важ
ными являются способ размножения и быстрота смен поколений.
Способ размножения не оказывает существенного влияния на натурализа
ционные возможности вида (табл. 9): к успешным колонизаторам относятся и
размножающиеся вегетативно подвижно Acorus и Elodea, и перекрестноопы
лители Acer negundo и Impatiens glandulifera, и самоопыляющиеся виды рода
Galinsoga. Однако способ размножения определяет механизм адаптаций. У пе
рекрестноопылителей адаптация осуществляется путем перекомбинации ге
нов и последующего естественного отбора, а у самоопылителей и апомиктов
Таблица 9
Особенности микроэволюции инвазионных видов
Амфимикты
Вегетативно размножающиеся
виды, самоопылители и
апомикты
Вид
Acer negundo, Echinocystis lobata,
Impatiens glandulifera
Acorus calamus, Amaranthus albus,
Ribes aureum, Aronia melanocarpa,
Impatiens parviflora, Epilobium
rubescens, Galinsoga parviflora,
G.quadriradiata
Способ
адапта-
ции
Адаптация осуществляется путем
перекомбинации генов и
последующего естественного отбора
В инициальной популяции
возникают микромутации
физиологических признаков,
связанные с расширением нормы
реакции, которые сохраняются в
чистых линиях, способных к
последующей натурализации
Форма
естес-
твенного
отбора
Движущий отбор
Стабилизирующий отбор
Характер
внутри-
видовой
измен-
чивости
В ходе натурализации вырабатываются
и генетически закрепляются важнейшие
адаптивные признаки: менее
продолжительный период роста и более
быстрое прохождение полного цикла
развития с юга на север. У Acer negundo
с юга на север увеличивается также
продолжительность периода покоя
семян и морозостойкость. При этом
наибольшее влияние на ритм роста и
развития у растений длинного дня
оказывают температурные условия и
режим осадков, а у растений короткого
дня -- длина дня.
Во вторичном ареале наблюдается
относительная константность
биоморфологических признаков.
Изменчивость ритма роста и развития
либо практически не выражена, либо
имеет беспорядочный
межпопуляционный (не клинальный)
характер
75
наиболее вероятны возникновение уже в инициальной популяции микрому
таций физиологических признаков, связанных с расширением нормы реак
ции, и сохранение их в чистых линиях, способных к последующей натурали
зации. В первом случае имеет место движущий отбор, а во втором -- стаби
лизирующий.
У перекрестноопылителей во вторичном ареале формируется клинальная
изменчивость фенологического ритма развития и ритма роста. В ходе натура
лизации вырабатываются и генетически закрепляются важнейшие адаптив
ные признаки: менее продолжительный период роста и более быстрое прохож
дение полного цикла развития с юга на север, что соответствует изменениям
климатических условий вдоль широтного профиля. При этом в комплексе кли
матических факторов наибольшее влияние на ритм роста и развития у расте
ний длинного дня оказывают температурные условия и режим осадков, а у
растений короткого дня -- длина дня.
У самоопыляющихся, апомиктных и вегетативно размножающихся видов
в их вторичном ареале наблюдается относительная константность биоморфо
логических признаков. Изменчивость ритма роста и развития либо практи
чески не выражена, либо имеет неупорядоченный межпопуляционный (не кли
нальный) характер, и это связано с тем, что отбор политолерантных линий
или клонов происходит на самых ранних этапах натурализации, а далее они
уже генетически почти не изменяются.
Во вторичном ареале могут иметь место и весьма значительные и констан
тные, сопоставимые с уровнем видообразования морфофизиологические пе
рестройки растений. Именно таков механизм возникновения Aronia mitschurinii.
76
Глава 5
ТАКСОНОМИЧЕСКИЕ ПРОБЛЕМЫ ИЗУЧЕНИЯ
ИНВАЗИОННЫХ ВИДОВ
Таксономическое распределение инвазионных видов флоры Средней Рос
сии таково: несомненный лидер -- семейство Compositae с 14 видами, на втором
месте Gramineae -- 7 видов, далее следуют семейства Cruciferae и Rosaceae -- по 4
вида и Onagraceae -- 3. Остальные семейства, среди которых Aceraceae, Acoraceae,
Amaranthaceae, Balsaminaceae, Boraginaceae, Chenopodiaceae, Cucurbitaceae,
Elaeagnaceae, Euphorbiaceae, Hydrocharitaceae, Juncaceae, Labiatae, Leguminosae,
Oleaceae, Oxalidaceae, Polygonaceae, Salicaceae и Umbelliferae, представлены 1--2
видами (рис. 16).
Отметим, что эта таксономическая структура соответствует пропорциям ад
вентивной флоры Европы, выявленной в результате паневропейского проекта
DAISIE [Lambdon et al., 2008]. Различия невелики: 3 е место в адвентивной фло
ре Европы занимает Rosaceae, заметно опережая семейство Cruciferae. Это от
личие легко объяснимо. Среди среднерусских адвентивных растений немало
широко распространенных видов розоцветных (Malus domestica, Prunus domestica,
Pyrus communis и др.), которые не являются инвазионными видами. Крестоцвет
ные, напротив, отличаются высокой активностью и широко расселяются из мест
заноса.
Интересно, что таксономический спектр адвентивного и инвазионного ком
понентов флоры Средней России имеют незначительные отличия, главным об
разом, количественные. В адвентивном компоненте первые пять семейств рас
полагаются следующим образом: Compositae (13%) -- Gramineae (13%) -- Cruciferae
(9%) -- Rosaceae (8%) -- Leguminosae (7%). Инвазионный компонент флоры Сред
ней России дает следующий таксономический спектр: Compositae (27%) --
Gramineae (10%) -- Cruciferae (8%) -- Rosaceae (6%) -- Onagraceae (6%). [Виног
радова, Майоров, Хорун, 2006]
Гораздо более существенные различия отмечены для таксономических спек
тров кенофитов в других регионах. Так, спектр кенофитов Украины: Compositae
(32%) -- Gramineae (8%) -- Cruciferae (8%) -- Umbelliferae (6%) -- Amaranthaceae
(6%), а спектр кенофитов Польши Compositae (24%) -- Cruciferae (9%) --
Leguminosae (8%) -- Gramineae (5%) -- Scrophulariaceae (5%) .
Впрочем, не стоит переоценивать важность таксономического распределе
ния для анализа парциальных флор при небольших выборках. При разнообраз
ных подсчетах следует учитывать статистический характер выделенных групп
и неизбежные разночтения или даже ошибки [Щербаков, 2001, 2003, 2006]. Бо
лее того, на таксономический спектр адвентивной флоры влияет степень ее вы
77
явления; в отличие от природной флоры, эти пропорции по мере накопления
данных изменяются [Щербаков, 2008]. Следует иметь в виду, что традиционно
в российских флористических исследованиях используют систему А. Энглера
(с изменениями!), западные флористы же постепенно переходят на систему APG
(the Angiosperm Phylogeny Group), в том числе и при исследовании адвентив
ной флоры [сравни: Маевский, 2007 и Lambdon et al., 2008].
Особую важность при работе с адвентивными растениями имеет надежная
идентификация видов, так как вся необходимая информация (происхождение,
тип ареала, экологические свойства и т. п.) «привязаны» к названию таксона.
Без точного определения инвазионных видов объединение усилий исследова
телей неэффективно [Pyšek, 2003; Pyšek et al., 2008]. Это определение необхо
димо для принятия ряда управленческих и экономических решений по контро
лю над инвазионными видами [Smith et al., 2008].
Точное определение заносных видов наталкивается на ряд объективных про
блем:
1) новые для данной территории адвентивные виды часто отсутствуют в
доступных региональных «Флорах» и определителях;
2) гербарные образцы из «иностранных» отделов Гербариев (MW, MHA и
даже LE) обычно немногочисленны и часто неоднородны из за заметного
числа неправильных определений;
3) адвентивные виды нередко представлены нетипичными, уклоняющими
ся формами;
4) адвентивные виды, особенно эргазиофиты, часто несут черты предка ос
нователя.
Все это существенно затрудняет надежную и точную идентификацию. По
этому в истории изучения адвентивной флоры немало случаев номенклатурной
и таксономической путаницы. Приведем несколько примеров.
Рис. 16. Таксономическое распределение инвазионных видов флоры
Средней России, вошедших в «Черную книгу»
78
Род бруннера (Brunnera Steven) на территории России представлен двумя
видами: кавказской бруннерой крупнолистной B. macrophylla I.M. Johnst. и брун
нерой сибирской B. sibirica Steven [Попов, 1953]. Как декоративное растение в
наших условиях выращивают преимущественно B. sibirica с более крупными
цветками и лучшими декоративными свойствами [Чуб, 2008]. Это неприхотли
вое растение подолгу удерживается на местах прежней культуры и встречается
как заносное у дачных поселков, по сорным местам. Однако во «Флоре евро
пейской части СССР» для нашей территории указывается лишь B. macrophylla
[Доброчаева, 1981]. Вслед за этим бруннера крупнолистная появилась в ряде
региональных работ [например, Цвелев, 2000; Григорьевская и др., 2004]. Таксо
номическое решение Д.Н. Доброчаевой основано, по видимому, на "Flora
Europaea" [Chater, 1972]. Рисунок в известном определителе Стейси подтверж
дает, что, по крайней мере, в Великобритании культивируется именно B.
macrophylla [Stace, 1997]. Среди адвентивных растений Европы значится толь
ко последний вид [Handbook..., 2009]. Однако описание бруннеры, данное Д. Н.
Доброчаевой [1981], соответствует B. sibirica у М. Г. Попова [1953].
Наши наблюдения за природными популяциями B. macrophylla на Кавказе, а
также за культивируемыми растениями в Ботаническом саду МГУ и подробное
изучение гербарных материалов с Кавказа и из Сибири (MW, MHA) показали,
что как декоративное растение культивируется именно бруннера сибирская. B.
macrophylla отличается от B. sibirica относительно мелкими цветками, наличием
эпигеогенных корневищ и более ранним цветением: цветки у нее раскрываются
еще до полного разворачивания крупных прикорневых листьев. Кроме того, у
B. macrophylla корневище сравнительно тонкое [Чуб, 2008]. Следует помнить,
что размер цветков не является надежным диагностическим признаком [Попов,
1953]. B. macrophylla отмечена в Подмосковье лишь в последние годы (личное
сообщение В.Д. Бочкина).
Таким образом, таксономическая обработка Brunnera в последней по време
ни крупной региональной сводке не описывает реальной ситуации с видами рода
на нашей территории. Поэтому без обращения к гербарным материалам литера
турные указания следует воспринимать как предварительные данные, посколь
ку не ясно точно, о каком именно виде идет речь.
Для викарных видов в пределах природного ареала нередко самым надеж
ным «признаком» является географическое положение популяции. Так, евро
пейско кавказский Acer tataricum L., среднеазиатский A. semenovii Regel et Herd.
и дальневосточный A. ginnala Maxim. аллопатричны и достаточно хорошо отли
чаются морфологически [Цвелев, 2000]. Однако, в последнее время многие ев
ропейские ботаники [см., например, Suomen..., 1992; Zander, 2002] объединяют
эти виды. Причиной такого мнения может быть как широкое распространение
в Европе сортовых форм с промежуточными признаками, так и недостаточно
точное морфологическое описание во «Флоре СССР» из за невнимания к гете
рофилии у этих видов на генеративных и вегетативных побегах [Пояркова, 1949].
A. tataricum и A. ginnala широко используются в озеленении, в последнее время
неоднократно отмечается самосев. Однако, как оценивать необычные формы
кленов из культуры с промежуточными признаками не ясно: либо это результат
сортового отбора уклоняющихся форм одного из видов, либо гибридизация близ
79
ких видов при вторичной симпатрии. Современные методы в ряде случаев спо
собны определить географическое происхождение инвазионных растений
[см., например: Milne, Abbott, 2000; Meekins et al., 2001; Durka et al., 2005; Kliber,
Eckert, 2005].
Особую проблему представляют гибридные комплексы. Так, в культуре ши
роко распространены близкородственные виды -- дальневосточная Spiraea
salicifolia L. с розовыми цветками и компактным цилиндрическим соцветием и
североамериканская S. alba L. с белыми цветками и пирамидальным соцветием.
Время от времени сообщается о находках этих спирей как заносных растений.
Кроме того, в культуре несколько реже встречается североамериканская S.
douglasii Hook. с интенсивно розовыми цветками, компактным соцветием и
обильно опушенными снизу листьями. Известны, описаны и широко культиви
руются гибриды между этими тремя видами (рис. 17). Следует учитывать, что
иногда крупные формы S. alba выделяют как S. latifolia Aiton, а в составе S.
douglasii известны и описаны гололистные растения-- это ssp. menziezii (Hook.)
Voss. Некоторые гибриды "S. latifolia" описаны как особые таксоны [Цвелев,
2000]. Очевидно, что в подобных случаях надежная идентификация гибридных
образцов едва ли возможна, поскольку ситуация осложняется селекцией высо
кодекоративных форм спирей, отбор направлен на окраску цветков и форму
соцветий -- диагностические признаки у родительских видов и гибридных форм.
Определение гибридных растений наталкивается на еще одну диагностичес
кую проблему. Нередко гибридизация происходит между растениями в услови
ях культуры вне природного ареала. Это, например, Reynoutria × bohemica, S. ×
uplandicum Nyman (S. officinale × S. asperum Lepechin), Aster × salignus и, вероят
но, Amelanchier spicata. В этом случае такие гибридные формы часто неизвестны
из мест симпатрии родительских видов (если эти ареалы пересекаются) и, есте
ственно, отсутствуют в региональных «Флорах...» и определителях.
Не всегда удается установить оба родительских вида для гибридного таксо
на. Так, ирония Мичурина, или черноплодная рябина, -- Aronia × mitschurinii
A.K.Skvortsov et Yu.K.Maitulina была получена при селекции A. melanocarpa
(Michx.) Nutt. et Elliott. Для этого И. В. Мичурин использовал гибридизацию с
другими видами, в том числе, вероятно, рябинами. К сожалению, история воз
Рис. 17. Спиреи S. salicifolia, S. alba, S. douglasii и их гибриды
80
никновения «черноплодки», несмотря на обширные публикации работ Мичу
рина, не ясна. Следует учитывать, что для гибридизации он нередко применял
смесь пыльцы разных растений, что, по мнению селекционера, должно было
повышать качество гибридного потомства. A. × mitschurinii отличается от A.
melanocarpa, с которой ее нередко путают, прежде всего большей мощностью
растения и качеством плодов [Скворцов, Майтулина, 1982]. Точное установле
ние второго родительского вида возможно только при применении методов
молекулярной генетики.
В 1970--80 е годы аронию широко культивировали в промышленных масш
табах как неприхотливое высокоурожайное ягодное растение, плоды которого,
среди прочего, выделяются высоким содержанием витамина Р. В последние годы
отмечено расселение аронии Мичурина по лесным опушкам и на вырубках, чаще
у дачных поселков или близ плантаций черноплодки. При этом она нередко
поселяется по сырым местам вдоль дорог, на вырубках, то есть в местообитани
ях аналогичных условиям произрастания A. melanocarpa в естественном ареале.
В некоторых случаях адвентивные растения показывают иной характер из
менчивости, чем в природном ареале. Так, кипреи из родства E.ciliatum Raf. пред
ставлены в Европейской России двумя видами: E. adenocaulon Hausskn. и E.
pseudorubescens A.Skvortsov. На территории Северной Америки E.ciliatum разде
ляют на большое число микровидов, не всегда хорошо различимых. В Евразии,
напротив, эти два кипрея ведут себя как независимые и вполне отчетливо раз
личимые виды. Первоначально E. pseudorubescens был определен как E. rubescens
Rydb., однако знакомство А.К. Скворцова с типовым материалом убедило его в
ошибочности такого решения. Обилие числа описанных видов и малая доступ
ность типового материала для кипреев не позволяет надежно сопоставить ад
вентивный белоцветковый кипрей и какой либо североамериканский микровид.
Это побудило А.К. Скворцова к описанию нового вида -- E. pseudorubescens. Не
исключено, что в дальнейшем удастся обнаружить исходный североамериканс
кий микровид из состава E. ciliatum [Скворцов, 1996].
В средней полосе Европейской части России среди обычного тростника
Phragmites australis (Cav.) Trin. ex Steud. как заносные отмечены очень крупные
растения с широкими листьями, мощными соцветиями, зацветающие поздно
осенью и размножающиеся, по видимому, только вегетативно [Папченков, 2008].
Эти растения получили название Ph. altissimus Benth. (Ph. isiaca Delile). В насто
ящее время в средней полосе они известны в Ярославской, Тверской, Московс
кой и Ульяновской областях, Мордовии [Папченков, 2008; MW]. Следует иметь
ввиду, что эта раса хорошо отличается от типичной лишь во вторичном ареале.
В обширном природном ареале наблюдается постепенный переход от растений
тростника привычного нам облика к огромным южным формам. Н.Н. Цвелев
эту форму выделяет в ранге подвида ssp. altissimus (Benth.) Clayton, в целом бо
лее южного, но симпатричного с типовым подвидом [Цвелев, 1974, 1976].
В ряде случаев идентификации трудно разграничимых видов помогает их
биологическое окружение. Так, в Северной Америке галловая мушка Eurosta
solidaginis предпочитает инфицировать Solidago altissima L., при этом близкий и
очень похожий S. canadensis ею практически не повреждается [Abrahamson et
al., 1989; Abrahamson; Weis, 1997]. Как это ни удивительно, наличие или отсут
81
ствие галлов -- один из лучших диагностических признаков для определения
этих близких видов золотарника [Semple, Cook, 2006]. Во вторичном ареале мы
естественным образом лишены этого удобного признака.
Нередко в случае с адвентивными видами имеет место таксономические раз
ночтения в трактовке тех или иных видов в зависимости от национальной шко
лы. Например, в англоязычных странах дурнишник, называемый в нашей стра
не X. albinum, принято ошибочно объединять с X. strumarium (см. статью ниже).
Или, американцы обычно называют адвентивный окопник из Европы Symphytum
officinale L., между тем, бульшая часть образцов принадлежит к S. Ч uplandicum --
US, NY, GH, A (определения С.Р. Майорова и Д.Д. Соколова). Несомненны по
добные случаи и в практике российских флористов. Примеры тому Cenchrus
echinatus L. и C. pauciflorus Benth., Phytolacca acinosa Roxb. и Ph. americana L.
Многие используемые в озеленении древесные растения поступают из спе
циализированных питомников или ботанических садов, где нередко близкие
виды культивируются совместно. При этом в ряде случаев спонтанно возника
ют межвидовые гибриды. Такая гибридизация характерна, например, для мно
гих розоцветных. Некоторые гибридные формы затем находят широкое приме
нение в озеленении, некоторые из таких таксонов способны дичать. В нашем
списке инвазионных видов к гибридогенным таксонам относятся Reynoutria ×
bohemica и Aster Ч salignus. По мнению некоторых ученых, такая гибридизация
может приводить к образованию инвазионных нотовидов [Abbott, 1992;
Ellstrand, Schierenbeck, 2000]. Поток генов между местными и инвазионными
видами неоднократно доказан на генетическом уровне и требует тщательного
изучения [Abbott et al., 2003; Petit, 2004; Bleeker, Matthies, 2005].
С другой стороны, гибридизация местных растений с адвентивными приво
дит к генетическому загрязнению, а интрогрессивная гибридизация может при
водить к исчезновению редких местных видов [Bleeker, Matthies, 2005; Bleeker
et al., 2007].
Следует отметить, что переход интродуцированного или заносного вида че
рез географические и биологические барьеры [Richardson et al., 2000] видоизме
няет инвазионный вид. Это было убедительно показано на гигантских борще
виках с использованием молекулярно генетических методов: инвазионные по
пуляции Heracleum mantegazzianum и H. sosnowskyi обнаружили отчетливые
отличия от природных [Jahodovб et al., 2007; Ecology..., 2007]. Этот переход тре
бует некоторого времени, в течение которого интродуцированный вид стано
вится способным к активному расселению. Это время в англоязычной литера
туре принято называть "lag phase". Для объяснения этого явления предложено
несколько гипотез [обзор см.: Pyšek, Hulme, 2005]. Нам ближе гипотеза о накоп
лении за это время генетических изменений, которые позволяют растению при
способиться к новым условиям и реализовать адаптивные преимущества
адвентивного вида [Hobbs, Humphries, 1995; Kowarik, 1995].
***
Точная таксономическая идентификация адвентивных видов растений дол
жна быть неотъемлемой частью любых работ, исследовательских или практи
ческих, с инвазионными видами [Smith et al., 2008]. Повысить точность опреде
ления можно лишь аккуратным изучением таксономической литературы, тща
82
тельным сравнением адвентивных растений с надежно определенными гер
барными образцами из мест естественного произрастания. Таксономичес
кие комментарии должны сопровождать публикации, посвященные адвен
тивным видам.
Остаются существенные проблемы в таксономии и диагностике адвентивных
видов в родах Aster s. l. (Symphyotrichum), Helianthus, Crataegus, Rosa и т.п.
В нашем списке к таксонам с окончательно не решенными таксономически
ми проблемами относятся Amelanchier spicata, Crataegus monogyna, Epilobium
adenocaulon, E. pseudorubescens, Solidago canadensis, Aster × salignus, Helianthus
tuberosum, Xanthium albinum. В очерках приводятся необходимые комментарии
по систематике этих видов.
С другой стороны, Cardaria draba в настоящее время чаще включают в со
став рода Lepidium, а Aster × salignus переносят в род Symphyotrichum. Мы сохра
нили привычные названия для этих растений, поскольку собственно границы
видов не изменяются.
83
Глава 6
БИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ
ИНВАЗИОННЫХ ВИДОВ ФЛОРЫ
СРЕДНЕЙ РОССИИ
СЕМЕЙСТВО ACERACEAE -- КЛЕНОВЫЕ
КЛЕН ЯСЕНЕЛИСТНЫЙ
Название Acer negundo L.
Синонимы: Negundo aceroides Moench
Морфологические признаки
Дерево высотой до 20 (25) м и диаметром 90 (100) см как в первичном, так и
во вторичном ареале [Hosia, 1969]. Его габитус сильно зависит от условий мес
тообитания. В лесах с умеренным увлажнением и ненарушенной почвой он имеет
форму прямостоячего дерева с одним стволом. Когда растение частично затене
но, например на опушке леса, его ветви принимают плакучую форму, а стволы
сильно наклоняются. В местообитаниях вне леса растение сильно ветвится и
редко достигает высоты более 12--15 м [Sutton, Johuson, 1974]. Листья непарно
перистосложные, число листочков изменяется от 1 у проростков до 5 и даже 7 в
старшем возрасте. Молодые побеги имеют зеленую кору, часто покрытую бело
ватыми волосками или восковым налётом. Имеется много культиваров с листь
ями различной формы и расцветки.
Выделяют шесть разновидностей A.negundo: var. arizonicum Sarg., var.
californicum (Torr et Gray) Sarg., var. interius (Britt.) Sarg., var. negundo, var. texanum
Pax, var. violaceum (Kirchn.) Jaeger.
Естественный ареал
Клен ясенелистный в природе произрастает в Северной Америке (от
Скалистых гор до Атлантического побережья и от Канады до Флориды). Не
прерывный (сплошной) ареал простирается от Нью Джерси и Нью Йорка к са
мому югу Онтарио, центру штата Мичиган, северу Миннесоты, центру Мани
тобы, центру Саскачевана и югу Альберты, центру штата Монтана и востоку
Вайоминга, Юты и Калифорнии; на юге граница проходит по югу штата Техас и
центру Флориды. Локальные популяции имеются также в Нью Гэмпшире, шта
тах Вермонт, Массачусетс, Коннектикут, Айдахо и Невада.
Географические расы A. negundo встречаются в горах Мексики (штаты Nuevo
Leon, San Luis Potosi и на юге Chihuahua), а также в Гватемале [Rosario, 1988].
Вторичный ареал
История интродукции и географическое распространение. A.negundo --
натурализовавшийся вид в штатах Мэн, Вашингтон и Орегон, в провинциях
Квебек, Нью Брансуик, Новая Шотландия, на о вах Принца Эдуарда.
Намеренно завезен в Европу вместе с многочисленными американскими
растениями в ХVII столетии. Первая достоверно известная дата -- 1688 г.; в то
84
время клен зарегистрирован в саду Fulham в Англии [Kowarik, 1992]. Позже он
интродуцирован в Голландию (1690 г.) и в Германию (1699 г.). В Польше впер
вые отмечен в 1808 г. [Szymanowski, 1960].
В России клен ясенелистный известен с конца ХVIII века. По сведениям
С.Я. Соколова [1957], в ботаническом саду Санкт Петербурга имелись уже взрос
лые экземпляры клена в 1796 году.
Первоначальные опыты разведения клена из семян в открытом грунте в Пе
тербурге были безуспешными -- сеянцы гибли от мороза, так как испытывались
образцы, полученные из южной части естественного ареала [Уханов, 1950]. По
беги взрослых деревьев при сильных холодах в значительной мере обмерзали, и
на зиму все деревья укутывали [Цигра, 1842], что дало повод считать A.negundo
непригодным для разведения в условиях Санкт Петербурга [Регель, 1879]. В
Москве сеянцы и взрослые деревья также сильно обмерзали, а потому клен ясе
нелистный рос в саду Демидова в холодной оранжерее [Паллас, 1781].
Опыты по интродукции вида в Прибалтике, относящиеся к началу ХIХ века
[Zigra, 1817; Вагнер, 1821], на первых порах также были неудачными: каждую
зиму растения сильно обмерзали, а в суровые зимы зимовали лишь под надеж
ным укрытием. Тем не менее клен ясенелистный выращивали во всех районах
Прибалтики [Klinge, 1883]; в частности, в Тартуском ботаническом саду -- с 1833 г.
[Tanavots, 1969].
Гораздо успешнее протекала интродукция клена ясенелистного на Украине,
куда семена вида попали еще в 1809 г. Их привез из США и Канады И.Н. Кара
зин и высеял в основанном им в том же году Основянском акклиматизацион
ном саду около Харькова [Кохно, 1970]. Деревья, выросшие из этих семян, от
личались быстрым ростом и сизыми от микроскопических волосков и специфи
ческого налета побегами. Американские авторы деревья со сходными
признаками относят к A.n.pseudocalifornicum Schwer. [Rehder, 1949].
Потомство, полученное от этих деревьев, использовали при посадках в са
дах и парках юга Средней России и на Украине. В 1816 г. клен ясенелистный
уже произрастал в Кременце [Besser, 1816], а с 1849 г. известен в Одессе (KW). В
1865 г. Графф собрал гербарные экземпляры клена в Великоанадольском лесниче
стве (KW). А.Воейков [1908] указывает, что деревья, обладающие быстрым ростом
и сизыми побегами, оказались значительно более выносливыми. Хотя в Петербур
ге и Москве их побеги не вызревали, в более южных районах они чувствовали себя
неплохо и лишь иногда теряли верхушечные почки, что при быстром росте было
почти незаметно. Деревья с данными морфологическими признаками в конце ХIХ
века часто встречались в лесничествах Тамбовской губернии и в Поволжье, где их
распространял питомник Н.П. Корбутовского [Воейков, 1908].
В конце ХIХ века Э.Л.Регель получил семена A.negundo из северных райо
нов Канады. Деревья, выросшие из этих семян, отличались более слабым рос
том, тонкими ветвями и густым слоем красного красящего вещества -- антоциа
на. Р.И. Шредер описал их под названием A.n.boreale, американскими авторами
деревья со сходными морфологическими признаками относятся к A.n.var.
violaceum (Kirchn.) Jaeg. [Rehder, 1949].
Потомство, полученное от разводимых Э.Л. Регелем деревьев, в условиях
Петербурга оказалось вполне выносливым, только семена вызревали не каж
85
дый год [Воейков, 1908]. Из семян, полученных из Петербурга, Р.И. Шредер
вырастил два экземпляра клена ясенелистного в Москве, которые каждый год
обильно плодоносили. В настоящее время деревья клена со слабыми красными
ветвями изредка встречаются почти по всей территории России в садах и пар
ках, но в инвазионных популяциях мы не находим деревьев со сходными мор
фологическими признаками.
В Прибалтике интродукция клена протекала сходным образом. В 1893 г.
М.Сиверс заложил в Скриверском дендропарке посадки клена ясенелистного
из семян, выписанных из провинции Манитоба (Канада), и в начале ХХ века
этот вид стал широко внедряться в декоративные посадки Латвии [Мауринь,
1970]. В Литве его начали культивировать в 1930 х г., а впервые он отмечен как
вид, сбежавший из культуры, в 1963 г. [Gudzinskas, 1998].
В 1897 г. A.negundo впервые появился на Урале. Он возделывался в питом
нике при Талицкой лесной школе, заложенном под руководством О.Г. Вронско
го. Интродукция вида в условиях Урала протекала весьма успешно. С 1909 г. его
стали массово высаживать в городские парки [Луговых, 1959].
Клен ясенелистный был среди первых древесных пород, высаженных в саду
Комиссарова, заложенного в 1896 г. в Западной Сибири на правом берегу Ир
тыша в 30 км южнее Омска [Крылов, Салатова, 1955].
В Крыму, в Никитском ботаническом саду этот вид разводится с 1814 г. [Де
ревья..., 1954].
В Среднюю Азию клен ясенелистный начали ввозить с 1870 х гг., после при
соединения этой территории к России. В 1891 г. вид собран в селении Гульча в
Киргизии (LE). В 1895 г. С.Коржинский (LE) собрал гербарные образцы клена
в Ашхабаде и Самарканде, а в 1899 г. -- в окрестностях г. Верный (ныне Алма
Ата). В 20--30 х гг. ХХ века во многих городах Средней Азии клен ясенелистный
высаживали массово. В 1922 г. посадки производились в г. Пишпек (Бишкек) при
создании зеленой зоны в Карагачевой роще, а затем в Нукусе, Турткуле, Кунгра
де, Иссык Кульской долине [Альжанов, Сагитов, 1963; Лысенко, 1973].
Таким образом, семена и саженцы A. negundo ввозили в Россию несколько
раз, они происходят из разных источников, а потому, естественно, выросшие из
них растения обладали различными биологическими признаками. А.Воейков [1908],
испытывавший в своем опытном саду в Симбирской губернии ряд образцов клена
ясенелистного, указывал, что "типичный" A.negundo мерзнет каждый год до снега,
A.n.pseudocalifornicum отличается пышным ростом и почти не страдает, а A.n.violaceum
растет хоть и не так сильно, зато без малейшего повреждения. Сходные результаты
получили и другие исследователи [Шредер, 1896; Вольф, 1915].
В настоящее время самая северная точка произрастания клена ясенелистно
го --Полярно альпийский ботанический сад, но там это растение обмерзает и не
плодоносит.
В Архангельске, где клен растет чаще в виде куста, он вполне вынослив, вер
хушки побегов обмерзают, но не ежегодно. Плодоносит нерегулярно, только в
наиболее благоприятные по погодным условиям годы [Уханов, 1950]. По нашим
наблюдениям, не дичает.
В Петрозаводске растет в большом количестве, но плодоносит нерегулярно.
В ряде пунктов города обнаружен самосев [Виноградова, 2006].
86
В Санкт Петербурге клен растет хорошо. В одиночных посадках и при со
блюдении элементарных мер ухода растет деревом, в иных условиях часто при
нимает кустообразную форму, что обусловлено способностью легко давать
обильную поросль. Вполне морозостоек. Плодоносит регулярно и обильно. По
нашим наблюдениям, на пустырях повсеместно размножается самосевом.
В Прибалтике клен распространен широко. Вполне зимостоек, иногда рас
тет в виде куста. Почти ежегодно обильно цветет и плодоносит. Во многих мес
тах мы отметили самосев. В Белоруссии растет в форме дерева. Вполне морозо
стоек, только в суровые зимы однолетние побеги повреждаются морозами [Не
стерович, 1955].
На Украине и в степной зоне России клен ясенелистный культивируют как
лесную породу. Зимует он хорошо, хотя в суровые зимы годичные побеги час
тично обмерзают. Засуху переносит хорошо. По нашим наблюдениям, всюду
размножается самосевом, особенно по берегам рек.
На юго востоке европейской части России при применении искусственного
орошения или по берегам рек достигает мощного развития. В Астрахани мы
видели деревья высотой до 20 м, обильно плодоносящие и с многочисленным
самосевом вокруг. В Махачкале растет в городских садах, но без полива совер
шенно не развивается и усыхает.
В Крыму клен хорошо растет и плодоносит. В Симферополе и окрестностях
размножается самосевом.
На Кавказе клен растет хорошо [Адо, 1934], но встречается лишь в крупных
городах. Самые старые экземпляры вида высажены в 20 х гг. ХХ века. Ежегод
но цветет и плодоносит. Местами размножается самосевом. Крона имеет не
правильную форму. Ствол искривленный, декоративность деревьев слабая. В
Азербайджане на бедных почвах Апшеронского полуострова при малом ороше
нии отмечается усиленное усыхание ветвей и пожелтение листьев среди лета.
Обильно плодоносит и дает многочисленный самосев. Самые старые экземпля
ры клена ясенелистного растут с 1900 г. [Стребкова, 1931].
Клен ясенелистный растет во всех крупных городах Западной Сибири вплоть
до Тюмени. Этот вид рекомендован для озеленения городов и рабочих поселков
этого региона, исключая самую северную его часть [Крылов, Салатова, 1955].
A.negundo встречается также в южных районах Восточной Сибири и на Даль
нем Востоке.
Во всех городах Средней Азии клен ясенелистный произрастает весьма ус
пешно. При морозе ниже 10°С отмечается частичное обмерзание побегов. Су
хим и жарким летом наблюдаются небольшие ожоги на листьях. Деревья в воз
расте 20 лет начинают суховершинить [Лысенко, 1973; Костелова, 1971]. По
нашим наблюдениям, самосев встречается на орошаемых территориях повсе
местно и во множестве.
По личному сообщению А.К. Скворцова, клен ясенелистный повсюду дича
ет и на территории Казахстана --от оз. Зайсан до Актюбинска.
Из вышеизложенного следует, что клен ясенелистный сформировал на тер
ритории стран бывшего СССР обширный ареал и во многих частях его вы
шел из культуры и внедрился в аборигенную флору, успешно размножаясь
самосевом.
87
Натурализация произошла в массовом масштабе в нарушенных антропоген
ных местообитаниях и по берегам рек во многих регионах Польши, Германии и
Литвы [Lohmeyer, Sukkopp, 1992; Kuusk et al., 1996; Gudzinskas, 1998]. В Шве
ции натурализовавшиеся растения встречаются редко. В Латвии клен растет в
старых парках, по берегам рек, на обочинах дорог и по железным дорогам
[Gavrilova, Sulcs, 1999]. В Дании этот вид используется в садах и парках, но не
натурализуется [Svart, Lyck, 1991]. В Эстонии A.negundo обычен и в культуре, и
в одичавшем виде [Kukk, 1999]. В Норвегии этот вид распространен не столь
широко, и в настоящее время его рекомендуют там как выносливое и декора
тивное дерево для посадки в садах и на крутых склонах. Только две локальные
популяции в окрестностях Осло, обе вблизи старых посаженных экземпляров,
рассматриваются как натурализовавшиеся.
Пути и способы заноса. Растение стало весьма популярным садовым дере
вом благодаря быстрому росту в первые годы жизни. В конце ХVIII века вид
был проверен в лесоводстве, но хорошего результата это не дало [Hegi, 1977;
von Schwerin, 1919]. Во второй половине XIX века вид был вновь предложен
для посадок в парках и по обочинам дорог. Его рекомендовали сажать как вет
розащитную и лесозащитную породу [Ehrendorfer, 1973; Tutin et al., 1968]. В
первой половине XX века он стал одним из самых обычных высаживаемых чу
жеродных растений. Культивары с белоокаймленными или желтыми листьями
до сих пор часто высаживают в садах. Они, по видимому, менее способны к ин
вазии, но необходимо учитывать инвазивность подвоя, на котором привиты эти
формы.
Этот вид пропагандировался также для культивирования на пасеках. A.
negundo -- один из наиболее ранних весенних источников пыльцы. Есть даже
специальный мед box elder honey, который продается в Польше в Беловежье.
Осенью и зимой семена вида переносятся ветром на далекие расстояния
(см. ниже).
Статус в регионе. Дичание A.negundo в Средней России началось во вто
рой половине XX века, и до настоящего времени клен занимает большое место
в зеленом строительстве. В Средней России он отличается буйным ростом и
регулярным плодоношением. Особо урожайные годы обычно бывают через два
года на третий. Всюду размножается самосевом. Сильные морозы и редкие пе
репады температур переносит хорошо: даже в очень суровую зиму 1978/79 час
тично обмерзли лишь однолетние побеги. Однако, хотя вид активно расселяет
ся во всех хозяйственно административных регионах Средней России, гербар
ных сборов его немного, так как A.negundo считают самым обычным и
малоинтересным растением (рис. 18).
Местообитания. A.negundo освоил многочисленные экологические ниши.
В естественном ареале этот вид входит в число различных фитоценозов (22 ас
социации; 13 типов экосистем), включая болота, пойменные леса, мезотрофные
лиственные леса, хвойные леса с соснами, елью и елью Дугласа; дубовые редко
лесья, чаппарали и различные виды прерий и полей [Rosario, 1988].
Чрезвычайно широкий диапазон местообитаний -- результат высокой толе
рантности к дефициту почвенной влаги и нехватке питательных веществ в по
чве. Отмечены различия между полом растений по их предпочтению к среде
88
обитания: к экстремальным условиям среды терпимы оба пола, однако женские
экземпляры лучше растут в более влажных и богатых питательными вещества
ми местообитаниях [Dawson, Ehleringer, 1993; Ward et al., 2002].
A. negundo умеренно стоек к подтоплению, то есть менее стоек, чем тополи и
ивы, но более устойчив, чем другие виды мезофитных лесов [Friedman, Auble,
1999]. В естественном ареале этот вид вдоль больших рек растет на надпоймен
ных террасах [Rosario, 1988; Everson, Boucher, 1998]. A. negundo захватил также
антропогенные местообитания, и его обычно называют деревом сорняком.
Местообитания, которые этот вид занимает в естественном и во вторичном
ареалах, весьма сходны. Он в изобилии населяет прибрежные фитоценозы (на
пример, пойменные леса) вдоль как мелких речушек, так и широких рек (рис. 19)
до высоты 1000 м над ур. м. Он найден в мезофитных дубравах, сосновых лесах,
особенно вдоль опушек. В отсутствие наводнений A. negundo в ходе сукцессии
обычно заменяется более теневыносливыми видами. Так же, как он внедряется
в естественные фитоценозы (леса и степи), A. negundo колонизирует и огром
ный диапазон полуестественных местообитаний -- заброшенные парки, посел
ки, заброшенные пахотные поля, обочины дорог, железнодорожное полотно,
свалки и даже крыши домов.
Так, в Мордовии вид успешно расселяется по долинам рек и лесным опуш
кам, конкурируя в древостое с ясенем, липой, кленом остролистным. Еще более
агрессивно он ведет себя в пойменных дубравах и ивняках, вытесняя из подро
ста местные виды. Активно расселяется A.negundo и на особо охраняемых при
Рис. 18. Расселение Acer negundo в Средней России
Рис. 19а. Пойменный лес с доминированием Acer negundo по Москве реке.
Рис. 19б. Заросли Acer negundo в заповеднике «Галичья Гора», участок
«Морозова Гора» (Липецкая область).
Рис. 20. Обильное плодоношение Acer negundo (Москва)
Рис. 21. Проростки Acer negundo на речном аллювии, р. Дон в Воронежской
области у с. Кривоборье
91
родных территориях: в Мордовском государственном заповеднике, в националь
ном парке "Смольный", природных парках и заказниках. Промышленную зону
вокруг поселков и в городах, равно как и разнообразные вторичные местооби
тания, вид уже завоевал [Бармин, 2006].
Размножение и жизненный цикл. A.negundo считают строго двудомным
видом, мужские растения зацветают чуть ранее женских. Изучение изменчиво
сти по половому признаку по профилю долины реки в естественном ареале по
казало, что в зависимости от пола деревья имеют предпочтения к тому или ино
му местообитанию [Ramp, Stephenson, 1988; Freeman, Klikoff, Harper, 1976].
Однако во многих случаях соотношение полов составляет 1:1 без какой либо
корреляции с условиями обитания [Medrzycki, 2002].
Цветение A.negundo начинается ранней весной до распускания листьев.
Пыльца разносится, главным образом, ветром. Имеются сообщения, что, по край
ней мере, дополнительное опыление производится пчелами, которые ранней вес
ной собирают много пыльцы. Плоды A.negundo -- двойные крылатки (рис. 20).
Возраст первого плодоношения зависит от наличия ресурсов -- на открытом
месте с нормальными почвами он может составлять 5 лет, а под пологом леса
может достигать 15 лет и более [Medrzycki, 2002]. Наличие света, по видимому,
влияет на урожайность -- женские экземпляры при лимите света производят в 5
раз меньше семян, чем растения на открытых местах. Максимальную продук
тивность оценить трудно: 100--500 тыс. крылаток на 1 дерево -- это еще не мак
симальное значение. Максимальный возраст деревьев ~100 лет.
Масса семян сопоставима с другими видами кленов, но необычно высока по
сравнению с иными ранневесеннецветущими древесными видами.
Расселение и распространение. Независимо от местообитания основной
способ рассеивания семян -- ветром. Крылатки могут разнестись по меньшей
мере на 50 м от родительского дерева [Sachse, 1991]. В некоторых местообита
ниях, например в долинах рек, семена могут распространяться и потоками воды.
Учитывая, что семена клёна ясенелистного способны выживать в воде в тече
ние по крайней мере 6 недель и прорастать еще до того, как попадут на слой
почвы, подобным способом семена могут распространяться на довольно дале
кие расстояния (рис. 21). Данные по инвазии A.negundo в 1900--2000 гг. в Бело
вежской Пуще показывают, что скорость расселения может составить от 0,6 до
1 м/год при распространении ветром и до 100 м/год при случайном распростра
нении автомобилями и поездами или при распространении по воде [Mеdrzycki,
2002].
Последствия внедрения
Воздействие на естественные фитоценозы и аборигенные виды. Влия
ние A.negundo на другие растения, по видимому, не отличается от воздействия
других лиственных деревьев. Его опад разлагается довольно хорошо, так что он
может ускорять минерализацию растительных остатков. Однако полностью этот
процесс не исследован. Эффект затенения других растений такой же, как у дру
гих кленов. Единственное специфическое влияние A.negundo -- его постоянный
высокий прирост биомассы, что ведет к доминированию вида в пойменных ле
сах. Типичное поведение A.negundo в Центральной Европе -- сильный наклон
или даже падение ствола, контакт с землей, образование придаточных корней и
92
формирование быстрорастущих побегов 2 ого порядка. Некоторые из горизон
тальных стволов могут неоднократно покрываться новыми речными отложени
ями. В результате образуется удивительно сложный организм, который начина
ет доминировать над более низким подлеском и почти полностью останавлива
ет возобновление ив и тополей [Kunstler, 1999]. Единственно возможный
лимитирующий фактор распространения A.negundo в речных долинах -- часто
та и продолжительность затопления.
В первичном ареале этот вид, кажется, склонен к поражениям различными
патогенами [White, Whitham, 2000]. Во вторичном ареале степень поражения
вида вредителями невысока.
Генетические изменения и вариабельность вида. Поскольку A. negundo во
вторичном ареале не имеет близкородственных (входящих в секцию Negundo)
видов, прямого риска гибридизации или интрогрессии нет.
Выявлено (см. главу 4), что во вторичном ареале внутривидовая изменчи
вость количественных признаков плодов не носит четко выраженного геогра
фического характера и является в основном внутрипопуляционной, тогда как в
естественном ареале имеет место клинальная изменчивость данных параметров.
Методом создания интродукционной популяции установлено, что за 9--11 смен
поколений у A.negundo уже выработалась генетически закрепленная географи
ческая изменчивость некоторых биологических признаков: с севера на юг досто
верно уменьшаются продолжительность периода покоя семян, морозостойкость
деревьев, годовой прирост диаметра ствола и биомассы растений, а у однолетних
сеянцев соответствующая пара листьев распускается на 2 недели позднее.
Влияние на здоровье человека. Пыльца A. negundo является аллергеном
[Ersh et al., 2001].
Экономическое и социальное (положительное и отрицательное) значе
ние. A.negundo не имеет прямой ценности для лесоводства. Как дикая форма,
так и многочисленные культивары в течение долгого времени использовались в
ландшафтном дизайне для парков, улиц и обсаживания обочин дорог в сельс
кой местности. В настоящее время дикая форма применяется редко, но все еще
высаживается в тех случаях, когда критериями выбора является дешевизна,
быстрый рост и толерантность к почвенным условиям, например для рекульти
вации земель.
В городах, где A.negundo высажен преднамеренно или где обычны спонтан
но занесенные растения, дерево приносит пользу, очищая воздух вследствие его
высокой скорости фотосинтеза, которая может превышать 25 мmol CO m 2‡ s 2
[Foster, 1992]. Однако изучение этого аспекта до сих пор не проводилось.
Меры борьбы
Предотвращение заноса может быть самым легким и самым дешевым спосо
бом управления инвазией A.negundo. Маловероятно, что этот вид будут выса
живать в местах спонтанного его распространения. Однако декоративные куль
тивары, которые часто прививаются на дикие подвои, до сих пор продаются во
многих питомниках и садоводческих центрах, и это дает мало шансов избежать
инвазии A.negundo. Желательно полностью прекратить посадки этого вида.
Около больших поселений сдерживание, то есть попытка ограничить появ
ление клена ясенелистного в пределах вторичного ареала, будет лучшим реше
93
нием. Из за относительно низкой скорости рассеивания семян ветром доста
точно сдерживать распространение семян в зоне 50--250 м от материнского ра
стения. Более далекое рассеивание возможно, но не в массовом масштабе (при
сильном снегопаде, шторме, буре и т.п.). Однако чем более связаны местообита
ния, тем больший шанс имеет случайный занос семян машинами и поездами
или потоками воды. При таких обстоятельствах скорость рассеивания может
быть даже в 100 раз большей, так что буферная зона 5--10 км будет вполне дос
таточной.
Уничтожение может быть реализовано, главным образом, путём механичес
кого удаления проростков и ювенильных экземпляров; химическая обработка
будет также весьма эффективной, поскольку A.negundo чувствителен ко мно
гим гербицидам, например к глифосфату.
В июле 2005 г. в окрестностях хут. Недвиговка Мясниковского р на Ростов
ской обл. нами отмечено массовое поражение A.negundo гусеницами американ
ской белой бабочки (Hyphantria cunea). Листья клена были практически полно
стью съедены, от них оставались лишь крупные жилки. Последствия этого по
ражения клена были заметны и в 2008 г.: на деревьях наблюдалось много усохших
ветвей. Однако, в свою очередь, американская белая бабочка является каран
тинным видом [Справочник..., 1995; Ижевский, 2002, 2008].
Обзор научных исследований и публикаций
В естественном ареале изучены многие аспекты биологии A.negundo. Фак
тически он стал модельным видом для изучения эволюционного процесса по
ловой дифференциации под влиянием среды обитания [Willson, 1986; Dawson,
Ehleringer, 1993; Ward, Dawson, Ehleringer, 2002]. Популяционная биология этого
вида изучена в естественном ареале в штате Висконсин [Sachse, 1991]. Изучен
масштаб инвазии A.negundo в зависимости от типа использования земель
[Medrzycki, 2002; Medrzycki, Pabjanek, 2001]. Описано поведение этого вида в
Варшаве и окрестностях [Kosim, 2005; Salapa, 2005; Banaszek, 2005; Caban, 2005].
Появление и воздействие A.negundo на пойменные леса изучены в долине р. Висла
[Kunstler, 1999]. Создана интересная ГИС база, моделирующая потенциальные
местообитания клена ясенелистного в Чехии [Hrazsky, 2005].
СЕМЕЙСТВО AMARANTHACEAE АМАРАНТОВЫЕ
ЩИРИЦА БЕЛАЯ
Название: Amaranthus albus L.
Морфологические признаки
Однолетнее растение высотой 15--60 см. Стебли прямостоячие, от основа
ния ветвистые (рис. 22). Листья голые, продолговато обратнояйцевидные, на
конце с шипиком; черешок в 2--3 раза короче пластинки. Цветки в пазухах лис
тьев. Прицветники шиловидно заострённые, в 1,5--2 раза длиннее околоцвет
ника. Околоцветник трехлистный, пленчатый, из яйцевидно ланцетных листоч
ков, чуть короче плода.
94
Естественный ареал
A.albus -- североамериканское растение. Естественный его ареал занимает
только прерии и район Великих Равнин США, а в остальных регионах североа
мериканского континента этот вид является заносным [Gleason, 1963].
Вторичный ареал
История интродукции и географическое распространение. Щирица белая
занесена в Средиземноморье в начале ХVIII века и впервые отмечена в 1723 г. в
Тоскане (Италия), а в 1797 г. -- в Алжире. В начале ХIХ века найдена в Турине и
Генуе (Италия), Испании, Португалии и Греции, а также отмечена среди других
адвентивных растений ботанического сада Монпелье. В Среднюю Европу (Гер
мания, Швейцария, Чехия) проникла в конце ХIХ века. В Польше впервые най
дена в 1907 г., но там она распространялась крайне медленно -- вторая находка
относится лишь к 1921 г. [Frey, 1974; Hegi, 1959; Boissier, 1879]. К середине ХХ
века A.albus натурализовалась практически по всей Европе (исключая Сканди
навию) и в Средиземноморье. Позднее отдельные ее местонахождения отмече
ны также в Юго Западной Азии и в Восточной Монголии [Tutin et al., 1964;
Aellen, 1972; Цэрэнбалжид, 1984].
По многочисленным литературным данным, занос A.albus в Россию произо
шел во второй половине ХIХ века. Однако в ботанических садах она росла и
ранее [Виноградова, 2002]. Первое упоминание о виде найдено в каталоге рас
тений сада А.Разумовского в Горенках, относящемся к 1812 г. В 1816 г. щирицу
белую выращивали в ботаническом саду Кременца, а в 1823--24 гг. -- в ботани
ческих садах Санкт Петербурга и Харькова [Fischer, 1812; Besser, 1816; Index ..,
1823; Фишер, 1824].
В 60 х гг. ХIХ века щирица белая с корабельным балластом была занесена в
порты Черного моря и начала распространяться по югу Российской империи. В
1869 г. она собрана в Одессе (LE), и до начала ХХ века обнаружена в Симферополе,
Евпатории, Массандре, Феодосии, на о. Куянлы, в Умани и Херсоне (LE, KW).
К 1911 г. образовались три первичных очага этого сорняка. Первый прости
рался от Одессы на запад и юго запад до устья Дуная, второй охватывал весь
Херсонский уезд, где A.albus стала обычным сорным растением на дорогах и в
посевах, а третьим очагом распространения щирицы стала ж/д от Крыма до
Мелитополя [Буянкин, 1974].
Ареал щирицы стремительно расширялся, охватывая все более северные районы.
В 1910 г. она найдена около Днепропетровска, в 1914 г. -- на окраинах Купянска, в
1916 г. -- на окраинах Киева (KW). Практически одновременно щирица белая рассе
лилась по Нижней Волге: в 1910 г. обнаружена в г. Покровск (ныне --Энгельс), в 1920
г. -- в окрестностях Царицына (ныне --Волгоград) и в Камышине. На север A.albus
распространялась, в основном, по железным дорогам. В 1921--1927 гг. она найдена в
Кременчуге, в Ворошиловградской обл., в Бортничах и Березани, в Лубнах, Белой
Церкви, Черкассах, Виннице [Протопопова,1973], а также в Харькове (LE) и в окре
стностях Минска и Витебска (картотека ВИРа). Все вышеперечисленные находки
сделаны или около ж/д вокзалов, или на полотне железной дороги.
Распространяясь в восточном направлении, A.albus в 1913 г. попала в Даге
стан, в 1916 г. -- в устье р. Кубань, в 1921 г. -- в окрестности Майкопа; в 1925--
1928 гг. стала массово встречаться на Дону и Северном Кавказе (LE).
95
По данным В.С. Ябровой Колаковской [1977], щирица появилась в Закав
казье в 1880 х гг., однако гербарными данными подтверждаются ее находки лишь
с 1924 г., когда вид обнаружен в Батуми и Ереване, а в 1928 г. -- в долине р. Арпа
(LE). В картотеке гербария ВИР имеются сведения о сборах щирицы в 1911 г. в
Тифлисской губернии. Имеются данные о находках вида в 1925 г. на Апшероне,
в 1926 г. -- в Кура Араксинской низменности и в 1931 г. в Ленкоранской низмен
ности [Буянкин, 1974; Мамедов, 1986].
В Средней Азии A.albus впервые найдена в 1914 г. в окрестностях Ташкента,
а в 1916 г. зарегистрирована на ж/д ст. Арысь (LE). В 1923 г. собрана в Чимкен
тском уезде на оз. Сары Куль у р. Сыр дарья (TASH). На западе Казахстана в
Уральской обл. собрана в 1924 г., а на востоке в окрестностях г. Семипалатинск --
в 1926 г. В 1927 г. обнаружена в Ашхабаде [Буянкин, 1974].
В 1926 г. В.Траншель собрал щирицу белую на Дальнем Востоке (LE), при
чем отметил, что там этот вид еще не указывался.
Таким образом, к началу 1930 х гг. северная граница ареала щирицы белой
проходила по линии Киев--Харьков--Саратов, а восточная граница находилась
в пределах Заволжья [Виноградова, 2002], и единичные находки отмечены в
Средней Азии и на Дальнем Востоке.
Ареал вида продолжал расширяться. За время Великой Отечественной вой
ны A.albus внедрилась в нарушенные местообитания по всему Казахстану и
Средней Азии (TASH, TAD). В 1942--1946 гг. щирица найдена во Львове и Бре
сте. В начале 1960 х гг. A.albus появилась в Забайкалье в окрестностях г. Улан
Удэ и стала массово встречаться в Приморье, Амурской обл. и в Хабаровском
крае [MHA; Буянкин, 1974; Котов, 1949; Фомина, 1965].
На север щирица белая продолжала поступать по железным дорогам. Еди
ничные ее находки отмечены в 1943 в г. Петрозаводск, в 1945 г. -- к северу от
Сыктывкара, в 1959 г. -- в Норильске. Большинство находок A.albus сделано
Ю.Д.Гусевым (LE) в 1961--1973 гг. Он обнаружил вид в г. Тарту (Эстония), в
Псковской обл. на песчаном берегу Чудского озера, на ж/д станциях Княжпо
гост, Сыктывкар и Помоздино (Коми АССР), в Ленинградской обл. близ Бело
острова, на станциях Тихвин и Молосковицы, а также на станции Сухона Во
логодской обл., в Могилеве (Белоруссия) и на ж/д станциях Киров, Юрья и
Фаленки (Кировская обл.).
В 1980 г. находки A.albus сделаны в Тандинском р не Тувы и на окраине Бар
наула [Зверева, 1981].
Таким образом, от первой находки A.albus в Италии (1723 г.) до начала ее
натурализации в Средней Европе (60 е гг. ХХ века) прошло 250 лет! В настоя
щее время A.albus является инвазионным видом практически по всей Европе,
на Кавказе, в Средней Азии и на Дальнем Востоке.
Пути и способы заноса. В регионах Средней России активно распространя
ется в основном по железным дорогам. В Московской обл. отмечены случаи рас
селения вида возле элеваторов.
Статус в регионе. На территории Средней России (г. Тверь) отмечена как
заносное растение в 1917 г., через год найдена в Саратове, а еще через 9 лет -- в
Москве [Буянкин, 1974]. В середине 1920 х гг. она стала уже самым обычным
сорняком от Ергеней до Саратова. В 1934 г. отмечена на окраинах Самары [Бу
96
янкин, 1974; Перечень семян..., 1934]. Распространение вида на север шло в ос
новном по железным дорогам. В 1942--1946 гг. щирица белая найдена в Улья
новской и Воронежской обл. В 1969--1975 гг. вид был собран в Смоленской и
Тверской обл. на ж/д станциях Духовская, Бологое, Лихославль и Ржев [LE;
Гусев, 1973; Малышева, 1979]. В 1975--1990 гг. новые находки A.albus сделаны в
г. Иваново, в Удмуртии (Ижевск, Сарапул, Каркалай), в Чувашии (Канаш), во
многих пунктах Рязанской обл. и в ряде мест по ж/дорогам Брянской обл. [MHA;
Шилова, 1984; Ильминских и др., 1981; Гущина, 1983].
Таким образом, за полвека A.albus стал массовым сорняком (рис. 23). Посколь
ку это растение однолетнее, можно считать, что период его натурализации в
Средней России насчитывает не меньше 40 смен поколений.
Местообитания. Места массового распространения A.albus связаны с песчаны
ми почвами. Однако вид отличается широкой эколого ценотической амплитудой
и встречается также в садах, рудеральных местах и посевах многих сельскохозяйствен
ных культур, а также вдоль дорог, железнодорожных путей, возле жилья (рис. 24). Об
ладает относительно высокой технотолерантностью и заселяет техногенные экото
пы южных степей и даже может входить в пионерные галофитные сообщества. В
Ростовской обл. произрастает на аллювиальных песках по берегам Дона.
Размножение и жизненный цикл. Однолетнее растение. Цветет с июля по
сентябрь, плодоносит с августа. Всхожесть семян сохраняется до 8 лет и состав
ляет в первый год до 85%. Для A.albus характерны растянутые сроки прораста
Рис. 23. Расселение Amaranthus albus в Средней России
Рис. 22. Amaranthus albus, общий вид
растения
Рис. 24. Растение Amaranthus albus
на ж/д насыпи, устойчивое к
гербицидам.
Рис. 25. Две формы
Amaranthus albus: слева ---
f. albus, справа --- f. minimus.
98
ния, что является приспособлением, направленным на сохранение вида и спо
собствующим накоплению запасов семян в почве.
Расселение и распространение. Мелкие семена, устилающие землю вокруг
материнского растения, разносятся дождевыми потоками и талыми водами на
большие расстояния. Так как растение образует форму "перекати поле", его мо
жет переносить ветер, и таким образом семена засоряют почву на значительной
площади. Семена легко прилипают вместе с грязью к ногам и колесам. Антропо
хорные способы заноса облегчаются тем, что растение массово произрастет на
рудеральных местах.
Последствия внедрения
Воздействие на естественные фитоценозы и аборигенные виды. Засоряет
посевы пропашных и зерновых культур, хлопчатника, кукурузы, сои, сахарной
свеклы, реже встречается на огородах, в виноградниках и садах.
Генетические изменения и вариабельность вида. Значительная внутривидо
вая изменчивость щирицы белой позволила выделить в ее вторичном ареале 12
форм [Hegi, 1959]: f.albus (типичная форма), f.minimus Priszter, f.umbrosus Polgar,
f.monosepalus Thell., f.puberulus Thell., f.viarum Priszter, f.prostrato ascendens
Thell., f.compactus Priszter, f.tenuis Priszter, f.scoparius Priszter, f.parviflorus Moq.
(= f.parvifolius Zimmermann), f.rubicundus Thell. У A.albus отмечена также [Ти
монин, 1984] внутрипопуляционная изменчивость анатомии побега (наличие/
отсутствие вздутых эпидермальных клеток).
Ни в одном из исследованных регионов [Виноградова, 2002] 100% ной реа
лизации фенофонда не наблюдалось. В Московской обл., где распространение
вида приурочено в основном к железным дорогам, степень реализации фено
фонда составила 25%. Чаще других встречается A.albus f.prostrato ascendens (ра
стения стелющиеся, раскидистые, площадь покрытия земли одним растением --
1--1,5 мІ, верхушки побегов приподнимающиеся). Типичная форма A.albus f.albus
встречается реже. Растения A.albus f.minimus (прямостоячие неветвящиеся по
беги высотой до 5 см с мелкими листьями длиной до 0,5 см) отмечаются в попу
ляциях только во второй половине лета.
В Волгоградской обл., в Калмыкии и Абхазии степень реализации фенофон
да выше и составляет 33%. Помимо вышеперечисленных, там произрастает
A.albus f.rubicundus (все растение, либо по крайней мере стебель, окрашено в
красный цвет).
В Средней Азии отмечена наиболее высокая степень реализации фенофонда --
42%. Преобладает A.albus f.viarum (растет на уплотненной почве; растения по
душкообразной формы диаметром 10--20 см, побеги сильно облиствены, листья
мелкие), A.albus f.parviflorus (листья длиной 0,5--1 см), а также упомянутые выше
распростертая A.albus f.prostrato ascendens и карликовая A.albus f.minimus фор
мы. Типичная A.albus f.albus растет только на клумбах при поливе [Виноградо
ва, 2002].
По литературным данным, в Польше степень реализации фенофонда A.albus
составляет 42%, в Словакии -- 67%, в Венгрии -- 25% [Frey, 1974; Grull, Priszter,
1967; Solymosi, 1983].
Экономическое и социальное (положительное/отрицательное) значение. В
южной части ареала A.albus -- главнейший засоритель посевов. Так, в Уральской
99
обл. засоренность посевов щирицей белой составляет 49% от общей засоренно
сти, а в Волго Ахтубинской пойме щирица белая засоряет 58% полей. Запас се
мян вида в почве в южных районах Украины составляет 53,4% запаса семян
остальных сорняков [Протопопова, 1973; Слесарев, Буянкин, 1971; Коблова,
1962]. Польза от щирицы белой крайне незначительна. Поскольку A.albus со
держит ~25% протеина, в Казахстане и в Болгарии ее используют для силосова
ния [Ковачев, 1966].
Меры борьбы
Защитные мероприятия: пожнивное лущение, междурядная обработка, хи
мическая прополка.
Обзор научных исследований и публикаций
Выявлено (см. главу 5), что описываемые в литературе формы A.albus явля
ются отражением модификационной изменчивости вида. Генетически закреп
лены лишь две формы: типичная A.albus f.albus, семена которой прорастают вес
ной и дают высокий, сильно разветвленный побег, и неотеничная A.albus
f.minimus, семена которой прорастают в середине лета и дают неветвящийся
побег высотой до 10 см (рис. 25). Методом создания интродукционных популя
ций выявлен очень низкий уровень генетической изменчивости А.albus во вто
ричном ареале, что объясняется обедненным генофондом первоначальных ма
лочисленных инвазионных популяций в сочетании с самоопыляемостью вида.
СЕМЕЙСТВО AMARANTHACEAE -- АМАРАНТОВЫЕ
ЩИРИЦА ЗАПРОКИНУТАЯ
Название: Amaranthus retroflexus L.
Морфологические признаки
Травянистый однолетник, большей частью бледно зеленый, иногда с
красноватым оттенком (рис. 26). Стебель прямостоячий, высотой 20--80 см.,
простой или ветвистый, сероватый от густых коротких волосков, слегка углова
тый. Листья довольно крупные, яйцевидно ромбические, на верхушке тупые или
несколько выемчатые, с небольшим шипиком. Верхняя сторона листа голая;
нижняя сторона, край и черешок коротко волосистые. Черешок по длине почти
равен листовой пластинке или чуть длиннее ее [Васильченко, 1936].
Цветки собраны на концах побегов в ложноколосовидные (очень плотные
метельчатые) соцветия, в нижней части ветвящиеся. Прицветники ланцетные,
заостренные, превышают околоцветник почти вдвое. Околоцветник состоит из
5 листочков. Растение однодомное; у мужских цветков доли околоцветника лан
цетные, у женских -- продолговато линейные; кверху лопатовидно расширен
ные, тупые или слегка выемчатые, с коротким острием на верхушке, беловатые,
пленчатые, с тонкой светло зеленой срединной жилкой, обычно не доходящей
до верхушки. Тычинок 5, пестик 1 с единственным семязачатком. Коробочка
односеменная, короче околоцветника, открывается крышечкой. Семена ~1 мм,
черные или черно коричневые, блестящие, с острой каймой по краю [Василь
ченко, 1936; Мосякин, 1996].
100
Семядоли продолговато линейные, длиной 8--10 мм, шириной 2 мм, на вер
хушке островатые, к основанию суженные, серовато зеленые. Первый лист ко
роткоовальный или яйцевидно ромбический, на верхушке выемчатый, к осно
ванию быстро сужается, длиной 8--10 мм и шириной 6--8 мм. Черешки и стебе
лек покрыты мелкими курчавыми волосками.
Естественный ареал
Естественный ареал A.retroflexus занимает только прерии и район Великих
Равнин США, в остальных районах Северной Америки этот вид является за
носным [www.efloras.org]. Вторичный ареал
История интродукции и географическое распространение. Как натура
лизовавшееся растение произрастает на территории Северной Америки от Аляс
ки и о вов Борден, Брок, Виктория, Маккензи Кинг, Принс Патрик, западной
части острова Мелвилл на севере и до провинции Британская Колумбия на юге
(отсутствует на территории штата Юкон, материковой части штата Ньюфаунд
ленд и северо востоке Канады) [www.plants.usda.gov].
О времени и месте заноса этого вида из Северной Америки в Старый Свет
существуют противоречивые мнения. Ряд исследователей полагали, что щири
ца запрокинутая занесена в XIV веке в Италию, а по другим сведениям, расте
ние занесено в виде семян из штата Пенсильвания в Ботанический сад Шве
Рис. 27. Расселение Amaranthus retroflexus в Средней России
Рис. 29. Всходы Amaranthus retroflexus в прибрежных сообществах по берегу р. Дон.
Рис. 26. Amaranthus retroflexus,
общий вид растения.
Рис. 28. Массовое засорение посевов
щирицей (Amaranthus retroflexus).
102
ции, где его в 1750 г. исследовал К.Линней, и откуда оно распространилось по
всей Европе [Буянкин, 1977]. Позднее, в 1783 г., оно встречалось во Франции, в
1785г. --вИталии,в1791г. --вВенгрии,в1794г. --вГермании,в1831г. -- в
Австрии [Буянкин, 1977].
Наиболее старые гербарные сборы щирицы сделаны на юге Европейской
России. В Гербарии МГУ хранится экземпляр щирицы запрокинутой, собран
ной Л.Ф. Гольдбахом в 1807 г. в Елизаветграде (ныне -- Кировоград, Украина). В
Гербарии БИНа также имеются ранние сборы сорняка, выполненные И.Д. Пре
скоттом в 1823 г. в Астраханской губернии и К.Ф. Ледебуром в 1818 г. -- в дельте
Волги. Там же находятся гербарные листы щирицы запрокинутой, собранные в
Екатеринославской губернии (ныне -- Днепропетровская обл.) неизвестными
авторами, датированные 1827 и 1832 гг. В это же время щирица запрокинутая
встречалась и на Кавказе. Так, К.А. Мейер собирал ее на побережье Каспийско
го моря еще в 1829--1830 гг. (LE). Позднее, в 1842 г., она найдена А.Демидовым
в Крыму в Симферополе.
Уже в первой половине XIX века щирица запрокинутая встречалась в раз
личных районах России и сопредельных государств. В 1838 г. вид отмечен в
Киеве, в 1840 г. найден Г.С. Карелиным и И.П. Кирилловым в Восточном Казах
стане у Бухтарминска, и Кюлевейном в 1846 г. -- в Санкт Петербурге. В 1840 х
гг. щирица запрокинутая встречалась в г. Ревель (Таллин) и некоторых других
пунктах Прибалтики. Ю. Шелл собирал ее в 1848 г. в Оренбурге (LE).
Во второй половине XIX века собрано много гербарных листов щирицы зап
рокинутой со всех губерний европейской части России, вплоть до 60° с.ш. В
БИНе (LE) хранится, например, несколько листов Н.Довнара, собранных им в
Могилевской провинции в 1857--1862 гг. Но наиболее часто вид отмечался опять
таки для южных районов страны. В 1854 г. в Новороссийске щирицу запроки
нутую собрал С.С. Щеглеев, в 1869 г. она впервые отмечена для Кишинева (LE).
В это время она в немалом количестве произрастала на территории всей Украи
ны, что подтверждается материалами гербариев. Так, в Одессе она зарегистри
рована в 1865 г., в Таганроге -- в 1875--1877 гг. (MW), в Харькове -- в 1877 г., в
Мелитополе -- в 1878 г., в Азове -- в 1867 г. В 1870 г. К.А. Гольде фиксирует ее для
Волынской флоры [Буянкин, 1977].
Одновременно продолжается распространение щирицы запрокинутой
на Кавказе. В 1865 г. она собрана в Бакинской губернии, в 1871 г. -- в Тифлисе
[LE; Буянкин, 1977]. Таким образом, к началу 1870 х гг. щирица запрокинутая
стала обычным сорняком в центральной и южной частях России. В это время
наблюдается расширение ее ареала на север, восток и юго восток страны. А.Бек
кер нашел ее в 1869 г. в Западном Казахстане на п ове Мангышлак.
В 1870--1880 гг. осуществлены сборы A.retroflexus в Красноярском крае, в 1884 г. --
в Омской губернии, в 1877 г. -- в Киргизии, в 1874 г. -- в Каракалпакии (LE,
MW). С.Коржинский в 1875 г. нашел щирицу запрокинутую в Казанской гу
бернии, Ю.Шелл -- в 1876--1879 гг. -- в Пермской и Виленской губерниях. На
рубеже XX века растение отмечено в окрестностях г. Ашхабад (1897 г.), г. Вер
ный (Алма Ата, 1894 г.), г. Кустанай (1898 г.), в Барнаульском округе (1890 г.), в
Прибайкалье (1891 г.) и на Дальнем Востоке в 1898 г. (LE). В 1896 и 1899 гг. оно
зарегистрировано в Псковской обл. [Пуринг, 1898, 1901].
103
Уже к началу XX века контуры ареала щирицы запрокинутой в основном
сложились. Это растение было широко распространено во всех земледельчес
ких губерниях европейской части России. Кроме того, имелись первичные оча
ги распространения в различных районах Казахстана, Средней Азии, Западной
и Восточной Сибири, а также на Дальнем Востоке [Буянкин, 1977].
В последующие годы наблюдались быстрое расселение сорняка из первич
ных очагов и слияние отдельных районов распространения в сплошной ареал.
К началу 30 х гг. ХХ века северной границей распространения щирицы запро
кинутой стала линия Санкт Петербург--Вологда--Вятка--Пермь--Тобольск--се
верная часть Байкала--устье Амура [Буянкин, 1977].
Многочисленные гербарные материалы различных авторов показывают, что
граница эта сохранилась и поныне. Однако есть сведения, что растение может
произрастать и в более северных районах, где она быстро натурализуется. Так,
щирица запрокинутая собрана в 1966 г. на юге Архангельской обл. у станции
Котлас, а в ВИРе хранятся 18 гербарных листов И.А. Иванова, собранных им в
1969--1971 гг. в г. Якутск. [Буянкин, 1977].
Пути и способы заноса. В посевы зерновых культур и кормовых трав попа
дает главным образом с семенами. Активно распространяется по железным до
рогам, часто встречается вблизи железнодорожных насыпей.
Статус в регионе. Самый ранний гербарный лист A.retroflexus собран М.А.
Максимовичем в 1824--1826 гг. в Москве (MW). Имеются сборы из Воронежс
кой (1838 г.) и Ярославской (1866 г.) губерний (LE). Активное расселение
A.retroflexus в губерниях Центрально Черноземной зоны и Нечерноземья прохо
дило во второй половине XIX века. В БИНе находится гербарный лист 1860 г.
неизвестного коллектора из Курской губернии, на котором указано, что расте
ние «произрастает в неимоверных количествах». В 1868 г. щирица запрокину
тая собрана в Калужской губернии (MW). В работе В.Я. Цингера [1885]
A.retroflexus приводится для Костромской (1867), Тверской, Смоленской, Вла
димирской, Нижегородской (1884), Рязанской (1851), Тульской (1880), Орловс
кой (1855), Тамбовской (1851), Саратовской (1851), Пензенской и Симбирской
(1854) губерний. Таким образом, уже к началу ХХ века ареал A.retroflexus на тер
ритории Средней России полностью сформировался (рис. 27). Отсутствие гер
барных сборов из Брянской обл. следует отнести к досадному недоразумению.
Местообитания. Встречается на полях, огородах, пустырях (рис. 28), план
тациях различных культур, на лугах и степных склонах, береговых обрывах, по
берегам водоемов, у дорог, в населенных пунктах [Васильченко, 1936]. Предпо
читает песчаные и супесчаные почвы [Бариева, 1997].
Размножение и жизненный цикл. Плод (коробочка) вскрывается поперечной
кольцевой щелью. На одном растении может быть до 117 400 семян [Radosevich et
al., 1997], или до 1 000 000 семян/м2 [Cousens, Mortimer, 1995]. Оптимальной для
прорастания является температура 26--36°С, минимальной -- 6--8°С [Баркинхоев,
2004]. Семена хорошо переносят отрицательные температуры, поздневсхожие, в
умеренных широтах дают всходы в мае -- начале июня [Бариева, 1997], сохраняют
всхожесть в течение 30--40 лет амбарного хранения [Ульянова, 2005].
Расселение и распространение происходит в основном с семенами посев
ных культур.
104
Последствия внедрения
Воздействие на естественные фитоценозы и аборигенные виды.
A.retroflexus хорошо приживается в нарушенных сообществах, например в раз
нотравно дерновинно злаковых (рис. 29), разнотравно типчаково ковыльных
и других степях, сильно пострадавших от чрезмерного выпаса [Сконникова,
1985], конкурируя с аборигенными видами.
Генетические изменения и вариабельность вида. A.retroflexus -- полимор
фный вид, для которого описано немало внутривидовых таксонов, большин
ство из которых не имеет серьезного систематического значения. На террито
рии Восточной Европы отмечены гибриды A. powellii ґ A. retroflexus (A. ґ
soproniensis Priszter et Karpati) и A. hybridus L.ґ A. retroflexus (A. ґ ozanonii
Thellung) [Мосякин, 1996].
Влияние на здоровье человека. Листья A.retroflexus содержат много каль
ция (~5,3% в пересчете на сухую массу) и целлюлозы, а потому могут использо
ваться в профилактике инфаркта миокарда и инсульта, а также при остеопорозе
и некоторых других заболеваниях [Чухно, 2007].
Экономическое и социальное (положительное/отрицательное) значе
ние. A.retroflexus -- злостный сегетальный сорняк пропашных культур (хлопчат
ник, кукуруза, соя и др.), реже зерновых и однолетних кормовых. Часто встре
чается в большом количестве в посадках картофеля [Бариева, 1997].
Семена можно использовать в качестве корма для птиц. Молодые отварен
ные листья употребляются в пищу. Молодые растения поедают свиньи. Щири
ца запрокинутая пригодна на силос [Васильченко, 1936].
Меры борьбы
Избавлению плантаций от A. retroflexus способствуют пары, пожнивное лу
щение, глубокая зяблевая вспашка, культивация в период появления всходов,
периодическая смена пропашных культур кормовыми и зерновыми [Ульянова,
2005]. Всходы и проростки легко уничтожаются поверхностными приемами об
работки почвы, повсходовыми и почвенными гербицидами [Баркинхоев, 2004].
По мнению Э.Р. Бариевой, одним из способов регулирования численности
щирицы запрокинутой является включение в севооборот многолетних трав, так
как многолетние травы способствуют уменьшению освещенности и уплотне
нию почвы, в результате чего изменяется аэрация верхнего слоя почвы, которая
воспринимается семенами как сигнал к прорастанию. Это приводит к мозаич
ному появлению всходов сорняка и к увеличению доли угнетенных особей, пре
вращая популяцию в депрессивную [Бариева, 1997].
Обзор научных исследований и публикаций
A.retroflexus -- факультативно короткодневный вид, то есть если родительс
кие растения цвели в условиях короткого светового дня (8 ч), семенам для про
растания нужна продолжительная ночь и температура ~30°С. Если же особи
цвели при длинном световом дне (16 ч), то семена прорастают при более низкой
температуре и более длинном дне [Kigel et al., 1979]. Таким образом, длина све
тового дня, необходимая для прорастания семян, зависит от места обитания
родительских растений, что является одной из причин хорошей приживаемос
ти этого сорняка на различных широтах.
105
СЕМЕЙСТВО APIACEAE (UMBELLIFERAE) ---
СЕЛЬДЕРЕЙНЫЕ (ЗОНТИЧНЫЕ)
БОРЩЕВИК СОСНОВСКОГО
Название: Heracleum sosnowskyi Mandenova
Морфологические признаки
Двулетнее или многолетнее монокарпическое растение. Стебель прямой,
округлый, бороздчатый, негусто опушенный. Прикорневые и нижние стебле
вые листья тройчатые, реже перистые; боковые сегменты широкояйцевидные
или округлые, неравнобокие, 3(5) лопастные или рассеченные; верхний сег
мент округлый, более или менее глубоко 3 надрезанный на широкояйцевид
ные, в свою очередь лопастные, доли. Верхние листья уменьшенные, с расши
ренным влагалищем и 3 лопастной пластинкой. Листья сверху голые, снизу --
мелко и оттопырено опушенные. Зонтики крупные, верхний нередко диамет
ром до 0,5 м; лучи зонтиков и зонтичков мелкошероховато опушенные; лис
точки оберток и оберточек линейно шиловидные с расширенным основани
ем. Зубцы чашечки хорошо заметные, треугольные, зеленые. Лепестки белые,
реже розовые, у краевых цветков в зонтике сильно увеличенные (рис. 30).
Плоды обратнояйцевидные или широкоовальные, длиной 10--12 мм и шири
ной 6--8 мм, со спинки сплюснутые.
Близкий вид H. mantegazzianum Somm. et Lev. -- Б. Мантегацци отличается
оттопырено опушенными лучами зонтиков и зонтичков и заостренными ко
нечными сегментами листьев [Манденова, 1951; Виноградова, 2004].
Естественный ареал
Восточная часть Большого Кавказа, Восточное и Юго Восточное Закавка
зье, северо восток Турции [Ecology..., 2007].
Вторичный ареал
История интродукции и географическое распространение. Как занос
ное растение, H.sosnowskyi встречается в России, Эстонии, Латвии, Литве,
Польше, Беларуси, на Украине, на востоке ФРГ (на территории бывш. ГДР), то
есть в СССР и странах бывш. социалистического лагеря [The Giant..., 2005;
Ecology..., 2007]. Недавние молекулярно генетические исследования выявили
этот борщевик в Дании [Jahodovб et al., 2007]. Борщевик Сосновского отнесен к
балтийскому типу1 распространения адвентивных растений Европы [Lambdon
et al., 2008].
Источником семян для посадок борщевика Сосновского были его кавказс
кие популяции (в частности, для диссертационного исследования А.А. Мар
ченко материал собран в Кабардино Балкарии) [Ecology..., 2007]. Следует учи
тывать, что на Кавказе растет несколько близких видов борщевика, при сборе
плодов в природе могли происходить ошибки. Так, для обширной программы
разведения борщевика Сосновского в Центральном ботаническом саду Бела
руси (Минск) материал собирали в окрестностях Красной Поляны близ Сочи,
где вид этот не растет [Ecology..., 2007].
1 Такое решение ошибочно принято без учета широкого распространения этого борще
вика в Европейской России, Беларуси и на Украине.
106
Поскольку H. sosnowskyi выносливое и холодоустойчивое растение, оно
широко культивировалось, особенно на северо западе Европейской России,
где впервые высажено в 1947 г. [Практическое..., 2005; Kabuce, 2006]. Борще
вик получил репутацию высокопродуктивной и устойчивой культуры [Иев
лев, 1983; Сацыперова, 1984; Тимофеев, 1993, 2002; Мишуров и др., 1999].
Однако постепенно выяснилось, что специфический запах борщевика сохра
няется в мясе и молоке животных, а при наличии других кормов коровы пло
хо поедали силос [Практическое..., 2005; The Giant..., 2005; Kabuce, 2006; соб
ственные данные]. К тому же стали очевидна опасность ожогов при контакте с
растениями борщевика. Поэтому, уже в 1980 е годы борщевик практически
перестали культивировать, но, по некоторым данным, на севере европейской
части России его культивируют до сих пор [Практическое..., 2005; The Giant...,
2005; Kabuce, 2006].
Для стран Балтии дичание отмечено во второй половине 1980 х гг.
[Gudћinskas, 1998; Kabuce, 2006]. К сожалению, в ходе международного про
екта, посвященного гигантским борщевикам (Giant Alien Project), который был
проведен в 2002--2005 гг. и в котором от России принимали участие БИН РАН
и ЗИН РАН, точное распространение именно Н. sosnowskyi осталось наименее
изученным [Практическое..., 2005; The giant..., 2005]. Известно лишь, что в
настоящее время борщевик широко расселился по Европейской России
[Виноградова, 2004].
Пути и способы заноса. Борщевик Сосновского внедрен в культуру как
высокопродуктивное силосное растение вскоре после Великой Отечествен
ной войны [Сацыперова, 1984]. Иногда годом интродукции называется 1947 й
[The Giant..., 2005; Ecology..., 2007]. Специалисты интродукторы рапортова
ли: «Борщевик Сосновского способен формировать надземную биомассу до
2000 ц/га»; «Непревзойденной силосной культурой является борщевик Со
сновского. Зеленая масса его богата сахарами, витаминами, содержит анти
бактериальные вещества, а самое главное, по продуктивности она в 3--4 раза
превосходит другие культуры», борщевик способен «обеспечивать продуктив
ность 11--7 тыс. л молока в год от фуражной коровы» [Иевлев, 1983; Сацыпе
рова, 1984; Тимофеев, 1993, 2002; Мишуров и др., 1999].
По данным Giant Alien Project [Практическое..., 2005; The Giant..., 2005],
наиболее вероятно появление борщевиков в следующих местах:
· находящихся на незначительном расстоянии от мест произрастания
Н.sosnowskyi и не отделенных преградой (возможен занос семян ветром);
· находящихся ниже по течению рек от занятых борщевиком мест;
· на обочинах дорог и ж/д полотна в том случае, если менее чем в 2 км про
израстают борщевики;
· на территориях, где поблизости борщевик специально выращивался.
Статус в регионе. Впервые в Средней России одичавшим H. sosnowskyi
был собран в 1948 г. Б.М. Кульковым в с. Рождествено Серпуховского р на
Московской обл. (МНА). В последующие годы борщевик дичал редко, встре
чаясь в основном близ мест культивирования [Игнатов и др., 1990], по край
ней мере в Московской обл. в 1960--е гг. этот вид не проявлял тенденции к
внедрению в естественный растительный покров [Скворцов, 1982]. В 1970 х
гг. дичание Н. sosnowskyi приобрело массовый характер [Игнатов и др., 1990;
собственные наблюдения]. Гербарные материалы, на основании которых можно
107
Рис. 30. Соцветия Heraсleum sosnowskyi с розовыми лепестками
Рис. 31. Расселение Heraсleum sosnowskyi в Средней России
Рис. 32. Heraсleum sosnowskyi,
общий вид цветущего растения.
Рис. 33. Внедрение Heraсleum
sosnowskyi в прибрежные
сообщества, р. Москва река ниже
с.Каринское в Одинцовском р не
Московской обл.
109
было бы изучить распространение борщевика, чрезвычайно ограничены. Это
связано, во первых, с нежеланием коллекторов иметь дело с опасным расте
нием, во вторых, с неудобством самого борщевика для гербаризации из за
крупных размеров растения. Из за этого проследить динамику расселения не
представляется возможным. Распространен во всех областях Средней России,
но гербарных образцов довольно мало (рис. 31) ввиду крупных размеров рас
тения и небезопасности его сбора.
Местообитания. На Кавказе борщевик Сосновского растет «в среднем и
верхнем лесном поясе, на лесных опушках, полянах и т.д.» [Манденова, 1951].
В европейских условиях борщевик предпочитает освещенные места, встреча
ется на опушках, но под полог леса практически не заходит. Чаще борщевик
поселяется на пустошах, залежах, обочинах дорог, берегам водоемов и т.п., в
то время как возделанные земли не являются привлекательными для данного
вида [Практическое..., 2005; The Giant..., 2005]. В целом, растение предпочита
ет солнечные места, мало используемые человеком, с влажной плодородной
почвой.
Размножение и жизненный цикл. Согласно И.П. Манденовой [1951], бор
щевик Сосновского является двулетним или многолетним растением. Близ
кий H.mantegazzianum2 в Чехии до цветения может расти 6--7 лет [Ecology...,
2007]. Н.sosnowskyi -- монокарпик [Ткаченко, 1989; The Giant..., 2005]. До цве
тения в фазе вегетативной розетки он пребывает 2--4 года, а в неблагоприят
ных условиях -- до 5--6 лет [Kabuce, 2006]. Цветет борщевик Сосновского в
конце июня -- в июле, поврежденные растения могут зацветать позднее. Это
насекомоопыляемое растение. Привлечению насекомых способствуют увели
ченные лепестки краевых цветков соцветия. Цветки посещают разнообразные
насекомые: двукрылые, перепончатокрылые, жуки и др. Как и многие зонтич
ные, борщевик имеет широкий круг опылителей, так как нектарники у него
открытые и крупные. По некоторым сведениям, борщевик способен к само
опылению [Практическое..., 2005; The Giant..., 2005]. Плодоношение обиль
ное. В условиях Ленинградской области одно растение борщевика дает
8836 плодов [Ткаченко, 1989].
Семена H. sosnowskyi нуждаются в стратификации холодом [Николаева и
др., 1985]. В лабораторных условиях достаточно снижения температуры до
+1--6°C. В природе семена прорастают ранней весной, летом семена не прора
стают [Ecology..., 2007]. Мерикарпии борщевиков образуют семенной банк. Око
ло 98% мерикарпиев H. sosnowskyi находятся в почве на глубине до 5 см, однако
небольшое их количество обнаружено и на глубине до 15 см [Moravcovб et al.,
2005, 2007]. Более 90 % семян прорастают в следующую весну. Таким образом,
семенной банк борщевика Сосновского относится к кратковременному
(transient) по классификации К.Томпсона с соавторами [Thompson et al., 1997].
Прорастание семян H. sosnowskyi в Литве после зимней стратификации мало
зависит от температуры и наблюдается даже в марте, ранее прочих видов бор
щевика [Ecology..., 2007]. В Коми в условиях интродукции зрелые листья от
личаются высокой морозостойкостью и способны сохраняться при кратков
ременном действии отрицательных температур до 8--10°С. Растения не по
вреждаются даже при сильных ранневесенних заморозках, доходящих до
12--14°С [Тимофеев, 2002].
2 Однако, по И.П. Манденовой [1951], H. mantegazzianum всегда многолетник!
110
Расселение и распространение. Как правило, мерикарпии борщевика па
дают в непосредственной близости от материнского растения. Когда высота
растения более 2 м, 60--90% мерикарпиев осыпаются на землю в радиусе 4 м.
Чем дальше от материнского растения, тем ниже число мерикарпиев, но от
дельные диаспоры могут оказаться довольно далеко, и таким образом проис
ходит захват новых территорий. По видимому, в долинах рек мерикарпии раз
носятся по течению, в том числе во время весеннего разлива. Зимой мерикар
пии разносятся ветром по замерзшей почве или насту [Практическое..., 2005;
The Giant..., 2005; Kabuce, 2006].
Авторы монографии "Ecology and management of giant hogweed (Heracleum
mantegazzianum)" считают, что граница расселения борщевика Сосновского в
бульшей степени определяется историей интродукции, чем давлением эколо
гических факторов [Ecology..., 2007].
Последствия внедрения
Воздействия на естественные фитоценозы и аборигенные виды. Появ
ление борщевика Сосновского вызывает обеднение видового состава окружа
ющей растительности, что, впрочем, характерно для всех видов зарослей бу
рьянного широкотравья. Крупные листья затеняют окружающие растения, а
сам борщевик способен расти в плотных зарослях [Практическое..., 2005; The
Giant..., 2005].
Генетические изменения и вариабельность вида. Недавние молекуляр
но генетические исследования инвазионных видов борщевика в Европе с ис
пользованием метода AFLP (Amplified Fragment Length Polymorphism) под
твердили наличие здесь видов (H. mantegazzianum, H. sosnowskyi и H. persicum
Desf.). Инвазионные популяции H. mantegazzianum и H. sosnowskyi обнаружи
ли отчетливые отличия от природных [Jahodovб et al., 2007; Ecology..., 2007].
Отметим, что образцы борщевика Сосновского для исследования были взяты
с территории Армении [Jahodovб et al., 2007]. Для интродукции же использо
вали растения Большого Кавказа [Сацыперова, 1984; Ecology..., 2007]. Обна
ружена популяция H. sosnowskyi в Бельгии, генетически отличающаяся от во
сточноевропейских, что свидетельствует о неоднократной интродукции этого
вида в Европу. Восточноевропейские образцы образуют единый кластер
[Jahodová et al., 2007; Ecology..., 2007].
Обращают на себя внимание различия в экологических предпочтениях ев
ропейских и кавказских растений борщевика. Кавказский H. sosnowskyi растет
в лесах и на полянах [Манденова, 1951], европейский предпочитает полное
освещение. Кавказский борщевик имеет высоту 1--1,5 м [Манденова, 1951],
инвазионные растения -- 2--4 м (рис. 32). Семена европейских растений замет
но крупнее: у борщевика из Литвы длина плодов 13,4±1,3 мм, а ширина 7,2±0,7
мм [Ecology..., 2007], по указаниям И.П. Манденовой [1951], их длина 7--9 мм,
ширина 4--6 мм.
У большинства растений венчик белый, розовоцветковые растения редки
[Практическое..., 2005; The Giant..., 2005]. Единичные растения с розовым вен
чиком встречены в окрестностях Звенигородской биостанции МГУ в долине
р. Москва (см. рис. 28).
Влияние на здоровье человека. Н. sosnowskyi представляет серьезную уг
розу здоровью человека. Растение содержит фотосенсибилизирующие соеди
нения фуранокумарины (фурокумарины). При контакте кожи с соком борще
111
вика и под воздействием ультрафиолетового излучения эти соединения вы
зывают ожоги. Содержание фуранокумаринов в разных частях растений раз
лично, реакция зависит от индивидуальной чувствительности, однако контак
тов с растением следует избегать в любом случае. Фототоксическая реакция
может проявляться под воздействием ультрафиолета уже через 15 мин, при
этом чувствительность кожи максимальна через 30--120 мин после контакта с
растением. Влага (например, пот) и тепло могут усилить кожную реакцию.
Через 24 часа кожа краснеет (эритрема), затем появляются волдыри с боль
шим скоплением жидкости, возможна воспалительная реакция. Приблизитель
но через неделю после заживления волдырей на пораженных участках появ
ляются красно розовые пятна и развивается необычное потемнение кожи (ги
перпигментация), которое сохраняется в течение нескольких месяцев, нередко
до года. Пораженные участки кожи могут быть чувствительны к ультрафио
лету в течение многих лет. К тому же некоторые фуранокумарины являются
канцерогенными и тератогенными веществами.
К группе риска относятся люди, вынужденные контактировать с растени
ем по работе (например, садовники, работники зеленого хозяйства), а также
дети, которые используют полые стебли как игрушки. Поскольку сам контакт
с растением практически безболезнен, контактировавшие могут в течение не
скольких часов не подозревать, какой опасности они подверглись.
Были отмечены случаи ожогов 3 й степени и летальные исходы для ма
леньких детей при обширных ожогах [Методические..., 2008].
Экономическое и социальное (положительное и отрицательное) зна
чение. Борщевик оказался опасным инвазионным видом. Он активно рассе
ляется вдоль дорог и, главное, вдоль малых и средних рек (рис. 33). Если не
организовать систему мониторинга и уничтожения борщевика Сосновского,
то, учитывая опасность фотохимических ожогов, через несколько десятиле
тий в летнее время реки окажутся закрытыми для рекреационного использо
вания. Методика расчета экономического ущерба от подобных инвазий не раз
работана. Необходимо планировать расходы на выявление популяций борще
вика, их уничтожение, систему мониторинга численности этого опасного
растения. Эти расходы на нашей территории могут быть весьма существенны.
Так, например, в 2007 г. в Эстонии Государственный центр охраны приро
ды боролся с борщевиком на площади 609 га, что обошлось в 4 млн эстонских
крон (то есть немногим менее 400 тыс. долларов США, или ~650 $/га). К на
стоящему времени в Эстонии остался 1041 участок распространения борще
вика общей площадью 1175 га, из них 123 участка появились в 2007 г. [Борь
ба..., 2007].
Меры борьбы
Для предотвращения дальнейшего расселения Н. sosnowskyi необходимо
уничтожить поля с культурой этого борщевика (если таковые еще остались!).
Для мониторинга за гигантскими борщевиками следует выявить места их
произрастания. В руководстве по борьбе с борщевиками [Практическое..., 2005;
The Giant..., 2005] рекомендуется обобщить информацию для планирования
работ на местном уровне и организовать контроль за их проведением, для чего
рекомендована специальная форма (табл. 10, 11).
3 Очевидно, что в этих экспериментах изучали поедаемость H. mantegazzianum,
с борщевиком Сосновского такие опыты, по видимому, не проводились.
112
В Чехии для учета популяций H. mantegazzianum используется аэрофотосъ
емка, что оказалось эффективным способом контроля [Müllerovб et al., 2005].
Оказалось, что в некоторых случаях заросли Н. sosnowskyi видны на космичес
ких снимках Европейской России, доступных пользователям GOOGLE Maps
(http://maps.google.com/).
Таблица 10
Обзор параметров, необходимых при создании карты и наблюдения за
территориями, занятыми борщевиком [Практическое..., 2005; The Giant..., 2005]
Цель
Параметры
Описание
Каждому участку присваивается
серийный номер для
идентификации его в поле
Определение
места
Местоположение
Описание точного местоположения
растений, нанесение сетки
Владение землей
Частное, государственное,
в ведении общественных
организаций или церкви
Регистрационный номер
участка
Факультативно
Состояние растения
Растет, цветет, плодоносит, умерло
Территория
Территория, занятая растениями, м2
Плотность
Число растений на 1 м2
Число растений
Территория, занятая растениями,
и плотность произрастания дают
основание для подсчета числа
растений
Использование земли
Сельскохозяйственное угодье,
берег реки, пустошь, др.
Описание
популяции
Доступ к территории
и качество земли
Расстояние до ближайшей дороги
и способность земли выдержать
тяжелую технику
Биологические свойства
территории
Описание растительного
сообщества, в частности видов,
которые находятся под угрозой из-
-за распространения борщевиков
Ценность как места
отдыха
Доступность зоны, близость
к жилищу, пригодность для
использования как места отдыха
Риск эрозии почвы
(особенно на бeрегах
водоемов)
Плотность насаждений и угол
наклона могут повлиять на
возникновение эрозии: уровень
высокий, средний или низкий
История борьбы
Ранее применявшиеся методы
борьбы, особые обстоятельства
Возможный
ущерб при
отсутствии
надлежащего
контроля
Методы борьбы
Принятые методы борьбы,
основывающиеся на собранных
данных
113
Для борьбы с популяциями борщевика Сосновского возможны различные под
ходы и способы, а также их сочетание [Практическое..., 2005; The giant..., 2005].
Механическое уничтожение. Растения борщевика удаляют вручную или с
помощью разнообразных косилок. Следует помнить, что скашивание назем
Таблица 11
Выбор способа борьбы с борщевиком [Практическое..., 2005; The Giant..., 2005].
Размер
популяции Действия
по контролю Затраченное время
Примечание
Уничтожение
корней
100 растений/час
(растения 2-го года) Трудоемкий, но весьма
эффективный метод
Скашивание
растений
100--200 растений
менее чем за 1 час при
использовании косы
Менее трудоемкий, но
и менее эффективный
метод
Отдельные
растения,
от5до100
Химический
контроль,
обработка
участка
100--200 растений/час Необходимо согласовы-
вать с государственными
правилами использования
гербицидов
Уничтожение
корней
100 растений/час
(растения 2-го года) Трудоемкий, но весьма
эффективный метод
Скашивание
растений
Механический
покос с помо-
щью сенокосилки
0,25--1 га/час
Требуется техника.
Тяжелая техника не
повсюду пройдет
Скашивание
косой
Высокая плотность:
1500 растений/час
Средняя плотность:
1000 растений/час
Небольшая плотность:
500 растений/час
Химический
контроль
300 м2/час
Ручное оборудование.
Необходимо согласовы-
вать с государственными
правилами использования
гербицидов
Небольшая
популяция,100
--1000 растений
Использование
под пастбище (см. ниже)
Возможно, если под
пастбище используются
соседние участки
Вспашка или
механический
покос
Механический покос с
помощью сенокосил-
ки: 0,25--1 га/час
Требуется техника.
Тяжелая техника не всюду
пройдет
Химический
контроль
0,5--1 га/час
Требуется техника.
Необходимо согласовы-
вать с государственными
правилами использования
гербицидов
Большая
популяция,
>1000 растений
Использование
под пастбище
1000 часов в год для
ежедневного наблюде-
ния и перемещения
170 овец, распределен-
ных по 10 различным
участкам
Стоимость зависит от
цены на изгородь и
расходов на содержание
животных
114
ной части не ведет к гибели растений. Они погибают лишь после 2--3 и более
скашиваний в течение вегетационного сезона на продолжении нескольких лет.
Только в этом случае истощаются запасы питательных веществ в корневище.
При ручном скашивании удобнее пользоваться не косой, а специальным инст
рументом, представляющим собой относительно короткое лезвие на длинной
рукоятке, длиннее, чем у привычной косы. Если плоды завязались, то растения
необходимо удалять для предотвращения их дозревания на срезанных экземп
лярах. Поэтому косить борщевик необходимо до начала массового цветения.
Необходимо и выкашивание буферной зоны шириной не менее 4 м от за
рослей борщевика [Методические..., 2008].
Более эффективным является удаление корневищ борщевика после ска
шивания. В таком случае можно удалить значительно быстрее. Однако это
существенно более трудоемкий способ, к тому же контакт с растениями до
вольно близкий, что повышает риск ожогов у людей.
Специалистами Института биологии Коми НЦ УрО РАН для борьбы с бор
щевиком предложено использовать черную полиэтиленовую пленку толщи
ной >100 мкм или иглопробивные геотекстильные полотна типа «Геоком Д»
или «Геоком ДТМ» с плотностью 100 г/м2 и более [Методические..., 2008].
Растения борщевика под черной пленкой погибают через 2--3 недели, а иглоп
робивное геополотно позволяет поместить грунт поверх пленки и провести
посевы луговых трав, то есть провести качественную рекультивацию [Мето
дические..., 2008]. Следует помнить, что только стоимость черной полимер
ной пленки на конец 2008 г. составила >3,5 руб. /м2, геополотна -- >15 руб./м2.
Использование мест произрастания борщевика под пастбище. Оказывает
ся, борщевик возможно использовать как пастбищное растение. Овцы, после
некоторого периода привыкания, охотно потребляют в пищу. Они предпочи
тают молодые свежие растения и быстро их уничтожают. Для этих целей луч
ше применять черных овец, поскольку у животных с пигментированной ко
жей риск фотохимических ожогов минимален3. Весной рекомендуется 20--30
овец на гектар, а летом численность можно сократить до 5--10 овец/га.
Гербициды. Для борьбы с борщевиком рекомендуется использовать раун
дап (глифосат). Достаточная концентрация -- 5 л/100 м2 стандартного раство
ра раундапа, то есть 360 г/л глифосата. Необходимо обработать 70--80% лис
товой поверхности растений борщевика [Методические..., 2008].
Первую обработку следует проводить весной, когда высота растений не
превышает 50 см, что позволяет безопасно проникнуть в центр его зарос
лей. Необходима также повторная обработка гербицидами после отраста
ния растений.
Правила работы с гербицидами, включая технику безопасности, описаны в
«Методическом пособии...» [2008].
Однако односезонная обработка раунапом не приводит к желаемому унич
тожению борщевика, на следующий год возникают массовые всходы Н.
sosnowskyi, через 2 года результатов обработки не заметно [Olukalns et al., 2005].
После уничтожения борщевика Сосновского необходима борьба с эрозией
почвы и его повторным заселением. Для этого желательны глубокая вспашка
(более чем на 25 см) и залужение обнажившейся почвы. Наиболее эффектив
ным является возвращение земель в сельскохозяйственный оборот [Практи
ческое..., 2005; The giant..., 2005; Olukalns et al., 2005].
115
СЕМЕЙСТВО ARACEAE АИРОВЫЕ
АИР ОБЫКНОВЕННЫЙ
Название: Acorus calamus L.
Морфологические признаки
Многолетнее травянистое корневищное растение (рис. 34). Листья мече
видные. Початок с длинным зеленым мечевидным покрывалом.
Естественный ареал
Родина аира Юго Восточная Азия, инвазионные популяции широко рас
пространены по всему Северному полушарию.
Вторичный ареал
История интродукции и географическое распространение. Считается, что в
Европу A.calamus интродуцирован из Малой Азии или Среднего Востока (ско
рее всего из Турции) в ХVI в. [Engler, 1905; Mucke, 1908; Schantz, 1959]. По дан
ным К.Wein [1940], к 1562 г. Acorus попал в Константинополь и Австрию, в 1579 г. --
вГессен,в1583г. --вИталию,в1587г. --воФранцию,в1599г. --вАнглиюив
1642 г. -- в Скандинавию. Аир довольно медленно завоевывал новые террито
рии. Так, например, следующая находка аира в Англии сделана почти полвека
спустя (в 1643 г.) в Западном Норфолке [Bennett, 1903]; третья находка аира в
графстве Чешир относится к 1711 г. [Britton, 1901].
Имеются данные о существовании в Польше и, возможно, в других райо
нах Балтии диплоидных форм A.calamus [Хромосомные числа..., 1969;
Kozlowski, 1960]. Эти формы, по мнению одних ученых, были занесены тата
рами [Mucke, 1908; Schantz, 1959], а по другим данным [Wulff, 1940--1941]
появились в ботаническом саду Копенгагена.
Первые документальные сведения о распространении аира в России отно
сятся к первой четверти ХIХ века: вид отмечен в каталогах ботанического сада
В.Бессера и ботанических садов Петербурга и Москвы [Besser, 1816; Index
plantarum..., 1824; Index seminum ..., 1826].
В Северной Америке встречаются ди и триплоидные формы, что явилось
причиной постановки вопроса, является аир аборигенным или инвазионным
растением. Еще в 1954 г. Merrill заявил, что обычный A.calamus занесен из Ев
ропы первыми колонистами, прибывшими в Джеймстаун в 1607 г. и в Плимут в
1620 г., или несколькими годами ранее французскими поселенцами в Восточ
ной Канаде [Packet, Ringius, 1984]. По данным, собранным R.Harper, впервые об
этом растении писал S.Elliott в 1821 г., который сообщил, что аир растет на юге
Калифорнии и в штате Джорджия; W.Darlington в 1837 г. отметил его в Пен
сильвании; J.Darby в 1855 г. в Оберне (неясно в котором из штатов Мэн или
Нью Йорк) и Алабаме; A.W. Chapman в 1860 г. нашел аир во Флориде; N.Britton
и A.Brown [1896] писали, что аир встречается от Новой Шотландии до Онта
рио, Миннесоты, Луизианы и Канзаса; Gattingen в 1901 г. отметил аир для фло
ры штата Теннесси [Harper, 1936].
Дальнейшие исследования показали, что в северных и центральных шта
тах Северной Америки встречаются естественные диплоидые популяции, об
разующие нормальные зрелые, успешно прорастающие семена [Хромосом
ные..., 1969; Löve, Löve, 1964; Jervis, Buell, 1964]. И, напротив, к востоку от
116
Аппалачей произрастают клоны, по видимому, интродуцированные из Евро
пы, которые являются триплоидами и не дают зрелых семян [Хромосомные ...,
1969; Jones, 1957; Thompson, 2007].
Ситуация с A.calamus, произрастающим в Канаде, стала вполне ясна после
исследований J.Packer и G.Ringius [1984], которые показали, что название
A.calamus относится к европейским инвазионным стерильным триплоидам,
тогда как американские природные фертильные диплоиды с более коротки
ми, чем у A.calamus, початками заслуживают видового статуса и называются
A. americanus (Raf) Raf.
A.americanus произрастает во всех провинциях Канады и в округе Маккен
зи Северо Западных территорий. A.calamus встречается реже и обнаружен всего
в 26 местонахождениях: 2 образца без точного места сбора (первое докумен
тальное подтверждение существования A.calamus в Канаде) относятся к 1855 г.,
в провинции Онтарио отмечено 6 местообитаний, самая ранняя находка в
1901 г.; в Квебеке -- 7; в Новой Шотландии -- 8; в Нью Брансуике -- 1, в 1878 г.;
на о ве Принца Эдуарда -- 2.
Пути и способы заноса. Заносится с речными паводками.
Статус в регионе. Аир обыкновенный в Восточной Европе представляет
собой стерильный, вегетативно размножающийся триплоид. Период произра
стания инвазионных популяций аира в России насчитывает, по видимому, >300
лет. Отмечен во всех регионах Средней России (рис. 35).
Местообитания. Аир обыкновенный -- гигрофит, типичный мегатроф, реже
мезотроф, предпочитает богатые аллювиальные отложения. Оптимальные
места его распространения -- неразмываемые берега водоемов, рек и ручьев со
стоячей или медленно текущей водой (рис. 36).
Размножение и жизненный цикл. В Средней России растения стерильны и
размножаются исключительно вегетативно. Отрастание листьев начинается в
середине апреля. Сначала развертываются 3--4 весенних листа, которые в се
редине мая постепенно начинают отмирать и окончательно засыхают в сере
дине июня.
Одновременно с отмиранием весенних листьев начинают отрастать летние
листья, развертывание которых завершается в конце июля. За вегетационный
период на одном растении образуется 6 7 летних листьев. Полное отмирание
летних листьев наблюдается в начале сентября, а за 2 недели до того, в середи
не августа, начинают расти листья (по 4 5 на побег), которые уходят под снег и
в следующем вегетационном сезоне развиваются как весенние листья.
Соцветия -- початки появляются в середине июня (рис. 37). На каждом ра
стении образуется только по 1 початку, средняя продолжительность цветения
составляет 3 недели. Все цветки стерильные. Длина покрывала всегда превы
шает длину листьев.
Расселение и распространение происходит с помощью кусочков корневи
ща, которые переносятся водными потоками. Существует легенда, что татары
принесли этот вид «на копытах коней» и специально бросали корневища в
водоемы для очистки воды.
Последствия внедрения
Воздействие на естественные фитоценозы и аборигенные виды. Конкури
рует с аборигенными прибрежно водными видами.
117
Рис. 34. Acorus calamus, общий вид соцветия
Рис. 35. Расселение Acorus calamus в Средней России
118
Рис. 37. Цветки Acorus calamus
Рис. 36. Acorus calamus по берегу р. Москва в Одинцовском р не Московской обл.
119
Генетические изменения и вариабельность вида. В литературе имеются до
вольно разноречивые сведения о кариологии A.calamus. Ряд исследователей
считали основным числом хромосом n=9, однако тщательный анализ показал,
что довольно мелкие хромосомы в метафазных пластинках в митозе имеют
тенденцию сливаться вместе, а в мейозе базовое число хромосом n=12 [Ammal
et al., 1967; Raquibuddowla et al., 1967; Wulff, 1940 1941; Хромосомные..., 1969;
Wieffering, 1972].
У A.calamus имеются диплоиды (2n=24), триплоиды (2n=36), тетрап
лоиды (2n=44--48) и гексаплоиды (2n=66). На родине в Южной и Восточной
Азии встречаются клоны со всеми 4 мя уровнями плоидности [Ammal et al.,
1967; Raquibuddowla et al., 1967; Wulff, 1940 1941; Хромосомные..., 1969; Wang
et al., 2001; Ito, 1942; Larsen, 1969].
Ди и тетраплоиды различаются по экологическим требованиям. Так, на
пример, в Северном Таиланде диплоиды приурочены к проточной воде и ка
менистому субстрату, в то время как тетраплоиды чаще встречаются в стоячей
воде на суглинке [Larsen, 1969]. В Европе обычны триплоидные популяции,
которые размножаются только вегетативно и семян не образуют. В мейозе у
этих форм имеется много уни и тривалентов [Wulff, 1940 1941; Palmgren, 1943;
Kozlowski, 1960; Sokolowska Kulczycka, 1961]. Аберрации в мейозе приводят к
образованию стерильной пыльцы.
А.Энглер выделял четыре разновидности A.calamus: vulgaris, verus,
angustatus, angustifolius. Однако эта классификация базировалась на морфо
логических признаках вегетативных органов, которые сильно варьируют, в том
числе под влиянием различных условий произрастания [Engler,1905]. Наибо
лее оптимальной представляется классификация А.Баранова [Baranov,1960]:
A.calamus L. var.americanus (Raf) Wulff (диплоид) = A. americanus (Raf) Raf.
A.calamus L. var.vulgaris Engl. (триплоид) = A.calamus L. var.сalamus
A.calamus L. var.spurius (Schott) Engl. (тетраплоид) = A.asiaticus Nakai
Внутривидовую фенотипическую изменчивость инвазионных популяций
аира изучали в 21 местообитании Словении [Rode et al., 1996]. Средняя длина
листа составляла 80±15,8 см, средний диаметр корневища -- 1,7±0,5 см. Сход
ные исследования проведены О. Г. Юкневичене [1971] в Литве. В зависимос
ти от экологических условий обитания длина листа составляла от 90,6 до 133,6
см; ширина -- от 1,48 до 1,72 см; длина стебля -- 42,3--70,3 см; диаметр стебля
0,92--1,15 см; длина покрывала 48,4--69,1 см; ширина покрывала 0,97--1,45 см;
длина соцветия 6,1--8,7 см; ширина соцветия 0,79--1,12 см. При этом мини
мальные значения параметров отмечались у растений, произрастающих на
влажной песчаной почве, а максимальные -- на влажной илистой почве в осо
ково аирных сообществах. Амплитуда изменчивости длины годового прирос
та корневища составляла 10,3--27,1 см, сырого веса годового прироста 9,3--68,2 г,
диаметр корневища 1,2--1,93 см. При этом минимальные параметры характер
ны для сухих суглинистых почв, а максимальные -- для влажных илистых почв.
Экономическое и социальное (положительное/отрицательное) значение. В
состав корневищ аира входит эфирное масло (1,3--8,0%), горький гликозид
акорин, алкалоид каламин, дубильные вещества, смолы, камедь, экстрактив
ные вещества, аскорбиновая кислота, витамин Е, каротин, крахмал. Количе
ство эфирного масла и содержащегося в нем активного вещества в азарона
варьируют в течение вегетационного сезона, максимумы наблюдаются в апре
120
ле, начале августа и в октябре. У различных образцов, собранных в одно и то
же время, содержание эфирного масла колебалось от 3,6 до 7,8 мл на 100 г
вещества в Литве и от 2,4 до 7,2 мл -- в Словении [Rode et al., 1996; Юкневиче
не, 1969]. При этом содержание эфирного масла являлось для одного образца
константной величиной и не менялось при пересадке образца в иные экологи
ческие условия.
Существенные различия в содержании азарона в эфирном масле аира от
мечены индийскими исследователями [Ammal et al., 1964]: при выращивании
в однородных условиях образцы аира из Кашмира содержали 5% азарона, а
образцы из Джамму -- >80% (!?-- ЮВ). Изучение химического состава
A.calamus из Монреаля и Копенгагена (2n), Киля (3n) и Ленинграда (4n) так
же показало наличие хеморас у этого вида [Wulff, Stahl, 1960].
В настоящее время промышленные заготовки аира осуществляются, в ос
новном, на Украине, где общая площадь зарослей составляет >300 га. Заготов
ки ведутся в таких масштабах, что аир оказался уже под угрозой исчезнове
ния, и требуется введение этого ценного лекарственного растения в широкую
культуру (ссылка). Поскольку количество и качество эфирного масла в кор
невищах аира является наследственным признаком, имеется возможность
выбирать для культивирования популяции с максимальным содержанием по
лезных веществ.
Меры борьбы
Специальные мероприятия не проводятся.
Обзор научных исследований и публикаций
Изучена генетическая закрепленность различных биологических призна
ков и амплитуда их внутривидовой изменчивости (см. главу. 4). Выявлено на
личие межпопуляционной изменчивости (не носящей клинального характера)
по темпам роста как весенних, так и летних листьев [Виноградова, 2004].
СЕМЕЙСТВО ASTERACEAE (COMPOSITAE) --
СЛОЖНОЦВЕТНЫЕ
АМБРОЗИЯ ПОЛЫННОЛИСТНАЯ
Название: Ambrosia artemisiifolia L.
Синонимы: A. elatior L., A. glandulosa Scheele; A. monophylla (Walter) Rydberg
Морфологические признаки
Однолетнее растение высотой 20 150 см с рассеянным опушением. Корень
стержневой, с мощным разветвлением, проникает в глубину до 4 м [Моска
ленко, 2001]. Листья дважды перисторассеченные, сверху темно зеленые, по
чти голые, снизу серо зеленые, коротковолосистые. Преимущественно одно
домное растение. Мужские цветки мелкие, диаметром 2--5 мм, желтого цвета,
собраны в корзинки по 5--25 цветков, последние собраны в колосовидные соц
ветия, расположенные на верхушке побегов. Женские цветки расположены в
пазухах листьев по 2--3, без венчика, заключены в сросшуюся обертку, в сред
ней части которой есть 5--8 шипиков. При созревании плодов обертка тверде
ет. Плод -- семянка обратнояйцевидной формы, гладкая, блестящая, оливко
121
Рис. 38. Ambrosia artemisiifolia, общий вид растения
Рис. 40. Ambrosia artemisiifolia по нарушенным местообитаниям у жилья
Рис. 45. Ambrosia artemisiifolia в пойменных лугах р. Дон
123
во серого или коричневого цвета, с небольшим выступом на вершине -- остат
ком столбика (рис. 38). Естественный ареал
Род Ambrosia насчитывает 21 вид, родина которых -- Северная Америка.
A.artemisiifolia естественно произрастает во всех провинциях Канады, в вос
точных и юго восточных штатах США. Популяции центральных и западных
регионов США рассматриваются как вторичные.
Вторичный ареал
История интродукции и географическое распространение. A. artemisii
folia -- наиболее инвазионный вид рода. Вид завезен в Германию в 1863 г., дву
мя годами позднее во Францию, быстро распространился в долине Роны, в
Бельгии и Нидерландах, в массе стал встречаться в Великобритании и про
двигаться на восток -- в Венгрию, на Балканы и на Украину.
В настоящее время A. artemisiifolia образует крупные популяции в долине
Роны, в Северной Италии и в Карпатском регионе [Sheppard et al., 2006]. Как
заносный вид встречается в Средней и Атлантической Европе, Средиземно
морье, в Молдове, на Украине, Кавказе, в Российской Федерации, Казахстане
и Средней Азии, Малоазиатском регионе, Иране, Японии, Китае, Южной Аме
рике, Австралии, Африке [Чужеродные виды на территории России, 2008].
Распространение A. artemisiifolia на территории бывшего СССР носило
характер экологического взрыва [Димитриев и др., 1994]. В 1918 г. вид впер
вые найден в окрестностях Ставрополя [Амелин, 1936], затем его обнаружили
в 1929 г. в окрестностях г. Орджоникидзе [Сорные..., 1935]. В 1960 е гг. амбро
зия полыннолистная была уже распространена в Ставропольском крае, Рос
товской обл., Северо Осетинской АССР, Волгоградской обл., Кабардино Бал
карской АССР, Чечено Ингушской АССР, Краснодарском крае [Клюшкин и
др., 1962]. В этот же время началось интенсивное продвижение этого вида в
центральные районы страны, Среднюю Азию, на юг Дальнего Востока [Кова
лев, 1989]. В равнинных и горных частях Ставрополья и на Северном Кавказе
за последние 40--45 лет A. artemisiifolia массово внедрилась в зону дерновинно
злаковых и луговых степей с черноземными почвами и продолжает проникать
на восток -- в сухие степи и на юг -- в горные долины, где поднимается на высо
ту до 1000 (1200) м над ур. м. Массовое распространение амброзии в этом ре
гионе отмечается там, где пашня составляет 80--90% от общей площади сель
хозугодий [Дзыбов, 1989]. В настоящее время амброзия полыннолистная рас
селилась почти во всех районах страны с периодом вегетации >150 дней
[Недолужко, 1984]. В качестве эфемерофита встречается и в районах с более
коротким периодом вегетации растений [Гусев, 1977].
Пути и способы заноса. Ксенофит. В Европе амброзия распространяется
вдоль транспортных осей, речных берегов, по эродированным почвам и оро
шаемым землям [Sheppard et al., 2006].
В Средней России A. аrtemisiifolia заносится с семенами сельскохозяйствен
ных культур, особенно поздних (подсолнечник, конопля, люцерна, овощные и т.д.),
уборка которых совпадает с созреванием сорняка (август сентябрь), а также с за
соренными отходами, сеном, при перегоне скота. Осенью и зимой семена амбро
зии с нескошенных растений разносятся ветром [Чужеродные виды..., 2008].
Статус в регионе. В Белгородской области A.аrtemisiifolia собрана в 2006 г.
в Вейделевском районе, в пос. Вейделевка, в тополевом парке (Н.Решетнико
124
ва, А.Мамонтов, МНА). В Воронежской обл. впервые найдена С.В. Голицы
ным в 1967 г. (VOR). Подробно расселение и распространение А. artemisiifolia
в этом регионе описаны в книге «Адвентивная флора Воронежской области»
[Григорьевская и др., 2004]. На первых этапах расселения А. artemisiifolia была
обнаружена в значительном количестве на лугах в южных районах области [Ба
рабаш и др., 1979], а в более северных районах спорадически появлялась и исче
зала [Александрова, Барабаш, 1987]. К настоящему времени встречается часто
единичными экземплярами и небольшими группами, однако на некоторых уча
стках, в том числе в пойме, образует заросли [Григорьевская и др., 2004], что
пока не отмечается севернее. В Воронеже -- единично [Григорьевская, 2000].
Первые известные сборы из Курской обл.: «г. Курск, пустырь у завода КЗТЗ,
23.08.1993, А.Полуянов» (КурМ); «Льговский р н, окрестности ж/д. ст. Бло
хино, на ж/д. путях, 2.08.1995, А.Полуянов» (КурМ, MOSP). А. artemisiifolia
известна из всех районов Липецкой обл. [Флора Липецкой области, 1996].
Впервые собрана в с. Хлевное на обочине дороги, 14.09.1981, М.В. Казаковой
(MW). В Орловской обл. вид встречается очень редко [Еленевский, Радыги
на, 1997]. А. artemisiifolia собрана в Орле, на свалке в районе СПЗ, 22.09.1982,
Л.А. Самсонова, В.И. Радыгина (OHHI). По нашим наблюдениям (2008 г.), на
юго востоке Орловской обл. вид остается редким, вырастает отдельными эк
земплярами на хлебоприемных пунктах и близ железной дороги. В Тульской
обл. впервые собрана в г. Ясногорск на сорном месте, 20.08.1979 Ю.Е. Алексе
евым [МW; Алексеев, Губанов, 1980]. В отличие от более южных регионов, на
территории Тульской обл. A. artemisiifolia довольно редка, хотя в последние
годы ее встречаемость заметно возросла.
В Московской обл. А. artemisiifolia появилась в 1970 е гг. [Октябрева и др.,
1978]. Впервые собрана в 1974 г. в Коломенском р не, близ ст. Голутвин [MW;
Игнатов и др., 1990]. В 1980--1990 гг. амброзия регулярно отмечалась на ж/д
насыпях, реже -- на сорных местах в Москве, всегда единичными особями. В те
годы амброзия в Московской обл. плоды формировала, но семена не вызревали
[Игнатов и др., 1990]. Во Владимирской обл. впервые собрана в Меленковском
р не, на ст. Бутылицы, 21.07.1976 г. А.В. Чичевым (MW). В последние годы A.
artemisiifolia отмечалась в разных р нах [Вахромеев, 2002], особенно часто в
Меленковском и Ковровском (А.П. Серегин, личное сообщение). В Смоленс
кой обл. единственный сбор сделан в г. Смоленск [Решетникова, 2004].
В Ивановской обл. A. аrtemisiifolia впервые обнаружена в 1985 г. в Кине
шемском р не [Борисова, 2007]. На крупных городских свалках Ивановской
обл. встречается ежегодно, нередко образует крупные заросли [MW; LE; ИУ;
Шилов, 1989; Борисова, 2005а]. В Нижегородской обл. амброзия полынноли
стная отмечена Т.Б. Силаевой в 1986--1987 гг. на ж/д. ст. Сергач. В гербарии
И.Л. Мининзона хранятся сборы A.artemisiifolia, сделанные с 1982 по 1991 г. в
Горьковском р не и г. Горький [Димитриев и др., 1994]. В Тверской обл. A.
аrtemisiifolia впервые собрана в г. Калинин (Тверь): «близ ст. Пролетарка, на
краю ж/д насыпи, 25.09.1978, В.Малышева» (MW).
В Калужской обл., по личному сообщению С.Р. Майорова, А. artemisiifolia
вырастает по ж/д насыпям редко, но местами в массе. Первый сбор в Рязанс
кой обл. сделан близ Рязани на обочине шоссе Рязань--Касимов в 1982 г. [Ка
закова, 2004].
125
В Тамбовской обл. A. аrtemisiifolia собрана в 1997 г. в Мичуринском р не, в
1 км западнее ж/д пл. Турмасово (А.Сухоруков, MW). В Пензенской обл. амб
розия встречется довольно редко, по ж/д путям [Васюков, 2004].
В Мордовии единичные экземпляры амброзии, находящиеся на разных ста
диях развития, обнаружены на ж/д станциях в 1982 г. [Силаева, Тихомиров,
1987]. Впоследствии вид периодически встречали на железных дорогах, пус
тырях, газонах в г. Саранск [Димитриев и др., 1994]. В Чувашии амброзия по
лыннолистная впервые обнаружена в середине 1950 х гг. в г. Чебоксары на
цветочной клумбе. С 1980 г. единичные экземпляры отмечаются на железных
дорогах республики, полях и территориях ферм [Димитриев и др., 1994].
Таким образом, к настоящему времени амброзия полыннолистная проникла
во многие регионы РФ и прочно вошла в состав адвентивной флоры (рис. 39).
Авторы, изучавшие распространение А.artemisiifolia в Волжско Камском ре
гионе [Димитриев и др., 1994], отмечают, что на север региона вид продвига
ется с трудом и ведет себя как эфемерофит, не образует жизнеспособных се
мян и не закрепляется в фитоценозах. В центре и на юге региона в лесостеп
ной и степной зонах А.artemisiifolia начинает проникать в агрофитоценозы.
Частота находок амброзии полыннолистной зависит от продолжительности
вегетационного периода в той или иной части территории, интенсивности гру
зооборота, степени нарушенности естественных биогеоценозов и ряда других
факторов [Димитриев и др., 1994].
Местообитания. В Северной Америке и Западной Европе A. artemisiifolia
в обилии растет вдоль обочин дорог, на сорных местах и по берегам рек
Рис. 39. Расселение Ambrosia artemisiifolia по Средней России
126
[Sheppard et al., 2006]. Временную динамику мест обитания A. artemisiifolia
исследовали в канадской провинции Квебек [Lavoie, 2007] по 707 гербарным
образцам, собранным с начала ХХ века по настоящее время. Выяснилось, что
распространение A. artemisiifolia в Квебеке можно разбить на 4 периода. Пер
вые находки приурочены к поймам рек. В агроценозы вид внедрился в 1920 х
гг., а вдоль железных и шоссейных дорог расселился в 1930 е гг. Авторы пола
гают, что на расселение вида в Квебеке в значительной степени повлияло раз
витие дорожной сети в 1930 е гг., и сделали вывод, что уничтожение амброзии
только на полях -- недостаточная мера борьбы с этим видом, так как обочины
дорог служат постоянными рефугиумами для заноса диаспор на поля.
В Средней России вид занимает весь спектр нарушенных мест обитания:
ж/д насыпи, пустыри (рис. 40), свалки, обочины шоссейных дорог, террито
рии хлебоприемных комбинатов, очистные сооружения, стройплощадки, бе
рега рек и прудов, засоренные луга, нарушенные участки в населенных пунк
тах [Борисова, 2007; Васюков, 2004; Вахромеев, 2002; Григорьевская, 2004;
Димитриев и др., 1994; Еленевский, Радыгина, 1997; Игнатов и др., 1990; Ка
закова, 2007; Маркелова, 2004; Полуянов, 2005; Силаева, Тихомиров, 1987;
Флора Липецкой области, 1996]. В последние годы из за массового примене
ния гербицидов амброзия на железных дорогах стала редка, а в других местах
обитания распространение вида не изменилось.
Размножение и жизненный цикл. Яровой однолетник, вегетирует с
мая по июль, соцветия появляются с середины лета до середины осени вплоть
до первых морозов [Sheppard et al., 2006]. Размножается семенами. Минималь
ная температура прорастания семян 6 8°C, оптимальная 22 24° C, максималь
ная глубина прорастания -- 6--8 см. Плодовитость -- до 88 тыс. семян с 1 расте
ния. Жизнеспособность семян в почве -- 40 лет [Нumangarden, 2008]. При бла
гоприятных условиях увлажнения всходы могут появляться в течение всего
вегетационного периода. При поздних всходах растения могут развиваться по
ускоренному циклу (неотения). Продолжительность вегетационного периода
составляет ~180 дней на юге Восточной Европы и ~150 дней в Приморском
крае [Москаленко, 2001]. Продвижение амброзии полыннолистной на север
лимитируют длина дня и температурный режим в период вегетации.
Расселение и распространение. В южных регионах Средней России
амброзия регулярно плодоносит и формирует в почве собственный банк се
мян, тогда как в северных областях расселение вида зависит от периодическо
го заноса диаспор. В центральных областях в отдельные годы семена
A.artemisiifolia могут вызревать; так, например, в Московской обл. в октябре
1989 г. были сделаны гербарные сборы растений с плодами.
Последствия внедрения
Воздействия на естественные фитоценозы и аборигенные виды. Не
смотря на обилие работ, посвященных экологии амброзии, вопросы ее взаимодей
ствия во вторичном ареале с видами природной флоры изучены недостаточно.
Генетические изменения и вариабельность вида. Во «Флоре Север
ной Америки» [Strother, 2006] выделены ряд разновидностей A. artemisiifolia:
A. artemisiifolia var. artemisiifolia; A. artemisiifolia var. elatior (L.) Descourtils; A.
artemisiifolia var. paniculata (Michx.) Blank. и гибридов: A. × intergradiens W.H.
Wagner -- гибрид между A. artemisiifolia и A. psilostachya DC., A. Ч helenae
Rouleau -- гибрид между A. artemisiifolia и A. trifida L.
127
Сравнение генетического разно
образия популяций A. artemisiifolia в
естественном (Северная Америка) и
вторичном (долина Роны, Франция)
ареалах проведено молекулярными
методами с использованием микро
сателлитов [Genton et al., 2005]. Воп
реки первоначальной гипотезе ока
залось, что уровень как внутри=, так
и межпопуляционной генетической
изменчивости ниже в естественном,
а не во вторичном ареале. Распреде
ление тестеров и редких аллелей
свидетельствует не в пользу проис
хождения французских популяций
из единого источника. Аллели, при
сущие различным североамериканс
ким популяциям, сосуществуют во
Франции в каждой из изученных популяций. Авторы сделали вывод, что фран
цузские популяции сформировались путем не одного, а «смеси» заносов, и даже
в области первичного заноса состоят из неродственных особей. Показано так
же, что донором формирования новых инвазионных популяций, удаленных
от области первичного заноса, являются не североамериканские, а местные
популяции.
Влияние на здоровье человека. В южных районах России пыльца амбро
зии -- один из наиболее сильных аллергенов (рис. 41). Настоящим бедствием
это растение стало в Ставропольском крае, в Ростовской и Волгоградской обл.,
где в период цветения амброзии численность страдающих поллинозами дос
тигает 40%. Пыльца амброзии циркулирует в воздухе и за пределами этих ре
гионов. До 2000 г. в аэропалинологическом спектре Москвы пыльцевые зерна
амброзии не отмечались. Впервые пыльца амброзии зарегистрирована в августе
2000 г. (суммарная концентрация 8 п.з./м3), и с того времени ее стали отмечать
ежегодно. В настоящее время концентрация пыльцы амброзии в Москве в 5--10
раз превышает пороговые значения и имеет стойкую тенденцию к возрастанию.
Как правило, кривая пыления амброзии прерывистая, периоды высокой концен
трации непродолжительные и не превышают 4--5 дней (рис. 42). (ссылка!)
Обнаружение пыльцевых зерен амброзии в составе спектра сопряжено с вет
ром южного или юго западного направления. Изменение направления ветра при
водит к быстрому (в течение нескольких часов) снижению концентрации пыль
цы. Отдельные пики концентрации пыльцы амброзии в 2006 г. (13--16 августа,
26--29 августа, 18--20 сентября) проанализированы методом обратных траекто
рий, позволяющим реконструировать возможные пути дальнего транспорта пыль
цы. Для всех исследованных эпизодов регионами -- потенциальными источника
ми пыльцы амброзии выступали южные районы России и Украина. Сопоставле
ние данных станций аэропалинологического мониторинга в Москве, Смоленске
и Запорожье показало, что время и продолжительность пиков концентрации прак
тически совпадают, что подтверждает предположение о возможном дальнем за
носе пыльцы амброзии в Среднюю Россию именно из южных регионов.
Рис. 41. Пыльцевое зерно Ambrosia
artemisiifolia (сканирующий
электронный микроскоп)
128
Рис. 42. Пыление амброзии в 2001--2006 гг. в Москве по данным аэропалиноло
гического мониторинга (по: Северова и др., 2008)
Рис. 44. Кривая пыления амброзии, иллюстрирующая дальний занос
(30.08.2003), по данным Е.Э. Северовой
Рис. 43. Кривая пыления амброзии, характеризующая локальное цветение
(27.08.2005), по данным Е.Э. Северовой
129
Общеизвестно, что для амброзии характерна четкая суточная ритмика пы
ления с максимумом между 9 и 13 ч. Анализ суточной динамики концентра
ции пыльцы амброзии в Москве показал, что, по видимому, часть регистриру
емой в составе спектра пыльцы локальна. Локальное цветение приходится на
конец августа --- начало сентября и характеризуется суточным подъемом кон
центрации в 9--12 ч. (рис. 43). Ночные пики концентрации, периодически на
блюдаемые в августе--сентябре, связаны, по видимому, с дальним транспор
том пыльцы амброзии (рис. 44).
Экономическое и социальное значение (положительное и отрицатель
ное). Развивая мощную надземную массу и корневую систему, амброзия по
лыннолистная сильно подавляет культурные растения, она расходует много
воды на образование единицы сухого вещества, что приводит к иссушению
почвы и значительному снижению ее плодородия. Наиболее часто страдают
яровые хлеба и пропашные культуры, особенно подсолнечник. При недоста
точном уходе за посевами этих культур амброзия полыннолистная перераста
ет их и сильно заглушает, что приводит к резкому снижению, а то и к полной
гибели урожая. На лугах и пастбищах амброзия вытесняет злаково бобовые
травы и снижает кормовые качества сена и выпасов, поскольку скот ее не по
едает из за содержания в ее листьях горьких эфирных масел (рис. 45 а или б).
Меры борьбы
Амброзия полыннолистная внесена в список карантинных растений,
ограниченно распространенных на территории Российской Федерации [Пе
речень..., 2003]; ввоз семян вида на территорию РФ запрещен.
Основными методами борьбы при очистке полей от амброзии являют
ся агротехнические [Москаленко, 2001]: правильное чередование культур в
севообороте, обработка почвы, уход за посевами, направленный на истощение
запасов семян сорняка в почве и предотвращение повторного засорения как
почвы, так и урожая сельскохозяйственных культур. На землях, сильно засо
ренных амброзией, нужно использовать чистые пары, что снижает засорен
ность на 70--80%. Сильно засоренные амброзией площади следует также отво
дить под бессменный (2--3 года) посев озимых зерновых с предшествующей
полупаровой обработкой почвы. На полях с легкими почвами, сильно засо
ренными семенами амброзии, не следует проводить предпосевную культива
цию зяби перед посевом ранних яровых зерновых культур. Такая культивация
создает благоприятные условия для прорастания семян амброзии и массового
появления ее всходов, подавляющих всходы ранних яровых. В этом случае
лучше ограничиться боронованием [Москаленко, 2001].
В посевах кормовых трав мероприятия по борьбе с амброзией сводят
ся к высококачественной обработке почвы, внесению удобрений, оптималь
ным срокам посева и т.д. Хорошо развитые травы в значительной степени по
давляют амброзию. В семенных посевах многолетних трав наиболее успеш
ное уничтожение сорняка достигается при летних посевах трав по чистым
парам, обработка которых в первой половине лета позволяет очистить почву
от амброзии [Москаленко, 2001].
Помимо агротехнических мер, применяются химические методы: диф
ференцированное использование гербицидов в зависимости от вида засорен
ной культуры [Чужеродные виды..., 2008].
130
Проекты по биологическому контролю численности A. artemisiifolia, реа
лизуемые в России, странах бывш. Югославии, Австралии и Китае, основаны
на использовании насекомых -- врагов A. artemisiifolia. Наиболее эффективны
Zygogramma suturalis и Epiblema strenuana. Причем E. strenuana имеет широкий
диапазон хозяев, включая другие виды рода Ambrosia и Xanthium, в том числе
европейские. Используются также Stobaera concinna, Trigonorhinus tomentosus,
Euaresta bella и Tarachidia candefacta, но эти виды требуют акклиматизации
[Sheppard et al., 2006].
Истории интродукции Zygogramma suturalis в СССР и Россию посвящено
значительное число публикаций специалистов ЗИН РАН [Reznik et al., 2004]:
жук листоед Zygogramma suturalis интродуцирован в СССР в 1978 г. О.В. Ко
валевым для борьбы с A.artemisiifolia в агроценозах с севооборотом. По дан
ным на 1983--1985 гг., жук прошел акклиматизацию и смог подавить амбро
зию по краям полей. Однако оценка плотности Z. suturalis, проведенная на пло
щади 25 тыс. га в 1988--2004 гг., показала очень низкую среднюю плотность
насекомых (~5 жуков на 1 кг амброзии), несмотря на наличие нескольких учас
тков, кишащих насекомыми. Статистический анализ показал, что при подоб
ной популяционной плотности Z.suturalis не может существенно влиять на чис
ленность особей A.artemisiifolia. На отдельных, ограниченных участках Z.suturalis
является причиной серьезного вреда амброзии. Однако локальные участки с
высокой плотностью Z.suturalis располагаются обычно по окраинам полей и не
играют серьезной роли в контроле за численностью амброзии в популяциях.
Таким образом, результат интродукции и акклиматизации Z.suturalis можно
рассматривать как умеренный биологический успех, но цели биологического
контроля за численностью A.artemisiifolia пока нельзя считать достигнутыми.
СЕМЕЙСТВО ASTERACEAE (COMPOSITAE) --
СЛОЖНОЦВЕТНЫЕ
АСТРА ИВОВАЯ
Название: Aster × salignus Willd. (A. lanceolatus Willd. × A. novi belgii L.)
Синонимы: Symphyotrichum1 × salignum (Willd.) G.L. Nesom [S. lanceolatum
(Willd.) G.L. Nesom × S. novi belgii (L.) G.L. Nesom], Aster salicifolius Schöller
(1787), non Lam. (1783).
Морфологические признаки
Многолетнее травянистое длиннокорневищное растение высотой 30--100 см
(рис. 46). Стебли одиночные, внизу округлые в сечении, вверху -- ребристые,
ветвление преимущественно в области рыхлого метельчатого соцветия из мно
1Морфологические и молекулярные данные несомненно подтверждает выде
ление североамериканских астр в особый род Symphyotrichum Nees [Корнiенко,
Мосякiн, 2006; Nesom, 1995; Noyes, Rieseberg, 1999]. Этот род принят во "Flora of
North America" [Brouillet et al., 2006]. Мы не считаем необходимым вводить эту
таксономическую новацию в настоящем издании и сохраняем привычное вклю
чение этих видов в род Aster s. l.
131
гочисленных корзинок. По стеблю от основания листьев низбегают две по
лоски коротких простых волосков, опушение в верхней части растения гуще.
Листья очередные, ланцетные или линейно ланцетные, голые, с клиновидным
основанием, у самого стебля немного расширенные, полустеблеобъемлющие,
с острой треугольной верхушкой, по краю шероховатые, в верхней части часто
зубчато пильчатые. Корзинки диаметром около 14--20 мм. Обертка цилинд
рическая, черепитчатая, листочки оберток голые или с короткими шерохова
тыми волосками, травянистые, в верхней половине без перепончатой каймы,
более или менее прижатые к обертке (рис. 47). Краевые цветки язычковые,
бледно лиловые, иногда белые, срединные цветки трубчатые, желтые, к кон
цу цветения красноватые.
Отметим, что в первоописании A. × salignus указано, что краевые цветки
белые, при отцветании голубые [Willdenow, 1804].
Считается, что A. × salignus является результатом гибридизации типовых
разновидностей A. lanceolatus и A. novi belgii [Brouillet et al., 2006]. Следует
иметь в виду, что само разграничение родительских видов на Атлантическом
побережье в зоне их совместного произрастания нередко затруднительно
[Brouillet et al., 2006].
A. lanceolatus отличается сравнительно узкими, обычно менее 1 см, листья
ми без стеблеобъемлющего основания, краевые цветки часто белые, а листоч
ки обертки длиной менее 5 мм. A. novi belgii имеет более рыхлую обертку, ее
внешние листочки в разной степени отогнутые [Игнатов, Макаров, 1984; Stace,
1997; Hаeupler, Muer, 2000, 2007].
Естественный ареал
A. × salignus -- возникший в Европе гибридогенный вид. Его произрастание
в Северной Америке пока не подтверждается, но возможны находки на вос
точном побережье, где пересекаются ареалы родительских видов [Brouillet et
al., 2006].
Вторичный ареал
История интродукции и географическое распространение. Широко
распространена в Европе и на юге Западной Сибири [Цвелев, 1994]. Вероят
но, есть на российском Дальнем Востоке [Баркалов, 1992], в Японии отмечена
как инвазионный вид [Müller, Okuda, 1998]. Известна более чем в половине
стран Европы, причем в 19 регионах является инвазионным видом [Lambdon
et al., 2008]. В Венгрии A. × salignus отнесена к видам трансформерам [Balogh
et al., 2004].
Эта астра отмечена во всех флористических районах центра Европейской
России [Цвелев, 1994]. Занесена в список инвазионных видов северо запада
Европейской России [Гельтман, 2003].
Астра ивовая культивируется как неприхотливое позднецветущее декора
тивное растение. Уже при первоописании этой астры К.Л. Вильденов указы
вал на достаточно широкое ее распространение в Центральной Европе: «Habitat
in Germania ad ripas Albis, et in Hungaria» [Willdenow, 1804].
Реальная история распространения астры ивовой в Европейской России
не вполне ясна. Долгое время почти все американские астры называли A. ×
salignus. М.С. Игнатов и В.В. Макаров обратили внимание на то, что в Мос
132
ковской обл. следует различать 4 вида: A. novi belgii L., A. lanceolatus, A. ×
salignus, A. versicolor (A. novi belgii × A. laevis Willd.) [Игнатов, Макаров, 1984;
Игнатов и др., 1990], позднее обнаружены некоторые новые дичающие севе
роамериканские астры. Это повлекло определение гербарных образцов занос
ных астр по формальным признакам строения обертки, особенно по форме
листочков обертки. В частных разговорах флористы признают сложность си
туации и проблемы в определении, тем не менее специального таксономичес
кого исследования группы A. lanceolatus -- A. novi belgii -- A. × salignus до сих
пор не предпринято, надежные диагностические признаки не найдены, точное
определение ряда форм затруднено [Hаeupler, Muer, 2000, 2007]. Так, предло
женный С.Стейси иллюстрированный ключ для определения североамерикан
ских астр, оставляющий хорошее впечатление, далеко не всегда позволяет оп
ределить каждое растение [Stace, 1997].
Тем не менее ясно, что активное расселение A. × salignus происходило в пер
вой половине XX века [MW, MHA; Тамамшян, 1959; Игнатов и др., 1990; Цве
лев, 1994]. Наибольшее распространение эта астра получила в лесных облас
тях Средней России.
Пути и способы заноса. Многолетние североамериканские астры в садо
вой культуре размножают делением куста. По видимому, корневища астр по
падают на сорные места при прополке дачных участков и цветников, когда
удаляют лишние побеги и избавляются от лишних растений.
Статус в регионе. Распространена довольно широко (рис. 48).
Местообитания. Астра иволистная встречается преимущественно по на
рушенным местообитаниям: на пустырях, вдоль дорог, у дачных поселков, на
залежах (рис. 49). В ряде случаев вид произрастает по берегам рек, в полосе
ивняков на прирусловом вале и по сыроватым лугам, как, например, вдоль р.
Клязьма во Владимирской обл. Такое распространение A. × salignus характер
но для регионов севернее Москвы. Аналогичная приуроченность астры отме
чена для Литвы [Priede, 2008]. В более южных областях эта астра встречается
преимущественно по рудеральным местообитаниям.
Размножение и жизненный цикл. Цветение у A. × salignus позднее, в авгу
сте--октябре. Семенное размножение для этой астры в Средней России пока
не отмечено, впрочем, специальных исследований семенной репродукции не
проводилось. A. × salignus -- самонесовместимое растение, образование семян
в Кембриджшире (Великобритания) не обнаружено, для успешного опыле
ния необходимо соседство растений из разных клонов [Briggs et al., 2006].
Впрочем, в Центральной Европе отмечено обильное плодоношение A. × salignus
[Jedliиka, Prach, 2006].
Расселение и распространение A. × salignus происходит фрагментами корне
вищ. Благодаря вегетативному размножению астра способна формировать обшир
ные одновидовые заросли. При благоприятных условиях в течение одного сезона
астра способна давать до сотни новых побегов [Jedliиka, Prach, 2006].
Последствия внедрения
Воздействия на естественные фитоценозы и аборигенные виды. Обшир
ные монотонные заросли A. × salignus, возникающие на месте заброшенной
пашни или огородов, могут замедлять процесс их естественного зарастания.
Генетические изменения и вариабельность вида. Между отдельными
клонами A. × salignus наблюдаются заметные различия. Растение могут отли
133
Рис. 46. Aster x salignus,
общий вид растения
Рис. 48. Расселение Aster x salignus в Средней России
Рис. 47. Корзинка
Aster x salignus
134
Рис. 49. Массовое размножение Aster Ч salignus на залежи в Калужской области
135
чаться размерами корзинок, интенсивностью окраски краевых цветков, высо
той побегов и мощностью соцветия, шириной листьев. Очевидно, что при куль
тивировании астры ивовый мог происходить отбор по этим признакам. Неиз
вестно, возникал ли этот гибрид единожды или многократно, его связи с аме
риканскими растениями не ясны. Специальные работы по изменчивости этой
астры нам не известны.
Экономическое и социальное (положительное и отрицательное) зна
чение. A. × salignus является неприхотливым позднецветущим декоративным
растением, популярным среди дачников. Но точный ассортимент американс
ких астр, используемых в озеленении, плохо изучен.
Меры борьбы
Специальные меры борьбы с A. × salignus не разработаны. Показано, что
быстрому вегетативному разрастанию A. lanceolatus и A. novi belgii препятству
ет местный ненарушенный растительный покров [Jedliиka, Prach, 2006].
СЕМЕЙСТВО ASTERACEAE (COMPOSITAE) --
СЛОЖНОЦВЕТНЫЕ
ЧЕРЕДА ОБЛИСТВЕННАЯ
Название: Bidens frondosa L.
Синонимы: B.melanocarpa Wieg.
Морфологические признаки
Однолетнее травянистое растение, побеги, прямостоячие, ветвящиеся, по
чти голые. Листья супротивные, 3--5 рассеченные. Боковые доли с черешками
длиной 3--5 мм. Конечная доля листа длиннее, чем боковые, с черешком дли
ной 10--15 мм. Доли листа ланцетовидные или продолговато ланцетовидные,
заостренные, крупнозубчатые (рис. 50).
Благодаря широкому ареалу и экологической пластичности B. frondosa
представляет собой полиморфный вид, или, вернее (поскольку это агамный
вид), комплекс мелких самоопыляющихся линий. Слабо выраженные микрови
ды столь многочисленны, что их таксономическая трактовка крайне затруднена.
Хотя по литературным данным различают 1 гибрид, 1 форму и ~6 разновиднос
тей череды облиственной [Sherff, 1937; Strother, Weedon, 2006], внутривидовой
полиморфизм в естественном ареале выражен столь сильно (табл. 12), что при
желании разновидностей или микровидов можно выделить еще больше.
Описаны: B. frondosa var. frondosa (доли листьев с широкотреугольными
зубчиками, внешние листочки обертки корзинки в 2 раза больше, чем внут
ренние); B. frondosa L. f. anomala (Porter ex Fernald) Fernald, которая отмечена
[Blake, 1925] в штатах Пенсильвания, Нью Джерси, Делавэр, Канзас Небрас
ка, Массачусетс, Колумбия и канадской провинции Новая Шотландия); B.
frondosa L. var. anomala Porter ex Fernald (отличается от типичной var. frondosa
вверх направленными зубцами на ости семянки); B. frondosa L. var. caudata
Sherff; B. frondosa L. var. pallida Wiegand; B. frondosa L. var. stenodonta Fernald
et H.St. John (имеет ланцетные, с оттянутой верхушкой зубчики на листочках
и внешние листочки обертки, в 4 раза большие, чем внутренние); B. frondosa
L. var. puberula Wiegand = B.vulgata Greene = B. frondosa Wiegand, not L.
136
(стебель опушенный, лист 3--5, иногда 7 раздельный; корзинка крупная, ди
аметром до 3 см; внешние листочки обертки реснитчатые; семянки оливко
вого, желтоватого или коричневого цвета); B.comosa (A.Gray) Wiegand × B.
frondosa L.
Коррелируют между собой только два морфологических признака: семян
ки коричневого цвета имеют прямые или чуть отклоненные ости, а семянки
желтоватого цвета снабжены сильно отклоненными остями. Остальные при
знаки не коррелируют и могут сочетаться разными способами; на гербарном
материале североамериканского происхождения (LE) и на основании изуче
ния литературных источников нами отмечено [Виноградова, 2003а] 9 вариан
тов их сочетания: чаще (39%) встречается вариант 000000, то есть набор при
знаков, характерный для B. frondosa L. var. frondosa. В 24% случаев отмечен
вариант 010000; далее по степени снижения частоты встречаемости следуют
варианты 000010 (8%), 211000 (8%), 010100 (5%), 110000 (5%), 110100 (5%),
011000 (3%) и 010101 (3%).
Помимо высокой амплитуды изменчивости морфологических признаков
семянок, в естественном ареале имеет место также изменчивость числа хро
мосом. В норме B. frondosa имеет 2n=48 хромосом. Однако в естественном аре
але отмечены отдельные экземпляры с 2n=24, 36 и 72 хромосомами [Lцve,
Lцve,1969; Scheerer, 1940; Crowe D.R., Parker, 1981].
Естественный ареал
На родине --- в Северной Америке --- B. frondosa распространена довольно
широко. Ее естественный ареал занимает юго восток Аляски, юг Канады и се
верные и центральные штаты США от 55 до 30є с. ш. и от Атлантического до
Тихоокеанского побережья [Scoggan, 1979].
Таблица 12
Варианты изменчивости морфологических признаков B. frondosa
в естественном ареале
Признак
0
1
2
Лист
тройчатосложный триждырассеченный рассеченный
на 5--7 долей
Семянки
бородавчатые
без бородавок
Семянки
коричневые
желтоватые или
оливковые
Ости семянок
прямые или чуть
отклоненные
сильноотклоненные
Опушение
побегов
отсутствует
имеется хотя бы
в верхней части
Внешние листочки
обертки
цельные
реснитчатые
Внешние листочки
обертки
по длине равны
корзинке или чуть
превышают ее
превышают длину
корзинки более чем
в 2 (иногда в 5!) раза
Рис. 51. Расселение Bidens frondosa в Средней России
Рис. 50. Bidens frondosa, общий вид растения
138
Рис. 53. Корзинка
Bidens frondosa
Рис. 52. Bidens frondosa вдоль р. Пехорка в Люберецком р не Московской обл.
139
Вторичный ареал
История интродукции и географическое распространение. Впервые B.
frondosa отмечена среди адвентивных растений ботанического сада Монпелье
(юг Франции) в 1762 г. Эти растения (LE) характеризовались комплексом при
знаков 010000 (с коричневыми семянками без бородавок). Именно эта форма в
течение >100 лет изредка встречалась в Западной Европе. По крайней мере, все
просмотренные нами гербарные листы, собранные в Европе в ХVIII--ХIХ ве
ках, имели только такое сочетание признаков. В 1777 г. B. frondosa обнаружили
в Польше на Одере близ Вроцлава [Lhotska, 1966]. В ХIХ веке она появилась в
Италии (на Сицилии в 1834 г. и во Флоренции в 1849 г.), в Португалии (с 1877 г.)
