/
































































































































































































































































































Текст
3. Н. КУДРЯШЕВА МИКОЛОГИЯ
С ОСНОВАМИ ФИТОПАТОЛОГИИ
Rebuild by Shadowlabs
ОТ АВТОРА
Учебное пособие «Микология с основами фитопатологии» предназначается для студентов биологических факультетов университетов и пединститутов, а также факультетов защиты растений сельскохозяйственных вузов.
При составлении данного учебного пособия автор стремился сделать его интересным как с микологической, так и с фитопатологической точки зрения. В кем дается общая характеристика грибов как особого отдела растений, в систематическом порядке описываются классы, а в качестве примеров приводятся фитопатогенные организмы—их циклы развития, распространение, вредоносность и меры борьбы.
В связи с тем что в имеющейся микологической литературе классу несовершенных грибов уделяется мало внимания, автор посвящает им значительный раздел в пособии. В этом разделе по системе Саккардо описаны порядки и семейства со ссылкой на главнейшие роды, представители которых вызывают инфекционные заболевания растений.
В настоящем учебном пособии дается общее представление о вирусных, бактериальных и непаразитарных болезнях растений, а также об иммунитете растений к инфекционным заболеваниям.
При составлении пособия автор старался использовать большой фактический материал, накопленный белорусскими микологами и фитопатологами.
Автор благодарен действительному члену АН БССР доктору сельскохозяйственных наук профессору Н. А. Дорожкину за научное редактирование рукописи, а также всем лицам, оказавшим помощь при составлении настоящего учебного пособия.
Глубокую благодарность автор выражает руководителю лаборатории микологии имени профессора А. А. Ячевского ВИЗР, доктору биологических наук профессору М. К. Хохрякову за внимательное рецензирование рукописи и ценные замечания.
Все критические замечания, которые возникнут при знакомстве с книгой, автор примет с благодарностью и постарается учесть в процессе работы со студентами.
Замечания направлять по адресу: г. Минск, Белорусский государственный университет имени В. И. Ленина, биологический факультет, кафедра систематики растений.
Минск, 1967 3. Кудряшева
ВВЕДЕНИЕ
Грибы известны человечеству с глубокой древности. Еще в IV веке до нашей эры древнегреческий ученый Теофраст, друг и ученик Аристотеля, в своих трудах упоминал о таких грибах, как шампиньоны, трюфели, сморчки. Позднее, в I веке нашей эры, знаменитый ученый-натуралист древнего Рима Плиний Старший обратил внимание на обилие трутовиков на стволах живых деревьев и на пнях, причем он совершенно правильно считал их грибами. Плиний отмечал их интересное свойство вызывать ночью свечение пней.
Первые попытки классификации макроскопических грибов принадлежат также Плинию. Основываясь на внешней форме и хозяйственном значении грибов, он делил их на съедобные и ядовитые.
В Риме среди съедобных грибов особенно высоко ценили кесарев гриб, который считался лакомым блюдом на императорских пирах. Римляне были хорошо осведомлены о ядовитых свойствах грибов и умело пользовались ими в тех случаях, когда надо было без особого шума удалить неугодного государственного деятеля или политического противника.
В древнем Риме, где было высоко развито земледелие, обращали внимание и на болезни сельскохозяйственных растений, такие, как головня, ржавчина, мучнистая роса. Однако природа этих болезней не была понятна, а появление их объясняли изменением частей тела самого растения под влиянием условий погоды и называли «сыпями».
Ботаники-натуралисты даже более позднего времени собирали и описывали макроскопические грибы, преимущест
венно шляпочные, но о жизни их еще ничего не знали. Обилие и разнообразие всевозможных грибов сбивало с толку натуралистов. Многие из них даже сомневались, считать ли грибы растениями или животными.
Большая заслуга в развитии науки о грибах принадлежит голландскому профессору ботаники Клаузиусу. В 1578 году вышла его замечательная коллекция цветных изображений 221 вида грибов, получившая название «Кодекса Клаузиуса». Этот альбом послужил пособием многим натуралистам для распознавания грибов.
Однако еще при жизни Клаузиуса альбом бесследно исчез. Через 100 лет в книге другого автора появились изображения грибов, перерисованные из этого альбома, а вскоре нашелся и знаменитый альбом. Кодекс Клаузиуса и поныне представляет большой интерес для науки и хранится в библиотеке Лейденского университета.
Истинная природа грибов не была выяснена и к началу XVIII столетия. Своеобразная форма, отличающая их ог остальных растений, массовое появление в определенное время года, отсутствие семян — все это вызывало сомнение и приводило к спорам, куда следует отнести грибы, к животным или растениям. Часто появление грибов объясняли всякими небылицами: то им приписывали божественное происхождение, то, наоборот, считали порождением дьявола. Известный французский ботаник Вейан, выступая в Париже на собрании ученых, характеризовал грибы как «проклятое племя, изобретение дьявола, придуманное им для того, чтобы нарушать гармонию остальной природы, созданной богом, смущать и приводить в отчаяние ботаников-исследователей».
Микроскопические грибы (головня, ржавчина и др.) либо продолжали считать сыпями, как результат изменения тела растений, либо объясняли «божьей карой».
Все ботаники в то время занимались главным образом цветковыми растениями, их легче было распознавать. Растения, не имеющие цветков, классифицировать было труднее, и многие исследователи относили к ним грибы. Так, например, профессор Парижского ботанического сада Турнефор в своих описаниях причислял грибы к травам и кустарникам без цветков и плодов.
Известный итальянский ботаник Микели считал, что и у грибов есть цветки с тычинками, но без околоцветника (чашечки и венчика). К такому выводу он пришел, исследуя нижнюю часть шляпки. Рассматривая под увеличительным стеклом тонкую пыль, высыпающуюся из грибных шляпок, он посчитал ее семенами. В дальнейшем, исследуя под микроскопом эту пыль и наблюдая за ее прорастанием, Микели сделал большое открытие о размножении грибов. Он доказал,
что грибы возникают в результате прорастания мельчайших крупинок, которые позднее были названы спорами. Своими исследованиями Микели положил конец всяким небылицам о возникновении шляпочных грибов. Исследования Микели пало считать зарождением микологии как науки.
Выдающийся натуралист XVIII века Линней вначале колебался, куда отнести грибы — к животным или к растениям. Он даже высказал предположение о том, что грибы следует считать животными, так как они имеют сходство с полипами. Однако позднее Линней изменил свою точку зрения и определил грибы как незеленые растения.
Строение макроскопических и особенно микроскопических грибов и в более позднее время не привлекало такого большого количества исследователей-ботаников, как строение высших растений. Это объяснялось их малой заметностью, а также сложностью микроскопического изучения их структуры. Поэтому научная систематизация грибов была разработана значительно позднее, чем систематизация высших растений.
Основоположники систематики грибов — голландский исследователь Персоон и профессор университета в Линде (Швеция) Е. Фриз. Работа, проделанная этими учеными, по своему значению приравнивается к работе Линнея по систематике высших растений. Исследования Персоона и Фриза завершают описательный период в развитии микологии, когда исследователи ставили своей задачей только сбор и описание новых форм грибов, а также их классификацию.
К середине XIX столетия микология стала радикально изменять свое направление. Развитие земледелия, его интенсификация предъявляли новые определенные требования к науке. Повышение урожайности сельскохозяйственных растений невозможно было без конкретных знаний причин появления различных болезней растений, поэтому внимание исследователей было направлено на изучение особенностей развития паразитных грибов. Тем самым были заложены научные основы фитопатологии как науки о болезнях растений.
Онтогенетическое направление в развитии всех отраслей ботаники имело большое значение и для микологии. Метод онтогенеза оказался особенно плодотворным при изучении грибов с их сложным циклом развития. При этом значительно изменился и круг объектов изучения. Вместо крупных шляпочных и трутовых грибов исследователи стали заниматься микроскопическими формами и главным образом грибами, паразитирующими на высших растениях.
Изучение грибного организма в процессе его онтогенеза показало, что в цикле развития гриб проходит ряд последовательно сменяющихся стадий, каждая из которых совершенно
не похожа на предшествующую. Экспериментальные исследования мучнисто-росяных, головневых, ржавчинных и других грибов, произведенные братьями Тюлян во Франции, позволили сделать вывод о наличии у грибов плеоморфизма (много-формия), т. е. образования одним видом гриба различных спороношений, настолько отличных между собой, что исследователи более раннего периода принимали их за разные виды.
Исключительно велика роль в развитии микологии выдающегося ученого Антона де Бари. Изучая многие паразитные грибы, он показал, что они являются не следствием болезней растений, а причиной. Де Бари определил и подробно изучил возбудителя болезни картофеля Phytofthora infestans\ различные головневые грибы; проследил цикл развития Puccinia graminis, установив при этом явление двухозяйственности. Большое значение имеют его исследования по сумчатым грибам.
Указанные работы, а также многие другие не только продвинули вперед развитие микологии, но имели и общебиологическое значение.
Де Бари не только талантливый исследователь. Его величайшей заслугой было создание большой школы микологов и фитопатологов, среди которых оказались виднейшие русские исследователи М. С. Воронин, И. Ф. «Шмальгаузен, А. С. Фа-минцин, И. Баранецкий, Ф. М. Каменский, С. Н. Виноградский и др.
Одним из наиболее близких учеников де Бари был М. С. Воронин (1838—1903). Почти три десятилетия продолжалась дружба и плодотворное сотрудничество двух великих биологов XIX века. Совместные труды учителя и ученика обогатили науку и, что самое главное, поставили микологию и фитопатологию в число экспериментальных наук.
Онтогенетическое направление в изучении фитопатогенных организмов стало основным, а исследования патогенеза болезней растений в их единстве со средой способствовали решению животрепещущих практических вопросов.
В более поздние годы М. С. Воронин изучил возбудителя ржавчины подсолнечника — Puccinia helianthi; паразитическую специализацию Plasmodiophora brassicae; открыл и подробно изучил Exobasidium vaccinii на бруснике. Причем Воронин с удивительной прозорливостью высказал мысль, что Е. vaccinii не единственный паразит этого рода на высших растениях. В 1895 году английский исследователь Масси, основываясь на исследованиях Воронина, описал возбудителя болезни чайного куста — паразитный гриб Exobasidium vexans.
Славным продолжателем дела М. С. Воронина был выда
ющийся ученый А. А. Ячевский (1863—1932). В России его деятельность как миколога и фитопатолога началась в 1894 году. В 1907 году им было создано в Петербурге Бюро но микологии и фитопатологии, завоевавшее вскоре репутацию крупного научного центра. Микологический гербарий в лаборатории А. А. Ячевского, по свидетельству крупнейших зарубежных ученых, считался одним из лучших в Европе.
Деятельность А. А. Ячевского как ученого весьма разно-стороняя. Общее количество опубликованных им работ свыше 600. Из них заслуживают быть отмеченными два выпуска: «Микологическая флора России» (т. I — Пероноспоровые, 1901; т. II — Слизевики, 1907), «Определители грибов» (т. I — Совершенные грибы, 1913; т. II — Несовершенные грибы, 1917). Общеизвестна его монография «Основы микологии». Ячевский был автором капитальных работ, посвященных заболеваниям отдельных культурных растений: «Ржавчина хлебных злаков» (1909), «Грибные и бактериальные болезни клевера» (1916), «Болезни хлопчатника» (1931) и др.
Громадное научное наследие Ячевского сыграло большую роль в подготовке кадров микологов и фитопатологов, а сам он по справедливости считается одним из основоположников дела защиты растений в СССР.
С А. А. Ячевским работал В. Г. Траншель (1868—1941) — выдающийся знаток ржавчинных грибов. Его исследования в этой области являются классическими, а капитальный труд «Обзор ржавчинных грибов СССР» служит настольной книгой каждого фитопатолога.
Сотрудник Ячевского видный миколог и фитопатолог Н. А. Наумов (1888—1859) написал свыше 100 работ, среди которых много оригинальных учебников, неоднократно издававшихся на различных языках народов СССР и стран народной демократии. Работы Н. А. Наумова в области систематики мукоровых грибов пользуются мировой известностью.
Одним из старейших русских микологов и фитопатологов является А. С. Бондарцев, начинавший свою работу еще у А. А. Ячевского. Произведенные им микологические и фитопатологические обследования громадных районов нашей Родины (Урал, Крым, Кавказ) дали возможность выявить районы распространения многих грибов. Им издано обширное руководство «Болезни культурных растений и меры борьбы с ними», неоднократно переиздававшееся и долгое время бывшее единственным учебником по фитопатологии. Громадное значение имеет капитальный труд А. С. Бондарцева «Трутовые грибы Европейской части СССР и Кавказа».
Для развития отечественной микологии большое значение шмели работы Л. И. Курсанова (1877—1954). Его учебник
«Микология» до настоящего времени является настольной книгой каждого миколога.
В настоящее время одним из крупнейших микологических центров продолжает оставаться лаборатория имени профессора А. А. Ячевского во Всесоюзном научно-исследовательском институте защиты растений в Ленинграде. Возглавляет лабораторию профессор М. К. Хохряков. Его перу принадлежит около 100 печатных работ по различным вопросам микологии и фитопатологии: токсономии грибов, проблеме вида и видообразования, гельминтоспориозам злаков, борьбе с увяданиями сельскохозяйственных культур и др.
Из ленинградских микологов и фитопатологов, известных своими исследованиями не только в Советском Союзе, но и за рубежом, следует назвать А. С. Бондарцева, Б. П. Василькова, Т. И. Федотову, П. Н. Головина, М. А. Литвинова, С. М. Тупиневича и др.
В Москве микологические и фитопатологические исследования осуществляются под руководством М. В. Горленко, М. С. Дунина, Н. А. Черемисинова и др.
Несмотря на обширность территории Советского Союза, исследования в области микологии и фитопатологии ведутся во всех его климатических зонах и республиках.
На Украине большие исследования проводились А. А. Потебней, Т. Д. Страховым, В. П. Муравьевым. В настоящее время ими руководят В. И. Билай, В. Ф. Пересыпкин, Н. М. Пидопличко, В. Н. Шевченко.
В Молдавской ССР Д. Д. Вердеревским и его учениками ведутся исследования по изучению возбудителей болезней винограда и плодовых.
В Армении большое внимание уделяется изучению гоммоза и вертициллеза хлопчатника, болезней табака, овощных и других культур. Исследовательская работа в республике возглавляется Д. Н. Тетеревниковой-Бабаян и А. А. Бабаян — учениками А. А. Ячевского.
В Грузии исследования по изучению микофлоры винограда, цитрусовых и других культур проводятся учеными во главе с Л. А. Канчавели.
В Азербайджанской ССР основное внимание уделяется болезням хлопчатника, косточковых культур, виноградной лозы. Руководит исследовательскими работами В. И. Ульянищев.
Большая исследовательская работа по изучению микофлоры и фитопатогенных организмов проводится в республиках Средней Азии.
В Прибалтийских республиках также имеются свои научные центры, где широко поставлены миколого-флористические и фитопатологические исследования. Ю. Смародс посвятил всю свою жизнь изучению микофлоры Латвии, а в настоящее
время начатую им работу продолжают многие ученые. А.Мин-гявичус занимает ведущее место среди фитопатологов Литвы. В Эстонии исследовательские работы проводятся под руководством А. Марланда, Э. Пармасто.
Для развития микологических знаний в Белоруссии большое значение имели труды М. И. Медиша и С. М. Тупиневи-ча. Начиная со второй четверти XX века и по настоящее время микологические и фитопатологические исследования возглавляются двумя крупнейшими учеными республики: членом-корреспондентом АН СССР В. Ф. Купревичем и действительным членом АН БССР Н. А. Дорожкиным.
В. Ф. Купревич и большой коллектив его учеников изучают базидиальные грибы, в частности ржавчинные, афиллофоро-вые, агариковые. Среди его многочисленных работ особого внимания заслуживают монографии «Болезни клевера и люцерны» (1954), «Ржавчинные грибы в СССР», т. 1 (сем. Ме-lampsoraceae), «Физиология больного растения в связи с общими вопросами паразитизма» (1947).
В результате многолетних фитопатологических исследований Н. А. До-рожкина и его учеников в условиях Белоруссии изучены грибные паразитные организмы различных сельскохозяйственных культур: зерновых, картофеля, люпина, овощных, плодово-ягодных. На основании изучения биологии возбудителей болезней разработан ряд новых оригинальных методов борьбы с ними. Н. А. Дорожкиным и его учениками опубликовано около 300 работ, из которых следует отметить «Порайонную характеристику заболеваний картофеля в БССР» (1933), «Головневые заболевания зерновых культур» (1934), «Порошистую паршу картофеля» (1936), «Болезни картофеля» (1955), «Вирусные болезни картофеля» (1964), «Болезни люпина» (1965) и др.
За годы Советской власти в Белоруссии подготовлено большое количество специалистов микологов и фитопатологов, которые плодотворно работают во всех уголках республики.
ГЛАВА ОБЩИЕ СВЕДЕНИЯ О СТРОЕНИИ,
ПИТАНИИ И РАЗМНОЖЕНИИ
] ГРИБОВ
МИЦЕЛИЙ И ЕГО ВИДОИЗМЕНЕНИЯ
Строение мицелия. Грибы (Fungi) — бесхлорофильные организмы-— принадлежат к низшим растениям (Tallophyta). Вегетативное тело их — мицелий, или грибница,— имеет нитчатое строение. Нити, или гифы, обладая вершинным ростом, могут быть простыми или разветвленными. Толщина гиф обычно незначительна-—от 1 до 10, реже 20 микрон, а длина их может достигать нескольких десятков сантиметров (в исключительных случаях она измеряется метрами).
Существенным признаком грибницы является отсутствие или наличие в гифах поперечных перегородок. Если гифы не имеют перегородок, такая грибница является по существу одноклетной. При наличии поперечных перегородок грибница называется септированной (многоклетной). Строение грибницы служит основным .признаком, согласно которому грибы делят на низшие '(грибница одноклетная) и высшие '(грибница многоклетная) (рис. 1). Рост грибницы происходит всегда в радиальном направлении по отношению к споре, из которой она получила развитие.
К низшим грибам принадлежат и примитивные формы, у которых мицелий не развит или имеется в зачаточном состоянии. Вегетативное тело таких грибов представляет собой голый комочек протоплазмы или же состоит из одноядерной клетки, от которой отходят зачаточные безъядерные гифы.
Грибы, не имея хлорофилла, питаются гетеротрофно, т. е. поглощают питательные вещества из того субстрата, на котором поселяются. Положение грибницы по отношению к питательному субстрату может быть поверхностным (экзогенная грибница), или же она развивается внутри субстрата (эндогенная грибница). Поглощение питательных веществ может
осуществляться всей поверхностью вегетативного тела гриба пли специализированными для этой цели гифами — гаустория* ми (присосками), которые представляют собой особые выросты, отходящие от гиф к источнику питания. При помощи I аустерий питательные вещества всасываются, а затем посту-
пают в грибницу. Форма и величина гаусторий различна у разных видов грибов (рис. 2).
Грибница весьма чувствительна к изменениям условий окружающей среды, а поэтому в природе встречается гораздо большее количество грибов со скрытой в том или ином субстрате грибницей. У таких грибов на поверхности находятся только органы спороношения.
По продолжительности жизни грибницы все грибы можно разделить на четыре группы:
1. Эфемерные грибы, у которых цикл развития завершается в несколько дней (например, водный гриб сапролегния).
2. Однолетние грибы, заканчивающие цикл своего развития в течение одного календарного года. К ним относятся некоторые виды головневых грибов (твердая головня пшеницы, стеблевая головня ржи).
3. Двулетние грибы. Это большая группа, в которую входят многие виды, паразитирующие на высших растениях. Как правило, они начинают цикл своего развития в начале вегетационного периода на живых тканях растений. Осенью грибница не отмирает, а питается сапрофитно на отмерших тканях в течение зимнего периода. К весне на ней развивается один из видов спороношения, которым заканчивается цикл развития гриба.
4. Многолетние грибы. Также весьма обширная группа. В нее входят многие почвенные сапрофитные грибы микори-зообразователи, большое количество паразитов высших многолетних растений: трутовые грибы, развивающиеся на стволах деревьев, а также паразиты, грибница которых может сохраняться в зимующих органах — корнях, корневищах, луковицах и др.
Рис. 2. Гаустории грибов.
По мере развития мицелия в нем можно наблюдать известную дифференцировку гиф. Одни из них, более тонкие и разветвленные, служат для всасывания питательных веществ из субстрата, а вторые, более толстые и почти не ветвящиеся,— для распространения по субстрату. У некоторых грибов (Rhizopus) такие стелющиеся по субстрату гифы напоминают собой стелющиеся по почве побеги высших растений (усы земляники). В определенных местах они образуют пучки тонких разветвленных гиф, внедряющихся в субстрат наподобие корней.
В процессе эволюционного развития у грибов выработа-
лась способность видоизменять вегетативное тело. Некоторые из таких видоизменений являются своеобразной стадией покоя, в которой гриб может пребывать продолжительное время. Образованию покоящейся стадии предшествует выделение и'з организма излишней воды, а это в свою очередь обеспечивает минимальную чувствительность к неблагоприятным условиям. Видоизменениями мицелия являются оидии, хламидоспоры, мицелиальные тяжи, ризоморфы и склероции.
Оидии. В некоторых случаях мицелий гриба способен распадаться на отдельные части округлой, цилиндрической или
Рис. 3. Видоизменения мицелия:
а —- опдии; б — хламидоспоры; в — мицелиальные тяжи; г — ризоморфы;
д — склероции.
эллипсоидальной формы. Распадению мицелия предшествует образование в нем большого количества поперечных перегородок с перетяжками в этих местах. По мере углубления перетяжек и происходит разделение на отдельные клетки — оидии (рис. 3, а).
Образование оидий свойственно многим грибам. У одних это явление связано с изменениями условий существования, у других пребывание в виде оидий является постоянным наследственным признаком. Такие грибы при любых условиях существования не развивают нормального мицелия. К последней группе относятся представители Saccharomycetales. Существуя в виде оидий, они приобрели способность вегетативно размножаться посредством .почкования, вследствие которого образуются веточки, состоящие из овальных или эллипсоидальных клеток. Такая разновидность вегетативного тела получила название почкующегося мицелия.
Хламидоспоры. Следующей формой видоизменения мицелия и одновременно стадией покоя являются хламидоспоры. Они могут возникать из одноклетного и многоклетного мицелия. При этом протоплазма в отдельных участках или отдельных клетках гифы начинает сгущаться. В местах сгущения появляются выпуклости. Постепенно сгущенные участки покрываются собственной плотной оболочкой, при этом, конечно, нарушается целостность гифы. Промежутки гиф между вновь образовавшимися клетками постепенно отмирают, и вместо обычного мицелия оказывается большое количество его отдельных частей, одетых плотной оболочкой. Хламидоспоры обладают свойством сохранять жизнеспособность в течение продолжительного времени и у некоторых грибов являются обязательной стадией в цикле развития (рис. 3, б).
Мицелиальные тяжи и ризоморфы. У многих представителей высших грибов гифы могут расти параллельно друг другу, склеиваясь между собой слизистым веществом, выделяемым оболочками клеток, или соединяясь посредством коротких анастомозов.1 В результате образуются мицелиальные тяжи ‘(рис. 3, в). В простейшем случае гифы, входящие в состав тяжа, однородны. Однако встречаются такие грибы, у которых наблюдается дифференцировка гиф в тяжах. Наружные, более тонкие гифы, отличаются окрашенными плотными оболочками и малыми просветами. Внутренние имеют больший диаметр, неравномерно утолщенные боковые стенки, отчасти разрушенные поперечные перегородки и, таким образом, представляют своеобразную проводящую систему (домовой гриб). Мицелиальные тяжи домового гриба могут достигать толщины карандаша и нескольких метров длины.
Ризоморфы отличаются более темной окраской поверхностных гиф и способностью ветвиться подобно корням высших растений. Они не чувствительны к изменению экологических условий, так как поверхностные гифы состоят из омертвелых
1 Анастомозы — короткие веточки мицелия, соединяющие гифы между собой.
и легок. Ризоморфы могут распространяться в почве, а также п.| корнях и в нижней части стволов деревьев между корой и древесиной (опенок) (рис. 3, г).
Склероции. Под этим названием А. А. Ячевский описывает «такое уплотнение грибных гиф, при котором получается нн'рдое тело различной формы и объема, состоящее из окрашенной (кутинизированной) оболочки и бесцветной сердцевины, клетки которой заполнены жиром». Согласно этому определению, склероции представляют собой чисто микологическое образование. Развиваются такие склероции из вегетативного мицелия вследствие переплетения и уплотнения его гиф, причем этот процесс сопровождается выделением излишней воды. В зрелом состоянии склероции обычно бедны водой (5—10%) п богаты питательными веществами, особенно жиром ?(до 30%). У одних грибов склероции приобретают более или менее постоянную форму (Claviceps purpurea), у других грибов форма их неопределенна (Sclerotinia libertiana) (рис. 3,<5). Величина склероциев может быть различной — от десятых долей миллиметра до 30 см в диаметре и до 20 кг веса (склероции тропических трутовиков).
У многих грибов в образовании склероциев принимают участие не только грибница, но и ткани субстрата, как например у паразитного гриба Monilinia fructigena, вызывающего плодовую гниль. Гифы гриба пронизывают ткани плода, при этом клетки мякоти съеживаются, подсыхают, или мумифицируются, и не загнивают. Грибница в таком склероции сохраняется в состоянии покоя, а при благоприятных условиях возобновляет свою жизнедеятельность. Продолжительность жизни склероциев, т. е. сохранение их грибницей жизнеспособности, у разных грибов различна. В отдельных случаях она может достигать нескольких лет, а в общем зависит в сильной степени от внешних условий.
СТРОЕНИЕ КЛЕТКИ
Оболочка. Только у самых простейших грибов вегетативное тело представлено голой протоплазмой, т. е. не имеет на своей поверхности твердой оболочки. У таких организмов защитную функцию выполняет поверхностный, уплотненный слой протоплазмы. У остальных грибов клетка покрыта твердой оболочкой.
Молодые клетки грибов, а у одноклетного мицелия растущие концы гиф имеют тонкие, бесцветные, однородные оболочки. По мере старения клетки оболочка утолщается и приобретает различную окраску. Утолщение может происходить двумя путями. В первом случае оболочка утолщается снаружи за счет появления на ее поверхности различных отложе-
2 3. Н. Кудряшева 17
ний в виде шипиков, бугорочков и т. п. Подобные отложения наблюдаются в тех случаях, когда клетка погружена в другую клетку и окружена ее протоплазмой. Эта протоплазма выделяет различные вещества, откладывающиеся на оболочке погруженной клетки. Такие изменения оболочки наблюдаются при формировании спор в спорангиях и сумках.
Во втором случае оболочка утолщается изнутри. Это происходит вследствие жизнедеятельности протоплазмы самой клетки, при этом оболочка часто приобретает слоистость. Нарастание слоев изнутри приводит к сужению полости клетки и наблюдается у грибов в тех частях тела, которым требуется особая прочность (мицелиальные тяжи домового гриба, ножки шляпочных грибов). Оболочка клетки может утолщаться еще вследствие разбухания и ослизнения ее наружных слоев, при этом гифы оказываются погруженными в слизистые чехлы, которые могут склеиваться между собой.
По химическому составу оболочка состоит в основном из углеводов: целлюлозы или ее видоизменения — метацеллюлозы, а также каллезы. У зигомицетов и высших грибов к углеводной основе оболочки присоединяются азотистые вещества, сходные с хитином насекомых. У многих грибов оболочка клетки по мере старения может кутинизироваться, а нередко инкрустироваться оксалатом кальция.
Оболочка спор грибов пропитывается жиром и воском, вследствие чего плохо смачивается водой.-
Протоплазма. Живая клетка содержит протоплазму и ядро. Молодые клетки, а также растущие концы гиф одноклетного мицелия сплошь заполнены зернистой протоплазмой, способной разбухать. По мере роста клетки в ней возникают мелкие вакуоли в виде капелек водного раствора различных веществ. С увеличением клетки увеличиваются и вакуоли, а протоплазма остается в виде тонкого постенного слоя с отходящими от него тяжами, как бы разграничивающими вакуоли. У простейших грибов, а также в зооспорах встречается особый вид вакуолей, получивший название пульсирующих, т. е. ритмически сокращающихся, а затем вновь расширяющихся.
Протоплазма в клетках грибов, так же как и в клетках высших растений, обладает способностью к движению. Ток ее идет всегда в акропетальном направлении. Особенно хорошо заметно движение протоплазмы к верхушечным молодым частям гиф одноклетного мицелия.
Клеточный сок вакуолей представляет запас различных питательных веществ, которые в дальнейшем потребляются протоплазмой. Кроме того, в клеточный сок могут выделяться всевозможные продукты, нерастворимые в воде, как капельки масла. Из включений в протоплазму следует отметить
хондриосомы в виде изогнутых палочек. Некоторые исследователи высказывают предположение, что в хондриосомах । рпбов откладываются тельца метахроматина или валютина, в дальнейшем переходящего в протоплазму. Валютин является одним из основных запасных продуктов в клетках не только грибов, но и некоторых водорослей и бактерий.
Запасными питательными продуктами являются также гликоген и жировые вещества, или липоиды. Гликоген откладывается в протоплазме в мелко раздробленном состоянии. От его присутствия протоплазма приобретает стекловатый вид. Особенно много гликогена содержится в протоплазме сумок. При созревании их гликоген ферментируется, превращаясь в сахар, при этом тургорное давление увеличивается, сумки раскрываются, а споры выбрасываются наружу. Липоиды откладываются в протоплазме в мелко распыленном состоянии, а также в виде более крупных жировых капелек в вакуолях. В протоплазме некоторых трутовых грибов откладываются смолистые вещества в виде бесформенных бурых телец или капель. У некоторых шляпочных грибов (рыжик) смолистые вещества находятся в мелко раздробленном состоянии в особых гифах, называемых млечниками. Их содержимое бывает горьким, острым, реже сладковатым.
Ядро. Грибам свойственны одноядерные и многоядерные клетки. У простейших представителей фикомицетов имеется только одно ядро. Более высоко организованные фикомице-ты, обладающие сильно развитым одноклетным мицелием, имеют очень большое количество мелких (1—1,5 мк) ядер. У высших грибов число ядер в клетках различно. У одних грибов наблюдается строгая одноядерность (мучнисто-росяные), у других первичная одноядерность заменяется двухъ-ядерностью в связи с переходом гриба из одной стадии развития в другую (базидиальные). Наконец, известно много видов, у которых клетки грибницы всегда многоядерные (большая часть сумчатых).
Размеры ядер варьируют от 1 до 10 мк в поперечнике. Наиболее крупные ядра наблюдаются в сумках и базидиях.
В клетках грибов, как и в клетках высших растений, происходит деление ядра: амитоз, митоз (кариокинез) и мейоз.
Амитоз у грибов довольно распространен и может происходить многократно в пределах одной клетки без последующего ее деления. В таких случаях возникают многоядерные клетки. Если деление ядра сопровождается делением всей клетки, то имеет место кариокинетическое деление ядра, обеспечивающее равномерное распределение хроматинового вещества между двумя дочерними клетками.
Мейоз, или редукционное деление ядра, при котором происходит уменьшение вдвое числа хромосом, предшествует
половому процессу, в котором слияние двух гаплоидных клеток (гамет) обеспечивает восстановление количества хроматинового вещества в зиготе.
ТКАНИ ГРИБОВ
Ткани грибов принято называть ложными, так как они образуются не в результате деления клеток во всех направлениях, а вследствие переплетения и срастания гиф мицелия. Однако поперечные срезы с массивных органов грибов — плодовых тел, склероциев и др.— при рассмотрении в микроскоп своим строением напоминают ткани высших растений. Ложная ткань грибов называется плектенхимой. В одних случаях она образована более или менее изодиаметрическими клетками, напоминает паренхиму и носит название параплектенхим ы. В других — клетки гиф, образующих ложную ткань, имеют вытянутую форму наподобие прозенхимных клеток высших растений, а поэтому она называется прозо-плектенхимой.
Представляя различные сочетания гиф, грибные ткани выполняют и различные физиологические функции, а в связи с этим могут быть разделены на такие же группы, как и ткани высших растений.
Образовательная ткань, или меристема. Функцию образовательной ткани выполняют растущие молодые кончики гиф, где происходит деление клеток.
Покровная ткань. Эта ткань у грибов хорошо развита на поверхности плодовых тел, склероциев. Гифы, образующие покровную ткань, могут иметь утолщенные и окрашенные оболочки, часто ослизняющиеся. У многих шляпочных грибов покровная ткань легко сдирается с поверхности шляпки (масленок, зеленка, сыроежка, рядовка и др.).
Механическая ткань. Состоит из параллельно идущих гиф с утолщенными стенками и узкими просветами. Такие гифы наблюдаются в осевых органах: в ножках шляпочных грибов, мицелиальных тяжах домового гриба и других образованиях.
Проводящие элементы. Они образованы маловетвящи-мися гифами с широкими просветами. Нередко боковые стенки их снабжены незначительными кольчатыми утолщениями, а живое содержимое редуцировано. Такие примитивные сосуды можно наблюдать в центральной части мицелиального тяжа домового гриба. В плодовых телах шляпочных грибов имеются гифы, проводящие органические вещества. Анатомической особенностью их является отсутствие поперечных перегородок и наличие на боковых стенках участков с ситовидными отверстиями. Последние обеспечивают поступление пи-
i .игольных и других веществ в проводящие элементы из соседних клеток. Гифы эти имеют живое содержимое, т. е. про-юнлазму и большое количество ядер, а клеточный »сок их бо-r.iT питательными веществами. Подходя к спороносной части шляпки плодового тела, они подносят необходимые для образования спор питательные вещества.
У некоторых шляпочных грибов содержимое таких гиф окрашено и имеет вид тонкой эмульсии из-за наличия мелких капелек смолы и жира (рыжик). Их называют млечными трубками, а содержимое—млечным соком.
ХИМИЧЕСКИЙ СОСТАВ
Отличительной чертой грибов является содержание большого количества воды. У некоторых видов шляпочных грибов оно достигает 90%. Несколько меньше воды в деревянистых плодовых телах трутовиков (60—80%). Наименьшим содержанием воды характеризуются склероции. Эта особенность позволяет им выдерживать неблагоприятные условия.
В состав мицелия грибов входят следующие элементы: углерод — 40%, кислород — 40%, азот 7—8%, водород —2— 3%. Содержание золы в грибах незначительно — в среднем 10% от сухого веса. Благодаря малому содержанию извести и магния большая часть золы растворяется в воде.
Примерно половину от всей массы золы составляет К2О, а затем Р2О5 (25—30%), однако соотношение этих составных частей золы может варьировать у разных грибов. В золе грибов имеются и такие вещества, как Na2O, CaO, SO2, а также незначительное количество магния, железа, алюминия и др.
Из органических веществ грибы содержат белки, углеводы, жиры, органические кислоты, пигменты, смолы, терпены, токсины, витамины.
Белковые вещества лучше изучены у дрожжей и шляпочных грибов. Последние содержат свыше 15% белка от сухого веса, а поэтому понятно значение грибов как пищевых продуктов. Однако далеко не все белки грибов одинаково удобоваримы. Исследования показали, что только около половины их переваривается желудочным соком и может быть усвоено организмом. Говоря о пищевом значении грибов, следует иметь в виду, что благодаря высокому содержанию белка при определенных условиях может происходить расщепление его с образованием ядовитых продуктов—птомаинов.
Из углеводов следует прежде всего отметить клетчатку как основную часть клеточной оболочки. Неудобоваримость грибов отчасти объясняется ее большим содержанием.
Грибы ие содержат крахмала, но имеют полисахарид— гликоген, пропитывающий протоплазму, отчего последняя
приобретает стекловатый вид. Наличие гликогена определяется также иодной реакцией (красно-бурая окраска).
Из сахаров у грибов встречается глюкоза, маннит и трегалоза, ее иначе называют микозой или грибным сахаром. Этот углевод обнаружен более чем у 250 видов грибов, главным образом у шляпочных, а также у спорыньи и мукоровых. Некоторые грибы обладают способностью выделять наружу сахаристые вещества в период конидиальных спороношений. Из таких грибов следует отметить спорынью с ее «медвяной росой», ржавчинные грибы, выделяющие сахаристую жидкость в пикнидиальной стадии.
Жиры весьма распространены у грибов как запасной питательный продукт, но содержатся они обычно в небольшом количестве (1—2%). Исключение составляют жировые дрожжи, у которых содержание жира достигает 30%, и рожки спорыньи с содержанием жира 30—35%. Являясь запасным питательным материалом, жиры откладываются в больших количествах в покоящихся органах и спорах в виде довольно крупных капель, окрашенных пигментами преимущественно в желто-оранжевый цвет. У шляпочных грибов наибольшее количество жира накапливается в гимениальном слое.
Органические кислоты у грибов весьма разнообразны. Среди них много жирных кислот: уксусная, масляная, стеариновая, олеиновая, молочная и др. Они входят в состав жиров. Грибам свойственны фумаровая, яблочная, янтарная, лимонная кислоты. Они встречаются в свободном состоянии и в виде солей. Из специфических кислот, свойственных только грибам, следует отметить гельвелловую, характерную для сморчковых грибов. Эта кислота ядовита, однако при варке плодовых тел сморчков она выщелачивается, и грибы обезвреживаются. Наличие органических кислот у грибов обусловливает кислую реакцию клеточного сока.
Пигменты содержатся у громадного большинства грибов (около 90%), и они довольно разнообразны. Но хлорофилла и антоциана, свойственных высшим растениям, грибы лишены. Особого физиологического значения пигменты грибов не имеют, и многие считают их продуктами отброса. В грибной клетке пигменты могут откладываться в оболочке и в полости самой клетки, а у некоторых видов способны выделяться наружу и окрашивать питательный субстрат, на котором обитает гриб (Fusarium).
Пигменты от зеленовато-желтого до красновато-оранжевого оттенка относятся к группе липохромов. Они отлагаются внутри клетки всегда в связи с капельками масла. Липохромы встречаются в эцидиоспорах и уредоспорах ржавчинных грибов и обусловливают их окраску. Группа красных пигментов встречается в кожице шляпок многих шляпочных грибов:
MWOMopa, сыроежки и др. Эти пигменты характеризуются не-cioiiкостью в водных растворах, но *в засушенном виде сохраняются долго. Бурые, темно-фиолетовые и черные пигменты обычно связаны с клеточными оболочками. Темно-фиолетовую окраску имеют поверхностные гифы рожков спорыньи. В плодовых телах рыжика совместно с желтыми пигментами в незначительном количестве присутствуют темно-фиолетовые.
У грибов широко распространены терпены (ароматические эфирные масла), обусловливающие так называемый грибной запах, свойственный сырым и сушеным грибам (особенно боровикам). Ароматические вещества, вырабатываемые грибами. могут быть строго специфическими для того или иного вида грибов. Так, например, твердая головня пшеницы издаст запах селедочного рассола; перечный запах свойствен лисичке; запахом падали обладает веселка, а запах сирени могут издавать некоторые виды фузариумов. Грибы могут обладать запахом чеснока, свежей муки, корицы, редьки и т. п. У многих грибов запахи усиливаются к периоду созревания спор. В этом случае следует отметить биологическое значение ароматических веществ, привлекающих насекомых и низших животных — переносчиков спор грибов.
Токсины, ядовитые вещества, также вырабатываются клетками грибов. У некоторых шляпочных грибов образующиеся токсины придают ядовитые свойства -плодовым телам и не могут быть извлечены кипячением в воде (бледная поганка, мухомор). У других грибов токсины сравнительно легко переходят в раствор при кипячении. Плодовые тела при этом обезвреживаются, а раствор приобретает токсические свойства (сморчковые).
Подробнее о токсических свойствах отдельных представителей сказано при описании соответствующих классов грибов.
ФЕРМЕНТЫ
Грибы обладают мощной системой ферментов, чем и объясняется приспособленность их к различным источникам питания.
Ферменты грибов имеют узкую специализацию и строго приурочены к действию на определенное вещество. Поскольку в клетках грибов могут происходить сложные процессы синтеза и расщепления веществ, они обладают несколькими ферментами, действующими в строгой последовательности.
Как правило, чем больше специализирован гриб по отношению к определенному субстрату, тем меньшее количество
ферментов имеется в его клетках. И, наоборот, всеядные грибы, способные поселяться на различных субстратах, вырабатывают разнообразный ассортимент ферментов.
Активность ферментов в значительной степени определяется условиями внешней среды: температурой, влажностью, количеством света.
У грибов обнаружены ферменты всех основных групп, т. е. гидролитические, окислительно-восстановительные и бродильные.
Гидролитические ферменты. Среди этой группы следует отметить протеолитические ферменты, наличие которых позволяет грибам использовать в качестве источника питания белковый субстрат. Действие этих ферментов приводит к расщеплению белка с образованием более простых соединений: альбумоз, пептонов и аминокислот. Последние в свою очередь расщепляются под влиянием других ферментов, и в конечном итоге получается мочевина и аммиак. К протеолитическим ферментам относят сычужный фермент, или химазу. Особенно много его у Pholiota mutabilis, Aspergillus oryzae и др.
Карбогидразы, или ферменты, .производящие гидролиз углеводов, также широко распространены среди грибов.
Жизнь грибов в основном связана с растительным субстратом. Основной органический материал, используемый грибами,— это целлюлоза оболочек неодревесневших клеток, пектиновые вещества, склеивающие клетки, и лигнин одревесневших оболочек. Расщепление целлюлозы осуществляется ферментом целлюлазой; пектиновые вещества гидролизуются пектиназой, а лигнин—гадромазой. Конечным продуктом гидролиза всех этих веществ являются простые сахара, потребляемые грибами.
Содержимое растительных клеток богато крахмалом. Гидролиз этого углевода осуществляется амилазой. Тростниковый сахар расщепляется на простые сахара под действием фермента инвертазы.
Но не все перечисленные ферменты в одинаковой степени распространены в разных группах грибов. Целлюлаза свойственна главным образом дереворазрушающим грибам (трутовым) и некоторым почвенным сапрофитам. Гадромазы характерны для домовых грибов. Дрожжевые грибы лишены амилазы, в связи с чем культура их связана с предварительным осахариванием питательного материала. Среди других грибов амилаза широко распространена.
Ферменты, производящие гидролиз жиров, или липазы, также свойственны грибам. Различают экто- и эндолипазы. Первые образуются у грибов, живущих преимущественно на субстратах, богатых жировыми веществами. Примером могут
служить энтомофторовые грибы, паразитирующие на насекомых и разлагающие в первую очередь жировые скопления их тел. Эндолипазы свойственны многим холобазидиальным 1 рпбам: шляпочным, рогатиковым, дождевикам. Под действием эндолипаз жиры превращаются в эмульсию и .потребляются протоплазмой.
Окислительно-восстановительные ферменты. Эти ферменты для грибов менее специфичны. Окисление субстрата может осуществляться присоединением кислорода. Этот процесс происходит при участии оксидаз, содержащихся у дереворазрушающих грибов. Отнятие водорода от субстрата также приводит к его окислению и осуществляется с помощью де-1 идраз, которые обнаружены у некоторых видов пенициллиу-ма и дрожжей. Восстановительные ферменты, или редуктазы, у грибов изучены довольно слабо.
Бродильные ферменты. Бродильные ферменты, или зимазы, свойственны дрожжевым, мукоровым и некоторым несовершенным грибам. Зимазы — это группа ферментов, каждый из которых участвует в определенной части процесса брожения. Лучше всего зимазы изучены у дрожжевых грибов. На их практическом применении основаны целые отрасли промышленности.
ПИТАНИЕ ГРИБОВ
Гетеротрофность грибов в основном выражается в их способности получать углерод из органических соединений. Азот и другие необходимые для питания элементы грибы могут получать как из органических, так и из неорганических соединений. В отношении приспособленности к источникам питания грибы делят на две группы.
Монофаги — приспособившиеся к строго определенным источникам питания. К этой группе принадлежат узко специализированные паразиты высших растений, приуроченные в своем паразитизме к определенным видам, а иногда и сортам растений. Примером таких грибов могут служить мучнисто-росяные, головневые, ржавчинные.
П олиф аги — грибы, обладающие широким диапазоном приспособления к различным источникам питания. Полифагами являются многочисленные сапрофитные формы, способные поселяться в почве и на различных растительных остатках.
Источники углерода. Для подавляющего большинства грибов наилучшим источником углерода является глюкоза или близкие к ней по атомному весу сахара. Питательная ценность других углеводов тем выше, чем легче они подвергаются гидролизу с образованием глюкозы или близких к ней са
харов. Известно, что процесс расщепления может идти только в присутствии ферментов. Если же гриб не обладает необходимыми ферментами, то даже при наличии в среде хорошего источника углеродистого питания, последний оказывается мало пригодным. Так, например, некоторые гименомицеты не могут использовать в качестве питательного материала тростниковый сахар вследствие того, что образуют мало инвертазы. Отсутствие у дрожжей амилазы делает непригодным для них крахмал. Грибы очень хорошо усваивают шестиатомные спирты (маннит), сходные по своему строению с сахарами, а также глицерин и хинную кислоту. На том же основании хорошим питательным материалом являются такие кислоты, как яблочная, винная и лимонная. Муравьиная, уксусная и щавелевая кислоты обладают малой питательной ценностью.
Азотистые органические соединения также могут служить источниками углерода, однако они менее приемлемы, так как при этом разлагается большое количество белка, что приводит к подщелачиванию среды.
Источники азота. В составе мицелия грибов азота примерно в 5—6 раз меньше, чем углерода, вместе с тем азотистое питание для грибов не менее важно, чем углеродное. Азот входит в состав хитина — составной части оболочки клетки, белков, нуклеиновых кислот, большинства ферментов и витаминов.
Источником азотистого питания для грибов могут служить органические (белки, пептоны, аминокислоты) и неорганические (соли аммония, нитриты, нитраты) соединения. Из органических азотистых соединений наибольшее значение имеют те, у которых легко происходит отщепление аммиака, из неорганических — соли аммония. Они легко проникают в клетки, идут на построение аминокислот, из которых образуется белок.
По способности усваивать азот из различных источников грибы делятся на четыре группы:
1. Атмосферный азот может усваиваться незначительным количеством грибов — некоторыми микоризообразователями, развивающими эндотрофную микоризу на брусничных и вересковых. Среди них следует указать роды Rhizoctonia, Pho-та. Способность фиксировать атмосферный азот впервые была обнаружена у паразита свеклы Phoina betae в 1901 году исследователем Саида.
2. Нитратный (NO3') и нитритный (NO2Z) азот способны усваивать не менее 90% грибов — многие сумчатые и несовершенные, некоторые базидиальные (дереворазрушающие). Нитраты и нитриты постепенно восстанавливаются в клетке до аммония. В некоторых случаях нитраты оказываются более подходящим источником азота, чем аммонийные соли.
3. Аммонийные соли усваиваются многими фикомицетами и базидиомицетами. Однако в некоторых случаях потребление азота из аммонийных солей может привести к накоплению аммиака в среде и вызвать отравление организма. Для предотвращения этото явления гриб должен одновременно активно использовать источник углерода, так как в этом случае образуются органические кислоты, которые связывают избыток аммиака.
4. В органических источниках азота нуждаются многие грибы, приспособившиеся к развитию на белковом субстрате (сапролегниевые, трихофитон и др.). Однако облигатно белковых грибов, т. е. таких, которые получали бы азот из белка без предварительного расщепления его, нет. Продукты расщепления — альбумозы, пептоны и особенно аминокислоты используются грибами в качестве источника азота. Грибы обладают избирательной способностью в отношении различных аминокислот.
Для многих грибов единственно хорошим источником азота является аспарагин. Для Aspergillus niger из 22 аминокислот лучшими источниками азота оказываются: аланин, аспарагиновая и глютаминовая кислоты и некоторые другие. У паразитных грибов набор аминокислот сказывается не только на росте грибов, но и на паразитических свойствах мицелия.
Витамины, гормоны, биос. Для нормального развития грибов естественный субстрат или искусственная среда должны содержать витамины и гормоны. С другой стороны, грибы способны самостоятельно синтезировать некоторые витамины и накапливать их в довольно больших количествах.
Растительные ткани всегда богаты витаминами и гормонами (ауксином), а поэтому рост и развитие грибов на этих естественных субстратах происходит значительно лучше, чем на питательных средах.
Для роста грибов наиболее необходимыми витаминами являются витамин Bi, аневрин или тиамин. Витамин содержится в большом количестве в хлебных дрожжах, которые применяются в лечебных целях как источник этого витамина.
Наличие витамина А, являющегося производным каротина, также характерно для грибов. Каротин содержится во многих грибах: он обусловливает цвет эцидио- и уредоспор ржавчинных, а также присутствует в гаметах многих других грибов.
Для грибов большое значение имеют вещества группы «биоса» — биологически активного начала, содержащегося в клетках дрожжей и способствующего их росту. Биос был открыт Вильдье в 1901 году. Впоследствии оказалось, что это
вещество представляет собой смесь водорастворимых витаминов группы В.
Минеральное питание. Кроме углерода, азота, водорода и кислорода, грибам необходим целый ряд элементов, составляющих так называемое минеральное питание. К ним относятся сера, фосфор, калий, магний, железо, цинк и др. В золе мицелия грибов можно обнаружить не менее 50 элементов, однако далеко не все из них необходимы для жизни грибов. В грибной клетке зольные элементы находятся в виде минеральных солей, либо входят в состав органических веществ.
Сера играет большую роль в жизни грибов, так как она входит в состав некоторых аминокислот, которые в свою очередь являются компонентами белков. Кроме того, сера входит в состав некоторых витаминов, а также служит стимулятором протеолитических ферментов.
Фосфор необходим для построения ядерных белков нукле-протеидов. В последнее время обнаружено, что фосфорные соединения входят в состав клеточной оболочки некоторых дрожжевых грибов.
Калий участвует в углеводном обмене. При недостатке его у некоторых грибов (аспергилловые) накапливается щавелевая кислота.
Микроэлементы—железо, медь, цинк, марганец, молибден и др.— также необходимы для нормального роста и развития грибов, так как участвуют в структуре ферментов.
Таким образом, грибы поглощают из окружающей среды все необходимые им вещества в той или иной форме. В клетках грибов при участии ферментов происходят сложные физиологические процессы ассимиляции новых веществ, из которых грибы строят свое тело. У них, как и у всех живых организмов, наряду с процессами ассимиляции происходят не менее важные процессы диссимиляции.
ДИССИМИЛЯЦИЯ У ГРИБОВ
В основе процесса диссимиляции лежит расщепление органических веществ, при котором происходит освобождение энергии. У грибов эта энергия потребляется на процессы роста и ассимиляции, а избыток ее может выделяться наружу.
Самым обычным и распространенным процессом диссимиляции является кислородное дыхание, при котором органические вещества в организме окисляются с выделением воды, углекислоты и энергии. В процессе дыхания грибы могут окислять разнообразные органические вещества, но наилучшим дыхательным материалом служат углеводы.
Грибы характеризуются высокой энергией дыхания. Последняя определяется количеством выделенной углекислоты
mi» «пюшению к собственному сухому весу. Обладая сравни-|с.чыю малым запасом балластных материалов, грибы имеют иг шачительный сухой вес.
Энергия дыхания находится в тесной связи с возрастом |рпба, условиями окружающей среды и характером дыха-i глиного материала.
Отношение объема выделенной углекислоты к объему по-। лощенного кислорода (СО2 : О2) называется д ы х а т е л ь-п м м коэффициентом. У грибов он обычно меньше единицы. Избыток кислорода, остающегося в свободном виде, идет на окисление различных веществ, накапливающихся в 1|)пбном организме (органических кислот и прежде всего щавелевой) .
Процесс дыхания ведет, как известно, к накоплению энер-i ни, которая выражается в повышении температуры и выделении тепла. Это можно заметить по нагреванию плодовых гел у шляпочных грибов.
Диссимиляционные процессы у грибов могут сопровождаться выделением света. Свечение свойственно главным образом базидиальным грибам (гименомицетам) и немногим сумчатым. Некоторые тропические шляпочные грибы излучают такой сильный свет, что ими пользуются как фонарями. У этих грибов свечение присуще главным образом гименофору (пластинкам).
В наших широтах свечение обнаружено на гнилых пнях, пронизанных мицелием сумчатого гриба Chlorosplenium, причем гнилушки излучают слабый синевато-зеленоватый свет. Светиться могут опавшие листья бука, клена, дуба, пронизанные грибницей базидиальных грибов Collibia, Marasmius и некоторых других. Наблюдается также свечение ризоморф опенка (Armillaria mellla), причем установлено, что светятся только самые молодые части ризоморф, излучая слабый белый свет. Остальные части тела опенка свечением не обладают. Свечение грибов является результатом процесса окисления особого органического вещества люциферина. Этот процесс может происходить при наличии фермента люциферазы, в присутствии воды и при достаточной температуре (оптимум 10—15°).
Грибам свойственно окислительное брожение — процесс неполного окисления органических веществ, при котором выделяется энергия, потребляемая в дальнейшем грибом. Чаще всего в результате окислительного брожения образуется щавелевая кислота, иногда лимонная. Процесс кислотообразо-вания у грибов широко распространен и дает им возможность успешно конкурировать с бактериями.
У некоторых грибов диссимиляционным процессом является настоящее брожение. Сюда следует прежде всего отнести
спиртовое брожение, которое могут вызывать многие грибы: дрожжевые (сем. Saccharomycetaceae), некоторые мукоро-вые, Manilla из несовершенных.
ВЛИЯНИЕ ВНЕШНИХ ФАКТОРОВ НА РОСТ И РАЗВИТИЕ ГРИБОВ
Для нормального развития грибов, кроме наличия питательного субстрата, требуется определенное сочетание факторов внешней среды — влажности, температуры, притока кислорода, освещения и т. п.
Вода. Известно, что грибы содержат большой процент воды, однако потребность в ней не у всех грибов одинакова. Водные грибы нормально развиваются при условии полного погружения мицелия в воду (Saprolegnia и др.). Размножение их также происходит в воде. Наземные формы грибов требуют значительной влажности субстрата в период развития вегетативного тела. Однако некоторые виды могут существовать при неполном насыщении субстрата водой. Примером может служить домовой гриб (Serpula lacrymans), развивающийся на древесине, влажность которой составляет 22—60% от ее сухого веса.
Значительно меньшую потребность в воде испытывают наземные грибы при формировании покоящейся стадии — склероцнев, а также в период спороношения. Вместе с тем влажность атмосферы всегда должна оставаться высокой, так как у грибов отсутствуют какие-либо приспособления, препятствующие испарению воды.
Быстрый рост плодовых тел шляпочных грибов также требует повышенной влажности воздуха. Следует отметить, что некоторые паразитные грибы (мучнисто-росяные) отличаются повышенной способностью к спорообразованию при сравнительной сухости атмосферы (относительная влажность — 55%). Интенсивное развитие гриба в этом случае объясняется понижением сопротивляемости растения, ослабленного засухой.
В пустынных районах существуют формы грибов, приспособившиеся к росту только во время дождливого периода.
Температура. Амплитуда колебаний температуры, при которой может идти развитие большинства грибов, имеет довольно широкие пределы: от 1 до 35°, при оптимуме 20—25°. Однако у отдельных видов оптимум может смещаться в ту или иную сторону. У головневых он лежит несколько ниже (10—20°); Macrosporium tomato — возбудитель коричневой пятнистости листьев томатов — является теплолюбивым организмом. Оптимум его развития находится между 25 и 30°
I 'pergillus fumigatus, участвующий в процессе самонагрева-ни*,। ссна и хлопка, выдерживает температуру 50°.
Температура может оказывать влияние на интенсивность прорастания спор грибов. Споры твердой головни пшеницы пнгснсивнее прорастают при температуре почвы 8—9°, а у । солевой головни температурный оптимум для прорастания । нор лежит значительно выше (19—21°). Нагревание спор Neurospora sitophila до 40—50° стимулирует их прорастание.
Характер прорастания спор некоторых грибов в известной < юнени также зависит от температуры. Спорангии Phytophtho-ni infestans прорастают сразу же с образованием мицелия при температуре 23°, а при 13° они дают зооспоры.
Кислород. Облигатных анаэробов среди грибов неизвестно. Однако некоторые из них могут выдерживать отсутствие кислорода сравнительно продолжительное время. Так, например, Fusarium oxysporum выживает в анаэробных условиях в течение 13 недель.
Большинство грибов лучше растет при достаточном доступе кислорода. Потребность в кислороде определяется качеством питательного субстрата, структурой почвы, а для паразитных грибов — физиологическими особенностями растения-хозяина.
Свет. Мицелий гриба может расти как на свету, так и в i емноте. Развитие органов спороношения гораздо успешнее идет при наличии света. Фототропическую реакцию спороносных органов можно наблюдать у Pilobolus crystallinus, у которого изгибание спорангиеносцев и «отстреливание» спорангиев идет в направлении источника света. У некоторых пире-номицетов хоботок перитеция также изгибается в сторону падающего света. Полное отсутствие света может вызвать стерильность грибницы Pilobolus, Coprinus и др. Известны виды грибов, приспособившиеся к развитию спороносных органов и в темноте: домовой гриб, шампиньон, аспергилло-вые и др.
Свет влияет на пигментообразование. Его действие можно проследить на плодовых телах некоторых шляпочных грибов, быстро приобретающих окраску при выходе на поверхность почвы.
Споры многих грибов отличаются темной окраской. Эту особенность следует рассматривать как приспособительную в отношении прямого действия солнечных лучей, которое может оказаться губительным во время переноса спор воздушными течениями.
Кислотность среды. Большинство грибов имеет оптимум развития при слабокислой реакции среды (pH 5,0—6,0). Однако1 некоторые из них приспособились к таким субстратам, которые представляют среду либо более кислую, либо щелоч
ную. Среди таких грибов следует отметить Penicillium, поражающий цитрусовые. Он успешно растет при очень высоком pH, равном 3,0.
Сапролегния, обитающая в воде на мертвых насекомых и рыбной икре, приспособилась переносить щелочную реакцию.
РАЗМНОЖЕНИЕ
У грибов известны все виды размножения: вегетативное, бесполое и половое.
Вегетативное размножение. У грибов широко распространено вегетативное размножение. В основе его лежит способность организма к регенерации. Вегетативное размножение может осуществляться отделенными кусочками грибницы, а также ее специализированными частями, как оидии и хламидоспоры.
Бесполое размножение. Оно происходит посредством особых спор. Процесс спорообразования требует много пластического материала, а поэтому может совершаться только на хорошо разросшемся мицелии. Такой мицелий дает резко обособленные специализированные гифы, на которых и происходит образование спор. Последние могут возникать эндогенно и экзогенно.
Эндогенные споры образуются в особых вместилищах — спорангиях, возникающих на концах специальных гиф — спорангиеносцах. Эндогенное развитие спор наблюдается у низших грибов и у некоторых несовершенных, а экзогенное свойственно немногим низшим и всем высшим грибам.
У наиболее примитивных форм низших грибов эндогенные споры представлены голым комочком протоплазмы с одним ядром и одним или двумя жгутиками, которые обеспечивают активное передвижение. Такие споры получили название зооспор (рис. 4, а).
Грибы с более высокой организацией, но принадлежащие к низшим, образуют неподвижные споры, представляющие большей частью многоядерные клетки, одетые твердой оболочкой, часто окрашенной. Эти споры называются спорангиоспорами (рис. 4 6).
Для распространения зооспор необходима водная среда, а поэтому они свойственны водным грибам или таким наземным формам, которые еще не потеряли окончательно связи с водной средой и в период своего размножения требуют наличия капельно-жидкой влаги в виде дождя или росы.
Спорангиоспоры образуются у типично наземных видов грибов. Распространение их осуществляется воздушными течениями после вскрывания спорангия.
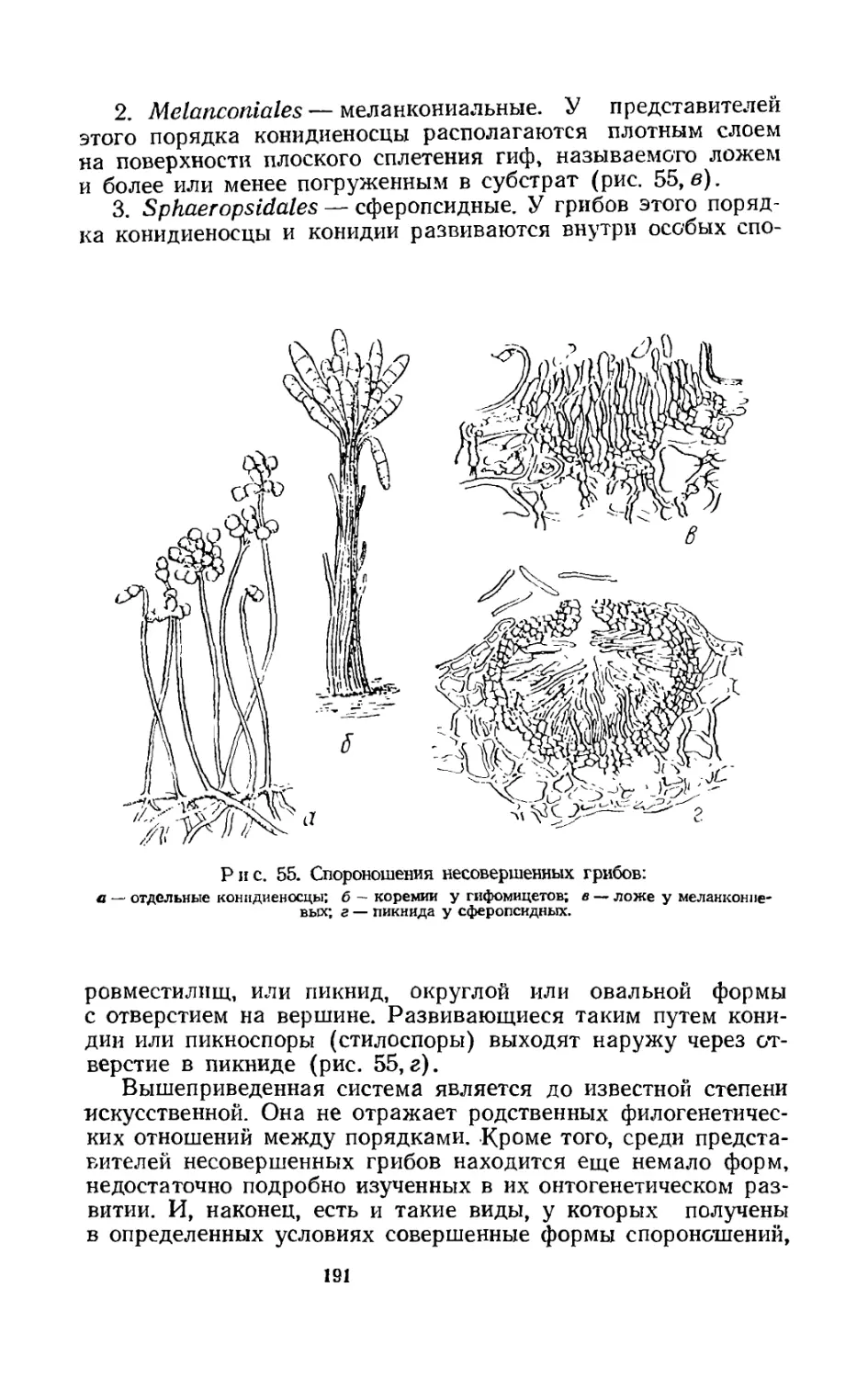
Экзогенное спороношение отличается исключительно большим разнообразием. Специальные гифы, или конидиеносцы, направлены, как правило, вверх; образующиеся на них споры, или конидии, оказываются в воздушной среде и легко рассеиваются после созревания. Конидиеносцы весьма разнообразны по своему строению, характеру ветвления и роста, а также окраске.
Рис. 4. Различные формы спороношений:
а — зооспоры; б — спорангиоспоры; в — конидии; г — сумкоспоры; д — базидиоспоры.
Еще большим разнообразием отличаются конидии (рис. 4, в). Они могут быть одноклетными и многоклетными и иметь самую разнообразную форму: округлую, овальную, вытянутую, веретеновидную, серповидную и т. п. Интенсивность окраски конидий также сильно варьирует. Встречаются конидии бесцветные и окрашенные в желтоватые, дымчатые, буроватые тона. У некоторых окраска настолько слаба, что единично взятые они кажутся бесцветными, а в массе имеют различную окраску — желтую, розовую, оливковую и т. п.
Образование конидий на концах конидиеносцев может идти двумя путями: базипетально и акропетально.
Базипетальное развитие конидий заключается в следующем: кончик конидиеносца разрастается, отделяется пере-
3 3. H. Кудряшева
33
тяжкой и принимает определенную форму, превращаясь в конидию. По мере ее роста конидиеносец образует под ней такую же вторую конидию, за ней третью и т. д. В результате возникает цепочка конидий, где самая зрелая находится на ее верхнем конце, а самая молодая — непосредственно на конидиеносце. Достигнув полного развития, верхняя конидия отпадает, а по мере того как идет созревание последующих, вершина конидиеносца образует все новые конидии. Базипетальное образование конидий широко распространено среди грибов (Penicillium, Aspergillus и др.).
Акропетальное развитие отличается тем, что первая конидия, образовавшаяся на конидиеносце, разрастается и на своей вершине дает вырост, развивающийся в следующую такую же конидию. На этой второй закладывается третья и т. д. Возникает цепочка, у которой самая старая и зрелая конидия находится внизу (Altennaria). Акропетальное развитие конидий у грибов распространено значительно реже.
Размножение при помощи конидий — весьма распространенное явление у высших грибов. На протяжении всего вегетационного периода размножение происходит только таким образом. Особенного многообразия конидиальное спороно-шениё достигает у класса сумчатых грибов. У базидиальных оно встречается реже, однако в некоторых порядках (Uredinales) в цикле развития одного вида гриба последовательно образуется несколько разнообразных конидиальных споро-ношений.
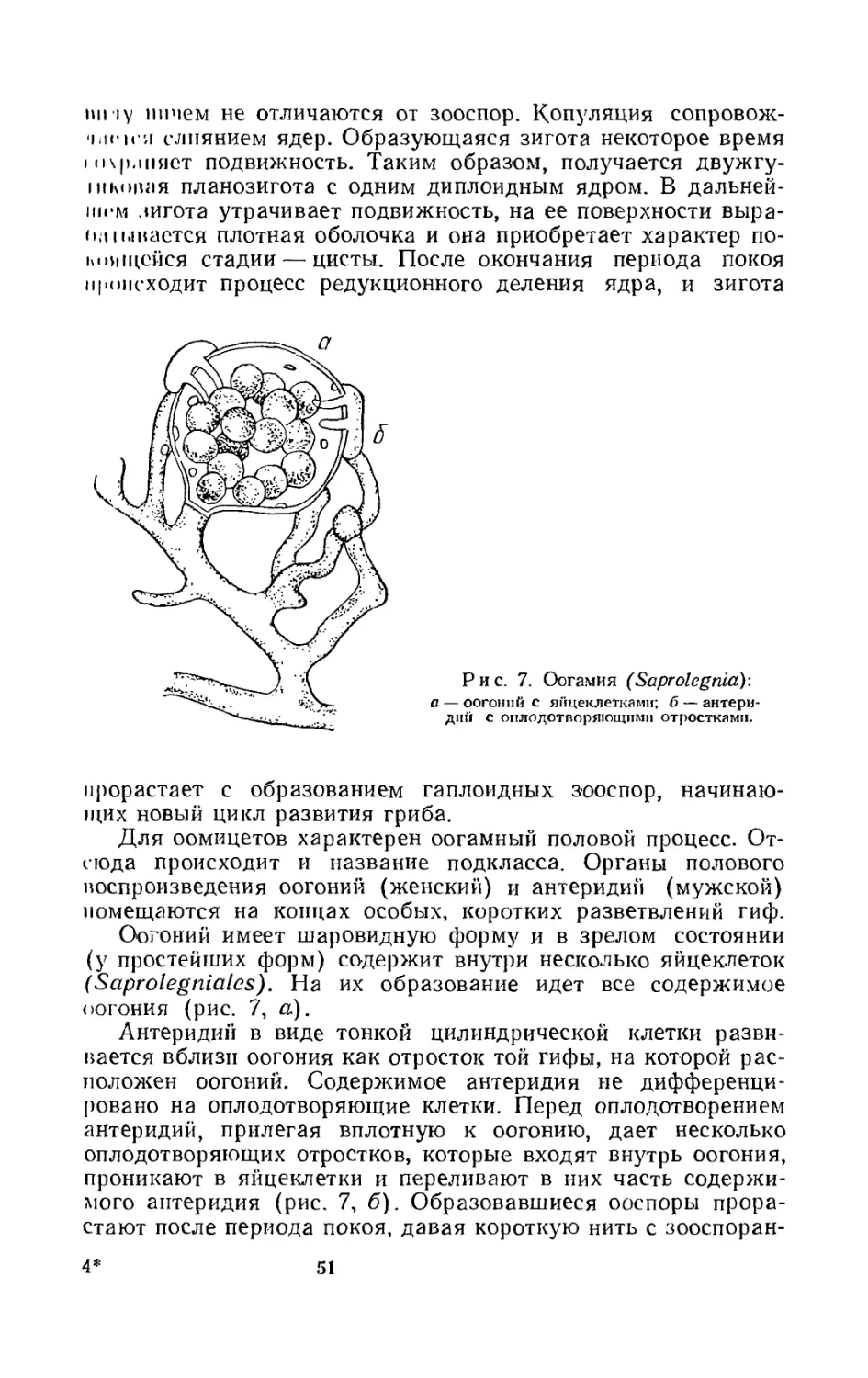
Половое размножение. Этот процесс, свойственный низшим и высшим грибам, осуществляется слиянием половых клеток, или гамет. У некоторых грибов может происходить слияние содержимого целых гаметангиев, не дифференцированных на гаметы. Продуктом слияния является зигота.