и в Германии (в Гамбурге с 1891 г.), причем предполагается, что и в Португалию,
и в Германию, и в Польшу вид был занесен непосредственно из Америки [Кор
нась, 1960; Hegi, 1975]. Известно [Besser, 1816; Index seminum ..., 1926; LE], что в
ботанических садах России вид культивировали в 1816 г. (Кременец), в 1825 г.
(Санкт Петербург) и в 1826 г. (Москва). Однако до начала ХХ века череда встре
чалась во вторичном ареале единично и не дичала.
В Петербургском ботаническом саду росли особи (LE) с набором призна
ков, не отмеченных в естественном ареале. Первый вариант 211001 (с неборо
давчатыми семянками и длинными листочками обертки) собран в 1825 г., а
второй 111000 (растения в некоторой степени похожие на B. frondosa var.
vulgata с трижды рассеченными листьями и желтовато зелеными голыми се
мянками) --- в 1851 г. Обе разновидности также никаких способностей к дича
нию не обнаружили.
И только в 1896 г. в Потсдаме зафиксирована разновидность 000000 с трой
чаторассеченными листьями, короткими листочками обертки и коричневыми
бородавчатыми семянками, которая явилась основателем инвазионной попу
ляции, проявившей большую склонность к дичанию и внедрению в естествен
ные прибрежно водные ценозы. В настоящее время в Евразии широко рас
пространенные инвазионные популяции B. frondosa физиономически едино
образны и представлены особями именно с таким набором признаков.
Объяснить этот феномен можно только тем, что лишь одна из разновиднос
тей, занесенных в Старый Свет, явилась основателем активно натурализую
щейся популяции. А поскольку B. frondosa --- самоопылитель, то современный
вторичный ареал вида в Европе сформирован из потомков этой инициальной
популяции.
Сходная картина отмечается и в других регионах. В 1950 г. во Владивосто
ке, а в 1990 г. в Батумском ботаническом саду найдены (MHA) растения с мел
кими густоопушенными семянками. В 1978 г. в Большехехцирском заповед
нике (MHA) отмечены особи с бородавчатыми семянками и длинными лис
точками обертки (B. frondosa var. stenodonta). Обе разновидности в массовом
количестве так и не распространились. Натурализовалась как в Колхиде, так
и в Приморском и Хабаровском краях, а также на о. Кунашир, где найдена в
1977 г. [Старченко, Дойко, 1981], опять таки только разновидность с тройча
торассеченными листьями, короткими листочками обертки и коричневыми
бородавчатыми семянками.
Исходя из вышеизложенного началом формирования инвазионных попу
ляций B. frondosa в Европе следует считать 1896 г., поскольку именно с этого
времени и началось фактическое расширение вторичного ареала череды об
140
лиственной. После Первой мировой войны места ее заноса в Европе стали
многочисленнее. Она массово произрастала на Рейне и Эльбе, в 1926 г. обна
ружена в Голландии, а позднее в Чехии. Однако до Второй мировой войны
натурализовалась B. frondosa только во Франции в долине р. Луара и в Ита
лии --- в Тоскане и на Сицилии [Anzalone, 1957; Hegi, 1975; Debray, 1942;
Trzcinska, 1961].
После Второй мировой войны началось быстрое расширение вторичного
ареала череды. В 1955 г. B. frondosa найдена Я.Корнасем в Бресте. По всей ве
роятности, она была занесена в Белоруссию из Западной Польши, где к тому
времени началась ее натурализация [Trzcinska, 1961]. Можно предположить,
что вид был распространен на территории европейской части России несколько
раньше, но просматривался из за габитуального сходства с B.tripartita, с кото
рой чаще всего произрастал совместно.
Через 15 лет B. frondosa обнаружена на Украине --- в 1970 г. в Каневе [Кор
нась, 1971], в 1972 г. --- в Мукачеве [Хртек, Шоуркова, 1977]. В европейскую
часть России череда проникла из Литвы и Белоруссии. Вполне натурализо
вавшаяся популяция найдена в 1972 г. Ю.Д. Гусевым [1980]. в Калининградс
кой обл. на Куршской косе. Динамика расселения вида в Восточной Европе
подробно описана в работах Е.А. Глазковой [2005, 2006], а потому мы приво
дим лишь две находки (МНА), которые не вошли в ее труды: в Самарской обл.
B. frondosa найдена в 1978 г. в пойме Волги в г.Тольятти; в Астраханской обл.
отмечена в Волго Ахтубинской пойме близ с. Селитренное Харабалинского
р на в 1993 г.
Помимо типичной формы, в Европе распространяется и B. frondosa var.
anomala, которая отличается от типичной B. frondosa var. frondosa вверх на
правленными зубцами на ости семянки. Появилась она во вторичном ареале
значительно позднее типичной формы. В 1954 г. экземпляры с подобными се
мянками найдены в Германии, а в 1960 г. --- в Польше [Trzcinska, 1961; Lhotska,
1965]. Позднее появились сведения о находках этой формы в Англии, Голлан
дии, Бельгии и Чехии. Отмечено, что для B. frondosa var. anomala характерно
появление в местностях с солоноватой водой. В настоящее время в Германии
эта форма распространена уже достаточно широко --- найдена в районе Берли
на, в мюнстерской гавани, на берегу Рейна в Гессене [Ludwig, 1994], а на бере
гу р. Рур и канала Рейн--Херпе B. frondosa var. anomala является преобладаю
щей [Keil, 1995]. До последнего времени наши целенаправленные поиски этой
формы в популяциях Московской и Рязанской обл., а также Волгоградской
обл. (где B. frondosa произрастает по берегам солоноватых водоемов) были
безуспешными. Но в 2003 г. А.К. Скворцов обнаружил B. frondosa var. anomala
(МНА) во дворе своего дома в Москве. По видимому, следует ожидать более
широкого распространения этой формы как в Центральной Европе, так и в
Средней России.
Таким образом, в настоящее время на территории бывшего СССР суще
ствуют три изолированных очага распространения B. frondosa: Приморье, куда
она занесена непосредственно из Америки не позднее 1950 г.; европейская часть
России, где вид стал расселяться с 1955 г., и Черноморское побережье Кавка
за, куда череда занесена в конце 1970 х гг. Во всех этих регионах череда актив
но натурализуется. В рудеральных популяциях через несколько лет числен
ность особей может существенно сокращаться, а порой популяции вообще
141
выпадают, даже в сырых местообитаниях. Подобное же сокращение числен
ности этого вида наблюдается и в некоторых районах Польши [Misiewicz, 1981].
Пути и способы заноса. Имеется два способа миграции вида [Корнась,
1960]: вдоль рек (по которым рассеивается путем гигрохории и эпизохории),
а также вдоль железных дорог (где мигрирует путем антропохории).
Статус в регионе. В 1975 г. вид был найден в Москве и Рязанской обл., а в
1979 г. обнаружен на границе Брянской и Сумской обл. в пойме Оки (MW). В
середине 1980 х гг. происходит взрывное расширение ареала череды: ее на
ходки сделаны в Воронежской (Новохоперский р н , Хоперский заповедник
1985. Печенюк . LE) и Ульяновской обл. (совхоз с. Архангельское Чердаклин
ского р на. Берег Куйбышевского водохранилища. 28.08.1991. Н.С. Раков.
MHA), а также в других регионах (рис. 51). Таким образом, через 20 смен по
колений череда облиственная в Средней России полностью натурализовалась,
а в Московской обл. практически полностью вытеснила аборигенную
B.tripartita.
Местообитания. Экологические условия, в которых обитает череда об
лиственная в естественном ареале, очень разнообразны. Чаще она произрас
тает по берегам водоемов (рис. 52), но встречается и на рудеральных местах, а
иногда как сорняк в полях [Scoggan, 1979]. Во вторичном ареале, так же как на
родине, B. frondosa более часто и обильно растет у берегов рек и на рудераль
ных местах, особенно у ж/д станций.
Размножение и жизненный цикл. B. frondosa --- однолетнее растение и
размножается только семенами. Всходы появляются в апреле--мае. Цветет и
плодоносит в августе--сентябре (рис. 53), на месяц полтора позднее, чем або
ригенная B.tripartita.
Расселение и распространение. Семена переносятся антропо и зоохор
но, поскольку ости семянки снабжены зубцами, с легкостью цепляющимися
за одежду или шерсть животных. Имеет место и расселение водным путем.
Последствия внедрения
Воздействия на естественные фитоценозы и аборигенные виды. Еще в
середине ХХ века отмечено [Trzcinska, 1961], что в Средней Чехии B. frondosa
вытесняет B.tripartita и со временем может заместить ее. Предположение ока
залось пророческим. По нашим наблюдениям, уже к 1999 г. по берегам водо
емов Московской обл. росла почти исключительно B. frondosa, а B.tripartita
встречалась крайне редко. В Ростовской обл. идет столь же заметное сокраще
ние ареала аборигенной B.cernua.
Проведена сравнительная оценка конкурентоспособности инвазионной B.
frondosa и аборигенной B.tripartita путем выращивания образцов различного
географического происхождения в однородных почвенно климатических ус
ловиях. В интродукционной популяции, заложенной в 2001 г. из семян, со
бранных в Ростовской обл. и в Москве [Виноградова, 2003б], по темпам про
растания семян, скорости роста, биомассы, особенностям размножения расте
ний (табл. 13) и другим показателям преимущество имела B. frondosa.
В 2003 г. в интродукционной популяции, заложенной из семян, собранных
в Ярославской и Московской обл., B. frondosa также показала заметное пре
имущество по сравнению с B.tripartita по всхожести семян (56% против 39) и
длине побегов (71,7±12,3 см против 37,6±11,4).
142
Для экспериментальной проверки конкурентного превосходства B. frondosa
осенью 2005 г. мы высеяли смесь семян обоих видов московской репродук
ции. Каждый из 4 образцов представлял собой смесь 60 семянок B. frondosa и
60 семянок B.tripartitа. Два образца высеяли на участок с низким содержани
ем гумуса и два --- на участок с высоким содержанием гумуса. Всхожесть
B.tripartitа составила 32,5% на бедной почве и 42,5% на богатой почве; всхо
жесть B. frondosa оказалась заметно выше и составила 51,7% на бедной почве
и 79,2% на богатой почве. Темп роста B. frondosa значительно превосходил темп
роста B.tripartitа и на богатой (64,4 см против 25,9), и на бедной почве (54,5 см
против 25,0). B.tripartitа образовала довольно мало корзинок (от 1 до 6, в среднем
1,8) с 8--10 семянками в корзинке. B. frondosa сформировала значительно боль
шее число корзинок (от 2 до 106, в среднем 8,2) с 10--14 семянками в корзинке.
Дальнейшие опыты подтвердили (см. главу 3), что B. frondosa имеет кон
курентное превосходство перед B.tripartitа по темпам роста и семенной про
дуктивности на почвах разного плодородия и водного режима (табл. 14). Эти
данные согласуются с результатами, полученными при изучении экологичес
ких и биологических особенностей видов рода Bidens в Центральной Европе
[Kock, 1988].
Генетические изменения и вариабельность вида. Сведения о гибридиза
ции B. frondosa с аборигенной B.tripartitа довольно противоречивы. Считает
ся, что эти два вида могут гибридизировать, в результате чего возникает B. ×
garumnae Jean. et Debr. [Hegi,1975]. Опираясь на многолетние наблюдения, В.Г.
Папченков [2003] отмечает высокую гибридогенную активность B. frondosa,
что, по его мнению, позволяет гибридному виду B. × garumnae постепенно вы
Таблица 13
Некоторые морфометрические показатели двух видов рода Bidens
в конце вегетационного периода (2002 г.)
Показатель
B. frondosa B.tripartita
Средняя высота, см
41
14
Среднее число метамеров
9
7
Низкое содержание
питательных веществ,
семена ростовского
происхождения
Среднее число корзинок
35
Нет
Средняя высота, см
92
85
Среднее число междоузлий
8
7
Высокое содержание
питательных веществ,
семена московского
происхождения
Среднее число корзинок
51
41
Таблица 14
Всхожесть семян двух видов череды в разных экологических условиях (%)
При поливе
Без полива
торф земля
супесь
торф земля
супесь
B. frondosa
13
19
17
31
38
14
B.tripartita
20
14
9
23
21
20
143
теснять B.tripartita из естественных ценозов в бассейне Волги. В водохрани
лищах Волги В.Г. Папченковым [2006] найдены и другие гибриды: B. frondosa
× B.radiata Thuill. и B.cernua × B. frondosa, а в последние годы и возвратный
гибрид B. frondosa × B. × garumnae [Васильева, Папченков, 2007]. Но посколь
ку для B. frondosa характерна агамоспермия, а нормальное оплодотворение на
блюдается крайне редко [Crowe, Parker, 1981], да к тому же B. frondosa зацве
тает в среднем на 3 недели позднее B.tripartita, то процесс гибридизации весь
ма затруднен. Большинство исследователей не отмечали гибридов B. frondosa.
Специальный поиск, осуществленный Н.Мосякиным [1988] в местах сплош
ного произрастания родительских видов на окраинах Киева, позволил выя
вить всего лишь несколько экземпляров B.garumnae из нескольких тысяч рас
тений череды. Да и у этих экземпляров (по крайней мере, у тех из них, кото
рые хранятся в МНА) признаки, считающиеся диагностическими для
B.garumnae (крылатые черешки листьев, неяснобородавчатые семянки и ме
нее выраженная зубчатость сегментов листа) практически укладываются в
амплитуду модификационной изменчивости аналогичных признаков B.
frondosa. Более того, на гербарном листе имеется детерминантка самого С.Мо
сякина, переопределившего растение двумя годами позднее в B. frondosa.
Во вторичном ареале обнаружены растения B. frondosa с мелкоопушенны
ми листьями, такие экземпляры найдены Н.М. Решетниковой (MHA) в 2007 г. в
Калужской обл.
Изучена вариабельность плодов B. frondosa во вторичном ареале. Внутри
и межпопуляционная изменчивость размеров и формы семянок, степени их
опушения и выраженности бородавок, а также угла отхода ости от семянок
довольно низкая [Виноградова, 2009]. Варьирование морфологических при
знаков идет в основном на индивидуальном уровне. Почти у каждой особи,
помимо типичных семянок с 2 остями, имеются 1--3% семянок с 3 остями, а
форма, размер и угол расхождения остей зависят от места расположения се
мянок относительно края корзинки. По краю корзинки располагаются округ
лые семянки с углом отхождения ости от оси семянки, равным 30--45º. Чем
ближе к центру корзинки, тем более удлиняются семянки и уменьшается угол
отстояния остей. В центре корзинки форма семянок практически цилиндри
ческая, а угол не превышает 5º. Отмечено существенное снижение полимор
физма в инвазионных популяциях по сравнению с первичным ареалом. Неко
торые аллели (например, отсутствие бородавок на плодах) исчезли вовсе, а
оставшиеся изменили частоту встречаемости.
Анализ изменчивости биологических признаков показал, что в европейс
кой части вторичного ареала общая генетическая изменчивость B. frondosa
очень низкая (см. главу 4).
Экономическое и социальное (положительное и отрицательное) зна
чение. В Америке B. frondosa используют как средство для лечения гинеколо
гических заболеваний, болезней горла и болей в грудной клетке [Mitich, 1994].
В России череда облиственная не входит в число лекарственных растений, а
вот аборигенная B.tripartita используется широко. В связи с тем, что по мор
фологическим признакам оба вида довольно похожи и не исключена возмож
ность случайного включения B. frondosa в лекарственные сборы, в Главном
ботаническом саду проведен сравнительный анализ инвазионной B. frondosa
и аборигенной B.tripartita по содержанию фенольных соединений и кремния
144
[Виноградова и др., 2007]. Оба вида не проявили существенных отличий, как
по содержанию фенольных соединений (табл. 15), так и по содержанию раз
личных форм кремния.
У обоих видов череды, независимо от места произрастания, содержание
простых полифенолов, фенолкарбоновых кислот и флавоноидов в ряду лис
тья '! корзинки '! стебли снижается, а содержание дубильных веществ, напро
тив, возрастает.
Сходные закономерности выявлены и относительно содержания кремния:
межвидовая и географическая изменчивость этого признака несущественны,
а превалирует изменчивость между отдельными органами растения. Наибо
лее ценный для организма человека органический кремний в наибольшей мере
(1,14%) представлен в листьях, в меньшей степени (0,73%) --- в корзинках, а
минимальное его количество (0,50%) отмечено в стеблях череды. Содержание
минерального растворимого кремния, напротив, в этом ряду возрастает (лис
тья --- 0,15; соцветия --- 0,20; стебли --- 0,22%). Возрастает также и содержание
малоусвояемого полимерного кремния (листья --- 0,21; соцветия --- 0,25; стеб
ли --- 0,49%). Таким образом, количество фенольных соединений и кремния в
растениях является низковариабельным признаком. Поскольку зависимости
содержания фенольных соединений и кремния от видовой принадлежности
образца не выявлено, можно прогнозировать успешность применения листь
ев и соцветий B. frondosa для фармацевтических целей.
Меры борьбы
Имеются сведения [Sharma, Singh, 2000] о применении глифосата для борь
бы с B. frondosa.
Таблица 15
Содержание веществ флавоноидного комплекса в двух видах рода Bidens
Кверцетин-
3-О-
глюкозид
Кемпфе-
рол-3-О-
глюкозид
Лютеолин-
7-О-
глюкозид
Диосме-
тин-7-О-
глюкозид
Апи-
генин
Апигенин-
7-О-
глюкозид
Σ
флаво-
ноидов
листья
B. frondosa 1,18
0,49
0,22
0,20
0,26
0,22
2,31
B.tripartita 1,33
0,42
0,21
0,21
0,23
0,20
2,60
соцветия
B. frondosa 0,55
0,45
0,20
0
0,18
0,17
1,55
B.tripartita 0,63
0,38
0,23
0
0,20
0,14
1,58
стебли
B. frondosa 0,21
0
0,20
0,21
0
0,16
0,77
B.tripartita 0,30
0
0,21
0,15
0
0,15
0,82
145
СЕМЕЙСТВО ASTERACEAE (COMPOSITAE) --
СЛОЖНОЦВЕТНЫЕ
РОМАШКА ДУШИСТАЯ
Название: Chamomilla suaveolens (Pursh) Rydb.
Синонимы: Santolina suaveolens Pursh, Artemisia matricarioides Less.,
Chamomilla discoidea (DC) J.Gay ex A.Br., Lepidotheca suaveolens (Pursh) Nutt.,
Matricaria discoidea DC., M. matricarioides (Less.) Porter, M. suaveolens (Pursh)
Buch., Chrysanthemum suaveolens Aschers., Cotula matricarioides Bong., Tanacetum
suaveolens Hook
Морфологические признаки
Однолетнее растение высотой 5--35 см. Стебель ветвистый, под корзинка
ми голый или опушенный. Листья дважды перисторассеченные. Цветоносы
длиной 5--15 мм, корзинки многочисленные, без краевых язычковых цветков,
диаметром 7--15 мм (рис. 54).
Естественный ареал
Естественно произрастает лишь в западной части Северной Америки от
Внутренней Аляски до Калифорнии.
Вторичный ареал
История интродукции и географическое распространение. Как заносное
растение отмечено в 1838 г. на пустырях вокруг г. Сент Луис и Миссури. В то
же время выращивалось в Ботаническом саду Филадельфии из семян, собран
ных на западе континента [Torrey, Gray, 1969].
В 1860 г. ромашка занесена на Беринговы острова, а позднее -- на Чукотку и
Камчатку. В этом регионе Ch.suaveolens считается именно адвентивным видом,
поскольку поначалу [Комаров, 1930] она встречалась только в селениях или у
реки вокруг мест, где пластают рыбу. Во Владивостоке отмечена в 1910 г.
(МНА).
Во флору европейского региона внедрилась как "беженец" из ботаничес
ких садов. В 1849 г. ромашка произрастала в Ботаническом саду Гельсингфор
са (Финляндия) и через 20 лет натурализовалась в этом регионе. В Великоб
ританию интродуцирована из Северной Америки в 1871 г. и вскоре стала здесь
одним из самых распространенных инвазионных видов. Из Краковского бота
нического сада "сбежала" в 1886 г. В период с 1850 по 1900 г. находки этого
вида были сделаны в Швеции (г. Упсала), Германии (Берлин, Кенигсберг),
Чехии (Прага), Норвегии, Англии, Франции, Польше (Варшава), Австрии
(Вена), Италии (Триест) и Швейцарии (Цюрих) [Kinchin, 1988; Гусев, 1964;
Белозеров, 1960; Witte, 1909; Ridley, 1930; Hegi, 1954; Протопопова, 1973;
Sudnik Wojcikowska, 1987; Шафер, 1956]. В настоящее время Ch.suaveolens
широко распространена во многих европейских странах, отсутствуя лишь в
Средиземноморье [Florae Europaea, 1976].
Помимо Европы, как заносное встречается с 1873 г. в Новой Зеландии, с
1877 г. -- в Палестине, а также в Чили (Вальдивия), Маньчжурии и Корее [Hegi,
1954; Крылов, 1949].
В 40 е гг. ХIХ века ромашку душистую разводили в Санкт Петербургском
ботаническом саду как редкость. Через 10 лет она уже росла в саду в большом
количестве, и семена ее указывались в обменном каталоге семян сада [Index
146
seminum, 1863], а гербарные экземпляры были собраны в 1860 г. на рудераль
ных местах в Парголово (MHA). Вскоре ромашка стала обычным растением в
Петербургской губернии [Гусев, 1964].
В Киеве ромашка душистая обнаружена в 1886 г. [Шмальгаузен, 1886]. Ука
зание В.В. Протопоповой на нахождение этого вида в том же году В.Я. Цинге
ром в Сарепте неверно, поскольку в этой работе отмечен вид с белыми языч
ковыми цветками --Matricaria chamomilla [Протопопова, 1973; Цингер, 1885].
Массовое распространение вида в России и на Украине, особенно по же
лезным дорогам, началось в 90 х гг. ХIХ столетия. В это время ромашка души
стая была отмечена и в Вологодской обл.
Особый очаг расселения ромашки душистой появился в Западной Сибири.
Около 1890 г. ее разводили в Ботаническом саду Томского университета из
семян, выписанных из Эрфута, а через 3--4 года она распространилась уже по
всей университетской усадьбе. В Омске этот вид указан с 1889 г., в Пермской
губернии -- с 1912 г., в Тюмени и Ишимском уезде -- с 1915 г., в окрестностях
Тобольска -- с 1916 г. После 1917 г. ромашка была найдена в Барнауле, в 1927 г. --
на Алтае [Крылов, 1949].
В 30 е гг. ХХ века ромашка попала в Башкирию, а также была отмечена как
распространенный сорняк г. Сыктывкара и ряда других мест Коми АССР [Бе
лозеров, 1960; Флора Юго Востока..., 1936]. После войны найдена на о. Иту
руп Курильской гряды и в Восточном Казахстане (МНА).
Таким образом, к середине ХХ века ромашка произрастала во всех районах
европейской части России, а северная граница ее распространения проходила
по западу Мурманской обл., Северной Карелии, по берегам Белого моря и
Северного Ледовитого океана, а также спорадично в Сибири и на Дальнем
Востоке. Широко была распространена на Украине и в Белоруссии, единич
ная находка вида отмечена в Прибалхашье (Казахстан) [Белозеров, 1960; Фло
ра СССР..., 1961].
За последние 50 лет ареал ромашки расширился незначительно, в основ
ном, благодаря отдельным ее находкам в Средней Азии. Она отмечена как за
носное в Катакургане Самаркандской обл., в Чуйской долине и в котловине о.
Иссык Куль, в окрестностях Душанбе, а также в ряде областей Казахстана.
Несколько находок вида сделано и в Астраханской обл. [Определитель расте
ний Средней Азии..., 1993; Сафонов, 1982].
Пути и способы заноса. В Европе Ch.suaveolens появилась в середине ХIХ
века и начала внедряться в нарушенные ценозы как "беженец" из ботаничес
ких садов. В Южной Финляндии и Санкт Петербурге этот процесс занял 20
лет, в Москве -- 10, в Томске -- 4 года. Бурная натурализация вида наблюда
лась в начале ХХ века. Следовательно, между первым появлением растения в
Европе и его натурализацией прошло не менее 20 смен поколений. При даль
нейшем расселении вида приспособление его к новым условиям шло все стре
мительнее, и вид начинал внедряться в рудеральные местообитания уже через
3 смены поколений. В настоящее время в России произрастает по меньшей
мере 100 е поколение ромашки душистой.
Статус в регионе. Первая находка вида в Московской обл. сделана в 1870 г.
(ст. Мытищи, на первой версте к Тарасовке. 20.06.1870. Д.Сырейщиков. MW).
В Москве найдена в 1887 г., а уже через 2 года росла "по сорным местам, всю
ду: Ростокино, Останкино и другие места". Спустя 8 лет растение было собра
147
Рис. 55. Расселение Chamomilla suaveolens в Средней России
Рис. 54. Облик Chamomilla suaveolens
Рис. 57. Начало
плодоношения Chamomilla
suaveolens
Рис. 56. Chamomilla
suaveolens вдоль тропинки
149
но в Подольском уезде, а еще через 10 лет Ch.suaveolens была уже широко рас
пространена в Московской обл. и местами даже вытеснила Matricaria
chamomilla [Кауфман, 1889; Бухгольц, 1897; Сырейщиков, 1910].
Массовое распространение этого вида, особенно по железным дорогам, на
чалось в конце ХIХ столетия. К этому времени относится гербарный сбор из
Курской губернии (Щигровский уезд. Усадьба. Аноним. 1898 г. MW). В 1890--
1895 гг. ромашка душистая проникла в Костромскую губернию. В 1898 г., пос
ле проведения Всероссийской выставки, ромашка появилась в окрестностях
Нижнего Новгорода. А к 1912 г. этот вид указывался уже для Тверской, Смо
ленской, Ярославской, Тульской, Владимирской, Орловской, Московской,
Калужской, Нижегородской обл. [Белозеров, 1960; Протопопова, 1973; Луки
на, 1989; Маевский, 1912]. Позднее ромашка появилась и в других местах:
«Брянская обл., г.Трубчевск. Городские луга. 15.07.1926 г. В.А. Арсеньев. MW»;
«г. Иваново Вознесенск. Дер. Сергельское. Близ озера. Старая порубь -- силь
ный выпас. 06.08.1924. Евгорова. MW». В настоящее время встречается во всех
областях Средней России (рис. 55).
Местообитания. Массово произрастает во дворах и по обочинам дорог, а также
в других рудеральных местах: на полях, лугах, по песчаным берегам рек (рис. 56).
Размножение и жизненный цикл. Ch.suaveolens -- яровой однолетник, но иног
да развивается как зимующий двулетник. Размножается семенами: одно рас
тение дает до 5300 семянок (рис. 57).
Расселение и распространение. Семянки разносятся ветром (на 2,5--3,5 м)
и дождевыми потоками. В дождливую погоду плоды ослизняются, легко при
липают к ногам и могут разноситься на дальние расстояния. Распространяет
ся и лошадьми, которые поедают это растение.
Последствия внедрения
Воздействие на естественные фитоценозы и аборигенные виды. В естествен
ных луговых ценозах нами найдена лишь однажды: растения достигали в вы
соту ~60 см и имели немногочисленные корзинки.
Генетические изменения и вариабельность вида. В инвазионных популяци
ях межпопуляционная изменчивость по количественным признакам практи
чески не проявляется [Виноградова, 2001], однако внутрипопуляционная из
менчивость довольно высока. Амплитуда изменчивости высоты растений со
ставляла 5--40 см, числа побегов 1--15, диаметра корзинки 7--15 мм, длины
цветоносов 5--15 мм. Диаметр корзинки и масса 100 соцветий варьировали на
среднем уровне (CV=20%); высота растений, длина цветоносов, количество
побегов и биомасса надземных органов варьировала на очень высоком уровне
изменчивости (CV >41%). Во всех популяциях встречались особи как с пря
мостоячими, так и с распростертыми побегами, а также с цветоносами как го
лыми, так и опушенными под корзинкой прижатыми волосками.
Сходные данные получены при изучении изменчивости этого вида в Киев
ской обл. [Четверня, 1987]. Коэффициент плодоношения ромашки душистой
варьировал там на низком уровне (CV=12%); число семян и семяпочек в соц
ветии, а также масса 1000 семян и высота растений -- на повышенном уровне
(CV=40%); урожайность соцветий, вегетативной массы и семян ромашки -- на
повышенном и очень высоком уровне изменчивости (CV=41%).
Экономическое и социальное (положительное/отрицательное) значение. В
надземной части ромашки душистой содержатся: эфирное масло, апиин, сали
150
циловая кислота, глицериды жирных кислот, аскорбиновая кислота, каротин,
горечи, слизи и камедь. Найдены также полисахариды, флавоноиды кверци
меритрин и лютеолин 7 глюкозид, аминоспирт холин, кумарин герниарин, а
также цис и трансбициклоэфиры [Протопопова, 1973; Mathe, Tyhak, 1962;
Просовский и др., 1985; Киселева, Кибальчич, 1970; Арак, Раал, 1987]. По ко
личественному содержанию эфирного масла она соответствует требованиям
государственной фармакопеи -- не менее 0,2% (ГОСТ 2237 75).
В эфирном масле ромашки душистой нет бизаболола и хамазулена, кото
рые являются основными носителями противовоспалительного действия, зато
в нем значительно выше, чем у ромашки аптечной, содержание фарнезена (об
ладает гранулирующим и эпителизирующим воздействием), фарнезола и цис
ен индициклоэфира (обладает фунгицидным и сильным спазмолитическим
воздействием). Оба вида содержат 19 общих фенольных соединений и 10 об
щих флавоноидных агликонов [Арак, Раал, 1987; Четверня, 1987], что предоп
ределяет использование ромашки душистой в качестве лекарственного расте
ния -- полноценного заменителя ромашки аптечной. Изучение качественного
состава эфирного масла ромашки душистой из 19 пунктов произрастания ее в
Эстонии [Арак, Раал, Таммеорг, 1981] показало, что существует несколько хе
мотипов данного вида. Аналогичные выводы сделаны и другими исследовате
лями.
Интересно, что в Канаде содержание эфирного масла у Ch.suaveolens край
не низкое, и химические компоненты его существенно иные [Просовский и
др., 1985]. Следовательно, содержание эфирного масла и его компонентный
состав зависят от места произрастания этого лекарственного растения, что
свидетельствует о необходимости тщательно изучать исходное сырье, приме
няемое для фармацевтических целей.
В ромашке душистой содержатся также фенольные соединения. Амплиту
да изменчивости содержания фенольных соединений определена [Виногра
дова, Колесников, 2005] в интродукционной популяции, сформированной из
семян, собранных в различных точках вторичного ареала вида. Изучено соот
ношение четырех групп фенольных соединений -- простых полифенолов и ок
сибензойных кислот, оксикоричных кислот и их слож-ных эфиров с хинной
кислотой, флавоноидов (включая полиметоксифлавоны), а также полимер
ных и конденсированных полифенолов. Семена, собранные в 2003 г. в 17 точ
ках вторичного ареала вида (в Латвии, Австрии, Карелии, Мурманской, Воло
годской, Псковской, Ярославской, Московской и Липецкой обл.) той же осе
нью были высеяны на экспериментальный участок ГБС РАН. В надземной
части ромашки душистой отмечено высокое общее содержание флавоноидов
(в среднем 3,10%). Наибольшее количество флавоноидов (3,44%) содержал
образец из Северной Карелии.
Определено содержание фенольных соединений в различных органах ро
машки душистой. Сумма флавоноидов в стеблях в 2 раза меньше, чем в соцве
тиях или листьях. Отсюда следует, что для лекарственных целей целесообраз
но брать не надземную часть растения в целом, а сбор листьев и соцветий.
При сравнении содержания фенольных соединений в спонтанных инвази
онных популяциях и в интродукционной популяции, выращенной в однород
ных условиях питомника, отмечено, что при интродукции растений из отдель
ной локальной популяции количественное содержание фенольных соединений
151
ни в соцветиях, ни в листьях, ни в стеблях растений практически не изме
няется.
Проведено сравнение содержания фенольных соединений в ромашке душис
той и модельных лекарственных растениях. По сумме конденсированных поли
фенолов (дубильных веществ) в листьях ромашка душистая (1,82%) превосхо
дит любой из ранее изученных по аналогичной методике 21 вида (от 0,41 до 1,23%).
Закономерностей изменчивости содержания фенольных соединений от
высоты над уровнем моря или широты географического пункта сбора семян
нами не отмечено. Следовательно, межпопуляционная изменчивость химичес
ких признаков, так же как морфологических и биологических, у самоопыляю
щегося вида Ch.suaveolens носит неупорядоченный, а не клинальный харак
тер. Это позволяет отбирать для культивирования в лекарственных целях об
разцы с наиболее высоким содержанием активных веществ и влечет
необходимость тщательно изучать исходное сырье, применяемое для фарма
цевтических целей. Дополнительным преимуществом Ch.suaveolens как ново
го лекарственного растения является то, что при культивировании количество
фенольных соединений в образце практически не меняется.
Меры борьбы
Специальных мероприятий по борьбе с ромашкой душистой не проводится.
Обзор научных исследований и публикаций
Изучена генетическая закрепленность различных биотипов Ch.suaveolens
и амплитуда внутривидовой изменчивости [Виноградова, 2001]. Выявлен су
щественный полиморфизм по феноритму и ряду количественных признаков
(см. главу 4).
По данным С.А. Четверни [1987], использовавшего аналогичную методи
ку, только в Киевской обл. насчитывается 6 биотипов ромашки душистой, раз
личающихся по высоте особей, числу корзинок, продолжительности вегета
ционного периода, длительности периода цветения, содержанию эфирного
масла и отдельных его компонентов, а также по содержанию флавоноидов.
СЕМЕЙСТВО ASTERACEAE (COMPOSITAE) --
СЛОЖНОЦВЕТНЫЕ
МЕЛКОЛЕПЕСТНИК КАНАДСКИЙ
Название: Conyza canadensis (L.) Cronquist
Синонимы: Erigeron canadensis L., Leptilon canadense (L.) Britton
Морфологические признаки
Однолетнее или озимое двулетнее растение (рис. 58). Корень стержневой,
короткий; сеянцы формируют розетку темно зеленых слабоопушенных лис
тьев, ширина которых редко превышает 1 см, с выраженным черешком и круп
нозубчатыми краями; по мере удлинения стебля базальная розетка усыхает.
Стебли вертикальные, высотой 10--180 см, опушенные, с многочисленными
мелкими цветоносными побегами в верхней части; стеблевые листья очеред
ные, многочисленные от ланцетных до линейных, почти цельнокрайные; са
мые верхние стеблевые листья имеют ширину всего 5 мм; корзинки мелкие,
152
диаметром 3--5 мм, многочисленные на коротких цветоножках; женские языч
ковые цветки белые, очень короткие, часто короче обертки корзинки. Семян
ки мелкие, длиной 1--2 мм длиной, прозрачные с паппусом длиной 3--5 мм.
Естественный ареал
Родина -- Северная Америка. C.canadensis найден к югу от 55є с.ш. во всех
регионах Канады, кроме о. Ньюфаундленд. C.Frankton и G.Mulligan [1987]
перечисляют его как обычный в Канаде вид, более редкий, однако, на Атлан
тическом побережье. Один образец зарегистрирован в Форт Смите (Северо
Западные территории). Естественный ареал занимает также большую часть
США, достигая на юге штатов Техас и Орегон [Torrey, Gray, 1969; Weaver, 2001].
Вторичный ареал
История интродукции и географическое распространение. В ХVII веке
мелколепестник завезен в ботанические сады Европы, по видимому, как ред
костное заморское растение; отмечен в каталоге Ботанического сада в окрест
ностях Нюрнберга в 1646 г. и в каталоге ботанического сада Блуа (Франция) в
1655 г. (в последнем случае под названием Aster canadensis annuus). В ХVIII веке
вид был уже довольно широко распространен в Средней Европе. В 1707 г. его
нашли в Пфальце, в 1710 г. -- в Бранденбурге, в 1715 г. -- в Гессене, в 1726 г.
--в
ЙенеиЛейпциге,в1728г. -- вЭльзасе,в1730г. --вВаршаве,в1747г. --в
Страсбурге, в 1753 г. -- в Геттингене и в 1762 г. -- в Вене [Hegi, 1954].
В настоящее время входит в состав синантропной флоры во всех странах
Европы, за исключением Ирландии и Исландии [Cronquist, 1976]. В конце
ХХ века обнаружен в Австралии и Японии [Weaver, 2001; Miyawaki, 1988].
В Российской империи этот вид появился, вероятно, в начале ХVIII века,
но сказать что либо определенное относительно места внедрения мелколепе
стника канадского в аборигенную флору не представляется возможным. Са
мый ранний источник сведений об этом адвентивном растении относится к
1753 г. -- так датирован гербарный лист мелколепестника, собранного в Одес
се (MW). В Каталоге растений сада П.Демидова в Москве C.canadensis упоми
нается уже как обычный сорняк [Паллас, 1781].
В первой половине ХIХ века мелколепестник канадский распространен уже
во многих районах европейской части Российской империи: с 1816 г. известен
в Кременце, в 1824 г. произрастал на рудеральных местах в окрестностях Санкт
Петербурга, а в 1826 г. --в Вологде. Гербарные образцы собраны в 1824 г. в Нов
город Северске, в 1832 г. -- в Днепропетровской обл. и в Оренбурге, в 1833 г. -- в
Риге, в 1837 г. -- в Киеве, в 1840--1848 гг. в Кировограде, Каунасе, Курляндии
(ныне южная часть Латвии и Литва), Самаре, Житомире, Астрахани [Besser,
1816; Weinmann, 1824; Фортунатов, 1826; LE, MW].
На Кавказе этот вид также встречался массово. Еще в 1829 г. мелколепест
ник найден в Карабахе, а позднее (1835--1843 гг.) -- в Иберии, Имеретии, Осе
тии, Малке и Екатеринограде (LE).
К первой половине ХIХ века (1841--1843 гг.) относятся также две находки
вида, сделанные в горах Алатау в Восточном Казахстане (KW).
В первой подробной сводке о флоре России мелколепестник канадский
приводится для Озилии (ныне -- о.Сааремаа), Ливонии, Куронии и Литвы
(т.е. для всей Прибалтики), Волыни, Курска, Тулы, Москвы, Пензы, Казани,
Симбирска, Саратова, Киева, Подолии, для Украины, Дона, а также для дру
Рис. 58. Conyza canadensis --- злостный сорняк посевов
Рис. 59. Расселение Conyza canadensis в Средней России
Рис. 60. Внедрение Conyza canadensis в прибрежные сообщества
Рис. 61. Розетка Conyza canadensis
155
гих мест в земле Донских казаков. Он произрастал в Таврии (ныне -- Крым),
на Волге, на Кавказе (Терек, Карабах, Рача, Ленкорань, Елизабетполь), оди
ночно в Уральской Сибири [Ledebour, 1845--1846].
Из литературных и гербарных источников середины ХIХ века известны
местонахождения мелколепестника между истоками Волги, Днепра, Западной
Двины и Ладожских вод, в Тамбове, Могилеве, в Бессарабии, в Харьковской
губернии и в Области войска Донского [Финк, 1854; LE, MW, KW]. В сводке о
флоре различных провинций России [Lindemann, 1860] C.canadensis приво
дится еще для ряда пунктов: Витебска, Гродно, Минска, Чернигова, Орла и
Полтавы. Таким образом, в середине ХIХ века мелколепестник канадский об
ладал уже обширным вторичным ареалом.
В 1860--1870 гг. в связи с присоединением к России Средней Азии число
местонахождений мелколепестника значительно возросло. В 1867 г. он обна
ружен в Туркестане, а в 1873--1874 гг. члены Аму Дарьинской экспедиции от
метили наличие этого вида в низовьях р. Аму--Дарья и в Хиве. C.canadensis
был встречен также в Ташкенте, Верном (Алма Ата), Андижане, Китабе (LE).
Множество находок вида в Средней Азии позволяет думать, что мелколепест
ник канадский, возможно, произрастал там задолго до того, как русские ис
следователи сделали первые его сборы.
В 1883 г. при обследовании Уфимской и Оренбургской губерний Ю.Шелль
[1885] нашел мелколепестник только "по эту сторону Уральских гор" в Ники
тине, Гатчине, Зеркле, Сакмарском городке и Оренбурге, но заметил, что "Лес
сингом это растение голословно указывается по ту сторону Уральских гор".
Временем заноса мелколепестника из Европейской России в Западную Си
бирь следует считать рубеж ХIХ ХХ вв. Впервые C.canadensis найден там в
Курганском уезде Тобольской губернии, а затем в Челябинской обл. и Тюмени
(LE). В 1928 г. А.А. Хребтов констатировал массовое распространение этого
вида в Шадринском уезде [Крылов, 1949].
Спонтанное расселение мелколепестника в Сибирь происходило, по всей
вероятности, двумя путями: из Европейской России и Восточного Казахста
на, причем занос мелколепестника из Восточного Казахстана в Алтайский край
случился значительно раньше. Это подтверждается находкой C.canadensis на
Алтае А.Н. Красновым еще в 1882 г. и более поздними (1895--1908 гг.) наход
ками этого вида в Минусинском округе Енисейской губернии, около Барнау
ла и в Томской губернии (LE). В 1911--1912 гг. мелколепестник канадский
произрастал уже во многих местах западной части Алтайского края. Самая
северная точка современного распространения мелколепестника в Сибири --
г. Маган близ Якутска [Макаров, Дубовик, 1977].
В начале ХХ века мелколепестник начал внедряться и во флору Дальнего
Востока. Уже в 1902 г. небольшая популяция вида обнаружена в с. Черниговс
кое Приморской обл. Позднее (1909--1911 гг.) он найден около Хабаровска, в
Иманском уезде и во Владивостоке. Через 10 лет мелколепестник канадский
стал обычным сорным растением Уссурийского р на и появился в Амурском
округе. В 1948 г. C.canadensis обосновалась и на Сахалине (LE).
Продвижение мелколепестника на север и северо восток европейской час
ти России шло более медленными темпами. К концу ХIХ века (1887--1889 гг.)
мелколепестник появился в Псковском уезде и в Твери (LE, MW).
156
По наблюдениям П.В. Сюзева [Крылов, 1949], в 1912 г. этот вид встречался
единично около Перми близ железной дороги. Для Архангельской губернии
C.canadensis впервые указан в 1912 г. В 1921 г. этот вид найден в Карелии на
побережье Сегозера, а в 1922 г. -- в Архангельске (LE). Для Вологодской обл.
первое указание на встречаемость этого растения появилось лишь в 1936 г.
[Белозеров, 1960]. Имеются также данные [Hulten, 1950] о единичных его на
ходках в окрестностях Лахденпохья и Оланги. В 60 х гг. ХХ века очень редко
встречался на юге Коми АССР [Определитель высших..., 1962] в Летском,
Прилузском и Сысольском р нах. В настоящее время изредка встречается как
полевой сорняк в Ленинградской, Новгородской, Вологодской и Костромс
кой обл. В более северных районах отмечены лишь единичные местонахожде
ния. Так, в Республике Коми вид встречается по железным дорогам на ст. Сык
тывкар, Микунь, Язель, Копты, Човью [Лавренко, Кустышева, 1990].
Таким образом, на территории России ареал С.canadensis состоит из двух
частей. Северная граница западной части ареала проходит на 10° севернее гра
ницы естественного ареала вида по линии Кировск--Архангельск --юг респуб
лики Коми и далее выклинивается по линии Тобольск--Красноярск--Мину
синск--Зайсан. Восточная часть ареала занимает Приморский край, юг Амур
ской обл. и южную часть о. Сахалин [Виноградова, 2005].
Пути и способы заноса. Приспособлением для рассеивания семян ветром
является паппус, который, по меньшей мере, в 2 раза длиннее семени.
Хотя высокие растения производят больше семян, чем низкие, D.Regehr и
F.Bazzaz [1979] выявили, что репродуктивное усилие (число семян на грамм
сухого веса надземной части растения) снижается с высотой растения. Они
предположили, что высокие растения имеют преимущество рассеивания, ко
торое является более важным, чем максимальное репродуктивное усилие. Они
сообщили о рассеивании семян на поле зерновых культур на расстояние до
122 м в подветренную сторону от края сплошных зарослей C.canadensis. Рас
сеивание семян может происходить также водным путем. Множество семян
C.canadensis, 80% из которых оказались жизнеспособными, найдено в иррига
ционных каналах и реках, граничащих с полевыми популяциями [Kelley, Bruns,
1975]. Y.Kasahara [Weaver, 2001] приписывал широкое распространение
C.canadensis в Японии способностью сеяться вдоль автомобильных и желез
ных дорог.
Статус в регионе. Впервые C.canadensis приводится Ледебуром [1845--
1846] для Курска, Тулы, Москвы, Пензы, Казани, Симбирска, Саратова. В се
редине ХIХ века отмечен в Тамбове и Орле. Для Костромской губернии впер
вые указан в 1912 г. В настоящее время встречается во всех областях Средней
России (рис. 59). Отмечено, что инвазионные популяции в России представ
лены по меньшей мере 200 м поколением вида. [Виноградова, 2005].
Местообитания. И в естественном, и во вторичном ареале обычно встре
чается на грубых почвах. Предпочитает каменные, песчаные (рис. 60) или пло
дородные суглинистые почвы и хорошо переносит засуху [Weaver, 2001]. Хотя
мелколепестник иногда колонизирует затапливаемые равнины, наводнений
он не терпит. Встречается также в ямах гравия, на утесах и скалистых берегах.
Чаще отмечается на рудеральных местах, вдоль дорог, на выгонах и пастби
щах. Отличается широкой эколого ценотической амплитудой приспособле
157
ния и относительно высокой технотолерантностью. В техногенных экотопах
образует иногда сплошные одновидовые заросли.
Размножение и жизненный цикл. Розеточная форма и озимый однолет
ний жизненный цикл позволяют C. canadensis обосновываться и расти без кон
куренции с яровыми однолетними сорняками. Размножается исключительно
семенным способом. Большинство сеянцев C.canadensis появляются с конца
августа до октября и формируют зимующие розетки (рис. 61). Часть пророст
ков появляется весной, с марта до начала мая. Стебли начинают удлиняться в
мае, и растение цветет в середине июля, а продуцирование семян достигает
пика в начале августа и в сентябре. Известно, что мелколепестник в более
южных широтах развивается как яровое растение, а на севере -- как озимое, то
есть осенью образует розетки и дает цветоносные побеги только на следую
щий год. В Москве мелколепестник растет преимущественно в озимой форме
[Виноградова, 2005].
C.canadensis -- самосовместимое растение [Mulligan, Findlay 1970]. Пыльца
выпускается прежде, чем корзинка полностью откроется, и это подразумева
ет, что вид является в основном самоопылителем, хотя можно наблюдать и
насекомых, посещающих открытые цветы. Используя устойчивость к герби
циду в качестве маркера, А.Smisek [Weaver, 2001] определил, что степень аут
кроссинга в популяции C.canadensis в графстве Эссекс составляет ~4% (от 1,2
до 14,5%).
Расселение и распространение. За вегетационный сезон на одном расте
нии формируется в среднем 50--300 корзинок, число семянок в каждой из них
составляет в среднем 30--40 [Галкина, Виноградова, 2008]. Лабораторная всхо
жесть семян -- 90--95%, полевая всхожесть ~80%. Семена созревают обычно
через 3 недели после опыления. Средний вес семени без паппуса 0,072 мг, из
которого 15% приходится на семенную оболочку и 85% -- на зародыш семени
[Fenner, 1983]. Число корзинок на 1 растение и общая семенная продуктив
ность пропорциональны высоте растения [Regehr, Bazzaz,1979]. Растение вы
сотой 40 см производит ~2000 семянок, в то время как растение высотой 1,5 м
производит ~230 тыс. семянок. Р.Bhowmik и M.Bekech [Weaver, 2001] сообщи
ли, что особи C.сanadensis, растущие при плотности 10 растений/м2 произво
дят ~200 тыс. семян/1 растение.
Последствия внедрения
Воздействие на естественные фитоценозы и аборигенные виды.
C.canadensis отмечена в разнообразных сообществах: встречается в садах, ви
ноградниках, старых полях, по обочинам дорог и железным дорогам, в нару
шенных местах естественных ценозов, например вдоль тропинок в лесистой
местности или в поймах рек, а также в городах. Это растение -- пионер на нео
брабатываемых сельскохозяйственных землях и лесных вырубках, особенно
после пожарищ, может сохраняться на заброшенных пастбищах. Значительно
увеличивает влияние как сорняк на тех пахотных полях, где обработка почвы
снижается или прекращается вовсе. Обычно встречается в сообществах с дру
гими озимыми одно= или двулетними сорными травами, которые поселяются
на нарушенных местообитаниях, такими как Capsella bursa pastoris, Lactuca,
Lepidium spp. и др.
Начинает проникать в естественные ценозы. В Северско Донецком при
родном комплексе встречается в борах, на суходольных лугах в формациях
158
полевицы и пырея, а в Липецкой обл. участвует в создании разнотравья в со
сновых лесах, встречается по песчаным берегам Дона и его притоков [Вьюко
ва, 1987].
Генетические изменения и вариабельность вида. Мелколепестник канад
ский --диплоид. Во всех исследованных популяциях (Северная Америка, Гер
мания, Польша, Узбекистан, Япония) 2n=18 [Bijok et al., 1972].
Данных по внутривидовой изменчивости канадских популяций не имеет
ся, подвидов и форм не описано [Weaver, 2001]. Спонтанные популяции вида
в Европе отличаются от популяций других инвазионных видов рода Conyza
(C.bonariensis, C.sumatrensis, C.blakei и C.floribunda) разнообразием морфоло
гических и электрофоретических характеристик. Европейские популяции де
монстрируют широкую амплитуду изменчивости как морфологических, так и
биологических признаков, несмотря на сравнительно низкую вариабельность
изоэнзимного комплекса.
Есть предположение, что в Европе происходит гибридизация между
C.canadensis и другими сорными видами рода Conyza, особенно C. sumatrensis
и C. bonariensis, поскольку они часто растут в смешанных популяциях, и име
ются промежуточные формы [Thebaud, Abbott, 1995]. Однако C.canadensis --
диплоид, в то время как другие виды являются аллополиплоидами, и авторы,
основываясь на морфологических и электрофоретических данных, предпола
гают, что эти 3 таксона генетически обособились намного ранее их интродук
ции в Европе [Thebaud, Abbott, 1995].
В Англии обнаружены два экотипа: при выращивании семян в условиях пи
томника первый (типичный) представляет собой растение с высоким (до 1 м)
стеблем и метельчатым соцветием; второй экотип, встречающийся на морских
побережьях, -- низкие (10--20 см) растения подушкообразной формы с много
численными боковыми побегами, идущими от основания стебля [Boyce, 1954].
Сравнительный анализ фенотипической изменчивости 16 количественных
признаков (высота растения, размеры листьев, длина соцветия и т.д.) 14 спон
танных популяций мелколепестника в Германии и на Украине выявил зависи
мость изученных признаков от экологических условий и в первую очередь от
степени антропогенной нарушенности территории [Тохтарь, 2002].
Анализ изменчивости биологических признаков во вторичном ареале, про
веденный с использованием метода создания интродукционных популяций
[Виноградова, 2005], показал, что в ходе натурализации у C.canadensis выра
ботались и генетически закрепились важнейшие адаптивные признаки: менее
продолжительный период роста и более быстрое прохождение полного цикла
развития по направлению с юга на север, что соответствует изменениям кли
матических условий вдоль широтного профиля. В комплексе климатических
факторов наибольшее влияние на ритм роста и развития C.canadensis оказы
вают температурные условия. Сумма эффективных температур, необходимая
для наступления соответствующей фенофазы, тем выше, чем южнее произра
стает популяция. Недостаточная сумма эффективных температур, накоплен
ная за вегетационный сезон на широте Москвы, не позволяет южным образ
цам мелколепестника пройти полный цикл развития. Мелколепестник канад
ский -- растение длинного дня, и чем далее к северу обитает его популяция,
тем более продолжительный период освещения требуюется входящим в нее
особям для полного прохождения жизненного цикла. Растения волгоградско
159
го и московского происхождения при коротком дне не способны к образова
нию цветоносного побега и вегетируют в виде розеток, а у более южных образ
цов наблюдаются морфологические уклонения: растения образуют сильно
рассеченную листовую пластинку, искривленный стебель и видоизмененное
соцветие с малочисленными крупными корзинками. У растений, произраста
ющих на южном пределе вторичного ареала (в Центральной Азии), генети
чески закреплено свойство ярового типа развития. Остальным образцам для
вступления в генеративный период развития требуется стратификация семян
(тогда они растут как однолетники). В противном случае наблюдается двух
летний цикл развития.
Экономическое и социальное (положительное/отрицательное) значе
ние. C.canadensis -- сорняк более 40 культур. В Северной Америке он навод
няет сады, виноградники, сенокосы, пастбища, посевы зерновых культур, сои
и хлопка. Этот вид -- обычный сорняк моркови и лука, особенно при отсут
ствии севооборота.
C.canadensis является растением -- хозяином Lygus lineolaris, который при
чиняет ущерб многим зерновым культурам. Это насекомое откладывает яйца
на цветках мелколепестника и кормится прежде всего плодами и частями цвет
ка, но также и листьями. C.canadensis также промежуточный хозяин
Adelphocoris lineolatus -- вредителя люцерны в Канаде, и микоплазменной бо
лезни, передающейся широкому диапазону растений -- хозяев вредителем
Macrosteles fascifrons Stal.
Имеются данные, что листья C. сanadensis могут вызывать у некоторых
людей раздражение кожи, а также раздражают ноздри лошадей при непосред
ственном контакте [Weaver, 2001].
С другой стороны, C.canadensis используется как лекарственное растение.
Масло мелколепестника, дистиллированное из листьев, продавалось в неко
торых штатах США как диуретик, тонизирующее и вяжущее средство. Масло
содержит 47 компонентов, >91% из них являются терпеноидами. Водный эк
стракт листьев используют как антигипертензивное средство [Weaver, 2001].
В растении содержатся танины, наибольшее количество которых найдено в
листьях (в период вегетации 0,2%, в период плодоношения 0,3%) и в цветках
(0,3%). Листья содержат целый ряд стеролов. В фазу цветения в свежем сырье
мелколепестника содержится до 1,08--1,45% эфирного масла [Kostecka
Madalska, Borowczak Niemczewska, 1968; Dubey, Gupta, 1988; Kostecka
Madalska, Polanowski, 1964].
В зависимости от географического местоположения популяции варь
ирует количественный выход эфирного масла, выделяемого из мелколепест
ника канадского и используемого в парфюмерной промышленности. Варьи
рует и качественный состав масла, основной компонент масла -- лимонен, его
содержание в эфирном масле достигает 83%.
В России использование C.canadensis в качестве лекарственного растения до
вольно ограничено. В старину он применялся "для ращения волос", в народной
медицине используется редко, в основном как кровоостанавливающее и закреп
ляющее средство. В связи с этим в Главном ботаническом саду проведен анализ
изменчивости содержания фенольных соединений и кремния в интродукцион
ной популяции C.canadensis, сформированной из семян, собранных в 25 различ
ных точках вторичного ареала вида [Виноградова, Колесников, 2007].
160
По содержанию фенольных соединений (3%) мелколепестник канадский
не уступает ряду широко распространенных лекарственных растений, 21 вид
из которых исследовался по аналогичной методике, а по содержанию апиге
нина во флавоноидном комплексе (1,03%) даже превосходит их. Закономер
ностей изменчивости содержания фенольных соединений от широты геогра
фического пункта сбора семян не отмечено.
В стеблях содержится значительно меньше полифенолов, флавоноидов и
оксикоричных кислот, чем в листьях и соцветиях, и при этом преобладают
конденсированные полифенолы. В связи с этим для лекарственных целей сле
дует брать не всю надземную часть растения, а только листья и соцветия.
Общее содержание кремния в растениях мелколепестника достаточно вы
сокое -- 1,23% абсолютно сухой массы. Амплитуда как внутрипопуляционной,
так и межпопуляционной изменчивости содержания различных форм крем
ния в интродукционной популяции очень низкая.
В соцветиях С.canadensis преобладает кремний органический, тогда как в
стеблях его количество в 2 раза ниже за счет повышения содержания поли
мерного и минерального кремния.
Анализ показывает, что многие лекарственные растения, обогащенные фла
воноидами, одновременно являются и кремниефильными. Возможно, их фар
макологическая значимость обусловлена как раз удачным сочетанием флаво
ноидого комплекса с органогенным кремнием, который также способствует
укреплению капилляров. И хотя по общему содержанию кремния (1,2%)
С.canadensis немного уступает другим видам, сочетание высокой доли веществ
флавоноидого комплекса и органического кремния позволяет рассматривать
этот вид в качестве перспективного лекарственного растения.
Предложено также включить C. canadensis в перечень полезных растений,
которые могут служить индикаторами содержания в почвах кадмия и хрома.
Обнаружено, что растения C. canadensis при произрастании на полевых участ
ках с высоким содержанием тяжелых металлов поглощают существенное ко
личество кадмия без симптомов повреждения. В тканях листьев и стеблей со
держалось до 47 мг /кг кадмия через 5 лет после того, как на поверхность уча
стка было вылито 11,6 кг/га кадмия в растворимой форме [Weaver, 2001].
Меры борьбы
Численность розеток C.canadensis можно эффективно контролировать не
специфическими гербицидами, содержащими глифосат или глюфосинат ам
мония. Применение параквата идет с переменным успехом: при дозе 300--
600 г/га растения повреждаются, но часто восстанавливаются [Bruce, Kells,
1990]. Применение гербицидов лучше проводить осенью или рано весной до
начала удлинения стебля, когда мелкие розетки активно растут.
В популяциях C.canadensis, растущих в посевах сои, наблюдалась изменчи
вость восприимчивости отдельных особей к ряду гербицидов (линурон, има
заквин, имазетапир и метрибузин), причем в большинстве случаев примене
ние этих препаратов результатов не дало [Bruce, Kells, 1990; Moseley, Hagood,
1990]. Имеются данные [Weaver, 2001], что на C.canadensis в посевах моркови
не действует линурон, а в посевах лука не действует оксифлуорфен, однако в
посевах озимого ячменя применение бромоксинила привело к сокращению его
популяции. В посевах зерновых культур для уничтожения C.canadensis при
меняют гербициды, содержащие дикамб. Имеются сообщения [Wiese et
161
al.,1995] об успешном уничтожении летом растений 30 см высотой атразином
с дикамба, хлорсульфуроном, метсульфуроном и тифенсульфуроном. На план
тациях сосны [South, 2000] C.canadensis можно контролировать путем приме
нения клопиралида.
К сожалению, применение химических способов борьбы с этим сорняком
привели к появлению ряда экотипов, устойчивых к тому или иному гербици
ду. Популяции C. canadensis выработали устойчивость к различным гербици
дам более чем в 10 странах. Устойчивые к параквату популяции отмечены в
нескольких садах в графстве Эссекс, в Онтарио, а также в Венгрии, Японии и
США. Атразин устойчивые популяции найдены в Великобритании, Франции,
Бельгии, Швейцарии, Польше, Чехословакии и Испании. Популяции
C.canadensis, стойкие к гербицидам, которые ингибируют синтез ацетолакта
та, отмечены в полях в штате Огайо и по железным дорогам в Польше. Попу
ляции, устойчивые к глифосату, найдены в штате Делавэр [Smisek et al., 1998;
Weaver, 2001]. Несколько популяций C.canadensis выработали устойчивость
сразу к ряду гербицидов с различными способами воздействия. Популяции,
стойкие и к параквату, и к атразину обнаружены в Венгрии. Популяции, ус
тойчивые и к атразину, и к хлорсульфурону, найдены в Израиле [Lehoczki et
al.,1984; Weaver, 2001].
Обзор научных исследований и публикаций
D.Regehr и F.Bazzaz [1976] исследовали фотосинтетическое поведение 5
озимых однолетних растений, включая C. canadensis. Они показали, что зиму
ющие розетки способны к существенной фиксации углерода и сохранению
энергии при низких температурах. При высокой солнечной радиации темпе
ратура розеточных листьев часто была на 10°C выше окружающей температу
ры. Оптимальная температура воздуха для фотосинтеза варьирует от 28°C
летом до 15°C зимой.
СЕМЕЙСТВО ASTERACEAE (COMPOSITAE) --
СЛОЖНОЦВЕТНЫЕ
ЦИКЛАХЕНА ДУРНИШНИКОЛИСТНАЯ
Название: Cyclachaena xanthiifolia (Nutt.) Fresen
Синонимы: Iva xanthiifolia Nuttall
Морфологические признаки
Однолетнее растение высотой от 30 до 200 см. Стебель прямостоячий, реб
ристый. Листья супротивные, черешковые, сердцевидно яйцевидные, по кра
ям пильчато зубчатые, сверху шероховатые от коротких щетинистых волос
ков, снизу сероватые от густого шелковисто войлочного опушения (рис. 62).
Корзинки многочисленные, поникающие, собраны в общее рыхлое метельча
тое соцветие. Краевые женские цветки в корзинке немногочисленные, с почти
редуцированным околоцветником; срединные --- обоеполые, трубчато коло
кольчатые, зеленовато белые, обычно бесплодные. Плоды --- темно коричне
вые семянки без хохолка [Губанов и др., 2004].
162
Естественный ареал
Вид природной флоры североамериканских прерий [Strother, 2006].
Вторичный ареал
История интродукции и географическое распространение. Явно адвен
тивный вид к востоку от Миссисипи и в западных штатах. Отмечен в Кали
форнии как засоритель производственных посевов моркови [Strother, 2006].
В Нью Йорке --- редкий заносный вид [New York Metropolitain..., 2008].
Занесен на все континенты, спорадически встречается в Средней и Атлан
тической Европе, Средиземноморье, на Кавказе, в Средней Азии и Сибири, на
Дальнем Востоке, в Японии, Китае, Южной Америке и Австралии [Куприя
нов, 1989; Красноборов, 2000; Ломоносова, Зыкова, 2003]. В Европе встреча
емость и обилие вида убывает с востока на запад. В Польше C.хanthiifolia
появилась в рудеральных экотопах во второй половине XIX века, распростра
нившись из ботанических садов [Alien species..., 2008]. Сейчас местонахожде
ний циклахены в Польше свыше 1 тыс., это массовый вид, образующий круп
ные заросли. C.xanthiifolia внесена в список видов, «наиболее инвазионных на
территории Европы и представляющих потенциальную опасность для Испа
нии» [Dana et al., 2008].
В 1870 х гг. (по другим источникам --- в начале XIX века) циклахену выра
стили в Киевском ботаническом саду, откуда в 1920 х гг. она распространи
лась по Украине, а затем по России. В MHA хранится сбор C.xanthiifolia, сде
ланный Н. Цингером «близ г. Киева, сорное по пустырям (в долине р. Лыбе
ди) близ города, 17.08. 1903 г.». Экземпляр, собранный в тот же день в Киеве,
хранится и в МГУ, причем на этикетке имеется любопытная приписка: «севе
роамериканское растение, по сведениям Роговича (Обозрен. раст. Киевск.
учебн. окр., 1869, с. 158) в 1842 г. распространилось из Ботанического сада
университета Св. Владимира и одичало на сорных местах в Киеве и его окрес
тностях. В настоящее время в ботаническом саду оно не растет, но на окраинах
Киева в качестве сорного, по видимому, прочно прижилось и в изобилии встре
чается в палисадниках, огородах, на пустырях и т.п., иногда покрывая сплошь
значительные пространства и образуя местами заросли значительно выше че
ловеческого роста. Н.Цингер». В гербарии МГУ (MW) имеется еще несколь
ко сборов из Киева и Киевской губернии --- 1885, 1887, 1927 и 1931 гг.
В Волгоградской обл. циклахена натурализовалась на сегетальных и руде
ральных местах [Сагалаев, Матвеев, 2000]. В Астраханской обл. обнаружена
во второй половине ХХ века, а к настоящему времени входит в ядро рудераль
ной флоры региона [Афанасьев, 2008]. Вид активно расселяется по террито
рии Республики Башкортостан, куда внедрилась с юга, из прилегающих райо
нов Оренбургской обл., где C.xanthiifolia зафиксирована значительно ранее
[Ануфриев, 2008]. В последние годы C.xanthiifolia распространяется в Запад
ной Сибири и на Алтае [Красноборов, 2000; Ломоносова, Сухоруков, 2000]. В
2002 г. циклахена впервые собрана в Новосибирске [Ломоносова, Зыкова,
2003]. В 2002 г. вид обнаружен в Иркутске на ж/д насыпи [Зарубин, 2008]. В
Приамурье впервые обнаружена в 2001 г. [Аистова, 2007].
Циклахена до сих пор остается относительно редким видом северных реги
онов России. Собрана в Заполярье: Мурманская обл., Кандалакшский р н, 3
км южнее ст. Ковда, на ж/д полотне, 26.08.1993 г., собр. Д.Д. Соколов, опр. С.Р.
Майоров (MW). В 2004 г. найдена в окрестностях Вологды, рядом с птице
Рис. 62. Вегетирующие растения Cyclachaena xanthiifolia
Рис. 63. Расселение Cyclachaena xanthiifolia в Средней России
64
Рис. 64 а. Внедрение Cyclachaena xanthiifolia в луговые сообщества
Рис. 64б. Cyclachaena xanthiifolia в посевах Рис. 65. Цветение Cyclachaena xanthiifolia
165
фабрикой, в массе, а в 2007 г. --- на железной дороге в Вологодской обл. в цве
тущем состоянии. В Ленинградской, Псковской, Новгородской обл.
C.xanthiifolia встречается редко, обычно не плодоносит [Цвелев, 2000]. Вид
внесен в информационную систему «Чужеродные виды Северо Запада Рос
сии» [2008] как инвазионный ксенофит.
Пути и способы заноса. Первичный источник заноса C.xanthiifolia в Сред
нюю Россию --- скорее всего, коллекции ботанических садов (см. выше). Кро
ме того, немаловажное значение в распространении вида имеют загрязненные
грузы, перемещаемые по железной дороге. Сходным образом вид завезен и
распространился в Европе. Например, в Болгарию циклахена попала предпо
ложительно с семенами кукурузы в период с 1994 по 1996 г. [Milanova, 2001].
Статус в регионе. В пределах Средней России встречаемость, разнообра
зие занимаемых экотопов и обилие вида закономерно уменьшаются с юга на
север (рис. 63). Первоначально C.xanthiifolia отмечена в Курской и Воронежс
кой обл. Ныне в черноземной полосе циклахена обычна и проникает севернее
по нарушенным местам, главным образом, вдоль шоссейных и ж/д путей
[Губанов и др., 2004].
В Белгородской обл. --- широко распространившийся заносный вид [Еле
невский и др., 2004]. В MW имеется сбор, произведенный в 1958 г. С.В. Голи
цыным в г. Новый Оскол, во дворе гостиницы.
В Воронежской обл. впервые найдена в окрестностях Воронежа на ж/д валу
в 1926---1928 гг. [Гроссет, Замятнин, 1935]. Один из первых гербарных сборов
сделан С.В. Голицыным в 1939 г. в Воронеже: «ж/д пути, по колее, группами»
(MW). Однако основной этап расселения вида пришелся на послевоенное вре
мя. В сводках XIX --- начала ХХ века для территории Курской обл. циклахена
не указана. Первые достоверные сборы известны с территории Стрелецкого
участка Центрально Черноземного заповедника: «Дедов лес, близ кордона,
1949 г., Г.П. Чувилин» [ЦЧЗ; Левицкий, 1957]. Сейчас произрастает во всех
районах Курской обл., часто массово [Полуянов, 2005].
В Липецкой обл. в 1967 и 1989 гг. собрана В.Н. Тихомировым и коллегами
в Задонском р не, окрестности с. Донское, на территории заповедника «Гали
чья гора» (MW). Ныне встречается по всей территории, в массе, часто образу
ет чистые заросли [Флора Липецкой области, 1996; Н.Ю. Хлызова, личное
сообщение]. История расселения вида в средней полосе России подробно рас
смотрена Н.А. Вьюковой [1987].
В Орловской обл. C.xanthiifolia собрана В.И. Радыгиной в Ливенском р не,
на берегу р. Сосна близ с. Успенское, 28.08.1983 г. (OHHI). По нашим наблю
дениям 2008 г., на юго востоке Орловской обл., циклахена, являясь массовым
видом, образует одновидовые заросли.
На границе Тульской обл. C.xanthiifolia собрана в 1979 г., на дамбе в пойме
Оки у шоссе Москва--Тула, 2.08.1979 г. [Ю.Алексеев, MW; Алексеев, Губанов,
1980]. По оценке вышеуказанных авторов, на тот период времени циклахена
была еще редким видом в приокских районах. Период ее массового распрост
ранения пришелся на 1980 е гг. Впервые в пределах Тульской обл. C.xanthiifolia
обнаружена И. Шереметьевой в 1987 г. в Туле, на пустыре у Московского вок
зала, 10.09.1987 г. (MW, ТГПУ). Затем последовали многочисленные находки
вблизи путей сообщения в разных районах области, а к концу 1980 х гг. отме
чено образование крупных одновидовых зарослей в Плавском и Ефремовс
166
ком р нах на пустырях вдали от ж/д путей [Хорун и др., 1995]. Сейчас --- впол
не обычный вид на территории области, однако роль ж/д мест обитания сни
жается, что связано с обработкой их гербицидами.
В Московской обл. C.xanthiifolia впервые обнаружена Т.Асеевой на ст. Вялки
Казанской железной дороги (MHA [этого сбора мы не обнаружили]), а затем
в 1969 г. А.К. Скворцовым на ст. Ржевская в Москве [1973б]. В 1957 г. в Моск
ве, на обочине Дмитровского шоссе, циклахену собрал В.Вехов (MW). В 1970
х гг. циклахена обнаружена еще около 10 раз, но то были находки единичных
растений на ж/д путях и на сорных местах в Москве. С 1980 х гг. встречае
мость циклахены значительно повысилась [Игнатов и др., 1990].
Для Владимирской обл., по данным И. В.Вахромеева [2002], имеется доре
волюционный сбор из г. Муром. Ряд современных сборов с территории Му
ромского, Гусь Хрустального, Ковровского, Суздальского, Собинского, Пету
шинского р нов сделан А.Серегиным в 2001--2006 гг. (MW) на ж/д полотне,
свалках, обочинах шоссейных дорог. По личному сообщению А.П. Серегина,
C.xanthiifolia в одном и том же месте несколько лет подряд не удерживается. В
начале 2000 х гг. этот вид в области встречался чаще, чем амброзия, теперь реже.
Для Тверской обл. впервые указана Ю.Д. Гусевым в 1969 г. на ст. Бологое
[LE; Гусев, 1973].
В Ивановской обл. C.xanthiifolia впервые отмечена в 1984 г. на ж/д насыпи
в г. Тейково (ИУ). В настоящее время обычно встречается одиночными экзем
плярами или небольшими группами, крупных зарослей не образует [MW, ИУ;
Шилов и др., 1989; Борисова, 2003б].
Вид указан для Смоленской обл. [MW; Решетникова, 2004] -- сбор из Гага
ринского р на Смоленской обл.: в 2 км западнее ж/д ст. Батюшково, по ж/д
полотну, 8.07.2001 г., А.Сухоруков (MW). В MHA и MW хранятся сборы 1975 г.
Ю.Е. Алексеева и В.В. Макарова из пос. Комаричи Брянской обл.: «обнаруже
но несколько хорошо развитых растений рядом друг с другом на привокзаль
ной территории. В самом поселке не замечено». В 1974 г. циклахену собирали
близ ж/д ст. Бочеп (П.З. Босек, MW).
В Калужской обл. впервые собрана в 1975 г. в Калуге на берегу р. Ока око
ло города, мусорное место. А.К. Скворцов (МНА). Растение встречается ред
ко, отдельными экземплярами (С.Майоров, личное сообщение).
В Рязанской обл. впервые собрана в 1983 г. (Васильев, РГУ), в настоящее
время распространена неравномерно: в Милославском, Новодеревенском,
Сараевском р нах нередко образует заросли в населенных пунктах, тогда как
в Рязанском и Спасском р нах встречается редко [Казакова, 2004]. В Мордо
вии циклахену собирали на территории Старошайговского р на, севернее с.
Богдановка, по обочине шоссе, 20.08.1980 г., Т.Силаева (MW) и в г. Саранск,
на ж/д насыпи 15.09.1990, С.Майоров (MW).
В Тамбовской обл. C.xanthiifolia впервые собрана в Мичуринском р не как
сорное на территории пос. Учхоза ТСХА им. Калинина (у Мановиц), по забо
рам, часто, 17.09.1966 г., В.В. Макаров (MHA).
Местообитания. В пределах естественного ареала встречается на забро
шенных полях, в понижениях рельефа, на затапливаемых пойменных землях
по берегам рек и других водоемов [Strother, 2006 ]. В Средней России растет в
населенных пунктах, на насыпях шоссейных дорог и ж/д полотна, на пусты
рях и залежах (рис. 64 а или б), в огородах, посевах, по улицам и сорным мес
167
там, свалкам, разного рода техногенным территориям, берегам водоемов [Ва
сюков, 2004; Еленевский, Радыгина, 1997; Еленевский и др., 2004; Казакова,
2004; Маркелова, 2004; Полуянов, 2005; Флора Липецкой области, 1996; Хо
рун, 1999; Григорьевская, 2000; Григорьевская и др., 2004].
Размножение и жизненный цикл. В южных регионах цветет в июле --- ок
тябре, семянки созревают в августе --- октябре [Губанов и др., 2004]. Севернее
(например, в Ивановской обл.) только цветет, но не успевает продуцировать
зрелые плоды [Борисова, 2007].
Изучению биологии циклахены в условиях вторичного ареала посвящено
значительное число работ. В 2004--2005 гг. Е.В. Аистовой определена всхо
жесть семян C.xanthiifolia в Амурской обл. Семена проращивали в чашках
Петри в двух температурных режимах --- 19 и 25°С. При 19°С семена прораста
ли через 7--10 дней, при 25°С прорастание не наблюдалось. При посеве семян
на поверхности почвы прорастание происходило быстрее, чем в чашках Пет
ри [Аистова, 2007]. В Республике Башкортостан определена общая семенная
продуктивность циклахены --- в среднем 5378 семянок на 1 растение. В пере
счете на 1 га этот показатель составляет 9,4 млрд семянок, что определяет до
минирование C.xanthiifolia в сообществах [Ануфриев, 2008].
Расселение и распространение. Будучи занесена на рудеральные участ
ки в урбаноэкосистемы, C.xanthiifolia активно распространяется на близлежа
щие культивируемые земли, заброшенные поля, берега рек и т.п. Это проис
ходит посредством антропо и зоохории, а также с помощью органических удоб
рений и ирригационных вод. Быстрое распространение C.xanthiifolia
обеспечивается благодаря ее высокой семенной продуктивности и конкурент
ной способности, а также некоторому аллелопатическому эффекту [Valkova,
Maneva, 2008]. При изучении спонтанного распространения в Центральной
Европе 3 сорных видов, в том числе циклахены, выявлено, что в первую оче
редь C.xanthiifolia захватывает нарушенные участки в естественной раститель
ности и открытые территории в антропогенных экотопах [Eliбљ, 1993]. Пер
воначально вид натурализуется в нескольких локальных центрах и формиру
ет богатый банк семян, что способствует быстрой последующей его экспансии
на новые территории.
Последствия внедрения
Воздействие на естественные фитоценозы и аборигенные виды. Цик
лахена формирует растительные группировки на начальных стадиях сукцес
сионного процесса, проходящих на нарушенных или незадерненных участках
(рис. 65). Особенно страдают техногенные территории, например отвалы уголь
ных шахт Донбасса [Жуков, 2003]. При сравнении локальных флор техноген
ных территорий Европы выявлено, что C.xanthiifolia распространяется на тех
ногенных территориях всех изученных областей России и Украины, однако в
Средней и Западной Европе этот процесс выражен значительно слабее [Тох
тарь и др., 2003].
В Беларуси C.xanthiifolia активно проникает на территории национальных
парков, в том числе и в Беловежскую Пущу [Джус, 2008]. На юге Самарской
обл. в результате неумеренного выпаса скота травяной покров побережий силь
но нарушился, в нем господствуют растительные группировки с доминирова
нием сорно рудеральных видов, в том числе циклахены [Петрова, Легоньких,
2008]. Аналогичные наблюдения сделаны при обследовании парковой расти
168
тельности г. Воронеж, включающей группировки с участием C.xanthiifolia [Гри
горьевская и др., 2002]. В Башкирском Предуралье циклахена встречается в
самых разнообразных фитоценозах --- от сильно нарушенных рудеральных
сообществ первой стадии восстановительной сукцессии до практически нена
рушенных человеком пойменных сообществ, и от сырых пойм с избыточным
увлажнением до сухих степей [Ануфриев, 2008]. Являясь агрессивным, высо
ко конкурентоспособным сорным растением, C.xanthiifolia при внедрении в
сообщества быстро становится доминантом с долей участия вида от 71 до 99%,
формируя большое число побегов (от 79 до 564 шт./м2) и надземную биомассу
(от 1,5 до 8,5 кг/ м2).
Болгарские исследователи [Valkova, Maneva, 2008] определяли конкурент
ную способность C.xanthiifolia в сравнении с другимии сорными видами:
Conium maculatum L., Xanthium strumarium L., Chenopodium album L., Sinapis
arvensis L. Показано, что C.xanthiifolia оказывает крайне негативное влияние
на рост и развитие Conium maculatum. Напротив, Xanthium strumarium по кон
курентной способности превосходит C.xanthiifolia. Присутствие Xanthium
strumarium и Sinapis arvensis уменьшает конкурентную способность
C.xanthiifolia, а присутствие Conium maculatum и Chenopodium album, наобо
рот, ее увеличивают.
Влияние на здоровье человека. C.xanthiifolia -- весьма аллергенный вид,
причина массовых поллинозов, в том числе с тяжелыми астматическими про
явлениями. В городах с большой концентрацией вредных производств такой
дополнительный неблагоприятный фактор может нанести серьезный урон
здоровью населения [Ануфриев, 2008]. Пыльца C.xanthiifolia находит приме
нение в медицине для диагностики аллергических заболеваний; ее этиологи
ческая роль подтверждена положительной реакцией у 38% больных поллино
зом при отрицательных результатах с другими видами аллергенов пыльцево
го происхождения [Андриеш, 2008].
Экономическое и социальное значение (положительное и отрицатель
ное). C.xanthiifolia -- злостный сорняк посевов подсолнечника, кукурузы, ово
щей, сахарной свеклы [Valkova, Maneva, 2008]. Циклахену не поедает скот, и
она вытесняет из растительных сообществ местные рудеральные виды.
Меры борьбы
C.xanthiifoliа является карантинным сорняком, и к ней применим перечень
превентивных мер, предусмотренных инструкцией карантинных инспекций
[Аистова, 2007, Ануфриев, 2008]:
1) карантинный фитосанитарный контроль импортируемых грузов и транс
портных средств;
2) мониторинг популяций, изучение биологических особенностей вида в
новых условиях обитания, разработка мер по сокращению его численности с
использованием химических, агротехнических и карантинных мероприятий;
3) обследование территорий с целью выявления новых местообитаний цик
лахены и очагов ее распространения, нуждающихся в контроле;
4) принятие мер по ликвидации первичных очагов C.xanthiifolia.
В 2004--2006 гг. на территории 4 районов Республики Башкортостан про
водились полевые опыты по определению эффективности использования для
борьбы с циклахеной гербицидов «Луварам», «Чисталан», «Торнадо», «Раун
дап», «Ураган» [Ануфриев, 2008]. Гербициды оказались достаточно эффектив
169
ными: через месяц доля погибших растений составила от 84 до 100%, а боль
шая часть выживших растений была повреждена. Эффект сохранялся до кон
ца вегетационного периода, повторного отрастания не наблюдалось. По срав
нению с июлем при обработке участка в августе процент гибели C.xanthiifolia
несколько снизился, что, по мнению автора, связано с тем, что к середине ав
густа растения значительно увеличиваются в размерах, прирост по высоте со
ставляет ~50%, а биомассы 30%, и норма внесения гербицида становится не
достаточной. В последующие годы наблюдается постепенное восстановление
популяций из семян, попадающих с соседних необработанных участков. Со
гласно выводам О.Ануфриева, это свидетельствует о хорошей восстановитель
ной способности растений и недостаточности однократного использования
гербицидов. В работе высоко оценивается эффективность подкашивания рас
тений в фазе бутонизации, указано на необходимость многократного ежегод
ного применения обоих методов [Ануфриев, 2008].
СЕМЕЙСТВО ASTERACEAE (COMPOSITAE) --
СЛОЖНОЦВЕТНЫЕ
МЕЛКОЛЕПЕСТНИК ОДНОЛЕТНИЙ
Название: Erigeron annuus (L.) Pers.
Синонимы: Erigeron strigosus Mьehl. ex Willd. var. septentrionalis (Fernald et
Wiegand) Frey, Baltisb. et Edwards, Phalacroloma annuum (L.) Dumort.,
Ph. septentrionale (Fernald et Wiegand) Tzvelev, Stenactis annua (L.) Less., S.
septentrionalis (Fernald et Wiegand) Holub.
Отнесение Erigeron strigosus Mьehl. ex Willd. var. septentrionalis в синонимы
мелколепестника однолетнего требует комментариев. Секция Phalacroloma
включает два вида -- E. annuus и E. strigosus [Nesom, 1989]1. Это однолетние
мелколепестники с относительно крупными язычковыми цветками и семян
ками с редуцированным внешним рядом щетинок хохолка [Цвелев, 1994; Noyes,
Allison, 2005]. В типичных формах E. annuus и E. strigosus хорошо отличаются
друг от друга [Illustraited..., 1998]. Разновидность var. septentrionalis занимает
среднее положение между двумя видами: тип опушения у нее как у E. annuus, а
характер листьев приближается к E. strigosus, и ее иногда выделяют в особый
вид [Цвелев, 1994; Cronquist, 1947]. Эта разновидность первоначально описа
на в составе E. ramosus (Walter) Britton, Sterns et Poggenb. (этот вид в настоя
щее время включают в состав E. strigosus [Nesom, 2006]), а затем перенесена в
состав E. strigosus (Fertnald, Wiegand, 1913; Fernald, 1942; Frey, 2003]. После
дняя комбинация принята во "Flora of North America" [Nesom, 2006]. Такое ре
шение заметно затрудняет разграничение E. annuus и E. strigosus, и, возможно,
потому в ряде флористических работ принято объединение этих видов в один
[например, Clement, Foster, 1994; Hаeupler, Muer, 2000, 2007]. Наконец, в 1965 г.
разновидность перенесена в состав E. annuus -- var. septentrionalis (Fernald et
Wiegand) Frey, Baltisb. et Edwards [Frey, 2003].
1 По видимому, в эту секцию следует включать E. tenuis Torrey et A. Gray и, вероятно,
E.geiseri Shinners [Noyes, Allison, 2005; Noyes, 2000b, 2006a]. Оба вида за пределами
Северной Америки неизвестны.
170
Накопились данные, поддерживающие такое решение:
-- E. annuus -- триплоидный (2n = 3x =27) апомиктический вид, E. strigosus
диплоидный (2n = 2x = 18) амфимиктический [Noyes, Rieseberg, 2000; Noyes,
Allison, 2005; Noyes, 2006a];
-- европейские и североамериканские растения var. septentrionalis -- трип
лодиды, 2n = 27 [Frey, 2003; Frey et al., 2003];
-- типовая разновидность и var. septentrionalis, без сомнения, имеют единый
геном [Edwards et al., 2006].
Мы принимаем точку зрения Д.Фрая и его коллег [Frey, 2003; Frey et al.,
2003] о включении var. septentrionalis в состав E. annuus.
E. strigosus s. str. отличается от E. annuus однородным опушением стебля
короткими прижатыми простыми волосками [Frey, 2003] и в средней полосе
Европейской России является редким заносным видом, известным лишь по
сборам 1968 и 1970 годов из Клепиковского и Касимовского р нов Рязанской
обл. (MW).
Морфологические признаки
Однолетник, яровой или озимый, высотой (30)50--100 см, с тонким кор
нем. Стебель прямой, в нижней части щитенисто опушенный отстоящими под
разными углами простыми волосками разной длины, часто также и коротки
ми прижатыми вниз простыми волосками, в соцветии опушение короче и вверх
прижатое. Листья разнообразной формы, от овальных до линейно ланцетных,
постепенно уменьшающиеся кверху, у var. annuus край листа зубчатый, ниж
ние листья черешковые, пластинка шириной до 20--40 мм, у var. septentrionalis
край листа от сглаженно зубчатого до цельного, пластинки в целом более уз
кие. Корзинки многочисленные, в рыхлом метельчатом соцветии (рис. 66).
Обертки чашевидные, диаметром 6--12 мм, листочки обертки многочисленные, в
2--3(--4) ряда, почти линейные, остроконечные, по краю пленчатые, голые или
по средней линии с длинными согнутыми многоклеточными волосками. Кра
евые цветки язычковые, чаще белые, но иногда сиреневые или бледно фиоле
товые, многочисленные, длиной 4--10 мм. Цветки диска трубчатые, желтые
или желто оранжевые. Семянки длиной 0,8--1 мм, бледные, хохолок в 2--2,5
раза длиннее семянки, из 15--25 белых щетинок, часто неравной длины.
Естественный ареал
По видимому, охватывает северные районы США и прилегающие терри
тории Канады; однако в настоящее время E. annuus широко распространен
практически по всей Северной Америке [Frey et al., 2003; Nesom, 2006].
Вторичный ареал
История интродукции и географическое распространение. E. annuus
интродуцирован в Европу как декоративное растение в 1635 г. в ботанический
сад в Париже, откуда он введен в другие ботанические сады [Edwards et al.,
2006]. На территории Швейцарии одичавшие растения впервые собраны в 1828 г.
[Frey, 2003]. В Чехии известен с 1884 г. [Pyљek et al., 2002]. По видимому, ак
тивное расселение мелколепестника в Европе началось в XX веке. Первые
указания для Австрии относятся к 1950--1960 м гг. [Essl, Rabitsch, 2002]. Для
Польши указывается лишь с конца XX века [Chmura, 2004]. В Испании E.
annuus отнесен к видам на начальной стадии инвазии [Sanz Elorza et al., 2001].
В Италии и Венгрии отнесен к инвазионным видам, в Швейцарии натурали
171
Рис. 66. Цветение Erigeron annuus
Рис. 67. Расселение Erigeron annuus в Средней России
172
Рис. 68. Внедрение Erigeron annuus
в луговые сообщества
Рис. 69. Корзинки Erigeron annuus
173
зовался [Invasive alien species in Hungary, 2003; Invasive alien species in
Switzerland, 2006]. Известен почти во всех странах Европы, в 27 регионах на
турализовался [Lambdon et al., 2008].
Вне Европы E. annuus указан для Японии [Mito, Uesugi, 2004], Кореи [Kil
et al., 2004], Новой Зеландии, где известен с 1929 г. [Given, 1984]. Обнару
жен на юге Дальнего Востока [Баркалов, 1992], но не отмечен для Сибири
[Королюк, 1997].
Пути и способы заноса. Первоначально E. annuus ввезен в Европу как де
коративное растение, однако позднейшее расселение, вероятно, непосредствен
но не связано с деятельностью человека как агента расселения. Для мелколе
пестника характерна анемохория [Edwards et al., 2006].
Статус в регионе. В Средней России E. annuus впервые обнаружен в окре
стностях Москвы: «Больш. Мытищи, на лугу близ жилья, 8.07.1902 г., 3288,
Д.Сырейщиков» (MW). Однако массовые сборы появляются заметно позднее,
во второй половине XX века (MW, MHA). К настоящему времени отмечен
практически во всех областях Средней России (рис. 67).
Местообитания. В пределах естественного ареала E. annuus обитает пре
имущественно по открытым нарушенным местообитаниям, особенно часто по
обочинам дорог, полям, пустошам [Nesom, 2006]. На западе США E. annuus --
один из пионерных видов залежей [Hancock, Wilson, 1976]. При зарастании
залежей мелколепестник удерживается дольше других однолетних сорных
видов [Bazzaz, 1996]. В Европе этот вид обитает в сходных условиях [Frey,
2003]. Спектр растительных ассоциаций, в которых обитает вид в Централь
ной Европе, довольно широк [Gregor, 2004]. Показано угнетающее аллелопа
тическое воздействие вытяжки из корней сосны на проростки E. annuus
[Allelopathy..., 1992]. По нашим наблюдениям, наибольшего обилия E. annuus
достигает на легких почвах (рис. 68), часто встречается на мелкоземе в овра
гах, карьерах или у подножия холмов.
Размножение и жизненный цикл. Традиционно считалось, что виды мел
колепестников секции Phalacroloma являются облигатно апомиктическими
[Cronquist, 1947]. Специальные исследования подтвердили, что E. annuus яв
ляется триплоидным (2n = 3x = 27) апомиктическим видом [Noyes, Rieseberg,
2000; Noyes, 2006a; Noyes et al., 2006]. Образование полиплоидности связыва
ют с нарушениями при мейозе с образованием нередуцированных гамет [Noyes,
2005, 2006a; Noyes et al., 2007]. Молекулярно генетические данные отвергают
облигатный апомиксис у E. annuus: генетический полиморфизм вида невоз
можно объяснить без предположения об изредка происходящем перекрестном
опылении. При этом в европейских популяциях амфимиксис происходит, ве
роятно, чаще, чем в североамериканских [Edwards et al., 2006]. Эксперимен
тально полученные гибриды между E. annuus и E. strigosus в первом поколении
высокостерильны [Noyes, 2000a].
E. annuus -- озимый однолетник [Tuyama, Murase, 1951; Werner, 1977; Regehr,
Bazzaz, 1976]. Впрочем, во «Flora of North America» сообщается как об ози
мых, так и яровых растениях [Nesom, 2006].
Расселение и распространение. Распределение генотипов позволяет пред
положить занос диаспор на дальние расстояния (long distance dispersal)
[Edwards et al., 2006].
174
Последствия внедрения
Воздействия на естественные фитоценозы и аборигенные виды мало
изучено.
Генетические изменения и вариабельность вида. Поскольку E. annuus --
апомиктический вид, можно было предполагать его высокую однородность.
Однако даже в пределах одного пункта мелколепестник обладает довольно
высоким полиморфизмом [Hancock, Wilson, 1976]. Наличие в европейских
популяциях амфимиксиса приводит к достаточно большой изменчивости ра
стений [Edwards et al., 2006].
На территории Средней России явно преобладает var. septentrionalis, типо
вая разновидность встречается заметно реже (рис. 69). Отметим, что среднерус
ские растения в мелких деталях отличаются от приводимого во «Flora of North
America» описания этого вида [Nesom, 2006]. Так, наши растения часто имеют
голые листочки обертки, в отличие от американских «sparsely villous or hirsuto
villous, minutely glandular» (E. annuus s. str.) или "strigose (hairs flattened, 0,5--1,2
mm)" (var. septentrionalis). Среди среднерусских растений не удалось найти об
разцы с железистым опушением или со сплюснутыми волосками (MW, MHA).
Экономическое и социальное (положительное и отрицательное) зна
чение. В традиционной медицине траву E. annuus использовали для лечения
диабета [Marles, Farnsworth, 1995; Traditional..., 2006].
Выявлена фугницидная активность штамма Gluconobacter sp. 71F против
Fusarium oxysporum f. sp. Lycopersici, вызывающего фузариоз томатов. Скри
нинг проведен для 41 вида растений, лишь с поверхности корней E. annuus
удалось выделить высокоактивный штамм Gluconobacter, что позволит разра
ботать эффективный фунгицидный препарат [Asante et al., 2008].
Меры борьбы
Поскольку E. annuus обычно не является сорняком селькохозяйственных
культур, специальных мер по борьбе с ним не разработано.
Обзор научных исследований и публикаций
Специальные исследования выявили 4 компонента, ингибирующих прора
стание семянок салата (Lactuca sativa). Это (5 butyl 3 oxo 2,3 dihydrofuran 2
yl) acetic acid, 3 hydroxy pyran 4 one, 4 hydroxycinnamic acid и 3,4
dihydroxycinnamic acid methyl ester, которые при концентрациях 2,13±0,03,
12,85±0,56, 4,97±0,24 и 4,87±0,25 mM вызывают 50% ное ингибирование про
растания семян -- IC50 [Oh et al., 2002].
СЕМЕЙСТВО ASTERACEAE (COMPOSITAE) --
СЛОЖНОЦВЕТНЫЕ
ГАЛИНЗОГА МЕЛКОЦВЕТКОВАЯ
Название: Galinsoga parviflora Cav.
Синонимы: G. laciniata Retz., G. semicalva (Gray) St. John and White
Морфологические признаки
Стебель прямостоячий сильноветвистый высотой 10--80 см, по большей
части голый, изредка в верхней части опушенный мелкими прижатыми во
175
лосками. Листья простые, супротивные длиной до 6 см и шириной 4 см, по
краю зубчатые. Корзинки диаметром менее 1 см, состоят из 4 или 5 белых трех
лопастных краевых цветков и многочисленных желтых трубчатых цветков
(рис. 70). Плод --- семянка длиной 1,5 мм, опушенная, сужающаяся от основа
ния к вершине, с белым паппусом, напоминающим корону. Чешуйки паппуса
трехлопастные, прицветники с округлой вершиной.
Естественный ареал
Естественный ареал G. parviflora --- горы Мексики.
Вторичный ареал
История интродукции и географическое распространение. Формирование
вторичного ареала G. parviflora началось в Европе не позднее 1785 г. К тому
времени относятся сведения о культивировании вида в Парижском ботаничес
ком саду из семян, присланных из Перу [Ridley, 1930], и в Ботаническом саду
Мадрида. В Англию интродуцирована как декоративный однолетник в 1796 г.
Расселение G. parviflora в Европе шло очень быстро [Klopfer, Schonfeld,
1986]: в 1798--1801 гг. она появилась в ботанических садах Германии (Берлин,
Карлсруэ, Бремен; причем в Бремен семена прислали именно из Парижского
ботанического сада), в 1801 г. --- в Швейцарии (Цюрих), в 1844 г. --- в Италии
(Больпано), в 1859 г. --- в Польше (Краков), в 1864 г. --- в Австрии (Вена), в
1872 г. --- в Дании (Хадерслев), в 1882 г. --- в Голландии (Багенинген).
Вне ботанических садов галинзогу стали находить: в Англии через 13 лет
после интродукции, в Бремене --- через 14, в Саксонии через 30 [Ridley, 1930;
Klopfer, Schonfeld, 1986; Schulz, 1987]. Однако подобные случаи отмечались
редко. В Англии, например, до начала ХХ века обнаружено только 4 спонтан
ные популяции этого вида [Ridley, 1930].
В обзоре распространения G. parviflora, приведенном в работе О.Барановсь
кого [1938], указаны более поздние даты появления ее в тех или иных странах,
но вывод однозначен: к началу ХХ века галинзога спорадически встречалась
по всей Средней Европе и постепенно внедрялась в рудеральные сообщества.
Помимо Европейского региона, к началу ХХ века G. parviflora встречалась
в Северной Америке (с 1882 г. в Нью Йорке, с 1883 г. в Чикаго), в Индии
(с 1840 г. в Западных Гималаях), на Яве (с 1881 г.), в Австралии (с 1873 г. в
окрестностях Сиднея и Мельбурна), в Южной Африке (с 1893 г. в Капской
колонии). В 1904 г. этот вид найден в Новой Зеландии, а в 1928 г. --- на Филип
пинах [Ridley, 1930; Барановський, 1938]. К середине ХХ века G. parviflora
встречалась уже в различных местообитаниях и агрофитоценозах Нового Све
та, Европы, Азии, Африки и Австралии на высотах от 40 до 3600 м. В настоя
щее время ее вторичный ареал продолжает медленно расширяться, охватывая
новые регионы [Canne, 1977; Husaini, Iwo, 1990; Folch, Abella, 1974], как с уме
ренным, так и с субтропическим климатом.
В Балтийских странах галинзога начала культивироваться с 1810 г. (в Дер
птском ботаническом саду, Тарту, Эстония). Так же как в Англии, проникла в
окрестности сада, но широкого распространения не получила. Через 115 лет в
качестве единственного местонахождения вида в Эстонии также указывался
только г. Тарту ("в садах, на грядках, редко"). Следовательно, за столь долгое
время она не только не вышла за пределы города, но и в черте города получила
ограниченное распространение [Круберг, 1955].
176
В Латвии наблюдалась аналогичная картина: G. parviflora найдена на руде
ральных местах в окрестностях Риги в 1893 г. (RGU), однако до 1930 г. встре
чалась довольно редко и только в пределах города. Позднее спорадические
местонахождения вида отмечены в Елгаве и Даугавпилсе [Круберг, 1955]. В
Литве обнаружена в 1894 г.
На Украине G. parviflora впервые отмечена в 1854 г. как очень редкое расте
ние, встречающееся в парке Александрия под Белой Церковью (LE). По ви
димому, она была занесена туда с семенами или рассадой. До начала ХХ века
выявлена в качестве редкого вида еще в 5 пунктах: в Львове (1866 г.), Кремен
це (1879 г.), Заболотове (1880 г.), окрестностях Ровно (1890 г.) и на окраинах
Харькова (1894 г.). В первой четверти ХХ века находки G. parviflora стали более
частыми: в 1905 г. обнаружена в Киеве, в 1919 г. --- в г. Богодухово, в 1920 г. --- в
Кировограде, в 1922 г. --- в Борисполе, 1937 г. --- в Одесской обл. Отдельные ее
местонахождения отмечены в Полтавской, Черкасской, Житомирской, Тер
нопольской, Ровенской, Кировоградской и Винницкой обл. В 1930 х гг. галин
зога мелкоцветковая стала на Украине довольно распространенным сорняком
[Протопопова, 1973].
В Белоруссии впервые отмечена в 1862 г. Единичные находки галинзоги в
конце ХIХ века сделаны в Люблинской, Виленской, Минской губерниях (LE).
В России G. parviflora выращивали в 1820 х гг. в Ботаническом саду Санкт
Петербурга. В 1831 г. ее наблюдали на рудеральных местообитаниях на Апте
карском острове в большом количестве, но уже через несколько лет она исчез
ла. Из обменных списков семян сада видно, что G. parviflora с 1834 г. часто
выращивали в саду, причем вводили в культуру в ХIХ --- ХХ веках. неоднок
ратно [Гусев, 1966].
В 1897 г. рудеральные популяции G. parviflora обнаружены в окрестностях
Владикавказа (LE), а через 25 лет вид стал на Северном Кавказе доминирующим
сорняком. В 1929 г. галинзога найдена в Аджарии на чайной плантации (LE).
Занесенная из Америки, G. parviflora стала во Владивостоке к середине ХХ
столетия одним из самых обычных растений пустырей, откосов и садов, но за
пределами города не встречалась. Через 30 лет вид не только завоевал все круп
ные города и поселки Приморского и Хабаровского краев, но и стал одним из
злейших засорителей полей [Воробьев, 1954].
После Великой Отечественной войны в Санкт Петербурге отмечено более
100 ее местообитаний галинзоги; единичные особи найдены также в 1958 г. в
Иван городе, а в 1965 г. --- в Луге [Гусев, 1966].
В 1960 е гг. галинзога мелкоцветковая обнаружена в Ижевском р не Уд
муртии и в Казани [Ефимова, Туганаев, 1964; Иванова, Туганаев, 1969].
В то же время зафиксированы находки G. parviflora в Средней Азии: в Таш
кенте (в 1958 г.) и Душанбе (в 1963 г.), а позднее (в 1978 г.) в Хороге (TASH,
TAD). Распространилась галинзога и в Армении [Понерт, 1971]: в Ереване (в
1955 г.), Дилижане и в Ноемберянском р не (в 1960 г.).
Взрывное расширение ареала галинзоги произошло в 1980 е гг. Растение
стало играть заметную роль в сложении рудеральных ценозов и доминиро
вать на клумбах и газонах в странах Балтии [Рамомавичюс, 1988; LATV], Да
гестане (MHA). В 1982 г. G. parviflora обнаружена нами лишь на одной клумбе
в г. Вологда [Майтулина, 1984]. Через 3 года это растение стало там массовым
сорняком в цветниках.
Рис. 70. Массовое цветение Galinsoga parviflora
Рис. 71. Расселение Galinsoga parviflora в Средней России
Рис. 72. Galinsoga parviflora --- злостный сорняк цветников
Рис. 73. Корзинки
Galinsoga parviflora
179
В 1983 г. G. parviflora собрана нами на Памире (Рушанский хребет в окрес
тностях кишлаков Сохгарв и Баргем на высотах от 1950 до 2600 м), а в 1985 г.
мы обнаружили ее во множестве в Фергане, Коканде и Фрунзе.
В последнее десятилетие ХХ века вид найден в Республиках Коми и Ма
рий Эл и массово произрастал практически во всех регионах Восточной Ев
ропы, а на западе Украины и в Молдавии стал злостным сорняком полей
[LE, Абрамов, Жиряков, 1989].
Пути и способы заноса. Большинство исследователей считают, что G.
parviflora «сбежала» из Ботанического сада Парижа, и именно оттуда ведут
происхождение большинство европейских популяций вида. Не исключено
также, что большую роль в проникновении вида в Европу играли морские пор
ты, через которые импортировалось американское зерно, засоренное семена
ми галинзоги мелкоцветковой [Барановський, 1938].
Статус в регионе. В средней полосе России впервые собрана в Нижнем
Новгороде [Маевский, 1917]. После Великой Отечественной войны вторич
ный ареал G. parviflora стал заметно расширяться. Она появилась в Москве
(MW, MHA), Ульяновске, Костроме [Голицын, 1945; Белозеров, 1960].
В 1960 е гг. G. parviflora стала обычным видом в Московской обл. В 1980 е гг.
галинзога мелкоцветковая стала доминантом рудеральных сообществ в Воро
неже, Чувашии (LE), Самарской и Липецкой обл. [Игошин и др., 1985; Вью
кова, 1987]. Найдена в Орловской обл. (г. Орел, газон близ Оки, 14.09.1984 г.
В.Радыгина. ОГУ) и в Ульяновской обл. (г.Инза, пункт первичной переработ
ки шерсти. На ж/д путях. 16.08.1989 г. Н.С. Раков. MHA).
В последнее десятилетие ХХ века вид занесен в Татарстан и Мордовию, а
также отмечен во флорах Ярославской, Владимирской и Ивановской обл.
[LE; Папченков, Дмитриев, 1989; Силаева, 1989; Шилов и др., 1989; Лавренко,
Кустышева, 1990]. Число новых местонахождений галинзоги мелкоцветковой
растет с каждым годом, и всюду отмечается массовый характер ее распростра
нения (рис. 71).
В ходе расширения вторичного ареала период от первого появления вида
до натурализации значительно сокращается. В Санкт Петербурге G. parviflora
начала дичать через 135 лет, в Латвии --- через 60 лет, на Украине --- через 55
лет, в Средней Азии --- через 25 лет, в Костромской обл. --- через 10 лет, в
г. Вологда --- всего через 3 года [Виноградова, 2002]. Поскольку G. parviflora
является однолетником, можно считать, что период натурализации вида в Рос
сии насчитывает не менее 40 смен поколений.
Местообитания. Галинзога предпочитает рыхлые гумусированные увлаж
ненные почвы. Во вторичном ареале она --- преимущественно сорняк цветни
ков, реже засоренных газонов (рис. 72), в последнее время стала встречаться
на ж/д насыпях. Обычна на культивируемых почвах и рудеральных участках
и является злостным сорняком зерновых культур и пашен
Размножение и жизненный цикл. Галинзога мелкоцветковая --- яровое одно
летнее растение, цветет с июня по сентябрь, плодоносит с июля до замороз
ков. Самоопылитель. Одно растение дает 5--30 тыс. семянок, средняя урожай
ность --- 600 тыс. семянок на 1 м2 ( ссылка). Семянки имеют растянутый пери
од вызревания, всхожесть семян сохраняется в течение 5 лет. У свежесозревших
семян всхожесть 28--49%. Прорастают с глубины не более 2--3 см. На прорас
тание большое влияние оказывает свет. Летнее осенние всходы не перезимо
вывают и гибнут при температуре 4°С.
180
Расселение и распространение. Семена распространяются ветром. В Юж
ной Африке галинзога распространяется водой по рекам и произрастает по
берегам [Ridley, 1930]. Последствия внедрения
Воздействие на естественные фитоценозы и аборигенные виды. G. parviflora
обладает высокой конкурентоспособностью и быстрым распространением, что по
зволяет виду становиться доминантом на нарушенных местообитаниях (рис. 73).
По данным ряда авторов, в некоторых районах Центрального Черноземья га
линзога мелкоцветковая внедрилась в лесные сообщества, в травостой донс
ких лугов и даже проникла на территорию Жигулевского заповедника [Алек
сандрова, Барабаш, 1987; Саксонов, 1989].
Генетические изменения и вариабельность вида. G. parviflora может гибри
дизировать с G.quadriradiata [Reinhardt et al., 2003]. У растений типовой раз
новидности во всех регионах первичного и вторичного ареалов выявлен дип
лоидный набор хромосом 2n=16. В естественном ареале выделена, однако, раз
новидность G. parviflora var. semicalva A.Gray, имеющая тетраплоидный набор
хромосом 2n=32, которая произрастает в Аризоне и Нью Мексико (США) и
отличается мелкими листьями (длиной обычно менее 5 см), колокольчатым
венчиком, чешуйками, более короткими, чем венчик, и заметно более трехраз
дельными, чем у типовой разновидности [Canne, 1977, 1983; Husaini, Iwo, 1990].
Во вторичном ареале у G. parviflora отмечено наличие межпопуляционной
изменчивости. В исследованных нами популяциях встречалась как типовая
разновидность с абсолютно голыми побегами, так и G. parviflora var. genuina
Thell. --- верхняя часть стебля, цветоложе и обертки соцветия покрыты про
стыми прижатыми волосками, иногда с небольшой примесью железистых во
лосков. При этом внутри отдельной микропопуляции изменчивости по при
знакам опушения не отмечалось. Небольшая амплитуда изменчивости наблю
далась по размеру листьев, степени ветвления и по степени надрезанности
чешуек в корзинке. Значительно варьирует число цветков и семян (от 4 до 100
корзинок на 1 растение).
Изучение семенной продуктивности вида по высотному профилю в Карпа
тах [Гамор, 1978] показало, что с увеличением высоты местности (от 150 до 700 м
над ур. м.) потенциальная и фактическая семенная продуктивность уменьшает
ся, а амплитуда внутрипопуляционной изменчивости увеличивается (коэффи
циент вариации числа корзинок на 1 растение возрастает с 47 до 133%).
Экономическое и социальное (положительное/отрицательное) значение. G.
parviflora сильно заглушает морковь, петрушку, свеклу, картофель, помидоры,
сорничает в посевах зерновых. Вид стал опасным сорняком в питомниках де
коративных растений.
Виды рода Galinsoga являются растениями --- хозяевами для многих насе
комых, вирусов и нематод, влияющих на урожай зерновых культур [Huffman,
2004].
Меры борьбы
Виды рода Galinsoga лучше контролируются в начале цветения, что позво
ляет предотвращать образование семян. Необходима глубокая обработка по
чвы, предпосевная культивация, своевременный посев, междурядные обработ
ки в посадках пропашных, соблюдение севооборота. Сорняк чувствителен к
большинству применяемых гербицидов.
181
Обзор научных исследований и публикаций
Изучена генетическая закрепленность различных биотипов G. parviflora
(см. главу 4). Отмечено, что популяции G. parviflora на всем протяжении вто
ричного ареала проявляют низкий уровень изменчивости по важнейшим адап
тивным признакам: ритму роста и фенологическому ритму развития, однако
наблюдается значительная межпопуляционная изменчивость по степени опу
шенности верхней части стебля, цветоложа и обертки.
СЕМЕЙСТВО ASTERACEAE (COMPOSITAE) --
СЛОЖНОЦВЕТНЫЕ
ГАЛИНЗОГА ЧЕТЫРЕХЛУЧЕВАЯ
Название: Galinsoga quadriradiata Ruiz et Pav.
Синонимы: Adventina ciliata Raf., Galinsoga urticifolia (Kunth) Benth., G. ciliata
(Raf.) S.F. Blake, G. hispida Benth., G. aristulata Bickn., G. bicolorata St.John and
White, G. caracasana (DC) Schultz Bip., Wilburgia urticifolia Kunth
Морфологические признаки
Стебель прямостоячий, сильноветвистый, высотой 10--80 см, покрыт жест
кими отстоящими волосками (рис. 74). Листья супротивные, темно зеленые,
длиной до 6 см и шириной 4 см, простые, oвальные, заостренные к верхушке,
наиболее широкие в основании; край листьев зубчатый; с обеих сторон листья
покрыты щетинистыми волосками. Соцветия диаметром ~1 см, состоят из 4
или 5 белых трехлопастных краевых цветков и многочисленных желтых сре
динных цветков. Плод -- семянка длиной 1,5 мм, опушенная, сужающаяся от
основания к вершине, с белым паппусом, напоминающим корону. Чешуйки
паппуса цельные, прицветники имеют очень узкую, острую вершину [Sulcs,
1998, Reinhardt et al., 2003]. Из за некоторого сходства этот вид часто путают
с G.parviflora, которая отличается более бледно зеленой окраской, короткими
волосками, трехлопастными чешуйками паппуса и прицветниками с более
широкой и округленной вершиной.
Естественный ареал
Естественный ареал G.quadriradiata охватывает часть Южной и Централь
ной Америки от Мексики до Чили, но благодаря действиям человека вид рас
пространился далеко за пределы естественного ареала.
Вторичный ареал
История интродукции и географическое распространение. G.quadriradiata --
сорняк многих зерновых культур, садов и пустырей в большинстве умерен
ных и субтропических регионах мира. За пределами естественного ареала ее
впервые обнаружили в 1836 г. в Филадельфии, где вид выращивали в ботани
ческом саду. Вскоре он довольно широко распространился в США, а в 1893 г.
достиг Канады (Монреаль). Через 30 лет G. quadriradiata стала в провинциях
Онтарио и Квебек обычным растением [Warwick, Sweet, 1983]. В 1970 х гг.
галинзога встречалась уже как адвентивная в Коста Рике, Доминиканской
Республике, Сальвадоре, Гондурасе, Гватемале, Никарагуа, Панаме, Пуэрто
Рико, Аргентине, Боливии, Бразилии, Чили, Колумбии, Эквадоре, Перу, Ве
182
несуэлле, в Индии, Японии, Непале, на Филиппинах, в Камеруне, Юго За
падной Африке, причем в Африке вид обилен [Canne, 1977].
В Европу G. quadriradiata занесена в середине ХIХ века. По одним данным,
это связано с усилением экспорта американского зерна [Schulz, 1987]. Однако
мы придерживаемся мнения, что вид "сбежал" из культуры, поскольку первые
упоминания о появлении его в Европе находим в каталогах ботанических са
дов. Расселение G.quadriradiata в Европе шло очень быстро [Klopfer, Schonfeld,
1986]: в 1850 г. она отмечена в Ботаническом саду Вены, в 1858 г. -- в Польше
(Вроцлав), в 1866 г. -- в Германии (Бойтценбург), в 1872 г. -- во Франции (ботани
ческий сад Гренобля), в 1901 г. -- в Чехии (Годонин), в 1914 г. -- Австрии (Инсб
рук), в 1917 г. -- в Швейцарии (Цюрих), в 1921 г. -- в Бельгии (Нинове), в 1934 г. --
в Голландии (Амстердам).
В Балтийских странах впервые отмечена в Эстонии. Гербарный образец,
неверно определенный как G.parviflora (RIG), собран в 1889 г. в Ботаничес
ком саду Тарту; в 1921 г. галинзога зарегистрирована уже там как «беженец»
из ботанического сада. В Литве собрана в окрестностях Вильнюса в 1924 г.
[Kuusk et al., 2003]. В Латвии впервые найдена в г. Лиепая в 1928 г. [Melderis,
1939]. В Скандинавских странах первые находки G.quadriradiata сделаны в
1900 г. в Норвегии [Lid, Lid, 2005], в 1926 г. в Швеции [Hylander, 1971], в 1927 г. в
Дании [Hansen, Pedersen, 1961], в 1928 г. в Финляндии [Saansalo Taubert, 1962].
В 70 х годах ХХ века встречалась как адвентивное в Нидерландах, Португа
лии и Англии [Canne, 1977]. Дальнейшее расширение ареала G.quadriradiata
практически прекратилось. До конца ХХ века вид был обнаружен только в
Испании и странах бывшей Югославии -- в Словении, Хорватии, Черногории
и Сербии [Trinajstic, 1983; Martinez, 1985]. Вид не встречен пока в Исландии,
Гренландии и на Фарерских о вах.
На территории СНГ галинзога впервые зафиксирована в Санкт Петербурге,
где ее культивировали в Ботаническом саду с 1842 г. [Гусев, 1966]. В 1905 г. найде
на в Любленской губернии (LE), в 1939 г. -- в Батумском ботаническом саду.
После Второй мировой войны G.quadriradiata появилась в западных обла
стях Украины, в Белоруссии, в Ленинградской и Калининградской обл. [Коз
ловская, 1962; LE]. Поначалу вид встречался еще очень редко, а целенаправ
ленные поиски, предпринятые, например, Ю.Д.Гусевым [1966] в Ленинградс
кой обл., часто были безуспешными: найти удалось лишь единичные
экземпляры в 1958 г. в Иван городе, а в 1961 г. в Ленинграде.
В конце 1960 х гг. началось массовое, взрывное распространение вида на
Восточно Европейской равнине и на Кавказе. G.quadriradiata обнаружена в
Пскове, Пермской обл., Нальчике, Пятигорске, Ростове на Дону, в Краснодар
ском крае и на территории Колхиды [Босек, 1983; Кушхов, 1980; Понерт, 1971].
На Украине отмечено продвижение вида в Черновицкую, Ровенскую и Сумс
кую обл. [Протопопова, 1973]. В 1980 х гг. ареал галинзоги волосистой про
должал расширяться. В 1982 г. мы нашли в Вологде на клумбе (вместе с
G.parviflora) всего 2 экз. вида [Майтулина,1984], а уже через 3 года он стал там
массовым сорняком. G.quadriradiata собрана также в Новгородской обл. и в
Кабардино Балкарии [Тихомиров, 1982; Кушхов, 1989]. В 1984 г. обнаружена
как сорняк цветников в Ленинабадском ботаническом саду, а в 1986 г. -- в Ду
шанбинском (TAD). Во время экспедиции 1985 г. в Ферганскую долину и в
Киргизию найдена нами в незначительном числе экземпляров в Андижане и
183
Рис. 75. Расселение Galinsoga quadriradiata в Средней России
Рис. 74. Характерное опушение побегов Galinsoga quadriradiata
Рис. 76. Клумба с Galinsoga quadriradiata в центре г. Кинешма, август 2008 г.
Рис. 77. Массовое цветение Galinsoga quadriradiata
185
Кара Куле и массово в Намангане, причем галинзоги мелкоцветковой в этих
пунктах не обнаружено. В Фергане встречались оба вида, но сорничали они на
разных клумбах, и в одной микропопуляции вместе не попадались.
В настоящее время вид натурализовался во многих европейских странах. В
Германии и Польше распространился по всей территории и особенно обычен
на культивируемых почвах и рудеральных участках [Reinhardt et al., 2003;
Tokarska Guzik, 2003]. В Польше это обычный полевой сорняк и рудеральное
растение, произрастающее в союзах Sisymbrion, Eu Polygonion и Panico Setarion.
Более редко он встречается в полуестественных местообитаниях -- по лесным
тропинкам, прогалинам и опушкам смешанных, хвойных и лиственных лесов
[Tokarska Guzik, 2005; Chmura, 2004]. В Балтийских странах вид обычен для
всей территории и является опасный сорняком зерновых культур и пашен
[Gudzinskas,1997; Kuusk et. al., 2003]. В Латвии и Литве G.quadriradiata -- ад
вентивный вид, который никогда не выращивали в ботанических садах; наи
более интенсивное распространение G.quadriradiata в Литве было отмечено в
1980 х гг. [Gudzinskas, 1997]. В последнее время вид очень быстро распростра
нился по всей Латвии и в некоторых местах гораздо более обычен и агресси
вен, чем G.parviflora. В настоящее время вид внедрился в естественные сооб
щества Эстонии, но натурализовался, главным образом, в южных районах.
Растет как надоедливый сорняк в полях и в садах, на ж/д насыпях, морских
побережьях и мусорных свалках [Kull, Kukk, 2005]. Вид обычен в Дании, но
не считается агрессивным [Hansen, Pedersen 1962; Mossberg, Stenberg, 1999].
В Финляндии G. quadriradiata -- довольно обычный сорняк теплиц и оранже
рей, но на открытых местах является редким и отмечен лишь в нескольких
городах на юге страны. Вследствие редкости и распространения исключительно
в искусственных местообитаниях вид не считается в Финляндии агрессивным.
В Норвегии вид является, главным образом, сорняком в городах и других ру
деральных местообитаниях. Довольно обычен в окрестностях Осло и имеет
рассеянные локальные популяции вдоль юго западного побережья. Считает
ся, что в Норвегии вид имеет возможность дальнейшего значительного рас
ширения ареала [Lid, Lid, 2005].
Хотя G.quadriradiata появилась на территории бывшего СССР позднее
G.parviflora, расселялась она быстрее, а в некоторых районах (например, на
Северном Кавказе, в Латвии и Колхиде) превалирует над G.parviflora. Неко
торые авторы [Козловская, 1962; Понерт, 1971] объясняют это различием в
биологических особенностях близких видов, указывая, что G.quadriradiata
предпочитает более влажные районы. Однако наблюдения, проведенные нами
в Средней Азии, свидетельствуют о том, что все не так однозначно: в Наманга
не, например, растет только G.quadriradiata, а в Коканде -- исключительно
G.parviflora.
Пути и способы заноса. Во многих европейских странах G.quadriradiata --
адвентивный вид, который прибыл из естественного ареала, транспортируе
мый с почвой, с зерном или с другими сельскохозяйственными продуктами.
G. quadriradiata выращивалась в европейских ботанических садах (с 1849 г.)
и, возможно, «сбежала» оттуда, подобно ситуации с близкородственной
G.parviflora. В Дании, вероятно, вид имел два основных пути непреднамерен
ного заноса: 1) через порты, в которые поступали товары; 2) вместе с семена
ми, закупленными для озеленения [Hansen, Pedersen, 1962]. В Германии
186
G.quadriradiata впервые обнаружена на помойке в кофейной куче, что позво
ляет предположить неумышленный завоз ее диаспор при импорте кофе
[Reinhardt et al., 2003]. В Финляндию, по видимому, вид занесен исключитель
но с импортируемыми семенами и рассадой декоративных растений.
Статус в регионе. В Москве G.quadriradiata появилась в 1922 г. (MW). В
конце 1960 х гг. как сорняк цветников галинзога отмечена в Смоленске, Кост
роме, Ульяновске, Нижнем Новгороде, Тверской и Брянской обл. (LE; Босек,
1983]. Позднее обнаружена в Орловской обл. (Дмитровский р н, г. Дмитровск,
сорное место, 25.07.1977 г. В.И. Радыгина. ОГУ). В 1980 х гг. галинзога четы
рехлучевая собрана в Саранске, в Рязанской, Липецкой и Калужской обл. [Ти
хомиров, 1982; Вьюкова, 1983; Силаева, 1989]. Натурализовалась во всех ре
гионах Средней России (рис. 75).
Местообитания. G.quadriradiata приспособлена к теплому климату и тя
желым, богатым азотом глинистым почвам. Преимущественно сорняк цвет
ников (рис. 76), но нередко встречается на огородах, улицах, во дворах, у ж/д
платформ (в последнем местообитании встречается значительно реже, чем
G.parviflora). Вид найден также в садах, оранжереях, зеленых насаждениях,
обочинах дорог и на мусорных местах, особенно в населенных пунктах
[Gudzinskas, 1997; Sulcs, 1998; Reinhardt et. al., 2003]. Эти местообитания сход
ны с теми, которые занимает G.quadriradiata в естественном ареале, где ее также
считают обычным сорняком. Проникновение G.quadriradiata в прибрежные
фитоценозы вдоль речных берегов отмечено в Литве [Gudzinskas, 1997].
Как показано американскими исследователями [Shontz, Shontz, 1970], в
Новой Англии, где G.quadriradiata произрастает в течение 70 лет, сформиро
вались 2 экотипа вида, различающихся по целому ряду морфологических и
физиологических признаков. Первый ("внешний") экотип произрастает на
нарушенных природных местообитаниях. Второй ("внутренний") экотип сор
ничает в оранжереях. Растения внешнего экотипа выше, могут (хотя и плохо)
расти в слабой тени, имеют более крупные семянки, прорастающие одинаково
быстро и при высокой, и при низкой температуре, и особенно быстро при су
точных колебаниях температуры от 10 до 24°С. Оранжерейный экотип не спо
собен расти в тени, в условиях короткого дня продуцирует семянки со слабой
скоростью прорастания, лучше прорастает при постоянной температуре 24°С.
При экспериментальном выращивании обоих экотипов в условиях оранже
реи слизни охотнее поедали растения "внешнего" экотипа.
Размножение и жизненный цикл. G.quadriradiata -- однолетнее растение,
распространяется только семенами, цветет с июня до поздней осени. Растение
является или самоопылителем, или его опыляют насекомые [Reinhardt et al.,
2003]. Семянки мелкие, длиной ~1,5 мм. Растение начинает цвести, когда высо
та его достигает 3 см, и продолжает цвести весь вегетационный сезон (рис. 77).
Растение в возрасте 8 9 недель может производить 3 тыс. соцветий и множе
ство (до 7500) семян [Kagima, 2000; Huffman, 2004]. Семена способны прорас
ти немедленно после контакта с теплой сырой почвой; поэтому растение при
благоприятных условиях (в США и Японии) может давать 2 4 поколения за
вегетационный сезон [Warwick, Sweet,1983; Reinhardt et al., 2003]. Однако вид
чувствителен к морозу, и для прорастания его семена требуют высоких темпе
ратур. Семена галинзоги остаются жизнеспособными в полевых условиях в
течение нескольких лет [Huffman, 2004].
187
Расселение и распространение. Семена разносятся ветром или их распрост
раняют животные [Reinhardt et al., 2003], а порой вода [Melderis, 1939]. Пло
ды цепляются за мех животных или одежду людей [Shontz, Shontz, 1970]. Глав
ную роль в распространении вида играют действия человека, связанные с озе
ленением (транспортировка садовых земель и растений) [Hansen, Pedersen,
1961]. В Финляндии, по крайней мере, наиболее эффективный вектор рассе
ления -- садовая земля, выброшенная после высадки рассады в грунт.
Последствия внедрения
Воздействие на естественные фитоценозы и аборигенные виды. G. quadri radiata
часто находят на пахотной земле и в садах. Вид конкурентоспособен и быстро
распространяется, что позволяет ему становиться доминантом. G.quadriradiata
наносит экономический ущерб только хлебным злакам (ссылка).
Генетические изменения и вариабельность вида. G.quadriradiata может гиб
ридизировать с G. parviflora [Reinhardt et al., 2003]. Основное число хромосом
х=8. Большинство популяций тетраплоиды (2n=32), однако кариологичес
кий анализ показал [Canne, 1977], что G.quadriradiata не является тетрапло
идным дериватом диплоидной G.parviflora. В Перу найден образец, имеющий
n=24, а одна популяция насчитывает n=32.
В естественном ареале различают 3 разновидности G.quadriradiata. Первая
распространена от севера Южной Америки до Канады. Имеет короткое кону
сообразное цветоложе с цельными чешуйками, железистые волоски и цвето
ножку длиной до 2 см. Вторая разновидность встречается от юга Мексики до
Колумбии, отличается высоким коническим цветоложем с неглубоко трехраз
дельными чешуйками, нежелезистыми волосками и цветоножками длиной 2--
3,5 см. Третья (розовоцветковая) разновидность из Мексики и Южной Аме
рики морфологически сходна с двумя предыдущими и имеет различное число
хромосом (n=16, n=24 и n=32). Во вторичном ареале в Европе встречаются
все 3 разновидности [Canne, 1977].
Исследованные нами на территории бывшего СССР популяции G. quadrira
diata показали очень широкую вариабельность всех этих признаков, практи
чески не позволяющую четко определить, к какой разновидности следует от
нести тот или иной образец. Нередко в одной корзинке имеются и цельные, и
неглубоко трехраздельные чешуйки. В городе на одной клумбе произрастают
растения с короткими, а на другой с длинными цветоножками. Форма цвето
ложа, степень надрезанности чешуек и наличие железистых/нежелезистых
волосков не коррелируют между собой. По всей видимости, наряду с само
опылением, которое обеспечивает существование изолированных инбридных
популяций, нередки и случаи перекрестного опыления. Это подтверждается
данными по изучению хромосом подобных промежуточных форм в мейозе,
которые показали, что промежуточные формы гетерозиготы по крайней мере
по двум хромосомным инверсиям [Canne, 1977]. Разновидность с розовыми
цветками мы не встречали.
Экономическое и социальное (положительное/отрицательное) значение.
G.quadriradiata постоянная проблема для многих озеленительных фирм, вклю
чая коммерческие оранжереи, но нет никаких данных относительно стоимос
ти контролирования этого сорного вида. G. quadriradiata отбирает питатель
ные вещества, необходимые для культивируемых растений, и ее присутствие
может на 10 50 % снижать урожаи на полях с овощными и зерновыми культу
188
рами [Kuniga, 1994]. Еще один ущерб заключается в относительно высокой
площади листовой поверхности G. quadriradiata, и ее листья могут затенять
культивируемые виды [Reinhardt et al., 2003].
Виды рода Galinsoga являются растениями хозяевами для многих насеко
мых, вирусов и нематод, которые сокращают урожай зерновых культур
[Huffman, 2004].
Меры борьбы
Виды рода Galinsoga лучше контролируются в начале зацветания, что по
зволяет предотвращать образование семян. Есть несколько довольно эффек
тивных для уничтожения G.quadriradiata гербицидов [Reinhardt et al., 2003],
но так как вид сорничает, главным образом, на полях овощных культур, ис
пользование гербицидов весьма ограничено или вовсе невозможно. Приори
тет отдается биологическим методам контролирования сорных видов
[Turakainena, 2001]. Необходимо применять севооборот например, менять
поля с зерновыми культурами плантациями помидоров [Kagima, 2000]. Яро
вые зерновые культуры Sorghum bicolor или S. sudanense подавляют прораста
ние семян G. quadriradiata. Ранняя обработка почвы может быть тоже полез
на, так как семена Galinsoga лучше прорастают в слегка нарушенной почве не
глубже чем от 2 до 3 см [Huffman, 2004]. Черная пластмассовая мульча -- хоро
шее средство контроля в интенсивном сельскохозяйственном производстве или
домашнем садоводстве. Один из возможных методов контроля использова
ние аллелопатии. Имеются сообщения об аллелопатическом воздействии ози
мой ржи [Turakainena, 2001]. Если Galinsoga появилась в саду недавно, наибо
лее успешна, вероятно, обработка участка мотыгой.
Обзор научных исследований и публикаций
У G.quadriradiata отмечена крайне низкая внутрипопуляционная и доволь
но значительная межпопуляционная изменчивость по степени вырезанности
чешуек, длине цветоножек, наличию или отсутствию железистых волосков и
другим морфологическим признакам (см. главу 4).
Исследование относительно влияния озимой ржи и озимого рапса на жиз
неспособность семян и развитие Galinsoga проводилось в 2001 г. в Сельскохо
зяйственном университете Латвии.
СЕМЕЙСТВО ASTERACEAE (COMPOSITAE) --
СЛОЖНОЦВЕТНЫЕ
ПОДСОЛНЕЧНИК КЛУБНЕНОСНЫЙ, ТОПИНАМБУР
Название: Helianthus tuberosus L.
Синонимы: H. subcanescens (A. Gray) E. Watson, H. Ч laetiflorus auct. non
Pers., p. p.
Морфологические признаки
Многолетнее травянистое растение высотой 0,5--2 м и более (рис. 78). Кор
невище удлиненное (в этом случае клубни удлиненные, диаметром менее 1 см)
или укороченное (культурные формы, клубни диаметром более 2 см, ветвис
тые); клубни формируются к концу вегетационного сезона. Стебель прямой,
шероховато колючий или волосистый. Листья супротивные или в верхней
Рис. 78. Массовое цветение Helianthus tuberosus
Рис. 80. Внедрение
Helianthus tuberosus в
прибрежные ивняки
на р. Оке в окрестностях
г. Калуга
Рис. 81. Корзинки
Helianthus tuberosus
191
части очередные, реже б. ч. очередные, черешковые, черешок длиной 2--8 см,
часто крылатый; листовые пластинки с 3 основными жилками, от яйцевид
ных до широколанцетных, 10--23x7--15 см, основание листовой пластинки от
широко-- до узкоклиновидного, край от пильчатого до цельного, сверху от ко
роткоопушённых или шероховатых до коротковойлочных с сидячими желез
ками, снизу --- грубо шершавые. Корзинки в рыхлом метельчатом соцветии из
(3)5--15(25) корзинок на ножках длиной 1--15 см. Обертка полушаровидная,
диаметром10--25 мм; листочки обертки в числе 22--35, темно зеленые, ланцет
ные, 8,5--15x2--4 мм, с отогнутой верхушкой, шероховатые или короткоопу
шенные, железисто точечные. Чешуи ложа корзинки длиной 8--9 мм, 3--зуб
чатые, на верхушке волосистые. Краевые цветки язычковые, длиной 25--40 мм,
желтые, в числе 10--20; трубчатых цветков 60 и более, длиной 6--7 мм, с жел
тым венчиком, тычинки от темно коричных до черных, с темным или желто
ватым надсвязником. Семянки длиной 5--7 мм, голые или в верхней половине
опушенные.
На облик растений топинамбура влияют условия произрастания. Это от
мечено нами в Калужской обл. в окрестностях Калуги на левобережье
р. Ока, ниже устья р. Угра. Здесь H. tuberosus высаживали по краю огородов, в
которых выращивали преимущественно картофель. Песчаные почвы быстро
истощаются, поэтому участки часто забрасывают, а топинамбур сохраняется.
Листья у растений на заброшенных участках относительно более узкие, яйце
видно ланцетные, желтовато зеленые и более мелкие, чем у растений на бога
тых суглинистых почвах. Листочки обертки корзинок по спинке почти голые,
Рис. 79. Расселение Helianthus tuberosus в Средней России
192
а по краю с длинными ресничками. Определить такие растения по "Flora of
North America" [Schilling, 2006] не удается.
Длиннокорневищные сорные растения с небольшими вытянутыми клубнями в
европейской части б. СССР традиционно относили к особому виду H. subcanescens
[см., например: Определитель растений Мещеры, 1987; Протопопова, 1994;
Mosyakin, Yavorska, 2002]. В связи с этим интересны наблюдения А.Я. Георгиевс
кой с соавторами. В Воронежском заповеднике в 1957--1959 годах для подкормки
оленей высаживали H. tuberosus s. str. В настоящее время на месте заброшенных
посадок произрастает типичный H. subcanescens [Григорьевская и др., 2004]. Похо
жую картину мы наблюдали в окрестностях Калуги на заброшенных огородах.
Тем не менее не исключено, что часть заносных растений относится к H. x
laetiflorus Pers. (H. tuberosus x H. pauciflorus Nutt.). Согласно «Flora of North
America», от H. tuberosus этот гибрид отличается резко неравными, обычно
прижатыми листочками обертки. У H. tuberosus эти листочки отогнутые, по
чти одинаковой длины. Кроме того, поскольку одним из родительских видов
H. x laetiflorus является H. pauciflorus, в целом листочки обертки более широ
кие --- от ланцетных до овальных [Schilling, 2006]. Только специальные гене
тические исследования могут подтвердить (или опровергнуть) наличие H. x
laetiflorus на территории Европейской России.
Естественный ареал
Растение Северной Америки, однако природный ареал установить едва ли
возможно, поскольку оно издавна использовалось индейцами как пищевое
[Жуковский, 1971; Schilling, 2006].
Вторичный ареал
История интродукции и географическое распространение. В Европу
топинамбур попал в начале XVI века, сначала во Францию; в Англии известен
с начала XVII века [Kays, Nottingham, 2007]. Со второй половины XIX века
известен широко и культивировался как пищевая и фуражная культура [Жу
ковский, 1971]. Так, в Чехии известен с 1885 г. [Pyљek et al., 2002; Pyљek, Prach,
2003]. Ныне H. tuberosus --- один из наиболее распространенных сорняков в
Европе [Weber, Gut, 2005; Lambdon et al., 2008]. В Австрии [Essl, Rabitsch, 2002],
Венгрии [Invasive..., 2003], Великобритании [Clement, Foster, 1994] подсол
нечник -- массово натурализовавшийся сорный вид. В Швейцарии он вклю
чен в надзорный лист (Watch Liste) инвазионных растений [Gigon, Weber,
2005]. В Центральной Италии --- один из наиболее распространенных инвази
онных видов [Pace, Tammaro, 2003]. EPPO (European and Mediterranean Plant
Protection Organization) предлагает уделять виду особое внимание как потен
циальному карантинному объекту [Identification..., 2005].
H. tuberosus натурализовался в Чили [Castro et al., 2005], известен как сор
ное растение в Австралии [Auld, Medd, 1992], Японии [Mito, Uesugi, 2004].
В России топинамбур разводили с начала XIX века [Эйхе, 1957; Жуковский,
1971]. В настоящее время H. tuberosus распространен широко: от северо запада ев
ропейской части России [Цвелев, 2000] до Сахалина [Баркалов, Таран, 2004]. К
сожалению, история распространения топинамбура как сорного растения плохо до
кументирована1 как гербарными материалами, так и литературными указаниями.
1 Впрочем, расселение новых адвентивных растений недостаточно документировано
даже в Швейцарии [Weber, 1999] --- стране с самой высокой в мире плотностью гербарных
сборов на 1 км2 территории.
193
Пути и способы заноса. H. tuberosus выращивался с начала XIX века как
пищевое растение [Жуковский, 1971]. Можно предположить, что именно эти
культурные растения стали источником для дичающих растений. Следует учи
тывать, что семенное размножение этого подсолнечника в условиях Средней
России либо отсутствует, либо подавлено. Обнаружить спелые семянки под
солнечника в благоприятные годы с теплой продолжительной осенью обнару
жить не удалось [Игнатов и др., 1990]. Даже в тех случаях, когда семянки за
вязывались, они оставались щуплыми (собственные наблюдения в окрестнос
тях г. Калуга). Очевидно, что существенную роль в образовании обширных
зарослей H. tuberosus в рудеральных местообитаниях, в том числе вдоль ж/д
насыпей, играет вегетативное размножение. По мнению В.О. Мокиевского
(личное сообщение), в распространении подсолнечника, особенно вдоль до
рог и насыпей, могут участвовать мышевидные грызуны, которые запасают
мелкие клубни подсолнечника. Рост H. tuberosus из клубней зависит от стадии
развития клубня, его размеров, регенерация с глубины по крайней мере до 30 см
вполне успешна [Swanton, Cavers, 1988].
Статус в регионе. Дичание H. tuberosus в Средней России впервые отмече
но А.К. Скворцовым [1973] для Московской обл. Видимо, ранее вид из культу
ры не распространялся, о чем свидетельствует отсутствие гербарных сборов из
Московской обл. до 1960 х гг. В соседних областях находки H. tuberosus в каче
стве занесенного растения появились только в 1970--1980 гг. (MW, MHA). К
1980 м гг. в Москве и несколько реже в Подмосковье вид стал обычным на пус
тырях, ж/д насыпях, по рудеральным местообитаниям [Игнатов и др., 1990].
Встречается во многих регионах Средней России и в последнее десятиле
тие активно расширяет вторичный ареал (рис. 79).
Местообитания. В Северной Америке встречается вдоль дорог, на полях,
по залежам и на пустырях [Schilling, 2006]. В пределах вторичного ареала
H. tuberosus растет по нарушенным антропогенным местообитаниям: пусты
рям, обочинам дорог, по краю полей (рис. 80). В Европе и Японии отмечена
приуроченность зарослей к долинам рек, в том числе малых [Miyawaki,
Washitani, 2004]. Предпочитает богатые почвы и высокую освещенность, но
при этом может расти и на бедных кислых почвах и в полутени.
Размножение и жизненный цикл. Опыление перекрестное, возможна гей
теногамия и самоопыление [Жуковский, 1971]. H. tuberosus -- высокополип
лодный вид, 2n=102 [Schilling, 2006; Kays, Nottingham, 2007]. При спорогене
зе наблюдаются многочисленные нарушения. Возможно, поэтому семенная
продуктивность этого подсолнечника незначительная [Жуковский, 1971]. В
Новой Зеландии, где H. tuberosus является инвазионным видом, его семенное
размножение не обнаружено [Esler, 1988].
H. tuberosus --- позднецветущее растение. В работах конца XIX --- начала XX
века [Кауфман, 1889; Сырейщиков, 1910] указано, что топинамбур зацветает
только поздней осенью. По видимому, за минувшее столетие время цветения
стало более ранним (рис. 81): в Подмосковье он обычно зацветает в конце ав
густа --- в начале сентября (но некоторые культурные формы зацветают поз
же!). В жаркие годы в Москве на склонах насыпей южной экспозиции отмече
но цветение H. tuberosus с конца июня [Игнатов и др., 1990].
Расселение и распространение. По видимому, расселение H. tuberosus в
Средней России происходит только вегетативными частями, а именно корне
194
вищами. Топинамбур дичает из посадок, в том числе дачных. В 1980--1990 х
годах топинамбур был популярной культурой, однако дачники часто не полу
чали желаемого урожая, и культура вышла из моды. При этом корневища выб
расывали за пределы дачного участка, на компостные кучи. Сейчас H. tuberosus
высаживают как позднецветущее декоративное растение, нередко у заборов
для создания кулис. Топинамбур нередко встречается как сорное растение у
дачных поселков, на сорных местах близ жилья.
В настоящее время H. tuberosus встречается также вдоль ж/д насыпей, где
порой образует обширные заросли. В подобных местообитаниях топинамбур
распространяется без непосредственного участия человека. По видимому, в
его расселении участвуют мышевидные грызуны.
Последствия внедрения
Воздействия на естественные фитоценозы и аборигенные виды. Взаи
модействие подсолнечника с растениями местной флоры не изучено, но, оче
видно, что в его зарослях не находится места аборигенным видам, как и в слу
чае других инвазионных видов бурьянного высокотравья.
Генетические изменения и вариабельность вида. H. tuberosus --- гексап
лоидный вид, 2n=102. Предполагается, что топинамбур может быть результа
том древней гибридизации 68 хромосомным и 34 хромосомным видами, сре
ди вероятных предков называются H. decapetalus L., H. hirsutus Raf. или H.
strumosus L. (2n=68) и H. giganteus L., H. grosseserratus Martens --- 2n=34 [Kays,
Nottingham, 2007]. Молекулярно генетические исследования не позволяют
пока однозначно выявить родительские виды [Timme et al., 2007].
Отмечен высокий уровень межпопуляционной изменчивости растений,
клоны отличаются по форме и размеру клубней, листьев, соцветий, семянок,
химическому составу клубней [Kays, Nottingham, 2007].
В начале XX века для Москвы указано очень позднее цветение топинамбура:
«у нас цвести не может, так как зацветает только в октябре» [Сырейщиков, 1910].
Уже к концу века отмечено «в жаркие годы (1981, 1982) на склонах насыпей
южной экспозиции в Москве... цветение H. tuberosus с конца июня» [Игнатов и
др., 1990]. Таким образом, очевидно изменение фенологии подсолнечника. От
метим, что по нашим наблюдениям, разные популяции H. tuberosus заметно от
личаются по срокам цветения: часть растений в условиях Подмосковья обычно
зацветает в конце августа --- начале сентября, другие же --- в начале октября.
Исследование H. tuberosus в Черногории показало большое морфологичес
кое сходство местных растений с североамериканскими. При этом выделено
две группы популяций, что позволяет предположить о двукратном заносе (за
возе) материала из Нового Света [Vasiж et al., 2002].
Экономическое и социальное (положительное и отрицательное) зна
чение. Топинамбур является пищевым и фуражным растением [Wiersema, Leуn,
1999]. В его клубнях высокое содержание инулина, и потому их используют для
диетического питания и как сырьевой источник для получения инулина [Эйхе,
1957; Kays, Nottingham, 2007]. Меры борьбы
Меры борьбы с подсолнечником клубненосным не разработаны. В специ
альных мерах пока нет необходимости, поскольку вид массово разрастается по
рудеральным местообитаниям. Но необходим мониторинг особенностей рассе
ления H. tuberosus, поскольку известны случаи его поселения в долинах рек.
195
CЕМЕЙСТВО ASTERACEAE (COMPOSITAE) ---
СЛОЖНОЦВЕТНЫЕ
КРЕСТОВНИК КЛЕЙКИЙ
Название: Senecio viscosus L.
Морфологические признаки
Однолетнее растение высотой 10--40(50) см, с обильным отстоящим лип
ким железистым опушением. Листья большей частью выемчато перистолопа
стные или неравномерно перисторассечённые, сидячие, с удлинённо обратно
яйцевидной пластинкой, клиновидно суженной к основанию (рис. 82). Кор
зинки в рыхлом общем соцветии, удлинённо яйцевидные, длиной 6--8(10) мм;
листочки внешнего ряда линейные, травянистые, разной длины; нередко нож
ка корзинок с мелкими линейными листьями. Язычковые цветки короткие,
едва длиннее обёртки, с плоским или отвёрнутым наружу отгибом, малоза
метные. Семянки веретеновидные, коричневые, с тонкими продольными рёб
рами, обычно голые, длиной 5--7 мм, с более длинным белым хохолком из про
стых волосков.
Естественный ареал
Европейский вид, распространенный в Западной и Центральной Европе, а
также на Кавказе [Шишкин, 1961; Meusel, Jäger, 1992a, b]. В европейской час
ти России этот крестовник в пределах естественного ареала встречается толь
ко на Северо Западе [Цвелев, 2000].
Вторичный ареал
История интродукции и географическое распространение. Продвиже
ние этого крестовника с запада на восток началось в XIX веке. И.Ф. Шмальга
узен [1897] сообщал, что вид встречается редко и только в западных р нах Рос
сии1. Образцы начала XIX века из гербария М.А. Максимовича (MW), по ви
димому, относятся к культивируемым растениям [Игнатов и др., 1990]. К
настоящему времени S. viscosus широко встречается, по видимому, на большей
части Европейской России, кроме самого юга территории [Конечная, 1994].
Заносное на Дальнем Востоке, в Северной Америке [Конечная, 1994; Барка
лов, Таран, 2004; Barkley, 2006].
Пути и способы заноса. В Европейской России, кроме Северо Запада,
S.viscosus появился сначала на железнодорожных насыпях, откуда позднее и
проник в естественные местообитания. В Ирландии расселение крестовника
также связывают с железными дорогами [Reynolds, 2002].
Статус в регионе. Первые находки крестовника клейкого приурочены к
Московской губернии, где в начале XX века его собирали неоднократно [MW;
Игнатов и др., 1990]. Первоначальное его расселение шло медленно. В.Н. Воро
шилов с соавт. [1966] все еще оценивали S. viscosus как редкий вид. С 1970 х
годов он становится массовым видом в Подмосковье [Игнатов и др., 1990], сле
дуют его многочисленные находки в других областях. К настоящему времени
S.viscosus встречается, по видимому, во всех регионах Средней России (рис. 83).
Местообитания. Крестовник клейкий относится к светолюбивым расте
ниям, обитающим в сухих местообитаниях на относительно бедных почвах, и
1 Следует помнить, что в это время в состав Российской империи входила Польша.
196
не выносит засоления, но в Великобритании этот вид предпочитает несколько
более влажные и плодородные почвы при широком диапазоне pH [Hill et al.,
1999]. По видимому, естественными местообитаниями для S.viscosus были пес
чаные холмы и прибрежные дюны [Meusel, Jäger, 1992a]. У растений, произра
стающих на угольных отвалах в Шотландии, обнаружена арбускулярная ми
кориза с Endogone sp. [Daft, Nicolson, 1974].
Крестовник клейкий встречается преимущественно по ж/д насыпям (рис. 84),
у дорог, реже возле жилья. В последние годы неоднократно встречен по сби
тым боровым пескам, вырубкам.
Размножение и жизненный цикл. По одним данным, S.viscosus --- озимый
однолетник [Meusel, Jäger, 1992a; Kadereit et al., 1995], по другим, --- яровой
[Harvie, 2004]. По нашим наблюдениям, в условиях Средней России S.viscosus
развивается исключительно по яровому типу. Крестовник посещается насеко
мыми [Gibson et al., 2006]. Тем не менее S.viscosus --- самоопыляемый вид, что
подтвердили опыты по изоляции корзинок [Akeroyd et al., 1978; Emig, Kadereit,
1993; Snell, Aarssen, 2005]. Цветение преимущественно в июле---августе (рис. 85).
В условиях Шотландии 1 растение производит от 30 до 900 семянок при по
тенциальной продуктивности до 2300--2400 семянок на 1 растение [Harvie,
2004]. Для S.viscosus характерно формирование почвенного семенного банка
[Harvie, 2004].
Расселение и распространение. Семянки S.viscosus распространяются вет
ром [Harvie, 2004].
Последствия внедрения
Воздействия на естественные фитоценозы и аборигенные виды. Крес
товник клейкий до недавнего времени был распространен почти исключитель
но на ж/д насыпях. В последние десятилетия он отмечен на вырубках, песча
ных пустырях, однако роль его в растительном покрове незначительна. В ес
тественных местообитаниях при зарастании нарушенных участков S. viscosus
легко вытесняют местные виды [Harvie, 2004].
В Европе известны гибриды с S.sylvaticus L. -- S. x viscidulus Scheele, эти
растения в основном стерильные [Stace, 1997]. Такие растения на территории
России неизвестны.
Генетические изменения и вариабельность вида. Для европейских попу
ляций S. viscosus указаны голые семянки [Stace, 1997; Hodálovб et al., 2004].
Однако среди наших растений изредка встречаются растения с явно опушен
ными семянками. Вероятно, это можно объяснить интрогрессивной гибриди
зацией с местными видами крестовника sect. Senecio: S.vulgaris L. или S.sylvaticus
L. Аналогичным образом объясняется возникновение разновидности S. vulgaris
с заметными краевыми цветками, так называемую var. hibernicus Syme, как ре
зультат гибридизации S. vulgaris и S. squalidus L. [Ingram et al., 1980; Abbott,
1992; Abbott et al., 1992; Lowe, Abbott, 2000]. Эта гипотеза, однако, требует
специальных доказательств, как в случае с var. hibernicus.
Специальные исследования показали очень низкий уровень межпопуля
ционной изменчивости английских и шведских популяций S. viscosus [Akeroyd
et al., 1978; Koniuszek, Verkicij, 1982]. Такой низкий уровень изменчивости не
редко считают результатом заносного характера локальных популяций [Webb,
1985]. Есть, правда, и иное возможное объяснение такому уровню изменчиво
сти: недавнее происхождение S. viscosus --- позднеплейстоценовое или даже
голоценовое [Kaderit et al., 1995].
Рис. 82. Клейкие листья Senecio viscosus
Рис. 83. Рассе
ление Senecio
viscosus в Сред
ней России
Рис. 84. Расселение Senecio
viscosus по железнодорожным
путям
Рис. 85. Корзинки
Senecio viscosus
199
Экономическое и социальное (положительное и отрицательное) зна
чение. S.viscosus как сорное растение не имеет серьезного экономического зна
чения.
Меры борьбы
S. viscosus уничтожается на железных дорогах при обработке гербицидами.
Встречаемость его на магистральных путях в последние годы стала заметна
меньше. При обработках следует уделять большее внимание сортировочным
и подъездным путям.
СЕМЕЙСТВО ASTERACEAE (COMPOSITAE) --
СЛОЖНОЦВЕТНЫЕ
ЗОЛОТАЯ РОЗГА КАНАДСКАЯ, или ЗОЛОТАРНИК КАНАДСКИЙ
Название: Solidago canadensis L.
Синонимы: S. altissima L., S. canadensis ssp. altissima (L). Bolos et Vigo, S.
canadensis var. scabra Torr. et A.Gray.
Solidago canadensis --- высоковариабельный вид. Таксономический статус
его не вполне ясен и труден для оценки. В естественном ареале в Северной
Америке в комплексе S. canadensis различают несколько таксономических
субъединиц, которым некоторые авторы предоставляют видовой статус [Weber,
2000]. В строгом смысле этот комплекс в Европе рассматривается как S. altissima
[Prieditis, 2002].