У низших грибов зигота некоторое время находится в состоянии покоя, а затем прорастает. Перед прорастанием происходит процесс редукционного деления ядра (или ядер). Зигота, прорастая, может дать сразу вегетативный мицелий (Monoblepharis) или, что бывает чаще, образует спорангий с большим количеством спор. Этот спорангий в отличие от спорангия, развивающегося при бесполом размножении, называется зародышевым (Mucor mucedo). Образующиеся в нем споры физиологически различны, т. е. часть их несет признаки одного пола ( + ), а часть другого ( —).
Почти у всех низших грибов вся жизнь протекает в гаплоидной фазе, диплоидной оказывается только зигота.
У высших грибов половой процесс может осуществляться переливанием содержимого мужского полового органа (антеридия) в женский (оогоний, архикарп). Непременным ус-
MiiiiicM -лому должно быть близкое расположение, обеспе-чни.пощсс соприкосновение обоих органов.
Оплодотворение женского органа может происходить iii’in 1ДВПЖНЫМИ клетками — спермациями, переносящимися шндушпыми течениями или насекомыми.
У ряда высших грибов произошло угасание полового про-пг<‘га. Оплодотворение совершается посредством переползания протопласта одной вегетативной клетки в другую (сома-шымия) или попарным соединением ядер в одной клетке (зигогамия). Характерной особенностью почти всех высших ipnooB является неодновременное слияние протоплазмы и и/ггр в момент оплодотворения. Обычно сначала наступает плазмогамия, т. е. слияние протоплазмы. Ядра разных половых знаков размножаются делением и размещаются парами, образуя дикарионы (дикариотическая фаза), после чего наступает кариогамия (половое слияние ядер). Каждое диплоидное ядро начинает делиться редукционно, а образующиеся гаплоидные ядра дают начало спорам полового размножения.
Таким образом, в цикле развития большинства высших грибов наблюдается закономерное чередование следующих грех фаз: гаплоидной, дикариотической (фаза парного рас-’ положения ядер) и диплоидной. Гаплоидная и дикариотическая фазы отличаются наибольшей продолжительностью, а диплоидная кратковременная, так как сразу же после образования диплоидных копуляционных ядер начинается процесс редукционного деления.
Количество спор, развивающихся после полового процесса, кратно 2, чаще всего бывает 8 или 4. Образуются они эндогенно в особых одноклетных вместилищах — асках, или с умка х, или же экзогенно на поверхности одноклетных или многоклетных выростов, называемых базидиями (рис. 4, а, д). В первом случае они называются аскоспорами, во втором — базидиоспорами. Аскоспоры отличаются большим разнообразием по сравнению с базидиоспорами. Они могут быть одиоклетные, двухклетные и многоклетные. Базидиоспоры всегда одноклетные.
Особенности полового процесса, свойственные отдельным группам грибов, излагаются при характеристике каждого класса.
В цикле развития грибов наблюдается закономерное чередование бесполого и полового размножений. Процесс бесполого образования спор совершается многократно в течение вегетационного периода, и таким образом происходит расселение гриба. Половой процесс наступает в конце вегетации. Образованием половых спор заканчивается цикл индивидуального развития каждого вида гриба.
ОБРАЗ ЖИЗНИ ГРИБОВ В ЕСТЕСТВЕННЫХ УСЛОВИЯХ
Не имея возможности синтезировать органические вещества, грибы нуждаются в готовом питательном материале, который они получают либо из органических остатков, либо из живых организмов. Соответственно этому определяется характер их питания — сапрофитный или паразитный.
Сапрофитизм. Это наиболее распространенное явление среди грибов. Оно характерно для 3/4 всех известных видов. В природе всегда имелось и имеется большое количество органических остатков, поэтому сапрофитные виды могут легко получать необходимые питательные вещества без особой специализации. В связи с этим сапрофитный способ питания следует считать первичным.
Остатки растительных организмов, а также субстраты растительного происхождения представляют наиболее благоприятную среду обитания по сравнению с животными. Сапрофитные грибы можно встретить в почве и на ее поверхности, где гниют всевозможные отмершие части растений — листья, стебли, плоды. Большое количество сапрофитных грибов развивается на различных пищевых и кормовых продуктах растительного происхождения, сопутствуя человеку в его повседневной жизни.
Предпочтение, оказываемое грибами растительному материалу перед животным, следует объяснять, во-первых, тем, что в отмерших частях тела растений и других растительных субстратах имеется много углеводов — наилучшего питательного материала для грибов. Во-вторых, кислая реакция растительных остатков подавляет развитие многих бактерий — антагонистов грибов. Попутно следует указать, что остатки животных организмов, а также субстраты животного происхождения мало доступны для грибов, ибо, являясь в основном белковыми продуктами, заселяются бактериями. Последние, разлагая белки, вызывают подщелачивание среды, которое большинство грибов не переносит.
Паразитизм грибов на растительных организмах более распространен, чем на животных. Переход от сапрофитизма к паразитному существованию у грибов осуществлялся постепенно в процессе их эволюции. Некоторые грибы, первоначально развивающиеся сапрофитно на мертвых тканях растений, постепенно приобрели возможность переходить на отмирающие, а затем и на живые части растений, поселяясь на них вначале в одной из стадий своего индивидуального развития. Вновь приобретенное свойство, оказавшись полезным, постепенно закрепилось в потомстве, и возникла категория грибов полусапрофитов, или фа культа-
। и в ii ы х паразитов. Примером таких грибов может । qу жить Botrytis, встречающийся на всходах некоторых рас-। г unit.
В процессе приспособления к условиям окружающей сре-1Ы и живым тканям обособилась и категория полупара-। и г о в, или факультативных сапрофитов. Это j рпбы, развивающиеся обычно на живых растениях, но при и\ отсутствии ведущие сапрофитный образ жизни. Такие ор-1.НП13МЫ с успехом растут и на искусственных питательных родах. К ним осносятся многие сумчатые, существующие в ьонидиальной стадии как паразиты, а в сумчатой как сапрофиты (Venturia).
Облигатный паразитизм. Это высший этап паразитного i уществования гриба, приспособившегося к определенному составу питательных веществ, доставляемых ему растением--нняином. Облигатные паразиты отличаются исключительно жой специализацией и в своем развитии бывают ограничены пределами вида, а иногда и сорта растения-хозяина. Приспособившись только к определенному кругу питающих растений с их качественным и количественным соотношением питательных веществ, облигатные паразиты не спо-гобны развиваться на искусственных питательных средах (Erysiphales, Uredinales).
Мицелий паразитных грибов в соответствии с условиями питания большей частью разрастается внутри тканей растений. Однако есть такие виды, у которых развитие мицелия происходит на поверхности того или иного органа. Соответственно различают грибы эндопаразиты и эктопаразиты.
Эндопаразитизм широко распространен и в свою очередь может быть представлен тремя типами. Одни паразитные грибы развиваются только внутри клеток растения. К ним относятся наиболее примитивные формы низших гринов, у которых вегетативное тело представляет собой комочек голой протоплазмы или оно в виде зачаточного мицелия (Myxochytridiales и Муcochyiridiales).
У других хорошо развитый одноклетный или многоклет-ный мицелий может распространяться как в межклетниках, гак и внутриклетно (Phytophthora, Kabatiella и др.).
Наконец, существуют такие паразиты, у которых гифы мицелия развиваются только в межклетных пространствах, а в клетки проникают гаустории в виде коротких и тонких выростов гиф, при помощи которых гриб извлекает питательные вещества (Cystopus и др.).
Эндопаразиты могут иметь мицелий диффузный и местный.
Диффузный мицелий свойствен облигатным паразитам, а также факультативным сапрофитам. Пронизывая органы,
целые побеги, а иногда и все растение, мицелий гриба может быть однолетним и многолетним. Однолетняя грибница начинает свое развитие вместе с прорастанием семени и, как правило, гибнет с отмиранием растения (многие головневые). Многолетняя грибница сохраняется из года в год в зимующих органах растений (корнях, корневищах, стеблях). Весною она начинает развиваться, пронизывая молодые, отрастающие органы (некоторые пероноспоровые, голосумчатые, ржавчинные).
Местный мицелий может быть распространен на незначительном расстоянии от первоначального места внедрения инфекции. В этом случае он обычно бывает однолетним (многие ржавчинные). У других паразитных грибов многолетний мицелий простирается на значительное расстояние и, ежегодно разрастаясь, захватывает все новые и новые участки тканей (трутовые грибы).
Эктопаразитизм встречается реже. Мицелий гриба распространяется по поверхности тела растения, образуя вначале паутинистый налет, впоследствии уплотняющийся. Питательные вещества из тела растения-хозяина гриб получает также при помощи гаусториев (Erysiphaceae).
Тот или иной вид паразитного гриба может встречаться на многих представителях одного и того же семейства. Например, стеблевая ржавчина Puccinia graminis поражает представителей семейства злаковых. Однако вид гриба как таксономическая единица может состоять из отдельных форм, которые, не отличаясь между собою морфологически, способны развиваться только на представителях определенного рода данного семейства. В таких случаях к названию гриба добавляют forma Speciales (сокращенно sp.) и родовое название растения в родительном падеже. Например, стеблевая ржавчина, развивающаяся на пшенице, будет называться Puccinia graminis f. sp. tritici, на овсе — P. grami-nis f. sp. avenae и т. д.
Еще более узкая специализация проявляется в способностях гриба развиваться на определенном сорте растений, что дает возможность выделять физиологические расы, а также биотипы. Для обозначения физиологических рас применяются порядковые номера, а для биотипов — буквы.
На животных организмах грибов-паразитов встречается значительно меньше, однако все же количество их равняется нескольким сотням. Среди низших грибов есть целое семейство Entomophthoraceae, представители которого паразитируют на насекомых, в том числе на комнатных мухах (Empusa muscae). Существуют паразиты низших позвоночных, например Saprolegnia, развивающаяся на рыбах. В зна-
чн гельной степени подвергаются нападению паразитных । рнбов птицы. В их дыхательных путях могут поселяться как низшие грибы (Мисог), так и высшие (Aspergillus, Peni-rillium). Однако больше всего паразитные грибы распространены на млекопитающих и в том числе на человеке. Болезнь, известная под названием «стригущий лишай», вызывается грибом Trichophyton. Множество других накожных полезней, называемых бластомикозами, обусловлены развитием грибов. В период лактации у грудных детей и молодых животных в полости рта развивается молочница — белый налег, образованный грибом Endomyces albicans. Ряд заболеваний внутренних органов человека (почки, печень) вызываются некоторыми видами рода Aspergillus.
Симбиоз. Среди грибов распространено сожительство с другими организмами, или симбиоз. Взаимоотношения между обоими компонентами чаще всего основаны на обоюдной пользе. Однако в некоторых случаях один из организмов своим поведением может нарушать установившееся равновесие и проявлять паразитические свойства.
Наиболее распространен симбиоз грибов с растительными организмами — водорослями и высшими растениями.
Симбиоз грибов с водорослями дает новый своеобразный тип живых организмов — лишайники. Вегетативное тело лишайника представляет слоевище, или таллом, и состоит из плотно сплетшихся грибных гиф, между которыми располагаются клетки пли нити водорослей (рис. 5). Обладая хлорофиллом, водоросль синтезирует из углекислоты и воды углеводы, которые использует и гриб. В свою очередь гриб снабжает водоросль водой и минеральными солями, а также защищает от высыхания и чрезмерной инсоляции.
Взаимоотношение гриба и водоросли в лишайнике часто называют мутуалистическим симбиозом, т. е. гармоническим сожительством двух организмов. Однако известно, что гриб для получения от водоросли питательных веществ способен направлять в ее клетки гаустории, т. е. проявлять паразитические свойства. В лишайнике гриб настолько приспособился к определенной водоросли, что потерял способность развиваться самостоятельно.
В создании лишайников принимают участие преимущественно сумчатые грибы (из пиреномицетов и дискомицетов), реже базидиальные. Водоросли, входящие в тело лишайника, принадлежат к типу зеленых (порядок протококковые), значительно реже встречаются представители типа сине-зеленых.
Симбиоз грибов с высшими растениями представляет исключительный интерес в теоретическом и практическом отношении. Семена плевела опьяняющего (Lolium temulen-
turn L.) в клетках оболочки у основания зародыша имеют грибницу. При прорастании семени гифы гриба проникают в зародыш, разрастаются одновременно с развивающимся растением, не нанося при этом ему никакого вреда. При образовании соцветия грибница также проникает в новые завязи. Плодоношений у гриба не бывает, а распространение
Рис. 5. Разрез слоевища лишайника: а — водоросли.
его идет исключительно вегетативно, т. е. заключенной в семенах грибницей. Это сожительство возникло в весьма отдаленные времена. Доказательством давности этого симбиоза служат семена плевела, обнаруженные в Египетских пирамидах, которым насчитывается более 4000 лет.
Пораженные грибом семена приобретают опьяняющие свойства, за что растение получило видовое название плевел опьяняющий. Гриб получает от растения необходимые ему углеводы, а плевел, имея в своих тканях грибницу, становится ядовитым, что является своеобразной защитой.
Еще больший интерес представляет сожительство грибов с корнями высших растений, известное под названием микоризы.
Первое глубокое исследование микоризы принадлежит профессору Одесского университета Ф. М. Каменскому и относится к 1881 году. Изучая анатомическое строение корней нодьельника (Monotropa hypopitus L.), он нашел, что окончания корней этого растения покрыты толстым слоем перепле-|.1К)|цихся гиф гриба. На основании этого Каменский сделал вывод о возможности симбиотических отношений между грином и корнями высшего растения. В дальнейшем при попытке вырастить растение в полевых условиях оказалось, что семена подъельника не развиваются при отсутствии гриба. Это пало возможность Каменскому утверждать, что подъельник является облигатным микотрофом.
Несколько позднее, в 1885 году, известный русский миколог М. С. Воронин высказал мысль о тесной связи некоторых шляпочных грибов с определенными лесными породами.
А. Б. Франк с 1882 года занимался изучением распространения трюфелей в природных условиях, нашел мицелий этих । рибов на корнях дуба и бука. Симбиотические отношения 1риба и корней высшего растения были им названы в 1885 году микоризой, что означает грибокорень.
В дальнейшем вопросами микотрофии у высших растений занимались многие исследователи. В настоящее время известно, что микориза широко распространена среди многолетних как семенных, так и архегониальных растений. У однолетних грав она встречается редко, а у водных растений вообще отсутствует. Следует отметить, что среди высших растений имеется относительно мало облигатных мпкотрофов (орхидные, вересковые), в большинстве же случаев наблюдается факультативная микотрофия, при которой гриб в значительной степени улучшает питание своего симбионта, обеспечивая ему нормальное развитие.
С другой стороны, для образования плодового тела у гриба, т. е. для развития совершенной формы спороношения, необходимо наличие соответствующего высшего растения, с которым бы гриб имел возможность вступить в симбиотические отношения, т. е. образовать микоризу. В противном случае грибница остается стерильной, и плодовое тело не развивается. Некоторые грибы проявляют большую специализацию п способны образовывать микоризу с определенными лесными породами. Так, например, масленок лиственничный связан исключительно с лиственницей; подберезовик — главным образом с березой; боровик — с сосной и елью. Другие виды менее разборчивы; рыжик образует микоризу с елью, сосной, пихтой и лиственницей, а мухомор — со многими лиственными и хвойными породами.
В зависимости от положения гриба-микоризообразователя относительно тканей корня различают три типа микоризы:
1. Эктотрофная микориза — наружная; гифы гриба снаружи густо оплетают кончики корня, образуя около него как бы чехол, а многочисленные свободные грибные гифы отходят в почву. Собственных корневых волосков корень при этом не развивает. С другой стороны, оплетающие кончик корня гифы заходят в межклетники первичной коры корешков. не проникая внутрь клеток (рис. 6, а).
Р и с. 6. Микориза: а — эктотрофная; б — эндотрофная.
2. Эндотрофная микориза — внутренняя; гифы располагаются главным образом внутри тканей — в клетках коровой паренхимы. Корни при этом несут нормальные корневые волоски (рис. 6,6).
3. Экт о- эндотрофная микориза является переходной между двумя первыми типами. Грибные гифы этой микоризы не только густо1 оплетают корень снаружи, но и заходят в большом количестве в коровую паренхиму.
Мицелий эндотрофной и экто-эндотрофной микоризы идет межклетно и внутриклетно. В последнем случае он обычно образует плотные клубки гиф или разветвления, напоминающие гаустории.
Каковы же физиологические взаимоотношения между обоими компонентами этого симбиоза?
Немецкий исследователь Мелин считает, что гриб привлекается и стимулируется в росте какими-то выделениями корня, которые способствуют также и прорастанию спор мико-
pH iiu.ix грибов. Вступая в тот или иной вид сожительства । корнем, гриб, как бесхлорофильный организм, получает от ni.H'iHero растения главным образом продукты его фотосинте-ы углеводы. С другой стороны, гриб своим густо разветв-loiiiihiM в почве мицелием, заменяя корню корневые волоски (при эктотрофной микоризе), усваивает из почвы не только поду п минеральные соли, но и сложные, главным образом, л юг истые органические вещества. Таким образом, высшее р.к'гение посредством гриба может использовать те органические вещества почвы, которые ему недоступны.
При эндотрофной микоризе иногда наблюдается отмирание и растворение в коровых клетках грибных гиф. При этом псе вещества усваиваются клетками корня. Это своеобразное внутриклетное переваривание сходно с фагоцитозом у жило гных.
Некоторые исследователи выделяют еще один тип микори-и>| — перитр о ф н у ю. Это специальная флора грибов, широко распространенная в ближайшем соседстве с корнями той или иной породы. Подобная грибная флора ризосферы корней может быть различна для разных видов, а также и для разных почв. Не будучи анатомически связаны с корнями высших растений, эти грибы привлекаются выделениями корней и в свою очередь, развиваясь в ризосфере, создают в ней соответствующую среду, оказывающую положительное влияние на рост и развитие растения.
Симбиотические отношения проявляются у грибов и с насекомыми, в частности с муравьями. Тропические муравьи (род Aita) устраивают в своих муравейниках так называемые «грибные сады», где разводят нужный им гриб, являющийся единственной их пищей. Для сдабривания почвы в «грибных садах» муравьи обгрызают паренхиму листьев, переносят ее в муравейники. Там этот растительный материал тщательно пережевывается ими и идет на удобрение. Грибные гифы пронизывают удобренные участки, а на их свободных концах образуются многочисленные скопления коротких и вздутых ветвей, собранные в головки и получившие название «кучки кольраби». Они-то и являются единственной пищей муравьев. Ухаживая за своим садом, муравьи не допускают развития спороношения у гриба. В случае удаления муравьев из муравейника на грибнице развивается конидиаль-ное спороношение.
Занос гриба в новый муравейник совершает самка. Вылетая из старого муравейника, она в своем теле уносит небольшой участок грибницы, сохраняющийся около слюнной железы. На новом месте эта грибница выбрасывается самкой наружу и поначалу сдабривается экскрементами. Дальнейшая работа проделывается рабочими муравьями.
ПУТИ И СПОСОБЫ РАСПРОСТРАНЕНИЯ ГРИБОВ
Большой общебиологпческий интерес имеет знание условий и способов распространения спор грибов. Научное обоснование методов защиты растений от различных болезнен невозможно без знания особенностей переноса спор того или иного возбудителя.
Споры грибов, развивающиеся в особых вместилищах — зооспорангиях, спорангиях, сумках, пикнидах, прежде всего должны быть освобождены от непосредственного контакта с клеткой или тканью, где произошло их образование. Для их освобождения основное значение имеет тургорное давление, увеличивающееся внутри спорангия, сумки к моменту созревания спор. Освободившиеся от материнского организма споры попадают в водную или воздушную среду, где и распространяются.
Зооспоры, обладая способностью к активному движению, могут распространяться только в водной среде.
У подавляющего большинства грибов, живущих на суше, споры, освободившиеся из споровместилищ или образовавшиеся открыто на конидиеносцах, могут распространяться с помощью ветра, дождя, мелких и крупных животных. Последние переносят споры на своем теле, либо выбрасывают их с экскрементами.
Существенным биологическим различием спор, определяющим пути распространения, является их состояние к периоду созревания. Одни из них сухие и могут распыляться, другие окружены слизистым веществом, выделяемым споровмести-лищем или конидиальным ложем. В связи с указанной биологической особенностью живущие на суше грибы можно разделить на две группы: «сухоспоровые» и «слизистоспоровые». Среди крупных сапрофитных грибов наиболее типичными сухоспоровыми являются дождевики из семейства Lycoperda-сеае. При окончательном созревании плодовых тел их оболочка (перидий) разрывается, и споры высыпаются в виде бурого порошка. Подхваченные воздушными течениями, они переносятся на большие расстояния.
К сухоспоровым относятся также шляпочные грибы (порядок Agaricales). Их споры, отрываясь от базидий, падают в свободное пространство под шляпкой и легко разносятся даже слабыми токами воздуха.
Таким же путем распространяются уредоспоры ржавчинных, споры пыльной головни и ряда других грибов.
Воздушные течения способны заносить споры на громадную высоту и распространять их на большие расстояния в горизонтальном направлении, при этом у спор сохраняется
। ши обность к прорастанию. Например, конидии Altemaria. । пбрапные с высоты 3000 м и уредоспоры Puccinia graminis, 11)бранные с высоты 2000 м, легко прорастали. Уредоспоры ржавчинных грибов могут переноситься на расстояние 900— ИKJO м от первоначальных очагов. Скорость распространения » пор по горизонтали зависит от силы ветра.
Если образование спор сопровождается выделением слизи, hi перенос их осуществляется насекомыми или брызгами кнкдя. В распространении спор грибов принимают участие • амые различные насекомые: мухи, жуки, бабочки, муравьи, пчелы и др.
Наиболее интересные примеры энтомофильных грибов можно найти среди представителей семейства Phallaceae, Широко распространенным видом является веселка обыкновенная (Phallus impudicus, см. рис. 54), у которой зрелое пло-човое тело имеет ножку и коническую небольшую шляпку, покрытую толстым слоем зеленовато-черной сахаристой слизи, । одержащей множество спор. Слизь издает отвратительный <апах падали, привлекающий мясных и навозных мух. За 2—3 часа они удаляют всю слизь, а с нею и споры, оставляя чистой белую ячеистую поверхность шляпки. Споры разносятся на 1сле и лапках насекомых, а также могут проходить через пищеварительный канал, не теряя при этом способности к прорастанию.
У австралийского гриба Asera rubra плодовое тело напоминает цветок. Ножка длиною 5—10 см на вершине несет ('порообразующую слизистую зону, окруженную своеобразным венчиком из красных стерильных лопастей, привлекающих своей окраской насекомых.
Интересным примером переноса спор насекомыми может служить конидиальная стадия паразита злаков — спорыньи. В период конидиального спороношения на пораженных завязях выделяется «медвяная роса» (Sphacelia), содержащая сахаристые вещества. Некоторые насекомые, питающиеся пыльцой злаков, привлекаются медвяной росой и на своих лапках переносят споры гриба.
У ржавчинных грибов перенос спермациев из одного спермогония на другой также совершается насекомыми, привлекающимися сахаристыми выделениями спермогониев.
Дождевые капли способствуют распространению спор, образующихся на слизистых конидиальных ложах, как например у представителей рода Kabatiella, Gloeosporium, Colle-totrichum и др.
Споры грибов могут распространяться крупными и мелкими животными, а также человеком. Корм травоядных животных всегда изобилует спорами различных грибов, которые, проходя через пищеварительный тракт животного, во многих
случаях не теряют способности к прорастанию. Если попробовать понаблюдать за свежим навозом травоядных животных, помещенным под стеклянным колпаком на свету при высокой влажности, то можно заметить как на нем будут развиваться различные грибы: Mucor, Pilobulus, Ascobolus, Arthro-botrys и др. С навозом травоядных могут распространяться покоящиеся споры возбудителя килы капусты (Plasmodio-phora brassicae), зимние зооспорангии возбудителя рака картофеля (Synchytrium endobioticum).
Интересным примером может служить гриб Basidiobolus ranarum, встречающийся на экскрементах лягушки. В его распространении участвуют два вида животных, стоящих далеко в систематическом положении: жук и лягушка. Мелкие жучки питаются спорангиями этого гриба, в свою очередь лягушка проглатывает жуков. В ее кишечнике происходит дозревание спорангиев и освобождение из каждого из них восьми спорангиоспор. Последние выделяются с экскрементами лягушки и прорастают, образуя мицелий.
Распространению шляпочных грибов помогают в значительной степени белки, мыши-полевки. Перетаскивая плодовые тела в свои гнезда и норы, они способствуют рассеиванию базидиоспор. Слизни, питаясь плодовыми телами шляпочных грибов, разносят споры на поверхности своего тела, а также выделяют вполне жизнеспособные споры с экскрементами.
Птицы также переносят споры. Известно, что дятлы и пищухи оказались виновниками катастрофического распространения заболевания коры американского каштана.
Водоплавающие птицы содействуют переносу водных грибов из одного изолированного водоема в другой.
В ряде случаев человек способствует распространению грибов сознательно, культивируя плесневые грибы, необходимые в медицине, сыроварении, разводя шампиньоны. Однако во многих случаях распространение спор грибов, причем очень часто патогенных, совершается по неведению или неосторожности. При этом часто патогенные грибы — возбудители серьезных заболеваний сельскохозяйственных и декоративных растений — попадают с одного континента на другой. Так, например, из Америки в Европу попала американская мучнистая роса крыжовника, а из Европы в Америку перекочевала ржавчина веймутовой сосны. Подобных примеров можно было бы привести великое множество. Резерваторами спор в таких случаях могут быть части тела растений, предназначенные для вегетативного размножения: черенки, луковицы, клубни и т. п., а также семена.
Следует остановиться еще на особой экологической группе грибов, плодовые тела которых образуются подземно,—
। рюфелсвых. В распространении их спор принимают участие 1рызуны, откапывающие плодовые тела для питания, и зей-,»i и пью черви, разносящие на своем теле освобожденные при l».i щушении плодовых тел споры.
В основе современной системы грибов как особого отдела р.«’гений лежит деление его на 4 класса:
I. Phycomycetes (фикомицеты).
2. Ascomycetes (аскомицеты).
3. Basidiomycetes (базидиомицеты).
4. Denteromycetes, или Fungi itnperfecli (несовершенные । рпбы).1
Первый класс представляет собой низшие грибы, харак-|сризующиеся более примитивной организацией. Остальные ip и —высшие грибы.
1 Классификация и построение филогенетической системы грибов относится к наиболее сложным теоретическим проблемам микологии. Этим можно объяснить, что в разное время были предложены зарубежными п нашими отечественными учеными различные схемы классификации грибов. (Прим, ред.)
ГЛАВА
I КЛАСС PHYCOMYCETES
(ФИКОМИЦЕТЫ)
ОБЩАЯ ХАРАКТЕРИСТИКА
Представители этого класса встречаются в водной среде и на суше. По образу жизни одни из них являются сапрофитами, другие приспособились к паразитному существованию на растительных и животных организмах.
У простейших видов вегетативное тело представлено зачаточным мицелием (ризомицелий), а у самых примитивных из них—голым комочком протоплазмы, покрывающимся оболочкой перед спорообразованием. Однако следует отметить, что простейших фикомицетов в природе встречается значительно меньше, чем более высокоорганизованных форм.
У большинства фикомицетов вегетативное тело представлено хорошо развитым одноклетным мицелием, гифы которого содержат большое количество ядер и обладают вершинным ростом.
Фикомицеты делятся на 3 подкласса:
1. Archimycetes (Архимицеты).
2. Oomycetes (Оомицеты).
3. Zygomycetes (Зигомицеты).
Размножаются фикомицеты вегетативным, бесполым и половым путями.
Вегетативный способ размножения наиболее высокоорганизованных представителей с хорошо развитым мицелием основан на способности кусочков гиф регенерировать и давать новые особи.
Бесполое размножение совершается посредством зооспор, спорангиоспор и конидий.
В зависимости от количества жгутиков, имеющихся у зо-<>< пор, или их отсутствия среди фикомицетов различают 3 эво-поционных ряда:
I. Uniciliata (одножгутиковые).
2. Bicillata (двужгутиковые).
3. Aciliata (безжгутиковые).
Зооспоры фикомицетов образуются в зооспорангиях, простейших при развитии зооспорангия расходуется все ве-и'гативное тело, и после образования и выхода зооспор существование данной особи прекращается (холокарпические формы). Наиболее примитивные формы фикомицетов могут образовывать спорангии двух видов: тонкостенные, созревающие в течение нескольких дней, и толстостенные, зимующие. Последние представляют собой покоящуюся стадию, или ци-ггу, и прорастают весной следующего года. У более высокоорганизованных форм зооспорангии развиваются на дифференцированных гифах мицелия, причем на их образование расходуется не все вегетативное тело, а только часть его (эукарпические формы). При размножении зооспорами необходимым условием является наличие водной среды или капельно-жидкой влаги в период размножения.
К одножгутиковому ряду принадлежат самые примитивные фикомицеты. Ряд одножгутиковых произошел от двужгутиковых форм, которые в своем развитии утратили один жгутик. Этот ряд в эволюционном отношении оказался ограниченным, не получил дальнейшего развития, а поэтому заканчивается слепо.
Двужгутиковые фикомицеты представляют основной эволюционный ряд. На его представителях можно проследить, как постепенно совершался переход от водного образа жизни к наземному существованию.
Наиболее низкоорганизованные двужгутиковые фикомицеты являются типичными водными обитателями (Saprolegnia-les). Другие, поселившись на суше, приспособились к паразитному образу жизни на высших растениях. Однако размножаться бесполым путем при помощи зооспор они могут только при наличии капельно-жидкой влаги. В противном случае зооспорангии у этих видов не дают зооспор, а прорастают только одним мицелиальным ростком, т. е. превращаются в конидию (род Phytophthora).
Третий эволюционный ряд — безжгутиковые фикомицеты — состоит из форм, утративших в своем развитии подвижную стадию. Этот ряд является наиболее прогрессивным среди фикомицетов. Его представители полностью приспособились к жизни в наземных условиях. Ряд безжгутиковых имеет продолжение в более высоко стоящих группах грибов — сумчатых, а возможно, и базидиальных.
4 3. H. Кудряшева
49
Бесполое размножение безжгутиковых осуществляется посредством спорангиоспор и конидий, которые с самого начала своего развития имеют хорошо оформленные клеточные оболочки.
Спорангиоспоры образуются в спорангиях—шаровидных вместилищах, расположенных на концах воздушных гиф. Количество спор может быть от одной до нескольких сотен. Освобождение их из спорангия происходит вследствие того, что вся оболочка его расплывается благодаря действию водяных паров, а также в результате разрыва оболочки. Распространение спор совершается с помощью ветра.
Спорангии по размерам и количеству содержащихся в них спор бывают двух типов:
1) главные спорангии или спорангии I рода обычно более крупные: от нескольких десятков до нескольких сотен микрон в диаметре, как правило, с большим количеством спор;
2) спорангии II рода не более 20 мк в поперечнике, с количеством спор от 1 до 10.
Постепенное превращение спорангия в конидию можно наблюдать и в этом ряду фикомицетов. Оно осуществлялось путем уменьшения количества спор в спорангиях II рода до одной и срастания ее оболочки с оболочкой спорангия. Так, например, у одного из представителей мукоровых грибов (Thamnidium), развивающегося на навозе, имеется два вида спорангиев: главный спорангий с большим количеством спор образуется на главной оси спорангиеносца, а на боковых ветвях развиваются спорангии II рода, содержащие менее 10 спор. При неблагоприятных условиях главный спорангий не развивается, а количество спор в спорангиях II рода не превышает одной, и они. таким образом, биологически превращаются в конидию, хотя срастание оболочки споры с оболочкой спорангия еще не наблюдается. У других мукоровых (Chaeto-cladium) в нормальных условиях существования образуются исключительно односпоровые спорангии II рода, причем оболочки споры и спорангия срастаются между собой так, что спорангий полностью превращается в конидию.
Имеющиеся сведения о бесполом размножении фикомицетов дают возможность считать исходным органом бесполого размножения зооспорангий; остальные две формы — спорангий и конидия — выработались путем эволюции зооспорангия в связи с переходом организмов от водного образа существования к жизни на суше.
Половое размножение у фикомицетов представлено хологамией, изогамией, оогамией и зигогамией.
У представителей подкласса Archimycetcs половой процесс заключается в копуляции двух вегетативных особей (хологамия) или двух пл аногамет (изогамия), которые по внешнему
пи чу ничем не отличаются от зооспор. Копуляция сопровож-ъпчгя слиянием ядер. Образующаяся зигота некоторое время । <।чроняет подвижность. Таким образом, получается двужгу-।пковая планозигота с одним диплоидным ядром. В дальнейшем зигота утрачивает подвижность, на ее поверхности выравнивается плотная оболочка и она приобретает характер по-кпнщсйся стадии — цисты. После окончания периода покоя п|кшеходит процесс редукционного деления ядра, и зигота
Рис. 7. Оогамия (Saprolegnia)\ а — оогоний с яйцеклетками; б — антеридий с оплодотворяющими отростками.
прорастает с образованием гаплоидных зооспор, начинающих новый цикл развития гриба.
Для оомицетов характерен оогамный половой процесс. Отсюда происходит и название подкласса. Органы полового воспроизведения оогонии (женский) и антеридий (мужской) помещаются на концах особых, коротких разветвлений гиф.
Оогоний имеет шаровидную форму и в зрелом состоянии (у простейших форм) содержит внутри несколько яйцеклеток (Saprolegnialcs). На их образование идет все содержимое оогония (рис. 7, а).
Антеридий в виде тонкой цилиндрической клетки развивается вблизи оогония как отросток той гифы, на которой расположен оогоний. Содержимое антеридия не дифференцировано на оплодотворяющие клетки. Перед оплодотворением антеридий, прилегая вплотную к оогонию, дает несколько оплодотворяющих отростков, которые входят внутрь оогония, проникают в яйцеклетки и переливают в них часть содержимого антеридия (рис. 7, б). Образовавшиеся ооспоры прорастают после периода покоя, давая короткую нить с зооспоран
гием на конце. При этом происходит процесс редукционного деления диплоидного ядра ооспоры.
Более высоко развитая форма оогамного полового процесса характерна для порядка Peronosporales. Оогоний в начале своего развития заполнен протоплазмой с большим количеством ядер. В дальнейшем протоплазма дифференцируется на периферическую более водянистую часть (периплазма), в которой сосредоточены многочисленные ядра оогония, и сгущенную зернистую центральную часть (ооплазма) с одним ядром в центре.
Антеридий развивается на той же гифе, на которой расположен оогоний, или на другой в виде булавовидного отрога, отделенного поперечной перегородкой от несущей его гифы. Антеридий представляет многоядерное образование, вначале тесно прилегающее к оогонию. Затем на нем образуется цилиндрический росток, называемый антеридиальным отростком, который проникает довольно глубоко в оогоний к центру ооплазмы. На вершине антеридиальный отросток разрывается, и часть протоплазмы антеридия с одним ядром вливается в ооплазму оогония. При этом происходит слияние мужской и женской протоплазмы (плазмогамия); ядра сближаются (но не сливаются), образуя дикарион.
Из периплазмы развивается многослойная оболочка ооспоры, причем все оставшиеся в ней ядра дегенерируют. После образования оболочки ооспоры наступает кариогамия, т. е. слияние мужского и женского ядер. Образовавшаяся ооспора обычно перезимовывает и начинает прорастать только весною, давая мицелиальный росток, развивающийся в новую грибницу. Перед прорастанием ооспоры происходит редукционное деление ее диплоидного ядра.
Таким образом, и у оомицетов диплоидная стадия представлена только зиготой (ооспорой). Вся остальная жизнь гриба проходит в гаплоидной стадии.
Для подкласса Zygomycetes характерен зигогамный половой процесс, который происходит следующим образом. Концы двух соседних гиф булавовидно вздуваются, отделяются поперечной перегородкой от остальной части гифы и в дальнейшем растут навстречу друг другу. При соприкосновении их происходит растворение оболочки и слияние внутреннего содержимого. Каждая из сливающихся клеток содержит большое количество ядер, а так как каждое ядро можно считать соответствующим гамете, то эти клетки часто называют гаметангиями. У некоторых представителей зигомицетов гаметангии несколько отличаются друг от друга и могут быть определены как мужской и женский гаметангий. Количество ядер в гаметангиях может уменьшаться иногда до одного. Такие гаметангии фактически содержат по одной гамете.
При слиянии содержимого двух гаметангиев вначале про-in ходит плазмогамия, ядра только попарно сближаются, а за-н'м уже наступает кариогамия. Какое бы количество ядер не содержалось в гаметангиях, результатом слияния является шгота, или зигоспора, имеющая характер покоящейся стадии, и.*|и покоящейся споры. Образуясь на месте слияния двух га-мсчангиев, зигоспора плотно срастается с их оболочками, чем
Рис. 8. Зигогамия (Мисог). Формирование зигоспоры и ее прорастание.
существенно отличается от ооспоры оомицетов, свободно лежащей в полости оогония. Зрелая зигоспора имеет двухслойную оболочку, внутренний слой которой — эндоспорий — бесцветный, наружный — экзоспорий — темноокрашенный и бугорчатый. Зигоспора поддерживается подвесками, или суспензорами,— остатками гиф, образовавших гаметангии.
Ко времени полного созревания диплоидные ядра зигоспоры делятся редукционным путем и, таким образом, внутри нее образуется большое количество гаплоидных ядер.
При прорастании зигоспора набухает, наружный окрашенный слой оболочки разрывается, а из внутреннего образуется слабоветвящаяся или неветвящаяся нить, заканчивающаяся спорангием, который носит название зародышевого (рис. 8).
Исследованиями Блэксли (1904—1920) установлено наличие гетероталлизма (раздельнополость) у некоторых зигоми-цетов, в частности у мукоровых. У гетероталличных форм
копулируют гаметангии, образовавшиеся на мицелии двух различных по половому знаку ( + ) или (—) особей, растущих рядом. «Соответственно и в зародышевом спорангии таких грибов споры также будут разного полового знака.
У гомоталличных, или гермофродитных, форм копуляция происходит между гаметангиями, возникшими на концах гиф, принадлежащих одному и тому же мицелию. Зародышевый спорангий, развившийся в результате прорастания зигоспоры гомоталличных организмов, содержит споры физиологически одинаковые.
ПОДКЛАСС ARCHIMYCETES
Архимицеты обитают в воде и на суше. По образу жизни это преимущественно внутриклетные паразиты водорослей, водных грибов и высших наземных растений. Некоторые могут жить сапрофитно в водной среде на разлагающихся растительных остатках.
Подкласс архимицетов делят на следующие порядки: 1) Plasmodiophorales; 2) Myxochytridiales, 3) Mycochytridia-les,
ПОРЯДОК PLASMODIOPHORALES
Сюда входит одно семейство Plasmodiophoraceae, включающее несколько родов внутриклетных паразитов высших растений, главнейшими из которых являются Plasmodiophora и Spongospora.
Сем. Plasmodiophoraceae
Род Plasmodiophora Wor. Его представители поражают корни растений из семейства крестоцветных. Наиболее изучено развитие Pl. brassicae Wor.— возбудителя заболевания корней капусты, известного под названием килы.
У заболевшего растения на корнях появляются наросты самых разнообразных форм и размеров, иногда достигающие величины кулака. Такие корни почти не ветвятся и мало всасывают воды.
Обнаруживается паразит при микроскопическом исследовании наростов. В сильно разросшихся паренхимных клетках содержится вегетативное тело паразита в виде густой протоплазматической массы, более зернистой, чем протоплазма клеток корня. Другие клетки, тоже увеличенные, могут быть заполнены мелкими шаровидными тельцами — это более поздняя стадия развития гриба, его споры (рис. 9).
Первым исследовал возбудителя болезни и подробно описал его развитие М. С. Воронин (1864).
Развитие гриба начинается весной после освобождения с|<» спор из сгнивших корней капусты. Перезимовавшие споры прорастают, давая по одной одножгутиковой зооспоре.1 Пере-мннгаясь в почвенной влаге, зооспоры проникают в корневые полоски и превращаются в миксамебы. В дальнейшем паразит
Р и с. 9. Plasmodiophora brassicae'. а — наросты па корнях; б — разрез корня.
из корневых волосков постепенно попадает в паренхимные клетки корня, в которых питается и делится. В одной клетке может оказаться несколько миксамеб. Затем отдельные микс-амебы сливаются в один сплошной плазмодий, заполняющий постепенно всю клетку. Образование плазмодия сопровождается попарным слиянием ядер. В зрелом плазмодии начинается процесс спорообразования, которому предшествует редук
1 В литературе высказываются разноречивые мнения относительно количества зооспор и жгутиков на них.
ционное деление всех диплоидных ядер. Плазмодий распадается на громадное количество одноядерных участков и дает гаплоидные зимующие споры.
Поскольку PL brassicae поражает только корни, болезнь с семенами передаваться не может. Однако ее проявление можно заметить на рассаде в парниках. Инфекция в виде зимующих спор может сохраняться в почве парников, а также и в поле при бессменной культуре капусты на одном и том же участке.
В связи с тем что килою поражаются только корни, симптомы болезни на надземных частях проявляются позднее. Растения отстают в росте. В начале болезни листья теряют тургор только в дневные, наиболее жаркие часы. Затем увядание становится все более и более продолжительным, и листья прежде времени отмирают.
Пораженные килою растения не завязывают кочана, так как продукты ассимиляции направляются в корни и служат материалом для образования гипертрофированных тканей. Боковые и придаточные корни недоразвиваются, больное растение имеет слабый контакт с почвой, и его можно легко вытащить из земли.
Plasmodiophora одинаково легко заражает растения в любом возрасте. Однако вредоносность болезни определяется возрастом растения в момент заражения. Если растения поражаются в молодом возрасте, то они погибают, не завязывая кочанов. Более позднее заражение может снизить урожай на 20—40%. Если же паразит встречает вполне сформировавшуюся корневую систему, он не успевает вызвать существенных изменений в корнях и не приносит большого вреда.
Экологические условия оказывают весьма существенное влияние на развитие килы. В более сильной степени болезнь развивается на суглинистых и супесчаных почвах, чем на почвах, богатых перегноем. Кислая реакция почвы способствует развитию килы.
Большое значение имеет степень влажности. Засушливые годы характеризуются незначительным распространением килы. Минимум влажности, при котором возможно развитие гриба, равен 50% от полной влагоемкости, оптимум — 75—90%). PL brassicae сильнее развивается на низких местах в поймах рек, особенно при бессменной культуре капусты на одном и том же участке.
Температура почвы также оказывает влияние на развитие гриба: минимальной температурой считается 12°, оптимальной — 18—24° и максимальной — 27°.
Относительно сохранения спорами гриба жизнеспособности в почве мнения расходятся. Американские исследователи считают, что споры в почве могут сохранять жизнеспособность
и м’чснис 10 и более лет (Дж. Уокер), а советские исследова-и in Б. А. Герасимов, Н. А. Наумов, Е. А. Осницкая пола-। инн, что их жизнеспособность ограничивается 4—5 годами.
Кила поражает различные виды капусты (белокочанную, грлсную, цветную, кольраби и др.), а также репу, брюкву, редис, редьку, турнепс. Из сорняков семейства крестоцветных поражаются пастушья сумка, дикая редька, ярутка и др.
Канадские исследователи установили 6 биотипов у PL bttissicae, поражающих белокочанную капусту. Из них первый и 1рстий развиваются также на брюкве и некоторых сорняках. Наименьшей агрессивностью обладают пятый и шестой био-НП1Ы.
Абсолютно иммунных к киле сортов капусты нет. Есть лишь сорта более или менее устойчивые к этому заболеванию. В Советском Союзе все устойчивые к киле сорта русско-io происхождения: Вальватьевка, Горбуновка, Каширка, Ку-оышка, Московская поздняя, Славянка и др.
Меры борьбы с килою капусты рекомендуются следующие:
1. Выращивание здоровой рассады в свежей незаражен-пой почве. В случае необходимости следует проводить химическое обеззараживание почвы парников 4%-ным раствором карбатиона (40%-ного) из расчета 2—2,5 л рабочей жидкости на 1 м2 с последующей перекопкой. Химическое обеззараживание производится осенью.
2. Подщелачивание почвы в парниках внесением свежегашеной извести перед посевом из расчета 1 —1,5 кг на раму.
3. Не допускать выращивания капусты по капусте. Возвращение капусты на прежнее место возможно после двух не-поражаемых килой культур.
4. Немедленное удаление с поля пораженных килой растений в период вегетации и кочерыг после уборки урожая.
Род Spongospora Brunch. Внутриклетные паразиты. Споры в клетке питающего растения не одиночные, а склеены в губчатые клубочки. Широко распространенным видом является Spongospora subterranea (Wallr.) Johnson, вызывающий заболевание клубней, столонов и корней картофеля — п о porn и с т у ю паршу. Наиболее характерно проявляется поражение клубней. На ранней стадии развития болезни под кожицей образуются мелкие бугорки (2—3 мм). В дальнейшем они разрываются, а кожица звездообразно подвертывается, обнажая темную порошкообразную массу, представляющую скопление спор гриба (рис. 10). Микроскопический анализ порошкообразной массы показывает, что споры не одиночные, а собраны в клубочки, или спорокучки, округлой или эллипсоидальной формы. Споры плотно прилегают одна к другой, отчего принимают несколько угловатые очертания. Попадая в почву, спорокучки зимуют, а весною распадаются на отдель
ные споры. Каждая спора дает одноядерную маленькую амебу, которая при неблагоприятных условиях (чрезмерная сухость почвы) покрывается толстой оболочкой, сохраняя под ней жизнеспособность. Если условия для заражения подходящи, амебы сливаются между собой в плазмодии, которые и заражают растущие клубни, столоны и корни. Попадая в ткани растения, плазмодии стимулируют усиленное деление кле-
Рис. 10. Spongospora subterranecr. а — пораженный клубень; б — клубочки спор в клетке клубня.
ток, в результате которого возникают бугорки. Их клетки заполнены плазмодиями. К концу вегетации образуются клубочки спор.
Развитию гриба способствует затяжная и холодная весна. Наибольшего’ распространения порошистая парша достигает на тяжелых суглинистых и заболоченных почвах.
В Белоруссии порошистая парша имеет повсеместное распространение преимущественно на приусадебных участках.
Меры борьбы с этим заболеванием следующие:
I. Отбор для посадки здоровых клубней и протравливание их формалином (1:200). Протравливание следует производить перед посадкой.
2. Выращивание картофеля в системе правильного севооборота так, чтобы картофель на старое место возвращался на 4—5-й год.
3. Введение в культуру устойчивых сортов. (Относительно устойчивыми сортами являются: Агрономический, Юбель, Остботе и др.).
ПОРЯДОК MVXOCHVTRIDIALES
Формы, объединяемые в порядок миксохитридиевых, являются внутриклетными паразитами высших растений и причиняют весьма существенный вред. Главнейшие семейства этого порядка: Olpidiaceae, Synchytriaceae.
Сем. Olpidiaceae
Род Olpidiutn А. Вг. Это основной род, насчитывающий около 20 видов. Наиболее широко распространен вид Olpi-diiini brassicae Dang., который поражает молодые растеньица капусты и вызывает болезнь, известную под названием черпая ножка капустной рассады. Возбудитель этой болезни впервые был изучен и описан М. С. Ворониным под названием Chytridium brassicae Wor.
Рис. 11. Olpiditim brassicae-.
a—поражение корневой шейки рассады; б—летние; в — зимние зооспорангии; г — зооспоры.
Симптомом болезни служит побурение тканей в области корневой шейки. Стебельки становятся тоненькими, в результате растение надламывается и засыхает.
Инфекция в виде одножгутиковой зооспоры проникает в растение у корневой шейки. Попадая в паренхимные клетки,
паразит принимает вид круглого протоплазменного комочка, разрастается, становится многоядерным, одевается оболочкой и превращается в летний зооспорангий. Его внутреннее содержимое распадается на большое количество зооспор. Для выхода их наружу на спорангии развивается более или менее длинная шейка, прорастающая через 3—4 ряда клеток, расположенных выше пораженной. При помощи такого выводного канала зооспоры выходят наружу и заражают новые растения (рис. 11).
У 01. brassicae имеются зимующие спорангии. Они залегают глубже летних, имеют звездчатую форму и толстые оболочки. Их образование происходит после копуляции двух зооспор, которые при определенных условиях функционируют как гаметы. Покоящиеся зооспорангии освобождаются после сгнивания тканей растения, а зооспоры образуются после периода покоя и редукционного деления диплоидного ядра.
В качестве мер борьбы рекомендуется:
1. Проветривать парники с целью уменьшения влажности.
2. Покрывать почву в парнике слоем песка в 2 см, что способствует образованию придаточных корешков выше места поражения.
3. При сильном развитии болезни следует менять почву в парниках перед посевом или дезинфицировать ее паром при 90—100° в течение 30 минут.
Сем. Synchytriaceae
Это семейство отличается от предыдущего более сложным развитием паразита в период вегетации растения-хозяина. Внедряясь в клетки растения, паразит проходит несколько стадий и в конечном итоге образует не один зооспорангий, а несколько (5—7).
Род Synchytrium de Вагу et Wor. Главный в семействе, насчитывает около 70 видов, из них наиболее распространен и вредоносен Synchytrium endobioticum (Schibb.) Pers.— возбудитель рака картофеля.
В Англии и Шотландии эта болезнь была известна с 1870 года, а в 1896 году ее обнаружили и описали в Венгрии. Постепенно распространяясь по странам Западной Европы, рак картофеля попал в Советский Союз после воссоединения западных областей Украины и Белоруссии с восточными в 1939—1940 годах.
В настоящее время граница распространения этой болезни в нашей стране простирается с юго-запада на северо-восток, где среднемесячное выпадение осадков составляет около 70 мм, а среднемесячная температура июля около 20°. В юж-
них засушливых районах рак картофеля отсутствует, в связи |<‘м что паразит требует сравнительно высокой влажности
1НН1И.1,
11о зарубежным данным, болезнь сильнее распространена и<> влажных горных и приморских районах.
Заболевание характеризуется образованием наростов на клубнях, столонах, а иногда на стеблях и листьях. Наросты представляют собой разросшуюся паренхимную ткань клубня, неприкрытую покровной, а в связи с этим доступную для це-'кно ряда других микроорганизмов и быстро загнивающую (рис. 12).
Возбудитель картофельного рака, кроме обычной формы поражения, может вызвать еще три: листовидную, паршеоб-ризную и гофрированную. При листовидной форме поражают-(я пазушные почки — глазки, при этом образуются уродливо разросшиеся листочки. Паршеобразная форма вызывает образование язвочек на поверхности клубня, а гофрированная — воеобразную морщинистость из наплывов и бороздок.
Исследования, проведенные в Белоруссии Н. А. Дорожкиным и С. В. Горленко, показали, что последние две формы рака картофеля могут быть вызваны неблагоприятными условиями для развития гриба.
При микроскопическом исследовании наростов присутствие паразита в клетках клубня обнаруживается легко. Пораженные клетки содержат зимние зооспорангии (цисты). Они зале-иют в большом количестве в паренхимной ткани по одному в клетке. Занимая весь объем клетки, зооспорангии легко заметны благодаря наличию толстой желто-бурой оболочки. В некоторых зооспорангиях бывает различимо довольно крупное диплоидное ядро.
При уборке пораженного картофеля наросты легко обламываются, а оставаясь в почве, сгнивают, освобождая зооспорангии паразита. С наступлением теплой погоды начинается их прорастание, которому, по-видимому, предшествует процесс редукционного деления ядра. Перед образованием зооспор гаплоидные ядра делятся, а содержимое зооспорангия распадается на соответствующее ядрам количество участков. Таким образом, происходит образование зооспор, а затем осво-оождение их из спорангия. Обладая длинным гибким жгутиком, зооспоры легко передвигаются в почвенной влаге и, встречая соответствующие органы растения (клубни, столоны), проникают в эпидермальные клетки. В период вегетативного роста в клетке хозяина гриб выделяет биологически активные вещества, способствующие ее разрастанию. Диффундируя в соседние клетки, эти же вещества вызывают беспорядочное деление их, приводящее к образованию раковых наростов.
Клетки эпидермиса, окружающие пораженную, делясь
Рис. 12. Synchytriutn endobioticunr:
a — пораженное растение; б — зооспоры; в — летняя циста; г — начало ее прорастания; д — сорус спорангиев; е — копуляция; ж — зимняя циста;
з — ее прорастание.
и разрастаясь, образуют своеобразную розетку, внутри которой оказывается клетка с паразитом. Вегетативное тело гриба растет, вырабатывает двухслойную оболочку, превращаясь в одноядерную летнюю цисту (просорус), занимающую не весь объем клетки хозяина. Летняя циста прорастает без периода
покоя. Прорастание заключается в том, что в свободное про-। ipaiicTBO клетки хозяина циста образует выпячивание, по-। pi.iгос только внутренней оболочкой (будущий сорус), в ко-юрос переходит протоплазма и ядро. Вначале многократно /шли гея ядро, а затем и содержимое на 5—7 многоядерных участков, имеющих до 200—300 ядер. Каждый участок покры-пасч'ся собственной оболочкой, и образуется сорус споран-। пев. В дальнейшем набухшие окружающие клетки выдавливают спорангии из их общей оболочки, а затем и из оболочки клетки хозяина. К этому моменту внутри каждого спорангия образуются зооспоры, содержащие по одному гаплоидному ядру. Цикл развития одного поколения гриба совершается на протяжении 12—14 дней. Образующиеся зооспоры способны вызывать новое заражение.
Установлено, что зооспоры, вышедшие из летних зооспорангиев, могут попарно копулировать, т. е. выполнять функцию гамет. Продуктом копуляции является двужгутиковая планозигота с одним диплоидным ядром. Производя также заражение эпидермальной клетки, паразит стимулирует ее деление в тангентальном направлении. В результате пораженная клетка оказывается в глубине ткани. Разрастаясь и покрываясь плотной оболочкой в клетке хозяина, паразит превращается в зимнюю цисту, которая прорастает весной сразу (не проходя стадии просоруса и соруса) и дает гаплоидные зооспоры.
Зимние зооспорангии, оставаясь в почве после разрушения наростов, долго сохраняют жизнеспособность (5—7 и более лет). Это значительно осложняет борьбу с заболеванием.
Процесс прорастания зимних зооспорангиев, как и всякий биологический процесс, требует наличия определенных условий. Таковыми являются влажность почвы (60—80% от полной влажности), температура (17—18°), достаточно хорошая аэрация почвы и pH, равное 7—8. Если прорастание зооспорангиев происходит при отсутствии поражаемых растений (картофель, паслен черный, паслен сладко-горький, белена черная и др.), зооспоры, не встретив подходящего хозяина, погибают и почва постепенно освобождается от инфекции.
В настоящее время более устойчивыми по отношению к возбудителю рака являются следующие сорта картофеля советской селекции: Агрономический, Зазерский, Кандидат, Темп, Камераз, Разваристый и др.
Меры борьбы с болезнью рекомендуются следующие:
1. Возделывание ракоустойчивых сортов.
2. Ограничение вывоза из зараженных районов не только картофеля, но и корнеплодов, луковиц, саженцев и т. п.
3. Обеззараживание очагов болезни 2—2,5%-ным раствором нитрафена (20 л на 1 м2).
4. Выращивание на зараженных участках в течение 5—7 лет культур, не поражаемых грибом.
5. Проведение систематического обследования участков для выявления очагов болезни.
ПОРЯДОК МУ СОСНУТ RIDI ALES
Вегетативное тело микохитридиевых с самого начала одето-оболочкой. У простейших оно состоит из одноядерной центральной клетки и отходящих от нее ризоидов. Зооспорангий образуется из центральной клетки после многократного деления ядра. Зооспоры снабжены одним длинным жгутиком, отходящим от заднего конца. Высшие микохитридиевые представляют формы с более развитым мицелием. Гифы его в некоторых местах вздуваются, образуя так называемые собирательные клетки, из которых впоследствии развиваются спорангии или покоящиеся цисты. Большинство микохитридпе-вых являются паразитами высших растений.
Микохитридиевые делят на несколько семейств. Наибольшего1 внимания заслуживает семейство Phyzodermaiaceae.
Сем. Phyzodermaiaceae
Род Phyzoderma Wallr. Его представители паразитируют на различных высших растениях. В качестве примера следует отметить Phyzoderma zeae Shaw., вызывающую бурую пятнистость листьев кукурузы. В южных штатах Америки, где болезнь сильно распространена, ее называют «оспой кукурузы». В Советском Союзе это заболевание встречается в Грузии.
Зимующей стадией паразита являются толстостенные округлые цисты, которые после разрушения пораженных тканей освобождаются и воздушными течениями переносятся на растения, вызывая новое заражение незадолго до выметывания метелок.
Прорастание цисты и образование зооспор прослежено в искусственных условиях в капле воды. В природе этот процесс совершается после того, как циста паразита попадает в пазухи листьев, где скопляются капельки воды. Оболочка цисты состоит из двух слоев. Наружный слой — экзоспорий, желтоватый, более плотный; внутренний — эндоспорий, тонкий и эластичный. От увеличивающегося тургорного давления внутри цисты на экзоспории появляется кольцевая трещина, отделяющая небольшую часть его в виде крышечки. Эндоспорий при этом вытягивается в виде сосочка на освободившемся участке (рис. 13). Из внутреннего содержимого цисты образуются зооспоры, выходящие наружу через маленькое от-
Таблица I. Claviceps purpurea.
а — колос ржи, пораженный спорыньей; б — проросший склероций с головчатыми стромами на иожках; в —разрез через строму, видны перитеции; г — перитеций с пучком сумок; д — сумка с нитевидными спорами; е — медвяная роса на колосе; ок — конидненосцы и конидни.
ш рстие на вершине сосочка. Попадая на листья и прилежании' участки стебля, зооспоры прорастают, образуя мицелий, который и проникает в паренхимную ткань. Внутри растения мицелий разрастается, на его гифах кое-где начинают формироваться собирательные клетки в виде небольших вздутий. Вначале эти клетки содержат одно ядро. Позднее они вытяги-п;потся, делятся на 3—4 одноядерные клетки, расположенные
Рис. 13. Phyzoderma геае\
а, б — прорастание цисты; в — развитие мицелия из зооспоры; г — прорастание собирательных клеток, вправо вегетативно, влево образованием цисты.
друг над другом по длине гифы. Обладая способностью прорастать, клетки вскоре дают отростки, одни из которых развиваются в новый мицелий, а другие, вздуваясь на конце, образуют многоядерные цисты. К концу вегетации нити мицелия исчезают, как и содержимое пораженных клеток хозяина. Симптомы болезни проявляются в виде бурых пятен па пораженных участках листьев и стебля.
Основной мерой борьбы с бурой пятнистостью кукурузы следует считать плодосмен.
ПОДКЛАСС OOMYCETES
Грибы, принадлежащие к подклассу оомицетов, обитают в водной среде и на суше. Они ведут как сапрофитный, так и паразитный образ жизни. Многие из паразитных форм причиняют значительный ущерб урожаю сельскохозяйственных растений.
Главнейшие порядки:!) Saprolegniales;2) Peronosporales.
5 3. Н. Кудряшева 65
ПОРЯДОК SAPROLEGNI ALES
Представители этого порядка — преимущественно водные сапрофиты, причем многие из них поселяются на трупах мелких животных. Вегетативное тело рассматриваемых грибов состоит из двух родов гиф: тонкие ризоиды внедряются в субстрат, а более толстые направляются от субстрата и несут зооспорангии, оогонии и антеридии.
Сем. Saprolegniaceae
Представители этого семейства почти исключительно водные сапрофиты, поселяющиеся на разлагающихся белковых телах.
Род Saprolegnia Nees von Esenb. Если взять воду из водоема, где всегда находятся зооспоры гриба, и бросить туда несколько мертвых мух, то через 3—4 дня вокруг тела мухи будет хорошо заметен мицелий Saprolegnia в виде белого пушка.
Тонкие гифы внедряются в тело мухи, а толстые и мало-ветвящиеся направляются от субстрата в воду. Концы их отделяются поперечной перегородкой и образуют зооспорангии. Их содержимое распадается на одноядерные участки, превращающиеся в зооспоры, которые плавают в окружающей их жидкости зооспорангия. Затем тургорное давление внутри зооспорангия повышается, он разрывается сверху, и зооспоры с силой выбрасываются в воду (см. рис. 4, а). Они имеют грушевидную форму, а на переднем конце клетки находятся два одинаковых жгутика. Эти зооспоры не способны производить новое заражение и развиваться в мицелий. Поплавав немного в воде, они останавливаются, одеваются оболочкой, втягивая жгуты. Через некоторое время оболочка сбрасывается, и в воду выходят зооспоры совершенно другой формы: они почковидны и имеют два жгутика сбоку. При движении один жгутик направлен вперед, другой — назад. При наличии подходящего субстрата зооспоры способны оседать на нем и развивать новый мицелий.
Виды рода Saprolegnia обоеполы. Оогонии и антеридии могут возникать как на одной и той же ветви мицелия, так и на разных его ветвях, для рода Achlya Nees von Esenb. известны случаи гетероталлизма (оогонии и антеридии развиваются на разных мицелиях).
Некоторые из представителей указанных выше родов могут паразитировать на рыбной молоди и икре, принося тем ^амым значительный ущерб рыбоводству.
ПОРЯДОК PERONOSPORALES
Этот порядок характеризуется очень большим количеством представителей (до нескольких сотен видов), которые весьма разнообразны по условиям существования. Есть формы чисто водные, но большей частью встречаются сухопутные виды, вполне приспособившиеся к наземному существованию.
По образу жизни пероноспоровые могут быть сапрофитами и 1паразитами. Последние распространены наиболее широко и отличаются высокой специализацией.
Все пероноспоровые имеют хорошо развитый неклет-пый мицелий, который у более низко организованных представителей слагается отчасти из свободно идущих гиф, отчасти из гиф, погруженных в субстрат (эндофитных). Последние проходят по межклетникам, либо же пронизывают и клетки, никогда не образуя при этом специальных гаусторий.
Органы бесполого размножения у одних пероноспоровых представлены зооспорангиями, а у других настоящими конидиями со всеми возможными переходами между этими крайними типами. Причем как у сапрофитных, так и у паразитных форм с эндофитным мицелием образование органов бесполого размножения происходит на специализированных гифах обычно вне субстрата.
Органы полового размножения оогонии и антеридии у паразитных форм закладываются на эндофитном мицелии, а ооспоры освобождаются и прорастают после сгнивания пораженных органов.
Среди представителей порядка Peronosporales есть формы, приносящие большой вред сельскому хозяйству. Являясь паразитами таких растений, как картофель, томаты, виноград, свекла, лук и др., они в благоприятные для своего развития годы значительно снижают урожай этих культур.
Порядок Peronosporales делится на 3 семейства: Pythia-свае; Peronosporaceae; A Ibuginaceae.
Сем. Pythiaceae
Простейшие из семейства питиевых являются типичными водными сапрофитами; более высокоорганизованные паразитируют на высших растениях и по своей организации сходны с представителями второго семейства Peronosporaceae.
Наибольшее значение в семействе питиевых грибов имеют роды Pythiutn и Phytophthora.
Род Pythium Pringsh. Большинство представителей — водные формы, сапрофиты или факультативные паразиты на во
дорослях. Некоторые паразитируют на наземных растениях, получая особенное развитие при повышенной влажности.
Наиболее важным и широко распространенным представителем рода Pythium является Р. de baryanum Hesse. Он может жить в почве сапрофитно, но вместе с тем при благоприятных условиях способен нападать на корни молодых сеянцев травянистых и древесных пород и вызывать их гибель.
Р de baryanum в паразитных условиях проявляет известную специализацию. В (пределах этого вида выявлено довольно значительное количество рас, отличающихся своими патогенными свойствами.
Из сельскохозяйственных растений этот вид приносит наибольший вред сеянцам сахарной свеклы и капусты.
Род Phytophthora de Вагу. Он широко распространен в различных климатических зонах. Его виды могут развиваться на самых различных растениях: древесных (хвойных и лиственных), кустарниках, травянистых (однолетних и многолетних). Паразитируя на растениях, представители рода Phytophthora вызывают в конечном итоге загнивание тканей. Вместе с тем для них характерна известная приспособленность к определенным органам растений. Например, Ph. infes-tans de Вагу поражает ботву и клубни картофеля, а у томатов от этого возбудителя особенно страдают плоды. Ph.cocto-rum Lebert., поражая орех, начинает свое развитие от корневой шейки, а в дальнейшем распространяется на корни. Ph. fragaria Nik. поражает проводящую ткань осевого цилиндра корня. Ph. phaseoli Thaxt., развиваясь на фасоли, поражает молодые побеги, листья, цветки, плоды и может проникнуть в семена.
Мицелий Phitophthora, как правило, эндофитный, развивается преимущественно по межклетникам, но может проходить и через клетки. У отдельных представителей (Ph. faseo-II) мицелий образуется и на поверхности пораженного органа (на плодах) в виде белого войлочного налета.
Наиболее важным и известным представителем является Ph. infestans de Вагу — возбудитель поздней гнили, или картофельной плесени, самого опасного заболевания ботвы и клубней картофеля. Это заболевание широко распространено во всех картофелепроизводящих странах мира.
Как показали исследования Н. А. Дорожкина, в Белоруссии вредоносность фитофторы выражается в снижении урожая на 30—50% в годы, благоприятные для развития гриба.
Впервые Phitophthora на картофеле появилась в Европе в 30-х годах XIX века и в дальнейшем широко распространилась по всем европейским странам. После сильного развития фитофторы в 1S45 году в Англии, Голландии и Германии на-
i । ушил голод, и картофелеводством в этих странах почти перестали заниматься. Эпифитотия фитофторы привлекла внимание как ученых специалистов, так и общественных дея-н'Лей.
Заслуга установления возбудителя гнили картофеля (/7/. infestans) и изучения его биологии принадлежит Антону /и* Бари. Его работа об особенностях развития гриба вышла в 1861 году. Исследования де Бари имели большое значение чля дальнейшего изучения Ph. infestans во всех странах, где возделывался картофель.
Ph. infestans поражает листья, стебли и клубни картофеля. Мицелий гриба распространяется в пораженных органах но межклетникам, а также проходит через клетки. Особенностью мицелия Phitophthora является то, что гифы его не образуют гаусторий (присосок). Грибница получает питание г помощью гиф, проходящих через клетки.
Гриб начинает свое развитие на листьях растения и проявляется в виде сероватых, а впоследствии буреющих пятен, расположенных чаще по краям листа. При влажной и теплой погоде, особенно в ранние утренние часы, на нижней стороне пораженных листьев можно видеть серовато-белый налет. г)тот налет состоит из разветвленных конидиеносцев, на концах которых развиваются конидии. Конидиеносцы выступают по одному или по несколько через устьица листа (рис. 14). Конидии — одноклетные, лимонообразной формы. Отпадающие конидии попадают на здоровые листья и, прорастая, заражают растение. Ветви конидиеносца Ph. infestans обладают способностью продолжать свой рост после отпадения конидий.
Конидии способны прорастать как в капле воды, так и при отсутствии капельно-жидкой влаги. При прорастании в капле воды конидия дает до 8 зооспор и, таким образом, функционирует как зооспорангий. Зооспоры имеют овальную форму и два жгута сбоку, направленные при движении один вперед, другой назад. Каждая зооспора способна производить заражение. В случае отсутствия капельно-жидкой влаги при прорастании конидии образуется росток в виде гифы, которая заражает здоровый орган.
Заражение клубней происходит в период вегетации и во время уборки картофеля. Если картофель плохо окучен и клубни находятся неглубоко, то они могут заражаться опавшими с листьев и прорастающими конидиями. Во время уборки клубни заражаются от соприкосновения с ботвой.
В цикле развития Ph. infestans имеется и половой процесс, протекающий по типу, обычному для порядка Регопо-sporales. Антеридии и оогонии могут развиваться как на гифах гриба в пораженных тканях растения после уборки, так
Рис. 14. Phytophthora infestans:
a — пораженный лист; б — разрез листа с выходящими через устьице конидиеносцами; в — конидия; г — прорастание ее зооспорангием; д — зооспора;
е — прорастание кониднн гифой.
и на мицелии, живущем сапрофитно в почве. При прорастании ооспоры образуется либо первичный зооспорангий, либо мицелий гриба. Возникновение первичной инфекции происходит главным образом gt слабозараженных клубней, из которых развиваются больные ростки. Образующиеся на них конидии заражают здоровые растения.
В последние годы как в зарубежной, так и в отечественной литературе появились указания на существование у Ph. in [estans различных рас. В настоящее время исследованиями ио определению биологической специализации Ph, infestans установлено 17 физиологических рас с разной степенью агрессивности. У более агрессивных рас конидии способны прорастать при температуре выше оптимума. Распространение их мицелия в тканях пораженного растения происходит быстрее. Выживаемость в хранилищах на пораженных клубнях значительно лучше при слабом поражении. Такие клубни не сгнивают, а, попадая в поле, являются очагами распространения инфекции.
В Белоруссии изучением степени агрессивности рас Ph. infestans и устойчивости сортов картофеля в полевых условиях занимается коллектив лаборатории иммунитета под руководством Н. А. Дорожкина в БНИИПОК. Выделены сорта, обладающие высокой степенью сопротивления к болезни (Лошиц-кий, Темп и др.). Практическая ценность таких сортов заключается в том, что они не могут служить массовым источником инфекции.
Против этого заболевания картофеля разработан целый ряд мероприятий, как агротехнических, так и химических.
Агротехническими мероприятиями являются:
1) просушивание клубней после выкопки;
2) яровизация картофеля;
3) глубокое окучивание;
4) скашивание ботвы перед уборкой и удаление ее с поля;
5) введение в культуру фитофтороустойчивых сортов (Камераз 1, 2, 3, Московский, Фитофтороустойчивый, Лошиц-кий, Темп и др.).
Из химических мер борьбы применяются следующие:
1. Авиаопрыскивание микродозами медного купороса. Первое опрыскивание производится через 7—8 дней после появления полных всходов. При 1%-ной концентрации раствора расходуется 100 л/га, при 2%-ной — 50 л/га. Второе опрыскивание проводится через 8—10 дней после первого. При тех же концентрациях количество раствора соответственно увеличивается: в первом случае 200 л/га, во втором 100 л/га.
2. Опрыскивание бордоской жидкостью: первое в период бутонизации; второе при проявлении первых признаков болезни; третье через 10—15 дней после второго. Концентрация раствора во всех случаях—1—2% при расходовании 300 л/га.
3. Опрыскивание хлорокисью меди. При авиаопрыскивании употребляется 1%-ная суспензия в количестве 300 л/га; при опрыскивании наземными машинами концентрация должна быть равна 5%, а количество раствора — 600 л/га.
Кроме картофеля, Ph. infestans в сильной степени поражает томаты. Особенно велика патогенность этого возбудителя к плодам томатов, которые, как правило, поражаются им чрезвычайно сильно. На листьях болезнь проявляется реже, даже у наиболее восприимчивых сортов томатов.
Ph. infestans поражает преимущественно еще зеленые плоды. Заболевание начинается чаще всего с вершины. Здесь образуется неправильной формы пятно, постепенно охватывающее весь плод. Пораженная ткань становится твердой. При микроскопическом анализе ее хорошо заметна эндофитная грибница Ph. infestans. Конидиальное спороношение гриба на пораженных плодах может развиваться только при повышенной влажности, поэтому влажная и теплая «погода способствует развитию болезни. Пораженные плоды гниют. Возбудитель болезни зимует в пораженных растительных остатках томатов и картофеля. Начало появления болезни наблюдается во вторую половину лета, недели через две после массового поражения картофеля.
Высокая восприимчивость томатов к Ph. infestans показывает, что данный паразит в природных условиях может легко переходить с картофеля на томаты. Это с особой остротой подчеркивает необходимость пространственной изоляции томатов от участков, занятых картофелем не только в текущем, но и в будущем году.
Из других мероприятий, направленных на борьбу с фито-фторозом томатов, большое значение имеют следующие: удаление с поля пораженных плодов; внесение повышенных доз калийных удобрений; опрыскивание 1%-но’й бордоской жидкостью (800—1000 л/га). В районах сильного распространения фитофтороза опрыскивание следует производить через каждые 10—15 дней, начиная с момента завязывания плодов, а в остальных районах со времени появления этой болезни на картофеле.
Phitophthora fragariae Nik. вызывает опасную болезнь земляники. Основным симптомом ее является красная окраска проводящей ткани осевого цилиндра корня. Около 1920 года это заболевание земляники появилось в Шотландии, а затем было обнаружено в 1930 году в США.
По сведениям многих зарубежных исследователей болезнь очень быстро распространяется и причиняет самый большой ущерб плантациям земляники в Северной Америке, Европе и Новой Зеландии.
В СССР пока это заболевание широко не распространено, но тем не менее представляет большую угрозу для культуры земляники.
Впервые в Советском Союзе фитофтороз земляники обнаружен в Ленинградской области Г. Ф. Маклаковой (1958),
.1 затем Н. Ф. Юровой. Весною 1962 года Ph. fragariae най-iriia на землянике в Краснодарском крае (Говорова, 1966). < .пмптомом болезни на надземной часта растений служит внезапное увядание всего растения, иногда только наружных листьев, а у молодых растений — цветоносов. При сильном поражении растения погибают во время жаркого периода. Слабое поражение проявляется в изменении окраски листьев, они становятся синевато-зеленоватого оттенка, теряют блеск. На краях старых листьев появляются желтые или буровато-красноватые пятна. Для точного диагноза болезни необходим анализ корневой системы. Развитие Ph. fragariae на корнях вызывает отмирание мелких разветвлений корней. Остаются оголенные, более крупные, суживающиеся книзу корни («крысиный хвост»). Центральный древесинный цилиндр этих корней приобретает красную окраску. Признаки фитофтороза особенно типичны у кустов на второй год поражения и обнаруживаются уже весной. Болезнь проявляется в запаздывании появления листьев. Листочки с серым налетом развиваются на укороченных черешках, а листовые пластинки мелкие, чашевидные.
В цикле развития гриба имеется грибница, зооспоры и ооспоры. Грибница растет по межклетникам. Оптимальная температура для ее роста 18—22°, при более высокой рост замедляется, а затем прекращается. Зооспорангии образуются на поверхности пораженных корней в сырой почве. Зооспоры сохраняют жизнеспособность в течение 3—4 часов. Оогонии и антеридии развиваются в центральном цилиндре корня, в коре боковых корешков и у вершины главных корней. В центральном цилиндре заболевших корней легко обнаружить ооспоры золотисто-бурого или кирпичного цвета.
Ph. fragariae не поражает дикорастущую землянику, даже в том случае, если последняя растет рядом с культурными сортами. Это говорит о резкой специализации гриба. Из коллекции сортов Майкопской опытной станции наиболее поражаемыми являются: Крупная, Мелитопольская, Поздняя из Загорья, Предгорная, Светлана и др. Устойчивы к фитофторе Консервная, Превосходная, Киевская № 2, Директор Юльке, Доктор Эфем, Елшанка и др.
Меры борьбы с фитофторозом земляники прежде всего должны быть направлены на тщательную выбраковку посадочного материала, с которым заболевание может попасть в новые районы. Большое значение имеет регулирование влажности на участках, занятых земляникой, применением дренажа и осушения. При сильном поражении растений участки распахивают. Возвращение на них земляники возможно не ранее чем через 6—8 лет.
Сем. Peronosporaceae
Представители семейства пероиоспоровых являются типичными облигатными паразитами исключительно цветковых растений и отличаются узкой специализацией. Они развивают всегда эндофитный мицелий, идущий ino межклетникам, а в клетки посылают гаустории (присоски), с помощью которых извлекают питательные вещества. В некоторых случаях на-
Рис. 15. Конидиеносцы Peronosporaceae: а — Plasmopara, б — Pcronospora.
блюдается диффузное распространение мицелия, т. е. он пронизывает все растение. Если растение многолетнее, мицелий зимует в корнях, а весною распространяется по надземным частям по мере их отрастания.
В основу классификации представителей семейства положено строение конидиеиосцев, которые выходят наружу через устьица. Конидиеносцы пероноспоровых не обладают свойством, как у Phytophthora продолжать свой рост после отпадения «конидий (рис. 15).
Наиболее широко распространены роды Plasmopara, Peronospora, Pseudoperonospora.
Род Plasmopara Schrot. Название рода (Plasmopara — рождающая плазму) обусловлено способностью конидий
выпускать наружу все содержимое в виде голой протоплазменной массы, которая затем одевается оболочкой и прорастает в нить мицелия. У некоторых представителей образуются зооспоры, т. е. конидия прорастает, как зооспорангий.
Важнейшим представителем данного рода является Plas-mopara viticola Berl. et de Toni, паразитирующий на вино-1раде. Этот гриб был занесен в Европу из Америки в 1878 го-1.у и распространился всюду, где возделывается виноград, выбывая болезнь его, известную под названием мильдью. Гриб поражает листья, усики и ягоды, а, кроме того, почки и ветви, где и перезимовывает грибница. Основным признаком болезни служит появление желтовато-зеленых маслянистых пятен на листьях.
В течение лета гриб размножается при помощи конидий. Они образуются в большом количестве на конидиеносцах, выходящих пучками через устьица на нижней поверхности листьев и переносятся ветром.
К осени на грибнице в пораженных органах (преимущественно в листьях) закладываются оогонии и антеридии, а образующиеся ооспоры освобождаются после сгнивания листьев. Весною каждая из них, прорастая, дает нонидиеносец с конидиями. Последние переносятся ветром на здоровые листья винограда и заражают их.
Однако не все ооспоры прорастают сразу. Этот процесс может продолжаться в течение 2—3 месяцев, а часть их вообще остается непроросшими до следующего года.
Р. viticola обладает явно выраженным генетическим консерватизмом и способностью к большой морфологической изменчивости в зависимости от условий среды (К. Рафаилэ). Эта особенность гриба послужила основанием для высказывания предположений о наличии у Р. viticola специализированных рас.
Из европейских сортов винограда сравнительно устойчивыми к мильдью являются Каберне-Савиньон, Рислинг рейнский, Алиготе и некоторые другие. Из восприимчивого европейского сорта Шасла методом индивидуального отбора получен новый сорт, высокоустойчивый к мильдью (Верде-ревский). Американские сорта винограда Рупестрис, Цине-рея, Рипария иммунны к мильдью, а поэтому используются в качестве подвоя для скрещивания с европейскими.
Мильдью приносит большой экономический ущерб. Так, по сообщению Д. Д. Вердеревского, урожай винограда в годы, благоприятные для развития болезни, снижается на 50%. Кроме того, сильное развитие болезни ослабляет виноградную лозу, что приводит к гибели центральных почек, а в связи с этим к значительному снижению урожая в следующем году.
В борьбе с мильдью рекомендуется следующее:
1. Уничтожать ооспоры, образующиеся в опавших листьях и представляющие зимующее инфекционное начало.
2. Не допускать возможности заражения растений в период вегетации путем опрыскивания и опыливания виноградников хлорокисью меди, динитророданбензолом совместно с хлорокисью меди, а также цинебом.
Первое опрыскивание (проводится при обнаруживании первых маслянистых пятен. Второе после прироста 3—5 новых листьев (через 8—10 дней).
3. Выводить устойчивые сорта путем межвидовой гибридизации.
Род Peronospora Cord а. Среди всех пероноспоровых этот род особенно обширен, насчитывает около 200 видов. Представители его наиболее специализированные паразиты, совершенно утратившие способность образовывать зооспоры. Мицелий у них развивается эндофитно, а конидиеносцы с конидиями образуются на поверхности пораженных листьев, вызывая явление, известное под названием ложной мучнистой росы.
Одним из наиболее распространенных паразитных грибов названного рода является Peronospora schleideni Ung.— возбудитель ложной мучнистой росы лука. Мицелий гриба зимует в луковицах, не вызывая их загнивания. Если зараженные луковицы высадить в почву, они дают больные растения. Грибница’ паразита переходит в листья (перо), пронизывая их ткани. Через 3—4 недели после посадки пораженные растения заметно отличаются от здоровых: листья становятся желтоватыми и вялыми, а на их поверхности появляется серовато-фиолетовый налет, состоящий из конидие-носцев и конидий паразита. Пораженные растения и являются первыми очагами, которые распространяют споры гриба. Болезнь особенно вредоносна для семенников. Пораженные стрелки желтеют, надламываются, семена получаются щуплые или не развиваются совсем.
При созревании лука грибница из листьев проникает в луковицу и сохраняется в ее чешуях в недеятельном состоянии. Влажная и теплая погода способствует развитию болезни.
Эффективными мероприятиями в борьбе с ложной мучнистой росой лука являются:
1. Прогревание посадочного материала.
2. Протравливание в день посадки гранозаном (300 г/ц)-Из других видов рода Peronospora следует отметить Per, schachtii Fuck, на свекле. Этот гриб распространен в районах свеклосеяния. Раньше всего пероноспороз проявляется на высадках, а затем переходит на свеклу первого года. Таким образом, первая вспышка болезни наблюдается в мае —
июне. Погодные условия этого периода способствуют развгь гню болезни. У высадков сначала заболевают молодые листья, а затем верхушки цветочных побегов. У свеклы первого года поражаются молодые центральные листья розетки. Нижняя сторона листьев покрывается серо-фиолетовым налетом, состоящим из конидиеносцев и конидий гриба. Листья оледнеют, утолщаются, скручиваются и в конце концов отмирают (рис. 16). По мере отрастания листья один за другим
Рис. 16. Pcronospora schachtir. а — пораженные листья; б — конпдиеносец с конидиями.
поражаются грибом. Распространение инфекции в течение вегетационного периода происходит конидиями. Если условия для развития конидиальной стадии неблагоприятны, болезнь может сохраняться в растении в скрытом состоянии, а затем давать новую вспышку. Зимует гриб главным образом в виде грибницы, сохраняющейся в тканях головки корнеплода маточной свеклы. Зимовка может происходить и ооспорами, образующимися в тканях пораженных листьев, остающихся на поверхности почвы, а также в семенах.
В борьбе с пероноспорозом свеклы рекомендуется:
1. Посевы, предназначенные для выращивания маточной свеклы, размещать на расстоянии не менее 1 км от высадков.
2. Тщательно выбраковывать во время уборки все пораженные растения и удалять их с поля.
3. При обнаружении весною очагов болезни производить опрыскивание бордоской жидкостью здоровых растений в радиусе не менее 5 м от очага болезни.
Род Pseudoperonospora Rostowzew. Он занимает промежуточное положение между двумя первыми родами и имеет черты, сходные как с первым, так и со вторым родом. При бесполом размножении у представителей данного рода образуются зооспоры, т. е. отпадающие от конидиеносцев конидии прорастают, как зооспорангии. По» этому признаку рассматриваемый род более сходен с родом Plasmopara; ветвление же конидиеносцев у Pseudoperonospora такое же, как и у Ре-ronospora. Род Pseudoperonospora довольно немногочисленный. Наиболее известны два его представителя: 1) Ps. cuben-sis В. et С.— паразит на листьях огурцов, вызывающий их преждевременное отмирание; 2) Ps. humuli Wils., поражающий побеги хмеля. Этот весьма вредоносный по своему действию паразит завезен из Японии. В Советском Союзе он получил особенно широкое распространение в Ленинградской области, в Белоруссии, на Северном Кавказе.
Сем. Albuginaceae
Все представители этого семейства ведут исключительно паразитный образ жизни. Характерной особенностью этих грибов является не только эндофитное развитие мицелия, но
Рис. 17. Albugo Candida. Разрез листа с конидиеносцами и конидиями под эпидермисом.
и образование органов бесполого размножения внутри субстрата. Они обычно прикрыты эпидермисом растения-хозяина и освобождаются лишь после его разрыва. Отпадающие спо
р.тгии прорастают только зооспорами. Органы полового размножения развиваются также эндофитно, а ооспоры освобож-|;потся после сгнивания тканей растения-хозяина. При их прорастании образуется спорангий, дающий зооспоры.
Наиболее широко распространен вид Albugo Candida (Pers.) Kze, паразитирующий на крестоцветных и особенно часто на пастушьей сумке. Он вызывает болезнь, известную под названием белой ржавчины. Пораженные органы растения покрываются как будто капельками белой масляной краски. Спороношение гриба, образовавшееся под эпидермисом, приподнимает его, а в дальнейшем разрывает (рис. 17). I Сраженные участки становятся матовыми и начинают порошить. Очень часто у больных растений можно наблюдать различные искривления и гипертрофию тканей.
ПОДКЛАСС ZYGOMYCETES
Зигомицеты — довольно обширный подкласс, насчитывающий около 400 видов почти исключительно сухопутных сапрофитных форм. Очень немногие из них вторично приспособились к водным условиям, а некоторые к паразитному существованию большей частью на животных организмах.
В соответствии с образом жизни в наземных условиях зи-гомицеты совершенно лишены подвижных жгутиковых стадий— зооспор, чем резко отличаются от оомицетов. Кроме различия в характере бесполого и полового размножения, между обоими подклассами есть и некоторые другие различия, а именно: у зигомицетов впервые появляется хитин в оболочках их гиф, который представлен и у высших наземных грибов. По своей химической (природе хитин представляет собой ацетилированный глюкоз амин и у зигомицетов сочетается в оболочке с пектиновыми веществами.
Подкласс зигомицетов делится на два порядка: 1) Мисо-rales; 2) Entomophthorales.
ПОРЯДОК MUCORALES
Порядок мукоровых грибов широко распространен в природе на самых разнообразных субстратах. Естественными местообитаниями для большинства служат почва, растительные остатки, экскременты травоядных животных.
Являясь сапрофитами — разрушителями органических веществ, они играют значительную роль в круговороте веществ в природе.
Многие мукоровые поселяются на различных пищевых продуктах растительного происхождения (хлебе, варенье, плодах, овощах), образуя плесени. Некоторые могут пора
жать семена в период их прорастания и тем самым значительно снижать процент всхожести.
Большое значение имеют мукоровые грибы в медицине и ветеринарии как паразиты животных (птиц, млекопитающих) и человека, вызывающие заболевания дыхательных путей и внутренних органов, известные под названием м у к о-ромикозов.
Все мукоровые имеют хорошо развитую белую или сероватую грибницу. Одна часть ее погружена в субстрат и представляет собой тонкие ризоиды, а другая в виде более толстых ветвящихся гиф, утончающихся к верхушке, направлена в воздушную среду. Воздушный мицелий многих мукоровых может давать столоны в виде дугообразно растущих гиф. Их образование происходит следующим образом: гифа сначала растет косо вверх, а затем плавно пригибается к субстрату. В месте соприкосновения с ним она образует пучок коротких ризоидов и продолжает расти дальше, таким же образом давая новую дугу. Иногда в старых гифах мукоровых могут появляться перегородки, а в некоторых случаях перегородки можно наблюдать в спороносных органах.
У немногих представителей мукоровых, как например у Mucor racemosus, наблюдается образование перегородок при определенных условиях развития и в молодом возрасте. Если культивировать этот гриб погруженным в жидкий субстрат, где недостаточная аэрация, то в гифах образуются перегородки, а в дальнейшем мицелий распадается по этим перегородкам на отдельные клетки. Образовавшиеся клетки по своему происхождению являются оидиями (видоизменение мицелия). Постепенно они приобретают шаровидную форму, размножаются почкованием и образуют так называемые му-корсвые дрожжи. Перенесенные на поверхность субстрата, они снова вырастают в обычный одноклетный мицелий.
Спорангиеносцы мукоровых приподнимаются над субстратом и обладают способностью ветвиться дихотомически, мо-ноподиальио и симподиально.
При развитии спорангия как органа бесполого размножения кончик спорангиеносца обычно вздувается, а затем у основания его образуется перегородка, которая при дальнейшем развитии спорангия куполообразно изгибается в полость последнего и очерчивает так называемую колонку. Колонка представляет собой как бы продолжение спорангиеносца. У некоторых видов имеется апофиза, представляющая расширение спорангиеносца под спорангием воронковидной или иной формы. Оболочка молодого спорангия по своим физическим и химическим свойствам ничем не отличается gt оболочки спорангиеносца или других гиф мицелия. По мере созревания в ней 'происходят изменения химического состава,
( называющиеся и на ее физических свойствах. У одних оболочка зрелого спорангия расплывается от воды и даже от влажного воздуха, причем это явление наблюдается только наверху спорангия. В его нижней части около колонки оболочка остается в виде воротничка (Mucor mucedo), У других при созревании спорангия оболочка разрывается на кусочки (Л4. racemosus). И, наконец, у некоторых, как например у Pi-lobolus, оболочка спорангия сильно кутинизирована и обладает значительной прочностью. Она не разрывается и спорангий не раскрывается, а отбрасывается от спорангиеносца развивающимся в нем тургорным давлением.
Результатом зигогамного полового процесса в нормальных условиях является зигоспора, или зигота, но наряду с нормальными зиготами могут закладываться азиготы, образующиеся без полового процесса из одного только гаметангия. Образование азигот обычно связано с изменением внешних условий, например с повышением температуры.
Порядок Mucorales включает несколько семейств, из которых наибольшее практическое значение имеет семейство Ми-согасеае.
Сем. Мисогасеае
Большинство представителей этого семейства — сапрофиты. Развиваясь на различных растительных пищевых продуктах, они образуют плесени. Наиболее распространены роды Mucor, Rhizopus, Pilobolus.
Род Mucor (L.) Fres. Основным и наиболее распространенным представителем этого рода является Mucor mucedo (L.) Fres. (рис. 18), образующий головчатую плесень, встречающуюся на хлебе, плохо сваренном варенье и других продуктах, хранящихся в сырых помещениях. Споры мукора находятся в большом количестве в воздухе и, попадая на благоприятный субстрат, при наличии необходимой влажности могут уже на 2—3-й день вызвать образование пышной грибницы.
Некоторые представители рода Mucor обладают бродильными свойствами, как например М. racemosus Fres., М. genevensis Lendn., М. javanicus Wehmer и др. Причем первые два бродят в состоянии мукоровых дрожжей, а другие — в виде нормального мицелия.
В Азии при приготовлении алкогольных напитков употребляют М. javanicus и М. rauxianus Wehmer, так называемые восточно-азиатские дрожжи. Они имеют некоторое преимущество перед настоящими дрожжами, так как сами осахаривают крахмал и не требуют предварительной обработки крахмалистых продуктов.
6 3. Н. Кудряшева
81
Некоторые представители рода Мисог паразитируют на животных, а также на человеке, вызывая поражение внутренних органов. Такими являются М, corymbifer Cohn и М. pusil-lus Hagem.
Род Rhizopus Ehrend. Один из видов этого рода Rh. oryzae Went et Pr. Geerl, как и М. javanicus, обладает способностью осахаривать крахмал и применяется в Японии для изготовления рисовой водки саке. Распространенный в Америке Rh. tri-tici Saito> поражает зерно, особенно -при повышенной влажности хранения.
а
Рис. 18. Мисог mucedo: а — мицелий со спорангиями, б — развитие спорангия.
Род Pilobolus Tode. На конском навозе развивается Pilo-bolus crystallinus Tode. Он обладает интересной особенностью отбрасывать спорангии целиком (стреляющие спорангии). Видовое название crystallinus объясняется способностью гриба выделять блестящие капельки жидкости на поверхности спорангиеносца.
ПОРЯДОК ENTOMOPHT MORALES
У представителей этого порядка мицелий рано получает перегородки и в зрелом состоянии оказывается уже многоклетным. Иногда он распадается на составляющие его клетки. У энтомофторовых спорангии полностью заменены конидиями,
р.ввивающимися на конидиеносцах. Зигогамный половой способ размножения известен только для некоторых представи-юлей.
В порядок Entomophthorales входит только одно семей-г гво Entomophthoraceae.
Сем. Entomophthoraceae
По образу жизни представители этого семейства распадаются на три группы:
1. Паразиты насекомых.
2. Кишечные паразиты низших позвоночных и сапрофиты па их экскрементах.
3. Паразиты и сапрофиты на растительных субстратах.
Здесь будут указаны только паразиты насекомых, представленные двумя родами: Empusa и Entomophthora.
Род Empusa Cohn. Гибель комнатных мух осенью обычно вызывается паразитным грибом Е. muscae Cohn.
Очень часто можно видеть мертвых мух, прикрепившихся к какому-либо предмету, например к оконному стеклу, которое на расстоянии 1—2 см как бы обсыпано беловатым порошком. На брюшной стороне тела мухи виден белый налет,
Рис. 19. Etnpusa muscaet а — пораженная муха; б — конидиеносцы.
представляющий собой неразветвленные конидиеносцы, выступающие наружу через дыхательные отверстия хитинового
покрова. На верхних концах конидиеносцев развиваются конидии, которые после созревания с силой отбрасываются и прилипают к стеклу (рис. 19). Попадая на здоровую муху, конидия дает ростковую трубочку, внедряющуюся внутрь тела насекомого и развивающуюся в жировом теле в мицелий. Последний распадается на отдельные клетки, которые разносятся по всему телу мухи и вызывают ее гибель. Конидиеносцы развиваются из клеток гриба, находящихся внутри тела.
В высохших мухах можно найти покоящиеся клетки гриба, которые одни исследователи считают хламидоспорами (видоизменение мицелия), а другие — азиготами. У Е. muscae настоящих зигоспор не обнаружено, а у других видов они известны.
Род Entomophthora Fres. Представители этого рода имеют разветвленные конидиеносцы. Цикл развития этих грибов в основном сходен с таковым у Е. muscae.
Паразитируя на комнатных мухах и других вредных насекомых, этот род, так же как и Empusa, может служить биологическим способом борьбы с общественно живущими формами насекомых.
ГЛАВА II КЛАСС ASCOMYCETES (СУМЧАТЫЕ ГРИБЫ)
ОБЩАЯ ХАРАКТЕРИСТИКА
Сумчатые грибы, или аскомицеты, представляют класс высших грибов. В настоящее время описано около 30 тысяч видов аскомицетов весьма различных как по образу жизни, так и по строению. Общим для всех этих видов является наличие сумки, или аска,— особого органа спороношения, в котором образуется определенное количество эндогенных спор, обычно равное восьми.
Аскомицеты отличаются от класса низших грибов — фикомицетов следующими основными чертами:
1. Мицелий у всех аскомицетов многоклетный, иногда способный распадаться на отдельные клетки. Клетки мицелия, как правило, многоядерные, но порой среди них могут встречаться представители сумчатых с одноядерными (Erysipha-сеае) или двухъядерными (Exoascales) клетками.
2. Клеточные оболочки аскомицетов содержат хитин с целлюлозой.
3. Бесполое размножение сумчатых грибов осуществляется исключительно при помощи конидий. Причем некоторые из них могут развивать несколько видов конидиального спороношения.
4. В период полового размножения аскомицетов образуется дикариотическая фаза, представляющая гифы, которые развиваются из оплодотворенного женского органа размножения. Клетки их содержат по паре ядер, где одно ядро является женским, другое — мужским.
5. У большинства сумчатых грибов полный цикл развития проходит в несколько фаз, каждая из которых имеет опреде
ленный внешний вид. Такое явление у грибов получило название плеоморфизма и было1 впервые описано в 40-х годах XIX столетия французским ученым Тюляиом.
6. Очень многие сумчатые грибы являются паразитами высших растений. Конидиальное спороношеиие гриба развивается в период вегетации растений. Период развития сумчатой стадии в отдельных группах грибов различен. У одних, как например у голосумчатых, сумки образуются в период вегетации растений; у других наблюдается закладывание сумчатых плодовых тел в конце вегетации, а полной зрелости они достигают только к началу вегетационного периода следующего года (у большинства мучнисто-росяных грибов). У некоторых сумчатая стадия может развиваться в результате прорастания перезимовавших склероциев. В этом случае плодовые тела созревают сравнительно быстро, сумкоспоры рассеиваются в первой половине вегетационного периода.
Как же развивается у сумчатых грибов их основной орган спороношения — сумка, или аск?
Наиболее простое развитие сумки у представителей порядка Protascales — первичносумчатых грибов. У некоторых организмов, обладающих почкующимся мицелием, как например у культурных пивных дрожжей (Saccharomyces cerevisiae), образование сумок идет партеногенетически — из одной клетки, без предварительной копуляции. При этом клетка несколько увеличивается и превращается в сумку. Одновременно происходит два, следующих одно за другим, деления ядра, и образовавшиеся четыре ядра дают начало сумкоспорам.
У дрожжей из рода Schizosaccharomyces перед возникновением сумки происходит копуляция двух свободно живущих клеток. При этом они образуют выросты навстречу друг другу, в которые переходят ядра из обеих клеток. Выросты и ядра сливаются, а соединенные клетки начинают разрастаться, образуя сумку. Копуляционное диплоидное ядро делится три раза, давая восемь ядер для будущих сумкоспор. Первое деление ядра является редукционным.
У представителей порядка Protascales, имеющих много-клетиый мицелий, сложенный из одноядерных клеток, как например у представителей рода Eremascus, сумки развиваются следующим образом. Две соседние клетки образуют отростки вблизи отделяющей их одну от другой перегородки. Ядра, находящиеся в клетках, делятся и по одному из двух образовавшихся после деления переходят в отростки. Последние растут навстречу друг другу и копулируют, а вскоре сливаются и ядра. Зигота отделяется перегородкой от гифы, на которой она возникла, и развивается в сумку с восемью сумкоспорами.
Описанные три случая образования сумок у первичносумчатых грибов показаны на рис. 20.
У наиболее высокоорганизованных сумчатых грибов Euas-comycetes, или плодосумчатых, образованию сумки предшествует развитие специальных органов размножения, затем происходит сложный половой процесс, при котором зигота никогда не превращается прямо в сумку, а дает выросты,
Р и с. 20. Образование сумок у первичносумчатых:
а — Saccharomyces cerevesiae (апогамно); б — Schizosaccharotnyces octosporus и е—Еге-inascus fertilis (половым путем).
называемые аскогенными гифами, и на их концах формируются сумки. При этом процессе слияния ядер, или кариогамия, несколько запаздывает. Мужские и женские ядра сближаются попарно, образуя так называемые дикарионы, а слияние их происходит только в развивающейся сумке.
Этот процесс «подробно был изучен в 1912 году Клауссеном на грибе Pyronema confluens Tul. Было установлено, что на мицелии гриба развиваются парами половые органы: женский архикарп и мужской антеридий, причем обычно образуется одновременно по нескольку пар, так что получаются своеобразные розетки органов размножения.
Архикарп состоит из двух клеток: нижняя — аскогон — представляет вздутую клетку, заполненную протоплазмой с большим количеством ядер; верхняя — трихогина — цилиндрической формы и только в начале развития имеет собственное живое содержимое (рис. 21).
Антеридий развивается в виде массивной цилиндрической клетки, также многоядерной. Перед оплодотворением вершина антеридия плотно срастается с кончиком трихогины, разделяющая их перегородка растворяется и антеридиалы-юе содержимое переливается в трихогину, у которой к настоящему моменту собственное живое содержимое уже дегенерировало.
В это же время в поперечной перегородке, разделяющей трихогину и аскогон, появляется отверстие, через которое антери-диальное содержимое поступает в аскогон, где происходит только слияние протоплазмы мужской и женской клеток. Мужские ядра сближаются попарно с женскими, образуя дикарионы. Периода покоя в данном случае не наступает, а аскогон начинает давать мешковидные выросты в количестве от 10 до 20. Эти выросты вытягиваются, ветвятся, приобретают клеточное строение, образуя так называемые аскогенные гифы. Нижние клетки аскогенных гиф, т. е. ближайшие к аскогону, содержат по нескольку пар ядер, верхние, или конечные,— по одной. Из них развиваются сумки (рис. 22).
Из рисунка видно, что конечная клетка аскогенной гифы начинает расти несколько необычно, загибаясь в сторону и образуя крючок (а). Дикарион ее при этом делится продольными осями и образуется четыре ядра, из которых одно заходит в крючок, другое, несестринское от его пары, остается в основании клетки, а пара передвигается в среднюю часть клетки (б). Следующая стадия представляет развитие двух перегородок: одной, отделяющей основание клетки с одним ядром, другой, отделяющей крючок с несестринским ядром. Образование двух перегородок отделяет клетку с дикарионом, которая и является материнской клеткой сумки (в). Крючок постепенно прирастает к основанию клетки. Разделяющая их оболочка растворяется, и ядро из крючка проходит в клетку,
восстанавливая в ней дикарион (г). Образовавшаяся двухъ-идсрная клетка вновь способна давать крючок, и процесс развития сумки может повторяться (д).
Материнская клетка сумки разрастается, диплоидное ядро ее делится последовательно три раза, причем первое деление является редукционным. В результате образуется
Рис. 22. Развитие сумки из конечной клетки аскогенной гифы.
восемь гаплоидных ядер, которые и дают начало восьми сум-коспорам (г, д, е). После полного созревания сумкоспоры выбрасываются и, попадая на благоприятный субстрат, прорастают, давая начало вегетативному гаплоидному мицелию.
Весьма существенным моментом при подобном половом процессе, -сопровождающем развитие сумок, является то, что процесс плазмогамии — слияние протоплазмы — и процесс кариогамии — слияние ядер — сильно раздвинуты, чего не наблюдается у низших грибов.
В цикле развития высших сумчатых грибов преобладает гаплоидная фаза. Она включает аскоспоры, вегетативный мицелий, конидиальное спороношеиие и сплетение гиф, образующих аскокарпы. Дикариотическую фазу составляют аскогенные гифы, клетки которых содержат дикарионы. Диплоидная, самая короткая, представлена материнской клеткой сумки до момента редукционного деления ее диплоидного ядра.
Этот процесс образования сумок является типичным для всех Euascomycetes — плодосумчатых грибов. Благодаря многоядерности половых органов происходит множественное образование дикарионов, а в дальнейшем — множественное оплодотворение. Подобная кариогамия имеется и у более низко организованных грибов, например у представителей порядка Mucorales из класса фикомицетов. При зигогам-ном половом процессе у них происходит слияние многоядерных гаметангиев, сопровождающееся множественной кариогамией.
У большинства аскомицетов строение половых органов, а также и половое воспроизведение сходны с таковыми у
Pyronema^ но встречаются и некоторые отличия, прежде всего в морфологическом строении архикарпа и антеридия. Например, у видов Ascobolus архикарп представляет собой многоклетную ветвь, в которой нижняя более широкая, многоядерная клетка выполняет функцию аскогона, а трихогина состоит из нескольких мелких клеток.
У некоторых плектасковых архикарп и антеридий представляют спирально -закрученные, большей частью много клетные ветви, а в других случаях обе половые ветви проходят более или менее прямо, не скручиваясь, но отличаясь от других гиф главным образом крупными размерами клеток. Такая половая ветвь, представляющая архикарп, получила название Воронинской гифы в честь русского миколога М. С. Воронина.
Известны и такие случаи, когда антеридий вообще не образуется, но аскогенные гифы из аскогона развиваются. В них, так же как и при оплодотворении, наблюдаются дикарионы в клетках. Образование дикарионов происходит благодаря соответствующей перегруппировке ядер внутри аскогона или путем перехода их в аскогон из вегетативных клеток мицелия, соприкасающихся с аскогоном.
Иногда вместо антеридия развиваются спермации, которые по своему строению сходны с конидиями. Они могут образовываться даже на значительном расстоянии от архикарпа, а на трихогину переносятся ветром или мелкими животными. Переливание содержимого и дальнейшее оплодотворение идут так же, как и в типичном случае.
В строении и форме сумок наблюдается большое разнообразие: наиболее типичная форма сумки представляет собой вытянутое тело с ножкой в нижнем конце (см. рис. 22, в). Споры обычно располагаются в верхнем конце сумки и плавают в ее водянистом содержимом — эпиплазме, заполняющей и нижнюю часть сумки. Эпиплазма содержит гликоген, который при созревании сумки ферментируется, превращаясь в сахар, отчего тургорное давление в сумке повышается, оболочка ее сначала растягивается, а затем разрывается и происходит активное выбрасывание сумкоспор. Освобождение сумкоспор может происходить и по причине общего ослизнения оболочки сумки.
Аскоспоры по своему внешнему строению не одинаковы. Чаще всего они имеют вид одноклетных округлых или овальных одноядерных образований. В одних случаях одноядерные вначале споры затем могут становиться многоядерными (например, Verpa из сморчковых); в других —• аскоспоры вытягиваются в виде нитевидных образований, оставаясь при этом одноклетными (Rhytlsma), или же в них могут возникать поперечные перегородки, тогда споры становятся многоклетны-
мп (Claviceps). Иногда перегородки могут быть не только поперечными, но и продольными (Cucurbitaria). Аскоспоры, в распространении которых принимают участие животные или насекомые, имеют на своей поверхности шипики или ребрышки (Tuber). У некоторых сумчатых грибов споры могу быть окружены сплошной слизистой обверткой (Ascobolus).
Систематическое деление класса сумчатых грибов основано на их способности развивать сумки прямо на мицелии или же внутри особых плодовых тел — аскокарпов. В связи с этой особенностью класс Ascomycetes делится на два подкласса:
1. Protascomycetes, или Gymnoascomycetes (первичносумчатые, или голосумчатые).
2. Euascomycetes, или Carpoascomycetes (собственносумчатые, или плодосумчатые).
ПОДКЛАСС PROTASCOMYCETES
Это сравнительно небольшой подкласс, насчитывающий около 400 видов. Его представители характеризуются образованием сумок одиночно или сплошным слоем прямо на мицелии и отсутствием плодовых тел.
Подкласс делится на порядки: 1) Protascales,i\mA Endoas-cales; 2) Exoascales, или Tafrinales.
ПОРЯДОК PPOTASCALES
Все представители этого порядка по условиям существования связаны с богатыми сахаром субстратами. В естественной обстановке в природе они встречаются в истечениях из поврежденных стволов деревьев, в нектаре цветов, на поверхности сочных и сладких плодов. Естественное брожение ягодных соков, наблюдающееся при приготовлении вина, особенно виноградного, объясняется присутствием на поверхности ягод дрожжевых грибов видов Saccharomyces и Schizosaccharomy-ces, попадающих на них из почвы.
Большая часть представителей порядка первичносумчатых ведет сапрофитный образ жизни. Среди сапрофитных видов наибольшее практическое значение имеют возбудители алкогольного брожения. Эта особенность свойственна многим представителям семейства Saccharomycetaceae.
Что касается паразитных форм, то необходимо отметить следующее: на пластинках шляпочных грибов, особенно опенка, иногда можно встретить беловатый налет, представляющий Endomyces decipiens. Этот организм является примером паразитизма одного гриба на другом. Второй представитель этого рода Endomyces albicans вызывает явление молочницы в по
лости рта у грудных детей и молодых животных. Есть среди первичносумчатых и паразиты насекомых, как Pericystis apis. В некоторых случаях дрожжевые грибы могут вызвать у человека заболевания кожи, называемые бластомикозами.
Первичносумчатые по некоторым чертам полового процесса ’занимают промежуточное положение между зигомице-тами и аскомицетами. Так, например, представители рода Pericystis своим половым процессом — слиянием многоядерных гаметангиев — напоминают мукоровые, а наличие многоклетного мицелия и способность зиготы прорастать без периода покоя в зародышевый спорангий представляют известный переход к Protascales, у которых сумка имеет по существу такое же происхождение и может быть гомологизиро-вана с зародышевым спорангием зигомицетов.
У Dipodascus albidus при образовании сумок также происходит половой процесс, напоминающий зигогамию мукоровых, но в отличие от последних здесь не происходит множественной кариогамии, а сливаются только два ядра, по одному из каждого гаметангия, остальные в дальнейшем отмирают. Копуляционное ядро многократно делится и около каждого формируется по аскоспоре.
У других первичносумчатых при формировании сумки наблюдается уже определенное число сумкоспор, и эту черту можно считать дальнейшим шагом эволюции.
Порядок Protascales включает четыре семейства: Dipodas-caceae, Endomycetaceae, Saccharomycetaceae, Schizosaccharo-mycetaceae.
Сем. Dipodascaceae
К нему относятся наиболее примитивные формы, стоящие на границе между зигомицетами и собственно аскомицетами. Из сапрофитных видов можно отметить Dipodascus albidus Lagerh., развивающийся в сахаристых истечениях стволов деревьев, особенно березы. Многоклетный минелий этого гриба обладает способностью распадаться на оидии, а при образовании сумок происходит половой процесс, напоминающий зигогамию мукоровых. Зигота без периода покоя развивается в многоспоровую сумку.
Из паразитных форм следует указать Pericystis apis Maassen, который развивается на личинках -пчел, вызывая у них заболевание под названием каменной или известковой детки. Гриб имеет многоклетный мицелий. При половом процессе развиваются два различных по величине гаметангия, содержимое которых не дифференцировано на отдельные гаметы. Более крупный — оогоний принимает содержимое меньшего—антеридия. Плазмогамия сопровожда-
я множественной кариогамией, копуляционные ядра де-гнся, а затем около образовавшихся гаплоидных ядер разминаются споры.
Сем. Endomycetaceae
К семейству Endomycetaceae относятся два рода Eremas-(/is и Endomyces.
Род Eremascus Eidam. Образование сумок у представи-|глей этого рода показано на рис. 20, в и описано при общей характеристике способов развития сумок у сумчатых грибов.
Род Endomyces Reess. У представителей этого рода сумки образуются чаще всего апогамически из отростков клеток пли из самих вегетативных клеток мицелия. Сумки содержат по четыре споры, довольно своеобразной формы в виде полу-шаровидных телец с утолщенным ободком. У представителей н ого рода широко распространено вегетативное размножение путем распадения мицелия на «почкующиеся клетки. Е, albicans Vuill.— паразит, вызывающий явление молочницы у грудных детей и молодых животных, а £. fibuliger Lindner — сапрофит, один из возбудителей алкогольного брожения. /'. vernalis Ludvig.— так называемые жировые дрожжи. Не обладая бродильной способностью, этот гриб в своих клетках может накапливать до 30% жира и представляет в этом отношении известный практический интерес как организм, перерабатывающий углеводы в жиры.
Во время империалистической войны в Германии жировые дрожжи разводились в широком масштабе, а получаемый жир находил применение и как пищевой продукт и особенно для технических целей (получение глицерина для взрывчатых веществ). У нас во время гражданской войны в Ленинграде также делались попытки получения жира из жировых дрожжей для питания.
Сем. Saccharomycetaceae 1
Характерным признаком вегетативного строения дрожжевых грибов является наличие почкующегося мицелия, образованного отдельными клетками или соединенными в цепочки и способного размножаться почкованием. Все дрожжи хо
1 В. И. Кудрявцев на основании своих исследований объединяет в новый порядок Vnicellomycetales все одноклетные немицелиальные и не являющиеся стадиями развития мицелиальных грибов дрожжевые организмы, независимо от их способности к образованию сумок со спорами. Вегетативное тело их в условиях постоянного обитания представляет одну нераз-ветвленную (в отличие от мукоровых) и окруженную оболочкой (в отличие от миксохитридиевых) клетку (Кудрявцев, 1954).
рошо развиваются на питательных средах, богатых углеводами. Правда, они могут использовать и углерод белковых веществ, но в очень слабой степени. Образующийся при этом в избытке аммиак быстро подщелачивает среду настолько, что дальнейшее развитие дрожжей становится невозможным.
Род Saccharomyces (Meyen) Reess. Среди дрожжевых грибов он имеет наибольшее распространение и практическое значение. Организмы, принадлежащие к этому роду, при обычных условиях не развиваются, например в пчелином меду, вареньях, джемах, потому что они не выносят имеющейся в этих субстратах исключительно высокой концентрации сахаров (до 80.% и больше). Достаточно, однако, понизить эту концентрацию до 25—30%, как сахаромицеты и особенно Sacch. vini оказываются господствующими организмами. Дрожжи обладают наиболее ярко выраженной способностью использовать различные сахара путем сбраживания и образования из них наибольшего количества спирта (от 10 до 19% по объему и больше). Таким образом, сахаромицеты характеризуются наибольшей стойкостью к ядовитому действию этого продукта углеводного обмена по сравнению с другими микроорганизмами. Эта биологическая особенность ставит дрожжи в более выгодное положение в борьбе с организмами других родов за места своего обитания.
Вегетативное размножение у сахаромицетов осуществляется почкованием. С наступлением неблагоприятных условий возникают более стойкие покоящиеся формы — артроспоры и сумки со спорами.
Артроспоры отличаются от вегетативных клеток более резко преломляющей свет плазмой, наличием в ней гликогена, капелек жира и утолщенной оболочкой. При попадании в благоприятные для развития условия артроспоры непосредственно превращаются в почкующиеся клетки. При этом плазма теряет свой блеск, капельки жира исчезают, оболочка становится тоньше и на одном из полюсов клетки появляется первая почка.
Сумки со спорами у дрожжей рода Saccharomyces возникают партеногенетически из вегетативных клеток, прекративших почкование (см. рис. 20,а). В одной сумке образуются 1—4 споры, очень редко 8. Освободившиеся из сумок споры разбухают, затем попарно копулируют или же копуляция наступает между первыми почками. В результате вегетативное поколение дрожжей сахаромицетов всегда бывает диплоидным. Гаплоидными оказываются только сумкоспоры.
Изучение сахаромицетов на фоне условий их жизни в природе и в хозяйстве человека дало возможность В. И. Кудрявцеву свести систематику этого рода к 18 видам против 220, описанных более ранними исследователями.
Наиболее широко распространенный вид Saccharomyces terevisiaea Meyen— пивные, или хлебные, дрожжи —живет исключительно в условиях бродильной промышленности, перерабатывающей хлебо-картофельное сырье. Вызывает брожение глюкозы, галактозы, сахарозы, мальтозы и простых декстринов. Согласно исследованиям В. И. Кудрявцева, в природных субстратах эти дрожжи не найдены.
Sacch. lactis Adametz, встречается только в кисломолочных продуктах, так как обладает способностью в отличие от других видов усваивать путем брожения молочный сахар, пли лактозу. В природных субстратах также пока не обнаружен.
Sacch. ellipsoideus Reess, или Sacch. vini Meyen, обнаруживается преимущественно в соках ягод и других плодов в условиях как промышленного, так и домашнего виноделия и обладает способностью сбраживать глюкозу, фруктозу, сахарозу, а также мальтозу с образованием этилового спирта до 19% и выше.
Sacch. oviformis Osterw. обитает в тех же субстратах, что и Sacch. vini. Разновидность этих дрожжей, известная под названием хересных, обладает способностью на поверхности ягодного сока вырабатывать пленку, состоящую из живых организмов дрожжей. Оказалось, что если виноградное вино покрывается такой дрожжевой пленкой, то оно не только не портится, как это бывает при развитии других микроорганизмов, образующих пленки на подобных субстратах, но и приобретает особый, так называемый «хересный тон». Отсюда соответствующие дрожжи получили название хересных. Они широко используются сейчас в промышленной практике, например при изготовлении шампанских вин.
Род Zygosaccharoinyces Barker. Он также принадлежит к семейству Saccharoinycetaceae и объединяет организмы, способные развиваться в субстратах с высокой концентрацией сахара — до 80% и выше. Такими субстратами являются нектар цветов, пчелиный мед, варенья, джемы. Кроме размножения почкованием, в цикл развития этих организмов входит образование покоящихся форм — сумок и артроспор, которые появляются с наступлением неблагоприятных для вегетативного развития условий.
Сумки возникают после копуляции вегетативных клеток, редко партеногенетически. Сумкоспоры при прорастании превращаются непосредственно в почкующиеся клетки. У этих организмов никогда не наблюдается копуляции сумкоспор или первых их почек. Поэтому вегетативное поколение дрожжей рода Zygosaccharomyces всегда является гаплоидным, чем они существенно отличаются от дрожжей рода сахаромицетов.
Сем. Schizosaccharoinycetaceae
Это семейство объединяет все дрожжи, размножающиеся не почкованием, а делением. Родиной их являются тропические и субтропические страны, где до недавнего времени они только и обнаруживались. В пользу этого говорит и их способность развиваться при более высоких температурах. Так, по наблюдениям, произведенным в Мексике на одном из спиртовых заводов, при температуре 30—42° в заторах господствовали шизосахаромицеты, а при понижении температуры до 25—30° они вытеснялись обычными почкующимися дрожжами. По степени сбраживания осахаренных крахмалистых субстратов делящиеся дрожжи несколько превосходят наши пивные.
По морфологическому строению клеток делящиеся дрожжи представляют палочковидные организмы. При наступлении неблагоприятных условий вегетативные клетки попарно копулируют, и зигота непосредственно превращается в сумку с 4 спорами. При прорастании споры набухают, разрывают оболочку сумки и превращаются в обычные вегетативные клетки.
Основным родом семейства является род S chizo sac char о-myces.
Род Schizosaccharoinyces Lindner. Среди видов этого рода наиболее широко распространен Schiz. pombe Lindner, впервые выделенный из негритянского пива, приготовляемого в Восточной Африке. Негритянское пиво представляет собой напиток, полученный путем сбраживания смеси сока бананов и разваренного проса.
Кроме этого, дрожжи Schiz. pombe употребляются при изготовлении крепких алкогольных напитков — аррака и рома.
ПОРЯДОК EXOASCALES
По условиям существования все грибы этого порядка являются паразитами высших растений и вызывают различные деформации пораженных органов. Образование сумок происходит прямо на мицелии, причем сумки располагаются не одиночно, а правильным тесным слоем, напоминающим гимений высших сумчатых грибов. К порядку Exoascales относятся два семейства: Exoascaceae, Protoinycetaceae.
Сем. Exoascaceae
В семействе Exoascaceae различают два рода: Exoascus и Taphrina.
Таблица II. Сморчковые грибы:
а — Morchella esculenta\ б — М. coniccr, в — Helvetia esculenta.
Род Exoascus Fuck. Его представители имеют многолетний мицелий, сохраняющийся из года в год в стеблях и поч-K.ix пораженных растений. Наиболее распространенным ви->1ом является Е. pruni TuL, развивающийся на черемухе и •ливах. Мицелий этого гриба зимует в ветках и почках дерена, распространяется по межклетникам пораженных органов. Весною через цветоножки гриб проникает в завязи и своим присутствием вызывает гипертрофию плода. Пораженные
Рис. 23. Exoascus pruni на сливе: а — ветка с одним здоровым и двумя пораженными плодами; б — слой сумок на поверхности плода.
плоды уже в половине июня представляют уродливые образования, по размерам в несколько раз более крупные, чем здоровые в это же время. Семя в таком плоде никогда не развивается, на его месте образуется полость. За эту особенность болезнь получила название дутые сливы или кармашки слив (рис. 23).
Характерной морфологической особенностью мицелия Е. pruni является то, что он состоит из коротких двухъядер-пых клеток. Перед образованием сумок ветви мицелия проникают в самые наружные слои плода между эпидермисом и кутикулой и здесь распадаются на двухъядерные клетки, в которых сразу же происходит кариогамия. После этого в верхней части клетки образуется вырост, куда переходит популяционное ядро. Вырост растет, пробивает кутикулу и превращается в сумку. Копуляционное ядро делится три раза и образуется восемь гаплоидных ядер, дающих начало сумкоспорам.
7 3. Н. Кудряшева 97
Последние, еще находясь в сумке, способны почковаться, а поэтому зрелая сумка Е. pruni содержит более восьми спор.
Нижняя часть клетки, образовавшей вырост, оказывается пустой, отделяется перегородкой и образует так называемую клетку-ножку. Пораженный плод покрывается беловатым налетом, представляющим многочисленные сумки гриба, расположенные сплошным гимениальным слоем. Из зрелых сумок сумкоспоры выбрасываются и, попадая на здоровые ветви дерева, прорастают. Пораженные плоды к концу июня начинают подсыхать и осыпаться с дерева. Количество больных плодов зависит от климатических условий. Так, например, замечено, что если во время цветения слив стоит холодная и сырая погода, то гриб получает большее развитие.
Е. pruni значительно снижает урожай плодов, так как, во-первых, пораженные завязи не получают нормального развития и осыпаются; кроме того, гриб, поселяясь в ветвях дерева, в значительной степени истощает его.
Борьба с этим грибом заключается в подрезке больных побегов и ветвей и в немедленном их сжигании. Пораженные плоды по возможности необходимо собирать и сжигать. Как профилактическую меру можно применить опрыскивание деревьев до распускания почек раствором железного купороса (400—800 г на 12 л воды или же 1%-ным раствором).
У некоторых видов этого рода доказано происхождение двухъядерного мицелия. Так, у Е. epiphy На Sa deb. гаплоидные сумкоспоры по выходе из сумок попадают на ветви дерева, но заражения не производят, а попарно копулируют, причем ядро одной переходит в другую, вырастающую в двухъядерный мицелий, который и производит заражение. Копуляция может наступать не только между сумкоспорами, но и между продуктами их почкования. Некоторые представители рода Taphrina являются гетероталличными формами — половина аскоспор в сумке несет признаки одного пола, а половина — другого.
Из других представителей рода Exoascus с зимующим мицелием необходимо отметить Е. cerasi Sadeb., вызывающую образование на вишне и черешне так называемых ведьминых метел.
Ведьмины метлы представляют множество густо растущих тонких, слаборазвитых ветвей с блестящими и утолщенными волнистыми листьями. Ведьмины метлы особенно хорошо заметны весной, так как они раньше покрываются листочками, чем здоровые ветви. Плодов на таких веточках никогда не образуется, но в то же время ведьмины метлы сильно истощают дерево, требуя огромного количества питательных веществ для своего развития. На нижней стороне листьев обычно развивается сероватый налет из сумок.
8 качестве основных мер борьбы с грибом можно рекомендовать обрезку и уничтожение ранней весной пораженных ветвей, а раны, наносимые дереву, замазывать садовой замазкой.
Е. deformans Tul. вызывает курчавость листьев персика, а Е. minor Sadeb.— курчавость листьев вишни.
Род Taphrina Fries. Паразитные грибы с однолетней грибницей, вызывающей местное заражение в виде пятен и искривлений на листьях.
На разных видах тополя паразитирует Т. aurea Fries, вызывающий образование вздутий желтого или коричневого цвета на верхней стороне листьев. На нижней стороне соответственно вздутиям образуются вдавленные участки, на которых развиваются сумки.
На листьях груши пузыревидные вздутия образуются вследствие поражения их Т. bullata Tul.
Сем. Protomycetaceae
Это семейство включает два рода: Protoinyces и Taphri-dium.
Роды Protoinyces Unger и Taphridium Link. Представители этих родов паразитируют исключительно на зонтичных. Под влиянием этих грибов на листовых черешках и жилках образуются пузыревидные опухоли. Мицелий состоит из многоядерных клеток, проходит по межклетникам. Некоторые клетки мицелия пузыревидно вздуваются, приобретают утолщенную оболочку и превращаются в хламидоспоры, которые освобождаются и прорастают следующей весной после сгнивания тканей растения-хозяина.
* # *
Относительно родственных связей Exoascales с другими группами сумчатых грибов существуют две точки зрения. Одни исследователи сближают их с Protascales на основании отсутствия у тех и других плодовых тел. Другие ученые утверждают, что между этими двумя группами мало общих черт, и основной упор делают на то, что у Protascales в цикле развития преобладает гаплоидная фаза, а у Exoascales — диплоидная. На основании такого признака, как расположение сумок правильным гимениальным слоем, сторонники последнего взгляда рассматривают экзоасковые грибы как редуцированные формы, произошедшие от наиболее высокоорганизованных сумчатых грибов-дискомицетов.
ПОДКЛАСС EUASCOMYCETES
Значительная часть сумчатых грибов принадлежит к этому подклассу. В целом он характеризуется наличием плодовых тел, внутри или на поверхности которых помещаются сумки. Процесс развития плодового тела идет одновременно с процессом полового воспроизведения. Вокруг половых органов и развивающихся после оплодотворения аскогона аскогенных гиф разрастаются ветви вегетативного мицелия гриба, которые, переплетаясь в плектенхиму (своеобразную грибную ткань), образуют плодовое тело, или аскокарп. Плодовое тело сумчатых грибов представляет образование двойственной морфологической природы. Главная масса его состоит из вегетативного мицелия и принадлежит к гаплоидной генерации, самая внутренняя часть — аскогенные гифы — представляет диплоидную генерацию.
У плодосумчатых грибов различают следующие типы плодовых тел.
1. Клейстокарпии — совершенно замкнутые плодовые тела обычно округлой формы. Сумки помещаются внутри плодового тела, часто без особого порядка. Освобождение сумок происходит вследствие общего разрушения оболочки (перидия) или ее разрыва, вызванного набуханием созревших сумок. Количество последних в плодовом теле может варьировать у разных видов грибов от одной до нескольких.
2. Перитеции — полузамкнутые плодовые тела в форме кувшина с узким отверстием (устьицем) наверху для выхода сумок. У одних форм перитеции имеют собственную стенку, представленную плотно сплетенными гифами, и поэтому могут быть свободно отделимы от подстилающего их вегетативного мицелия (Venturia inaequalis). У других представителей перитеции в виде бутылкообразных полостей в общей строме плотно сплетенных грибных гиф (Epichide typhina, Claviceps purpurea). Сумки в перитециях вытянутой формы, располагаются на дне в виде пучка. По мере созревания они отрываются, поднимаясь кверху, высовываются своим верхним концом через выводное отверстие плодового тела и активно выбрасывают сумкоспоры.
3. Апотеции — открытые плодовые тела в большинстве блюдцевидной формы. Сумки располагаются на верхней свободной стороне слоем, называемым гимением. Между сумками находятся бесплодные гифы — парафизы. Они придают упругость гимению, предохраняют сумки от слипания и высыхания. Форма сумок обычно цилиндрическая, и расположены они вертикально и параллельно друг другу. По мере созревания они выбрасывают сумкоспоры через верхний конец, несколько высовывающийся над общей поверхностью гимения.
Из трех типов плодовых тел сумчатых грибов наиболее примитивны по своей организации клейстокарпии, так как они менее приспособлены к функции рассеивания спор. Плодовое тело раскрывается один раз, освобождая все имеющиеся там сумки, из которых и рассеиваются сразу все сумко-споры.
Перитеции, имея на вершине устьице для высовывания сумок по мере их созревания, более приспособлены к активному разбрасыванию сумкоспор, причем период созревания сумок и освобождения сумкоспор бывает более растянутый.
Наиболее высокоорганизованной формой плодового тела является апотеций. Гимений, расположенный открыто и широким слоем на поверхности плодового тела, обеспечивает максимальную продукцию спор и их активное выбрасывание. Последнему способствуют парафизы, поддерживающие с боков оболочку сумки, не давая ей раздуваться от увеличивающегося тургорного давления. Его сила направлена на верхний свободный конец сумки, где помещаются сумкоспоры, которые и выбрасываются на расстояние 10 см и даже более. Пустая сумка спадает, на ее место снизу врастает новая.
Опасность повреждения гимения в процессе развития апотеция устраняется тем, что вначале он закладывается только из одних парафиз, а сумки продвигаются снизу по мере созревания. Кроме этого, в некоторых случаях гимений защищен сплетением вершин парафиз, образующих так называемый эпитеций. И, наконец, у многих грибов защитную функцию выполняет особый кроющий слой, образованный разрастающимися стенками самого плодового тела, отчего последнее в молодости бывает почти замкнутым.
Основным признаком, положенным в основу классификации плодосумчатых грибов, является форма плодового тела. Подкласс Euascoinycetes делят на следующие порядки: 1) Plectascales, 2) Erysiphales (плодовые тела клейстокарпии); 3) Hypocreales, 4) Dothidiales, 5) Sphaeriales, 6) Не-mis phaer tails, 7) Laboulbeniales (плодовые тела перитеции); 8) Hysteriales, 9) Phacidiales, 10) Pezizales, 11) Elelotia-les, 12) Tuberales (плодовые тела апотеции). Порядки 3—7 объединяются в группу порядков Pyrenomycetes. Порядки 8—12 объединяются в группу порядков Dyscomycetes.
ПОРЯДОК PLECTASCALES
У простейших представителей этого порядка плодовые тела зачаточные, у более совершенных видов клейстокарпии характеризуются беспорядочным расположением сумок внутри плодового тела.
В порядке Plectascales следует отметить семейства: Asper-dillaceae, Terfeziaceae, Elaphomycetaceae, Onygenaceae.
Сем. Aspergillaceae
Среди плектасковых наибольшее практическое значение представляют грибы, принадлежащие к семейству Aspergillaceae, насчитывающему более 200 видов, среди которых будут отмечены некоторые виды родов Aspergillus, Penicillium.
Род Aspergillus Mich. Этот род включает около сотни видов преимущественно сапрофитных форм, распространенных
Рис. 24. Aspergillus.
в почве и в виде плесени на пищевых продуктах, богатых углеводами. Некоторые виды Aspergillus известны как возбудители заболеваний (микозов) человека и высших животных.
Aspergillus oryzae, обладающий чрезвычайно сильным диастатическим действием, употребляется в Японии для осахаривания риса при приготовлении водки.
Представители рода Aspergillus встречаются преимущественно в конидиальных стадиях, построенных чрезвычайно характерно. Конидиеносцы не ветвятся, а на их вздутой вершине радиально располагаются стеригмы. На свободных концах стеригм отшнуровываются базипетальные цепочки конидий (рис. 24). Сумчатые спороношения известны только у немногих видов.
Род Penicillium Link. Этот род также широко распространен в природе. У его представителей конидиеносцы на конце ? -3 раза ветвятся, а конечные клетки разветвлений функционируют как стеригмы и также отшнуровывают базипетальные цепочки конидий. Своим расположением на разветвленных концах конидиеносцев они напоминают микроскопическую
Р и с. 25. Penicillium.
кисточку, откуда п произошло русское название гриба — кистевик (рис. 25).
Представители рода Penicillium имеют большое практическое значение. Они очень обильно представлены в почве и играют там немалую роль при разрушении органических остатков. Весьма обильны они и как плесени на хлебе, варенье, коже, обоях. Известны виды, поражающие плоды в период хранения, например на плодах цитрусовых получают развитие Р. italicum Wehmer и Р. digitatuin £ас.
Некоторые виды Penicillium обладают способностью вырабатывать вещества, убивающие или подавляющие развитие многих других микроорганизмов, в том числе и патогенных. Вытяжки из этих грибов под названием пенициллина применяются при лечении гнойных ран и воспалительных процессов.
Для придания сыру аромата и вкуса в сыроварении применяется Р. roquefortii Thom, и др.
У большинства видов названных грибов конидии зеленоватые или сизые, вследствие чего первоначально бесцветный мицелий в состоянии спороношения приобретает зеленый цвет от покрывающих его конидий (зеленая плесень).
Сем. Terfeziaceae и Elaphomycetaceae
Принадлежащие к этим семействам грибы характеризуются крупными плодовыми телами, развивающимися под землей и похожими на трюфели. Они отличаются от последних беспорядочным расположением сумок внутри плодового тела. Примерами их могут служить съедобный закавказский трюфель Terfezia transcaucasica W. Tichomirov, плодовое тело которого достигает величины кулака, и олений трюфель Elaphoinyces cervinus (Pers.) Schrot., имеющий плодовое тело величиной с грецкий орех, несъедобный для человека, но поедаемый грызунами (например, белками). Он встречается у нас в средней полосе.
Сем. Onygenaceac
Интересно семейство Onygenaceac, представители которого отличаются тем, что поселяются на довольно своеобразных субстратах — животных остатках, содержащих роговые вещества: рога, копыта, перья. Своими выделениями эти грибы вызывают гидролиз рогового вещества.
ПОРЯДОК ERVSIPHALES
Грибы, принадлежащие к порядку Erysiphales, имеют замкнутые плодовые тела (клейстокарпии). Сумки обычно в небольшом числе, но располагаются они не беспорядочно, как у плектасковых, а соединены в пучок, поднимающийся от нижней части плодового тела. Форма сумок большей частью овальная, парафизы в плодовых телах отсутствуют. Большинство организмов, относящихся к этому порядку, является эктопаразитами высших растений. В порядок входят два семейства: Erysiphaceae, Perisрогiaceae.
Сем. Erysiphaceae
Это высокоспециализированная группа облигатных паразитов высших растений, приспособившихся к необычному характеру паразитизма посредством поверхностного мицелия. Внутренний мицелий у большинства их совершенно отсутствует. Поселяются эти грибы обычно на листьях и других молодых частях различных растений, где их поверхностно развитый мицелий имеет вид беловатого пушка, очень скоро покрывающегося как бы мучнистым налетом от развивающихся конидий (явление известное под именем мучнистой росы, отчего грибы называют мучнисто-росяными).
У некоторых представителей мицелий имеет сероватый и даже буроватый оттенок и может несколько уплотняться, ста-
повиться ватообразным, почти войлочным, как например у возбудителя мучнистой росы злаков.
У большинства мучнисто-росяных грибов, паразитирующих на однолетних растениях и на растениях с опадающими на зиму листьями, грибница однолетняя. У некоторых вегетативная грибница может зимовать на поверхности зимующих листьев и других органов, как например у Erysiphe graminis на озимой пшенице, у Sphaerotheca pannosa на розах и персике. Возможность зимовки отмечена и для таких видов, у которых клейстокарпии редко развиваются. Грибница в течение зимы у таких грибов может сохраняться в почках и трещинах коры (Uncinula necator — мучнистая роса винограда, Podosphaera leucotricha — мучнистая роса яблони).
Вначале грибница развивается на верхней стороне листьев, ио со временем может переходить и на нижнюю. Прикрепление грибницы к органам питающего растения осуществляется следующим образом. Рыхло стелющийся мицелий на поверхности эпидермиса в некоторых местах более плотно прикладывается к эпидермальным клеткам, образуя здесь особые, иногда лопастные расширения, называемые аппрессориями. От основания последних отходят очень тонкие ветви грибницы, прободающие стенку эпидермальной клетки и внедряющиеся внутрь, где они вздуваются на концах и превращаются в особые специализированные органы питания — гаустории, или присоски. В отдельных случаях гаустории приобретают форму лопастных образований, занимающих почти всю полость клетки хозяина. У некоторых видов мучнисто-росяных грибов гаустории развиваются нс в эпидермальных клетках, а глубже, в клетках мезофилла. У немногих представителей (род Phyllactinia) поверхностный мицелий не развивает гаусторий, а образует короткие выросты, проникающие внутрь листа через устьица, а на конечных клетках этих выростов образуются гаустории. Часть грибницы оказывается эндофитной.
Размножение мучнисто-росяных грибов осуществляется двумя способами — бесполым и половым.
Бесполое размножение происходит при помощи конидий, образующихся в большом количестве и в нескольких генерациях в течение вегетационного периода. Конидии легко распространяются воздушными течениями, заражают новые растения и таким образом идет массовое распространение гриба в период вегетации растений. Конидиальная стадия мучнисторосяных грибов получила название Oidiuin, Одноклетные короткие конидиеносцы поднимаются вверх от поверхностной грибницы и на своих концах развивают базипетальные цепочки одноклстных бесцветных конидий боченкообразной формы.
По своим биологическим особенностям конидии характе
ризуются недолговременной сохраняемостью. Они гибнут обычно через неделю от высыхания, а также от высоких или низких температур. В отличие от спор большинства грибов конидии мучнисто-росяных способны прорастать при относительно небольшом насыщении воздуха влагой.
Половой способ размножения мучнисто-росяных грибов осуществляется при помощи сумкоспор, возникающих в сумках, заключенных в плодовых телах — клейстокарпиях. Развитию последних предшествует половой процесс. Клейстокарпии закладываются, как правило, в конце вегетационного периода на поверхностной грибнице. Они имеют вид мелких, едва видимых простым глазом шарообразных телец, вначале желтоватых, затем постепенно темнеющих и, наконец, черных. Оболочка клейстокарпия, или перидий, состоит из более или менее ясно выраженных клеток с побуревшими и утолщенными стенками. Внутренняя часть, или так называемое ядро плодового тела, образовано нежными, бесцветными клетками псевдопаренхиматического характера. Среди них и располагается пучок сумок (или одна сумка). Некоторые клетки оболочки клейстокарпия дают выросты, называемые придатками или подвесками. Особенности их строения являются весьма важным систематическим признаком и принимаются во внимание при характеристике родов и видов мучнисто-росяных грибов. Они различаются по расположению на плодовом теле, по форме, строению, длине, окраске и другим признакам. Придатки плодового тела имеют очень большое биологическое значение, способствуя удерживанию клейстокарпия на субстрате и распространению видов.
Клейстокарпии представляют зимующую стадию гриба и служат для сохранения вида в течение неблагоприятного периода года. Несмотря на то что закладываются они на мицелии осенью, полного созревания обычно достигают только весной, т. е. к началу следующего вегетационного периода. Весеннее возобновление мучнисто-росяных грибов, как правило, осуществляется при заражении растений спорами, вышедшими из сумок.
Мучнисто-росяные грибы являются чрезвычайно специализированными облигатными паразитами. В пределах вида существуют еще специализированные формы, приспособленные к узкому кругу питающих растений. В естественной обстановке развитию мучнисто-росяных грибов способствуют сухость атмосферы и яркое освещение. Некоторые авторы указывают, что в сухом воздухе образование конидий происходит обильнее и всхожесть их больше. Непосредственное наблюдение также показывает, что весьма сильное развитие эти грибы получают в сухое лето, особенно в странах с сухим климатом (Средняя Азия). Существовало даже мнение, что
мучнисто-росяные грибы — особый тип сухолюбивых грибов. Но детальное исследование биологии этих паразитов показало, что причиной наиболее интенсивного развития их в засушливые годы служит то, что в условиях засухи растения-хозяева становятся наиболее восприимчивыми. Сухость атмосферы и повышенная температура понижают устойчивость растений по отношению к возбудителям мучнистой росы. Очень сильное развитие это заболевание получает также в оранжереях и вегетационных домиках.
Мучнисто-росяные грибы, поражая многие культурные растения как в открытом, так и в закрытом грунте, а также многие полезные дикие растения, приносят громадный вред. Развитие гриба на листьях значительно уменьшает ассимилирующую поверхность, а поглощение грибом питательных веществ при помощи гаусторий истощает растение. Листья, пораженные мучнистой росой, преждевременно засыхают.
Известны и такие случаи, когда мучнисто-росяные грибы, поселяясь на кормовых растениях, сообщают им ядовитые свойства. При скармливании таких растений скоту наступают кормовые отравления. По указаниям М. Полянского и П. Бе-гучева, ядовитыми свойствами обладает розовый клевер, пораженный мучнистой росой.
Потери в урожае, наблюдаемые при поражении растений мучнистой росой, достигают крупных размеров. Например, недобор урожая винограда вследствие развития на нем мучнистой росы для республик Средней Азии определяется как минимум 5%, а для Южного Крыма — 10%. В годы с особенно благоприятными условиями для развития гриба потери урожая могут достигать 25—30%. Очень сильно страдают от мучнистой росы тыквенные, особенно дыни, а в закрытом грунте огурцы. Листья преждевременно стареют и засыхают, а плоды развиваются в небольшом количестве и плохого качества.