Морфологические признаки
В Европе побеги S. canadensis достигают высоты 70--210 см, в основании
они голые, а выше, по крайней мере в верхней половине, опушенные, имеют
40--110 листьев, несколько уменьшающихся в размере снизу вверх. Стебле
вые листья ланцетные, суживающиеся, снизу опушенные, с зубчатыми края
ми, с 2 заметными боковыми жилками. Соцветие представляет собой широко
пирамидальную метелку с центральной осью и отогнутыми боковыми ветвя
ми. Листочки обертки соцветия линейные, тупые или заостренные. Цветки
желтые, собраны в многочисленные мелкие корзинки. Число корзинок на од
ном побеге -- от 41 до 4600 (в среднем 1400). Внутренних трубчатых цветков
обычно меньше, чем краевых язычковых, венчик длиной 2,4--2,8 мм. Плод --
семянка длиной 0,9--1,2 мм, короткоопушенная, с паппусом длиной 2--2,5 мм.
От близкого вида S.gigantea Ait. отличается опушенным стеблем (рис. 86),
опушенной нижней стороной листа и резко зазубренным краем по всей длине
листа [Квитка, Виноградова, 2006].
Основное число хромосом S. canadensis х = 9, и в естественном ареале этот
вид наиболее часто представляет собой гексаплоид (2n = 54), однако встреча
ются и триплоидные (2n = 27), и тетраплоидные (2n = 36) цитотипы. В Евро
пе найдены только диплоидные растения (2n = 18) [Raven, 1960; Tutin et al.,
1976; Weber, 2000].
Естественный ареал
S. canadensis естественно произрастает в Северной Америке от 26 до 45° с. ш.,
достигая 65° с. ш. на западе Канады и на Аляске. В США вид найден от
штата Северная Дакота до штатов Флорида, Техас и Аризона на юге, а в
200
Канаде --- от п ова Новая Шотландия до провинции Онтарио [Hegi, 1979;
Weber, 2000].
Вторичный ареал
История интродукции и географическое распространение. S. canadensis
--- одно из самых старых декоративных растений, интродуцированных в Ев
ропу из Северной Америки. В Англии вид известен с 1645 г. [Kowarik, 2003].
Сначала золотую розгу культивировали в ботанических садах и распростра
няли по питомникам. Поскольку вид весьма привлекателен и легко выращи
вается, его широко использовали садоводы.
Даты начала регистрации S. сanadensis, как натурализовавшегося вида в
Европе: 1857 г. --- в Германии, 1864 г. --- в Швеции, 1866 г. --- в Дании [Weber,
1998], 1872 г. --- в Польше [Tokarska Guzik, 2003], 1887 г. --- в Норвегии [Weber,
1998], 1907 г. --- в Латвии [Kupffer, 1907], 1910 г. --- в Финляндии [Jalas, 1980]
и 1983 г. --- в Литве [Gudzinskas, 1997]. В Эстонии S. canadensis интродуциро
вана в 1807 г. в Ботаническом саду Тарту.
В начале XX века виды рода Solidago стали рассматривать не только как
декоративные, но и как каучуконосные растения. Н.И. Вавиловым в Россию
были завезены семена наиболее перспективного каучуконосного вида S.
leavenworthii Torr. et Gray, а также семена других золотарников, в том числе 16
образцов S. canadensis. В 1937 г. на участках Интродукционного питомника
субтропических культур в г. Сухуми было представлено уже 50 видов Solidago
[Волховская, 1937]. Для производства каучука все эти виды оказались непри
годны, но успешно акклиматизировались, благодаря нетребовательности к
условиям произрастания.
В настоящее время S. canadensis встречается от юга Скандинавии до Север
ной Италии. Помимо Европы, S. canadensis натурализовалась в Австралии,
Японии и Новой Зеландии, на Tайване, в Закавказье и Сибири [Weber, 2000].
Рассеянные популяции S. canadensis довольно обычны на севере Централь
ной Норвегии до 63о с. ш., а одна находка сделана в графстве Nordland (~65,5о
с. ш.); на юго востоке Норвегии S. canadensis широко распространена и быст
ро расселяется [Sunding, 1989]. Вид обилен в Польше, особенно в южных вое
водствах [Weber, 2000; Solarz et al., 2005]. В Финляндии S. canadensis натура
лизуется почти до 63о с. ш., но более или менее обычен только в южных, плот
но населенных регионах, подобно пригороду Хельсинки --- Hameenlinna [H;
Kurtto, Ulynranta, 1998]. В Эстонии S. canadensis распространена спорадичес
ки. В Латвии и Литве натурализовалась во всех р нах [Kuusk et al., 2003;
Gudzinskas, 1997]. В Исландии S. canadensis выращивается в Ботаническом
саду Рейкьявика с 1967 г. и культивируется как декоративный многолетник с
начала 1990 х гг. В Исландии цветет поздно, и его семена не вызревают
[D.Jakobsdottir, личное сообщение).
Пути и способы заноса. S. canadensis интродуцирована из Северной Аме
рики в Европу как декоративное растение [Weber, 1998], и многие из натура
лизовавшихся популяций вида являются результатом «бегства из культуры»
из ботанических садов, питомников декоративных растений или из старых
усадеб [Kurtto, Helynranta, 1998].
Статус в регионе. Как интродуцированное растение отмечено в Московс
кой обл. в 1863 г., в Тульской обл. --- в 1880 г., в Костромской обл. --- в 1882 г.
(MW). В настоящее время как декоративное и изредка дичающее растение
201
Рис. 87. Расселение Solidago сanadensis в Средней России
Рис. 86. Побеги Solidago
сanadensis (вверху)
и Solidago gigantea
(внизу)
Рис. 88. Внедрение Solidago
сanadensis в луговые сообщества,
Московская обл., пойма
р. Клязьма
Рис. 89. Плодоношение
Solidago сanadensis
203
встречается по всей Средней России (рис. 87). В естественные ценозы актив
но внедряется с 2000 х гг.
Местообитания. В естественном ареале S. canadensis обнаружена, глав
ным образом, на опушках леса и по обочинам дорог, на заброшенных полях и
других неконтролируемых местообитаниях, которые вид колонизирует сразу
же, как только их перестают обрабатывать (рис. 88). S. canadensis может встре
чаться на полях зерновых культур, но ее не считают серьезным сорняком од
нолетних зерновых культур, так как она может удаляться при вспашке. Одна
ко вид вторгается на заброшенные пастбища и может также быть серьезным
сорняком в лесных питомниках и среди многолетних зерновых культур и са
довых растений [Werner et al., 1980], потому что S. canadensis распространяет
ся очень быстро, а главное, в широком диапазоне почвенных условий, как по
структуре, так и по плодородию почвы [Weber, 2000].
Во вторичном ареале S. canadensis благодаря высокой адаптационной спо
собности может произрастать на почвах различного механического состава ---
от легких песчаных до тяжелых глинистых. Она найдена на многих нарушен
ных участках: вдоль ж/д путей, по обочинам дорог, на заброшенных полях,
опушках леса, нарушенных лесах и по берегам рек [Weber, 2000; Prieditis, 2002].
Местообитания не всегда только нарушенные, поскольку вид присутствует и
в естественных прибрежных фитоценозах.
Размножение и жизненный цикл. S. canadensis --- корневищный гемик
риптофит. Отдельные клоны живут подолгу и достигают возраста 100 лет.
Отдельные невысокие побеги могут оставаться вегетативными в течение 1--3
лет, но при хороших условиях растения способны к репродукции уже на 1 м
году жизни [Weber, 2000]. Цветение начинается в конце июля, но пик наблю
дается между серединой августа и концом сентября, а порой может продол
жаться весь октябрь [Weber, 2000].
S. canadensis опыляют насекомые. Частые посетители золотой розги в
Европе --- представители Phalacridae, Muscidae, Syrphidae, Apidae, Formicidae,
Sphecidae и Panorpidae [Weber, 2000]. Биология цветения изучена недостаточ
но. По одним данным, цветки S. canadensis протерандричные, самостерильные,
а потому вид относят к строгим перекрестноопылителям [Melville, Morton,
1982]. По другим данным, пыльцевые зерна S. canadensis стерильные, поэтому
данный вид не способен к нормальному формированию семян и размножает
ся вегетативно [Pullaiah, 1978].
Расселение и распространение. S. canadensis распространяется семенами
и фрагментами корневищ. Семена продуцируются в большом количестве --- в
Европе один генеративный побег (рис. 89) может продуцировать >10 тыс. се
мян [Weber, 2000]. Семена рассеиваются на дальние расстояния, что обеспе
чивает колонизацию незанятых участков. Рассеиванию ветром способствуют
небольшие размеры семян и низкая скорость их падения на почву [Weber,
2000]. Семена, падающие с высоты 1 м над уровнем почвы, при скорости ветра
до 5 м/сек, рассеиваются в среднем на 0,3 м (0,6--2,4 м).
Распространение на близкое расстояние возможно с помощью фрагментов
корневищ. Популяция разрастается, главным образом, в результате роста кло
нов [Weber, 2000].
Случайная интродукция возможна как побочный результат деятельности
человека --- например, сбору соцветий на букеты, которые после отцветания
204
выбрасывают в кучи мусора. Осенью общепринято срезать в садах отцветшие
побеги и также удалять их в кучи мусора вне садов; если рядом с местами скоп
ления мусора протекают ручьи или речушки, их потоки могут распространять
побеги вместе с семенами по течению. Таким же образом фрагменты корневищ
могут образовывать новые популяции вдоль рек и ручьев. Семена и корневища
могут распространяться и в результате транспортировки земли в ходе строитель
ных работ, при прокладывании автодорог и железных полотен [CABI, 2004].
Последствия внедрения
Воздействия на естественные фитоценозы и аборигенные виды. Боль
шой вторичный ареал S. canadensis --- результат неправильного управления
использованием земли, что позволяет виду обосновываться на новом месте и
конкурировать с аборигенными растениями, ведя к серьезным изменениям и
отрицательно воздействуя на флору так, что характерные для отдельных пун
ктов виды могут исчезнуть полностью.
Маловидовые сообщества S. canadensis преобразуют естественные ландшаф
ты [CABI, 2004]. Внедрившись в то или иное местообитание, S. canadensis мо
жет оставаться доминантом в течение длительного времени. Благодаря кло
нальному росту S. canadensis может создавать густые заросли, плотность кото
рых достигает 309 побегов/м2 [Weber, 2000; Kowarik, 2003].
Генетические изменения и вариабельность вида. В естественном ареале
в пределах популяции могут существовать генетические вариации вида, ус
тойчивые к гербицидам. В Европе отмечена широкая генетическая как внут
ри , так и межпопуляционная изменчивость по фенологическим и морфоло
гическим признакам [Weber, 2000]. S. canadensis иногда гибридизирует с або
ригенной S.virgaurea, по крайней мере в Фенноскандии [Nilsson, 1976; Sunding,
1989; Mossberg, Stenberg, 2003]. Однако виды столь сильно различаются по
времени цветения (S. canadensis цветёт почти на месяц позднее), что малове
роятно, чтобы подобная гибридизация происходила в крупном масштабе.
Влияние на здоровье человека. Вопреки сложившемуся мнению, золотар
ники играют весьма незначительную роль в проявлении сенной лихорадки.
Период цветения этих растений совпадает с сезоном самой высокой вспышки
сенной лихорадки, и, поскольку вид наиболее заметен, его подозревали ви
новником заболевания. Пыльца золотарников провоцирует, конечно, сенную
лихорадку, но обычно тяжелая липкая пыльца вида переносится насекомыми
или падает на землю недалеко от растений. Лишь изредка в сухую и ветреную
погоду пыльца золотарников, рассеянная в воздухе, может причинить вред
чувствительным людям.
Экономическое и социальное значение (положительное и отрицатель
ное). S. canadensis --- альтернативный хозяин для насекомых, которые могут
быть переносчиками патогенов зерновых культур, однако специального изу
чения этого явления не проводилось [CABI, 2004]. S. canadensis выращивает
ся в Европе как декоративное растение. Регионы с доминированием S.
canadensis пригодны для производства меда. Венгерские исследователи пред
положили, что ацетоновый экстракт из S. canadensis может влиять на другие
сорняки как аллелопатическое средство [Solymosi, 1994].
S. canadensis --- лекарственное растение; используется в европейской фи
тотерапии как урологическое и противовоспалительное средство [Apati et al.,
2003]. Надземная часть содержит ряд действующих веществ: эфирное масло,
205
дубильные вещества, горечи, сапонины и флавоноиды. Государственная служ
ба здравоохранения Германии рекомендует S.саnadensis для промывания при
воспалении мочевыводящих путей и мочекаменной болезни. Кроме того,
S.саnadensis может оказывать стимулирующее воздействие на обмен веществ,
эффективен при кожных болезнях, а также при заболеваниях печени. Экст
ракт надземной части S.саnadensis входит в состав российского препарата «Про
станорм», используемого при лечении заболеваний предстательной железы.
Изучена амплитуда изменчивости содержания кремния и 4 групп феноль
ных соединений в инвазионных популяциях S.саnadensis в различных точках
вторичного ареала вида [Акимова и др., 2008]. Уровень изменчивости по об
щему содержанию полифенолов между образцами разных видов из различ
ных местообитаний очень низкий. Однако содержание полифенолов в разных
органах растений (стебель, лист, соцветие) сильно варьирует (табл. 16). В ли
стьях золотарников суммарное количество полифенолов в среднем составля
ет от 5,39 до 6,29%. Содержание простых полифенолов и оксибензойных кис
лот варьирует от 0,34 до 0,55%, фракции оксикоричных кислот --- от 0,12 до
0,17%, конденсированных полифенолов (дубильные вещества) --- от 1,35 до
1,72%. Содержание веществ флавоноидного комплекса изменяется в диапазо
не от 3,45 до 3,94%. В состав флавоноидного комплекса входят: кверцетин 3
О глюкозид (1,32%), кверцетин 3 О рамнозид (0,1%), рутин (0,43%), кемп
ферол (0,4%), кемпферол 3 О глюкозид (1,13%), лютеолин 7 О глюкозид
(0,22%), апигенин 7 О глюкозид (0,09%).
В соцветиях золотарников общее содержание полифенолов снижается на
20% и в среднем составляет 4,75% абсолютно сухой массы. При этом содержа
ние наиболее ценных веществ --- флавоноидов снижается на 30% по сравне
нию с листьями, а содержание наименее ценных дубильных веществ повыша
ется на 10%. Содержание простых полифенолов и оксибензойных кислот ва
рьирует от 0,32 до 0,47%, фракции оксикоричных кислот --- от 0,10 до 0,14%,
конденсированных полифенолов --- от 1,64 до 1,84%. Содержание веществ
флавоноидного комплекса изменяется в диапазоне от 2,4 до 2,7%. В состав
флавоноидного комплекса входят: кверцетин 3 О глюкозид (1,15%), кверце
тин 3 О рамнозид (0,06%), рутин (0,08%), изорамнетин 3 О глюкозид (0,07%),
кемпферол 3 О глюкозид (0,3%), кемпферол 3 О рамнозид (0,03%), лютео
лин 7 О глюкозид (0,3 %), апигенин --- 7 О глюкозид (0,03%).
Таблица 16
Фракционный состав полифенолов (ПФ) в различных органах золотарников
(% абс. сухой массы)
Простые ПФ и фенол-
карбоновые кислоты
Дифенил-
пропаноиды
Орган Общая
сумма
ПФ простые ПФ
и оксибензой-
ные кислоты
Оксико-
ричные
кислоты
сумма
флаво-
ноидов
апи-
ге-
нин
Конденси-
рованные
ПФ (ду-
бильные
вещества)
Листья
5,86
0,47
0,15
3,69 0,1
1,55
Соцветия 4,75
0,39
0,11
2,51 0,3
1,73
Стебли
3,25
0,26
0,06 0,61
0
2,32
206
В стеблях общее содержание полифенолов минимальное, почти на 50%
ниже, чем в листьях (от 3,04 до 3,5%), при этом количество конденсирован
ных полифенолов максимальное (на 50% больше, чем в листьях). Содержание
простых полифенолов и оксибензойных кислот варьирует от 0,22 до 0,3%,
фракции оксикоричных кислот --- от 0,05 до 0,09%, конденсированных поли
фенолов --- от 2,18 до 2,44%. Общее содержание веществ флавоноидного ком
плекса --- 0,53--0,71%; это кверцетин (0,12%), кверцетин 3 О глюкозид (0,25%),
лютеолин 7 О глюкозид (0,24%), остальные флавоноиды в стеблях золотар
ников не обнаружены.
Для золотарников характерно довольно значительное содержание общего
кремния: до 1,56% абс. сухой массы. Наибольшее количество суммарного крем
ния содержится в листьях (на 15% больше, чем в стеблях и соцветиях). В наи
большей мере (1,23%) представлен в листьях и наиболее ценный для организ
ма человека органический кремний. В несколько меньшей степени (0,74%) он
имеется в корзинках, а минимальное его количество (0,45%) отмечено в стеб
лях. Содержание минерального растворимого кремния, напротив, в этом ряду
возрастает (листья --- 0,12%; соцветия --- 0,17%; стебли --- 0,23%). Подобным
образом возрастает и содержание малоусваиваемого полимерного кремния
(листья --- 0,21%; соцветия --- 0,25%; стебли --- 0,50%).
Таким образом, у всех видов золотарников, независимо от места произрас
тания, содержание простых полифенолов, фенолкарбоновых кислот и флаво
ноидов в ряду листья---корзинки---стебли снижается, а содержание дубильных
веществ, напротив, возрастает.
Сходные закономерности выявлены и относительно содержания кремния:
межвидовая и географическая изменчивость этого признака несущественны,
а превалирует изменчивость между отдельными органами растений.
Существенных отличий по содержанию фенольных соединений и кремния
между инвазионными золотарниками и лекарственной S.virgaurea не выявле
но. В связи с этим можно прогнозировать успешность применения листьев и
соцветий инвазионных видов рода Solidago для фармацевтических целей.
Меры борьбы
S. canadensis внесен в «Список инвазионных видов ЕРРО», в котором пе
речислены виды, наносящие серьезный ущерб аборигенным видам растений,
окружающей среде и биологическому разнообразию. Странам, в которых про
израстает вид, рекомендовано принять меры, чтобы предотвратить его даль
нейшую интродукцию и распространение.
Имеется несколько механических способов удаления зарослей S. canadensis.
Один из эффективных способов избавления от видов рода Solidago --- коше
ние дважды в год (в мае и августе) в течение нескольких лет или перекапыва
ние почвы в течение лета при сухих погодных условиях. После кошения золо
тарник можно накрыть легкой непроницаемой полимерной пленкой, что так
же снижает рост растений. Однако этот метод уничтожает всю растительность.
Применимость различных мер зависит от состояния участка (влажный, су
хой), степени насыщенности региона зарослями золотарника и представлен
ности других таксонов --- например, наличия редких видов, находящихся под
охраной [Weber 2000].
В естественном ареале расселение S. canadensis находится под биологичес
ким контролем, поскольку на распределение биомассы влияют травоядные
207
животные. В Европе влияние травоядных невелико. Улитки и мелкие грызу
ны изредка питаются стеблями и листьями вида. В Швейцарии известно 18
насекомых фитофагов, питающихся S. canadensis [Weber, 2000]. Нет данных о
выпасе скота, а это могло бы стать хорошим методом контролирования чис
ленности S. canadensis после механического удаления вида.
Молодые растения могут контролироваться химическими методами, пото
му что они чувствительны к гербицидам, но ближе к концу вегетационного
сезона почвенные гербициды становятся малоэффективными. Глифосат и не
которые другие контактные гербициды применяются для обработки S.
canadensis при высоте растений 10--15 см [CABI, 2004].
Обзор научных исследований и публикаций
S. canadensis продолжает использоваться как декоративное до сих пор и
имеется в каталогах и на веб сайтах коммерческих питомников и ботаничес
ких садов, так что вполне вероятна и дальнейшая его интродукция. Есть не
сколько информационных листов о биологии, экологии и распространении S.
Canadensis --- например, немецкие веб страницы NeoFlora и Crop Protection
Compedium.
Виды рода Solidago интенсивно изучались по химическому составу. У
Solidago отмечено наличие дитерпенов, которые могут применяться в качестве
средств от поедания насекомыми и как ингибиторы роста [Weber, 2000].
СЕМЕЙСТВО ASTERACEAE (COMPOSITAE) --
СЛОЖНОЦВЕТНЫЕ
ЗОЛОТАЯ РОЗГА ГИГАНТСКАЯ,
или ЗОЛОТАРНИК ГИГАНТСКИЙ
Название: Solidago gigantea Ait.
Синонимы: Aster latissimifolius (Miller) Kuntze var. serotinus Kuntze; Doria
dumetorum (Lunell) Lunell; D. pitcheri (Nuttall) Lunell; Solidago cleliae DC.; S.
dumetorum Lunell; S. gigantea var. leiophylla Fernald; S. gigantea var. pitcheri
(Nuttall) Shinners; S. gigantea subsp. serotina (Kuntze) McNeill; S. gigantea var.
serotina (Kuntze) Cronq.; S. gigantea var. shinnersii Beaudry; S. pitcheri Nuttall; S.
serotina Ait., 1789, not Retz. 1781; S. serotina var. gigantea (Ait.) A. Gray; S. serotina
var. minor Hooker; S. serotinoides Б.Lцve et D.Lцve; S. shinnersii (Beaudry) Beaudry;
S. somesii Rydb.
Морфологические признаки
Solidago gigantea --- высоковариабельный вид с неясным и трудным для оцен
ки таксономическим статусом. Это прямостоячее многолетнее длиннокорне
вищное растение. Надземные побеги достигают от 30 до 280 см в высоту и от 5
до 11 мм в диаметре. Стебли ветвятся только в соцветиях, которые составля
ют в среднем 1/3 общей высоты побега, хотя такое соотношение довольно силь
но варьирует (10--60%) и зависит от популяции, географического положения
и времени цветения. Более высокие растения имеют тенденцию к позднему
сроку цветения. Побеги до начала соцветия гладкие, часто пурпурные и по
крыты беловатым восковым налетом.
208
Листья S.gigantea простые, очередные, продолговато ланцетные, длиной 80--
180 мм и шириной 10--30 мм. В течение вегетационного сезона образуется до
90 листьев. Более крупные листья образуются в середине стебля, листья мель
че --- в верхней и нижней частях стебля и в соцветиях. Листья с тремя более
или менее заметными главными жилками; центральная жилка выделяется
более явно, нежели латеральные жилки. Край листовой пластинки слабозуб
чатый вдоль всего листа, размер зубчиков варьирует в популяции весьма зна
чительно. Листовые пластинки обычно гладкие сверху и снизу, но изредка
могут быть опушенными на нижней стороне по главной жилке.
В верхнем слое (10--20 см) почвы S.gigantea, в отличие от S.canadensis, обра
зует длинные плагиотропные фиолетовые или красноватые корневища (рис. 90).
Одно растение формирует за год от 3 до 50 корневищ. При неблагоприятном
воздействии на растение число образующихся корневищ возрастает. Корне
вища достигают длины до 90 см и диаметра 1 см, часто ветвятся. На корневи
щах образуются почки (до 2,2 на 1 см), наибольшая их густота отмечается у
основания стебля и на конце корневища.
Форма соцветий --- пирамидальная метелка. Обычно серповидно изогну
тые веточки (рис. 91) содержат многочисленные (1200 190) корзинки [Schmid
et al., 1988]. Корзинка диаметром 2--3 мм, обертка соцветия имеет длину 3--4
мм. В корзинке имеется два типа цветков: краевые ярко желтые язычковые
женские и трубчатые обоеполые. Трубчатые цветки длиной 4--8 мм. Плоды
длиной 1--1,8 мм с паппусом длиной 2,5--4 мм, опушенные длинными волос
ками, которые способствуют распространению семянок с помощью ветра
[Weber, Jakobs, 2005].
От близкого вида, S.canadensis, хорошо отличается наличием корневищ,
голым стеблем с беловатым налетом, неопушенной нижней стороной листа,
более плотной архитектурой соцветия и коричневатым оттенком паппуса
[Квитка, Виноградова, 2006].
Основное число хромосом Solidago gigantea х = 9, в естественном ареале у
вида выделено три цитотипа: диплоид 2n=18, тетраплоид 2n=36, гексаплоид
2n=54. J.Beaudry рассматривал цитотипы 2n=18 и 2n=36 как 2 отдельных вида:
S.gigantea и S.serotina Ait. Позднее он выделил в пределах S.gigantea s.l. и тре
тий цитотип --- S.shinnersii, имеющий 2n=54 [Melville, Morton,1982].
Ранее считалось [Weber, Schmid,1998], что в Европе натурализовалась толь
ко тетраплоидная форма S.gigantea. Однако в последнее время в Европе были
обнаружены все три цитотипа [Weber, Jakobs, 2005], поэтому имеет смысл рас
сматривать S.gigantea как комплекс:
S.gigantea 2n = 2x =18 (опушение по главной жилке с нижней стороны
листа);
S.serotina 2n = 4x =36 (гладкие узкие листья);
S.shinnersii 2n = 6x = 54 (гладкие широкие листья).
Естественный ареал
S. gigantea естественно произрастает по всей Северной Америке южнее 55°с.
ш., за исключением штата Аризона. Существует определенный характер в рас
пределении трех цитотипов: диплоидная форма тяготеет к востоку гор Аппа
лачи, тетраплоидная форма распространена по восточной лесной территории,
гексаплоидная форма тяготеет к прериям [www.eFlora.org].
Рис. 90. Корневища
Solidago сanadensis
(вверху) и Solidago
gigantea (внизу)
Рис. 91. Соцветия
Solidago gigantea
(слева) и Solidago
сanadensis
(справа)
Рис. 93. Внедрение Solidago gigantea в прибрежные сообщества
Рис. 92. Расселение Solidago gigantea в Средней России
211
Вторичный ареал
История интродукции и географическое распространение. Впервые как
декоративное интродуцированное растение S.gigantea зарегистрирована в Бо
таническом саду Лондона в 1758 г. [Weber, 1998]. Вскоре она была отмечена в
садах и питомниках континентальной Европы, но происхождение этих ини
циальных популяций остается неизвестным. В первой половине ХIХ века
S.gigantea зарегистрирована во Франции, в Швейцарии и Германии [Weber,
Jakobs, 2005]. Вид стал инвазионным через ~100 лет: первое упоминание о само
произвольном распространении в Германии датируется 1832 г., в Австрии ---
1857 г. [Wagenitz, 1979]. Между 1850 и 1880 гг. описано много новых местона
хождений, и растение начало распространяться по всей Центральной Европе
[Wagenitz, 1979]. На основании гербарных образцов и литературных источни
ков, можно утверждать, что скорость экспансии составляла ~910 км2/год
[Weber, 1998]. В 1950 г. S.gigantea уже распространилась по всей Европе от
севера Испании до Восточной Европы и от Северной Италии до юга Сканди
навии [Tutin et al., 1964]. В настоящее время ареал вида в Европе простирает
ся от 42 до 63°с.ш. и ожидается дальнейшее расширение его границ в восточ
ном направлении [Meusel, Jager, 1992; Weber, 2001]. S.gigantea встречается,
главным образом, ниже 1200 м над ур. м., но может иногда наблюдаться на
больших высотах [Weber, Jakobs, 2005].
В европейской части ареала отмечено локальное разнообразие фенотипов:
внутривидовые формы широко распространены в Центральной и Восточной
Европе, но значительно реже встречаются в Скандинавии и Северной Герма
нии. По мнению ряда ученых [Botta Dukát, Dancza, 2001a], именно высокий
уровень изменчивости морфологических признаков обеспечил столь высокий
инвазионный потенциал вида в Европе.
Помимо Европы, S. gigantea также натурализовалась в Японии, Корее
[Enomoto, 2000; Tsuyuzaki, 2002] и на Дальнем Востоке РФ [МНА; Баркалов,
1992], в Новой Зеландии, Австралии, на Азорских и на Гавайских о вах, а так
же в Мексике [Weber, Jakobs, 2005].
Пути и способы заноса. S.gigantea интродуцирована в Европу как декора
тивное растение и пользовалась большим успехом у садовников из за легкос
ти выращивания. Культивирование золотой розги на приусадебных участках
до сих пор является основным путем заноса вида на новые территории.
При натурализации S.gigantea вначале захватывает рудеральные нарушен
ные территории --- берега рек и обочины автомобильных дорог и ж/д полотна.
С конца ХIХ века S.gigantea также закрепилась в полуестественных и есте
ственных местообитаниях [Guzikowa, Maycock, 1986]. Урбанизация, строитель
ство автомобильных дорог и ж/д путей, заброшенные поля способствуют рас
пространению вида. Избавление от садового мусора (удаление растений с дач
ных участков) и выращивание золотой розги как медоносного растения также
содействуют его инвазии в новые места обитания [Weber, Jakobs, 2005]. По
сравнению с другими видами золотарника, интродуцироваными в Европе, ско
рость распространения S. gigantea оказалась наиболее высокой [Guzikowa,
Maycock,1986; Weber, 1998], и сегодня вид обоснованно относится к одним из
наиболее агрессивных инвазионных растений Европы.
В 1937 г. на участках Интродукционного питомника субтропических куль
тур в г. Сухуми, куда Н.И. Вавиловым были завезены в Россию вместе с перс
212
пективным каучуконосом S. leavenworthii Torr. et Gray семена других золотар
ников, было представлено уже 50 видов Solidago L., в том числе 1 образец
S.gigantea [Волховская,1937].
Статус в регионе. Как интродуцированное растение отмечено в Воронеж
ской обл. в 1868 г. [MW]. В настоящее время как декоративное и изредка ди
чающее растение встречается по всей Средней России (рис. 92). В естествен
ные ценозы активно внедряется в последнее десятилетие. Массовые заросли
отмечены в Калужской, Курской и Московской обл.
Местообитания. В естественном ареале S.gigantea является частью сооб
ществ прерий и заливных лугов. Это высокотолерантный вид, не требователь
ный к свету, влажности почвы, содержанию питательных веществ, температу
ре и рН фактору [Weber, Jakobs, 2005]. Хотя S. gigantea предпочитает богатые
и достаточно увлажненные почвы, она встречается на различных по структу
ре грунтах и на разных типах почв. Наилучшего развития S. gigantea достигает
в прибрежных местообитаниях и на топких местах, но растет также и в сухих
местах, таких как обочины автомобильных дорог и ж/д насыпи.
Во вторичном ареале в Европе S.gigantea занимает схожие экологические
ниши. Вид произрастает, главным образом, в местообитаниях, богатых азотом,
но также найден на относительно бедных питательным веществом участках ---
пустырях и обочинах дорог. На увеличение питательных веществ в почве вид
отвечает значительным увеличением размеров, особенно размера соцветий и
числа произведенных семян. В сырых и влажных местообитаниях ее жизнен
ность выше, чем в сухих местах, и часто этот вид образует плотные одновидо
вые насаждения (рис. 93). Однако продолжительного затопления этот вид не
выносит. В более сухих местах S. gigantea часто сосуществует с другими вида
ми [Botta Dukát, Dancza, 2001a], и в таких условиях вид менее конкурентос
пособен. S.gigantea реагирует на фактор сухости уменьшением листовой по
верхности и редукцией общей биомассы. Механическое удаление корневищ
может сократить доминирование S.gigantea в таких местообитаниях, посколь
ку скорость побегообразования замедляется [Botta Dukát, Dancza, 2001b].
Размножение и жизненный цикл. S.gigantea --- геофит, внутри клонов прак
тически не наблюдается сеянцев, а потому исследователи предполагают, что
семена служат, главным образом, для распространения на дальние расстояния
и заселения новых мест. У близкородственного вида S.altissima внутри клонов
прорастание семян ингибируется дитерпеноидами, выделяемыми растения
ми, подобное может иметь место и в случае S.gigantea [Werner et al., 1980].
Иногда при наличии открытых участков сеянцы могут появляться после ве
сеннего покоса [Weber, Jakobs, 2005]. Однако размер популяции обычно уве
личивается за счет клонального роста. Клон растет, главным образом, в тече
ние первых лет после возникновения, а по мере его старения роль полового
размножения возрастает.
Семеношение наблюдается каждый год, хотя невысокие побеги внутри кло
на могут оставаться вегетативными. С другой стороны, у изолированно расту
щих растений в неблагоприятных условиях могут цвести даже небольшие по
беги высотой до 25 см. В благоприятных условиях почти все растения цветут
и плодоносят на втором году жизни. S.gigantea цветет поздно, с июля по но
ябрь, с пиком цветения в августе--сентябре. Вид является перекрестноопыли
телем: соцветия привлекают опылителей яркими желтыми цветами и боль
213
шим количеством пыльцы и нектара. Частые посетители цветов в Европе ---
виды таких семейств, как Phalacridae, Muscidae, Syrphidae, Apidae, Formicidae,
Sphecidae и Panorpidae [Weber, Jakobs, 2005].
Отрастание побегов начинается весной, и высота их увеличивается почти
линейно до конца июля. Растения заканчивают рост только после формирова
ния соцветия и раскрывания первых цветков, обычно между августом и сен
тябрем. В благоприятных условиях отдельный генеративный побег продуци
рует до 19 тыс. семянок. Семена легко разносит ветер, но большинство семя
нок остаются на стебле в течение зимы. Семянки прорастают весной, но период
прорастания довольно растянут и продолжается до начала лета. Всхожесть
семян S.gigantea может составлять 100%, однако этот параметр зависит от по
чвенных условий: на глинистой почве и на торфе всхожесть составила 47%,
тогда как на фильтровальной бумаге в течение 3 недель проросло 99% семян
[Weber, Jakobs, 2005]. В условиях ограниченных ресурсов половое размноже
ние сокращается, а преобладать начинает вегетативное размножение.
Расселение и распространение. S.gigantea может увеличивать захвачен
ные площади как с помощью семян, которые ветер разносит на далекие рассто
яния, так с помощью фрагментов корневищ, которые переносятся водными по
токами и легко укореняются. Распространение вдоль водного пути часто может
быть заметным; например, популяция золотой розги легко распространилась
вдоль реки Тальяменто на севере Италии [Weber, Jakobs, 2005].
Имеется отрицательная зависимость между массой семян и их численнос
тью, и соотношение этих величин зависит от конкретных почвенно климати
ческих условий. В менее благоприятных условиях и вытаптываемых (часто на
рушаемых) местах семян формируется больше, и они легче, что гарантирует их
перенос на дальние расстояния; там, где условия благоприятнее, семян меньше,
но они крупнее, а это повышает вероятность образования сеянцев в близлежа
щих местах с сомкнутым растительным покровом [Werner, Platt, 1976].
Корневища растут в направлении от центра клона. Корневища довольно
длинные и сильно ветвятся, таким образом, популяции состоят из нескольких
или многих смешивающихся генет. Соотношение генеративных и вегетатив
ных побегов существенно меняется в зависимости от почвы и условий осве
щенности. Отмечено [Weber, Jakobs, 2005], что число генеративных побегов
отрицательно коррелирует с плотностью побегов в популяции. В центре за
рослей сопутствующие виды практически отсутствуют, по направлению к кра
ям число сопутствующих видов повышается [Lukac, 1988]. На некосимом за
ливном лугу в Швейцарии площадь популяции за счет роста клонов увеличи
валась на 0,3--0,8 м/год [Weber, Jakobs, 2005]. В одной рамете за вегетационный
сезон формируется от 5 до 55 корневищ. Большое число почек на корневищах
позволяет растениям быстро повторно отрастать после повреждений. Число
ростовых почек на корневищах у S. gigantea в 10 раз выше, чем у S.canadensis
[Weber, Jakobs, 2005], и это в какой то мере объясняет успешность колониза
ции S. gigantea. Кроме того, корневища S.gigantea в среднем намного более
длинные; таким образом, вид может покрыть большие площади и расширить
размер участка, захваченный популяцией, за относительно более короткий
промежуток времени [Schmid et al., 1988]. При отсутствии травоядных живот
ных корневища остаются в виде неделимой системы в течение нескольких лет.
Вероятно, отсутствие травоядных животных во вторичном ареале может быть
214
одной из причин более высокого числа корневищ, отмечаемых в инвазионных
популяциях. Размер площади заросли S. gigantea зависит от количества осен
них осадков [Weber, Jakobs, 2005]. В норме длина корневищ составляет до 90
см. В благоприятных условиях корневища в среднем короче, и тогда клоны
более плотные; более длинные корневища, которые развиваются в менее бла
гоприятных условиях, имеют больше шансов достигнуть подходящих участ
ков. На недавно заброшенных полях индивидуальные клоны часто легко раз
личимы и достигают диаметра 2--5 м. Крупные старые клоны могут состоять
из 1 тыс. рамет. Иногда клоны формируют «ведьмины круги» вокруг началь
ного центра генеты.
Последствия внедрения
Воздействие на естественные фитоценозы и аборигенные виды. Еди
ножды обосновавшись на новой территории, S. gigantea очень скоро стано
вится доминантом благодаря быстрому росту клонов и высокой конкурентос
пособности. Плотность побегов достигает 30--170 шт/м2, а размер локальных
популяций варьирует от нескольких до 50 тыс. м2 [Weber, Jakobs, 2005].
В результате инвазии вида в фитоценозы аборигенные виды, как правило,
вытесняются, а образуются монодоминантные растительные сообщества раз
личной видовой насыщенности. S.gigantea обычно образует ассоциации с
Achillea millefolium, Aegopodium podagraria, Arrhenatherum elatius, Artemisia
vulgaris , Calystegia sepium, Cirsium arvense, Dactylis glomerata, Elymus repens,
Galium album, G.aparine, Heracleum sphondylium, Symphytum officinale, Tanacetum
vulgare и Urtica dioica. Вид вторгается в полуестественные сообщества, такие
как Equiseto Alnetum, Salicion и Convolvulion, а также во вторичные леса из
Robinia pseudoacacia и в урбанофитоценозы [Weber, Jakobs, 2005].
Рудеральные местообитания обычно явно сокращают видовое разнообра
зие после внедрения в них золотой розги. На богатых видами суходольных
лугах экологическое воздействие может быть еще более серьезным. На лугах с
золотарником в среднем отмечается 12,5 вида, заметно ниже, чем в таких же
местообитаниях без золотарников, где среднее число видов 22,9 [Weber, Jakobs,
2005]. В первую очередь исчезают такие виды, как Lysimachia vulgaris, Potentilla
erecta, Betonica officinalis, Filipendula ulmaria, Scabiosa columbaria, Centaurea
scabiosa, Campanula glomerata и Succisa pratensis. Чем выше численность вида,
тем меньше он реагирует на вселение S.gigantea. В речных долинах, плотные
заросли золотой розги могут подавлять омоложение пойменной растительно
сти. Там S.gigantea часто сосуществует с другими инвазионными видами.
В Западной Европе S.gigantea редко произрастает совместно с другими ви
дами рода Solidago, тогда как в Восточной Европе часто встречаются ассоциа
ции, где произрастают и S. gigantea, и S.canadensis.
Генетические изменения и вариабельность вида. Согласно литературным
данным, S.gigantea и S.canadensis не скрещиваются между собой. Лишь при
искусственном опылении, когда в качестве материнского растения при скре
щивании была взята тетраплоидная форма S.gigantea, получено небольшое
число (10,5%) сформированных семянок, которые, однако, оказались не спо
собными к прорастанию [Melville, Morton,1982].
Экономическое и социальное значение (положительное и отрицатель
ное). S.gigantea выращивают в Европе как декоративное растение. Этот вид
славится как медонос и пергонос, а когда то ценился также красильщиками,
извлекавшими из травы и цветков желтую и коричневую краски.
215
Траву эту игнорируют домашние животные, хотя дикие ее как будто поеда
ют. Имеются данные о поедании S.gigantea в Германии кроликами, которые,
однако, не оказывают существенного влияния на распространение вида. Часть
источников указывает на токсичность растения.
S. gigantea -- лекарственное растение; надземная часть которого содержит
ценные действующие вещества: эфирное масло, дубильные вещества, горечи,
сапонины и флавоноиды. Используется для лечения болезней мочевого пузы
ря и почек. Кроме того, S.gigantea может оказывать стимулирующее воздей
ствие на обмен веществ, быть эффективной при кожных болезнях, а также при
заболеваниях печени. В многочисленные чаи (особенно от ревматизма, подаг
ры, а также так называемые кровоочистительные) как компонент входит и зо
лотая розга.
По количественному и качественному составу фенольных соединений и
кремния S. gigantea и S.canadensis достоверно не различаются [Акимова, Ви
ноградова, Колесников, 2008]. У обоих видов, независимо от места произрас
тания, содержание простых полифенолов, фенолкарбоновых кислот и флаво
ноидов в ряду листья--корзинки--стебли снижается, а содержание дубильных
веществ, напротив, возрастает.
Для обоих видов характерно довольно значительное содержание общего
кремния: до 1,56% абс. сухой массы. Наиболее ценный для организма человека
органический кремний в наивысшей степени (1,23%) представлен в листьях, в
меньшей степени (0,74%) --- в корзинках, а минимальное его количество (0,45%)
отмечено в стеблях золотарника. Содержание минерального растворимого крем
ния, напротив, в этом ряду возрастает (листья -- 0,12%; соцветия -- 0,17%; стеб
ли -- 0,23%). Подобным образом возрастает и содержание малоусвояемого по
лимерного кремния (листья --0,21%; соцветия -- 0,25%; стебли -- 0,50%).
Существенных различий по содержанию фенольных соединений и крем
ния между инвазионными золотарниками и аборигенной S. virgaurea, входя
щей в Государственный реестр лекарственных средств, не выявлено, что по
зволяет прогнозировать успешность применения S. gigantea для фармацевти
ческих целей.
Негативные эффекты касаются, главным образом, экологии и защиты ес
тественной среды. В лесных питомниках и при лесовосстановлении S. gigantea
наносит серьезный экономический ущерб [Weber, Jakobs, 2005].
Меры борьбы
S. gigantea, наряду с S.canadensis, внесена в «Список активно распространя
ющихся чужеродных видов Европейской и Средиземноморской организации по
защите и карантину растений» [EPPO List...], в котором перечислены виды
растений, которые представляют угрозу для благополучия растений, окружа
ющей среды и биологического разнообразия на территории Европы.
В естественном ареале S.gigantea сильно повреждают тля, а также другие
насекомые вредители: Eurosta solidaginis, Rhopalomyia ssp., Trirhabda ssp.
[Abrahamson et al., 1991, 2001], которые контролируют распространение дан
ного вида. Напротив, во вторичном ареале у S. gigantea вредителей практичес
ки не наблюдается.
Имеется несколько механических способов удаления зарослей S. gigantea.
Один из эффективных способов -- кошение дважды в год (в мае и августе) в
течение нескольких лет, или перекапывание почвы в течение лета при сухих
216
погодных условиях. После сенокошения для контролирования роста зарос
лей S. gigantea следует высевать травосмесь, в результате чего значительно
снизится густота побегов растений. Эффективные мероприятия по удалению
плотных зарослей и подавлению жизнеспособности включают удаление раз в
год с мульчированием, или удаление два раза в год без мульчирования в тече
ние 3 летнего периода. Однако подобные механические воздействия могут
нанести также вред естественной растительности, таким образом, способ этот
рекомендовать нельзя. S.gigantea реагирует на удаление уменьшением диаметра
стеблей и стимуляцией сильного повторного отрастания из корневищ. Коше
ние раз в год ненамного снижает густоту побегов и даже провоцирует проти
воположный эффект -- увеличение плотности популяции на следующий год.
Мульчирование подавляет рост сильнее, чем просто удаление, возможно по
тому, что срезанная трава содержит ингибиторы роста [Weber, Jakobs, 2005].
В Европе выясняют возможность препятствовать внедрению S.gigantea в
леса посредством формирования вокруг них густых одновидовых зарослей
аборигенных видов [Weber, Jakobs, 2005]. Контролировать расселение
S.gigantea сложно и дорого. Искоренение вида нереально, и управление долж
но быть ограничено особыми площадями специальных экологических пред
приятий.
Обзор научных исследований и публикаций
Обнаружено [Schmid et al.,1988], что фотосинтетическая активность
S.gigantea в начале вегетационного сезона высокая (19 ± 2мmol CO2/m2/s), а
позднее снижается (16 ± 2 мmol CO2/m2/s).
СЕМЕЙСТВО ASTERACEAE (COMPOSITAE) --
СЛОЖНОЦВЕТНЫЕ
ДУРНИШНИК ЭЛЬБСКИЙ
Название: Xanthium albinum (Widd.) H. Scholz
Синонимы: X. californicum Greene, X. italicum Moretti, X. palustre Greene,
X. riparium Itzigs. et Hertsch, X. ripicola Holub, X. strumarium auct., non L. p. p.
Полный список синонимов огромен. Так, во «Flora of North America» приво
дится 16 названий [Strother, 2006], а на сайте «The Atlas of Florida Vascular Plants» --
86 синонимов! Выбор точного названия для растений из Средней России без спе
циальных номенклатурных и таксономических изысканий затруднителен. Мы
принимаем наиболее привычное и традиционное для флористических работ
последних 20--30 лет, понимая некоторую условность этого решения1.
Морфологические признаки
Однолетнее травянистое растение высотой до 80(120) см. Стебель прямос
тоячий, у крупных растений ветвистый, с жестким шероховатым опушением,
часто с красноватыми или красно бурыми пятнами. Листья очередные, череш
ковые, яйцевидные или широкояйцевидные, нередко трехлопастные, шерохо
вато опушенные, по краю неправильно дваждызубчатые. Корзинки однопо
1 По видимому, номенклатурно корректным названием для нашего дурнишника будет
X.orintale L. [Hаeupler, Muer, 2000, 2007; Parsons, Cuthbertson, 2001].
217
лые, собранные в колосовидные соцветия на верхушке стебля и в пазухах ли
стьев. Тычиночные корзинки находятся в верхних частях таких соцветий, ша
ровидные, с однорядной оберткой из свободных листочков. Тычиночные цвет
ки с пятизубчатым венчиком и свободными тычинками. Пестичные корзинки
со сросшимися листочками обертки, с 2 пестичными цветками, на верхушке с
2 клювовидными или прямыми шипами, часто неравной длины. При созрева
нии обертка твердеет и образует соплодия с 2 семянками. Соплодия длиной
15--25(30) мм, густо покрыты крючковидными шипами до самой верхушки,
основание шипов с жесткими щетинками (редко в небольшом числе), иногда
заканчивающихся железкой, с более или менее обильным железистым опуше
нием. Зрелые соплодия от соломенных до красновато бурых (рис. 94).
В Средней России, по нашему мнению, произрастают всего два вида дур
нишника из секции Xanthium: X. albinum и X. strumarium L. Уверены, вслед за
В.Н. Тихомировым, что все эти названия, под которыми дурнишник появля
ется в публикациях [например, Дугуян, 1960; Цвелев, 1988; Протопопова, 1994;
Еленевский и др., 2000], относятся к одному виду [Тихомиров, 1979; Опреде
литель растений Мещеры, 1987]. Нередко даже на одном растении соплодия
настолько разные, что если строго следовать ключу В.В. Протопоповой [1994],
то их следовало бы относить к разным видам при дробном понимании видов
дурнишника. К югу преобладают растения с более крупными соплодиями, ко
торые при созревании приобретают коричневато бронзовую окраску; север
нее --- соплодия мельче, созревшие соломенные. Изменчивость этого дурниш
ника не позволяет разделять его на несколько видов. Подтверждение тому
найдено при наблюдении за дурнишниками в европейской части России, в
Крыму, на Кавказе, в Турции, при просмотре американских материалов в US,
NY, GH, A. Однако значительная часть англоязычных ботаников объединяют
X. albinum и X. strumarium [Löve, Dansereau, 1959; Löve, 1976; Clement, Foster,
1994; Stace, 1997; Maggi, Ahles, 1999; Synthesis..., 1999], что, по нашему мнению,
ошибочно. Исследования североамериканских дурнишников показывают их
существенное генетическое и морфологическое сходство [Lцve, Dansereau, 1959;
Tranel, Wassom, 2000]. Однако не исключено, что при их изучении авторы име
ли дело лишь с одним видом [Lцve, Dansereau, 1959; Tranel, Wassom, 2000;
Wassom et al., 2002], поскольку X. strumarium s. str. в Северной Америке редок и
встречается в основном на Восточном побережье (US, NY, GH, A).
Различия между двумя видами можно отразить в следующем диагности
ческом ключе [Майоров, 2006]:
1. Соплодия длиной 15--25(30) мм, густо покрыты крючковидными шипа
ми до самой верхушки, основание шипов с жесткими щетинками (редко в не
большом числе), заканчивающихся иногда железкой, с более или менее обиль
ным железистым опушением, практически без простых волосков. Зрелые со
плодия от соломенных до красновато бурых. Средние стеблевые листья
яйцевидные или широкояйцевидные, иногда трехлопастные. Стебель и череш
ки листьев покрыты жесткими щетинистыми волосками X. albinum.
--- Соплодия длиной 10--15 мм, крючковидные шипы на верхушке сопло
дий обычно отсутствуют, без жестких щетинистых волосков, но с более или
менее обильными светлыми простыми волосками (поэтому соплодия серова
тые), железистое опушение слабое. Средние стеблевые листья широкояйце
видные, трехлопастные. Стебли и черешки листьев шероховатые, покрыты
сравнительно мягкими волосками X. strumarium.
218
Не исключено, что X. strumarium --- вид Старого Света [Opravil, 1974, 1983].
Лектотип X. strumarium (LINN 1113.1)2, выделенный во «Flora Iranica» [1989],
вполне соответствует этому виду в нашем понимании. Следует обратить вни
мание, что наряду с типовой формой К. Линней выделял американскую:
в. Xanthium elatius & majus americanum, fructii spinulis aduncis armato [Linnaeus,
1753]. Таким образом, совершенно справедливо K.Рехингер выделил лекто
тип из «европейского» материала [Flora Iranica, 1989].
Американский вид дурнишника --- X. albinum в нашем понимании --- разде
ляют на две разновидности: var. canadense (P. Miller) Torrey et Gray, которая
широко натурализовалась в Европе, и var. glabratum (DC.) Cronq., в Европей
ской России не встреченная3. Естественный ареал
X. albinum --- американское растение, широко распространенное в Север
ной, Центральной и Южной Америке [Strother, 2006]. Полагают, что естествен
ный ареал дурнишника приурочен к Центральной или Южной Америке, от
куда он расселился сначала по Америке, а затем по странам Старого Света
[Löve, Dansereau, 1959].
Вторичный ареал
История интродукции и географическое распространение. К сожале
нию, из за номенклатурной путаницы литературные указания о распростра
нении X. albinum не всегда точны. Так, в статье, посвященной X. strumarium из
весьма качественной серии «The biology of canadian weeds», изображен несом
ненный X. albinum [Weaver, Lechowicz, 1982]. Тем не менее можно утверждать,
что к настоящему времени дурнишник широко расселился по Евразии: от Ве
ликобритании [Clement, Foster, 1994] до Кореи [Kil et al., 2004]. В Чехии он
известен уже с 1872 г. [Pyšek et al., 2002]. Как сорное растение широко распро
странен в восточной части Австралии [Parsons, Cuthbertson, 2001].
История расселения неясна. В конце XIX века этот дурнишник (под назва
нием X. italicum) отмечался «на песчаных берегах, только в Польше» [Шмаль
гаузен, 1897]. С 1900 г. известен в окрестностях Риги (LE, MW). В Сибири
этот дурнишник был собран в 1915 г. в Тюмени [Эбель, 2003].
Пути и способы заноса. Соплодия дурнишника хорошо адаптированы к
разнообразным способам расселения. По видимому, его проникновение на
нашу территорию произошло из вторичного ареала в Западной или Централь
ной Европе, а затем этот вид расселялся без прямого вмешательства человека.
Статус в регионе. Первый раз в Средней России был собран Г.Э. Гроссе
том: «Окр. Пензы. Лев. берег р. Суры у слияния с р. Пензой, 1922.XI.7, Г.Гр.
опр. В. Бочкин» (MW). До 1960 х гг., когда он уже широко расселился, на него
не обращали внимания, путая с X. strumarium. В настоящее время X. albinum
широко распространен по всей Средней России (рис. 95).
Любопытны сведения о сборах X. albinum и X. strumarium в Европейской Рос
сии (рис. 96). На первый взгляд мы видим типичную картину вселения инвази
онного вида (X. albinum), который вытесняет местный вид (X. strumarium).
2 http://www.nhm.ac.uk/research curation/projects/linnaean typification/ --- The Linnaean
Plant Name Typification Project.
3 На первый взгляд, эта разновидность достаточно резко отличается от var. canadense
и, возможно, представляет собой «хороший» вид. Но для подтверждения этой точки зрения
необходимы исследования изменчивости природных популяций.
Рис. 94. Соплодия Xanthium albinum
Рис. 95. Расселение Xanthium albinum в Средней России
Рис. 97. Внедрение
Xanthium albinum
в прибрежные
сообщества, р. Ока
в Калужской обл.
Рис. 98. Соплодия
трех Xanthium abinum
(слева), X. strumarium
(в центре), X. spinosum
(справа)
221
Последний почти исчезает, однако затем восстанавливает численность на мень
шем уровне. «Запаздывание» кривой X. albinum объясняется отсутствием вни
мания флористов на ранних этапах инвазии. Однако наблюдаемые изменения
численности на самом деле представляют собой сплошные артефакты, свя
занные с неоднородностью накопления гербарных материалов. Так, пик X.
strumarium в конце XIX века связан с деятельностью В.Я. Цингера. Минимум
в 40 е годы объясняется Великой Отечественной войной. У X. albinum пик в
70 е годы --- результат массовых сборов во время Мещерской экспедиции под
руководством В.Н. Тихомирова. При существующей плотности гербарных сбо
ров в Средней России от 0,2 до 3,3 гербарного листа на 1 км2 [Щербаков, Сере
гин, 2002] детальная реконструкция инвазии, увы, невозможна.
Местообитания. X. albinum часто растет на свежем речном аллювии
(рис. 97), при этом явно предпочитает песчаные отложения [собственные
наблюдения; Weaver, Lechowicz, 1983]. Нередко встречается по рудеральным
местообитаниям, вдоль дорог, в городских условиях.
Размножение и жизненный цикл. Преимущественно самоопыляемое рас
тение [Lцve, Dansereau, 1959; Weaver, Lechowicz, 1982; Dinelli et al., 2003]. Од
нако при этом дурнишник обладает довольно высокой внутрипопуляционной
изменчивостью, что, по мнению итальянских исследователей, обеспечивает
экологическую пластичность вида [Dinelli et al., 2003]. Апомиксис у дурниш
ника не обнаружен [McMillan, 1974a, b, 1975]. Наличие апомиксиса докумен
тально подтверждено лишь у аномальной формы X. strumarium (? --- СМ) с
многочисленными, до 25, семянками в одном соплодии, при этом у нормаль
ной формы апомиксис не обнаружен [Abbas et al., 1999]. Цветение в июле ---
августе. Соплодия созревают к концу лета --- началу осени. На одном растении
образуется 500--5400 соплодий в зависимости от мощности растения (рис. 98).
В густых зарослях дурнишника число соплодий уменьшается до 70--580
[Weaver, Lechowicz, 1983]. Они способны частично удерживаться на материн
ском растении до следующего вегетационного сезона.
Расселение и распространение. Соплодия X. albinum, зацепившиеся за
шерсть животных или за одежду людей, могут быть перенесены на значитель
Рис. 96. Частота гербарных сборов Xanthium strumarium и X. albinum по данным
MW, по горизонтали --- десятилетия, по вертикали --- число гербарных образцов
дурнишника за десятилетие.
222
ные расстояния. Высохшие соплодия обладают высокой плавучестью и раз
носятся речными водами [Löve, Dansereau, 1959; Weaver, Lechowicz, 1982].
Поскольку часть соплодий остается на сухих растениях до лета, то вполне ве
роятно их распространение с весенним половодьем. Кроме того, речной песок
нередко используется в строительстве, что также может способствовать рас
селению дурнишника. Последствия внедрения
Воздействия на естественные фитоценозы и аборигенные виды. На
песчаных пляжах в долинах среднерусских рек дурнишник нередко образует
обширные заросли, где может конкурировать с аборигенными видами. Неред
ко считают, что американский X. albinum вытеснил местный X. strumarium,
который в последние десятилетия стал редким видом4 [Тихомиров, 1979; Оп
ределитель растений Мещеры, 1987]. Следует обратить внимание, что еще И.Ф.
Шмальгаузен [1897] указывал X. strumarium только для сорных мест, при этом
X. albinum --- прежде всего аллювиальный вид, произрастающий также на сор
ных местах с бедными почвами. Таким образом, эти виды занимали (и занима
ют!) в значительной мере разные экологические ниши, поэтому их реальная
конкуренция не столь остра. Причины современной редкости X. strumarium,
по видимому, иные.
Генетические изменения и вариабельность вида. При исследовании дур
нишника из рудеральных и естественных местообитаний показано, что попу
ляции заметно отличаются по многим признакам, но их изменчивость не кор
релирует с характером местообитания [Blais, Lechowicz, 1989].
Влияние на здоровье человека. У некоторых людей возможно появление
контактного дерматита [Weaver, Lechowicz, 1983].
Экономическое и социальное (положительное и отрицательное) зна
чение. Дурнишник ухудшают качество речных пляжей: его заросли из грубой
шероховатой травы затягивают открытый песок, а соплодия колючие и спо
собны проколоть кожу на подошве. X. albinum выходит на поля как сорное
растение, но массовым сорняком на полях в Европейской России дурнишник
пока не является. Для Северной Америки отмечено существенное (до 60%!)
снижение урожая сои на полях, засоренным дурнишником. Кроме того, на
блюдается заметное ухудшение качества шерсти овец на засоренных пастби
щах [Weaver, Lechowicz, 1982; Parsons, Cuthbertson, 2001].
Меры борьбы
В посевах численность дурнишника вполне хорошо контролируется гер
бицидами для двудольных [Weaver, Lechowicz, 1983]. В США разрабатывают
ся меры биологического контроля [Logarzo et al., 2002].
Обзор научных исследований и публикаций
Исследования итальянских ботаников [Baldoni et al., 2000; Dinelli et al.,
2003] показали неоднородность комплекса X. strumarium s. l. в Италии. Доста
точно надежно выделяются 3 группы: Xanthium italicum, X. strumarium и X.
orientale. Напротив, американские популяции весьма однородны [Löve,
Dansereau, 1959; Tranel, Wassom, 2000; Wassom et al., 2002].
4 В последние десятилетия X. strumarium чаще удается встретить на почти сельских
улицах старых уездных городков, реже на сельских пустырях.
223
СЕМЕЙСТВО BALSAMINACEAE --
БАЛЬЗАМИНОВЫЕ
НЕДОТРОГА ЖЕЛЕЗКОНОСНАЯ
Название: Impatiens glandulifera Royle
Синонимы: I.royle Walp.
Морфологические признаки
Высокое однолетнее растение с гладким, обычно пустотелым и членистым
стеблем, который легко ломается. Стебель может достигать высоты 3 м, его
диаметр доходит до нескольких сантиметров. Листья супротивные или в трех
членной мутовке, голые, от ланцетных до эллиптических, длиной 5--18 см и
шириной 2,5--7 см. Соцветие рацемозное из 2--14 цветков длиной 25--40 мм.
Цветки зигоморфные, самый нижний чашелистик мешковидно вздут и закан
чивается прямой шпорой. Окраска цветков варьирует от белого до розового и
фиолетового, растения цветут с июня по октябрь (рис. 99). Коробочки длиной
1,5--3,5 см и шириной до 1,5 см. Коробочка содержит до 16 семян, каждый дли
ной 4--7 мм и шириной 2--4 мм со средней воздушно сухой массой 2,35 мг
[Beerling, Perrins, 1993]. Естественный ареал
Родина I.glandulifera -- западные Гималаи -- центр происхождения рода
Impatiens, где обитает еще ~400 видов рода. I.glandulifera произрастает здесь
на высотах от 1800 до 4000 м над ур.м. [Gupta, 1989].
Вторичный ареал
История интродукции и географическое распространение. Вторичный аре
ал вида очень широк и охватывает Северную Америку и Евразию. В Европе
(Англия) I.glandulifera впервые появилась в 1838 г. Семена с 3 растений этого
вида с лиловыми цветами и с 1 растения var. candida с белыми цветами при
слали в то время в Кью и начали разводить как декоративное растение. По
зднее, в 1898 г., в Англию были завезены растения var. pallidiflora, которые
встречаются в Сиккимских Гималаях [Valentine, 1971]. Упоминания о дича
нии растения в Британии долгое время были крайне редкими: один случай
зафиксирован в 1855 г. в Девоншире [Valentine, 1971], другой -- в 1859 г. около
Манчестера, затем находки участились -- в 1864 г. I.glandulifera одичала в Лан
кашире, в 1895 г. в графстве Суррей, в 1898 г. -- в Вустершире [Britten, 1900].
В Германии I.glandulifera появилась, по одним данным, одновременно с по
явлением ее в Англии [Valentine, 1971], а по другим -- в 1897 г. в окрестностях
Гамбурга [Hegi, 1923]. Около 100 лет этот однолетник изредка культивирова
ли в парках, на кладбищах, в сельской местности, порой возделывали как ме
донос. На юго западе Германии в некоторых местах недотрога была довольно
обычна уже в 1920 х гг., распространившись из швейцарских популяций (где
вид натурализовался в 1904 г.) по Рейну. Однако к 1923 г. недотрога встреча
лась не более чем в 50 пунктах Средней Европы. Отмечалось, что I.glandulifera
плохо закрепляется на одном месте, поскольку семена ее полностью не вызре
вают [Hegi, 1923].
Вид был интродуцирован Ботаническим садом Хельсинки в конце 1800 х
гг. [Kurtto, 1992]. Первые натурализовавшиеся популяции наблюдались в
224
Швеции в конце 1920 х гг., в Норвегии -- в конце 1930 х гг, а в Финляндии -- в
1947 г. [Kurtto, 1996]. В Дании вид отмечен впервые в 1888 г., в 1956 г. зареги
стрировано уже ~40 находок, главным образом, в восточной части страны
[Pedersen, 1956], а в 1988 г. I.glandulifera произрастала уже по всей Дании
[Hansen, 1991]. В Польше вид интродуцирован в Судетские горы в 1890 г. В
Латвии I.glandulifera интродуцирована как садовое растение, а первый гербар
ный сбор датирован 1898 г. (RIG).
Расширение ареала I.glandulifera началось, по видимому, после Второй ми
ровой войны. В 1957 г. недотрога найдена на берегу водоема в Амьене, Фран
ция [Wintrebert, 1957]. В 1968 г. она отмечена в Хорватии, до 1989 г. ее обнару
жили только 15 раз, а экспансия вида началась лишь в последнее десятилетие
прошлого века [Lukac, 1989]. В 1979 г. I.glandulifera отмечена близ г. Павия в
качестве нового для флоры Ломбардии вида [Soldano, 1979]. В начале 1960 х
гг. началась бурная экспансия вида в Средней Европе, и по сведениям
E.Daumann [1967], в некоторых местах на юге Богемии I.glandulifera вытесни
ла аборигенную I.noli tangere. В Литве вид как беглец из культуры зарегист
рирован в 1959 г. [Gudzinskas, 1998]. Первые популяции в естественных пой
менных лесах по рекам Польши отмечены в начале 1960 х гг.[Jasnowski,1961;
Dajdok, Aniol Kwiatkowska, 1998].
Теперь I.glandulifera широко распространена в 18 странах Европы между
30 и 64° с. ш.: в Ирландии, Англии, на юге и на восточном побережье Швеции,
юге Норвегии и Финляндии, в Дании и Германии. Продолжается инвазия вида
в Чехии и Словакии [Pyšek, Prach, 1995], а также в Швейцарии, где вид широ
ко распространился уже в поясе до 800 м над ур.м. Как обычное растение не
дотрога железконосная отмечена во Французских Пиренеях, в Голландии,
Австрии (особенно в Альпах), Польше, Венгрии, бывш. Югославии [Beerling,
Perrins, 1993].
Спонтанные популяции обнаружены нами в 2003 г. в Австрийских Альпах
на берегу горной речки Мелль на высоте 900 м над ур.м., а на юге Австрии вид
рос в придорожных канавах на протяжении 3--5 км, достигая в высоту 3 м. Во
время экспедиционной поездки в Польские Татры (2005 г.) мы наблюдали за
росли недотроги железконосной, тянущиеся на километры. Обилие недотроги
в странах Европы с каждым годом увеличивается, несмотря на проводимые гид
ромелиоративные мероприятия и применение гербицидов [Mountford, 1994].
В Северную Америку (Канада) недотрога попала, по видимому, в 1906 г.,
изредка ее культивировали, а в 1960 е гг. в ряде штатов США (Калифорния,
Айдахо, Массачусетс, Мэн, Мичиган, Монтана, Нью Йорк, Орегон, Вермонт,
Вашингтон) и в ряде провинций Канады (Британская Колумбия, Онтарио,
Квебек, Нью Брансуик и Новая Шотландия) недотрога начала внедряться в
естественную растительность по берегам рек и по влажным местообитаниям
[Gleason,1963; Scoggan, 1978].
Как интродуцированное растение отмечена во флоре Ирана [Grey Wilson, 1979].
На территорию России I.glandulifera попала в конце ХIХ века. Известно,
что в Петербургском ботаническом саду недотрогу культивировали в 1895 г.,
поскольку вид упомянут в списке семян сада [Delectus seminum..., 1895]. Сбо
ры I.glandulifera в Курляндии сделаны в 1898 г. (RIG). Случаи дичания вида
на территории бывшего СССР были в то время, так же как в Средней Европе,
крайне редки. К 1941 г. известны лишь 2 местонахождения: окрестности оз.
225
Рис. 100. Расселение Impatiens glandulifera в Средней России
Рис. 99. Внедрение Impatiens glandulifera в прибрежные сообщества,
Московская обл., р. Москва в Одинцовском р не
Рис. 101. Массовое
цветение Impatiens
glandulifera в
Москве, в овраге в
Битцевском
лесопарке
Рис. 102. Розовая
окраска венчика
Impatiens glandulifera
227
Селигер [Маевский, 1940] и околицы г. Каменец Подольский (Украина), где
недотрога найдена в 1939 г. [Берко, 1963]. Во Флоре СССР указано [Победи
мова, 1949] еще одно место сбора -- Эстония. Однако и там вид встречался
дико только в двух местообитаниях вблизи г. Тарту рядом с ботаническими
садами, где недотрога в изобилии культивировалась [Talts, 1959].
После войны зарегистрировано еще несколько местообитаний недотроги:
в Литве в 1959 г. в одичалом состоянии [Gudzinskas, 1995] и на Украине: в
1953 г. -- в Прикарпатье, г. Трускавец, в цветнике (LE); в районе г. Ковель Во
лынской обл. [Карнаух, 1955] и в 1962 г. в с. Верхняя Билка Львовской обл. в
запущенном парке [Берко, 1963]. В 1956 г. растение обнаружено в одичалом
виде в Ленинграде [Гусев, 1973].
В 1960--1970 х гг. бурная натурализация недотроги началась и в европейс
кой части СССР: в 1967 г. она отмечена в г. Приморск Ленинградской обл., а
ранее указана и для Петродворца [Гусев, 1973]. В 1974 г. крупная популяция
одичавшей недотроги отмечена в г. Рига (LATV). В 1978 г. недотрога найдена
Н.Н. Цвелевым на левом берегу р.Уда на юго западной окраине Харькова (LE).
Маccoвoe дичание I.glandulifera наблюдалось и в Удмуртии: А.Н. Пузырев
[1985] отмечает, что на небольшой речке у ст. Можга цветущие растения обра
зовали заросли по берегам на протяжении нескольких метров. В 1986 г. найде
на в Псковской обл.
Еще во второй половине 1920 х гг. недотрога железконосная отмечена в
г. Орджоникидзе и окрестностях [Горбачев, 1932]. Но здесь она не распростра
нялась, а возможно, и исчезла вовсе. Была приведена для окрестностей Кис
ловодска как первое указание для Кавказа в 1979 г. [Котов, 1979]. В 80 х гг.
прошлого века найдена в регионе еще в нескольких пунктах на рудеральных
местообитаниях: единично на окраине пос. Мизур в Алагирском ущ. в 1983 г.,
в окрестностях г. Орджоникидзе в 1984 г., по склонам г. Лысая в Лесистом хр.
в 1987 г. [Комжа, Попов, 1990].
Довольно редки случаи разведения недотроги в Сибири. В 1988 г. она от
мечена В.В. Штаркером на приусадебных участках в районе заповедника Стол
бы. Через год найдена по берегам рек Танзыбей и Оя в Ермаковском р не Крас
ноярского края [Степанов, 1990]. В 90 х гг. прошлого века I.glandulifera стала
дичать из культуры в Томской обл. [Пяк, 1994].
Во Владивостокском р не недотрога собрана В.Н. Ворошиловым как сор
ное растение в 1960 г. (MHA). В 1974 г. найдена Т.Н. Ульяновой в Южно Са
халинске у забора (MHA) и в Корсаковском р не в сорных местах близ ж/д
насыпей [Воробьев и др., 1974]. К 1980 г. недотрога уже широко распростра
нилась по огородам, садам и паркам г. Хабаровск [Шлотгауэр, Небайкин, 1984],
а в начале 90 х гг. прошлого века натурализовалась на о. Кунашир [Нечаева,
Петрова, 1995].
В настоящее время недотрога широко известна как агрессивный чужерод
ный вид в умеренных областях в Европе, Азии, Северной Америке и Новой
Зеландии.
В более или менее естественных местообитаниях отмечается повсюду в
Балтийском регионе [Kuusk et al., 1996]. В Финляндии с 1990 х гг. расшире
ние ареала I.glandulifera ускорилось, и теперь этот вид весьма обычен в плотно
населенных р нах на юге и в центре страны, а наиболее северные популяции
достигли Северного полярного круга.
228
В Швеции недотрогу считают одним из пяти наиболее агрессивных видов
растений. Она найдена по всей Швеции, кроме Лапландии [Larsson, Martinsson,
1998]. В Норвегии вид быстро распространяется, занимая естественные и по
луестественные местообитания -- сырые леса, сырые луга, затопляемые зем
ли, канавы, побережья, речные потоки и различные типы сырых рудеральных
участков; найден на юго востоке Норвегии и по северу побережья до 69,5° с.ш.
[Aim, 2002; Lid, Lid, 2005]. В обоих ботанических садах Исландии вид отсут
ствует, и неизвестно, натурализовался ли он где либо в Исландии [Nobanis, 2009].
В Литве I.glandulifera наиболее интенсивно распространяется по речным
берегам, однако произрастает и во многих других типах естественных и антро
погенных местообитаниях [Gudzinskas, Sinkeviciene, 1995; Gudzinskas, 1998],
и ее считают инвазионным видом.
Информации о расселении I.glandulifera в Латвии мало, и существует нео
днозначное мнение о ее агрессивности. Этот вид вторгся по берегам рек, най
ден в старых парках, садах, обочинах дорог и мусорных местах. Реки -- глав
ный путь распространения недотроги в Латвии [Nobanis, 2009].
В настоящее время I.glandulifera распространилась по всей Польше, но чаще
встречается на юге, это один из 20 самых агрессивных инвазионных видов в
стране [Tokarska Guzik, 2003, 2005]. В Германии растение является наиболее
широко распространенным чужеземным видом и растет по берегам рек и дру
гим влажным, богатым питательными веществами естественным местообита
ниям. В Германии вопрос относительно агрессивности I.glandulifera дискус
сионный, потому что нет доказанных случаев, когда опасности был бы под
вергнут аборигенный вид. Расселение I.glandulifera ведет к сокращению
природных видов, однако в высокодинамичных местообитаниях пойменных
лесов всегда есть новые постоянно возникающие местообитания, и популя
ции I.glandulifera через какое то время исчезают (например, в годы с неподхо
дящими погодными условиями или при затоплении). Кроме того, вид преиму
щественно замещает Urtica dioica, которая не является редким или находящим
ся под угрозой исчезновения видом [Kasperek, 2004].
Пути распространения. Основной способ интродукции -- «бегство» из час
тных садов и пасек. В Финляндии вид распространился, главным образом, при
ввозе семян из за границы и частном обмене семенным материалом. Часто
распространяется также вокруг населенных мест из заброшенных садов или с
почвой [Kurtto, 1996].
Статус в регионе. В одичалом состоянии недотрога впервые встречена близ
Сенежского озера в Клинском уезде Московской губернии в большом коли
честве [Сырейщиков, 1914]. К 1941 г. известно лишь еще одно ее местонахож
дение в окрестностях оз. Селигер [Маевский, 1940]. В Мocквe и Калуге оди
чавшие экземпляры недотроги собраны в 1955 г. (МНА). По личным сообще
ниям садоводов любителей известно, что в конце 1950 х гг. семена I.glandulifera,
как декоративного растения, пользовались в Москве большим спросом.
В 1970 х гг. началась бурная натурализация недотроги во всех областях
Средней России. В 1971 г. I.glandulifera собрана Ю.Д. Гусевым в Московской
обл. на ст. Дмитров (LE), в 1973 г. -- в Рязанской обл. (г. Касимов по склону
оврага, в бурьяне. 1973. В.Тихомиров, Н.Октябрева. MW), где уже через 2 года
отмечена как компонент флоры Рязанской Мещеры [Тихомиров, 1975]. В 1976 г.
недотрога найдена в Брянской обл. (МНА). При изучении флоры Калининс
229
кой обл. I.glandulifera найдена на пустырях, особенно во влажных местах
(в канавах, около прудов, в понижениях рельефа) в Калинине, Торжке, Стари
це, Кувшинове, Нелидове, Западной Двине, Торопце [Малышева, 1979]. В 1980
г. заросли цветущих растений найдены в г. Чаплыгин Липецкой обл. в пойме
р. Становая Ряса, годом позднее в г. Липецк также в большом количестве, а в
1987 г. -- в поймах р. Воронеж и Дон [Вьюкова, 1983, 1987]. К 1982 г. относятся
сборы в Орловской обл. (г.Орел, стройка у сараев. 11.08.1982 г. В.И. Радыги
на. ОГУ). В 1983 г. недотрога встречена во многих районах и пригородах Ряза
ни [Гущина, 1983], а в 1988 г. -- в Ульяновской обл. (МНА). В 1989 г.
I.glandulifera обнаружена во влажных местообитаниях на берегу Воронежско
го водохранилища [Барабаш, Камаева, 1989] и занесена в Марийскую АССР
[Абрамов, Киряков, 1989]. В настоящее время широко распространена во всех
областях Средней России (рис. 100).
Местообитания. В природе I.glandulifera растет на выcоте 1800--3000 м,
часто по берегам рек, по краям полей и в нарушенных влажных лесах [Valentine,
1971], а в Пакистане -- до 4300 м над ур.м. (Nasir, 1980).
В пределах вторичного ареала произрастает в широком диапазоне место
обитаний, но лучше всего растет на сырых местах, богатых питательными ве
ществами, особенно по берегам озер, по рекам и ручьям (рис. 101). Часто вид
также можно обнаружить в антропогенно нарушенных и искусственных место
обитаниях -- на полях, в кустарниках, канавах, обочинах дорог и живых изгоро
дях [Kurtto, 1996]. Наиболее подходящими для I.glandulifera являются биото
пы, подверженные антропогенному воздействию [Garkaje, 2006]. В таких био
топах I.glandulifera обычно растет вместе с растениями, которые нуждаются в
запасе азота в почве -- Aegopodium podagraria, Urtica dioica, Calystegia septum,
Deschampsia caespitosa, Stellaria nemorum и Galium aparine. Вид вторгается в мно
голетнюю травянистую растительность речных берегов, светлые пойменные леса
и влажные луга и найден на разнообразных типах почвы [Kowarik, 2003].
В Европе растения I.glandulifera всех возрастов нехолодостойки. Обычно
все взрослые растения погибают осенью при первых морозах, а проростки по
гибают при поздних весенних заморозках [Sebald et al., 1998]. Вид также не
выносит засухи и быстро увядает, и растения могут выживать, только если
период засухи короток [Beerling, Perrins, 1993].
Недотрога железконосная -- теневыносливое растение. Рост ее, однако, ухуд
шается, когда мощность светового потока становится менее 30% полного днев
ного света [Beerling, Perrins, 1993]. В качестве факторов, которые ограничива
ют инвазию I.glandulifera, могут выступать требовательность к высокой влаж
ности почвы и слабая морозоустойчивость. Этот вид отмечен как на равнинных
участках, так и на крутых (>40°) склонах, при условии умеренного затенения,
способен произрастать как на тонком, так и на грубом аллювии, морской галь
ке, хоро-шо дренируемых минеральных почвах, отвалах каменного угля и тор
фах [Beerling, Perrins, 1993]. Недотрога железконосная обычно растет на бога
тых минеральным азотом почвах, предпочитает почвы с нейтральной реакци
ей, но может расти на кислых и щелочных (рН от 4,5 до 7,3--7,7). Обитает на
почвах от влажных, не просыхающих, до сырых, часто плохо аэрируемых. Для
успешного выживания проростков требуются умеренная нарушенность рас
тительного покрова (вывалы деревьев, упавшие ветки деревьев, зимнее под
топление берегов) и наличие голых участков грунта [Марков и др., 1997].
230
В Московской обл. I.glandulifera встречается на берегу р. Москва на протя
жении десятков километров в Можайском, Рузском и Одинцовском р нах.
Местами заходит на ключевые болота. В ряде р нов вид образует обширные
заросли по заболоченным лугам у водоемов [Игнатов и др., 1990]. Следова
тельно, во вторичном ареале I.glandulifera занимает те же местообитания, что
и на родине.
Габитус растений меняется в зависимости от влажности местообитания.
Высокие, маловетвистые экземпляры обитают в довольно влажных условиях,
низкие растения с обильно ветвящимися стеблями, короткими междоузлия
ми и мелкими листьями характерны для нарушенных лесных участков с непо
стоянным увлажнением [Марков и др., 1997]. Плотность популяций в сред
нем составляет 40 растений/м2, но может возрастать и до 70 особей [Wadsworth
et al., 2000]. Достижение высокого роста растений при низком уровне освеще
ния в плотных популяциях обеспечивается физиологическим процессом на
копления в клетках стебля аниона NO3 [Andrews et al., 2005].
Размножение и жизненный цикл. Цветки I.glandulifera протандричные, по
этому опыление их облигатно перекрестное (рис. 102). Однако может иметь
место опыление между цветками с одного и того же растения. Вид энтомо
фильный, опыляется шмелями.
I.glandulifera -- однолетнее растение. В зависимости от плотности популя
ции и от характеристики места произрастания одна особь может продуциро
вать от 95 до 390 плодов и от 500 до 2500 семян [Марков и др., 1997]. Обычно
семена выбрасываются из коробочки на расстояние до 5 м, максимальное чис
ло семян падает на почву в радиусе 1--2 м от материнского растения. Подсчи
тано, что скорость распространения популяции в одном местообитании состав
ляет 2,47 м в год [Beerling, Perrins, 1993]. В переувлажненных местах набух
шие семена I.glandulifera погружаются в воду и способны прорастать под водой
[Lhotska, Kopecky, 1966]. Более высокие растения образуют большее число
плодов и большее число семян в одном плоде. Масса семян не зависит от чис
ла плодов на растении или от числа семян в плоде. Масса 100 семян варьирует
незначительно [Виноградова, 2008] и составляет в среднем от 1,05 г (Санкт
Петербург) до 1,59 г (Вологда). На высокопитательных почвах I.glandulifera
продуцирует большее количество семян, а не более крупные семена [Willis,
Hulme, 2004].
В Англии семена могут прорастать иногда через 18 месяцев. Без холодной
стратификации семян большинство сеянцев появляется в течение 4 недель
[Beerling, Perrins, 1993]. По нашим данным, семена I.glandulifera теряют всхо
жесть уже через 1 год сухого хранения.
I.glandulifera имеет хорошую регенерационную способность, и на повреж
денных стеблях формируются новые ветви и цветки. Мелкие растения также
могут образовывать цветки и семена.
В Германии время от прорастания до начала цветения -- 13 недель, а цвете
ние продолжается в течение следующих 12 недель [Sebald et al., 1998].
Расселение и распространение. I.glandulifera распространяется только се
менами. Распространение семян происходит гидро , антропо и автохорно.
Когда коробочка созревает, она взрывается и выбрасывает семена. Имеются
сообщения, что семена рассеиваются от материнского растения на расстояние
до 7 м. Отдельное растение может производить более 4 тыс. семян, и в чистых
231
зарослях образуется до 32 тыс. семян/м2 [Koenies, Glavac, 1979]. Экспансия
вида в речных системах, в частности, обусловлена способностью семян к рас
пространению по водным путям, поскольку они могут переноситься и плавно
текущей водой (в осадке), и сухими плавучими семенами. Вместе с наносами
сезонных паводков се-мена могут переноситься вдоль рек на большие рассто
яния. Мелкие семена легко переносятся в новые местообитания с почвой или
на обуви. Семена, вероятно, также распространяются муравьями (мирмеко
хория). Плодоносящие экземпляры или их фрагменты также переносятся с
почвой или потоками воды [Kurtto, 1993]. Для Великобритании рассчитана
способность к расселению, которая составляет 2,6--5 км в год (ссылка).
Репродуктивная стратегия основана на активном распространении семян
и на их высокой всхожести, которая составляет до 80% [Grime, 1987]. Расте
ния конкурируют на речных берегах путем синхронного прорастания множе
ства семян, быстро вырабатывают достаточную биомассу и подавляют соседние
виды. I.glandulifera растет довольно быстро и формирует плотные заросли.
Последствия внедрения
Воздействие на естественные фитоценозы и аборигенные виды. Хотя в ка
честве декоративного растения I.glandulifera выращивали в садах Европы с
середины ХIХ века, а в Северной Америке -- с начала ХХ века, первые случаи
дичания вида отмечены через 20 смен поколений, внедрение в естественные
ценозы -- через 60, а бурная экспансия -- через 100 смен поколений.
I. glandulifera встречается на побережьях озер и морей, на лугах, по рекам и
ручьям, в зарослях черной ольхи и в лесах около селений [Kurtto, 1996]. Отме
чено, что во влажных лесах вид занимает «новые» местообитания, то есть явля
ется пионерным. В Швеции заросли I. glandulifera сохранялись в течение по
крайней мере 70 лет [Larsson, Martinsson, 1998]. При некоторых обстоятельствах
вид только сокращает численность аборигенных видов, которые все же находят
достаточно мест для произрастания, например в высоко динамичном поймен
ном лесу [Kasperek, 2004]. Но в иных ситуациях растение формирует плотные
заросли, которые покрывают всю почву и «душат» другую растительность, при
чем I.glandulifera конкурирует не только с однолетними, но и с многолетними
видами растений [Larsson, Martinsson, 1998]. В окрестностях Осло (Норвегия)
отмечалось, что распространение I.glandulifera уменьшило видовое разнообра
зие в некоторых локальных ценозах. В речных долинах I. glandulifera формиру
ет плотные заросли собственного сообщества, которое классифицируется фи
тосоциологами как ассоциация Impatienti Calystegium [Moor,1958; Soo, 1971].
Способность I. glandulifera к конкуренции с природной флорой и ее предсказу
емая экспансия по водным путям указывают, что в будущем вид может стать
более серьезной угрозой сохранению естественного биоразнообразия [Prach,
1994; Pyšek, Prach, 1995]. Известно, что I. glandulifera предотвращает возобнов
ление лесов в сырых полузатененных местообитаниях [Lhotska, Kopecky, 1966].
Поскольку I. glandulifera, по видимому, положительно реагирует на увели
чение CO2 и повышение температуры, потенциально вид может стать в усло
виях изменяющегося климата еще более агрессивным захватчиком.
I. glandulifera привлекает опылителей, и вид, по видимому, способен кон
курировать с другими видами Impatiens за опылителей [Daumann, 1967]. Пу
тем конкуренции за опылителей I.glandulifera может заместить и некоторые
другие природные виды [Chittka, Schurkens, 2001].
232
Генетические изменения и вариабельность вида. В естественном ареале в
Кашмире и Джамму 2n=20 [Khoshoo, 1957]. Во вторичном ареале вид суще
ствует по меньшей мере в двух формах, имеющих 2n=18 и 2n=20, причем пер
вая считается производной, возникшей на новой родине [Khoshoo, 1957]. По
пуляция растений с 2n=18 и n=9 обнаружена в Манчестере [Valentine, 1971].
А в Кью обе формы произрастают вместе в одной и той жe микропопуляции
[Jones, Smith, 1966].
И в естественном, и во вторичном ареале отмечен полиморфизм по окраске
цветков I.glandulifera. Основные цвета: лиловый, бледно розовый и белый. Уже
при первичной интродукции в Англию в 1838 г. завезена не только лилово , но
и белоцветковая форма. Однако натурализовавшиеся растения с белыми цве
тами появились в Британии только в 1921 г. [Valentine, 1971]. В континен
тальной Европе эта форма встречается только в культуре: отмечена в Цюрихе
в 1914 г. [Hegi, 1923] и на юге Польше [Valentine, 1971]. Выращивается форма и
в канадской провинции Онтарио [Scoggan, 1978]. В СССР форма с белыми цвет
ками также отмечена только в культуре -- собрана А.К. Скворцовым в г. Мука
чево (Закарпатье) и Д.Басаргиным [1989] на Дальнем Востоке.
Бледно розовые цветки встречаются редко. В Канаде такая форма отмечена
в провинциях Новый Брансуик, Британская Колумбия, Онтарио и Новая Шот
ландия [Scoggan, 1978]. На Дальнем Востоке такая форма встречается только в
садах, а в европейской части России -- и в спонтанных популяциях.
По данным D.Valentine [1971], только одна популяция натурализовавшей
ся I.glandulifera около г. Манчестер и одна -- на севере Голландии имеют боль
ший процент светло розовых и белых цветков, чем темно пурпурных. Во всех
остальных европейских популяциях наблюдается обратное соотношение.
Одной из причин узкого распространения белых цветков считалось то, что
бледноцветковые растения продуцируют меньше нектара и реже посещаются
шмелями и пчелами, чем темные цветы в соотношении 1:2 или 1:3 [Valentine,
1971]. Однако на экспериментальном участке ГБС РАН в интродукционной
популяции, созданной из семян, собранных в различных точках вторичного
ареала вида, все цветовые формы насекомые посещали одинаково часто, а бе
лоцветковые формы завязали такое же количество семян, что и лиловоокра
шенные. При свободном переопылении различноокрашенных форм в первой
генерации произошло расщепление у потомства семян, собранных с белоцвет
ковых форм, в соотношении 2 пурпурных : 2 розовых : 1 белоцветковое расте
ние; в последующих генерациях число особей с белыми цветками еще более
сократилось [Виноградова, 2008].
Таким образом, в Англии и Западной Европе, где I.glandulifera натурализо
валась ранее, наблюдается 5 вариаций окраски лепестков, причем голубые
цветки возникли, по видимому, уже во вторичном ареале. В районах, где
I.glandulifera появилась недавно, форм с голубыми цветками не найдено вов
се, белые и светло розовые имеются только в садах, а дичают экземпляры с
темно пурпурными и розовыми цветками.
Анализ дискретных признаков эстивации (взаимного расположения лепе
стков зигоморфного цветка) показал слабое внутрипопуляционное разнооб
разие [Басаргин,1989].
Экономическое и социальное (положительное/отрицательное) значение.
Уничтожение зарослей -- дорогостоящий и долговременный процесс из за
233
высокой способности вида к восстановлению и распространению. Агентство
по охране окружающей среды Великобритании подсчитало, что уничтожение
I. glandulifera только в Англии и Уэльсе будет стоить от 210 до 240 млн евро.
Растение популярно среди коллекционеров бабочек, так как вид является
важным источником нектара и пыльцы. Цветы часто посещают пчелы и шме
ли [Beerling, Perrins, 1993]. Кроме того, I. glandulifera считается красивым де
коративным растением; его семена продолжают высевать, и это играет важ
ную роль в распространении вида.
Меры борьбы
Удаление инвазионного чужеродного вида должно начаться как можно рань
ше, до того как проблема станет слишком серьезной. Для успеха действий об
щественность должна быть проинформирована о негативном влиянии расте
ния и его быстром распространении по водным путям. Особо настойчиво сле
дует информировать об особенностях вида пасечников и владельцев
питомников декоративных растений.
Естественные враги для ограничения распространения вида не известны, а
биологические методы контроля для I.glandulifera пока не разработаны
[Sheppard et al. 2006].
Уничтожение и меры контроля включают как удаление растений, так и пре
дотвращение формирования и распространения семян. Наиболее важен вы
бор времени для уничтожения. Если удалить растения слишком рано, они вос
становятся, а если сделать это слишком поздно, сформируются зрелые семе
на, которые прорастут на следующий год. Лучшее время -- конец июля, когда
появляются первые цветы. Из за способности вида к регенерации (см. выше)
чрезвычайно важно удалить весь растительный материал. I.glandulifera может
быть легко удалена путем прополки, кошения или обрезки. Удаление должно
продолжаться до тех пор, пока в течение по крайней мере 2--3 лет не будут
появляться новые растения. Использования химических веществ следует из
бегать, и во многих регионах это запрещено, особенно по берегам водоемов.
Имеются рекомендации по использованию пастбищных животных для
уничтожения вида [Larsson, Martinsson 1998].
Обзор научных исследований и публикаций
Методом создания интродукционной популяции выявлено (см. главу 4),
что у I.glandulifera выработалась генетически закрепленная географическая
изменчивость некоторых биологических признаков: с севера на юг достовер
но возрастают высота и число метамеров главного побега, увеличиваются и
продолжительность периода роста растений и биомасса надземной части, а
также позднее наступает соответствующая фенофаза. В ряду последующих
поколений микроэволюция адаптивных признаков идет в сторону сближения
с соответствующими параметрами московских экземпляров; нивелирование
различий между образцами различного географического происхождения на
блюдается уже через 5 смен поколений [Виноградова, 2007, 2008].
Подобная прямая зависимость размеров растений от климатических усло
вий места их произрастания установлена на северо востоке Англии [Willis,
Hulme, 2002; Willis, Hulme, 2004]. Семена I.glandulifera в течение 2 лет высева
ли на пробных площадках по высотному градиенту -- от 10 до 600 м над ур. м.
Отмечено, что с увеличением высоты над уровнем моря уменьшались разме
234
ры растений и сокращалось число продуцируемых семян, однако масса семян
возрастала.
СЕМЕЙСТВО BALSAMINACEAE -- БАЛЬЗАМИНОВЫЕ
НЕДОТРОГА МЕЛКОЦВЕТКОВАЯ
Название: Impatiens parviflora DC
Морфологические признаки
Травянистое растение высотой 80--90 см с голыми ланцетными листьями.
Цветоносы формируются в пазухах верхних листьев и на 6--8 см возвышают
ся над верхушкой побега (рис. 103). Соцветие кистевидное из 4--12 цветков
[Марков и др., 1997]. В естественном ареале произрастают две цветовые формы
недотроги -- с желтыми и лиловыми цветками, ранее расссматриваемые [Побе
димова, 1949] в качестве отдельных видов. Однако нами установлено, что в Сред
ней Азии I. nevskii Pobed. не отличается от недотроги мелкоцветковой ни по од
ному из изученных нами признаков: ареалу, экологии, морфологии цветков, ка
честву семян, фенологии растений в природе и при интродукции [Майтулина,
1988]. По нашему мнению, недотрогу Невского нельзя рассматривать в каче
стве самостоятельного вида и следует считать ее лиловоцветковой формой I.
parviflora DC. Окраска лиловых лепестков недотроги по направлению к восточ
ной границе ареала постепенно бледнеет. Так, в ущельях хр. Кугитанг лепестки
интенсивно лилового цвета, в Душанбинском р не -- бледно лилового, а в г. Ру
шане на Памире -- практически белого с лиловым оттенком.
И у желто , и у лиловоцветковой недотроги даже в одной микропопуля
ции форма лепестков сильно варьирует: они имеют различную степень надре
занности -- и двулопастные, и двулопастные с неглубоким вырезом одной из
лопастей, и трехлопастные. Более того, даже у одного и того же цветка правые
и левые боковые лепестки могут различаться по степени надрезанности. Дли
на шпорца лепестковидного чашелистика также довольно изменчива. В одной
и той же популяции у обеих форм встречаются и короткие (3 мм) ширококли
новидные шпорцы, и длинные (до 9 мм) узкоклиновидные шпорцы, иногда
булавовидные на конце.
Естественный ареал
Центральноазиатский вид с естественным ареалом, расположенным в ос
новном на территории республик Средней Азии.
Вторичный ареал
История интродукции и географическое распространение. В Европе недо
трога впервые появилась в 1831 г. в Ботаническом саду Женевы, в 1837 г. ее
культивировали в Дрездене. В течение следующих нескольких десятилетий
отмечались единичные находки во многих пунктах в Европе, обычно возле
ботанических садов: Женева, Прага, Тарту, Кенигсберг, Берлин, Дрезден
[Coombe, 1956]. В гербарии Львовского университета хранятся образцы из
парков и садов Вены (1860 г.), Кракова (1861 г.), Дублян (1895 г.).
В Англии первые находки вида не связаны с ботаническими садами. Впер
вые I. parviflora обнаружена в Чешире в 1848 г., следующие находки датирова
235
Рис. 103. Цветение Impatiens parviflora
Рис. 104. Расселение Impatiens parviflora в Средней России
Рис. 105. Внедрение Impatiens
parviflora в лесные сообщества
Рис. 106. Форма Impatiens parviflora
с лиловой окраской венчика
237
ны 1851 и 1856 гг. В 1864 г. недотрога отмечена в Шотландии. К началу ХХ
века вид встречался в Англии довольно часто, но в Ирландии отсутствовал
[Coombe, 1956].
В середине ХХ века ареал I. parviflora в Европе стал стремительно расши
ряться. С 1947 г. она известна в Южной Швеции [Riska, 1957--1958]. В то же
время началась натурализация вида в Дании, на юге Финляндии, Польше,
Чехословакии, Германии, Венгрии, Австрии, Голландии, Бельгии, на севере и
востоке Франции. Одна находка сделана в Норвегии. В Швейцарии обычна
до высоты 760 м над ур. м., но поднимается и выше: максимальная высота, на
которой недотрога обнаружена в Альпах, -- 1860 м [Coombe, 1956]. В конце
ХХ века вид найден в Италии [Gredaro, Pirola, 1987] и в Болгарии [Чешмед
жиев, 1988].
В Северной Америке отмечена как заносное в Британской Колумбии (Ка
нада). Найдена впервые в Ванкувере в 1949 г., а в 1972 г. собрана там же, а
также в 15 милях к северу от предыдущего местообитания [Barkworth, 1973].
Интродуцирована на острове Принца Эдуарда в Квебеке [Coombe, 1956]. Есть
данные [Zinov'eva Stahovitch, Grant, 1984] о натурализации ее в Монреале.
В конце ХIХ в. недотрога мелкоцветковая выращивалась в Российской
империи в садах и парках Львова (герб. Львовского ун та, 1871, сборы
Iarolem'a), С. Петербурга, а также в трех ботанических садах Прибалтики
(RIG): Тарту (1893), Риге (1907) и Пярну (1909).
Вторичный ареал вида расширялся довольно медленно. В середине ХХ века
I. parviflora собрана в Латвии (LATV) в Валмиере (1962 г.), Огре (1972 г.),
Валке (1974 г.), Сигулде (1976 г.), Алуксне (1977 г.), Стучке (1981 г.), Бауске
(1983 г.), Мадоне (1984 г.). Позднее недотрога мелкоцветковая была собрана в
одном пункте Карелии [Раменская, 1983] и отмечена в качестве нового адвен
тивного вида в Белорусском Поозерье [Мартыненко, Сюборова, 1984]. На
Украине и в Молдавии I. parviflora буквально за несколько лет не только по
явилась на сорных местах, но и начала внедряться в естественные фитоценозы.
Самые северные точки ее вторичного ареала находятся на ст. Язель в Коми АССР
[Лавренко, Кустышева, 1990], а также в Вологде, где мы обнаружили недотрогу
в 1982 г. на немногих рудеральных местообитаниях, но в довольно большом ко
личестве. Растения цвели и плодоносили [Майтулина, 1984]. Южная граница
вторичного ареала -- Северная Осетия, где недотрога собрана в 1980 г. во Влади
кавказе и в 1983 г. -- в г.Алагир [Комжа, Попов, 1990]. При этом сообщается, что
в последнем местообитании популяция I. parviflora существует несколько лет,
но распространение ее на ближайшую территорию не наблюдается.
На Дальнем Востоке, в дендрарии г. Хабаровск, впервые собрана в 1964 г.
В.Н. Ворошиловым (MHA). Через 20 лет собрана в том же месте, но других
находок вблизи первого заноса не обнаружено [Шлотгауэр, 1984], в конце ХХ
века начала интенсивно распространяться в регионе [Ворошилов, 1982].
Пути и способы заноса. Несомненно, началом формирования вторичного
ареала недотроги в континентальной Европе следует считать ее «бегство» из
ботанических садов. В Англии первые находки сделаны на землях крупных
сельских поместий, что дало основание предположить, что семена недотроги
были занесены сюда с гречихой, используемой на корм фазанам.
Статус в регионе. Первый гербарный экземпляр недотроги, собранный
А.Петунниковым в Москве, относится к 1884 г. (MW). Н.Кауфман [1889] со
238
общал, что вид встречается во многих садах и парках. Число местонахожде
ний недотроги в Москве увеличивалось крайне медленно, и только через 40
лет Д.Сырейщиков [1927] отметил быстрое распространение вида по сорным
местам близ жилья и по садам.
Первая находка в Калужской обл. сделана в начале прошлого века (г.Жиз
дра, занесена 30.08.1906 г. В.Н. Хитрово. ОГУ), единичные находки отмеча
лись в Тульской обл. [Маевский, 1912]. Однако расширение ареала недотроги
началось лишь с 1970 х гг., то есть через 90 лет после первого появления в
Европейской России. Она найдена Н.А.Вьюковой [1987] в цветнике в г.Елец,
а также в ивняках и ольшаниках в поймах р. Воронеж, Дон, Становая Ряса в
Липецкой обл.
В настоящее время произрастает во всех областях Средней России (рис. 104).
Местообитания. В естественном ареале растет в узких глубоких лощинах,
ущельях, под кроной деревьев. В горных районах поднимается на высоту 2500 м
над ур. м.
Во вторичном ареале произрастает в посадках, по берегам рек, в садах, влаж
ных тенистых местах, лесопарках и нарушенных лесных массивах (рис. 105).
Усиленно внедряется в широколиственные леса Европы, вытесняя абориген
ную I.noli tangere [Csontos, 1986].
Литературный обзор по экологии I. parviflora сделан в «Биологической
флоре Московской области» [Марков и др., 1997]. Большинство местообита
ний, где произрастает недотрога мелкоцветковая, характеризуется той или
иной степенью затенения. Зависимость роста растений от освещенности, фо
топериодизма, температурных условий, условий минерального питания нео
днократно изучена по темпам накопления биомассы [Evans, Hughes, 1961;
Hughes, Evans, 1962; Hughes, 1966; Rackham, 1966; Peace, Grubb, 1982]. В це
лом, недотрога обнаруживает чрезвычайно высокую пластичность по отноше
нию к затенению. Ухудшение роста наблюдается при освещенности около 10%
полного света.
Наблюдения над недотрогой мелкоцветковой, проведенные в Главном бо
таническом саду РАН в.Москве, позволяют считать вид холодостойким: он не
повреждается морозами и заморозками [Карписонова, 1985].
Растет на самых разных, но достаточно увлажненных, почвах при условии,
что они хорошо аэрируются и не подтопляются. Предпочитает почвы, уме
ренно или очень богатые основаниями (но необязательно карбонатами), с рН
корнеобитаемого слоя 4,5--7,6. Почвы местообитаний недотроги обычно бога
ты мяг-ким гумусом. Однако при достаточно высокой освещенности недотро
га может длительно существовать и на очень бедной почве, занимая свобод
ные от других растений участки [Hughes, 1966].
Недотрога мелкоцветковая обычно растет на нарушенных почвах. Это пре
имущественно полуестественные лесные сообщества, парки или отмели рек с
облесенными берегами. Например, она встречается в донских приречных ив
няках [Тихомиров, 1987]. На подзолистых почвах произрастает исключитель
но по нарушенным местам [Coombe, 1956].
Размножение и жизненный цикл. Однолетнее растение. Цветы протандрич
ные. Рыльце созревает после растрескивания и опадания тычинок. Насеко
мые опылители до начала июля появляются редко, а во влажные и холодные
дни не появляются вовсе, тем не менее практически каждый цветок семена
239
завязывает. Превалирует хазмогамия, очень редко наблюдается клейстогамия
[Coombe, 1956]. Случаев перекрестного опыления и образования гибридов не
зафиксировано. Семенная продуктивность сильно варьирует. В лесах на сла
боизвестковых почвах в коробочке содержится обычно 1 семя, а на одном рас
тении за вегетационный сезон созревает 1--5 семян. На богатых увлажненных
почвах в коробочке содержится 4--5 семян, а одно растение дает до 10 тыс.
семян. Диссеминация может продолжаться до 3 месяцев.
Практически ни по одному показателю качества семян (масса, продолжи
тельность периода покоя, лабораторная всхожесть, энергия прорастания и др.)
не наблюдается никаких существенных различий между желто и лиловоц
ветковой формами I. parviflora, за исключением более высокой лабораторной
всхожести последней [Виноградова, 1992]. Средняя масса 100 семян у обеих
форм -- 0,5 г. У желтоцветковой формы лабораторная всхожесть семян в сред
нем составила 48%, у лиловоцветковой -- 60%.
Расселение и распространение. Семена разлетаются на 1--2 м, могут рас
пространяться по рекам. Последствия внедрения
Воздействие на естественные фитоценозы и аборигенные виды. В Централь
ной Европе недотрога типична для ши-роколиственных буковых лесов. В Че
хии может быть доминантом в травяном покрове ассоциации Querco
Populetum. В Западной Европе распространена в лесном экотоне Querco
petraeae -- Carpinetum pannonicum и Mercuriali Tilietum, положительно реаги
руя на сокращение площа-ди, занятой покровом из геофитов. В этих условиях
(например, при снижении покрытия аборигенных видов до 10% и ниже) недо
трога может стать доминантом и сохранять лидирующее положение, по край
ней мере в течение лета. Недотрога типична для нитрофильной растительнос
ти на нарушенных богатых почвах в кустарниковых зарослях, в парках, на пу
стырях, произрастая вместе с Chelidonium majus, Poa nemoralis, Solidago gigantea,
Urtica dioica, Glechoma hederacea, Geranium robertianum, Scrophularia nodosa
[Csontos, 1986; Марков и др., 1997].
Один из немногочисленных примеров вытеснения адвентивным видом або
ригенного -- вытеснение I. parviflora аборигенной I.nolitangere. Недотрога мел
коцветковая может существенно потеснить недотрогу обыкновенную в мес
тах совместного произрастания: во влажных лесах, в лесопарках и пойменных
фитоценозах около населенных пунктов. В лесопарках и нарушенных лесных
массивах нередко образует сплошные заросли, вытесняя естественную расти
тельность. На Украине и в Молдавии I. parviflora начала внедряться в есте
ственные фитоценозы широколиственных лесов, в том числе заповедные. Ра
стет в заповеднике Дунайские Плавни и в Каневском заповеднике, где в тра
вяном покрове преобладают Aegopodium podagraria и Galeobdolon luteum
[Дубына, Протопопова, 1985; Любченко, 1986; Пынзарь, 1988].
Недотрога мелкоцветковая более конкурентоспособна, чем недотрога обык
новенная, так как имеет более широкую экологическую амплитуду [Weise, 1968;
Csontos, 1986]. Это можно объяснить также морфологическими особенностя
ми вида и, в частности архитектурой корневой системы в ее онтогенетической
динамике [Марков, 1994]. Недотрога мелкоцветковая, начиная с конца июня,
не так резко снижает относительный вклад биомассы в корневую систему, чем
недотрога обыкновенная.
240
Генетические изменения и вариабельность вида. Данные по числу хромо
сом неоднозначные: 2n=24 у материала, собранного в Германии, Швеции и
Англии, есть также сообщения о 2n=20 и 2n=26 у европейских популяций
[Coombe,1956], в Монреале 2n=26 [Zinov'eva Stahоvitch, Grant, 1984].
Популяции вида во вторичном ареале обнаруживают довольно низкую измен
чивость. В Британии [Coombe, 1956] отмечена фенотипическая вариабельность
интенсивности окраски желтого околоцветника. В московских, петербургских и
вологодских популяциях интенсивностъ желтой окраски также варьирует.
Лиловоцветковые особи (рис. 106) стали дичать с 1990 х гг. только непода
леку от нашего экспериментального участка, где их долгое время выращива
ли. В других пунктах вторичного ареала лиловоцветковых растений не нахо
дили вплоть до начала 2000 х гг., когда Н.Н. Цвелев обнаружил эту форму в
Ленинградской обл. (LE).
Экономическое и социальное (положительное/отрицательное) значение. В
Средней Азии жители используют недотрогу в качестве красителя [Марков и
др., 1997]. Ранее ее разводили как декоративное растение в садах и парках Ев
ропы, несмотря на невзрачные цветки.
Меры борьбы
Исследования Р.А. Карписоновой [1985], проведенные в Главном бо
таническом саду РАН в Москве показали, что недотрога мелкоцветковая бо
лезнями и насекомыми не повреждается. Известно, что вид выступает в каче
стве одного из промежуточных хозяев паразитического ржавчинного гриба
Puccinia komarovii, который образует эцидии исключительно на нижней части
стебля, иногда на семядолях, но никогда на листьях, тогда как уредоспоры об
разуются на листьях. В случае произрастания недотроги на бедных почвах на
ее корнях развивает микоризу гриб типа Endogone [Peace, Grubb, 1982].
Обзор научных исследований и публикаций
Желто и лиловоцветковая формы сравнивали по скорости испарения воды
всеми частями растения при высыхании, что является косвенным показате
лем засухоустойчивости растений -- важнейшего свойства среднеазиатских
видов [Майтулина, 1988]. Установлена зависимость интенсивности испарения
двух форм недотроги от их возрастного состояния. У всходов, ювенильных и
имматурных особей наблюдается более быстрое испарение воды стеблями ра
стений по сравнению с семядолями или листьями. У особей в генеративном
состоянии листья и стебли испаряют воду с одинаковой скоростью, а цветы и
завязи плодов намного быстрее.
Резких различий в степени испарения воды между желто и лиловоцветко
вой формами недотроги не прослеживается.
СЕМЕЙСТВО BORAGINACEAE -- БУРАЧНИКОВЫЕ
ОКОПНИК КАВКАЗСКИЙ
Название: Symphytum caucasicum Bieb.
Морфологические признаки
Многолетнее корнеотпрысковое травянистое растение с укороченным ве
ретеновидным корневищем. Все растение с обильным сероватым опушением
241
из простых волосков, которое в целом мягче, чем у других крупных окопни
ков средней полосы России. Волоски двух типов: с расширенным коническим
многоклеточным основанием и обычные, без такого расширения. Стебель пря
мой, опушенный волосками обоих типов, в верхней части опушение гуще, вы
сотой до 100--150 см. Листья простые, от яйцевидно ланцетных до ланцетных,
нижние -- черешковые, верхние сидячие, с низбегающим на стебель основани
ем, отчего стебель вверху узкокрылатый (в сухом виде крылья малозаметны!).
Листовая пластинка сверху с вдавленной сетью жилок, с однородным опуше
нием из длинных волосков с расширенным основанием, опушение снизу во
лоски без расширенного основания, разной длины, по жилкам волоски длин
ные, между ними -- преимущественно короткие, направленные в разные сто
роны. Соцветия -- боковые завитки с многочисленными цветками; цветение
продолжительное, поэтому к его окончанию соцветие насчитывает до 40--45
отцветших цветков. Цветок типичного для бурачниковых строения, пятичлен
ный. Чашечка узко колокольчатая, надрезана на 1/3--1/2, с треугольными зуб
цами, с обильным опушением из простых волосков, нередко дуговидно изог
нутых (рис.107). Венчик голубой, однотонный, до раскрытия розовый, була
вовидный, в зеве с 5 полыми треугольными чешуйками, закрывающими вход
в трубку венчика. При созревании плодов чашечка увеличивается. Эремы пря
мые, не изогнутые.
Поскольку при определении окопников нередки ошибки, считаем необхо
димым привести ключ для их определения, в значительной степени основан
ный на опубликованном в «Новостях систематики высших растений» [Тихоми
ров и др., 1998]. В поле S. caucasicum сравнительно легко узнаваем по голубым
венчикам, относительно неглубоко рассеченной чашечки, многоцветковому соц
ветию и мягкому сероватому опушению. Однако предварительное полевое оп
ределение желательно подтвердить с использованием более «тонких» призна
ков. Следует учитывать, что крылатость стеблей на гербарном материале вид
на плохо.
1. Пыльники на верхушке с надсвязником (иногда несколько раздвоенным!)
или со связником, доходящим до верхушки гнезд; гнезда в верхней части па
раллельны друг другу. Эремы бугорчатые или гладкие, матовые или блестя
щие ........................................................................................................................................... 2
--- Связник заметно короче гнезд, поэтому пыльники на верхушке раздво
енные, с расходящимися гнездами. Эремы всегда бугорчатые, матовые .... 4
2. Эремы гладкие, блестящие. Свободные (т.е. не приросшие к венчику) час
ти тычиночных нитей короче пыльников или равны им по длине, а в нижней
части равны им по ширине. Венчик фиолетовый или розовато фиолетовый, без
синих или голубых тонов, реже белый или желтоватый .......... S. officinale L.
--- Эремы бугорчатые, матовые, реже блестящие. Свободные части тычи
ночных нитей обычно более длинные и узкие. Венчик разнообразной окраски,
обычно с синими или голубыми тонами, иногда белый или желтоватый ... 3
3. При плодах чашечка обычно полураскрытая из за того, что зубцы более или
менее параллельны друг другу; зубцы чашечки примерно равны по длине трубке ча
шечки, короче или длиннее ее. Венчик розово голубой, обычно с неоднородной ок
раской: розовые и голубые продольные полоски чередуются друг с другом; редко вен
чик белый или желтоватый. Стебли всегда крылатые......................S. × mosquense
(см. также ст. 6)
242
--- При плодах (до опадения эремов!) чашечка замкнутая, поскольку зубцы
сходятся друг с другом; зубцы чашечки всегда заметно длиннее трубки. Венчик
фиолетовый, розовато фиолетовый, грязно фиолетовый, голубоватый, редко бе
лый или желтоватый. Стебли крылатые или бескрылые..S. ×uplandicum Nyman
(см. также ст. 7)
4(1). Венчик желтоватый, иногда почти белый. Стеблевые листья, по крайней
мере нижние, черешковые, черешки обычно не крылатые. Стебли не крылатые.
На цветоносном побеге ко времени цветения нередко сохраняется розетка зеле
ных, обычно некрупных листьев. Обычно невысокие (до 50 см) растения лесов и
зарослей кустарников, цветущие весной или в самом начале лета....S. tauricum
Willd.
--- Венчик голубой, синий, фиолетовый или пурпурный. Стеблевые листья
сидячие или с крылатыми черешками. Розетки листьев, если они развиты во
время цветения, расположены на отдельных побегах, крупные. Обычно более
высокие растения, приуроченные ко вторичным местообитаниям ................ 5
5. Чашечка при плодах полураскрытая из за того, что ее зубцы более или
менее параллельны друг другу, после отделения эремов остающаяся полурас
крытой, надрезанная на 1/3--2/3, в 2--3 раза короче венчика. Стебли крыла
тые. Листья розеток, если они развиты, с клиновидным или округло клино
видным основанием ........................................................................................................... 6
--- Чашечка при плодах (до опадения эремов!) замкнутая, поскольку зубцы
сходятся друг с другом, после отделения эремов широко раскрывающаяся,
надрезанная на 2/3 или почти до основания, в 2--4 раза короче венчика. Стеб
ли крылатые или бескрылые. Листья розеток, если они развиты, с клиновид
ным или сердцевидным основанием ............................................................................ 7
6. Венчик голубой. Чашечка надрезана на 1/3--1/2. Эремы «прямые», то
есть их внутреннее, обращенное к столбику, ребро направлено почти перпен
дикулярно плоскости прикрепления эрема .................................. S. caucasicum
--- Венчик розово голубой, обычно с неоднородной окраской: розовые и голу
бые продольные полоски чередуются друг с другом. Чашечка надрезана на 1/3--
3/4. Эремы обычно изогнуты внутрь, то есть их внутреннее ребро направлено под
значительным углом к плоскости прикрепления эрема..................S. × mosquense
(см. также ст. 3)
7(5). Венчик синий или сине фиолетовый, обычно более или менее однотон
ный. Стебель не крылатый. Чашечка в 3,5--5 раз короче венчика. Листья розеток,
если развиты, с усеченным или сердцевидным основанием .............. S. asperum
--- Венчик грязно фиолетовый или розовато фиолетовый, обычно неодно
тонный, часто продольно полосатый. Стебли крылатые или бескрылые. Ча
шечка в 2--3 раза короче венчика. Листья розеток, если развиты, с усеченным
основанием ............................................................ S. × uplandicum (см. также ст. 3)
В Ботаническом саду МГУ им. М.В. Ломоносова на Воробьевых горах сорни
чает еще один окопник -- S. azureum Van Hall, который, видимо, для флоры России
ранее не указывался. Этот малоизвестный вид, отсутствующий в большинстве
«Флор», неплохо известен в Европе специалистам озеленителям [Borchardt,
2008]. Он напоминает S. caucasicum -- венчик голубой и опушение сравнительно
мягкое, но растения невысокие, обычно не выше 50(60) см, чашечка узкая, по
чти трубчатая, цветение раннее, преимущественно в мае. Указание на его сибир
ское происхождение [Borchardt, 2008], очевидно, ошибочно, по видимому,
243
Рис. 108. Расселение Symphytum caucasicum в Средней России
Рис. 107. Цветки Symphytum caucasicum с характерной колокольчатой чашечкой
Рис. 109. Расселение Symphytum
caucasicum по железной дороге,
Московская обл., близ платформы
«Тарасовка»
Рис. 110. Плодоношение Symphytum
caucasicum
245
S.azureum -- культигенный вид, описанный из ботанического сада Гронингена,
Нидерланды (Van Hall, 1849).Естественный ареал
S. caucasicum -- широко распространенный кавказский вид, естественный
ареал которого сходен с ареалом S. asperum Lepechin, но в целом он встречает
ся реже [Попов, 1953; Гвиниашвили, 1976].
Вторичный ареал
История интродукции и географическое распространение. В Западной Ев
ропе вид выращивают давно: в Англию S. caucasicum завезен в 1820 г., в Герма
нию -- в 1816 г., но он известен гораздо меньше, чем S. asperum и S. × uplandicum
Nyman [Bucknall, 1913; Hegi, 1927]. В настоящее время широко известен как де
коративное растение [Bennett, 2003]. В Великобритании отмечены немногочис
ленные случаи дичания S. caucasicum из культуры1 [Clement, Foster, 1994]. Во
«Флоре европейской части СССР» вид указан для Крыма [Доброчаева, 1981].
Пути и способы заноса. Выращивается как теневыносливая неприхотли
вая декоративная культура [Карписонова, 1999; Robinson, 1987]. Устойчив в
местах прежних посадок, где сохраняется без малейшего ухода. Дичает и в
последнее время встречается на пустырях, у жилья, на ж/д насыпях. Его рас
пространение связано в основном с культивированием.
Статус в регионе. В Средней России S. caucasicum культивируют как деко
ративное растение в городах, деревенских палисадниках, на дачных участках,
особенно часто -- в Москве. Иногда окопник ускользает из культуры. Сведе
ния о распространении S. caucasicum в Средней России практически отсутству
ют; первое указание относится к Москве [Куваев и др., 1992]. В гербариях MW
и MHA хранятся сборы окопника из Владимирской, Калужской, Московской
и Тульской обл., а также из Мордовии. Первые гербарные сборы S. caucasicum
в Средней России находятся в MHA: «Москва, Останкино, 30.06.1956 г., А.Хох
ряков», «Московская область, Мытищи, 19.07.1976 г., М.Шумова, В.Макаров».
Долгое время S. caucasicum не был замечен флористами, его путали с S.
asperum [Тихомиров и др., 1998]. Например, указание М.С.Игнатова с соавт.
[1990] на широкое распространение S. asperum в Московской обл. относится,
очевидно, в основном к S. caucasicum. Дополнительную путаницу вносит то,
что в сельскохозяйственной практике «кавказским окопником» часто назы
вают S. asperum [Астахов, 1951, 1953].
В настоящее время вид отмечен в большинстве регионов Средней России
(рис. 108).
Местообитания. В пределах естественного ареала растет по лесным опуш
кам, полянам, у рек и ручьев от лесного до субальпийского пояса, встречается
как сорняк на выгонах, паровых полях и огородах, сорных местах [Малеев,
1934; Гвиниашвили, 1976]. В пределах вторичного ареала приурочен в основ
ном ко вторичным местообитаниям у жилья: пустырям, обочинам дорог, опуш
кам у дачных поселков (рис. 109).
Размножение и жизненный цикл. При повреждении корней даже небольшие
куски корня могут дать новое растение, поэтому S. caucasicum нередко образует
обширные заросли [Малеев, 1934]. По видимому, в Средней России размножа
ется преимущественно вегетативно, так как, по крайней мере в пределах конк
ретного местонахождения, плодов завязывается крайне мало (рис. 110). Мож
1 В Германии дичание кавказского окопника пока не отмечено [Hаeupler, Muer, 2000, 2007].
246
но предположить, что, подобно S. officinale [Goulson et al., 1998], S. caucasicum
является самонесовместимым перекрестноопыляемым растением. Однако не
большое число эремов тем не менее завязывается. Растение зацветает на вто
рой год после прорастания семян [Малеев, 1934].
Расселение и распространение. Нередко дачники высаживают S. caucasicum
вдоль заборов или выбрасывают лишние растения на компостные кучи, где он
приживается и разрастается. Такие группы растений можно обнаружить на опуш
ках у дачных поселков, вдоль дорог, по сельским улицам, сорным местам.
Последствия внедрения
Воздействия на естественные фитоценозы и аборигенные виды. В настоя
щее время происходит лишь начальная стадия натурализации S. caucasicum.
Однако в Калужской обл. в окрестностях Калуги у устья р. Калужка вид обна
ружен в прибрежных ивняках (MW). Таким образом, расселение окопника из
антропогенных местообитаний в естественные возможно.
Генетические изменения и вариабельность вида. М.Г. Попов указывал на
наличие железистого опушения на чашечке S. caucasicum [Попов, 1953]. Од
нако у дичающих растений такое опушение отсутствует, волоски на чашечке
простые, разного строения, но железистых среди них нет (MW, MHA).
Среди гербарных образцов одичавших растений S. caucasicum изредка встре
чаются экземпляры, у которых чашечка рассечена глубже обычного (MW,
MHA), при этом форма листьев, тип опушения характерны именно для кав
казского окопника. Возможно, это результат интрогрессивной гибридизации,
но для правильной оценки таких растений необходимо изучение изменчивос
ти в природных популяциях.
Отмечена гибридизация S. caucasicum с местным S. officinale. Гибрид получил
название S. × mosquense S.R. Majorov et D.D. Sokoloff по месту первой находки
[Тихомиров и др., 1998]. К настоящему времени гибрид обнаружен в Москве,
Московской и Калужской обл. [Маевский, 2006; собственные наблюдения].
Экономическое и социальное (положительное и отрицательное) значение.
Окопник -- трудноискоренимый сорняк садов и дачных участков. «Разраста
ется моментально, особенно если начинаешь его срезать и копать. Чуть про
смотришь, и буквально за неделю розетки шершавых листьев отрастают, как
лопухи. И все норовят занять соседние территории» -- из комментария посе
тителя «Дачного форума» [2008].
Окопник кавказский, как и другие виды рода, является источником биоло
гически активных соединений, которые в последние годы активно изучаются.
Это, прежде всего, пирролизидиновые алкалоиды [Барбакадзе и др., 2002; Зу
зук и др., 2004; Rizk, 1990; Gomes et al., 2007].
Меры борьбы
В настоящее время S. caucasicum рекламируется как чрезвычайно удобное
растение для тенистых участков сада [например, Карписонова, 1999]. Инте
рес к окопнику подогревается многочисленными сообщениями в Интернете о
лекарственных свойствах видов этого рода. В пособиях для садоводов необхо
димо предупреждать цветоводов, что окопник кавказский является обреме
нительным сорняком, а также об опасности использования окопника как ле
карственного растения [Rode, 2002].
247
CЕМЕЙСТВО CRUCIFERAE (BRASSICACEAE) --
КРЕСТОЦВЕТНЫЕ
КАРДАРИЯ ОБЫКНОВЕННАЯ
Название: Cardaria draba (L.) Desv.
Синонимы: Lepidium draba L.
Морфологические признаки
Многолетнее травянистое корнеотпрысковое растение с длинным прочным
стержневым корнем и горизонтальными ветвистыми боковыми корнями. Цве
тоносные побеги прямостоячие, высотой 20--60(80) см, голые или с прижаты
ми простыми короткими волосками, олиственные, в поперечном сечении уг
ловатые. Листья от салатно зеленых до серо зеленых из за прижатого корот
кого простого опушения, овальные до ланцетных, нижние черешковые, с
клиновидным основанием, обычно ко времени цветения высыхающие, сред
ние и верхние -- сидячие, полустеблеобъемлющие, с ушками при основании,
край листа от цельного до зубчато лопастного. Соцветие ветвистое, с 2--4 бо
ковыми кистями (рис. 111). Цветки диаметром 4--6(8) мм, типичного для кре
стоцветных строения. Лепестки белые, вдвое длиннее чашелистиков. Стручоч
ки невскрывающиеся, широкосердцевидные или округлые, килеватые, длиной
около 4 мм, с сохраняющимся столбиком, созревшие --- с сетчатыми или глад
кими сворками. Семян обычно 2, но иногда развивается только 1 или же 4.
Европейские и американские специалисты различают два подвида: spp.
draba и ssp. chalepensis (L.) O.E. Shulz [Буш, 1939а; Scurfield, 1962; Stace, 1997].
При этом считается, что типовой подвид встречается в основном на юге Евро
пы и Сибири, а ssp. chalepensis приурочен к азиатской части ареала: Кавказу,
Малой и Средней Азии.
Следует иметь в виду, что в последние годы в Саратовской обл. (MW) и в
Москве (MHA) обнаружена габитуально похожая кардария опушенная ---
C. pubescens (C.A. Meyer) Jarm. (Hymenophysa pubescens C.A. Meyer, Lepidium
pubescens auct., non Desv.), отличающаяся прежде всего опушенными стручоч
ками [Буш, 1939б; Scurfield, 1962; Taiyan et al., 2001]. Соцветие этого вида бо
лее рыхлое, несколько напоминающее соцветие L. campestre R. Br.
Закавказский вид кардарии C. propinqua (Fisch. et C.A. Meyer) N.Busch так
же нередко синонимизируют с C. chalepensis (L.) Hand. Mazz. [ssp. chalepensis
(L.) O.E. Schulz] [Jafri, 1973; Taiyan et al., 2001]. По видимому, для этого есть
основания, так лектотип C. chalepensis [LINN No. 824.20; http://www.nhm.ac.uk/
research curation/research/projects/linnaean typification/) весьма напоминает
закавказские растения (MW, MHA). Однако типовой материал Фишера и
Мейера нами не изучен. Следует иметь в виду, что нередко под названием C.
chalepensis часто приводят растения C. draba с округлым или ширококлино
видным (а не сердцевидным) основанием стручочков [Sell, 1993]. Следует
иметь в виду, что типичная структура стручочков формируется только у прак
тически созревших плодов, что может приводить к ошибкам при определе
нии, о чем свидетельствуют многочисленные фотографии кардарии на амери
канских интернет сайтах.
Сложнее ситуация с C. repens (Schrenk) Jarm. [C. draba var. repens (Schrenk)
O.E. Shulz, Lepidium repens (Schrenk) Boiss.]. Зарубежные ботаники обычно
248
включают этот вид в состав C. chalepensis. [например: Jafri, 1973; Rollins, 1993].
Однако просмотр гербарных образцов из Средней Азии в MW и MHA пока
зал неоднозначность такого решения. Очевидно, что виды кардарии нужда
ются в дополнительном изучении, при этом необходимо исследование кавказ
ских и среднеазиатских природных популяций.
К сожалению, определительные ключи А.В. Ярмоленко [1934] в «Сорных
растениях СССР» и Н.А. Буша [1939а] во «Флоре СССР» не позволяют, по
нашему мнению, составить определенное представление о различиях этих так
сонов. Предлагаем поэтому оригинальный вариант ключа для различения этих
кардарий (рис. 112).
1. Стручочки опушенные, с округлым или даже клиновидным основанием,
нередко вверху с антоциановым окрашиванием, створки без заметных жилок, но
совершенно созревшие оголяющиеся и мелкоячеистые. Верхние стеблевые лис
тья выемчато зубчатые, ланцетные или линейно ланцетные .......... C. pubescens
--- Стручочки голые, без антоцианового окрашивания. Верхние стеблевые
листья обычно более широкие, от ланцетных до яйцевидных, цельнокрайные
или городчато зубчатые, реже выемчато зубчатые. ............................................... 2
2. Средние и верхние стеблевые листья с клиновидным основанием, голые
(оголяющиеся?), немного кожистые (как у Lepidium latifolium). Стручочки с
округлым или усеченным основанием, мелкоячеистые (если отдельные жил
ки и видны, то жилки не образуют ячеек). ........................................ C. chalepensis1
--- По крайней мере, средние стеблевые листья полустеблеобъемлющие, с
отчетливыми ушками при основании, с обильным сероватым коротким опу
шением, травянистые. Стручочки с сердцевидным или округлым основанием,
ячеисто сетчатые, реже гладкие. ..................................................................................... 3
3. Зрелые стручочки сильно вздутые, почти шаровидные, без жилок, шири
ной до 5--6 мм, с округлым или усеченным основанием, боковой киль не выра
жен; при гербаризации недозревшие стручочки сминаются. Стеблевые листья
овальные или яйцевидно овальные, с округлой верхушкой, цельнокрайные или
сглажено зубчатые. ............................................................................................... C. repens
--- Зрелые стручочки менее вздутые, шириной обычно менее 5 мм, с боко
вым килем; при гербаризации стручочки не сминаются; створки сетчатые из
за выступающих жилок (реже --- почти гладкие). Стеблевые листья обычно с
треугольной верхушкой, разной ширины: от ланцетных до яйцевидных; край
листа от цельного до зубчатого. ...................................................................... C. draba
Мнения авторов таксономических обработок крестоцветных о распростра
нении видов Cardaria не совпадают [Ярмоленко, 1934; Буш, 1939а; Котов, 1979;
Дорофеев, 2002]. А.В. Ярмоленко [1934] расширял ареал C. repens до Нижней
Волги, что не подтверждается гербарными сборами2. Для Москвы М.И. Наза
ров указывал C. repens, которую отмечал в 1919--1920 гг. на путях Малой ок
ружной ж/д, однако соответствующих гербарных сборов обнаружить не уда
лось [Назаров, 1927; Игнатов, 1990]. Не исключена, впрочем, номенклатурная
путаница. Следует отметить однородность заносных растений в Средней Рос
сии, которые относятся к C. draba s. str. (MW, MHA).
1 Включая С. propinqua.
2 Оригинальных детерминаток А. В. Ярмоленко мы не видели (LE, MW, MHA), что не
позволяет вынести определенного решения о северной границе распространении этого вида.
249
Рис. 112. Плоды трех видов рода Cardaria: C. draba --- слева, C. repens --- в центре,
C. pubescens --- справа
Рис. 111. Цветение
Cardaria draba
Рис. 114. Обо
чина дороги с
Cardaria draba
Рис. 113. Рассе
ление Cardaria
draba в Средней
России
251
Род Cardaria Desv. очень близок к Lepidium L., от которого отличается взду
тыми невскрывающимися плодами и сливающимися нектарниками [Taiyan et
al., 2001]. Впрочем, эти признаки изредка встречаются и у клоповников
[Bowman et al., 1999]. В последнее время, особенно на Западе, род Cardaria
нередко не выделяют из состава Lepidium [Al Shehbaz et al., 2002]. Молеку
лярно генетическое исследование рода подтверждает эту точку зрения, выде
ление особого рода Cardaria не подтверждается. Принимаемый авторами ис
следования таксономический ранг этой группы --- секция: Cardaria (Desv.) DC.
Ближайшей родственной группой оказывается секция Lepia DC. с L. campestre
[Mummenhoff et al., 2001]. Таким образом, оказывается, что сходство облика
C. pubescens и L. campestre имеет глубокую генетическую основу.
Последствием объединения Lepidium и Cardaria, которое необходимо учи
тывать, является образование нового названия для C. pubescens --- L. appelianum
Al Shehbaz, поскольку в роде клоповник уже есть вид с таким название, это L.
pubescens Desv. [Al Shehbaz et al., 2002].
По видимому, объединение этих родов скоро войдет в повседневный обиход,
однако в данном издании мы принимает традиционное и привычное их разделе
ние, широко принятое в том числе и в литературе по инвазионным видам.
Естественный ареал
Природный ареал C. draba приурочен к Старому Свету. Он охватывает юг
Европы, Северную Африку, Западную и Центральную Азию, юг Сибири [Буш,
1939а; Котов, 1979; Дорофеев, 2002; Scurfield, 1962; Jafri, 1973; Ball, 1993; Taiyan
et al., 2001].
Вторичный ареал
История интродукции и географическое распространение. В настоящее вре
мя C. draba широко распространена во внетропических областях земного шара
в Европе, Азии, Северной и Южной Америке, Австралии, Южной Африке
[Scurfield, 1962; Jafri, 1973; Ball, 1993; Clement, Foster, 1994; Taiyan et al., 2001;
Auld et al., 2003].
В Европе расселение кардарии началось, по видимому, в начале XIX века
[Hegi, 1919; Szafer, 1966]. Подробная история расселения рассмотрена в «Illustrierte
Flora von Mittel Europa» [Hegi, 1919]. Ныне C. draba встречается в большинстве
стран Европы, в 17 регионах натурализовалась [Lambdon et al., 2008].
Впервые в США кардария собрана в штате Калифорния в 1876 г., в Канаде
в провинции Онтарио в 1878 г. [Mulligan, Findlay, 1974]. В настоящее время
известна во всех штатах США и провинциях Канады, местами представляет
собой злостный сорняк посевов [Anderson, 1999]. В настоящее время C. draba
известна в Южной Америке [Castro et al., 2005], Австралии [Parsons,
Cuthbertson, 2001], Японии [Auld et al., 2003]. Активное расселение кардарии
произошло в конце XIX --- первой половине XX века.
В России кардария распространилась широко: от Калиниградской обл. до
Сахалина [Конспект сосудистых..., 1999; Баркалов, Таран, 2004]. На Кавказе
C. draba распространена очень широко3 [Гроссгейм, 1950]. В Сибири известна
в южных районах или как заносное растение [Никифорова, 1994]. На Даль
нем Востоке C. draba известна пока только на Сахалине [Беркутенко, 1988;
Баркалов, Таран, 2004].
3 C. chalepensis произрастает только в Закавказье [Гроссгейм, 1950].
252
Пути и способы заноса. Кардария относится к непреднамеренно занесен
ным растениям, которая оказывается вне естественного ареала разнообразны
ми способами. Семена кардарии могут разноситься с зерном, кормами, занос
ные растения при этом обнаруживаются у дорог, на железнодорожных насы
пях. Появление C. draba в Северной Америке связывают с люцерной из Азии,
но не исключен ее занос с судовым балластом [Mulligan, Findlay, 1974]. В рай
онах с поливным земледелием плоды могут разноситься по системе поливных
каналов и дренажа [Scurfield, 1962].
Статус в регионе. В Средней России кардария обнаружена впервые, по
видимому, в Москве: «Москва, в Александровском саду, 27 мая 1881 г., одичав
шее с выставки 1872 г, В.Я. Цингер» [MW; Цингер, 1885]. В Подмосковье ре
гулярно собиралась на ж/д насыпях с 20--х гг. XX века [MW; Игнатов и др.,
1990]. За первую половину XX века расселилась по всей Средней России. В
настоящее время отмечена во многих регионах Средней России (рис. 113). В
южных районах массовый обременительный сорняк, в северных как заносное
растение, обычно колонофит или эфемерофит, устойчивых популяций не фор
мирует [Дорофеев, 2002, 2006].
Н.А. Буш считал, что в Средней России в Волжско Донском флористичес
ком районе вид находится на северном пределе естественного ареала [Буш,
1939а]. Однако М.И. Котов [1979] признавал все среднерусские местонахож
дения заносными.
Местообитания. Засухоустойчивое и солевыносливое растение. Обитает
на различных почвах, но предпочитает слабощелочные [Qasem, 2004]. На юге
и юго востоке Европейской России C. draba растет на солонцах, меловых скло
нах, вдоль дорог, в полях, на газонах, в лесополосах (рис. 114), иногда в пой
мах, на севере --- заносное вдоль транспортных магистралей, в населенных пун
ктах, предпочитает засоленные местообитания [Дорофеев, 2002; Dicot..., 1988].
Размножение и жизненный цикл. Цветет в апреле --- июне. По видимому,
кардария --- самоопыляемое растение [Scurfield, 1962]. Однако, по нашим на
блюдениям, цветущую C. draba активно посещают разнообразные насекомые:
мухи, жуки, в меньшем числе перепончатокрылые. Плодоносит в июле. Семен
ная продуктивность оценивается от 850 плодов на цветоносный побег [Scurfield,
1962] до 1200--4800 плодов на растение [Ярмоленко, 1934; Parsons, Cuthbertson,
2001; Wilson, Littlefield, 2006]. Семена обладают высокой жизнеспособностью --
до 80% [Parsons, Cuthbertson, 2001]. Вес 1000 семян -- 1,8 г [Ярмоленко, 1934].
Всходы появляются следующей весной --- в начале лета. Минимальная темпе
ратура прорастания семян + 3--4°C, оптимальная + 15--25°С. Зацветает на 2 й
год [Ярмоленко, 1934]. Семена прорастают в почве с глубины не более 5--7 см,
сохраняют жизнеспособность не менее 3 лет [Scurfield, 1962]. Однако, по дру
гим данным, на 3 й год всхожесть семян незначительна [Anderson, 1999]. Ак
тивно размножается вегетативно, образуя вегетативные обширные клоны [Яр
моленко, 1934]. За год прирост зарослей может достигать 60--70 см [Anderson,
1999]. Способна регенерировать из отрезка корня длиной 4--5 см [Scurfield, 1962].
Всходы из семян и побеги от корневых почек (с глубины до 80 см) появляются
в апреле --- мае и летом. Наземные органы повреждаются при -- 5--6°С.
Расселение и распространение. Скорость распространения C. draba в Евро
пе оценивается от 2,7 км/год для Великобритании до 0,5 км/год для Ирлан
дии [Williamson et al., 2005].
253
Последствия внедрения
Воздействия на естественные фитоценозы и аборигенные виды. C. draba
является активным аллелопатическим растением, ее выделения подавляют
всходы других растений, чем объясняется формирование обширных зарослей
кардарии в Новом Свете [Kiemnec, Mcinnis, 2002; Qasem, 2004].
Генетические изменения и вариабельность вида. Специальное исследование
не выявило существенных различий между популяциями Старого и Нового
Света по конкурентной мощности [McKenney et al., 2007]. Выделены микро
сателитные маркеры для дальнейшего исследования генетики инвазионных
популяций [Bon et al., 2005].
Экономическое и социальное (положительное и отрицательное) значение. В
южных областях кардария является злостным сорняком. Однако в Средней
России C. draba такой активностью пока не обладает.
Меры борьбы
Как показали наблюдения последних лет в Москве, кардария оказалась
неустойчивой к регулярному выкашиванию газонов. Многие ранее устойчи
вые ее колонии выпали после 2--3 лет выкашивания.
В США рассматривается возможность применения для биологического
контроля галловых клещей Aceria drabae (Eriophyidae) и жуков Ceutorhynchus
spp. (Coleoptera, Curculionidae), Psylliodes isatidis (Coleoptera, Chrysomelidae).
Однако при этом возможен ущерб для американских крестоцветных, в том
числе охраняемым видам, так как выискать строгих стенофагов среди вреди
телей Cruciferae едва ли возможно [Wilson, Littlefield, 2006].
До сих пор наиболее приемлемым гербицидом для борьбы с C. draba оста
ется 2,4--D, при этом требуется неоднократная обработка для уничтожения
регенерирующих растений [Peachey et al., 2007].
CЕМЕЙСТВО CRUCIFERAE (BRASSICACEAE) ---
КРЕСТОЦВЕТНЫЕ
РОГАЧКА ФРАНЦУЗСКАЯ
Название: Erucastrum gallicum (Willd.) O.E. Schulz.
Синонимы: Sisymbrium gallicum Willd.; Erucastrum pollichii K.F. Schimp. et
Spenn.; Brassica erucastrum L.; Kibera gallica (Willd.) V.I. Dorof.
Морфологические признаки
Однолетнее или двулетнее растение высотой до 60 см, с опушенным стеб
лем. Листья перисторассеченные, с продолговатыми, зубчато надрезанными
долями. Цветки собраны в облиственные кисти. Чашелистики косо вверх тор
чащие. Лепестки бледно желтые, редко желтые, длиной 6--8 мм (рис. 115).
Цветоножки в 3--4 раза короче косо вверх торчащих стручков. Плоды -- почти
четырехгранные стручки длиной 2--4 см, сидячие (без плодоножек); носик
стручков длиной 1,5--4 мм, линейно цилиндрический [Губанов и др., 2003].
Естественный ареал
Европейский по происхождению вид. Область его сплошного природного
распространения охватывает центральные и южные регионы Европы [Flora of
254
NW Europe, 2008]. Средняя Европа: Австрия, Чехия, Словакия, Германия,
Бельгия, Венгрия, Нидерланды, Люксембург, Швейцария; Южная Европа:
Италия, страны бывшей Югославии, Франция, Испания [Germplasm Resources
Information (GRIN), 2008]. Вторичный ареал
История интродукции и географическое распространение. История интро
дукции рогачки в Северную Америку подробно описана в работе J.Luken с со
авторами [1993]. Вид впервые обнаружен в США в штате Висконсин в 1903 г.
[Robinson, 1911]. С 1911 по 1920 гг. сборы E. gallicum сделаны еще в 7 штатах.
В Канаде первая находка датирована 1932 г. [Groh, 1933]. Вскоре E.gallicum
распространилась по ж/д путям в Канаде и США, и в настоящее время отме
чена по всей Северной Америке, в центральных и западных штатах США -- в
массе [Warwick, Wall, 1998]. Встречается рогачка и в Мексике [Germplasm
Resources Information (GRIN), 2008].
В регионах Азии E. gallicum указывается как натурализовавшаяся в Япо
нии, Корее, (где впервые обнаружена в 1992 г.), а также в Индонезии, Иорда
нии, и на о. Бали [Alien Plants of Korea, 2008; Zipcodezoo.com, 2008].
В Северной и Восточной Европе E. gallicum -- заносное растение. В Ислан
дии, Великобритании, Финляндии указывается как эфемерофит, в Норвегии,
Австрии, Германии, Чехии, Беларуси -- как натурализовавшийся вид [GCW
(Global Compendium of Weeds), 2008]. Во многих регионах Восточной Европы
вид отмечен относительно недавно и, вероятно, продолжает продвигаться к
востоку и северо востоку. Указана для Турковского и Сколевского р нов Львов
ской обл. как новый вид флоры Украинских Карпат. Предполагается, что рас
тение занесено в Карпаты с востока Украины, однако существует вероятность
заноса из Румынии, Венгрии или Словакии [Данилюк, 2007].
Продолжает расселяться в Европейской России. Указывается как новый
вид для Ульяновской обл. [Силаева, 2008]. В Волгоградской обл. E. gallicum
встречается по берегам рек, распространяется водным транспортом [Матвеев,
Сагалаев, 2000]. На северо западе -- в Ленинградской, Новгородской и Псков
ской обл. встречается у дорог, преимущественно на ж/д путях, на песках мор
ского побережья, в населенных пунктах, нередко [Цвелев, 2000].
Пути и способы заноса. В середине 1980 х гг. (рис. 116) рогачка французская
стала доминирующим видом в растительном покрове щебневых карьеров в Туль
ской и Калужской обл. [Крылов, 2008]. Учитывая экологическую приуроченность
вида в естественном ареале, можно предположить, что именно из карьеров
E.gallicum расселяется по другим типам мест обитания -- обочинам дорог, сорным
местам, свалкам, пустырям, газонам, известняковым обнажениям, берегам рек,
кучам строительного мусора, территориям промышленных предприятий.
Статус в регионе. На юге Воронежской обл. E.gallicum встречается по от
крытым местам обитания [Камышев, Хмелев, 1976]. Севернее вид единично
отмечен в г. Воронеж, на мусорной куче в ботаническом саду ВГУ [Григорьев
ская, 2000]. «Возможно, встречается чаще, но не фиксируется коллекторами»
[Григорьевская и др., 2004].
В Белгородской обл. вид собран 20.09.1956 г. в окрестностях Алексеевки,
на меловых склонах (MW, фамилия коллектора неразборчива). В настоящее
время -- обыкновенно по мелам, мусорным местам, нарушенным степным скло
нам [Еленевский и др., 2004].
Рис. 115. Цветение
Erucastrum gallicum
Рис. 116. Расселение
Erucastrum gallicum
в Средней России
Рис. 117. Внедрение
Erucastrum gallicum в
естественные
сообщества,
Воронежская обл.,
меловое обнажение
у с. Мастюгино в
долине р. Девица
Рис. 118. Стручки
Erucastrum gallicum
с прицветниками
при плодоножках
257
В Курской обл. рогачка впервые собрана Л.В. Тишиной в Касторенском р
не, в окрестностях д. Горяиново, в 1994 г. (КурМ). На востоке Курской обл. E.
gallicum встречается редко на меловых обнажениях и по склонам оврагов и
балок, на остальной территории области возможны находки по ж/д [Полу
янов, 2005].
В Орловской обл. лишь однажды собрана в г. Орел, на ж/д у завода «Дор
маш», 7.08.1995 г., А. Сотников (OHHI). Несмотря на специальные поиски и
просмотр подходящих мест обитания, вид не был отмечен нами на юго восто
ке Орловской обл. в июле 2008 г.
К 1996 г. для Липецкой обл. было известно лишь два сбора в Добринском
р не на ж/д полотне и в Липецке на газоне [1979, MW; Флора Липецкой обла
сти, 1996]. Впервые в Тульской обл. E. gallicum собрана Ю.Е. Алексеевым в Яс
ногорском р не между шпалами и на откосах ж/д в 1980 г. (MW). Затем после
довали многочисленные сборы и наблюдения E. gallicum на рудеральных местах
обитания в Тульской обл. [Шереметьева, 1989], и к настоящему времени рогач
ка французская -- обычный вид нарушенных, незадерненных участков.
В Московской губернии E. gallicum появилась в конце XIX века [Горожан
кин, 1888], но распространилась лишь локально в окрестностях Петровской
академии. В 1970 е гг. рогачка появилась на ж/д путях [Октябрева и др., 1978]:
«Московская обл., Люберецкий р н, между пл. Яничкино и ст. им. Дзержин
ского на ж/д путях, 9.08.1974 г., Н. Октябрева, А. Чичев» (MW). В начале 1980 х
годов E. gallicum перешла с ж/д местообитаний на газоны и щебневые покры
тия [Игнатов и др., 1990], а впоследствии вид распространился в массе.
В 1978 г. E. gallicum впервые собрана в Конаковском, Кимрском и Калининском
р нах Тверской обл. Один из первых сборов: «Калининская обл., г. Кимры, близ ст.
Савелово, ж.д. насыпь, собр. В. Малышева, опр. Ю. Гусев, 11.07.1978 г.» (MW). К
настоящему времени вид отмечен во всех р нах области [Маркелова, 2004].
Для Владимирской обл. указывается Е.А. Борисовой [Борисова, 2006а], на
основании следующего сбора: «Владимирская обл., окр. г. Александров, у ж.д.
моста через р. Серая, склон ж.д. насыпи, 14.08.2005 г., Е. Борисова» (MW).
Указывается в ряде населенных пунктов Ивановской обл. [Борисова, 2007].
E. gallicum собрана Е. Борисовой в Ивановской обл., в г. Вичуга, близ ж/д вок
зала, 16.06.1991 г. (MW). В г. Приволжск растет по известняковой насыпи уз
коколейной железной дороги [М. Голубева, личное сообщение].
Собрана в Смоленске на ж/д насыпи [Третьяков, 1996].
Собрана А.К. Скворцовым в 1973 г. на склоне ж/д насыпи ст. Шемякино
Калужской обл. (МНА). В настоящее время в Калужской обл. рогачка фран
цузская встречается по ж/д насыпям и в большом числе по грунтовым доро
гам, по которым возят известняк [С.Р. Майоров, личное сообщение].
Во флоре Рязанской обл. вид не отмечен [Казакова, 2004] и, несмотря на
наши активные поиски в 2004 г., обнаружить рогачку французскую в Рязанс
кой обл. нам не удалось.
В Пензенской обл. -- очень редко, по данным В.М. Васюкова, имеются сбо
ры конца ХХ века с севера Никольского р на [Васюков, 2004].
В Тамбовской обл. вид собран А. Сухоруковым в Тамбовском р не, на ж/д
пл. Челнавская, по ж/д полотну, 20.08.2005 г. (MW).
Местообитания. Все ранние сборы E.gallicum в США сделаны по ж/д на
сыпям [Luken et al., 1993]. В Канаде спектр первоначальных мест обитания
258
был более широким: вид собирали на ж/д полотне, обрабатываемых полях,
близ элеваторов, мельниц и в портах [Warwick, Wall, 1998]. Сейчас в Канаде
рогачка растет вдоль автомобильных и железных дорог, на мусорных кучах,
пустырях, полях, в том числе паровых, в огородах и садах, на песчаных нано
сах по берегам рек и ручьев, на сухих лугах, каменистых склонах, в известня
ковых карьерах (рис. 117). Успешно развивается на разных типах почв -- от
меловых и песчаных до глинистых [Warwick, Wall, 1998].
В пределах вторичного ареала в Центральной и Северной Европе E.gallicum
встречается на засоренных и пахотных землях [Flora of NW Europe, 2008].
В условиях Средней России вид растет на песках, мелах, нарушенных степ
ных склонах, ж/д насыпях, в щебневых карьерах, посевах, на газонах и сорных
местах, в долинах рек и по дорогам.
Размножение и жизненный цикл. В Средней России E.gallicum -- однолетнее
растение, цветет с июня до октября, плодоносит с июля [Губанов и др., 2003].
В США и Канаде E. gallicum также является однолетником, а в южных р нах
Северной Америки и в Южной Европе вид может вести себя как двулетник
[Hanf, 1983; Rich, 1991; Rollins, 1993]. Наиболее благоприятными для роста и
развития растения являются температуры от 25 до 27°С., при 10 и 35°С расте
ния развиваются слабо.
Большинство проростков появляется в апреле или начале мая. Пиков мас
сового прорастания не выявлено. Прорастание может продолжаться до конца
сентября [Chepil, 1946]. В засушливые годы всходы не появляются до дождя.
Отдельные особи способны переживать зиму в вегетативном состоянии
[Klemow, 1984]. В условиях Канады (провинция Манитоба) цветение E. gallicum
начинается через 5 недель после прорастания и продолжается с начала июля
до начала августа. [Wall, 1997]. Плоды появляются в начале августа (рис. 118).
Плодоношение продолжается до морозов [Chepil, 1946; McDonald, 1994].
E. gallicum -- самоопылитель, и может давать богатый урожай семян при
отсутствии опылителей [Warwick, Wall, 1998]. У растений, выращенных в оран
жерее, масса 1000 семян составляет в среднем 0,36 г, а в полевых условиях --
0,30--0,35 г [Wall, 1997]. Стручок в среднем содержит 21 семя. Среднее коли
чество стручков на растении -- 351 (7065 семян). Отдельные особи могут проду
цировать до 430 стручков и 10886 семян [Wall, 1995]. В условиях пониженной
влажности растения формируют меньшее число семян в стручке, но семена при
этом более крупные. На паровых землях семенная продуктивность рогачки выше
по сравнению с обрабатываемыми полями. Наименьшая семенная продуктив
ность наблюдается на полях льна и пшеницы. На поле льна, например, 1 расте
ние продуцирует в среднем 233 семени при засушливых условиях и 426 -- при
нормальных. На поле пшеницы -- еще меньше: 17 и 92 семени соответственно
[Warwick, Wall, 1998]. При уборке урожая семена E. gallicum формируют почвен
ный семенной банк и примешиваются к урожаю, что создает возможности для
дальнейшего расселения вида [Chepil, 1946; Warwick, Wall, 1998].