Значительный вред наносит мучнистая роса плодовым, ягодным, зерновым, кормовым и другим культурам. Немалый вред причиняет она и лесам, поражая дуб, березу и другие породы.
Вредоносность мучнистой росы вызывает необходимость борьбы с заболеванием с целью устранения паразита и повышения урожая. Борьба с мучнистой росой, как и вообще с болезнями растений, требует применения как специальных мероприятий химического характера, так и косвенных, направленных на создание условий, способствующих защите растений от заражения, повышению их сопротивляемости и выносливости.
Из специальных мероприятий, направленных на борьбу с мучнистой росой, следует указать на эффективность при
менения препаратов, содержащих серу. Сера может быть использована в чистом виде (серный цвет) для опыливания растений. Коллоидная сера применяется в виде 0,2 %-него раствора для опрыскивания винограда, крыжовника, роз и др.
Из предупредительных мероприятий в первую очередь необходимо указать на полное уничтожение всего того, что может нести на себе инфекцию (остатки урожая, опавшие листья, сучья, осыпавшиеся плоды и др.). Необходимо тщательно осматривать посадочный материал—саженцы, черенки, так как на них также может сохраняться инфекция. Наконец, в числе общих мероприятий огромное значение имеют подбор и выведение устойчивых к мучнистой росе сортов культурных растений.
Классификация представителей семейства Erysiphaceae основана на особенностях строения клейстокарпиев: количестве содержащихся в них сумок, форме придатков, строении оболочки и других признаках. Главные роды следующие:
I. Клейстокарпии с одной сумкой
I. Sphaerotheca. Придатки нитевидные, переплетающиеся с грибницей (рис. 26, а).
2. Podosphaera. Придатки прямые, направленные вверх, вильчато разветвленные на концах (рис. 26,6).
II. Клейстокарпии с несколькими сумками
1. Erysiphe. Придатки нитевидные, короткие, переплетающиеся с грибницей (рис. 26,в).
2. Leveillula. Придатки, как у Erysiphe. Основное отличие этого рода от предыдущего заключается в характере грибницы. Вначале она эндофитная, позднее из нее развивается эпифитная грибница, на которой и образуются плодовыетела.
3. Microsphaera. Придатки свободные, располагаются в верхней части плодового тела, вильчато разветвленные (рис. 26,а).
4. Uncinula. Придатки свободные на концах, закрученные спирально или крючком (рис. 26,6).
5. Phyllactinia. Придатки двух родов: одни — шиловидные со вздутием у основания, располагаются экваториально; другие— разветвленные кистевидно, клейкие, расположены на вершине (рис. 26,6).
Род Sphaeroiheca Lev. Грибница вначале паутинистая, а затем уплотняющаяся и в старости чернеющая. Уплотнение мицелиального налета и его потемнение объясняется образованием так называемого вторичного мицелия в виде приподнимающихся кверху или изогнутых гиф с утолщенными стенками и более или менее интенсивной окраской. Этот вторич-
Р и с. 26. Клейстокарпии мучнисто-росяных грибов:
а — Sphacrotheca", б — Podosphacrax в — Ery siphex г — Micros phacrax
д — Uncinula и Phyllactirda.
ный мицелии и образует войлочное сплетение, в которое погружены клейстокарпии. Обычно развитие вторичного мицелия наблюдается у тех видов, у которых слабо развиты придатки на плодовых телах, и, таким образом, вторичный мицелий принимает на себя функцию придатков. Образование вторичного мицелия и развитие придатков стоят в тесной связи со степенью опушенности пораженных органов. На сильно опушенных листьях вторичный мицелий не развивается.
Наиболее широко распространен вид Sphaerotheca mors-uvae (Schwein) Berk, et Curt — возбудитель мучнистой росы крыжовника. Этот вид североамериканского происхождения. Он был завезен в Европу в 1900 году. Впервые его обнаружили в Ирландии и почти одновременно на территории России.
Мучнистая роса крыжовника не только значительно понижает урожай, но может за короткое время погубить целые плантации. Белый налет гриба после своего появления настолько увеличивается в размерах, что обволакивает почти все ягоды и концы молодых побегов, затем он постепенно темнеет и уплотняется. Ягоды прекращают рост, часто растрескиваются или же ссыхаются. Заболевшие кусты почти не дают прироста, слабеют и при сильном поражении гибнут.
Плодовые тела заметны даже невооруженным глазом в виде разбросанных черных точек, погруженных в бурый налет. Их придатки переплетаются с грибницей. Плодовое тело содержит одну сумку с восемью спорами. Сумкоспоры дозревают в течение зимы, а весною выбрасываются из сумок и производят заражение. Первые признаки болезни в виде паутинистого мицелия на листьях и молодых веточках наблюдаются через 2—3 недели после заражения.
Устойчивыми сортами крыжовника являются: Зеленый бутыльчатый, Пятилетка, Московский № 17, мичуринские сорта— Черный мавр, Штамбовый и др. Сильно поражаемые сорта — Виноградный, Финик.
Осенью после уборки полезно опрыскивать кусты 3%-ным полисульфидом кальция, а весною—арсенатом натрия (20 г на 12 л воды) или кальция (4 г на 12 л).
Кроме крыжовника, американская мучнистая роса замечена на красной и черной смородине.
Sphaerotheca paiinosa Lev. var. rosae Woronich. повсеместно распространена па дикорастущих и культурных видах розы. Вызывает уродливость листьев, искривление побегов, а при сильном поражении бутонов приводит к их гибели. Клейстокарпии развиваются почти исключительно на побегах и ягодах.
Род Podosphaera Kunze. Грибница белая, слаборазвитая, иногда исчезающая ко времени созревания клейстокарпиев. Конидии малочисленные. Клейстокарпии содержат одну сумку. Придатки их резко отличаются от грибницы, расположены узким веерообразным пучком на вершине плодового тела или по его экватору радиально. На концах несколько раз дихотомически разветвленные. Виды рода Podosphaera паразитируют на груше, сливе, березе, чернике и др.
Род Erysiphe Link. Это наиболее обширный род по количеству специализированных форм, приспособившихся к раз
витию на определенных видах растений. Грибница поверхностная, весьма разнообразная по плотности и окраске. Конидирльная стадия очень обильная. Придатки клейстокар-пиев не отличаются от грибницы, с которой обычно переплетаются. Иногда они очень короткие, располагаются в нижней части клейстокарпия. Сумки яйцевидные или грушевидные, в большинстве случаев многочисленные. Количество спор в сумке от 2 до 8. Чрезвычайно распространенным видом является Erysiphe graminis D. С. — мучнистая роса з л а-к о в, паразитирующая как на культурных, так и на дикорастущих видах. Этот вид характеризуется распространением грибницы с конидиальным плодоношением на обеих сторонах листа. По мере старения грибница уплотняется и приобретает вид выпуклых подушечек грязно-серого цвета, в которые погружены клейстокарпии. Придатки их обычно короткие, часто очень слаборазвитые. Сумки многочисленны — от 9 до 30 в одном плодовом теле. Спор — от 4 до 8 в сумке.
В настоящее время в пределах этого вида различают более 30 специализированных форм, как например Er. graminis f. tritici Marchal на пшенице; f. secalis Marchal на ржи; f. fes-tucae Jacz. на видах овсяницы и др.
Культурные злаки, главным образом пшеница и рожь, поражаются повсеместно, и чем больше загущен посев, тем поражение сильнее.
Потери от мучнистой росы на культурных злаках достаточно велики, так как пораженные растения дают щуплое, неполновесное зерно.
Свое развитие мучнистая роса начинает весною, располагаясь вначале у основания стеблей, затем переходит на листья, листовые влагалища, а при сильном поражении грибницу можно видеть и на колосковых чешуях и остях.
Цикл развития формы, специализированной на пшенице, детально изучен М. В. Горленко. Им установлено, что зимовка паразита может происходить в стадии вегетативного мицелия на зимующих зеленых частях растений (озими). Клейстокарпии в этом случае служат для сохранения гриба в течение лета, для перенесения неблагоприятных условий в период между уборкой урожая и развитием озимых. Таким образом, выход сумкоспор и заражение озимых происходят осенью.
Исследования Е. Г. Гулецкой в Белоруссии в 1952— 1953 годах также показали, что в годы с благоприятными климатическими условиями возбудитель мучнистой росы на озимой ржи и пшенице может проходить весь цикл развития с образованием и «полным созреванием сумчатой ‘стадии в течение одного вегетационного периода. Зимовка гриба в данном случае происходит в виде мицелия.
Учитывая узкую специализацию паразита, основным мероприятием в борьбе с мучнистой росой злаков следует считать правильный севооборот. Кроме того, рекомендуются рбщеги-гиенические мероприятия в виде хорошей обработки почвы с запашкой пожнивных остатков, на которых может сохраняться инфекция.
Род Phyllactinia Lev. Имеет нежную, паутинистую грибницу на нижней поверхности листа, проникающую через устьица в межклетники. Таким образом, у этого рода часть грибницы является эндофитной. Конидиеносцы удлиненные, с поперечными перегородками, в большинстве случаев имеют на конце по одной крупной, булавовидной или веретеновид-ноп, конидии.
Клейстокарпии несут придатки двух родов: одни в небольшом числе, жесткие, прямые, утончающиеся к концам и с шаровидно вздутым гигроскопическим основанием, расположены радиально и отходят от экватора клейстокарпия; вторые многочисленные, слабозаметные, ослизняющиеся, расположены на вершине плодового тела (см. рис. 26, е).
Оболочка шаровидных вздутий вследствие неодинаковой толщины при понижении влажности в нижней части сморщивается. При этом придатки приподнимаются и упираются своими острыми концами в субстрат, приподнимая клейстокарпий и отрывая его от грибницы. Плодовое тело оказывается стоящим на своих придатках, как на ножках, легко отрывается и переносится воздушными течениями на значительное расстояние. При падении вниз клейстокарпий поворачивается основанием кверху и приклеивается короткими ослизняющимися придатками к новому субстрату.
Это морфологическая особенность строения придатков объясняет ряд указаний на нахождение плодовых тел видов Phyllactinia на самых разнообразных субстратах, вплоть до шляпок некоторых гименомицетов, тогда как развиваются эти грибы только на деревьях и кустарниках.
Сем. Perisporiaceae
В отличие от Erysiphaceae это семейство ограничено весьма неясно и представляет некоторый переход от плекта-сковых к пиреномицетам. Сюда относятся преимущественно субтропические и тропические формы, сапрофиты и эктопаразиты. Поверхностный мицелий темноокрашенный, а внутренний — бесцветный. Плодовые тела развиваются на поверхностном мицелии и по своему строению несколько напоминают клейстокарпии Erysiphaceae. Семейство Perisporiaceae, несомненно, должно считаться более примитивным в порядке
как по строению плодовых тел, так и по характеру паразитизма. В то время как Erysiphaceae представляют высокоспециализированную группу облигатных паразитов, приспособившихся к паразитизму через посредство поверхностного мицелия, среди Perisporiaceae имеется большое число сапрофитов, а паразитные формы малоспециализированы.
ГРУППА ПОРЯДКОВ pyRENOMVCETES
Пиреномицеты объединяют несколько порядков плодосумчатых грибов, у которых плодовые тела построены по типу перитециев. Это группа огромная, насчитывает более 10 000 видов, среди которых сравнительно мало форм с детально изученной историей развития.
ПОРЯДОК НУ РОС REALES
Этот порядок содержит около 1000 видов. Мицелий обычно бесцветный, плодовые тела светлоокрашенные, мягкие. Они могут располагаться у одних форм свободно, у других — погружены в мягкую строму. Сумки всегда расположены пучком па дне плодового тела.
В порядке Hypocreales следует отметить семейства Нуро-сгеасеае и Nectriaceae.
Сем. Нуросгеасеае
Род Claviceps Tul. Содержит около десяти видов паразитных грибов. Главным из них является С. purpurea Tul. — спорынья, особенно часто встречается на ржи, но поражает и другие злаки (табл. I).
В цикле развития спорыньи имеется покоящаяся стадия — склероций. Упавшие на землю из колосьев пли высеянные вместе с рожью склероции способны переносить самые неблагоприятные условия. Ранним летом склероций прорастает. Черно-фиолетовая кора его лопается, и наружу выступают до 20—30 красноватых стром, состоящих из длинной ножки и шаровидной головки на ее конце. Через 15—20 дней головки становятся бугристыми, а каждый бугорок имеет отверстие (устьице), ведущее в грушевидную полость — перитеций. У всех представителей рода Claviceps перитеций не имеет дифференцированной собственной стенки. Со дна перитеция поднимается пучок вытянутых сумок, парафизы отсутствуют.
Ко времени цветения ржи споры созревают. Сумки поодиночке отрываются от основания плодового тела, поднимаясь кверху, высовываются своими верхушками из отверстия перитеция, лопаются, и споры, подхваченные воздушными тече
ниями, рассеиваются. Попадая на цветок, споры прорастают. Нежная, тонкая грибница проникает в завязь и постепенно вытесняет ее ткани. Через четыре дня после заражения на грибнице развивается конидиальное спороношение, известное под именем Sphacelia. Конидиеносцы короткие, на своих концах отчленяют одноклетные бесцветные конидии; одновременно с этим выделяется сахаристый сок, привлекающий на-секомых (медвяная роса). Насекомые разносят конидии на другие цветы, и таким образом распространяется заражение. Ко времени созревания ржи выделение медвяной росы прекращается; грибные гифы, совершенно разрушив завязь, сплетаются более плотно, образуя покоящуюся стадию грибницы — склероций, окрашенный в черно-фиолетовый цвет. Достигая у ржи нескольких сантиметров в длину, склероций высовывается из цветочных и кроющих чешуи наружу, а на верхнем конце его можно видеть -остатки засохшей кони ди альной стадии. Созревший склероций падает на землю, где и зимует, а весною прорастает.
Рожь больше других злаков поражается спорыньей, потому что, являясь перекрестноопыляющимся растением, имеет открытое цветение, а поэтому ее завязь легко доступна для заражения. Большинство же других хлебных злаков — самоопылители и имеют закрытое цветение.
Спорынья наносит очень большой вред. Он заключается в основном не в снижении урожая, так как процент пораженных цветков ржи не так уж велик, а в ядовитых свойствах этого гриба. Ядовитые вещества спорыньи — корнутин, эрготинин и другие — вызывают сокращение гладкой мускулатуры и при больших дозах дают тяжелое, иногда смертельное заболевание, известное под названием злые корчи, или эрготизм. Это заболевание наблюдается ;при употреблении в пищу хлеба из муки, полученной из недостаточно отсортированного зерна.
Особенно сильные вспышки этого заболевания наблюдались в Западной Европе в период X—XII веков. В те времена болезнь именовалась «огонь св. Антония» (в честь ордена святого Антония, посвятившего себя уходу за больными). При систематическом потреблении хлеба с примесью рожков спорыньи эрготизм может проявляться в виде конвульсивных сведений рук и ног, а также образования гангренозных участков на конечностях. С другой стороны, спорынья находит применение в медицине. Благодаря способности ее вызывать сокращение гладкой мускулатуры настоем спорыньи пользуются в гинекологической и акушерской практике. В аптеках она известна под названием Secale cornutum (рогатая рожь).
Cl. purpurea может расти и в искусственной сапрофитной культуре, развивая мицелий и конидиальное спороношение.
Вытяжка из такой культуры содержит те же вещества, как и естественный продукт.
До недавнего времени в микологической литературе можно было встретить утверждения, что спорынья с крупными рожками, заражающая завязь таких крупнозерновых злаков, как рожь, ячмень и др., относится к виду Cl. purpurea, злаки же с мелкими завязями (тимофеевка, лисохвост, мятлик и др.) поражаются другим видом спорыньи — Cl. microcephala, для которого характерно образование мелких склероциев.
Отсюда делался вывод, что гриб не может переходить с дикорастущих злаков на культурные. Однако исследования Л. И. Пшедецкой (1953) по перекрестному заражению набора злаков разными формами спорыньи показали, что Cl. purpurea одинаково легко заражает любой злак, меняя в зависимости от величины его завязи размеры своих склероциев. И, таким образом, оказалось, что все виды спорыньи на злаках — это синонимы, т. е. формы одного вида, имеющего широкий круг хозяев. Поэтому борьба с этим паразитом должна проводиться не только на хлебных злаках, но и на дикорастущих злаковых сорняках.
Если склероции ржаной спорыньи распространяются благодаря хозяйственной деятельности человека, то у других видов распространению могут способствовать вода, воздушные течения, животные. Склероции этих видов спорыньи имеют соответствующие приспособления.
Раньше спорынья широко встречалась в России, особенно в северных районах культуры ржи. Заготовки ее как лекарственного сырья были обширны и покрывали потребности не только внутри страны, но в большом количестве спорынья шла и на экспорт. Укрепление колхозного строя в СССР повлекло за собой повышение агротехники, контроль за семен-ным материалом и привело к значительному сокращению зараженности спорыньей посевов ржи.
Род Epichide Fries. Наиболее распространенным представителем его является Е. typhina (Pers.) Winter — паразит на луговых злаках, особенно на тимофеевке и еже. Развивается на верхушках стеблей еще до цветения растений, в виде сначала беловатого, а затем желтеющего плотного войлока, напоминающего как бы чехол, надетый на конец стебля, за что болезнь получила название чехловидной. В итоге рост стебля прекращается и соцветие не развивается. Налет этот состоит из плотно сплетенных грибных гиф, на свободных концах которых отчленяются мелкие бесцветные конидии. Плодовые тела развиваются позднее и имеют вид бутылкообразных полостей в общей войлочной строме. Сумки вытянутые. Сумкоспоры вначале одноклетные, а затем становятся многоклетными. Гриб способен зимовать в виде мицелия в
почках, образующихся с осени. С целью предотвращения заражения почек рекомендуется возможно ранний и повторный укос пораженных растений.
Сем. Nectriaceae
Род Nectria Fries. Содержит около 400 видов. Наиболее известным и широко распространенным видом этого рода является Nectria galligena Brea.— факультативный паразит, вызывающий заболевание плодовых деревьев, известное под названием обыкновенного рака или по месту его наибольшего распространения европейского рака.
При этом заболевании поражаются главным образом стволы деревьев, развилки скелетных ветвей и ветви первого порядка. Поражение развилок ветвей — наиболее опасный случай заболевания, так как поврежденные ветви обычно не могут противостоять ветру и обламываются.
Первым симптомом появления болезни является потемнение коры, а затем ее отмирание более или менее правильными концентрическими кругами. В результате развития болезни на пораженных частях дерева образуются глубокие раны, затрагивающие не только кору, но и древесину. Как результат сопротивления дерева развитию паразита вокруг пораженных участков возникают наплывы в виде каллюса (рис. 27). При сильном поражении раком яблонь урожайность снижается более чем в два раза.
Грибница, внедряясь в ткань пораженного растения, ветвится в разных направлениях, поглощая органические вещества, за счет которых происходят дальнейший рост и развитие паразита. Мицелий, достигая древесины, распространяется вдоль органа, а затем проникает в новые участки коры, вызывая при этом новые очаги поражения. Способность мицелия N. galligena проникать глубоко в ткани коры и древесины предохраняет его от влияния неблагоприятных климатических условий в течение зимнего периода, и, таким об-
Р и с. 28. Конидии возбудителя рака яблони.
разом, вегетативная грибница служит для перезимовки возбудителя.
В цикле развития N. galligena имеются конидиальиая и сумчатая стадии.
В начале весны непосредственно на мицелии или на мицелиальных стромах, имеющих вид белых подушечек, гриб образует разнородные конидии. Одни из них мелкие од-ноклетные, так называемые микроконидии; другие—более крупные, слегка согнутые многоклетяые— макроконидии (рис. 28). Оболочка конидий очень тонкая, к перезимовке они не способны. Основная роль конидий сводится к размножению паразита, особенно во влажные периоды года.
К осени на тех же стромах образуются сумчатые плодовые тела — перитеции. Н. В. Виноградова (1956) установила, что в климатических условиях Белоруссии сумчатое спороношение следует считать преобладающим, так как оно имеет превосходство в численности, продолжительности образования спор на протяжении года и стойкости их к неблагоприятным условиям.
Наиболее иммунными сортами яблони являются: Антоновка обыкновенная, Польское сахарное (Каштеля), Пепин литовский; сильнопоражаемыми — Титовка, Папировка, Ана
насный ранет; среднепоражаемыми— Боровинка, Штрейф-линг, Малиновка, Цыганка.
Наиболее эффективной мерой борьбы с раком яблони является применение комбинированных лечебных замазок с предварительным зачищением раковых ран. Таковы, например, нигроловые замазки, содержащие 60% нигрола, 20% канифоли, 20% парафина и один из фунгицидов (0,3% паранитрофенола или 0,3% трихлорфенолата натрия).
Большое значение имеют и агротехнические мероприятия: осенне-весенняя вспашка междурядий; обработка приствольных кругов; осенняя и весенняя побелка стволов со своевременным замазыванием лечебной замазкой всех случайных поранений; удаление и уничтожение всевозможных пораженных частей дерева: коры ветвей, мумифицированных плодов и др.
ПОРЯДОК SPHAERIALES
Этот обширный порядок насчитывает более 7000 видов, преимущественно сапрофитов на различных растительных субстратах. Плодовые тела у большинства форм имеют собственную оболочку черного цвета, большей частью твердой углистой консистенции. Огромное большинство этой обширной группы грибов еще недостаточно изучено.
Наибольшего внимания заслуживают семейства: Ceratos-tomaceae, Mycosphaerellaceae, Pleosporaceae.
Сем. Ceratoslomaceae
Для этого семейства весьма характерно строение перитециев. Они очень мелкие, черные и оттянутые на вершине в длинную шейку, по размерам часто превышающую в несколько раз диаметр перитеция (рис. 29). Активного выбрасывания спор через шейку не происходит. Когда созревают сумки, их оболочки разрушаются, а споры выдвигаются разбухающей слизью из полости перитеция через канал шейки и скопляются в виде шапочки на ее вершине.
Род Ceratostomella Saccardo. Некоторые его виды имеют практическое значение. С. ulmi Schwarz широко распространен в Западной Европе и вызывает массовое отмирание вязов.
Виды С. picea Munch., С, pint Munch., С. coerulea Munch, хотя это и сапрофитные формы, вызывают широко распространенное явление синевы хвойной древесины. Изменение цвета представляет чисто оптическое явление рассеивания световых лучей между бесцветными стенками клеток древесины и бурыми гифами гриба. Рыночная ценность такой древесины значительно снижается.
Сем. Mycosphaerellaceae
Представлено главным образом паразитными формами. У большинства этих грибов в период вегетации образуется только конидиальная стадия, сумчатая же—-развивается после зимовки в отмершей и полуразрушенной ткани. Пери-
Р и с. 29. Ceratostoinella ulnii\ перитеций.
теции у них одиночные, без стромы, погруженные в ткань растения и высовывающиеся только своим устьицем.
Род Venturia de Not. Широко распространен и приносит огромный вред плодовым культурам. Представлен двумя видами: V. inaequalis Aderh. с конидиальной стадией Fusicla-diutn dendriticum (Waller) Fuck, паразитирует на яблоне и
pirina Aderh. с конидиальной стадией F. pirinum (Libert.) :k. поражает грушу. Заболевание, вызываемое этими грили, известно под названием парши.
Р и с. 30. Venturia inaequalis:
а — пораженный лист; б — спороношение гриба; в — перитеций с сумками; г — сумка со спорами.
На яблоне болезнь развивается главным образом на тьях и плодах, вызывая образования округлых, зеленовато-вковых бархатистых пятен (рис. 30). На листьях они зна-ельно крупнее, чем на плодах, и менее резко очерчены.
Мицелий и конидиеносцы развиваются в местах пятен под кожицей, а также и в клетках кожицы. Под напором образующихся конидиеносцев кожица разрывается, причем наружные стенки кожицы вместе с кутикулой отворачиваются. Конидиеносцы парши яблони гладкие и прямые. Конидии яйцевидные или обратно-булавовидные, в молодости одноклетные, а затем двухклетные, 20—30x7—9 мк. Конидии переносятся ветром и насекомыми и, попадая на здоровые листья и плоды, прорастают, причем росток может пробуравливать кожицу и развивать новую грибницу. При благоприятных условиях, когда дождливые дни сменяются солнечными, развитие парши яблони происходит очень быстро. Конидии очень устойчивы и в некоторых случаях сохраняют способность к прорастанию до весны.
При поражении плодов под пятном образуется пробковая ткань, препятствующая мицелию гриба проникать глубоко в мякоть плода.
Кроме листовых пластинок и плодов, паршой могут поражаться черешки листьев, плодоножки и даже цветки. В некоторых районах (Сибирь, Казахстан, Северный Кавказ) отмечено образование парши на побегах яблони, главным образом дикой. Л. Д. Казенас (1953) отмечает, что в 1949 году в горах Заилийского Алатау на дикой яблоне поражение побегов паршой наблюдалось у 95,3% деревьев (по Дементьевой). Проявляется парша на побегах в виде небольших вздутий, со временем разрывающихся, откуда выступает конидиальное спороношенне. В этих случаях возбудитель парши зимует в конидиальной стадии в ветвях.
В большинстве же других климатических зон гриб зимует в опавших листьях, образуя весной сумчатую стадию, а в районах, где отмечено поражение ветвей, зимовка гриба может проходить при помощи мицелия, на котором весной развивается конидиальное спороношение.
Вредоносность парши велика. Она снижает количество и качество урожая. Образование пятен на листьях уменьшает ассимилирующую поверхность, а разрыв кожицы повышает транспирацию. Все это способствует преждевременному опадению листьев, ослаблению дерева, понижению морозоустойчивости.
Пораженные плоды растут уродливыми, а образующиеся трещины в местах развития пробки способствуют проникновению микроорганизмов внутрь плода и его загниванию в период хранения.
Парша груши в общих чертах похожа на паршу яблони, однако имеются и некоторые различия. Конидиальное спороношение развивается преимущественно на нижней стороне листьев. Конидиеносцы искривленные, с неровной бугристой
поверхностью от зубчиков, которые служили основанием уже отпавших конидий. Гриб в сильной степени поражает ветви и побеги, особенно молодые. Они чернеют, искривляются, растрескиваются и отпадают. Прироста дерева не наблюдается, а ослабленные паршой молодые деревца подвержены вымерзанию. В пораженных веточках мицелий гриба перезимовывает, а весною на нем развивается спороношеиие, которое обусловливает первичное заражение паршой. Трещины, возникающие на ветвях, являются убежищем для насекомых, а также служат путями проникновения различной инфекции в глубь тканей дерева.
При развитии парши на плодах груши не вырабатывается пробка. Грибница, не имея никаких барьеров, проникает в плод, пронизывая всю мякоть. Плод становится жестким, теряет вкусовые качества и совершенно не годен к употреблению.
Сумчатая стадия Venturia развивается только к весне на □павших листьях. На поверхности прошлогодних листьев можно видеть невооруженным глазом шаровидные величиной булавочную головку плодовые тела — перитеции, почти погруженные в ткань листа. Внутри помещаются булавовидные уумки с восемью зеленоватыми двухклетными спорами грушевидной формы (см. рис. 30). Споры освобождаются с наступ-пением теплых весенних дней. Выбрасываются они обычно после дождя, разносятся воздушными течениями по всему :аду и заражают молодые листья. Выбрасывание спор повторяется после каждого дождя и продолжается до середины пета, пока не сгниют все опавшие прошлогодние листья. Через 10—16 дней после заражения на листьях появляются характерные для парши пятна.
Не все сорта яблони и груши поражаются паршой в оди-шковой степени. Кроме того, различная восприимчивость юртов зависит и от экологических условий.
Практически устойчивыми сортами яблони являются: Петин шафранный, Пармен зимний золотой, Ренет шампанский т др. Сильно поражаемые сорта: Боровинка, Бельфлер-ки-айка, Анис серый, Ренет Симиренко и др.
Наиболее устойчивые сорта груши: Бере Лигеля, Бере Слержо, Бере Боек, Кюре и др. Сильно поражаемые сорта: Тесная красавица, Бере Диль, Тонковетка, Любимица Клап-та и др. (по Дементьевой).
В целях борьбы с паршой в первую очередь рекомендуется 'ничтожение очагов зимующей инфекции и опрыскивание деревьев бордоской жидкостью. Первое опрыскивание проводится в апреле — мае в фазу «зеленого конуса» 1%-ным раствором; второе — в период порозовения бутонов; третье — юсле опадения лепестков.
Сем. Pleosporaceae
Большинство принадлежащих сюда грибов ведут сапрофитный образ жизни, поселяясь на перезимовавших стеблях различных растений. Однако имеются и паразитные формы на корнях, стеблях и листьях. Конидиальное спороношение у них может быть пикнидиального (Phoma) или гифомицет-ного типа (Alternaria, Macrosporium и др.). Перитеции мелкие, плотной консистенции, вначале всегда погруженные, а затем постепенно освобождающиеся вследствие разрушения тканей субстрата. Сумки цилиндрические с парафизами, несколько расширенные к вершине, содержат по 8 бесцветных или слабо окрашенных спор цилиндрической или нитевидной формы.
Из этого семейства следует отметить два рода: Ophiobolus и Pleospora.
Род Ophiobolus Riess. Характеризуется окрашенным мицелием, перитеции шарообразные или шарообразно-конические, погруженные. Устьице хорошо выражено, иногда несколько вытянуто. Сумки длинно-цилиндрические. Споры со многими перегородками нитевидно вытянутые. Нередко одна из конечных клеток споры вздута, что делает ее похожей на тело змеи (отсюда название рода).
В северо-западной зоне Европейской части СССР и в предгорьях Кавказа широко распространен Ophiobolus graminis Saccardo. Этот гриб поражает озимую пшеницу, озимый ячмень и сорняки: костер, щетинник и др. Развиваясь на корнях всходов, гриб вызывает корневые гнили, а поражая основание стеблей способствует их полеганию и пустоколосости. На пораженных стеблях развивается мицелий, конидиальное и сумчатое спороношение. Поверхностное заспорение семян злаков возможно при обмолоте. Перезимовывает гриб на растительных остатках злаковых культур и сорняков, где и происходит развитие сумчатого спороношения.
Род Pleospora Robenhorst. Характеризуется шарообразноприплюснутыми, иногда шаровидно-коническими перитециями с очень плотной, почти деревянистой оболочкой и хорошо развитым устьицем. Расположены они одиночно или скученно. Сумки булавовидные содержат по 8 спор эллиптической формы с перетяжкой по середине. Споры окрашены в желтые и бурые тона, снабжены многими поперечными и несколькими продольными перегородками. Pl. chartarum Fuck, может развиваться на бумаге, поражая книги в библиотеках и хранилищах.
PI. vulgaris Niessl — широко распространенный вид на различных травянистых растениях: вике, лютике, колокольчике. погремке и многих других.
ПОРЯДОК D0THID1ALES
Преимущественно паразиты на листьях и стеблях растений. Порядок включает около 400 видов. Для всех их характерно, что перитеции погружены в ткань темноокрашенных стром и не имеют собственной оболочки. Большая часть грибов этого порядка является тропическими формами. У нас довольно распространены представители рода Phyllachora Nitschke.
ПОРЯДОК Н EMI SPHAERI ALES
Эта группа грибов охватывает тропические формы, еще недостаточно изученные. Мицелий разрастается радиально, более или менее равномерно во все стороны из общего центра. Наиболее характерной чертой Hemisphaeriales является строение плодового тела. Оно имеет вид приплюснутого образования, прикрытого сверху особым диском из темных кутинизи-рованных клеток. Очень часто сумки располагаются в виде гимения. Эта особенность представляет некоторый переход к дискомицетам. Представители порядка паразитируют на живых листьях растений (реже на ветвях).
ГРУППА ПОРЯДКОВ DVSCOMVCETES
Дискомицеты включают грибы, у которых сумчатое плодовое тело представляет апотеций — блюдцевидное образование. Верхняя сторона его несет гимений, состоящий из сумок и парафиз. Дискомицеты связаны с пиреномицетами постепенными переходами. Такие порядки дискомицетов, как Hysteriales и Phacidiales, включают организмы, у которых наиболее отчетливо выступает связь с предыдущей группой.
ПОРЯДОК HYSTERIALES
По строению сумчатых плодовых тел и характеру их раскрывания порядок Hysteriales занимает промежуточное положение между пиреномицетами и дискомицетами. Плодовые тела вытянутые, погруженные в субстрат, снаружи черного цвета. Сверху прикрыты плотным сплетением гиф, под которым развивается гимений. При созревании сумок кроющий слой разрывается продольной щелью, причем гифы этого слоя весьма чувствительны к изменению влажности: во влажную погоду они расходятся, щель становится шире и обнажается гимений, а в сухую — снова сближаются и гимений закрывается.
Наиболее широко распространенным представителем ггого порядка является Lophodermiutn pinastri (Schrad.) Chev., паразитирующий на хвое сосны. При сильном развитии гриб вызывает отмирание и опадение хвои и наносит значительный ущерб сосновым сеянцам в питомниках.
В паразитных условиях на второй год после заражения развивается конидиальная стадия на живой хвое, а сумчатая — после ее отмирания. Среди немецких лесоводов эта болезнь получила название Schutte.
ПОРЯДОК PHACIDIALES
К порядку Phacidiales относится около 600 видов преимущественно сапрофитных форм. Встречаются и паразитные формы. Среди последних лучше изучен паразит на листьях клена — Rhytisma acerinym Fr., образующий черные блестящие пятна (рис. 31).
Р и с. 31. Rhytisma acerinym на листе клена.
Гифы гриба распространяются преимущественно в клетках верхнего эпидермиса и, сплетаясь между собой, образуют сплошное плоское ложе, так как боковые стенки клеток эпидермиса со временем разрушаются.
Конидиеносцы развиваются на мицелиальном ложе тесными группами, а отшнуровывающиеся конидии прорывают наружную стенку эпидермальных клеток и, таким образом, выступают наружу. Во второй половине лета стромы утолщаются и чернеют, приобретая склероциальный характер. Осенью в них закладываются апотеции, созревающие только весной. Аскоспоры заражают молодые листья клена, и цикл развития гриба возобновляется.
ПОРЯДОК PEZIZALES
Апотеции грибов из порядка Pezizales образуются прямо на мицелии. В раннем периоде они полузамкнутые, позднее при созревании разворачиваются. Зрелые плодовые тела мягкой, нередко почти сочной консистенции, яркой или бурой окраски, сидящие на более или менее длинной ножке.
Пецициевые представляют большой интерес в теоретическом отношении. Среди представителей этого порядка лучше всего можно проследить постепенную утрату половой функции и половых органов. Простейшие представители Pezizales, как Pyronema confluens, имеют типично развитый и функционирующий половой аппарат. С другой стороны, наиболее специализированные семейства, как Helvellaceae, представляют пример далеко зашедшей редукции половой .фУнкиии*
В промежуточных семействах, как Ascobolaceae и Pezizaceae наряду с типично половыми формами встречаются и такие, у которых сумки развиваются апогамно.
Наибольшего внимания в порядке Pezizales заслуживают семейства Pyronemaceae, Pezizaceae, Helvellaceae.
Сем. Ругопетасеае
Насчитывает не более 25 видов, но представляет интерес в том отношении, что к нему принадлежит Pyronema confluens Tul.— сапрофитный гриб, послуживший в свое время Клауссену для изучения нормального полового процесса у сумчатых грибов. Pyronema встречается на местах бывших костров, так как прогретая и отчасти стерилизованная почва исключает возможность подавления развития мицелия Ругопепга другими почвенными грибами, а также бактериями. Апотеции красноватые, мелкие (до 1 мм в диаметре), но скученные группами, а поэтому легко заметные.
Сем. Pezizaceae
Обширнее предыдущего, насчитывает около 800 видов главным образом сапрофитных форм, широко распространенных на почве, древесине и навозе травоядных. Плодовые тела представителей семейства Pezizaceae обычно служат демонстрационным объектом для лабораторных занятий по изучению типичного строения апотеция.
На рис. 32 изображены плодовые тела представителей сем. Pezizaceae в натуральную величину.
Сем. Helvellaceae
Семейство Helvellaceae — сморчковые грибы. 1\ нему относится около 100 видов сапрофитных форм, растущих на почве. Плодовые тела большей частью крупные, состоят из ножки и шляпки. Последняя несет на своей ячеистой или складчатой поверхности гимений.
Рис. 32. Плодовые тела представителей семейства Pezlzaceae’. а — Lachnca; б — Huinaria\ в — Macropodia,
В семействе сморчковых различают следующие важнейшие роды: Morchella (сморчки), Gyromita, Verpa и Helvetia (строчки).
Род Morchella Dillen. Наиболее распространены виды М. esculenia и М. conica.
М. esculenia Pers. — сморчок обыкновенный имеет яйцевидной формы шляпку светло-коричневого цвета с ямчатой поверхностью и выступающими ребрами (табл. II, а). Поверхность шляпки напоминает соты, только ямки выстланы гимением, а ребра стерильны. Споры бесцветные, эллиптические.
М. conica Pers.— сморчок конический (табл. II. б), отличается от первого вида конической формой шляпки, цвет ее бурый или оливковый. Поверхность шляпки и расположение гимения такие же, как и у М. esculenia. Растут оба вида сморчков преимущественно в хвойных лесах и на старых лесных пожарищах с апреля до конца мая. Все исследователи считают сморчки съедобными и безвредными грибами.
Род Helvetia Linne. Helvetia esculenia Pers,— строчок обыкновенный, имеет шляпку неправильно шаровидной формы, чаще темно-коричневого цвета, иногда она окрашена в более светлые тона (табл. II, в). Поверхность шляпки покрыта складками, напоминающими извилины мозга. Гимений развивается сплошь по всей поверхности шляпки. Споры бесцвет-
ные или желтоватые, эллиптические; растет по хвойным или смешанным лесам, преимущественно на песчаных почвах, с апреля до конца мая.
Строчки по многовековой ошибке считаются в быту съедобными грибами и допущены к открытой продаже во всех странах. В некоторых местах, например в Латвии, ядовитое действие их приписывается не самим строчкам, а паразитам, живущим на них. Население, продавцы и большинство врачей обычно смешивают виды сморчков и строчков, несмотря на то что у них есть определенные морфологические отличия, на которые не обращают достаточного внимания.
Некоторые авторы считают строчки очень ядовитыми грибами. Так, еще в 1885 году Бему и Кюльцу удалось выделить из спиртового настоя, а также из водного горячего отвара строчков гельвелловую кислоту (С12Н20О7). Токсичность ее была проверена на животных. У отравленных собак наблюдалась желтуха, а затем разрушение эритроцитов крови.
Указанными авторами в дальнейшем была установлена токсичность отвара строчков и практическая безвредность самих отваренных грибов. Н. И. Орлов обращает внимание на неодинаковое действие строчков на разные организмы людей: в одних случаях отравление может привести к гибели, в других случаях люди, употреблявшие в пищу строчки, остаются здоровыми или получают легкое отравление. Оказывается, степень отравления зависит от состояния печени. У лиц со здоровой печенью имеется относительный иммунитет к яду строчков. Люди, не имеющие острого или хронического заболевания кишечного тракта, также относительно иммунны.
Кроме того, токсичность строчков колеблется в зависимости от различных факторов: погоды, времени, места сбора, условий вегетации в отдельные годы, а также, возможно, и возраста грибов. Согласно наблюдениям, строчки, собранные в дождливую и теплую погоду, а также старые грибы содержат больше ядовитого начала.
Обезвреживание строчков достигается двумя способами: отвариванием с обязательным удалением отвара или сушкой. При отваривании яд переходит в воду, а обезвреживание сушеных строчков наступает через 3—4 недели после сушки. Причем сушку необходимо производить только на воздухе. Ядовитое начало в данном случае, по-видимому, подвергается разрушению благодаря окислению кислородом воздуха.
ПОРЯДОК HELOTIALES
Этот порядок насчитывает около 300 родов, объединяемых различными авторами в разное количество семейств.
Таблица HI. Amanita phcilloides.
Наибольшего внимания заслуживает семейство Sclerotiniaceae.
Сем. Sclerotiniaceae
Грибы, объединяемые в семейство Sclerotiniaceae, развиваются на плодовых, овощных и технических культурах, а также на сорных растениях. Цикл развития гриба начинается на живых тканях растения, а заканчивается на растительных остатках. Для всех видов характерно образование склероция.
Большое практическое значение имеют представители родов Sclerotinia и Monilinia.
Род Sclerotinia Fuck. Склероции неопределенной формы, образованы плотно сплетшимися гифами гриба. Размеры их варьируют от нескольких миллиметров до 2 см в поперечнике. В некоторых случаях склероции образуются значительно больших размеров, принимают очертания полостей или промежутков между частями растений. Из наиболее распространенных видов следует отметить S. libertiana Fuck — возбудителя белой гнили.
Этот гриб обладает широкой избирательной способностью,, поражает культурные растения и сорняки в период вегетации, а также корнеплоды в период хранения.
Особенный вред S. libertiana приносит подсолнечнику, развиваясь во всех районах его возделывания. Поражает все части растения в любую фазу развития—от всходов до созревания корзинок.
Цикл развития S. libertiana протекает в виде чередования двух основных стадий — мицелия и образующихся на нем склероциев. Конидиального спороношения этот вид не образует, однако на его мицелии развиваются микроконидии, роль которых не вполне выяснена (есть предположение, что они участвуют в процессе оплодотворения). Склероции могут прорастать в мицелий, но в более типичных случаях образуют апотеции.
Грибница местного характера. В сухую погоду она развивается внутри пораженных тканей, а в сырую — образует на поверхности белый ватообразный налет. Распространение гриба в период вегетации совершается кусочками подсохшей грибницы.
На всходах подсолнечника поражается подсемядольное колено, в результате чего растения гибнут. Пораженные стебли взрослых растений покрываются бурыми пятнами, на которых впоследствии развивается белый налет грибницы. На корзинках в начале развития болезни также появляются бурые мокнущие пятна, покрывающиеся затем грибницей. Пора
женные части растений загнивают и разрушаются, а на грибнице образуются склероции. Развиваясь в паренхимной ткани корзинки, гриб проникает внутрь семян, где также могут образовываться склероции.
Непосредственно в почве гриб не живет и не размножается. Зимует мицелием и склероциями на растительных остатках и в семенах. Весною перезимовавшая грибница подсыхает, разносится воздушными течениями и служит одним из источников распространения болезни.
Прорастающие весной склероции образуют плодовые тела— апотеции. При созревании сумок споры выбрасываются на несколько сантиметров, переносятся воздушными течениями и заражают растения. Выбрасывание спор сумками одного апотеция может совершаться в течение 2—3 декад. В почве склероции сохраняют жизнеспособность до трех лет, что значительно затрудняет борьбу с возбудителем болезни.
Для борьбы с белой гнилью подсолнечника рекомендуются следующие мероприятия:
1. Посевы производить после непоражаемых белой гнилью культур с возвращением подсолнечника на прежнее место не ранее чем через 3 года.
2. С целью уничтожения семенной инфекции протравливать семена гранозаном (1,5 кг/т) или ТМТД (4—5 кг/т).
3. Удалять с поля и уничтожать больные растения, а также послеуборочные остатки.
В значительной степени S. libertiana поражает клубни и корнеплоды в хранилищах. Пораженные органы загнивают и превращаются в кашицеобразную массу. Очень скоро грибница выделяет капельки воды и постепенно уплотняется, образуя склероции. Внешние очертания их весьма изменчивы, а размеры могут достигать горошины и даже больше. Склероции могут прорастать сразу же, но при отсутствии необходимой влажности очень долго сохраняются, не теряя способности к прорастанию. Апотеции мелкие, чашеобразной формы, на длинных ножках. Споры, освобождающиеся из сумок, производят заражение здоровых органов растений. Распространение гриба в хранилище может происходить и вегетативно, кусочками грибницы.
Борьба с белой гнилью овощей должна прежде всего заключаться в уничтожении очагов заразы как в хранилищах, так и в почве, где могут сохраняться склероции гриба. Хранилища перед закладкой овощей должны быть продезинфицированы. Овощи рекомендуется закладывать в просушенном виде и, кроме того, пересыпать сухим песком. После уборки •овощей следует проводить зяблевую вспашку, строго придерживаться плодосмена и не высевать семена от больных растений.
S. libertiana широко распространена на различных культурных растениях и в Белоруссии. В 1929 году она была обнаружена впервые на люпине С. М. Тупиневичем. Как показали более поздние исследования Н. А. Дорожкина и А. И. Кустовой (1956) и Н. А. Дорожкина и Н. И. Чекалин-ской (1965), развитие белой гнили на желтом безалкалоидном люпине приводит к значительному снижению урожая семян и зеленой массы. На льне впервые в республике белая гниль найдена О. Я. Стрельской (опытная станция «Устье» Витебской обл., 1961), а Н. А. Дорожкиным она обнаружена и на картофеле (Ганусовская опытная станция Минской обл., 1948).
В последние десятилетия все шире и шире ставится вопрос о применении биологических методов борьбы с различными патогенными микроорганизмами. Известно, что многие микроорганизмы, сапрофитно живущие в почве, обладают антагонистическими свойствами по отношению к паразитам — возбудителям инфекционных заболеваний растений.
Установлено, что почвенный сапрофитный гриб Trichoder-та lignorum подавляет развитие многих паразитных грибов и в том числе S. libertiana (Федоринчик, 1951).
Trichoderma lignorum Harz, по своему систематическому положению относится к классу несовершенных грибов — Deuteromycetes, порядку Hyphomycetales, семейству Mucedi-пасеае. Этот гриб обладает способностью выделять ферменты, растворяющие не только вегетативный мицелий гриба, но и его видоизменение — склероции, и употреблять их в процессе питания.
Принимая во внимание то, что склероции возбудителя белой гнили в большом количестве находятся в почве, а при прорастании служат источником инфекции, значение Trichoderma совершенно очевидно.
В настоящее время биопрепарат «триходермин-3», представляющий культуру Trich. lignorum, искусственно выращенную и в массе накопленную на прогретом торфе, применяется в борьбе с различными возбудителями болезней растений и в том числе с S. libertiana.
Исследования А. А. Сапоговой (1960) в Белоруссии также подтверждают эффективность применения триходермина в борьбе с возбудителем белой гнили огурцов в закрытом грунте.
Род Monilinia Honey. Представители этого рода широко распространены на семечковых и косточковых культурах, а в последние годы в условиях Дальнего Востока обнаружены на черноплодной рябине и боярышнике (Хохрякова, 1965).
В европейской части СССР особенно вредоносным является возбудитель плодовой гнили Monilinia fructigena (Schrot)
Honey1, поражающий плоды яблони, груши, айвы. Конидиаль-ная стадия этого возбудителя относится к роду Monilia Pers, и характеризуется цепочками лимонообразных бесцветных конидий (см. гл. V. Несовершенные грибы).
Склероции М. fructigena представляют мумифицированные пораженные плоды, в которых грибница сохраняет жизнедеятельность до весны. В 1931 году в окрестностях Ленинграда было отмечено прорастание склероция плодовыми телами — апотециями (Солкина). Другими авторами сумчатая стадия нигде не обнаружена.
На перезимовавшем мумифицированном плоде обычно развивается конидиальное спороношение, дающее начало новому циклу развития гриба.
Исследования показывают, что различные сорта яблонь и груш обладают разной степенью устойчивости к заболеванию.
Яблоня. Сорта менее устойчивые: Апорт, Антоновка, Анис, Боровинка, Белый налив. Более устойчивыми сортами являются: Пепин шафрановый, Пепин литовский, Борсдорф-китайка, Украинка, Титовка.
Груша. Сорта менее устойчивые: Сапежанка, Бергамот; более устойчивые: Бере зимняя Мичурина, Октябрьская, Аврора.
Степень устойчивости зависит от многих внешних условий, в частности от особенностей местности. Сорт, менее поражаемый в одной местности, может оказаться более поражаемым в другой.
Широкое распространение плодовой гнили и значительный ущерб, причиняемый ею в садах, а также в период зимнего хранения плодов, настоятельно требуют проведения определенных мер борьбы с этим заболеванием.
Прежде всего необходимо тщательно убирать и уничтожать все загнившие плоды, как падалицы, так и висящие на деревьях. Рекомендуется также опрыскивать деревья перед цветением 1%-ным раствором бордоской жидкости, а после цветения опрыскивание повторять в те же сроки, что и против парши, добавляя инсектицид для борьбы с вредителями сада.
ПОРЯДОК tuberales
Порядок Tuberales, или трюфелевые грибы, включает около сотни видов с подземными плодовыми телами в виде мясистых, довольно крупных клубней, иногда весом до 1 кг.
1 М. fructigena (Schrot) Honey в отечественной литературе известна под названием Sclerotinia fructigena SchrotfStromatinia fructigena Aderhold.) -
Во взрослом состоянии они совершенно не похожи на апотеции дискомицетов, но история развития их показывает, что вначале эти плодовые тела закладываются как апотеции. В дальнейшем вследствие давления почвы на развивающееся плодовое тело края его заворачиваются и смыкаются, оставляя узкое отверстие, и, таким образом, сумки оказываются внутри клубнеобразного плодового тела.
Рис. 33. Трюфель:
а — разрез плодового тела; б — сумки при большом увеличении.
Зрелое плодовое тело трюфеля одето бурой многослойной оболочкой — перидием, а внутренняя мясистая часть имеет своеобразный мраморный рисунок из чередующихся светлых и темных прослоек (рис. 33). Светлые прослойки или внутренние вены, представляют складки гимения, который не мог свободно развиваться благодаря давлению почвы и постепенному смыканию краев апотеция. В начале своего развития гимений состоит из одних парафиз. В дальнейшем парафизы сильно разрастаются и переплетаются с парафизами противолежащей складки, образуя слой наиболее темной и рыхлой ткани — наружные вены. Сумки развиваются на границе между наружными и внутренними венами. Споры образуются в количестве от 2 до 4 и имеют мелкошиповатую оболочку. Освобождение спор из сумок и плодового тела происходит или при общем его разрушении, или при поедании животными, особенно роющими грызунами.
Трюфелевые имеют ограниченные области распространения и являются обязательными микоризообразователями лиственных пород. Важнейшими представителями Tuberales являются виды Tuber — съедобные грибы, имеющие высокую ценность. Особенно ценен черный, или французский, трюфель
T brumale var. melanosporum Vittadini. Встречается он во Франции в дубовых лесах на известковой почве и имеет большое промышленное значение.
Доволен ценен также Т aestivum Vittadini, произрастающий у нас в Закавказье. Менее ценен, но съедобен белый трюфель Choiromyces meandriformis Vittadini, встречающийся под Москвой. Его плодовые тела напоминают по внешнему виду картофель. При созревании они слегка выступают из земли.
Эволюция аскомицетов выразилась в образовании аскогенных гиф после полового процесса, увеличении числа сумок и развитии плодовых тел.
Формирование плодовых тел шло в двух направлениях. Одно от замкнутых клейстокарпиев с беспорядочным расположением сумок привело к перитециям с узким отверстием на вершине для выдвижения сумок и активного выбрасывания сумкоспор. Другое направление выразилось в развитии апотециев, содержащих гораздо большее число сумок и приспособленных к активному выбрасыванию сумкоспор сразу многими сумками.
ГЛАВА HI КЛАСС BASIDIOMYCETES (БАЗИДИАЛЬНЫЕ ГРИБЫ)
4
* ОБЩАЯ ХАРАКТЕРИСТИКА
Обширный класс базидиальных грибов включает организмы, имеющие большое практическое значение. К нему относятся грибы шляпочные, съедобные и ядовитые; разрушающие древесину, а также паразиты сельскохозяйственных растений, вызывающие их болезни — головню и ржавчину.
Базидиальные грибы, как и сумчатые, относятся к высшим грибам, но отличаются от сумчатых циклом развития, характером спороношения и другими признаками. Существенной особенностью этого класса является наличие двух типов мицелия в цикле развития. При прорастании споры базидиаль-ного гриба образуется одна или несколько ростковых трубочек, которые делятся поперечными перегородками и дают первичный, или гаплоидный, мицелий, который может размножаться бесполым путем (оидиями и конидиями), но обычно отличается кратковременным существованием. Дальнейшее развитие гриба возможно только в том случае, если на гаплоидном мицелии разовьется дикариотический, или вторичный, мицелий.
У гомоталличных базидиомицетов переход от гаплоидной к дикариотической фазе осуществляется в пределах одного мицелия, а у гетероталличных — при соединении двух разнородных в половом отношении гаплоидных мицелиев. В обоих случаях этот переход осуществляется соприкосновением двух близлежащих клеток, между которыми образуются короткие перемычки, называемые анастомозами. Через них ядро или все содержимое из одной клетки перемещается в другую. При этом получаются первые двухъядерные клетки вторичного, или дикариотического, мицелия (рис. 34,а). Ядра в клетках
не сливаются, а только сближаются, образуя дикарионы. При делении этих клеток ядра делятся одновременно и параллельно друг другу.
У гетероталличных форм есть еще один путь диплоидиза-ции мицелия — перенесение гаплоидных оидии или конидий
Рис. 34. Развитие вторичного мицелия у гетероталличных базидио-мицетов (а) и образование пряжек (б, в).
с мицелия одного пола на мицелий другого пола. При соприкосновении двух клеток ядро конидий перемещается в мицелиальную клетку и возникает двухъядерная клетка.
На вторичном мицелии базидиальных грибов нередко встречаются особые анастомозы, имеющие вид маленьких дугообразных клеток. Они лежат сбоку на гифах против поперечных перегородок и носят название пряжек. Образование последних происходит при делении клеток следующим образом. Сначала на гифе замечается небольшой боковой вырост, после чего оба ядра делятся одновременно и параллельно, причем оси деления располагаются вдоль клетки. Из полученных четырех ядер два отходят в верхний конец клетки, одно — в нижний, а одно—в боковой вырост. Затем сама клетка делится поперечной перегородкой, расположенной немного ниже бокового выроста. В результате верхняя отделившаяся клетка получает два несестринских ядра, образуя дикарион, нижняя восстанавливает дикарион благодаря переходу ядра из бокового выроста, который к этому времени прирастает к стенке клетки пониже поперечной перегородки. Переход ядра осуществляется вследствие растворения оболочки в месте срастания кончика выроста с боковой стенкой клетки (рис, 34, б, в).
Основным органом спороношения базидиальных грибов является базидия, образующаяся только на дикариотическом мицелии. Возникает она апогамно из конечной клетки гифы мицелия (рис. 35). Два ядра ее сливаются в одно диплоидное, которое затем делится дважды. При этом происходит ре-
Рис. 35. Развитие базидии из конечной
клетки мицелия.
дукция числа хромосом, так что в молодой базидии получается четыре гаплоидных ядра. Одновременно на вершине базидии появляются четыре выроста — стеригмы. Последние, вздуваясь на конце, дают начало будущим базидиоспорам. Стеригмы сообщаются узкими каналами с полостью базидии, через которые проталкивается по одному гаплоидному ядру. По созревании базидиоспоры отбрасываются от своих стеригм благодаря повышению тургорного давления внутри базидии. Канал стеригмы узкий, поэтому сила тургорного давления весьма незначительна, и базидиоспоры отбрасываются не более как на несколько миллиметров. Расположение базидиоспор на базидии может быть двух типов. У большинства представителей базидиоспоры расположены на вершине базидии и на одном уровне. Такие базидии получили название акроспоровых. В другом случае базидиоспоры развиваются на боковой стороне базидии. Эти базидии называются плев-роспоровыми (рис. 36).
Базидиоспоры всегда одноклетны и большей частью одноядерны. Форма их округлая, овальная или нитевидно вытянутая. Наиболее типичное число базидиоспор на базидии — четыре, однако известны двух-, восьми- и односпоровые базидии.
У одних грибов базидии остаются одноклетными и назы
ваются холобазидиями. У других грибов вслед за делением диплоидного ядра сама базидия делится поперечными или продольными перегородками на четыре клетки, а каждая из них развивает по одной базидиоспоре. Подобные базидии называются фрагмю базидиям и.
У некоторых представителей базидиальных грибов базидии образуются непосредственно на дикариотическом мицелии
Рис. 36. Типы базидий.
или берут начало из спор, которые в свою очередь возникают из мицелия (склеробазидии головневых и ржавчинных). У большинства же базидии образуются на особых плодовых телах, аналогичных плодовым телам сумчатых грибов, но не гомологичных им. Если у аскомицетов плодовые тела имеют двойственную морфологическую природу — снаружи они состоят из вегетативных гаплоидных гиф и только внутренняя часть образована гифами из двухъядерных клеток, то у базидиальных грибов плодоносны целиком образованы дикарио-тическим мицелием. Плодовые тела у базидиомицетов могут иметь мясистую или деревянистую консистенцию и чрезвычайно разнообразны по форме.
В простейшем случае плодовое тело представляет плоское гифенное сплетение, расположенное непосредственно на субстрате и нарастающее своими краями. При этом базидии располагаются на верхней свободной поверхности. Такие плодовые тела называются ресупинатными (Serpula, Corticum, Hypochtius).
У более высокоорганизованных базидиомицетов плодовые тела несколько приподнимаются над субстратом, оставаясь плоскими, а иногда приобретают коническую или воронковидную форму, базидии развиваются на боковой стороне (Cantharellus).
Еще более совершенна копытообразная форма плодовых тел, свойственная многим трутовым грибам. Подобные плодовые тела боком прирастают к субстрату, и базидии располагаются на нижней стороне.
Наивысшего развития плодовое тело достигает у шляпочных грибов, где оно обычно дифференцировано на ножку или пенек, и шляпку, несущую базидии на нижней поверхности.
Какова бы ни была форма плодового тела, оно всегда состоит из бесплодной ткани, или трамы, и плодущей части — гименофора. Гименофор в свою очередь может иметь различную форму: складок, лабиринтовидных ходов, шипов, трубочек и, наконец, пластинок. Все эти особенности строения гименофора значительно увеличивают его поверхность. Последняя покрыта гимением — плотным слоем булавовидных, цилиндрических или мешковидных базидий, между которыми располагаются парафизы. В молодости парафизы содержат по два ядра, как и базидии, но эти ядра не сливаются и поэтому парафизы остаются бесплодными.
Рассматривая строение гимения под микроскопом, можно видеть, что базидии и парафизы выходят из плотного сплетения гиф так называемого субгимениального слоя, берущего свое начало в мякоти гименофора. Парафизы как сочные тургесцентные образования придают упругость всему гиме-ниальному слою, поддерживая и отделяя базидии друг от друга, предохраняют сидящие на них базидиоспоры от слипания. У некоторых базидиальных грибов в гимениальном слое развиваются еще особые образования — цистиды — различной формы и окраски и более крупные, чем парафизы. Цистиды обычно возвышаются своими концами над общим уровнем гимения. Они играют защитную функцию, предохраняя молодой гимений от соприкосновения с другими поверхностями, что может повредить развивающиеся базидиоспоры.
Плодовые тела некоторых шляпочных грибов (гриб-зонтик, опенок, масленок и др.) в молодости имеют особое пленчатое образование на нижней поверхности шляпки, называемое частным покрывалом (velum partiaale). Его функция
защитная—прикрывать гимений в период развития. Впоследствии покрывало разрывается и повисает на ножке в виде кольца (гриб-зонтик, опенок) или в виде бахромок на краю шляпки (волнушка). Иногда покрывало может быть паутинистым и даже слизистым. Такие плодовые тела называются гемиангиокарпными в отличие от гимнокарп-н ы х, где базидии развиваются открыто на поверхности плодового тела.
Кроме частного покрывала, некоторые грибы из высших шляпочных в молодости имеют общее покрывало, или вольву (velum universale). При дальнейшем росте гриба оно разрывается, образуя при основании ножки мешковидное влагалище (бледная поганка). В то же время на шляпке остаются хлопьевидные лоскутки, так хорошо заметные у мухомора. Среди базидиальных грибов встречаются и такие, у которых плодовые тела совершенно замкнутые — это так называемые дождевики. У них базидии образуются внутри плодового тела и совсем не обнажаются. После окончательного созревания плодового тела происходит разрыв или разрушение общей плодовой оболочки и освобождение базидий и базидиоспор. Подобные замкнутые плодовые тела называются а н г и о-к а р п н ы м и.
В основе систематического деления базидиальных грибов лежит: во-первых, строение базидии, а во-вторых, плодового тела. Таким образом, установлено два подкласса базидиальных грибов:
1) Holobasidiomycetes; 2) Phragmobasidiomycetes.
ПОДКЛАСС HOLOBASIDIOMYCETES
Систематическое деление холобазидиальных грибов можно представить несколькими порядками и группами порядков: 1) Dacryomycetales; 2) Exobasidiales; 3) Ну шепоту cetes; 4) Gastromycetes (3 и 4 объединяют группы порядков).
ПОРЯДОК DACRYOMYCETALES
Этот порядок очень небольшой, насчитывающий около 30 видов сапрофитных грибов со студенистыми, часто окрашенными плодовыми телами в виде корочек, морщинистых подушечек или коралловидных разветвленных образований. Характерной особенностью их является наличие вильчато разветвленных базидий и способность базидиоспор делиться на несколько клеток при прорастании. Представители этого порядка встречаются как сапрофиты на гнилой древесине.
ПОРЯДОК EXOBASIDIALES
Порядок экзобазидиальных грибов с сем. Exobasidiaceae был установлен русским микологом и фитопатологом М. С. Ворониным в 1867 году.
Сем. Exobasidiaceae
Род Exobasidium Wor. Несколько лет подряд Воронин находил на бруснике (Vaccinium vitis ideae L.) гипертрофированные листья, стебли, а иногда и цветки. Для выяснения сущности обнаруженного явления он исследовал много тысяч деформированных растений брусники и установил, что гипертрофия органов вызывается высшим грибом, который развивает базидии прямо на мицелии.
Патологические явления, наблюдаемые на бруснике, характеризуются следующими признаками: гриб поражает листья, стебли и реже цветки. На верхней стороне листьев появляются вогнутые участки чаще карминно-красной окраски. Нижняя поверхность этих участков листа становится выпуклой и приобретает снежно-белый цвет. На стеблях болезнь проявляется в виде беловато-розоватых вздутий, которые распространяются по всей окружности стебля и могут захватывать несколько междоузлий. Цветки поражаются реже. У больных цветков чашечка и венчик ненормально разрастаются, превращаясь в мясистые образования слабо розовой окраски и во много раз по размерам превосходящие соответствующие части нормальных цветков (рис. 37, а, б, в). В пораженных завязях развитие семян подавляется.
Тщательное исследование нижней снежно-белой поверхности пораженных листьев позволило установить, что гриб образует булавовидные базидии, располагающиеся сплошным гимениальным слоем. Вырастая, они разрывают эпидермис и высовываются наружу, а затем на них появляются базидиоспоры по 4—5 на каждой базидии (рис. 37, г).
Базидиоспоры вначале одноклетные, а при прорастании делятся на 3—5 клеток. Причем только некоторые споры содержат во всех своих клетках мелкозернистую плазму. У большинства спор она имеется лишь в двух крайних клетках, каждая из которых прорастает только одной ростковой трубкой (рис. 37, д, е).
В дальнейшем, если прорастание идет в воде, ростковые трубки отшнуровываются от спор в виде одноклетных образований или в виде разветвленных членистых гиф, при этом отшнуровывание происходит неоднократно.
При прорастании спор на молодых листьях брусники ростковые трубки, не отшнуровываясь от спор, проникают сразу
Рис. 37. Exobastdium vaccinu\
а, б, e — поражение листьев, стебля и цветка; г — базидии гриба между клетками эпидермиса; д, е — базидиоспоры и их прорастание.
внутрь листа через устьица или сквозь стенки эпидермальных клеток.
Впервые открытый и подробно изученный организм, вызывающий болезнь брусники, М. С. Воронин назвал Exobasidium vaccinii Wor.
Результаты исследований М. С. Воронина сыграли очень важную роль при изучении экзобазидиоза чайного куста. В 1895 году английский миколог и фитопатолог Масси, основываясь на этих открытиях, определил, что давно известная болезнь чайного куста под английским названием «блистэр
блайт» (Blister blight) вызывается грибом, относящимся к установленному М. С. Ворониным роду Exobasidium. Масси описал «чайный» экзобазидиум под названием Exobasidium vexans Mass.
На чайных плантациях в Советском Союзе экзобазидиоз не встречается. В целях предотвращения заноса инфекции применяется ряд карантинных мероприятий.
М. С. Дунин указывает на чрезвычайно сильное распространение экзобазидиоза чайного куста во многих районах Индии. Наблюдая за развитием болезни на чайных плантациях в период своего путешествия по Индии в 1950 году, Дунин заметил, что Е. vexans развивается по тому же типу и вызывает такие же изменения в зараженных растениях чая, как Е. vaccinii, изученный М. С. Ворониным на бруснике, а затем на голубике.
Исследования грибов рода Exobasidium, выполненные М. С. Ворониным, способствовали выяснению биологических особенностей и обоснованию мероприятий против заболевания чайного куста на плантациях Индии.
Кроме брусники, голубики и чайного куста, виды рода Exobasidium могут поражать лавр, рододендрон и некоторые другие растения.
ГРУППА ПОРЯДКОВ HYMENOMYCETES
По практической значимости и по количеству видов (около 16 000) гименомицеты занимают первое место среди холо-базидиальных грибов.
В теоретическом отношении эта обширная группа грибов также очень интересна, так как на ее представителях можно проследить всю эволюцию плодовых тел, от ресупинатных плоских до вертикально стоящих, дифференцированных на ножку и шляпку.
По образу жизни большинство гименомицетов относится к почвенным макрофитам, причем многие из них являются обязательными микоризообразователями. Значительная часть гименомицетов живет в качестве сапрофитов на древесине и паразитов на стволах деревьев.
Большинство микологов в настоящее время делят гименомицеты на следующие порядки: I) Aphyllophorales; 2) Agaricales.
ПОРЯДОК APHYLLOPHORALES
У представителей этого порядка гименофор имеет вид шипов, складок или трубочек, причем последние не отделяются от мякоти плодового тела. Плодовые тела различной формы:
ресупинатные, отстающие от субстрата в виде пластинок, иногда приподнимающиеся в виде воронок, копытообразные, и лишь у немногих в виде шляпки на ножке. По консистенции они обычно плотные, не мягкомясистые и не загнивающие в старости.
Порядок афиллофоровых делят на следующие семейства: Thelephoraceae, Clavariaceae, Hydnaceae, Polyporaceae.
Сем, Thelephoraceae
К семейству телефоровых относится более 500 видов грибов, главным образом живущих сапрофитно на древесных остатках и на почве. В этом семействе следует отметить роды Hypochnusy Coniophora, Stereum, Thelephora, Craterel-lus и Cantharellus.
Род Hypochnus Fries. Простейшие представители этого рода еще не имеют плодового тела, а базидии развиваются из конечных клеток рыхлого паутинистого сплетения гиф. Ясно выраженного гимения также не наблюдается. Весьма широко распространен вид Нypochnus solani Р. et D., развивающийся на нижних частях стеблей картофеля, покрывая их беловатым паутинистым войлочком, на котором и развиваются базидии. Гриб может развиваться и на клубнях картофеля, образуя шероховатые выпуклые корочки. В молодой стадии они имеют беловатый цвет и состоят из рыхлосплетенных грибных гиф, позднее чернеют, становятся твердыми, напоминая частицы почвы, приставшие к клубню. Эта бесплодная стадия гриба (Rhisoctonia solani Kuhn), известная под названием черной парши.
Как факультативный паразит гриб существует во всякой почве и оттуда попадает па растение. Если поражаются молодые ростки, то присутствие гриба может привести к их гибели. Развитие гриба на взрослых растениях не вызывает никаких патологических явлений.
Род Coniophora D. С. Представители этого рода имеют ресупинатные плодовые тела. Весьма распространенным видом является Coniophora cerebella (Pers.) Schrot. — пленчатый домовой гриб. Плодовые тела, распростертые, перепончатые, мясистые, легко отделяются от субстрата, вначале беловатые, а затем темнеющие.
Пленчатый гриб распространен на мертвой древесине хвойных и лиственных пород, поселяется в погребах, на шпалах, столбах и т. п. Мицелий развивается в древесине, разрушает ее, вызывая образование мелких и частых трещин. Кроме мицелия и плодовых тел, Coniophora образует слаборазвитые желтые пленки и тонкие, ветвистые коричневые шнуры. Максимальная температура, убивающая гриб, равна
80° при действии в течение 20 минут, минимальная — ниже —30°.
М. Н. Родигин указывает на способность пленчатого гриба поражать клубни картофеля в период хранения в подвалах. При этом пораженные клубни покрываются светло-серым паутинистым, а местами довольно плотным налетом. Поскольку пленчатый домовой гриб поражает только кожицу, разрушение мякоти клубня впоследствии идет за счет других микроорганизмов.
Для предупреждения заболевания клубней необходима тщательная очистка и дезинфекция подвалов перед засыпкой картофеля на хранение.
Род Stereum Fries. Сильно распространен в лиственных и еловых древостоях. Плодовые тела у его представителей, хотя и ресупинатны, приподнимаются по краям, верхняя поверхность при этом имеет концентрические круги. У некоторых видов плодовые тела тонкокожистые в виде черепитчато расположенных шляпок с приподнятым краем. Поселяясь на живом дереве, гриб вызывает бурую центральную гниль, начинающуюся с вершины. Распространению гриба способствуют различные повреждения и поражения дерева, как например подсочка.
Род Thelephora Ehernb. Плодовые тела кожистые, ресупи-натные или же приподнимающиеся. В этом случае они снабжены ножкой, причем гимений расположен на наружной (морфологически нижней) поверхности.
Роды Craterellus Pers, и Cantharellus Fries. В пределах семейства телефоровых наибольшего развития плодовое тело достигло у Craterellus (вороночник) и Cantharellus (лисичка). Оно имеет воронковидное строение с гимением, развивающимся на продольных складках нижней поверхности, весьма сходных с гименофором агариковых. Лисички являются почвенными сапрофитами, широко распространенными в лиственных и смешанных лесах. Они вполне съедобны, но по вкусовым качествам относятся к третьей категории грибов.
Сем. Clavariaceae
Большинство представителей семейства рогатиковых являются лесными почвенными сапрофитами. Их плодовые тела обычно мясистые, со всех сторон покрытые гимением.
Род Clavaria Vaillant. Этот род представлен наиболее обширно (300 видов). Его представители имеют плодовые тела, неветвящиеся, булавовидные или коралловидно разветвленные— Cl. cristata Schaff. (рис. 38). Рогатиковые — весьма распространенные представители грибной флоры хвойных и
10 3. Н. Кудряшева 145
смешанных лесов. В молодом возрасте плодовые тела их могут употребляться в пищу.
Род Typhula Pers. У его представителей плодовое тело развивается при прорастании склероциев и имеет форму булавы с утолщением на вершине (рис. 39), где и располагаются базидии. Многие представители этого рода являются
Рис. 38. Clavaria (ориг.).
Рис. 39. Typhula trifolii— проросший склероций.
паразитами сельскохозяйственных растений: злаков, свеклы, клевера. Так, Typhula trifolii Rostr. на клевере особенно сильно развивается на втором году жизни и может образовывать до 1500 склероциев на 1 м2.
Сем. Hydnaceae
Ежевиковые в большинстве случаев — сапрофиты на древесных субстратах или почве, но не исключены и случаи паразитизма. На представителях этого семейства можно проследить эволюцию плодовых тел от ресупинатных до прямостоящих, высоко организованных, дифференцированных на ножку и шляпку с гименофором на нижней стороне. Строение
гименофора своеобразно — он имеет вид бородавок, шипов, зубцов.
Род Mucronella Fries. Включает в себя весьма своеобразные примитивные формы, встречающиеся на гнилой древесине. Плодового тела по существу не образуется, а гименофор развивается прямо на субстрате и имеет вид сравнительно длинных шипов, собранных в пучки.
Р и с. 40. Hudnutn repandutn и его гименофор (ориг.).
Более высоко организованные формы имеют ресупинатные плодовые тела с гименофором в виде бугорков или бородавочек на верхней стороне.
Род Hydnum L. В большинстве случаев его представители имеют вертикально стоящие плодовые тела, дифференцированные на ножку и шляпку. Нижняя поверхность последней несет шипы, ориентированные более или менее вертикально. Некоторые виды съедобны, например Hydnum repandum L. (рис. 40).
Паразитные виды рода Hydnum развиваются на древесных породах — дубе, сосне, яблоне и вызывают разрушение древесины.
Сем. Polyporaceae
Трутовые грибы имеют очень большое научное и практическое значение. Одни из них участвуют в процессе круговорота веществ, другие составляют большую группу разрушителей живой и мертвой древесины. Плодовые тела у трутовых ресупинатные, кожистые, а также более или менее твердые, копытообразные, чаще многолетние, в старости не загнивающие. Гименофор в виде неопределенных складок, а у большинства в виде трубочек, не отрывающихся от бесплодной части плодового тела и расположенных на его нижней стороне.
Всестороннему изучению трутовых грибов — разрушителей древесины— многие годы своей научной деятельности посвятил один из крупнейших советских микологов А. С. Бон-дарцев.
Трутовые грибы обладают способностью быстро разрушать древесину, как живую на корню, так и мертвую. Проникая в древесину, они оказывают более или менее сильное воздействие на стенки ее клеток и являются причиной различного типа гнилей. В мицелии и плодовых телах этих грибов содержится большое количество ферментов, принадлежащих к гидролитическим. Поэтому гидролиз целлюлозы, геммицеллюлозы и лигнина клеточных стенок древесины происходит быстро. Если не проводить никаких профилактических мероприятий, гибель дерева, а также разрушение мертвой древесины может наступить в сравнительно короткий промежуток времени.
Плодовые тела трутовых грибов, разрушающих древесину, могут быть то распростертыми по субстрату (Serpula lacry-mans), то с отвороченными в виде шляпки краями (Fomi-iopsis annosa), то копытообразными, боковой стороной приросшими к субстрату (Fames fomenlarius).
Плодовое тело обычно развивается на вегетативном мицелии, но только тогда, когда гниение древесины достигло определенной стадии.
Наблюдения показывают, что плодовые тела у трутовых грибов образуются на деревьях там, где впервые произошло заражение. Дерево инфицируется через поврежденные места (морозобоины, ожоги, повреждение насекомыми). Развитие мицелия после заражения идет обычно снизу вверх, но иногда и вниз по ядровой части дерева. Окраска плодовых тел весьма разнообразна — от белой, желтой, оранжевой до красной, коричневой, почти черной. По консистенции они могут быть рыхлые, почти ватообразные, кожистые, хрящеватые, мясистые и даже деревянистые. Однолетние и двулетние плодовые тела обычно более мягкой консистенции, а многолетние — деревянистые с наличием концентрической бороздча-тости на верхней стороне. У многолетних плодовых тел трубчатый гименофор нарастает каждый год заново в виде одного слоя трубочек. По числу слоев, заметных на разрезе многолетнего плодового тела, можно более или менее точно определить его возраст.
Род Serpula Karst. Домовой гриб (Serpula lacrymans Schum.), разрушающий деловую древесину, по вредоносности занимает первое место. Как и большая часть домовых грибов S. lacrymans встречается на древесине построек, в сырых, плохо проветриваемых помещениях. Домовой гриб может успешно развиваться при влажности древесины от 22 до 60%.
Вызываемое им гниение древесины сопровождается выделением из нее воды, а поэтому внедрившийся в древесину гриб может развиваться даже при отсутствии влажности.
При благоприятных условиях роста грибница достигает больших размеров и имеет вид пушистых ватообразных скоплений, выделяющих капли водянистой жидкости. За это грибу дано видовое название lacrymans— слезоточивый.
Рис. 41. Ser pula lacrymans — мицелиальный гяж.
Характерной особенностью домового гриба является образование мицелиальных шнуров или тяжей. Своеобразное строение последних заключается в том, что гифы их дифференцируются по выполняемой функции, соответственно чему изменяется и их строение. Так, у S. lacrymans имеется три разновидности гиф: одни из них очень нежные и тонкие (гифы основной ткани), другие с более толстыми стенками и узким просветом (механические гифы) и, наконец, третьи — большого диаметра (до 50 мк) с очень широким просветом, неравномерно утолщенными внутренними стенками и отчасти разрушенными поперечными перегородками (сосудовидныегифы) (рис. 41). Шнуры S. lacrymans могут достигать толщины карандаша и нескольких метров длины, благодаря чему гриб может перекидываться на значительное расстояние по субстрату, не способному питать грибницу (например, из одного этажа на другой по кирпичной стене). При встрече вновь
подходящих условий от тяжа образуются тонкие веточки, развивающиеся в пышную грибницу.
Плодовое тело распростертое, мягкое (рис. 42). Гименофор на верхней поверхности охристо-желтый или коричневый, образует извилистые складки. Споры эллиптические, желтовато-коричневые.
S. lacrymans является наиболее опасным домовым грибом. Действие его на древесину относится к типу деструктивного разрушения. Древесина темнеет, постепенно растрескивается
и с. 42. Ser pula lacrymans — плодовое тело.
продольными и поперечными щелями на кубические участки. Поскольку гриб разрушает целлюлозу и лигниновые вещества 43 клеточных стенок, то древесина, потерявшая прочность, застирается между пальцами в порошок. Древесина заражается спорами, которые образуются в громадном количестве <а плодовых телах, легко переносятся воздухом и животными. Возможно инфицирование грибницей при соприкосновении ее со здоровой древесиной. Легче всего домовой гриб разрушает древесину хвойных пород. Древесина некоторых лиственных, 1апример дуба, поражается слабо, так как гриб весьма чувствителен к таннинам, находящимся в древесине дуба.
Меры борьбы с домовыми грибами:
1. Для построек рекомендуется применять древесину тща-ельно высушенную. Опыты показали, что домовой гриб не ложет развиваться на древесине, имеющей влажность ниже 8%.
2. Обеспечивать нормальную вентиляцию в подпольях.
3. Деревянные части здания должны быть предохранены >т попадания влажности из фундамента, штукатурки. А все асти деревянных конструкций, например концы балок, должны быть пропитаны специальными антисептиками.
4. Рекомендуется применение следующих антисептиков: пористого натрия в виде 3%-ного раствора и кремнефтори-того натрия в виде 2—2,5%-ного водного раствора; камен
ноугольного креозотового масла, торфяного креозота, березового дегтя (каждый из них в чистом виде).
Из трутовых грибов, развивающихся на живых деревьях, следует указать корневую губку, ложный трутовик, березовую губку, серно-желтый трутовик.
Род Fomes Gillet. Корневая губка (Fomes annosus (Fr.) Cke. syn. Fomitopsis annosa (Fr.) Bond et Sinq.) приносит большой вред хвойным деревьям: ели, сосне, лиственнице, пихте, кедру (рис. 43). Она растет на пнях и корнях,
Р и с. 43. Femes annosus.
Р и с. 44. Fomes igniarius.
выступающих из почвы. Дерево инфицируется в результате попадания спор на поверхность ранок на коре у основания ствола, а также грибницей при соприкосновении больных корней со здоровыми. Способность гриба распространяться через корни приводит к образованию целых очагов больных деревьев. Мицелий постепенно проникает в ствол дерева и в конечном итоге может привести к образованию дупла. У сосны и кедра гниль ограничивается только комлевой частью ствола и выше 2 м обычно не поднимается, а у ели и пихты, как пород менее богатых смолою, она поднимается значительно выше — до 6—8 м, а в некоторых случаях до 15—20 м. Гриб повреждает как 2—5-летние, так и старые деревья. Деревья, ослабленные болезнью, подвергаются ветровалу и бурелому, что приводит к изреженности древостоя.
Плодовые тела гриба образуются на нижней стороне корней и в пустотах между ними. Гриб избегает света, и на
верхней поверхности корней плодовые тела образуются только в густых древостоях.
Плодовые тела имеют очень разнообразную и часто неправильную форму от 5 до 15 см и до 3,5 см толщины, распростерто-отогнутые, часто срастающиеся по нескольку. Вследствие того что плодовые тела F. annosus развиваются на корнях или у основания ствола, т. е. близко от поверхности почвы, ветер не играет существенной роли в распространении спор. Некоторые исследователи высказывают предположения, что основными переносчиками спор являются мыши, кроты и другие роющие животные.
Ложный трутовик (Fomes igniarius (L). Gill. syn. Phellinus igniarius (L. ex Fr.) Quel) встречается во всех странах света, широко распространен и в СССР не только в лесах, но в парках и садах (рис. 44). Он развивается в течение всего года на живых деревьях лиственных пород: березе, осине, ольхе, иве и других, а также на отмерших пнях. Гриб является опасным паразитом, вызывающим белую или слегка желтоватую гниль, наблюдаемую на деревьях разного возраста. Загнивание распространяется, как и при всякой внутренней гнили, от внутренних слоев древесины к наружным. На отломах пораженной древесины можно наблюдать скопления рыжеватой грибницы. В конечной стадии гниения нередко образуется дупло.
Плодовые тела этого гриба многолетние, достигают иногда 30—35-летнего возраста. Их форма довольно изменчива. Чаще встречаются копытообразные плодовые тела, реже — ресупинатные. Верхняя поверхность с концентрическими бороздками, большей частью темная, иногда с поперечными трещинками.
А. С. Бондарцев установил несколько форм у этого гриба: f. tremulae* f. betulae, f. ain't и др. На березе часто развивается стерильная, без спор, форма, известная под названием «чага», применяющаяся издавна в народной медицине как средство против рака. Последнее время вводится в научную медицину.
Примером трутовиков с однолетними плодовыми телами служит большой род Polyporus, причем форма плодового тела чрезвычайно разнообразна.
Род Polyporus Fries. Березовая губка Polyporus betulinus (Bull) Fr. имеет подушковидное или почковидное плодовое тело.
У серно-желтого трутовика (Polyporus sulfurets (Bull) Fr.) плодовые тела обычно собраны в группы. Шляпки их со слегка загнутыми краями постепенно суживаются к месту прикрепления их к стволу дерева. Верхняя поверхность шляпок светло-желтая или оранжевая. Мягкие
в начале развития плодовые тела впоследствии отвердевают. Род Polyporus развивается на стволах живых деревьев лиственных и реже хвойных пород. Грибница проникает через ранку в древесину и вызывает центральную гниль. Со временем древесина приобретает бурую окраску.
Как было отмечено выше, трутовые грибы, развиваясь на живых деревьях, срубленных и 'заготовленных лесоматериалах, дровах, в деревянных постройках и вызывая гниение древесины, причиняют колоссальный вред. Однако многие трутовики в определенный период своего развития приносят и известную пользу. Так, в молодом возрасте плодовые тела некоторых трутовиков, как Р. squamosus Huds ex Fr. и другие, могут употребляться в пищу. Еще в недавнее время в отдаленных северных районах из плодовых тел трутовых грибов из рода Fomes приготовлялся трут для добывания огня. А из мягкой массы плодового тела трутовиков в ряде стран изготовляют перчатки, сумочки, рамки. Плодовые тела некоторых трутовиков содержат красящие вещества и в настоящее время находят применение в Средней Азии в качестве красителей тканей.
Одним из полезных свойств трутовых грибов является их способность давать декоративную древесину. Это объясняется тем, что некоторые трутовые грибы на начальных стадиях своего роста, еще почти не нарушая прочности древесины, отлагают в ней различные пигменты, что приводит к появлению цветных пятен, полос, а иногда и характерного муарового рисунка. Подобная древесина после полировки становится особенно красивой, поэтому она находит применение в строительстве при различных столярных работах. Очень ценной является древесина ореха из Кахетии и Гурии, древесина ясеня из западной Грузии. Древесина клена, пораженная в начальной стадии одним из видов трутовых грибов, употребляется для изготовления музыкальных струнных инструментов (балалайки, гитары).
В процессе минерализации древесных остатков большую положительную роль играют сапрофитные трутовые грибы. Они являются своеобразными санитарами леса.
Однако отрицательное действие паразитных трутовых грибов настолько велико, что для предотвращения громадных потерь в лесном хозяйстве необходимо изучать местную флору трутовых грибов. Это позволит установить видовой состав и географическое распространение отдельных видов, а также разработать необходимую систему мероприятий для успешной борьбы с трутовыми грибами.
Большая заслуга по изучению флоры трутовых грибов в БССР принадлежит Э. П. Комаровой. При обследовании лесничеств Минской, Брестской, Гомельской, Могилевской
и Витебской областей обнаружены 109 видов трутовых грибов. Причем 19 видов ранее не были известны для Белоруссии, а трутовый гриб Abortiporus fractipes (Berk et Curt) Bond, до сего времени не был известен на территории СССР и Европы. Этот вид найден в Брестской и Минской областях. Кроме того, Э. П. Комарова обнаружила и описала три овые для науки формы гриба рода Tyromyces.
В борьбе с трутовыми грибами рекомендуется:
1. Выращивание смешанных насаждений. Это значительно снижает распространение трутовых грибов, способных поражать одну или близкие породы.
2. Вырубка и удаление из леса зараженных деревьев. Причем для предохранения заражения поросли через пень следует срезать деревья возможно ниже.
3. Охрана стволов и корней деревьев от механических повреждений.
ПОРЯДОК AGARICALES
Плодовые тела агариковых грибов, дифференцированные ia ножку и шляпку, обычно мягкомясистые, у большинства
Рис. 45. Гименофор Agaricales-. с — трубчатый (поперечный разрез); б — пластинчатый (тангентальный разрез).
старости загнивающие. Гименофор расположен на нижней тороне шляпки в виде трубочек или пластинок (рис. 45).
У многих видов агариковых (масленок, шампиньон, осеней опенок) гименофор вначале прикрыт особой пленкой
из рыхлого сплетения гиф — частным покрывалом. Позднее оно разрывается и остается в виде кольца на ножке.
У других, кроме частного покрывала, развивается еще общее, или вольва, одевающее в молодости все плодовое тело. При разрастании плодового тела оно также разрывается, нижняя часть его остается вокруг ножки в виде чашечки, а верхняя в виде лоскутков на шляпке (мухомор). Порядок агариковых делят на два семейства: Boletaceae, Agaricaceae.
Сем. Boletaceae
Плодовые тела у болетусовых мясистые и скоро загнивающие. Раньше эти грибы относили к трутовым, в настоящее время их выделяют в самостоятельное семейство порядка ага-риковых.
От трутовых они отличаются формой плодового тела и особенностью трубчатого гименофора. Трубочки легко отделимы от мякоти шляпки, чего нет у трутовых. С пластинчатыми их связывают переходные формы, имеющие гименофор в виде радиально расположенных, сетчатоальвеолярных складок (Phy Порог us). Наиболее крупным родом в семействе является род Boletus, включающий много съедобных грибов.
Род Boletus Pat. Среди грибов этого рода первое место принадлежит боровику — Boletus edulus Bull (Fr.). По высоким вкусовым качествам он не имеет себе равных. Шляпка в молодости полушаровидная, трубчатый гименофор белый. С возрастом шляпка становится более плоской, а цвет гименофора изменяется до желтого и желто-зеленого. Окраска поверхности шляпки по мере старения плодового тела почти не изменяется, но может варьировать от светло-буроватой (почти белой) до темно-бурой и даже фиолетово-бурой в зависимости от условий произрастания. Боровик растет в различных лесах — хвойных и лиственных.
К семейству Boletaceae относится подосиновик (Кгот-bholzia aurantiaca (Roques) Gilb.), подберезовик (Kr. scabra (Fr.) Karst.), масленок поздний (Ixocotnus lutens (Fr.) Quel.). Они, хотя и уступают по вкусовым качествам боровику, являются также очень ценными грибами.
Среди многочисленных представителей Boletaceae есть несъедобные грибы.
Желчный гриб (Tylopilus felleus (Fr.) Karst.) внешне очень похож на боровик. Основным отличием его является розоватый оттенок гименофора и слабое порозовение мякоти на изломе. Встречается желчный гриб в тех же местах, где и белый. Он обладает горьким вкусом и должен быть отнесен к несъедобным грибам.
По многим указаниям ядовитым считается сатанинский гриб (Boletus satanas Lentz.), способный вызывать головокружение, рвоту. Однако в ряде местностей его употребляют в пищу.
Сем. Agaricaceae
Семейство пластинчатых грибов следует рассматривать как высшее среди гименомицетов. По количеству видов (более 7000) оно также превосходит болетусовые. Большинство его представителей является почвенными сапрофитами, а у некоторых (опенок) образ жизни связан с древесным субстратом.
Плодовые тела у пластинчатых грибов достигают наиболее высокой степени организации: состоят из центральной ножки и шляпки, почти всегда мясистые, однолетние. Гименофор на нижней стороне шляпки имеет вид пластинок, радиально расходящихся от центра к ее краю. У большинства представителей в гимениальном слое отмершие старые базидии заменяются новыми, которые закладываются в субгимениальном слое, а затем выдвигаются до общего уровня гимения. У немногих видов в гимении развивается только одно поколение базидий.
Плодовое тело у пластинчатых грибов закладывается в виде округлого сплетения гиф, в котором затем начинает дифференцироваться шляпка и ножка. На ранних стадиях развития уже намечается гименофор. Он может закладываться открыто по перетяжке, отделяющей шляпку от ножки. В дальнейшем, когда шляпка разрастается и края ее несколько заворачиваются вниз, гименофор оказывается в кольцевой щели, образовавшейся между шляпкой и ножкой. Полное обнажение гименофора происходит при созревании шляпки.
У большинства пластинчатых щель, в которой закладывается гименофор, бывает прикрыта частным покрывалом. Наличие общего покрывала также характерно для многих представителей этого семейства. В семействе Agaricaceae следует отметить роды Amanita, Armillaria, Hypholoma, Lada-rius, Coprinus.
Род Amanita Fries. Кесарев или ц e з a p с к и й гриб (Amanita caesarea (Scop) Fr.) за отличные вкусовые качества высоко ценился еще в древнем Риме. Растет в дубовых лесах, в южных широтах.
Бледная поганка (A. phalliodes (Vaill.) Fr.) (табл. Ill) — самый ядовитый гриб. В Белоруссии встречается в смешанных лесах. В молодости плодовое тело заключено в общее покрывало, которое впоследствии остается в виде мешковидного влагалища у основания ножки и хлопьевидных лоскутков на поверхности шляпки. Частное покрывало при росте шляпки открывается и повисает в виде кольца на ножке. Токсические свойства бледной поганки настолько сильны, что
достаточно одному плодовому телу попасть среди партии хороших грибов, чтобы вызвать тяжелое отравление при употреблении в пищу. Оно сопровождается сильнейшими страданиями и заканчивается смертельно. Ядовитым веществом бледной поганки является токсин фаллин.
Чтобы отравить кошку, достаточно 4 мг этого вещества, 25 мг смертельно действует на собаку, а для отравления человека требуется всего лишь 30 мг. Первые признаки отравления бледной поганкой обнаруживаются через 10—12 часов после принятия пищи, т. е. тогда, когда практически невозможно оказать помощь пострадавшему, так как пища попала в тонкий кишечник и токсин уже проник в кровь. Симптомы отравления: головная боль, головокружение, нарушение нормального зрения, а затем сильнейшие боли в желудке и печени, особенно при надавливании; судороги в конечностях, при этом пульс резко падает. Через некоторое время состояние больного улучшается, а затем приступы возобновляются, наступает потеря сознания. Индивидуальные свойства больного и количество усвоенного токсина сказываются на характере течения болезни, но так или иначе выздоровление почти никогда не наступает. От действия фаллина у больного наступает разрушение тканей печени, почек, селезенки, сердечных мышц, кровоизлияние в гемолиз, т. е. разложение красных кровяных телец. По данным некоторых авторов, фаллин не теряет свою активность при варке, при температуре 100° и не растворяется в воде, сохраняясь в грибных тканях.
Род Armillaria Fries. Опенок (Armillaria mellia (Vail.) Quel.) (табл. IV, а) растет на пнях и живых деревьях, принося им существенный вред. Мицелий гриба дает ризоморфы, благодаря которым он и распространяется от дерева к дереву. Шляпка обычно сухая, чешуйчатая, серовато-буроватая. У молодых шляпок гименофор закрыт частным покрывалом в виде рыхлого белого сплетения гиф. Ножка снабжена кольцом — остатком частного покрывала. Опенок съедобен в свежем, соленом и маринованном виде, но относится к грибам третьей категории.
Род Hypholoma Fries. Ложноопенок сер но-ж ел-т ы й (Hypholoma fasciculare (Huds.) Quel.) (табл. IV, б) часто путают с настоящим опенком. Шляпка его более ярко окрашенная, желтоватая, а в центре кирпично-красная, гладкая. Пластинки сначала серо-желтые, потом зеленовато-оливковые. Ножка его ровная или суженная к основанию. Гриб обладает горьким вкусом и считается ядовитым. Он растет всюду на пнях и около них, а иногда и на живых деревьях.
Род Lactarius Fries. Рыжик (Lactarius deliciosus Fr.). Шляпка гомогенная с ножкой и от нее неотделима. В плодовом теле отсутствует частное и общее покрывало. Особен
ностью рыжика является наличие хорошо развитых млечных трубок, в которых содержатся запасные питательные вещества, имеющие вид тонкой эмульсии от присутствия мелких капель распыленной смолы и жира, окрашенных пигментами. Млечный сок рыжика оранжево-желтый, сладковатый, немного острый, со смолистым запахом, на воздухе зеленеющий.
Рыжик растет в различных хвойных лесах. Различают две формы: первая оранжево-желтая, ножка более короткая — обитает в сосновых борах; вторая приурочена к еловым насаждениям, имеет зеленовато-желтую шляпку более конической формы. Рыжик считается первосортным съедобным грибом, употребляется в соленом, маринованном и свежем виде.
Род Coprinus Pers. Коп р и н у с (рис. 46) включает более 200 видов, относится к экологической группе копрофильных,.
Рис. 46. Coprinus.
или навозных, грибов. Виды его селятся на навозе травоядных, хорошо удобренных почвах, а также на пнях и на гниющих растительных остатках. Плодовое тело белое или сероватое, состоит из центральной тонкой полой ножки и колоколовидной шляпки. Гименофор представлен тонкими пластинками, покрытыми чрезвычайно правильно построенным гимением, образующим только одно поколение базидий, в
связи с чем гименофор и плодовое тело в целом у некоторых видов отличается весьма эфемерным существованием.
Созревание базидий начинается по нижнему краю шляпки и продвигается постепенно кверху. После созревания базидиоспор гимений, несущая его часть пластинки и прилегающая часть шляпки подвергаются автолизу, превращаясь в быстро чернеющую жидкость, стекающую каплями вниз. Это приводит к постепенному укорачиванию шляпки.
Капельки автолизированного гименофора раньше добавлялись к чернилам и туши, употреблявшимся для подписей особо важных государственных бумаг. Наличие или отсутствие прилипших с чернилами спор гриба давало возможность обнаруживать подделку. За эту особенность гриб получил название чернильного.
Шляпочные грибы порядка Agaricales широко распространены и известны в качестве продуктов питания с незапамятных времен. На территории Советского Союза они встречаются повсюду, от самых крайних пределов растительности на севере до субтропических лесов Закавказья. По их количеству и разнообразию видов особенно выделяется лесная зона, дающая обильные урожаи первосортных съедобных грибов.
Изучением особенностей строения плодовых тел шляпочных грибов и их графическим распространением в Советском Союзе много занимались Л. А. Лебедева, Б. П. Васильков и др. Большое значение для изучения географического распространения различных видов Agaricales имеют флористические сборы и описания грибов в отдельных республиках и районах СССР.
В Белоруссии такая работа проделана Г. И. Сержаниной. Ею собрано в разных районах БССР 384 вида грибов, из них 192 обнаружено впервые в республике, а 17 являются новыми и для СССР. Из собранных в Белоруссии грибов порядка Agaricales 202 вида съедобных и 38 ядовитых. Остальные виды еще недостаточно изучены в пищевом отношении.
ГРУППА ПОРЯДКОВ GASTROMYCETES
Особенностью гастромицетов является образование совершенно замкнутых ангиокарпных плодовых тел. Базидии закладываются внутри плодового тела, и там же происходит отделение базидиоспор, а освобождение их совершается позднее вследствие разрыва или общего разрушения оболочки плодового тела.
По образу жизни гастромицеты почти исключительно почвенные сапрофиты. Количество их видов составляет около 1000. Практическое значение гастромицетов невелико: плодовые тела лишь некоторых видов в молодости съедобны.
Однако среди представителей гастромицетов известны виды с весьма своеобразной и наиболее сложной формой плодового тела. Мицелий гастромицетов в почве может образовывать довольно мощные мицелиальные тяжи. На них подземно закладываются плодовые тела, которые затем выдвигаются на поверхность.
Форма плодового тела в большинстве случаев округлая, снаружи имеет покров, называемый перидием и состоящий из бесплодных гиф, часто окрашенных.
Внутренняя часть плодового тела — глеба — в молодости представляет бесцветную рыхлую ткань, в которой затем образуются различной формы полости, называемые камерами. Из окружающей ткани в них вдаются базидии, на которых образуются базидиоспоры большей частью темноокрашенные. Окраска базидиоспор сообщает цвет глебе при созревании плодового тела. Между камерами остаются прослойки ткани, или трамы, превращающиеся к моменту созревания плодового тела в волокнистое образование — к а п ил л и ц ий. Нити капиллиция обладают гигроскопическими свойствами, а поэтому при разрывании плодового тела способствуют рассеиванию спор.
У других видов трамы сохраняются не измененными, поэтому внутреннее содержимое плодового тела остается плотным и часто имеет мраморный рисунок из-за чередующихся темноокрашенных от базидиоспор камер и бесцветных прослоек трам.
Базидии гастромицетов несколько отличаются своей формой (часто расширены) и расположением базидиоспор: они могут быть почти сидячими или на очень длинных стеригмах. Базидиоспоры во многих случаях располагаются на разной высоте базидии и ориентированы не вверх, а вбок.
Разные авторы делят гастромицеты на различное количество порядков и семейств. Самые интересные семейства Ly-coperdaceae, наиболее обширное, и Phallaceae с плодовыми телами, достигающими наивысшей степени организации.
Сем. Lycoperdaceae
Большинство грибов, принадлежащих к этому семейству, имеют надземные плодовые тела. В зрелом состоянии их содержимое представлено рыхлой бурой массой, образованной нитями капиллиция и массой спор между ними.
На лесных опушках и лугах часто встречаются грибы, называемые дождевиками. Среди них наиболее распространены два рода: Bovisia и Lycoperdon.
Роды Bovista Pers, и Lycoperdon Tournefort. Они весьма близки друг к другу, но отличаются тем, что в плодовом теле
Таблица IV. Armillaria mellea (a); Hypholoma fasciculare (6).
первого вся глеба плодущая, а у второго нижняя часть его стерильная и несколько вытягивается, образуя зачаточную ножку (рис. 47).
Некоторые виды дождевиков обладают крупными плодовыми телами. Так, например, L. giganteum Batsch. имеет пло
Рис. 47. Bovista (a); Lycoperdon (б).
довое тело величиной с большой арбуз. Продукция спор его исчисляется примерно в 7,5 триллиона. Раньше этот гриб считали самым плодовитым, однако в Северной Америке был обнаружен дождевик (Calvaiia maxima) еще большего размера. Его плодовое тело имеет форму лепешки длиною 1,6 м, шириной 1,35 м и высотой в 24 см. Подсчитано, что такое плодовое тело может дать около 160 триллионов спор.
Сем. Phallaceae
Наиболее сложное строение плодовых тел известно у представителей семейства Phallaceae, Залагаются плодовые тела подземно на мицелиальных тяжах, как и у многих других гастромицетов. В дальнейшем они выступают на поверхности почвы в виде крупных яйцевидных тел, одетых плотной белой оболочкой — вольвой, под которой находится глеба. Последняя состоит из стерильной и плодущей части. При созревании плодового тела вольва разрывается, стерильная часть глебы (рецептакулум) сильно вытягивается, вынося кверху плодущую, которая принимает вид колокола, очень часто ярко окрашенного, а при полном созревании ослизняющегося.
Род Phallus Pers. Веселка обыкновенная Phallus impudicus Fries, (рис. 48) довольно распространена в широко-
лиственных лесах. Плодовое тело этого гриба имеет особый плодоносен, состоящий из длинной губчатой ножки (стерильная глеба) с ячеистой шляпкой на конце, покрытой буро-зеленой слизью и издающей запах падали. Споры находятся в этой слизи и переносятся мухами.
Рис. 48. Phallus impudicus.
Рис. 49. Dictyophora phalloides.
Род Dictyophora Е. Fischer. Близкий к веселке тропический вид Dictyophora phalloides (рис. 49) имеет еще более интересное и изящное плодовое тело. Оно снабжено особым покрывалом — индузием, которое в виде опрокинутой сетчатой воронки прикреплено к вершине ножки под глебой.
За яркую окраску и красивую форму плодовых тел представители семейства Phallaceae получили название грибов-цветков (Pilzblumen).
ПОДКЛАСС PHRAGMOBASIDIOMYCETES
Подкласс фрагмобазидиальных грибов характеризуется наличием клеточной базидии и включает четыре порядка: 1) Aurlculariales\ 2) Tremellales; 3) Ustilaginales; 4) Uredi-nales.
Представители первых двух порядков (Auriculariales и Tremellales) развиваются главным образом на древесных сучьях, пнях и других подобных субстратах и характеризуются наличием примитивных плодовых тел обычно студенистой консистенции. Базидии располагаются по поверхности плодового тела под общим покровом слизи довольно правильным слоем, который можно сравнить с гимением. При высыхании такие плодовые тела превращаются в мало заметные корочки.
Практическое значение обоих семейств невелико. Развиваясь в большинстве случаев на мертвом древесном субстрате, они вызывают разрушение древесины.
ПОРЯДОК USTILAGINALES
В настоящее время известно около 900 видов головневых грибов — паразитов высших растений, вызывающих заболевание их, известное под названием головни.
Организация грибов, входящих в эту группу, проста и представлена мицелием, спорами (хламидоспорами), базидиями, базидиоспорами и конидиями. Других образований, в том числе и плодовых тел, в цикле развития не наблюдается.
Простота строения головневых грибов и отсутствие у них плодовых тел тесно связаны с паразитическим образом жизни.
Мицелий головневых грибов представлен, как у всех базидиальных, двумя видами: гаплоидным (первичным), весьма слабо развитым и неспособным заражать растение, и дика-риотическим (вторичным) — паразитирующим. Развивается он чаще по межклетникам, изредка внутриклетно и обладает, как правило, гаусториями. В зависимости от особенности того или иного вида головневого гриба мицелий может пронизывать различные органы растения: корни, стебли, листья, соцветия. Гифы гриба, развиваясь в пораженных органах, образуют плотные сплетения, а затем в их клетках под старой оболочкой образуется новая, более плотная и темная. Старые оболочки постепенно разрушаются, и гриб дает массу округлых двухъядерных головневых спор, называемых хламидоспорами. Их оболочка может быть гладкой, сетчатой, щетинистой или бородавчатой. Масса головневых спор, образующая
ся из мицелия гриба, представляет более или менее распыляющийся черный, фиолетовый или красноватый порошок, внутри пораженных тканей питающего растения. Очень часто появле-
Рис. 50. Головня:
a — U^tllago б—Tiltetia triiiri\ в — Urocystes occulia\ & — Ustilago
panicd — Ustilago zcae.
нию спор предшествует изменение внешнего вида пораженного органа: гипертрофия, искривление, скручивание и т. п. Все эти признаки настолько характерны, что головневые грибы легко распознаются (рис. 50).
У некоторых головневых грибов образуются не одиночные споры, а спорокучки, состоящие из большего или меньшего количества спор, не разъединяющихся при созревании. У одних родов спорокучки образованы одинаковыми спорами, у других же в состав спорокучки входят клетки двух видов: одни заполнены живым содержимым и способны прорастать; другие лишены содержимого, наполнены воздухом и способствуют переносу спорокучки воздушными течениями.
Рис. 51. Прорастание хламидоспор:
а — у возбудителя пыльной головни пшеницы; б —твердой головни пшеницы; в — пузырчатой головни кукурузы; г — стеблевой головни ржи.
Размеры хламидоспор головневых грибов микроскопические— от 4 до 30 мк. Споровая масса твердой головни, заключенная в оболочке зерновки пшеницы, состоит из 8—10 млн. спор, а в желваке пузырчатой головни кукурузы их содержатся миллиарды.
У одних видов хламидоспоры могут прорастать сразу же после созревания (пыльная головня пшеницы), у других они требуют некоторого периода покоя (карликовая головня пшеницы) и могут долго сохраняться жизнеспособными (2—3 года, в отдельных случаях — 7—8 лет). Споры, заключенные в оболочку зерна или желвака, дольше сохраняют способность к прорастанию, чем свободные. Споры, находящиеся на поверхности почвы, обычно прорастают через 1—2 месяца в зависимости от внешних условий.
Исследуя жизнеспособность головневых спор по гербарным образцам, немецкий ученый Фишер в 1936 году получил очень интересные данные. Споры твердой головни пшеницы Tilleiia levis сохраняли жизнеспособность в течение 25 лет, споры твердой головни ячменя Ustilago hordei — 23 года.
При прорастании хламидоспоры образуется базидия (рис. 51). У одних видов базидия снабжена поперечными пе
регородками и является типичной фрагмобазидией обычно четырехклетной. На каждой клетке базидии развивается по одной базидиоспоре. Однако есть виды головневых грибов, не образующие базидиоспор на фрагмобазидии,— это пыльная головня пшеницы (U. tritici) и пыльная головня ячменя (U. nuda).
У других головневых базидия представлена одной клеткой. В этом случае на ее вершине развивается 4—8 базидиоспор. Базидиоспоры некоторых головневых грибов обладают способностью почковаться, находясь еще на базидии. Отпочковывающиеся клетки являются конидиями и, почкуясь, в свою очередь придают базидии вид разветвленного деревца (базидия возбудителя пузырчатой головни кукурузы).
Многие виды головневых грибов — гетероталличные организмы. Раздельнополость проявляется уже у отдельных клеток фрагмобазидии, а образующиеся на них базидиоспоры также несут признаки того или иного пола.
В цикле развития головневых грибов наблюдается чередование трех стадий, или фаз: гаплоидной, дикариотической и диплоидной.
Гаплоидная стадия представляет базидиоспоры и первичный мицелий. Поскольку первичный мицелий, как правило, не способен заражать растение, эта стадия кратковременна.
Дикариотическая стадия включает вторичный мицелий, клетки которого содержат по два ядра, а также молодые хламидоспоры. Возникновение дикариотической стадии у головневых грибов происходит после весьма примитивного полового процесса, заключающегося в копуляции двух базидиоспор разного знака или их отпочковавшегося потомства. У таких видов как пыльная головня пшеницы и ячменя, не образующих базидиоспор, копулируют две соседние клетки фрагмобазидии. В обоих случаях содержимое одной клетки переходит в другую. Опустевшая клетка вскоре отмирает, а другая, имеющая два ядра, прорастает в дикариотический мицелий, способный заражать растение. Дикариотическая стадия наиболее продолжительна в цикле развития головневого гриба.
Диплоидная стадия весьма короткая. Она наступает после процесса кариогамии, происходящего в созревающей хламидоспоре. Таким образом у головневых диплоидны только зрелые хламидоспоры.
Прорастание спор головневых грибов сопровождается процессом редукционного деления ядра. Каждая клетка базидии получает по одному гаплоидному ядру, которые в дальнейшем переходят в базидиоспоры. У головневых, имеющих одноклетную базидию (твердая головня пшеницы), диплоидное ядро также делится сначала редукционно, а затем простым делением. В результате этого в базидии образуется 4—8 гаплоид
ных ядер, которые и переходят в базидиоспоры. Количество их равно количеству ядер.
Возникновение дикариотической стадии несколько иначе происходит у пузырчатой головни кукурузы. Копуляции между базидиоспорами или продуктами их почкования не наблюдается. Заражение растения может произойти только тогда, когда в тканях его встречаются два гаплоидных мицелия разного полового знака, клетки которых и копулируют. Для совершения этого процесса необходимо, чтобы базидиоспоры или отпочковавшиеся от них клетки воздушными течениями были перенесены на растение. При благоприятных условиях возможно их прорастание и проникновение первичного мицелия в ткани растения.
Биологическая особенность гриба давать почкующиеся базидиоспоры значительно увеличивает выход споровой продукции и обеспечивает его дальнейшее развитие, т. е. возможность встречи двух первичных мицелиев в тканях растения и копуляцию их клеток.
Головневые грибы встречаются главным образом на травянистых растениях, но иногда развиваются и на древесных породах, как например головня, паразитирующая на стволах эвкалипта.
Головневые грибы прошли большой путь эволюции, и их паразитизм достиг высшей степени совершенства. Зараженное растение ими не убивается, а долгое время вегетирует, и только к концу его жизненного цикла обнаруживается поражение заболеванием. Паразитизм головневых характеризуется весьма высокой степенью приспособленности. Она выражается в том, что один ботанический вид гриба состоит из нескольких рас, морфологически не отличимых между собой, но способных поражать только определенные сорта культурного растения.
Различные способы инфекции растений и характер воздействия гриба на питающее растение также свидетельствует об исключительной приспособленности паразита.
Способы заражения питающих растений головневыми грибами могут быть различными в зависимости от вида паразита. Установлены следующие три основных типа заражения.
Первый тип. Пыльная головня пшеницы Ustila-go tritici Jens, и пыльная головня ячменя Ustilago nuda Jens.
Распыление головневых спор происходит в фазу цветения растений, так как к этому периоду пораженные растения имеют совершенно зрелые споры. Хламидоспора при помощи ветра заносится на рыльце пестика и здесь прорастает в четырехклетную базидию, не образующую базидиоспор. После копуляции соседних клеток фрагмобазидии начинает разви
ваться дикариотический мицелий. Его гифы растут по пути, проложенному пыльцевыми трубками, и, достигая завязи внедряются в семяпочку. Последняя не погибает, а развивается в семя, зародыш которого (под почечкой и в щитке) содержит гифы гриба.
Мицелий в зерне находится в покоящемся состоянии до тех пор, пока зерно не начинает прорастать. Присутствие гриба не мешает нормальному прорастанию семян. Одновременно начинает расти и мицелий гриба, проникает в конус нарастания, разрастаясь особенно сильно в тканях, содержащих наибольшее количество питательного материала,— в колосе. В конце концов происходит разрушение тканей колоса, а мицелий гриба распадается на большое количество спор.
На первых этапах развития пораженное растение ничем не отличается от здорового, но во взрослом состоянии оно несколько ниже и обильнее кустится.
Присутствие гриба в тканях растения повышает его физиологические функции. По данным А. Л. Курсанова, больные растения в начале колошения испаряют воды на 20—25% больше, чем здоровые. А энергия дыхания у больных растений в любой момент вегетации обычно на 20—30% выше, чем у здоровых.
На основании данных о биологии этих видов головневых грибов разработаны и соответствующие мероприятия по борьбе с ними. В данном случае, когда возбудитель болезни находится не на поверхности зерна, а внутри его в виде покоящегося мицелия, обработка семян химическими препаратами не эффективна. В настоящее время находит широкое применение термический метод обеззараживания посевного материала путем прогревания зерна в горячей воде. Термическое обеззараживание производится двумя способами: двухфазным и однофазным. Двухфазный заключается в том, что сначала семена предварительно увлажняются в воде при температуре 28—32° в течение 3—5 часов, причем нельзя допускать наклевывания семян (1-я фаза). Цель этой фазы вызвать набухание мицелия гриба. 2-я фаза—активная обработка семян в горячей воде при температуре 52° в течение 8 минут, а при 53° — в течение 7 минут. Во время второй фазы набухший мицелий погибает. После активного прогревания производится воздушное охлаждение семян. Семена рассыпают тонким слоем (5—8 см) и часто перелопачивают. При однофазном способе семена замачивают в воде, подогретой до 45°, в течение 3—4 часов, а при температуре воды 47° время замачивания сокращается до 2 часов. По истечении установленного срока семена вынимают и просушивают.
По мнению К. Я- Калашникова, эффект термического обеззараживания заключается в том, что в тканях семени, размяг-
ценных намачиванием, под воздействием температуры активизируются физиологические процессы. В изменившихся условиях нарушаются взаимоотношения между паразитом и растением-хозяином. При этом развитие гриба сначала тормозится, а затем вообще прекращается.
П. В. Пак отмечает, что термическое обеззараживание положительно сказывается и на урожае семян последующих поколений, повышая их жизнеспособность. Оздоровленный один раз термическим способом семенной материал в течение 3—4 лет может быть свободен от пыльной головни.
Второй тип. Головня проса — Ustilago panici-miliacei Bui., твердая головня пшеницы — Tilletia tritici Wint., стеблевая головня ржи — Urocystis occulata Wint., карликовая головня пшеницы — Tilletia соп-troversa Kuhn, и др.
Головневые споры этих грибов во время обмолота прилипают к семенам, а затем вместе с ними попадают в почву. Здесь они прорастают в типичную четырехклетную базидию, образующую базидиоспоры. Последние могут копулировать сразу же или предварительно почкуются, в последнем случае копулируют продукты почкования. После копуляции двух гаплоидных клеток происходит развитие дикариотического мицелия. К этому же времени прорастает зерно, и мицелий проникает в росток, доходит до точки роста и растет вместе с ней.
В формирующемся соцветии мицелий очень быстро разрастается, разрушает его ткани, а сам распадается на хламидоспоры. При выколашивании выдвигается пораженное соцветие.
Заражение овса пыльной головней Ustilago avenae Jens, может происходить и по вышеописанному типу, и в период цветения (первый тип). В последнем случае головневая спора прорастает на цветке, дает базидию с базидиоспорами, после их копуляции мицелий не проникает в завязь, а разрастается преимущественно на внутренней поверхности кроющих чешуй. Очень скоро мицелий дает покоящиеся клетки — геммы. При посеве такого зерпа в почву одновременно с его прорастанием трогаются в рост и геммы, образуя грибницу, достигающую проростка.
В целях борьбы с головневыми грибами, споры которых находятся на поверхности семян или в наружных тканях, применяется химическое протравливание семенного материала. Известны три способа химической обработки семян: сухое, полусухое и влажное протравливание. Наиболее широко в качестве протравителей применяются следующие препараты: гранозан, меркуран, агронал, ТМТД, формалин (в разведении 1 : 300 с последующим томлением 2 часа).
Третий тип. Пузырчатая головня кукурузы — Ustilago zeae Unger.
Этот вид головни отличается тем, что, заражая растение в любом возрасте, он поражает молодые ткани: нижние части междоузлий стебля, листья, цветки и даже корни. Головневые споры, прорастая в почве, образуют фрагмобазидии с базидиоспорами, которые обладают способностью очень интенсивно почковаться. Базидия с почкующимися базидиоспорами имеет вид разветвленного деревца (см. рис. 51, в). Отпочковавшиеся клетки могут переноситься на растения воздушными течениями и насекомыми, где и прорастают в гаплоидный мицелий. Однако этот мицелий не может заразить растение. Оно инфицируется в том случае, если встречаются два гаплоидных мицелия и происходит диплоидизация, т. е. две клетки гаплоидных мицелиев сливаются и образуют одну двухъядерную. Из нее развивается росток, заражающий растение. Мицелий пузырчатой головни местный, так что количество пораженных головнею участков равно количеству заражений.
Развитие мицелия гриба сопровождается притоком питательных веществ. В результате получаются желваки, наросты или вздутия различных размеров от едва заметных бугорков до крупных пузырей, достигающих 15 см и больше в диаметре. Наиболее крупные вздутия развиваются на початках и стеблях. От момента заражения до образования заметного глазом головневого пятнышка проходит 7—12 дней, а формирование вздутия совершается в течение 7—15 дней в зависимости от погодных условий и места заражения.
В начале развития вздутия образованы паренхимными клетками растения и только пронизаны гифами гриба. Позднее мицелий сильно разрастается, разрушает полностью ткани растения, кроме наружных покровов, и, наконец, распадается на хламидоспоры. Молодые вздутия покрыты белой пленкой, которая придает им серебристый оттенок. По мере созревания пленка подсыхает, лопается и споры попадают в почву, где могут прорастать без периода покоя. Во время уборки споры попадают на початки, а затем на обмолоченные семена. Последние, загрязненные спорами, служат источником распространения инфекции из зараженных районов в незараженные.
Есть предположение, что пузырчатая головня кукурузы была завезена в Европу из Америки с семенным материалом. В Московской области до 1951 года пузырчатая головня не наблюдалась. В 1951 году были обнаружены единичные случаи поражения ею растений, а в последующие годы она сильно распространилась. В 1955 году на некоторых посевах количество пораженных растений уже составляло 30%.
Споры, распыленные в почве, сохраняют жизнедеятельность недолго, около двух месяцев, а во вздутиях — до двух
лет. При уборке урожая пораженные головней части растений, попадая в почву, являются основными источниками инфекции на будущий год.
Из агротехнических приемов в борьбе с пузырчатой головней большое значение имеет комбайновая уборка кукурузы. Головневые вздутия, попадая в комбайн и проходя через различные части машины, подвергаются деформации, разрушению и споры распыляются.
По данным Ф. Е. Немлиенко и И. Е. Сиденко, около 96% головневых вздутий попадает в комбайн и только 4%, расположенные ниже места среза, остаются в поле. При ручной уборке крупные вздутия отламываются от початков и остаются в поле. Это и приводит к сохранению инфекции до середины следующего вегетационного периода.
Наблюдения показали, что на участках, где в течение ряда лет кукуруза убиралась комбайном, поражение пузырчатой головней было в 5—7 раз меньше, чем при уборке вручную. Из других агротехнических мер борьбы с пузырчатой головней рекомендуется избегать посевов кукурузы на зараженной спорами почве; обрывать вздутия первичного заражения до распыления спор; уничтожать или запахивать осенью послеуборочные остатки.
В Советском Союзе головневые грибы распространены повсеместно. Они встречаются на культурных и дикорастущих злаках, на осоках, а также на двудольных растениях (семейства гвоздичных, фиалковых, гречишных и некоторых других растений).
Поражение культурных злаков отдельными видами головни зависит от многих причин: природных условий, особенности возделываемых сортов, качества проводимых мероприятий по борьбе с этими заболеваниями и др.
Наиболее распространенными видами головни являются: пыльная и твердая головня пшеницы, стеблевая головня ржи, пыльная головня овса, пузырчатая головня кукурузы.
Обнаруживаются и новые, ранее неизвестные для тех или иных районов головневые грибы. Так, в 1947 году в Армении были обнаружены первые очаги к ар ликов ой головни пшеницы Tilletia controversa Kuhn. В последующие годы эта болезнь появилась и в других районах страны (Алма-Атинской и Закарпатской областях, Молдавии, Азербайджане и др.).
Карликовая головня пшеницы по внешним признакам напоминает твердую головню. Поэтому вполне возможно, что ее ареалы еще не вполне выявлены. Впервые карликовая головня была обнаружена немецким микологом Кюном в 1874 году на соцветиях пырея. Оказалось, что гриб может зимовать в корневищах растения.
В Германии ее обнаружили на пшенице в 1930 году, а в США карликовая головня как самостоятельный тип заболевания пшеницы описана несколько раньше (1925).
Название заболевания вполне соответствует диагностическим признакам. Карликовая головня вызывает сильную задержку роста стеблей, а иногда и чрезмерное кущение пораженных растений. Пораженные растения могут быть настолько низкорослыми, что остаются несрезанными во время уборки урожая. Больные растения имеют обычно более плотный колос и увеличенное число пораженных завязей в колосках по сравнению с растениями, пораженными твердой головней. Важной биологической особенностью гриба является повышенная жизнеспособность спор. В почве они не теряют способности к прорастанию до 7 лет (по зарубежным данным). Лабораторные исследования хламидоспор карликовой головни показали, что в первый период после созревания они либо совсем не прорастают, либо прорастают в единичных случаях, т. е. не обладают достаточной физиологической зрелостью.
Карликовая головня, кроме озимой пшеницы, может поражать другие культурные, а также дикорастущие злаки: озимую рожь, озимый ячмень, виды пырея, костер, лисохвост и др.
В настоящее время для борьбы с этим видом головни рекомендуются следующие мероприятия:
1. Протравливание семян 50%-ным гекса хлорбензолом (2 кг/т).
2. Опыливание посевов 50%-ным гексахлорбензолом через 2—3 недели после всходов.
3. Возвращение пшеницы на прежнее поле не ранее чем через 3—4 года.
В Белоруссии головневые грибы также широко распространены и наносят значительный ущерб зерновым культурам.
Изучение видового состава головневых грибов, их распространения и вредоносности начато в 30-х годах.
Согласно данным Н. А. Дорожкина, в 1927 году в Горец-ком районе поражение овса пыльной головней доходило до 40—50%, а в 1929 году повреждение посевов ржи стеблевой головней в Борисовском районе составляло 10%. Позднее этим же исследователем отмечено быстрое распространение пузырчатой головни на посевах кукурузы и ее вредоносность, которая проявляется не только в потере урожая зеленой массы и початков, но и понижении качества силоса.
Наиболее распространены в Белоруссии: на пшенице — пыльная головня, меньше — твердая; на овсе — пыльная, на кукурузе — пузырчатая. Пузырчатая головня особенно распространена в южных районах республики, где в отдельных хозяйствах поражение посевов кукурузы достигает 15%.
Много внимания в республике уделяется вопросам изучения степени восприимчивости сельскохозяйственных культур к грибным заболеваниям, в том числе злаковых к головне. Различие физиологических особенностей того или иного сорта, а также условия его произрастания определяют большую или меньшую его восприимчивость к заболеваниям. Наблюдения П. В. Пак показали, что организмы, временно ослабленные, например перегреванием семян, в фазе проростков в большей степени подвергаются заражению твердой головней, чем проростки, выросшие из неперегретых семян.
ПОРЯДОК UREDINALES
Явления, вызываемые у высших растений ржавчинными грибами, известны человечеству с давних времен. Еще до нашей эры у римлян, египтян, греков упоминается о болезненных признаках (мучнистая роса, ржавчина), проявляющихся у сельскохозяйственных растений. Однако эти явления в те далекие времена толковались как сыпи, возникающие вследствие изменения частей тела растений под влиянием избыточной влажности. Истинная природа ржавчины, ее причины выяснены значительно позднее.
В- 1767 году были опубликованы результаты исследований итальянского ученого Ф. Фонтана, который впервые обстоятельно описал стеблевую ржавчину пшеницы и зарисовал спо-роношения. В настоящее время известно, что все ржавчинные грибы являются облигатными паразитами. Все они чрезвычайно строго специализированы и приурочены к развитию на определенных видах, разновидностях, а часто и сортах растений. Известно также, что эти паразиты способны развиваться только на вполне здоровых и упитанных растениях и с трудом заражают ослабленные.
Ржавчинные грибы имеют большой научный и практический интерес. Во-первых, они обладают резко выраженным свойством плеоморфизма, т. е. имеют различные формы споро-ношений, которые называются стадиями развития гриба. Они чередуются в строгой закономерности и наследственно закреплены. Во-вторых, среди ржавчинных грибов широко распространено явление разнодомности или двухозяйности, т. е. для прохождения полного цикла развития гриб требует последовательной смены двух растений-хозяев.
Ржавчинные грибы наносят большой вред культурным растениям, особенно хлебным злакам. В 1932 году в СССР потери зерна от ржавчины составляли 25 млн. ц, а в 1933 году — свыше 25,5 млн. ц. Поэтому неудивительно, что большое число исследований посвящено ржавчинным грибам. А. А. Ячевский, В. Г. Траншель, Н. А. Наумов, А. Ф. Русаков, В. И. Ульяни
щев, В. Ф. Купревич, Н. А. Дорожкин и многие другие советские ученые занимались изучением ржавчинных грибов.
Цикл развития ржавчинного гриба начинается от прорастания базидиоспор. Проникновение ростка происходит через устьице или через наружную стенку эпидермальной клетки. Грибница распространяется почти исключительно в межклетниках, а в клетки направляет гаустории, при помощи которых и получает питательные вещества. В большинстве случаев у ржавчинников грибница местного характера и занимает сравнительно небольшой участок тканей в месте проникновения ростка. Иногда грибница диффузная. Диффузное поражение нередко вызывает деформацию пораженных органов.
Характер расположения спороношений гриба на местной и диффузной грибнице несколько различен. При местном поражении спороносные пустулы вначале единичны. По мере того как возникают новые очаги поражения, пустулы могут сливаться между собой. При диффузном поражении спороносные участки рассеяны равномерно по всему пораженному органу.
Местная грибница обычно однолетняя, диффузная же большей частью многолетняя, зимующая в стеблях, почках, корнях.
В цикле развития ржавчинных грибов имеются следующие стадии: базидии (IV), спермогонии (0), эцидии (I), уредопу-стулы (II), телейтопустулы (HI) L В каждой стадии развивается характерное для нее спороношение: базидиоспоры, спермации, эцидиоспоры, уредоспоры, телейтоспоры. Ржавчинные грибы отличаются циклами развития. Если имеются все пять стадий, такой цикл считается полным. При отсутствии той или иной стадии в развитии гриба цикл будет неполным. Для их обозначения пользуются прибавлением соответствующей приставки к родовому названию.
Ей — приставка перед родовым названием, например Еи-Puccinia. Имеются все формы спороношения 0, I, II, III, IV.
Нету — приставка перед родовым названием, например Hemy-Puccinia. Отсутствуют спермогонии и эцидии; имеются спороношения II, III, IV.
Brachy — приставка перед родовым названием, например Brachy-Puccinia. Отсутствуют эцидии; имеются спороношения 0, II, III, IV.
Opsis — приставка после родового названия, например Pucciniopsis. Отсутствует уредо-, имеются спороношения 0, I, III, IV.
Micro — приставка перед родовым названием, например Micro-Puccinia. Отсутствуют эцидии и уредо-, иногда испер-
1 Римскими цифрами, указанными в скобках, пользуются для сокращенного обозначения стадий развития ржавчинных грибов.
могонии, имеются спороношения 0, III, IV. Телейтоспоры прорастают после зимовки.
Lepto — приставка перед родовым названием, например Lepto-Puccinia. Отсутствуют эцидии и уредо-, имеются спороношения 0, III, IV. Телейтоспоры прорастают сразу после образования.
Спермогонии всегда развиваются на мицелии, образовавшемся в результате прорастания базидиоспоры, и располагаются на верхней поверхности листа. От внутренних стенок спермогония в его полость врастают короткие конидиеносцы, отчленяющие шаровидные или яйцевидные спермации. Наиболее часто встречающаяся форма спермогониев — кувшинообразная, с выводным отверстием — устьем на вершине, через которое выступают парафизы, или устьичные нити (сем. Pucci-niaceae). Реже встречаются спермогонии плоской формы (сем. М elamp soraceae).
Спермогоний обладает способностью выделять сладкую и душистую жидкость, привлекающую насекомых. Вместе с жидкостью наружу выступают спермации, которые и переносятся насекомыми на другие спермогонии. Нового заражения спермации вызвать не могут. Но для дальнейшего развития гриба, для перехода его к следующей форме спороношения такой перенос спермациев необходим. Подробно о его значении будет сказано ниже.
Эцидии закладываются в виде шаровидных или плоских тел на мицелии, развившемся из базидиоспоры (первичный мицелий) в ткани растения. Дальнейшее развитие их возможно только после диплоидизации мицелия.
В зависимости от отсутствия или наличия на поверхности эцидия особого покрывала — перидия различают следующие типы эцидиев: 1) Саеота (без перидия), 2) Aecidium, 3) Roestelia, 4) Peridermium (с перидием различной формы).
Саеота закладывается под эпидермисом в виде плоского сплетения гиф. Образующиеся на нем цепочки эцидиоспор давят на эпидермис, он разрывается и эцидиоспоры освобождаются. Цеомы представлены у родов Melampsora и Phrag-midium. У видов Phragmidium по краю эцидиоспор располагаются венцом булавовидно расширенные парафизы.
Aecidium закладывается в глубине тканей в виде шаровидного сплетения и прикрыт перидием, образованным одним слоем клеток. По мере роста эцидия перидий выступает на поверхность и к моменту созревания эцидиоспор разрывается зубчатым краем.
Roestelia имеет перидий, вытянутый в длинную трубочку, отчего спороношеиие принимает форму рожка, выступающего из субстрата. Раскрывание перидия происходит отверстием на вершине трубочки, либо расщеплением продольными лентами
сверху вниз или боковыми щелями, при этом вершина остается замкнутой.
Peridermium имеет пузыревидно вздутый перидий. При созревании эцидиоспор он разрывается в разных направлениях.
Перед образованием эцидиоспор базальные клетки эци-дия отчленяют цепочки двухъядерных материнских клеток эцидиоспор. Каждая такая клетка делится на две: верхнюю, боЛее крупную — эцидиоспору и нижнюю маленькую— промежуточную, позднее разрушающуюся. Зрелые цепочки эцидиоспор легко разъединяются вследствие разрушения промежуточных клеток, и споры высыпаются из раскрытого эцидия.
.Оболочка эцидиоспор обычно бесцветная, густобородавчатая, а содержимое окрашено в желтый цвет. В большинстве случаев эцидиоспоры обладают способностью прорастать сразу же после образования. Только у некоторых видов Gymnos-porangium прорастание возможно после периода покоя.
Уредопустулы ржавой окраски представляют скопление уредоспор гриба. У громадного большинства ржавчинников они не имеют собственных покровов и только до созревания прикрыты эпидермисом растения-хозяина. У некоторых видов уредопустулы окружены парафизами, заменяющими перидий. Уредоспоры одноклетные, округлой или эллипсоидальной формы. Имеют бесцветную, либо окрашенную в желтый или бурый цвет, густо бородавчатую или шиповатую оболочку. Содержимое уредоспоры окрашено. Его цвет зависит от окрашенных пигментами капелек масла. У большинства ржавчинных грибов уредоспоры образуются на длинных ножках, а по созревании отпадают от них.
В распространении ржавчинных грибов уредоспоры имеют основное значение, так как каждая из них, попадая на здоровый участок ткани растения, может вызвать новое заражение, а через несколько дней (7—9) образование новой уредопустулы.
Т е л е й т о п у с т у л ы бурой, почти черной окраски развиваются в конце вегетации растения-хозяина и представляют скопления телейтоспор гриба. Телейтоспоры в отличие от уредоспор очень разнообразны. Их форма, количество клеток, длина ножки или ее отсутствие (сем. Melampsoraceae) является систематическим признаком ржавчинных грибов. Оболочка телейтоспор большей частью утолщенная, бурая, иногда почти черная. У некоторых заметны ростковые поры (по одной в клетке— Urotnyces, Puccinia; по нескольку — Phraginiditun, Gymnosporangium). Содержимое телейтоспор бурое, мелкозернистое. Часто хорошо различимо одно крупное (диплоидное) ядро в каждой клетке. У многих ржавчинных грибов телейтоспоры прорастают только после зимовки, хотя имеется
немало видов, у которых телейтоспоры способны прорастать вскоре после образования.
Базидии ржавчинных грибов развиваются при прорастании телейтоспор. Каждая клетка телейтоспоры дает одну четырехклетную базидию (см. табл. V, ж). Клетки базидии имеют по одному гаплоидному ядру и по одной короткой и тонкой боковой веточке — стеригме. Вершина стеригмы вздувается, сюда переходит ядро и ютшнуровывается базидиоспора. Форма базидиоспор может быть округлая, эллипсоидальная, почковидная. Базидиоспоры отбрасываются от стеригм с некоторой силой. Однако она гораздо меньше силы, выбрасывающей сумкоспоры из сумок. Узкий канал стеригмы значительно снижает силу тургорного давления внутри клетки базидии и базидиоспоры отбрасываются на незначительное расстояние.
Цитологические особенности ржавчинных грибов можно охарактеризовать тремя последовательно сменяющимися фазами в цикле развития.
Г аплоидная фаза представлена базидием с базидиоспорами, первичным мицелием, спермогониями со сперма-циями.
Дикариотическая фаза включает вторичный мицелий, эцидии с эцидиоспорами, всю уредостадию и частично телейтостадию (мицелий и молодые телейтоспоры).
Диплоидная фаза — зрелые телейтоспоры с одним диплоидным ядром.
Как показали исследования канадского миколога и фитопатолога Крэги (1927), ржавчинные грибы являются формами раздельнополыми (гетероталличными). На одной базидии развиваются две базидиоспоры одного полового знака ( + ) и две другого (—). Их прорастание дает начало гаплоидным грибницам соответствующего полового знака.
Проявление гетероталлизма у ржавчинных грибов прежде всего наблюдается при развитии эцидиальной стадии.
На гаплоидной грибнице, развившейся в результате прорастания базидиоспоры, образуются спермогонии и зачатки эцидпев. Развитие их и образование эцидиоспор возможны только в том случае, если на спермогоний одного пола будут перенесены спер-мации другого пола. Перенесенные спермации копулируют с парафизами спермогония, образуя двухъядерную клетку, или прорастают в грибницу, проникающую к основанию эцидия. Там ее клетки копулируют с клетками первоначальной грибницы. Образующиеся двухъядерные клетки дают начало дикариотической фазе, а эцидии является первой формой спороношения вторичного мицелия.
Эцидиоспоры содержат по два ядра. При их прорастании образуется грибница, состоящая также из двухъядерных кле
ток. Через некоторое время на ней развиваются двухъядер-ные уредоспоры, а затем и телейтоспоры, В клетках телейтоспор ядра очень скоро сливаются в одно диплоидное ядро, которое делится редукционно при прорастании телейтоспоры и образовании базидии.
По современной систематике ржавчинные грибы делят на два семейства; Melampsoraceae, Pucciniaceae.
Сем. Melampsoraceae
Мелямпсоровые характеризуются следующими признаками: у одних видов телейтоспоры без ножек соединены в подушечки или корочки, прикрытые эпидермисом растения-хозяина (Melampsora). У других телейтоспоры образуют длинные роговидные колонки, выступающие над поверхностью разорванной кожицы (Cronartium ribicola).
Большая часть Melampsoraceae является разнохозяйными паразитами, причем гаплоидная стадия развивается на различных видах голосемянных растений. Основным родом является род Melampsora.
Род Melampsora Cast. Из наиболее распространенных видов необходимо отметить ржавчину льна Melampsora Uni (Pers.) Desm. Это одна из наиболее вредоносных и распространенных болезней льна. Паразит однохозяйный, с полным циклом развития (рис. 52).
В июне на листьях растений появляются довольно мелкие, бледные многочисленные спермогонии шаровидной формы и без устьичных нитей, расположенные под эпидермисом листа. Эцидии развиваются в конце июня преимущественно на нижней стороне листьев -и не имеют перидия. Эцидиальная стадия Melampsora Uni не отличается обильным спороношением и часто проходит незаметно для наблюдателя.
В июле начинают появляться уредопустулы. Массовое развитие их совпадает с фазой цветения льна. Уредопустулы образуются на обеих сторонах листьев, а также на стеблях. Вначале они прикрыты эпидермисом. Позднее он разрывается и наружу выступает масса уредоспор, между которыми развиваются головчатые или булавовидные парафизы. Уредо-споры ярко-оранжевые или желтые с мелкошиповатой оболочкой. Рассеиваясь, они вызывают новое заражение растений. Уредоспоры ржавчины льна не теряют способности к быстрому прорастанию в течение трех месяцев.
К концу вегетации льна на листьях и преимущественно на стеблях появляются блестящие в виде корочек телейтопу-стулы, прикрытые эпидермисом. Телейтоспоры призматические, одноклетные, без ножек.
В пораженных местах стебель льна утолщается, а лубяные волокна разрываются. Телейтопустулы прочно прикрепляются к волокну, отчего оно становится грубым и приобре-
Р и с. 52. Melampsora Uni:
а — уредопустулы на листьях; б — уредоспора; в — телейтопустулы на стеблях; г — разрез через телейтопустулу; д — волокно с телейтопустулами.
тает бурую окраску. Под влиянием гриба волокно трудно отделяется от древесины, оно как бы спрессовывается, отчего болезнь получила название «присухи».
12*
179
Ржавчина снижает не только качество волокна, но и ассимилирующую способность растения. Наличие большого количества уредопустул на листьях растений уменьшает ассимилирующую поверхность, а разрыв эпидермиса приводит к повышенной транспирации. Все это снижает урожай семян на 8—10%.
Гриб зимует в виде телейтоспор на растительных остатках. Прорастание телейтоспор происходит весной, а образующиеся базидиоспоры обусловливают весеннее заражение растений.
В борьбе с ржавчиной льна проводятся мероприятия:
1. Тщательная очистка семенного материала от остатков вегетативных органов, на которых могут сохраняться телепто-пустулы гриба.
2. Удаление растительных остатков с поля как источников возобновления инфекции.
3. Выведение устойчивых к ржавчине сортов льна.
Род Cronartiurn Fries. Из двухозяйных грибов, принадлежащих к семейству Melampsoraceae, следует отметить ржавчину смородины (Cronartiurn ribicola Dietr.). Эцидиальная стадия этого гриба развивается на веймутовой сосне и сибирском кедре. Гриб поражает ветви, образуя под корой спермогонии. Через трещинки в ко;ре выделяются капельки сахаристой жидкости, а с -нею вместе спермации, которые и переносятся насекомыми. Эцидии в большом количестве выступают из-под коры и имеют вид пузырей довольно крупных размеров: 2—7 мм длины и 2—3 мм ширины. Это заболевание получило название пузырчатой ржавчины сосны.
Вторым питающим растением этого гриба являются виды смородины и крыжовника. Эцидиоспоры переносятся на листья и вызывают заражение растений, а в середине лета на нижней поверхности появляются оранжевые пустулы уре-доспор округлой или яйцевидной формы. Уредоспоры могут вызывать повторное заражение.
К концу лета среди уредоспор появляются роговидносогнутые столбики до двух миллиметров длиной, состоящие из склеенных одноклетных светло-бурых телейтоспор (рис. 53), Они способны прорастать осенью без периода пбкоя.
Интересной биологической особенностью' гриба является продолжительный период инкубации. После заражения дерева осенью эцидии появляются только через два года. Старая грибница, сохраняющаяся в ветвях дерева, ежегодно может развивать эцидии. Пораженные ветви сосны деформируются и в конце концов отмирают.
Исследованиями А. С. Бондарцева установлено, что Сго-nartium ribicola высоко специализированный паразит и может
заражать только определенные виды смородины или крыжов-ника.
Постоянными очагами ежегодного возобновления инфекции являются пораженные ветви, поэтому рекомендуется их срезать .и сжигать.
Рис. 53. Cronartium ribico-la — столбик телейтоспор: а — телейтоспора проросла фраг-мобазидией с четырьмя базидиоспорами.
Р и с. 54. Periderniiuni pint — эцидиальная стадия па веточке сосиы (ориг.).
На ветвях обыкновенной сосны также может встречаться пузырчатая ржавчина, но она вызывается другими возбудителями: Cronartium asclepiadeum Fr. и Peridermium pint Kleb.
Эцидиоспоры первого прорастают на пионах, мытнике, бальзаминах; для второго пока неизвестны уредо- и телейто-стадии, а эцидии способны непосредственно заражать сосну, причем инкубационный период может длиться до 3 лет.
Пузырчатая ржавчина на обыкновенной сосне встречается и в Белоруссии и в отдельные годы достигает большого распространения. Нам приходилось наблюдать сильное развитие ее в 1957 г. в Мядельском районе Минской области (леса окрестностей озера Нарочь) (рис. 54).
Сем. Pucciniaceae
Семейство Pucciniaceae более обширное, включает одно-и двухозяйных паразитов. Телейтоспоры на ножках отличаются большим разнообразием 'морфологического строения. Они могут быть одноклетные, двухклетные и многоклетные. Форма и величина клеток также варьируют.
Наиболее крупными родами в семействе являются: Puccinia и Uromyces.