По мнению W. Chepil [1946], период покоя семян E. gallicum превышает 3
года. Однако другие авторы [Warwick, Wall, 1998] полагают, что покой семян
для этого вида не обязателен, и свежие семена успешно прорастают как в оран
жерее, так и в полевых условиях.
Размножение и жизненный цикл E. gallicum в зависимости от многолетней
динамики увлажнения изучался в штате Нью Йорк на территории щебневого
карьера [Klemow, Raynal, 1979; 1983].
259
Расселение и распространение. Распространение и сохранение устой
чивых популяций E. gallicum возможно благодаря высокой семенной продук
тивности, растянутому периоду прорастания семян, созданию почвенного се
менного банка и тому, что жизнеспособные семена сохраняются в почве как
минимум год [Klemow, Raynal, 1983]. В условиях вторичного ареала вид рас
селяется при перевозке щебневого субстрата по карьерам, ж/д насыпям, стро
ительным площадкам, а впоследствии занимает и другие нарушенные места
обитания.
Последствия внедрения
Воздействия на естественные фитоценозы и аборигенные виды. E.gallicum --
растение нарушенных субстратов, имеет низкую конкурентную способность,
и при задернении участков вытесняется другими видами [Klemow, Raynal, 1983;
Milberg, 1991].
В литературе активно обсуждается вопрос о возможных генетических
последствиях скрещивания сорного вида E.gallicum c культивируемыми вида
ми рода Brassica: B.napus L., B.rapa L., B.juncea (L.) Czern. [Lefol et al., 1997]. В
лабораторных условиях получены гибриды B.rapa × E.gallicum и B.napus ×
E.gallicum. Гибрид между E.gallicum и B.juncea получить не удалось. Гибрид
B.rapa × E.gallicum оказался весьма жизнеспособным и фертильным, прекрас
но рос в естественных местах обитания, отличался хорошей семенной продук
тивностью и успешно возвратно гибридизировал с B.rapa и немного менее -- с
E.gallicum. Гибрид B.napus × E.gallicum оказался менее жизнеспособным, но
также фертильным и возвратно гибридизировал с E.gallicum. Наличие такого
рода гибридов может иметь отдаленные, но весьма заметные последствия, так
как имеются опасения, что гены, отвечающие за устойчивость к гербицидам,
могут проникать из культивируемых трансгенных модификаций B.rapa и
B.napus в E.gallicum, которая впоследствии будет засорять посевы. C целью
проверки этой гипотезы в Канаде проведена оценка потока генов от B.napus к
четырем близким видам: B.rapa, Raphanus raphanistrum L., Sinapis arvensis L. и
E.gallicum как в условиях оранжереи, так и в агрокультуре. Во всех вариантах
эксперимента, при всестороннем исследовании более 60 тыс. проростков, гиб
ридов между B. napus и E. gallicum не обнаружено. Согласно выводам авторов,
эти факты говорят о том, что возможность потока генов от трансгенной B.napus
к E.gallicum крайне низка (< 2--5x10 5), однако исключать ее полностью нельзя
[Hall, 2008].
Генетические изменения и вариабельность вида. Число хромосом n = 15; 2n =
30 как в Канаде [Mulligan, 1957, 1984; Warwick, Anderson, 1993], так и в евро
пейских популяциях в Австрии, Чехии, Дании и Словакии [Manton, 1932;
Harberd, 1972; Vachova, Ferakova, 1978, 1980; Sikka, Scharma, 1979; Kirschner
et al., 1982]. E. gallicum -- аллополиплоидный или амфиплоидный таксон (n =
1+8), родителями которого могут быть Erucastrum nasturtiifolium (Poir.) O.E.
Schulz (n = 8) и Diplotaxis erucoides (L.) DC. (n = 7) [Harberd, Me Arthur, 1980;
Takahata, Hinata, 1983; Warwick, Black, 1993; Anderson, 1995]. Филогенетичес
кие исследования рода Brassica и близких к нему родов указывают на наличие
2 групп таксонов: 1) Brassica rapa/Brassica oleracea и 2) Brassica nigra. Вместе с
Diplotaxis erucoides E. gallicum входит в первую группу; вероятно, эти виды
имеют общего предка. Результаты подтверждают генетическую близость
E.gallicum к видам рода Brassica и возможность обмена генами между ними
260
[Warwick, Black, 1993]. Популяции E.gallicum, как в Старом Свете, так и на
североамериканском континенте, могут служить потенциальным источником
ценных признаков, повышающих урожайность B.napus, например, устойчи
вость к болезням и вредителям [Warwick, Wall, 1998].
Экономическое и социальное (положительное и отрицательное) значение.
Сорное. В Канаде E.gallicum занимает 27 место в списке наиболее распростра
ненных сорняков зерновых и масличных культур [Thomas, Wise, 1984]. В про
винции Саскачеван вид отмечен на 0,9% осмотренных в 1976--1979 гг. полей, в
1986 г. эта цифра составила 0,5%, а в 1995 г. -- 2,2% [Thomas, Wise, 1983, 1984;
Thomas et al., 1996]. Установлено, что растение менее часто встречается в по
севах зерновых и льна по сравнению с масличными культурами из рода Brassica
[Thomas, Wise, 1984, 1988].
Меры борьбы
По данным канадских авторов, E.gallicum имеет низкую плотность популя
ций в посевах льна и зерновых и на таких полях в специальных мерах контро
ля не нуждается. Кроме того, при очистке семенного материала семена рогач
ки хорошо отделяются и не вызывают беспокойства как загрязнители указан
ных выше культур [Wall, 1997].
Однако в посевах масличных культур рода Brassica рогачка требует жест
ких мер контроля, так как здесь популяции E.gallicum имеют бульшую плот
ность и существует опасность гибридизации с культурными растениями и при
обретения гибридами генов устойчивости к гербицидам, что впоследствии се
рьезно затруднит их уничтожение [Warwick, Wall, 1998].
В современных работах перечисляется большой набор гербицидов, пред
лагаемых разными авторами для уничтожения E.gallicum [Warwick, Wall, 1998].
Однако в их использовании имеется ряд ограничений, например, чувствитель
ность близких к E.gallicum видов к одним и тем же гербицидам, а также необ
ходимость постоянно изменять препараты и дозы для получения желаемого
эффекта.
CЕМЕЙСТВО CRUCIFERAE (BRASSICACEAE) ---
КРЕСТОЦВЕТНЫЕ
КЛОПОВНИК ГУСТОЦВЕТКОВЫЙ
Название: Lepidium densiflorum Schrader
Синонимы: L.apetalum sensu N.Busch, non Willd., L.neglectum Thell., L.virgi
nicum L. ssp. neglectum (Thell.) P.Fourn.
Морфологические признаки
Яровой однолетник высотой (5)10--45(60) см (рис. 119). Стебель прямой,
жесткий, вверху разветвленный, с более или менее густым опушением из ко
ротких цилиндрических простых волосков. Нижние листья в прикорневой
розетке, рано отмирающие, обратноланцетные, клиновидно суженные в чере
шок, пластинка от зубчато лопастной до перисторассеченной, обычно длиной
до 5--8 см. Стеблевые листья уменьшающиеся, от ланцетных до линейных, край
пластинки от зубчатого до почти цельного. Листья сверху голые, снизу обыч
но опушенные лишь в нижней части или по жилкам. Соцветие многоцветко
Рис. 120. Расселение Lepidium
densiflorum в Средней России
Рис. 119. Плодоношение
Lepidium densiflorum
Рис. 121. Внедрение
Lepidium densiflorum в
естественные сообщества
Рис. 122. Песчаные почвы
с Lepidium densiflorum
263
вое, густое. Цветки с 4 чашелистиками длиной менее 1 мм, по краю белоплен
чатыми, у верхушки обычно с пучком простых волосков. Лепестки отсутству
ют. Тычинок 2. Стручочки двусемянные, обратнояйцевидные, реже овальные,
длиной до 2,5--3 мм, с небольшой (0,2--0,4 мм) выемкой; рыльце сидячее или
на столбике 0,1--0,2 мм длины. Плодоножка превышает длину стручочка ме
нее чем в 1,5 раза. Семена овальные, 1,0--1,3 мм, бурые или рыжевато бурые,
мелко папиллозные.
А.В. Ярмоленко [1934] и Н.А. Буш [1939] указывали для европейской час
ти как заносное растение близкий азиатский вид -- L. apetalum Willd.1. Нам не
удалось обнаружить ни одного образца этого вида из Европейской России
(MW, MHA). Не указывается он и В.И. Дорофеевым [2002]. В гербарных сбо
рах L. densiflorum из Европы нами найден единственный сбор L. apetalum2:
«Planta Poloniae Exsicatae. 424. Lepidium densiflorum Schrader. Warszawa: Ruiny
w okolicach Zelezney Bramy. In ruinai prope "Zelezna Brama". 1946. Lg.
R.Kobendza» (MHA).
Прежде всего, L. apetalum отличается формой волосков, которые у него бу
лавовидные или головчатые, а у L. densiflorum -- папилловидные или цилинд
рические [Jafri, 1973; Taiyan et al., 2001]. Кроме того, L. densiflorum -- яровой
однолетник, который чаще ветвится в верхних 1/2--2/3 стебля, а L. apetalum --
двулетнее растение (или озимый однолетник?), обычно ветвящееся от осно
вания, стручочки с наибольшей шириной около середины, часто с небольши
ми, но отчетливыми треугольными или округло треугольными крыльями на
верхушке плода, выемка на верхушке обычно V образная, в то время как у
L.densiflorum U образная. У L. apetalum нередко видны остатки лепестков, а
чашелистики, как правило, голые (MW, MHA). А.Л. Эбель указывает, что
L. apetalum, подобно L. ruderale, обладает неприятным запахом, но более сла
бым [Эбель, 2001]. Обнаружены заметные отличия этих видов в нуклетидной
последовательности интрона PI [Lee et al., 2002].
Совершенно неожиданно в «Сосудистых растениях советского Дальнего
Востока» [Беркутенко, 1988] и «Флоре Сибири» [Никифорова, 1994]
"L. apetalum auct., non Willd."3 отнесен в синонимы L. densiflorum, при этом соб
ственно L. apetalum для Сибири и Дальнего Востока не указываются. Однако
L. apetalum описан из Сибири: "Habitat in Sibiria" [Willdenow, 1800; Буш, 1939].
В.И. Дорофеев [1998] также поддерживает гипотезу об азиатском происхож
дении L. densiflorum (то есть о синонимичности L. apetalum и L. densiflorum).
Клоповник густоцветковый появился в Сибири сравнительно недавно [Эбель,
2001], в MW и MHA нет ни одного образца L. densiflorum из Сибири, с таким
определением хранятся только образцы L. apetalum. Среди дальневосточных
растений есть и L. apetalum, и L. densiflorum (MHA). Таким образом, при ука
заниях L. densiflorum для Азиатской России имеет место досадная номенкла
турная путаница.
1 Это название иногда отвергают, так как оно имеет более ранний омоним --- Lepidium
apetalos Gilib., 1782, Fl. Lit. Inch., II: 54 (= L. ruderale L.), и заменяют на L. incisum Roth,
1802, Neue Beitr., I: 224. Однако "Flora lituanica inchoata", включено в Приложение VI Ко
деска ботанической номенклатуры и, согласно правилу 32.9, название L. apetalos Gilib. не
является действительно обнародованным [International..., 2006].
2 По видимому, для адвентивной флоры Европы L. apetalum не ранее приводился
[Handbook..., 2009].
3 То есть как у М. И. Котова [1979] для европейской части СССР!
264
Изредка L. densiflorum наряду с L. apetalum и L. neglectum включают в со
став L. virginianum L. [Clement, Foster, 1994].
Естественный ареал
Североамериканский вид, естественно произрастающий в США, Канаде,
Мексике [Synthesis..., 1999].
Вторичный ареал
История интродукции и географическое распространение. Широко на
турализовался в Европе [Akeroyd, Rich, 1993]. Однако время появления вида
в Европе неясно. В Чехии известен с 1904 г. [Pyљek et al., 2002], в Австрии -- c
1948 г. [Essl, Rabitsch, 2002]. Самый ранний известный нам гербарный сбор сде
лан на Украине: «Herbarium Florae Rossicae. 558 (как L. apetalum -- СМ). Черни
говск. губ., Остерск. у., края дороги бл. Бровары, 25.05.1898 г., Н. Цингер» (MHA).
Чуть ранее И. Шмальгаузен [1895] указывал этот вид (как L. apetalum)4 для ок
рестностей Киева, Крыма и Астраханской губернии, уточняя, что киевские рас
тения видел лично. В Европе в настоящее время известен более чем в половине
регионов, в трети из них натурализовался [Lambdon et al., 2008].
L. densiflorum известен в Азии: Корее, Монголии, Китае [Taiyan et al., 2001] и
Японии [Mito, Uesugi, 2004]. Вид обнаружен в Австралии [Jacobs, Pickard, 1981]
и с 1948 г. в Новой Зеландии [Garnock Jones, 1979]. Недавно появились сооб
щения о находке L. densiflorum в Южной Америке -- в Аргентине [Prina, 1997].
Распространение вида в России требует дополнительных исследований.
L. densiflorum указан для Северного Кавказа [Дорофеев, 2002; Зернов, 2006].
Нам не удалось найти растение среди кавказских гербарных сборов. Все не
многочисленные кавказские образцы клоповника в MW, определенные как
L. densiflorum и на которые ссылается А.С. Зернов [2006], являются L.virginicum.
В Сибири L. densiflorum достоверно известен из Омской, Кемеровской, Ново
сибирской и Томской обл., Алтайского края, Республики Алтай, Хакасии
[Эбель, 2001]. Вид, несомненно, есть на Дальнем Востоке (MHA).
Пути и способы заноса. По мнению Э.Егера, расселение L. densiflorum про
исходило с востока на запад вдоль путей сообщения [Jдger, 1980]. В.Н. Тихо
миров склонялся к тому же мнению [Тихомиров, 19**]. По видимому, это не
совсем так. Документированные находки этого вида не позволяют выявить
направленное и закономерное расселение по Европейской России.
Статус в регионе. По видимому, первые находки L. densiflorum в Евро
пейской России сделаны в Нижегородской обл. «ст. Игумново Московско
Курской железной дороги, 07.07.1927 г., Н.Покровский», «Нижний Новгород,
близ городского парка, 22.07.1932 г., он же» (LE; Дорофеев, 1998]. В Московс
кой обл. собран заметно позднее: «Ленинский р н, близ с. Михайловка, на заб
рошенной ж/д насыпи, 07.07.1946 г., В.Н. Ворошилов» (МНА). Вскоре этот
клоповник указан в 8 м изд. «Флоры средней полосы европейской части
СССР» П.Ф. Маевского [1954]. Г.Э. Гроссет собрал его в «окрестностях г. Пен
зы, у дороги в сосновом лесу близ Ахун, на песчаной почве, 04.06.1962 г., Г.Грос
сет» [MW] и, по видимому, обратил на него бульшее внимание московских
флористов. Через 10 лет Гуго Эдгарович собрал этот клоповник уже на не
скольких московских улицах, сопроводив один из образцов комментарием:
«наблюдался в течение ряда лет в различных местах Киевского района [Мос
4 Речь идет именно о L. densiflorum, поскольку в описании указаны признаки этого вида.
265
квы]» (MW). В 1970 х гг. этот вид собран во многих областях и республиках
Средней России. Уже к 1975 г. L. densiflorum стал массовым видом в Московс
кой, Рязанской, Владимирской и Тульской обл. [Октябрева и др., 1978], и, ве
роятно, в других регионах Европейской России.
В настоящее время широко распространен во всех областях Средней Рос
сии [Дорофеев, 2002], однако в южных районах обнаруживает явное тяготе
ние к песчаным почвам и встречается реже (рис. 120).
Местообитания. Клоповник густоцветковый произрастает по нарушен
ным местообитаниям с несомкнутым растительным покровом, при этом оче
видно предпочитает почвы механического состава, а также на субстратах с
хорошим дренажем -- щебенистые насыпи, кучи шлака и т.п. [Игнатов и др.,
1990]. L. densiflorum нередко растет в долинах рек, но не собственно по бере
гам у воды, а на песчаных отложениях в пойме и на террасах, особенно часто
вдоль дорог (рис. 121).
Размножение и жизненный цикл. L. densiflorum -- яровое однолетнее рас
тение. В литературе немало указаний на то, что этот клоповник может быть и
двулетником [Беркутенко, 1988; Никифорова, 1994]. По видимому, эти сведе
ния -- результат номенклатурной путаницы (см. выше). По нашим наблюде
ниям, L. densiflorum развивается по яровому типу, по крайней мере, в Средней
России. Клоповник густоцветковый -- самоопыляемое растение, обычно все
цветки образуют завязь (рис. 122). Однако у отдельных экземпляров в начале
плодоношения стручочки не завязываются вовсе. После 30--60 плодоножек
без стручочков фертильность восстанавливается и выше все плодоножки не
сут стручочки. Причина такого явления неясна. В каждом стручочке 2 семени.
На Аляске 1 растение L. densiflorum может произвести до 5 тыс. семян [Royer,
Dickinson, 1999]. Наряду с крупными растениями клоповника на бедных суб
стратах вырастают экземпляры, несущие всего 10--20 плодов. Семена доволь
но быстро теряют всхожесть: при хранении в темноте через 4 месяца при тем
пературе 15°С всхожесть падает более чем на 50% [Toole et al., 1955].
Расселение и распространение. Способы распространения неясны. Для
Аляски указано, что L. densiflorum может расселяться как перекати поле. Едва
ли этот способ распространения представлен на нашей территории.
Последствия внедрения
Воздействия на естественные фитоценозы и аборигенные виды. По
некоторым данным, L. densiflorum вытесняет L. ruderale [Тихомиров, 19**]. Так,
по материалам гербария Калужского гос. пед. университета в Калужской обл.
после появления клоповника густоцветкового число сборов клоповника сор
ного заметно сократилось [Воронкина и др., 2005].
Генетические изменения и вариабельность вида. В Северной Америке
различают несколько разновидностей L. densiflorum. У var. densiflorum стру
чочки длиной не более 2,5 мм, голые, плодоножки не уплощенные. Прочие
разновидности имеют стручочки длиной около 3 мм и более, плодоножки от
четливо сплюснутые: var. macrocarpum Mulligan отличается голыми стручоч
ками, у var. pubicarpum (A. Nelson) Thell. стручочки опушены по всей поверх
ности, у var. elongatum (Rydb.) Thell. -- только по краю [Rollins. 1993;
Illustrated..., 1998]. У var. ramosum (A. Nelson) Thell. стручочки более плоские,
чем у типовой разновидности: их ширина примерно вдвое превышает толщи
ну [The Jepson..., 1993]. Типовая разновидность распространена широко, дру
266
гие приурочены в основном к западу США [Synthesis..., 1999]. Следует отме
тить, что растения нетиповой разновидности из западных частей США могут
иметь мезотоническое ветвление, габитуально напоминая L.ruderale. В Евро
пейской России встречаются только типовая разновидность клоповника.
Экономическое и социальное (положительное и отрицательное) зна
чение. Молодые растения L. densiflorum могут использоваться как пряная зе
лень, подобно L.sativum [Seidemann, 2005]. Но трава этого клоповника быстро
грубеет и становится малосъедобной. В традиционной медицине индейцев
Северной Америки клоповник использовался как анальгетик и успокоитель
ное средство [Moerman, 2009]. Меры борьбы
Клоповник густоцветковый не является обременительным сорняком. Спе
циальных мер борьбы с ним не разработано, и, видимо, явной необходимости
в них нет.
CЕМЕЙСТВО CRUCIFERAE (BRASSICACEAE) ---
КРЕСТОЦВЕТНЫЕ
ГУЛЯВНИК ВОЛЖСКИЙ
Название: Sisymbrium volgense Bieb. ex Fourn.
Синонимы: Есть разногласия по поводу того, как правильно писать видо
вой эпитет: wolgense или volgense. В литературе встречаются оба варианта на
писания.
Морфологические признаки
Многолетнее растение высотой 40--60 см, голое, сизоватое, в нижней части
опушенное мелкими волосками. Стебель ветвистый. Листья утолщенные, ниж
ние -- копьевидные или струговидно раздельные, с длинными черешками, вер
хние -- мелкие, продолговатые, зубчатые, у зубчиков мозолисто утолщенные.
Чашелистики длиной 3,5--4 мм. Лепестки желтые, длиной 7--9 мм. Стручки
длиной 4--6 см, голые, с очень коротким столбиком, сидящие на утолщенных
кверху ножках длиной 5--10 мм (рис. 123). Семена овально складчатые, блестя
щие, желтовато коричневые, у основания более темные, длиной 0,75--1,25 мм,
шириной 0,5--0,75 мм и толщиной 0,5 мм. Масса 1000 семян 0,1--0,3 г. [Губанов
и др., 2003].
Естественный ареал
Европейский вид, распространенный на юго востоке европейской части
России, в зоне степей. Эндемик Нижней Волги и Нижнего Дона, известный
здесь как сорный вид [Губанов и др., 2003].
Вторичный ареал
История интродукции и географическое распространение. S.volgense
натурализовался во многих странах Европы, включен в список заносных ви
дов Польши, натурализовался в Беларуси [Œwies, Wrresieс, 2004; Germplasm,
2008]. Включен в Список карантинных видов Республики Словения [2008].
Распространение вида началось в 1880 г., когда его нашли на мельнице в
местечке Vaasa в Финляндии [Jehlík, 1981]. К 1971 г. в стране известно уже 22
267
местонахождения вида -- главным образом, вокруг мельниц и на ж/д насыпях;
везде вид был устойчив и размножался вегетативно [Suominen, 1979]. В 1887
г. S.volgense обнаружили в Германии, близ Берлина, на паровом поле у зерно
хранилища. В ХХ веке число мест нахождения возросло, особенно вдоль авто
мобильных дорог, по ж/д насыпям и в портах; вероятно, вид завезен в Герма
нию с российским зерном [Jehlík, 1981]. В Швеции S.volgense обнаружен в 1891
г. в Maльме [Hylander, 1971], впоследствии число мест нахождения непрерыв
но росло, главным образом в портах [Jehlík, 1981]. В 1896 г. вид зарегистриро
вали в Польше, в окрестностях Варшавы, на ж/д насыпи [Majewski, 1971]. К
1980 м гг. известно уже 16 местонахождений вида, преимущественно на ж/д
станциях [Jehlík, 1981]. Однако до сих пор в Польше имеются лишь единич
ные местонахождения гулявника, в каждом из которых встречено не более 10
особей [Alien species..., 2008], возможно, это связано с прекращением поставок
российского зерна на запад. В Великобритании к 1980 м гг. зарегистрировано
7 местонахождений, первое из которых -- в 1896 г.; все точки -- близ пшенич
ных мельниц и доков, где сгружалось зерно, во всех местах обитания вид ус
тойчив [Clement, 1979]. В Дании гулявник впервые найден в 1899 г., и до 1958
г. обнаружено 6 его местообитаний в портовых городах, близ мельниц и на
обочинах дорог [Pedersen, 1958]. В Нидерландах вид обнаружен впервые око
ло 1900 г. [Hock, 1900]. К началу 1980 х гг. во Франции известно лишь одно
местонахождение -- близ Парижа, указанное в 1902 г. [Thellung, 1919]. В Нор
вегии вид обнаружен в 1911 г. на зерновой мельнице [Jörgensen, Ouren, 1969].
Позднее S.volgense обнаруживали неоднократно; очень стойкий вид во всех
местах нахождения, образует зрелые семена [Jehlík, 1981]. На территории Бель
гии вид обнаружен в 1953 г. на ж/д станции [Lawalrée, 1957]. Первые указания
о новом адвентивном виде в Чехословакии опубликованы в 1968 и 1971 гг.
[Houda, 1968; Jehlík, 1971]: вид найден в 1960 г. в Пардубице, на разгрузочной
ж/д станции. До 1980 г. в Чехословакии обнаружено 51 местообитание вида,
главным образом на ж/д станциях, и S.volgense стал устойчивым видом флоры
Чехии и Словакии [Jehlík, 1981]. В Австрии вид зарегистрирован в 1963 г. в
Вене [Melzer, 1964].
Собран в Риге около 1894 г. [Behrendsen, 1896; Lehmann, 1896] Это место
нахождение имеет особое значение, так как в прошлом Рига была важнейшим
портом, специализировавшимся на экспорте зерна из Российской империи, и
конечным пунктом железной дороги Царицын (ныне -- Волгоград) -- Рига. Со
времен Первой мировой войны эта важнейшая магистраль соединяла юго во
сток Европейской России с Балтией [Jehlík, 1981].
Уже в первой половине ХХ века S.volgense зафиксирован на Украине как
вид, растущий вдоль железных дорог и внедряющийся в поля [Котов, 1926]. К
середине прошлого века вид стал на Украине обременительным сорняком по
лей и в массе распространился по рудеральным местам обитания [Протопопо
ва, 1966; Котов, 1970].
История расселения S.volgense в Европе и в Нечерноземье описана в конс
пекте флоры адвентивных растений Московской обл. По мнению авторов, рас
селение S.volgense в России связано с массовым переселением крестьян после
отмены крепостного права [Игнатов и др., 1990].
В Ленинграде вид собран в 1921 г., а в середине ХХ века продолжал актив
но распространяться по железным дорогам Северо Запада [Гусев, 1971]. Не
268
редко встречается в Псковской и Новгородской обл. [Цвелев, 2000], в каче
стве ксенофита указывается в информационной системе «Чужеродные виды
Северо Запада России» [2008]. Небольшие колонии S.volgense обнаружены на
шоссейных дорогах Удмуртии, первая находка сделана в 1984 г. [Пузырев,
2006]. В Сибири указывается для г. Курган и с.Утятское на юге Томской и
Иркутской обл. Обнаружен в Новосибирске [Ломоносова, Зыкова, 2003]. От
мечен в Троицком, Первомайском, Косихинском и Михайловском р нах Ал
тайского края и в Барнауле, собран Е.Комовой в 1997 г. в г. Камень на Оби
[Терехина, Копытина, 1999; Ломоносова, Зыкова, 2003]. Распространился по
Средней Азии [Терехина, Копытина, 1999; Ломоносова, Зыкова, 2003]. Вне
сен в список инвазионных видов Японии [Invasive Plants Database, 2008]. (Гу
банов и др., 2003)v/cgi bin/npgs/html/taxon/pl?401849).а, 1999).
За пределами Европы S.volgense впервые собран на Дальнем Востоке, на
ж/д ст. Первая Речка во Владивостоке. Растение цвело и образовывало плоды
[Нечаева, 1978].
Пути и способы заноса. Первичный способ заноса S.volgense на новые тер
ритории связан с импортом российского и советского зерна в европейские стра
ны в XIX и XX веках [Jehlík, 1981]. Упоминаются случаи завоза S.volgense че
рез порты Черного моря в порты Германии, где разгружали российское зерно
[Bonte, 1930]. Импорт зерна из СССР в страны Центральной и Восточной
Европы был очень велик -- например, экспорт Чехословакией с 1955 по 1978 г.
российской пшеницы составлял до 74% . Помимо пшеницы, из СССР везли
ячмень на фураж, кукурузу и рожь, хотя и в значительно меньших объемах.
Большинство мест обитания гулявника в Европе сконцентрировано вблизи
ж/д станций, зернохранилищ, мельниц и портов, где разгружали и хранили
импортное зерно [Jehlнk, 1981]. Впоследствии S.volgense распространялся во
вторичном ареале вегетативным и семенным способом уже из этих первона
чальных популяций.
По железным дорогам с транспортом зерна S.volgense занесен также в се
верную и северо западную часть Европейской России и на Украину. Расселе
ние S. volgense именно по железной дороге отразилось во включении вида в
группу «железнодорожных растений» [Голицын, 1947].
Статус в регионе. Известен во всех областях Средней России [Маевский,
2006]. Природный ареал вида охватывает только Саратовскую обл., в осталь
ных регионах S.volgense является заносным растением (рис. 124).
В Белгородской обл. S.volgense -- прогрессирующий адвентивный вид на
пустырях, обочинах дорог и меловых склонах (рис. 125). Значительные попу
ляции отмечены в Белгороде и Старом Осколе [Еленевский и др., 2004]. В
Воронежской обл. впервые обнаружен Г.Э. Гроссетом в 1927 г. на ст. Сомово
[Григорьевская и др., 2004]. Позднее, в 1941 и 1946 гг., вид неоднократно соби
рали на территории Воронежского заповедника, а в 1980 х гг. и позднее -- ре
гулярно наблюдали на железных дорогах области и в населенных пунктах. В
Воронеже встречается по рудеральным местам [Григорьевская, 2000].
Для территории Курской обл. в сводках XIX и первой половины XX века
не указан. Первый сбор: [Дмитриевский р н], окр. Дмитриева, ж/д насыпь,
9.06.1961 г., С.В. Голицын (VU). В настоящее время встречается в области спо
радически, а по ж/д полотне юго восточного направления является массовым
видом [Полуянов, 2005].
Рис. 124. Расселение
Sisymbrium volgense
в Средней России
Рис. 123. Sisymbrium
volgense, общий вид
растения
Рис. 125. Внедрение
Sisymbrium volgense в
естественные
сообщества
Рис. 126. Гербарный
лист Sisymbrium volgense
271
Первый сбор для Липецкой обл. сделан в Грязинском р не, г. Грязи, на пра
вом берегу р. Матыра, на пустыре у ж.д., 12.07.1984 г., В.Тихомировым, Т.Тро
ицкой, и Т.Кумаховой (MW). К настоящему времени встречается по всей тер
ритории Липецкой обл., местами в массе, быстро расселяется и распространя
ется [Флора Липецкой области, 1996]. Собран в Орловской обл. (г. Орел, по
ул. Мопра, у железной дороги Орел--Брянск, 19.06.1982 г.. В.И. Радыгина,
А.Г. Еленевский. OHHI).
В Тульской обл. гулявник волжский впервые собран А.И. Алюшиным в 1976 г.
[ТГПУ; Алюшин, 1982]. К настоящему времени S. volgense -- нередкое в облас
ти растение, успешно расселяется и прочно удерживает позиции даже на вы
шедших из активного употребления землях.
Собран во Владимирской обл., Муромском р не, в 10 км от г. Муром, по
дороге во Владимир, обочина шоссе, довольно обильно, 21.06.1969 г., В.Тихо
миров и др. (MW). Есть также гербарные сборы 1971, 1972 гг. (MW; LE) из
Вязниковского и Гусь Хрустального р нов. По личному сообщению А.П. Се
регина, вид в области встречается довольно редко, но регулярно, причем мес
та обитания уже не ограничиваются ж/д полотном, как это было несколько
лет назад: S.volgense произрастает и по обочинам шоссе.
В Московской обл. S. volgense впервые собран на ст. Быково Казанской же
лезной дороги в конце XIX века [1897?, Сырейщиков, 1907]. Во время Граж
данской войны неоднократно заносился по железным дорогам: М.И. Назаров
[1927] сообщил о находках S.volgense на 5 из 11 обследованных станций. Вплоть
до начала 1960 х гг. гулявник волжский оставался в Московской обл. редким
видом, однако к началу 1970 х гг. ситуация изменилась, о чем свидетельству
ют многочисленные гербарные сборы (MW, рис. 126). По свидетельству А.К.
Скворцова [1973], в Московской обл. гулявник «встречается не только почти
по всем железнодорожным насыпям, но уже нередко и по городским улицам...,
и кое где и по откосам шоссейных дорог». Сходным образом вел себя вид и
позднее [Игнатов и др., 1990], однако в последнее десятилетие, благодаря ис
пользованию гербицидов, численность особей S.volgense на ж/д насыпях су
щественно сократилась (С.Майоров, личное сообщение).
В Калининской обл. (ныне -- Тверской) вид собран Ю.Д. Гусевым между
ст. Санаторий и Тверца, на юго западном склоне ж/д насыпи, 30.08.1971 г.
(MW). В Ивановской обл. встречается на ж/д насыпях дорог всех направле
ний, по олуговелым склонам берегов рек близ железной дороги в г. Иваново,
Кинешма [Шилов, 1989; Шилов и др., 1992; Борисова, 1995а, 2007]. Старых сбо
ров обнаружить не удалось. В 1991 г. S.volgense собран Е.Борисовой в г. Иваново
(MW). В Костроме вид собран по краю ж/д насыпи, 29.07.1979 г., И.Гусев (MW).
Есть сборы из Брянской обл. -- один из первых сделан в 1920 е гг.: «Черниговс
кая губ., Новозыбковский уезд, 1920--1922 гг., собр. К.Марусяк, опр. Ю.Алексе
ев (MW)», позднее последовали многочисленные сборы 1970 х гг. (МНА).
Указан для г. Смоленска [Решетникова, 2004]. Cобран в Хиславичском р не
Смоленской обл., в окрестностях пос. Хиславичи, в пойме р. Сож, на обочине
дороги, 05.07.2006 г., Н.Решетникова, И.Фадеева (MHA). В Калужской обл.
встречается редко, но в местах произрастания образует обширные колонии по
железным дорогам (С.Майоров, личное сообщение). Собран в Козельском р
не, в 3 км. к востоку от г. Козельск, на ж/д насыпи, 6.06.1984 г., собр. С.Майо
ров, опр. В.Тихомиров (MW).
272
Для Рязанской обл. впервые отмечен в Сасовском р не, на ж/д насыпи ст.
Пичкиряево, 10.06.1979 г., собр. Е.Гущина, Н.Кошелева, опр. В. Тихомиров
(MW). Собран в Большеберезниковском р не Республики Мордовия, в 3 км
северо западнее с. Паракино, на обочине шоссе, 9.06.1980 г., В.Тихомиров
(MW). Нередко произрастает в Пензенской обл. [Васюков, 2004].
Собран в Тамбовской обл., Рассказовском р не, в 2 км западнее ж/д ст. Рас
сказово, на полосе отчуждения ж/д полотна, часто, 25.05.1996 г., А.Сухоруков
(MW). В Саратовской обл. собран в Ртищевском р не, в 1 км восточнее ж/д ст.
Ртищево 1, на запасных путях, 25.06.1999 г., А.Сухоруков (MW).
Местообитания. В естественном ареале является частью сорной флоры
степной зоны, растет как сорное по обочинам дорог и в бурьяне, на каменис
тых и рудеральных участках [Schulz, 1924; Walter, 1974; Котов, 1979]. В Казах
стане встречается на каменистых местах в зоне полупустынь вокруг Каспийс
кого моря [Васильева, 1961].
В степной зоне Сибири известен как сорное в посевах, а в лесостепной --
как заносное по дорогам и мусорным местам [Игнатов, Игнатова, 1982].
В условиях вторичного ареала в Центральной и Западной Европе вид рас
тет на ж/д станциях и полотне, вокруг мельниц и зернохранилищ, пунктов
разгрузки зерна, в портах, на улицах городов, рудеральных местах обитания,
на свалках, по берегам прудов близ утиных ферм [Jehlík, 1981].
В Средней России основные места обитания вида -- территории ж/д стан
ций и ж/д насыпи, обочины автомобильных дорог и рудеральные местообита
ния, шахтные отвалы, территории предприятий, свалки, пустыри [Васюков,
2004; Казакова, 2004].
В Курской обл. гулявник растет по щебнистым откосам ж/д насыпей, обо
чинам шоссейных дорог, дамбам, карьерам, прирусловым валам [Полуянов,
2005]. В Мордовии -- по разбитым придорожным пескам, редко [Тихомиров,
Силаева, 1990].
В Тульской обл. успешно расселяется и прочно удерживает позиции даже
на вышедших из активного употребления землях. Растет на свалках, пусты
рях и обочинах шоссе, но явно преобладает на шлаковых и щебневых ж/д на
сыпях, где образует крупные колонии. На юго востоке области мы наблюдали
большие заросли S.volgense на слабозадерненных шахтных отвалах и неисполь
зуемых подъездных путях закрытых шахт.
Размножение и жизненный цикл. Цветет в мае июне, плодоносит в июле
сентябре. Всходы появляются в марте мае. Семена прорастают с глубины не
более 1,5 2 см. Свежесозревшие семена не имеют биологического покоя, и че
рез 8 дней после созревания всхожесть составляет 99,3% [Список растений...,
2008]. Существенное значение имеет размножение корневыми отпрысками,
благодаря чему в степной и полупустынной зонах на рудеральных местах и
полях S.volgense является трудноискоренимым сорняком [Губанов и др., 2003].
Во вторичном ареале вид чаще размножается вегетативно, хотя известно и се
менное размножение [Jehlík, 1981].
Во вторичных местообитаниях вид ведет себя как эпекофит, реже -- как
эфемерофит. S.volgense принадлежит к типичным синантропным растениям,
которые образуют множество семян и имеют высокую жизненность. В 1970 г.
фертильность S.volgense изучалась в порту Holeљovice в Праге. Растение, име
ющее один побег, в среднем продуцирует 1099 плодов, каждый из которых со
273
держит в среднем 29 семян, что составляет 32040 семян/особь. Собранные в
этом местообитании семена, выдержанные в течение 3 месяцев, имели всхо
жесть 51 100% [Hejný et al., 1973].
Расселение и распространение. В Средней России S.volgense устойчив и
имеет тенденцию к активному распространению. Предпочитает бедные щеб
невые и шлаковые субстраты техногенного происхождения, расширение ко
торых способствует его расселению: ж/д насыпи, территории предприятий,
шахтные отвалы.
Существует вероятность, что в настоящее время вид попадает в европейс
кую часть России уже из вторичного ареала Центральной Европы. В частно
сти, по мнению А.Н. Пузырева [2006], в Удмуртию S.volgense попадает с им
портным зерном.
Последствия внедрения
Воздействие на естественные фитоценозы и аборигенные виды.
S.volgense, как правило, растет с обычными сорными и рудеральными видами
в составе пионерных группировок. Со временем, при увеличении размеров
популяций посредством вегетативного размножения, вид способен образовы
вать чистые заросли и занимать значительные пространства на более или ме
нее обнаженных субстратах.
В 1980 е гг. S.volgense был наиболее активен в Чехии, Словакии, Польше,
Германии и Финляндии. Западнее встречался реже, и его распространение ог
раничивалось немногочисленными местонахождениями. Сделано предполо
жение, что в будущем в Европе гулявник станет сорняком полей [Jehlík, 1981].
Однако во многих европейских странах, куда вид занесен в середине и конце
XIX века, он так и остался относительно нечастым видом рудеральных мест
обитания и существенного влияния на природные сообщества не оказывает
[Alien species..., 2008].
Генетические изменения и вариабельность вида. Специальные исследо
вания генетической изменчивости вида нам неизвестны.
Экономическое и социальное значение (положительное и отрицательное).
Трудноискоренимое сорное растение [Germplasm, 2008]. Внесено в списки каран
тинных сорных видов стран Восточной и Центральной Европы [Hejný et al., 1973].
Меры борьбы
В большинстве европейских стран в контроле не нуждается ввиду мало
численности [Alien species..., 2008]. Колонии S.volgense, формирующиеся на ж/д
полотнах Средней России, как правило, уничтожаются с помощью гербици
дов, а за пределами железных дорог вид пока не агрессивен, и специальные
меры борьбы с ним нам неизвестны.
CЕМЕЙСТВО CHENOPODIACEAE --- МАРЕВЫЕ
ЛЕБЕДА ТАТАРСКАЯ
Название: Atriplex tatarica L.
Синонимы: A.laciniata L.
Морфологические признаки
Яровой однолетник высотой 15--50 см. Корень стержневой, крепкий. Се
мядоли длиной 12--24 мм, шириной 2--6 мм, линейно яйцевидные. Эпикотиль
274
невысокий. Гипокотиль сверху слегка розовато зеленый. Стебель прямостоя
чий или приподнимающийся, голый, при плодоношении часто краснеющий.
Листья первой пары черешковые, длиной 12--25 мм, шириной 8--12 мм, яйце
видные, реже эллиптические, по краю неравноволнисто зубчатые. Сверху и
особенно снизу пластинки покрыты серебристым мучнистым налетом. Лис
тья серовато зеленые, с нижней стороны более светлые, удлиненно яйцевид
ные, треугольно яйцевидные, реже ланцетные или копьевидные, длиной 1,5--
8 см, шириной 0,5--5 см, по краю неравнозубчатые, редко почти цельнокрай
ние. Верхние листья покрыты серебристыми чешуйками, особенно густо по
нижней поверхности (рис. 127). Цветки в клубочках в пазухах листьев в об
щих колосовидных, при основании облиственных, соцветиях. Прицветнички
до половины сросшиеся, ромбические, б.м. трехлопастные, с выступающими
жилками, при созревании плодов твердеющие, на спинке с более или менее
выраженными выростами, по краю -- зубчатые. Плод округло сдавленный,
соломенно желтый или белесовато желтый мелкоточечный блестящий оре
шек, в околоцветнике, длиной 2--2,5, шириной 3--4, толщиной 1--1,5 мм. Семе
на диаметром 2,5--3 мм [Ломоносова, 1992].
Не все авторы считают A.laciniata синонимом A.tatarica. Во «Флоре Европы»,
например, A.tatarica и A.laciniata разделяют по строению соцветий. У A. tatarica
соцветия безлистные, верхушечные, часто длинные, тогда как у A. laciniata соцве
тия олиственные. Кроме того, эти виды различают по размерам: A.laciniata -- кар
ликовое растение высотой до 30 см с мелкими листьями, в то время как A.tatarica
-- крупная лебеда высотой до 150 см с крупными листьями [Aellen, 1964].
Естественный ареал
Естественный ареал включает следующие районы: Северная Африка (Ал
жир, Египет, Марокко, Тунис), Западная Азия (Афганистан, Иран, Израиль,
Иордания, Турция), Закавказье (Армения, Азербайджан), РФ (Кавказ, Дагес
тан, Алтай, Западная Сибирь, юг европейской части), Центральная Азия (Ка
захстан, Кыргызстан, Таджикистан, Туркменистан, Узбекистан), Монголия,
Китай, Пакистан, Центральная Европа (Австрия, Чехия, Словакия, Германия,
Венгрия, Польша), Восточная Европа (Беларусь, Украина), юг Европы (Ал
бания, Болгария, Греция, Италия, Румыния, страны бывшей Югославии, Фран
ция, Испания) [Germplasm Resources..., 2008].
Вторичный ареал
История интродукции и географическое распространение. Из области
естественного распространения вид расселился в северные, северо западные
и центральные районы европейской части России, в Западную и Северную
Европу, на Дальний Восток и на восточное побережье США, исключая севе
ро восточные штаты [Aellen, 1960; Welsh, 2003].
Граница между естественным и вторичным ареалом вида проходит в Цент
ральной Европе, в частности в Чешской Республике [Jalas, Suominen, 1988].
В Средней России A.tatarica -- обычный вид черноземной полосы, где встре
чается во всевозможных вторичных местообитаниях. В Нечерноземье -- рас
пространенное «железнодорожное» растение, иногда отмечается по обочинам
автомобильных дорог, особенно в крупных населенных пунктах [Маевский,
2006]. К северо западу России встречаемость лебеды татарской резко падает,
в Ленинградской и сопредельных областях она встречается как заносное у до
рог, в населенных пунктах, довольно редко [Цвелев, 2000].
Рис. 127. Листья
Atriplex tatarica
покрыты белым
налетом
Рис. 128. Расселение
Atriplex tatarica в
Средней России
Рис. 129. Внедрение
Atriplex tatarica
в луговые сообщества
Рис. 130. Цветение
Atriplex tatarica
277
Пути и способы заноса. Вероятно, заносится по путям сообщения, пре
имущественно по ж/д насыпям с щебневым и шлаковым покрытием.
Статус в регионе. В Воронежской обл. в конце 1950 х гг. A. tatarica отме
чалась единично, но по всей территории [Камышев, 1959а]. В МНА хранится
сбор В.Н. Ворошилова: «Новоусманский р н, близ с. Рогачевка, сорное, сбоку
проселочной дороги, 1.08.1945 г.». Имеется гербарный сбор, сделанный в 1903
г. на склоне меловой горы под Белгородом (МНА). Во флористических свод
ках XIX века для Курской губернии вид не приведен, но к началу ХХ века A.
tatarica, вероятно, была там уже широко распространена. Первый сбор сделан
в окрестностях Курска: «...на улицах, пустырях, в изобилии; у сл. Ямской -- по
насыпям ж/д», 1902, 1905 гг., В.Алехин (MW). В настоящее время в Курской
обл. лебеда татарская встречается по всей территории, часто. В городах и по
селках один из самых массовых видов [Полуянов, 2005]. Один из первых сбо
ров для Орловской флоры сделан в г. Болхов, 15.08.1927 г., В.Н. Хитрово
(OHHI). В Тульской обл. A. tatarica впервые собрана А.И. Алюшиным в Туле,
на территории Московского ж/д вокзала в 1975 г. [ТГПУ; Хорун, 1998]. По
зднее, в 1979 г., обнаружена Ю.Е. Алексеевым в г. Ясногорск (MW). В 1980--
1990 х гг. A.tatarica стала доминирующим видом ж/д полотна и насыпей.
В Московской обл. A.tatarica впервые собрана в 1904 г. Д.П. Сырейщико
вым на ст. Быково Рязанской железной дороги (MW). Во время Первой ми
ровой войны лебеду многократно собирали на железных дорогах [Назаров,
1927], но, судя по этикеткам, ранние сборы регистрировали как A.laciniata L. и
переименовали в A.tatarica позднее. В настоящее время лебеда татарская ре
гулярно встречается на железных дорогах и изредка в карьерах и на рудераль
ных местах, особенно в южной части области. Нередко образует значитель
ные колонии [Игнатов и др., 1990]. В самые последние годы из за активного
использования гербицидов на железных дорогах Московской обл. A.tatarica
встречаться стала реже.
Во Владимирской обл. A.tatarica известна из всех районов, нечасто [Вахро
меев, 2002]. В MW имеется сбор из Вязниковского р на: «окр. д. Коурково, по
ж/д насыпи, 1.07.1971 г., В. Новиков, М. Шабалина», а также хранятся много
численные сборы А.Серегина 2001--2007 гг. В настоящее время, по личному
сообщению А.П. Серегина, на железной дороге вид встречается значительно
реже, чем в середине 1990 х гг., что связано с использованием гербицидов.
Ранние сборы A.tatarica из Тверской обл. сделаны М.Назаровым (MW): «близ
г. Твери, сорное на линии, 8.09.1917; г. Тверь, по полотну ж/д, 9.06.1917 г.».
В Ивановской обл. вид впервые отмечен в: «г. Иваново, хутор Митрофа
новский, у ж/д переезда, на ж/д полотне, несколько экземпляров, 8.08.1991 г.,
Е.Борисова, опр. А.Сухоруков» (MW). В настоящее время встречается еди
нично вдоль ж/д путей на ст. Иваново, Сортировочная, Кохма, Тейково и еже
годно отмечается в массе на ст. Шуя [Борисова, 2000, 2007].
В МНА и MW имеется ряд сборов из Брянской обл., сделанных в 1975--
1976 гг. Ю.Е. Алексеевым и В.В. Макаровым. Во всех случаях цветущие и пло
доносящие растения собраны по щебневому ж/д полотну или у привокзаль
ных строений. В MW хранится множество сборов A.tatarica из разных райо
нов Калужской обл.; наиболее ранний сделан Л.Ф. Волосновой в 1979 г. в
Сухиничском р не. Один из первых сборов в Рязанской губернии сделан
29.08.1904 г. И.В. Новопокровским (MW).
278
Имеется старый сбор с территории, предположительно, Тамбовской обл: «кн.
Вяземский, Тамбовская губ., 1868--1869 гг.» (MW). В МНА хранится сбор из
Мичуринского р на Тамбовской обл.: «близ с. Мановицы, на залежи, массово,
21.09.1966 г., В.В. Макаров». В Смоленской обл. не отмечена [Решетникова, 2004].
Местообитания. В естественном ареале A. tatarica растет на солончаках,
по берегам рек, каналов, арыков, на солонцеватых лугах, каменистых склонах,
а также по мусорным местам, встречается на высоте до 4500 м над ур. м., обра
зует заросли [Лекарственные растения..., 2008].
В черноземной части Средней России, ближе к границам естественного аре
ала, A.tatarica занимает широкий спектр экотопов -- растет в населенных пунк
тах, у дорог и троп, на засоренных лугах и лесных полянах, по окраинам полей,
берегам водоемов, всевозможным сорным местам. В нечерноземной части пред
почитает щебневые насыпи железных дорог, где растет в массе, образуя чистые
одновидовые, иногда многокилометровые заросли вдоль ж/д полотна. Вне же
лезных дорог встречается нечасто и в меньшем количестве -- на мусорных местах,
пустырях, окраинах полей, обочинах дорог [Васюков, 2004; Григорьевская, 2000;
Григорьевская и др., 2004; Еленевский, Радыгина, 1997; Казакова, 2004].
Во вторичном ареале растет в экотопах, сходных по экологическим харак
теристикам с естественными местообитаниями -- на бедных засоленных по
чвах или по участкам с щелочной реакцией субстрата. Предпочитает светлые
и умеренно глинистые почвы, нуждается в хорошем дренаже. Не выносит тень.
Устойчива к засухе [Plants for a future, 2008].
Размножение и жизненный цикл. Судя по гербарным сборам, в Средней
России (рис. 128) A. tatarica успешно плодоносит и возобновляется семенным
путем. Минимальная температура прорастания семян 3--4°С, оптимальная 18--
22°С, максимальная глубина прорастания 4--5 см [Humangarden, 2008]. Всхо
ды появляются в марте--мае. Цветет в июле--сентябре. Плодоносит в сентяб
ре--октябре (рис. 129).
Вид обладает гетерокарпией, описаны два типа плодов: B и С. Тип В -- ма
ленькие, черные, блестящие, гладкие, тип С -- коричневые и весьма крупные
орешки. В то время как семена из плодов типа В находятся в состоянии покоя,
формируют банк семян и для успешного прорастания нуждаются в стратифи
кации, семена с плодов типа С не впадают в состояние покоя и при оптималь
ных условиях быстро прорастают [Mandгk, 2003b]. Таким образом, семена из
плодов типа В характеризуются поздним прорастанием и высокой выживае
мостью проростков, а семена из плодов типа С, напротив, отличаются ранним
прорастанием и низкой выживаемостью проростков. Эта приспособительная
особенность делает понятным широкое расселение вида и дает возможность
популяциям уцелеть при сильной нарушенности мест обитания и переносить
неблагоприятные сезоны [Mandгk, 2003b].
Расселение и распространение. В Средней России распространяется по
путям сообщения, в первую очередь по железным дорогам, а впоследствии за
нимает подходящие места обитания на участках с нарушенным растительным
покровом (рис. 130).
Последствия внедрения
Воздействия на естественные фитоценозы и аборигенные виды.
A.tatarica -- вид ранних сукцессионных стадий, в первые 2--3 года часто доми
нирует и конкурирует с местными сорными видами [Mandгk, 2003b].
279
Генетические изменения и вариабельность вида. 2n = 18 [Цвелев, 2000].
Во «Флоре СССР» [Ильин, 1936] описано 4 разновидности A. tatarica, разли
чающиеся по морфологии стебля (лежачий/прямостоячий) и листа: var.
concolor Fenzl, var. diffusa (Ten.) Gьrke, var. virgata Boiss., var. discolor (K.Koch)
Graebn. Во «Флоре Китая» выделяются еще две разновидности A. tatarica:
var. tatarica и var. pamiriса, различающиеся по форме верхних листьев [Gelin
et al., 2003].
Опираясь на особенности размножения A.tatarica, коллектив авторов в Чеш
ской Республике посвятил целую серию работ выявлению морфологических,
карпологических, экологических и генетических признаков, определяющих
высокую инвазионную способность вида на границе естественного распрост
ранения. Работы посвящены выяснению следующих вопросов: 1) как семена,
собранные с разных типов плодов, прорастают в различных экологических
условиях; 2) каким образом генотип семян влияет на параметры прорастания;
3) каким образом генетические различия между популяциями зависят от раз
ных типов плодов и как эти различия наследуются; 4) связано ли число семян
у отдельной особи с генетическими различиями между популяциями.
В ранних публикациях изучались условия прорастания инвазионных
(A.sagittata Borkh., A.tatarica L.) и неинвазионных (A.hortensis L., A. rosea L.)
видов рода Atriplex в Чешской Республике [Mandгk, 2003]. Их местообитания --
преимущественно урбанизированные экотопы, обочины дорог, свалки и ж/д
насыпи. Сделана попытка найти различия условий прорастания видов с уче
том сравнительно экологических данных и исследовать признаки, определяю
щие степень инвазионности. Виды оценивались по наличию/отсутствию покоя
семян и условиям прорастания при различном уровне нитратов и засоления.
Выявлено, что все изученные виды обладают выраженной гетерокарпией, одна
ко существенно различаются биологией семян. Семена инвазионной A.sagittata
имеют длительный период прорастания и сохраняют всхожесть несколько лет.
В паре A.tatarica (инвазионный вид) -- A.rosea (неинвазионный вид) растения
по длительности периода покоя семян и дружности прорастания существенно
не отличались, несмотря на различную частоту встречаемости в Чешской Рес
публике. Это объясняется высокой конкурентной способностью A.tatarica, эф
фективным распространением диаспор и устойчивостью к вредителям.
Генетическое разнообразие A. tatarica изучали в 25 природных популяциях
на западной границе ареала вида, в Чешской Республике [Mandгk et al., 2005].
Согласно выводам авторов, A. tatarica проявляет умеренный уровень генети
ческого разнообразия по сравнению с другими травянистыми растениями, так
же растущими в маргинальных популяциях. Географической изоляции между
популяциями на изучаемой территории выявлено не было. Генетическое раз
нообразие на границе распространения вида cнижается несущественно, что
показывает частичную необоснованность гипотезы относительно cнижения
внутрипопуляционного генетического разнообразия на границе ареала.
Тот же коллектив авторов [Mandak et al., 2006] изучали генетическую струк
туру экспериментальных популяций и репродуктивную активность A. tatarica.
Выявляли корреляцию между разными типами плодов и интенсивностью
прорастания сформировавшихся в них семян при различных температурных
условиях и степени засоленности. Из семян, прораставших при оптимальных
условиях, вырастал больший процент гетерозиготных растений, тогда как из
280
семян, прораставших в субоптимальных условиях, вырастало больше гомози
готных растений.
Определено влияние возрастного состояния популяций на уровень внутрипо
пуляционной изменчивости. Показано, что с увеличением возраста популяции
(превалированием генеративных особей над имматурными и ювенильными) про
исходит рост числа гетерозигот, уменьшение уровня инбридинга и рост генети
ческого разнообразия внутри популяций. Таким образом, благодаря высокому
уровню генетического разнообразия растений, выросших из семян почвенного
банка, в популяциях реализуется микроэволюционный потенциал и повышается
способность A. tatarica к активному расселению [Mandak et al., 2006].
Влияние на здоровье человека. Из пыльцы лебеды татарской производят
иммунобиологический диагностический препарат «Аллерген». Растение со
держит алкалоиды, кумарины, флавоноиды (0,48--0,5%). В семенах обнару
жены сапонины. С лечебными целями используется трава (стебли, листья,
цветки), семена. Отвар семян обладает мочегонным и рвотным действием. В
Туркмении отвар листьев лебеды татарской применяют при желтухе и как мо
чегонное средство [Лекарственные растения, 2008]. В г. Астрахань специально
изучали гиперчувствительность к пыльцевым аллергенам у детей в зависимос
ти от района проживания [Райский и др., 2008]. По мнению авторов, наиболее
частой причиной сезонной пыльцевой аллергии у астраханских детей является
гиперчувствительность к пыльце Atriplex tatarica и Artemisia absintium.
Экономическое и социальное значение (положительное и отрицатель
ное). Виды рода Atriplex не содержат токсины и имеют более или менее съе
добные листья. Однако при росте на искусственно удобренных субстратах они
могут накапливать в листьях высокий уровень нитратов [Plants for a future,
2008]. Лебеда татарская используется как пищевое и кормовое растение, лис
тья пригодны для маринадов, щей, овощных блюд, квашения [Лекарственные
растения, 2008]. При крайней необходимости растения можно есть сырыми.
Семена используют в пищу и применяют как загуститель при приготовлении
хлеба [Plants for a future, 2008]. Растение служит кормом для верблюдов и овец.
Сорное, засоряет посевы, особенно пропашных культур [Энциклопедия сель
ского хозяйства, 2008].
Меры борьбы
К превентивным методам следует отнести очистку семенного материала,
раннюю зяблевую вспашку с предварительным лущением, своевременную
предпосевную обработку и рыхление междурядий. С целью уничтожения по
пуляций A. tatarica применяются гербициды: 2,4 Д в посевах зерновых, 2М
4Х -- льна, симазин -- в посевах кукурузы, трефлан -- на плантациях хлопчат
ника, проводится уничтожение сорняков на необрабатываемых землях [Эн
циклопедия сельского хозяйства, 2008].
СЕМЕЙСТВО CUCURBITACEAE -- ТЫКВЕННЫЕ
ЭХИНОЦИСТИС ШИПОВАТЫЙ
Название: Echinocystis lobata (Mich.) Torr. et Gray
Синонимы: E. echinata (Muhl.) Vass.; Micrampelis lobata (Mich.) Greene.
281
Морфологические признаки
Однолетнее вьющееся растение (рис. 131). Стебель голый с 3--4 раздель
ными усиками. Листья пятилопастные. Венчик желтовато белый, с 6 узкими
долями. Пестичные цветки одиночные, с опушенной завязью. Тычиночные
цветки собраны в многоцветковое метельчатое соцветие. В естественном аре
але подвидов и форм не имеется. Отмечается высокая внутрипопуляционная
изменчивость по качественным признакам семян. Семена, находящиеся в пло
дах, собранных с одной особи, имеют форму, цвет и рисунок семенной кожу
ры, присущие именно этой особи, прямо таки как отпечатки пальцев у людей,
варьируют лишь масса семян и их абсолютные размеры. В 2005 г. нами [Ви
ноградова, 2006] определена фенетическая гетерогенность семян E.lobata в
естественной популяции в окрестностях Национального арборетума Минне
соты (45° с. ш.; 93° з. д.). Выявлено 17 дискретных вариаций, или фенов: тек
стура семенной кожуры -- гладкая/морщинистая, с рисунком/без рисунка;
форма семени -- округлая (l/d < 2)/овальная (2d" l/d d"3)/ланцетная (l/d >
3); окраска семени -- черная/коричневая/серая, с боковой полосой/без поло
сы; рисунок семенной кожуры -- мраморный/пятнистый; пятна -- бесформен
ные /треугольные/ округлые.
В естественной популяции преобладают растения с семенами морщинисты
ми (57%), без рисунка (64%), овальной формы (71%), коричневого цвета (50%),
без полосы (79%). Округлая форма семян отмечена лишь у одной особи, а лан
цетная -- у трех. Черные семена собраны с трех особей, серые -- с четырех.
Естественный ареал
Естественный ареал -- Северная Америка. В Канаде в природных место
обитаниях встречается в провинциях: Саскачеван (северная граница прохо
дит по 52°с. ш.), Манитоба (самое северное местообитание -- оз. Уэкуско, 55°с.
ш.), Онтарио (самая северная находка -- р. Капескейсин, 49°с. ш.), г. Квебек
(п ов Гаспе), о. Принца Эдуарда, провинции Нью Брансуик и Новая Шот
ландия [Flora of Canada, 1979]. В США встречается повсеместно, исключая
крайние юго западные и юго восточные штаты.
Вторичный ареал
История интродукции и географическое распространение. В качестве инт
родуцированного вида отмечен в провинциях Британская Колумбия и Аль
берта [Flora of Canada, 1979].
В Европе впервые найден в 1904 г. в Румынии, однако широкого распрост
ранения он в то время не получил. Были отмечены лишь отдельные находки в
1911 г. в Чехии, в 1933 и 1936 гг. -- в Эльзас Лотарингии, в 1937 г. -- в Венгрии,
в 1939 г. -- в Мюнхене, в 1940 г. -- в Лейпциге. Имеются предположения, что
американскими войсками во время Второй мировой войны эхиноцистис был
занесен в Европу вторично, а, кроме того, он «сбежал» из культуры из некото
рых европейских ботанических садов. Хотя с полной уверенностью утверж
дать о независимой и неоднократной интродукции вида нельзя, тем не менее,
после 1946 г. эхиноцистис найден на юге Польши, в 1952 г. обнаружен в Наум
бурге, а годом позднее -- в окрестностях Галле. В 1945 г. E.lobata впервые от
мечен в Словении, в 1956 г. -- в Хорватии. В Боснии и Герцеговине известен по
двум находкам 1964 и 1967 гг. В 1982 г. как одичавшее растение обнаружен в
Вестфалии. В 1983 г. отмечено значительное расширение ареала эхиноцисти
282
са в юго восточной Сербии [Cecconelli, 1976; Chrtkova, 1983; Duty, 1960;
Abadzic, 1987; Sonneborn, Sonneborn, 1986; Randelovic et al., 1984].
Формирование вторичного ареала E.lobata в бывшем СССР шло из разных
районов доноров, причем практически одновременно и в европейской, и в ази
атской частях страны [Виноградова, 2006]. Первым литературным источни
ком, указывающим на распространение вида в России, является «Определитель
растений Дальневосточного края» [Комаров, Клобукова Алисова, 1931--1932 гг.],
где отмечено, что «Micrampelis lobata разводится в садах по окраинам Владивос
тока». С 1960 г. E.lobata использовался для вертикального озеленения в г. Улан
Удэ. В Амурскую обл. попал в 1959--1964 гг. и уже через 10 лет дико произрас
тал на окраинах многих населенных пунктов, в то же время начал сорничать и в
Иркутске [Спицын, 1965; Дымина, Еремеева, 1979; Тельпуховская, 1970].
В европейской части ареала эхиноцистис отмечен впервые (причем сразу
как натурализовавшееся растение) в 1946 г. на Украине в Закарпатье, и 30 лет
инвазионные популяции этого вида встречались только здесь. В 1950--1960 х
гг. E.lobata изредка отмечался на Украине, в Литве, Белоруссии и Московской
обл., но только как культивируемый в садах [Виноградова, 2006].
В начале 1970 х гг. произошло прямо таки взрывное расселение эхиноцис
тиса по Восточно Европейской равнине. В 1971 г. E.lobata уже разводился
[Захаренкова, 1971] во многих населенных пунктах Белоруссии (в Орше, Ста
рых Дорогах, Вилейке, Ошмянах, Волковичах). Вид стал изредка встречаться
как заносное и одичавшее растение в Молдавии [Гейдеман, 1975]. Во Флоре
Литвы [1971] указано, что вид широко распространен во всех районах респуб
лики, где культивируется в декоративных целях.
Большая заслуга в выявлении новых местонахождений E.lobata принадле
жит Ю.Д. Гусеву. В 1974 г. в г. Ростов на берегу оз. Неро на свалке он обнару
жил 1 экземпляр эхиноцистиса, изредка наблюдал вид в этом же городе в куль
туре. Ю.Д. Гусев отметил, что эхиноцистис дичает в Новосокольниках Псков
ской обл., в Санкт Петербурге и в Павловске Ленинградской обл. В 1975 г.
эхиноцистис обнаружен Ю.Д. Гусевым на заросшем кустами пустыре в Витеб
ске, на ж/д насыпи в Полоцке, на сорных местах возле ст. Фомино и Орша
Восточная. В 1976 г. Ю.Д. Гусев собрал эхиноцистис в Ижевске в 1 км север
нее вокзала на склонах ж/д насыпи, одичавшие растения отмечены им в 2 ме
стах между ст. Заводская и Ижевск. Ему же принадлежит ряд находок
эхиноцистиса в Львовской обл. и в Литве (LE).
В 1972 г. эхиноцистис обнаружен в окрестностях Махачкалы [Львов, 1979].
Повторно вид найден П.Львовым на свалке по берегу канала в Махачкале в
1976 г. Наши наблюдения 1984 г. показали, однако, что устойчивой, пусть даже
культивируемой, популяции эхиноцистиса в г. Махачкала в то время не было,
а несколько жителей разводят вид на балконах и на приусадебных участках из
семян, купленных или присланных из более северных регионов. П.Д. Львов
сообщил, что он также не встречал более одичавшего эхиноцистиса в том мес
те, где он произрастал в 1976 г. Натурализовался эхиноцистис в прибрежных
сообществах в Северной Осетии близ г. Орджоникидзе и Алагир только к 1990 г.
[Комжа, Попов, 1990].
E.Jager в 1977 г. собрал эхиноцистис в кустарниках на берегу р. Белая в Уфе
[Jager, 1980]. Нами гербарные сборы одичавшего E.lobata сделаны в 1978 г. на
сорных местах в Волгограде. В 1982 г. мы собрали семена эхиноцистиса с куль
283
Рис. 132. Расселение Echinocystis lobata в Средней России
Рис. 131. Массовое цветение Echinocystis lobata
Рис. 133. Внедрение Echinocystis lobata в приречные сообщества
Рис. 134. Плоды Echinocystis lobata
285
тивируемых растений в г. Таганрог; в 1983 г. нами отмечены крупные популя
ции одичавшего эхиноцистиса в Ленинградской, Новгородской и Тверской обл.
В Вологде в 1982 г. вид отмечен нами на сорных местах недалеко от реки, при
чем наблюдаемые популяции сохранились в 1983 и 1984 гг., так что их можно
считать устойчивыми, хотя в природные ценозы -- ивняки по берегам р. Во
логда -- E.lobata к тому времени еще не внедрился; в 100 км к северо западу в г.
Кириллов Вологодской обл. вид отмечен только в культуре. В 1983 г. эхиноцис
тис спорадически встречался возле домов и изредка дичал на мусорных местах
в районе Кунгурской лесостепи Пермской обл., но в г. Пермь не был замечен
(личное сообщение А.Овеснова). В 1984 г. вид (личное сообщение Л.Гречушки
ной Сухоруковой) культивировался в садах на окраинах Ставрополя.
В 1970--1980 е гг. E.lobata бурно расселялся и в азиатской части вторично
го ареала. Р.В. Камелин [1971] среди адвентивных растений Средней Азии на
зывал эхиноцистис, встреченный им в усадьбах г. Джамбул (сорно) и в искус
ственной роще на берегу р. Талас выше г. Гродеково. Г.Жалалов и А.Дзевал
товский [1973] изучали эмбриологию эхиноцистиса, используя в работе
растения, культивируемые в г. Ташкент. В работе по адвентивной флоре Бар
наула [Игнатов, Игнатова, 1972] эхиноцистис отмечен как культивируемый и
одичавший в ряде мест в окрестностях города. Н.И. Золотухин [1983] сообща
ет об устойчивости популяции эхиноцистиса в Алтайском заповеднике: этот
вид изредка встречается в пос. Яймс и на кордонах. В 1980 г. собран автором
на кордоне Беле, где эхиноцистис произрастал у заборов и на мусорных мес
тах. E.lobata повсеместно дичал, распространяясь по заброшенным садам, пу
стырям и обочинам троп в южной части Хабаровского края [Шлотгауэр, Не
байкин, 1984]. В 1987 г. эхиноцистис собран на хр. Кулумыс в Красноярском
крае [Назимова, Степанов, 1988] и стал обычным сорняком на нижнем Амуре
и в Большехехцирском заповеднике [Мельникова, 1989; Шага, 1989]. Во вре
мя экспедиций ГБС по Енисею, Алтаю и Казахстану в 1983--1984 гг. А.Кукли
на обнаружила культивируемые экземпляры эхиноцистиса в с. Верхнеимбат
ском Туруханского р на Красноярского края, нашла 1 экз. в вегетативном со
стоянии на клумбе в г. Игарка, отметила, что этот вид разводится в садах на
окраинах Усть Каменогорска, Горно Алтайска, Ончудая и Шебалино. В 1984
г. мы получили от С.Горожанкиной семена эхиноцистиса, собранные в г. Крас
ноярск и в с. Назимово Енисейского р на Красноярского края. В 1988 г. со
бран в Ононском р не Читинской обл. [MHA]. В 2002 г. отмечен в качестве
адвентивного вида на севере Хакасии [Эбель, 2002].
Таким образом, в Евразии имеется два изолированных очага расселения
эхиноцистиса -- Центральная Европа и Приморский край. На Дальний Вос
ток E.lobata, надо полагать, был занесен непосредственно из Северной Америки
в 20 х гг. прошлого века. В Центральной Европе эхиноцистис появился в нача
ле ХХ века, но до Второй мировой войны имел лишь точечные местонахожде
ния и только начиная с 1946 г. стал постепенно расширять ареал и продвигаться
на северо восток. Последние 25 лет ознаменовались дальнейшим расширением
и европейской, и азиатской частей вторичного ареала [Виноградова, 2006]. В
настоящее время граница вторичного ареала E.lobata в России проходит на 5°
севернее границы его естественного ареала по линии Санкт Петербург--Волог
да--Пермь--Красноярск--Иркутск--Тында--Комсомольск на Амуре.
Пути и способы заноса. Площадь обширного вторичного ареала в умерен
ном поясе Евразии c каждым годом увеличивается. Новые территории E.lobata
286
осваивает преимущественно благодаря разведению его в качестве декоратив
ного растения. Дичает довольно быстро, поселяясь в местах с богатой влаж
ной почвой: огородах, помойках, сорных местах. Конечная фаза натурализа
ции эхиноцистиса -- внедрение его в приречные кустарниковые фитоценозы,
свойственные виду в естественном ареале [Виноградова, 2007а].
Статус в регионе. Впервые отмечен в культуре в Московской обл. в 1963 г.,
через 5 лет стал дичать. К началу 1970 х гг. эхиноцистис обнаружен в есте
ственных фитоценозах во многих пунктах Средней России: в Московской,
Калужской, Рязанской и Брянской обл., начал культивироваться в Липецкой
обл. и в Чувашии (MHA, MW), стал распространяться на пустырях, свалках и
других рудеральных местах в Твери [Малышева, 1979].
В 1980 е гг. площадь вторичного ареала E.lobata в Средней России продол
жала увеличиваться. Этот вид стал спорадически встречаться во всех районах
Ульяновской обл. как одичавший вид по паркам, садам, сорным местам, пой
менным ивнякам. В Саратовской обл. стал наиболее распространенным ад
вентивным растением. В 1983 г. эхиноцистис впервые обнаружен в Воронежс
кой обл. Полностью натурализовался в Рязанской обл., стал дичать в Марий
ской АССР, Чувашии и Татарстане [Маевский, Иванов, 1984; Гущина, Казакова,
1989; Папченков, Дмитриев, 1984]. В настоящее время встречается во всех,
без исключения, районах Средней России, во многих местах массово (рис. 132).
Местообитания. Растет вдоль берегов рек, обвивая своими плетями при
речные кустарники (рис. 133).
Изучена зависимость роста и фенологии E.lobata от экологических усло
вий обитания [Виноградова, 2008б]. Семена, собранные с одной особи, высе
вали на свету и в тени. Различия между световыми и теневыми экземплярами
по средней длине главной оси, среднему числу метамеров на главной оси и
порядковому номеру листа, в пазухе которых завязываются мужские соцве
тия, были небольшими. Значительные различия отмечены по общей длине всех
побегов: в зависимости от пункта сбора семян на свету растения были в 2, 3 и
даже (у образца, собранного в г. Мценск) в 7 раз длиннее, чем в тени, и имели
в 2--6 раз больше метамеров. По числу побегов 1 го порядка экземпляры, вы
росшие на свету, превосходят теневые в 3--6 раз, а у образца из Белгорода -- в
16 раз! Различие в порядковом номере листа, в пазухе которого развивается
первый плод, составляет у разных образцов от 2 до 16.
Таким образом, экспериментально установлено, что экземпляры, произра
стающие на свету, независимо от пункта сбора семян образуют более длинные
побеги с большим числом метамеров, сильнее ветвятся и формируют плоды в
пазухе более раннего листа, и это позволяет сделать заключение о том, что
E.lobata -- светолюбивое растение.
Размножение и жизненный цикл. Эхиноцистис -- однодомное растение; в
каждом узле образуется одиночный невзрачный женский цветок и многочис
ленные мужские цветки, собранные в сложную метелку. Всходы появляются
в начале мая, цветет в июне -- начале июля, семеношение начинается в конце
августа--сентябре.
За вегетационный сезон одно растение может образовать от 10 до 40 пло
дов (рис. 134), в каждом из которых развивается по 4 семени. Лабораторная
всхожесть семян 90--95%, полевая всхожесть ~70%. Терофит. Вегетативно не
размножается.
287
Расселение и распространение. Распространяется в основном при помощи
человека, дичая из культуры по окраинам небольших населенных пунктов.
Последствия внедрения
Воздействие на естественные фитоценозы и аборигенные виды. Инвазион
ные популяции в Московской обл. очень устойчивы, хотя флуктуация чис
ленности особей и имеет место. Так, в конце 1970 х гг. наблюдалась вспышка чис
ленности вида, и приречные ивняки были столь сильно увиты эхиноцистисом,
что издали напоминали стога и копны сена. После суровой зимы 1978/79 в тех же
пунктах наблюдались лишь единичные особи, а через 4 года численность популя
ций восстановилась. E.lobata активно вытесняет аборигенные виды из естествен
ных приречных фитоценозов, создавая густую тень, в которой не способны про
израстать свойственные этим местообитаниям травянистые растения.
Генетические изменения и вариабельность вида. Высокая внутрипопуляци
онная изменчивость по качественным признакам семян, свойственная виду на
родине, сохраняется и во вторичном ареале. В 16 обследованных инвазионных
популяциях [Виноградова, 2006] в Закарпатье, Благовещенске и в Европейс
кой России от Вологды до Донецка выявлены все 17 дискретных вариаций.
Во всех популяциях, так же как в естественном ареале, превалировали рас
тения с морщинистыми семенами (не менее 53%), но, в отличие от естествен
ного ареала, с рисунком (не менее 60%). Фен наличия полосы на боку семян
преобладал во всех закарпатских популяциях и в популяциях на р. Нара и в г.
Мценск. Во всех популяциях, за исключением популяции г. Крестцы Новго
родской обл., преобладают семена овальной формы, однако присутствуют и
особи с семенами округлой формы, тогда как семена ланцетной формы отме
чены лишь в 6 популяциях. В 6 популяциях отмечено наибольшее число ко
ричневых семян. В остальных популяциях число черных, коричневых и серых
семян оказалось примерно равным. Корреляционной зависимости между вы
деленными фенами не наблюдается. При анализе географического распреде
ления фенов в пределах вторичного ареала E.lobata клинальной изменчивости
частоты встречаемости дискретных вариаций не отмечено. И в недавно сфор
мированных инвазионных популяциях, и в популяциях, произрастающих уже
длительное время, и даже в г. Благовещенск (этот семенной материал, по всей
вероятности, имеет совершенно другой источник заноса -- непосредственно
из Америки) фены комбинировались случайным образом в самые разнообраз
ные сочетания. Каждая популяция характеризуется различной степенью реа
лизации фенофонда, то есть числом вариаций, найденных в конкретных попу
ляциях и выраженных в процентах к числу фенов, выделенных для вида в це
лом. У эхиноцистиса степень реализации фенофонда очень высока и в 14
изученных популяциях составляет 100%. В двух популяциях на северном пре
деле распространения, куда E.lobata занесен не так давно и где встречаемость
его еще довольно низка, а популяции немногочисленны -- в г. Вологда и Нов
городской обл., -- степень реализации фенофонда составляет 65%.
Следует, однако, отметить, что фенетическая гомогенность семян вовсе не
предполагает генетической гомогенности: растения, выросшие из семян, со
бранных с одной особи, в однородных условиях питомника показывают неко
торые различия по темпам роста и фенологическому ритму развития.
Поскольку эхиноцистис является перекрестноопыляемым растением, се
мена дочерних растений первого поколения F1 по форме и окраске также сильно
288
варьируют на внутрипопуляционном уровне и могут вовсе не походить на се
мена материнских растений.
Таким образом, в инвазионных популяциях перекрестноопыляющихся видов,
расширяющих вторичный ареал (к которым, несомненно, относится E.lobata),
сохраняется высокий уровень генетического разнообразия, и именно это обеспе
чивает их успешную адаптацию к новым условиям произрастания.
Экономическое и социальное (положительное/отрицательное) значение.
Инвазия эхиноцистиса в прибрежные фитоценозы приводит к резкому сокра
щению биологического разнообразия.
Меры борьбы
Прекращение выращивания в декоративных целях [Виноградова, 2008а]
приведет к заметному сокращению инвазионных популяций. Эхиноцистис
легко выпалывается, и на ограниченных площадях может быть быстро уничто
жен. По поймам рек, по видимому, меры борьбы нереальны. В естественном аре
але популяции сильно поражаются болезнями и вредителями, что обусловли
вает мозаичность распространения вида. Во вторичном ареале исследования
практически не проводились, имеются лишь сведения о выявлении в Югославии
паразитирующего на эхиноцистисе гриба из рода Oidium [Rankovic, 1989].
Обзор научных исследований и публикаций
Изучены изменчивость инвазионных популяций эхиноцистиса [Виногра
дова, 2006, 2007а, 2007б, 2008б].
СЕМЕЙСТВО ELAEAGNACEAE -- ЛОХОВЫЕ
ЛОХ УЗКОЛИСТНЫЙ
Название: Elaeagnus angustifolia L.
Синонимы: E. hortensis Bieb., E. orientalis L., ? E. oxycarpa Schlecht.
Объем вида до сих пор является предметом дискуссий: отечественные бо
таники принимают несколько «мелких» видов [Горшкова, 1949; Цвелев, 2003,
2004], которые за рубежом обычно объединяют в один вид [Nasir, 1975;
Fernandez Arias, Castrovieho, 1997; Haining, Gilbert, 2007].
Морфологические признаки
Листопадное дерево с низкой широкой кроной, реже кустарник, высотой
до 5--8(10) м. Молодые ветви серебристые от белых чешуек, 2--3 летние ---
красно бурые или коричневые, нередко колючие. Листья очередные, яйцевид
но ланцетные, удлиненно овальные, ланцетные (или более узкие), цельнок
райные, сверху серовато зеленые со светлыми жилками, снизу серебристо бе
лые от обильных чешуек. Лох цветет после распускания листьев в конце вес
ны --- начале лета. Растение полигамное, с обоеполыми и тычиночными
цветками на одном растении, расположенными по 2--4(6) в пазухах листьев
на однолетних побегах. Околоцветник простой, трубчато колокольчатый, с 4
треугольными лопастями, снаружи серебристо серовато белый от покрываю
щих его звездчатых чешуек, внутри ярко жёлтый; цветки с сильным прият
ным запахом. Плод --- эллипсоидная или яйцевидная ложная костянка, или
сфалерокарпий [Каден, Кондорская, 1967], длиной 7--15 мм, мучнистая, бу
рая или коричневая, незрелая --- серебристая от многочисленных чешуек; кос
289
Рис. 135. Побеги
Elaeagnus
angustifolia
Рис. 136. Рассе
ление Elaeagnus
angustifolia в
Средней России
Рис. 137. Внедре
ние Elaeagnus
angustifolia в
естественные
сообщества
Рис. 138. Плоды
Elaeagnus
angustifolia
291
точка светло коричневая с 8--10 тонкими продольными бороздками; образо
ван орешком, формирующимся из завязи, и разросшейся мясистой нижней
частью околоцветника (рис. 135).
Естественный ареал
E. angustifolia издревле широко культивируется, поэтому точно установить
его естественный ареал едва ли возможно. Происхождение лоха обычно свя
зывают с Малой и Средней Азией [Fernandez Arias, Castrovieho, 1997]. Так, в
«Species plantarum» Карл Линней описывал распространение следующим об
разом: Habitat in Bohemia, Hispania, Syria, Cappadocia [Linnaeus, 1753].
Вторичный ареал
История интродукции и географическое распространение. Лох узколис
тный широко культивируется как устойчивое декоративное растение, особенно
ценное для безлесных аридных регионов. Во многих странах отмечено его дича
ние. В Европе E. angustifolia признан инвазионным видом в Венгрии [Invasive...,
2003; Balogh et al., 2004] и Испании [Sanz Elorza et al., 2001], отмечен как занос
ное растение в Австрии [Essl, Rabitsch, 2002] и Чехии [Pyљek et al., 2002]
В США лох интродуцирован во второй половине XIX века как декоратив
ное и медоносное растение и для защиты песчаных почв от эрозии, где он ши
роко одичал, особенно на западе страны [Hayes, 1976; Knopf, Olson, 1984]. До
1980--1990 х гг. посадки лоха субсидировали из федерального и местных бюд
жетов [Olson, Knopf, 1986]. Однако в то же время отмечена его широкая нату
рализация на востоке США [Katz, Shafroth, 2003]. В настоящее время E.
angustifolia в США считается опасным инвазионным видом [Miller, 2003;
Stannard et al., 2002; Katz, Shafroth, 2003; Williamson, 2003]. Лох известен в
аридных регионах Южной Америки [Klich, 2000].
В Европейской России E. angustifolia разводится как декоративное расте
ние [Горшкова, 1949; Цвелев, 2003, 2004]. Широко используется в лесомелио
рации в южных аридных областях как засухо и солеустойчивая культура.
Пути и способы заноса. Лох распространяется из мест культивирования
[Katz, Shafroth, 2003].
Статус в регионе. На территории средней полосы Европейской России
лох узколистный является инвазионным видом лишь на юге территории, осо
бенно в юго восточных областях. В северной части вид известен преимуще
ственно в культуре, но изредка встречается как заносное растение (рис. 136).
Местообитания. Лох способен расти в разнообразных местообитаниях,
но предпочитает песчаные равнины, а также речные долины (рис. 137). На юге
Европейской России обнаруживает тяготение к засоленным почвам. E.
angustifolia обычно поселяется в местах достаточного увлажнения, хотя бы
временного, однако может расти и при существенном дефиците влаги [Katz,
Shafroth, 2003]. Лох образует арбускулярно везикулярную микоризу [Riffle,
1977] и клубеньки на корнях с азотфиксирующим актиномицетом из рода
Frankia [Zitzer, Dawson, 1992; Frankia..., 2003; Nalin et al., 2006], что делает лох
нетребовательным к содержанию в почве азота [Kohls, Baker, 1989]. Не выдер
живает кислых почв с pH <6,0 [Tu, 2003]. Лох устойчив к засолению и спосо
бен выдерживать концентрацию NaCl до 10--15 г/л [Katz, Shafroth, 2003].
Размножение и жизненный цикл. E. angustifolia размножается семенами.
При благоприятных условиях роста лох способен плодоносить на 3--5 й год.
Плодоношение регулярное и обильное (рис. 138). Есть сведения о вегетатив
292
ном размножении корневыми отпрысками [Stannard et al., 2002]. Эти отпрыс
ки образуются при повреждении горизонтальных корней, которые залегают
близко к поверхности почвы.
Расселение и распространение. Семена лоха разносят птицы, которые
питаются его плодами [Pearce, Smith, 2001]. Внешние слои семени непрони
цаемы для желудочного сока, что позволяет им сохранить всхожесть при про
хождении через желудочно кишечный тракт [Shafroth et al., 1995]. Однако
плоды могут разноситься и потоками воды, и во время весеннего ледохода
[Pearce, Smith, 2001]. В почве семена сохраняют жизнеспособность в течение
3 лет [Shafroth et al., 1995; Pearce, Smith, 2001].
Последствия внедрения
Воздействия на естественные фитоценозы и аборигенные виды. Посе
ляясь в речных долинах, лох способен достаточно быстро расселяться, вытес
няя местные виды и образуя одновидовые заросли [Pearce, Smith, 2001].
В штате Монтана, США, E. angustifolia вытесняет из долинных лесов топо
ля, поскольку бобры значительно меньше повреждают деревья лоха [Lesica,
Miles, 1999]. Кроме того, появление E. angustifolia уменьшает эрозию почвы,
что, в свою очередь, уменьшает число сеянцев тополя, приспособленных к ро
сту на обнаженном субстрате [Katz et al., 2001]. Влияние инвазии лоха на жи
вотное население сложнее. В зарослях E. angustifolia общее богатство фауны
птиц и мелких млекопитающих может и не меняться, но разнообразие насеко
мых заметно уменьшается [Katz, Shafroth, 2003].
Генетические изменения и вариабельность вида. При изучении изменчиво
сти листьев E. angustifolia следует учитывать, что их размер, форма, анатомичес
кие параметры зависят от положения листьев в кроне дерева [Klich, 2000].
Экономическое и социальное (положительное и отрицательное) зна
чение. В аридных районах Азии и отчасти Европы лох широко культивирует
ся как полезное засухоустойчивое растение. Древесина используется в хозяй
стве как строительный материал, для поделок и как топливо, листья и кора
являются прекрасными дубителями, а само дерево используется в полезащит
ном лесоразведении. Лох -- хороший медонос. Плоды, известные под названи
ем «джида», съедобны, а недозревшие используются в медицине как вяжущее
средство [Горшкова, 1949; Nasir, 1975; Hussain et al., 2007; Khan, Khatoon, 2007].
Лох можно использовать как индикатор загрязнения среды тяжелыми метал
лами, в частности цинка [Aksoy, Єahin, 1999].
Меры борьбы
Необходим контроль расселения E. angustifolia из мест культуры, следует
прекратить его посадки в полезащитном лесоразведении. В США для борьбы
с лохом используются гербициды, в частности, гарлон--4 --- Garlon 4, гарлон--
3А --- Garlon 3А и арсенал --- Arsenal [Miller, 2003; Stannard et al., 2002; Parker,
Williamson, 2003]. Этот способ уничтожения существенно эффективнее меха
нического истребления или выжигания, поскольку лох может возобновлять
ся корневыми отпрысками [Stannard et al., 2002].
В Западном Прикаспии временами наблюдается усыхание лоха под влия
нием заболонника Ярошевского (Scolytus jaroschewskii Schevyrew) и патоген
ной микрофлоры [Кузьмичев, Петров, 1991; Петров, Кузьмичев, 1994].
293
СЕМЕЙСТВО ELAEAGNACEAE --- ЛОХОВЫЕ
ОБЛЕПИХА КРУШИНОВИДНАЯ
Название: Hippophaл rhamnoides L. Латинизированная форма греческого
названия растения hippophaes происходит от hippos -- лошадь и phaos -- блеск.
Морфологические признаки
Деревце или крупный кустарник высотой 4--5, редко 6--8 м, с острыми креп
кими колючками длиной 2--7 см (рис. 139). Молодые побеги серебристые от
покрывающих их чешуек и волосков. Листья очередные, линейно ланцетные,
длиной 2--8 см и шириной ~0,5 см. Растение двудомное. Цветки мелкие, раз
дельнополые, расположены на побегах прошлого года. Мужские цветки с дву
раздельным околоцветником, собраны в короткие соцветия -- колосья, женс
кие цветки -- с двулопастным околоцветником, расположены по 2--5 в пазухах
веток и колючек. Костянки сочные, оранжевые или красные, шарообразные
или эллипсоидальные, длиной 0,5 см и диаметром 0,3 0,5 (0,8) см, с блестящи
ми темно коричневыми косточками [Вехов и др., 1978]. Косточки продолго
вато округлые, в поперечном сечении эллипсовидные, длиной 4--7 мм и ши
риной 2,5--3,5 мм. Окраска косточек от серовато бурой до темно коричневой,
почти черной, зависит от содержания дубильных веществ и пигментов в по
кровах косточек. Абсолютная масса 1000 косточек в воздушно сухом состоя
нии 14--19 г. Из 1 кг плодов западноевропейских и восточносибирских попу
ляций получают 30--100 г косточек [Ермаков, Фаустов, 1983].
Естественный ареал
В диком виде растет в приатлантических и южноскандинавских районах
Западной Европы, на Балканах, в Турции, Иране, Афганистане, Пакистане,
гималайских районах Индии, Китае, в том числе в Тибете, Монголии, почти
во всех горных районах по южной границе России (на Кавказе, в предгорной и
горной частях Средней Азии, Казахстане), а также на юге Сибири и в Забай
калье [Вехов и др., 1978]. Вид подходит к западной границе Европейской Рос
сии -- встречается от островов в Балтийском море на севере до Черного моря
на юге. На Украине растет дико в устье Дуная [Петрова, 1987].
История формирования природного ареала облепихи в Западной Европе
восстановлена с помощью молекулярных методов исследования [Bartish et al.,
2006]. Ядерные и цитоплазматические маркеры указывают, что облепиха была
распространена в Юго Восточной Европе и именно с этой территории нача
лась колонизация ею Центральной Европы и Скандинавии. В районе Север
ных Альп проходила контактная зона между популяциями из Альп и популя
циями, происходящими из Скандинавии, Восточной и Центральной Европы.
В ходе исследования были определены по крайней мере 4 важнейших эпизода
увеличения размеров популяций, случившихся за последние 40 тыс. лет. В
частности, синхронное расширение популяций в разных частях ареала в Цен
тральной, Восточной Европе и Малой Азии произошло 13 11,6 тыс. лет назад,
что, вероятно, было связано с глобальным похолоданием.
Вторичный ареал
История интродукции и географическое распространение. H.rhamnoides
занесена в Северную Америку. Расширению ареала вида способствует его вы
сокая устойчивость к экстремальным условиям, включая температуру от 43
294
до +40°С, засуху, затопление, большие высоты, засоленность и щелочность
грунтов [Ercisli et al., 2008].
Благодаря декоративности и лечебным свойствам плодов облепиха полу
чила значительное распространение в лесной и лесостепной зоне России, куда
она интродуцирована в начале XIX века. В декоративное озеленение облепиха
была введена Санкт Петербургским ботаническим садом из растений и косто
чек, привезенных в начале XIX века с Горного Алтая. В начале ХХ века облепи
хой заинтересовались уже как плодовым растением, и ее начали разводить в
европейской части России. Однако особо популярной облепиха стала в 1960 е
гг. [Петрова, 1987]. В Вологодской обл. облепиха дичает по ж/д путям и свал
кам, на улицах встречается редко. В северо западных областях России облепи
ха известна как интродуцированный вид, сведения о дичании не приводятся
[Цвелев, 2000]. Южнее, в областях Средней России, облепиха становится обыч
ным видом улиц городов, местами в массе заполняет прибрежный аллювий.
Пути и способы заноса. Вероятно, изначально преобладает преднамерен
ный занос вида как закрепителя песков, декоративного, пищевого и маслич
ного растения. Занос птицами и вегетативное размножение корневыми отпрыс
ками служит способом распространения H. rhamnoides уже в пределах вторич
ного ареала.
Статус в регионе. За последние годы облепиха как ягодная культура ши
роко распространилась по всем областям Средней России (рис. 140). Как ди
чающее интродуцированное растение отмечена в Ивановской, Тверской, Ни
жегородской, Московской, Владимирской, Тульской, Липецкой, Курской, Во
ронежской, Калужской, Рязанской, Пензенской, Тамбовской и Ульяновской
обл. [Борисова, 2007; Васюков, 2004; Григорьевская и др., 2004; Игнатов и др.,
1990; Казакова, 2004; Маевский, 2006; Маркелова, 2004; Мининзон, 2007; По
луянов, 2005; Флора Липецкой области, 1996]. Вид, однако, собирают мало, и
гербарные коллекции содержат скудный материал по облепихе. Старых сбо
ров, как правило, обнаружить не удается.
В Воронежской обл. широко культивируется, дичает [Григорьевская и др.,
2004]. В восточной части Липецкой обл. спорадически отмечается одичавшей
(Н.Ю. Хлызова, личное сообщение). Первые сведения о культивировании вида
в Курской обл. содержатся в работе С.И. Машкина [1971]; ныне разводится в
садах и лесополосах и дичает [Полуянов, 2005].
В старых ботанических работах по Тульской обл. сведения о разведении
облепихи отсутствуют, вид введен в культуру как ценное плодовое и лекар
ственное растение только в середине ХХ века. Сборы, сделанные в Щекинс
ком р не Тульской обл., хранятся в MW: лесхоз «Тульские засеки», Ярцевское
лесничество, посадки по опушкам дубравы, 25.05.1960 г., Смирнова; лесхоз
«Тульские засеки», вблизи с. Ярцево, 29.05.1960 г., Ганюшкина. В том же райо
не сделаны еще 3 сбора в 1961 1967 гг., однако неизвестно, были это посажен
ные или дичающие особи. Это интересно знать, так как в литературных источ
никах 1980 х гг. [Алексеев, Губанов, 1980; Алюшин, 1982] вид отсутствует. В
настоящее время встречается регулярно и в большом числе. По нашему мне
нию, массовое внедрение облепихи во флору области произошло недавно, в
течение последних 5 6 лет.
В Московской обл. к настоящему времени -- обычный вид. Один из первых
сборов сделан А.Н. Петунниковым: Московская губерния, университетский
Рис. 139. Hippophae
rhamnoides между
бетонными плитами
Рис. 140. Расселение
Hippophae rhamnoides
в Средней России
Рис. 141. Внедрение
Hippophae rhamnoides
в естественные
сообщества
Рис. 142. Цветы
Hippophae rhamnoides
297
ботанический сад [в г. Москве, на просп. Мира], 12.07.1885 г. (MW). Разведе
ние облепихи в Подмосковье отмечал Д.П. Сырейщиков [1907], однако попу
лярность этой культуры в 1980 х гг. сильно возросла [MW; Игнатов и др., 1990].
Во Владимирской обл. собран в Юрьев Польском р не, в 3 км. к западу от
г. Юрьев Польский, на залежи у дороги на с. Горки, вегетативно разросся на
площади более 10 мІ, 23.08.2007 г., А.Серегин (MW). Теперь H.rhamnoides встре
чается по всей области, особенно часто в ополье. Долгое время облепиху мож
но было наблюдать вдоль заборов дачных поселков, а в последние 3 года она
стала выходить на залежи (А.П. Серегин, личное сообщение).
В Ивановской обл. облепиха встречается единичными экземплярами и
крупными зарослями [Борисова, 2007]; собрана в окрестностях г. Иваново, на
ст. Текстильная близ ж/д переезда, 1 куст между ж/д путями, 5.09.1991 г., Е.Бо
рисова (MW).
На территории Тверской обл. облепиху культивировали уже в первой по
ловине XIX века [Покровский, 1879]. В одичавшем состоянии собрана А.П.
Хохряковым в 1996 г. в Калининском р не (TVBG). К настоящему времени
облепиха прочно удерживается в местах заноса, наблюдается активное раз
множение посредством корневых отпрысков [Маркелова, 2004].
В Калужской обл. вид местами массовый (С.Майоров, личное сообщение).
Впервые собран в 1966 г. в Калуге, в сквере им. Пушкина (Н.Н. Гурова, КОКМ).
Собран в Рязани, на южной окраине, по краю городской свалки, несколько
экземпляров, 20.07.2004 г., Л.Хорун, В.Сычев, Д.Ламзов (MW). В настоящее
время, по нашим наблюдениям, проявляет склонность к инвазии.
В Тамбовской обл. H.rhamnoides собрана в Мичуринском р не, между ж/д
пл. Каменка и электродепо, на песке у ж.д. близ моста через р. Каменка,
15.06.2007 г., А.Сухоруков (MW).
В Смоленской обл. дичает по ж/д насыпям, но встречается не часто [Ре
шетникова, 2004].
Местообитания. В естественном ареале на территории России H.rhamnoides
образует сплошные массивы по отмелям и на песчаных берегах водоемов, рек,
морей, озер, ручьев, в тугаях, тальниково тополевых лесах, на равнинах и в го
рах, преимущественно в нижнем и среднем поясе гор [Петрова, 1987]. В Европе
вид заселяет песчаные дюны в Великобритании и на Атлантическом побережье
континента, берега фьордов в Норвегии, гравийные речные берега в долине р.
Рона, гравийные берега альпийских рек [Binggeli et al., 2008].
В процессе дичания в Средней России вид концентрируется в местах, сход
ных по условиям с природными -- на прибрежных песках и наносах (рис. 141),
золоотвалах, в стыках бетонных плит на набережных, а также успешно осваи
вает придорожные и сорные места обитания: обочины шоссейных дорог и ж/д
путей, улицы, пустыри, дворы, скверы, лесополосы, окрестности садоводчес
ких товариществ.
Размножение и жизненный цикл. Облепиха размножается семенами, ко
торые сохраняют высокую всхожесть (до 96%) в течение 2 лет. Однако цен
ные сортовые качества наследуются лишь при вегетативном размножении че
ренками, отводками, отпрысками. Быстро образует придаточные корни при
засыпании землей или песком. Способна формировать нескольких ярусов
горизонтальных придаточных корней. В естественных условиях корневые от
прыски появляются на горизонтальных корнях материнского растения и об
298
разуют собственную систему придаточных корней, однако связи с материнс
кими растениями, как правило, не теряют. Диаметр корневой системы высо
корослых деревьев облепихи превышает диаметр их кроны в 1,5 3 раза [Ерма
ков, Фаустов, 1983]. Морозо и жароустойчивая культура. Вследствие повер
хностного расположения корневой системы предпочитает дренированные, но
достаточно влажные почвы, богатые кальцием; не нуждается в азотном удоб
рении, так как на ее корнях живут азотфиксирующие актиномицеты. Хорошо
отзывается на фосфорные удобрения. На плотных почвах с недостатком воз
духа в корнеобитаемом слое чувствует себя угнетенно. Растет довольно мед
ленно. Начинает плодоносить с 3--6 лет. Цветет в апреле--мае, до распускания
листьев или одновременно с ним (рис. 142). Плоды созревают с конца августа
по октябрь. От начала цветения до полного созревания плодов проходит 12--
15 недель. Одно дерево дает 7--9 (16) кг плодов, а некоторые сорта >20 кг, уро
жайность плантации -- до 120 ц/га. Семена высевают осенью или весной после
3 х месячной стратификации. Можно размножать отсадкой корневых отпрыс
ков, зелеными и одревесневшими черенками. Продолжительность жизни в
естественных условиях -- 80 лет и более, в культуре срок использования -- 15--
20 лет [Вехов и др., 1978; Петрова, 1987].
В последнее время выявлено >30 видов вредителей, повреждающих облепиху.
Из насекомых постоянный вред наносят облепиховые муха, моль и зеленая тля.
Если первые два вредителя паразитируют на дикорастущих растениях, то облепи
ховая зеленая тля встречается и в районах интродукции [Ермаков, Фаустов, 1983].
Расселение и распространение. Семена распространяют птицы, но
значительный вклад в распространение облепихи вносит вегетативное размно
жение корневыми отпрысками.
Последствия внедрения
Воздействие на естественные фитоценозы и аборигенные виды. Зна
чительное внимание вопросам влияния облепихи на природные экосистемы
уделяется в Западной Европе. В Северной Ирландии, например, изучается
воздействие H.rhamnoides на экосистемы песчаных дюн [Binggeli et al., 2008].
H.rhamnoides интродуцирована в Ирландию в 1838 г. и растет на больших пло
щадях песчаных дюн с начала ХХ века изначально как задернитель дюн, а за
тем и в одичалом состоянии. Песчаные дюны Северной Ирландии являются
частью охраняемых природных территорий и, как и большинство незадернен
ных пространств, имеют слабую устойчивость к биологическим инвазиям. На
их территорию попадает значительное число адвентивных видов растений,
однако H. rhamnoides, по мнению авторов, является самым агрессивным из них.
С 1949 по 1989 г. площадь дюн Северной Ирландии, покрытая облепихой, воз
росла с 1,6 до 13%. К настоящему времени облепиха представляет серьезную
угрозу для флоры и фауны дюн. Показано, что присутствие H. rhamnoides на
песчаных дюнах несколько изменяет уровень карбоната кальция и кислотность
грунта, но не оказывает влияния на содержание органического вещества и
соединения азота. Наблюдаются различия в видовом составе сопутствующей
флоры, мезофауны и сообществах почвенных организмов. Под влиянием об
лепихи в зависимости от степени сукцессии происходит сдвиг в сторону фор
мирования типичных лесных сообществ.
Китайские авторы, напротив, указывают на положительное влияние обле
пихи на обедненные экосистемы лессовых плоскогорий Китая [Ruan Chengjiang,
299
Li Daiqiong, 2002]. Одним из главных ограничивающих факторов развития тех
территорий является серьезная почвенная и водная эрозия. H. rhamnoides являет
ся «пионерным» растением на лессовых плоскогорьях и играет существенную роль
в восстановлении растительности на эродированных участках. Облепиха облада
ет высокой приспособляемостью, быстрым ростом: через 4--5 лет уже формирует
кустарниковые сообщества с устойчивой структурой и высокой продуктивнос
тью. По мнению авторов, вид играет защитную роль против ветра и движения
песков, сохраняет почву и воду, регулирует микроклимат, улучшает биологичес
кое разнообразие и восстанавливает стабильность экосистем плоскогорий. Фи
томасса H. rhamnoides служит хорошим кормом, удобрением и топливом.
Генетические изменения и вариабельность вида. Облепиха -- доволь
но вариабельный вид, имеющий в Европе ряд подвидов, 6 из которых приво
дятся для России и сопредельных государств: H. rhamnoides subsp. carpatica
Rousi, H. rhamnoides subsp. caucasica Rousi, H. rhamnoides subsp. mongolica Rousi,
H. rhamnoides subsp. pamiroalaica Avdeev, H. rhamnoides subsp. salicifolia (D. Don)
Servettaz, H. rhamnoides subsp. turkestanica Rousi [Черепанов, 1995].
В Великобритании экземпляры H. rhamnoides достигают высоты 3 м, а в
Норвегии -- 1 м [Binggeli et al., 2008].
Изменчивость биохимических признаков облепихи изучена в Финляндии
[Tütinen et al., 2006], Пакистане [Sabir et al., 2005] и Польше [Kawecki et al.,
2006]. Исследование внутривидовой изменчивости молекулярно генетичес
кими методами с использованием RAPD анализа проводилась в Китае [Sheng
et al., 2006] и Индии [Panjii Singh et al., 2006]. Популяции облепихи из турец
кой провинции Анталья послужили материалом для комплексного исследо
вания состава жирных кислот в мезокарпе плодов [Ercisli et al., 2008]. Соотно
шение 7 жирных кислот и последующий RAPD анализ позволил выделить в
турецких популяциях облепихи 10 различных генотипов.
Применение RAPD анализа показало также более высокий генетический
полиморфизм российских популяций облепихи по сравнению с китайскими
[Liu et al., 2007].
Природные популяции облепихи, произрастающие в экстремальных усло
виях Гималаев, демонстрируют высокую изменчивость по цвету, форме, разме
рам плодов, листьев, колючек и т.д., что позволило отобрать из них 17 морфоти
пов [Panjii Singh et al., 2006]. Последующий анализ этих морфотипов с использо
ванием RAPD маркеров подтвердил наличие 5 генетических групп, при этом ни
один из морфотипов не показывает сходство >25%, что указывает на высокий
уровень генетического разнообразия естественных популяций. По нашему мне
нию, такой высокий уровень генетического разнообразия помогает виду реализо
вывать адаптационный потенциал в ходе формирования вторичного ареала.
Влияние на здоровье человека. Плоды содержат витамины А (до 11
мг%), В1 (до 0,035 мг%), В2 (до 0,56 мг%), В6 (до 0,79 мг%), С (до 316 мг%), Е
(до 8 мг%), а также жирное масло (до 9% в мякоти плодов и до 12% -- в косточ
ках), которое применяется в медицине. В масле содержится до 350 мг% каро
тина и каротиноидов, до 165 мг% витамина Е и значительное количество ви
тамина F, регулирующего обмен веществ кожи. Облепиховое масло обладает
ранозаживляющими и болеутоляющими свойствами, применяется для лече
ния ожогов, обморожений, экзем, плохо заживающих наружных язв, язвен
ной болезни желудка и двенадцатиперстной кишки, лучевых поражений орга
300
низма, а также в онкологической практике и в качестве профилактического
средства для уменьшения дегенеративных изменений слизистой оболочки
пищевода при лучевой терапии рака [Вехов и др., 1978].
Экономическое и социальное значение (положительное и отрицатель
ное). Плоды облепихи имеют пищевое значение. В их мякоти, помимо жирно
го масла и большого количества витаминов, содержатся сахара (до 6,6%) и
органические кислоты (до 2,5%). Плоды облепихи называют «сибирским ана
насом», едят свежими и варят из них варенье, кисели, желе, пастилу, готовят
соки, вино, настойки и наливки, примешивают к муке для изготовления осо
бого хлеба. Листья облепихи содержат до 10% дубильных веществ, их можно
использовать для дубления и окраски кож. Плодами раньше красили ткани:
молодые побеги с солями железа дают черно бурую, а плоды -- желтую пище
вую краску. Древесина мелкослойная, плотная, красивого желтоватого и жел
товато коричневого цвета, с довольно отчетливой штриховой текстурой, хо
рошо полируется, идет на мелкие поделки. H.rhamnoides разводят одиночны
ми экземплярами и группами, в виде живых стриженых изгородей, используют
для укрепления песков, берегов, откосов, ж/д выемок и каналов и в защитных
полосах от снежных заносов. Сообщества облепихи дают убежище и пищу
многочисленным птицам и мелким грызунам. Облепиха декоративна, может
быть использована в одиночных и групповых посадках садов и парков, но с
этой целью ее разводят относительно редко [Вехов и др., 1978; Петрова, 1987].
Значению H.rhamnoides для лесообразовательного процесса на нарушенных
землях и закономерностям процесса формирования популяций облепихи на
Урале посвящено исследование А.П. Кожевникова [2003].
В научно исследовательских учреждениях России проводится работа по
выведению и районированию столовых, технических и декоративных сортов.
Особое внимание уделяется продуктивности, крупноплодности, легкости от
деления ягод при сборе, высокому содержанию масла, витаминов, приспособ
ляемости к разным условиям выращивания, прочности и долговечности кро
ны, отсутствию колючек, срокам созревания плодов [Петрова, 1987]. Есть и
неординарные тенденции в селекции. Например, созданы карликовые формы
облепихи, для сбора плодов путем скашивания [Ермаков, Фаустов, 1983].
Меры борьбы
В литературе упоминаются, скорее, меры охраны природных популяций H.
rhamnoides. В частности, вид внесен в Красную книгу МНР, Республики Буря
тия, числится в списке редких и исчезающих растений Тывы. На сайте «Крас
ная книга растений Иркутской области» H. rhamnoides рассматривается как вид
с сокращающейся численностью и ареалом. В качестве лимитирующих факто
ров приводятся срубание и срывание ветвей при заготовке плодов, хозяйствен
ное использование территорий, весенние палы [Ляхова, 2008].
СЕМЕЙСТВО EUPHORBIACEAE --- МОЛОЧАЙНЫЕ
МОЛОЧАЙ БУТЕРЛАКОВЫЙ
Название: Euphorbia peplus L.
Синонимы: Euphorbia hyrcana Grossh. Вид не следует путать со сходным по
написанию E. peplis L.
301
Морфологические признаки
Однолетнее растение высотой до 25 см, ветвистое от основания, с тонким
корнем. Листья черешковые, нижние почти округлые, верхние -- обратнояй
цевидные, тупые. Зонтик трехлучевой, цветоносы на конце 2--5 раз вильчато
разветвленные; листочки обертки с черешками длиной ~3 мм, широкообрат
нояйцевидные, листочки оберточек в числе двух, яйцевидные или треуголь
но ромбические (рис.143). Плод приплюснуто шаровидный, длиной ~2 мм и
несколько большей ширины. Семена пепельно серые, шестигранные, с бороз
дками и ямками, на конце с придатком [Губанов и др., 2003].
Естественный ареал
Вид распространен в Европе (кроме северных районов), Северной Африке
и в ряде регионов Западной Азии [Губанов и др., 2003]. В пределах Восточной
Европы: южные регионы европейской части России, Краснодарский край,
Украина [Humangarden, 2008].Вторичный ареал
История интродукции и географическое распространение. За предела
ми естественного ареала вид широко распространился во всех умеренных и
субтропических регионах мира. E.peplus натурализовался в Канаде и обнару
жен в 35 штатах США, включая Гавайи и Аляску [Poindexter, 2008]. Занесен в
Южную Америку: в Чили произрастает в разных условиях увлажнения, встре
чается как в горных центральных областях на высотах 500 2000 м над ур. м,
так и в прибрежных районах от 0 до 500 м над ур. м. [Флора Чили, 2008]. Ши
роко известен в Австралии и Новой Зеландии, куда в начале XIX века завезен
европейскими поселенцами в медицинских целях [Discover Life, 2008].
В России E.peplus спорадически встречается в европейской части, в том
числе по всей территории Средней России, но чаще в черноземной полосе [Гу
банов и др., 2003]. В Ленинградской обл. известен как редкий заносный вид в
садах и огородах, у дорог; собран в Санкт Петербурге и у ст. Можайская [Цве
лев, 2000]. На севере, например в Вологодской обл., не обнаружен.
Пути и способы заноса. Многие авторы считают, что E.peplus заносится в
новые места обитания с семенами газонных трав и цветочно декоративных
культур [Полуянов, 2005; Письмаркина и др., 2006].
Статус в регионе. Произрастает во многих регионах Средней России (рис.
144). Наиболее ранний сбор в Воронежской обл. сделан в Воронежском уезде,
с. Гололобово, 16.07.1917 г., подпись коллектора неразборчива (MW, из герба
рия М.И. Назарова). Начиная с 1970 х гг. находки E.peplus регистрировались
в г. Воронеж неоднократно [Григорьевская, 2000; Григорьевская и др., 2004]. В
Белгородской обл. E.peplus -- редкое растение [Еленевский и др., 2004]. Со
бран в Белгороде: «луговина на пустыре перед жилищным комплексом,
27.09.1997 г., Н.Н. Чаадаева, М.А. Полежаева, А.Г. Еленевский» (OHHI). В
Курске собран в 1999 г. как сорное на цветочной клумбе (MW).
В Орловской обл. E.peplus собран В.И. Радыгиной в г. Орел, сорняк в ого
роде, 1.08.1983 г. (OHHI). В настоящее время в Орле -- обычный сорный вид;
в июле 2008 г. мы наблюдали его в массе на огородах. В юго восточных р нах
Орловской обл. вид редок, встречен единожды у забора элеватора в пос. Вер
ховье (Л.Хорун, А.Сотников, MW). В Липецке впервые отмечен в 1921 г., по
зднее, с 1979 г., последовали регулярные его находки в разных р нах области
[Александрова и др., 1996].
302
Согласно литературным данным, в конце XIX -- начале ХХ века E.peplus
встречался на территории Тульской губернии более или менее регулярно [Ро
зен, 1893]. К 1916 г. вид обнаружен в Туле и Ефремове [Розен, 1916]. В после
дующие 80 лет его не отмечали. После значительного перерыва, имевшего ме
сто не только в Тульской обл., но и в соседних регионах, E.peplus был найден
сразу в нескольких областях Средней России: Московской [А.Шретер, 1985,
MW]; Рязанской [М.Казакова, 1987, MW]; Липецкой [К.Александрова, 1989,
MW] и Тульской [Л.Хорун, 1996, MW]. К настоящему времени в Тульской
обл. вид значительно увеличил численность.
В Московской обл. молочай бутерлаковый известен с начала ботанических
исследований [Stephan, 1792; Martius, 1817; Максимович, 1826; (MW); Дви
губский, 1828]. В конце XIX века и первой половине XX века до 1942 г. его
находили часто, а затем, после значительного перерыва, обнаружили в посел
ке ВИЛАР на юге Москвы в 1985 г. [Игнатов и др., 1990]. Для Владимирской
обл. указан в сборнике В.Я. Цингера [1885]. Гербарные сборы неизвестны.
Для Тверской обл. вид указан еще в конце XVIII века [Pallas, 1784 1788].
Собран М.Л. Невским в 1947 г. в окрестностях д. Красный Бор Калининского
р на. Во второй половине XX -- начале XXI века на территории области вид
ни разу не отмечали [Маркелова, 2004]. Для Ивановской обл. приведен Е.А.
Борисовой [2007] по указаниям нескольких изданий флоры П.Ф. Маевского
[Маевский, 1933, 1940, 1964] как редкий огородный сорняк. В последние годы
не отмечен, возможно, исчез [Борисова, 2007]. В Нижегородской обл. E.peplus
собран В.В. Макаровым: г. Арзамас, сорное вдоль заборов и на газонах по ул.
Ленина, в посадках кустарников у музея Гайдара, 31.08.1986 г. (MHA).
В Калуге E.peplus впервые собран на огороде, 7.08.1882 г., П.Саницкий
(MW). До сих пор встречается только в Калуге по газонам, на ограниченной
территории (С.Р. Майоров, личное сообщение) и в других пунктах области не
отмечен. В Смоленской обл. молочай собрала Н.М. Решетникова в Демидовс
ком р не, в д. Городище, у забора, 28.07.1999 г. (MW). В Рязанской обл. -- сор
ный вид, места обитания -- газоны, огороды. Собран в Рязани в 1987 г. М.В.
Казаковой на газоне у дома [MW; Казакова, 2004]. В Мордовии E.peplus -- ред
кий сорный вид, приводится на основании гербарного сбора 1968 г. из с. Коч
курово [GMU; Письмаркина и др., 2006], позднее собран в г. Ардатов, на клум
бе около здания районной администрации, 19.08.2004 г., Е. Письмаркина
(GMU). В Пензенской обл. -- в г. Пенза, очень редко. Сборы датированы 1993 г.
[Васюков, 2004]. В Тамбовской обл. собран в г. Мичуринск, по ул. Советской,
в частном огороде, в массе, 26.07.2003 г., А. Сухоруков (MW).
Местообитания. Встречается как сорное в населенных пунктах (рис. 145).
Растет на обрабатываемых незадерненных участках, в садах, огородах, пали
садниках, по мусорным кучам и нарушенным местам. Предпочитает ровные,
хорошо освещенные участки с дренированной почвой.
Размножение и жизненный цикл. E.peplus -- однолетник, обладающий
быстрым ростом. Жизненный цикл особи составляет 12--14 недель. Цветет с
мая до осени, плодоносит с июня. Семена очень мелкие (в среднем 1700 семян
в 1 г), их всхожесть составляет не менее 70% [Davis, Parsons, 2002]. Биологию
семян E.peplus изучли в 5 летнем эксперименте. Выявлено, что семена прора
стают преимущественно весной. Большая часть всходов E.peplus появляется в
первый год, в последующие годы число проростков закономерно уменьшает
ся [Roberts, Boddrell, 1983].
Рис. 143. Euphorbia
peplus, общий вид
Рис. 144. Расселение
Euphorbia peplus в
Средней России
Рис. 145. Euphorbia
peplus в трещинах
асфальта
Рис. 146. Внедрение
Euphorbia peplus в
естественные
сообщества
305
Расселение и распространение. В благоприятные годы вид успешно рас
селяется по обработанной почве. Распространение E.peplus в регионах Сред
ней России неравномерно и локально -- процесс расселения подвержен дли
тельным флуктуациям. В XIX -- начале XX века E.peplus отмечен в большин
стве губерний Средней России, в середине XX века находки стали редкими и
спорадическими, но в конце XX века ситуация вновь изменилась: молочай
бутерлаковый стал встречаться местами, но, как правило, в массе на возделан
ной почве. По нашим наблюдениям, в конце 1990 -- начале 2000 х гг. в Туле
E.peplus в массе произрастал на огородах, цветниках, обочинах дорог, улицах
частного сектора (рис. 146). В последние 5 лет, однако, численность вида за
метно уменьшилась: нужно пройти несколько улиц, чтобы увидеть небольшую
популяцию.
Последствия внедрения
Воздействия на естественные фитоценозы и аборигенные виды. E.
peplus растет на участках с обнаженной почвой. Малоустойчив и не выдержи
вает конкуренции с аборигенными растениями, поэтому при задернении уча
стков выпадает.
Генетические изменения и вариабельность вида. 2n = 16 [Цвелев, 2000].
Влияние на здоровье человека. Млечный сок растения токсичен. В тибет
ской медицине применяется для удаления бородавок, папиллом и сухих мозо
лей [Буддизм в Украине, 2008]. В народной австралийской медицине сок
E.peplus традиционно использовали при солнечных ожогах и заболеваниях
кожи, в том числе онкологических [Green, Beardmore, 1988]. Австралийские
ученые подтверждают эффективность лечения некоторых раковых заболева
ний кожи с помощью E.peplus: в 88% случаев его применение помогло пациен
там с базальной и сквамозной клеточной карциномами. Активный компонент
E.peplus запатентован австралийской кампанией Peplin Biotech [Hohmann et
al., 2000]. Разработана методика лабораторного получения и очистки компо
нентов из свежих или высушенных растений. Урожай составляет 1--1,5 кг/мІ
при содержании полезных компонентов 100--300 мг/кг [Davis, Parsons, 2002].
Экономическое и социальное значение (положительное и отрицатель
ное). Сорный вид, может засорять большие участки возделанной почвы, за
нимать огороды и палисадники. Так же, как и у других молочаев, сок E.peplus
содержит ядовитые смолы [Hohmann et al., 2000].
Меры борьбы
Австралийские авторы считают, что E. peplus не является опасным сорняком и
конкурентом полевых культур, так как не распространяется за пределы теневых
участков [Davis, Parsons, 2002]. Как сорняк E. peplus уничтожают прополкой до
созревания семян. Для химического контроля за E. peplus предлагается использо
вать селективный гербицид, активным компонентом которого является оксадиа
зон [Гербицид Chipco Ronstar®C. Инструкция к применению, 2008].
306
СЕМЕЙСТВО FABACEAE -- БОБОВЫЕ
ЛЮПИН МНОГОЛИСТНЫЙ
Название: Lupinus polyphyllus Lindley
Синоним: L.macrophyllus Beniham
Морфологические признаки
Травянистый двулетник или короткоживущий многолетник высотой 50--
150 см. Стебли мощные, прямые, почти голые (слабоопушенные). Листья оче
редные, пальчатосложные, с длинным черешком и 10--15 узко ланцетными
листочками шириной 1--2 см. Стебли и черешки с довольно редкими, корот
кими волосками. Прилистники шиловидные, обычно на ѕ сросшиеся с череш
ком, явственно опушенные. Цветоносы, чашечка и молодые листочки имеют
серебристое опушение. Соцветие терминальное длинное, обычно рыхлое, с
многочисленными цветками. Венчик в 3 раза длиннее чашечки, разной окрас
ки, чаще всего фиолетовый, но встречаются формы с синими, голубоватыми,
розовыми и белыми венчиками (рис. 147). Лодочка голая. Цветоножки длин
ные, почти равные длине цветков. Губы чашечки цельные или слабозубчатые.
Прицветник рано опадающий, удлиненно ланцетовидный, не длиннее чашеч
ки, опушенный. Бобы многочисленные, плоские, удлиненные, в зрелом состо
янии черные, густо опушенные седыми прижатыми волосками. Семена более
или менее овальные, слабосдавленные, чаще темно коричневые, почти черные
[Палибин, 1945; Майсурян, Атабекова, 1974].
L.polyphyllus отличается от садового гибрида (или гибридного комплекса)
L. × regalis (L.аrboreus × L.polyphyllus) слабоветвистым стеблем и менее поли
морфной окраской венчика [Clement, Foster,1994; http://nlbif.eti.uva.nl/bis/
flora.php?menuentry=soorten&id=2993].
Естественный ареал
Западные части Северной Америки с океаническим климатом: Канада (про
винция Британская Колумбия), США (штат Аляска, запад штата Орегон, за
пад штата Вашингтон, северная часть штата Калифорния).
Вторичный ареал
История интродукции и географическое распространение. L.polyphyllus
интродуцирован в восточных частях Северной Америки и является культиви
руемым и натурализовавшимся видом во многих странах Европы. Во «Flora
Europaea» [Franco, da Silva, 1968] вид отмечен в Дании, Финляндии, Герма
нии, Норвегии, Польше, центральной части Eвропейской России и в Швеции,
однако не приводится для Великобритании, хотя в действительности уже >100
лет произрастает и в Англии, и в Шотландии [Clement, Foster,1994]. В настоя
щее время в список стран, в которых наблюдается инвазия люпина, следует
добавить Эстонию, Латвию и Литву (и Беларусь? Козловская).
L.polyphyllus привезен в Англию в 1826 г. известным шотландским «охот
ником за растениями» Дэвидом Дугласом и вскоре был интродуцирован в
Центральную Европу как садовое растение [Krausch, 2003]. Уже в 1840 х гг.
питомники и ботанические сады предлагали разнообразные цветовые формы
вида. К 1900 г. культивирование и коммерческое распространение вида воз
росло, и в то же время он одичал в Великобритании [Preston et al., 2002]. Де
Рис. 147. Соцветия
Lupinus polyphyllus
Рис. 148. Расселение
Lupinus polyphyllus в
Средней России
Рис. 149. Плоды Lupinus polyphyllus
Рис. 150. Внедрение Lupinus polyphyllus в луговые сообщества
309
коративный гибрид Lupinus × regalis был представлен на Европейском рынке
в 1935 г.
В Швеции L.polyphyllus впервые отмечен в списке «беглецов» из сада в 1870 г.
[Hylander, 1971]. В Норвегии вид упомянут в списке растений в 1831 г., но
зарегистрирован как «беглец из культуры» только в 1940 е гг. [Nordhagen,1940;
Elven, Fremstad, 2000]. В Финляндии люпин впервые зарегистрирован как
«беглец из культуры» в 1895 г. В Германии первые дичающие заросли наблю
дались в Баварии в 1890 г. [Hegi, l964].
L.polyphyllus интродуцирован в Эстонии, в Ботаническом саду Тарту в 1807 г.
В Литве люпин стали культивировать в начале XX века, а в 1931 г. он впервые
зарегистрирован как «беженец из ботанического сада» в окрестностях Кауна
са [Gudzinskas, 1999, 2005]. В Латвии первая регистрация L. polyphyllus как
натурализовавшегося растения сделана в 1921 г. В Польше люпин интродуци
рован около 1877 г. и стал инвазионным видом с 1950 г. [Tokarska Guzik, 2003].
В Великобритании L.polyphyllus в настоящее время растет в садах реже, чем
ранее, и заменен L. × regalis. «Беглецы из культуры» обычно регистрируются
как L. polyphyllus, но наиболее вероятно, что это L. × regalis, кроме Шотландии,
где L. polyphyllus, по видимому, более распространен [Stace, 1997]. Неясно,
показывает ли карта ареала [Preston et al., 2002] распространение в Великоб
ритании и Ирландии только L.polyphyllus, или в нее включен и L. × regalis.
В Норвегии культивирование L. × regalis в садах увеличивается, но
L.polyphyllus в садах все еще обычен, и «беглецом из сада» является именно
L.polyphyllus. L. × regalis, по видимому, менее вынослив и пока не прижился
вне садов [Lid, Lid, 2005]. L.polyphyllus обычен на юге Норвегии, более редко
встречается на севере, но зарегистрирован во всех округах (то есть до 70° с.
ш.); это прочно обосновавшийся и обычный чужеземный вид в стране. В Шве
ции L.polyphyllus широко распространен вплоть до северных берегов Ботни
ческого залива [Mossberg, Stenberg, 2004]. В Финляндии L.polyphyllus -- обыч
ный прочно обосновавшийся чужеземный вид на юге страны, но изредка встре
чается и в Аландском архипелаге, и в регионах севернее Северного полярного
круга [Hamet Ahti et al., 1998].
В Эстонии L.polyphyllus встречается локально, является довольно обычным
на северо востоке страны и очень редким или вовсе отсутствующим на запад
ных островах [Kukk, Kull, 2005]. В Эстонии нет никаких проблем с «бегством
из садов». Имеет место «бегство с культурных полей» на обочины дорог и в от
крытые сосновые леса, где растения оказывают неблагоприятное влияние на
аборигенные виды. Некоторые из эстонских растений, возможно, распростра
нились из Латвии, где вид найден по всей территории [Kuusk et al., 1996]. В
Литве является агрессивным сорным растением, занимает большие площади и
включен в список видов Литвы, подлежщих уничтожению [Gudzinskas, 2005].
В Польше L.polyphyllus найден по всей стране, более чем в 1000 местооби
таний [Tokarska Guzik, 2003; Solarz et al., 2005]. В Германии растение прочно
обосновалось, довольно обычно и является одним из 15 наиболее широко рас
пространенных чужеземных растений на кислых и слабокислых почвах и ред
ким только на известковых почвах и в районах с низким уровнем летних осад
ков. В Рейкьявике (Исландия) L.polyphyllus найден в нескольких пунктах как
реликт оставленных садов или в рудеральных местах; одна его находка сдела
на на обочине дороги в Северной Исландии, где вид произрастал совместно с
L.nootkatensis.
310
Пути и способы заноса. Во всех северных европейских странах L.polyphyllus
был изначально преднамеренно интродуцирован, прежде всего как садовое рас
тение. Позднее культивировался также для других целей, особенно для улуч
шения и закрепления почв и как фураж для домашних и диких животных.
Этот вид «сбегает» из с/х культуры, что является основной причиной его
широкого распространения в странах Европы, гораздо более важной, чем ди
чание из садовой культуры. Даже если прекратить использование L. polyphyllus
в качестве растения, закрепляющего почву, случаи его дичания, время от вре
мени, вероятно, неизбежны.
L.polyphyllus растет в симбиозе с азотфиксирующей бактерией Bradyrhizobium
sp., которая вызывает рост корневых клубеньков и фиксирует молекулярный
азот из атмосферы. Почвы, где растет L polyphyllus, обогащены азотом, который
могут использовать также другие растения. Способность увеличивать изоби
лие почвы -- главная причина разнообразного и широкого использования рас
тения. В Германии основным путем его интродукции было использование в ка
честве вида, улучшающего почву и препятствующего ее засолению, особенно
для бедных, кислотных почв в горных регионах. Этот вид и до сих пор выса
живается для укрепления открытых почв после строительных дорожных ра
бот или после вырубки деревьев. L.polyphyllus также высаживается как «зеле
ное удобрение» (промежуточная культура) на культурных полях и как фураж.
Этот вид использовался в лесозащитных полосах в Литве [Gudzinskas,1999].
Статус в регионе. Гербарный сбор L.polyphyllus из культуры относится к
началу ХХ века -- «Москва, Петровское Разумовское. Ботанический сад.
02.07.1914 г. А.Соловьева. MOSP», но натурализация вида началась позднее.
Да и «убежал» люпин не из садов, а из сельскохозяйственных угодий (рис.
148). Впервые отмечен в качестве натурализующегося растения в Ярославс
кой обл. в 1921 г. С 1952 г. стал дичать в Московской обл. В 1957 г. собран в
Калужской обл., «близ д.Заборово. В поле по краю посевов ржи. 20.06.1957.
М.Камурова. MOSP». Первые сборы во Владимирской обл. сделаны в 1969 г.,
в Тверской -- в 1980 г. В Калужской обл. впервые отмечена инвазионная попу
ляция с полиморфной окраской цветков «близ с. Митин Двор в 6 км к Ю от
Перемышля. Сырой лужок. Обильно возобновляется. 2 экз. с белыми цветка
ми, 1 с пестрыми бело сиреневыми. 02.07.1974. В.Макаров, М.Баженова.
MHA». К 1976 г. относится первая находка из Брянской обл. «в 3 4 км южнее
г.Стародуба, по обочине шоссе. Встречен 1 экз. на песчаных местах. 24.07.1976.
Ю.Алексеев, В.Макаров. MHA».
В настоящее время на заброшенных угодьях в Смоленской и Калужской
обл. сформировались обширные инвазионные популяции L.polyphyllus, пред
ставляющие собой одновидовые заросли, уходящие за горизонт. Активно рас
селяется в Рязанской, Московской, Тверской, Владимирской и Ивановской
областях.
Местообитания. В естественном ареале L.polyphyllus растет на берегах рек,
на лугах и обочинах дорог и в других нарушенных местообитаниях; вид явля
ется сорняком даже в естественном ареале [Scoggan, 1978]. Природные место
обитания характеризуются как «теневые, умеренно сухие с почвами от песча
ных до суглинистых».
Сходные экологические условия люпин занимает и во вторичном ареале:
он широко натурализовался по рекам, автомобильным дорогам и ж/д путям;
311
рудеральным местам [Franco, da Silva, 1968; Stace, 1997; Fremstad, Elven 2004].
В Польше растет в лесах, на опушках леса в ассоциации Epilobietea, а в Скан
динавии -- на бедных питательными веществами почвах: обочинах автомобиль
ных дорог и ж/д насыпях, карьерах для добычи гравия, рудеральных местах
[Mossberg, Stenberg, 2004]. Вид нередок в Норвегии, где растет в зарослях вы
сокой травы по обочинам дорог. В Германии вид проникает в полуестествен
ные ценозы -- поля с бедными кислыми почвами и высокотравье [Otte et
al.,2002; Otte, Maul, 2005].
В Балтии L.polyphyllus растет в сосновых лесах на песчаном подзоле, на су
хих лугах с кислой почвой, обрабатываемых полях и захватывает разнообраз
ные рудеральные местообитания: обочины полей и дорог, откосы железнодо
рожных путей, карьеры, канавы и т.д., а также в антропогенно нарушенные
леса. В Латвии L. polyphyllus часто культивируется и «сбегает» в леса и на обо
чины дорог и в другие заброшенные места [Kuusk et al., 1996].
Размножение и жизненный цикл. L.polyphyllus цветет в мае -- июне (в бо
лее северных регионах -- в июле). В Литве среднее число цветков в соцветии --
120, а максимальное -- 200 [Gudzinskas, 2005]. Семена вызревают в конце лета
или в начале осени. Сильно опушенные бобы содержат 4--10(12) семян, кото
рые распространяются на близкое расстояние от материнского растения при
растрескивании бобов (рис. 149). Lupinus polyphyllus -- гемикриптофит, но не
редко может распространяться и посредством подземных корневищ. Растения
могут быть размножены делением или выращены из семян. Долговечность
семян может достигать >50 лет [Otte et al., 2002].
Изучена семенная продуктивность L. polyphyllus на 5 пробных площадках в
Смоленской и Московской обл. [Ткачева, Виноградова, 2009]. В Сафоновс
ком р не Смоленской обл. на 1 генеративном побеге формируется от 25 до 61
плода (в среднем 40,4±11,6). В каждом плоде содержится от 1 до 10 зрелых
семян (в среднем 4,6±1,9). Таким образом, на 1 генеративном побеге форми
руется 185,6±59,5 зрелых семян (амплитуда изменчивости признака от 109 до
290 семян). Индивидуальная изменчивость растений по числу семян в плоде
высокая и очень высокая, коэффициент вариации составляет до 43%. В Егорь
евском р не Московской обл. на 1 генеративном побеге формируется от 31 до
88 плодов (в среднем 49,6±18,8). В каждом плоде содержится от 1 до 9 зрелых
семян (в среднем 4,4±0,9). Генеративный побег несет в среднем 224,4±105,3
зрелых семян (амплитуда изменчивости признака от 110 до 477 семян). Ин
дивидуальная изменчивость растений по числу семян в плоде высокая и очень
высокая, коэффициент вариации составляет до 46%.
Расселение и распространение. L.polyphyllus распространяется, главным
образом, семенами. Помимо культивирования, семена могут переноситься
транспортными средствами или при транспортировке почвы. В тех местах, где
L. polyphyllus встречается во множестве (например, на юге Норвегии, Elven,
Fremstad, 2000] этот вид высеян преднамеренно для стабилизации почвенно
го субстрата.
Последствия внедрения
Воздействие на естественные фитоценозы и аборигенные виды. Отме
чен в широком диапазоне местообитаний, имеющих, однако, общие черты: до
статочно легкие, бедные питательными веществами почвы и ограниченную
конкуренцию с другими видами. Влияние на аборигенные виды наиболее оче
312
видно в случае образования обширных, довольно плотных зарослей, которые
подавляют естественную растительность (рис. 150). L.polyphyllus может кон
курировать с местными видами, встречающимися на обочинах дорог, в руде
ральных местах, каменистых берегах рек и других открытых местообитаниях.
Хотя большинство видов, растущих в подобных местообитаниях, не вхо
дит в группу охраняемых, некоторые редкие или находящиеся под угрозой
исчезновения (по крайней мере, в местном масштабе) неконкурентоспособ
ные аборигенные виды могут быть обнаружены даже и здесь. Благодаря азот
фиксирующим бактериям L.polyphyllus изменяет химию почвы в пользу ви
дов, требовательных к азоту. Таким образом, L.polyphyllus изменяет плодоро
дие почвы и в конечном счете естественные фитоценозы. Эвтрофикация
бедных питательными веществами участков и последующие изменения в
структуре сообществ и биоразнообразии -- главная проблема вторжения
L.polyphyllus в тот или иной регион.
В нескольких странах L.polyphyllus вообще не рассматривается как пробле
ма, хотя произрастать может массово. В Польше вид, по видимому, не причи
няет серьезных проблем, хотя недооценка его влияния на лесные травы [Solarz
et al., 2005], место имеет. Но проблема с L.polyphyllus может стать очевидной
при изменении режима землепользования, которое позволит виду продуци
ровать и распространять зрелые семена. В Литве L.polyphyllus быстро распро
страняется и угрожает биологическому разнообразию лугов и песков
[Gudzinskas, 1999]. В течение последних десятилетий L.polyphyllus вторгся в
состоящие из гравия речные берега (аллювиальные участки) на нескольких
из самых больших рек Центральной Норвегии.
В окрестностях Хельсинки, по нашим наблюдениям, формирует крупные
одновидовые заросли плотностью 23 генеративных побега/м2 с практически
полным отсутствием сопутствующих аборигенных растений.
Генетические изменения и вариабельность вида. В Восточной Европе не
имеется естественных видов этого рода. Следовательно, нет никакой опаснос
ти гибридизации L.polyphyllus с природным видом. В Шотландии [Stace, 1997],
Норвегии [Lid, Lid,2005], Швеции [Karlsson, 1981] и Исландии отмечен гиб
рид L. nootkatensis × L. polyphyllus.
Влияние на здоровье человека. Нет никаких данных об опасностях для
здоровья человека, связанных с L.polyphyllus, обработка растений вручную так
же не вредна. Растение и его семена содержат алкалоиды, потребление кото
рых может быть вредно для овец и крупного рогатого скота. Содержание алка
лоидов может варьировать от 3,5% в семенах до 2% в вегетативных органах.
Основной алкалоид -- люпанин, C15H24N2O. Выведены линии с низким содер
жанием алкалоидов, культивируемые для получения низкоалкалоидных се
мян, которые используются для выращивания фуража; безалкалоидные семе
на также могут использоваться человеком [Santen, Hill, 2004].
Экономическое и социальное (положительное и отрицательное) зна
чение. L.polyphyllus -- важное сельскохозяйственное растение с широким раз
нообразием культиваров, различающихся по химическим признакам (содер
жание аминокислот, белков, масел и алкалоидов и т.д.), темпам роста и ис
пользованию. Сено, собранное с полей, на которых рос L.polyphyllus, может
быть менее ценным из за содержания в растении алкалоидов.
313
Меры борьбы
Едва ли реалистично запретить культивирование L. polyphyllus как декора
тивного растения, однако этот источник распространения, по всей видимости,
менее проблематичен. Самые большие заросли вида возникают из за массово
го его высева дорожными организациями и их субподрядчиками, а также сель
скохозяйственными предприятиями. В Норвегии для защиты аборигенных
видов запрещен высев L.polyphyllus по обочинам дорог. Немецкое федераль
ное агентство по охране природы рекомендует не высевать L. polyphyllus для
закрепления и улучшения почвы возле местообитаний, представляющих ин
терес для сохранения биоразнообразия.
Во многих местообитаниях уничтожение L.polyphyllus не является необхо
димостью. Отдельные экземпляры могут легко быть удалены при прополке. В
тех местообитаниях, в которых L.polyphyllus произрастает массово и угрожает
естественной растительности и где ставится цель полного уничтожения вида,
лучшие и самые быстрые результаты дает применение раундапа, однако гер
бицид нельзя применять в массовом масштабе ввиду его влияния и на есте
ственные ценозы.
Пастьба обширных стад овец также дает довольно хорошие результаты при
условии пастьбы скота в течение 2 лет. Правильное кошение или пастьба
(в зависимости от того, является угодье пастбищем или лугом) постепенно
уменьшает число популяций и отдельных особей. Рекомендуется сочетать ко
шение с пастьбой. Кошение должно проводиться дважды в год -- перед цвете
нием и 2 месяцами позднее, в течение 3--5 лет. В дальнейшем кошение может
быть сокращено до 1 раза в год перед цветением или, по крайней мере, перед
созреваниям семян. В этом случае на некоторых участках конкуренция
L.polyphyllus с аборигенными видами снизится, однако, по всей вероятности,
полностью вид не исчезнет [Otte et al., 2002].
Власти, отвечающие за охрану природы, должны сообщать о потенциаль
ном отрицательном влиянии вида тем учреждениям (сельское хозяйство, стро
ительство дорог), которые в массовом масштабе используют и распространя
ют L.polyphyllus.
СЕМЕЙСТВО HYDROCHARITACEAE --- ВОДОКРАСОВЫЕ
ЭЛОДЕЯ КАНАДСКАЯ
Название: Elodea canadensis Michx.
Синонимы: Anacharis canadensis (Michx.) Planch.; Udora canadensis (Michx.) Nutt.
Морфологические признаки
Многолетнее водное растение, ветвящиеся побеги которого могут дости
гать в длину 30--100 см. Стебли ломкие, у основания укореняющиеся, обычно
несут еще и длинные плавающие в воде придаточные корни. Листья в мутов
ках по 3--4(6), продолговатые или линейно ланцетные, коротко заостренные,
с очень мелкими зубчиками по краю; в пазухах листьев имеются 2 цельнок
райные чешуйки. Растения двудомные; цветки пазушные, одиночные, на длин
ных цветоносах, выходящих из двураздельного пленчатого яйцевидного или
линейного покрывала; концы покрывала заостренные. Чашечка с удлиненной
314
нитевидной трубкой и маленьким трехраздельным отгибом, красноватых ча
шелистиков 3; лепестков также 3, они белые, округлые. Женские цветки несут
пестик с 3 двунадрезанными пурпурными рыльцами; мужские --- 9 тычинок.
Пыльники во время цветения взрывообразно вскрываются и высыпают не сма
чиваемую водой пыльцу. Пыльца, плавая на поверхности воды, вступает в кон
такт с рыльцами, обеспечивая опыление. В континентальную Евразию занесе
ны только женские экземпляры, которые здесь и распространились (рис. 151).
Хромосомные числа 2n = 24, 48 [Федченко, 1934; Цвелев, 1982].
Система рода Elodea весьма запутанна. Сент Джон описывал 17 видов, Д. и
Г. Кореллы для территории США приводили всего 4 вида (Elodea bifoliata St
John, E. longivaginata St. John, E. canadensis и E. nuttallii (Planch.) H. St. John)
[Corell, Corell 1975], а Кук и Урми Кёниг [Cook, Urmi Kцnig 1985] --- 5 видов:
(E. canadensis, E. potamogeton (Bert) Espinosa, E. bifoliata, E. nuttallii и E.
callitrichoides Casp.), признавая при этом наличие гибридов между E. сanadensis
и E. nuttallii. Они же создали ключи для диагностики видов в вегетативном и
генеративном состоянии, один из которых мы приводим в упрощенной форме
[цит. по: Bowmer et al., 1995]:
1. Большинство листьев шириной 1,75 мм или более ................................... 2
--- Большинство листьев шириной менее 1,75 мм ........................................... 4
2. По направлению к верхушке стебля листья обычно налегают друг на
друга, образуя ряды, вытянутые вдоль стебля. Листья продолговатые или
овальные ...................................................................................................... E. canadensis
--- По направлению к верхушке стебля л. обычно не налегают друг на
друга, располагаясь неправильно и рассеянно. Листья обычно ланцетные
или от эллиптических до линейных ......................................................................... 3
3. Листья, как правило, в мутовках по 4, от ланцетных до эллиптичес
ких, верхушка листа тупая или заостренная ............................. E. potamogeton
--- Листья, как правило, супротивные, линейные, верхушка листа ост
рая ... ................................................................................................................... E. bifoliata
4(1). Листья обычно вдоль сложенные, с волнистым краем, не более 10 мм
длины; их верхушки несколько завернуты к основанию .................... E. nuttallii
--- Л. обычно плоские, с ровными краями и более длинные ... E. callitrichoides
Естественный ареал
Элодея канадская --- североамериканское плюризональное водное растение,
характерное для большинства умеренных районов США и Канады [Bowmer
et al., 1995].
Вторичный ареал
История интродукции и географическое распространение. Элодея ка
надская --- одно из немногих растений, распространение которого за предела
ми естественного ареала, в частности, в Северной Евразии, прослежено отно
сительно хорошо. Этому способствовали общая немногочисленность видов
собственно водных растений, хорошая узнаваемость растения в природе и
обычно «взрывной» характер заселения им водоемов.
Впервые достоверно элодея была занесена в Европу в 1836 г., позднее по
явилась в Азии, Африке и Австралии [Игнатов и др., 1990]. Натурализовалась
в Европе, Северной и Южной Африке, Северной и Восточной Азии, на юго
востоке и севере Австралии, в Новой Зеландии, Мексике, Центральной и
Южной Америке, на Гавайских островах [Germplasm..., 2008].
315
Рис. 151. Цветущая Elodea canadensis
Рис. 152. Расселение Elodea canadensis в Средней России
Рис. 153. Внедрение Elodea canadensis в естественные сообщества
Рис. 154. Побеги Elodea canadensis
317
Интерес к распространению E. сanadensis в пределах Российской империи
появился уже в конце XIX --- начале XX века. [Артари, 1886; Стемпинский,
1909; Вернер, 1910]. В Польше, в бассейнах Вислы и Одера, вид замечен в 1884 г.,
в этом же году элодея канадская отмечена и в бассейне р. Неман. В Прибалти
ке элодея появилась немногим ранее: в 1882 г. она завезена в Гельсингфорс
(Хельсинки), а затем распространилась по всей приморской Финляндии [Стем
пинский, 1909]. В бассейне Невы вид впервые обнаружен в 1881--1882 гг., и за
последующие 10 лет широко распространился по всей Петербургской губер
нии. Вероятно, в это же время произошло вселение и натурализация элодеи и
в Невской губе [Жакова, 2008].
К 1950 м гг. элодея канадская распространилась практически по всей Вос
точной Европе [Дексбах, 1951], а к середине 1970 х гг. встречалась на ее севе
ре в бассейне Северной Двины и к западу от нее [Флора северо востока..., 1974].
В бассейне р. Волга вид впервые обнаружен в 1885 г. в Московской губер
нии, в бассейне р. Днепр зафиксирован в 1894 г. в районе Херсона [Стемпинс
кий, 1909], откуда через непродолжительное время проник в Одессу [Шесте
риков, 1912].
На Урал вид занесен в 1889 г. и в течение нескольких десятилетий рассе
лился на обширном участке бассейна Оби в пределах современных Свердлов
ской, Челябинской, Курганской и Тюменской обл. [Свириденко, 1986].
В частности, в пределах современной Тюменской обл. первые сведения о
появлении элодеи датируются началом XX века, когда она была обнаружена в
протоках р. Пышма. В настоящее время E. сanadensis входит в состав расти
тельного покрова 41% подтаежных и 75% пресноводных озер лесостепной ча
сти этого региона [Бабушкин, 2003, цит. по: Токарь, 2004]. В настоящее время
здесь она распространилась примерно до 60° с. ш. [Доронькин, 2003].
В Восточной Сибири элодея канадская впервые отмечена на р. Енисей у
Красноярска, а также в Иркутском вдхр. [Паутова, Галимулин, 1980]. В Бай
кале вид впервые отмечен в 1980 г. на Селенгинском мелководье [Паутова,
Галимулин, 1980].
В Северном Казахстане появление вида зафиксировано относительно по
здно, в 1982--1985 гг. [Свириденко, 1986], что, вероятно, связано с тяготением
E. canadensis к водоемам лесной полосы и избеганием засоленных экотопов.
Вероятно, именно по этой же причине лишь в последние годы вид проник
на Северный Кавказ в Адыгею [Зернов, 2006], а в Закавказье элодею до сих
пор никто не видел (А.С. Зернов, личное сообщение).
Из за особенностей экономик КНР и КНДР в первую страну элодея канадс
кая проникла совсем недавно, где была, в частности, зарегистрирована близ Уха
ня [Qio et al., 1997] при туманном прогнозе дальнейшего распространения. На
российском Дальнем Востоке до сих пор не отмечена [Флора российского..., 2006].
В середине XX века в водоемы Европы и Японии вторглась E. nuttalii, мес
тами заместив E. сanadensis [Cook, Urmi Kцnig, 1985]. Распространение E.
callitrichoides, имеющей более короткий жизненный цикл, ограничено в основ
ном приокеаническими районами: впервые этот вид обнаружен за пределами
естественного ареала в 1940 х гг. Есть данные о его натурализации в Великоб
ритании [Sheppard et al., 2006].
Пути и способы заноса. Первоначальными агентами заноса вида, вероят
но, являются аквариумисты и ботанические сады. Впоследствии распростра
318
нение элодеи канадской было связано преимущественно с водным транспор
том [Дексбах, 1951; Свириденко, 1986]: попадание обрывков стеблей в кожу
хи колесных пароходов и щели деревянных барж, перевозки речного песка и
щебня. На локальном уровне из водоема в водоем диаспоры элодеи могут пе
реноситься на сетевых орудиях лова, лапах водоплавающих и околоводных
птиц, конечностях крупных зверей.
Статус в регионе. В Средней России, в бассейне Волги, вид впервые об
наружен А.Н. Петунниковым в 1885 г. (рис. 152): на р. Оке у Голутвинского
ж/д моста в Коломне [MOSP; Артари, 1886; Петунников, 1886], а также близ
с. Коробчеево (MW).
На территории современной Белгородской обл. элодея обнаружена отно
сительно поздно: не позднее 1940 г. [Маевский, 1940]. В конце 1970 х гг. Н.С.
Камышев оценивал здесь элодею как редкое растение, имеющее встречаемость
от 20 до 40% по оригинальной, разработанной им, шкале [Камышев, 1978]. В
настоящее время на северо западе области растение является довольно обыч
ным, на юго западе и в центральном р нах становится нередким, а на юго вос
токе и юге встречается редко (Н.Н. Чаадаева, личное сообщение).
В Воронежской обл. E. сanadensis впервые отмечена в р. Воронеж в 1910 г.
[Камышев, Хмелев, 1976]. В настоящее время отмечена в р. Дон и во всех его
притоках [Григорьевская и др., 2004], однако на восток области вид попал от
носительно недавно, и встречаемость его здесь невелика (Н.Ю. Хлызова, лич
ное сообщение).
В Саратовской обл. элодея канадская зарегистрирована до 1940 г. [Маевский,
1940]. В настоящее время встречается по всей территории изредка [Еленевский и
др., 2008], явно тяготея к долинам крупных рек [Флора Нижнего..., 2006].
В Курской обл. E. canadensis также зарегистрирована не позднее 1940 г.
[Маевский, 1940]. На современной территории региона распространение вида
оценивается А. В. Полуяновым [2005] как частое и равномерное, хотя Н.Ю.
Хлызова (личное сообщение), обоснованно считает, что в бассейне р. Сосна
элодея встречается заметно реже.
В Орловском регионе элодея появилась не ранее середины 1920 х гг., по
скольку она отсутствует в сборах В.Н. Хитрово, но и не позднее начала 1930 х гг.
[Маевский, 1933]. В настоящее время здесь наблюдается уменьшение встречае
мости этого вида с северо запада на юго восток. Если в пределах Днепровско
Деснянского Полесья она встречается часто, то в бассейне Верхней Оки --- неред
ко, а в бассейне р. Сосна -- нечасто.
В Липецкой обл. вид распространен по всей территории региона, но чаще --
в водоемах бассейна р. Воронеж, а также в крупных реках на западе области:
Дон, Сосна, Красивая Меча. Если в центре и на востоке области вид довольно
обычен [Флора Липецкой..., 1996], то к западу от Дона элодея встречается
нечасто (Н.Ю. Хлызова, личное сообщение).
В Тамбовский регион элодея канадская, вероятно, проникла не позднее на
чала ХХ века, когда одноименная губерния выходила на Оку. Вероятно, имен
но к приокской части губернии относится указание на произрастание здесь
элодеи в 4 м издании «Флоры...» П.Ф. Маевского [1912]. Однако в начале 1920 х
гг. близ Тамбова экспедицией П.А. Смирнова этот вид не регистрировался, а
после классических исследований Д.И. Литвинова [1886--1888] систематичес
ких флористических работ на современной территории области не велось
319
[Флора Средней России..., 1998]. В настоящее время в бассейне рек Цна и Во
ронеж вид относительно обычен, тогда как в бассейне р. Ворона встречается
заметно реже (А.П. Сухоруков и Н.Ю. Хлызова, личные сообщения).
В.И. Матвеев [1983] считает, что в Пензенской обл. элодея канадская по
явилась после 1903 г. К сожалению, специально водная флора данного регио
на не изучалась, а в общефлористической работе В.М. Васюкова [2004] рас
пространение этого растения оценивается как «нередко, местами довольно
часто и в массе».