Род Puccinia Pers. Самый многочисленный по количеству видов (свыше 2000) и самый вредоносный. Его виды поражают растения из различных семейств, но особенный вред они причиняют злаковым культура-м. Одни представители рода Puccinia имеют полный цикл развитая с обязательным участием эцидиальной стадии, развивающейся на соответствующем промежуточном хозяине. Сюда относятся стеблевая ржавчина злаков, корончатая ржавчина овса и др.
Другие виды могут развиваться по неполному циклу -{без участия эцидиальной стадии) и зимовать в стадии уредоми-целия и уредопустул. К этой группе относятся бурая листовая ржавчина пшеницы (в Европейской части Союза), желтая ржавчина злаков, а в Западной Сибири и линейная ржавчина. На хлебных злаках наиболее распространены следующие виды ржавчины: линейная или стеблевая ржавчина (Р. graminis Pers.), бурая ржавчина пшеницы (Р. iriticina Erikss.), бурая ржавчина ржи (Р. dispersa Erikss. et Henn.), желтая ржавчина злаков (Р. glumarum (Schm.) Erikss. et Henn.), корончатая ржавчина овса (Р. coronifera Kleb.).
Большее или меньшее проявление того или иного вида ржавчины определяется зональными особенностями района.
Линейная, или стеблевая, ржавчина (Puccinia graminis Pers.) распространена на всех континентах: в Европе, Азии, Африке, Северной и Южной Америке, Австралии. Этот гриб встречается в низинных местах и в горах; в посевах — на культурных злаках и сорняках; на лугах и опушках леса.
Р. graminis — двухозяйный вид. Спермогонии и эцидии развиваются на видах барбариса (Berberis L.) и магонии (Mahonia L.) а уредо- и телейтостадия—на злаках (табл. V).
Пораженные участки листьев промежуточного хозяина изменяют окраску, становятся оранжево-красными. На верх
ней стороне, а иногда и на нижней развиваются спермогонии небольшими группами или одиночные, шаровидной формы. Эцидии образуются на нижней стороне листьев, черешках, молодых побегах, даже на завязях. Форма эцидиев обычно чашевидная, они покрыты перидием, который по созреванию эцмдия разрывается, края его отгибаются и споры рассеиваются. Эцидиоспоры чаще всего шаровидные, имеют бесцветную оболочку, покрытую бородавочками и окрашенное в желтый цвет содержимое.
Уредопустулы образуются на стеблях, листовых влагалищах и пластинках листа, а при сильном поражении — на колосковых чешуях и остях. Форма уредоспор овальная; оболочка— мелкошиповатая; окраска желтовато-бурая.
Телейтопустулы появляются на тех же частях тела растения. но по размерам они значительно больше уредопустул, так как могут сливаться между собой и достигать 22 мм длины. Вначале они прикрыты эпидермисом и очень плотные. Телейтоспоры чаще двухклетные, на длинной ножке, иногда одноклетные. Оболочка клеток гладкая, желтовато-бурая. Перезимовывает линейная ржавчина телейтоспорами. При прорастании телейтоспоры каждая клетка образует четырехклетную фрагмобазидию с базидиоспорами. Есть указания, что только те телейтоспоры способны весною прорастать, которые сохранялись на вертикально стоящих соломинах. Телейтоспоры на разбросанной по полю соломе или в скирдах не жизнеспособны. В некоторых климатических зонах, где не произрастает барбарис (Западная Сибирь, Северный Казахстан), возбудитель линейной ржавчины приспособился зимовать в виде уредомицелия в тканях пырея ползучего (Agro-pyron repens L.). Весною на перезимовавшем мицелии развивается уредоспороношение, способное заражать рожь и ячмень. Пшеница не заражается уредоспорами с пырея и пока не выяснено, на каком растении в указанных условиях зимует возбудитель линейной ржавчины пшеницы.
В настоящее время известно несколько специализированных форм Р. graminis, состоящих из большего или меньшего числа биотипов.
Наиболее изученной формой является Р. graminis f. sp, tritici Erikss. et Henn., поражающая пшеницу, ячмень, овсяницу гигантскую и др. Для этой формы ржавчины выявлено свыше 150 биотипов, имеющих строгую специализацию, приуроченную к определенным сортам.
Основными мерами борьбы со стеблевой ржавчиной является:
1. Введение в культуру ржавчиноустойчивых сортов.
2. Повышение уровня агротехники (лущение стерни, вспашка под зябь, ранние сроки сева).
3. Уничтожение промежуточных хозяев — барбариса и ма-гонии.
Бурая листовая ржавчина пшеницы (Р. tri-iicina Erikss.) широко распространена в районах возделывания пшеницы и наиболее вредоносна. Бурая листовая ржавчина на территории Советского Союза существует в двух формах. Одна форма, широко распространенная на всей европейской части СССР, может заражать в качестве промежуточного растения-хозяина василистник (Thalicirutn flavum L.). Однако в естественных условиях в природе цикл развития гриба проходит без эцидиальной стадии и представлен только уредо- и телейтостадией. Телейтостадия хотя и развивается, но не имеет значения в цикле развития гриба в силу того, что не вызывает заражения никаких растений. Основное значение принадлежит уредостадии, в которой гриб способен повсюду зимовать.
Другая форма бурой ржавчины распространена в Сибири, имеет активного промежуточного хозяина — лещицу (Isopy-rum futnarioides L.). Эцидиальная стадия на лещице может развиваться с осени, но массовое появление ее наблюдается в последней декаде мая. Характерной особенностью сибирской формы Р. triticina является растянутость периода прорастания телейтоспор. Часть их прорастает еще осенью (чем и объясняется появление в этот период эцидиальной стадии на лещице), основная масса — после перезимовки.
Наиболее характерные симптомы поражения пшеницы бурой листовой ржавчиной следующие. На листовых пластинках, преимущественно на верхней стороне, а также на влагалищах появляются мелкие округлые или эллиптические уредопустулы буро-охряного оттенка. При благоприятных для гриба условиях перезимовки сохранившиеся па озимых уредопустулы можно обнаружить сразу после схода снега. Новое поколение уредоспор появляется в мае, а затем инфекция постепенно нарастает за счет все новых и новых генераций уредоспор. Образование уредопустул сопровождается развитием хлороза тканей пораженных участков. К периоду молочной спелости бурая ржавчина достигает максимального развития. В это же время па тех же органах появляются черные глянцевитые телейтопустулы. Уредоспоры гриба одноклетные, округлые, эллипсоидальные, мелкошиповатые. Телейтоспоры двухклетные, верхняя клетка иногда усеченная, на короткой ножке.
Р. triticina отличается от Р. graminis большей экологической пластичностью. Она может развиваться в широких температурных пределах. Минимальной температурой, при которой возможно прорастание уредоспор, является 2,5°, максимальной— 31° тепла. Уредоспоры сравнительно долго не теряют
способности к прорастанию. Например, при температуре 5° и относительной влажности воздуха они остаются жизнеспособными в течение двух месяцев.
В южных районах между уборкой урожая и посевом озимых проходит продолжительное время. В сохранении бурой ржавчины на полях в это время большое значение имеет самосев, являющийся резерватором болезни и источником ее распространения на озимые. Р. iriticina обладает очень узкой специализацией. Биотипы ее способны паразитировать далеко не на всех сортах.
В борьбе с бурой ржавчиной пшеницы основное значение имеют агротехнические и организационно-хозяйственные мероприятия:
1. Посев и уборка пшеницы в сжатые, рекомендуемые для данной местности сроки.
2. Пожнивное лущение стерни и глубокая зяблевая вспашка до появления всходов озимых.
3. Боронование озимых весной.
4. Посев устойчивых сортов (Безостая 1, Белоцерковская 198, Мироновская 264 и др.).
Бурая ржавчина ржи (Puccinia dispersa Erikss. et Henn.) также широко распространена.
Эцидиальной стадией поражаются виды воловика Anchu-sa L. и кривоцвета Lycopsis L. Эцидии могут развиваться уже во второй половине лета, под осень. Это связано со способностью телейтоспор прорастать сразу же после их образования и заражать указанные выше растения из семейства бурачниковых. В Белоруссии пораженные воловик и особенно крпвоцвет можно встретить в конце июля — начале августа.
Уредоспоры появляются на всходах озимой ржи, которые могут заражаться через самосев, заражающийся в свою очередь от растительных остатков после уборки урожая.
На озимых гриб может зимовать в стадии уредомицелия. Весною, с началом вегетации озимых, молодой уредомицелий отрастает вокруг прошлогодних пустул и продуцирует уредоспоры, которые и заражают отрастающие листья.
Таким образом, гриб, появившийся однажды на посевах, может расселяться с одного поля на другое в стадии уредо-спороношения и обходиться без промежуточного хозяина. Однако необходимо искоренять на полях воловик и криво-цвет, так как первыми источниками распространения инфекции в поле могут быть и эти растения.
Основным мероприятием, направленным на борьбу с бурой ржавчиной, является выведение устойчивых сортов ржи.
Род Uromyces Link. Насчитывает около 600 видов однохозяйных и двухозяйных ржавчинных грибов, способных поражать почти все бобовые, дикорастущие и культурные.
Очень распространенный вид Urotnyces pisi (Pers.) Schot. развивается на горохе и некоторых чинах. Это двудомный паразит, в эцидиальной стадии поражает различные виды молочая, а уредо- и телейтоспоры развиваются на горохе.
Мицелий гриба зимует в подземных частях молочая, а весною диффузно распространяется в отрастающих побегах. Пораженные побеги отличаются отсутствием ветвления, сильно вытянутыми междоузлиями и мелкими, плотными желтовато-зелеными листьями. Эцидии густо покрывают нижнюю поверхность листьев, дают большое количество эцидиоспор, которые и заражают горох.
К началу цветения гороха на листьях и стеблях появляются оранжево-коричневые уредопустулы. Уредоспоры окрашены в ржаво-коричневый цвет, одноклетные, неправильно-округлые с тонкошиповатой оболочкой.
К периоду созревания гороха развиваются телейтоспоры на коротких ножках, шаровидно-овальной формы. Оболочка их снабжена небольшими шипиками и имеет утолщение наверху в виде сосочка. Весною телейтоспоры, зимовавшие с остатками урожая, прорастают, а образующиеся базидиоспоры ранней весной заражают молочай. Под влиянием паразита листья гороха желтеют, растения сильно отстают в росте и дают значительно пониженный урожай.
Ржавчина гороха широко распространена в Белоруссии. Н. А. Дорожкин отмечал сильное поражение посевов в Оршанском округе еще в 1929 году. Из видов молочая, произрастающих в республике, особенно сильно поражается эцидиальной стадией молочай прутьевидный (Euphorbia virgata W. et К.). В 1951 году нам приходилось наблюдать массовое поражение этого вида молочая по опушкам леса, у дорог и по межам в Бегомльском районе Минской области (Березинский заповедник).
Из наиболее действенных мер борьбы с ржавчиной гороха рекомендуются следующие:
1. Уничтожение молочая как промежуточного хозяина.
2. Тщательная уборка с поля растительных остатков гороха, содержащих зимующую стадию гриба.
Выше указывалось, что ржавчинные грибы являются паразитными организмами, весьма приспособленными к определенным растениям-хозяевам. Им свойственно образование физиологических рас, а также биотипов, обладающих узкой специализацией. Возникновение новых рас в природной обстановке происходит в связи с присущей ржавчинным грибам раздельнополостью и переносом спермациев с одного гаплоидного мицелия на другой.
В результате такой гибридизации и возникают организмы, обладающие очень тонкими индивидуальными особенностями,
несмотря на то что они принадлежат к одному и тому же виду и даже к одной и той же форме.
Расообразовательными процессами, происходящими у ржавчинных грибов в естественных условиях, и объясняется потеря через некоторое время устойчивости к паразиту многими сортами.
Возникающие новые расы приспосабливаются к физиологическим и биохимическим особенностям клеток растений ранее устойчивых сортов, и через некоторое время сорт, считавшийся несколько* лет устойчивым к ржавчине, поражается ею.
В связи с этим основная работа по борьбе с этими паразитами должна быть направлена на выведение иммунных к ржавчине сортов, к изысканию способов поддержания устойчивости их в течение длительного периода.
В заключение необходимо остановиться на филогенетических связях базидиомицетов.
В настоящее время большинство микологов признает общее происхождение сумчатых и базидиальных грибов от зигомицетов. Однако происхождение сумки из зародышевого спорангия зигомицетов более очевидно, чем происхождение базидии. Предполагается, что на определенном этапе эволюции зародышевый спорангий зигомицетов получил возможность вырабатывать споры, расположенные экзогенно, и, таким образом, произошло превращение его в холобазидию, которая и должна рассматриваться как первичная.
Наиболее примитивными формами из холобазидиальных грибов следует считать те, которые не имеют плодового тела. От них эволюция пошла в двух направлениях: в сторону развития паразитизма при отсутствии плодовых тел (экзобази-диальные) и в сторону возникновения и дальнейшего усложнения плодового тела в группах порядков как гимено- так и гастромицетов. Причем высказывается мнение, что гемиан-гиокарпные плодовые тела пластинчатых произошли от ан-гиокарпных плодовых тел гастромицетов. Покрывало, имеющееся на плодовом теле у многих агариковых, следует рассматривать как рудимент перидия их гастральных предков.
Подкласс фрагмобазидиальных грибов следует рассматривать как ветвь, берущую начало от наиболее примитивных хо-лобазидиомицетов, но сильно уклонившуюся в сторону. Эволюция этого ряда грибов в основе своей имела особенности развития базидии. В этом случае готовая базидия состоит как бы из двух частей. Нижняя из них, или .гипобазидия, соответствует холобазидии и зародышевому спорангию фико-
мицетов, а верхняя — эпибазидия вырастает и представляет разделенную на клетки фрагмобазидию.
Развитие эпибазидии шло постепенно. У дакриомицетов она представлена в виде двух толстых ветвей на вершине базидии. У дрожалковых ветви подобного рода срастаются между собой и дают продольно разделенную фрагмобазидию, а у аврикуляриевых в базидии появляются поперечные перегородки.
Паразитный образ жизни головневых и ржавчинных грибов явился причиной утраты ими плодовых тел, а цикл развития, при котором гриб имеет стадию покоя, привел к превращению гипобазидии в покоящуюся спору (хламидоспору, телейтоспору), на которой при определенных условиях вырастает эпибазидия, обычно клеточного строения.
ГЛАВА IV КЛАСС. DEUTEROMYCETES
(НЕСОВЕРШЕННЫЕ ГРИБЫ)
ОБЩАЯ ХАРАКТЕРИСТИКА
В микологической учебной литературе несовершенным грибам уделяется мало внимания. Между тем этот обширный и разнообразный класс включает большое количество видов (до 25 тыс.), имеющих громадное значение в хозяйственной деятельности человека. Для его представителей характерно наличие клеточного мицелия и бесполого размножения при помощи конидий. Экспериментальные исследования отдельных представителей этого класса грибов показали, что некоторые из них могут давать сумчатое спороношеиие. Бази-диальное развивается в более редких случаях. Известны также и такие формы, у которых грибница остается совершенно стерильной.
Образ жизни несовершенных грибов может протекать в самых различных условиях. Они широко представлены в почве как сапрофитно живущие организмы, а у некоторых обнаружено свойство хищничества. Громадное количество грибов этого класса ведет паразитный образ жизни, развиваясь преимущественно на растениях. Очень многие несовершенные грпбы прекрасно развиваются на различных питательных средах в лабораторных условиях.
Значение несовершенных грибов в природе и хозяйственной деятельности человека может быть оценено двояко. Многие из них являются организмами не только полезными, но и, безусловно, необходимыми, как например, почвенные сапрофиты, принимающие участие в процессе круговорота веществ, и формы, живущие в почве и обладающие способностью улавливать нематод (так называемые «хищные грибы»), среди которых имеется много видов, опасных для человека, животных и растений. Некоторые несовершенные грибы могут вы-
делять ферменты, используемые человеком в ряде производственных процессов. Например, ферменты, вырабатываемые Aspergillus oryzae и A. niger, применяются в производстве натуральных фруктовых соков для их осветления, а ферменты Botrytis citierera используются для придания прозрачности некоторым сортам вина.
Среди несовершенных грибов немало и таких организмов, которые в процессе своей жизнедеятельности выделяют антибиотические вещества, имеющие широкое применение в медицине, ветеринарии и фитопатологии. Такими организмами являются некоторые виды Penicilliuin, Aspergillus, Tricho-derma и др.
Однако несовершенные грибы широко известны и с отрицательной стороны. Многие из них вырабатывают токсины, ядовитые для человека и животных (некоторые виды Fusarium, Dendrodochiutn и др.). Громадное количество видов несовершенных грибов является паразитами высших растений. Следует отметить, что форма паразитизма несовершенных грибов на высших растениях является факультативной. Однако, несмотря на это, несовершенные грибы приносят существенный вред культурным сельскохозяйственным и декоративным растениям. В связи с этим изучение их биологических особенностей, а также экологических условий развития имеет очень большое значение для разработки рациональных мероприятий в борьбе за сохранение урожая.
Преобладающие внешние признаки, с которыми по большей части бывает связано поражение высших растений несовершенными грибами,— это увядание растения, начиная с верхушки, образование различных пятнистостей на наземных вегетативных органах, цветках, плодах, а также всевозможные гнили. В первом случае заболевания растений вызываются явлением трахеомикоза, т. е. закупориванием гифами гриба элементов ксилемы, проводящих воду; во втором — отмиранием тканей под воздействием локализованного в этих участках мицелия гриба и продуктов его жизнедеятельности; в третьем — загниванием того или иного органа.
Для обширного класса несовершенных грибов в настоящее время общепринята система, предложенная итальянским исследователем Саккардо. В ее основу положены разнообразные формы конидиальных спороношений, свойственные несовершенным грибам.
Согласно этому принципу, Саккардо делит все несовершенные грибы на три порядка:
1. Hyphomycetales — гифомицеты. Конидиальное спороношение у них образуется на вегетативной грибнице в виде отдельных конидиеносцев или же последние собраны в пучки (коремии) или в подушечки (спородохии) (рис. 55, а, б).
2. Melanconiales — меланкониальные. У представителей этого порядка конидиеносцы располагаются плотным слоем на поверхности плоского сплетения гиф, называемого ложем и более или менее погруженным в субстрат (рис. 55,в).
3. Sphaeropsidales — сферопсидные. У грибов этого порядка конидиеносцы и конидии развиваются внутри особых спо-
Рис. 55. Спороношения несовершенных грибов:
в —' отдельные конидиеносцы; б — коремии у гифомицетов; в — ложе у меланконне-вых; г — пикнида у сферопсидных.
ровместилищ, или пикнид, округлой или овальной формы с отверстием на вершине. Развивающиеся таким путем конидии или пикноспоры (стилоспоры) выходят наружу через отверстие в пикниде (рис. 55,г).
Вышеприведенная система является до известной степени искусственной. Она не отражает родственных филогенетических отношений между порядками. Кроме того, среди представителей несовершенных грибов находится еще немало форм, недостаточно подробно изученных в их онтогенетическом развитии. И, наконец, есть и такие виды, у которых получены в определенных условиях совершенные формы споронотпений,
в связи с чем явилась возможность отнести их в соответствующие классы высших грибов.
Однако система Саккардо обладает тем преимуществом перед другими системами (А. А. Потебня, Fr. Hoehnel), что она проста и удобна для целей определения несовершенных грибов.
Перечисленные порядки системы Саккардо подразделяются на семейства. У гифомицетов в основу деления принимается строение и окраска конидиеносцев и конидий, у сферопсид-ных — форма, окраска, характер залегания пикнид, а также форма и окраска пикноспор. Меланкониевые включают только одно семейство.
ПОРЯДОК НУ PHOMVCETALES
Гифомицеты могут жить как в сапрофитных, так и в паразитных условиях. В последнем случае они являются возбудителями весьма вредоносных заболеваний сельскохозяйственных и декоративных растений из самых различных семейств. В порядке гифомицетов отмечены три семейства: Mucedina-ceae, Dematiaceae, Tuberculariaceae.
Сем. Mucedinaceae
Мицелий, конидиеносцы и конидии грибов из этого семейства обычно бесцветные, реже сероватой или буроватой окраски. Конидиеносцы прямые или ветвящиеся (ветвление древовидное, кистевидное, мутовчатое и других типов), одиночные или расположенные группами, нс тогда не сросшиеся между собой. Конидии располагаются по одной на разветвлениях конидиеносцев или же образуют цепочки, гроздья или головки. В цикле развития грибов образуются различной формы склероции, при помощи которых организмы долгое время сохраняют жизнеспособность.
Наиболее широко распространенными представителями семейства Mucedinaceae, вызывающими болезни растений, являются грибы из родов: Botrytis, Monilia, Verticillium и др.
Род Botrytis Micheli. Виды рода Botrytis могут паразитировать на самых различных сельскохозяйственных и декоративных растениях. Они поражают все вегетативные органы: корни, стебли и листья, а также части цветка, плоды. У одних видов грибница способна зимовать в корнях растения, а весною при отрастании стеблей распространяться по всему растению, давая диффузный мицелий, на котором развивается спороношеиие. У других в цикле развития гриба образуются склероции, которые после перезимовки прорастают и образуют новый молодой мицелий с конидиеносцами и конидиями.
Таблица V. Puccinia graminis-.
a — эцидии на листьях и ягодах барбариса; б — поперечный разрез через спермогонии и эцидии; а — уредопустулы на стебле овса; г — уредоспоры; д — телейтопу-стулы на стебле овса; е — телейтоспоры; ж — прорастание телейтоспоры.
Конидиеносцы на конце древовидно ветвящиеся, конидии — одноклетные, овальные и располагаются на концах разветвлений конидиеносца на очень коротких ножках. Грибница, конидиеносцы и конидии Botrytis слегка дымчатой или буроватой окраски (иногда бесцветные).
Представители рода Botrytis обладают способностью вырабатывать различные ферменты и в качестве источника питания использовать различные субстраты. Одни развиваются в сапрофитных условиях на лесной подстилке, гниющих веточках и листьях. Другие паразитируют на многих однодольных и двудольных сельскохозяйственных и декоративных растениях.
Botrytis развивается в широких температурных пределах. Грибы этого рода можно найти в период вегетации растений в поле, оранжереях, а также в хранилищах на корнеплодах, луковицах, клубнелуковицах, кочанах и т. п.
Очень широко распространен вид В. cinerea Pers. Этот гриб в литературе известен как факультативный паразит многих высших растений. Его относят к гидрофильным грибам, так как наибольшего развития он достигает в условиях высокой влажности. Развитие В. cinerea вызывает заболевание растений, известное под названием серой гнили.
Серая гниль земляники весьма распространенное заболевание. Гриб поражает листья, бутоны, цветки, плодоножки и ягоды. На листьях болезнь проявляется в виде крупных расплывчатых темно-серых пятен, на которых образуется легкий пушок, состоящий из редко расположенных ко-нидиеносцев с конидиями.
Наиболее типично серой гнилью поражаются плоды, где первым симптомом заболевания служит появление сначала бурого мокнущего пятна, а затем образование на нем серого войлочного налета с пылящими конидиями (табл. VI, а). Пятно и налет быстро разрастаются, покрывая весь плод, который в дальнейшем съеживается, высыхает — мумифицируется. Конидиеносцы гриба расположены довольно густо и на концах древовидно разветвлены. Конидии обладают свойством очень скоро прорастать, что способствует быстрому распространению болезни. В сухом виде конидии не теряют способности к прорастанию в течение 1—2 лет. Однако для успешного развития и прорастания конидий нужна капельножидкая влага. Диапазон температур для развития серой гнили земляники очень широк-— от 2 до 35°. Оптимальной температурой является 22—25°.
Распространение гриба в течение периода вегетации осуществляется при помощи конидий, а перезимовка — склеро-циями. Последние образуются вследствие уплотнения мицелия как на поверхности, так и внутри пораженных органов. Скле~
роции обычно неправильной формы от 2 до 5 мм в диаметре.
В годы, благоприятные для своего развития, серая гниль может вызвать гибель 50% урожая плодов, а если учесть, что болезнь развивается на листьях, вследствие чего уменьшается ассимилирующая поверхность, а также на цветоножках и цветках, где она мало заметна, то вредоносность серой гнили окажется еще большей.
Мероприятия, направленные на борьбу с серой гнилью, носят профилактический характер и заключаются в очистке плантаций земляники от мертвых остатков растений, так как они могут служить очагами возникновения болезни вследствие прорастания склероциев; правильном уходе за растениями в период вегетации.
В. cinerea может вызывать и серую гниль ягод винограда. Развиваясь при различных условиях погоды, гриб оказывает двоякое физиологическое воздействие на ягоды. Если гриб развивается при хорошей, теплой погоде на уже созревших кистях винограда, то под его влиянием в мякоти ягод сильно возрастает содержание сахара. Это явление положительно сказывается при выработке вина. Кроме того, вино, получаемое из таких ягод, обладает всегда особым ароматом, отличающим, например рейнские и бордоские вина. Развитие гнили при подобных условиях весьма желательно, и она получила название «благородной гнили» винограда.
Однако, если во время созревания винограда сухая, теплая погода сменяется сырой и дождливой, ягоды винограда, покрывшиеся серым пушком гриба, начинают загнивать и приобретают неблагоприятные свойства. Полученное из такого винограда вино имеет неприятный привкус плесени, причем красные вина еще и изменяют окраску.
Весьма существенный ущерб наносит развитие серой гнили капусте, свекле, моркови, брюкве в период их зимнего хранения. Вредоносность гриба выражается не только в уничтожении овощей и корнеплодов, предназначенных для пищевого и кормового использования, но и в том, что серая гниль является одной из основных причин выпадения семенников при их весенней высадке.
На капусте болезнь проявляется в том, что поверхность кочанов покрывается пушистым серым налетом из конидиеносцев и конидий, пораженные листья ослизняются и сгнивают. Гриб распространяется в хранилище за счет конидий, а сохраняет жизнеспособность благодаря образованию склероциев, которые очень скоро становятся заметными, особенно на главных жилках листьев кочана. При недостаточно тщательной очистке и дезинфекции хранилищ склероции могут служить источником' инфекции для последующих партий закладываемых овощей.
По мере развития гнили гриб добирается до кочерыги и поражает ее верхушечную почку, т. е. наиболее жизненную часть, обеспечивающую рост. Такие кочерыги, высаженные весной в качестве маточных растений, не развивают цветоноса из основной верхушечной почки, а образуют их из боковых, более слабых и менее устойчивых по отношению к другим возбудителям болезней. Таким образом, серая гниль, развиваясь на кочанах в период хранения, может стать причиной гибели семенников.
Корнеплоды — свекла, морковь и брюква — также поражаются серой гнилью. В. cinerea развивается преимущественно на ослабленной ткани корнеплода. Проникновению гриба внутрь корнеплода способствуют различные травмы на поверхности. Болезнь, как правило, начинается с кончика корнеплода, ткани которого прежде всего теряют тургор и становятся более восприимчивыми.
Если температура и влажность хранилища способствуют развитию серой гнили, гриб распространяется по всему корнеплоду. Позднее мицелий гриба уплотняется, образуя склероции. При сильном поражении корнеплод сгнивает. В борьбе с болезнями овощных во время их хранения большое значение имеет дезинфекция хранилищ.
Незначительно пораженные корнеплоды свеклы при недостаточно тщательной переборке весной могут попадать в поле, и тогда гриб развивается на высадках, причем поражается верхушка побегов и происходит недоразвитие клубочков.
В борьбе с серой гнилью на высадках рекомендуется опрыскивание растений бордоской жидкостью.
Шейковая гниль лука, вызываемая В. allii Munn., признана наиболее вредоносной болезнью луковиц в период зимнего хранения. Заражение луковиц происходит еще в поле, а проявление болезни наблюдается через 1,5—2 месяца после уборки. Первые симптомы заболевания проявляются в размягчении чешуй сначала в области шейки луковицы, затем поражение распространяется на всю поверхность чешуй, и они приобретают вид вареных. Через некоторое время на пораженных участках появляется сероватый налет из конидиенос-цев и конидий гриба. Конидиеносцы довольно длинные — около 1 мм, конидии 7—16X4—6 мк (Бондарцев). Конидии, попадая на здоровые луковицы, распространяют болезнь. Через некоторое время на пораженных участках возникают склероции, которые, сливаясь между собой, образуют сплошные корочки. В тех случаях, когда поражены многие чешуи, вся луковица мумифицируется.
Здоровые луковицы, взятые из хранилища, где была распространена шейковая гниль, на своей поверхности несут споры гриба. Использованные в качестве посадочного материала,.
они заносят в почву инфекцию. Споры прорастают и мицелий вначале развивается на отмирающих чешуях материнской луковицы и тут же образует спороношение. Конидии попадают вместе с частицами почвы в пазухи листьев. Однако в период активного роста болезнь развивается медленно, так как большое количество вырабатываемых растением фитонцидов в этот период затормаживает развитие гриба. Понижение физиологической активности клеток растения к концу вегетации обусловливает быстрое развитие мицелия в листьях лука, а затем и проникновение его из листьев в шейку луковицы. Если листья лука срезают зелеными, то шейка луковицы служит местом внедрения гриба. Шейковой гнилью могут поражаться стрелки и семенные головки, а поэтому болезнь может передаваться семенами.
Б. А. Герасимов и Е. А. Осницкая указывают, что при благоприятных условиях шейковая гниль может уничтожить до 80% лука в период зимнего хранения. Все сорта лука подвержены заболеванию, однако существует различие между ними по степени заражения. Сладкие сорта лука, независимо от окраски чешуи, являются более восприимчивыми, чем острые. Из острых сортов наиболее легко заражаются белые; желтые и красные более устойчивы.
Для предотвращения развития шейковой гнили лук должен убираться совершенно зрелым. Очень важным профилактическим мероприятием является тщательная просушка луковиц после уборки в течение 8—10 суток при температуре 30—35° и последующая обрезка.
В хранилище должна поддерживаться влажность не выше 70—75%, а температура — 0—2°. Рекомендуется также проводить предпосадочное протравливание лука гранозаном. Это снижает поражение шейковой гнилью в 2—3 раза.
Русский миколог и фитопатолог А. С. Бондарцев 50 лет тому назад впервые открыл, изучил и описал развитие цветочной плесени красного клевера. Новому виду гриба было дано название Botrytis anthophila Bond.
Цветочная плесень проявляется на тычинках цветков красного клевера. Конидиеносцы развиваются очень обильно на пыльниках, и конидии в виде пыльцевого облачка поднимаются при раскрывании цветка. Форма конидиеносцев В. anthophila довольно разнообразна: они могут быть прямые, колен-чато изогнутые, вильчато разветвленные, с расширенными концами и короткими зубчиками, на которых прикрепляются продолговато-овальные бесцветные конидии (14—22X3,8—6 мк, табл. VI, б). Пораженные пыльники не развивают пыльцы.
Зрелые конидии прорастают только на рыльце пестика. Переносу конидий с больных головок на здоровые в значительной степени способствуют насекомые. Росток мицелия прони
кает в завязь, не мешая развитию семени. Покоящаяся грибница сохраняется под кожурой. Пораженные семена выглядят более щуплыми по сравнению со здоровыми.
При прорастании зараженных семян гриб растет вместе с растением и образует диффузную грибницу, проходящую по межклетникам тканей стебля. Поднимаясь до цветочной головки, мицелий пронизывает все части цветка, но особенного развития он достигает в пыльниках, на поверхности которых и образуется спороношеиие. В головке клевера обычно поражаются все цветки. Они отличаются более светлой окраской.
В. anthophila имеет многолетнюю грибницу, зимующую в корнях. Весною при отрастании молодых стеблей грибница развивается и болезнь возобновляется.
Цветочная плесень клевера распространена почти во всех районах клеверосеяния, но чаще встречается в северных областях Советского Союза. Развитию цветочной плесени способствует влажная и теплая погода в период цветения клевера, а также чрезмерная загущенность посевов.
Поскольку болезнь передается семенами, необходимо проводить отвеивание щуплых семян. Рекомендуется также предпосевное протравливание семенного материала гранозаном (2 г на 1 кг семян), ТМТД (3 кг на 1 т).
Гладиолусы, лилии и тюльпаны — красивейшие цветы, пользующиеся большой популярностью у населения всего земного шара, также подвержены нападению Botrytis.
На гладиолусах развивается В. gladiolorum Timm, и вызывает заболевание, известное под названием коричневой сердцевинной гнили. Во второй половине вегетационного периода развития растения можно обнаружить у основания листьев большие продолговатые коричневые пятна. Отсюда болезнь перемещается на верхние части листовых пластинок, цветоносы и сердцевину клубнелуковиц. На листовых пластинках появляются пятна светло-коричневого оттенка с красноватыми краями. В сухую и теплую погоду их можно заметить только на нижней поверхности листа, притом они ржаво-бурые и очень мелкие. Такие же пятна обнаруживаются и на цветоносах. Во влажную погоду очень восприимчивыми к заражению оказываются также и лепестки цветков, на которых образуются водянистые пятна.
На клубнелуковицах, прикрытых чешуей, часто можно не заметить признаков болезни, но при нажиме на донце обнаруживается, что сердцевина сгнила. Заражение сердцевины клубнелуковицы может происходить как на стеблевом конце, так и у донца. В первом случае инфекция проникла от больного остатка цветоноса, а поражение у донца является результатом перехода болезни от материнской клубнелуковицы, пораженной еще в предыдущем сезоне.
Во влажных условиях хранения вся клубнелуковица превращается в мягкую гниющую массу, а на ее поверхности образуются спороношения и склероции гриба. На вегетативных органах в поле спороношение образуется на засохших листьях в нижней части (табл. VI, 3). Заражение клубнелуковиц может происходить еще в поле, а также в период сушки образовавшимися на отмерших листьях конидиями.
Инфекция в клубнелуковицах может сохраняться из года в год, не проявляя себя на вегетирующих растениях. Появление болезни в текущем году в поле происходит за счет прорастающих в почве склероциев и образования спор. Склероции могут развиваться на гниющих растительных остатках и сохраняться жизнеспособными в течение многих лет.
С коричневой сердцевинной гнилью гладиолусов рекомендуются следующие меры борьбы:
1. Возвращение гладиолусов на прежнее место не раньше чем через 5 лет.
2. Сжигание сильно зараженных клубнелуковиц.
3. Выкапывание и сжигание всех тех клубнелуковиц, у которых в период вегетации развивается искривленная ботва и пожелтевшие листья.
4. Клубнелуковицы без видимых следов поражения, но могущие содержать гриб на своей поверхности, тщательно просушивать при температуре 25—30° сразу же после уборки.
5. При хранении клубнелуковиц температура должна быть 4—6°, а относительная влажность воздуха не выше 70%.
6. Опыливание сухим гранозаном из расчета 3—5 г на 1 кг клубнелуковиц.
7. Опрыскивание растений не менее 2 раз в месяц в течение всего периода вегетации 1—2 %-ной бордоской жидкостью или 0,7—0,75%-ным раствором цинеба.
Серая плесень, или гниль, лилий давно известна и распространена на всем земном шаре. Некоторые виды, как например Liliutn candidtun L., могут быть уничтожены целиком этой болезнью, если условия для ее развития благоприятны. Возбудителем является В. elliptica (Веге.) Cooke. Гриб поражает растения во всех фазах развития, появившиеся всходы могут оказаться больными, при этом поражается точка роста и прекращается развитие побега.
Наиболее известным симптомом является пятнистость листьев. Пятна вначале небольшие до 2 мм в диаметре, округлой или удлиненной формы со светлой срединой и красновато-коричневым окаймлением. При благоприятных условиях— холодной и влажной погоде — пятна увеличиваются, сливаются, достигая 1 см и больше в диаметре.
Гриб может поразить стебли вблизи поверхности почвы. При сильном поражении желтеют все выше расположенные
листья. Бурые пятна могут появляться на бутонах и цветках. В прохладную и влажную погоду болезнь очень быстро развивается и цветки превращаются в мокрую, ослизненную массу, покрытую налетом спор. Размеры спор 20—28X13—28 мк. Сухая и теплая погода затормаживает развитие болезни и пятна подсыхают.
Американский исследователь К. Гулд указывает, что оптимальной температурой для заражения листьев является 15,5°, но после того, как гриб проник в растение, он лучше развивается при температуре 21,1°. После попадания споры на лист растения при благоприятных условиях можно наблюдать образование водянистого пятна через 10 часов, а через несколько дней развивается спороношение, обеспечивающее новое заражение. Таким образом, в течение вегетационного периода цикл развития гриба повторяется несколько раз.
В это же время на пораженных листьях растения образуются склероции округлой, эллиптической, а иногда неправильной формы, длиною 1—5 мм, шириной 0,5—1 мм. Они прорастают следующей весной и образуют конидии.
Гриб может перезимовывать не только в форме склероциев, но и как сапрофит на растительных остатках. У некоторых видов лилий он перезимовывает в прикорневых розетках листьев.
Наиболее восприимчивым к заболеванию видом считают Lilium candidum L., менее восприимчив L. longifloruin Thnb., а среди устойчивых видов отмечают L. giganteum Wall.
Меры борьбы с заболеванием рекомендуются следующие:
1. Не высаживать растения на пониженных и сырых участках.
2. При сильном развитии болезни следует сменить участок, а луковицы после выкопки опыливать серой.
3. При разведении культуры в оранжереях следует проветривать их, поддерживая ровную температуру, чтобы влага не скапливалась на листьях.
4. Опрыскивать растения и почву препаратами, содержащими медь и серу.
Гниль тюльпанов, возбудителем которой является В. tulipae (Lib.) Lind, передается через больные луковицы или зараженную почву. На растительных остатках, а также на шелухе луковиц гриб перезимовывает в стадии склероциев, которые при прорастании весной образуют спороношение. Если молодые ростки развиваются на здоровых луковицах, то, пробиваясь через зараженную почву, они соприкасаются с грибом и поражаются заболеванием. Слабо пораженные луковицы также дают больные ростки. В любом случае всходы тюльпанов слабо развиты, искривлены и засыхают в сухую погоду. При избытке влажности больные ростки превращают-
’Ся в бесформенную массу серовато-бурого цвета, покрытую спорами гриба. От первоначальных очагов инфекция может распространяться на другие растения. В первую очередь ею поражаются листья. На них образуются сначала мелкие, а затем увеличивающиеся пятна бурой окраски с более темным водянистым краем. Вскоре в центре пятна развивается спо-роношение. Споры бесцветные или светло-серые 12—24X8— —20 мк. Крупные пятна, находящиеся у основания листа и на стебле, могут стать причиной переломов.
Очень восприимчивы к заболеванию цветки. На них также образуются бурые пятна с серым налетом спор.
На луковицах болезнь проявляется в образовании вдавленных желтых или бурых воронкообразных углублений на наружных мясистых чешуях. В углублениях на наружных покровах луковиц, а также на пятнах в нижней части листьев и стеблей развиваются мелкие склероции, обеспечивающие перезимовку гриба на растительных остатках (табл. VI, 4).
Несмотря на то что В. tulipae может поражать здоровые растения, проникновению инфекции способствуют различные повреждения кожицы (орудиями ухода, градом, морозом).
В борьбе с болезнью следует применять все агротехнические приемы в сочетании с химическими мерами борьбы:
1. Возвращать тюльпаны на прежнее место не раньше чем через 3 года.
2. Уничтожать больные луковицы и удалять с поля пораженные растения.
3. Избегать загущенных посадок.
4. Опыливать луковицы перед посадкой препаратом АБ, а во время вегетации опрыскивать растения коллоидной серой.
Род Monilia Pers. Широко распространен на косточковых и семечковых плодовых породах. Некоторые виды имеют многолетнюю грибницу, зимующую в ветвях дерева, у других грибница сохраняется жизнеспособной в склероциях. Кони-диальное спороношеиие может развиваться с ранней весны до осени на вегетирующей грибнице, а также при прорастании склероциев. Конидиеносцы расположены группами, выступают через разорванную кожицу пораженного органа и несут на своих концах цепочки бесцветных овальных конидий. У разных видов размеры конидий варьируют в пределах 9—25Х Хб—15 мк.
Монилиальный ожог косточковых и семечковых пород сильно распространен на вишнях и сливах в условиях влажного климата. Возбудителем его является Monilia cinerea Schrot. Зимующая грибница сохраняется в ветвях дерева и в мумифицированных плодах.
Болезнь обнаруживается весной в период цветения вишни и сливы. Ее первые симптомы — побурение и засыхание цвет
ков. Вслед за цветками засыхают листья, молодые веточки, а также однолетние побеги. Пораженные части растения, в том числе и цветки, не опадают, а остаются на дереве, даже до следующего года. При сильном развитии болезни дерево выглядит опаленным. Отсюда и произошло название болезни мо ни л иальный ожог. Завязи, пораженные в начале развития, буреют, засыхают и не опадают. Поражение плодов в период созревания проявляется в образовании небольших темных пятен, быстро разрастающихся по всему плоду. Мякоть плода буреет, а поверхность покрывается мелкими, а иногда сливающимися подушечками спороношения гриба серого цвета. Эта фаза проявления монилиоза обусловила второе название болезни — серая гниль. Под влиянием гриба плоды засыхают, мумифицируются, сохраняя грибницу до следующего года.
Спороношение гриба может развиваться не только на пораженных плодах в период их налива, но также на черешках листьев, цветоножках и частях цветка. Таким образом, запас инфекции все время увеличивается и болезнь все больше распространяется в насаждениях вишни и сливы. Конидии М. cinerea мельче, чем у других видов,— 9,5—12X6—9.
Распространению конидий способствуют насекомые. Конидии легко прорастают на рыльцах пестиков и пыльниках, откуда мицелий гриба распространяется в завязи, цветоножку и веточки. Все пораженные части буреют и засыхают. Однако грибница сохраняет жизнеспособность в течение зимнего периода, и весной образуется обильное спороношение на всех засохших и оставшихся частях растения. Инфекция попадает на распускающиеся цветки и листья, и болезнь возобновляется.
Влажная и теплая погода в период цветения деревьев способствует образованию конидиальных подушечек, прорастанию конидий и распространению болезни. Поражению созревающих плодов способствует различное повреждение кожицы вредителями сада, птицами, градом, а также простое соприкосновение больных плодов со здоровыми.
Меры борьбы с монилиальным ожогом косточковых должны быть прежде всего направлены на уничтожение всех пораженных частей растения в текущем вегетационном периоде. Рекомендуется также опрыскивание деревьев бордоской жидкостью. Первое опрыскивание проводится по «розовому бутону» раствором 3—4 %-ной концентрации (так называемое «голубое опрыскивание»), второе — после цветения, третье — через месяц после второго. Второе и третье опрыскивания производят 1%-ным раствором бордоской жидкости с добавлением парижской зелени для одновременной борьбы с казаркой и плодожоркой.
На Дальнем Востоке и в некоторых других районах Советского Союза (Адыгейская автономная область) обнаружен монилиальный ожог яблони. Заболевание наносит существенный вред яблоне весною, так как гриб поражает цветки завязи и молодые веточки. В дальнейшем на уже оформившихся плодах вплоть до их созревания гриб не получает развития.
В борьбе с монилиальным ожогом яблони рекомендуется проводить опрыскивание деревьев до цветения 3%-ным раствором медного купороса, а после цветения—1%-ным раствором.
Монилиоз, или плодовая гниль, губит значительную часть плодов яблони и груши как в период их созревания, так и при хранении. Вызывается заболевание грибом Monilia fructigena Pers. Начинается болезнь появлением на плоде бурого пятна, вскоре распространяющегося на всю поверхность. Мякоть плода размягчается, становится губчатой, теряя вкусовые качества. На поверхности таких плодов вскоре развивается спороношение гриба в виде серовато-желтых подушечек, расположенных правильными концентрическими кругами (рис. 56). Конидии М. fructigena крупнее, чем у
Рис. 56. Monilia fructigena'.
а — пораженный плод; б — разрез через конидиальнукэ подушечку.
М. cinerea,— 17,5—25X11—15 мк и расположены также цепочками на коротких конидиеносцах. Правильное расположение подушечек на поверхности пораженных плодов объясняется тем, что развитие мицелия от места первоначального заражения идет равномерно во все стороны. По мере того как грибница достигает физиологической зрелости, на ней появляется спороношение.
Пораженные плоды, оставшиеся висеть на дереве, мумифицируются, сохраняя жизнедеятельную грибницу. Весною плоды покрываются конидиальными подушечками и являются источником первичной инфекции. Наиболее быстрого развития
плодовая гниль достигает при достаточной сырости и температуре 24—28°. Конидии прорастают через 12 часов, а инкубационный период равен 3—5 дням. В зараженных плодах, положенных на хранение, болезнь развивается медленнее, так как условия хранения плодов характеризуются относительно низкими температурами. Спороношеиие не развивается, плоды буреют, а потом чернеют.
Проникновение мицелиального ростка в плод возможно только в случае повреждения кожицы насекомыми, градом и другими причинами или же при длительном соприкосновении больных плодов со здоровыми.
Распространению плодовой гнили в значительной степени способствуют вредители сада, например казарка. Этот жук питается плодами, пораженными грибом. Вместе с мякотью жук поглощает и конидии, которые, проходя через его кишечник, не теряют способности к прорастанию. Откладывая яйца в здоровые плоды, самка замазывает ямку, в которой находится яйцо, экскрементами и таким образом заносит конидии непосредственно в мякоть плода. Личинки жука казарки могут жить только в плодах, пораженных грибом.
Мероприятия по борьбе с плодовой гнилью должны быть направлены одновременно и на уничтожение вредителей сада.
Рекомендуется проводить трехкратное опрыскивание сада 1%-ной бордоской жидкостью: первое, предупреждающее,до цветения по «розовому бутону», второе — при появлении первой падалицы, третье — через 10 дней после второго. При втором и третьем опрыскивании к бордоской жидкости следует добавить парижскую зелень.
Из санитарно-профилактических мероприятий рекомендуется тщательная уборка падалицы, а осенью — сбор и уничтожение мумифицированных плодов.
Род Verticillium Nees. Вертициллиум — почвенный гриб. Он может вести сапрофитный образ жизни на различных растительных остатках и вместе с тем является патогенным организмом для растений самых различных семейств. Поражая растения, гриб проникает в сосудистую систему, развивая мицелий в сосудах, проводящих воду, т. е. вызывает трахео-микозное заболевание, проявляющееся в увядании растений. Конидиеносцы Verticillium мутовчато-разветвленные, с заостренными концами. Конидии шаровидные или яйцевидные. Окраска конидиального спороношения беловато-сероватая или ржавого оттенка. Спороношеиие гриба крайне редко развивается на поверхности пораженных органов. Обычно его можно видеть во влажной камере на кусочках пораженных частей стебля, корня и листьев. В пораженных тканях растения гриб может образовывать микросклероции, обладающие высокой стойкостью (сохраняются в почве 3—4 года).
Вертициллиозное увядание картофеля и томатов вызывается Verticillium albo-atrum Rke et Berth. Болезнь одинаково протекает на обеих культурах. Влияние гриба особенно сильно сказывается в засушливые периоды.
Болезнь начинает проявляться в фазе цветения. Прежде всего у растений понижается тургор (вначале только днем). Позднее привядание держится более продолжительное время. Затем начинают желтеть листья, на них появляются темные пятна с желтоватой каймой, и листья со временем отмирают. Мицелий гриба проникает в растение из почвы и развивается в элементах ксилемы, постепенно заполняя сосуды корня, стебля, черешков листа, жилок, вызывая их потемнение. Конидиальное спороношение развивается также внутри сосудов. Конидии бесцветные, продолговато-овальные 5—12x3 мк. Гриб поражает и глазки клубней, они чернеют и сгнивают. На месте погибших глазков образуются впадинки, в которых развивается спороношение. При слабом поражении высаженные клубни дают ростки, на которых со временем все же проявляется болезнь.
V albo-atrum — многоядный организм и способен заражать многие растения: огурцы, дыню, тыкву, хлопчатник, табак, землянику и др.
Специальных мер для борьбы с вертициллиозным увяданием картофеля и томатов не разработано. Рекомендуется проводить общепрофилактические мероприятия и прежде всего удалять растительные остатки, на которых гриб зимует в виде микросклероциев.
Увядание, или вертициллиозный вилт, малины распространено в Западной Европе и Северной Америке. Так как возбудителем болезни является все тот же V. albo-atrum, не исключена возможность ее появления и в Советском Союзе. Вертициллиозное увядание малины в Америке называется «голубым стеблем» (Джефферс, 1956). Сильно пораженные молодые побеги приобретают голубую окраску. При меньшей степени поражения голубая окраска распространяется только на отдельные сосудисто-волокнистые пучки, отчего стебель кажется полосатым. Листья на таких побегах желтеют и погибают. В более тяжелых случаях болезнь может вызвать гибель старых побегов, а также загнивание и гибель корней. Наиболее острая форма болезни наблюдается на тяжелых, плохо дренируемых почвах. Черная малина отличается большей восприимчивостью к вер-тициллиозному увяданию, чем красная. В борьбе с этой болезнью рекомендуется не производить посадок малины на тех участках, где произрастали культуры, поражаемые вер-тициллиозом.
На зараженных вертициллиозных участках можно производить новые посадки малины только в том случае, если почва была продезинфицирована фунгицидом, содержащим медь, или раствором формалина (1 часть 40%-ного формалина на 50 частей воды). В этом случае посадка может быть произведена не раньше чем через месяц.
В последнее время появилось сообщение Злотина (1964) о развитии вертициллиоза на яблоне в Таджикской ССР, проявляющемся в виде ожога. Возбудителем болезни является Verticillium sp. Болезнь обнаруживается уже во время цветения. Чашелистики, лепестки и тычинки буреют и гибнут, иногда соцветия вообще недоразвиваются. В период распускания почек на молодых листочках обнаруживаются мелкие разбросанные бурые пятна, на которых развивается спороношение гриба, заметное только под микроскопом. Через некоторое время плодоножки буреют, а плоды усыхают, что сопровождается образованием бурых вдавленных пятен с концентрическими кругами, постепенно разрастающимися. Заражение боковой поверхности плода происходит только в местах ранений. Со временем окраска плода становится темно-бурой с фиолетовым оттенком. Часть плодов постепенно опадает, другая мумифицируется и остается висеть на дереве до следующего года.
В течение вегетационного периода болезнь на листьях не развивается, однако конидиальное спороношение, образовавшееся на некоторых молодых листочках, может способствовать развитию болезни на плодах. В борьбе с вертициллиоз-ным ожогом яблони рекомендуется опрыскивание деревьев 8%-ным раствором карболинеума в зимний период и 1%-ной бордоской жидкостью в период распускания почек.
Вертициллиозный вилт хлопчатника встречается во всех зонах, где возделывается хлопчатник. При этом потери урожая весьма значительны. Обследования посевов, проведенные А. А. Бабаяном в 1928 г. в Армении, показали, что 60,8%) всех площадей поражены этим заболеванием. Каждый больной куст сравнительно устойчивых сортов теряет 30 %) урожая; у сильно восприимчивых потери достигают 60% с каждого куста.
На вредоносность вертициллиозного увядания указывает и Дж. Пресли. В Калифорнии недобор урожая вследствие развития вертициллиозного увядания может достигать 50%), а потери, равные 10—15%), считаются обычным явлением.
Возбудителем увядания является Verticillium albo-atrum Rke et Berth., живущий в почве.
Некоторые исследователи — Ван дер Меер, В. В. Лунд-брук, Е. 3. Окнина — считают, что возбудителем вертициллиозного увядания может быть второй вид V. dahliae, способ
ный давать микросклероции. А. А. Бабаян на основании изучения вертициллиозного увядания хлопчатника в Армении пришел к заключению, что V. dahliae и V. albo-atrum один и тот же вид, но обладающий большой изменчивостью свойств в зависимости от условий существования питающих растений. Род Verticillium в природе широко распространен и является многоядным, однако, развиваясь на хлопчатнике, становится разборчивым. Поражая в сильной степени сорта Gassypium hirsu-turn, слабо поражает сорта G. barbadense. Такая «избирательная способность» в отношении выбора растений-хозяев отмечается не только в Советском Союзе, но и в США (Дунин, 1946).
Вертициллиозное увядание — это болезнь в основном взрослых растений. В силу своих физиологических способностей они обладают большей восприимчивостью. Кроме того, их корни находятся в большем контакте с имеющейся в почве инфекцией.
Гриб проникает в корневую систему через прорывы коры, которые образуются благодаря выходу боковых корней. Инфекция может попадать также и при ранении коры орудиями ухода и всевозможными вредителями. Гифы гриба проникают в сосуды, паренхимную ткань, в волокна либроформа. В сосудах гриб проходит все стадии развития — образует мицелий, конидиальное спороношеиие, а под конец вегетации микросклероции и хламидоспоры. Конидии токами воды разносятся по растению и вызывают возникновение новых очагов болезни, в том числе и в листьях. В результате пораженные грибом участки можно найти на разных ярусах стебля. Симптомы болезни в начале ее развития появляются на нижних листьях, а затем на средних и верхних. Края листьев и участки пластинок между жилками желтеют, листья становятся пятнистыми. Сильно зараженные растения теряют все листья и большую часть коробочек. Развитию болезни благоприятствует температура 23—25° От общего количества образующейся новой вертициллиозной инфекции, ежегодно поступающей в почву хлопковых полей, до 70% падает на долю зараженных листьев, в которых образуется масса микроскле-роциев. В связи с этим основным мероприятием в борьбе с вертициллиозным вилтом хлопчатника является удаление с поля зараженных растений вместе с корнями после уборки урожая.
Сем. Dematiaceae
Грибы, принадлежащие к этому семейству, характеризуются наличием прямых или коленчато-изогнутых конидиеносцев, всегда окрашенных, собранных пучком, но друг с другом не
сросшихся. Конидиеносцы отходят непосредственно от мицелия или от сплетения его гиф. Конидии могут быть окрашенные и бесцветные, иногда одноклетные, либо двухклетные или многоклетные. Наиболее распространенными родами, имеющими практическое значение, как возбудители болезней растений являются: Fusicladium, Helminthosporium, Cercospora, Macrosporium и др.
Род Fusicladium Bonord. Как указывают Н. И. Васильевский и Б. П. Каракулин (1937), вегетативный мицелий Fusicladium разрастается главным образом под кутикулой или в клетках кожицы, образуя древовидноразветвленные тяжи. Конидиеносцы выступают наружу через разрыв кожицы, одиночные, небольшими пучками или сплошными дерновинками, но никогда не срастаются между собой. Обычно неразветв-ленные прямые, иногда коленчато изогнутые с небольшими рубчиками на боковой поверхности — местами прикрепления конидий. Конидии окрашенные, одиночные, образующиеся на вершине или с боков, овальные, яйцевидные или обратно-булавовидные. Вначале одноклетные, затем с одной-двумя перегородками. Наиболее распространенным видом является F. dendriticum (Wallr.) Fuck. (см. гл. Ill Ascomycetes).
Род Helminthosporium Link. Представители этого обширного рода паразитируют преимущественно на культурных и дикорастущих злаках. В одних случаях грибница может пронизывать отдельные органы растения (листья) и носит характер диффузной; в других— она местная, развивается на небольших участках тканей растения. В обоих случаях на грибнице развивается конидиальное спороношение. У некоторых возбудителей гельминтоспориоза грибница может быть покоящейся. Получая незначительное развитие в семенах в текущем вегетационном периоде, она сохраняется до весны следующего года.
Конидиеносцы гриба прямые, узловатые, слабо разветвленные, темной окраски, выходят пучками, конидии цилиндрические или удлиненно булавовидные, со многими перегородками, в большинстве случаев такой же окраски, как и конидиеносцы. Конидии обладают большой стойкостью, сохраняют жизнеспособность в разных условиях: в почве, на растительных остатках и на поверхности семян.
Гельм интоспориозы злаков. На злаках могут паразитировать различные виды рода Helminthosporium, но наиболее распространенными являются Н. gramineum Ro-benh., Н, teres Sacc., H. sativum R.K. et В. Развитие этих видов на злаках проявляется в разных формах заболевания: пятна на листьях, корневая гниль, почернение зародыша.
Широко распространенным является Н. gramineum, вызывающий полосатую пятнистость листьев ячменя.
В связи, с тем что инфекция может передаваться через почву и с семенным материалом, болезнь проявляется очень рано и в дальнейшем развивается вплоть до уборки ячменя. Мицелий Н. gramiiieum поражает всходы и сосредоточивается вблизи точки роста, а по мере развития растения прони-
Р и с. 57. Нelminihosporlum grarnineunv.
а — пораженный лист; б — спороношение гриба и прорастание конидий.
кает в листья. Развитие гриба в тканях листьев вызывает образование на их поверхности удлиненных пятен, вначале бледных, а затем окаймленных темным ободком (рис. 57). В дальнейшем пятна сливаются по длине листа, образуя полосы, нередко тянущиеся от верхушки до основания. В результате ткань на пораженных участках отмирает и листья расщепляются на продольные полосы. С пластинок болезнь передается и на влагалища. На отмершей ткани во влажных
условиях развивается спороношеиие гриба. Короткие конидиеносцы выступают из устьиц и несут многоклетные вытянутые конидии (80—110X12—23 мк). В период цветения споры гриба попадают на цветки и прорастают. Мицелий проникает в завязь, где и сохраняется в покоящемся состоянии, не оказывая существенного влияния на развитие семени.
Во время обмолота споры гриба могут прилипать к поверхности здоровых семян и сохранять способность к прорастанию до весны. Таким образом, возбудитель полосатого гельминтоспориоза ячменя способен сохраняться в виде внутренней и наружной инфекции на семенах.
Гриб может образовывать склероции на пораженных тканях растения и зимует в этой стадии на растительных остатках в поле. Как показали исследования Дресклера еще в 1923 году, на склероциях весной развиваются конидиеносцы с конидиями.
Большая вредоносность полосатого гельминтоспориоза ячменя очевидна. Поражение ассимилирующей поверхности листьев сказывается отрицательно на продуктивности фотосинтеза, а это, в свою очередь, влечет недоразвитие, щуплость зерна. Всхожесть пораженного ячменя низкая, а общее количество урожая иногда снижается на 50% и более.
Еще два возбудителя гельминтоспориоза Н. teres и Н. sativum в отличие от Н. gramineum не способны давать диффузно-распространенную в листьях грибницу. Каждое пятно на листе является результатом местного заражения и возникает благодаря прорастанию конидии.
Н. teres поражает ячмень. Пятна на листьях небольшие, по краю желтоватые, продолговатые, с сетчатым рисунком, расщепления листовых пластинок не наблюдается. Цилиндрические конидии у этого вида могут быть несколько длиннее — 30— 175 X15—22 мк.
Третий возбудитель гельминтоспориоза 77. sativum, помимо ячменя, больше всего поражает пшеницу. Кроме того, этот вид широко распространен на таких дикорастущих злаках, как тимофеевка, лисохвост, костер безостый, ежа сборная и др. Все они могут являться резерваторами инфекции гельминтоспориоза.
Грибы из рода Helminthosporium могут вызывать корневые гнили. Пораженные зерна или заразившиеся из почвы при прорастании дают ослабленные растения с побуревшей корневой системой, которая очень скоро теряет способность к росту и развитию. При высокой агротехнике, обеспечивающей благоприятные условия для развития узла кущения, растения выправляются, в противном случае погибают.
Существует еще одна форма проявления гельминтоспориоза— образование черного зародыша. Наиболее
восприимчивой к этой форме является пшеница. Проявление болезни заключается в образовании на зерне вдавленностп и почернения в области зародыша. В этой части зерна развивается грибница. Заражение происходит еще в поле в период молочной и даже восковой спелости зерна. Влияние гриба на всхожесть зерна различно. При сильном развитии мицелия в области зародыша (а это зависит от погодных условий во время заражения) зерно теряет способность к прорастанию. Если грибница развита незначительно, зерна могут прорасти, но проростки часто бывают уродливыми.
Меры борьбы с гельминтоспориозом рекомендуются следующие: предпосевная очистка семян от щуплых зерен; протравливание семян ртутными препаратами (гранозан, мерку-ран); термическое обеззараживание как и против пыльной головни пшеницы и ячменя; уничтожение сорняков; лущение стерни с целью уничтожения растительных остатков, на которых может происходить зимовка паразита.
Исследования Л. Р. Войтовой (1963) биологических особенностей возбудителей гельминтоспориозов злаков в Белоруссии, а также путей их распространения показали, что возбудитель полосатого гельминтоспориоза ячменя в цикле развития имеет склероции, которые после перезимовки могут давать, кроме конидиального спороношения, еще и сумчатое. Таким образом, перезимовка полосатого гельминтоспориоза происходит главным образом на растительных остатках, хотя не исключена возможность передачи заболевания и с семенным материалом. Н. sativum также в значительной степени передается с семенами.
Углубленные экспериментальные исследования полиморфного рода Helminthosporium японскими микологами Ниси-кадо (1929) и Ито (1930) показали, что по целому ряду признаков, в том числе по способности давать определенный род сумчатого спороношения (Pyrenophora), некоторые виды должны быть исключены из рода Helminthosporium и отнесены к роду Drechslera Ito.
Позднейшие исследования советского миколога М. К. Хохрякова (1953) морфологических и культурных признаков, а также специализации и циклов развития представителей рода Helminthosporium подтвердили основные положения японских микологов. К выделенному роду Drechslera относят Н. gramineum, называя его Drechslera graminea Ito и Н. teres — D. teres Ito.
Род Cercospora Fresen. Род Cercospora выделен в 1863 году Фрезениусом. К настоящему времени известно около 900 видов этого рода, причем все они являются паразитами высших растений из самых разнообразных семейств.
Паразитизм видов Cercospora главным образом эндофитный, но изредка может возникать и наружная форма. Развивается церкоспороз преимущественно на листовых пластинках, иногда поражаются и другие части растений: черешки, стебли, плоды и семена. Поражение листьев приводит к образованию пятнистости. Вегетативный мицелий распространяется в тканях растений, а также может скопляться в виде плотных клубочков в подустьичных полостях. От мицелия отходят пучки конидиеносцев, выходящие через устьица. В более редких случаях они выступают наружу, прорывая эпидермис. Конидиеносцы Cercospora окрашенные, прямостоячие или располагаются под углом, а в некоторых случаях стелются по субстрату; на поверхности имеют небольшие рубчики — следы отпавших конидий. Конидии бесцветные или окрашенные, цилиндрической, обратно-булавовидной или червеобразной формы, кверху утончающиеся, с несколькими перегородками. Размеры конидиеносцев и конидий, а также число перегородок в них могут в сильной степени варьировать в зависимости от погодных условий. Так, во влажные периоды размеры конидиеносцев и конидий значительно больше, чем в сухую погоду. Количество перегородок также больше у конидий, образовавшихся при повышенной влажности.
Некоторые представители рода Cercospora не отличаются резко выраженной специализацией. Такое свойство грибов сказывается и на морфологических особенностях их кониди-ального аппарата. Один и тот же вид, развиваясь на разных растениях-хозяевах, может иметь различие в размерах и форме конидиеносцев и конидий. Некоторые виды Cercospora образуют склероции, а для очень немногих установлена связь с сумчатой стадией из семейства Mycosphaerellaceae порядка Sphaeriales.
На сахарной и столовой свекле паразитирует Cercospora beticola Sacc. Особенно опасен этот гриб для посевов сахарной свеклы, так как его развитие значительно снижает урожай и сахаристость корнеплодов, а также их лежкость в буртах.
Внешние признаки церкоспороза свеклы в период вегетации растений заключаются в появлении на листовых пластинках своеобразных округлых белых или пепельно-серых пятен с красновато-бурым ободком (рис. 58). Величина и расположение их значительно варьируют в зависимости от сорта и метеорологических условий.
На черешках листьев и стеблях также возникают подобные пятна, только форма их соответственно органам вытянутая. Со временем в центре пятен развиваются бурые конидиеносцы, несущие бесцветные конидии.
Исследованиями В. К- Горовец в Белоруссии установлено, что морфологические признаки гриба — длина конидиеносцев и конидий — могут изменяться в зависимости от погодных условий. Понижение температуры и относительной влажности воздуха приводит к уменьшению размеров конидий и количества поперечных перегородок в них.
Рис. 58. С er corpora beticolcn
а — пораженный лист; б — спороношеиие гриба.
Зимует гриб при помощи мицелия в растительных остатках и в поверхностных слоях почвы. Весною на перезимовавшем мицелии образуются конидии, способные производить заражение новых посевов, причем инфекция проникает через устьица при наличии капельно-жидкой влаги. Средние листья растений оказываются наиболее подходящими для попадания инфекции, а поэтому на них-то и развивается гриб. Дальнейшее расселение паразита происходит за счет образовав
шихся на листьях конидий, в распространении которых основное значение имеют брызги дождя. Одиночные конидии могут переноситься и воздушными течениями.
Cercospora beticola обладает широкой избирательной способностью и может развиваться на целом ряде растений из различных семейств: щавеле, мальве, лебеде, подорожнике, люцерне, горохе и др.
Основное мероприятие в борьбе с церкоспорозом свеклы— это опрыскивание 1%-ной бордоской жидкостью или 1 %-ной суспензией цинеба. Большое значение имеют и такие профилактические приемы, как удаление с поля растительных остатков (листья) и борьба с сорняками, на остатках которых паразит может перезимовывать.
Род Macrosporium Fries. Виды Macrosporium могут развиваться как паразиты главным образом на картофеле и томатах, реже — на баклажанах, а как сапрофиты — на различных растительных остатках. Как паразитные организмы эти грибы вызывают образование характерных бурых пятен главным образом на листьях растений, на которых со временем появляется конидиальное спороношение.
Конидиеносцы чаще расположены группами. Конидии — обратно-булавовидной формы, с большим вытянутым придатком, с несколькими поперечными и одной-двумя продольными перегородками. Паразитные виды Macrosporium — теплолюбивые организмы, развивающиеся наиболее интенсивно при температуре 25—30°.
Сухая, или коричневая, пятнистость листьев картофеля и томатов вызывается Macrosporium solani Ell. et Mart, на картофеле и томатах и М. tomato Ckl. на плодах томата. Наибольшего развития оба представителя достигают в южных районах, а в средней полосе — в годы с жарким летом.
Уже в конце июня на нижних листьях куста можно обнаружить округлые, иногда несколько угловатые, темно-коричневые пятна с легко заметной зональностью (рис. 59). Стечением времени при благоприятных условиях развития болезни пятна могут сливаться, захватывая большую часть пластинки листа, что приводит к значительному уменьшению ассимилирующей поверхности. Конидиеносцы гриба выступают одиночно или пучками на поверхности пятен. Конидии булавовидные, буроватые с многими поперечными и 2—3 продольными перегородками, 120—200X14—20 мк.
Распространение заболевания происходит путем переноса конидий брызгами дождя и насекомыми. Зимует гриб на растительных остатках, а весною заражает сначала нижние листья растения, а затем заболеванием могут быть охвачены и выше расположенные листья.
На томатах макроспориозом поражаются и плоды. На них появляются вдавленные округлые пятна, под которыми ткань темнеет, делая плод совершенно несъедобным. Мицелий гриба проникает до семян и заражает их, поверхность плода покры
Рис. 59. Macrosporium solani'. а — пораженный лист; б — конидии гриба.
вается черным бархатистым налетом спор. В связи с тем что инфекция может сохраняться в семенах, признаки макроспо-риоза обнаруживаются на рассаде в парниках.
По сообщениям Е. П. Охова, время проявления болезни в поле зависит от предшественника. Если рассада высажена после помидоров или картофеля, первые признаки макроспорио-за обнаруживаются через 10—15 дней, а по другим предшественникам — на 9—10 дней позднее.
Из рекомендуемых мер борьбы против макроспориоза картофеля и томатов следует упомянуть глубокую зяблевую вспашку (для томатов); опрыскивание 1%-ной бордоской жидкостью (для обоих культур); севооборот.
Сем. Tuberculariaceae
Грибы из данного семейства обладают тонкой бесцветной или окрашенной грибницей. Конидиеносцы короткие, иногда ветвящиеся, собраны в более или менее рыхлые подушечки. Конидии — одноклетные или многоклетные. Некоторые виды могут образовывать хламидоспоры. По образу жизни сапро-фиты и паразиты. Из родов, принадлежащих к этому семейству, наибольший интерес представляет род Fusarium.
Род Fusarium Link. Грибы из этого рода широко распространены в природе. Обладая большим набором ферментов, они могут поселяться на самых разнообразных субстратах. Большинство представителей рода Fusarium по образу жизни являются сапрофитами, обитая в почве и на различных растительных остатках, вместе с тем они способны при определенных условиях проявлять паразитические свойства и развиваться на различных культурных и диких растениях. Грибы этого рода хорошо растут на питательных средах. В фитопатологическом отношении род Fusarium по широте распространения является важнейшим из несовершенных грибов. Однако тут не отмечается строго выраженной специализации.
Патологические явления, вызываемые этими грибами, отличаются большим разнообразием. Это могут быть гнили семян, сеянцев, корней, корне- и клубнеплодов, увядание растений, задержка в росте и т. п.
Отдельные виды рода Fusarium обладают способностью выделять токсины, действующие на растительные и животные организмы. Употребление в пищу человеком или в качестве корма животным растительных продуктов, пораженных токсическими грибами, вызывает заболевания известные под названием микотоксикозы. Токсины грибов из рода Fusarium вызывают различные по клиническим проявлениям заболевания у человека и животных. Они могут поражать нервную систему и в первую очередь центральную; влиять на слизистую оболочку полости рта и горла, вызывая алиментарно-токсическую алейкию (септическую ангину у людей) и др.
У растений действие токсических продуктов вызывает увядание, а развитие гриба в проводящей системе растения приводит к закупорке сосудов, или трахеомикозу.
В результате многолетних исследований советский миколог В. И. Билай (1955) предложила новую систематику этого рода, согласно которой род Fusarium представлен 9 секциями, 26 видами и 29 разновидностями. Такое систематическое деление значительно проще ранее существовавшей систематики рода и облегчает определение видов.
Конидиальное спороношеиие Fusarium весьма разнообразно и по морфологии конидий, и по способу их образования.