Точное время проникновения E. canadensis в Самарский регион неизвест
но, но произошло это не позднее начала 1930 х гг. [Маевский, 1933]. Совре
менные исследователи флоры области оценивают данное растение как весьма
обычное [Матвеев, 1969; Плаксина, 2001],
Во флору Брянской обл. элодея канадская внедрилась не позже 1939 г. [Ма
евский, 1940], а, вероятнее всего, значительно раньше. В настоящее время
встречается здесь часто и по всей территории [Булохов, Величкин, 1998].
Калужский регион --- одно из первых мест появления элодеи в Средней
России. Не учитывая находку вида на Протве, на границе с Московской гу
бернией, в 1886 г., в том же году она собрана И.Н. Горожанкиным на той же
реке близ д. Кислино Тарусского уезда (MW). В связи с этим не исключено,
что первым среднерусским регионом, где появилась элодея, была именно Ка
лужская губерния. В пользу этой гипотезы говорит то, что выше устья Про
твы данный вид был обнаружен почти на 10 лет позже, близ Калуги [Литви
нов, 1895]. Против данного предположения работает то, что в те годы бассейн
Протвы представлял собой оторванное от путей сообщения --- железных до
рог и водных путей (в отличие от Серпухова и Коломны) захолустье, куда за
нос данного вида был значительно менее вероятен. В настоящее время E.
canadensis в Калужской обл. является одним из самых массовых сосудистых
водных растений, встречаясь здесь практически повсеместно и в большом ко
личестве (А.А. Шмытов, личное сообщение).
Хотя один из ведущих исследователей флоры Тульской губернии, В.В. Ро
зен [1916] на основании сбора из бывшего Каширского уезда считает, что эло
дея канадская появилась в этом регионе в середине 1900 х гг., он отодвигает
это событие по меньшей мере на 20 лет, поскольку на границе Тульской, Ка
лужской и Московской губерний она была обнаружена в 1886 г., а близ Ко
ломны -- годом ранее. Несомненно, в середине 1880 х гг., это растение должно
было встречаться в Оке, по которой тогда проходила граница между Москов
ской и Тульской губерниями. Однако В. Голицын [1905--1907] не приводит
этот вид для восточной части Епифанского уезда Тульской губернии. Вид не
был обнаружен и на сопредельных участках Рязанской губернии в конце 1920 х
гг. А. Ипатовой. А.И. Алюшин [1982] отмечал E. canadensis в северной части
области -- в Суворовском, Ясногорском и Алексинском р нах, везде обильно.
Сейчас вид к северу от Тульских засек и по Оке встречается очень часто, но к
юго востоку области его встречаемость заметно уменьшается: в верховьях ок
ских притоков элодея встречается нечасто, а в бассейнах рек Дон и Красивая
Меча -- изредка [Щербаков, 1999].
Первый гербарный сбор элодеи в Рязанской обл. сделан в 1963 г. в пос. Гусь
Железный Касимовского р на (RSU), но растение появилось здесь значительно
раньше. Дело в том, что в XIX веке правый берег Оки против Коломны и оба
320
ее берега чуть ниже Коробчеева относились к Рязанской губернии. Учитывая
интенсивность судоходства по Оке и активное рыболовство, можно ожидать,
что в пределы современной Рязанской обл. вид проник не позднее середины
1890 х гг. Отсутствие же документальных подтверждений этому связано с тем,
что после экспедиции В. Мешаева ботанические работы здесь были возобнов
лены только в середине ХХ века. На эту же версию работают и находки элодеи
ниже по Оке: близ Мурома и Велетьмы [Флёров, 1902]. В настоящее время
элодея здесь встречается часто и равномерно по всей территории [Казакова,
2004], кроме небольшого участка на крайнем юго западе области, где встреча
емость вида заметно ниже.
В Республике Мордовия первая достоверная находка вида датирована 1961 г.
(GMU), хотя, несомненно, учитывая судоходство по р. Мокша, вид должен
был проникнуть сюда значительно раньше. В настоящее время растение встре
чается здесь часто и повсеместно, местами образуя заросли (Е.В. Варгот, лич
ное сообщение).
Время инвазии элодеи канадской в Ульяновскую обл. не установлено, но это
произошло не позднее начала 1930 х гг. [Маевский, 1933]. Сейчас по оценкам
местных ботаников, элодея встречается по всей области часто, местами образуя
сплошные подводные заросли [Определитель растений Среднего..., 1984].
По данным Я.Я. Алексеева [1961], на Верхнем Днепре, то есть на современ
ной территории Смоленской обл., E. canadensis появилась в 1916--1917 гг. В
настоящее время отмечена на всей территории области [Решетникова, 2004]..
В 1885 г. в Московской губернии, на Оке близ ж/д моста в Коломне элодею
канадскую в массе обнаружил А.Н. Петунников [1886]. К началу ХХ века это
растение стало в губернии практически повсеместным [Сырейщиков, 1906]. В
настоящее время вид равномерно распространен по всей территории области,
обнаружен во всех типах водоемов, причем по частоте встречаемости лидиру
ет среди сосудистых водных растений и уступает только Lemna minor L. и
Spirodela polyrhisa (L.) Sсhleid. [Щербаков, 1990]
Самый ранний сбор элодеи во Владимирской губернии сделан А.Ф. Флё
ровым на Оке близ Мурома в 1900 г. (MW) и на р. Клязьма близ Владимира в
1901 г. (LE). Годом позднее Флёров указывал это растение для окрестностей
Мурома, близ сел Варех, Шайск и Волосово, а также для некоторых других
мест [Флёров, 1902]. Сейчас E. сanadensis распространена во всех районах,
очень часто [Вахромеев, 2002], примерно так же, как и в Московской обл. (А.П.
Серегин, личное сообщение).
Поскольку в настоящее время граница между Владимирской и Нижегород
ской обл. проходит по Оке, указание А. Ф. Флёрова [1902] на произрастание
E. canadensis близ Мурома относится и к последнему региону, тем более что в
цитируемой работе в качестве одного из мест встречи этого вида фигурирует
Велетьма, входившая тогда в Муромский уезд. К середине 1930 х гг. растение
стало в регионе очень обычным по Оке и Волге [Аверкиев, 1938], а в дальней
шем распространилось и по всей области [Аверкиев, Аверкиев, 1985].
Данные по флоре Чувашии довольно скудны [Флора Средней России...,
1988], и в ботанической литературе первые указания на проникновение сюда
вида относятся к началу 1930 х гг. [Маевский, 1933]. Однако нам представля
ется, что элодея появилась здесь никак не позднее 1910 г., учитывая ее наход
ки в конце XIX -- начале ХХ века. под Муромом и Казанью [Флёров, 1902;
321
Стемпинский, 1909], а также интенсивность тогдашнего судоходства и рыбо
ловства на Волге и нижней Оке. В середине ХХ века растение часто встреча
лось в озерах и старицах Волги, Суры и Цивиля [Куданова, 1965].
Первая находка элодеи канадской в Казанской губернии датируется 1896 г.
[Стемпинский, 1909]. В настоящее время, по оценкам казанских ботаников
[Бакин и др., 2000], растение встречается здесь часто и во всех флористичес
ких районах.
Впервые в Тверской обл. вид, по видимому, отмечен в 1912 г. А.П. Ильинс
ким [Ильинский, 1924] близ с. Белый Городок бывшего Калязинского уезда, а
также в 1913 г. в озерах и канавах у д. Дымовка. К настоящему времени расте
ние встречается здесь часто и во всех типах водоемов, причем в прудах, карье
рах и канавах иногда наблюдается массовое цветение [Маркелова, 2004].
В Ярославской обл. элодея появилась в первом десятилетии ХХ века [Оп
ределитель растений Ярославской..., 1961]. В настоящее время является здесь
обыкновенным и встречается по всей территории видом [Определитель выс
ших растений Ярославской..., 1986].
Указание Е.А. Борисовой [2007] на проникновение E. canadensis в пределы
современной Ивановской обл. в 1902 г. не совсем правомерно, поскольку, ве
роятнее всего, оно основано на цитированном выше сборе А.Ф. Флёрова из
окрестностей Владимира. Впервые на современной территории области вид
собран А.Е. Жадовским: в береговом озере на левом берегу Волги против впа
дения р. Томна, близ г. Кинешма, в 1914 г. (MW); но, не исключено, что на
пограничный с Владимирской обл. участок Клязьмы это растение могло про
никнуть из под Владимира и раньше. К настоящему времени встречается во
всех типах водоемов, часто, местами в массе [Борисова, 2007].
В Костромской губернии впервые указана в начале ХХ века Н.Л. Скалозу
бовым [Белозеров, 2008], что отражено в 5 м издании «Флоры...» П.Ф. Маев
ского [1917]. В середине ХХ века она широко распространилась по всему ре
гиону, где была отмечена во многих водоемах; проникла отсюда в Вологодс
кую обл. [Белозеров, 1956, 2008].
Местообитания. Вид с широчайшей экологической амплитудой, встреча
ющийся в разнообразных водоемах и водотоках: реках, прудах, материковых
и пойменных озерах, ручьях, каналах, а также в головных частях заливов во
дохранилищ и даже в мочажинах болот (рис. 153). Обычно предпочитает пес
чаные или илистые грунты; плохо переносит волнобой [Флора СССР, 1934].
По материалам, полученным при изучении североказахстанских и уральских
популяций, экологический ареал E. canadensis включает гидрокарбонатно натри
евые и хлоридно натриевые воды, имеющие минерализацию 0,30--0,68 г/л, об
щую жесткость 1,69--5,70, кальциевую жесткость 0,97--1,07 мг экв./л, рН 7,20--
8,39 [Дексбах, 1951]; иногда встречается в водоемах с рН 10,4--10,6; прозрач
ность в течение вегетационного периода 0,5--3,8 м [Свириденко, 1986]. Элодея
канадская особенно требовательна к концентрации в воде солей калия и каль
ция, является типичным кальцефилом, обычно гибнет при концентрации хло
рида натрия более 0,5% [Дексбах, 1951], хотя может произрастать и в олиго и
дистрофных водоемах с умеренно кислыми водами.
E. сanadensis на всей территории России и сопредельных стран демонстри
рует сходное поведение. Уже через 3--4 года после вселения растение может
образовать в водоеме обширные заросли и становиться важнейшим продуцен
322
том органического вещества [Свириденко, 1986]. Конкурентоспособность эло
деи канадской в местах ее массового распространения большинством специа
листов оценивается как очень высокая. Максимального обилия вид обычно
достигает к осени, особенно в умеренно загрязненных органическими соеди
нениями водоемах. Продуктивность, состав и распространение сообществ в
разные годы могут колебаться: от полного исчезновения вида до обильного
разрастания [Постовалова, 1966: цит. по: Флора северо востока..., 1974; Пап
ченков и др., 1988; Strand, Weisner, 2001; Жакова, 2008], максимальная фито
масса в зарослях составляет 50--90 г/м2 [Жакова, 2008].
Размножение и жизненный цикл. Растение быстро размножается вегета
тивно, причем даже относительно короткий обрывок стебля способен образо
вать новую особь [Энциклопедия..., 2008]. На родине представители рода Elodea
зимуют в виде семян и зимующих почек, или турионов; в нашей флоре семен
ное размножение отсутствует.
Активный рост при температуре воды более + 15°С ведет к быстрому нараста
нию биомассы, так что к концу лета побеги элодеи могут заполнить всю толщу
воды. Длинные горизонтальные побеги захватывают новые участки дна водоема
и закрепляются на нем с помощью придаточных корней [Sheppard et al., 2006].
Скорость распространения в водоеме обломков побегов E. canadensis и их
закрепления на дне специально изучалась во Франции в условиях 10 недель
ного модельного эксперимента [Barrat Segretain et al., 1998]. Было показано,
что фрагменты элодеи развивали множество отростков, способных к распрос
транению, но при этом в грунте не закреплялись.
Расселение и распространение. Занос элодеи в другие страны, как прави
ло, связан с аквариумной культурой или деятельностью ботанических садов,
из которых растения случайно попадают в водоемы. Если природные условия
благоприятны для роста элодеи, а данный клон в достаточной степени к ним
приспособлен, дальнейшее распространение растения идет путем переноса
частей растения (обрывков стеблей либо турионов) в пределах одного водо
ема или между ними.
Обрывки стеблей и турионы цепляются за выступающие части корпусов и
ходовой части судов или попадают в щели их обшивки (особенно благоприят
ны в этом отношении суда с деревянным наборным корпусом, а также колес
ные пароходы), вместе с песком оказываются на шаландах при производстве
дноуглубительных работ, перевозятся с навалочными грузами водным транс
портом. Все это способствует переносу диаспор элодеи в пределах судоход
ных участков рек, а также из одного речного бассейна в другой, если они со
единены судоходными каналами. Турионы в качестве агентов переноса на даль
ние расстояния, возможно, даже более эффективны, чем стеблевые фрагменты.
Кроме того, турионы элодеи и небольшие обрывки ее стеблей могут прили
пать к лапам крупных водоплавающих и околоводных птиц или к конечнос
тям диких и домашних животных и таким образом переноситься из одного
смежного водоема в другой. Подобный же перенос имеет место и на сетевых
орудиях лова.
О находках зрелых семян элодеи сообщается крайне редко, и распростра
нение семенами на территориях, где элодея канадская стала сорным растени
ем, большого значения не имеет. Тем более, что в известных местах заноса на
блюдаются только однополые популяции.
323
Интересный прогноз распространения элодеи канадской в Европе в случае
глобального потепления дан в работе, посвященной распространению адвен
тивных водных видов растений в Швеции [Larson, 2007]. Согласно смодели
рованному сценарию, этот вид расширяет площади распространения даже силь
нее, чем это прогнозировалось. Из 13 рассмотренных в этом исследовании ви
дов сходную картину дала только Alisma gramineum.
Во всех случаях основными ограничителями для расселения E. canadensis
являются условия среды: температурный, световой, гидрологический и гид
рохимический режимы водных объектов.
Последствия внедрения
Воздействия на естественные фитоценозы и аборигенные виды. Явля
ясь крайне мощным конкурентом в условиях водоемов умеренной полосы,
элодея канадская при внедрении в них способная активно вытеснять абори
генные виды, формируя на больших площадях чистые заросли. Этому же спо
собствует и аллелопатическое влияние выделений элодеи на некоторые виды
водных растений.
В частности, при специальных исследованиях распространения и условий
обитания этой элодеи в Чивыркуйском зал. оз. Байкал было установлено, что ее
сообщества особенно хорошо чувствовали себя на илистых грунтах на глубинах
до 3 м, практически вытеснив все природные растительные сообщества с затиш
ных и илистых участков [Базарова, Пронин, 2008]. В настоящее время продолжа
ется широкое распространение этого растения по мелководьям Байкала, связан
ное в том числе с эвтрофикацией отдельных участков озера [Азовский, 2006].
В оз. Финьясьон в Швеции после первоначального роста числа видов вод
ных растений при улучшении условий их обитания, затем в течение несколь
ких лет их численность уменьшилась, что было приписано конкурентному
вытеснению со стороны элодеи канадской. Однако это уменьшение продол
жилось и после практически полного исчезновения элодеи на исследуемых
участках озера в 1997 г. В связи с этим, авторы работы подчеркивают, что на
видовое богатство оказывают влияние и другие факторы, а не только конку
ренция со стороны E. сanadensis [Strand, Weisner, 2001].
Считается, что сообщества с богатым видовым составом способны лучше
сопротивляться инвазиям [Kennedy et al., 2002; Davis et al., 2005]. С целью
проверки этой гипотезы было изучено 275 озер Центральной Швеции. Озера
с популяциями E. canadensis и E. nuttallii сравнивались с озерами, где занос
ные водные растения отсутствовали. В ходе работ было выявлено, что число
природных видов растений в озерах положительно кореллирует с инвазибель
ностью этих водоемов. В озерах, содержащих заносные виды, в целом насчи
тывалось больше видов, чем в озерах, где заносных видов не было. Более того,
даже в пределах одного озера участки, заселенные заносными видами, имели
большее видовое разнообразие, чем участки, где такие виды отсутствовали.
Одно из возможных объяснений этого явления, по мнению авторов, состоит в
том, что озера, где встречались заносные виды, имеют большую гетерогенность
среды и, следовательно, больше ниш для заселения, которые и осваиваются не
только аборигенными, но и адвентивными видами [Larson, 2007].
Генетические изменения и вариабельность вида. Во флоре Северной
Америки выделяют 2 разновидности: E. canadensis var. angustifolia Farw. и var.
planchonii Farw. [Haynes, 2000].
324
Экономическое и социальное (положительное и отрицательное)зна
чение. Заросли E. сanadensis служат субстратом, на котором поселяются мел
кие животные и их личинки, защитой для икры и молоди рыб, хотя по этому
показателю они уступают большинству видов аборигенных погруженных сосу
дистых водных растений. Вегетативная масса может быть использована на корм
скоту, в частности, свиней, и в качестве органического удобрения [Флора СССР,
1934; Рычин, Сергеева, 1939; Определитель растений Татарской..., 1979], одна
ко на практике с подобными фактами нам встречаться не доводилось.
Наряду с другими водными и прибрежными видами, элодея, имея боль
шую биомассу и значительную поверхность ассимилирующих органов, выпол
няет важную роль в самоочищении рек и других водоемов и водотоков. В днев
ные часы растение хорошо насыщает воду кислородом, однако в теплые авгу
стовские ночи его заросли в стоячем водоеме могут стать причиной летнего
замора рыбы, а зимой ее разлагающаяся биомасса часто вызывает здесь же и
зимние заморы.
Элодея канадская используется как неприхотливое водное растение для хо
лодноводных и умеренно теплых аквариумов, и упоминание о ней можно встре
тить практически в любом руководстве по аквариумистике (рис. 154). При этом
обращается внимание, что сок элодеи для некоторых видов рыб является ток
сичным, а потому ее черенкование лучше проводить в отдельной посуде.
Растение широко используется для биомониторинга и рекультивации вод
ных экосистем. Исследованиями Института биохимии и физиологии расте
ний и микроорганизмов РАН была выявлена способность E. сanadensis разру
шать сырую нефть, фенол, толуол, нафталин, бензол; причем в случае толуола
активность элодеи по этому показателю превышала таковую для микроорга
низмов. Показано, что ферментативной активностью в отношении исследо
ванных загрязнителей обладают экссудаты элодеи и экстракты из клеток это
го растения [Отчет..., 2007, с. 115].
Элодея канадская способна поглощать из грунта ионы тяжелых металлов, в
том числе таких высокотоксичных и радиоактивных, как медь, цинк, никель, а
также плутоний и америций 241. Так, в золе элодеи концентрация америция 241
оказалась в 5 раз выше, чем в тканях, а в целом его концентрация в растении ока
залась в 2000 раз выше, чем в окружающей среде [Болсуновский и др., 2004].
В акватории Енисея, подвергшейся радиоактивному загрязнению Горнохи
мическим комбинатом, проводилось цитогенетическое изучение E. сanadensis,.
У растений при этом были выявлены различные хромосомные расы: 2n = 24, 32,
40, 48, 56, 64, 72 и 96, а также миксоплоидия и анеуплоидия. Предполагается,
что изменчивость хромосомных чисел у элодеи канадской в исследуемом райо
не объясняется радиоактивным загрязнением, а также загрязнением воды соеди
нениями тяжелых металлов [Болсуновский и др., 1999; Муратова и др. 2008].
Это растение широко использовалась в качестве тест объекта в цитогене
тических исследованиях, а также как учебный объект на занятиях по физио
логии растений.
При буйном развитии элодея канадская вытесняет из гидрофитоценозов
другие виды водных растений, ее заросли могут стать препятствием для пла
вания маломерных и мелкосидящих судов, а также для рыболовства. В зимнее
время разлагающиеся растения, как отмечалось выше, вызывают заморы рыбы,
отрицательно влияют на качество питьевой воды. Массовое развитие E.
325
сanadensis уменьшает прозрачность воды, влияет на ее температуру, содержа
ние кислорода и кислотность. Это оказывает влияние на все аспекты суще
ствования и функционирования водных сообществ, включая рыбопродуктив
ность [Sheppard et al., 2006]. Меры борьбы
Во многих районах мира E. canadensis является трудноискоренимым сор
ным растением. Особенно сильно ее негативное влияние отмечается в ороси
тельных системах юго востока Австралии. Для контроля за этим видом здесь
разработаны экономичные, безопасные и эффективные методы: осушение ка
налов, их прочистка и фильтрация воды, применение гербицидов и, где это
возможно, разведение растительноядных рыб [Bowmer et al., 1995].
В ряде стран мира для уничтожения E. canadensis в 1980 е гг. использова
лись гербициды, в частности --- растворы акролеина и солей эндотала [Bowmer,
Smith, 1984]. Однако к настоящему времени из за токсичности этих соедине
ний и их малой эффективности применение данного метода весьма ограниче
но. Подробный обзор использования гербицидов для борьбы с элодеей приве
ден в статье Боумера и соавторов [Bowmer et al.,1995].
Существует обширная литература по биологической борьбе с погружен
ными водными растениями с использованием травоядных рыб, например, бе
лого амура [Bonar et al., 1987; Pauley et al., 1987]. Некоторые авторы полагают,
что элодея, наряду с другими видами растений, обладает для этих рыб прият
ным вкусом и они нередко предпочитают ее другим видам. Однако большин
ство исследователей [Bowers et al., 1987] считают, что виды рода Elodea по их
аппетитности для белого амура располагаются ближе к концу списка кормовых
объектов. Кроме того, запуск растительноядных рыб в озера со смешанным на
бором видов растений может привести к росту биомассы «невкусных» видов и
росту расходов на увеличение количества «вкусных» [Fowler, Robson, 1978].
Механические методы борьбы с элодеей (срезка, боронование и т.п.) оказа
лись крайне неэффективными и обычно не рекомендуются, так как они спо
собствуют еще более сильному распространению этого вида. Подобные мето
ды могут быть применены лишь в крайнем случае и там, где E. canadensis уже
расселилась [Bowmer et al., 1995].
Понижение уровня воды в эвтрофных водоемах как способ борьбы с вод
ными макрофитами [Cooke, 1980] также не получил широкого применения,
поскольку эти растения, в том числе и элодея, быстро восстанавливают свою
численность при повторном затоплении. Тем не менее, установлено [Hejny,
Husák, 1978], что летний и зимний сброс воды, ведущие к промораживанию
дна водоемов в холодных районах или к его обсыханию --- в засушливых, в
Чехословакии оказалось эффективным для противодействия расселению E.
canadensis. Но подобный прием требует длительного времени и на крупных
водоемах и водотоках возможен далеко не всегда [Bowmer et al., 1995]. Так, в
1997 г. в оз. Финьясьон (Швеция) был резко понижен уровень воды, после
чего E. сanadensis здесь практически исчезла. Однако при восстановлении уровня
в течение короткого времени ее популяции за счет интенсивного вегетативного
размножения восстановила свою численность [Strand, Weisner, 2001].
326
СЕМЕЙСТВО JUNCACEAE -- СИТНИКОВЫЕ
СИТНИК ТОНКИЙ
Название: Juncus tenuis Willd.
Синонимы: J. macer S.F. Gray
Морфологические признаки
Многолетнее короткокорневищное растение, образующее плотные дерно
вины (рис. 155). Стебли высотой до 40--50 см, прямостоячие, реже поникаю
щие, диаметром 0,6--1,2 мм. Побег с 7--9 листьями, из них 2--3 чешуевидные,
бурые, остальные уплощенно желобчатые, шириной (0,5)1--1,5 мм и длиной
до 15 см; ушки пленчатые, длиной 2--4 мм, заостренные. Соцветие верхушеч
ное, рыхлое, щитковидно метельчатое; цветки одиночные или частично в не
многоцветковых группах; нижние (1) 2 (3) прицветника листовидные, часто
длиннее соцветия. Листочки околоцветника более или менее равной длины
(внешние чуть длиннее), зеленоватые (по средней жилке травянистые, с края
светлые, пленчатые), ланцетные, заостренные, длиной 3,5--4,4 мм. Тычинок 6,
тычиночные нити длиной 0,5--1,1 мм, пыльники -- 0,4--0,7 мм, обычно в 2--3
раза короче нити. Коробочка треугольно овальная, одногнездная или ложно
трехгнездная, длиной 3,8--4,7 мм, на верхушке короткозаостренная или тупая.
Семена 0,3--0,5 мм длиной, красновато коричневые, тонкосетчатые.
Естественный ареал
J. tenuis -- североамериканский вид, известный от Ванкувера и Ньюфаундлен
да до Мексики. Наиболее обычен на северо востоке США и востоке Канады, но
встречается также южнее и западнее [Brooks, Clemants, 2000; Juncaceae..., 2002].
Вторичный ареал
В Европе ситник тонкий впервые обнаружен в 1795 г. или 1796 г. в Вели
кобритании. В 1824 г. найден в Бельгии, затем в 1825 г. в Голландии и в 1834 г.
в Германии [Новиков, 1978; Richards, 1943]. В Ирландии и на юго западе Ве
ликобритании J. tenuis встречается разреженно по солнцеватым приморским бо
лотам, что позволило предположить о естественном характере вида в Великобри
тании [Scully, 1889; Praeger, 1934, цит. по: Richards, 1943; Rose, 1989]. В Чехии
известен с 1851 г. [Pyšek et al., 2002], а в Польше -- лишь с 1901 г. [Biological...,
1990]. На территории Украины впервые собран И.Ф. Шмальгаузеном: «Вышне
вец, Волынская губ., 29.06.1885» (KW). В настоящее время ситник натурализо
вался более чем в половине регионов Европы [Lambdon et al., 2008].
Широко натурализовался в Турции [Byfield, Baytop, 1998]. Как заносное
растение известен с 1875 г. в Новой Зеландии [Cheeseman, 1878], позднее -- в
Австралии и Южной Америке [Richards, 1943]. В настоящее время J. tenuis
известен во многих внетропических странах обоих полушарий [Brooks,
Clemants, 2000; Guofang, Clemants, 2000; Juncaceae..., 2002].
На Кавказе, по указанию А.А. Гроссгейма [1940], известен раньше, чем в
Европейской России: «Сочи, Красная Поляна, 30.07.1904 г., И.Я. Акинфеев»1.
В Сибири впервые собран Н.И. Кузнецовым (LE): «Долина р. Лена, на лугу
между селениями Головновка и Тутура, 17.06.1910» (совр. Читинская обл.).
1 Гербарный образец, подтверждающий находку, нами не обнаружен.
Рис. 155. Juncus
tenuis, общий вид
Рис. 156. Рассе
ление Juncus
tenuis в Средней
России
Рис. 157. Внедрение
Juncus tenuis
в естественные сообщества
Рис. 158. Плодоношение
Juncus tenuis
329
Ныне ситник тонкий в Сибири редок [Ковтонюк, 1987], на Дальнем Востоке
встречается в южной части региона [Новиков, 1985].
Пути и способы заноса. J. tenuis распространился вне явной связи с же
лезной дорогой и другими транспортными путями. Ситники обладают весьма
эффективным способом распространения семян (см. ниже). Деятельность че
ловека создала подходящую сеть местообитаний, что создало предпосылки для
быстрого расселения вида.
Статус в регионе. В Средней России J. tenuis впервые обнаружен в 1922--
1923 гг. в г. Сергиев Посад (Московская обл.) и окрестностях [MW, Иванова,
1927]. Долгое время на ситник не обращали внимание, и за несколько десяти
летий он стремительно расселился [Скворцов, 1959; Тихомиров и др., 1960]. В
1970 е гг. J. tenuis неоднократно собран в ряде областей Средней России, что
свидетельствует о его широком расселении (рис. 156).
Источник появления ситника на нашей территории неясен. С одной сторо
ны, вид культивировали в ботаническом саду в Горенках: «Ex hort. Gorenki.
Cult. com. Steven, 1810 г.» (LE) и «1827 г.. Culta. Gorenki» (MW, гербарий Бош
няка; LE); с другой -- в окрестностях сада в начале XX века проводились мас
совые гербарные сборы, в которых ситник отсутствует, что скорее свидетель
ствует о появлении J. tenuis с территории Европы -- из центра вторичного рас
пространения [Игнатов и др., 1990].
Местообитания. J. tenuis -- растение открытых местообитаний, предпочи
тающий умеренно закисленные влажные почвы, бедные или умеренно плодо
родные (4--5 я ступень по шкале Элленберга), при засолении отсутствует [Hill
et al., 1999].
Ситник тонкий встречается по лесным и низкотравным пустошным лугам,
вырубкам, разработанным торфяникам, лесным и проселочным дорогам, кра
ям лесных болот. Чаще встречается на участках с нарушенным растительным
покровом, предпочитает песчаные и торфянистые почвы (рис. 157). J. tenuis ---
один из сравнительно немногих адвентивных видов Европы, освоивших раз
нообразные места обитания [Pyšek et al., 1998].
Размножение и жизненный цикл. Ветроопыляемое растение [Richards,
1943; Michalski, Durka, 2007]. Обычно цветки раскрываются между 7 и 8 ч утра,
а пыльники вскрываются между 9 и 10 ч, к полудню листочки околоцветника
смыкаются [Новиков, 1978]. Отмечено одновременное зацветание растений в
пределах популяции [Michalski, Durka, 2007].
По видимому, J. tenuis может зацветать в первый год (рис. 158). Семена
мелкие, 0,014--0,017 мг. В одной коробочке образуется 60--450 семян, в сред
нем ~350. Среднее число цветков на генеративном побеге ~25, но их число ко
леблется: от 5 до 60. Развитое растение имеет 15--20 генеративных побегов,
таким образом, одна развитая дерновинка производит 150--200 тыс. семян
[Новиков, 1978; Richards, 1943].
Способен образовывать почвенный банк семян [Rabinowitz, 1981; Bigwood,
Inouye, 1988; Sullivan, Ellison, 2006]. Сухие семена, хранимые в лабораторных
условиях, сохраняют всхожесть более 2 лет [Richards, 1943].
Семена прорастают весной, и к началу осени при благоприятных условиях
растения ситника достигают высоты 6--10 см и имеют 7--10 побегов. Новые
побеги закладываются до глубокой осени. В тенистых и влажных местах рас
тения J. tenuis часто отмирают на 2 й год. По видимому, на более светлых и
сухих местах возраст растений все же не превышает 5--6 лет, однако точное
330
определение возраста затруднено, так как отмирающие части быстро разруша
ются [Новиков, 1978].
Расселение и распространение. Для ситников характерен своеобразный
специализированный способ распространения семян. При избыточном увлаж
нении после дождя или при росе семена выделяют слизь, вместе с которой легко
прилипают к ногам человека или шерсти животных. При подсыхании слизь
образует пленку с семенами, и в таком виде они могут переноситься на дальние
расстояния [собственные наблюдения; Новиков, 1978; Richards, 1943]. Считает
ся, что в Северной Америке агентом расселения на дальние расстояния являет
ся белохвостый олень --- Odocoileus virginianus [Myers et al., 2004], а в Европе
кабан --- Sus scrofa [Heinken et al., 2006]. Кроме того, возможно распростране
ние семян водой и ветром [Новиков, 1978].
Последствия внедрения
Воздействия на естественные фитоценозы и аборигенные виды. Яв
ных признаков воздействия на местные растения не обнаружено.
Генетические изменения и вариабельность вида. В Европе извесен толь
ко один экотип изменчивого J. tenuis [Richards, 1943]. Следует, однако, отметить,
что ушки при основании листовой пластинки растений Средней России мень
ше 3--6 мм, как это указывается для американских растений [Brooks, Clemants,
2000; Juncaceae..., 2002].
Экономическое и социальное (положительное и отрицательное) зна
чение. В лесной зоне ситник тонкий является одним из пионерных видов зара
стания обнаженного грунта на вырубках, по грунтовым лесным дорогам; при
этом он наиболее обилен в условиях небольшого избыточного увлажнения. При
этом он неизбежно конкурирует с видами местной флоры. На низкотравных
пустошных лесных лугах часто обилен, и долго не уступает место аборигенным
растениям.
Меры борьбы
Поскольку J. tenuis не является сельскохозяйственным сорняком, специаль
ные меры борьбы с ним не разработаны и едва ли необходимы.
СЕМЕЙСТВО LABIATAE -- ГУБОЦВЕТНЫЕ
ЭЛЬШОЛЬЦИЯ РЕСНИТЧАТАЯ или ПАТРЕНА
Название: Elsholtzia ciliata (Thunb.) Hyl. Родовое название происходит от
имени Иоганна Сигизмунда Эльшольца (1623--1688), немецкого лекаря, бота
ника и натуралиста XVII века. По В.Далю, несколько видов пряных растений
называли в России шандрой, возможно, от слова «шантить», то есть вонять.
Шандрой, или собачьей мятой, называют и эльшольцию реснитчатую, так как
она имеет насыщенный запах [Гостева, 2008].
Синонимы: Sideritis ciliata Thunb., Elsholtzia patrinii (Lepech.) Garcke, E. patrinii
var. ramosa Nakai, E. cristata Willd., E. formosana Hayata, E. minina Nakai, E.
pseudocristata H.Lйveillй et Vaniot, Hyssopus ocymifolius Lam., H. bracteatus Gmel.
ex Steud., Mentha baicalensis Georgi, M.cristata Buch, M. ovata Cavanilles, M. patrinii
Lepech., Perilla polystachya D. Don.
331
Морфологические признаки
Однолетнее травянистое растение высотой 30--50 см., опушенное редкими
волосками. Побеги прямые, ветвистые. Листья на длинных черешках, с яйце
видной, по краям городчато зубчатой пластинкой, вытянутые от 2 до 8,5 см и
шириной от 0,8 до 2,5 см. Цветки собраны в густое однобокое колосовидное соц
ветие длиной 1--5 см (рис. 159). Прицветники травянистые, налегающие друг
на друга, коротко заостренные, по краям мелко реснитчатые. Лепестки бледно
лиловые, длиной 3--5 мм, с бахромчатыми лопастями. Плодики яйцевидные,
темно бурые [Доронькин, 1997].
Естественный ареал
Центром происхождения и видообразования видов рода Elsholtzia является
Китай, где E.ciliata растет во всех провинциях. Есть данные о нахождении E.ciliata в
Непале на высотах от 1500 до 3400 м над ур. м. Широкое распространение E.ciliata
получила по всей Восточной Азии, Индокитаю и Японии. Вид известен в боль
шинстве районов Индии, а также в Малайзии, Монголии [Hsi wen, Hedge, 1994].
В пределах России ареал E.ciliata охватывает Западную и Восточную Си
бирь до 54° с.ш. [Доронькин, 1997].
Вторичный ареал
История интродукции и географическое распространение. Как сорное
E.ciliata впервые отмечена в 1887 г. в городке Нотр Дам дю Лак (канадская
провинция Квебек) в местообитании, где культивирование вида вряд ли было
возможно. Следующая находка сделана в 1909 г. в городе New Limerick (штат
Мэн, США) в 100 милях к югу от первого сбора. В 1938 г. E.ciliata найдена в
палисаднике г. Челси (штат Вермонт), а в 1943 г. -- в Reverc Beach (штат Масса
чусетс), разросшейся вдоль тротуара [Invasive Plant Atlas of New England
(IPANE), 2008]. Специальная работа посвящена распространению E.ciliata в
Нью Йорке [Monachino, 1958]. В настоящее время вид обнаружена в штатах
Северная Дакота, Миннесота, Висконсин, Нью Йорк, Пенсильвания, Нью
Джерси, Западная Виргиния, Северная Каролина, Мэн, Вермонт, Массачусетс
и Коннектикут [Invasive Plant Atlas of New England (IPANE), 2008].
В странах Европы вид культивируется как декоративное и эфиромаслич
ное растение и успешно дичает.
Пути и способы заноса. Как правило, изначально вид попадает на новую
территорию в качестве эргазиофита, но через несколько лет начинает размно
жаться семенным способом и занимает подходящие экотопы.
Статус в регионе. В Средней России вид распространен относительно рав
номерно, однако находки вне населенных пунктов до сих пор большая редкость
(рис. 160).
Впервые для Воронежской обл. вид отмечен в 1926--1928 гг. [Гроссет, Замят
нин, 1935; Григорьевская и др., 2004]. В 1930 е гг. наблюдалось массовое рассе
ление E.ciliata на северо западе области, а также указывалось, что к юго восто
ку вид быстро исчезал [Попов, 1933]. В настоящее время эльшольция реснитча
тая встречается по всей территории часто, иногда в массе [Григорьевская и др.,
2004]. Для флоры Белгородской обл. приведены два сбора: 1) Белгород, р н
Сокол, 27.09.1997 г. и 2) Грайворонский р н, заповедник «Лес на Ворскле», доро
га на Красный Куток, 4.09.1974 г., Евминова. WORSKL [Еленевский и др., 2004].
E.ciliata указана для Курской губернии начиная с первых флористических
сводок. В.М. Черняев [1836] приводит ее под названием «пчелиная трава» в списке
332
медоносов, высеваемых рядом с пасеками. А.Мизгер [1869] указывает ее под на
званием «французская мята». Один из первых сборов E.ciliata сделан в 1897 г.:
Курская губерния и уезд. Дачи Рудакова. Берег Сейма, 1897 г. Подпись коллекто
ра неразборчива (MW). В 1902 г. E.ciliata собрана в окрестностях Курска В.Але
хиным (MW). В настоящее время в Курской обл. встречается по всей террито
рии, нередко во множестве [Полуянов, 2005]. Первый сбор в Липецкой обл. сде
лан в 1873 г. Грунером (MW). Сейчас E.ciliata отмечена во всех р нах области,
но неравномерно и спорадически, хотя местами развивается в массе [Флора
Липецкой области, 1996]. В Новосильском уезде Тульской губернии (ныне --
Орловская обл.) E.ciliata собрали в 1885 г. (MW). Сейчас в Орловской обл. обыч
ный вид [Еленевский, Радыгина, 1997].
В гербарии М.И. Голенкина и С.Н. Милютина, относящегося к Калужской
губернии, есть сбор, сделанный в 1887 г. в г. Лихвин (ныне -- г. Чекалин) Суво
ровского р на Тульской обл. (MW). Вероятно, его и следует считать первым
для территории современной Тульской обл. В 1954--1955 гг. E.ciliata собрал В.И.
Соболевский в Заокском р не: «в д. Бехово, близ Поленово, на окраине деревни.
Очень часто и много в Бехово и других деревнях» (МНА). Однако начало мас
сового распространения вида следует отнести к 1970 м гг. [Алексеев, Губанов,
1980; Алюшин, 1982]. К настоящему времени E.ciliata принадлежит к самым
распространенным сорным растениям тульской флоры.
Впервые в Московской губернии E.ciliata собрана Г.Борхманом в 1840 х гг.:
«по пригоркам у р. Москвы, редко» (MW). Позднее вид распространился в ок
рестностях Серпухова, что подтверждается многочисленными сборами 1872 г. и
последующих лет (MW). До 1960 х гг. E.ciliata оставалась сравнительно ред
ким видом; с 1970 х гг. вид стал регулярно встречаться на железных дорогах, а
затем на сорных местах в Москве [Игнатов и др., 1990].
Впервые в Ивановской обл. E.ciliata обнаружена на пустыре в г. Иваново в
начале 1980 х гг. В настоящее время распространение вида охватывает не всю
территорию области, а ряд городов: Иваново, Кинешма, Шуя, Тейково, Гаври
лов Посад, Пучеж. [Борисова, 2007]. Во Владимирской обл. E.ciliata встречается
во всех р нах, но довольно редко [Вахромеев, 2002]. Вид собран всего раз как
сорное на клумбе в национальном парке «Мещера» (А.Серегин, 2002, MW),
более ранних сборов обнаружить не удалось.
В Брянской обл. E.ciliata активно собирали в 70 е гг. ХХ века. В MHA хра
нятся сборы 1973--1979 гг. из Комаричского, Трубчевского, Климовского, Жу
ковского р нов.
Первый сбор в Смоленской обл. сделан в 1894 г. в окрестностях Смоленска
(аноним, опр. В.Цингер, MW). Один из более поздних сборов сделан вне насе
ленного пункта и указывает на возможность дичания вида: «Рудненский район,
сырой березово ольховый лес в 14 км южнее Понизовья, 30.07.1974 г., Н.Беля
нина, Г.Проскурякова» (MHA). В настоящее время отмечена во многих насе
ленных пунктах области [Решетникова, 2004].
Вероятно, одним из первых вид собрал в окрестностях Калуги Д.И. Литви
нов, 19.08.1894 г., (MHA). В 1953--55 гг. в Калужской обл. E.ciliata несколько раз
собрал В.И. Соболевский. По указанию на этикетках, вид рос «по улицам, са
дам, огородам, дворам г. Тарусы и прилегающим деревням» (MHA). Позднее, в
70--80 гг. ХХ века E.ciliata собирали в населенных пунктах Угодско Заводского,
Ферзиковского и Юхновского р нов Калужской обл. (MHA). Сейчас в г. Калуга
Рис. 160. Расселение
Elsholzia ciliata
в Средней России
Рис. 159. Цветение
Elsholzia ciliata
334
и населенных пунктах Калужской обл., по данным С.Р. Майорова, E.ciliata рас
пространена повсеместно, однако вне жилья встречается редко.
В 1915 г. E.ciliata собирал М.Назаров в г. Касимов Рязанской губернии (MW).
В настоящее время в Рязанской обл. эльшольция встречается по всей терри
тории, изредка, в местах произрастания порой в массе [Казакова, 2004]. Первое
указание для Пензенской флоры сделал В.Цингер [1885], сейчас в Пензенской
обл. -- редко, приведены всего 3 местонахождения вида [Васюков, 2004]. В гер
барии МГУ имеется старый сбор E.ciliata, сделанный князем Вяземским в Там
бовской губернии: 1866--1869 гг., без указания точного места находки (MW).
Местообитания. В пределах естественного ареала, в Китае, E.ciliata растет
на холмах, пустырях, солнечных газонах, берегах рек, в лесах [His wen, Hedge,
1994]. В Корее и Японии -- на горных луговинах [Plant For a Future, 2008]. В
Сибири -- на лугах, приречных галечниках, по лесным опушкам, обочинам до
рог, огородам [Доронькин, 1997].
В Европе как заносный вид встречается в рудеральных местах, по берегам и
отмелям рек. В США растет на сорных местах, вдоль дорог, на старых полях и
опушках лесов [Гостева, 2008].
В Средней России во множестве разрастается в палисадниках, огородах, са
дах, на влажных сорных местах, вдоль пешеходных дорожек, по берегам рек и
ручьев. За пределами населенных пунктов встречается значительно реже и мо
жет совсем отсутствовать.
Размножение и жизненный цикл. Растение предпочитает солнечные мес
та, умеренно суглинистые и тяжелые глинистые почвы. Почва обязательно дол
жна быть влажной. Растение успешно размножается семенами [Plants For a
Future database, 2008]. Биологические особенности E.ciliata в Московской обл.
изучала М.В. Гринева. Выявлено, что семена сохраняют высокую всхожесть в
течение 3 лет, но при этом энергия прорастания снижается уже после 2 го года
хранения. В условиях Московской обл. всходы появляются в среднем через 15
дней после посева. Фаза бутонизации наступает в 1 ой декаде августа, массовое
цветение отмечается в 3 ей декаде августа, массовое плодоношение -- во 2 ой
декаде сентября. Вегетационный период в среднем составляет 120 дней. Расте
ние проходит весь цикл роста и развития. Оптимальными сроками посева в
Московской обл. являются конец апреля -- начало мая [Гринева, 2007].
Расселение и распространение. Растение распространяется с помощью
мелких семян, имеющих высокий процент всхожести [Invasive Plant Atlas of
New England (IPANE), 2008].
Последствия внедрения
Воздействие на естественные фитоценозы и аборигенные виды. Явля
ясь компонентом пионерной растительности, E.ciliata не выдерживает конку
ренции со стороны многолетних видов травянистых растений, заселяющих
местообитание на более поздних сукцессионных стадиях. Например, в природ
ных сообществах Китая, на альпийских лугах в ходе ранних стадий сукцессии в
сообществе доминируют пионерные виды, размножающиеся семенами, в том
числе E.ciliata. По мере развития сукцессионных процессов в сообществе возра
стает роль видов с вегетативным типом размножения, при этом виды первой
группы позиции теряют [Jiang et al., 2004]. В японской префектуре Нагано, на
заброшенном рисовом поле, где вид является компонентом природной флоры,
подлежащим охране, изучалась встречаемость эльшольции на участках поля,
335
различающихся по условиям увлажнения. Плотность локальных популяций
оказалась выше на откосах мелководных гряд, по сравнению с их вершинами,
что объясняется положительной корреляцией между обилием вида и уровнем
почвенного увлажнения [Shizuka et al., 2001].
Генетические изменения и вариабельность вида. Во «Флоре Китая» вы
делены 4 разновидности E.ciliata: var. brevipes С.Y. Wu et S.C. Huang, var.
depauperata C.Y. Wu et S.C. Huang, var. ramosa (Nakai) С.Y. Wu et H.W. Li, var.
remota C.Y. Wu et S.C. Huang [Hsi wen, Hedge, 1994]. Во «Флоре СССР» [Волко
ва, 1954] выделены 2 формы, описанные с Дальнего Востока: f. saxatilis Kom. и f.
ruderalis Kom.
Отмечены различия по биохимическому составу растительного сырья
E.ciliata в различных регионах мира. Выявлено, что в зависимости от климати
ческих или экологических условиий в составе эфирного масла преобладают либо
эльшольция кетон, либо его производное -- дегидроэльшольция кетон. Обна
ружен и третий компонент аромата -- розафуран. В состав вьетнамского типа
эльшольции реснитчатой входит также лимонен и соединения гераниольного
типа. Основными компонентами эфирного масла сибирского типа E.ciliata яв
ляются эльшольция кетон (19%), дегидроэльшольция кетон (68%), периллен
(4%), хумулен (3%), дегидроэльшольцион (2%). Основными компонентами даль
невосточного типа являются розафуран (42%), цитраль (14,9%), лимонен (6,1%),
б хумулен (6,4%), гермакрен Д (6,2%), причем в цветках сосредоточен в основ
ном розафуран, в листьях и стеблях -- цитраль [Гостева, 2008]. Образцы E.ciliata,
собранные на сорных местах в пригороде Новосибирска, содержали следую
щие основные компоненты: периллен (2,1--3,9%), эльшольция кетон (3,3--19,3%),
б дегидро эльшольцион (2,0 5,7%), дегидро эльшольция кетон (66,1--72,4%) и
хумулен (1,5--3,8%) [Korolyuk et al., 2002]. При изучении компонентного соста
ва эфирного масла E.ciliata, выращенной в Московской обл., установлено, что
основными компонентами являются нагина кетон (71,8%), а содержание эль
шольция кетона составляет всего 22,6% [Гринева, 2007].
Экономическое и социальное (положительное и отрицательное) зна
чение. Лекарственным сырьем у E.ciliata являются побеги, собранные в фазе
цветения. Сырье содержит эфирные масла (до 0,6%), горькое вещество марубин
(дитерпеновый лактон), алкалоиды (бетоницин, турицин, этахидрин), дубиль
ные вещества (~7%), слизи. Компоненты эфирного масла оказывают желчегон
ное, спазмолитическое действие, облегчают отхаркивание, нормализуют сер
дечную деятельность, возбуждают аппетит, стимулируют деятельность кишеч
ника. В тибетской и японской медицине траву E.ciliata применяют в качестве
мочегонного и стимулирующего пищеварение средства, а также для лечения
дыхательной системы и в онкологии [Гостева, 2008]. Компоненты E.ciliata вхо
дят в состав вьетнамских препаратов для комплексного лечения органов дыха
ния и используются при онкологических заболеваниях. В народной медицине
эльшольцию используют как антибактериальное, жаропонижающее, противо
вирусное, вяжущее, ветрогонное, диуретическое средство. Эфирное масло
E.ciliata обладает противогрибковым действием, ускоряет заживление ран, сти
мулирует репаративные процессы [Гринева, 2007].
Молодые листья можно использовать как пряную приправу для супов и
салатов. Во Вьетнаме листья используют в пищу в сыром виде. Их режут, до
бавляют к салатам или используют как зелень. E.ciliata подходит ко многим
336
блюдам, богатым протеинами -- яйцам, птице, рыбе, фасоли и гороху [Гостева,
2008], а также к вегетарианским блюдам. Порошок семян используют как при
праву для придания блюдам и продуктам аромата [Plants For a Future, 2008].
Растение медоносное.
Меры борьбы
Е.ciliata -- потенциально инвазионный вид на северо востоке США и вклю
чен в местные региональные списки инвазионных видов [Invasive Exotic Plants
of the Northeast, 2008]. В штате Коннектикут, например, законодательно, под
угрозой штрафа, запрещено распространять E.ciliata [An Act Concerning
Revisions..., 2008].
Однако, так как E.ciliata является видом пионерных сообществ, она не нуж
дается в специальных мерах контроля, по крайней мере в тех местообитаниях,
где идет процесс задернения и не остается увлажненных обнаженных участков.
Кроме того, по оценкам экспертов, в США вид занимает около 10% площади
подходящих для него экотопов и не представляет большой инвазионной опас
ности [Invasive Non native plant Management during 2002, 2008]. В Средней Рос
сии, вероятно, картина сходная -- вид не требует пока специальных мер по кон
тролю и уничтожению.
CЕМЕЙСТВО OLEACEAE -- МАСЛИННЫЕ
ЯСЕНЬ ПЕНСИЛЬВАНСКИЙ
Название: Fraxinus pennsylvanica Marsh.
Синонимы: F. lanceolata Borkh., F. americana auct. Fl. Ross. p. p., non L., F.
pubescens Lam.
Морфологические признаки
Прямоствольное высокое дерево с сетчато трещиноватой серой корой
(рис. 161). Молодые ветви с рыжеватым опушением или, реже, голые. Листья
супротивные, непарноперистосложные, о 5--7(9) листочках; листочки от яйце
видных до ланцетных, на коротких черешочках, по краю от сглажено пильча
тых до цельнокрайных, сверху темно зеленые, практически голые, снизу свет
лее, с простыми волосками, хотя бы вдоль главных жилок, иногда почти голые.
Молодые листья, помимо обильного опушения простыми волосками, с рассеян
ными сидячими желёзками. Растения двудомные. Соцветия метельчатые, па
зушные. Пестичные цветки длиной около 4 мм, околоцветник простой, яйце
видный, вверху неправильно зубчатый; завязь скрыта в околоцветнике; рыль
ца 2 лопастные, грязно вишневые, длиной около 2,5 мм; оси соцветия почти
голые. Тычиночные цветки с воронковидным околоцветником длиной менее 0,5
мм; тычинок 2--3, тычиночная нить длиной около 0,5 мм, пыльники -- 2--3 мм,
желтые, с коротким надсвязником; оси тычиночного соцветия обильно опушен
ные. Плоды -- узколопатчатые крылатки длиной 2--5 см при ширине 3--5 мм, с
остатками чашечки, сначала светло желтовато зеленые, затем светло бурые;
семенная камера составляет 1/3--2/5 длины крылатки, крыло плоское.
В России нередко указывают на использование в озеленении F. americana L.,
наряду с местным F. excelsior L. [см., например: Васильев, 1952; Головач, 1960;
Цвелев, 2004]. При этом, однако, авторами признается разделение F. pennsylvanica
337
на несколько видов, но важные диагностические признаки опускаются -- на
пример, двудомность этих американских ясеней [Rehder, 1949]. Поэтому счита
ем необходимым привести определительный ключ для этих трех видов.
1. Почки бархатисто черные (реже на жировых побегах бурые). Молодые
веточки голые. Листья из (7)9--11(15) почти сидячих листочков, удлиненно эл
липтических или ланцетных, голых или снизу по главной жилке с рассеянными
волосками. Растение однодомное. Цветки голые, обоеполые и тычиночные в од
ном соцветии. Крылатки почти до основания крылатые, с закругленным основа
нием без остатков чашечки, часто винтовидно скрученные . ........... F. excelsior
--- Почки коричневые, иногда рыжеватые. Молодые веточки опушённые или
голые. Листья о 5--7(9) листочках на коротких черешочках, от яйцевидных до
ланцетных. Растение обычно двудомное. Цветки с небольшой зеленой чашеч
кой. Крылатки узкообратноланцетные, практически плоские, семенное гнездо
не крылатое, с узко клиновидным основанием ...................................................... 2
2. Листовой рубец полулунный, с явственно вогнутой верхней кромкой.
Молодые побеги и листья снизу голые ............................................. F. americana
--- Листовой рубец в форме буквы D, верхний его край выгнутый или почти
плоский. Молодые побеги и листья снизу в разной степени опушенные, редко
голые ...................................................................................................... F. pennsylvanica
При составлении ключа использовались преимущественно американские
интернет ресурсы (в этой ситуации на редкость единодушные), а также сборы в
MW, MHA1. Используется ли реально F. americana в озеленении на территории
России, нам неизвестно. По крайней мере, в гербарных сборах из Европейской
России в Гербариях Москвы он отсутствует. Впрочем, следует учитывать, что
сборы американских видов ясеня крайне ограничены.
Естественный ареал
Североамериканский вид ясеня с самым обширным ареалом. Его естествен
ный ареал охватывает восточные и центральные районы Северной Америки:
от центра провинции Саскачеван и юга Квебека до востока штата Техас и севе
ра Флориды, на западе до штатов Монтана, Юта и Нью Мексико [Gleason,
Cronquist, 1991; Burns, Honkala, 1990]. В области естественного распростране
ния широко культивируется как быстрорастущая устойчивая культура.
Вторичный ареал
История интродукции и географическое распространение. В Европе в
культуре с 1723 г. Популярным стал с первой половины XX века, наряду с Acer
negundo, A. saccharinum, гибридными тополями [Mкdrzycki, 2007]. Сведения о
дичании пенсильванского ясеня появляются во второй половине XX века. В
настоящее время есть сведения о той или иной степени натурализации в Авст
рии [Essl, Rabitsch, 2002], Венгрии [Invasive..., 2003], Германии [Doppelbaur, 1963;
Schaffrath, 2001; Hаeupler, Muer, 2007], Польше [Kosiсski, Bednorz, 2003; Urbisz,
Urbisz, 2006], Чехии [Pyšek et al., 2002], Хорватии [Kremer et al., 2006], на терри
тории бывшего СССР [Цвелев, 2004]. С другой стороны, в Великобритании
ясень известен в единичных пунктах [Clement, Foster, 1994]. Есть данные о нату
рализации F. pennsylvanica в Японии [Mьller, Okuda, 1998], Аргентине [Achinelli,
Delucchi, 2000]. В СССР широко использовался для создания полезащитных
1 В MHA образцы F. americana достаточно многочисленны, в том числе оригинальные
сборы А.К. Скворцова, которого интересовали различия ясеней американского проис
хождения.
338
лесонасаждений, особенно на юго востоке Европейской России [Васильев, 1952;
Головач, 1960].
Пути и способы заноса. F. pennsylvanica широко используется в городском
озеленении. В лесостепных и степных областях Европейской России применя
ется для создания полезащитных лесополос. Плодоношение ясеня пенсильван
ского обильное (рис. 162).
Крылатки облетают с материнских растений как осенью, так и зимой. При
этом возможно их распространение не только ветром на сравнительно корот
кие расстояния, но дальний занос с талыми водами [Burns, Honkala, 1990]. Од
носемянные плоды обладают высокой всхожестью. Они прорастают на участ
ках без сомкнутого растительного покрова: на свежем аллювии, на вторичных
местообитаниях.
Статус в регионе. В культуре широко распространен по всей Средней Рос
сии. В южных областях высоко активен как инвазионный вид, на севере терри
тории дичание отмечено преимущественно в пределах городов и поселков, реже
в пригородных лесах. Натурализация отмечена во всех регионах средней поло
сы. В Средней России впервые2 собран в Московской обл., «Серпуховской р н,
бл. с. Лужки, пойма р. Оки, в ивняке по Можайки, 1 дерево в 4 (или 7? -- СМ) м,
6.08.1971 г., П.Смирнов (как F. excelsior), опр. С.Майоров» (MW). К сожалению,
самостоятельное расселение крайне скудно документировано гербарными ма
териалами. В крупнейших гербариях России сборы F. pennsylvanica не из куль
тивируемого материала ограничены первыми десятками гербарных листов (LE,
MW, MHA). Активное расселение началось, вероятно, в 1970--1980 гг.. В 1966 г.
в «Определителе растений Московской области» было написано: «видимо, у
нас не дичает» [Ворошилов и др., 1966] (рис. 163).
Местообитания. В пределах естественного ареала F. pennsylvanica растет
по речным долинам, реже на болотах. Оптимальные условия для ясеня -- плодо
родные слабокислые супесчаные почвы речных долин. Поселяется как пионер
ный вид на аллювиальных почвах вместе с Populus deltoides, P. tremuloides, Salix
nigra. На склонах речных долин ясень растет вместе с красным кленом (Acer
rubrum), американским вязом (Ulmus americana) и показывает высокие прирос
ты [Burns, Honkala, 1990].
В районах вторичного ареала приурочен к рудеральным местообитаниям,
растет на городских пустырях, в оврагах, на дорогах и обочинах, выемках грунта, у
фундаментов. В последние годы все чаще отмечается в речных долинах [Müller,
Okuda, 1998; Kremer et al., 2006], в том числе и в Средней России (собственные
наблюдения). Ясень отмечен в нарушенных пригородных лесах [Борисова, 2006].
Он активно расселяется по меловым обнажениям Среднерусской возвышенности,
где растет преимущественно на шлейфах выноса на мелкоземе, в карьерах, но от
дельные растения встречаются и на степных и меловых склонах (собственные на
блюдения). Особенно активно F. pennsylvanica внедряется в пойменные леса Ниж
ней Волги, где вытесняет местные виды (личное сообщение И.А. Шанцера).
Размножение и жизненный цикл. Ясень зацветает одновременно с нача
лом распускания листьев во второй половине апреля -- начале мая. Цветение
продолжается 1--2 недели, а отдельное дерево цветет 3--4 дня (рис. 164). Цвете
2 В MW есть более ранний сбор F. pennsylvanica: «сов. Знаменское Московск. у., лес,
8.06.1925 г., В.Ворошилов (как F.ecxelsior), опр. С.Майоров» (MW). Однако из этой этикет
ки неясно, собран образец с деревьев культивируемых или нет.
Рис. 161. Внедрение
Fraxinus pennsylvanica в
естественные сообщества
Рис. 162. Плодоношение
Fraxinus pennsylvanica
3
Рис. 164. Цветение
Fraxinus pennsylvanica
Рис. 165. Усыхание Fraxinus
pennsylvanica в насаждениях
г. Москвы
Рис. 166. Изумрудная златка --
вредитель Fraxinus pennsylvanica
341
ние ежегодное. Полного созревания плоды достигают к концу лета -- началу
осени, при этом цвет крылаток изменяется с зеленого на бурый.
Прорастание надземное. В 1 ый год сеянцы не ветвятся и могут достигать
высоты 30--40 см. В Пенсильвании (США) при благоприятных условиях ясень
достигает высоты 20--30 м при диаметре ствола 20--30 см. Деревья пенсильванско
го ясеня способны к цветению при диаметре 8--10 см и высоте 6--8 м. На Великих
Равнинах достигает стадии плодоношения на 6--7 год [Burns, Honkala, 1990].
Расселение и распространение. Широко используется в городском озеле
нении, так и при создании лесополос, особенно в южных областях Европейской
России [Головач, 1960]. Нередко дает массовый самосев и в последние десятиле
тия стал внедряться в пойменные и нарушенные пригородные леса, чаще в ок
рестностях городов [Овчаренко, Золотухин, 2003]. Впервые самосев отмечен Р.
Регелем: «остр. Валаам (Kl) сорным, по дор. у монаст. у забора III, 1,
27.06(10.07)1917; опр. Н.Цвелев, 2001» (LE). Тем не менее в литературе на дича
ние ясеня указано существенно позднее для Ростовской обл. [Головач, 1960].
Последствия внедрения
Воздействия на естественные фитоценозы и аборигенные виды. Наи
большую активность ясень проявляет на пригородных территориях. Отмечено
внедрение ясеня в прибрежные местообитания, в том числе и на особо охраняе
мых природных территориях [Овчаренко, Золотухин, 2003]. В пределах есте
ственного ареала отмечено угнетающее влияние F. pennsylvanica на местные
Рис. 163. Расселение Fraxinus pennsylvanica в Средней России
342
древесные породы, прежде всего на виды дуба -- Quercus pagoda Raf., Q. phellos L.,
Q. nigra L., Q. texana Buckley, Q. nuttallii Palmer [Oliver et al., 2005]. Подобного
воздействия следует ожидать и в наших условиях.
Генетические изменения и вариабельность вида. Так называемый зеле
ный ясень [var. subintegerrima (Vahl) Fern. -- F. lanceolata, 'green ash'] в крайних
формах хорошо отличается от типичной разновидности (красный ясень -- 'red
ash') более узкими ланцетными листочками, при этом нередко пильчатыми, а
также голыми листьями и молодыми побегами [Preston, 1966; Maggi, Ahles, 1999].
Однако обилие переходных форм и отсутствие резких границ позволяют счи
тать его лишь формой F. pennsylvanica [Rehder, 1949; Preston, 1966; Maggi, Ahles,
1999]. Это противоречит российской традиции дробного понимания видов [Ва
сильев, 1952; Головач, 1960; Цвелев, 2002, 2004].
В Москве на территории 4 й городской больницы отмечено однодомное ра
стение с обоеполыми соцветиями, что отнюдь не характерно для видов этой
секции [Rehder, 1949].
В США на территории Великих Равнин выделено 3 экотипа. Популяции с
засушливой северо западной части более засухоустойчивы, экотип северных
штатов растет медленнее, но обладает большей морозостойкостью, чем популя
ции Восточного побережья. Однако распределение экотипов по всему ареалу не
прослежено [Burns, Honkala, 1990].
Влияние на здоровье человека. В некоторых случаях пыльца ясеня может
вызывать поллиноз [Клиническая..., 2000; Nelson, 1975]. В Москве в пик цвете
ния ясеня (середина мая) концентрация пыльцы может достигать **** пыльце
вых зерен на 1 м3, большую часть этой пыльцы составляет именно
F. pennsylvanica. (********************).
Экономическое и социальное (положительное и отрицательное) зна
чение. Ясень пенсильванский оказался устойчивым к местным условиям ви
дом, широко востребованным в зеленом строительстве. До недавнего времени
он самовозобновлялся преимущественно в городских условиях. При этом не
редко самосев ясеня развивается в массе вдоль дорог, в нарушенных парках,
часто на фундаментах зданий. Экономические затраты на удаление самосева не
подсчитаны.
Его начавшееся внедрение в пойменные ценозы может представлять угрозу
для местной биоты. К тому же в Москве F. pennsylvanica оказался базой для вне
дрения опасного вредителя -- Agrilus planipennis, подлежащего жесткому конт
ролю. Экономический ущерб от повреждения ясеня пока не подсчитан, но огро
мен (см. ниже).
Меры борьбы
Следует исключить пенсильванский ясень из списка деревьев для городско
го озеленения и лесополос. В настоящее время многие питомники предлагают
породу для озеленения.
В 2003 г. в Москве отмечена ясеневая изумрудная узкотелая златка Agrilus
planipennis Fairmaire (A. feretriue Obenberger, A. marcopoli Obenberger) [Волко
вич, 2007; Ижевский, 2007; Мозолевская, 2007; Шанхиза, 2007; Baranchikov at
al., 2008]. Насекомое распространено в Восточной Азии: на Корейском п ове, в
Северо Восточном Китае, Японии, Монголии, на Тайване, в России известно
на Дальнем Востоке в Приморском и Хабаровском краях [Волкович, 2007; Ижев
ский, 2007; Baranchikov at al., 2008]. В пределах естественного ареала узкотелая
343
златка заселяет ясени (Fraxinus chinensis Roxb., F. japonica Blume ex Koch, F.
lanuginosa Koidz., F. mandshurica Rupr, F. rhynchophylla Hance) и некоторые дру
гие породы [Juglans mandshurica, Pterocarya rhoifolia, Ulmus davidiana, U. propinqua)
[Ижевский, 2007; Baranchikov at al., 2008]. В Хабаровске отмечено катастрофи
ческое поражение посадок F. pennsylvanica [Baranchikov at al., 2008].
В пределах вторичного ареала в Северной Америке личинки златки обнару
жены только на ясенях [Baranchikov at al., 2008]. В Москве она также поражает
ясени (рис. 165). Личинки питаются камбиальным слоем, что вызывает усыха
ние деревьев [Волкович, 2007]. «На ранних стадиях (1 й год) заражение злат
кой установить довольно трудно. Основные симптомы заражения -- характер
ные летные отверстия на стволах и главных ветвях -- появляются позднее
(рис. 166). Для обнаружения ходов необходимо удалить кору. Зараженные дере
вья обычно имеют разреженную крону, листья раньше желтеют, вдоль старых
ходов появляются вздутия и трещины, развиваются вторичные побеги вдоль
ствола и главных ветвей. На 3 й год заражения многие ветви отмирают, кроны
сильно разреживаются, имеются многочисленные летные отверстия, стволы
покрыты трещинами» [Волкович, 2007]. Летные отверстия имеют характер
ную D образную форму диаметром до 3--4 мм (рис. 159, 160) [Ижевский, 2007;
Ижевский, Мозолевская, 2008; Haack et al., 2002].
Распространение златки в Москве стремительное. Так, на территории 4 й
городской больницы 2--3 года назад признаков повреждения заметно не было,
но в 2007 г. повреждены и усыхают все растения F. pennsylvanica и 2/3 деревьев
F. excelsior. К настоящему времени A. planipennis широко расселилась не только
по Москве, но и активно распространяется по области. В 2006 г. она найдена в
30 км от МКАД по Рижскому направлению [Шанхиза, 2007]. В последние годы
отмечено резкое уменьшение количества пыльцы ясеня в московском воздухе
(личное сообщение Е.Э. Северовой).
В 1990 х годах в США отмечено усыхание ясеней (F. pensylvanica, F. americana
L., F. velutina Torr., F. nigra Marsh.). Причина стала ясна в 2002 г., когда златка
была достоверно выявлена [Haack et al., 2002; New Threats..., 2003]. В штатах
Мичиган и Огайо заражено уже >2000 миль2 (то есть более 500 тыс. га) ясене
вых лесов и лесопосадок [New Threats..., 2003]. Златка быстро расселяется в
Новом Свете, инвазия приурочена к району Великих озер и с момента первого
обнаружения уничтожено свыше 20 млн деревьев ясеня [Волкович, 2007; Poland,
McCullough, 2006; Baranchikov at al., 2008]. По данным на 7 июля 2007 г., златка
выявлена в США (Огайо, Индиана, Мичиган, Иллинойс, Мэриленд) и Канаде
(Онтарио) (http://www.emeraldashborer.info/files/MultiState_EABpos.pdf). В
США и Канаде A. planipennis отнесен к карантинным видам [Kimoto, Duthie
Holt, 2006]. Только для штата Огайо ущерб оценивается в 1,8--7,6 млн долларов
[Sydnor et al., 2007]. Подробная информация об инвазии размещена на сайте
(http://www.emeraldashborer.info). Для борьбы с вредителем предприняты се
рьезные усилия, в том числе изучение биологии златки и ее паразитов [см., на
пример, Liu et al., 2003; Gates et al., 2005; Yang et al., 2005; Muirhead et al., 2006;
Rodriguez Saona et al., 2006]. Однако остановить златку пока не удается. Ведут
ся даже работы по созданию генетически модифицированных растений
F. pennsylvanica, устойчивых к A. planipennis [Du, Pijut, 2005, 2007]. О важности
для США проблемы инвазии узкотелой златки свидетельствует хотя бы тот
факт, что на сайте американского конгрессмена от Иллинойса М.С. Кирка есть
344
регулярно обновляемая страница о вторжении к A. planipennis (http://
www.house.gov/kirk/Emerald.Ash.Borer2.shtml).
EPPO (European and Mediterranean Plant Protection Organization) предла
гает усилить карантинный контроль для предотвращения проникновения A.
planipennis в Европу (http://www.eppo.org/).
По мнению С.С. Ижевского [2007], «занос (инвазия) A. planipennis в Москов
ский регион, скорее всего, произошел в начале 1990 х годов с посадочным мате
риалом из Северной Америки». Однако это точка зрения кажется нам ошибоч
ной. Более вероятным путем представляется занос, как и в США, с упаковоч
ным лесом -- горбылем или деревянной тарой [Волкович, 2007].
В Москве и России меры карантина не приняты3. Значительная часть ясене
вых насаждений Москвы поражены златкой [Шанхиза, 2007; собственные на
блюдения]. Поражен также местный F. excelsior. Нам не удалось обнаружить ни
одного массива ясеня без златки, значительная часть пенсильванского ясеня
суховершинит. Лишь несколько внешне здоровых деревьев F. pennsylvanica най
дено по 1 му Подольскому переулку. Экономический ущерб от инвазии может
быть огромен. При этом вдоль московских проспектов на место выпавшей липы
в настоящее время активно высаживают пенсильванский ясень.
Необходимо удалить пораженные деревья, вывезти и уничтожить раститель
ные остатки, заменить ясень в городских посадках [Мозолевская, Исмаилов,
2007]. Златка уже выбралась за пределы Москвы. Ее может остановить только
спорадическое распространение F. excelsior на широте Москвы, а массовые по
садки ясеня пенсильванского приурочены в основном к городам--спутникам
Москвы. В пределах естественного ареала узкотелая златка не является стено
фагом [Ижевский, 2007; Ижевский, Масляков, 2008; Ижевский, Мозолевская,
2008], что увеличивает опасность ее дальнейшего расселения. Следует учесть,
что в более южных областях России значительная часть лесополос состоит из
F. pennsylvanica. Необходимы безотлагательные меры по изучению реальной
картины заражения A. planipennis [Ижевский, 2008]. «Но уже сейчас ясно, что
картина угрожающая» [Шанхиза, 2007].
СЕМЕЙСТВО OENOTHERACEAE -- ОСЛИННИКОВЫЕ
КИПРЕЙ ЖЕЛЕЗИСТОСТЕБЕЛЬНЫЙ
Название: Epilobium adenocaulon Hausskn.
Синонимы: E. ciliatum Raf. subsp. ciliatum Hoch et Raven, E. dominii M. Popov,
E. graebneri Rubner. Морфологические признаки
В Северной Америке различают несколько разновидностей, рассматривае
мых иногда в качестве отдельных видов [Gleason, 1963; Scoggan, 1979]: var.
adenocaulon -- стебли прямостоячие, часто высокие, листья, главным образом,
ланцетные, внизу округлые, сидячие или с очень коротким черешком; var.
3 Департамент природопользования и охраны окружающей среды г. Москвы в неявном
виде признает наличие златки на территории Москвы и необходимость контроля ее чис
ленности (http://www.moseco.ru/ru/showArticle/atlID/65).
345
occidentale Trel. -- в западной части ареала, листья узколанцетные; var. perplexans
Trel.(=E. americanum Rydb.) -- в северо западной части ареала, стебли слабые,
прямостоячие или приподнимающиеся (восходящие), высотой 1--3 дюйма, с
редкими железками, листья в основании сужены в заметный черешок (рис. 167).
Белоцветковая форма -- E. adenocaulon f. albiflorum G.Bernard, найденная в
провинции Квебек [Bernard, 1968], по видимому, должна быть отнесена к E.
rubescens.
Естественный ареал
Родина E. adenocaulon -- Северная Америка, где его ареал простирается от
Ньюфаундленда и Аляски до Калифорнии и Огайо [Scoggan, 1979].
Вторичный ареал
История интродукции и географическое распространение. E. adenocaulon впер
вые собран в Британии в Лестершире в 1891 г. [Ash, 1953] и произрастал там
более или менее стабильно в течение 40 лет, не расширяя своего ареала. Широ
кое распространение на юге Англии началось лишь в 1930 е гг., когда вид нату
рализовался в графстве Суррей. С того времени ареал E. adenocaulon стал стре
мительно расширяться, и распространение вида приобрело в 1950 е гг. взрыв
ной характер. К 1942 г. он достиг Уэльса, к 1957 г. -- Шотландии, к 1958 г. --
Ирландии [Preston, 1988], к 1990 г. спонтанно появился на шотландском о. Mull
[Clark, Jermy,1990].
В 1904 г. вид собран на юге Швеции [Скворцов, 1995], но там тоже долго не
расширял ареал, по крайней мере, в 1970 е гг. отмечался как довольно редкий, и
на юго востоке Швеции был найден всего в 4 местообитаниях [Nylander, 1973].
В 1910 г. E. adenocaulon обнаружен в Финляндии; до 1920 г. в регионе сделана
еще лишь одна находка кипрея, а с конца 1920 х гг. его ареал в Восточной Скан
динавии стал бурно расширяться [Eklund, 1927--1928; Piispala, 1964].
В 1917 г. E. adenocaulon зарегистрирован в Польше, в Беловежской Пуще
[Hegi, 1906 1931]; в 1947 г. -- в Вестфалии [Wittig, Pott, 1980], в 1937 г. [Holub,
Vicol, 1971] -- в Румынии (но очень редко -- до 1960 х гг. обнаружен только 6
раз), в 1952 г. -- во Франции [Ash,1953]. В 1950 х гг. вид встречался уже в Бель
гии, Чехии, Словакии, Дании, Голландии и Норвегии [Raven, 1968].
Помимо Европейского региона, E. adenocaulon обосновался на Гавайских о
вах, в Австралии и Новой Зеландии [Raven, Raven, 1976]. В Новую Зеландию
растение занесено из Британии (а не из Северной Америки) не позднее 1896 г.,
когда оно было собрано на Северном о ве, где широко распространилось уже
через 11 лет. В 1912 г. E. adenocaulon собран на Южном о ве и продвинулся до его
южной оконечности к 1950 м гг., когда его зарегистрировали и на о ве Стьюарт.
В австралийском регионе E. adenocaulon обильнее всего в Тасмании, где он
впервые найден в 1918 г. На австралийский материк, в штат Виктория, он по
пал в 1964 г. Кроме того, он известен как сорняк садов в окрестностях Сиднея,
Канберры, Аделаиды и Брисбена [Raven, Raven, 1976].
В России вид впервые собран в самом конце XIX века в Псковской губернии,
но не был как следует распознан до 1927 г., когда приобрел уже довольно значи
тельное распространение [Скворцов, 2005].
В 1918 г. вид найден в Риге [Hegi, 1906--1931], затем последовали единичные
его находки в Лиепае и Усме [Bumbure, 1957. p. 342]. В Эстонии E. adenocaulon
встречался «рассеянно по всей республике» [Talts, 1959. p. 412]. Спорадически
встречался по всей Ленинградской обл. [Флоровская, 1961]. В Литве к 1971 г.
346
достоверных его находок не было. Имелось лишь указание Рубнера, что вид
растет в лесу между г. Паланга и Лиепая [Natkeviиaite Ivanauskiene, 1971. p.765].
Изредка вид встречался в Белоруссии (собран в 1917 г. в Себежском уезде
Витебской губернии, МНА) и в лесных районах Украины [Доброчаева, 1955].
В Карелии отмечен в Приладожском, Межозерском, Шокшинском, Заонеж
ском р нах и на Ругозере [Раменская, 1960]. Две находки сделаны в Мурманс
кой обл.: Миддендорфом в районе Кольского залива и Шляковым в Апатитах
[Шляков, 1959].
C середины 1960 х гг. E. adenocaulon начал стремительно завоевывать новые
территории. Собран А.К. Скворцовым на Урале (MW). Многочисленные наход
ки отмечены на северо востоке [Скворцов, 1976]: Соловецкие о ва, Онега (Засо
лье, Вага, Пинега, Мезень, Усогорск), Вычегда, Ухта, Печора (ст. Козлавом).
В 1970--1980 х гг. E. adenocaulon найден в окрестностях Перми [Овеснов,
Шилова, 1975], встречен в Марийской АССР и на северо западе Оренбургской
обл. [Папченков, 1984], отмечен С.В. Баландиным [1989] в заповеднике «Басеги»
на Среднем Урале. Увеличился ареал вида в Литве: в 1984 г. его местонахождения
отмечены в Лаздийском р не, в Вильнюсе и Шяуляе [Матекайтите, 1985].
Стремительно расширялся ареал вида на Вологодчине: в 1982 г. кипрей же
лезистостебельный обнаружен нами только в г. Вологда [Майтулина, 1984], а
через 3--4 года он встречался уже по всей области [Орлова, 1989].
В 1995 г. отмечен как компонент флоры Алтая [Терехина, 1995].
В настоящее время вид распространен по всей Восточной Европе, кроме
Крайнего Севера. Редко встречается на Урале и в Сибири; рассеянно -- на юге
Дальнего Востока, где часто смешивается с местным E.maximowiczii [Скворцов,
2005]. В Финляндии стал бурно расширять ареал через 20 лет, а в Европейской
России -- только через 50 лет после первого появления.
Пути и способы заноса. Семена разносятся ветром.
Статус в регионе. Впервые собран в Московской обл. «близ дер. Подушкино
Звенигородского уезда Московской губернии. В сырой придорожной канаве в
значительном числе особей. 6.08.1909 г. А.А.Хорошков. MW». В последующем
обнаружен во Владимирской обл. «дер. Жердево Покровскаго у. Владимирской
г., в кустарнике. 06.07.1912. MW» и в окрестностях Тамбова «на болоте по р.
Цна. 29.06.1921 г. П.Смирнов. MW». В 1924 г. находки E. adenocaulon сделаны
З.Снятиновской в Ивановской обл. «Иваново Вознесенский у. и губ. д. Сергель
ское, по разреженному смешанному лесу. MW» и П.Смирновым в Тульской обл.
«Тульская губ., Веневский у. Близ дер. Гурьево. Сырой известковый берег р.
Осетра. MW».
В 1920 е гг. гербарные сборы кипрея железистостебельного сделаны на тер
ритории Москвы и Московской обл. 9 раз [Игнатов и др., 1990]. До середины
ХХ века вид дважды собирали в Калининской обл. [Невский, 1952] и отмечали
для Ярославской обл. [Определитель..., 1961]. В Рязанской обл. найден в 1953 г.
«Ерахтурский р н. В 3 км на юг от с. Ерахтур. Сосново брезовый молодой лесок
на песчаной террасе. 14.07.1953 г.. Собр. П.Жудова, опр. В.Тихомиров. MW».
C начала 1970 х гг. E. adenocaulon начал стремительно завоевывать новые
территории (рис. 168): найден в Орловской обл. [Еленевский, Радыгина,1984]
и на Верхней Волге [Лисицына, Экзерцева, 1984]. В 1969--80 гг. E. adenocaulon
впервые встречен на Приволжской возвышенности в Чувашской и Татарской
АССР, Ульяновской и Куйбышевской обл. [MW; Папченков, 1984]. Распростра
Рис. 167. Цветение
Epilobium adenocaulon
Рис. 168. Расселение
Epilobium adenocaulon
в Средней России
Рис. 169. Epilobium adenocaulon
в трещинах асфальта
348
Рис. 170. Лесная вырубка с зарослями Epilobium adenocaulon,
окрестности Звенигорода
349
нился кипрей и в Смоленской (1974 г., МНА), Калужской (1973 г., МНА) и
Брянской (1973 г., МНА) областях.
В 1983 г. обнаружен в Саратовской обл. [Березуцкий, Дайковский, 1988], от
мечен в Липецкой обл. в ивняках и ольшаниках в поймах р. Воронеж, Дон, Ста
новая Ряса [Вьюкова, 1987] и в Воронежской обл. [Григорьевская, 1987].
Местообитания. Фенотипическая изменчивость помогает кипрею занимать
широкую амплитуду местообитаний -- вид растет на легких и тяжелых, на изве
стковых и неизвестковых почвах.
Во вторичном ареале проявляет такую же широкую пластичность, как на
родине: растет на почвах различной степени кислотности и разного механичес
кого состава. Чаще всего E. adenocaulon распространен в сообществах открыто
го типа, на илистых грунтах по берегам водоемов и в лесах. Встречается также
на различных вторичных, антропогенных местообитаниях: у дорог, канав, на
ж/д откосах, а нередко и как сорное растение в садах и огородах (рис. 169). Интен
сивно растет на лесных трелевочных дорогах, где уничтожен травяной и моховой
покровы и перемешаны почвы с оголением подзолистого горизонта (рис. 170). В
Австралии кипрей расширяет ареал вдоль дорог [Raven, Raven, 1976].
Размножение и жизненный цикл. E. adenocaulon -- самоопылитель с высокой
степенью гомозиготности, однако имеется слабая способность к перекрестному
опылению. Двулетник, но у раннелетних проростков семена образуются уже в
первый год жизни. Осенние проростки образуют розетку и цветут на 2 ой год.
Вегетативный способ размножения осуществляется дочерними розетками,
которые образуются во время цветения на коротком корневище у основания
побега и перезимовывают, давая на следующий год новые побеги.
Расселение и распространение. Кипрей железистостебельный распространя
ется в основном семенами. Средний вес семени 0,062 мг [Myerscough, Whitehead,
1967]. С учетом способности вида давать семена в 1 ый год жизни, можно утвер
ждать, что кипрей приспосабливался к новым условиям существования в тече
ние 50 смен поколений, и со времени широкого его распространения в Европей
ской России прошло не менее 30 смен поколений.
Последствия внедрения
Воздействие на естественные фитоценозы и аборигенные виды. Интересны
проведенные в Британии исследования биологических особенностей E.
adenocaulon в сравнении с другими видами, заселяющими пионерные местооби
тания -- Tussilago farfara, Chamenerion angustifolium и Epilobium montanum
[Myerscough, Whitehead, 1966, 1967]. Оказалось, E. adenocaulon превосходит эти
виды по способности прорастать при низких (10оС) температурах, низком уров
не освещения, а также в засушливых условиях.
В фитосоциологическом отношении E. adenocaulon лежит в пределах поряд
ка Convolvuletalia sepii. В Западной Европе он встречается, главным образом, в
сообществах с господством Bidens, Alopecurus aequale, Ranunculus scleratus, но
также в зарослях тростника, Eleocharis palustris, Typha latifolia, крупноосочни
ках и других сообществах открытого типа [Wittig, Pott, 1980].
Генетические изменения и вариабельность вида. Диплоид, 2n=36 [Raven, Raven,
1976]. Может образовывать искусственные гибриды, способные давать генера
цию F2 с 14 аборигенными новозеландскими таксонами рода Epilobium. Разли
чие в хромосомном наборе между E. adenocaulon и этими видами (1 реципрокт
ная транслокация) не влияет на фертильность [Raven, Raven, 1976].
350
Более того, во вторичном ареале в районе Австралии и на прилегающих
островах обнаружены единичные растения, являющиеся естественными гиб
ридами между E. adenocaulon и 6 аборигенными видами. При этом один из гиб
ридов -- E. adenocaulon × E.brunnescens -- начал распространяться, размножаясь
как вегетативным путем, так и (очень редко) семенами [Raven, Raven, 1976].
В Скандинавии найден естественный гибрид E. adenocaulon × E.obscurum
[Oredsson, 1964], в Чехословакии -- E. adenocaulon × E.hirsutum [Smejkal, 1974]. В
России также отмечены естественные гибриды E. adenocaulon с E.montanum,
E.roseum и E.palustre [Скворцов, 1995].
П.А. Смирнов описал во вторичном ареале форму pumila P.Smirnov высотой
1--25 см [Штейнберг, 1949].
Экономическое и социальное (положительное/отрицательное) значение.
Сокращает биоразнообразие естественных сообществ.
Меры борьбы
Сорные экземпляры в огородах и на дачных участках легко удаляются при
прополке. На лесных просеках меры борьбы не проводят. Сведениями о приме
нении химических способов воздействия на этот инвазионный вид авторы не
располагают. Обзор научных исследований и публикаций
Генетическая закрепленность морфологических признаков E. adenocaulon и
амплитуда их внутривидовой изменчивости изучена в интродукционной попу
ляции, заложенной в однородных условиях из семян различного географичес
кого происхождения [Виноградова, 1992]. Выявлено, что во вторичном ареале в
Европе E. adenocaulon имеет очень низкую генетическую вариабельность: раз
личий по феноритму не наблюдается, а изменчивость ритма роста выражена не
вполне отчетливо. При выращивании E. adenocaulon в однородных условиях в
теплицах также отмечена малая генетическая вариабельность [Myerscough,
Whitehead, 1966], которую авторы объясняют особенностями биологии цвете
ния видов рода кипрей.
СЕМЕЙСТВО OENOTHERACEAE -- ОСЛИННИКОВЫЕ
КИПРЕЙ ЛОЖНОКРАСНЕЮЩИЙ
Название: Epilobium pseudorubescens A. Skvorts.
Синонимы: E. rubescens auctt. non Rydb., E ciliatum Raf. subsp. ciliatum Hoch et
Raven.
Морфологические признаки
Очень близок к E.adenocaulon, и рядом авторов рассматривался ранее как
его белоцветковая форма [Raven,1968; Фатаре, 1979]. Однако исследованиями
А.К. Скворцова [1973] установлено, что E. pseudorubescens отличается более уз
кими ланцетными листьями, белыми цветками с более узкими лепестками, пре
обладанием самоопыления и в среднем более крупными семенами (рис. 171).
Долгое время этот вид принимался за E. rubescens Rydb. Однако сравнение с
типом последнего (NY) показало ошибочность такого определения; поэтому не
исключена возможность недавнего происхождения E. pseudorubescens из амери
канских корней -- но в Европе [Скворцов, 1995].
351
Рис. 172. Расселение Epilobium pseudorubescens в Средней России
Рис. 171. Цветение Epilobium
pseudorubescens
Рис. 173. Epilobium pseudorubescens --
огородный сорняк
352
Рис. 174. Внедрение Epilobium pseudorubescens в естественные сообщества
353
Естественный ареал
Предковые формы имеют естественный ареал в Северной Америке.
Вторичный ареал
История интродукции и географическое распространение. Впервые собран в
Финляндии в 1915 г. [Piispala, 1964]; к 1930 г. в Южной Финляндии известно
только 4 местонахождения вида. Постепенное расширение его ареала началось
через 15--20 лет после первой находки.
В Эстонии известен с 1925 г. [Гусев, 1980], но встречался очень редко [Talts,
1959. p. 418]. Единичные находки отмечены в Латвии [Bumbure, 1957. p. 345]. На
остальной территории СССР до Великой Отечественной войны, по видимому,
отсутствовал [Скворцов, 1973].
К 1950 г. найден в Карелии в г. Сортавала и Петрозаводск [Раменская, 1960, 1983].
Сведения о дальнейшем расширении ареала вида стали публиковаться только
после идентификации его А.К. Скворцовым [1973]. До того E. pseudorubescens,
по видимому, не отграничивали от E.adenocaulon. Ю.Д. Гусев собрал E.
pseudorubescens в 1967 г. в Пушкине и Приморске, а в 1978 г. -- между ст. Ульянка
и Броневая Ленинградской обл. [Гусев, 1980]. В 1980 г. E. pseudorubescens приве
ден для флоры Налибокской Пущи в Белоруссии [Сауткина, 1983]. Нами най
ден в 1982 г. в Вологде [Майтулина, 1984].
Ареал вида расширяется к востоку России: в 1988 г. он отмечен экспедицией
ГБС в окрестностях Читы и на Амуре между Хабаровском и Комсомольском
(МНА).
Пути и способы заноса. Вероятно, заносится с садовой землей. Семена разно
сит ветер.
Статус в регионе. В 1959 г. собран А.К. Скворцовым на юго западе Москвы и
в пос. Жаворонки Московской обл.. За последующие 10--15 лет вид распростра
нился по всей Средней России (рис. 172). Историю его расселения можно про
следить по нижеперечисленным гербарным сборам:
Рязанская обл., Спасский р н. Окский гос. заповедник, кв. 14. Сырой черно
ольшаник. 07.07.1967 г. В.Тихомиров, В.Артеменко (MW);
Владимирская обл., Ковровский р н. Ольшаник в пойме р. Нерехта выше
Бабенк. моста. 19.07.1967 г. А.Бутряков. (MW);
Тульская обл., Заокский р н. Выходы грунтовых вод в долине р. Скнижка у д.
Выдумки. 14.06.1973 г.. Ю.Алексеев (MW);
Калужская обл., Жуковский р н., луг 3 км юго западнее дер. Дроздово.
3.07.1975 г. В.Васильков, Н.Голубева (MW);
Брянская обл. Придорожная канава близ пос. Батагово. 16.08.1976 г. Ю.Алек
сеев (MW);
Липецкая обл., Данковский р н, правый берег р. Дон против с. Ново Ни
кольское. У воды. 30.06.1984 г.. В.Тихомиров, В.Никифорова, А.Белов (MW);
Ивановская обл., 1991 много сборов Е.Борисовой (MW);
Костромская обл. Павинский р н. 7 км к СЗ от с. Павино. Левый берег р.
Пызмас. Вырубка на месте елового леса. 18.06.1991 г.. Н.Г. Прилепский (MW);
Нижегородская обл. Борский р н. В 1 км к югу от пос. Рустай. Заповедник
Керженский, кв.128. Вырубка. 21.07.1997. Н.М. Решетникова, С.П. Урбанавиуте
(MW);
Смоленская обл., Демидовский р н. В д. Покровское. В огороде. 09.07.1999 г.
Н.Решетникова (MW);
354
Курская обл., Железногорский р н. 0,5 км к востоку от д.Гнань. Ключевое
болотце у подножия склона долины р. Свапа. 20.07.2004 г. А.Полуянов (MW);
В настоящее время встречается во всех регионах Средней России.
Местообитания. В Московской обл. E. pseudorubescens -- обычный вид, рас
тет, главным образом, в более или менее сухих рудеральных местообитаниях,
на насыпях, в городах, в трещинах асфальта [Игнатов и др., 1990]. Встречается
и на других нарушенных местообитаниях: по берегам водоемов, в засоренных
лесах, в садах, на сорных местах (рис. 173, 174). По сравнению с предыдущим
видом заметно более теневынослив.
В Рязанской обл. уже с начала 1980 х гг. имел тенденцию к активному про
никновению в естественные фитоценозы [Тихомиров, 1982; Гущина, Казакова,
1989]. В Калужской и Брянской обл. отмечен как на сорных местах, так и в
составе естественных сырых лугов [Алексеев, Макаров, 1981]. В Липецкой обл.
встречается в заболоченных дубравах и по берегам водоемов [Вьюкова, 1987].
Размножение и жизненный цикл. Кипрей краснеющий -- облигатный само
опылитель. Двулетник, однако раннелетние проростки цветут и дают семена
уже на 1 ом году. Может размножаться вегетативно дочерними розетками так
же, как E.adenocaulon.
Расселение и распространение. В лесах расселяется вдоль тропинок, треле
вочных дорог и по вырубкам.
Последствия внедрения
Воздействие на естественные фитоценозы и аборигенные виды. На вырубках
соседствует и конкурирует с аборигенными растениями -- пионерами откры
тых растительных сообществ.
Генетические изменения и вариабельность вида. При стабильном генотипе E.
pseudorubescens обладает значительной фенотипической изменчивостью, помо
гающей ему занимать широкую амплитуду местообитаний.
Экономическое и социальное (положительное/отрицательное) значение. Та к
же как E.adenocaulon, является сорным растением. Сокращает биоразнообразие
естественных ценозов.
Меры борьбы
Поскольку серьезного экономического ущерба вид не наносит, борьба с ним
ведется только в локальных масштабах -- на дачных участках и огородах, где E.
pseudorubescens удаляется при прополке.
Обзор научных исследований и публикаций
Амплитуда внутривидовой изменчивости изучена в интродукционной по
пуляции, заложенной в однородных условиях на экспериментальном участке
ГБС РАН из семян, собранных в Москве и Вологде [Виноградова, 1992]. При
осеннем посеве не отмечено достоверных отличий между образцами ни по тем
пам роста, ни по фенологическому ритму развития. Растения зацвели одновре
менно в конце июля, через 4 недели образовали семена и оставили зимующие
розетки, которые на следующий год так же дружно вступили в генеративный
период развития и погибли. Таким образом, кипрей ложнокраснеющий являет
ся двулетником, формируя розетки при коротком дне и цветоносные побеги
при длинном дне.
355
СЕМЕЙСТВО OENOTHERACEAE -- ОСЛИННИКОВЫЕ
ЭНОТЕРА ДВУЛЕТНЯЯ или ОСЛИННИК ДВУЛЕТНИЙ
Название: Oenothera biennis L.
Синонимы: Onagra biennis Scop., Onagra vulgaris Spach,
Морфологические признаки
Двулетнее растение со стержневым толстым корнем и прямостоячим стеб
лем высотой до 1,5 м. Стебель обычно простой, иногда ветвится в верхней части;
негусто опушенный (как чашечка и коробочка) короткими железистыми и бо
лее длинными простыми светлыми волосками, густо олиственный, в нижней
части иногда краснеет или имеет красные матовые пятна. Прикорневые листья
длиной 10--20 см, шириной 2--6 см, длинночерешковые, продолговато яйцевид
ные или эллиптические, слабовыемчато зубчатые, почти цельнокрайные, за
канчиваются острием. Стеблевые листья очередные, на коротких черешках,
верхние почти сидячие, продолговато ланцетные, в основании клиновидные,
тонкозубчатые, покрытые с верхней и нижней сторон прижатыми мелкими во
лосками. Cредняя жилка красная. Длина стеблевых листьев составляет 5--7 см,
ширина ~2 см.
Соцветие -- длинная кисть с крупными желтыми актиноморфными цветка
ми в пазухах мелких прицветных листьев (рис. 175). Лепестки широкояйцевид
но треугольные, слегка выемчатые, краями налегающие друг на друга; длиной
до 20 мм, длиннее тычинок. При цветении доли чашечки отогнуты вниз. Рыльце
четырехраздельное. Плод -- локулицидная 4 гнездная коробочка длиной 2--4
см, в нижней части более толстая, чем в верхней. Семена многочисленные, гори
зонтальносидящие, неправильноугловатые, мелкие, с кожистой оторочкой, от
темно серого до темно коричневого цвета, длиной ~1,5 мм [Штейнберг, 1949;
Маевский, 2006].
Естественный ареал
Широко встречается на территории Северной Америки от 60°с. ш., кроме
штатов Аризона, Юта, Колорадо, Айдахо, Вайоминг [www.plants.usda.gov], од
нако преобладает на востоке материка [Dietrich et al., 1997].
Вторичный ареал
История интродукции и естественное распространение. O.biennis за
везена в Европу из Северной Америки в 1614 г. Растение культивировалось в
садах из за ароматных цветков, а затем широко распространилось как сорное
[Dietrich, Wagner, 1988]. Из Западной Европы O.biennis проникла в европейс
кую часть России и распространилась как сорняк в южных и центральных рай
онах [Тохтарь, 2003]. Севернее встречается крайне редко [Скворцов, 1996]: пос
ле первой находки в Вологодской обл. в 1928 г. долгое время никаких упомина
ний о виде не было, вторая находка нескольких цветущих экземпляров сделана
в Вологде лишь в 1982 г. [Майтулина, 1984].
Пути и способы заноса. Распространению энотеры способствовало разве
дение в садах. Распространяется по ж/д путям (перевозится с грузами и т.п.),
может расти на ж\д насыпях [Тохтарь, 2003].
Статус в регионе. Первая находка в Московской обл. относится к 1824--
1826 гг. (MW). В конце XIX столетия энотера отмечалась в Белгородской, Во
356
ронежской, Курской, Липецкой, Пензенской, Саратовской, Тамбовской Орлов
ской и Ивановской обл. (LE, MHA, MW). Неясны причины гораздо более по
здних сборов энотеры в Смоленской и Калужской обл.: «Смоленская обл. у с.
Слобода (Пржевальское) в бору на песчаной прогалине. 13.07.1962. А.Сквор
цов. MHA», «Калужская обл., Ферзиковский р н. с. Кольцово у Оки. Песок в
долине Оки. 04.09.1971. А.К. Скворцов. MHA». В настоящее время встречается
по всей Средней России, в южных регионах массово (рис. 176).
Местообитания. На открытых песках, особенно в долинах крупных рек, а
также иногда у дорог, на насыпях и изредка как сорное [Маевский, 2006]. Зак
репляет почву, являясь пионерным растением техногенных экотопов.
Размножение и жизненный цикл. Двулетний, реже многолетний травяни
стый монокарпик (рис. 177). В первый год образует только вегетативную ро
зетку, прилегающую к почве. В средней полосе России цветет в июне--июле
[Штейнберг, 1949; Маевский, 2006]. Плодоношение наступает в августе--сен
тябре, массовое созревание семян в сентябре--октябре (рис. 178). В среднем на 1
генеративном побеге формируется 84±16 плодов, 3--7% из которых не успевают
созреть [Климахин и др., 2005].
Для вида отмечены и анемогамия, и энтомогамия, и автогамия [Rostaсski et
al., 2004].
Расселение и распространение. Плод содержит многочисленные мелкие
семена. Масса семян в одной коробочке колеблется от 0,05 до 0,17 г. Установлена
достаточно высокая семенная продуктивность -- 8,1±1,7 г на 1 побег. Семенам ос
Рис. 176. Расселение Oenothera biennis в Средней России
Рис. 175. Цветение Oenothera biennis
Рис. 178. Плодоношение
Oenothera biennis
Рис. 177. Oenothera biennis
358
линника свойственно состояние органического покоя [Климахин и др., 2001]. Семе
на распространяются посредством анемохории, барохории и эндозоохории.
Последствия внедрения
Воздействие на естественные фитоценозы и аборигенные виды. Вне
дряется во вторичные местообитания, и не отмечен в нетронутых лесных или
луговых сообществах [Ульянова, 2005].
Генетические изменения и вариабельность вида. Диплоидный набор хро
мосом 2n = 14, 28, 42, 56 [Dietrich et al., 1988].
Вступает в гибридизацию с другими видами рода Oenothera, чаще с видами
секции Oenothera. В Северной Америке встречается O.villosa subsp. strigosa, ге
ном которого схож с геномом O.biennis [Dietrich, 1997].
Морфологические признаки в различных популяциях довольно сильно ва
рьируют. При этом популяции, произрастающие в Германии и в Польше, раз
личаются между собой меньше, чем популяции из Польши и степной зоны Рос
сии и Украины (табл. 17), что может быть объяснено бульшим расстоянием,
разделяющим последнюю пару географических р нов. Предварительный ана
лиз z трансформированных корреляционных матриц исследованных призна
ков в различных географических популяциях O.biennis свидетельствует о буль
шем влиянии на их корреляционную структуру экологических, нежели геогра
фических, факторов [Тохтарь, 2003; Rostaсski et al., 2004].
Отмечено, что O.biennis, характеризующаяся средними размерами цветка,
способна произрастать в различных условиях и обладает более широкой амп
литудой изменчивости, чем другие виды энотер как с крупными, так и с мелки
ми цветками [Тохтарь, 2003].
Влияние на здоровье человека. Лекарственным сырьем являются семена
энотеры (Semen Oenotherae, Semen Onagrae), которое заготовляют осенью пос
ле 2 ого года вегетации, в период созревания плодов. В связи с неравномерным
созреванием семян сначала заготавливают спелые, полуспелые и зеленоватые
плоды, которые потом высушивают естественным способом или в сушильнях
при температуре не более 50°С. Затем высушенные семена отделяют от плодов
и при необходимости досушивают. Из сухих семян энотеры методом экстрак
ции органическими растворителями или методом горячего прессования полу
чают жирное масло золотистого цвета, которое используют для изготовления
лекарственных средств. Зрелые семена содержат от 10 до 20% жирного масла,
Таблица 17
Амплитуда изменчивости таксономических признаков генеративных органов
O.biennis в различных инвазионных популяциях (Ч -- Чехия; Р -- Россия;
П -- Польша; Г -- Германия (по данным В.Тохтаря, неопублик.)
Длина
лепестка,
мм
Ширина
лепестка,
мм
Длина
гипантия Длина
пыльника
Длина
нити
пыльника
Длина
верхушек
чашелистиков
14--29 (Ч)
20--30 (Р,
П)
15--30 (П)
12--25 (Г)
16--37 (Ч)
14--27 (Г)
24--35 (П)
24--38 (Ч)
25--40 (Г)
25--40 (П)
7--10 (Р)
5--12 (Ч)
5--10 (П)
9--17 (Ч) 2,5--7,5 (Ч)
2,5--3 (Г)
359
состав которого зависит от климатических и агрономических условий выращи
вания растения. Благодаря содержанию г линоленовой кислоты его рекомен
дуют при заболеваниях, сопровождающихся сниженным содержанием кисло
ты и ее метаболитов (дигомо г линоленовой и арахидоновой кислот) в плазме
крови и жировой ткани [Куцик, Зузук, 2005].
Экономическое и социальное (положительное/отрицательное) значе
ние. Изредка культивируется как декоративное растение [Маевский, 2006].
Отварные корни съедобны. Также используются цветки (для приготовления
салатов) и, по некоторым источникам, молодые побеги [Facciola, 1990].
Меры борьбы
O.biennis практически не переходит из рудеральных местообитаний на поля
и, как правило, растет в условиях, где может составить конкуренцию лишь псам
мофитам, а не культурным растениям, поэтому специальных мер по борьбе с
этим видом пока не предложено.
Обзор научных исследований и публикаций
Имеются данные о существовании вида в условиях повышенного радиаци
онного фона [Дощечкина, 1989].
СЕМЕЙСТВО OXALIDACEAE -- КИСЛИЧНЫЕ
КИСЛИЦА ПРЯМОСТОЯЧАЯ
Название: Oxalis stricta L.
Синонимы: O. fontana Bunge, O. europaea Jordan, Xanthoxalis stricta (L.) Small,
X. fontana (Bunge) Holub.
Морфологические признаки
Растение более или менее опушенное, с редкими белыми прижатыми волос
ками; корневище тонкое, ползучее (рис. 179). Стебель прямостоячий, простой
или ветвистый, высотой 15--45 см, красноватый или более или менее багрянис
тый, большей частью опушенный. Листья очередные, тройчатосложные, без
прилистников, черешки прямые, в основании с сочлененим, от почти голых до
густо опушенных многоклеточными волосками, длиной 2,5--6(8) см. Листочки
обратносердцевидные, голые или иногда волосистые, по краю реснитчатые с
маленькими, длиной 0,3 мм, опушенными черешочками. Цветочная стрелка па
зушная, в основании сочлененная, опушенная, длиной 4,5--5 см с ланцетными
прицветниками длиной 1,5--2 мм и шириной 0,3 мм, расположенными в основа
нии 3--6(8) слабоопушенных цветоножек длиной 4--6 мм. Чашечка длиной 4 мм,
почти в 2 раза короче венчика, чшелистики ланцетные, длиной 3--3,5 мм и ши
риной 0,7 мм, с белыми прямыми волосками. Венчик почти колокольчатый, ле
пестки желтые, длиной 5,5--8 мм и шириной 1,5--3 мм, ноготок прямой, пластин
ка лепестка продолговато обратнояйцевидная; тычиночные нити без зубцов, в
основании сросшиеся, внутренние почти голые или вверху более или менее ред
коволосистые, на 1/3 длиннее наружных, голых; пыльники продолговатые, жел
тые, налегающие; коробочка продолговатая, ребристая, длиной 12--20 мм, по
крытая отстоящими волосками. Семена яйцевидные, сплюснутые, длиной 1--
1,3 мм и шириной 0,7 мм, коричневые, поперек ребристые [Горшкова, 1949].
360
К O. stricta морфологически и экологически близки два вида: O. dillenii Jacq. ---
кислица Дилениуса и O. corniculata L. --- кислица рогатая, с которыми ее нередко
путают. От O.stricta их можно отличить по опушению стебля и коробочек. У
O.stricta коробочки с редкими отстоящими многоклеточными волосками или
почти голые, стебель чаще голый, а иногда опушен оттопыренными многокле
точными волосками (f. villicaulis Weig.). У O.dillenii и O.corniculata коробочки,
как правило, с густыми прижатыми одноклеточными волосками, а стебель опу
шен прижатыми волосками [Маевский, 2006; Eiten,1963; Lovett et al., 1985].
Естественный ареал
Северная и Центральная Америка. Повсеместно распространена в Канаде и
восточных районах США [Горшкова, 1949].
Вторичный ареал
История интродукции и географическое распространение. Отмечена
как чужеродный вид в европейской части России, на Украине, в Беларуси и
странах Балтии, в Западной Сибири, на Дальнем Востоке, в Средней и Южной
Европе, Средиземноморье, Японии, Корее, Китае [Горшкова, 1949; Quanru,
Watson, 2008].
В северо западных регионах России -- пока относительно редкий вид: извес
тен лишь из окрестностей Санкт Петербурга и Луги как заносный в садах и
парках, у дорог, в культуре редко. Есть сорта с красными листьями [Цвелев,
2000]. В Вологодской обл. во множестве растет по цветникам.
Пути и способы заноса. Вероятнее всего, первичный занос сделан с семена
ми декоративных растений закрытого грунта и почвой для теплиц. В качестве
декоративного растения кислицу выращивают в цветниках.
Статус в регионе. В Белгородской обл. редко, имеются сборы из Грайво
ронского (MW) и Шебекинского р нов [Еленевский и др., 2004].
В Воронежской области единичные экземпляры O.stricta впервые отмечены
С.В. Голицыным [1961] в 1946--1947 гг. в дубраве Воронежского заповедника.
Впоследствии вид стал распространяться на заповедной территории Усманс
кого бора по обочинам дорог и просекам, тогда как в незаповедной части лесно
го массива O. stricta является сорняком огородов на песчаной почве [Григорьев
ская и др., 2004]. В Воронеже вид отмечен впервые сравнительно недавно [Ба
рабаш, Камаева, 1989], но быстро распространяется по городской территории
по газонам и рудеральным местам [Григорьевская, 2000]. В последнее время
вошли в культуру сорта с темно красными листьями, которые быстро дичают и
легко образуют гибридные формы [Хлызова, Агафонов, 2003].
Первые указания для Курской обл. относятся к 1970 м гг.: «Глушковский р н,
с. Теткино, школьный огород»; «с. Карыж, саженый сосняк на надпойменной
террасе р. Сейм, 1972 г.»; «г. Курск, на сорных местах, 1972 г.». Отмечен также в
Курском и Льговском р нах [Кузнецова, Макаренко, 1976; Полуянов, 2005]. В
Курске на ул. Пирогова А.В. Полуяновым 7.06.2002 г. одновременно собраны и
зеленолистная, и краснолистная формы вида (MW).
Для Орловской обл. один из первых сборов сделан В.Н. Хитрово: Хотынец,
13.06.1920 г. (OHHI). Впоследствии кислицу находили в Дмитровском и Ор
ловском р нах [Еленевский, Радыгина, 1997]. По нашим наблюдениям, в июле
2008 г. на юго востоке Орловской обл. O.stricta -- редкий вид, высаживается на
цветниках, из посадок, как правило, не выходит, преобладает краснолистная
361
Рис. 179. Цветение
Oxalis stricta
Рис. 180. Расселение Oxalis stricta в Средней России
Рис. 181. Плодоношение Oxalis stricta
Рис. 182. Oxalis stricta -- огородный сорняк
363
форма. Зеленолистную форму мы нашли лишь на территории старого кладби
ща в г. Ливны.
Во «Флоре Липецкой области» [1996] O.stricta указывается для трех пунк
тов: Липецк, Добринка, Усмань (сборы К.И. Александровой и Н.А. Вьюковой;
1980, 1981: MW, MOSP, ГЛПИ). Сейчас обычный вид в городах, крупных насе
ленных пунктах, преобладает краснолистная форма (Н.Ю. Хлызова, личное
сообщение).
В Тульской обл. вид впервые собран на окраине г. Ефремов, 1894 г., Н.В.
Цингер (MW), зеленая форма. Собран А.И. Алюшиным в 1980 г. в Туле, в рай
оне Мясново (ТГПУ). В 1990 е гг. O.stricta отмечалась И.Шереметьевой на цвет
нике в г. Тула и нами -- на территории МУП «Декоративные культуры», из
парникового хозяйства которого вид предположительно распространялся по
городу и области. Еще в конце 1990 х гг. мы встречали лишь отдельные одичав
шие экземпляры вида [Хорун, 1999], однако к настоящему времени натурализа
ция носит массовый характер. В отличие от других регионов, в Тульской обл.
доминирует форма с красными листьями. Зеленолистная форма отмечена только
раз -- на окраине д. Краинка Суворовского р на, у источника минеральных вод.
В Московской обл. O.stricta впервые обнаружил Арефьев в 1868--1870 гг. близ
Серпухова в огородах как одичалое (MW [этого листа в 2008 г. мы не видели]), но
тот сбор, как отмечал А.Н. Петунников [1896], сделан на самом деле в Москве, на
огороде Новодевичьего монастыря. Почти все образцы из Московской обл., храня
щиеся в MW, имеют зеленые листья, и только один гербарный лист -- красные:
«пос. Тарасовка, 55°57' с.ш. и 37°51' в.д., на улице дачного поселка, сорное, живые
растения интенсивно пурпурно фиолетовые, 27.08.2004 г., С. Майоров (MW)».
В Тверской обл. O.stricta отмечена в конце 1980 х гг. на территории старого пар
ка в д. Бреднево Калининского р на под вязами и вдоль грунтовой дороги. В конце
1990 х гг., вид встречен в окрестностях нескольких усадебных парков, на пустырях
и набережной в г. Тверь, в поселках, на свалках, ж/д насыпях [Маркелова, 2004].
Во Владимирской обл. форму с зелеными листьями собрал А.Серегин: «Суз
дальский район, сырой сбитый газон и сорное на грядках в дачном поселке на
коренном левом склоне долины р. Клязьмы в пределах черты г. Владимира, в 1
км. к северо востоку от ТЭЦ, 13.08.2000. MW». Краснолистная форма собрана в
Судогодском р не: «лесное кладбище дер. Соколово, сосняк на песке, дичает меж
ду могил, 27.07.2004 г., А. Серегин. MW». По личному сообщению А.П. Сереги
на, на территории области кислица встречается редко. Краснолистная форма
преобладает на кладбищах, зеленая, как сорное -- во Владимире и Муроме.
В Ивановской обл. O.stricta впервые отмечена в 1922 г., в г. Иваново Возне
сенск [ОКМ; Хорошков, 1925]. Изредка встречалась и позднее [Маевский, 1964;
Алявдина, Виноградова, 1972; Шилов, 1989]. В последние годы наблюдается по
всеместное дичание краснолистной формы, ускользающей из культуры. Форма с
зелеными листьями используется в декоративном озеленении [Борисова, 2007].
Один из первых сборов в Брянской обл. сделан в пос. Злынка Гомельской
губернии [ныне -- Брянская обл.], в саду, 20.08.1920 г. (MW, зеленолистная фор
ма). Форму с зелеными листьями собрал и Ю.Е. Алексеев «близ г. Стародуб, в
лесополосе, 15.06.1976 г., MHA». Там же найдена и краснолистная форма: «сорное
в г. Стародубе, 23.06.1976 г., Ю.Алексеев, В.Макаров» (MHA). В 1988 г. зеленоли
стная форма O.stricta собрана в парке с. Гринево Богарского р на. 25.08.1988 г.,
Л.Дейстфельдт (MHA).
364
В Калужской обл. идет активное расселение вида, чаще встречается форма с
зелеными листьями, растения с красными листьями изредка вырастают на га
зонах (С.Майоров, личное сообщение). Гербарные образцы с зелеными листья
ми собраны на северо восточной окраине г. Калуга, на гравийной дорожке дачно
го участка, 30.07.1995 г., С.Майоров, Е.Дараган (MW) и в Юхновском р не, в
окрестности д. Александровка, в долине Угры, на залежи, рассеянно, 14.08.2003 г.,
С.Полевова, Н.Решетникова (MHA).
В Рязани и окрестностях -- обычный вид в садах и огородах [Казакова, 2004].
В Пензенской обл. O.stricta -- редкий вид парковых газонов [Васюков, 2004].
Имеется сбор зеленолистной формы из Тамбовского р на Тамбовской обл., сде
ланный на окраине пос. СПТУ 10, на свалке, 27.07.2004 г., А.Сухоруков (MW).
В Смоленской обл. не отмечена [Решетникова, 2004].
Отметим, что старые сборы в Средней России представлены, в основном,
зеленолистной формой вида, в то время как сейчас встречаются обе формы, и
численно преобладает краснолистная (рис. 180). Не исключено, что эти две
формы были занесены в разное время и из разных источников, этот вопрос
требует дальнейшего изучения.
Местообитания. Встречается по запущенным паркам, садам, огородам,
тенистым сорным местам, реже -- на газонах [Еленевский, Радыгина, 1997; Ма
евский, 2006]. В Московской обл. найден в рудеральных местообитаниях, на ж/
д путях, в цветниках, парках [Игнатов и др., 1990]. Частый сорняк теплиц, обра
ботанных почв, цветочных контейнеров, палисадников (рис. 181). Отмечен на
полях, песках и галечниках речных долин. В Курской обл. найден вдали от жи
лья в саженых сосняках [Кузнецова, Макаренко, 1976; Полуянов, 2005].
В Китае растение встречается в лесах и оврагах, на высотах до 1500 м [Quanru,
Watson, 2008].
Предпочитает дренированные, влажные, щелочные почвы, солнечные мес
та. Может расти на бедных, глинистых почвах. Не очень хорошо растет в тени.
Не выносит задернения участка, особенно краснолистная форма.
Размножение и жизненный цикл. Цветет с июля по сентябрь. Прекрасно
размножается семенами. Зрелые коробочки растрескиваются при любом каса
нии и с силой разбрасывают семена, которые легко прилипают к любой повер
хности (собственные наблюдения). Таким образом, семена O.stricta могут раз
носиться человеком или животными. Как показало специальное исследование
в Оклахоме (США), среди диаспор растений (семена, плоды) на шерсти бизона
(Bison bison) семена кислицы довольно обычны (Rosas et al., 2008).
Расселение и распространение. Вид активно распространяется семенным
путем по возделанным и нарушенным участкам.
Последствия внедрения
Воздействие на естественные фитоценозы и аборигенные виды. Силь
ное задернение служит серьезным препятствием для расселения вида. В Сред
ней России в обработанных цветниках и палисадниках O.stricta часто образует
сплошной ковер, а за пределами участков, на луговине, растений нет или они
единичные (рис. 182).
В высокотравных североамериканских прериях, которые страдают от пожа
ров и от выпаса бизонов, встречаемость O.stricta на участках, вытаптываемых
бизонами, значительно выше, чем на участках, на которых выпас отсутствует
[Hartnett et al., 1996].
365
Генетические изменения и вариабельность вида. O. stricta --- изменчивый
вид, в пределах которого выделяют ряд разновидностей (var. europaea R.Knuth
, var. decumbens Bitter, var. europaea (Jord. ex F.Schultz) R.Knuth, var. piletocarpa
Wiegand , var. rufa Farw., var. villicaulis (Wiegand) Farw. var. viridiflora Hus), разли
чающихся характером опушения, формой роста и пигментацией растения. Точ
ная их номенклатура сложна и запутана. В Средней России чаще других встреча
ется var. rufa с выраженной красно вишневой пигментацией, реже растет var.
decumbens с распростертым стеблем. Растения с обильным железистым опушени
ем (var. villicaulis) на нашей территории достоверно не отмечены (MHA, MW; Цве
лев, 2000), но известны южнее, на территории Украины (MW; Цвелев, 2000).
Влияние на здоровье человека. O.stricta содержит витамин С и помогает
предотвращать цингу; применяется при гипертонии, диабете, ознобе. Настойку
кислицы применяют для снятия жара, при желудочных спазмах и тошноте. В
связи с большим количеством аскорбиновой и яблочной кислоты в листьях, упот
ребление кислицы следует ограничить людям, страдающим ревматизмом и арт
ритом. Припарки из растения помогают избежать припухлостей при ушибах.
Экономическое и социальное (положительное и отрицательное) зна
чение. Все части растения съедобны, с выраженным кислым вкусом, как и у
других видов рода Oxalis. Листья, цветки и плоды используются для приготов
ления салатов, для украшения и придания вкуса блюдам. Все части растения
можно жевать для утоления жажды. Из листьев готовят напиток, по вкусу на
поминающий лимонад. Все растение заваривают как чай, используют как аро
матическую приправу при приготовлении блюд из зеленой фасоли. Путем ки
пячения из растения получают оранжевую краску.
Меры борьбы
В США O.stricta внесена в списки видов, которые распространяются глав
ным образом по нарушенным местам обитания, с трудом проникая в естествен
ные сообщества и являются сорняками сельскохозяйственных культур
[Kentucky Exotic Pest Plant Council, 2008]. O.stricta -- трудноискоренимый сорняк
кукурузных полей США и Канады. Он отмечен, например, на 50% обследованных
участках в посадках кукурузы в канадской провинции Квебек. Для уничтожения
сорняка на полях кукурузы используется гербицид mesotrione; при правильно по
добранной концентрации которого уничтожается до 100% O.stricta [Proccedings...,
2008]. На промышленных, транспортных и других несельскохозяйственных
участках в США используют гербицид «Арсенал» [Hartnett et al., 1996].
СЕМЕЙСТВО POACEAE (GRAMINEAE) -- ЗЛАКИ
КОСТЕР КРОВЕЛЬНЫЙ
Название: Anisantha tectorum (L.) Nevski
Синонимы: Bromus tectorum L., Schedonorus tectorum (L.) Fries, Bromus
setaceus Buckl., Zerna tectorum (L.) Lindm., Anisantha pontica C.Koch
Морфологические признаки
Озимый монокарпик или яровой однолетник. У яровых особей насчитыва
ется от 1 до 5 побегов, у озимых -- до 24 побегов. Длина побегов 20--50 см, число
366
листьев на одном побеге варьирует от 3 до 7. Соцветие -- односторонняя метел
ка длиной 5--10 см с 14--25 тонкими, поникающими, дуговидно изогнутыми ве
точками (рис. 183).
В естественном ареале листья могут быть опушены смесью длинных и ко
ротких волосков, только короткими волосками или, реже, быть голыми; в неко
торых популяциях встречаются особи со всеми тремя вариациями опушения
листьев. Во вторичном ареале отмечены только особи с опушенными листьями.
Основание метелки голое или опушенное; в долинных популяциях выше
частота встречаемости неопушенного основания оси метелки, а в высокогор
ных опушенного основания [Виноградова, 2000а].
Нижняя цветковая чешуя длиной 20--30 мм, ланцетная или широколанцет
ная, с широким пленчатым краем, наверху слегка притупленная и с длинной
остью. В естественном ареале преобладают растения, у которых верхняя цвет
ковая чешуя полностью или хотя бы в нижней части голая. Ниже частота встре
чаемости растений с цветковой чешуей, слабоопушенной короткими волоска
ми; и очень редко встречаются единичные особи с цветковой чешуей, сильно опу
щенной смесью коротких и длинных волосков. Во вторичном ареале преобладают
особи с сильноопушенными цветковыми чешуями, а неопушенные цветковые че
шуи вовсе не отмечены. Такая же закономерность прослеживается по степени
опушения колосковых чешуй: в естественном ареале значительно преобладают
особи с голыми колосковыми чешуями, а во вторичном ареале -- с опушенной
колосковой чешуей [Виноградова, 2000а]. В Северной Америке pастения с колос
ками «скорее голыми, чем опушенными» рассматриваются как А.tectorum var.
glabratus Spenner. Встречается эта разновидность очень редко: в Канаде, напри
мер, найдена лишь в 4 пунктах в провинции Британская Колумбия, в 1 пункте в
провинции Альберта и 1 экземпляр -- в провинции Онтарио [Upadhyaya, 1986].
Наименьшее число цветков в колоске отмечено в естественном ареале у рас
тений из равнинных популяций. У растений горных популяций среднее число
цветков в колоске увеличивается до 4,1--4,4. Во вторичном ареале цветков в
колоске еще больше -- до 5--8. Следовательно, на северном и высотном пределах
ареала колосок состоит из большего числа цветков, что позволяет несколько
увеличить число образовавшихся семян.
Во вторичном ареле в Волгоградской и Московской обл. встречаются целые
популяции костра с побегами пурпурного цвета. В естественном ареале подоб
ную окраску можно наблюдать только у единичных особей.
Естественный ареал
Родина А.tectorum -- южные районы Восточной Европы и Западной Азии,
Центральная Азия и Средиземноморье.
Вторичный ареал
История интродукции и географическое распространение. Вторичный
ареал костра кровельного на североамериканском континенте по площади не
уступает естественному ареалу. Первые особи нового пришельца появились в
восточных штатах в 1861 г., а в западных -- в 1889 г. Медленное расширение
начальной инвазионной популяции длилось ~15 лет, после чего последовало
быстрое, взрывообразное расселение вида. A.tectorum сначала прочно поселил
ся на сильно урбанизированных территориях, затем проник в менее нарушен
ные биотопы и, наконец (через 25 лет в США и 50 лет в Канаде) освоил природ
367
Рис. 184. Расселение Anisantha tectorum в Средней России
Рис. 183. Метелка Anisantha tectorum
Рис. 185. Плодоно
шение Anisantha
tectorum
Рис. 186. Anisantha
tectorum на ж/д на
сыпи, ст. Тарасовка
Московской обл.
369
ные биотопы -- климаксовые степные сообщества. Затем наступил период ста
билизации, который продолжается и в настоящее время и сопровождается мед
ленным проникновением отдельных спонтанных популяций вида как в новые
регионы, так и в новые экологические ниши [Mack, 1981; Klemedson, Smith,1964;
Anderson, 1984].
В середине ХIХ века костер стал проникать в северные районы Европы. По
меньшей мере 10 инвазионных популяций A.tectorum были отмечены в 1909 г. в
Швеции [Witte, 1909]. В Латвии вид собран в 1919 г. на ст.Тукумс, а в 1924 г. -- на
cт. Елгaвa. В 1970 х гг. обнаружен еще в 3 населенных пунктах Латвии [Фатаре,
1977]. В Литве найден в 1956 г. на песчаном берегу р. Рожничанка близ г. Друски
нинкай (MHA). По данным Л.Сапегина [1985], в качестве элемента синантропной
флоры костер кровельный встречается в поймах рек Белорусского Полесья.
В северных районах европейской части России встречается довольно редко.
Найден в 1930 г. на ст. Апатиты. В 1960 е гг. произрастал по железным дорогам
в 4 пунктах Ленинградской обл. В 1968 г. обнаружен на ст. Беломорск в Каре
лии; позднее отмечен в г. Шуя и г. Кондопога [Гусев, 1968, 1973; Раменская, 1983].
В 1957 г. костер кровельный был собран на ж/д ст. Кизнер в Удмуртии [Ефимо
ва, 1961]. Произрастал этот вид также около Ухты и по Печоре [Определитель
Коми ACCP, 1962].
В 1974 г. костер собран в морском порту Владивостока. Еще 2 находки вида в
окрестностях Владивостока сделаны в 1979 и 1981 гг. [Нечаева,1985].
В настоящее время костер кровельный в качестве заносного отмечен в Се
верной Америке, Исландии, Гренландии, Абиссинии, Южной Африке, Японии,
на острове Чеджудо (Корея), в Китае, на Гавайских островах, в Австралии и
Новой Зеландии [Виноградова, 2000а].
Пути и способы заноса. Зерновки опадают в течение недели после созрева
ния, и ветер рассеивает их на короткие расстояния. Обычно плоды, подгоняе
мые ветром, катятся по поверхности почвы и не поднимаются в воздух. Но ос
новной способ распространения -- засоренное семенами зерно, выброшенная ис
пользованная подстилка для рогатого скота и солома из упаковочных ящиков.
Животные также способствуют распространению семян, так как длинная ость ко
стра кровельного цепляется к их шкуре и шерсти [Mitich,1999]. В посадки зерно
вых культур и кормовых трав попадает, главным образом, с семенами. Также ак
тивно распространяется по ж/д путям, часто встречается вблизи ж/д насыпей.
Статус в регионе. Процесс проникновения вида в Среднюю Россию на
чался в середине ХIХ века. В Московской обл. костер отмечали еще в 1817 г., но
встречался он очень редко -- в ХIХ веке его нашли только 4 раза, первый гербар
ный сбор относится к 1863 г. (MW). В других регионах находки тоже были неча
сты: имеются отдельные гербарные сборы 1870 г. из Воронежской обл., 1879 г. -- из
Орловской обл., 1887 г. -- из Курской обл. (рис. 184). Вид собрали в 1897 г. на ст.
Бологое в Тверской губернии и дважды (1917 и 1927 гг.) -- в окрестностях Твери
[Малышева, 1979].
В ХХ веке стал встречаться в Московской обл. регулярно и отмечен (МНА)
по железным дорогам в Клинском уезде (1922 г.), в Опалихе (1927 г.), Москве
(1946 г.), Ромашково (1947 г.), Кашире (1962 г.), а также по песчаным склонам
рек -- Москвы (1927 г.) и Оки (1956 г.).
С середины ХХ века костер кровельный распространился по всем областям
Средней России. Поскольку A.tectorum -- однолетник или озимый двулетник,
370
можно считать, что спонтанные популяции вида существуют здесь не менее 30
поколений. В Московском регионе отмечены крупные популяции A.tectorum,
устойчивые в течение многих лет.
Местообитания. На родине произрастает на закрепленных песках в сухих
степях и полупустынях и на щебнистых склонах гор до высоты 3000 м. Во вто
ричном ареале в Средней России популяции костра кровельного встречаются
как по железным дорогам, так и по песчаным берегам рек и рудеральным место
обитаниям; отмечен также по выпасаемым степям и лугам, залежам, пустырям,
полям, обнажениям коренных пород, обочинам дорог (рис. 185, 186).
Во вторичном ареале в Америке костер кровельный типичен для степной
зоны, где выпадает от 150 до 560 мм осадков. Растет в широком диапазоне по
чвенных условий -- от сухих, песчаных щелочных до плодородных перегнойных
почв. Очень редко, но встречается и на засоленных почвах. Cпорадически про
израстает в горных лесах до высоты 2700 м [Pierson, Mack, 1990].
Размножение и жизненный цикл. Костер кровельный -- в высокой степени
самофертильный клейстогамный вид [Mack,1981; МсKone,1985] с очень низ
кой, если не совсем невозможной, способностью к перекрестному опылению.
Ни один исследователь никогда не видел цветков с тычинками, полностью вы
совывающимся из цветка, и лишь однажды в популяции на железной дороге ст.
Вербилки Московской обл. мы видели растения с тычинкой, частично высовы
вающейся между цветковыми чешуями.
Расселение и распространение. A.tectorum формирует множество семян,
причем даже в годы с неблагоприятными климатическими условиями. При хоро
ших условиях роста урожай семян может достигать 2,6 млрд. зерновок/га. От
дельная особь в густых популяциях может произвести ~25 зерновок каждая. Од
нако изолированно растущее растение с многочисленными веточками в метелке
на солнечном участке при обильной влажности может сформировать до 5 тыс. зер
новок. В условиях природных пастбищ незаделанные в почву семена костра кро
вельного обычно остаются жизнеспособными не дольше 1 года [Mitich,1999].
Последствия внедрения
Воздействие на естественные фитоценозы и аборигенные виды. В ес
тественном ареале плотность популяций невысока: не превышает 200 растений
на 1 м2 [Виноградова, 2000б]. Во вторичном ареале в Америке этот показатель
намного выше. Средняя плотность обычно составляет от 50 до 2000 растений на
1 м2. При этом общая биомасса составляет 400 600 г/м2 Американскими учены
ми отмечено, что при повышении плотности популяции уменьшается вес расте
ний, вес 1 побега, число зерновок на 1 побеге и на всем растении и число побегов
на растении: при плотности популяции 5382 растения на 1 м2 все они были с
единственным побегом [Thill, Beck, Callihan, 1984].
Кроме того, изобилие сухих побегов А.tectorum летом приводит к повыше
нию частоты пожаров в степях с доминированием видов рода Artemisia. В ре
зультате естественные заросли кустарниковой полыни резко сократились
[DiTomaso, 2000].
В регионах, где должным образом регулируется выпас скота и предотвра
щается пал, костер кровельный в конечном итоге естественно замещается более
ценными и устойчивыми многолетними видами [Mitich,1999].
Генетические изменения и вариабельность вида. Во всех изученных по
пуляциях в первичном и вторичном ареале 2n=14 [Naganowska, Krzakowa, 1984;
371
Mc Kone, 1985]. Гибридизация в природных условиях (если она вообще случа
ется) происходит в исключительных случаях. Помимо специфического строе
ния цветка, гибридизация невозможна и потому, что A.tectorum зацветает на 2
недели раньше других однолетних видов костра.
Гибрид между A.tectorum и Bromus mollis не дает семян [Knowles, 1944].
При скрещивании А. tectorum (2n =14) и B.carinatus (2n=56) произошла аберра
ция хромосом, и гибрид содержал 2n=35. Гибрид Bromus × laagei=A. tectorum
×B.squarrosus получен искусственным путем [De Cugnac, Camus, 1944], но и он
оказался стерильным. Сообщение [Scholz, 1989] о находке гибрида Bromus seriseus
ssp. fallax на Синайском п ове (A. tectorum × Bromus sericeus ssp. sericeus) представ
ляется нам сомнительным, поскольку эти так называемые «гибриды» собраны в
пункте, значительно удаленном от основного ареала Bromus sericeus ssp. sericeus.
В естественном ареале популяции костра кровельного полиморфны: встре
чаются особи с тремя дискретными вариациями опушения листьев и цветковых
чешуй, с двумя вариациями опушения колосковых чешуй и основания оси ме
телки и двумя вариациями окраски побегов. Однако преобладают особи с зеле
ными побегами и неопушенными органами.
Во вторичном ареале как в Европейской России, так и в Северной Америке,
наоборот, популяции более однородны, причем преобладают опушенные экземп
ляры с побегами пурпурного цвета. В Америке костер отличается более крупными
размерами и большим числом колосков на 1 растении [Виноградова, 2000а].
Экономическое и социальное (положительное/отрицательное) значе
ние. Дали положительные результаты опыты [George et al.,1981] по использо
ванию костра кровельного для зарастания истощенных нефтяных разработок,
субстрат которых имеет рH=9.
В Америке костер используется как кормовое растение, которое особенно
ценится из за быстрого весеннего отрастания листьев, когда традиционные
многолетние травы еще не вегетируют. Ежедневный прирост биомассы костра
составляет 2,9 г/м2 [Morrow, Stahlman, 1984], а при применении азотных удоб
рений этот показатель может быть увеличен в 4 раза. По содержанию азота и
белка зеленая масса костра сравнима с лучшими кормовыми травами, а уро
жайность составляет 370 г/м2 сухой массы [Klemedson, Smith, 1964].
Однако животные, пасущиеся в популяциях костра в период созревания се
мян, или поедающие зрелые семена с сеном, часто травмируются. Зрелые длин
ные жесткие ости легко прокалывают слизистую оболочку во рту, горле и ки
шечнике домашнего скота, вызывая раны и инфекцию. Кроме того, высохшие
растения сильно увеличивают пожароопасность природных пастбищ, посколь
ку костер кровельный очень быстро воспламеняется [Mitich, 1999].
Более известен костер кровельный как злостный сорняк. При засорении по
лей костром в количестве до 110 растений на 1 м2 урожайность пшеницы сокра
щается на 40%, а при плотности костра до 538 растений на 1 м2, урожай пшени
цы сокращается до 90% [Wicks, 1984]. В некоторых штатах вид распространен
очень широко: на юге Айдахо, например, он доминирует на 4 млн акров и со
ставляет до 95% биомассы. В Орегоне доминирует на 10 млн акров [Klemedson,
Smith, 1964].
Меры борьбы
В посевах озимой пшеницы в Америке с костром кровельным борются
путем весенней (март) и осенней (ноябрь) обработок посевов метрибузином в
372
дозе 0,42 кг/га и метрибузином + тербутрином (0,28 + 0,67 кг/га). Лучшие ре
зультаты получены при осенней обработке метрибузином + тербутрином: сор
няк уничтожается на 93%, а урожайность пшеницы повышается на 36% [Swan,
Whitesides, 1988].
В качестве агента биологической борьбы с костром кровельным использует
ся штамм D7 ризобактерии Pseudomonas fluorescens [Kennedy et al., 2001].
Обзор научных исследований и публикаций
Отмечено, что при формировании вторичного ареала костра кровельно
го происходит снижение полиморфизма инвазионных популяций по призна
кам опушения. Некоторые аллели исчезают вовсе, а оставшиеся демонстриру
ют изменение частоты встречаемости (в естественном ареале преобладают осо
би с неопушенными органами, а во вторичном с опушенными). Проведено
сравнение характера опушения верхней цветковой чешуи с данными L.Hulbert
[1955], полученными на обширном материале, включающем 119 образцов из 21
пункта сбора в четырех штатах США [Виноградова, 2000б].
Отмечено значительное варьирование количественные признаки костра
кровельного в естественных и инвазионных популяциях (табл. 18).
Имеются и другие данные, касающихся гетерогенности костра кровельного.
Так, при исследовании изменчивости 19 польских популяций этого вида и 6 по
пуляций из Португалии, Италии, Чехии, Словакии, Венгрии и Швеции [Krzakowa,
Kraupe, 1981] в отношении систем ферментов установлено, что изученные попу
ляции мономорфны по двум системам ферментов и полиморфны по двум другим
системам ферментов эстеразе и глютамат оксалат трансаминазе.
Сходная работа [Novak, Mack, 1989] проводилась с использованием элект
рофореза в крахмальном геле для определения генетической вариабельности
24 популяций костра кровельного из его естественного ареала. Всe популяции
проанализированы на изоферментную вариабельность по 15 ферментам, ко
дируемым 25 генетическими локусами. Полученные результаты сравнили с дан
Таблица 18
Сравнительная характеристика некоторых количественных признаков костра
кровельного в естественном и вторичном ареалах
Средние значения по всем популяциям
Изученные
признаки
естественный
ареал
вторичный ареал
Таджикистан
Россия
Северная Америка
Число цветков
на 1 растении
292,5
(135--941)
365,0
(238--531) 327,5
[Mc Kone, 1985]
Число колосков
на 1 растении
81,3
(31--277)
70,2
(52--91) 108,5
[Mc Kone, 1985]
Число цветков
в колоске
3,6 (2--7)
5,2 (4--8) 3,0 [Mc Kone, 1985]
Длина верхней
цветковой чешуи 11,53
(10,7--14,4)
12,52
(9,7--13,8) 11,58
[Mc Kone, 1989]
Средняя высота,
см
26,3
(21,4--41,0)
32,8
(23,5--52,0) 70,4 [Mc Kone, 1985]
49,3 (35--65) [Hulbert, 1955]
373
ными, приводимыми ранее для североамериканских популяций. Обнаружена
незначительная дифференциация по уровню полиморфизма внутри популя
ций костра кровельного и зафиксировано исчезновение некоторых редких ал
лелей при внедрении вида в Северную Америку. В евразийских популяциях
52% локусов были полиморфными, а в североамериканских только 28%.
Впоследствии по той же методике электрофореза в крахмальном геле был
проведен [Novak, Mack, Soltis, 1991] анализ генетической вариабельности по 25
локусам 2141 экз. костра кровельного, выращенных из семян, собранных в 60
популяциях в 4 регионах вторичного североамериканского ареала: 1) восточ
ная часть Скалистых гор (14 популяций); 2) штаты Невада и Калифорния
(14 популяций); 3) Великая равнина (22 популяции); 4) канадская провинция
Британская Колумбия (10 популяций). Общая генетическая изменчивость
А.tectorum по сравнению с другими цветковыми растениями оказалась низкой.
Полиморфность в пределах одной популяции составила 4,6%, охватывая 1,05
аллели на локус при средней ожидаемой гетерозиготности 0,012. Фактически
гетерозиготность не выявлена ни у одной особи и реально составила 0,000. Гене
тическое сходство между популяциями равно 0,980. Авторы сделали вывод, что
для А.tectorum, так же как для других самоопыляющихся видов, характерны
низкая генетическая изменчивость и существенная генетическая дифференци
ация между популяциями.
В дальнейший анализ включено [Bartlett et al., 2002] 38 популяций
А.tectorum из восточных штатов США для оценки их генетического родства с
популяциями западных штатов. Генетическая изменчивость восточных попу
ляций низкая: среднее число аллелей на локус -- 1,01% полиморфных локусов в
популяции -- 1,05 %, гетерозигот не обнаружено. Тринадцать популяций в вос
точной и западной части Северной Америки отличаются по локусу Pgm 1a и
Pgm 2a, в то время как 8 популяций отличаются по локусу Mdh 2b и Mdh 3b.
Однако несколько аллелей, обнаруженные в западной части Северной Амери
ки (Got 4c, Got 4d и Pgi 2b), не были найдены в восточных популяциях. Следо
вательно, инвазия А.tectorum в Северную Америку произошло на обоих побере
жьях путем многократной интродукции. Исторические и генетические данные
позволяют предположить, что восточные популяции возникли в результате как
минимум двух независимых заносов. Распространение А.tectorum с востока на
запад, вероятно, произошло в ХIХ столетии.
СЕМЕЙСТВО POACEAE (GRAMINEAE) -- ЗЛАКИ
ОВСЯНИЦА ШЕРШАВОЛИСТНАЯ
Название: Festuca trachyphylla (Hackel) Krajina, Gramineae (Poaceae)
Синонимы: F. duriuscula auct., F. brevipila R. Tracey1
1 Использование названия F. trachyphylla дискуссионно. С одной стороны, М. Вилкин
сон и К. Стайси показали высокое сходство F. ovina subsp. euovina var. duriuscula subvar.
trachyphylla Hack. и F. brevipila, но название F. trachyphylla Hackel ex Druce на видовом
уровне ранее уже использовано [Wilkinson, Stace, 1988]. С другой стороны, А. К. Скворцов
[20**] отстаивал идентичность F. trachyphylla и F. macutrensis Zapaі., последний вид описан
раньше, чем F. brevipila. В данной работе мы используем название F. trachyphylla, как широ
ко используемое флористами Европы, Азии и Америки [Цвелев; 1974; Markgraf Dannenberg,
1980; Aiken et al., 2005; Lu et al., 2006], понимая возможную номенклатурную некоррект
ность такого решения.
374
Морфологические признаки
Плотнодерновинный злак высотой (20)30--40(50) см. Возобновление побе
гов внутривлагалищное, горизонтальные удлиненные корневища отсутству
ют. Основание побегов с пурпурным оттенком. Нередко сохраняются серова
тые влагалища прошлогодних листьев. Листья обычно темно зеленые, длиной
8--25 см, жесткие, на верхней (внутренней) поверхности с волосками, на ниж
ней (внешней) гладкие или чаще шероховатые от коротких волосков
(рис. 187). Язычок длиной 0,1--0,3 мм, с ресничками. Листовые пластинки вдоль
сложенные, шириной 0,4--0,6 мм, а развернутые часто шириной более 1 мм, с 3--
5 ребрами и с (5)7 жилками, склеренхима на нижней поверхности листа в 3
тяжах, реже с перемычками между тяжами (рис 188).
Соцветия --- многоколосковые метелки, длиной 5--15 см, на тонких длинных
соломинах. Колоски 4--7(8) цветковые, длиной 5,5--10 мм. Цветковые чешуи 4--
5 мм длиной с остями 0,5--2,5 мм. Пыльники 2--3(3,5) мм. Вершина завязи голая.
Зерновка длиной 2,5--3,5 мм.
Естественный ареал
Природный ареал F. trachyphylla охватывает северные районы Централь
ной Европы [Markgraf Dannenberg, 1980; Wilkinson, Stace, 1988]
Вторичный ареал
История интродукции и географическое распространение. Из за но
менклатурных и таксономических проблем восстановление истории расселе
ния F. trachyphylla наталкивается на недостаток информации. Трудно предста
вить специальную интродукцию этой овсяницы в Европе, как это произошло в
США. В Великобритании считается заносным видом [Stace, 1997]. Восточная
граница распространения овсяницы не определена, ее произрастание в Китае
достоверно не подтверждено [Lu et al., 2006].
В США овсяница широко интродуцирована как фуражная культура и рас
тение для газонов [Aiken et al., 2005; Darke, 2007]. Cледует иметь в виду, что
североамериканская овсяница часто имеет почти непрерывное (но неравномер
ное) кольцо склеренхимы на абаксиальной поверхности листа [Aiken et al., 2005],
что не характерно для европейских растений, а потому нуждается в дополни
тельном таксономическом исследовании.
Пути и способы заноса. По видимому, для F. trachyphylla необходимы учас
тки с нарушенным растительным покровом, которые являются подходящими
местообитаниями для вида. Овсяница предпочитает обширные песчаные мас
сивы, как флювиогляциальные, так и пески речных долин (но не пойменные!).
По видимому, зрелые колоски овсяницы цепляются за одежду людей и шерсть
животных, и таким образом зерновки разносятся.
Статус в регионе. В европейской части России расселение овсяницы про
исходило в середине XX века. В.И. Кречетович и Е.Г. Бобров [1934] приводили
овсяницу только для Верхне Днепровского и северо запада Верхне Волжского
флористических р нов с вопросом для Курска. Н.Н. Цвелев [1974] указывал вид
для Верхне Днепровского, всего Верхне Волжского, запада Волжско Донского
флористических р нов. Авторами «Атласа экономически значимых растений и
вредных объектов России и сопредельных государств» составлен ареал2 совре
2 Следует иметь в виду, что овсяница распространилась восточнее, чем это указано на
карте. Эта неточность объясняется тем, что карта из «Атласа...» составлена на на основе
литературных источников и материалов LE, без учета сборов из московских гербариев.
375
Рис. 187. Гербарный лист Festuca trachyphylla
Рис. 190. Внедрение
Festuca trachyphylla в
естественные сообщества
Рис. 191. Плодоношение
Festuca trachyphylla
377
менного распространения F. trachyphylla в России (http://www.agroatlas.spb.ru/
related/Metadata/Meta_Festuca_trachyphylla_ru.htm). Следует иметь в виду, что
овсяница распространилась восточнее, чем указано на карте. Неточность объяс
няется, видимо, тем, что составлена на основе литературных источников и ма
териалов LE, без учета сборов в московских и региональных гербариях. На во
сток в Европейской России F. trachyphylla распространилась до Удмуртии
[Пузырев, 2008].
Рис. 188. Изменчивость анатомической структуры листьев F. trachyphylla
[по: Wilkinson, Stace, 1988, с изменениями]
(пр.п. --- проводящие пучки, скл. --- склеренхима).
Рис. 189. Расселение Festuca trachyphylla в Средней России
378
F. trachyphylla в настоящее время известна из большинства областей Сред
ней России, кроме, возможно, самых восточных территорий (рис. 189).
Местообитания. В Великобритании предпочитает кислые хорошо дрени
рованные почвы [Stace, 1997]. Однако в США для F. trachyphylla оптимальны
ми считаются суглинистые почвы и хорошее освещение, но овсяница способна
приоизрастать и на песчаных почвах при широком диапозоне pH почвы, вы
держивает засоление [Darke, 2007; Pessarakli, Pessarakli, 2007]. Обладает высо
кой засухоустойчивостью.
В Московской обл. встречается на песчаных участках на склонах к рекам и
во вторичных местообитаниях, весьма обычна на ж/д насыпях. Кроме того, она
активно расселилась по свежим наслоениям песка на уличных газонах [Игна
тов и др., 1990].
Размножение и жизненный цикл. F. trachyphylla плотнодерновинный мно
голетний злак (рис. 190, 191). Опыляется ветром. Распространяется только зер
новками, вегетативное расселение практически отсутствует.
Расселение и распространение. Есть сведения, что F. trachyphylla анемо
хорное растение. По нашему мнению, более вероятна зоохория и антропохория.
Последствия внедрения
Воздействия на естественные фитоценозы и аборигенные виды. Мес
тами образует обширные заросли, как, например, в окрестностях дер. Ивановка
Темниковского р на Республики Мордовия (MW), которые, по нашим наблю
дениям, на открытых песках занимают несколько гектаров. В подобных случа
ях овсяница конкурирует с местными пионерными псаммофитными видами.
Генетические изменения и вариабельность вида. Известны садовые фор
мы с сизыми, почти голубыми листьями [Aiken et al., 2005].
Экономическое и социальное (положительное и отрицательное) зна
чение. Экономическое значение F. trachyphylla минимально. В США она исполь
зуется как фуражная и газонная культура [Aiken et al., 2005; Darke, 2007].
Меры борьбы
F. trachyphylla конкурирует, возможно, лишь с псаммофитными видами, об
ширные ее заросли известны в редких случаях. По мере зарастания песков она
обычно исчезает. Поэтому в специальных мерах борьбы овсяница не нуждается.
Обзор научных исследований и публикаций
Недавние исследования популяционной изменчивости комплекса F. valesiaca
s. l. показали континуальную изменчивость по 72 морфологическим призна
кам, что не позволяет однозначно выделять из этого комплекса некоторые «мел
кие» виды [Беднарська, 2007].
СЕМЕЙСТВО POACEAE (GRAMINEAE) -- ЗЛАКИ
ЯЧМЕНЬ ГРИВАСТЫЙ
Название: Hordeum jubatum L.
Синонимы: Critesion jubatum (L.) Nevski, C. geniculatum Rafin.
Морфологические признаки
Однолетнее или многолетнее плотнодерновинное голое растение. Генера
тивные побеги прямостоячие или восходящие, высотой 25--50 см, диаметром
379
около 2 мм, с 3--5 узлами. Листовые пластинки плоские, шириной 1,5--4 мм при
длине до 10--12 см. Колосья зеленые или серовато зеленые, поникающие, дли
ной 5--10 см (включая ости), мягкие, ось колоса ломкая (рис. 192). Колоски в
группах по три: боковые колоски редуцированы до 1--3 остей или, реже, тычи
ночные, центральный колосок обоеполый, нижняя и верхняя цветковые чешуи
равной длины 5--6(8) мм c остью длиной до 7 см. При созревании зерновок ости
отогнутые, ось колоса разламывается.
Естественный ареал
Охватывает северную часть Северной Америки (кроме арктических райо
нов) и северо восток Азии.
Вторичный ареал
История интродукции и географическое распространение. Как занос
ное растение встречается во многих странах Европы [Lambdon et al., 2008], в
Чехии известен с 1880 г. [Williamson et al., 2005].
В бывшем СССР ячмень гривастый был известен как заносное растение в
европейской части России, Западной Сибири, Средней Азии [Невский, 1934] и,
возможно, в Приморье.
Пути и способы заноса. H. jubatum -- анемохорный вид, этому способствуют
длинная ость и относительно мелкие зерновки [von Bothmer et al., 1995]. Одна
ко большая часть гербарных образцов, по крайней мере самые ранние для реги
она находки, приурочена к ж/д насыпям.
Пути заноса и распространения, очевидно, двояки. H. jubatum -- обычное
«железнодорожное» растение. С другой стороны, культивируется на цветниках
в качестве декоративного, и первые его находки связаны именно с такой культу
рой. Выращивается дачниками как декоративное растение, семена можно при
обрести в специализированных магазинах.
Для вида отмечены случаи так называемого «дальнего расселения» с ветром
и птицами [Royer, Dickinson, 1999].
Статус в регионе. Расселение ячменя в Средней России произошло во вто
рой половине XX века. Впервые отмечен Д.П. Сырейщиковым [1914] в Подмос
ковье: «Химки, около дач, в большом числе особей (Слуцкий)» и собран в Пет
ровско Разумовском, «у канавы среди полей, 29.6.1918 г., В.Милованов» (MW).
С.А. Невский сообщал только о 3 известных ему точках для всей Европейской
России1, в Средней России -- только для Рыбинска [Невский, 1934]. H. jubatum
регулярно указывался как «редкое заносное растение» [Сырейщиков, 1927; Во
рошилов и др., 1966]. Н.Н. Цвелев [1974] указывал этот вид как заносное расте
ние уже для Верхне Волжского и Волжско Донского флористических районов
[Цвелев, 1974]. Массовое расселение ячменя гривастого произошло, по види
мому, в 1950--1960 е гг. В настоящее время H. jubatum распространен по всей
Средней России (рис. 193). В последние годы из за широкого использования
гербицидов на железных дорогах ячмень встречается заметно реже.
Местообитания. В Северной Америке H. jubatum произрастает преимуще
ственно на лугах и пастбищах на равнинах и в нижнем горном поясе, широко
распространен по обочинам дорог, пустырям, на полях [North..., 2003]. Этот вид
принадлежит к растениям, которые первыми поселяются после нарушения ра
стительности и могут доминировать в растительном покрове [Bakker, Wilson,
1 Отметим, что К. Ледебур сообщал о произрастании H. jubatum у Астрахани [Ledebour, 1853].
380
2004]. На более поздних стадиях сукцессии ячмень не обладает высокой числен
ностью. Ячмень гривастый предпочитает незасоленные местообитания, но при
этом обладает существенной солеустойчивостью, до 10 или даже 15 г/л [Badger,
Ungar, 1991; Wang, Redmann, 1996]. В пределах естественного ареала локаль
ные популяции адаптированы к локальному уровню засоления [Wang,
Redmann, 1996].
Во вторичном ареале ячмень гривастый встречается на ж/д путях, иногда
образует колонии на прилегающих пустырях [Игнатов и др., 1990]. В Мещере
отмечено расселение H. jubatum по песчаным пустырям у жилья [Определитель
растений Мещеры, 1986].