Основным фактором, влияющим на изменчивость конидиаль-ного аппарата, являются условия окружающей среды.
Фузариумы могут давать макро- и микроконидии. Форма макроконидий бывает серповидная, веретеновидно-серповид-ная с различной степенью изогнутости. При классификации этих грибов учитывается особенность строения конидий: количество перегородок, изогнутость, форма полярных клеток и т. п.
Макроконидии образуются на простых или разветвленных конидиеносцах, обычно собранных в спородохии или пионноты.
Спородохии представляют собой подушечки, покрытые с поверхности короткими конидиеносцами, а снизу образованные более или менее плотным плектенхиматическим сплетением гиф. Макроконидии дают слизистые скопления. У разных видов в массе они могут быть окрашены в оранжевый, лососевый, синий или фиолетовый тона.
Пионнот ы отличаются более рыхлым сплетением гиф, не имеющим плектенхиматического характера. Макроконидии также образуют слизистый слой. Окраска пионнот такая же.
В некоторых случаях макроконидии образуются прямо на воздушном мицелии.
Независимо от характера образования, макроконидии фузариумов всегда многоклетные. Количество клеток чаще от 4 до 6, они обычно бесцветные.
В том случае, когда макроконидии образуются в споро-дохиях и пионнотах, оболочка их выделяет слизь, которая в массе окрашена в различные цвета. У некоторых видов пигментация наблюдается в клетках мицелия. Прорастание макроконидий происходит как полярными, так и центральными клетками.
Микроконидии бывают одноклетными или двухклетными, овальной или яйцевидной формы. Характер образования микроконидий также разнообразен: они могут возникать как на более или менее разветвленных конидиеносцах, так и на коротких веточках мицелия и располагаться одиночно, цепочками или головками. В некоторых случаях они образуются в таком большом количестве, что дают порошковидные скопления на поверхности мицелия.
Значительное большинство видов Fusarium способно давать хламидоспоры в период интенсивного роста мицелия. Отличаются хламидоспоры от остальных клеток грибницы более толстой, иногда двойной оболочкой — гладкой, шиповатой или мелкобугорчатой, округлой формой и большим диаметром. В старых культурах хламидоспоры могут отделяться, и в то время как остальные клетки мицелия отмира
ют, хламидоспоры остаются в виде округлых телец, размером 12X12 мк. Они бывают бесцветными и окрашенными в охряно-бурые тона. В некоторых случаях хламидоспоры образуются на первых коротких веточках прорастающих конидий. Прорастание хламидоспор происходит одной или двумя ростковыми гифами.
Как в культурах, так и в естественных условиях существования на развитие гриба во всех его стадиях (рост мицелия, образование конидий и хламидоспор) большое влияние оказывают условия окружающей среды.
Для некоторых видов Fusarium известно сумчатое спороношение, относящееся к родам Gibberella, Nectria, Colonectria, Hypomyces.
Фузариозы хлебных злаков настолько распространены, что вред, причиняемый ими, не меньший, чем от головни и ржавчины. Вызываются фузариозы различными видами гриба. Все они мало требовательны к условиям окружающей среды, чрезвычайно пластичны, а поэтому широко распространены и занимают большие области.
Снежная плесень получила свое название за то, что поражение растений замечается сразу же после таяния снега, а развитие болезни зависит от толщины снегового покрова и интенсивности его таяния. Больше других злаков подвержены этому заболеванию озимые рожь и пшеница. Снежная плесень вызывается многими видами Fusarium, чаще же всего F. nivale Ces.
Ранней весной, сразу же после таяния снега, можно наблюдать на озимых гибель большого количества растений вследствие развития на их листьях рыхлого паутинистого мицелия. Поражение охватывает большие участки и на посевах образуются плеши как будто выпревших растений, за что эту болезнь называют еще выпреванием озимых.
Постепенно болезнь охватывает и листовые влагалища, а у основания стеблей вскоре становятся заметны розоватые подушечки спороношения гриба. Конидии с тремя перегородками 20,6—25,2x3,7—4,3 мк.
Гриб имеет сумчатую стадию (Colonectria graminicola (В. et Вг.) Wr., которая принадлежит к порядку Hypocreales. Развивается она на отмерших листьях.
F. nivale может жить сапрофитно в почве и при благоприятных условиях заражать озимые еще с осени. Если снег выпал на незамерзшую землю, болезнь развивается и под снегом. Кроме такого способа инфекции растений, снежная плесень может распространяться весной за счет перезимовавших на остатках листьев хламидоспор гриба. Семенной материал также может оказаться зараженным грибом. Развитие конидиального спороношения F. nivale можно наблю
дать на остатках погибших растений вплоть до цветения и налива зерна. Конидии гриба, попадая на развивающиеся зерновки, прорастают. Мицелиальный росток при этом внедряется в зерно, задерживая его нормальный налив. Такие щуплые зерна, находясь в посевном материале, дают пораженные проростки, которые и служат очагами инфекции.
Вредоносность болезни зависит от погодных условий. Если таяние снега идет медленно, болезнь быстро распространяется, и при поражении грибом области узла кущения растения погибают. Если же снег тает быстро и стоит солнечная и ветреная погода, развитие болезни прекращается и растения выправляются.
Снежная плесень встречается во всех районах возделывания озимой ржи и пшеницы: в нечерноземной полосе, Сибири, на Дальнем Востоке, а также во многих зарубежных странах.
В борьбе со снежной плесенью большое значение имеет весеннее боронование и подкормка озимых, очистка семенного материала от щуплого и легковесного зерна, а также мокрое и сухое протравливание ртутными препаратами.
Пьяный хлеб—вид фузариоза, вызываемый F. gra-minearum Schw.
Гриб поражает рожь, пшеницу, овес, кукурузу, ячмень, причем не только в период вегетации, но и после уборки, а также во время хранения. Развитию гриба благоприятствует влажная погода.
Внешний вид зараженного зерна зависит от степени его поражения. Сильно пораженное фузариозом зерно легковесное, морщинистое, без блеска, часто трухлявое. Окраска такого зерна от бледно-розового до темно-розового цвета.
На посевах первые признаки фузариоза обнаруживаются к моменту вызревания хлебов. Первым симптомом болезни бывает появление на некоторых колосках розоватого налета — спороношения гриба. Сами же колоски отличаются при этом более бледным цветом и кажутся как бы выцветшими. При более сильном развитии болезни конидиальные подушечки появляются не только на колосковых и цветочных чешуйках, но и на зерне (рожь), а также на влагалищах листьев и узлах. Со временем отдельные подушечки на колосках сливаются в сплошной налет, особенно в верхней части колоса. Конидиальное спороношеиие у F. graminearum в виде спородохиев. Грибница рыхлая, различной окраски — от бело-розовой и золотисто-желтой до пурпурной.
По исследованиям Н. А. Наумова, грибница может встречаться во всех частях растения, пронизывая все ткани, за исключением проводящей. Конидии гриба серповидно изогнутые с заостренной верхней клеткой и ножкой у основания,
чаще с 3—5 перегородками, размером 3,3—7,0X30—96 мк. В отдельности конидии бесцветные, но в массе дают розоватую, иногда желтовато-охряную окраску.
Конидии распространяются с помощью дождя, ветра, насекомых. Заражение растений может произойти в любой момент, даже после уборки и в период хранения.
В конце вегетации на колосках, а также и после уборки в снопах на пораженных участках развивается сумчатое спороношение Gibbirella saubinetti (Mont.) Sacc. (принадлежит к порядку Hypocrealles). Конидии и сумкоспоры не теряют способности к прорастанию в течение года и могут вызвать заражение проростков.
Развиваясь на зерновых культурах, гриб не ограничивается их поверхностным поражением. Мицелий гриба проникает в зерно. При слабом поражении грибница обнаруживается только в покровах, при сильном — она проникает в эндосперм как в алейроновый слой, так и в крахмалоносную часть. Наконец, грибница может добираться и до зародыша, нередко лишая его всхожести. В пораженном зерне мицелий гриба сохраняет жизнеспособность до 3 лет. Проникая в эндосперм семени, гриб оказывает глубокое воздействие на запасные питательные вещества, резко изменяя их химический состав. Как указывает Н. А. Наумов, развитие гриба в алейроновом слое семени приводит к разложению белковых веществ с выделением аммиака и других токсических продуктов. Пораженное фузариозом зерно приобретает ядовитые свойства. Хлеб, приготовленный из токсического зерна, получил название «пьяного» хлеба. Употребление такого хлеба человеком, равно как скармливание скоту, вызывает отравление.
Признаки отравления у человека заключаются в слабости, чувстве тяжести в конечностях, связанности походки, головокружении. Больной не способен ни к физической, ни к умственной работе. Наступает расстройство пищеварения, а зачастую и обморочное состояние. На следующий день состояние больного как после тяжелого опьянения. Длительное питание таким хлебом может отразиться на психике больных, особенно детей. При этом наступает резкое истощение и расстройство зрения.
Из животных наиболее восприимчивы к отравлению зерном лошади. Крупный рогатый скот относительно устойчив к этому роду заболевания. У животных отмечается инстинктивная самозащита в отношении болезнетворных кормов. Поевши пораженный корм, животное в дальнейшем может длительно голодать, но не прикасаться к токсическому корму.
Заболевание, возникающее от приема в пищу пьяного хлеба, впервые было обнаружено на Дальнем Востоке в
1882 году Н. А. Пальчевским. В 1891 году им была опубликована монография о пьяном хлебе и вызываемом им заболевании. Однако старожилам южных районов Приморья эта болезнь давно была известна. Китайцы называли ее «ми-чун», а пораженное зерно узнавали по розовому налету. Эта болезнь известна в Швеции, Финляндии, Германии, Северной Америке и других зарубежных странах.
В настоящее время разработана целая система мероприятий против этого вида фузариоза: тщательное отделение щуплых семян из посевного материала; протравливание семян гранозаном (1—2 кг/т); своевременная уборка урожая, не допускающая перестаивания на корню, тщательная просушка снопов перед скирдованием, засыпка зерна на хранение с влажностью, не превышающей 13—14 % •
На перезимовавших в поле по тем или иным причинам сельскохозяйственных растениях (рожь, пшеница, ячмень, овес, горох, гречка) развиваются токсические грибы F. sporotrichioides Sherb. и F. роае (Peck.) Wr. Употребление продуктов, полученных из таких растений, в пищу или на корм скоту также вызывает тяжелое заболевание — септическую ангину.
F. sporotrichioides широко распространен в Европе, Азии и Северной Америке, встречается на картофеле, хлебных злаках, горохе, косточковых, сеянцах хвойных -пород и других растениях.
В Советском Союзе в 1932 году в Казахстане наблюдалось отравление продуктами от перезимовавших в поле зерновых культур.
С 1942 года большая работа по изучению токсических свойств F. sporotrichioides проводится во Всесоюзной научно-исследовательской лаборатории по изучению ядовитых грибов под руководством А. X. Саркисова. Гриб обнаружен в образцах перезимовавших в поле злаков из Западной Сибири, центральных районов европейской части СССР, Башкирской и Татарской АССР.
Исследованиями В. И. Билай установлено, что отличительным микроскопическим признаком этого вида является наличие лимонообразных микроконидий. Ею также отмечается токсическое действие гриба на растительные ткани и целые растения. При поражении зерна мицелий гриба проникает в зародыш, вызывая его отмирание. Нанесение экстракта гриба на поверхность листьев и стеблей проростков вегетирующих растений вызывает некроз клеток и тканей растений.
Употребление в пищу человеком продуктов из зерна, пораженного F. sporotrichioides, вызывает септическую ангину (алиментарно-токсическая алейкия). Симптомом этого забо
левания являются: кровоизлияние в слизистых оболочках ротовой полости и горла, лейкопения (уменьшение числа лейкоцитов в крови). Заболевание этого же типа наблюдается и у животных. При этом часты смертельные случаи.
Другой вид токсического гриба F. роае был обнаружен Н. М. Пидопличко в 1949 году в Сумской области в соломе, скармливание которой лошадям вызвало их заболевание.
Развитие различных видов фузариумов в проводящей ткани растений вызывает явление увядания, или в и л т. Относимые сюда виды обычно живут сапрофитно в почве, откуда грибница проникает в корни, распространяется в сосудах ксилемы, закупоривает их и препятствует поступлению воды в вышележащие части.
Фузариоз льна вызывается Fusarium Uni Boll, и др. Гриб распространен во всех районах возделывания льна. Вредоносность болезни особенно велика, так как заражение может происходить во всех фазах развития растений. Проникновение гриба в ткани растения происходит главным образом через корневую систему, однако не исключена возможность попадания инфекции и через надземные вегетативные органы.
Пораженные молодые растения очень быстро увядают, образуя бурые пятна в посевах. Наибольшего развития болезнь достигает тогда, когда лен находится в фазе «елочки», т. е. имеет до 10 пар листьев. По времени это совпадает с концом июня. Лен, пораженный в этой фазе фузариозом, всегда имеет поникшую верхушку побега. Больные растения легко выдергиваются из земли, так как их корни загнивают. В сырую погоду у основания стеблей увядших растений можно заметить плесневидный налет розоватого оттенка—это мицелий и спороношеиие гриба. Конидии F. lini четырехклетные, серповидные, 27—38x3—3,5 мк, одиночно взятые— бесцветные, в массе дают розовую окраску (рис. 60).
Растения, заболевшие в более позднем возрасте, не погибают, но их стебли дают короткое, потемневшее волокно со значительно пониженной номерностью. Кроме того, гриб из стеблей проникает в коробочки, а оттуда в семена. Пораженные фузариозом семена щуплые, матовые, с пониженной всхожестью. Масло, приготовленное из фузариозных семян, обладает такими же токсическими свойствами, как и пьяный хлеб. Развитию заболевания благоприятствует повышенная влажность почвы и воздуха, а также температура 16—32°.
Первоисточником фузариоза льна может служить почва, где гриб живет сапрофитно, и семена, полученные из пораженных коробочек.
В борьбе с увяданием льна рекомендуется протравливать семена перед посевом гранозаном, возвращать лен на то же
поле не ранее чем через 5—6 лет, сеять лен после клевера п других бобовых, удалять с поля растительные остатки.
Некоторые виды рода Fusarium развиваются на картофеле. Одни из них, как например F. oxysporum Schl., вызывают
Рис. 60. Fusarium Uni'.
а — пораженное растение ; б—конидии гриба и грибница на разрезе стебля.
явление увядания, другие, как F. solani (Mart) Арр. et Wr,.— гниль клубней.
Фузариозное увядание картофеля в Советском Союзе особенно распространено в южных районах. Испытывая по
стоянный недостаток воды в почве и иссушающее действие ветров, картофель становится особенно восприимчив к этому заболеванию. Возбудитель болезни обычно обитает в почве и попадает в растение через корневые волоски. Оттуда он продвигается до подземной части стебля и развивается в сосудах ксилемы. Сосуды буреют, основание стебля при этом загнивает. Заражение картофеля происходит в июне, а видимые признаки увядания проявляются к фазе цветения картофеля, т. е. во второй половине июля. Постоянный недостаток воды в растении приводит к уменьшению урожая клубней в 2—3 раза, к значительному (до 4%) понижению содержания крахмала.
F. solani вызывает сухую гниль клубней. Симптомы этой болезни проявляются в период хранения. Вначале на клубнях появляются вдавленные места с несколько сморщенной кожурой. Затем обнаруживается спороношение гриба в виде спородохий, окрашенных или бесцветных. Конидии гриба слабо изогнуты с 1—5 перегородками. Дальнейшее развитие гнили зависит от условий хранения. При избыточной влаге развивается мокрая гниль. Мякоть клубня буреет, мокнет и сгнивает.
В условиях сухого хранения пораженные фузариозом клубни сморщиваются, на поверхности их появляется зональная складчатость, а вся мякоть пронизывается грибницей. Грибница Fusarium не разрушает крахмальных зерен, и клубни превращаются в плотные комки.
Основной мерой борьбы с заболеванием является тщательная просушка клубней перед закладкой на хранение и соблюдение необходимого режима влажности и температуры в хранилище.
Фузариозное увядание хлопчатника, вызываемое F. oxysporum Schl f. vasinfectutn Bilai, опасное и вредоносное заболевание. Оно может проявляться в любой фазе развития растений, начиная со всходов, при этом буреют семядоли, первые листочки и проростки погибают.
Первые признаки болезни более взрослых растений — остановка роста, пожелтение, увядание и опадение листьев. Изменение окраски листьев начинается с краев листовых пластинок, около жилок. На стеблях наблюдается почернение тканей древесины, а на поперечном разрезе заметно кольцеобразное побурение тканей вокруг сосудистых пучков. В случае сильного развития болезни побурение распространяется по всему стеблю вплоть до плодоножек и коробочек, откуда гриб может проникать в семена. На больных растениях коробочки раскрываются до созревания.
Гриб живет в почве, образуя нитевидную грибницу, конидии и хламидоспоры. По данным А. Смита, он может сохра
няться в почве до 25 лет даже при отсутствии хлопчатника. Проникает гриб в растения через корневые волоски. Попаданию инфекции в корни особенно благоприятствует повреждение их нематодами. Однако при особенно обильном наличии гриба в почве заражение может происходить и без предварительного повреждения корней.
Для борьбы с этим заболеванием хлопчатника прежде всего следует не допускать накопления инфекции в почве — производить тщательную уборку растительных остатков, с которых гриб может переходить на сапрофитное существование в почву. Весьма эффективно введение в севооборот растений, не поражаемых фузариозным увяданием (люцерны, злаков и др.).
Возбудителем фузариозного увядания астр является F oxysporum f. callestephi Beach. Этот вид гриба отличается узкой специализацией, и астра является его единственным растением-хозяином.
Симптомы заболевания можно наблюдать на любой фазе развития растения. У пораженных всходов болезнь проявляется в полегании растений, вызванном загниванием стеблей у поверхности почвы. Чем теплее почва, тем скорее гибнет рассада. У больных растений, достигших высоты девяти и более сантиметров, обнаруживается полосатчатость стебля в соответствии с пораженными сосудистыми пучками. Иногда такие полосы появляются на одной стороне стебля, а листья и боковые побеги, находящиеся на этой стороне, отмирают и растение становится ассиметричным. Во влажную погоду у больных растений на стеблях у поверхности почвы появляется розоватый налет мицелия со спороношением гриба. Конидии образуются в виде слизистых скоплений и переносятся водой. Во влажных условиях прорастание их возможно сразу же после образования, а при сухой погоде они сохраняют всхожесть в течение нескольких месяцев.
Гриб способен образовывать хламидоспоры, обладающие гораздо большей устойчивостью к высыханию и обработке фунгицидами, чем мицелий и конидии. В связи с этим гриб может сохраняться в почве даже при отсутствии астр около 5 лет. Перенос инфекции может осуществляться инвентарем и обувью обслуживающего персонала.
Астры восприимчивы к фузариозному увяданию, начиная от прорастания семян и до полной зрелости. Заражение возможно через совершенно здоровые корни, так как возбудитель проникает между клетками корневого чехлика. Кроме того, проникновение гриба в ткани растения возможно через ранки на стебле, поэтому следует ухаживать за растениями осторожно, избегая поранения поверхности стебля.
У устойчивых сортов астр наблюдается слабый рост мице-
Таблица VI. Развитие Botrytis на различных растениях:
1 — В. cinerea на землянике; 2 — В. anthophila на цветке клевера (а — пораженный цветок; б — конидиеносцы и конидии); 3 — В. gladiolorum на гладиолусах (засохшее растение с налетом спороношений); 4 — В. tall рае склероции на пораженной луковице тюльпана.*
лия в кончике корня. Он локализуется в виде крошечных пятнышек в местах первоначального внедрения, но жизнеспособности при этом не теряет, У восприимчивых сортов грибница проникает из кончика корня в сосуды ксилемы. Однако пышного развития не получает, сосудов не закупоривает и подача воды не задерживается. Гибель растений обусловлена действием токсинов, выделяемых грибом, которые разносятся током воды по растению.
Оптимальная температура почвы, при которой увядание принимает широкие размеры,— 25—26°, максимальный предел— 32° При 12,3° растения мало страдают от фузариоза, а при 3,9° рост гриба прекращается (по Бейке).
Для борьбы с заболеванием следует:
1. Удалять и уничтожать растительные остатки.
2. Возвращать астры на старое место не ранее чем через четыре года.
3. Известковать почву, протравливать ее за 15 дней до посадки растений раствором формалина (250 см3 на ведро воды) при количестве раствора 10 л на 1 м2.
4. Протравливать семена водной суспензией гранозана (1 г препарата на 1 л воды при экспозиции 25 мин).
Весьма распространен фузариоз гладиолусов — излюбленных декоративных растений во многих странах. Современные сорта их представляют результат 125-летней упорной работы цветоводов по скрещиванию различных видов этого рода — выходцев из Южной Африки. В настоящее время десятки миллионов клубнелуковиц высаживаются в самых разнообразных климатических поясах. С целью обмена посадочным материалом клубнелуковицы гладиолусов транспортируются из одних зон в другие, а это способствует распространению вредителей и болезней культуры.
Одной из наиболее опасных болезней гладиолусов является фузариоз, вызываемый Fusarium oxysporum f. gladioli (Mass.) Snyder et Hansen. Гриб внедряется из почвы в сосудистую систему корней клубнелуковиц и в основания листьев. Фузари-озная инфекция проявляется в виде бурой гнили клубнелуковиц, а также пожелтения и деформации надземных вегетативных органов и цветоноса. Заражение клубнелуковиц происходит еще в почве, и болезнь быстро развивается в хранилищах, если клубнелуковицы перед закладкой были недостаточно хорошо просушены.
Признаки гнили клубнелуковиц проявляются различно у разных сортов, причем в первую очередь загниванию подвергаются поврежденные при копке или перевозке клубнелуковицы. Загнивание может начаться в любой точке, но чаще всего у донца, при этом пораженная ткань темнеет. Гниль может распространяться вплоть до самой центральной части, или же,
наоборот, поражение охватывает периферическую часть клубнелуковицы, образуя округлые или овальные пятна, обычно вдавленные и хорошо заметные снаружи. Поверхность клубнелуковицы приобретает концентрическую складчатость с расположенными кругами подушечками спор гриба розовато-белого цвета. В период хранения болезнь может переноситься при помощи спор гриба на здоровые клубнелуковицы. При сильном поражении клубнелуковицы ссыхаются, мумифицируются и стучат как камни. При незначительном поражении болезнь может быть не обнаружена, но после высадки такие клубнелуковицы быстро загнивают. Фузариозная инфекция может сохраняться в клубнелуковицах в скрытом состоянии и передаваться из года в год молодым клубнелуковицам. Такое скрытое заражение обычно и является источником распространения инфекции.
Этот тип фузариоза представляет собой сосудистое заболе-гание, а поэтому поражение клубнелуковиц сопровождается патологическими явлениями и на наземных частях растения. Прежде всего наблюдается пожелтение, а затем побурение нижних листьев. Молодые листья у более взрослых растений деформируются. Цветонос становится более мягким, тонким, часто искривленным. У пораженных растений может происходить изменение формы, окраски и величины цветков, причем иногда только на одной стороне соцветия.
Фузариоз гладиолусов широко распространен во всех зонах возделывания. Американский исследователь Р. Мейджи указывает, что в 1943 году в штате Флорида это заболевание поразило такое большое количество растений, что в следующем году потребовался завоз 200 млн. клубнелуковиц сорта Пикарди взамен сгнивших.
В борьбе с фузариозом гладиолусов рекомендуются следующие мероприятия:
1. Возвращать гладиолусы на то же место не ранее чем через 4 года.
2. Тщательно выбраковывать больные клубнелуковицы перед посадкой.
3. Удалять больные растения с участка сразу после их обнаружения.
ПОРЯДОК MELANCON I ALES
Порядок меланкониальных грибов имеет всего лишь одно семейство Melanconiaceae. Для представителей этого порядка характерно наличие погруженного в субстрат мицелиального ложа, состоящего из плотного сплетения гиф, на котором развивается конидиальное спороношение. Если субстратом служат ткани растений, то вначале ложе прикрыто покровной тканью
растения. При развитии конидиеносцев и конидий это покрытие прорывается. У большинства представителей на поверхность выступают только конидии, а у некоторых и конидиеносцы.
Конидиеносцы и конидии могут быть бесцветными и окрашенными. Конидии одноклетные или с перегородками, разнообразные по форме. У большинства видов конидии развиваются на вершине конидиеносцев, но у некоторых сбоку. Большей частью они образуются по одной на конидиеносце, но в некоторых родах — по нескольку, причем иногда на более или менее -заметных стеригмах.
Почти все меланкониевые характеризуются слизистым образованием конидий, распространяющихся поэтому при помощи воды или насекомых, к телу которых они прилипают.
У некоторых представителей в конидиальных ложах имеются темноокрашенные щетинки, значительно превышающие своими размерами конидиеносцы с конидиями. Они могут быть прямыми или изогнутыми, прямостоящими или наклоненными, одноклетными или с перегородками.
Среди меланкониевых очень много паразитных форм, которые, развиваясь на сельскохозяйственных и других полезных растениях, наносят значительный ущерб урожаю, снижая количество и качество получаемой продукции. Меланкониевые грибы, развиваясь на растениях, вызывают появление пятен, а затем язв на вегетативных органах и плодах. На пораженных участках можно обнаружить конидиальные ложа гриба в виде маленьких подушечек от светлоокрашенных до грязно-бурых, почти черных, студенистой, восковой или роговой консистенции. *
Сем. Melanconiaceae
В данном семействе следует отметить роды Kabatiella, Colletotrichum и Gloeosporium.
Болезни многих сельскохозяйственных растений, вызываемые этими грибами, получили название антракнозов.
Род Kabatiella Bub. Представители данного рода развиваются как паразиты на листьях, стеблях, плодах и семенах различных растений. Конидиальные ложа, образующиеся под эпидермисом, обычно мелкие, реже — простирающиеся более широко, скоро становятся слизистыми. Конидиеносцы — тесно сплоченные, булавовидные, чаще бесцветные. Конидии —> одноклетные, продолговатые, овальные, веретеновидные, слегка изогнутые. Отчленяются они по нескольку на вершине конидие-носца и располагаются на едва заметных стеригмочках. Характерная особенность конидий — способность к почкованию еще до отчленения от конидиеносцев — является причиной интенсивного размножения гриба.
Возбудитель антракноза красного клевера — Kabatiella caulivora (Kirchn.) Karak.— поражает все надземные части растения и получил широкое распространение во всех районах клеверосеяния. Название болезни происходит от греческого anthrax — уголь; американцы называют эту болезнь Scorch—обожженное место, а немцы — Stengelbrenner, что означает «ожог стеблей». Действительно, пораженное антракнозом клеверное поле имеет вид обожженного. Растения поражаются на всех фазах развития, начиная с проростков. Гриб образует местную грибницу, локализованную только в местах поражения. Распространение болезни происходит при помощи спор, образующихся на пораженных частях растения (листовых пластинках, черешках, стеблях и головках). Размеры спор— 18—25,4X3,3—5,9 мк.
В течение ряда лет антракноз красного клевера изучался нами в условиях Белоруссии. Выяснено, что, несмотря на незначительный процент поражения семян (1), болезнь проявляется на проростках в виде поражения подсемядольного колена. Следствием этого является гибель проростка и возникновение небольшого очага болезни, от которого происходит распространение спор гриба.
Следующий этап развития болезни проявляется на листовых пластинках и наблюдать его проще. Симптомом служит образование буроватых пятен с последующим разрывом ткани углом по направлению к главной жилке. С листовых пластинок гриб переходит на черешки, вызывая петлеобразное изгибание их (рис. 61).
По мере отрастания стеблей возбудитель антракноза поражает междоузлия. На них возникают желтовато-коричневые вытянутые пятна, окаймленные черным ободком. Через некоторое время в центре пятна можно видеть даже невооруженным глазом беловато-розоватые подушечки спор гриба сначала матовые, а затем несколько слизистые. Основная масса спор гриба образуется на больных стеблях и распространяется с брызгами дождя. Затем начинается разрушение тканей стебля под пятном, образуется трещина, которая бывает настолько глубоком, что стебель переламывается. На одном междоузлии может образоваться несколько трещин. Размеры их варьируют от 0,5 до 3,5 см. О. М. Миняева установила, что антракнозом могут поражаться не только вегетативные органы растения, но и цветочные головки. В Белоруссии поражение антракнозом цветочных головок не наблюдалось.
Интересной биологической особенностью возбудителя антракноза является большая жизнеспособность грибницы. В пораженных стеблях клевера она остается жизнедеятельной и развивает спороношение после длительного и самого разнообразного хранения.
На перезимовавшей в естественных условиях грибнице образуются своеобразные плодовые тела — псевдопикниды темней окраски, несколько погруженные в ткань стебля. Созревание псевдопикнид и освобождение из них пикноспор в условиях Минской области начинается в конце третьей декады апреля.
Антракноз является самой вредоносной болезнью клевера. Его развитие приводит к преждевременному засыханию лис-
Рис. 61. Kabaiiella caulivora:
а — петлеобразное изгибание черешка листа; б — пятна и трещины на стебле; в — конидии гриба (орнг.).
тьев, разрушению проводящей ткани стеблей и как следствие к недоразвитию головок. При сильном поражении стебли растений переламываются.
В Белоруссии вредоносность антракноза сказывается сильнее на одноукосном клевере. Это следует объяснить многократным заражением и длительным развитием гриба на одном и том же травостое.
Вредоносность заболевания сказывается не только на уменьшении урожая. Под влиянием антракноза значительно снижается его качество, так как развивающийся в тканях гриб вызывает уменьшение содержания в растении сахара (глюкозы) и общего азота, что приводит к понижению питательности клевера как кормового растения.
В борьбе с антракнозом красного клевера большое значение имеет удаление с поля растительных остатков, в которых
сохраняется жизнедеятельная грибница. Весьма эффективна весенняя подкормка калийными удобрениями, внесенными в повышенных дозировках (90 кг/га и 120 кг/га К2О). Она значительно повышает устойчивость клевера к антракнозу.
Из других видов рода Kabatiella следует отметить Kabatiella Uni (Laff.) Karak., который развивается на льне во всех районах культуры и поражает семядоли, стебли, чашелистики и коробочки. Исследователь этого заболевания Лафферти (1921) отнес его возбудителя к роду Polyspora (Р. lini Laff.). С того времени заболевание получило название полиспо-роза льна, сохранившееся до настоящего времени, несмотря на то, что исследованиями Б. П. Каракулина (1950) установлено, что возбудителем болезни является Kabatiella lini.
В Белоруссии это заболевание льна детально изучалось Н. Н. Лучиной (1961). Полиспороз на льне проявляется в течение всего вегетационного периода и начинается побурением семядолей. В дальнейшем болезнь переходит на стебли, вызывая образование бурых пятен и изломов в местах поражения. Пораженные стебли дают короткое волокно желтой окраски, непригодное для текстильных изделий. К фазе цветения бурые пятна появляются и на листьях, они поникают, прилипая к стеблю. В дальнейшем болезнь проявляется на чашелистиках, коробочках и, в конце концов, гриб поражает семена.
В период вегетации на семядолях, листьях и чешелистиках развивается спороношение гриба. Спороносные ложа мелкие, конидиеносцы слабо вздутые на вершине, конидии бесцветные, округлые, овальные или слегка продолговатые 4—20x4—8 мк.
В условиях Белоруссии перезимовка К. Uni происходит хламидоспорами (Лучина, 1961). Последние образуются из клеток мицелия, развившегося в стеблях и семенах. Весною при высеве пораженных семян хламидоспоры прорастают, заражая проростки. Всходы льна могут заражаться прорастающими хламидоспорами из больных растительных остатков.
В Белоруссии все районированные сорта льна-долгунца поражаются полиспорозом, но в разной степени. Наиболее устойчивыми являются Светоч и Вайжгантас.
В борьбе с полиспорозом льна рекомендуются следующие мероприятия:
1. Тщательно сортировать семенной материал.
2. Заблаговременно протравливать семена гранозаном (1,5—2 кг/т) или ТМТД (3 кг/т).
3. Возвращать лен на прежнее место не раньше чем через 5 лет.
4. Опрыскивать посевы в фазу бутонизации и массового цветения 1%-ным раствором хлорокиси меди (800 л/га).
Род Colletotrichum Sacc. Почти все виды Colletotrichum (около 200) являются паразитами высших растений. Однако
наиболее интенсивно они поражают ослабленные растения или менее активные в жизненном отношении их части, главным образ-ом плоды.
При поражении вегетативных органов распространение грибницы всегда бывает местным, а в случае поражения плодов мицелий может пронизывать весь плод. Конидиальные ложа развиваются в местах поражения, обычно в клетках эпидермиса, реже — под кожицей и после ее разрыва становятся заметны невооруженным глазом. Окраска их может варьировать у разных видов от розовато-желтоватой до грязно-бурой. Поверхность ложа слизистая или воскоподобная от массы образующихся конидий, склеенных студенистым веществом. Конидиеносцы развиваются плотным слоем. Вначале, они бесцветные, а у основания более или менее окрашенные, цилиндрические или слегка булавовидные, у основания часто разветвленные, одноклетные или с перегородками. Между ко-нидиеносцами обычно образуются окрашенные щетинки, располагающиеся по краям ложа или по всей его поверхности. Щетинки бывают одноклетными или снабжены перегородками. Значение щетинок для гриба заключается в том, что они, разрывая и приподнимая верхнюю стенку эпидермиса или весь эпидермис, облегчают конидиям выход наружу. Конидии образуются на вершине конидиеносца по одной. Они одноклетные, яйцевидные, продолговатые или овальные, иногда слабо согнутые. Одиночные конидии бесцветные, а в массе розоватые, оранжево-желтые, а иногда коричневые, размером 10—25x3,5—6 мк.
Сравнительно немногие представители этого рода имеют сумчатую стадию типа Glomerella. Однако эта стадия играет незначительную роль в сохранении и распространении того или иного вида.
Гриб обычно зимует ложами или мицелием, а иногда — склероциальными подушечками и пикнидами.
Сельскому хозяйству виды рода Colletoiriclium наносят большой ущерб, так как многие из них поражают и губят вегетативные органы и плоды овощных, плодовых и технических культур.
Антракноз фасоли вызывается Colletoirichum Нп-demuihianuin (Sacc. et Magn.) Bl. et Cav. и широко распространен в Америке и многих странах Европы, в том числе и в Советском Союзе. Впервые антракноз фасоли был обнаружен в Италии Линдемутом, откуда и происходит видовое название возбудителя болезни.
Заболеванием поражаются все органы растения, даже корни, наибольшего развития болезнь достигает на плодах.
Как указывает Д. Н. Тетеревникова-Бабаян (1965), признаки антракноза можно наблюдать уже на сеянцах фасоли,
а именно на семядолях и подсемядольном колене. По мере роста растений болезнь переходит на вегетативные органы, а затем на цветки, бобы и семена (рис. 62). На пораженных органах растений появляются красновато-коричневые пятна. На
листьях они угловатые, а на осевых органах несколько вытянутые. На пятнах спороносные ложа гриба располагаются концентрическими кругами. В зрелом состоянии ложа покрыты буроватой слизью, состоящей из массы конидий. Щетинки разбросаны по всему ложу или только по его краям. Они слегка изогнуты, кверху утончаются, имеют 1—4 перегородки. Конидиеносцы бесцветные и только в нижней части слабо окрашенные. Конидии главным образом продолговато-цилиндрические, прямые или согнутые, размером в среднем 17X48 мк.
Особенно интенсивно болезнь развивается на плодах. В местах поражения образуются мелкие, а затем укрупняющиеся пятна. Иногда они сливаются между собой, и стенки плода буреют. Если плоды находятся в ранней стадии развития, то отмирание ткани идет через всю толщу стенки плода и гриб может перейти на семена. При слабом поражении гриб распространяется только в оболочке семени, образуя бурые
пятна. При сильном заражении мицелий проникает до зародыша и семядолей. Больные семена твердеют, сморщиваются и часто теряют всхожесть, а если и прорастают, то дают больные проростки.
Возбудитель антракноза перезимовывает посредством мицелия в семенах и в растительных остатках, главным образом в стеблях и стенках плодов, которые как более грубые органы труднее поддаются действию почвенных микроорганизмов и сохраняются до весны не разложившимися. Конидии гриба зимовать не способны.
Colletoirichum lindemuthianum имеет много физиологических рас, отличающихся друг от друга неодинаковым отношением к различным сортам фасоли. Желтоплодные сорта менее устойчивы к заболеванию.
Для борьбы с антракнозом фасоли прежде всего необходимо проводить оздоровление посевного материала. Для этой цели рекомендуется погружать семена в воду. При этом щуплые семена всплывают и могут быть легко отделены. Имеются данные о положительном действии прогревания семян в воде при температуре 47—48° в течение 25 минут. Такому прогреванию должно предшествовать замачивание семян в воде с температурой 18—22° в продолжение 15 часов. Действие этого метода основано на том, что предварительное замачивание позволяет мицелию гриба тронуться в рост, а температура 47— 48° при втором этапе обработки убивает мицелий.
К профилактическим мероприятиям относится соблюдение плодосмена с возвращением фасоли на поле не ранее чем через 3 года. Необходимо также тщательно очищать поле от растительных остатков после уборки урожая.
Первые сведения о возбудителе антракноза льна Colletotrichum lini Boll, появились в литературе в 1910 г. и принадлежат американскому исследователю Боллею. Несколько позднее описание болезни приводится у различных авторов как зарубежных, так и отечественных. В настоящее время антракноз льна является широко известной распространенной болезнью во всех льноводческих районах.
Особенность этого заболевания заключается в том, что его возбудитель может развиваться на растениях, начиная от всходов вплоть до фазы созревания семян, при этом поражает все вегетативные органы, а также цветки, коробочки и семена. С. lini проявляет себя как узкоспециализированный паразит, приспособившийся только к развитию на различных видах льна.
Отрицательное действие антракноза прежде всего сказывается на семенной продукции. Под влиянием гриба семена льна либо теряют всхожесть, либо дают больные проростки. Наблюдения японского исследователя Хиура показали, что
интенсивность развития гриба в семенах определяется степенью зрелости семян в момент проникновения в них инфекции. Если семена заражаются грибом, когда пигментный слой в них еще не образовался, грибница проникает далеко в зародыш, он погибает и, следовательно, семена теряют всхожесть. При более позднем заражении, после образования пигментного слоя, развитие грибницы происходит только в самом верхнем ослизняющемся слое семенной оболочки. Такие семена прорастают, но дают больные проростки.
Исследования О. Я. Стрельской в Белоруссии показали, что грибница возбудителя антракноза льна может сохраняться жизнеспособной в кожуре семени в течение продолжительного времени (до 8 лет) в том случае, если семена не теряют всхожесть. В противном случае жизнедеятельность грибницы значительно понижается.
На проростках антракноз проявляется на корешках, подсемядольном колене и семядолях. На корешках и подсемядольном колене образуются ржавые, водянисто-слюдяные, удлиненные пятна, превращающиеся в язвочки. Этот симптом болезни на подсемядольном колене является самым характерным диагностическим признаком. На семядолях пятна округлые, желтоватые, постепенно увеличивающиеся, захватывающие всю поверхность семядоли. Гибель всходов изреживает посевы, инфицирует почву, способствуя дальнейшему распространению болезни.
Если болезнью поражается часть проростка, находящаяся под почвой, то растение может развивать добавочные корни выше зараженного участка и благодаря этому выживает, однако в дальнейшем оно отстает в росте, приобретает много-ярусность, снижающую качество волокна.
При развитии болезни на молодых листьях растений они становятся пятнистыми, засыхают. На стеблях в период их роста так же появляются буроватые пятна, обусловливающие так называемую «мраморную пятнистость», впоследствии на них возникают удлиненные трещинки. Коробочки также поражаются грибом. Мицелий через оболочки проникает в семена и заражает их.
Спороносные ложа гриба образуются на всех пораженных участках. Они мелкие, слегка розоватые, щетинки у основания бурые, а на вершине почти бесцветные, с 2—3 перегородками. Конидиеносцы короткие, бесцветные, конидии также лишены окраски, имеют продолговато-цилиндрическую форму, слегка изогнуты, размером 14,3—21,4x2,9—5,7 мк.
Влажная погода способствует распространению болезни в поле, так как расселение конидий происходит с каплями дождя. Насекомые так же переносят конидии, прилипающие к их телу.
Из всего сказанного видно, что вредоносность заболевания очень велика. Кроме снижения урожая семян и гибели сеянцев, значительно падает качество волокна, оно получается более коротким, обработка его сопряжена с трудностями.
Основные мероприятия в борьбе с антракнозом льна:
1. Механическая очистка семенного материала от щуплых, легковесных семян.
2. Химическая обработка — протравливание гранозаном и ТМТД.
3. Возвращение льна на то же поле севооборота не ранее чем через 5 лет.
4. Посев льна в оптимальные сроки и хорошо прогретую почву.
5. Соблюдение правильной нормы высева без загущения посевов.
Род Gloeosporium Desm. et Mont. Род.Gloeosporium впервые описали в 1849 году Демазьер и Монтань. Согласно этим авторам, к данному роду относятся грибы, паразитирующие преимущественно на листьях, реже на стеблях, ветвях и плодах растений. Конидиальные ложа очень мелкие, расположены под эпидермисом и имеют вид блестящих бугорков. На тонких листьях они особенно хорошо заметны на свет. Кони-диеносцы — тесно сплоченные, иногда у основания дихотомически расщепленные. В качестве наиболее характерного признака, по которому было дано само название рода («глоеос» по греч. клейкий), отмечается склеивание конидий и выход их наружу после разрыва кожицы слизистыми массами. Форма конидий в основном эллипсоидальная или цилиндрическая, иногда серповидная. Совместно с типичными конидиями (макроконидиями) иногда в отдельных ложах могут развиваться микроконидии. Они обычно значительно меньше макроконидий и имеют палочковидную форму. Макро- и микроконидии видов Gloeosporium одноклетные. Некоторые представители рода Gloeosporium в цикле развития имеют совершенную стадию, принадлежащую к родам Gnomonia и Pseudopeziza класса сумчатых грибов.
Атракноз смородины и крыжовника вызывается грибом Gloeosporium ribes (Lib.) Mont, et Desm. Поражая листья всех видов смородины и крыжовника, гриб обладает биологической специализацией, образуя следующие формы: G, ribis f. nigri — на черной смородине; G. ribis f. rubri — на красной и белой; G. ribis f. grossulariae — на крыжовнике.
Заболевание широко распространено и сопровождает эти культуры во всех зонах возделывания. В Советском Союзе особенно вредоносен антракноз в северной и средней частях.
На листьях растений образуются очень мелкие, несколько выпуклые пятнышки с блестящей поверхностью, в которых
и формируются спороносные ложа гриба (рис. 63). Есть указания, что количество этих пятен на одном листе может до-стигаты 200. Распространение мицелия не ограничивается отдельными пятнышками, гифы пронизывают и здоровые участки листа. Созревающие споры, склеенные слизистым веществом, прорывают покровную ткань и выступают наружу в виде беловатых крупинок. Конидиеносцы короткие, палочко-
Рис. 63. Gloeosporium г ibis: а — пораженный лист; б — конидии.
видные, наверху утончающиеся. В конидиальных ложах гриба образуются одноклетные, серповидно-изогнутые макроконидии, в среднем имеющие 23,4x6,3 мк. Вместе с макрокони-диями, а иногда на отдельных ложах образуются палочковидные микроконидии 5,7X1,5 мк. Биологическое значение мик-роконидий не выяснено. Есть предположение, что их развитие связано с половым процессом, в котором они участвуют в качестве спермациев.
Поражение листьев влечет за собой их засыхание. У черной смородины засохшие листья долго остаются висеть на кусте, а у красной — сразу же опадают. При сильном поражении можно наблюдать в середине июля совершенно голые кусты, на концах ветвей которых начинают развиваться из спящих почек молодые листочки.
Гриб поражает черешки, зеленые побеги, плодоножки, вызывая образование мелких бурых язвочек. Изредка поражаются плоды.
Для G. ribis известна совершенная стадия, принадлежащая к дискомицетам — Pseudopeziza ribis Kleb.
Эта стадия развивается в сапрофитных условиях на опавших листьях. К весне формируются плодовые тела — апотеции. Созревание их в средней зоне происходит в основном во второй половине мая. Сумкоспоры являются источником первичной инфекции, хотя наряду с этим гриб может зимовать мицелием и конидиями.
Присутствие паразита весьма отрицательно сказывается на росте и развитии растений. Опадение листвы среди лета истощает кусты и они почти не плодоносят, одревеснение побегов происходит недостаточно, а развитие молодых листочков из спящих почек во второй половине лета еще больше ослабляет растения. Все это значительно снижает урожай не только в текущем, но и в будущем году.
Наиболее рациональной мерой борьбы следует считать осеннюю уборку листвы и последующее закапывание ее в землю. Эта мера исключает возможность развития совершенной стадии гриба и тем самым предотвращает заражение весной молодых листьев аскоспорами.
Рекомендуется также трехкратное опрыскивание растений 1%-ным раствором бордоской жидкости в следующие сроки: первое — сразу же после цветения; второе — через 10 дней после первого и третье — после уборки урожая.
ПОРЯДОК SPHAEROPSI DALES
По системе Саккардо, принятой для несовершенных грибов, сферопсидные представляют собой третий порядок. Их особенностью является образование пикнид — своеобразных плодовых тел, в которых развиваются конидиеносцы и конидии.
Сферопсидные грибы широко распространены в природе как на культурных, так и на дикорастущих растениях. Паразитные формы развиваются главным образом на листьях растений, вызывая образование пятен, но могут поражать и другие органы, стебли, плоды. Характер пятен очень разнообразен и зависит от возбудителя болезни и свойств растения-хозяина. В одних случаях пятна резко очерчены более темной каймой. Иногда такое поражение ткани может сопровождаться ее выпадением, и листья становятся продырявленными. В других случаях наблюдается постепенный переход от пораженного участка к здоровой ткани. Пятна могут сливаться между собой, постепенно захватывая большую часть пластинки листа. Спустя некоторое время после образования пятен на их верхней или нижней стороне можно заметить черные точечки — пикниды гриба. Количество пикнид на одном пятне и характер их расположения бывают различными. У одних грибов пикниды сгруппированы в центре пятна, у других —
рассеяны по всей его поверхности, у третьих—расположены концентрическими кругами на пораженном участке.
Морфологические признаки пикнид довольно разнообразны и определяются различными факторами: характером пятен, глубиной залегания, особенностями тканей растения-хозяина. По форме пикниды могут быть округлыми, несколько сплюснутыми или же вытянутыми подобно перитецию сумчатых грибов. Они окрашены в более или менее темный цвет, что делает их хорошо заметными на поверхности пятен. Открываются пикниды на вершине округлым или щелевидным устьицем. Иногда вершина пикниды оттянута в виде сосочка или хоботка, на котором находится устьице. Стенка пикниды образована сплетением гиф, клетки которых имеют параплектенхимное или прозоплектенхимное строение. У некоторых представителей она более плотная, склероциального характера. Внутренняя полость покрыта короткими конидие-носцами, отчленяющими конидии (пикноспоры или стилло-споры), весьма разнообразные по форме, величине и количеству клеток. У многих представителей порядка, имеющих мелкие конидии, последние выходят через устьице пикниды в виде слизистого извивающегося тяжа. Это объясняется наличием особого склеивающего вещества, выделяющегося в полости пикниды. Конидии сферопсидных грибов распространяются каплями дождя или росы, воздушными течениями, насекомыми. Многие сферопсидные грибы представляют ко-нидиальную стадию сумчатых из группы пиреномицетов. Зимуют такие виды в сапрофитных условиях на растительных остатках, формируя к весне сумчатое спороношение. Некоторые способны перезимовывать конидиями, удерживающимися на зимующих органах растений.
Наиболее обширным по количеству видов, паразитирующих на высших растениях, является семейство Sphaerioida-сеае.
Сем. Sphaerioidaceae
Из принадлежащих к этому семейству родов наибольшее практическое значение имеют Ascochyta, Septoria и Phoma.
Род Ascochyta Libert. Включает несколько сот видов, паразитирующих на стеблях, листьях и плодах многих культурных и дикорастущих растений. Болезни, вызываемые представителями рода Ascochyta, получили общее название аско-хитозов.
Аскох птоз гороха широко распространенное заболевание гороха во всех странах Европы, Азии, Америки. Особенный ущерб этой культуре болезнь наносит в местностях с большим увлажнением. Вызывается аскохитоз несколькими
возбудителями: Asc. pisi Lib., Asc. pinodes Jones., Asc. pino-della Jones., Asc. pseudopinodella Bond., Mont, et Was.
Аскохитоз может развиваться на всех надземных органах растения, за исключением цветков, но больше всего подвержены заболеванию листья и бобы. Кроме того, второй и третий возбудители поражают еще и корневую шейку.
Рис. 64. Ascochita pisi: а — поражение листьев и бобов; б — разрез через пикниду, в — пикноспоры.
В связи с тем что есть некоторые различия в симптомах болезни, определяющиеся характером возбудителя, в проявлении аскохитоза различают два типа поражения.
Первый тип поражения вызывается Asc. pisi, который впервые был описан в 1830 году Либертом и долгое время считался единственным возбудителем аскохитоза гороха. Гриб развивается на листьях, стеблях и бобах, вызывая образование светлоокрашенных пятен с темно-коричневым ободком (рис. 64). На листьях и бобах пятна более или менее округлые, на стеблях вытянутые. На стеблях и особенно на бобах они постепенно становятся вдавленными. Через некоторое время в центре пятен на всех пораженных органах образуются группы пикнид. Пораженные грибом листья быстро засыхают, а стебли становятся хрупкими и легко ломаются.
При более позднем поражении бобов пятна на них не успевают образоваться. Мицелий гриба распространяется более или менее равномерно по створкам боба, вызывая их общее пожелтение. В этом случае пикниды развиваются рассеянно по всей створке. Со створок гриб переходит на семена, однако пикнид на созревающих семенах почти никогда не образуется. Обильное развитие их на семенах наступает в период набухания и прорастания последних. Пораженные семена можно легко отличить от здоровых: они мелкие, сморщенные. Мицелий гриба в большинстве случаев находится в оболочке, а иногда поражает зародыш.
Второй тип поражения обусловлен развитием главным образом Asa pinodes и Asc. pseudopinodella. В этом случае пятна темно-коричневые, на листьях быстро сливающиеся, на бобах — не вдавленные, а выпуклые, часто мелкие. Иногда поражается корневая шейка, где пятна .постепенно переходят в язвы. Такие растения обычно погибают.
Asc. pinodella развивается главным образом на корнях и корневой шейке. Вредоносность этого возбудителя особенно велика в связи с его способностью сохраняться в почве более двух лет. Остальные возбудители такими свойствами не обладают. После разложения растительных остатков они погибают под влиянием антагонистических микроорганизмов. Размер пикнид варьирует в пределах 70—185 мк в диаметре. Конидии в начале развития не имеют перегородок, затем появляется одна перегородка и небольшая перетяжка около нее. У Asc. pisi конидии крупнее, чем у других видов, и в среднем достигают И — 14x3,5—3,7 мк.
Для Asc. pinodes известна сумчатая стадия — Didimella pinodes Berk, et Blox., развивающаяся на растительных остатках. У других возбудителей аскохитоза сумчатое спороношение не обнаружено.
Из вышеизложенного видно, что первичным источником распространения аскохитоза гороха могут быть семена и растительные остатки. В связи с этим должны быть организованы и меры борьбы:
1. Посев следует производить здоровыми семенами. Предварительная экспертиза семян путем проращивания позволяет установить степень их пораженности, а затем соответственно применить химическую обработку.
2. Трехкратное опрыскивание 1%-ной бордоской жидкостью (первое — перед цветением, второе — через десять дней после первого, третье — через 20 дней после второго).
3. Протравливание семян гранозаном (3 кг на 1 т) или ТМТД (4—6 кг на 1 т).
4. Агротехнические мероприятия: запахивание растительных остатков; посев гороха в смеси с другими растениями:
горчицей, рыжиком, бобами. Это предохраняет горох от полегания, а следовательно, и ют переувлажнения надземных органов, подверженных заболеванию.
Род Septoria Fries. Представители его распространены в самых разнообразных экологических условиях. В настоящее время науке известно 1500 видов. В Эстонской ССР их насчитывается около 273 видов (Марланд), а в Армении —150 (Тетеревникова-Бабаян). Всего для Советского Союза известно около 500 видов представителей этого рода.
По приуроченности к питающим растениям эти грибы отличаются большим разнообразием. Их можно встретить на дикорастущих и культурных злаках, на древесно-кустарниковых породах и на декоративных цветочных растениях.
Основным диагностическим признаком септориозов растений является образование характерных пятен главным образом на листьях. Форма и окраска пятен может варьировать в зависимости от питающего растения. Чаще они бывают округлые, со светлой серединой и буро-коричневым ободком. На светлом участке пятна пикниды образуются рассеянно или более или менее скученно. Размер пикнид от 50 до 200 мк в диаметре. Конидии сильно вытянуты, одноклетные или с несколькими (до 5) перегородками, прямые или изогнутые, цилиндрические или же на нижнем конце расширенные, а на верхнем — заостренно-вытянутые. Размер их чрезвычайно варьирует у разных видов, особенно по длине — от 10 до И0 мк. В ширине наблюдаются меньшие разнообразия — от 0,5 до 6 мк.
Септориоз томатов — наиболее широко распространенная болезнь, вызываемая грибом Septoria lycopersici Speg. Паразит может развиваться на томатах, выращиваемых в парниках, теплицах и в открытом грунте, причем поражает растения любого возраста.
Как указывает Е. А. Осницкая, первые септориозные пятна могут появляться на семядолях, на которые инфекция в виде перезимовавших конидий попадает с растительных остатков (с семенами болезнь не передается). Установлено, что от момента проникновения инфекции до образования пикнид со зрелыми пикноспорами достаточно 14 дней. С семядолей гриб переходит на листья, вызывая появление на их поверхности большого количества водянистых пятен неправильно-округлой формы. Центр пятна — серой окраски, а края окаймлены более темной каймой.
При дальнейшем развитии болезни в центре пятен образуются мелкие черные пикниды с широким устьицем для выхода конидий. Последние —нитевидноцилиндрические, несколько утолщены на одном конце, прямые или слабо изогнутые,со многими перегородками, 65—123x2—Змк(рис.65).
В условиях влажной погоды конидии выступают на поверхность пятен и переносятся на здоровые листья дождем и сельскохозяйственным инвентарем при уходе за растениями. Пораженные листья засыхают и при сильном развитии болезни остается только несколько самых верхних листьев.
М. С. Дунин считает, что возбудителю септориоза томатов свойственна резко выраженная онтогенетическая специа-
Рис. 65. Septoria lycopersici. а — пораженный лист;
б — разрез через пикниду;
в — пикноспоры.
лизация, проявляющаяся в том, что заражению наиболее подвержены нижние листья, физиологически более старые.
Развитию болезни благоприятствует высокая влажность воздуха и обилие осадков, так как конидии выходят из пикнид во время дождя или росы.
Некоторые авторы наблюдали развитие гриба на стеблях, цветоножках, чашечках и плодах томатов. Есть указание, что гриб может паразитировать и на других пасленовых: картофеле, баклажанах, черном паслене и дурмане.
В качестве профилактических мероприятий рекомендуется тщательно удалять с поля все остатки больных растений, чтобы исключить возможность перезимовки гриба в виде конидий в пикнидах. Если болезнь появилась в парниках, то при высадке рассады следует осторожно обрывать пораженные листочки, чтобы предотвратить занос инфекции в поле.
По мнению М. С. Дунина, необходимо применять калийные подкормки и вносить микроэлементы под культуру поми
доров, так как эти мероприятия задерживают физиологическое старение листьев и делают их наиболее устойчивыми. Рекомендуется также опрыскивание растений 2%-ным раствором бордоской жидкости не менее трех раз за лето. Е. А. Осниц-кая рекомендует опрыскивать рассаду 1%-ным раствором бордоской жидкости за две недели до высадки в грунт.
Многолетний люпин в сильной степени поражается Septaria lupini Kazn. В Белоруссии септориоз люпина считается наиболее в-редоносным. Как указывают Н. А. Дорожкин и Н. И. Чекалинская, пораженность посевов люпина усиливается с возрастом культуры, так как инфекция накапливается в поле за счет неубранных растительных остатков, и ко второму году выращивания поражение посевов достигает 38,3%.
В середине мая обнаруживаются пораженные грибом листья. На их дольках образуются довольно крупные, неправильно-округлой формы пятна с беловатой серединой и темно-коричневым ободком. Со временем на светлом фоне пятен на верхней стороне листочков становятся хорошо заметны пикниды диаметром 120—200 мк. Конидии на обоих концах закругленные и слегка изогнутые, 40—120X3—4 мк.
Из однолетних видов люпина, культивируемых в Белоруссии, в слабой степени поражается только желтый люпин, причем, по данным выше приведенных авторов, желтый люпин заражается от многолетнего в случае отсутствия пространственной изоляции между посевами.
Уничтожение послеуборочных остатков является основным мероприятием в борьбе с септоризом люпина.
Род Phoma Fr. Виды рода Phoma разнообразны по образу жизни: одни из них могут существовать сапрофитно на растительных -остатках, другие являются весьма вредоносными паразитами культурных растений, причем развиваются на различных частях: корнях, корнеплодах, стеблях, листьях и семенах. Развитие гриба можно наблюдать на растениях в самом молодом возрасте, еще до выхода всходов на поверхность почвы. Это приводит к сильному изреживанию посевов. На стеблях представители рода Phoma могут вызывать образование глубоких пятен с поражением сосудистой системы. Такие растения увядают или надламываются. На листьях гриб образует пепельно-серые пятна, часто с ясно выраженной зональностью. Со временем пятна покрываются большим количеством черных точек — пикнид гриба с очень мелкими одноклетными конидиями.
Всходы всех видов свеклы поражаются болезнью, которая получила название корнееда. Проявляется она в загнивании всходов еще до выхода их на поверхность почвы. Заболевание корнеедом вызывается целым комплексом возбудите
лей, в числе которых весьма обычным является и Phoma betae Frank.
При развитии на всходах свеклы Ph. betae симптомом болезни является поражение подземной части проростка: корешок поражается при выходе из клубочка, а подсемядольное колено — у корневой шейки. Образуется светло-бурое продольное пятнышко, очень скоро чернеющее и распространяющееся по окружности органа. На пораженном участке можно при помощи лупы заметить пикниды гриба 100—180 мк. Конидии мелкие, размером 5—7X3—4 мк (Тетеревникова-Бабаян).
При благоприятных условиях развития болезни поражение заходит глубоко и захватывает сосудистые пучки. Такие растения погибают. Если же условия для развития гриба неблагоприятны, больные растения могут оправиться, образуя придаточные корешки выше места поражения. Однако такие растения заметно отстают в росте и дают деформированные корни.
Корнеед, вызываемый Ph. betae, передастся семенами. Пикниды гриба обнаруживаются на клубочках и на оболочках семян.
В борьбе с этим видом корнееда большое значение имеет получение здорового семенного материала. Для этого требуется тщательно отбирать корнеплоды при высадке их в грунт как маточные растения. Рекомендуется также маточные корни выращивать из семян, полученных от главных цветоносов, так как растения, выращенные из таких семян, оказываются более устойчивыми, жизнеспособными и дают значительную прибавку в урожае корнеплодов. Для уничтожения инфекции, имеющейся на поверхности клубочков, следует проводить заблаговременное протравливание семян гранозаном (2—4 кг на 1 т) или ТМТД (5—6 кг на 1 т).
Ф о м о з распространен на всех видах свеклы, относится к числу наиболее вредоносных. Паразитизм Ph. betae на вегетативных органах взрослого растения отличается большим многообразием. В первый год жизни поражаются листья, корнеплоды, а при их высадке на второй год — стебли, листья и семенные клубочки. Наконец, гриб участвует в сложном комплексном явлении — кагатной гнили.
Развитие гриба на листьях вызывает появление некротических сероватых пятен с концентрическими кругами (зональная пятнистость). Заболеванию подвержены главным образом листья с пониженной жизнеспособностью (нижние подсыхающие или пораженные церкоспорозом). Однако образовавшиеся на них пикниды способствуют расселению гриба и проявлению других форм болезни. При благоприятных условиях может наблюдаться отмирание точки роста и внутренних листочков розетки. В дальнейшем болезнь переходит на корнеплод, вызывая сердцевинную гниль (гниль сердечка).
Развитию сердцевинной гнили способствует сухая погода и недостаток солей бора в почве. В области шейки ткань корнеплода пронизывается гифами гриба, темнеет, а на поверхности возникают сероватые пятна, на которых могут образовываться пикниды.
Больные корнеплоды, попадая в хранилище, дают типичную форму кагатной гнили, которая представляет результат комплексного воздействия нескольких микроорганизмов, в том числе Ph. betae, на пораженный корнеплод.
У семенников поражаются стебли, листья, цветоносы и семенные клубочки.
В конце июля можно наблюдать на стеблях появление коричневых некротических пятен, которые вскоре в центре приобретают серую окраску и покрываются пикнидами гриба. На цветоносах и клубочках пятен не обнаруживается, но появляется характерная точечность, обусловленная развитием большого количества пикнид.
В борьбе с многочисленными формами проявления паразита рекомендуются следующие мероприятия:
1. Улучшение физических свойств почвы с задержанием по возможности в ней влаги.
2. Уничтожение послеуборочных остатков, особенно на высадочных полях.
3. Возвращение свеклы на то же поле не ранее чем через 4 года.
4. Быстрая уборка и обмолот семян.
5. Протравливание семян гранозаном (2—4 г на I кг).
6. Поддержание в хранилищах пониженной температуры (около 1°) и относительной влажности воздуха (85%).
ГЛАВА ВИРУСНЫЕ, БАКТЕРИАЛЬНЫЕ
И НЕИНФЕКЦИОННЫЕ БОЛЕЗНИ
U ВИРУСНЫЕ БОЛЕЗНИ
Природа вирусов. Инфекционные болезни растений, животных и человека могут вызываться вирусами, характеризующимися ультрамикроскопической величиной, отсутствием клеточного строения и способностью к размножению в живых клетках восприимчивого организма. Вирус — слово латинское и в переводе означает яд. Вирусные болезни существуют в течение многих веков, однако природа этих возбудителей долгое время не была выяснена. Одни исследователи высказывали предположение, что вирусы — это ультрамикроскопи-ческие организмы, способные проходить через самые тонкие фильтры. Другие считали, что это сложные вещества со свойствами простейших существ.
В настоящее время наука о вирусах — вирусология — продвинулась значительно вперед. Исследования физико-химических и биологических свойств вирусов показали, что это живые организмы со всеми присущими им свойствами: они способны размножаться, различным образом реагировать на внешние условия и т. п.
Вирусология как наука зародилась в нашей стране. Выдающийся русский микробиолог Н. Ф. Гамалея еще в 1886 году обнаружил, что возбудитель чумы крупного рогатого скота способен проходить через те фильтры, которые удерживают микробов. В 1892 году известный русский ученый Д. И. Ивановский, изучая мозаичную болезнь табака, установил, что ее возбудителем также является вирус. За способность вирусов проходить через поры бактериальных фильтров их стали называть фильтрующимися вирусами.
Эти открытия имели громадное научное и практическое значение для дальнейших исследований природы многих инфекционных болезней растений, животных и человека.
За первую половину текущего столетия открыто свыше 200 новых только фитопатогенных вирусов. Они существовали в отдельных локализованных точках земного шара. Местные культурные и дикорастущие растения вследствие длительного совместного существования приобрели определенную устойчивость к этим формам вирусов, а поэтому и не особенно страдали при поражении. Интродукция культурных растений в новые районы способствовала обнаружению вирусных болезней и установлению их возбудителей, так как не приспособившиеся новые виды сильно страдали от поражения, а симптомы болезни выступали отчетливей.
Форма, величина и химический состав вирусов. Современные методы исследования вирусов при помощи центрифугирования, электронного микроскопа и т. д. дали возможность установить различие вирусов по величине, форме и химическому составу.
Величина вирусов исчисляется миллимикронами. Форма их может быть разнообразна: палочковидная, нитевидная, угловатая, сферическая. Палочковидный вирус табачной мозаики имеет размеры 15X300 ммк. Сферической формы вирус мозаики люцерны около 15 ммк в диаметре.
Фитопатогенные вирусы, выделенные до настоящего времени, имеют сходный химический состав. Основными компонентами являются нуклеиновая кислота и протеин. Рибонуклеиновая кислота (РНК) характерна для всех фитопатогенных вирусов. Изучение вируса табачной мозаики показало, что белок лишен инфекционных свойств, а нуклеиновая кислота, будучи изолирована, способна вызвать вирусное заболевание.
Размножение и некоторые другие свойства вирусов. Фитопатогенные вирусы обладают способностью к размножению в живых тканях растения-хозяина, при этом они проходят через определенный цикл развития. Так, например, цикл развития вируса табачной мозаики имеет две фазы. Первая вегетативная фаза, когда собственно и происходит его размножение, характеризуется слабой инфекционностью или полным отсутствием инфекционных свойств. Вторая — фаза покоя, когда заканчивается процесс размножения и формирования вирусных частиц. В этой фазе вирус обладает высокой инфекционностью. Размножение вирусов может так же происходить в изолированных тканях растений, произрастающих на питательных средах.
Жизнеспособность вирусов в мертвых тканях растений не одинакова и часто определяется условиями окружающей сре
ды. Наиболее изученный по физиологическим свойствам вирус табачной мозаики может сохранять жизнеспособность в сухом табаке до 50 лет, а в больных сухих листьях и стеблях в поле активность его пропадает через 3—4 года. Этот же вирус, находясь в соке, выделенном из растения, инактивируется через несколько месяцев.
Все фитопатогенные вирусы легко переносят низкую температуру. К воздействию высоких температур некоторые вирусы очень чувствительны. Например, вирус желтухи персиков инактивируется при 35°. Другие, наоборот, проявляют большую стойкость к высоким температурам. Инактивация вируса курчавости листьев сахарной свеклы наступает при 75—80°, а табачной мозаики — при 90°
Весьма важным свойством вирусов является их способность сохранять патогенные свойства при сильном разведении. Вирус мозаики табака, будучи разведен водой в отношении 1 : 10 9, способен вызвать заражение. Вирусы хорошо противостоят действию препаратов, убивающих бактерии. Карболовая кислота, спирт, формальдегид не вызывают инактивации вирусов. Исключение представляет вирус табачной мозаики, легко инактивирующийся спиртом.
Многие вирусы, выделенные в чистом виде из сока растений, обладают способностью образовывать кристаллы, которые можно видеть при сравнительно слабых увеличениях микроскопа. Каждому типу вируса свойственна определенная форма кристаллов. Вирус табачной мозаики выделяется в виде игловидных кристаллов, а вирус некроза табака дает кристаллы ромбической формы.
Выделенные в кристаллической форме вирусы сохраняют свои патогенные свойства и при введении в здоровые растения вызывают такие же заболевания.
Симптомы и типы поражений. Вирусные болезни широко распространены в природе и наносят большой вред сельскохозяйственным растениям. Они могут вызвать быструю гибель растений, но чаще приводят к значительному снижению урожая.
По типу симптомов вирусные болезни делят на две группы: мозаики и желтухи.
Мозаика характеризуется неравномерной окраской листьев. Патологические явления наблюдаются, главным образом, в хлорофиллоносных тканях. При этом происходит распад хлоропластов. Хлоротичные или желтоватые участки различной величины и формы чередуются с нормально окрашенной тканью.
Проявление мозаичной болезни может быть и на лепестках цветков в виде крапчатости, полосчатости, пятнистости. Так выглядят многие пестрые формы тюльпанов, гладиолу
сов, левкоев. Вирусные болезни типа мозаик вызывают появление некротических пятен на листьях, стеблях и плодах (бронзовость томатов).
Однако, кроме изменения окраски, может изменяться и форма листовой пластинки; появляется морщинистость, курчавость. Эти симптомы обусловлены задержкой в росте жилок листа, в то время как мякоть листа продолжает разрастаться. К этому типу болезней относятся: мозаика табака, морщинистая и полосчатая мозаика картофеля, мозаика томатов, свеклы, малины и т. д.
Желтуха, как показывает название, вызывает общее пожелтение листьев, т. е. общий хлороз. При этом типе болезни происходит поражение флоэмы (ситовидных трубок), задерживается отток продуктов фотосинтеза, вследствие чего в клетках скопляется большое количество крахмала, листья теряют мягкость и легко ломаются.
Могут наблюдаться и другие симптомы: скручивание листьев (у картофеля, пораженного вирусом скручивания); гипертрофия отдельных частей тела растений (при столбуре томатов — гипертрофия чашелистиков); карликовость растений (у овса, пораженного вирусом закукливания) и т. п.
Пути передачи и распространения вирусов. Перенос вирусов от больного растения к здоровому может осуществляться различными путями: насекомыми, соком больных растений, больными частями и органами растений при вегетативном размножении, семенами, через почву.
Большинство фитопатогенных вирусов распространяется сосущими насекомыми: тлями, цикадками, червецами, щитовками, трипсами и др.
Насекомые могут быть механическими переносчиками вирусов. Питаясь на больном растении, они загрязняют свои ротовые части мельчайшими частицами растительной ткани, содержащей вирус, а затем, посещая здоровые, заносят вирус в его клетки.
В другом случае насекомые являются биологическими переносчиками и передают вирус при помощи кровеносной системы и слюнных желез. Питаясь на больном растении, они через некоторое время становятся инфекционными. Вирус, попав в тело насекомого, должен пройти инкубационный период, в течение которого он попадает в кровеносную систему и внутренние органы насекомого.