Размножение и жизненный цикл. Ячмень гривастый размножается семе
нами (рис. 194). Каждое растение способно произвести более 170 зерновок [Royer,
Dickinson, 1999]. Прорастание весеннее и в конце вегетационного периода
[Badger, Ungar, 1991]. Семена способны прорастать с глубины до 7 см [Royer,
Dickinson, 1999]. Прорастание семян подавляется высокими летними темпера
турами [Badger, Ungar, 1989]. Образует семенной банк [Badger, Ungar, 1994;
Ungar, 1995]. Семена сохраняют жизнеспособность до 7 лет [Royer, Dickinson,
1999]. В зависимости от уровня засоления H. jubatum может развиваться как
однолетник или многолетник [Badger, Ungar, 1991]. Засоление 20 г/л и более
резко уменьшает прорастание семян ячменя [Keiffer, Ungar, 1997].
Расселение и распространение. Скорость расселения H. jubatum в Чехии
оценивается в 2,1 км/год [Williamson et al., 2005].
Последствия внедрения
Воздействия на естественные фитоценозы и аборигенные виды. Влия
ние H.jubatum на растения местной флоры, вероятно, незначительно.
Генетические изменения и вариабельность вида. Дальневосточные рас
тения в северной части ареала резко отличаются распростертыми побегами и
красновато малиновой окраской колосьев, растения Приморья и Хабаровско
го края имеют прямостоячие стебли и зеленые колосья [Пробатова, 1985].
Из состава H. jubatum нередко выделяют H. caespitosum Scribn. ex Pammel
[например, Baum, Bailey, 1994]. Этот ячмень указан для Москвы [Алексеев, 2006].
Однако в последние годы его включают в состав ssp. intermedium Bowden (ssp.
breviaristatum Bowden × ssp. jubatum) [Synthesis..., 1999].
Экономическое и социальное (положительное и отрицательное) значение.
H.jubatum может быть обременительным сорняком посевов [von Bothmer et al., 1995].
Наличие ячменя гривастого в кормах для лошадей и других животных может вызы
вать у скота беспокойство, раздражение пищеварительного тракта и даже язвы [Lewis
et al., 1996; Royer, Dickinson, 1999]. H. jubatum самый декоративный среди ячменей
(рис. 195) и используется для сухих букетов [Darke, 1999; Quattrocchi, 2006].
В пределах природного ареала может использоваться для рекультивации
земель [Quattrocchi, 2006].
Меры борьбы
Для борьбы с H. jubatum на территории США, где он местами является обре
менительным сорняком, используется обработка полей гербицидами [Conn,
Deck, 1995; von Bothmer et al., 1995; Blackshaw et al., 1998, 1999].
Необходим мониторинг распространения ячменя гривастого, который по
тенциально может быть сорняком посевов, особенно в районах широкого рас
пространения песчаных почв.
Рис. 193. Расселение
Hordeum jubatum
в Средней России
Рис. 192. Колосья
Hordeum jubatum
Рис. 194. Плодоношение
Hordeum jubatum
Рис. 195. Разведение
Hordeum jubatum в
декоративных целях
383
СЕМЕЙСТВО POACEAE (GRAMINEAE) -- ЗЛАКИ
МЯТЛИК ПРИЗЕМИСТЫЙ
Название: Poa supina Schrader
Морфологические признаки
Многолетнее рыхлодерновинное растение (рис. 196). Стебли стелющиеся,
длиной 5--15(25) см. Соломина цилиндрическая, гладкая. Листья в основном
прикорневые, плоские или вдоль сложенные, длиной 2--7 см и шириной 2--3 мм,
внезапно заостренные. Язычок пленчатый, тупой, его высота у верхнего листа
генеративного побега 0,1--1,3 мм [Чертопруд, Соколов, 2001; Соколов, Чертоп
руд, 2002]. Соцветие рыхлая метелка длиной 2--4 см. Ветви соцветия гладкие
(рис. 197). Колоски одиночные, 4--6 цветковые, овально ромбические, сжатые с
боков, длиной 3,5--5 мм. Ось колоска и каллус при основании нижней цветковой
чешуи гладкие. Колосковые чешуи длиной около 1,5 мм, килеватые, с 3 жилка
ми. Нижняя цветковая чешуя длиной 2,5--3,5 мм, с 5 жилками, средняя жилка с
ресничками. Верхняя цветковая чешуя немного короче нижней, кожистая, по
килям с ресничками. Пыльники длиной 1,2--2,0 мм мм [Чертопруд, Соколов,
2001; Соколов, Чертопруд, 2002].
Естественный ареал
Естественный ареал охватывает горные районы Европы, а также большую
часть горных систем внетропической Азии, вплоть до Дальнего Востока [Про
батова, 1985; Олонова, 1990; Cope, 1982; Guanghua et al., 2006]. В Европе произ
растает в горах Центральной и Юго Западной Европы, Феноскандии, на Апен
нинах и Северо Западе России [Цвелев, 2000; Edmonson, 1980].
Вторичный ареал
История интродукции и географическое распространение. Появление и
расселение изучены недостаточно. Известны находки на Северо Западе России
[Скворцов, 1973; Цвелев, 1974, 1976]. Долгое время P. supina просто не замечали, а
гербарные сборы других видов мятлика, с которым его путали, ограничены.
Пути и способы заноса. Не исключено, что изначально P. supina мог по
явиться в парках Средней России вместе с посадочным материалом из Европы
или как газонный злак. Позднее он распространился на нарушенные местооби
тания, как и другие европейские злаки с похожей историей расселения.
Статус в регионе. В Средней Росии P. supina впервые собран в Московской
обл.: «Богородский уезд, с. Шевелкино, 8.05.1891 г. Аноним» (MW). В 1960 х гг.
Р. supina отмечен уже во многих районах Подмосковья [Скворцов, 1973]. Вскоре
обнаружен Тверской области в Центрально Лесном заповеднике (Миняев, Ко
нечная, 1976). Ранние этапы расселения мятлика плохо документированы, по
скольку внешне он очень похож на P. annua или P. pratensis (рис. 198). Можно
предположить, что P. supina культивировался в ботаническом саду в Горенках,
поскольку сейчас вдоль тропинок и по сбитым лужкам это массовый и обычный
для этого парка мятлик (впрочем, не исключен и более поздний занос P. supina в
Горенки). В настоящее время мятлик, по видимому, широко распространен в
лесных областях. Его активное расселение происходило во второй половине
XX века. Находки этого мятлика пришлись на середину 1970 х гг., поскольку в
«Злаках СССР» Н.Н. Цвелев [1976] не указывает P. supina среди адвентивных
384
злаков средней полосы, а во «Флоре европейской части СССР» сообщает о его
находках в Верхне Волжском районе [Цвелев, 1974]. Он легко просматривает
ся, если заранее не быть готовым к его находкам. Нередко этот мятлик находят
на хорошо изученных местах, и часто в большом количестве. Так, в Калужской
обл. на жиздринском участке национального парка «Угра» найден в 1990 х гг.,
местами в массе, а 10--15 годами ранее на этой территории мятлик отмечен не
был [MW, MHA; Решетникова и др., 2005]. На Звенигородской биостаниции
МГУ (Московская обл.) обнаружен всего около 10 лет назад. Оказалось, что P.
supina обычен на территории усадьбы биостанции (Нижние и Верхние Дачи) и в
окрестностях; чаще всего растет вдоль проселочных лесных дорог и на просеках
[Чертопруд, Соколов, 2001; Соколов, Чертопруд, 2002; Учебное..., 2008]. Очевид
но, что здесь мятлик растет давно, прочно натурализовался и просто просматри
вался (и это при ежегодных интенсивных ботанических экскурсиях!).
Местообитания. P. supina встречается преимущественно вдоль лесных до
рожек, иногда образует сплошной покров на лесных лужках и полянах [Сквор
цов, 1973; Игнатов и др., 1990].
Размножение и жизненный цикл. P. supina многолетник с ползучими кор
невищами и вегетативными побегами [Скворцов, 1973; Чертопруд, Соколов,
2001; Соколов, Чертопруд, 2002]. По другим данным, двулетник [Олонова, 1990].
Цветение в мае --- начале лета, чем отличается от P. annua c его почти непрерыв
ным цветением [Скворцов, 1973; Чертопруд, Соколов, 2001]. Очевидно, P. supina
сочетает вегетативное и семенное размножение, что позволяет ему активно рас
Рис. 198. Расселение Poa supinа в Средней России
Рис. 197. Метелка
Poa supinа
Рис. 196. Корневища Poa supinа
Рис. 199. Расселение Poa supinа вдоль лесных тропинок
386
селяться вдоль дорожек и по тропинкам. Семннная продуктивность мятлика,
однако, невысока [Casler, Duncan, 2003].
Расселение и распространение. Зрелые колоски мятлика (рис. 199) и их
части легко цепляются за одежду и обувь, а зерновки вместе с землей прилипа
ют к обуви.
Последствия внедрения
Воздействия на естественные фитоценозы и аборигенные виды. Не
изучено.
Генетические изменения и вариабельность вида. Азиатские растения P.
supina отличаются более плотной дерновиной и обедненным соцветием и опи
саны как особый вид P. ustulata S.E. Frцhner [P. supina ssp. ustulata (S.E. Fröhner)
Б. et D. Löve]. Но отличия эти нестабильны, и выделение отдельного вида едва
ли оправдано [Цвелев, 1974, 1976; Cope, 1982; Guanghua et al., 2006].
Экономическое и социальное (положительное и отрицательное) зна
чение. В Германии, Австрии, Великобритании и США P. supina приобретает
популярность как газонное растение для легкоатлетических стадионов, полей
для гольфа и отчасти для тенниса. Он вымокает меньше других злаков, устой
чив к низким температурам и перспективен для спортивных полей Северной
Европы [Puhalla et al., 1999; Casler, Duncan, 2003]. Однако его широкому приме
нению препятствует низкая семенная продуктивность [Casler, Duncan, 2003].
Поскольку P. supina относительно теневынослив, его рекомендуют для устрой
ства газонов в полутени [Vernon et al., 2009].
Меры борьбы
Специальные меры борьбы с P. supina не требуются, поскольку он пока не
является злостным сорняком. Необходим мониторинг за его расселением.
Обзор научных исследований и публикаций
По видимому, P. annua L. представляет собой гибридогенный вид, возник
ший при естественном скрещивании P. supina и P. infirma Kunth. [Vargas, Turgeon,
2003].
На Звенигородской биостаниции МГУ (далее --- ЗБС) изучена изменчивость
морфологических признаков P. supina и P. annua, обычно используемых для ди
агностики этих видов [Цвелев, 1974, 1976]. Оказалось, что у P. annua длина пыль
ников изменяется от 0,5 до1,25 мм, а длина язычка верхнего листа генеративно
го побега составляет 1,2--3,0 мм. У P. supina длина пыльника 1,2--2,0 мм, длина
язычка листа --- 0,1--1,3 мм. Таким образом, каждый из этих двух признаков не
позволяет различить виды, хиатус установить не удалось. Однако совместный
анализ двух признаков показал возможность четкого разграничения этих мят
ликов в условиях ЗБС. Сравнение по длине пыльников в пределах растения и
внутрипопуляционной изменчивости показало их значительное сходство. Для
надежного определения P. supina необходимо использовать комплекс призна
ков, наиболее важными из которых служат длина пыльников и длина язычка,
далее по степени значимости следуют наличие вегетативных побегов, корневи
ща, число опушенных жилок на нижней цветковой чешуе [Чертопруд, Соколов,
2001; Соколов, Чертопруд, 2002].
387
СЕМЕЙСТВО POACEAE (GRAMINEAE) -- ЗЛАКИ
БЕСКИЛЬНИЦА РАССТАВЛЕННАЯ
Название: Puccinellia distans (Jacq.) Parl.
Синонимы: Poa distans Jacq., Atropis distans (Jacq.) Griseb., Phippsia distans
(Jacq.) A. et D. Lцve, Puccinellia pseudoconvoluta Klok., P. sachalinensis Ohwi.
Морфологические признаки
Дерновинный многолетник. Стебли высотой 15--50 (75) см, часто внизу
коленчато восходящие. Листовые пластинки шириной 2--5 мм, плоские или
реже свернутые, сверху и по краям шероховатые, язычок длиной 1--2,5 мм.
Метелка длиной 5 15(20) см, многоколосковая, с нижними веточками, откло
ненными горизонтально или вниз. Колоски длиной 3--5 мм, 3--6 цветковые,
бледно зеленые или реже фиолетовые (рис. 200). Нижняя цветковая чешуя
длиной 1,8--2,2 мм, с золотистой оторочкой, при основании голая. Пыльники
длиной 0,5--0,9 мм [Губанов и др., 2002].
Естественный ареал
Как вид природной флоры, P.distans известна на юге Европы, на Кавказе, в
Средней Азии [Цвелев, 1974]. В России вид распространен в Предкавказье, в
степной зоне европейской части -- Поволжье и Ростовской обл., на Южном
Урале и в Западной Сибири -- в Тюменской, Курганской, Омской и Томской
обл. [Белый и др., 2008; Лысенко, 2005; Моисеев, 1998; Бубнова, 1990].
Характер ареала в Скандинавии, Центральной Европе, Средиземноморье
и ряде регионов Азии не совсем ясен, этот вопрос требует дополнительного
изучения.
Вторичный ареал
История интродукции и географическое распространение. P. distans
занесена в Северную Америку [Cusick, 1983; Vincent, Cusick, 1998], обнаруже
на в Новой Зеландии [Edgar, 1996], быстро распространяется по обочинам
дорог в европейских странах: в Польше [Mirek, Trzcinska Tacik, 1982], Бель
гии [Vanden Berghen, 1994], Люксембурге [Steichen, 1995] и других.
В пределах азиатской части России занесена в Восточную Сибирь, где
произрастает в Новосибирской, Кемеровской и Иркутской обл., Алтайском и
Красноярском крае, в Хакасии, Туве, Бурятии и Якутии [Бубнова, 1990], и на
Дальний Восток [Губанов и др., 2002]. Отмечен во многих районах о. Сахалин
[Баркалов, Таран, 2008]; на северо западе Камчатки -- редкий вид у дорог и
жилья [Чернягина, Якубов, 2008].
Вполне обычный адвентивный вид для Северо Западного региона Рос
сии: часто встречается в Ленинградской, Псковской и Новгородской обл. [Цве
лев, 2000]. Внесен в список инвазионных видов Северо Запада России в каче
стве ксенофита [Чужеродные виды северо западной России, 2008]. Обычный
заносный вид в Волжско Камском регионе [Пузырев, 2006].
Пути и способы заноса. Первичный занос возможен по путям сооб
щения, в том числе с перевозимой сельхозпродукцией (зерном, фуражом, фрук
тами), а также с гравием и другими материалами, используемыми для строи
тельства насыпей [Пузырев, 2006].
Статус в регионе. Природный ареал включает Саратовскую, Воронежс
кую и Белгородскую обл., южные районы Пензенской, Тамбовской, Курской и
388
Липецкой обл. [Васюков, 2004; Еленевский и др., 2004; Григорьевская, 2000;
Полуянов, 2005]. В остальных регионах Средней России P. distans -- заносное
растение (рис. 201).
В Курске является обычным видом, часто образует большие скопления
вдоль бордюров тротуаров [Полуянов, 2005]. Собран в Тамбовской губернии
[ныне -- Липецкая обл.]: «Усманский уезд, по берегу р. Байгора, в окр. с. Коро
бовка, 23.08.1917 г., П.А. Смирнов» (MW). Один из первых сборов для Орлов
ской обл. сделан в г. Орел: «левый берег Оки, 8.07.1980 г., Тетерина, Ступина,
Ивлева, Малахова» (OHHI), позднее инвазионные популяции отмечены в
Дмитровском, Знаменском, Колпнянском, Ливенском, Новодеревеньковском
и Орловском р нах [Еленевский, Радыгина, 1997].
В Тульской обл. P. distans впервые отмечена В.Голицыным [1905] для фло
ры Епифанского уезда: «...у опушки Мурина леса, очень редко». На дальней
шее распространение P. distans повлияла Гражданская война. Многочислен
ная популяция бескильницы обнаружена М.Назаровым (1920 г., MW) на ст.
Тула. Вплоть до 1980 хх гг. в области вид встречался нечасто [Алексеев, Губа
нов, 1980; Алюшин, 1982]. Известны лишь сборы Т.Вернандер 1933 г. из со
временного Щекинского р на (MW). К настоящему времени P. distans прочно
перешла в группу эпекофитов, освоив увеличившиеся по площади засолен
ные места обитания.
Впервые в Москве вид собран в начале XIX века (MW, гербарий московс
кой флоры М.А. Максимовича, 1824--1826 гг.), и вплоть до Первой мировой
войны оставался крайне редким. Начиная с 1916 года, сделано множество сбо
Рис. 201. Расселение Puccinelia distans в Средней России
Рис. 200. Гербарный лист
Puccinelia distans
Рис. 203. Кущение
Puccinelia distans
Рис. 202. Внедрение
Puccinelia distans в
рудеральные сообщества
(Москва, Битцевский
лесопарк).
391
ров с ж/д насыпей и сорных мест. В настоящее время P.distans -- массовый вид
в населенных пунктах. Повышение численности вида приходится, вероятно,
на 1970 е гг., поскольку в середине 1960 х гг. бескильницу расставленную еще
не относили к обычным видам [Ворошилов и др., 1966; Игнатов и др., 1990]. В
Тверской обл. P. distans, по видимому, появилась во второй половине XIX века
[Малышева, 1988б, Бакунин, 1879]. Один из первых сборов сделан близ Тве
ри, на ж/д полотне 13.06.1917 г., М.И. Назаров (MW). В настоящее время вид
широко распространился во всех р нах области [Маркелова, 2004].
Один из первых сборов P. distans во Владимирской обл: «г. Меленки, в при
дорожной канаве, 18.06.1920 г., М. Назаров» (MW). Встречается во всех р
нах, спорадически, нечасто [Вахромеев, 2002]. В Ярославской губернии, Рос
товском уезде, близ г. Ростов, на берегу о. Неро бескильницу собрал А.Жадов
ский, 12.08.1929 г. (MW). Дальнейшие сборы сделаны лишь в 1970--х гг.
В Ивановской обл. вид впервые отмечен в 1920 г. [ОКМ; Хорошков, 1923]. В
«Определителе...» [Алявдина, Виноградова, 1972] бескильница приведена в ка
честве редкого вида. К настоящему времени обычна [MW; ИУ; PLES; Борисова,
1996б; 2007]. В Костроме cобрана в 1882 г. П. и Е.Жадовскими (MW); следую
щие сборы в Костроме сделаны в 1889 и 1920 гг. (MW), а более поздние находки
в Костромской обл. датированы уже 1997--2001 гг. (Е.А. Борисова, MW).
Указана для Смоленской обл. в качестве адвентивного вида [MW; Решет
никова, 2004]. Собрана в Демидовском р не, в 1 км к западу от о. Баклановс
кое, на обочине дороги у песчаного карьера, 13.07.1999 г., Н.М. Решетникова
(MW). Обнаружена Д.И. Литвиновым [1895] в г. Калуга. Позднее А.Ф. Фле
ров [1912] указывал P.distans для ряда р нов Калужской обл. Один из первых
сборов сделан в Мещевском уезде, в г. Серпейск: «канава у «городка», очень
редко, 12.08.1925 г., М. Караваев» (MW). В настоящее время растет по всей
территории, нередко (С.Майоров, личное сообщение). Собрана в Брянской
обл., на загрязненной пустоши в г. Карачев, 19.08.1976 г., Ю. Алексеев (MW).
В Рязанской обл. одним из первых можно считать сбор, сделанный В.Н.
Тихомировым и Н.Б. Октябревой: «Рязанская область и район, пос. Варские,
у полотна узкоколейной ж/д, 28.06.1970 г.». Ныне встречается по всей терри
тории повсеместно [Казакова, 2004].
C территории современной Мордовии есть сбор из Краснослободского уез
да, сделанный К.А. Космовским в 1888 г. (MW). В Нижегородской губернии
вид собран в Лукояновском уезде: «близ с. Протасово, внизу оврага при доро
ге, 18.06.1925 г. Собр. В.В. Алехин, И.Г. Басов, К.В. Доброхотова» (MW). В
1925--1927 гг. P. distans собирали также в Сергачском, Арзамасском, Павловс
ком, Выксунском уездах Нижегородской губернии (MW). На территории
Чувашии P. distans собирали в Порецком р не: «в 5 6 км северо восточнее с.
Митрополье Горьковской обл., на заболоченном берегу р. Киша, 3.07.1983 г.,
В.Тихомиров, Л.Зелинская» (MW).
Местообитания. В природном ареале растет на засоленных лугах, при
речных песках, по берегам водоемов, по грунтовым дорогам, сорным местам,
ж/д насыпям [Бубнова, 1990; Губанов и др., 2002]. В Средней России P. distans
находится на границе ареала: в южных областях встречается по солонцеватым
лугам и солодям, поймам рек и ручьев, сырым придорожным кюветам и обо
чинам, железным дорогам, сырым местам в населенных пунктах. В северных
регионах -- заносное растение, интенсивно расселяется по вторичным антро
392
погенным местообитаниям, на участках с выраженным засолением иногда об
разует сплошные заросли (рис. 202). В массе -- по ж/д насыпям и крупным
шоссейным дорогам, по краям газонов, на пустырях, сорных местах, террито
риях промышленных предприятий, складов и баз.
Размножение и жизненный цикл. В северных регионах цветение вида
продолжительное за счет образования новых генеративных побегов в июле и
августе (рис. 203). Плодоношение может продолжаться до октября. В южных
районах цветение заканчивается в начале июня, плодоносит в июле--августе
[Губанов и др., 2002].
Расселение и распространение. P.distans интенсивно распространяется в
городах, особенно в Москве [Губанов и др., 2002]. Резкое увеличение числен
ности вида связано с применением солевых противогололедных смесей. Эта
особенность отмечена во многих странах Западной Европы [Игнатов и др.,
1990; Mirek, Trzcinska Tacik, 1982; Vanden Berghen, 1994; Steichen, 1995]. Та
же закономерность выявлена и в США: вид быстро распространяется вдоль
скоростных шоссе, особенно обилен на тех участках, где зимой применяют соль
[Cusick, 1983; Vincent, Cusick, 1998]. Вероятно, городские и междугородние
магистрали с засоленными обочинами являются «коридорами», связывающи
ми популяции вида в естественном и вторичном ареале.
Последствия внедрения
Воздействия на естественные фитоценозы и аборигенные виды. Име
ется ряд работ, посвященных взаимодействию P. distans с другими галофита
ми в условиях природного ареала [Moravcovб, 2001; Piernik, 2006; Tarasoff et
al., 2007]. Во вторичном ареале этой проблеме уделялось значительно меньше
внимания. Указано, что вид доминирует в растительных сообществах на тер
ритории химических предприятий в Башкирском Предуралье [Зейферт, 2008].
Сообщества с доминированием P. distans изучали на индустриальных терри
ториях Рура в Германии [Dettmar, 1993]. Можно предположить, что степень
влияния бескильницы расставленной на растительные сообщества во вторич
ном ареале не слишком высока, так как она сосредоточена на открытых засо
ленных участках, которые способны занимать немногие виды.
Генетические изменения и вариабельность вида. Н.Н. Цвелев выделяет
в составе P.distans 3 подвида: subsp. distans; subsp. glauca (Regel) Tzvel.; subsp.
sevangensis (Grossh.) Tzvel. 2n = 28 [Цвелев, 1976, с. 505].
Экономическое и социальное (положительное и отрицательное) зна
чение. В районах естественного произрастания растение используют как кор
мовой злак [Белый и др., 2008].
Меры борьбы
P. distans -- массовый вид на территориях предприятий, откосах желез
ных и шоссейных дорог, стройплощадках и т.п. Для этой категории несельско
хозяйственных объектов предлагается применять гербицид «Арсенал», успеш
но апробированный для уничтожения травянистой и древесной растительно
сти [Спиридонов, Шестаков, 2008].
393
СЕМЕЙСТВО POLYGONACEAE --- ГРЕЧИШНЫЕ
РЕЙНУТРИЯ ЯПОНСКАЯ
Название: Reynoutria japonica Houtt.
Синонимы: Polygonum cuspidatum Siebold et Zucc., non Willd. ex Sprengel;
Fallopia japonica (Houtt.) Ronce Decr.; Pleuropterus cuspidatus (Siebold et Zucc.)
H. Gross
Морфологические признаки
Многолетнее травянистое растение с мощным ветвистым подземным кор
невищем. Стебли многочисленные, высотой 1--3 м, полые, прямостоячие, ввер
ху ветвистые и часто дуговидно изогнутые, внизу обычно с красноватыми пят
нами или штрихами. Раструбы косоворонковидные, разрушающиеся. Листья
черешковые, листовая пластинка яйцевидная или овальная, голая или снизу
по жилкам папиллозная, сверху темно зеленая, снизу заметно светлее, во вто
рой половине вегетационного сезона кожистая, с оттянутой треугольной вер
хушкой, основание усеченное или широко клиновидное, часто несколько не
равнобокое (рис.204). Длина листьев на главном побеге до 15--20(30) см, ши
рина примерно в 1,5 раза меньше, на боковых побегах листья мельче. Соцветия
пазушые, метельчатые, со скошенными трубчатыми раструбами, в пазухах каж
дого по 2--4(6) цветков (рис.205). Цветоножки длиной 2--4 мм, тонкие, с со
членением посредине. Околоцветник белый, бледно желтоватый или едва ро
зоватый, с 5 долями; при плодах в обоеполых цветках 3 доли увеличиваются и
смыкаются вокруг плода, на внешней поверхности крылатым килем. Обоепо
лые цветки с 8 тычинками длиннее околоцветника; пестичные цветки с 3 ло
пастным бахромчатым рыльцем и зачатками тычинок (рис. 206).
Помимо R. japonica, в Средней России встречается также R. × bohemica
Chrtek et Chrtkovб --- Р. богемская. Это растение у нас до сих пор малоизвест
но, обычно его не отличают с R. japonica [Майоров, 2002]. Впрочем, это харак
терно для многих регионов мира [Zika, Jacobson, 2003]. Например, в руковод
стве по биологическому контролю за инвазионными растениями востока США
(весьма качественная сводка! -- СМ) под названием R. japonica помещена фо
тография R. × bohemica [Shaw, Seiger, 2002].
Третий вид рейнутрии флоры Средней России --- R. sachalinensis (Fr. Schmidt
ex Maxim.) Nakai [Polygonum sachalinense Fr. Schmidt ex Maxim., Fallopia
sachalinensis (Fr. Schmidt ex Maxim.) Ronce Decr.] --- Рейнутрия сахалинская ---
встречается сравнительно редко и преимущественно в культуре.
Предлагаем определительный ключ для видов рода Reynoutria, произрас
тающих в Средней России (рис. 207).
1. Листья снизу с длинными (часто более 2 мм) многоклеточными волосками.
Листья длиной до 20--45 см, с явственно сердцевидным основанием . .R. sachalinensis
--- Листья снизу голые либо с короткими, менее 1 мм, одноклеточными, реже
3 клеточными волосками. Основание листьев от усеченного до клиновидно
го, реже сердцевидное, длина листьев обычно менее 20--25 см ...................... 2
2. Листья снизу хотя бы по жилкам с густыми острыми коническими
волосками. Листья с усеченным, реже сердцевидным или ширококлиновид
ным основанием ......................................................................................... R. × bohemica
394
--- Листья снизу совершенно голые или по жилкам папиллозные. Листья с
усеченным или клиновидным основанием ................................... R. japonica --- 3
3. Развитые растения высотой 1--4 м. Самые крупные листья длиной до 20--
25 см. Околоцветник белые, едва розоватый или желтоватый . ...... var. japonica
--- Развитые растения высотой до 1 м. Самые крупные листья длиной до
10 см. Околоцветник розовый или красноватый .......................... var. сompacta
Естественный ареал
Ареал R. japonica охватывает юг Приморья, Южный Сахалин, Южные Ку
рилы, Японию, Корею, бульшую часть Китая, Тайвань [Цвелев, 1989; Jäger,
1995; Anjen, Park, 2003b].
Естественный ареал R. japonica нуждается в дополнительном изучении
[Mandбk et al., 2004]. Дело в том, что корейские и японские растения описаны
под названием R. forbesii (Hance) Yamazaki [Fallopia forbesii (Hance) Yonekura
et H.Ohashi]. Исследованиями корейских ботаников подтверждено морфоло
гическое своеобразие корейских популяций: листья R. forbesii относительно
более широкие, почти овальные [Kim, Park, 2000]. Однако, во «Flora of China»
R. forbesii включена в состав рода Fallopia, и ареал ее ограничен Кореей и Ки
таем [Anjen, Park, 2003a]. Таким образом, не вполне ясно, произрастает ли R.
japonica s. str. в Корее и каково распространение вида в Китае.
Вторичный ареал
История интродукции и географическое распространение. В Европу вид
интродуцирован в 1825 г. Лондонским сельскохозяйственным обществом
(London Horticultural Society) из Китая. Питомник Зибольда в Лейдене с 1840
г. рассылал посадочный материал. Растение стало популярным и в 1847 г. по
лучило золотую медаль The Society of Agriculture & Horticulture в Утрехте за
замечательные садовые качества. Примерно в то же время, в середине XIX века,
в Великобританию интродуцирована и var. compacta [Conolly, 1977; Child,
Wade, 2000; Bailey, Conolly, 2001; Mandбk et al., 2004; Weston et al., 2005]. В
Феноскандии в культуре известна с 1880 х годов [Neilsen, Handeland, 2000].
В Европе расселение рейнутрии вне культуры произошло в XIX веке. В
Чехии рейнутрия известна с 1883 г. [Mandбk et al., 2004]. Массовое расселе
ние R. japonica в Германии произошло позднее, с 1950 х годов [Биоиндика
ция..., 1988]. В настоящее время R. japonica широко распространена в странах
Центральной, Западной и Северной Европы1 [Atlas..., 1979; Neilsen, Handeland,
2000]. Следует иметь в виду, что часть указаний «Атласа» относится к R. ×
bohemica [Bailey et al., 1996]. По видимому, распространение вида в Европе
определяется климатическими факторами. Так, северная граница определяет
ся среднегодовой изотермой +5°С [Beerling et al., 1995]. В настоящее время
вид известен почти во всех странах Европы [Lambdon et al., 2008].
В США натурализация R. japonica отмечена с 1880 х гг. вдоль автострад,
проселочных дорог, в прибрежных местообитаниях вдоль водотоков [Child,
Wade, 2000; Weston et al., 2005; Barney, 2006]. Вид известен в Австралии и Но
вой Зеландии [Barney et al., 2006].
Во вторичном ареале R. japonica является активным инвазионным видом.
Так, в Великобритании она известна более чем в половине квадратов 10 км2
1 К сожалению, в 1979 г. советские ботаники не принимали участие в проекте "Atlas
Florae Europaeae", поэтому данные о распространении вида на территории Восточной Ев
ропы практически отсутствуют.
Рис. 204. Побег Reynoutria japonica,
Московская обл., ж. д. ст. Тарасовка
Рис. 205. Массовое цветение
Reynoutria japonica
Рис. 206. Тычиночные цветки
Reynoutria japonica
396
Рис. 207. Форма листь
ев европейских видов Rey
noutria: 1 --- R. sachalinensis,
2a --- R. japonica var. japoni
ca, 2b --- R. japonica var.
compacta, 3 --- R. × bohemica
[по: Bailey et al., 1996, с из
менениями]
Рис. 208. Расселение Reynoutria japonica в Средней России
Biological Records Centre mapping system [Hollingsworth, Bailey, 2000]. В Швей
царии R. japonica включена в черный список (Schwarze Liste) инвазионных
растений [Gigon, Weber, 2005]. Вид включен IUCN (the International Union for
Conservation of Nature) в сотню наиболее вредных инвазионных видов мира
[Lowe et al., 2003].
397
Любопытно, как оценивал В.Л. Комаров [1936] возможность распростра
нения вида: «Способно дичать, что отмечено уже во Владивостоке. Вообще
годно для районов с непромерзающей зимой почвой».
Пути и способы заноса. R. japonica распространяется из мест культивиро
вания или вторичных местообитаний вегетативно: фрагментами корневища
или стеблей с почвой, перевозимой человеком, или вдоль водотоков с талыми
или ливневыми водами [Conolly, 1977; Beerling et al., 1994; Bailey et al., 1996;
Pyšek et al., 2003; Zika, Jacobson, 2003; Weston et al., 2005; Barney, 2006]. Рассе
ление вида в Германии связывают с долинами рек [Биоиндикация..., 1988] (при
веденная карта -- рис. 2.52 -- свидетельствует скорее о приуроченности рей
нутрии к городским агломерациям -- СМ). Впрочем, и в Великобритании R.
japonica считается связанной преимущественно с долинами рек [Beerling et
al., 1994; Hollingsworth et al., 1998].
В последние годы появились сведения о наличии семенного размножения
у R. japonica на территории США [Forman, Kesseli, 2003; Wymer et al., 2007].
Статус в регионе. В Средней России первыми обратили внимание на по
явление рейнутрии А.К. Скворцов [1973], И.А. Губанов и Ю.Е. Алексеев [1975].
Первый гербарный образец относится к 1932 г. и собран в Смоленской обл
(MW). К сожалению, расселение плохо документировано гербарными матери
алами (рис. 208), что чрезвычайно важно в данном случае. Так, первые указания
[Скворцов, 1973; Игнатов и др., 1990] на сборы R. japonica в 1924 и 1964 гг. отно
сятся на самом деле к R. × bohemica (MW, MHA).
Местообитания. В Японии рейнутрия -- пионерный вид зарастания вул
канических пустынь, произрастающий от уровня моря до высоты 1500 м, ко
торый растет также по открытым участкам с обнаженной почвой, вдоль дорог
[Adachi et al., 1996]. В Китае рейнутрия растет среди кустарников по горным
склонам, долинам и краю полей [Anjen, Park, 2003b]. Рейнутрия является для
вулканических субстратов естественным фитомелиоратором, осваивающим
безжизненные вулканические отложения и делающим его пригодным для дру
гих растений [del Moral, Grishin, 1999]. Для популяций г. Фудзи показано на
личие арбускулярной микоризы [Wu et al., 2004; Fujiyoshi et al., 2005].
В пределах вторичного ареала R. japonica является преимущественно руде
ральным растением, произрастающим на пустырях, ж/д насыпях, вдоль ав
Рис. 209. Схема основных направлений гибридизации в роде Reynoutria для
Европы (по: Bailey et al., 2007, с изменениями)
398
томобильных дорог, на сорных местах. В Центральной и Западной Европе, в США
отмечено расселение вида по берегам рек и ручьев [Beerling et al., 1994; Brabec, Pyšek,
2000; Shaw, Seiger, 2002; Pyšek et al., 2003; Mandák et al., 2004; Barney et al., 2006]. В
Бельгии ~70% местонахождений рейнутрии приурочено к полосе шириной всего
10 м вдоль автомобильных и железных дорог, а также рек [Tiébré et al., 2008].
На территории России случаи внедрения рейнутрии в прибрежные место
обитания пока единичны. В Москве она нередко произрастает в оврагах у вре
менных водотоков, откуда корневища могут быть перенесены ливневыми сто
ками в долины малых рек.
Размножение и жизненный цикл. В Японии рейнутрия распространяется как
вегетативно, так и семенным путем. Семена прорастают весной после зимних хо
лодов [Maruta, 1983; Nishitani, Masuzawa, 1996]. Но семена не нуждаются в обяза
тельной стратификации [Huebner et al., 2005]. Оптимальная температура для
развития сеянцев 20--25°С [Nishitani, Masuzawa, 1996]. Образование новых дер
новин происходит преимущественно семенным путем [Zhou et al., 2003].
В Центральной и Западной Европе R. japonica представлена преимуществен
но одним половым типом: это пестичные растения, тычинки в цветках недо
развиваются [Beerling et al., 1994; Bailey et al., 1996; Hollingsworth, Bailey, 2000;
Pyšek et al., 2003; Grimsby et al., 2007]. Более того, значительная часть расте
ний принадлежит одному клону. Это предположение подтверждается молеку
лярно генетическими исследованиями [Pyšek et al., 2003]. В Северной Америке
известны обе половые формы [Forman, Kesseli, 2003; Barney et al., 2006]. В Рос
сии половая структура популяций растений этого рода не исследована. Осенью
2006 г. близ ж/д ст. Тарасовка в Московской обл. нами встречены растения с
длинными тычинками (MW), что характерно для обоеполых цветков [Barney
et al., 2006]. Наличие жизнеспособной пыльцы у этих растений не проверено.
В пределах вторичного ареала в Европе R. japonica размножается преимуще
ственно вегетативно. Рейнутрия обладает высокой способностью к вегетатив
ному возобновлению [Wade at al., 1996; Brabec, Pyšek, 2000; Bímová et al., 2003].
Отрезок корневища весом 5 г способен к регенерации. Корневище может прора
стать с глубины 1 м, а побеги способны взломать 5 см слой асфальта [Shaw, Seiger,
2002]. При эксперименте в теплице до 25% весенних зеленых черенков длиной
всего 40 мм после 6 дней культивирования давали почки [De Waal, 2001].
В США, напротив, семенное размножение распространено довольно ши
роко [Forman, Kesseli, 2003; Bram, McNair, 2004; Barney et al., 2006]. Отметим,
что жизнеспособные плоды собраны в штате Филадельфия в декабре [Forman,
Kesseli, 2003], а в Пенсильвании -- в октябре [Bram, McNair, 2004].
Расселение и распространение. Распространению R. japonica способство
вало широкое внедрение вида в культуру как экзотического декоративного рас
тения [Bailey, Conolly, 2001]. На территории США распространение рейнутрии
описывается логистической кривой, лаг фаза составила ~50 лет [Barney, 2006].
Из мест культивирования растения стало расселяться по вторичным местооби
таниям вегетативно благодаря высокой способности к регенерации. В Бель
гии за год площадь заросли увеличивается более чем на 30% [Tiébré et al., 2008].
Последствия внедрения
Воздействия на естественные фитоценозы и аборигенные виды. Мощ
ные заросли R. japonica затеняют местные растения, что приводит к уменьше
нию биоразнообразия [Bímová et al., 2004; Shaw, Seiger, 2002]. Местные расте
399
ния в ее густых зарослях практически отсутствуют [Barney et al., 2006]. Пока
зано, что разнообразие почвенных жуков среди рейнутрии ниже, чем в есте
ственной рудеральной растительности [Topp et al., 2008]. Аналогичные резуль
таты получены для других групп беспозвоночных [Kappes et al., 2007]. Сооб
щества речных гифомицетов и бентосных беспозвоночных довольно
эффективно перерабатывают биомассу R. japonica, тем не менее наличие при
брежных зарослей приводит к уменьшению биоразнообразия [Lacerf et al.,
2007]. В США зеленая лягушка (Rana clamitans) в зарослях рейнутрии добы
вает меньше пищи, чем в естественных растительных сообществах [Maerz et
al., 2005]. Оказалось, что успешность вселения рейнутрии в новом пункте не
зависит от исходного уровня биоразнообразия [Bímová et al., 2004].
Генетические изменения и вариабельность вида. В Корее у R. japonica
var. japonica выявлено 3 уровня плоидности: тетраплоидный -- 2n = 44, гексап
лоидный --2n = 66 и октоплоидный -- 2n = 88 [Kim, Park, 2000], Китае -- только
тетраплоидный [Anjen, Park, 2003b]. В адвентивных популяциях Великобри
тании и Чехии выявлен только октоплоидный цитотип, обладающий мужс
кой стерильностью, культивируемые растения var. compacta обладали тетрап
лоидным цитотипом [Bailey, Stace, 1992; Mandák et al, 2003]. В последние годы
в Европе выявлены растения как с обоеполыми цветками, так и только с пес
тичными [Bailey, Conolly, 2001]. Генетические исследования в США показали
заметные различия среди североамериканских популяций R. japonica [Weston
et al., 2005; Grimsby et al., 2007].
Разновидность var. compacta Hook. fil. отличается меньшей (до 1 м) высотой,
относительно более широкими листьями (их ширина примерно равна длине) и
красноватыми соцветиями [Stace, 1997]. Но в пределах естественного ареала
вида эта разновидность морфологически не выделяется [Mandák et al., 2004].
Считалось, что на территории Европы (Великобритания, Чехия, Германия,
Франция) и США R. japonica представлена единственным генотипом
[Hollingsworth, Bailey, 2000]. Впрочем, позднейшие исследования в США и в
Европе не подтвердили это утверждение [Bailey, Conolly, 2001; Forman, Kesseli,
2003; Bram, McNair, 2004].
Гибридизация с Fallopia baldschuanica (Regel) Holub (Bilderdykia bald
schuanica (Regel) D.A. Webb, Reynoutria baldschuanica (Regel) Moldenke)
[Tiйbrй et al., 2007] может приводить к образованию жизнеспособных семян,
что важно для дальнейшей эволюции всего комплекса Reynoutria в Европе
[Bailey et al., 2007], рис. 209.
Экономическое и социальное (положительное и отрицательное)значе
ние. Рейнутрия, поселяясь в городах и поселках, способна разрушать асфальто
вое покрытие, повреждать фундаменты и легкие постройки. Заросли R. japonica
увеличивают эрозию почвы при весеннем половодье, так как местные дерно
винные виды вытесняются, а почва оказывается не защищенной от смыва. Кор
невища могут повреждать гидротехнические сооружения [Barney et al., 2006].
В Великобритании стоимость обработки раундапом (glyphosate) зарослей
рейнутрии оценивается в 1,6 $/м2. Считается, что наличие зарослей на строи
тельной площадке увеличивает общий бюджет подготовки к строительству на
10%. Известен случай, когда при строительстве автомобильной стоянки круп
ный супермаркет потратил 600 тыс. долларов США для уничтожения зарос
лей R. japonica [Shaw, Seiger, 2002].
400
Экстракт из рейнутрии, выпускаемый под торговой маркой Milsana®, ока
зался эффективным средством борьбы с мучнистой росой томатов [Konstan
tinidou Doltsinis et al., 2006].
Меры борьбы
До сих пор в России нет контроля за состоянием популяций рейнутрии.
Опыт европейских стран и США показывает, что неконтролируемый рост вида
может приводить к существенным экономическим потерям. Необходимы раз
работка мер борьбы с распространением R. japonica с учетом местной специ
фики и контроль за состоянием существующих популяций.
Механическое скашивание не является эффективным методом контроля
за R. japonica. Ежемесячное скашивание рост зарослей останавливает, но не
уничтожает [Weber, 2003; Barney et al., 2006]. По видимому, в настоящее вре
мя самым эффективным способом уничтожения зарослей является неоднок
ратная обработка раундапом, пиклорамом или имазапиром, однако такая об
работка малоприемлема для прибрежных зарослей [Shaw, Seiger, 2002; Weston
et al., 2005; Barney et al., 2006].
Успешная борьба с рейнутрией требует кропотливой работы и сочетания
разных методов борьбы с этим инвазионным видом [Wilson, 2007].
В США и Европе разрабатываются биологические меры борьбы с R. japonica
[Kurose et al., 2007]. Для этих целей рекомендовано использовать жуков
Gallerucida nigromaculata Baly и ржавчинный гриб Puccinia polygoni weyrichii
Miyabe [Shaw, Seiger, 2002; DAISIE..., 2009]. Исследования взаимодействия
фитофагов и рейгутрии в пределах природного ареала активно продолжаются
[Kawano et al., 1999]. Однако действительно эффективных мер борьбы до сих
пор не разработано [Barney et al., 2006].
СЕМЕЙСТВО POLYGONACEAE --- ГРЕЧИШНЫЕ
РЕЙНУТРИЯ БОГЕМСКАЯ
Название: Reynoutria × bohemica Chrtek et Chrtková (R. japonica × R.
sachalinense), Polygonaceae
Синонимы: Fallopia × bohemica (Chrtek et Chrtková) J. P. Bailey; Polygonum
Ч bohemicum (Chrtek et Chrtková) P. F. Zika et A. L. Jacobson.
Морфологические признаки
По облику близка к R. japonica, но основание листьев обычно неглубоко
сердцевидное, иногда усеченное (рис. 210). Их форма обычно позволяет отли
чить рейнутрию богемскую с расстояния в несколько метров. Листья снизу по
жилкам с острыми коническими 1--3 клеточными волосками. У теневых рас
тений листья иногда почти голые, но конические волоски сохраняются хотя
бы в основании листовой пластинки. В начале мая при развертывании листо
вых пластинок, по крайней мере у части растений, с обеих сторон листа хоро
шо заметны более длинные простые волоски длиной 1--2 мм (наблюдения в
Москве в 2008--2009 гг.). Соцветия R. × bohemica обычно более густые, чем у
R. japonica (рис. 211).
Рис. 210. Побеги Reynoutria × bohemica
Рис. 211. Массовое цветение Reynoutria × bohemica
Рис. 212. Расселение
Reynoutria × bohemica
в Средней России
Рис. 213. Внедрение
Reynoutria × bohemica
в естественные
сообщества
403
Естественный ареал
Рейнутрия богемская описана сравнительно недавно, в 1983 г. [Иas. Nár.
Mus. Ser. Natur., 152: 120 -- n. v.; цит. по: Chrtek, 1990]. По видимому, культи
генный гибрид центральноевропейского происхождения [Neilsen, Handeland,
2000; Bailey, Conolly, 2001]. Вторичный ареал
История интродукции и географическое распространение. В Европе
гибрид достоверно известен с конца XIX века [Bailey, Conolly, 2001]. В насто
ящее время широко распространен в Западной, Центральной и Северной Ев
ропе [Neilsen, Handeland, 2000; Bailey, Conolly, 2001; Muller, 2004].
R. × bohemica не вошла во «Флору Восточной Европы» [Цвелев, 1996]. Впер
вые для России указана в «Определителе сосудистых растений Северо Запад
ной России» [Цвелев, 2000], а затем обнаружена в средней полосе Европейс
кой России [Майоров, 2002].
Пути и способы заноса. R. Ч bohemica распространяется так же, как R.
japonica: фрагментами корневища или стеблей с почвой, или вдоль водотоков
с талыми или ливневыми водами [Mandák et al., 2004; Zika, Jacobson, 2003].
На территории Чехии распространяется быстрее, чем R. japonica [Mandák
et al., 2004]. Однако в Бельгии эта закономерность не прослеживается [Tiébré
et al., 2008].
Статус в регионе. Достоверно известна из немногих областей Средней
России, но, несомненно, просматривается, поскольку отсутствует в большин
стве определителей (рис. 212).
Местообитания. R. × bohemica произрастает по городским пустырям, ж/д
насыпям, сорным местам, вдоль дорог.
В Бельгии ~70% местонахождений R. × bohemica, как и R. japonica, приуро
чено к узкой полосе вдоль дорог и рек [Tiébré et al., 2008]. В Европе она неред
ко произрастает по берегам рек, дренажным и оросительным каналам. В евро
пейской части России рейнутрия обитает преимущественно по рудеральным
местообитаниям, но в Московской обл. отмечена нами по берегам р. Пехорка
(наблюдения 2008 г.).
Размножение и жизненный цикл. Размножается вегетативно. Регенера
ция из частей корневища или зеленых побегов у R. × bohemica несколько выше,
чем у родительских видов [Bímová et al., 2004]. В Великобритании у R. ×
bohemica отмечено образование небольшого количества плодов [Bailey et al.,
1996]. В Москве нами также обнаружены завязавшиеся плоды, которые, одна
ко, не вызрели из за осенних заморозков.
Расселение и распространение. Рейнутрия богемская обладает большой
мощностью и активным ростом (рис. 213). В Бельгии за год площадь заросли
увеличивается более чем на 30% [Tiébré et al., 2008].
Последствия внедрения
Воздействия на естественные фитоценозы и аборигенные виды. Воз
действие R. × bohemica на местные виды аналогично влиянию R. japonica и
приводит к заметному снижению биоразнообразия [Barney et al., 2006].
Генетические изменения и вариабельность вида. Для территории Евро
пыотмечено3уровняплоидности:2n=4x=44,2n=6x=66,2n=8x=88при
преобладании гексаплоидов [Bailey, Stace, 1992; Mandák et al, 2003; Tiйbrй et
al., 2007b]. Основной путь возникновния R. Ч bohemica -- гибридизация R.
404
japonica var. japonica (2n = 88) и R. sachalinense (2n = 44). Тетраплоиды образо
вались при гибридизации R. japonica var. compacta (2n = 44) и R. sachalinense.
Образование октоплоидов не вполне ясно [Mandák et al., 2003; Bailey et al.,
2007]. Возможно, часть плодов образуется при гибридизации с Fallopia
baldschuanica (Regel) Holub -- широко распространенной в Центральной Ев
ропе декоративной кустарниковой лианы [Tiébré et al., 2007a, b].
Экономическое и социальное (положительное или отрицательное) зна
чение. В европейских программах по биоэнергетике предполагалось исполь
зовать R. × bohemica как источник биомассы, учитывая высокую продуктив
ность растения. Однако из за инвазионной активности рейнутрии от такого
предложения рекомендовано отказаться [Pude, Franken, 2001].
В остальном ущерб от R. × bohemica аналогичен таковому от рейнутрии
японской.
Меры борьбы
Меры борьбы с зарослями R. × bohemica те же, что и для R. japonica.
СЕМЕЙСТВО ROSACEAE -- РОЗОЦВЕТНЫЕ
ИРГА КОЛОСИСТАЯ
Название: Amelanchier spicata (Lam.). K.Коch.
Синонимы: Crataegus spicata Lam. (1783), Pyrus ovalis Willd. (1796),
Amelanchier ovalis (Willd.) Pers. (1803), non Medik., A.oblongifolia Blanchard
(1834), A. canadensis (L.) Medik. var. spicata Sargent (1892), A.saxatilis Blanchard
(1907), A. humilis Wiegand (1912), A. stolonifera Wiegand (1912), A.spicata
Wollman (1914), A. botryapiumTorrey var. ovalis Farwell (1916), A. botryapium
var. micropetala Farwell (1916), A. botryapium var. conferta Farwell (1916), A.
austromontana Ashe (1919), A. stolonifera var. lucida Fernald (1922), A. mucronata
Nielsen (1939), A. canadensis var. micropetala Rehder (1945).
Высказана гипотеза [Fernald, 1946; Franco, 1968], что A. spicata появилась в
результате естественной гибридизации между европейским видом A. ovalis
Medik. и завезенной в Европу A. сanadensis (L.) Medik. По другой гипотезе
[Rehder, 1949; Schroeder, 1972; Цвелев, 2001], A. spicata возникла при гибри
дизации A. сanadensis и A. stolonifera Weig., последняя является низкорослым
североамериканским кустарником, едва достигающим высоты 2 м, и по цело
му ряду признаков сходна с иргой колосистой.
Морфологические признаки
Кустарник высотой до 4--8 м с прямостоячими стволиками и мощной сис
темой гипогеогенных корневищ (рис. 214). Вопреки указаниям многочислен
ных литературных источников, корневую поросль не образует! Почки кону
сообразные, острые, красновато коричневые. Верхушечная почка длиной
6--12 мм, боковые почки мельче и отклонены вбок. Почечные чешуи неопу
шенные, с золотисто коричневой пленчатой каймой, по краю с длинными бе
лыми реснитчатыми волосками. Листья широкоэллиптические, яйцевидные
или овальные, длиной 2,5--6 см, шириной 2--3,5 см. Верхушка листа коротко
заостренная, основание округленное или сердцевидное. Молодые листья олив
ково зеленого цвета. Листья с нижней стороны в начале вегетации войлоч
Рис. 214. Amelanchier
spicata, общий вид
растения
Рис. 215. Расселение
Amelanchier spicata в
Средней России
Рис. 216. Внедрение Amelanchier
spicata в сосновый лес
Рис. 217. Цветение
и плодоношение
Amelanchier spicata
407
ные, позднее голые. По краю листа мелкие острые зубчики, не доходящие до
основания пластинки, обычно 5--7 зубцов на 1 см длины. Листья без заметной
осенней окраски. Прилистники линейные, опушенные, быстро опадающие.
Диаметр цветка 16--24 мм. Чашелистики острые, продолговато треугольные,
длиной 2--3 мм. Лепестки белые, длиной 6--10 мм, продолговато овальные. В
цветке 20 тычинок с голыми нитями, 5 столбиков пестиков срастаются при
мерно до половины. Короткая и узкая кисть направлена вертикально, длиной
8--12 см с 4--10 цветками, напоминает колос. Плоды пурпурно черные с си
зым налетом, при плодах сохраняются прямостоячие чашелистики, верхушка
завязи густошерстистая. Естественный ареал
Естественный ареал предка A. spicata занимает восточную часть Канады и
США: от о. Ньюфаундленд до штата Алабама, на запад до Миссури и Минне
соты, на юг до Южной Каролины [Jones, 1946; Артюшенко, 1959].
Вторичный ареал
История интродукции и географическое распространение.
В конце XVI века (1590 г.) из французского поселения канадской провин
ции Квебек Жан и Веспасиан Робины привезли в Париж для Королевского
ботанического сада (Jardin des Plantes) иргу канадскую A. сanadensis (L.) Medik.
[Schroeder, 1972]. В тот же период, в 1596 г., в культуру введена ирга овально
листная A. ovalis Medik. -- единственный вид с европейским ареалом [Rehder,
1949]. За 100--200 лет интродукции A. сanadensis успешно обосновалась в Цен
тральной и Северной Европе. Вероятно, в результате естественной гибриди
зации во второй половине XVIII века в Европе сформировался новый вид -- A.
spicata [Куклина, 2006а]. Показано, что в Северной Америке такой вид есте
ственно не произрастал [Fernald, 1946]; однако, несмотря на это, виды с похо
жими признаками, но значительно меньшего размера, во многих изданиях при
водятся для Северной Америки как A. spicata [Jones, 1946].
В Дании, Норвегии и Финляндии A. spicata известна с начала 1800 х гг., в
Польше --с 1820 г, в Латвии -- с 1896 г. [Kabuce, 2005].
В России A. spicata стала известна с конца XIX века. В Санкт Петербургс
ком ботаническом саду имелись «разновидности» ирги, среди которых ирга
канадская, названная «?. canadensis из Канады», как самая распространенная
в садах, и разновидность (?. rotundifolia) с более округлыми листьями (веро
ятно, A. spicata), похожая на A. сanadensis [Регель, 1874]. В Москве в конце
XIX века в дендрарии сельскохозяйственного института выращивали иргу
колосистую, под названием A. ovalis Sargent.(= A. botryapium Torrey), A.
macrocarpa hort., -- как «более позднее видоизменение» [Шредер, 1899]. Рас
тения были устойчивы и давали обильную поросль. Э.Регель [1874] пишет о
появлении в садах одного из лучших кустарников для климата России -- A.
vulgaris в. canadensis, называя ее канадской разновидностью и считая синони
мом A. spicata.
Согласно гербарным сборам, в конце XIX века иргу колосистую выращи
вали в Киеве (1893 г., LE), Вильнюсе (1897 г., LE), в г. Новосильск Тульской
обл. (1890 г., LE). В XX веке A. spicata начала распространяться шире и отмече
на в Ленинградской обл., г. Петергоф (1905 г., LE); Пензе (1905 г., LE); Рязанс
кой обл., с. Галенчино (1910 г., LE); Псковской обл., г. Новоржевск (1913 г., LE);
Тверской обл., Новоторжокский уезд (1914 г., LE); Самарской обл., г. Бугурус
408
лан (1916 г., LE). Ярославской обл, пос. Крест Багор (1924, LE) и на Урале: г.
Уфа (1923, LE), г. Уральск (1924, LE), г. Стерлитамак (1928, LE).
Вид обладает высокими адаптационными свойствами и натурализовался в
Австрии, Бельгии, Чехии, Голландии, Германии, Дании, Эстонии, на юге Фин
ляндии и Норвегии [Schroeder, 1972]. Вид растет по всей европейской части
России, вплоть до Архангельска и Мурманска, известен на Урале, в Западной
Сибири, на Алтае, в Забайкалье (Иркутск) и на Дальнем Востоке. Везде зимо
стоек и плодоносит. Степень распространенности вида в регионах различна,
но сведения порой отсутствуют.
Пути и способы заноса. Сначала A. spicata ценили за неприхотливость и
декоративные качества, а позднее ею заинтересовались как устойчивой пло
довой культурой. В Енисейской губернии (Минусинск) И.П. Бедро выращи
вал ее с 1910 г. для получения плодов [Куминов, 1996]. В промышленных мас
штабах разведение ирги начато в 1939 г. в Пермской обл. (Кудымкар). Культи
вировали, вероятно, два вида -- A. spicata и A. alnifolia. Эти кустарники обладают
исключительной зимостойкостью и выдерживали суровые зимы в районах до
60є с. ш. В период 1949--1962 гг. было получено 170 тыс. саженцев ирги, кото
рые рассылались в 114 регионов, включая среднюю полосу России, Урал, При
морский край и Северный Казахстан [Овчинников, 1963].
Статус в регионе. Вид отмечен почти во всех областях Средней России
(рис. 215): Пензенской (Конторы, 1921 г., LE; Поймский р н, 1955 г., LE); Во
ронежской (Березовка, 1951 г., LE); Саратовской (Калининск, 1968 г., LE);
Калужской (окр. Козельска, 1970 г., KLH; Мосальск, 1981 г., KLH; Мещовск,
1986 г., KLH; Юхнов, 1991 г., MW); Курской (Короча, 1998 г., LE); Орловской
(Хотынец, 1999 г., MW) и Владимирской (Гусь Хрустальный, 2000 г., MW;
Покров, 2001 г., MW). Она регулярно встречается в Московской обл. -- Горенки
(1924 г., LE), Барвиха (1930 г., LE), Подольск (1946 г., MW), Поречье (1947 г.,
MW), Серебряный Бор (1948 г., MW), Волоколамск (1958 г., LE), Воскресенск
(1975 г., MW), Солнечногорск (1975 г., MW), Битца, (1993 г., MW), Коломен
ское (1998 г., MW), Дмитров (1999 г., MW).
Местообитания. В Канаде вид, сходный по морфологическим признакам
с A. spicata, произрастает на каменистых и покрытых гравием прибрежных
участках, по берегам рек, крутым обрывам, скалистым выступам, в лесах, су
хих сосняках, на песчаниках и известняках. Он растет совместно с видами ро
дов Sorbus и Crataegus, Prunus pensilvanica и P. serotina [Cinq Mars, 1971].
Во вторичном ареале в Европе вид приурочен к сухим сосновым лесам
(рис. 216), реже встречается совместно с елью. Отмечается устойчивость ирги
как на известковых, так и на кислых почвах. На северо западе России A. spicata
произрастает чаще всего во влажных и заболоченных сосняках. В подобном
местообитании в Ленинградской обл. почвенные условия характеризовались
кислой средой (pH 4), невысоким содержанием гумуса (7,8%), азота (NO3=0,89
мг/100 г), фосфора (P2O5=3,1 мг/100 г), калия (K2O=4 мг/100 г) (ссылка)
Размножение и жизненный цикл. По многолетним наблюдениям в Мос
кве вегетация A. spicata начинается в среднем 17 апреля и длится ~166 дней
[Щербацевич, 1975]. Ирга начинает цвести раньше многих плодовых культур:
в Москве -- в мае, в Волгограде -- в конце апреля, в Томске -- в мае июне. Цве
тение длится 7--12 дней и совпадает с ростом побегов, который заканчивается
в августе. Ежегодный прирост побегов составляет 30--60 см.
409
У A. spicata типичные обоеполые цветки. Устройство цветка и его медонос
ные свойства указывают на энтомофилию. Близость пыльцы к рыльцу цветка
предполагает наличие самоплодности и отсутствие перекрестного опыления.
Посещается насекомыми, чаще всего пчелами. У близкородственных видов ирги
(A. canadensis, A. stolonifera, A. laevis) обнаружен апомиксис, при котором тре
буется лишь стимулирующее воздействие своей или чужеродной пыльцы [Oslon,
1984]. Жизнеспособность пыльцы 95--98% [Некрасов и др., 1973; Семенютина,
1985]. Согласно нашим опытам, проведенным в 2006--2007 гг. в ГБС РАН, пло
ды A. spicata в изолированных соцветиях полностью завязываются.
Время созревания плодов ирги зависит от метеорологических условий и
района выращивания. В Волгограде плоды созревают уже в июне, в Москве --
в конце июля, начале августа. На Урале (в Пермском крае) плоды поспевают к
20--25 июля, а в годы с холодным летом -- в середине августа.
Плодоносит с 3--4 лет, долговечность плодоносящего куста -- 50--70 лет
(рис. 217). Плоды округлой формы диаметром 6--10 мм, массой 0,27--0,38 г, с
тонкой кожицей, сочные, сладковатые, съедобные. В одном плоде формируется
2--5 коричневых серповидно изогнутых семян длиной 5 мм и шириной 3 мм.
Вес 1000 семян 4,3--9 г. Жизнеспособность семян -- 100% [Некрасов и др., 1973].
Лучшие результаты прорастания наблюдаются у свежесобранных семян.
Изучение корневой системы показало, что ирга колосистая вегетативно размно
жается не корневой порослью (как указано в многочисленных работах, посвящен
ных культивированию кустарника), а образует систему корневищ, так что нередко
заросль представляет собой единственный клон (С.Майоров, не опубликовано).
Расселение и распространение. Семена A. spicata распространяют мно
гие виды птиц, мелкие млекопитающие и медведи [Kabuce, 2005]. По наблю
дениям ботаников ГБС РАН, A. spicata натурализуется в Московской и Свер
дловской обл., в Пермском и Краснодарском крае -- по опушкам смешанных
лесов; в Тульской и Орловской обл. -- вблизи лесозащитных полос; в Псковс
кой и Новгородской обл. -- в заболоченных сосняках, в Смоленской, Влади
мирской, Калужской обл. и в Мордовии -- в сухих сосняках, в Белгородской
обл. -- в сосняке по меловым выходам [Куклина, 2006 в].
Последствия внедрения
Воздействие на естественные фитоценозы и аборигенные виды. В
Средней России расселяется умеренно, образуя единичные вкрапления в под
лесок лесного ценоза и лишь изредка формируя обширные заросли. Такие за
росли, например, отмечены в Усманском бору в северной части Воронежской
обл. В них ирга подавляет возобновление не только аборигенных древесных
пород, но даже и видов травянистого яруса.
В странах Балтии [Kabuce, 2005] вид вторгается в прибрежные области,
занимая лесистые дюны. Быстрое распространение A. spicata может привести
к угнетению роста сосны и изменению всего пейзажа. В Германии и Дании A.
spicata встречается вдоль железных дорог, на обочинах шоссе, по опушкам ду
бовых лесов, на бедных сухих почвах.
Генетические изменения и вариабельность вида. A.spicata, вероятно,
имеет гибридное происхождение. Встречаются экземпляры, обладающие на
бором признаков, свойственных и A.spicata, и A. alnifolia. Ирга колосистая легко
вступает в гибридизацию с другими видами ирги [Kabuce, 2005]. Хромосом
ный набор 2n=68.
410
Морфометрические признаки в инвазионных популяциях варьируют не
значительно (табл. 19). Средняя длина листа 3,2--5,2 см, средняя ширина 2,2--
4,0 см. Значение индекса листа (соотношение длины листовой пластинки и ее
ширины = l/d) для европейских популяций составляет от 1,14 до 1,51, отра
жая невысокий уровень межпопуляционной изменчивости. Относительно ста
бильны степень зазубренности края листа, форма основания и вершины лис
товой пластинки.
Признаки генеративных органов также отличаются определенной ста
бильностью: отсутствует изогнутость лепестков, постоянны характер и сте
пень опушенности частей цветка, степень срастания столбиков (на Ѕ длины).
Амплитуда изменчивости количественных признаков (длины цветоножки,
длины кисти, числа цветков в кисти, длины чашелистиков, ширины основа
Таблица 19
Изменчивость морфометрических параметров листовой пластинки
A. spicata [Куклина, 2006 б]
NoNo
Популяция
Индекс ли-
ста (l/d),
М±м
Средняя
длина
листа, см
Средняя
ширина
листа, см
1 Канада, провинция Онтарио
1,87±0,13 5,37
2,88
2 США, штат Висконсин
1,68±0,24 3,81
2,30
3 Финляндия, г. Хельсинки
1,36±0,15 3,25
2,34
4 Финляндия, г. Оулу
1,51±0,16 3,63
2,30
5 Финляндия, г. Хяменлинна
1,44±0,15 3,69
2,56
6 Германия, г. Кёльн
1,51±0,21 3,14
2,16
7 Латвия, г. Мадона
1,36±0,08 3,66
2,71
8 Ленинградская обл., г. Волхов
1,18±0,13 4,20
3,58
9 Ленинградская обл., г. Ораниенбаум 1,38±0,01 4,30
3,14
10 Ленинградская обл., г. Елизаветино 1,19±0,06 4,14
3,46
11 Ленинградская обл., г. Контори
1,50±0,07 3,42
2,28
12 Московская обл., ст. Битца
1,23±0,18 4,10
3,37
13 Московская обл., с. Поречье
1,52±0,19 3,87
2,55
14 Московская обл., г. Солнечногорск 1,30±0,18 3,26
2,52
15 Московская обл., г. Звенигород
1,39±0,15 3,74
2,69
16 Московская обл., г. Подольск
1,24±0,15 4,22
3,52
17 Московская обл., г. Воскресенск
1,36±0,16 5,34
3,93
18 Московская обл., г. Дмитров
1,29±0,07 3,03
2,35
19 Москва, Серебряный Бор
1,29±0,11 4,94
3,85
20 Москва, Коломенское
1,14±0,17 3,38
2,99
21 Калужская обл., г. Кондрово
1,30±0,10 5,28
4,01
22 Владимирская обл., г. Покров
1,38±0,16 4,32
3,13
23 Владимирская обл., г. Гусь-
Хрустальный
1,25±0,15 3,97
3,43
24 Тульская обл., г. Алексин
1,26±0,08 3,58
2,84
25 Тульская обл., г. Богородск
1,25±0,10 4,32
3,46
26 Липецкая обл., г. Елец
1,36±0,18 4,16
3,04
27 Орловская обл., г. Хотынск
1,19±0,10 3,68
2,84
411
ния чашелистиков, длины и ширины лепестков, числа тычинок и столбиков,
длины тычинок и столбиков) также невысока [Лунева, Бурмистров, 2001].
Экономическое и социальное (положительное и отрицательное) зна
чение. A. spicata ценится как плодовая культура. Плоды, содержащие витами
ны и другие полезные вещества, используют в пищу в свежем виде и в каче
стве переработанной продукции -- желе, джем, варенье, пастила, сиропы и вино.
Благодаря высокой зимостойкости, эта культура пригодна для садоводства
даже северных районов России. Она долговечна и урожайна.
A. spicata -- декоративное и медоносное растение. Применяется в основном
ассортименте древесных и кустарниковых пород при закладке новых парков,
а также при озеленении населенных пунктов. Используется при создании ле
созащитных, снегозащитных полос, а в степных районах -- как опушечный вид
в полезащитных полосах почвозащитного и противоэрозионного значения.
Плодами ирги кормятся несколько видов птиц -- дрозды, воробьи и малинов
ки, которые уничтожают значительную часть урожая [Куклина, 2007а, б].
Плотная и крепкая древесина ирги ценится за прочность, красивый цвет,
отлично полируется, используется в художественном промысле для изготов
ления мелких поделок.
Меры борьбы
Необходимость в устранении вида из биоценозов вторичного ареала не воз
никала. Очагов массового произвольного заселения окультуренных ландшаф
тов пока не отмечено. Достаточно отслеживать темпы распространения вида
на новых территориях.
Вид довольно устойчив к антропогенному влиянию. После обрезки куста
наблюдается сильное вегетативное возобновление. В случае вырубки вида на
территории культурного ландшафта также происходит его восстановление за
счет обильной поросли. Даже полная выкорчевка клона вряд ли возможна без
использования сильнодействующих гербицидов.
В результате мониторинга 2004--2006 гг. обнаружено, что вид устойчив к
болезням, от вредителей страдает не сильно. Листья и плоды повреждаются
насекомыми, часть из которых развиваются на яблоне: пяденица обдирало
(Erannis defoliaria Cl.), зимняя пяденица (Operophthera brumata L.), розанная
(Cacoecia rosana L.) и смородинная кривоусая листовертка (Pandemis ribeana
Hb.), зеленая яблонная тля (Aphis pomi Deg.), яблонная моль (Yponomeuta
malinellus.). Плодами питается яблонная плодожорка (Laspeyresia pomonella L.)
и боярышница (Aporia crataegi L.). Древесину ослабленных кустов ирги по
вреждает морщинистый заболонник (Scolytus rugulosus Ratz.).
СЕМЕЙСТВО ROSACEAE -- РОЗОЦВЕТНЫЕ
ИРГА ОЛЬХОЛИСТНАЯ
Название Amelanchier alnifolia (Nutt.). Nutt. (1834)
Синонимы: Aronia alnifolia Nutt. (1818), Pyrus alnifolia (Nutt.) Sprengel
(1825), Amelanchier ovalis Hooker (1834), A. canadensis var. alnifolia (Nutt.) Torrey
et Gray (1840), A. montana Hort. (1894), A. cuneata Piper (1900), A. florida Piper
(1906), A. utahensis Piper (1906), A. alnifolia var. typica Schneider (1906), A.
412
alnifolia var. pumila Schneider (1906), A. macrocarpa Lundl. (1913), A. leptodendron
Lundl. (1918), A. carrii Rudberg (1931), A. humilis Raup (1936), A. alnifolia subsp.
florida (Lundl.) Hulten (1973).
Морфологические признаки
Многоствольный кустарник высотой 4 м и более с гладкой темно серой
корой и рыхлой кроной (рис. 218). Почки овальные или яйцевидно овальные,
верхние чешуи с редким опушением. Листья эллиптические, почти округлые
шириной до 4 см. Вершина листа вытянутая или закругленная, основание
овальное или слегка округлое, иногда сердцевидное. Край листа грубопильча
тый или до половины зубчатый, с каждой стороны листа имеется по 5--10 круп
ных зубцов (2--5 шт. на 1 см). Лист плотный, в начале вегетации снизу войлоч
ный, затем голый. Цветки белые, ароматные диаметром 12--20 мм, собраны по
10--15 в прямые густые кисти длиной 3--6 см. Лепестки обратнояйцевидные
или яйцевидные, тупые, длиной 6--10 мм и шириной 2--3 мм. У основания ле
пестка на коротком ноготке имеется маленький хохолок из волосков. В цветке
20 тычинок с голыми тычиночными нитями, 5 столбиков (редко 4) почти до
половины сросшихся. Чашелистики ланцетные длиной 1,5--3 мм, в начале цве
тения внутри опушенные, позднее голые. Верхушка завязи всегда войлочная.
Плоды шаровидной и обратногрушевидной формы диаметром 10--15 мме, мас
сой до 0,41--0,65 г. Плоды пурпурные или почти черные, сладкие, съедобные.
Широко распространен подвид ssp. florida (Lindl.) Hulten, который одни
систематики считают синонимом A. alnifolia [Index..., 1974; Kartesz, 1980], а
другие выделяют в качестве самостоятельного вида [Jones, 1946; Rehder, 1949;
Цвелев, 2001]. Его ареал сосредоточен на западной прибрежной территории
США и частично перекрывает ареал основного вида. Морфологически под
вид выделяется более крупными цветками (диаметр 20--30 мм) с лепестками
длиной 12--15 мм, без хохолка у основания. Кустарник высотой 8 м с серо
коричневой корой. Почки опушенные, пурпурные. Лист яйцевидный с серд
цевидным основанием и заостренной верхушкой, длиной 3--4 см. Зубцов по
краю листа больше, чем у A. alnifolia, и они доходят почти до основания листа.
Естественный ареал
Обширный ареал вида занимает западную часть Северной Америки: в Ка
наде -- провинции Юкон и Британская Колумбия, в США -- штат Аляска и
регионы Великих Равнин и Скалистых гор от штатов Вашингтон и Орегон до
Небраски и Колорадо.
Вторичный ареал
История интродукции и географическое распространение. В США
в качестве плодовой винодельческой культуры иргу выращивают приблизи
тельно с 1800 г., отобраны промышленные сорта [Rehder, 1949]. В XIX веке
вид культивировали в садах и парках Европы, в некоторых местностях Вели
кобритании он одичал [Schroeder, 1970].
В Москве в конце XIX века в дендрарии Сельскохозяйственного институ
та выращивали A. alnifolia, которая отличалась устойчивостью и давала по
росль [Шредер, 1899]. В 1921 г. вид был высажен П.Н. Крыловым в Томске в
дендрарии Сибирского ботанического сада [Осипова, 1970]. Вид культивиро
вали в Москве, Санкт Петербурге, Киеве, Минске, Таллине, Владивостоке как
декоративное и плодовое растение, пригодное для лесозащитных насаждений
[Артюшенко, 1954].
Рис. 218. Побеги
Amelanchier alnifolia
Рис. 219. Расселение
Amelanchier alnifolia в
Средней России
Рис. 220. Цветение
Amelanchier alnifolia
Рис. 221. Плодо
ношение
Amelanchier alnifolia
415
В настоящее время растет в Балтии и Центральном районе Восточной Ев
ропы, натурализовался в пос. Громово Ленинградской обл. [Цвелев, 2001]. По
наблюдениям А.Г. Куклиной [2007, 2007а], натурализовался в Ленинградской,
Орловской обл. и Пермском крае (Кудымкар, Соликамск). Из за того, что вид
не всегда правильно определяют и часто относят к A. spicata, точных сведений
о его натурализации совсем мало.
Пути и способы заноса. В промышленных масштабах разведение ирги в
России начато в 1939 г. в Пермской обл. (Кудымкар). Вероятно, помимо A.
spicata, там культивировали также A. alnifolia, многочисленный разновозраст
ный самосев которой найден летом 2006 г. при посещении питомника в Ку
дымкаре [Куклина, 2006а, б]. Этот кустарник обладает исключительной зимо
стойкостью и выдерживает суровые зимы в районах до 60є с. ш. В период 1949--
1962 гг. получены многочисленные саженцы (в том числе ирги ольхолистной),
которые были разосланы в 114 областей России, на Урал, в Приморский край
и Северный Казахстан [Овчинников, 1963].
Статус в регионе. Ирга ольхолистная отмечена в качестве натурализо
вавшегося вида в Московской, Тверской, Калужской, Смоленской, Ивановс
кой и Ярославской обл. [Цвелев, 2001]. Обнаружена под пологом естествен
ных лесных фитоценозов в Орловской обл. [Куклина, 2007, 2007а]. По види
мому, распространена в Средней России гораздо шире, но при сборах
принимается за A.spicata (рис. 219).
Местообитания. В Северной Америке образует обширные заросли, иног
да за счет разрастания корневищ, по берегам рек и ручьев, на влажных скло
нах холмов, в лесах, на откосах каньонов [Scoggan, 1978], на осветленных и
тенистых участках лиственных лесов. В Канаде вид обитает совместно с Populus
tremuloides, Prunus virginiana, Shepherdia argentea, Eleagnus commutata [Pierre,
Steeves, 1990].
Во вторичном ареале произрастает на опушках и освещенных участках в
садах, парках и лесах среди кустарникового подлеска совместно с различны
ми аборигенными видами (рис. 220).
Размножение и жизненный цикл. В Москве вегетация A. alnifolia начина
ется в среднем со второй половины апреля, цветение происходит в конце мая
или начале июня. Цветение длится 7--12 дней. Плодоношение начинается кон
це июля -- начале августа и растянуто почти на месяц, так как плоды в кисти
созревают неодновременно (рис. 221). Массовый листопад наблюдается во
второй половине сентября [Древесные растения..., 2005]. В Сибири (Томск)
ирга также цветет в мае--июне, плоды начинают созревать в начале июля, а
массовое плодоношение наступает во второй половине июля и длится ~2 не
дель [Осипова, 1970]. В Краснодаре и Волгограде цветение и созревание пло
дов отмечается на месяц раньше: цветение -- 2 мая, созревание плодов -- в на
чале июня [Корунчикова, 1994; Семенютина, 1985].
У A. alnifolia типичные обоеполые энтомофильные цветки, их органогенез
детально изучен [Oslon, 1984; Steeves, Steeves, 1991]. Жизнеспособность пыль
цы высокая -- до 90--94% [Некрасов и др., 1973; Семенютина, 1985]. Согласно
нашим опытам в ГБС РАН, проведенным в 2006 г., плоды завязываются в изо
лированных соцветиях, что подтверждает возможность самоопыления и апо
миксиса. Биология развития плодов и влияние гиббереллина на степень их
созревания изучены канадскими исследователями [Sumner & Co, 1999].
416
Плоды A. alnifolia содержат 2--3 коричневых семени (длиной 4--5 мм, ши
риной 3 мм), они асимметрично овальные, плоские, гладкие. Жизнеспособ
ность семян 100% [Некрасов и др., 1973]. Масса 1000 семян -- 6--9,2 г. После
посева всходы появляются на 10--12 день. Лучшие результаты прорастания
наблюдаются у свежесобранных семян.
Вегетативное размножение происходит при разрастании корневищ.
Расселение и распространение. Семена A. alnifolia распространяют мно
гие виды птиц.
Последствия внедрения
Воздействие на естественные фитоценозы и аборигенные виды. Встре
чается единично в подлеске лесных фитоценозов. По видимому, пока не явля
ется сильным конкурентом для местных видов, но детально вопрос не изучен.
Генетические изменения и вариабельность вида. Хромосомный набор
2n=68. Вид склонен к гибридизации. Гибриды A. alnifolia с A. spicata обнару
жены в Московской обл. и Пермском крае.
В 1966 г. описана A. confuse Hylander, являющаяся гибридом между A.
alnifolia и A. laevis Wieg. Ирга конфузная как декоративное растение культи
вируется с 1830 г. в Южной и Центральной Швеции и там натурализовалась
[Schroeder, 1970, 1972]. Схожее по морфологическим признакам растение об
наружено Н.Н. Цвелевым в Ленинградской обл. [LE].
Cуществует разновидность var. cusickii (Fern.) C.L.Hitchc. с ареалом в Ка
наде (провинция Британская Колумбия) и США (штаты Вашингтон, Орегон,
Айдахо, Монтана) [Index..., 1974]. Иногда ее описывают в ранге самостоятель
ного вида -- A. cusickii Fern. [Jones, 1946]. Кустарник высотой до 10 м, с продол
говатыми листьями длиной 5 см, с 7--10 парами жилок. По краю листа 3--6
зубцов на 1 см. Цветки очень крупные, диаметром до 50 мм. Лепестки длиной
25 мм и шириной 8,5 мм. Кисти короткие, из 3--6 цветков.
Изредка в ботанических садах культивируется описанный в 1925 г. межро
довой гибрид A. alnifolia с Sorbus scopulina Greene -- Amelosorbus jackii Rehder
[Jones, 1946].
Изучение гербарных образцов, собранных в Канаде и США [LE, MHA],
показало, что форма листовой пластинки и форма плодов обладают опреде
ленной межпопуляционной изменчивостью, при этом диапазон вариабельно
сти не слишком широк [Куклина, 2007].
Вариабельность генеративных признаков у A. alnifolia исследована на рас
тениях, культивируемых в Ленинградской обл. [Лунева, Бурмистров, 2001].
Определена амплитуда изменчивости количественных признаков -- длины
цветоножки, длины кисти, числа цветков в кисти, длины чашелистиков, ши
рины основания чашелистиков, длины и ширины лепестков, числа тычинок и
столбиков, длины тычинок и столбиков: наибольшая стабильность отмечена
по яйцевидной форме лепестков с ноготком в основании. Лепестки опушены
в различной степени, чашечка и цветоножка густо опушены. Столбики чаще
сросшиеся на 2/3 длины, реже до 1/2 длины.
Экономическое и социальное (положительное и отрицательное) зна
чение. Ценится как неприхотливая зимостойкая плодовая культура. Плоды
используются в пищу в свежем и переработанном виде. Декоративное и медо
носное растение. Применяется при создании лесозащитных полос.
417
Меры борьбы
Поскольку инвазия вида в природные ценозы выражена слабо, вполне дос
таточно отслеживание темпов его расселения. Птицы (в основном дрозды ря
бинники) уничтожают часть урожая. Листья повреждаются той же группой
листогрызущих насекомых, что и A. spicata:Erannis defoliaria Cl., Operophthera
brumata L., Cacoecia rosana L., Pandemis ribeana Hb., Aphis pomi Deg., Yponomeuta
malinellus.
СЕМЕЙСТВО ROSACEAE -- РОЗОЦВЕТНЫЕ
БОЯРЫШНИК ОДНОПЕСТИЧНЫЙ
Название: Crataegus monogyna Jacq.
Синонимы: Mespilus monogyna (Jacq.) All., M. oxyacantha (L.) Scop. var.
monogyna (Jacq.) Schmalh., Crataegus oxyacantha (L.) var. intermixta Wenz., C.
curvisepala auct. non Lindem., C. oxyacantha var. paulii (Rehd.) Rehd.
Морфологические признаки
Небольшое деревце высотой до 5 м, с округлой кроной, нередко растущее
кустообразно, особенно на пастбищах, а также в садах или парках (рис. 222).
Колючки длиной до 1 см, немногочисленные, нередко вовсе отсутствуют. Ли
стья слегка кожистые, темно зеленые сверху и светлые снизу, 3--5 раздель
ные, с часто и несимметрично расположенными лопастями, туповатыми, цель
нокрайными или с немногими зубцами близ вершины. Соцветия прямостоя
чие, диаметром не более 5 см, из 5--25 цветков. Цветки диаметром 1--1,5 см, с 5
белыми лепестками, многочисленными красными тычинками и 1 пестиком.
Гипантий голый. Чашелистики не длиннее половины гипантия, широкооваль
ные, оттянутые в притупленный кончик. Плоды широкоэллипсоидальные,
коричневато красные, ~1 см длиной, односеменные [Губанов и др., 2003].
Естественный ареал.
Как вид природной флоры, произрастает в Западной Европе, кроме север
ных и восточных регионов. В Великобритании -- дико, за исключением Север
ной Шотландии. [Royal Botanic Gardens, Kew, 2009]. В России встречается в Ка
лининградской обл., в черноземной полосе европейской части (Воронежская,
Белгородская обл. и южнее) и в Предкавказье [Ареалы..., 1980]. Растет в средней
и южной части Молдовы и Украины, в Крыму и Западном Закавказье [Агроэко
логический атлас ..., 2008]. Встречается в Западной Азии и Северной Африке.
Вторичный ареал
История интродукции и географическое распространение. Вид интро
дуцирован в континентальных районах США [IT IS Report, 2008]. В Беларуси
разводится в парках и питомниках в качестве декоративного растения [Пояр
кова, 1950]. Вполне натурализовавшиеся популяции встречаются в централь
ных и северных регионах Средней России [Губанов и др., 2003]. На Северо
Западе России в Ленинградской, Псковской, Новгородской обл. C. monogyna
изредка культивируется в садах и парках, на улицах населенных пунктов, у
дорог, иногда дичает [Цвелев, 2000].
Пути и способы заноса. Заносится в основном как эргазиофит. Для деко
ративного и защитного озеленения саженцы привозят из питомников и садо
418
вых центров. Таким же способом завозят и гибридные таксоны с участием C.
monogyna.
Статус в регионе. По нашему мнению, на территории Средней России в
природе встречается C. curvisepala Lindem. (C. rhipidophylla Gand.) и C.
monogyna. Однако бедность известных нам гербарных сборов последнего вида
и сомнения в достоверности определения не позволяют с достаточной степе
нью подробности описать распространение указанных таксонов в Средней
России (рис. 223). Возможности достоверного определения C. monogyna объек
тивно ограничены, так как морфологические признаки чрезвычайно вариабель
ны, и, например, при проработке всей листовой серии отдельного растения
можно обнаружить признаки сразу нескольких таксонов. Приведенные во
«Флоре Чехии...» [Holub, 19..] иллюстрации (рис. 224) хорошего качества, од
нако при работе с природным и гербарным материалом их, разумеется, недо
статочно. Более западный вид C. laevigata (Poir.) DC. и его гибриды в нашей
флоре могут быть встречены как заносные. Требует проверки частота встреча
емости в Средней России гибрида C. curvisepala × C. monogyna (C. × kyrtostyla
Fingerh.). Этот гибридный таксон встречается в культуре, но достоверно оп
ределенных образцов, собранных в природе, мы не видели.
В Курской обл. первый сбор C. monogyna сделан С.В. Голицыным в Дмит
риевском р не, в окрестностях Дмитриева, на защитной полосе ж/д, 9.06.1961г.
(VU). Позднее вид собирали в Глушковском, Обоянском, Пристенском р нах
[VU, ЦЧЗ; Полуянов, 2005]. На Зоринском участке Центрально Черноземно
го заповедника отмечены активное расселение вида в естественных сообще
ствах и его гибридизация с C. rhipidophylla [Золотухин, Золотухина, Полуянов,
2001]. Для Липецкой обл. C. monogyna не указан, приведен вид природной
флоры C. curvisepala Lindem. со следующим комментарием: «указания на на
хождение в области в диком виде C. monogyna следует относить к C. curvisepala»
[Флора Липецкой области, 1996]. Однако в MHA есть сбор из Липецкой обл.,
определенный как C. monogyna: «Склон террасы долины р. Дон, луговая степь,
11.06.2006 г., М.А. Беспанод». В Тульской обл. C. monogyna собран нами в Яс
ногорском р не, на ст. Ясногорск, вдоль ж/д пути в 1992 г. (ТГПУ).
По личному сообщению А.П. Серегина, на территории Владимирской обл.
C.monogyna встречается нечасто. Взрослые растения очень редки. Отмечена
мелкая поросль у лесного питомника. Кроме того, для Владимирской обл. при
водится C.rhipidophyllа [Вахромеев, 2002]. В Ивановской обл. C. monogyna
собран Е.А. Борисовой в сосновом лесу в окрестностях г. Иваново и на склоне
ж/д насыпи на ст. Строково (ИУ; Борисова, 2007].олго удерживается в местах
культуры, ежегодно
Указан для г. Смоленск [Решетникова, 2004], но на юге области дичает
С.curvisepala.
Для Рязанской обл. C. monogyna не указан, приведен вид природной фло
ры С.curvisepala, находящийся на северной границе ареала [Казакова, 2004]. В
Пензенской обл. C.monogyna собран одичавшим в Тамалинском р не [LE; Ва
сюков, 2004].
Местообитания. В пределах естественного ареала растет одиночно или
группами на каменистых склонах среди степных кустарников, на опушках, в
сухих дубравах, реже в сосново лиственных лесах [Ареалы..., 1980]. Во вто
ричном ареале C. monogyna -- обычный вид в населенных пунктах, где растет в
культуре и нередко дает самосев.
Рис. 222. Побеги
Crataegus monogyna
Рис. 223. Расселение
Crataegus monogyna
в Средней России
Рис. 225. Деревце Crataegus monogyna
421
Встречается одиночными деревьями по склонам речных берегов и балок,
среди кустарников, по опушкам лиственных лесов, иногда в глубине лесного
массива. Вырастает по насыпям железных дорог и обочинам шоссе. Часто фор
мирует кустарниковый ярус в лесополосах.
Размножение и жизненный цикл. Опыляется насекомыми. Цветет в июне,
плоды созревают в сентябре [Губанов и др., 2003]. Возраст жизни отдельных
особей достигает 250 лет и более.
Рис. 224. Рисунок
из книжки
Crataegus monogyna
Рис. 226. Схема отношений
C. monogyna и близких видов
422
Светолюбивый вид, мезофит, микротерм, мезотроф [Ареалы..., 1980]. Об
ладает быстрым ростом, особенно в первые 15 лет. Вид нетребователен к по
чвам и температурному режиму, что может являться одной из причин его ус
пешного расселения и распространения во многих странах [Weed US, 2008]. В
Европе уделяют значительное внимание борьбе с вредителями и болезнями
C. monogyna [Wilson et al., 1990].
Расселение и распространение. В пределах вторичного ареала C. monogyna
возобновляется на ж/д насыпях, обочинах дорог, по улицам и пустырям, вблизи
мест культивирования. C. monogyna -- орнито и зоохор [Ареалы..., 1980]. Пло
ды хорошо поедают птицы, особенно дрозды, распространяя таким образом
боярышник по лесным территориям [Губанов и др., 2003]. Известны случаи
распространения C. monogyna животными. Например, в Австралии поссум
Trichosurus vulpecula, поедая плоды C.monogyna, оставляет экскременты на рас
стоянии до 50 м от растения. Всхожесть семян, прошедших кишечник поссу
ма, довольно высока -- 73%. Всхожесть свежих семян составляет 87,6%. Авто
ры исследования отмечают, что распространение семян C. monogyna поссумом
часто недооценивается, а это важный фактор инвазии боярышника в лесные
территории Австралии [Bass, 1990].
Последствия внедрения
Воздействия на естественные фитоценозы и аборигенные виды. Вид
образует ассоциации с Prunus spinosa, Euonymus europaea, Rosa afzeliana,
Rhamnus cathartica, Ulmus carpinifolia, Cerasus fruticosa, степными видами спи
рей и т.д. C. monogyna -- ассектатор кустарниковых зарослей и, реже, подлеска
[Ареалы..., 1980]. На территории Средней России C. monogyna, по нашим оцен
кам, не является пока видом, угрожающим целостности сообществ (рис. 225).
Отдельные особи встречаются в лесах, чаще пригородных, но больших зарос
лей нам наблюдать не приходилось.
В 1970--1980 гг. C. monogyna указывали среди опасных сорных видов, угро
жающих агрофитоценозам Австралии. Его распространение, наряду с други
ми сорными видами, изучали в 10 сельскохозяйственных провинциях в Вик
тории (Австралия). Основная масса особей C. monogyna сосредоточена на обо
чинах дорог и рядом с фермами. Присутствие C.monogyna оказывало
негативное влияние на растительный покров пастбищ и мешало задернению
обочин [Lanea, 1979]. Распространение C.monogyna представляет опасность и
для лесных территорий, а также лугов и выгонов Австралии [Bass, 1990]. Опас
ность вида на континенте не ограничивается только внедрением его в природ
ные сообщества. Он представляет угрозу для садоводства, так как является
пристанищем большого числа насекомых--вредителей плодовых деревьев и
кустарников [SGA: Sustainable Gardening Australia, 2008].
Вид создает серьезные проблемы для природных экосистем в Калифор
нии, включен в список опасных инвазионных видов как таксон, требующий
мер по сокращению его численности [Calflora, 2008]. Опасность вида отмече
на не только для Калифорнии, но для всей территории США. C. monogyna
включен в базу данных Weed US [2008] как вид, активно внедряющийся в при
родные сообщества.
С другой стороны, в пределах естественного ареала вид страдает от ряда
факторов, например, чрезмерного выпаса овец. Этот аспект исследован на тер
ритории Северного Уэльса [Good et al., 1990].
Рис. 227. Генератив
ные побеги Sorbaria
sorbifolia
Рис. 228. Расселение
Sorbaria sorbifolia в
Средней России
Рис. 229. Внедрение
Sorbaria sorbifolia в
опушечные сообщества
Рис. 230. Обильное
цветение Sorbaria sorbifolia
425
Генетические изменения и вариабельность вида. 2n = 32, 34 [Агроэколо
гический атлас ..., 2008]. Род Crataegus обоснованно труден для систематичес
ких исследований вследствие гибридизации, полиплоидии и апомиксиса внут
ри рода. C. monogyna --полиморфный вид, разбиваемый в настоящее время на
несколько рас, понимаемых многими как самостоятельные виды. J.Franco вы
деляет 6 подвидов C. monogyna: subsp. nordica Franco (С. orientobaltica Cin.); subsp.
monogyna; subsp. leiomonogyna (Klokov) Franco; subsp. brevispina (G. Kunze) Franco;
subsp. aegeica (Pojark.) Franco; subsp. azarella (Griseb.) Franco [Franco, 1968].
Г.Крюссман приводит 4 подвида C. monogyna: ssp. nordica Franco, ssp. monogyna,
ssp. brevispina (G. Kuntze) Franco, ssp. azarella (Griseb.) Franco. При этом Крюс
сман, в отличие от Франко, не признает виды М.И. Клокова и А.И. Поярковой
[Krьssmann, 1976]. Недавно обозначены лектотипы для C. monogyna f. subdigyna и
C.monogyna var. ronnigeri [Christensen, Janjic, 2004].
C.monogyna часто гибридизирует как с видами природной флоры в местах
совместного произрастания, так и с декоративными интродуцированными
видами. Один из самых распространенных гибридов -- C. × kyrtostyla Fingerh.
(= C. rhipidophylla × C.monogyna s. l.; C. × heterodonta Pojark.; C. monogyna var.
rubra Pall., C. monogyna auct. plur. fl. Ross. non Jacq., C. monogyna var. kyrtostyla
Lange, C. oxyacantha var. monogyna Schmalh. ex parte; C. curvisepala Lindm.) -- Б.
согнутостолбиковый [Пояркова, 1950; Цвелев, 2000]. В дикорастущем состо
янии вид встречается в южной части Республики Беларусь. По мнению А.И.
Поярковой, это «единственный дикорастущий вид в БССР; все указания C.
oxyacantha и C. monogyna для БССР относятся к этому виду» [Пояркова, 1950,
с. 62]. От сходного, распространенного западнее C.laevigata (Poiret) DC.,
C.monogyna отличается формой листовой пластинки: у C.monogyna листья с
более глубокими лопастями, цветки имеют 1 пестик, а не 2--3. Однако хорошо
различимы только типичные формы, а не гибриды. Гибрид C. monogyna × C.
laevigata (C. × macrocarpa Hegetschw., syn. C. × media Bechst.) часто использу
ется как садовый кустарник со множеством форм, из которых в Европе весьма
популярен 'Paul's Scarlet' с темно розовыми махровыми цветками. Однако во
«Флоре Европы...» и «Флоре Чехии...» [Holub, 19..] C. × macrocarpa Hegetschw.
и C. × media Bechst. не синонимы, а два самостоятельных таксона [Franco,
1968, c. 75; ...].
Схему отношений C. monogyna и близких видов можно представить следу
ющим образом (рис. 226).
Crataegus curvisepala Lindm., C. laevigata (Poiret) DC. и C. monogyna Jacq.
формируют гибридные комплексы в результате интрогрессии в Северной Ев
ропе. В исследованиях показано, что ряд ранее описанных видов на самом деле
являются гибридами этих трех таксонов. Например, C.palmstruchii Lindm. яв
ляется синонимом C.laevigata. В свою очередь, C.eremitagensis Raunk.,
C.raavadensis Raunk. и С.shumacheri Raunk. представляют собой гибрид С.
сurvisepala × С. laevigata. Вместе с тем, разграничение гибридов С.сurvisepala
× С.laevigata и С.monogyna × С.laevigata представляет значительные трудно
сти [Christensen, 1983]. Датский систематик K.Христенсен проводил сравни
тельное изучение греческого, югославского и датского материала C.monogyna,
С.сurvisepala и их гибридов с применением дискриминантного анализа. Он
пришел к выводу о целесообразности выделения C. × albanica Pojark.
(C.monogyna × C.orientalis) и C.×kyrtostyla Fingerh. (С.сurvisepala × C.monogyna)
426
[Christensen, 1984]. C целью разграничения C.monogyna, C. pentagyna и
С.laevigata было проведено хемосистематическое исследование, направлен
ное на получение маркеров для точной идентификации этих видов, широко
применяемых в европейской фитотерапии. Выявлены и описаны различия в
составе флавоноидов трех видов [Prinz et al., 2007]. Гибридные формы с учас
тием C. monogyna известны по всему обширному ареалу вида. Например, было
проведено молекулярное и морфологическое исследование с участием образ
цов из Южной Сирии: Crataegus monogyna Jacq., C. azarolus var. aronia L. и C. ×
sinaica Boiss. ssp. Sinaica и показано, что образцы С. × sinaica Boiss. ssp. sinaica
отличаются высокой степенью изменчивости признаков и по данным секве
нирования, скорее всего имеют гибридное происхождение с участием C.azarolus
и C.monogyna [Albarouki, Peterson, 2007].
В Северо Западной Европе изучали фенотипическую изменчивость лис
товой пластинки C. monogyna с целью выяснения уровня гибридизации осо
бей этого вида в популяциях. Проанализирована изменчивость размеров и
формы листовых пластинок, взятых с побегов в одном клоне и имеющих, та
ким образом, одинаковый генотип. Оказалось, что только один морфологи
ческий признак листовой пластинки из 9 изученных зависит от условий по
чвенного питания [Gosler et al., 1994].
Влияние на здоровье человека. Плоды содержат виннокаменную, лимон
ную и кофейную кислоты, ацетилхолин, изобутиламин, сахара, флавоновые
гликозиды, тритерпеновые кислоты, дубильные вещества и витамин С. В цвет
ках содержится эфирное масло, кверцетин, холин, гиперозид и другие соеди
нения. В медицинской практике используются цветки и плоды. Цветки соби
рают вполне распустившимися, а плоды заготавливают осенью. Препараты
растения обладают болеутоляющим и успокаивающим действием, их приме
няют при гипертонии, сердечных неврозах и нервных заболеваниях. В науч
ной медицине при функциональных расстройствах сердечной деятельности
используют жидкий экстракт (Extractum Crataegi fluidum) из плодов или на
стойку (Tinctura Crataegi) из цветков. В народной медицине настой из пло
дов и цветков используют для повышения работоспособности и уменьшения
возбудимости ЦНС [Лекарственные растения, 2008].
Экономическое и социальное значение (положительное и отрицатель
ное). Издавна разводится в садах и парках, особенно в живых изгородях, так
как прекрасно поддается стрижке и обладает большой побегообразующей спо
собностью, к тому же оказывается почти непроницаемым барьером и хорошо
изолирует территории усадеб. Широко распространены различные декоратив
ные формы -- с махровыми белыми, розовыми и красными цветками, с пира
мидальной и плакучей кроной, со скрученными ветками, скученными в пучки
колючками и пр. Г.Крюссманн приводит серию садовых форм C. monogyna,
замечая, что формы часто неверно относят к C. laevigata [Krьssmann, 1976, р.
433]: 'Aurea' -- с желтыми плодами; 'Bicolor' (C. monogyna gumperi Voss) -- лепе
стки белые с розовыми краями; 'Biflora' -- цветет дважды за лето; 'Compacta'
(C.monogyna compacta Spдth) -- с короткими толстыми ветвями, без колючек;
'Ferox' ('Horrida') -- с мелкими цветками и многочисленными крепкими ко
лючками; 'Fissa' -- листья до 7 см, глубоко рассеченные; 'Flexuosa' (C. monogyna
tortuosa hort.) -- с повислыми ветвями; 'Granatensis' -- ветви повислые, колю
чек очень мало, происходит из Испании; 'Inermis' -- ветви густые, колючки
427
отсутствуют, происходит из Америки; 'Lutescens' -- листья желто зеленые; C.
monogyna f. microphylla Chenev. -- листья очень мелкие; 'Pendula' -- ветви по
вислые; 'Pteridiflora' (C. monogyna filicifolia hort.) -- листья рассеченные, вееро
видные; 'Rosea' -- цветки розовые; 'Semperflorens' (C. monogyna Bruanti hort.) --
цветет с мая по август; 'Stricta' (C. monogyna fascigiata и C. monogyna pyramidalis
hort.) -- с прямыми длинными ветвями.
Молодые листья боярышника однопестичного используются для приготов
ления салатов. Плоды идут на вино, желе, ими обогащают вкус бренди, ис
пользуют для изготовления суррогата кофе и чая, муку из плодов домешива
ют в хлебобулочные изделия. В Китае сушеные плоды и особенно хлопья из
них употребляют в пищу. Желе или хлопья, приготовленные из плодов боя
рышника, используют для поддержки пищеварения после тяжелых мясных
блюд в китайской медицине. Сеянцы C. monogyna обычно используют в каче
стве подвоев [Агроэкологический атлас..., 2008].
Меры борьбы
В Австралии C. monogyna является карантинным сорняком и запрещен к
ввозу в пределы страны. На австралийском континенте предлагается усилить
контроль за обочинами дорог с целью уничтожения C. monogyna и предотвра
щения его внедрения в другие типы ценозов. Однако на практике вырубание
зарослей C. monogyna сильно затруднено из за наличия колючек [Lanea, 1979;
SGA: Sustainable Gardening Australia, 2008].
В других регионах мира принимать подобные меры борьбы с C. monogyna
пока не требуется, так как пока распространение вида носит ограниченный
характер.
СЕМЕЙСТВО ROSACEAE -- РОЗОЦВЕТНЫЕ
РЯБИННИК РЯБИНОЛИСТНЫЙ
Название: Sorbaria sorbifolia (L.) A. Br.
Синонимы: Spiraea sorbifolia L., S. pinnata Moench, Basilima sorbifolia (L.)
Raf., Schizonotus sorbifolius (L.) Lindl., Sorbaria stellipila (Maxim.) C.K. Schneider,
S. assurgens hort. ex C. K. Schn.
Вид описан К.Линнеем как Spiraea sorbifolia, род Sorbaria выделен А.Брау
ном в 1860 г. В современном понимании род состоит из нескольких видов, иног
да культивируемых в садах и часто встречающихся в специализированных
коллекциях, дендрариях или ботанических садах. Некоторые таксоны описа
ны неоднократно и под разными наименованиями, что являлось причиной
номенклатурных недоразумений [Tomaszewski, 2001]. Согласно последней ре
визии [Rahn, 1989], род Sorbaria включает 4 вида: Sorbaria sorbifolia (L.) A. Br.
(включая S.stellipila C. K. Schn.), S.grandiflora (Sweet) Maxim. [=S. alpina (Pall.)
Dippel, S.pallasii (G. Don) A. Pojark., S.rhoifolia Komarov], S.kirilowii (Regel)
Maxim. (включая S.assurgens Vilm. et Bois, S.arborea C.K. Schn.) и S.tomentosa
(Lindl.) Rehder [= S.lindleyana (Lindl.) Maxim., включая S.olgae Zinscrl. и
S.gilgitensis] с var. angustifolia, comb. nov. (=S. altchisonii).
В садах и дендрариях виды рода Sorbaria часто сажают совместно, что при
водит к их гибридизации. Так, в ботанических садах Познани и Варшавы об
наружены гибридные особи между S.sorbifolia, S.kirilowii и S.tomentosa
428
[Tomaszewski, 2001]. Эти растения имели простые волоски на нижней стороне
листочков (как у S.kirilowii и S.tomentosa) и опушенные листовки (как у
S.sorbifolia и S.grandiflora). Процент жизнеспособной пыльцы у гибридов ниже,
чем у видовых растений (24 87 и 72 100% соответственно). В культуре отмече
ны также гибриды между S.sorbifolia и S.grandiflora и между S.sorbifolia и
S.kirilowii [Rahn, 1989].
Приводим ключ для определения трех видов рода, культивируемых в
Польше [Tomaszewski, 2001]:
1.Плоды всегда опушенные. Тычинок обычно 30 или более. Нижняя сторона
листочков без простых волосков. Кустарники до 3 м высотой .......... S. sorbifolia
--- Плоды голые или редко едва опушенные. Тычинок обычно 20. Нижняя
сторона листочков хотя бы частично с простыми волосками, иногда скрытыми
под звездчатыми. Кустарники до 7 м высотой, обычно -- более 3 м ................... 2
2. Основание столбика находится значительно ниже верхушки листовки.
Нижняя сторона листочков по жилкам более или менее опушена звездчатыми
и простыми волосками. Соцветие очень рыхлое, его ветви более или менее ото
гнутые ..................................................................................................................... S. kirilowii
--- Столбик загнут назад, его основание находится над или около верхушки
листовки. Нижняя сторона листочков с простыми волосками, расположенны
ми вдоль главной жилки или голая, звездчатые волоски обычно отсуствуют,
соцветие густое, плотное ............................................................................. S. tomentosa
При просмотре весьма скудных гербарных материалов по этому роду мы
убедились, что все образцы, собранные в Средней России, относятся к
S.sorbifolia, по крайней мере те, которые собраны вне культуры. Гибридных
растений мы не обнаружили.
Морфологические признаки
Кустарник высотой 1--3 м, с тонко опушенными, редко голыми побегами.
Листья непарноперистосложные, продолговато эллиптические, длиной 12--25
см и шириной 6--13 см, на черешках длиной 2--5 см, с 9--21, а чаще -- с 17, лан
цетовидными, заостренными листочками, по краю двоякопильчатыми, дли
ной 2,5--8 см, шириной 0,8--2,5 см, голыми или реже снизу по жилкам с про
стыми или разветвленными желтыми волосками. Прилистники от широко
сердцевидных до линейных, зубчатые или цельнокрайние. Метелки длиной
12--30 см и шириной 5--12 см. Оси соцветия, цветоножки, а часто и нижняя
часть гипантия снаружи обычно тонко пушистые с примесью короткого желе
зистого опушения или более или менее густо покрытые кустистыми рыжева
тыми волосками; в последнем случае обычно и нижняя сторона листьев по
жилкам с таким же опушением. Цветки диаметром 7--11 мм с почти округлы
ми лепестками и тычинками, вдвое превышающими лепестки. Листовки опу
шенные, 5 мм длиной (рис. 227). Цветет со второй половины июня до начала
сентября, плодоносит с августа [Флора СССР, 1939].
Естественный ареал
Западная Сибирь: Томская обл., Красноярский край, Восточная Сибирь:
Иркутская, Читинская обл., Бурятия, Республика Саха (Якутия), Дальний
Восток, Камчатка, Северная Монголия, Япония, Корея, Китай, Гималаи, Цен
тральная Азия [Флора СССР, 1939; Флора Сибири, 1988].
429
Вторичный ареал
История интродукции и географическое распространение. Рябинник
интродуцирован в Европу в середине XVIII века и к настоящему времени
широко распространился в культуре [Bulygin, Firsov, 1998; Hegi, 1922;
Shipchinskii, 1954; Tomaszewski, 2001]. В качестве заносного вида отмечен в
Центральной и Северной Европе [Interactive flora of NW Europe, 2008], Польше
[Tomaszewski, 2001]. Интродуцирован в Северную Америку, где не является
массовым видом и лишь изредка формирует большие, плотные заросли. Рас
пространен на северо востоке США, в восточных и центральных провинциях
Канады; на Западном побережье встречается только на Аляске и в одном из
штатов США [USDA plants, 2008].
Культивируется в населенных пунктах и разрастается в местах посадок в
Вологодской, Ленинградской, Псковской и Новгородской обл. [Цвелев, 2000].
Пути и способы заноса. На новую территорию вид попадает исключитель
но как эргазиофит, так как является чрезвычайно популярным декоративным
растением.
Статус в регионе. В Средней России широко используется в озеленении,
долго сохраняется на месте старых посадок, местами дичает и иногда образует
обширные заросли [Маевский, 2006].
В Воронежской обл. S.sorbifolia культивируется, в одичавшем состоянии
отмечен на окраине кладбища г. Воронеж [Григорьевская, 2000; Григорьевс
кая и др., 2004]. В Курской обл. дичание вида впервые отмечено А.В. Полу
яновым (личное сообщение) в окрестностях Курска, близ д. Моква, у ручья
Моква близ дамбы Моковского пруда, в конце 1980 х гг. Для Орловской фло
ры один из первых сборов сделан в Болховском уезде Орловской губернии, в
с. Муратово, в роще, 17.07.1926 г., О. Хитрово (MW).
Впервые в Московской обл. собран, вероятно, А.Н. Петунниковым в Кузь
минках [г. Москва], в парке, 19.06.1887 г. (MW). Отмечен Н.Н. Кауфманом
[1889], как разводимый и дичающий вид. В Ярославской обл. собран в Неко
узском р не: «Борок, парк, 4.06.1960 г., Шорохова, Полякова» (MW). В Тверс
кой обл. вид найден в 1915 г. в качестве одичавшего растения в запущенном
парке у имения Конюшино Новоторжского уезда и в окрестностях д. Дымов
ка Кашинского уезда [Ильинский, 1924]. Во Владимирской обл. известен во
всех районах, но редко. Собран А. Серегиным в Гусь Хрустальном р не, на тер
ритории национального парка «Мещера», в 4 км. к северо востоку от пос. Ур
шельский, в разрушенной деревне Мочаловка, на обочине дороги у р. Бужа,
8.07.2002 г. (MW). Более ранние сборы с территории Владимирской обл. нам
неизвестны. Cобран в 1880 г. в Костромском уезде Костромской губернии,
1.07.1880 г., И.Ф. Мейснер (MW), с припиской на этикетке: «в садах разво
дится как декоративное».
Вид указывается для Смоленской обл. [Решетникова, 2004] как вид, дича
ющий из посадок, но не обширных зарослей не образующий. Собран в Деми
довском р не, в одичалом парке на месте помещичьей усадьбы у о. Баклановс
кое, поросль под пологом леса, в тени, много, 2.08.1974 г., В. Макаров, М. Ба
женова (MHA). В Калужской обл. распространение локальное, есть сплошные
заросли в лесах, на месте бывших посадок (С. Майоров, личное сообщение).
Вид собран в окрестностях г. Таруса, «в боковом темном сыром овражке, близ
Ильинского, 18.08.1954 г., В.И. Соболевский» (MHA); «Юхновский район, в 5
430
км к юго западу от д. Александровка. Национальный парк «Угра». По левому
берегу р. Собжи, около 4 км выше ее устья (впадения в р. Угру), в березняке с
сосной и в ольшанике, на протяжении нескольких километров, 12.08.2004 г.,
Н.Решетникова, С.Полевова» (MHA).
Собран А.Сухоруковым в Уметском р не Тамбовской обл. в 1 км восточнее
д. Натальевка, в широколиственной роще на просеке, 17.07.1996 г. (MW).
Таким образом, S. sorbifolia дичает во всех областях Средней России (рис. 228),
однако недостаток гербарного материала по ряду областей не всегда позволя
ет составить детальную картину истории расселения вида на исследуемой тер
ритории.
Местообитания. В природе S.sorbifolia растет по берегам горных и лес
ных ручьев и речек, местами образуя очень густые и большие заросли, по не
густым хвойным, смешанным и лиственным лесам и опушкам, окраинам бо
лот [Флора СССР, 1939].
В условиях вторичного ареала культивируется как декоративный кустар
ник в населенных пунктах и быстро разрастается благодаря образованию кор
невой поросли. Встречается одичавшим в парках, в том числе старинных, у
заборов, в палисадниках, вдоль дорог, на влажных сорных местах, вдоль же
лезных и шоссейных дорог, на пустырях, вблизи дачных участков, в пригород
ных лесах и лесопарковых зонах (рис. 229). Вид, вероятно, устойчив к засоле
нию почвенного покрова, так как высаживается и успешно разрастается вок
руг городских свалок (например, в г. Узловая Тульской обл.). Во Владимирской
обл. образует большие заросли под пологом леса близ брошенных деревень
(А.П. Серегин, личное сообщение). В Ивановской обл. присутствует в старых
усадебных парках в Ивановском и Приволжском р нах [Борисова, 2007; Ва
сюков, 2004; Вахромеев, 2002; Григорьевская и др., 2004; Еленевский, Радыги
на, 1997; Игнатов и др., 1990; Казакова, 2004; Маркелова, 2004; Полуянов, 2005;
Флора Липецкой области, 1996]. В местах бывшего культивирования активно
разрастается, формируя обширные заросли, однако сплошного покрытия вто
ричного ареала в условиях Средней России пока не наблюдается вследствие
малой вероятности семенного размножения.
Случаи самосева вида отмечены лишь в Ивановской области, в дендрарии
Ботанического сада Ивановского университета [Борисова, 2007].
Размножение и жизненный цикл. Рябинник рябинолистный дает обиль
ные корневые отпрыски, образуя густые, очень эффектные заросли. Растет
быстро. Обладает высокой побегообразующей способностью. К почве нетре
бователен, но лучше растет на дренированной и влажной. Выносит небольшое
затенение. Обильно и продолжительно цветет: цветение может длиться до 30
дней (рис. 230). В культуре размножается семенами, а также корневыми от
прысками, одревесневшими черенками и отводками. Семена высеваются вес
ной без предварительной обработки на увлажненную почву, 1 2 месячная стра
тификация улучшает прорастание. Обильно цветет на протяжении 30 дней.
Зимостоек. Требователен к богатству и влажности почвы. Переносит времен
ное затопление, хуже --- засуху. [Аксенов, Аксенова, 2000; Энциклопедия де
коративных садовых растений, 2008].
Расселение и распространение. В природе виды рода Sorbaria занимают
берега рек и ручьев, их легкие семена перемещаются по поверхности воды и
быстро прорастают [Tomaszewski, 2001]. В культуре семенами размножается с
431
трудом, дичает посредством разрастания, образуя обильные корневые отпрыс
ки, самосев редок.
Последствия внедрения
Воздействия на естественные фитоценозы и аборигенные виды. В на
стоящее время в условиях Средней России S. sorbifolia способен образовы
вать сплошные обширные заросли под пологом леса и вытеснять местные виды,
что негативно сказывается на динамике состава и структуры лесных сообществ.
Генетические изменения и вариабельность вида. 2n = 36. На юге Куриль
ских островов встречаются растения S.sorbifolia f. stellipila с более опушенны
ми цветоножками, гипантием и нижней стороной листьев. Иногда эту расу
считают отдельным видом [Алексеев и др., 1997]. В современных обработках
S.stellipila не выделяют как самостоятельный вид и включают в S.sorbifolia
[Rahn, 1989].
Влияние на здоровье человека. Вид известен как лекарственное растение,
активно применяется в китайской и японской медицине [Kim, Zee, 2000; Xiao
Dong et al., 2002; Zhang et al., 2007].
Экономическое и социальное (положительное и отрицательное) зна
чение. S.sorbifolia --- один из наиболее распространенных декоративных кус
тарников в садах и парках и по берегам водоемов. Живописно выглядит в пе
риод массового цветения и осенней окраски листьев. Используется в одиноч
ных и групповых посадках, для создания опушек и подлеска, живых изгородей,
закрепления откосов и оврагов. Все рябинники обладают высокими фитон
цидными свойствами. [Аксенов, Аксенова, 2000; Энциклопедия декоративных
садовых растений, 2008].
Меры борьбы
Меры по уничтожению и предотвращению распространения S.sorbifolia вне
культуры не разработаны, так как активное расселение вида и образование им
обширных зарослей отмечено только в последние годы.
СЕМЕЙСТВО SALICACEAE -- ИВОВЫЕ
ТОПОЛЬ БЕЛЫЙ
Название: Populus alba L.
Синонимы: Populus alba в. nivea Ait., P. bachofenii Wierzb. ex Rochel, P. bolleana
Lauche, P. nivea (Ait.) Willd., P. pseudonivea Grossh., P. alba subsp. major (Mill.)
R. Kam nom. invalid. (P. major Mill.), P. triloba Dode, P. treyviana Dode.
Морфологические признаки
Дерево высотой 18--35 м, с широкой кроной; кора очень светлая, в молодо
сти гладкая, у старых деревьев зеленовато серая с неглубокими трещинами
(рис. 231). Молодые ветви беловойлочные, зимние почки опушенные, реже
голые с реснитчатыми краями. Листья удлиненных побегов дельтовидные с
прямым основанием и двумя маленькими лопастями на углах, выше зубча
тые, снизу с обильным, хлопьевидным опушением, белые; листья укорочен
ных побегов округлые, со слегка усеченным основанием и острыми зубцами,
несимметричные, сохраняющие серое хлопьевидное опушение до осени. Пыль
никовые сережки толстые, длиной 3--7 см, прицветники ржавого цвета, от эл
432
липтических до клиновидно эллиптических, неравномерно зубчатые или же
почти цельнокрайние, умеренно мохнато ресничатые. Тычинок 6--8 (3--10) с
пыльниками сначала светло пурпурными, позднее желтыми. Плодущие сереж
ки длиной 10--12 см, оси их с редкими, мягкими волосками; завязи продолго
вато конусовидные, на коротких ножках; рыльца 2, розово красные, лопасти
их узкие, расположены крестообразно; коробочки голые. Цветет весной одно
временно с распусканием листьев [Комаров, 1936].
Естественный ареал
Юг Европейской части России, Крым, Кавказ (Предкавказье), Западная
Сибирь, Средняя Азия, Южная часть Средней и Западной Европы, Средизем
номорье, Балканы, Малая Азия. В Европе ареал P. alba тянется от Испании до
Марокко через всю Центральную Европу, доходит на север до Германии и
Польши, а затем переходит на юг Европейской России. Самый северный пункт
произрастания культивируемых деревьев 68° с. ш. в Норвегии, а дико расту
щих -- 58°15' у с. Ново Ильинского на р. Обь южнее устья р. Кеть. Самым вос
точным местом обитания вида является 58° в. д. на р. Чулым, а самым южным --
Китайская Джунгария, р. Урунгу [Комаров, 1936].
Вторичный ареал
В Северную Америку P. alba интродуцирован первыми поселенцами ранее
1785 г. и широко распространился вдоль восточного побережья США и Кана
ды, оказавшись весьма конкурентоспособным [Spies, Barnes, 1981]. В Северной
Америке вид встречается в массе от 40° с.ш. до 30° ю.ш. Несмотря на то, что вид
присутствует на северо американском континенте более 200 лет, его нельзя счи
тать натурализовавшимся, так как преобладает вегетативное размножение, осо
би образуют клоны, и почти все из них -- женские [Spies, Barnes, 1981]. Вид яв
ляется инвазионным сорняком в южной Африке. В западной Австралии фор
мирует плотные заросли по речным долинам [Weed identification, 2008].
Разграничение природного и вторичного ареала P.alba в Европе сопряже
но с серьезными трудностями. В рамках европейского проекта «Биоразнооб
разие природных и культивируемых тополей» проведены молекулярные ис
следования образцов P.alba и достоверно выявлены прямые и возвратные гиб
риды с P.tremula, а также показано природное происхождение популяций вида
в северной Италии [Fossati et al., 2004]. Определено происхождения популя
ций P.alba на о. Сардиния, произрастающих там, согласно отрывочным палео
ботаническим данным, с античного периода, но достоверно зарегистрирован
ных с середины XIX века [Zapelli et al., 2005]. На севере Сардинии в популя
циях P. alba выявлено лишь 3 генотипа, распространенных локальными
группами. Авторы склоняются к гипотезе о природном происхождении попу
ляций вида на острове и делают предположение, что все особи из современ
ных популяций Сардинии, произошли путем вегетативного размножения от
этих трех предковых генотипов.
История интродукции и географическое распространение. На севере
России, в Вологодской обл., P.alba изредка встречается в посадках и не прояв
ляет склонности к дичанию. На Северо Западе России вид культивируется в
населенных пунктах, садах и парках, у дорог, по берегам водоемов, иногда ди
чает [Цвелев, 2000].
Пути и способы заноса. Первичный способ заноса P.alba -- намеренная
интродукцию, то есть вид является эргазиофитом. Исключение составляет
Рис. 231. Пойменный
лес из Populus alba
Рис. 232. Расселение
Populus alba в
Средней России
Рис. 233. Клон Populus alba
Рис. 234. Сережка Populus alba
435
территория, по которой проходит граница ареала. Например, в Курской обл.
соседствуют как природные, так и заносные популяции P. alba, и происхожде
ние каждой из них определить затруднительно [Золотухина, Золотухин, По
луянов, 2001]. Трудно также выяснить, был ли вид занесен на конкретный уча
сток человеком или появился там случайно.
Статус в регионе. Во всех областях Средней России часто разводится в
садах и парках и нередко встречается одичалым на месте уже не существую
щих селений и усадеб (рис. 232). В южных областях иногда разводится пира
мидальная форма P. bolleana Lauche (Тополь Болле или самаркандский), ве
роятно, среднеазиатского происхождения [Маевский, 2006]. В ряде сводок
P.bolleana имеет статус вида [Комаров, 1936], однако в Европе и Северной
Америке его считают формой P. alba 'Pyramidalis'. В Средней России P.bolleana
нам встречать не приходилось; вдоль улиц, дорог и в парках обычно высажи
вают пирамидальную форму P.nigrа.
Во всех областях Средней России тополь белый известен с XIХ столетия.
В этот период вид уже широко использовался в качестве декоративного расте
ния, и авторы отмечали его способность к вегетативному размножению. Се
менное возобновление P.alba за пределами природного ареала в Орловской,
Тульской и Московской обл. отмечено в 1980 е гг. Вероятно, именно наличие
или отсутствие семенного возобновления определяет характер и скорость рас
пространения P.alba в Средней России. Если в Ивановской обл. в большин
стве случаев встречаются лишь клоны, то в Московской, Тульской обл. и юж
нее P.alba прекрасно возобновляется самосевом. Ниже приводятся данные об
истории и современном распространении P.alba в областях Средней России
по градиенту юг север.
Для Курской губернии вид приводится в литературе, начиная с первых
флористических сводок [Черняев, 1836; Мизгер, 1869]. Первая гербарная на
ходка тополя сделана в Тимском уезде Курской губернии, с. Никольское, Д.Д.
Санцовым и Ф.А. Гриневским 16.07.1887 г. (MW). В XIX веке P. alba собирали
в Орловской губернии: 1) Болховский уезд, 23 апреля 18**, Герминьяр (MW);
2) г. Орел, 1.05.188* [фамилия коллектора выцвела, из гербария Цингера],
(MW). В 1907 г. в с. Муратово Орловской губернии P. alba собрал В.Н. Хитро
во (OHHI). В Тульской обл. P.alba разводится давно [Цингер, 1885]. В «Денд
рологическом списке...» В.В. Розена [1913] P. alba причисляется к древесным
интродуцентам, способным в наших условиях разрастаться вегетативно. В MW
хранится сбор начала XIX века, сделанный в Московской губернии: Zarizina,
26.07.1814 г., д р Виаль. Во Владимирской губернии P.alba собран М.Назаро
вым в г. Меленки, 2.05.1910 г. (MW). Один из первых сборов в Калужской
губернии сделан в Жиздринском уезде княгиней Е.Л. Голицыной, 188*, [из
гербария Н.Я. Цингера], (MW); а позднее вид собран В.И. Соболевским в ок
рестностях Тарусы, в старинной усадьбе, 24.08.1954 г. (MHA). Вероятно, од
ним из первых сборов с территории современной Рязанской обл. можно счи
тать следующий: Елатомский уезд, в самом г. Елатьма, госп. Орлов, 18**, из
гербария Н.Я. Цингера (MW). В 1950--1970 гг. P. alba активно собирали в раз
ных районах Рязанской обл. (MW).
Учитывая недостаток данных, можно сделать несколько замечаний о со
временном распространении и характере возобновления P.alba. По современ
ной Курской обл. проходит граница естественного ареала вида. P.alba произ
436
растает здесь в пойменных, реже водораздельных, дубравах, ольшаниках, по
склонам речных долин, оврагов и балок. В настоящее время по всей области
встречается нередко, но рассеянно, больших скоплений не образует. Повсеме
стно разводится в населенных пунктах и лесополосах и легко дичает [Полу
янов, 2005]. Границу между природными и одичавшими популяциями вида
провести трудно, как минимум некоторые из них имеют естественное проис
хождение [Золотухин, Золотухина, Полуянов, 2001].
В Воронежской и Белгородской обл. тополь распространен сходным обра
зом: природные популяции сочетаются с широким применением вида в куль
туре. Например, в Воронежской обл. вид произрастает в поймах крупных рек,
а в г. Воронеж --культивируется и встречается в парках и лесопарках, садах,
скверах, на бульварах [Григорьевская, 2000]. В Белгородской обл. встречается
нечасто, имеются несколько сборов из Старооскольского, Прохоровского и
Грайворонского р нов (LECB, ОГУ), сделанных у дорог, в поймах рек и водо
хранилищ [Еленевский и др., 2004].
В Липецкой обл. тополь высаживают вдоль дорог, в населенных пунктах;
нередко вид дичает, встречается по опушкам и долинам рек [Флора Липецкой
области, 1996]. Дичание P. alba в Тамбовской губ. отмечал Д. Литвинов во вто
рой половине XIX века: Кирсановский уезд, поселяется в долине р. Ворона,
близ с. Пущино, дико, июль 1879 г., Д.Литвинов (MW), в 1889 г. вид собран в
Тамбове (MW). В Орловской обл. P.alba разводится в городах и поселках, из
редка встречается одичалым [Еленевский, Радыгина, 1997]. В июле 2008 г. в
юго восточных р нах Орловской обл. мы наблюдали массовое вегетативное и
семенное возобновление P.alba.
Данных о распространении P. alba в юго восточных регионах Средней Рос
сии недостаточно: в Пензенской обл. часто культивируется и встречается оди
чавшим по берегам водоемов и у дорог [Васюков, 2004]. Имеется старый сбор
с территории современной Самарской обл.: Симбирская губ., Сызранский уезд,
культурное в парке техникума в с. Усолье, лето 1919 г., (MW, фамилия коллек
тора отсутствует).
В Центральных областях Средней России, по современным данным, наблю
дается семенное возобновление P. alba.
В Брянской обл. вид собран в Климовском р не, на днище песчаного карье
ра, 5.09.1975 г., В.Макаров (MHA). Указывается для Смоленской обл. в каче
стве адвентивного вида [Решетникова, 2004]; собран в Демидовском р не, в
1,5 км. к юго востоку от д. Городище, на месте бывшей помещичьей усадьбы у
р. Гобза, 30.07.1999 г., Н.Решетникова.
В Калужской обл. P. alba встречается по старым усадьбам, в пригородных
лесах (С.Майоров, личное сообщение).
В Тульской обл. тополь белый дает обильный самосев (семенное возобнов
ление отмечено И.Шереметьевой в 1980 е гг.), активно расселяется вдоль до
рог, по свалкам и сорным местам, дичает в старых парках и усадьбах. Мы
наблюдали очень старые деревья в Заокском р не, в лесном массиве на терри
тории музея усадьбы А.Т. Болотова. Под кронами тополей сформировался
сплошной покров из разновозрастных отпрысковых растений. Нельзя исклю
чать, что посадки P.alba могли быть сделаны самим А.Болотовым в конце XVIII
века. Молодые растения тополя белого нередки на ж/д насыпях, однако здесь,
как правило, подрост уничтожается при очистке путей. В последнее время вид
активно перемещается в пригородные леса.
437
В Московской обл. широко используется в озеленении и часто распростра
няется и порослью, и самосевом по ж/д и рудеральным местам обитания. В 51
из 74 обследованных старых парков отмечено возобновление P.alba, из них в
27 -- при помощи самосева [Игнатов и др., 1990].
В Рязанской обл. высаживается в населенных пунктах, в парках, дичает. В
долине Оки растет на песчаных дюнах и в междюнных понижениях; предпо
ложительно, некогда его высаживали здесь для закрепления песков [Казакова,
2004]. По нашим наблюдениям, вид является весьма обычным компонентом фло
ры населенных пунктов Рязанской обл. и их окрестностей. С территории Мордо
вии в MW хранится сбор, сделанный в Теньгушевском р не, в с. Белораменье, на
месте бывшего кордона, 16.07.1979 г., Т.Силаева (MW).
На севере Средней России семенное возобновления тополя отсутствует. Во
Владимирской обл. вид легко дичает, однако размножается корневыми отпрыс
ками, самосев отсутствует и, как следствие, гибридные особи не отмечены
(А.П. Серегин, личное сообщение).
В Ивановской обл. образует крупные заросли по берегам Волги в городах
Пучеж и Кинешма, на песках в Южском районе (ИУ, ОКМ). Активно форми
рует корневую поросль, размножается вегетативно. Часто выращивается в го
родах и поселках, у жилья, в парках, дичает [Борисова, 2007]. Семенное раз
множение в пределах области не отмечено, крупные заросли на волжских пес
ках формируются, как правило, в местах бывших посадок (М. Голубева, личное
сообщение). По Нижегородской обл. данных недостаточно: вид собирали в
Нижегородской губ., Сергачском уезде, с. Малое Казариново, на проселочной
дороге, 12.08.1926 г. собр. Н.Туранов, К.Ковалев, опр. А.Уранов (MW).
Местообитания. В условиях природного ареала вид встречается в пой
менных лесах, среди пойменных лугов одиночно или рощами. Культивирует
ся повсеместно как садово парковое дерево и активно дичает.
Размножение и жизненный цикл. На севере Средней России вид размно
жается вегетативно, южнее -- семенным способом, и экземпляры семенного
происхождения встречаются в прибрежных лесах, вдали от жилья и мест куль
тивирования. Дает обильную корневую поросль, размножается черенками,
корневыми отпрысками (рис. 233). Корневая поросль растет от боковых кор
ней, иногда на 20--30 м от ствола и часто формирует клоны. В естественном
ареале такие колонии могут занимать значительные пространства. P.alba --
двудомное дерево. Цветение происходит рано весной (рис. 234), созревание
семян -- поздней весной или ранним летом. Большинство культивируемых в
северной Европе деревьев -- женские экземпляры. Вид нуждается в хорошем
освещении и питании, хорошо переносит паводок, любит слабокислые почвы,
устойчив к засолению.
В Европе, где P.alba выращивают для декоративных и технических целей,
разработаны специальные методики выращивания деревьев. Например, для
ускорения роста на плантациях используют углекислый газ, что приводит к
повышению биомассы корневой системы и микоризообразованию [Lukac et
al., 2003; Wittig et al., 2005].
В конце 1970 х гг. обнаружено поражение особей P.alba «американской
ржавчиной» Melampsora medusae Thьm. [Sharma, Heather, 1977]. В 1980 гг. бо
лезнь «Witches' Broom» наблюдалась у P.alba в Париже и его окрестностях по
обочинам магистральных дорог. Болезнь вызвана присутствием в больных
438
образцах микоплазматических организмов [Sharma, Cousin, 1986]. Эта же бо
лезнь в 1990 гг. обнаружена у P.alba в Венгрии и Германии [Berges et al., 1997].
Расселение и распространение. Первоначальным способом распростра
нения тополя белого в Средней России было его введение в культуру как
неприхотливого декоративного дерева, а также закрепителя песчаных дюн,
в дальнейшем расселение вида зависело от природных условий конкретных
территорий.
Последствия внедрения
Воздействия на естественные фитоценозы и аборигенные виды. В рай
онах совместного произрастания P.alba и P.tremula встречается их межвидо
вой гибрид P. × canescens Smith (P.hybrida Bieb.) -- Т. сереющий, имеющий про
межуточные признаки. Это высокое дерево с беловато серой корой, листья
удлиненных побегов почти сердцевидные, слегка лопастные, с зубчиками, ко
торые по краю образуют неправильную пильчатость. Крона сравнительно то
щая, почти плакучая, светло зеленая, листья укороченных побегов округлые,
неправильно выемчатые, по краю зубчатые с тонким хлопьевидным опушени
ем, рыльца карминово красные и приподнятые параллельно длинной оси за
вязей. III IV. [Комаров, 1936]. В гербариях образцы P. × canescens представле
ны очень скудно, но заметно, что верхняя сторона листовой пластинки у них
голая или почти голая, как у осины, а нижняя сторона имеет совершенно не
характерное для осины плотное бело серое опушение. Большинство особей P.
× canescens в культуре -- мужские экземпляры, женские встречаются значи
тельно реже. Отличить P. ×canescens от его родителей относительно несложно,
однако при возвратном скрещивании внутри гибридных зон морфологичес
кие признаки малоэффективны. Специальные исследования посвящены изу
чению процессов формирования микоризы у P. × canescens в условиях гибрид
ной зоны [Gafur et al., 2003].
P. ×canescens растет дико в пойменных лесах на юге Европейской части Рос
сии и на Кавказе. В Европе, где природные ареалы родительских видов пере
крываются, гибрид встречается часто. В Средней России редок, однако отдель
ные его экземпляры находят регулярно. В Москве, например, отмечено 2--3
дерева P. × canescens (С.Майоров, личное сообщение). В августе 2008 г. в окре
стностях д. Федяшево Белевского р на Тульской обл. И.С. Шереметьевой и
М.В. Казаковой отмечен 1 экземпляр P. × canescens (личное сообщение).
Во вторичном ареале в Северной Америке тополь белый гибридизиру
ет с местными видами рода Populus -- P.grandidentata Michaux и P.tremuloides
Michaux. Гибриды между P.alba и P.grandidentata (=P. ×rouleauiana Boivin) ши
роко распространены в природе и хорошо отличаются от родительских видов
по морфологическим признакам. Значительно реже встречается P. × heimburgeri
Boivin -- гибрид между P.alba и P.tremuloides [Spies, Barnes, 1981].
Генетические изменения и вариабельность вида. Наличие в Европе про
тяженных природных гибридогенных зон между P.alba и P.tremula побуждает
европейских исследователей использовать этот модельный объект для изуче
ния микроэволюционных процессов. В работе К.Лексер с соавт. [Lexer et al.,
2005] приведены данные о размерах и направленности интрогрессии между
P.alba и P.tremula вдоль большой симпатрической зоны в долине Дуная. Для
генетического исследования взяты образцы 93 гибридных морфотипов и ро
дительских особей из 4 популяций как внутри, так и за пределами зоны сим
439
патрии. Результаты показали, что интрогрессия направлена, в основном, от
P.tremula к P. alba. Однонаправленная модель обусловлена высоким уровнем
распространения пыльцы P.tremula и непосредственной близостью особей P.
alba в условиях пойменного леса, несмотря на относительно небольшие раз
меры популяций обоих родительских видов.
Изучена внутренняя структура генов, потенциально обеспечивающих эко
логическую дивергенцию между P.alba и P.tremula. Обнаружены маркеры, ко
торые могут быть использованы для исследования геномных барьеров потока
генов между экологически дивергентными видами [Joseph, Lexer, 2007]. Вни
мание уделяется генетическому изучению не только родительских, но и гиб
ридных таксонов. Проводилось исследование пространственной генетической
структуры (SGS) и клоновости (клональности) P. × canescens и его симпатри
ческого родителя P. alba в Центральноевропейской гибридной зоне [Loo et al.,
2007]. Авторы отмечают, что мультилокусную SGS в гибридной зоне изучают
редко, хотя она является ключом к пониманию явления клональности (кло
новости) для естественных гибридов между двумя диплоидными, экологически
дивергентными европейскими древесными видами со смешанным семенным
и вегетативным размножением. Для изучения клональности, SGS и расстоя
ний, на которые исторически мог происходить поток генов, были использова
ны ядерные микросателлиты с 407 деревьев P. × canescens и P. alba большой
Центральноевропейской гибридной зоны. Авторами выдвинута предваритель
ная гипотеза о превалировании семенной рекомбинации обеих таксонов в при
роде, однако P. × canescens имеет тенденцию создавать гораздо бульшие по
площади клоны, чем P.alba. Эта ситуация лучше объясняется наличием более
устойчивой (в 3--4 раза по сравнению с P. alba) генетической структурой P. ×
canescens, способствующей наличию межвидового потока генов, выборочно
му скрещиванию и увеличению клональности (клоновости) у гибридов.
Экономическое и социальное (положительное и отрицательное) зна
чение. Древесина P.alba крупнослойная, лоснящаяся, очень мягкая, хорошо
колющаяся, легкая. Используется для получения целлюлозы, идет на доски
для обшивки помещений, двери, окна, паркет, поделки, ящики. Листья исполь
зуются на корм скоту, кора является дубителем [Комаров, 1936]. Поскольку
вид дает корневую поросль и устойчив к засолению, его используют для зак
репления береговых песчаных дюн. Особенно велико значение P.alba в куль
туре: зелено белые листья и декоративная крона делают его очень эффектным
деревом, используемом в озеленении в условиях природного ареала и за его
пределами.
Меры борьбы
В литературе мы не обнаружили данных о методах и практическом
опыте уничтожения особей P.alba. Вероятно, это объясняется отсутствием или
незначительным уровнем ущерба, который приносит этот вид. Например, в
Канаде P.alba распространен локально, и в списке канадских инвазионных
видов отнесен в группу «Minor Invasive aliens» [Canadian Wildlife service, 2008].
В США также не имеет склонности к инвазии [Spies, Barnes, 1981]. Кроме того,
положительные качества растения, высокая декоративность и хозяйственная
ценность, не позволяют пока рассматривать его как источник угрозы.
440
ЗАКЛЮЧЕНИЕ
ПРОБЛЕМЫ ВЕДЕНИЯ
РЕГИОНАЛЬНЫХ «ЧЕРНЫХ КНИГ»
Мы хорошо понимаем, что список инвазионных видов Средней
России, представленный в книге, во многом субъективный и, разумеется,
далеко не полный. Однако поднимать проблему растительных инвазий надо
было незамедлительно, и мы начали с тех видов, которые представлялись
для всех троих авторов наиболее проблематичными.
Приятно отметить, что наша работа послужила точкой отсчета для начала
создания региональных «Черных книг». Региональные «Черные книги» при
званы решать следующие задачи [Крылов, 2008]:
-- изучение исторического и современного распространения адвентивных
видов, их обилия и численности на территории региона;
-- создание основы для мониторинга, изучения динамики популяций ад
вентивных видов;
-- выявление путей и способов заноса;
-- изучение эколого биологических особенностей адвентивных видов;
-- оценка экономического ущерба;
-- разработка конкретных мер, препятствующих внедрению чужеродных
видов в растительный покров региона;
-- привлечение внимания административных органов;
-- популяризация знаний и привлечение общественных организаций к изу
чению инвазионных видов.
Одна из задач региональных «Черных книг» -- это выявление местных ин
вазий адвентивных видов. Так, например, в список инвазионных растений
Калужской обл. [Крылов, 2008] включены виды, не попавшие в наш список,
но которые активно внедряются в естественные и полуестественные фитоце
нозы конкретного региона и на которые рекомендовано обратить внимание на
сопредельных территориях: Arrhenatherum elatius, Festuca arundinacea, Trisetum
flavescens, Aronia mitchurinii, Sambucus racemosa. И наоборот, ряду видов из спис
ка инвазионных растений Средней России включения в «Черную книгу» Ка
лужской области пока не требуется: Hordeum jubatum, Populus alba, Reynoutria
× bohemica, Cardaria draba, Erucastrum gallicum, Sisymbrium wolgense, Crataegus
monogina, Euphorbia peplus, Elaeagnus angustifolia, Symphytum caucasicum,
Ambrosia artemisiifolia и Cyclachaena xanthiifolia.
В региональные «Черные книги» [Виноградова и др., 2006; Крылов и др.,
2007; Крылов, 2008] мы предлагаем включать растения со степенью натурали
зации N7, N8 и N9, то есть виды, которые способны к активному возобновлению,
расселению и внедрению в естественные и полуестественные местообитания,
и виды, изменяющие сложившиеся фитоценозы. Разная степень выраженнос
ти этих характеристик может быть отражена в разном статусе видов «Черной
книги» по аналогии с «краснокнижными» растениями. Таким образом, пред
лагается выделять 3 категории видов «Черной книги» с разным статусом: 3 я
441
категория -- это растения со степенью натурализации N7 (адвентивные виды, в
настоящее время активно расселяющиеся и натурализующиеся по нарушен
ным местообитаниям); 2 ая категория -- N8 (адвентивные растения, расселяю
щиеся и натурализующиеся по нарушенным, полуестественным и естествен
ным местообитаниям); 1 ая категория -- N9 «трансформеры» [Richardson et
al., 2000] -- активно внедряются во вторичные естественные и полуестествен
ные сообщества, изменяют облик и природу экосистем, нарушают сукцесси
онные связи, становятся эдификаторами, образуют одновидовые заросли, пре
пятствуя возобновлению видов природной флоры.
Надеемся, что вслед за «Черной книгой» Калужской области подобные ра
боты будут проводиться и в других регионах.
Помимо этого, выполняя рекомендации Стратегии по инвазионным видам
Европы, согласно которой в каждом регионе необходимо вести контролиро
вание расселения 100 наиболее вредоносных чужеродных видов, в последую
щие «Черные книги» Средней России предстоит включить еще 48 видов. Ду
мается, что в этот список должны попасть те заносные растения, которые фор
мируют инвазионные популяции хотя бы в одном из регионов Средней России.
В качестве проекта «black list» мы предлагаем обсудить нижеследующий спи
сок растений, находящихся большей частью на первых стадиях инвазионного
процесса и проявляющих тенденцию к активному расширению ареала.
1. Acer ginnala
2. Adenocaulon adhаerеscens
3. Amaranthus blitoides
4. Ambrosia trifida
5. Amorpha fruticosa
6. Aronia x mitchurinii
7. Arrhenatherum elatius
8. Artemisia dubia
9. Artemisia sieversiana
10. Asclepias syriaca
11. Aster novi belgii
12. Bellis perennis
13. Brunnera sibirica
14. Calystegia inflata
15. Caragana arborescens
16. Corispermum declinatum
17. Cornus albus
18. Cotoneaster lucidus
19. Cuscuta campestris
20. Erigeron droebachensis
21. Gailardia aristata
22. Galega orientalis
23. Geum macrophyllum
24. Juglans manshurica
25. Kochia scoparia
26. Lemna minuta
27. Lonicera tatarica
28. Oenothera rubricaulis
29. Parthenocissus inserta
30. Petasites hybridus
31. Physocarpus opulifolius
32. Polygonon weyrichii
33. Prunus virginiana
34. Quercus rubra
35. Ribes aureum
36. Robinia pseudacacia
37. Rosa rugosa
38. Rudbeckia hirta
39. Salsola tragus
40. Sambucus racemosa
41. Symphytum Ч uplandicum
42. Teleckia speciosa
43. Thladiantha dubia
44. Ulmus pumila
45. Veronica filiformis
46. Veronica persica
47. Vinca minor
48. Zizania latifolia
Надеемся, что дискуссия ботаников и экологов, а также широкий обмен
информацией о случаях натурализации чужеродных видов на той или иной
территории приведет к формированию перечня видов растений, которые тре
буют первоочередного изучения и мониторинга в целях предотвращения био
логических инвазий.
442
ЛИТЕРАТУРА
Абрамов Н.В., Жиряков А.З. 1989. Адвентивные и синантропные виды во флоре Марийской АССР и
их классификация // Проблемы изучения адвентивной флоры СССР. М.: Наука. С. 49--51.
Аверкиев Д.С. 1938. Определитель растений Горьковской области. -- Горький: Облгиз. 360 с.
Аверкиев Д.С., Аверкиев В.Д. 1985. Определитель растений Горьковской области. -- 2-е изд., испр.
и доп. -- Горький: Волго-Вят. кн. изд-во. 320 с.
Агроэкологический атлас России и сопредельных стран [Electronic resource.] Mode of access:
http://www.agroatlas.spb.ru/ru/content/related/Crataegus_monogyna/. Accessed 30 Mar. 2008.
Адо М.И. 1934. Экзоты черноморского побережья. М,: Академия ком.х-ва при СНК РСФСР.
117 c.
Азовский М.Г. 2006. Особенности распространения высшей водной растительности вдоль за-
падного берега озера Байкал // География и природные ресурсы. № 3. С. 62--68.
Аистова Е.В. 2007. Адвентивная флора Амурской области / Автореф. дисс. ... канд. биол. наук. --
Владивосток.
Акимова А.О., Виноградова Ю.К., Колесников М.П. 2008. Содержание фенольных соединений и
кремния в некоторых видах рода Solidago L. // Интродукция нетрадиционных и редких расте-
ний. Мат-лы VIII Международной конф. Мичуринск-наукоград. Изд-во МичГАУ. С.13--15.
Аксенов Е.С., Аксенова Н.А. 2000. Декоративные растения. Т. 1. (Деревья и кустарники). Изд.
2-е. Энциклопедия природы России. М.: АБФ/ABF. 560 c.
Александрова К.И., Барабаш Г.И. 1987. Сорная флора донских лугов // Растительный покров
Центрального Черноземья и его охрана: Межвуз. сб. науч. тр. Воронеж. С. 20--23.
Александрова К.И., Казакова М.В., Новиков В.С., Ржевуская Н.А., Тихомиров В.Н. 1996. Флора
Липецкой области -- М.: Аргуc. 376 c.
Алексеев Ю. Е. 2006. Gramineae Juss. [Poaceae (R. Br.) Barnh.] -- Злаки // Маевский П. Ф. Флора
средней полосы европейской части России. С. 63--120.
Алексеев Ю.Е., Губанов И.А. 1980. Флора окрестностей Пущина-на-Оке. -- М.: Изд-во МГУ. 101 с.
Алексеев Ю.Е., Жмылев П.Ю., Карпухина Е.А. 1997. Деревья и кустарники. Энциклопедия при-
роды России. М.: -- 592 с.
Алексеев Ю.Е., Макаров В.В. 1981. Дополнение к флоре Брянской и Калужской областей // На-
уч. докл. высшей школы. Биол. науки. Т. 9. С.73--77.
Алексеев Я.Я. 1961. Определитель растений Смоленской и смежных с ней областей. -- 3-е изд. --
Смоленск: Кн. изд-во. 415 с.
Альжанов М., Сагитов С. 1963. Из опыта интродукции видов клена в ботаническом саду//
Вестник Каракалпакского филиала АН УзССР. № 4. С. 99.
Алюшин А.И. 1982. Растения Тульского края (очерки растительного покрова). -- Изд. 2-е. -- Ту-
ла: Приокск. кн. изд-во. 144 с.
Алюшин А.И. 1982. Растения Тульского края (очерки растительного покрова). -- Изд. 2-е. -- Ту-
ла: Приокск. кн. изд-во. 144 с.
Алявдина К.П., Виноградова В.П. 1972. Определитель растений / Под ред. О.Н. Шалыгановой. --
Ярославль: Верх.-Волж. кн. изд-во. 399 с.
Амелин И.С. 1936. Сорная растительность Ставропольских полей. - Ставрополь.
Андриеш Л.П. 2008. Способ диагностики поллиноза, [Electronic resource.] Mode of access:
http://ru-patent.info/20/10-14/2011385.html. Accessed 1 June 2008.
Ануфриев О.Н. 2008. Инвазивные виды семейства Asteraceae Dumort. в Башкирском Предура-
лье: распространение, биология и контроль численности / Автореф. дисс. ... канд. биол.
наук. Оренбург. -- 18 с.
Арак Э.Х., Раал А.Э. 1987. О возможности использования надземной части Matricaria discoidea
DC. в качестве лекарственного сырья // Растительные ресурсы. Т. 23. № 4. С. 584--590.
443
Арак Э.Х., Раал А.Э., Таммеорг Й.К. 1981. Изучение качественного состава эфирного масла
ромашки душистой эстонского происхождения // Тез. докл. 2-го съезда фармацевтов ЭССР.
Таллинн. С. 83--84.
Ареалы деревьев и кустарников СССР. 1980. - Том 2. Гречишные-Розоцветные. Л., Наука, С. 72
(Авторы: Соколов С.Я., Связева О.А., Кубли В.А.).
Артари А. 1886. Заметка по поводу нахождения «водяной заразы» (Elodea canadensis Rich.) в
Средней России // Сад и огород. № 22. С. 226.
Артюшенко З.Т. 1954. Amelanchier Medik. // Деревья и кустарники СССР. М.-Л.: Изд-во АН
СССР. Т. 3. С. 495--507.
Астахов И.И. 1951. Кавказский комфрей (окопник) как кормовая культура // Сб. тр. Ленингр.
обл. с.-х. опытной станции. 1925--1950. Л. С. 111--123.
Астахов И.И. 1953. Кавказский комфрей (окопник) // Достижения науки -- сельскохозяйствен-
ному производству. Полеводство. Л. С. 252--255.
Афанасьев В.Е. 2008. Адвентивная флора Астраханской области / Автореф. дисс.
... канд.
биол. наук. Саратов. 20 с.
Базарова Б.Б., Пронин Н.М. 2006. Элодея канадская в Чивыркуйском заливе озера Байкал //
География и природные ресурсы. № 1. С. 59--62.
Бакин О.В., Рогова Т.В., Ситников А.П. 2000. Сосудистые растения Татарстана. -- Казань: Изд-
во Казан. ун-та. 496 с.
Баландин С.В. 1989. Адвентивная флора заповедника "Басеги" // Проблемы изучения адвен-
тивной флоры СССР. М. С. 56--57.
Барабаш Г.И., Камаева Г.М. 1989. Новые адвентивные растения во флоре Воронежа // Проблемы
изучения адвентивной флоры СССР: мат-лы совещ., 1-3 февраля 1989 г. М. С. 46-47.
Барабаш Г.И., Камаева Г.М., Карташева Н.М. 1979. О карантинных сорняках Воронежской
области // Защита растений. № 10. С. 48.
Барановський О.Л. 1938. Бiологiя буряну Galinsoga та засоби боротьби з ним. - Киiв. - 95 с.
Барбакадзе В.В., Кемертелидзе Э.П., Таргамадзе И.Л., Шашков A.С., Усов A.И. 2002. Новый
биологически активный полимер 3-(3,4-дигидроксифенил)--глицериновой кислоты из двух
видов окопника Symphytum asperum и S. caucasicum (Boraginaceae) // Биоогр. химия. Т.28.
№ 4. С. 362--366.
Бариева Э.Р. 1997. Эколого-популяционный анализ и структура щирицы запрокинутой (Ama-
ranthus retroflexus L.) в различных агроэкологических условиях / Автореф. дисс. ... канд.
биол. наук. Удмуртский гос. ун-т. Ижевск. 24 с.
Баринхоев М.М. 2004. Сорные растения. «Эль-Фа». Нальчик. 335 с.
Баркалов В.Ю., Таран А.А. 2004. Список видов сосудистых растений острова Сахалин // Расти-
тельный и животный мир острова Сахалин (Мат-лы Международного сахалинского проек-
та). Владивосток. Ч. 1. С. 39--66.
Баркалов В.Ю. 1992. Астра -- Aster L. // Сосудистые растения советского Дальнего Востока. Т.
6. С. 54--64.
Баркалов В.Ю. 1992. Золотарник -- Solidago L. // Сосудистые растения советского Дальнего
Востока. Т.6. С. 45--53.
Баркалов В.Ю. 1992. Фалакролома -- Phalacroloma Cass. // Сосудистые растения советского
Дальнего Востока. Т. 6. С. 78--82.
Баркалов В.Ю., Таран А.А. 2008. Список видов сосудистых растений острова Сахалин. [Electronic
resource.] Mode of access: http://www.biosoil.ru/files/00000309/pdf. Accessed 23 May 2008.
Бармин Н.А. 2006. Адвентивная флора республики Мордовия / Автореф. дисс. ... канд. биол. наук.
Бармин Н.А. 2000. Адвентивная флора Республики Мордовия. - Дисс... канд. биол. наук. Москва.
Басаргин Д.Д. 1989. Недотрога железконосная на Советском Дальнем Востоке // Проблемы
изучения адвентивной флоры СССР. М.: Наука. С. 115--116.
Беднарьска I.O. 2007. Рiд Festuca L. (Poaceae) у флорi захiдних регioнiв Украïни. / Автореф.
дисс. ... канд. биол. наук. Киïв. 21 с.
Белозеров П.И. 1957. О распространении сорняков из семейства амарантовых на север евро-
пейской части СССР // Ботан. журн. Т. 42. № 3. С. 454--456.
Белозеров П.И. 1960. О распространении некоторых видов сорных растений на северо-востоке
европейской части СССР // Ботан. журн. Т. 45. № 8. С. 1227--1232.
444
Белозеров П.И. 1956. К вопросу о распространении на север элодеи канадской // Бот. журн. -- Т.
41, № 2. С. 262--263.
Белозеров П.И. 2008. Флора Костромской области: Монография.
-- Кострома: Изд-во Костр.
гос. технол. ун-та. 197 с.
Белый В.В., Белая А.М., Матецкая А.Ю., Утянская С.В., Ханин М.В. 2008. «Виртуальный гер-
барий Ростовской области», [Electronic resource.] Mode of access: http://rspu.edu.ru/ pro-
jects/plants/puccinel.html. Accessed 23 May 2008.
Березуцкий М.А. 1999. Антропогенная трансформация флоры // Ботан. журн. Т. 84. № 6. С. 8--19.
Березуцкий М.А. 2000. Антропогенная трансформация флоры южной части Приволжской воз-
вышенности / Дисс... докт. биол. наук. Воронеж.
Березуцкий М.А., Дайковский В.С. 1988. Дополнение к Конспекту флоры Саратовской области
// Вопросы ботаники Юго-Востока. Саратов. № 6. С. 89--92.
Берко О.М. 1963. Проiзростання розрiв-трави (Impatiens roylei) пiд Львовом // Укр. ботан.
журн. Т. 20. № 1. С. 102--103.
Беркутенко А.Н. 1988. Капустовые, или Крестоцветные -- Brassicaceae Burnett (Cruciferae Juss.)
// Сосудистые растения советского Дальнего Востока. Т. 3. С. 38--115.
Биоиндикация загрязнения наземных экосистем. 1988. Под ред. Р. Шуберта. - М.: Мир. - 350 с.
Болсуновский А.Я., Зотина Т.А., Бондарева Л.Г., Дегерменджи А.Г. 2004. Оценка интенсивно-
сти накопления трансуранового элемента америция-241 водным растением Elodea canaden-
sis // ДАН Т. 399, № 4. С. 130--133.
Борисова Е.А. 1993. Адвентивная флора Ивановской области. Дисс... канд. биол. наук. Москва.
Борисова Е.А. 2003. Характеристика адвентивного компонента флоры г. Иваново // Проблемы
изучения адвентивной и синантропной флоры в регионах СНГ / Мат-лы науч. конф. / Под
ред. В.С. Новикова и А.В. Щербакова. -- М.: Ботан. сад МГУ; Тула: Гриф и К. С. 22--24.
Борисова Е.А. 2005. Род Ambrosia L. (Asteraceae) во флоре бассейна Верхней Волги // Природа
и человек / Мат-лы IV науч.-практ. конф. (Иваново, 23-24 нояб. 2005 г.) /Под ред. В.А.
Исаева. -- Иваново: Иван. гос. ун-т. С. 46--48.
Борисова Е.А. 2006. Новые и редкие адвентивные виды Ивановской, Владимирской и Костром-
ской областей // Бюл. МОИП. Отд. биол. Т. 111, вып. 6. С. 63--67.
Борисова Е.А. 2006. Флористическое загрязнение пригородных лесов г. Иваново // Экология.
№ 3. С. 168--172.
Борисова Е.А. 2007. Адвентивная флора Ивановской области. -- Иваново: Иван. гос. ун-т. 188 с.
Борисова Е.А. 2008. Адвентивная флора Верхневолжского региона (современное состояние,
динамические тенденции, направленность процессов формирования) / Дисс... докт. биол.
наук. -- Москва.
Борисова М.А. 2002. Флора транспортных путей Ярославской области / Дисс. ... канд.. биол.
Наук. -- Ярославль.
Борьба с борщевиком обошлась в 4 миллиона // http://rus.postimees.ee/291007/glavnaja/estonija/
24583.php (23.11.2007).
Босек П.З. 1983. Новые материалы по адвентивной флоре Брянской области // Ботан. журн. Т. 68.
№ 5. С. 672--674.
Бочанцев В.П. 1959. Мелколепестник -- Erigeron L. // Флора СССР. - Т. 25. - С. 191--288.
Бочкин В.Д. 1991. Растения сем. Poaceae Barnhart во флоре железных дорог Москвы // Систе-
матика и эволюция злаков / Мат-лы совещ. 10--14 сентября 1991 г.- Краснодар.- С. 15--17.
Бубнова С.В. Флора Сибири. Т. 2. 1990, «Энциклопедия растений Сибири», [Electronic
resource.] Mode of access: http://skazka.nsk.ru/atlas/id1919. Accessed 25 May 2008.
Буддизм в Украине [Electronic resource.] Mode of access: http://ningma.org.ua/index.php?option=
com.content. Accessed 15 November 2008.
Булохов А.Д., Величкин Э.М. 1998. Определитель растений Юго-Западного Нечерноземья
(Брянская, Калужская, Смоленская области). -- 2-е изд., перераб. и доп. -- Брянск: Изд-во
Брянск. пед. ун-та. -- 380 с.
Бухгольц Ф.В. 1897. Гербарий. Список семенных и высших споровых растений. -- М.: Кушне-
ревиК.29с.
Буш Н.А. 1939. Крестоцветные -- Cruciferae B. Juss. // Флора СССР. Т. 8. С. 14--629.
Буш Н.А. 1939а. Клоповник -- Lepidium L. // Флора СССР. Т. 8. С. 501--524.
Буш Н.А. 1939б. Двугнездка -- Hymenophysa C. A. M. // Флора СССР. Т. 8. С. 535--537.
445
Буянкин В.И. 1974. Распространение Amaranthus albus L. в СССР // Ботан. журн. Т.59. № 6.
С. 864--866.
Буянкин В.И. 1977. История распространения и ареал Amaranthus retroflexus L. и A. blitoides S.
Wats. (Amaranthaceae) в СССР // Ботан. журн. Т. 62. № 12. С. 1779--1785.
Вагнер К.И. 1821 Каталог плодовых дерев. -- Рига: тип.Мюллера; 46 с.
Васильев В.Н. 1952. Ясень -- Fraxinus L. // Флора СССР. Т. 18. С. 485--502.
Васильева Н.В., Папченков В.Г. 2007. Распространение в бассейне Волги и биоэкологические
особенности Bidens frondosa L. // Естественные и инвазийные процессы формирования
биоразнообразия водных и наземных экосистем / Тезисы докладов международной науч-
ной конференции. -- Ростов-на-Дону. Южный научный центр РАН. С. 64--65.
Васильченко И.Т. 1936. Amaranthaceae // Флора СССР. Т. VI. изд-во АН СССР, Ленинград, Мо-
сква. C. 354--369.
Васюков В.М. 2004. Растения Пензенской области (конспект флоры). -- Пенза: Изд-во Пенз. гос.
ун-та. 184 с.
Вахромеев И.В. 2002. Определитель сосудистых растений Владимирской области. -- Владимир.
314 с.
Вернер Э. 1910. К вопросу о распространении Elodea canadensis Michx. в России // Тр. Бот. са-
да Юрьев. ун-та. -- Юрьев. Т. 2. С. 128.
Виноградова В.М. 2004. Apiaceae Lindl. (Umbelliferae Juss.) -- Сельдереевые (Зонтичные) [excl.
Trinia, Rumia, Pimpinella] // Флора Вост. Европы. Т. 11. С. 315--437.
Виноградова Ю.К. 1992. Процессы микроэволюции у адвентивных и интродуцированных рас-
тений / Автореф. дисс. ... докт. биол. наук. М. 40 с.
Виноградова Ю.К. 2000а. Внутривидовая изменчивость костра кровельного в естественных и
спонтанных интродукционных популяциях // Бюлл. Гл. ботан. сада. Вып. 179. С. 37--45.
Виноградова Ю.К. 2000б. Анализ амплитуды внутривидовой изменчивости костра кровельного
в естественном и вторичном ареалах // Бюлл. Гл. ботан. сада. Вып. 180. С. 47--54.
Виноградова Ю.К. 2001. Натурализация, биологические особенности и внутривидовая измен-
чивость ромашки душистой // Бюлл. Гл. ботан. сада. Вып. 182. С.7--15.
Виноградова Ю.К. 2002а. Внутривидовая изменчивость щирицы белой во вторичном ареале //
Бюлл. Гл. ботан.сада. Вып.183. С. 8--18.
Виноградова Ю.К. 2002б. Формирование вторичного ареала и внутривидовая изменчивость галин-
зоги мелкоцветковой (Galinsoga parviflora Cav.) // Бюлл. Гл. ботан. сада. Вып. 184. С. 24--31.
Виноградова Ю.К. 2003а. Роль эффекта основателя при формировании вторичного ареала
Bidens frondosa L. // Тез. докладов Х1-го международного совещания по филогении расте-
ний. -- М. Изд-во Центра охраны дикой природы. С. 31--32
Виноградова Ю.К. 2003б. Экспериментальное изучение растительных инвазий (на примере ро-
да Bidens) // Проблемы изучения адвентивной и синантропной флоры в регионах СНГ /
Мат-лы научн. конф. -- Изд-во ботан. сада МГУ; Тула: Гриф и К°. С. 31--33.
Виноградова Ю.К. 2003в. Внутривидовая изменчивость галинзоги волосистой (Galinsoga
ciliata) в естественном и вторичном ареалах // Бюлл. Гл. ботан. сада. Вып. 185. С. 63--69.
Виноградова Ю.К. 2004. Изменчивость биологических признаков аира обыкновенного (Acorus
calamus L.) в естественных и инвазионных популяциях // Бюлл. Гл. ботан. сада. Вып. 187.
С. 23--31.
Виноградова Ю.К. 2005. Экспериментальное изучение инвазионных популяций мелколепестника
канадского (Сonyza canadensis (L.) Cronquist // Бюлл. Гл. ботан. сада. Вып. 189. С. 53--76.
Виноградова Ю.К. 2006. Формирование вторичного ареала и изменчивость инвазионных попу-
ляций клена ясенелистного (Acer negundo L.) // Бюлл. Гл. ботан. сада. Вып. 190. С. 25--47.
Виноградова Ю.К. 2007а. Биоразнообразие Impatiens glandulifera Royle в естественном и во
вторичном ареале // Биологическое разнообразие. Интродукция растений (Мат-лы 4 Меж-
дународной научн. конф.). -- С.-Петербург. БИН РАН. С. 26--28.
Виноградова Ю.К. 2007б. Этапы формирования вторичного ареала и изменчивость инвазион-
ных популяций эхиноцистиса шиповатого // Бюлл. Гл. ботан. сада. Вып. 192. С. 8--23.
Виноградова Ю.К. 2008. Микроэволюция недотроги железконосной (Impatiens glandulifera
Royle) в процессе формирования вторичного ареала // Бюлл. Гл. ботан. сада. Вып. 194.
Виноградова Ю.К. 2009. Изменчивость и конкурентоспособность череды облиственной Bidens
frondosa во вторичном ареале // Бюлл. Гл. ботан. сада. -- Вып. 196.
446
Виноградова Ю.К., Возна Л.И. 2008. Инвазибельность естественных фитоценозов и конку-
рентные отношения между аборигенными и инвазионными видами // Биоразнообразие:
проблемы и перспективы сохранения. Ч. 1. Пенза. С. 17--19.
Виноградова Ю.К., Колесников М.П. 2005. Фенольные соединения в ромашке душистой
[Lepidotheca suaveolens (Pursh) Nutt.] // Ботанические сады как центры сохранения биораз-
нообразия и рационального использования растительных ресурсов. -- Москва. С. 89--91.
Виноградова Ю.К., Колесников М.П. 2007. Содержание фенольных соединений и кремния в
растениях мелколепестника канадского (Сonyza canadensis (L.) Cronquist // Бюлл. Гл. ботан.
сада. Вып. 193. С. 117--127.
Виноградова Ю.К., Майоров С.Р., Хорун Л.В. 2006. Первые итоги реализации проекта «Черная
книга» Средней России // Флористические исследования в Средней России. Москва. Това-
рищество научных изданий КМК. С. 45--48.
Виноградова Ю.К., Рыхликова А.А., Колесников М.П. 2007. Сравнительный анализ некоторых
видов рода Bidens L. по содержанию фенольных соединений и кремния // Новые и нетра-
диционные растения и перспективы их использования / Мат-лы VII Международного сим-
позиума. Москва. Т. 2. С. 72--74.
Воейков А.Д. 1908. О натурализации древесных пород // Лесной журн.
Волкова Е.В. 1954. Род Elsholtzia // Флора СССР. Ленинград: изд-во АН СССР.
-- Т. XXI.
С. 634--636.
Волкович М. Г. 2007. Узкотелая златка Agrilus planipennis -- новый опаснейший вредитель ясеней в ев-
ропейской части России // http://www.zin.ru/ANIMALIA/COLEOPTERA/rus/eab_2007.htm. 2007.
Волховская У.В. 1937. Золотарник. (Итоги интродукции) // Труды интродукционого питомника
субтропических культур. -- Сухуми. Вып. 8. 38 с.
Вольф Э.Л. 1915. Декоративные кустарники и деревья для садов и парков. Петроград. 461 с.
Воробьев Д.П. 1954. К вопросу о заносных и сорных растениях в Приморском крае // Комаров-
ские чтения. Владивосток. Вып. 4. С. 3--22.
Воробьев Д.П., Ворошилов В.Н., Гурзенков Н.Н. и др. 1974. Определитель высших растений
Сахалина и Курильских островов. -- Л.: Наука.372 с.
Ворошилов В.Н., Скворцов А.К., Тихомиров В.Н. 1966. Определитель растений Московской об-
ласти. -- М.: Наука. 368 с.
Ворошилов В.Н. 1982. Определитель растений советского Дальнего Востока. -- М. Наука. 672 с.
Вьюкова Н.А. 1983. Новые материалы к характеристике адвентивной флоры Липецкой области
// Бюлл. МОИП. Отд. биол. Т. 88. Вып .1. С. 128--133.
Вьюкова Н.А. 1985. Адвентивная флора Липецкой и сопредельных областей /Автореф. дисс. ...
канд. биол. наук. -- М. 16 с.
Вьюкова Н.А. 1987. Приуроченность адвентивных растений к фитоценозам Липецкой области
// Растительный покров Центрального Черноземья и его охрана. Воронеж. С. 29.
Вьюкова Н.А. 1987. Формирование комплекса адвентивных растений -- новый этап в развитии
флоры Липецкой области // Природа малых охраняемых территорий. -- Воронеж. -- С. 78--84.
Галанин А.В. 1991. Флора и ландшафтно-экологическая структура растительного покрова.
--
Владивосток: ДВО АН СССР. 272 с.
Галкина М.А., Виноградова Ю.К. 2008. Сравнительный анализ биоморфологических признаков
Conyza canadensis и C.bonariensis -- инвазионных видов флоры юга России // Биоразнообра-
зие: проблемы и перспективы сохранения. Пенза. Ч. 1. С. 25--28.
Гамор Ф.Д. 1978. Плодоношення Galinsoga parviflora у рiзних агрофiтоценозах Закарпаття //
Укр. ботан. журн. Т. 35. № 4. С. 362--366.
Гвиниашвили Ц.Н. 1976. Кавказские представители рода Symphytum L. (Boraginaceae Juss.). --
Тбилиси: Мецниереба. 146 с.
Гельтман Д.В. 2003. Понятие «инвазивный вид» и необходимость изучения этого явления //
Проблемы изучения адвентивной и синантропной флоры в регионах СНГ / Мат-лы науч.
конф. под ред. В.С. Новикова, А.В. Щербакова. М.: Изд-во ботан. сада МГУ; Тула, Гриф и
К°. С. 35--36.
Гербицид Chipco Ronstar®C. Инструкция к применению, [Electronic resource.] Mode of access:
http://www.auburn.nsw.gov.au/reploaded/Files/AuburnWeb/Residents/Ronstar/2520. Accessed
17 November 2008.
447
Глазкова Е.А. 2005. Bidens frondosa (Asteraceae) -- новый адвентивный вид флоры северо-
запада России и история его расселения в Восточной Европе // Ботан. журн. Т. 90. № 10.
С. 1525--1540.
Глазкова Е.А. 2006. Новые данные о распространении Bidens frondosa (Asteraceae) в России //
Ботан. журн. Т. 91. № 11. С. 1749--1752.
Голицын В.М. 1905. Очерк флоры Епифанского уезда Тульской губернии // Тр. Ботан. сада
Юрьев. ун-та. -- Т. 5 (1904 г.) Вып. 4. С. 230--243.
Голицын В.М. 1906. Очерк флоры Епифанского уезда Тульской губернии [окончание] // Тр.
бот. сада Юрьев. ун-та. -- Юрьев. -- Т. 6 [1905 г.], вып. 1. -- С. 20--31.
Голицын В.М. 1907. Первое дополнение к очерку флоры Епифанского уезда Тульской губернии
// Тр. бот. сада Юрьев. ун-та. -- Юрьев, -- Т. 7 [1906 г.], вып. 4. -- С. 231--233.
Голицын С.В. 1945. К вопросу об антропохорных миграциях растений // Советская ботаника.
Т.13. № 6. С. 19--29.
Голицын С.В. 1961. Список растений Воронежского государственного заповедника // Тр. Воро-
неж. гос. зап. -- Воронеж: Изд-во Воронеж. гос. ун-та. -- Вып. 10. -- 101 с.
Головач А.Г. 1960. Ясень -- Fraxinus L. // Деревья и кустарники СССР. Т. 5. С. 406--430.
Головкин Б.Н. 1988. Культигенный ареал растений. -- М.: Наука. 165 с.
Горбачев Б.Н. 1932. Сорно-полевая растительность и меры борьбы с ней в зоне ГЗОС // Тр.
Горской зональной опытной станции. Cер. 1. № 3. С.1--51.
Горожанкин И.Н. 1888. Материалы для флоры Московской губернии // Bull. Soc. Nature. Mos-
cow. Т. 64. № 2. P. 349--372.
Горчаковский П.Л. 1979. Тенденции антропогенных изменений растительного покрова Земли //
Ботан. журн. Т. 64. № 12. С. 1697--1713.
Горчаковский П.Л., Козлова Е.В. 1998. Синантропизация растительного покрова в условиях за-
поведного режима // Экология. № 3. С. 171--177.
Горшкова С.Г. 1949. Лоховые -- Elaeagnaceae Lindl. // Флора СССР. - Т. 15. - С. 515--525.
Горшкова С.Г. 1949. Род Oxalis / Флора СССР. - T. XIV, М.-Л.: Изд-во АН СССР. - С.78-79.
Гостева Г. Энциклопедия трав, специй и пряностей, [Electronic resource.] Mode of access:
http://worldspice.sitecity.ru/index.phtml. Accessed 25 Oct. 2008.
Григорьевская А.Я., Стародубцева Е.А., Хлызова Н.Ю., Агафонов В.А. 2004. Адвентивная фло-
ра Воронежской области: исторический, биогеографический, экологический аспекты. - Во-
ронеж: Изд-во Воронежск. ун-та. 320 с.
Григорьевская А.Я. 1987. Сорная флора заповедника "Галичья гора" // Растительный покров
Центрального Черноземья и его охрана. -- Воронеж. С. 41--51.
Григорьевская А.Я. 2000. Флора города Воронежа. -- Воронеж: изд-во Воронеж. гос. ун-та. --
200 с.
Григорьевская А.Я. 2003. Антропогенная трансформация растительного покрова среднерус-
ской лесостепи / Дисс... докт. геогр. наук. -- Воронеж.
Григорьевская А.Я., Хрипякова В.Я., Прохорова О.В. 2002. Экоиндикационные ряды раститель-
ности массивов и урочищ г. Воронежа // Геоэкология. Вестник ВГУ, серия география и
геоэкология. №1. С. 74--94.
Гринева М.В. 2007. Интродукционные исследования Котовника кошачьего Nepeta cataria L. и
Эльсгольции реснитчатой Elsholtzia ciliata (Lep) (Garcke) в Московской области / Авто-
реф...канд. биол. наук. -- М. 24 с.
Гроссгейм А.А. 1940. Флора Кавказа. - Т. 2. Баку: Изд. Аз ФАН. 284 c.
Гроссгейм А.А. 1950. Флора Кавказа. 2-е изд. Т. 4. М., Л.: Изд-во АН СССР. xxii, 311 с.
Гроссет Г.Э., Замятнин Б.Н. 1935. Новые материалы по инвентаризации флоры окрестностей
г. Воронежа // Тр. Воронеж. гос. ун-та. -- Воронеж, Т. 7. С. 147--152.
Губанов И.А., Алексеев Ю.Е. 1975. Новые местонахождения редких и интересных растений в
Московской области // Бюлл. МОИП. Отд. биол. Т. 80. Вып. 3. С. 128--130.
Губанов И.А., Киселева К.В., Новиков В.С., Тихомиров В.Н. 2002. Иллюстрированный опреде-
литель растений Средней России. Т. 1: папоротники, хвощи, плауны, голосеменные, по-
крытосеменные (однодольные). -- М.: Товарищество научных изданий КМК. 526 с.
Губанов И.А., Киселева К.В., Новиков В.С., Тихомиров В.Н. 2003. Иллюстрированный опреде-
литель растений Средней России. Том 2: Покрытосеменные (Двудольные: раздельнолепе-
стные). Москва: Товарищество научных изданий КМК. -- 665 с.
448
Губанов И.А., Киселева К.В., Новиков В.С., Тихомиров В.Н. 2004. Иллюстрированный опреде-
литель растений Средней России. Том 3: Покрытосеменные (двудольные: раздельнолепе-
стные). -- М.: Товарищество научных изданий КМК. -- 520 с.
Гусев Ю.Д. 1966. Расселение видов Galinsoga в Ленинградской области // Ботан. журн. Т.51.
№ 4. С. 577--579.
Гусев Ю.Д. 1968. Новые и редкие адвентивные растения ленинградской и карельской флоры //
Ботан.журн. Т.53. № 2. С. 267--269.
Гусев Ю.Д. 1973. Дополнения к адвентивной флоре северо-западных областей европейской
России // Ботан. журн. Т. 58. № 6. С. 904--909.
Гусев Ю.Д. 1977. Материалы по адвентивной флоре Удмуртии // Ботан. журн. Т. 62. № 5.
С. 691--694.
Гусев Ю.Д. 1980. Новые сведения по адвентивной флоре разных областей таежной зоны
Европейской части СССР // Ботан.журн. Т. 65. № 2. С. 249--255.
Гусев Ю.Д. 2005. Натурализация американских растений в бассейне Финского залива // Ботан.
журн. 1964. Т. 49. № 9. С. 1262--1271.
Гущина Е.Г. 1983. Адвентивные растения г.Рязани и его окрестностей // МОИП Докл.
Теоретические и прикладные аспекты результатов изучения растений и животных. -- М.,
Наука. С. 149--150.
Гущина Е.Г., Казакова М.В. 1989. Состояние изученности адвентивной флоры Рязанской об-
ласти // Проблемы изучения адвентивной флоры СССР. -- М.: Наука. С. 41--42.
Данилюк Е.Н. 2007. Erucastrum gallicum (Willd.) O.E. Schulz (Cruciferae) -- новый адвентивный
вид во флоре Украинских Карпат // Украïнський ботанiчний журнал, № 4б.
Дачный форум [Электронный ресурс]. 2008 // Наша дача. [web-сайт]. (12.10.2008).
http://dacha.wcb.ru/index.php?showforum=2.
Двигубский И. 1828. Московская флора или описание растений, дикорастущих в Московской
губернии. М. -- 576 с.
Дгебуадзе Ю.Ю. 2002. Проблемы инвазий чужеродных организмов // Экологическая безопас-
ность и инвазии чужеродных организмов / Сб. мат-лов круглого стола Всерос- С. конф. по
экологической безопасности России (4--5 июня 2002 г.). -М.: ИПЭЭ им. А.Н. Северцева,
IUCN (МСОП). C. 11--14.
Дексбах Н.К. 1951. Элодея канадская в водоемах Среднего Урала и Зауралья // Тр. Всесоюз.
гидробиол. о-ва. -- М., Т. 3. С. 204--216.
Деревья и кустарники СССР. 1954. -- М.: Изд-во АН СССР. Т. 3. 671 с.
Джус М.А. Дополнения к списку флоры сосудистых растений ГНП «Беловежская Пуща» и НП
«Припятский», с сайта «Беловежская Пуща. XXI век», [Electronic resource.] Mode of access:
http://bp21.org.by/ru/books/celebo75.html. Accessed 3 June 2008.
Дзыбов Д.С. 1989. Фитоценотический метод борьбы с амброзией полыннолистной --Ambrosia
artemisiifolia L. // Теоретические основы биологической борьбы с амброзией. Тр. зоол. ин-
та. -- Л. С. 227--229.
Димитриев А.В., Абрамов Н.В., Мининзон И.Л., Папченков В.Г., Пузырев А.Н., Раков Н.С., Си-
лаева Т.Б. 1994. О распространении Ambrosia artemisiifolia (Asteraceae) в Волжско-Камском
регионе // Ботан. журн. Т. 79. № 1. С. 79--83.
Доброчаева Д.М. 1955. Onagraceae Lindl. //Флора УРСР. Киiв. Изд-во АН УРСР. Т. 7. C. 410--445.
Доброчаева Д.Н. 1981. Сем. Boraginaceae Juss. -- Бурачниковые // Флора европ. части СССР. Т. 5.
С. 113--179.
Дорогостайская Е.В. 1972. Сорные растения Крайнего Севера СССР. -- Л.: Наука. -- 172 c.
Доронькин В.М. 1997. Род Elsholtzia // Флора Сибири. Т. 11. Новосибирск, Наука. С. 224--225.
Доронькин В.М. 2003. Том1. Lycopodiaceae -- Hydrocharitaceae // Флора Сибири. Т. 14. Допол-
нения и исправления. Алфавитные указатели. С. 11--19.
Дорофеев В.И. 1998. Новые находки крестоцветных -- Brassicaceae (Cruciferae) в Нижнегород-
ской области // Turczaninowia. Vol. 1. № 4. С. 28--32.
Дорофеев В.И. 2002. Крестоцветные (Cruciferae Juss.) Европейской России // Turczaninowia. -
Т. 5. Вып. 3. С. 5--114.
Дорофеев В.И. 2006. Cruciferae B. Juss., nom. alter. (Brassicaceae Burnett) -- Крестоцветные // Фло-
ра средней полосы европейской части России. Изд. 10, испр. и доп. М.: КМК. С. 256--283.
449
Дорофеев В.И. 2007. Конспект семейства крестоцветные -- Cruciferae B. Juss. Нижнего Дона:
supertribus brassicidiinae V. Avet. / Turczaninovia. Т. 10. № 2. С. 38--49.
Древесные растения Главного ботанического сада им. Н.В.Цицина РАН. 2005. М: Наука. 586 с.
Дубына Д.В., Протопопова В.В. 1985. Анализ адвентивной флоры заповедника "Дунайские
плавни"// Биологические науки. № 10. С. 68.
Дугуян Д.К. 1960. О видах рода Xanthium L. флоры СССР // Тр. Рост. отд. ВБО. Вып. 1. С. 42--57.
Еленевский А. Г., Радыгина В.И., Буланый Ю.И. 2000. Растения Саратовского Правобережья
(конспект флоры). -- Саратов: Изд-во Саратовского пед. ин--та. 101 с.
Еленевский А.Г., Буланый Ю.И., Радыгина В.И. 2008. Конспект флоры Саратовской области. --
Саратов: Изд. центр «Наука» -- 232 с.
Еленевский А.Г., Радыгина В.И. 1984. Флора Орловской области и перспективы ее исследова-
ния // Состояние и перспективы исследования флоры средней полосы Европейской части
СССР. М. С. 69--70.
Еленевский А.Г., Радыгина В.И. 1997. Определитель сосудистых растений Орловской области.
Орел: Труд. -- 208 с.
Еленевский А.Г., Радыгина В.И., Чаадаева Н.Н. 2004. Растения Белгородской области (кон-
спект флоры). -- Москва. -- 120 с.
Ефимова Т.И., Туганаев В.В. 1964. О некоторых редких и новых для флоры Удмуртии видах
растений // Ботан. журн. Т.49. № 12. С. 1797--1798.
Ефимова Т.П. 1961. Адвентивные растения в Удмуртии // Ботан. журн. Т. 46. № 7. С. 1045.
Жадовский А.Е. 1918. Рецензия на «Список растений, найденных в Тульской губернии до 1916 г.
В.В. Розен»// Тр. по прикл. бот. и сел. Пг. -- XI. №5/6. -- С. 49--50.
Жакова Л.В. Чужеродные виды в водных и прибрежно-водных фитоценозах Невской губы. --
Электр. ресурс. Режим доступа: http://www.zin.ru/conferences/rtable2007/PDF/doklad
Zhakova.pdf. -- Acc. 2 Nov. 2008.
Жуков С.П. 2003. Взаимоотношения растений на первой стадии сукцессии растительности от-
валов угольных шахт Донбасса // Промышленная ботаника. -- Вып. 3. -- С. 25--28.
Жуковский П.М. 1971. Культурные растения и их сородичи. Систематика, география, цитоге-
нетика, иммунитет, экология, присхождение, использование. 3 изд. Л.: Колос. 751 с.
Зарубин А.М. Материалы к флоре Байкальской Сибири: сборник научных статей. Вып. 1, - С.
141, [Electronic resource.] Mode of access: http://window.edu.ru/window_catalog/pdf2txt?p_id=
27035&p_page=15. Accessed 20 June 2008.
Зверева Г.А. 1981. Новые и редкие виды высших растений во флоре Тувы // Ботан. журн. Т.66.
№ 8. С. 1215--1218.
Зейферт Д.В. Растительные сообщества и почвенная мезофауна территорий химических пред-
приятий в степной зоне Башкирского Предуралья, [Electronic resource.] Mode of access: http:
// www.oglib.ru/cgi-bin/cgibf.exe. Accessed 10 May 2008.
Зернов А.С. 2006. Флора Северо-Западного Кавказа. - М.: КМК. 664 с.
Золотухин Н.И., Золотухина И.Б., Полуянов А.В. 2001. Сосудистые растения Зоринского уча-
стка Центрально-Черноземного заповедника // Природные условия и биологическое разно-
образие Зоринского заповедного участка в Курской области. Тр. Центрально-Черноземного
государственного заповедника. Вып. 17. -- Тула. -- С. 41--84.
Зузук Б.М., Куцик Р.В., Костюк И.Р., Мельничук Г.Г., Гайдук Р.Й. 2004. Окопник лекарствен-
ный // Провизор. № 17. № 18. № 19. (http://www.provisor.com.ua/index.php)
Зыкова Е.Ю. Database of plants and fungi of Altai-Sayan Ecoregion, [Electronic resource.] Mode of
access: http://www.bioaltai-sayan.ru/regnum/endem.php. Accessed 26 Oct. 2008.
Иванова Н.А. 1927. Растительность Сергиевского уезда. -- М.: Новая Москва. 80 с.
Иванова Р.Г., Туганаев В.В. 1969. О распространении некоторых видов адвентивных растений
на юге вятско-камского бассейна // Ботан. журн. Т. 54. № 4. С. 605.
Игнатов М.С. 1989. Об особенностях расселения адвентивных растений // Проблемы изучения
адвентивной флоры СССР/ Мат-лы совещ. 1--3 февраля 1989 г. С. 15--17.
Игнатов М.С., Макаров В.В. 1984. Новости адвентивной флоры Московской области // Бюлл.
Гл. ботан. сада АН СССР. Вып. 132. С. 49--51.
Игнатов М.С., Макаров В.В., Чичев А.В. 1990. Конспект флоры адвентивных растений Москов-
ской области // Флористические исследования в Московской области. -- М.: Наука. С. 5--105.
450
Игошин Г.К., Киселев О.Б., Мозговая О.А. и др. 1985. Флора сосудистых растений городов Куй-
бышевской области // Проблемы изучения синантропной флоры СССР. -- М.: Наука. С. 56--59.
Иевлев Н.И. 1983. Кормовые растения на торфяных почвах европейского Севера. -- Л.: Наука.
152 с.
Ижевский С.С. 2002. О возможности вывода американской белой бабочки из числа карантин-
ных объектов // Защита и карантин растений. № 12. С. 14--17.
Ижевский С.С. 2008. Инвазия чужеземных насекомых -- угроза экологической и экономиче-
ской безопасности России // АгроXXI. № 4--6. C. 34--36.
Ижевский С.С. Угрожающие находки ясеневой изумрудной узкотелой златки Agrilus
planipennis в Московском регионе // http://www.zin.ru/ANIMALIA/COLEOPTERA/ rus/agr-
plaiz.htm, 2007.
Ижевский С.С., Масляков В.Ю. 2008. Новые инвазии чужеземных насекомых в европейскую
Россию // Российский Журнал Биологических Инвазий. № 2 (http://www.sevin.ru/invasjour/
issues/2008_2.html).
Ижевский С.С., Мозолевская Е. Г. 2008. Изумрудная узкотелая златка (Agrilus planipennis
Fairmaire) на московских ясенях // Российский Журнал Биологических Инвазий. № 1
(http://www.sevin.ru/invasjour/issues/2008_1.html).
Изучение флоры Восточной Европы: достижения и перспективы. Тезисы докладов междуна-
родной конференции (Санкт-Петербург, 23--28 мая 2005 г.) / Под ред. А.Н. Сенникова и
Д.В. Гельтмана. М., СПб.: Товарищество научных изданий КМК, 2005. 111 с.
Ильинский А.П. 1924. Материалы к флоре Тверской губернии. II. Впервые указываемые и ред-
кие растения Тверской флоры // Журн. Рус. Ботан. об-ва. Т. 7 (1922 г.). С. 193--197.
Ильминских Н.Г. 1982. Анализ городской флоры (на примере флоры города Казани). - Дисс...
канд. биол. наук. -- Ленинград.
Ильминских Н.Г. 1986. Флорогенез в условиях урбанизированной среды (на примере городов
Вятско-Камского края) / Дисс... докт. биол. наук. Ленинград, и их динамика за 60-80 лет. --
Дисс... канд. биол. наук. Днепропетровск, 1992.
Ильминских Н.Г. 1988. Увеличивается ли сходство флоры городов в ходе их исторического
развития? // Агрофитоценозы и экологические пути повышения их стабильности и продук-
тивности/ Мат-лы Всесоюзн. совещ. -- Ижевск. С. 134--136.
Ильминских Н.Г., Дмитриев А.В., Мильчаков Л.В. 1981. О некоторых редких и новых адвентив-
ных растениях во флоре Волжско-Камского края // Ботан. журн. Т.66. № 8. С. 1221--1224.
Ишбирдина Л.М., Ишбирдин А.Р. 1992. Урбанизация как фактор антропогенной эволюции
флоры и растительности // Журн. общ. биол. Т. 53. № 2. С. 211--224.
Каден Н.Н., Кондорская В.Р. 1967. Морфология цветка и плода лоховых // Морфология расте-
ний. Сб. ст., посвященных памяти проф. К. И. Мейера. С. 102--118.
Казакова М.В. 2004. Флора Рязанской области. -- Рязань: Русское слово.-- 388 с.
Камышев Н.С. 1959. К классификации антропохоров // Ботан. журн. Т. 44. № 11. С. 1613--1615.
Камышев Н.С. 1978. Флора Центрального Черноземья и ее анализ. -- Воронеж: Изд-во Воро-
неж. ун-та. -- 116 с.
Камышев Н.С., Хмелев К.Ф. 1976. Растительный покров Воронежской области и его охрана. --
Воронеж: изд-во Воронеж. гос. ун-та. -- 184 с.
Карнаух Е.Д. 1955. Balsaminaceae // Флора УРСР. -- Киiв: Изд-во АН УРСР. Т. 7. С. 231--235.
Карписонова Р. 1999. Сад в тени. -- М.: Культура и традиции. 200 с.
Карписонова Р.А. 1985. Травянистые растения широколиственных лесов СССР. -- М. Наука. 205 с.
Кауфман Н.Н. 1889. Московская флора или описание высших растений и ботанико-
географический обзор Московской губернии. 2-е изд. -- М. -- 798 с.
Квитка А.Ю., Виноградова Ю.К. 2006. Инвазионные виды рода Solidago во флоре Средней
России // Флористические исследования в Средней России. Москва. Товарищество науч-
ных изданий КМК. С. 74--78.
Киселева Е.Я., Кибальчич П.Н. 1970. О ромашке душистой // Фармация. № 3. С. 70--71.
Клиническая иммунология и аллергология. / Под ред. Г. Лолора, Т. Фишера, Д. Эйдельмана.
Пер. с англ. под ред. Е.Н. Образцовой, В.М. Нечушкиной, А.С. Апта. -- М.: Практика, 2000.
806 с.
Клюшкин Т.К., Тихонов Н.П., Никонова К.А. 1962. Карантинные сорные растения и борьба с
ними. -- М.: Изд-во МСХ РСФСР.
451
Коблова М.Н. 1962. Сорная растительность Волго-Ахтубинской поймы // Уч. зап. Кабардино-
Балкарского ун-та. -- Вып. 12. С. 91--102.
Ковалев О.В. 1989. Расселение адвентивных растений трибы амброзиевых в Евразии и разра-
ботка биологической борьбы с сорняками рода Ambrosia L. (Ambrosieae, Asteraceae) // Тео-
ретические основы биологической борьбы с амброзией. -- Л.: Наука. С. 7--23.
Ковачев И.Г. 1966. Принос върху изясняване химичния състав на няколко вида от рода
Amaranthus // Научни тр. Висш. селскостомански ин-т "Васил Коларов". Т.15, № 1. С. 63--65.
Ковтонюк Н.К. 1987. Семейство // Флора Сибири. Araceae -- Orchidaceae. С. 16--43.
Козловская Н.В. 1962. Некоторые заносные растения Белоруссии // Ботан. журн. Т.47. № 11.
С. 1684--1686.
Козловская Н.В. 1978. Адвентивные и прогрессирующие виды во флоре Белоруссии // Ботани-
ка: исследования. -- Минск: Наука и техника. Вып. 20. С. 88--95.
Комаров В.Л. 1936. Горец -- Polygonum L. // Флора СССР. -- Т. 5. С. 594--661, 672--701.
Комаров В.Л. 1930. Флора полуострова Камчатки. -- Л.: Изд-во АН СССР. -- Т.3. С. 140--141.
Комаров В.Л. 1936. Род Populus / Флора СССР. -- Т. V. Москва-Ленинград: Изд-во АН СССР.
С. 224.
Комжа А.Л., Попов К.П. 1990. Новые данные об адвентивной флоре Северной Осетии // Ботан.
журн. -- Т.75. № 1. С. 109.
Конвенция о биологическом разнообразии. 1995. Текст и приложения. UNEP/CBD. 34 с.
Конечная Г.Ю. 1994. Senecioneae Cass. // Флора европ. ч. СССР. -- Т. 7. С. 52--77.
Конспект сосудистых растений Калининградской области: Справочное пособие / Губарева И.
Ю., Дедков В. П., Напреенко М. Г., Петрова Н. Г., Соколов А. А. Под ред. В.П. Дедкова. -
Калининград: издание Калинингр. Ун-та, 1999. -- 107 с.
Конспект флоры Рязанской Мещеры / Водолазская Н.Н., Губанов И.А., Киселева К.В. и др.;
Под ред. В.Н.Тихомирова. 1975. -- М: Изд-во Лесная пром-сть. -- 328 с.
Корнiенко O.M., Мосякiн С.Л. 2006. Номенклатура культивпваних та здичавiлих в Украïнi
пiвнiчноамериканських «Айстр» з точки зору делiмiтацiï родiв у трибi Astereae (Asreraceae)
// Укр. ботан. журн. -- Т. 63. № 2. С. 159--165.
Корнась Ян. 1960. О находке Bidens frondosa в Бресте // Ботан. мат-лы гербария БИНа. Т. 20. С.
337--339.
Корнась Ян. 1971. Сучаснi антропогеннi змiни у флорi Польшi // Укр. ботан. журн. Т. 28. № 2.
С. 167--173.
Королюк Е.А. 1997. Erigeron L.
-- Мелколепестник // Флора Сибири.
-- Т. 13: Asteraceae
(Compositae). С. 37--43.
Корунчикова В.В. 1994. Эколого-ботаническое изучение рода ирги // Бюлл. ботан. сада им.
И.С.Косенко. № 1. С. 21--27.
Костелова Г.С. 1971. Опыт интродукции видов рода Клен в условия бот. сада АН УзССР
г.Ташкента / Автореф. дисс. ... канд. биол. наук. -- Ташкент. 21 с.
Котов М.И. 1979. Brassicaceae Burnett (Cruciferae Juss. nom. altern.) -- Крестоцветные // Флора
европ. ч. СССР. -- Т. 4. С. 30--148.
Котов М.И. 1949. Адвентивные растения в УССР// Укр. ботан. журн. - Т.6. - № 1. - С.74-78.
Котов М.И. 1979. Растительность и флора Кисловодска и его окрестностей // Актуальные во-
просы современной ботаники. -- Киев: Наук. думка. С. 96--102.
Кохно Н.А. 1970. Итоги и теоретические основы интродкции на Украине видов рода Клен
/Автореф. дисс. ... докт. биол. наук. -- Киев. 47 с.
Красноборов Н.М. 2000. По поводу некоторых видов во флоре Алтайского края // Turczani-
nowia. --Т.3. --Вып.1. --С.56--57.
Краткий словарь ботанических терминов. 1993. Coст. Буланая М.В., Буланый Ю.И., Еленев-
ский А.Г., Черепанова Л.А. / Под ред. А.Г. Еленевского. -- Саратов: Изд-во Саратов. пед.
ин-та. -- 152 с.
Кречетович В.И., Бобров Е.Г. 1934. Овсяница -- Festuca L. s. str. // Флора СССР. Т. 2. С. 497--535.
Круберг Ю.К. 1955. О появлении во флоре Ленинграда галинзоги // Уч. зап. Ленинград. пед.
ин-та. -- Т.109. С. 239--245.
Крылов А.В. 2008. Адвентивный компонент флоры калужской области: динамика и натурали-
зация видов / Дисс... канд. биол. наук. -- М. -- 208 с.
452
Крылов Г.В., Салатова Н.Г. 1955. Озеленение городов и рабочих поселков Западной Сибири. --
Новосибирск: ВНИТОЛес. 56 с.
Крылов П.Н. 1949. Флора Западной Сибири. -- Томск: Изд-во ТГУ. № 11. С. 2734--2735.
Куваев В.Б., Шелгунова М.Л., Константинов Л.К. 1992. Флора окрестностей Знаменского.
Опыт долговременного мониторинга и сохранения урбанизируемой флоры Подмосковья.
М.: Наука. 358 с.
Куданова З.М. 1965. Определитель высших растений Чувашской АССР. -- Чебоксары: Чуваш.
кн. изд-во. -- 345 с.
Кузнецова Е.А., Макаренко Л.С. 1976. О некоторых новых и редких во флоре Курской области
растениях // Науч. тр. Курск. гос. педагогич. ин-та. -- Т. 69 (162). -- С. 21--31.
Кузьмичев Е.П., Петров А.В. 1991. Сосудистый микоз лоха и роль заболонника Ярошевского в
распространении болезни // Мат-лы Второй Всесоюзной Научно-технической конф. Охра-
на лесных экосистем и рациональное использование лесных ресурсов. Тезисы докладов. Ч.
1. М.: МЛТИ. С. 145--147.
Куклина А.Г. 2005. Основные этапы интродукции североамериканских видов ирги в Евразию //
Мат-лы VI Международного симпозиума «Новые и нетрадиционные растения и перспекти-
вы их использования». М: РУДН. -- Т. 1. С. 74--76.
Куклина А.Г. 2006. Распространение североамериканских видов ирги в Средней России // Фло-
ристические исследования в Средней России. Мат-лы VI совещания по флоре Средней
России. (Тверь, 15-16 апреля 2006 г.) М: Товарищество научных изданий КМК. -- С. 91--94.
Куклина А.Г. 2006а. Возможные пути происхождения некоторых видов ирги // Нетрадицион-
ные и редкие растения, природные соединения и перспективы их использования. Т. 1. Бел-
город. -- С. 38--40.
Куклина А.Г. 2006б. Изменчивость вегетативных признаков ирги колосистой (Amelanchier spi-
cata (Lam.) K. Koch) в европейских инвазионных популяциях // Бюлл. Гл. ботан. сада. --
Вып. 191.
Куклина А.Г. 2007. Инвазионные популяции ирги ольхолистной (Amelanchier alnifolia (Nutt.)
Nutt.) в России //Бюлл. Гл. ботан. сада. Вып. 198?.
Куклина А.Г. 2007а. Жимолость, ирга. Пособие для садоводов-любителей. - М: Изд-во «Ниола-
пресс»; Издательский дом «ЮНИОН-паблик». 204 с.
Куклина А.Г. 2007б. Калина, ирга. -- М: Кладезь-Букс. 96 с.
Куклина А.Г. 2007в. Изучение биоразнообразия видов рода Amelanchier Medik. // Биологиче-
ское разнообразие. Интродукция растений /Мат-лы IV Междунар. научн. конф. 5-8 июня
2007 г., Санкт-Петербург. СПб: БИН. С. 47--48.
Куминов Е.П. 1994. Ирга // Нетрадиционные садовые культуры. Мичуринск: ВНИИС. С. 149--161.
Куприянов А.Н. 1989. Новые и редкие растения для Центрального Казахстана // Ботан. журн. --
Т. 74. № 4. С. 545--547.
Кушхов А.Х. 1980. Некоторые новые для флоры КБАССР и СОАССР растения // Вопросы бо-
таники. -- Нальчик: Кабардино-Балкарский гос. ун-т. С. 163--166.
Кушхов А.Х. 1989. Новинки адвентивной флоры для КБ АССР и сопредельных территорий //
Проблемы изучения адвентивной флоры СССР. -- М.: Наука. С. 77--78.
Лавренко А.Н., Кустышева А.А. 1990. Новые и редкие для Коми АССР виды адвентивной фло-
ры // Ботан. журн. -- Т.75. № 2. С. 268.
Левицкий С.С. 1957. Список сосудистых растений Центрально-Черноземного государственного
заповедника // Тр. Центр.-Чернозем. гос. заповедника им. проф. В.В. Алехина. ---- Вып. 4. --
С. 110--178.
Лекарственные растения»: http://lekar-trava.ru/serdechno_sosud-3.html
Лисицына Л.И., Экзерцева В.А. 1984. О редких видах водных и прибрежно-водных растений
водоемов Волжского бассейна // Состояние и перспективы исследования флоры средней
полосы Европейской части СССР. -- М. С. 42--43.
Литвинов Д.И. 1895. Список видов, дикорастущих в Калужской губернии, с указанием полезных
и вредных. -- Калуга: Изд. Калуж. о-ва сел. хоз-ва, -- VIII, 112 с.
Литвинов Д.И. 1886--1888. Список растений, дикорастущих в Тамбовской губернии // Bull. Soc.
Nat.Mosc. --1886. --T.61(1885г.),№3. --P.1--49;1887. --T.62(1886г.),№4. --P.277--
295; 1887. -- Nouv.ser. T. 1, № 3. -- P. 789--812; 1888. -- Nouv.ser. T. 2, № 1. -- P. 96--118;
1888. -- Nouv.ser. T. 2, № 2. -- P. 220--260.
453
Ломоносова М.Н., Зыкова Е.Ю. 2003. Флористические находки в городе Новосибирске //
Turczaninowia. -- Т. 6. -- Вып. 1. С. 63--66.
Ломоносова М.Н., Сухоруков А.П. 2000. Флористические находки в Южной Сибири //
Turczaninowia. -- Т. 3. Вып. 4. С. 64--66.
Луговых П.В. 1959. Акклиматизация древесных и кустарниковых растений на Урале // Бюлл.
Гл. ботан. сада. Вып. 34. С. 24--29.
Лукина Е.В. 1989. Об адвентивных видах флоры Горьковской области // Проблемы изучения
адвентивной флоры СССР. -- М.: Наука. С. 42--44.
Лунева Н.Н., Бурмистров Л. А. 2001. Оценка внутривидового генетического разнообразия ирги
(Amelanchier Medik.) // Проблемы интродукции и систематики культурных растений и их
диких сородичей. Труды по прикл. ботанике, генетике и селекции. -- Т. 154. С. 123--127.
Лысенко З.Е. 1973. Биологические особенности видов рода Клен, интродуцированных в Се-
верную Киргизию / Автореф. дисс. ... канд. биол. наук. -- Фрунзе. -- 17 с.
Лысенко Т.М. 2005. Галофитная растительность Среднего Поволжья // Поволжский экологиче-
ский журн. -- № 3. С. 214--217.
Любченко В.М. 1986. Недотрога мелкоцветковая в фитоценозах Каневского заповедника //
Бюлл. Гл. ботан. сада. --Вып. 143. С. 39--43.
Маевский П.Ф. 1912. Флора Средней России: Иллюстрированное руководство к определению
среднерусских цветковых растений. -- 4-е изд., испр. и доп. / Под ред. Д.И. Литвинова. -- М.:
Изд. Сабашниковых. -- XXXVI, 731 с.
Маевский П.Ф. 1917. Флора Средней России: Иллюстрированное руководство к определению
среднерусских цветковых растений. -- 5-е изд., испр. и доп./ Под ред. Д.И. Литвинова. -- М.:
Изд. Сабашниковых. -- XXXII, 910 с.
Маевский П.Ф. 1933. Флора Средней России: Иллюстрированное руководство к определению
среднерусских семенных и сосудистых споровых растений центральных областей европейской
части СССР. -- 6-е изд., испр. и доп./ Под ред. Б.А. Федченко. -- М.; Л.: Сельхозгиз. -- 748 с.
Маевский П.Ф. 1940. Флора средней полосы Европейской части СССР. 7-е изд., испр. и доп.
М.-Л. Сельхозгиз. 824 с.
Маевский П.Ф. 1954. Флора средней полосы европейской части СССР. 8-е изд., испр. и доп. /
Под ред. Б.К.Шишкина. -- М.; Л.: Сельхозгиз. 912 с.
Маевский П.Ф. 1964. Флора Средней России. 9-е изд. -- М. 880 с.
Маевский П.Ф. 2006. Флора средней полосы европейской части России. 10-е изд., испр. и доп. -
М. : Товарищество научных изданий КМК. -- 600 с.
Маевский П.Ф. Флора Средней России. -- 5-е изд. -- 1917. -- 910 с.; 6-е изд. -- 1933. -- 760 с.; 7-е
изд. -- 1940. -- 824 с.; 8-е изд. -- 1954. -- 912 с., 9-ое изд. -- 1964. -- 880 с.;
Майоров С.Р. 2002. Новые сведения к флоре европейской России // Бюлл. Моск. о-ва испыт.
прир. Отд. биол. -- Т. 107. Вып. 6. С. 62--64.
Майоров С.Р. 2006. Xanthium L. -- Дурнишник // Маевский П. Ф. Флора средней полосы евро-
пейской части России. Изд. 10-е, испр. и перераб. -- С. 494--495.
Майсурян Н.А., Атабекова А.И. 1974. Люпин. -- М.: «Колос». 463 с.
Майтулина Ю.К. 1980. О морфологии и прорастании семян клена ясенелистного из различных
географических пунктов // Бюлл. Гл. ботан. сада. -- Вып. 117. С. 85--89.
Майтулина Ю.К. 1984. Новые и редкие адвентивные растения Вологодской области // Бюлл.
Гл. ботан. сада. Вып. 132. С. 45--46.
Майтулина Ю.К. 1988. К биологии и систематике среднеазиатских видов рода Недотрога //
Бюлл. Гл. ботан. сада. -- Вып. 150. С. 59--64.
Майтулина Ю.К. 1990. Влияние фотопериодической реакции растений на результаты их ин-
тродукции // Адаптационная изменчивость растений при интродукции.
-- Рига. Зинатне.
С. 87--90.
Макаров А.А., Дубовик Л.А. 1977. Материалы по флавоноидоносности растений Центральной
Якутии // Материалы к изучению лекарственной флоры Якутии. -- Якутск. С. 53--62.
Максимович М. 1826. Список растений Московской флоры // М. -- Ч. 2. № 3. С. 203--224.
Малеев В.П. 1934. Boraginaceae -- Бурачниковые // Сорные растения СССР. Т. 3.
Малышев Л.И. 1981. Изменение флор земного шара под влиянием антропогенного давления //
Науч. докл. высшей школы. Биол. науки. -- № 3. С. 5--20.
454
Малышева В.Г. 1988. Пути и способы формирования адвентивной флоры Калининской облас-
ти // Флора и растительность южной тайги. -- Калинин: изд-во КГУ. С. 25--31.
Малышева В.Г. 1979. К адвентивной флоре Калининской области // Экология растений южной
тайги. -- Калинин. С. 48--54.
Малышева В.Г. 1979. Новые и редкие адвентивные растения Калининской области // Ботан.
журн. -- Т.64. №3. С. 438--441.
Малышева В.Г. 1980. Адвентивная флора Калининской области / Дисс... канд. биол. наук. --
Ленинград.
Мамедов Н.А. 1986. Щирицы -- адвентивные сорняки низменностей Азербайджана // Изв. АН
АзССР. Сер. биол. № 6. С. 26--30.
Манденова И.П. 1951. Борщевик -- Heracleum L. // Флора СССР. -- Т. 17. С. 223--259.
Маркелова Н.Р. 2004. Динамика состава и структуры адвентивной флоры Тверской области /
Дисс... канд. биол. наук. -- М. 282 с.
Марков М.В. 1994. Сравнительная характеристика морфологии корневых систем у двух видов
Недотроги Impatiens noli-tangere L. и I. рarviflora DC. (Balsaminaceae) на ранних этапах
развития // Флора и растительность Тверской области. -- Тверь. С. 64-- 68.
Марков М.В., Уланова Н.Г., Чубатова Н.В. 1997. Род Недотрога // Биологическая флора Мос-
ковской области. -- Вып.13. -- М.: Изд-во Полиэкс. С. 128--168.
Мартыненко В.П., Сюборова С.Ф. 1984. Новые виды для флоры Белорусского Поозерья. -Деп.
Ред. ж. Изв. АН БССР Сер. биол. науки.
-- Минск, 13 с. Рукопись деп. в ВИНИТИ
26.01.1984 № 492-84 Деп.
Матвеев В.И. 1983. Динамика флоры Белого озера за последние 80 лет // Сложение и динамика
растительного покрова: Межвуз. сб. науч. тр. -- Куйбышев, -- С. 62--71.
Матвеев В.И. 1969. Флора водоемов Средней Волги и ее притоков // Уч. зап. Куйбышев. пед.
ин-та. -- Вып. 68: Бот. и сел. хоз-во. -- С. 30--78.
Матвеев Д.Е. Конспект адвентивных видов Волгоградской области, ...26 С.
Матекайтите В.П. 1985. О новых и редких рудеральных видах в Литовской ССР // Ботан.
журн. -- Т. 70. № 10.С. 1410--1414.
Мауринь А.М. 1970. Опыт интродукции древесных растений в Латвийской ССР. -- Рига: Зинат-
не. 257 с.
Методические рекомендации по борьбе с неконтролируемым распространением растений
борщевика Сосновского / Составители Далькэ И.В., Чадин И.Ф. -- Сыктывкар. 2008. - 28 с.
Мизгер А. 1869.Конспект растений, дикорастущих и разводимых в Курской губернии. -- Курск. --
115 с.
Миняев Н.А., Конечная Г.Ю. 1976. Флора Центрально-Лесного государственного заповедника.
Л. 104 с.
Миркин Б.М., Наумова Л.Г. 2001. Адвентизация растительности: инвазивные виды и инвази-
бельность сообществ // Успехи совр. биологии. -- Т. 121. № :. -- С. 550--562.
Мишуров В.П., Волкова Г.А., Портнягина Н.В. 1999. Интродукция полезных растений в подзо-
не средней тайги Республики Коми. -- СПб.: Наука. 216 с.
Мозолевская Е. 2007. Златка угрожает // Российская лесная газета. № 7--8 (189--190) от
25.02.2007.
Мозолевская Е.Г., Исмаилов А.И. 2007. Ясеневая изумрудная узкотелая златка в городских на-
саждениях Москвы // Лесной бюллетень. -- № 35. С. 17--20.
Моисеев Д.А. 1998. Аннотированный список высших растений музея-заповедника «Аркаим» и
его окрестностей. - Челябинск: -- 72 с.
Морозова О.В. 2003а. Участие адвентивных видов в формировании разнообразия и структуры
флор Восточной Европы // Известия РАН. Серия геогр. -- № 3. -- С. 63--71.
Морозова О.В. 2003б. База данных по адвентивным видам растений (AliS) и перспективы соз-
дания единого банка информации // Проблемы изучения адвентивной и синантропной фло-
ры в регионах СНГ / Мат-лы междунар. науч. конф. / Под ред. В.С. Новикова и А.В. Щер-
бакова. -- М.: Узд. Бот. сада МГУ; Тула: Гриф и К°. -- С. 72--73.
Москаленко Г.П. 2001. Карантинные сорные растения России. -- 278 с.
Мосяк Н.С. 1988. Рiд Bidens у флорi УРСР // Укр. ботан. журн. -- Т. 45. № 6. С. 63--64.
Мосякин С.Л. 1996. Amaranthus // Флора Восточной Европы. т. IX. «Мир и семья -- 95», --
Санкт-Петербург. С. 11--18.
455
Муратова Е.Н., Корнилова М.Г., Пименов А.В., Седельникова Т.С., Владимирова О.С. 2006. Ци-
тогенетические эффекты влияния горно-химического комбината на клетки элодеи канад-
ской (Elodea canadensis Michx.) // Вестник Красноярск. гос. аграрного ун-та. -- Красноярск, --
Вып. 14. -- С. 159--163.
Назаров М.И. 1927. Адвентивная флора средней и северной части РСФСР за время войны и ре-
волюции // Изв. Гл. ботан. сада СССР. - Т. 26. - Вып. 3. С. 223--234.
Название! [Electronic resource.] Mode of access: http://trifoly.by.ru/wight/Str_w_1/stranisi/4.html.
Accessed 20 Mar. 2008.
Невский М. 1952. Флора Калининской области. -- Калинин. Обл. книжное изд-во; -- Ч. 2. 1033 с.
Невский С.А. 1934. Ячменевые -- Hordeeae Benth. // Флора СССР. -- Т. 2. С. 390--728.
Недолужко В.А. 1984. Распространение Ambrosia artemisiifolia (Asteraceae) в Приморском крае
// Ботан. журн. Т. 69. № 4. С. 527--529.
Некрасов В.И., Князева О.М., Смирнова Н.Г. 1973. Семенная продуктивность интродуцентов
дендрария Главного ботанического сада // Опыт интродукции древесных растений (Сбор-
ник научных статей). -- М.: ГБС АН СССР. С. 205--226.
Нестерович Н.Д. 1955. Плодоношение интродуцируемых древесных растений и перспективы
разведения их в БССР. -- Минск: АН БССР 384 с.
Нечаева Т.И. 1985. Дополнения к флоре Владивостока и его окрестностей// Новости
систематики высших растений. -- Т. 22. С. 257.
Нечаева Т.И., Петрова О.В. 1995. Находки адвентивных растений на острове Кунашир // Бо-
тан. журн. -- Т.80. № 8. С. 118.
Никифорова О.Д. 1994. Cardaria Desv. -- Кардария (Сердечница) // Флора Сибири. -- Т. 7. Ber-
beridaceae -- Grossulariaceae. С. 144--145.
Никифорова О.Д. 1994. Lepidium L. -- Клоповник // Флора Сибири. -- Т. 7. Berberidaceae -- Gros-
sulariaceae. С. 138--144.
Николаева М.Г., Разумова М.В., Гладкова Н.В. 1985. Справочник по проращиванию покоящих-
ся семян. -- Л.: Наука, 348 с.
Новиков В.С. 1978. Род Ситник // Биологическая флора Московской области. - Вып. 4. Под ред.
Т. А. Работнова. С. 3--51.
Новиков В.С. 1985. Ситниковые -- Juncaceae Juss. // Сосудистые растения советского Дальнего
Востока. Т. 1. C. 57--88.
Нотов А.А. 1999. Материалы к флоре Тверской области. Ч.1: Высшие растения. 2-е изд., пере-
раб. и дополн. Тверь. 119 с.
Нотов А.А. 2000. Материалы к флоре Тверской области. Ч.1: Высшие растения: (Электрон. ре-
сурс). 3-я версия, перераб. и доп.
-- Тверь. Режим доступа: http://garden.tversu.ru/
publ/flora/spisok_2.
Нотов А.А., Маркелова Н.Р. 2005. Адвентивный компонент флоры Тверской области. -- Тверь:
Изд-во «ГЕРС».
Овеснов А.М., Шилова С.И. 1975. О некоторых новых адвентивных и редких видах Централь-
ных районов Пермской области // Экология опыления. № 1. С. 135--137.
Овчаренко А.А., Золотухин А.И. 2003. О распространении Acer negundo L. и Fraxinus pennsyl-
vanica Marsh. в пойменных лесах степной зоны Балашовского района (Саратовская об-
ласть) // Проблемы изучения адвентивной и синантропной флоры в регионах СНГ. / Мат-
лы научной конференции. С. 75--76.
Овчинников И.Ф. 1974. Ирга. Кудымкар: Перм. кн. изд-во. 32 с.
Октябрева Н.Б., Тихомиров В.Н., Чичев А.В. 1978. Новые материалы к характеристике адвентив-
ной флоры Московской области // Биол. науки. -- № 12. С. 89--94.
Олонова М.В. 1990. Poa L. -- Мятлик // Флора Сибири. -- Т. 2. С. 163--186.
Определитель высших растений Коми ACCP. 1962. -- М.-Л.: Изд-во АН СССР. 359 с.
Определитель растений Мещеры. 1986. Ч. 1 / Под ред. В.Н. Тихомирова. -- М.: Изд-во Моск.
ун-та. 240 с.
Определитель растений Мещеры. 1987. Ч. 2 / Под ред. В. Н. Тихомирова. -- М.: Изд-во Моск.
ун-та. 224 с.
Определитель растений Среднего Поволжья / В.В. Благовещенский, Ю.А. Пчёлкин, Н.С. Раков и
др. -- Л.: Наука, 1984. -- 392 с.
Определитель растений Средней Азии. 1993. Ташкент: ФАН, Т.10. С. 620--621.
456
Определитель растений Татарской АССР / Н.Г. Афанасьева, Р.Г. Иванова, З.И. Макарова и др. --
Казань: Изд-во Казан. ун-та, 1979. -- 371 с.
Определитель растений Ярославской области / Н.М. Беловашина, В.К. Богачев, М.М. Прозорова и
др. -- Ярославль: Кн. изд-во, 1961. -- 500 с.
Определитель сосудистых растений Орловской области / А.Г. Еленевский, В.И. Радыгина. --
Орел: Труд. 1997. -- 208 с.
Орлова Н.И. 1989. Обзор видов семейства кипрейных флоры Вологодской области // Вестник
ЛГУ. -- Сер. 3. № 1. С. 32--38.
Осипова В.Д. 1970. Ирга в Томске // Бюлл. Сиб. ботан. сада. Вып. 7. С. 89.
Отчет о деятельности Российской Академии Наук в 2006 году. Основные результаты в области
естественных, технических, гуманитарных и общественных наук. М.: Наука., 2007. 215 с.
Палибин И.В. 1945. Люпин // Флора СССР. -- М.-Л.: АН СССР. Т. 11. С. 48--51.
Паллас П.С. 1781. Каталог растениям находящимся в Москве в саду его превосходительства,
действительнаго статскаго советника и Императорскаго Воспитательнаго дома знаменита-
го благодетеля, Прокопия Акинфиевича Демидова. 163 с.
Панасенко Н.Н. 2002. Урбанофлора Юго-Западного Нечерноземья России (на примере городов
Брянской области). -- Дисс... канд. биол. наук. -- Брянск.
Папченков В.Г. 1984. Флора рек Среднего Поволжья // Состояние и перспективы исследования
флоры средней полосы Европейской части СССР. -- М. С. 44--45.
Папченков В.Г. 2003. К определению сложных групп водных растений и их гибридов // Гидро-
ботаника. -- Рыбинск. Рыбинский дом печати. - С.82-91.
Папченков В.Г. 2003. Растения-вселенцы и их воздействие на мелководные экосистемы басей-
на Волги Проблемы изучения адвентивной и синантропной флоры в регионах СНГ / Мат-
лы междунар. науч. конф. / Под ред. В.С. Новикова и А.В. Щербакова. -- М.: Изд. Бот. сада
МГУ; Тула: Гриф и К°. -- С. 79--81.
Папченков В.Г. 2006. Гибриды водных растений и особенности их определения // Мат-лы VI
Всероссийской школы-конференции по водным макрофитам. -- Рыбинск. С. 49--57.
Папченков В.Г. 2008. О распространении Phragmites altissimus (Benth.) Nabille (Poaceae) [Элек-
тронный ресурс] // Российский Журнал Биологических Инвазий. 2008. № 1. <http://
www.sevin.ru/invasjour/issues/2008_1/Papchenkov_08_1.pdf> (01.08.2009).
Папченков В.Г., Голубева И.Д., Шпак Т.Л. 1988. Флора мелководий Куйбышевского водохрани-
лища // Вторая Всесоюз. конф. по высшим водным и прибрежно-водным раст.: (Тезисы
докл.). -- Борок. -- С. 33--34.
Папченков В.Г., Дмитриев А.В. 1989. Об адвентивной флоре автономных республик Среднего
Поволжья // Проблемы изучения адвентивной флоры СССР. М.: Наука. С. 47--49.
Паутова В.Н., Галимулин М.Г. 1980. О находках редких для Восточной Сибири видов высших
водных растений // Бот. журн. -- Т. 65, № 7. С. 1020--1022.
Перечень вредителей, возбудителей болезней растений, сорняков, имеющих карантинное зна-
чение для Российской Федерации. МСХ, 2003.
Перечень семян ботанического сада Самары. 1934.
Петров А. В., Кузьмичев Е. П. 1994. Усыхание лоха на Западном побережье Каспия под влия-
нием заболонника Ярошевского и патогенной микрофлоры // Лесоведение. № 3. С. 48--53.
Петрова А.Ю., Легоньких О.А. Особенности растительности водоемов юга Самарской области,
[Electronic resource.] Mode of access: http://www.ksu.ru/conf/botan200/p244.rtf. Accessed 12
June 2008.
Петунников А.Н. 1896. Критический обзор Московской флоры: в 3 ч. Ч. 1 Dialypetalae // Ботан.
зап. (изд. При Ботан. саде Спб. ун-та). Т. 6. Вып. 13. С. 1--121.
Петунников А.Н. 1886. Водяная зараза (Elodea canadensis Rich.) // Сад и огород.
--№19.
С. 163--165.
Письмаркина Е.В., Силаева Т.Б., Агеева А.М., Бармин Н.А. / Приволжский экологический жур-
нал, 2006. №1. С. 87--90, [Electronic resource.] Mode of access: http://www.sevin.ru/volecomag/
issues/ 2006_1/PEJ_2006_1_87_90.pdf. Accessed 15 November 2008.
Плаксина Т.И. 2001. Конспект флоры Волго-Уральского региона. -- Самара: Изд. Самар. ун-та, --
387 с.
Победимова Е.Г. 1949. Balsaminaceae // Флора СССР. -- М.-Л. Изд-во АН СССР. Т. 14. С. 624--634.
Полуянов А.В. 2005.Флора Курской области. -- Курск: изд. Курск. гос. ун-та. 264 с.
457
Понерт Й. 1971. Критические заметки о флоре Колхиды // Новости систематики высших рас-
тений. № 8. С. 292--294.
Попов В.И. 2000. Адвентивный компонент синантропной флоры Санкт-Петербурга. Автореф.
дис...канд. биол. наук / БИН РАН. СПб. - 18 с.
Попов М.Г. 1953. Сем. Бурачниковые -- Boraginaceae G. Don // Флора СССР. Т. 19. С. 97--691.
Попов Т.И. 1933. Лекарственные и душистые растения ЦЧО. -- Воронеж. 107 с.
Пояркова А.И. 1949. Кленовые -- Aceraceae Lindl. // Флора СССР. Т. 14. С. 580--622.
Пояркова А.И. 1950. Род 293. Боярышник -- Crataegus L. // Флора БССР. Том III. -- Минск: изд-
во АН БССР. -- С. 52--65.
Практическое пособие по борьбе с гигантскими борщевиками (на основе европейского опыта
борьбы с инвазивными сорняками). Ред. Ш. Нильсен, Г. П. Равн, В. Нентвинг, М. Вэйд.
Hoersholm: Forest & Landscape Denmark, 2005. 44 с.
Пробатова Н.С. 1985. Мятликовые, или Злаки -- Poaceae Barnh. (Gramineae Juss.) // Сосуди-
стые растения советского Дальнего Востока. - Т. 1. C. 89--382.
Просовский М.А., Рыбалко К.С., Шейченко В.И. и др. 1985. Химический состав ромашки души-
стой // Хим.-фармацевтич. журн. - Т.19. - № 8. - С. 981-984.
Протопопова В.В. 1994. Heliantheae Cass. // Флора европ. ч. СССР. - Т. 7. - С. 25--52.
Протопопова В.В. 1994. Дурнишник -- Xanthium L. // Флора европ. ч. СССР. - Т. 7. - С. 48--52.
Протопопова В.В., Мосякін С.Л., Шевера М.В. 2002. Фітоінвазії в Україні як загроза
біорізноманіттю: сучасний стан і завдання на майбутнє. -- Киiв: Ін. Ботан. ім. М. Г. Холод-
ного НАНУ. -- 32 с.
Протопопова В.В. 1973. Адвентивнi рослини лiсостепу I степу Украiни. -- Киев. Наук. думка.
192 с.
Протопопова В.В. 1991. Синантропная флора Украины и пути ее развития. -- Киев: Наук. Дум-
ка. --204с.
Пузырев А.Н. 1985. Дополнение к адвентивной флоре Удмуртии // Ботан. журн. Т.70. № 2.
С. 268--271.
Пузырев А.Н. 2006. О находках адвентивных видов растений на шоссейных дорогах Удмуртии
// Вестник Удмуртского университета. Сер. Биология. № 10. С. 25--36.
Пузырев А.Н. 2008. Дополнение к адвентивной флоре шоссейных дорог Удмуртии // Вестник
Удмуртского университета Сер. Биология. Науки о Земле. - Вып. 2. С. 139--150.
Пынзарь П.Я. 1988. Новые для Молдавии адвентивные растения // Изв. АН МССР. Сер. биол. и
хим. наук. № 5. С. 68--69.
Пяк А.И. 1994. Адвентивные растения Томской области // Ботан. журн. Т.79. 11. С. 45--51.
Раков Н.С. 2003. Флора города Ульяновска и его окрестностей. -- Ульяновск. -- 216 с.
Раменская М.Л. 1960. Определитель высших растений Карелии. -- Петрозаводск: изд-во Ка-
рельской АССР. 485 с.
Раменская М.Л. 1983. Анализ флоры Мурманской области и Карелии. -- Л.: Наука. 215 с.
Рамомавичюс В.А. 1988. Сегетальные сообщества Литовской ССР // Тр. АН ЛитССР. Сер.В. №
1/101. С. 27--37.
Регель Э. 1874. Русская дендрология. Спб: Типография В. Грацианского. Вып. IV. 473 с.
Регель Э. 1879. Русская дендрология. № 5. С. 355--473.
Решетникова Н.М. 2003. Активность адвентивних видов сосудистых растений на территории
национального парка «Смоленское поозерье» // Проблемы изучения адвентивной и синант-
ропной флоры в регионах СНГ / Мат-лы междунар. науч. конф. / Под ред. В.С. Новикова и
А.В. Щербакова. -- М.: Узд. Бот. сада МГУ; Тула: Гриф и К°. -- С. 89--91.
Решетникова Н.М. 2004. Материалы к флоре Смоленской области // Бюлл. Гл. ботан. сада. --
Вып. 188. С. 70--102.
Решетникова Н.М., Скворцов А.К., Майоров С.Р. 2005. Сосудистые растения национального
парка «Угра». [Флора и фауна национальных парков. Вып. 6] -- 143 с.
Род Matricaria // Флора СССР. М-Л.: Изд-во АН СССР, 1961. Т. 26. С. 150--151.
Род Matricaria // Флора Юго-Востока Европейской части СССР. Л.: Изд-во АН СССР, 1936.
Вып. 6. -- С. 342.
Розен В.В. 1893. Список растений Веневского уезда Тульской губернии // Памятная книжка
Тульской губ. за 1893 г. -- Тула. -- Отд. 8. С. 1--183.
458
Розен В.В. 1913. Дендрологический список растений, культура которых возможна в пределах
тульской губернии // Изв. Тул. об-ва любит. Естествознания. ---- Вып. 2. -- С. 3-63; 81-83.
Розен В.В. 1916. Список растений, найденных в Тульской губернии до 1916 года // Изв. Тул.
об-ва любит. естествознания. ---- Вып. 4. - С. 1- 282.
Рычин Ю.В., Сергеева П.В. 1939. Водная и прибрежная флора: Краткий определитель по веге-
тативным признакам прибрежных и водных сосудистых растений центральной части евро-
пейской территории СССР. -- М.: Учпедгиз,-- 184 с.
Сагалаев В.А., Матвеев Д.Е. 2000. Флористические находки на Среднем Дону // Ботан. журн. -
Т. 85. № 10. -- С. 115--119.
Саксонов С.В. 1989. Очаги концентрации синантропных видов в Жигулевском заповеднике //
Проблемы изучения синантропной флоры СССР. -- М.: Наука. С. 72--74.
Сапегин Л.М. 1985. Синантропная флора травяных сообществ поймы Белорусского полесья //
Проблемы изучения синантропной флоры СССР. -- М.: Наука. С. 34--35.
Сауткина Т.А., Сауткина Н.И. 1983. Род Epilobium во флоре Белоруссии // Вестник Белорус-
ского ун-та. сер.2. -- №3. С. 23--26.
Сафонов Г.Е. 1982. Новые адвентивные растения во флоре Астраханской области // Бюлл. Гл.
Ботан. сада. -- Вып. 124. -- С. 48.
Сацыперова И.Ф. 1984. Борщевики флоры СССР -- новые кормовые растения. -- Л. Наука. 223 с.
Свириденко Б.Ф. 1986. Находки Elodea canadensis (Hydrocharitaceae) в Северном Казахстане //
Бот. журн. -- Т. 71, № 12. -- С. 1686--1688.
Северова Е., Siljamo Р., Skjoth C.A. 2008. Дальнезаносная пыльца в аэропалинологическом
спектре Москвы // Палинология: стратиграфия и геоэкология. Сборник научных трудов 12
Всероссийской Палинологической конференции. СПб. T. 1. С. 185--190.
Семенютина А.В. 1985. Интродукция видов рода Amelanchier Medik в условиях Нижнего По-
волжья //Селекционные основы повышения устойчивости защитных лесных насаждений в
аридной зоне. Волгоград: ВНИИ агромелиорации. -- С. 92--94.
Серегин А.П., Щербаков А.В. 2002. Основные гербарные фонды по флоре Средней России //
Флора Середней России: Аннотированная библиография. Первое дополнение / И.А. Губа-
нов, И.М. Калиниченко, А.В. Щербаков. М.: Изд-во Центра охраны дикой природы.
--
C. 60--71.
Силаева Т.Б. 1989. О новых адвентивных растениях флоры Мордовии // Проблемы изучения
адвентивной флоры СССР. -- М.: Наука. С. 44--45.
Силаева Т.Б. Флора бассейна реки Суры [Electronic resource.] Mode of access: http:// www.ksu.ru/
conf/botan200/p181.rtf. Accessed 23 Mar. 2008.
Силаева Т.Б., Тихомиров В.Н. 1987. Изменение флоры Мордовской АССР под влиянием антро-
погенных факторов // Генетические основы интродукции и селекции растений. - Саранск:
Изд-во Мордовск. гос. ун-та. - С. 120-127.
Скворцов А.К. 1959. Новые данные о распространении в Московской области некоторых ред-
ких и интересных растений // Биол. науки. - № 3. - С.164--167.
Скворцов А.К. 1973. Новые данные об адвентивной флоре Московской области. 2 // Бюлл. Гл.
ботан. сада. Вып. 88. С. 31--35.
Скворцов А.К. 1973. Новые данные об адвентивной флоре Московской области. 1 // Бюлл. Гл.
бот. сада. Вып. 87. С. 3--11.
Скворцов А.К. 1976. Род Epilobium // Флора северо-востока Европейской части СССР. Т. 3.
С. 211--216.
Скворцов А.К. 1982. Новые данные об адвентивной флоре Московской области. III // Бюлл. Гл.
ботан. сада. -- Вып. 124. С. 43--48.
Скворцов А.К. 1995. К систематике и номенклатуре адвентивных видов рода Epilobium
(Onagraceae) во флоре России // Бюлл. Моск. о-ва испытат. природы. Отд. биол. Т. 100. №
1. С. 74--78.
Скворцов А.К. 1996. Onagraceae Juss. -- Ослинниковые, Кипрейные // Флора Восточной Евро-
пы. Т. 9. С. 299--316.
Скворцов А.К. 2005. Конспект рода Кипрей (Epilobium L.s.str) в России и сопредельных странах
// Бюлл. Гл. ботан. сада. -- Вып. 189. С. 90--104.
Скворцов А.К., Майтулина Ю.К. 1982. Об отличиях культурной черноплодной аронии от ее
диких родоначальников // Бюлл. Гл. ботан. сада. - Вып. 126. С. 35--40.
459
Скворцов А.К., Майтулина Ю.К. 1989. Адвентивные растения как модель для изучения микро-
эволюционных процессов // Проблемы изучения адвентивной флоры СССР / Мат-лы со-
вещ. 1--3 февраля 1989 г. С. 6--8.
Сконникова В.В. 1985. Растительный мир Урала и его антропогенные изменения. -- Свердловск:
УНЦ АН СССР. -- С. 106--121.
Слесарев В.Н., Буянкин В.И. 1971. Биологические особенности щирицы белой и меры борьбы с
ней // Тр. Зап.-Казахстан. с.-х. ин-та. -- Т. 1. С. 29--31.
Соколов Д.Д., Чертопруд Е.С. 2002. Диагностические признаки Poa supina Schrad. (Gramineae)
по материалам из Московской области // Бюл. МОИП. Отд. биол. Т. 107. Вып. 4. С. 87--90.
Соколов С.Я. 1957. Современное состояние теории акклиматизации и интродукции растений //
Тр. БИНа. -- Сер. 6. № 5. С. 10--32.
Сорные растения СССР. 1935. Руководство к определению сорных растений СССР. -- Т. IV. М.;
Л.: Изд-во АН СССР. -- 414 с.
Сосудистые растения Татарстана / О.В. Бакин, Т.В. Рогова, А.П. Ситников. Изд-во Казан. ун-
та. 2000. -- 496 с.
Спиридонов Ю.Я., Шестаков В.Г. Применение гербицида Арсенал, ВК (250 г/л имазапира)
кампании БАСФ на объектах несельскохозяйственного пользования (рекомендации), [Elec-
tronic resource.] Mode of access: http: // www.basf.ru/files/ArsenalBooklet_final.pdf. Accessed
10 May 2008.
Справочник по вредителям, болезням растений и сорнякам, имеющим карантинное значение
для территории Российской Федерации. 1995. Нижний Новгород: Арника. -- 231 с.
Стародубцева Е.А. 1995. Антропогенные изменения флоры и растительности Усманского бора
/ Дисс... канд. биол. наук. -- Воронеж.
Старченко В.М., Дойко Э.В. 1981. Новые местонахождения некоторых заносных и редких на
Дальнем Востоке // Ботан. журн. -- Т. 66. № 1. С. 82.
Стемпинский Л. 1909. История распространения Elodea canadensis Michx. в России // Тр. Бот.
сада Юрьев. ун-та. -- Юрьев. -- Т. 10, вып. 4. -- С. 214--229.
Степанов Н.В. 1990. Флористические находки в Красноярском крае // Ботан. журн. -- Т. 75. №
5. С. 725--729.
Стребкова А. 1931. Культуры древесных пород на Апшеронском полуострове // Тр. по прикл.
бот., ген., селекции. -- Т. 27. № 3. С. 206--228.
Сырейщиков Д.П. 1906. Иллюстрированная флора Московской губернии: В 4 ч. -- Ч. 1: [Сосу-
дистые споровые, голосеменные, однодольные]. -- М., -- IV, 274 с.
Сырейщиков Д.П. 1910. Иллюстрированная флора Московской губернии. -- Ч. 3. М.: Изд. книж.
маг. Торг. Дома Лахтин, Сырейщиков и Ко. 397 с.
Сырейщиков Д.П. 1927. Определитель растений Московской губернии. -- М.: Об-во изучения
Московской губернии. 294 с.
Тамамшян С.Г. 1959. Астра -- Aster L. // Флора СССР. Т. 25. С. 77--110.
Терехина Т.А. 1995. Конспект флоры Кания-на-Оби // Ботан. исслед. Сибири и Казахстана. -
Барнаул, изд-во Алтайского гос. ун-та. С. 150--163.
Тимонин А.К. 1984. Некоторые особенности опушения стеблей видов Amaranthus в связи с их
диагностикой // Науч. докл. высш. школы. Биол. науки. -- Вып.1. С. 61--67.
Тимофеев Н.П. 1993. Некоторые практические итоги внедрения нетрадиционного кормопроиз-
водства // Мат-лы VIII Всероссийского симпозиума по новым кормовым растениям. - Сык-
тывкар. С. 152--153.
Тимофеев Н.П. 2002. Протеиновая ценность новых культур в условиях Севера (Теоретическое
обоснование и практическая реализация) // Нетрадиционные природные ресурсы, иннова-
ционные технологии и продукты. Сб. научн. тр. -- Вып. 6. С. 115--139.
Тихомиров В.Н. 1979. 5896. Xanthium riparium Itz. et Hertsch. // Список растений Гербария фло-
ры СССР. -- Т. 22. Вып. 118. С. 97.
Тихомиров В.Н. 1982. Три новых вида для флоры Рязанской области // Науч. докл. высшей
школы. Биол. науки. -- № 12. С. 52--53.
Тихомиров В.Н. 1987. Новые материалы к флоре Воронежской области // Биол. науки. Т. 6.
С. 74--78.
Тихомиров В.Н., Загородняя Г.Ю., Старобогатов Я.И., Шведчикова Н.К. 1960. Juncus macer
S.F. Gray в Московской области // Биол. науки. -- № 2. С. 121--124.
460
Тихомиров В.Н., Майоров С.Р., Соколов Д.Д. 1998 (1999). О роде Symphytum L. (Boraginaceae) в
Средней России // Новости систематики высших растений. -- Т. 31. С. 231--248.
Тишков А.А. 2005. Экологические последствия вступления России во всемирную торговую ор-
ганизацию (ВТО) // Экономическая политика. http://trade.ecoaccord.org/docs/tishkov.htm
Ткачева Е.В., Виноградова Ю.К. 2009. Сравнительный анализ семенной продуктивности таксо-
нов рода Люпин - Lupinus polyphyllus и L. × regalis // Тезисы в Белгороде.
Ткаченко К.Г. 1989. Особенности цветения и семенная продуктивность некоторых видов
Heracleum L., выращенных в Ленинградской области. // Растит. Ресурсы. Т. 25. Вып. 1.
С. 52--61.
Токарь О.Е. Elodea canadensis Michx. как интродуцируемый вид в Тюменской области // II мо-
лод. конф. «Исслед. молодых ботаников Сибири», 24--27 февр. 2004 г., Новосибирск,
Центр. Сиб. бот. сад СО РАН. -- Электронный ресурс. Режим доступа: http://www.ict.nsc.ru/
ws/ show_abstract.dhtml?ru+98+6788. -- Acc. 13 Nov. 2008.
Тохтарь В.К. 2001. Исследование структуры количественных морфологических признаков
различных географических популяций Conyza сanadensis // Промышленная ботаника. Вып.
1. С. 126--129.
Тохтарь В.К., Хархота А.И., Ростаньски А., Виттиг Р. 2003. Сравнение локальных флор тех-
ногенных территорий Европы // Промышленная ботаника, -- Вып. 3. С. 3--11.
Третьяков Д.И. 1996. О новых и редких видах адвентивных растений для города Смоленска,
Ленинградской области и Краснодарского края // Бот. журн. -- Т. 81. № 5. С. 82--90.
Туганаев В.В., Пузырев А.Н. 1988. Гемерофиты Вятско-Камского междуречья.-- Свердловск:
Изд-во Уральск. ун-та. -- 125 с.
Ульянова Т.Н. 2005. Сорные растения во флоре России и сопредельных государств. Алтайский
гос. ун-т, Южно-Сибирский Ботанический Сад. «Азбука», -- Барнаул. 295 с.
Усадебные парки русской провинции: проблемы сохранения и использования / Мат-лы Всерос.
науч. конф. / Великий Новгород. 2003. -- 260 с.
Уханов В.В. 1950. Клены Северной Америки в районе Ленинграда и возможности их культуры
в Европейской части СССР // Тр.БИНа. -- Сер 4. № 1. С. 20--57.
Учебное пособие к летней практике по ботанике: Флора высших растений Звенигородской
биостанции МГУ и ее окрестностей / Ю.Е. Алексеев, П.Ю. Жмылев, З.А. Слука, Е.А. Кар-
пухина, С.Р. Майоров, Н.Г. Прилепский, А.К. Тимонин. Тула: Гриф и К., 2008. 76 с.
Фатаре И.Я. 1977. Род Bromus в Латвийской ССР// Флора и растительность Латв.ССР. Кур-
земский геоботанический р-н. -- Рига: Зинатне. С. 121--132.
Фатаре И.Я. 1979. Род Epilobium в Латвийской ССР// Флора и растительность Латв. ССР. --
Рига. C. 128--132.
Федченко Б.А. 1934. Водокрасовые -- Hydrocharitaceae Aschers. // Флора СССР. Т. 1. С. 293--298.
Финк Э. 1854. Список растениям, дикорастущим и оклиматизованным на возвышенности меж-
ду источниками рек: Волги, Днепра, Западной Двины и Ладожских вод. -- Санкт-Петербург.
Фишер Ф.Б. Index plantarum anno 1824 in Horto botanico Imperiali Petropolitano vigentium.
-
Petropoli.
Флёров А.Ф. Флора Владимирской губернии // Тр. О-ва естествоиспытателей при Юрьев. ун-те.
-- Юрьев, 1902. -- Т. 10. -- С. 3--338.
Флора Липецкой области / К.И. Александрова, М.В. Казакова, В.С. Новиков и др. -- М.: Аргус,
1996. -- 376 с.
Флора Нижнего Поволжья.
-- Т. 1: Споровые, голосеменные, однодольные / Ю.Е. Алексеев,
Г.Ю. Клинкова, А.П. Лактионов и др.; Под общ. ред. А.К. Скворцова. -- М.: Т-во науч. изд.
КМК, 2006. -- 435 с.
Флора российского Дальнего Востока: Дополнения и изменения к изданию «Сосудистые рас-
тения советского Дальнего Востока». Т. 1--8 (1985--1996) / Отв. ред. А.Е. Кожевников и
Н.С. Пробатова. -- Владивосток: Дальнаука, 2006. -- 456 с.
Флора северо-востока европейской части СССР: В 4 т.
-- Т 1. Семейства Polypodiaceae --
Gramineae. -- Л.: Наука, 1974. -- 275 с.
Флора Сибири, 1988, с сайта: http://skazka.nsk.ru/atlas/id.3731
Флора Средней России: Аннотированная библиография / В.Н. Тихомиров, И.А. Губанов, И.М.
Калиниченко, Р.А. Лозарь; Под ред. В.Н. Тихомирова. -- М.: Рус. университет, 1998. -- 199 с.
Флора СССР. Т. IX. Москва-Ленинград: Изд-во АН СССР, 1939, С. 313.
461
Флора Чехии...
Флора Чили, [Electronic resource.] Mode of access: http://www.chileflora.com/Florachilena/ Flora-
Russian/ HighResPages/RH0549.htm. Accessed 15 November 2008.
Флоровская Е.Ф. 1961. Семейство Кипрейные // Флора Ленинградской области. Изд-во Ленин-
градского ун-та. -- Т. 3. С. 201--210.
Фомина З.В. 1965. О новых видах сорных растений в Улан-Удэнском районе Бурятской АССР
// Тр. Бурятского СХИ. 18 с.
Фортунатов А. 1826. Исчисление растений, дикорастущих в Вологодском уезде //Новый Ма-
газин Естественной Истории Двигубскаго. -- Ч. 3. № 3. С. 207--215.
Хлызова Н.Ю., Агафонов В.А. 2003. Особенности новейшего этапа формирования адвентивно-
го компонента флоры Воронежской области // Проблемы изучения адвентивной и синан-
тропной флоры в регионах СНГ / Мат-лы науч. конф. -- М.; Тула. -- С. 115--116.
Хмелев К.Ф., Березуцкий М.А. 2001. Состояние и тенденции развития флоры антропогенно-
трансформированных экосистем // Журн. Общ. Биол. -- Т. 62. - № 4. -- С. 339--351.
Хорошков А.А. 1925. Материалы для флоры Иваново-Вознесенской губернии // Бюлл. МОИП.
Отд. биол. -- Т. 33. Вып. 3--4. С. 244--257.
Хорун Л.В. 1999. Адвентивная флора Тульской области / Дисс... канд. биол. наук. - Москва,
368 с.
Хорун Л.В., Шереметьева И.С., Щербаков А.В. 1995. Некоторые новые данные о распростра-
нении адвентивных растений в Тульской области // Бюлл. МОИП. Отд. биол. -- Т. 100. Вып.
5. С. 90--93.
Хромосомные числа цветковых растений. 1969. Л.: Наука. 926 с.
Хртек И., Шоуркова М. 1977. Дополнение к флоре западных областей Украинской ССР // Бо-
тан. журн. Т. 62. № 1. С. 90--92.
Цвелев Н.Н. 1972. Род Festuca в СССР // Новости сист. высш. раст. Т. 9. С. 15--46.
Цвелев Н.Н. 1974. Poaceae Barnh. (Gramineae Juss., nom. altern.) -- Злаки // Флора европ. ч.
СССР. Т. 1. С. 117--368.
Цвелев Н.Н. 1976. Злаки СССР. М.: Наука. -- 788 с.
Цвелев Н.Н. 1976. Злаки СССР. Ленинград: Наука. -- 788 с.
Цвелев Н.Н. 1982. Семейство Водокрасовые (Hydrocharitaceae) // Жизнь растений. В 6 т. Т. 6.
С. 17--24.
Цвелев Н.Н. 1988. Флора Хоперского государственного заповедника. Л.: Наука. 191 с.
Цвелев Н.Н. 1989. Гречиховые -- Polygonaceae Juss. // Флора советского Дальнего Востока. Т. 4.
С. 25--122.
Цвелев Н.Н. 1994. Astereae Cass. // Флора европ. ч. СССР. Т. 7. С. 174--206.
Цвелев Н.Н. 1996. Polygonaceae Juss. -- Гречишные [excl. Rumex] // Флора Вост. Европы. Т. 9. С.
98--101, 119--157.
Цвелев Н.Н. 2000. Определитель сосудистых растений Северо-Западной России (Ленинградская,
Псковская и Новгородская области). - СПб.: Изд-во СПХФА. 781 с.
Цвелев Н.Н. 2001. Amelanchier Medik. // Флора Восточной Европы. СПб.: Мир и семья. Т. 10. С.
552--555.
Цвелев Н.Н. 2002. К систематике семейства Oleaceae в Восточной Европе // Новости система-
тики высших растений. Т. 34. С. 138--136.
Цвелев Н.Н. 2003. О родах Elaeagnus и Hippophaë (Elaeagnaceae) в России и сопредельных го-
сударствах // Ботан. журн. Т. 87. № 11. С. 74--86.
Цвелев Н.Н. 2004. Elaeagnaceae Adans. Лоховые // Флора Восточной Европы. Т. 11. С. 478--485.
Цвелев Н.Н. 2004. Oleaceae Hoffmgg. et Link -- Маслиновые // Флора Восточной Европы. Т. 11.
С. 451--478.
Цигра И. 1842. Древесная флора. Санкт-Петербург. 926 с.
Цингер В.Я. 1885. Сборник сведений о флоре Средней России. М. Изд-во МГУ. 520 с.
Цэрэнбалжид Г. 1984. Монгол орны ургамлын аймагт зэегдмел ишнэ зуйл // Бот. хурээлэнгийн
эрдэмшинжилгээнит бутээл. БНМАУ шинжлэх ухааны акад. № 2. С. 100--104.
Чернягина О.А., Якубов В.В. Аннотированный список флоры Тигильского района Корякского А.О. /
«Труды» КФ ТИГ ДВО РАН. Вып. V. Материалы к флоре северо-западной Камчатки, [Electronic
resource.] Mode of access: http://www.terrakamchatka.org/ publications/trudy/trudy5/14.htm. Ac-
cessed 15 May 2008.
462
Черняев В. 1836. О произведениях растительного царства Курской губернии // Журн. Мин-ва
внут. дел. Ч. 22. № 12. С. 505--514.
Чертопруд Е.С., Соколов Д.Д. 2001. Poa supina Schrad. (Gramineae) в окрестностях Звениго-
родской биостанции МГУ // Роль биостанций в сохранении биоразнообразия России. Ма-
териалы конф., посвящ. 250-летию МГУ им. М.В. Ломоносова и 90-летию Звенигородской
биологической станции им. С.Н. Скадовского. М. С. 182--183.
Четверня С.А. 1987. Биологические особенности и сравнительная оценка действующих ве-
ществ ромашки аптечной и ромашки безъязычковой // Автореф. дисс. ... канд. биол. наук.
Киев. 17 с.
Чешмеджиев И.В. 1988. Нови материали и хронологичны дани за флората на Българиа // Фи-
тология. Т.35. С. 58--63.
Чичев А.В. 1985. Адвентивная флора железных дорог Московской области / Автореф. дисс. ...
канд. биол. наук. М. 24 с.
Чуб В.В. 2008. Многолетники для ландшафтного сада. М.: Эксмо. 576 с.
Чужеродные виды на территории России»: [Electronic resource.] Mode of access: http://
www.sevin.ru/ invasive/invasion/plants/ragweed.html. Accessed 10 Nov. 2008.
Чужеродные виды северо-запада России, [Electronic resource.] Mode of access: http://
www.zin.ru/projects/invasions/rus/plants/nw_list.htm. Accessed 21 June 2008.
Чухно Т. 2007. Лекарственные растения, иллюстрированный словарь. М., «Эксмо», 768 с.
Шанхиза Е.В. Инвазия узкотелой златки Agrilus planipennis в Московском регионе //
http://www.zin.ru/ANIMALIA/COLEOPTERA/rus/fraxxx.htm, 2007.
Шафер В. 1956. Основы общей географии растений. -- М.: Изд-во иностр. лит-ры, -- 140 с.
Швецов А.Н. 2008. Кандидатская диссертация.
Шелль Ю. 1885. Материалы для ботанической географии Уфимской и Оренбургской губерний
цветковых растений // Тр. об-ва естествоиспытателей при Казанском ун-те. Т. 12. №4-5.
122 с.
Шереметьева И.С. 1989. Находки адвентивных видов в Тульской области // Проблемы изуче-
ния адвентивной флоры СССР (Материалы совещ. 1-3 февр. 1989 г.). -- М., Наука, С. 38--39.
Шестериков П.С. 1912. Определитель растений окрестностей Одессы. -- Одесса: Сапожников.--
120 с.
Шилов М.П. 1989. Местная флора: Учеб. пособие. -- Иваново: Иван. гос. ун-т. -- 96 с.
Шилов М.П., Силаева Т.Б., Борисова Е.А. 1990. Новые и редкие для флоры Ивановского Повол-
жья растения // Проблемы изучения Плеса / Мат-лы III науч. конф. -- Плес. -- С. 28--29.
Шилов М.П., Шилова Т.Н., Богданова М.А. 1989. Новые заносные растения во флоре Ивановской
области // Проблемы изучения адвентивной флоры СССР. - М.: Наука. - С. 28-29.
Шилова Т.Н. 1984. О флоре г. Иваново // Состояние и перспективы исследования флоры сред-
ней полосы... - М. МГУ. - С. 24-26.
Шишкин Б. К. 1961. Крестовник -- Senecio L. // Флора СССР. -- Т. 26. С. 699--788.
Шлотгауэр С.Д., Небайкин В.Д. 1984. К познанию адвентивной флоры южной части Хабаров-
ского края // Бюлл. Гл. ботан. сада. - Вып. 133. - С.42-45.
Шляков Р.Н. 1959. Семейство Кипрейные // Флора Мурманской области. -- М.-Л., АН СССР. Т.
4. С. 211--225.
Шмальгаузен И. 1886. Флора юго-западной России. -- Киев. 783 с.
Шмальгаузен И.Ф. 1895. Флора средней и южной России, Крыма и Северного Кавказа. Руко-
водство для определения семенных и высших споровых растений. -- Т. 1. Киев: т-во И.Н.
Кушнеров и Ко. XXXI. 468 с.
Шмальгаузен И.Ф. 1897. Флора средней и южной России, Крыма и Северного Кавказа. Руко-
водство для определения семенных и высших споровых растений. Т. 2. Киев. XXX. 752 с.
Шредер Р.И. 1896. Список зимующих в открытом грунте древесных пород в средних и отчасти
в северных губерниях. -- Санкт-Петербург. 256 с.
Шредер Р.И. 1899. Указатель растений дендрологического сада Московского сельскохозяйст-
венного института. М: Товарищество И.Н.Кушнерев.
Штейнберг Е.И. 1949. Семейство Кипрейные -- Onagraceae Lindl. // Флора СССР. М.-Л., АН
СССР. Т. 15. С. 565--637.
Щербаков А.В. 1990. Конспект флоры водоемов Московской области // Флористич. исслед. в
Моск. обл. -- М., -- С. 106--120.
463
Щербаков А.В. 1999. Атлас флоры водоемов Тульской области. -- М.: Изд-во Рус. университет, 45 с.
Щербаков А.В. 2001. Некоторые размышления по поводу анализа региональных флор // Фло-
ристич. исслед. в Центральной России на рубеже веков / Мат-лы научн. совещ. -Рязань.
С. 165--166.
Щербаков А.В. 2003. Изучение и анализ региональных флор водоемов // Гидроботаника: мето-
дология, методы / Мат-лы школы по гидроботанике. -- Рыбинск. С. 56--69.
Щербаков А.В. 2006. Региональная водная флора как модельный объект для флористического
анализа // Мат-лы VI Всерос. школы-конференции по водным макрофитам «Гидроботаника --
2005». пос. Борок, 11--16 октября 2005 г. -- Рыбинск. С. 34--48.
Щербаков А.В. 2008. Влияние изученности адвентивной флоры на систематический спектр //
Мат-лы междунар. научн. конф. «Биоразнообразие: проблемы и перспективы сохранения».
Пенза. Ч. 1. С. 351--353.
Щербацевич В.Д. 1973. Фенологические группы североамериканских лиственных деревьев и
кустарников // Опыт интродукции древесных растений (Сборник науч. статей). М.: ГБС АН
СССР. С. 125--152.
Эбель А. Л. 2001. О двух видах рода Lepidium L. (секция Dileptium DC.) во флоре Сибири // Ис-
следования молодых ботаников Сибири / Тезисы докладов (20--22 февраля 2001). - Новоси-
бирск, ЦСБС СО РАН). [Электронный ресурс] http://www.sbras.ru/ws/ show_abstract.
dhtml?ru+ 18+1285. Дата доступа 23.04.2009.
Эбель А. Л. 2003. О распространении некоторых эукенофитов на юге Сибири // Проблемы изучения
адвентивной и синантропной флоры в регионах СНГ / Мат-лы науч. конф. С. 128--130.
Эйхе Э.П. 1957. Топинамбур или земляная груша: (основы возделывания и нар.-хоз. значение).
М., Л.: Изд. АН СССР. 190 с.
Элтон Ч.С. 1960. Экология нашествий животных и растений. -- М.: Изд-во иностр. лит-ры. --
231 с.
Энциклопедия декоративных садовых растений.
-- Электронный ресурс. Режим доступа:
http://flower.onego.ru/voda/elodea/html. -- Acc. 17 Dec. 2008.
Энциклопедия декоративных садовых растений» , [Electronic resource.] Mode of access:
http://flower.onego.ru/kustar/sorbaria.html. Accessed 9 Feb. 2008.
Энциклопедия растений Сибири, [Electronic resource.] Mode of access: http://skazka.nsk.ru/atlas/
id.4100. Accessed 20 Oct. 2008.
Юкневичене О-Г.К. 1969. Некоторые вопросы изучения биологии аира и вахты трехлистной в
Литовской ССР. Автореф. дисс. ... канд. биол. наук. -- Вильнюс. 28 с.
Юкневичене О-Г.К. 1971. Рост и развитие аира // Тр. АН ЛитССР, сер.В. Биол. науки. Т. 3.
С. 35--41.
Юрцев Б.А., Камелин Р.В. 1991. Основные понятия и термины флористики / Учебное пособие
по спецкурсу. -- Пермь: Изд. Пермск. ун-та. 81 с.
Яброва-Колаковская В.С. 1977. Адвентивная флора Абхазии. -- Тбилиси: Мецниереба. 64 с.
Ярмоленко А.В. 1934. Cruciferae Juss. -- Крестоцветные // Сорные растения СССР. Т. 3. С. 107.
Abbas H.K., Pantone D.J., Paul R.N. 1999. Characteristics of multiple-seeded cocklebur: a biotype of
common cocklebur (Xanthium strumarium L.) // Weed Technology. - Vol. 13. - Is. 2. - P. 257--263.
Abbott R. J. 1992. Plant invasions, interspecific hybridization and the evolution of new plant taxa //
Trends Ecol. Evol. - Vol. 7. - Is. 12. - P. 401--405.
Abbott R.J., Ashton P.A., Forbes. D.G. 1992. Introgressive origin of the radiate groundsel, Senecio
vulgaris L. var. hibernicus Syme: Aat-3 evidence // Heredity. -- Vol. 68. P. 425--435.
Abbott R.J., James J.K., Milne R.I., Gillies A.C. M. 2003. Plant introductions, hybridization and gene
flow // Phil. Trans. R. Soc. Lond. B. - Vol. 358. P. 1123--1132.
Abrahamson W.G., Eubanks M.D., Blair C.P., Whipple A.V. 2001. Gall flies, inquilines, and goldenrods: a
model for host-race formation and sympatric speciation // Am. Zool. - Vol. 41. - P. 928--938.
Abrahamson W.G., McCrea K.D., Anderson S.S. 1989. Host preference and recognition by the gold-
enrod ball gallmaker Eurosta solidaginis (Diptera: Tephritidae) // Amer. Midland Natur. - Vol.
121. - № 2. - P. 322--330.
Abrahamson W.G., McCrea K.D., Whitwell A.J., Vernieri L.A. 1991. The role of phenolics in golden-
rod ball gall resistance and formation // Biochem. Syst. Ecol. - Vol.19. - P. 615--622.
Abrahamson W.G., Weis A.E. 1997. Evolutionary ecology across three trophic levels: goldenrods,
gallmakers and natural enemies. - Princeton: Univ. Press. - 456 p.
464
Achinelli F.G., Delucchi G. 2000. El "fresno americano" naturalizado en la Provincia de Buenos
Aires (Argentina): Fraxinus pennsylvanica Marshall (Oleaceae) // Rev. Mus. La Plata, Sect. Bot. -
Vol. 14. - №. 110-113. - P. 477--482.
Adachi N., Terashima I., Takahashi M. 1996. Central die-back of monoclonal stands of Reynoutria
japonica in an early stage of primary succession on mount Fuji // Annals of Botany. - Vol. 77. - P.
477--486.
Aellen P. 1972. Amaranthaceae // Flora Iranica. - Wien: Akad. Druck-u.Verlags-anstalt; - Vol. 91. -
P.1--19.
Aiken S.G., Dallwitz M.J., McJannet C.L., Consaul L.L. 1996 onwards. Festuca of North America:
descriptions, illustrations, identification, and information retrieval. Version: 19th October 2005 //
http://delta-intkey.com/festuca/
Aiken S.G., Picard R.R. 1980. The influence og substrate on the growth and morphology of
Myriophyllum exalbescens and M. spicatum // Can. J. Bot. - Vol.58. - №. 9. - Р. 1111--1118.
Aim T. 2002. Kjempespringfra (Impatiens glandulifera) - en fremmed art i rask spredning i Nord-
Norge // Polarflokken. - Tromso. - Bd 26. - № 2. - S. 123--131.
Akeroyd J.R., Rich T. C.G. 1993. Lepidium L. // Flora Europaea. 2nd ed. - Vol. 1. - P. 398--402.
Akeroyd J.R., Warwick S.I., Briggs D. 1978. Variation in four populations of Senecio viscosus L. as
revealed by a cultivation experiment // New Phytologist. - Vol. 81. - №. 2. - P. 391--400.
Aksoy A., Şahin U. 1999. Elaeagnus angustifolia L. as a biomonitor of heavy metal pollution // Tr. J.
of Botany. - Vol. 23. - P. 83--87.
Albarouki E., Peterson A., 2007. Molecular and morphological characterization of Crataegus L.
species (Rosaceae) in Southern Syria / Botanical Journalof the Linnean Society. - Vol. 153, -
Issue 3. - P. 255-263.
Alien plants in Korea [Electronic resource.] Mode of access: http://alienplant.nier.go.kr/eng/html/
search_detail.isp?id=47. Accessed 25 Mar. 2008.
Alien species in Poland", [Electronic resource.] Mode of access: http://www.iop.krakow.pl/ids/
species.asp?108. Accessed 14 June 2008.
Allard R.W. 1965. Genetic systems associated with colonizing ability in predominatly self-pollinated
species // Genetics of colonizing species. - New-York-London: Hafner publishing company. - P.
49-75.
Allard R.W. 1975. The mating system and microevolution // Genetica. - Vol. 79. -- Suppl. -P. 115--
126.
Allelopathy: basic and applied aspects. 1992. / Ed. V.Rizvi - Springer. - 480 p.
Al-Shehbaz I. A., Mummenhoff K., Appel O. 2002. Cardaria, Coronopus, and Stroganowia are
united with Lepidium (Brassicaceae) // Novon: A Journal for Botanical Nomenclature. - Vol. 12. -
№1. -P.5--11.
Ammal E.K. et al. 1964. The interrelationship between polyploidy, altitude and chemical composition
in Acorus calamus // Current Sci. - Vol. 33. - № 16. - Р.500.
An Act Concerning Revisions to the Invasive Plant Provisions, [Electronic resource.] Mode of access:
http://www.cga.ct.gov/2005/fc/2005SB-00590-R000154-FC.htm. Accessed 21 Oct. 2008.
An inventory of alien species and their threat to biodiversity and economy in Switzerland / Ed. by
R.Wittenberg. 2005. CABI Bioscience Switzerland Centre report to the Swiss Agency for
Environment, Forests and Landscape. - 416 p.
Anderson L.C. 1984. Noteworthy plants from North Florida // Sida Contrib. Bot. - Vol. 10. - № 4. -
P.295-297.
Anderson W. P. 1999. Perennial Weeds: Characteristics and identification of selected herbaceous
species. - Las Cruces: New Mexico State Univ. - 244 p.
Anderson J.K. 1995. Systematic implications of isozyme number variation on tribe Brassiceae
(Brassicaceae). Master's Thesis, University of Ottawa, Ottawa, ON. 139 p.
Andrews M., Maule H.G., Raven J.A., Mistry A. 2005. Extension Growth of Impatiens glandulifera at
low irradiance: importance of nitrate and potassium accumulation // Annals of Botany. - Vol. 95. -
№ 4. - Р.641-648.
Anjen L., Park Ch.-W. 2003a. Fallopia Adanson // Flora of China. - Vol. 5. - P. 315--318.
Anjen L., Park Ch.-W. 2003b. Reynoutria Houttuyn // Flora of China. - Vol. 5. - P. 319.
Anzalone B. 1957. Nuove localita italiane di Bidens frondosa L. // Nuovo qiorn. bot. ital. ns. - Vol.
64. - № 4. - P.738-739.
465
Apati P., Kristo T.Z., Szoke E., Kery A., Szentmihalyi K., Vinkler P. 2003. Comprehensive evaluation of
different Solidaginis herba extracts // Acta Hort. (ISHS) 597. - P. 69-73.
Asante A., Hashidoko Y., Deora A., Tahara S. 2008. Antagonistic Gluconobacter sp. induces
abnormal morphodifferentiation to Fusarium oxysporum f. sp. lycopersici hyphae // Journal of
Pesticide Science. - Vol. 33. - № 2. - P. 138--145.
Ash G.M. 1953. Epilobium adenocaulon in Britain // The changing flora of Britain. Arbroath.
-
P.168--170.
Atlas Florae Europaeae. Distribution of Vascular Plants in Europe. 4. Polygonaceae. 1979. /Eds.
J.Jalas, J.Suominen // Helsinki: The Committee for Mapping the Flora of Europe & Societas
Biologica Fennica Vanamo. - 71 p.
Auld B. A., Medd R. W. 1992. Weeds. An illustrated botanical guide to the weeds of Australia.
Melbourne: Inkata Press. - 255 p.
Auld B., Morita H., Nishida T., Ito M., Michael P. 2003. Shared exotica: Plant invasions of Japan and
south eastern Australia // Cunninghamia. - Vol. 8. - №. 1. - P. 147--152.
Azuma Shizuka, Nakashima Atsushi, Yamada Hiroyuki, Yabu Shinobu. 2001. A Study of
Conservation of the Growing Condition of Japanese Native Fragrance Herbs: Mentha arvensis
var. piperascens and Elsholtzia ciliata / Environmental Systems Research. - Vol. 29. - P. 179-184.
Badger K. S., Ungar I. A. 1989. The effects of salinity and temperature on the germination of the
inland halophyte Hordeum jubatum // Can. J. Bot. - Vol. 67. - №. 5. - P. 1420--1425.
Badger K. S., Ungar I. A. 1991. Life history and population dynamics of Hordeum jubatum along a
soil salinity gradient // Can. J. Bot. - Vol. 69. - №. 2. - P. 384--393.
Badger K. S., Ungar I. A. 1994. Seed bank dynamics in an inland salt marsh, with special emphasis
on the halophyte Hordeum jubatum L. // International Journal of Plant Sciences. - Vol. 155. - №.
1. - P. 66--72.
Bailey J. P., Bímová K., Mandák B. 2007. The potential role of polyploidy and hybridisation in the further
evolution of the highly invasive Fallopia taxa in Europe // Ecol. Res. - Vol. 22. - P. 920--928.
Bailey J. P., Child L. E., Conolly A. P. 1996. A survey of the distribution of Fallopia × bohemica
(Chrtek and Chrtkova) J. Bailey (Polygonaceae) in the British Isles // Watsonia. - Vol. 21. - P.
187--198.
Bailey J. P., Conolly A. P. 2001. Prize-winners to pariahs -- a history of japanese knotweed s. l.
(Polygonaceae ) in the British Isles // Watsonia. - Vol. 23. - P. 93--110.
Bailey J. P., Stace C. A. 1992. Chromosome number, morphology, pairing, and DNA values of
species and hybrids in the genus Fallopia (Polygonaceae) // Pl. Syst. Evol. - Vol. 180. - P. 29--52.
Bailey J., Wisskirchen R. 2004. The distribution and origins of Fallopia × bohemica (Polygonaceae)
in Europe // Nordic Journal of Botany. - Vol. 24. - Is. 2. - P. 173--199.
Bais H.P., Vepachedu R., Gilroy S., Callaway R.M., Vivanco J.M. 2003. Allelopathy and exotic plants:
from genes to invasion // Science. - Vol. 301. - Is. 5638. - P. 1377--1380.
Baker H.G. 1959. The contribution of autecological end genecological studies to our knowledge of
the past migrations of plants // Amer. Natur. - Vol. 93. - № 871. - P. 255--272.
Baker H.G. 1974. The evolution of weeds // Ann. Rev. Ecol. Syst. - Vol. 5. - P. 1--24.
Bakker J. D., Wilson S. D. 2004. Using ecological restoration to constrain biological invasion //
Journal of Applied Ecology. - Vol. 41. - P. 1058--1064.
Baldoni G., Viggiani P., Bonetti A., Dinelli G., Catizone G. 2000. Classification of Italian of
Xanthium strumarium complex based on biological triats, electrophoretic analysis and response to
maize interference // Weed Research. - Vol. 40. - P. 191--204.
Ball P. W. 1993. Cardaria Desv. // Flora Europaea. 2nd ed. - Vol. 1. - P. 68.
Balogh L., Dancza I., Király G. 2004. Actual list of neophytes in Hungary and their classification
according to their success // Biological invasions in Hungary. Invasive plants. / Ed. B.Mihály,
Z.Botta-Dukát - Budapest: TermészetBÚVÁR Alapítvány Kiadó. - P. 61--92.
Banaszek E. 2005. Problemy pielejnacyjne wynikajace z cech wzrostu i reprodukcji osobnikow klonu
jesionolistnego Acer negundo L. na wybranych typach obiektow architektury krajobrazu w
Wolominie w roku 2004. /Diploma Thesis at the Faculty of Architecture. Univ. of Ecology and
Management in Warsaw.
Baranchikov Y., Mozolevskaya E., Yurchenko G., Kenis M. 2008. Occurrence of the emerald ash
borer, Agrilus planipennis in Russia and its potential impact on European forestry // EPPO
Bulletin. - Vol. 38. - № 2. - P. 233--238.
466
Baranov A. 1960. On the case of sprouting of the seeds of Acorus calamus in North Manchuria //
Phyton. - Vol. 9, - № 1-2. - P. 21-23.
Barkley Th. M. 2006. Senecio // Flora of North America. - Vol. 20. - P. 544--570.
Barkworth M.E. 1973. Impatiens parviflora in British Columbia // Madrono. - Vol. 22. - №1. - P. 24.
Barney J. 2006. North american history of two invasive plant species: phytogeographic distribution,
dispersal vectors, and multiple introductions // Biological Invasions. - Vol. 8. - № 4. - P. 703--717.
Barney J. N., Tharayil N., DiTommaso A., Bhowmik P. C. 2006. The Biology of Invasive Alien
Plants in Canada. 5. Polygonum cuspidatum Sieb. & Zucc. [= Fallopia japonica (Houtt.) Ronse
Decr.] // Can. J. Plant Sci. - Vol. 86. - P. 887--905.
Barrat-Segretain M.-H, Bornette G., Hering-Vilas-Bôas A. 1998. Comparative abilities of vegetative
regeneration among aquatic plants growing in disturbed habitats// Aquat. Bot. Vol. 60, № 3.
P. 201--211.
Bartlett E., Novak S.J., Mack R.N. 2002. Genetic variation in Bromus tectorum (Poaceae):
differentiation in the eastern United States // Amer.J.Bot. - Vol. 89. - № 4. - Р.602-612.
Bass D.A. 1990. Dispersal of an introduced shrub (Crataegus monogyna) by the Brush-tailed Possum
(Trichosurus vulpecula) / Austral Ecology. - Vol. 15. - Issue 2. - P. 227-229.
Baum B.R., Bailey L.G. 1994. Taxonomy of Hordeum caespitosum, H. jubatum, and H. lechleri
(Poaceae: Triticeae) // Plant Syst. Evol. - Vol. 190. - Is. 1. - P. 97--111.
Bazzaz F.A. 1996. Plants in changing environments: linking physiological, population, and
community ecology. -- Cambridge: Univ. Press. - 320 p.
Beerling D. J., Bailey J. P., Conolly A. P. 1994. Biological Flora of the British Isles. № 183 Fallopia
japonica // J. Ecol. - Vol. 82. - № 4. - P. 959--979.
Beerling D. J., Huntley B., Bailey J. P. 1995. Climate and the distribution of Fallopia japonica: use of
an introduced species to test the predictive capacity of response surfaces // Journal of Vegetation
Science. - Vol. 6. - № 2. - P. 269--282.
Beerling D.J., Perrins J.M. 1993. Impatiens glandulifera Royle (Impatiens roylei Walp.) // The
Journal of Ecology. - Vol. 81. - № 2. - Р. 367-382.
Bennett A. 1903. Acorus calamus in England // Jour.of Bot. - Vol. 41. - P. 23-24.
Bennett M. 2003. Pulmonarias and the Borage Family. - Portland: Timber Press. - 240 p.
Berges R., Cousin M.-T., Roux J., Mäurer R., Seemüller E. 1997. Detection of phytoplasma
infections in declining Populus nigra "Italica" trees and molecular differentiation of the aster
yellows phytoplasmas identified in various Populus species / European Journal of Forest
Pathology. - Vol. 27. - Issue 1. - P. 33-43.
Bernard F.G. 1968. Forme blanche d'epilobium adenocaulon // Naturaliste Canad. - Vol. 95. - № 1. -
P.225.
Besser W. 1816. Catalogus plantarum in Horto botanico gymnasii volhyniensis cremeneci cultarum. -
Cremeneci.
Bigwood D. W., Inouye D. W. 1988. Spatial pattern analysis of seed banks: an improved method and
optimized sampling // Ecology. - Vol. 69. - № 2. - P. 497--507.
Bijok K., Krenska B., Pawlak T. 1972. Karyological investigation on Erigeron canadensis and
E.annuus from natural stands in Poland // Acta Soc.bot.Polon. -- Bd 41. - № 3. - S. 433-438.
Bímová K., Mandák B., Kašparová I. 2004. How does Reynoutria invasion fit the various theories of
invasibility? // Journal of Vegetation Science. - Vol. 15. - Is. 4. - P. 495--504.
Bímová K., Mandák B., Pyšek P. 2003. Experemental study of vegetative regenaration in four
invasive Reynoutria taxa (Polygonaceae) // Plant Ecology. - Vol. 166. - P. 1--11.
Binimelis R., Born W., Monterroso I., Rodríguez-Lapajos B. 2007. Socio-economic impacts and
assessment of biological invasions // Biological invasions. Ed. W.Nentwig - Berlin: Springer. - P.
331--347. (Ecological Studies. Vol. 193).
Biodiversity Portrait of the St. Lawrence [Electronic resource.] Mode of access: http://
www.gc.ec.gc.ca/faune/biodiv/en/recherche/especes/map.asp?T=PV&id=652. Accessed 2 April,
2008.
Biological invasions in Europe and the Mediterranean basin. 1990. Ed. E.Di Castri, A.J.Hansen,
M.Debussche - Dordrecht (Netherlands): Kluwer Academic. - ix, - 447 p.
Blackshaw R. E., Semach G., Entz T. 1998. Postemergence control of foxtail barley (Hordeum
jubatum) seedlings in spring wheat (Triticum aestivum) and flax (Linum usitatissimum) // Weed
Technology. - Vol. 12. - Is. 4. - P. 610--616.
467
Blackshaw R. E., Semach G., Xiangju Li, O'donovan J. T., Harker K. N. 1999. An integrated weed
management approach to managing foxtail barley (Hordeum jubatum) in conservation tillage
systems // Weed technology. - Vol. 13. - № 2. - P. 347--353.
Blais P. A., Lechowicz M. J. 1989. Variation among populations of Xanthium strumarium L.
(Compositae) from natural and ruderal habitats // Amer. J. Bot. - Vol. 76. - № 6. - P. 901--908.
Blake S.F. 1925. Records of Bidens frondosa var. anomala Porter // Rhodora. - Vol. 27. - P.34-35.
Bleeker W. 2003. Hybridization and Rorippa austriaca invasion in Germany // Mol. Ecol. -Vol. 12. -
Is. 7. - P. 1831--1841.
Bleeker W., Matthies A. 2005. Hybrid zones between invasive Rorippa austriaca and native R.
sylvestris (Brassicaceae) in Germany: ploidy levels and patterns of fitness in the field // Heredity.
- Vol. 94. - P. 664--670.
Bleeker W., Schmitz U., Ristow M. 2007. Interspecific hybridisation between alien and native plant
species in Germany and its consequences for native biodiversity // Biol. Conserv. - Vol. 137. - Is.
2. - P. 248--253.
Boissier E. 1879. Amaranthus L. // Flora orientalis. - Genevae: Bibliopolam. - Vol. 4. - P.990-991.
Bon M.-C., Hurard C., Gaskin J., Risterucci A.-M. 2005. Polymorphic microsatellite markers in
polyploid Lepidium draba L. ssp. draba (Brassicaceae) and cross-species amplification in closely
related taxa // Molecular Ecology Notes. - Vol. 5. - № 1. - P. 68--70.
Bonar S.A., Thomas G.L., Pauley G.B. 1987. Estimation of triploid White Amur stocking densities
for aquatic plant control for Devils Lake, Oregon // Proc. 21st Ann. Meeting Aquatic Plant Control
Res. Progr. -- Vicksburg (Mississippi). -- P. 122--132.
Borchardt W. 2008. PflanzenLexikon fur ausbildung und berif. Garten- und landschaftsbau,
landschaftsarchitektur. Lernhilfe Wissensspeicher. - 319 p.
Botta-Dukát Z., Dancza I. 2001a. Effect of weather conditions on the growth of Solidago gigantea //
In: Brundu, G., etal. (Eds.), Plant Invasions: Species Ecology and Ecosystem Management.
--
Leiden: Backhuys Publ. - P. 185--197.
Botta-Dukát Z., Dancza I. 2001b. Morphological plasticity in the rhizome system of Solidago
gigantea: comparison of wet and dry habitats // Sixth International Conference on the Ecology
and Management of Invasive Plants, Emapi, Loughborough, UK.
Bowers K.L., Pauley G.B., Thomas G.L. 1987. Feeding preference on Pacific Northwest aquatic plant
species by diploid and triploid Grass Carp (Ctenopharyngodon idella) // Proc. 21st Ann. Meeting
Aquatic Plant Control Res. Progr. -- Vicksburg (Mississippi), -- P. 133--140.
Bowman J. L., Brüggemann H., Lee J.-Y., Mummenhoff K. 1999. Evolutionary changes in floral
structure within Lepidium L. (Brassicaceae) // Int. J. Plant Sci. - Vol. 160. - № 5. - P. 917--929.
Bowmer K.H., Sainty G.R., Smith G.H, Shaw K. 1979. Management of Elodea in Australian
irrigation systems // J. Aquat. Plant Management. ---- Vol. 17. -- P. 4--12.
Bowmer K.H., Smith G.H. 1984. Herbicides for injection into flowing water: acrolein and endothal-
amin // Weed Res. Vol. 24, № 3. P. 201--211.
Boyce S.G. 1954. The salt spray community // Ecological monographs. - Vol. 24. - №1. - P. 29--67.
Brabec J., Pyšek P. 2000. Establishment and survival of three invasive taxa of the genus Reynoutria
(Polygonaceae) in mesic mown meadows: a field experimental study // Folia Geobotanica - Vol.
35. - P. 27--42.
Bram M. R., McNair J. N. 2004. Seed germinability and its seasonal onset of Japanese knotweed
(Polygonum cuspidatum) // Weed Science. - Vol. 52. - P. 759--767.
Briggs D., Block M., Jennings S. 2006. The possibility of determining the age of colonies of clonally
propagating herbaceous species from historic records: the case of Aster novi-belgii L. (first
recorded as A. salignus Willd.) at Wicken fen nature reserve, Cambridgeshire, England // New
Phytologist. - Vol. 112. - № 4. - P. 577--584.
Britten J. 1900. Impatiens Roylei in England // J. Bot. - № 38. - P. 50--51.
Britton J. 1901. Acorus in Cheshire // J. Bot. - Vol. 39. - P. 38.
Britton N., Brown A. 1896. Acorus // An illustrated flora of Northern United States, Canada and
British possessions. - Vol. 1. - P.364.
Brooks R. E., Clemants S. E. 2000. Juncaceae // Flora of North America. - Vol. 22. - P. 211--267.
Brouillet L., Semple J.C., Allen G.A., Chambers K., Sundberg S. 2006. Symphyotrichum Nees // Flora
of North America. - Vol. 20. Asteraceae/ - Part 2. Astereae and Senecioneae. - P. 465--539.
468
Brown A. 1878. Plants introduced with ballast and on made land // Bull. Torrey Bot. Club. - Vol. 6. -
P. 255--258.
Bruce J. A., Kells J. J. 1990. Horseweed (Conyza canadensis) control in no-tillage soybeans (Glycine
max) with preplant and preemergence herbicides // Weed Technol. - Vol. 4. - P.642-647.
Buchner R., Weber M., 2000. PalDat - a palynological database: Descriptions, illustrations,
identification, and information retrieval // http://www.paldat.org
Bucknall C. 1913. A revision of the genus Symphytum Tourn. // J. Linn. Soc. London (Bot.). - Vol.
41. - P. 491--556.
Bulygin N.E., Firsov G.A., 1998. Малоизвестные материалы I. Fal´ka о времени введения в
культуру некоторых древесных видов флоры России // Бот. журн. 8: 85-89.
Bumbure M. 1957. Onagraceae // Latvijas PSR flora. Riga: Latvijas valsts izdevnieciba. Т. 3. P. 333-352.
Burdon J.J., Marshall D.R., Groves R.H. 1980. Izozyme variation in Chondrilla juncea in Australia //
Austral. J. Bot. - Vol. 28. , № 2. -P. 193-198.
Burns R. M., Honkala B. H. 1990. Silvics of North America.
- Vol. 2. Hardwoods. Agriculture
Handbook 654. Washington (D. C.): U. S. Dept. of Agriculture, Forest Service. - viii, 877 p.
Byfield A. J., Baytop A. 1998. Three alien species new to the flora of Turkey // Tr. J. of Botany. -
Vol. 22. - Is. 3. - P. 205--208.
Caban M. 2005 Inwazja klonu jesionolistnego a uzytkowanie ziemi na przykladzie Piaseczna k.
Warszawy w roku 2005. / Master Thesis at the Faculty of Ecology. Univ. Ecology and
Management in Warsaw.
CABI, 2004. Solidago canadensis L. data sheet. Crop Protection Compendium, Wallingford, UK: CAB
International (www.cabcompendium.org/cpc)
Calflora, [Electronic resource.] Mode of access: http://www.calflora.org/cgi-bin/species_query.cgi?
where-calrecnum= 8429. Accessed 18 Mar. 2008.
Canne J.M. 1977. A revision of the genus Galinsoga // Rhodora. - Vol. 79. - № 819. - P. 319-389.
Canne J.M. 1983. Cytological and morphological observations in Galinsoga and related genera //
Rhodora. - Vol. 85. - № 843. - P. 355-366.
Carilli J.P., Fairbrothers D.E. 1988. Allozyme variation in three Eastern United States species of
Aeschynomene, including the rare A.virginica // Syst. Bot. - Vol. 13. - № 4. - P. 559--566.
Casler M.D., Duncan R.R. 2003. Turfgrass biology, genetics, and breeding. Hoboken (NJ): John
Wiley & Sons. 384 p.
Castro S. A., Figueroa J. A., Muñoz-Schick M., Jaksic F. M. 2005. Minimum residence time,
biogeographical origin, and life cycle as determinants of the geographical extent of naturalized
plants in continental Chile // Divers. Distribut. - Vol. 11. - Is. 3. - P. 183--191.
Chater A. O. 1972. Brunnera // Flora Europaea. - Vol. 3. - P. 106.
Cheeseman T. F. 1878. Art. LXVII.--Notice of the Occurence of Juncus tenuis, Willd., in New
Zealand // Transactions and Proceedings of the Royal Society of New Zealand.
- Vol. 11.
-
P. 433--434.
Chepil W.S. 1946. Germination of weed seeds. I. Longevity, periodicity of germination, and vitality
of seeds in cultivated soil. Sci. Agric. 26: 307-346.
Child L., Wade M. 2000. The japanese knotweed manual. Chichester (UK): Packard Publishing Ltd. -
123 p.
Chittka L., Schürkens S. 2001. Succesful invasion of a floral market: an exotic plant has moved in on
Europe's river banks by bribing pollinators // Nature. - Vol. 411. - P. 653-655.
Chmura D. 2004. Penetration and naturalisation of invasive alien plant species (neophytes) in
woodlands of the Silesian upland (Southern Poland) // Nature Conservation. - Vol. 60. - P. 3--11.
Christensen K.L.B. 1983. A biometric study of some hybridizing Crataegus populations in Denmark
// Nordic J. Bot. - Vol. 2. -- Is. 6. - P. 537-548.
Christensen K.L.B. 1984. The morphological variation of some Crataegus populations (Rosaceae) in
Greec and Yugoslavia // Nordic J. Bot. - Vol. 4. -- Is. 5. - P. 585-595.
Christensen K.L.B., Janjic N. 2004. Taxonomic notes on European taxa of Crataegus (Rosaceae). //
Nordic J. Bot. - Vol. 24. -- Is. 2. - P. 143-147.
Chrtek J. 1990. Reynoutria Houtt. -- křídlatka // Květena České republiky. - Vol. 2. - P. 362--366.
Chytrý M., Jarošík V., Pyšek P., Hájek O., Knollová I., Tichý L., Danihelka J. 2008a. Separating
habitat invasibility by alien plants from the actual level of invasion // Ecology. - Vol. 89. - № 6. -
P. 1541--1553.
469
Chytrý M., Maskell L., Pino J., Pyšek P., Vila M., Font X., Smart S. 2008b. Habitat invasions by alien
plants: a quantitative comparison between Mediterranean, subcontinental and oceanic regions of
Europe // J. Appl. Ecol. - Vol. 45. - № 2. - P. 448--458.
Chytrý M., Pyšek P., Tichý L., Knollová I., Danihelka J. 2005. Invasions by alien plants in the Czech
Republic: a quantitative assessment across habitats // Preslia. - Vol. 77. - № 4. - P. 339--354.
Cinq-Mars L. 1971. Le genre Amelanchier au Quebec // Naturaliste Canadien. - Vol. 98. - № 3. - P.
329-346.
Clark J.W., Jermy A.C. 1990. Additions to the flora of Mull and adjacent small islands // Watsonia. -
Vol. 18. - № 2. - P.179-187.
Clement E.J., Foster M.C. 1994. Alien plants of the British Isles. A provisional catalogue of vascular
plants (excluding grasses). London: Botanical Society of the British Isles. 590 p.
Cocks P.S., Boyce K.G., Kloot P.M. 1976. The Hordeum murinum complex in Australia // Austral. J.
Bot. - Vol. 24. - № 5. - P.651-662.
Conn J. S., Deck R. E. 1995. Optimal glyphosate application time for control of foxtail barley
(Hordeum jubatum) // Weed technology. - Vol. 9. - № 2. - P. 267--269.
Conolly A. P. 1977. The distribution history in the British Isles of some alien species of Polygonum
and Reynoutria // Watsonia. - Vol. 11. - P. 291--311.
Considine M.L. 1986. Finding a pattern to plant invasions // Ecos. - № 47. - P. 30--31.
Cook C.D.K., Urmi-König K. 1985. A version of the genus Elodea (Hydrocharitaceae) // Aquat. Bot.
Vol. 21. -- P. 111--156.
Cooke G.D. 1980. Lake level drawdown as a macrophyte control technique // J. of the Amer. Water
Resources Association. Vol. 16, № 2. -- P. 317--322.
Coombe D.C. 1956. Biological flora of the British Isles: Impatiens parviflora // J. Ecol. - Vol. 44. - P.701.
Cope T. A. 1982. Poaceae // Flora of Pakistan. - № 143. - P. 1--678.
Corell D.C., Corell H.B. 1975. Aquatic and Wetland Plants of South-Western United States.
--:
Stanford (California): Univ. Рress. 167 p.
Council of Europe. 2003. European strategy on invasive alien species: Final Version [Electronic
resourse] / Document prepared by Genovesi P., Shine C. // NOBASS. North European and Baltic
Network on Invasive Alien Species [web-сайт]. (14.05.2009). http://www.nobanis.org/files/
eu_strategy_inva.pdf.
Cousens R., Mortimer M. 1995. Dynamics of weed populations. -- Cambridge: Univ. Press. - 332 p.
Credaro V., Pirola A. 1987. Alcuni reperti interessanti per la flora Lombarda // Atti. Ist. bot. Lab.
Crittog. Univ. Pavia. - Vol. 6. - P.51--59.
Cronquist A. 1947. Revision of the North American species of Erigeron, north of Mexico // Brittonia.
- Vol. 6. - P. 121--302.
Cronquist A. 1976. Conyza // Florae europaea. - Cambridge. Univ.press. - Vol. 4. - P.120.
Crowe D.R., Parker W.H. 1981. Hybridization and agamospermy of Bidens in north-western Ontario
// Taxon. - Vol.30. - № 4. - P.749-760.
Csontos P. 1986. Phytosociological description of a hilly country stand of Impatiens parviflora //
Stud. bot. hung. - Vol. 19. - P. 115-118.
Cullen J.M., Moore A.D. 1983. The influence of three populations of Aceria chondrillae on three
forms of Chondrilla juncea // J. Appl. Ecol. - Vol. 20/ - № 1. - P. 235-243.
Cusick A.W. 1983. Puccinellia distans (Jacq.) Parl.: a halophytic grass rapidly spreading in Ohio.
Sida 9. - P. 360-363.
CWS (Canadian Wildlife service), [Electronic resource.] Mode of access: http://www.cws-
scf.ec.gc.ca/PUBLICATIONS/NV/14_e.cfm. Accessed 5 October 2008.
Daehler C.C. 1998. The taxonomic distribution of invasive angiosperm plants: ecological insights and
comparison to agricultural weeds // Biol. Conserv. - Vol. 84. - № 2. - P. 167--180.
Daft M. J., Nicolson T. H. 1974. Arbuscular mycorrhizas in plants colonizing coal wastes in Scotland
// New Phytologist. - Vol. 73. - № 6. - P. 1129--1138.
DAISIE. Handbook of Alien Species in Europe / Ed. by Pyšek P., Lambdon P.W., Arianoutsou M.,
Kiihn I., Pino J., Winter M. Alien Vascular Plants of Europe. Springer. 2009. xxviii, 400 p.
(Invading Nature -- Springer Series in Invasion Ecology. Vol. 3).
Dajdok Z., Aniol-Kwiatkowska A. 1998. Wstejme wyniki badan nad ekspansja, niecierpka
gruczolowatego (Impatiens glandulifera Royle) w dolinie Odry // Bad. Fizjogr. Pol Zach. - Ser. B. --
Bd 47. - S.195-204.
470
Dana E.D., Sanz-Eloiza M., Sobrino E. Plant invaders in Spain [check-list], "The unwanted citizens",
[Electronic resource.] Mode of access: http://www.ual.es/personal/edana/alienplants/checklist.pdf.
Accessed 15 June 2008.
Darbyshire S. J. Inventory of Canadian Agricultural Weeds [Electronic resource.] Mode of access:
http://dsp-psd.pwgsc.gc.ca/Collection/A42-100-2003E.pdf. Accessed 23 April, 2008.
Darbyshire S.J. 1988. Inventory of Canadian agricultural weeds. -- Ottawa.
Darke R. 1999. The color encyclopedia of ornamental grasses: sedges, rushes, restios, cat-tails, and
selected bamboos. - Timber Press. - 325 p.
Darke R. 2007. The Encyclopedia of grasses for livable landscapes - London: Timber Press. - 487 p.
Daumann E. 1967. Zur Bestaubungs- und Verbreitungsokologie dreier Impatiens-Arten // Preslia
(Praha). - Bd 39. - № 1. - S.43-58.
Davis Craig; Parsons Peter, 2002. Growing Milkweed, a plant with prospective anti-cancer properties:
a review of the medicinal potential of members of the Euphorbiaceae family / A report for the
Rural Industries Research and Development Corporation. Janiary. 16 p., http: //
www.rirdc.gov.au/reports/EOI/02-001.pdf
Davis M.A., Thompson K., Grime J.P. 2005. Invasibility: the local mechanism driving community
assembly and species diversity // Ecography. Vol. 28. P. 696--704.
Dawson Т.Е., Ehleringer J.R. 1993. Gender-specific physiology, carbon isotope discrimination, and
habitat distribution in boxelder, Acer negundo // Ecology. - Vol. 74. - P. 798--815.
De Candolle A.P. 1855. Géographie Botanique Raisonné. - Vol. 2. V. Masson. - Paris.
De Cugnac A., Camus A. 1944. Recherches phyletiques sur le genre Bromus // Bull. Soc. Bot. Fr. - №
91. - P.172-174.
De Waal L. C. 2001. A viability study of Fallopia japonica stem tissue // Weed Research. - Vol. 41. -
Is. 5. - P. 447--460.
Debray M. 1942. Bidens frondosa et Paspalum longipilum dans la valee de la Loire // Bull. Soc. bot.
France. - Vol. 89. - P. 43-44.
del Moral R., Grishin S. Yu. 1999. Volcanic Disturbances and Ecosystem Recovery // Ecosystems of
Disturbed Ground. /Ed. L. R. Walker. - P. 137--160.
Delectus seminum quae Hortus botanicus imperialis Petropolitanus. 1895. - Cпб.: Тип. В.Демакова.
--42с.
Derick B. Poindexter, 2008. Silene flos-cuculi ssp. flos-cuculi (Caryophyllaceae) and Euphorbia
peplus (Euphorbiaceae) new to North Carolina // Journal of the Botanical Research Institute of
Texas -- Vol. 2. - № 1. P. 649-650.
Dicot weed. 1988. 1 / Ed. W.Püntener. - Basel: CIBA -- GEIGY. - 335 p.
Dinelli G., Bonetti A., Viggiani P. 2003. Genetic structure and mating system of Italian Xanthium
strumarium complex // Weed Sc. - Vol. 51. - Is. 1. - P. 69--77.
Discover Life, [Electronic resource.] Mode of access: http: // www.discoverlife.org/20/q?search
=Euphorbia+peplus. Accessed 16 November 2008.
DiTomaso J.M. 2000. Invasive weeds in rangelands: species, impacts, and management // Weed Sc. -
Vol. 48. - № 2. - P. 255-265.
Doppelbaur H. 1963. Fraxinus pennsylvanica Marsh. in Bayern // Ber. Bayer. Bot. Ges. - Bd 36. -
S. 67--68.
Dreyer G.D., Baird L.M., Fickler Ch. 1987. Celastrus scandens and C. orbiculatus: comparisons of
reproductive potential between a native and an introduced woody vine // Bull. Torrey Bot. Club. -
Vol. 114. - № 3. - P. 260--264.
Du N., Pijut P. M. Genetic transformation and regeneration of green ash (Fraxinus pennsylvanica) for
resistance to the emerald ash borer. 2005. (http://www.agriculture.purdue.edu/fnr/HTIRC/
documents/ningxia_000.pdf).
Du N., Pijut P. M. Regeneration and genetic transformation of green ash for resistance tot he emerald
ash borer. 2007. (http://www.agriculture.purdue.edu/fnr/HTIRC/documents/ningxia_003.pdf).
Dubey S., Gupta K.C. 1988. Sterol composition of leaves of Erigeron canadensis // Fitoterapia.- Vol.
59. -№5. -P.428--429.
Durka W., Bossdorf O., Prati D., Auge H. 2005. Molecular evidence for multiple introductions of
garlic mustard (Alliaria petiolata, Brassicaceae) to North America // Mol. Ecol. - Vol. 14. - № 6. -
P. 1697--1706.
471
Ecology and management of giant hogweed (Heracleum mantegazzianum) 2007. / Ed. P. Pyšek , M.
J. W. Cock, H. P. Ravn, W. Nentwig Totnes (UK): CABI Publ. xvii, 324 pp.
Edgar E., 1996. Puccinellia Parl. (Gramineae: Poeae) in New Zealand / New Zealand J. Bot. -- Vol.
34. - P.17-32.
Edmonson J.R. 1980. Poa L. // Flora Europaea. Vol. 5. P. 159--167.
Edwards P. J., Frey D., Bailer H., Baltisberger M. 2006. Genetic variation in native and invasive
populations of Erigeron annuus as assessed by RAPD markers // Int. J. Plant Sci. - Vol. 167. - №
1. - P. 93--101.
Ehrendorfer F. 1973. Liste der GefaBpflanzen Mitteleuropas. - Berlin, Hamburg.
Eiten G. Taxonomy and regional variation of Oxalis section Corniculatae. I. Introduction, keys and
synopsis of the species // Amer. Midland Naturalist. - Vol. 69. - № 2 (apr. 1963). - P. 257--309.
Eklund O. 1927-28. Epilobium adenocaulon Hausskn fur Karelia australis neu // Memoranda
societatis pro fauna et flora fennica. - Bd. 4. - S.10-11.
Eliáš P. 1993. Invasive behavior of alien annuals / 2-nd International Workshop "Plant Invasions --
Theory and Applications", September 16-19, 1993. Kostelec n. Cernými lesy, Czech republic.
Abstracts. С сайта: http: //www.lboro.ac.uk/research/cens/invasives/2iceap_abstracts.htm
Ellstrand N. C., Schierenbeck K. A. 2000. Hybridization as a stimulus for the evolution of
invasiveness in plants? // PNAS. - Vol. 97. - № 13. - P. 7043--7050.
Elven R., Fremstad E. 2000. Fremmede planter i Norge. Flerârige arter av slekten lupin Lupinus L. // Blyttia. --
Bd 58. - S. 10--22.
Emig W., Kadereit J. W. 1993. The comparative biology of the closely related Senecio nebrodensis
and S. viscosus, a narrow endemic and a widespread ruderal // Nord. J. Bot. - Vol. 13. - № 4. -
P. 369--375.
Engler A. 1905. Das pflanzenreich. - Leipzig: Auftrage der Preuss. Akademie der Wissenschaften. -
S. 308-313.
Enomoto T. Weeds of Japan. 2000. http://rib.okayamau.ac.jp/wild/zassou/z_table.html
EPPO List of invasive alien plants (Европейская и Средиземноморская Организация по Защите и
Карантину Растений) http://www.eppo.org/QUARANTINE/ias_plants.htm
Erkamo V. 1946. Beobachtungen über die mit deutschen Truppen im Jahre 1944 nach Helsinki
eingeschleppten pflanzearten // Ann. Bot. Soc. Zool. Bot. Fenn. Vanamo. - Bd. 21.
Esch R.E., Hartsell C.J., Creenshaw R., Jacobson R.S. 2001. Common Allergenic Pollens, Fungi, Animals,
and Arthropods. // Clinical Reviews in Allergy and Immunology. - Vol. 21. - P. 261-292.
Esler A.E. 1988. The naturalisation of plants in urban Auckland, New Zealand. 4. The nature of the
naturalised species // New Zealand J. Bot. - Vol. 26. - № 3. - P. 345--385.
Essl F., Rabitsch W. 2002. Neobiota in Österreich. - Wien: Umweltbundesamt. - 432 p.
Europe's environment: The Dobris assessment. 1995. / Ed. D. Stanners, P.Bourdeau -Copenhagen:
Europ. Envir. Agency. - 676 p.
European Commission. 1998. A European community biodiversity strategy [Electronic resourse] //
European Commission [web-сайт]. (14.05.2009). http://ec.europa.eu/environment/docum/pdf/
com_98_42_en.pdf
European Commission. 2002. Thematic report on alien invasive species (based on the questionnaire
provided by the CBD secretariat ). 2nd report of the European Community to the Convention on
Biological Diversity [Electronic resourse] // Convention on Biological Diversity [web-сайт].
(14.05.2009). http://www.cbd.int/doc/world/eur/eur-nr-ais-en.doc
European Commission. 2004. Alien species and nature conservation in the EU. The role of the LIFE
program / R.Scalera, D.Zaghi [Electronic resourse] // European Commission [web-сайт].
(14.05.2009). http://ec.europa.eu/environment/life/publications/lifepublications/lifefocus/
documents/ alienspecies_en.pdf
Europe's environment: the second assessment 1998. / Europ. Envir. Agency. Publ. Europ. Commun.
Oxford (United Kingdom): Elsevier. - 293 pp.
Europe's environment: the third assessment 2003. / Europ. Envir. Agency. Office for Luxembourg:
Official Publ. Europ. Commun. 341 pp. (Envir. Assessment Report № 10).
Evans G.C., Hughes A.P. 1961. Plant growth and aerial environment. I. Effect of artificial shading on
Impatiens parviflora DC. // New Phytol. - Vol. 60. - № 2. - P. 150--180.
Everson D.A., Boucher D.H. 1998. Tree species-richness and topographic complexity alon the
riparian edge of the Potomac River // Forest Ecol. and Management. - Vol. 109. - P. 305-314.
472
Fenner M. 1983. Relationships between seed weight, ash content and seedling growth in twenty-four
species of Compositae // New Phytol. - Vol. 95. - P. 697-706.
Fernald M.L. 1946. Amelanchier spicata not an American species // Rhodora. - Vol. 48. - P. 125-134.
Fernandez Arias M. I., Castrovieho S. 1997. Elaeagnus L.// Flora Iberica. - Vol. 8. - P. 140--142.
Fertnald M. L. 1942. Three transfers in the Compositae // Rhodora. - Vol. 44. - № 525. - P. 340.
Fertnald M. L., Wiegand K. M. 1913. A northern variery of Erigeron ramosus // Rhodora. - Vol. 15. -
№ 169. - P. 59--61.
Fischer F. 1812. Catalogue du jardin des plantes de A.Razoumoffsky a Gorenki. - M.
Flora Iranica. № 164. Compositae, VII. / Ed. K. H. Rechinger. Graz (Austria): Akademische Druck-
u. Verlagsanstalt, 1989. - 200 p.
Flora of California [Electronic resource.] Mode of access: http://www.calflora.org/cgi-
bin/species_query.cgi?where-calrecnum=3455. Accessed 17 Ahril, 2008.
Flora of China, Vol. 11, p. 3-4, [Electronic resource.] Mode of access: http://www.efloras.org.
Accessed 4 April 2008.
Flora of Helsinki [Electronic resource.] Mode of access: http://www.helsinki.fi/lehdet/uh/299b.htm.
Accessed 18 April, 2008.
Flora of North America. http://www.efloras.org/flora_page.aspx?flora_id=1
Florae europaea -Cambridge: Univ.press. 1964-1972. Vol.1-5 (надо 1968)
Florae europaea. 1976. Cambridge: Univ.press. Vol.4. P.165
Folch R., Abella C. 1974. Galinsoga parviflora y Guizotia abyssinica, dos adventicias nuevas para la
flora Catalana // Collect. Bot. - Vol. 9. - № 7. - Р.183-189.
Forman J., Kesseli R. V. 2003. Sexual reproduction in the invasive species Fallopia japonica
(Polygonaceae) // Amer. Jour. Bot. - Vol. 90. - № 4. - P. 586--592.
Fossati T., Patrignani G., Zapelli I., Sabatti M., Sala f., Castiglione S. 2004. Development of
molecular markers to assess the level of introgression of Populus tremula into Populus alba
natural populations / Plant Breeding. - Vol. 123. - Issue 4. - P. 382-385.
Foster J. R. 1992. Photosynthesis and water relations of the floodplain tree, boxelder (Acer negundo
L.)// Tree Physiol. - Vol.11. - P.133-149.