Промежуток времени, необходимый насекомому для поглощения вируса при питании на больном растении, может иметь протяженность от 5 минут до 6 часов; период инкубации— от нескольких часов до 14 дней. Например, вирус скручивания листьев картофеля может быть передан тлей здоровому растению через 24—28 часов после его поступле
ния в тело тли. Для передачи вируса в растение насекомым требуется всего несколько минут питания на нем. Насосавшись сока больного растения, насекомое сохраняет способность заражать здоровые растения на протяжении периода времени от 7 дней до 3—4 месяцев, не пополняя запасы вируса.
Известны случаи, когда один и тот же вид тли переносит один вирус чисто механическим путем, а для другого является биологическим переносчиком. Например, персиковая тля для многих вирусов является механическим переносчиком, а вирусами мозаики гороха, желтухи свеклы и скручивания листьев картофеля может заражать здоровые растения только после определенного инкубационного периода.
Некоторые вирусы, распространяющиеся насекомыми, могут передаваться и их потомству. Большинство же вирусов этой способностью не обладает, и каждое новое поколение насекомых-переносчиков должно инфицироваться на больном растении.
Перенос вирусной инфекции может осуществляться посредством сока больных растений. Всевозможные ранения вегетативных органов во время ухода за растениями в течение вегетационного периода способствуют механической передаче вируса здоровым растениям.
Механическим путем многие вирусы легко передаются при различных способах вегетативного размножения: прививками, клубнями, луковицами и т. п. (картофель, малина, земляника, тюльпаны, гладиолусы и др.).
Несмотря на то что в пораженных растениях вирус может накапливаться во всех органах, в том числе и в цветках, семенами большинство вирусов не передается. У большинства растений в созревающих семенах образуются вещества, инактивирующие вирусы.
Многие вирусы могут передаваться через почву, в которой сохраняются зараженные растительные остатки. Некоторые вирусы обладают способностью сохраняться в почве в активной форме до 6 лет (вирус мозаики озимой пшеницы). Из почвы в растения вирусы проникают через поранения корневой системы, наносимые орудиями обработки и почвенными насекомыми.
Меры борьбы с вирусными болезнями. В борьбе с вирусными болезнями основное значение имеют агротехнические профилактические мероприятия:
1. Удаление с поля больных растений.
2. Тщательное уничтожение сорняков, являющихся часто источником вирусной инфекции.
3. Отбор и оздоровление семенного и посадочного материала.
4. Уничтожение насекомых — переносчиков вирусных болезней.
5. Введение в культуру устойчивых сортов.
БАКТЕРИАЛЬНЫЕ БОЛЕЗНИ РАСТЕНИЙ
Общие сведения о бактериях. Болезни растений, вызываемые бактериями, или бактериозы, имеют широкое географическое распространение и значительно снижают количество и качество урожая сельскохозяйственных растений. Особенно большой ущерб бактериозы причиняют в период хранения картофеля и овощей.
Бактерии представляют мельчайшие бесхлорофилльные организмы весьма примитивной организации. Тело бактерий состоит из одной клетки, покрытой плотной оболочкой из гемицеллюлозы и пектина. Иногда в состав оболочки входят белковые вещества. Наружные слои оболочки могут набухать и ослизняться, образуя слизистые капсулы, нередко превосходящие размеры самой клетки. Полость клетки заполнена протоплазмой; у стареющих клеток появляются вакуоли.
У громадного большинства бактерий нет дифференцированного ядра, а ядерные вещества находятся в диффузном состоянии. Только у немногих есть вполне оформленное ядро.
Большинство бактерий бесцветны, однако у некоторых в протоплазме имеются пигменты — бактериопурпурин, состоящий из бактериохлорина и бактериоэритрина, и бактериохлорофилл.
Запасными продуктами бактерий являются жир, гликоген, валютин; крахмала никогда не образуется.
По форме различают следующие основные группы бактерий: кокки — шаровидные; палочки (бациллы) — палочковидные; вибрионы — в форме запятой; спириллы — спирально закрученные.
Многие бактерии обладают способностью к активному движению благодаря наличию жгутиков — длинных тончайших выростов протоплазмы, видимых только после окрашивания и фиксации. В зависимости от характера расположения жгутиков бактерии делятся на три группы: монотри-х и, с одним полярным жгутиком; лофотрихи, с пучком жгутиков на одном конце клетки; перитрихи, с большим количеством жгутиков, расположенных по всей поверхности клетки.
По характеру питания большинство бактерий — гетеротрофные организмы и поглощают питательные вещества осмотическим путем через клеточную оболочку.
Автотрофных бактерий в природе значительно меньше. Они в свою очередь делятся на ф ото а в тотр оф н ы е
и хемоавтотрофные, первые синтезируют органическое вещество за счет световой, вторые — за счет химической энергии.
По способу дыхания бактерии делятся на две группы: аэробные и анаэробные. Аэробные могут развиваться только при наличии свободного кислорода в окружающей среде. Анаэробные бактерии живут при отсутствии кислорода. Необходимую для жизни энергию они получают при расщеплении сложных органических веществ на более простые.
Размножаются бактерии путем простого деления клетки на две части. При благоприятных условиях процесс деления идет чрезвычайно быстро. Некоторые бактерии могут делиться через каждые 20—30 минут.
По образу жизни бактерии бывают сапрофитами и паразитами. Сапрофитные бактерии живут на самых разнообразных субстратах, преимущественно в почве. Среди паразитных форм имеется значительное количество фитопатогенных, строго приуроченных к определенным растениям-хозяевам. Вместе с тем среди фитопатогенных бактерий совершенно отсутствуют облигатные паразиты.
По степени паразитической активности среди фитопатогенных бактерий следует различать две группы:
1) факультативные сапрофиты, вполне приспособившиеся к паразитному существованию на растениях, однако при отсутствии последних способные питаться сапро-фитно. к этой группе принадлежат такие широко распространенные бактерии как Pseudomonas tumefaciens Stevens — возбудитель корневого рака многих растений, Pectobacterium phytophthorum Dowson, вызывающий черную ножку картофеля;
2) факультативные паразиты, обычно живущие сапрофитно, но при известных условиях способные поражать живые растения и вызывать их заболевания. Примером этой группы является Xanthomonas heteroceae Savulescu — возбудитель пятнистостей надземных вегетативных органов многих растений.
Пути проникновения бактерий в растения и способы их распространения. Большинство фитопатогенных бактерий не вырабатывают ферментов, растворяющих клетчатку. В связи с этим они могут проникать внутрь растительных тканей либо через природные отверстия (устьица, гидатоды, чечевички и т. п.), либо через повреждения кожицы, которые наносят животные, а также человек во время ухода за растениями. Передвижению бактерий по поверхности тела растений и проникновению внутрь через естественные отверстия или поврежденные участки способствует наличие капельно-жидкой влаги. Дальнейшее развитие бактериозов также в значительной
степени зависит от условий влажности. Например, кольцевая гниль клубней картофеля (Corynebacterium sepedonicum Scapt. et Burkh.) развивается сильнее в тех районах, где в период уборки преобладает влажная погода (северная и средня полоса РСФСР, Белоруссия).
Проникнув в растение, возбудитель бактериоза может сохраняться в некоторых его частях до следующего вегетационного периода. Семена, клубни, луковицы и другой посадочный материал является местом сохранения многих видов фи-топатогенных бактерий, а также источником распространения бактериозов в новые районы.
Заражение семян бывает поверхностным и внутренним. На поверхность семян бактерии попадают при извлечении их из больных плодов. Внутренняя инфекция происходит во время их развития. При этом бактерии проникают в формирующиеся семена по проводящей системе.
Распространению бактерии с посевным и посадочным материалом в новые районы в значительной степени способствует хозяйственная деятельность человека. При перевозке культурных растений из одной страны в другую, как правило, перевозятся и различные возбудители болезней. Так, в 1911 году из Японии во Флориду был завезен возбудитель рака цитрусовых.
Человек может разносить некоторые бактериозы, пользуясь специальными приемами возделывания культурных растений. Например, при вершковании махорки может переноситься бактериальная рябуха табака; при пасынковании томатов — бактериальный рак и т. п.
Источником заражения растений бактериальными болезнями могут быть и насекомые. Ростковая муха способна переносить возбудителя черной ножки картофеля — Р ectob ас terium phytophthorum; в кишечнике жука-листоеда зимует бактерия, вызывающая увядание огурцов,— Bacterium tracheiphi-lum Burgw. и т. д.
Бактериальная инфекция может сохраняться в течение зимнего периода в растительных остатках: листьях, стеблях, покровах плодов, а весною распространяться различными способами: насекомыми, водой, ветром и др.
В почве фитопатогенные бактерии сохраняться долго не могут и погибают вследствие наличия там других микроорганизмов — антагонистов. Поэтому почва может служить источником инфекции лишь некоторое время.
Симптомы и типы поражения растений бактериями. Следует различать два вида поражения растений бактериозами: общий и местный. Общее поражение характеризуется проявлением заболевания на всем растении и происходит обычно* в том случае, когда бактериальная инфекция попадает в про
водящую систему растения и таким путем разносится по всему телу. Общие бактериозы отличаются большой вредоносностью, они обычно вызывают гибель растений.
При местном поражении бактерии внедряются в отдельные участки тела растений и тоже наносят значительный ущерб, хотя и не вызывают полной гибели растений.
Внешние признаки (симптомы) бактериальных заболеваний, так же как и характер течения болезней, отличаются большим разнообразием. Бактериозы могут проявляться в виде наростов, ожогов, некротических участков, увядания растений и гнилей.
Наросты обычно возникают на корнях и имеют вид опухолей. В пораженных участках клетки начинают усиленно делиться (явление гиперплазии) и ткань разрастается. Различают два типа опухолей: раковые и туберкулезные. Раковые опухоли не имеют полостей внутри; в туберкулезных вследствие распада ткани образуются полости или каверны, заполненные бактериальной слизью. К этому типу бактериозов относится корневой рак многих растений (Pseudomonas tume-faciens) и туберкулез корня свеклы (Xanthomonas beticola Dowson).
Ожоги характеризуются почернением и отмиранием пораженных участков. Они могут проявляться на цветочных и листовых почках, на лепестках цветка и листьях молодых побегов, а также на коре деревьев. Например, бактериальный ожог косточковых, вызываемый Pseudomonas cerasi Griff., широко распространен на Украине, им поражаются цветки, отдельные ветви и кора дерева абрикосов, черешен, слив.
Некрозы, или бактериальные пятнистости, также весьма распространенный тип бактериоза, проявляющийся в образовании небольших пятен со светлым окаймлением. Бактериальные пятнистости могут возникать на всех надземных органах растений, особенно на листьях. Ткань в пораженных участках отмирает и может выпадать, при этом листья становятся продырявленными. Примером является угловая пятнистость огурцов (Pseudomanas lachrymans Sm. et Br.).
Увядание растений обусловлено проникновением бактерий в сосудистую систему. Быстро размножаясь, они закупоривают проводящие воду сосуды и вызывают задержку тока воды. Кроме этого, бактерии в процессе своей жизнедеятельности выделяют токсические вещества, вызывающие отмирание и других тканей. Иногда одним из симптомов бактериального увядания является потемнение жилок, как например у капусты, пораженной сосудистым бактериозом (Xantho-monas campestris Dowson).
Гнили чаще всего наблюдаются на мясистых частях растений, богатых углеводами и азотистыми веществами:
клубнях, луковицах, плодах и т. п. Пораженные органы размягчаются, превращаясь в кашицеобразную бесформенную массу вследствие того, что под воздействием фермента пектиназы, выделяемого бактериями, происходит разрушение межклетного вещества. Примером этого типа бактериозов может служить мокрая гниль моркови и лука (Рectobacterium carotovorum Dowson) и мокрая гниль клубней картофеля (Pseudomonas xanthochlora Stapp.).
Меры борьбы с бактериозами растений. В борьбе с бактериозами основное значение имеют профилактические мероприятия:
1. В связи с тем что многие бактериозы передаются с семенами, необходимо производить протравливание семенного материала различными химическими препаратами. Например, в борьбе с бактериозами фасоли (Xanthomonas phpaseoli Dowson) и гороха (Pseudomonas pisi Stapp.) достаточно эффективным оказывается протравливание семян гранозаном (3 кг/т) или ТМТД (4—8 кг/т).
2. В тех случаях, когда возбудители бактериозов сохраняются в трудно сгниваемых послеуборочных остатках (возбудители сосудистого и слизистого бактериозов капусты и др.), следует своевременно удалять с поля после уборки урожая все пораженные части растении.
3. К профилактическим мероприятиям следует отнести также и борьбу с насекомыми — передатчиками бактериальной инфекции (мухами — капустной, луковой, морковной; жуками— листоедами, хлебными клопами и др.).
4. Большое значение имеет соблюдение общегигиенических условий, как например дезинфекция почвы и помещений при выращивании растений в закрытом грунте; удаление и уничтожение пораженных растений еще в период вегетации и т. п.
5. Во многих случаях большую роль играет применение бактериофага. Под его влиянием прекращается патологический процесс в растении, так как у бактерий резко изменяются морфологические и физиологические свойства.
6. Наконец, исключительное значение в борьбе с фитопа-тогенными бактериями имеет введение в культуру устойчивых к бактериозам сортов растений.
НЕИНФЕКЦИОННЫЕ БОЛЕЗНИ РАСТЕНИЙ
В основе любого патологического процесса лежит нарушение обмена веществ. В растениях обмен веществ может быть нарушен не только проникновением в их клетки и ткани фитопатогенных микроорганизмов. Всякое нарушение жизненных условий, необходимых для нормального развития растений, может вызвать их угнетение.
Болезни растений, возникающие под воздействием неблагоприятных абиотических факторов, относятся к группе неинфекционных или непаразитарных заболеваний. Основными абиотическими факторами являются почвенные и метеорологические условия.
Болезни, вызываемые неблагоприятными почвенными условиями. Для нормального роста и развития растений требуется определенное количество питательных веществ и влаги в почве. Недостаток питательных веществ вообще или относительный недостаток того или иного элемента минерального питания (азота, фосфора, калия, железа, микроэлементов) может явиться причиной различных заболеваний растений, называемых голоданием.
Наиболее распространенным типом минерального голодания является азотное. Азот входит в состав белка, а поэтому необходим для роста и развития всех тканей. Недостаток его в почве сразу же вызывает у растений симптомы азотного голодания. Прежде всего листья теряют нормальную зеленую окраску. Пожелтение листовых пластинок (хлороз) начинается между жилками. В конце концов листья отмирают, начиная с верхушки к основанию.
Из других симптомов азотного голодания следует указать на ожог, приостановку в росте, опадение завязей.
При калийном голодании также может наблюдаться хлороз, но только на листьях более старых, причем пожелтение тканей в первую очередь наблюдается около жилок (кукуруза). Признаками калийного голодания служат также некротические пятна, появляющиеся на нижних листьях (сахарная свекла) сначала по краям листовых пластинок, а затем в центре. Такие листья постепенно отмирают. Недостаток калия в почве может вызвать на листьях появление желтовато-белой крапчатости (хлопок).
Фосфорное голодание характеризуется прежде всего отставанием в росте, а также изменением окраски листьев. У картофеля нижние листья приобретают фиолетовую окраску; у сахарной свеклы — темно-бурую, у кукурузы — красноватую и т. д. Симптомами фосфорного голодания служат образование многочисленных боковых почек и слабое ветвление корней.
Недостаток железа в почве вызывает у растений хлороз. Железо необходимо для образования хлорофилла. Если этого элемента недостаточно, хлорофилл в хлоропластах не образуется или его количество значительно снижается.
Хлороз может проявляться в виде общего пожелтения растений — диффузный хлороз или в виде появления хлоротичных пятен на листовых пластинках — местный хлороз.
Отсутствие в почве микроэлементов — бора, марганца,
меди, молибдена, цинка и др.— также может нарушить обмен веществ у растений и вызвать патологические процессы. Например, недостаток бора в почве приводит к отмиранию точек роста стеблей и корней, общей задержке роста, отсутствию цветения. Отмирание верхушки стеблей табака, гниль сердечка свеклы, побурение внутренних тканей у капусты — все это болезни, вызываемые недостатком бора в почве.
Для нормального течения физиологических процессов в растении необходима почвенная влага. Однако избыток или недостаток ее в почве может существенно нарушить обмен веществ у растений.
Переувлажнение почвы, хотя бы и временное, затрудняет доступ воздуха к корням, они задыхаются и не могут нормально подавать воду надземным органам, а это приводит к ослаблению растений. Избыток влаги вызывает вымокание озимых вследствие застоя воды на поверхности почвы. Это явление особенно наблюдается на пониженных участках, а также па тяжелых глинистых почвах.
Если периоду избыточного увлажнения почвы предшествовала засуха, то происходит растрескивание плодов (вишня, слива, груша, яблоня) и корнеплодов (морковь, свекла, редис, петрушка и др.). Засуха способствует формированию механических элементов, и ткани теряют способность к растяжению. Поступление в растение большого количества воды ведет к быстрому увеличению отдельных органов, а наружные слои, не способные к растяжению, не выдерживая давления изнутри, растрескиваются, обнажая мясистые и сочные внутренние ткани.
Недостаток влаги в почве вызывает у травянистых растений карликовый рост и преждевременное созревание. У злаковых культур недостаток почвенной влаги препятствует нормальному наливу зерна, оно получается щуплым, мелким, быстро созревающим.
У древесных пород сухость почвы может вызвать длительное завядание, опадение завязей, отмирание корневых волосков и мелких корней. Последнее особенно опасно, так как приводит к засыханию верхушек деревьев (суховершинность).
Сухость почвы способствует образованию склеренхимных клеток у плодов груши, айвы, что значительно снижает их вкусовые качества.
Болезни, вызываемые неблагоприятными метеорологическими условиями. Среди метеорологических условий, вызывающих патологические процессы у растений, на первое место следует поставить высокие и низкие температуры.
Высокие температуры у растений могут вызвать ожоги надземных вегетативных органов. Особенно страдают листья, на них появляются желтые или бурые пятна неправильной
формы. При сильных повреждениях тканей листья засыхают и опадают.
Действие высокой температуры нарушает нормальную работу устьичного аппарата. Устьица теряют способность своевременно закрываться, растение чрезмерно испаряет влагу, а это также приводит к засыханию и опадению листьев.
У семечковых и косточковых пород в летний период возникают солнечные ожоги коры. Высокие дневные температуры вызывают образование на коре штамба и скелетных сучьев, красноватых, вначале пузыревидных, а затем западающих пятен различной величины преимущественно с юго-западной стороны. Особенно сильные ожоги наблюдаются у сортов с темной окраской коры. Пораженная солнечным ожогом кора растрескивается и отстает от древесины.
Низкие температуры вызывают замерзание целых растений или их отдельных частей. Сущность замерзания заключается в том, что вода из клеток поступает в межклетные пространства, где и замерзает, превращаясь в кристаллики льда, механически разрушающие клеточные оболочки. Обезвоженная протоплазма не способна впитывать воду даже, если ткани оттаивают. Клетки теряют тургор и погибают.
Чем больше воды содержится в растении, тем более оно подвержено замерзанию. Поэтому молодые побеги, листья, а также цветки особенно чувствительны к действию низких температур. Даже ранние осенние заморозки могут вызвать их гибель. Части растения, вступившие в период покоя, бедны водой и отличаются большей выносливостью.
Резкое падение температуры в зимнее время ведет к образованию морозобоин. Древесина обладает плохой теплопроводностью, вследствие чего наружные слои ее охлаждаются быстрее внутренних, сокращаются больше и не могут охватывать центральную часть дерева. В результате получаются разрывы наружных частей по радиусу. Такие разрывы и называются морозобоинами.
Если после больших морозов температура резко повышается, наружные части ствола нагреваются и расширяются сильнее внутренних, образуется кольцевая трещина между годичными слоями древесины или между корой и древесиной. Эти разрывы называются отлупами.
Повреждения растений вызываются различными атмосферными явлениями — молнией, градом, затяжными дождями, ветрами и т. п.
Молния может вызвать расщепление ствола дерева или повредить отдельные ветви, а у травянистых растений убить ткани стебля и листьев.
Град наносит сильные повреждения и опасен в любой период вегетации. Сильный град пробивает листья, сбивает
цветки, повреждает плоды, а в некоторых случаях вызывает полную гибель посевов, садов, виноградников. Кроме того, поврежденные градом растения становятся более доступными для фитопатогенных микроорганизмов.
Затяжные дожди в период налива зерна у хлебных злаков могут вызвать «стекание зерна». Нормальные функции ферментов нарушаются и запасные питательные вещества, отложенные в эндосперме семян, вновь переходят в растворимое состояние. В результате резкого увеличения осмотического давления внутри зерен и под влиянием выщелачивания качество зерна значительно снижается.
Ливни, сопровождаемые сильными ветрами, приводят к полеганию колосовых хлебов, что в дальнейше^м значительно затрудняет их уборку. В садах и лесах ливневые бури вызывают полом ветвей, сучьев, а иногда и стволов деревьев, так называемые буреломы. Через поврежденные участки коры и древесины проникают возбудители гнилей и других заболеваний.
ГЛАВА ИММУНИТЕТ РАСТЕНИЙ
К ИНФЕКЦИОННЫМ
У ЗАБОЛЕВАНИЯМ
ОБЩИЕ СВЕДЕНИЯ
ОБ ИММУНИТЕТЕ РАСТЕНИЙ
Иммунитетом растений называют невосприимчивость, проявляемую организмом при непосредственном контакте с паразитным агентом, способным вызвать заболевание при наличии необходимых условий для заражения. Иммунитет в переводе с латинского (immunis) означает освобождение от чего-либо.
В практике возделывания сельскохозяйственных растений давно было подмечено, что растения одного вида, произрастающие в одинаковых условиях, проявляют различное отношение к возбудителям болезней. Одни поражаются сильно, другие слабо, а третьи остаются здоровыми. Подмечено было также, что непоражаемые растения лучше растут и дают более высокий урожай. Естественно, что практики-растениеводы отбирали растения устойчивые к тому или иному заболеванию. Со временем накопилось немало сведений о существовании невосприимчивости у растений к возбудителям болезней. В конце XIX века начинаются первые исследования по этому вопросу и возникают различные теории относительно природы иммунитета у растений.
Невосприимчивость растений связывали с анатомо-морфологическими особенностями строения растений; качественным составом клеточного сока (количество органических кислот, дубильных веществ, антоциана). Высказывались предположения, что невосприимчивость растений к инфекционным заболеваниям зависит от величины осмотического давления клеточного сока и тургора клеток.
Среди теорий, пытавшихся объяснить явление иммунитета у растений, особое место занимала фагоцитарная теория, пред
ложенная И. И. Мечниковым. Согласно этой теории, иммунитет рассматривался как защитный приспособительный признак высшего организма в борьбе с патогенными микроорганизмами, проявляющийся в способности клеток хозяина захватывать и переваривать клетки паразита (фагоцитоз).
В последние годы Д. Д. Вердеревский выдвинул теорию, согласно которой невосприимчивость растений зависит от их фитонцидных свойств.
Все эти теории односторонне объясняют невосприимчивость растений к патогенным микроорганизмам. Иммунитет растений — явление весьма сложное и многообразное, и, по-видимому, еще нет достаточного количества экспериментальных данных для создания единой теории иммунитета. Растения различно реагируют на внедрение каждого паразита. Например, при поражении облигатными паразитами причинами устойчивости являются одни свойства растений, а в случае поражения факультативными паразитами — другие. При поражении разных сортов одной и той же болезнью защитные реакции одного сорта будут отличаться от таковых у другого. Например, И. Н. Найденовой установлено, что у одних миль-дьюустойчивых сортов винограда защитным свойством являются некротические реакции, а у других — фитонцидность клеточного сока.
Учение об иммунитете растений — фитоиммунология — представляет часть фитопатологии, в которой разбираются взаимоотношения паразитов и растений-хозяев в определенных условиях окружающей среды, а также вопросы повышения устойчивости к болезням у имеющихся сортов и выведения новых, обладающих комплексным иммунитетом сразу к нескольким возбудителям болезней.
По происхождению и проявлению различают два типа иммунитета: врожденный и приобретенный.
ВРОЖДЕННЫЙ ИММУНИТЕТ
Под врожденным или естественным иммунитетом понимают присущее данному виду или сорту растений свойство не поражаться тем или иным заболеванием. Это свойство является таким же наследственным признаком вида или сорта, как морфологические особенности, и способно изменяться под влиянием условий внешней среды.
Врожденный иммунитет в свою очередь разделяется на две категории: пассивный и активный.
Пассивный иммунитет — это совокупность факторов или свойств, имеющихся у растений, которые препятствуют проникновению в их ткани (и развитию) возбудителей болезней. Свойства эти выработались у растений вне связи с защитными реакциями против паразитов.
Главнейшие из них следующие:
1. Особенности строения покровных тканей (толщина ку-тикуляторного покрова, опушенность листьев, количество устьиц и т. п.). Известно, что устойчивость видов барбариса к заражению базидиоспорами стеблевой ржавчины зависит от толщины кутикулы, а более устойчивые к мильдью сорта винограда имеют меньшее количество устьиц на единицу поверхности.
2. Особенности прорастания семян. У устойчивых к твердой головне сортов яровых пшениц энергия прорастания семян выше, чем у восприимчивых. Поскольку период заражения твердой головней короткий (от прорастания семян до появления всходов), возможность заражения у первых меньше, чем у вторых.
3. Характер цветения. Сорта пшениц, у которых большинство цветков цветет закрыто или открывается на короткий период, меньше поражаются пыльной головней, чем сорта с более длительным периодом открытого цветения.
4. Движение устьичного аппарата. Прорастающая на поверхности листа спора гриба дает ростковую нить, которая в большинстве случаев попадает во внутренние ткани через устьица. Поэтому возможность заражения выше у тех растений, устьица которых открываются на продолжительное время. Например, сорта пшениц с устьицами, открывающимися на короткий период, менее поражаются ржавчиной по сравнению с теми, у которых они открыты целый день.
5. Качественный состав клеточного сока: содержание свободных кислот, антоцианов, фенольных соединений, фитонцидов, дубильных и ростовых веществ и других химических соединений.
Известно, что в клеточном соке устойчивых к мильдью сортов винограда содержится от 6,2 до 10,3% свободных кислот, а у восприимчивых — от 0,5 до 1,9%.
Возбудителем шейковой гнили лука (Botrytis allii) сильнее поражаются сорта с белой чешуей, чем с темноокрашенной. Это объясняется тем, что последние содержат антоциан и фенольные соединения, обладающие фунгицидным действием.
Активный и м м у н и т с т— способность растений активно реагировать на внедрение и распространение в тканях паразитных организмов. Проявляется это свойство только в случаях нападения паразита.
Устойчивые к болезням растения способны развивать защитные реакции, сущность которых заключается в том, что они направлены либо на локализацию паразита, либо на его уничтожение вырабатывающимися антитоксинами.
Активными реакциями растений, или факторами активного иммунитета, являются следующие:
1. Образование защитных некрозов — своеобразных барьеров, ограничивающих распространение паразита. Внешне защитные некрозы представляют собой отмершие участки ткани, в которых облигатный паразит не может развиваться и погибает. Защитный некроз может проявляться опробковением клеток вокруг пораженного участка, что также является препятствием для распространения паразита. Использовав все питательные вещества на изолированном участке, паразит быстро погибает.
Сорта хлебных злаков, устойчивые к ржавчине, могут служить хорошим примером образования защитных некрозов. При проникновении гифы гриба в ткани образуется некротическое или хлоротическое пятно, или кольцо вокруг пораженной ткани. При крайней степени устойчивости уредопустулы совсем не развиваются и болезнь проявляется только образованием некротических пятен в тех местах, где произошло внедрение паразита.
Появление точечных некрозов на листьях клевера свидетельствует о его защитной реакции против возбудителя мучнистой росы (Erysiphe polygoni). Как только гриб проникает через оболочку в клетку, содержимое ее начинает отмирать, а вместе с ним отмирает и внедрившаяся туда гифа гриба.
2. Антитоксические реакции растений. В образовании активных защитных реакций растений большое значение принадлежит окислительным ферментам. У устойчивых сортов имеется мощная система окислительных ферментов, стойких к токсинам грибов и активизирующихся под влиянием этих же токсинов. У восприимчивых сортов окислительные ферменты представлены значительно слабее.
Токсины грибов содержат аммиак, амины, органические кислоты и т. п. Окисление этих веществ сопровождается их разрушением, а следовательно, и потерей токсичности. Если гриб питается за счет тканей, предварительно им убитых, как например Botrytis cinerea. то защитные реакции растения в таком случае направлены на разрушение выделяемого грибом токсина. Устойчивость растений к таким паразитам объясняется окислением токсических веществ и превращением их в безвредные соединения. У устойчивых к болезням растений активизируются и синтетические процессы, парализующие деятельность гидролитических ферментов паразита.
ПРИОБРЕТЕННЫЙ ИММУНИТЕТ
Способность растений приобретать устойчивость к заболеваниям в процессе роста и развития называется приобретенным иммунитетом. В некоторых случаях это свойство может передаваться по наследству. Приобретенный иммунитет может быть неинфекционным и инфекционным.
Неинфекционный приобретенный иммунитет создается человеком. Выращивая растения и применяя различные приемы обработки почвы, семян, ухаживая за растениями в период вегетации, можно повысить их устойчивость к различным заболеваниям.
Еще в 1903 году русские ученые И. Я- Шевырев и С. А. Мокржецкий разработали метод внекорневого питания растений минеральными соединениями и отметили его благотворное действие на повышение у них невосприимчивости к заболеваниям и повреждению насекомыми.
В 1932 году Н. А. Наумов на основании своих исследований также сделал вывод, что воздействуя на растение в период его онтогенеза различными факторами, в том числе и химическими, можно добиться значительного повышения невосприимчивости к болезням. Основные приемы, при помощи которых можно создать у растений неинфскционный иммунитет, следующие: вакцинация растений; химическая иммунизация; антибиотические вещества.
Вакцинация растений — это введение внутрь тканей убитых культур или экстрактов возбудителей болезней. Практически вакцинация растений производится путем обработки семян, опрыскиванием растений и т. п. П. М. Жуковский путем опрыскивания пшеницы настоем конидий гриба Erysiphe graminis создавал у растений иммунитет к мучнистой росе.
Оказалось возможным создать устойчивость пшеницы к Helminthosporium sativum путем проращивания семян в экстрактах гриба. Обработка всходов фасоли фильтратом Botrytis cinerea способствовала выработке устойчивости у них к этому грибу.
Химическая иммунизация — это обработка растений различными химическими веществами (химическими иммунизаторами) для повышения их устойчивости к возбудителям болезней. Химическая иммунизация может осуществляться следующими приемами: предпосевной обработкой семян химическими соединениями; внесением химикатов в почву в сухом или жидком виде; внекорневой подкормкой растений путем опрыскивания их в период вегетации слабыми растворами химических веществ.
И. М. Поляков провел большие исследования по предпосевной обработке семян, установив, что из семян пшеницы, обработанных роданом, вырастают растения, обладающие устойчивостью к трем важнейшим заболеваниям: пыльной головне, стеблевой ржавчине и мучнистой росе, причем эта устойчивость передается следующему поколению.
В качестве химических иммунизаторов применяются этил-меркурхлорид, этилмеркурфосфат, медный купорос.
В 1922 году Т. Д. Страхов установил, что, регулируя режим питания, т. е. внося в почву те или иные минеральные соединения, можно повысить болезнеустойчивость растений. К настоящему времени получено много данных относительно значения вносимых в почву веществ. Калийные удобрения, как правило, повышают устойчивость растений, азотные же, наоборот, усиливают восприимчивость.
А. А. Ячевский еще в 1910 году указывал, что в целях повышения устойчивости растений к заболеваниям следует применять микроэлементы: бор, марганец, медь, цинк и др. Позднее обстоятельные исследования в этом направлении были проведены Т. Д. Страховым и его учениками. В настоящее время микроэлементы широко используются в практике растениеводства как средство повышающее болезнеустойчивость растений. Микроэлементы применяются при предпосевной обработке семян, их можно вносить в почву в чистом виде и в сочетании с полным минеральным удобрением, в качестве внекорневной подкормки растений.
В Белоруссии широко применяется разработанный Н. А. Дорожкиным метод внекорневой подкормки картофеля 0,1—0,2%-ным раствором сернокислой меди с целью повышения устойчивости к фитофторозу.
М. С. Дунин считает, что все болезни растений можно разделить на две группы, если за основу принять возраст поражаемых тканей: 1) болезни, развивающиеся на молодых тканях (I период вегетации) и 2) болезни, поражающие стареющие ткани (II период вегетации). В случае, если нападению паразитов подвергаются молодые ткани, для повышения их болезнеустойчивости целесообразно практиковать такие приемы, которые увеличивают темп роста молодых органов и тем самым укорачивают период наиболее уязвимой фазы. Например, применение обрезки деревьев и кустарников задерживает начало вегетации, а затем, наоборот, вызывает быстрый темп роста и развития побегов, что приводит к укорачиванию наиболе благоприятного для заражения периода. Этот способ позволил повысить устойчивость абрикосов к мо-нилиозу и клястероспорозу.
Повысить устойчивость растений к болезням второй группы можно такими приемами, которые задерживают старение тканей, например подкашивание многолетних трав. Известно, что этот прием вызывает устойчивость клевера к антракнозу, а люцерны к мучнистой росе.
Инфекционый иммунитет возникает у растений вследствие перенесения какой-либо болезни. Инфекционный иммунитет — явление специфичное. Растение приобретает устойчивость только к тому возбудителю, который вызвал заболевание.
Переболевшее растение может остаться носителем инфекции и представлять опасность для окружающих здоровых растений. Такой инфекционный иммунитет называется нестерильным.
Если у выздоровевшего растения не сохраняется возбудитель инфекции, оно не опасно для окружающих здоровых растений. Повторное заражение его тем же возбудителем также не вызывает заболевания. Такой инфекционный иммунитет носит название стерильного.
* *
Выше указывалось, что иммунитет растений — явление весьма сложное и многообразное, определяющееся сочетанием свойств растения-хозяина и паразита — возбудителя болезни в определенных условиях внешней среды. Принимая во внимание, что каждый из указанных компонентов обладает большим разнообразием и изменчивостью свойств, степень устойчивости иммунитета не является постоянной.
Период сохранения устойчивости к заболеваниям у возделываемых сортов растений различен. Известны сорта многих культурных растений, десятки лет сохраняющих устойчивость к возбудителям болезней. С другой стороны, имеются сорта, которые некоторое время считались болезнеустойчивыми, а затем теряли устойчивость.
На изменчивость иммунитета могут влиять половая и вегетативная гибридизация среди растений-хозяев, адаптация паразита к новым сортам, условия окружающей среды и. что, пожалуй, самое главное, способность паразита давать новые физиологические расы, обладающие более агрессивными свойствами. Последнее особенно имеет место при половой гибридизации у возбудителей болезни. Вследствие этого сорта, обладавшие какое-то время устойчивостью, начинают поражаться, и многолетние работы по выведению устойчивых сортов сводятся на нет.
Потерянную сортами устойчивость можно восстановить путем отбора непоражаемых растений, особенно на фоне естественного и искусственного заражения. Инфекционный фон дает возможность выявить иммунные растения.
Борьба с потерей болезнеустойчивости сортов должна быть направлена также на тщательное изучение расового состава возбудителей болезней.
ЛИТЕРАТУРА
Амбросов А. Л. Вирусные болезни картофеля и методы выращивания здорового семенного материала в Белорусской ССР. Минск, 1965.
Бондарцев А. С. Болезни культурных растений и меры борьбы с ними. М.—Л., 1931.
— Трутовые грибы Европейской части СССР и Кавказа. М.—Л., 1953.
Б и л а й В. И. Фузарпп. Киев, 1955.
— Ядовитые грибы иа зерне хлебных злаков. Киев, 1953.
Бабаян А. А. Краткие итоги изучения вертпцпллезиого увядания хлоп-чат ника в Армении.— Сб. научи, тр. Научи.-псслед. ин-та земледелия. 1961.
Беккер 3. Э. Физиология грибов и их практическое использование. М., 1963.
Васильков Б. П. Очерк географического распространения шляпочных грибов в СССР. М.—Л., 1955.
Васильевский Н. И., К а р а к у л и н Б. П. Паразитные несовершенные грибы. Ч. 1. Гифомицеты. М.— Л., 1950; ч. II Мсланкониальные. М.— Л., 1950.
Воронин М. С. Избранные произведения. М., 1961.
Виноградова Н. В. Биологические особенности возбудителя рака яблони (Nectria galligena Brea.) и разработка мер борьбы с ним в условиях Белоруссии. Дисс. Гродно, 1956.
Вердеревский Д. Д. Иммунитет растений к паразитарным болезням. М., 1959.
— О некоторых теоретических вопросах иммунитета растений.— Тезисы докладов IV Всесоюзного совещания по иммунитету сельскохозяйственных растений, 1965.
Горленко М. В. Болезни пшеницы. М., 1951.
Краткий курс иммунитета растений к инфекционным болезням. М., 1959.
Г у т н е р А. С. Головневые грибы. Л., 1941.
Гулд К. Гнили лилий и тюльпанов.— Болезни растений. Под ред. М. С. Дунина. М., 1956.
Головин П. Н. Мучнисто-росяные грибы, паразитирующие на культурных и полезных растениях. М.—Л., 1960.
Головин П. Н. Принципы систематики низших грибов.— Тр. ВИЗР, вып. 23, 1964.
Г у л е ц к а я Е. Г. Некоторые данные относительно мучнистой росы злаков.— Уч. зап. Белорусок, ун-та, вып. 33, сер. биол. Минск, 1957.
Г о р о в е ц В. К. Биология и экология возбудителя церксспороза сахарной свеклы и меры борьбы с ним. Минск, 1954.
Грушевой С. Е. Сельскохозяйственная фитопатология. М., 1965.
Дарожк1н М. А. Пералж 1ржавых хвароб на раслшах Аршанскай акруп i ашсаине хвароб культурных с-г. раслш Бранскай губерж.— За-niCKi БДАН. Т. 6. 1929.
— Хваробы збожжа, некаторых бабовых культур i меры барацьбы з iMi. Минск, 1935.
— Грыбныя хваробы Гласных культур у БССР.— Савецкая Kpaina, 1932, № 1.
Дорожкин Н. А. Рак картофеля и меры борьбы с ним. Минск, 1948. — Болезни картофеля. Минск, 1955.
Дорожкин Н. А., Кустова А. И. Опыты и наблюдения над склеротинией в БССР.— Уч. зап. Белорусск. ун-та, вып. 26, сер. биол., 1956.
Дорожкин Н. А. Биологическое обоснование агротехнических способов борьбы с возбудителем рака картофеля Synehylrum endobivtieum (Schibb.) Pers.— Rostlinn^ vyroba. Прага, 1959.
Дорожкин H. А., Ч e к а л и н с к а я Н. И. Болезни люпина. Минск, 1965.
Дементьева М. И. Болезни плодовых культур. М., 1962.
Д о б р о з д р а к о в а Т. Л., Летова М. Ф., Степанов К- М., Хохряков М. К- Определитель болезней растений. М.—Л., 1956.
Дуни н М. С. По Афганистану, Пакистану и Индии. М., 1954.
3 о р г е в и ц А. Гладиолусы. Рига, 1961.
Инголд Ц. Пути и способы распространения грибов. М., 1957.
К у р с а н о в Л. И. Микология. М., 1940.
К у р с а н о в Л. И., К о м а р и и ц к п й Н. А. Курс систематики низших растений. Л., 1945.
Комарницкий Н. А., Кудряшев Л. В., Ураков А. А. Систематика растений. М., 1962.
Кудрявцев В. И. Систематика дрожжей. М., 1954.
Калашников К- Я. Головня зерновых культур. М.—Л., 1962.
Келли А. Микотрофия у растений. М., 1952.
Комарова Э. П. Рет кие виды трутовых грибов, обнаруженные на территории БССР.—Изв. АН БССР, 1966, № 2.
Купревич В. Ф. Физиология больного растения в связи с общими вопросами паразитизма. М.—Л., 1947.
Купревич В. Ф., Транше ль В. Г. Флора споровых растений. Т. IV. Мелампсоровыс. М.—Л., 1957.
Кудряшева 3. Н. Антракноз красного клевера в Белоруссии п значение режима питания в борьбе с ним.— Болезни сельскохозяйственных культур в БССР. Минск, 1958.
— Фпкомицеты (Phycomycetes). Минск, 1960.
— Сумчатые грибы (Ascomycetes). Минск, 1962.
Л у ч и и а Н. Н. Биологические особенности развития возбудителя поли-спороза льна (Kabatiella lini (Laff.) Кагак.) в условиях БССР и меры борьбы с ним. Минск, 1961.
Мейджн Р. Некоторые грибные болезни гладиолусов/—Болезни растений. Под ред. М. С. Дунина. М., 1956.
Наумов Н. А. Определитель мукоровых. М.—Л., 1935.
— Ржавчина хлебных злаков СССР. М., 1939.
— Болезни сельскохозяйственных растений. М., 1952.
— Фитофтора картофеля. Л—М., 1961.
Натальина О. Б. Болезни ягодников. М., 1963.
— Определитель низших растений. Под ред. Л. И. Курсакова. Т. 3, 4. Грибы. М., 1954.
Орлов Н. И. Съедобные и ядовитые грибы, грибные отравления и их профилактика. М., 1953.
Пак П. В. Г ельминтоспориозы хлебных злаков и меры борьбы с внутренней инфекцией семян.— Селекция и семеноводство. Минск, 1950.
Проценко Е. П. и Проценко А. Е. Краткий атлас болезней декоративных растений. М., 1961.
С а п р у и о в Ф. Ф. Хищные грибы — гифомицеты и их применение в борьбе с патогенными нематодами. Ашхабад, 1958.
Саркисов А. X. Микотоксикозы. М., 1954.
Сержанина Г. И. Съедобные и ядовитые грибы порядка Agaricales и их хозяйственное значение. Минск, 1962.
Стрельская О. Я- Антракноз льна (Colletoi richuni lini Boll.) в БССР и обоснование мер борьбы с ним. Минск, 1961.
Те т ер ев н и к ов а - Б аба я и Д. Н. Обзор грибов из рода Septoria. Ереван, 1962.
— Болезни овоще-бахчсвых культур в Армении и меры борьбы с ними. Ереван, 1964.
Траншель В. Г. Обзор ржавчинных грибов СССР. М.—Л., 1939.
Ульянищев В. П. Микофлора Азербайджана. Т. I—III, Баку, 1952— 1960.
Ф е д о р и н ч и к И. С. Роль антагонистов в подавлении заразного начала возбудителей болезней, передающихся через почву.— Тр. ВИЗР, вып. 3, 1951.
Хохряков М. К- Морфолого-биологическое обоснование систематики грибов рода Helminthosporium на злаках. Л., 1953.
— Вредные и полезные грибы. Л.—М., 1961.
— Проблема вида и видообразования у грибов.— Тр. ВПЗР, вып. 23, 1964.
— К 100-летию со дня рождения профессора А. А. Ячевского.— Тр. ВИЗР, вып. 23, 1964.
Хохрякова Т. М. Монилиозы плодовых культур Дальнего Востока. Л, 1966.
Ч е р е м и с и и о в Н. А. Общая патология растений. М., 1965.
Я ч ев с кин А. А. Справочник фитопатологических наблюдений. Л., 1930. — Основы микологии. М.— Л., 1933.
19 3. Н. Кудряшева
УКАЗАТЕЛЬ РУССКИХ НАЗВАНИЙ ГРИБОВ И БОЛЕЗНЕЙ, ВЫЗЫВАЕМЫХ ИМИ
А
Аврнкуляриевые 188
Агариковые 145, 154, 155
Алиментарно-токсическая алим-ппя 215
Антракнос 227, 232
— красного клевера 228, 229
— -крыжовника 235
— льна 233
— смородины 235
— подсолнуха 231, 233
Архимицеты 48, 54
Аскомицеты 47, 85, 89, 92, 134
Аскохнтос 238, 239
— гороха 238, 239, 240
Аспергилловые 28, 31
Афнллофоровые 144
Б
Базидиальные грибы 19, 26, 29. 34.
39, 49, 135, 136, 137, 138, 187
Базидиомнцеты 46, 138, 139, 140, 187
Без жгутиковые 49, 50
Березовая губка 151, 152
Бластомикозы 39, 92
Бледная поганка 23, 140, 156, 157
Болетусовые 155
Боровик 23, 41. 155
В
Ведьмины метлы 98
Веселка обыкновенная 23, 45, 161
Вертпциллиоз яблони 205
Вилт 221
— вертициллиозный малины 204
---- хлопчатника 205
Волнушка 140
Вороночник 145
Выпревание озимы 217
Высшие грибы 19 32, 34, 35, 39.
46, 135
Г
Г а ст ром и петы 159, 160, 187
Гельминтоспорпоз 207, 209, 210
— злаков 9, 207
г— полосатый ячменя 209
Гидрофильные грибы 193
Гифомицеты 190, 192
Гниль 190
— белая 129, 130. 131
— кагатная 242, 245
— коричневая сердцевинная гладиолусов 197, 198
— мокрая клубней картофеля 223
— поздняя------68
— серая 193, 201
---- винограда 194
----земляники 193
---- лилий 198
----овощных культур 194
----тюльпанов 199
— сердцевинная свеклы (гниль сердечка) 244
— сухая клубней картофеля 223
— шейковая лука 195
Головневые грибы 7, 25, 38, 163,
165. 166, 167, 169, 171, 172, 188
Головня 4, 5, 135, 163, 164= 173
Головня карликовая пшеницы 165, 169, 171, 172
— проса 169
— пыльная 44
---- овса 169, 172
----пшеницы 165. 166, 167, 172
----ячменя 166, 167
— стеблевая ржи 14, 169
— твердая пшеницы 14, 23, 165,
166, 169, 172, 173
---- ячменя 165
Голосумчатые грибы 38, 86, 91
Гриб-зонтик 139, 140
д
Дакриомнцеты 188
Двужгутиковые 49
Дереворазрушающие грибы 24, 25, 26
Дискомицеты 39, 99, 124, 236
Дождевики 25, 160, 161
Домовой гриб 16, 18, 20, 24, 30,
148, 149, 150
— — пленчатый 144, 145
Дрожалковые 188
Дрожжевые грибы 24, 25, 30, 31, 93, 94
Дрожжи 25, 82, 86, 93
— восточно-азиатские 82
— жировые 22, 93
— мукоровые 82
— пивные .или хлебные 86, 95
— хересные 95
Дутые сливы 97
Е
Ежевпковые 146
Ж
Желчный гриб 155
3
Зеленка 20
Зигом инеты 18, 48, 53, 79, 92, 187
К
Каменная или известковая детка 92
Кармашки слив 97
Кесарев гриб 4, 156
Кила капусты 46, 54, 56, 57
Кистевик 103
Копринус 158
Копрофплъные грибы 158
Корневая губка 151
Корнеед свеклы 243, 244
Курчавость листьев вишни 99 ---- персика 99
Л
Лисичка 23, 145
Ложная мучниста роса 76
------ лука 76
Ложноопенок серно-желтый 157
М
Макроспормоз 214
Масленок 20, 139, 154 — лиственничный 41 — поздний 155
Меланкониальные 191, 192, 226
Мелямпсоровые 178
Микозы 102
.Микотоксикозы 215
Микохитридпевые 64
Миксохитридиевые 58, 93
Мильдью 75
Молочница * детей 39, 91, 93
Монилпальный ожог 201
----косточковых 200, 201
— — семечковых 200
----яблони 202
Молилиоз 201, 202
Мукаровые грибы 8, 22 25, 30, 50, 53, 79, 80, 92, 93
Мукоромикозы 80
Мучнистая роса 4, 104, 107
---- винограда 105
---- злаков 111, 115
----- крыжовника 46, ПО
---- яблонь 105
Мучнисто-росяные грибы 7, 19, 25, 30, 86, 104, 105, 106, 107
Мухомор 23, 41, 155
Н
Несовершенные грибы 8, 25, 30, 32, 46, 131, 132, 189, 190, 191*, 192, 215, 237
Низшие грибы 32, 37, 39, 46,
О
Огонь св. Антония 114
Одножгутиковые 49
Оомицеты 48, 51, 65, 79
Опенок 17, 91, 139, 140, 157 — осенний 154
П
Парша 120, 122
— груши 121
— порошистая картофеля 10, 57, 58
— черная— 144
— яблони 121
Пенициллиум 25
Первичносумчатыс 86, 87*, 91, 92
Пероноспоровые 8, 38, 67, 74, 76
Пероноспороз 76
— свеклы 77
Пиреномицеты 31, 39, 113, 124
Питиевые 67
Пластинчатые 156
Плектасковые 90, 102
Плесень 79
— головчатая 82
— зеленая 103
— картофельная 68
— снежная 217, 218
— цветочная клевера 196, 197
Плодосумчатые 89, 91, 100. 101
Подберезовик 41, 155
Подосиновик 155
Полиспороз льна 230
Присуха 179
Пьяный хлеб 218, 219
Пятнистость 190
— бурая листьев кукурузы 64, 65
— коричневая—картофеля 213
------томатов 30, 213
— полосатая 208
Р
Рак европейский 116
— картофеля 60, 61, 63
— обыкновенный 116
— яблони 118
Ржавчина 4, 5, 135, 173
— белая 79
— бурая пшеницы 182, 183
---ржи 182, 185
— гороха 186
— желтая пшеницы 182
— корончатая овса 182
— линейная злаков 182, 183
— льна 178, 180
— подсолнечника 7
— пузырчатая сосны 180. 181
— смородины 180
Ржавчинные грибы 7, 8, 22, 25, 38, 44, 173, 174, 176, 177, 178, 186, 187, 188
Рогатиковые 25, 145, 146
Рыжик 19, 21, 23, 41, 157, 158
Рядовка 20
С
Сапролегниевые 27
Сапролегния 14, 32
Сатанинский гриб 156
Сахаромицеты 94. 95
Септическая ангина 215, 220
Септориоз 241
— люпина 243
— томатов 241, 242
Синева древесины 118
Сморчки 4, 22, 127, 128
Сморчковые грибы 22, 23, 90, 127
Сморчок конический 127
— обыкновенный 127
Спорынья 22, 23, 113, 114, 115
Стригущий лишай 39
Строчки 127, 128
Строчок обыкновенный 127
Сумчатые грибы 7, 19, 34, 37. 49, 85, 87, 89, 91, 99, 100'; 101, 135, 187, 235
Сферопсидные 191, 192, 232
Сыроежка 20, 23
Т
Телефоровые 144. 145
Трахеомикоз 215
Трихофптон 27
Трутовик ложный 151, 152
— серпо-желтый 151, 152
Трутовики 17
Трутовые грибы 6, 14, 24, 38, 46,
147, 148, 151, 153, 155
Трюфелевые грибы 132, 133
Трюфели 4, 104
Трюфель 133
— белый 134
— закавказский 104
— оленин 104
— черный 133
У
Увядание 190, 221
— вертиипллиозное 205. 206
— — картофеля 204
—• — томатов 204
— фузариозное астр 224
---картофеля 222
•-- хлопчатника 223
ф
Фикомицеты 19, 47, 48, 49, 50, 90, 188
Фитофтора 68, 69
Фитофтороз 72, 73
— земляники 72
— картофеля 72
— томатов 72
Фомоз 244
Фрагмобазндпальные грибы 163, 187
Фузариоз 218, 219, 220
— гладиолусов 225, 226
— льна 221
— хлебных злаков 217
Фузариум 23, 216
X
Холобазидпальные грибы 140, 187
ц
Цезаре кий гриб 156
Церкоспороз 211
— свеклы 211, 213
Ч
Чага 152
Черный зародыш 210
Черная ножка капусты 59
Чехловидная болезнь злаков 115
Ш
Шампиньон 4, 31, 154
Шпзосахаромицеты 96
Шляпочные грибы 6, 18, 20, 21, 22, 23, 25, 29, 30, 31, 41, 44, 91. 135, 139, 140, 159
Э
Экзоасковые 99
Экзобазидиальные 141, 187
Экзобазидпоз чайного куста 142,
143
Экзобазидиум 143
Энтомофторовые 25, 82
Эрготизм 114
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ ГРИБОВ
А
Abortiporus fractipes 154
Achlya 66
Aciliata 49
Agaricaceae 155, 156
Agaricales 44, 143, 154 *
Albuginaceae 67, 78
Albugo Candida 79
Alternaria 34, 44, 126
Amanita 156
— caesarea 156
— phalloides 156
Aphyllophorales 143
Archimycetes 48, 50
Armillaria 156, 157
— mellea 29, 157
Arthrobotrys 46
Ascobolaceae 126
Ascobolus 46, 90, 91
Ascochyta 238
— pinodella 239, 240
— pinodes 239, 240
— pi si 239 :1:
— pseudopinodella 239, 240
Ascomycetes 47, 85, 91, 207
Asera rubra 45
Aspergillaceac 102
Aspergillus 34, 39, 102 *, 190
— fumigatus 31
— niger 27, 190
— oryzae 24, 102, 190
Aunculariales 163
В
Basidiobolus ranarum 46
Basidiomycetes 47, 135
Biciliata 49
Boletaceae 155
Boletus 155
— edulus 155
— satanas 156
Botrytis 192, 193
— allii 195
— anthophila 196, 197
— cinerea 190, 193, 194, 195
— elliptica 198
— gladiolorum 197
— tulipae 199, 200
Bovista 160, 161
C
Calvatia maxima 161
Cantharelhis 139, 144, 145
Carpoascomycetes 92
Ceratostomaceae 118
Ceratostomella 118
— coerulea 118
— picea 118
— pini 118
— ulmi 118, 119 *
Cercospora 207, 210, 211
— beticola 211, 212*, 213
Chaetocladium 50
Chlorosplenium 29
Choiromyces meandriformis 134
Chytridium brassicae 59
Clavaria 145, 146*
— cristata 146
Clavariaceae 144, 145
Claviceps 91, 113
— microcephala 115
— purpurea 17, JOO, 113, 114, 115
Colletotrichum 45, 227, 230, 231 — lindemuthianum 231, 232*, 233 — lini 233 Collybia 29 Colonectria 217 — graminicola 217 Coniophora 144 — cerebella 144 Coprinus 31, 153, 158 * Corti cum 139 Craterellus 144, 145 Cronartium 180 — asclepiadeum 181 — ribicola 178, 180, 181 Cucurbitaria 91 Cystopus 37
D
Dacryomycetales 140
Dematiaceae 192, 206
Dendrodochium 190
Deuteromycetes 47, 131, 189
Dictiophora 162 — phalloides 162* Didimella pinodes 240 Dipodascaceae 92 Dipodascus albidus 92 Discomycetes 101, 124 Dothidiales 101, 124 Drechslera 210 — graminea 210 — teres 210
E
Elaphomyces cervinus 104
Elaphomycetaceae 102, 104
Empusa 83, 84
— muscae 38, 83 *, 84
Endoascales 91
Endomyces 93
— albicans 39, 91, 93
— decipiens 91
— fibuliger 93
— vernalis 93
Endomycetaceae 92, 93
Entomophthora 83, 84
Entomophthoraceae 38, 83
Entomophthorales 79, 82, 83
Epichloe 115
— typhina 100, 115
Eremascus 86, 93
— fertilis 87 *
Erysiphaceae 38, 85, 104, 112, 113
Erysiphales 37, 101, 104
Erysiphe 108, 109 , 110, 111
— graminis 105, 111
-----f. festicae 111
Erysiphe graminis f. secalis 111
----f. tritici 111
Euascomycetes 87, 89, 91, 100, 101
Exoascaceae 85, 96, 141
Exoascales 91, 96, 99
Exoascus 96, 97, 98
— cerasi 98
— epiphylla 98
— deformans 99
— minor 99
— pruni 97 *, 99
Exobasidiales 140, 141
Exobasidium 141, 143
— vaccinii 7, 142 *, 143
— vexans 7, 143
F
Fomes 151
— annosus 151 *
— fomentarius 148
— igniarius 152
----f. alni 152
----f. betula 152
----f. tremulae 152
Fomitopsis annosa 148, 151
Fungi 12
— imperfecii 47
Fusarium 22, 190, 215, 217
— graminiarum 218
— lini 221, 222 *
— nivale 217
— oxysporum 31, 222, 223
----f. vasinfectum 223
----L colestephi 224
----L gladioli 225
— poae 220
— sporotrichioides 220
Fusicladium 207
— dcndriticum 119, 207
— pirinum 119
G
Gastromycetes 140, 159
Gibberella 217
— saubinetti 219
Gloeosporium 45, 227, 235
— ribis 235, 236
----f. grossularia 235
----f. nigri 235
----f. rubri 235
Glomerella 231
Gnomonia 235
Gymnoascomycetes 91
Gymnosporangium 176
Gyromitra 127
н
Helminthosporium 207, 209, 210 — gramineum 207, 208* 209, 210 — sativum 207, 209 — teres 207, 209, 210 Helotiales 101, 124, 128 Helvella 127 — esculenta 127 Helvellaceae 126
Hemisphaeriales 101, 124
Holobasidioniycetes 140
Humaria 128*
Hydnaceae 144, 146
Hydnum 147
— repandum 147 *
Hymenomycetes 140, 143
Hypholoma 156, 157
— fasciculare 157
Hyphomycetales 131, 190, 192
Hypochnus 139, 144
— solani 144
Hypocreaceae 113
Hypocreales 101, 113, 217, 219
Hypomyces 217
Hysteriales 101, 124
I
Ixocomus luteus 155
К
Kabatiella 36, 45, 227, 230
— caulivora 228, 229 *
— lini 230
Krombholzia aurantiaea 155
— scabra 155
L
Laboulbeniales 101
Lachnea 127 *
Lactarius 156, 157
— delic iosus 157
Leveillulla 108
Lophodermium pinastri 125
Lycoperdaceae 44, 160
Lycoperdon 160, 161 * — giganteum 161
M
Macropodia 127 *
Macrosporium 123, 213
— solani 213, 214 *
— tomato 30, 213
Marasmius 29
Melampsora 175, 178
— lini 178, 179 *
Melampsoraceae 10, 176, 178
Melanconiaceae 227
Melanconiales 191, 226 ;Microsphaera 108, 109* Monilia 30, 132, 192, 200 — cinerea 200, 202 — fructigena 202 Monilinia 129, 131 — fructrigena 17, 131, 132
Monoblepharis 34
M orchell a 127
conica 127
— esculenta 127
AAucedinaceae 131, 192
Mucor 39, 46, 53 *, 82
— corimbifer 82
— genevensis 82
— javanicus 82
— mucedo 34, 81 , 82
— pusillus 82
— racemosus 80, 81, 82
— rauxianus 82
Mucoraceae 81
Mu corales 79, 81. 89
Mucronella 147
Mycochytridiales 37, 54, 64
Mycosphaerell aceae 118, 119, 211
Myxochytridiales 37, 54
N
Nectria 217
— galligena 116, 117
Nectriaceae 113, 116
О
Oidium 105
Olpidiaceae 58, 59
Olpidium 59
— brassicae 59 *, 60
On ygen aceae 102, 104
Oomycetes 48, 65
Ophiobolus 123
— graminis 123
P
Penicillium 32, 34, 39, 102, 103 *, 190
— digitatum 103
— italicurn 103
Penicillium roquefortii 103
Pericystis apis 92
Peridermium 181
Perisporiaceae 104, 112, 113 Peronospora 74, 75 — schachlii 75, 77 * — schleideni 75 Peronosporaceae 64, 74 * Peronosporales 52, 65, 67, 69 Pezizaceae 126, 127 551 Pezizales 101, 124, 126 Phacidiales 101, 124, 125 Phallaceae 45, 160, 161, 162 Phallus 161 — impudicus 45, 161, 162* Phellinus igniarius 152 Pholiota mutabilis 24 Phoma 26, 238, 243 — betae 26, 243, 244, 245 Phragmidium 175, 176 Phragmobasidiomycetus 140 Phycomycetes 47, 48 Phyllachora 124 Phyllactinia 105, 108, 109 112
Phvlloporus 155 Phytophthora 37, 49, 67, 68, 69, 74 — fragaria 68, 72, 73 — infestans 7, 31, 68, 69, 70 , 71, 72 — phaseoli 68 Physoderma 64 — zeae 64, 65 * Physodermataceae 64 Pilobolus 31, 46, 82 - - crystallinus 31, 82 Plasmodiophora 54, 56 — brassicae 7. 46, 54, 55 56
Plasmodiophoraceae 54 Plasmodiophorales 54 Plasmopara 74, 78 — vilicola 75 Plectascales 101, 102 Pleospora 123 — chartarum 123 — vulgaris 123 Pleosporaceae 118, 123 Podosphaera 108, 109*, 110 — leucotrucha 105 Polyporaceae 144, 147 Polyporus 152, 154, — betulinus 152 — squamosus 153 — sulfureus 152 Polyspora 230 — lini 230 Protascales 86. 91, 92, 99 Protascomycetes 91 Protomyces 99 Protomycetaceae 96, 99 Pseudoperonospora 74, 78 — cubensis 78 — humuli 78 Pseudopeziza 235
— ribis 236
Puccinia 176, 182
— coronifera 182
— dispersa 182, 185
— graminis 7, 38, 182, 183, 184
----f. avenae 38
----f. tritici 38, 183
— glum arum 182
— helianthi 7
— triticina 182, 184, 185
Pucci niaceae 178, 182
Pyrenomycetes 101, 113
Pyrenophora 210
Pyronema 90
— confluens 87, 88 *, 126
Pyronemaceae 126
Pythiaceae 67
Pythium 67, 68
— de baryanum 68
R
Rhizoctonia 26 — solani 144
Rhizopus 14, 82
— oryzae 82
— tritici 82
Rhytisma 90 — acerinum 125 *
S
Saccharomyces 91, 93
— cerevisiae 86, 87 *, 95
— ellipsoideus 95
— lactis 95
— oviform is 95
— vini 93, 95
Saccharomycetaceae 30, 91, 92, 93
Saccharomycetales 16
Saprolegnia 30, 38, 51 *. 66
Saprolegniaceae 66
Saprolegniales 49, 51, 65, 66
Schizosaccharomyces 86, 91, 96
— octosporus 87 *
— pombe 96
Schizosaceharomycetaceae 92, 96
Sclerotinia 129
— fructigena 132
— libertiana 17, 129, 130, 131
Sclerotiniaceae 129
Septoria 238, 241
— lupini 243
— lycopersici 241, 242
Serpula 139, 148
— lacrymans 30, 148, 149 150 *
Sphacelia 45, 114
Sphaeriales 101, 118,211 Sphaerioidaceae 238 Sphaeropsidales 18, 237 Sphaerotheca 108, 109* — mors-uvae 110 — pannosa 105, 110 Spongospora 54, 57 — subterranea 57, 58 Stereum 144, 145 Stromatinia fructigena 132 Synchytriaceae 58, 60 Synchytrium 60 — endobioticum 46, 60, 62
T
Taphridium 99 Taphrina 96, 98, 99 — aurea 99 — bullata 99 Taphrinales 91 Terfezia transcaucasica 104 Terfeziaceae 102, 104 Thamnidium 51 Thelephora 144, 145 Thelephoraceae 144
Tilletia controversa 169, 171 — levis 165 — tritici 164*, 169 Tremellales 163 Trichoderma 131, 190 — lignorum 131
Trichophyton 39
Tuber 91, 133 — aestivum 134 — brumale 134
Tuberales 101, 124, 132, 133 Tuberculariaceae 192, 215 Tylopilus fellens 155 Typhula 146 — trifolii 146 *
Tyromyces 154
11
Uncinula 108, 109
— necator 105 Unicellomycetales 93 Uniciliata 34, 37, 163, 173 Urocystis occulta 164*, 169 Uromyces 176, 182, 185 — pisi 185
Ustilaginales 163
Ustilago avenae 169
— hordei 165
— nuda 166, 167
— panici-miliacei 164 *, 169 — tritici 164*, 166, 167
— zeae 164 *, 170
V
Venturia 119, 122
— inaequalis 100, 119
— pirina 119
Verpa 90
Verticillium 192, 203, 206
— albo-atrum 203, 205, 206
— dahliae 206
— sp. 205.
Z
Zygomycetes 48, 52, 79 Zygosaceharomyces 95
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ ВОЗБУДИТЕЛЕЙ БАКТЕРИОЗОВ
Bacterium tracheiphilum 253
Corynebacterium sepedonicum 253
Pectobacterium carotovorum 255
— phytophthorum 252, 253
Pseudomonas cerasi 254
— lachrymans 254
— pisi 255
— tumefaciens 252, 254
— xanthochlora 255
Xanthomonas beticola 254
— campestris 254
— heteroceae 252
— phaseoli 255
ОГЛАВЛЕНИЕ
Введение 5
Глава 1. Общие сведения о строении, питании и размножении грибов 12
Мицелий и его видоизменения 12
Строение клетки 17
Ткани грибов 20
Химический состав 21
Ферменты . . 23
Питание грибов ... 25
Диссимиляция у грибов ... 28
Влияние внешних факторов на рост и развитие грибов 30
Размножение .... . 32
Образ жизни грибов в естественных условиях 36
Пути и способы распространения грибов 44
Глава 2. I класс Phycomycetcs (фикомицеты) 48
Общая характеристика 48
Подкласс Archimycetes 54
Порядок Plasmodiophora! es 54
Сем. Plasmodiophoraceae . 54
Порядок Myxochytridiales 58
Сем. Olpidiaceae . 59
Сем. Synchyiriaceae . . 60
Порядок Mycochytridiales 64
Сем. Phyzodermataceae 64
Подкласс Oomycetes . 65
Порядок Saprolegniales 66
Сем. Saprolegniaceae . 66
Порядок Peronosporales Ы
Сем. Pythiaceae . 67
Сем. Peronosporaceae 74
Сем. Albuginaceae . 76
Подкласс Zygomycetes 79
Порядок Mucorales 79
Сем. Mucoraceae .... 81
Порядок Entomophthorales 82
Сем. Entomophthoraceae 83
Глава 3. 11 класс Ascomycetes (сумчатые грибы) 85
Общая характеристика 85
Подкласс Protascomycetes 91
Порядок Protascales 91
Сем. Dipodascaceae 92
Сем. Endomycetaceae 93
Сем. Saccharomycetaceae . 93
Сем. Schizosaccharomycetaceae 96
Порядок Exoascales 96
Сем. Exoascaceae 96
Сем. Protomycetaceae . . 99
Подкласс Euascomycetes 100
Порядок Plectascales 101
Сем. Aspergillaceae................................................... 102
Сем. Terfeziaceae и Elaphomycetaceae 104
Сем. Onygenaceae . 104
Порядок Erysiphales 104
Сем. Erysiphaceae 104
Сем. Perisporiaceae .... 112
Группа порядков Pyrenomycetes ИЗ
Порядок Hypocreates 113
Сем. Hypocreaceae 113
Сем. Nectriaceae . 116
Порядок Sphaertales 118
Сем. Ceratostoinaceae 118
Сем. Mycosphaerellaceae 119
Сем. Pleosporaceae . 123
Порядок Dothidiales . 124
Порядок Hemisphacriales . . 124
Группа порядков Dyscomycetes 124
Порядок Hysteriales 124
Порядок Phacidiales 125
Порядок Pezizales 126
Сем. Ругопепгасеае 126
Сем. Pezizaceae 126
Сем. Helvellaceae 127
Порядок Helotiales 128
Сем. Sclerotiniaceae 129
Порядок Tuberales 132
Глава 4. III класс Basidioiiiycetes (базидиальные грибы) 135
Общая характеристика 135
Подкласс Holobasidiomycetes 140
Порядок Dacryomycetales 140
Порядок Exobasidiales 141
Сем. Exobasidiaceae . 141
Группа порядков Hymenomycetes 143
Порядок Aphyllophorales 143
Сем. Telephoraceae 144
Сем. Clavariaceae 145
Сем. Nydnaceae 146
Сем. Polyporaceae 117
Порядок Agaricales 154
Сем. Boletaceae 155
Сем. Agaricaceae................................................... 156
Группа порядков Gastromycetes 159
Сем. Lycoperdaceae 160
Сем. Phallaceae ................ 161
Подкласс Phragmobasidiomycetes 163
Порядок Ustilaginales 163
Порядок Ur edinales 173
Сем. Melampsoraceae 178
Сем. Pucciniaceae 182
Глава 5. IV класс Gcuteromycetes (несовершенные грибы) 189
Общая характеристика 189
Порядок Н yphomycetales 192
Сем. Mucedinaceae 192
Сем. Dematiaceae . 206
Сем. Tuberculcuiaceae 215
Порядок Melanconiales 226
Сем. Melanconiaceae . 227
Порядок Sphaeropsidales 237
Сем. Sphaerioldaceae 238
Глава 6. Вирусные, бактериальные и неинфекционные болезни 246
Вирусные болезни................................................. 246
Бактериальные болезни растений . 251
Неинфекционные болезни растений 255
Глава 7. Иммунитет растений к инфекционным заболеваниям 260
Общие сведения об иммунитете растений 260
Врожденный иммунитет 261
Приобретенный иммунитет 263
Литература 267
Указатель 270
Кудряшева Зинаида Никандровна
МИКОЛОГИЯ С ОСНОВАМИ ФИТОПАТОЛОГИИ
Под ред. Н. А. Дорожкина. Минск, «Вышэйшая школа», 1968.
283 стр. с илл. 582
Редактор С. Шердюкова
Оформление Г. Г рака
Худож. редактор И. Андрианов
Техн, редактор М. Кислякова
Корректор Г. Вагабова
АТ 00415. Сдано в набор 12/IV 1967 г. Подписано в печать 4/XI 1967 г. Бумага 60х901/1б- типогр. № 1. Печ. л. 17,75+6 вкл. Уч.-изд. л. 16,63. Изд. № 66—82. Тип. Зак. 242. Тираж 4000 экз. Цена 95 коп.
Издательство «Вышэйшая школа» Государственного Комитета Совета Министров БССР по печати. Редакция литературы по естественным наукам. Тем. план 1967 г., № 10. Минск, ул. Кирова, 24.
Типография издательства «Звязда». Минск, Ленинский пр., 79.