Fowler M.C., Robson T.O. 1978. The effects of the food preferences and stocking rates of Grass Carp
(Ctenopharyngodon idella Val.) on mixed plant communities // Aquat. Bot. ---- Vol. 5, № 2. -- P.
261--276.
Franco А.J., da Silva Р.A.R. 1968. 28. Lupinus L. P. 105-106 in Tutin T.G. et al. (eds) Flora Europaea. 2.
Rosaceae to Umbelliferae. - Cambridge: Univ. Press.
Franco А.J. 1968. Crataegus L. // Flora Europaea. Vol. 2: Rosaceae to Umbelliferae. -- Cambridge:
Univ. Press. -- P. 73-77.
Frankia Symbiosis. Proceedings of the 12th meeting on Frankia and actinorhizal plants. Carry-le-
Rouet, France, June 2001 / Ed. P.Normand, K.Pawlowski, J. O. Dawson 2003. [Developments in
Plant and Soil Sciences. Vol. 100]. Norwell, MA: Kluwer Academic Publishers. - 252 p.
Frankton C. 1963. Weeds of Canada. Ottawa, Canada, Canada department of agriculture. - 196 p.
Frankton C., Mulligan G.A. 1987. Weeds of Canada. - Toronto. - Ministry of Supply and Services
Canada. NC Press Limited. - 217 pp.
Freeman B.C., Klikoff L.G., Harper K.T. 1976. Differential resource utilization by sexes of dioecious
plants // Science. - № 193. - P. 597-599.
Fremstad E., Elven R. 2004. Perennial lupins in Fennoscandia // Santen E., Hill G.D. (eds) Wild and
cultivated lupins from thé tropics to thé pôles. Proceedings of thé 10* International Lupin Conférence,
Laugarvatn, Iceland, 19-24 June 2002. International Lupin Association, Canterbury, New Zealand. - P.
178-183.
Frey A. 1974. Rodzaj Amaranthus L. w Polsce // Fragm. Flor. Geobot. - Bd 20. - № 2. - S.143-201.
Frey D. 2003. Patterns of variation within the Erigeron annuus complex in the United States and
Europe. A dissertation submitted to the Swiss Federal Institute of Technology Zurich for the
degree of doctor of natural science. Diss. ETH № 15405 // http://e-collection.ethbib.ethz.ch/
view/eth:27343.
Frey D., Baltisberger M., Edwards P. 2003. Cytology of Erigeron annuus s.l. and its consequences in
Europe // Botanica Helvetica. - Vol. 113. - № 1. - P. 1--14.
Friedman J.M., Auble G.T. 1999. Mortality of riparian boxelder from sediment mobilization and
extended inundation // Regulated rivers-research i Management. - № 15.
473
Fujiyoshi M., Kagawa A., Nakatsubo T., Masuzawa T. 2005. Successional changes in mycorrhizal
type in the pioneer plant communities of a subalpine volcanic desert on Mt. Fuji, Japan // Polar
Biosci. - Vol. 18. - P. 60--72.
Gafur A., Schützendübel A., Langenfeld-Heyser R., Fritz E., Polle A. 2003. Compatible and
incompetent Paxillus involutus isolates for ectomycorrhiza with poplar (Populus x canescens)
differ in H2O2 production / Plant biology. - Vol. 6. -- Is. 1. P. 91-99.
Galenieks P. 1959. Latvijas PSR flora 4. Latvijas Valsts izdevnieciba. - Riga. - 524 p.
Garkaje A. 2006. Impatiens glandulifera (Royle) impact of the river - banks in Latvia. Bachelor work.
-Riga. -59p.
Garnock-Jones P. J. 1979. Checklist of dicotyledons naturalised in New Zealand. 4. Rhoeadales //
New Zealand J. Bot. - Vol. 17. - P. 303--310.
Gates M. W., Liu H., Bauer L. S., Schauff M. E. 2005. A new species of Pediobius (Hymenoptera:
Eulophidae) parasitizing Chyliza apicalis (Diptera: Psilidae) in ash trees attacked by Agrilus
planipennis (Coleoptera: Buprestidae) // The great lakes entomologist. Vol. 38. № 1--2. P. 76--82.
Gavrilova G., Sulcs V. 1999. Latvijas vaskularo augu flora: Taksonu saraksts. - Riga: Latv. Akad. b-
ka. -- 1361 pp.
GCW (Global Compendium of Weeds) [Electronic resource.] Mode of access: http://
www.hear.org/gcw/species/erucastrum_gallicum. Accessed 12 April 2008.
Genton B.J., Shykoff J.A., Giraud T. 2005. High genetic diversity in French invasive populations of
common ragweed Ambrosia artemisiifolia, as a result of multiple sources of introduction //
Molecular Ecology. -- Vol. 14. -- Is. 14. -- P. 4275-4285.
George M.R., Mc Kell C. M., Richardson S.G. 1981. The Establishment of Cheatgrass (Bromus tectorum
L.) on Spent Oil Shale from the Paraho Process // J. Environ. Qual. - Vol. 10. - P.166 - 169.
Germplasm Resources Information Network» [GRIN].
-- Режим доступа: http://www.ars-
grin.gov//cgi-bin/npgs/html/taxon.pl?400047. -- Acc. 17 Dec. 2008.
Gibson R. H., Nelson I. L., Hopkins G. W., Hamlett B. J., Memmott J. 2006. Pollinator webs, plant
communities and the conservation of rare plants: arable weeds as a case study // J. Appl. Ecol. -
Vol. 43. - P. 246--257.
Gigon A., Weber E. 2005. Invasive Neophyten in der Schweiz: Lagebericht und Handlungsbedarf.
Bericht zu Handen des BUWAL. Bern. - 44 s.
Given D. R. 1984. Checklist of dicotyledons naturalised in New Zealand. 16. Compositae -- tribes
Vernonieae, Eupatorieae, Astereae, Inuleae, Heliantheae, Tageteae, Calenduleae, and Arctoteae
// New Zealand J. Bot. - Vol. 22. - P. 183--190.
Gleason H. 1963. Amaranthus L. // The new Britton and Brown illustrated flora of Northeastern
United States and adjacent Canada. New York Botan. garden. - Vol. 2. - P.102.
Gleason H. A., Cronquist A. 1991. Manual of the vascular plants of northeastern United States and
adjacent Canada. 2nd ed. New York: New York Botanical Garden. XXV, 910 p.
Gleason H.A. 1963. The New Britton and Brown Illustrated Flora of Northeastern United States and
Adjacent Canada. N.Y.: Hafner Publ.Co.; Vol.2. 655p.
Gleason H.A. The New Britton and Brown Illustrated Flora of Northeastern United States and
Adjacent Canada. New York: Hafner Publ. Co. 1963. Vol.3. P.114 ТАКОЙ КНИГИ НЕТ
Gomes M. F. P. L., Massoco C. O., Xavier J. G., Bonamin L. V. 2007. Comfrey (Symphytum
officinale L.) and experimental hepatic carcinogenesis: a short-term carcinogenesis model study //
Evidence-based Complementary and Alternative Medicine (eCAM) December 26
Good J.E.G., Bryant R., Carlill P. 1990. Distribution, longevity and survival of upland hawthorn
(Crataegus monogyna) scrub in North Wales in relation to sheep grazing // J. Appl. Ecol. - Vol.
27. -№1. -P.272-283.
Gosler A.G., Kelly C.K., Blakey J.K. 1994. Phenotypic plasticity in leaf morphology of Crataegus
monogyna (Rosaceae): an experimental study with taxonomic implications // Botanical Journal of
the Linnean Soc. - Vol. 115. -- Is. 3. P. 211-219.
Goulson D., Stout J. C., Hawson S. A., Allen J. A. 1998. Floral display size in comfrey, Symphytum
officinale L. (Boraginaceae): relationships with visitation by three bumblebee species and
subsequent seed set // Oecologia. - Vol. 113. - P. 502--508.
Green A.C., Beardmore G.L. 1988. Home treatment of skin cancer and solar keratoses // Australasian
Journal of Dermatology. - Vol. 29. -- Is. 3. - P. 127-130.
474
Gregor Th. 2004. Apomicts in the Vegetation of Central Europe // http://www.uni-vechta.de/
ifd/biologie/upload/Gregor_Thomas/Forschungsprojekte/Apomikten/apomictsinthevegetationofce
ntraleurope.pdf (06.05.2008)
Grey-Wilson C. 1979. Balsaminaceae // Flora Iranica. - № 143. -- 15 р.
Grigulis K., Lavorel S., Davies I.D., Dossantos A., Lloret F., Vilà M. 2005. Landscape-scale positive
feedbacks between fire and expansion of the large tussock grass, Ampelodesmos mauritanica in
Catalan shrublands // Glob. Change Biol. - Vol. 11. - № 7. - P. 1042--1053. (нет этой статьи в
этом номере!)
Grime J.P. 1987. Dominant and subordinate components of plant communities: implications for
succession, stability and diversity // Colonization, Succession, Stability and Diversity (eds AJ.
Gray, M.J. Crawley, P.J. Edwards) P. 413-428. - Symposia of the British Ecological Society, 26.
Blackwell Scientific Publications, Oxford.
Grimsby J. L., Tsirelson D., Gammon M. A., Kesseli R. 2007. Genetic diversity and clonal vs. sexual
reproduction in Fallopia spp. (Polygonaceae) // Amer. Jour. Bot. - Vol. 94. - № 4. - P. 957--964.
Groh H. 1933. Some recently noticed mustards. Sci. Agric. Ottawa 13: 722-727.
Grull F., Priszter S. 1967. Die Variabilitat und Verbreitung einiger Amaranthus-Arten auf den
Ablagerungsplatzen und Schutthalden der Stadt Brno // Preslia. -- Bd 39. - № 3. - S. 306-311.
Guanghua Z., Liang L., Soreng R.J., Olonova M.V. 2006. Poa // Flora of China. - Vol. 22. - P. 257--
309.
Gudzinskas Z. 1997. Conspectus of alien plant species of Lithuania. 4. Asteraceae // Botanica Lithuanica. -
Vol.3. -№4. -P.335-366.
Gudzinskas Z. 1998. Conspectus of alien plant species of Lithuania. 8. Aceraceae, Balsaminaceae,
Elaeagnaceae,Geraniaceae, Hippocastanaceae, Linaceae, Lythraceae, Onagraceae,
Oxalidaceae, Rutaceae, and Vitaceae // Bot. Lithuan. - Vol. 4. - № 4. - P. 363-377.
Gudžinskas Z. 1998. Conspectus of alien plant species of Lithuania. 7. Apiaceae, Apocynaceae,
Asclepiadaceae, Caprifoliaceae, Dipsacaceae, Oleaceae,Sambucaceae, Viburnaceae, and
Valerianaceae // Botanica Lithuanica. - Vol. 4. - № 3. - P. 249--265.
Gudzinskas Z. 1999. Conspectus ofalien plant species of Lithuania. 10. Fabaceae // Botanica Lithuanica. -
Vol.5. -№2. -P.103-114.
Gudzinskas Z. 2005. Fecundity of Lupinus polyphyllus in relation to habitats and age of individuals //
Proceedings of 8th International Conference on the Ecology and Management of Alien Plant
Invasions. Univ. of Silesia. Katowice, Poland. - P. 10.
Gudzinskas Z., Sinkevicene Z. 1995. Distribution, biology and naturalization of Impatiens
glandulifera Royle (Balsaminaceae) in Lithuania // Bot. Lithuan. - Vol. 1. - № 1. - Р.21-33.
Guénégou M.C., Levasseur J.E. 1993. La nouvelle espèce amhiploïde Spartina anglica C.E. Hubbard:
son origine, argumentation et implications // Biogeographica. - Vol. 69. - P. 125--133.
Guide to wild Germplasm of Brassica and Allied Crops (Tribe Brassiceae, Brassicaceae): Part III.
Interspecific and Intergeneric Hybridization Data [Electronic resource.] Mode of access:
http://www.brassica.info/resources/crucifer_genetics/brass05.pdf. Accessed 14 April, 2008.
Guofang W., Clemants S. E. 2000. Juncaceae // Flora of China. - Vol. 24. - P. 44--69.
Gupta R.K. 1989. The Living Himalayas. Vol. 2. Aspects of Plant Exploration and Phytogeography.
Today and Tomorrow Printers and Publlishers. New Delhi.
Guzikowa M., Maycock P.P. 1986. The invasion and expansion of three North American species of
goldenrod (Solidago canadensis L. sensu lato, Solidago gigantea Ait. and Solidago graminifolia
(L.) Salisb.) in Poland // Acta Societatis Bot. Poloniae. - Vol. 55. - P. 367-384.
Haack R. A., Jendek E., Liu H., Marchant K.R., Petrice T.R., Poland T.M., Ye H. 2002. The Emerald
Ash Borer: A New Exotic Pest in North America // Newsletter of the Michigan Entomological
Soc. -Vol.47. -№3--4. -P.1--5.
Haining Q., Gilbert M. G. 2007. Elaeagnaceae // Flora of China. - Vol. 13. - P. 251--273.
Hall L. Gene flow in herbicide resistant Brassica napus (oil seed rape) in Canada [Electronic
resource.] Mode of access: http://www.gmsciencedebate.org.uk/meetings/pdf/110203-
speakers.pdf. Accessed 20 April, 2008.
Hâmet-Ahti L., Suominen J., Ulvinen T., Uotila P. (eds) 1998. Retkeilykasvio.
- Luonnontieteellinen
keskusmuseo, Kasvimuseo. - Helsinki. - 656 s.
Hancock Jr. F., Wilson R. E. 1976. Biotype selection in erigeron annuus during old field succession //
Bull. Torr. Bot. Club. - Vol. 103. -№ 3. - P. 122--125.
475
Handbook of Alien Species in Europe. DAISIE. Springer, 2009. XXVIII, 400 p. [Series: Invading
Nature -- Springer Series in Invasion Ecology. Vol. 3].
Hanf M. 1983. The arable weeds of Europe. BASF United Kingdom Limited. 494 pp.
Hansen A., Pedersen A. 1961. Noter om danske flora og vegetation 18-21. Flora og Fauna. - Bd 76.
S. 129-144.
Hansen K. 1991. (ed.) Dansk Feltflora. Gyldendal.
Harberd D. J., McArthur E.D. 1980. Meiotic analysis of some species and genus hybrids in the
Brassiceae. Pages 65-87 in S. Tsunoda, K. Hinata, and C. Gomez-Campo, eds. Brassica crops and
wild allies. Japan Scientific Societies Press, Tokyo, Japan.
Harberd D.J. 1972. A contribution to the cyto-taxonomy of Brassica (Cruciferae) and its allies. Bot. J.
Linn. Soc. 65: 1-23.
Harland, 1947
Harper R.M. 1936. Is Acorus calamus native in the United States? // Torreya. - Vol. 36. - № 6. - Р.
143-147.
Hartnett D.C., Hickman K.R., Walter L.E. 1996. Effects of bison grazing, fire, and topography on
floristic diversity in tallgrass prairie / J. Range Manag. --Vol. 49. - № 5. P. 413-420.
Harvie B. 2004. The mechanisms and processes of vegetation dynamics on oil-shale spoil bings in
West Lothian, Scotland. The West Lothian question. Doctoral dissertation (PHD(R)). University
of Edinburgh, College of science and engineering, School of geoscience. 248 p. [Electronic
resourse] // Atmospheric and Environmental Science PhD thesis collection [web-сайт].
(23.4.2009). (http://www.ifremer.fr/avano/notices/00096/857.htm).
Hayes B. 1976. Planting the Elaeagnus russian and autumn olive for nectar // Amer. Bee J. - Vol. 116.
-P.74,82.
Haynes R.R. 2000. Hydrocharitaceae // Flora of North America. Vol. 22. P. 26--38.
Hegi G. 1906--1931. Illustrierte Flora von Mittel-Europa. Mit besonderer Berücksichtigung von
Deutschland, Oesterreich und der Schweiz, zum Gebrauche in den Schulen und zum
Selbstunterricht. Bd. I--VII. München: Lehmann. Bd. 1: CLVIII, 402 s. Bd II: 405 s. Bd III: 607 s.
Bd IV/1, 2, 3: 1748 s. Bd V/1, 2: 1562 s. Band VI/1: 544 s., Band VII: 561 s.
Hegi G. 1935--1979. Illustrierte Flora von Mittel-Europa. Mit besonderer Berücksichtigung von
Deutschland, Österreich und der Schweiz. 2 Aufl. 6 Bände in 15 Bänden (I/1, I/2, II/1, III/1, III/2,
III/3, IV/1, IV/2A, IV/3, V/1, V/2, V/3, V/4, VI/1,VI/3). Berlin u. Hamburg: Paul Parey.
Heidel B., Christy E.J., Gilmartin A.J. 1982. Numerical phenetic analysis of variation in populations
of Poa secunda and Bromus japonicus // Northwest Science. - Vol. 56. - № 2. - P. 108--120.
Heinken T., Schmidt M., von Oheimb G., Kriebitzsch W.-U., Ellenberg H. 2006. Soil seed banks near
rubbing trees indicate dispersal of plant species into forests by wild boar // Basic and Applied
Ecol. -Vol.7. -Is.1. -P.31--44.
Hejda M, Pyšek P. 2006. What is the impact of Impatiens glandulifem on species diversity of invaded
riparian vegetation? // Biol. Conserv. - Vol. 132. - № 2. - P. 143--152.
Hejny S., Husák S. 1978. Higher plant communities // Pond Littoral Ecosystems Struct. and
Functioning / D. Dykyjová and J. Kvet, eds. -- Berlin. -- P. 36--37.
Herbicide "Arsenal", [Electronic resource.] Mode of access: http://www.plunketts.net/arsenal-l.pdf.
Accessed 5 April 2008.
Hickey R.J., Vincent M.A., Guttman S.L. 1991. Genetic variation in running buffalo clover
(Trifoflium stofoniferum, Fabaceae) // Conserv. Biol. - Vol. 5. - № 3. - Р. 309--316.
Hierro J.L., Maron J.L, Callaway R.M. 2005. A biogeographical approach to plant invasions: the
importance of studying exotics in their introduced and native range // J. Ecol. - Vol. 93. - № 1. -
P. 5--15.
Hill M.O., Mountford J.O., Roy D. B., Bunce R. G. H. 1999. Ellenberg's indicator values for British
Plants // ECOFACT. - Vol. 2a. Technical Annex. - 46 p.
Hobbs R. J., Humphries S. E. 1995. An integrated approach to the ecology and management of plant
invasions // Conserv. Biol. - Vol. 9. - № 4. - P. 761--770.
Hodálová I., Ťavoda O., Grulich V. 2004. Rozšírenie Senecio sylvaticus (Compositae) na Slovensku //
Bull. Slov. Bot. Spoločn., Bratislava. - Vol. 26. - P. 111 -- 116.
Hohmann J., Evanics F., Berta L., Bartók T., 2000. Diterpenoids from Euphorbia peplus // Planta
Med. -- Vol. 66. -- P. 291-294.
476
Hollingsworth M. L., Bailey J. P. 2000. Evidence for massive clonal growth in the invasive weed
Fallopia japonica ( Japanese Knotweed) // Bot. J. Linnean Soc. - Vol. 133. - P. 463--472.
Hollingsworth M. L.., Hollingsworth P. M., Jemkins G. I., Bailey G. I., Ferris C. 1998. The use of
molecular markers to study patterns of genotypical diversity in some invasive alien Fallopia spp.
(Polygonaceae) // Molecular Ecol. - Vol. 7. - P. 1681--1691.
Holub J., Jirásek V. 1967. Zur Vereinheitlichung der Terminologie in der Phytogeographie // Folia
Geobot. Phytotax. - Bd. 2. - S. 69--113.
Holub J., Vicol E.C. 1971. Epilobium adenocaulon Hausskn - specie noua in flora Romaniei //
Contrib.Bot. - P.105-110.
Hosie R.C. 1969. Native trees of Canada. Canadian Forestry Service, Department of Fisheries.
-
Ottawa. - 380 p.
Hrazsky Z. 2005. Potential distribution modeling of the invasive tree Acer negundo in the Czech
Republic. / Poster presented at the 8th Internationl conference on the Ecology and Management of
Alien Plant Invasions. Katowice. Poland.
Hsi-wen L., Hedge I.C. 1994. Lamiaceae / Flora of China. - Vol. 17. - P. 50-299.
http://nlbif.eti.uva.nl/bis/flora.php?menuentry=soorten&id=2993
http://www. agra.com.ua Большая агрономическая энциклопедия болезней и вредителей.
http://www. Plantarium.ru/page/taxonomy/taxon/11895.html
http://www.biodat.ru/doc/lib/WTO.htm)
http://www.efloras.org
http://www.plants.usda.gov A PLANTS profile of Amaranthus retroflexus (redroot amaranth) from
the USDA PLANTS database
Huebner C.D., Olson C., Smith H.C. 2005. Invasive plants field and reference guide: an ecological
perspective of plant invaders of forests and woodlands. - USDA Forest Service, State and Private
Forestry, Northeastern Area. - vii, 79 p.
Huffman L. 2004. Problem Weed of the Month: Hairy Galinsoga. - Ontario Ministry of Agriculture, Food
and Rural Affairs.
Hughes A.P, Evans G.C. 1962. Plant growth and the aerial environment. II. Effects of light intensity
on Impatiens parviflora DC. // New Phytol. - Vol. 61. - № 2. - P. 154--174.
Hughes A.P. 1966. The importance of light compared with other factors affecting plant growth //
Light as an ecological factor. - Oxford: Blackwell. -- P. 121--146.
Hulbert L.C. 1955. Ecological studies of Bromus tectorum and other annual bromegrasses // Ecol.
Monographs. - Vol. 25. - № 2. - Р.181-213.
Hull J., Groves R.H. 1973. Variation in Chondrilla juncea in South-Eastern Australia // Austral. J.
Bot.- Vol. 21. - № 1. P. 113-135.
Hulme Ph.E. 2007. Biological invasions in Europe: drivers, pressures, states, impacts and responses //
Biodiversity under threat / Ed. R.M.Harrison, R.E. Hester. - Cambridge: Royal Soc. Chem. - P.
56--80. (Issues in Envir. Sci. Technol. - Vol. 25).
Hulme Ph.E., Bacher S., Kenis M., Klotz S., Kühn I., Minchin D., Nentwig W., Olenin S., Panov V.,
Pergl J., Pyšek P., Roques A., Sol D., Solarz W., Vilà M. 2008. Grasping at the routes of
biological invasions: a framework for integrating pathways into policy // J. Appl. Ecol. - Vol. 45.
-№2. -P.403--414.
Hulme Ph.E., Bremner E.T. 2006. Assessing the impact of Impatiens glandulifera on riparian
habitats: partitioning diversity components following species removal // J. Appl. Ecol. - Vol. 43. -
№1. -P.43--50.
Hulme Ph.E., Roy D.B., Cunha T., Larsson T.-B. 2009. A pan-European inventory of alien species:
rationale, implementation and implications for managing biological invasions // DAISIE,
Handbook of Alien Species in Europe. - P. 1--14.
Hulten E. 1950. Atlas over vaxternas utbredning i Norden. Stockholm: «Generalstabens litografiska
anstalts forl.». 119, 512s.
Humangarden [Electronic resource.] Mode of access: http://humangarden.ru/sofia/
findnewdb.php?rast=14200. Accessed 14 June 2008.
Hume L., Cavers P.B. 1983. Differences in the flowering requirements of widespread populations of
Rumex crispus // Can. J. Bot. -- Vol. 61. - № 10. P. 2760-2762.
Husaini S.W.H., Iwo G.A. 1990. Cytology of some weedy species of the family Compositae from Jos
Plateau, Nigeria // Feddes repert. - Vol.101. - № 1-2. - P. 49-62.
477
Hussain F., Shah S. M., Sher H. 2007. Sher traditionnal resource evaluation of some plants of Mastuj,
district Chitral, Pakistan // Pak. J. Bot. - Vol. 39. - № 2. - P. 339--354.
Hylander N. 1971. Prima loca plantarum vascularium sueciae. Plantae subspontaneae vel in tempore
recentiore adventitiae // Svensk botanisk Tidskrift. - Bd 64. - Suppl. 332 s.
Hаeupler H., Muer T., 2000. Bildatlas der Farn- und Bluttenpflazen Deutschlands. - Stuttgart: Ulmer.
-759s.
Hаeupler H., Muer T., 2007. Bildatlas der Farn- und Bluttenpflazen Deutschlands. 2 Aufl. Stuttgart:
Ulmer. - 789 s.
Identification of risks and management of invasive alien species using the IPPC framework.
Proceedings of the workshop on invasive alien species and the International Plant Protection
Convention, Braunschweig, Germany, 22--26 September 2003. Rome: FAO, 2005. xii + 301 p.
Illustrated Flora of British Columbia. Vol. 1: Gymnosperms and Dicotyledons (Aceraceae through
Asteraceae). 1998. Ed. by Douglas G.W., Straley G.B., Meidinger D.V., Pojar J. Victoria: B.C.
Ministry of Environment, Lands & Parks and B.C. Ministry of Forests. - 436 p.
Illustrated flora of British Columbia. Vol. 2: Dicotyledons (Balsaminaceae through Cucurbitaceae).
1998. Ed. by Douglas G. W., Straley G. B., Meidinger D. V., Pojar J. Victoria (B. C.): B. C.
Ministry of Environment, Lands & Parks and B. C. Ministry of Forests. - 401 p.
Index of Garden plants. 1974. - London, Basingstoke; MakMillan Press. Ltd. - 1234 p.
Index plantarum anno 1824 in Horto botanico Imperiali Petropolitano vigentium. 1824. Petropoli. --
75 p.
Index plantarum horti botanici Univers. Caes. Charcoviensis. 1823. Charcov.
Index seminum et plantarum horti botanici Universitatic Caesereao Mosquensis. 1826. -Mosquae.
Index seminum et plantarum horti botanici Universitatis Caesareae Mosquensis. Mosquae. Typis
Universitatis. 1826. - 82 p.
Index seminum... 1863. Petersburg. -- 40 c.
Ingram R., Weir J., Abbott R. J. 1980. New evidence concerning the origin of inland radiate
groundsel, S. vulgaris L. var. hibernicus Syme// New Phytol. - Vol. 84. - P. 543--546.
Interactive Flora of NW Europe [Electronic resource.] Mode of access: http://ip30.eti.uva.nl/bis/
flora.php?selected. Accessed 15 Mar. 2008.
Interactive flora of NW Europe, [Electronic resource.] Mode of access: http://nlbif.eti.uva.nl/bis/
flora.php?menuentry=soorten&id=2577. Accessed 3 Mar. 2008.
International Code of Botanical Nomenclature (Vienna Code) adopted by the Seventeenth
International Botanical Congress Vienna, Austria, July 2005. Ed. by McNeill J., Barrie F. R., Burdet
H. M., Demoulin V., Hawksworth D. L., Marhold K., Nicolson D. H., Prado J., Silva P. C., Skog J.
E., Wiersema, J. H., Turland, N. J. // Regnum Vegetabile. Vol. 146. 2006. XVIII, 568 p.
Invasive alien species in Hungary. 2003. // National ecological network (Budapest). - № 6. - 20 p.
Invasive alien species in Switzerland. An inventory of alien species and their threat to biodiversityand
economy in Switzerland. 2006. Bern: Federal Office for the Environment FOEN. - 155 p.
Invasive alien species: a toolkit for best prevention and management practices / Ed. R.Wittenberg,
M.J.W. Cock 2001. - Wallingford: CABI. - 228 p.
Invasive Exotic Plants of the Northeast, [Electronic resource.] Mode of access:
http://arboretum.conncoll.edu/salt/invasives.html. Accessed 20 Oct. 2008.
Invasive Non-native Plant Management during 2002 by Miles Falck and Steve Garske, administrative
Report 02-12, February 10, 2003, Great Lakes Indian Fish & Wildlife Comission Biological
Services Division p.o. Box 9, Odanah, WI54861 (715)682-6619, [Electronic resource.] Mode of
access: http://www.glifwc-maps.org/pdf/Administrative%20Report%2002-12.pdf. Accessed 14
Oct. 2008.
Invasive Plant Atlas of New England (IPANE), [Electronic resource.] Mode of access:
http://invasives.eeb.uconn.edu/alphaipane/ctcouncil/4-16-04htm. Accessed 12 Oct. 2008.
IT IS Report, [Electronic resource.] Mode of access: http://www.itis.gov/servlet/SingleRpt/
SingleRpt?search...Crataegus+ monogyna@search. Accessed 20 Mar. 2008.
Ito I. 1942. Chromosomen und sexualitat von den Araceae. 1. Somatische Chromosomenzahlen
einiger Arten // Cytologia. -- Bd 12. - № 2-3. - S. 313-325.
IUCN guidelines for the prevention of biodiversity loss caused by alien invasive species. Information
Paper. 2000 [Electronic resourse] // Invasive Species Specialist Group [web-сайт]. (14.05.2009).
http://www.issg.org/infpaper_invasive.pdf
478
Jacobs S. W. L., Pickard J. 1981. Plants in New South Wales.
- Sydney: New South Wales
Government Printer. - 227 p.
Jafri S. M. H. 1973. Brassicaceae // Flora of West Pakistan. - Vol. 55. - P. 1--308.
Jäger E. J. 1980. Floristische Neufunde in der Baschkirischen ASSR und Bemerkungen zur
Ausbreitungsgeschichte von Lepidium densiflorum, Echinocystis lobata und Collomia linearis //
Wiss. Zeitschr. Martin-Luther-Univ. Halle-Wittenberg, Biowiss. - № 29, math.-nat. Reihe. - H. 4.
- S. 117--124. (publ. 1981).
Jäger E. J. 1988. Möglichkeiten der Prognose synanthroper Pflanzenausbrietungen // Flora. -Bd. 180.
- № 1--2. - S. 101--131.
Jäger E. J. 1995. Die Gesamtareale von Reynoutria japonica Houtt. und R. sachalinensis (F. Schmidt)
Nakai, ihre klimatische Interpretation und Daten zur Ausbreitungsgeschichte // Schr.-R. f.
Vegetationskde., Sukopp-Festschrift. - H. 27. - S. 395--403.
Jahodová Š., Trybush S., Pyšek P., Wade M., Karp A. 2007. Invasive species of Heracleum in
Europe: an insight into genetic relationships and invasion history // Diver. Distrib. - Vol. 13. - №
1. - P. 99--114.
Jain S.F., Marshall D.R. 1967. Population studies in predominantly self-pollinating species. X.
Variation in natural populations of Avena fatua and A. barbata // Amer. Natur.
- Vol. 101. -
P. 19--33.
Jalas J. (ed.) 1980. Suuri kasvikirja III. Otava, Helsinki. - 944 p.
Jalas J. 1955. Hemerobe und hemerochore Pflanzenarten: ein terminologischen Reformversuch //
Acta Soc. Fauna et Flora Fenn. - Bd. 72. - № 11. - S.1--15.
Jasnowski M. 1961. Impatiens Roylei - nowa roslina w Polsce // Fragm. florist. et geobot. - Vol.7. - №
1. - P. 77-80.
Jedlička J., Prach K. 2006. A comparison of two North-American asters invading in central Europe //
Flora. Morphology, Distribution, Functional Ecology of Plants. - Vol. 201. - Is. 8. - P. 652--657.
Jervis R.A., Buell M.F. 1964. Acorus calamus in New Jersey // Bull. Torrey Bot. Club. - Vol. 91. - №
4. - Р.335-336.
Jiang X., Zhang W., Yang Z., Du G. 2004. Plant diversity variations in zokor-mound communities
along a successional stage / Ying Yong Sheng Tai Xue Bao.; 15(5): 814-8 (ISSN: 1001-9332).
Jones G.E. 1957. Chromosome numbers and phylogenetic relationships in the Araceae // Ann.
Arbor. - № 1.
Jones G.N. 1946. American species of Amelanchier // Illinois biol. monographs. - Vol. 20. - № 2. -
P. 1-126.
Jones K., Smith B. 1966. The cytogeography of Impatiens // Kew Bull. - Vol. 20. - № 1. - P.63-72
Joseph A.J., Lexer Ch. 2007. A set of novel DNA polymorphisms within candidate genes potentially
involved in ecological divergence between Populus alba and P. tremula, two hybridizing
European forest trees / Molecular Ecol. Resources. - Vol. 8. -- Is. 1. -- P. 188-192.
Juncaceae. 3: Juncus subg. Agathryon. Species Plantarum: Flora of the World. 8. Compailed by J.
Kirschner. Canberra, 2002. - 192 p.
Kabuce N. 2005. Invasive Alien species fact sheet Amelanchier spicata. Nobanis.
(www.nobanis.org.).
Kabuce N. 2006. NOBANIS -- Invasive Alien Species Fact Sheet. Heracleum sosnowskyi // Online
Database of the North European and Baltic Network on Invasive Alien Species -- NOBANIS
www.nobanis.org, Date of access x/x/200x.
Kadereit J. W., Comes H. P., Curnow D. J., Irwin J. A., Abbott R. J. 1995. Chloroplast DNA and
isozyme analysis of the progenitor-derivative species relationship between Senecio nebrodensis
and S. viscosus (Asteraceae) // Amer. J. Bot. - Vol. 82. - № 9. - P. 1179--1185.
Kagima D. 2000. Bibliography and Biology of Galinsoga spp. /The ISU Weed Biology Library. - 17 pp.
Kappes H., Lay R., Topp W. 2007. Changes in different trophic levels of litter-dwelling macrofauna
associated with giant knotweed invasion // Ecosystems. - Vol. 10. - P. 734--744.
Karlsson T. 1981. Den gammaldags lupinen i Sunnerbo // Svensk botanisk Tidskrift. - Bd 75. - S. 265--278.
Kartesz J.T., Kartesz R.A 1980. Synonymized checklist of the vascular flora of the United States,
Canada, and Greenland. - Chapel Hill: Univ. North Carolina Press. -- 500 p.
Kasperek G. 2004. Fluctuations in numbers of neophytes, especially Impatiens glandulifera, in
permanent plots in a west German floodplain during 13 years // Neobiota. - Vol. 3. - P. 27-37.
479
Katz G. L., Friedman J. M., Beatty S. W. 2001. Effects of physical disturbance and granivory on
establishment of native and alien riparian trees in Colorado, U.S.A. // Diversity and Distributions.
-Vol.7. -№1/2. -P.1--14.
Katz G. L., Shafroth P. B. 2003. Biology, ecology and management of Elaeagnus angustifolia L.
(Russian olive) in western North America // Wetlands. - Vol. 23. - № 4. - P. 763--777.
Kawano S., Azuma H., Ito M., Suzuki K. 1999. Extrafloral nectaries and chemical signals of Fallopia
japonica and Fallopia sachalinensis (Polygonaceae), and their roles as defense systems against
insect herbivory // Plant Species Biology. - Vol. 14. - № 2. - P. 167--178.
Kays S. J., Nottingham S. 2007. Biology and chemistry of Jeruslaem artichoke: Helianthus tuberosus
L. CRC Press. - 478 p.
Keiffer C. H., Ungar I. A. 1997. The effect of extended exposure to hypersaline conditions on the
germination of five inland halophyte species //Am. J. Bot. - Vol. 84. - № 1. - P. 104--111.
Keil P. 1995. Haufigkeit und Verbreitung von Bidens frondosa var. anomala am Beispiel des unteren
Ruhrtales im westlichen Ruhrgebiet // Florist. Rundbriefe. -- Bd 29. - № 2. - S.129-134
Kellock A.W., Stubbs L.L., Parbery D.G. 1978. Seedborne Fusarium species on subterranean clover
and other pasture legumes // Austral. J. Agricult. Res. -- Vol. 29. - № 5. P. 975-982.
Kelly A., Bruns V. 1975. Dissemination of weed seeds by irrigation water // Weed Sci. - Vol. 23. -
P. 486-493.
Kennedy A.C., Johnson B.N., Stubbs T.L. 2001. Host range of a deleterious rhizobacterium for
biological control of downy brome // Weed Sc. - Vol. 49. - № 6. - P. 792-797.
Kennedy T.A., Naeem S., Howe K.M., Knops J.M.H., Tilman D., Reich P. 2002. Biodiversity as a
barrier to ecological invasion // Nature. -- № 417. -- Р. 636--638.
Kentucky Exotic Pest Plant Council (KY-EPPC), [Electronic resource.] Mode of access:
http://www.se-eppc/org/ky/list-htm. Accessed 5 April 2008.
Khan S. W., Khatoon S. 2007. Ethnobotanical studies on useful trees and shrubs of Haramosh and
Bugrote valleys, in Gilgit northern areas of Pakistan // Pak. J. Bot. - Vol. 39. - № 3. - P. 699--710.
Khoshoo T.N. 1957. Cytology of some Impatiens species // Caryologia. - Vol. 10. - P. 55-74.
Kiemnec G. L., Mcinnis M. L. 2002. Hoary cress (Cardaria draba) root extract reduces germination
and root growth of five plant species // Weed Technology. - Vol. 16. - Is. 1. - P. 231--234.
Kigel J., Gibly A., Negbi M. 1979. Seed germination in Amaranthus retroflexus L. as affected by the
photoperiod and age during flower induction of the parent plants// J. Experimental Bot. - Vol. 30.
-№5. -Р.997--1002.
KilJ.H.,ShimK.C.,ParkS.H.,KohK.S.,SuhM.H.,KuY.B.,SuhS.U.,OhH.K.,KongH.Y.
2004. Distributions of Naturalized Alien Plants in South Korea // Weed Technology. - Vol. 18. -
P. 1493--1495.
Kim D.K., Zee O.P. 2000. A new cyanogenic glycoside from Sorbaria sorbifolia var. stellipila //
Chem. Pharm. Bull. - Vol. 48. - № 11. P. 1766-1767.
Kim J. Y., Park Ch.-W. 2000. Morphological and chromosomal variation in Fallopia section
Reynoutria (Polygonaceae) in Korea // Brittonia. - Vol. 52. - № 1. - P. 34--48.
Kimoto T., Duthie-Holt M. 2006. Exotic Forest Insect Guidebook. Canadian Food Inspection Agency.
-128p.
Kinchin J.M. 1988. Ecology of the mural flora // J. Biol. Educ. - Vol. 22. - № 4. - P.263-266.
Kirschner J., Stepanek J., Stepankova J. 1982. A. Love, ed. IOPB chromosome number reports.
LXXVI. Taxon 31: 574.
Klemedson J.O., Smith J.G. 1964. Cheatgrass (Bromus tectorum) // Bot. rev. - Vol. 30.
-№2. -
P. 226-262.
Klemow K.M., Raynal D.J. 1979. The population ecology of Erucastrum gallicum (Cruciferae).
Contributed paper: Ecological Society of America meeting. Abstract: Bulletin of the Ecological
Society of America 60:112.
Klemow K.M., Raynal D.J. 1983. Population Biology of an Annual Plant in a Temporally Variable
Habitat // The Journal of Ecology, Vol. 71, № 3: 691-703.
Klemow К.М. 1984. Plant community development in an abandoned limestone quarry: A
demographic assessment. Pages 195-210 in T. N. Veziroglu, ed. The biosphere: Problems and
solutions. Elsevier Science Publ., Amsterdam, The Netherlands.
Kliber A., Eckert Ch.G. 2005. Interaction between founder effect and selection during biological
invasion in an aquatic plant // Evolution. - Vol. 59. - № 9. - P. 1900--1913.
480
Klich M. G. 2000. Leaf variations in Elaeagnus angustifolia related to environmental heterogeneity //
Environmental and experimental botany. - Vol. 44. - № 3. - P. 171--183.
Klinge J. 1883. Die Holzgewachse von Est-, Liv- und Kurland.-Dorpat: Mattiesen. -- 290 s.
Kloot P.M. 1987. The naturalized flora of South Australia. 1. The documentation of its development //
J. Adelaide Bot. Gardens. - Vol. 10. - P. 81--90.
Klopfer K., Schonfeld S. 1986. Zur Verbreitungsgeschichte der Galinsoga - Arten in Mitteleuropa //
Wiss. Z. Pad. Hochsch. K. Lubknrecht Potsdam. -- Bd 30. - № 1. - S. 81--94.
Klotz S., Kühn I., Durka W. 2003. BIOLFLOR -- Eine Datenbank mit biologisch-ökologischen
Merkmalen zur Flora von Deutschland. Landwirtschaftsvlg Münster. - xvi, 334 S., mit einer CD--
ROM (Schriftenreihe für Vegetationskunde. - Bd 38).
Knopf F. L., Olson Th. E. 1984. Naturalization of russian-olive: implications to Rocky Mountain
wildlife // Wildlife Society Bulletin. - Vol. 12. - № 3. - P. 289--298.
Knowles P.F. 1944. Interspecific hybridization of Bromus//Genetics. - № 29. - Р. 128--140.
Kock U.V. 1988. Okologische Aspekte der Ausbreitung von Bidens frondosa in Mitteleuropa.
Verdrangt er Bidens tripartita? // Flora. - Bd 180. - № 3-4. - S. 177-190.
Koenies H., Glavac V. 1979. Uber die Konkurrenzfahigkeit des Indischen Springkrauts (Impatiens
glandulifera Royle) am Fuldaufer bei Kassel.// Philippia. - Bd 4. - S. 47--59.
Kohls S. J., Baker D. D. 1989. Effects of substrate nitrate concentration on symbiotic nodule
formation in actinorhizal plants // Plant and Soil. - Vol. 118. - P. 171--179.
Koniuszek J.W.J., Verkicij J.A.C. 1982. Genetical variation in two related annual Senecio species
occurring on the same habitat // Genetica. - Vol. 59. - № 2. - P. 133--137.
Konstantinidou-Doltsinis S., Markellou E., Kasselaki A.-M., Fanouraki M. N., Koumaki C. M.,
Schmitt A., Liopa-Tsakalidis A., Malathrakis N. E. 2006. Efficacy of Milsana®, a formulated
plant extract from Reynoutria sachalinensis, against powdery mildew of tomato (Leveillula
taurica) // BioControl. - Vol. 51. - P. 375--392.
Kornás J. 1968. A geographical-historical classification of synanthropic plants // Mater. Zakl. Fitosoc.
Stos. VW. -- Warszawa-Bialowiera. - № 25. - P. 33--41.
Kornás J. 1982. Man's impact upon the flora: proceses and effects // Memorabilia Zool. - Vol. 37. -
P. 11--30.
Korolyuk E.A., Konig W., Tkachev A.V. 2002. Composition of essential oil of Elsholtzia ciliata
(Thunb.) Hyl. from the Novosibirsk region, Russia // Химия растительного сырья. № 1. P. 31-36.
Kosim B. 2005. Problemy pielejnacyjne wynikaja_ce z cech wzrostu i reprodukcji osobnikow klonu
jesionolistnego Acer negundo L. na terenie wybranych parkow miejskich w Warszawie w roku
2004. /Diploma Thesis at the Faculty of Architecture. Univ. of Ecology and Management in
Warsaw.
Kosiński P., Bednorz L. 2003. Trees and shrubs of the Polish part of the Eastern Sudety Mts. //
Dendrobiology. - Vol. 49. - P. 31--42.
Kostecka-Madalska O.I., Borowczak-Niemczewska E. 1968. The tannines in Erigeron canadensis L.
// Acta Soc. bot. Polon. - Vol.37. - № 4. - P.657-664.
Kostecka-Madalska O.I., Polanowski A. 1964. Olejek lotny z krajowego ziela Erigeron canadensis //
Acta Poloniae pharmac. -- Bd 21. - № 3. - S. 275-279.
Kowarik I. 1992. Einfuhrung und Ausbreitug nichteinheimischer Geholzarten in Berlin und
Brandenburg -- Verb. Bot. Ver., - Berlin, Brandenburg. Beheft 3. - 188 s.
Kowarik I. 1995. Time lags in biological invasions with regard to the success and failure of alien
species //. Plant invasions: general aspects and special problems / Ed. P.Pyšek, K.Prach,
M.Rejmanek, M.Wade Amsterdam: SPB Acad. Publ. - P 15--38.
Kowarik I. 2003. Biologische Invasionen: Neophyten und Neozoen in Mitteleuropa. - Ulmer, Stuttgart. - 380 s.
Kozlowski J. 1960. Kariotyby tataraku (Acorus calamus) na terenie Polski // Biul. Inst. Roslin
Leczniczych. - № 6. - S.65-70.
Krausch H.-D. 2003. Kaiserkron und Pâonien rot...' Entdeckung und Einfuhrung unserer Gartenblumen.
Dôlling und Galitz Verlag, - Hamburg - 536 s.
Kremer D., Pernar R., Ančić M. 2006. Distribution of North American ash species in the Drava river
basin and Danube basin (Croatia) // Acta Bot. Croat. - Vol. 65. - № 1. - P. 57--66.
Krüssmann G., 1976. Hardbuch der Lanbgehöbze.
- Band 1, A-D. 2 auflage.
-- Berlin, Hamburg:
Parey -- 486 s.
481
Krzakowa М., Kraupe А. 1981. Isozyme investigation of naturel populations of the cheatgrass
(Bromus tectorum L.) // Bot. Jahrb. Syst. Pflanzengesch und Pflanzengeogr. - Vol.102. - № 1-4. -
S. 393-399.
Kühn I., Klotz S. 2003. The alien flora of Germany -- basics from a new German database // Plant
invasions: ecological threats and management solutions. Child L.E., Brock J.H., Brundu G., Prach
K., Pyšek P., Wade P.M., Williamson M. Leiden: Backhuys. P. 89--100.
Kühn I., May R., Brandl R., Klotz S. 2003. Plant distribution patterns in Germany -- Will aliens match
natives? // Feddes Repert. - Vol. 114. - № 7--8. - P. 559--573.
Kukk T. 1999. Eesti taimestik. Tartu-Tallinn, (in Estonian, Flora of Estonia).
Kukk T., Kull T. (eds) 2005. Atlas of thé Estonian flora. - Tartu. - 528 p.
Kull T., Kukk T. 2005. Invasiivsed voorliigid Eestis. Keskkonnaministeerium. - Tallinn.
Kuniga I. 1994. SIkziedu galinsogas (Galinsoga parviflora L.) biologijas izpete, tas ietekme uz dazu
kuluraugu razu. Bakalaura darbs. - Latvijas Lauksaimniecibas Universitate, Jelgava. -- P. 1--67.
Kunstler P. 1999. The role of Acer negundo L. in the structure of floodplain forests in the middle
course of the Vistula river // Proceedings of the 5th International Conference on the Ecology of
the Invasive Alien Plants. - La Maddalena, Sardinia, Italy.
Kupffer K.R. 1907. Kleine Notizen. Korr.-Bl. d. Naturf.-Ver. Zu Riga, Jg. 50. - S. 180-210.
Kurose D., Renals T., Shaw R., Furuya N., Takagi M., Evans H. 2007. Fallopia japonica, an
increasingly intractable weed problem in the UK: Can fungi help cut through this Gordian knot? //
Mycologist. - Vol. 20. - Is. 4. - P. 126--129.
Kurtto A. 1992. Jattipalsami (Impatiens glandulifera) - kuriton, mutta kiinnostava // Lutukka. --Bd 8. -
S.14--29.
Kurtto A. 1996. Impatiens glandulifera (Balsaminaceae) as an ornamental and escape in Finland, with
notes on the other Nordic countries //Acta Universitatis Upsaliensis, Symbolae Botanicae
Upsalienses. -- Bd 31. - № 3. - Р. 221-228.
Kurtto A., Helynranta L. 1998. Helsingin kasvit. Kukkivilta kivilta metsan syliin. [Flora of Helsinki. From
Flowering Stones to Forest Floor]. City of Helsinki Environment Centre and Helsinki University Press,
Helsinki. - 400 p.
Kuusk V., Plotniece M., Jankeviciene R. 1996. Balsaminacea A. Rich. // Flora of the Baltic Countries
II (eds. V.Kuusk, L.Tabaka, R.Jankeviciene). Eesti Loodusfoto AS. - 177 p.
Kuusk V., Tabaka L., Jankeviciene R. (eds) 1996. Flora of thé Baltic countries. Compendium of vascular
plants. II. Eesti Loodusfoto AS, - Tartu. - 372 p.
Kuusk V., Tabaka L., Jankeviciene R. 2003. Flora of the Baltic countries 3. - Tartu, Estonian Agricultural
University, Institute of Zoology and Botany. - 406 p.
Lambdon P.W., Pyšek P., Basnou C., Hejda M., Arianoutsou M., Essl F., Jarošík V., Pergl J., Winter
M., Anastasiu P., Andriopoulos P., Bazos I., Brundu G., Celesti-Grapow L., Chassot P.,
Delipetrou P., Josefsson M., Kark S., Klotz S., Kokkoris Y., Kühn I., Marchante H., Perglová I.,
Pino J., Vila M., Zikos A., Roy D., Hulme Ph. E. 2008. Alien flora of Europe: species diversity,
temporal trends, geographical patterns and research needs // Preslia. - Vol. 80. - № 2. - P. 101--
149.
Lanea D. 1979. The significance of noxious weeds on roadsides in agricultural areas of Victoria,
Australia / Weed Research - Vol. 19. - Issue 3. - P. 151--156.
Larsen K. 1969. Studies in the flora of Thailand. 54. Cytology of vascular plants. III. A study of Thai
Aroids // Dansk Bot. Ark. - Vol. 27. - № 1. - Р. 39-59.
Larson D. 2007. Non-indigenous Freshwater Plants: Patterns, Processes and Risk Evaluation: Doct.
thesis. -- Uppsala: Swed. Univ. of Agric. Sciences, -- [Acta Univ. Agric. Suec., 2007; Bd. 15].
Larsson C., Martinsson K. 1998. Jattebalsamin Impatiens glandulifera i Sverige - invasionsart eller
harmlos tradgardsflykting? // Svensk Botanisk Tidskrift. -- Bd 92. - S. 329-345.
Laubhan M. K., Shaffer T. L. 2006. Seed germination of Cirsium arvense and Lepidium latifolium:
implications for management of Montane wetlands // Wetlands. - Vol. 26. - Is. 1. - P. 69--78.
Lavoie Cl., Jodoin Yv., Goursand de Merlis A. 2007. How did common ragweed (Ambrosia
artemisiifolia L.) spread in Quebec? A historical analysis using herbarium records // Journal of
Biogeography -- Vol. 34. -- Is. 10. -- P. 1751-1761.
Lecerf A., Patfield D., Boiché A., Chauvet E., Dobson M. 2007. Stream ecosystems respond to
riparian invasion by Japanese knotweed (Fallopia jaonica) // Can. J. Fish. Aquat. Sci. - Vol. -
P. 1273--1283.
482
Ledebour C.F. 1845-1846. Flora Rossica. Stuttgartise: Schweizerbart. -- Bd 2. - № 2. - S.463--937.
Ledebour С. F. 1853. Flora Rossica; sive, Enumeratio plantarum in totius Imperii Rossici provinciis
Europaeis, Asiaticis et Americanis hucusque observatarum, auctore Carolo Friderico a Ledebour.
-Vol.4. -741p.
Lee J.-Y., Mummenhoff K., Bowman J. L. 2002. Allopolyploidization and evolution of species with
reduced floral structures in Lepidium L. (Brassicaceae) // PNAS. - Vol. 99. - № 26. - P. 16835--
16840.
Lefol E., Séguin-Swartz G., Downey R.K. 1997. Sexual hybridization in crosses of cultivated
Brassica species with the crucifers Erucastrum gallicum and Raphanus raphanistrum: potential
for gene introgression // Euphytica, Volume 95, № 2: 127-139.
Lehoczki E., Laskay G., Polos E., Mikulas J. 1984. Resistance to triazine herbicides in horseweed
(Conyza canadensis) // Weed Sci. - Vol. 32. - P. 669-674.
Lesica P., Miles S. 1999. Russian olive invasion into cottonwood forests along a regulated river in
north-central Montana // Can. J. Bot. - Vol. 77. - № 8. - P. 1077--1083.
Levine J.M., D'Antonio C.M. 2003. Forecasting biological invasions with increasing international
trade // Conserv. Biol. - Vol. 17. - № 1. - P. 322--326.
Lewis L., Knight A., Lewis B., Lewis C. 1996. Feeding and care of the horse. 2nd ed. Oxford, etc.
Blackwell Publishing. - 446 p.
Lexer C., Fay M.F., Joseph J.A., Nica M.-S., Heinze B. 2005. Barrier of gene flow between two
ecologically divergent Populus species, P. alba (White Poplar) and P. tremula (European aspen):
the role of ecology and life history in gene introgression / Molecular Ecology. - Vol. 14. -- Is. 4. P.
1045-1057.
Lhotska M. 1965. Bidens frondosa var. anomala v ceskych zemich. // Preslia. - Bd. 37. - № 2. - S.
218-219
Lhotska M. 1966. Der alteste Fund der Art Bidens frondosa in Europa // Folia geobot. et phytotaxon
bohemoslov. -- Bd 1. - № 2. - S. 186-189.
Lhotska М., Kopecky К. 1966. Zur Verbreitungsbiologie und Phytozonologie von Impatiens
glandulifera Royle an den Flusssystemen der Svitava, Svratka und oberen Odra // Preslia (Praha).
-Bd38. -№2. -S.376--385.
Li, Xiao-Dong; Wu, Li-Jun; Zang, Xiao-Yan; Zheng, Jian, 2002. Studies on chemical constituents of
Sorbaria sorbifolia // Zhongguo Zhongyao Zazhi - Vol. 27. - № 11. - P. 841--843.
Lid J. 1944. Norsk flora. Det Norske Samlaget, Oslo. - 637 p.
Lid J., Lid D.T. 2005. Norsk flora. 7. utg. Red.: Reidar Elven. Det Norske Samlaget, Oslo. - 1230 p.
Lindemann E. 1860. Index plantarum quas in variis Rossiae provinciis hucus que invenit et
observavit. // Bull. Soc. Nat. Mosc. - Vol. 33. - № 3. - Р.77--190.
Linnaeus C. 1753. Species Plantarum, exhibentes plantas rite cognitas, ad genera relatas, cum
differentiis specificis, nominibus trivialibus, synonymis selectis, locis natalibus, secundum
systema sexuale digestas. T. 1. Stockholm: L. Salvius. xii, 560 p.
Linnaeus C. 1753. Species Plantarum, exhibentes plantas rite cognitas, ad genera relatas, cum
differentiis specificis, nominibus trivialibus, synonymis selectis, locis natalibus, secundum
systema sexuale digestas. T. 2. Stockholm: L. Salvius. P.ii, 561-1200.
Liu H., Bauer L. S., Gao R., Zhao T., Petrice T. R., Haack R. A. 2003. Exploratory survey for the
emerald ash borer, Agrilus planipennis (Coleoptera: Buprestidae), and its natural enemies in
China // The great lakes entomologist. - Vol. 36. - № 3--4. - P. 191--204.
Logarzo G., Gandolfo D., Cordo H. 2002. Biology of Apagomerella versicolor (Boheman)
(Coleoptera: Cerambycidae) in Argentina, a candidate for biological control of cocklebur
(Xanthium spp.) // Biological Control. - Vol. 25. - Is. 1. - P. 22--29.
Lohmeyer W., Sukopp H. 1992. Agriophyten in der Vegetation Mitteleuropas // Schriftenreiche fur
Vegetationskunde - Bd 25. - S. 1-185.
Lonsdale M. 1999. Global patterns of plant invasions and the concept of invasibility // Ecology. - Vol.
80. - № 5. - P. 1522--1536.
Loo M., Joseph J.A., Heinze B., Fay M.F., Lexer Ch. 2007. Clonality and spatial genetic structure in
Populus x canescens and its sympatric parent P. alba in a Central European hybrid zone / New
Phytologist - Vol. 177. -- Is. 2. - P. 506-516.
Lousley J.E. 1953. The recent influx of aliens into the British flora // The changing flora of Britain /
Ed. J.E. Lousley - P. 140--160.
483
Löve A., Löve D. 1961. Chromosome number of Central and Nordwest Europaean Plant spesies.
Stockholm. -- 581 p.
Löve A., Löve D. 1964. Chromosome number reports // Taxon. - Vol. 13. - № 3. - Р. 99-110.
Löve D. 1976. Xanthium L. // Flora Europaea. - Vol. 4. - P. 143.
Löve D., Dansereau P. 1959. Biosystematic studies on Xanthium: taxonomic appraisal and ecological
status // Can. J. Bot. - Vol. 37. - P. 173--208.
Lovett D.L., Mackinnon A. 1985. Biology of Canadian weeds: 71. Oxalis stricta, Oxalis corniculata,
Oxalis dillenii ssp. dillenii and Oxalis dillenii ssp. filipes. // Can. J. Plant Science -- Vol. 65. - №
3. -- P. 691-710.
Löwe A. J., Abbott R. J. 2000. Routes of origin of two recently evolved hybrid taxa: Senecio vulgaris
var. hibernicus and york radiate groundsel (Asteraceae) // American Journal of Botany - Vol. 87. -
№ 8. - P. 1159--1167.
Löwe S., Browne M., Boudjelas S. 2003. 100 of the World's Worst Invasive Alien Species. Published
by ISSG. Auckland (New Zealand): ISSG office. - 11 p.
Lu Sh., Chen X., Aiken S. G. 2006. Festuca // Flora of China. - Vol. 22 - P. 225--242.
Ludwig W. 1994. Bidens frondosa var. anomala in Hessen// Hess. florist. Briefe. 43. - Bd 2. - S. 17--19.
Lukac G. 1988. Some structural characteristics of the stands of the species Solidago gigantea Ait. and
Helianthus tuberosus L. and their ornithocenoses in northwest Croatia (Yugoslavia)// Acta Bot.
Croatica. - Vol. 47. - P.63--76.
Lukac G. 1989. Nova nalazista vrste Impatiens glandulifera u Hrvatskoj // Acta bot. croat. -- Bd 48. -
S.165--168.
Lukac M., Calfapietra C., Godbold D.L. 2003. Production, turnover and mycorrhizal colonization of
root systems of three Populus species grown under elevated CO2 (POPFACE) // Global Change
Biology - Vol. 9. -- Is. 6. - P. 838--848.
Luken J.O., Thieret J.W., Kartesz J.R. 1993. Erucastrum gallicum (Brassiceae): invasion and spread
in NorthAmerica. SID A Contributions to Botany 15: 569-582.
Mack R.N. 1981. Invasion of Bromus tectorum into Western North America an ecological chronicle //
Agro-Ecosystems. - Vol. 7. - № 2. - P. 145--165.
Mack R.N. 2000. Cultivation fosters plant naturalization by reducing environmental stochasticity //
Biol. Invasions. - Vol. 2. - № 2. - P. 111--122.
Maerz J. C., Blossey B., Nuzzo V. 2005. Green frogs show reduced foraging success in habitats
invaded by Japanese knotweed // Biodiversity and Conservation. - Vol. 14. - P. 2901--2911.
Maggi D. W., Ahles H. E. 1999. Flora of the Northeast. A Manual of Vascular Flora of New England
and Adjacent New York. Drawings by A. Rorer. - Amherst: University of Massacusetts Press. -
XXXI + 1213 p.
Mandák B., Pyšek P., Bímová K. 2004. History of the invasion and distribution of Reynoutria taxa the
Czech Republic: a hybrid spreading faster than its parents // Preslia (Praha). - Vol. 76. - P. 15--64.
Mandák B., Pyšek P., Lysák M., Suda J., Krahulcová A., Bímová K. 2003. Variation in DNA-ploidy
levels of Reynoutria taxa in the Czech Republic // Ann. Bot. - Vol. 96. - № 2. - P. 265--272.
Manton I. 1932. Introduction to the general cytology of the Cruciferae. Ann. Bot. (bond.) 46: 509-556.
Markgraf-Dannenberg I. 1980. Festuca L. // Flora Europaea. - Vol. 5. - P. 125--133.
Marles J.R., Farnsworth N.R. 1995. Antidiabetic plants and their active constituents // Phytomedicine.
-Vol.2. -№2. -P.123--189.
Martin R.J., Carnahan J.A. 1983. The effect of field storage and laboratory conditions on germination
of five Xanthium species // Austral. J. Agricul. Res. - Vol. 34. - № 3. - P. 249--260.
Martindale I.S. 1876. The introduction of foreign plants // Bot. Caz. - Vol. 2. - № 1. - P. 55--58.
Martinez X. 1985. Algunas aportaciones a la flora Gallega // Anales del Jardin botanico de Madrid -
Vol. 42. - № 1. - P.191-196.
Martins P.S., Jain S.K. 1980. Interpopulation variation in rose clover: a recently introduced species in
California rangelands // J. Heredity. - Vol.71. - № 1. - P. 29--32.
Martius H. 1817. Prodromus Florae mosquensis . 2 ed. Lipsiae. - 304 p.
Maruta E. 1983. Growth and survival of current-year seedlings of Polygonum cuspidatum at the upper
distribution limit on Mt. Fuji // Oecologia. - Vol. 60. - № 3. - P. 316--320.
Mathe I., Tyhak E. 1962. Adatok a Matricaria matricarioides hazai elterjedesehez es hatoanyag
vizsga latahoz. // Acta pharm. hungarica. - Vol. 32. - № 1. - P.43-48.
484
Mc Kone M.J. 1985. Reproductive biology of several bromegrass (Bromus): breeding system, pattern
of fruit maturation, and seed set // Amer. J. Bot. - Vol. 72. - № 9. - P.1334--1339.
Mc Kone M.J. 1989. Intraspecific variation in pollen yield in bromegrass (Bromus) // Amer. J. Bot. -
Vol. 76. - № 2. - Р.231--237.
McDonald R. 1994. Outcrossing frequency of glufosinate resistant canola HCN92 with Diplotaxis
muralis, Raphanus raphanistrum, Brassica rapa and Erucastrum gallicum under field conditions.
Report ACI94-08, AgrEvo Canada Inc., Regina, SK. 15p.
McKenney J. L., Cripps M. G., Price W. J., Hinz H. L., Schwarzländer M. 2007. No difference in
competitive ability between invasive North American and native European Lepidium draba
populations // Plant Ecology. - Vol. 193. - № 2. - P. 293--303.
McMillan C. 1974a. Photoperiodic adaptation of Xanthium strumarium in Europe, Asia Minor, and
northern Africa // Can. J. Bot. - Vol. 52. - № 8. - P. 1779--1791.
McMillan C. 1974b. Photoperiodic responses in experimental hybrids of cockleburs // Nature - Vol.
249. - № 5453. - P. 183--186.
McMillan C. 1975. Experimental hybridization of Xanthium strumarium (Compositae) from Asia and
America. I. Responses of F1 hybrids to photoperiod and temperature // American Journal of
Botany. - Vol. 62. - № 1. - P. 41--47.
McVean D.N. 1966. Ecology of Chondrilla juncea in south-eastern Australia // J. Ecol. - Vol. 54. -- P.
345--365.
Mędrzycki P. 2002. Inwazja amerykanskiego klonu Acer negundo L. a uzytkowanie ziemi w Puszczy
Biatowieskiej / PhD Thesis, Faculty of Biology, Warsaw University.
Mędrzycki P. 2007. NOBANIS -- Invasive Alien Species Fact Sheet -- Acer negundo // Online
Database of the North European and Baltic Network on Invasive Alien Species -- NOBANIS
www.nobanis.org, Date of access 21/10/2007.
Mędrzycki P., Pabjanek P. 2001. Linking land use and invading species features: a case study of Acer
negundo L. in Bialowieza village (NE Poland) // G. Brundu, J. Brock, I. Camarda, L. Child, M.
Wade. (red.). Plant invasions: Species Ecology and Ecosystem Management. Backhuys
Pubblishers. - S.123--132.
Meekins J.F., Ballard H.E. Jr., McCarthy B.C. 2001. Genetic variation and molecular biogeography
of a north american invasive plant species (Alliaria petiolata, Brassicaceae) // Int. J. Plant Sci. -
Vol. 162. - № 1. - P. 161--169.
Melderis A. 1939. Jauns ienacejs ziedaugs Latvijas flora - Galinsoga quadriradiata Ruiz and Pavon. Daba un
Zinatne, - Nr. 4. -- P. 142 -- 146.
Melville M.R., Morton J.K. 1982. A biosystematic study of the Solidago canadensis (Compositae)
complex. I. The Ontario populations // Can. J. Bot. - Vol. 60. - P. 976--997.
Meusel H., Jäger E. J. 1992a. Vergleichende Chorologie der Zentraleuropäischen Flora. Text. Band
III. -- Jena etc.: Gustav Fischer. -- 333 p.
Meusel H., Jäger E. J. 1992b. Vergleichende Chorologie der Zentraleuropäischen Flora. Karten,
Literatur, Register. Band III. -- Jena etc.: Gustav Fischer. -- 267 p.
Meyer A., Schmid B. 1991. Experimentelle Demography von Pflanzen: Solidago altissima.
Meyerson L.A., Mooney H.A. 2007. Invasive alien species in an era of globalization // Front. Ecol.
Environ. - Vol. 5. - № 4. - P. 199--208.
Michalski S. G., Durka W. 2007. Synchronous pulsed flowering: analysis of the flowering phenology
in Juncus (Juncaceae) // Ann. Bot. - Vol. 100. - Is. 6. - P. 1271--1285.
Milanova S.D. 2001. Some morphological and bioecological characteristics of Iva xanthifolia (Nutt.).
Bulgarian Journal of Agricultural Science -- Vol. 7. -- P. 141--146.
Milberg P. 1991. Erucastrum gallicum, a vanishing plant at Lake Takern. Svensk. Bot. Tidskr. 85:
105-109.
Miller C., Kettunen M., Shine C. 2006. Scope options for EU action on invasive aline species (IAS)
Final report for the European Commission.
- Brussels: Institute for European Environmental
Policy (IEEP). - 109 pp.
Miller J. H. 2003. Nonnative invasive plants of southern forests: a field guide for identification and
control. Gen. Tech. Rep. SRS--62. Asheville, NC: U.S. Department of Agriculture, Forest Service,
Southern Research Station. - 93 p.
Milne R. I., Abbott R. J. 2000. Origin and evolution of invasive naturalized material of
Rhododendron ponticum L. in the British Isles // Mol. Ecol. - Vol. 9. - P. 541--556.
485
Mirek Z., Trzcinska-Tacik H. 1982. Spreading of Puccinellia distans along the roads in Southern
Poland / Ekologia polska - Bd 29. -- S. 343--352.
Misiewicz J. et al. 1981. Flora synantropijna swinoujscia I okolicy // Fragm. floristica et geobot. - Bd.
27. -№4. -S.591-605.
Mitich L.W. 1994. Beggarticks // Weed Technology - Vol. 8. - № 1. - P.172--175.
Mitich L.W. 1999. Downy brome, Bromus tectorum L. // Weed Technol. - Vol.13. - № 3. - P. 665--
668.
Mito T., Uesugi T. 2004. Invasive alien species in Japan: the status quo and the new regulation for
prevention of their adverse effects // Global Environmental Research. - Vol. 8. - № 2. - P. 171--
191.
Miyawaki A. 1988. Die veranderung innerhalb der japanischen anthropogenen Vegetation // Flora. --
Bd 180. - № 3-4. - S. 191-201.
Miyawaki Sh., Washitani I. 2004. Invasive alien plant species in riparian areas of Japan: the
contribution of agricultural weeds, revegetation species and aquacultural species // Global
Environmental Research. - Vol. 8. - Is. 1. - P. 89--101.
Moerman D. E. 2009. Native american medicinal plants: an ethnobotanical dictionary. Timber Press. -
800 p.
Monachino J. 1958. Elsholtzia ciliata in New York // Bulletin of the Torrey Botanical Club - Vol. 85.
-№2. --P.139-140.
Moravcová L., Gudžinskas Z., Pyšek P., Pergl J., Perglová I. 2007. Seed ecology of Heracleum
mantegazzianum and H. sosnowskyi, two invasive species with different distributions in Europe //
Ecology and Management of Giant Hogweed (Heracleum mantegazzianum) / Ed. P.Pyšek,
M.J.WCock., H.P.Ravn, W.Nentwig - P. 157--169.
Moravcová L., Jarolímova V., Zákravský P. 2001. Morphological differences and chromosome
numbers in Puccinellia distans and P. limosa populations from Central Europe // Preslia, Praha -
Vol. 73. - № 2. -- P. 161--172.
Moravcová L., Perglová I., Pyšek P., Jarosík V., Pergl J. 2005. Effects of fruit position on fruit mass
and seed germination in the alien species Heracleum mantegazzianum (Apiaceae) and the
implications for its invasion // Acta Oecol. - Vol. 28. - № 1. - P. 1--10.
Morley F.H.W., Katznelson J. 1965. Colonization in Australia by Trifolium subterraneum L. // The
Genetics of colonizing species. - New-York-London: Hafner publishing company. -- P. 269--282.
Morrow L., Stahlman Ph. 1984. The history and distribution of Bromus tectorum in North America //
Weed Science. - Vol. 32. - suppl.1. - P. 2--6.
Moseley C.M., Hagood E.S.Jr. 1990. Horseweed (Conyza canadensis) control in full-season no-till
soybeans (Glycine max) // Weed Technol. - Vol. 4. - P. 814--818.
Mossberg B., Stenberg L. 1999. Den Store Nordiske Flora. Oversat af Jon Feilberg og Bernt Lejtnant. G.E.C.
Gads Forlag, Kоbenhavn. - 710 p.
Mossberg B., Stenberg L. 2003. The Nordic Flora. Den nordiska floran. Wahlstrom and Widstrand.
Mossberg B., Stenberg L. 2004. Den nya nordiska floran. Wahlstrôm and Widstrand. - 928 p.
Mosyakin S.L., Yavorska O.G. 2002. The nonnative flora of the Kiev (Kyiv) urban area, Ukraine: a
checklist and brief analysis // Urban Habitats. - Vol. 1. - № 1. - P. 45--65.
Mountford J.O. 1994. Floristic change in English grazing marshes: The impact of 150 years of
drainage and land-use change // Watsonia. Vol. 20. - № 1. - P. 3--24.
Mucke M. 1908. Uber der Bau und die Entwicklung der Fruchte und uber die Herkunft von Acorus
calamus // Bot. zeit. -- Bd 66. - S. 1--23.
Müller N., Okuda Sh. 1998. Invasion of alien plants in floodplains -- a comparison of Europe and
Japan // Plant Invasions: Ecological Mechanisms and Human Responses / Eds.: U.Starfinger,
K.Edwards, I.Kowarik, M.Williainson - Leiden (Netherlands): Backhzlys Publ. - P. 321--332.
Müllerová J., Pyšek P., Jarošík V., Pergl J. 2005. Aerial photographs as a tool for assessing the
regional dynamics of the invasive plant species Heracleum mantegazzianum // J. Appl. Ecol. -
Vol. 42. - P. 1042--1053.
Muirhead J.R., Leung B., van Overdijk C., Kelly D.W., Kanavillil N., Marchant K.R., MacIsaac H.J.
2006. Modeling local and long-distance dispersal of invasive emerald ash borer Agrilus
planipennis (Coleoptera) in North America. // Diversity and distributions. - Vol. 12. - P.71--79.
Muller S. 2004. Plantes Invasives en France. Paris: Muséum National d'Histoire Naturelle. - 168 p.
(Сollection patrimoines naturels. 62).
486
Mulligan G. A., Findlay J. N. 1970. Reproductive systems and colonization in Canadian weeds // Can.
J. Bot. - Vol. 48. - P. 859-860.
Mulligan G.A. 1957. Chromosome numbers of Canadian weeds. I. Can. J. Bot. 35: 779-789.
Mulligan G.A. 1984. Chromosome numbers of some plants native and naturalized in Canada.
Naturaliste Can. (Rev. Ecol. Syst.) Ill: 447-449.
Mulligan H.A., Findlay J. N. 1974. The Biology of Canadian Weeds. 3. Cardaria draba, C.
chalepensis and C. pubescens // Can. J. Plant Science. - Vol. 54. - P. 149--160.
Mummenhoff K., Bruggemann H., Bowman J. L. 2001. Chloroplast DNA phylogeny and
biogeography of Lepidium (Brassicaceae) // Am. J. Bot. - Vol. 88. - № 11. - P. 2051--2063.
Myers J. A., Vellend M., Gardescu S., Marks P. L. 2004. Seed dispersal by white-tailed deer:
implications for long-distance dispersal, invasion, and migration of plants in eastern North
America // Oecologia. - Vol. 139. - № 1. - P. 35--44.
Myerscough P.J., Whitehead F.H. 1966-1967. Comparative biology of Tussilago farfara,
Chamenerion angustifolium, Epilobium montanum and E.adenocaulon // New Phytologist. - Vol.
65. -№2. -P.192-210;Vol.66. -№4. -P.785-823.
Naganowska B., Krzakowa M. 1984. Karyotype analysis of cheatgrass (Bromus tectorum) // Bull.
Soc. amis sci et lett. Poznan. - Д. 24. - S.105-108.
Nalin R., Normand Ph., Domenach A.-M. 2006. Distribution and N2-fixing activity of Frankia strains
in relation to soil depth // Physiologia Plantarum. - Vol. 99. - № 4. - P. 732--738.
Nasir Y. 1975. Elaeagnaceae // Flora of West Pakistan. - № 85 / Ed. E.Nasir, S.I.Ali - P. 1--6.
Nasir Y.J. 1980. Balsaminaceae // Flora of Pakistan. - № 133 / Ed. E.Nasir, S.I.Ali - P. 1--17.
Natkevičaite-Ivanauskiene M. 1971. Epilobium // Lietuvos TSR flora. Vilnius: Mokslas. Vol. 4.
P. 755-766.
Neilsen H., Handeland S. 2000. Fallopia Adans. // Flora Nordica. - Vol. 1. - P. 273--278.
Nelson R. 1975. Pollen guide for allergy. Spokane (WA): Hollister-Stier Laboratories. - 24 p.
Nesom G. L. 1989. Infrageneric taxonomy of New World Erigeron (Compositae: Astereae) //
Phytologia. - Vol. 67. - P. 67--93.
Nesom G. L. 1995(1994). Review of the taxonomy of Aster sensu lato (Asteraceae: Astereae),
emphasizing the New World species // Phytologia. - Vol. 77. - № 3. - P. 141-297.
Nesom G. L. 2006. Erigeron Linnaeus // Flora of North America. - Vol. 20. - P. 256.
New Threats to North American Forests. A summary of a forum and workshop exploring the impact
of Asian Longhorned Beetle and Emerald Ash Borer on forests and forest-based economies.
February 27, 2003. / Ed. D.A.Hobson, M. L.Tyrrell, A.E.Camp // YFFReview. - Vol. 6. - № 1. -
40 p.
New York Metropolitain Flora Projects, [Electronic resource.] Mode of access:
http://nymf.bbg.org/species/2931. Accessed 17 June 2008.
Nilsen E.T., Muller W.H. 1980. A comparision of the relative naturalization ability of two Schinus
species in Southern California // Bull. Torr. Bot. Club. - Vol. 107. - № 1. - P. 51--56.
Nilsson A. 1976. Spontana gullrishybrider (Solidago canadensis × virgaurea) i Sverige och Denmark.
//Svensk Bot. Tidskr. - Bd 70. - S.7-16.
Nishitani S., Masuzawa T. 1996. Germination characteristics of two species of Polygonum in relation
to their altitudinal distribution on mt. Fuji, Japan // Arctic and Alpine Research. - Vol. 28. - № 1. -
P. 104--110.
Nobanis, 2009. (http://www.nobanis.org/Factsheets.asp). 29.05.2009
Nordhagen R. 1940. Norsk flora. H. Aschehoug and Co., Oslo. 766 p.
North American Wildland Plants: A Field Guide. 2003 / Stubbendieck J. L., Hatch S. L., Landholt L.
M., Rhodes Hays K. L. - Lincoln: Univ. of Nebraska Press. - 501 p.
Novak S., Mack R. 1989. Electrophoretic variation in native range population of Bromus tectorum:
evidence for genetic bottlenecks following introduction // Amer. J. Bot. - Vol.76. - № 6. - Suppl. -
P.151.
Novak S., Mack R., Soltis D. 1991. Genetic variation in Bromus tectorum. Population differentiation
in its North American range // Amer. J. Bot. - Vol. 78. - № 8. - P. 1150-1161.
Noyes R. D. 2000a. Diplospory and parthenogenesis in sexual × agamospermous (apomictic)
Erigeron (Asteraceae) hybrids // Int. J. Plant Sci. - Vol. 161. - № 1. - P. 1--12.
Noyes R. D. 2000b. Biogeographical and evolutionary insights on Erigeron and allies (Asteraceae)
from ITS sequence data // Plant Syst. Evol. - Vol. 220. - P. 93-114.
487
Noyes R. D. 2005. Inheritance of apomeiosis (diplospory) in fleabanes (Erigeron, Asteraceae) //
Heredity. - Vol. 94. - P. 193--198.
Noyes R. D. 2006a. Intraspecific nuclear ribosomal DNA divergence and reticulation in sexual
diploid Erigeron strigosus (Asteraceae) // Am. J. Bot. - Vol. 93. - № 3. - P. 470--479.
Noyes R. D. 2006b. Apomixis via recombination of genome regions for apomeiosis (diplospory) and
parthenogenesis in Erigeron (daisy fleabane, Asteraceae) // Sex. Plant Reprod. - Vol. 19. - P. 7--18.
Noyes R. D., Allison J. R. 2005. Cytology, ovule development, and pollen quality in sexual Erigeron
strigosus (Asteraceae) // Int. J. Plant Sci. - Vol. 166. - № 1. - P. 49--59.
Noyes R. D., Baker R., Mai B. 2007. Mendelian segregation for two-factor apomixis in Erigeron
annuus (Asteraceae) // Heredity - Vol. 98. - P. 92--98.
Noyes R. D., Gerling H., Vandervoort C. 2006. Sexual and apomictic prairie fleabane (Erigeron
strigosus) in Texas: geographic analysis and a new combination (Erigeron strigosus var. traversii,
Asteraceae) // Sida. - Vol. 22. - № 1. - P. 265--276.
Noyes R. D., Rieseberg L.H. 1999. ITS sequence data support a single origin for north american
Astereae (Asteraceae) and reflect deep geographic divisions in Aster s. l. // Amer. J. Bot. - Vol.
86. -№3. -P.398--412.
Noyes R. D., Rieseberg L.H. 2000. Two independent loci control agamospermy (apomixis) in the
triploid flowering plant Erigeron annuus // Genetics. - Vol. 155. - P. 379--390.
Nylander C.-E. 1973. Fynd av Epilobium adenocaulon I Blekinge // Svensk. Bot. Tidskr. - Bd. 67. -
№1. -S.68.
Oh H., Lee S., Lee H. S., Lee D. H., Lee S. Y., Chung H. T., Kim T. S., Kwon T. O. 2002. Germination
inhibitory constituents from Erigeron annuus // Phytochemistry. - Vol. 61. - № 2. - P. 175-179.
Oka H.-I. 1983. Life history characteristics and colonizing success in plants // Amer. Zool. - Vol. 23. -
№ 1. - P. 99--109.
Olenin S., Didziulis V. 2009. Introduction to the species list // DAISIE, Handbook of alien species in
Europe. - Dordrecht: Springer. - P. 129--132.
Oliver Ch.D., Burkhardt E.C., Skojac D.A. 2005. The increasing scarcity of red oaks in Mississippi
River floodplain forests: Influence of the residual overstory // Forest Ecology and Management. -
Vol. 210. - P. 393--414.
Olson Th. E., Knopf F. L. 1986. Agency subsidization of a rapidly spreading exotic // Wildlife
Society Bulletin. - Vol. 14. - № 4. - P. 492--493.
Olukalns A., Berzins A., Lapins D., Lejins A., Sprincina A. 2005. Studies on hogweed (Heracleum)
restriction in Latvia in 2002--2004 // Agronomijas Vēstis (Latvian Journal of Agronomy). - № 8. -
P. 228--232.
Opravil E. 1974. Xanthium strumarium L. -- evropsky archeofyt? // Acta Inst. Bot. Acad. Sci. Slov. -
Ser. A. Tax. Geobot. - Vol. 1. - P. 83--87.
Opravil E. 1983. Xanthium strumarium L.-- ein europäischer Archäophyt? // Flora. - Vol. 173. - P. 71-79.
Oredsson A. 1964. Epilobium adenocaulon, Epilobium adenocaulon x obscurum och E.rubescens i
Matterods socken // Smarre uppsatser och meddelanden. - S.425-426.
Oslon A.R. 1984. Structural aspects of pollination in Amelanchier alnifolia (Maloideae) // Canad. J.
Bot. -Vol.62. -№4. -P.858-864.
Otte A., Maul P. 2005.Verbreitungsschwerpunkte und strukturelle Einnischung der Stauden-Lupine (Lupinus
polyphyllus Lindl.) in der Bergwiesen der Rhôn // Tuexenia. - Vol.25. - P.151-182.
Otte A., Obert S., Volz H., Weigand E. 2002. Effekte von Beweidung auf Lupinuspolyphyllus Lindl. in
Bergwiesen des Biosphârenreservates Rhôn // Neobiota. - Vol. 1. - P.101-133.
Pace L., Tammaro F. 2003. The Main Invasive Alien Plants in the Protected Areas in Central Italy
(Abruzzo) // Advances in Global Change Research. 9. Global Change and Protected Areas / Ed.
G.Visconti, M.Beniston, E.Iannorelli, D.Barba - P. 495--504.
Packet J.G., Ringius G.S. 1984. The distribution and status of Acorus in Canada // Can. J. Bot. - Vol.
62. - № 11. - Р. 2248-2252.
Pallas P.S. 1784-1788. Flora Rossica seu Imperii Rossici per Europam et Asiam indigenarum
descriptioneseticones. --Petropoli. -T.1. --P.1. -80f.;-T.1. --P.2. -114f.
Palmgren O. 1943. Chromosome number in angiospermous plants // Bot. Not. - Р. 348-352.
Parker D., Williamson M. 2003. Low-impact, selective herbicide application for control of exotic
trees in riparian areas: saltcedar, russian-olive and siberian elm. A preliminary field guide.
-
Washington, D.C.: USDA -- FS Southwestern region. ii, - 6 p.
488
Parsons W. T., Cuthbertson E. G. 2001. Noxious weeds of Australia. 2nd ed. - Collingwood: CSIRO
Publishing. - 712 p.
Pauley G.B., Thomas G.L., Thiesfeld S.L., Bonar S.A., Bowers K.L. 1987. An overview of the use of
triploid Grass Carp (Ctenopharyngodon idella) as a biological control of aquatic macrophytes in
Devils Lake, Oregon // Proc. 21st Ann. Meeting Aquatic Plant Control Res. Progr. -- Vicksburg
(Mississippi) -- P. 115-121.
Peace W. J.H., Grubb P. J. 1982. Interaction of light and mineral nutrient supply in the growth of
lmpatiens parviflora DC. // New Phytol. - Vol. 90. - № 1. - P. 127--150.
Peachey E., Ball D., Parker R., Yenish J. P., Miller T. W., Morishita D. W., Hutchinson P. J. S. 2007.
Pacific Northwest Weed Control Handbook. - Corvallis: Oregon State Univ. - 463 p.
Pearce C. M., Smith D. G. 2001. Plains cottonwood's last stand: can it survive invasion of Russian
olive onto the Milk River, Montana floodplain? // Environmental Management. - Vol. 28. - № 5. -
P. 623--637.
Peden D.G. 1982. Factors associated with growth of wild rice in northern Saskatchewan // Arctic. -
Vol. 35. - № 2. - P. 307--311.
Pedersen A. 1956. Rubiaceernes, Polygalaceernes, Linaceernes, Oxalidaceernes og Balsaminaceernes
udbredelse I Danmark //Botanisk tidsskrift -- Bd 47. - P.373--480.
Pernes J. 1987. Incidences du mode de reproduction sur la domestication des cereales // Actes Colloq.
biol. populat. - Lyon. - P. 80--83.
Pessarakli P., Pessarakli M. 2007. Handbook of Turfgrass Management and Physiology. - Tucson:
CRC Press. - 720 p.
Pétillon J., Ysnel F., Canard A., Lefeuvre J.-C. 2005. Impact of an invasive plant (Elymus athericus)
on the conservation value of tidal salt marshes in western France and implications for
management: responses of spider populations // Biol. Conserv. - Vol. 126. - № 1. - P. 103--117.
Petit R. J. 2004. Biological invasions at the gene level // Divers. Distribut. - Vol. 10. - Is. 3. - P. 159--165.
Piernik A. 2006. Growth of three meadow species along a salinity gradient in an inland saline habitat:
transplant experiment // Polish journal of ecology. - Vol. 54. - № 1. - P. 117-126.
Pierre St.R.G., Steeves T.A. 1990. Observations on soot morphology, anthesis, flower namber, and
seed production in the saskatoon, Amelanchier alnifolia. (Rosaceae) // Canad. Field-Naturalist. -
Vol. 104. - № 3. - P. 379-386.
Pierson E.A., Mack R.N. 1990. The population biology of Bromus tectorum in forests// Oecologia. -
Vol. 84. - № 4. - Р. 519-525.
Piispala E. 1964. Epilobium adenocaulon und E.rubescens in Ostfennoskandien // Ann. Bot. Fenn. --
Bd1. -№1. -S.36-46.
Pilate-Andre S., Sandmeier M., Toure A., Pernes J. 1987. Evaluation du polymorphisme enzymatique
des mils penicillaires de l'Afrique de l'Quest // Actes Colloq. biol. populat. Lyon. - P. 352--356.
Pimentel D., Lach L., Zuniga R., Morrison D. 2001. Environmental and economic costs of nonindigenous
species in the United States // BioScience. - Vol. 50. - № 1. - Р. 53--65.
Plant For a Future Database, [Electronic resource.] Mode of access: http://www.ibiblio.org/pfaf/cgi-
bin/arr_html?Elsholtzia+ciliata. 1 Oct. 2008.
Poland Th. M., McCullough D. G. 2006. Emerald ash borer: invasion of the urban forest and the
threat to North America's ash resource // Journal of Forestry. - P. 118--124.
Prach K. 1994. Seasonal dynamics of Impatiens glandulifera in two riparian habitats in central
England // Ecology and management of invasive riverside plants. / eds. L.C.Waal, L.E.Child,
M.Wade - John Wiley and Sons.
Preston C.D. 1988. The spread of Epilobium ciliatum Raf. in the British Isles // Watsonia. - Vol.17. -
P. 279-288.
Preston Ch.D., Pearman D.A., Dines T.D. (eds) 2002. New atlas of the British and Irish flora.
-
Oxford: Univ. Press. - 912 pp.
Preston Ch.D., Pearman D.A., Hall A.R. 2004. Archaeophytes in Britain // Bot. J. Linn. Soc. - Vol.
145. - № 3. - P. 257--294.
Preston R. J. Ju. 1966. North American Trees (exclusive of Mexico and Tropical United States). Rev.
ed. - Cambridge (USA, Mass.): The M.T.I. Press - 395 p.
Priede A. 2008. Distribution of some invasive alien plant species in riparian habitats in Latvia //
Botanica Lithuanica. - Vol. 14. - № 3. - P. 137--150.
Prieditis N. 2002. Latvijas daba (http://www.latvijasdaba.lW)
489
Prina A. O. 1997. Lepidium densiflorum Schrad. (Brassicaceae) in Argentine // Hickenia. - Vol. 2. -
№ 47. - P. 233--235.
Prinz S., Ringi A., Huefner A., Pemp E., Kopp B. 2007. 4'''-Acetylvitexin-2''-C-rhamnoside,
Isoorientin, Orientin, and 8-Methoxylaemp...glucoside as markers for the differentiation of
Crataegus monogyna and Crataegus pentagyna from Crataegus laevigata (Rosaceae) // Chemistry
& Biodiversity - Vol. 4. -- Is. 12. P. 2920--2931.
Proceedings of the Sixieth Annual Meeting of the Northeastern Weed Science Society, [Electronic
resource.] Mode of access: http://www.newss.org/proceedings/proceedings_2006.pdf. Accessed 5
April 2008.
Pude R., Franken H. 2001. Reynoutria bohemica -- eine Alternative zu Miscanthus × giganteus? // Die
Bodenkultur. - Vol. 52. - № 1. - S. 19--27.
Puhalla J., Krans J., Goatley M. 1999. Sports fields: a manual for design, construction, and
maintenance. Hoboken (NJ): John Wiley & Sons. 464 p.
Pullaiah T. 1978. Studies in the Embryology of Compositae III. The Tribe-Astereae // Bot. Mag.
Tokyo. - Vol. 91. - № 1023. - P.197--205.
Pyke D.A. 1990. Comparative demography of co-occurring introduced and native tussock grasses:
persistence and potential expansion // Oecologia. - Vol. 82. - № 4. - P. 537--543.
Pyšek P. 1998. Is there a taxonomic pattern to plant invasions? // Oikos (Copenhagen). - Vol. 82. - №
2. - P. 282--294.
Pyšek P. 2003. How reliable are data on alien species in Flora Europaea? // Flora. - Vol. 198. - № 6. -
P. 499--507.
Pyšek P., Brock J. H., Bímová K. Mandák B., Jarošik V., Koukolíková I., Pergl J., Štĕpánek J. 2003.
Vegetative regenaration in invasive Reynoutria (Polygonaceae) taxa: the determinant of
invasibility at the genotype level // Amer. Jour. Bot. - Vol. 90. - № 10. - P. 1487--1495.
Pyšek P., Hulme P. E. 2005. Spatio-temporal dynamics of plant invasions: linking pattern to process //
Ecoscience. - Vol. 12. - № 3. - P. 302--315.
Pyšek P., Prach K. 1995. Invasion dynamics of Impatiens glandulifera -- a century of spreading
reconstructed // Biol. Conserv. - Vol.74. - № 1. - P. 41--48.
Pyšek P., Prach K., Mandák B. 1998. Invasions of alien plants into habitats of Central European
landscape: an historical pattern // Plant invasions: Ecological mechanisms and human responses /
Ed. U.Starfinger, K.Edwards, I.Kowarik, M.Williamson - Leiden: Backhuys Publishers. - P. 23--32.
Pyšek P., Richardson D. M., Rejmánek M., Webster G. L., Willamson M., Kirschner J. 2004. Alien
plant in checklist and floras: towards better communication between taxonomists and ecologists //
Taxon. - Vol. 53. - № 1. - P. 131--143.
Pyšek P., Richardson D.M., Jarošík V. 2006. Who cites who in the invasion zoo: insights from an analysis
of the most highly cited papers in invasion ecology // Preslia. -- Vol. 78. - № 4. - P. 437--468.
Pyšek P., Richardson D.M., Pergl J., Jarošík V., Sixtová Z., Weber E. 2008. Geographical and
taxonomic biases in invasion ecology // Trends Ecol. Evol. - Vol. 23. - № 5. - P. 237--244
Pyšek P., Sádlo J., Mandák B. 2002. Catalogue of alien plants of the Czech Republic // Preslia. - Vol.
74. -№2. -P.97--186.
Pyšek P., Sádlo J., Mandák B., Jarošík V. 2003. Czech alien flora and the historical pattern of its
formation: what came first to Central Europe? // Oecologia. - Vol. 135. - P. 122--130.
Qasem J. R. 2004. Allelopathic plants: 11. Cardaria draba (L.) Desv. // Allelopathy Journal. - Vol.
13. -№2. -P.165--172.
Qiu D., Wu Z., Yan G., Li Y., Zhov Y. 1997. Study of the ecological restoration of aquatic
macrophytes in a euthrophic shallow like // China J. Oceanol. Limnol. -- Vol. 15, № 1. -- P. 52--60.
Quanru L., Watson M.F. 2008. Oxalidaceae // Flora of China. Vol. 11. P. 1--6.
Quattrocchi U. 2006. CRC world dictionary of grasses: common names, scientific names, eponyms,
synonyms, and etymology. - Vol. 1. A--D. CRC Press. - 2408 p.
Rabinowitz D. 1981. Buried viable seeds in a north american tall-grass prairie: the resemblance of
their abundance and composition to dispersing seeds // Oikos. - Vol. 36. - № 2. - P. 191--195.
Rackham O. 1966. Radiation, transpiration and growth in a woodland annual //Light as an ecological
factor. - Oxford: Blackwell. - P. 167--185.
Radosevich S. R., Holt J. S., Ghersa C. 1997. Weed ecology: implications for management. 2nd ed.
John Wiley and Sons. - 589 p.
490
Rahn K. 1989. A survey of the genus Sorbaria (Rosaceae) // Nordic Journal of Botany -- Vol. 8. --
P. 557--563.
Ramp P.P., Stephenson S.N. 1988. Gender dimorphism in growth and mass partitioning by boxelder
(Acer negundo) //American Midland Naturalist. - Vol.119. - P. 420--430.
Raquibuddowla M.M., Siddiqueullah R.S., Dewan 1967. A. Studies on the solvent extraction of oil
from Acorus calamus // Sci. Res. Dacca. - № 4. - Р.234--239.
Raven P.H. 1968. Epilobium // Florae europaea. Cambridge: Univ. press. - Vol.2. - 455 p.
Raven P.H. et al. 1960. Chromosome numbers in Compositae. I. Astereae //Am. J. Bot. - Vol. 47. - №
2. - P. 124--132.
Raven P.H., Raven T.E. 1976. The genus Epilobium in Australasia: a systematic and evolutionary
study. - Christchurrch: N.Z. Dep. of scientific and industr. research -- 321 p.
Regehr D. L., Bazzaz F. A. 1976. Low temperature photosynthesis in successional winter annuals //
Ecology. - Vol. 57. - № 6. - P. 1297--1303.
Regehr D. L., Bazzaz F. A. 1979. The population dynamics of Erigeron canadensis, a successional
winter annual. // J. Ecol. - Vol. 67. - P. 923-933.
Rehder A. 1949. Manual of cultivated trees and shrubs hardy in North America. 2nd ed. NY: - The
Macmillan Company. - 996 p.
Reinhard F., Herle M., Bastiansen F., Streit B. 2003. Economic Impact of the Spread of Alien species in
Germany. - Berlin - 229 p.
Reynolds S. C. P. 2002. A catalogue of alien plants in Ireland // Glasra. Occasional Papers. - № 14. --
414 p.
Reznik S. Ya., Volkovitsh M.G., Dolgovskaya M. Yu., Cristofaro M. 2004. Introduction of the
ragweed leaf beetle Zygogramma suturalis (Coleoptera, Chrysomelidae) into Russia for biological
control of common ragweed Ambrosia artemisiifolia (Asteraceae) as a model of biological
invasion / Abstr. of Neobiota, 3 International Conference on Biological Invasions (Bern,
Switzerland, 30 September -- 1 October 2004). - P. 111.
Rich Т.С.G. 1991. Crucifers of Great Britain and Ireland. Botanical Society of the British Isles,
London, UK. Handbook No. 6, 336 p.
Richards P. W. 1943. Juncus macer S. F. Gray (Juncus tenuis auct. mult. non Willd.) / Biological
flora of British Isles // Journal of Ecology. - Vol. 31. - № 1. - P. 51--59.
Richardson D.M., Pyšek P. 2006. Plant invasions: merging the concepts of species invasiveness and
community invasibility // Progr. Phys. Geogr. - Vol. 30. - № 3. - P. 409--431.
Richardson D.M., Pyšek P., Rejmánek M., Barbour M.G., Panetta F.D., West C.J. 2000.
Naturalization and invasion of alien plants: concepts and definitions // Divers. Distribut. - Vol. 6.
-№2. -P.93--107.
Ridley H.N. 1930. The dispersal of plants throughout the world. - Ashford (Kent): Reeve. xx, 744 p.
Rieseberg L.H., Sollis D.F., Sollis P.S. 1988. Genetic variation in Helianthus annuus and H.bolanderi
// Biochem. Syst. Ecol. - Vol. 16. - № 4. - P. 393--399.
Riffle J. W. 1977. First report of vesicular-arbuscular mycorrhizae on Elaeagnus angustifolia //
Mycologia. - Vol. 69. - № 6. - P. 1200--1203.
Rikli M. 1903. Die Anthropochoren und der Formenkrcis der Nasturtium palustre D.C. // Ber.
Zürcher. Botan. Ges. - Bd 8. - S. 71--82.
Riska S. 1957. Impatiens parviflora I Grankulla // Mem. Soc. fauna et flora fenn. - Vol. 58. - P.34--49.
Rizk A. M. 1990. Naturally occurring pyrrolizidine alkaloids. - London -- NY: CRC Press. - 240 p.
Roberts H.A., Boddrell J.E. Seed survival and periodicity of seedling emergence in ten species of
annual weeds / Annals of Applied Biology - Vol. 102. -- Is. 3. - P. 523--532.
Robinson B.L. 1911. Erucastrum pollichii adventive in America. Rhodora 13: 10-12.
Robinson W. 1987. The wild garden or the naturalization and natural grouping of hardy exotic plants
with a chapter on the garden of british wild flowers. - NY: Sterling Pub. Co. - 320 p.
Rode D. 2002. Comfrey toxicity revisited // Trends in Pharmacological Sciences. - Vol. 23. - Is. 11. -
P. 497--499.
Rode J.,Mastnak-Culk C.,Wagner T. 1996. Acorus calamus L. in Slovenia // Beitr. Zuchtungsforsch. -
Jg.2. -№1. -S.88--91.
Rodriguez-Saona C., Poland Th.M., Miller J.R., Stelinski L.L., Grant G.G., de Groot P., Buchan L.,
MacDonald L. 2006. Behavioral and electrophysiological responses of the emerald ash borer,
491
Agrilus planipennis, to induced volatiles of Manchurian ash, Fraxinus mandshurica //
Chemoecology. - Vol. 16. - P. 75--86.
Rollins R.C. 1993. The Cruciferae of Continental North America. Stanford University Press,
Stanford, CA. pp. 471-472.
Rollins R.C. 1993. The Cruciferae of continental North America: Systematics of the mustard family
from the Arctic to Panama. - Stanford University Press. - 976 p.
Rosario L. C. 1988. Acer negundo // Fire Effects Information System, [Online]. U.S.
Rosas C.A., Engle D.M., Shaw J.H., Palmer M.W. 2008. Seed dispersal by Bison bison in a tallgrass
prairie // Journal of Vegetation Science. Vol. 19. P. 769--778.
Rose F. 1989. Colour identification guide to the grasses, sedges, rushes and ferns of the British Isles
and north-western Europe. - London: Viking. - 240 p.
Royal Botanic Gardens, Kew [Electronic resource.] Mode of access: http://apps.kew.org/trees/
?page_id=120. Accessed 12 Jan. 2009.
Royer F., Dickinson R. 1999. Weeds of the Northern U. S. and Canada. - Edmonton: The University
of Alberta press. - 434 p.
Russell J., Levin D.A. 1988. Competitive relationships of Oenothera species with different
recombination systems // Amer. J. Bot. - Vol. 75. - № 8. - P. 1175--1180.
Saarisalo-Taubert A. 1962. Kotiutuvia tulokaskasvejamme. Luonnon Tutkija - № 66. P. 22--24.
Sachse U. 1991. Die Populationsbiologie von Acer negundo L., einem agressivem Neophyten in
Eurasien /Postdoktorandenstipendium SA 445/1-1. DFG, - Berlin.
Sala O.E., Chapin F.S. III, Armesto J.J., Berlow E., Bloomfield J., Dirzo R., Huber-Sanwald E.,
Huenneke L.F., Jackson R.B., Kinzig A., Leemans R., Lodge D.M., Mooney H.A., Oesterheld M.,
LeRoy P.N., Sykes M.T., Walker B.H., Walker M., Wall D.H. 2000. Global biodiversity
scenarios for the year 2100 // Science. - Vol. 287. - № 5459. - P. 1770--1774.
Salapa K. 2005. Uprawa alejowa i odnawianie sie, klonu jesionolistaego Acer negundo L. w
zachodniej cze_sci dzielnicy Praga Pohidnie w Warszawie w roku 2004 /Diploma Thesis at the
Faculty of Architecture. University of Ecology and Management in Warsaw.
Santen E., Hill G.D. (eds) 2002. Wild and cultivated lupins from thé tropics to thé pôles // Proceedings of thé
10* International Lupin Conférence, Laugarvatn, Iceland, 19-24 June 2002. International Lupin
Association, Canterbury, New Zealand.
Sanz-Elorza M., Dana E., Sobrino E. 2001. Listado de plantas alóctonas invasoras reales y potenciales
en España. // Lazaroa. - Vol. 22. - P. 121--131.
Schaffrath J. 2001. Vorkommen und spontane Ausbreitung der Rotesche (Fraxinus pennsylvanica
Marshall) in Ost-Brandenburg // Naturschutz und landschaftspflege in Brandenburg. - Bd 10. - №
4. - S. 134--139.
Schantz M. 1959. Uber die Verbreitung und Herkunft von Acorus calamus // Medd. Norsk Farm.
Selskap. -- Bd 21. - S.13--21.
Scheerer H. 1940. Chromosomenzahlen aus der Schleswig-Holsteinischen Flora // Planta. - Bd 30. -
№ 5. - S.716--725.
Schilling E. E. 2006. Helianthus Linnaeus // Flora of North America. - Vol. 21. P. ***--***.
Schlichting C.D., Levin D.A. 1986. Phenotypic plasticity: an evolving plant character // Biol. J. Linn.
Soc. -Vol.29. -№1. -P.37--47.
Schmid В., Puttick G.M., Burgess K.H., Bazzaz F.A. 1988. Correlations between genet architecture
and some life-history features in three species of Solidago // Oecologia - Vol. 75. - P. 459--464.
Scholz H. 1989. Bromus sericeus subsp. fallax, eine neue unterart aus dem vorderen grient (Sinai) //
Willdenowia - № 19. - S. 133--136.
Schroeder F.-G. 1969. Zur Klassifizierung der Anthropochoren // Vegetatio. - Bd 16 - S. 225--238.
Schroeder F.-G. 1970. Exotic Amelanchier species naturalised in Europe and their occurrence in
Great Britain // Watsonia. - Vol. 8. - № 2. - P. 155--162.
Schroeder F.-G. 1972. Amelanchier-Arten als Neophyten in Europa // Abhandlungen
Naturwissenschaftlichen Verein Bremen. - Bd 37. - Heft 3/3. - S. 287--419.
Schulz D. 1987. Zur Ausbreitung der Galinsoga - Arten in Sachsen // Ber. Arbeitsgemeinsch sachs.
Bot. -- Bd 13. - S.65--72.
Schwaegerle K.E., Schaal B.A. 1979. Genetic variability and founder effect in the pitcher plant
Sarracenia purpurea // Evolution (USA). - Vol. 33. - № 4. - P. 1210--1218.
492
Scoggan H.J. 1978. Amelanchier Medik. // The Flora of Canada - Ottawa: National Museum of
Natural Sciences. - Part 3. - P. 904--908.
Scoggan H.J. 1978. Balsaminaceae // The Flora of Canada - Ottawa: Nat. Mus. of Nat. Sci., Nat.
Mus. of Canada - Part 3. - P.1079--1080.
Scoggan H.J. 1978. The Flora of Canada. 3. Dicotyledonae (Saussuraceae to Violaceae). National Muséum
of Natural Sciences, Publications in Botany 7-3. - 1115 p.
Scoggan H.J. 1979. The Flora of Canada. Ottawa: Nat. Mus. of Nat. Sci., Nat. Mus. of Canada; Part 4.
- P. 1131--1136.
Scurfield G. 1962. Cardaria draba (L.) Desv. [Biological flora of British Isles] // Jour. Ecol. - Vol.
50. -№2. -P.489--499.
Sebald O., Seybold S., Philippi G., Worz A. 1998. Die Farn- und Blutenpflanzen Baden-
Wurttembergs. - Ulmer, Stuttgart.
Seidemann J. 2005. World spice plants. 4th ed. Springer. - 591 p.
Sell P. W. 1993. Cardaria Desv. // Flora Europaea. 2nd ed. - Vol. 1. - P. 402.
Semple J. C., Cook R. E. 2006. Solidago L. // Flora North Amer. - Vol. 20. - P. 107--166.
SGA: Sustainable Gardening Australia, [Electronic resource.] Mode of access:
http://www.sgaonline.org.au/invasive_crtaegusmonogyna.html). Accessed 19 Mar. 2008.
Shafroth P. B., Auble G. T., Scott M. L. 1995. Germination and establishment of the native plains
cottonwood (Populus deltoides Marshall subsp. monilifera) and the exotic russian-olive
(Elaeagnus angustifolia L.) // Conservation Biology. - Vol. 9. - № 5. - P. 1169--1175.
Sharma A.K., Cousin M.-T. 1986. Mycoplasmalike Organisms (MLOs) Associated with the
Witches'Broom Diseas / Journal of Phytopathology - Vol. 117. -- Is. 4. - P. 349--356.
Sharma J.K., Heather W.A. 1977. Infection of Populus alba var. hickeliana by Melampsora medusae
Thüm. / European Journal of Forest Pathology. - Vol. 7. -- Is. 2. - P. 119--124.
Sharma S.D., Singh M. 2000. Optimizing foliar activity of glyphosate on Bidens frondosa and
Panicum maximum with different adjuvant types // Weed Res. - Vol. 40. - P.523--533.
Shaw R. H., Seiger L. A. 2002. Japanese knotweed // Biological control of invasive plants in the
Eastern United States / Technical coordinators Bargeron Ch. T., Moorhead D. J., Douce G. K.,
Reardon R. C., Miller A. E. The University of Georgia, USDA APHIS PPQ and USDA Forest
Service, Forest Health Technology Enterprise Team. - P. 159--166.
Sheppard A.W., Shaw R.H., Sforza R. -- 2006. Top 20 environmental weeds for classical biological
control in Europe: a review of opportunities, regulations and other barriers to adoption // Weed
Research.-- Vol. 46. -- P. 93--117.
Sheppard A.W., Shaw R.H., Sforza, R. 2006. Top 20 environmental weeds for classical biological
control in Europe: a review of opportunities, regulations and other barriers to adoption // Weed
Research. - Vol. 46. - P. 93--117.
Sherff E.E. 1937. The genus Bidens // Publications of field museum of natural history. Botany. -
Chicago, U.S.A. - Vol. XVI. P. 17--73.
Shontz N.N., Shontz J.P. 1970. Galinsoga ciliata: its arrival and spread in the northeastern United
States // Rhodora. - Vol.72. P. 386--392.
Shouliang Ch., Guanghua Z. 2006. Tribe Triticeae // Flora of China. - Vol. 22. - P. 386--444.
Sikka K., Sharma A.K. 1979. Chromosome evolution in certain genera of Brassiceae. Cytologia 44:
467--477.
Simpson D.A. 1990. Displacement of Elodea canadensis by Elodea nuttallii in the British Isles //
Watsonia. - Vol. 18. - № 2. - Р. 173--177.
Smejkal M. 1974. Epilobium x novae-civitatis hybr. nova (Epilobium adenocaulon x hirsutum), ein
neuer Bastard // Preslia. - Vol. 46. - № 1. - P.64--66.
Smisek A., Doucet C., Jones M., Weaver S. 1998. Paraquat resistance in horseweed (Conyza
canadensis) and Virginia pepperweed (Lepidium virginiciim) from Essex County, Ontario //
Weed Sci. - Vol. 46. - P. 200--204.
Smith A.H. 1867. On colonies of plants observed near Philadelphia // Proc. Acad. Sci. Philadelphia. -
P. 15--24.
Smith I.M.B. 1981. Colonist ability, altitudinal range and origins of the flora of Mt Field, Tasmania //
J. Biogeogr. - Vol. 8. - № 3. - P. 249--261.
Smith R. D., Aradottir G. I., Taylor A., Lyal C. 2008. Invasive species management -- what taxonomic
support is needed? - Nairobi (Kenya): Global Invasive Spec. Progr. - vi, 44 p.
493
Snell R., Aarssen L.W. 2005. Life history traits in selfing versus outcrossing annuals: exploring the
'time-limitation' hypothesis for the fitness benefit of self-pollination // BMC Ecology. - Vol. 5. -
Is. 2. - P. 2--15.
Sokolowska-Kulczycka A. 1961. Further additions to chromosom numbers of Polish Angiosperms //
Acta Soc. Bot. Pol. - Vol. 30. - № 3. - Р. 463--489.
Solarz W., Chmura D., Krol W., Cierlik G. 2005. Alien species in Poland: an online database // Proc.
of 8th Int. Conf. on the Ecology and Management of Alien Plant Invasions. Katowice (Poland):
Univ. Silesia. - P. 81.
Solarz W., Tokarska-Guzik B., Zajac K., Chmura D., Cierlik G., Krôl W. 2005. Principles of management of
alien species in native fauna and flora. - Institute of Nature Conservation, Krakôw. - 118 p.
Soldano A. 1978-1979(80). Naturalizzazione nel pavese di Amaranthus bouchonii e di altre sette
esotiche nuove per la Lombardia // Atti. Ist. bot. Univ. Lab. Crittog. Pavia. - Vol. 13. - P. 137--143.
Solymosi P. 1983. Study of distribution of some intraspecific Amaranthus taxa in Hungary // Bot.
Kozl. - Vol. 70. - № 1-2. - P. 43--54.
Solymosi P. 1994. Crude plant extracts as weed biocontrol agents // Acta Phytopathol. Entomol. Hungarica. -
Vol. 29. - № 3--4. - P. 361--370.
South D. B. 2000. Tolerance of southern pine seedlings to clopyralid. // Southern J. Appl. For. - Vol.
24. - P. 51--56.
Spies T.A., Barnes B.V. 1981. A Morphological Analysis of Populus alba, P. grandidentata and their
Natural Hybrids in Southeastern Michigan / Silvae Genetica -- Vol. 30. - № 2-3. P. 102--106.
Stace C. 1997. New Flora of the British isles. Second edition. - Cambridge: Univ. Press. - xxvii, 918 p.
Stannard M., Ogle D., Holzworth L., Scianna J., Sunleaf E. 2002. History, biology, ecology,
suppression and revegetation of Russian-olive sites (Elaeagnus angustifolia L.) // USDA--
National Resources Conservation Service − Montana. Plant Materials. № MT--43, Technical
Notes. - P. 1--14.
Steeves T.A., Steeves M.W. 1991. Flower development in Amelanchier alnifolia (Maloideae) //
Canad. J. Bot. - Vol. 69. - № 4. - P. 844-857.
Steichen J. 1995. On the distribution of Puccinellia distans (L.) Parl. on roadsides in Luxembourg //
Bulletin de la societe des Naturalistes Luxembourgeois, 0, 3-8.
Stephan F. 1792. Enumeratio stirpium agri Mosquensis. - Moscuae - 69 p.
Strand J.A., Weisner S.E.B. 2001. Dynamics of submerged macrophyte populations in response to
biomanipulation // Freshwater Biology. ---- Vol. 46, № 10. -- P. 1397--1408.
Strother J. L. 2006. Xanthium Linnaeus // Flora of North America. - Vol. 21. - P. 19--20.
Strother J., Weedon R. 2006. Bidens // Flora of North America. New York: Oxford University Press. -
Vol. 21. - P. 205--218.
Strother J.L. 2006. Ambrosia // Flora of North America. - Vol. 21. www.eFloras.org
Strother J.L. 2006. Cyclachaena // Flora of North America - Vol. 21. P. 27-28 [Electronic resource.]
Mode of access: http://www.eFloras.org. Accessed 16 June 2008.
Sudnik-Wojcikowska B. 1987. Dynamik der Warschauer Flora in den letrten 150 jahren //
Gleditschia. -- Bd 15. - № 1. - S. 7--23.
Sulcs V. 1998. Sikgalvites, galinsogas. //Enciklopedija Latvijas Daba, 5. sej. - Preses names - P. 88 -- 89.
Sullivan K. A., Ellison A. M. 2006. The seed bank of hemlock forests: implications for forest
regeneration following hemlock decline // Journal of the Torrey Botanical Society. - Vol. 133. -
№ 3. - P. 393--402.
Sumner M.J., Remphrey W.R., Martin R. 1999. Pollen development in relation to phenological stages
of inflorescence expansion in Amelanchier alnifolia (saskatoon), with a comparison with buds
forced out of dormancy // Can. J. Bot. / Rev. Can. Bot. - Vol. 77. - № 2. - P. 262--268.
Sunding P. 1989. Naturaliserte Solidago - garter i Norge // Blyttia. - Vol. 47. - P. 23--27.
Suominen puu- ja penskaskvio. 1992. / Eds. L.Hämet-Ahti, A.Palmén, P.Alanko, P.M.A.Tigerstedt -
Helsinki: Dendrologian Seura -- Dendrologiska Sallskaper r. y. - 373 pp.
Sutton R. F., Johnson C. W. 1974. Landscape plants from Utah's mountains. EC-368. Logan, UT. -
135 s.
Svart H.E., Lyck G. 1991. Introducerede planter - forvildede og adventive arter. Second edition.
Skov- og Naturstyrelsen.
Swan D.G., Whitesides R.E. 1988. Downy brome (Bromus tectorum) control in winter wheat // Weed
Technol. - Vol. 2. - № 4. - Р.481--485.
494
Swanton C. J., Cavers P. B. 1988. Regenerative capacity of rhizomes and tubers from two populations
of Helianthus tuberosus L. (Jerusalem artichoke) // Weed Research. - Vol. 28. - Is. 5. - P. 339--
345.
Sydnor T. D., Bumgardner M., Todd A. 2007. The Potential economic impacts of emerald ash borer
(Agrilus planipennis) on Ohio, U.S., Communities // Arboriculture & Urban Forestry. - Vol. 33. -
№1. -P.48--54.
Synthesis of North American Flora. Version 1.0. 1999. Kartesz J. T. -- Scientific Data, Meacham Ch.
A. -- Software Program. Published by the North Carolina Botanical Garden. Pentium 90 MHz
processor; 32 Mb RAM; SVGA display (800 x 600 pixel resolution) with 16 color; Microsoft
Window 3.1, NT, 95, 98 or 2000 operating system; CD-ROM drive for instalation; mouse or
pointing device; keyboard.
Szafer W. 1966. The Vegetation of Poland. Oxford, New York [etc.], Warszawa: Pergamon, PWN.
xxiii, - 738 p.
Szymanowski T. 1960. Kiedy zostaly wprowadzone obce gatunki drzew do uprawy w Polsce?
//Rocznik Dendrologiczny. - Bd14. - S.81--99.
Taiyan Zh., Lianli L., Guang Y., Al-Shehbaz I. A. 2001. Brassicaceae (Cruciferae) // Flora of China.
- Vol. 8. - P. 1--193.
Takahata Y., Hinata K. 1983. Studies on cytodemes in sub-tribe Brassicinae (Cruciferae). Tohoku J.
Agric. Res. 33: 111--124.
Talts S. 1959. Balsaminaceae. // Eesti NSV floora. - Tallinn: Valgus. - Т. 3. - P. 468--476.
Talts S. 1959. Onagraceae // Eesti NSV flora. - Tallinn: Valgus. - Т. 3. - P. 392-432.
Tanavots J. 1969. Lehtpuuliikide introduktsiooni tulemusi TRU botaanikasias. / Diplomitoo. --Tartu. --
93 s.
Tarasoff C.S., Mallory-Smith C.A., Ball D.A. 2007. Comparative plant responses of Puccinellia
distans and Puccinellia nuttalliana to sodic versus normal soil types // Journal of arid
environments. - № 3. -- P.. 403--417.
Taylor S.M.O. 1972. Ecological and genetic isolation of Fraxinus americana and Fraxinus
pennsylvanica / A dissertation submitted in partial fulfillment of the requirements for the degree
of Doctor of Philosophy (Botany) in the University of Michigan.
- 161 p. (http://
www.emeraldashborer.info/files/taylor.pdf).
The Atlas of Florida Vascular Plants // http://www.plantatlas.usf.edu/default.asp
The Giant Hogweed Best Practice Manual. Guidelines for the management and control of an invasive
weed in Europe / Ed. C.Nielsen, H.P.Ravn, W.Nentwig, M.Wade. 2005. Hoersholm (Denmark):
Forest & Landscape. - 44 p.
The Jepson Manual. Higher Plants of California / Ed. James C. Hickman. 1993. Berkeley: Univ. of
California Press. - 1424 p.
Thebaud C., Abbott R. J. 1995. Characterization of invasive Conyza species (Asteraceae) in Europe:
Quantitative trait and isozyme analysis // Am. J. Bot. - Vol. 82. - P. 360-368.
Thellung A. 1905. Einteilung der Ruderal- und Adventivflora in Genetische Gruppen // Die Flora des
Kanton Zürich. - Teil. 1. - Die Ruderal- und Adventivflora des Kanton Zürich / Eds. O.Naegeli,
A.Thellung - Vjschr. Naturforsh. Ges. Kanton Zürich - № 50. - S. 232--236.
Thellung A. 1915. Pflanzenwanderungen unter dem Einfluss des Menschen. //Bot. Jahrb. Syst. Biebl.
Leipzig. - Bd 53 - S. 37--66.
Thellung A. 1918--1919. Zur Terminologie der Adventiv- und Ruderalfloristik. // Allg. Bot. Zeitschr.
-- Bd 24/25 -- S. 36--42.
Thill D.C., Beck G., Callihan R.H. 1984. The Biology of Downy Brome (Bromus tectorum)//Weed
Science - Vol. 32. - suppl.1. - P.7-12.
Thomas A.G., Wise R.F. 1983. Weed survey of Saskatchewan cereal and oilseed crops from 1976 to
1979. Weed Survey Series, Publ. 83-6, Agriculture Canada, Regina, SK. 260 pp.
Thomas A.G., Wise R.F. 1984. Weed survey of Manitoba cereal and oilseed crops 1978, 1979 and
1981. Weed Survey Series, Publ. 84-1, Agriculture Canada, Regina, SK. 230 pp.
Thomas A.G., Wise R.F. 1988. Weed survey of Manitoba cereal and oilseed crops 1986. Weed
Survey Series, Publ. 88-1, Agriculture Canada, Regina, SK. 201 pp.
Thomas A.G., Wise R.F., Frick B.L., Juras L.T. 1996. Saskatchewan weed survey of cereal, oilsed,
and pulse crops in 1995. Weed Survey Series. Agriculture and Agri-Food Canada, Saskatoon, SK.
Publ. 96-1, 419 p.
495
Thompson K., Bakker J. P., Bekker R. M. 1997. The soil seed banks of North West Europe
methodology, density and longevity (with disk). - Cambridge: Univ. Press. - 288 p.
Thompson Sue. Acoraceae // Flora of North America. - New York: Oxford Univ. Press. - Vol. 22.
Tiébré M.-S., Bizoux J.-P., Hardy O. J., Bailey J. P., Mahy G. 2007. Hybridization and
morphogenetic variation in the invasive alien Fallopia (Polygonaceae) complex in Belgium //
American Journal of Botany. - Vol. 94. - № 11. - P. 1900--1910.
Tiébré M.-S., Saad L., Mahy G. 2008. Landscape dynamics and habitat selection by the alien invasive
Fallopia (Polygonaceae) in Belgium // Biodivers. Conserv. - Vol. 117.
Tiébré M.-S., Vanderhoeven S., Saad L., Mahy G. 2007. Hybridization and sexual reproduction in the
invasive alien Fallopia (Polygonaceae) complex in Belgium // Annals of Botany. - Vol. 99. - №
1. - P. 193--203.
Timme R.E., Simpson B. B., Linder C.R. 2007. High-resolution phylogeny for Helianthus
(Asteraceae) using the 18S-26S ribosomal DNA external transcribed spacer // Am. J. Botany. -
Vol. 94. - № 11. - P. 1837--1852.
Tokarska - Guzik B. 2005. The establishment and spread of alien plant species (kenophytes) in the
flora of Poland. Wydawnictwo Uniwersytetu Slaskiego. - Katowice. - 192 p.
Tokarska-Guzik В. 2003. The expansion of some alien plant species (neophytes) in Poland // Plant
invasions: Ecological treats and management solutions. -- Leiden: Backhuys Publishers. - P. 147--167.
Tomaszewski D. 2001. Sorbaria species cultivated in Poland / Dendrobiology - Vol. 46. -- P. 59--64.
Toole E. H., Toole V. K., Borthwick H. A., Hendricks S. B. 1955. Photocontrol of Lepidium seed
germination // Plant Physiology. - Vol. 30. - № 1. - P. 15--21.
Topp W., Kappes H., Rogers F. 2008. Response of ground-dwelling beetle (Coleoptera) assemblages
to giant knotweed (Reynoutria spp.) invasion // Biol. Invasions. - Vol. 10. - P. 381--390.
Torrey J.M.D., Gray A.M.D. 1969. A Flora of North America. - New-York-London: Hafner publ.
comp.; - Vol.2. - P. 413
Torrey J.M.D., Gray A.M.D. 1969. Erigeron //A Flora of North America.
- New-York-London:
Hafner publ. comp. - Vol. 2. - P. 167.
Touchy S. 1857. I'herborisation faite le 10 juimiaux bois de la moure et anport juvenal, et dirigreepar
M.M. Chation et Touchy. Bull. Soc. Fr. - Vol. IV. - P. 592--593.
Traditional medicines for modern times: antidiabetic plants / Ed.: A.Soumyanath 2006. CRC Press. -
314 p.
Tranel P. J., Wassom J. J. 2000. Genetic relationships of common cocklebur accessions from the
United States // Weed Science. - Vol. 49. - № 3. - P. 318--325.
Traveset A., Moragues E. 2004. Effect of Carpobrotus spp. on the pollination success of native plant
species of the Balearic Islands // Biol. Conserv. - Vol. 122. - № 4. - P. 611--619.
Trinajstic I. 1983. Novi podaci a rasprostranjenosti adventivne vrste Galinsoga ciliata u Jugoslaviji //
Fragm. herbol. Jugosl. -- Bd 13.- № 2. - S.63--66.
Trist P.J.O., butler, J.K. 1995. Puccinellia distans (Jacq.) Parl. subsp. borealis (O. Holmb.) W.E.
Hughes (Poaceae) in mainland Scotland and the outer isles. / Watsonia. -- Vol. 20. -- P. 391--396.
Trzcinska H. 1961. Badania nad zasiegami roslin synantropijnych. 1. Bidens melanocarpus w Polsce
// Fragm. floristica et geobot. - Bd. 7. - № 1. - S. 161--168.
Tsuyuzaki S. 2002. Vegetation development patterns on skislopes in lowland Hokkaido, northern
Japan //Biol. Conserv. - Vol. 108. - P. 239--246.
Tu M. 2003. Element stewardship abstract for Elaeagnus angustifolia L. // http:// tncweeds.
ucdavis.edu/esadocs/documnts/elaeang.pdf
Turakainena A. 2001. Galinsogu (Galinsoga spp.) izplatibas biologiskas ierobezosanas petljumi / Magistra
darbs. - Latvijas Lauksaimniecibas Universitate, Jelgava. - P. 1-- 57.
Tutin T.G. 1964. Amaranthaceae // Florae Europaea. Cambridge: Univ.press. Vol.1. P.108--111
Tutin T.G. et al. 1964. Flora Europaea. Vol. 4. Cambridge University Press, Cambridge.
Tutin T.O, Heywood VH, Surges NA, Moore DM, Valentine DH, Walters SM, Webb DA (eds)
(1964-1980) Flora Europaea. Vols. 1-5. Cambridge University Press, Cambridge
Tutin Т.О., Heywood V. H., Burges N. A., Moore D. M., Valentine D. H., Walters S. M., Webb D. A.
1968. Acer negundo Flora Europaea. Vol 2. Cambridge.
Tutin Т.О., Heywood V.H., Burges N.A., Moore D.M., Valentine D.H., Walters S.M., Weeb D.A. (Ed.)
1976. Flora Europaea. Vol. 4. Cambridge University Press. 505p.
496
Tuyama T., Murase A. 1951. On the polymorphism of Erigeron annuus Pers. and E. strigosus Muhl.
II : Situation of rosettes in their life history // Natural science report of the Ochanomizu
University. - Vol. 1. - P. 95--101.
Ungar I. A. 1995. Seed germination and seed-bank ecology in halophytes // Seed development and
germination / Ed. J.Kigel, G.Galili. - P. 599--628.
Upadhyaya M.K. 1986. The biology of Canadian weeds. Bromus tectorum.// Canad. J. Plant Sc. - Vol.
66. -№3. -Р.689--709.
Urbisz A., Urbisz A. 2006. Invasive vascular plant species in the south-western part of Silesian
Upland (south Poland) // Biodiversity Research and Conservation - Vol. 1. - № 2. - P. 143--146.
USDA plants", [Electronic resource.] Mode of access: http://plants.usda.gov/java/ profile?symbol=
sos02. Accessed 15 April 2008.
Vachova M., Ferakova V. 1978. In A. Love, ed. IOPB chromosome number reports. LXI. Taxon
27: 382.
Vachova M., Ferakova V. 1980. In A. Love, ed. IOPB chromosome number reports. LXIX. Taxon
29: 722.
Valentine D.H. 1971. Flower-colour polymorphism in Impatiens glandulifera Royle // Boissiera. -
Vol. 19. - P. 339--343.
Valkova М., Maneva S. 2008. Interactions between Iva xanthifolia (Nutt.) and other weed species /
Herbologia (An international journal of weed research and control). - Vol. 9. - № 1. - P. 11--21.
Van Hall H.C. 1849. Botanische aanteckeningen // Tijdschr. Wetensch. II. P. 126--132.
Vanden Berghen C. 1994. A galonitrophilous plant community with Puccinellia distans along the
roads in Belgium / Naturalistes Belges -- Vol. 75. - P. 41--46.
Vargas J.M., Turgeon A.J. 2003. Poa annua: physiology, culture, and control of annual bluegrass. -
Hoboken: John Wiley and Sons. - 176 p.
Vasić D., Miladinović J., Marjanović-Jeromela A., Škorić D. 2002. Variability between Helianthus
tuberosus accessions collected in the USA and Montenegro // Helia. - Vol. 25. - № 37. - P. 79--84.
Vernon S., Garmory N., Tennant R. 2009. Landscape Architect's Pocket Book. Oxford (UK):
Architectural Press. 299 p.
Vilà M., Corbin J. D., Dukes J. S., Pino J., Smith S. D. 2006. Linking plant invasions to
environmental change // Terrestrial ecosystems in a changing world / Ed. J.Canadell, D.Pataki,
L.Pitelka - Berlin: Springer. - P. 115--124.
Vilà M., Lloret F., Ogheri E., Terradas J. 2001. Positive fire-grass feedback in Mediterranean basin
woodlands // Forest Ecol. Manag. - Vol. 147. - № 1. - P. 3--14.
Vilà M., Tessier M., Suehs C.M., Brundu G., Carta L., Galanidis A., Lambdon P., Manca M., Medail
F., Moragues E., Traveset A., Troumbis A.Y., Hulme Ph.E. 2006. Local and regional assessment
of the impacts of plant invaders on vegetation structure and soil properties of Mediterranean
islands // J. Biogeogr. - Vol. 33. - № 5. - P. 853--861.
Vilà M., Weber E., D'Antonio C.M. 2000. Conservation implications of invasion by plant
hybridization // Biol. Invasions. - Vol. 2. - P. 207--217.
Vincent M.A., Cusick A.W. 1998. New records of alien species in the Ohio vascular flora // Ohio J.
Sci. --Vol.98. -№2. --P.10-17.
von Bothmer R., Jacobsen N., Baden C., Jørgensen R. B., Linde-Laursen I. 1995. An ecogeographical
study of the genus Hordeum. 2nd ed. [Systematic and Ecogeographic Studies on Crop Genepools.
7.] International Plant Genetic Resources Institute. - Rome: IBPGR. - 129 p.
von Schwerin F. 1919. Zur Ehrenrettung des Acer negundo// Mittilungen der Deutschen
Dendrologischen Gesellschaft. -- Bd 28. - S.146--150.
Wade M., Child L., Adachi N. 1996. Japanese knotweed -- a cultivated coloniser // Biological
Sciences Review. - Vol. 8. - P. 31--33.
Wadsworth R.A., Collingham Y.C., Willis S.G., Huntley B., Hulme P.E. 2000. Simulating the spread
and management of alien riparian weeds: are they out of control? // J. Appl.Ecol. - Vol. 37. -
Suppl. 1. - P. 28--38.
Wagenitz G. 1979. Solidago L. // Hegi G. (Ed.). Illustrierte Flora von Mitteleuropa. - Carl Hanser
Verlag, Munchen. P. 16--29.
Wall D.A. 1995. Comparative analysis of three cruciferous weeds: growth, development, and
competitiveness. Weed Sci. 43: 75-80.
497
Wall D.A. 1997. Dog mustard (Erucastrum gallicum) response to crop competition. Weed Sci.
45: 397-403.
Wang H., Li W.-L., Gu Z.-J., Chen Y.-Y. 2001. Cytological study on Acorus in Southwestern China with
some cytogeographical notes on A.calamus // Acta bot. Sinica. - Vol. 43. - № 4. - Р. 354--358.
Wang X. Y., Redmann R. E. 1996. Adaptation to salinity in Hordeum jubatum L. populations studied
using reciprocal transplants // Plant Ecology - Vol. 123. - № 1. - P. 65--71.
Ward J. K., Dawson T. E., Ehleringer J. R. 2002. Responses of Acer negundo genders to Wein K.
1931. Die Einfuhrung nordamerikanischer Geholze in Europa II. // Mittelungen des Deutschen
Dendrologischen Gesellschaft. - Bd 43. - S. 95--154.
Warwick S.I., Anderson J.K. 1993. Guide to the wild germplasm of Brassica and allied crops. Part II.
Chromosome numbers in the tribe Brassiceae (Cruciferae). Agriculture Canada Research Branch
Tech. Bull. Ottawa, ON. 1993-15E, 22 pp.
Warwick S.I., Black L.D. 1993a. Molecular relationships in subtribe Brassicinae (Cruciferae, tribe
Brassiceae). Can. J. Bot. 71:906-918.
Warwick S.I., Sweet R.D. 1983. The biology of canadian weeds. 58. Galinsoga // Can. J. Plant Sci. -
Vol. 63. - P. 695--709.
Warwick S.I., Wall D.A., 1998. The biology of Canadian weeds. Erucastrum gallicum (Willd.) O.E.
Schulz // Canadian Journal of Plant Science, 78(1): 155-165.
Wassom J. J., Tranel P. J., Wax L.M. 2002. Variation among U. S. accessions of common cocklebur
(Xanthium strumarium) // Weed Technology. - Vol. 16. - Is. 1. - P. 171--179.
Watson H.S. 1870. Compendium of the Cybele Britannica, or, British plants in their geographical
relations. - London: Longmans, Green, Reader et Dyer. - vi, 651pp.
Weaver S. 2001. The biology of Canadian weeds. 115. Conyza сanadensis // Can. J. Plant Sci. - Vol.
81. -№4. -P.867--875.
Weaver S. E., Lechowicz M. J. 1982. The Biology of Canadian weeds. 56. Xanthium strumarium L. //
Can. J. Plant Sci. - Vol. 63. - P. 211--225.
Webb D. A. 1985. What are the criteria for presuming native status? // Watsonia. Vol. 15. P. 231--236.
Weber E. 1998. The dynamics of plant invasions: a case study of three exotic goldenrod species
(Solidago L.) in Europe // J. Biogeogr. - Vol. 25. - P. 147-154.
Weber E. 1999. Gebietsfremde Arten der Schweizer Flora -- Ausmass und Bedeutung // Bauhinia. -
Vol. 13. - S. 1--10.
Weber E. 2000. Biological flora of Central Europe: Solidago altissima L. // Flora. Switzerland. - Vol. 195. -
P. 123--134.
Weber E. 2001. Current and potential ranges of three exotic goldenrods (Solidago) in Europe //
Conserv. Biol. - Vol. 15. - P. 122--128.
Weber E. 2003. Invasive plant species of the world: a reference guide to environmental weeds. -
Wallingford (UK): CABI Publ. - 548 pp.
Weber E., Gut D. 2005. A survey of weeds that are increasingly spreading in Europe // Agron.
Sustain. Dev. - Vol. 25. - № 1. - P. 109--121.
Weber E., Jakobs J. 2005. Biological flora of central Europe: Solidago gigantea Aiton // Flora.
Switzerland.
- Vol. 200.
- P. 109--118. http://www.botany.hawaii.edu/staffpages/staff_pubs/
WeberJakobs2004.pdf
Weber E., Schmid В. 1998. Latitudinal population differentiation in two species of Solidago
(Asteraceae) introduced into Europe // Am. J. Bot. - Vol. 85. - P. 1110--1121.
Weber E.F. 1997. The alien flora of Europe: a taxonomic and biogeographic overview // J. Veg. Sci. -
Vol. 8. - P. 565--572.
Webster L. 1992. Origin and distribution of adventive alien flowering plants in Hawai'I // Alien Plant
Invasions in Native Ecosystems of Hawai'i: Management and Research/ Eds С.P.Stone,
C.W.Smith, J.T. Tunison - Honolulu: Univ. Hawaii, Coop. Nat. Park Resources Studies Unit. -
P. 99--154.
Weed identification", [Electronic resource.] Mode of access: http://www.weeds.org.au. Accessed 6
October 2008.
Weed US, Database of Plants, Invading Natural Areas in the United States [electronic resource.] Mode of
access: http://www.invasive.org/weedus/subject:html?sub=5392. Accessed 15 May 2008.
Wein K. 1940. Die alteske Einfuhrungs u Ausbreitungs geschichte von Acorus calamus // Hercynia. --
Bd 3. - S.71--128.
498
Weinmann J.A. 1824. Elenchus plantarum Horti imperialis pawlowskensis et agri petropolitani.
-
Petropoli. -- 472 p.
Weise B. 1966--1967(1968). Untersuchungen uber die Konkurrenzbeziehungen von Impatiens parviflora
und Impatiens noli-tangere // Ber. Arbeitsgemeinsch. Sachsisch. Bot. - Bd 8. - S. 101--122.
Werner P. A. 1977. Colonization success of a "biennial" plant species: experimental field studies of
species cohabitation and replacement // Ecology. - Vol. 58. - № 4. - P. 840--849.
Werner P.A., Bradbury I.K., Gross R.S. 1980. The biology of Canadian weeds. 45. Solidago canadensis L.
//Canadian Journal of Plant Science. - Vol. 60. - № 4. - P. 1393--1409.
Werner P.A., Platt W.J. 1976. Ecological relationships of co-occurring goldenrods (Solidago -
Compositae) // Am. Nat. - Vol. 110. - P. 959--971.
Weston L. A., Barney J. N., DiTommaso A. 2005. A review of the biology and ecology of three
invasive perennials in New York State: Japanese knotweed (Polygonum cuspidatum), mugwort
(Artemisia vulgaris) and pale swallow-wort (Vincetoxicum rossicum) // Plant and Soil. - Vol. 277.
P. 53--69.
Whalley R.D.B., Burfitt J.M. 1972. Ecotypic variation in Avena fatua, A. sterilis and A. barbata in New
South Wales and southern Queensland // Austral. J. Agric. Res. - Vol. 23. - № 5. - P. 799--810.
White J.A., Whitham T.G. 2000. Associational susceptability of cottonwood to a box elder herbivore
// Ecology. - Vol. 81. - P. 1795--1803.
Wicks G. 1984. Integrated Systems for Control and Management of Bromus tectorum // Weed
Science. - Vol. 32. - suppl.1. - P. 26--31.
Wieffering I.H. 1972. Some notes on the diploid chromosome number of the genus Acorus //Acta bot.
Neerl. - Vol. 21. - № 5. - P. 555--559.
Wiersema J. H., León B. 1999. World economic plants: a standard reference. - Boca Raton, Florida:
CRC Press. xxxv, - 749 p.
Wiese A. F., Salisbury C. D., Bean B.W. 1995. Downy brome (Bromus tectorum), jointed goatgrass
(Aegilops cylindrica) and horseweed (Conyza canadensis) control in fallow //Weed Technol. -
Vol. 9. - P. 249--254.
Wilkinson M. J., Stace C. A. 1988. The taxonomic relationships and typification of Festuca brevipila
Tracey and F. lemanii Bastard (Poaceae) // Watsonia. - Vol. 17. - P. 289--299.
Willdenow C. L. 1800. Caroli a Linné Species plantarum: exhibentes plantas rite cognitas, ad genera
relatas, cum differentiis specificis, nominibus trivialibus, synonymis selectis, locis natalibus,
secundum systema sexuale digestas. - T. 3. - Pars 1. - Berlin. - 847 p.
Willdenow C. L. 1804. Species plantarum, exhibentes plantas rite cognitas ad genera relatas cum
differentiis specificis, nominibus trivialibus, synonymis selectis, locis natalibus secundum
systema sexuale digestas. Olim curante Carolo Ludovico Willdenow. - T. 3. - Pars 3. Berolini.
Impensis G. C. Nauk. - P. 1477--2409.
Williams R.D., Winstead J.E. 1972. Populational variation in weights and analysis of caloric content
in fruit of Acer negundo L. // Castanea. - Vol. 37. - № 2. - P. 125--130.
Williamson M. 1996. Biological Invasions. - London: Chapman & Hill. - 244 pp.
Williamson M., Fitter A. 1996. The varying success of invaders // Ecology. Vol. 77. № 6. P. 1661--1666.
Williamson M., Pyšek P., Jarošík V., Prach K. 2005. On the rates and patterns of spread of alien
plants in the Czech Republic, Britain and Ireland // Ecoscience. - Vol. 12. - № 3. - P. 424--433.
Williamson M.H. 2002. Alien plants in the British Isles // Biological invasions: economic and
environmental costs of alien plant, animal and microbe species / Ed. D.Pimentel - Boca Raton:
CRC Press. - P. 91--112.
Willis S. G., Hulme P. E. 2004. Environmental severity and variation in the reproductive traits of
Impatiens glandulifera // Functional Ecology. - Vol. 18. - № 6. - P. 887--898.
Willis S.G., Hulme P.E. 2002. Does temperature limit the invasion of Impatiens glandulifera and
Heracleum mantegazzianum in the UK? // Functional Ecol. - Vol.1 6. - № 4. - Р. 530--539.
Willson M. F. 1986. On the costs of reproduction in plants: Acer negundo. Amer. Midi.
Wilson L. M. 2007. Key to Identification of Invasive Knotweeds in British Columbia. Kamploops (B.
C.): B. C. Min. For. & Range, For. Prac. Br. - 10 p.
Wilson L., Littlefield J. 2006. Proposed host specificity test plant list for potential biological control
agents of hoary cresses Lepidium draba, and L. appelianum, perennial pepperweed, L. latifolium,
and dyer's woad, Isatis tinctoria (Brassicaceae), in the United States and Canada. For submission
499
to USDA -- APHIS -- Technical Advisory Group (TAG) for review. Bozeman (MT): Montana
State University. - 48 p.
Wilson M., Epton H.A.S., Sigee D.C. 1994. Biological control of the blight of hawthorn (Crataegus
monogyna) with Erwinia... under protected conditions / Plant Pathology - Vol. 39.
--Is.2.
-
P. 301--308.
Wintrebert J. 1957. Sur un peuplement d'Impatiens roylei en Haute-Saone // Ann. Scient. Univ.
Besancon. - Ser. 2. - Bd 9. - S. 185.
Witte H. 1909. Nagra bidrag till Kannedomen om vegetationen pa vara ruderatplatser // Svensk
Botanisk Tidskift. - Bd 3. - h. 2. - S.174--182.
Wittig R., Pott R. 1980. Zur Verbreitung, Vergesellschaften und zum Status des Epilobium
adenocaulon in der Westfalischen Bucht // Natur und Heimat. - Bd. 40. - № 3. - S. 83--87.
Wittig V.E., Bernacchi C.J., Xin-Guang Zhu, Calfapietra C., Сeulemans R., Deangelis P., Miglietta
F., Morgan P.B., Long S.P. 2005. Gross primary production is stimulated for three Populus
species grown under free-air CO2 enrichment from planting through canopy closure // Global
Change Biology. - Vol. 11. -- Is. 4. - P. 644--656.
Woodward, Morley,1974 -- клевер в Австралии],
Wu B., Isobe K., Ishii R. 2004. Arbuscular mycorrhizal colonization of the dominant plant species in
primary successional volcanic deserts on the southeast slope of Mount Fuji // Mycorrhiza. - Vol.
14. - P. 391--395.
Wulff H.D. 1940-1941. Uber die Ursache der Sterilitat des Kalmus (Acorus calamus) // Planta. -- Bd
31. -№3. -S.478--491.
Wulff H.D., Stahl E. 1960. Chemische Rassen bei Acorus calamus // Naturwissenschaften. - Bd. 47. -
№5. -S.114.
Wymer C. L., Gardner J., Steinberger Z., Peyton D. K. 2007. Polygonum cuspidatum (Polygonaceae)
genetic diversity in a small region of eastern Kentucky // Journal of the Kentucky Academy of
Science. - Vol. 68. - Is. 1. - P. 89--95.
Yang Z., Strazanac J. S., Marsh P. M., Van Achterberg C., Choi W. 2005. First recorded parasitoid
from China of Agrilus planipennis: a new species of Spathius (Hymenoptera: Braconidae:
Doryctinae) // Annals of the Entomological Society of America. - Vol. 98. - № 5. - P. 636--642.
Zander: Handwörterbuch der Pflanzennamen. 2002 / Eds W.Erardt, E.Götz, N.Bödeker, S.Seybold 17
Auf. - Stuttgart: Eugen Ulmer. - 990 s.
Zapelli I., Fossati T., Patrignani G., Brundi G., Camarda I., Sala F., Castiglione S. 2005. AFLPs to
Assess the Controversial Status of Populus Alba L. of Sardinia // The Role of Biotechnology -
Villa Gualino, Turin, Italy. - P. 165--166.
Zhang X.W., Gui C.X., Chen Ly. 2007. Inhibition of Sorbaria sorbifolia on proliferation of hepatoma
HepG-2 cell line // Zhong Yao Cai -- Vol. 30. - № 6. -- P. 681--684.
Zhou Zh., Miwa M., Nara K., Wu B., Nakaya H., Lian Ch., Miyashita N., Oishi R., Maruta E.,
Hogetsu T. 2003. Patch establishment and development of a clonal plant, Polygonum cuspidatum,
on Mount Fuji // Mol. Ecol. - Vol. 12. - № 6. - P. 1361--1373.
Zigra J. 1817. Ausfuhrliches verzeichnis derjenigen Pflanzen, Baume und straucher, welche in Riga
in dem Garten von J.H.Zigra gezogen werden. -- Riga. -- 62 s.
Zika P.F., Jacobson A.L. 2003. An overlooking hybrid japense knotweed (Polygonum cuspidatum x
sachalinense; Polygonaceae) // Rhodora. - Vol. 105. - № 922. - P. 143--152.
Zinov'eva-Stahevitch A.E., Grant W.F. 1984. Chromosome numbers in Impatiens // Can. J. Bot. -
Vol. 62. - № 12. - Р. 2630--2635.
Zipcodezoo.com [Electronic resource.] Mode of access: http://zipcodezoo.com/plants/E/
Erucastrum_gallicum.asp. Accessed 16 April, 2008.
Zitzer S.F., Dawson J.O. 1992. Soil properties and actinorhizal vegetation influence nodulation of
Alnus glutinosa and Elaeagnus angustifolia by Frankia. // Plant and Soil - Vol. 140. - P. 197--204.
500
ОГЛАВЛЕНИЕ
Предисловие Ю.Ю.Дгебуадзе ........................................................................................ 3
Введение. Экспансия чужеродных видов .................................................................. 6
Глава 1. Понятие «инвазионный вид» и история его формирования .......... 17
Глава 2. Чужеродные и инвазионные виды в Европе ........................................ 28
Глава 3. Гипотезы успешности инвазионных видов ............................................ 44
Глава 4. Микроэволюция инвазионных видов ....................................................... 50
Глава 5. Таксономические проблемы изучения инвазионных видов .......... 76
Глава 6. Биологические особенности инвазионных видов флоры
Средней России ................................................................................................................... 83
Семейство Aceraceae ........................................................................................ 83
Acer negundo L. ............................................................................... 83
Семейство Amaranthaceae ............................................................................. 93
Amaranthus albus L. ...................................................................... 93
Amaranthus retroflexus L. ............................................................ 99
Семейство Apiaceae ........................................................................................ 105
Heraсleum sosnowskyi Manden. ............................................... 105
Семейство Araceae .......................................................................................... 115
Acorus calamus L. ........................................................................ 115
Семейство Asteraceae .................................................................................... 120
Ambrosia artemisiifolia L. .......................................................... 120
Aster × salignus Willd. .............................................................. 130
Bidens frondosa L. ........................................................................ 135
Chamomilla suaveolens (Pursh.) Rydb. ................................. 145
Conyza canadensis (L.) Cronq. ............................................... 151
Cyclachaena xanthiifolia (Nutt.) Fresen. ............................. 161
Erigeron annuus (L.) Pers. ......................................................... 169
Galinsoga parviflora Cav. ........................................................... 174
Galinsoga quadriradiata Ruiz et Pav. ..................................... 181
Helianthus tuberosus L. .............................................................. 188
Senecio viscosus L. ........................................................................ 195
Solidago сanadensis L. ................................................................ 199
Solidago gigantea Ait. ............................................................... 207
Xanthium albinum (Widd) H.Scholz. ..................................... 216
Семейство Balsaminaceae ............................................................................. 223
Impatiens glandulifera Royle. ................................................... 223
Impatiens parviflora DC. ........................................................... 234
Семейство Boraginaceae ............................................................................... 240
Symphytum caucasicum Bieb. ................................................... 240
Семейство Brassicaceae ................................................................................. 247
Cardaria draba (L.) Desv. .......................................................... 247
Erucastrum gallicum (Willd.) O.E. Schulz ........................... 253
Lepidium densiflorum Schrad. ................................................... 260
501
Sisymbrium volgense Bier. ex Fourn. ....................................... 266
Семейство Chenopodiaceae ......................................................................... 273
Atriplex tatarica L. ....................................................................... 273
Семейство Cucurbitaceae ............................................................................. 280
Echinocystis lobata (Michx.) Torr. et Gray ........................... 280
Семейство Elaeagnaceae ............................................................................... 288
Elaeagnus angustifolia L. ........................................................... 288
Hippophae rhamnoides L. ........................................................... 293
Семейство Euphorbiaceae ............................................................................ 300
Euphorbia peplus L. ...................................................................... 300
Семейство Fabaceae ....................................................................................... 306
. Lupinus polyphyllus Lindl. ......................................................... 306
Семейство Hydrocharitaceae ...................................................................... 313
. Elodea canadensis Michx. ........................................................ 313
Семейство Juncaceae ...................................................................................... 326
. Juncus tenuis Willd. .................................................................... 326
Семейство Lamiaceae ..................................................................................... 330
. Elsholzia ciliata (Thunb.) Hyl. ................................................ 330
Семейство Oleaceae ........................................................................................ 336
. Fraxinus pennsylvanica Marsh. ................................................ 336
Семейство Onagraceae .................................................................................. 344
. Epilobium adenocaulon Hausskn. ............................................ 344
. Epilobium pseudorubescens A.Skvortsov ............................... 350
. Oenothera biennis L. .................................................................... 355
Семейство Oxalidaceae ................................................................................. 359
. Oxalis stricta L. ............................................................................. 359
Семейство Poaceae ......................................................................................... 365
. Anisantha tectorum (L.) Nevski ............................................... 365
. Festuca trachyphylla (Hack.) Krajina .................................... 373
. Hordeum jubatum L. .................................................................... 378
. Poa supinа Schrad. ....................................................................... 383
. Puccinelia distans (L.) Parl. ....................................................... 387
Семейство Polygonaceae .............................................................................. 393
. Reynoutria japonica Houtt. ....................................................... 393
. Reynoutria × bohemica Chrtek et Chrtkova ........................ 400
Семейство Rosaceae ....................................................................................... 404
. Amelanchier spicata (Lam.) C.Koch ....................................... 404
. Amelanchier alnifolia (Nutt.) Nutt ......................................... 411
. Crataegus monogyna Jacq. ......................................................... 417
. Sorbaria sorbifolia (L.) A.Br. .................................................... 427
Семейство Salicaceae ..................................................................................... 431
. Populus alba L. .............................................................................. 431
Заключение. Ведение «Черных книг» региональных флор ....................... 440
Литература ......................................................................................................................... 442
502
Научное издание
Юлия Константиновна Виноградова
Сергей Робертович Майоров
Людмила Викторовна Хорун
ЧЕРНАЯ КНИГА
ФЛОРЫ СРЕДНЕЙ РОССИИ
(ЧУЖЕРОДНЫЕ ВИДЫ РАСТЕНИЙ
В ЭКОСИСТЕМАХ СРЕДНЕЙ РОССИИ)
Редактор издательства Н.Б. Пурганская
Макет Н.П. Кураленко
Подписано к печати 08.12.2009
Формат 70х100 1/16. Бумага офсет 1, 80 г/м
Гарнитура Таймс. Печать офсетная. Уч. изд. л. 50,0 п.л.
Тираж 800 экз.
Тип. ВТИИ. Москва. Зак.
.
Издательство ГЕОС
125315, Москва, 1 й Амбулаторный пр., 7/3 114.
Тел./факс: (095) 152 19 14, тел. 230 80 92
E mail: geos@ginras.ru
http://www.geos books.ru